Embed Size (px)
Citation preview

i
UNIVERSIDADE FEDERAL DO CEARÁ UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
MORFOFISIOLOGIA DO DOSSEL E DESEMPENHO DE OVINOS EM CAPIM-TIFTON 85
ANA PATRÍCIA ALMEIDA BEZERRA
FORTALEZA - CE
Março de 2011

ii
UNIVERSIDADE FEDERAL DO CEARÁ UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
MORFOFISIOLOGIA DO DOSSEL E DESEMPENHO DE OVINOS EM CAPIM-TIFTON 85
ANA PATRÍCIA ALMEIDA BEZERRA
Engenheira Agrônoma
FORTALEZA - CE
Março de 2011

Dados Internacionais de Catalogação na Publicação Universidade Federal do Ceará
Biblioteca de Ciências e Tecnologia B469 Bezerra, Ana Patrícia Almeida.
Morfofisilogia do dossel e desempenho de ovinos em capim-tifton 85. /Ana Patrícia Almeida Bezerra. – 2011.
173 f. : il., color. Tese (doutorado) – Universidade Federal do Ceará, Centro de Ciências Agrárias,
Departamento de Zootecnia, Programa de Doutorado Integrado em Zootecnia, Fortaleza, 2011. Área de Concentração: Forragicultura e Pastagens. Orientação: Prof. Dr. Magno José Duarte Cândido. Coorientação: Dr. Tadeu Vinhas Voltolini. 1. Morfogênese. 2. Pastagens. I. Título.
CDD 636.08

ii
ANA PATRÍCIA ALMEIDA BEZERRA
MORFOFISIOLOGIA DO DOSSEL E DESEMPENHO DE OVINOS EM CAPIM-TIFTON 85
Tese submetida à Coordenação do Curso de Pós-Graduação em Zootecnia, da Universidade Federal do Ceará, como requisito parcial para obtenção do grau de Doutora em Zootecnia.
Área de Concentração: Forragicultura e Pastagens
Orientação
Prof. D. Sc. Magno José Duarte Cândido
Coorientação
D. Sc. Tadeu Vinhas Voltolini
FORTALEZA - CE
Março de 2011

iii
ANA PATRÍCIA ALMEIDA BEZERRA
MORFOFISIOLOGIA DO DOSSEL E DESEMPENHO DE OVINOS EM CAPIM-TIFTON 85
Tese defendida e aprovada pela comissão examinadora em 14 de março de 2011.
Comissão Examinadora:
___________________________________________
Prof. D.Sc. Magno José Duarte Cândido (Orientador)
Universidade Federal do Ceará Departamento de Zootecnia-DZ/UFC
___________________________________________
D.Sc. Tadeu Vinhas Voltolini (Coorientador)
Empresa Brasileira de Pesquisa Agropecuária - Centro de Pesquisa Agropecuária do Trópico Semiárido (CPATSA)
___________________________________________
Profª. D.Sc. Maria Socorro de Souza Carneiro (Membro)
Universidade Federal do Ceará Departamento de Zootecnia-DZ/UFC
___________________________________________
Profª. D.Sc. Maria Fernanda Soares Queiroz (Membro)
Universidade Federal da Paraíba Departamento de Agropecuária-DAP/UFPB
___________________________________________
Prof. D.Sc. Claudio Mistura (Membro)
Universidade Estadual da Bahia Departamento de Tecnologia e Ciências Sociais-DTCS/UNEB

iv
DADOS CURRICULARES DA AUTORA
Ana Patrícia Almeida Bezerra, filha de Maria Zenir de Almeida Bezerra e
Francisco Airton Bezerra, nasceu em 29 de junho de 1975, em Quixadá - Ceará.
Em janeiro de 2000, recebeu o título de Engenheira Agrônoma pela
Universidade Federal da Paraíba - UFPB.
Em março de 2004, recebeu o título de Mestre em Agronomia, pelo
Departamento de Fitotecnia da Universidade Federal do Ceará - UFC.
De 2005 e 2006 trabalhou como professora nos Cursos de Agronomia e
Veterinária na Universidade Estadual do Maranhão – UEMA e nos Cursos de Zootecnia
e Farmácia na Faculdade de Imperatriz – FACIMP.
Em março de 2007, iniciou o curso de Doutorado em Zootecnia, área de
concentração em Forragicultura e Pastagens, no Departamento de Zootecnia da
Universidade Federal do Ceará - UFC.
Realizou o experimento de campo na Empresa Brasileira de Pesquisa
Agropecuária - Centro de Pesquisa Agropecuária do Trópico Semiárido (CPATSA).
Em agosto de 2010, foi aprovada no concurso público para professor adjunto no
Curso de Agroecologia, na área de produção animal na Universidade Federal da Paraíba
- UFPB.

v
DEDICO
Aos meus pais Maria Zenir de Almeida Bezerra (in memoriam) e Francisco Airton Bezerra
Ao meu esposo e amor Leonardo Augusto Fonseca Pascoal
A minha irmã Antônia Silvana Carla Almeida Bezerra
Que sempre estiveram comigo em todas as conquistas, que me apoiaram nos momentos
difíceis, que me ajudaram a perseverar em meus anseios e a vencer a distância e
saudade...
OFEREÇO
A Maria Zenir de Almeida Bezerra (in memoriam), em reconhecimento de uma vida
integra e repleta de sabedoria, que me serve de exemplo e orgulho, através de suas
palavras: “o único legado e o mais importante, que os pais são capazes de deixar aos
filhos é o estudo”.

vi
“Os teus caminhos posso não entender Senhor, mas sei que tudo é visando o meu
crescer, se lutas e tribulações eu tenho que passar te peço forças para continuar.”
(Jó)

vii
AGRADECIMENTOS
À Deus, acima de tudo.
À Universidade Federal de Ceará – UFC e ao Departamento de Zootecnia, pela
oportunidade de realização do curso.
À Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico -
FUNCAP, pela concessão da bolsa.
À Empresa Brasileira de Pesquisa Agropecuária - Centro de Pesquisa
Agropecuária do Trópico Semiárido (CPATSA), pelo o desenvolvimento do
experimento de tese.
Ao Instituto Federal de Educação, Ciência e Tecnologia do Sertão de
Pernambuco (IF Sertão-PE) pela viabilização do abate dos animais.
Ao professor Magno José Duarte Cândido, pela orientação, pelos ensinamentos e
por ter acreditado na realização desse trabalho.
Ao pesquisador Tadeu Vinhas Voltolini por viabilizar a execução do projeto na
Embrapa Semiárido, por todo empenho, coorientação, confiança, amizade e por ter me
engajado na carreira científica.
À professora Maria Socorro de Souza Carneiro pela amizade e ensinamentos,
durante o curso.
À pesquisadora Salete Alves de Moraes pela amizade e valiosos comentários
para a elaboração desse trabalho.
À professora Maria Fernanda pela amizade e correções dos capítulos.
Ao professor Claudio Mistura, pela participação na Banca Examinadora e
valiosa contribuição.
Ao meu esposo, Leonardo, pela ajuda com a análise estatística.
À coordenação do Programa de Pós-Graduação em Zootecnia, nas pessoas do
Coordenador Arlindo de Alencar Araripe e da Secretária Francisca.
À todos os professores que contribuíram para minha formação profissional.
Aos Funcionários da Embrapa Semiárido, especialmente os senhores: José e
Telito, pela convivência e auxílio nos trabalhos de campo.

viii
Aos Funcionários do Laboratório de Nutrição Animal da Embrapa: Benedito e
Alcides, pela atenção dedicada a mim, sempre que busquei ajuda e o café da manhã (ô,
riqueza!).
Aos amigos, Kaio Victor, Ernandes, Fabrícia e Rafael, pela amizade, sem essas
pessoas esse experimento jamais sairia do papel.
Aos estagiários: Iranildo (Bolinha), Elida, Samires, Elane, Isabelle, Camila,
Getúlio, Moara e Daniel (Osama), inestimável contribuição para a realização desse
experimento.
A galera do “Bode Assado”: Greicy, Genilson, Rafael e Kaio, nossa única fonte
de energia nos finais de semana.
Aos colegas de orientação: Patrícia Lima, Marieta, Suely, Roberto, Cutrim,
Gregório e Liandro, nunca serão esquecidos.
Aos novos amigos, Marcílio Costa, Marquinhos, Luiz Neto, Labib, Marieta,
Igor, Carol, Guilherme, Rildson, Gilson, Mirlânda, Katiane, Paty e Liandro. E a todos
os colegas de Pós-Graduação pela convivência.
Em fim a todos que contribuíram para a realização deste trabalho,
OBRIGADA!

ix
SUMÁRIO
Página
LISTA DE TABELAS ...................................................................................... xii
LISTA DE FIGURAS ....................................................................................... xv
RESUMO GERAL ............................................................................................ xvii
ABSTRACT ....................................................................................................... xix
Considerações Iniciais ...................................................................................... 2
Capítulo 1. Referencial Teórico ....................................................................... 5
1. As pastagens no Brasil e o gênero Cynodon sp .............................................. 5
2. A dinâmica de produção de forragem ............................................................. 6
2.1. Características morfogênicas ....................................................................... 7
2.2. Características estruturais ............................................................................ 9
3. Estratégias de manejo: período de descanso e resíduo pós-pastejo ................ 12
4. Ambiente de pastoril: planta–animal .............................................................. 15
REFERÊNCIAS ................................................................................................. 16
Capítulo 2. Crescimento e componentes da biomassa inicial do dossel do
capim-tifton 85 ..................................................................................................
23
RESUMO ............................................................................................................ 23
ABSTRACT ....................................................................................................... 24
INTRODUÇÃO .................................................................................................. 25
MATERIAL E MÉTODOS ................................................................................ 26
RESULTADOS E DISCUSSÃO ........................................................................ 29
CONCLUSÕES .................................................................................................. 37
REFERÊNCIAS ................................................................................................. 38
Capítulo 3. Características morfogênicas do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo ............................
42
RESUMO ............................................................................................................ 42
ABSTRACT ....................................................................................................... 43
INTRODUÇÃO .................................................................................................. 44

x
MATERIAL E MÉTODOS ................................................................................ 45
RESULTADOS E DISCUSSÃO ........................................................................ 49
CONCLUSÕES .................................................................................................. 58
REFERÊNCIAS .................................................................................................
58
Capítulo 4. Componentes da biomassa e características estruturais do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo .........................................................................................................
62
RESUMO ............................................................................................................ 62
ABSTRACT ....................................................................................................... 63
INTRODUÇÃO .................................................................................................. 64
MATERIAL E MÉTODOS ................................................................................ 65
RESULTADOS E DISCUSSÃO ........................................................................ 69
CONCLUSÕES .................................................................................................. 81
REFERÊNCIAS ................................................................................................. 81
Capítulo 5. Composição química e digestibilidade in vitro da matéria seca de lâminas foliares e colmos do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo ..........................................
86 RESUMO ............................................................................................................ 86
ABSTRACT ....................................................................................................... 87
INTRODUÇÃO .................................................................................................. 88
MATERIAL E MÉTODOS ................................................................................ 89
RESULTADOS E DISCUSSÃO ........................................................................ 92
CONCLUSÕES .................................................................................................. 103
REFERÊNCIAS ................................................................................................. 103
Capítulo 6. Comportamento e desempenho de ovinos terminados em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo .........................................................................................................
108
RESUMO ............................................................................................................ 108
ABSTRACT ....................................................................................................... 109
INTRODUÇÃO .................................................................................................. 110

xi
MATERIAL E MÉTODOS ................................................................................ 111
RESULTADOS E DISCUSSÃO ........................................................................ 116
CONCLUSÕES .................................................................................................. 129
REFERÊNCIAS ................................................................................................. 130
Capítulo 7. Características da carcaça, componentes não carcaça e composição centesimal do músculo Longíssimus dorsi de ovinos terminados em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo ..................................................................................
134 RESUMO ............................................................................................................ 134
ABSTRACT ....................................................................................................... 135
INTRODUÇÃO .................................................................................................. 136
MATERIAL E MÉTODOS ................................................................................ 137
RESULTADOS E DISCUSSÃO ........................................................................ 142
CONCLUSÕES .................................................................................................. 149
REFERÊNCIAS ................................................................................................. 150
Considerações Finais e Implicações ................................................................ 153

xii
LISTA DE TABELAS
Capítulo 2
Páginas
Tabela 1. Dados climáticos observados durante o período experimental ............... 26
Capítulo 3
Tabela 1. Dados climáticos observados durante o período experimental ............... 45
Tabela 2. Características químicas do solo da área experimental em duas
profundidades .......................................................................................... 46
Tabela 3. Características morfogênicas, taxa de produção e acúmulo de forragem
em pastagem de capim-tifton 85 manejado sob três períodos de
descanso e dois resíduos pós-pastejo ....................................................... 51
Tabela 4. Efeito dos ciclos de pastejo sobre as características morfogênicas em
pastagem de capim-tifton 85 sob três períodos de descanso e dois
resíduos pós-pastejo ................................................................................ 55
Tabela 5. Efeito dos ciclos de pastejo sobre as características morfogênicas em
pastagem de capim-tifton 85 sob três períodos de descanso e dois
resíduos pós-pastejo ................................................................................ 56
Tabela 6. Efeito dos ciclos de pastejo sobre as características morfogênicas em
pastagem de capim-tifton 85 sob três períodos de descanso e dois
resíduos pós-pastejo ................................................................................ 57
Capítulo 4
Tabela 1. Dados climáticos observados durante o período experimental ............... 65
Tabela 2. Características químicas do solo da área experimental em duas
profundidades .......................................................................................... 66
Tabela 3. Componentes da biomassa e características estruturais do capim-tifton
85 manejado sob três períodos de descanso e dois resíduos pós-pastejo 70
Tabela 4. Efeito dos ciclos de pastejo sobre os componentes da biomassa e as
características estruturais em pastagem de capim-tifton 85 sob três
períodos de descanso e dois resíduos pós-pastejo ..................................
74

xiii
Tabela 5. Componentes da biomassa e características estruturais do capim-tifton
85 manejado sob três períodos de descanso e dois resíduos pós-pastejo 78
Tabela 6. Efeito dos ciclos de pastejo sobre os componentes da biomassa e as
características estruturais em pastagem de capim-tifton 85 sob três
períodos de descanso e dois resíduos pós-pastejo ..................................
80
Capítulo 5
Tabela 1. Dados climáticos observados durante o período experimental ............... 89
Tabela 2. Características químicas do solo da área experimental em duas
profundidades .......................................................................................... 90
Tabela 3. Composição química e digestibilidade in vitro da matéria seca da
lâmina foliar do capim-tifton 85 manejado sob três períodos de
descanso e dois resíduos pós-pastejo ....................................................... 93
Tabela 4. Efeito dos ciclos de pastejo sobre a composição química e
digestibilidade in vitro da matéria seca da fração lâmina foliar do
capim-tifton 85 manejado sob três períodos de descanso e dois
resíduo pós-pastejo ................................................................................ 97
Tabela 5. Composição química e digestibilidade in vitro da matéria seca da
fração colmo do capim-tifton 85 manejado sob três períodos de
descanso e dois resíduos pós-pastejo ....................................................... 99
Tabela 6. Efeito dos ciclos de pastejo sobre a composição química e
digestibilidade in vitro da fração colmo do capim-tifton 85 manejado
sob três períodos de descanso e dois resíduos pós-pastejo ...................... 101
Tabela 7. Efeito dos ciclos de pastejo sobre a composição química e
digestibilidade in vitro da fração colmo do capim-tifton 85 manejado
sob três períodos de descanso e dois resíduos pós-pastejo ...................... 102
Capítulo 6
Tabela 1. Dados climáticos observados durante o período experimental ............... 112
Tabela 2. Características químicas do solo da área experimental em duas
profundidades .......................................................................................... 114

xiv
Tabela 3. Atividades contínuas e pontuais de ovinos em capim-tifton 85
manejado sob três períodos de descanso e dois resíduos pós-pastejo .....
120
Tabela 4. Taxa de bocado, número diário de bocados e massa do bocado de
ovinos em capim-tifton 85 manejado sob três períodos de descanso e
dois resíduos pós-pastejo ........................................................................ 121
Tabela 5. Atividades contínuas e pontuais nas 24 horas do 1º e 4º dia de pastejo
de ovinos em capim-tifton 85 manejado sob três períodos de descanso e
dois resíduos pós-pastejo ..................................................................... 121
Tabela 6. Taxa de bocado, número diário de bocados e massa do bocado nas 24
horas do 1º e 4º dia de pastejo de ovinos em capim-tifton 85 manejado
sob três períodos de descanso e dois resíduos pós-pastejo ...................... 124
Tabela 7. Composição químico-bromatologica e DIVMS das amostras obtidas
através da simulação de pastejo no 1º e 4º dia de ocupação do capim-
tifton 85 manejado sob três períodos de descanso e dois resíduos pós-
pastejo........................................................................................................ 127
Tabela 8. Desempenho produtivo e consumo de matéria seca de ovinos em
capim-tifton 85 manejado sob três períodos de descanso e dois resíduos
pós-pastejo ............................................................................................... 128
Tabela 9. Taxa de lotação de ovinos em capim-tifton 85 manejado sob três
períodos de descanso e dois resíduos pós-pastejo .................................. 129
Tabela 10. Consumo de matéria seca de ovinos no 1º e 4º dia de ocupação do
capim-tifton 85 manejado sob três períodos de descanso e dois
resíduos pós-pastejo ............................................................................... 131
Capítulo 7
Tabela 1. Características da carcaça, espessura de gordura e área de olho de lombo
de ovinos em pastagem de capim-tifton 85 manejado sob três períodos
de descanso e dois resíduos pós-pastejo ....................................................
144
Tabela 2. Pesos (kg) dos não componentes da carcaça de ovinos em capim-tifton
85 manejado sob três períodos de descanso e dois resíduos pós-pastejo.... 146
Tabela 3. Pesos (kg) dos cortes comerciais de ovinos em capim-tifton 85 manejado
sob três períodos de descanso e dois resíduos pós-pastejo ........................
147

xv
Tabela 4. Composição centesimal do músculo longíssimus dorsi de ovinos em
capim-tifton 85 manejado sob três períodos de descanso e dois resíduos
pós-pastejo ................................................................................................. 149
LISTA DE FIGURAS
Capítulo 2
Páginas
Figura 1. Analisador LAI 2000 da LI COR Biocienses ........................................... 27
Figura 2. Evolução do índice de área foliar (IAF) e interceptação da radiação
fotosinteticamente ativa (RFA) em dossel de capim-tifton 85(Cynodon
sp), em diferentes idades de rebrotação .................................................. 30
Figura 3. Componentes da biomassa inicial (g/m²): Massa seca de forragem total
(MSFT), massa seca de forragem verde (MSFV), massa seca de lâmina
verde (MSLV), massa seca de colmo verde (MSCV) e massa seca de
forragem morta (MSFM) em dossel de capim-tifton 85 (Cynodon sp),
em diferentes idades de rebrotação ......................................................... 31
Figura 4. Taxa de crescimento relativo (TCR) em dossel de capim-tifton 85
(Cynodon sp), em diferentes idades de rebrotação................................. 32
Figura 5. Taxa de assimilação liquida (TAL) em dossel de capim-tifton 85
(Cynodon sp), em diferentes idades de rebrotação.................................. 33
Figura 6. Razão de área foliar (RAF) em dossel de capim-tifton 85 (Cynodon
sp), durante a avaliação de crescimento, em diferentes idades de
rebrotação................................................................................................. 34
Figura 7. Taxa de acúmulo de forragem (TAF) em dossel de capim-tifton 85
(Cynodon sp), em diferentes idades de rebrotação................................... 35
Figura 8. Densidade populacional de perfilhos (perfilhos/m2) em em dossel de
capim-tifton 85 (Cynodon sp), em diferentes idades de rebrotação......... 36
Figura 9. Número de folhas vivas/perfilh em em dossel de capim-tifton 85
(Cynodon sp), em diferentes idades de rebrotação...................................
37

xvi
Capítulo 4
Figura 1. Condição de pré-pastejo no pasto manejado sob PD de 8,5 fol/perf ....... 67
Figura 2. 2º dia de pastejo ....................................................................................... 67
Figura 3. 3º dia de pastejo ....................................................................................... 67
Figura 4. Condição residual de 10 cm .................................................................... 67
Capítulo 6
Figura 1. Animais no 1º dia de pastejo ................................................................... 113
Figura 2. Animais no 4º dia de pastejo ................................................................... 113
Figura 3. Preparo da balança no centro de manejo ................................................ 114
Figura 4. Pesagem dos animais .............................................................................. 114
Figura 5. Preparo dos animais para coleta de fezes ................................................ 115
Figura 6. Animal com bolsa coletora de fezes e arreio .......................................... 115
Capítulo 7
Figura 1. Carcaça de ovinos acondicionadas em sacos plásticos ........................... 139
Figura 2. Carcaças de ovinos na câmara fria a 5ºC ................................................ 139
Figura 3. Meia carcaça foi dividida em seis cortes comerciais: pescoço, serrote,
costilhar, paleta, lombo e pernil .............................................................. 140
Figura 4. Corte longitudinal da carcaça ................................................................. 141
Figura 5. Traçado da área de olho de lombo em película transparente .................. 141
Figura 6. Medição da espessura de gordura, com o paquímetro digital ................. 141
Figura 7. Longíssimus dorsi ................................................................................... 141

xvii
RESUMO GERAL
MORFOFISIOLOGIA DO DOSSEL E DESEMPENHO DE OVINOS EM
CAPIM-TIFTON 85
Objetivou-se avaliar as características morfofisiológicas do dossel e o
desempenho de ovinos em capim-tifton 85. Os três períodos de descanso utilizados
foram 4,5, 6,5 e 8,5 novas folhas formadas após a saída dos animais. A condição
residual preconizada para a saída dos animais foi de uma altura de 5 e 10 cm. O
delineamento experimental foi inteiramente casualização em arranjo fatorial 3x2 com
quatro repetições (piquetes), os dados foram analisados inicialmente o efeito dos
períodos de descanso e dos resíduos pós-pastejo, em seguida, o efeito dos ciclos
sucessivos de pastejo. Utilizou-se o método de pastejo sob lotação intermitente, com
lotação variável. Foram utilizados 30 ovinos de prova e um número variável de animais
de reguladores, da raça Santa Inês, com peso corporal de 20 kg e, em grupos de cinco,
distribuídos por sorteio em seis tratamentos. Na avaliação de crescimento inicial, o
aumento da massa seca de colmo verde pode estar relacionado com o aumento do índice
de área foliar ao longo da avaliação e ter sido o responsável na redução da massa seca
de lâmina verde. Foram observadas reduções nos valores de taxa de crescimento
relativo, assimilação liquida e razão de área foliar com o avanço da idade de rebrotação.
A taxa de acúmulo de forragem aumento ao longo da avaliação. Foram verificadas
reduções na densidade populacional de perfilhos e no número de folhas vivas. As
características morfogênicas foram afetadas pelos períodos de descanso (PDs) e os
resíduos pós-pastejo (RPPs) e efeito dos ciclos de pastejo. Os PDs superiores ao
necessário para formação de 6,5 fol/perf tornam todas as características morfogênicas
estudadas indesejáveis do ponto de vista forrageiro. Os componentes da biomassa no
pré e pós-pastejo, foram afetados pela interação PD x RPP e pelos ciclos de pastejo. O
prolongamento do PD comprometeu a estrutura do dossel. O uso do capim-tifton 85 não
deve exceder ao PD de 6,5 fol/perf para evitar perdas de forragem por senescência,
sendo possível utilizar uma altura residual de 5 cm sem comprometer a perenidade da
pastagem. O teor de proteína bruta (PB) das duas frações foi reduzido com o aumento
do PD. Os coeficientes de DIVMS da fração lâmina foliar, encontram-se adequados.
Observou-se que a variação dos valores de DIVMS da fração colmo dos pastos

xviii
manejados sob os três PDs e os dois RPPs foi de pequena magnitude. Os ciclos de
pastejo afetaram a composição química e a DIVMS das duas frações. Os pastos
manejados sob PD de 8,5 fol/perf e resíduo de 10 cm proporcionaram maior tempo de
pastejo e ruminação. O 4º dia de pastejo proporcionou maior tempo em pastejo e
ruminação. O consumo de sal e água foi bastante reduzido. A maior taxa de bocados foi
verificada nos pastos sob PD de 4,5 fol/perf. O 4º dia de pastejo proporcionou aumento
da taxa e número de bocados e redução da massa do bocado. Os PDs correspondentes a
4,5 e 6,5 fol/perf apresentaram melhores desempenho animal. A maior capacidade de
suporte, em unidades animal, observada para os pastos manejados sob PD de 8,5
fol/perf. O maior consumo de matéria seca foi verificado 1º dia de pastejo. As
estratégias de manejo afetaram as características da carcaça. O PD de 8,5 fol/perf foi
responsável pelo maior peso da pele. O RPP mais elevado proporcionou maior
rendimento biológico. Os cortes não foram afetados pelas estratégias de manejo. Os
teores de umidade e extrato etéreo foram influenciados pelo PD.
Termos para indexação: estrutura do pasto, morfogênese, valor nutritivo, carcaça.

xix
ABSTRACT
MORPHOPHYSIOLOGY OF FORAGE AND SHEEP PERFORMANCE ON
TIFTON 85 GRASS
The aim of the study was to evaluate morphophysiological characteristics of forage and
sheep performance on Tifton 85 Bermudagrass. Three periods of rest were used with
4.5, 6.5 and 8.5 new leaves per tiller after stocking. The residual condition to delimited
the end of stocking was forage height of 5 and 10 cm. It was used a completely
randomized design arranged in a 3x2 fatorial scheme, with four replicates and two post-
grazing periods (5 and 10 cm). Data of effect of periods of rest (PR) and post-grazing
(PG) periods were initially analyzed and then the effect of continually grazing cycles. It
was used the intermittent stocking grazing method, with variable stocking. A total of 30
Santa Ines sheep and variable number of regulating animals, with 20 kg of body weight,
were distributed in groups of 5 animals among 6 treatments. At evaluation of initial
growth, an increase of green culm dry matter maybe related to the increase of leaves
area index throughout the evaluation and being responsible to reduce the green blade
dry matter. Reductions in the values of relative growth rate, net assimilation and leaves
area ratio with increasing the age of regrowth were observed. The rate of forage
accumulation increased throughout the evaluation. Reductions were observed in tiller
density and number of green leaves. The morphogenetic characteristics were affected by
periods of rest (PR) and post-grazing residue (PG) and the effect of grazing cycles.
Unnecessary higher PR to formation 6.5 leaves/tiller became morphogenical
characteristics undesirable. The components of biomass in pre and post-grazing were
affected by the interaction between PR x PG and the grazing season. The extension of
the PR affect the forage structure. The use of Tifton 85 Bermudagrass should not exceed
the PR of 6,5 leaves/tiller to avoid loss of forage due to senescence, using a residual
height of 5 cm without compromising the sustainability of the pasture. The crude
protein (CP) of both fractions was reduced with increasing PR. The coefficients of
DIVMS of leaves fraction were adequate. It was observed that the variation of DIVMS
of culm fraction of pastures grazed under three PR and two PR had low
magnitude. Grazing cycles affected the chemical composition and DIVMS of two
fractions. The pastures grazed under PR of 8,5 leaves/tiller and residue of 10 cm
provided higher grazing time and ruminating. The 4th day of grazing provided more

xx
time of grazing and ruminating. The salt and water intakes were reduced. The highest
bite rate was found in pastures under PD of 4,5 leaves/tiller. The 4th day of grazing
provided an increase int the rate and number of bites and reduction at mass bite. PR
corresponding to 4.5 and 6.5 leaves/tiller provided better performance. The higher
support capacity, in animal units, was observed to pastures grazed under PD of 8.5
leaves/tiller. The higher dry matter intake was recorded at the 1st day of grazing. The
management strategies affect carcass characteristics. The PD of 8.5 leaves/tiller was
responsible for the greater weight of skin. The PR showed higher biological yield. The
cuts were not affected by management strategies. The levels of humidity and lipids were
affected by PR.
Index terms: forage structure, morphogenesis, nutritional value, carcass

1
CONSIDERAÇÕES INICIAIS
____________________________________________________________

2
CONSIDERAÇÕES INICIAIS
A pecuária é uma das principais atividades econômicas do Brasil e se baseia
quase exclusivamente no uso de pastagens nativas ou cultivadas como suprimento
alimentar dos animais domésticos, principalmente, os ruminantes. Dentre os métodos de
utilização das culturas forrageiras, o pastejo é o mais econômico, uma vez que a colheita
da forragem é feita pelo próprio animal, reduzindo os custos com mão-de-obra. Com
base nisso, conclui-se que a planta forrageira é um componente chave do sistema de
produção de ruminantes a pasto no Brasil.
Os sistemas de produção no Nordeste não diferem muito dos demais sistemas de
produção no restante do país, que baseiam-se principalmente em pastagens. As
pastagens constituem a base do sistema produtivo sustentável e econômico de ovinos,
desde que manejada racionalmente, considerando os aspectos da escolha das plantas
forrageiras, incremento da fertilidade do solo, ajuste da pressão de pastejo e controle
parasitário, alcançando boas produções por animal e por área, aumentando a
rentabilidade do empreendimento.
A produção de forragem é um processo contínuo, regulado por variáveis do
ambiente e por características da pastagem. À medida que tecidos foliares crescem nas
plantas, esses estão sujeitos ao envelhecimento e à senescência, responsáveis pela
redução no acúmulo de massa seca e pelas perdas quantitativas e qualitativas da
forragem produzida. Portanto o sucesso na produção de forragem em pastagens depende
não só da disponibilidade de água, mas de um conjunto de fatores, como a compreensão
dos mecanismos morfofisiológicos e de sua interação com o ambiente, ponto
fundamental para suportar tanto o crescimento quanto a manutenção da capacidade
produtiva do pasto.
Os estudos de fluxo de biomassa vêm se constituindo em importante ferramenta
para avaliação da dinâmica de folhas e perfilhos em comunidades de plantas forrageiras.
Nesses termos, os pastos podem ser considerados como sistemas dinâmicos, onde
alterações na morfogênese determinam modificações na estrutura do dossel, resultando
em alterações no índice de área foliar (IAF) e, conseqüentemente, na eficiência de
intercepção e de utilização da luz. Por sua vez, a eficiência de utilização desses fatores
abióticos determinará a quantidade e a qualidade da forragem produzida redundando,

3
em última análise, no desempenho individual do animal, na capacidade de suporte da
pastagem e na produtividade do ecossistema pastoril.
O conhecimento das relações ambiente-planta-animal torna-se de suma
importância, uma vez que, conhecidas as variáveis determinantes da otimização do uso
da pastagem, se possa criar ambientes através do manejo que não venham a limitar o
animal no emprego de suas estratégias de pastejo e também não prejudiquem a
sustentabilidade da pastagem. As estratégias de manejo para as plantas forrageiras
tropicais têm sido aplicados com bastante sucesso na região Sudeste do Brasil, contudo,
para o Semiárido brasileiro, apesar de relevantes, são escassas essas informações, que
podem se constituir em importantes indicativos ao manejo das pastagens.
Considerando a importância dos estudos sobre o manejo de pastagens e das
inter-relações solo-planta-animal, esta pesquisa foi conduzida com o objetivo de avaliar
as respostas da pastagem e do rebanho ovino ao manejo do capim-tifton 85 sob efeito
dos três períodos de descanso 4,5; 6,5 e 8,5 (número de folhas/perfilho) e dois resíduos
pós-pastejo 5 e 10 cm, nas condições do Nordeste do Brasil.

4
CAPÍTULO 1
____________________________________________________________
Referencial teórico

5
1. As pastagens no Brasil e o gênero Cynodon
As pastagens totalizam aproximadamente ¼ da superfície terrestre
(MARCELINO et al., 2006). No Brasil a área de pastagens é de 25 %, o que representa
mais de 172 milhões de hectares (ANUALPEC, 2008), realçando a grande contribuição
desse recurso como base alimentar da produção pecuária nacional. A produção de
ruminantes baseada em pastagens e a forma mais econômica de se obter produtos de
alto valor biológico a baixo custo, uma vez que o próprio animal se encarrega de
selecionar e colher o próprio alimento. As pastagens assumem dois aspectos
fundamentais, o primeiro é que viabilizam a competitividade brasileira, e a segunda, é o
fato de possibilitarem o atendimento da grande demanda mundial por alimento
produzido de forma natural, com respeito ao ambiente e aos animais (EUCLIDES,
2005).
Nas duas últimas décadas, foram introduzidas no Brasil, gramíneas tropicais
melhoradas e adaptadas, bastante produtivas quando utilizadas práticas de manejo
adequadas. Dentre as forrageiras utilizadas no estabelecimento de pastagens no Brasil,
destacam-se as do gênero Cynodon. As gramíneas deste gênero são capazes de produzir
grandes quantidades de matéria seca e elevada relação lâmina foliar/colmo, resultando
em um adequado valor nutritivo. Essas forrageiras são bastante flexíveis quanto à forma
de utilização nos sistemas produtivos, pois se adaptam à colheita mecânica, bem como
ao pastejo; podem ser utilizadas para a produção de feno ou silagem; em condições de
pastejo, podem ser manejadas em lotação contínua ou em lotação intermitente.
Os capins do gênero Cynodon dividem-se em dois grupos: as “gramas” ou
“bermudas” e as “estrelas”. São pertencentes ao grupo das “bermudas” os capins
Swannee, Costal Bermuda, Florarkik, Coastcross e Tifton e pertencem ao grupo das
“estrelas” os capins Ona, Florico, Florona e Estrelas. As plantas do primeiro grupo
apresentam rizomas e estolões, enquanto as do segundo grupo possuem apenas estolões
(GARCIA et al., 2004). De modo geral, são forrageiras, exigentes em fertilidade do solo
e com grande potencial de produção. Atualmente, as cultivares mais comumente
utilizadas no Brasil como recurso forrageiro são: Estrela roxa, Florico, Florona,
Florakirk, Tifton 68, Tifton 78, Coastcross e Tifton 85 (PEDREIRA & TONATO,
2006).

6
O Tifton 85, uma das espécies do gênero Cynodon, possui boas características de
produção e elevada capacidade de crescimento, apresentando grande potencial de uso
como forrageira nas condições subtropicais e tropicais. Seu dossel atinge até um 1 m de
altura e é resistente a cortes freqüentes. A matéria seca produzida, em boas condições de
manejo e adubação, é de boa qualidade, permitindo bom desempenho animal na
produção de leite e carne (MATOS et al., 2008). Essa planta destacou-se nos Estados
Unidos e posteriormente no Brasil por sua alta produção de matéria seca e teor de
proteína bruta elevado (14 t/ha/ano e 15,2%) respectivamente, em experimentos de
corte, Alvim et al. (1997). Fagundes (1999) ressaltou as altas taxas de acúmulo de
forragem e Carnevalli et al. (2001), o valor nutritivo, digestibilidade, desempenho
animal e a capacidade de suporte dessa forrageira.
2. A dinâmica de produção de forragem
A utilização de variáveis arbitrárias tais como taxa de lotação, pressão de pastejo
e duração do período de descanso não podem ser consideradas como determinantes
primários da produção de forragem ou do desempenho animal, uma vez que seus efeitos
são mediados por características estruturais do dossel, que determinam a condição e
estrutura do pasto (HODGSON, 1985). Essas variáveis tornam-se, então, parte de uma
estratégia de ação, cujo objetivo é a manutenção do pasto em condições de estrutura do
dossel tidas como ideais para determinado sistema de produção (HODGSON & Da
SILVA, 2002).
Avanços significativos na busca do entendimento da dinâmica do ecossistema de
pastagem só foram possíveis quando foi reconhecido que o uso de plantas forrageiras
em pastagens deveria ser considerado dentro de um contexto sistêmico, no qual a
adoção de técnicas e estratégias de manejo em pontos isolados do sistema dificilmente
resultaria em mudanças significativas na eficiência global final do processo produtivo.
Para tanto segundo Nabinger e Pontes (2001), tornaram-se relevantes os estudos de
dinâmica da produção primária das forrageiras, realizados por meio de avaliações das
características morfogênicas e estruturais das mesmas. A maximização da eficiência de
produção e colheita, por meio das avaliações morfogênicas e estruturais, permite gerar
informações, que levam ao desenvolvimento de estratégias de manejo baseadas nas

7
condições do pasto com metas de manejo definidas em termos de altura do dossel
(HODGSON, 1990) ou massa de forragem (MATTHEWS et al., 1999).
2.1. Características morfogênicas
A produtividade de uma gramínea decorre da contínua emissão de folhas e
perfilhos, processo importante após o corte ou pastejo para restaurar a área foliar da
planta e permitir a perenidade do pasto. O entendimento de características morfogênicas
permite ao técnico uma visualização da curva de produção, acúmulo de forragem e uma
estimativa da qualidade do pasto (GOMIDE, 1997a), e uma possibilidade de
recomendação de práticas de manejo diferenciadas (GOMIDE, 1998).
Segundo Lemaire e Chapman (1996), a morfogênese pode ser definida como a
dinâmica de geração e expansão da forma da planta no espaço. Embora seja
geneticamente programada essa dinâmica pode ser fortemente influenciada por
variáveis ambientais tais como temperatura, disponibilidade hídrica e de nutrientes. Esta
pode descrita por três características principais como: taxa de aparecimento de folhas
(TApF), taxa de alongamento de folhas (TAlF) e duração de vida da folha (DVF). Em
algumas espécies de plantas tropicais, particularmente aquelas de crescimento ereto,
existe um outro componente importante do crescimento que interfere significativamente
na estrutura do pasto e nos equilíbrios dos processos de competição por luz, a taxa de
alongamento de colmos (SBRISSIA & Da SILVA, 2001; CÂNDIDO (2003).
A Taxa de aparecimento de folhas (TApF) é a característica de maior destaque,
devido à sua influência direta sobre cada um dos componentes estruturais do pasto
(LEMAIRE & CHAPMAN, 1996). Essa variável, por ser central dentro do programa
morfogênico das gramíneas, é a última a ser penalizada pela planta quando de um
estresse (CHAPMAN & LEMAIRE, 1993). Esta característica é expressa como o
número médio de folhas surgidas em um perfilho por unidade de tempo (ANSLOW,
1966), sendo o seu inverso o filocrono. Oliveira et al. (2000) estudando a morfogênese
de capim-tifton 85 (Cynodon sp.) em diferentes idades de rebrotação, concluíram que
ocorre redução na taxa de aparecimento de folhas individuais entre as idades de 14 e 70
dias, correspondente a variações no filocrono de 1,6 e 4,1 dias/folha.

8
De acordo com Skinner e Nelson (1994) o aumento do filocrono (dias/folha)
com a idade na fase vegetativa ocorre em razão do aumento do tempo necessário para a
folha percorrer a distância entre o meristema apical e a extremidade do pseudocolmo
formado pelas bainhas das folhas mais velhas, enquanto na fase reprodutiva ocorre o
oposto deste fenômeno. Valores de filocronos para o capim-tifton 85, foram relatados
por Pinto et al. (2001) correspondendo a 3,0 a 5,0 dias/folha, sendo menores daqueles
relatados por Carnevalli e Da Silva (1999), que variaram de 3,0 a 5,8 dias/folha. Há uma
relação direta da TApF com a densidade populacional de perfilhos, o que determina o
potencial de perfilhamento de dado genótipo, pois cada folha formada traz na sua axila
uma gema, representando potencialmente o surgimento de um novo perfilho.
A TApF é ainda, passível de ser utilizada no processo de seleção de cultivares,
tornando-o mais preciso no que se refere à descrição da adaptação da planta ao meio
ambiente e ao potencial de perfilhamento, fatores que estão ligados de forma direta à
estabilidade das plantas na área, permitindo informações a respeito da persistência e/ ou
adaptação da planta forrageira ao corte ou ao pastejo.
Das características morfogênicas, a taxa de alongamento de folhas (TAlF)
apresenta a maior correlação com a produção, no caso das gramíneas do tipo C3 durante
toda sua fase vegetativa (HORST et al., 1978) e durante parte desta fase, no caso das
gramíneas do tipo C4 (CÂNDIDO, 2003). A TAlF se correlaciona positivamente com a
massa de forragem dos pastos (HORST et al., 1978) e com o crescimento por perfilho
(NELSON et al., 1977), no entanto, negativamente com o número de perfilhos por
planta (JONES et al., 1979). A TAlF é uma das características mais importantes no
desenvolvimento de plantas forrageiras; entretanto, maiores taxas não significam,
necessariamente, maior acúmulo de biomassa, pois, nesse caso, a planta pode estar
comprometendo o surgimento de novas folhas, com aumentos na taxa de senescência e,
ou, incrementando o tamanho das lâminas foliares, porém diminuindo o número dessas
(FARIA, 2009).
A outra característica morfogênica de grande importância é a taxa de senescência
foliar (TSF) ou o seu inverso, duração de vida da folha (TVF), em razão do menor valor
forrageiro dos tecidos senescentes e por representar perda de forragem produzida.
Segundo Lemaire (1997) a duração de vida das folhas representa o período durante o
qual há acúmulo de folhas no perfilho sem que haja detecção de qualquer perda por

9
senescência. O conhecimento da duração de vida das folhas é fundamental no manejo da
pastagem, pois, de um lado indica o teto potencial de rendimento da espécie (máxima
quantidade de material vivo por área) e, por outro lado, é um indicador fundamental
para a determinação da intensidade de pastejo com lotação contínua ou da freqüência do
pastejo em lotação rotacionada que permita manter índices de área foliar próximos da
maior eficiência de intercepção e máximas taxas de crescimento.
Em plantas tropicais e subtropicais cespitosas, o alongamento das hastes assume
grande importância relativa como característica morfogênica e determina as
características estruturais como relação lâmina foliar/colmo (SBRISSIA & DA SILVA,
2001). O sombreamento mútuo reduz a relação vermelho/vermelho extremo (V/Ve) da
luz, transmitida ao longo do dossel, sendo percebida pelo aparato de fitocromos (TAIZ
& ZEIGER, 2004), desencadeando processos de mudanças na estrutura do dossel por
meio de alongamento das colmos (bainhas e colmos). Esse alongamento do colmo é fato
de grande relevância na produção animal nos trópicos, por ser característica marcante
em forrageiras de clima tropical, tanto que Sbrissia e Da Silva (2001) e Cândido (2003)
propuseram a inclusão dessa característica ao esquema de Chapman e Lemaire (1993).
Cândido (2005a), avaliando o capim-mombaça sob estratégias de pastejo rotativo,
observaram um aumento crescente na proporção de colmos, ocasionando uma acentuada
redução na relação lâmina foliar/colmo na forragem acumulada, com o prolongamento
do período de descanso.
2.2. Características estruturais
A estrutura do dossel forrageiro é definida por um conjunto de características
genéticas da espécie, denominadas morfogênicas, que são condicionadas por fatores de
ambiente como luz, temperatura, umidade e outros (LEMAIRE & CHAPMAN, 1996).
A combinação das características morfogênicas determina as três principais
características estruturais do dossel forrageiro: comprimento final da folha (CFF);
Densidade populacional de perfilhos (DPP); Número de folhas vivas por perfilhos
(NFV) e relação lâmina foliar/colmo.
Os fatores determinantes do tamanho final da folha (TFF) são TApF e TAlF,
uma vez que, para dado genótipo, o período de alongamento da folha é uma fração

10
constante no intervalo do aparecimento de folhas sucessivas (DALE, 1982). O
comprimento da folha como foi relatado a pouco é determinado pela relação taxa de
alongamento/taxa de aparecimento de folhas. A taxa de aparecimento de folhas é
inversamente correlacionada com o tamanho final da folha (NABINGER & PONTES,
2001).
Dessa forma, variações na TAlF e TApF por meio de práticas de manejo
(intensidade de pastejo, freqüência de desfolhação, fertilização) ou flutuações climáticas
podem ocasionar variações no comprimento da folha. Enquanto a TAlF está diretamente
correlacionada com o comprimento da folha, folhas de menor tamanho são associadas a
valores elevados de TApF (NABINGER & PONTES, 2001). Outros fatores que
apresentam influência no tamanho da folha são o comprimento da bainha – que é
determinado pela altura de corte ou pastejo –, o nível de inserção da folha e o tipo de
perfilho. Logo, o comprimento da lâmina foliar é uma característica vegetal plástica
responsiva à intensidade de desfolhação, considerada por Lemaire e Chapman (1996)
como um mecanismo de escape capaz de conferir à planta graus variáveis de tolerância
ao pastejo. Assim, sua mensuração e análise podem auxiliar a identificação das
diferentes respostas morfofisiológicas desencadeadas pelas plantas em resposta à forma
como os fatores abióticos interagem entre si.
A densidade populacional de perfilhos em comunidades de plantas forrageiras é
função do equilíbrio entre as taxas de aparecimento e morte de perfilhos (LEMAIRE &
CHAPMAN, 1996). Em outras palavras, o número de perfilhos vivos por planta, ou por
unidade de área, é determinado pela relação entre a periodicidade de aparecimento de
novos perfilhos e a longevidade dos mesmos. O perfilhamento é influenciado ainda por
vários fatores relacionados ao ambiente e ao manejo adotado. Assim é que nutrição
mineral, manejo de cortes ou pastejo e fatores de ambiente, como luz, temperatura,
fotoperíodo e disponibilidade hídrica terão grande efeito sobre o perfilhamento da
planta (OLIVEIRA, 1999). A melhor estrutura no dossel para se otimizar a
produtividade e a qualidade da forragem é aquela que mantém o equilíbrio entre
perfilhos reprodutivos e vegetativos. Os perfilhos reprodutivos garantem taxa
fotossintética elevada e perfilhos mais pesados enquanto os perfilhos vegetativos
propiciam rápida recuperação após o corte e elevada qualidade do material pastejado,
que seria composto principalmente de folhas (CORSI, 1994).

11
Os perfilhos originados das gemas axilares (perfilhos aéreos) não alongam as
colmos tão rapidamente como os perfilhos que se originam das gemas situadas na coroa
das touceiras (perfilhos basais). Difante et al. (2008), avaliando a dinâmica do
perfilhamento do capim-marandu submetido a duas alturas (15 e 30 cm) e três intervalos
de corte (aparecimento de três, quatro e cinco folhas por perfilho), verificaram que a
produção de perfilhos aéreos representou proporção elevada da população total de
perfilhos (34 a 56%). Entretanto, esses perfilhos são pequenos e de vida curta, surgindo
em época do ano não favorável ao crescimento e, apesar de sua alta participação na
população, pouco contribuem para a produção de forragem.
O número de folhas vivas por perfilho (NFV) é geneticamente determinado,
sofrendo pouca influência dos fatores externos. Nabinger e Pontes (2001) afirmaram
que o número de folhas vivas por perfilho é uma constante genotípica bastante estável
na ausência de deficiências hídricas ou nutricionais. Binnie e Chestnutt (1994) não
observaram variação no número de folhas vivas em relvados de azevém e azevém/trevo
ou mesmo entre as diferentes alturas avaliadas. Lemaire e Chapman (1996) citaram que
longevidade das folhas, determina a produção teto de um pasto, mais que o número de
folhas vivas por perfilho. Pinto (2000) mencionou um número de folhas vivas/perfilho
para o Tifton 85 de 5,4, valor inferior ao encontrado por Oliveira (1999) que foi de 9,5.
A relação lâmina foliar/colmo está diretamente relacionada com a qualidade da
forragem e apresenta grande relevância, uma vez que é responsável por expressar a
quantidade de matéria seca das lâminas foliares em relação à quantidade de matéria seca
das colmos. Da sua diminuição, depreende-se que, do total da matéria seca de forragem
produzida, houve crescimento percentual da participação da massa seca de colmo verde
na massa de forragem total. Com isso, há diminuição de qualidade na massa de
forragem ofertada ao animal.
Para as gramíneas C4, essa relação toma maior importância, visto que o processo
de alongamento das colmos verifica-se ainda na fase vegetativa. Após o dossel atingir o
IAF crítico, há desencadeamento do processo de alongamento de colmo em pastagens
formadas por gramíneas C4, como discutido anteriormente. Desse processo, resulta o
estreitamento da relação lâmina foliar/haste. Chapman e Lemaire (1993) relataram que a
manutenção do pasto com altura e IAF elevados ocasionam diminuição da relação
lâmina foliar/haste, fato esse que influencia na capacidade fotossintética do dossel

12
quando na rebrotação, podendo elevar, mas comprometendo a taxa de ingestão por parte
dos animais em pastejo. Cândido (2003) enfatizou que a relação lâmina foliar/colmo é a
principal característica ligada à estrutura do pasto no referente ao desempenho animal
para gramíneas do tipo C4. Neste mesmo estudo, verificou-se redução na relação lâmina
foliar/colmo em pastos de capim-mombaça, com o prolongamento do período de
descanso.
3. Estratégias de manejo: período de descanso e resíduo pós-pastejo
O homem exerce maior controle do ambiente pastoril, a partir do controle da
densidade e, ou, tempo de permanência na área, do tamanho, das espécies e das
categorias de animais utilizadas. Fundamentalmente, isto significa manejar o grau de
desfolhação das plantas forrageiras. O conhecimento da reação da comunidade de
plantas à desfolhação é básico para se conceber uma estratégia de manejo que propicie a
maximização da produtividade de forragem com elevado valor nutritivo e que permita o
ajuste de seu crescimento às necessidades dos animais sem comprometer a perenidade
da pastagem (RODRIGUES & RODRIGUES, 1987). Informações nesse sentido só
podem ser geradas se características estruturais do dossel forrageiro forem controladas
com o objetivo de se manter um balanço entre altos valores de eficiência de utilização e
de desempenho animal.
O rendimento forrageiro após corte ou pastejo é proporcional à área foliar
existente, intensificando-se à medida que aumenta a intercepção da radiação luminosa
em decorrência do crescimento do IAF do dossel. Imediatamente após a saída dos
animais do piquete, o IAF do dossel é baixo, apresentando predominância de folhas
velhas; contudo, após os primeiros dias do período de descanso verifica-se nele maior
proporção de folhas emergentes e, subseqüentemente, de folhas recém expandidas.
Consequentemente, a taxa de fotossíntese bruta do dossel cresce curvilinearmente
durante o período de descanso, refletindo o crescimento do IAF (KING et al, 1984;
PARSONS et al, 1988) e o avanço da idade de suas folhas. O rendimento forrageiro se
aproxima do máximo quando o dossel intercepta 95% da radiação incidente, momento
em que seu IAF se aproxima de seu valor máximo. Entretanto, este critério não é de
aplicação prática no dia a dia do manejo da pastagem.

