Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
INSTITUTO DE QUÍMICA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA DE ALIMENTOS
Análise da diversidade microbiana de grãos de kefir, caracterização da bebida
fermentada e potencial probiótico das estirpes isoladas
Aluna: Analy Machado de Oliveira Leite
Orientadores: Vânia Margareth Flosi Paschoalin
Joab Trajano Silva
Co-orientador: Marco Antônio Lemos Miguel
Rio de Janeiro
2012

ANALY MACHADO DE OLIVEIRA LEITE
Análise da diversidade microbiana de grãos de kefir, caracterização da
bebida fermentada e potencial probiótico das estirpes isoladas
Tese de Doutorado apresentada ao Programa de Pós-
Graduação em Ciência de Alimentos, Instituto de Química,
Universidade Federal do Rio de Janeiro, como parte dos
requisitos necessários à obtenção do título de Doutor em
Ciência de Alimentos.
Orientadores: Prof. Dra Vânia M. Flosi Paschoalin
Prof. Dr. Joab Trajano Silva
Co-orientador: Prof. Dr. Marco Antonio Lemos Miguel
Rio de Janeiro
2012

Leite, Analy Machado de Oliveira.
Análise da diversidade microbiana de grãos de kefir,
caracterização da bebida fermentada e potencial probiótico das
estirpes isoladas/ Analy Machado de Oliveira Leite - Rio de Janeiro:
UFRJ, 2012. 236 f.: Orientadora: Vânia Margareth Flosi Paschoalin Orientador: Joab Trajano Silva Co-orientador: Marco Antônio Lemos Miguel Tese (Doutorado) – Universidade Federal do Rio de Janeiro,
Instituto de Química, 2012.
1. Grãos de kefir 2. kefir 3.Comunidade microbiana 4. Potencial probiótico – Teses. I.Paschoalin, Vânia M. Flosi (Orient.). II. Silva, Joab Trajano. III Miguel,Marco Antonio Lemos. IV Universidade Federal do Rio de Janeiro. Instituto de Química. Programa de Pós-Graduação em Ciência de Alimentos. V. Título.

Analy Machado de Oliveira Leite
Análise da diversidade microbiana de grãos de kefir, caracterização da bebida fermentada e potencial probiótico
das estirpes isoladas
Aprovada em
_______________________________________
(Vânia Margaret Flosi Paschoalin, Doutora – UFRJ/
Joab Trajano Silva, Doutor - UFRJ)
___________________________________________
(Marco Antônio Lemos Miguel, Doutor - UFRJ)
_________________________________________
(Selma Gomes Ferreira Leite, Doutora , UFRJ)
__________________________________________
(Raquel Regina Bonelli, Doutora - UFRJ)
__________________________________________
(Marcia Barreto Feijó, Doutora - UFF)
______________________________________________________
(Robson Maia Franco, Doutor – UFF)

Agradecimentos
A Deus, por me dar oportunidade de realizar este trabalho, proporcionar toda esta experiência
e estar sempre ao meu lado, guiando-me.
À minha família, em especial, minha mãe, por estar presente em todos os momentos, me
apoiando, incentivando, acreditando, comemorando cada conquista, como se fosse sua. Aos
meus tios Cleide e Renato que sempre estiveram próximos, me incentivando e apoiando. A
minha irmã, pela ajuda na revisão deste trabalho, por sempre torcer por mim e acreditar na
minha escolha. Ao meu pai que mesmo distante, sei que torce e se orgulha de mim. A minha
avó, que onde quer que esteja, estará sempre presente.
Aos meus orientadores, profa. Dr
a. Vânia Paschoalin e prof. Dr. Joab Trajano Silva, por
confiar em mim, apoiar minhas idéias e me dar à oportunidade de desenvolver este trabalho.
Obrigada pela paciência e disponibilidade da discussão dos resultados, pelo incentivo na
realização do doutorado sanduíche, pela orientação, dedicação nos momentos mais difíceis
desta tese e ótima convivência durante todos esses anos.
Ao meu co-orientador e amigo prof. Dr. Marco Miguel por sua companhia maravilhosa, por
estar sempre presente, por nossas longas conversas, por me fazer enxergar o mundo de uma
outra forma, pelos ensinamentos, por estar sempre disposto a ajudar, aconselhar, conversar,
apoiar, incentivar e discutir meus resultados.
À profa. Dr
a. Raquel Silva Peixoto e ao prof. Dr. Alexandre Soares Rosado por me receberem
e me acolherem no Laboratório de Ecologia Microbiana Molecular (LEMM), como se fosse
aluna do grupo. Obrigada por todo apoio, conversas, conselhos e oportunidade em adquirir
novas experiências.
Aos pesquisadores Dr. Baltasar Mayo e Dra. Susana Delgada pela oportunidade dada em
trabalhar no Instituto de Productos Lacteos de Asturias (IPLA), por todos os ensinamentos,
pelo apoio, pela atenção e ótima recepção na Espanha.

