Embed Size (px)
Citation preview

VIROLOGIA
1. Introdução aos Vírus
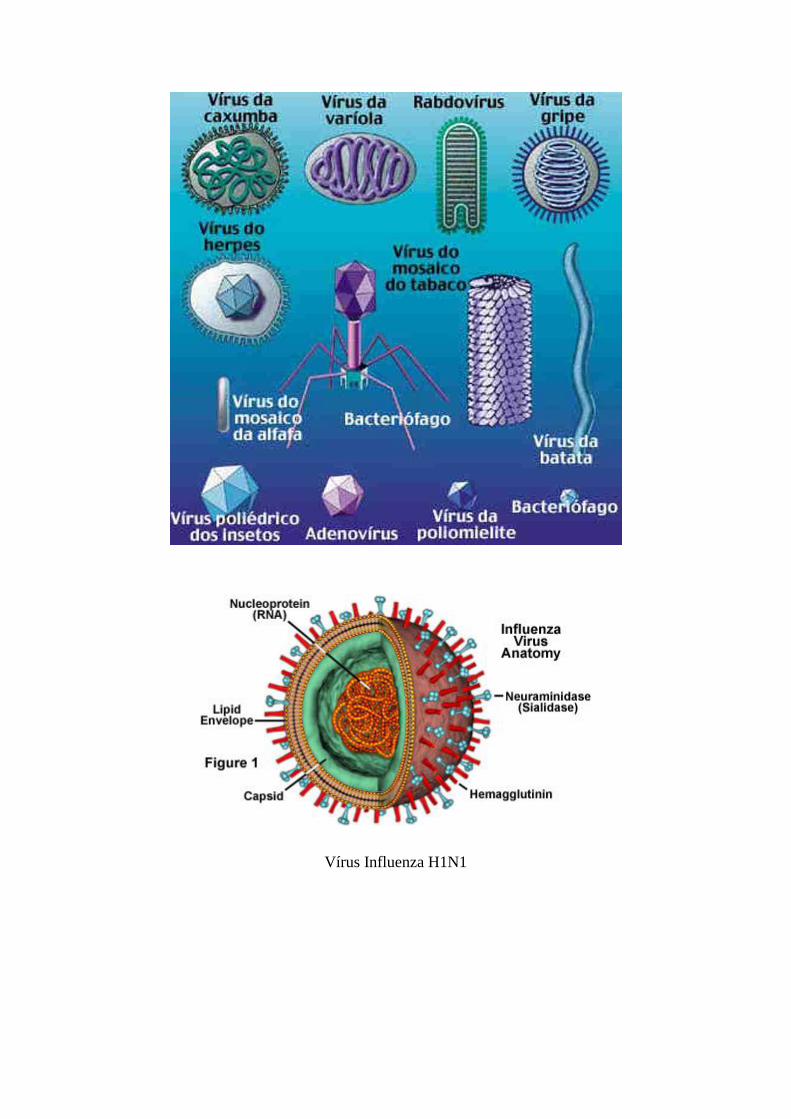
Vírus são entidades acelulares compostas basicamente por um invólucro
protéico denominado capsídio que confina o genoma viral, constituído
exclusivamente de DNA ou RNA que podem ser de cadeia única ou dupla e
estarem presentes como uma única molécula contendo todo o genoma ou
em múltiplas moléculas nas quais se distribui o genoma.
A partícula viral individual completa é denominada vírion.
A complexidade estrutural tanto do capsídio quanto do genoma varia
grandemente conforme a família viral, existindo vários tipos morfológicos
que variam em tamanho e complexidade.
Vírus Influenza H1N1

Vírus da AIDS
Vírus HPV

Vírus da Hepatite b
Vírus da Hepatite c
As dimensões da maioria dos vírus conhecidos variam entre 10 a 250 nm e
seus genomas variam de mil pares de base a 400 mil pares de base.
Tais partículas virais só podem ser visualizadas através de microscopia
eletrônica.
Os vírus podem ser encontrados em todos os ambientes do planeta, com
grande importância nos oceanos, onde compõem o reservatório da maior
parte da diversidade genética.
Os vírus ocorrem em todos os domínios da vida havendo vírus específicos
para arqueas, bactérias, fungos, protozoários, algas, plantas, animais e
humanos.

