Embed Size (px)
Citation preview

UFRRJ
INSTITUTO DE ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ZOOTECNIA
DISSERTAÇÃO
Suplementação de Betaína em Rações de Frangos de Corte
Infectados Experimentalmente com Eimeria acervulina
(TYZZER, 1929)
Tania Marcia Soares Niang
2005

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO INSTITUTO DE ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ZOOTECNIA
SUPLEMENTAÇÃO DE BETAÍNA EM RAÇÕES DE FRANGOS DE CORTE INFECTADOS EXPERIMENTALMENTE COM Eimeria
acervulina (TYZZER, 1929)
TANIA MARCIA SOARES NIANG
Sob a Orientação do Professor Augusto Vidal da Costa Gomes
e Co-orientação do Professor Fernando Augusto Curvello
Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Ciências em Zootecnia, Área de Concentração em Produção Animal.
Seropédica, RJ Agosto de 2005

636.5084 N577s T
Niang, Tania Marcia Soares, 1976- Suplementação de betaína em rações de frangos de corte infectados experimentalmente com Eimeria acervulina (Tyzzer, 1929) / Tania Marcia Soares Niang. – 2005. 76 f. : il. Orientador: Augusto Vidal da Costa Gomes. Dissertação (mestrado) – Universidade Federal Rural do Rio de Janeiro, Instituto de Zootecnia. Inclui bibliografia. 1. Frango de corte – Alimentação e rações - Teses. 2. Aminoácidos na nutrição animal – Teses. 3. Dieta de alto teor protéico – Teses. 4. Frango de corte – Parasito - Teses. 5. Eimeria – Teses. 6. Coccidiose em ave – Teses. I. Gomes, Augusto Vidal da Costa. II. Universidade Federal Rural do Rio de Janeiro. Instituto de Zootecnia. III. Título.

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO INSTITUTO DE ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM ZOOTECNIA
TANIA MARCIA SOARES NIANG Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Ciências no Programa de Pós-Graduação em Zootecnia, área de Concentração em Produção Animal, DISSERTAÇÃO APROVADA EM 03/08/2005
Geraldo Luiz Colnago. Dr. UFF
Cristina Amorim Ribeiro de Lima. Dra. UFRRJ
Augusto Vidal da Costa Gomes. Dr. UFRRJ (Orientador)

DEDICO
A DEUS: pelo amor incondicional que me foi dado em todos os momentos da vida.
Por ter me dado força para vencer obstáculos e por ter permitido que muitos dos meus
sonhos se tornassem realidade!
A Minha Mãe Marcilene e a Minha Avó Francisca: pelas lições de vida e perseverança
que me ensinaram e por toda dedicação e amor!
Aos Meus Irmãos: Tonio, Glebson, Jeogan, Michele e Larissa, pelo amor, carinho e
amizade, e por entenderem que a distância foi um mal necessário em nossas vidas!
A Todos os Meus Parentes e Amigos: que sempre torceram por mim!
Aos Meus Amigos: Simone Moreira Bastos, Eduardo Fernandes e Janete Bastos pela
confiança e amizade!
Ao Meu Namorado e Marido Amadou: pelo amor e pelo imenso companheirismo que
sempre me dedicou nestes ótimos anos de relacionamento!
“Uma vida sem amor é como árvores sem flores e sem frutos.
E um amor sem beleza é como flores sem perfume.
Vida, amor, beleza: eis a minha trindade”.

AGRADECIMENTOS
A Universidade Federal Rural do Rio de Janeiro: “Que como um oásis, imponente,
recebe, abriga e protege seus filhos, saciando-lhes à imediata sede de sabedoria e
preparando-os para desafios futuros”.
A Coordenação do Programa de Pós-Graduação em Zootecnia: pelo apoio e competência
com qual me orientaram no decorrer das atividades curriculares.
A CAPES: pela concessão de bolsa de estudo.
A Btech Tecnologias Agropecuárias e Comércio Ltda: pelo auxilio e pela concessão do
produto testado.
Ao Dr. Carlos Alberto Fagonde Costa da Embrapa Suínos e Aves: que gentilmente cedeu
o inóculo contendo os oocistos de Eimeria acervulina que foram usados neste
experimento.
Ao Professor e Ex-Reitor da UFRRJ José Antônio de Souza Veiga: pela amizade e pelo
apoio, um grande ser humano, que sempre tratou com respeito e atenção todos os alunos
desta instituição.
Aos Professores e/ou Pesquisadores: Augusto Vidal da Costa Gomes, Carlos Wilson
Gomes Lopes, Celso Guimarães Barbosa, Cristina Amorim Ribeiro de Lima, Fernando
Augusto Curvello, Francisco Gerson Araújo, Geraldo Luiz Colnago, Jorge Jacob Neto,
Marcos Antônio da Silva Batista, Mauricio Ballesteiro, Mauro Portela Piña Rodrigues,
Nelson Jorge Moraes Matos, Pedro A. Muniz Malafaia e Zelson Giacomo Loss: pelos
ensinamentos e apoio.
Aos Amigos e Grandes Colaboradores na Execução do Experimento: Ana Carla Silva,
Arley Alves de Oliveira, Luciana dos Santos da Silva, Marcel Teixeira, Nelson Alves
Junior, Renata Nunes, Rosane Vaz, Saul Emmanuel de Mattos Quintans e Viviane
Aparecida Carli Costa: pela dedicação, competência e amizade.

Aos Funcionários do Instituto de Zootecnia: Emanuel, Evandro, Fátima, Marcus Pessoa,
Pedro Timótheo e Valdeci: pela colaboração e carinho.
A Toda Equipe do Laboratório de Ecologia de Peixes: André Pessanha, André, Alejandra,
Amilton, Ana Paula, Bianca, Carla Caetano, Francisco Guimarães, Leonardo, Luciano,
Magna, Márcia, Márcio, Marcus Costa, Rafaela, Rosana Milagre: pelas trocas de
conhecimento e ajuda mútua que aconteceram no período de estágio durante a graduação.
As Amigas e Companheiras do Alojamento da Graduação: Aline, Bianca, Edimeia,
Fernanda, Gilda, Giliane, Katiúscia, Maria Celestina, Mônica, Tatiana e Vera: pela
amizade e companheirismo e pela incomensurável troca de conhecimento e alegria.
Aos Amigos e Colegas do Alojamento da Pós-Graduação: Alzira, Anilce, Antonieta,
Daniela, Denise, Elisângela, Eloíza, Érica, Fabiana, José Dias, Kênia, Luciana Helena,
Luciana, Luziane, Mariela, Marli, Michele, Natali, Nilza, Renata, Rosana, Rose, Sandra e
Silvana: por tornarem os momentos cotidianos mais agradáveis e alegres.
Aos Amigos: Bárbara Werneck, Claudson Brito e José Amilton: pela força e amizade.
A secretaria de alojamento da graduação, mais especificamente ao senhor Antônio: por
todo o auxilio prestado durante os momentos de conserto no alojamento da graduação,
assim como, no alojamento da pós-graduação e, mais ainda, pela atenção e amizade!

“Contar com a companhia dos amigos nas estradas da vida é fundamental.
Ajuda a manter o rumo e a não se perder na estrada.
Quem caminha sozinho pode até chegar mais rápido.
Mas aquele que vai acompanhado, com certeza chegará mais longe.
Que bom que vocês fizeram parte desta caminhada!”
Obrigada!

RESUMO
NIANG, Tania Marcia Soares. Suplementação de betaína em rações de frangos de corte infectados experimentalmente com Eimeria acervulina (TYZZER, 1929). 2005. 90p. Dissertação (Mestrado em Zootecnia). Instituto de Zootecnia, Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2007.
Este trabalho teve como objetivo estudar a utilização da betaína em substituição ao uso dos aditivos tradicionais em dietas para frangos de corte infectados com Eimeria acervulina. A betaína como osmoprotetora deverá preparar as estruturas da mucosa intestinal das aves para uma futura infecção por coccídios e aumentar a disponibilidade dos principais compostos utilizados na síntese de proteína para o crescimento muscular. Um experimento foi realizado a fim de avaliar a digestibilidade aparente da ração, a estrutura da mucosa intestinal, o desempenho e as características da carcaça dos frangos de corte infectados experimentalmente com E. acervulina. Foram utilizados 420 pintos de corte machos Cobb, de um dia de idade, distribuídos e alojados em baterias metálicas de três andares num delineamento de blocos ao acaso com 5 tratamentos e 6 repetições com 14 aves/repetição, onde cada andar representava um bloco. As dietas foram fornecidas de 1 a 21 e 22 a 42 dias de idade. Os tratamentos foram: 1) controle positivo (CP) = dieta basal mais promotor de produção e coccidiostático; 2) 0,0% de betaína = dieta basal sem betaína (0,0%); 3) 0,05% betaína = dieta basal mais 0,05% de betaína; 4) 0,10% betaína = dieta basal mais 0,10% de betaína; 5) 0,15% betaína = dieta basal mais 0,15% de betaína. No décimo quarto dia de idade, as aves foram inoculadas individualmente com aproximadamente 2x105 oocistos esporulados de E. acervulina. Foram realizados dois ensaios de digestibilidade, no primeiro, a betaína afetou positivamente a digestibilidade do nitrogênio (N) e os valores de energia metabolizável (EM) da ração, não diferindo do promotor de produção e coccidiostático, porém, não afetou positivamente a digestibilidade da matéria seca (MS) nesta fase. Já no segundo ensaio, a betaína proporcionou efeitos positivos sobre a digestibilidade da MS, mas não sobre os valores de EM da ração. A estrutura da mucosa intestinal foi estudada com aves necropsiadas no 14º, 18°, 21º e 28° dia de vida. Mesmo não tendo sido observado diferença significativa (p>0,05) entre os tratamentos para altura de vilosidades aos 14, 18 e 28 dias, os níveis de betaína suplementados na dieta sempre produziram aumentos nas alturas das vilosidades. Aos 21 dias, o nível de 0,05% de betaína e o promotor de produção e coccidiostático proporcionaram as maiores (p<0,05) altura de vilosidades em relação aos demais tratamentos. Já a profundidade das criptas e a relação vilo:cripta não foram afetadas pelos diferentes tratamentos (p>0,05). Os índices de desempenho foram analisados no período de 1 a 14, 14 a 21, 1 a 21, 22 a 28, 28 a 42, 22 a 42 e 1 a 42 dias de idade das aves. Aos 43 dias de idade, duas aves de cada repetição, totalizando 12 aves por tratamento, foram abatidas para estudo das características de carcaça. A betaína afetou positivamente o desempenho e as características da carcaça dos frangos, tendo muitas vezes não diferido estatisticamente do promotor de produção e coccidiostático.
Palavras chave: Coccidiose, Osmoproteção, Síntese protéica.

ABSTRACT
NIANG, Tania Marcia Soares. Betaine Supplementation in diets of broiler chicks infected experimentally with Eimeria acervulina (TYZZER, 1929). 2005. 90p. Dissertation (Magister Scientiae in Animal Science). Instituto de Zootecnia, Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2007.
This work had as aims to study the use of betaine to replace traditional drugs in diet of broiler chicken Eimeria acervulina-infected. Since betaine acts as osmoprotector it should prepare the structure of chicken intestinal mucosal against a coccidian future infection and increase the availability of main compounds used in protein synthesis to muscular growth. An experiment was carried out to assess diet’s apparent digestibility, intestinal mucosal structure, growth performance and carcass characteristics of chicken infected experimentally with E. acervulina. A total of 420 day-old males Cobb broiler chicks were distributed in cage battery of three floors in a randomized block experimental design with 5 treatments and 6 replicates with 14 birds per replicate, where each floor represented a block. Diets were supplied from 1 to 21 and 22 to 42 days. The treatments were as follow: 1) Positive control (PC) = basal diet plus a production promoter and a coccidiostatic; 2) 0.0% of betaine = basal diet without betaine (0.0%); 3) 0.05% betaine = basal diet plus 0.05% betaine; 4) 0.10% betaine = basal diet plus 0.10% betaine; 5) 0.15% betaine = basal diet plus 0.15% de betaine. At fourteen-day of age the birds were individually inoculated with approximately 2x105 sporulated oocysts of E. acervulina. Two digestibility assays were performed, in the first, the betaine affected positively the nitrogen digestibility and the metabolisable energy diet, not differing from the PC diet but not affecting positively the dry matter digestibility. In the second assay, the betaine showed positives effects on dry matter digestibility, but not on the value of metabolizable energy of diet. The intestinal mucosal structure was studied in birds killed at, 14º, 18°, 21º and 28° day of life. Although no significant differences (P>0.05) were shown among the treatment for villus height at 14, 18 and 28 days, the levels of betaine added to the diet always showed a trend to increase the villus height. At 21 days, the level of 0.05% betaine and the PC diet showed the highest villus height (p<0.05) compared with the remains treatments. The crypts depth and the ratio of villus:crypt were not affected by different treatments (p>0.05). Performance was assessed from 1 to 14, 14 to 21, 1 to 21, 22 to 28, 28 to 42, 22 to 42 and 1 to 42 days of age. At 43 day of age, two chicks from each replicate, totalizing 12 chicks for treatment, were slaughtered for assessing the carcass characteristics. The betaine affected positively the performance and carcass characteristics; showing in several cases no significant difference from the PC diet.
Key words: Coccidiose. Osmoprotectant. Protein synthesis.

ÍNDICE DE ILUSTRAÇÕES
CAPITULO I Influência da Betaína na Digestibilidade da Ração de Frangos de Corte
Infectados Experimentalmente com Eimeria acervulina (TIZZER, 1929)
TABELAS TABELA 1. Composição das dietas basais 17TABELA 2. Digestibilidade aparente da matéria seca (%MS), do nitrogênio metabolizável
(%N) e da energia metabolizável (EM) da ração, expressa na base da matéria seca, por fase, em função dos diferentes tratamentos
20 FUGURAS
FIGURA 1. Influência da suplementação de betaína nos valores de EM aparente da ração em frangos de corte infectados experimentalmente com E. acervulina no primeiro ensaio de digestibilidade............................... .............................................................................................
21
FIGURA 2. Influência da suplementação de betaína na digestibilidade aparente da MS da ração em frangos de corte infectados com E. acervulina no segundo ensaio de digestibilidade
22CAPÍTULO II Influência da Betaína no Desenvolvimento de Vilos e Criptas do
Intestino Delgado de Frangos de Corte Infectados Experimentalmente com Eimeria acervulina (TYZZER, 1929)
TABELAS TABELA 1. Médias de altura dos vilos (µm) do duodeno de frangos de corte necropsiados
aos 14, 18, 21 e 28 dias de idade 44
TABELA 2. Médias de profundidade das criptas (µm) do duodeno de frangos de corte
necropsiados aos 14, 18, 21 e 28 dias de idade 46
TABELA 3. Médias da relação vilo:cripta (µm) do duodeno de frangos de corte
necropsiados aos 14, 18, 21 e 28 dias de idade 48
FIGURAS FIGURA 1. Corte histológico com demonstração das medidas tomadas para alturas das
vilosidades (A) e profundidades das criptas (C) do intestino delgado de frangos de corte
41
FIGURA 2. Lesões na mucosa intestinal do duodeno de aves infectadas experimentalmente com E. acervulina aos quatro (A), sete (B) e quatorze (C) dias após a inoculação
43
FIGURA 3. Influência dos aditivos dietéticos sobre a altura das vilosidades intestinais dos frangos de corte submetidos à infecção experimental com E. acervulina aos 14 dias de idade
45
FIGURA 4. Influência dos aditivos dietéticos sobre a profundidade das criptas intestinais dos frangos de corte submetidos à infecção experimental com E. acervulina aos 14 dias de idade.............................................
47
FIGURA 5. Influência dos aditivos dietéticos sobre a relação vilo:cripta do intestino de frangos de corte submetidos à infecção experimental com E. acervulina aos 14 dias de idade
49

CAPÍTULO III Influência da Betaína no Desempenho e nas Características da Carcaça de Frangos de Corte Infectados Experimentalmente com Eimeria acervulina (TYZZER, 1929)
TABELAS TABELA 1. Total de frangos utilizados no ensaio de desempenho de acordo com as fases
de retiradas destes para estudos de necropsia 64
TABELA 2. Efeito da betaína sobre o desempenho de pintos de corte no período de 1 a 14 dias de idade........................................................................
67
TABELA 3. Efeito da betaína e infecção com Eimeria acervulina sobre o desempenho de frangos de corte no período de 14 a 21 dias de idade
69
TABELA 4. Efeito da betaína sobre o desempenho de frangos de corte no período de 1 a 21 dias de idade
70
TABELA 5. Efeito da betaína e inoculação de Eimeria sobre o desempenho de frangos de corte no período de 22 a 28 dias de idade
72
TABELA 6. Efeito da betaína e inoculação de Eimeria sobre o desempenho de frangos de corte no período de 28 a 42 dias de idade
72
TABELA 7. Efeito da betaína sobre o desempenho de frangos de corte no período de 22 a 42 dias de idade
74
TABELA 8. Efeito da betaína sobre o desempenho de frangos de corte no período de 1 a 42 dias de idade
76
TABELA 9. Características da carcaça de frangos de corte machos no período de 1 a 43 dias de idade
79
FIGURAS FIGURA 1. Efeito da suplementação dietética de betaína no ganho de peso médio de
frangos de corte no período de 1 a 14 dias de idade 68
FIGURA 2. Efeito da suplementação dietética de betaína no consumo médio de ração dos frangos de corte no período de 14 a 21 dias de idade
70
FIGURA 3. Efeito da suplementação dietética de betaína no consumo médio de ração dos frangos de corte no período de 28 a 42 dias de idade
73
FIGURA 4. Efeito da suplementação dietética de betaína no ganho de peso médio dos frangos de corte no período de 28 a 42 dias de idade
74
FIGURA 5. Efeito da suplementação dietética de betaína no consumo médio de ração dos frangos de corte no período de 22 a 42 dias de idade
75
FIGURA 6. Efeito da suplementação dietética de betaína no consumo médio de ração de frangos de corte no período de 1 a 42 dias de idade
77
FIGURA 7. Efeito da suplementação dietética de betaína no ganho de peso médio dos frangos de corte no período de 1 a 42 dias de idade
78
FIGURA 8. Efeito da suplementação dietética de betaína no peso da carcaça resfriada de frangos de corte
80

SUMÁRIO
INTRODUÇÃO.............................................................................................................. 01 RESUMO......................................................................................................................... 02 ABSTRACT.................................................................................................................... 03
CAPITULO I INFLUÊNCIA DA BETAÍNA NA DIGESTIBILIDADE DA RAÇÃO DE FRANGOS DE CORTE INFECTADOS EXPERIMENTALMENTE COM EIMERIA ACERVULINA (TYZZER, 1929)................................................................
04
RESUMO......................................................................................................................... 05 ABSTRACT.................................................................................................................... 06 1 INTRODUÇÃO........................................................................................................... 07 2 REVISÃO DE LITERATURA................................................................................... 08 2.1 Digestão...................................................................................................................... 08 2.2 Absorção..................................................................................................................... 08 2.3 Eimeria...................................................................................................................... 08 2.3.1 Ciclo de vida do coccídio....................................................................................... 10 2.4 Eimeria Acervulina 11 2.5 A Coccidiose e sua Interferência nos Processos de Absorção................................... 12 2.6 A Coccidiose em Operações Comerciais de Frangos de Corte.................................. 13 2.7 Betaína........................................................................................................................ 14
3 MATERIAL E MÉTODOS........................................................................................ 16
3.1 Local........................................................................................................................... 16 3.2 Animais e Manejo....................................................................................................... 16 3.3 Dietas Experimentais................................................................................................. 16 3.4 Material Infectante...................................................................................................... 17 3.5 Ensaio de Digestibilidade........................................................................................... 18 3.6 Análise Estatística...................................................................................................... 19
4 RESULTADOS E DISCUSSÃO................................................................................ 20 5 CONCLUSÕES............................................................................................................ 24 6 REFERÊNCIAS BIBLIOGRÁFICAS...................................................................... 25
CAPÍTULO II
INFLUÊNCIA DA BETAÍNA NO DESENVOLVIMENTO DE VILOS E CRIPTAS DO INTESTINO DELGADO DE FRANGOS DE CORTE INFECTADOS EXPERIMENTALMENTE COM EIMERIA ACERVULINA (TYZZER, 1929) ...........................................................................................................
32
RESUMO........................................................................................................................ 33 ABSTRACT.................................................................................................................... 34 1 INTRODUÇÃO........................................................................................................... 35

2 REVISÃO DE LITERATURA.................................................................................. 34 2.1 Mudanças Fisiológicas na Mucosa Intestinal das Aves............................................. 37 2.2 Nutrição e Coccidiose................................................................................................. 38 3 MATERIAL E MÉTODOS........................................................................................ 40 3.1 Local........................................................................................................................... 40 3.2 Metodologia Utilizada Anteriormente ao Preparo Histológico.................................. 40 3.3 Preparo Histológico.................................................................................................... 40 3.4 Avaliação da Relação Vilo/Cripta.............................................................................. 40 3.5 Análise Estatística...................................................................................................... 41 4 RESULTADOS E DISCUSSÃO................................................................................ 43 4.1 Altura das Vilosidades Intestinais.............................................................................. 43 4.2 Profundidade das Criptas Intestinais.......................................................................... 46 4.3 Relação Vilo:Cripta Intestinais................................................................................... 48 5 CONCLUSÕES............................................................................................................ 50 6 REFERÊNCIAS BIBLIOGRÁFICAS...................................................................... 51
CAPÍTULO III INFLUÊNCIA DA BETAÍNA NO DESEMPENHO E NAS CARACTERÍSTICAS DA CARCAÇA DE FRANGOS DE CORTE INFECTADOS EXPERIMENTALMENTE COM EIMERIA ACERVULINA (TYZZER, 1929) ...........................................................................................................
56
RESUMO........................................................................................................................ 57 ABSTRACT.................................................................................................................... 58 1 INTRODUÇÃO........................................................................................................... 59 2 REVISÃO DE LITERATURA.................................................................................. 61 2.1 A importância da Água no Organismo Vivo.............................................................. 61 2.2 Impacto da Eimeria acervulina no Organismo das Aves........................................... 61 2.3 Betaína como Doadora de Grupamentos Metila......................................................... 62 2.4 Betaína como Suplemento Nutricional....................................................................... 62 3 MATERIAL E MÉTODOS........................................................................................ 64 3.1 Local........................................................................................................................... 64 3.2 Metodologia Utilizada Anteriormente ao Ensaio de Desempenho............................ 64 3.3 Ensaio de Desempenho............................................................................................... 64 3.4 Características da Carcaça.......................................................................................... 65 3.5 Análise Estatística....................................................................................................... 65 4 RESULTADOS E DISCUSSÃO................................................................................ 67 4.1 Desempenho............................................................................................................... 67 4.1.1 Desempenho dos frangos de corte no período de 1 a 14 dias de idade................... 67 4.1.2 Desempenho dos frangos de corte no período de 14 a 21 dias de idade................. 69 4.1.3 Desempenho dos frangos de corte no período de 1 a 21 dias de idade................... 70 4.1.4 Desempenho dos frangos de corte no período de 22 a 28 dias de idade................. 71 4.1.5 Desempenho dos frangos de corte no período de 28 a 42 dias de idade................. 72 4.1.6 Desempenho dos frangos de corte no período de 22 a 42 dias de idade................. 74

4.1.7 Desempenho dos frangos de corte no período de 1 a 42 dias de idade................... 75 4.2 Características da Carcaça.......................................................................................... 78 5 CONCLUSÕES............................................................................................................ 83 6 REFERÊNCIAS BIBLIOGRÁFICAS...................................................................... 84 7 CONCLUSÕES GERAIS........................................................................................... 90

- 1 -
INTRODUÇÃO
Atualmente a produção de carne de aves tem se expandido significativamente em relação há várias décadas atrás. A indústria avícola tem evoluído de plantéis de fundo quintal a um engenhoso sistema de produção alimentar. O crescimento e o rendimento das aves foram aumentados com a ajuda de dietas altamente especializadas e pelo uso da genética.
O enfoque primário da indústria avícola é maximizar os lucros, promover máximo rendimento e manter a saúde e o bem-estar das aves. Qualquer interferência à saúde do animal diminuirá a rentabilidade. Melhorias em tecnologias relativas a vacinas ou a nutrição poderiam economizar dinheiro às indústrias e permitir que elas operem mais eficazmente com rendas crescentes e custos globais diminuídos. Uma das principais despesas enfrentadas pela indústria é a perda de produção associada a doenças avícolas, incluindo os custos com a vacinação, prevenção e tratamento das doenças; perdas no ganho de peso e mortalidade.
A coccidiose é uma doença parasitária responsável por perdas que ocorrem na produção animal mundialmente. A incidência da coccidiose aviária ocorre devido as altas densidades e práticas de manejo mais intensivas. Assim, há necessidade de pesquisas que visem trazer informações para ajudar no tratamento e prevenção da doença.
Uma alternativa poderia ser o uso de ingredientes naturais na ração de frangos de corte. A betaína ou trimetilglicina, que é um produto natural, sintetizado por muitos organismos vivos, tais como as plantas pertencentes à família Chenopodiaceae, da qual a beterraba é a mais comum, e está entre as melhores acumuladoras dessa substância que tem recebido muita atenção por parte dos pesquisadores por atuar de forma benéfica no organismo do animal quando este está sendo parasitado por espécies do gênero Eimeria.
O interesse dos pesquisadores na utilização da betaína em dietas de frango de corte tem sido despertado devido às funções que esta apresenta como osmólito orgânico e como doadora de grupamentos metil. A natureza bipolar da betaína é importante para a osmorregulação porque ajuda no balanço de água dentro das células que é gerada pela mudança na concentração de partículas que ficam carregadas na solução. Como doadora de grupo metil, a betaína pode contribuir para a síntese de importantes compostos, incluindo proteína, DNA/RNA e aumentar a disponibilidade de metionina para síntese de proteína e permitir o crescimento muscular. Além disso, é também conhecido que no metabolismo da betaína, há produção de glicina, um aminoácido também importante na síntese de proteína que colabora para o desenvolvimento muscular.
Muitas pesquisas têm elucidado que a betaína causa efeitos positivos sobre o desempenho de frangos de corte, melhora as características de carcaça e a digestibilidade dos nutrientes em aves infectadas ou não com coccídios. E que seus efeitos como osmólito têm resultado em melhorias significativas na resistência do epitélio intestinal contra infecções coccídianas em frangos.
O presente experimento teve como objetivo estudar a influência da suplementação de betaína na digestibilidade da ração, no desenvolvimento de vilos e criptas do intestino delgado, no desempenho e nas características da carcaça de frangos de corte infectados experimentalmente com E. acervulina.

- 2 -
RESUMO
NIANG, Tania Marcia Soares. Suplementação de betaína em rações de frangos de corte infectados experimentalmente com Eimeria acervulina (TYZZER, 1929). 2005. 76p. Dissertação (Mestrado em Zootecnia). Instituto de Zootecnia, Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2005.
Este trabalho teve como objetivo estudar a utilização da betaína em substituição ao uso dos aditivos tradicionais em dietas para frangos de corte infectados com Eimeria acervulina. A betaína como osmoprotetora deverá preparar as estruturas da mucosa intestinal das aves para uma futura infecção por coccídios e aumentar a disponibilidade dos principais compostos utilizados na síntese de proteína para o crescimento muscular. Um experimento foi realizado a fim de avaliar a digestibilidade aparente da ração, a estrutura da mucosa intestinal, o desempenho e as características da carcaça dos frangos de corte infectados experimentalmente com E. acervulina. Foram utilizados 420 pintos de corte machos Cobb, de um dia de idade, distribuídos e alojados em baterias metálicas de três andares num delineamento de blocos ao acaso com 5 tratamentos e 6 repetições com 14 aves/repetição, onde cada andar representava um bloco. As dietas foram fornecidas de 1 a 21 e 22 a 42 dias de idade. Os tratamentos foram: 1) controle positivo (CP) = dieta basal mais promotor de produção e coccidiostático; 2) 0,0% de betaína = dieta basal sem betaína (0,0%); 3) 0,05% betaína = dieta basal mais 0,05% de betaína; 4) 0,10% betaína = dieta basal mais 0,10% de betaína; 5) 0,15% betaína = dieta basal mais 0,15% de betaína. No décimo quarto dia de idade, as aves foram inoculadas individualmente com aproximadamente 2x105 oocistos esporulados de E. acervulina. Foram realizados dois ensaios de digestibilidade, no primeiro, a betaína afetou positivamente a digestibilidade do nitrogênio (N) e os valores de energia metabolizável (EM) da ração, não diferindo do promotor de produção e coccidiostático, porém, não afetou positivamente a digestibilidade da matéria seca (MS) nesta fase. Já no segundo ensaio, a betaína proporcionou efeitos positivos sobre a digestibilidade da MS, mas não sobre os valores de EM da ração. A estrutura da mucosa intestinal foi estudada com aves necropsiadas no 14º, 18°, 21º e 28° dia de vida. Mesmo não tendo sido observado diferença significativa (p>0,05) entre os tratamentos para altura de vilosidades aos 14, 18 e 28 dias, os níveis de betaína suplementados na dieta sempre produziram aumentos nas alturas das vilosidades. Aos 21 dias, o nível de 0,05% de betaína e o promotor de produção e coccidiostático proporcionaram as maiores (p<0,05) altura de vilosidades em relação aos demais tratamentos. Já a profundidade das criptas e a relação vilo:cripta não foram afetadas pelos diferentes tratamentos (p>0,05). Os índices de desempenho foram analisados no período de 1 a 14, 14 a 21, 1 a 21, 22 a 28, 28 a 42, 22 a 42 e 1 a 42 dias de idade das aves. Aos 43 dias de idade, duas aves de cada repetição, totalizando 12 aves por tratamento, foram abatidas para estudo das características de carcaça. A betaína afetou positivamente o desempenho e as características da carcaça dos frangos, tendo muitas vezes não diferido estatisticamente do promotor de produção e coccidiostático.
Palavras chave: Coccidiose, Osmoproteção, Síntese protéica.

