Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE SANTA CATARINA
CENTRO DE CIÊNCIAS BIOLÓGICAS
GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
Vanessa Miranda
AVALIAÇÃO DA FENOLOGIA E MATURAÇÃO DE VARIEDADES DE
VIDEIRA POTENCIAIS PARA OS VALES DA UVA GOETHE
Florianópolis
2017

Vanessa Miranda
AVALIAÇÃO DA FENOLOGIA E MATURAÇÃO DE VARIEDADES DE
VIDEIRA POTENCIAIS PARA OS VALES DA UVA GOETHE
Trabalho de Conclusão de Curso
apresentado ao Curso de Graduação em
Ciências Biológicas da Universidade
Federal de Santa Catarina, como requisito
para a obtenção do grau de Bacharel em
Ciências Biológicas.
Orientador
Prof. Dr. Aparecido Lima da Silva
Coorientador
Me. Tiago Camponogara Tomazetti
Florianópolis
2017


Vanessa Miranda
AVALIAÇÃO DA FENOLOGIA E MATURAÇÃO DE VARIEDADES DE
VIDEIRA POTENCIAIS PARA OS VALES DA UVA GOETHE
Este Trabalho de Conclusão de Curso foi julgado adequado para a obtenção do Grau de
Bacharel em Ciências Biológicas e aprovado em sua forma final pelo Curso de Ciências
Biológicas.
Florianópolis, 4 de julho de 2017.
Prof. Dr. Carlos Roberto Zanetti
Coordenador do Curso de Ciências Biológicas
Banca Examinadora:
Me. Tiago Camponogara Tomazetti
Presidente
Prof. Dr. José Afonso Voltolini
Membro Titular
Dr. Marcelo Borghezan
Membro Titular

Este trabalho é dedicado às minhas avós,
Eulália e Dalmira (in memoriam), que me
ensinaram o amanho da terra em seus jardins.

AGRADECIMENTOS
À minha família, por permitir que eu chegasse até esse momento.
Ao professor Aparecido Lima da Silva, por aceitar me orientar e por sempre dizer “tudo
vai dar certo” mesmo nas horas de dificuldade. Por todas as conversas, ensinamentos e
pela amizade sincera.
Ao meu coorientador Tiago Camponogara Tomazetti, que esteve ao meu lado em todas
as partes desse trabalho, ensinando e ajudando. Pelas viagens até Urussanga e todo o
serviço de campo. Pela paciência durante os experimentos no laboratório e redação de
trabalhos.
À Márcia Denise Rossarolla, por ter me aconselhado em alguns momentos de desânimo
e pelo auxílio durante os experimentos.
À Angela Costa, por ter sido de extrema ajuda no último dia de análises.
A todos do NEUVIN que de alguma forma me auxiliaram durante esse trabalho, pelas
risadas, viagens e aprendizados.
À Estação Experimental da Epagri de Urussanga, em especial ao técnico Olívio que foi
fundamental na realização dessa pesquisa.
Ao meu parceiro Matheus Henrique Stofela Sarolli, por ter estado ao meu lado durante
todo este percurso. Pelo acalento nos momentos difíceis. Pelas risadas sem fim. Pela
companhia, pelas conversas, pelas aventuras e pelo amor.
Aos meus colegas do curso de Ciências Biológicas, pela parceria nos seminários, saídas
de campo e peripécias sem fim. À panelinha, pela amizade, por serem os melhores.
A todos os grandes professores do curso de Ciências Biológicas, por todo o conhecimento
passado com paciência e maestria.
Obrigada.

“Il y a plus de philosophie dans une
bouteille de vin que dans tous les livres. ”
(Louis Pasteur)

RESUMO
A vitivinicultura nos “Vales da Uva Goethe” se inicia com a imigração italiana no
final do século XIX. A região, nessa época, produzia vinhos para consumo local e que
seriam também levados para grandes cidades brasileiras. Atualmente a viticultura avança
na região, mesmo com o gargalo do míldio, doença responsável por prejuízos
significativos na produção vitícola do Brasil e do mundo. Visando coibir essas perdas,
alguns programas de melhoramento genético da videira buscam gerar genótipos com
resistência ao míldio, oídio e outras doenças. Para a consolidação de novas variedades
resistentes, estudos devem ser realizados para observar sua adaptação em diferentes
regiões e climas. Neste contexto, o objetivo com este trabalho foi avaliar aspectos
produtivos e fenológicos de 11 genótipos de videira resistentes ao míldio e oídio na região
de Urussanga, SC, durante o ciclo 2016/2017. O estudo foi efetuado na Estação
Experimental da Epagri de Urussanga (altitude 49 m). Durante todo o ciclo foi realizado
o monitoramento das variáveis climáticas de temperatura do ar, radiação solar global,
precipitação pluviométrica e umidade relativa do ar. Foram calculados os índices
bioclimáticos de Winkler e Huglin, além do requerimento térmico. O número de nós por
sarmento foi contado quinzenalmente, gerando o plastocrono para 5 genótipos
selecionados. Os estágios de brotação, floração, véraison e maturação foram
determinados durante o ciclo semanalmente. Durante a maturação das uvas foi observada
a evolução dos sólidos solúveis totais, pH, acidez total titulável, antocianinas e polifenóis
totais. A região de Urussanga foi classificada como “Região V” pelo Índice de Winkler e
“Região muito quente” pelo Índice de Huglin. O genótipo GF-6 apresentou brotação mais
precoce e ciclo mais longo, totalizando 121 dias e acumulando 1190 °C dia. “Helios”
apresentou o ciclo mais curto, totalizando 106 dias e acumulando 1080,7 °C dia. A
variedade Baron apresentou o maior requerimento térmico, acumulando 1198,9 °C dia.
Os valores de plastocrono ficaram entre 39,5 °C dia nó-1 para ‘Regent’ e 73,5 °C dia nó-1
para ‘Bronner’. O número total de folhas por planta variou entre 115 folhas para ‘Regent'
e 230,6 folhas para o genótipo GF-6. Fatores climáticos apresentaram forte influência
sobre a maturação das uvas, reduzindo a concentração de sólidos solúveis totais e
compostos fenólicos. Considerando estes resultados, a região apresenta potencial para a
produção de vinhos jovens, que não exijam grande período de envelhecimento.
Palavras-chave: viticultura, resistência a patógenos, adaptação de variedades, fenologia,
plastocrono, maturação.

ABSTRACT
The viticulture in the “Vales da Uva Goethe” begins with the Italian immigration
by the end of the 19th century. This region, at the time, produced wine for local
consumption and also for big Brazilian cities. Nowadays the viticulture spreads
throughout the region, even with the harm brought by downy mildew, a disease
responsible for a significant damage inflicted upon the winegrowing in Brazil and across
the globe. In order to cover these losses, some grapevine genetic improvement programs
objective the generation of genotypes resistant to downy and powdery mildew as well as
other diseases. For the consolidation of new resistant varieties, studies must be conducted
to observe their adaptive capability in different regions and climates. In this context, the
goal of this work was to evaluate the productive and phenological aspects of 11 grapevine
genotypes resistant to downy and powdery mildew in the city of Urussanga, SC, on the
2016/2017 cycle. The research took place at Epagri’s Experimental Station in Urussanga
(49 mamsl). During the whole cycle, there was a monitoring of climatic variables, such
as air temperature, global solar radiation, rainfall and relative humidity. Winkler’s and
Huglin’s bioclimatic indexes were calculated, in addition to the thermal requirement. The
number of nodes per cane was counted once every two weeks providing the plastochron
index for 5 selected genotypes. The bud break, flowering, véraison and harvest stages
were determined during the vegetative cycle on a weekly basis. During the maturation,
the evolution of the total soluble solids, pH, titratable acidity, total anthocyanins and
polyphenols were observed. The Urussanga was classified as "Region V" by Winkler's
index and "Very warm region" by Huglin's index. The GF-6 genotype had the earliest bud
break and the longest cycle, totaling 121 days and accumulating 1190 °C day. "Helios"
showed the shortest cycle, with a total of 106 days and accumulating 1080,7 °C day. The
Baron variety showed the highest thermal requirement, accumulating 1198,9 °C day. The
plastochron index were between 39,5 °C day per node for ‘Regent’ and 73,5 °C day per
node for ‘Bronner’. The total number of leaves per plant varied between 115 leaves for
‘Regent' and 230,6 leaves for the GF-6 genotype. Climatic factors had a strong influence
on the maturation of the grapes, reducing the concentration of total soluble solids and
phenolic compounds. Considering these results, the region presentes good potential for
the production of young wines, wich does not require aging time.
Keywords: viticulture, resistance to pathogens, varieties adaptation, phenology,
plastochron index, maturation.

LISTA DE FIGURAS
Figura 1. Localização de Urussanga no estado de Santa Catarina .................................. 4
Figura 2. Genealogia da variedade Bronner .................................................................... 6
Figura 3. Genealogia da variedade Baron ....................................................................... 7
Figura 4. Genealogia da variedade Regent ...................................................................... 8
Figura 5. Genealogia da variedade Calandro ................................................................... 8
Figura 6. Genealogia da variedade Helios ....................................................................... 9
Figura 7. Genealogia da variedade Prior ........................................................................ 10
Figura 8. Genealogia da variedade Felicia ..................................................................... 10
Figura 9. Genealogia da variedade Aromera .................................................................. 11
Figura 10. Cachos das variedades Baron (A), Calandro (B), Prior (C), Regent (D),
Aromera (E), Bronner (F), Felicia (G) e Helios (H) ....................................................... 12
Figura 11. Diagrama mostrando o aumento de volume do fruto a partir da antese ....... 15
Figura 12. Estágio de ponta verde (A), inflorescências surgindo junto às folhas (B), queda
da caliptra (C), floração plena (D), crescimento dos frutos (E), véraison (F) e maturação
(G) .................................................................................................................................. 16
Figura 13. Somatório da precipitação pluviométrica mensal (mm) e médias mensais da
umidade relativa do ar (%), na região de Urussanga – SC, ciclo 2016/2017 .................. 24
Figura 14. Valores médios por hora da radiação solar global (W m-2) durante o período
de maturação dos genótipos no município de Urussanga-SC, Brasil, ciclo 2016/2017
........................................................................................................................... 25
Figura 15. Duração (dias) de cada estágio fenológico para cada genótipo, na região de
Urussanga, SC, ciclo 2016/2017 ............................................................................ 29
Figura 16. Requerimento térmico (°C dia) através do Método 3.3 para cada genótipo, em
cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017 ...................... 29

LISTA DE TABELAS
Tabela 1. Temperaturas médias, mínimas e máximas do ar (°C), amplitude térmica média
(°C), precipitação (mm) e umidade relativa do ar (%) de cada variedade (B – Brotação, F
– Floração, V – Véraison, M – Maturação), em cada estágio fenológico, na região de
Urussanga, SC, ciclo 2016/2017 ............................................................................ 23
Tabela 2. Requerimento térmico (°C dia) através do Índice de Winkler para cada
variedade, em cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017 .. 26
Tabela 3. Requerimento térmico (°C dia) através do Índice de Huglin para cada genótipo,
em cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017 ................. 27
Tabela 4. Data de ocorrências dos estágios fenológicos dos genótipos estudados na região
de Urussanga, SC, ciclo 2016/2017 ........................................................................ 28
Tabela 5. Requerimento térmico (°C dia) através do Método 3.3 para cada genótipo, em
cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017 ...................... 28
Tabela 6. Plastocrono (°C dia nó-1), coeficiente de determinação (R2), número de
sarmentos por planta e número de nós por sarmento dos genótipos selecionados, região
de Urussanga, SC, ciclo 2016/2017......................................................................... 30
Tabela 7. Valores de SST, pH e ATT de cada genótipo, durante o período de maturação,
na região de Urussanga, SC, ciclo 2016/2017 .......................................................... 32
Tabela 8. Valores de PT de cada genótipo e AMT das variedades tintas, durante o período
de maturação, na região de Urussanga, SC, ciclo 2016/2017 ..................................... 38

LISTA DE ABREVIATURAS E SIGLAS
= Absortividade Molar
°Brix = Grau Brix
AT = Absorbância Total
ATT = Acidez Total Titulável
CIRAM = Centro de Informações de Recursos Ambientais e de Hidrometeorologia de
Santa Catarina
DF = Fator de Diluição
Epagri = Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina
h = Hora
HCl = Ácido Clorídrico
km = Quilômetro
m = Metro
M = Molar
meq = Miliequivalente
mL = Mililitro
mm = Milímetro
N = Normal
NaOH = Hidróxido de Sódio
nm = Nanômetro
O = Oeste
ºC = Grau Celsius
PM = Peso Molecular
PT = Polifenóis Totais
r = Coeficiente de Correlação
R² = Coeficiente de Determinação
S = Sul
SC = Santa Catarina
SST = Sólidos Solúveis Totais
STd = Soma Térmica Diárias
Tb = Temperatura de Base Inferior
TB = Temperatura de Base Superior
Tmax = Temperatura Máxima
Tmean = Tméd = Temperatura Média
Tmin = Temperatura Mínima
Tot = Temperatura Ótima
UFSC = Universidade Federal de Santa Catarina
W = Watt Σ = Somatório