Ainda não foram encontrados vírus de procariotos termófilos ou
hipertermófilos, mas talvez seja apenas uma questão de tempo.
Recentemente, foi descoberto um vírus que infecta a ameba Acanthamoeba
polyphaga, cujo capsídio tem um diâmetro de 400 nm envolvido por longas
fibrilas de 80 nm de comprimento.
O seu genoma, composto de uma molécula de DNA de cadeia dupla
circular de 1,2 milhões de pares de base, contendo pelo menos 911 genes é
o maior de todos os vírus caracterizados até então e é mais extenso que o
genoma de algumas bactérias.
Esse vírus foi denominado Mimivirus e parece ser membro de uma nova
família viral.
Outros vírus de DNA de cadeia dupla com capsídios variando entre 200 nm
e 600 nm com genomas variando de 300 mil pares de base a 1,2 milhões de
pares de base estão sendo descobertos em números cada vez maiores em
amostras de água do mar e água doce, associados a microrganismos
planctônicos.
Os maiores vírus conhecidos têm dimensões aproximadas às das menores
bactérias e podem inclusive serem visualizados com microscopia óptica.
O capsídio tem duas funções básicas:
(1) proteger o ácido nucléico viral contra injúrias causadas por fatores
ambientais e

(2) permitir a adsorção do vírion à membrana da célula hospedeira, o
primeiro passo para a infecção viral.
A maioria dos genomas virais é muito pequena e geneticamente limitada.
Os genomas dos menores vírus podem conter de 3 a 7 genes, enquanto que
os genomas dos vírus maiores podem conter entre 150 e 911.
Os genes virais contêm informação exclusiva para a síntese de vírions
completos, incluindo algumas enzimas virais que possibilitam sua entrada e
saída da célula hospedeira e, também, para programar a maquinaria
sintética da célula hospedeira para a replicação de componentes do vírus.
Por causa de sua limitação genética, os vírus não apresentam uma estrutura
celular com sistema enzimático próprio para síntese de proteínas e
obtenção de energia e, portanto, não realizam nenhuma das funções
metabólicas usuais das células vivas, ou seja, são metabolicamente inertes e
não se reproduzem.
Os vírus são parasitas intracelulares obrigatórios e essencialmente
infecciosos, pois sua multiplicação e continuidade dependem exclusiva e
inteiramente da maquinaria enzimática de uma célula viva que lhes sirva de
hospedeira.
São, também, denominados "parasitas informacionais" por transportarem
informação genética para sua replicação.
Pelo fato dos vírus serem metabolicamente inertes e não se reproduzirem,
as partes constituintes do capsídio e do genoma são sintetizados pela célula
hospedeira e depois montados em vírions completos que são liberados da
célula.
Considerar os vírus como seres vivos representando os mais simples
microrganismos ou matéria inanimada na forma das mais complexas
entidades moleculares é objeto de controvérsia e depende do conceito de
vida adotado. Alguns vírus podem ser cristalizados sem perder seu poder
infeccioso.
2 – Importância médica dos vírus