- 3 -
ABSTRACT
NIANG, Tania Marcia Soares. Betaine Supplementation in diets of broiler chicks infected experimentally with Eimeria acervulina (TYZZER, 1929). 2005. 76p. Dissertation (Magister Scientiae in Animal Science). Instituto de Zootecnia, Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2005.
This work had as aims to study the use of betaine to replace traditional drugs in diet of broiler chicken Eimeria acervulina-infected. Since betaine acts as osmoprotector it should prepare the structure of chicken intestinal mucosal against a coccidian future infection and increase the availability of main compounds used in protein synthesis to muscular growth. An experiment was carried out to assess diet’s apparent digestibility, intestinal mucosal structure, growth performance and carcass characteristics of chicken infected experimentally with E. acervulina. A total of 420 day-old males Cobb broiler chicks were distributed in cage battery of three floors in a randomized block experimental design with 5 treatments and 6 replicates with 14 birds per replicate, where each floor represented a block. Diets were supplied from 1 to 21 and 22 to 42 days. The treatments were as follow: 1) Positive control (PC) = basal diet plus a production promoter and a coccidiostatic; 2) 0.0% of betaine = basal diet without betaine (0.0%); 3) 0.05% betaine = basal diet plus 0.05% betaine; 4) 0.10% betaine = basal diet plus 0.10% betaine; 5) 0.15% betaine = basal diet plus 0.15% de betaine. At fourteen-day of age the birds were individually inoculated with approximately 2x105 sporulated oocysts of E. acervulina. Two digestibility assays were performed, in the first, the betaine affected positively the nitrogen digestibility and the metabolisable energy diet, not differing from the PC diet but not affecting positively the dry matter digestibility. In the second assay, the betaine showed positives effects on dry matter digestibility, but not on the value of metabolizable energy of diet. The intestinal mucosal structure was studied in birds killed at, 14º, 18°, 21º and 28° day of life. Although no significant differences (P>0.05) were shown among the treatment for villus height at 14, 18 and 28 days, the levels of betaine added to the diet always showed a trend to increase the villus height. At 21 days, the level of 0.05% betaine and the PC diet showed the highest villus height (p<0.05) compared with the remains treatments. The crypts depth and the ratio of villus:crypt were not affected by different treatments (p>0.05). Performance was assessed from 1 to 14, 14 to 21, 1 to 21, 22 to 28, 28 to 42, 22 to 42 and 1 to 42 days of age. At 43 day of age, two chicks from each replicate, totalizing 12 chicks for treatment, were slaughtered for assessing the carcass characteristics. The betaine affected positively the performance and carcass characteristics; showing in several cases no significant difference from the PC diet.
Key words: Coccidiose, Osmoprotectant, Protein synthesis.

- 4 -
CAPÍTULO I
INFLUÊNCIA DA BETAÍNA NA DIGESTIBILIDADE DA RAÇÃO DE FRANGOS DE CORTE INFECTADOS EXPERIMENTALMENTE COM
EIMERIA ACERVULINA (TYZZER, 1929)

- 5 -
RESUMO
NIANG, Tania Marcia Soares. Influência da betaína na digestibilidade da ração de frangos de corte infectados experimentalmente com Eimeria acervulina (TYZZER, 1929).
O objetivo deste estudo foi avaliar o efeito da suplementação de betaína sobre a digestibilidade aparente da matéria seca (MS), do nitrogênio metabolizável (N) e da energia metabolizável (EM) da ração de frangos de corte machos submetidos à infecção experimental com Eimeria acervulina em dois ensaios de digestibilidade. Foram utilizados 420 pintos de corte Cobb, de um dia de idade, distribuídos num delineamento de blocos ao acaso com 5 tratamentos e 6 repetições, alojados em baterias metálicas de três andares, onde cada andar representava um bloco. As dietas foram fornecidas de 1 a 21 e 22 a 42 dias de idade. Os tratamentos foram: 1) controle positivo (CP) = dieta basal mais promotor de produção e coccidiostático; 2) 0,0% de betaína = dieta basal sem betaína (0,0%); 3) 0,05% betaína = dieta basal mais 0,05% de betaína; 4) 0,10% betaína = dieta basal mais 0,10% de betaína; 5) 0,15% betaína = dieta basal mais 0,15% de betaína. No décimo quarto dia de idade as aves foram inoculadas individualmente com aproximadamente 2x105 oocistos esporulados de E. acervulina. O primeiro ensaio de digestibilidade compreendeu a primeira fase da ração e o período em que as aves estavam passando por um desafio-coccídio (17 a 21 dias de idade) e o segundo ensaio abrangeu a segunda fase da ração e o período de uma provável recuperação (27 a 31 dias de idade). A digestibilidade da MS da ração no primeiro ensaio foi superior (p<0,05) nas aves que receberam o alimento contendo promotor de produção e coccidiostático em relação às aves que receberam os demais tratamentos. Já a digestibilidade do N da ração foi maior (p<0,05) nas aves que consumiram a ração com promotor de produção e coccidiostático e naquelas que consumiram a ração contendo 0,05% de betaína em relação às aves que não foram suplementadas com betaína (0,0%) ou que foram suplementadas com os níveis de 0,10 e 0,15%. Os maiores valores de EM da ração (p<0,05) foi observado nos frangos suplementados com 0,10% de betaína e com a ração contendo promotor de produção e coccidiostático em relação aos frangos que não receberam a suplementação de betaína (0,0%) ou que receberam o nível de 0,15%. No segundo ensaio, não houve diferença (p>0,05) entre os tratamentos para digestibilidade da MS e do N da ração. Os maiores valores de energia metabolizável da ração (p<0,05) foram observados nos frangos que consumiram a ração contendo promotor de produção e coccidiostático em relação aos frangos que consumiram as demais dietas. Palavras chave: coccidiose aviária, metabolismo, nutriente alternativo.

- 6 -
ABSTRACT
NIANG, Tania Marcia Soares. Influence of betaine on digestibility of the diet of broiler chickens infected experimentally with Eimeria acervulina (TIZZER, 1929).
The objective of this study was to evaluate the betaine supplementation on apparent digestibility of dry matter (DM), metabolizable nitrogen (N) and metabolizable energy (ME) of males broiler chicken diets submitted to experimental infection with Eimeria acervulina in two-digestibility assay. A total of 420 day-old males Cobb broiler chicks were distributed in battery cage of three floors in a randomized block experimental design with 5 treatments and 6 experimental units per treatment, where each floor represented a block. Diets were supplied from 1 to 21 and 22 to 42 days. The treatments were as follow: 1) Positive control (PC) = basal diet plus a growth production promoter and a coccidiostatic; 2) 0.0% of betaine = basal diet without betaine (0.0%); 3) 0.05% betaine = basal diet plus 0.05% betaine; 4) 0.10% betaine = basal diet plus 0.10% betaine; 5) 0.15% betaine = basal diet plus 0.15% de betaine. At fourteen day of age, the birds were individually inoculated with approximately 2x105 sporulated oocysts of E. acervulina. The first assay comprised the 17-21 days feeding period and the time when the chickens were under coccidian challenge. The second assay comprised the 27-31 days feeding period and the infection recover period. DM digestibility diet in the first assay was higher (p<0.05) in the chickens that received the PC diet in relation to those in the other treatments. The digestibility of N was higher (p<0.05) for the chicks consuming the PC diet and in those consuming the diet containing 0.05% betaine in relation to the chickens that did not receive betaine (0.0%) or those receiving the diet supplemented with the 0.10 e 0.15% betaine levels. The highest (p<0.05) digestibility values of ME diet was found in the chickens supplemented with 0.10% betaine and with the PC diet in relation to chickens that did not receive betaine supplementation (0.0%) or those which received the 0.15% betaine level. In the second assay, there was no difference (p>0.05) among the treatments for DM digestibility and metabolizable N. The highest digestibility values (p<0.05) of ME was observed in the chickens that consumed the PC diet in relation to chicken that consumed the remaining diets. Key words: alternative nutrients, avian coccidiosis, metabolism.

- 7 -
1 INTRODUÇÃO Desde os primórdios da avicultura industrial, no início do século XX, a coccidiose
sempre foi uma preocupação onipresente das pessoas que estudavam a alimentação e a nutrição das aves. Uma das primeiras pesquisas publicadas que faz menção à relação entre a coccidiose e a alimentação e nutrição das aves foi publicada em 1925 (BEACH & CORL, 1925). Com o desenvolvimento que se sucedeu na avicultura nas décadas seguintes, até o advento dos agentes anticoccidianos na década de 40, alterações na composição dos ingredientes da fórmula de ração e manipulações nutricionais foram as principais ferramentas empregadas pelos avicultores para auxiliar no controle da doença.
Por ser um parasita que ataca a área do organismo das aves mais diretamente associada ao processo de digestão e absorção dos nutrientes, a coccidiose aviária foi e continua sendo a doença parasitária que mais traz preocupação e desafios para o nutricionista de aves. A princípio, ela pode interferir no processo nutricional das aves através da competição por nutrientes com o organismo invadido, pela mobilização destes nutrientes pelos mecanismos de defesa, pela redução na digestão e absorção dos nutrientes e pela própria recuperação das células e tecidos atingidos pelo parasita no período pós-infecção. Então, a coccidiose em espécies avícolas é uma doença entérica, e a infecção está associada com desordens osmóticas e iônicas (CROMPTON, 1976; VIRTANEN, 1995). Estas desordens são provavelmente causadas por desidratação e diarréia que são características de infecção coccidiana.
Vários nutrientes têm sido conhecidos por mudar a severidade das infecções por coccídios, dentre estes, apresenta-se a betaína, que é amplamente encontrada na natureza e sintetizada por uma variedade de plantas e organismos (BOCH et al., 1994). O acúmulo de betaína protege as células do estresse osmótico e lhes permite continuar a regular as atividades metabólicas em condições nas quais as células seriam normalmente inativadas (RUDOLPH et al., 1986; PETRONINI et al., 1992; KO et al., 1994). Os efeitos da suplementação de betaína incluem melhor absorção de proteína, lisina e metionina em aves submetidas ao estresse (REMUS, 2001). Assim, substâncias que tenham ação trófica sobre a mucosa intestinal, aumentando a sua capacidade funcional, poderão propiciar melhor digestibilidade às aves por apresentar uma maior capacidade de digerir e absorver os nutrientes da dieta. Nesse sentido, a suplementação de betaína parece ser uma alternativa para melhorar a mucosa intestinal dos frangos de corte submetidos a um desafio por Eimeria.
Dessa forma, o presente estudo foi desenvolvido com o objetivo de avaliar a influência da suplementação de betaína sobre a digestibilidade aparente da matéria seca, do nitrogênio e da energia metabolizáveis da dieta de frangos de corte submetidos a uma infecção experimental com E. acervulina.

- 8 -
2 REVISÃO DA LITERATURA
2.1 Digestão
Conforme descreve ANDRIGUETTO et al. (1999), os alimentos, em sua quase totalidade, se apresentam em formas combinadas de macromoléculas, que para serem utilizadas pelo organismo precisam ser transformadas em formas menores e absorvíveis. Ao conjunto de processos que sofrem os alimentos desde a ingestão, transformação ao longo do tubo digestivo e eliminação dos resíduos não absorvidos, denomina-se digestão.
Durante o processo digestivo, as substâncias alimentícias sofrem uma degradação que as transformam em produtos de peso molecular mais baixo, perdendo assim, suas características específicas originais, com a finalidade de deixar à disposição do organismo os nutrientes que as compõem. A digestão constitui uma função vital composta por processos físicos e químicos, intimamente relacionados aos sistemas nervoso e humoral.
2.2 Absorção
Entende-se por absorção a passagem de substâncias vindo do meio externo para o interior do organismo animal. Na absorção digestiva entende-se que certos produtos derivados dos alimentos passam através da mucosa intestinal para o sangue e linfa. A absorção digestiva é mais representativa no intestino delgado, sendo menos intensa no intestino grosso, onde é absorvida principalmente a água. Em todas as espécies animais, é ao nível do intestino delgado que se encontram reunidas às condições mais favoráveis a absorção de nutrientes. O epitélio da mucosa intestinal representa uma superfície considerável que se avalia em m2. Esta superfície é constituída por duas formações: as válvulas coniventes e as vilosidades. A primeira corresponde a dobras dispostas transversalmente, não ocupando mais que uma parte da superfície interior do intestino, onde elas duplicam ou triplicam a superfície. A segunda, apresenta capilares sanguíneos, e em seu centro, um capilar linfático. As vilosidades apresentam movimentos que facilitam a absorção dos princípios nutritivos. As vias de coleta dos nutrientes são as vias sanguínea e linfática (ANDRIGUETTO et. al., 1999).
2.3 Eimeria
Coccidiose é uma doença infecciosa e normalmente auto-restrita a área digestiva, é causada por hospedeiros específicos, parasitas protozoários intracelulares do gênero Eimeria. Os coccídios são classificados segundo o sub-reino Protozoa do filo Apicomplexia (JEURISSEN et al., 1996; LILLEHOJ & TROUT, 1996). Como um grupo, os coccídios do gênero Eimeria causam o problema de saúde mais difundido na indústria de frangos, sendo uma das doenças

- 9 -
mais caras da produção avícola comercial (EDGAR, 1992; HENKEN et al., 1994a,b; YUN et al., 2000).
As aves infectadas com oocistos coccídios não se desenvolvem tão bem quanto às aves não infetadas, como resultado de moderado a severo danos na mucosa intestinal, apresentam redução no ganho de peso, aumentam a conversão alimentar, e em alguns casos, elas podem parecer assintomática, porém, sua habilidade para maximizar a eficiência do alimento estará limitada. De acordo com EDGAR (1992), só basta um oocisto viável para estabelecer a presença do coccídio num aviário. Isto é possível devido ao alto índice de reprodução do parasita. A ingestão de um oocisto de E. acervulina no estado infectivo tem a capacidade de gerar aproximadamente 72,000 oocistos em um ciclo de vida completo (HENKEN et al., 1994a,b).
A prevalência da coccidiose é mundial e pode ser encontrada em quase todo o plantel avícola comercial (McDOUGALD & REID, 1997; COX, 1998). Na avicultura, existem sete espécies do gênero Eimeria que infectam as aves (SHIRLEY, 1986; McDOUGALD & REID, 1997). Todos os coccídios de aves são patogênicos, entretanto, algumas espécies produzem efeitos mais severos que outras, tais como morbidez severa e mortalidade. Este microscópico parasita unicelular invade o seu hospedeiro pela rota fecal-oral e a imunidade é alcançada uma vez que o parasita completa seu ciclo de vida no hospedeiro (BRACKETT & BLIZNICK, 1950; McDOUGALD & REID, 1997). Frangos de todas as idades são suscetíveis a coccidiose, porém, aves com três a cinco semanas de idade são mais vulneráveis (EDGAR, 1992; YUN et al., 2000).
As espécies do gênero Eimeria têm um ciclo de vida extremamente complexo que inclui ambos estágios, interno e externo no hospedeiro. A infecção ocorre através da ingestão de oocistos coccídios esporulados encontrados na cama, no solo, no alimento ou na água contaminada. Após a ingestão, os protozoários passam por uma série de estágios intracelular, extracelular, assexuais e sexuais para produzir oocistos viáveis que são excretados nas fezes (ROSE, 1987). Após um breve período fora do hospedeiro, os oocistos se tornam novamente infectivos pelo processo de esporulação e o ciclo de vida se completa.
Os impactos da coccidiose são significativos, porém, a erradicação é impraticável devido aos mecanismos protetores dos oocistos. As espécies do gênero Eimeria possuem uma parede exterior espessa que age como uma barreira protetora que aumenta a chance de sobrevivência sob condições adversas. Os oocistos são capazes de permanecerem infectivos fora do hospedeiro por longos períodos, e suas propriedades protetoras lhes permitem serem resistentes a muitas substâncias químicas severas e a desinfetantes (ESCHENBACHER et al., 1996; JEURISSEN et al., 1996; YUN et al., 2000).

- 10 -
2.3.1 Ciclo de vida do coccídio
Como a espécie do gênero Eimeria tende a ser muito específica na região intestinal a qual ela invade, seu ciclo de vida é semelhante com o grau de especificidade da espécie. Eimeria completa seu ciclo de vida em três fases distintas, incluindo esporogonias, merogonias, esquizogonias e gametogonias, porém, as durações destas fases são únicas para as espécies (YUN et al., 2000).
O ciclo de vida seguinte é adaptado de EDGAR (1992). A coccidiose, causada pela ingestão de oocistos microscópicos, é facilmente transmissível através de meios mecânicos como calçados e equipamentos contaminados, ou pode ser encontrada na cama, na terra, no alimento ou na água contaminada (CONAWAY & McKENZIE, 1991; McDOUGALD & REID, 1997).
As aves podem ser infectadas pelo coccídio uma vez que os oocistos se desenvolvam em uma fase infectiva fora do hospedeiro. Esporogonia é o processo de um zigoto unicelular dentro do oocisto que sofre uma série de divisões para formar esporozoítos que está contido dentro dos esporocistos. Somente oocistos que sofrem este processo é que podem causar a doença.
Oocistos esporulados contêm quatro esporocistos, e cada esporocisto, contém dois esporozoítos. A ação mecânica da moela e enzimas pancreáticas, tais como a tripsina e os sais biliares, causam a destruição da parede exterior dos oocistos, na qual lança os esporocistos dentro do trato digestivo. Os esporocistos são então, mais adiante, excisados pela tripsina e sais biliares que estão presentes no intestino. Os esporozoítos invadem as células epiteliais dos vilos, ao longo de localizações específicas por toda o trato digestivo, dependendo da espécie do gênero Eimeria. Algumas espécies viajam dentro da mucosa, através da lâmina própria para as células epiteliais da cripta. Uma vez dentro das células dos vilos ou das criptas, o processo de merogonia toma lugar.
O esporozoíto se desenvolve dentro de um corpo arredondado chamado de trofozoíto, e então, dentro de uma primeira geração de esquizonte (meronte) assexualmente reprodutiva. O esquizonte cresce e se divide rapidamente para produzir muitas primeiras-gerações de merozoítos. Os corpos rompem e lançam centenas de merozoítos de primeira-geração que procuram e invadem outras células epiteliais.
A segunda-geração de trofozoítos se desenvolve dentro da segunda-geração de esquizontes. Quando a segunda-geração de esquizontes amadurecem e rompem, são lançadas altas quantidades de merozoítos invasores, causando infecção difundida. O número de estágios assexuais e o tempo requerido para cada um, depende da espécie do gênero Eimeria envolvida, porém, a maioria das espécies terão menos de quatro gerações assexuais reprodutivas (ROSE, 1987; EDGAR, 1992).
Os merozoítos produzidos pelas gerações de esquizontes posteriores desenvolvem-se em formas sexuais chamada gametócitos, alguns machos e outras

- 11 -
fêmeas. Esta fase de reprodução sexual é denominada gametogonia. O gametócito fêmea amadurece em um macrogameta e o gametócito macho amadurece e rompe lançando um grande número de microgametas móveis, biflagelados. Os microgametas penetram no macrogameta fêmea maduro e a fertilização acontece. Seguinte a fertilização, é formada uma parede protetora espessa ao redor do zigoto. Neste estágio, o zigoto é considerado um oocisto imaturo.
Quando amadurecem, os oocistos rompem a célula hospedeira, entram no lúmen, e são expelidos nas fezes. Os sinais clínicos estão associados com destruição de tecido, devido o lançamento dos merozoítos e oocistos maduros da superfície da mucosa durante as últimas gerações de merogonias e ao longo das gametogonias. Em infecções severas, grande parte do epitélio mucosal é modificado e a absorção de nutrientes é comprometida (JEURISSEN et al., 1996; McDOUGALD & REID, 1997; YUN et al., 2000).
Os oocistos excretados das aves permanecem no ambiente e tem potencial para infectar outras aves. Sob condições ambientais favoráveis, a esporulação dos oocistos ocorre num período de 24 a 48 horas, e o ciclo continuará (EDGAR, 1992; GRAAT et al., 1994; McDOUGALD & REID, 1997). De acordo com EDGAR (1992), uma vez que o oocisto é esporulado, pode permanecer infectivo para as aves por vários meses, um ou dois anos, caso se proteja de condições muito quentes, secas ou geladas. Os oocistos coccídios têm características rígidas, mas não é totalmente indestrutível. De acordo com LEE & SHIH (1988), oocistos não esporulados são mais suscetíveis aos agentes físicos e químicos que os oocistos esporulados, talvez devido a um estado metabólico altamente sensível.
Outros fatores, tais como amônia e condições anaeróbicas, são também letais aos oocistos no ambiente. O desenvolvimento cessa quando os oocistos são expostos a altos níveis de CO2 ou NH4, ou quando são expostos a sais, cianeto e cloreto de mercúrio, devido à habilidade destes agentes para penetrar a parede dos oocistos (KHEYSIN, 1972). O período pré-patente, tempo que leva para os oocistos serem vistos nas fezes após a ingestão, é de aproximadamente quatro a sete dias para as espécies do gênero Eimeria (HENKEN et al., 1994a,b; JEURISSEN et al., 1996; McDOUGALD & REID, 1997).
2.4 Eimeria Acervulina
Na indústria avícola, nove espécies do gênero Eimeria têm sido descritas em frangos, porém, atualmente, só sete destas espécies são conhecidas por existir e causar efeitos patogênicos (CONAWAY & MCKENZIE 1991; WILLIAMS, 1998). As espécies de coccídios são classificadas baseado em atributos como: 1) localização da infecção intestinal, 2) aparecimento de lesões intestinais, 3) tamanho, forma e cor dos oocistos, 4) média de duração do período pré-patente, 5) tamanho dos esporozoítos e merozoítos invasores e 6) tipo de células parasitadas e localização dentro das células epiteliais. Baseado nestas características, E.

- 12 -
acervulina, E. brunetti, E. maxima, E. mitis, E. necatrix, E. praecox e E. tenella são consideradas patogênicas para os frangos.
Das sete espécies, E. acervulina é um dos coccídios parasitas freqüentemente encontrado que infectam as aves em operações comerciais (HENKEN et al., 1994b; McDOUGALD & REID, 1997). HENKEN et al. (1994a) assume que todo o plantel é infectado com coccídios parasitas, e E. acervulina é responsável pela maioria destes casos. Dentro da indústria comercial de frangos a infecção com E. acervulina gera impacto importante na produção. Estes impactos estão relacionados como o hospedeiro reage à infecção do parasita. No entanto, há outros fatores afetando a severidade da doença, tais como, a quantidade de oocistos viáveis de ingeridos e a patogenicidade dos isolados, como também o estado genético, nutricional, e imune das aves (McDOUGALD & REID, 1997).
Uma infecção moderada com E. acervulina poderia produzir efeitos sub-clínicos, tais como, redução na utilização dos alimentos e aumento no tempo de passagem do alimento, que pode aumentar os custos alimentares e a taxa global de conversão alimentar. Entretanto, uma infecção moderada a severa pode produzir perda de apetite, redução no consumo de água, redução no ganho de peso, perda de pigmentação da pele, diarréia, letargia, piora na conversão alimentar e lesões branca-acinzentada da mucosa intestinal. As infecções severas podem causar lesões para coalescer e pode ocorrer a morte (EDGAR, 1992; McDOUGALD & REID, 1997).
Enquanto cada espécie do gênero Eimeria tende a ter um local específico, E. acervulina invade a porção anterior do intestino delgado, especificamente a alça duodenal, e em infecções severas, a porção anterior do jejuno. Especula-se que as moléculas secretadas dos locais invadidos e as moléculas localizadas na superfície do parasita poderiam ser responsáveis por esta especificidade de local (LILLEHOJ & TROUT, 1993; JEURISSEN et al., 1996).
2.5 A Coccidiose e sua Interferência nos Processos de Absorção
A coccidiose tem sido responsabilizada por diminuir a absorção de alguns minerais nas aves em vários estudos realizados por TAKHAR & FARREL (1979a); WILLIS & BAKER (1981); GIRALDO et al. (1987); WATKINS et al. (1989); WARD et al. (1990), e também por aumentar a absorção de outros minerais (SOUTHERN & BAKER, 1982, 1983a,b; CZARNECKI & BAKER, 1984; BROWN & SOUTHERN, 1985).
A coccidiose não afeta somente a absorção de minerais, mas também têm efeito sobre a absorção de aminoácidos e vitaminas, bem como na utilização de energia. De acordo com KOUWENHOVEN & VAN DER HORST (1972) a infecção coccidiana diminui a absorção de vitamina A. TAKHAR & FARREL (1979a,b) relataram que a utilização da energia metabolizável e a retenção de nitrogênio foram diminuídas em frangos infectados com coccídios.

- 13 -
2.6 A Coccidiose em Operações Comerciais de Frangos de Corte
A coccidiose tem atingido a indústria avícola desde o inicio de 1900, mas somente nos anos quarenta, foram coletadas informações consideráveis sobre a doença. A produção avícola em larga escala foi desenvolvida, e com isso, houve a necessidade de se estudar o controle de doenças impactantes (BRACKETT & BLIZNICK, 1950). Atualmente, muito conhecimento tem sido gerado a respeito da coccidiose, mas ela continua tendo um impacto econômico muito grande na indústria avícola comercial.
Segundo redação da AVICULTURA INDUSTRIAL (2003), o Brasil tem um prejuízo de aproximadamente sessenta milhões de dólares em perdas anuais relativo a infecção coccidiana, tratamento e prevenção. Porém, é difícil calcular com precisão o total das perdas monetárias sofridas pela indústria avícola no mundo, que é o resultado da coccidiose, sua prevenção ou controle (DANFORTH, 1998; WILLIAMS, 1999), porque a coccidiose infecta qualquer tipo de ave com muita facilidade, sendo de ocorrência mundial (McDOUGALD & REID, 1997).
Espécies de coccídios de frangos são altamente específicas. As aves podem abrigar a doença e disseminá-la após a infecção (LEE & SHIH, 1988; WILLIAMS, 1998). A maioria das infecções são moderadas devido à ingestão de poucos oocistos, e dessa forma, a doença não seria noticiada. Com tais infecções, a eficiência alimentar e a utilização dos alimentos pelas aves só serão diminuídos ligeiramente. No entanto, a ingestão de milhões de oocistos pode causar infecções severas e poderia acontecer uma erupção danosa no epitélio intestinal.
Plantéis infectados, como resultado de moderada a severa exposição, normalmente apresentam uma acentuada redução no consumo de alimentos e água, e as aves tornam-se deprimidas e tendem a amontoar-se. A redução no ganho de peso acontece como resultado de transtorno na mucosa intestinal onde está ocorrendo a mínima absorção. A diarréia pode ser resultante da tentativa que o organismo do hospedeiro encontra para eliminar a água do corpo, que pode levar a desidratação. As lesões na mucosa intestinal e a perda de pigmentação também podem tornar-se aparentes durante as fases posteriores a infecção (CONAWAY & MCKENZIE, 1991; EDGAR, 1992; LILLEHOJ & TROUT, 1993; McDOUGALD & REID, 1997). A mortalidade resultante poderia ser devido à falta do consumo adequado de nutrientes, infecções secundárias e outros estressores contínuos associados com o estado da doença.
Desde os anos cinqüenta, o controle da coccidiose tem sido alcançado por compostos anti-coccídios administrados no alimento que previne ou reduz as infecções para um nível sub-clínico (DANFORTH, 1998). Quando usados corretamente, estes compostos provêem o controle adequado da doença. Porém, as práticas de manejo atual, como o confinamento intensivo, encorajam a severidade e a transmissão da coccidiose. Com o uso contínuo da aplicação de anti-coccídios no alimento, os coccídios de aves têm se tornado resistente às drogas, reduzindo assim, a eficácia destes medicamentos. A mais séria limitação da terapia anticoccídiana é o aumento da tolerância do parasita (McDOUGALD & REID, 1997).