SUMÁRIO
1. Introdução ................................................................................................................ 1
2. Objetivos .................................................................................................................. 3
2.1. Objetivo Geral ......................................................................................................... 3
2.2. Objetivos Específicos .............................................................................................. 3
3. Revisão Bibliográfica .............................................................................................. 4
3.1. Características geográficas e histórico vitivinícola da região de Urussanga ........... 4
3.2. Variedades de videiras resistentes ........................................................................... 5
3.2.1. Variedade Bronner .............................................................................................. 5
3.2.2. Variedade Baron ................................................................................................. 6
3.2.3. Variedade Regent ............................................................................................... 7
3.2.4. Variedade Calandro ............................................................................................ 8
3.2.5. Variedade Helios ................................................................................................ 9
3.2.6. Variedade Prior ................................................................................................... 9
3.2.7. Variedade Felicia .............................................................................................. 10
3.2.8. Variedade Aromera ........................................................................................... 11
3.3. Caracterização fenológica ...................................................................................... 12
3.3.1. Índices Bioclimáticos e Requerimento Térmico ............................................... 16
4. Material e Métodos ................................................................................................ 19
5. Resultados e Discussão .......................................................................................... 22
5.1. Variáveis Climáticas .............................................................................................. 22
5.1.1. Temperatura do Ar e Amplitude Térmica ......................................................... 22
5.1.2. Precipitação Pluviométrica e Umidade Relativa .............................................. 24
5.1.3. Radiação Solar .................................................................................................. 24
5.2. Índices Bioclimáticos ............................................................................................ 25
5.3. Caracterização Fenológica e Requerimento Térmico ............................................ 27
5.3.1. Plastocrono ....................................................................................................... 30
5.4. Avaliação da Maturação ....................................................................................... 30
5.4.1. Sólidos Solúveis Totais ................................................................................... 30
5.4.2. pH .................................................................................................................... 31
5.4.3. Acidez Total Titulável ..................................................................................... 31
5.4.4. Antocianinas Monoméricas Totais .................................................................. 32
5.4.5. Polifenóis Totais .............................................................................................. 32
6. Conclusões ................................................................................................................ 34
REFERÊNCIAS ........................................................................................................... 35
ANEXOS ........................................................................................................... 41

1
1. Introdução
A videira é uma planta trepadeira com gavinhas, caule lenhoso e porte arbustivo.
Suas folhas são alternas, pecioladas, cordiformes, com cinco lóbulos sinuados dentados,
glabras na parte superior e tomentosas na parte inferior. As flores são pequenas e de cor
branco esverdeado, dispostas em racimos (POMMER, 2003). O cacho é composto por
uma parte lenhosa e pelas bagas. O formato, cor e consistência das bagas variam de acordo
com a cultivar (AMARANTE, 2015).
O gênero Vitis, ao qual pertence a videira, está inserido na família Vitaceae, que
possui 910 espécies dispostas em 14 gêneros (CHRISTENHUSZ; BYNG, 2016).
Majoritariamente composta por trepadeiras, a família Vitaceae apresenta distribuição
cosmopolita, tendo como principais centros de diversidade as Américas, a África e o
sudeste asiático (STEVENS, 2017).
A uva é considerada um dos frutos de domesticação mais antiga que se tem
conhecimento, devido ao registro de muitas civilizações (SOUSA, 1996), tendo surgido
no início do período Terciário, provavelmente na Groenlândia (ALVARENGA et al.,
1998). A videira foi inicialmente cultivada há cerca de 11.000 anos, na área conhecida
como Crescente Fértil do Mediterrâneo Oriental, onde hoje se localizam os países do
Líbano, Síria, Turquia, Irã, Jordânia, Iraque e Israel (RAVEN; EVERT; EICHHORN,
2014).
Com o início das Grandes Navegações, a viticultura que antes se concentrava na
Europa se espalhou para os demais continentes. No Brasil, as primeiras plantas foram
introduzidas no Sudeste por Martim Afonso de Souza, em 1532. Porém, o cultivo só
obteve sucesso quando os jesuítas a levaram para o sul, onde as características climáticas
eram mais adequadas para o desenvolvimento da videira (DALL'AGNOL, 2007).
O estado de Santa Catarina produziu 34.116 toneladas de uvas em 2016
(CARVALHO et al., 2016). Em extensão de área colhida, é classificado em 4º lugar entre
os estados brasileiros, com 4.700 hectares colhidos em 2016 (EPAGRI, 2016).
A vitivinicultura no sul catarinense, principalmente na região de Urussanga, se
confunde com a história da imigração italiana no final do século XIX. Colonizada a partir
de 1877, a região produzia vinhos para consumo local e que seriam também levados para
grandes cidades brasileiras, como Rio de Janeiro e São Paulo (VALES DA UVA
GOETHE, 2013).

2
O avanço da viticultura na região, enfrenta como importante gargalo as condições
de exposição ao oomiceto Plasmopara viticola, responsável por prejuízos significativos
na vitivinicultura do sul do Brasil, bem como nas demais áreas vitícolas do mundo
(CZERMAINSKI; SÔNEGO, 2004). Este patógeno pode causar perdas de até 100% da
produção se as condições climáticas estiverem favoráveis ao seu desenvolvimento:
temperatura entre 20 e 25 ºC e a umidade relativa do ar elevada, que pode ocasionar a
presença de água livre na superfície dos tecidos vegetais.
Além de afetar toda a parte vegetativa da videira, o P. viticola também age sobre
as inflorescências e frutos. Os primeiros sintomas são o aparecimento de manchas
amareladas e translúcidas (manchas de óleo) na face adaxial das folhas, podendo ocorrer
esporulação de cor branca na face inferior, seguida de necrose do tecido foliar e possível
desfolha precoce da planta (NAVES et al., 2005).
Outra doença que traz grandes problemas para a vitivinicultura é o oídio, causado
pelo fungo Uncinula necator. Ao contrário do míldio, o oídio requer umidade relativa do
ar baixa para o seu perfeito desenvolvimento; entretanto, é necessária temperatura
elevada, entre 20 e 27 ºC. O fungo ataca todos os órgãos da videira, exibindo manchas
difusas e esporulação branca característica. As folhas infectadas apresentam queda na
taxa fotossintética e frequentemente sofrem senescência prematura seguida de abscisão.
Outros sintomas da doença incluem flores e frutos secos que caem e/ou rachaduras nas
bagas (NAVES et al., 2005; JONES et al., 2014).
O míldio e o oídio, como outras doenças e pragas, foram levados da América do
Norte para a Europa durante o século XIX. Desde então, há um grande esforço na criação
de variedades resistentes a essas doenças. Espécies fontes de resistência têm sido
extensamente investigadas para utilização em programas de melhoramento, visando
introduzir, via retrocruzamentos, genes de resistência a estas doenças em variedades
europeias (Vitis vinifera); os exemplares mais promissores para esse objetivo são as
videiras nativas americanas e asiáticas (STAUDT; KASSEMEYER, 1995).
Além do controle de patógenos, para uma boa produtividade é fundamental que
sejam feitos estudos sobre o comportamento da cultura em relação aos fatores ambientais,
sendo este estudo chamado de fenologia. O conhecimento da fenologia de uma planta
possibilita a redução de tratamentos fitossanitários, melhoria na qualidade dos frutos e
economia de insumos (MURAKAMI et al., 2002).
As análises fenológicas, na vitivinicultura, buscam caracterizar a duração das
diferentes fases de desenvolvimento da planta e sua relação com o clima. Este

3
conhecimento desempenha um papel importante pois, por meio dele, é possível
interpretar o potencial climático de uma região e demonstrar o efeito das variações
estacionais sobre o genótipo (TERRA; PIRES; NOGUEIRA, 1998).
O processo de caracterização fenológica é especialmente necessário em
variedades introduzidas. Se forem utilizados índices bioclimáticos analisados
externamente ao local de plantio e produção, os resultados gerados podem não
corresponder às expectativas do produtor (MANDELLI, 1984).
2. Objetivos
2.1. Objetivo Geral
Avaliar aspectos fenológicos de genótipos de videira resistentes ao míldio e oídio,
cultivados na Estação Experimental da Epagri, no município de Urussanga, SC.
2.2.Objetivos Específicos
• Acompanhar as variáveis climáticas durante a safra 2016/2017 na Estação
Experimental de Urussanga;
• Acompanhar a fenologia e o requerimento térmico de 11 genótipos resistentes ao
míldio e oídio;
• Caracterizar o plastocrono de 5 desses genótipos;
• Avaliar as curvas de maturação tecnológica e fenólica.

4
3. Revisão Bibliográfica
3.1. Características geográficas e histórico vitivinícola da região de Urussanga
O município de Urussanga localiza-se na região Sul do estado de Santa Catarina,
na latitude 28° 31' 04'' S, longitude 49° 19' 15'' O e altitude de 49 m (Figura 1). Segundo
a metodologia de Köppen, o clima do município é classificado como Cfa - subtropical
úmido, sem estação seca e com verão quente. A temperatura média anual é de 19,2 ºC, a
precipitação média anual é de 1.540 mm e a umidade relativa do ar média é de 81,5%.
O solo da região apresenta frações de podzólico vermelho-amarelo álico e
cambissolo álico, com textura média à argilosa, podendo ser cascalhenta ou pedregosa
(POTTER, 2004).
Figura 1. Localização de Urussanga no estado de Santa Catarina.
Fonte: Raphael Lorenzeto de Abreu.
Santa Catarina tem a vitivinicultura presente em três regiões principais, divididas
de acordo com suas características e tradição de produção: região Tradicional, região
Nova e região Super Nova. A região Super Nova, ou de altitude, compreende as cidades
produtoras do planalto serrano. A região Nova é composta pelos municípios de Nova

5
Trento e Rodeio, além de cidades da região Oeste. Por fim, a região Tradicional abrange
duas áreas distintas dentro do estado: o Vale do Rio do Peixe e a região Carbonífera, no
Sul, composta pelos municípios de Urussanga, Pedras Grandes, Braço do Norte, Nova
Veneza e Morro da Fumaça (ROSIER, 2005). O nome “Carbonífera” se deve a extração
de carvão mineral, que é a principal atividade econômica da região há décadas.
O Núcleo Colonial de Urussanga foi fundado em 26 de maio de 1878 pelo
engenheiro Joaquim Vieira Ferreira, data em que ocorreu a chegada dos primeiros
colonizadores, em sua maioria oriundos da Comune de Longarone, província de Belluno,
na região de Vêneto, norte da Itália (BALDIN, 1999).
A região Carbonífera possui colonização predominantemente italiana. Os
imigrantes, tendo uma cultura intimamente relacionada ao vinho e a videira, trouxeram
consigo mudas de Vitis vinifera para cultivo na nova terra. O vinho produzido apresentava
baixa qualidade, pois a videira europeia não se adaptava às características climáticas
locais. A solução para manter a bebida na dieta tradicional foi o plantio de variedades
americanas, em sua maioria pertencentes à espécie V. labrusca, bem como variedades
híbridas, obtidas no cruzamento entre videiras europeias e americanas, que melhor
resistiam às doenças e pragas facilitadas pelo clima quente e úmido da região
(MARZANO, 1985).
Dessas novas variedades introduzidas, a ‘Goethe', uma variedade híbrida
americana, se tornou a uva típica da região: apresentou boa adaptação às características
climáticas e ao solo, além de produzir um vinho que agradava a população. Em 2011, o
vinho de Goethe da região de Urussanga recebeu o registro IP, tornando-se a primeira
Indicação Geográfica do estado de Santa Catarina, consolidando a identidade dos “Vales
da Uva Goethe” (PROGOETHE, 2012).
3.2. Variedades de Videira Resistentes
3.2.1. Variedade Bronner
A uva ‘Bronner’ é uma variedade branca híbrida (Figura 10), desenvolvida em
1975 por Norbert Becker, no Instituto Nacional de Viticultura em Freiburg, Alemanha.
No cruzamento foram utilizadas a variedade Merzling e o genótipo GM 6494 (JKI, 2017).
A variedade recebeu esse nome em homenagem ao farmacêutico e pioneiro
viticultor alemão, Johann Philipp Bronner (1792-1864) (WEIN-PLUS, 2016), e apresenta

6
boa resistência ao míldio e ao Botrytis, além de alta resistência ao oídio, exigindo menor
número de tratamentos com fungicidas em condições normais de produção. Essa
resistência se deve a presença de videiras selvagens americanas e asiáticas em sua
genealogia (Figura 2), como Vitis rupestris, V. lincecumii, e V. amurensis
(BEURSKENS; CRONE; HOUBEN, 2014).
A variedade ainda não é amplamente conhecida na vitivinicultura mundial por ser
consideravelmente nova, porém, é cultivada em vinhedos na Alemanha, Holanda e Itália.
Seu vinho apresenta forte acidez e aroma frutado, lembrando os vinhos de ‘Pinot Blanc’
e ‘Pinot Gris’ (SIBBUS, 2011).
A videira ‘Bronner’ foi utilizada como parental nos cruzamentos que deram
origem às variedades ‘Baron’, ‘Cabernet Carbon’ e ‘Souvignier Gris’ (todas ‘Cabernet
Sauvignon’ x ‘Bronner’) e ‘Divico’ (‘Gamaret’ x ‘Bronner’) (JKI, 2017).
Figura 2. Genealogia da variedade Bronner.
Fonte: Vanessa Miranda, adaptado de JKI (2017).
3.2.2. Variedade Baron
A uva ‘Baron’ é uma variedade tinta híbrida (Figura 10), criada em 1983 por
Norbert Becker, no Instituto Nacional de Viticultura em Freiburg, Alemanha. O
cruzamento teve como parentais as variedades Cabernet Sauvignon e Bronner (JKI,
2017).
A variedade apresenta alta resistência ao míldio e boa resistência ao oídio, devido
a sua genealogia híbrida advinda da parental ‘Bronner’ (Figura 3). O vinho de ‘Baron’
apresenta coloração tinta intensa, com alta concentração de compostos fenólicos, aroma
frutado e de especiarias (WINE PLANT, 2016).