A todos da Faculdade de Farmácia da UFF, especialmente do Departamento de Bromatologia
de Alimentos, pela ótima convivência durante o meu contrato de professor Substituto, por
apoiarem e incentivarem o meu estágio sanduíche na Espanha.
Ao prof. Dr. Sergio Seabra, ao técnico Eliandro Lima e ao aluno Laidson Paes Gomes pela
ajuda técnica e apoio nas análise de Microscopia Eletrônica de Varredura dos grãos.
A todos os amigos do laboratório 545, novos e antigos Eduardo Eustáquio, Bruna Bellei,
Giselli Asensi, Laidson Paes (Antunes), Patricia, Fernanda Pinhati, Luciana, Raquel, Thiago
Alvarez, Sabrina, Filipe, Felipe, Tiyoko, Karine, Carlos, Nathalia, Ana Letícia, Rodrigo, Ana
Carolina, Claudius, ao técnico Ricardo Bretas e ao primo Rafael Luiz por conviverem comigo
durante todos esses anos da melhor forma possível, pelas trocas de informações, pelas
conversas, desabafos, almoços, experimentos, por me apoiarem e me ouvirem nos momentos
difíceis. À minha querida aluna de iniciação científica Bianca Fernandes Arruda, por me
ajudar nos momentos mais estressantes deste trabalho, por estar sempre disposta a ajudar,
contribuindo em grande parte dos resultados. Ao Dr. Eduardo Mere por toda paciência, por
todas as brigas, momentos de descontração, orientação, discussões, correções e ajuda.
Aos amigos do laboratório de Microbiologia de Alimentos, Renata, Priscila, Leonisa, Carol,
Marcela, Juliana, Aline, Bia, Duda, Ana, Henrique, Max, Tayná por toda ajuda, carinho,
almoços, conversas, e ótima convivência durante todos esses anos. Em especial ao técnico
Antonio por toda disponibilidade, paciência, apoio, carinho, ensinamentos e ajuda nos meus
experimentos.
Aos amigos do LEMM pela recepção, por todas as conversas, almoços, churrascos,
aprendizado e companheirismo. Em especial gostaria de agradecer a minha prima postiça
Deborah Leite por sua disposição em me ajudar desde o início, por sua simpatia, sua
dignidade, pelos experimentos, idéias, discussão dos resultados, carinho e apoio em todos os
momentos. Ao aluno Caio Rachid por toda paciência, por todo apoio, conversas,
ensinamentos, ajudas nos experimentos e análises bioinformática, pelas discussões dos
resultados e por todos os ótimos momentos de convivência. A Regina Macedo por suas sábias
palavras nos momentos mais desesperadores, a Luiza Lessa por sua companhia maravilhosa e
pela amizade construída em tão pouco tempo.

A todos do IPLA pela recepção maravilhosa, por toda convivência nos sete meses na
Espanha, por toda preocupação, por me deixarem tão à vontade, por toda atenção e
companhia. Em especial a técnica Alicia Noriega pela amizade, companheirismo, pelos
jantares, calimochos, saídas, conversas e ajudas nos experimentos. Aos alunos Luca Losurdo,
Marutpong Panya, Ángel Alegría, Elena Fernandes pela ótima convivência no laboratório e
pela ajuda, amizade e paciência.
A todas as minhas amigas maravilhosas Fernanda Vasconcellos, Nádia Vidal, Daniele Fasano,
Manuela Dias, Bruna Bellei, Giselli Asensi, Thaiz Rangel, Patrícia Baptista, Daniela Catarina,
Ana Vater, Erica Castro, Veronica Catão, Thais Lin, Flavia Sousa, Clara Slade e Juliana Sá
que estiveram sempre ao meu lado, me apoiando e incentivando. Em especial, à Luciana
Romão, por todos os conselhos, conversas, ajudas e revisões, sempre com muita dedicação e
boa vontade.
A Capes pela bolsa de doutorado e doutorado sanduíche concedida (BEX 5019/10-9).
Ao CNPq e FAPERJ pela apoio financeiro.
A todos que de alguma forma contribuíram para realização deste trabalho. Muito obrigada!!

“Tu te tornas eternamente responsável
por aquilo que cativas”
“Mas o vaidoso não ouviu.
Os vaidosos só ouvem os elogios”
O Pequeno Príncipe
Antoine de Saint Exùpèry

LEITE, Analy Machado de Oliveira. ANÁLISE DA DIVERSIDADE MICROBIANA DE
GRÃOS DE KEFIR, CARACTERIZAÇÃO DA BEBIDA FERMENTADA E POTENCIAL
PROBIÓTICO DAS ESTIRPES ISOLADAS. Rio de Janeiro, 2012. Tese (Doutorado em
Ciência de Alimentos) - Instituto de Química, Universidade Federal do Rio de Janeiro, Rio de
Janeiro, 2012
Resumo
Kefir é uma bebida láctea fermentada por uma comunidade microbiana mista presente no grão
de kefir. A microbiota de grãos brasileiros e da bebida fermentada foi analisada por PCR-
DGGE (denaturing gradient gel electrophoresis), pirosequenciamento, e ARDRA (amplified
ribosomal DNA restriction analysis); os metabólitos da fermentação foram analisados por
cromatografia líquida de alta eficiência; além da avaliação do potencial probiótico das cepas
isoladas. Lactobacillus kefiranofaciens, L. kefiri e Saccharomyces cerevisiae foram
encontradas majoritariamente em todos os grãos. Membros do gênero Lactobacillus
totalizaram 96% das espécies bacterianas, ao contrário de Lactococcus spp, Leuconostoc spp,
e Acetobacter spp, presentes em baixos níveis. Na bebida fermentada (24 h), Bactérias Ácido
Láticas (BAL), Bactérias Ácido Acéticas (BAA) e leveduras foram encontradas em 10, 7,8 e 6
log de UFC/mL, respectivamente. Durante a estocagem do kefir (28 dias/4°C), as contagens
de BAL e leveduras mantiveram-se constante, mas a contagem de BAA foi reduzida para 7,2
log UFC/mL. Os isolados do kefir foram identificados como L. lactis cremoris (45%), L.
lactis lactis (5%), L. mesenteroides (29%), A. lovaniensis (10%) e S. cerevisiae (11%).
Entretanto, as espécies predominantes identificadas a partir do sequenciamento das bandas
isoladas do PCR-DGGE foram L. kefiranofaciens, L. kefiri, L. parakefiri e S. cerevisiae,
indicando que a combinação de técnicas independentes e dependentes de cultivo foi
importante para a completa caracterização da microbiota. Durante a fermentação e estocagem
do kefir o pH (6,55-4,31), o teor de lactose (46,3/8,4 mg/mL) e ácido cítrico (4,18/2,89
mg/mL) foram reduzidos, enquanto que glicose (0,18/0,37 mg/mL), galactose (0,46/0,88
mg/mL), ácidos lático (0/9,53 mg/mL), acético (0/1,15 mg/mL), butírico (0/0,63 mg/mL),
propiônico (0,44/0,96 mg/mL) e etanol (0/1,36 mg/mL) foram aumentados. 33 BAL isoladas
dos grãos e classificadas como L. mesenteroides (52%), L. casei/paracasei (15%), L. lactis
lactis (9%) e L. lactis cremoris (24%) foram tolerantes ao pH ácido e a sais biliares, capazes
de inibir a proliferação de patógenos, sem apresentar atividade hemolítica ou enzimática
indesejável, e foram sensíveis à maioria dos antibióticos testados. Cinco isolados produziram
bacteriocinas e dois foram capazes de aderir às células Caco-2 de forma comparável ao
controle. Sendo assim, duas cepas de L. casei/paracasei mostraram-se adequadas para uso
como probiótico.
Palavras-chave: grãos de kefir, kefir, comunidade microbiana, potencial probiótico