- 14 -
Produtos anticoccídios medicamentosos inovadores são pouco prováveis de serem comercializados devido aos custos elevados de se aprovar tais componentes para alimentação animal e as perspectivas do consumidor relativo a esses aditivos alimentares (DANFORTH, 1998; VERMEULEN et al., 2001). Em tais circunstâncias, os produtores e os pesquisadores tentam identificar substâncias alternativas para o controle da coccidiose, seja através da melhoria da sanitização; como a limpeza da cama, seleção de reprodutores para melhorar a imunidade, programas de vacinação e ingredientes alternativos nas rações para frangos de corte (WILLIAMS, 1998). Em se tratando de ingrediente alternativo, a betaína tem sido muito estuda como tal, para auxiliar em melhorias no organismo das aves.
2.7 Betaína
Os efeitos significativos da aplicação de betaína na nutrição avícola têm sido apresentados desde meados dos anos quarenta (Du VIGNEUAUD et al., 1939; McGINNIS et al., 1942; ALMQUIST & GRAU, 1943, 1944). Essa glicina betaína, trimetilglicina ou oxineurina pode ser naturalmente sintetizada por uma ampla variedade de espécies animais e vegetais (BUDAVARI, 1989; BOCH et al., 1994). Concomitantemente, a maioria dos organismos animais e vegetais usam a betaína como uma fonte de grupamentos metil em certas reações metabólicas (ex.: sínteses de fosfatidilcolina, DNA, RNA, carnitina, creatina, etc.) e como osmoprotetora (ex.: estresses iônicos/osmóticos).
Algumas plantas da família da beterraba (Chenopodiaceae), a maioria dos microrganismos e uma série de invertebrados marinhos têm a capacidade de acumular betaína intracelular (KONOSU & HAYASHI, 1975). A principal razão pela qual a betaína se acumula nestes organismos é para proteger as células do estresse osmótico em condições de seca ou de alta salinidade. O acúmulo de betaína dentro das células e organelas celulares expostas ao estresse osmótico é para repor íons inorgânicos e proteger as macromoléculas da inativação iônica.
As aves mantêm sua concentração de água intracelular (homeostasia) a partir da osmorregulação em nível de rins, intestino delgado, ceco e cloaca (SHOEMAKER, 1972). Osmorregulação é a habilidade da célula em manter sua estrutura/volume e funcionalidade através da regulação do movimento de água intra e extracelular. Osmólito é uma substância que afeta o movimento da água, acumulando-a rapidamente em nível intracelular, sem alterar o metabolismo celular/mitocôndria.
Diarréias causadas por bactérias (ex: Clostridium, Escherichia ou Salmonella) e vírus (ex: Adenovírus, coronavírus, parvovírus ou rotavírus) estimulam a adenil ciclase e o AMPc, os quais elevam a atividade secretória das células das criptas intestinais e reduz a absorção de sódio e cloro por parte de seus vilos (FONDACARO, 1986). Como resultado, ocorre a formação de uma solução hiperosmótica em nível de lúmen intestinal (CANTEY, 1993) que estimula o

- 15 -
acúmulo de betaína nas células epiteliais do intestino, onde exerce seus efeitos protetores e mantedores da integridade e funções das mesmas (KIDD et al., 1997).
Como descrito acima, a presença da coccidiose em aves, causada por protozoários do gênero Eimeria, pode resultar em diminuição na absorção intestinal, enterites e eventualmente, altas mortalidades. Perdas de fluídos como resposta a coccidiose cecal pode causar desbalanços iônicos, que ativam a ação osmoprotetora da betaína, se presente naquele local (enterócitos). VIRTANEN & ROSI (1995) constataram que a betaína reduz os escores de lesão nas aves expostas a E. tenella, E. acervulina e E. máxima, desde que a presença do coccídio na célula intestinal crie algum tipo de estresse/desafio. A habilidade da betaína em manter o controle do balanço hídrico pode ser um importante mecanismo que permite a célula enfrentar esse tipo de condição.
A betaína melhora a retenção de água corporal. MOONEY et al. (1998) constataram que os frangos tratados com a betaína perderam menores quantidades de água do que os frangos controle, quando expostos a estresse cíclicos de temperatura e/ou desafio por coccidiose. Dessa forma, betaína, o osmólito, pode influenciar no balanço hídrico das células, dos tecidos, e de um modo geral, de todo o organismo animal.
Melhorias significativas na digestibilidade aparente da lisina, proteína, gordura e carotenóides foram observadas em frangos tratados com betaína sob condições desafiadoras de coccidiose (REMUS et al, 1995). AUGUSTINE & DANFORTH (1999) também relataram que o uso da betaína em dietas de frangos que foram submetidos a um desafio coccídio apresentaram um aumento na digestibilidade dos nutrientes.
Resultados de alguns estudos têm demonstrado que suínos em crescimento usam uma quantidade surpreendente da energia de mantença (40 a 50%) para conservar o equilíbrio osmótico (movimento de água) no intestino e em outras vísceras. A betaína dietética tem sido responsável por poupar parte desta energia que poderá ser então, usada para o crescimento e deposição de carne magra em suínos que recebem um suprimento adequado de aminoácidos. Assim, ao reduzir a necessidade de requerimento da energia de mantença dos animais através da melhoria da osmoproteção, a betaína poderá ajudar na absorção de nutrientes e contribuir para o melhor desempenho dos suínos (PARTRIDGE, 2002; 2003).

- 16 -
3 MATERIAL E MÉTODOS
3.1 Local
O experimento foi conduzido no Departamento de Nutrição Animal e Pastagens do Instituto de Zootecnia da Universidade Federal Rural do Rio de Janeiro, no município de Seropédica, RJ, Brasil, no período de 29/06 a 29/07 de 2004. As análises laboratoriais foram realizadas no laboratório de análises bromatológicas do referido departamento.
3.2 Animais e Manejo
Foram usados 420 pintos de corte machos da linhagem comercial Cobb, de um dia de idade, os quais foram distribuídos em um delineamento de blocos ao acaso com cinco tratamentos e seis repetições de 14 aves/repetição . Os pintos foram alojados em baterias metálicas de três andares com medidas de 96 x 80 x 39cm, providas de bebedouros tipo pressão e comedouros tipo prato. Cada andar foi equipado com uma lâmpada incandescente de 150w para fornecer calor nas duas primeiras semanas, sendo substituído por lâmpada de 60w na metade da segunda semana, suficientes para manter a temperatura aproximadamente em 32°C e 29°C respectivamente. Os equipamentos de primeira idade foram gradativamente sendo trocados por comedouros e bebedouros tipo calha a partir do 11º dia de vida das aves.
Os pintainhos foram vacinados contra a doença de Marek no incubatório e no 7º dia de vida receberam a vacina de Newcastle (New-Vacin La Sota), aplicada por via ocular.
3.3 Dietas Experimentais
As dietas basais (Tabela 1) foram fornecidas em um programa de alimentação com duas fases (1 a 21 e 22 a 42 dias de idade), tendo sido formuladas de acordo com recomendações de ROSTAGNO et al. (2000). A ração e a água foram fornecidas ad libitum.
A suplementação de betaína (Betafin® 96% de pureza) foi feita nas duas fases em substituição ao ingrediente inerte (caulim) da ração. As dietas contendo níveis de betaína foram comparadas com um tratamento que não teve a inclusão de qualquer tipo de aditivo (0% de betaína) e também com um tratamento controle positivo (CP), contendo promotor de produção: penicilina G potássica (2.000mg/kg do produto) e um coccidiostático: salinomicina (20.000mg/kg do produto), sem a presença de betaína. Os tratamentos foram designados da seguinte forma:
1) Controle positivo (CP) = dieta basal + promotor de produção + coccidiostático; 2) 0,00% de Betaína = dieta basal; 3) 0,05% de Betaína = dieta basal + 0,05% de betaína; 4) 0,10% de Betaína = dieta basal + 0,10% de betaína; 5) 0,15% de Betaína = dieta basal + 0,15% de betaína;
- 17 -
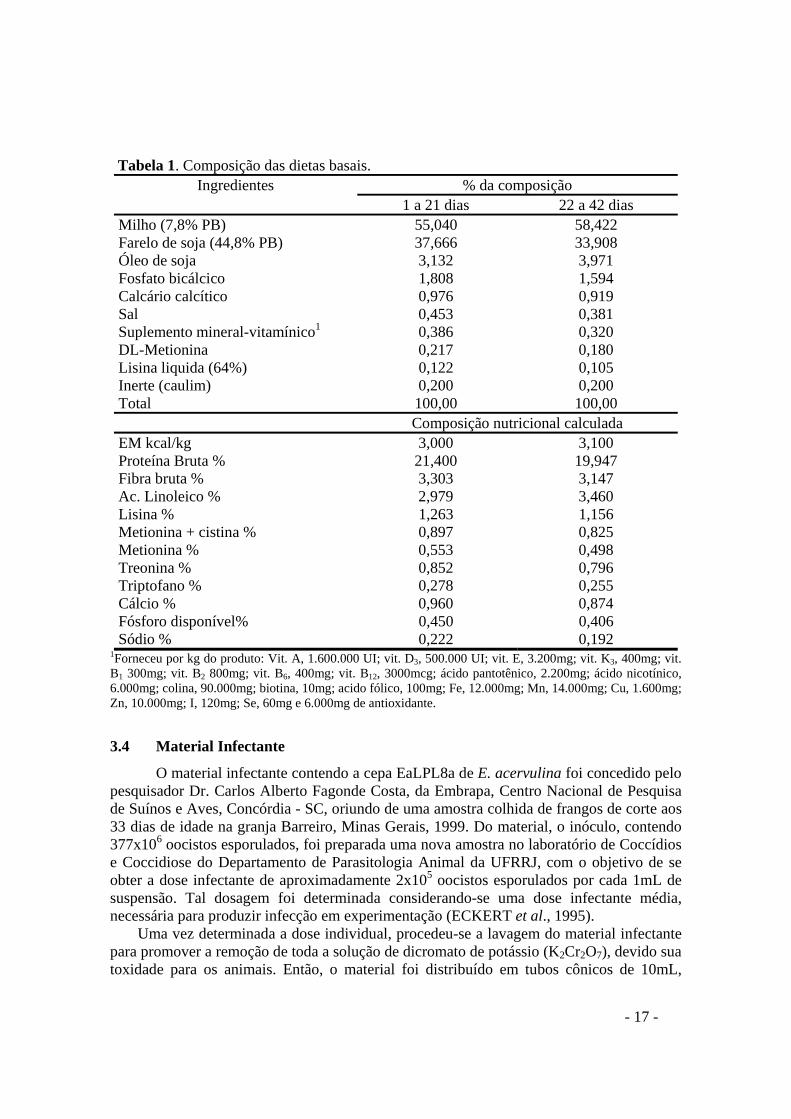
Tabela 1. Composição das dietas basais.
Ingredientes % da composição 1 a 21 dias 22 a 42 dias
Milho (7,8% PB) 55,040 58,422 Farelo de soja (44,8% PB) 37,666 33,908 Óleo de soja 3,132 3,971 Fosfato bicálcico 1,808 1,594 Calcário calcítico 0,976 0,919 Sal 0,453 0,381 Suplemento mineral-vitamínico1 0,386 0,320 DL-Metionina 0,217 0,180 Lisina liquida (64%) 0,122 0,105 Inerte (caulim) 0,200 0,200 Total 100,00 100,00 Composição nutricional calculada EM kcal/kg 3,000 3,100 Proteína Bruta % 21,400 19,947 Fibra bruta % 3,303 3,147 Ac. Linoleico % 2,979 3,460 Lisina % 1,263 1,156 Metionina + cistina % 0,897 0,825 Metionina % 0,553 0,498 Treonina % 0,852 0,796 Triptofano % 0,278 0,255 Cálcio % 0,960 0,874 Fósforo disponível% 0,450 0,406 Sódio % 0,222 0,192
1Forneceu por kg do produto: Vit. A, 1.600.000 UI; vit. D3, 500.000 UI; vit. E, 3.200mg; vit. K3, 400mg; vit. B1 300mg; vit. B2 800mg; vit. B6, 400mg; vit. B12, 3000mcg; ácido pantotênico, 2.200mg; ácido nicotínico, 6.000mg; colina, 90.000mg; biotina, 10mg; acido fólico, 100mg; Fe, 12.000mg; Mn, 14.000mg; Cu, 1.600mg; Zn, 10.000mg; I, 120mg; Se, 60mg e 6.000mg de antioxidante.
3.4 Material Infectante
O material infectante contendo a cepa EaLPL8a de E. acervulina foi concedido pelo pesquisador Dr. Carlos Alberto Fagonde Costa, da Embrapa, Centro Nacional de Pesquisa de Suínos e Aves, Concórdia - SC, oriundo de uma amostra colhida de frangos de corte aos 33 dias de idade na granja Barreiro, Minas Gerais, 1999. Do material, o inóculo, contendo 377x106 oocistos esporulados, foi preparada uma nova amostra no laboratório de Coccídios e Coccidiose do Departamento de Parasitologia Animal da UFRRJ, com o objetivo de se obter a dose infectante de aproximadamente 2x105 oocistos esporulados por cada 1mL de suspensão. Tal dosagem foi determinada considerando-se uma dose infectante média, necessária para produzir infecção em experimentação (ECKERT et al., 1995).
Uma vez determinada a dose individual, procedeu-se a lavagem do material infectante para promover a remoção de toda a solução de dicromato de potássio (K2Cr2O7), devido sua toxidade para os animais. Então, o material foi distribuído em tubos cônicos de 10mL,

- 18 -
centrifugados a 3000 rpm (2200g) por 10 minutos, e a seguir, o sedimento foi suspenso em solução salina 0,9% NaCl. Tal operação foi repetida por mais duas vezes até que os oocistos estivessem livres da solução de dicromato de potássio. O sedimento final contendo os oocistos foi suspenso em nova solução salina e armazenado sob refrigeração de 4-8 °C. Após a lavagem procedeu-se a quantificação dos oocistos presentes no material infectante com auxílio da câmara de Neubauer, de acordo com o procedimento descrito no CURSO DE PRÁCTICAS DE BIOLOGÍA CELULAR – UNIVERSITAT DE BARCELONA (2004-2005). Por meio de regra de três, foi calculada a quantidade total de diluente (solução salina) necessária para se obter doses suficientes do inóculo para a infecção das aves do experimento.
No décimo quarto dia de idade, todas as aves do experimento foram inoculadas individualmente com oocistos esporulados de E. acervulina por via oral com 1mL do inóculo preparado. Foram utilizadas pipetas do tipo Ependorff® de 1,25mL introduzidas pelo canal oral objetivando-se aproximar ao inglúvio.
3.5 Ensaio de Digestibilidade
O ensaio foi conduzido em duas fases, a primeira correspondendo a fase inicial de criação (17 a 21 dias de idade), período em que as aves passavam pela fase mais crítica do desafio, e a segunda fase correspondendo a fase final de criação (27 a 31 dias de idade), período de uma provável recuperação das mesmas. Na primeira fase foram utilizadas onze aves por unidade experimental, totalizando 330 animais, e na segunda, nove aves, sendo 270 animais no total. A redução do número de aves nas unidades experimentais no decorrer do experimento foi devido à retirada destas para necropsia e preparo de lâminas para análise histológica.
O período de colheita das excretas em cada ensaio foi de cinco dias, totalizando dez dias para os dois ensaios. Foi colocado sob cada compartimento da bateria, bandeja metálica forrada com lona plástica para evitar contaminação do material. As excretas foram colhidas duas vezes ao dia com intervalo de 12 horas, acondicionadas em sacos plásticos identificados de acordo com o respectivo tratamento e repetição, e congeladas (-18°C) para análises posteriores. Após o término do experimento, as excretas foram descongeladas e homogeneizadas, e retirou-se uma amostra de 500g de cada repetição para pré-secagem em estufa ventilada a 55°C por 48 horas; após secagem, as excretas foram expostas ao ar até que atingissem a temperatura ambiente para serem pesadas, moídas e armazenadas em potes plásticos devidamente identificados para análises posteriores.
Foram registrados o consumo das dietas experimentais e a quantidade de excreta produzida por cada unidade experimental, as quais foram submetidas a análise de matéria seca (MS) e nitrogênio metabolizável (N), segundo a AOAC (1984). A energia bruta (EB) foi determinada usando a bomba calorimétrica PARR, de acordo com a técnica descrita por HARRIS (1970).
A determinação dos coeficientes de digestibilidade aparente das rações foi baseada no consumo de MS da ração e na excreção da MS e nos teores de N e EB das rações e das excretas, de acordo com a fórmula descrita por SCHINEIDER & FLAT (1975).
Digestibilidade Aparente = Nutriente ingerido(g)-Nutriente excretado(g) x 100 Nutriente ingerido (g)

- 19 -
3.6 Análise Estatística
O delineamento experimental usado foi o de blocos ao acaso, onde se utilizaram baterias metálicas compostas de três andares, cujo bloco foi representado por cada andar, totalizando três blocos por bateria. O modelo estatístico adotado foi escolhido pensando-se nas condições de conforto térmico que poderiam ser diferentes em cada um dos andares, uma vez que o animal experimental é muito sensível ao fator temperatura. Foram usadas dez baterias metálicas compostas de três blocos (superior, médio e inferior) onde cada bateria foi ocupada por aves que estavam recebendo o mesmo tratamento, cujo objetivo foi evitar possíveis contaminações dos tratamentos entre os blocos. Em cada bloco, um único tratamento foi repetido por duas vezes, totalizando seis repetições por tratamento (duas no bloco superior, duas no bloco médio e duas no bloco inferior), então, o mesmo tratamento foi distribuído em duas baterias metálicas diferentes. Como havia cinco tratamentos, foi necessário utilizar dois pares de cinco baterias, que somado as seis repetições de cada tratamento, deu um total de 30 observações.
As análises de variância foram realizadas utilizando-se o pacote computacional Sisvar, descrito por FERREIRA (2000). As variáveis de cada tratamento tiveram suas médias comparadas pelo teste de Student-Newman-Keuls (SNK) a um nível de significância de 5% (H0:p<0,05), assim como as análises de regressão foram calculadas por meio da técnica de polinômios ortogonais.
O modelo estatístico adotado para análise do ensaio de digestibilidade para o teste
de comparação de médias foi:
ebty ijjiij+++= µ
Em que: yij
é o valor observado da característica estudada, no tratamento i ( i = 1,2,3,4,5,6), no
bloco j (j = 1,2,3) µ é a média geral paramétrica
tié o efeito do tratamento i
bj é o efeito do bloco j
eijé o erro associado à observação yij
, ou efeito dos fatores não controlados sobre a
observação yij.
O modelo estatístico adotado para análise de regressão do ensaio de digestibilidade
foi: ipipiii e+Χ++Χ+Χ+=Υ ββββ ...22110
em que: iΥ é a variável dependente, obtida na observação i
pββββ ,...,,, 210 são os parâmetros da regressão
piii ΧΧΧ ,...,, 21 são variáveis independentes, na observação i
ie é o erro associado a observação iΥ .

- 20 -
4 RESULTADOS E DISCUSSÃO
Os resultados da digestibilidade aparente da matéria seca (MS) e nitrogênio metabolizável (N) e os valores de energia metabolizável (EM) da ração dos frangos de corte suplementados com betaína durante o primeiro e o segundo ensaio de digestibilidade estão apresentados na Tabela 2 e Figuras 1 e 2.
A digestibilidade da MS da ração no primeiro ensaio foi superior (p<0,05) nas aves que receberam o alimento contendo promotor de produção e coccidiostático em relação às aves que receberam a ração sem a suplementação de betaína (0,0%) e as rações contendo 0,05; 0,10 e 0,15% de betaína (Tabela 2). Este resultado difere daquilo que foi encontrado por VIRTANEN (1995), o qual relata que aves oralmente inoculadas com uma mistura de três espécies do gênero Eimeria aos 14 dias de idade, recebendo uma dieta contendo 0,15% de betaína, apresentaram maior digestibilidade da matéria seca do que aquelas que receberam a dieta controle (sem suplementação de betaína).
De acordo com vários autores (DE BLAS, 1984; FULLER, 1989; KERMAUNER & STRUCKLEC, 1996; BEDFORD, 2000), aditivos utilizados em dietas, como antibióticos e ácidos orgânicos atuam controlando a proliferação de microrganismos patogênicos, favorecendo a proliferação de microrganismos desejáveis ou atuando sobre o pH do trato digestivo, e como resultado, diminuem os distúrbios gastrintestinais melhorando o aproveitamento dos nutrientes.
Tabela 2. Digestibilidade aparente da matéria seca (%MS), do nitrogênio metabolizável (%N) e dos valores de energia metabolizável (EM) da ração expressa na base da matéria seca, por fase, em função dos diferentes tratamentos.
Tratamentos
Primeiro ensaio de digestibilidade: 17 a 21 dias de idade das aves, 3dpi
Segundo ensaio de digestibilidade: 27 a 31 dias de idade das aves, 13 dpi
%MS %N EM/kg MS MS %N EM/kg MS CP 65,6a 53,1a 2652,34a 70,7a 57,8a 3013,77a 0,00% Bet. 60,5b 47,7b 2422,18b 70,7a 61,1a 2865,97b 0,05% Bet. 60,5b 50,1ab 2576,03ab 73,1a 62,2a 2864,03b 0,10% Bet. 58,8b 47,2b 2692,29a 73,5a 61,9a 2795,13b 0,15% Bet. 58,9b 45,4b 2476,17b 73,8a 59,6a 2886,48b CV Regressão
4,23 ns
6,89 ns
4,24 Q
3,16 L
6,30 ns
2,97 ns
dpi = dias após a inoculação das aves. a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). ns = não significativo para análise de regressão (p>0,05). L = Efeito linear (p<0,05) Q = Efeito quadrático (p<0,05)
A digestibilidade do N metabolizável da ração no primeiro ensaio foi maior (p<0,05) nas aves que consumiram a ração com promotor de produção e coccidiostático e naquelas que consumiram a ração contendo 0,05% de betaína do que nas aves que não

- 21 -
foram suplementadas com betaína (0,0%) ou que foram suplementadas com os níveis de 0,10 e 0,15% (Tabela 2). VIRTANEN (1995) encontrou diferença significativa para a digestibilidade da proteína na ração suplementada com betaína em relação à dieta controle de aves inoculadas, tendo sido utilizado o nível de 0,15% de betaína. Segundo resultados de trabalhos conduzidos no PARC Institute nos EUA, frangos inoculados com Eimeria, que receberam dietas suplementadas com 0,15% de betaína, também apresentaram melhor digestibilidade da proteína que os controles (sem betaína), no entanto, as respectivas dietas estavam suplementadas com um coccidiostático ionóforo (HRUBY et al., 2001).
Foram observados os maiores valores de EM da ração (p<0,05) nos frangos suplementados com 0,10% de betaína e com a ração contendo coccidiostático e promotor de produção em relação aos frangos que não receberam a suplementação de betaína (0,0%) ou que receberam o nível de 0,15% (Tabela 2). Além disso, observou-se que não houve diferença (p>0,05) nos valores da EM da ração dos frangos que receberam o nível de 0,10% e o nível de 0,05% de betaína (Tabela 2). Em experimentos realizados com suínos, foi comprovado o conceito de que a betaína aumenta o valor de energia/nutriente da dieta quando fornecida a um nível de 0,10 ou 0,125% na ração (CASARIN et al., 1997; MATTHEWS et al., 1997).
Quanto à análise de regressão, observou-se no primeiro ensaio um efeito quadrático (y = -36998x2 + 6106,1x + 2407,4) para os valores de EM da ração, onde se pode constatar um aumento da energia quando se adicionou betaína até o nível de 0,082%. À partir da adição de níveis mais altos que este valor ocorreu um decréscimo na EM da ração (Figura 1).
Figura 1. Influência da suplementação de betaína nos valores de EM aparente da ração em frangos de corte infectados experimentalmente com E. acervulina no primeiro ensaio de digestibilidade.
Ainda de acordo com os dados apresentados na Tabela 2 no segundo ensaio, foi observado que não houve diferença significativa (p>0,05) entre os tratamentos para
y = -36998x2 + 6106.1x + 2407.4R2 = 0.8976
2400.00
2460.00
2520.00
2580.00
2640.00
2700.00
0.00 0.05 0.10 0.15Níveis de betaína (%)
Ene
rgia
Met
abol
izáv
el (k
cal/k
g)
0,082%
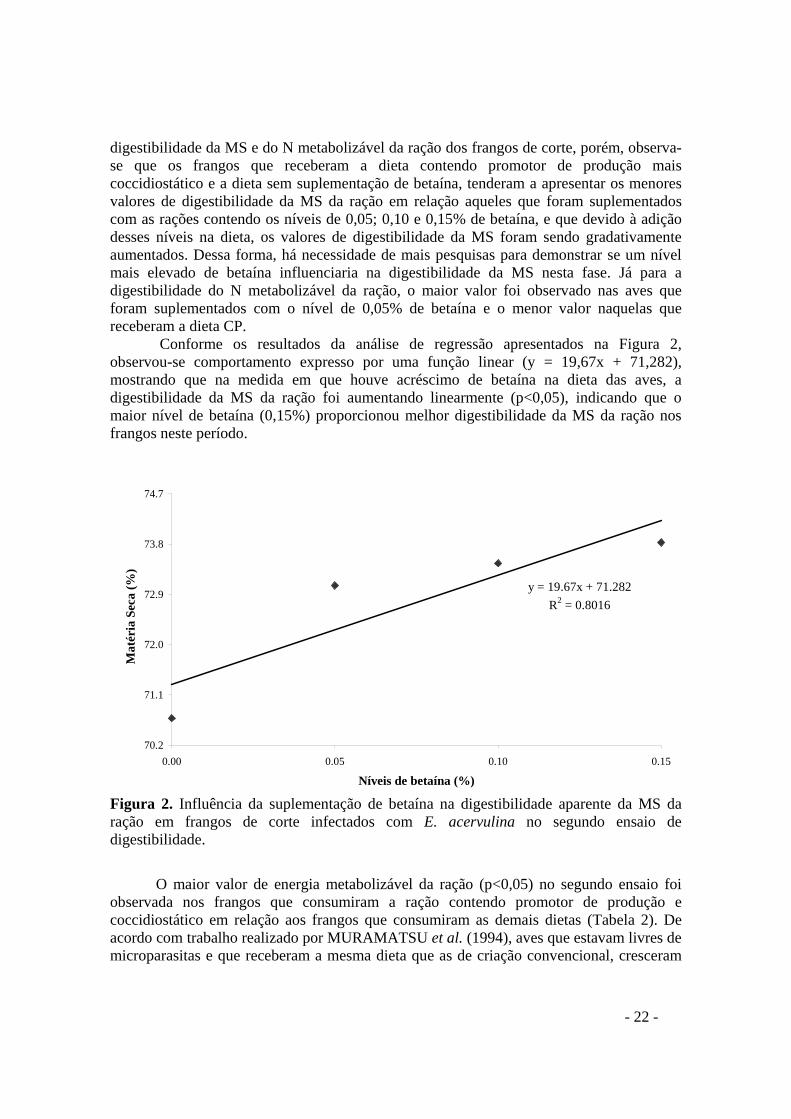
- 22 -
digestibilidade da MS e do N metabolizável da ração dos frangos de corte, porém, observa-se que os frangos que receberam a dieta contendo promotor de produção mais coccidiostático e a dieta sem suplementação de betaína, tenderam a apresentar os menores valores de digestibilidade da MS da ração em relação aqueles que foram suplementados com as rações contendo os níveis de 0,05; 0,10 e 0,15% de betaína, e que devido à adição desses níveis na dieta, os valores de digestibilidade da MS foram sendo gradativamente aumentados. Dessa forma, há necessidade de mais pesquisas para demonstrar se um nível mais elevado de betaína influenciaria na digestibilidade da MS nesta fase. Já para a digestibilidade do N metabolizável da ração, o maior valor foi observado nas aves que foram suplementados com o nível de 0,05% de betaína e o menor valor naquelas que receberam a dieta CP.
Conforme os resultados da análise de regressão apresentados na Figura 2, observou-se comportamento expresso por uma função linear (y = 19,67x + 71,282), mostrando que na medida em que houve acréscimo de betaína na dieta das aves, a digestibilidade da MS da ração foi aumentando linearmente (p<0,05), indicando que o maior nível de betaína (0,15%) proporcionou melhor digestibilidade da MS da ração nos frangos neste período.
Figura 2. Influência da suplementação de betaína na digestibilidade aparente da MS da ração em frangos de corte infectados com E. acervulina no segundo ensaio de digestibilidade.
O maior valor de energia metabolizável da ração (p<0,05) no segundo ensaio foi observada nos frangos que consumiram a ração contendo promotor de produção e coccidiostático em relação aos frangos que consumiram as demais dietas (Tabela 2). De acordo com trabalho realizado por MURAMATSU et al. (1994), aves que estavam livres de microparasitas e que receberam a mesma dieta que as de criação convencional, cresceram
y = 19.67x + 71.282R2 = 0.8016
70.2
71.1
72.0
72.9
73.8
74.7
0.00 0.05 0.10 0.15
Níveis de betaína (%)
Mat
éria
Sec
a (%
)

- 23 -
substancialmente mais rápido e pareceram absorver menos energia metabolizável da dieta. O autor explica ainda que esta discrepância na absorção de energia é uma conseqüência da microflora que extrai uma quantidade significativa de energia da dieta, energia que não estará disponível para a ave. No caso do experimento em questão, supõe-se que as aves que receberam a ração com promotor de produção e coccidiostático poderiam estar com os microparasitas controlados, e dessa forma, a energia metabolizável foi mais bem aproveitada pelo organismo do animal, e como conseqüência, ocorreu uma melhoria na capacidade de digestão dos frangos.
O primeiro ensaio de digestibilidade foi realizado objetivando-se acompanhar o período crítico da coccidiose, admitindo-se que o período pré-patente de Eimeria acervulina varia em torno de 4 a 9 dias após a infecção, conforme descrito por ALLEN & FETTERER (2002). E de fato, como pode ser observado na Tabela 2, houve uma drástica redução dos valores de digestibilidade durante o primeiro ensaio. TAKHAR & FARREL (1979a,b) também observaram que a utilização da energia metabolizável e a retenção de nitrogênio foram diminuídas em frangos infectados com coccídios.
Após a fase crítica, no segundo ensaio (Tabela 2), observa-se que os valores de digestibilidade aumentaram substancialmente. Observa-se ainda que a betaína teve efeitos positivos sobre a digestibilidade do N metabolizável e da EM da ração durante o primeiro ensaio. No segundo ensaio, a betaína foi tão efetiva ou até mesmo tendeu a ser mais efetiva que o promotor de produção e coccidiostático para aumento da digestibilidade aparente da MS e do N metabolizável da ração, porém, a mesma afirmativa não pode ser feita para os valores de EM nesta fase, pois o promotor de produção e coccidiostático foram bem mais eficazes em termos de aumento dos valores de EM, entretanto, pode ser observado que o maior nível de betaína tendeu à aumentar os valores da EM da ração.
Para o aumento da digestibilidade que ocorreu no segundo ensaio, após a fase crítica da coccidiose, pode-se sugerir a hipótese da compensação na capacidade de absorção que acontece no intestino das aves após um período de infecção com coccídio. TURK (1978) relatou que no intestino delgado das aves, foi observado que nas áreas mais infectadas ocorreu uma redução na capacidade de absorção dos aminoácidos, mas, por outro lado, áreas menos prejudicadas aumentaram sua taxa de absorção.

- 24 -
5 CONCLUSÕES
Considerando-se a digestibilidade do nitrogênio e os valores de energia metabolizável da ração durante a fase crítica da coccidiose, foi positiva a suplementação de betaína em rações de frangos de corte infectados experimentalmente com Eimeria acervulina.
Baseando-se na digestibilidade da MS e do nitrogênio metabolizável da ração, foi positiva a suplementação de betaína em rações de frangos de corte, após período critico da coccidiose.
A betaína pode ser de utilidade, em conjunto com outras estratégias nutricionais, para a viabilização da retirada dos antibióticos como promotores de produção em dietas de frangos de corte.

