Embed Size (px)
DESCRIPTION
Curso - Fisiologia Geral
Citation preview

1
Programa de Educação Continuada a Distância
Curso de
Fisiologia Geral
Aluno:
EAD - Educação a Distância Parceria entre Portal Educação e Sites Associados

Curso de Fisiologia Geral
MÓDULO I Atenção: O material deste módulo está disponível apenas como parâmetro de estudos para este Programa de Educação Continuada, é proibida qualquer forma de comercialização do mesmo. Os créditos do conteúdo aqui contido são dados aos seus respectivos autores descritos na Bibliografia Consultada.
2 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

3
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
MÓDULO I
1. INTRODUÇÃO À FISIOLOGIA
1.1. A célula e seu funcionamento Não há vida sem as células. Esses pequenos compartimentos, limitados por uma
membrana e preenchidos por uma substância aquosa repleta de compostos químicos (o
citoplasma), desempenham em miniatura todas as funções vitais. A célula move-se,
cresce, reage a estímulos, defende-se e se reproduz. Para manter rotina tão variada, a
célula eucariota utiliza-se de um conjunto bem organizado de estruturas que lembram
um pequeno complexo industrial. Cada estrutura, ou organela, tem funções definidas.
1.2. O transporte através da Membrana Celular
Muitas das substâncias (gases, íons, açúcares, etc.) dissolvidas em nosso
compartimento intracelular ou extracelular podem atravessar a membrana celular e
passar de um compartimento a outro.
Existem várias formas através das quais as diversas substâncias podem
atravessar a membrana celular. As principais e mais bem conhecidas são: Difusão
simples, difusão facilitada e transporte ativo.
Na difusão simples, a substância passa de um meio a outro (do intracelular para o
extracelular ou do extracelular para o intracelular) simplesmente devido ao movimento
aleatório e contínuo da substância nos líquidos corporais, devido a uma energia cinética
da própria matéria. Em tal meio de transporte não ocorre gasto de ATP(adenosina
trifosfato) intracelular nem ajuda de carreadores.
Podemos citar um exemplo de difusão simples: gases como oxigênio ou dióxido
de carbono atravessam a membrana celular com grande facilidade, simplesmente se
dissolvendo na matriz lipídica desta membrana (oxigênio e dióxido de carbono são
lipossolúveis).
Na difusão facilitada, a substância se utiliza também de seus movimentos
aleatórios e contínuos nos líquidos corporais e passa também de um lado a outro da
membrana celular. Porém, por ser insolúvel na matriz lipídica (não lipossolúvel) e de

tamanho molecular grande demais para passar através dos diminutos "poros" que se
encontram na membrana celular, a substância apenas se dissolve e passa através da
membrana celular ligada a uma proteína carreadora específica para tal substância,
encontrada na membrana celular (Figura1). Em tal transporte também não há gasto de
ATP intracelular.
Podemos citar um exemplo de difusão facilitada: A glicose é um importante
monossacarídeo, que atravessa a membrana celular de fora para dentro da célula (do
meio de maior concentração para o meio de menor concentração de glicose) ligada a
uma proteína carreadora específica para glicose.
FIGURA 1 – Difusão simples e difusão facilitada
No transporte ativo, a substância é levada de um meio a outro através da
membrana celular por uma proteína carreadora que é capaz, inclusive, de transportar
esta substância contra um gradiente de concentração, de pressão ou elétrico (a
4 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

substância pode, por exemplo, ser transportada de um meio de baixa concentração para
um meio de alta concentração da mesma). Para tanto, O carreador liga-se quimicamente
à substância a ser transportada através da utilização de enzima específica, que
catalizaria tal reação. Além disso há um consumo de ATP intracelular para transportar a
substância contra um gradiente de concentração.
Podemos citar um exemplo de transporte ativo: Bomba de Sódio e Potássio que
transporta constantemente, nas células excitáveis, através da membrana, íon sódio de
dentro para fora e íon potássio de fora para dentro da célula. Ambos os íons são
transportados contra um gradiente de concentração, isto é, de um meio menos
concentrado para um mais concentrado do mesmo íon (Figura 2).
FIGURA 2 – Transporte ativo
2. FISIOLOGIA DOS MÚSCULOS E DOS NERVOS
2.1. Potenciais de membrana e Potenciais de ação
A eletricidade é um processo natural em nosso organismo e está envolvida na
função específica de certas células especiais no cérebro e nos músculos estriados e
lisos. Cada padrão de luz, som, calor, dor, cada piscar de olhos, estalar de dedos, cada
pensamento, se traduz em uma sequência de pulsos elétricos.
As células nervosas possuem propriedades similares às outras células em muitos
aspectos: elas se alimentam, respiram, passam por processos de difusão e osmose em
suas membranas, etc., mas diferem em um aspecto importante: elas processam
informação. A habilidade das células nervosas processarem informação depende de
5 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

6
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
propriedades especiais da membrana do neurônio, a qual controla o fluxo de substâncias
ao lado interno da célula (íons sódio, cálcio, potássio, etc).
Os neurônios não existem isoladamente: eles também se conectam uns aos
outros formando as chamadas cadeias neuronais, as quais transmitem informações a
outros neurônios ou músculos. Por essas cadeias caminham os impulsos nervosos. Dois
tipos de fenômenos estão envolvidos no processamento do impulso nervoso: elétrico e
químico. Os eventos elétricos propagam um sinal dentro de um neurônio, e o químico
transmite o sinal de um neurônio a outro ou a uma célula muscular. O "engate" ou junção
entre um neurônio e outro, é denominado sinapse.
Um impulso nervoso é a transmissão de uma alteração elétrica ao longo da
membrana do neurônio a partir do ponto em que ele foi estimulado. A direção normal do
impulso no organismo é do corpo celular para o axônio. Esse impulso nervoso, ou
potencial de ação, é uma alteração brusca e rápida da diferença de potencial
transmembrana. Normalmente, a membrana do neurônio é polarizada em repouso,
sendo que o potencial é negativo (-70 mV). O potencial de ação consiste de uma
redução rápida da negatividade da membrana até 0mV e inversão deste potencial até
valores de cerca de +30mV, seguido de um retorno também rápido até valores um pouco
mais negativos que o potencial de repouso de -70mV.
O mais importante exemplo de transporte ativo presente na membrana das
células excitáveis é a Bomba de Sódio e Potássio. Tal bomba transporta, ativamente e
constantemente, íons sódio de dentro para fora da célula e, ao mesmo tempo, íons
potássio em sentido contrário, isto é, de fora para dentro das células. Mas os íons (sódio
e potássio) não são transportados com a mesma velocidade: A Bomba de Sódio e
Potássio transporta mais rapidamente íons Sódio (de dentro para fora) do que íons
Potássio (de fora para dentro).
Para cada cerca de 3 íons sódio transportados (para fora), 2 íons potássios são
transportados em sentido inverso (para dentro). Isso acaba criando uma diferença de
cargas positivas entre o exterior e o interior da célula, pois ambos os íons transportados
pela bomba (sódio e potássio) são cátions (com 1 valência positiva), e a Bomba de Sódio
e Potássio transporta, portanto, mais carga positiva de dentro para fora do que de fora
para dentro da célula.

7
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
Cria-se assim um gradiente elétrico na membrana celular: No seu lado externo
acaba se formando um excesso de cargas positivas enquanto que no seu lado interno
ocorre o contrário, isto é, uma falta de cargas positivas faz com que o líquido intracelular
fique com mais cargas negativas do que positivas.
O gradiente elétrico então formado é conhecido como Potencial de Membrana
Celular. Na maioria das células nervosas tal potencial equivale a algo em torno de 90mv.
Quando a membrana de uma célula excitável realmente se excita, uma sucessão
de eventos fisiológicos ocorrem através da tal membrana. Tais fenômenos, em conjunto,
produzem aquilo que chamamos de Potencial de Ação.
Geralmente a excitação ocorre no momento em que a membrana recebe um
determinado estímulo. Alguns estímulos como: calor, frio, solução salina hipertônica ou
hipotônica, ácidos, bases, corrente elétrica, pressão, etc.
Algumas células desencadeiam o Potencial de Ação sem a necessidade de
receberem estímulos, devido a uma alta excitabilidade que as mesmas apresentam. Tais
células são denominadas auto-excitáveis, e os potenciais por elas gerados são
denominados de potenciais espontâneos.
Um típico potencial de ação em uma típica célula excitável dura apenas alguns
poucos milésimos de segundo, e pode ser dividido nas seguintes fases: despolarização e
repolarização.
A despolarização é a primeira fase do potencial de ação. Durante esta fase ocorre
um significativo aumento na permeabilidade aos íons sódio na membrana celular. Isso
propicia um grande fluxo de íons sódio de fora para dentro da célula através de sua
membrana, por um processo de difusão simples.
Como resultado do fenômeno citado acima, o líquido intracelular passa a
apresentar uma grande quantidade de íons de carga positiva (cátions) e a membrana
celular passa a apresentar agora um potencial inverso daquele encontrado nas
condições de repouso da célula: Mais cargas positivas no interior da célula e mais
cargas negativas no seu exterior.
O potencial de membrana neste período passa a ser, portanto, positivo (algo em
torno de +45 mv).
A repolarização é a segunda fase do potencial de ação e ocorre logo em seguida
à despolarização.

8
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
Durante este curtíssimo período, a permeabilidade na membrana celular aos íons
sódio retorna ao normal e, simultaneamente, ocorre agora um significativo aumento na
permeabilidade aos íons potássio. Isso provoca um grande fluxo de íons potássio de
dentro para fora da célula (devido ao excesso de cargas positivas encontradas neste
período no interior da célula e à maior concentração de potássio dentro do que fora da
célula). Enquanto isso, os íons sódio (cátions) que estavam em grande quantidade no
interior da célula, vão sendo transportados ativamente para o exterior da mesma, pela
bomba de sódio-potássio. Tudo isso faz com que o potencial na membrana celular volte
a ser negativo (mais cargas negativas no interior da célula e mais cargas positivas no
exterior da mesma).
O potencial de membrana neste período passa a ser algo em torno de -95 mv.
(ligeiramente mais negativo do que o potencial membrana em estado de repouso da
célula).
O repouso é a terceira e a última fase deste processo. É o retorno às condições
normais de repouso encontradas na membrana celular antes da mesma ser excitada e
despolarizada. Nesta fase a permeabilidade aos íons potássio retorna ao normal e a
célula rapidamente retorna às suas condições normais. O potencial de membrana celular
retorna ao seu valor de repouso (cerca de -90 mv.).
Todo o processo descrito acima dura, aproximadamente, 2 a 3 milésimos de
segundo na grande maioria das células excitáveis encontradas em nosso corpo.
Mas algumas células excitáveis apresentam um potencial bem mais longo do que o
descrito acima. As células musculares cardíacas, por exemplo, apresentam potenciais de
ação que chegam a durar 0,15 a 0,3 segundos (e não alguns milésimos de segundo,
como nas outras células). Tais potenciais, mais longos, apresentam um período durante
o qual a membrana celular permanece despolarizada, bastante prolongado. Estes
potenciais são denominados Potenciais em Platô.

FIGURA 3 – O gráfico mostra o comportamento do Potencial de ação na célula.
Na letra A da figura mostra uma visão esquemática do potencial de ação
idealizado. Ilustra as suas várias fases à medida que ele percorre um único ponto da
membrana plasmática. Na letra B do esquema mostramos registros reais de potenciais
de ação que são comumente distorcidos em comparação às visões esquemáticas devido
a variações nas técnicas eletrofisiológicas de registro.
A velocidade de propagação do potencial de ação pode ser variada ao se variar o
tempo de duração de alguma das duas fases da propagação. Contudo, a fase ativa
costuma ser constante nas células, durando em torno de 4ms. Deste modo, a célula
9 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

10
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
varia a duração da fase passiva, havendo dois modos básicos:
Aumento ou diminuição do calibre do axônio ou célula.
Maior ou menor isolamento da membrana (ao variar a espessura da mielina, se
houver).
O aumento do calibre do axônio ou célula provoca um aumento da velocidade de
propagação do potencial de ação, pois há diminuição da resistência longitudinal,
provocada por uma maior área de secção transversal.
Em alguns axônios do polvo Atlântico, a velocidade de propagação do potencial
de ação alcança velocidades superiores a 100 m/s, em virtude do calibre elevado e da
mielina espessa.
Outro fator fundamental para desencadear um potencial de ação é o tipo de
bainha de mielina. A bainha de mielina é uma membrana lipídica modificada e
espessada. Ela pode ser sintetizada por duas células: oligodendrócitos, no sistema
nervoso central, e células de Schwann, no sistema nervoso periférico. A espessura da
bainha de mielina é de acordo com o número de voltas que a membrana das células de
Schwann ou dos oligodendrócitos dão em torno do axônio. Em axônios de calibre
pequeno, não há mielina envolvendo; já em axônios de calibre grande, a mielina é mais
espessada que os outros menores que a possuem.
A bainha de mielina fornece um aumento do isolamento celular (aumento da
resistência de membrana), em virtude de não haver canais de vazamento de membrana
onde há mielina, deste modo, a fase passiva perde menos íons, o que aumenta a chance
do potencial de ação ter sucesso. Além de não haver canais de vazamento de
membrana, não há também praticamente nenhum tipo de canal de membrana quando há
bainha de mielina (ex.: bombas de sódio e potássio), o que provoca para a célula uma
menor necessidade de síntese protéica, ou seja, menos gasto energético.
A bainha de mielina permite uma maior velocidade da fase passiva da propagação
do potencial de ação (diminui a capacitância de membrana e aumenta a resistência de
membrana). Além disso, diminui o número de fases ativas da propagação do potencial
de ação, tornando a propagação mais veloz ainda. As fases ativas da propagação
ocorrem em máculas da bainha de mielina, os nódulos da Ranvier. Neles,
diferentemente da zona cercada por bainha de mielina, há abundância de canais de íon
sódio voltagem-dependentes (densidade até quatro ordens de magnitude a mais que nas

membranas amielínicas), o que permite a ocorrência do potencial de ação, que
corresponde à fase ativa da propagação do potencial de ação. A distância entre os
nódulos de Ranvier deve ser muito bem calculada pelas células, de modo que o
potencial passivo chegue com íons suficientes para provocar o potencial de ação.
A conseqüência da bainha de mielina queimar etapas na propagação, ao diminuir
o número de potenciais ativos, são os movimentos saltatórios, que possuem este nome
em virtude de haver a impressão de que os potenciais de ação saltam de nódulo em
nódulo.
FIGURA 4 – Estrutura e condução na bainha de mielina
De acordo com a Figura acima, os cátions à esquerda, dentro da célula, são
conseguidos a partir de um potencial de ação. Passivamente, eles se difundem para
outro nódulo de Ranvier, onde gerarão um novo potencial de ação.
2.2. Contração do Músculo Estriado e Contração do Músculo Liso
Para que um músculo, esquelético ou visceral, se ponha em ação, isto é, se
contraia, deve ser excitado. Experimentalmente o músculo responde a diversos tipos de
excitação:
- excitações mecânicas, como são as determinadas por uma pancada, uma
picada, um esmagamento etc.;
- excitações térmicas, como o aumento de temperatura;
11 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

12
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
- excitações elétricas; este tipo de excitação é o ideal porque o experimentador
pode fazer variar a intensidade e o grau de excitabilidade do próprio músculo.
No ser vivo, a excitação chega ao músculo através dos nervos motores.
O músculo excitado responde ao estímulo contraindo-se. A contratibilidade é a
característica essencial do músculo. O músculo excitado se deforma, se encolhe,
aumenta de espessura, mas o seu volume total não muda. Diversa é a contração nos
músculos estriados e nos músculos lisos. Os primeiros se contraem muito mais
rapidamente do que os segundos. Uma vez contraído, o músculo se afrouxa, voltando à
sua forma primitiva. O músculo é, portanto, dotado de elasticidade. Isto se pode
constatar distendendo um músculo pelas suas extremidades: observa-se que o músculo
retorna ao seu primitivo comprimento uma vez cessada a tração, com a condição de que
esta não tenha sido muito forte ou muito violenta.
A elasticidade do músculo é indispensável. O músculo deve, na verdade, voltar à
sua forma primitiva para poder contrair-se de novo. Além disso, nos músculos
considerados antagônicos, isto é, que desempenham funções opostas, têm lugar,
contemporaneamente, dois fenômenos contrários: quando um deles se contrai o outro se
afrouxa.
Assim, quando dobramos o antebraço sobre o braço, temos a contração do
bíceps e, ao mesmo tempo, o afrouxamento do tríceps, o músculo antagônico.
Contraindo-se, os músculos esqueléticos agem sobre os ossos, que constituem
verdadeiras "alavancas". Quando levantamos um peso com a mão, dobrando o cotovelo,
o antebraço constitui a alavanca, a articulação do cotovelo é o ponto de apoio, a força
desenvolvida pelos bíceps constitui a força motora e o peso a resistência. Os músculos
realizam sempre um "trabalho". Em física, define-se o "trabalho" como o produto de uma
força pelo deslocamento do ponto de aplicação dessa força. Mas os músculos realizam
um trabalho mesmo sem deslocamento das alavancas ósseas. Para manter na
respectiva posição a cabeça, o tronco e os membros, é necessária uma harmônica
contração de diversos grupos musculares. A manutenção da posição ereta é, sob este
ponto de vista, qualquer coisa de maravilhoso, porque a base da figura constituída pelo
corpo humano é muito pequena e o centro de gravidade está situado muito no alto. O
corpo tende a cair ora para diante e ora para trás, tanto para a direita como para a
esquerda; apesar da nossa aparente imobilidade, somos constrangidos, para evitar a

queda, a contrair, de momento a momento e no tempo oportuno, diversos grupos
musculares. Que tudo impõe trabalho muscular fica demonstrado pelo fato que basta
perder, mesmo por um único instante, a consciência, para cair no chão.
O trabalho necessário para manter o equilíbrio se chama "trabalho estático" e é
comparado ao trabalho fornecido pelo músculo para manter um peso a uma determinada
altura.
Para entendermos as características particulares da contração cardíaca é
relevante recordarmos os mecanismos responsáveis pela capacidade contrátil do
músculo estriado. No sarcômero, unidade contrátil do músculo encontra-se filamentos
grossos e finos intercalados e dispostos como mostrado a seguir:
FIGURA 5 – Filamentos finos e filamentos grossos
O filamento grosso é composto por aproximadamente duzentas moléculas de
miosina. A miosina é uma proteína formada por duas cadeias polipeptidicas pesadas e
quatro leves; as cadeias pesadas possuem uma estrutura globular em suas
extremidades denominada cabeça da miosina, e as duas cadeias pesadas formam uma
dupla hélice deixando as cabeças livres na extremidade. As quatro cadeias leves se
localizam na cabeça da miosina, duas em cada cabeça. Os corpos das moléculas de
miosina formam a cauda do filamento grosso e dela saem proeminências da porção
helicoidal da molécula, mantendo a cabeça longe do corpo: é o braço da molécula. O
conjunto formado chama-se ponte cruzada.
13 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 6 – Ponte Cruzada
O filamento fino é composto por três proteínas, a actina, a troponina e a
tropomiosina. A actina é a molécula central, que polimeralizada forma uma dupla
hélice e contém os sítios de ligação com a miosina. A tropomiosina é uma molécula
presa à actina de forma espiralada sobre a dupla hélice. A tropomiosina impede a
ligação actina/miosina bloqueando o sítio de ligação. A troponina fica presa à
molécula de tropomiosina, e possui três subunidades: uma com afinidade à actina,
outra a tropomiosina e uma última ao Ca2+, a troponina regula o bloqueio do sítio de
ligação feito pela tropomiosina.
FIGURA 7 – Estrutura da actina, troponina e tropomiosina
Os filamentos de actina e miosina têm uma grande afinidade e ligam-se
facilmente sem a presença do complexo troponina/tropomiosina. Nota-se que esse
complexo impede a ligação na ausência de Ca2+. O mecanismo de liberação do sítio
de ligação actina/miosina começa com a chegada do potencial de ação à membrana
do músculo, promovendo a entrada maciça de íons Ca2+ . Estes íons ligam-se à
troponina C, causando uma mudança conformacional da mesma que se reflete na
molécula de tropomiosina, que libera então os sítios da actina que estavam
bloqueados. A interação actina/miosina se dá imediatamente desde que haja ATP e
magnésio (ambos presentes em condições normais). A contração ocorre à medida
que os filamentos finos deslizam sobre os grossos, encurtando o sarcômero:
14 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

15
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
• A cabeça da miosina possui um sítio onde se liga uma molécula de ATP a
ser hidrolisada em ADP e Pi, que permanecem fixos à cabeça, ocupando o
sítio. Este estado permite que a cabeça se estenda em direção ao filamento
fino;
• Assim que o Ca2+ se liga à troponina C e o complexo troponina-tropomiosina
libera o sítio de ligação actina/miosina, a ligação entre os filamentos ocorre;
• Segue-se então o chamado movimento de tensão, que ocorre como
decorrência da energia acumulada na mudança conformacional da cabeça
da miosina em direção ao filamento de actina e da nova alteração
conformacional da cabeça que se curva em direção do braço da miosina;
• Este movimento provoca o deslizamento do filamento fino sobre o filamento
grosso e permite a liberação do ADP e do Pi armazenados na cabeça da
miosina;
• O sítio é então ocupado por uma nova molécula de ATP e a cabeça se solta
do filamento de actina; lembremo-nos que a cabeça só se ligou à actina
devido à hidrólise do ATP e à mudança conformacional. Com a entrada de
um ATP a molécula retorna à sua conformação original e promove a quebra
do ATP em ADP e Pi para recomeçar o ciclo.
No músculo estriado esquelético, a força da contração é determinada não só pela
quantidade de Ca2+ disponível como também pela quantidade de fibras motoras
ativadas. Cada fibra muscular é inervada por um neurônio, porém um mesmo neurônio
inerva mais do que uma fibra. Este conjunto (neurônio + fibras por ele inervadas) é
denominado unidade motora. A relação fibras/neurônio varia em cada tipo de músculo;
em músculos cujos movimentos devem ser precisos a relação chega a ser de duas a três
fibras por neurônio. Já em músculos cuja precisão não é tão necessária a relação é de
centenas de fibras inervada por cada neurônio.
A força da contração cardíaca não se deve à quantidade fibras ativadas
primeiramente porque o coração não recebe inervação motora do sistema nervoso
central, pois possui um sistema especializado de excitabilidade e condutibilidade: a fibra
cardíaca é formada por muitas células individuais separadas entre si por discos
intercalares que possuem uma resistência elétrica muito baixa em relação à membrana

16
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
sarcoplasmática normal. A baixa resistência elétrica permite que o músculo cardíaco se
comporte como um sincício onde as células são interligadas, o que implica na chamada
lei do tudo ou nada - essa lei determina que uma vez que tenha chegado à membrana
de uma miocélula um potencial de ação que se propaga por todas as demais, e logo
todas irão se contrair quase que ao mesmo tempo e com mesma intensidade. O coração
contrai de uma só vez ou simplesmente não contrai.
3. FISIOLOGIA DO CORAÇÃO
O coração é um órgão oco e musculoso, especializado em bombear sangue para
os pulmões, órgãos e tecidos. Localiza-se anatomicamente um pouco à esquerda do
centro do tórax, no sentido antero-posterior, sendo que o ápice do coração é
acentuadamente deslocado para a esquerda.
A bomba cardíaca é, na realidade, formada por duas bombas distintas: o coração
direito e esquerdo. Ou seja, constitui uma bomba muscular dupla e auto-reguladora.
Cada uma dessas partes, as quais impelem sangue para os pulmões e para os tecidos,
é formada por um átrio e um ventrículo. O átrio, segundo (Guytonet al., 1997), funciona
como uma bomba de escorva para o ventrículo. Esse, por sua vez, é o grande
responsável por impulsionar o sangue para a circulação pulmonar e sistêmica. O átrio
direito recebe sangue venoso (carregado de dióxido de carbono CO2) pelas veias cavas
superiores e inferior. Deve-se observar que a denominação “veia” corresponde a todo
vaso cujo sangue chega ao coração, proveniente da circulação sistêmica. Normalmente,
75% do sangue que chega ao átrio direito flui diretamente para o ventrículo direito,
mesmo antes que o átrio se contraia. Portanto, apenas um quarto do sangue é
bombeado para o ventrículo pela contração atrial, sendo, por isso, os átrios considerados
como bomba de escorva. O ventrículo direito, então, bombeia o sangue venoso, através
da artéria pulmonar. Deve-se observar que a denominação “artéria”.

FIGURA 8 – Vista anterior do coração
FIGURA 9 – Vista lateral do coração
3.1. Características do Músculo Cardíaco
O músculo cardíaco é encontrado nas paredes do tubo cardíaco embrionário e no
coração do adulto e é derivado de uma massa restrita do mesênquima esplâncnico. As
17 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

18
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
fibras são longas, ramificadas e apresentam um ou dois núcleos localizados
centralmente na célula. O sarcoplasma próximo aos pólos nucleares contém muitas
mitocôndrias que se localizam em cadeias entre os miofilamentos e os grânulos de
glicogênio. A disposição dos miofilamentos forma estriações semelhantes às do músculo
esquelético. O retículo sarcoplasmático no músculo cardíaco é menos organizado que o
do músculo esquelético. Os túbulos T cardíacos ocorrem ao nível da linha Z. Na maioria
das células, os túbulos T se associam com uma cisterna única e expandida do retículo
sarcoplasmático, formando díades ao invés de tríades. As células musculares cardíacas
formam áreas juncionais altamente especializadas nas extremidades celulares,
conhecidas como discos intercalares. Eles favorecem o acoplamento elétrico entre fibras
musculares cardíacas adjacentes e transmitem o estímulo para a contração de célula a
célula.
Há dois tipos de fibras musculares cardíacas. As fibras musculares cardíacas
atriais são pequenas e possuem menos túbulos T que as fibras ventriculares. Elas
contêm pequenos grânulos com um precursor do fator natriurético atrial, um hormônio
secretado em resposta ao aumento do volume sanguíneo e que atua sobre os rins
causando perda de sódio e água. As fibras musculares cardíacas ventriculares são
maiores, contém mais túbulos T e não apresentam grânulos.
As fibras musculares cardíacas se contraem espontaneamente com um ritmo
intrínseco. O coração recebe inervação autônoma através de axônios que terminam
próximos às fibras, mas nunca formam sinapses com as células musculares cardíacas.
Os estímulos autônomos não podem iniciar a contração, mas podem acelerar ou retardar
os batimentos intrínsecos. O estímulo que inicia a contração é gerado por um conjunto
de células musculares cardíacas especializadas localizadas no nódulo sinoatrial e
conduzido por outras células especializadas denominadas células de Purkinje para
outras células musculares cardíacas. O estímulo é passado entre células adjacentes
através de junções gap que estabelecem uma continuidade iônica entre fibras
musculares cardíacas e que permite que elas trabalhem juntas como se fossem um
sincício funcional.

19
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
3.2. O Ritmo Cardíaco
Há mais de 200 anos, Guyton et al., 1997, demonstraram que fenômenos elétricos
estavam intimamente ligados às contrações rítmicas e espontâneas do coração. De fato,
as células cardíacas são altamente excitáveis, ou seja, são capazes de gerar
rapidamente, em suas membranas, variações nos impulsos eletroquímicos, os quais
podem ser utilizados para a transmissão d sinais ao longo das membranas dessas
células.
Em condições normais, os potenciais de ação só podem ser conduzidos do
sincício atrial para o sincício ventricular por meio de um sistema especializado de
condução, o feixe atrioventricular (feixe AV), que é um feixe de fibras condutoras
especializadas.
A bomba cardíaca é dotada de um sistema especializado em gerar e conduzir
impulsos elétricos, que compreendem desde de nodos, os quais são responsáveis pela
origem desses impulsos, capazes de promover a contração de forma rítmica do músculo
cardíaco, até feixes e vias, os quais, por sua vez, são especializados em conduzir os
impulsos para todo o coração.
O sistema rítmico e condutor do coração pode ser lesado em doenças cardíacas,
em especial pela isquemia dos tecidos cardíacos, resultante do fluxo sangüíneo
coronário insuficiente, o chamado infarto do miocárdio.
A maioria das fibras cardíacas são dotadas da capacidade de auto-excitação,
processo que pode provocar descarga e contrações automáticas e rítmicas. Todavia, o
nodo sinusal (NSA) é o responsável por controlar, normalmente, a freqüência de
batimento de todo o coração, por possuir uma auto-excitação em maior grau, e, portanto,
gerar os impulsos elétricos que ocasionam toda a excitabilidade do coração.
O nodo sinusal, segundo Guyton et al., 1997, é uma estrutura pequena, formada
por músculo especializado em forma de elipse, com cerca de 3 mm de largura, 15 mm
de comprimento e 1 mm de espessura. O NSA está situado na parede lateral superior do
átrio direito, imediatamente abaixo ou quase ao lado do orifício da veia cava superior.