Os vírus causam diversas doenças em humanos variando de infecções
brandas e benignas como o resfriado comum, herpes simples e gripes até
enfermidades graves e potencialmente fatais como hepatite, febre amarela,
dengue, poliomielite e varíola.
Pela vacinação em massa da população mundial, doenças virais tais como a
poliomielite e a varíola foram erradicadas.
Vírus da varíola Vírus da Polimielite
Alguns vírus causam quadros clínicos agudos facilmente reconhecidos
como gripes e resfriados; outros causam doença e parecem desaparecer,
permanecendo latentes por muitos anos, e então se manifestam causando
formas mais severas da doença como acontece com o vírus varicela-zoster
que, em um primeiro momento, na infância, causa a varicela (popularmente
conhecida como catapora) e anos após a cura da doença pode reaparecer na
forma de herpes zoster.
Alguns vírus, denominados oncovírus, não matam suas células hospedeiras,
mas induzem sua multiplicação descontrolada originando tumores
cancerosos.
Estudos epidemiológicos mostram que em países desenvolvidos as
infecções virais são a causa mais comum de doenças agudas de evolução
benigna e que normalmente não requerem hospitalização.
Por outro lado, em países em desenvolvimento, as doenças virais são
importantes causas de mortalidade e invalidez permanente, notadamente
entre crianças.
Enquanto que infecções por bactérias e por protozoários podem ser
relativamente bem controladas, as infecções virais representam uma
ameaça relativamente maior e menos controlável à saúde humana.
Infecções virais emergentes e re-emergentes, tais como as causadas por
linhagens altamente virulentas do vírus Influenza (vírus da gripe), ocorrem
periodicamente e representam sério risco à saúde pública.
Os vírus são altamente específicos quanto aos seus hospedeiros. Uma
espécie ou grupo de vírus pode ser capaz de infectar e causar doença em
uma única espécie de hospedeiro como o vírus da varíola, que só ataca
humanos ou em um grupo de espécies relacionadas como o vírus da raiva
que ataca mamíferos.
Os vírus de eucariotos multicelulares infectam tecidos específicos de um
determinado hospedeiro, fenômeno denominado de tropismo de tecido.
Essa especificidade de hospedeiros advém do fato de os vírus terem que se
aderir à superfície da célula hospedeira para poder infectá-la.
Quadro 1: Algumas viroses humanas com agente causador, as formas de infecção e de
prevenção.
VIROSE VÍRUS CARACTERÍSTICAS INFECÇÃO PREVENÇÃO
Catapora ou
varicela
Varicella zoster virus (VZV)
Envelopado, DNA
cadeia dupla
Vias respiratórias
DNA incubado
gânglios nervosos
espinhais
Vacina e
Evitar
contato
Herpes labial Herpes simplex
tipo 1 (HSV-1)
Envelopado, DNA
cadeia dupla
Por contato
DNA viral incubado
gânglios do nervo
trigêmeo
Evitar
contato e
Higiene
Herpes
genital
Herpes simplex
tipo 2 (HSV-2)
Envelopado, DNA
cadeia dupla
Por contato
DNA viral incubado
gânglios do nervo
sacral
Evitar
contato e
Higiene
Rubéola Rubella virus Envelopado, RNA
cadeia simples(+)
Vias respiratórias
ou gotículas de saliva
Vacina
tríplice e
Evitar
contato
Sarampo Morbillivírus Envelopado, RNA
cadeia simples(-)
Vias respiratórias ou
gotículas de saliva
Vacina
tríplice e
Evitar

contato
Varíola Orthopoxvirus
viriolae
Envelopado, DNA
cadeia dupla
Vias respiratórias ou
gotículas de saliva
Vacina e
Evitar
contato
Poliomielite Enterovirus
Grupo Poliovírus (3 sorotipos).
Não-envelopado,
RNA cadeia
simples(+)
Ingestão de
alimentos, água
contaminados com
fezes e pela saliva de
portadores
Vacina e
Evitar
contato
Raiva Lyssavirus Envelopado, RNA
cadeia simples(-)
Pelo contato com
saliva de animal
infectado na mordida
ou em ferimentos
Vacina de
animais
domésticos
e Evitar
contato
Dengue Flavivirus
(arbovírus)
Envelopado, RNA
cadeia simples(-)
Picada de mosquito
Aedes aegyoti
Eliminar
criadouros
do mosquito
Febre
amarela
Flavivirus
(arbovírus)
Envelopado, RNA
cadeia simples(+)
Picada de mosquito
Aedes aegyoti
Eliminar
criadouros
do mosquito
Gripe Influenza A virus (H1N1)
Envelopado, com 8
moléculas de RNA
cadeia simples(-)
Vias respiratórias ou
gotículas de saliva
Vacina e
Evitar
contato
Resfriado
comum
Rhinivirus e
Coronavirus
Não-envelopado,
RNA cadeia
simples(+) e
Envelopado, RNA
cadeia simples(+)
Contato com
secreção nasal ou em
ambientes
contaminados
Evitar
contato
Caxumba Paramyxovirus Envelopado, RNA
cadeia simples(-)
Vias respiratórias ou
gotículas de saliva
Vacina
tríplice e
Evitar
contato
Gastrenterite
rotaviral
Rotavirus Não-envelopado,
com 11 moléculas
RNA cadeia dupla
Ingestão de
alimentos, água
contaminados com
fezes
Higiene
Saneamento
básico
Hepatite A e
E
Hepatitis A
(HAV) e
Hepatitis E
(HEV)
Não-envelopado,
RNA cadeia
simples(+)
Ingestão de
alimentos, água
contaminados com
fezes
Vacina
hepatite A e
Evitar
contato
Higiene
Saneamento
básico
Hepatite B Hepadnavirus Envelopado, DNA
cadeia dupla
Transfusão de sangue
. gestação, parto,
amamentação e
relações sexuais.
Contato com fluidos
corporais (saliva,
Vacina
hepatite B e
Evitar
contato