13
A correta definição do período de descanso contribui para o sucesso desse
sistema de manejo. Períodos de descanso de adequada duração propiciam à restauração
da área foliar, a intercepção da luz, a produção de forragem, a restauração das reservas
orgânicas, mas períodos de descanso muito longos comprometem o valor nutritivo da
forragem, a estrutura do dossel forrageiro, assim como o consumo de forragem e o
rendimento animal na pastagem. A literatura relata diversos experimentos sobre
critérios pertinentes à definição do período de descanso, como restauração das reservas
orgânicas (FULKERSON & DONAGHY, 2001), área foliar (BROUGHAM, 1956),
intercepção da radiação luminosa (KORTE et al.,1982; PARSONS et al., 1983), taxa
média de acúmulo de massa de forragem (ROBSON,1973; PARSONS & PENNING,
1988) e, número constante de folhas vivas por perfilho (GRANT et al.,1988;
FULKERSON & DONAGHY, 2001). Este último critério, fundamentado na
morfogênese da forrageira, visa prevenir as perdas de biomassa por senescência e morte
de folhas e perfilhos, de modo a otimizar a eficiência de utilização da forragem
produzida.
O número de folhas vivas por perfilho, constante a partir do momento em que a
taxa de senescência foliar iguala a taxa de aparecimento, constitui critério objetivo e
prático para definição da duração do período de descanso, ou seja, o momento da
introdução dos animais no piquete (FULKERSON & SLACK, 1995; GRANT et al,
1988). Para uma pastagem com um filocrono de 10 dias, os animais voltariam ao
piquete, quando o número de folhas/perfilho fosse de 3,5, o corresponderia a 35 dias. A
partir deste ponto o prolongamento deste período de descanso, resultaria em perdas por
senescência, redução da relação lâmina foliar/colmo e do valor nutritivo.
Um ferramenta de manejo que pode ser adotada é a altura, que devido a
facilidade da medição de altura do dossel faz com que esta seja potencial no manejo do
pastejo e de fácil aplicação no campo. Embora a altura do dossel não possa ser tomada
como variável determinante dos processos de crescimento e utilização da forragem em
pastagens tropicais, a facilidade de sua medição, conforme inúmeros trabalhos já
relatados (CAVALCANTE, 2001; GOMIDE et al., 2003; CÂNDIDO, 2003), sugere
que uma vez determinada a estratégia de manejo, com base nos parâmetros
ecofisiológicos, como a interceptação da luz, índices morfogênicos e características
estruturais, possa ser adotada uma altura prática de manejo na entrada e outra na saída

14
dos animais do piquete. A altura do resíduo pós-pastejo é um dos fatores condicionantes
das taxas de rebrotação de pastagens de gramíneas forrageiras tropicais (GOMIDE et
al., 1979). De acordo com Brâncio et al. (2003) o resíduo após pastejo pode estar
associado à camada de folhas ou, ainda, à barreira formada por material morto e às
bainhas das folhas na parte basal da planta
Carvalho et al. (2005) verificaram que as duas alturas de resíduo pós-pastejo de
100 ou 50 cm, não influenciaram as variáveis morfogênicas e produtivas do capim-
elefante. A altura de resíduo influencia com maior intensidade a produção de folhas, o
comprimento internodal e o número de nós por perfilho do capim-elefante que o
intervalo de desfolhação (BUTT et al.,1993). Contudo, estudos mais recentes
desenvolvidos em pastagens de gramíneas forrageiras tropicais de crescimento
cespitoso, avaliados sob pastejo, têm revelado pouca influência da altura do resíduo
sobre as variáveis morfogenéticas e estruturais da pastagem (BARBOSA et al., 2002;
GARCEZ NETO et al., 2002; CASAGRANDE et al., 2003; LAMBERTUCCI et al.,
2003).
A altura do resíduo pós-pastejo exerce um importante efeito sobre a qualidade da
pastagem. Forbes e Coleman (1993) afirmaram que o resíduo determina mudanças na
estrutura da pastagem, isto é, na participação dos componentes: lâmina foliar, colmo e
material morto e, como conseqüência altera a qualidade da forragem disponível para o
pastejo animal. Bianchini (1998) comparou o efeito de quatro alturas de manejo em
pastagem de coast cross no desempenho com ovinos. Alturas da forragem remanescente
após o pastejo foram: T1= 4 a 7 cm, T2= 10 a 13 cm, T3= 16 a 19 cm e T4= 22 a 25 cm.
A produção animal por área foi de 460, 436, 254 e 264 kg PV/ha, para T1, T2, T3 e T4,
respectivamente. As duas maiores alturas de manejo mostrou redução no teor de
proteína bruta e digestibilidade in vitro da matéria seca, bem como, menor ganho de
peso vivo por área, comparando com as menores alturas. No entanto, o autor não
correlacionou a densidade de forragem com a disponibilidade de forragem (relação
lâmina foliar/haste).

15
4. Ambiente de pastoril: planta–animal
A produção animal obtida em pastagens é o resultado do processo fotossintético
das plantas, que utilizam a energia solar para formação de biomassa que deverá ser
consumida pelo pastejo e convertida em produto animal. A desfolhação, por meio do
pastejo, necessita ser controlada, pois se de um lado as plantas crescem utilizando
energia solar, água e nutrientes fornecidos pelo solo, de outro lado o animal influencia o
seu crescimento pelo pastejo seletivo, além do pisoteio e das dejeções.
O animal exerce influência sobre a produção de forragem, principalmente devido
à remoção de material fotossinteticamente ativo e por alterar a interceptação luminosa
do dossel forrageiro, uma vez que pelo pastejo ocorre a desfolhação que condiciona as
respostas fisiológicas e morfológicas por parte da planta, as quais resultam em alteração
de sua forma e função, ou seja, a estrutura do dossel forrageiro (SBRISSIA & Da
SILVA, 2001). E ao mesmo tempo a estrutura do dossel forrageiro é fundamental tanto
para desempenho animal como de plantas (MAXWELL & TREACHER, 1987). A
definição de estratégias de manejo do pastejo devem ser baseadas no controle e
manipulação de características específicas do pasto num estado de equilíbrio dinâmico
(HODGSON, 1985).
A estrutura do dossel é determinante do valor nutritivo da forragem ofertada e
consumida pelos animais. A qualidade de uma planta forrageira é definida como sendo
sua capacidade em gerar desempenho animal, compreendendo, portanto sua composição
química, digestibilidade, consumo voluntário e interação de fatores hereditários e do
ambiente (MOTT, 1970; MOORE, 1994). Em ecossistemas de pastoris o valor nutritivo
da forragem está consistentemente associado à maturidade da planta forrageira. Quando
períodos de descanso muito prolongados são utilizados, ocorrem tanto modificações nas
proporções dos componentes da planta (incremento na proporção de colmos e material
morto e redução na proporção de lâminas foliares) quanto modificações (redução) na
digestibilidade desses componentes (TERRY & TILLEY, 1964).
A relação lâmina foliar/colmo também tem influência no consumo devido à
preferência dos animais pelas lâminas foliares (FORBES & HODGSON, 1985), que
apresentam maior facilidade de apreensão e maior valor nutritivo. Ainda que o animal
possa compensar o menor valor nutritivo do pasto pela sua seletividade (CHACON &

16
STOBBS, 1976), ou aumento no tempo de pastejo (CHACON & STOBBS, 1976), o
consumo será afetado pela redução na taxa de bocado (FORBES & HODGSON, 1985).
REFERÊNCIAS
ALVIM, M.J.; RESENDE, H.; BOTREL, M.A. Efeito da freqüência de cortes e de níveis de nitrogênio sobre a produção e qualidade da matéria seca do “coastcross”. In: WORKSHOP SOBRE O POTENCIAL FORRAGEIRO DO GÊNERO CYNODON, 1996, Juiz de Fora. Anais... Juiz de Fora: EMBRAPA CNPGL, 1996. p.45-55. ANSLOW, R.C. The rate of appearance of leaves on tillers of the gramineae. Herb. Abstr., v.36, n.3, p.149-155, 1966. ANUALPEC: Anuário da pecuária brasileira. FNP CONSULTORIA & COMÉRCIO. São Paulo, 2008. 380p. BARBOSA, R.A., NASCIMENTO JR, D. EUCLIDES, V.P.B. et al. Características morfogênicas e acúmulo de forragem do Capim- Tanzânia (Panicum maximum Jacq. cv. Tanzânia) em dois resíduos forrageiros pós-pastejo. Revista Brasileira de Zootecnia, v.31, n.2, p.583-593, 2002. BIANCHINI, D. Efeito de quatro alturas de manejo sobre as características quantitativas e qualitativas em pastagem de coast cross no desempenho de cordeiros em fase de terminação. Jaboticabal, 1998. 135p. Tese (Doutorado) – Universidade Estadual Paulista, Faculdade de Ciências Agrárias e Veterinárias, 1998. BINNIE, R.C., CHESTNUTT, D.M.B. Effects of continuous stocking by sheep at four sward heights on herbage mass, herbage quality and tissue turnover on grass/clover and nitrogen-fertilized grass swards. Grass and Forage Science, v.49, n.2, p.192-202, 1994. BRÂNCIO, P. A. et al. Avaliação de três cultivares de Panicum maximum Jacq. sob pastejo: Composição da dieta, consumo de matéria seca e ganho de peso animal. Revista Brasileira de Zootecnia, v. 32, n. 5, p.1037-1044, 2003.
BROUGHAM, R. W. Effect of intensity of defoliation on regrowth of pasture. Australian Journal of Agricultural Research, v.7, p.377-387, 1956. BUTT, N.M.; DONART, G.B.; SOUTHWARD, M.G.; PIEPER, R.D.; MOHAMMAD, N. Effect of defoliation on plant growth of Napier grass. Tropical Science, v.33, p.111-120, 1993.

17
CÂNDIDO, M.J.D.; GOMIDE, C.A.M.; ALEXANDRINO, E. et al. Morfofisiologia do dossel de Panicum maximum cv. Mombaça sob lotação intermitente com três períodos de descanso. Revista Brasileira de Zootecnia, v.34, n.2, p.406-415, 2005a. CÂNDIDO, M.J.D. Morfofisiologia e crescimento do dossel e desempenho animal em Panicum maximum cv. Mombaça sob lotação intermitente com três períodos de descanso. Viçosa: UFV, 2003. 134p. Tese (Doutorado em Zootecnia) – Universidade Federal de Viçosa, 2003. CARNEVALLI, R. A.; SILVA, S.C.; CARVALHO, C.A.B. et al. Desempenho de ovinos e respostas de pastagens de Coastcross submetidas a regimes de desfolha sob lotação contínua. Pesquisa Agropecuária Brasileira., v.36, n.6, p.919-927, 2001. CARNEVALLI, R.A.; DA SILVA, S.C. Validação de técnicas experimentais para avaliação de características agronômicas e ecológicas de pastagens de Cynodon dactylon cv. Coastcross-1. Scientia Agrícola, v.56, p.489-499, 1999. CARVALHO, A. B. ed; PACIULLO, D. S. C.; ROSSIELLO, R. O. P. et al Morfogênese do capim-elefante manejado sob duas alturas de resíduo pós-pastejo. B. Indústr.anim., N. Odessa,v.62, n.2, p.101-109, 2005. CASAGRANDE, D.R.; MISTURA, D.; FONSECA, D.M. et al. Efeito de alturas de resíduo pós-pastejo na taxa de acúmulo de forragem em pastagem de capim-elefante. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 40, 2003, Santa Maria. Anais... Santa Maria: SBZ. 2003, CD ROM. CAVALCANTE, M.A.B. Características morfogênicas e acúmulo de forragem em relvado de Brachiaria decumbens cv. Basilisk sob pastejo, a diferentes alturas. Viçosa, MG: Universidade Federal de Viçosa, 2001. 64p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Viçosa, 2001. CHACON, E. e STOBBS, T.H. Influence of progressive defoliation of a grass sward on the eating behaviour of cattle. Australian Journal of Agricultural Research, v.27, n.5, p.709-727, 1976. CHAPMAN, D.F., LEMAIRE, G. Morphogenetic and structural determinants of plant regrowth after defoliation. In: BAKER, M.J. ed. Grasslands for our world. Wellington: SIR Publishing, p.55-64, 1993. CORSI, M. Uréia como fertilizante na produção de forragem. In: SIMPÓSIO SOBRE NUTRIÇÃO DE BOVINOS: URÉIA PARA RUMINANTES, 2, 1984, Piracicaba. Anais... Piracicaba: FEALQ, 1994. p. 239-268. DALE, J. E. The growth of leaves. London: Edward Arnold, 1982. 60 p. (Studies in biology, 137).

18
DIFANTE, G. dos S.; NASCIMENTO JÚNIOR, D.; da SILVA, S. C. et al. Dinâmica do perfilhamento do capim-marandu cultivado em duas alturas e três intervalos de corte. Revista Brasileira de Zootecnia, v. 37, n.2, p.189-196, 2008. EUCLIDES, V. P. B. Manejo de Pastagem e Valor Nutritivo das Forrageiras. In: Simpósio sobre Manejo Sustentável em Pastagens. CECATO, U; JOBIM, C. C. et al. Anais... Maringá – PR. 2005. FAGUNDES, J. L.; SILVA, S. C.; PEDREIRA, C. G. S. Light interception and herbage accumulation in Tifton-85 swards grazed by sheep under continuous stocking. In: SIMPÓSIO INTERNACIONAL GRASSLAND ECOPHYSIOLOGY AND GRAZING ECOLOGY, 1999, Curitiba. Anais… Curitiba: UFPR, 1999. p. 339-343. FARIA, D. J. G. Características morfogênicas e estruturais dos pastos e desempenho de novilhos em capim-braquiária sob diferentes alturas. Tese (Doutorado) – Universidade Federal de Viçosa.Viçosa/MG, 2009. 148 f. 2009. FORBES, T.D.A., COLEMAN, S.W. Forage intake and ingestive behaviour of catle grazing old world bluestems. Agronomy of Journal, Madison, v.85,n.4,p.808-16, 1993. FORBES, T. D. A., HODGSON, J. Comparative studies of fhe influence of sward conditions on the ingestive behaviour of cows and sheep. Grass and Forage Science, v.40, p.69-77, 1985. FULKERSON, W. J.; DONAGHY, D.J. Plant soluble carbohydrate reserves and senescence – key criteria for developing an effective grazing management system for ryegrass based pasture: a review. Australian Journal Experimental Agriculture, v. 41, p.261-275, 2001. FULKERSON, W.J., SLACK, K. Leaves number as a criterion for determining defoliation time for Lolium perenne. 2. Effect of defoliation frequency and height. Grass Forage Sci., v.50, n.1, p.16-20, 1995. GARCIA, R. ROCHA, F.C.; BERNARDINO, F.S. et al. Forrageiras utilizadas no sistema integrado agricultura-pecuária. In: ENCONTRO SOBRE MANEJO INTEGRADO: INTEGRAÇÃO AGRICULTURA-PECUÁRIA, 6., 2004. Anais... Viçosa: UFV, 2004, p.331-351. GOMIDE, J. A.; CÂNDIDO, M. J. D.; ALEXANDRINO, E. As interfaces solo, planta e animal da exploração da pastagem. In: FORRAGICULTURA E PASTAGENS: TEMAS EM EVIDÊNCIA – SUSTENTABILIDADE, 4., 2003, Lavras. Anais... Lavras: Editora da UFLA, 2003. p. 75-116. GOMIDE, J.A Capim-elefante Mott, uma variedade anã de Pennisetum purpureum. Informe Agropecuário, 19(192): 62-65, 1998. GOMIDE, J.A. SIMPÓSIO INTERNACIONAL SOBRE PRODUÇÃO ANIMAL EM PASTEJO. Viçosa: UFV, 1997, 471p.

19
GOMIDE, J.A. et al. Introdução de leguminosas tropicais em pastagens de gramíneas. Ver. Sociedade Brasileira de Zootecnia., 8 (4), 1979. GRANT, S.A.; BARTHRAM, G.T.; TORVELL. L. et al. Comparison of herbage production under continuous stocking and intermittent grazing. Grass and Forage Science, v. 43, n. 1, p. 29-39, 1988. HODGSON, J.; DA SILVA, S. C. Options in tropical pasture management. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39.; SIMPÓSIO INTERNACIONAL DE FORRAGICULTURA, 2002, Recife, PE. Anais... Recife: SBZ, 2002. p. 180-202. HODGSON, J. Grazing management: Science into practice. Longman Scientific and technical, Longman group, 1990. HODGSON, J. The control of herbage intake in the grazing ruminant. Proceedings of the Nutrition Society, v. 44, p. 339-346. 1985. HORST, G.L., NELSON, C.J., ASAY, K.H. Relationship of leaves elongation to forage yield of tall fescue genotypes. Crop Science., v.18, p.715-719, 1978. KORTE, C.J.; WATKIN, B.R.; HARRIS, W. Use of residual leaves area index and light interception as criteria for spring-grazing management of ryegrass dominant pasture. New Zealand Journal of Agricultural Research, v.25, p.309-319, 1982. JONES, R.J.; NELSON, C.J.; SLEPER, D.A. Seedling selection for morphological characters associated with yield of tall fescue. Crop Science, v.19, p.367-372, 1979. KING, J.; SIM, E.M. e GRANT, S.A. Photosynthetic rate and carbon balance of grazed ryegrass pastures. Grass Forage Sci., v.39, n.1, p.81-92, 1984. LAMBERTUCCI, D.; MISTURA, C.; FONSECA, D.M. et al. Estrutura do pasto na disponibilidade de matéria seca em pastagem de capim-elefante em diferentes alturas de resíduo pós-pastejo. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 40, 2003, Santa Maria. Anais... Santa Maria: SBZ. 2003, CD ROM. LEMAIRE, G. The physiology of grass growth under grazing: tissue turnover. In: SIMPÓSIO INTERNACIONAL DE PRODUÇÃO ANIMAL EM PASTEJO, 1997, Viçosa. Anais... Viçosa: UFV, 1997. p.117-144. LEMAIRE, G., CHAPMAN, D.F. Tissue flows in grazed plant communities. In: HODGSON, J., ILLIUS, A.W. (Ed.). Ecology and management of grazing systems. Wallingford: CAB International, 1996. p.3-36. MARCELINO, K.R.A., NASCIMENTO JR, D., Da SILVA, S.C. et al. Características morfogênicas e estruturais e produção de forragem do capim marandu submetido a intensidades e freqüências de desfolhação. Revista Brasileira de Zootecnia., 35, 2243-2252,2006 (in Portuguese, English abstract).

20
MAXWELL, T.J.; TREACHER, T.T. Decision rules for grassland management. In: efficient sheep production form grass. POLLOTT, G.E. (Ed.). BRITISH GRASSLAND SOCIETY OCCASONAL SYMPOSIUM, Hurley, Berkshire, U.K, 1987. Proceedings. UK: British Grassland Society, 1987. p.67-78. MATTHEW, C; ASSUERO, S.G.; BLACK, C. K. et al. Tiller dynamics of grazed swards, In: Simpósio Internacional “Grassland Ecophysiology and grazing ecology”. Curitiba, 1999. Anais… Curitiba: UFRP/ UFRGS, p. 109-133, 1999. MATOS, A. T.; ABRAHÃO, S. S.; PEREIRA, O. G. Desempenho agronômico de capim-tifton 85 (Cynodon sp) cultivado em sistemas alagados construídos utilizados no tratamento de água residuária de laticínios. Ambi-Água, Taubaté, v. 3, n. 1, p. 43-53, 2008. MOORE, J.E. Forage quality, evaluation and utilization. In: NATIONAL CONFERENCE ON FORAGE QUALITY, EVALUATION AND UTILIZATION. Lincoln, 1994. Proceedings. Lincoln: University of Nebraska, 1994, p.967-998: cap.24: Forage Quality Índices: Development and application. MOTT, G.O. Evaluacion de la produccion de forajes. In: HUGHES, H.D.; HEATH, M.E.; METCALFE, D.S. (Ed.). Forrajes – la ciencia de la agricultura baseada en la procuccion de pastos. México: CECSA, 1970. p.131-141. NABINGER, C., PONTES, L.S. Morfogênese de plantas forrageiras e estrutura do pasto. In: SIMPÓSIO SOBRE A PRODUÇÃO ANIMAL NA VISÃO DOS BRASILEIROS /REUNIÃO DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38, 2001, Piracicaba. Anais... MATTOS, W.R.S. et al. (EE.). Piracicaba: FEALQ, 2001. p. 755-771. NELSON, C. J.; ASAY, K. H.; SLEPER, D. A. Mechanisms of canopy development of tall fescue genotypes. Crop Science, v. 17, p. 449-452, 1977. NETO, A.F.G.; NASCIMENTO JUNIOR, D.; REGAZZI, A.J. et al. Respostas morfogênicas e estruturais de Panicum maximum cv. Mombaça sob diferentes níveis de adubação nitrogenada e alturas de corte. Revista Brasileira de Zootecnia, v.31, n.5, p.1890-1900, 2002. OLIVEIRA, M.A., PEREIRA, O.G., HUAMAN, C.A.M. et al. Características Morfogênicas e Estruturais do Capim-Bermuda „Tifton 85‟ (Cynodon spp.) em Diferentes Idades de Rebrota. Revista Brasileira de Zootecnia, v.29, n.6, supl.1, p.1939-1948, 2000. OLIVEIRA, M.A. Morfogênese, análise de crescimento e valor nutritivo do capim-tifton 85 (Cynodom sp) em diferentes idades de rebrota. Viçosa: UFV, 1999. 94p. Dissertação (Mestrado em Zootecnia) – Universidade Federal de Viçosa, 1999.

21
PACIULLO, D. S. C.; DERES, F.; AROEIRA, L. J. M. et al. Morfogênese e acúmulo de biomassa foliar em pastagem de capim-elefante avaliada em diferentes épocas do ano. Pesquisa Agropecuária Brasileira, Brasília, v. 38, n. 7, p. 881-887, 2003. PARSONS, A.J.; PENNING, P.D. The effect of duration of regrowth on photosynthesis, leaves death and the average rate of growth in a rotationally grazed sward. Grass and Forage Science, v.43, p.15-27, 1988. PARSONS, A.J.; LEAVESE, E.L.; COLLETT, B. et al. The physiology of grass production under grazing. II – Photosynthesis, crop growth and animal intake of continuously grazed sward. Journal of Applied Ecology, v.20, n.1, p.127- 139, 1983. PEDREIRA, C.G.S.; TONATO, F. Bases ecofisiológicas para o manejo de gramíneas do gênero Cynodon. In: SIMPÓSIO SOBRE MANEJO ESTRATÉGICO DA PASTAGEM, 3., 2006, Viçosa. Anais... Viçosa: UFV, 2006. p.93-115. PINTO, L. F. M.; SILVA, S. C.; SBRISSIA, A. F. et al. Dinâmica do acúmulo de matéria seca em pastagens de Tifton 85 sob pastejo. Scientia Agricola, v. 58, n. 3, p. 439-447, 2001. PINTO, L.F.M. Dinâmica do acúmulo de matéria seca em pastagens de Cynodon
spp. Dissertação (Mestrado) Piracicaba, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 2000, 124p. RODRIGUES, L.R.A., RODRIGUES, T.J.D. Ecofisiologia de plantas forrageiras. In: Ecofisiologia da Produção Agrícola Piracicaba: Potafos, 1987. p. 47-68. ROBSON, M.J. The growth and development of simulated swards of perennial ryegrass. I. Leaves growth and dry weight change as related to the ceiling yield of a seedling sward. Annals of Botany, v.37, p.487-500, 1973. SBRISSIA, A.F., SILVA, S.C. O ecossistema de pastagens e a produção animal. In: SIMPÓSIO SOBRE A PRODUÇÃO ANIMAL NA VISÃO DOS BRASILEIROS /REUNIÃO DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38, 2001, Piracicaba. Anais... MATTOS, W.R.S. et al. (EE.). Piracicaba: FEALQ, 2001. p. 731-754. SKINNER, R.H.; NELSON, C.J. Role of leaves appearance rate and the coleoptile tiller in regulating tiller production. Crop Science, Madisnon, v.34, n.1, p.71-75, 1994. TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 3ª Edição – Porto Alegre: Artmed, p. 719; 2004. TILLEY, J.M.A.; TERRY.R.A. The digestibility of the leaves and stems of perennial ryegrass, coksfoot, timoty, tall fescue, Lucerne and sain foin, as measured by in vitro procedure. Journal Britain Grassland Society 19:p. 363-372, 1964.

22
CAPÍTULO 2
____________________________________________________________
Crescimento e componentes da biomassa inicial do dossel do capim-tifton 85

23
Crescimento e componentes da biomassa inicial do dossel do capim-tifton 85
RESUMO: A avaliação de crescimento foi conduzida numa área de 0,82 ha, após o
corte de uniformização. Avaliaram-se os índices de crescimento, componentes da
biomassa inicial e as características estruturais do capim-tifton 85 (Cynodon sp) em sete
idades de rebrotação (0; 5; 10; 15; 20; 25 e 30 dias). Os dados foram ajustados às
equações que melhor estimassem as variáveis avaliadas, e apresentados de modo
descritivo, através de curvas em função da idade de rebrotação da gramínea. Os gráficos
foram plotados através do programa computacional Excel. O índice de área foliar (IAF)
e a interceptação da radiação fotossinteticamente ativa aumentaram com o avanço da
idade de rebrotação. A massa seca de forragem total (MSFT) elevou-se ao longo da
idade de rebrotação, fato também observado para a massa seca de forragem verde
(MSFV). O aumento da massa seca de forragem morta (MSFM), ao longo da avaliação
de rebrotação não foi suficiente para comprometer o aumento da (MSFV). O aumento
da massa seca de colmo verde (MSCV) pode estar relacionado com o aumento do IAF
ao longo da avaliação e ter sido o responsável na redução da massa seca de lâmina verde
(MSLV). Foram observadas reduções nos valores de taxa de crescimento relativo
(TCR), assimilação liquida (TAL) e razão de área foliar (RAF) com o avanço da idade
de rebrotação. A taxa de acúmulo de forragem (TAF) aumento ao longo da avaliação.
Foram verificadas reduções na densidade populacional de perfilhos e no número de
folhas vivas ao longo do período de avaliação.
Termos para indexação: índice de área foliar, interceptação da radiação
fotossinteticamente ativa, índices de crescimento, rebrotação

24
Growth and components of the initial biomass of the canopy of tifton 85 grass
ABSTRACT: The growth evaluation was lead in an area of 0,82 ha, after the
uniformização cut. The indices of growth, the structural components of the initial
biomass and characteristics of tifton 85 grass (Cynodon sp) in seven ages of regrowth
had been evaluated (0; 5; 10; 15; 20; 25 and 30 days). The data had been adjusted to the
equations that better esteem the 0 variable evaluated, and presented in descriptive way,
through curves in function of the age of regrowth of the grassy one. The graphs had
been located through the computational program Excel. The leaf area index (IAF) and
the interception of the fotossinteticamente active radiation had increased with the
advance of the age of regrowth. The dry mass of total fodder plant (MSFT) was raised
throughout the age of regrowth, fact also observed for the dry mass of green fodder
plant (MSFV). The increase of the dry mass of fodder plant deceased (MSFM),
throughout the regrowth evaluation was not enough to compromise the increase of
(MSFV). The increase of green the dry mass of stem (MSCV) can be related with the
increase of the IAF throughout the evaluation and to have been the responsible one in
the reduction of the dry mass of green blade (MSLV). Reductions in the values of rate
of relative growth had been observed (TCR), net assimilation rate (TAL) and leaf area
ratio (RAF) with the advance of the age of regrowth. The rate of accumulation forage
(TAF) increase throughout the evaluation. Reductions in the population density of tiller
and the number of alive leaves alive had been verified throughout the period of
evaluation.
Index-terms: leaf area index, interception of the fotossinteticamente active radiation,
growth indexes, regrowth

25
INTRODUÇÃO
O crescimento das plantas segue uma curva sigmóide, com uma fase inicial
lenta, seguida de uma de rápido crescimento ou fase logarítmica, e, posteriormente,
outra em que o crescimento é lento ou ausente. Finalmente, a curva toma a direção
decrescente, coincidindo com a senescência das plantas. O fundamento da análise de
crescimento é a medida seqüencial da acumulação de matéria orgânica e sua
determinação é realizada por meio da avaliação do acúmulo de massa seca da planta e
do crescimento da área de folhas (SANTOS JR. et al., 2004). Permite obter informações
dos efeitos do ambiente sobre as diferenças entre espécies (ANDRADE et al., 2004),
avaliando o crescimento final da planta como um todo e a contribuição dos diferentes
órgãos no crescimento total. A partir dos dados de crescimento pode-se inferir atividade
fisiológica, isto é, estimar-se, de forma bastante precisa, as causas de variações de
crescimento entre plantas geneticamente diferentes ou entre plantas crescendo em
ambientes diferentes.
A avaliação do crescimento constitui uma parte da fisiologia vegetal em que se
faz uso de fórmulas e modelos matemáticos para avaliar índices de crescimento das
plantas, sendo muitos deles relacionados com a atividade fotossintética (BENINCASA,
2004). Como o crescimento é avaliado por meio de variações de tamanho de algum
aspecto da planta, geralmente morfológico, em função do acúmulo de material
resultante da fotossíntese líquida, esta passa a ser o aspecto fisiológico de maior
importância para a avaliação de crescimento.
Segundo Radford (1967), as principais características de crescimento são as
seguintes: taxa de crescimento da cultura; taxa de crescimento relativo; taxa
assimilatória líquida; razão de área foliar, que se decompõe em área foliar específica e
razão de peso foliar; índice de área foliar e duração da área foliar.
A pesquisa foi realizada com o objetivo de avaliar o crescimento, as
características do crescimento e os componentes da biomassa inicial do capim-tifton 85
(Cynodon sp), nas condições do Semiárido Nordestino, em diferentes idades de rebrotação (0;
5; 10; 15; 20; 25 e 30 dias).

26
MATERIAL E MÉTODOS
A pesquisa foi desenvolvida no Setor de Produção de Ovinos do Campo
Experimental de Bebedouro, pertencente à Embrapa Semiárido, localizado no
Município de Petrolina-Pernambuco. O período de avaliação teve início em agosto e se
estendeu até de setembro do ano de 2008. O clima da região é classificado como quente
e seco, sendo caracterizada por duas estações bem definidas, uma chuvosa, de janeiro a
maio, e outra seca, de junho a dezembro. Os dados sobre as condições de temperaturas
máxima e mínima do ar e precipitação pluvial durante o período de avaliação foram
obtidos da Estação Agrometeorologia de Bebedouro, Petrolina-Pernambuco e são
apresentados na Tabela 1.
Tabela 1. Dados climáticos observados durante o período experimental
Mês Precipitação (mm) Temperatura (ºC)
Máxima Mínima Média
agosto 1,2 31,5 19,5 24,9 setembro 0,0 34,4 20,7 26,7 Dados obtidos da Estação Agrometeorologia do Campo Experimental de Bebedouro, Embrapa Semiárido, Petrolina-Pernambuco.
A avaliação de crescimento foi conduzida na área de 0,82 ha, em pastagem
cultivada de capim-tifton 85 (Cynodon sp), no crescimento em rebrotação após o corte
de uniformização de 5 cm, sem influência de nenhuma estratégia de manejo. Foram
sorteados três pontos para cada data de avaliação, a intervalos de cinco dias. Portanto,
foram demarcados 21 pontos {[(30 dias de crescimento 5 dias de intervalo)+1] x 3
pontos por data de avaliação}.
As avaliações foram realizadas nas seguintes idades de rebrotação após o corte
de uniformização (0; 5; 10; 15; 20; 25 e 30 dias). Em cada uma destas datas, foram
realizadas as seguintes avaliações:
a) Leitura da intercepção de radiação fotossinteticamente ativa (RFA) e do
índice de área foliar, utilizando-se o Analisador LAI 2000 da LI-COR Bioscienses
(Figura 1), posicionado o analisador no topo do dossel e ao nível do solo;

27
Figura 1. Analisador LAI 2000 da LI-COR Biocienses.
b) Coleta da massa de forragem em cada um dos pontos, utilizando-se uma
moldura em formato quadrado de 0,25 m², padronizando-se o corte a 5 cm do solo. Tais
amostras foram levadas ao laboratório, sendo então separado o material vivo do material
morto e em seguida as lâminas foliares dos colmos do material vivo, os quais foram
secos em estufa de ventilação forçada a 55º C e pesados, para a quantificação dos
componentes da biomassa: massas secas de forragem total (MSFT), de forragem verde
(MSFV), de forragem morta (MSFM), de lâmina foliar verde (MSLV), de colmo verde
(MSCV), além das relações material vivo/material morto e relação lâmina foliar/haste.
c) Determinação dos índices de crescimento, tomando-se os dados de MSFV e
área foliar (AF), onde foram ajustados modelos polinomiais de segundo grau em função
da idade do dossel, ao tempo das amostragens. Em seguida, a partir da derivada
primeira do modelo quadrático da MSFV em função da idade, foram estimados os
valores instantâneos para a taxa de crescimento relativo (TCR) e taxa de assimilação
líquida (TAL), conforme Radford (1967), pelas fórmulas:
TCR = (1/W) x (dW/dt)
TAL = (1/AF) x (dW/dt)
onde,
W = valor instantâneo da massa seca de forragem verde estimado a partir da respectiva
equação quadrática (g/m2);

28
AF = valor instantâneo da área foliar estimado a partir da respectiva equação quadrática
(m2fol/m2);
dW/dt = equação derivada primeira da respectiva equação quadrática da massa seca de
forragem verde em função do tempo de rebrotação (g/m2 x dia).
O valor instantâneo da razão de área foliar (RAF) foi obtido pelo quociente entre
os valores estimados de AF e MSFV para cada idade. De posse das estimativas de TCR,
TAL e RAF, foram elaborados gráficos ligando-se os pontos obtidos para cada
estimativa.
d) Acompanhamento da morfogênese do capim-tifton 85, registrando-se o
aparecimento, comprimento de segmentos verde e/ou senescente de lâminas foliares,
conforme Davies (1993) e o de alongamento das hastes, a fim de se estimar as
respectivas taxas. Em cada um dos pontos anteriormente marcados, foram escolhidos ao
acaso 5 perfilhos, identificados com anéis coloridos de fio telefônico, aos quais foram
atadas fitas coloridas da mesma cor para facilitar a localização. Nos referidos perfilhos,
registrou-se a cada cinco dias o comprimento total e também da porção verde de todas
as lâminas foliares que não estivessem completamente mortas a partir da lígula da
própria folha - quando já expandida, ou da lígula da folha mais recentemente expandida
- em se tratando de folha emergente. O comprimento da porção senescente foi obtido
pela diferença entre o comprimento total da lâmina foliar e da sua porção ainda verde.
Estimou-se, ainda, o alongamento dos colmos, como a diferença de altura da lígula mais
distante da base do colmo por ocasião da primeira e última leitura, durante o período de
descanso.
Para expressar o crescimento linear em crescimento ponderal, estimou-se o
respectivo fator de conversão. Foram coletados aleatoriamente trinta perfilhos em cada
repetição, os quais foram levados ao laboratório e separados em colmo, lâmina de folha
expandida e lâmina de folha emergente. Cada uma dessas frações teve o comprimento
de seus componentes registrado, sendo então submetidas à secagem em estufa de
ventilação forçada a 55ºC, durante 72 horas e pesadas, obtendo-se um índice de peso
por unidade de comprimento para lâmina foliar emergente (α1), um índice de peso por
unidade de comprimento para lâmina foliar adulta (α2) e um índice de peso por unidade
de comprimento para colmos (β). Assim, foi possível estimar a taxa de acúmulo de
forragem (TAF) durante o período de descanso, a partir das taxas de alongamento

29
(TAlF) e senescência (TSF) de lâmina foliar, da taxa de alongamento das hastes (TAlH)
e da densidade de perfilhos (DPP), conforme a equação:
TAF = {[(TAlF x α1) - (TSF x α2)] + (TAlH x β)} x DPP
onde,
TAF = taxa de acúmulo de forragem durante o período de rebrotação (kg MS/ha);
TAlF = taxa de alongamento de lâmina foliar (cm/dia x perfilho);
α1 = índice de peso por unidade de comprimento para lâmina foliar emergente (g/cm);
TSF = taxa de senescência de lâmina foliar (cm/dia x perfilho);
α2 = índice de peso por unidade de comprimento para lâmina foliar adulta (g/cm);
TAlH = taxa de alongamento da haste (cm/dia x perfilho);
β = índice de peso por unidade de comprimento para colmo (g/cm);
DPP = densidade populacional de perfilhos no período de rebrotação (perfilhos/ha).
e) Algumas medidas estruturais do pasto também foram tomadas a cada idade de
rebrotação, tais como: densidade populacional de perfilhos, contando-se o número de
perfilhos vivos presentes na moldura de 0,5 x 0,5 m, antes de ser realizado o corte da
biomassa.
Os dados foram ajustados às equações que melhor estimassem as variáveis
avaliadas, e apresentados de modo descritivo, através de curvas de em função da idade
de rebrotação da gramínea. Os gráficos foram plotados através do programa
computacional Excel.
RESULTADOS E DISCUSSÃO
Pode-se observar uma relação entre o índice de área foliar (IAF) e a
interceptação da radiação fotossinteticamente ativa (IRFA), uma vez que, à medida IAF
do dossel eleva-se (Figura 2), há uma tendência deste interceptar mais luz em
decorrência da quantidade de folhas com intensa atividade fotossintética, permitindo,
portanto, crescimento da IRFA, em função do aumento do IAF.
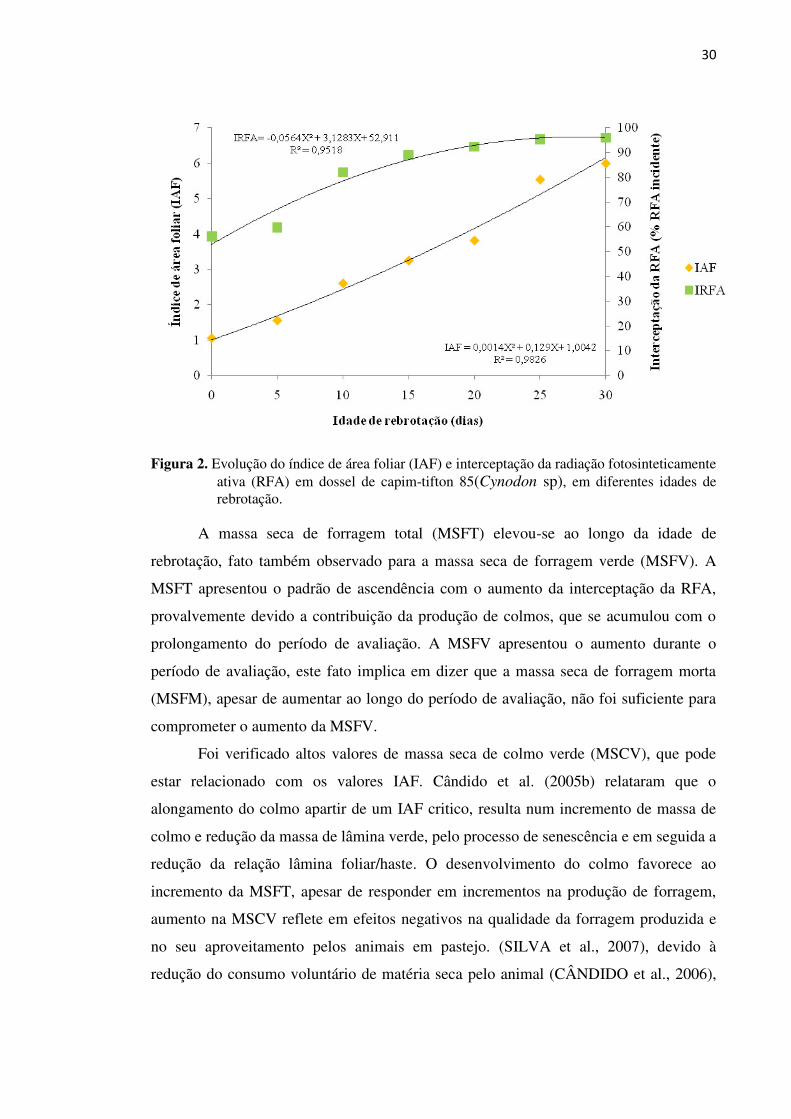
30
Figura 2. Evolução do índice de área foliar (IAF) e interceptação da radiação fotosinteticamente
ativa (RFA) em dossel de capim-tifton 85(Cynodon sp), em diferentes idades de rebrotação.
A massa seca de forragem total (MSFT) elevou-se ao longo da idade de
rebrotação, fato também observado para a massa seca de forragem verde (MSFV). A
MSFT apresentou o padrão de ascendência com o aumento da interceptação da RFA,
provalvemente devido a contribuição da produção de colmos, que se acumulou com o
prolongamento do período de avaliação. A MSFV apresentou o aumento durante o
período de avaliação, este fato implica em dizer que a massa seca de forragem morta
(MSFM), apesar de aumentar ao longo do período de avaliação, não foi suficiente para
comprometer o aumento da MSFV.
Foi verificado altos valores de massa seca de colmo verde (MSCV), que pode
estar relacionado com os valores IAF. Cândido et al. (2005b) relataram que o
alongamento do colmo apartir de um IAF critico, resulta num incremento de massa de
colmo e redução da massa de lâmina verde, pelo processo de senescência e em seguida a
redução da relação lâmina foliar/haste. O desenvolvimento do colmo favorece ao
incremento da MSFT, apesar de responder em incrementos na produção de forragem,
aumento na MSCV reflete em efeitos negativos na qualidade da forragem produzida e
no seu aproveitamento pelos animais em pastejo. (SILVA et al., 2007), devido à
redução do consumo voluntário de matéria seca pelo animal (CÂNDIDO et al., 2006),

31
uma vez que este alongamento é acompanhado de uma lignificação acentuada, que
resulta na queda do valor nutritivo da forragem e repletirá negativamente sobre o
desempenho animal (FARIA, 2009).
Figura 3. Componentes da biomassa inicial (g/m²): Massa seca de forragem total (MSFT),
massa seca de forragem verde (MSFV), massa seca de lâmina verde (MSLV), massa seca de colmo verde (MSCV) e massa seca de forragem morta (MSFM) em dossel de capim-tifton 85 (Cynodon sp), em diferentes idades de rebrotação.
Os valores dos índices de crescimento, taxa de crescimento relativo (TCR), taxa
de assimilação liquida (TAL) e razão de área foliar (RAF), durante o período de
avaliação, encontram-se nas Figuras 4, 5 e 6. Observa-se em todos eles a natureza
decrescente, semelhante à encontrada para capim-tifton 85, cultivado em parcelas
(OLIVEIRA et al., 2000).
A TCR reduziu com o avanço da idade de rebrotação (Figura 4). Sabe-se que a
TCR apresenta valores mais altos nos primeiros dias de rebrotação, reduzindo
posteriormente. Valores obtidos no presente estudo encontram-se dentro do esperado,
uma vez que, de acordo com Beadle (1993), os valores de TCR para a maioria das
espécies de plantas C4, encontram-se entre o intervalo de 0,02 a 0,5 g/g x dia. Oliveira et
al. (2000) observaram variação exponencial, com valores de TCR de 0,12 a 0,026 g/g x
dia, dos 14 aos 70 dias, apresentando queda mais acentuada até a idade de 35 dias. Em
trabalhos conduzidos por Gomide (1996), com cinco cultivares do gênero Cynodon,
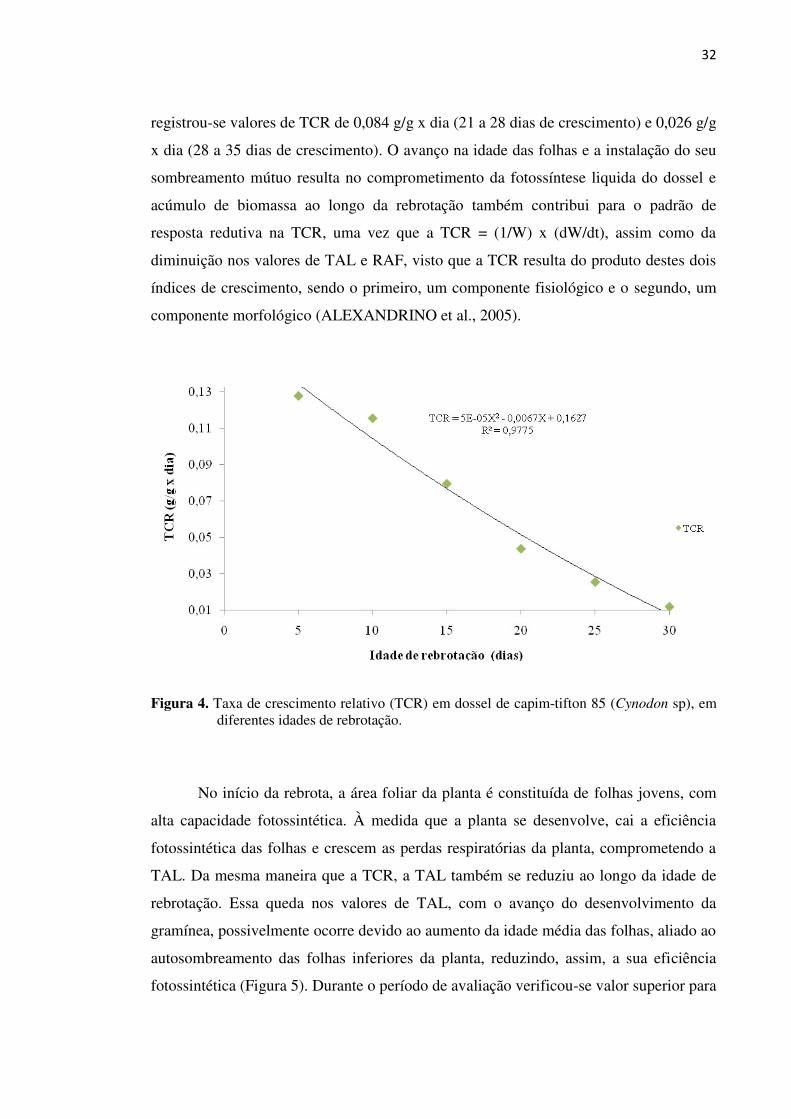
32
registrou-se valores de TCR de 0,084 g/g x dia (21 a 28 dias de crescimento) e 0,026 g/g
x dia (28 a 35 dias de crescimento). O avanço na idade das folhas e a instalação do seu
sombreamento mútuo resulta no comprometimento da fotossíntese liquida do dossel e
acúmulo de biomassa ao longo da rebrotação também contribui para o padrão de
resposta redutiva na TCR, uma vez que a TCR = (1/W) x (dW/dt), assim como da
diminuição nos valores de TAL e RAF, visto que a TCR resulta do produto destes dois
índices de crescimento, sendo o primeiro, um componente fisiológico e o segundo, um
componente morfológico (ALEXANDRINO et al., 2005).
Figura 4. Taxa de crescimento relativo (TCR) em dossel de capim-tifton 85 (Cynodon sp), em diferentes idades de rebrotação.
No início da rebrota, a área foliar da planta é constituída de folhas jovens, com
alta capacidade fotossintética. À medida que a planta se desenvolve, cai a eficiência
fotossintética das folhas e crescem as perdas respiratórias da planta, comprometendo a
TAL. Da mesma maneira que a TCR, a TAL também se reduziu ao longo da idade de
rebrotação. Essa queda nos valores de TAL, com o avanço do desenvolvimento da
gramínea, possivelmente ocorre devido ao aumento da idade média das folhas, aliado ao
autosombreamento das folhas inferiores da planta, reduzindo, assim, a sua eficiência
fotossintética (Figura 5). Durante o período de avaliação verificou-se valor superior para