7
Figura 3. Genealogia da variedade Baron.
Fonte: Vanessa Miranda, adaptado de JKI (2017).
3.2.3. Variedade Regent
A uva ‘Regent’ é uma variedade tinta (Figura 10), sendo um híbrido complexo
desenvolvido em 1967 por Gerhardt Alleweldt, no Instituto Julius-Kühn, Alemanha. No
cruzamento foram utilizadas as variedades Diana e Chambourcin (JKI, 2017). A
variedade foi nomeada como ‘Regent’ em alusão ao diamante “Le Régent”, um diamante
azulado de 140,64 quilates encontrado na Índia em 1698, atualmente parte do Tesouro
Real da França (PRITCHARD, 2016).
Existem amplas áreas de cultivo na Europa, principalmente na Alemanha (sendo
a 12ª variedade mais cultivada), Suíça, Polônia, Reino Unido e Bélgica, além do norte
dos Estados Unidos. O vinho de ‘Regent’ apresenta alta qualidade, sendo um vinho
encorpado de cor tinta intensa e acidez moderada, taninos presentes e aromas de cereja,
groselha e cassis (ROBINSON; HARDING; VOUILLAMOZ, 2012).
A videira ‘Regent’ apresenta brotação precoce e adaptação às regiões de frio
intenso, possui média resistência ao Botrytis e alta resistência ao oídio e ao míldio,
podendo reduzir em até 80% os tratamentos com fungicidas. Considerada como a uva
tinta de maior resistência à patógenos, a ‘Regent’ possui genealogia (Figura 4) com
múltiplos cruzamentos com videiras selvagens americanas, como Vitis rupestris, V.
riparia, V. lincecumii, V. aestivalis, V. cinerea, V. berlandieri e V. labrusca (EIBACH;
TÖPFER, 2003).
Esta variedade foi utilizada como parental nos cruzamentos que originaram
‘Calandro’ (‘Domina’ x ‘Regent’), ‘Julius’ (‘Regent’ x ‘Kozma 20-3’) e ‘Reberger’
(‘Regent’ x ‘Blaufraenkisch’) (JKI, 2017).

8
Figura 4. Genealogia da variedade Regent.
Fonte: Vanessa Miranda, adaptado de JKI (2017).
3.2.4. Variedade Calandro
A uva ‘Calandro’ é uma variedade tinta híbrida (Figura 10), desenvolvida em
1984 por Rudolf Eibach e Reinhard Töpfer, no Instituto Julius-Kühn, Alemanha. O
cruzamento que originou ‘Calandro’ utilizou as variedades Domina e Regent como
parentais (JKI, 2017).
A variedade é altamente resistente ao míldio, ao oídio e ao Botrytys; sua
resistência se deve a forte genealogia americana da parental ‘Regent’ (Figura 5). Apesar
de não ser amplamente conhecida, a ‘Calandro’ é cultivada em vinhedos experimentais
na Alemanha desde 2005. O vinho produzido é encorpado e tânico, com aromas
defumados e de frutas vermelhas, além de bom potencial de guarda (ROBINSON;
HARDING; VOUILLAMOZ, 2012).
Figura 5. Genealogia da variedade Calandro.
Fonte: Vanessa Miranda, adaptado de JKI (2017).

9
3.2.5. Variedade Helios
A uva ‘Helios’ é uma variedade branca híbrida (Figura 10), criada em 1973 por
Norbert Becker, no Instituto Nacional de Viticultura em Freiburg, Alemanha. As videiras
parentais utilizadas no cruzamento foram ‘Merzling’ e ‘FR 986-60’ (JKI, 2017). A
variedade foi nomeada ‘Helios’ em alusão ao titã grego “Helius”, a personificação do Sol.
A videira ‘Helios’ apresenta boa resistência ao míldio, ao oídio e ao Botrytys. A
parental ‘Merzling’ possui cruzamentos com videiras americanas em sua genealogia
(Figura 6), como as espécies Vitis rupestris, V. riparia, V. lincecumii e V. berlandieri
(STRIEM, 2010).
Figura 6. Genealogia da variedade Helios.
Fonte: Vanessa Miranda, adaptado de JKI (2017).
3.2.6. Variedade Prior
A uva ‘Prior’ é uma variedade tinta híbrida complexa (Figura 10), criada em 1987
por Norbert Becker, no Instituto Nacional de Viticultura em Freiburg, Alemanha. No
cruzamento foram utilizados os genótipos FR 4-61 e FR 236-75R. Destes, o FR 236-75R
apresenta os mesmos parentais de ‘Bronner’ (‘Merzling’ X ‘GM 6494’) (JKI, 2017).
A variedade foi nomeada ‘Prior’, do latim “primeira, antiga”, sugerindo uma
qualidade superior da uva e aludindo à sua relativamente precoce maturação. A ‘Prior’ é
cultivada em áreas limitadas na Suíça, Itália e Alemanha (ROBINSON; HARDING;
VOUILLAMOZ, 2012). O vinho de ‘Prior’ é rico em taninos e compostos fenólicos,
apresentando coloração vermelha intensa com aromas leves e frutados, lembrando os
vinhos de ‘Pinot Noir’ (PRITCHARD, 2016).
Apresenta alta resistência ao míldio, porém é suscetível ao Botrytis e ao oídio,
sendo necessária a aplicação de tratamentos fungicidas para uma boa produção. A

10
resistência ao míldio da ‘Prior’ se deve à sua complexa genealogia (Figura 7),
envolvendo cruzamentos naturais e deliberados entre 70 parentais distintos, incluindo
espécies selvagens americanas e asiáticas, como Vitis rupestris, V. riparia, V. lincecumii,
V. aestivalis, V. cinerea, V. berlandieri, V. amurensis e V. labrusca (ROBINSON;
HARDING; VOUILLAMOZ, 2012).
Figura 7. Genealogia da variedade Prior.
Fonte: Vanessa Miranda, adaptado de JKI (2017).
3.2.7. Variedade Felicia
A ‘Felicia’ é uma variedade branca híbrida (Figura 10), criada em 1984 por
Rudolf Eibach e Reinhard Töpfer, no Instituto Julius-Kühn, Alemanha. As parentais
utilizadas no cruzamento são as variedades Sirius e Vidal Blanc (JKI, 2017).
Esta variedade apresenta alta resistência ao oídio e média resistência ao míldio,
pois compartilha parte de sua genealogia (Figura 8) com as variedades Prior e Helios,
onde estão presentes videiras americanas e asiáticas. O vinho de ‘Felicia’ apresenta
aromas florais e acidez equilibrada, lembrando os vinhos de uvas moscatel
(PRITCHARD, 2016).
Figura 8. Genealogia da variedade Felicia.
Fonte: Vanessa Miranda, adaptado de JKI (2017).

11
3.2.8. Variedade Aromera
A ‘Aromera’ é uma variedade branca híbrida (Figura 10), criada pela InnoVitis,
na Itália. As parentais utilizadas no cruzamento são as variedades Muscat Ottonel e Eger
2 (JKI, 2017).
‘Aromera’ apresenta alta resistência ao míldio e ao Botrytis e média resistência ao
oídio, sendo necessário o uso de tratamentos fungicidas. Uma das parentais dessa
variedade, ‘Eger 2’, apresenta na sua genealogia (Figura 9) as espécies americanas Vitis
rupestris, V. lincecumii, V. berlandieri e V. labrusca (STRIEM, 2010). O vinho de
‘Aromera’ é aromático, traz frutas cítricas e rosas no paladar, lembrando os vinhos
moscatel (INNOVITIS, 2013).
Figura 9. Genealogia da variedade Aromera.
Fonte: Vanessa Miranda, adaptado de JKI (2017).

12
Figura 10. Cachos das variedades Baron (A), Calandro (B), Prior (C), Regent (D),
Aromera (E), Bronner (F), Felicia (G) e Helios (H).
Fonte: Gyula Gyukli (A), Doris Schneider/JKI (B, D, G), Ursula Brühl/JKI (C) InnoVitis (E),
Wolfgang Renner (F), Vanessa Miranda (H).
3.3. Caracterização Fenológica
A fenologia é a área que analisa os eventos anuais e periódicos dos seres vivos e
sua relação com as variações do clima. No caso dos vegetais, as etapas geralmente
observadas são o desenvolvimento de folhas, a floração e a frutificação (BLOESCH;
VIRET, 2008).
Na viticultura existem três principais sistemas de estágios fenológicos que são
referência para a videira, são eles o sistema ‘Baggiolini’ (subdividido em 16 estágios, de
A a P), o sistema ‘Eichhorn e Lorenz’ (subdividido em 22 estágios, de 01 a 47) e o sistema
‘BBCH’ (subdividido em 31 estágios, de 00 a 97) (BAILLOD; BAGGIOLINI, 1993).
A fenologia desempenha papel fundamental na introdução de novas variedades,
desvendando como o clima local e as variações sazonais vão interagir com a planta,

13
possibilitando determinar a adaptação da variedade à região de cultivo, além de planejar
atividades de manejo e estimar a data da colheita. Considerando que a fenologia é
dependente do genótipo e das condições geográficas da região produtora, faz-se
necessário uma nova análise fenológica a cada diferente variedade em novo local de
cultivo (JONES; DAVIS, 2000; NAGATA et al., 2000).
O ciclo vegetativo da videira ocorre anualmente e compreende o intervalo de
tempo entre o início da brotação e a queda das folhas. Entre os ciclos, durante o inverno,
a videira atravessa um período de dormência no qual ocorre uma drástica redução das
atividades metabólicas. Esse período de frio é fundamental para que a brotação ocorra de
forma homogênea no vinhedo, pois a intensidade e o tempo de exposição da videira às
baixas temperaturas determina sua capacidade de brotação (VIANA, 2009).
O mecanismo de indução da dormência, apesar de não estar completamente
conhecido, é diretamente relacionado com o aumento da concentração de ácido abscísico
na planta. Durante o inverno, a videira sofre um estresse hídrico, pois a água se torna
menos disponível em áreas de pouca chuva ou congelamento. Sob estas condições, o
tecido vegetal produz muito ácido abscísico, que estimula o fechamento dos estômatos,
evitando a perda de água (RAVEN; EVERT; EICHHORN, 2014; ASSIS; LIMA FILHO;
LIMA, 2004).
Normalmente, existe um equilíbrio entre as concentrações de citocininas, auxinas,
etileno e ácido abscísico, que regulam o metabolismo da planta, porém, em situações de
estresse, o desbalanço desses hormônios pode gerar um estágio de dormência
(KERBAUY, 2012). Nesse estágio, a umidade da gema cai de 80% para 50%, visando
preservar os tecidos vegetais contra um possível congelamento; com os estômatos
fechados, a respiração celular se torna extremamente baixa, não há sinais de mitose nos
tecidos e o crescimento dos ramos é inibido (JACKSON, 2008).
Chegando ao fim do período de estresse hídrico, as citocininas e auxinas voltam a
ser produzidas em maior quantidade e passam a estimular a divisão celular, o crescimento
das gemas e a expansão foliar, levando a quebra da dormência (TAIZ; ZEIGER, 2013).
A brotação começa quando as gemas intumescem e abrem suas escamas,
chegando ao estágio de ponta verde (Figura 12). Após o aparecimento de folhas
rudimentares, o caule herbáceo se expande e as folhas jovens se desenvolvem
rapidamente. As inflorescências surgem junto às folhas (Figura 12) e com o passar do
tempo os botões florais se individualizam, abandonando a estrutura aglomerada inicial
(BLOESCH; VIRET, 2008).