leite, sêmen)
contaminados
Hepatite C Hepatitis C
(HCV)
Envelopado, RNA
cadeia simples(+)
Transfusão de sangue
. gestação, parto,
amamentação e
relações sexuais.
Contato com fluidos
corporais (saliva,
leite, sêmen)
contaminados
Evitar
contato
Hepatite D Hepatitis D
(HDV) ou vírus
delta
Sem envoltório
protéico associado
ao HBV
Transfusão de sangue
. gestação, parto,
amamentação e
relações sexuais.
Contato com fluidos
corporais (saliva,
leite, sêmen)
contaminados
Vacina
hepatite B e
Evitar
contato
Condiloma Papillomavirus
(HPV)
Não-Envelopado,
DNA cadeia dupla
Relações sexuais,
parto e objetos
contaminados
Vacina e
evitar
contato
Hantavirose Hantavitus Envelopado, RNA
cadeia simples(-) inalação de aerossóis da urina, da saliva e das fezes de ratos silvestres
Evitar
contato
Higiene
Saneamento
e higiene
Ebola Ebolavirus Envelopado, RNA
cadeia simples (-) Contato com pessoa infectada
AIDS Lentivirus (HIV-1 e HIV-2)
Envelopado, RNA
cadeia simples (-) Contaminação por relações sexuais e pelo sangue.
HIV ataca
Linfócito T CD4+
(T4)
Sexo seguro
Evitar
contato
com sangue
3 – Importância ambiental dos vírus
Os vírus podem ter representado – e ainda representar – um importante
fator de seleção natural de espécies animais.
Um exemplo clássico é a seleção natural de coelhos resistentes ao vírus do
Mixoma durante epidemias induzidas como forma de controle biológico
das populações de coelhos da Austrália.

Evidências indiretas sugerem que a mesma ação seletiva foi desempenhada
pelo vírus da varíola em populações humanas.
Nos oceanos, os vírus – de DNA e de RNA – constituem parte significativa
da biomassa das águas costeiras em números de até 50 milhões de
partículas por mililitro, atingindo um total estimado de 25 a 270
megatoneladas. A maioria dos vírus oceânicos não foi caracterizada.
As infecções por vírus oceânicos são principal causa de mortalidade desde
microrganismos a grandes mamíferos.
Como resultado, influencia as composições das comunidades marinhas e os
ciclos biogeoquímicos.
Os microrganismos compõem mais de 90% da biomassa viva dos mares,
sendo importantes componentes do fitoplanctôn e do bacterioplâncton.
Estima-se que os vírus matem cerca de 20% dessa biomassa por dia.
Portanto, os vírus desempenham a principal forma de controle das
populações de microplâncton e, conseqüentemente, sobre a produção de
oxigênio e de dimetilssulfeto atmosférico, um fator importante na
regulação climática.
4- Importância Científica
Os vírus representaram e ainda representam instrumentos importantes para
o desenvolvimento da pesquisa científica, tanto acadêmica quanto aplicada.
Os bacteriófagos, podem, ao serem liberados de uma célula bacteriana
infectada, transportar fragmentos do cromossomo dessa bactéria para outra,
a qual, se sobreviver à infecção viral (fase lisogênica) poderá recombinar
seu material genético com o fragmento de DNA trazido pelo vírus.
Este fenômeno, conhecido como transdução, foi amplamente utilizado
experimentalmente nos primeiros estudos de genética de bactérias.