33
TAL na fase inicial de rebrotação de 14,8 g/m² fol x dia. Além disso, com o avanço da
maturidade da planta, ocorre maior aporte de assimilados para o desenvolvimento do
colmo e de outras partes, aumentando ainda mais as perdas respiratórias da planta, o que
leva à redução da TAL (OLIVEIRA et al., 2000). Segundo Gomide e Gomide (1999), a
redução na TAL ao longo do período de avaliação se deve à competição por luz, ao
avanço na idade média da área foliar da planta e ao incremento das demandas
respiratórias.
Figura 5. Taxa de assimilação liquida (TAL) em dossel de capim-tifton 85 (Cynodon sp), em
diferentes idades de rebrotação.
Normalmente a RAF é reduzida com o aumento da idade de rebrotação. Segundo
Benincasa (1988), a RAF representa a unidade de área foliar que está sendo usada pela
planta para produzir uma unidade de peso de matéria seca. Assim, durante o estádio de
crescimento vegetativo, há maior aporte de assimilados para a produção de folhas, a fim
de aumentar a captura da luz incidente e promover rápido crescimento. Nesse período,
observam-se, então, valores altos de razão de área foliar, devido ao crescente aumento
relativo no aparelho assimilatório. O alto valor de RAF pode ser verificado o inicio do
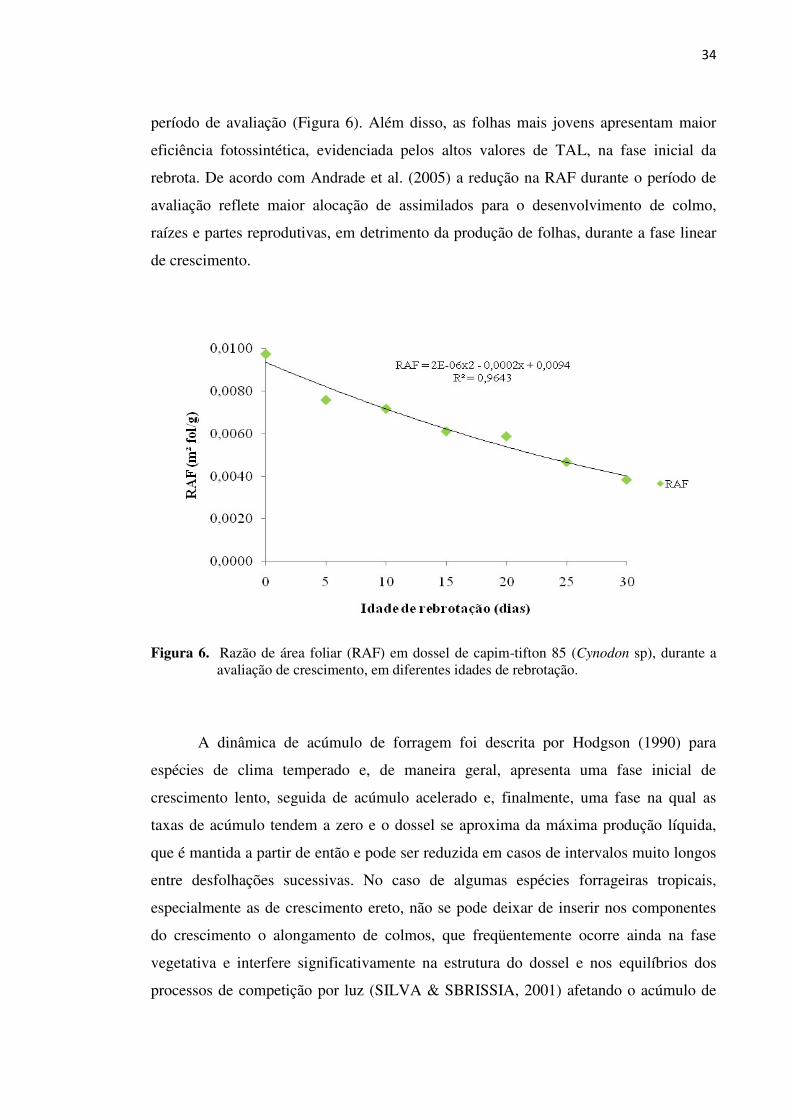
34
período de avaliação (Figura 6). Além disso, as folhas mais jovens apresentam maior
eficiência fotossintética, evidenciada pelos altos valores de TAL, na fase inicial da
rebrota. De acordo com Andrade et al. (2005) a redução na RAF durante o período de
avaliação reflete maior alocação de assimilados para o desenvolvimento de colmo,
raízes e partes reprodutivas, em detrimento da produção de folhas, durante a fase linear
de crescimento.
Figura 6. Razão de área foliar (RAF) em dossel de capim-tifton 85 (Cynodon sp), durante a
avaliação de crescimento, em diferentes idades de rebrotação.
A dinâmica de acúmulo de forragem foi descrita por Hodgson (1990) para
espécies de clima temperado e, de maneira geral, apresenta uma fase inicial de
crescimento lento, seguida de acúmulo acelerado e, finalmente, uma fase na qual as
taxas de acúmulo tendem a zero e o dossel se aproxima da máxima produção líquida,
que é mantida a partir de então e pode ser reduzida em casos de intervalos muito longos
entre desfolhações sucessivas. No caso de algumas espécies forrageiras tropicais,
especialmente as de crescimento ereto, não se pode deixar de inserir nos componentes
do crescimento o alongamento de colmos, que freqüentemente ocorre ainda na fase
vegetativa e interfere significativamente na estrutura do dossel e nos equilíbrios dos
processos de competição por luz (SILVA & SBRISSIA, 2001) afetando o acúmulo de
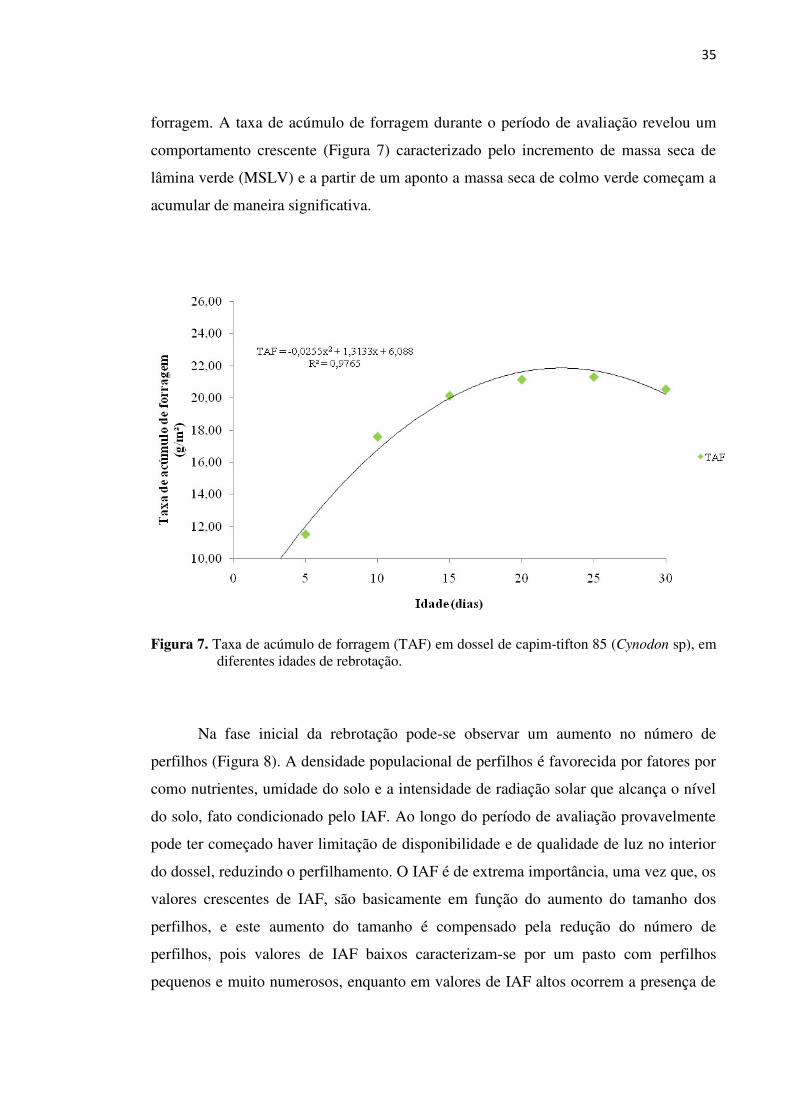
35
forragem. A taxa de acúmulo de forragem durante o período de avaliação revelou um
comportamento crescente (Figura 7) caracterizado pelo incremento de massa seca de
lâmina verde (MSLV) e a partir de um aponto a massa seca de colmo verde começam a
acumular de maneira significativa.
Figura 7. Taxa de acúmulo de forragem (TAF) em dossel de capim-tifton 85 (Cynodon sp), em
diferentes idades de rebrotação.
Na fase inicial da rebrotação pode-se observar um aumento no número de
perfilhos (Figura 8). A densidade populacional de perfilhos é favorecida por fatores por
como nutrientes, umidade do solo e a intensidade de radiação solar que alcança o nível
do solo, fato condicionado pelo IAF. Ao longo do período de avaliação provavelmente
pode ter começado haver limitação de disponibilidade e de qualidade de luz no interior
do dossel, reduzindo o perfilhamento. O IAF é de extrema importância, uma vez que, os
valores crescentes de IAF, são basicamente em função do aumento do tamanho dos
perfilhos, e este aumento do tamanho é compensado pela redução do número de
perfilhos, pois valores de IAF baixos caracterizam-se por um pasto com perfilhos
pequenos e muito numerosos, enquanto em valores de IAF altos ocorrem a presença de

36
perfilhos grandes e pouco numerosos. Este comportamento pode ter sido responsável
pela redução do número de perfilhos. Se o período de rebrotação se estender muito
além, a mortalidade de perfilhos aumenta, podendo vir a comprometer a capacidade de
recuperação da área foliar e a habilidade competitiva da planta.
Figura 8. Densidade populacional de perfilhos (perfilhos/m2) em dossel de capim-tifton
85 (Cynodon sp), em diferentes idades de rebrotação.
Segundo Oliveira et al. (1998), o perfilho apresenta aumento crescente no
número de folhas vivas no início da rebrotação, atingindo o máximo, mas estabiliza-se
com menor número de folhas vivas/perfilho, graças à presença da senescência foliar.
O número total de folhas vivas (NFV) por perfilho expressa o potencial de
assimilação de carbono e de produção de forragem ao nível de perfilho. Essa variável
apresenta ainda grande importância na avaliação e manejo das plantas forrageiras, por
tratar-se do componente da biomassa com melhor atributo qualitativo, sendo a fração
mais selecionada pelos animais em pastejo e também pela sua aplicação como critério
de definição prático para a determinação do período de descanso numa área de pastejo
(GRANT et al., 1988; FULKERSON & DONAGHY, 2001). À medida que o dossel

37
aumenta sua capacidade de interceptação da RFA, este tende a produzir mais folhas,
aumentando o número de folhas vivas, sendo esta característica estrutural uma
ferramenta de manejo que pode ser utilizada para determinar a condição de entrada dos
animais no pasto (Figura 9). Segundo Gomide e Gomide (2000), durante o
desenvolvimento da forrageira, o NFV cresce enquanto não se instala o processo de
senescência e morte de folhas. O NFV obtido no presente estudo foi em média de 5,
apresentando semelhança para o número de folhas vivas/perfilho para o Tifton 85
relatado por Pinto (2000) de 5,4 e inferior para o valor encontrado por Oliveira (1999),
que foi de 9,5.
Figura 9. Número de folhas vivas/perfilho em dossel de capim-tifton 85 (Cynodon sp), em
diferentes idades de rebrotação.
CONCLUSÕES
O índice de área foliar e a interceptação da radiação fotossinteticamente ativa se
elevaram o avanço da idade de rebrotação.
Foi observado incremento da massa seca de forragem total e forragem verde
com o aumento da idade de rebrotação.

38
Os valores dos índices de crescimento TCR, TAL e RAF se reduziram com o
avanço da idade de rebrotação.
A densidade populacional de perfilhos e o número de folhas/perfilho se
reduziram com o avanço da idade de rebrotação.
REFERÊNCIAS
ALEXANDRINO, E.; NASCIMENTO JÚNIOR, D.; REGAZZI, A. J. et al. Características morfogênicas e estruturais da Brachiaria brizantha cv. Marandu submetida a diferentes doses de nitrogênio e freqüências de cortes. Acta Scientiarum Agronomy, Maringá, v. 27, n. 1, p. 17-24, 2005. ALVIM, M.J.; VILELA, D.; LOPES, R.S. Efeitos de dois níveis de concentrado sobre a produção de leite de vacas da raça Holandesa em pastagem de coast-cross (Cynodon
dactilum (L.) Pers). Revista Brasileira de Zootecnia, v.26, p.967-975, 1997. ANDRADE, A. C.; FONSECA, D. M.; LOPES, R. S. et al. Características morfogênicas do capim-elefante Napier adubado e irrigado. Ciência e Agrotecnologia, Lavras, v. 29, n. 1, p. 150-159, 2005. BEADLE, C. L. Growth analysis. In: HALL, D. O.; BOLHARNORDENKAMPF, H. R.; LEEGOOD, R. C.; LONG, S. P. (Eds.). Photosynthesis and production in a changing environment: a field and laboratory manual. London: [s.n.], 1993. p. 36-46. BENICASA, M. M. P. Análise de Crescimento de Plantas (noções básicas ). Jaboticabal. FUNEP. 2004. 42p. BENINCASA, M. M. P. Análise de crescimento de plantas: noções básicas. Jaboticabal: FUNEP, 1988. 41p CÂNDIDO, M.J.D.; SILVA, R.G.; NEIVA, J.N.M. et al. Fluxo de biomassa em capimtanzânia pastejado por ovinos sob três períodos de descanso. Revista Brasileira de Zootecnia, v. 35, n. 6, p. 2234-2242, 2006. CÂNDIDO, M.J.D.; GOMIDE, C.A.M.; ALEXANDRINO, E. et al. Morfofisiologia do dossel de Panicum maximum cv. Mombaça sob lotação intermitente com três períodos de descanso. Revista Brasileira de Zootecnia, v.34, n.2, p.406-415, 2005. CARNEVALLI, R. A.; SILVA, S. C. da; CARVALHO, C. A. B. et al. Desempenho de ovinos e respostas de pastagens de Coastcross (Cynodon spp.) submetidas a regimes de desfolha sob lotação contínua. Pesquisa Agropecuária Brasileira, Brasília, v. 36, n. 6, p. 919-927, jun. 2001a.

39
DAVIES, A. Tissue turnover in the sward. In: DAVIES, R.D. et al. eds. Sward measurement Handbook, 2nd ed., Reading, p.183-216, 1993. FAGUNDES, J.L.; SILVA, S.C. da; PEDREIRA, C.G.S. et al. Índice de área foliar, interceptação luminosa e acúmulo de forragem em pastagens de Cynodon spp. sob diferentes intensidades de pastejo. Scientia Agricola, v.56, p.1141-1150, 1999. FULKERSON, W.J.; DONAGHY, D.J. Plant soluble carbohydrate reserves and senescence – Key criteria for developing an effective grazing management system for ryegrass based pastures: a review. Australian Journal of Experimental Agriculture, v.41, n.2, p.261-275, 2001. GOMIDE, C.A.M.; GOMIDE, J.A. Morfogênese de cultivares de Panicum maximum
Jacq. Revista Brasileira de Zootecnia, v.29, n.2, p.341-348, 2000. GOMIDE, C.A.M.; GOMIDE, J.A. Análise de crescimento de cultivares de Panicum
maximum Jacq. Revista Brasileira de Zootecnia, v.28, n.4, p.675-680, 1999. GOMIDE, C. C. C. Algumas características fisiológicas e químicas de cinco cultivares de Cynodon. 1996. 100 f. Dissertação (Mestrado em Zootecnia) – Universidade Estadual Paulista, Jaboticabal, 1996. GRANT, S.A.; BARTHRAM, G.T.; KING, L.T.J. et al. Comparison of herbage production under continuous stocking and intermittent grazing. Grass and Forage Science, v.43, n.1, p.29-39, 1988. HODGSON, J. Grazing management: science into practice. New York: Wiley; Burnt Mill, Harlow, Essex: Longman Scientific and Technical, 1990. 203p. OLIVEIRA, M.A.; PEREIRA, O.G.; GOMIDE, J.A. et al. Análise de crescimento do capim-bermuda „Tifton 85‟ (Cynodon sp). Revista Brasileira de Zootecnia, v.29, n.6, p.1930-1938, 2000 (supl.1). OLIVEIRA, M.A. Morfogênese e análise de crescimento e valor nutritivo do capim Tifton 85 (Cynodon sp) em diferentes idades de rebrota. Dissertação de Mestrado, Viçosa, Universidade Federal de Viçosa, 1999, 94p., Orientador: Prof. Odilon Gomes Pereira. OLIVEIRA, M.A.; PEREIRA, O.G.; GARCIA, R. et al. Morfogênese de folhas do tifton 85 (Cynodon sp) em diferentes idades de rebrota. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, 1998. v.2, p.302-303. PINTO, L.F.M. Dinâmica do acúmulo de matéria seca em pastagens de Cynodon
spp. Dissertação (Mestrado) Piracicaba, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 2000, 124p.

40
RADFORD, P.J. Growth analysis formulae – their use and abuse. Crop Science, v.7, n.3, p.171-175, 1967. SANTOS JÚNIOR, J. D. G.; MONTEIRO, F. A.; JUNIOR, J. L. Análise de Crescimento do Capim-marandu Submetido a Doses de Nitrogênio. Revista Brasileira de Zootecnia, v. 33, n. 6, p. 1985-1991, 2004. SILVA, M. J. D.; CÂNDIDO, M. J. D.; NEIVA, J. N. M. et al. Características estruturais do dossel de pastagens de capim-tanzânia mantidas sob três períodos de descanso com ovinos. Revista Brasileira de Zootecnia, v. 36, n. 5, p. 1255-1265, 2007 a. SILVA, S.C. da; SBRISSIA, A.F. A planta forrageira no sistema de produção. In: SIMPÓSIO SOBRE MANEJO DA PASTAGEM, 17., 2001, Piracicaba. Anais. Piracicaba: Fealq, 2001. p.71-88.

41
CAPÍTULO 3
____________________________________________________________
Características morfogênicas do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo

42
Características morfogênicas do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
RESUMO: Foram avaliadas as características morfogênicas e o acúmulo de forragem
do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
em diferentes ciclos de pastejo. Os tratamentos corresponderam a combinações de três
condições de pré-pastejo (número de folhas/perfilho: 4,5; 6,5 e 8,5) e duas condições
pós-pastejo (altura residual: 10 e 5 cm), distribuídos em um delineamento experimental
inteiramente casualizado em arranjo fatorial 3x2, com quatro repetições (piquetes). As
avaliações foram realizadas logo após o pastejo e em intervalos variados de acordo com
o período de descanso. Não foi verificado efeito do período de descanso (PD), resíduo
pós-pastejo (RPP), da interação PD x RPP e efeito dos ciclos (P 0,05), sobre a razão
entre a taxa de alongamento da folha 1 e 2 (TAlF1/TAlF2). O PD de 8,5 fol/perf
apresentou menor taxa de alongamento foliar (TAlF). O resíduo de 10 cm, apresentou
maior TAlF. As maiores taxas de alongamento das hastes (TAlH), foram observadas
para os pastos com PD de 6,5 e 8,5 fol/perf. Houve efeito da interação PD x RPP
(P 0,05) para taxa de senescência anterior (TSFa) e os pastos com 4,5 fol/perf, em
ambos os RPP, apresentaram valores inferiores para esta variável. Os maiores valores de
taxa de senescência posterior (TSFp) e senescência total (TST), são observados nos
pastos com PD de 8,5 fol/perf. As maiores taxas de aparecimento foliar (TApF) foram
observadas nos pastos com PD de 4,5 e 6,5 fol/perf e resíduos de 10 e 5 cm, que
resultaram em elevadas taxas de produção de forragem (TPF) e acúmulo de forragem
(TAF). Os ciclos de pastejo causaram efeito sobre as TAlF e TApF, apresentando
valores elevados, ao longo do período experimental. As TSFa e TST em função dos
ciclos, se elevaram nos pastos sob o PD 8,5 fol/perf e resíduos de 10 e 5 cm. A TSFp,
foi elevada ao longo dos ciclos de pastejo, para com PD de 4,5 e 8,5 fol/per e resíduo de
5 e 10 cm, respectivamente. As características morfogênicas foram afetadas pelos PDs,
RPPs e efeito dos ciclos de pastejo. Os PDs superiores ao necessário para formação de
6,5 fol/perf tornam todas as características morfogênicas estudadas indesejáveis do
ponto de vista forrageiro.
Termos para indexação: acúmulo de forragem, perfilho, taxa de aparecimento foliar

43
Morphogenic characteristics of the tifton 85 grass managed under three periods of
rest and two post-grazing residues
Abstract: The morphogenic characteristics and the accumulation of fodder plant of
tifton 85 grass managed under three periods of rest had been evaluated and two residues
after-pastejo in different cycles of pastejo. The treatments had corresponded the
combinations of three conditions of daily pay-pastejo (leaves number/tiller: 4,5; 6,5 and
8,5) and two conditions after-pastejo (residual height: 10 and 5 cm), distributed in an
experimental delineation entirely casualizado in factorial arrangement 3x2, with four
repetitions (plots). The evaluations had been carried through soon after pastejo and in
varied intervals in accordance with the period of rest. Effect of the period of rest was
not verified (PD), residue after-pastejo (RPP), of interaction PD x RPP and effect of the
cycles (P 0,05), on the reason the tax of allonge of leaves 1 and 2 enters
(TAlF1/TAlF2). The 8,5 PD of leaves/tiller presented minor tax of foliar allonge
(TAlF). The residue of 10 cm, presented TAlF greater. The biggest rates of allonge of
the connecting steam (TAlH), had been observed for the grass with 8,5 PD of 6,5 and
leaves/tiller. It had effect of interaction PD x RPP (P 0,05) for rate of previous
senescence (TSFa) and the grass with 4,5 leaves/tiller, in both the RPP, had presented
inferior values for this 0 variable. The biggest values of rate of posterior senescence
(TSFp) and total senescência (TST), are observed in the grass with 8,5 PD of
leaves/tiller. The biggest rates of foliar appearance (TApF) had been observed in the
grass with 6,5 PD of 4,5 and leaves/tiller and residues of 10 and 5 cm, that had resulted
in high taxes of production of fodder plant (TPF) and accumulation of fodder plant
(TAF). The cycles of pastejo had caused effect on the TAlF and TApF, having
presented raised values, throughout the experimental period. TSFa and TST in function
of the cycles, if had raised in the grass under 8,5 PD leaves/tiller and residues of 10 and
5 cm the TSFp, was raised throughout the cycles of pastejo, stops with PD of 4,5 and
8,5 leaves/tiller and residue of 5 and 10 cm, respectively. The morphogenic
characteristics had been affected by the PDs, RPPs and effect of the cycles of pastejo.
The PDs superior to the necessary one for formation of 6, 5 leaves/tiller becomes all the
undesirable studied morphogenic characteristics of the forrageiro point of view.
Index-terms: accumulation of forage, tiller, rates of foliar appearance

44
INTRODUÇÃO
A área de pastagens no território brasileiro corresponde a 22 %, o que representa
mais de 172 milhões de hectares (ANUALPEC, 2008), realçando a grande contribuição
desse recurso na exploração pecuária e na economia do país. A produção de ruminantes
baseada em pastagens é a forma mais econômica de se obter produtos de origem animal
de alto valor biológico.
A produção de forragem é um processo contínuo, regulado por variáveis do
ambiente e por características da pastagem. À medida que tecidos foliares crescem nas
plantas, esses estão sujeitos ao envelhecimento e à senescência, responsáveis pela
redução no acúmulo de massa seca e pelas perdas quantitativas e qualitativas da
forragem produzida. Avanços significativos na busca do entendimento da dinâmica do
ecossistema de pastagem só foram possíveis quando foi reconhecido que o uso de
plantas forrageiras em pastagens deveria ser considerado dentro de um contexto
sistêmico, no qual a adoção de técnicas e estratégias de manejo em pontos isolados do
sistema dificilmente resultaria em mudanças significativas na eficiência global final do
processo produtivo. Para tanto segundo Nabinger e Pontes (2001), tornaram-se
relevantes os estudos de dinâmica da produção primária das forrageiras, realizados por
meio de avaliações das características morfogênicas e estruturais das mesmas.
De acordo com Nascimento Júnior et al. (2002), a taxa de aparecimento, a taxa
de alongamento e a duração de vida das folhas são o referencial morfogênico que
permite integrar diferentes características estruturais do pasto, com a finalidade de
atingir os principais objetivos da produção animal em pastagens. As características
morfogênicas apresentam estreita correlação com o rendimento forrageiro (HORST et
al., 1978) e podem ser usadas como critério de seleção em trabalhos de melhoramento
genético e em estudos para avaliar efeitos dos fatores do meio sobre a produtividade das
gramíneas (GRANT & MARRIOT, 1994).
Segundo Pontes et al. (2003), diferentes alturas de manejo proporcionam
diferenças na estrutura da pastagem e, conseqüentemente, na morfogênese da planta,
uma vez que alteram a dinâmica do processo de desfolhação efetuado pelo animal.
Diante do exposto, esta pesquisa foi conduzida com o objetivo de avaliar o efeito
dos três períodos de descanso 4,5; 6,5 e 8,5 (folhas/perfilho) e dois resíduos pós-pastejo

45
5 e 10 cm, sobre a morfogênese do capim-tifton 85, nas condições do Semiárido
Nordestino.
MATERIAL E MÉTODOS
A pesquisa foi desenvolvida no Setor de Produção de Ovinos do Campo
Experimental de Bebedouro, pertencente à Embrapa Semiárido, localizado no
Município de Petrolina-Pernambuco. Durante o mês de outubro foi efetuada a divisão
da área em piquetes e ajuste da metodologia de trabalho. A coleta de dados ocorreu
entre os meses de novembro de 2008 a junho de 2009, totalizando nove meses.
O clima da região é classificado como quente e seco, sendo caracterizada por
duas estações bem definidas, uma chuvosa, de janeiro a maio, e outra seca, de junho a
dezembro. Os dados sobre as condições de temperaturas máxima e mínima do ar e
precipitação pluviométrica durante o período experimental foram obtidos da Estação
Agrometeorologia de Bebedouro, Petrolina-Pernambuco e são apresentados na Tabela
1.
Tabela 1. Dados climáticos observados durante o período experimental
Mês Precipitação (mm) Temperatura (ºC)
Máxima Mínima Média
outubro/2008 0,0 33,7 21,7 27,7 novembro/2008 0,0 34,8 23,2 28,7 dezembro/2008 54,6 33,7 22,6 27,1 janeiro/2009 45,7 34,3 23,0 27,5 fevereiro/2009 127,0 32,4 22,1 26,1 março/2009 152,9 33,0 22,2 26,3 abril/2009 223,6 31,1 22,3 25,5 maio/2009 85,4 28,8 21,2 24,2 Junho/2009 14,5 29,7 19,5 23,6 Dados obtidos da Estação Agrometeorologia de Bebedouro, Embrapa Semiárido, Petrolina/Pernambuco.

46
Foram avaliados três períodos de descanso 4,5; 6,5 e 8,5 (número de
folhas/perfilho) e dois resíduos pós-pastejo 5 e 10 cm, em um delineamento
experimental inteiramente casualizado em arranjo fatorial 3x2 com quatro repetições
(piquetes). A área experimental constou de 0,82 ha de pastagem cultivada com capim-
tifton 85 irrigada por aspersão com canhões com a distribuição da água durante o dia.
Utilizou-se o método de pastejo sob lotação intermitente, com lotação variável,
mantendo-se animais prova nos piquetes e também animais reguladores para ajuste da
taxa de lotação, segundo a técnica da taxa de lotação variável. A pastagem foi
subdivida, com tela campestre, em 37 piquetes de 180 m2 cada, destes piquetes 24 foram
considerados amostrais. Foram utilizados 30 ovinos de prova e um número variável de
animais de equilíbrio, da raça Santa Inês, machos castrados, com peso corporal (PC)
médio inicial de ± 20 kg, os quais foram previamente identificados, vermifugados e, em
grupos de cinco, distribuídos por sorteio em seis tratamentos. Foram efetuadas
adubações de reposição com nitrogênio, em cada piquete, no momento da saída dos
animais, nas seguintes doses: 0,971; 1,110; 1,320; 1,475; 1,666 e 1,800 g, de acordo
com o período de descanso.
O solo da área experimental é classificado como Podzólico Amarelo Eutrófico
Latossólico (EMBRAPA, 1999), teve suas características químicas avaliadas (Tabela 2)
em outubro de 2008. A área foi adubada em função das necessidades da análise do solo.
A adubação com fósforo e potássio foi realizada a lanço e em cobertura, na forma de
superfosfato simples e cloreto de potássio, respectivamente, em 02 de novembro de
2008 em uma aplicação com 120 kg/ha de P2O5 e 100 kg/ ha de K2O. A adubação
nitrogenada (600 kg de N/ha) foi fracionada em aplicações, ao dos ciclos de pastejo,
após a saída dos animais dos piquetes, na forma de uréia, de acordo com os períodos de
descanso.
Tabela 2. Características químicas do solo da área experimental em duas profundidades
Profundidade MO pH:H2O P K Al Ca Mg H + Al CTC SB -----(cm)---- g/Kg-1 1: 2,5 mg/dm3 ----Cmolc dm-3---- %
0 – 5 24,14 6,18 41,51 0,21 0,05 2,45 0,94 1,83 5,48 66,41 5 - 20 7,10 6,09 22,67 0,14 0,05 1,33 0,59 1,16 3,25 63,51
Nos piquetes amostrais, foram marcadas duas hastes após a saída dos animais,
mudadas a cada novo ciclo de pastejo. Em cada uma das hastes, quatro perfilhos foram

47
identificados aleatoriamente com anéis coloridos de fio telefônico e fitas coloridas da
mesma cor, atadas para facilitar sua localização. A escolha de perfilhos para marcação
foi feita procurando-se evitar plantas debilitadas ou injuriadas por qualquer razão. Nos
referidos perfilhos, registrou-se, depois de dois dias da saída dos animais e também a
cada quatro dias, o comprimento total das lâminas expandidas, das lâminas emergentes
e a altura do colmo. O comprimento das lâminas expandidas foi feito por meio da
medição da distância da sua lígula até o ápice da folha. O comprimento da lâmina
emergente foi obtido por meio da medição da distância do ápice dessa folha até a lígula
da folha expandida imediatamente anterior. Já o comprimento do colmo foi obtido pela
medição da distância da última lígula exposta até a base do perfilho. Fez-se a estimativa
do ângulo do perfilho em relação ao solo, com medição do comprimento do colmo no
sentido horizontal (do solo até a lígula mais alta, de modo que fosse seguida a
inclinação do perfilho) e vertical (do solo até a lígula mais alta na posição vertical). O
comprimento da porção senescente foi obtido pela diferença entre o comprimento total
da lâmina foliar e o comprimento de sua porção ainda verde.
Através das medidas dos comprimentos das lâminas foliares e do colmo, foram
estimados os seguintes índices: a) taxa de alongamento foliar (TAlF) (referente ao
alongamento médio diário das lâminas foliares de todo o perfilho durante o período de
descanso); b) taxa de alongamento da haste (TAlH) (referente ao alongamento médio
diário do colmo de todo o perfilho durante o período de descanso); c) razão entre as
taxas de alongamento de lâminas foliares da primeira e da segunda folha produzida no
início da rebrotação de cada perfilho (TAlF1/TAlF2); d) TSFa e a TSFp, que expressam,
respectivamente, a taxa de senescência de lâminas foliares formadas anteriormente ao
período de descanso em avaliação e a taxa de senescência de lâminas foliares formadas
o período de descanso em avaliação; e) TST, que corresponde à taxa de senescência
foliar total ao final de um determinado período de descanso e representa a soma das
TSFa e TSFp; f) taxa de aparecimento foliar (TApF), que mede a velocidade de
surgimento da folha, expressa, em dias.
Foram determinados os índices gravimétricos. Para tanto, ao final de cada
período de descanso, foram colhidos aproximadamente 30 perfilhos por piquete
amostral, levados ao laboratório e separados em colmos, lâminas foliares expandidas e
lâminas foliares emergentes. Cada uma dessas frações teve seu comprimento total

48
registrado, para serem, então, submetidas à secagem em estufa de ventilação forçada a
65°C, durante 72 horas e pesadas, para a determinação posterior do índice de peso por
unidade de comprimento da lâmina foliar emergente (α1), da lâmina foliar expandida
(α2) e das hastes (β).
De posse das variáveis morfogênicas, foi possível estimar também a taxa de
acúmulo de forragem (TAF) e a taxa de produção de forragem (TPF) durante o período
de descanso, a partir das taxas de alongamento (TAlF) e senescência (TSF) de lâmina
foliar, da taxa de alongamento das hastes (TAlH), dos índices gravimétricos e da
densidade populacional de perfilhos (DPP), conforme as equações, adaptada de Davies
(1993):
TPF = {[(TAlF x α1)] + (TAlH x β)} x DPP
TAF = {[(TAlF x α1) - (TSF x α2)] + (TAlH x β)} x DPP
onde:
TPF = taxa de produção de forragem durante o período de descanso i (kg MS/ha x dia);
TAF = taxa de acúmulo de forragem durante o período de descanso i (kg MS/ha x dia);
TAlF = taxa de alongamento de lâmina foliar (cm/dia x perfilho);
α1 = índice de peso por unidade de comprimento para lâmina foliar emergente (g/cm);
TSF = taxa de senescência de lâmina foliar (cm/dia x perfilho);
α2 = índice de peso por unidade de comprimento para lâmina foliar adulta (g/cm);
TAlH = taxa de alongamento do colmo (cm/dia x perfilho);
β = índice de peso por unidade de comprimento para colmo (g/cm);
DPP = densidade populacional de perfilhos no início do período de descanso
(perfilhos/ha).
Os dados foram submetidos análise de variância e teste de comparação de
médias. Para comparar o efeito dos tratamentos, foi efetuada análise de variância do
efeito dos períodos de descanso e dos resíduos pós-pastejo, na média de todos os ciclos,
em arranjo fatorial completo, de modo que fosse desdobrada a interação quando
significativa ao nível de 5% de probabilidade. As médias foram comparadas por meio
do teste t de Tukey, ao nível de 5% de probabilidade. Para avaliar o efeito dos ciclos
sucessivos de pastejo, foi efetuada análise de variância dos ciclos dentro de cada um dos
seis tratamentos. As médias dos ciclos foram comparadas por meio do teste t de Tukey,
ao nível de 5% de probabilidade. Como ferramenta de auxílio às análises estatísticas,

49
utilizou-se o procedimento GLM do programa estatístico SAS (SAS INSTITUTE,
2003).
RESULTADOS E DISCUSSÃO
Não foi verificado efeito do nem PD (P 0,05), nem dos RPP (P 0,05) e nem
houve interação entre PD x RPP (P 0,05) para a razão entre a taxa de alongamento da
primeira e segunda folha (TAlF1/TAlF2) (Tabela 3), com médias de 0,74, 0,79 e 0,83 para
PD de 4,5; 6,5 e 8,5 fol/perf e 0,74 e 0,78 para RPP de 10 e 5 cm. Estes valores estão
adequados ao manejo, uma vez que valores próximos de 1, sugerem uma menor
mobilização de reservas orgânicas para rebrotação. Os valores encontrados neste estudo
estão próximos aos encontrados por Cutrim Junior et al. (2010) trabalhando com capim-
Tanzânia, sob três frequências de desfolhação e dois resíduos pós-pastejo. Não houve
efeito dos ciclos de pastejo, sobre a razão TAlF1/TAlF2 (Dados não apresentados).
Houve efeito do PD (P 0,05) e do RPP (P 0,05) sobre a taxa de alongamento
foliar (TAlF) (Tabela 3). Foram obtidos valores de 3,04; 2,23 e 1,79 cm/perf x dia para
o PD de 4,5; 6,5 e 8,5 fol/perf, respectivamente e 2,51 e 2,19 cm/perf x dia para o RPP
de 10 e 5 cm de altura residual. A TAlF reduziu com o aumento do PD, ou seja, com o
avanço da maturidade do capim, ocorreu maior competição por fotoassimilados, que são
mobilizados para o desenvolvimento de perfilhos aéreos e de estruturas reprodutivas
que surgem no meristema apical, o que provavelmente contribuiu para a redução na
TAlF. Em relação aos valores de TAlF para as alturas residuais, pode-se perceber que o
resíduo de 5 cm, apresentou menor TAlF, uma vez que, valores de TAlF baixos são
decorrentes de um pastejo mais intenso, promovendo um habito de crescimento mais
prostrado do capim, o que ocasiona um maior sombreamento e menor alongamento
foliar. Carnevalli (2003) e Barbosa et al. (2007) que verificaram maior TAlF quando
manejaram o pasto com alturas residuais mais elevadas. A TAlF variou (P 0,05) de
acordo com os ciclos de pastejo nos pastos com 4,5 fol/per com 10 e 5 cm de resíduo;
6,5 fol/perf com 5cm de resíduo e 8,5 fol/perf com 10 de resíduo (Tabelas 4, 5 e 6). Nos
últimos ciclos foi constatada redução no alongamento de folhas, indicando que as
condições ambientais, principalmente a precipitação, não foram ideais nesse período.

50
Houve efeito do PD (P 0,05) e do RPP (P 0,05) sobre a taxa de alongamento
das hastes (TAlH) (Tabela 3). Nos pastos com 6,5 e 8,5 fol/perf, foram maiores quando
comparadas à TAlC do pasto com 4,5 fol/perf, este comportamento resulta do fato de
que o pasto com 4,5 fol/perf, foi capaz de manter o alongamento da haste com menor
valor e que o processo de desencadeamento das colmos somente ocorreu no momento
da expansão da sexta lâmina produzida na rebrotação, que resultou num processo de
alongamento mais tardio. Oliveira et al. (2010) verificaram que o alongamento da haste
aumenta a produção de massa seca de forragem total, porem traz prejuízos para a
qualidade da forragem ofertada e, portanto, deve ser controlado. Outro fato observado
neste estudo é que o maior resíduo foi responsável pela maior TAlH, esta resposta pode
ser relacionada as alterações na estrutura do dossel promovidas por uma baixa
intensidade de pastejo, resultando no alongamento das hastes. A taxa de alongamento da
haste (TAlH) foi afetada pelo efeito dos ciclos de pastejo (P 0,05), sendo mais elevada
nos primeiros ciclos de pastejo e reduzindo ao longo das avaliações, para os pastos com
4,5 fol/perf e resíduo de 10 cm (Tabela 4), possivelmente a altura maior do resíduo,
tenha influenciado para esta condição. Contudo, os pastos com 4,5 fol/perf e 5 cm de
resíduo, 6,5 fol/perf e resíduo de 10 cm e 8,5 fol/perf com resíduo de 10 e 5 cm (Tabela
4, 5 e 6) apresentaram um comportamento diferente, onde a TAlH foi maior nos últimos
ciclos de pastejo. Este comportamento pode ser explicado pelo fato da planta modificar
sua estrutura com o objetivo de elevar o meristema apical e, desta maneira, evitar que
este seja decapitado pelo corte ou pastejo, evitando assim a morte do perfilho.
Ocorreu efeito da interação PD x RPP (P 0,05) sobre a taxa de senescência
foliar anterior (TSFa) (Tabela 3), no qual os pastos com 4,5 fol/perf, com resíduo de 10
e 5 cm, apresentaram menores valores. A TSFa pode ser usada como indicativo da
intensidade de pastejo adotada. Intensidades de pastejo alta tendem a diminuir a TSFa,
pois, com menor resíduo pós-pastejo, uma menor quantidade de folhas iniciará o
processo de senescência devido ao menor sombreamento que a forragem remanescente
promove, o que favorece a senescência. Contudo, não houve efeito do resíduo pós-
pastejo sobre a TSFa (P>0,05). De acordo com Cutrim Junior et al. (2010) frequências
de pastejo baixa promovem um aumento na TSFa de senescência, pois as primeiras
folhas produzidas durante o PD entram em senescência e serão rejeitadas pelo animal,
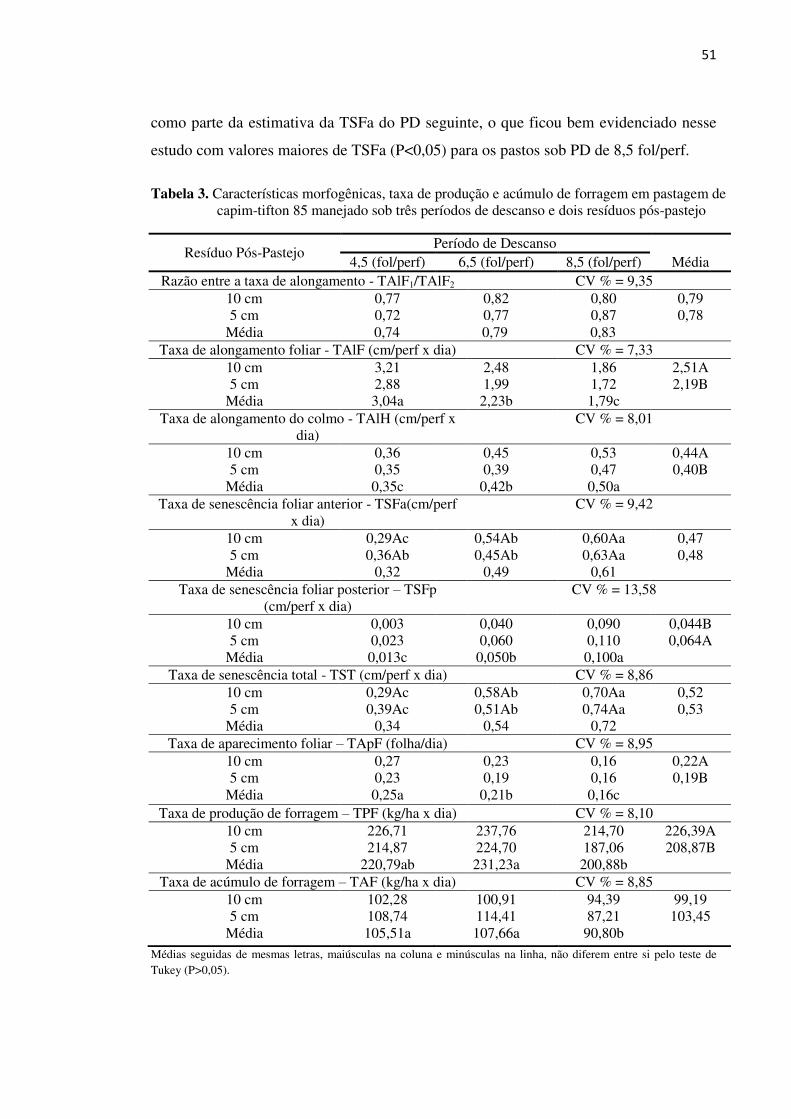
51
como parte da estimativa da TSFa do PD seguinte, o que ficou bem evidenciado nesse
estudo com valores maiores de TSFa (P<0,05) para os pastos sob PD de 8,5 fol/perf.
Tabela 3. Características morfogênicas, taxa de produção e acúmulo de forragem em pastagem de
capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de mesmas letras, maiúsculas na coluna e minúsculas na linha, não diferem entre si pelo teste de Tukey (P>0,05).
Resíduo Pós-Pastejo Período de Descanso
4,5 (fol/perf) 6,5 (fol/perf) 8,5 (fol/perf) Média Razão entre a taxa de alongamento - TAlF1/TAlF2 CV % = 9,35
10 cm 0,77 0,82 0,80 0,79 5 cm 0,72 0,77 0,87 0,78
Média 0,74 0,79 0,83 Taxa de alongamento foliar - TAlF (cm/perf x dia) CV % = 7,33
10 cm 3,21 2,48 1,86 2,51A 5 cm 2,88 1,99 1,72 2,19B
Média 3,04a 2,23b 1,79c Taxa de alongamento do colmo - TAlH (cm/perf x
dia) CV % = 8,01
10 cm 0,36 0,45 0,53 0,44A 5 cm 0,35 0,39 0,47 0,40B
Média 0,35c 0,42b 0,50a Taxa de senescência foliar anterior - TSFa(cm/perf
x dia) CV % = 9,42
10 cm 0,29Ac 0,54Ab 0,60Aa 0,47 5 cm 0,36Ab 0,45Ab 0,63Aa 0,48
Média 0,32 0,49 0,61 Taxa de senescência foliar posterior – TSFp
(cm/perf x dia) CV % = 13,58
10 cm 0,003 0,040 0,090 0,044B 5 cm 0,023 0,060 0,110 0,064A
Média 0,013c 0,050b 0,100a Taxa de senescência total - TST (cm/perf x dia) CV % = 8,86
10 cm 0,29Ac 0,58Ab 0,70Aa 0,52 5 cm 0,39Ac 0,51Ab 0,74Aa 0,53
Média 0,34 0,54 0,72 Taxa de aparecimento foliar – TApF (folha/dia) CV % = 8,95
10 cm 0,27 0,23 0,16 0,22A 5 cm 0,23 0,19 0,16 0,19B
Média 0,25a 0,21b 0,16c Taxa de produção de forragem – TPF (kg/ha x dia) CV % = 8,10
10 cm 226,71 237,76 214,70 226,39A 5 cm 214,87 224,70 187,06 208,87B
Média 220,79ab 231,23a 200,88b Taxa de acúmulo de forragem – TAF (kg/ha x dia) CV % = 8,85
10 cm 102,28 100,91 94,39 99,19 5 cm 108,74 114,41 87,21 103,45
Média 105,51a 107,66a 90,80b

52
A taxa de senescência foliar posterior (TSFp) apresentou efeito da PD x RPP
(P 0,05) e do RPP (P<0,05). Os valores verificados para os pastos sob PD de 4,5; 6,5 e
8,5 fol/perf foram de 0,01; 0,05 e 0,10 cm/perfilho x dia, respectivamente, e para os
RPP de 10 e 5 cm foram de 0,04 e 0,06 cm/perfilho x dia, respectivamente (Tabela 3).
Os maiores valores para esta variável são observados nos pastos com PD de 8,5 fol/perf,
resultando em grande perda de massa de forragem, diminuindo a eficiência de seu uso,
sendo prejudicial ao consumo e desempenho dos animais. Este resultado era esperado,
pois, a primeira folha formada na rebrotação só entra em senescência quando a quarta
folha está quase completamente expandida. Estes valores elevados para TSFp nos
pastos mantidos sob maior PD, também foram verificados por Cândido et al. (2006) em
capim-tânzania.
A TST foi afetada pela interação PD x RPP (P 0,05) (Tabela 3), apresentando
maiores valores para os pastos com 8,5 fol/perf e resíduos de 10 e 5 cm. Esse resultado
mostra uma elevação da TST prolongamento do PD, fato bem documentado (HUNT,
1965; CÂNDIDO et al., 2005), mas mostra um resultado inesperado em relação ao RPP,
uma vez que, o esperado seria de haver maior senescência nos pastos mantidos com
maior RPP (PARSONS et al., 1983; MAZZANTI et al., 1994). Provavelmente o maior
PD, ocorrido nos pastos manejados com menor RPP tenha proporcionado um dossel
com maior quantidade de folhas. Com isso, o maior sombreamento dessas na base do
perfilho acarreta uma maior perda de forragem por senescência. Com maior PD, a
planta deva ter ultrapassado seu ponto ótimo de uso, o que acarreta naturalmente uma
senescência das folhas basais. Isso demonstra a efetividade da adoção de PDs curtos em
minimizar as perdas de forragem por senescência (CUTRIM JUNIOR et al., 2010).
Quando se observa a TSFa e TSFT, em função dos ciclos, verifica-se que ambas
as taxas se elevaram nos pastos sob o PD 8,5 fol/perf e resíduos de 10 e 5 cm (Tabela
6), esta elevação pode ser atribuída a uma tendência do maior PD nos dois resíduos pós-
pastejo, proporcionando senescência das folhas que não foram pastejadas no ciclo
anterior, visto que a TSFa é afetada principalmente pela condição residual, e pode ser
também influenciada pela duração do período de descanso anterior ao último pastejo.
Para os pastos sob PD de 4,5 e 6,5 fol/perf e resíduos de 10 e 5 cm, as taxas se elevaram
do início ao meio do período experimental, com posterior queda ao final do mesmo,
essa redução podendo ser justificada pelo pastejo mais intenso, resultando em menor

53
tamanho residual das lâminas, que pode ter retardado o processo de senescência das
mesmas. Para a TSFp, observou-se efeito apenas para os pastos com PD de 4,5 fol/per e
resíduo de 5 cm e 8,5 fol/perf e resíduo de 10 cm, onde houve elevação desta taxa ao
final das avaliações dos ciclos de pastejo.
Os valores médios de taxa de aparecimento foliar (TApF) são verificados na
Tabela 3. Houve efeito do PD (P 0,05) e do RPP (P 0,05), para esta característica. Os
pastos com PD de 4,5 fol/perf e altura residual de 10 e 5 cm apresentaram as maiores
TApF, com valores de 0,27 e 0,23 folha/dia. O fato que pode ter elevado a TApF nos
pastos com PD de 4,5 fol/perf e resíduo de 10 e 5 cm pode ter sido o esgotamento maior
das reservas da planta, que atrasaram sua rebrotação, demandando maior tempo para a
completa expansão de cada folha. Observa-se ainda que o prolongamento do PD
promoveu redução da TApF, como foi observado para os pastos mantidos com 6,5 e 8,5
fol/perf, uma vez que esta redução é associada ao maior comprimento da bainha em
níveis de inserção cada vez mais altos, ou seja, a folha percorre um maior trajeto entre
seu ponto de conexão com o meristema e a extremidade do pseudocolmo formado pelas
bainhas das folhas mais velhas (ANDRADE et al., 2008). Oliveira et al. (2000)
estudando a morfogênese de capim-tifton 85 em diferentes idades de rebrotação,
concluíram que ocorre redução na TApF individuais entre as idades de 14 e 70 dias.
A taxa de produção da forragem (TPF) foi influenciada pelo PD (P 0,05) e do
RPP (P 0,05) (Tabela 3), esta variável é influenciada pela TApF, fato este que pode ter
sido responsável por maiores valores nos pastos que apresentaram maiores valores de
TApF, que pode ser observado nos pastos com 4,5 e 6,5 fol/perf. Paciullo et al. (2005)
verificaram TPF de 53,0 a 125,7 kg/ ha/dia de MS para o capim-coastcross. Não houve
efeito dos ciclos de pastejo, sobre a taxa de produção de forragem (TPF), para os pastos
com PD de 4,5 fol/perf e resíduo de 10 cm, 6,5 fol/perf e resíduo de 5 cm e 8,5 fol/perf
e resíduo de 10 cm (Dados não apresentados). A TPF foi reduzida ao longo dos ciclos
de pastejo para os pastos com PD de 6,5 fol/perf e resíduo de 10 e 8,5 fol/perf e resíduo
de 5 cm (Tabelas 5 e 6), no entanto observou-se elevação desta taxa para os pastos com
PD de 6,5 fol/perf e resíduo de 5 cm.
A taxa de acúmulo de forragem, determinada pela diferença entre os processos
de crescimento e senescência (HODGSON, 1990), é característica importante para o
manejo das pastagens, pois permite que o equilíbrio entre oferta e demanda de forragem

54
seja alcançado sem prejudicar o desempenho dos animais. A taxa de acúmulo de
forragem (TAF) foi influenciada apenas pelo PD (P<0,05). Os pastos sob PD de 4,5 e
6,5 fol/perf apresentaram os maiores valores de TAF (105, 51 e 107,66 kg MS/ha x dia)
respectivamente, decrescentes com o aumento do PD, de forma que chegaram a 90,08
kg MS/ha x dia para os pastos manejados sob PD de 8,5 fol/perf (Tabela 3). Tal fato
ocorreu em virtude da grande influência da TAlF sobre a TAF, em que essa se
apresentou superior para nos pastos manejados com menor PD, o que relata um
incremento apenas de lâmina foliar, haja vista que em menores interceptações ainda não
há alongamento do colmo. Na literatura são encontradas taxas de acúmulo de forragem
de espécies de Cynodon variando de 15,7 a 105,0 kg/ ha/dia de MS (CARNEVALLI &
SILVA, 1999; FAGUNDES et al., 1999; CARVALHO et al., 2001; PINTO et al.,
2001). A taxa de acúmulo de forragem (TAF) foi reduzida nos últimos ciclos de pastejo
para os pastos com PD de 4,5 e 6,5 fol/perf e resíduos de 5 cm, período que corresponde
a menor TAlF para estes pastos.