- 25 -
6 REFERÊNCIAS BIBLIOGRÁFICAS
ALLEN, P.C.; FETTERER, R.H. Recent advances in biology and immunobiology of Eimeria species and in diagnosis and control of infection with these coccidian parasites of poultry. Clinical Microbiology Reviews, v.15, n.1, p.58-65, 2002.
ALMQUIST, H.J.; GRAU, C.R. Growth-promoting activity of betaine in chick. Journal of Biology Chemistry, v.149, p.575-576, 1943.
ALMQUIST, H.J.; GRAU, C.R. Interrelatioship of methionine, choline, betaine and arsenocholine in the chick. Journal of Nutrition, v.27, p.263-269, 1944.
ANDRIGUETTO, J.M.; PERLY, L.; MINARDI, I.; GAMAEL, A.; FLEMMING, J.S.; SOUZA, G.A.; BONA FILHO, A. Nutrição Animal: as bases e os fundamentos da nutrição animal. 6. ed. São Paulo: Nobel, 1999. v.1, 395p.
ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS (AOAC). Official methods of analysis. 14º ed.: Washington, D.C., 1984, 1141p.
AUGUSTINE P.C.; DANFORTH, H.D Influences of betaine and salinomycin on intestinal absorption of methionine and glucose and on the ultrastructure of the intestinal cells and parasite developmental stage in chicks infected with Eimeria acervulina. Avian Disease, v.43, p.89-97, 1999. AVICULTURA INDUSTRIAL. Seqüência de aminoácidos. Redação AI 23/06/2003. Disponível em: <http://www.aviculturaindustrial.com.br>. Acesso em: 08 jan. 2004. BEACH, J.R.; CORL, J.C. Studies in the control of avian coccidiosis. Poultry Science, v. 4, p. 83-93, 1925. BEDFORD, M. Removal of antibiotic growth promoters from poultry diets: implications and strategies to minimize subsequent problems. World’s Poultry Science Journal, v.56, p.347-365, 2000. BOCH, J.; KEMPF, B.; BREMER, E. Osmoregulation in Bacillus subtilis: synthesis of the osmoprotectant glycine betaína from exogenously provided choline. Journal of Bacteriology, v.176, p.5364-5371, 1994. BRACKETT, S.; BLIZNICK, A. The occurrence and economic importance of coccidiosis in chickens. Lederle Laboratories Division, American Cyanamid Company. 1950.

- 26 -
BROWN, D.R.; SOUTHERN, L.L. Effect of Eimeria acervulina infection in chicks fed excess dietary cobalt and/or manganese. Journal of Nutrition, v.115, p.347-351, 1985. BUDAVARI, S. The Merck Index. In: RAHWAY, N.J. Merck and Co. Inc. USA, 1989, p.1201-1202. CANTEY J.R. Escherichia coli diarrhea. Gastroenterology Clinics of North America, v.22, p.609-622, 1993. CASARIN, A.; FORAT, M.; ZABARAS-KRICK, B.J. Interrelationships between betaine (Betafin-BCR) and level of feed intake on the performance parameters and carcass characteristics of growing-finishing pigs. Journal of Animal Science, v.75 (suppl. 1), n.75 (Abstr.), 1997. CONAWAY, D.P.; MCKENZIE, E. Poultry coccidiosis: diagnostic and testing procedures. Pfizer Incorporated. Studio Zaba, New York, NY, 1991.
COX, F.E.G. Control of coccidiosis: lessons from other sporozoa. International Journal for Parasitology. v.28, p.165-179, 1998.
CROMPTON, D.W.T. Malfunctioning of the gut: parasitism. In: BOORMAN, K.N. & FREEDMAN, B.M. Digestion in the fowl. Edinburgh, UK: Editor British Poultry Science Ltda, 1976, p.193-245. CZARNECKI, G.L.; BAKER, D.H. Feed additive interactions in the chickens: reduction of tissue copper deposition by dietary roxarsone in healthy and in Eimeria acervulina-infected or Eimeria tenella-infected chicks. Poultry Science, v.63, p.1412-1418, 1984. DANFORTH, H.D. Use of live oocyst vaccines in the control of avian coccidiosis: experimental studies and field trials. International Journal for Parasitology, v.28, p.1099-1109, 1998. De BLAS, C. Alimentación del conejo. Madri: Mundi-Prensa, 1984, 215p. DU VIGNEAUD, V.; CHANDLER, J.P.; MOYER, A.W.; KEPPEL, D.M. The effect of choline on the ability of homocystine to replace methionine in the diet. Journal of Biological Chemistry, v.131, p.57-76, 1939. ECKERT, J.; TAYLOR, M.; LICOIS, D.; COUDERT, P.; CATCHPOLE, J.; BUCKLAR, H. Identification of Eimeria and Isospora species and strains. Morphological and biological characteristics. In: ECKERT, J.; BRAUN, R.; SHIRLEY, M.W.; COUDERT, P. Biotechnology. Guidelines on Techniques in Coccidioisis Research. Luxembourg, 1995, p. 103-119. EDGAR, S.A. Field diagnosis of coccidiosis in chickens. Agri-Bio Corporation, 1992.

- 27 -
ESCHENBACHER, K.H., EGGLI, P.; WALLACH, M.; BRAUN, R. Characterization of a 14kDa oocyst wall protein of Eimeria tenella and E. acervulina. Parasitology, v.112, p.169-176, 1996.
FERREIRA, D.F. SISVAR – Sistema de Análise Estatística. Lavras, UFLA, 2000. Comunicação pessoal.
FONDACARO, J.D. Intestinal ion transport and diarrheal disease. American Journal of Physiology, 250 (Gastrointestinal Liver Physiology 13) G1-G8, 1986.
FULLER, R. Probiotics in man and animals: a review. Journal of Applied Bacteriology, n.66, p.365-378, 1989.
GIRALDO, C.A.; BROWN, D.R.; WATKINS, K.L.; SOUTHERN, L.L. Responses to excess dietary magnesium as affected by experimental Eimeria acervulina infection or by dietary ammonium chloride ingestion in the chick. Journal of Nutrition, v.117, p.1053-1059, 1987.
GRAAT, E.A.M.; HENKEN, A.M.; PLOEGER, H.W.; NOORDHUIZEN, J.P.T.M.; VERTOMMEN, M.H. Rate and course of sporulation of oocysts of Eimeria acervulina under different environmental conditions. Parasitology, v.108, p.497-502, 1994.
HARRIS, L.E. Os métodos químicos e bioquímicos empregados na análise de alimentos. Gainesville, Universidade da Flórida, 1970, paginação irregular.
HENKEN, A.M.; PLOEGER, H.W.; GRAAT, E.A.M.; CARPENTER, T.E. Description of a simulation model for the population dynamics of Eimeria acervulina infection in broilers. Parasitology, v.108, p.503-512, 1994a.
HENKEN, A.M.; GRAAT, E.A.M.; PLOEGER, H.W.; CARPENTER, T.E. Description of a model to simulate effects of Eimeria acervulina infection on broiler production. Parasitology, v.108, p.513-518, 1994b.
HRUBY, M.; BARRATT, A.; REMUS, J. Betaine improves poultry carcass characteristics. Poultry Digest, 2001. JEURISSEN, S.H.M.; JANSE, E.M.; VERMEULEN, A.N.; VERVELDE, L. Eimeria tenella infections in chickens: aspects of host-parasite interaction. Veterinary Immunology and Immunopathology, v.54, p.231-238, 1996.

- 28 -
KERMAUNER, A.; STRUCKLEC, M. Addition of probiotic to feeds with different energy and ADF content in rabbits. 1. Effect on the digestive organs. World Rabbit Science, v.4, n.4, p.187-193, 1996. KHEYSIN, Y.M. Life cycles of coccidia of domestic animals. University Park Press. 1972.
KIDD, M.T.; FERKET, P.R.; GARLICH, J.D. Nutritional and osmoregulatory functions of betaine. World’s Poultry Science Journal, v.53, p.125-139, 1997.
KO, R.; SMITH, L.T.; SMITH, G.M. Glycine betaine confers enhanced osmotolerance and cryotolerance on Listeria monocytogenes. Journal of Bacteriology, v.176, p.426-431, 1994. KONOSU, S.; HAYASHY, T. Determination of beta-alanine betaine and glycine betaine in some marine invertebrates. Bulleetin Jp. Society Science Fish, v.41, p.743-746, 1975. KOUWENHOVEN, B.; VAN DER HORST, C.J.G. Disturbed intestinal absorption of vitamin A and carotenes and the effect of a low pH during Eimeria acervulina infection in the domestic fowl (Gallus domesticus). Zoology Parasitenked, v.38, p.152-161, 1972.
LEE, M.R.; SHIH, J.C.H. Effect of anaerobic digestion on oocysts of the protozoan Eimeria tenella. Applied and Environmental Microbiology, v.54, p.2335-2341, 1988.
LILLEHOJ, H.S.; TROUT, J.M. Coccidia: A review of recent advances on immunity and vaccine development. Avian Pathology, v.22, p.3-31, 1993.
LILLEHOJ, H.S.; TROUT, J.M. Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clinical Microbiology Reviews, July, p.349-360, 1996.
MATTEWS, J.O.; SOUTHERN, L.L.; BIDNER, T.D. Interactive effects of betaine (Betafin-BCR), crude protein, and net energy on growth, carcass traits, and serum metabolites of gilts. Journal of Animal Science, v.75 (Suppl. 1), n.62 (Abstr.), 1997.
McDOUGALD, L.R.; REID, W.M. Coccidiosis. In: CALNEK, B.W. Diseases of Poultry. 10th ed. Ames, IA: Editor Iowa State University Press, 1997, p.865-883.
McGINNIS, J.; NORRIS, L.C.; HEUSER, G.F. Effect of ethanolamine and betaine on perosis in chicks. Experimental Biology and Medicine, v.51, p.293-294, 1942.

- 29 -
MOONEY, M.; BELAY, T.; TEETER, R.G.; VIRTANEN, E.; REMUS, J. Effect of betaine on loss of corporal water. In: SOUTHERN POULTRY SCIENCE SOCIETY AND SOUTHERN CONFERENCE ON AVIAN DISEASES, 1998, Atlanta, GA, Proceedings… Atlanta: jan.19-20, 1998.
MURAMATSU, T.; NAKAJIMA, S.; OKUMURA, J. Modification of energy metabolism by the presence of the gut microflora in the chickens. British Journal of Nutrition, v.71, p.709-717, 1994.
PARTRIDGE, G. Betaine from sugarbeet gives an energy boost. Pig International, v.32, n.1, p.21, 2002.
PARTRIDGE, G. Betaine’s dual role. Pig Progress, v.19, n.10, 2003.
PETRONINI, P.G.; DEANGELIS, E.M.; BORGHETTI, P.; BORGHETTI, A.F.; WHEELER, K.P. Modulation by betaine of cellular responses to osmotic stress. Biochemistry Journal, v.282, p.69-73, 1992.
REMUS, J.; VIRTANEN, E.; ROSI, L.; McNAUGHTON, J. Effect of betaína on nutrient utilization of 21 day-old broilers during coccidiosis. In: EUROPEAN SYMPOSIUM ON POULTRY NUTRITION, 10, 1995, Antalya, Turkey. Proceedings… Antalya, 1995, p. 371-372.
REMUS, J.C. Betaine for increased breast meat yield in turkeys. World Poultry, v.17, n.2, 2001.
ROSE, M.E. Immunity to Eimeria infections. Veterinary Immunology and Immunopathology, v.17, p.333-343, 1987.
ROSTAGNO, H.S.; SILVA, D.S.; COSTA, P.M.A.; FONSECA, J.B.; SOARES, P.R.; PEREIRA, J.A.A.; SILVA, M.A. Composição de alimentos e exigências nutricionais de aves e suínos (Tabelas Brasileiras). Viçosa, UFV, Imprensa Universitária, 2000, 69p.
RUDOLPH, A.S.; CROWE, J.H.; CROWE, L.M. Effects of three stabilizing agents-proline, betaine and trehalose- on membrane phospholipids. Archives of Biochemistry and Biophysics, v.245, p.134-143, 1986.
SCHNEIDER, B.A.; FLATT, W.P. The evaluation of feeds through digestibility experiments. Athens, The University of Georgia, 1975, 423p.

- 30 -
SHIRLEY, M.W. New methods for the identification of species and strains of Eimeria. In: McDOUGALD, L.R.; JOYNER, L.P.; LONG, P.L. Research in Avian Coccidiosis. University of Georgia, Athens, Georgia, 1986, p.13-35. SHOEMAKER, V.H. Osmoregulation and excretion in birds. In: FARNER, D.S. KING, J.R.; PARKERS, K.C. Avian Biology, v.2, Academic Press, 1972, p.527-574.
SOUTHERN, L.L.; BAKER, D.H. Eimeria acervulina infection in chicks fed cobalt in the presence or absence of excess dietary methionine. Journal of Nutrition, v.112, p.1220-1223, 1982.
SOUTHERN, L.L.; BAKER, D.H. Eimeria acervulina infection and the zinc-copper interrelationship in the chick. Poultry Science, v.62, p.401-404, 1983a.
SOUTHERN, L.L.; BAKER, D.H. Eimeria acervulina infection in chicks fed deficient or excess levels of manganese. Journal of Nutrition, v.113, p.172-177, 1983b. TAKHAR, B.S.; FARREL, D.J. Energy and nitrogen metabolism of chickens infected with either Eimeria acervulina or Eimeria tenella. British Poultry Science, v.20, p.197-211, 1979a. TAKHAR, B.S.; FARREL, D.J. Energy and nitrogen metabolism of chickens subjected to infection and reinfection with Eimeria acervulina. British Poultry Science, v.20, p.213-224, 1979b. TURK, D.E. The effects of coccidiosis on intestinal function and gut microflora. In: LONG, P.L.; BOORMAN, K.N.; FREEMAN, B.M. Avian Coccidiosis. Edinburg, Scotland: Editor British Poultry Science Ltda, 1978, p.227-267. UNIVERSITAT DE BARCELONA. Departament de Biología Cel.lular. Prácticas de biología celular: curso 2004-2005. Barcelona, 2004, 40p. Disponível em: <http://www.ub.es. Acesso em: 2004. VERMEULEN, A.N.; SCHAAP, D.C.; SCHETTERS, Th.P.M. Control of coccidiosis in chickens by vaccination. Veterinary Parasitology, v.100, p.13-20, 2001. VIRTANEN, E. Piecing together the betaine puzzle. Feed Mix, v.3, n.1, p.12-17, 1995.
VIRTANEN, E.; ROSI, L. Effects of betaine on methionine requirements of broilers under various environmental conditions. In: AUSTRALIAN POUTRY SCIENCE SYMPOSIUM, 1995, Sidney. Proceedings…Sidney: University of Sidney, Australia, 1995, v.7, p.88-98.
WARD, T.L.; WATKINS, K.L.; SOUTHERN, L.L. Interactive effects of sodium zeolite A (Ethacal®) and monesin in uninfected and Eimeria acervulina-infected chicks. Poultry Science, v.69, p.276-280, 1990.

- 31 -
WATKINS, K.L.; VAGNONI, D.B.; SOUTHERN, L.L. Effect of dietary sodium zeolite A and excess calcium on growth and tibia calcium and phosphorus concentration in uninfected and Eimeria acervulina-infected chicks. Poultry Science, v.68, p.1236-1240, 1989.
WILLIAMS, R.B. Epidemiological aspects of the use of live anticoccidial vaccines for chickens. International Journal for Parasitology, v.28, p.1089-1098, 1998.
WILLIAMS, R.B. A Compartmentalized model for the estimation of the cost of coccidiosis to the world’s chicken production industry. International Journal for Parasitology, v.29, p.1209-1229, 1999.
WILLIS, G.M.; BAKER, D.H. Phosphorus utilization during Eimeria acervulina infection in the chick. Poultry Science, v.60, p.1960-1962, 1981.
YUN, C.H.; LILLEHOJ, H.S.; LILLEHOJ, E.P. Intestinal immune responses to coccidiosis. Developmental and Comparative Immunology, v.24, p.303-324, 2000.

- 32 -
CAPÍTULO II
INFLUÊNCIA DA BETAÍNA NO DESENVOLVIMENTO DE VILOS
E CRIPTAS DO INTESTINO DELGADO DE FRANGOS DE
CORTE INFECTADOS EXPERIMENTALMENTE COM EIMERIA
ACERVULINA (TIZZER, 1929)

- 33 -
RESUMO
NIANG, Tania Marcia Soares. Influência da betaína no desenvolvimento de vilos e criptas do intestino delgado de frangos de corte infectados experimentalmente com Eimeria acervulina (TIZZER, 1929).
Foi avaliado o efeito da suplementação de betaína sobre a estrutura da mucosa intestinal de frangos de corte machos submetidos à infecção experimental com Eimeria acervulina. Foram utilizados 420 pintos de corte Cobb, de um dia de idade, distribuídos num delineamento de blocos ao acaso com 5 tratamentos e 6 repetições, alojados em baterias metálicas de três andares, onde cada andar representava um bloco. As dietas foram fornecidas de 1 a 21 e 22 a 42 dias de idade. Os tratamentos foram: 1) controle positivo (CP) = dieta basal mais promotor de produção e coccidiostático; 2) 0,0% de betaína = dieta basal sem betaína (0,0%); 3) 0,05% betaína = dieta basal mais 0,05% de betaína; 4) 0,10% betaína = dieta basal mais 0,10% de betaína; 5) 0,15% betaína = dieta basal mais 0,15% de betaína. No décimo quarto dia de idade, as aves foram inoculadas individualmente com aproxidamente 2x105 oocistos esporulados de E. acervulina. Uma ave de cada unidade experimental, totalizando 30 aves por amostragem, foi sacrificada por deslocamento da coluna cervical no 14º, 18°, 21º e 28° dia de vida para que fossem coletadas amostras de tecidos da alça duodenal para preparo histológico. Foi observado que não houve diferença (p>0,05) entre os tratamentos para altura das vilosidades aos 14 dias de idade, entretanto, observou-se que na medida em que se elevaram os níveis de betaína na dieta, houve aumento na altura dos vilos. Aos 18 dias de idade, não houve diferença (p>0,05) entre os tratamentos para altura dos vilos, porém, observou-se que o maior valor para altura das vilosidades foi encontrado nas aves suplementadas com 0,05% de betaína na ração. Já aos 21 dias, os frangos que receberam a dieta contendo 0,05% de betaína e a dieta CP apresentaram as maiores alturas de vilosidades (p<0,05) em relação aos frangos suplementados com 0,10% de betaína na ração. Aos 28 dias de idade, observou-se que mesmo não havendo diferença (p>0,05) entre os tratamentos para altura dos vilos, foi possível averiguar que na medida em que a betaína foi acrescida a dieta, houve aumento dos mesmos, e que o maior valor para altura de vilosidades foi encontrado nos frangos suplementados com o maior nível de betaína (0,15%). A profundidade das criptas e a relação vilo:cripta não foram afetadas (p>0,05) pelos diferentes tratamentos aos 14, 18, 21 e 28 dias de idade dos frangos.
Palavras chave: Coccidiose. Mucosa intestinal. Suplemento dietético.

- 34 -
ABSTRACT
NIANG, Tania Marcia Soares. Influence of betaine on the villus and crypts development of small intestinal of broiler chickens infected experimentally with Eimeria acervulina (TIZZER, 1929).
The effect of the betaine supplementation on the intestinal mucosal structure of males broiler chickens submitted to the experimental infection with Eimeria acervulina was evaluated. A total of 420 day-old males Cobb broiler chicks were distributed in battery cage of three floors in a randomized block experimental design with 5 treatments and 6 experimental units per treatment, where each floor represented a block. Diets were supplied from 1 to 21 and 22 to 42 days. The treatments were as follow: 1) Positive control (PC) = basal diet plus a production growth promoter and a coccidiostatic; 2) 0.0% of betaine = basal diet without betaine (0.0%); 3) 0.05% betaine = basal diet plus 0.05% betaine; 4) 0.10% betaine = basal diet plus 0.10% betaine; 5) 0.15% betaine = basal diet plus 0.15% de betaine. At fourteen day of age, the birds were individually inoculated with approximately 2x105 sporulated oocysts of E. acervulina. One bird from each experimental unit, total 30 birds per sampling, was sacrificed by displacement of the cervical column at the 14, 18, 21 and 28 day of life so that samples of tissue of the duodenal loop were collected for histologyc exams. It was observed that there was not difference (p>0.05) among the treatments for villus height at the 14 days of age, however, it was observed that as the betaine levels increased, the villus heights tended to increase. At 18 days of age, there was not difference (p>0.05) among the treatments for villus height, however, it is observed that the largest value for villus height was found in the birds supplemented with 0.05% of betaine in the ration. At 21 days, the chickens that received the diet with 0.05% of betaine and the PC diet had the largest villus height (p<0.05) in relation to the chickens supplemented with 0.10% of betaine in the ration. At 28 days of age, it was observed that there was no difference (p>0.05) among the treatments for villus height, but it was possible to observe that as the levels of the betaine was added to diet increased, the villus height was increased, and that the largest value for villus height was found in the chickens supplemented with the largest betaine level (0.15%). The crypts depth and the villus:crypt ratio were not affected (p>0.05) by the different treatments at the 14, 18, 21 and 28 days of age of the chickens.
Key words: Coccidiosis. Dietary supplement. Intestinal mucosal.

- 35 -
1 INTRODUÇÃO
O estudo da mucosa intestinal reflete um importante aspecto da fisiologia da digestão, pois ela apresenta uma extensa área de exposição a agentes exógenos que estão presentes nesta região a partir do início da ingestão, digestão e absorção dos nutrientes. Seu desenvolvimento é decorrente de dois eventos citológicos primários associados: renovação celular (proliferação e diferenciação), resultante das divisões mitóticas sofridas por células totepotentes localizadas na cripta e ao longo dos vilos (UNI et al., 1998; UNI et al., 2000) e a perda de células por descamação, que ocorre naturalmente no ápice dos vilos. O equilíbrio entre esses dois processos é determinado por uma taxa de renovação constante, e, portanto, também a capacidade digestiva e de absorção intestinal.
De acordo com MACARI (1999) o número de vilosidades e seu tamanho, bem como o de microvilos, em cada segmento do intestino delgado, confere a eles características próprias, sendo que na presença de nutrientes a capacidade absortiva do segmento será diretamente proporcional ao número de vilosidades ali presentes, tamanho dos vilos e área de superfície disponível para absorção.
Normalmente, o epitélio intestinal age como uma barreira natural contra bactérias patogênicas e substâncias tóxicas presentes na luz intestinal. Distúrbios na microflora normal ou nas células epiteliais intestinais, causadas por algum tipo de estresse, patógenos, substâncias químicas e radiação, podem alterar a permeabilidade desta barreira natural, facilitando a invasão de patógenos e outras substâncias nocivas, modificando o metabolismo, a capacidade de digestão e absorção de nutrientes e podendo ainda causar inflamações crônicas na mucosa intestinal (HOFSTAD, 1972; PODOLSKY, 1993; OLIVEIRA, 1998). Conseqüentemente ocorre diminuição das vilosidades, aumento do turnover celular e diminuição da atividade digestiva e absortiva (VISEK, 1978).
Microparasitas intracelulares do gênero Eimeria, causam uma doença conhecida como coccidiose, uma infecção intestinal de grande importância e que está associada à mudanças metabólicas e estruturais que ocorrem na mucosa intestinal do animal hospedeiro (RUFF & ALLEN, 1990). A doença causa diarréia, que pode levar à desidratação e a mudanças osmóticas (CHADWICK et al., 1985). Suplementos alimentares como a betaína têm sido estudados por contribuir diretamente para um melhor desempenho de frangos infectados com coccídios e por inibir parcialmente a invasão e o desenvolvimento destes, e indiretamente, por apoiar a estrutura e a função intestinal na presença de uma infecção coccídiana, apesar de seu modo de ação não estar completamente entendido (AUGUSTINE et al., 1997).
A betaína ou trimetilglicina está presente em todos os organismos vivos, embora em quantidades altamente variáveis. É acumulada para osmoproteção contra grandes gradientes de concentração em células renais de mamíferos (BAGNASCO et al., 1986; KEMPSON, 1998), brânquias de mexilhão marinho (WRIGHT et al., 1992), e muitas espécie de bactérias (CSONKA, 1989). Ela também tem um papel na osmoproteção intestinal das aves (KIDD et al., 1997; AUGUSTINE & DANFORTH, 1999).
Um grande número de células animais, vegetais e procarióticas adaptam-se ao estresse osmótico externo através do acúmulo de osmólitos orgânicos de baixo peso

- 36 -
molecular, tais como aminas metiladas (betaína), certos aminoácidos (lisina, taurina, prolina, glutamina e arginina) e álcoois de açúcares (sorbitol e mio-inositol) ou inorgânicos, sendo os mais comuns o potássio, o magnésio e o fósforo/fosfato (WUNZ & WRIGHT, 1993).
Como osmólito, a betaína possui a habilidade de estabilizar a função metabólica celular sob diferentes condições de estresse osmótico (YANCEY et al. 1982; DRAGOLOVICH, 1994), a um baixo custo energético. As propriedades osmoprotetoras que cabem à betaína relacionam-se à sua característica bipolar e à sua alta solubilidade em água (CHAMBERS & KUNIN, 1985).
O propósito deste trabalho foi determinar a influência da suplementação de betaína sobre a estrutura da mucosa intestinal de frangos de corte infectados experimentalmente por E. acervulina.

- 37 -
2 REVISÃO DA LITERATURA
2.1 Mudanças Fisiológicas na Mucosa Intestinal das Aves
A resposta ao parasita do gênero Eimeria na mucosa intestinal pode ser mediada diretamente pelo parasita ou pela resposta imunoinflamatória do hospedeiro, que poderá resultar em mudanças na morfologia da mucosa intestinal (BARKER, 1993). As mudanças fisiológicas relacionadas à infecção foram bem documentadas em experimentos com frangos infectados com E. acervulina. Os resultados têm indicado que ao nível de mucosa intestinal, respostas locais e sistêmicas aos coccídios são mediadas de modos diferentes.
FERNANDO & McCRAW (1973) medindo a resposta intestinal a uma única dose de E. acervulina, observaram que as aves infectadas mostraram um acentuado aumento na espessura da mucosa, redução na altura dos vilos e aumento na profundidade das criptas no duodeno e em menor extensão, no jejuno anterior. As alterações da mucosa foram mais severas no período crítico da infecção, e foi observado ainda, um acentuado aumento na taxa de substituição das células epiteliais intestinais, além de um aumento no turnover celular do epitélio duodenal.
ALLEN (1983) observou um aumento no metabolismo das células da mucosa no intestino delgado, onde estas poderiam aumentar o crescimento compensatório e ajudar a ave a superar os efeitos negativos da infecção em relação aos parâmetros de produção. Outras mudanças morfológicas foram evidentes, tais como, aumento do comprimento do intestino e aumento da umidade do tecido (edema) no intestino, que não foram associados com a fome, mas ao único efeito de desafio. Se as mudanças no ambiente intestinal forem causadas devido ao desafio, pode ocorrer ainda redução do pH na área infectada (RUFF & WILKINS, 1984) e aumento no tempo de passagem do alimento (STEPHENS et al., 1974; McKENZIE et al., 1987). Essas mudanças na homeostase intestinal podem alterar o consumo de alimentos, que leva a diminuição da digestão e absorção de nutrientes, mudanças no metabolismo e diminuições globais no ganho de peso (ADAMS et al., 1996).
De maneira interessante, os pesquisadores têm contrastado resultados em experimentos onde o ganho de peso está sendo mensurado. STEPHENS et al. (1974) e ADAMS et al. (1996) relataram redução no ganho de peso associados com desafio de E. acervulina, enquanto OGBUOKIRI & EDGAR (1985) e McKENZIE et al. (1987) não encontraram diferença significativa no ganho de peso quando comparado aos controles não desafiados. Esta diferença nos resultados poderia estar associada a vários fatores, tais como a genética das aves, rota de administração do desafio e quantidade de oocistos infectivos administrados para o desafio de E. acervulina (DANFORTH & AUGUSTINE, 1989).
Estudos adicionais têm demonstrado que o contato inicial dos esporozoítos com a mucosa intestinal produz uma resposta inflamatória incluindo marcante infiltração celular no local da infecção (ROSE et al., 1979). Esta infiltração consiste em múltiplas subpopulações de leucócitos, macrófagos, morte celular natural, granulócitos e linfócitos que poderiam modular e aumentar a resposta imune; portanto, alterando a absorção de nutrientes e diminuindo o ganho de peso (JEURISSEN et al., 1996).
Os mecanismos específicos pelo qual cada espécie de coccídio causa a doença não estão claros, contudo, o estabelecimento de infecções secundárias devido à alteração na mucosa intestinal pode ser responsável por uma resposta diferencial do hospedeiro (BARKER, 1993). Isto é possível porque os coccídios são capazes de interagir com outros

- 38 -
patógenos, tais como bactérias e vírus que podem ampliar os efeitos observados (RUFF, 1993). Como descrito acima, a habilidade das aves para controlar a severidade da infecção e desenvolver imunidade protetora aos parasitas coccídios depende de inúmeros fatores. A resposta do hospedeiro ao parasita envolve uma série complexa de fatores internos que são dependentes do estágio de desenvolvimento do parasita, estado imune das aves e espécies do parasita do gênero Eimeria.
2.2 Nutrição e Coccidiose
A influência de fatores dietéticos sobre a coccidiose avícola tem sido estudada por diversos autores (BEACH & CORL, 1925; ALLEN, 1932; BECKER, 1937; MAN, 1947; ERASMUS et al., 1960; BRITTON et al., 1964; KOLVEIT, 1969; MURILLO et al., 1976; WILLIS & BAKER, 1981; COLNAGO et al., 1984a,b,c; AUGUSTINE et al., 1997; KLASING et al., 2002).
A influência da dieta no hospedeiro e no desenvolvimento da infecção por coccídio foi estudada por BEACH & CORL (1925) e TYZZER (1929). Estes autores reportaram que a severidade da infecção com Eimeria tenella foi reduzida quando se adicionou leite em pó desnatado a dieta das aves. Antes do uso de drogas anticoccídianas foram recomendadas formulações dietéticas que melhorassem a severidade da infecção por coccídio (BEACH & CORL, 1925; BECKER, 1937).
Vários nutrientes são conhecidos por mudar a severidade das infecções coccídianas. Estes incluem a vitamina A (ERASMUS et al., 1960; PANDA et al., 1964), vitamina K (BRITTON et al., 1964), cálcio (HOLMES et al., 1937), selênio (JENSEN et al., 1978) e betaína (VIRTANEN, 1995).
A betaína é extraída do açúcar de beterraba e é subseqüentemente purificada. Suas funções no corpo como osmólito ajudam a defender contra desidratações. Osmólito é uma substância que afeta o movimento da água, acumulando-a rapidamente em nível intracelular, sem alterar o metabolismo celular/mitocôndria. Em trabalhos de pesquisa a betaína tem sido relatada por reduzir escores de lesão (VIRTANEN et al., 1996; REMUS et al., 1996) e por causar resistência à tensão intestinal (REMUS & QUARLES, 2000) em frangos com um histórico de desafio coccídio.
A suplementação dietética de betaína tem mostrado efeitos positivos no balanço de água no organismo de frangos de corte estressados por alta temperatura ambiental ou por coccidiose (AUGUSTINE & DANFORTH, 1999; TEETER et al., 1999). Em trabalho conduzido por AUGUSTINE et al. (1997) verificaram que in vitro, exceto em altas concentrações, a betaína não foi tóxica para os esporozoítos de E. acervulina, e teve pouco efeito sobre a invasão e o desenvolvimento destes na cultura de células. No entanto, in vivo, a invasão por esporozoítos de E. acervulina foi significativamente reduzida em todas as aves alimentadas com dietas contendo betaína ou salinomicina quando comparada com aves que receberam o tratamento controle.
A relação cripta/vilos foi diminuída através da suplementação dietética de betaína em frangos saudáveis e infectados, sugerindo que a betaína dietética protege a vilosidade jejunal contra infecção coccídiana e também estabiliza a estrutura da mucosa em frangos de corte saudáveis (KETTUNEN et al., 2001).
KLASING et al. (2002) examinando a relação entre betaína, osmolaridade e coccidiose em aves que receberam uma dieta a base de milho e soja contendo 0,0; 0,05 ou 0,10% de betaína, observaram que a betaína controlou a osmolaridade no duodeno,

- 39 -
especialmente em aves desafiadas com coccídios E. acervulina. A altura dos vilos foi diminuída pela coccidiose e, foi então, melhorada com a adição de 0,10% de betaína na dieta. A coccidiose aumentou o número de leucócitos na lâmina própria duodenal, especialmente na presença de altos níveis de betaína (0,05% e 0,10%).