Anteriormente à descoberta e o amplo uso dos antibióticos na terapia das
doenças causadas por bactérias cogitou-se na administração de
bacteriófagos na prevenção e tratamento das infecções bacterianas.
Com a descoberta dos antibióticos e de sua eficácia no tratamento dessas
infecções, os estudos clínicos utilizando bacteriófagos como agentes
antibacterianos não tiveram continuidade no Ocidente, mas esses vírus
continuaram sendo utilizados na ex-União Soviética e na Europa Oriental.
A emergência de bactérias patogênicas resistentes a múltiplos antibióticos é
um dos mais críticos problemas da prática médica moderna,
particularmente por causa do concomitante aumento de indivíduos
imunossuprimidos.
O risco de a humanidade voltar à era pré-antibióticos é real e o
desenvolvimento de formas alternativas de tratamento de infecções
bacterianas tornou-se uma das maiores prioridades da medicina moderna e
da biotecnologia.
A “bacteriofagoterapia” ressurge no campo da saúde pública como uma
dessas alternativas.
Outros vírus são extensivamente utilizados na engenharia genética para
transferir material genético entre diferentes espécies de hospedeiros. Por
exemplo, genes virais têm sido utilizados na obtenção de alimentos
transgênicos.
Vírus que parasitam insetos têm grande potencial para o controle biológico
de pragas agrícolas, atuando como inseticidas biológicos.
Uma vez que os vírus são específicos para seus hospedeiros, o uso de vírus
como controladores biológicos é considerado seguro para o ecossistema e
para o homem.
Por exemplo, os vírus do gênero Nucleopolyhedrovirus (família
Baculoviridae) são utilizados no controle biológico da lagarta da soja
(Anticarsia gemmatalis).

A origem dos vírus é uma incógnita. Os vírus não representam a forma de
vida mais primitiva por dependerem de células vivas para sua replicação.
Não há registro fóssil de vírus.
2. Estrutura viral
Um vírus, durante seu ciclo infeccioso, assume várias formas e
manifestações tais como uma partícula viral completa ou um ácido nucléico
em replicação no hospedeiro.
A partícula viral individual ou vírion é caracterizada por propriedades
intrínsecas tais como forma, massa, composição química, tipo de
genoma e subunidades protéicas, entre outras.
A complexidade estrutural de um vírion varia conforme a família viral.
Existem famílias cujas partículas virais apresentam estruturas simples
como o vírus da poliomielite e há aquelas com vírions exibindo uma ultra-
estrutura complexa como acontece com o vírus da imunodeficiência
humana (HIV).
O tamanho e a forma de um vírion, o tipo de ácido nucléico que transporta
e a presença ou não de envelope, são características constantes e utilizadas
na classificação dos vírus.

2.1 Capsídio
2.1.1 Organização do capsídio
O invólucro protéico, denominado “capsídio”, pode ser simples ou
complexo, envolvido ou não por um envelope lipídico.

O capsídio determina a forma do vírion e é formado pela agregação
espontânea e organizada de múltiplas subunidades protéicas denominadas
“capsômeros”.
O capsídio pode ser constituído por um único tipo de proteína ou por várias
proteínas diferentes.
O número de proteínas e o arranjo espacial dos capsômeros são
características específicas de cada família viral e são utilizadas na
identificação e classificação dos vírus.
O capsídio tem duas funções principais:
(1) proteger o ácido nucléico viral contra injúrias causadas por fatores
ambientais e
(2) permitir a ligação do vírion a receptores específicos presentes na
superfície da célula hospedeira, no processo da infecção viral.
Sem a proteção do capsídio, a carga negativa da superfície da membrana
plasmática repeliria o ácido nucléico que também possui carga negativa.
A organização do capsídio em subunidades protéicas repetitivas (estrutura
polimérica) decorre da quantidade exígua de material genético disponível
nos vírus e é um meio de resistência a mutações, pois algumas subunidades
sintetizadas erroneamente não são incorporadas ao vírion evitando a
montagem de partículas defeituosas.
A natureza particulada dos vírus reflete duas grandes propriedades:
(1) Especificidade: composição química bem definida capaz de determinar
respostas imunológicas identificáveis;
(2) Plasticidade: capacidade de sofrer alterações antigênicas durante sua
passagem de um hospedeiro para outro.
2.2 Envelope viral