LEITE, Analy Machado de Oliveira. MICROBIAL DIVERSITY ANALYSIS OF KEFIR
GRAINS, CHARACTERIZATION OF THE FERMENTED BEVERAGE AND
PROBIOTIC POTENTIAL OF ISOLATED STRAINS. Rio de Janeiro, 2012. Thesis
(Doctorate in Food Science) – Chemistry Institute, Universidade Federal do Rio de Janeiro,
Rio de Janeiro, 2012
Abstract
Kefir is a milk beverage fermented by a mixed microbial community found in kefir grains.
The microbiota of Brazilian grains and the fermented beverage were assessed by PCR-DGGE
(denaturing gradient gel electrophoresis), pyrosequencing, and ARDRA (amplified ribosomal
DNA restriction analysis); fermentation metabolites were analyzed by high performance
liquid chromatography, and the evaluation of the probiotic potential of isolated strains was
conducted. Lactobacillus kefiranofaciens, L. kefiri and Saccharomyces cerevisiae were the
predominant species found in all grains. Members of the Lactobacillus genus amounted to
96% of the bacterial species, while Lactococcus spp, Leuconostoc spp and Acetobacter spp
were present in low levels. In the fermented beverage (fermented for 24 h), lactic acid bacteria
(LAB), acetic acid bacteria (AAB) and yeast were found in 10, 7.8 and 6 log CFU/ml,
respectively. During kefir storage (28 days at 4 °C), the LAB and yeast counts remained
constant, but the BAA count was reduced to 7.2 log CFU/ml. The isolated strains were
identified as L. lactis cremoris (45%), L. lactis lactis (5%), L. mesenteroides (29%), A.
lovaniensis (10%) and S. cerevisiae (11%). However, the predominant species identified by
DGGE band sequencing were L. kefiranofaciens, L. kefiri, L. parakefiri and S. cerevisiae,
indicating that the application of a polyphasic approach, using culture-dependent and -
independent techniques, was important for a complete microbiota characterization. During
fermentation and kefir storage pH (6.55 to 4.31), lactose (46.3 - 8.4 mg / ml) and citric acid
(4.18 - 2.89 mg / ml) content were reduced, while glucose (0.18 - 0.37 mg / mL), galactose
(0.46 - 0.88 mg / ml), lactic (0/9, 53 mg / ml), acetic (0.00 -1.15 mg / ml), butyric (0.00 - 0, 63
mg / ml) and propionic acids (0.44 / 0.96 mg / ml) and ethanol (0.00 - 1.36 mg / ml) were
increased. 33 LAB isolated from kefir grains and identified as L. mesenteroides (52%), L.
casei / paracasei (15%), L. lactis lactis (9%) and L. lactis cremoris (24%) were tolerant to
acidic pH and bile salts, capable of inhibiting pathogen proliferation without introducing
undesirable hemolytic or enzyme activity and were sensitive to most of the tested antibiotics.
Five isolates produced bacteriocins and two were able to adhere to the Caco-2 cells in a
manner comparable to the control. Thus, two L. casei/paracasei strains proved suitable for
use as probiotics.
Keywords: Kefir grains, kefir, microbial community, probiotic potential.