FIGURA 10 – Vista lateral do ventrículo esquerdo
As fibras do nodo sinusal possuem seu diâmetro, em média, 3 ou 4 vezes
menores que as das fibras atriais circundantes, as quais estão conectadas diretamente
às fibras sinusais. Assim sendo, qualquer impulso elétrico gerado pelo NSA se propaga
imediatamente para todas as regiões dos átrios, em uma velocidade de
aproximadamente 1m/s. Uma via especial, a faixa miocárdica interatrial anterior, conduz
o impulso do NSA diretamente para o átrio esquerdo. A onda de impulsos elétricos,
responsáveis pela excitação, que se prossegue inferiormente através do átrio direito
finalmente alcança, por meio da vias internodais, o nodo átrio ventricular (NVA),
considerada normalmente a única via de condução entre os átrios e os ventrículos.
20 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 11 – Vista lateral do átrio esquerdo
Já o nodo átrio ventricular, segundo Berne et al., 2000, tem comprimento de
aproximadamente 22 mm, 10 mm de largura e espessura de 3 mm. Esse nodo situa-se
posteriormente na face direita do septo interatrial, próximo ao óstio do seio coronário.
Também mostra os intervalos de tempos, em frações de segundo, desde a geração do
impulso cardíaco no NSA até sua passagem pelo septo ventricular. Nota-se um retardo
de tempo na passagem do impulso dos átrios para os ventrículos, permitindo que os
átrios esvaziem seu conteúdo sangüíneo nos ventrículos antes que comece a contração
rítmica ventricular. O NVA continua como feixe de His, que dá origem a um ramo
esquerdo no ápice do septo interventricular e continua como ramo direito. O ramo direito
se divide em fascículos anterior e posterior. Cada ramo se divide progressivamente em
ramos cada vez mais finos, os quais cursam pelas paredes internas das cavidades
21 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

ventriculares, retornado em direção à base do coração. Os terminais dessa complexa
rede, que se espalham pelas superfícies subendocárdicas de ambos os ventrículos, são
chamados de fibras de Purkinje, cujas fibras se disseminam por todas as regiões do
miocárdio ventricular.
FIGURA 12 – Esquema do sistema de condução cardíaca
Segundo Berne et al.,2000, as fibras de Purkinje são as células mais largas do
coração, tendo diâmetro de 70 a 80 micrômetros, o que lhes permitem condução rápida
dos impulsos cardíacos, chegando a uma velocidade de condução de até 4 m/s. Essa
alta velocidade permite a ativação rápida de toda a superfície endocárdica dos
ventrículos.
A Figura mostra a representação esquemática do potencial de ação cardíaco.
Nela pode-se observar a presença dos platôs, tanto no músculo atrial, quanto no
músculo ventricular, o que permite que a contração muscular seja mais prolongada.
Durante o platô, a permeabilidade ao potássio é reduzida devido ao influxo
excessivo do cálcio pelos canais específicos desse íon, retardando dessa maneira a
volta do potencial ao seu valor de repouso. Quando os canais lentos de cálcio e sódio
terminam por se fechar, ocorre o aumento instantâneo e rápido da permeabilidade da
membrana ao potássio. A perda rápida de potássio pela fibra faz com que o potencial de
membrana retorne ao seu valor de repouso, e é, assim, finalizado o potencial de ação.
22 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 13– Esquema de um potencial de ação
Em termos do sistema cardiovascular, existem dois tipos principais de potenciais
de ação: os potenciais de ação rápida e de ação lenta.
O potencial de resposta rápida, que também se inicia com grande velocidade a
partir do valor mais negativo de polarização da célula, é característico de células
musculares atriais e ventriculares normais e das fibras de Purkinje. Nessas fibras, o
potencial de membrana em repouso é entre –80 a –90 mV, sendo a velocidade de
ascensão do potencial de ação em torno de 150 mV/s e uma velocidade de condução
muito rápida. A rápida despolarização celular até o ponto de potencial limiar se deve,
basicamente, pelo aumento extremamente rápido da permeabilidade de íons sódio (Na+)
para o interior celular. Essa permeabilidade é resultado da abertura abrupta das
comportas iônicas na membrana, que é representada pelo movimento através dos
canais rápidos de sódio. Nesse período também ocorre a redução espontânea e rápida
de íons potássio (K+).
O potencial de ação lento é característico das células nodais sinusais e
atrioventriculares normais, nas quais o potencial de repouso é de –40 a –70 mV, a
velocidade de ascensão do potencial de ação é no máximo de 10 V/s e a velocidade de
condução do impulso é muito baixa. A despolarização lenta dessas células é devido às
correntes lentas de influxo de Na+ e Ca++. A velocidade de despolarização é em torno
de 5 MV/s.
23 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 14 – Sistema de condução do coração e os potenciais de ação.
Sabe-se que, normalmente, o impulso cardíaco origina-se no NSA. As fibras do
nodo átrio ventricular, quando não são estimuladas por alguma fonte externa,
apresentam atividade rítmica na freqüência intrínseca de 40 a 60 batimentos/minuto
(bat/min) e as fibras de Purkinje apresentam ritmicidade entre 10 a 40 bat/min. Essas
freqüências contrastam com a freqüência normal do nodo sinusal, que é de 70 a 80
bat/min. Isto se deve ao fato de que quando o NSA gera um impulso elétrico, a
freqüência da descarga é bem maior que a do NAV ou da fibras de Purkinje. Cada vez
que o NSA entra em atividade, seu impulso é conduzido para o NAV e para as fibras de
Purkinje, descarregando suas membranas excitáveis. O NSA, o NAV e as fibras de
Purkinje recuperam-se do potencial de ação, ficando hiperpolarizados quase que ao
mesmo tempo. Mas o NSA se despolariza, liberando sua carga, muito mais rapidamente
que qualquer das outras duas estruturas mencionadas. Dessa maneira, o nodo sinusal
produz um novo impulso cardíaco antes que o nodo átrio ventricular ou as fibras de
Purkinje possam atingir seus limiares para auto-excitação. Esse ciclo continua
indefinidamente durante todo funcionamento do coração, com o nodo sinusal sempre
excitando esses outros tecidos, potencialmente autoexcitáveis.
24 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 15 – Estruturas responsáveis pela harmonia do ritmo cardíaco
---------- FIM DO MÓDULO I ----------
25 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Curso de
Fisiologia Geral
MÓDULO II
Atenção: O material deste módulo está disponível apenas como parâmetro de estudos para este Programa de Educação Continuada, é proibida qualquer forma de comercialização do mesmo. Os créditos do conteúdo aqui contido são dados aos seus respectivos autores descritos na Bibliografia Consultada.

27
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
MÓDULO II
1.1.
roeletrolíticos), ressalvando-se que algumas anomalias cardíacas não
alteram
es patológicas, ressalvando-se que algumas anomalias
cardía
l conectadas a um fio condutor)
dispos
xo QRS é comumente formado por três ondas distintas, a onda Q,
a onda
nto a onda P quanto os
compo
do estado de despolarização. Esse processo normalmente ocorre no músculo ventricular
Algumas características do Eletrocardiograma O coração apresenta atividade elétrica por variação na quantidade relativa de ions
presentes dentro e fora das células do miocárdio. Esta variação cíclica gera diferença de
concentração dos referidos íons na periferia do corpo. Eletrodos sensíveis colocados em
pontos específicos do corpo registram esta diferença elétrica. Eletrocardiograma (ECG) é
o registro dos fenômenos elétricos que se originam durante a atividade cardíaca, auxiliar
valioso no diagnóstico de grande número de cardiopatias e outras condições patológicas
(ex.distúrbios hid
o ECG.
Eletrocardiograma (ECG) é o registro dos fenômenos elétricos que se originam
durante a atividade cardíaca, auxiliar valioso no diagnóstico de grande número de
cardiopatias e outras condiçõ
cas não alteram o ECG.
Esse registro é realizado através de um aparelho denominado eletrocardiógrafo. O
eletrocardiógrafo nada mais é do que um galvanômetro (aparelho que mede a diferença
de potencial entre 2 pontos) que mede pequenas intensidades de corrente que recolhe a
partir de dois eletrodos (pequenas placas de meta
tos em determinados pontos do corpo humano.
O eletrocardiograma normal é composto por uma onda P, um complexo QRS e
uma onda T. O comple
R e a onda S.
A onda P é produzida por potenciais elétricos gerados à medida que os átrios se
despolarizam, antes de contrair-se. O complexo QRS se deve aos potenciais gerados
quando os ventrículos se despolarizam, antes de contrair-se, isto é, conforme a onda de
despolarização se propaga através dos ventrículos. Assim ta
nentes do complexo QRS são ondas de despolarização.
A onda T é devida aos potenciais gerados durante a recuperação dos ventrículos

em 0,25 a 0,30 segundo após a despolarização, sendo após a despolarização, sendo
essa onda conhecida como onda de repolarização.
FIGURA 1 – Esquema das ondas eletrocardiográficas
FIGURA 2 – Variações do complexo QRS
1. FISIOLOGIA DA CIRCULAÇÃO
Os vasos sanguíneos são responsáveis pelo transporte do sangue, o qual contém
gases, nutrientes e resíduos. Na circulação sanguínea, o coração lança o sangue a
pressões elevadas através das artérias e este é transportado até chegar ao nível de
capilares, onde ocorrem as trocas de substâncias. O leito capilar vai ser drenado por
elementos venosos que fazem com que o sangue retorne ao coração. Os vasos
sanguíneos são constituídos por três camadas de tecidos: túnica íntima, túnica média e
28 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

túnica adventícia. Estas camadas são mais definidas nas artérias e são ausentes nos
capilares, onde distingüe-se apenas um endotélio.
A Túnica íntima é constituída de células endoteliais pavimentosas simples que
revestem a luz do vaso e um tecido conjuntivo subendotelial. A Túnica média é composta
por células musculares lisas de disposição circular e de tecido conjuntivo fibroelastico. A
Túnica média é mais proeminente nas artérias e pouco distinta nas veias. A Túnica
adventícia é camada a mais externa, sendo constituída de tecido conjuntivo e pode conter
músculo liso. É a camada mais desenvolvida nas veias.
FIGURA 3– O esquema mostra as túnicas das veias e artérias
29 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 4 – Representação do lúmen e túnicas íntima, média e adventícia.
1.1. Funções especiais das Artérias
As artérias são os vasos que, partindo dos ventrículos cardíacos, levam o sangue
do coração a todas as partes do nosso corpo, dividindo-se em vasos mais finos, os
capilares.
A camada interna das artérias tem o nome de endotélio ou túnica interna. As
paredes das artérias, ao contrário das veias, têm alguma resistência, fazendo com que
mesmo quando não contêm sangue mantêm a sua forma tubular. O calibre (tamanho) das
artérias pode ser maior ou menor, e a sua constituição também varia. No entanto, as
artérias grossas e internas possuem fibras elásticas, como a aorta, que é a artéria mais
grossa de todo o corpo. As artérias finas e superficiais têm uma quantidade menor de
fibras elásticas, tendo assim maior quantidade de fibras de tipo muscular. É devido à
elasticidade das artérias e ao bombeamento propulsor efetuado pelo coração que o
30 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

sangue circula continuamente. É também devido a este fator que consegue-se determinar
o número exato de pulsações por unidade de tempo.
FIGURA 5 – Estrutura das artérias
FIGURA 6 – Tipos de camadas arteriais
31 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 7 – Imagem gerada o microscópio de uma artéria
1.2. Funções especiais das Veias
Ao contrário das artérias, as veias são os vasos que levam o sangue de todas as
partes do corpo até ao coração, chegando à aurícula direita. Em outras palavras veia é a
designação genérica dos vasos sangüíneos que conduzem ao coração o sangue
distribuído pelas artérias em todas as partes do corpo. Têm paredes delgadas que
aumentam gradativamente de calibre até formar os grossos troncos que terminam no
coração. As paredes das veias são menos resistentes e mais delgadas do que as das
artérias embora apresentem três camadas. Visto que possuem uma quantidade menor de
fibras elásticas e musculares, no entanto, quando não transportam sangue perdem a sua
forma de cilindro. Mas, a superfície interna das veias possui uma série de pregas duplas
que agem como verdadeiras válvulas que controlam, conforme o fecho e a abertura, para
que o sangue no interior da veia siga a direção certa.
32 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Figura 8 – Estrutura das veias
As veias levam o sangue de todos os órgãos até ao coração. Os problemas
principais das veias são a inflamação, a coagulação e os defeitos que conduzem à
dilatação e às varizes. Os membros inferiores contêm dois grupos principais de veias: as
superficiais, localizadas na camada gorda por debaixo da pele, e as profundas,
localizadas nos músculos. Existem veias curtas que ligam as superficiais com as
profundas. Normalmente, a pressão do sangue em todas as veias é baixa; e nos
membros inferiores, esta pressão baixa pode representar um problema. Quando uma
pessoa está de pé, o sangue deve circular das veias dos membros inferiores para cima
até chegar ao coração. As veias profundas desempenham um papel crucial na propulsão
do sangue para cima, uma vez que ao estarem localizadas dentro dos poderosos
músculos da panturrilha, estas veias são profundamente comprimidas em cada passada.
Estas veias transportam 90 % ou mais do sangue que vai dos membros inferiores para o
coração.
33 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
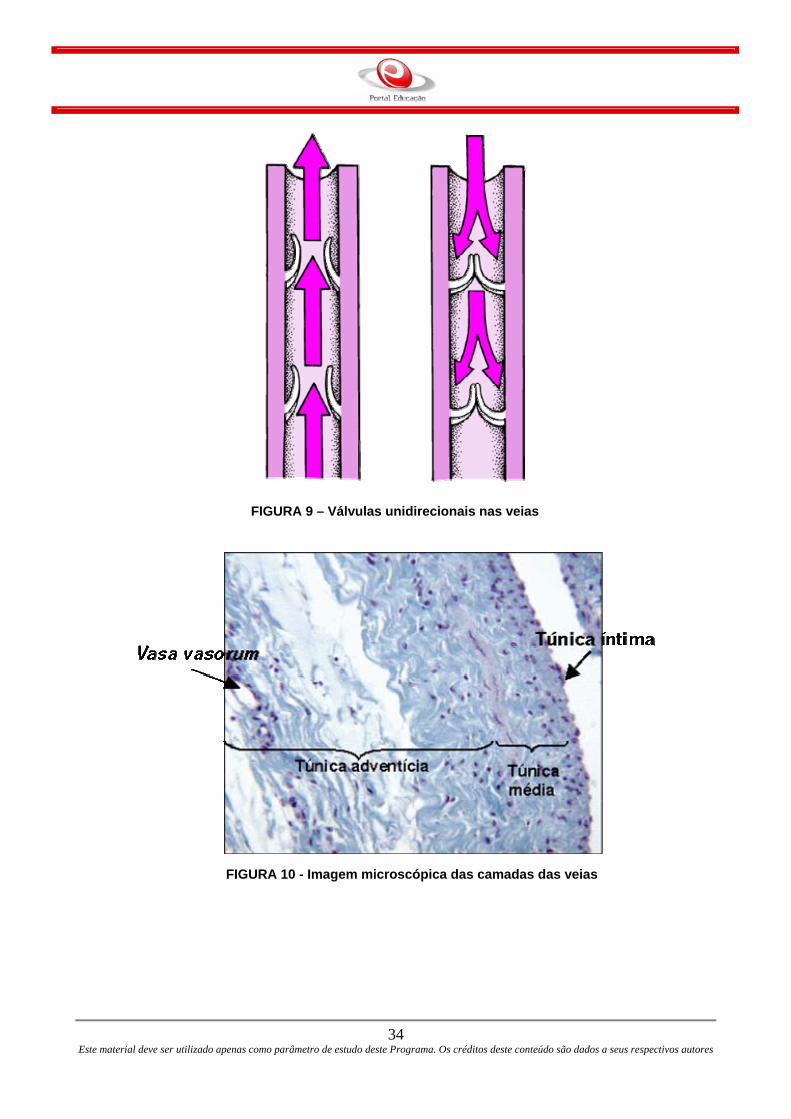
FIGURA 9 – Válvulas unidirecionais nas veias
FIGURA 10 - Imagem microscópica das camadas das veias
34 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

1.3. Funções especiais dos Capilares
Os capilares sanguíneos, ou vasos capilares são órgãos do sistema circulatório
com forma de tubos de pequeníssimo calibre (tamanho). Constituem a rede de
distribuição e recolhimento do sangue nas células. Estes vasos estão em comunicação,
por um lado, com ramificações originárias das artérias e, por outro lado, com as veias de
menor dimensão. Os capilares existem em grande quantidade no nosso corpo. Podem
deformar-se com muita facilidade, impedindo assim a passagem de glóbulos vermelhos. A
parede dos capilares é constituída por uma única camada de células que é a túnica
interna das artérias. É nas paredes dos capilares que ocorrem as trocas dos gases. As artérias, chegando à periferia do corpo humano, isto é, nos músculos, na pele e
em todos os órgãos, se dividem em artérias sempre menores (arteríolas) até que o seu
calibre se torna microscópico: é a este nível que têm lugar as trocas entre sangue e
células. Estes vasos microscópicos chamam-se capilares e se formam nos órgãos e nos
tecidos uma vasta rede. Os capilares confluem para pequenas veias (vênulas) que aos
poucos se vão unindo umas com outras, tornam-se veias verdadeiras e trazem de volta o
sangue ao coração. Do coração partem duas grandes artérias: a artéria pulmonar e a
artéria aorta.
Figura 11 – Representação da estrutura capilar sanguínea
35 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 12 – A figura mostra a estrutura dos capilares sanguíneos
FIGURA 13 – Estrutura capilar
36 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

37
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
1.4. O papel da Regulação Humoral
A regulação humoral da circulação significa a regulação por substâncias
secretadas ou absorvidas para os líquidos corporais como hormônios, íons e assim por
diante. Algumas dessas substâncias são formadas por glândulas especiais e, então,
transportadas no sangue por todo o corpo. Outras são formadas em áreas localizadas de
tecido em resposta a condições locais ou são liberadas por nervos excitados. Causam
então efeitos circulatórios locais. Entre os fatores humorais mais importantes que afetam
a função circulatória estão os seguintes: agentes vasoconstritores e agentes
vasodilatadores.
Os agentes vasoconstritores mais conhecidos são norepinefrina, epinefrina,
angiotensina, vasopressina e a endotelina.
A norepinefrina é um hormônio vasoconstritor especialmente poderoso. A
epinefrina é menos e, em algumas instâncias, até causa discreta vasodilatação. Quando o
sistema nervoso simpático é estimulado durante o estresse ou o exercício, as terminações
nervosas simpáticas liberam norepinefrina, que excita o coração, as veias e as arteríolas.
Os nervos também fazem com que as medulas adrenais secretem tanto norepinefrina
quanto epinefrina no sangue. Esses hormônios, então circulam no sangue e causam
quase os mesmos efeitos excitatórios sobre a circulação que a estimulação simpática
direta, fornecendo assim um duplo sistema de controle.
A angiotensina é uma das substâncias vasoconstritoras mais poderosas das que
são conhecidas. Uma quantidade tão pequena como um milionésimo de grama pode
aumentar a pressão arterial (PA) de uma pessoa por até 50 ou mais mmHg. O efeito da
angiotensina é provocar constrição muito intensa das pequenas arteríolas. A verdadeira
importância da angiotensina no sangue é que ela normalmente atua de forma simultânea
sobre todas as arteríolas do corpo para aumentar a resistência periférica total,
aumentando assim a pressão arterial. Por causa disso e de mais vários efeitos
estimulatórios renais e adrenocorticais da angiotensina, este hormônio desempenha um
papel integral na regulação da PA.
A vasopressina, também chamada de hormônio antidiurético, é formada no
hipotálamo, mas é transportada para baixo, ao longo do centro de axônios nervosos, para
a glândula hipófise posterior, onde é finalmente secretada no sangue. A vasopressina é

38
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
ainda mais poderosa que a angiotensina como vasoconstritor, que talvez a torne assim a
substância constritora mais potente do corpo. Normalmente apenas quantidades muito
diminutas de vasopressina são secretadas. Entretanto, após a hemorragia grave que
cause uma grande baixa da PA, a concentração de vasopressina pode subir o suficiente
para aumentar a PA até 60 mmHg, em muitos casos, isto pode, por si só, trazer a PA
quase de volta ao normal.
A endotelina é um grande peptídeo com 21 aminoácidos que com pequenas
quantidades pode causar uma vasoconstrição poderosa. Essa substância está presente
nas células endoteliais de todos ou quase todos os vasos sanguíneos do corpo. O
estímulo usual para a liberação é a lesão do endotélio, como a causada pelo
esmagamento dos tecidos ou pela injeção de uma substância química traumatizante
dentro do vaso sanguíneo. Após lesão grave do vaso sanguíneo, são provavelmente a
liberação de endotelina local e a vasoconstrição subseqüente que impedem o
sangramento profuso das artérias de até 5 mm de diâmetro, que foram abertas para lesão
do esmagamento.
Os agentes vasodilatadores mais importantes na regulação humoral são:
bradicinina, histamina e prostaglandinas.
Várias substâncias chamadas de cininas que podem causar vasodilatação
potente são formadas no sangue e nos líquidos teciduais de alguns órgãos. Uma dessas
substâncias é a bradicinina. As cininas são pequenos clivados por enzimas proteolíticas a
partir de globulina alfa 2 no plasma ou nos líquidos teciduais. Uma enzima proteolítica de
particular importância é a calicreína, que está presente no sangue e nos líquidos teciduais
numa forma inativa. A calicreína é ativada pela maceração do sangue, pela inflamação do
tecido e outros efeitos químicos e físicos semelhantes sobre o sangue ou os tecidos.
Quando a calicreína é ativada, atua imediatamente sobre a globulina alfa 2 para liberar
uma cinina chamada calidina, que é então convertida pelas enzimas dos tecidos em
bradicinina. Uma vez formada, a bradicinina persiste por apenas alguns minutos porque é
inativada pela enzima carboxipeptidase ou pela enzima de conversão, uma enzima que
também desempenha um papel essencial na ativação da angiotensina. A bradicinina
causa dilatação arteriolar muito potente e também permeabilidade capilar aumentada. A
injeção, por exemplo, de 1 micrograma de bradicinina na artéria braquial de uma pessoa
aumenta o fluxo sanguíneo do braço até seis vezes, e mesmo pequenas quantidades

39
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
ainda menores, injetadas localmente nos tecidos, podem causar edema acentuado por
causa do aumento do tamanho dos poros dos capilares.
Há razões para se acreditar que as cininas desempenhem papéis especiais na
regulação do fluxo sanguíneo e no vazamento capilar de líquidos nos tecidos inflamados.
Acredita-se também que a bradicinina participa da regulação do fluxo sanguíneo na pele e
nas glândulas salivares e gastrointestinais.
A histamina é liberada essencialmente em cada tecido do corpo quando este é
lesado, inflamado ou sujeito a uma reação alérgica. A maior parte da histamina é derivada
dos mastócitos nos tecidos acometidos e dos basófilos no sangue. A histamina tem
poderoso efeito vasodilatador sobre as arteríolas e, como a bradicinina, também tem a
capacidade de aumentar muito a porosidade capilar, permitindo o extravasamento tanto
de líquido quanto de proteínas plasmáticas para dentro dos tecidos. Em muitas condições
patológicas, a intensa dilatação arteriolar e a porosidade capilar aumentada, causadas
pela histamina motivam o vazamento de tremendas quantidades de líquidos para fora da
circulação e para dentro dos tecidos, induzindo ao edema. Os efeitos vasodilatadores e
produtores de edema locais da histamina são especialmente proeminentes nas reações
alérgicas.
Quase todos os tecidos do corpo contêm quantidades pequenas a moderada de
várias substâncias químicas aparentadas chamadas de prostaglandinas. Estas
substâncias têm efeitos intracelulares especialmente importantes, mas, além disso,
algumas delas também são liberadas nos líquidos teciduais locais e no sangue circulante
em condições fisiológicas e patológicas. Apesar de algumas prostaglandinas causarem
vasoconstrição, a maioria das mais importantes parece ser, sobretudo das agentes
vasodilatadores.
1.5. O papel dos Rins na regulação da Pressão Arterial A elevação da pressão nas artérias pode ocorrer de várias maneiras. Por exemplo,
o coração pode bombear com mais força, ejetando mais sangue a cada minuto. Outra
possibilidade é as artérias de maior calibre perderem sua flexibilidade normal e tornarem-
se rígidas, de modo que elas não conseguem expandir para permitir a passagem do
sangue bombeado pelo coração.

40
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
Assim, o sangue ejetado em cada batimento cardíaco é forçado através de um
espaço menor que o normal e a pressão arterial aumenta. É isto o que ocorre em pessoas
idosas cujas paredes arteriais se tornaram espessadas e rígidas por causa da
arteriosclerose. De modo similar, a pressão arterial eleva em casos de vasoconstrição,
quando artérias muito finas (arteríolas) se contraem temporariamente devido à
estimulação nervosa ou por hormônios presentes no sangue.
Uma terceira forma de elevação da pressão arterial é através do aumento do
aporte líquido ao sistema. Isto ocorre quando os rins funcionam mal e são incapazes de
remover a quantidade adequada de sal e água do organismo. O volume de sangue no
corpo aumenta e a pressão arterial também. Por outro lado, se a função de bombeamento
de sangue do coração diminui, se as artérias dilatarem ou se houver perda de líquido do
sistema, a pressão arterial é reduzida.
Os ajustes desses fatores são regidos por alterações da função renal e do sistema
nervoso autônomo (parte do sistema nervoso que regula automaticamente muitas funções
do organismo). O sistema nervoso simpático, o qual faz parte do sistema nervoso
autônomo, aumenta temporariamente a pressão arterial durante a resposta de “luta ou
fuga” (reação física diante de uma ameaça).
O sistema nervoso simpático aumenta tanto a freqüência quanto a força dos
batimentos cardíacos. Ele também produz uma contração da maioria das arteríolas, mas
expande as arteríolas de determinadas áreas, como na musculatura esquelética, onde é
necessária uma maior irrigação sangüínea. Além disso, o sistema nervoso simpático
diminui a excreção renal de sal e água, aumentando assim o volume sangüíneo do corpo.
O sistema nervoso simpático também libera os hormônios epinefrina (adrenalina) e
norepinefrina (noradrenalina), os quais estimulam o coração e os vasos sangüíneos. Os
rins controlam a pressão arterial de vários modos. Se a pressão aumenta, os rins
aumentam a excreção de sal e água, o que reduz o volume sangüíneo e faz a pressão
retornar ao normal. Por outro lado, se a pressão cai, os rins diminuem a excreção de sal e
água e, conseqüentemente, o volume sangüíneo aumenta e a pressão retorna ao normal.
Os rins também podem elevar a pressão arterial secretando a enzima renina, a
qual estimula a produção do hormônio angiotensina, o qual, por sua vez, desencadeia a
liberação do hormônio aldosterona. Devido ao importante papel dos rins no controle da
pressão arterial, muitas doenças e anomalias renais podem causar o aumento da pressão

41
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
arterial. Por exemplo, o estreitamento da artéria que irriga um dos rins (estenose da
artéria renal) pode causar hipertensão. Da mesma forma, inflamações renais de diversos
tipos e a lesão renal uni ou bilateral também podem provocar aumento da pressão arterial.
Sempre que uma alteração provoca a elevação da pressão arterial, é
desencadeado um mecanismo de compensação que procura compensar esse aumento e
manter a pressão em níveis normais. Assim, um aumento no volume do sangue
bombeado pelo coração, o qual tende a aumentar a pressão arterial, faz com que os
vasos sangüíneos dilatem e que os rins aumentem a excreção de sal e água, o que tende
a reduzir a pressão arterial. Entretanto, a arteriosclerose produz enrijecimento das
artérias, impedindo sua dilatação, a qual auxiliaria na redução da pressão arterial aos
seus níveis normais. Alterações arterioscleróticas renais podem comprometer a
capacidade dos rins de excretar sal e água, o que contribui para a elevação da pressão
arterial.
Uma queda na pressão arterial (1) provoca a liberação de renina, uma enzima
renal. Por sua vez, a renina (2) ativa a angiotensina (3), um hormônio que provoca
contração das paredes musculares das pequenas artérias (arteríolas), aumentando a
pressão arterial. A angiotensina também desencadeia a liberação do hormônio
aldosterona pelas glândulas adrenais (4), provocando a retenção de sal (sódio) e a
excreção de potássio. O sódio promove a retenção de água e, dessa forma, provoca a
expansão do volume sangüíneo e o aumento da pressão arterial.

FIGURA 14 – Esquema do controle da pressão arterial
------ FIM MÓDULO II -----
FIGURA 15 – Metabolismo e regulação dos agentes vasoconstritores e vasodilatadores
42 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 16 – Metabolismo da angiotensina e a influência na excreção de água e sódio
FIGURA 17 – Ciclo da regulação da PA desde a pró-renina até angiotensina II
---------- FIM DO MÓDULO II ----------
43 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Curso de
Fisiologia Geral
MÓDULO III
Atenção: O material deste módulo está disponível apenas como parâmetro de estudos para este Programa de Educação Continuada, é proibida qualquer forma de comercialização do mesmo. Os créditos do conteúdo aqui contido são dados aos seus respectivos autores descritos na Bibliografia Consultada.