55
Tabela 4. Efeito dos ciclos de pastejo sobre as características morfogênicas em pastagem de capim-tifton 85 sob três períodos de descanso e dois resíduos pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
Período de descanso 4,5 e resíduo pós-pastejo 10 cm TAlF 3,80a 3,40ab 2,72bc 3,34ab 3,47ab 3,37ab 3,12abc 3,11abc 3,45ab 2,34c 3,21 10,54
TAlC 0,66a 0,53ab 0,60ab 0,50bc 0,59ab 0,59ab 0,46bc 0,47bc 0,39bc 0,48bc 0,52 11,38
TApF 0,28a 0,22b 0,29a 0,26ab 0,29a 0,28a 0,26ab 0,28a 0,28a 0,25ab 0,26 8,44
TSFa 0,23bc 0,52ab 0,31abc 0,31abc 0,27abc 0,55a 0,13c 0,14c 0,21bc 0,23abc 0,29 44,75
TST 0,26abc 0,52a 0,31abc 0,31abc 0,27abc 0,56a 0,14c 0,14c 0,21bc 0,23bc 0,32 44,49
Período de descanso 4,5 e resíduo pós-pastejo 5 cm
TAlF 3,20a 3,05a 2,79ab 2,97a 3,22a 2,98a 3,09a 2,52bc 2,10 - 2,88 6,5
TAlC 0,39bc 0,51abc 0,57ab 0,36bc 0,57ab 0,67a 0,50abc 0,31c 0,35bc - 0,47 20,84
TApF 0,24a 0,20ab 0,23a 0,23a 0,23a 0,23a 0,23a 0,23a 0,24a - 0,22 6,24
TSFa 0,32bc 0,25bc 0,70a 0,34bc 0,51ab 0,73a 0,15c 0,14c 0,13c - 0,36 38,40
TSFp 0,005c 0,014abc 0,009bc 0,034abc 0,013abc 0,013abc 0,030ab 0,042ab 0,047a - 0,023 61,90
TST 0,33dc 0,27dc 0,70ab 0,37bcd 0,53abc 0,77a 0,20dc 0,17d 0,14d - 0,38 37,46
TAF 114,05abc 119,62ab 136,29a 114,08abc 136,20a 115,06abc 66,60c 92,58abc 83,88bc - 108,70 19,15
Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

56
Tabela 5. Efeito dos ciclos de pastejo sobre as características morfogênicas em pastagem de capim-tifton 85 sob três períodos de descanso e dois resíduos pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
Período de descanso 6,5 e resíduo pós-pastejo 10 cm TAlC 0,38ab 0,54ab 0,55a 0,35b 0,35b 0,50ab 0,43ab 0,51ab - - 0,45 17,39
TSFa 0,71ab 0,35cd 0,62ab 0,47bc 0,69ab 0,85a 0,47bc 0,15d - - 0,53 23,56
TST 0,76ab 0,40cd 0,66ab 0,54bc 0,71ab 0,88a 0,52bc 0,23d - - 0,58 20,69
TPF 207,22b 224,73ab 231,56ab 207,15b 267,22ab 223,67 340,34a 191,71b - - 236,7 22,64
Período de descanso 6,5 e resíduo pós-pastejo 5 cm
TAlF 2,11a 2,03a 2,03a 2,41a 2,32a 1,53b 1,53b - - - 1,99 10,45
TApF 0,18ab 0,16b 0,17ab 0,21a 0,21a 0,21a 0,19ab - - - 0,19 8,79
TSFa 0,34ab 0,23b 0,61a 0,41ab 0,42ab 0,65a 0,46ab - - - 0,44 30,09
TST 0,39ab 0,30b 0,69a 0,47ab 0,47ab 0,71a 0,53ab - - - 0,57 28,58
TAF 131,96ab 156,97a 92,97b 130,47ab 107,63ab 78,14b 102,63ab - - - 114,39 20,94
TPF 214,39ab 211,28ab 210,73ab 176,76b 246,07ab 230,40ab 292,29a - - - 226,06 21,66
Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

57
Tabela 6. Efeito dos ciclos de pastejo sobre as características morfogênicas em pastagem de capim-tifton 85 sob três períodos de descanso e dois resíduos pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
Período de descanso 8,5 e resíduo pós-pastejo 10 cm TAlF 1,88ab 1,79ab 1,82ab 2,41a 1,82ab 1,47b - - - - 1,86 15,26
TApF 0,16abc 0,15bc 0,17ab 0,14c 0,18a 0,18a - - - - 0,16 6,77
TAlC 0,30c 0,30c 0,30c 0,37bc 0,40ab 0,46a - - - - 0,35 10,56
TSFa 0,43c 0,49bc 0,68b 0,52bc 0,57bc 0,93a - - - - 0,60 17,24
TSFp 0,09ab 0,12a 0,10ab 0,10ab 0,06b 0,08ab - - - - 0,09 25,54
TST 0,53c 0,61bc 0,79b 0,63bc 0,63bc 1,02a - - - - 0,70 13,35
Período de descanso 8,5 e resíduo pós-pastejo 5 cm
TAlC 0,25c 0,29bc 0,32abc 0,42ab 0,46a - - - - - 0,34 17,24
TApF 0,15bc 0,14c 0,16abc 0,17ab 0,18a - - - - - 0,16 8,65
TSFa 0,44c 0,61abc 0,83a 0,55bc 0,71ab - - - - - 0,62 18,49
TST 0,58b 0,74ab 0,94a 0,66b 0,78ab - - - - - 0,74 16,46
TPF 101,72c 220,47a 142,04bc 265,70a 205,42ab - - - - - 187,07 17,71
Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

58
CONCLUSÕES
As características morfogênicas do capim-tifton 85 são afetadas pelos períodos
de descanso, pelos resíduos pós-pastejo, e também pelos dos ciclos de pastejo.
A razão entre as taxas alongamento de lâminas foliares da primeira e da segunda
folha produzida na rebrotação de cada perfilho, foi a única característica morfogênica,
não influenciada pelos, períodos de descanso, resíduo pós-pastejo e ciclos de pastejo.
Períodos de descanso superiores ao necessário para formação de 6,5 fol/perf
tornam todas as características morfogênicas estudadas indesejáveis do ponto de vista
forrageiro.
REFERÊNCIAS
ANDRADE, A. C.; RODRIGUES, B. H. N.; MACHADO, D. M. et al. Características Morfológicas da Canarana-Ereta-Lisa (Echinochloa pyramidalis Lam.) em Diferentes Idades de Rebrotação. Revista Ciência e Produção Animal. v. 10, n. 1, p. 37- 49, 2008. ANUALPEC: Anuário da pecuária brasileira. FNP CONSULTORIA & COMÉRCIO. São Paulo, 2008. 380p. BARBOSA, R.A.; NASCIMENTO JÚNIOR, D.; EUCLIDES, V. P. B. et al. Capim-tanzânia submetido a combinações entre intensidade e freqüência de pastejo. Pesquisa Agropecuária Brasileira, Brasília, v.42, n.3, p.329-340, 2007. CÂNDIDO, M. J. D.; Da SILVA, R. G.; NEIVA, J. N. M. et al. Fluxo de biomassa em capim-tanzânia pastejado por ovinos sob três períodos de descanso. Revista Brasileira de Zootecnia, v.35, n.6, p.2234-2242, 2006. CÂNDIDO, M.J.D.; GOMIDE, C.A.M.; ALEXANDRINO, E. et al. Morfofisiologia do dossel de Panicum maximum cv. Mombaça sob lotação intermitente com três períodos de descanso. Revista Brasileira de Zootecnia, v.34, n.2, p.338-347, 2005b. CARNEVALLI, R. A. Dinâmica da rebrotação de pastos de capim Mombaça submetidos a regimes de desfolhação intermitente. Piracicaba, 2003. 136 p. Tese (Doutorado) – Escola Superior de Agricultura Luiz de Queiroz. Universidade de São Paulo. CARNEVALLI, R.A.; SILVA, S.C. da. Validação de técnicas experimentais para avaliação de características agronômicas e ecológicas de pastagens de Cynodon
dactylon cv. Coast-cross-1. Scientia Agricola, v. 56, n. 2, p. 489-499, 1999.

59
CARVALHO, P.C.F.; RIBEIRO FILHO, H.M.N.; POLI, C.H.E.C.; MORAES, A.; DELAGARDE, R. Importância da estrutura da pastagem na ingestão e seleção de dietas pelo animal em pastejo. In: A PRODUÇÃO ANIMAL NA VISÃO DOS BRASILEIROS, Piracicaba, 2001, Anais... Piracicaba: FEALQ, 2001. p. 853-871. CUTRIM JÚNIOR, J. A. A.; CÂNDIDO, M. J. D.; VALENTE, B. S. M. et al. Fluxo de biomassa em capim-tanzânia sob três frequências de desfolhação e dois resíduos pós-pastejo. Revista Brasileira Saúde Produção Animal, v.11, n.3, p.618-629, 2010. DAVIES, A. Tissue turnover in the sward. In: DAVIES, A.; BAKER, R.D.; GRANT, S.A. et al. (Eds.). Sward measurement handbook. London: British Grassland Society, 1993. p.183-216. EMBRAPA, Centro Nacional de Pesquisa de Solos. Sistema Brasileiro de Classificação de Solos. Brasília: EMBRAPA/ CNPS, 1999. 412p. il. (Documentos, 5). FAGUNDES, J.L.; SILVA, S.C. da; PEDREIRA, C.G.S. et al. Índice de área foliar, interceptação luminosa e acúmulo de forragem em pastagens de Cynodon sp. Sob diferentes intensidades de pastejo. Scientia Agricola, v. 56, n. 4, p. 1141-1150, 1999. GRANT, S.A.; MARRIOT, C.A. Detailed studies of grazed sward-tecniques and conclusions. Journal Agriculture Science, v. 122, n. 1, p. 1-6, 1994. HODGSON, J. Grazing management: science into pratice. Unit Kingdom: Longman Scientific and Technical, Logman Group, 1990. 203 p. HORST, G.L., NELSON, C.J.; ASAY, K.H. Relationship of leaves elongation to forage yield of tall fescue genotypes. Crop Sci., v.18, p.715-719, 1978. HUNT, L. A. Some implications of death and decay in pasture production. Journal of the British Grassland Society, v.20, p.27-31, 1965. MAZZANTI, A.; LEMAIRE, G.; GASTAL, F. The effect of nitrogen fertilization upon herbage production of tall fescue sward continuously grazed with sheep. 1- Herbage growth dinamics. Grass and forage Science, v. 49, n. 2, p. 111-120, 1994. NASCIMENTO Jr., D.; GARCEZ NETO, A.F.; BARBOSA, R.A. et al. Fundamentos para o manejo de pastagens: evolução e atualidade. In: OBEID, J.A.; PEREIRA, O.G.; FONSECA, D.M.; NASCIMENTO Jr., D. (Eds.). SIMPÓSIO SOBRE MANEJO ESTRATÉGICO DA PASTAGEM, 1., 2002, Viçosa. Anais... Viçosa: UFV, 2002. p.149-196. NABINGER, C., PONTES, L.S. Morfogênese de plantas forrageiras e estrutura do pasto. In: SIMPÓSIO SOBRE A PRODUÇÃO ANIMAL NA VISÃO DOS BRASILEIROS /REUNIÃO DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38, 2001, Piracicaba. Anais... MATTOS, W.R.S. et al. (EE.). Piracicaba: FEALQ, 2001. p. 755-771.

60
OLIVEIRA, L. E. V.; CAVALCANTE, A. C.R.; CUTRIM JÚNIOR, J. A. et al. Efeito de diferentes intensidades de manejo sobre componentes do fluxo de biomassa do capim-tanzânia (Panicum maximum cv. Tanzânia). 47a Reunião Anual da Sociedade Brasileira de Zootecnia Salvador, BA – UFBA, Anais... 27 a 30 de julho de 2010 OLIVEIRA, M.A; PEREIRA, O.G.; MARTINEZ Y HUAMAN, C.A. et al. Características morfogênicas e estruturais do capim-bermuda „Tifton 85‟ (Cynodon
spp.) em diferentes idades de rebrota. Revista Brasileira de Zootecnia, v. 29, n. 6, p. 1939-1948, 2000. PACIULLO, D. S. C.; AROEIRA, L. J. M.; HEINEMANNI, A. B. Morfogênese, características estruturais e acúmulo de forragem em pastagem de Cynodon dactylon, em diferentes estações do ano. Ciência Animal Brasileira v. 6, n. 4, p. 233-241, 2005. PARSONS, A.J.; LEAVES, E.L.; COLLETT, B.; STILES, W. The physiology of grass production under grazing. 1. Characteristics of leaves and carropy photosynthesis of continuously grazed swards. Journal of Applieo Ecology, v. 20, p. 117-126, 1983. PINTO, L. F. M.; SILVA, S. C.; SBRISSIA, A. F.; CARVALHO, C. A. B. et al. Dinâmica do acúmulo de matéria seca em pastagens de Tifton 85 sob pastejo. Scientia Agricola, v. 58, n. 3, p. 439-447, 2001. PONTES, L. S.; NABINGER, C.; CARVALHO, P. C. F. et al. Variáveis morfogênicas e estruturais de Azevém Anual (Lolium multiflorum Lam.) manejado em diferentes alturas. Revista Brasileira de Zootecnia, v.32, n.4, p.814-820, 2003 SAS INSTITUTE. SAS system for Windows. Version9. 1. Cary: SAS Institute Inc. 2003. 2 CD – ROMs.

61
CAPÍTULO 4
____________________________________________________________
Componentes da biomassa e características estruturais do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo

62
Componentes da biomassa e características estruturais do capim-tifton 85 sob três períodos de descanso e dois resíduos pós-pastejo
RESUMO: Avaliaram-se as características estruturais do capim-tifton 85 sob três
períodos de descanso e dois resíduos pós-pastejo em diferentes ciclos de pastejo. Os
tratamentos corresponderam a combinações de três condições de pré-pastejo, período de
descanso (PD) 4,5; 6,5 e 8,5 (folhas/perfilho) e duas condições de pós-pastejo, resíduo
pós-pastejo de 10 e 5 cm, distribuídos em um delineamento experimental inteiramente
casualizado em arranjo fatorial 3x2, com quatro repetições (piquetes). Estimaram-se as
produções de massa seca de forragem total (MSFT), forragem morta (MSFM), forragem
verde (MSFV), lâmina foliar verde (MSLV) e colmo verde (MSCV), a altura (ALT) e o
número de folhas/perfilho (F/P); as relações folha/colmo (F/C) e material vivo/morto
(MV/MM) no pré e pós-pastejo; a densidade populacional de perfilhos (DPP).
Observou-se, aumento na MSFT, com o prolongamento do período de descanso (PD). O
aumento da MSFV deveu-se a elevações da MSLV e MSCV, principalmente nos pastos
sob PD de 8,5 fol/perf e resíduos de 10 e 5 cm. A menor MSFM foi observada nos
pastos com PD de 4,5 fol/perf e resíduos de 10 e 5 cm. Os pastos sob maior PD
apresentaram menor relação MV/MM. Houve aumento na altura do dossel com o
decorrer dos ciclos de pastejo. A produção de colmos ocasionou a redução da relação
F/C. Os pastos com o maior PD apresentaram maior número de folhas/perfilho (F/P).
Houve redução no número de perfilhos/m² com o prolongamento do PD. Os
componentes da biomassa residual e a relação MV/MM apresentaram comportamento
semelhante ao da condição de pré-pastejo. A relação (F/Cres) do pasto sob o PD 8,5
fol/perf foi inferior aos demais. Os componentes da biomassa no pré e pós-pastejo,
foram afetados pela interação PD x RPP e pelos ciclos de pastejo. O prolongamento do
PD compromete a estrutura do dossel. O resíduo pós-pastejo afetou a massa seca de
forragem total e a altura no pré-pastejo e massa seca de lâmina verde e altura no pós-
pastejo. O uso do capim-tifton 85 não deve exceder ao período de descanso de 6,5
fol/perf para evitar perdas de forragem por senescência, sendo possível utilizar uma
altura residual de 5 cm sem comprometer a perenidade da pastagem.
Termos para indexação: altura, perfilhos, massa seca de lâmina foliar verde, Cynodon
sp, relação lâmina foliar/haste

63
Structural components of the biomass and characteristics of the tifton 85 grass
managed under three periods of rest and two post-grazing residues
ABSTRACT: The structural characteristics of capim-tifton 85 under three periods of
rest and two residues after-pastejo in different cycles of pastejo had been evaluated. The
treatments had corresponded the combinations of three conditions of daily pay-pastejo,
period of rest (PD) 4,5; 6,5 and 8,5 (leaves/ tiller) and two conditions of after-pastejo,
residue after-pastejo of 10 and 5 cm, distributed in an experimental delineation entirely
casualizado in factorial arrangement 3x2, with four repetitions (poles). Total herbage
dry mass (MSFT), death herbage dry mass (MSFM), green herbage dry mass (MSFV),
green leaves blade dry mass (MSLV), green stem dry mass (MSCV), canopy height
(ALT); number of live leaves per tiller (F/P), leaves/stem (F/C) and live/death material
(MV/MM) ratios, pre- and post-grazing, as well as the tiller populational density (DPP)
pre-grazing were determined. It was observed, increase in the MSFT, with the
prolongation of the period of rest (PD). The increase of the MSFV had it rises of the
MSLV and MSCV, mainly in the grass under 8,5 PD of leaves/ tiller and residues of 10
and 5 cm minor MSFM was observed in the grass with 4,5 PD of leaves/ tiller and
residues of 10 and 5 cm. The grass under bigger PD had presented minor relation
MV/MM. It had increase in the height of the canopy with elapsing of the cycles of
pastejo. The production of stem caused the reduction of relation F/C. The grass with the
biggest PD had presented greater leaves number/tiller (F/P). with the prolongation of the
PD had reduction in the number of tiller. The components of the residual biomass and
relation MV/MM had presented similar behavior to the one of the daily pay-pastejo
condition. The relation (F/Cres) of the grass under 8,5 PD leaves/ tiller was inferior to
excessively. The components of the biomass in the daily pay and after-pastejo, had been
affected by interaction PD x RPP and the cycles of pastejo. The prolongation of the PD
compromises the structure of the canopy. The residue after-pastejo affected the dry
mass of total fodder plant and the height in the daily pay-pastejo and dry mass of green
blade and height in the one after-pastejo. The use of capim-tifton 85 does not have to
exceed to the period of rest of 6,5 leaves/ tiller to prevent losses of fodder plant for
senescência, being possible to use a residual height of 5 cm without compromising the
perenidade of the pasture.
Index-terms: height, tiller, green leaves blade dry mass, Cynodon sp, leaves/stem ratio

64
INTRODUÇÃO
A estrutura do dossel forrageiro é definida como a distribuição e o arranjo das
partes da planta, ou a quantidade e organização de seus componentes, dentro de sua
comunidade, sobre o solo (LACA & LEMAIRE, 2000). Características como altura da
planta, relação lâmina foliar/haste, taxa de crescimento, dinâmica de perfilhamento,
remoção de meristemas apicais, expansão foliar, entre outras, apresentam uma relação
direta com a produtividade e a qualidade da forragem em oferta, além de subsidiarem a
adoção de práticas de manejo mais adequadas. As plantas forrageiras variam,
naturalmente, quanto à estrutura da vegetação, em função da arquitetura da planta, do
estádio de desenvolvimento, das condições edafoclimáticas e do manejo que lhes é
imposto.
Sob pastejo com lotação intermitente, a duração do intervalo de pastejos
sucessivos – período de descanso – é o que determina a recuperação do índice de área
foliar do dossel (IAF) e, conseqüentemente, maximiza a produção de forragem em cada
ciclo de pastejo (GRANT et al., 1988). O resíduo na pastagem exerce efeito importante
na qualidade da forragem, pois, segundo Forbes e Coleman (1993), a estrutura do pasto
é modificada com a participação de seus componentes folha, haste e material morto. De
acordo com Janusckiewicz et al. (2010) o sistema de pastejo que combina períodos de
descanso e alturas de resíduo pós-pastejo adequados torna-se imprescindível para que
ocorra rebrota mais efetiva da planta e, conseqüentemente, melhor expressão do seu
potencial genético. A desfolhação interfere diretamente na estrutura do pasto e no
acúmulo de matéria seca, bem como no valor nutritivo da forragem.
Nos últimos anos as plantas do gênero Cynodon despertaram grande interesse e
ganharam popularidade devido à sua facilidade de cultivo, alta produção de forragem
(20 a 25 t MS. ha/ano) e considerável valor nutritivo (11 a 13% de proteína bruta e 58 a
65% de digestibilidade) (PEDREIRA, 1996). Apresenta também alta taxa de
crescimento, resistência ao pastejo e elevada capacidade de suporte (LIMA & VILELA,
2005). Misley e Pate (1996) destacam ainda como vantagem o rápido estabelecimento,
ressaltando, porém, que a digestibilidade varia entre os cultivares. O cultivar Tifton 85
(Cynodon sp.) é o resultado do cruzamento do Tifton 68 com um acesso proveniente da
África do Sul. Apresenta colmos grandes, folhas finas de cor verde escura e rizomas

65
bem desenvolvidos. Fagundes et al. (1999) que ao avaliarem a produção de forragem de
capins do gênero Cynodon sp., incluindo o Tifton 85, observaram produções que
variaram de 3.740 a 8.110 kg de matéria seca ha-1 em quatro alturas de manejo (5, 10,
15 e 20 cm) e 30 dias de intervalos de cortes.
Considerando o que foi relatado acima, esta pesquisa foi conduzida com o
objetivo de avaliar os componentes da biomassa e as características estruturais do
capim-tifton 85 manejado sob três períodos de descanso 4,5; 6,5 e 8,5 (folhas/perfilho) e
dois resíduos pós-pastejo 10 e 5 cm.
MATERIAL E MÉTODOS
A pesquisa foi desenvolvida no Setor de Produção de Ovinos do Campo
Experimental de Bebedouro, pertencente à Embrapa Semiárido, localizado no
Município de Petrolina-Pernambuco. Durante o mês de outubro foi efetuada a divisão
da área em piquetes e ajuste da metodologia de trabalho. A coleta de dados ocorreu
entre os meses de novembro de 2008 a junho de 2009, totalizando nove meses. O clima
da região é classificado como quente e seco, sendo caracterizada por duas estações bem
definidas, uma chuvosa, de janeiro a maio, e outra seca, de junho a dezembro. Os dados
sobre as condições de temperaturas máxima e mínima do ar e precipitação pluvial
durante o período experimental foram obtidos da Estação Agrometeorologia de
Bebedouro, Petrolina-Pernambuco e são apresentados na Tabela 1.
Tabela 1. Dados climáticos observados durante o período experimental
Mês Precipitação (mm) Temperatura (ºC)
Máxima Mínima Média
outubro/2008 0,0 33,7 21,7 27,7 novembro/2008 0,0 34,8 23,2 28,7 dezembro/2008 54,6 33,7 22,6 27,1 janeiro/2009 45,7 34,3 23,0 27,5 fevereiro/2009 127,0 32,4 22,1 26,1 março/2009 152,9 33,0 22,2 26,3 abril/2009 223,6 31,1 22,3 25,5 maio/2009 85,4 28,8 21,2 24,2 Junho/2009 14,5 29,7 19,5 23,6 Dados obtidos da Estação Agrometeorologia de Bebedouro, Embrapa Semiárido, Petrolina-Pernambuco.

66
Foram avaliados três períodos de descanso 4,5; 6,5 e 8,5 (número de
folhas/perfilho) e dois resíduos pós-pastejo 5 e 10 cm, em um delineamento
experimental inteiramente casualizado em arranjo fatorial 3x2 com quatro repetições
(piquetes). A área experimental foi composta de 0,82 ha de pastagem cultivada com
capim-tifton 85 irrigada por aspersão com canhões com a distribuição da água durante o
dia. Utilizou-se o método de pastejo sob lotação intermitente, com lotação variável,
mantendo-se animais prova nos piquetes e também animais reguladores para ajuste da
taxa de lotação, segundo a técnica da taxa de lotação variável. A pastagem foi
subdivida, com tela campestre, em 37 piquetes de 180 m2 cada, destes piquetes 24 foram
considerados amostrais. Foram utilizados 30 ovinos de prova e um número variável de
animais de equilíbrio, da raça Santa Inês, machos castrados, com peso corporal inicial
(PC) em torno de ± 20 kg, os quais foram previamente identificados, vermifugados e,
em grupos de cinco, distribuídos por sorteio em seis tratamentos. Foram efetuadas
adubações de reposição com nitrogênio, em cada piquete, no momento da saída dos
animais, nas seguintes doses: 0,971; 1,110; 1,320; 1,475; 1,666 e 1,800 g, de acordo
com o período de descanso.
O solo da área experimental é classificado como Podzólico Amarelo Eutrófico
Latossólico (EMBRAPA, 1999), teve suas características químicas avaliadas (Tabela 2)
em outubro de 2008. A área foi adubada em função das necessidades da análise do solo.
A adubação com fósforo e potássio foi realizada a lanço e em cobertura, na forma de
superfosfato simples e cloreto de potássio, respectivamente, em 02 de novembro de
2008 em uma aplicação com 120 kg/ha de P2O5 e 100 kg/ ha de K2O. A adubação
nitrogenada (600 kg de N/ha) foi fracionada em aplicações, ao dos ciclos de pastejo,
após a saída dos animais dos piquetes, na forma de uréia, de acordo com os períodos de
descanso.
Tabela 2. Características químicas do solo da área experimental em duas profundidades
Profundidade MO pH:H2O P K Al Ca Mg H + Al CTC SB -----(cm)---- g/Kg-1 1: 2,5 mg/dm3 ----Cmolc dm-3---- %
0 – 5 24,14 6,18 41,51 0,21 0,05 2,45 0,94 1,83 5,48 66,41 5 - 20 7,10 6,09 22,67 0,14 0,05 1,33 0,59 1,16 3,25 63,51

67
Ao final de cada período de descanso - anteriormente à entrada dos animais, e
também imediatamente após o período de pastejo, foram feitas as seguintes avaliações
nos piquetes pertinentes, correspondendo nas condições de pré e pós-pastejo (Figuras 1;
2; 3 e 4).
Figuras 1; 2; 3 e 4. (1) Condição de pré-pastejo no pasto manejado sob PD de 8,5 fol/perf; (2)
2º dia de pastejo; (3) 3º dia de pastejo e (4) condição residual de 10 cm.
a) A altura do dossel em pré e pós-pastejo foi determinada com o auxílio de uma
bengala (cano PVC de ½) graduada em centímetros, medindo-se a planta desde o nível
do solo até a curvatura da última folha completamente expandida. Foram tomadas 30
medidas de altura por piquete para o pré e pós-pastejo.
b) Quantificação dos componentes da biomassa, por meio da colheita da massa
de forragem a 5 cm do solo em cada um dos pontos, usando molduras de 0,5 x 0,5 m.
As amostras foram levadas ao laboratório, sendo então separados o material vivo do
material morto e em seguida as lâminas foliares dos colmos do material vivo, os quais
foram secos em estufa de ventilação forçada a 55º C e pesados, para a quantificação dos
componentes da biomassa: massas secas de forragem total (MSFT), de forragem verde
(MSFV), de forragem morta (MSFM), de lâmina foliar verde (MSLV), de colmo verde
(MSCV), além das relações material vivo/material morto e relação lâmina foliar/haste.
1 2
3 4

68
Além das medidas anteriores, foi avaliado em cada um dos piquetes, apenas no
pré-pastejo o número de novas folhas por perfilho (F/P), essa avaliação foi realizada
pela contagem de todas as folhas produzidas na rebrotação, atribuindo o valor 0,5
àquelas folhas cuja lígula ainda não estivesse exposta; após cinco dias do pastejo a
densidade populacional de perfilhos (DPP), foi avaliada pela contagem do número de
perfilhos vivos presentes na moldura de 0,5 x 0,5 m, antes de ser feito o corte da
biomassa.
Os dados foram analisados por meio de análise de variância e teste de
comparação de médias. Para comparar o efeito dos tratamentos, foi efetuada análise de
variância do efeito dos períodos de descanso e dos resíduos pós-pastejo, na média de
todos os ciclos, no fatorial completo, de modo que fosse desdobrada a interação quando
significativa ao nível de 5% de probabilidade. As médias foram comparadas por meio
do teste Tukey de, ao nível de 5% de probabilidade. Para avaliar o efeito dos ciclos
sucessivos de pastejo, foi efetuada análise de variância dos ciclos dentro de cada um dos
seis tratamentos. As médias dos ciclos foram comparadas por meio do teste de Tukey,
ao nível de 5% de probabilidade. Como ferramenta de auxílio às análises estatísticas,
utilizou-se o procedimento GLM do programa estatístico SAS (SAS INSTITUTE,
2003).
RESULTADOS E DISCUSSÃO
Componentes da biomassa e características estruturais no pré-pastejo
A massa seca de forragem total (MSFT) foi influenciada pelo PD (P 0,05) e o
RPP (P 0,05), com valores de 3685,89; 5014,11 e 6403,17 kg/ha para o PD de 4,5; 6,5
e 8,5 fol/perf, respectivamente e 4785,49 e 5283,29 kg/ha para o RPP de 10 e 5 cm
(Tabela 3). Estes valores de MSFT encontram-se próximos aos relatados por Souza et
al. (2010) para a mesma espécie deste estudo. Fagundes et al. (1999) observaram
produções que variaram de 3.740 a 8.110 kg de matéria seca/ha em quatro alturas de
manejo (5, 10, 15 e 20 cm) e 30 dias de intervalos de cortes. Observa-se, uma
ascendência na MSFT, com o aumento do PD, provavelmente devido o aumento na

69
produção de colmos (GOMIDE et al., 2007) relataram aumento da biomassa forrageira
com o prolongamento do PD, assim como o observado neste estudo. De acordo com
Barbosa et al. (2007) a elevação da MSFT nos pastos sob PD mais longos não implica
em maior produtividade, uma vez que foram obtidos à custas de longos PD, reduzindo,
portanto, o número de ciclos de pastejo, promovendo alterações na estrutura do dossel
em função do elevado alongamento dos colmos e a quantidade de material morto.
Barbosa et al. (2007) avaliaram as características estruturais do capim-tanzânia
submetido a diferentes combinações de intensidades e freqüências de pastejo e
concluíram que em relação aos resíduos, o maior acúmulo de forragem foi registrado
nos tratamentos de 25 cm de resíduo, em relação àqueles de 50 cm. Comportamento
similar para o acúmulo de forragem foi observado no presente estudo. Não houve efeito
dos ciclos de pastejo (P 0,05) sobre a MSFT (Dados não apresentados).
Houve efeito da interação PD x RPP (P 0,05) sobre a massa seca de forragem
(MSFV) (Tabela 3). Sendo menor para os pastos manejados sob PD de 4,5 fol/perf e
ambos os resíduos e maior para os pastos manejados sob PD de 8,5 fol/perf e resíduo de
5 cm. Isso ocorreu devido a menor produção de forragem verde e um menor incremento
de colmos na MSFV para os pastos com PD de 4,5 fol/perf, ocorrendo o inverso para os
pastos com PD de 6,5 e 8,5 fol/perf. Comportamento semelhante foi observado por
Cândido et al. (2006) manejando o capim-tânzania com diferentes PDs. Os valores de
MSFV estão acima dos preconizados por Minson (1990) onde valores de 2000 kg/ha
MSFV são adequados para que não ocorra redução na ingestão, em decorrência da
diminuição do tamanho do bocado e aumento do tempo de pastejo. O efeito dos ciclos
de pastejo (P 0,05) sobre a MSFV foi observado, apenas para os pastos com PD de 6,5
fol/perf e resíduo de 5 cm (Tabela 4), apresentando valores homogêneos ao longo do
período experimental a partir do segundo ciclo de pastejo.
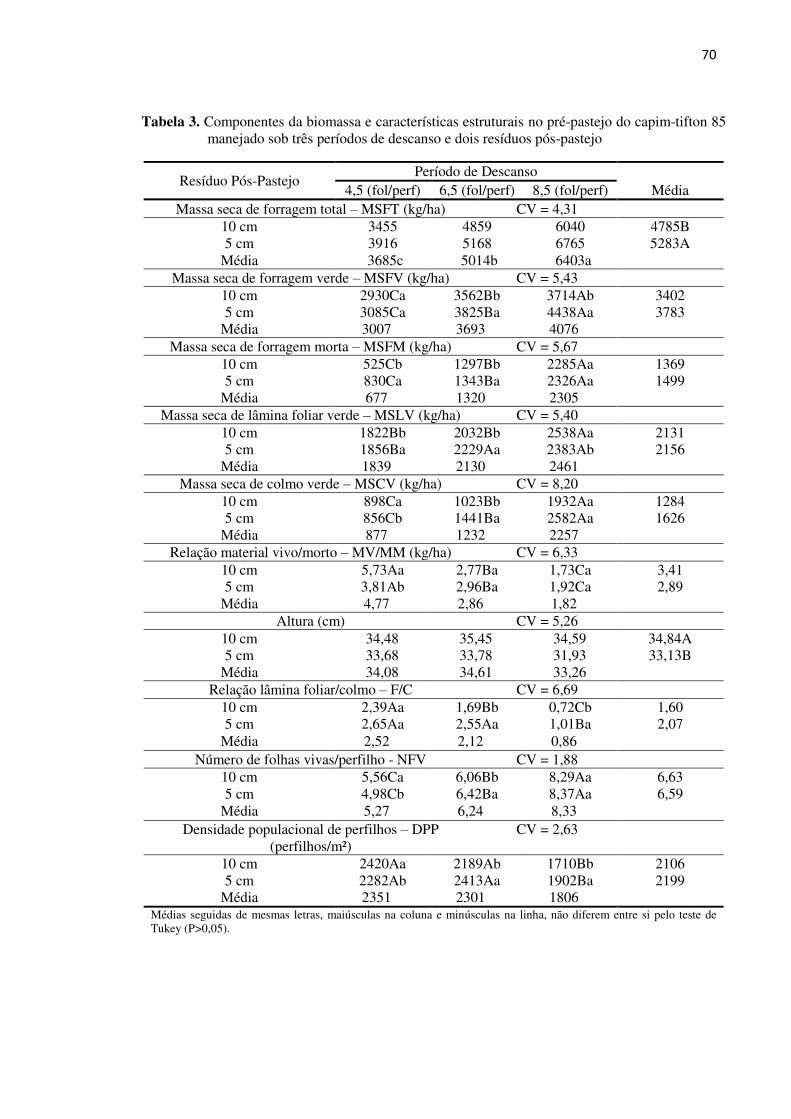
70
Tabela 3. Componentes da biomassa e características estruturais no pré-pastejo do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Resíduo Pós-Pastejo Período de Descanso
4,5 (fol/perf) 6,5 (fol/perf) 8,5 (fol/perf) Média Massa seca de forragem total – MSFT (kg/ha) CV = 4,31
10 cm 3455 4859 6040 4785B 5 cm 3916 5168 6765 5283A
Média 3685c 5014b 6403a Massa seca de forragem verde – MSFV (kg/ha) CV = 5,43
10 cm 2930Ca 3562Bb 3714Ab 3402 5 cm 3085Ca 3825Ba 4438Aa 3783
Média 3007 3693 4076 Massa seca de forragem morta – MSFM (kg/ha) CV = 5,67
10 cm 525Cb 1297Bb 2285Aa 1369 5 cm 830Ca 1343Ba 2326Aa 1499
Média 677 1320 2305 Massa seca de lâmina foliar verde – MSLV (kg/ha) CV = 5,40
10 cm 1822Bb 2032Bb 2538Aa 2131 5 cm 1856Ba 2229Aa 2383Ab 2156
Média 1839 2130 2461 Massa seca de colmo verde – MSCV (kg/ha) CV = 8,20
10 cm 898Ca 1023Bb 1932Aa 1284 5 cm 856Cb 1441Ba 2582Aa 1626
Média 877 1232 2257 Relação material vivo/morto – MV/MM (kg/ha) CV = 6,33
10 cm 5,73Aa 2,77Ba 1,73Ca 3,41 5 cm 3,81Ab 2,96Ba 1,92Ca 2,89
Média 4,77 2,86 1,82 Altura (cm) CV = 5,26
10 cm 34,48 35,45 34,59 34,84A 5 cm 33,68 33,78 31,93 33,13B
Média 34,08 34,61 33,26 Relação lâmina foliar/colmo – F/C CV = 6,69
10 cm 2,39Aa 1,69Bb 0,72Cb 1,60 5 cm 2,65Aa 2,55Aa 1,01Ba 2,07
Média 2,52 2,12 0,86 Número de folhas vivas/perfilho - NFV CV = 1,88
10 cm 5,56Ca 6,06Bb 8,29Aa 6,63 5 cm 4,98Cb 6,42Ba 8,37Aa 6,59
Média 5,27 6,24 8,33 Densidade populacional de perfilhos – DPP
(perfilhos/m²) CV = 2,63
10 cm 2420Aa 2189Ab 1710Bb 2106 5 cm 2282Ab 2413Aa 1902Ba 2199
Média 2351 2301 1806 Médias seguidas de mesmas letras, maiúsculas na coluna e minúsculas na linha, não diferem entre si pelo teste de Tukey (P>0,05).

71
A massa seca de forragem morta (MSFM) foi influenciada pela interação PD x
RPP (P 0,05) (Tabela 3), onde a menor MSFM foi observada nos pastos com PD de 4,5
fol/perf e resíduos de 10 e 5 cm (Tabela 3). O comportamento verificado para a MSFM
era esperado uma vez que, pastos submetidos a PDs mais curtos, apresentam menores
perdas por senescência. O efeito dos ciclos de pastejo sobre a MSFM foi verificado para
os pastos sob PD 4,5 fol/perf e resíduo de 10 cm (Tabela 4), pode-se observar uma
pequena oscilação entre os ciclos, porém com a maior MSFM é produzida no oitavo
ciclo de pastejo. As flutuações na proporção de material morto no dossel ao longo do
período experimental acompanharam as flutuações em massa de forragem seca de
forragem total, fato esse bastante sugestivo e indicativo da contribuição desse
componente para os elevados valores de massa de forragem seca de forragem total
encontrados.
A massa seca de lâmina verde (MSLV) foi afetada pela interação PD x RPP
(P 0,05), apresentando maiores médias para os pastos submetidos a PD de 8,5 fol/perf e
resíduos de 10 e 5 cm (Tabela 3). Embora isso possa sugerir maior qualidade dos pastos
manejados sob PDs mais longos, o aumento na massa seca de colmo verde (MSCV)
nestes pastos sugere dificuldades para o animal em pastejo, pois essa maior massa de
folhas possivelmente se localiza num horizonte de pastejo menos acessível, uma vez
que os pastos com PDs mais longos atingiram uma maior elevação dos colmos.
Cândido et al. (2005b) ao avaliarem o capim-mombaça também observaram aumento da
MSLV com o aumento do PD. Foi verificada aumento da MSLV ao longo dos ciclos de
pastejo (P 0,05), para os pastos com PD de 6,5 fol/perf e resíduo de 5 cm (Tabela 4).
Foi verificado efeito da interação PD x RPP (P 0,05) sobre a massa seca de
colmo verde (MSCV), apresentando um aumento com o prolongamento do PD (Tabela
3), resposta esperada uma vez que o aumento do PD resulta na elevação dos colmos,
que por sua vez é responsável pela redução da qualidade da forragem. De acordo com
Faria (2009) o alongamento de colmo é acompanhado de uma lignificação acentuada,
devido à necessidade de sustentação do órgão; havendo, nesse caso, queda do valor
nutritivo da forragem disponível, o que refletirá negativamente sobre o desempenho
animal. Além da restrição no consumo pelo animal devido a redução no valor nutritivo
da forragem, o alongamento do colmo também promove restrição física à ingestão de
forragem, pois o comportamento ingestivo dos animais é sensível a variações na

72
estrutura do dossel forrageiro (PALHANO et al., 2007). A MSCV foi afetada pelos
ciclos de pastejo (P 0,05) para os pastos com PD de 4,5 e 6,5 fol/perf, ambos com
resíduo de 5 cm, no entanto o PD com 4,5 fol/perf, apresentou a MSCV no primeiro
ciclo de pastejo, tornado-se posteriormente constante nos demais ciclos. Para o pasto
submetido ao PD com 6,5 fol/perf, foi observado maior MSCV (Tabela 4). Ao contrário
do observado no presente estudo, Fagundes et al. (1999) verificaram aumento na
porcentagem de colmos para os cultivares de Cynodon sp à medida que avançou o
período experimental.
Houve efeito da interação PD x RPP (P 0,05) sobre a relação entre o material
vivo/morto (MV/MM), onde os pastos com PD de 8,5 fol/perf e resíduo de 10 e 5 cm,
apresentaram os menores valores para esta relação (Tabela 3). Esse comportamento é
atribuído à maior elevação na MSFM nos pastos com PD de 8,5 fol/perf e resíduo de 10
e 5 cm, ocasionando diminuição na MV/MM, especialmente nos PDs mais longos, onde
o processo de senescência se intensifica e nos maiores resíduos pós-pastejo, devido ao
menor uso da forragem produzida. Os ciclos de pastejo influenciaram a relação
MV/MM (P 0,05) para os pastos submetidos ao PD de 6,5 fol/perf e resíduo de 5 cm
(Tabela 4). Este comportamento é decorrente do acúmulo de material morto, fração
rejeitada pelos animais em pastejo, ao longo dos ciclos sucessivos de pastejo, onde o
processo de senescência foi menos intenso a partir do segundo ciclo de pastejo,
resultando num maior eficiência de utilização de forragem nestes pastos. Isso denota
maior efeito da MSFV na relação MV/MM, que comportou- se de maneira similar.
A altura pré-pastejo do dossel foi afetada pelo RPP (P 0,05). Os pastos com
RPP de 10 cm apresentaram maior altura pré-pastejo do dossel (Tabela 3). A altura do
dossel também foi afetada pelos ciclos de pastejo (P 0,05) para os pastos com PD de
4,5 e 8,5 fol/perf, em ambas as alturas e também nos pastos com 6,5 fol/perf resíduo de
5 cm (Tabela 4). Com o decorrer dos ciclos de pastejo foi observado aumento na altura
do dossel com o decorrer dos ciclos de pastejo. O alongamento do colmo ao longo dos
ciclos de pastejo refletiu em aumento da altura dos pastos nos ciclos subseqüentes, em
decorrência da rejeição desse componente pelos animais em pastejo. De acordo
Casagrande et al. (2010) quando ocorre aumento na proporção de colmos na parte
superior do dossel, dificilmente a altura do pasto reduz, visto que esses colmos se

73
tornam um impedimento físico ao pastejo, pela dificuldade em que os animais
encontram para consumi-los.
Foi observado efeito da interação PD x RPP (P 0,05) sobre a relação lâmina
foliar/colmo (F/C). Para os pastos sob PD de 8,5 fol/perf e resíduos de 10 e 5 cm foram
estimados valores inferiores a 1, o que pode acarretar comprometimento da qualidade da
forragem (Tabela 3). Possivelmente a mudança no hábito de crescimento da planta
tenha elevado a produção das colmos, onde o aumento dessa produção em detrimento a
de MSLV, ocasionou na redução da relação F/C. Os valores para relação F/C no
presente estudo, foram superiores aos relatados por Oliveira et al. (2000) ao observarem
redução da relação F/C, com o avanço da idade da planta, com valores de 1,39 e 0,45
aos 14 e 70 dias de idade, respectivamente. O progressivo alongamento dos colmos,
resulta na redução da relação F/C (P<0,05) com a sucessão dos ciclos de pastejo (Tabela
4) para os pastos com PD de 4,5 e 6,5 fol/perf e resíduos de 5 cm. O controle do
alongamento dos colmos é um grande desafio a ser solucionado pelo manejo.