LISTA DE ILUSTRAÇÕES
Figura 1 – Grãos de Kefir 39
Figura 2 - Micrografia eletrônica de grão de kefir mostrando bactérias e leveduras na matriz
de polissacarídeo e proteína. (foto do autor) 42
Figura 3 – Esquema ilustrativo do sequenciamento na plataforma 454 55
Figura 4 – Desenho experimental 58
Figura 5 – Estrutura macroscópica de grãos de kefir 60
Figura 6 - Microscopia eletrônica de varredura da microbiota de grãos de kefir brasileiros. A,
C, E : porções externas; B,D,F: porções internas. Setas: micrografia A: cocos; micrografia
D:material fibrilar – polissacarídeo kefiran; micrografia E: seta 1- material granular – proteína
coagulada, seta 2 – diferentes leveduras 63
Figura 7 - Produção artesanal do kefir. (1) Grãos de kefir são adicionados ao leite (2), e
mantidos a temperatura ambiente por 18-24 h (3); Após a fermentação, são coados (4) e o
leite fermentado (kefir) está pronto para consumo (5) e os grãos (1), podem ser reutilizados 66
Figura 8 - Fluxograma de produção da bebida kefir. (A) Método russo; (B) Método com
culturas puras (bebida “tipo kefir”). 67
Figura 9 - Perfis do DGGE da comunidade microbiana de três grãos de kefir brasileiros
(amostras AR, AD e AV). Painel A: Perfil de DGGE do gene codificador do RNAr 16S
bacteriano amplificado com iniciadores universais (1) Lactobacillus kefiranofaciens; (2)
Lactobacillus kefiranofaciens; (3) Lactococcus lactis; (4) Lactobacillus kefiri; (5)
Lactobacillus kefiranofaciens. Painel B: Perfil de DGGE do domínio eucarioto D1 do gene
codificador do RNAr 26S (6) Saccharomyces cerevisiae; (7) Kazachstania unispora. Painel C:
Perfil DGGE do gene codificador do RNAr 16S bacteriano amplificado com iniciadores
específico para grupo de lactobacilos (8) Lactobacillus kefiranofaciens; (9) Lactobacillus
kefiri. Painel D: Perfil de DGGE do gene codificador do RNAr 16S amplificado com
iniciadores específicos para grupo lactococos (10) Lactococcus lactis 77
Figura 10 - Curvas de rarefação das sequências parciais do gene rrs de grãos de kefir
brasileiros (AD, AR e AV) a 97% de similaridade. (A) e curvas de rarefação normalizadas
com relação ao menor número de sequências das amostra (B) 78
Figura 11 - Diagrama de Venn com disposição das OTU comuns e específicas em grãos de
kefir brasileiros AD, AR e AV, e o percentural de ocorrência das sequências totais (entre
parênteses) 79
Figura 12 - Abundância relativa em nível de família, baseada na classificação parcial das
sequencias parciais do gene rrs bacteriano de grãos de kefir brasileiros AD, AR e AV, usando
o RDP-Classifier. 80
Figura 13 - Árvore filogenética bacteriana mostrando leituras representativas da análise de
pirosequenciamento. A árvore foi construída utilizando o algoritmo neighbor joining com
sequências representativas para cada OTU selecionda pelo programa MOTHUR. Números
nos nós indicam valor de bootstrap (expressos como percentual de 1000 repetições). Valores
nos parênteses representam o número de sequências encontrada para cada OTU. Símbolos e
cores representam o grupo de amostras que contribuíram para cada OTU. 81
Figura 14 - Enumeração microbiana (log10 ufc/mL) presuntiva de BAL (M17 and MRS), de
BAA (AAB) e levedura (YGC) na bebida kefir brasileira ao longo dos períodos de
fermentação e estocagem (tempo 0 = após inoculação dos grãos). Dados são a média de três
produções da bebida kefir, barras de erro mostram o desvio padrão. (*) indica diferença
significativa (p < 0.05) entre a contagem de células viáveis durante a fermentação e
estocagem. 90

Figura 15 - Perfis obtidos com a técnica de PCR-DGGE da comunidade microbiana das
amostras de leite, grão de kefir e bebidas kefir coletadas durante diferentes períodos de
fermentação e estocagem. Painel A: Dendrogramas de similaridade gerados pela análise de
perfis de bandas obtidos pelo DGGE da comunidade de bactéria (A.1) e de leveduras (A.2),
usando a correlação de Dice e agrupamento UPGMA. A barra de escala indica o percentual de
similaridade. Painel B: Perfis de bandas obtidos com os produtos de PCR analisadas da região
V3 do gene codificador do RNAr 16S para bactéria (B.1) e da região D1 do gene codificador
da RNAr 26S para leveduras (B.2). A identificação das bandas está descrita na tabela 2. 92
Figura 16 - Perfis de bandas obtidos por PCR-DGGE (DNA) e RT-PCR-DGGE (RNA) da
região codificadora do RNAr 16S de bactérias (A) e do RNAr 26S de leveduras (B) do grão
de kefir utilizado como inóculo para fabricação da bebida kefir e das amostras da bebida kefir
durante diferentes períodos de fermentação e estocagem. A identificação das bandas está
descrita na tabela 2. 93
Figura 17 - Atividade antagônica e sensibilidade à proteólise de estirpes de L. lactis isoladas
de grãos de kefir. Painel A: Inibição de E. coli ATCC 25922 pelas estirpes L. lactis subsp.
cremoris MRS26 (1), MRS20 (2) e M171 (3). Painel B: Atividade antimicrobiana da estirpe
MRS26 contra o indicador Lb. sakei CECT 906 e sua sensibilidade à proteólise. Poços: 1 –
sobrenadante livre de células neutralizado da estirpe L. lactis subsp. cremoris MRS26 ; PK -
proteinase K (20mg/ml) e PN – pronase (20 mg/ml). 110
Figura 18 - Dendrograma de similaridade construído a partir da combinação dos perfis
obtidos com as técnicas rep-PCR e RAPD-PCR, usando os iniciadores BOXA1R, BOXA2R e
M13. 111
Figura 19 - Percentual de adesão (UFC/mL de bacteria aderida em relação à UFC/mL de
bacteria adicionada) dos quatro isolados de Lactobacillus case/paracasei à linhagem de célula
intestinal Caco-2. Colunas que não compartilham a mesma letra são estatisticamente
diferentes, de acordo com o teste de comparação de médias LSD (least significant differences)
(p<0,05). Lactobacillus rhamnosus GG foi utilizado como cepa de referência para avaliação
da capacidade de adesão. 112