- 40 -
3 MATERIAL E MÉTODOS 3.1 Local
O experimento foi conduzido no Departamento de Nutrição Animal e Pastagens do Instituto de Zootecnia da Universidade Federal Rural do Rio de Janeiro, no município de Seropédica, RJ, Brasil, no período de 29/06 a 26/07 de 2004. As análises laboratoriais foram realizadas no laboratório de coccídios e coccidiose do Departamento de Parasitologia Animal do Instituto de Veterinária da UFRRJ.
3.2 Metodologia Utilizada Anteriormente Ao Preparo Histológico
A metodologia aplicada para animais e manejo, dietas experimentais e material infectante é a mesma descrita no capitulo i, por isso, a mesma não foi repetida neste capítulo.
3.3 Preparo Histológico Para realizar o preparo histológico retirou-se uma ave com o peso médio do lote de
cada unidade experimental, sendo seis aves por tratamento e trinta aves no total. Posteriormente, as mesmas foram sacrificadas por deslocamento da coluna cervical no 14º, 18°, 21º e 28° dia de vida. As aves necropsiadas aos 14 dias não foram infectadas por E. acervulina, portanto, serviram como controle em relação àquelas que foram infectadas e retiradas nos períodos subseqüentes. As demais necropsias foram realizadas para estudar o período pós-infecção (4, 7 e 14 dpi). O intestino delgado das aves foi cortado e medido com o auxílio de um paquímetro em milímetros. Cortes transversais de aproximadamente 30mm da alça duodenal foram coletados e armazenados em vidros com capacidade de 50mL contendo solução de paraformaldeido a 10% para preparo posterior.
Os tecidos obtidos foram processados por técnica de inclusão em parafina ou Histosec®(Merck), cortados em 5µm e corados pela hematoxilina e eosina (HE) ou PAS, conforme BEHMER et al. (1976).
3.4 Avaliação da Relação Vilo/Cripta
A avaliação das lâminas histológicas do duodeno constituiu-se na análise de três medidas: altura das vilosidades, profundidade das criptas e razão entre esses dois valores. Tal procedimento teve a finalidade de complementar a avaliação dos possíveis danos causados à estrutura da mucosa intestinal pelo parasitismo mediante aos tratamentos aplicados. Para tal, utilizou-se um microscópio binocular Carl Zeiss em objetiva de 4x acoplada de ocular micrométrica K-15x-PZO (Polônia) para mensuração. Foram efetuadas medidas de vilosidades e criptas (Figura 1) dentre as repetições de cada tratamento (n=50), totalizando-se 1500 mensurações. Os valores foram expressos em micrômetros (µm) tanto para altura das vilosidades como para profundidade das criptas.

- 41 -
Figura 1. Corte histológico com demonstração das medidas tomadas para alturas das vilosidades (A) e profundidades das criptas (C) do intestino delgado de frangos de corte.
3.5 Análise Estatística
O delineamento experimental usado foi o de blocos ao acaso, onde se utilizaram baterias metálicas compostas de três andares, cujo bloco foi representado por cada andar, totalizando três blocos por bateria. O modelo estatístico adotado foi escolhido pensando-se nas condições de conforto térmico que poderiam ser diferentes em cada um dos andares, uma vez que o animal experimental é muito sensível ao fator temperatura. Foram usadas dez baterias metálicas compostas de três blocos (superior, médio e inferior), onde cada bateria foi ocupada por aves que estavam recebendo o mesmo tratamento, cujo objetivo foi evitar possíveis contaminações dos tratamentos entre os blocos. Em cada bloco, um único tratamento foi repetido por duas vezes, totalizando seis repetições por tratamento (duas no bloco superior, duas no bloco médio e duas no bloco inferior), então, o mesmo tratamento foi distribuído em duas baterias metálicas diferentes. Como havia cinco tratamentos, foi necessário utilizar dois pares de cinco baterias, que somado as seis repetições de cada tratamento, deu um total de 30 observações.
As análises de variância foram realizadas utilizando-se o pacote computacional Sisvar, descrito por FERREIRA (2000). As variáveis de cada tratamento tiveram suas médias comparadas pelo teste de Student-Newman-Keuls (SNK) a um nível de significância de 5% (H0:p<0,05), assim como as análises de regressão foram calculadas por meio da técnica de polinômios ortogonais.
O modelo estatístico adotado para análise de altura das vilosidades, profundidade das criptas e a relação vilo/cripta para o teste de comparação de médias foi:
ebty ijjiijm +++=
em que:

- 42 -
yij é o valor observado da característica estudada, no tratamento i ( i = 1,2,3,4,5,6), no
bloco j (j = 1,2,3) m é a média geral de todas as observações do experimento ti
é o efeito do tratamento i
bj é o efeito do bloco j
eij é o erro associado à observação yij, ou efeito dos fatores não controlados sobre a
observação yij.
O modelo estatístico adotado para análise de regressão para altura das vilosidades, profundidade das criptas e a relação vilo/cripta foi:
ipipiii e+Χ++Χ+Χ+=Υ ββββ ...22110 em que:
iΥ é a variável dependente, obtida na observação i
pββββ ,...,,, 210 são os parâmetros da regressão
piii ΧΧΧ ,...,, 21 são variáveis independentes, na observação i
ie é o erro associado a observação iΥ .

- 43 -
4 RESULTADOS E DISCUSSÃO
4.1 Altura das Vilosidades Intestinais
As lesões na mucosa intestinal do duodeno das aves foram claramente aparentes em todos os tratamentos analisados após a infecção com Eimeria acervulina, aos quatro (A), sete (B) e quatorze (C) dias após a inoculação (dpi), onde é possível observar placas coalescentes na parede do intestino com espessura aumentada (Figura 2A e B), porém estas placas foram menos expressivas aos 14 dpi (Figura 2C).
Figura 2. Lesões na mucosa intestinal do duodeno das aves infectadas experimentalmente por E. acervulina aos quatro (A), sete (B) e quatorze (C) dias após a inoculação.
De acordo com o que foi descrito por ALLEN & FETTERER (2002) o período pré-
patente das espécies de Eimeria varia de quatro a cinco dias após a infecção. E a maior produção de oocistos varia de seis a nove dpi. No presente trabalho, foi observado que a infecção coccídiana causada por E. acervulina foi mais pronunciada aos 21 dias de idade das aves, ou seja, aos sete dpi, com redução tanto da altura das vilosidades quanto da relação vilo:cripta, e aumento da profundidade das criptas, (Tabelas 2, 3 e 4).
Analisando-se os resultados apresentados na Tabela 2, pode-se observar que não houve diferença (p>0,05) entre os tratamentos para altura das vilosidades intestinais dos frangos aos 14 dias de idade. Entretanto, observa-se que na medida em que se elevam os níveis de betaína na dieta, a altura dos vilos tende a aumentar. É importante salientar que aos 14 dias de idade as aves não estavam infectadas com Eimeria, portanto, isto explica o fato de não ter havido diferença significativa entre os tratamentos.
A B C

- 44 -
Tabela 1. Médias de altura dos vilos (µm) do duodeno de frangos de corte necropsiados aos 14, 18, 21 e 28 dias de idade.
Altura dos Vilos (µm) Idade em dias
Tratamentos
14 18 21 28
Controle positivo 1212,13a 1349,80a 1193,68a 1334,47a 0,00% de betaína 1175,33a 1257,55a 1121,62ab 1560,44a 0,05% de betaína 1263,68a 1423,57a 1245,85a 1635,48a 0,10% de betaína 1268,12a 1384,57a 962,98b 1412,64a 0,15% de betaína 1325,63a 1391,11a 1100,43ab 1641,24a CV (%) 13,59 12,96 12,19 13,02 a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0.05) pelo teste de Student Newman Keuls (SNK).
Os resultados apresentados na tabela 2 mostram que aos 18 dias de idade, quatro dias após a infecção com E. acervulina (dpi) nos frangos, não houve diferença (p>0,05) entre os tratamentos para altura dos vilos, porém, observa-se que o maior valor para altura das vilosidades foi encontrado nas aves que foram suplementadas com o nível de 0,05% de betaína na ração. Aos 21 dias de idade, ou seja, sete dpi, observa-se uma drástica redução na altura dos vilos, e também os frangos que receberam a dieta contendo 0,05% de betaína e a dieta com promotor de produção mais coccidiostático (CP) apresentaram as maiores alturas de vilosidades (p<0,05) em relação aos frangos que foram suplementadas com 0,10% de betaína na ração nesta idade. Estes resultados diferem em alguns aspectos dos resultados encontrados por KLASING et al. (2002), que ao inocular oralmente as aves com uma dose de 4,6x104 oocistos esporulados de E. acervulina aos 14 dias de idade, relataram que E. acervulina diminuiu a altura do vilos dos quatro até os sete dias após a infecção das aves, e que esta redução foi mais pronunciada nos animais que consumiram a dieta não suplementada com betaína, enquanto o encurtamento das vilosidades foi menos severo nas aves que receberam 0,10% de betaína na ração, aos sete dias após inoculação.
No período em que já é esperada a recuperação dos frangos, aos 28 dias de idade (Tabela 2), observa-se que mesmo não havendo diferença significativa (p>0,05) entre os tratamentos para altura de vilos, é possível observar que na medida em que a betaína é acrescida a dieta, a altura dos mesmos é aumentada, e que o maior valor para altura de vilosidades é encontrado nos frangos que foram suplementados com o maior nível de betaína (0,15%), por isso, cabe ressaltar que mais pesquisas são necessárias para dizerem se níveis maiores que 0,15% de betaína proporcionariam diferença significativa entre os tratamentos para este período. É importante também frisar que aos 28 dias, a altura dos vilos foi substancialmente aumentada em relação ao período critico da coccidiose, aos 21 dias de idade.
Os resultados obtidos por meio da análise de regressão para altura das vilosidades intestinais dos frangos de corte, levam em consideração o efeito dos tratamentos no decorrer das idades, elucidando de forma concisa o comportamento da doença para cada característica estudada.

- 45 -
As aves que receberam a dieta contendo promotor de produção e coccidiostático não apresentaram diferença (p>0,05) para alturas das vilosidades no período de 14 a 28 dias de idade (Figura 3). Já as aves que não foram suplementadas com betaína na dieta (0,0%) apresentaram como resposta para altura dos vilos comportamento expresso por uma função quadrática (y = 89.155x2 - 343.84x + 1469.7), onde se pode observar uma leve redução na altura dos vilos dos 14 aos 18 dias e um crescente aumento após os 19 dias, pressupondo-se que os animais deste tratamento apresentaram pouca variação na altura dos vilos no decorrer deste período, o que torna difícil uma explicação para tal efeito.
Os frangos que foram suplementados com os níveis de 0,05; 0,10 e 0,15% apresentaram como resposta para altura dos vilos comportamento expresso por uma função cúbica (y = 150.82x3 - 1073.7x2 + 2325.3x - 138.69; y = 234.87x3 - 1678.2x2 + 3506.9x - 795.52 e y = 197.93x3 - 1365.6x2 + 2776.9x - 283.52), na qual observa-se aumento na altura dos vilos dos 14 até os 16 dias, com pouca variação entre os tratamentos, e subseqüentemente, uma acentuada redução dos mesmos dos 17 até os 24 dias, onde aumenta a variação entre os tratamentos, que pode ser explicado pelo fato dos frangos terem passando pela fase critica da coccidiose, porém, após os 24 dias observa-se um aumento na altura dos vilos, tendo evidente melhora aos 28 dias, quando as aves já estão no período de recuperação da doença. Neste caso, o nível 0,05% de betaína foi o que proporcionou menor oscilação na altura dos vilos em relação aos maiores níveis (Figura 3).
Em suma, os frangos que foram suplementados com promotor de produção mais coccidiostático (CP) e com a dieta sem betaína (0,00%) apresentaram menos oscilação na altura das vilosidades do que aqueles que receberam os demais tratamentos.
Figura 3. Influência dos aditivos dietéticos sobre a altura das vilosidades intestinais dos frangos de corte submetidos a infecção experimental com E. acervulina, representadas pelas seguintes equações de regressão:
600.0
800.0
1000.0
1200.0
1400.0
1600.0
1800.0
14 16 18 20 22 24 26 28 30Idade (dias)
Altu
ra d
os v
ilos (
µm)
0.00% Bet
0.05% Bet
0.10% Bet
0.15% Bet

- 46 -
0,00% BetQ (y = 3.5832x2 - 125.98x + 2266.5, R2 = 0.85);
0,05% BetC (y = 1.833x3 - 111.32x2 + 2187.1x – 12567, R2 = 1);
0,10% BetC (y = 3.1934x3 - 193.48x2 + 3755.3x – 22146, R2 = 1 ) e
0,15% BetC (y = 2.3995x3 - 143.35x2 + 2751.3x – 15679, R2 = 1). Onde, Q = efeito quadrático e C = efeito
cúbico.
4.2 Profundidade das Criptas Intestinais
A profundidade das criptas não foi influenciada (p>0,05) pelos diferentes tratamentos aos 14, 18, 21 e 28 dias de idade dos frangos (Tabela 3). KLASING et al. (2002) também não encontraram diferença significativa para profundidade das criptas no duodeno das aves que receberam os tratamentos contendo 0,0; 0,05 e 0,10% de betaína na dieta aos 2, 4 e 7 dpi. Ainda de acordo com os resultados apresentados na Tabela 3 é possível averiguar que aos 14 e aos 18 dias de idade foram encontrados os menores valores para profundidades de criptas, período em que as aves não estavam infectadas ou que a coccidiose ainda não havia produzido efeitos, entretanto, aos 21 dias, período crítico da doença, esses valores aumentaram consideravelmente, voltando a se normalizar aos 28 dias quando as mesmas já passavam pela fase de recuperação da doença.
Tabela 2. Médias de profundidade das criptas (µm) do duodeno de frangos de corte necropsiados aos 14, 18, 21 e 28 dias de idade.
Profundidade das Criptas (µm) Idade em dias
Tratamentos
14 18 21 28
Controle positivo 153,97a 139,97a 282,10a 154,65a 0,00% de betaína 134,50a 144,08a 306,84a 206,45a 0,05% de betaína 137,53a 147,85a 310,54a 191,75a 0,10% de betaína 176,52a 168,42a 290,64a 170,87a 0,15% de betaína 151,95a 162,27a 295,65a 194,70a CV (%) 33,57 20,70 15,63 20,28 a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0.05) pelo teste de Student Newman Keuls (SNK).
Conforme os dados da análise de regressão apresentados na Figura 4, todos os tratamentos apresentaram como resposta para altura dos vilos comportamento expresso por uma função cúbica (p<0,05), cujas equações foram as seguintes: y = -0.9877x3 + 59.614x2 - 1148.7x + 7261.4 para os frangos que receberam as rações suplementadas com promotor de produção mais coccidiostático, y = -1.0189x3 + 61.408x2 - 1176.1x + 7359.3 para os que receberam as rações sem a suplementação de betaína, y = -1.0354x3 + 62.253x2 - 1190.2x

- 47 -
+ 7439.7 para aqueles suplementados com 0,05% de betaína, y = -0.8497x3 + 51.143x2 - 982.65x + 6241.1 para aqueles suplementados com 0,10% de betaína e y = -0.848x3 + 50.927x2 - 972.43x + 6111.2 para os que receberam 0,15% de betaína no período de 14 a 28 dias de idade, onde se é observado os menores valores para profundidade das criptas no período de 14 a 16 dias, e um eminente aumento das mesmas após os 16 dias, com as maiores profundidades ocorrendo aos 24 dias, voltando a decrescer após este período. Neste caso, nenhum dos tratamentos analisados no presente estudo interferiram de forma efetiva nas oscilações que ocorreram nas profundidades das criptas dos frangos infectados no período estudado. Sendo assim, observa-se que até mesmo as medicações profiláticas não tiveram efeitos positivos sobre os coccídios, impedindo que estes alterassem a profundidade das criptas durante o período da infecção. Dessa forma, há necessidade em buscar meios que possam auxiliar o controle dos parasitas e interferência destes nas profundidades das criptas duodenais.
Figura 4. Influência dos aditivos dietéticos sobre a profundidade das criptas intestinais dos frangos de corte submetidos a infecção com coccídios, representadas pelas seguintes equações de regressões: CPC (y = -0.9877x3 + 59.614x2 - 1148.7x + 7261.4, R2 = 1);
0,00% BetC (y = -1.0189x3 + 61.408x2 - 1176.1x + 7359.3, R2 = 1);
0,05% BetC (y = -1.0354x3 + 62.253x2 - 1190.2x + 7439.7, R2 = 1);
0,10% BetC (y = -0.8497x3 + 51.143x2 - 982.65x + 6241.1, R2 = 1) e
0,15% BetC (y = -0.848x3 + 50.927x2 - 972.43x + 6111.2, R2 = 1). Onde, C = efeito cúbico.
80.0
130.0
180.0
230.0
280.0
330.0
380.0
430.0
480.0
14 16 18 20 22 24 26 28Idade (dias)
Prof
undi
dade
das
cri
ptas
(µm
)
CP
0.00% Bet
0.05% Bet
0.10% Bet
0.15% Bet

- 48 -
4.3. Relação Vilo:Cripta Intestinais
Tabela 3. Médias da relação vilo:cripta (µm) do duodeno de frangos de corte necropsiados aos 14, 18, 21 e 28 dias de idade.
Relação Vilo/Cripta (µm) Idade em dias
Tratamentos
14 18 21 28
Controle positivo 8,45a 10,62a 4,30a 9,33a 0,00% de betaína 9,65a 9,37a 3,81a 7,92a 0,05% de betaína 9,44a 9,95a 4,18a 9,05a 0,10% de betaína 8,60a 8,78a 3,41a 8,48a 0,15% de betaína 9,60a 8,88a 4,11a 9,20a CV (%) 26,85 21,94 20,24 25,18 a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0.05) pelo teste de Student Newman Keuls (SNK).
Segundo os resultados obtidos com a análise de regressão (Figura 5), a resposta para relação vilo/cripta teve comportamento expresso por uma função cúbica (p<0,05) em todos os tratamentos, no período de 14 a 28 dias de idade. A relação vilo/cripta dos frangos que receberam a dieta contendo promotor de produção mais coccidiostático (y = 0.0472x3
- 2.8785x2 + 56.247x - 344.25); a dieta sem a suplementação de betaína (y = 0.0355x3 - 2.137x2 + 40.88x - 241.32) e as dietas contendo os níveis de 0,05% (y = 0.0398x3 - 2.4016x2 + 46.286x - 276.95); 0,10% (y = 0.0367x3 - 2.2087x2 + 42.382x - 252.59) e 0,15% (y = 0.0311x3 - 1.8511x2 + 35.059x - 203.75) de betaína foram totalmente influenciadas no período critico da coccidiose, o que era de se esperar devido ao desenvolvimento de criptas mais profundas, que em associação com o encurtamento dos vilos, resultou numa significativa redução desta relação.
Os maiores valores da relação vilo/cripta foram observados dos 14 aos 16 dias, sendo abruptamente reduzidos após os 16 dias, e persistindo em queda até os 24 dias, acompanhando a fase crítica da doença, e voltando a aumentar consideravelmente após os 24 dias, mantendo-se normalizada aos 28 dias (Figura 5).

- 49 -
Figura 5. Influência dos aditivos dietéticos sobre a relação vilo:cripta do intestino de frangos de corte
submetidos a infecção com coccídio, representadas pelas seguintes equações de regressão:
CPC (y = 0.0472x3 - 2.8785x2 + 56.247x - 344.25, R2 = 1);
0,00% BetC (y = 0.0355x3 - 2.137x2 + 40.88x - 241.32, R2 = 1);
0,05% BetC (y = 0.0398x3 - 2.4016x2 + 46.286x - 276.95, R2 = 1);
0,10% BetC (y = 0.0367x3 - 2.2087x2 + 42.382x - 252.59, R2 = 1) e
0,15% BetC (y = 0.0311x3 - 1.8511x2 + 35.059x - 203.75, R2 = 1). Onde, C = efeito cúbico.
Consoante os dados obtidos no presente estudo, é possível sugerir que durante a infecção, o aumento na profundidade das criptas estava diretamente relacionado ao aumento da substituição ou ao turnover das células epiteliais, sugerindo ainda que os mecanismos de repostas intestinais do hospedeiro estavam tentando compensar os efeitos da infecção causados por E. acervulina. Estas mudanças foram evidentes na relação vilo/cripta, que está correlacionada a quantidade de vilos atrofiados e hiperplasia das criptas durante a infecção. Respostas similares tem sido reportadas em outros estudos com aves (FERANDO & MCCRAW, 1973). Desta forma, constatou-se que nenhum dos tratamentos testados neste estudo foi capaz de amenizar o impacto da E. acervulina nas células intestinais, contrariando os resultados obtidos por KETTUNEN et al., 2001.
-2.0
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
14 16 18 20 22 24 26 28
Idade (dias)
Rel
ação
vilo
/cri
pta
(µm
)
CP
0.00% Bet
0.05% Bet
0.10% Bet
0.15% Bet

- 50 -
5 CONCLUSÕES
Sugere-se que para amenizar a infecção intestinal causada por E. acervulina na altura das vilosidades do duodeno dos frangos de corte, pode-se utilizar como recurso nutricional o nível de 0,05% betaína, uma vez que neste estudo ele produziu os mesmos efeitos que o promotor de produção mais coccidiostático.
Todos os tratamentos testados neste estudo não interferiram de forma positiva na profundidade das criptas e na relação vilo/cripta do duodeno de frangos de corte infectados experimentalmente por E. acervulina nos períodos estudados.
A betaína pode ser de utilidade, em conjunto com outras estratégias nutricionais, para a viabilização da retirada dos antibióticos como promotores de produção das dietas de frangos de corte.

- 51 -
6 REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, C.; VAHL, H.A.; VELDMAN, A. Interaction between nutrition and Eimeria acervulina infection in broiler chickens: development of an experimental infection model. British Journal of Nutrition, v.75, p.867-873, 1996.
ALLEN, E.A. The influence of diet on the development of experimental coccidiosis of chickens kept under sanitary conditions. American Journal of Hyg., v.15, p.163-185, 1932.
ALLEN, P.C. Physiological responses of chicken gut tissue to infection with Eimeria acervulina. Avian Diseases, v.28, p.868-876, 1983.
ALLEN, P.C.; FETTERER, R.H. Recent advances in biology and immunobiology of Eimeria species and in diagnosis and control of infection with these coccidian parasites of poultry. Clinical Microbiology Reviews, v.15, n.1, p.58-65, 2002.
AUGUSTINE, P.C.; McNAUGHTON, J.L.; VIRTANEN, E.; ROSI, L. Effect of betaine on the growth performance of chicks inoculated with mixed cultures of avian Eimeria species and on invasion and development of Eimeria tenella and Eimeria acervulina in vitro and in vivo. Poultry Science, v.76, p.802-809, 1997.
AUGUSTINE P.C.; DANFORTH, H.D Influences of betaine and salinomycin on intestinal absorption of methionine and glucose and on the ultrastructure of the intestinal cells and parasite developmental stage in chicks infected with Eimeria acervulina. Avian Disease, v.43, p.89-97, 1999.
BAGNASCO, S.; BALABAN, R.; FALES, H.M.; YANG, Y.; BURG, M. Predominant osmotically active organic solutes in rat and rabbit renal medullas. Journal of Biological Chemistry, v.261, n.13, p.5872-5877, 1986.
BARKER, I. K. Pathological processes associated with coccidiosis. In: INTERNATIONAL COCCIDIOSIS CONFERENCE, 6, 1993, Guelph. Proceedings… Guelph - Canada, 1993, p. 81-94.
BEACH, J.R.; CORL, J.C. Studies in the control of avian coccidiosis. Poultry Science, v.4, p.83-93, 1925. BECKER, E.R. Dietary control in experimental coccidiosis. Science, v.86, p.403-404, 1937.

- 52 -
BEHMER, O.A.; TOLOSA, E.M.C.; FREITAS NETO, A.G. Manual de técnicas para histologia normal e patológica. São Paulo: EDART – São Paulo Livraria Editora, Editora da Universidade de São Paulo, 1976. 241p. BRITTON, W.M.; HILL, C.H.; BARBER, C.W. A mechanism of interaction between dietary protein levels and coccidiosis in chicks. Journal of Nutrition, v.82, p.306-310, 1964.
CHADWICK, A.; RAPSON, E.B.; CARLOS, G.M.; LEE, D.L. Circulating prolactin concentrations in chickens infected with Eimeria tenella. British Poultry Science, v.26, p.17-23, 1985.
CHAMBERS, S.T.; KUNIN, C.M. The osmoprotective proprieties of urine for bacteria: the protective effect of betaine and human urine against low pH and high concentrations of electrolytes, sugars, and urea. Journal of Infectious Diseases, v.152, p.1308-1316, 1985. COLNAGO, G.L.; JENSEN, L.S.; LONG, P.L. The effect of feeding a semipurified diet to chickens infected with coccidia. Poultry Science, v.63, p.636-646, 1984a. COLNAGO, G.L.; JENSEN, L.S.; LONG, P.L. Effect of selenium and vitamin E on the development of immunity to coccidiosis in chickens. Poultry Science, v.63, p.1136-1143, 1984b. COLNAGO, G.L.; JENSEN, L.S.; LONG, P.L. Effect of natural feedstuffs added to a semi-purified diet on Eimeria tenella. Poultry Science, v.63, p.2145-2152, 1984c. CSONKA, L.N. Physiological and genetic responses of bacteria to osmotic stress. Microbiology Rev., v.53, p.121-147, 1989. DANFORTH, H.D.; AUGUSTINE, P.C. Coccidia vaccines. Veterinary Protozoan and Hemoparasite Vaccines. 1989, chapter 8, p.165-175.
DRAGOLOVICH, J. Dealing with salt stress in animal cells: the role and regulation of glycine betaine concentrations. Journal of Experimental Zoology, v.168, p.139-144, 1994. ERASMUS, J.; SCOTT, M.L.; LEVINE, P.P. Relationship between coccidiosis and vitamin A nutrition in chicks. Poultry Science, v.39, p.565-572, 1960. FERNANDO, M.A.; McCRAW, B.M. Mucosal morphology and cellular renewal in the intestine of chickens following a single infection of Eimeria acervulina. The Journal of Parasitology, v.59, p.493-501, 1973. FERREIRA, D.F. SISVAR – Sistema de Análise Estatística. Lavras, UFLA, 2000. Comunicação pessoal.