45
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
MÓDULO III
1.1. Líquidos Extracelular e Intracelular
imentos do
organi o de
ua e pelos eletrólitos é fundamental para a
preven
do gorduroso tem um baixo
teor de
lar corresponde à
água do plasma sanguíneo (4%) e à água do líquido intersticial (16%).
1. FISIOLOGIA DOS RINS
A troca de nutrientes e dejetos entre o sangue e os tecidos é realizada por uma
extensão de capilares, equivalente a aproximadamente 700 metros quadrados. Aquelas
trocas requerem a presença da água, como o meio nobre em que as células vivem e
realizam as suas funções; a permanência da água nos diferentes compart
sm pende da presença de um teor adequado de diversos eletrólitos.
As alterações da distribuição da água e dos eletrólitos são bastante comuns
e podem levar à complicações de extrema gravidade, ou mesmo determinar a morte do
indivíduo. A circulação extracorpórea pode produzir distúrbios da composição hídrica e
eletrolítica do organismo, capazes de gerar numerosas complicações. O reconhecimento
das principais funções desempenhadas pela ág
ção das complicações e suas seqüelas.
A água corresponde à maior parte do peso dos indivíduos. Em um neonato, a água
corresponde a cerca de 75 a 80% do peso. Aos 12 meses de idade o teor de água do
organismo é de 65% e na adolescência alcança o valor de 60% no sexo masculino e 55%
no feminino, que se mantém na vida adulta. Essa pequena diferença se deve à maior
quantidade de tecido gorduroso no organismo feminino. O teci
água em relação aos músculos e aos órgãos internos.
A água do organismo está distribuída em dois grandes compartimentos: o
intracelular e o extracelular. A água do interior das células (líquido ou compartimento
intracelular), corresponde a cerca de 40% do total do peso do indivíduo, enquanto a água
do líquido extracelular corresponde a 20%. O compartimento extracelu
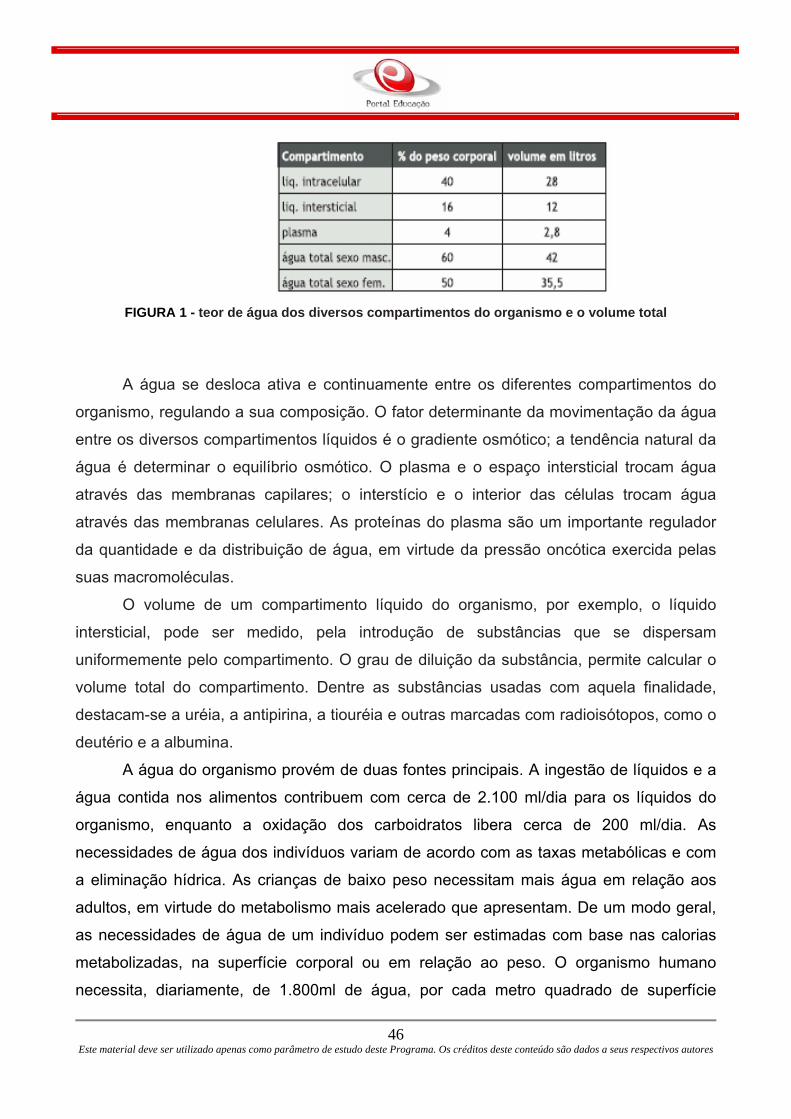
FIGURA 1 - teor de água dos diversos compartimentos do organismo e o volume total
A água se desloca ativa e continuamente entre os diferentes compartimentos do
organismo, regulando a sua composição. O fator determinante da movimentação da água
entre os diversos compartimentos líquidos é o gradiente osmótico; a tendência natural da
água é determinar o equilíbrio osmótico. O plasma e o espaço intersticial trocam água
através das membranas capilares; o interstício e o interior das células trocam água
através das membranas celulares. As proteínas do plasma são um importante regulador
da quantidade e da distribuição de água, em virtude da pressão oncótica exercida pelas
suas macromoléculas.
O volume de um compartimento líquido do organismo, por exemplo, o líquido
intersticial, pode ser medido, pela introdução de substâncias que se dispersam
uniformemente pelo compartimento. O grau de diluição da substância, permite calcular o
volume total do compartimento. Dentre as substâncias usadas com aquela finalidade,
destacam-se a uréia, a antipirina, a tiouréia e outras marcadas com radioisótopos, como o
deutério e a albumina.
A água do organismo provém de duas fontes principais. A ingestão de líquidos e a
água contida nos alimentos contribuem com cerca de 2.100 ml/dia para os líquidos do
organismo, enquanto a oxidação dos carboidratos libera cerca de 200 ml/dia. As
necessidades de água dos indivíduos variam de acordo com as taxas metabólicas e com
a eliminação hídrica. As crianças de baixo peso necessitam mais água em relação aos
adultos, em virtude do metabolismo mais acelerado que apresentam. De um modo geral,
as necessidades de água de um indivíduo podem ser estimadas com base nas calorias
metabolizadas, na superfície corporal ou em relação ao peso. O organismo humano
necessita, diariamente, de 1.800ml de água, por cada metro quadrado de superfície
46 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

corporal. As necessidades de água dos diferentes indivíduos estão relacionadas na tabela
7.2, conforme o peso corporal. Aqueles valores referem-se à indivíduos sadios, sem
disfunção renal, cardiovascular ou metabólica e, portanto, sem restrições à ingestão
normal de água. As alterações da água consistem, principalmente, de desidratação,
quando há perda excessiva de líquidos do organismo ou, ao contrário, hiperidratação,
quando há oferta excessiva de líquidos ao organismo.
FIGURA 2 - Intercâmbio líquido entre os diferentes compartimentos do organismo
Na circulação extracorpórea, principalmente em crianças, não é rara a ocorrência
de hiperidratação, causada pelo excesso de soluções cristalóides no perfusato. Devemos
considerar que durante um procedimento cirúrgico, a administração de água e eletrólitos é
feita pelo perfusionista através o perfusato; pelo anestesista, através das soluções
venosas administradas durante a operação e pelo cirurgião, através da administração das
soluções cardioplégicas, principalmente a cardioplegia cristalóide. Sem controle
adequado, a soma dos volumes infundidos pode ultrapassar em muito, as necessidades
diárias dos pacientes que, além de tudo, receberão mais líquidos no pós-operatório
imediato.
47 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

48
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A hiperidratação pode também ocorrer em pacientes com quantidades de proteínas
abaixo do normal. A pressão oncótica do plasma fica reduzida e permite o
extravasamento de líquidos do plasma para o espaço intersticial, especialmente se a
oferta líquida não for adequadamente dimensionada. Quando há perda excessiva ou
insuficiente administração de sódio, também pode ocorrer hiperidratação. A causa é a
redução da pressão osmótica do líquido extracelular, em relação ao interior das células. A
água passa do interstício para o líquido intracelular, para refazer o equilíbrio osmótico. O
paciente hiperidratado pode apresentar edema de face ou generalizado, ascite, derrame
pleural, insuficiência respiratória, astenia, desorientação, delírio e convulsões ou outras
manifesta manifestações neurológicas. A migração da água entre os diferentes
compartimentos, depende da concentração dos eletrólitos, para que o equilíbrio hídrico do
organismo seja mantido.
Os eletrólitos, quando em uma solução aquosa, comportam-se como íons. Os íons
são a menor porção de um elemento químico que conserva as suas propriedades. Os
cátions são os íons que tem carga elétrica positiva, como o sódio (Na+) e o potássio (K+).
Os anions são os íons que tem carga elétrica negativa, como o cloro (Cl-) ou o
bicarbonato (HCO2).
O equilíbrio químico de uma solução significa a existência de igual número de
cátions e anions. Os eletrólitos são quantificados em miliequivalentes, que correspondem
à milésima parte de um equivalente grama, ou simplesmente equivalente. O equivalente
de uma substância é a menor porção da substância, capaz de reagir quimicamente e,
corresponde ao peso atômico ou ao peso molecular, dividido pela valência. Em geral, nos
líquidos do organismo, os eletrólitos são considerados em termos de miliequivalentes por
litro (mEq/l).
Os líquidos orgânicos têm uma composição semelhante, sob o ponto de vista da
atividade química e das pressões osmóticas. A natureza dos íons, contudo, difere entre os
compartimentos intracelular e extracelular.
O líquido extracelular inclui o liquido intersticial e o plasma sanguíneo. O liquido
extracelular tem grandes quantidades de sódio e cloreto. O sódio é o cátion predominante
do líquido extracelular, enquanto o potássio é o cátion predominante no líquido
intracelular. Aproximadamente 95% do potássio existente no organismo estão situados no

49
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
interior das células. A distribuição do magnésio, como o potássio, também é
predominantemente intracelular.
Os principais eletrólitos celulares são o potássio, magnésio, fosfato, sulfato,
bicarbonato e quantidades menores de sódio, cloreto e cálcio. O liquido intracelular possui
grande quantidade de potássio e pequena quantidade de sódio e de cloreto. As grandes
proteínas e alguns tipos de ácidos orgânicos ionizáveis existem exclusivamente no líquido
intracelular; não existem no plasma e no líquido intersticial.
As diferenças de composição entre os líquidos intracelular e extracelular são muito
importantes, para o desempenho adequado das funções celulares. O liquido extracelular
inclui ainda a linfa, o liquor, o liquido ocular e outros líquidos especiais do organismo,
menos importantes em relação à regulação hídrica e eletrolítica. O plasma e o líquido
intersticial são os grandes responsáveis pela regulação da água do organismo; a sua
composição eletrolítica é praticamente a mesma, exceto pela presença das proteínas no
plasma. Os íons presentes nos líquidos orgânicos desempenham funções essenciais à
manutenção do perfeito equilíbrio funcional celular.
O sódio é o cátion mais abundante no líquido extracelular; é fundamental na
manutenção do equilíbrio hídrico. A perda de sódio causa redução da pressão osmótica
do líquido extracelular, que resulta na migração de água para o interior das células. O
aumento da concentração do sódio no líquido extracelular, ao contrário, aumenta a sua
pressão osmótica e favorece o acúmulo de água no interstício, produzindo edema. O
sódio também é importante na produção
do impulso para a condução cardíaca e para a contração muscular. Um mecanismo
especial chamado de bomba de sódio controla o fluxo de sódio e potássio através da
membrana celular, mantendo o sódio no exterior e o potássio no interior das células. A
concentração do sódio é controlada pelos rins, pela secreção de aldosterona e pela
secreção do hormônio antidiurético.
O potássio é o cátion intracelular mais importante; é transportado para o interior
das células pelo mecanismo da bomba de sódio e tem ação fundamental na condução do
impulso elétrico e na contração muscular. O acúmulo excessivo de potássio no líquido
extracelular (hiperpotassemia) pode causar redução da condução elétrica e da potência

50
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
da contração miocárdica, levando à parada cardíaca em assistolia. Esse efeito do
potássio é o princípio fundamental da sua utilização nas soluções cardioplégicas.
O cálcio é essencial à formação dos dentes, ossos e diversos outros tecidos. É
também um fator importante na coagulação do sangue. A presença de pequenas
quantidades de cálcio é essencial à manutenção do tônus e da contração muscular,
inclusive miocárdica; a deficiência do cálcio (hipocalcemia) pode produzir efeitos
semelhantes aos do excesso de potássio.
O magnésio é um íon importante na função de numerosas enzimas e participa
ativamente no metabolismo da glicose, de diversos outros hidratos de carbono e das
proteínas. Participa também, ativamente, nos processos da contração e irritabilidade
neuromuscular; o seu excesso (hipermagnesemia) pode produzir relaxamento muscular,
inclusive miocárdico, além de alterações da condução elétrica cardíaca.
O anion cloro (cloreto) é predominante no líquido extracelular; sua função principal
é a manutenção do equilíbrio químico com os cátions presentes. O cloro participa ainda
nos efeitos tampão do sangue em intercâmbio com o bicarbonato.
A função mais importante do íon bicarbonato é a regulação do equilíbrio ácido-
básico, em que participa com o ácido carbônico (dióxido de carbono + água ), formando o
principal sistema tampão do organismo.
Para que ocorra o intercâmbio de água por osmose, através da membrana capilar
ou celular, é necessário que haja diferença na concentração total de solutos nos dois
lados da membrana. As membranas celulares e capilares são permeáveis à água e aos
solutos dos líquidos orgânicos e não são permeáveis às proteínas.
Um soluto é uma substância, como o cloreto de sódio, cloreto de potássio, glicose,
ou proteína, que pode ser dissolvida em um solvente, para formar uma solução; a solução
salina, por exemplo, tem o cloreto de sódio como soluto e a água como o solvente.
Na prática, as soluções podem ser classificadas conforme o tamanho das
partículas do soluto ou conforme a sua natureza. Uma solução cristalóide é aquela que
contém partículas homogeneamente dispersas no solvente até que ocorra a passagem de
uma corrente elétrica ou a sua mistura com outra solução. Os solutos das soluções
cristalóides, ou simplesmente cristalóides, são pequenos íons, ácidos e bases simples,
aminoácidos, pequenas moléculas orgânicas, como glicose e frutose, pequenas

51
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
moléculas nitrogenadas, como uréia e creatinina ou pequenas cadeias de polipeptídeos.
O limite superior para o tamanho das partículas cristalóides está em torno de 50.000
Dalton. Uma solução coloidal ou, simplesmente, colóide, contém partículas que quando
deixadas em repouso por um tempo prolongado, tendem a depositar, perdendo a
homogeneidade; o processo de deposição pode ser acelerado por centrifugação e outros
meios físico-químicos. As partículas que formam as soluções coloidais têm peso
molecular maior que os solutos cristalóides, acima de 50.000 Daltons.
As membranas biológicas, membrana capilar e membrana celular, não permitem a
passagem dos colóides e permitem a livre passagem de água e dos cristalóides. Se
colocarmos uma solução de cloreto de sódio (NaCl) em um lado de uma membrana
permeável à água e ao sal, e colocarmos água pura no outro lado da membrana, as
moléculas de sódio, cloro e água, vão passar livremente através dos dois lados da
membrana, até que a concentração de sódio e cloro nos dois lados seja a mesma. A
passagem da água e dos eletrólitos Na+ e Cl - para o lado da membrana, onde a sua
concentração é menor, ocorre pelo fenômeno da osmose.
A pressão osmótica corresponde à pressão exercida pelas partículas ou íons de
soluto em uma determinada solução. A pressão osmótica é medida em osmol ou
miliosmol (mOsm). Uma molécula de cloreto de sódio, por exemplo, se dissocia em dois
íons, Na+ e Cl-; portanto, a solução de uma molécula de cloreto de sódio exercerá uma
pressão osmótica de 2 osmol/litro de água ou por Kg de água (1litro de água = 1 Kg).
O intercâmbio de água entre os diferentes compartimentos é governado pela
osmose. As membranas celulares e capilares são muito permeáveis à água e o
intercâmbio diário é enorme, entre os compartimentos líquidos do organismo. Quando a
pressão osmótica se altera, a água se move através das membranas, para restabelecer o
equilíbrio e manter o estado isosmótico.
A tonicidade compara as diferentes soluções em termos da pressão osmótica que
exercem. Duas soluções com o mesmo número de partículas dissolvidas por unidade de
volume, têm a mesma pressão osmótica e são chamadas, soluções isotônicas. Quando
uma solução tem um número maior de partículas, é dita hipertônica em relação à outra e,
finalmente, se o número de partículas de uma solução é menor que a solução de
comparação, diz-se que ela é hipotônica. O padrão de comparação que nos interessa, é o

52
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
plasma sanguíneo. As soluções que serão misturadas ao plasma devem ser isotônicas, a
fim de evitar alterações significativas da pressão osmótica. As soluções hipertônicas, se
necessário, podem apenas ser administradas em pequenos volumes, para corrigir déficits
de algum eletrólito específico.
As moléculas de colóides, em geral, são adicionadas às soluções para acrescentar
pressão oncótica. As soluções coloidais são o plasma sanguíneo, as soluções de
albumina. A pressão osmótica de uma solução depende do número de partículas ou
moléculas na solução. Quanto menor o peso da molécula de uma substância, mais
moléculas existirão, em um determinado peso da substância. Dessa forma, 1 grama de
cloreto de sódio conterá um número infinitamente maior de moléculas do que 1 grama de
albumina; o peso da molécula de cloreto de sódio é 58,5 enquanto o peso da molécula de
albumina é 80.000. Podemos, portanto, afirmar que 1 grama de cloreto de sódio exerce
uma pressão osmótica muito maior que 1 grama de albumina. Quando em uma solução,
adicionamos um soluto como a albumina, cuja molécula é de elevado peso, confinada por
uma membrana impermeável à albumina, esta exercerá uma grande pressão oncótica (ou
coloido-osmótica). A adição de grandes moléculas, como albumina, dextran e outras,
aumentam a pressão oncótica da solução. Contudo, como o número de moléculas na
solução é pequeno, o seu efeito sobre a pressão osmótica é negligível. A pressão
oncótica é expressa em milímetros de mercúrio (mmHg) e tem grande importância na
manutenção da água do plasma e na captação da água do líquido intersticial. Quando a
pressão oncótica do plasma está reduzida a água tende a migrar para o líquido intersticial.
O organismo normal mantém o equilíbrio entre o ganho e a perda diária de água,
regulando a diurese, o suor e as perdas insensíveis. Qualquer interferência nos
mecanismos normais da regulação pode gerar distúrbios do equilíbrio dos líquidos e de
eletrólitos. Durante a circulação extracorpórea, a oferta excessiva de líquidos ou de
eletrólitos através do perfusato, pode romper aquele equilíbrio e produzir complicações. A
perda diária de água corresponde à eliminação pela urina, pelas fezes, pela evaporação
nos pulmões, durante a respiração (perda insensível), e pela formação do suor,
dependendo da temperatura ambiente e do grau de atividade física. A perda total diária de
um indivíduo adulto é de aproximadamente 2.400 à 2.900 m

FIGURA 3 – Perdas diárias de água de um adulto, pelas diversas vias de eliminação
O adequado equilíbrio da água e dos eletrólitos do organismo deve ser lembrado
na preparação da perfusão, na escolha dos componentes do perfusato e nos volumes
necessários ao procedimento. As soluções para o perfusato devem ter a composição
química e a pressão osmótica idênticas ao plasma, para minimizar a possibilidade de
produzir distúrbios hídricos e eletrolíticos.
A liberação de radicais livres e de numerosas citoquinas e outros agentes pró-
inflamatórios durante a circulação extracorpórea altera a permeabilidade das membranas
capilares e celulares e contribui substancialmente para alterar os volumes de água
contidos nos diferentes compartimentos do organismo. Esse processo é parte importante
da reação inflamatória sistêmica do organismo e, quando intenso, pode produzir
complicações difíceis de controlar ou reverter.
1.2. Formação da Urina
O sistema urinário é responsável pela formação da urina, ou seja, pela eliminação
de grande quantidade de excretas e também do excesso de água e de outras substâncias
do organismo. É composto por: rins, ureteres, bexiga urinária e uretra.
Os rins são órgãos com formato de grão de feijão com aproximadamente 10 cm de
comprimento localizados na parte posterior da cavidade abdominal de ambos os lados da
coluna vertebral. São órgãos filtradores do sangue e formadores da urina; o sangue chega
aos rins pela artéria renal e sai dos mesmos pelas veias renais, que despejam o sangue,
já filtrado, na veia cava; a urina formada segue para os ureteres.
53 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Os rins podem excretar diariamente cerca de 50mEq. de íons hidrogênio (H+) e
reabsorver 5.000 mEq. de íon bicarbonato (HCO3-). Os rins eliminam material não volátil
que os pulmões não tem capacidade de eliminar. A eliminação renal é de início mais
lento, torna-se efetiva após algumas horas e demora alguns dias para compensar as
alterações existentes. A eliminação de bases e seus cátions é feita exclusivamente pelos
rins. Os rins tem a capacidade de reabsorver o sódio (Na+) e o potássio (K+) filtrados
para a urina, eliminando o íon hidrogênio (H+) em seu lugar; o sódio reabsorvido pode ser
usado para produzir mais bicarbonato e reconstituir a reserva de bases do organismo.
Além de influir na restauração do equilíbrio ácido-base, os rins reagem à desidratação, à
hipotensão, aos distúrbios da osmolaridade e eliminam ácidos fixos. Os rins
desempenham fundamentalmente duas funções no organismo: a eliminação de produtos
terminais do metabolismo, como uréia, creatinina e ácido úrico e; controle das
concentrações da água e de outros constituintes dos líquidos do organismo como sódio,
potássio, hidrogênio, cloro, bicarbonato e fosfatos.
FIGURA 4 – Vista em corte longitudinal do rim esquerdo
As vias urinárias são condutos por onde a urina segue após sair dos rins até ser
eliminada. Os ureteres são dois tubos musculares que coletam a urina nos rins e a
despejam na bexiga urinária, que é um órgão muscular em forma de bolsa onde
desembocam os ureteres; sua função é armazenar a urina constantemente produzida nos
rins para posterior eliminação.
54 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Por fim, temos a uretra, que é um canal muscular que conduz a urina ao meio
externo.
FIGURA 5 – Ilustração do sistema urinário humano
FIGURA 6 – Vista anterior e lateral de um rim
55 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

56
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A formação da urina começa nos rins, nos néfrons, que são as unidades funcionais
dos rins, onde a urina é realmente formada; cada rim possui aproximadamente um milhão
de néfron que são formados por túbulos contorcidos e microscópicos.
No processo de formação de urina, a arteríola aferente é um ramo da artéria renal
e numa região envolvida pela cápsula de Bowman, se enrola formando o glomérulo.
Quando o sangue passa pelo glomérulo uma parte do plasma extravasa através da
cápsula de Bowman, ocorrendo o processo de filtração. O filtrado glomerular possui
tantas substâncias úteis ao organismo, como glicose, água sais minerais, aminoácidos e
vitaminas, quanto excretas inúteis ao organismo. As substâncias úteis precisam ser
reabsorvidas, ou seja, passar dos túbulos do néfron para os capilares que os envolvem
por meio de mecanismos especiais que as células dos túbulos possuem, que é o
processo de reabsorção. Ao longo do trajeto pelos túbulos do néfron também ocorre a
passagem de algumas substâncias dos capilares para os túbulos, que é o processo de
secreção. Desta maneira pode-se dizer que na formação da urina fazem parte os
processos de filtração, reabsorção e secreção de substâncias, e desses processos resta
nos túbulos do néfron as excretas (principalmente uréia) e o excesso de sais minerais e
de água. A urina segue para o túbulo coletor e deste sai dos rins através dos ureteres e é
armazenada na bexiga urinária e é eliminada para o meio exterior através da uretra.
Existe um hormônio indispensável no processo de controle da urina que é o
hormônio anti-diurético (ADH). O ADH é produzido no hipotálamo e atua no túbulo
contorcido distal dos néfrons estimulando a reabsorção passiva de água, em outras
palavras, diminui a quantidade de urina. O papel do ADH é importantíssimo, sem ele a
diurese pode chegar a 20 litros, quando o normal está na faixa de 1,5 litros. A
desidratação e a sede tornam-se intensa. A ingestão de bebidas alcoólicas inibe a
produção de ADH, desta maneira aumentando a diurese.
1.3. Funcionamento da Micção
A fisiologia bem como a neurofisiologia da micção não estão completamente
compreendidas. O fenômeno simples e quase inconsciente da micção envolve complexos
mecanismos e interações neurais que têm sido objeto de inúmeros estudos nas últimas

57
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
décadas. O desenvolvimento de técnicas histoquímicas especiais, estudos com
estimulação elétrica nervosa em raízes sacrais e principalmente a maior difusão e
padronização de estudos urodinâmicos têm permitido esclarecimentos de alguns pontos
fundamentais para sua compreensão.
A uretra e a bexiga mantêm entre si continuidade anatômica e guardam relação
funcional bastante íntima. A parede vesical no corpo da bexiga é composta de
musculatura lisa que se distribui em todos os sentidos. Próximo ao colo vesical, organiza-
se em três camadas anatomicamente distintas. A camada mais interna orienta-se no
sentido longitudinal prolongando-se com a camada longitudinal interna da uretra. A
camada muscular média, mais espessa e evidente a este nível, interrompe-se no colo
vesical, não se prolongando até a uretra. A camada muscular externa tem sentido oblíquo
nos mais variados graus de inclinação; tem, de modo geral, orientação espiralada,
continuando-se com a camada externa uretral.
Existem fibras musculares estriadas que envolvem a uretra: nos homens, entre o
verumontanum e uretra bulbar;nas mulheres, envolvem principalmente a porção média.
A uretra posterior masculina (compreendendo a uretra prostática e a uretra
membranosa) corresponde a praticamente toda uretra feminina, tendo a mesma origem
embriológica.
No homem adulto, o parênquima prostático localiza-se na porção supramontanal;
envolvendo a uretra por todos os lados, o que dificulta a identificação das camadas
musculares uretrais e leva a confundir suas fibras musculares lisas que envolvem os
ácinos prostáticos com as da musculatura uretral.
A musculatura vesico-uretral tem papel fundamental na função de armazenamento
e esvaziamento vesical. Durante a fase de esvaziamento, é necessário não apenas que a
musculatura vesical se contraia, mas também que musculatura uretral se relaxe.A
contração vesical ocorre basicamente por um estímulo parasimpático. Um arco reflexo,
poderia dar-nos uma idéia simplista do funcionamento vesical. Fibras sensitivas partindo
dos proprioceptores da parede vesical atingem os nervos pré-sacrais (não existe um
nervo sensitivo específico, mas sim um verdadeiro plexo nervoso que se localiza
anteriormente ao sacro). Este plexo organiza-se ao nível dos forames sacrais S2,S3 e S4
fazendo parte das raízes nervosas sacrais S2,S3 e S4, atingindo o cônus medular através

58
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
de ramos da cauda eqüina, fazendo aí sinapse. Deste nível partem fibras motoras
parasimpáticas que, também através das raizes sacrais S2,S3 e S4, passam pelas fibras
do plexo pré-sacral e atingem a parede vesical, estabelecendo-se sinapse nos gânglios
intramurais, partindo daí as fibras motoras vesicais pós-sinápticas. Este arco reflexo
também está sob influência direta cortical, com mecanismo facilitatórios e inibidores. A
sensibilidade da distensão vesical através da medula também é informada ao córtex
cerebral, tomando-se consciência da situação da distensão vesical. São estes
mecanismos que permitem ao indivíduo adulto urinar ou não, ao ser informado pelos
proprioceptores da situação de distensão vesical.
Como já dissemos, para que ocorra a micção não basta que a contratação vesical ocorra,
mas também a resistência uretral deve diminuir, ocorrendo relaxamento esfincteriano. A
inervação da musculatura estriada periuretral é feita por fibras que também trafegam
pelos ramos S2 a S4 e compõem o nervo pudendo. Impulsos nervosos contínuos,
transportados pelo nervo pudendo, atingem o "esfíncter estriado" e o mantêm sob
contração involuntária durante o enchimento vesical. O aumento involuntário desta
contração, acompanhando o enchimento vesical, é um fato normalmente observado.
Quando ocorre a contração vesical, existe uma inibição reflexa desse tônus e - o que, por
sua vez, causa o relaxamento esfincteriano. É interessante observa-se que esta interação
depende de mecanismos neurológicos mais altos situados ao nível da ponte (a conexão
entre o encéfalo e a medula). Essa interação entre o conus medular e a ponte é que
permite também que o reflexo miccional ocorra até o completo esvaziamento vesical. Nos
bebês, esta interação pontino medular está íntegra não tendo as crianças controle por
falta de integração cortical; em um paciente com lesão medular acima do cônus medular,
interrompe-se esta via, deixando de haver essa interação. Têm, assim, muito
freqüentemente, contrações vesicais reflexas com contrações esfincterianas durante a
contração vesical (a chamada dissinergia vesico-esfincteriana) e contrações vesicais de
duração insuficiente. Apresentam, portanto, micção de alta pressão, com elevado volume
de resíduo pós-miccional.
A musculatura uretral, pelo seu tônus, exerce força constrictiva sobre a luz uretral,
ocluindo-a mantendo os níveis pressóricos mais elevados na uretra do que na bexiga, não
ocorrendo perda urinária. A atividade muscular uretral é composta de dois elementos

59
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
básicos: o esfíncter muscular liso,aceito genericamente como esfíncter interno, distribuído
por todo o comprimento da uretra feminina e pela uretra prostática masculina; e o
esfíncter voluntário, estriado, de localização preferencial no terço médio da uretra
feminina, e próximo à uretra membranosa masculina. A atividade do esfíncter voluntário e
do esfincter interno se sobrepõem em razoável trajeto uretral. Se o indivíduo se submete
a um esforço, ocorre um aumento da pressão abdominal que se transmite à bexiga, e o
mecanismo esfincteriano responde: em parte aumentando sua eficiência, através de
reflexo neurológico que contrai a musculatura estriada; e em parte sofrendo transmissão
direta da pressão abdominal. O gradiente de pressão uretral mantém-se maior que a
pressão vesical, não ocorrendo, portanto, perda de urina.Necessária e fundamental para a
continência urinária é além da integridade dos mecanismos esfincterianos, a acomodação
vesical durante a fase de seu enchimento.
A bexiga tem a capacidade de receber significativo volume de urina, sem que se
verifique expressiva elevação pressórica. Mesmo quando se atinge a capacidade vesical
máxima e o desejo miccional se torna imperioso, os níveis pressóricos da bexiga mantêm-
se baixos; e assim, mesmo em tais condições extremas, consegue-se inibir sua
contração.
Os baixos níveis pressóricos vesicais durante a fase de enchimento da bexiga são
fundamentais para a continência. Pacientes nos quais esse fator não se verifique em
decorrência de cirurgia ou por alteração da constituição da parede vesical apresentam-se
com polaciúria intensa, comportando-se clinicamente incontinentes, ainda que o
mecanismo esfincteriano se mostre normal.
Quando a distensão vesical atinge volume ao redor de 150 mL, começamos a
sentir o desejo miccional que atinge seu máximo quando o volume acumulado se iguala à
capacidade vesical máxima (cerca de 500 mL). A musculatura vesical constitui-se
provavelmente no único músculo liso do corpo humano sujeito a algum controle voluntário
cortical. Possuímos a capacidade voluntária de inibir e de iniciar a contração vesical.
Imediatamente antes da contração vesical, ocorre relaxamento esfincteriano e do
assoalho pélvico, o que permite a descida do colo vesical, sendo este um provável
estímulo para a contratação vesical. A contração da musculatura longitudinal interna da
uretra e concomitantemente com a da bexiga leva ao encurtamento uretral e ao