Dependendo da família viral, os vírions podem possuir um envoltório
membranoso, denominado “envelope”, constituído por uma porção de
membrana plasmática que o envolve ao emergir da célula hospedeira em
um processo denominado “exocitose” ou “brotamento”.
Na estrutura do envelope encontram-se tanto proteínas codificadas pelo
genoma da célula hospedeira quanto proteínas codificadas pelo genoma
viral.
O envelope protege o vírion da ação do sistema imunitário por ter
composição semelhante às membranas celulares do hospedeiro e auxilia na
infecção por conter as proteínas de ligação virais que reconhecem
receptores específicos na superfície celular.
Em alguns vírus, como o HIV, por exemplo, além de o envelope conter
proteínas de ligação, este se funde à membrana plasmática da célula
hospedeira introduzindo o capsídio em seu interior.
A presença ou ausência de envelope classifica os vírions em “envelopados”
e “não-envelopados”, respectivamente, e está associada a aspectos
epidemiológicos relacionados com a transmissão dos vírus (uma de suas
propriedades relacionais) e, neste contexto, o termo vírus pode ser
empregado, como nos exemplos: os vírus envelopados são transmitidos por
via respiratória ou contato direto e os não-envelopados são transmitidos por
via oral.
Estas características estão relacionadas com a termolabilidade e fraca
resistência do envelope às enzimas do trato digestivo.
Os vírions envelopados podem ser inativados por quaisquer agentes
químicos e físicos que danifiquem membranas celulares como cloro,
peróxido de hidrogênio, fenol, solventes lipídicos (detergentes e álcool),
luz ultravioleta, temperatura (calor, congelamento e descongelamento) e
pH (menor que 6 e maior que 8).
Por outro lado, os não-envelopados são mais resistentes às condições
ambientais, detergentes e desinfetantes.
A presença de um envelope é uma característica comum nos vírus de
animais, mas incomum em vírus de plantas.

2.3 Proteínas de ligação
Todos os vírions contêm em sua superfície externa, um ou mais tipos de
proteínas ou glicoproteínas, denominadas proteínas de ligação ou ligantes
que interagem com receptores específicos presentes na superfície da célula
hospedeira e que propiciam sua anexação à mesma.
A aderência de um vírion à superfície da célula hospedeira é denominada
“adsorção” e é o primeiro passo tanto para a sua penetração na célula
hospedeira quanto para o estabelecimento de uma infecção viral.
Algumas proteínas de aderência são denominadas “espículas” por se
projetarem diretamente da superfície do capsídio ou do envelope,
facilitando o contato com os receptores celulares.
Nos vírions envelopados, as espículas são chamadas de “peplômeros”.
Pelo fato de serem específicos para seus receptores celulares, todos os vírus
têm um espectro limitado de hospedeiros.
Mesmo dentro de um hospedeiro particular, os vírions se adsorvem e
invadem apenas os tipos celulares que possuam receptores específicos para
as suas proteínas de aderência.
Por exemplo, o vírus da varíola e o HIV atacam apenas humanos, o vírus
da gripe aviária infecta aves em geral, mas potencialmente humanos, os
vírus da raiva ataca mamíferos, enquanto que os bacteriófagos atacam
unicamente procariotos.
2.4 Morfologia

A estrutura da maioria das partículas virais reflete duas propriedades
fundamentais do processo de organização morfológica:
(1) as subunidades protéicas do capsídio, codificadas pelo genoma viral,
correspondem a cópias idênticas de um número reduzido de proteínas e
(2) a organização morfológica final dos vírus mais complexos resulta da
organização de cada um dos componentes do vírion.
Os vírus apresentam uma ampla gama de tipos morfológicos sendo a
grande maioria de morfologia regular com seus elementos organizados
segundo estruturas helicoidais ou icosaédricas.
Nos helicoidais, os capsômeros organizam-se segundo simetria helicoidal e
o ácido nucléico viral associa-se à parte interna das subunidades protéicas
como acontece, por exemplo, no vírus do mosaico do tabaco (TMV).
Nos icosaédricos, os capsômeros organizam-se formando um corpo
simétrico de vinte faces triangulares eqüiláteras, 12 vértices e 30 arestas,
como ocorre, por exemplo, nos vírus da Família Adenoviridae.