- 53 -
HOFSTAD, M.S. Diseases of poultry. 6. ed. The Iowa State University Press: Editor Ames, 1972, 1176p.
HOLMES, E.E.; HERRICK, C.A.; OTT, G.L. Studies in coccidiosis in chickens: calcium carbonate additions and coccidian. Poultry Science, v.16, p.335-339, 1937.
JENSEN, L.S.; JOHNSON, J.; RUFF, M.D. Selenium status and response of broiler chicks to coccidial infection. Poultry Science, v.57, n.1147 (Abstr.), 1978.
JEURISSEN, S.H.M.; JANSE, E.M.; VERMEULEN, A.N.; VERVELDE, L. Eimeria tenella infections in chickens: aspects of host-parasite interaction. Veterinary Immunology and Immunopathology, v.54, p.231-238, 1996.
KEMPSON, S.A. Differential activation of system A and betaína/GABA transport in MDCK cell membranes by hypertonic stress. Biochimica et Biophysica Acta, v.1372, p.117-123, 1998.
KETTUNEN, H.; TIIHONEN, K.; PEURANEN, S.; SAARINEN, M.T.; REMUS, J.C. Dietary betaína accumulates in the liver and intestinal tissue and stabilizes the intestinal epithelial structure in healthy and coccidian-infected broiler chicks. Comparative Biochemistry and Physiology Part A, v.130, p.759-769, 2001.
KIDD, M.T.; FERKET, P.R.; GARLICH, J.D. Nutritional and osmoregulatory functions of betaine. World’s Poultry Science Journal, v.53, p.125-139, 1997.
KLASING, K.C.; ADLER, K.L.; REMUS, J.C.; CALVERT, C.C. Dietary betaína increases intraepithelial lymphocytes in the duodenum of coccidian-infected chicks and increases functional properties of phagocytes. Journal of Nutrition, v.132, p.2274-2282, 2002.
KOLVEIT, A.J. Effect on mortality of semi-purified diets versus natural feedstuffs fed to chickens infected with E. tenella. Avian Disease, v.13, p.288-296, 1969.
MACARI, M. Fisiologia do sistema digestivo das aves (I). Aves e ovos, 1999, p. 2-20.
MANN, T.B. Roughage and protein as dietary factors influencing coccidiosis in chickens, with notes on the limitations of sulphamezathine in the control of coccidiosis. Journal of Agriculture Science, v.37, p.145-151, 1947.
McKENZIE M.E.; COLNAGO, G.L.; LEE, S.R.; LONG, P.L. Gut stasis in chickens infected with Eimeria. Poultry Science, v.66, p.264-269, 1987.

- 54 -
MURILLO, M.G.; JENSEN, L.S.; RUFF, M.D.; RAHN, A.P. Effect of dietary methionine status on response of chicks to coccidial infection. Poultry Science, v.55, p.642-649, 1976.
OGBUOKIRI U.D.E.; EDGAR, S.A. Effect of mild infections with six species of Eimeria on skin pigmentation of broiler chickens. Poultry Science, v.65, p.1816-1818, 1985.
OLIVEIRA, P.B. Influência de fatores antinutricionais de alguns alimentos sobre o epitélio intestinal e o desempenho de frangos de corte. Maringá, SP, 1998. 42p. Dissertação (Mestrado em Zootecnia) – Universidade Estadual de Maringá.
PANDA, B.; COMBS, G.F.; DEVOLT, H.M. Studies on coccidiosis and vitamin A nutrition of broilers. Poultry Science, v.43, p.154-164, 1964.
PODOLSKY, D.K. Regulation of intestinal epithelial proliferation: a few answers, many questions. Animal Journal Physiologic, v.264, p.G179-G186, 1993. REMUS, J.; VIRTANEN E.; ROSI, L. McNAUGHTON, J. In: THE WORLD’S POULTRY CONGRESS, 1996. Proceedings… 1996, v.4, n.336, p.2-5.
REMUS, J.C.; QUARLES, C.L. Poultry Science, v.79 (suppl. 1), n.118, 2000.
ROSE, M.E.; HESKETH, P.; OGILVIE, B.M. Peripheral blood leukocyte response to coccidial infection: a comparison of the response in rats and chickens and its correlation with resistance to reinfection. Immunology, v.36, p.71-79, 1979.
RUFF M.D.; WILKINS, G.C. Intestinal absorption following challenge of immune chicks with Eimeria acervulina. Avian Pathology, v.13, p.25-35, 1984.
RUFF, M.D.; ALLEN, P.C. Pathophysiology of coccidial infections. In: LONG, P.L. Coccidiosis of man and domestic animals. CRC Press: Editor Boca Raton, 1990, p.263-280.
RUFF M.D. Factors affecting coccidiosis. In: INTERNATIONAL COCCIDIOSIS CONFERENCE, 6, 1993, Guelph. Proceedings… Guelph - Canadá, 1993, p.73-79.
STEPHENS, J.F.; BORST, W.J.; BARNETT, B.D. Some physiological effects of Eimeria acervulina, E. brunetti, and E. mivati infections in young chickens. Poultry Science, v.53, p.1735-1742, 1974.

- 55 -
TEETER, R.G.; REMUS, J.C.; BELAY, T.; MOONEY, M. VITANEN, E.; AUGUSTINE, P. The effects of betaine on water balance and performance in broilers reared under differing environmental conditions. In: AUSTRALIAN POULTRY SCIENCE SYMPOSIUM, 1999, Australia. Proceedings… Australia, 1999, p. 165.
TYZZER, E.E. Coccidiosis in gallinaceous birds. American of Journal Hyg., v.10, p.269-383, 1929.
UNI, Z.; GANOT, S.; SJLAN, D. Posthach development of mucosal function in the broiler small intestine. Poultry Science, v.77, p.75-82, 1998.
UNI, Z. Vitamin A deficiency interferes with proliferation and maturation of cells in the chickens small intestine. British Poultry Science, v.41, p.410-415, 2000.
VIRTANEN, E. Piecing together the betaine puzzle. Feed Mix, v.3, n.1, p.12-17, 1995.
VIRTANEN, E.; REMUS, J.; ROSI, L.; McNAUGHTON, J.; AUGUSTINE, P. The effect of betaine and salinomycin during coccidiosis in broilers. Poultry Science, v.75 (suppl. 1), n.149, 1996. VISEK, W.J. The mode of growth promotion by antibiotics. Journal of Animal Science, v.46, p.1447-1469, 1978. WILLIS, G.M.; BAKER, D.H. Eimeria acervulina infection in the chickens:sulfur amino acid requirement of the chick during acute coccidiosis. Poultry Science, v.60, p.1892-1897, 1981. WRIGHT, S.H.; WUNZ, T.M.; SILVA, A.L. Betaine transport in the gill of a marine mussel, Mytilus californianus. American Journal of Physiology, v.263 (Regulatory Integrative Comparative Physiology, v.32), p.R226-R232, 1992. WUNZ, M.; WRIGHT, S. Betaine transport in rabbit renal brush-border membrane vesicles. American Journal of Physiology, v.264. p.F948-F955, 1993. YANCEY, P.H.; CLARK, M.E.; HAND, S.C.; BOWLUS, R.D.; SOMERO, G.N. Living with water stress: evolution of osmolyte systems. Science, v.217, p.1214-1223, 1982.

- 56 -
CAPÍTULO III
INFLUÊNCIA DA BETAÍNA NO DESEMPENHO E NAS
CARACTERÍSTICAS DA CARCAÇA DE FRANGOS DE CORTE
INFECTADOS EXPERIMENTALMENTE COM EIMERIA
ACERVULINA (TIZZER, 1929)

- 57 -
RESUMO
NIANG, Tania Marcia Soares. Influência da betaína no desempenho e nas características da carcaça de frangos de corte infectados experimentalmente com Eimeria acervulina (TIZZER, 1929).
Investigou-se o efeito da suplementação de betaína no desempenho e nas características da carcaça de frangos de corte machos submetidos à infecção com Eimeria acervulina. Foram utilizados 420 pintos de corte Cobb, de um dia de idade, distribuídos num delineamento de blocos ao acaso com 5 tratamentos e 6 repetições, alojados em baterias metálicas de três andares, onde cada andar representava um bloco. As dietas foram fornecidas de 1 a 21 e de 22 a 42 dias de idade. Os tratamentos foram: 1) controle positivo (CP) = dieta basal mais promotor de produção e coccidiostático; 2) 0,0% de betaína = dieta basal sem betaína (0,0%); 3) 0,05% betaína = dieta basal mais 0,05% de betaína; 4) 0,10% betaína = dieta basal mais 0,10% de betaína; 5) 0,15% betaína = dieta basal mais 0,15% de betaína. No décimo quarto dia de idade, as aves foram inoculadas individualmente com 2 x 105 oocistos esporulados de Eimeria acervulina. Os índices de desempenho foram analisados no período de 1 a 14, 14 a 21, 1 a 21, 22 a 28, 28 a 42, 22 a 42 e 1 a 42 dias de idade das aves. Aos 43 dias de idade, duas aves de cada repetição, totalizando 12 aves por tratamento, foram abatidas para estudo das características de carcaça. No período de 1 a 14 dias os frangos que foram alimentados com a ração contendo os níveis de betaína apresentaram o mesmo (p>0,05) ganho de peso que aqueles que receberam a dieta contendo promotor de produção mais coccidiostático. Não houve diferença (p>0,05) entre os tratamentos para consumo de ração e conversão alimentar neste período. Para os períodos de 14 a 21, 1 a 21 e 22 a 28 dias, os frangos que consumiram o alimento suplementado com 0,10% de betaína apresentaram menor consumo (p<0,05) de ração. Nos períodos de 28 a 42 e 22 a 42 dias, não houve diferença (p>0,05) entre os tratamentos para consumo de ração, ganho de peso e conversão alimentar. Entretanto, no período de 1 a 42 dias, os frangos que receberam a ração contendo os níveis de 0,10 e 0,15% de betaína não apresentaram diferença (p>0,05) no ganho de peso e na conversão alimentar em relação àqueles que foram suplementados com promotor de produção e coccidiostático. Ademais, não foi observado diferença (p>0,05) entre os tratamentos para consumo de ração. A equação de regressão foi significativa (p<0,05) para peso da carcaça resfriada, apresentando comportamento linear. As aves que receberam a ração contendo 0,10% de betaína apresentaram maior rendimento de carcaça (p<0,05) em relação àquelas que receberam a ração com 0,05% de betaína. Para peso do peito e rendimento de peito, não foi observado diferença entre os tratamentos (p>0,05). Os frangos alimentados com a ração contendo promotor de produção mais coccidiostático e com a ração que não foi suplementada com betaína (0,0%) apresentaram maior conteúdo de gordura abdominal (p<0,05) em relação àqueles que receberam 0,05% de betaína na dieta.
Palavras chave: Aves. Coccidiose. Nutrição.

- 58 -
ABSTRACT
NIANG, Tania Marcia Soares. Influence of betaine on growth performance and carcass characteristics in broiler chickens infected experimentally with Eimeria acervulina (TIZZER, 1929).
The effect of the betaine supplementation in the growth performance and carcass characteristics of male broilers chickens submitted to experimental infection with Eimeria acervulina was investigated. A total of 420 day-old males Cobb broiler chicks were distributed in battery cage of three floors in a randomized block experimental design with 5 treatments and 6 experimental units per treatment, where each floor represented a block. Diets were supplied from 1 to 21 and 22 to 42 days. The treatments were as follow: 1) Positive control (PC) = basal diet plus a production growth promoter and a coccidiostatic; 2) 0.0% of betaine = basal diet without betaine (0.0%); 3) 0.05% betaine = basal diet plus 0.05% betaine; 4) 0.10% betaine = basal diet plus 0.10% betaine; 5) 0.15% betaine = basal diet plus 0.15% de betaine. At fourteen day of age, the birds were individually inoculated with approximately 2x105 sporulated oocysts of E. acervulina. The performance was studied in the period from 1 to 14, 14 to 21, 1 to 21, 22 to 28, 28 to 42, 22 to 42 and 1 to 42 days of age of the birds. At 43 days of age, two birds of each repetition, total 12 birds per treatment, were killed for studied the carcass characteristics. In the period from 1 to 14 days the chickens that were fed the diets with betaine levels had the same (p>0.05) weight gain that those receiving the diet with production growth promoter plus coccidiostatic. There was not difference (p>0.05) among the treatments for feed intake and feed conversion in this period. For the periods from 14 to 21, 1 to 21 and 22 to 28 days, the chickens that consumed the food supplemented with 0.10% of betaine had reduced feed intake (p <0.05). In the periods from 28 to 42 and 22 to 42 days, there was not difference (p>0.05) among treatments for feed intake, weight gain and feed conversion. However, in the period from 1 to 42 days, the chickens that received the diet with the levels of 0.10 and 0.15% of betaine showed no difference (p>0.05) on weight gain and feed conversion in comparison to those that were supplemented with production growth promoter plus coccidiostatic. Besides, difference was not observed (p>0.05) among the treatments for feed intake. There was a linear effect (p <0.05) of betaine levels on chilled carcass weight, birds that received the diet with 0.10% of betaine had larger carcass yield (p <0.05) in relation to those that received the diet with 0.05% of betaine. For breast weight and breast yield, differences were not observed among the treatments (p>0.05). The chickens fed diet with production growth promoter plus coccidiostatic and with the diet without betaine supplementation (0.0%) had larger percentage of abdominal fat (p<0.05) in relation to those that received 0.05% of betaine in the diet.
Key words: Coccidiosis. Nutrition. Poultry.

- 59 -
1 INTRODUÇÃO
A coccidiose é uma doença comum em aves domésticas, principalmente quando as mesmas são criadas em condições de intenso confinamento, onde as perdas podem ser potencialmente grandes. A doença resulta na redução do crescimento e na piora da eficiência alimentar, causadas por desordens iônicas e osmóticas no intestino, que se tornam evidentes através do baixo pH duodenal (RUFF & REID, 1975; FOX & SOUTHERN, 1987) e da reduzida capacidade de absorção dos nutrientes (TURK & STEPHENS, 1970; RUFF et al., 1976).
O rápido desenvolvimento de resistência às drogas pela Eimeria, o aumento nos preços dos medicamentos e a desconfiança do público consumidor sobre a carne de animais tratados com drogas, têm levado a uma busca por aditivos naturais alternativos. Produtos naturais com propriedades medicinais também podem ter efeitos benéficos no controle da coccidiose. Componentes da dieta ou suas formas de apresentação podem agir diretamente sobre o desenvolvimento do parasita, fortalecer a defesa do hospedeiro ou favorecer sua recuperação.
Para maximizar o potencial genético das aves, os nutricionistas têm desenvolvido dietas de alto valor nutritivo. Os níveis dos nutrientes, tais como a proteína bruta e os aminoácidos essenciais nas rações podem afetar o crescimento dos frangos de corte. Alguns suplementos alimentares tais como a betaína, podem melhorar estas características de desempenho e rendimento da carcaça de frangos infectados ou não infectados por Eimeria sp (VIRTANEN et al., 1993; AUGUSTINE et al., 1997; TEETER et al. 1999; WALDENSTEDT et al., 1999; SIVERSIDES et al., 1999; REMUS, 2001; HRUBY et al., 2001; HRUBY, 2002).
A Betaína (trimetilglicina) é um aminoácido metilado, que apresenta dupla função como osmólito e como doadora de grupo metil em vertebrados. É uma pró-vitamina obtida comercialmente da beterraba processada. É um osmólito com ação conhecida em rins de mamíferos (WUNZ & WRIGHT, 1993; FERRARIS et al. 1996), macrófagos hepáticos (ZHANG et al. 1996) e fibroblastos embriônicos de frangos (PETRONINI et al. 1992). Células da medula renal acumulam osmólitos orgânicos como betaína, sorbitol e inositol para proteger as células contra a resistência osmótica causada pela uréia e cloreto de sódio (BURG, 1995). A betaína é também apropriada para desenvolver uma função osmolítica no intestino de frangos (KIDD et al. 1997).
Como osmólito, a betaína possui a capacidade de estabilizar a função metabólica celular sob diferentes condições de estresse osmótico (YANCEY et al. 1982; DRAGOLOVICH, 1994), a um baixo custo energético. As propriedades osmoprotetoras da betaína relacionam-se à sua característica bipolar e a alta solubilidade em água (CHAMBERS & KUNIN, 1985).
Como doadora de grupo metil a betaína pode aumentar o rendimento de carne ao contribuir para a síntese de importantes compostos, incluindo proteína e DNA/RNA e aumentar a disponibilidade de metionina para síntese de proteína e permitir o crescimento muscular. Além disso, o metabolismo da betaína produz glicina, um aminoácido também importante na síntese de proteína que colabora para o crescimento muscular (HRUBY, 2002).

- 60 -
O presente trabalho teve como objetivo avaliar a influência da suplementação de betaína no desempenho produtivo e nas características da carcaça de frangos de corte machos submetidos a uma infecção experimental com E. acervulina.

- 61 -
2 REVISÃO DA LITERATURA
2.1 A Importância da Àgua no Organismo Vivo
A água apresenta uma função básica em todos os tipos de células, por ser a matriz que banha e apóia as organelas celulares. É o solvente “chave” que regula o metabolismo celular (HAUSSINGER, 1996). Uma grande porção de energia dentro do corpo é usada para manter um volume de água relativamente constante no interior das células. Estima-se que 30 a 60% da energia de mantença utilizada dentro dos órgãos viscerais está associada com a bomba de sódio-potássio ATPase (McBRIDE & MILLIGAN, 1985; McBRIDE & KELLY, 1990). Esta bomba ajuda a manter o balanço de água nas células e é particularmente importante na ausência de osmólitos como a betaína. Músculo de frangos de corte e perus com idade de abate contém aproximadamente 74% a 75% água (USDA, 1999).
A vantagem do osmólito betaína versus a bomba de sódio-potássio ATPase é que menos energia é necessária para acumular betaína dentro da célula. Além disso, é seguro para a célula acumular níveis altos de betaína, embora altos níveis de eletrólitos possam afetar adversamente a atividade metabólica da mesma (YANCEY et al., 1982). Especificamente a betaína tem sido estudada por aliviar a desidratação celular durante um estresse osmótico (BAGNASCO et al., 1986; PETRONINI et al., 1993). Em frangos de corte, tem sido conhecida por aumentar a retenção de água no corpo durante um estresse calórico e por ajudar no desempenho desde que ocorra um desafio coccídio (VIRTANEN & ROSI, 1995; VIRTANEN et al., 1996; REMUS et al., 1996).
As células animais tentam reter água acumulando íons como o potássio. Altas concentrações destes íons geralmente têm um efeito negativo no processo metabólico dentro das células. A betaína age estabilizando as enzimas celulares e mantendo o processo metabólico. Isto, somado ao efeito de doador metil conduz a um aumento no rendimento de peito em produções avícolas comerciais (LUNDEEN, 2001).
2.2 Impacto da Eimeria acervulina no Organismo das Aves
Eimeria acervulina, também conhecida como um coccídio duodenal, é um protozoário parasita que invade as células epiteliais do duodeno das aves (BEAVER et al., 1984). Uma infecção coccídiana aguda pode levar à diminuição do desempenho (McDONALD et al., 1982; SOUTHERN & BAKER, 1983; MATTHEWS & SOUTHERN, 2000).
KOUWENHOVEN (1972) reportou perda de peso durante uma infecção aguda de coccidiose, como também anorexia e diarréia nas aves. A infecção coccídiana aguda pode levar a um aumento da morbidade (HEIN, 1968), lesões na mucosa epitelial (KOUWENHOVEN & VAN DER HORST, 1972; MATTEWS & SOUTHERN, 2000), distensão dos segmentos do intestino e aumento de edema (ALLEN, 1983), redução das vilosidades e das enzimas da mucosa em forma de borda de escova (ALLEN, 1987) e enterites necróticas (STEPHENS et al., 1974). Já a coccidiose crônica resulta em completa anorexia, diarréia e morte (MOREHOUSE & McGUIRE, 1958; HEIN, 1968).
Em experimentos realizados por McKENZIE et al. (1987), observaram que o tempo de passagem do alimento pelo intestino de frangos de corte infectados com 1x104 ou 5x105

- 62 -
oocistos de E. acervulina aumentou significativamente aos sete e dez dias após a infecção. Estes autores citaram AYLOTT et al. (1986) os quais observaram que os efeitos das infecções com coccídio sobre o tempo de passagem pelo intestino provavelmente não estavam diretamente relacionados aos efeitos da coccidiose sobre a absorção de nutrientes. Entretanto, McKENZIE et. al. (1987) relataram que a diminuição da motilidade no intestino pode predispor algumas aves a outras infecções, especialmente a enterites necróticas.
2.3 Betaína como Doadora de Grupamentos Metila
Betaína, metionina e colina, junto com a vitamina B6, ácido fólico e vitamina B12, são as principais fontes de grupos metil na dieta de animais e humanos. Os grupos metil são necessários para sintetizar vários compostos fisiologicamente essenciais, tais como a metionina, carnitina, creatina, fosfolípideos, hormônios adrenais, RNA e DNA (BAKER, 1984; FRIEDEL et al., 1989; SMOLIN & BENEVENGA, 1989; FRONTIERA et al., 1994). Embora nenhuma recomendação nutricional sobre os grupo metil na dieta de animais foi estabelecida por causa do papel estratégico da metilação nos sistemas nervosos, imunes, renais e cardiovasculares. Acredita-se que tanto animais em crescimento quanto adultos requerem um suprimento constante de grupos metil (DU VIGNEAUD, 1952; NEWBERNE, 1993).
A betaína tem sido classificada como uma metilamônia em virtude de seus três grupamentos metila quimicamente reativos (lábeis) unidos ao átomo de nitrogênio de uma molécula de glicina (YANCEY et al., 1982 & KIDD et al., 1997).
Necessariamente, a metionina deve ser adenosilada/ATP a S-adenosilmetionina (forma ativa desse componente) para que possa doar seu grupamento metila disponível (SMOLIN & BENEVENGA, 1989). Já a colina deve ser convertida; duplamente oxidada a forma de betaína na mitocôndria celular, para exercer sua função de doadora de metila no citosol celular (MANN et al., 1938; TYLER, 1977; DRAGOLOVICH, 1994).
Resultados de pesquisa indicam que a eficiência da colina para reações de metilação é altamente variável (MOLITORIS & BAKER, 1976; BAKER, 1984; LOWRY et al., 1987). Tem se estabelecido desde o passado que a colina é ineficiente, comparada a betaína e a metionina nas reações de metilação de homocisteína e de creatina. STEKOL et al., (1953) reportaram que no metabolismo de frangos de corte, a betaína é três vezes mais eficiente que a colina na metilação de homocisteína a metionina.
É também provável que essa ineficiência na conversão da colina à betaína seja exarcebada com a presença de anticoccidianos ionóforos nessas dietas (TYLER, 1977). Esses produtos interferem no transporte da colina pela parede mitocondrial. Ademais, como grande parte da colina dietética é incorporada à lecitina e esfingomielina quando emerge do trato gastrintestinal, somente uma pequena quantidade de colina livre será disponibilizada para a oxidação à forma de betaína. Então, a betaína, como uma doadora direta de grupo metil pode ser mais efetiva (VIRTANEN, 1995).
2.4 Betaína como Suplemento Dietético Numa série de experimentos conduzidos por WALDENSTEDT et al. (2000), para
estudar os efeitos dos diferentes fatores da alimentação sobre a performance de frangos

- 63 -
durante as infecções coccídianas, estes autores observaram que uma dieta à base de trigo em combinação com as vitaminas A, D3, E, C, K3 e selênio não influenciaram na resistência das aves contra a infecção, mas que ao adicionar betaína na dieta, houve uma influência positiva sobre o ganho de peso em aves inoculadas e não inoculadas com Eimeria.
CREVIEU & NACIRI (2001) observaram que o desenvolvimento da coccidiose pode ser alterado pelo uso de uma dieta balanceada a base de grãos e cereais, que pode agir indiretamente na modificação da fisiologia digestiva. Uma elevada porcentagem de proteína na dieta aumenta a secreção pancreática e favorece o desenvolvimento do parasita. A recuperação da infecção pode ser beneficiada pela vitamina A, C ou K e também por betaína, que modifica os efeitos da Eimeria no tecido epitelial intestinal.
SCHUTTE et al. (1997) reportaram que a betaína não afetou o crescimento, mas aumentou o rendimento de peito em frangos de corte.
Trabalhos realizados nos EUA, México e Canadá segundo REMUS (2001), resultaram num aumento de 2% em média, no rendimento de peito de frangos de corte que foram alimentados com dietas contendo betaína. Cita ainda que a adição de 0,10% de betaína em uma dieta balanceada para perus proporcionou um aumento de 3% no rendimento de peito.
O fornecimento de betaína (0,10 e 0,15%) na dieta de aves submetidas a desafio ambiental como o estresse calórico e/ou coccídio, proporcionou melhoria na conversão alimentar e aumentou o rendimento de peito (HRUBY, 2002).
Trabalhos conduzidos no centro de pesquisa aviária da Geórgia para investigar o efeito da suplementação de betaína sobre o ambiente intestinal de frangos jovens, que foram infectados com coccídio via dieta aos 15 dias de idade, mostraram que houve uma redução do peso quando as aves receberam alimento deficiente em metionina, levando a uma piora da conversão alimentar. Porém, a suplementação, tanto com metionina quanto betaína, resultou em aumento do peso e melhorou significativamente a conversão alimentar aos 42 dias de idade (SILVERSIDES et al., 1999).
Ainda de acordo com SILVERSIDES et al. (1999) em trabalho conduzido no Colorado Quality Research (EUA), a betaína foi capaz de compensar os baixos níveis de metionina para o desempenho quando não houve um desafio coccídio, enquanto na presença de um desafio coccídio, a betaína obteve melhor desempenho que a metionina. Neste trabalho, estavam incluídos nas dietas dois coccidiostáticos e um promotor de produção.
Dois trabalhos realizados no Colorado Quality Research (EUA) investigaram o papel das propriedades osmoprotectantes da betaína sobre os efeitos da coccidiose no desempenho de frangos de corte recebendo betaína, colina ou metionina, com ou sem um coccidiostático e com um desafio moderado ou alto de coccídio, o resultado mostrou que a integridade intestinal das aves alimentadas com betaína foi melhor em todos os casos, especialmente sob condições de alto desafio coccídio (SILVERSIDES et al., 1999).
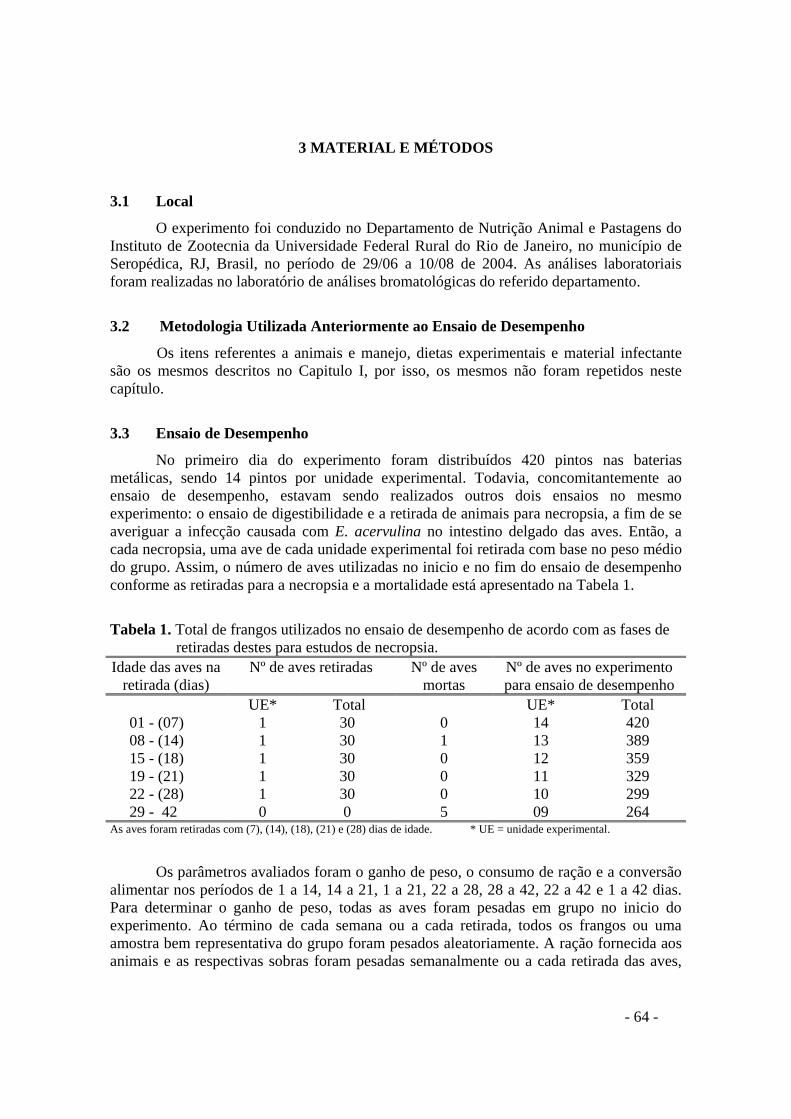
- 64 -
3 MATERIAL E MÉTODOS 3.1 Local
O experimento foi conduzido no Departamento de Nutrição Animal e Pastagens do Instituto de Zootecnia da Universidade Federal Rural do Rio de Janeiro, no município de Seropédica, RJ, Brasil, no período de 29/06 a 10/08 de 2004. As análises laboratoriais foram realizadas no laboratório de análises bromatológicas do referido departamento.
3.2 Metodologia Utilizada Anteriormente ao Ensaio de Desempenho
Os itens referentes a animais e manejo, dietas experimentais e material infectante são os mesmos descritos no Capitulo I, por isso, os mesmos não foram repetidos neste capítulo.
3.3 Ensaio de Desempenho
No primeiro dia do experimento foram distribuídos 420 pintos nas baterias metálicas, sendo 14 pintos por unidade experimental. Todavia, concomitantemente ao ensaio de desempenho, estavam sendo realizados outros dois ensaios no mesmo experimento: o ensaio de digestibilidade e a retirada de animais para necropsia, a fim de se averiguar a infecção causada com E. acervulina no intestino delgado das aves. Então, a cada necropsia, uma ave de cada unidade experimental foi retirada com base no peso médio do grupo. Assim, o número de aves utilizadas no inicio e no fim do ensaio de desempenho conforme as retiradas para a necropsia e a mortalidade está apresentado na Tabela 1.
Tabela 1. Total de frangos utilizados no ensaio de desempenho de acordo com as fases de retiradas destes para estudos de necropsia.
Idade das aves na retirada (dias)
Nº de aves retiradas Nº de aves mortas
Nº de aves no experimento para ensaio de desempenho
UE* Total UE* Total 01 - (07) 1 30 0 14 420 08 - (14) 1 30 1 13 389 15 - (18) 1 30 0 12 359 19 - (21) 1 30 0 11 329 22 - (28) 1 30 0 10 299 29 - 42 0 0 5 09 264 As aves foram retiradas com (7), (14), (18), (21) e (28) dias de idade. * UE = unidade experimental.
Os parâmetros avaliados foram o ganho de peso, o consumo de ração e a conversão alimentar nos períodos de 1 a 14, 14 a 21, 1 a 21, 22 a 28, 28 a 42, 22 a 42 e 1 a 42 dias. Para determinar o ganho de peso, todas as aves foram pesadas em grupo no inicio do experimento. Ao término de cada semana ou a cada retirada, todos os frangos ou uma amostra bem representativa do grupo foram pesados aleatoriamente. A ração fornecida aos animais e as respectivas sobras foram pesadas semanalmente ou a cada retirada das aves,