60
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
afunilamento do colo vesical, contribuindo para o direcionamento da força vesical e a
diminuição da resistência uretral. A micção ocorre com baixa resistência uretral, e a
pressão dentro da bexiga mantém-se em níveis baixos (ao redor de 10 - 15 cm H2O). A
pressão uretral mantém-se baixa durante toda a micção, permitindo um fluxo contínuo (da
ordem de 15 - 25 mL/s), que varia com o volume urinado, o sexo e a idade. Somente ao
término do esvaziamento da bexiga, a contração vesical cessa, e o tônus uretral volta aos
níveis basais. Quando desejamos interromper voluntariamente a micção antes do total
esvaziamento vesical, realizamos (através do nervo pudendo) a contração tanto das fibras
estriadas periuretrais, quanto da musculatura perineal resultando no aumento da
resistência uretral e na conseqüente interrupção do fluxo. A musculatura detrusora
mantém-se contraída ainda por alguns poucos segundos, relaxando-se reflexamente à
seguir. Portanto, não interrompemos diretamente o arco reflexo miccional, mas sim, de
forma voluntária, o fluxo urinário interrompendo-se, reflexamente, a contração vesical.
Sabe-se bastante sobre a ação simpática na continência, porém sua ação na micção é
questionável. Alguns autores, através de técnicas histoquímicas mostram que a inervação
do esfíncter estriado é feita por fibras simpáticas, parasimpáticas e somáticas. A ação
simpática também é evidente na ejaculação.
A estimulação simpática promove contração das fibras que envolvem os ácinos
prostáticos, provocando a expulsão da secreção acumulada anteriormente, para a luz
uretral. A contração simultânea de todo o parênquima prostático, por sua localização
preferencial entre o verumontanum e o colo vesical, se traduzirá por constricção mais
acentuada desta porção, não permitindo a ejaculação retrógrada. Receptores beta-
adrenérgicos, que têm ação de relaxamento de fibras lisas, foram encontrados em grande
número na parede vesical sendo provavelmente sua ação de relaxamento o que, atuando
com a falta de ação parassimpática na fase de enchimento, permite que a acomodação
vesical ocorra a baixa pressão.
As disfunções neurológicas podem levar a alterações das funções vesico-uretrais,
sendo conhecidas como bexiga neurogênica. Disto pode resultar o comprometimento das
fibras sensitivas vesicais, como acontece, por exemplo, no diabetes, situação na qual as
fibras sensitivas, por serem as mais finas, são as primeiras acometidas. Como
consequência deste acometimento, os pacientes passam inicialmente a apresentar o

61
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
primeiro desejo somente com grandes distensões vesicais. Quando solicitado, o paciente
consegue urinar e urina, então, grandes volumes, pois a capacidade vesical se encontra
bastante aumentada. Essa distensão vesical crônica acarreta lesão da própria
musculatura detrusora, o que por sua vez, impede o bom esvaziamento vesical; disto
resulta a formação de resíduo pós-miccional, que progressivamente se acumula, levando
à retenção urinária e a suas repercussões no trato urinário superior. Ao lado disto, a
progressão da lesão neurológica causa interrupção do arco reflexo miccional.
Quando a lesão compromete as fibras motoras, tem-se o quadro de bexiga
neurogênica paralítico-motora, como o que se verifica na poliomielite e no trauma ou no
tumor medular. Nesta situação, a sensibilidade está preservada e o paciente percebe o
grau de distensão vesical, porém não consegue desencadear o reflexo miccional. A
Bexiga neurogênica paralítico-motora é uma situação patológica bastante rara de
encontrarmos na prática clínica.
Quando há comprometimento tanto das fibras sensitivas quanto das motoras, ou
ainda do próprio cônus medular, tem-se a chamada bexiga neurogênica autônoma. Por
ser no cônus medular, como sabemos, que ocorre o fechamento do arco reflexo vesical,
lesões do cônus ou das vias aferentes e eferentes vesico-uretrais levam à incapacidade
de se obter reflexamente o arco reflexo. Como estas vias são também responsáveis por
outros reflexos, o reflexo bulbo-cavernoso e o reflexo cutâneo anal estarão de iguais
modos comprometidos.
O grau de comprometimento do esfíncter vai estar relacionado com o grau de
comprometimento neurológico e poderá haver pacientes que, apesar de não terem
contração vesical, serão incontinentes por falta de atividade esfincteriana. Por outro lado,
a retenção pode ser o achado clínico neste tipo de lesão nos casos onde não existe
contração vesical, porém o esfíncter é ativo. Devemos lembrar que um paciente
retencionista pode apresentar incontinência clínica, pois à medida que vai ocorrendo o
enchimento vesical, a pressão intravesical vai se elevando, até o momento que vence a
resistência uretral, ocorrendo, a partir daí, perda constante de urina (é a chamada
incontinência paradoxal). Portanto, para a correta avaliação se um paciente é
retencionista ou incontinente devemos verificar o grau de esvaziamento vesical, e não
somente se o paciente apresenta saída involuntária de urina pela uretra. A bexiga

62
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
autônoma pode ser encontrada em portadores de tumores medulares, trauma ou
malformações congênitas como mielomeningocele ou agenesia sacral.
Quando a lesão ocorre acima do cônus medular que no adulto está localizado ao
nível "ósseo" T12 - L1, o arco reflexo está liberado, ocorrendo contração vesical reflexa à
distensão vesical.A contração vesical é involuntária e sem sensibilidade. Como já foi dito
anteriormente, nesta situação pode ocorrer contração esfincteriana simultânea à
contração vesical e o paciente tem micções de altíssima pressão, levando a repercussões
graves do trato urinário. É o tipo de comportamento vesical (bexiga neurogênica reflexa)
encontrado no trauma medular, na mielomeningocele e na esclerose múltipla, dentre
outras.
Nesta situação, além do reflexo miccional, outros reflexos abaixo da lesão (como
bulbo cavernoso e cutâneo anal) estão também liberados. Cabe registrar um aspecto
freqüentemente observado em lesões agudas, como as verificadas logo após o trauma
medular: o fato de todos os reflexos abaixo da lesão encontrarem-se bloqueados. Este
"silêncio medular" abaixo da lesão pode durar de horas a meses (fase de choque
medular), evoluindo na situação crônica para a liberação dos reflexos.
Outro tipo de comportamento vesica encontrado, como exemplo típico, está na
Moléstia de Parkinson, em que o paciente apresenta o arco reflexo normal, com
sensibilidade e relaxamento esfincteriano, porém as fibras responsáveis pela inibição do
arco reflexo são as comprometidas. Nesta situação, o paciente apresenta incapacidade
de inibir o arco reflexo, configurando-se um quadro clínico de urgência miccional com
incontinência por urgência -- ou seja, no momento em que tem o desejo miccional, ocorre
o arco reflexo e o paciente é incapaz de inibir a micção. Basicamente o que ocorre é uma
desconexão entre a córtex cerebral e a ponte, perdendo-se a capacidade de inibir o
reflexo miccional. Preserva-se a função pontina, e a micção ocorre coordenada, sem
dissinergia.
Como vimos, as disfunções neurológicas podem levar a disfunções miccionais
graves. Sabemos que o trato urinário mantém níveis pressóricos baixos -- e que o
armazenamento, o transporte e a eliminação da urina se fazem com níveis pressóricos
baixos. Elevações pressóricas intravesicais acima de 35 cm H2O causam dificuldade de
drenagem do ureter, acarretando dilatações ureterais.

Com o progressivo aumento da pressão intravesical, esta se transmite ao ureter,
resultando em aumento da pressão intrapiélica e conseqüentemente às repercussões
renais. Ao lado disto, dificuldade de drenagem vesical pode promover alterações da
própria parede vesical -- alterações anatômica essas que podem resultar no aparecimento
de refluxo vesico-ureteral ou ainda levar diretamente a obstrução ureteral na passagem
do ureter para a bexiga (Hiato Ureteral).
Com as alterações da parede vesical -- persistindo da obstrução --, a própria
parede vesical (músculo Detrusor) entra em falência, propiciando o aparecimento do
resíduo pós-miccional que causa infecções urinária de difícil controle. Assim os
mecanismos que levam a disfunção vesical de causa neurológica a repercussões diretas
da função renal são muitos; e pacientes com bexiga neurogênica requerem
acompanhamento e tratamentos urológicos de longo prazo.
FIGURA 7 – A ilustração mostra a inervação no processo de micção
63 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

64
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
2. FISIOLOGIA DO SANGUE
Todas as células do nosso organismo têm necessidade, para manter-se em vida e
desempenhar as suas funções, de receber oxigênio e materiais nutritivos. A tarefa de
transportar a elas esses elementos cabe ao sangue, o qual, por sua vez, recebe das
células as substâncias de rejeição. Para realizar esse refornecimento, o sangue tem
necessidade de "circular" continuamente: o canal dentro dos quais o sangue circula são
os vasos sangüíneos, enquanto o coração é a bomba que dá ao sangue o seu impulso
para circulação. Coração e vasos constituem, no seu conjunto, o aparelho circulatório. O
sangue humano é constituído por um líquido amarelado, o plasma, e por células e
pedaços de células, genericamente denominados elementos figurados.
2.1. Hemácias e Leucócitos
Os glóbulos vermelhos são células também conhecidas como eritrócitos, hemácias
ou células vermelhas que estão presentes no sangue em número de cerca de 5 milhões
por milímetro cúbico, em condições normais. São constituídas basicamente por globulina
e hemoglobina, composta de 4 moléculas protéicas e 1 grupamento heme, que contém o
ferro, e a sua função é transportar o oxigênio(principalmente) e o gás carbônico (em
menor quantidade) aos tecidos.
A cor vermelha se deve à alta concentração da molécula de transporte de oxigênio
dentro das células, a hemoglobina. Há cerca de 5 milhões de eritrócitos em um milímetro
cúbico de sangue humano; eles são produzidos numa velocidade de 2 milhões por
segundo por um tecido especial que se localiza na medula óssea, o tecido
hematopoiético, e as células velhas são destruídas e removidas pelo baço. As baixas
tensões de oxigênio nas grandes altitudes estimulam maior produção de hemácias para
que o transporte de oxigênio seja facilitado.
Quando colocadas em solução hipotônica (menos concentrada), as hemácias
sofrem hemólise, ou seja, se rompem. Em meio hipertônico (mais concentrado), perdem
água e murcham, ocorrendo plasmólise. Quando os eritrócitos se rompem, liberam a

hemoglobina, que é convertida em bilirrubina e eliminada pela vesícula biliar ao sistema
gastrintestinal.
Os leucócitos, também conhecidos por glóbulos brancos, são células sanguíneas
com a função de defender o organismo de agressões externas através da fagocitose. Os
leucócitos são divididos nos seguintes tipos: neutrófilos, basófilos, eosinófilos, que são
granulócitos (divididos em lóbulos), apresentam grânulos em seu citoplasma, com núcleo
arredondado e formados pelas células mães da medula óssea; linfócitos e monócitos, que
são agranulócitos (não são divididos em lóbulos), Não apresentam grânulos no seu
citoplasma. Os neutrófilos e monócitos são responsáveis pela fagocitose, ou seja,
envolvimento e digestão de corpos invasores, que penetram no nosso corpo, como
espinhos, cacos de vidro, bactérias. Os basófilos e os eosinófilos combatem processos
alérgicos.
FIGURA 8 – Neutrófilos granulócitos
FIGURA 9 – Eosinófilos granulócitos
65 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 10 – Basófilos granulócitos
FIGURA 11 – Linfócito agranulócito
66 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 12 – Monócito agranulócito
2.2. Grupos Sanguíneos
Os tipos de Grupos Sanguíneos foram descobertos no início do século XX, quando
o cientista austríaco Karl Landsteiner dedicou-se a comprovar que havia diferenças no
sangue de diversos indivíduos. Ele colheu amostras de sangue de diversas pessoas,
isolou os glóbulos vermelhos e fez diferentes combinações entre plasma e glóbulos
vermelhos, tendo como resultado a aglutinação dos glóbulos em alguns casos, formando
grânulos,e em outros não. Landsteiner explicou então por que algumas pessoas morriam
depois de transfusões de sangue e outras não. Em 1930 ele ganhou o Prêmio Nobel por
esse trabalho.
Os resultados dos experimentos realizados por Landsteiner o levaram a sugerir o
Sistema ABO. Ele considerou que havia três tipos de sangue: A, B e O (Doador
Universal). Outros cientistas identificaram um quarto tipo, nomeado AB (Receptor
Universal). A diferença entre esse grupo de sangue deve-se à presença, nas hemácias,
de uma substância chamada aglutinogênio A e B. Dependendo dessa substância na
hemácia, existe no plasma uma substância chamada aglutinina, que pode ser Anti-A e
67 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Anti-B. Atualmente sabe-se que existem outros antígenos na superfície das hemácias que
também podem estar implicados em reações hemolíticas transfusionais, exemplos destes
são: Fator Rh, Antígeno Kell, Antígeno Duffy, Antígeno Kidd e o Antígeno Lewis.
FIGURA 13 – A tabela mostra a compatibilidade entre os tipos de sangue
A distribuição dos grupos sanguíneos na população humana não é uniforme. O
grupo sanguíneo mais comum é O+, enquanto que que o mais raro é o AB-.
Em 1940, Karl Landsteiner e Alexander Solomon Wiener realizaram experiências
com o sangue de macaco. Ao injetar sangue em cobaias, perceberam que elas produziam
anticorpos, gradativamente. Concluíram que havia nas hemácias do sangue do macaco
um antígeno que foi denominado de fator RH. O anticorpo produzido no sangue da cobaia
foi denominado de anti-Rh. Os indivíduos que apresentam o fator Rh são conhecidos
como Rh+, apresentando os genótipos RR ou Rr. Os indivíduos que não apresentam o
fator Rh são denominados Rhֿ e apresentam o genótipo rr, sendo geneticamente
recessivos.
Quando se procede a uma transfusão sanguínea é necessário verificar se o
receptor tem Rh-. Se assim for, o paciente só poderá receber sangue também Rh-, já que
se receber sangue Rh+ o sistema imunológico poderá reagir, causando hemólise. O
contrário, contudo, o paciente com Rh+ podem receber sangue Rh-, já que este último
não traz consigo os antígenos que provocam a reação imunológica.
68 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

69
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
2.3. Coagulação do Sangue
O organismo conta com um mecanismo vital contra as perdas excessivas de
sangue denominado coagulação que auxilia na interrupção das hemorragias fechando os
vasos sangüíneos abertos e, portanto, impedindo que o sangue extravase. Desde que os
organismos estão sujeitos a sofrer traumatismos que podem romper vasos sangüíneos, o
mecanismo de coagulação pode ser considerado como um fator de defesa natural.
Por outro lado, quando ocorrem perturbações no mecanismo da coagulação,
mesmo lesões pequenas como um corte superficial num dedo ou uma simples extração
dentária podem provocar sangramentos intensos que duram horas ou dias, chegando a
comprometer seriamente a vida do indivíduo.
Esses distúrbios no mecanismo de coagulação poderão ocorrer de forma inversa,
ou seja, provocando coagulação anormal no interior dos vasos sangüíneos (trombose),
fechando-os. Conseqüentemente, os tecidos servidos pelos vasos sangüíneos fechados
sofrem falta de irrigação sangüínea e acabam por apresentar necrose tissular (morte do
tecido).
Outra possibilidade é a de o coágulo, ou parte dele, destacar-se do local de sua
formação, indo obstruir vasos sangüíneos situados em regiões mais distantes do
organismo (fenômeno que caracteriza a chamada embolia), provocando nesses locais
distúrbios circulatórios que, freqüentemente, levam o doente à morte.
Normalmente, o sangue em circulação é líquido, coagulando-se somente quando
transborda dos vasos sangüíneos.
A fluidez do sangue no organismo depende dás propriedades físicas especiais do
endotélio vascular (camada celular que reveste o interior dos vasos), da velocidade do
fluxo sangüíneo, do número de células sangüíneas e da presença de anticoagulantes
naturais, como a heparina, por exemplo.
Quando retirado do interior dos vasos, o sangue perde rapidamente sua fluidez,
tornando-se inicialmente viscoso e adquirindo gradativamente consistência gelatinosa. Se
uma pequena quantidade de sangue extravasar, em pouco tempo haverá formação de um
coágulo semi-sólido.
De maneira simplificada, admite-se que o mecanismo de coagulação do sangue
consiste em uma extensa reação em cadeia, na qual interferem diversas substâncias

70
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
sangüíneas e celulares que agem umas sobre as outras, levando à formação de uma
proteína especial, a fibrina, responsável final pelo processo de coagulação.
Apesar de o mecanismo da coagulação não ser completamente conhecido, existe
uma teoria bastante difundida que atribui à coagulação a ação de doze fatores, indicados
em algarismos romanos por convenção internacional.
A fase final da coagulação é determinada pela formação de fibrina, que se deposita
sob a forma de um emaranhado de fios microscópicos, os quais acabam por aprisionar
completamente as células sangüíneas. Os fios recém-formados aderem uns aos outros,
às células do sangue, aos tecidos e à superfície alterada do revestimento interno dos
vasos - está formado o coágulo. O sangue extravasado transforma-se numa massa
gelatinosa, interrompendo a hemorragia. No entanto, esse é o final do processo de
coagulação; para a formação de fibrina é necessário que todos os outros fatores tenham
exercido sua atividade.
A fibrina resulta da transformação do fibrinogênio, proteína diluída no plasma (parte
líquida do sangue) sangüíneo. Mas, para que o fibrinogênio se transforme, é necessária a
intervenção da tromba que, por sua vez, é o resultado da transformação da protrombina,
uma proteína (globulina) formada no ligado.
A responsável pela transformação da protrombina é a tromboplastina, substância
presente nos tecidos e no interior das plaquetas (pequenos fragmentos celulares que se
originam de grandes células da medula vermelha dos ossos, os megacariócitos). Quando
a tromboplastina é liberada, inicia-se o processo de coagulação. Quando o sangue sai
dos vasos sangüíneos, através de algum ferimento, as plaquetas se aglutinam, libertando
a tromboplastina. Por outro lado, quando os tecidos também são atingidos pelo
traumatismo, liberam a mesma substância.
Essa liberação de tromboplastina sofre a interferência do fator íons-cálcio, fator
anti-hemofilico A e fator anti-hemofilico B. É o início da reação em cadeia. É por esse
motivo que, quando se prende o dedo em uma porta, por exemplo, o sangue poderá
coagular mesmo por baixo da pele. Os tecidos lesados liberam a tromboplastina, que vai
agir sobre a protrombina e assim por diante, desencadeando a coagulação do sangue.
Como a formação de protrombina é realizada no fígado, quando esse órgão se encontra
lesado por alguma afecçao grave sua produção é reduzida. Além disso, sua formação

depende de um suprimento e absorção adequados de vitamina K. Nos casos de doenças
grave do fígado ou de carência de vitamina K, existe urna falta relativa de protrombina e,
portanto, forte tendência a hemorragias. A transformação da protrombina em trombina
pela tromboplastina depende basicamente da participação dos íons-cálcio e é acelerada
pela presença do fator pró-acelerina (fator acelerador).
FIGURA 14 – Mecanismo de formação de um coágulo
3. FISIOLOGIA RESPIRATÓRIA
3.1. A Ventilação e a Circulação Pulmonar
Nossas células necessitam, enquanto vivas e desempenhando suas funções, de
um suprimento contínuo de oxigênio para que, num processo químico de respiração
celular, possam gerar a energia necessária para seu perfeito funcionamento e produção
de trabalho.
Da mesma forma que um motor de automóvel necessita, para produzir seu trabalho
mecânico, além da fonte de energia orgânica fornecida pelo combustível (gasolina, álcool
ou diesel), de fornecimento constante de oxigênio; da mesma forma que uma chama num
palito de fósforo, para permanecer acesa necessita além da matéria orgânica presente na
madeira do palito, também de oxigênio, nossas células também, para manterem seu
perfeito funcionamento necessitam, além da fonte de energia proporcionada pelos
diversos alimentos, de um fornecimento constante de oxigênio.
71 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

72
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
O oxigênio existe em abundância em nossa atmosfera. E para captá-lo
necessitamos de nosso aparelho respiratório. Através deste, parte do oxigênio da
atmosfera se difunde através de uma membrana respiratória e atinge a nossa corrente
sanguínea, é transportado pelo nosso sangue e levado às diversas células presentes nos
diversos tecidos. As células, após utilizarem o oxigênio, liberam gás carbônico que, após
ser transportado pela mesma corrente sanguínea, é eliminado na atmosfera também pelo
mesmo aparelho respiratório.
Para que seja possível uma adequada difusão de gases através da membrana
respiratória, oxigênio passando do interior dos alvéolos para o sangue presente nos
capilares pulmonares e o gás carbônico se difundindo em sentido contrário, é necessário
um processo constante de ventilação pulmonar.
A ventilação pulmonar consiste numa renovação contínua do ar presente no interior
dos alvéolos. Para que isso ocorra é necessário que, durante o tempo todo, ocorram
movimentos que proporcionem insuflação e desinsuflação de todos ou quase todos os
alvéolos. Isso provoca, no interior dos alvéolos, uma pressão ligeiramente, ora mais
negativa, ora mais positiva do que aquela presente na atmosfera.
Durante a inspiração, devido a uma pressão intra-alveolar de aproximadamente 3
mmHg mais negativa do que a atmosférica, uma certa quantidade de ar atmosférico é
inalado pelo aparelho respiratório; durante a expiração, devido a uma pressão intra-
alveolar de aproximadamente 3 mmHg mais positiva do que a atmosférica, a mesma
quantidade de ar é devolvida para a atmosfera.
Para que possamos insuflar e desinsuflar nossos alvéolos, devemos inflar e
desinflar nossos pulmões. Isso é possível através de movimentos que acarretem aumento
e redução do volume no interior da nossa caixa torácica, onde nossos pulmões estão
localizados.
Podemos expandir o volume de nossa caixa torácica levantando nossas costelas e
contraindo o nosso músculo diafragma. Para retrairmos o volume da caixa torácica
fazemos exatamente o contrário: rebaixamos nossas costelas enquanto relaxamos o
nosso diafragma.
A inspiração, que promove a entrada de ar nos pulmões, dá-se pela contração da
musculatura do diafragma e dos músculos intercostais. O diafragma abaixa e as costelas

elevam-se, promovendo o aumento da caixa torácica, com conseqüente redução da
pressão interna (em relação à externa), forçando o ar a entrar nos pulmões.
A expiração, que promove a saída de ar dos pulmões, dá-se pelo relaxamento da
musculatura do diafragma e dos músculos intercostais. O diafragma eleva-se e as
costelas abaixam, o que diminui o volume da caixa torácica, com conseqüente aumento
da pressão interna, forçando o ar a sair dos pulmões.
FIGURA 15 – Fisiologia da Mecânica Respiratória
Portanto temos diversos músculos que nos são bastante importantes durante
nossa respiração:
Os principais músculos utilizados na inspiração são: diafragma,
esternocleidomastoideos, intercostais externos, escalenos e serráteis anteriores.
Os músculos utilizados na expiração são intercostais internos, retos abdominais e
demais músculos localizados na parede anterior do abdômen.
73 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 16 – Atuação dos músculos expiratórios e inspiratórios
Durante a inspiração e durante a expiração, o ar passa por diversos e diferentes
segmentos que fazem parte do aparelho respiratório:
O Nariz é o primeiro segmento por onde, de preferência, passa o ar durante a
inspiração. Ao passar pelo nariz, o ar é filtrado, umidificado e aquecido. Na
impossibilidade eventual da passagem do ar pelo nariz, tal passagem pode acontecer por
um atalho, a boca. Mas infelizmente, quando isso acontece, o ar não sofre as importantes
modificações descritas acima.
Após a passagem pelo nariz, antes de atingir a laringe, o ar deve passar pela
faringe, segmento que também serve de passagem para os alimentos.
A Laringe, normalmente, permite apenas a passagem de ar. Durante a deglutição
de algum alimento, uma pequena membrana (epiglote) obstrui a abertura da laringe, o
que dificulta a passagem de fragmentos que não sejam ar para as vias respiratórias
74 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

75
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
inferiores. Na laringe localizam-se também as cordas vocais, responsáveis para produção
de nossa voz.
A Traquéia é um tubo cartilaginoso que liga as vias respiratórias superiores às
inferiores, logo abaixo. Em seguida, a traquéia se ramifica em Brônquios que são
numerosos e ramificam-se também numerosamente, como galhos de árvore e permitem
a passagem do ar em direção aos alvéolos. Os Bronquíolos respiratórios e terminais
estão entre os brônquios e os sacos alveolares, de onde saem os alvéolos.
Por toda a mucosa respiratória, desde o nariz até os bronquíolos, existem
numerosas células ciliadas, com cílios móveis, e grande produção de muco. Tudo isso
ajuda bastante na constante limpeza do ar que flui através das vias respiratórias.
Os alvéolos apresentam certa tendência ao colabamento. Tal colabamento
somente não ocorre normalmente devido à pressão mais negativa presente no espaço
pleural, o que força os pulmões a se manterem expandidos. O grande fator responsável
pela tendência de colabamento dos alvéolos é um fenômeno chamado Tensão
Superficial. A Tensão Superficial ocorre no interior dos alvéolos devido a grande
quantidade de moléculas de água ali presente e revestindo, inclusive, toda a parede
interna dos alvéolos. A Tensão Superficial no interior dos alvéolos certamente seria bem
maior do que já é se não fosse a presença, nos líquidos que revestem os alvéolos, de
uma substância chamada surfactante pulmonar. O surfactante pulmonar é formado
basicamente de fosfolipídios (dipalmitoil lecitina) por células presentes no epitélio alveolar.
A grande importância do surfactante pulmonar é sua capacidade de reduzir
significativamente a tensão superficial dos líquidos que revestem o interior dos aléolos e
demais vias respiratórias.

FIGURA 17 – Vista anterior do sistema respiratório
Quanto à circulação pulmonar, esta é chamada de pequena circulação. A artéria
pulmonar parte do ventrículo direito e se bifurca logo em artéria pulmonar direita e artéria
pulmonar esquerda, que vão aos respectivos pulmões. Uma vez entradas nos pulmões,
ambas se dividem em tantos ramos quantos são os lobos pulmonares; depois uma ulterior
subdivisão ao nível dos lóbulos pulmonares, estes se resolvem na rede pulmonar. As
paredes dos capilares são delgadíssimas e os gases respiratórios podem atravessá-las
facilmente: o oxigênio do ar pode assim passar dos ácinos pulmonares para o sangue; ao
contrário, o anidrido carbônico abandona o sangue e entra nos ácinos pulmonares, para
ser depois lançado para fora. Aos capilares fazem seguimento as vênulas que se reúnem
entre si até formarem as veias pulmonares. Estas seguem o percurso das artérias e se
lançam na aurícula esquerda. A artéria pulmonar contém sangue escuro, sobrecarregado
de anidrido carbônico (sangue venoso). As veias pulmonares contêm, contrariamente,
sangue que abandonou o anidrido carbônico e se carregou de oxigênio, tomando a cor
vermelha (sangue arterial).
76 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

77
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
3.2. O Transporte de Oxigênio e Dióxido de Carbono
O termo "transporte do O2" engloba todos os processos envolvidos na mobilização
do oxigênio desde o ar inspirado até a mitocôndria.
Um fornecimento contínuo de oxigênio é necessário para a manutenção da
integridade e função normal das diversas células do organismo; esse oxigênio serve como
aceptor final de elétrons na cadeia respiratória mitocondrial, processo acoplado à
fosforilação oxidativa que gera ATP. Essa é a principal via metabólica pela qual o
organismo consome oxigênio.
O movimento das moléculas de O2 da atmosfera às mitocôndrias requer o
funcionamento integrado dos pulmões, coração, sangue e rede vascular que compõem
um complexo sistema de transporte de massa capaz de suportar variações no consumo
de oxigênio (VO2) tão grandes quanto 15 a 20 vezes.
É por difusão que as moléculas de O2 se movem do gás alveolar para o sangue
que percorre os capilares pulmonares; é também por difusão que este gás se move do
capilar sistêmico até às mitocôndrias, nos diversos órgãos sistêmicos. O movimento do
CO2 é no sentido oposto mas também se processa por difusão. Ambos os gases sofrem
reações na corrente sangüínea no início e no fim de suas jornadas entre os pulmões e os
tecidos periféricos.