74
Tabela 4. Efeito dos ciclos de pastejo sobre os componentes da biomassa e as características estruturais em pastagem de capim-tifton 85 sob três períodos de descanso e dois resíduos pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
Período de descanso 4,5 e resíduo de 10 cm MSFM (kg/ha) 399b 548ab 500ab 486ab 586ab 544ab 464ab 643a 580ab 503ab 525 19,15 Altura (cm) 19,86c 29,50b 31,40ab 34,82ab 35,85ab 36,22ab 36,22ab 39,29a 40,55a 40,52a 34,42 11,04 F/P 4,50ef 4,25f 4,87def 4,75def 5,25de 5,50cd 8,75a 6,37bc 6,60b 4,87def 5,57 6,61 DPP (perf/m²) 1396c 1764ab 1532bc 1928a 1788ab 1700ab 1636bc 1940a 1784ab 1640bc 1711 6,45
Período de descanso 4,5 e resíduo de 5 cm MSCV (kg/ha) 1129a 841ab 851ab 781b 900ab 807b 736b 849ab 811b - 856 14,31 Altura (cm) 19,87d 27,97cd 30,85bc 32,75abc 35,45abc 35,45abc 41,27a 38,00abc 41,07ab - 33,63 12,91 F/P 4,50b 4,50b 5,00b 4,87b 4,75b 4,75b 4,75b 4,75b 6,62a - 4,94 8,89 F/C 2,84ab 2,64ab 2,48ab 2,78ab 2,34b 2,85ab 3,37a 2,39b 2,18b - 2,65 14,82 DPP 1760bc 1984ab 1668c 1800bc 1968ab 1920abc 1932ab 2084a 2004ab - 1902 5,68
Período de descanso 6,5 e resíduo de 10 cm F/P 6,25a 6,12ab 6,37a 6,62a 6,37a 6,50a 5,12b 5,12b - - 6,05 7.43 DPP (perf/m²) 2296ab 2156bc 1968c 2296ab 2208abc 2452a 1976c 2160bc - - 2189 4,80
Período de descanso 6,5 e resíduo de 5 cm MSFV (kg/ha) 3317c 3961abc 4189ab 3576abc 3935abc 3491bc 4245a - - - 3816 8,34 MSLV (kg/ha) 1826b 2330ab 2536a 2328ab 2472a 2349ab 2840a - - - 2383 10,98 MSCV (kg/ha) 1490abc 1630ab 1652a 1247bc 1523abc 1141c 1405abc - - - 1441 12,12 MV/MM 2,27b 3,36ab 3,33ab 2,54ab 3,10ab 2,56ab 3,53a - - - 2,84 17,54 Altura (cm) 28,50c 28,55c 31,30bc 34,10abc 37,00ab 37,00ab 40,70a - - - 33,87 16,59 F/C 2,06ab 2,09a 1,54abc 1,87ab 1,65abc 1,22c 1,43bc - - - 1,69 9,19
Período de descanso 8,5 e resíduo de 10 cm Altura (cm) 23,14b 27,02b 34,57a 40,12a 41,35a 41,35a - - - - 34,59 9,18 DPP (perf/m²) 2132b 2936a 2344b 2372b 2372b 2368b - - - - 2420 7,97
Período de descanso 8,5 e resíduo de 5 cm Altura (cm) 21,60b 24,22b 36,15a 37,77a 39,92a - - - - - 31,93 11,66 DPP (perf/m²) 1816d 2744a 2284bc 2524ab 2044cd - - - - - 2282 8,78
Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

75
O número de folhas vivas por perfilho (F/P) foi afetado pela interação PD x RPP
(P 0,05) (Tabela 3). Obteve-se um valor médio de 5,27; 6,24 e 8,33 folhas/perfilho para
os pastos manejados sob PD de 4,5; 6,5 e 8,5 fol/perf, respectivamente. Á medida que
ocorre o prolongamento do PD, este tende a produzir mais folhas, sendo tal
característica estrutural uma ferramenta de manejo que pode ser adotada para o controle
da entrada dos animais no pasto, essa maior produção de folhas, no entanto, não implica
em melhor qualidade do pasto, pois no PD prolongado ocorre o aumento dos colmos,
que contribui para a redução da qualidade do pasto, além, do fato de que as folhas
produzidas podem estar menos acessível ao animal em pastejo. A maior produção de
folhas nos pastos manejados com menor RPP pode ser atribuída a interferência da luz,
que atingiu o dossel aumentando a eficiência de uso da radiação, pois. Houve efeito dos
ciclos de pastejo (P 0,05), para os pastos com PD de 4,5 fol/perf com resíduos de 10 e 5
cm e 6,5 fol/perf com resíduo de 10 cm (Tabela 4).
A densidade populacional de perfilhos (DPP) foi afetada pela interação PD x
RPP (P 0,05) (Tabela 3). Houve uma redução no número de perfilhos/m² com o
prolongamento do PD. Nos pastos manejados com PD de 8,5 fol/perf e resíduo de 10
cm, foi verificado menor número de perfilhos, quando comparado aos demais pastos.
Simon e Lemaire (1987) afirmaram que com o aumento do período de descanso da
pastagem, ocorre à diminuição da DPP, pois com o aumento do Índice de área foliar,
aumenta o sombreamento mútuo, que inibe a brotação das gemas, devido à menor
qualidade de luz transmitida no interior do dossel, o que pode explicar as diferenças
ocorridas entre os períodos de descanso no presente estudo. Em PDs curtos, observa-se
perfilhos numerosos e pequenos, enquanto em pastos com PD longos são encontrados
pefilhos grandes e em menor quantidade (CARVALHO et al., 2000; SBRISSIA et al.,
2003). À media que o PD se prolonga e os pastos são manejados a uma altura residual
mais elevada, estes tendem a perfilhar menos. Alguns estudos mostraram a existência de
relação inversa entre a altura do pasto e a densidade populacional de perfilhos
(BIRCHAM & HODGSON, 1983; CARVALHO et al., 2000, 2001). Tal relação deve
se a um mecanismo de compensação tamanho/densidade populacional de perfilhos,
existente em comunidades de plantas, em que os pastos mantidos baixos apresentam
maior população de perfilhos menores em relação aos pastos mantidos altos
(MATTHEW et al., 1995). Houve efeito dos ciclos de pastejo (P 0,05) para quase todos

76
os tratamentos com exceção do pasto com PD de 6,5 fol/perf e resíduo de 5 cm (Dados
não apresentados), no entanto para os demais pastos observou-se uma oscilação da DPP
ao longo do período experimental (Tabela 4).
Componentes da biomassa e características estruturais no pós-pastejo
A massa seca de forragem total residual (MSFTres) foi afetada pela interação
PD x RPP (P 0,05) (Tabela 5). A maior MSFTres foi verificada para os pastos
manejados sob PD de 8,5 fol/perf e resíduos de 10 e 5 cm. Provavelmente o
prolongamento do PD, tenha favorecido aos maiores acúmulos de MSFMres e
MSCVres observados nestes pastos, uma vez que o componente com maior participação
na MSFTres foi o colmo, seguido do material morto e das lâminas foliares. De acordo
com Azar (2007) ideal é manter uma oferta de forragem de 4000 kg de MS/ha e resíduo
pós-pastejo de 2000 kg de MS/ha para garantir vigor de rebrota da pastagem após a
desfolha, portanto, o resíduo pós-pastejo obtido nesta pesquisa encontra-se dentro do
recomendado, pelo autor supracitado. Foi verificado efeito dos ciclos de pastejo sobre a
MSFTres (P 0,05) (Tabela 6), para os pastos com PD de 4,5; 6,5 e 8,5 fol/perf e
resíduos de 5; 10 e 5 cm, respectivamente. As maiores MSFTres foram observadas no
terceiro ciclo de pastejo para os pastos com PD 4,5 e 8,5 fol/perf e resíduo de 5 cm e no
sexto ciclo de pastejo para os pastos com 6,5 fol/perf e resíduo de 10 cm (Tabela 6).
A massa seca de forragem verde residual (MSFVres) foi influenciada do PD
(P 0,05) (Tabela 5). Os pastos com PD de 8,5 fol/perf apresentaram maiores MSFVres,
possivelmente devido ao maior acúmulo de colmo observado para estes pastos durante o
período experimental. Houve efeito dos ciclos de pastejo sobre a MSFVres (P 0,05)
(Tabela 6) para os pastos sob PD de 4,5; 6,5 e 8,5 fol/perf e resíduos de 5, 10 e 5,
respectivamente. A MSFVres para estes pastos foi bastante variável ao longo do
período experimental.
A massa seca de forragem morta residual (MSFMres) foi afetada apenas pela
interação PD x RPP (P 0,05) (Tabela 5). A maior MSFMres foi verificada nos pastos
sob PD de 8,5 fol/perf e resíduos de 10 e 5 cm e a menor 4,5 fol/perf e ambos os
resíduos. Este comportamento esperado, uma vez que o prolongamento do PD favorece
ao acúmulo de material morto, o que reflete em perda de forragem, que deixa de ser

77
consumida. A predominância de material morto no resíduo de um pasto pode indicar
grande seletividade por parte dos animais e um baixo índice de utilização da forragem
(CARNEVALLI & SILVA, 1999). Os ciclos de pastejo afetaram a MSFMres (P 0,05)
(Tabela 6) para os pastos sob PD de 6,5 e 8,5 fol/perf e resíduos de 10 e 5 cm,
respectivamente. Percebe-se uma redução da MSFMres ao longo dos ciclos de pastejo
nos pastos manejados sob PD de 8,5 fol/perf, representado uma condição favorável, o
que mostra uma eficiência no manejo destes pastos. Nos demais PD e RPP a MSFMres
permaneceu constante ao longo dos ciclos de pastejo.
Foi verificado efeito do PD e do RPP (P 0,05) sobre a massa seca de lâmina
verde residual (MSLVres). Os pastos sob PD de 4,5 fol/perf e resíduo de 10 e 5 cm,
apresentaram menores MSLVres. A MSLVres é fundamental na rebrotação do pasto,
pois a área remanescente após o pastejo é diretamente proporcional à taxa de
fotossíntese líquida do pasto (GOMIDE et al., 2002). A maior quantidade de lâminas
remanescentes favorece a menor mobilização das reservas orgânicas, permitindo um
crescimento mais rápido da planta. Houve efeito dos ciclos de pastejo (P 0,05) sobre a
MSLVres (Tabela 6) para os pastos com PD de 4,5 fol/perf e resíduos de 10 e 5 cm e
8,5 fol/perf e resíduo de 5 cm. MSLVres permaneceu praticamente constante ao longo
do ciclos de pastejo.
A massa seca de colmo verde residual (MSCVres) foi afetada apenas pelo PD
(P 0,05) sendo mais elevada para os pastos sob PD de 8,5 fol/perf e resíduos de 10 e 5
cm (Tabela 5), fato este que contribuiu para o aumento da MSFTres e MSFVres para
estes pastos. Cândido et al. (2005b) verificou aumento na MSCVres com o
prolongamento do PD para o capim-mombaça. Houve efeito dos ciclos de pastejo
(P 0,05) sobre a MSCVres nos pastos sob PD de 4,5 fol/perf e resíduo de 5 cm; 6,5
fol/perf e resíduos de 10 e 5 cm e 8,5 fol/perf e resíduo de 5 cm (Tabela 6).
A relação material vivo/morto residual (MV/MMres) sofreu efeito da interação
PD x RPP (P 0,05). A maior relação foi verificada para os pastos sob PD de 4,5 fol/perf
e resíduo de 10 cm. Esse comportamento é atribuído à menor MSFM residual no pasto
sob o PD 4,5 fol/perf, ocasionando aumento na MV/MMres.
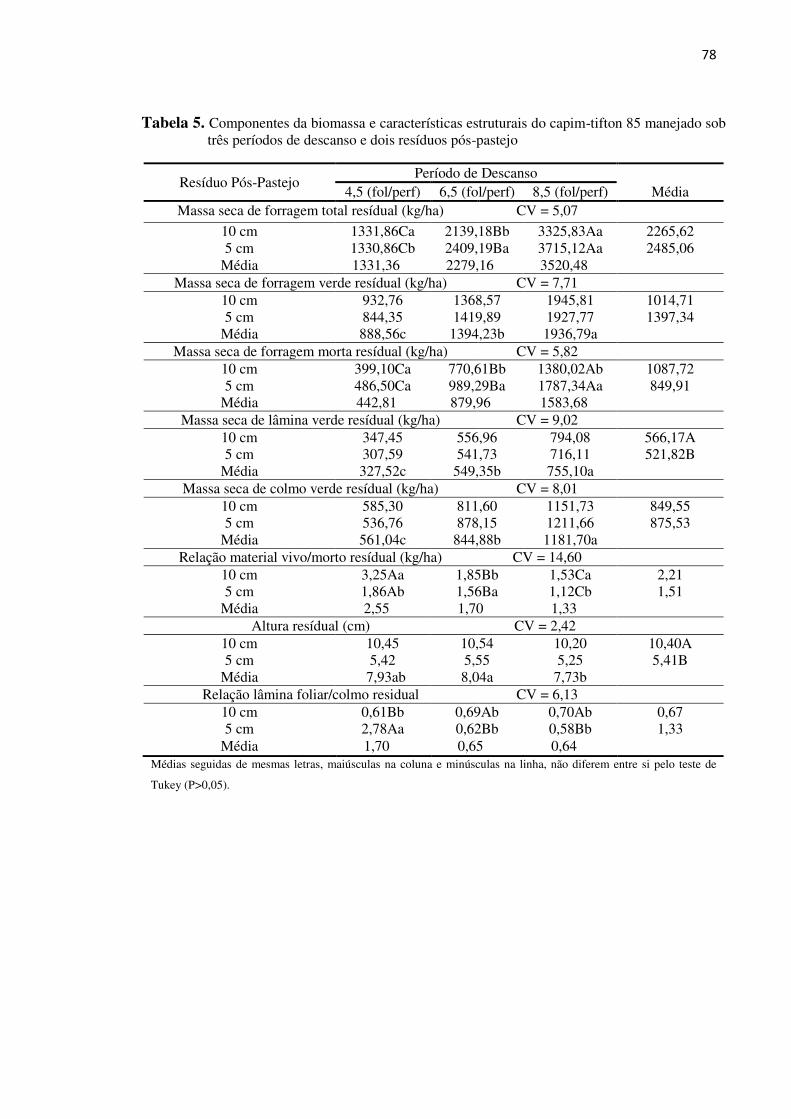
78
Tabela 5. Componentes da biomassa e características estruturais do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de mesmas letras, maiúsculas na coluna e minúsculas na linha, não diferem entre si pelo teste de
Tukey (P>0,05).
Resíduo Pós-Pastejo Período de Descanso
4,5 (fol/perf) 6,5 (fol/perf) 8,5 (fol/perf) Média Massa seca de forragem total resídual (kg/ha) CV = 5,07
10 cm 1331,86Ca 2139,18Bb 3325,83Aa 2265,62 5 cm 1330,86Cb 2409,19Ba 3715,12Aa 2485,06
Média 1331,36 2279,16 3520,48 Massa seca de forragem verde resídual (kg/ha) CV = 7,71
10 cm 932,76 1368,57 1945,81 1014,71 5 cm 844,35 1419,89 1927,77 1397,34
Média 888,56c 1394,23b 1936,79a Massa seca de forragem morta resídual (kg/ha) CV = 5,82
10 cm 399,10Ca 770,61Bb 1380,02Ab 1087,72 5 cm 486,50Ca 989,29Ba 1787,34Aa 849,91
Média 442,81 879,96 1583,68 Massa seca de lâmina verde resídual (kg/ha) CV = 9,02
10 cm 347,45 556,96 794,08 566,17A 5 cm 307,59 541,73 716,11 521,82B
Média 327,52c 549,35b 755,10a Massa seca de colmo verde resídual (kg/ha) CV = 8,01
10 cm 585,30 811,60 1151,73 849,55 5 cm 536,76 878,15 1211,66 875,53
Média 561,04c 844,88b 1181,70a Relação material vivo/morto resídual (kg/ha) CV = 14,60
10 cm 3,25Aa 1,85Bb 1,53Ca 2,21 5 cm 1,86Ab 1,56Ba 1,12Cb 1,51
Média 2,55 1,70 1,33 Altura resídual (cm) CV = 2,42
10 cm 10,45 10,54 10,20 10,40A 5 cm 5,42 5,55 5,25 5,41B
Média 7,93ab 8,04a 7,73b Relação lâmina foliar/colmo residual CV = 6,13
10 cm 0,61Bb 0,69Ab 0,70Ab 0,67 5 cm 2,78Aa 0,62Bb 0,58Bb 1,33
Média 1,70 0,65 0,64

79
Houve efeito (P 0,05) do PD e RPP sobre a altura residual (alturares), com
valores de 7,93; 8,04 e 7,73 cm para os pastos sob PD de 4,5; 6,5 e 8,5 fol/perf
respectivamente (Tabela 5). As alturas do dossel no pós-pastejo foram inferiores às
relatadas por Voltolini et al. (2009) que verificaram alturares de 11,86 cm, para o
mesmo capim, deste estudo. Em relação ao RPP, os pastos com maiores resíduos em
decorrência da maior quantidade de material remanescente, apresentaram maior altura
resídual. A alturares elevou-se até o sétimo ciclo de pastejo para os pastos sob PD de
4,5 fol/perf e 10 cm de resíduo, havendo posteriormente uma leve redução.
Comportamento semelhante foi verificado para os pastos sob PD de 6,5 fol/perf e
resíduo de 10 cm.
A relação lâmina foliar/colmo residual (F/Cres) do pasto sob o PD 8,5 fol/perf
foi inferior (P<0,05) aos demais. A redução da relação F/Cres foi devido à maior
presença de colmos no pasto sob o PD 8,5 fol/perf em relação aos demais (Tabela 5).
Esse mesmo comportamento foi verificado por Gomide (2001) e Cândido (2003),
estudando capim-mombaça (P. maximum) sob três períodos de descanso, onde o maior
acúmulo de colmo foi verificado no maior período de descanso, processo esse inerente
às espécies de gramíneas C4, ainda mais quando submetidas à baixa freqüência de
desfolhação. Houve efeito dos ciclos de pastejo (P<0,05) para os pastos sob PD de 4,5
fol/perf e resíduo de 10 cm (Tabela 6), pode-se observar maiores relações no primeiro e
segundo ciclos de pastejo.

80
Tabela 6. Efeito dos ciclos de pastejo sobre os componentes da biomassa e as características estruturais em pastagem de capim-tifton 85 sob três períodos de descanso e dois resíduos pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
Período de descanso 4,5 e resíduo de 10 cm MSLV (kg/ha) 321ab 452a 208b 306ab 375ab 442a 297ab 393ab 383ab 292ab 347 23,09 Altura (cm) 9,52b 9,97ab 10,52ab 10,75ab 10,22ab 10,37ab 11,12a 10,75ab 10,95ab 10,37ab 10,45 5,41 F/C 0,62ab 0,98a 0,34b 0,46b 0,56b 0,67ab 0,60ab 0,65ab 0,74ab 0,48b 0,61 28,44
Período de descanso 4,5 e resíduo de 5 cm MSFT (kg/ha) 1330ab 1205b 1934a 1267b 1218b 1332ab 1075b 1065b 1548ab - 1330 20,54 MSFV (kg/ha) 858ab 842b 1226a 877ab 768b 805b 689b 644b 886ab - 844 18,61 MSLV (kg/ha) 274b 323ab 446a 336ab 275b 314ab 248b 222b 326ab - 307 22,28 MSCV (kg/ha) 584ab 518b 780a 541ab 493b 491b 440b 421b 559ab - 536 20,41
Período de descanso 6,5 e resíduo de 10 cm MSFT (kg/ha) 2016ab 2024ab 1880b 2136ab 2214ab 2501a 2050ab 2288ab - - 2139 10,27 MSFV (kg/ha) 1329ab 1286ab 1081b 1466ab 1408ab 1557a 1428ab 1390ab - - 1368 14,29 MSFM (kg/ha) 687ab 738ab 798ab 670ab 806ab 944a 621b 897ab - - 770 16,93 MSCV (kg/ha) 823ab 713ab 634b 832ab 841ab 888ab 923a 826ab - - 810 14,48 Altura (cm) 9,80b 9,86ab 10,42ab 10,77ab 11,05a 10,87ab 10,95ab 10,60ab - - 10,54 4,91
Período de descanso 6,5 e resíduo de 5 cm MSFV (kg/ha) 1129b 1611ab 1539ab 1381ab 1720a 1311ab 1245ab - - - 1419 17,03 MSCV (kg/ha) 699b 973ab 1011ab 833ab 1047a 819ab 762ab - - - 878 16,05
Período de descanso 8,5 e resíduo de 5 cm MSFT (kg/ha) 3671ab 3351b 4217a 3887ab 3446ab - - - - - 3715 10,30 MSFV (kg/ha) 1734bc 1577c 2070ab 2286a 1970abc - - - - - 1927 9,82 MSFM (kg/ha) 1937ab 1774ab 2147a 1601ab 1476b - - - - - 1787 15,03 MSLV (kg/ha) 612bc 548c 787ab 880a 751ab - - - - - 716 12,92 MSCV (kg/ha) 1121b 1029b 1283ab 1405a 1218ab - - - - - 1211 10,15 MV/MM(kg/ha) 0,89c 0,89c 0,97bc 1,48a 1,38ab - - - - - 1,12 19,68
Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

81
CONCLUÇÕES
Os componentes da biomassa no pré e pós-pastejo, são afetados, tanto pela
interação entre os períodos de descanso e resíduos pós-pastejo e pelos ciclos de pastejo.
O prolongamento do período de descanso compromete a estrutura do dossel.
O resíduo pós-pastejo afetou a massa seca de forragem total e a altura no pré-
pastejo e massa seca de lâmina verde e altura no pós-pastejo.
O uso do capim-tifton 85 não deve exceder ao período de descanso de 6,5
fol/perf para evitar perdas de forragem por senescência, sendo possível utilizar uma
altura residual de 5 cm sem comprometer a perenidade da pastagem.
REFERÊNCIAS AZAR, G. S. Avaliação de cultivares de Cynodon nos períodos seco e chuvoso, na região norte do Piauí /Teresina, 2007. 48 f. Dissertação (Mestrado em Ciência Animal) - Universidade Federal do Piauí. 2007. BARBOSA, R.A.; NASCIMENTO JR., D.; EUCLIDES, V.P.B. et al. Capim-tanzânia submetido a combinações entre intensidade e frequência de pastejo. Pesquisa Agropecuária Brasileira, v.42, p.329-340, 2007. BIRCHAM, J.S.; HODGSON, J. The influence of sward condition on rates of herbage growth and senescence in mixed sward under continuous stocking management. Grass and Forage Science, v. 38, n. 4, p. 323-331, 1983. CÂNDIDO, M. J. D.; Da SILVA, R. G.; NEIVA, J. N. M. et al. Fluxo de biomassa em capim-tanzânia pastejado por ovinos sob três períodos de descanso. Revista Brasileira Zootecnia, v.35, n.6, p.2234-2242, 2006. CÂNDIDO, M.J.D.; GOMIDE, C.A.M.; ALEXANDRINO, E. et al. Morfofisiologia do dossel de Panicum maximum cv. Mombaça sob lotação intermitente com três períodos de descanso. Revista Brasileira de Zootecnia, v.34, n.2, p.338-347, 2005b. CÂNDIDO, M.J.D. Morfofisiologia e crescimento do dossel e desempenho animal em Panicum maximum cv. Mombaça sob lotação intermitente com três períodos de descanso. Viçosa: UFV, 2003. 134p. Tese (Doutorado em Zootecnia) – Universidade Federal de Viçosa, 2003. CARNEVALLI, R.A. Dinâmica da rebrotação de pastos de capim- Mombaça submetidos a regimes de desfolhação intermitente. Tese (Doutorado em Agronomia - Ciência Animal e Pastagens), Piracicaba, ESALQ, 2003. Orientador: Prof. Dr. Sila Carneiro da Silva.

82
CARNEVALLI, R.A.; DA SILVA, S.C. Validação de técnicas experimentais para avaliação de características agronômicas e ecológicas de pastagens de Cynodon dactylon cv. Coastcross-1. Scientia Agricola, v.56, p.489-499, 1999. CARVALHO, P. C. F.; RIBEIRO FILHO, H. M. N.; POLI, C. H. E. C. et al. Importância da estrutura da pastagem na ingestão e seleção de dietas pelo animal em pastejo. In: REUNIÀO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38, 2001, Piracicaba, Anais... Piracicaba: FEALQ, p. 853-871, 2001.
CARVALHO, C.A.B. Padrões demográficos de perfilhamento e acúmulo de forragem em pastagens de Cynodon spp. manejadas em quatro intensidades de pastejo. 2000. 96f. Dissertação (Mestrado) – ESALQ/USP, Piracicaba, SP. CASAGRANDE, D. R.; RUGGIERI, A. C.; JANUSCKIEWICZ, E. R. et al. Características morfogênicas e estruturais do capim-marandu manejado sob pastejo intermitente com diferentes ofertas de forragem. Revista Brasileira de Zootecnia, v.39, n.10, p.2108-2115, 2010. EMBRAPA - EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistema brasileiro de classificação de solos. Brasília: EMBRAPA, 1999. 412p. (Produção de Informação). FAGUNDES, J.L.; SILVA, S.C. da; PEDREIRA, C.G.S. et al. Índice de área foliar, interceptação luminosa e acúmulo de forragem em pastagens de Cynodon spp. Sob diferentes intensidades de pastejo. Scientia Agricola, v. 56, n. 4, p. 1141-1150, 1999. FARIA, D. J. G. Características morfogênicas e estruturais dos pastos e desempenho de novilhos em capim-braquiária sob diferentes alturas. Tese (Doutorado) – Universidade Federal de Viçosa. Viçosa/MG, 2009. 148 f. 2009. FORBES, T.D.A., COLEMAN, S.W. Forage intake and ingestive behaviour of catle grazing old world bluestems. Agron. Journal, Madison, v.85,n.4,p.808-16, 1993. GOMIDE, C.A.M.; GOMIDE, J.A.; ALEXANDRINO, E. Características estruturais e produção de forragem em pastos de capim-mombaça submetidos a períodos de descanso. Pesquisa Agropecuária Brasileira, v.42, p.1487-1494, 2007. GOMIDE, C.A.M. Características morfofisiológicas associadas ao manejo do capim-mombaça (Panicum maximum Jacq.). 2001. 107 f. Tese (Doutorado) – Universidade Federal de Viçosa. Viçosa/MG, 2001. GRANT, S.A.; BARTHRAM, G.T.; TORVEL, L. et al. Comparison of herbage production under continuous stocking and intermittent grazing. Grass and Forage Science, v.43, n.1, p.29-39, 1988. JANUSCKIEWICZ, E. R.; MAGALHÂES, M. A.; RUGGIERE, A. C. et al. Massa de forragem, composição morfológica e química de capim-tanzânia sob diferentes dias de

83
descanso e alturas de resíduo pós-pastejo. Bioscience Journal, Uberlândia, v. 26, n. 2, p. 161-172, Mar./Apr. 2010 LACA, E. A.; LEMAIRE, G. Measuring sward structure. In: T‟MANNETJE, L.; JONES, R. M. (Eds.). Field and laboratory methods for grassland and animal production research. New York: CABI, 2000. p. 103-122. LIMA, J.A. de; VILELA, D. Formação e manejo de pastagens de Cynodon. IN: Cynodon: Forrageiras que estão revolucionando a pecuária brasileira. Juiz de Fora: EMBRAPA GADO DE LEITE, 2005. p.11-32. MATTHEW, C.; LEMAIRE, G.; SACKVILLE HAMILTON, N. R. et al. A modified self-thinning equation do describe size/ density relationships for defoliated swards. Annals of Botany, v. 76, p. 579-587, 1995. MINSON, D.J. Forage in ruminant nutrition. San Diego: Academic Press, 1990. 483p. MISLEY, P.; PATE, F.M. Establishment, management, and utilization of Cynodon
grasses in Florida. In: WORKSHOP SOBRE O POTENCIAL FORRAGEIRO DO GÊNERO CYNODON, 1996, Juiz de Fora. Anais... Juiz de Fora: Embrapa-CNPGL, 1996. p.128-138. OLIVEIRA, M.A; PEREIRA, O.G.; MARTINEZ Y HUAMAN, C.A. et al. Características morfogênicas e estruturais do capim-bermuda „Tifton 85‟ (Cynodon
spp.) em diferentes idades de rebrota. Revista Brasileira de Zootecnia, v. 29, n. 6, p. 1939-1948, 2000. PALHANO, A. L.; CARVALHO, P. C. F.; DITTRICH, J. R. et al. Características do processo de ingestão de forragem por novilhas holandesas em pastagens de capim-mombaça. Revista Brasileira de Zootecnia, v. 36, p. 1014-1021, 2007. PEDREIRA, C.G.S. Avaliação de novas gramíneas de gênero Cynodon para a pecuária dos Estados Unidos, In.: WORKSHOP SOBRE O POTENCIAL FORRAGEIRO DO GÊNERO CYNODON, Juiz de Fora, 1996. Anais... Juiz de Fora, EMBRAPA-CNPGL, 1996, p. 111-125. SAS INSTITUTE. SAS system for Windows. Version9. 1. Cary: SAS Institute Inc. 2003. 2 CD – ROMs. SBRISSIA, A.F., Da SILVA, S.C., MATTHEW, C. et al. Tiller size/density compensation in grazed Tifton 85 bermuda grasss wards. Pesquisa Agropecuária Brasileira, Brasília, v.38, n.12, p.1459-1468, 2003. SILVA, R.G. Morfofisiologia do dossel e desempenho produtivo de ovinos em Panicum maximum (Jacq.) cv. Tanzânia sob três períodos de descanso. Fortaleza: Universidade Federal do Ceará, 2004. 114p. Dissertação (Mestrado em Zootecnia) - Universidade Federal do Ceará, 2004.

84
SIMON, J.C. & LEMAIRE, G. Tillering and leaves area index in grasses in the vegetative phase. Grass and Forage Science, v. 42, p. 373-380, 1987. SOUZA, R. A.; VOLTOLINI, T. V.; PEREIRA, L. G. R. P. et al. Desempenho produtivo e parâmetros de carcaça de cordeiros mantidos em pastos irrigados e suplementados com doses crescentes de concentrado. Acta Scientiarum. Animal Sciences. Maringá, v. 32, n. 3, p. 323-329, 2010. VOLTOLINI, T. V.; MOREIRA, J. N.; NOGUEIRA, D. M. et al. Fontes protéicas no suplemento concentrado de ovinos em pastejo. Acta Scientiarum. Animal Sciences, v. 31, n. 1, p. 61-67, 2009.

85
CAPÍTULO 5
____________________________________________________________
Composição químico-bromatológica e digestilibidade in vitro da matéria seca de lâminas foliares e colmos do capim-tifton 85 manejado
sob três períodos de descanso e dois resíduos pós-pastejo

86
Composição químico-bromatológica e digestilibidade in vitro da matéria seca de lâminas foliares e colmos do capim-tifton 85 manejado sob três períodos de
descanso e dois resíduos pós-pastejo
RESUMO: Foram avaliadas a composição química e a digestibilidade in vitro da
matéria seca de lâminas foliares e colmos do capim-tifton 85 sob três períodos de
descanso e dois resíduos pós-pastejo em diferentes ciclos de pastejo. Os tratamentos
corresponderam a combinações de três condições de pré-pastejo 4,5; 6,5 e 8,5 (número
de folhas/perfilho) e duas condições de pós-pastejo 10 e 5 cm, distribuídos em um
delineamento experimental inteiramente casualizado em arranjo fatorial 3x2, com
quatro repetições (piquetes). O aumento do período de descanso do capim-tifton 85
determinou acréscimos nos teores de matéria seca, extrato etéreo, fibra insolúvel em
detergente neutro, fibra insolúvel em detergente ácido e hemicelulose da fração lâmina
foliar. A interação afetou apenas a matéria mineral da fração lâmina foliar. O aumento
no período de descanso apesar de produzir acréscimos nos teores de matéria de seca,
fibra insolúvel em detergente neutro e fibra insolúvel em detergente ácido do capim-
tifton 85, não reduziu a digestibilidade in vitro da matéria seca na fração lâmina foliar.
Na altura residual de 10 cm foi verificada aumento nas concentrações de fibra insolúvel
em detergente neutro, fibra insolúvel em detergente ácido e lignina na fração lâmina
foliar. O teor de proteína bruta das duas frações foi reduzido com o aumento do período
de descanso.
Termos para indexação: componentes estruturais, maturidade da planta, valor
nutritivo.

87
Bromatologic-chemical composition and digestilibidade in vitro of the dry matter of leaf blades and stems of the tifton 85 grass managed under three periods of rest
and two post-grazing residues
Index-terms: The chemical composition and the digestibilidade had been evaluated in
vitro of the dry matter of leaf blades and stems of the tifton 85 grass managed under
three periods of rest and two post-grazing residues in different cycles of pastejo. The
treatments had corresponded the combinations of three conditions of daily pay-pastejo
4,5; 6,5 and 8,5 (leaf number/tiller) and two conditions of after-pastejo 10 and 5 cm,
distributed in an experimental delineation entirely casualizado in factorial arrangement
3x2, with four repetitions. The increase of the period of rest of tifton 85 grass
determined additions in texts of dry matter, etéreo extract, insoluble neutral detergent
fiber, insoluble acid detergent fiber and hemicelulose of the fraction foliar blade. The
interaction affected only the mineral substance of the fraction foliar blade. The increase
in the period of rest although to produce additions in texts of drought substance,
insoluble fiber in neutral detergent and insoluble fiber in acid detergent of tifton 85
grass, in did not reduce the digestibilidade in vitro of the dry matter in the fraction leaf
blade. In the residual height of 10 cm increase in the insoluble fiber concentrations was
verified in neutral detergent, insoluble in acid detergent and lignin fiber in the fraction
leaf blade. The crude protein text of the two fractions was reduced with the increase of
the period of rest.
Index-terms: structural components, maturity of the forage, nutritive value.

88
INTRODUÇÃO
O valor nutritivo de uma forragem é caracterizado pela sua composição química,
digestibilidade e natureza dos produtos digestíveis, enquanto a qualidade da forragem
envolve uma avaliação integrada de seu valor nutritivo e do nível de consumo de
matéria seca pelo animal (CROWDER & CHHEDA, 1982). Assim, seu baixo valor
nutritivo é determinado pelo reduzido teor de proteína bruta e mineral, pelo alto
conteúdo de fibra e pela baixa digestibilidade (EUCLIDES, 1995).
O valor nutritivo das plantas é afetado por fatores fisiológicos, morfológicos,
ambientais e por diferenças entre espécies, sendo que, no caso das plantas forrageiras, o
declínio do valor nutritivo associado ao aumento da idade, normalmente é explicado
como o resultado da maturidade da planta. Isto ocorre porque, com o envelhecimento
do pasto e conseqüente diminuição da taxa de alongação e aparecimento de folhas, o
conteúdo da parede celular nas plantas aumenta, indicando desenvolvimento de tecidos
estruturais em folhas e colmos (WILSON, 1994).
De acordo com Reis et al. (2005) a medida que a idade fisiológica da planta
avança, aumentam as porcentagens de celulose, hemicelulose e lignina, reduzindo a
proporção dos nutrientes potencialmente digestíveis (carboidratos solúveis, proteínas,
minerais e vitaminas), produzindo uma queda acentuada na digestibilidade.
Em um pasto, a presença de material verde indica tecido novo, de maior valor
nutritivo, significando do ponto de vista fisiológico o vigor de rebrota, e o potencial
para a realização de fotossíntese. O material seco indica partes mortas ou senescentes,
representando a porção com menor valor nutritivo e baixa aceitabilidade. A relação
entre material vivo/material morto, além de ser influenciada pelo manejo da pastagem, é
decorrente do potencial genético das plantas e do grau em que as condições ambientais
favorecem o crescimento. O resíduo de forragem na pastagem exerce efeito importante
na qualidade da forragem, pois, segundo Forbes e Coleman (1993), a estrutura do pasto
é modificada com a participação de seus componentes folha, colmo e material morto.
Nos últimos anos as plantas do gênero Cynodon despertaram grande interesse e
ganharam popularidade devido à sua facilidade de cultivo, alta produção de forragem
(20 a 25 t MS/ ha/ano) e bom valor nutritivo (11 a 13% de proteína bruta e 58 a 65% de
digestibilidade) (PEDREIRA, 1996).

89
Considerando o que foi relatado acima, esta pesquisa foi conduzida com o
objetivo de avaliar a composição químico-bromatologica e a digestibilidade in vitro da
matéria seca do capim-tifton 85 manejado sob três períodos de descanso 4,5; 6,5 e 8,5
(número de folhas/perfilho) e dois resíduos pós-pastejo 10 e 5 cm, na condição de pré-
pastejo.
MATERIAL E MÉTODOS
A pesquisa foi desenvolvida no Setor de Produção de Ovinos do Campo
Experimental de Bebedouro, pertencente à Embrapa Semiárido, localizado no
Município de Petrolina-Pernambuco. Durante o mês de outubro foi efetuada a divisão
da área em piquetes e ajuste da metodologia de trabalho. A coleta de dados ocorreu
entre os meses de novembro de 2008 a junho de 2009, totalizando nove meses.
O clima da região é classificado como quente e seco, sendo caracterizada por
duas estações bem definidas, uma chuvosa, de janeiro a maio, e outra seca, de junho a
dezembro. Os dados sobre as condições de temperaturas máxima e mínima do ar e
precipitação pluviométrica durante o período experimental foram obtidos da Estação
Agrometeorologia de Bebedouro, Petrolina-Pernambuco e são apresentados na Tabela
1.
Tabela 1. Dados climáticos observados durante o período experimental
Mês Precipitação (mm) Temperatura (ºC)
Máxima Mínima Média
outubro/2008 0,0 33,7 21,7 27,7 novembro/2008 0,0 34,8 23,2 28,7 dezembro/2008 54,6 33,7 22,6 27,1 janeiro/2009 45,7 34,3 23,0 27,5 fevereiro/2009 127,0 32,4 22,1 26,1 março/2009 152,9 33,0 22,2 26,3 abril/2009 223,6 31,1 22,3 25,5 maio/2009 85,4 28,8 21,2 24,2 Junho/2009 14,5 29,7 19,5 23,6 Dados obtidos da Estação Agrometeorologia de Bebedouro, Embrapa Semiárido, Petrolina-Pernambuco.

90
Foram avaliados três períodos de descanso 4,5; 6,5 e 8,5 (número de
folhas/perfilho) e dois resíduos pós-pastejo 5 e 10 cm, em um delineamento
experimental inteiramente casualizado em arranjo fatorial 3x2 com quatro repetições
(piquetes). A área experimental foi composta de 0,82 ha de pastagem cultivada com
capim-tifton 85 irrigada por aspersão com canhões com a distribuição da água durante o
dia. Utilizou-se o método de pastejo sob lotação intermitente, com lotação variável,
mantendo-se animais prova nos piquetes e também animais reguladores para ajuste da
taxa de lotação, segundo a técnica da taxa de lotação variável. A pastagem foi
subdivida, com tela campestre, em 37 piquetes de 180 m2 cada, destes piquetes 24 foram
considerados amostrais. Foram utilizados 30 ovinos de prova e um número variável de
animais de equilíbrio, da raça Santa Inês, machos castrados, com peso corporal inicial
(PC) em torno de 20 kg, os quais foram previamente identificados, vermifugados e, em
grupos de cinco, distribuídos por sorteio em seis tratamentos. Foram efetuadas
adubações de reposição com nitrogênio, em cada piquete, no momento da saída dos
animais, nas seguintes doses: 0,971; 1,110; 1,320; 1,475; 1,666 e 1,800 g, de acordo
com o período de descanso.
O solo da área experimental é classificado como Podzólico Amarelo Eutrófico
Latossólico (EMBRAPA, 1999), teve suas características químicas avaliadas (Tabela 2)
em outubro de 2008. A área foi adubada em função das necessidades da análise do solo.
A adubação com fósforo e potássio foi realizada a lanço e em cobertura, na forma de
superfosfato simples e cloreto de potássio, respectivamente, em 02 de novembro de
2008 em uma aplicação com 120 kg/ha de P2O5 e 100 kg/ ha de K2O. A adubação
nitrogenada (600 kg de N/ha) foi fracionada em aplicações, ao dos ciclos de pastejo,
após a saída dos animais dos piquetes, na forma de uréia, de acordo com os períodos de
descanso.
Tabela 2. Características químicas do solo da área experimental em duas profundidades
Profundidade MO pH:H2O P K Al Ca Mg H +
Al CTC SB
-----(cm)---- g/Kg-
1 1: 2,5 mg/dm3 ----Cmolc dm-3---- %
0 – 5 24,14 6,18 41,51 0,21 0,05 2,45 0,94 1,83 5,48 66,41 5 - 20 7,10 6,09 22,67 0,14 0,05 1,33 0,59 1,16 3,25 63,51

91
Foram analisadas as amostras das frações folha e colmo, obtidas na avaliação da
estrutura do dossel no pré-pastejo. A composição químico-bromatológica das amostras
das frações folha e colmo foram determinadas no Laboratório de Nutrição Animal,
pertencente a Embrapa Semiárido, no Município de Petrolina-Pernanbuco. Após a
secagem das amostras em estufa de ventilação forçada a 55° C por 72 horas, as mesmas
foram trituradas em moinho de facas tipo “Thomas Wiley”, utilizando-se peneira com
malha de 1,0 mm e acondicionadas em sacos fechados. Foram determinados os teores
de fibra insolúvel em detergente neutro (FDN), fibra insolúvel em detergente ácido
(FDA), lignina (LIG), proteína bruta (PB), segundo metodologias descritas em Silva e
Queiroz (2002). O estrato etéreo foi determinado por meio do extrator de gordura,
modelo ANKOM® XT – 10. A Lignina foi analisada com a adição de ácido sulfúrico
72% no resíduo insolúvel da determinação da FDA. Os teores de Hemicelulose foram
obtidos pela diferença entre os teores de FDN e FDA. Para a determinação da
digestibilidade in vitro da matéria seca (DIVMS), foi utilizado o fermentador artificial
Daisy II® (ANKOM - Technology Corporation).
Os dados foram analisados por meio de análise de variância e teste de
comparação de médias. Para comparar o efeito dos tratamentos, foi efetuada análise de
variância do efeito dos períodos de descanso e dos resíduos pós-pastejo, na média de
todos os ciclos, no fatorial completo. Para avaliar o efeito dos ciclos sucessivos de
pastejo, foi efetuada análise de variância dos ciclos dentro de cada um dos seis
tratamentos. As médias dos ciclos foram comparadas por meio do teste de Tukey, ao
nível de 5% de probabilidade. Como ferramenta de auxílio às análises estatísticas,
utilizou-se o procedimento GLM do programa estatístico SAS (SAS INSTITUTE,
2003).

92
RESULTADOS E DISCUSSÃO
Composição química e digestibilidade in vitro da matéria seca da fração lâmina foliar
A matéria seca da lâmina foliar (MS) foi afetada apenas pelos PDs (P 0,05),
com valores de 24,49; 26,17 e 26,75 kg/ha para os pastos manejados sob PD de 4,5; 6,5
e 8,5 fol/perf, respectivamente (Tabela 3). Os pastos sob PD de 4,5 fol/perf e resíduos
de 10 e 5 cm, apresentaram menor teor de MS, que é justificado pelo fato da planta
quando nova apresentar altos teores de água, e, quanto mais próximo da maturidade esse
teor é reduzido e ocorrendo um aumento nos teores de MS. De acordo com Drudi e
Favoreto (1987) à medida que se prolonga o intervalo de pastejo, o teor de MS de
forragem tende a aumentar. Comportamento semelhante foi observado por Oliveira et
al. (2000) avaliando o valor nutritivo do capim-tifton 85 em diferentes idades de
rebrota, com valores semelhantes aos encontrados nesta pesquisa, para a fração lâmina
foliar. O efeito dos ciclos de pastejo (P 0,05) sobre o teor de matéria seca foi
observado para os pastos sob PD de 4,5 fol/perf e resíduos de 10 e 5 cm; 6,5 e 8,5
fol/perf e resíduo de 10 cm (Tabela 4) onde os maiores teores de matéria seca,
corresponderam aos meses de maior precipitação.
Houve efeito da interação PD x RPP (P 0,05) para a matéria mineral (MM) da
lâmina foliar (Tabela 3). Os pastos manejados sob com PD de 6,5 fol/perf e resíduo de
10 e 5 cm, apresentaram maior teor de MM, quando comparado aos demais pastos. O
efeito dos ciclos de pastejo (P 0,05) sobre o teor de matéria mineral foi observado para
os pastos sob PD de 4,5 fol/perf e resíduos de 10 e 5 cm; 6,5 e 8,5 fol/perf e resíduo de
10 cm (Tabela 4) no qual, para este componente, foi verificado valores variáveis ao
longo do período experimental.