14
O fenômeno da brotação parece estar associado com o aumento da concentração
de peróxido de hidrogênio e o consequente estresse oxidativo gerado pelos baixos níveis
de catalase. A brotação inicia com a mobilização de reservas que foram acumuladas nos
ramos e raízes durante o período de dormência. Essa translocação de nutrientes é
estimulada pelo aumento da temperatura do ambiente e pela maior incidência de luz solar
sobre a planta. As reservas são utilizadas até que os novos tecidos sejam capazes de se
sustentar através da fotossíntese (JACKSON, 2008).
O início da floração é marcado pela queda da caliptra ou antese (Figura 12),
momento em que os estames da flor são expostos. Na plena floração (Figura 12), 50%
das flores já perderam a sua caliptra. A velocidade de crescimento das gemas fica reduzida
a partir da floração, pois há uma competição pelos nutrientes nas diferentes partes em
desenvolvimento na planta (BLOESCH; VIRET, 2008; GIOVANNI, 1999). Após a
fecundação, as peças florais caem e começa o crescimento dos frutos, um processo
dividido em três estágios, como proposto por Coombe (1992) (Figura 11).
No estágio I, as paredes do ovário se desenvolvem formando o pericarpo do fruto.
As bagas crescem progressivamente (Figura 12), no início aumentado o número de
células e depois através da expansão celular, por meio do acúmulo de solutos. Ao término
desse estágio, as sementes atingem seu tamanho final (KENNEDY, MATTHEWS,
WATERHOUSE, 2000).
No primeiro estágio, a clorofila é o pigmento principal e a auxina é o hormônio
em maior concentração. A atividade metabólica dentro da baga é intensa: a taxa de
respiração celular é alta e uma série de ácidos, principalmente ácido tartárico e málico, se
acumulam nos tecidos do fruto (RIBÉREAU-GAYON et al., 2006). Este é um estágio
sensível do desenvolvimento, pois nele se formam a maior parte dos minerais,
aminoácidos e compostos aromáticos presentes na uva, trazendo consequências diretas
para a qualidade do mosto e do vinho produzido posteriormente (CONDE et al., 2007).
No estágio II, os embriões já estão formados e as sementes podem ser
consideradas maduras. O segundo estágio é caracterizado como uma fase de latência: há
queda na concentração de clorofila e auxinas, as atividades metabólicas são reduzidas e a
baga praticamente não aumenta seu peso. Ao fim do estágio II, os ácidos acumulados
começam a ser transformados em açúcares, dando início a maturação (CONDE et al.,
2007; MALINOVSKI, 2013).

15
Figura 11. Diagrama mostrando o aumento de volume do fruto a partir da antese.
Fonte: Vanessa Miranda, adaptado de Coombe (1992).
Iniciado pelo véraison (Figura 12), o estágio III compreende a maturação da baga.
A entrada no terceiro estágio pode ser percebida visualmente pela mudança de coloração
na epiderme dos frutos. Há alta concentração de ácido abscísico, que inibe o processo
mitótico e sinaliza a baga como órgão senil. O crescimento dos ramos praticamente cessa,
pois, os esforços energéticos da planta são direcionados para os frutos (MOTA et al.,
2006).
A baga aproximadamente dobra seu tamanho entre o véraison e a colheita. As
substâncias produzidas no primeiro estágio, como ácidos, taninos e compostos aromáticos
reduzem sua concentração, pois são diluídas no açúcar e água que agora fazem parte da
baga (KENNEDY, 2002).
A principal característica do período de maturação é o acúmulo de açúcares, etapa
fundamental para originar polifenóis, antocianinas e aromas, além do álcool
posteriormente produzido na fermentação. Os açúcares podem ser procedentes de raízes,
troncos, sarmentos e folhas, migrando para o fruto durante o amadurecimento, mas
também podem ser produzidos dentro da baga através da transformação do ácido málico
em glicose (MOTA et al., 2006).

16
Progressivamente a acidez diminui, o pH aumenta e pigmentos se acumulam na
casca, dando a cor final da variedade. O terceiro estágio de crescimento muda
drasticamente as características dos frutos: antes pequenas, rígidas, ácidas e com pouco
açúcar, ao fim da maturação (Figura 12) as uvas se tornam grandes, tenras, doces e menos
ácidas, com maior intensidade de sabor e coloração (CONDE et al., 2007).
Figura 12. Estágio de ponta verde (A), inflorescências surgindo junto às folhas (B),
queda da caliptra (C), floração plena (D), crescimento dos frutos (E), véraison (F) e
maturação (G).
Fonte: Dane Cellars (A), Vanessa Miranda (B, D, E, F), Pancrat (C), Ursula Brühl/JKI (G).
3.3.1. Índices Bioclimáticos e Requerimento Térmico
O período de tempo que a videira demora para completar cada fase fenológica
durante seu ciclo pode ser relacionado com variáveis climáticas, principalmente a
temperatura (TONIETTO; CARBONNEAU, 2004). O requerimento térmico ou soma
térmica quantifica essa passagem do tempo e é expresso em °C dia.

17
Índices bioclimáticos são utilizados na viticultura para classificar regiões de
cultivo de acordo com suas temperaturas e, frequentemente, seu uso é extrapolado na
caracterização do requerimento térmico de variedades de videira. Os principais índices
utilizados com esses fins são o Índice de Winkler e o Índice de Huglin.
O Índice de Winkler (WINKLER et al., 1974) considera em sua equação as
temperaturas mínimas (Tmin) e máximas (Tmax), além da temperatura de base (10 °C). O
somatório é referente aos dias entre os meses de outubro e abril no hemisfério sul (HALL;
JONES, 2010):
Esse método classifica as regiões de cultivo em sete categorias:
• Região muito fria: < 850 °C dia
• Região I: 850 – 1389 °C dia
• Região II: 1389 – 1667 °C dia
• Região III: 1667 – 1944 °C dia
• Região IV: 1944 – 2222 °C dia
• Região V: 2222 – 2700 °C dia
• Região muito quente: >2700 °C dia
O Índice de Huglin (HUGLIN, 1978) considera em sua equação as temperaturas
médias (Tmean) e máximas (Tmax), além da temperatura de base (10 °C) e do coeficiente K
(varia de acordo com a latitude do local, corrigindo a equação conforme o comprimento
do dia). O somatório é referente aos dias entre os meses de outubro e março no hemisfério
sul (HALL; JONES, 2010):
Esse método classifica as regiões de cultivo em oito categorias:
• Região extremamente fria: <1200 °C dia
• Região muito fria: 1200 – 1500 °C dia
• Região fria: 1500 – 1800 °C dia
• Região temperada: 1800 – 2100 °C dia

18
• Região temperada quente: 2100 – 2400 °C dia
• Região quente: 2400 – 2700 °C dia
• Região muito quente: 2700 – 3000 °C dia
• Região extremamente quente: >3000 °C dia
Tanto o Índice de Winkler quanto o de Huglin apresentam uma série de limitações
no cálculo de requerimento térmico de variedades de videira. A temperatura de base
normalmente utilizada é de 10 °C, porém algumas variedades de videira apresentam
temperaturas tão baixas quanto 5,1 °C para quebrar a dormência e iniciar a brotação
(NENDEL, 2009).
A temperatura tem um papel fundamental no desenvolvimento da videira, atuando
diretamente sobre a quebra de dormência, crescimento vegetativo, floração e
desenvolvimento dos frutos. A temperatura influencia o processo de maturação da uva,
gerando variações na concentração de açúcares, ácidos e compostos fenólicos
(COOMBE, 1987).
Segundo Jackson (2008), o aumento da temperatura (até um limite variável)
favorece a maioria dos processos bioquímicos que ocorrem na planta, acelerando o seu
desenvolvimento. Durante a fase de maturação, para que as bagas atinjam níveis
satisfatórios de açúcares e ácido málico, Jackson (2008) propõe uma faixa de temperatura
ótima entre 20 e 25 °C.
A principal limitação desses índices bioclimáticos é a ausência de uma
temperatura de base superior. Apesar de terem uma temperatura de base inferior, abaixo
da qual presumivelmente não ocorre desenvolvimento vegetativo, ela não é suficiente
para demonstrar o real acúmulo térmico da videira, já que temperaturas acima de 32 °C
diminuem a capacidade fotossintética e a qualidade da fertilização (JACKSON, 2008).
Considerando estas limitações, Tomazetti et al. (2015) propuseram um método de
soma térmica (Método 3.3), que apresenta em sua equação as temperaturas mínimas
(Tmín), médias (Tméd) e máximas (Tmáx), além da temperatura de base inferior (Tb=10 °C),
temperatura ótima (Tot=25 °C) e temperatura de base superior (TB=35 °C). O método de
cálculo da soma térmica diária (STd) é descrito da seguinte forma:
• STd = [(Tmáx - Tb)0,5] ×1 dia, quando Tméd<Tot e Tmín<Tb;
• STd = (0) ×1 dia, quando Tmáx<Tb;
• STd = (Tméd - Tb) ×1 dia, quando Tméd<Tot e Tmín>Tb;

19
• STd = {(Tot - Tb).[(TB - Tméd)/(TB - Tot)]} ×1 dia, quando Tméd>Tot e Tmáx<TB;
• STd = {(TB - Tot)×[(TB - Tméd)/(TB - Tot)]} ×1 dia, quando Tméd>Tot e Tmáx>TB,
se Tmáx>TB, então Tmáx=TB.
Em climas quentes como o do sul catarinense e de outras regiões do Brasil,
métodos que considerem as três temperaturas cardinais apresentam maior precisão no
requerimento térmico. Principalmente no período entre véraison e maturação, onde as
temperaturas são mais elevadas e a planta incorre em maior estresse pelo calor
(TOMAZETTI et al, 2015).
O requerimento térmico pode ser utilizado também para quantificar o plastocrono,
que é o período de tempo necessário para o aparecimento de um novo nó no caule da
planta (no caso da videira, um novo nó no sarmento) e tem como unidade °C dia nó-1. O
número de nós é utilizado como parâmetro de desenvolvimento vegetativo em diferentes
espécies de plantas, pois está relacionado com o surgimento de folhas e inflorescências.
Logo, a taxa de aparecimento de nós nos sarmentos da videira é um componente
importante em modelos matemáticos de simulação do crescimento (BAKER; REDDY,
2001; STRECK et al., 2008).
4. Material e Métodos
O experimento foi realizado na Unidade de Pesquisa da Estação Experimental da
Epagri (Anexo A), no município de Urussanga, SC, a 28º 32’ 00,73” S e 49º 18’ 58,20”
O, em altitude de 49 m.
Foram avaliados 11 genótipos de videira resistentes ao míldio e ao oídio, sendo
eles: Aromera, Baron, Bronner, Calandro, Felicia, GF-6, GF-24, GF-15, Helios, Prior e
Regent; enxertados sobre ‘Paulsen 1103’, conduzidos em sistema de espaldeira, contendo
50 plantas por genótipo, espaçamento de 3 m entre filas e 1 m entre plantas. Foi realizada
poda curta no dia 6 de setembro de 2016, deixando 2-3 gemas por esporão. Quando a
poda foi executada, muitas plantas já haviam iniciado o período de brotação, estando entre
os estágios A e F de BAILLOD e BAGGIOLINI (1993), dependendo da variedade. As
avaliações iniciaram em setembro de 2016, a partir da poda de frutificação e se
estenderam até a colheita, em janeiro de 2017.

20
Durante o período do experimento, foram utilizados tratamentos fitossanitários
somente para antracnose e o manejo de plantas daninhas foi realizado através de
herbicidas (linha) e roçada (entrelinha).
Para observação das variáveis climáticas da região foram coletados os dados da
Estação Meteorológica Automática Telemétrica da Epagri/CIRAM/UFSC de Urussanga.
Os parâmetros climáticos observados foram: temperatura máxima, média e mínima do ar,
umidade relativa do ar, precipitação e radiação solar global.
O acompanhamento semanal da fenologia foi realizado por um técnico da Epagri
de Urussanga, incluso ao acompanhamento quinzenal dos pesquisadores. Nas visitas,
foram observados os estágios fenológicos que as plantas se encontravam nas respectivas
datas. Os estágios fenológicos foram adaptados de BAILLOD e BAGGIOLINI (1993) da
seguinte forma: brotação (quando mais de 50% das gemas atingiram o estágio de
emergência das folhas), floração (quando mais de 50% das flores estavam abertas),
véraison (quando as bagas iniciaram a mudança de coloração) e maturação (quando todas
as bagas mudaram a coloração e/ou estavam tenras ao toque). Os estágios foram
demarcados quando mais de 75% dos indivíduos de cada variedade avaliada se
encontravam dentro da descrição.
Para a quantificação do plastocrono, cinco plantas de cinco genótipos de maior
interesse foram marcadas. Estes genótipos foram escolhidos por conta de sua alta
resistência ao míldio e ao oídio (GF-6, GF-15, GF-24) e por seu maior reconhecimento
na vitivinicultura (Regent e Bronner). As plantas avaliadas foram sorteadas em
delineamento inteiramente casualizado. Nas visitas quinzenais, foram contabilizados os
nós pertencentes aos sarmentos separadamente.
A avaliação da maturação foi realizada em delineamento inteiramente casualizado
em fileiras marcadas aleatoriamente. Foram coletadas 50 bagas por variedade há cada
duas semanas, a partir do dia 12 de dezembro, quando três variedades já haviam iniciado
o véraison. O mosto extraído e as cascas das uvas foram utilizados para avaliação da
maturação tecnológica e da maturação fenólica.
As análises de maturação foram realizadas no Laboratório de Morfogênese e
Bioquímica Vegetal e no Laboratório de Fisiologia do Desenvolvimento e Genético
Vegetal, ambos localizados do Centro de Ciências Agrárias da UFSC.
Para avaliação da maturação tecnológica, foi acompanhada a evolução no teor de
sólidos solúveis totais, acidez total titulável e o pH do mosto, como proposto por
AMERINE e OUGH (1974):