FIGURA 18 – O esquema mostra o transporte da hemoglobina
A eficiência do processo de difusão simples pela membrana alvéolo-capilar é
grandemente aumentada pela rápida renovação circulatória do sangue em contato com os
capilares dos alvéolos pulmonares e dos tecidos, pelas rápidas reações químicas que
sofrem os gases respiratórios assim que atingem o sangue; as ligações químicas
reversíveis da hemoglobina com o O2 e o CO2 são complementares e colabam
consideravelmente para a capacidade de transporte do sangue, pela presença de
enzimas específicas que aceleram a captação de O2 nas células transportadoras, bem
como a combinação de CO2 com a água na corrente sangüínea e pela adequação da
circulação sangüínea com o volume de ventilação alveolar em função de exigências
metabólicas.
Diariamente, cerca de 1 Kg de cada um dos gases respiratórios é trocado pela
atmosfera com os tecidos. A despeito da intensa troca gasosa, a composição do ar
alveolar persiste admiravelmente constante, sem alterações bruscas na PaO2 ou PaCO2.
Isso ocorre principalmente porque cerca de 10 % do ar alveolar é renovado a cada ciclo
78 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

79
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
respiratório, pois o volume corrente é adicionado a uma CRF (Capacidade Residual
Funcional) quase dez vezes maior.
A velocidade do fluxo aéreo decresce enormemente na medida em que nos
aproximamos dos alvéolos, pois as sucessivas ramificações da árvore traqueobrônquica
aumentam muito a área de secção transversal; ao nível das unidades de trocas gasosas,
não há fluxo mensurável, sendo a renovação do ar alveolar decorrente do processo de
difusão molecular. Como a chegada de o2 é superior ao seu consumo pelo sangue
venoso que se arterializa, essa difusão da fase gasosa não limita, em condições normais,
a quantidade de O2 que atravessa a membrana e se combina com a hemoglobina.
A próxima fase é a difusão através da membrana alvéolo-capilar, onde o sangue
venoso é exposto ao gás alveolar que contém mais O2 (PaO2 = 105 mmHg) e muito
menos CO2 (PaCO2 = 40 mmHg) que ele próprio (PVO2 = 40 mmHg e PVCO2 = 45 mmHg).
Em função dessas diferenças de pressão, o CO2 difunde-se do capilar para o alvéolo e o
O2 difunde-se em sentido oposto.
O equilíbrio entre as pressões dos gases alveolares e capilares é atingido tão
rápido quanto em 0.25 s ou 1/3 do tempo de trânsito de cada hemácia (0.75 s).
No exato momento em que as moléculas de O2 atravessam a membrana alvéolo-
capilar e penetram no plasma, forma-se uma diferença de PO2 entre plasma e
hemoglobina contida no citoplasma da hemácia; isso desencadeia a "difusão na fase
sangüínea" em que o O2 migra do plasma a hemoglobina. Embora a ligação de O2 seja
extremamente rápida, ela se constitui no fator limitante da difusão alvéolo-capilar.
O O2 oferecido pelos alvéolos é captado pela hemoglobina, que, assim, se
converte de seu estado reduzido para a forma oxigenada (oxidada). A oxi-Hemoglobina é
um ácido mais forte que a hemoglobina reduzida; passa a neutralizar radicais alcalinos
antes neutralizados por outros ânions (Cl-, HCO3-) presentes nos eritrócitos.
Para equilibrar esta captação de cátions pela oxi-Hemoglobina, um número
correspondente de íons cloreto se difunde para o exterior dos eritrócitos, enquanto que
íons bicarbonato penetram nos eritrócitos, numa troca. Este bicarbonato é convertido em
CO2 molecular e H2O numa reação acelerada pela anidrase carbônica, presente no
interior dos eritrócitos. O CO2 assim liberado se difunde para o plasma e daí para o ar
alveolar.

80
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A oxidação da hemoglobina também provoca uma liberação de CO2 de grupos
amínicos, com os quais o CO2 se combina quando a molécula se encontra reduzida; os
grupos amínicos são incapazes de manter esta combinação quando a molécula se torna
mais ácida em virtude da oxigenação. Esta fração de CO2 também difunde para o plasma
e em seguida para os alvéolos. À medida que o CO2 dissolvido no plasma se difunde para
o ar alveolar, a tensão de CO2 do plasma cai e quantidades adicionais de CO2 são
liberadas de combinações químicas.
A perda de CO2 do sangue torna-se mais alcalino, o que permite à hemoglobina
combinar-se com mais oxigênio do que seria possível se a reação do sangue
permanecesse inalterada. Assim, a captação de O2 expulsa CO2 do sangue, enquanto
que a perda de CO2 permite ao sangue absorver mais O2 .
Nos tecidos, esta relação se inverte completamente. O CO2 produzido pela engenharia
metabólica dos diferentes tecidos reage com o sangue contido nos capilares sistêmicos,
sendo transportados até os pulmões principalmente na forma de bicarbonato de sódio,
onde se difunde para alvéolos pulmonares; o O2 presente no ar alveolar interage com a
hemoglobina, após atravessar a membrana alvéolo-capilar, sendo transportado até à
intimidade dos tecidos sistêmicos.
Os tecidos requerem uma pressão de oxigênio surpreendentemente pequena para
manter funcionando adequadamente suas mitocôndrias. Uma pressão de 5 mmHg no
citoplasma, e de apenas 1 mmHg na mitocôndria é mais do que suficiente para que as
mitocôndrias trabalhem a plena carga.
O sistema de transporte de oxigênio é capaz de fornecer todo o O2 necessário ao
metabolismo celular em repouso e de aumentar esse fornecimento quando se elevam as
demandas metabólicas, como se dá no exercício.
Quando cessa o transporte de O2, como ocorre na parada cardíaca, a morte sobrevem
em 3 a 10 minutos.
Um adulto normal de 70 Kg de peso corporal transporta aproximadamente 1050 ml
de O2 por minuto, dos quais os tecidos consomem 250 ml/min (3 a 5 ml/Kg/min); para
tanto os pulmões movimentam cerca de 10 000 litros de ar e recebem cerca de 8 mil litros
de sangue venoso para ser arterializado, diariamente, "quer chova ou faça sol".

O ar contém oxigênio a uma pressão que é aproximadamente igual a 159 mmHg;
desde o ar ambiente até a mitoc6ondria, o oxigênio passa de um compartimento para
outro por diferença de pressão parcial; assim é que, no alvéolo, sua pressão é de 105
mmHg, no sangue arterial em torno de 95 mmHg e no capilar sistêmico, próximo de 45
mmHg, se difundindo daí em direção aos tecidos, onde a pressão é inferior a 10 mmHg.
FIGURA 19 - Eliminação de CO2 pelo alvéolo
O transporte de gás carbônico pelo sangue tem em comum com o transporte de O2
o fato de que a maior parte ocorre após reações químicas reversíveis envolvendo a
hemoglobina, mais que dissolvido no plasma. O CO2 é produto do metabolismo celular
tanto aeróbio quanto anaeróbio; uma vez formado difunde-se para o plasma sangüíneo
obedecendo a diferença de concentração entre a célula e o capilar. 10% do CO2
produzido é transportado como gás dissolvido no plasma, enquanto os 90% restantes
estão relacionados com a hemoglobina
O transporte de gás carbônico está descrito como "relacionado à Hb" porque o CO2
que se liga quimicamente à hemoglobina forma carbamino-compostos, os quais
correspondem a apenas 10 a 20% do total do transporte de gás carbônico; entretanto o
81 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

82
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
restante do transporte "viaja" na forma de bicarbonato também depende da hemoglobina
para que se processe.
Em células metabolicamente ativas, a PCO2 tecidual é muito mais elevada que a
PCO2 do sangue que flui pelos capilares sistêmicos, permitindo sua difusão desde a
mitocôndria (onde ocorrem as descarboxilações que geram CO2) até os capilares. O
transporte do CO2 se inicia no local de formação no interior da célula ativa ou da
mitocôndria. Nesta altura, não existe fluxo de líquido para carregar o metabólito para fora
e além disso a membrana celular impede a passagem de íons bicarbonato. Portanto, todo
o CO2 produzido deve deixar a célula por difusão de moléculas gasosas dissolvidas sem
carga elétrica, que se movimentam de regiões de PCO2 alta, no interior das células, para
as regiões de pressões parciais inferiores, presentes nos capilares.
As distâncias percorridas nesse processo de difusão são finitas, mas em virtude do
alto coeficiente de difusão do CO2 nos tecidos (mais de 20 vezes que o observado para o
O2) o gradiente de pCO2 de célula capilar não supera 1 a 2 mmHg.Assim que uma
molécula de CO2 penetra num capilar sangüíneo, o trabalho de transporte até os pulmões
depende do coração. No entanto, a eficiência do transporte de CO2 depende de uma série
de características importantes do próprio sangue, relacionando-se intimamente com a
função de transporte de O2.
Uma parte do gás carbônico presente no sangue venoso dissolve no plasma, e
essa quantidade corresponde 10% do total transportado a qualquer instante. Outra
quantidade verdadeiramente desprezível combina-se com a água formando ácido
carbônico; entretanto essa reação processa-se muito lentamente, pois não há anidrase
carbônica no plasma humano. Uma terceira parte gera carbamino-compostos, reagindo
com o grupamento – NH2 de resíduos aminoácidos das proteínas plasmáticas.
Embora o gás carbônico seja 20 vezes mais solúvel no plasma que o O2, somente
10% do total produzido pode ser transportado dissolvido no plasma; somente 1,5 ml de
CO2 estão dissolvidos em 100 ml de sangue. A maior parte de CO2 presente no plasma,
se encontra combinado de 3 formas: compostos carbamínicos, ácido carbônico e
bicarbonato. A combinação de CO2 com o grupamento amina das proteínas plasmáticas
gera carbamino-compostos, que também ocorre em pequenas quantidades, contribuindo
muito pouco para o transporte do CO2.

O ácido carbônico resulta da hidratação do anidrido carbônico conforme a equação
CO2 + H2O -------- H2CO3 . Essa combinação entre CO2 e H2O é molecular. A reação se
processa para a direita, no sentido da hidratação, ao nível dos capilares teciduais, e para
esquerda (de hidratação) nos capilares pulmonares.
O ácido carbônico rapidamente sofre dissociação iônica, formando o bicarbonato:
H2CO3 ----------H+ + HCO3- . O ácido carbônico é fraco pouco dissociado de modo que
somente se forma uma quantidade muito pequena de bicarbonato. As proteínas
plasmáticas e os fosfatos plasmáticos podem aceitar H+, permitindo pelo aumento na
formação de HCO3-. Entretanto a maior parte do bicarbonato presente no plasma é
oriundo da hemácia.
FIGURA 20 – Formação de dióxido de carbono
A maior parte do CO2 retido nos tecidos é transportado até os alvéolos pulmonares
graças à hemoglobina contida nas hemácias. O CO2 interage com a hemoglobina de três
maneiras: uma parte pequena permanece como CO2 dissolvido no citoplasma da
hemácia; uma quantidade pouco maior reage com o grupamento amino dos resíduos
aminoácidos da hemoglobina formando carbamino-compostos: Trata-se de uma reação
muito rápida que não requer catalisador especial; e A maior parte (80%) combina-se com
água em decorrência da riqueza de anidrase carbônica, uma enzima que acelera a reação
de hidratação do anidrido carbônico em até 100 000 vezes.
O ácido carbônico formado dissocia liberando H+ e HCO3-. O hidrogenionte é
tamponado pela Hb, com alteração mínima no pH (reações isoídricas); tal reação é
favorecida pela simultânea perda de oxigênio pela Hb, que assim torna-se um ácido mais
fraco capaz de assimilar com mais facilidade o H+ oriundo do ácido carbônico.
83 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

84
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A membrana plasmática das hemácias é permeável aos ânions de pequeno peso
molecular, de modo que o bicarbonato difunde-se para fora da célula, fenômeno descrito
por Zuntz. A perda de carga negativa cria alteração no gradiente elétrico da célula,
resultando na entrada de outras cargas negativas que substituam o bicarbonato: quem
entra para compensar é o cloreto dado ao processo; esse constitui o "fenômeno de
Hambúrguer". Portanto, para manter a neutralidade elétrica do plasma e a negatividade
intracelular das hemácias, íons cloreto (Cl-) se difundem para o interior das hemácias,
fenômeno descrito por Hambúrguer como "troca de cloretos" ou "desvio de cloretos".
Ao nível dos capilares pulmonares, ao invés de o sangue ser exposto a um
ambiente rico em CO2, entra em contato com o ar alveolar, que tem pouco gás carbônico;
isso inverte o equilíbrio destas reações bioquímicas que passam a se processar no
sentido oposto. Assim, embora o plasma contenha mais de 8% de todo o CO2 retirado dos
tecidos, são as reações que ocorrem no interior da hemácia quem fornece praticamente
todo o bicarbonato transportado no plasma. O carregamento de CO2 e o desgarramento
de O2 são processos que auxiliam mutuamente; um aumento na pCO2 capilar (e
decréscimo no pH) facilita a liberação de O2 (efeito Bohr), enquanto que a liberação de O2
facilita a captação de CO2 (efeito Haldane). A hemoglobina participa do transporte de CO2
exatamente nesse ponto na medida em que retira o H+, pois o tamponamento deste
próton permite que a reação de hidratação do gás carbônico continue.
É através do processo acima descrito que 90% do CO2 produzido é transportado
dos tecidos até os alvéolos pulmonares e daí à atmosfera. Cada litro de sangue arterial
(pH 7,40) que penetra nos tecidos já carrega cerca de 22 milimoles de CO2. A presença
dessa quantidade de CO2 depende da existência no sangue de bases de efeitos
tamponantes do H2CO2 e da pressão parcial de CO2 imposta ao sangue arterial pelo
equilíbrio entre ventilação alveolar e velocidade de produção de CO2
No sangue arterial, o CO2 se distribui de maneira desigual entre plasma e as
hemácias. No estado de equilíbrio, o CO2 existe em três formas, tanto no plasma quanto
nos eritrócitos: CO2 fisicamente dissolvido, compostos carbamino e íons bicarbonato. O
plasma contém as maiores quantidades de CO2. A maior parte do CO2 plasmático
apresenta-se na forma de íons bicarbonato de modo que a relação entre bicarbonato e
CO2 fisicamente é em geral de 20:1. Uma parte do CO2 fisicamente dissolvido se hidrata

85
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
de acordo com a reação de equilíbrio CO2 + H2O ------------ H2CO3 no qual a proporção de
CO2 molecular para H2CO3 é de aproximadamente 1000:1.
No plasma, a hidratação do CO2 não é acelerada pela ação da anidrase carbônica,
a exemplo do que ocorre no eritrócito onde essa enzima é encontrada. A quantidade de
CO2 combinada com um grupo –NH2, de proteínas plasmáticas para formar complexo
carbamino-protéico é muito pequena. O O2 e o CO2 ligam-se a pontos quimicamente
distintos da molécula de hemoglobina. Entretanto, ao se ligarem, cada um influencia a
reação de hemoglobina com o outro. A quantidade de CO2 presente na forma de íon
bicarbonato no interior das hemácias é consideravelmente menor que aquela carregada
na forma de bicarbonato no plasma arterial; a diferença depende da concentração da
hemoglobina.
As alterações produzidas no sangue em sua transição do estado arterial ao venoso
ao passar pelos capilares tissulares, dependem da quantidade de CO2 acrescentada a
cada litro de sangue. Isto, por sua vez, depende de: fluxo sangüíneo por unidade de
tecido metabolicamente ativo e nível de atividade metabólica no tecido perfundido. Por
esses motivos, a composição em CO2 do sangue venoso difere entre os tecidos.
Alterações ao nível dos capilares tissulares O aparecimento de CO2 molecular no
sangue capilar coincide com o aumento da taxa de difusão de O2 para os tecidos e, deste
modo, com a desoxigenação de hemoglobina. Cerca de 2,0 mmol de CO2 penetram em
1litro de sangue em sua passagem através dos capilares, elevando a PCO2 de 41 para 46
mmHg. Este aumento de 5 mmHg na pCO2 representa uma elevação de apenas 0,1 mmol
de CO2 fisicamente dissolvido, mas esse aumento tem grande significação fisiológica.
Uma quantidade igualmente pequena de CO2, cerca de 0,1 mmol, permanece no
plasma na forma de íons bicarbonato formados no plasma e tamponados pelas proteínas
plasmáticas. O restante passa para o interior da hemácia, onde têm lugar etapas
adicionais de trocas gasosas intracapilares. Deste modo, aproximadamente 1,8/2,0 mmol,
ou 90% do CO2 absorvido por um litro de sangue, penetram nas hemácias e dependem
de fatores intra-eritrocitários para seu transporte.
Quando a hemoglobina se desoxigena nos tecidos, ela passa a fornecer dois
mecanismos para o transporte de CO2. O primeiro é combinação do CO2 com os grupos
amino das cadeias polipeptídicas da hemoglobina, para formar carbamino-hemoglobina.

86
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
O segundo mecanismo, que se desenrola rapidamente, envolve o papel da hemoglobina
como tampão e como fator dominante no transporte de CO2.
3.3. A Regulação Respiratória
Durante uma situação de repouso inspiramos e expiramos aproximadamente 500
ml de ar a cada ciclo. Em repouso executamos aproximadamente 12 ciclos a cada minuto.
Portanto, aproximadamente 6.000 ml de ar entram e saem de nossas vias aéreas durante
1 minuto.
Quando executamos uma atividade física aumentada, nossas células produzem
uma quantidade bem maior de gás carbônico e consomem também quantidade bem
maior de oxigênio. Por isso devemos aumentar também bastante nossa ventilação
pulmonar pois, caso isso não ocorra, teremos no nosso sangue uma situação de
hipercapnia e hipóxia. Tanto a hipercapnia quanto a hipóxia podem nos levar a um estado
de acidose. A acidose, se não tratada, pode nos levar a um estado de coma e,
posteriormente, à morte.
Tudo isso normalmente é evitado graças a um mecanismo automático que regula,
a cada momento, nossa respiração, de acordo com a nossa necessidade a cada instante.
No tronco cerebral, na base do cérebro, possuímos um conjunto de neurônios
encarregados de controlar a cada instante a nossa respiração: Trata-se do Centro
Respiratório. O Centro Respiratório é dividido em várias áreas ou zonas com funções
específicas cada uma:
A Zona Inspiratória é a zona responsável por nossa inspiração. Apresenta células
auto-excitáveis que, a cada 5 segundos aproximadamente, se excitam e fazem com que,
durante aproximadamente 2 segundos nos inspiremos. A partir desta zona parte um
conjunto de fibras (via inspiratória) que descem através da medula e se dirigem a diversos
neurônios motores responsáveis pelo controle dos nossos diversos músculos da
inspiração.
A Zona Expiratória é a zona que quando ativada, emite impulsos que descem
através de uma via expiratória e que se dirigem a diversos neurônios motores

87
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
responsáveis pelos nossos músculos da expiração. Através de um mecanismo de inibição
recíproca, quando esta zona entra em atividade, a zona inspiratória entra em repouso, e
vice-versa. Durante uma respiração em repouso a zona expiratória permanece
constantemente em repouso, mesmo durante a expiração. Acontece que, em repouso,
não necessitamos utilizar nossos músculos da expiração, apenas relaxamos os músculos
da inspiração e a expiração acontece passivamente.
A Zona Pneumotáxica, constantemente em atividade, tem como função principal
inibir (ou limitar) a inspiração. Emite impulsos inibitórios à zona inspiratória e, dessa
forma, limita a duração da inspiração. Portanto, quando em atividade aumentada, a
inspiração torna-se mais curta e a freqüência respiratória, consequentemente, aumenta.
A Zona Quimiossensível, situada entre as zonas inspiratória e expiratória, controla
a atividade de ambas. Quanto maior a atividade da zona quimiossensível, maior será a
ventilação pulmonar. Esta zona aumenta sua atividade especialmente quando certas
alterações gasométricas ocorrem: Aumento de Gás Carbônico, Aumento de íons
Hidrogênio livres (redução de pH) e, em menor grau, redução de Oxigênio.
O fator que provoca maior excitação na zona quimiossensível, na verdade, é o
aumento na concentração de íons Hidrogênio livres no meio, isto é, uma situação de
acidose. Mas acontece que, na prática, verificamos que um aumento de gás carbônico no
sangue (hipercapnia) provoca muito mais o aumento na atividade da zona
quimiossensível do que um aumento na concentração de Hidrogênio em igual proporção
no sangue. Isso ocorre porque o gás carbônico apresenta uma solubilidade muitas vezes
maior do que a do hidrogênio e, com isso, atravessa a membrana das células nervosas
com muito mais facilidade. No interior das células da zona quimiossensível, o gás
carbônico reage com a água lá presente e, graças à enzima anidrase carbônica,
rapidamente forma ácido carbônico. Este, então, se dissocia formando íon bicarbonato
mais íon Hidrogênio livre, sendo este último exatamente o que mais excita a zona
quimiossensível.

FIGURA 21– Regulação respiratória do PH
A hipóxia também excita a zona quimiossensível, mas de uma outra maneira bem
diferente: Na croça da aorta e nos seios carotídeos existem receptores muito sensíveis a
uma queda na concentração de oxigênio no sangue: os quimioceptores (aórticos e
carotídeos). Quando a concentração de oxigênio no sangue se torna mais baixo do que a
desejável, estes receptores se excitam mais intensamente e enviam sinais à zona
quimiossensível aumentando a excitabilidade desta e, com isso, aumentando a ventilação
pulmonar.
Um aumento da atividade física também provoca aumento na ventilação pulmonar
de outras formas. Impulsos provenientes da área motora cortical, responsável pelo
comando consciente de nossa atividade motora, ao se dirigirem para baixo, em direção à
medula, passam pelo tronco cerebral (além de outras áreas) e fazem conecções com
alguns neurônios desta região. Isso pode provocar aumento na ventilação pulmonar,
muitas vezes mesmo antes que as alterações gasométricas (hipercapnia, hipóxia ou
acidose) aconteçam.
Movimentos passivos também podem aumentar a ventilação pulmonar: Na
profundidade de nossos músculos esqueléticos, nos tendões e mesmo no interior de
muitas das nossas cápsulas articulares, possuímos receptores que se excitam a cada
movimento dessas estruturas. Ao se excitarem, enviam impulsos que se dirigem à medula
e também, muitas vezes, ao encéfalo, passando pelo tronco cerebral e fazendo conexões
com neurônios do Centro Respiratório.
88 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

89
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A concentração de íons hidrogênio do sangue ou, em outras palavras, o pH do
sangue, modifica a ventilação alveolar, através do centro respiratório. Esta estrutura do
sistema nervoso central se comporta como um "sensor" do pH do sangue. Quando a
concentração de íons hidrogênio do sangue está elevada (pH baixo) o centro respiratório
aumenta a freqüência dos estímulos respiratórios, produzindo taquipneia. Com o aumento
da freqüência respiratória, aumenta a eliminação do CO2 do sangue; a redução dos níveis
sanguíneos do CO2 eleva o pH. A concentração de H+ no sangue é permanentemente
acompanhada pelo centro respiratório, que regula seus estímulos de acordo com ela,
conforme demonstra o diagrama da figura 9. Ao contrário, quando a concentração de íons
hidrogênio (H+) está baixa (pH elevado), o centro respiratório diminui a freqüência dos
estímulos à respiração e ocorre bradipneia, que reduz a eliminação do CO2 tentando
corrigir o pH do sangue.
Na realidade, a regulação respiratória do pH, por estímulos do centro respiratório,
não normaliza o pH do sangue, porque, à medida que a concentração do íon hidrogênio
se aproxima do normal, o estímulo que modifica a atividade respiratória vai
desaparecendo. Apesar disso, a compensação respiratória é extremamente eficaz para
impedir grandes oscilações do pH.
---------- FIM MÓDULO III ----------

Curso de
Fisiologia Geral
MÓDULO IV
Atenção: O material deste módulo está disponível apenas como parâmetro de estudos para este Programa de Educação Continuada, é proibida qualquer forma de comercialização do mesmo. Os créditos do conteúdo aqui contido são dados aos seus respectivos autores descritos na Bibliografia Consultada.

91
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
MÓDULO IV
1. NEUROFISIOLOGIA 1.1.
recebe no corpo
celular
pré-sináptica, a zona de aposição, ou fenda
sináptica, e a membrana pós-sináptica. Quanto ao processo de transmissão de sinais, as
Funções Básicas das Sinapses e dos Neurotransmissores
A expressão sinapse foi introduzida por Sherrington, no final do século XIX, para
designar a região de aposição de neurônios, anteriormente descrita por Ramón y Cajal. A
transmissão química foi descoberta em 1920, por Loewi. Cada neurônio
e nos dendritos algo em torno de 1000 sinapses. Como são 10¹¹ neurônios ao
todo, são, em números aproximados, 1014 sinapses no sistema nervoso.
Constituem a sinapse a terminação
sinapses podem ser químicas ou elétricas.
FIGURA 1 – Representação de um neurônio, axônios e dendritos.
uma célula para a célula seguinte. As sinapses elétricas ocorrem em locais especializados
A maioria das sinapses dos mamíferos são sinapses químicas, mas existe uma
forma simples de sinapse elétrica que permite a transferência direta da corrente iônica de

92
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
as células glandulares, glia, etc. Elas também são comuns em vários
inverte
élulas do músculo esquelético, miocárdio, células da glândula pineal) que eles
inerva
de sua membrana, e quase que
invaria
e ligar-se aos receptores (proteínas específicas) na membrana de
um neurônio vizinho.
chamados junções. Elas formam canais que permitem que os íons passem diretamente
do citoplasma de uma célula para o citoplasma da outra. A transmissão nas sinapses
elétricas é muito rápida; assim, um potencial de ação no neurônio pré-sináptico, pode
produzir quase que instantaneamente um potencial de ação no neurônio pós-sináptico.
Sinapses elétricas no sistema nervoso central de mamíferos são encontradas
principalmente em locais especiais onde funções normais exigem que a atividade dos
neurônios vizinhos seja altamente sincronizada. Embora as junções sejam relativamente
raras entre os neurônios de mamíferos adultos, eles são muito comuns em uma grande
variedade de células não neurais, inclusive as células do músculo liso cardíaco, células
epiteliais, algum
brados.
Na sinapse química, o sinal de entrada é transmitido quando um neurônio libera um
neurotransmissor na fenda sináptica, o qual é detectado pelo segundo neurônio através
da ativação de receptores situados do lado oposto ao sítio de liberação. Os
neurotransmissores são substâncias químicas produzidas pelos neurônios e utilizadas por
eles para transmitir sinais para outros neurônios ou para células não-neuronais (por
exemplo, c
m.
A ligação química do neurotransmissor aos receptores causa uma série de
mudanças fisiológicas no segundo neurônio que constituem o sinal. Normalmente a
liberação do primeiro neurônio (chamado pré-sináptico) é causado por uma série de
eventos intracelulares evocados por uma despolarização
velmente quando um potencial de ação é gerado.
“Resumindo sinapse, podemos definir como um mecanismo que ocorre quando um
impulso elétrico ao viajar para a “cauda” da célula, chamado axônio”, chega a seu
término, disparando vesículas que contêm um neurotransmissor as quais se movem em
direção a membrana terminal. As vesículas se fundem com a membrana terminal para
liberar seus conteúdos. Uma vez na fenda sináptica (o espaço entre dois neurônios) o
neurotransmissor pod

FIGURA 2 – Representação de uma sinapse química
Existem dois tipos de sinapse química: sinapses inibitórias e sinapses excitatórias.
As Sinapses excitatórias causam uma mudança elétrica excitatória no potencial pós-
sináptico (EPSP). Isso acontece quando o efeito líquido da liberação do transmissor é
para despolarizar a membrana, levando-o a um valor mais próximo do limiar elétrico para
disparar um potencial de ação. Esse efeito é tipicamente mediado pela abertura dos
canais da membrana (tipos de poros que atravessam as membranas celulares para os
íons cálcio e potássio. As sinapses inibitórias causam um potencial pós-sináptico inibitório
(IPSP), porque o efeito líquido da liberação do transmissor é para hiperpolarizar a
membrana, tornando mais difícil alcançar o potencial de limiar elétrico. Esse tipo de
sinapse inibitória funciona graças à abertura de diferentes canais de íons nas membranas:
tipicamente os canais cloreto (Cl-) ou potássio (K+).
Quimicamente, os neurotransmissores são moléculas relativamente pequenas e
simples. Diferentes tipos de células secretam diferentes Neurotransmissores. Cada
substância química cerebral funciona em áreas bastante espalhadas, mas muito
específicas do cérebro e podem ter efeitos diferentes dependendo do local de ativação.
Cerca de 60 neurotransmissores foram identificados e podem ser classificados, em geral
em uma das quatro categorias: colinas, das quais a acetilcolina é a mais importante; as
aminas biogênica, se destacando a serotonina, a histamina, e as catecolaminas; os
93 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

94
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
aminoácidos como o glutamato e o aspartato que são os transmissores excitatórios bem
conhecidos, enquanto que o ácido gama-aminobutírico (GABA), a glicina e a taurine são
neurotransmissores inibidores; e neuropeptídeos, que são formados por cadeias mais
longas de aminoácidos (como uma pequena molécula de proteína). Sabe-se que mais de
50 deles ocorrem no cérebro e muitos deles têm sido implicados na modulação ou na
transmissão de informação neural.
1.2. Fisiologia da Dor
A dor nociceptiva ocorre como o resultado da ativação de nociceptores em tecidos
cutâneos e profundos. Os receptores sensoriais, preferencialmente sensíveis a estímulos
nocivos ou potencialmente nocivos, encontram-se na pele, músculos, tecidos conjuntivos
e vísceras torácicas e abdominais. Estas unidades têm aparência morfológica bem
definida à microscopia óptica e eletrônica e, fisiologicamente, caracterizam-se pelos seus
padrões de reações a estímulos cutâneos, mecânicos, térmicos e químicos. Uma vez
ativados, os nociceptores conduzem impulsos via fibras aferentes mielínicas finas A -
delta, ou pelas fibras não-mielínicas C.
Estudos fisiológicos têm demonstrado que os nociceptores não são ativos
espontaneamente, mas podem ser sensibilizados, particularmente após o dano térmico da
pele. A sensibilização manifesta-se como uma diminuição do limiar de ativação após o
dano, intensidade aumentada da reação a um dano prejudicial ou pelo aparecimento de
atividade espontânea. A sensibilização de nociceptores pode ocorrer dentro de minutos
após um dano térmico e podem durar horas. Tem-se especulado que isto possa ser a
relação fisiológica da hiperpatia, que ocorre depois de um dano térmico na pele, e,
também, um mecanismo de dor persistente no homem. A sensibilização dos nociceptores
pode ser decorrente de substâncias químicas tais como o íon potássio, bradicinina e
prostaglandinas liberadas como resultado de dano tecidual.
A ativação de nociceptores viscerais decorre da irritação das superfícies mucosa e
serosa, torção ou tração do mesentério, distensão ou contração de uma víscera oca e
resultado do impacto direto, tais como ocorre nos traumatismo. Estímulos similares são
necessários para provocar dor na bexiga, ureter ou uretra. Os nociceptores gastro-