93
Tabela 3. Composição química e digestibilidade in vitro da matéria seca da lâmina foliar do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de letras maiúsculas na coluna e minúsculas na linha diferem entre si (P<0,05) pelo teste de Tukey.
Os valores de proteína bruta (PB) foram reduzidos com o aumento no período de
descanso PD (P 0,05) (Tabela 3). Os menores teores foram observados nos pastos
Resíduo Pós-Pastejo Período de Descanso
4,5 (fol/perf)
6,5 (fol/perf)
8,5 (fol/perf)
Média
Matéria seca (%) CV = 4,69 10 cm 23,90 25,62 27,37 25,63 5 cm 25,08 26,72 26,13 25,98
Média 24,49b 26,17a 26,75a Matéria mineral (% MS) CV = 5,43
10 cm 6,90Ba 7,19Ab 8,07Aa 7,39 5 cm 7,33Ba 8,02Aa 6,95Bb 7,43
Média 7,12 7,60 7,51 Proteína bruta (%MS) CV = 4,37
10 cm 13,85 14,63 12,09 13,52 5 cm 13,57 14,02 11,75 13,11
Média 13,71a 14,33a 11,92b Extrato etéreo (% MS) CV = 5,65
10 cm 2,65 2,60 3,00 2,75 5 cm 2,61 2,70 3,01 2,83
Média 2,63b 2,68b 3,05a Fibra insolúvel em detergente neutro (% MS) CV = 1,26
10 cm 66,31 70,80 73,78 70,28A 5 cm 64,53 69,22 73,21 68,99B
Média 65,42c 70,01b 73,48a Fibra insolúvel em detergente ácido (% MS) CV = 3,97
10 cm 27,49 29,68 31,45 29,54A 5 cm 25,71 28,48 31,39 28,53B
Média 26,60c 29,08b 31,42a Hemicelulose (% MS) CV = 2,31
10 cm 37,69 39,88 42,20 39,92 5 cm 37,83 41,71 42,26 40,60
Média 37,76c 40,79b 42,23a Lignina (% MS) CV = 10,02
10 cm 2,18 2,06 2,42 2,22A 5 cm 1,97 2,00 2,10 2,02B
Média 2,07 2,03 2,26 Digestibilidade in vitro da matéria seca (% MS) CV = 4,01
10 cm 71,86 69,24 70,64 70,58 5 cm 69,98 71,37 68,44 69,91
Média 70,92 70,30 69,54

94
manejados sob PD de 8,5 fol/perf (11,92 %) os quais diferiram dos (13,71 e 14,33 %)
encontrados nas frações de lâmina foliar dos pastos sob PD de 4,5 e 6,5 fol/perf, sendo
que entre estes dois últimos não se evidenciaram (P 0,05) diferenças. Os teores
observados para PB nos pastos com menor PD, onde a forragem foi colhida em estágio
vegetativo inicial, provavelmente, devem-se às elevadas percentagens de folhas e baixas
porcetagens de colmos, além da conseqüente elevada relação lâmina foliar/haste.
Segundo El-Memari Neto et al. (2002) as folhas são notadamente mais digestíveis e
nutricionalmente mais ricas. Castro et al. (2004) trabalhando com Brachiaria brizantha
cv. Marandu com idade de corte de 28, 56, 84 e 112 dias, verificaram que os valores de
PB diminuíram com o aumento da idade de corte. O teor de PB aumentou ao longo dos
ciclos de pastejo (P 0,05) para os pastos sob PD de 4,5; 6,5 e 8,5 fol/per e resíduo de 10
cm (Tabela 4), este aumento provavelmente foi devido ao manejo da adubação em
associação a maior precipitação pluvial ocorrida ao longo do período experimental.
O extrato etéreo da lâmina foliar teve efeito do PD (P 0,05) para lâmina foliar
(Tabela 3). Os pastos sob PD de 8,5 fol/perf e resíduos de 10 e 5 cm, apresentaram
maiores teores de extrato etéreo. Gonçalves (2006) avaliando o efeito de três períodos de
descanso (1,5; 2,5 e 3,5 fol/perf) para capim-tanzânia, verificou teores de extrato etéreo
na ordem de 2,6; 2,5 e 2,8. O teor de extrato etéreo reduziu-se ao longo dos ciclos de
pastejo para os pastos sob PD de 4,5 fol/perf e resíduo de 5 cm e 6,5 e 8,5 fol/perf e
resíduo de 10 cm, onde se observa uma elevação até a metade do período experimental
(Tabela 4).
Balsalobre et al. (2001) relatam que as maiores mudanças que ocorrem na
composição das plantas forrageiras são aquelas decorrentes de sua maturidade. A
maioria das espécies forrageiras sofre declínio no seu valor nutritivo com o aumento da
idade, resultando da menor relação lâmina foliar/colmo combinada com a crescente
lignificação da parede celular.
Foi verificado efeito do PD e RPP (P 0,05) para a fibra insolúvel em detergente
neutro (FDN) da fração laminar foliar (Tabela 3). Os pastos sob PD 8,5 fol/perf e
resíduo de 5 e 10 cm, apresentaram maiores teores. O aumento no percentual de FDN
com o avanço na idade de corte indica redução na proporção do capim passível de ser
digerido pelo animal. Esse fato pode ser atribuído, provavelmente, ao espessamento e
lignificação da parede celular vegetal com o avanço da idade da planta, conforme

95
determinado por Bhering (2006). A maioria dos trabalhos com capim do gênero
Cynodon tem mostrado aumento no teor de FDN com a maturidade da planta
(HERRERA & HERNANDEZ, 1988; PALHANO, 1990; GOMIDE, 1996; RIBEIRO et
al., 1998a; CASTRO et al., 1998). Maiores teores de FDN foram também observados
por Carnevalli et al. (1999) quando pastagens de Tifton-85 foram manejadas à maiores
alturas. Observou-se maior concentração de FDN nos pastos manejados com resíduo
mais alto, provavelmente devido a maior quantidade de parede celular existente nas
lâminas foliares que não foram colhidas pelos animais em pastejo. Houve efeito dos
ciclos de pastejo sobre o teor de fibra insolúvel em detergente neutro (P 0,05).
Observou-se menor concentração de FDN nos últimos ciclos de pastejo (Tabela 4), para
os pastos sob PD de 4,5 fol/perf e resíduos de 10 e 5 cm e 6,5 fol/perf e resíduo de 10
cm, talvez o período de maior precipitação pluvial, tenha promovido redução nos
componentes fibrosos da parede celular e, conseqüentemente, redução no teor de fibra
insolúvel em detergente neutro.
Foi verificado efeito do PD e RPP (P 0,05) para a fibra insolúvel em detergente
ácido (FDA) da fração laminar foliar (Tabela 3). Os pastos sob PD 8,5 fol/perf e resíduo
de 5 cm, apresentaram maiores teores. As maiores mudanças que ocorrem na
composição química das plantas forrageiras são aquelas que acompanham a maturação.
À medida que a planta envelhece, a proporção dos componentes digestíveis tende a
diminuir e a fibra, aumentar. As variações observadas nas frações folha dos PD 4,5 e 8,5
fol/perf estão associadas ao avanço no estádio de maturação das plantas, que passaram a
ter seus tecidos mais lignificados, reduzindo assim a sua qualidade e facilidade de
fragmentação e digestão. Em relação à altura do resíduo, podemos observar que os
pastos mantidos a um resíduo de 5 cm, resultaram a menores teores de fibra insolúvel
em detergente neutro, quando comparado aos pastos com altura residual de 10 cm. Silva
et al. (1994) relataram que pastos manejados com baixas pressões de pastejo
apresentavam teores de FDA de lâminas foliares significativamente superiores àquelas
manejadas com média e alta pressão de pastejo. Foi verificado efeito dos ciclos de
pastejo (P 0,05) para os pastos sob PD de 6,5 e 8,5 fol/perf e resíduo de 10 cm (Tabela
4), com comportamento semelhante à fibra insolúvel em detergente neutro,
provavelmente devido a precipitação pluvial.

96
O teor de hemicelulose para a lâmina foliar foi afetado apenas pelo PD (P 0,05)
para os pastos sob PD de 8,5 fol/perf que apresentaram maiores valores (Tabela 3),
resultado do maior avanço ocorrido na maturação dos tecidos da planta, em comparação
aos teores encontrados nos pastos submetidos ao menor PD, resultado semelhante foi
encontrado por Velásquez et al. (2010) ao avaliarem a composição químico-
bromatológica de forrageiras tropicais em diferentes idades de corte, incluindo capim-
tifton 85, relataram resultados semelhantes aos observados neste experimento. Os ciclos
de pastejo afetaram o teor de hemicelulose (P 0,05), nos pastos com PD de 4,5 fol/perf
e resíduos de 10 e 5 cm (Tabela 4), que foi mais elevado apenas no sexto ciclo de
pastejo.
Foi verificado efeito do RPP (P 0,05) para o teor de lignina da Lâmina foliar
(Tabela 3). Os maiores teores de lignina foram observados para os pastos com maior
RPP. Janusckiewicz et al. (2010) também verificaram maior teor de lignina em função
do maior resíduo pós-pastejo. Mesmo que não tenha sido verificado diferença
significativa entre os PDs nos quais os pastos foram submetidos, o teor de lignina é
bastante relacionado com o grau de maturidade da planta, pois, com o avanço da idade,
aumenta o grau de lignificação dos tecidos. Os teores de lignina encontram-se bem
abaixo dos relatados por Oliveira et al. (2000) que obtiveram teores que variaram de
4,10 a 9,23, aos 14 e 70 dias de rebrota, para o Tifton 85. Foi verificado efeito dos ciclos
de pastejo (P 0,05) apenas para os pastos sob PD de 4,5 e 5 cm de resíduo, onde pode-
se observar uma redução do teor de lignina ao longo do período experimental (Tabela
4).
Também não foi verificado efeito dos PD, RPP e da interação (P 0,05) para o
coeficiente de digestibilidade in vitro da matéria seca (DIVMS) (Tabela 3). No entanto
pode-se inferir que a faixa de PD de máxima digestibilidade in vitro correu nos PDs de
4,5 e 6,5 fol/perf. Os valores de digestibilidade in vitro encontrados nesta pesquisa,
estão próximos aos encontrados por Oliveira et al. (2000). Do ponto de vista de nutrição
animal, os coeficientes de DIVMS encontram-se adequados, pois, de acordo com
Hamilton et al. (1970) valores acima de 65% para a DIVMS são indicativos de
adequado valor nutritivo, permitindo bom consumo de energia digestível pelo animal.
Não houve efeito dos ciclos de pastejo (P 0,05) (Dados não apresentados).

97
Tabela 4. Efeito dos ciclos de pastejo sobre a composição química e digestibilidade in vitro da matéria seca da fração lâmina foliar do capim-tifton 85 manejado sob três períodos de descanso e dois resíduo pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
4,5 fol/perf e resíduo de 10 cm Matéria seca 19,67d 21,83cd 21,34cd 20,50d 23,82bc 25,34ab 26,43ab 26,22ab 26,56ab 27,30a 23,90 5,18 M. mineral 7,91a 6,28bc 7,69ab 5,60c 5,35c 6,43abc 7,45ab 7,45ab 7,39ab 7,45ab 6,9 9,30
Proteína bruta 12,54f 12,76ef 14,59b 14,04bc 14,10bc 13,98bc 13,64cd 13,88bcd 13,19de 16,10a 1388 2,33 FDN 63,98b 63,65b 64,10b 64,80b 63,36b 72,24a 63,03b 63,03b 63,30b 63,30b 64,42 1,64
Hemicelulose 36,93b 37,96b 37,59b 39,35b 37,64b 47,50a 25,90c 38,24b 37,47b 38,28b 37,68 7,46 4,5fol/perf e resíduo de 5 cm
Matéria seca 20,54d 23,44bcd 21,99d 23,03cd 27,69a 27,44a 26,41abc 26,66ab 28,56a - 25,08 5,89 M. mineral 7,20abc 8,29ab 7,88abc 5,91c 6,16bc 6,40bc 7,71abc 7,71abc 8,76a - 6,47 13,38
Extrato etéreo 2,48bc 2,85ab 2,82ab 3,49a 2,29bc 2,60b 2,69b 1,78c 2,53b - 2,61 11,34 FDN 72,36a 66,46b 66,30bc 66,30bc 66,30bc 65,70bc 65,83bc 64,92bc 62,83bc - 66,31 2,36
Hemicelulose 39,31b 38,16b 39,59b 36,20b 39,02b 46,19a 26,70c 40,09ab 35,24b - 37,83 7,03 Lignina 2,77a 2,73a 2,76a 1,98ab 1,86ab 2,12ab 1,53b 2,00ab 1,87ab - 2,18 18,89
6,5fol/perf e resíduo de 10 cm Matéria seca 23,09cd 23,98bc 21,91d 22,72d 28,44a 30,60a 27,54ab 26,98ab - - 25,65 6,99 M. mineral 8,34a 6,06b 6,06b 7,29ab 7,29ab 7,67a 7,39ab 7,45ab - - 7,19 9,32
Proteína bruta 11,37b 15,84a 13,29b 12,73b 13,02b 16,89a 16,93a 16,97a - - 14,63 6,34 Extrato etéreo 2,54ab 2,64ab 2,49ab 3,15a 3,19a 2,66ab 1,91b 2,25b - - 2,60 12,86
FDN 72,29ab 73,09a 71,91ab 71,91ab 70,46bc 68,50c 66,01d 59,57e - - 69,21 1,42 FDA 30,18a 26,69a 27,42a 29,86a 28,92a 32,44a 28,36a 23,98b - - 28,48 10,52
8,5fol/perf e resíduo de 5 cm Matéria seca 23,74c 23,02c 27,35bc 21,97c 32,89a 35,26a - - - - 27,37 9,38 M. mineral 8,50ab 9,68a 9,68a 6,36c 6,36c 7,83bc - - - - 8,06 9,69
Proteína bruta 10,12c 11,42bc 11,91bc 11,80bc 13,93a 13,37ab - - - - 12,09 7,56 Extrato etéreo 3,45a 3,64a 3,20ab 2,83bc 2,49c 2,36c - - - - 2,99 7,84
FDA 34,31a 33,72a 32,50ab 30,29ab 30,25ab 27,28b - - - - 31,39 8,14 Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

98
Composição química e digestibilidade in vitro da matéria seca da fração colmo
Não foi observado efeito dos PDs e dos RPPs na composição da matéria seca
(MS) da fração colmo, sendo afetada pela interação PD x RPP (P 0,05), para os pastos
sob PD de 8,5 fol/perf e resíduo de 10 cm (Tabela 5). O efeito dos ciclos de pastejo
(P 0,05) sobre o teor de MS foi observado para os pastos sob PD de 4,5 fol/perf e
resíduos de 10 e 5 cm; 6,5 e 8,5 fol/perf e resíduo de 10 cm e 8,5 fol/perf e resíduos de
10 e 5 cm (Tabelas 5 e 6).
Houve efeito da interação PD x RPP (P 0,05) para a matéria mineral (MM) (Tabela
5), os pastos com PD de 8,5 fol/perf e resíduo de 10 cm, apresentaram maior teor de
matéria mineral quando comparado aos demais pastos. O efeito dos ciclos de pastejo
(P 0,05) sobre o teor de MM foi observado para os pastos sob PD de 6,5 fol/perf e
resíduo de 10 cm e 8,5 fol/perf e resíduo de 10 cm (Tabelas 6).
Os teores de PB observados dos colmos foram bastante inferiores aos
observados na fração lâmina folia, estando de acordo com a literatura (SANTOS, 1997;
BRÂNCIO et al., 1997, DIOGO, et al., 1995). Foi verificado efeito do PD (P 0,05) para
o teor de proteína bruta (PB) (Tabela 5). Os menores teores nos pastos sob PD de 8,5
fol/perf (5,85%) os quais diferiram dos (7,17 e 6,13 %) encontrados nos colmos dos
pastos sob PD de 4,5 e 6,5 fol/perf, sendo que entre estes dois últimos não se
evidenciaram (P 0,05) diferenças. A redução nos teores de PB com o avanço da
maturidade das plantas provavelmente deve-se ao efeito de diluição destes na matéria
seca produzida (GOMIDE, 1996). O teor de proteína bruta aumentou ao longo dos
ciclos de pastejo (P 0,05) para os pastos sob PD de 4,5 fol/per e resíduo de 10 e 5 cm e
8,5 fol/perf e 5 cm de resíduo (Tabelas 5 e 6). O aumento no teor de PB ao longo dos
ciclos de pastejo, provavelmente, foi devido ao manejo da adubação nitrogenada
(BALSALOBRE et al., 2003).
O extrato etéreo (EE) não foi afetado pelo PD, RPP e da interação PD x RPP (P 0,05)
(Tabela 5). O teor de extrato etéreo se reduziu ao longo dos ciclos de pastejo para os
pastos sob PD de 4,5 fol/perf e resíduos de 10 e 5 cm e 6,5 fol/perf e resíduo de 10 cm e
8,5 fol/perf e resíduos de 10 e 5 cm (Tabela 6).

99
Tabela 5. Composição química e digestibilidade in vitro da matéria seca da fração colmo do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de letras maiúsculas na coluna e minúsculas na linha diferem entre si (P<0,05) pelo teste de Tukey.
Resíduo Pós-Pastejo Período de Descanso
4,5 (fol/perf)
6,5 (fol/perf)
8,5 (fol/perf)
Média
Matéria seca (%) CV = 4,69 10 cm 20,70Ba 18,26Cb 23,69Aa 20,89 5 cm 19,76BA 22,88Aa 18,71Cb 20,45
Média 20,23 20,57 21,20 Matéria mineral (% MS) CV = 5,43
10 cm 7,64Ba 7,26Ba 8,33Aa 7,74 5 cm 6,97Ba 7,63BA 7,63Aa 7,41
Média 7,31 7,45 7,98 Proteína bruta (%MS) CV = 4,37
10 cm 6,65 6,04 5,70 6,13 5 cm 7,68 6,23 5,99 6,64
Média 7,17a 6,13ab 5,85b Extrato etéreo (% MS) CV = 5,65
10 cm 2,34 2,34 2,44 2,37 5 cm 2,37 2,23 2,35 2,32
Média 2,35 2,29 2,39 Fibra insolúvel em detergente neutro (% MS) CV = 1,26
10 cm 76,06 79,67 81,34 79,02 5 cm 75,95 77,51 76,51 76,65
Média 76,01b 78,59a 78,93a Fibra insolúvel em detergente ácido (% MS) CV = 3,97
10 cm 35,92 39,78 41,28 38,99 5 cm 34,71 38,24 39,69 37,54
Média 35,31b 39,01a 40,48a Hemicelulose (% MS) CV = 2,31
10 cm 39,23 40,66 39,29 39,72 5 cm 39,59 38,34 38,85 39,92
Média 39,07 39,41 39,50 Lignina (% MS) CV = 10,02
10 cm 3,84 3,74 3,93 3,94 5 cm 3,67 4,04 4,12 3,84
Média 3,75 3,89 4,03 Digestibilidade in vitro da matéria seca (% MS) CV = 2,92
10 cm 61,96 63,56 60,96 62,16 5 cm 62,61 61,95 62,51 62,36
Média 62,29 62,75 61,73

100
O avanço no PD da fração colmo resultou em aumentos (P<0,05) dos teores de
FDN e FDA (Tabela 4). Este comportamento pode ser atribuído à intensa deposição de
lignina na parede celular. Enquanto os teores de hemicelulose não aumentaram com o
aumento do PD (P 0,05) (Tabela 4). Os teores de lignina da fração colmo mesmo não
tendo sido afetados pelos PD, RPP e da interação PD x RPP (P 0,05) se elevaram (Tabela
5). Mais altos teores de lignina em segmentos de colmo foram observados nos colmos
dos pastos com maior PD de 6,5 e 8,5 fol/perf.
Os teores de FDA e FDN da fração colmo apresentaram aumento ao longo dos
ciclos de pastejo (P<0,05). nos pastos manejados com PDs de 4,5; 6,5 e 8,5 fol/perf e
resíduos de 10 e 5 cm. Isto ocorreu devido ao envelhecimento do pasto e conseqüente
diminuição da taxa de alongação e aparecimento de folhas, com aumento no conteúdo
da parede celular nas plantas aumenta, indicando desenvolvimento de tecidos estruturais
em colmos (WILSON, 1994).
A digestibilidade da fração colmo não foi afetada pelos PD, RPP e da interação
PD x RPP (P 0,05) (Tabela 5). Observou-se que a variação dos valores de DIVMS dos
pastos manejados sob os três PDs e os dois RPPs foi de pequena magnitude. Esperava-
se mais intensa redução na digestibilidade com o desenvolvimento do colmo,
principalmente nos pastos sob maior PD, uma vez que, a idade mais avançada do colmo
apresentou maiores concentrações dos componentes estruturais (FDN, FDA e lignina) e
menores teores de PB. Foi verificado efeito dos ciclos de pastejo (P 0,05) sobre a
DIVMS apenas nos pastos manejados sob PD de 6,5 fol/perf e resíduo pós-pastejo de 10
cm, onde podemos observar aumento do coeficiente de DIVMS a partir do segundo
ciclo de pastejo, que corresponde a redução dos componentes estruturais FDN e FDA, o
que resultou na melhoria deste coeficiente.
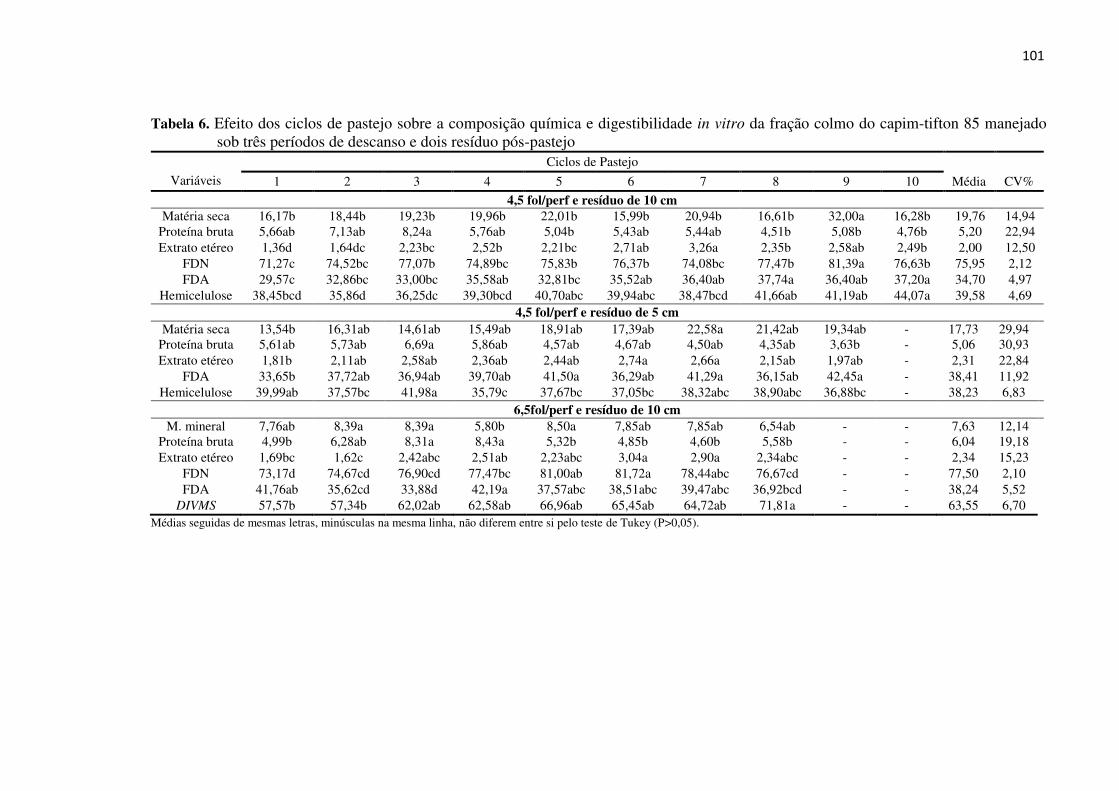
101
Tabela 6. Efeito dos ciclos de pastejo sobre a composição química e digestibilidade in vitro da fração colmo do capim-tifton 85 manejado sob três períodos de descanso e dois resíduo pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
4,5 fol/perf e resíduo de 10 cm Matéria seca 16,17b 18,44b 19,23b 19,96b 22,01b 15,99b 20,94b 16,61b 32,00a 16,28b 19,76 14,94
Proteína bruta 5,66ab 7,13ab 8,24a 5,76ab 5,04b 5,43ab 5,44ab 4,51b 5,08b 4,76b 5,20 22,94 Extrato etéreo 1,36d 1,64dc 2,23bc 2,52b 2,21bc 2,71ab 3,26a 2,35b 2,58ab 2,49b 2,00 12,50
FDN 71,27c 74,52bc 77,07b 74,89bc 75,83b 76,37b 74,08bc 77,47b 81,39a 76,63b 75,95 2,12 FDA 29,57c 32,86bc 33,00bc 35,58ab 32,81bc 35,52ab 36,40ab 37,74a 36,40ab 37,20a 34,70 4,97
Hemicelulose 38,45bcd 35,86d 36,25dc 39,30bcd 40,70abc 39,94abc 38,47bcd 41,66ab 41,19ab 44,07a 39,58 4,69 4,5 fol/perf e resíduo de 5 cm
Matéria seca 13,54b 16,31ab 14,61ab 15,49ab 18,91ab 17,39ab 22,58a 21,42ab 19,34ab - 17,73 29,94 Proteína bruta 5,61ab 5,73ab 6,69a 5,86ab 4,57ab 4,67ab 4,50ab 4,35ab 3,63b - 5,06 30,93 Extrato etéreo 1,81b 2,11ab 2,58ab 2,36ab 2,44ab 2,74a 2,66a 2,15ab 1,97ab - 2,31 22,84
FDA 33,65b 37,72ab 36,94ab 39,70ab 41,50a 36,29ab 41,29a 36,15ab 42,45a - 38,41 11,92 Hemicelulose 39,99ab 37,57bc 41,98a 35,79c 37,67bc 37,05bc 38,32abc 38,90abc 36,88bc - 38,23 6,83
6,5fol/perf e resíduo de 10 cm M. mineral 7,76ab 8,39a 8,39a 5,80b 8,50a 7,85ab 7,85ab 6,54ab - - 7,63 12,14
Proteína bruta 4,99b 6,28ab 8,31a 8,43a 5,32b 4,85b 4,60b 5,58b - - 6,04 19,18 Extrato etéreo 1,69bc 1,62c 2,42abc 2,51ab 2,23abc 3,04a 2,90a 2,34abc - - 2,34 15,23
FDN 73,17d 74,67cd 76,90cd 77,47bc 81,00ab 81,72a 78,44abc 76,67cd - - 77,50 2,10 FDA 41,76ab 35,62cd 33,88d 42,19a 37,57abc 38,51abc 39,47abc 36,92bcd - - 38,24 5,52
DIVMS 57,57b 57,34b 62,02ab 62,58ab 66,96ab 65,45ab 64,72ab 71,81a - - 63,55 6,70 Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

102
Tabela 7. Efeito dos ciclos de pastejo sobre a composição química e digestibilidade in vitro da fração colmo do capim-tifton 85 manejado sob três períodos de descanso e dois resíduo pós-pastejo
Ciclos de Pastejo
Variáveis 1 2 3 4 5 6 7 8 9 10 Média CV%
8,5 fol/perf e resíduo pós-pastejo de 10 cm Matéria seca 20,18ab 25,20a 21,08ab 20,77ab 11,66c 13,bc - - - - 18,71 18,96 M. mineral 8,04ab 8,38a 8,38a 7,40ab 7,30ab 6,29b - - - - 7,63 11,93
Proteína bruta 4,64b 7,99a 8,14a 8,13a 6,06ab 4,95b - - - - 6,65 14,39 Extrato etéreo 1,83b 1,92b 2,66ab 2,62ab 2,91a 2,68ab - - - - 2,43 16,83
FDN 81,16ab 81,35ab 80,51b 81,72ab 80,34b 82,98a - - - - 81,34 1,33
FDA 39,01b 41,58ab 41,29ab 41,55ab 41,69ab 42,55a - - - - 41,27 3,26
Hemicelulose 42,15a 39,77a 39,22a 40,17a 39,83a 34,60b - - - - 39,29 3,51
8,5 fol/perf e resíduo pós-pastejo de 5 cm Matéria seca 25,44b 20,68c 30,49a 20,89c 20,97c - - - - - 23,69 6,76
Extrato etéreo 1,88b 2,12ab 2,80a 2,29ab 2,67a - - - - - 2,35 14,26 FDN 72,90b 76m41b 76,06b 74,85b 82,33a - - - - - 76,51 2,58
Hemicelulose 35,14b 38,49ab 41,25ab 36,09ab 43,29a - - - - - 38,85 8,81 Lignina 3,05b 3,93ab 4,01ab 4,86a 4,37ab - - - - - 4,04 18,18
Médias seguidas de mesmas letras, minúsculas na mesma linha, não diferem entre si pelo teste de Tukey (P>0,05).

103
CONCLUSSÔES
O aumento no período de descanso apesar de produzir acréscimos nos teores de
matéria de seca, fibra insolúvel em detergente neutro e fibra insolúvel em detergente
ácido do capim-tifton 85, não reduziu a digestibilidade in vitro da matéria seca na fração
lâmina foliar.
Na altura residual de 10 cm foi verificada aumento nas concentrações de fibra
insolúvel em detergente neutro, fibra insolúvel em detergente ácido e lignina na fração
lâmina foliar.
O teor de proteína bruta das duas frações foi reduzido com o aumento do período
de descanso.
REFERÊNCIAS
ALEXANDRINO, E.; NASCIMENTO JUNIOR, D.; MOSQUIM, P. R. et al. Características morfogênicas e estruturais na rebrotação da Brachiaria brizantha cv. Marandú submetida a três doses de nitrogênio. Revista Brasileira de Zootecnia, v. 33, n. 6, p. 1372-1379, 2004. BALSALOBRE, M.A.A.; CORSI, M.; SANTOS, P.M. et al. Composição química e fracionamento do nitrogênio e dos carboidratos do capim-Tanzânia irrigado sob três níveis de resíduo pós-pastejo. Revista Brasileira de Zootecnia, v.32, n. 3, p. 519-528, 2003. BALSALOBRE, M. A. A.; NUSSIO, L. G.; MARTHA JÚNIOR, G. B. Controle de perdas na produção de silagem de gramíneas tropicais. In: SIMPÓSIO SOBRE MANEJO DA PASTAGEM, 2001, Piracicaba. Anais... Piracicaba: FEALQ, 2001. p. 890-911. BHERING, M. Produção e valor nutritivo do capim-elefante cv. Roxo (Pennisetum
purpureum Schum.) em diferentes idades de corte. 2006. 86f. Dissertação (Mestrado em Agricultura Tropical) - FAMEV – UFMT, Cuiabá. BRÂNCIO, P.A.; NASCIMENTO Jr., D.; MORAES, E.A. et al. Avaliação de pastagem nativa dos cerrados submetida à queima anual. 2. qualidade da dieta de bovinos. Revista Brasileira de Zootecnia, v.26, n.3, p.438-442, 1997. CARNEVALLI, R.A.; SILVA, S.C.; PEDREIRA, C.G.S. et al. Pasture and animal responses of Tifton-85 swards grazed by sheep under continuous stocking. In: INTERNATIONAL “GRASSLAND ECOPHYSIOLOGY AND GRAZING

104
ECOLOGY”, 1., 1999, Curitiba, PR. Proceedings... Curitiba: Universidade Federal do Paraná, 1999. p.357-361. CASTRO, G. H. F.; GRAÇA, D. S.; GONÇALVES, L. C.; BORGE. et al. Degradabilidade in situ da matéria seca e proteína bruta da Brachiaria brizantha cv. Marandu em quatro diferentes idades de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41, 2004, Campo Grande. Anais... Campo Grande: SBZ, 2004. CD-ROM. CASTRO, F.G.F.; HADDAD, C.M.; VIEIRA, A.C. et al. Efeito da idade e valor nutritivo de Cynodon nlemfluensis vanderyst var. nlemfluensis cv. Florico. In: Reunião Anual da Sociedade Brasileira de Zootecnia, 35, Botucatu, 1998. Anais... Botucatu, SBZ, 1998. P.578-579. CROWDER, L.V.; CHHEDA, H.R. Tropical grassland husbandry. New York: Longman, (Tropical Agriculture Series) 561p. 1982. DIOGO, J.M.S.; NASCIMENTO Jr., D.; TORREGROSA, S.L.J. Composição botânica da dieta selecionada por novilhos em pastagem natural de Viçosa-MG. Revista da Sociedade Brasileira de Zootecnia, v.24, n.6, p.884-895, 1995. DRUDI, A.; FAVORETTO, V. Influência da freqüência, época e altura do corte na produção e na composição química do capim-andropógon. Pesquisa Agropecuária Brasileira, Brasília, v. 22, n. 12, p. 1287-1292, 1987. EL-MEMARI NETO, A. C.; ZEOULA, L. M.; CECATO, U. Avaliação produtiva e química da Brachiaria brizantha no inverno e primavera. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: SBZ, 2002. CD-ROM. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Centro Nacional de Pesquisa de Solos. Sistema Brasileiro de Classificação de Solos. Brasília: EMBRAPA, 1999., 412p. EUCLIDES, V.P.B. Valor alimentício de espécies forrageiras do gênero Panicum. In: SIMPOSIO SOBRE PASTAGEM, 12., Piracicaba, 1995. Anais... Piracicaba: Fundação de Estudos Agrários Luiz de Queiroz, 1995. p.245-273. FORBES, T.D.A., COLEMAN, S.W. Forage intake and ingestive behaviour of catle grazing old world bluestems. Agronomy Journal, Madison, v.85,n.4,p.808-16, 1993. GOMIDE, C. C. C. Algumas características fisiológicas e químicas de cinco cultivares de Cynodon. 1996. Dissertação (Mestrado em Zootecnia) Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, 1996. GONÇALVES, J.S. Composição química e fracionamento dos carboidratos da biomassa de Panicum maximum cv. Tanzânia sob três períodos de descanso. Fortaleza–CE: UFC, 2006. 82f. Dissertação (Mestrado em Zootecnia), Universidade Federal do Ceará, 2006.

105
HAMILTON, R.I., LAMBOURNE, L.J., ROE, R. et al. Quality of tropical grasses for milk production. In: INTERNATIONAL GRASSLAND CONGRESS, 11, 1970, Surfers Paradise. Proceedings... Surfers Paradise: s.ed., 1970. p.860-864. HERRERA, R.S., HERNANDÉZ, Y. Efecto de la edad Del rebrote en algunos indicadores de la calidad de la bermuda cruzada-1. II. Componentes estructurales y digestibilidad de la massa seca. Pastos y Forrajes, v.11, n.2, p.177 182, 1988. JANUSCKIEWICZ, E. R.; MAGALHÃES, M. A.; RUGGIERI, A. C. et al. Massa de forragem, composição morfológica e química de capim-tanzânia sob diferentes dias de descanso e alturas de resíduo pós-pastejo. Bioscicence Journal, Uberlândia, v. 26, n. 2, p. 161-172, 2010 OLIVEIRA, M.A; PEREIRA, O.G.; MARTINEZ Y HUAMAN, C.A. et al. Características morfogênicas e estruturais do capim-bermuda „Tifton 85‟ (Cynodon sp.) em diferentes idades de rebrota. Revista Brasileira de Zootecnia, v. 29, n. 6, p. 1939-1948, 2000. PALHANO, A.L. Recrutamento de nutrientes e valor nutritivo de Cynodon
dactylon (L.) Pers. cv. Coastcross N1. Piracicaba: Escola Superior de Agricultura “Luis de Queiroz”, 1990. 122p. Dissertação (Mestrado em Zootecnia) - Escola Superior de Agricultura “Luis de Queiroz”, 1990. PEDREIRA, C.G.S. Avaliação de novas gramíneas de gênero Cynodon para a pecuária dos Estados Unidos, In.: WORKSHOP SOBRE O POTENCIAL FORRAGEIRO DO GÊNERO CYNODON, Juiz de Fora, 1996. Anais... Juiz de Fora, EMBRAPA-CNPGL, 1996, p. 111-125. REIS, R.A.; MELO G.M.P.; BERTIPAGLIA L.M.A. et al. Otimização da utilização da forragem disponível através da suplementação estratégica. In: REIS R.A.; SIQUEIRA, G.R.; BERTIPAGLIA, L.M.A. et al. (Eds.). Volumosos na produção de ruminantes. Jaboticabal: Funep, 2005. p.187-238. RIBEIRO, K.G.; PEREIRA, O.G.; GARCIA, R. et al Rendimento forrageiro e valor nutritivo do capim-tifton-85, em três freqüências de corte, sob diferentes doses de nitrogênio. In: Reunião Anual da Sociedade Brasileira de Zootecnia, 35, Botucatu, 1998. Anais... Botucatu, SBZ, 1998. p.542-544.
SANTOS, M.V.F. Métodos agronômicos para estimativa de consumo e de disponibilidade de forragem na Zona da Mata, Viçosa - MG. Viçosa, MG: Universidade Federal de Viçosa, 1997. 155p. Tese (Doutorado em Zootecnia) – Universidade Federal de Viçosa, 1997. SAS INSTITUTE. SAS system for Windows. Version9. 1. Cary: SAS Institute Inc. 2003. 2 CD – ROMs. SILVA, D. J.; QUEIROZ, A. C. Análise de alimentos: métodos químicos e biológicos. 3. ed. Viçosa: UFV, 2002. 235 p.

106
SILVA, D.S.; GOMIDE, J.A.; QUEIROZ, A.C. Pressão de pastejo em pastagem de capim-elefante anão (Pennisetum purpureum, Schum. cv. Mott): Efeito sobre o valor nutritivo, consumo de pasto e produção de leite. Revista da Sociedade Brasileira de Zootecnia, v.23, n.3, p.453-464, 1994b. VELÁSQUEZ, P. A. T.; BERCHIELLI, T. T.; REIS, R. A. et al. Composição química, fracionamento de carboidratos e proteínas e digestibilidade in vitro de forrageiras tropicais em diferentes idades de corte. Revista Brasileira de Zootecnia, v.39, n.6, p.1206-1213, 2010.
WILSON, J.R. Cell wall characteristcs in relation to forage by ruminantes. Journal Agric. Sci., 122:173-182, 1994.

107
CAPÍTULO 6
____________________________________________________________
Comportamento e desempenho de ovinos terminados em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo

108
Comportamento e desempenho de ovinos terminados em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
RESUMO: Este experimento foi conduzido com o objetivo de avaliar a resposta de
ovinos ao pastejo em capim-tifton 85. Foram utilizados 30 ovinos prova e número
variável de animais reguladores, com peso corporal (PC) de 20 kg, os quais foram
distribuídos em um delineamento experimental inteiramente casualizado em arranjo
fatorial 3x2, sendo três períodos de descanso (PD): 4,5; 6,5 e 8,5 (nº de folhas/perfilho)
e dois resíduos pós-pastejo (RPP): 5 e 10 cm. Para determinação da composição
químico-bromatológica e da digestiblidade in vitro da matéria seca foi realizada
simulação de pastejo no 1º e 4º dia de ocupação. Os PDs e os RPPs influenciaram o
tempo de pastejo dos animais. Os pastos manejados sob PD de 8,5 fol/perf e resíduo de
10 cm proporcionaram maior tempo de pastejo e ruminação aos animais comparados
aos demais pastos. O 4º dia de pastejo proporcionou maior tempo em pastejo e
ruminação. O consumo de sal e água foi bastante reduzido. As médias de taxa de
bocados verificadas nos pastos sob PD de 4,5 fol/perf demonstram que a mais baixa
biomassa obrigou os animais a aumentarem o número de bocados como forma de
otimizar o consumo de forragem. O 4º dia de pastejo proporcionou aumento da taxa e
número de bocados e redução da massa do bocado. Os PDs correspondentes a 4,5 e 6,5
fol/perf proporcionaram melhores desempenho animal. A maior capacidade de suporte,
em unidade animal, foi observada para os pastos manejados sob PD de 8,5 fol/perf.
Termos para indexação: consumo, pastejo, ruminação, valor nutritivo

109
Behavior and performance of sheep finished in tifton 85 grass managed under
three periods of rest and two post-grazing residues
ABSTRACT: This experiment was lead with the objective to evaluate the reply of
sheep to pastejo in capim-tifton 85. Sheep ones had been used 30 prove and changeable
number of regulating animals, with initial weight corporal (PC) of 20 kg, which had
been distributed in an experimental delineation entirely casualizado in factorial
arrangement 3x2, being three periods of rest (PD): 4,5; 6,5 and 8,5 (leaves/tiller) and
two post-grazing residues (RPP): 5 and 10 cm. For determination of the bromatologic-
chemical composition and the in vitro digestibility of the dry substance was carried
through simulation of pastejo in 1º and 4º day of occupation. The PDs and the RPPs had
influenced the time of pastejo of the animals. The grass managed under 8,5 PD of
leaves/tiller and residue of 10 cm had provided to greater time of grazing and
ruminating to the comparative animals to the too much grass. 4º day of grazing provided
to greater time in grazing and ruminating. The intake of salt and water sufficiently was
reduced. The averages of tax of bits verified in the grass under 4,5 PD of leaves/tiller
demonstrate that the lowest biomass compelled the animals to increase the number of
bits as form to optimize the fodder plant intake. 4º day of pastejo provided to increase of
the tax and number of bits and reduction of the mass of the bit. Better the 6,5 PDs
corresponding 4,5 and leaves/tiller had provided animal performance. The biggest
capacity of support, in animal unit, was observed for the grass managed under 8,5 PD of
leaves/tiller.
Index-terms: intake, grazing, ruminating, nutritive value.

110
INTRODUÇÃO
Para a eficiente exploração da pastagem é necessário o conhecimento das
relações existentes na interface planta-animal, o que envolve o estudo de como as
condições de pastejo interferem no comportamento ingestivo animal e no seu
desempenho, de forma a identificar condições de manejo adequadas à categoria animal
e ao sistema de produção adotado. A disponibilidade de forragem é um importante fator
no consumo, porém, a habilidade física do animal em colher a forragem e os efeitos da
estrutura do pasto no comportamento ingestivo podem ser determinantes no controle da
ingestão (PENNING et al., 1991). As práticas de manejo interferem no ritmo de
atividade dos animais em pastejo, tornando fundamental a avaliação do padrão diário de
variação do ritmo de atividade dos ovinos para a redução das interferências no
comportamento natural de pastejo (MEDEIROS et al., 2007).
A quantidade de alimento consumida em determinado período de tempo é
afetada conforme o número de refeições, a duração e a velocidade de ingestão, além de,
como especificado por Pedroso et al. (2004), variações no tempo de pastejo, na taxa de
bocados, no peso do bocado e na qualidade da forragem ingerida. Os diferentes estádios
fenológicos das forrageiras também fazem com que os animais utilizem diversas
estratégias para otimizar o consumo, podendo variar o peso e/ou freqüência do bocado
ou o tempo de pastejo, influenciando também o período de ruminação dos animais.
A avaliação do comportamento ingestivo constitui-se em ferramenta valiosa na
avaliação das dietas, possibilitando ajustar o manejo alimentar dos animais, que poderá
refletir em melhoria do desempenho produtivo. Nesse sentido, o conhecimento do
comportamento animal torna-se importante, pois possibilita promover o melhor
ajustamento da produção animal (GRANDIN, 2000). De acordo com Raslan (2008) os
conhecimentos gerados em função de investigações nos comportamentos dos animais
têm sido utilizados com uma freqüência cada vez maior, para melhorar as estratégias de
manejo e ajudar no desenvolvimento de novas técnicas de produção.
As atividades diárias dos ovinos compreendem períodos alternados de pastejo,
ruminação e ócio, sendo que a duração e distribuição dessas atividades são
influenciadas pelas características da pastagem, manejo, condição climática, categoria e
atividades dos animais em grupo (FISCHER et al., 1998).

111
Em pastejo intermitente, em especial sob pressões de pastejo médio e alta, ao
longo do período de ocupação do piquete, há redução na disponibilidade de forragem e
mudanças na estrutura das plantas, principalmente na relação lâmina foliar/colmo, que
pode afetar de forma severa o comportamento ingestivo e, conseqüentemente, a
produção animal (CHACON & STOBBS, 1976).
Considerando a hipótese de que a estrutura do pasto pode interferir no consumo
de forragem, no desempenho animal produtivo e no comportamento de ovinos, este
experimento foi conduzido com o objetivo de avaliar, a resposta de ovinos ao pastejo
em capim-tifton 85 manejados sob três períodos de descanso e dois resíduos pós-
pastejo.
MATERIAL E MÉTODOS
A pesquisa foi desenvolvida no Setor de Produção de Ovinos do Campo
Experimental de Bebedouro, pertencente à Embrapa Semiárido, localizado no
Município de Petrolina-Pernambuco. Durante o mês de outubro foi efetuada a divisão
da área em piquetes e ajuste da metodologia de trabalho. A coleta de dados ocorreu
entre os meses de novembro de 2008 a junho de 2009, totalizando nove meses.
O clima da região é classificado como quente e seco, sendo caracterizada por
duas estações bem definidas, uma chuvosa, de janeiro a maio, e outra seca, de junho a
dezembro. Os dados sobre as condições de temperaturas máxima e mínima do ar e
precipitação pluvial durante o período experimental foram obtidos da Estação
Agrometeorologia de Bebedouro, Petrolina-Pernambuco e são apresentados na Tabela
1.

112
Tabela 1. Dados climáticos observados durante o período experimental
Mês Precipitação (mm) Temperatura (ºC)
Máxima Mínima Média
outubro/2008 0,0 33,7 21,7 27,7 novembro/2008 0,0 34,8 23,2 28,7 dezembro/2008 54,6 33,7 22,6 27,1 janeiro/2009 45,7 34,3 23,0 27,5 fevereiro/2009 127,0 32,4 22,1 26,1 março/2009 152,9 33,0 22,2 26,3 abril/2009 223,6 31,1 22,3 25,5 maio/2009 85,4 28,8 21,2 24,2 Junho/2009 14,5 29,7 19,5 23,6 Dados obtidos da Estação Agrometeorologia de Bebedouro, Embrapa Semiárido, Petrolina-Pernambuco.
Foram avaliados três períodos de descanso 4,5; 6,5 e 8,5 (número de
folhas/perfilho) e dois resíduos pós-pastejo 5 e 10 cm, em um delineamento
experimental inteiramente casualizado em arranjo fatorial 3x2 com cinco repetições
(animais). A área experimental foi composta de 0,82 ha de pastagem cultivada com
capim-tifton 85 irrigada por aspersão com canhões com distribuição de água durante o
dia. Utilizou-se o método de pastejo sob lotação intermitente, com lotação variável,
mantendo-se animais prova nos piquetes e também animais reguladores para ajuste da
taxa de lotação, segundo a técnica da taxa de lotação variável. A pastagem foi
subdivida, com tela campestre, em 37 piquetes de 180 m2 cada, destes piquetes 24 foram
considerados amostrais. Foram utilizados 30 ovinos de prova e um número variável de
animais de equilíbrio, da raça Santa Inês, machos castrados, com peso corporal (PC)
inicial de ± 20 kg, os quais foram previamente identificados, vermifugados e, em grupos
de cinco, distribuídos por sorteio em seis tratamentos. Foram efetuadas adubações de
reposição com nitrogênio, em cada piquete, no momento da saída dos animais, nas
seguintes doses: 0,971; 1,110; 1,320; 1,475; 1,666 e 1,800 g, de acordo com o período
de descanso.
Após alguns ciclos de pastejo, com a intensificação do efeito dos tratamentos
sobre a estrutura do pasto, foi conduzido o ensaio de comportamento ingestivo, ao longo
de 24 horas, nos dias 30/01 e 02/02; 26/05 e 29/05 de 2009 que correspondem ao 1º e 4º
dia de pastejo (em cada tratamento) (Figuras 1 e 2), avaliados simultaneamente. Os
animais prova foram devidamente identificados com colares coloridos para facilitar a

113
visualização à distância. Para a realização da avaliação de comportamento utilizaram-se
observadores treinados, cada um responsável pela observação dos cinco animais de
prova de cada tratamento.
Figuras 1 e 2. (1): animais no 1º dia de pastejo e (2): animais no 4º dia de pastejo.
As anotações foram feitas a cada dez minutos, por meio de observação visual,
sendo registradas as atividades de pastejo, ruminação e ócio. O tempo de pastejo
representou o período em que o animal apreendeu ou selecionou a forragem. O tempo
de ruminação foi o período em que o animal não pastejou e mastigou o bolo alimentar
que retornou do rúmen, no qual foram observados movimentos mandibulares do animal
(TREVISAN et al., 2004). Denominou-se ócio como o período em que os animais não
realizaram nenhuma destas atividades, isto é, o período em que os animais não estão
comendo, ruminando ou ingerindo água (COSTA et al., 1983). Também foram tomados
os dados referentes à taxa de bocados dos animais (bocados/minuto), contando-se o
número de bocados por animal durante o processo de pastejo, dividindo-se esse número
pelo tempo transcorrido em segundos e multiplicando-se o resultado por 60, a fim de se
obter a taxa em bocados por minuto (FORBES & HODGSON, 1985). Multiplicando-se
a taxa de bocados pelo tempo de pastejo, obteve-se o número diário de bocados (NDB).
Dividindo-se o consumo de forragem estimado nos dias de avaliação do comportamento
ingestivo (em g MS) pelo NDB, obteve-se a massa estimada de cada bocado, em g
MS/bocado (FORBES, 1988).
Os animais de prova foram pesados a cada 20 dias para estimativa do desempenho
e os reguladores foram pesados mensalmente para acompanhamento do ganho de peso
(Figuras 3 e 4). As pesagens foram realizadas após jejum de alimento e água de 16
1 2

114
horas (COATES & PENNING, 2000). O ganho de peso total (GPT) foi determinado de
acordo com a equação: GPT = PCF – PCI, sendo: PCF = peso corporal final e PCI =
peso corporal inicial. Já o ganho médio diário (GMD) foi obtido dividindo-se o valor de
GPT pelo período em que os animais permaneceram no sistema. O peso corporal final
foi o peso obtido na última pesagem.
Figuras 3 e 4. (3): Preparo da balança, no centro de manejo e (4): pesagem dos animais.
Nos dias correspondentes à entrada e saída dos animais nos piquetes (1º e 4º dias
de pastejo), foi coletada a massa de forragem, aparentemente consumida pelos animais
prova, em vinte pontos diferentes do piquete por meio da técnica de simulação de
pastejo (GIBB & TREACHER, 1976). Foram colhidos 400g de massa fresca de
forragem, que foram pesados e submetidos à pré-secagem em estufa de ventilação
forçada a uma temperatura de 550C, até atingir o peso constante. Em seguida, as
amostras foram moídas em peneira com crivos de 1mm e armazenadas em sacos
plásticos para a realização das análises químico-bromatológicas.
A estimativa do consumo de matéria seca foi efetuada com a coleta total de fezes,
compreendendo três dias de coleta ao fim de cada ciclo de pastejo. As fezes foram
coletadas e pesadas diariamente (às 8h:30). Após a homogeneização do material,
retirou-se uma alíquota diária de 10% para preparação de uma amostra composta por
animal, que foi armazenada em freezer a -5°C. Ao término do ensaio as amostras foram
descongeladas à temperatura ambiente em seguida, procedeu-se à pré-secagem das fezes
em estufa de ventilação forçada a 55oC, até atingir o peso constante. As amostras foram
3 4

115
trituradas em moinho tipo Wiley, em peneira com crivos de 1 mm para determinação
dos teores de matéria seca (MS), conforme descrito por Silva e Queiroz (2002).
Figuras 5 e 6. (5): preparo dos animais para coleta de fezes e (6): animal com bolsa coletora de
fezes e arreio.
A determinação do consumo de forragem pelos animais procedeu-se com base nas
estimativas da produção fecal do animal e da digestibilidade in vitro da forragem,
conforme indicado por Astigarraga (1997):
Consumo (kg/animal. dia) = produção fecal total (kg MS/dia)
(1 - DIVMS)
Na análise dos aspectos comportamentais foi adotada inicialmente análise de
variância no fatorial completo, formado pelos três períodos de descanso e dois resíduos
pós-pastejo. O desdobramento da interação no fatorial completo foi efetuado quando
esta foi significativa ao nível de 5% de probabilidade. Na comparação das médias foi
realizado o teste de Tukey, ao nível de 5% de probabilidade. Em seguida, avaliado o
efeito do dia de pastejo para cada variável comportamental, na média dos seis
tratamentos, definindo-se a diferença estatística entre o primeiro e último dia de pastejo
por meio do teste F, ao nível de 5% de probabilidade. O desempenho animal, a
composição químico-bromatológica e a digestibilidade in vitro da matéria seca, foram
submetidos a análise de variância e teste de comparação de médias. O desdobramento da
interação no fatorial completo foi efetuado quando esta foi significativa ao nível de 5% de
5 6

116
probabilidade. A comparação de médias foi efetuada utilizando-se o teste de Tukey, ao
nível de 5% de probabilidade. Como ferramenta de auxílio às análises estatísticas, utilizou-
se o procedimento GLM, do programa estatístico SAS (SAS INSTITUTE, 2003).
RESULTADOS E DISCUSSÃO
O tempo de pastejo (min/dia) foi influenciado pela interação PD x RPP (P 0,05)
(Tabela 2). Os ovinos mantidos nos pastos sob PD de 6,5 fol/perf e resíduo de 5 cm
despenderam menor tempo diário pastejando, ao passo que os animais mantidos nos
pastos sob o PD 8,5 fol/perf tenderam a apresentar maior tempo diário de pastejo,
comportamento semelhante ao ocorrido nos pastos manejados sob o PD 4,5 fol/perf.
Sabe-se que os animais em pastejo apresentam um requerimento diário de nutrientes,
recorrendo a essa atividade até saciá-lo. Os ovinos nos pasto manejados sob PD 6,5
fol/perf, despenderam menor tempo com pastejo, provavelmente devido ao pasto
apresentar elevado valor nutritivo e estrutura adequada ao pastejo (alta densidade de
perfilhos e relação lâmina foliar/colmo). De acordo com Carvalho e Moraes (2005) um
pasto com uma massa de forragem adequadamente dimensionada, com alta proporção
de folhas jovens e, ou, uma massa de forragem residual pós-pastejo com elevada
proporção de folhas, representa um ambiente confortável e adequado do ponto de vista
alimentar para os animais em pastejo. Na situação contrária, os ovinos no pasto sob o
PD 8,5 fol/perf necessitaram de maior tempo de pastejo, a fim de atingir seus
requerimentos diários, já que o valor nutritivo do pasto sob PD mais longo não foi
elevado (Capítulo 5) e a estrutura (Capítulo 4) também não favoreceu o consumo (baixa
relação lâmina foliar/colmo) e por haver maior dispersão da massa de forragem ao
longo do dossel.
O tempo de ruminação (min/dia) foi influenciado pelo PD (P 0,05) (Tabela 2).
Esta atividade foi fortemente influenciada pelo tempo de pastejo e a qualidade da dieta,
ou seja, os pastos sob PD de 4,5 e 8,5 fol/perf, despenderam maior tempo em
ruminação, como consequência do maior tempo gasto com a atividade de pastejo. Como
as atividades do comportamento ingestivo são excludentes, a maior permanência dos
animais em pastejo, nos pastos sob PD de 4,5 e 8,5 fol/perf e resíduos de 10 cm,
implicou redução do tempo de ócio total, que diferiu entre o RPP (P<0,05) (Tabela 2).