21
• Sólidos solúveis totais: leitura direta do mosto através de um refratômetro digital de
bancada.
• Acidez total titulável: foi preparada uma solução com 10 mL de mosto para 70 mL
de água destilada; essa mistura foi colocada sob agitação e a titulação foi realizada
adicionando-se lentamente uma solução de NaOH [0,1 N] até ocorrer a virada do pH
da mistura.
• pH: leitura do mosto em pHmetro.
Para avaliação da maturação fenólica, foram analisados os valores de antocianinas
monoméricas totais e índice de polifenóis totais obtidos a partir do extrato metanólico das
cascas das uvas. Para isso, as cascas foram separadas da polpa, secas, pesadas e
adicionadas de metanol acidificado [HCl 1%] na proporção de 1:5; os extratos foram
mantidos no escuro a 4,0 ± 1º C por 24 h (LEES; FRANCIS, 1972).
Após esse período, os extratos foram filtrados com papel filtro sobre um funil de
vidro e transferidos para frascos âmbar, posteriormente vedados e mantidos em
temperatura de -18º C até a realização das análises (FERNANDES, 2014):
• Antocianinas monoméricas totais: foi utilizado o método do pH diferencial, onde o
extrato metanólico das uvas tintas foi diluído na proporção de 1:60 em tampão de
cloreto de potássio [0,025 M] de pH 1, e 1:60 em tampão de acetato de sódio [0,4 M]
de pH 4,5; as soluções foram mantidas em repouso por 15 minutos, ao abrigo da luz,
e então foi realizada a leitura da absorbância em cada tampão nos comprimentos de
onda de 520 e 700 nm em espectofotômetro. Os resultados foram baseados nas
seguintes equações, descritas por GIUSTI e WROLSTAD (2001):
AT = (A520 – A700)pH 1 – (A520 – A700)pH 4,5
Sendo AT = absorbância total, A520 = absorbância em 520 nm e A700 =
absorbância em 700 nm.
AMT (mg L-1) = (AT. PM.DF.1000)/( .1)
Sendo AMT= antocianinas monoméricas totais, AT = absorbância total, PM
= peso molecular da antocinanina majoritária (malvidina 3-glicosídeo, PM = 529),
DF = fator de diluição (60) e = absortividade molar da antocianina majoritária
(malvidina 3-glicosídeo, = 28000).

22
• Polifenóis totais: foi utilizado o método espectrofotométrico de Folin-Ciocalteu
como descrito por SINGLETON e ROSSI (1965), no qual em um tubo colocou-se
7,9 mL de água destilada, 0,1 mL do extrato metanólico diluído em água destilada
(proporção de 1:6 em uvas tintas e 1:3 em uvas brancas) e 0,5 mL do reagente de
Folin-Ciocalteu. Após 3 minutos adicionou-se 1,5 mL de carbonato de sódio 20% e
as soluções foram mantidas no escuro por 2 h. Foi feita uma curva de calibração a
partir de uma diluição seriada de ácido gálico. As leituras da absorbância de cada
amostra foram realizadas no comprimento de onda de 760 nm em espectofotômetro.
O valor de polifenóis totais foi obtido inserindo a absorbância de cada amostra na
equação da curva de calibração de ácido gálico (y = 0,1843x + 0,002).
5. Resultados e Discussão
5.1. Variáveis Climáticas
Os dados climáticos foram observados entre a brotação e a colheita, durante o
ciclo 2016/2017, compreendendo os meses de setembro a janeiro, no município de
Urussanga, sul catarinense.
5.1.1. Temperatura do Ar e Amplitude Térmica
Durante o período de maturação, as temperaturas médias em Urussanga variaram
entre 24,8 e 25,7 °C, já a amplitude térmica diária oscilou entre 10,5 e 11,2 °C (Tabela
1). As altas temperaturas aumentam a concentração de açúcares na baga e diminuem a
concentração de ácidos, dificultando também a produção de antocianinas nas cascas dos
frutos (COOMBE, 1987), contudo, o equilíbrio açúcares/ácidos pode ser mantido se o dia
quente for seguido por temperaturas baixas durante a noite, característica atestada pela
amplitude térmica do local. O acúmulo de compostos fenólicos é estimulado pela
amplitude térmica, desde que a mesma não ultrapasse os 15 °C (MORI; SUGAYA;
GEMMA, 2005).

23
Tabela 1. Temperaturas médias, mínimas e máximas do ar (°C), amplitude térmica média
(°C), precipitação (mm) e umidade relativa do ar (%) de cada variedade (B – Brotação, F
– Floração, V – Véraison, M – Maturação), em cada estágio fenológico, na região de
Urussanga, SC, ciclo 2016/2017.
Genótipo Estágio
Fenológico Temperatura
Média (°C) Temperatura Mínima (°C)
Temperatura Máxima (°C)
Amplitude Térmica (°C)
Precipitação (mm)
Umidade Relativa (%)
GF-6
B - F 18,9 11,4 27,0 15,7 2,8 63,6
F - V 19,9 14,5 26,5 12,0 350,6 72,4
V - M 24,8 20,4 31,5 11,2 113,8 74,0
B - M 20,7 15,3 27,4 12,1 467,2 72,3
GF-15
B - F 18,2 13,7 24,2 10,5 139,8 75,9
F - V 21,5 15,8 28,3 12,6 218,0 70,5
V - M 25,7 21,9 32,4 10,5 258,2 77,8
B - M 21,4 16,4 27,9 11,6 613,0 73,4
GF-24
B - F 18,2 13,7 24,2 10,5 139,8 75,9
F - V 21,5 15,8 28,3 12,5 218,0 70,5
V - M 25,7 21,9 32,4 10,5 258,2 77,8
B - M 21,4 16,4 27,9 11,5 613,0 73,4
Regent
B - F 16,6 11,8 22,7 10,9 49,0 74,8
F - V 21,3 15,8 28,0 12,1 301,0 72,1
V - M 25,0 20,7 31,7 11,0 222,6 75,2
B - M 21,4 16,4 27,9 11,5 613,0 73,4
Bronner
B - F 16,6 11,8 22,7 10,9 49,0 74,8
F - V 21,3 15,8 28,0 12,2 301,0 72,1
V - M 25,0 20,7 31,7 11,0 222,6 75,2
B - M 21,4 16,4 27,9 11,5 613,0 73,4
Felicia
B - F 16,7 11,3 23,1 11,8 49,6 73,1
F - V 21,3 15,9 28,0 12,1 305,8 72,0
V - M 24,8 20,6 31,4 10,8 110,0 74,4
B - M 20,8 15,6 27,4 11,8 464,4 72,7
Calandro
B - F 16,5 11,4 22,8 11,4 12,8 73,4
F - V 20,9 15,6 27,5 11,8 343,4 72,7
V - M 25,0 20,9 31,6 10,7 259,2 75,5
B - M 21,4 16,4 27,9 11,5 613,0 73,4
Aromera
B - F 18,2 13,7 24,2 10,5 139,8 75,9
F - V 21,5 15,8 28,3 12,5 218,0 70,5
V - M 25,7 21,9 32,4 10,5 258,2 77,8
B - M 21,4 16,4 27,9 11,5 613,0 73,4
Baron
B - F 16,7 11,3 23,1 11,8 49,6 73,1
F - V 21,3 15,9 28,0 12,1 305,8 72,0
V - M 25,0 20,9 31,6 10,7 259,2 75,5
B - M 21,1 16,0 27,7 11,7 613,6 73,1
Prior
B - F 16,6 10,9 23,3 12,3 13,4 71,7
F - V 20,9 15,6 27,5 11,9 343,4 72,7
V - M 25,0 20,9 31,6 10,7 259,2 75,5
B - M 21,1 16,0 27,7 11,7 613,6 73,1
Helios
B - F 16,5 11,4 22,8 11,4 12,8 73,4
F - V 20,9 15,6 27,5 11,9 343,4 72,7
V - M 24,8 20,6 31,4 10,8 110,0 74,4
B - M 21,1 16,0 27,6 11,6 463,8 73,0

24
5.1.2. Precipitação Pluviométrica e Umidade Relativa
Na Figura 13, encontram-se a precipitação pluviométrica e a umidade relativa do
ar durante os meses do ciclo 2016/2017.
Figura 13. Somatório da precipitação pluviométrica mensal (mm) e médias mensais da
umidade relativa do ar (%), na região de Urussanga – SC, ciclo 2016/2017.
Entre a brotação e a maturação, a precipitação acumulada em Urussanga foi de
616,4 mm e a umidade relativa média de 72,7%. Durante o período de amadurecimento
dos frutos, a precipitação acumulada foi de 263 mm e a umidade relativa média de 75,2%.
A precipitação em excesso durante o ciclo pode acarretar em prejuízos para a
floração, diminuindo a polinização pela dificuldade de queda da caliptra
(DOKOOZLIAN, 2000), e para a maturação, reduzindo a qualidade da uva através da
diluição de seus compostos ou da predisposição a doenças, como o míldio. A baixa
precipitação, por sua vez, favorece a concentração de açúcares e melhora a qualidade do
vinho (JACKSON, 2008).
5.1.3. Radiação Solar
O maior valor médio horário de radiação global registrado no período de
maturação foi de 784,9 W.m-2 (Figura 14). A quantidade de radiação varia de acordo
74,0
153,4
52,4
257,8
293,6
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
90,0
100,0
0,0
50,0
100,0
150,0
200,0
250,0
300,0
Setembro Outubro Novembro Dezembro Janeiro
Um
idad
e R
elat
iva
(%)
Pre
cip
itaç
ão (
mm
)
Precipitação Umidade

25
com a latitude, longitude, altitude, umidade do ar e nebulosidade do local, além do horário
e mês da análise (PANDOLFO, 2010).
Figura 14. Valores médios por hora da radiação solar global (W m-2) durante o período
de maturação dos genótipos no município de Urussanga-SC, Brasil, ciclo 2016/2017.
O resultado encontrado pode ser explicado pela proximidade entre Urussanga e o
oceano (aproximadamente 33 km em linha reta), já que grandes corpos d’água geram
maior quantidade de vapor, formando nuvens. A nebulosidade gera menores valores de
radiação global, pois a mesma é refletida. Em baixas altitudes, como na região (49 m), a
radiação também precisa atravessar uma maior coluna de ar para chegar a superfície
terrestre, reduzindo sua intensidade (AYOADE, 1996).
5.2. Índices Bioclimáticos
Considerando o Índice de Winkler, a região de Urussanga teve um somatório de
2651,8 °C dia entre os meses de outubro de 2016 e abril de 2017. Dessa forma, Urussanga
é classificada como “Região V” pelo Índice de Winkler. Jones et al. (2010) colocam que
em áreas classificadas como Região V são tipicamente produzidas apenas uvas e vinhos
de mesa, devido às altas temperaturas. Porém, Pirie (2007) destaca que as ilhas gregas, a
região de Jerez na Espanha, e as ilhas de Sardenha e Sicília na Itália se classificam
0
100
200
300
400
500
600
700
800
900
00:00 02:00 04:00 06:00 08:00 10:00 12:00 14:00 16:00 18:00 20:00 22:00
Rad
iação
Glo
bal (W
m-2)

26
também como Região V e são produtoras de uvas finas, à exemplo da ‘Primitivo’ e ‘Nero
D’Avola’.
Durante o período de brotação à maturação (setembro a janeiro), as variedades
cultivadas durante o experimento acumularam entre 1167,3 °C dia (Helios) e 1336 °C dia
(Baron), como observado na Tabela 2.
Tabela 2. Requerimento térmico (°C dia) através do Índice de Winkler para cada
variedade, em cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017.
Índice de Winkler
Genótipo B - F F - V V - M B - M
GF-6 75,4 894,6 329,1 1299,1
GF-15 245,5 701,8 345,3 1292,6
GF-24 245,5 700,5 345,8 1291,8
Regent 141,1 718,1 437,5 1296,7
Bronner 141,1 718,1 437,5 1296,7
Felicia 187,8 737,7 303,7 1229,2
Calandro 94,4 779,0 412,1 1285,5
Aromera 245,5 700,5 345,8 1291,8
Baron 187,8 736,1 412,1 1336,0
Prior 141,1 776,0 406,2 1323,3
Helios 94,4 776,0 296,9 1167,3
Para o Índice de Huglin, a região de Urussanga teve um somatório de 2843,6 °C
dia entre os meses de outubro de 2016 e março de 2017. Dessa forma, Urussanga é
classificada como “Região muito quente” pelo Índice de Huglin. Regiões com essa
classificação excedem o requerimento térmico necessário para o amadurecimento da
maioria das variedades, podendo gerar estresse na planta pelas altas temperaturas. No
entanto, regiões muito quentes podem ser indicadas para produção de alguns vinhos
fortificados, como ocorre em Jerez, na Espanha (TONIETTO; CARBONNEAU, 2004).
Durante o período de brotação à maturação (setembro a janeiro), as variedades
cultivadas durante o experimento acumularam entre 1473,1 (Helios) e 1676,4 °C dia
(Baron), como observado na Tabela 3.