95
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
intestinais reagem mais intensamente à dilatação excessiva ou à contração do intestino
e/ou torção do mesentério. Nociceptores polimodais suprem o canal anal, que é sensível
à dor em toda a sua extensão e tem inervação que se assemelha mais às estruturas
cutâneas do que vicerais.
A dor visceral é profunda, dolorosa. Mal localizada e, frequentemente, relacionada
a um ponto cutânea, relacionada a um ponto cutâneo, que pode ser sensível. Assim,
afecções pancreáticas e endometriais podem provocar dor referida nas costas; hepatoma
ou metástases no fígado podem gerar dor no ombro direito; neoplasias de próstata
desencadeiam dor no abdome e coxa. O mecanismo da dor referida não é totalmente
compreendido, mas pode ser relacionado, mas pode ser relacionado à convergência de
impulso sensorial cutâneo e visceral em células do trato espinotalâmico na medula
espinhal. A dor é relacionada à pele porque áreas encefálicas interpretam “mal”o impulso,
ou porque algumas fibras aferentes inervam estruturas somáticas e viscerais.
A dor nociceptiva, tal como ocorre na infiltração tumoral ou no processo
inflamatório, promove a liberação associada de mediadores químicos algiogênicos ou
pele, osso e vísceras, que ativam e sensibilizam os nociceptores. Este fato gera atividade
espontânea e sensibiliza fibras nociceptivas, resultando em dor. Os impulsos são
conduzidos ao sistema nervoso central (SNC) através das fibras A-delta ou fibras C. Estas
fibras penetram na medula, lateralmente na raiz dorsal, e fazem sinapse nas lâminas
superficiais (I e II) e profundas (V) do corno posterior, onde ativam sistemas nociceptivos
ascendentes, tais como o trato espinotalâmico, espinocervical e espinorreticular, podendo
gerara percepção consciente de dor.
A integridade das vias nociceptivas no sistema nervoso periférico (SNP) e sistema
nervoso central (SNC), bem como dos centros de processamento e de modulação da dor
no SNC, é necessária para que o fenômeno nococeptivo se processe normalmente.
Quando há lesão das fibras nervosas, surgem, imediatamente, potenciais de
grande amplitude nos aferentes primários durante alguns segundos. Os cotos proximais
dos axônios secionados são, a seguir, selados e a bainha de mielina adjacente, bem
como os axônios, degeneram na extensão de alguns milímetros. Após algum tempo,
grupos de axônios emergem dos bulbos terminais e, sob condições adequadas, alcançam
as terminações nervosas nos tecidos. As fibras nervosas em crescimento geram
potenciais de ação espontaneamente e são sensíveis a estímulos mecânicos e a ação da

96
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
adrenalina. Após certo tempo certo tempo, ocorre restauração da função normal dos
receptores nociceptivos. Quando o crescimento do nervo é bloqueado, ocorre formação
dos neuromas. Quando a lesão é parcial e a regeneração é bloqueada, a diferentes
intervalos, surgem microneuromas. Foram registrados potenciais espontâneos oriundos
dos neuromas nas fibras A-delta e C que alcançam o corno posterior da medula espinhal
(CPME). A atividade ectópica surge alguns dias após a lesão, aumenta a primeira semana
e decai progressiva,mente a seguir. Os potenciais ectópicos dos neuromas são mais
abundantes quando há isquemia tecidual, elevação da concentração do potássio
extracelular e acúmulo de peptídeos. A atividade ectópica dos neuromas das fibras
mielinizadas, reduz-se com o calor e eleva-se com o frio. Nas neuropatias periféricas, há
regeneração e degeneração disseminadas ao longo das fibras, do que resulta a
ocorrência freqüente de dor espontânea. A atividade elétrica dos gânglios sensítivos
aumenta em situações em que ocorre lesão do nervo periférico e constitui fonte adicional
significante de potenciais anormais, que se somam àqueles produzidos pelos neuromas.
O acúmulo de potássio e de outros íons na região do neuroma pode gerar despolarização
nas fibras nervosas vizinhas. Correntes efáticas parecem ocorrer nos neuromas de
amputação e nas fibras nervosas em degeneração. Potenciais ectópicos podem, também,
surgir quando os potenciais de ação dos aferentes primários prolongam-se além do
período refratário absoluto, gerando re-excitação da membrana neuronal. A lesão dos
nervos periféricos gera modificações anatômicas nos núcleos das células ganglionares do
nervos sensitivos e nas projeções centrais das raízes nervosas.
A hiperexcitabilidade neuronal é devida ao surgimento de marca-passos nos
nervos periféricos em regeneração e resulta da modificação da permeabilidade da
membrana axonal e do número, distribuição e cinética dos canais de sódio, cálcio e
potássio.
Estudos microneurográficos demonstraram haver número excessivo de potenciais
excitatórios nos nervos periféricos, associado à sensação de parestesias em seres
humanos. Foi observada hiperatividade neuronal na lâmina V do CPME, após rizotomia
espinal e avulsão de raízes espinais; o mesmo ocorre no núcleo do trato espinal do nervo
trigêmeo, após a rizotomia trigeminal. Hiperatividade neuronal talâmica contralateral à
lesão das raízes nervosas foi, também, observada tardiamente em animais de

97
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
experimentação. A desinibição das vias nociceptivas e a excitação das sinapses centrais
estão envolvidas no mecanismo de hiperatividade neuronal nas células do CPME.
Em casos de avulsão das raízes nervosas, o campo receptivo das células do
CPME amplia-se em grau menor do que em casos de rizotomia, talvez em decorrência da
lesão das estruturas do trato de Lissauer que exercem atividade facilitatória sobre as
unidades sensitivas da medula espinal.
Observou-se que as unidades celulares, predominantemente excitadas por
estímulos nociceptivos,, passam a responder, também, a estímulos de baixa intensidade
em casos de desaferentação. Os aferentes A-delta e C mantêm a capacidade de exitação
das células do CPME, mas há redução do mecanismo inibitório segmentar, alteração do
padrão de organização da chegada dos potenciais de ação ao CPME, aumento do
número de potenciais ectópicos, degeneração das projeções centrais dos aferentes
primários e, provavelmente, das células de origem dos tratos de projeção ascendentes,
podendo modificar a quantidade de neurotransmissores liberados pelas terminações
nervosas proximais.
A lesão das raízes sensitivas espinais e trigeminais resulta em degeneração dos
axônios e das suas projeções no SNC e, após alguns meses, em aumento da distribuição
dos aferentados. Este fenômeno é atribuído ao mecanismos de brotamento.
As lesões do SNP acarretam alterações funcionais nos neurônios de Segunda
ordem do CPME. A lesão parcial do CPME resulta em hipersensibilidade neuronal,
algumas horas após o traumatismo. Tardiamente, ocorrem anormalidades funcionais no
tálamo. Após a rizotomia, há queda da concentração de substância P nas lâminas I, II e
V,, seguida de retorno aos níveis progressos em cerca de 4 semanas. Em casos de
avulsão de raízes, ocorre redução da beta-encefalina e da substância P nas terminações
das lâminas I e II e da somatostatina na lâmina II, fatos que se acentuam na semana
subseqüente, associadamente ao desaparecimento quase completo da substância P nas
lâminas I e V, mas não da encefalina nas lâminas I, II e V. Estes achados sugerem que a
atividade excitatória da substância P e a ausências da atividade inibitória da encefalina,
que atuam pré-sinapticamente nas lâminas I e II e pós-sinapticamente na lâmina do
CPME, aliadas à queda da atividade da somatostatna, com ação inibitória nas lâminas II e
V, causam hiperatividade por desenervação observada nas lâminas I, II e V ao CPME, de
onde emergem as fibras dos tratos de projeção supra-segmentares.

98
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
As alterações nas concentrações dos neurotransmissores do CPME são, em parte,
revertidas pela aplicação do fator de regeneração nervosa na extremidade proximal da
estrutura nervosa secionada. Em casos de ganglietomia, a atividade da colecistoquimina
e da substância P recupera-se, não sendo, o mesmo, observado em relação à
somatostatina.
Há evidências de que a hiperatividade neuronal das células do CPME seja
reduzida pela hiperatividade da estimulação da cápsula interna, núcleos talâmicos do
complexo vetrobasal e do córtex cerebral. Observou-se que, em casos de secção
transversal da medula espinal, ocorre expansão dos campos receptivos e hiperatividade
das células do CPME. Nesta eventualidade, existe maior influência de aferências
inibitórias contralaterais, sejam influenciadas por vias decendentes.
As lesão dos funículos posteriores provocam surtos de atividades espontânea no
núcleo grácil e a lesão destes núcleo resulta em aumento do campo receptivo das
unidades celulares desaferentadas do complexo ventrobasal do tálamo.
As zonas de gatilho, que se observam em doentes com paraplegia e avulsão de
raízes, parecem ser devidas ao aumento da atividade excitatória prolongada, originada
em áreas distantes com inervação normal. Esse fato pode indicar que existe
reorganização e aumento da área de projeção dos aferentes nociceptivos nessas
unidades neuronais.
No ser humano, a síndrome de dor por desaferentação é conhecida há várias
décadas. Segundo Tasker e Emmers, dor por desaferentação é aquela resultante das
lesões de estruturas nervosas. É descrita como queimação, pontada, dormência,
formigamento ou sensações bizarras e é, geralmente, de início tardio. Como a dor não se
manifesta em todos os doentes com lesões similares, admite-se que haja fatores
genéticos envolvidos na sua ocorrência. A fisiopatologia da dor por desaferentação ainda
não foi completamente esclarecida. A função dos neurônios das vias sensitivas é de
codificar, conduzir e transferir informações aos diferentes níveis do SNC, envolvendo alto
grau de especialização regional. As terminações nervosas são especializadas em
codificar a informação sensitiva e em veicular informações sensitivas até o CPME, sem
modificações qualitativas e qualificativas. As propriedades funcionais dos axônios e das
unidades centrais precisam ser mantidas íntegras para que o processamento da
informação sensitiva ocorra de modo adequado. Havendo modificações na função das

99
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
terminações nervosas periféricas, troncos nervosos periféricos, ou das vias de condução
e de processamento central da informação nociceptiva, dor espontânea ou gerada por
estímulos não-nóxicos costuma manifestar-se.
A sensibilização dos receptores, a ocorrência de focos ectópicos de potencial de
ação nas fibras nervosas periféricas e nas vias centrais, bem como a atividade anormal
das unidades de processamento central das aferências sensitivas são, seguramente, os
mecanismos mais importantes na gênese da dor por desaferentação.
Na dor rápida, a dor é sentida em cerca de 0,1 s depois que o estimulo doloroso é
aplicado. Este tipo de dor é sentido frente a um traumatismo agudo, geralmente térmico
ou mecânico.
Na dor lenta, a dor é sentida após 1s ou mais, e aumenta lentamente , durante
muitos segundos até minutos. Este tipo de dor está geralmente associado a destruição de
tecidos, podendo ser um estimulo químico,físico ou mecânico. Pode levar a um sofrimento
insuportável e prolongado. Pode ocorrer tanto na pele como nos tecidos mais profundos
do corpo.
1.3. Neurofisiologia da Visão
Desde os rudimentares fotorreceptores de certos organismos unicelulares ao
complexo olho humano, a evolução percorreu um longo caminho para dotar os seres
vivos de instrumentos eficazes para a percepção dos objetos e a experiência do espaço
em grande parte, aos órgãos da visão.
Visão é o processo fisiológico por meio do qual se distinguem as formas e as cores
dos objetos. Em linhas gerais, o olho funciona como uma câmara fotográfica que projeta
uma imagem invertida do mundo exterior em sua porção interna posterior, onde existe um
revestimento fotossensível, a retina, que envia informações codificadas ao sistema
nervoso central, dando ao indivíduo a sensação da visão.
A visão se produz em nível molecular graças a substâncias fotossensíveis, os
fotopigmentos, que sofrem transformações químicas sob a ação da luz. Essas
transformações produzem estímulos em células e fibras sensoriais, que são transmitidos
aos centros nervosos correspondentes. Entre os vertebrados, o olho mais perfeito e

100
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
desenvolvido é o dos mamíferos, que corresponde, em linhas gerais, à configuração do
olho humano. No homem, os dois globos oculares estão alojados no interior das
cavidades orbitárias e se unem às paredes ósseas graças aos chamados músculos
extrínsecos, de natureza estriada e contração voluntária, responsáveis pela
movimentação dos olhos. Os músculos retos -- superior, inferior, externo e interno --
possibilitam a movimentação do globo ocular para cima, para baixo, para a direita e para
a esquerda. Outros dois, o oblíquo maior e menor, permite ao olho deslocar-se em todas
as direções.
O olho humano é constituído de três camadas. A mais externa, fibrosa, tem função
protetora e é chamada esclerótica. Em sua porção anterior, a esclerótica é transparente e
recebe o nome de córnea. Na parte posterior e lateral, é opaca. A camada intermediária é
abundante em vasos sangüínea e formada pela coróide, pelo corpo ciliar e pela íris. A
camada interna é a retina, onde se localizam as células fotorreceptoras. A córnea é
recoberta pela conjuntiva, fina membrana que se estende também pela face interna das
pálpebras.
Chama-se trato uveal o conjunto de coróide, corpo ciliar e íris. Muito vascularizada,
a coróide abastece de nutrientes e oxigênio os tecidos oculares. Nela situam-se também
células pigmentares, cuja função é absorver luz, como a pintura preta do interior das
câmaras fotográficas, para evitar que reflexos prejudiquem a qualidade da imagem
projetada na retina.

FIGURA 3 – Estrutura de um globo ocular
O corpo ciliar é o prolongamento anterior da coróide, formado pelos chamados
processos ciliares e pelo músculo ciliar. Os processos ciliares são ligamentos que unem
ao músculo ciliar o cristalino, estrutura transparente, com forma de lente biconvexa, que
permite focalizar os objetos situados a diversas distâncias. O músculo ciliar, cuja
contração é produzida pelo sistema parassimpático, controla o grau de curvatura do
cristalino e permite ajustar o foco.
Na porção anterior do cristalino, a íris controla, como um diafragma, a quantidade
de luz que atinge a retina. Pigmentos na íris lhe dão cor característica, que varia do negro
ao azul. As musculaturas lisas radial e circular da íris abrem e fecham seu orifício central,
a pupila. O espaço entre a córnea e o cristalino, dita câmara anterior, é preenchido pelo
humor aquoso, que mantém constante a pressão interna do globo ocular. Já a cavidade
entre o cristalino e a retina, a câmara posterior, contém uma substância gelatinosa
chamada como humor vítreo.
101 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
FIGURA 4 – Estruturas responsáveis pelo funcionamento da visão
Na retina estão situadas as células encarregadas de registrar as impressões
luminosas e transmiti-las ao cérebro por intermédio do nervo óptico, que sai da parte
posterior do globo ocular. As células fotorreceptoras são chamadas cones e bastonetes,
em virtude da forma de seus prolongamentos. Os cones dispõem-se na região central da
102 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

103
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
retina e são responsáveis pela visão colorida, enquanto os bastonetes, mais abundantes
nas regiões periféricas, processam uma visão de contornos, de contraste claro-escuro,
em condições de baixa luminosidade. A região de onde parte o nervo óptico é chamada
ponto cego, por ser insensível à luz. Já a região chamada fóvea, composta apenas de
cones e situada acima do ponto cego, é a área da retina onde a visão é mais nítida.
Alguns órgãos anexos ou acessórios protegem o globo ocular: as pálpebras,
dobras de pele que recobrem a parte anterior do olho de forma a impedir seu
ressecamento, movem-se em conseqüência da ação dos músculos palpebrais (elevador,
para abrir, e orbicular, para fechar); as sobrancelhas, aglomeração cutâneo-muscular
coberta de pêlos, estão localizadas acima de cada olho e evitam que neles penetre o
suor; os cílios são uns conjuntos de pêlos que nascem nas bordas das pálpebras e
protegem os olhos contra a entrada de pó e de outras partículas; e o aparelho lacrimal
produz as lágrimas (líquido aquoso e salino que também contém lisozimas, moléculas que
destroem microrganismos).
Para a formação da imagem do mundo exterior sobre a retina, o olho dispõe de um
conjunto de elementos refratores, constituídos pela córnea, humor aquoso, cristalino e
humor vítreo. As propriedades ópticas das superfícies refratoras estão relacionadas com
seu grau de curvatura e com o índice de refração dos meios que ela separa. A unidade
empregada para se medir o poder de refração é a dioptria, que é a recíproca da distância
focal, tomada em metros.
A face anterior da córnea é a principal superfície refratora do olho: pequenas
irregularidades que nela se verifiquem podem redundar em graves problemas para a
visão perfeita. A principal função do cristalino, segundo elemento refrator de importância
no sistema óptico do olho, está relacionada com sua capacidade de acomodação, ou seja,
com a propriedade de, mudando de forma (graças à contração e ao relaxamento da
musculatura ciliar), variar seu poder refrator. O cristalino permite, dessa maneira, uma
focalização perfeita da imagem sobre a fóvea, funcionando como o ajuste de foco de uma
máquina fotográfica.
Distinguem-se na retina três camadas de células, em que os corpos celulares dos
neurônios se agrupam densamente, separadas por duas camadas sinápticas, em que se
misturam prolongamentos de dendritos e axônios. A primeira camada é formada pelas
células fotorreceptoras, os cones e bastonetes. Os axônios da primeira camada fazem

104
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
sinapse com dendritos de células da segunda camada, as células bipolares. Os axônios
da segunda camada, por sua vez, fazem sinapse com os dendritos das células
ganglionares, da terceira camada celular.
Uma vez formada a imagem sobre a retina, essa luz estimulará os cones e os
bastonetes, da mesma forma que sensibilizaria a emulsão de um filme ou os elementos
fotossensíveis de uma câmara de televisão. Os elementos fotossensíveis da retina
contêm um pigmento, que, no caso dos bastonetes, é a rodopsina, proteína dotada de um
grupamento cromatóforo. Estimulada pela luz, essa substância desencadeia um complexo
de reações químicas que culminará com a despolarização da célula receptora, a ativação
das células bipolares e ganglionares e o aparecimento de uma informação, no nervo
óptico, sob a forma de impulso nervoso.
Os campos visuais de cada olho são diferentes, mas se superpõem em parte. A retina
divide-se em zonas, a interna (nasal) e a externa (temporal), em função do trajeto das vias
ópticas que se dirigem para o córtex cerebral. As fibras nervosas das duas zonas
temporais (olhos direito e esquerdo) passam para o córtex cerebral do hemisfério
correspondente, enquanto as das regiões nasais cruzam-se no quiasma óptico, indo para
o córtex cerebral do hemisfério oposto.
A luz que emana de um objeto visualizado atinge a zona temporal de um globo
ocular e a zona nasal do outro. Em função da disposição das vias ópticas, a atividade
nervosa resultante vai para o mesmo hemisfério cerebral. A superposição de campos
visuais permite ao cérebro uma interpretação estereoscópica, com percepção de altura,
largura e profundidade.
A capacidade de discriminação de cores pelo olho está relacionada com a
existência de três tipos de cones caracterizados pela presença de três diferentes
fotopigmentos. Ao que parece, esses fotopigmentos são sensíveis principalmente aos
comprimentos de luz azul, verde e vermelha. A teoria de Young-Helmholtz da visão a
cores explica a sensação das diversas cores como uma decorrência do fato de que cada
um desses pigmentos seria estimulado de forma diferente pela radiação luminosa e que
suas mensagens seriam interpretadas no cérebro de forma combinada. Uma luz amarela,
por exemplo, estimularia os receptores de vermelho e verde, mas exerceria pouca
influência no de azul. A sensação combinada seria, portanto, a do amarelo.

No estudo da fisiologia da visão, é importante o conceito de acuidade visual, que
consiste na capacidade do olho de reconhecer dois pontos no espaço, medida pelo
ângulo formado pelos dois pontos e o olho do observador. Dois pontos situados num
ângulo menor do que um certo limite aparecerão ao observador como um ponto único no
espaço. Normalmente, considera-se como valor médio da acuidade visual no homem um
ângulo de um minuto. O fenômeno é complexo e dele participam inúmeros fatores, entre
os quais: fatores ópticos, fatores retinianos (como o estado dos cones) e fatores
relacionados com o estímulo, como brilho, iluminação de fundo, duração etc.
Sempre que as imagens se formam corretamente na mancha amarela, a visão é
nítida, e o olho é considerado emetrope ou normal. Quando isso não ocorre, dizemos que
há defeito de visão. Dentre esses defeitos destacam-se a miopia, a hipermetropia, o
astigmatismo, o estrabismo e a presbiopia. Outros problemas de visão são o daltonismo,
a catarata e a conjuntivite. Na miopia a formação da imagem ocorre antes da retina,
porque o olho é anormalmente longo, os míopes enxergam mal de longe. Corrige-se esse
defeito com o uso de lentes (óculos ou lentes de contato) divergentes. Atualmente, já há
tratamento cirúrgico para olhos para míopes.
FIGURA 5 – Formação de uma imagem na miopia
105 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Na hipermetropia a formação da imagem ocorre, teoricamente, atrás da retina,
porque o olho é curto demais. Os hipermétropes enxergam mal de perto. O defeito é
corrigido com lentes convergentes.
FIGURA 6 – Formação de uma imagem na hipermetropia
O astigmatismo consiste em defeito na curvatura da córnea e mais raramente, do
cristalino. Em conseqüência, o olho não é capaz de distinguir, ao mesmo tempo, com a
mesma nitidez, linhas verticais e horizontais. Essa anomalia pode se somar à miopia ou à
hipermetropia.
FIGURA 7 – Formação de uma imagem na astigmatismo
106 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

107
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
1.4. Neurofisiologia das Sensações Térmicas
Em repouso, a exposição ao frio provoca duas respostas fisiológicas essenciais
que contribuem para a manutenção do equilíbrio térmico. Essas respostas são a
termogênese e a vasoconstrição periférica: estas levam ao aumento da produção de calor
e à diminuição da perda de calor, respectivamente. O aumento da termogênese durante a
exposição a baixas temperaturas é resultante do tremor, resposta fisiológica que aumenta
significativamente a taxa metabólica. Toda energia despendida pelo tremor é convertida
em calor.
O tremor pode aumentar a taxa metabólica em três vezes ou mais que o nível em
repouso. A resposta do tremor ao frio é controlada pelo centro termorregulador, localizado
no hipotálamo. Termorreceptores situados na pele, na medula espinhal e no cérebro
respondem a quedas tanto de temperatura cutânea como da interna e transmitem essas
informações ao hipotálamo que, por sua vez, emite a resposta apropriada. Em seres
humanos, a queda da temperatura central estimula muito mais o tremor que a queda da
temperatura cutânea. A diminuição na temperatura cutânea e interna também é
responsável pela vasoconstrição periférica, outra importante resposta do organismo ao
frio. A constrição dos vasos sangüíneos cutâneos provoca uma diminuição no seu fluxo à
pele, reduzindo o envio de calor para a pele e aumentando o efeito isolante dos tecidos
corporais. Além disso, o desvio de sangue para tecidos mais profundos ajuda a conservar
o calor, uma vez que as veias mais profundas dos membros estão próximas e paralelas
às artérias. Este é chamado "mecanismo contracorrente de troca de calor" que ajuda a
conservar o calor, pois este calor do sangue arterial mais quente é transferido ao sangue
venoso mais frio que está retornando à circulação central. É interessante notar que a
vasoconstrição periférica ocorre na maioria das partes do corpo, mas não nos vasos
superficiais da cabeça. Isto significa que grande parte do calor dissipado para o ambiente
durante a exposição ao frio é perdido através da cabeça, talvez 25% da perda total de
calor. As alterações fisiológicas que permitem aclimatarmo-nos ao frio costumam não ser
tão bem compreendidas como as alterações que acompanham exposição ao calor e a
grandes altitudes. Um exemplo da capacidade de aclimatação dos seres humanos à

108
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
exposição prolongada ao frio é a assim chamada "aclimatação isolante", observada nos
aborígenes australianos. Tornaram-se mais tolerantes ao dormirem nus ao relento,
aumentando a capacidade de vasoconstrição periférica muito mais de que pessoas não
aclimatadas, levando à queda da temperatura cutânea. Outro exemplo de aclimatação ao
frio é um maior isolamento secundário devido a um espessamento da pele. Além disso,
pode ocorrer uma "aclimatação metabólica", em que a produção de calor eleva-se após
exposição prolongada ao frio. Isso pode ser causado por um aumento na liberação de
hormônios da tiróide e/ou sensibilidade mais acentuada dos tecidos à norepinefrina; esses
dois tipos de hormônio elevam o consumo de oxigênio mitocondrial e a produção de calor.
Por último, sabe-se que roedores são capazes de aumentar a produção de calor através
de uma termogênese sem tremor, em que o tecido adiposo marrom, ao ser estimulado
pela norepinefrina, gera uma alta taxa de calor. Entretanto, não há provas concretas que
confirmem esse mecanismo em seres humanos adultos.
1.5. Neurofisiologia da Audição
O som é produzido por ondas de compressão e descompressão alternadas do ar.
As ondas sonoras propagam-se através do ar exatamente da mesma forma que as ondas
propagam-se na superfície da água. Assim, a compressão do ar adjacente de uma corda
de violino cria uma pressão extra nessa região, e isso, por sua vez, faz com que o ar um
pouco mais afastado se torne pressionado também. A pressão nessa segunda região
comprime o ar ainda mais distante, e esse processo repete-se continuamente até que a
onda finalmente alcança a orelha.
A orelha humana é um órgão altamente sensível que nos capacita a perceber e
interpretar ondas sonoras em uma gama muito ampla de freqüências (16 a 20.000 Hz -
Hertz ou ondas por segundo). A captação do som até sua percepção e interpretação é
uma seqüência de transformações de energia, iniciando pela sonora, passando pela
mecânica, hidráulica e finalizando com a energia elétrica dos impulsos nervosos que
chegam ao cérebro.
O pavilhão auditivo capta e canaliza as ondas para o canal auditivo e para o
tímpano. Já o canal auditivo serve como proteção e como amplificador de pressão
Quando se choca com a membrana timpânica, a pressão e a descompressão
alternadas do ar adjacente à membrana provocam o deslocamento do tímpano para trás e
para frente.
O centro da membrana timpânica conecta-se com o cabo do martelo. Este, por sua
vez, conecta-se com a bigorna, e a bigorna com o estribo. Essas estruturas, como já
mencionado anteriormente (anatomia da orelha média), encontram-se suspensas através
de ligamentos, razão pela qual oscilam para trás e para frente.
A movimentação do cabo do martelo determina também, no estribo, um movimento
de vaivém, de encontro à janela oval da cóclea, transmitindo assim o som para o líquido
coclear. Dessa forma, a energia mecânica é convertida em energia hidráulica.
FIGURA 8 – Estruturas que participam do mecanismo da audição
Os ossículos funcionam como alavancas, aumentando a força das vibrações
mecânicas e por isso, agindo como amplificadores das vibrações da onda sonora. Se as
ondas sonoras dessem diretamente na janela oval, não teriam pressão suficiente para
mover o líquido coclear para frente e para trás, a fim de produzir a audição adequada,
pois o líquido possui inércia muito maior que o ar, e uma intensidade maior de pressão
seria necessária para movimentá-lo. A membrana timpânica e o sistema ossicular
convertem a pressão das ondas sonoras em uma forma útil, da seguinte maneira: as
ondas sonoras são coletadas pelo tímpano, cuja área é 22 vezes maior que a área da
109 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

janela oval. Portanto, uma energia 22 vezes maior do que aquela que a janela oval
coletaria sozinha é captada e transmitida, através dos ossículos, à janela oval. Da mesma
forma, a pressão de movimento da base do estribo apresenta-se 22 vezes maior do que
aquela que seria obtida aplicando-se ondas sonoras diretamente à janela oval. Essa
pressão é, então, suficiente para mover o líquido coclear para frente e para trás.
À medida que cada vibração sonora penetra na cóclea, a janela oval move-se para
dentro, lançando o líquido da escala vestibular numa profundidade maior dentro da
cóclea. A pressão aumentada na escala vestibular desloca a membrana basilar para
dentro da escala timpânica; isso faz com que o líquido dessa câmara seja empurrado na
direção da janela oval, provocando, por sua vez, o arqueamento dela para fora. Assim,
quando as vibrações sonoras provocam a movimentação do estribo para trás, o processo
é invertido, e o líquido, então, move-se na direção oposta através do mesmo caminho, e a
membrana basilar desloca-se para dentro da escala vestibular.
FIGURA 9 – Movimento de líquido na cóclea
A vibração da membrana basilar faz com que as células ciliares do órgão de Corti
se agitem para frente e para trás; isso flexiona os cílios nos pontos de contato com a
membrana tectórica (tectorial). A flexão dos cílios excita as células sensoriais e gera
impulsos nas pequenas terminações nervosas filamentares da cóclea que enlaçam essas
células. Esses impulsos são então transmitidos através do nervo coclear até os centros
auditivos do tronco encefálico e córtex cerebral. Dessa forma, a energia hidráulica é
convertida em energia elétrica.
110 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