117
Estes resultados similares aos verificados por Manzano et al. (2007) ao avaliarem o
comportamento ingestivo de novilhos sob suplementação em pastagens de capim-
tanzânia sob diferentes intensidades, verificaram que o maior tempo em ócio dos
animais poderia ser explicado pelos menores tempos de pastejo e ruminação.
Em relação às atividades consumo de sal e água, o período de permanência ou o
número de vezes, em que foram ao saleiro e ao cocho, foi pequeno, ocorrido devido à
idade do pasto, horário das avaliações e também pelos requerimentos dos animais. Foi
verificado efeito da interação PDxRPP (P 0,05) para o consumo de sal. Nos pastos sob
PD de 6,5 fol/perf e resíduo de 10 cm e 8,5 fol/perf e resíduos de 10 e 5 cm, foi
observado os menores consumo de sal (Tabela 3). A ingestão de água foi influenciada
pelo RPP (P 0,05) (Tabela 3). Os animais mantidos nos pastos manejados sob RPP de
10 cm foram mais vezes a procura de água, este comportamento pode estar associado ao
maior consumo de matéria seca (MS), fato observado para este resíduo (Tabela 6). A
freqüência de ingestão de água depende da temperatura (MENDES et al., 1976) e
também da natureza da dieta. A ingestão de água observada nos animais deste estudo foi
reduzida, provavelmente em decorrência da temperatura amena observada nos dias de
avaliação, associada a forragem verde, resultando em baixa ingestão.

118
Tabela 2. Atividades contínuas e pontuais de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Resíduo pós-pastejo
Períodos de Descanso (folhas/perfilho)
Média 4,5 6,5 8,5
Pastejo (min/dia)
10 cm 577,00Aa 520,00Aa 592,00Ba 563,00
5 cm 543,00Ab 514,50Bb 553,00Aa 536,83
Média 560,00 517,25 572,5 -
Ruminação (min/dia)
10 cm 525,00 517,50 537,00 513,50
5 cm 443,00 416,00 474,50 444,50
Média 484,00a 466,75b 505,75a -
Ócio (min/dia)
10 cm 338,00 403,00 350,00 363,66B
5 cm 454,00 510,50 373,50 446,00A
Média 396,00 456,75 361,75 -
Ingestão de Sal (nº de vezes/ovino x dia)
10 cm 1,20Aa 0,75Bb 0,70Ba 0,88
5 cm 1,15Aa 1,65Aa 0,50Bb 1,10
Média 1,17 1,20 0,60 -
Ingestão de Água (nº de vezes/ovino x dia)
10 cm 1,80 1,30 0,90 1,33A
5 cm 0,85 1,05 0,95 0,95B
Média 1,32 1,17 0,92 -
Médias seguidas de mesmas letras, maiúscula na coluna e minúscula na linha, não diferem entre si pelo teste de Tukey (P>0,05).
Houve influência do PD (P<0,05) para a taxa de bocado (Tabela 3). Nos pastos
manejados sob PD de 4,5 fol/perf foram verificados maiores taxas quando comparados
aos pastos manejados sob PD de 6,5 fol/perf. O animal em pastejo procura ajustar seu
comportamento ingestivo de modo a manter seu consumo mesmo com a variação na

119
estrutura da pastagem. Acredita-se que a superioridade da taxa de bocados dos ovinos
pastejando nestes pastos tenha sido em função da menor disponibilidade de forragem e
menor número de perfilhos, estando à massa de forragem mais dispersa ao longo do
perfil horizontal da pastagem. As médias de taxa de bocados verificadas nos pastos sob
PD de 4,5 fol/perf demonstraram que a baixa biomassa obrigou os animais a
aumentarem o número de bocados como forma de otimizar o consumo de forragem,
tendo em vista o suprimento das suas necessidades diárias. Silva et al. (2007)
verificaram que quanto menor a oferta de pasto e o número de perfilhos, maior a taxa de
bocados indicando que os animais tentam compensar à disponibilidade limitada. Para os
ovinos submetidos ao pastejo nos pastos manejados sob o PD 8,5 fol/perf a menor taxa
de bocados provavelmente ocorreu em função da maior dispersão da forragem ao longo
do perfil vertical da pastagem, além da ocorrência de lâminas mais resistentes à
apreensão pelo animal. Em condições em que a quantidade de forragem apreendida em
cada bocado é prejudicada a compensação pode ser feita aumentando o tempo de
pastejo ou alterando a taxa de bocados (CARVALHO et al., 2001). Neste estudo, os
ovinos utilizaram a taxa de bocados como principal estratégia para manter seu consumo
diante da variação na estrutura da pastagem.
Observou-se efeito da interação PDxRPP (P 0,05) para o número diário de
bocados (Tabela 3). Nos pastos manejados com PD de 6,5 fol/perf e ambos os resíduos
foram verificados menor número de bocados por dia.
Houve efeito da interação PDxRPP (P 0,05) para a massa do bocado (Tabela 3).
Com a variação na taxa de bocados era esperada uma variação na massa de bocados,
como forma de compensação para manter o consumo (CARVALHO et al., 2001;
AGREIL et al., 2005). Nos pastos manejados sob PD de 6,5 fol/perf e ambos os
resíduos foram verificados maiores valores de massa do bocado. Este comportamento
era esperado, visto que nestes pastos a taxa de bocado foi menor. A associação entre
massa do bocado e tempo do bocado, geralmente é negativa, pois, quanto maior for a
massa de forragem apreendida em um bocado, menor será o número de bocados
efetuados em determinado período de tempo.
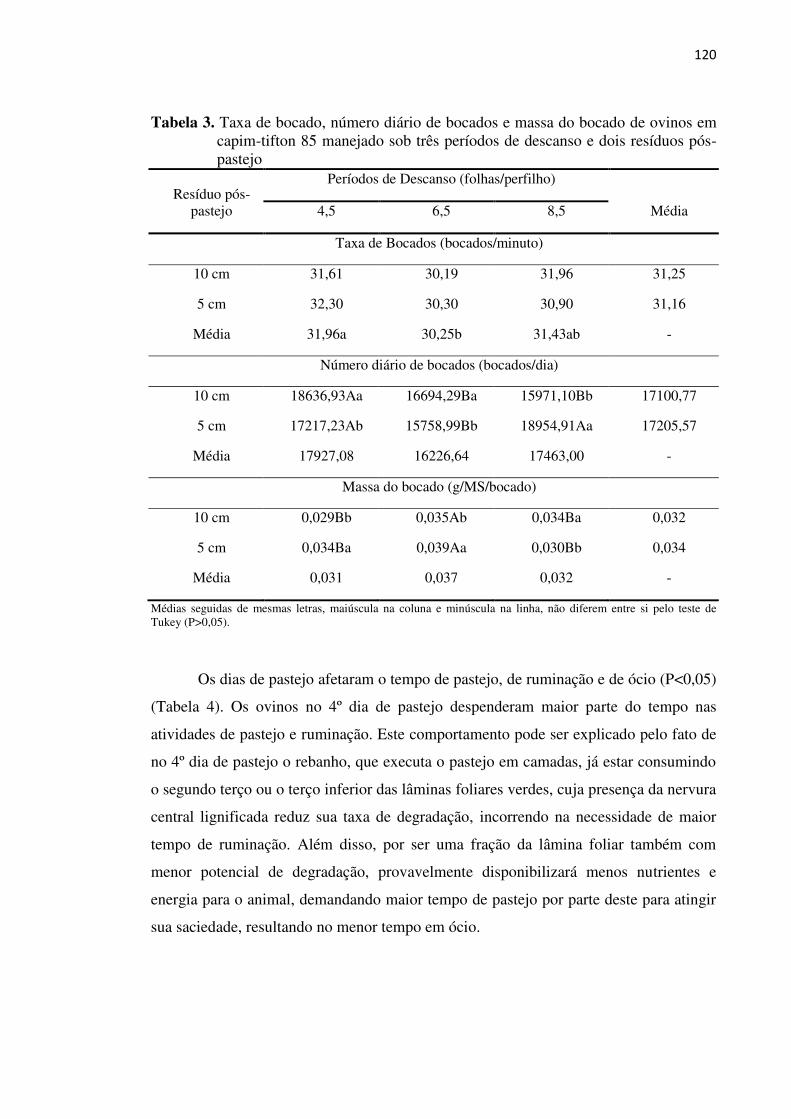
120
Tabela 3. Taxa de bocado, número diário de bocados e massa do bocado de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Resíduo pós-pastejo
Períodos de Descanso (folhas/perfilho)
Média 4,5 6,5 8,5
Taxa de Bocados (bocados/minuto)
10 cm 31,61 30,19 31,96 31,25
5 cm 32,30 30,30 30,90 31,16
Média 31,96a 30,25b 31,43ab -
Número diário de bocados (bocados/dia)
10 cm 18636,93Aa 16694,29Ba 15971,10Bb 17100,77
5 cm 17217,23Ab 15758,99Bb 18954,91Aa 17205,57
Média 17927,08 16226,64 17463,00 -
Massa do bocado (g/MS/bocado)
10 cm 0,029Bb 0,035Ab 0,034Ba 0,032
5 cm 0,034Ba 0,039Aa 0,030Bb 0,034
Média 0,031 0,037 0,032 -
Médias seguidas de mesmas letras, maiúscula na coluna e minúscula na linha, não diferem entre si pelo teste de Tukey (P>0,05).
Os dias de pastejo afetaram o tempo de pastejo, de ruminação e de ócio (P<0,05)
(Tabela 4). Os ovinos no 4º dia de pastejo despenderam maior parte do tempo nas
atividades de pastejo e ruminação. Este comportamento pode ser explicado pelo fato de
no 4º dia de pastejo o rebanho, que executa o pastejo em camadas, já estar consumindo
o segundo terço ou o terço inferior das lâminas foliares verdes, cuja presença da nervura
central lignificada reduz sua taxa de degradação, incorrendo na necessidade de maior
tempo de ruminação. Além disso, por ser uma fração da lâmina foliar também com
menor potencial de degradação, provavelmente disponibilizará menos nutrientes e
energia para o animal, demandando maior tempo de pastejo por parte deste para atingir
sua saciedade, resultando no menor tempo em ócio.
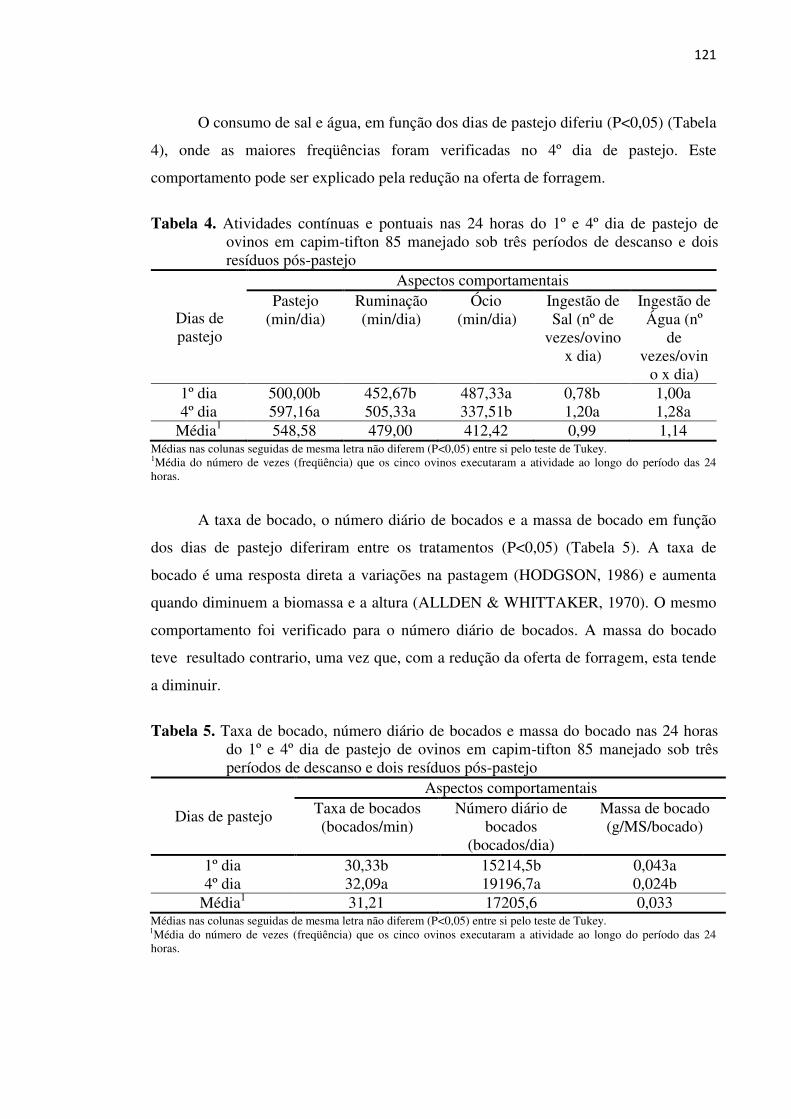
121
O consumo de sal e água, em função dos dias de pastejo diferiu (P<0,05) (Tabela
4), onde as maiores freqüências foram verificadas no 4º dia de pastejo. Este
comportamento pode ser explicado pela redução na oferta de forragem.
Tabela 4. Atividades contínuas e pontuais nas 24 horas do 1º e 4º dia de pastejo de
ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Dias de pastejo
Aspectos comportamentais Pastejo
(min/dia) Ruminação (min/dia)
Ócio (min/dia)
Ingestão de Sal (nº de
vezes/ovino x dia)
Ingestão de Água (nº
de vezes/ovin
o x dia) 1º dia 500,00b 452,67b 487,33a 0,78b 1,00a 4º dia 597,16a 505,33a 337,51b 1,20a 1,28a
Média1 548,58 479,00 412,42 0,99 1,14 Médias nas colunas seguidas de mesma letra não diferem (P<0,05) entre si pelo teste de Tukey. 1Média do número de vezes (freqüência) que os cinco ovinos executaram a atividade ao longo do período das 24 horas.
A taxa de bocado, o número diário de bocados e a massa de bocado em função
dos dias de pastejo diferiram entre os tratamentos (P<0,05) (Tabela 5). A taxa de
bocado é uma resposta direta a variações na pastagem (HODGSON, 1986) e aumenta
quando diminuem a biomassa e a altura (ALLDEN & WHITTAKER, 1970). O mesmo
comportamento foi verificado para o número diário de bocados. A massa do bocado
teve resultado contrario, uma vez que, com a redução da oferta de forragem, esta tende
a diminuir.
Tabela 5. Taxa de bocado, número diário de bocados e massa do bocado nas 24 horas
do 1º e 4º dia de pastejo de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Dias de pastejo
Aspectos comportamentais Taxa de bocados (bocados/min)
Número diário de bocados
(bocados/dia)
Massa de bocado (g/MS/bocado)
1º dia 30,33b 15214,5b 0,043a 4º dia 32,09a 19196,7a 0,024b
Média1 31,21 17205,6 0,033 Médias nas colunas seguidas de mesma letra não diferem (P<0,05) entre si pelo teste de Tukey. 1Média do número de vezes (freqüência) que os cinco ovinos executaram a atividade ao longo do período das 24
horas.

122
A matéria seca (MS) das amostras obtidas através da simulação de pastejo foi
influenciada pelos dias de pastejo (P<0,05) (Tabela 6). O maior teor de MS foi
verificado no 1º dia de pastejo.
Os teores de proteína bruta (PB) foram influenciados pelos períodos de descanso
(PD) e resíduos pós-pastejo (RPP) (P<0,05) (Tabela 6). As amostras obtidas nos pastos
manejados sob PD de 4,5 fol/perf e RPP de 5 cm apresentaram maior teor de PB no 1º
dia de pastejo, provavelmente pelo menor PD e conseqüentemente uma menor idade
fisiológica, resultando melhor qualidade da forragem ofertada para o animal. Carnevalli
et al. (2001), avaliando o desempenho de ovinos e as características do dossel forrageiro
de Tifton 85 em pastos mantidos em diferentes alturas, também observaram maior teor
de PB para os pastos mantidos mais baixos. Isso demonstra a importância de se manter
os pastos em idade mais jovem, apresentando menor massa de forragem. Voltolini et al.
(2009) em pesquisa conduzida com capim-tifton 85 em ciclos de pastejo de 24 dias,
verificaram teor de proteína bruta de 10,86%, inferior aos teores obtidos nesta pesquisa
no 1º dia de pastejo, indicando que a forragem utilizada no 1º dia de pastejo apresentou
elevado valor nutritivo.
Os teores de extrato etéreo (EE) em função dos PD; RPP e dias de pastejo, não
apresentaram diferença (P 0,05) (Tabela 6).
O teor de fibra insolúvel em detergente neutro (FDN) em função dos dias de
pastejo diferiu (P<0,05). Os teores no 4º dia de pastejo foram superiores em todos os
pastos em comparação aos teores no 1º dia de pastejo (Tabela 6). Esta superioridade
pode ser explicada pela condição estrutural do pasto, pois no 1º dia de pastejo o pasto
contém maior proporção de lâminas foliares verdes e conseqüentemente, melhor
qualidade, e com o pastejo seletivo feito pelo animal sobre a fração lâmina foliar nos
primeiros dias de pastejo a relação lâmina foliar/colmo diminui, aumentando o teor de
FDN do pasto ofertado e, conseqüentemente, da dieta selecionada pelos ovinos nos
últimos dias de pastejo. De acordo com Barthram e Grant (1984) essa resposta também
é conseqüência do pastejo em camadas realizado pelos ruminantes, ingerindo a
forragem contida no ápice do dossel nos primeiros dias, restando à forragem mais velha
para ser consumida nos últimos dias de pastejo. Cândido et al. (2005) avaliando o valor
nutritivo da dieta amostrada por pastejo simulado em piquetes de Panicum maximum cv.

123
Mombaça ao longo do período de pastejo verificou teores de FDN para o 1º dia de pastejo
em média de 66,1% e no 5º dia de pastejo 70,5% de FDN na dieta amostrada.
O teor de fibra insolúvel em detergente ácido (FDA) em função dos dias de
pastejo diferiu (P<0,05). Os teores de FDA no 4º dia de pastejo foram superiores em
todos os pastos em relação aos teores no 1º dia de pastejo (Tabela 6). Estas diferenças
estão associadas à mudança da composição da estrutura do pasto no decorrer dos dias de
pastejo, conforme explicado anteriormente, nos manejados sob RPP de 5 cm foi
verificado que os teores de FDA foram inferiores, quando comparado aos pastos com
RPP de 10 cm. Os menores teores de FDA no RPP de 5 cm podem ser explicados
devido à constante renovação de folhas em pastos manejados mais baixos, contribuindo
para a menor proporção de constituintes da parede celular. Avaliando o teor de FDA da
dieta amostrada por pastejo simulado em piquetes de Panicum maximum cv. Mombaça
sob três períodos de descanso, Cândido et al. (2005) obteve médias em função dos dias
de pastejo médias de 31,5% para o 1º dia de pastejo e 35,9% para o 5º dia de pastejo
Os dias de pastejo afetaram a digestibilidade in vitro da matéria seca (DIVMS)
da dieta consumida pelos ovinos (P<0,05) (Tabela 6). No 1º dia de pastejo, as médias
observadas para a digestibilidade aparente da forragem foram superiores (P<0,05), em
todos os pastos, em relação às medias observadas nos pastos no 4º dia de pastejo. O
decréscimo da DIVMS foi mais destacado à medida que avançou o período de pastejo do
1º para o 4º dia de pastejo, especialmente nos maiores períodos de descanso (6,5 e 8,5
fol/perf), reafirmando que a qualidade da forragem diminui com o suceder dos dias do
período de pastejo, decorrente da seleção dos ovinos das frações de maior valor
nutritivo nos primeiros dias, reduzindo essa oferta nos últimos dias de pastejo. Do ponto
de vista de nutrição animal, Hamilton et al. (1970) relataram que valores acima de 65%
para a DIVMS são indicativos de bom valor nutritivo, o que permite bom consumo de
energia digestível.
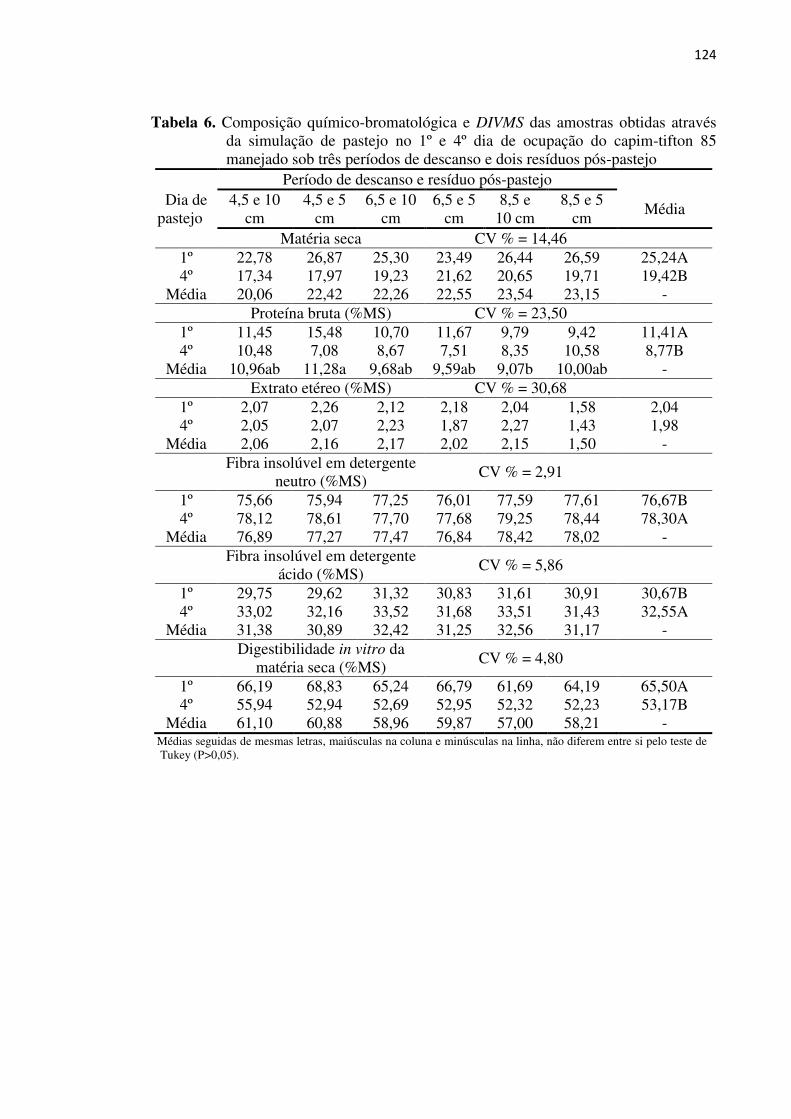
124
Tabela 6. Composição químico-bromatológica e DIVMS das amostras obtidas através da simulação de pastejo no 1º e 4º dia de ocupação do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Dia de pastejo
Período de descanso e resíduo pós-pastejo 4,5 e 10
cm 4,5 e 5
cm 6,5 e 10
cm 6,5 e 5
cm 8,5 e
10 cm 8,5 e 5
cm Média
Matéria seca CV % = 14,46 1º 22,78 26,87 25,30 23,49 26,44 26,59 25,24A 4º 17,34 17,97 19,23 21,62 20,65 19,71 19,42B
Média 20,06 22,42 22,26 22,55 23,54 23,15 - Proteína bruta (%MS) CV % = 23,50
1º 11,45 15,48 10,70 11,67 9,79 9,42 11,41A 4º 10,48 7,08 8,67 7,51 8,35 10,58 8,77B
Média 10,96ab 11,28a 9,68ab 9,59ab 9,07b 10,00ab - Extrato etéreo (%MS) CV % = 30,68
1º 2,07 2,26 2,12 2,18 2,04 1,58 2,04 4º 2,05 2,07 2,23 1,87 2,27 1,43 1,98
Média 2,06 2,16 2,17 2,02 2,15 1,50 -
Fibra insolúvel em detergente
neutro (%MS) CV % = 2,91
1º 75,66 75,94 77,25 76,01 77,59 77,61 76,67B 4º 78,12 78,61 77,70 77,68 79,25 78,44 78,30A
Média 76,89 77,27 77,47 76,84 78,42 78,02 -
Fibra insolúvel em detergente
ácido (%MS) CV % = 5,86
1º 29,75 29,62 31,32 30,83 31,61 30,91 30,67B 4º 33,02 32,16 33,52 31,68 33,51 31,43 32,55A
Média 31,38 30,89 32,42 31,25 32,56 31,17 -
Digestibilidade in vitro da
matéria seca (%MS) CV % = 4,80
1º 66,19 68,83 65,24 66,79 61,69 64,19 65,50A 4º 55,94 52,94 52,69 52,95 52,32 52,23 53,17B
Média 61,10 60,88 58,96 59,87 57,00 58,21 - Médias seguidas de mesmas letras, maiúsculas na coluna e minúsculas na linha, não diferem entre si pelo teste de
Tukey (P>0,05).

125
O peso corporal final (PC) dos animais foi influenciado pelo PD (P<0,05)
(Tabela 7), e os ovinos mantidos nos pastos manejados sob PD de 4,5 fol/perf
apresentaram maior PC final quando comparados aos ovinos dos demais tratamentos,
provavelmente o maior PC, deve ser resultado do consumo de pastos de melhor
qualidade e que tenham suprido as exigências nutricionais dos animais.
O ganho de peso total (GPT) foi influenciado pelo PD (P<0,05) (Tabela 7). Os
ovinos mantidos nos pastos manejados sob PD de 4,5 fol/perf em ambas as alturas
residuais, apresentaram maior GPT. Isso é resultado do menor período de descanso ao
qual a gramínea foi submetida, propiciando uma forragem de melhor qualidade, o que
corrobora o maior ganho médio diário GMD observado nos ovinos mantidos nestes
pastos. Neste estudo o maior acúmulo de forragem foi verificado nos pastos manejados
sob maior PD, no entanto isso não resultou em maiores ganhos de peso para os animais,
pode-se deduzir, portanto, que ocorreu menor eficiência de utilização da forragem
produzida nos pastos sob PD de 8,5 fol/perf, uma vez que, a maior eficiência de
utilização é conseguida quando o manejo associa período de descanso e resíduo pós-
pastejo de forma a minimizar as perdas de forragem. Em experimento avaliando a
resposta animal em relação à altura do dossel em pastagem de Tifton 85, numa
amplitude de 5 a 20 cm, Carnevalli et al. (2001)verificaram maiores ganhos individuais
de ovinos e produção por área nas maiores alturas do pasto, onde também foram
verificadas as maiores disponibilidades de forragem.
O GMD foi afetado pelo PD e pelo RPP (P<0,05) (Tabela 7), observou-se que os
ovinos mantidos nos pastos manejados sob PD de 4,5 fol/perf apresentaram
desempenho superior aos mantidos nos pastos sob PD de 6,5 fol/perf, que por sua vez,
foram superiores àqueles mantidos nos pastos sob PD de 8,5 fol/perf. Os maiores GMD,
observados nos PDs de 4,5 e 6,5 fol/perf, podem ser justificados pela melhor qualidade
da forragem disponível, uma vez que folhas mais jovens apresentam valor nutritivo
superior e permitem maiores incrementos diários de peso vivo. O comprometimento do
desempenho animal, nos pastos manejados com maiores PDs, pode ser consequência da
redução do consumo de forragem, devido as características estruturais, principalmente a
redução da relação lâmina foliar/colmo. Os pastos manejados com o maior resíduo
foram diferentes (P<0,05) dos pastos com menor resíduo. Essa resposta era esperada,
uma vez que, um resíduo mais alto favorece a seletividade do animal, permitindo o

126
consumo de uma forragem de melhor qualidade, e, por consequência um desempenho
superior. Bianchini (1998) comparou o efeito de quatro alturas de manejo em pastagem
de coast cross no desempenho de ovinos, onde as alturas da forragem remanescente pós-
pastejo foram: 4 a 6; 10 a 13; 16 a 19 e 22 a 25 cm, onde as duas maiores alturas de
manejo, mostraram redução na proteína e disgetibilidade da matéria seca, bom como,
menor ganho de peso vivo por área. Os valores de GMD dos ovinos no presente estudo
foram similares aos relatados por Carnevalli et at. (2001), que, estudando o desempenho
de ovinos em pastagem de Tifton 85 sob lotação contínua, relataram ganhos variando de
31,3 a 46,8 g/ovino x dia.
O consumo de matéria seca foi influenciado pelo RPP (P<0,05) (Tabela 7), os
pastos manejados sob RPP de 10 cm obtiveram melhor resposta no consumo de matéria
seca, com médias diárias de 0,604 (g/ovino), em relação a menor altura residual de
pastejo. Esta resposta no consumo de matéria seca, nos pastos com maior altura
residual, é reflexo da maior oferta de forragem, permitindo o pastejo seletivo e o
consumo de forragem de melhor qualidade, propiciando ao animal uma melhor
performance. Andrade (2003) observou em bovinos que de uma forma geral, pastos
mantidos mais baixos resultaram em menor massa de bocado, o que levou a um menor
consumo de forragem e, conseqüentemente, desempenho animal, apesar da tentativa dos
animais compensarem essa redução por meio do aumento do número de bocados
realizados por unidade de tempo e aumento do tempo gasto com a atividade de pastejo.
No entanto o consumo de matéria seca dos ovinos neste estudo foi inferior ao relatado
na literatura, e provavelmente tenha sido comprometido pela ocorrência de chuvas ao
longo do período experimental.

127
Tabela 7. Desempenho produtivo e consumo de matéria seca de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Resíduo pós-pastejo
Períodos de Descanso (folhas/perfilho)
Média 4,5 6,5 8,5
Peso corporal final (kg) CV% = 10,25
10 cm 32,92 31,90 31,50 32,10
5 cm 31,30 31,14 30,26 30,90
Média 32,11a 31,52b 30,88b -
Ganho de peso total (kg) CV% = 26,40
10 cm 8,18 9,08 6,58 7,94
5 cm 8,98 6,48 5,68 7,04
Média 8,58a 7,78ab 6,13b -
Ganho médio diário (g/animal x dia) CV% = 26,62
10 cm 46,21 34,81 45,40 42,14A
5 cm 50,33 29,58 31,76 37,22B
Média 48,27a 38,58ab 32,19b -
Consumo de matéria seca (g/ovino x dia) CV% = 16,15
10 cm 0,620 0,669 0,633 0,640A
5 cm 0,511 0,530 0,555 0,532B
Média 0,565 0,599 0,594 -
Médias seguidas de mesmas letras, maiúsculas na coluna e minúsculas na linha, não diferem entre si pelo teste de Tukey (P>0,05).
A taxa de lotação de animais por hectare, foi influenciada somente pelos PDs
(P<0,05) as menores taxas foram verificadas nos pastos manejados sob PD de 4,5 e 6,5
fol/perf em relação àqueles manejados sob PD 8,5 fol/perf (Tabela 8). A taxa de lotação
em unidades animal por hectare teve o mesmo comportamento que os pastos manejados
sob PD 8,5 fol/perf foram superiores (P<0,05) (Tabela 8) aos pastos sob PDs 4,5 e 6,5
fol/perf e que este último tendeu a ser superior aos piquetes sob PD mais curto. As
diferenças entre as duas taxas de lotação entre os pastos são reflexos, em grande parte,
do maior tempo de crescimento dado aos pastos sob o PD 8,5 fol/perf em relação aos
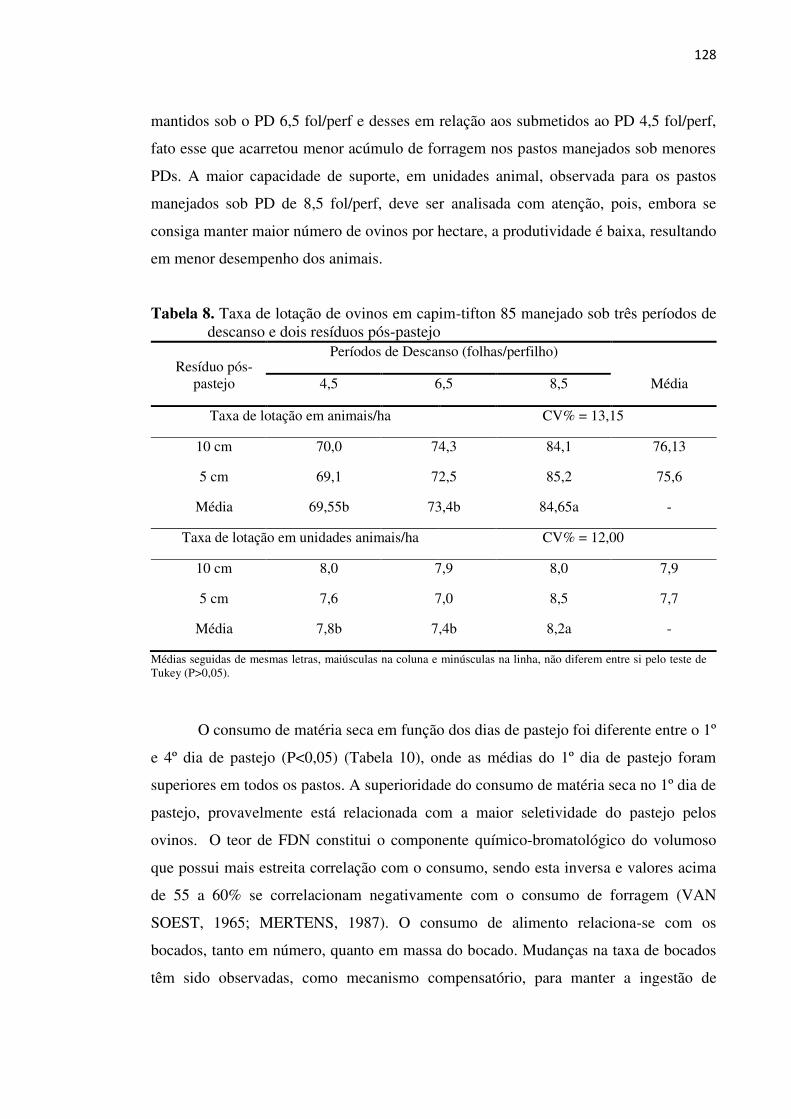
128
mantidos sob o PD 6,5 fol/perf e desses em relação aos submetidos ao PD 4,5 fol/perf,
fato esse que acarretou menor acúmulo de forragem nos pastos manejados sob menores
PDs. A maior capacidade de suporte, em unidades animal, observada para os pastos
manejados sob PD de 8,5 fol/perf, deve ser analisada com atenção, pois, embora se
consiga manter maior número de ovinos por hectare, a produtividade é baixa, resultando
em menor desempenho dos animais.
Tabela 8. Taxa de lotação de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Resíduo pós-pastejo
Períodos de Descanso (folhas/perfilho)
Média 4,5 6,5 8,5
Taxa de lotação em animais/ha CV% = 13,15
10 cm 70,0 74,3 84,1 76,13
5 cm 69,1 72,5 85,2 75,6
Média 69,55b 73,4b 84,65a -
Taxa de lotação em unidades animais/ha CV% = 12,00
10 cm 8,0 7,9 8,0 7,9
5 cm 7,6 7,0 8,5 7,7
Média 7,8b 7,4b 8,2a -
Médias seguidas de mesmas letras, maiúsculas na coluna e minúsculas na linha, não diferem entre si pelo teste de Tukey (P>0,05).
O consumo de matéria seca em função dos dias de pastejo foi diferente entre o 1º
e 4º dia de pastejo (P<0,05) (Tabela 10), onde as médias do 1º dia de pastejo foram
superiores em todos os pastos. A superioridade do consumo de matéria seca no 1º dia de
pastejo, provavelmente está relacionada com a maior seletividade do pastejo pelos
ovinos. O teor de FDN constitui o componente químico-bromatológico do volumoso
que possui mais estreita correlação com o consumo, sendo esta inversa e valores acima
de 55 a 60% se correlacionam negativamente com o consumo de forragem (VAN
SOEST, 1965; MERTENS, 1987). O consumo de alimento relaciona-se com os
bocados, tanto em número, quanto em massa do bocado. Mudanças na taxa de bocados
têm sido observadas, como mecanismo compensatório, para manter a ingestão de

129
matéria seca relativamente constante, frente a variações na massa de bocados em
situação de baixa disponibilidade de forragem (CHACON et al., 1976; PENNING et al.,
1991).
Tabela 10. Consumo de matéria seca de ovinos no 1º e 4º dia de ocupação do capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Dia de pastejo
Período de descanso e resíduo pós-pastejo 4,5 e 10
cm 4,5 e 5
cm 6,5 e 10
cm 6,5 e 5
cm 8,5 e 10
cm 8,5 e 5
cm Méd
ia Consumo de matéria seca
(g/ovino x dia) CV % = 17,24
1º 0,712 0,573 0,802 0,615 0,731 0,641 0,679A
4º 0,528 0,449 0,536 0,445 0,535 0,468 0,494B
Média 0,620ab 0,511b 0,669a 0,530ab 0,633ab 0,554ab - Médias seguidas de mesmas letras, maiúsculas na coluna e minúsculas na linha, não diferem entre si pelo teste de Tukey (P>0,05).
CONCLUSÕES
O comportamento ingestivo de animais em pastejo é sensível a variações na
estrutura do dossel forrageiro
Os ovinos mantidos nos pastos sob PD de 4,5 e 8,5 fol/perf em ambos os
resíduos pós-pastejo despederam maior tempo em pastejo.
O maior consumo de matéria seca foi obtido no 1º dia de pastejo.
Os ovinos mantidos nos pastos manejados a uma altura residual de 10 cm
apresentaram maior ganho médio diário e maior consumo de matéria seca.
Os pastos manejados sob períodos de descanso de 4,5 e 6,5 fol/perf
proporcionaram melhor desempenho animal.

130
REFERÊNCIAS
AGREIL, C.; FRITZ, H.; MEURET, M. Maintenance of daily intake through bite mass diversity adjustment in sheep grazing on heterogeneous and variable vegetation. Applied Animal Behaviour Science, v.91, p.35-36, 2005. ALLDEN, W. G.; WHITTAKER, I. A. M. The determination of herbage intake by grazing sheep: the interrelationship of factors influencing herbage intake and availability. Australian Journal of Agricultural Research, v. 21, n. 1, p. 755-766, 1970. ANDRADE, F.M.E. Valor nutritivo da forragem e desempenho de bovinos de corte em pastos de Brachiaria brizantha cv Marandu submetidos a regimes de lotação contínua. Piracicaba, 2003. Dissertação (Mestrado) – Escola Superior de Agricultura “Luiz de Queiroz” 2003. ASTIGARRAGA, L. Técnicas para la medición del consumo de rumiantes en pastoreo. In: SIMPÓSIO SOBRE AVALIAÇÃO DE PASTAGENS COM ANIMAIS, 1997, Maringá. Anais... Maringá: UEM, 1997. p. 1-23. BARTHRAM, G.T.; GRANT, S. Defoliation of ryegrass-dominated swards by sheep. Grass Forage Sci., v.39, n.3, p.211-219, 1984. BIANCHINI, D. Efeito de quatro alturas de manejo sobre as características quantitativas e qualitativas em pastagem de coast cross no desempenho de cordeiros em fase de terminação. Jaboticabal, 1998. 135p. Tese (Doutorado) – Universidade Estadual Paulista, Faculdade de Ciências Agrárias e Veterinárias, 1998. CÂNDIDO, M. J. D.; ALEXANDRINO, E.; GOMIDE, C. A. de M. et al. Período de Descanso, Valor Nutritivo e Desempenho Animal em Pastagem de Panicum maximum
cv. Mombaça sob Lotação Intermitente. Revista Brasileira de Zootecnia, v.34, n.5, p.1459-1467, 2005. CARVALHO, P.C.F.; RIBEIRO FILHO, H.M.N.; POLI, C.H.E.C. et al. Importância da estrutura da pastagem na ingestão e seleção de dietas pelo animal em pastejo. A produção animal na visão dos brasileiros. Piracicaba: Fundação de Estudos Agrários Luiz de Queiroz, 2001. p.853-871. CARVALHO, P.C. de F.; MORAES, A. Comportamento ingestivo de ruminantes: bases para o manejo sustentável do pasto. In: Simpósio sobre Manejo Sustentável das Pastagens, 2005, Maringá. Anais. Maringá: UEM, 2005. CD-ROM. CARNEVALLI, R. A.; SILVA, S. C. da; FAGUNDES, J. L. et al. Desempenho de ovinos e respostas de pastagens de Tifton 85 (Cynodon sp.) sob lotação contínua. Scientia Agricola, v.58, n.1, p.7-15, 2001.

131
CHACON, E.; STOBBS, T.H.; SANDLAND, R.L. Estimation of herbage consumption by cattle using measurements of eating behaviour. Journal of British Grassland Society, v.31, p.81-87, 1976. COATES, D.B.; PENNING, P. Measuring animal performance. In: MANNETJE, L.; JONES, R.M. (Eds.). Field and laboratory methods for grassland and animal production research. Wallingford: CAB International, p.353-402, 2000. COSTA, M.J.R.P.; MESQUITA, J.C.; JUNQUEIRA FILHO, A.A. Comportamento de vacas Holandesas em pastagem. In: Encontro Paulista de Etologia, 1, Jaboticabal, 1983. Anais... Jaboticabal-SP: UNESP/FCAVJ, 1983, 251p. EMPRESA BRASILEIRA DE PESQUISA E AGROPECUÁRIA - EMBRAPA. Sistema brasileiro de classificação de solos. Brasília: Centro Nacional de Pesquisa de Solos, 1999. 412p. FISCHER, V. et al. Padrões Nictemerais do Comportamento Ingestivo de Ovinos. Revista Brasileira de Zootecnia, v.27, p.362-369, 1998. FORBES, T.D.A. Researching the plant-animal interface: The investigation of ingestive behaviour of cows and sheep. Journal of Animal Science, v.66, p.2369-2379, 1988. FORBES, T.D.A.; HODGSON, J. Comparative studies on the influence of swards conditions on the ingestive behaviour of cows and sheep. Grass and Forage Science, v.40, p.69-77, 1985. GIBB, M. J.; TREACHER, T.T. The effect of herbage allowance on lambs grazing perennial ryegrass and red clover swards. Journal of Agricultural Science, v.86, p.355-365, 1976. GRANDIN, T. Princípios de comportamento animal para o manejo de bovinos e outros herbívoros em condições extensivas. Wallingford, Oxon (Reino Unido), cap.5, p.63-85, 2000. HAMILTON, R.I., LAMBOURNE, L.J., ROE, R. et al. Quality of tropical grasses for milk production. In: INTERNATIONAL GRASSLAND CONGRESS, 11, 1970, Surfers Paradise. Proceedings... Surfers Paradise: s.ed., p.860-864, 1970 MANZANO, R. P.; NUSSIO, L. G.; CAMPOS, F. P.; ANDREUCCIS, M. P.; COSTAS, R. Z. M. Comportamento ingestivo de novilhos sob suplementação em pastagens de capim-tanzânia sob diferentes intensidades de desfolhação. Revista Brasileira de Zootecnia, v.36, n.3, p.550-557, 2007. MEDEIROS, R.B.; PEDROSO, C.E.S.; JORNADA, J.B.J et al. Comportamento ingestivo de ovinos no período diurno em pastagem de azevém anual em diferentes estádios fenológicos. Revista Brasileira de Zootecnia, v.36, n.1, p.198-204, 2007.