27
Tabela 3. Requerimento térmico (°C dia) através do Índice de Huglin para cada
genótipo, em cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017.
Índice de Huglin
Genótipo B - F F - V V - M B - M
GF-6 113,3 1152,7 388,3 1654,3
GF-15 326,4 884,3 407,3 1618,0
GF-24 326,4 884,3 407,3 1618,0
Regent 197,2 895,1 516,8 1609,0
Bronner 197,2 895,1 516,8 1609,0
Felicia 268,9 927,2 351,8 1547,9
Calandro 140,4 980,9 480,3 1601,6
Aromera 326,4 884,3 407,3 1618,0
Baron 268,9 927,2 480,3 1676,4
Prior 212,2 980,9 480,3 1673,3
Helios 140,4 980,9 351,8 1473,1
5.3. Caracterização Fenológica e Requerimento Térmico
Na Tabela 4 encontram-se as datas de início dos estágios fenológicos observados
em cada variedade durante o ciclo 2016/2017. O genótipo mais precoce foi o GF-6, tendo
iniciado sua brotação alguns dias antes da poda, em 6 de setembro. A maioria das
variedades analisadas iniciaram a brotação duas semanas após a poda. A precocidade da
variedade GF-6 continuou durante a floração, iniciando já uma semana após a brotação e
três semanas antes de todas as outras variedades. O período do véraison apresentou uma
diferença máxima de uma semana entre as variedades.
O genótipo de ciclo mais longo foi o GF-6, totalizando 121 dias e acumulando
1190 °C dia de requerimento térmico (Tabela 5), seguido pelas variedades Baron e Prior,
ambas com 120 dias. A variedade com ciclo mais curto foi a “Helios”, totalizando 106
dias e acumulando 1080,7 °C dia.
A variedade Baron, apesar de não ter o ciclo mais longo, apresentou o maior
requerimento térmico, acumulando 1198,9 °C dia entre a brotação e a maturação. As
demais variedades apresentaram ciclo de 113 dias (Figura 15).
O período entre brotação e floração mais curto foi do GF-6, com 9 dias de duração
e 75,1 °C dia acumulados (Figura 16), e o mais longo da variedade Aromera e dos
genótipos GF-24 e GF-15 com 30 dias de duração e 228,7 °C dia acumulados. O período
entre floração e véraison mais curto foi da variedade Aromera e dos genótipos GF-24 e
GF-15, com 61 dias de duração e 669,3 °C dia acumulados e o mais longo do GF-6, com
90 dias de duração e 850,5 °C dia acumulados. O período entre véraison e maturação

28
mais curto foi das variedades Felicia e Helios, com 20 dias de duração e 243,4 °C dia
acumulados e o mais longo das variedades Bronner e Regent, com 29 dias de duração e
344 °C dia acumulados.
A data de brotação do genótipo GF-6 é muito próxima da observada
historicamente para a variedade Goethe na mesma região, 8 de setembro. A floração de
‘Goethe’ também está dentro do mesmo período observado para os genótipos do
experimento, ocorrendo próxima ao dia 12 de outubro. Já a colheita, geralmente ocorre
mais tardiamente, próxima ao dia 17 de janeiro (DELLA BRUNA; ARCARI; PETRY,
2016).
Tabela 4. Data de ocorrências dos estágios fenológicos dos genótipos estudados na
região de Urussanga, SC, ciclo 2016/2017.
Genótipo Início da
Brotação
Início da
Floração
Início do
Véraison Maturação
GF-6 06/set 14/set 12/dez 02/jan
GF-15 21/set 20/out 19/dez 09/jan
GF-24 21/set 20/out 19/dez 09/jan
Regent 21/set 11/out 12/dez 09/jan
Bronner 21/set 11/out 12/dez 09/jan
Felicia 14/set 11/out 14/dez 02/jan
Calandro 21/set 05/out 14/dez 09/jan
Aromera 21/set 20/out 19/dez 09/jan
Baron 14/set 11/out 14/dez 09/jan
Prior 14/set 05/out 14/dez 09/jan
Helios 21/set 05/out 14/dez 02/jan
Tabela 5. Requerimento térmico (°C dia) através do Método 3.3 para cada genótipo, em
cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017.
Genótipo B - F F - V V - M B - M
GF-6 75,1 850,5 264,4 1190,0
GF-15 228,7 669,3 265,4 1163,4
GF-24 228,7 669,3 265,4 1163,4
Regent 128,7 692,2 344,0 1165,0
Bronner 128,7 692,2 344,0 1165,0
Felicia 165,9 709,9 243,4 1119,3
Calandro 87,4 749,8 323,1 1160,3
Aromera 228,7 669,3 265,4 1163,4
Baron 165,9 709,9 323,1 1198,9
Prior 124,6 749,8 323,1 1197,5
Helios 87,4 749,8 243,4 1080,7

29
Figura 15. Duração (dias) de cada estágio fenológico para cada genótipo, na região de
Urussanga, SC, ciclo 2016/2017.
Figura 16. Requerimento térmico (°C dia) através do Método 3.3 para cada genótipo,
em cada estágio fenológico, na região de Urussanga, SC, ciclo 2016/2017.
9
30
30
21
21
28
15
30
28
22
15
90
61
61
63
63
65
71
61
65
71
71
22
22
22
29
29
20
27
22
27
27
20
0 10 20 30 40 50 60 70 80 90 100 110 120
GF-6
GF-15
GF-24
Regent
Bronner
Felicia
Calandro
Aromera
Baron
Prior
Helios
Dias após a brotação
B - F F - V V - M
75,1
228,7
228,7
128,7
128,7
165,9
87,4
228,7
165,9
124,6
87,4
850,5
669,3
669,3
692,2
692,2
709,9
749,8
669,3
709,9
749,8
749,8
264,4
265,4
265,4
344,0
344,0
243,4
323,1
265,4
323,1
323,1
243,4
0 200 400 600 800 1000 1200
GF-6
GF-15
GF-24
Regent
Bronner
Felicia
Calandro
Aromera
Baron
Prior
Helios
Requerimento Térmico (°C dia)
B - F F - V V - M

30
5.3.1. Plastocrono
Os valores de plastocrono observados (Tabela 6) ficaram entre 39,5 °C dia nó-1
(Regent) e 73,5 °C dia nó-1 (Bronner). Dessa forma, a variedade Regent precisa de 34 °C
dia nó-¹ a menos para produzir um nó, quando comparada com a variedade Bronner,
estando ambas expostas a mesma temperatura e outras condições climáticas.
Considerando a presença de uma folha a cada nó do sarmento, o número total de folhas
por planta variou entre 115 (Regent) e 230,6 folhas (GF-6).
As regressões lineares das equações do plastocrono apresentaram uma alta relação
(R2≥0,88) entre o número de nós por sarmento e o requerimento térmico. A linearidade
demonstrada entre esses dois fatores sugere que a temperatura é a principal condição que
atua sobre o aparecimento de nós no caule da planta (ZEIST et al., 2017).
Tabela 6. Plastocrono (°C dia nó-1), coeficiente de determinação (R2), número de
sarmentos por planta e número de nós por sarmento dos genótipos selecionados, região
de Urussanga, SC, ciclo 2016/2017.
Genótipo Plastocrono
(°C dia/nó-¹) R²
Número de
sarmentos por planta
Número de nós
por sarmento Número de folhas
por planta
GF-24 63,6 0,88 15,9 14,5 230,6
GF-15 58,1 0,96 10,9 15,6 170,0
GF-6 62,5 0,93 19,5 13,2 257,4
Regent 39,5 0,99 7,1 16,2 115,0
Bronner 73,5 0,88 11,3 12,1 136,7
Média 59,4 0,92 12,9 14,3 181,9
CV (%) 21,02 - 37,33 11,80 33,35
5.4. Avaliação da Maturação
5.4.1. Sólidos Solúveis Totais
Na Tabela 7 é observada a evolução nos níveis de sólidos solúveis totais (SST)
das variedades, durante o ciclo 2016/2017. O genótipo GF-24 não apresentou mosto
suficiente para a realização das análises na primeira coleta (12/12/16). As variedades
Felicia e Aromera e o genótipo GF-15 não foram analisadas após a colheita.

31
Os níveis de SST aumentaram de forma gradual, porém com queda na coleta do
dia 6 de janeiro, provavelmente devido a intensa precipitação pluviométrica ocorrida
nesta data e consequente diluição dos componentes presentes na baga. Os SST variaram
entre 15,0 °Brix (GF-6) e 17,6 °Brix (GF-24) nas análises pós-colheita.
Para a elaboração da maioria dos vinhos finos de qualidade é recomendado que os
SST se encontrem acima de 18 °Brix (JACKSON, 2008). Dessa forma, é possível afirmar
que nenhuma variedade foi colhida com SST na faixa ideal para elaboração de vinhos
finos. Apesar disso, historicamente na região dos Vales da Uva Goethe, a variedade
Goethe é colhida com SST em torno de 14,5 °Brix (DELLA BRUNA; ARCARI; PETRY,
2016).
5.4.2. pH
Na Tabela 7, é possível observar a evolução dos valores de pH em cada variedade,
durante o ciclo 2016/2017. Os valores subiram de forma gradual, porém também houve
queda na coleta do dia 6 de janeiro, provavelmente influenciado pelo mesmo fator
supracitado. O pH variou entre 3,3 (Helios) e 3,49 (GF-24).
Para produção de vinhos finos, é recomendável o pH do mosto entre 3,3 e 3,8
(INSTITUT RHODANIEN, 1998); portanto as uvas apresentaram valores ótimos de pH
quando colhidas.
5.4.3. Acidez Total Titulável
Na Tabela 7, pode ser observada a evolução dos valores de acidez total titulável
(ATT) em cada variedade, durante o ciclo 2016/2017. Não foi feita análise de ATT após
a colheita. A maioria das variedades apresentou uma diminuição dos valores ao longo do
ciclo. A ATT variou entre 87,1 meq L-1 (Baron) e 486,1 meq L-1 (GF-24) na última
análise.
Os valores desejáveis de ATT para produção de vinhos finos ficam entre 90 e 120
meq L-1, quando este valor é ultrapassado, pode ser necessário o uso de fermentação
malolática ou ajuste da acidez do mosto antes do processo de vinificação (MILLER, 2009;
ALLEBRANDT, 2012).