- 65 -
assim, o cálculo do consumo de ração foi obtido com base no consumo médio por unidade experimental, dentro de cada período. Os dados de conversão alimentar foram obtidos com base no consumo médio de ração e no ganho de peso médio por período experimental. A mortalidade foi registrada diariamente.
3.4 Características da Carcaça e dos Cortes
Aos 43 dias de idade as aves passaram por um jejum de 8 horas, sendo então retiradas duas aves de cada repetição próximas do peso médio do grupo, totalizando 12 aves por tratamento, que foram pesadas e identificadas individualmente nas patas por meio de esparadrapos cirúrgicos escritos com canetas marcador permanente que indicavam o respectivo tratamento e repetição.
Os frangos foram abatidos por sangria na veia jugular, escaldados, depenados e eviscerados no abatedouro do Instituto de Zootecnia para avaliação das características de carcaça. Após este processo, as carcaças foram levadas ao chiller e permaneceram no mesmo, até que atingissem a temperatura entre 2 e 4°C. Em seguida, foi realizado o gotejamento a fim de reduzir o excesso de água absorvida na etapa anterior. Posteriormente, as carcaças foram pesadas e armazenadas em sacos plásticos lacrados e identificados. A seguir, foram estocadas em câmera fria permanecendo por 24 horas até a avaliação das características de carcaça.
Para os cálculos dos rendimentos de carcaças, foram tomados como base o peso vivo após o jejum, imediatamente antes do abate e o peso da carcaça resfriada, sem cabeça e sem pés. Os rendimentos dos cortes foram calculados com base no peso da carcaça resfriada, sendo que a pesagem das carcaças, dos cortes e das gorduras abdominais foram feitas em balança com precisão de 1g.
Foi avaliado o peso vivo, o peso da carcaça e das partes resfriadas, bem como o rendimento da carcaça e das partes resfriadas, e a gordura abdominal. A gordura abdominal foi constituída pelo tecido adiposo presente ao redor da cloaca, da Bursa de Fabricius e dos músculos abdominais adjacentes, conforme metodologia descrita por SMITH (1993).
3.5 Análise Estatística
O delineamento experimental usado foi o de blocos ao acaso, onde se utilizaram baterias metálicas compostas de três andares, cujo bloco foi representado por cada andar, totalizando três blocos por bateria. O modelo estatístico adotado foi escolhido pensando-se nas condições de conforto térmico que poderiam ser diferentes em cada um dos andares, uma vez que o animal experimental é muito sensível ao fator temperatura. Foram usadas dez baterias metálicas compostas de três blocos (superior, médio e inferior), onde cada bateria foi ocupada por aves que estavam recebendo o mesmo tratamento, cujo objetivo foi evitar possíveis contaminações dos tratamentos entre os blocos. Em cada bloco, um único tratamento foi repetido por duas vezes, totalizando seis repetições por tratamento (duas no bloco superior, duas no bloco médio e duas no bloco inferior), então, o mesmo tratamento foi distribuído em duas baterias metálicas diferentes. Como havia cinco tratamentos, foi necessário utilizar dois pares de cinco baterias, que somado as seis repetições de cada tratamento, deu um total de 30 observações.
As análises de variância foram realizadas utilizando-se o pacote computacional Sisvar, descrito por FERREIRA (2000). As variáveis de cada tratamento tiveram suas

- 66 -
médias comparadas pelo teste de Student-Newman-Keuls (SNK) a um nível de significância de 5% (H0:p<0,05), assim como as análises de regressão foram calculadas por meio da técnica de polinômios ortogonais.
O modelo estatístico adotado para análise do ensaio de desempenho e características da carcaça para o teste de comparação de médias foi:
ebty ijjiijm +++=
onde: yij
é o valor observado da característica estudada, no tratamento i ( i = 1,2,3,4,5), no
bloco j (j = 1,2,3) m é a média geral de todas as observações do experimento ti
é o efeito do tratamento i
bj é o efeito do bloco j
eijé o erro associado à observação yij
, ou efeito dos fatores não controlados sobre a
observação yij.
O modelo estatístico adotado para analise de regressão do ensaio de desempenho e característica da carcaça foi:
ipipiii e+Χ++Χ+Χ+=Υ ββββ ...22110 em que:
iΥ é a variável dependente, obtida na observação i
pββββ ,...,,, 210 são os parâmetros da regressão
piii ΧΧΧ ,...,, 21 são variáveis independentes, na observação i
ie é o erro associado a observação iΥ .

- 67 -
4 RESULTADOS E DISCUSSÃO
4.1 Desempenho
4.1.1 Desempenho dos frangos de corte no período de 1 a 14 dias de idade
Os valores de desempenho das duas primeiras semanas de vida dos pintos de corte estão relacionados na Tabela 2. Analisando-se as médias pelo teste de Student Newman Keuls, os animais que receberam a ração com o promotor de produção e coccidiostático (CP) apresentaram maior ganho de peso (p<0,05) em relação aos que receberam a ração com 0% de betaína.
Tabela 2. Efeito da betaína sobre o desempenho de pintos de corte no período de 1 a 14 dias de idade.
Tratamentos2 Consumo (kg) Ganho de peso (kg) Conversão alimentar
Controle positivo 0,545a 0,427a 1,28a 0,00% de betaína 0,537a 0,404b 1,33a 0,05% de betaína 0,543a 0,418ab 1,30a 0,10% de betaína 0,540a 0,418ab 1,29a 0,15% de betaína 0,555a 0,421ab 1,32a CV (%) 3,20 2,97 4,12 Análise de regressão ns L ns a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). L = Efeito linear (p<0,05). ns = não significativo para análise de regressão (p>0,05). 2Cada tratamento consistiu de 6 repetições com 14 aves/repetição na 1ª semana e 13 aves na 2ª semana.
Ainda de acordo com os dados apresentados na Tabela 2, os frangos que foram alimentados com a ração contendo os níveis de 0,05; 0,10 e 0,15% de betaína, não apresentaram ganho de peso diferente entre si e nem daqueles que receberam a dieta contendo promotor de produção mais coccidiostático e a dieta sem adição de betaína (p>0,05), entretanto, observou-se uma tendência de aumento no ganho de peso das aves que foram suplementadas com os níveis de 0,05; 0,10 e 0,15% de betaína na dieta. Além disso, não houve diferença (p>0,05) entre os tratamentos para consumo de ração e conversão alimentar neste período.
Todavia, para os resultados obtidos por meio da análise de regressão (y = 0,1025x + 0,4076) no período de 1 a 14 dias, pode-se constatar que o ganho de peso das aves aumentou linearmente (p<0,05), na medida em que se elevou o nível de betaína na dieta, ilustrando que o maior nível (0,15% de betaína) foi o que proporcionou o maior ganho de peso aos frangos (Figura 1). Estes dados não estão em concordância com o que foi encontrado por KLASING et al. (2002), os quais, ao testar níveis de 0,0; 0,05 e 0,10% de betaína numa dieta a base de milho e soja, antes da infecção com E. acervulina em

- 68 -
frangos de corte, observaram que a betaína dietética não afetou o ganho de peso, o consumo de ração e a eficiência alimentar no período de 0 a 14 dias de idade. WALDENSTEDT et al. (1999) também não encontraram diferença significativa entre os tratamentos para peso vivo, consumo de ração e conversão alimentar dos frangos de corte, que receberam dietas a base de trigo e soja contendo os diferentes suplementos dietéticos: 0,10% de betaína; 0,10% de betaína mais 70ppm de narasin; 70ppm de narasin e nenhum aditivo, aos 10 dias de idade das aves, antes da infecção com uma mistura de espécies do gênero Eimeria.
Figura 1. Efeito da suplementação dietética de betaína no ganho de peso médio de frangos de corte no período de 1 a 14 dias de idade.
No presente estudo, o aumento no ganho de peso dos frangos atribuído ao acréscimo de betaína na dieta no período de 1 a 14 dias de idade, antes da infecção, pode ser comparado com os resultados obtidos por alguns autores (WALDENSTEDT et al., 1999; FETTERER et al., 2003), os quais observaram melhor desempenho em aves não infectadas recebendo a suplementação de betaína na ração quando comparadas com aquelas que foram infectadas com espécies do gênero Eimeria e que também receberam a suplementação dietética de betaína. Admitindo-se então, que as aves saudáveis são capazes de aproveitar da melhor forma possível os benefícios que a betaína apresenta como suplemento dietético, mesmo que não ocorra nenhum tipo de desafio, e como conseqüência, ocasiona melhorias no desempenho.
MATTHEWS et al. (1997), observaram que o nível de 0,10% de betaína na dieta dos frangos de corte aos 14 dias de idade, proporcionou aumento no ganho de peso das aves, porém, as mesmas estavam infectadas com 5x105 oocistos esporulados de E. acervulina. Em outro trabalho realizado por MATTHEWS & SOUTHERN (2000), observaram que houve um aumento no ganho de peso médio total dos frangos que receberam a dieta suplementada com 0,075% de betaína mediante uma infecção crônica de 2,5x105 oocistos esporulados de E. acervulina.
y = 0.1025x + 0.4076R2 = 0.73
0.402
0.408
0.414
0.420
0.426
0.00 0.05 0.10 0.15
Níveis de Betaína (%)
Gan
ho d
e Pe
so (k
g)

- 69 -
4.1.2 Desempenho dos frangos de corte no período de 14 a 21 dias de idade
Os frangos que consumiram a ração suplementada com 0,10% de betaína apresentaram menor consumo (p<0,05) em relação àqueles que receberam a ração com promotor de produção e coccidiostático, ração sem a suplementação de betaína (0,0%) e com 0,05% de betaína (Tabela 3). Já aqueles que tiveram a dieta suplementada com 0,15% de betaína não apresentaram consumo diferente (p>0,05) dos que receberam 0,10% de betaína na ração. Para as demais variáveis analisadas: ganho de peso e conversão alimentar, não houve diferença significativa (p>0,05) entre os tratamentos. Porém, neste período considerado crítico para as aves devido aos efeitos da coccidiose, observa-se que mesmo não havendo diferença entre os tratamentos para conversão alimentar, a mesma apresenta valores muito altos em relação aos períodos apresentados em seguida. Portanto, nesta fase, a coccidiose pode ter sido responsável pela piora na conversão alimentar dos frangos de corte.
Tabela 3. Efeito da betaína e infecção com Eimeria acervulina1 sobre o desempenho de frangos de corte no período de 14 a 21 dias de idade.
Tratamentos2 Consumo (kg) Ganho de peso (kg) Conversão alimentar
Controle positivo 0,744a 0,403a 1,85a 0,00% de betaína 0,742a 0,393a 1,91a 0,05% de betaína 0,742a 0,375a 1,98a 0,10% de betaína 0,712b 0,383a 1,87a 0,15% de betaína 0,722ab 0,384a 1,88a CV (%) Análise de regressão
2,56 L
6,95 ns
6,63 ns
a-bValores de média na coluna sem subscrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). 1Eimeria acervulina com dose infectante 2x105 oocistos/ml de suspensão. 2Cada tratamento consistiu de 6 repetições com 11 aves em cada repetição.
KLASING et al. (2002) observaram em seus estudos que os frangos desafiados com E. acervulina (4,6x104oocistos esporulados) aos 14 dias tiveram as mais baixas taxas de ganho de peso durante a semana subseqüente a infecção (14-21dias) quando comparado aos frangos não desafiados, e que estes tenderam a ter uma diminuição no consumo de ração e na eficiência alimentar. No presente estudo também houve redução no consumo de ração para as aves suplementadas com o nível de 0,10 e 0,15% de betaína e piora na conversão alimentar.
A análise de regressão neste período foi significativa (p<0,05) para consumo de ração, apresentado efeito linear (y = -0,1816x + 0,7431), a qual ilustra que os frangos que consumiram os maiores níveis de betaína (0,10 e 0,15%) apresentaram os menores consumo de ração (Figura 2).

- 70 -
Figura 2. Efeito da suplementação dietética de betaína no consumo médio de ração dos frangos de corte no período de 14 a 21 dias de idade.
4.1.3 Desempenho dos frangos de corte no período de 1 a 21 dias de idade
Conforme os resultados apresentados na Tabela 4, pode-se verificar que os frangos que foram alimentados com a dieta contendo 0,10% de betaína apresentaram menor consumo de ração (p<0,05) em relação àqueles que receberam as demais dietas. Entretanto, não houve diferença (p>0,05) entre os tratamentos para ganho de peso e conversão alimentar neste período. Ainda assim, é possível observar que as aves suplementadas com 0,10% de betaína tenderam a apresentar melhorias no ganho de peso e na conversão alimentar, ao contrário daquelas suplementadas com o nível de 0,05% .
Tabela 4. Efeito da betaína sobre o desempenho de frangos de corte no período de 1 a 21 dias de idade.
Tratamentos Consumo (kg) Ganho de peso (kg) Conversão alimentar
Controle positivo 1,289a 0,830a 1,55a 0,00% betaína 1,279a 0,797a 1,61a 0,05% betaína 1,285a 0,793a 1,62a 0,10% betaína 1,252b 0,801a 1,56a 0,15% betaína 1,278a 0,805a 1,59a CV (%) Análise de regressão
1,46 ns
3,69 ns
3,32 ns
a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). ns = não significativo para análise de regressão (p>0,05).
y = -0.1816x + 0.7431R2 = 0.593
0.711
0.719
0.727
0.735
0.743
0.00 0.05 0.10 0.15Niveis de betaína (%)
Con
sum
o (k
g)

- 71 -
Os resultados descritos acima estão coerentes com aqueles encontrados por KLASING et al. (2002), que verificaram que as aves infectadas com E. acervulina apresentaram uma tendência de redução no consumo de ração aos 21 dias de idade, porém, estes autores não citam qual foi o nível de betaína que proporcionou tal tendência.
FETTERER et al. (2003) trabalhando com dietas basais, não suplementadas ou suplementadas com 0,15% de betaína para frangos de corte infectados aos 15 dias de idade com três espécies do gênero Eimeria (E. acervulina, E. máxima ou E. tenella), observaram que aves infectadas com E. máxima e que receberam 0,15% de betaína tiveram aumento no ganho de peso quando comparadas com aves que receberam a dieta não suplementada com betaína. Já aquelas que foram infectadas com as três espécies de coccídios e que receberam a dieta sem a suplementação de betaína, apresentaram redução no ganho de peso. Citam ainda que a suplementação com 0,15% de betaína foi eficaz em proporcionar melhorias no ganho de peso das aves infectadas com E. tenella quando comparadas com aves infectadas com E. máxima que receberam a dieta sem a suplementação de betaína, mas que teve pouco efeito sobre o ganho de peso dos frangos que foram infectados com E. acervulina. Estes dados estão de acordo com o que foi observado no presente experimento, em que, o ganho de peso dos frangos infectados com E. acervulina não foi afetado pela suplementação de betaína no período estudado.
Trabalhos conduzidos por AUGUSTINE et al. (1997) com frangos de corte criados em baterias metálicas de 1 até os 21 dias de idade e inoculados aos 14 dias com uma mistura de E. acervulina, E. máxima e E. tenella, mostraram que a betaína (0,15%) contribuiu para melhorar o peso vivo e a conversão alimentar, porém, estas variáveis foram bem mais expressivas quando a betaína foi dada em combinação com a salinomicina. Ainda de acordo com AUGUSTINE et al. (1997), o resultado obtido para aves criadas em boxes, foi semelhante aquele observado para as aves criadas em baterias metálicas.
ESTEVE-GARCIA & MACK (2000) ao testarem os efeitos da DL-metionina (0,0; 0,06 e 0,12%) e betaína (0,0 e 0,05%) no desempenho de frangos de corte sadios, aos 21 dias de idade, observaram que houve resposta significativa a suplementação de metionina em termos de peso final e conversão alimentar, mas que os efeitos da betaína foram pequenos e não significativos, embora pudesse ser notado que eles foram na direção de melhorar a conversão alimentar neste período. Estes dados não estão condizentes com o que foi observado no presente estudo, porém, vale ressaltar que o nível de betaína suplementado na ração foi muito baixo em relação aos níveis de metionina testados, é possível que isto explique a diferença encontrada no desempenho dos frangos suplementados com metionina e aqueles suplementados com betaína no trabalho dos autores citado acima.
Resumidamente, foi possível observar neste estudo que o consumo de ração foi altamente afetado no período mais abrangente, de 1 a 21 dias, e, sobretudo, nas aves que foram infectadas. No entanto, melhorias no ganho de peso só foram observadas nas aves sadias, antes da infecção, não apresentando resultados significativos nos períodos pós-infecção. Vale ainda ressaltar que a conversão alimentar não foi melhorada por nenhum dos tratamentos testados aqui, em nenhum dos períodos estudados acima.
4.1.4 Desempenho dos frangos de corte no período de 22 a 28 dias de idade
No período de 22 a 28 dias (Tabela 5) os frangos que receberam a ração contendo promotor de produção e coccidiostático apresentaram maior consumo (p<0,05) em relação àqueles que receberam a dieta contendo 0,10% de betaína. No entanto, não houve diferença

- 72 -
significativa (p>0,05) no consumo dos frangos que receberam as rações suplementadas com os níveis 0,00; 0,05 e 0,15% de betaína. Para ganho de peso e conversão alimentar também não houve diferença significativa (p>0,05) entre os tratamentos.
Tabela 5. Efeito da betaína e inoculação de Eimeria1 sobre o desempenho de frangos de corte no período de 22 a 28 dias de idade.
Tratamentos2 Consumo (kg) Ganho de peso (kg) Conversão alimentar
Controle positivo 0,930a 0,665a 1,40a 0,00% betaína 0,877ab 0,587a 1,52a 0,05% betaína 0,890ab 0,634a 1,40a 0,10% betaína 0,853b 0,656a 1,31a 0,15% betaína 0,893ab 0,636a 1,41a CV (%) Análise de regressão
4,60 ns
7,32 ns
9,88 ns
a-bValores de média na coluna sem subscrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). 1Eimeria acervulina com dose infectante 2 x 105 oocistos/ml de suspensão aos 14d. 2Cada tratamento consistiu de 6 repetições com 10 aves cada neste período.
4.1.5 Desempenho dos frangos de corte no período de 28 a 42 dias de idade
Os dados apresentados na Tabela 6 resume o desempenho das aves no período de 28 a 42 dias, na qual observa-se que não houve diferença significativa (p>0,05) entre os tratamentos para consumo de ração, ganho de peso e conversão alimentar. Neste período já havia passado os 15 dias após a inoculação com E. acervulina, sendo que as aves já se encontravam em processo de recuperação da doença, o que de certa forma explique a falta de resultados com significância para o teste de médias.
Tabela 6. Efeito da betaína e inoculação de Eimeria1 sobre o desempenho de frangos de corte no período de 28 a 42 dias de idade.
Tratamentos2 Consumo (kg) Ganho de peso (kg) Conversão alimentar
Controle positivo 3,444a 1,989 a 1,73 a 0,00% betaína 3,465 a 1,892 a 1,84 a 0,05% betaína 3,379 a 1,912 a 1,77 a 0,10% betaína 3,369 a 1,928 a 1,75 a 0,15% betaína 3,477 a 1,975 a 1,76 a CV (%) Análise de regressão
2,64 Q
4,22 Q
4,29 ns
a-bValores de média na coluna sem subscrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). 1Eimeria acervulina com dose infectante 2 x 105 oocistos/ml de suspensão aos 14d. 2Cada tratamento consistiu de 6 repetições com 09 aves cada neste período. Q = Efeito quadrático.

- 73 -
A análise de regressão realizada neste período para consumo de ração apresentou
comportamento expresso por uma função quadrática (y = 16,629x2 – 2,4672x + 2,5838), onde é possível observar redução no consumo quando baixos níveis de betaína (0,00 a 0,074%) são adicionados à dieta, porém, na medida em que esses níveis são elevados acima de 0,074%, o consumo de ração volta a aumentar (Figura 3).
Figura 3. Efeito da suplementação dietética de betaína no consumo médio de ração dos frangos de corte no período de 28 a 42 dias de idade.
A análise de regressão apresentada na Figura 4 para este estudo exibe o comportamento quadrático (y = 11.76x2 - 1.431x + 1.3095) encontrado para ganho de peso neste período, onde é possível observar redução no ganho de peso das aves quando estas consomem a dieta com baixos níveis de betaína (0,00 a 0,06%), contudo, o baixo desempenho é superado quando níveis acima de 0,06% de betaína são adicionados a dieta.
y = 16.629x2 - 2.4672x + 2.5838R2 = 0.95
2.489
2.519
2.549
2.579
0.00 0.05 0.10 0.15
Niveis de betaína (%)
Con
sum
o (k
g)
0,074

- 74 -
Figura 4. Efeito da suplementação dietética de betaína no ganho de peso médio dos frangos de corte no período de 28 a 42 dias de idade.
4.1.6 Desempenho dos frangos de corte no período de 22 a 42 dias de idade
De acordo com os dados apresentados na Tabela 7, com base no teste de médias, não foi observada diferença significativa (p>0,05) entre os tratamentos para as variáveis consumo de ração, ganho de peso e conversão alimentar no período de 22 a 42 dias de idade das aves.
Tabela 7. Efeito da betaína sobre o desempenho de frangos de corte no período de 22 a 42
dias de idade. Tratamentos Consumo (kg) Ganho de peso (kg) Conversão alimentar
Controle positivo 3,444a 1,989a 1,73a
0,00% betaína 3,465a 1,892a 1,84a 0,05% betaína 3,379a 1,912a 1,77a 0,10% betaína 3,369a 1,928a 1,75a 0,15% betaína 3,477a 1,975a 1,76a CV (%) Análise de regressão
2,64 Q
4,22 ns
4,29 ns
a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). ns = não significativo para análise de regressão (p>0,05). Q = Efeito quadrático (p<0,05).
Consoante a análise de regressão, os frangos que receberam os níveis de betaína acrescidos à dieta neste período, apresentaram como resposta para consumo de ração
y = 11.76x2 - 1.431x + 1.3095R2 = 0.9435
1.250
1.280
1.310
1.340
1.370
0.00 0.05 0.10 0.15Níveis de betaína
Gan
ho d
e pe
so (k
g)
0,06
(%)

- 75 -
comportamento expresso por uma função quadrática (y = 19.281x2 - 2.8405x + 3.4668), na qual observou-se uma redução no consumo de ração das aves quando a dieta oferecida conteve os níveis de betaína variando de 0,0 até 0,074%, seguido de um aumento no consumo quando o nível de betaína foi maior que 0,074% (Figura 5).
Figura 5. Efeito da suplementação dietética de betaína no consumo médio de ração dos frangos de corte no período de 22 a 42 dias de idade.
Em resumo, o período mais abrangente (22 a 42 dias) analisado neste estudo refletiu
que o consumo de ração das aves foi significativamente influenciado pelos níveis de betaína acrescidos à dieta, embora o mesmo não tenha sido observado para o ganho de peso, exceto no período de 28 a 42 dias, e para a conversão alimentar, que mais uma vez não foi melhorada por nenhum dos tratamentos em todos os períodos estudados acima.
4.1.7 Desempenho dos frangos de corte no período de 1 a 42 dias de idade
Segundo os dados apresentados na Tabela 8, os frangos que foram alimentados com a dieta contendo promotor de produção e coccidiostático apresentaram maior ganho de peso (p<0,05) em relação aqueles que receberam a ração sem a suplementação de betaína (0,0%) e a ração com 0,05% de betaína. Contudo, as aves que receberam a dieta contendo os
y = 19.281x2 - 2.8405x + 3.4668R2 = 0.9909
3.360
3.390
3.420
3.450
3.480
0.00 0.05 0.10 0.15
Niveis de betaína (%)
Con
sum
o de
raç
ão (k
g)
0,074

- 76 -
diferentes níveis de betaína (0,0; 0,05; 0,10 e 0,15%), não apresentaram diferença (p>0,05) entre elas para ganho de peso. Tabela 8. Efeito da betaína sobre o desempenho de frangos de corte no período de 1 a 42 dias de idade.
Tratamentos Consumo (kg) Ganho de peso (kg) Conversão alimentar
Controle positivo 4,734a 2,819a 1,68b 0,00% betaína 4,744a 2,689b 1,77a 0,05% betaína 4,665a 2,705b 1,72ab 0,10% betaína 4,621a 2,729ab 1,70ab 0,15% betaína 4,755a 2,779ab 1,71ab CV (%) Análise de regressão
2,14 Q
2,55 L
2,62 ns
a-bValores de média na mesma coluna sem sobrescrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). ns = não significativo para análise de regressão (p>0,05). L= efeito linear e Q = Efeito quadrático
Os frangos que receberam a ração contendo os níveis de 0,10 e 0,15% de betaína (Tabela 8) não apresentaram diferença no ganho de peso (p>0,05) em relação aqueles que foram suplementados com promotor de produção e coccidiostático. Além disso, foi observada uma tendência de aumento no ganho de peso dos frangos na medida em que se adicionou betaína a dieta neste período. WALDENSTEDT et al. (1999), descreveram que a betaína como um simples suplemento alimentar (0,10%), aumentou significativamente o peso vivo das aves com coccidiose em 5,7; 5,4 e 5,6% aos 22, 29 e 36 dias de idade respectivamente, comparada com aves que receberam a ração controle (sem betaína).
AUGUSTINE et al. (1997) também observaram que a suplementação de 0,15% de betaína na ração de frangos de corte no período de 1 a 45 dias de idade aumentou o peso vivo das aves em relação aquelas que não receberam a suplementação de betaína, porém, o peso vivo foi consideravelmente maior quando houve adição de betaína junto com salinomicina na dieta.
A conversão alimentar foi significativamente melhor (p<0,05) nos animais que receberam a dieta com promotor de produção e coccidiostático em relação àqueles que receberam a ração sem a suplementação de betaína (0,0%). Porém, as aves que receberam o alimento contendo os níveis de 0,05; 0,10 e 0,15% de betaína apresentaram a mesma conversão alimentar (p>0,05) que as aves suplementadas com promotor de produção e coccidiostático (Tabela 8).
Mesmo não tendo sido observada diferença (p>0,05) para conversão alimentar dos frangos que receberam os diferentes níveis de betaína na dieta (Tabela 8), houve uma tendência de melhoria da mesma quando foi adicionado o nível de 0,05; 0,10 e 0,15% de betaína na ração, corroborando que mais trabalhos são necessários para responder se um nível mais alto de betaína ocasionaria melhorias significativas no desempenho das aves. Ademais, não foi observada diferença significativa (p>0,05) entre os tratamentos para consumo de ração no período de 1 a 42 dias.
WALDENSTED et al. (1999) não observaram diferença significativa para consumo de ração e conversão alimentar das aves suplementadas com 0,10% de betaína
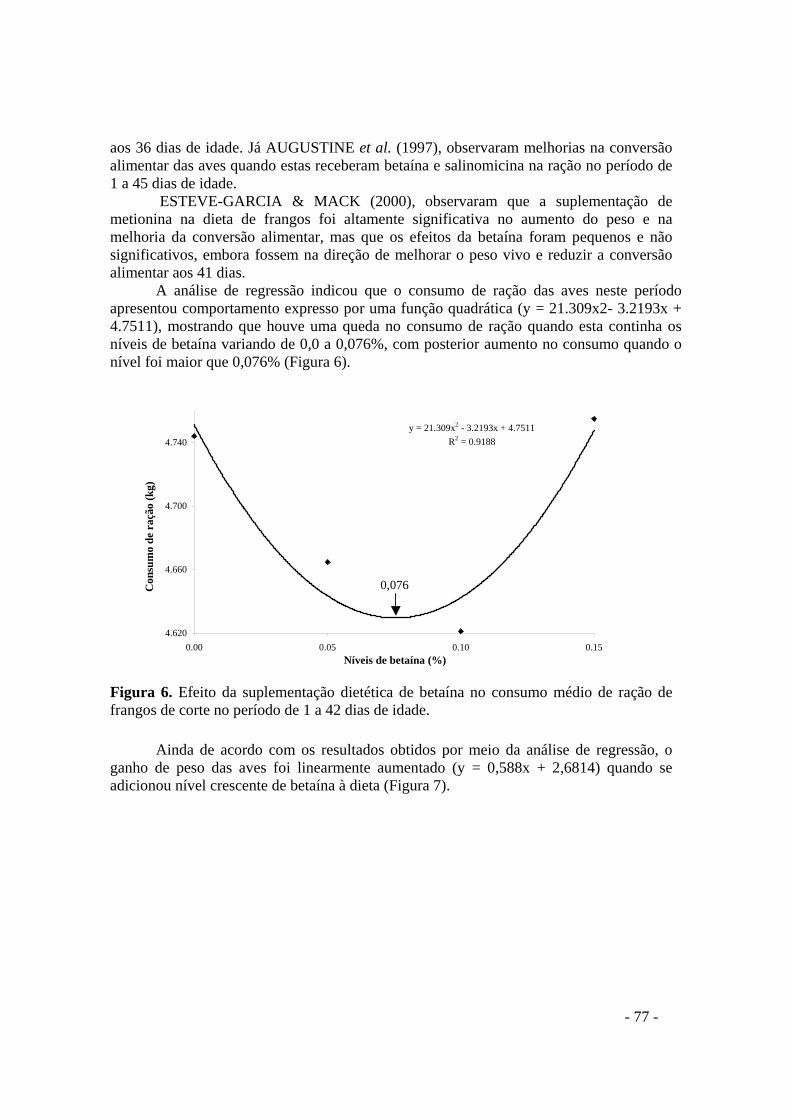
- 77 -
aos 36 dias de idade. Já AUGUSTINE et al. (1997), observaram melhorias na conversão alimentar das aves quando estas receberam betaína e salinomicina na ração no período de 1 a 45 dias de idade.
ESTEVE-GARCIA & MACK (2000), observaram que a suplementação de metionina na dieta de frangos foi altamente significativa no aumento do peso e na melhoria da conversão alimentar, mas que os efeitos da betaína foram pequenos e não significativos, embora fossem na direção de melhorar o peso vivo e reduzir a conversão alimentar aos 41 dias.
A análise de regressão indicou que o consumo de ração das aves neste período apresentou comportamento expresso por uma função quadrática (y = 21.309x2- 3.2193x + 4.7511), mostrando que houve uma queda no consumo de ração quando esta continha os níveis de betaína variando de 0,0 a 0,076%, com posterior aumento no consumo quando o nível foi maior que 0,076% (Figura 6).
Figura 6. Efeito da suplementação dietética de betaína no consumo médio de ração de frangos de corte no período de 1 a 42 dias de idade.
Ainda de acordo com os resultados obtidos por meio da análise de regressão, o ganho de peso das aves foi linearmente aumentado (y = 0,588x + 2,6814) quando se adicionou nível crescente de betaína à dieta (Figura 7).
y = 21.309x2 - 3.2193x + 4.7511R2 = 0.9188
4.620
4.660
4.700
4.740
0.00 0.05 0.10 0.15Níveis de betaína (%)
Con
sum
o de
raç
ão (k
g)
0,076