LISTA DE QUADROS
Quadro 1 - Culturas de microrganismos utilizados na fabricação de alguns leites fermentados
.................................................................................................................................................. 22
Quadro 2 - Diferenciação dos principais gêneros de BAL ...................................................... 23 Quadro 3– Potenciais efeitos benéficos de alimentos funcionais contendo probiótico. ......... 29 Quadro 4 - Alegação de propriedade funcional e de saúde aprovada pela ANVISA para
produtos probióticos ................................................................................................................. 36 Quadro 5- Requisitos físico-químicos e microbiológicos do kefir ......................................... 37
Quadro 6 - Meios de cultura comumente empregados para isolamento e identificação de
bactérias de interesse presentes nos grãos e no kefir ................................................................ 43 Quadro 7 - Meios de cultura comumente empregados para isolamento e identificação de
leveduras de interesse presentes nos grãos e no kefir ............................................................... 43

LISTA DE TABELAS
Tabela 1 - Riqueza de OTU estimada, cobertura da amostragem e índice de diversidade das
bibliotecas do gene rrs de amostras de grãos de kefir brasileiros ............................................ 78 Tabela 2 – Identificação dos microrganismos baseados na comparação com as sequências
disponíveis no banco de dados do Genbank, após clonagem e sequenciamento das bandas
selecionadas do PCR e RT-PCR-DGGE, utilizando iniciadores universais para bactéria
(RNAr 16S) e leveduras (RNAr 26S). ...................................................................................... 91 Tabela 3 – Compostos químicos quantificados por CLAE durante a fermentação do kefir e
estocagem da bebida a 4°C. ...................................................................................................... 95 Tabela 4 - Caracterização de BAL isolada de grãos de kefir brasileiro ................................ 109
Tabela 5 - Espécies e origem das cepas de BAL isoladas de grãos de kefir brasileiros ........ 110 Tabela 6 - Atividades enzimáticas das estirpes de Lactobacillus casei/paracasei mensurada
com sistema API-ZYM (bioMérieux) .................................................................................... 112 Tabela 7 - Concentração minima inibitória (MIC) em µg/mL de vários antibióticos frente a
estirpes de Lactobacillus por microdiluição em caldo. .......................................................... 113

LISTA DE SIGLAS E ABREVIATURAS
µL: Microlitros
16S rRNA: Subunidade 16S do RNA ribossomal
Ácido lático D(-): Ácido lático dextrógiro
Ácido lático L(+): Ácido lático levógiro
ANOVA: Analysis of Variance, Análise de variância
ANVISA: Agência Nacional de Vigilância Sanitária
APM: Acetobacter peroxydans medium
APS: Adenosine 5'- Phosphosulfate, Adenosina 5’- Fosfosulfato
ARDRA: Amplified Ribosomal DNA Restriction Analysis, Análise de restrição do DNA
ribosomal amplficado
ATP: Adenosine Triphosphate, Adenosina trifosfato
BAA: Bactéria Ácido Acética ou AAB: Acetic Acid Bacteria
BAL: Bactéria Ácido Lática ou LAB: Lactic Acid Bacteria
BHI: Brain Heart Infusion, infusão cérebro e coração
CCD: Charge-Coupled Device, dispositivo de carga acoplado
CLAE: Cromografia Líquida de Alta Eficiência
CO2: Dióxido de carbono
d.C.: depois de Cristo
DGGE: Denaturing Gradient Gel Eletrophoresis, Eletroforese em Gel de Gradiente
Desnaturante .
DNA : Deoxyribonucleic Acid , Ácido Desoxirribonucléico
dNTP: Deoxynucleotide Triphosphate, Desoxiribonucleotídeo Tri-fosfato
ECA: Enzima Conversora de Angiotensina-I
EDTA: Ethylenediamine Tetraacetic Acid, Ácido Etilenodiamino Tetra-Acético.
EFSA: European Food Safety Authority
EPS: Exopolissacarídeo
ERIC: Enterobacterial Repetitive Intergenic Consensus, consenso intergênico repetitivo
FAO: Food and Agriculture Organization
FEEDAP: Panel on Additives and Products or Substances used in Animal Feed
GC: conteúdo de Guanina e Citosina
GOS: Galacto-Oligossacarídeos
h: horas
IPTG: isopropyl-β-D-1-thiogalactopyranoside;
ITS: Internal Transcribed Spacer, espaçador não-codificante interno
KCA: Potassium Carboxymethyl Cellulose Agar
kDa: kilo dalton
KPL: Kefir grain polysaccharide-producing lactobacilli
L: litro
LAW: Lactic Acid Whey
log: unidade logarítmica
MAPA: Ministério da Agricultura Pecuária e Abastecimento
MEA: Malt Extract Agar
MEV: Microscopia Eletrônica de Varredura
mg/mL: Miligramas por Mililitro
MgCl2: Cloreto de Magnésio
MIC: Minimum Inhibitory Concentration
mL: mililitros