32
Tabela 7. Valores de SST, pH e ATT de cada genótipo, durante o período de
maturação, na região de Urussanga, SC, ciclo 2016/2017.
Genótipo SST (°Brix) pH
ATT (meq L-¹)
12/12 29/12 06/01 12/01 12/12 29/12 06/01 12/01 12/12 29/12 06/01
GF-6 8,3 14,3 12,7 15 2,25 3,59 3,42 3,38 214,6 119,3 126,9
GF-15 3,7 6,5 6,7 - 2,21 3,15 3,33 - 265,3 317,1 362,4
GF-24 - 10,1 5,9 17,6 - 3,25 3,13 3,49 - 366,9 486,1
Regent 10,8 13,5 13,2 15,8 2,55 3,65 3,66 3,45 152,3 120,6 114,0
Bronner 8,5 13,5 14 15,6 2,34 3,35 3,49 3,28 250,4 160,2 127,6
Felicia 9,9 12,4 10,5 - 2,53 3,42 3,45 - 145,3 132,8 112,9
Calandro 7,4 11,7 11,8 16,2 2,31 3,58 3,17 3,37 221,8 110,1 162,8
Aromera 3,8 4,1 11,6 - 2,28 2,67 3,16 - 288,3 386,7 183,0
Baron 11,3 16,4 13,9 15,1 2,38 3,45 3,75 3,43 185,4 85,5 87,1
Prior 11,6 16,2 13,3 17,5 2,22 3,42 3,34 3,31 179,6 185,2 133,0
Helios 10,9 10,8 12,7 15,6 2,44 2,88 3,66 3,3 109,0 108,0 127,0
5.4.4. Antocianinas Monoméricas Totais
Na Tabela 8, observamos a evolução dos valores de antocianinas monoméricas
totais (AMT) em cada uma das variedades tintas, durante o ciclo 2016/2017. Não foi feita
análise de AMT após a colheita. As variedades apresentaram aumento gradual da
concentração de antocianinas ao longo do tempo, como esperado durante a maturação,
porém, houve queda na coleta do dia 6 de janeiro, provavelmente devido aos motivos
supracitados. As AMT variaram entre 643,9 mg L-1 (Baron) e 1191,4 mg L-1 (Prior) na
última análise.
5.4.5. Polifenóis Totais
Na Tabela 8, observamos a evolução dos valores de polifenóis totais (PT) em
cada variedade, durante o ciclo 2016/2017. Não foi realizada análise dos PT após a
colheita. As variedades apresentaram (em sua maioria) diminuição da concentração de
polifenóis ao longo do tempo. Os PT variaram entre 180,7 mg L-¹ (Aromera) e 953,9 mg
L-¹ (Regent) na última análise.
É esperado que as uvas acumulem maior quantidade de polifenóis ao longo da
maturação, porém esse fator é altamente dependente de condições climáticas (COOMBE,
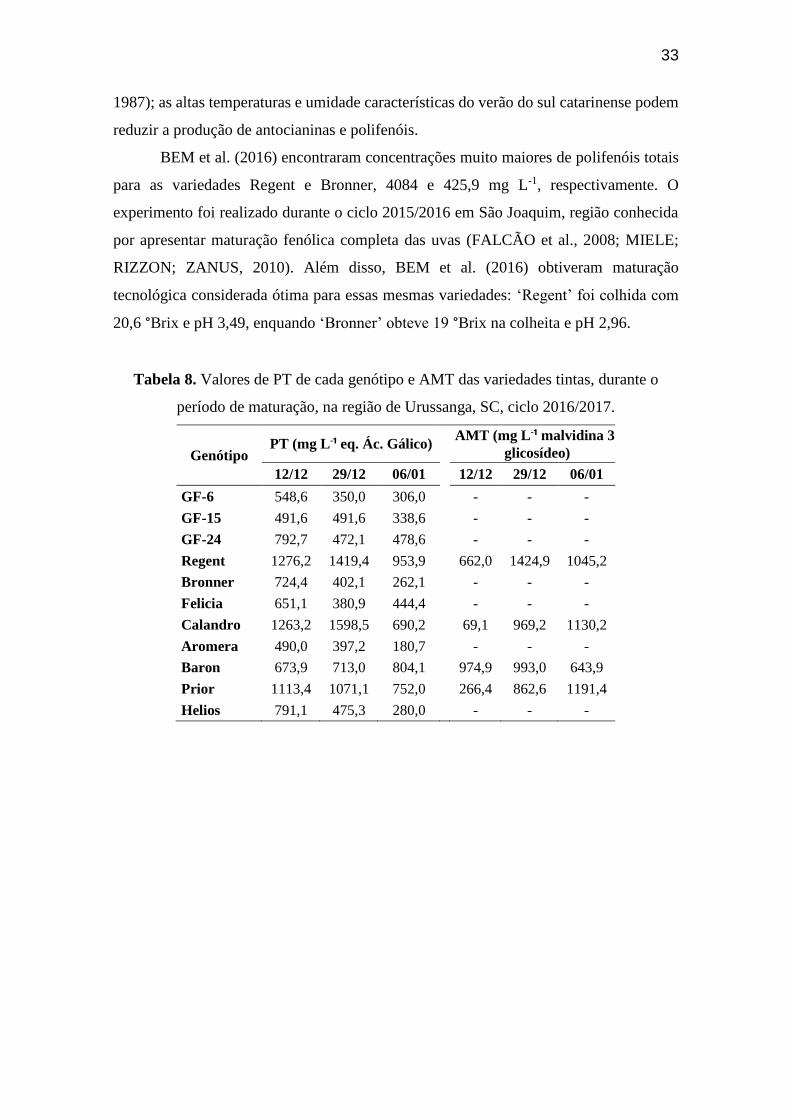
33
1987); as altas temperaturas e umidade características do verão do sul catarinense podem
reduzir a produção de antocianinas e polifenóis.
BEM et al. (2016) encontraram concentrações muito maiores de polifenóis totais
para as variedades Regent e Bronner, 4084 e 425,9 mg L-1, respectivamente. O
experimento foi realizado durante o ciclo 2015/2016 em São Joaquim, região conhecida
por apresentar maturação fenólica completa das uvas (FALCÃO et al., 2008; MIELE;
RIZZON; ZANUS, 2010). Além disso, BEM et al. (2016) obtiveram maturação
tecnológica considerada ótima para essas mesmas variedades: ‘Regent’ foi colhida com
20,6 °Brix e pH 3,49, enquando ‘Bronner’ obteve 19 °Brix na colheita e pH 2,96.
Tabela 8. Valores de PT de cada genótipo e AMT das variedades tintas, durante o
período de maturação, na região de Urussanga, SC, ciclo 2016/2017.
Genótipo PT (mg L-¹ eq. Ác. Gálico) AMT (mg L-¹ malvidina 3-
glicosídeo)
12/12 29/12 06/01 12/12 29/12 06/01
GF-6 548,6 350,0 306,0 - - -
GF-15 491,6 491,6 338,6 - - -
GF-24 792,7 472,1 478,6 - - -
Regent 1276,2 1419,4 953,9 662,0 1424,9 1045,2
Bronner 724,4 402,1 262,1 - - -
Felicia 651,1 380,9 444,4 - - -
Calandro 1263,2 1598,5 690,2 69,1 969,2 1130,2
Aromera 490,0 397,2 180,7 - - -
Baron 673,9 713,0 804,1 974,9 993,0 643,9
Prior 1113,4 1071,1 752,0 266,4 862,6 1191,4
Helios 791,1 475,3 280,0 - - -

34
6. Conclusões
As variáveis climáticas da região de Urussanga demonstraram alta disponibilidade
térmica para o desenvolvimento fenológico dos genótipos estudados, podendo gerar um
ciclo com menor tempo de duração quando comparado com as áreas de origem das
variedades (Alemanha e Itália).
Fatores como a temperatura do ar, amplitude térmica e precipitação pluviométrica
apresentaram forte influência sobre a maturação das uvas, reduzindo a concentração de
sólidos solúveis totais e compostos fenólicos. Considerando estes resultados, a região
apresenta potencial para a produção de vinhos jovens, que não exijam grande período de
envelhecimento.
Para os índices bioclimáticos, a região de Urussanga foi classificada no ciclo
2016/2017 como ‘Região V’ para o Índice de Winkler e ‘Região muito quente’ para o
Índice de Huglin.
O genótipo com brotação mais precoce e ciclo mais longo foi o GF-6, totalizando
121 dias e acumulando 1190 °C dia de requerimento térmico. A variedade com ciclo mais
curto foi a “Helios”, totalizando 106 dias e acumulando 1080,7 °C dia. A variedade
Baron, apresentou o maior requerimento térmico, acumulando 1198,9 °C dia entre a
brotação e a maturação. As demais variedades apresentaram ciclo de 113 dias
Os valores de plastocrono observados ficaram entre 39,5 °C dia nó-1 para ‘Regent’
e 73,5 °C dia nó-1 para ‘Bronner’. Considerando a presença de uma folha a cada nó do
sarmento, o número total de folhas por planta variou entre 115 para ‘Regent’ e 230,6
folhas para ‘GF-6’.
Os genótipos não apresentaram maturação tecnológica e fenólica dentro da faixa
considerada ideal para a produção de vinhos finos de qualidade.

35
REFERÊNCIAS
ALLEBRANDT, R. Caracterização da maturação e composição das uvas ‘Cabernet
Sauvignon’ e ‘Merlot’ produzidas em São Joaquim-SC. 2012. Trabalho de Conclusão
de Curso (Graduação em Agronomia) – Centro de Ciências Agrárias, Universidade
Federal de Santa Catarina, Florianópolis. Disponível em: <https://repositorio.ufsc.br/
bitstream/handle/123456789/79185/Ricardo%20Allebrandt.pdf?sequence=1&isAllowed
=y>. Acesso em: 25 novembro 2016.
ALVARENGA, A. A. et al. Origem e classificação botânica da videira. Informe
Agropecuário, Belo Horizonte, v. 19, n. 194, p. 5-8, 1998.
AMARANTE, J. O. A. Os segredos do vinho para iniciantes e iniciados. 4. ed. São
Paulo: Mescla, 2015. 624 p.
AMERINE, M. A.; OUGH, C. S. Wine and must analysis. Hoboken: John Wiley and
Sons, 1974.
ASSIS, J. S.; LIMA FILHO, J. M. P.; LIMA M. A. C. Fisiologia da Videira. Embrapa
Semiárido. In: FENAGRI, Petrolina, 2004, 26 p. Disponível em: < https://ainfo.cnptia.
embrapa.br/digital/bitstream/CPATSA/34234/1/OPB705.pdf>. Acesso em: 18 junho
2017.
AYOADE, J. O. Introdução a climatologia para os trópicos. 4. ed. Rio de Janeiro:
Bertrand Brasil, 1996. 332 p.
BAILLOD, M., BAGGIOLINI, M. Les stades repères de la vigne. Revue Suisse de
Viticulture, Arboriculture, Horticulture, v. 38, n. 1, p. 7–9, 1993.
BAKER, J. T.; REDDY, V. R. Temperature effects on phenological development and
yield of muskmelon. Annals of Botany, Oxford, v. 87, p. 605-613, 2001.
BALDIN, N. Tão fortes quanto a vontade, história da imigração italiana no Brasil: os
vênetos em Santa Catarina. Florianópolis: Insular/Editora da UFSC, 1999. 279 p.
BEM, B. et al. Downy mildew intensity in tolerant grapes varieties in highlands of
southern Brazil. In: Congresso Mundial da Uva e do Vinho, 39., 2016, Bento Gonçalves.
Anais... Bento Gonçalves, 2016.
BEURSKENS, S.; CRONE, S.; HOUBEN, G. The Development of Grape Varieties:
Towards a more sustainable viticulture. 2014. 30 p. Disponível em:
<http://brabantsewijnbouwers.nl/starnet/media/downloads/20141220%20-The%20
Development%20of%20Grape%20Varieties%20-%205.pdf>. Acesso em: 13 fevereiro
2017.
BLOESCH, B.; VIRET, O. Stades phénologiques repères de la vigne. Revue Suisse de
Viticulture, Arboriculture, Horticulture, Nyon, v. 40, n. 6, p. I-IV, 2008. Disponível
em: <http://www.agrometeo.ch/sites/default/files/documents/ stades_pheno_vigne.pdf>.
Acesso em: 16 fevereiro 2017.

36
CARVALHO, C. et al. Anuário Brasileiro da Uva 2016. Santa Cruz do Sul: Gazeta
Santa Cruz, 2016. 64 p. Disponível em: <http://www.editoragazeta.com.br/flip/anuario-
uva-2016/files/assets/common/downloads/publication.pdf>. Acesso em: 25 março 2016.
CHRISTENHUSZ, M. J. M.; BYNG, J. W. The number of known plants species in the
world and its annual increase. Phytotaxa, v. 261, n. 3, p. 201–217, 2016. Disponível em:
<https://www.biotaxa.org/Phytotaxa/article/view/phytotaxa.261.3.1/20598>. Acesso em:
13 maio 2017.
CONDE, C. et al. Biochemical changes throughout grape berry development and fruit
and wine quality. Food, v. 1, n. 1, p. 1-22, 2007.
COOMBE, B. G. Influence of temperature on composition and quality of grapes. Acta
Horticulturae, n. 206, p. 25–35, 1987.
COOMBE, B. G. Research on development and ripening of the grape berry. American
Journal of Enology and Viticulture, v. 43, n. 1, p. 101-110, 1992.
CZERMAINSKI, A. B. C.; SÔNEGO, O. R. Influência das condições climáticas sobre a
eficácia de fungicidas empregados para o controle do míldio em Vitis vinifera. Ciência
Rural, Santa Maria, v. 34, n. 1, p. 5-11, 2004. Disponível em: <http://www.scielo.br/pdf/
cr/v34n1/a02v34n1.pdf>. Acesso em: 16 abril 2016.
DALL'AGNOL, I. Origem Histórica e Dispersão da Videira. Confraria do Vinho:
Bento Gonçalves, 2007. Disponível em: <http://www.confrariadovinho-bg.com.
br/artigos.php?action=mostrar&id=16>. Acesso em: 22 junho 2016.
DELLA BRUNA, E.; ARCARI, S. G.; PETRY, H. B. A videira ‘Goethe’ e seus clones
nos Vales da Uva Goethe. Florianópolis: Revista Agropecuária Catarinense, v. 29, n. 2,
p. 53-57, 2016.
DOKOOZLIAN, N. K. Grape Berry Growth and Development. In: CHRISTENSEN, L.
P. Raisin Production Manual. Davis: University of California, 2000. p. 30-37.
Disponível em: <http://iv.ucdavis.edu/files/24467.pdf>. Acesso em: 03 junho 2017.
EIBACH, R.; TÖPFER, R. Success in resistance breeding: “Regent” and its steps into the
Market. ISHS Acta Horticulturae 603: VIII International Conference on Grape Genetics
and Breeding, p. 687-691, 2003.
EPAGRI. Síntese Anual da Agricultura de Santa Catarina 2015/2016. Florianópolis:
Epagri/Cepa, 2016, 188 p. Disponível em: <http://docweb.epagri.sc.gov.br/website_cepa/
publicacoes/Sintese_2016.pdf>. Acesso em: 25 março 2016.
FALCÃO, L. D. et al. Maturity of Cabernet Sauvignon berries from grapevines grown
with two different training systems in a new grape growing region in Brazil. Ciencia e
Investigación Agraria, v. 35, n. 3, p. 271-282, 2008.
FERNANDES, G. V. Ecofisiologia da videira (Vitis vinifera L.) sob cultivo protegido,
variedades Sauvignon Blanc, Cabernet Sauvignon e Merlot. 2014. 135 p. Dissertação
(Mestrado em Recursos Genéticos Vegetais) – Centro de Ciências Agrárias, Universidade