A flexão dos cílios nos pontos de contato com a membrana tectórica excita as
células sensoriais, gerando impulsos nervosos nas pequenas terminações nervosas
filamentares da cóclea que enlaçam essas células.
Um fenômeno chamado ressonância ocorre na cóclea para permitir que cada
freqüência sonora faça vibrar uma secção diferente da membrana basilar. Essas
vibrações são semelhantes àquelas que ocorrem em instrumentos musicais de corda.
Quando a corda de um violino, por exemplo, é puxada para um lado, fica um pouco mais
esticada do que o normal e esse estiramento faz com que se mova de volta na direção
oposta, o que faz com que a corda se torne esticada mais uma vez, mas agora na
direção oposta, voltando então à primeira posição. Esse ciclo repete-se várias vezes,
razão pela qual uma vez que a corda começa a vibrar, assim permanece por algum
tempo.
FIGURA 10 – Percepção da altura de um som
Quando sons de alta freqüência penetram na janela oval, sua propagação faz-se
apenas num pequeno trecho da membrana basilar, antes que um ponto de ressonância
seja alcançado. Como resultado, a membrana move-se forçosamente nesse ponto,
enquanto o movimento de vibração é mínimo por toda a membrana. Quando uma
freqüência média sonora penetra na janela oval, a onda propaga-se numa maior extensão
ao longo da membrana basilar antes da área de ressonância ser atingida. Finalmente,
uma baixa freqüência sonora propaga-se ao longo de quase toda a membrana antes de
111 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

112
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
atingir seu ponto de ressonância. Dessa forma, quando as células ciliares próximas à
base da cóclea são estimuladas, o cérebro interpreta o som como sendo de alta
freqüência (agudo), quando as células da porção média da cóclea são estimuladas, o
cérebro interpreta o som como de altura intermediária, e a estimulação da porção superior
da cóclea é interpretada como som grave.
1.6. Neurofisiologia da Olfação
O olfato humano é pouco desenvolvido se comparado ao de outros mamíferos. O
epitélio olfativo humano contém cerca de 20 milhões de células sensoriais, cada qual com
seis pêlos sensoriais (um cachorro tem mais de 100 milhões de células sensoriais, cada
qual com pelo menos 100 pêlos sensoriais). Os receptores olfativos são neurônios
genuínos, com receptores próprios que penetram no sistema nervoso central.
A cavidade nasal, que começa a partir das janelas do nariz, está situada em cima
da boca e debaixo da caixa craniana. Contém os órgãos do sentido do olfato, e é forrada
por um epitélio secretor de muco. Ao circular pela cavidade nasal, o ar se purifica,
umedece e esquenta. O órgão olfativo é a mucosa que forra a parte superior das fossas
nasais - chamada mucosa olfativa ou amarela, para distingui-la da vermelha - que cobre a
parte inferior.
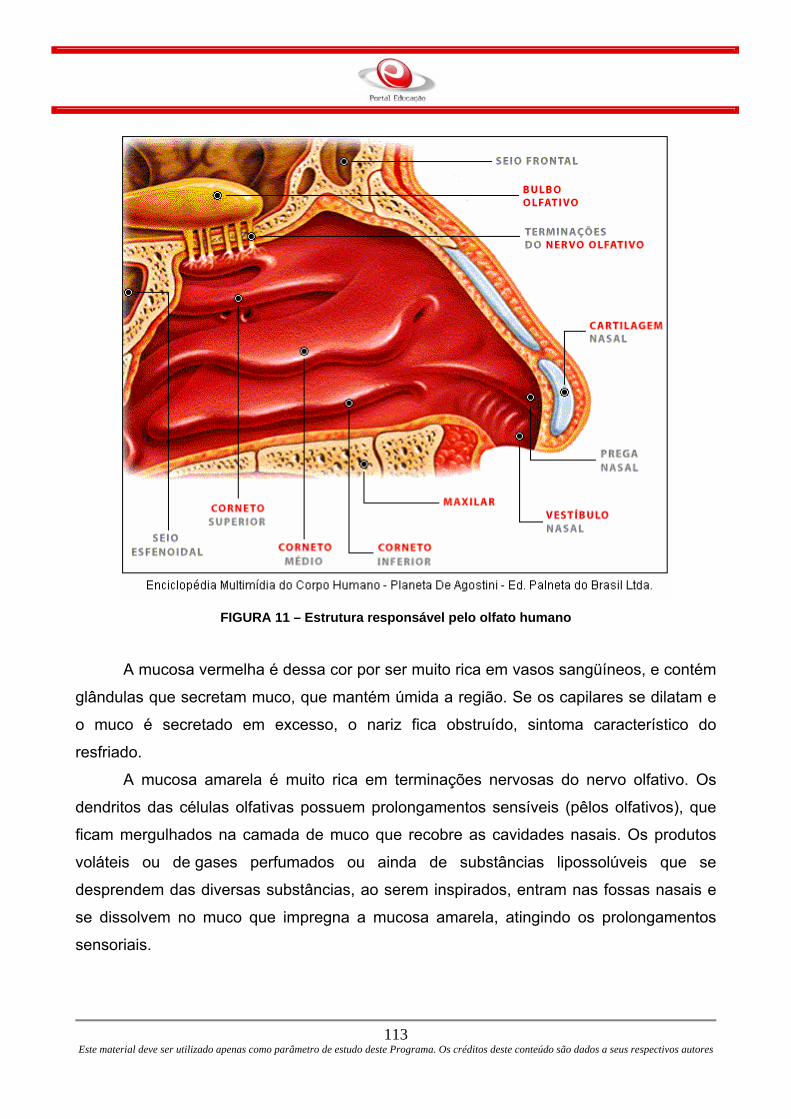
FIGURA 11 – Estrutura responsável pelo olfato humano
A mucosa vermelha é dessa cor por ser muito rica em vasos sangüíneos, e contém
glândulas que secretam muco, que mantém úmida a região. Se os capilares se dilatam e
o muco é secretado em excesso, o nariz fica obstruído, sintoma característico do
resfriado.
A mucosa amarela é muito rica em terminações nervosas do nervo olfativo. Os
dendritos das células olfativas possuem prolongamentos sensíveis (pêlos olfativos), que
ficam mergulhados na camada de muco que recobre as cavidades nasais. Os produtos
voláteis ou de gases perfumados ou ainda de substâncias lipossolúveis que se
desprendem das diversas substâncias, ao serem inspirados, entram nas fossas nasais e
se dissolvem no muco que impregna a mucosa amarela, atingindo os prolongamentos
sensoriais.
113 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

Dessa forma, geram impulsos nervosos, que são conduzidos até o corpo celular
das células olfativas, de onde atingem os axônios, que se comunicam com o bulbo
olfativo. Os axônios se agrupam de 10-100 e penetram no osso etmóide para chegar ao
bulbo olfatório, onde convergem para formar estruturas sinápticas chamadas glomérulos.
Estas se conectam em grupos que convergem para as células mitrais. Fisiologicamente
essa convergência aumenta a sensibilidade olfatória que é enviada ao Sistema Nervoso
Central (SNC), onde o processo de sinalização é interpretado e decodificado.
Aceita-se a hipótese de que existem alguns tipos básicos de células do olfato, cada
uma com receptores para um tipo de odor. Os milhares de tipos diferentes de cheiros que
uma pessoa consegue distinguir resultariam da integração de impulsos gerados por uns
cinqüenta estímulos básicos, no máximo. A integração desses estímulos seria feita numa
região localizada em áreas laterais do córtex cerebral, que constituem o centro olfativo.
FIGURA 12 – Região olfatória
A mucosa olfativa é tão sensível que poucas moléculas são suficientes para
estimulá-la, produzindo a sensação de odor. A sensação será tanto mais intensa quanto
maior for a quantidade de receptores estimulados, o que depende da concentração da
substância odorífera no ar.
O olfato tem importante papel na distinção dos alimentos. Enquanto mastigamos,
sentimos simultaneamente o paladar e o cheiro. Do ponto de vista adaptativo, o olfato tem
uma nítida vantagem em relação ao paladar: não necessita do contato direto com o objeto
114 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
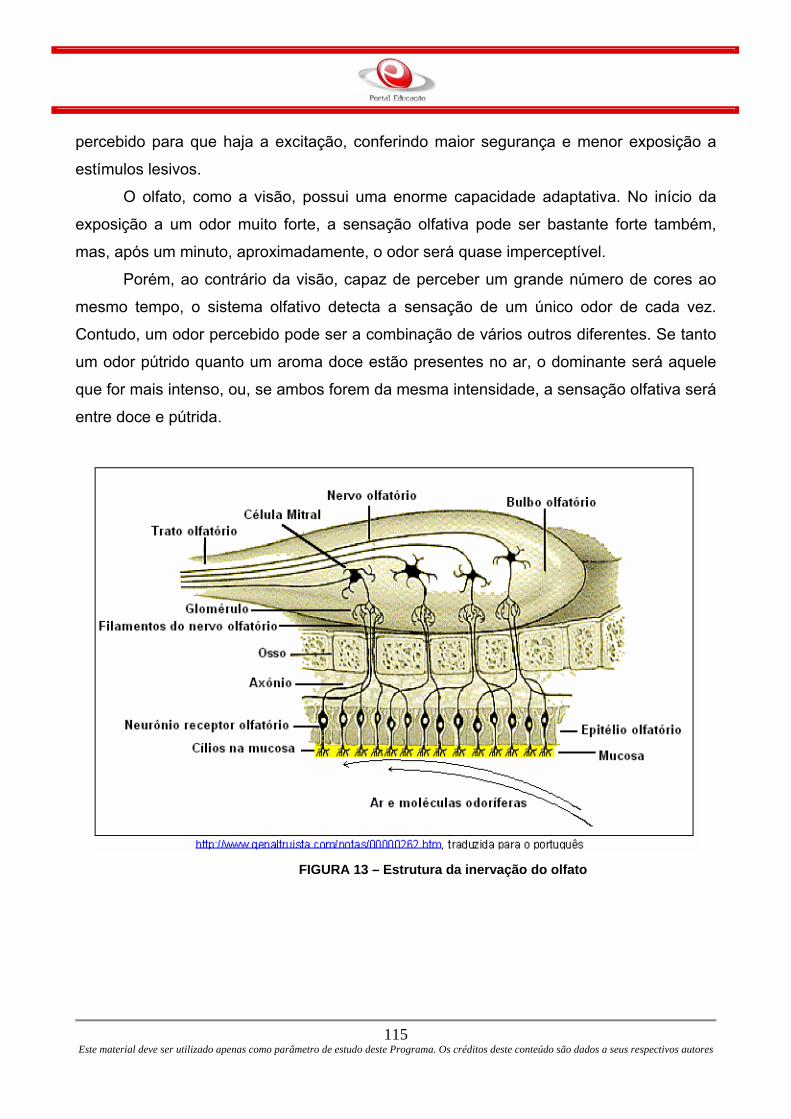
percebido para que haja a excitação, conferindo maior segurança e menor exposição a
estímulos lesivos.
O olfato, como a visão, possui uma enorme capacidade adaptativa. No início da
exposição a um odor muito forte, a sensação olfativa pode ser bastante forte também,
mas, após um minuto, aproximadamente, o odor será quase imperceptível.
Porém, ao contrário da visão, capaz de perceber um grande número de cores ao
mesmo tempo, o sistema olfativo detecta a sensação de um único odor de cada vez.
Contudo, um odor percebido pode ser a combinação de vários outros diferentes. Se tanto
um odor pútrido quanto um aroma doce estão presentes no ar, o dominante será aquele
que for mais intenso, ou, se ambos forem da mesma intensidade, a sensação olfativa será
entre doce e pútrida.
FIGURA 13 – Estrutura da inervação do olfato
115 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

1.7. Reflexos da Medula Espinhal
Nossa medula espinhal tem a forma de um cordão com aproximadamente 40 cm
de comprimento. Ocupa o canal vertebral, desde a região do atlas - primeira vértebra - até
o nível da segunda vértebra lombar. A medula funciona como centro nervoso de atos
involuntários e, também, como veículo condutor de impulsos nervosos.
Da medula partem 31 pares de nervos raquidianos que se ramificam. Por meio
dessa rede de nervos, a medula se conecta com as várias partes do corpo, recebendo
mensagens e vários pontos e enviando-as para o cérebro e recebendo mensagens do
cérebro e transmitindo-as para as várias partes do corpo. A medula possui dois sistemas
de neurônios: o sistema descendente controla funções motoras dos músculos, regula
funções como pressão e temperatura e transporta sinais originados no cérebro até seu
destino; o sistema ascendente transporta sinais sensoriais das extremidades do corpo até
a medula e de lá para o cérebro.
FIGURA 14 – Estrutura da medula espinhal no canal vertebral
Os corpos celulares dos neurônios se concentram no cerne da medula – na massa
cinzenta. Os axônios ascendentes e descendentes, na área adjacente – a massa branca.
As duas regiões também abrigam células da Glia. Dessa forma, na medula espinhal a
116 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

massa cinzenta localiza-se internamente e a massa branca, externamente (o contrário do
que se observa no encéfalo).
FIGURA 15 – Substância branca e substância cinzenta
Durante uma fratura ou deslocamento da coluna, as vértebras que normalmente
protegem a medula podem matar ou danificar as células. Teoricamente, se o dano for
confinado à massa cinzenta, os distúrbios musculares e sensoriais poderão estar apenas
nos tecidos que recebem e mandam sinais aos neurônios “residentes” no nível da fratura.
Por exemplo, se a massa cinzenta do segmento da medula onde os nervos rotulados C8
for lesada, o paciente só sofrerá paralisia das mãos, sem perder a capacidade de andar
ou o controle sobre as funções intestinais e urinárias. Nesse caso, os axônios levando
sinais para “cima e para baixo” através da área branca adjacente continuariam
trabalhando. Em comparação, se a área branca for lesada, o trânsito dos sinais será
interrompido até o ponto da fratura.
Infelizmente, a lesão original é só o começo. Os danos mecânicos promovem
rompimento de pequenos vasos sangüíneos, impedindo a entrega de oxigênio e
nutrientes para as células não afetadas diretamente, que acabam morrendo; as células
lesadas extravasam componentes citoplasmáticos e tóxicos, que afetam células vizinhas,
117 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

118
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
antes intactas; células do sistema imunológico iniciam um quadro inflamatório no local da
lesão; Células da Glia proliferam criando grumos e uma espécie de cicatriz, que impedem
os axônios lesados de crescerem e reconectarem. O vírus da poliomielite causa lesões na
raiz ventral dos nervos espinhais, o que leva à paralisia e atrofia dos músculos.
1.8. Sistema Nervoso Autônomo
O conjunto de nervos cranianos e raquidianos forma o sistema nervoso periférico.
Com base na sua estrutura e função, o sistema nervoso periférico pode subdividir-se em
duas partes: o sistema nervoso somático e o sistema nervoso autônomo ou de vida
vegetativa.
As ações voluntárias resultam da contração de músculos estriados esqueléticos,
que estão sob o controle do sistema nervoso periférico voluntário ou somático. Já as
ações involuntárias resultam da contração das musculaturas lisa e cardíaca, controladas
pelo sistema nervoso periférico autônomo, também chamado involuntário ou visceral.
O SNP Voluntário ou Somático tem por função reagir a estímulos provenientes do
ambiente externo. Ele é constituído por fibras motoras que conduzem impulsos do
sistema nervoso central aos músculos esqueléticos. O corpo celular de uma fibra motora
do SNP voluntário fica localizado dentro do SNC e o axônio vai diretamente do encéfalo
ou da medula até o órgão que inerva.
O SNP Autônomo ou Visceral, como o próprio nome diz, funciona
independentemente de nossa vontade e tem por função regular o ambiente interno do
corpo, controlando a atividade dos sistemas digestório, cardiovascular, excretor e
endócrino. Ele contém fibras nervosas que conduzem impulsos do sistema nervoso
central aos músculos lisos das vísceras e à musculatura do coração. Um nervo motor do
SNP autônomo difere de um nervo motor do SNP voluntário pelo fato de conter dois tipos
de neurônios, um neurônio pré-ganglionar e outro pós-ganglionar. O corpo celular do
neurônio pré-ganglionar fica localizado dentro do SNC e seu axônio vai até um gânglio,
onde o impulso nervoso é transmitido sinapticamente ao neurônio pós-ganglionar. O
corpo celular do neurônio pós-ganglionar fica no interior do gânglio nervoso e seu axônio

conduz o estímulo nervoso até o órgão efetuador, que pode ser um músculo liso ou
cardíaco.
O sistema nervoso autônomo compõe-se de três partes:
O Sistema Nervoso Autônomo (SNA) é composto por duas porções distintas:
Simpático e Parassimpático, cujas ações são antagônicas. Estas duas vertentes atuam
normalmente em simultâneo sendo do equilíbrio entre a força de ação de cada uma delas
(tônus) que nasce a extrema capacidade regulatória do SNA, essas ações estendem-se a
diversos domínios biofisiológicos do nosso organismo, incluindo o débito sanguíneo pelos
tecidos.
FIGURA 16 – Sistema nervoso simpático e parassimpático
O sistema nervoso autônomo divide-se em: sistema nervoso simpático e sistema
nervoso parassimpático. De modo geral, esses dois sistemas têm funções contrárias
(antagônicas). Um corrige os excessos do outro. Por exemplo, se o sistema simpático
acelera demasiadamente as batidas do coração, o sistema parassimpático entra em ação,
diminuindo o ritmo cardíaco. Se o sistema simpático acelera o trabalho do estômago e
dos intestinos, o parassimpático entra em ação para diminuir as contrações desses
órgãos.
119 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 17 – Comunicação das fibras simpáticas e parassimpáticas com o sistema nervoso central
O SN autônomo simpático, de modo geral, estimula ações que mobilizam energia,
permitindo ao organismo responder a situações de estresse. Por exemplo, o sistema
simpático é responsável pela aceleração dos batimentos cardíacos, pelo aumento da
pressão arterial, da concentração de açúcar no sangue e pela ativação do metabolismo
geral do corpo. O Simpático tem ação essencialmente vasoconstritora, mediante a
libertação do neurotransmissor norepinefrina (vasoconstritor) pelos seus botões terminais,
ao contrário do Parassimpático.
Já o SN autônomo parassimpático estimula principalmente atividades relaxantes,
como as reduções do ritmo cardíaco e da pressão arterial, entre outras do Parassimpático
que tem ação vasodilatadora mediante a libertação de acetilcolina.
1.9. Córtex Cerebral
A palavra córtex vem do latim para "casca". Isto porque o córtex é a camada mais
externa do cérebro. A espessura do córtex cerebral varia de 2 à 6 mm. O lado esquerdo e
direito do córtex cerebral são ligados por um feixe grosso de fibras nervosas chamado de
120 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

corpo caloso. Em mamíferos superiores (como humanos, primatas e cetáceos), o cérebro
tem uma superfície irregular, cheia de áreas mais protuberantes (chamadas de giros),
intercaladas com pequenos vales (chamados de sulcos) e que dão à aparência de
"enrugado" ao cérebro. O cérebro de mamíferos menos desenvolvidos como
camundongos e ratos tem poucos giros e sulcos, sendo a superfície cerebral mais lisa. As
principais funções são: pensamento, movimento voluntário, linguagem, julgamento e
percepção.
FIGURA 18 – Vista em corte transverso do córtex cerebral
O córtex cerebral, parte externa dos hemisférios, com somente poucos
milímetros de espessura, é composto de substância cinzenta, em contraste com o interior
do encéfalo, constituído parcialmente de substância branca. A substância cinzenta é
formada principalmente dos corpos das células nervosas e gliais, enquanto a substância
branca consiste predominantemente dos processos ou fibras dessas células.
O interior dos hemisférios cerebrais, incluindo o diencéfalo, contém não
somente substância branca mas também massas bem delimitadas de substância
cinzenta, coletivamente conhecidas como gânglios da base. Os mais destacados são os
núcleos caudado e lentiforme, e os tálamos. O interior do encéfalo encerra também
cavidades denominadas ventrículos, repletas de líquido cerebrospinal.
121 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

FIGURA 19 – Vista externa e interna do córtex cerebral
A substância cinzenta da superfície de cada hemisfério é denominada córtex
cerebral. Ela se encontra pregueada ou convolucionada em giros, que estão separados
uns dos outros pelos sulcos. O padrão é variável, e é necessário remover a pia-aracnóide
para a identificação individual dos giros e sulcos.
Uma porção do córtex cerebral denominada ínsula localiza-se profundamente ao
sulco lateral. A área do córtex imediatamente anterior ao sulco central é conhecida como
área motora e está relacionada com a atividade muscular, sobretudo a metade oposta do
corpo. O controle contralateral pode ser demonstrado por estimulação artificial desta
área, particularmente da região conhecida como giro pré-central ou área 4, tendo como
resultado movimentos na metade oposta do corpo. Além disso, o corpo está representado
numa posição invertida na área motora, isto é, a estimulação da parte superior da área
motora dá origem, predominantemente, a movimentos do membro superior, enquanto a
estimulação da parte inferior, dá origem a movimentos do membro inferior oposto e a
estimulação da parte média, a movimentos da cabeça e pescoço.
122 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

123
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A área do córtex imediatamente atrás do sulco central (o giro pós-central) é uma
área receptiva primária importante, para a qual se projetam vias aferentes por meio de
sinapses no tálamo. O córtex de cada hemisfério cerebral está dividido arbitrariamente em
lobos frontal, parietal, occipital e temporal.
2. FISIOLOGIA DO TRATO GASTROINTESTINAL 2.1. Controle Nervoso do Trato Gastrointestinal
O sistema digestivo inferior consiste do intestino delgado, do intestino grosso, do
reto, dos esfíncteres anais interno e externo e do canal anal. O intestino grosso armazena
o material residual. A intervalos regulares, não controlados, este resíduo é empurrado do
intestino grosso para o reto por uma série de ondas e contrações conhecidas como
movimentos peristálticos. O esfíncter anal interno, que é o músculo localizado abaixo do
reto, se abre automaticamente quando sente que existem fezes no reto. Nervos
localizados no canal anal mandam uma mensagem para o cérebro indicando que
devemos evacuar.

FIGURA 20 – Representação do sistema digestivo
O ato de defecar não ocorre naquele momento particular por causa do esfíncter
externo, um músculo que geralmente aprendemos a controlar entre as idades de um e
três anos. Quando é conveniente (por exemplo, quando estamos sentados no vaso
sanitário), este músculo se relaxa e uma combinação de contrações do reto e
revezamento dos músculos do estômago empurra as fezes para fora do corpo.
Num sistema digestivo inferior que funciona normalmente o esfíncter externo se
contrai quando o reto está cheio e retém as fezes no canal anal. No entanto, algumas
patologias como espinha bífida, uma vez que existe pouco ou nenhum controle sobre o
esfíncter anal externo, as fezes com freqüência saem em ocasiões inadequadas.
Sensibilidade limitada afeta a capacidade de perceber quando o reto está cheio e nervos
danificados impedem o cérebro de receber a mensagem para esvaziar o intestino. Se não
é percebido que o intestino está cheio e não existe controle do esfíncter externo então o
sistema digestivo inferior pode se abrir quando menos se espera.
124 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

125
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
2.2. Digestão e Absorção no Trato Gastrointestinal
O epitélio de revestimento do trato gastrointestinal é uma das interfaces do
organismo que, delimitando o meio externo do meio interno, permite intercâmbios
seletivos ao longo de toda sua extensão. No intestino, essas trocas se processam através
do seu epitélio transportador, por meio de fluxos resultantes de água e de solutos nos dois
sentidos: da luz intestinal (meio externo) para interstício e vasos (meio interno e vice-
versa. Os primeiros denominam-se fluxos absortivos, os segundos, secretores). O
delgado absorve os produtos da digestão protéica, glicídica e lipídica além de vitaminas. A
absorção e secreção de água e íons são geralmente acopladas aos processos de
absorção dos nutrientes orgânicos neste segmento intestinal. A digestão das proteínas
começa no estômago pela ação das pepsinas. As proteases pancreáticas rapidamente
clivam as proteínas no duodeno e jejuno para oligopeptídeos.
As enzimas clivadoras de peptídeos da borda-em-escova da membrana reduzem
oligopeptídeos para aminoácidos individuais e para dipeptídeos e tripeptídeos. Os
aminoácidos são levados para o enterócito por um conjunto de proteínas transportadoras
de aminoácidos na borda-em-escova da membrana. Dipeptídeos e tripeptídeos são
captados por uma proteína de transporte de peptídeos da borda-em-escova, com ampla
especificação. As α-amilases da saliva e do suco pancreático clivam o amido ramificado
em maltose, maltotriose e dextrinas α-limite.
Esses produtos da digestão são então reduzidos a moléculas de glicose por
glicoamilase e isomaltase, enzimas que ingerem carboidrato na borda-em-escova da
membrana plasmática. A borda-em-escova também contém as dissacaridases sucrase e
lactase, que clivam sucrose e lactose em monossacarídeos. Estes podem ser
transportados para os enterócitos pelas proteínas de transporte de monossacarídeos da
borda-em-escova da membrana. Um humano típico ingere 2L de água por dia, e cerca de
7L entram no aparelho gastrintestinal nas secreções gastrintestinais. Cerca de 99% da
água introduzida no aparelho gastrintestinal são absorvidos; aproximadamente 100mL de
água escapam nas fezes por dia. A absorção de água é potenciada pela absorção de íons
e nutrientes, predominantemente no intestino delgado. As células epiteliais maduras nas

126
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
pontas das vilosidades do intestino delgado são ativas na absorção de água e eletrólitos.
A absorção efetiva que normalmente ocorre no intestino delgado é o resultado de fluxos
secretórios e absortivos muito maiores
3. FISIOLOGIA DOS HORMÔNIOS
Dá-se o nome de sistema endócrino ao conjunto de órgãos que apresentam como
atividade característica a produção de secreções denominadas hormônios, que são
lançados na corrente sangüínea e irão atuar em outra parte do organismo, controlando ou
auxiliando o controle de sua função. Os órgãos que têm sua função controlada e/ou
regulada pelos hormônios são denominados órgãos-alvo.
Os tecidos epiteliais de secreção ou epitélios glandulares formam as glândulas, que
podem ser uni ou pluricelulares. As glândulas pluricelulares não são apenas aglomerados
de células que desempenham as mesmas funções básicas e têm a mesma morfologia
geral e origem embrionária - o que caracteriza um tecido. São na verdade órgãos
definidos com arquitetura ordenada. Elas estão envolvidas por uma cápsula conjuntiva
que emite septos, dividindo-as em lobos. Vasos sangüíneos e nervos penetram nas
glândulas, fornecendo alimento e estímulo nervoso para as suas funções.
Os hormônios influenciam praticamente todas as funções dos demais sistemas
corporais. Freqüentemente o sistema endócrino interage com o sistema nervoso,
formando mecanismos reguladores bastante precisos. O sistema nervoso pode fornecer
ao endócrino a informação sobre o meio externo, ao passo que o sistema endócrino
regula a resposta interna do organismo a esta informação. Dessa forma, o sistema
endócrino, juntamente com o sistema nervoso, atuam na coordenação e regulação das
funções corporais.
3.1. Hormônios da Hipófise
Situa-se na base do encéfalo, em uma cavidade do osso esfenóide chamada tela
túrcica. Nos seres humanos tem o tamanho aproximado de um grão de ervilha e possui
duas partes: o lobo anterior (ou adeno-hipófise) e o lobo posterior (ou neuro-hipófise).

127
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
Além de exercerem efeitos sobre órgãos não-endócrinos, alguns hormônios,
produzidos pela hipófise são denominados trópicos (ou tróficos) porque atuam sobre
outras glândulas endócrinas, comandando a secreção de outros hormônios. São eles:
Tireotrópicos: atuam sobre a glândula endócrina tireóide;
Adrenocorticotrópicos: atuam sobre o córtex da glândula endócrina adrenal
(supra-renal);
Gonadotrópicos: atuam sobre as gônadas masculinas e femininas;
Somatotrófico: atua no crescimento, promovendo o alongamento dos ossos e
estimulando a síntese de proteínas e o desenvolvimento da massa muscular.
Também aumenta a utilização de gorduras e inibe a captação de glicose plasmática
pelas células, aumentando a concentração de glicose no sangue (inibe a produção
de insulina pelo pâncreas, predispondo ao diabetes).
Localizado no cérebro diretamente acima da hipófise, o hipotálamo é conhecido por
exercer controle sobre ela por meios de conexões neurais e substâncias semelhantes a
hormônios chamados fatores desencadeadores (ou de liberação), o meio pelo qual o
sistema nervoso controla o comportamento sexual via sistema endócrino.
O hipotálamo estimula a glândula hipófise a liberar os hormônios gonadotróficos
(FSH e LH), que atuam sobre as gônadas, estimulando a liberação de hormônios
gonadais na corrente sanguínea. Na mulher a glândula-alvo do hormônio gonadotrófico é
o ovário; no homem, são os testículos. Os hormônios gonadais são detectados pela
pituitária e pelo hipotálamo, inibindo a liberação de mais hormônio pituitário, por feed-
back.

FIGURA 21 – Controle do hipotálamo sobre a hipófise
Como a hipófise secreta hormônios que controlam outras glândulas e está
subordinada, por sua vez, ao sistema nervoso, pode-se dizer que o sistema endócrino é
subordinado ao nervoso e que o hipotálamo é o mediador entre esses dois sistemas.
FIGURA 22 – Controle hormonal no organismo
128 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

129
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
O hipotálamo também produz outros fatores de liberação que atuam sobre a
adeno-hipófise, estimulando ou inibindo suas secreções. Produz também os hormônios
ocitocina e ADH (antidiurético), armazenados e secretados pela neuro-hipófise.
3.2. Hormônios da Tireóide
Localiza-se no pescoço, estando apoiada sobre as cartilagens da laringe e da
traquéia. Seus dois hormônios, triiodotironina (T3) e tiroxina (T4), aumentam a velocidade
dos processos de oxidação e de liberação de energia nas células do corpo, elevando a
taxa metabólica e a geração de calor. Estimulam ainda a produção de RNA e a síntese de
proteínas, estando relacionados ao crescimento, maturação e desenvolvimento. A
calcitonina, outro hormônio secretado pela tireóide, participa do controle da concentração
sangüínea de cálcio, inibindo a remoção do cálcio dos ossos e a saída dele para o plasma
sangüíneo, estimulando sua incorporação pelos ossos.
As células foliculares tireoideanas sintetizam, durante todo o tempo, uma proteína,
na qual se formam e armazenam os hormônios tireoideanos. Esta proteína se chama
tireoglobulina e é formada por uma cadeia de aminoácidos tirosina. Mas para que se
formem os hormônios tireoideanos não basta uma normal produção de tireoglobulina.
Também é de fundamental importância uma captação de íon iodeto, necessário à
formação dos hormônios.
A captação de iodeto se faz através de um transporte ativo (bomba de iodeto), que
bombeia contantemente estes íons do exterior para o interior das células foliculares,
armazenando uma concentração cerca de 40 vezes maior no líquido intracelular. Mas os
íons iodetos devem ser transformados na forma elementar de iodo no interior das células,
para que possam ser utilizados na formação dos hormônios. Isso se faz com a importante
ajuda de uma enzima denominada peroxidase.
Na medida em que as moléculas de tireoglobulina vão sendo produzidas,
moléculas de iodo vão se ligando quimicamente aos radicais tirosina das proteínas. Mas
para que as moléculas de iodo se liguem com a devida rapidez e em quantidade
satisfatória, se faz necessário a presença de uma enzima, a iodinase, que cataliza a
reação do iodo com os radicais tirosina das tireoglobulinas.