mM: milimolar
MRS: Man, Rogosa and Sharpe
MSE: Mayex, Sandine and Elliker
m/v: massa/volume
Nm: Nanômetros oC: Graus Celsius
OGYE: Oxytetracicline GlucoseYeast Extract
OTU: Operational Taxonomic Unit, Unidade Taxonômica Operacional
p/p: peso/peso
pb ou bp: pares de base, base pairs
PBS: Phosphate Buffered Saline, Solução tampão fosfato
PCR: Polimerase Chain Reaction , Reação em Cadeia de Polimerase.
pH: Potencial de Hidrogênio
PPi: pirofosfato
QPS: Qualified Presumption Safety, qualificado presumidamente como seguro
RAPD: Random Amplification of Polymorphic DNA, amplificação aleatória de DNA
polimórfico
REP: Repetitive Extragenic Palindromic Elements, elementos palindrômicos extragênicos
repetitivos
rep-PCR: repetitive element sequence based PCR, PCR de seqüências repetitivas de DNA
RNA: ácido ribonucleico, ribonucleic acid
RNAr: ácido desoxirribonucléico ribossomal, ribossomal deoxyribonucleic acid
rrs: gene que codifica a subunidade 16S do RNA ribossomal
RTIQ: Regulamento Técnico de Identidade e Qualidade
RT-PCR: reverse transcription PCR, PCR por transcrição reversa
TAE: Tris Acetato EDTA
TAE: Tris, ácido acético e EDTA
TBE: Tris Borato EDTA
TGI: Trato Gastrintestinal
T-RFLP: Terminal Restriction Fragment Length Polymorphism, polimorfismo de
comprimento de fragmentos de restrição terminal
UBA: Universal Beer Agar
UFC: Unidade Formadora de Colônia
UFLA: Universidade Federal de Lavras
UFRJ: Universidade Federal do Rio de Janeiro
UFV: Universidade Federal de Viçosa
UHT: Ultra-High-Temperature
UNIFAL: Univesidade Federal de Alfenas
UPGMA: Unweighted Pair Group Method with Arithmetic Mean, médias aritméticas entre
pares não ponderados.
UV: ultravioleta
WHO: World Health Organization
X-Gal: 5-bromo-4-chloro-3-indolyl-β-D-galacto-pyranoside
YEC: Yeast Extract Chloramphenicol
YELN: Yeast Extract Lactate + Naladixic acid
YEPG: Yeast Extract Peptone Glucose)
YGC: Yeast Glucose Chloramfenicol

SUMÁRIO
1 INTRODUÇÃO 19
2 REVISÃO BIBLIOGRÁFICA 22
2.1 LEITES FERMENTADOS 22
2.1.1 Papel dos microrganismos em alimentos fermentados 23
2.1.1.1 Bactérias Ácido Láticas (BAL) 23
2.1.1.1.1 Gênero Lactobacillus 24
2.1.1.1.2 Gênero Lactococcus 25
2.1.1.1.3 Gênero Leuconostoc 25
2.1.1.2 Bactérias Ácido Acéticas (BAA) 26
2.1.1.3 Leveduras 27
2.2 ALIMENTOS FUNCIONAIS 27
2.2.1 Probióticos 28
2.2.2 Efeitos benéficos dos probióticos 28
2.2.3 Critérios de seleção de bactérias probióticas 30
2.2.3.1 Aspectos funcionais 31
2.2.3.2 Aspectos de segurança 32
2.2.3.3 Aspectos tecnológicos 35
2.3 KEFIR 37
2.3.1 Definição 37
2.3.2 Aspectos gerais 37
2.3.3 Grãos de kefir 39
2.3.4 Aspectos microbiológicos dos grãos e do kefir 40
2.3.4.1 Bactérias 44
2.3.4.2 Leveduras 45
2.3.4.3 Interações entre microrganismos no kefir 45
2.3.5 Características do kefir 46
2.3.5.1 Químicas 46
2.3.5.2 Nutricionais 47
2.3.5.3 Compostos bioativos no kefir 48
2.3.5.4 Terapêuticas 49
2.4 MÉTODOS DEPENDENTES E INDEPENDENTES DE CULTIVO EMPREGADOS
NO ESTUDO DE COMUNIDADES MICROBIANAS 50
2.4.1 Métodos tradicionais 50
2.4.2 Métodos moleculares 50
2.4.2.1 Técnicas de fingerprinting de comunidades microbianas 51
2.4.2.2 Técnicas de sequenciamento dos membros da comunidade microbiana 53
4 OBJETIVOS 57
4.1 OBJETIVO GERAL 57
4.2 OBJETIVOS ESPECÍFICOS 57
5 ESTRATÉGIA EXPERIMENTAL 58
6 CAPÍTULOS 59
6.1 CAPÍTULO I - KEFIR: UMA BEBIDA PROBIÓTICA NATURAL 59
6.2 CAPÍTULO II - AVALIAÇÃO DA DIVERSIDADE MICROBIANA DE GRÃOS DE
KEFIR2 73
6.2.1 Material e Métodos 73
6.2.1.1 Amostras de grãos de kefir 73
6.2.1.2 Isolamento do DNA microbiano total 73
6.2.1.3 Análise de DGGE dos grãos de kefir 73