A organização icosaédrica é possível pela existência de hexâmeros (grupos
de seis capsômeros) que ocupam as faces triangulares ou arestas e de
pentâmeros (grupos de cinco capsômeros) que ocupam os vértices.
Dentre os menores vírus icosaédricos conhecidos estão os da Família
Picornaviridae cujos capsídios são constituídos por doze capsômeros e
dentre os maiores encontra-se o vírus do díptero Tipula iridescent, cujos
capsídios são formados por 1500 capsômeros.
As vantagens das estruturas helicoidais e icosaédricas residem no fato de as
subunidades se auto-agruparem sem a participação de enzimas ou outros
agentes.
Nos vírus de estrutura mais complexa (por exemplo, os bacteriófagos) há
síntese prévia de enzimas que participam do reagrupamento das
subunidades, mas o processo de automontagem é condição básica para o
reagrupamento enzimático.
A grande maioria dos vírions conhecidos tem dimensões entre 10 e 250 nm
podendo ser visualizados somente pela microscopia eletrônica.
Dentre os vírus humanos, os da família Picornaviridae (por exemplo, o
vírus da poliomielite e o rinovírus) estão entre o menores vírions
conhecidos com capsídios icosaédricos de 27 a 30 nm de diâmetro.
Até recentemente, os maiores vírions conhecidos eram os pertencentes à
família Poxviridae (por exemplo, o vírus da varíola humana) com
morfologia complexa, ovalada ou assemelhando-se a um paralelepípedo de
140 a 260 nm de diâmetro e de 220 a 450 nm de comprimento.
Mas, em 2003, um grupo de pesquisadores franceses isolou e caracterizou
vírions encontrados na ameba de vida livre Acanthamoeba polyphaga, cujo
capsídio – aparentemente icosaédrico – tem um diâmetro de cerca de 400
nm envolvido por longas fibrilas de 80 nm de comprimento.
Esse novo vírus foi denominado Mimivirus e parece ser membro de uma
nova família viral.
Outros vírus com capsídios variando entre 200 nm e 600 nm estão sendo
descobertos em números cada vez maiores em amostras de água do mar e
água doce, associados a microrganismos planctônicos.

Os maiores vírus conhecidos têm dimensões aproximadas às das menores
bactérias e podem inclusive serem visualizados com microscopia óptica.
2.5 Genomas virais
Os genomas virais são extremamente diversos. Os vários grupos de vírus
possuem tipos característicos e específicos de ácidos nucléicos:
A) DNA de cadeia dupla,
B) DNA de cadeia única,
C) RNA de cadeia dupla ou
D) RNA de cadeia única.
Quando o genoma é de RNA de cadeia simples, este pode ser “positivo” ou
“negativo”.
Um RNA genômico de cadeia simples positiva tem uma seqüência de
nucleotídios idêntica àquela do RNA mensageiro viral e um RNA
genômico de cadeia simples negativa tem uma seqüência de nucleotídios
complementar à do RNA mensageiro viral.
O genoma viral pode ser composto por uma única molécula do respectivo
ácido nucléico (genoma não-segmentado) ou por mais de uma molécula
(genoma segmentado).
Quando o genoma é segmentado, cada molécula de ácido nucléico contém
apenas uma parte dos genes virais e o genoma se distribui nas múltiplas
moléculas.
A maioria dos vírus conhecidos contém genomas compostos por moléculas
lineares, mas alguns vírus possuem genomas compostos por moléculas
circulares.
O tipo de genoma transportado por um vírion reflete o modo de o vírus
transmitir sua informação genética de uma geração para outra e seu modo
de expressar essa informação na célula hospedeira.