As moléculas de tireoglobulina, conforme vão sendo produzidas, vão saindo da
célula e armazenando-se no interior dos folículos, submersas num líquido gelatinoso
denominado colóide.
Cada molécula de tireoglobulina carrega, portanto, vários radicais tirosina
impregnados com molécula de iodo. 2 radicais tirosina, ligados entre sí, com 2 íons
iodetos em cada uma de suas moléculas, reagem-se entre sí formando uma molécula de
tiroxina (T4); 2 radicais tirosina, ligados entre sí, sendo um com 2 íons iodeto e outro com
apenas 1 íon iodeto, reagem-se também entre sí formando uma molécula de
triiodotironina (T3).
Diante do exposto acima, podemos então imaginar que cada molécula de
tireoglobulina carrega vários hormônios tireoideanos (a maioria T4) em sua fórmula.
Portanto, podemos dizer que os hormônios tireoideanos armazenam-se no interior dos
folículos tireoideanos na forma de tireoglobulina.
FIGURA 23 – Influência do hipotálmo sobre os hormônios da tireóide
130 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

A face voltada para o interior do folículo, da célula folicular tireoideana, faz
constantemente o processo de pinocitose. Através da pinocitose, constantemente,
diversas moléculas de tireoglobulina acabam retornando para o citoplasma da célula,
desta vez carregando diversas moléculas de hormônio tireoideano em sua estrutura. No
interior da célula, a tireoglobulina sofre ação de enzimas proteolíticas. Como
conseqüência, a tireoglobulina se fragmenta em numerosos pedaços pequenos, liberando
os hormônios tireoideanos (T3 e T4) na circulação, através da outra face celular. Os
hormônios, então, ligam-se a proteínas plasmáticas e assim circulam em nossa rede
vascular, atingindo quase todas as células de nosso corpo.
São raros os tecidos que não sofrem uma ação direta ou mesmo indireta dos
hormônios tireoideanos. Sob seu estímulo, as células aumentam seu trabalho, sintetizam
mais proteínas, consomem mais nutrientes e oxigênio, produz mais gaz carbônico, etc.
3.3. Hormônios Cortiço-Supra-Renais
São duas glândulas localizadas sobre os rins, divididas em duas partes
independentes – medula e córtex - secretoras de hormônios diferentes, comportando-se
como duas glândulas. O córtex secreta três tipos de hormônios: os glicocorticóides, os
mineralocorticóides e os androgênicos.
FIGURA 24 –Localização da glândula adrenal
131 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

132
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A supra-renal (ou adrenal), localizada acima de cada rim, com dimensões
aproximadas de 5 cm. por 1 cm., apresenta 2 tecidos histologicamente e fisiologicamente
bem distintos: medula e córtex. A medula secreta adrenalina e nor-adrenalina e faz parte
do sistema nervoso autônomo (simpático).
Já a córtex, importante glândula endócrina, produz e secreta dezenas de
hormônios. Todos os hormônios secretados por este tecido são sintetizados a partir do
colesterol e pertencem, portanto, ao grupo dos hormônios esteróides.
Os diversos hormônios produzidos pela córtex da adrenal, de acordo com seus efeitos,
são divididos em grupos:
• Mineralocorticóides: atuam no metabolismo de minerais, principalmente no
controle dos íons sódio e potássio. O principal mineralocorticóide, responsável por
pelo menos 95% da função mineralocorticóide da supra-renal, é o hormônio
aldosterona. Outros mineralocorticóides bem menos importantes são:
desoxicorticosterona e corticosterona.
• Glicocorticóides: atuam no metabolismo dos carboidratos, proteínas e
gorduras. O principal hormônio deste grupo é o cortisol.
• Androgênios: produzem efeitos masculinizantes, semelhantes àqueles
produzidos pela testosterona, secretada em grande quantidade pelas gônadas
masculinas.
A córtex da adrenal é dividida em 3 camadas: zona glomerulosa, zona fasciculada
e zona reticular.
A aldosterona é produzida na zona glomerulosa; as zonas fasciculada e retitular
produzem cortisol e androgênios. É o principal mineralocorticóide, controla os níveis
plasmáticos dos íons sódio e potássio. Exerce seu efeito no túbulo contornado distal e no
ducto coletor do nefron, aumentando a reabsorção de sódio e a excreção de potássio.
Como este transporte é mais efetivo ao sódio do que ao potássio, mais cátions são
reabsorvidos do que excretados nestes segmentos distais do nefron. A reabsorção de
sódio provoca, por atração iônica, reabsorção também de cloretos. A reabsorção de sal
(NaCl), por sua vez, reabsorve água (por osmose). Portanto, um aumento na secreção de
aldosterona, pela supra-renal, promove nos túbulos renais um aumento na reabsorção de

133
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
sal e água. Um aumento na reabsorção de sal e água promove, como conseqüência, um
aumento no volume do líquido no compartimento extra-celular. Isto faz com que ocorra um
aumento no volume sanguíneo e no débito cardíaco. Como conseqüência ocorre também
um aumento na pressão arterial.
Existem diversos fatores que influenciam a secreção da aldosterona. Os principais são:
• Potássio: Um aumento no nível plasmático deste íon estimula a zona glomerulosa
a aumentar a secreção de aldosterona;
• Angiotensina: Também exerce um importante efeito estimulante na secreção de
aldosterona;
• Sódio: Quanto menor sua concentração no líquido extra-celular, maior é a
secreção de aldosterona;
• ACTH: Estimula principalmente a secreção de cortisol, mas exerce também um
pequeno efeito estimulador de aldosterona.
O cortisol exerce importantes efeitos no metabolismo dos carboidratos, proteínas e
gorduras. Além disso, estabiliza membrana de lisossomos, reduz a utilização da glicose
pelas células, reduz a glicogênese e aumenta a glicogenólise. Como conseqüência
aumenta a glicemia.
O cortisol faz com que as células, de um modo geral, reduzam a síntese de
proteínas e aumentem a lise das mesmas: Isso promove uma redução das proteínas e um
aumento na quantidade de aminoácidos circulantes. No fígado ocorre o contrário:
aumento na síntese e redução na lise protéica. Como conseqüência, aumento na
quantidade de proteínas plasmáticas. O cortisol aumenta a mobilização de ácidos graxos
dos tecidos adiposos e a utilização das gorduras pelas células para produção de energia
e estabiliza a membrana dos lisossomas, dificultando seu rompimento durante uma lesão
tecidual.
Os hormônios androgênicos são bastante semelhantes ao hormônio masculino
testosterona e são secretados, felizmente, em quantidades bastante baixas. Quando
ocorre uma hiper-secreção anormal destes hormônios em crianças ou em mulheres,
efeitos masculinizantes como puberdade precoce, pilificação, calvície em pessoas
hereditariamente predispostas, voz mais grave, aumento nas dimensões do pênis ou
clítoris, além de outras alterações podem ser observadas.

3.4. Hormônios do Pâncreas
O pâncreas é uma glândula mista ou anfícrina – apresenta determinadas regiões
endócrinas e determinadas regiões exócrinas (da porção secretora partem dutos que
lançam as secreções para o interior da cavidade intestinal) ao mesmo tempo. As
chamadas ilhotas de Langerhans é a porção endócrina, onde estão as células que
secretam os dois hormônios: insulina e glucagon, que atuam no metabolismo da glicose.
FIGURA 25 – Metabolismo do pâncreas
A Insulina é o hormônio responsável pela redução da glicemia (taxa de glicose no
sangue), ao promover o ingresso de glicose nas células. Ela também é essencial no
consumo de carboidratos, na síntese de proteínas e no armazenamento de lipídios
(gorduras).
134 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

135
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
É produzida nas ilhotas de Langerhans, células do pâncreas endócrino. Ela age em
uma grande parte das células do organismo, como as células presentes em músculos e
no tecido adiposo, apesar de não agir em células particulares como as células nervosas.
Quando a produção de insulina é deficiente, a glicose se acumula no sangue e na
urina, matando as células de fome: é a diabetes mellitus. Para pacientes nessa condição,
a insulina é provida através de injeções, ou bombas de insulina. Recentemente foi
aprovado o uso de insulina inalada. Ancia de saude britanica não recomenda seu uso.
A insulina é um polipeptídeo de estrutura química plenamente conhecida, e pode
ser sintetizada a partir de diversos animais. Mais recentemente, surgiram os
medicamentos análogos de insulina, que não são propriamente a insulina em si, mas
moléculas de insulina modificadas em laboratório
A insulina é sintetizada nos humanos e em outros mamíferos dentro das células-
beta das ilhotas de Langerhans, no pâncreas. Um a três milhões de ilhotas de Langerhans
formam a parte endócrina do pâncreas, que é principalmente uma glândula exócrina. A
parte endócrina totaliza apenas 2% da massa total do órgão. Dentro das ilhotas de
Langerhans, as células-beta constituem 60-80% do todo. As ações da insulina no
metabolismo humano como um todo incluem: Controle da quantidade de certas
substâncias que entra nas células, principalmente glicose nos tecidos muscular e adiposo
(que são aproximadamente 2/3 das células do organismo); Aumento da replicação de
DNA e de síntese de proteínas via o controle de fornecimento de aminoácidos; e
Modificação da atividade de inúmeras enzimas (controle alostérico)
O Glucagon é um hormônio polipeptídeo produzido nas células alfa das ilhotas de
Langerhans do pâncreas e também em células espalhadas pelo trato gastrointestinal. São
conhecidas inúmeras formas de glucagon, sendo que a forma biologicamente ativa tem 29
aminoácidos.
A palavra glucagon deriva de gluco, glucose (glicose) e agon, agonista, ou agonista
para a glicose. Sua ação mais conhecida é aumentar a glicemia, contrapondo-se aos
efeitos da insulina. O glucagon age na conversão do ATP (trifosfato de adenosina) a
AMP-cíclico, composto importante na iniciação da glicogenólise, com imediata produção e
liberação de glicose pelo fígado.
Em condições normais, a ingestão de glicose suprime a secreção de glucagon. Há
aumento dos níveis séricos de glucagon durante o jejum. A secreção de glucagon é

estimulada por aminoácidos e alguns peptídeos gastrointestinais; sua secreção é inibida
pela somatostatina e por ácidos graxos livres.
3.5. Hormônios da Paratireóide
São pequenas glândulas, geralmente em número de quatro, localizadas na região
posterior da tireóide. Secretam o paratormônio, que estimula a remoção de cálcio da
matriz óssea (o qual passa para o plasma sangüíneo), a absorção de cálcio dos alimentos
pelo intestino e a reabsorção de cálcio pelos túbulos renais, aumentando a concentração
de cálcio no sangue. Neste contexto, o cálcio é importante na contração muscular, na
coagulação sangüínea e na excitabilidade das células nervosas.
O paratormônio é o hormônio da paratireóide que regula a taxa de cálcio,
estimulando a remoção de cálcio da matriz óssea (o qual passa para o plasma
sangüíneo), a absorção de cálcio dos alimentos pelo intestino e a reabsorção de cálcio
pelos túbulos renais, aumentando a concentração de cálcio no plasma.
FIGURA 26 – Metabolismo da tireóide e paratireóide
136 Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores

137
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
4. FISIOLOGIA DA REPRODUÇÃO
4.1. Hormônios Masculinos
Os testículos da criança permanecem inativos até que são estimulados entre 10 e
14 anos pelos hormônios gonadotróficos da glândula hipófise (pituitária). O Hipotálamo
libera fatores liberadores dos hormônios gonadotróficos que fazem a hipófise liberar fsh
(hormônio folículo estimulante) e lh (hormônio luteinizante).
O FSH estimula a espermatogênese pelas células dos túbulos seminíferos e o LH
estimula a produção de testosterona pelas células intersticiais dos testículos à
características sexuais secundárias, elevação do desejo sexual.
A testosterona faz com que os testículos cresçam. Ela deve estar presente,
também, junto com o folículo estimulante, antes que a espermatogênese se complete.
Depois que um feto começa a se desenvolver no útero materno, seus testículos começam
a secretar testosterona, quando tem poucas semanas de vida apenas. Essa testosterona,
então, auxilia o feto a desenvolver órgãos sexuais masculinos e características
secundárias masculinas. Isto é, acelera a formação do pênis, da bolsa escrotal, da
próstata, das vesículas seminais, dos ductos deferentes e dos outros órgãos sexuais
masculinos. Além disso, a testosterona faz com que os testículos desçam da cavidade
abdominal para a bolsa escrotal; se a produção de testosterona pelo feto é insuficiente, os
testículos não conseguem descer; permanecem na cavidade abdominal. A secreção da
testosterona pelos testículos fetais é estimulada por um hormônio chamado gonadotrofina
coriônica, formado na placenta durante a gravidez. Imediatamente após o nascimento da
criança, a perda de conexão com a placenta remove esse feito estimulador, de modo que
os testículos deixam de secretar testosterona. Em conseqüência, as características
sexuais interrompem seu desenvolvimento desde o nascimento até à puberdade. Na
puberdade, o reaparecimento da secreção de testosterona induz os órgãos sexuais
masculinos a retomar o crescimento. Os testículos, a bolsa escrotal e o pênis crescem,
então, aproximadamente mais 10 vezes.
Além dos efeitos sobre os órgãos genitais, a testosterona exerce outros efeitos
gerais por todo o organismo para dar ao homem adulto suas características distintivas.
Faz com que os pêlos cresçam na face, ao longo da linha média do abdome, no púbis e

138
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
no tórax. Origina, porém, a calvície nos homens que tenham predisposição hereditária
para ela. Estimula o crescimento da laringe, de maneira que o homem, após a puberdade
fica com a voz mais grave. Estimula um aumento na deposição de proteína nos músculos,
pele, ossos e em outras partes do corpo, de maneira que o adolescente do sexo
masculino se torna geralmente maior e mais musculoso do que a mulher, nessa fase.
Algumas vezes, a testosterona também promove uma secreção anormal das glândulas
sebáceas da pele, fazendo com que se desenvolva a acne pós-puberdade na face.
Na ausência de testosterona, as características sexuais secundárias não se
desenvolvem e o indivíduo mantém um aspecto sexualmente infantil.
4.2. Hormônios Femininos
A pituitária (hipófise) anterior das meninas, como a dos meninos, não secreta
praticamente nenhum hormônio gonadotrópico até à idade de 10 a 14 anos. Entretanto,
por essa época, começa a secretar dois hormônios gonadotrópicos. No inicio, secreta
principalmente o hormônio foliculo-estimulante (FSH), que inicia a vida sexual na menina
em crescimento; mais tarde, secreta o harmônio luteinizante (LH), que auxilia no controle
do ciclo menstrual.
O Hormônio Folículo-Estimulante causa a proliferação das células foliculares
ovarianas e estimula a secreção de estrógeno, levando as cavidades foliculares a
desenvolverem-se e a crescer. Já o Hormônio Luteinizante aumenta ainda mais a
secreção das células foliculares, estimulando a ovulação.
Os dois hormônios ovarianos, o estrogênio e a progesterona, são responsáveis
pelo desenvolvimento sexual da mulher e pelo ciclo menstrual. Esses hormônios, como os
hormônios adrenocorticais e o hormônio masculino testosterona, são ambos compostos
esteróides, formados, principalmente, de um lipídio, o colesterol. Os estrogênios são,
realmente, vários hormônios diferentes chamados estradiol, estriol e estrona, mas que
têm funções idênticas e estruturas químicas muito semelhantes. Por esse motivo, são
considerados juntos, como um único hormônio.
O estrogênio induz as células de muitos locais do organismo, a proliferar, isto é, a
aumentar em número. Por exemplo, a musculatura lisa do útero, aumenta tanto que o

139
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
órgão, após a puberdade, chega a duplicar ou, mesmo, a triplicar de tamanho. O
estrogênio também provoca o aumento da vagina e o desenvolvimento dos lábios que a
circundam, faz o púbis se cobrir de pêlos, os quadris se alargarem e o estreito pélvico
assumir a forma ovóide, em vez de afunilada como no homem; provoca o
desenvolvimento das mamas e a proliferação dos seus elementos glandulares, e,
finalmente, leva o tecido adiposo a concentrar-se, na mulher, em áreas como os quadris e
coxas, dando-lhes o arredondamento típico do sexo. Em resumo, todas as características
que distinguem a mulher do homem são devido ao estrogênio e a razão básica para o
desenvolvimento dessas características é o estímulo à proliferação dos elementos
celulares em certas regiões do corpo.
O estrogênio também estimula o crescimento de todos os ossos logo após a
puberdade, mas promove rápida calcificação óssea, fazendo com que as partes dos
ossos que crescem se "extingam" dentro de poucos anos, de forma que o crescimento,
então, pára. A mulher, nessa fase, cresce mais rapidamente que o homem, mas pára
após os primeiros anos da puberdade; já o homem tem um crescimento menos rápido,
porém mais prolongado, de modo que ele assume uma estatura maior que a da mulher, e,
nesse ponto, também se diferencia os dois sexos.
O estrogênio tem, outrossim, efeitos muito importantes no revestimento interno do
útero, o endométrio, no ciclo menstrual.
a progesterona tem pouco a ver com o desenvolvimento dos caracteres sexuais
femininos; está principalmente relacionada com a preparação do útero para a aceitação
do embrião e à preparação das mamas para a secreção láctea. Em geral, a progesterona
aumenta o grau da atividade secretória das glândulas mamárias e, também, das células
que revestem a parede uterina, acentuando o espessamento do endométrio e fazendo
com que ele seja intensamente invadido por vasos sangüíneos; determina, ainda, o
surgimento de numerosas glândulas produtoras de glicogênio. Finalmente, a progesterona
inibe as contrações do útero e impede a expulsão do embrião que se está implantando ou
do feto em desenvolvimento.
O ciclo menstrual na mulher é causado pela secreção alternada dos hormônios
folículo-estimulante e luteinizante, pela pituitária (hipófise) anterior (adenohipófise), e dos
estrogênios e progesterona, pelos ovários. O ciclo de fenômenos que induzem essa
alternância tem a seguinte explicação:

140
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
1. No começo do ciclo menstrual, isto é, quando a menstruação se inicia, a pituitária
anterior secreta maiores quantidades de hormônio folículo-estimulante juntamente
com pequenas quantidades de hormônio luteinizante. Juntos, esses hormônios
promovem o crescimento de diversos folículos nos ovários e acarretam uma
secreção considerável de estrogênio (estrógeno);
2. Acredita-se que o estrogênio tenha, então, dois efeitos seqüenciais sobre a
secreção da pituitária anterior. Primeiro, inibiria a secreção dos hormônios folículo-
estimulante e luteinizante, fazendo com que suas taxas declinassem a um mínimo
por volta do décimo dia do ciclo. Depois, subitamente a pituitária anterior começaria
a secretar quantidades muito elevadas de ambos os hormônios, mas principalmente
do hormônio luteinizante. É essa fase de aumento súbito da secreção que provoca o
rápido desenvolvimento final de um dos folículos ovarianos e a sua ruptura dentro de
cerca de dois dias;
3. O processo de ovulação, que ocorre por volta do décimo quarto dia de um ciclo
normal de 28 dias, conduz ao desenvolvimento do corpo lúteo ou corpo amarelo, que
secreta quantidade elevada de progesterona e quantidades consideráveis de
estrogênio;
4. O estrogênio e a progesterona secretados pelo corpo lúteo inibe novamente a
pituitária anterior, diminuindo a taxa de secreção dos hormônios folículo-estimulante
e luteinizante. Sem esses hormônios para estimulá-lo, o corpo lúteo involui, de modo
que a secreção de estrogênio e progesterona cai para níveis muito baixos. É nesse
momento que a menstruação se inicia, provocada por esse súbito declínio na
secreção de ambos os hormônios;
5. Nessa ocasião, a pituitária anterior, que estava inibida pelo estrogênio e pela
progesterona, começa a secretar outra vez grandes quantidades de hormônio
folículo-estimulante, iniciando um novo ciclo. Esse processo continua durante toda a
vida reprodutiva da mulher.

141
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
5. FISIOLOGIA DO EXERCÍCIO
5.1. Sistema Anaeróbio Alático
O sistema Anaeróbio alático, também conhecido como sistema fosfagênio ou
sistema ATP-CP, representa uma fonte imediata de energia para o músculo ativo.
Atividades que exigem altos índices de energia durante breve período de tempo
dependem basicamente, da geração de ATP a partir das reações enzimáticas do sistema.
O ATP necessário à contração do músculo está disponível tão rapidamente, porque esse
processo de geração de energia requer poucas reações químicas, não requer oxigênio e
o ATP e o PC estão armazenados e disponíveis no músculo. Este é o processo menos
complicado de gerar ATP.
A fosfocreatina (PC) tem uma cadeia de fosfato de alta energia, como a do ATP,
que também é chamada fosfagênio (daí o nome " sistema fosfagênio" ). O PC se
decompõe na presença da enzima creatina fosfoquinase e a energia liberada é utilizada
para formar o ATP, a partir do ADP.
PC ------------------ C + P + Energia P + Energia + ADP------------ ATP
Esta reação enzimática " ligada bioquimicamente " continuará até que se esgotem
os depósitos de fosfocreatina do músculo. O sistema ATP-CP fornece energia para as
contrações, durante os primeiros segundos do exercício.
5.2. Sistema Anaeróbio Lático
Esse sistema metabólico gera o ATP para necessidades energéticas
intermediárias; ou seja, as que duram de 45 -90 segundos, tendo como exemplo
atividades tipo: corridas de 400-800 m. , provas de natação de 100-200 m., também
proporcionando energia para piques de alta intensidade no futebol, róquei no gelo,
basquetebol, voleibol, tênis, badmington e outros esportes. O denominador comum
dessas atividades é a sustentação de esforço de alta intensidade e não ultrapassam os
dois minutos.

142
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
A glicólise anaeróbica, assim como o sistema ATP-CP, não requer oxigênio e
envolve a quebra incompleta do carboidrato em ácido lático. O corpo transforma os
carboidratos em açúcares simples, a "glicose", usada imediatamente ou depositada no
fígado e no músculo, como glicogênio. A glicose anaeróbia refere-se à quebra do
glicogênio na ausência do oxigênio. Esse processo é mais complicado quimicamente do
que o sistema ATP-CP e requer uma série mais longa de reações químicas. O sistema
ácido lático talvez seja bem mais lento do que o sistema fosfagênio, porém produz
quantidades mais altas de ATP (3 contra 1 do sistema fosfagênio), com a formação do
ácido lático, produto desse sistema, a produção pode nem chegar a 3. Quando o ácido
lático chega ao músculo e ao sangue, provoca a fadiga ou, até, uma falência muscular.
O sistema de ácido lático, ou glicose anaeróbia, não requer oxigênio; gera como
subproduto o ácido lático, que causa fadiga muscular; usa somente carboidratos; e libera
aproximadamente duas vezes mais ATP do que o sistema fosfagênico.
O sistema ácido lático também proporciona uma fonte rápida de energia, a glicose.
Ele é a primeira fonte para sustentar exercícios de alta intensidade. O principal fator
limitante na capacidade do sistema não é a depleção de energia, mas o acúmulo de
lactato no sangue. A maior capacidade de resistência ao ácido lático de um indivíduo é
determinado pela habilidade de tolerar esse ácido. A principal fonte de energia desse
sistema é o carboidrato.
5.3. Sistema Aeróbio
Este sistema fornece uma quantidade substancial de ATP, utiliza o oxigênio para
gerar o ATP e é ativado para produzir energia, durante períodos mais longos do
exercício. Fornece energia para exercícios de intensidade baixa para moderada.
Atividades como dormir, descansar, sentar, andar e outros. Quando a atividade vai se
tornando um pouco mais intensa a produção de ATP fica por parte do sistema ácido lático
e ATP-CP. Atividades mais intensas como caminhada, ciclismo,fazer compras e trabalho
em escritório também são supridas em parte pelo sistema aeróbico, até que a intensidade
atinja o nível moderado-alto (acima de 75%-85% da Frequência Cardíaca Máxima),
depois é recrutado para suprir energia suplementar.

143
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
Os melhores exemplos de exercícios que recrutam o sistema aeróbio são: aulas de
aeróbica e hidroginástica de 40-60 min., corridas mais longas que 5000 m., natação (mais
que 1500 m.), ciclismo (mais que 10 km.), caminhada e triathlon. Qualquer atividade
sustentada continuamente em um mínimo de 5 min. pode ser considerada aeróbia.
O ATP liberado da quebra da glicose e/ou dos ácidos graxos, em presença de O²,
custa centenas de reações químicas complexas, que envolvem centenas de enzimas. A
quebra ocorre num compartimento especializado da célula muscular, a mitocôndria. As
mitocôndrias são consideradas as "usinas energéticas" da célula e são capazes de
fornecer grandes quantidades de ATP para alimentar as contrações musculares.
O sistema aeróbio possui 3 fases. A quebra do glicogênio na presença do O², ou
glicólise anaeróbia, discutida acima, e a glicólise aeróbia é que o O² evita o acúmulo de
ácido lático. O glicogênio e os ácidos graxos são duas principais fontes de combustível
utilizadas no sistema metabólico aeróbio. Ocasionalmente a proteína pode ser também
usada como fonte de combustível metabólico, mas ocorre quando o corpo está
fisiologicamente desgastado por excessos, por dietas ou por níveis extremamente baixos
de gordura e glicogênio. Em suma, o O² ou sistema metabólico aeróbio requer grande
quantidade de O² para aconverter o glicogênio em 39 moléculas de ATP e os ácidos
graxos, em 130 moléculas de ATP. O ácido graxo ou glicogênio são quebrados e
preparados par ao ciclo de Krebs e o transporte de elétrons e, como resultado do
processo, temos CO², H²O e energia. O CO² evapora; a água é eliminada através da
evaporação e da radiação; e a energia é usada na segunda parte da reação ligada, para
sintetizar o ATP.
---------- FIM MÓDULO IV ----------
----------FIM DO CURSO ----------

144
Este material deve ser utilizado apenas como parâmetro de estudo deste Programa. Os créditos deste conteúdo são dados a seus respectivos autores
BIBLIOGRAFIA CONSULTADA 1. Anthony, C.P.; Thibodeau, G.A. – Textbook of Anatomy and Physiology. C.V. Mosby Co. St. Louis,1979. 2. Assumpção, C.R. – Súmula de Anatomo-Fisiologia. Coração, sangue e pulmões. Introdução à Circulação Extracorpórea. Módulo Teórico No 1. SBCEC, Rio de Janeiro, 1985. 3. Barner, H.B. – Cardiovascular Function andPhysiology. Glenn, W.W.L.; Baue, A.E.; Geha, A.S.;Hammond, G.L; Laks, H. – Thoracic and Cardiovascular Surgery. 4th. ed. Appleton-Century-Crofts, Norwalk, 1983. 4. Berger, E.C. – What is sequestration ? J. Can. Society of Extracorporeal Technology. 3, 20-25, 1974. 5. Berger, E.C. – The physiology of Adequate Perfusion. The C.V. Mosby Co., St. Louis, 1979. 6. Best, C.H. – Best and Taylor’s Phyisiological Basis of Medical Practice. Williams & Wilkins, Baltimore, 1979. 7. De Wall, R.A.; Lillehei, C.W.; Sellers, R.D. – Hemodilution perfusions for open-heart surgery. N. Engl. J. Med. 266, 1078-1084, 1962. 8. De Wall, R.A. – What constitutes adequate perfusion? J. Thorac. Cardiovasc. Surg. 16, 458-466, 1975. 9. Doty, D.B. – Venous Compliance DuringCardiopulmonary Bypass. In Utley, J.R. –Pathophysiology and Techniques of Cardiopulmonary bypass. Vol. II. Williams &Wilkins, Baltimore, 1983. 10. Fulton, G.P.; Zweifach, B.W. – Factors Regulating Blood Flow. American Physiological Society. Washington, 1958. 11. Galletti, P.M.; Brecher, G.A. – Heart-Lung Bypass. Principles and Techniques of Extracorporeal Circulation. Grune & Stratton, New York, 1962. 12. Gollan, F. – Physiology of Cardiac Surgery. Charles C. Thomas, Springfield, 1959. 13. 20. Good, A.E. – The cellular aspect of shock. J. Extracorp. Technol. 4, 18-23, 1972. 14. Gordon, R.J.; Ravin, M.; Rawitscher, R.E.; Daicoff, G.R. – Changes in arterial pressure, viscosity, and resistance during cardiopulmonary bypass. J. Thorac. Cardiovasc. Surg. 69, 552-558, 1975. 15. Grant, B. – Grant’s Atlas of Anatomy. 5th. edition.























![EAD-Enfermagem a Distância-Material Do Curso[Fisiologia Geral]](https://img.document.onl/doc/110x75/577c86d71a28abe054c2cad9/ead-enfermagem-a-distancia-material-do-cursofisiologia-geral.jpg)