6.2.1.3.1 Amplificação por PCR de fragmentos do gene codificador do RNAr 16S e RNAr 26S
73
6.2.1.3.2 Condições de eletroforese e identificação das bandas 74
6.2.1.4 Análise de pirosequenciamento dos grãos de kefir 74
6.2.1.4.1 Iniciadores e condições de amplificação do gene rrs 74
6.2.1.4.2 Tratamento das sequências e análise de bioinformática 75
6.2.2 Resultados 76
6.2.2.1 Análise por PCR-DGGE das comunidades bacterianas e de leveduras 76
6.2.2.2 Composição bacteriana e estrutura da comunidade determinada por
pirosequenciamento 77
6.2.3 Discussão 82
6.3 CAPÍTULO III - CARACTERÍSTICAS MICROBIOLÓGICAS E QUÍMICAS DA
BEBIDA KEFIR DURANTE A FERMENTAÇÃO E ESTOCAGEM 84
6.3.1 Material e Métodos 84
6.3.1.1 Obtenção do grão de kefir 84
6.3.1.2 Produção do kefir 84
6.3.1.3 Análises microbiológicas 84
6.3.1.4 Análise de restrição do DNA ribossomal amplificado (ARDRA) 85
6.3.1.5 Extração de DNA e RNA da bebida kefir e grãos 86
6.3.1.6 Protocolo do PCR e RT-PCR 86
6.3.1.7 Análise do kefir por DGGE 87
6.3.1.8 Análise dos compostos químicos por CLAE 88
6.3.1.9 Análises estatísticas 89
6.3.2 Resultados 89
6.3.2.1 Enumeração microbiana durante a fermentação e estocagem da bebida kefir 89
6.3.2.2 Identificação dos isolados microbianos 90
6.3.2.3 Estrutura da comunidade microbiana 90
6.3.2.4 Análise dos metabólitos durante a fermentação e estocagem da bebida kefir 93
6.3.3 Discussão 96
6.4 CAPÍTULO IV - POTENCIAL PROBIÓTICO DAS ESTIRPES BACTERIANAS
ISOLADAS DE GRÃOS DE KEFIR 101
6.4.1 Material e métodos 101
6.4.1.1 Isolamento de bactérias 101
6.4.1.2 Sobrevivência dos isolados em pH ácido 101
6.4.1.3 Tolerância à bile das culturas 102
6.4.1.4 Identificação de BAL 102
6.4.1.5 Análise de tipificação molecular 103
6.4.1.6 Propriedades antimicrobianas 103
6.4.1.6.1 Inibição de patógenos 103
6.4.1.6.2 Produção de bacteriocina 104
6.4.1.7 Ensaio de adesão 105
6.4.1.8 Avaliação das propriedades de segurança 106
6.4.1.8.1 Produção de hemolisinas 106
6.4.1.8.2 Susceptibilidade a antimicrobianos 106
6.4.1.9 Atividades enzimáticas 106
6.4.2 Resultados 107
6.4.2.1 Tolerância ao pH e sais biliares 107
6.4.2.2 Identificação de BAL 107
6.4.2.3 Propriedades antimicrobianas 108
6.4.2.3.1 Atividade antimicrobiana frente a patógenos 108

6.4.2.3.2 Produção de substância semelhante à bacteriocina 108
6.4.2.4 Tipificação das estirpes de Lactobacillus casei/paracasei 111
6.4.2.5 Atividades enzimáticas 111
6.4.2.6 Teste de adesão 112
6.4.2.7 Resistência antimicrobiana e produção de hemolisina 113
6.4.3 Discussão 113
7 CONCLUSÕES FINAIS 118
REFERÊNCIAS 119
APÊNDICES 143

19
1 INTRODUÇÃO
O processo de fermentação foi descoberto acidentalmente milhares de anos atrás. A
primeira fermentação provavelmente ocorreu com a estocagem de excedentes de leite,
resultando em um produto fermentado 1. A origem das bebidas lácteas fermentadas é antiga, e
remontam à domesticação de certos mamíferos, como vaca, cabra e ovelha. Pode-se assumir
que o processo de fermentação ocorreu de forma acidental, uma vez que os leites eram
estocados à temperatura ambiente. Devido a variações climáticas e condições ambientais de
diferentes partes do mundo, estirpes específicas de microrganismos tornaram-se dominantes
nestes produtos e, como consequência, tipos específicos de fermentações evoluíram e
tornaram-se características de uma determinada região 2.
Há relatos do consumo de leites fermentados desde antes da era Fenício, no Oriente
Médio. Posteriormente, a tradição do leite fermentado foi disseminada por todo o leste
Europeu e Rússia pelos tártaros, hunos e mongóis durante suas conquistas. Isso ocorria, tanto
pelo potencial conservante do alimento, quanto pela variedade de sabores e características
sensoriais dos produtos 1. Como consequência, vários produtos populares ainda existem
nessas regiões e afirma-se que alguns, como o iogurte e o kefir, são originários dos Balcãs e
Europa Oriental 3.
A modulação da composição da microbiota intestinal para melhora da saúde tem sido
realizada empiricamente desde tempos remotos, com relatos do emprego de leite fermentado,
para o tratamento de infecções gastrointestinais, já no ano 76 d.C.4. No início do século XX,
foi sugerido que a humanidade não só havia feito uso de uma mistura de microrganismos para
elaboração e/ou conservação de uma parte substancial do seu abastecimento de alimentos,
como também existiam algumas bactérias que exerciam efeitos benéficos para a saúde dos
hospedeiros, especialmente quando estes sofriam de infecções intestinais. Então, Metchnikoff
3; 5, ganhador do prêmio Nobel em 1908, publicou o livro Prolongation of Life no qual
postulava que as bactérias que participavam da fermentação do iogurte, contribuíam para a
manutenção da saúde mediante a supressão das “fermentações putrefativas” da microbiota
intestinal, e que esta era a causa da longevidade dos camponeses búlgaros, grandes
consumidores de iogurte 3; 6
. Atualmente, sabe-se que somente iogurtes e leites fermentados
contendo microrganismos probióticos podem ser considerados alimentos funcionais 7; 8
.
Alimentos funcionais são aqueles que, além de fornecerem a nutrição básica, promovem a
saúde. Esses alimentos possuem potencial para promover a saúde por mecanismos não