132
MENDES, M. A.; LEÃO, M. I.; SILVA, J. F. C. et al. Efeito da temperatura ambiente e do teor de energia da ração sobre os consumos de alimentos e de água e algumas variáveis fisiológicas de ovinos. Revista Brasileira de Zootecnia, Viçosa, v. 5, n. 2, p. 173-187, 1976. MERTENS, D.R. Predicting intake and digestibility using mathematical models of ruminal fuction. Journal of Animal Science., 64(8):1548-1558, 1987. PEDROSO, C. E. da S.; MEDEIROS, R.B. de; SILVA, M.A. da. et al. Produção de Ovinos em Gestação e Lactação sob Pastejo em Diferentes Estádios Fenologicos de Azevém Anual Revista Brasileira de Zootecnia, v.33, n.5, p.1345-1350, 2004. PENNING, P.D.; PARSONS, A.J.; ORR, R.J.; TREACHER, T.T. Intake and behaviour responses by sheep to changes in sward characteristics under continous stocking. Grass and Forage Science, v.46, p.15-28, 1991. RASLAN, L. S. A. Aspectos comportamentais e fisiológicos de ovino SRD sob pastejo com e sem sombreamento. Dissertação (Mestrado) Universidade Estadual do Sudoeste da Bahia - UESB – Itapetinga-BA: UESB, 2008, 99p. SILVA, R. G. da; NEIVA, J. N. M.; CANDIDO, M. J. D. et al. Aspectos comportamentais e desempenho produtivo de ovinos mantidos em pastagens de capim-tanzânia manejado sob lotação intermitente. Ciência Animal Brasileira, v. 8, n. 4, p. 609-620, out./dez. 2007. SILVA, D.J.; QUEIROZ, A.C. Análises de alimentos (métodos químicos e biológicos). 3.ed. Viçosa, MG: Editora UFV, 2002. 235p. SAS INSTITUTE. SAS system for Windows. Version9. 1. Cary: SAS Institute Inc. 2003. 2 CD – ROMs. TREVISAN, N.B.; QUADROS, F.L.F.; CORADINI, F.S. et al. Comportamento ingestivo de novilhos de corte em pastagem de aveia preta e azevém com níveis distintos de folhas verdes. Ciência Rural, v.34, n.5, p.1543-1548, 2004. VOLTOLINI, T. V.; MOREIRA, J. N.; NOGUEIRA, D. M. et al. Fontes protéicas no suplemento concentrado de ovinos em pastejo. Acta Scientiarum. Animal Sciences, v. 31, n. 1, p. 61-67, 2009.

133
CAPÍTULO 7
____________________________________________________________
Características de carcaça, componentes não-carcaça e composição centesimal do músculo Longissimus dorsi de ovinos terminados em capim-
tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo

134
Características de carcaça, componentes não-carcaça e composição centesimal do músculo Longissimus dorsi de ovinos em capim-tifton 85 manejado sob três
períodos de descanso e dois resíduos pós-pastejo
RESUMO: Avaliaram-se as características da carcaça de ovinos da raça Santa Inês,
submetidos a diferentes estratégias de manejo, em pastagem de capim-tifton 85. Foram
utilizados 30 ovinos prova e número variável de animais reguladores, com peso corporal
(PC) inicial de 20 kg, os quais foram distribuídos em um delineamento experimental
inteiramente casualizado em arranjo fatorial 3x2, sendo três períodos de descanso (PD):
4,5; 6,5 e 8,5 (nº de folhas/perfilho) e dois resíduos pós-pastejo (RPP): 5 e 10 cm. Após
alguns ciclos de pastejo, com a intensificação do efeito dos tratamentos sobre o ganho
de peso dos animais, foi realizado o abate, quando os animais atingiram uma média de
32 kg, pelo grupo de animais de prova de tratamento, sendo que, neste momento, todos
os animais de prova de todos os tratamentos foram abatidos no mesmo dia. Períodos de
descanso do pasto de 4,5; 6,5 e 8,5 fol/perf não alteram o rendimento comercial, a
espessura de gordura e a área de olho de lombo em ovinos pastejando em capim-tifton
85. O resíduo pós-pastejo mais elevado proporcionou maior rendimento biológico.
Termos para indexação: cortes, espessura de gordura, manejo, rendimento de carcaça.

135
Characteristics of carcass, components not-carcass and centesimal composition of the Longissimus muscle dorsi of sheep of the tifton 85 grass managed under three
periods of rest and two post-grazing residues
Abstract: The characteristics of the carcass of sheep of the race had been evaluated
Saint Ines, submitted the different strategies of handling, in pasture of capim-tifton 85.
Sheep ones had been used 30 prove and changeable number of regulating animals, with
weight corporal (PC) initial of 20 kg, which had been distributed in an experimental
delineation entirely casualizado in factorial arrangement 3x2, being three periods of rest
(PD): 4,5; 6,5 and 8,5 (leaves/tiller) and two residues after-pastejo (RPP): 5 and 10 cm
After some cycles of pastejo, with the intensification of the effect of the treatments on
the profit of weight of the animals, abate were carried through it, when the animals had
reached a of 32 kg, for the group of animals of test of treatment, being that, at this
moment, all the animals of test of all the treatments had been abated in the same day.
Periods of rest of the grass of 4,5; 6,5 and 8,5 leaves/tiller do not modify the
commercial income, the thickness of fat and the loin eye area in sheep grazing in
capim-tifton 85. The raised post-grazing residues more provided to greater biological
income.
Index-terms: cuts, thickness of fat, handling, income of carcass.

136
INTRODUÇÃO
A criação de pequenos ruminantes no Nordeste brasileiro é uma atividade básica
e generalizada, sendo praticada por grande número de produtores rurais, representando
uma atividade de grande importância sócio-econômica. A exploração de ovinos na
região Nordeste é conduzida na maioria, sob forma extensiva, apresentando pouca
produtividade, em razão ao baixo nível tecnológico aplicado ao manejo animal e do
potencial genético dos rebanhos, que muitas vezes só permite que o animal adquira peso
adequado ao abate ao atingir idade elevada, encarecendo, portanto, o produto final
(SIQUEIRA, 1999; REIS, 2001).
O rebanho ovino do Brasil é de 14.027.271 cabeças, sendo que na Região
Nordeste encontra-se 8.001.613 cabeças (ANULPEC, 2008). O país ocupa o 16º lugar
no ranking mundial em número de cabeças, sendo que a China está em 1º lugar com
366.641.000 cabeças. O consumo de carne ovina no Brasil ainda é baixo, em torno de
800 g/per capita/ano, no entanto, o setor está em fase de crescimento, pois estima-se
que a produção atingirá 100 milhões de cabeças nos próximos 10 anos e o segmento se
firmará como boa alternativa de investimento. A carne apresenta-se com uma alternativa
viável economicamente para o setor da ovinocultura. O mercado da carne ovina está em
franca ascensão em todo o país. Os preços hoje praticados no âmbito de unidade
produtiva representam bem mais do que o preço pago pela carne bovina nas mesmas
condições.
A produção de carne representa uma alternativa econômica para o Nordeste,
principalmente pela a adaptação dos animais as condições climáticas. Entretanto nota-se
que há escassez de informações sobre as características quantitativas da carcaça. Estas
informações são de grande importância, uma vez que, torna-se um ponto chave para
obter carcaças magras com peso e idade de abate adequados, em virtude de um mercado
consumidor cada vez mais exigente em produtos saudáveis. A preferência pela carne
ovina apresenta aspectos comuns, como busca por carne macia com pouca gordura e
muito músculo, comercializada a preços acessíveis (SILVA SOBRINHO, 2001).
A carne ovina, esta é uma fonte de proteína de alto valor biológico, presente na
dieta das populações de quase todos os países (ALMEIDA, 1990). Contudo, devido à
atenção que o consumidor tem dado para a relação entre dieta e saúde, há uma crescente

137
preocupação com o conteúdo de gordura e colesterol dos produtos de origem animal.
Atualmente, os profissionais da saúde recomendam dietas com baixas calorias, baixo
teor de gorduras saturadas e baixo colesterol a fim de reduzir o risco à saúde. Devido a
estes aspectos, o conhecimento da composição tecidual da composição centesimal e do
teor de colesterol da carne ovina adquire grande importância, fazendo-se necessários
estudos que identifiquem produtos mais saudáveis e que atendam a demanda dos
consumidores.
Alguns aspectos são fundamentais, nesta fase de plena expansão da atividade de
produção de carne, como: adoção de estratégias de manejo e abate adequado, com o
objetivo de se obter um produto de boa qualidade. De acordo com Sañudo e Sierra
(1993), a alimentação pode alterar as características da carcaça. A utilização de
forrageiras de alta qualidade e elevado valor nutritivo é considerada uma alternativa nos
sistemas de produção de carne ovina, o que reflete de maneira significativa no ganho de
peso do animal. Outro aspecto de extrema relevância no sistema de produção é a
exploração de raças adaptadas as condições da Região Nordestina. A raça Santa Inês é a
que apresenta precocidade aliada a uma resistência as parasitoses, fertilidade e
prolificidade, mostrando-se promissora para a produção de carne.
A pesquisa foi realizada com o objetivo de avaliar as características da carcaça e
composição centesimal da carcaça de ovinos da raça Santa Inês, terminados em
pastagem de capim-tifton 85 submetidos a três períodos de descanso e dois resíduos
pós-pastejo, no Nordeste brasileiro.
MATERIAL E MÉTODOS
A pesquisa foi desenvolvida no Setor de Produção de Ovinos do Campo
Experimental de Bebedouro, pertencente à Embrapa Semiárido, localizado no
Município de Petrolina-Pernambuco. Durante o mês de outubro foi efetuada a divisão
da área em piquetes e ajuste da metodologia de trabalho. A coleta de dados ocorreu
entre os meses de novembro de 2008 a junho de 2009, totalizando nove meses. O clima
da região é classificado como quente e seco, sendo caracterizada por duas estações bem
definidas, uma chuvosa, de janeiro a maio, e outra seca, de junho a dezembro.

138
Foram avaliados três períodos de descanso 4,5; 6,5 e 8,5 (número de
folhas/perfilho) e dois resíduos pós-pastejo 5 e 10 cm, em um delineamento
experimental inteiramente casualizado em arranjo fatorial 3x2 com cinco repetições
(animais). A área experimental foi composta de 0,82 ha de pastagem cultivada com
capim-tifton 85 irrigada por aspersão com canhões com distribuição de água durante o
dia. Utilizou-se o método de pastejo sob lotação intermitente, com lotação variável,
mantendo-se animais prova nos piquetes e também animais reguladores para ajuste da
taxa de lotação, segundo a técnica da taxa de lotação variável. A pastagem foi
subdivida, com tela campestre, em 37 piquetes de 180 m2 cada, destes piquetes 24 foram
considerados amostrais. Foram utilizados 30 ovinos de prova e um número variável de
animais de equilíbrio, da raça Santa Inês, machos castrados, com peso corporal (PC)
inicial de ± 20 kg, os quais foram previamente identificados, vermifugados e, em grupos
de cinco, distribuídos por sorteio em seis tratamentos. Foram efetuadas adubações de
reposição com nitrogênio, em cada piquete, no momento da saída dos animais, nas
seguintes doses: 0,971; 1,110; 1,320; 1,475; 1,666 e 1,800 g, de acordo com o período
de descanso.
O abate foi realizado no dia 2 de junho de 2009, no Abatedouro Escola do
Instituto Federal de Educação, Ciência e Tecnologia do Sertão de Pernambuco (IF
Sertão PE). O critério utilizado para determinar o abate dos animais foi o alcance de um
peso corporal de ± 32 kg pelo grupo de animais de prova de um tratamento, sendo que a
neste momento, todos os animais de prova de todos os tratamentos foram abatidos no
mesmo dia. Na véspera do abate, os ovinos foram pesados e mantidos em jejum
alimentar por 16 horas (COATES & PENNING, 2000). O abate foi realizado através de
atordoamento e sangria da veia jugular e artéria carótida, com coleta e pesagem do
sangue. Em seguida foi feita a esfola e a evisceração. O trato gastrintestinal (TGI), a
bexiga e a vesícula biliar foram esvaziados e limpos para a obtenção do peso do corpo
vazio (PCV), que foi obtido subtraindo-se do peso ao abate (PA), os pesos referentes ao
conteúdo do TGI e aos líquidos contidos na bexiga e na vesícula biliar. Todos os não
componentes da carcaça foram separados e pesados, sendo: vísceras, conteúdo do trato
gastrintestinal cheio, conteúdo do trato gastrintestinal vazio e pele (OSÓRIO, 1992).
Após a separação dos não componentes da carcaça foi obtido o peso da carcaça
quente (PCQ), que foi utilizado para estimar o rendimento de carcaça quente (RCQ =

139
PCQ/PA x 100). O rendimento biológico (RB) foi obtido a partir da razão entre o peso
da carcaça quente e o peso de corpo vazio (RB = PCQ/PCV x 100). Em seguida, as
carcaças foram acondicionadas em sacos plásticos e transportadas para uma câmara fria
a 5ºC, onde permaneceram penduradas pelos tendões da perna com ganchos de metal
por um período de 24 horas (Figuras 1 e 2). Ao final desse período as carcaças foram
pesadas para a obtenção do peso da carcaça fria (PCF), que foi utilizado para estimar o
rendimento da carcaça fria (RCF = PCF/PA x 100). O porcentual de perda de peso da
carcaça pelo resfriamento foi obtido pela fórmula PPR= ((PCF/ PCQ) x 100).
Figuras 1 e 2. (1): carcaças de ovinos acondicionadas em sacos plásticos e (2): Carcaças de
ovinos na câmara fria a 5ºC.
As carcaças foram divididas longitudinalmente (Figura 4), em seguida, a meia
carcaça direita foi dividida em seis cortes comerciais: pescoço, serrote, costilhar, paleta,
lombo e pernil, os quais foram pesados para cálculo dos seus rendimentos em relação ao
peso da carcaça (Figura 3). Entre a 12ª e a 13ª vértebras torácicas, foi realizado um corte
para expor a secção transversal do músculo Longíssimus dorsi, sobre o qual foi traçada
a área de olho do lombo (AOL) em película transparente (Figura 5) e em seguida foram
efetuadas mensurações para a obtenção da referida área, com o auxilio do programa
AutoCAD. Ainda no músculo longissimus dorsi, foi obtido à espessura de gordura, com
o paquímetro (Figura 6) e as amostras do músculo (Figura 7) foram em seguida
1 2

140
homogeneizadas em multiprocessador para serem determinados os teores de umidade,
matéria mineral e proteína bruta, conforme procedimentos descritos em Carvalho e Jong
(2002). O estrato etéreo foi determinado por meio do extrator de gordura, modelo
ANKOM® XT – 10.
Figura 3. Meia carcaça foi dividida em seis cortes comerciais: pescoço, serrote, costilhar, paleta, lombo e pernil.
Pescoço
Paleta Pernil
Lombo
Serrote
Costilhar
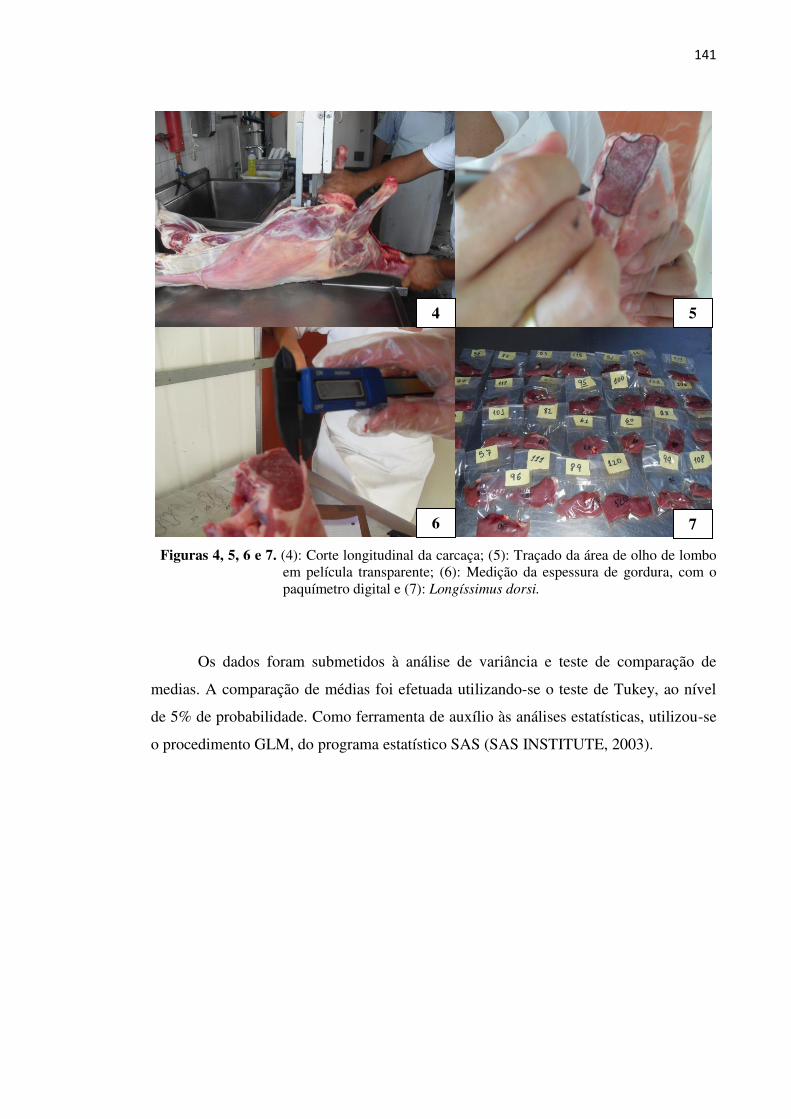
141
Figuras 4, 5, 6 e 7. (4): Corte longitudinal da carcaça; (5): Traçado da área de olho de lombo em película transparente; (6): Medição da espessura de gordura, com o paquímetro digital e (7): Longíssimus dorsi.
Os dados foram submetidos à análise de variância e teste de comparação de
medias. A comparação de médias foi efetuada utilizando-se o teste de Tukey, ao nível
de 5% de probabilidade. Como ferramenta de auxílio às análises estatísticas, utilizou-se
o procedimento GLM, do programa estatístico SAS (SAS INSTITUTE, 2003).
4 5
6 7

142
RESULTADOS E DISCUSSÃO
O peso vivo de abate (PVA) e peso da carcaça fria (PCF) não foram
influenciados pelos PDs (P 0,05), nem pelos RPPs (P 0,05) e nem pela PDxRPP
(Tabela 1). Apesar de não ter sido verificado diferenças significativas entre os ovinos
mantidos nos pastos manejados sob PDs e RPPs diferentes, os terminados nos pastos
sob PD 4,5 fol/perf e resíduo de 10 cm apresentaram maior valor numérico nas médias
para o PVA.
O peso da carcaça quente (PCQ) foi afetado pelo PD (P 0,05) (Tabela 1). Os
ovinos terminados nos pastos manejados com PD 4,5 fol/perf apresentaram maiores
médias. Os PCF e PCQ encontrados neste experimento foram considerados aceitáveis
para o comércio, pois de acordo com Pires et al. (1999) o consumidor prefere carcaças
com peso entre 12 e 15 kg. O maior PCQ correspondeu ao peso mais elevado de abate,
igualmente ao relatado por Motta et al. (2001). Essa clássica relação positiva, entre o
peso vivo e o peso da carcaça, sugere o uso do peso vivo pelos produtores para a
realização da seleção de animais para o abate, sejam de uma mesma raça ou
geneticamente semelhantes. O peso máximo de abate foi obtido nos pastos manejados
sob o menor PD. Destaca-se o fato de que pequenas mudanças na altura ou oferta de
lâminas foliares resultaram em grandes alterações no peso final dos ovinos.
Os rendimentos apresentados neste experimento foram inferiores aos obtidos
para borregos Santa Inês que são especializados para carne (ALVES et al. 2003). Esses
menores rendimentos podem ser atribuídos aos animais com grau de acabamento de
carcaça inferior. Houve efeito do PD (P<0,05) para rendimento verdadeiro (RV) onde
os maiores rendimentos foram obtidos nos pastos manejados sob PD de 4,5 fol/perf
(Tabela 1).
O período de descanso (P<0,05) e a altura de manejo do pasto (P<0,05)
interferiram no rendimento biológico (RB). Os ovinos mantidos os pastos manejados
sob PD de 4,5 fol/perf e resíduo de 10 cm apresentaram maior RB. O menor período de
descanso proporcionou aos ovinos uma forragem de melhor valor nutritivo, atendendo
as necessidades nutricionais. A altura de manejo determinou, indiretamente, a
quantidade de nutrientes disponíveis para serem convertidos em produto animal. Em
baixas alturas residuais, como nos pastos a 5 cm de altura, o desempenho dos animais

143
foi restringido tanto pela oferta de forragem como pela sua dificuldade de apreensão
(POPPI et al., 1987).
A diferença verificada com relação ao rendimento de carcaça é um dado
importante do ponto de vista de mercado, pois, de acordo com Carvalho et al. (2005b), o
menor rendimento de carcaça faz com que uma menor proporção de material comestível
resultante do abate esteja disponível para o consumidor, o que acaba prejudicando o
resultado final obtido pelo sistema produtivo utilizado.
Não foi verificado efeito do PD (P 0,05), do RPP (P 0,05) e nem da interação
PDxRPP (P 0,05) para a espessura de gordura (EG) e a área de olho de lombo (AOL).
A EG quando em excesso, pode ser restritiva na escolha do consumidor. O valor médio
para esta variável neste trabalho foi de 1,48 mm, podendo ser considerado baixo, não
proporcionando boa cobertura de gordura nas carcaças. De acordo com Silva Sobrinho
(2001) a baixa espessura de gordura (EG) pode estar relacionada à idade dos animais ou
processo de retirada do couro, uma vez que parte dessa gordura pode ter sido removida
com a pele, pode-se ainda citar a alimentação a base exclusivamente de pastagem como
um dos fatores para o resultado esperado. Para a variável AOL, o valor médio para
todos os animais foi 10,65 cm², sendo considerado que a quantidade de músculo da
carcaça produzida, é que a parte comercialmente mais valorizada.

144
Tabela 1. Características da carcaça, espessura de gordura e área de olho de lombo de ovinos em pastagem de capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de mesmas letras maiúsculas na coluna e minúsculas na linha diferem entre si (P<0,05) pelo teste de Tukey.
De acordo com Osório et al. (1996b) o não-aproveitamento adequado dos
componentes de não-carcaça representa uma perda de alimento e matéria-prima, que
Resíduo Pós-Pastejo Período de Descanso (n0 de folhas/perfilho)
4,5 6,5 8,5 Média Peso vivo de abate – PVA (kg) CV = 8,40
10 cm 32,91 29,16 31,39 31,15 5 cm 31,42 30,26 31,71 31,13
Média 32,16 29,71 31,55
Peso da carcaça quente – PCQ (kg) CV = 11,05
10 cm 14,34 12,10 14,03 13,49 5 cm 14,18 11,99 13,88 12,86
Média 14,26a 12,04b 13,95ab
Peso da carcaça fria – PCF (kg) CV = 12,56
10 cm 13,75 11,83 13,48 13,02 5 cm 12,09 11,75 12,41 12,57
Média 12,92 11,79 12,94
Rendimento verdadeiro – RV (%) CV = 6,33
10 cm 45,74 41,45 42,57 43,25 5 cm 44,80 39,69 39,35 41,28
Média 45,27a 40,57b 40,94b
Rendimento biológico – RB (%) CV = 6,32
10 cm 50,93 46,86 48,23 48,67A 5 cm 49,95 44,37 44,68 46,33B
Média 50,44a 45,61b 46,45b
Rendimento comercial – RC (%) CV = 8,42
10 cm 42,96 40,48 41,72 41,72 5 cm 43,68 38,88 38,32 40,29
Média 43,32 39,68 40,02
Espessura de gordura – EG (mm) CV = 27,92
10 cm 1,86 1,30 1,34 1,50 5 cm 1,29 1,56 1,57 1,47
Média 1,57 1,43 1,45
Área de olho do lombo – AOL (cm) CV = 20,78
10 cm 10,68 11,39 10,12 10,73 5 cm 10,89 10,63 10,20 10,57
Média 10,78 11,01 10,16

145
poderiam colaborar para diminuir o preço dos produtos para os consumidores e
melhorar o nível de vida das populações de baixo poder aquisitivo. Além do que,
conforme Silva Sobrinho (2001), a comercialização de constituintes de não-carcaça
também pode trazer benefícios econômicos para os produtores de cordeiros, agregando
valor ao produto. De acordo com Delfa et al. (1991) o fígado, o coração e os rins
apresentam elevada demanda pelos consumidores, pois são os mais atrativos e de fácil
digestão (HOPKINS, 1981), o que os tornam mais valorizados comercialmente, quando
comparados com as demais vísceras comestíveis, como exemplo, os pulmões, traquéia,
língua, entre outros.
Foi verificado efeito do PD (P 0,05) para o peso (kg) do conteúdo do trato
gastrointestinal cheio (Tabela 2). Os ovinos terminados nos pastos sob PD de 8,5
fol/perf apresentaram maior peso para este componente, quando comparado aos ovinos
dos demais pastos. O aumento do peso do conteúdo do trato gastrointestinal cheio com
o avanço do período de descanso pode ser explicado pela necessidade dos animais em
ingerir uma maior quantidade de pasto visando atender a necessidade energética.
Segundo MERTENS (1992), alimentos volumosos apresentam uma baixa densidade
energética quando comparados a alimentos concentrados, o que leva a uma maior
necessidade de ingestão. Outra justificativa pode estar relacionado ao aumento dos
níveis de fibra da dieta reduzindo a digestibilidade e o consumo de MS (capítulo 6), o
que resultou no enchimento do trato gastrointestinal, fazendo com que o alimento
permanecesse por mais tempo no rúmen.
Os pesos (kg) do conteúdo do trato gastrointestinal vazio, coração, fígado,
pulmão, rins e sangue não diferiram entre as estratégias de manejo impostas aos pastos
(P>0,05) (Tabela 2). Os resultados obtidos podem ser justificados pelo fato dos animais
apresentarem idade e peso corporal próximos, podendo-se inferir que o
desenvolvimento dos órgãos também está ligado ao tamanho do animal. Mahendrakar
et al. (1988) e Fimbres et al. (2001), quando avaliaram características de carcaça de
cordeiros, alimentados em pastagens e concluíram que os pesos dos órgãos são mais
relacionados ao tamanho dos animais.
O peso (kg) da pele foi afetado pelo PD (P 0,05). Foram verificados maiores
pesos de pele nos ovinos terminados nos pastos sob PD de 4,5 e 8,5 fol/perf (Tabela 2).
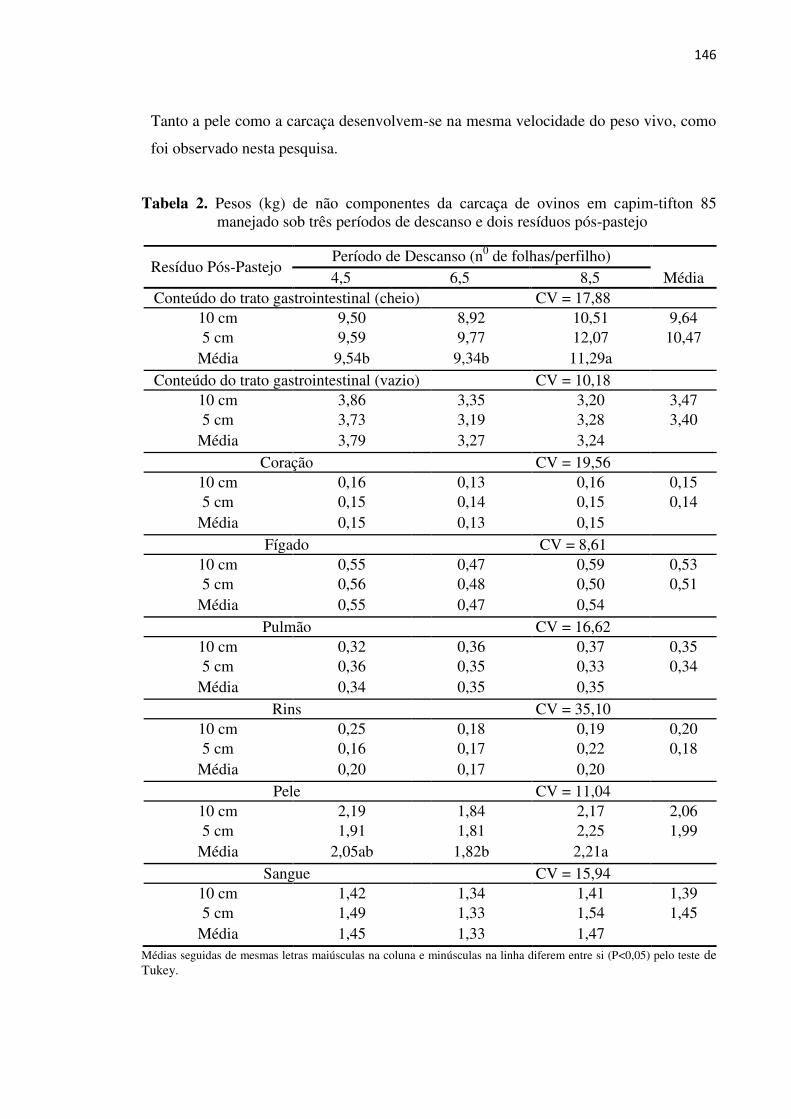
146
Tanto a pele como a carcaça desenvolvem-se na mesma velocidade do peso vivo, como
foi observado nesta pesquisa.
Tabela 2. Pesos (kg) de não componentes da carcaça de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de mesmas letras maiúsculas na coluna e minúsculas na linha diferem entre si (P<0,05) pelo teste de Tukey.
Resíduo Pós-Pastejo Período de Descanso (n0 de folhas/perfilho) 4,5 6,5 8,5 Média
Conteúdo do trato gastrointestinal (cheio) CV = 17,88 10 cm 9,50 8,92 10,51 9,64 5 cm 9,59 9,77 12,07 10,47
Média 9,54b 9,34b 11,29a Conteúdo do trato gastrointestinal (vazio) CV = 10,18
10 cm 3,86 3,35 3,20 3,47 5 cm 3,73 3,19 3,28 3,40
Média 3,79 3,27 3,24 Coração CV = 19,56
10 cm 0,16 0,13 0,16 0,15 5 cm 0,15 0,14 0,15 0,14
Média 0,15 0,13 0,15 Fígado CV = 8,61
10 cm 0,55 0,47 0,59 0,53 5 cm 0,56 0,48 0,50 0,51
Média 0,55 0,47 0,54 Pulmão CV = 16,62
10 cm 0,32 0,36 0,37 0,35 5 cm 0,36 0,35 0,33 0,34
Média 0,34 0,35 0,35 Rins CV = 35,10
10 cm 0,25 0,18 0,19 0,20 5 cm 0,16 0,17 0,22 0,18
Média 0,20 0,17 0,20 Pele CV = 11,04
10 cm 2,19 1,84 2,17 2,06 5 cm 1,91 1,81 2,25 1,99
Média 2,05ab 1,82b 2,21a Sangue CV = 15,94
10 cm 1,42 1,34 1,41 1,39 5 cm 1,49 1,33 1,54 1,45
Média 1,45 1,33 1,47

147
Não foi verificado efeito dos PDs (P 0,05), nem dos RPPs (P 0,05) e nem pela
PDxRPP para o peso (kg) do pescoço, serrote, costilhar, paleta, lombo e pernil (Tabela
3). Isso possivelmente devido à proporcionalidade de crescimento das distintas regiões
da carcaça, ocorrendo maior peso dos cortes com o aumento da massa corporal e
reforçando a lei da harmonia anatômica (BOCCARD & DUMONT, 1960), segundo a
qual carcaças de pesos e quantidades de gordura similares apresentam proporções
semelhantes das diferentes regiões anatômicas, independentemente da conformação dos
genótipos considerados.
Tabela 3. Pesos (kg) dos cortes comerciais de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de mesmas letras maiúsculas na coluna e minúsculas na linha diferem entre si (P<0,05) pelo teste de Tukey.
Resíduo Pós-Pastejo Período de Descanso (n0 de folhas/perfilho) 4,5 6,5 8,5 Média
Pescoço CV = 15,38 10 cm 0,57 0,48 0,58 0,54 5 cm 0,43 0,54 0,58 0,51
Média 0,50 0,51 0,58 Serrote CV = 17,36
10 cm 0,57 0,54 0,71 0,60 5 cm 0,51 0,63 0,67 0,60
Média 0,54 0,58 0,69 Costilhar CV = 14,31
10 cm 1,07 1,04 1,02 1,04 5 cm 0,96 1,05 1,01 1,00
Média 1,01 1,04 1,01 Paleta CV = 12,43
10 cm 1,23 1,05 1,20 1,16 5 cm 1,10 1,11 1,18 1,13
Média 1,16 1,08 1,19 Lombo CV = 18,50
10 cm 0,60 0,55 0,59 0,58 5 cm 0,51 0,57 0,67 0,58
Média 0,55 0,56 0,63 Pernil CV = 14,56
10 cm 2,11 1,93 2,16 2,06 5 cm 1,94 2,05 2,19 2,06
Média 2,02 1,99 2,17

148
De acordo com Prata (1999), a composição centesimal da carne ovina apresenta
valores médios de 75% de umidade. Neste experimento, os resultados observados
estiveram dentro da média relatada pelo referido autor. A carne dos ovinos terminados
nos pastos manejados sob os maiores PDs apresentou o maior teor de umidade (P<0,05)
(Tabela 4). O aumento do teor de umidade da carne em relação aos PDs foi
provavelmente devido ao alto teor de gordura. Almeida Jr. et al. (2004) relataram que à
medida que o teor de gordura aumenta, também aumenta o teor de umidade.
Não foi verificado efeito dos PDs (P 0,05), nem dos RPPs (P 0,05), e nem
PDxRPP (Tabela 4). Os valores de matéria mineral variam muito pouco, mesmo
comparando-se com trabalhos em confinamento (SOUZA et al., 2002).
Diversas pesquisas realizadas têm observado que a proteína é o componente que
menos varia nas carnes. O teor de proteína não foi influenciado pelos PDs (P 0,05),
nem pelos RPPs (P 0,05) e nem PDxRPP (Tabela 4). Gutler el al. (1987) afirmaram que
a proteína é o principal componente orgânico do músculo, apresentando teores em torno
de 20%, valores similares aos obtidos neste experimento para os ovinos terminados nos
pastos sob PD de 4,5; 6,5 e 8,5 fol/perf em ambos os resíduos. Os ovinos em pastejo
apresentam alta atividade metabólica, e direcionam os nutrientes em quantidade
adequada para formação muscular resultando em elevado valor de proteína bruta.
Houve efeito somente dos PDs (P<0,05) para o teor de extrato etéreo da carne.
Maiores teores foram verificados nos ovinos terminados nos pastos manejados sob PD
de 8,5 fol/perf (Tabela 4). Contudo, o teor de extrato etéreo obtido nesta pesquisa é
bastante inferior aos relatos da literatura. Kolb et al. (1984) afirmaram que as carnes
com conteúdo de gordura menor que 5% são consideradas magras. Diaz et al. (2002)
mencionaram que cordeiros terminados em pastejo, geralmente, têm menos gordura
que cordeiros terminados em confinamento, isso porque animais em pastejo tem
maiores exigências de energia que animais confinados em razão de um aumento no
metabolismo basal devido à presença de fibra na sua dieta e aumento da atividade
associada ao pastejo.
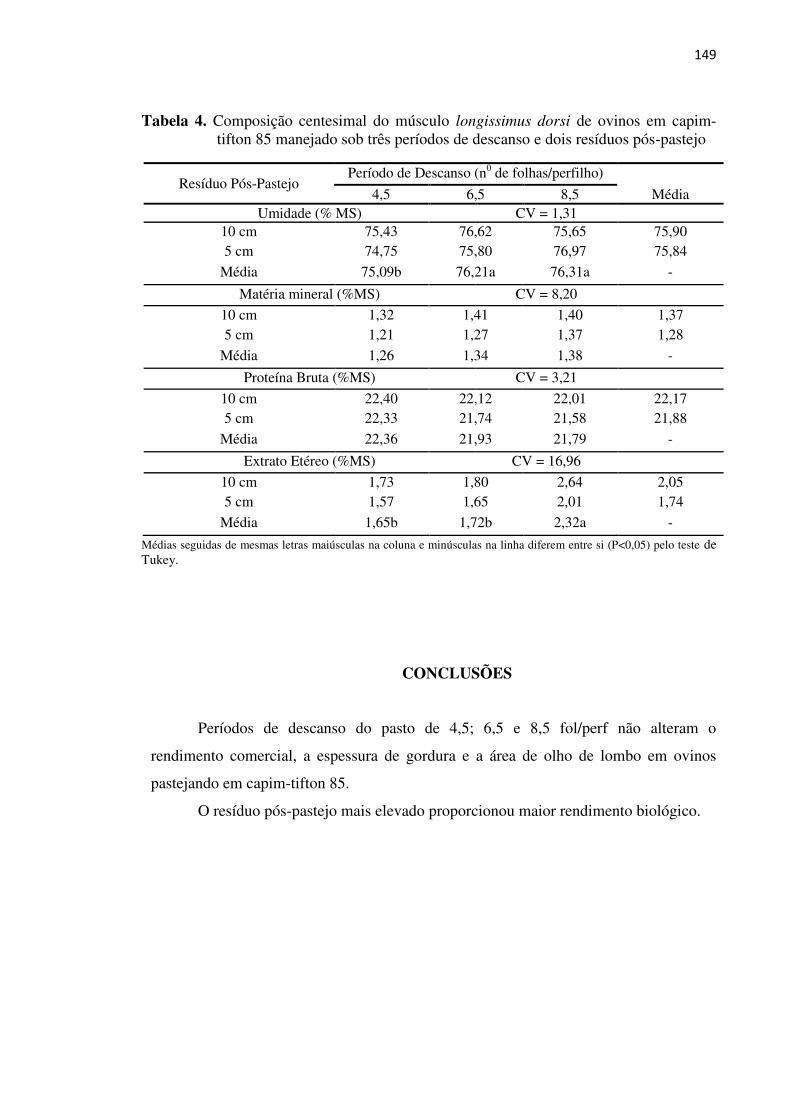
149
Tabela 4. Composição centesimal do músculo longissimus dorsi de ovinos em capim-tifton 85 manejado sob três períodos de descanso e dois resíduos pós-pastejo
Médias seguidas de mesmas letras maiúsculas na coluna e minúsculas na linha diferem entre si (P<0,05) pelo teste de Tukey.
CONCLUSÕES
Períodos de descanso do pasto de 4,5; 6,5 e 8,5 fol/perf não alteram o
rendimento comercial, a espessura de gordura e a área de olho de lombo em ovinos
pastejando em capim-tifton 85.
O resíduo pós-pastejo mais elevado proporcionou maior rendimento biológico.
Resíduo Pós-Pastejo Período de Descanso (n0 de folhas/perfilho)
4,5 6,5 8,5 Média Umidade (% MS) CV = 1,31
10 cm 75,43 76,62 75,65 75,90 5 cm 74,75 75,80 76,97 75,84
Média 75,09b 76,21a 76,31a -
Matéria mineral (%MS) CV = 8,20
10 cm 1,32 1,41 1,40 1,37 5 cm 1,21 1,27 1,37 1,28
Média 1,26 1,34 1,38 -
Proteína Bruta (%MS) CV = 3,21
10 cm 22,40 22,12 22,01 22,17 5 cm 22,33 21,74 21,58 21,88
Média 22,36 21,93 21,79 -
Extrato Etéreo (%MS) CV = 16,96
10 cm 1,73 1,80 2,64 2,05 5 cm 1,57 1,65 2,01 1,74
Média 1,65b 1,72b 2,32a -

150
REFERÊNCIAS
ALMEIDA JR., G.A; COSTA, C.; MONTEIRO, A.L.G. et al. Qualidade da carne de cordeiros, criados em creep feeding com silagem de grãos úmidos de milho. Revista Brasileira de Zootecnia, v.33, n.4, p.1039-1047, 2004. ALMEIDA, M.M.M. Estudo da composição química das carnes de caprinos e ovinos criados no sertão do Ceará. 1990. 78f. Dissertação (Mestrado em Tecnologia de Alimentos) – Curso de Pós-graduação em Tecnologia de Alimentos, Universidade Federal do Ceará. ALVES, K.S.; CARVALHO, F.F.R.; FERREIRA, M.A. et al. Níveis de energia em dietas para ovinos Santa Inês: características de carcaça e constituintes corporais. Revista Brasileira de Zootecnia, v.32, n.6, p.1927-1936, 2003 (supl. 2). ANUALPEC – Anuário da Pecuária Brasileira. São Paulo, SP: Instituto FNP/AGRA FNP Pesquisas Ltda, Consultoria & Comércio, 2008. p. 291. BOCCARRD, R.; DUMONT, B.L. Etude de la production de La viande chez les ovins. II variation de I importance relative dês differents régions corporelles de I agneau de boucherie. Annales de Zootechnie, v.9, n.4, p.355-365, 1960. CARVALHO, S. et al. Avaliação da suplementação concentrada em pastagem de Tifton-85 sobre os componentes não carcaça de cordeiros. Ciência Rural, Santa Maria, v.35, n.2, p.435- 439, 2005. CARVALHO, H.H.; JONG, E.V. Alimentos – métodos físicos e químicos de análise. Porto Alegre: UFRGS, 2002. 180p. COATES, D.B.; PENNING, P. Measuring animal performance. In: MANNETJE, L.; JONES, R.M. (Eds.). Field and laboratory methods for grassland and animal production research. Wallingford: CAB International, p.353-402, 2000. DELFA, R.; GONZALES, C; TEIXEIRA, A. El quinto quarto. Revista Ovis, v.17, p.49-66, 1991.
DIAZ, M.T.; VELASCO, S.; CAÑEQUE, V. et al. Use of concentrate or pasture for fattening lambs and its effect on carcass and meat quality. Small Ruminant Research. v.43, p.257-268, 2002. FIMBRES, H; HERNÁNDEZ-VIDALA, G; PICÓN-RUBIOA, J. F. et al. Productive performance and carcass characteristics of lambs fed finishing ration containing various forage levels. Small Ruminant Research, v.43, n.3, Março 2001, pages 283-288. KOLB, E.; GUTLER, H.; KETZ, A.; SCHRODER, L.; SEIDEL, H. Kolb: Fisiologia veterinária. Editoria de Erich Kolb, traduzido sob supervisão de Gandolfi. 4 ed. Rio de Janeiro: Guanabara Koogan, 1984. 612p.

151
HOPKINS, D.T. Effects of variation in protein digestibility. In: Protein Quality in Humans: Assessment and in Vitro Estimation (Bodwell, C.E.; Adkins, J.S. and Hopkins, D.T. Eds.). Publishing, Westport, CT. p.169.1981. MAHENDRAKAR, N.S; KHABADE, V.S.; DANI, N.P. Studies on the effect of fattening on carcass characteriastics and quality of meat from Bannur lambs. Journal Food Science Technology, v.25, n.4, p. 228-231, 1988. MERTENS, D.R. Análise da fibra e sua utilização na avaliação de alimentos e formulação de rações. In: SIMPÓSIO INTERNACIONAL DE RUMINANTES, 1992, Lavras, MG. Anais... Lavras : SBZ, 1992. p.188- 219. MOTTA, O.S.; PIRES, C.C.; SILVA, J.H.S.; ROSA, G.T.; FÜLBER, M. Avaliação da carcaça de cordeiros da raça Texel sob diferentes métodos de alimentação e pesos de abate. Ciência Rural, v.31, p.1051-1056, 2001. OSÓRIO, J. C. S. et al. Produção de carne em ovinos de cinco genótipos. Ciência Rural, v. 26, p. 447- 481, 1996b. OSÓRIO, J.C.S. Estudio de la calidad de canales comercializadas em el tipo ternasco segun la procedencia: bases para la mejora de dicha calidad em Brasil. 1992. 335p. Tese (Doutorado em Veterinaria) – Facultad de Veterinaria, Universidad de Zaragoza, Zaragoza, 1992. PIRES, C.C.; CARVALHO, S.; GRANDI, A. et al. Características quantitativas e composição tecidual de carcaça de cordeiros terminados em confinamento. Ciência Rural, v.29, n.3. p.539-543. 1999. POPPI, D.P.; HUGHES, T.P.; L‟HUILLIER, P.J. Intake of pasture by grazing ruminants. In: NICOL, A.M. (Ed.). Livestock feeding on pasture. Hamilton: New Zealand Society of Animal Production, 1987. p.55-64. PRATA, L.F. Higiene e inspeção de carnes, pescado e derivados. Jaboticabal: FUNEP. 1999. 217p. REIS, W. et al. Características da carcaça de cordeiros alimentados com dietas contendo grãos de milho conservados em diferentes formas. Revista Brasileira de Zootecnia, v. 30, p. 1308-1315, 2001. SAS INSTITUTE. SAS system for Windows. Version9. 1. Cary: SAS Institute Inc. 2003. 2 CD – ROMs. SAÑUDO, C., SIERRA, I. Calidad de la canal y de la carne en la especie ovina. Ovino y Caprino. Monografias del Consejo General de Colegios Veterinários. Madrid, España, p. 207-254, 1993.

152
SILVA SOBRINHO, A.G. Aspectos quantitativos e qualitativos da produção de carne ovina. In: MATTOS, W.R.S. (Ed.). A produção animal na visão dos brasileiros. Piracicaba: FEALQ, 2001. p.425-446. SIQUEIRA, E. R. Potencialidades da ovinocultura de corte. Tecnologia e Treinamento. Ano 3, nº 10, 1999. SOUZA, R.X.; PEREZ, J.R.O.; BRESSAN, M.C. et al. Composição centesimal do músculo Bíceps Femoris de cordeiros em crescimento. Ciencia e agrotecnologia, Lavras. Edição especial, p. 1507-1513, 2002.

153
CONSIDERAÇÕES FINAIS E IMPLICAÇÕES
A produção sustentável de pequenos ruminantes a pasto deve ter como base o
manejo da pastagem, permitindo a maximização da produção. Este manejo deve levar
em consideração o período de descanso e o resíduo pós-pastejo que possam favorecer a
rebrotação da planta. O menor período de descanso associado a uma menor altura
residual mostrou-se mais eficiente na redução da perda excessiva da massa seca de
forragem verde por senescência e no controle do alongamento do colmo, sem resultar na
degradação da pastagem, em virtude da maior frequência e intensidade de pastejo.
As mudanças nas características estruturais do pasto, juntamente com a
composição químico-bromatologica do mesmo, são de fundamentais importância no
momento de se avaliar o valor nutritivo dos pastos, com a finalidade de se obter
informações mais concretas a respeito do valor nutritivo dos mesmos além do
conhecimento do vigor da rebrotação do pasto, persistência e produtividade animal por
área. Sendo assim, o pasto manejado sob o período de descanso de 4,5 fol/perf e resíduo
pós-pastejo de 5 cm apresentou a melhor composição químico-bromatologica dentre os
que foram avaliados neste experimento.
Os resultados obtidos confirmam que a adoção de práticas ideais de manejo para
gramíneas, como as do gênero Cynodon sp, possibilitam altas taxas de produção animal.
Apesar de, na realidade do produtor rural, ser difícil praticar a lotação variável, em
virtude principalmente da dificuldade de se manejar parte dos animais tão
constantemente. Um segundo problema é o ajuste da taxa de lotação, pela
impossibilidade de conseguir ou retirar animais em tempo hábil para manter a estrutura
do pasto no alvo desejado.
Vale ainda salientar que esses resultados foram relativos ao pasto adubado e
irrigado e que, portanto, devem ser planejadas outras estratégias de manejo, a fim de
atender à demanda dos animais por alimento nas condições do Semiárido Brasileiro,
garantindo o suprimento de nutrientes o ano inteiro, de acordo com as metas planejadas
e,ou, idealizadas para o sistema de produção. Considera-se ainda que a produção animal
e a pecuária competitiva devem ser economicamente sustentáveis, respeitando o meio
ambiente e ainda atendendo às necessidades do mercado consumidor.

















![Forragicultura e Manejo de Pastajens [Zootecnia] Www.azootecnia.blogspot.com](https://img.document.onl/doc/110x75/54e1b0f54a7959d4418b48a7/forragicultura-e-manejo-de-pastajens-zootecnia-wwwazootecniablogspotcom.jpg)