37
Federal de Santa Catarina, Florianópolis. Disponível em:
<https://repositorio.ufsc.br/bitstream/handle/123456789/123373/326997.pdf?sequence=
1&isAllowed=y>. Acesso em: 01 junho 2016.
GIOVANNINI, E. Produção de uvas para vinho, suco e mesa. Porto Alegre:
Renascença, 1999. 364 p.
GIUSTI, M. M.; WROLSTAD, R. E. Characterization and measurement of anthocyanins
by UV-visible spectroscopy. Current Protocols in Food Analytical Chemistry, 2001.
HALL, A.; JONES, G. V. Spatial analysis of climate in winegrape-growing regions in
Australia. Australian Journal of Grape and Wine Research, v. 16, p. 389-404, 2010.
HUGLIN, M. P. Nouveau mode d'évaluation des possibilités héliothermiques d'un milieu
viticole. Comptes Rendus de l'Académie d'Agriculture de France, v. 64, p. 1117-
1126, 1978.
INNOVITIS. Resistant varieties: Aromera. 2013. Disponível em: <http://www.
innovitis.eu/tl_files/InnoVitis/pdf/Datenblaetter%20englisch/Aromera.pdf>. Acesso em:
16 fevereiro 2017.
INSTITUT RHODANIEN. Pour une bonne maîtrise de l'acidité des vins. Point de
Repère, n. 5, 1998.
JACKSON, R. S. Wine Science: Principles and Applications. Burlington: Academic
Press-Elsevier, 2008. 770 p.
JKI – Julius Kühn-Institut. Vitis International Variety Catalogue (VIVC). 2017.
Disponível em: <http://www.vivc.de/index.php?r=site%2Findex>. Acesso em: 13
fevereiro 2017.
JONES, G. V. et al. Spatial analysis of climate in winegrape growing regions in the
western United States. American Journal of Enology and Viticulture, v. 61, n. 3, p.
313-326, 2010.
JONES, G. V.; DAVIS, R. E. Climate influences on grapevine phenology, grape
composition, and wine production and quality for Bordeaux, France. American Journal
of Enology and Viticulture, v. 51, n. 3, p. 249-261, 2000. Disponível em:
<https://inside.sou.edu/assets/envirostudies/gjones_docs/JonesDavisAJEV.pdf>. Acesso
em: 27 fevereiro 2017.
JONES, L. et al. Adaptive genomic structural variation in the grape powdery
mildew pathogen, Erysiphe necator. BMC Genomics, 2014. Disponível em:
<http://bmcgenomics.biomedcentral.com/articles/10.1186/1471-2164-15-1081>.
Acesso em: 16 abril 2016.
KENNEDY, J. A.; MATTHEWS, M. A.; WATERHOUSE, A. L. Changes in grape seed
polyphenols during fruit ripening. Phytochemistry, v. 55, n. 1, p. 77-85, 2000.
Disponível em: <http://matthews.ucdavis.edu/publications-1/Kennedy-%2 02000.pdf>.
Acesso em: 27 fevereiro 2017.

38
KENNEDY, J. Understanding grape berry development. Practical Winery & Vineyard,
San Rafael, July/August, p. 14-23, 2002. Disponível em: <https://www.practicalwinery.
com/JulyAugust02/julaug02p14.htm>. Acesso em: 27 fevereiro 2017.
KERBAUY, G. B. Fisiologia Vegetal. 2. ed. Rio de Janeiro: Guanabara Koogan, 2012,
431 p.
LEES, D. H.; FRANCIS, F. J. Standardization of pigment analyses in cranberries.
HortScience, Alexandria, v. 7, n. 1, p. 83-84, 1972.
MALINOVSKI, L. I. Comportamento viti-enológico da videira (Vitis vinifera L.) de
variedades autóctones italianas na região dos Campos de Palmas em Água Doce –
SC – Brasil. 2013. 255 p. Tese (Doutorado em Ciências, área de concentração Recursos
Genéticos Vegetais) – Centro de Ciências Agrárias, Universidade Federal de Santa
Catarina, Florianópolis. Disponível em: <https://repositorio.ufsc.br/bitstream/handle/
123456789/107585/318102.pdf?sequence=1&isAllowed=y>. Acesso em: 13 fevereiro
2017.
MANDELLI, F. Comportamento fenológico das principais cultivares de Vitis vinifera
L. para a região de Bento Gonçalves, RS. 1984. 125 p. Dissertação (Mestrado em
Agronomia, área de concentração Física do Ambiente Agrícola) – Escola Superior de
Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba.
MARZANO, L. Colonos e missionários italianos na floresta do Brasil. Florianópolis:
Editora da UFSC, 1985. 200 p.
MIELE, A.; RIZZON, L. A.; ZANUS, M. C. Discrimination of Brazilian red wines
according to the viticultural region, varietal, and winery origin. Ciência e Tecnologia
de Alimentos, v. 30, n. 1, p. 268-275, 2010.
MILLER, M. Wine from de inside out. Genco Winemakers: Sonoma County, 2009.
Disponível em: < http://www.gencowinemakers.com/docs/ Acids%20Presentation.pdf>.
Acesso em: 11 junho 2017.
MORI, K.; SUGAYA, S.; GEMMA, H. Decreased anthocyanin in grape berries grown
under elevated night temperature condition. Scientia Horticulturae, v. 105, n. 3, p. 319-
330, 2005.
MOTA, R. V. et al. Fatores que afetam a maturação e a qualidade da uva para vinificação.
Informe Agropecuário, Belo Horizonte, v. 27, n. 234, p. 56-64, 2006.
MURAKAMI, K. R. N. et al. Caracterização fenológica da videira cv. Itália (Vitis
vinifera L.) sob diferentes épocas de poda na região norte do estado do Rio de Janeiro.
Revista Brasileira de Fruticultura, Jaboticabal, v. 24, n. 3, p. 615-617, 2002.
Disponível em: <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-2945
2002000300008>. Acesso em: 22 junho 2016.

39
NAGATA, K. R. et al. Temperatura-base e soma térmica (graus-dia) para videiras 'Brasil'
e 'Benitaka'. Revista Brasileira de Fruticultura, Cruz das Almas, v. 22, n. 3, p. 329-
333, 2000.
NAVES, R. L. et al. Doenças e seu controle. In: Sistema de Produção de Uva de Mesa
no Norte do Paraná. Embrapa Uva e Vinho, 2005. Disponível em:
<https://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Uva/MesaNorteParana/do
encas.htm>. Acesso em: 16 abril 2016.
NENDEL, C. Grapevine bud break prediction for cool winter climates. International
Journal of Biometeorology, v. 54, p. 231-241, 2009.
PANDOLFO, C. Sistemas atmosféricos, variáveis meteorológicas e mudanças
climáticas na potencialidade do cultivo da videira (Vitis vinifera L.) no estado de
Santa Catarina. 2010. 174 p. Tese (Doutorado em Ciências, área de concentração
Recursos Genéticos Vegetais) – Centro de Ciências Agrárias, Universidade Federal de
Santa Catarina, Florianópolis. Disponível em: <https://repositorio.ufsc.br/bitstream/
handle/123456789/103325/280813.pdf?sequence=1&isAllowed=y>. Acesso em: 06
junho 2017.
PIRIE, A. Defining Cool Climate. Stratford: Brave New World, 2007, London.
POMMER, C. V. Uva: tecnologia de produção, pós-colheita, mercado. Porto Alegre:
Cinco Continentes, 2003, 777 p.
POTTER, R. O. et al. Solos do Estado de Santa Catarina. Boletim de Pesquisa e
Desenvolvimento, n. 46. Rio de Janeiro: Embrapa Solos, 2004.
PRITCHARD, D. Vitis Varieties. Minehead: Winegrowers Supplies. 2016. Disponível
em: <http://www.winegrowers.info/varieties/Vine_varieties/>. Acesso em: 13 fevereiro
2017.
PROGOETHE. Histórico. Disponível em: <http://www.progoethe.com.br/historico.php
?id=1>. Acesso em: 10 maio 2016.
RAVEN, P. H.; EVERT, R. F.; EICHHORN, S. E. Biologia Vegetal. 8. ed. Rio de
Janeiro: Guanabara Koogan, 2014. 876 p.
RIBÉREAU-GAYON, P. et al. Handbook of Enology: The Microbiology of Wine and
Vinifications. 2. ed. Chichester: John Wiley & Sons Ldt., 2006. v. 1. 497 p.
ROBINSON, J.; HARDING, J.; VOUILLAMOZ, J. Wine Grapes: A complete guide to
1.368 vine varieties, including their origins and flavours. New York: Ecco Press, 2012.
1859 p.
ROSIER, J. P. Vitivinicultura em Santa Catarina. In: BRDE – Banco Regional de
Desenvolvimento do Extremo Sul. Vitivinicultura em Santa Catarina: Situação atual e
perspectivas. Florianópolis: BRDE, 2005. p. 28-35. Disponível em: < http://www.brde.
com.br/media/brde.com.br/doc/estudos_e_pub/Vitivinicultura%20em%20Santa%20Cat
arina.pdf>. Acesso em: 12 fevereiro 2017.

40
SIBBUS, R. White Wine Varieties: Bronner. 2011. Disponível em:
<http://www.sibbus.com/en/vine-nursery/varieties/white-wine-varieties/bronner.html>.
Acesso em: 13 fevereiro 2017.
SINGLETON, V. L.; ROSSI, J. A. Colorimetry of total phenolics with phosphomolybdic-
phosphotungstic acid reagent. American Journal of Enology and Viticulture, v. 16, p.
144-158, 1965.
SOUSA, J. S. I. Uvas para o Brasil. 2. ed. Piracicaba: FEALQ, 1996.
STAUDT, G.; KASSEMEYER, H. H. Evaluation of downy mildew resistance in various
accessions of wild Vitis species. Vitis, v. 34, n. 4, p. 225-228, 1995. Disponível em:
<http://www.vitis-vea.de/admin/volltext/ e035589.pdf>. Acesso em: 20 abril 2016.
STEVENS, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 13. Vitales.
2017. Disponível em: <http://www.mobot.org/MOBOT/Research/APweb/orders/
vitalesweb.html#Vitales>. Acesso em: 13 maio 2017.
STRECK, N. A., et al. Estimativa do plastocrono em cultivares de soja. Bragantia,
Campinas, v. 67, n. 1, p. 67-73, 2008. Disponível em: < http://www.iac.sp.gov.br/
publicacoesbragantia/volumesonline.php>. Acesso em: 29 maio 2017.
STRIEM, M. Grape hybrid varieties and accession’s parentage and their genetic
percent of Vitis species. 2010. 18 p. Disponível em:
<http://michaelstriem.com/files/Species.pdf>. Acesso em: 14 fevereiro 2017.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 5. ed. Porto Alegre: Artmed, 2013, 954 p.
TERRA, M. M., PIRES, E. J. P., NOGUEIRA, N. A. M. Tecnologia para produção de
uva 'Itália' na região Noroeste do Estado de São Paulo. Campinas: CATI, 1998. 51 p.
(Documento Técnico, 97).
TOMAZETTI, T. C. et al. Fenologia e acúmulo térmico em videiras viníferas na região
da Fronteira Oeste do Rio Grande do Sul. Pesquisa Agropecuária Brasileira, v. 50, n.
11, p. 1033-1041, 2015.
TONIETTO, J.; CARBONNEAU, A. A multicriteria climatic classification system for
grape-growing regions worldwide. Agricultural and Forest Meteorology, v. 124, p. 81-
97, 2004.
VALES DA UVA GOETHE. Histórico. Disponível em: <http://www.valesdauvagoethe.
com.br/historico.php?id=1>. Acesso em: 10 maio 2016.
VIANA, L. H. Fenologia e quebra de dormência da videira Niagara Rosada cultivada
na região norte fluminense em diferentes épocas de poda. 2009. 74 p. Tese (Doutorado
em Produção Vegetal) – Centro de Ciências e Tecnologias Agropecuárias, Universidade
Estadual do Norte Fluminense Darcy Ribeiro, Campos dos Goytacazes. Disponível em:
<http://uenf.br/pos-graduacao/producao-vegetal/files/2014/10/Leandro-Viana.pdf>.
Acesso em: 27 fevereiro 2017.

41
WEIN-PLUS. Glossary: Bronner. 2016. Disponível em: <http://www.wein-plus.eu/en/
Bronner+%28Rebsorte%29_3.0.3733.html>. Acesso em: 13 fevereiro 2017.
WINE PLANT. Resistant red wine grapes. 2016. Disponível em:
<http://www.wineplant.it/en/resistant-red-wine-grapes.html>. Acesso em: 14 fevereiro
2017.
WINE-SEARCHER. Regent Wine. 2015. Disponível em: <http://www.wine-
searcher.com/grape-981-regent>. Acesso em: 13 fevereiro 2017.
WINKLER, A. J. et al. General Viticulture. Berkeley: University of California, 1974.
710 p.
ZEIST, A. R. Estimativa do plastocrono das videiras 'Cabernet Sauvignon' e 'Chardonnay'
na Fronteira Oeste do Rio Grande do Sul. Pesquisa Agropecuária Brasileira, v. 52, n.
4, p. 244-251, 2017. Acesso em: < https://seer.sct.embrapa.br/index.php/pab/article/view/
24821/13733>. Acesso em: 11 junho 2017.
ANEXOS
Anexo A. Localização do vinhedo experimental (destacado em amarelo) na Estação
Experimental da Epagri de Urussanga, SC.
Fonte: Google Earth





