A diversidade observada nos genomas virais e nas suas formas de
replicação pode refletir as possíveis múltiplas origens dos vírus durante a
evolução.
A estratégia de replicação de um vírus depende da natureza de seu genoma.
Os genomas dos menores vírus são constituído por cerca de mil bases
podendo conter de três a sete genes, enquanto que os genomas dos vírus
maiores podem conter entre 400 mil e 1,2 milhões de bases contendo de
150 a mais de 900 genes.
Por exemplo, o genoma do Mimivirus, composto por 911 genes é o maior
de todos os genomas virais caracterizados até então e é mais extenso que o
genoma de algumas bactérias.
Os genes virais contêm informação exclusiva para a síntese de vírions
completos, incluindo algumas enzimas virais que possibilitam sua entrada e
saída da célula hospedeira e, também, para programar a maquinaria
sintética da célula hospedeira para a replicação de componentes do vírus.
Por causa de sua limitação genética, os vírus não apresentam uma estrutura
celular com sistema enzimático próprio para síntese de proteínas e
obtenção de energia e, portanto, não realizam nenhuma das funções
metabólicas usuais das células vivas, ou seja, são metabolicamente inertes e
não se reproduzem. Isto lhes condiciona o parasitismo intracelular
obrigatório.
Multiplicação Viral
a) Adsorção
A infecção de uma célula viva por um vírus ocorre após a interação de
proteínas do capsídio viral com receptores específicos presentes na
superfície celular, fenômeno denominado "adsorção" e que constitui o
primeiro e crucial evento da infecção viral.

A interação do capsídio viral com receptores celulares é altamente
específica e por isso todos os vírus têm um espectro limitado de
hospedeiros.
Mesmo dentro de um hospedeiro particular, os vírus só reconhecem e
invadem tipos celulares nos quais encontrem receptores específicos.
b) Penetração
Após a adsorção, ou o vírion insere seu genoma através da membrana
plasmática, permanecendo o capsídio vazio e sem função sobre a superfície
celular ou o vírion penetra inteiro na célula.
c) Desnudação
No último caso, em seguida à penetração do vírion, dá-se a exposição do
genoma viral pela remoção do capsídio.
d) Transcrição
Em ambos os casos, o genoma viral passa a dominar as funções normais da
célula.
Ocorre a síntese de m-RNA, os mecanismos de biossíntese da célula
hospedeira (energia, moléculas precursoras e enzimas) são utilizados pelos
vírus, iniciando a síntese viral. Uma célula infectada produz mais proteínas
e ácidos nucléicos virais que seus próprios produtos.
e) Tradução
O m-RNA viral liga-se aos ribossomos da célula, codificando a síntese das
proteínas virais.
f) Replicação
Os vários componentes do vírion são sintetizados separadamente pela
célula hospedeira e então montados para formar novas partículas que são
liberados por lise da célula ou por brotamento de vírions completos através
da membrana plasmática.

Os novos vírions estão prontos para infectar novas células.
A replicação por montagem de componentes pré-formados é exclusiva dos
vírus e os distingue de todas as outras formas de parasitas intracelulares.
Alguns vírus podem permanecer latentes em suas células hospedeiras por
longos períodos de tempo sem causar nenhuma alteração óbvia nas funções
celulares. Este estado é conhecido como "fase lisogênica".
Quando um vírus latente é ativado, este inicia a "fase lítica", com a
formação e liberação de novos vírions como acontece com os
bacteriófagos, o vírus da herpes e o vírus da imunodeficiência humana
(HIV).

A replicação viral no interior de células hospedeiras depende, também de
outros fatores, tais como a temperatura.
Por exemplo, os rinovírus, causadores do resfriado comum, requerem uma
temperatura que não exceda a 34ºC, o que restringe seu crescimento apenas
em células da camada mais fria da mucosa nasal evitando sua disseminação
para tecidos mais profundos onde as temperaturas são maiores.
O conhecimento das propriedades dos vírus e das relações que estabelecem
com seus hospedeiros é crucial para uma investigação bem sucedida e para
uma administração clínica dos processos patológicos.
Do ponto de vista evolutivo, os vírus representam uma modo de uma
determinada quantidade de informação genética, na forma de moléculas de
DNA ou RNA, garantir sua perpetuação.
Por isso a maioria dos vírus não matam seus hospedeiros.
O fato de sua manutenção ser absolutamente dependente da sobrevivência
de seu hospedeiro cria um dilema evolucionário para os vírus que matam
seus hospedeiros, tais como o HIV, o Ebola, o Hantavirus, o vírus rábico e
o vírus da varíola.
A multiplicação viral causa injúria e destruição das célula hospedeiras e
como os vírus delas dependem inteiramente, estes tendem a estabelecer
infecções brandas nas quais a morte do hospedeiro é mais uma exceção que
uma regra.





