- 78 -
Figura 7. Efeito da suplementação dietética de betaína no ganho de peso médio dos frangos de corte no período de 1 a 42 dias de idade.
De uma forma geral, o período de 1 a 42 dias retratou de maneira abrangente e explicita o quanto os níveis de betaína influenciaram o consumo de ração, o ganho de peso e a conversão alimentar das aves. elucidando ainda, que as aves suplementadas com a dieta contendo promotor de produção e coccidiostático apresentaram a mesma equivalência estatística que as aves suplementadas com os maiores níveis de betaína. cabe ainda ressaltar, que neste período, tanto o consumo de ração, quanto o ganho de peso e a conversão alimentar foram melhorados, sugerindo que as aves possam ter desenvolvido um mecanismo compensatório para melhorar o desempenho após a doença. essa explicação se baseia nas pesquisas de turk (1978) e a melhoria no desempenho das aves, nos dados de produção comercial de frangos de corte aos 42 dias comparados aos dados de desempenho obtidos neste experimento, visto que não havia um controle não infectado no período de 1 a 42 dias para fazer tal comparação.
4.2 Características Da Carcaça Os dados para análise dos componentes da carcaça estão ilustrados na Tabela 9 e
Figura 8. Para as variáveis peso vivo dos frangos em jejum imediatamente antes do abate e peso da carcaça resfriada, não foram observadas diferença significativa (p>0,05) entre os tratamentos.
y = 0.588x + 2.6814R2 = 0.934
2.660
2.690
2.720
2.750
2.780
0.00 0.05 0.10 0.15
Níveis de betaína (%)
Gan
ho d
e pe
so (k
g)

- 79 -
Tabela 9. Características da carcaça de frangos de corte machos no período de 1 a 43 dias de idade1.
Variáveis CP 0,0% Bet.
0,05% Bet.
0,10% Bet.
0,15% Bet.
CV (%)
Peso vivo ao abate, kg 2,811a 2,708a 2,732a 2,734a 2,794a 3.35Peso carcaça resfriada, kgL 1,941a 1,860a 1,820a 1,922a 1,929a 4.13% carcaça 69,0ab 68,6ab 66,8b 70,3a 69,0ab 2.71Peso do peito com osso, kg 0,662a 0,645a 0,653a 0,702a 0,690a 7.17% peito2 34,0a 34,6a 35,9a 36,5a 35,7a 4.39Peso coxa + sobrecoxa, kg 0,651a 0,614ab 0,602b 0,612ab 0,629ab 4.67% coxa + sobrecoxa2 33,5a 33,0a 33,0a 31,9a 32,7a 3.32Peso do dorso, kg 0,340a 0,332ab 0,311b 0,334ab 0,327ab 4.80% do dorso2 17,5a 17,9a 17,1a 17,4a 17,0a 3.56Peso da asa, kg 0,213a 0,205a 0,197a 0,208a 0,212a 5.00% da asa2 11,0a 11,0a 10,8a 10,8a 11,0a 4.36Gordura abdominal, kg 0,49a 0,46aa 0,35b 0,44ab 0,42ab 17.3% gordura abdominal2 2,48a 2,48a 1,91a 2,23a 2,15a 16.8a-bValores de média na mesma linha sem sobrescrito comum, diferem significativamente (p< 0,05) pelo teste de Student Newman Keuls (SNK). L = Efeito linear 1 = média de 12 frangos por tratamento 2 = O rendimento das partes foi calculado em relação ao peso da carcaça resfriada. Bet. = Betaína.
Todavia, os resultados obtidos com a análise de regressão (y = 0,6193x + 1,836) foram significativos (p<0,05) para peso da carcaça resfriada, apresentado comportamento expresso por uma função linear, mostrando que houve um aumento no peso da carcaça na medida em que a betaína foi adicionada a dieta, e que o maior nível de betaína (0,15%) proporcionou este aumento no peso da carcaça resfriada (Figura 8).
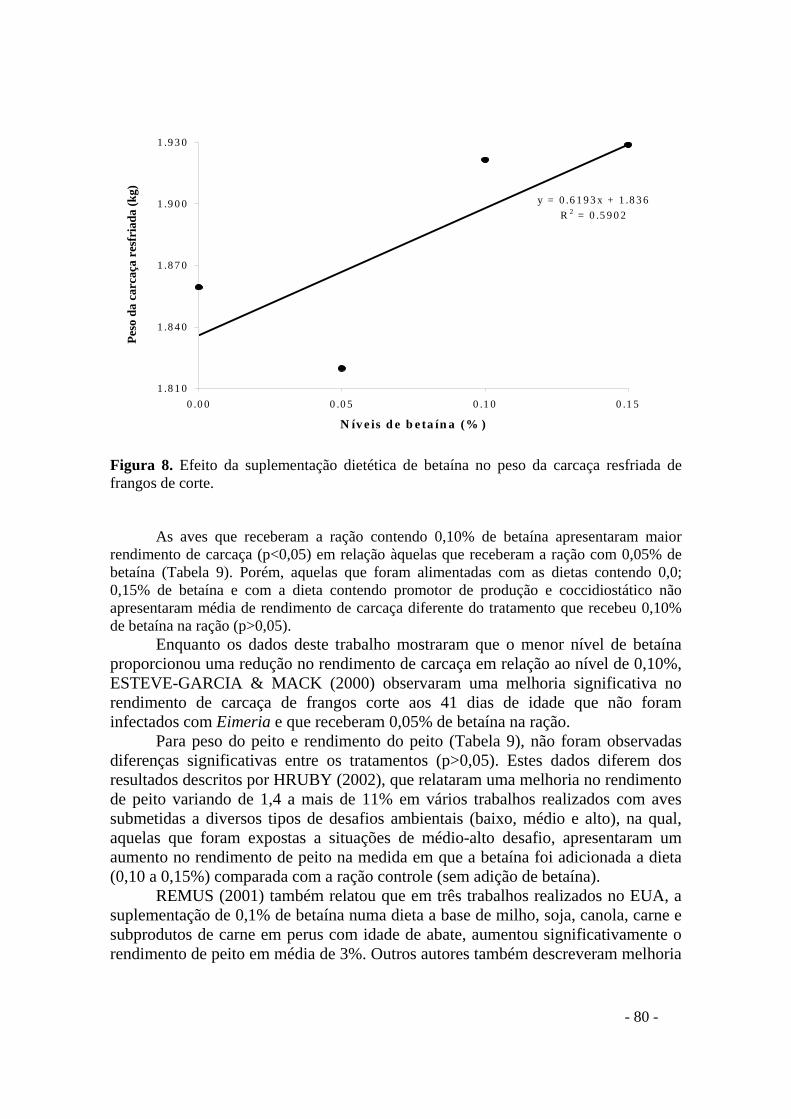
- 80 -
Figura 8. Efeito da suplementação dietética de betaína no peso da carcaça resfriada de frangos de corte.
As aves que receberam a ração contendo 0,10% de betaína apresentaram maior
rendimento de carcaça (p<0,05) em relação àquelas que receberam a ração com 0,05% de betaína (Tabela 9). Porém, aquelas que foram alimentadas com as dietas contendo 0,0; 0,15% de betaína e com a dieta contendo promotor de produção e coccidiostático não apresentaram média de rendimento de carcaça diferente do tratamento que recebeu 0,10% de betaína na ração (p>0,05).
Enquanto os dados deste trabalho mostraram que o menor nível de betaína proporcionou uma redução no rendimento de carcaça em relação ao nível de 0,10%, ESTEVE-GARCIA & MACK (2000) observaram uma melhoria significativa no rendimento de carcaça de frangos corte aos 41 dias de idade que não foram infectados com Eimeria e que receberam 0,05% de betaína na ração.
Para peso do peito e rendimento do peito (Tabela 9), não foram observadas diferenças significativas entre os tratamentos (p>0,05). Estes dados diferem dos resultados descritos por HRUBY (2002), que relataram uma melhoria no rendimento de peito variando de 1,4 a mais de 11% em vários trabalhos realizados com aves submetidas a diversos tipos de desafios ambientais (baixo, médio e alto), na qual, aquelas que foram expostas a situações de médio-alto desafio, apresentaram um aumento no rendimento de peito na medida em que a betaína foi adicionada a dieta (0,10 a 0,15%) comparada com a ração controle (sem adição de betaína).
REMUS (2001) também relatou que em três trabalhos realizados no EUA, a suplementação de 0,1% de betaína numa dieta a base de milho, soja, canola, carne e subprodutos de carne em perus com idade de abate, aumentou significativamente o rendimento de peito em média de 3%. Outros autores também descreveram melhoria
y = 0 .6 1 9 3 x + 1 .8 3 6R 2 = 0 .5 9 0 2
1 .8 1 0
1 .8 4 0
1 .8 7 0
1 .9 0 0
1 .9 3 0
0 .0 0 0 .0 5 0 .1 0 0 .1 5
N ív e is d e b e ta ín a (% )
Peso
da
carc
aça
resf
riad
a (k
g)

- 81 -
no rendimento de peito de frangos e perus com a suplementação de betaína na dieta comparada com o controle, sem betaína (VIRTANEN & RUMSEY, 1996; HRUBY et al., 2001; LUNDEEN, 2001).
As aves que receberam a dieta com promotor de produção e coccidiostático na ração (Tabela 9) apresentaram maior peso de coxa mais sobrecoxa e de dorso (p<0,05) em relação àquelas que receberam a dieta suplementada com 0,05% de betaína. No entanto, as aves alimentadas com os níveis de 0,0; 0,10 e 0,15% de betaína não diferiram entre si (p>0,05) e nem daquelas suplementadas com promotor de produção e coccidiostático (p>0,05). Porém, houve uma tendência de aumento no peso da coxa mais sobrecoxa dos frangos ao se adicionar os níveis de 0,10 e 0,15% de betaína na ração.
Conforme os resultados apresentados na Tabela 9, não houve diferença estatística entre os tratamentos (p>0,05) para rendimento da coxa mais sobrecoxa, rendimento do dorso, rendimento da asa e peso da asa. Dados sobre a suplementação dietética de betaína para peso da carcaça resfriada, peso do peito, peso e rendimento de: (coxa mais sobrecoxa, dorso e asa), em frangos de corte são ainda escassos na literatura.
Segundo os dados apresentados na Tabela 9, os frangos alimentados com a ração contendo promotor de produção mais coccidiostático e com a ração sem suplementação de betaína (0,0%), apresentaram maior conteúdo de gordura abdominal (p<0,05) em detrimento aqueles que receberam 0,05% de betaína na dieta. Contudo, não houve diferença no teor de gordura abdominal (p>0,05) das aves que receberam os níveis de 0,05; 0,10 e 0,15% de betaína na ração.
A percentagem de gordura abdominal não foi afetada (p>0,05) pelos diferentes tratamentos (Tabela 9). Apesar de não ter sido encontrada diferença significativa para rendimento de gordura abdominal, é possível observar no presente estudo que na medida em que o nível de betaína é aumentado, há uma tendência de redução tanto do peso, quanto da percentagem de gordura abdominal das aves, onde o nível de 0,05%, mostra-se mais efetivo com relação à redução da gordura, podendo-se entender que a betaína tem um papel mais efetivo no metabolismo dos lipídeos que o promotor de produção com o coccidiostático, cuja finalidade pode estar na redução da gordura abdominal da carcaça de frangos de corte.
KIDD et al. (1997) ao fazer comentários das aplicações da betaína no metabolismo dos lipídeos, comenta que SAUNDERSON & MACKINLAY (1990) observaram uma redução de gordura na carcaça das aves alimentadas com a dieta suplementada com a combinação de betaína e metionina, comparada com a dieta suplementada com a combinação de colina e metionina. Outros autores também observaram redução do rendimento de gordura abdominal de frangos de corte aos 45 dias de idade na medida em que foram adicionados níveis crescentes de betaína na dieta: 0,0, 0,05, 0,10 e 0,15% ( HRUBY et al., 2001).
Analisando-se os resultados das características da carcaça de uma maneira geral, foi possível observar que o nível de 0,10% de betaína foi efetivo na melhoria do rendimento de carcaça, e que, tanto a suplementação de betaína quanto à adição de promotor de produção e coccidistático na dieta, apresentaram o mesmo efeito nas

- 82 -
características da carcaça das aves, sugerindo que a betaína pode ser uma possível alternativa ao promotor de produção nas rações formuladas para frangos de corte.
O rendimento da carcaça, o peso da coxa mais sobrecoxa e o peso do dorso foram reduzidos nas aves alimentadas com 0,05% de betaína na ração, sendo este, portanto, considerado um nível não indicado para estas características, exceto, para a redução do teor de gordura abdominal, que foi tido como o melhor nível.

- 83 -
5 CONCLUSÕES
O desempenho dos frangos submetidos a coccidiose foi positivamente afetado pela suplementação de betaína na ração aos 42 dias de idade; O nível 0,10% de betaína na dieta melhorou o rendimento de carcaça dos frangos aos 43 dias de idade; O nível 0,05% de betaína não foi indicado para melhorar as características de carcaça dos frangos de corte aos 43 dias de idade, exceto, para a redução do teor de gordura abdominal.
A betaína pode ser de utilidade, em conjunto com outras estratégias nutricionais, para viabilizar a retirada dos antibióticos como produtores de produção das dietas de frangos de corte.

- 84 -
6 REFERÊNCIAS BIBLIOGRÁFICAS
ALLEN, P.C. Physiological responses of chicken gut tissue to infection with Eimeria acervulina. Avian Disease, v. 28, p. 868-876, 1983.
ALLEN, P.C. Physiological responses of chicken gut tissue to coccidial infections: comparative effects of Eimeria acervulina and Eimeria mitis on mucosal mass, carotenoid content, and brush border enzyme activity. Poultry Science, v. 66, p. 1306-1315, 1987.
AUGUSTINE, P.C.; McNAUGHTON, J.L.; VIRTANEN, E.; ROSI, L. Effect of betaine on the growth performance of chicks inoculated with mixed cultures of avian Eimeria species and on invasion and development of Eimeria tenella and Eimeria acervulina in vitro and in vivo. Poultry Science, v. 76, p.802-809, 1997.
AYLOTT, M.V.; VESTAL, O.H.; STEPHENS, J.F.; TURK, D.E. Effect of coccidial infection upon passage rates of digestive tract contents of chicks. Poultry Science, v. 47, p. 900-904, 1968.
BAGNASCO, S.; BALABAN, R.; FALES, H.M.; YANG, Y.; BURG, M. Predominant osmotically active organic solutes in rat and rabbit renal medullas. Journal of Biological Chemistry, v. 261, n. 13, p. 5872-5877, 1986.
BAKER, D.H. Amino acid interactions with vitamins, minerals and drugs. In:, NUTRITION INSTITUTE ON AMINO ACIDS, 1984, West Des Moines. Handbook… West Des Moines, IA, USA: National Feed Ingredients Association, 1984, p. 1-9.
BEAVER, P.C.; JUNG, R.C.; CUPP, E.W. Clinical Parasitology. 9th ed. Philadelphia, PA, ed. Lea and Febiger, 1984.
BURG, M.B. Molecular basis of osmotic regulation. American Journal of Physiology, v. 268, p. f983-f996, 1995.
CHAMBERS, S.T.; KUNIN, C.M. The osmoprotective proprieties of urine for bacteria: the protective effect of betaine and human urine against low pH and high concentrations of electrolytes, sugars, and urea. Journal of Infectious Diseases, v. 152, p. 1308-1316, 1985. CREVIEW, G.I.; NACIRI, M. Dietary effect on chichen coccidiosis. Productions – Animales, v. 14, n. 4, p. 231-246, 2001.

- 85 -
DRAGOLOVICH, J. Dealing with salt stress in animal cells: the role and regulation of glycine betaine concentrations. Journal of Experimental Zoology, v. 168, p. 139-144, 1994. DU VIGNEAUD, V. Dynamic processing of neuropeptides. Universities Cornell Press, 1952, p. 59. ESTEVE-GARCIA, E.; MACK, S. The effect of DL-methionine and betaine on growth performance and carcass characteristics in broilers. Animal Feed Science and Technology, v. 87, p. 85-93, 2000. FERRARIS, J.D.; BURG, M.B.; WILLIAMS, C.K.; PETERS, E.M.; GARCÍA, P. Betaine transporter cDNA cloning and effect of osmolytes on its mRNA induction. American Journal of Phisiology, v. 39, p. C650-C654, 1996.
FERREIRA, D.F. SISVAR – Sistema de Análise Estatística. Lavras: UFLA, 2000. Comunicação pessoal.
FETTERER, R.H.; AUGUSTINE, P.C.; ALLEN, P.C.; BARFIELD, R.C. The effect of dietary betaína on intestinal and plasma levels of betaína in uninfected and coccidian-infected broiler chicks. Parasitol Res, v. 90, p. 343-348, 2003.
FOX, M.C.; SOUTHERN, L.L. Effect of ammonium chloride ingestion on Eimeria acervulina-infected chicks fed excess copper. Poultry Science, v. 66, p. 1019-1022, 1987.
FRIEDEL H.A.; GOA K.L.; BENFIELD P. S-adenosyl-L-methionine: a review of its pharmacological properties and therapeutic potential in liver dysfunction and affective disorders in relation to its physiological role in cell metabolism. Drugs, v. 38, n.3, p. 389-416, 1989.
FRONTIERA, M.S.; STABLER, S.P.; KOLHOUSE, J.F.; ALLEN, R.H. Regulation of methionine metabolism: effect of nitrous oxide and excess methionine. Journal of Nutritional Biochemistry, v. 5, p. 28-38, 1994.
HAUSSINGER, D. The role of cellular hydration in the regulation of cell function: review article. Biochemistry Journal, Great Britain, v. 313, p. 697-710, 1996.
HEIN, H. The pathogenic effects of Eimeria acervulina in young chicks. Experimental Parasitology, v. 22, p. 1-11, 1968. HRUBY, M.; BARRATT, A.; REMUS, J. Betaine improves poultry carcass characteristics. Poultry Digest, 2001.

- 86 -
HRUBY, M. Increasing breast meat yield in broilers and turkeys. Watt Publishing Company, Poultry International, 2002.
KIDD, M.T.; FERKET, P.R.; GARLICH, J.D. Nutritional and osmoregulatory functions of betaine. World’s Poultry Science Journal, v. 53, p. 125-139, 1997.
KLASING, K.C.; ADLER, K.L.; REMUS, J.C.; CALVERT, C.C. Dietary betaína increases intraepithelial lymphocytes in the duodenum of coccidian-infected chicks and increases functional properties of phagocytes. Journal of Nutrition, v. 132, p. 2274-2282, 2002.
KOUWENHOVEN, B. The significance of infection dose, body weight and age of the host on the pathogenic effect of Eimeria acervulina in the fowl. Zoology Parasitenked, v. 39, p. 255-268, 1972.
KOUWENHOVEN, B.; VAN DER HORST, C.J.G. Disturbed intestinal absorption of vitamin A and carotenes and the effect of a low pH during Eimeria acervulina infection in the domestic fowl (Gallus domesticus). Zoology Parasitenked, v. 38, p. 152-161, 1972.
LUNDEEN, T. Methionine, betaína supplementation improves turkey breast meat yield. Feedstuffs, v. 1, 2001.
LOWRY, K.R.; IZQUIERDO, Q.A.; BAKER, D.H. Efficacy of betaína relative to choline as a methyl donor. Poultry Science, v. 55 (suppl.), n 135 (abstr), 1987.
MANN, P.J.G.; WOODWARD, H.E.; QUASTEL, J.H. Hepatic oxidation on choline and arsenocholine. Biochemistry Journal, v. 32, p. 1024-1032, 1938.
MANUAL DE FRANGO DE CORTE. Uberlândia: granja Planalto Ltda, jan., 2003, 34p.
MATTHEWS, J.O.; WARD, T.L.; SOUTHERN, L.L. Interactive effects of betaína and monensin in uninfected and Eimeria acervulina-infected chicks. Poultry Science, v. 76, p. 1014-1019, 1997.
MATTHEWS, J.O.; SOUTHERN, L.L. The effect of dietary betaína in Eimeria acervulina-infected chicks. Poultry Science, v. 79, p. 60-65, 2000.
McBRIDE, B.W.; MILLIGAN, L.P. Journal of Biochemistry, v. 17, n. 43, 1985.
McBRIDE, B.W.; KELLY, J.M. Energy cost of absorption and metabolism in the ruminant portal-drained viscera and liver: a review. Journal of Animal Science, v. 68, p. 2997-3010, 1990.

- 87 -
McDONALD, V.; BALLINGAL, S.; SHIRLEY, M.W. A preliminary study of the nature of infection and immunity in chickens given an attenuated line of Eimeria acervulina. Parasitology, v. 84, p. 21-30, 1982.
McKENZIE, M.E.; COLNAGO, G.L.; LEE, S.R.; LONG, P.L. Gut stasis in chickens infected with Eimeria. Poultry Science, v. 66, p. 264-269, 1987.
MOLITORIS, B.A.; BAKER, D.H. Choline utilization in the chick as influence by dietary protein and methionine. Journal of Nutrition, v.106, p. 412-418, 1976.
MOREHOUSE, N.F.; McGUIRE The pathogenicity of Eimeria acervulina. Poultry Science, v. 37, p. 665-672, 1958.
NEWBERNE, P. M. The methyl-deficiency model: history, characteristics and research directions. Journal of Nutrition Biochemistry, v. 4, p. 618-624, 1993.
PETRONINI, P.G.; De ANGELIS, E.M.; BORGHETTI, P.; BORGHETTI, A.F.; WHEELER, K.P. Modulation by betaine of cellular responses to osmotic stress. Biochemistry Journal, v. 282, p. 69-73, 1992.
PETRONINI, P. G.; De ANGELIS, E. M.; BORGHETTI, A. F.; WHEELER, K. P. Effect of betaine on HSP70 expression and cell survival during adaptation to osmotic stress. Biochemical Journal, v. 293, p. 553-558, 1993. REMUS, J.; VIRTANEN E.; ROSI, L.; McNAUGHTON, J. In: WORLD’S POULTRY CONGRESS, 1996. Procedings…1996, v. 4, n. 336, p. 2-5.
REMUS, J.C. Betaine for increased breast meat yield in turkeys. Elsevier, World Poultry, v. 17, n. 2, 2001.
RUFF, M.D.; REID, W.M. Coccidiosis and intestinal pH in chickens. Avian Disease, v. 19, p. 52-58, 1975.
RUFF, M.D.; WITLOCK, D.R.; SMITH, R.R. Eimeria acervulina and E. tenella: Effect on methionine absorption by the avian intestine. Experimental Parasitology, v. 39, p. 244-251, 1976.
SAUNDERSON, C.L.; MACKINLAY, J. Changes in body-weight, composition and hepatic enzymes activities in response to dietary methionine, betaine and choline levels in growing chicks. British Journal of Nutrition, v. 63, p. 339-349, 1990.

- 88 -
SCHUTTE, J.B.; DE JONG, J.; SMINK, W.; PACK, M. Replacement value of betaína for DL-methionine in male broiler chicks. Poultry Science, v. 76, p. 321-325, 1997. SILVERSIDES, F.G.; NEWCOMBE, M.; REMUS, J. Betaine, coccidiosis and coccidiostats in poultry. Elsevier, World Poultry Special, 1999. SMITH, M.O. Parts yield of broiler reared under cycling high temperatures. Poultry Science, v. 72, p. 1146-1150, 1993. SMOLIN, L.A.; BENEVEGA, N.J. Methionine, homocysteine, cysteine-metabolic interrelationships. In: MENDEL FRIEDMAN: Editor. Absorption and utilization of amino acids. CRC Press, Inc., Boca Raton, 1989, v. 1, p. 155-187. SOUTHERN, L.L.; BAKER, D.H. Eimeria acervulina infection and the zinc-copper interrelationship in the chick. Poultry Science, v. 62, p. 401-404, 1983.
STEKOL, J.A.; HSU, P.T.; WEISS, S.; SMITH, P. Labile methyl group and its synthesis de novo in relation to growth in chicks. Journal of Biological Chemistry, v. 203, p. 763-773, 1953.
STEPHENS, J.F.; BORST, W.J.; BARNETT, B.D. Some physiological effects of Eimeria acervulina, E. brunetti, and E. mivati infections in young chickens. Poultry Science, v. 53, p. 1735-1742, 1974.
TEETER, R.G.; REMUS, J.C.; BELAY, T.; MOONEY, M. VITANEN, E.; AUGUSTINE, P. The effects of betaine on water balance and performance in broilers reared under differing environmental conditions. In: AUSTRALIAN POULTRY SCIENCE SYMPOSIUM, 1999, Australia. Proceding… Australia,1999, p.165.
TURK, D.E.; STEPHENS, J.F. Effects of serial inoculations with Eimeria acervulina or Eimeria necatrix upon zinc and oleic acid absorption in chicks. Poultry Science, v. 49, p. 523-526, 1970.
TURK, D.E. The effects of coccidiosis on intestinal function and gut microflora. In: LONG, P.L.; BOORMAN, K.N.; FREEMAN, B.M. Avian Coccidiosis. Edinburg, Scotland: Editor British Poultry Science Ltda, 1978, p. 227-267.
TYLER, D.D. Transport and oxidation of choline by liver mitochondria. Biochemistry Journal, v. 166, p. 571-581, 1977.
USDA NUTRIENTE DATABASE FOR STANDARD REFERENCE, release 13, 1999. Disponível em: http://www.nal.usda.gov/fnic/foodcomp.

- 89 -
VIRTANEN, E.; McNAUGHTON, J.; ROSI, L.; MAY, D. Effects of betaine supplementation on intestinal lesions, mortality, and performance of coccidian-challenged broiler chicks. In: EUROPEAN SYMPOSIUM IN POULTRY NUTRITION, 9, 1993, Jelenia Gora. Proceedings… Jelenia Gora - Poland, 1993. VIRTANEN, E. Piecing together the betaine puzzle. Feed Mix, v. 3, n. 1, p. 12-17, 1995.
VIRTANEN, E.; ROSI, L. Effects of betaine on methionine requirements of broilers under various environmental conditions. In: THE AUSTRALIAN POUTRY SCIENCE SYMPOSIUM, 1995, Sidney. Proceedings… Sidney – Australia: University of Sidney, 1995, v. 7, p. 88-98. VIRTANEN, E.; RUMSEY, G. Betaine supplementation can optimize use of methionine, choline in diets. Feedstuffs, v. 68, n. 42, p. 12-13, 1996. VIRTANEN, E.; REMUS, J.; ROSI, L.; McNAUGHTON, J.; AUGUSTINE, P. The effect of betaína and salinomycin during coccidiosis in broilers. Poultry Science, v. 75 (suppl. 1), n. 149, 1996. WALDENSTEDT, L.; ELWINGER, K.; THEBO, P.; UGGLA, A. Effect of betaína supplement on broiler performance during a n experimental coccidial infection. Poultry Science, v. 78, p. 182-189, 1999.
WALDENSTEDT, L.; THEBO, P.; LUDEN, A. Broiler production without coccidiostats. Svensk – Veterinartidning, v. 52, n. 2, p. 77-84, 2000.
WUNZ, M.; WRIGHT, S. Betaine transport in rabbit renal brush-border membrane vesicles. American Journal of Physiology, v. 264. p. F948-F955, 1993.
YANCEY, P.H.; CLARK, M.E.; HAND, S.C.; BOWLUS, R.D.; SOMERO, G.N. Living with water stress: evolution of osmolyte systems. Science, v.217, p. 1214-1223, 1982.
ZHANG, F.; WARSKLAT, U.; HAUSSINGER, D. Modulation of tumor necrosis factor alfa release by anisoosmolarity and betaine in rat liver macrophages (Kupffer cells). FEBS Lett., v. 391, p. 239-296, 1996.

- 90 -
7 CONCLUSÕES
Considerando-se o os parâmetros de desempenho aos 42 dias de idade, as características de carcaça, a digestibilidade da ração e a altura das vilosidades, a suplementação de betaína foi considerada positiva em rações de frangos de corte expostos a E. acervulina;
A betaína pode ser de utilidade, em conjunto com outras estratégias nutricionais, para a viabilização da retirada dos antibióticos como promotores de produção das dietas de frangos de corte;





