20
previstos na nutrição convencional, devendo ser salientado que esse efeito restringe-se à
promoção da saúde e não à cura de doenças 9; 10
. São consumidos em dietas convencionais,
mas possuem capacidade de regular funções corporais de forma a auxiliar na proteção contra
doenças como hipertensão, diabetes, câncer, osteoporose e coronariopatias 11
. Uma subclasse
de alimentos funcionais são os alimentos probióticos que, segundo a Fao/Who 12
, são
microrganismos vivos que, quando ingeridos em quantidades adequadas, conferem benefícios
à saúde do hospedeiro.
Vários fatores têm contribuído para o desenvolvimento dos alimentos funcionais, entre os
quais o aumento da consciência dos consumidores que desejam melhorar a qualidade de vida
e optam por hábitos mais saudáveis 11
. Em função disso, as indústrias de produtos lácteos
estão respondendo com o desenvolvimento de novos produtos funcionais. Cerca de 70% dos
lançamentos de iogurte na Europa tem posicionamento funcional, sendo o LC1 da Nestlé e o
Actimel da Danone os mais consumidos 13
. Desde 1986, a Lifeway Foods Inc. lançou no
mercado americano, vários produtos lácteos à base de kefir contendo dez microrganismos
vivos originários do kefir tradicional, fermentadores do leite e duas culturas probióticas,
clinicamente comprovadas, alegando propriedades funcionais benéficas como, por exemplo, o
equilíbrio do ecossistema corporal, apoio à saúde digestiva e imunidade 14; 15
.
Kefir é um leite fermentado de sabor ácido, viscoso, levemente carbonatado e de baixo
teor alcoólico 16
, consumido tradicionalmente na Europa Oriental, Rússia e sudoeste asiático
15. Atualmente, o consumo desse alimento está se expandindo devido a suas propriedades
sensoriais únicas e sua longa história associada aos efeitos benéficos à saúde humana. Por
esse motivo, é considerado um alimento funcional probiótico, e muitos o designam como o
iogurte do século XXI 16; 17
.
Ao comparar o kefir com iogurte tradicional, o primeiro contém maior diversidade de
microrganismos, que participam do processo de fermentação, além de ser menos viscoso,
formar um coágulo menor, ser menos resistente e mais facilmente digerível, e conter etanol e
gás carbônico, além do ácido lático 8; 15; 18
.
A produção artesanal do kefir fundamenta-se na tradição dos povos do Cáucaso,
difundida a partir do final do século passado, para outras partes do mundo, integrando suas
indicações nutricionais e terapêuticas ao cotidiano alimentar atual. No Brasil, a bebida vem
sendo divulgada há pouco tempo e sua fabricação é exclusivamente artesanal, obtida pela
fermentação do grão em leite ou em água contendo açúcar mascavo. O produto é

21
desconhecido pela maior parte da população brasileira, bem como os possíveis benefícios da
inclusão desse alimento probiótico na dieta 8.
Apesar do valor nutricional e terapêutico do produto, o método tradicional de obtenção
do kefir, a partir de fermentações sucessivas com a reinoculação dos grãos, gera produtos não
padronizados. A composição da microbiota pode variar de uma produção para outra, tornando
difícil ou mesmo impossível a manutenção do padrão de qualidade do produto 8; 19
. Dessa
forma, estudos da composição microbiana e das características dos grãos de kefir e de seus
produtos fermentados disponíveis no Brasil, podem contribuir para fornecer base científica
para maiores aplicações do produto, de forma padronizada, facilitando a introdução desse
alimento no mercado consumidor brasileiro, em escala comercial.

22
2 REVISÃO BIBLIOGRÁFICA
2.1 LEITES FERMENTADOS
Segundo o Regulamento Técnico de Identidade e Qualidade (RTQI) de Leites
Fermentados 20
, entendem-se por leites fermentados os produtos adicionados ou não de outras
substâncias alimentícias, obtidas por coagulação e diminuição do pH do leite, in natura ou
reconstituído, adicionado ou não de outros produtos lácteos, por fermentação lática mediante
ação de cultivos de microrganismos específicos. Estes microrganismos devem ser viáveis,
ativos e abundantes no produto final durante seu prazo comercial (Quadro 1).
Segundo o Codex Alimentarius 21
, leite fermentado é um produto lácteo obtido por
meio da fermentação do leite, que pode ter sido elaborado a partir de produtos obtidos do
leite, com ou sem modificações na sua composição, por meio da ação de microrganismos
adequados e tendo como resultado a redução do pH, com ou sem coagulação (precipitação
isoelétrica). Estes cultivos de microrganismos deverão ser viáveis, ativos e abundantes no
produto até a validade comercial. Se o produto é tratado termicamente após a fermentação,
não se aplica o requisito de microrganismos viáveis.
Certos leites fermentados caracterizam-se por um cultivo específico (ou cultivos
específicos) utilizado para a fermentação, conforme exposto no quadro 1.
Quadro 1 - Culturas de microrganismos utilizados na fabricação de alguns leites fermentados
Iogurte Cultura simbiótica de Streptococcus thermophilus e Lactobacillus
delbrueckii subsp. bulgaricus
Iogurte a base de
culturas alternativas
Culturas de Streptococcus thermophilus e toda espécie de
Lactobacillus.
Leite acidófilo Lactobacillus acidophilus
Kefir Cultura preparada a partir de grãos de kefir, Lactobacillus kefiri,
espécies do gênero Leuconostoc, Lactococcus e Acetobacter que
crescem em uma estreita relação específica.
Kumys Lactobacillus delbrueckii subsp. bulgaricus e Kluyveromyces
marxianus Fonte: Brasil
20

23
2.1.1 Papel dos microrganismos em alimentos fermentados
2.1.1.1 Bactérias Ácido Láticas (BAL)
O grupo das Bactérias Ácido Láticas (BAL) é o principal representante dos probióticos
em alimentos, pertencentes ao filo Firmicutes, classe Bacilli. As BAL, como grupo de
microrganismos apresentam alguns fenótipos típicos, tais como: são Gram-positivos,
geralmente imóveis, não produzem esporos, não produzem a enzima catalase, são anaeróbios
aerotolerantes, fastidiosos, ácido-tolerantes e estritamente fermentadores, sendo o ácido lático,
o principal produto final da fermentação do açúcar. Em geral, estas bactérias são mesófilas,
mas podem crescer em baixas temperaturas como 10°C, ou altas como 45°C, em situações
específicas; são pouco proteolíticas e lipolíticas e requerem aminoácidos, vitaminas do
complexo B e bases púricas e pirimídicas para o crescimento22
. Geralmente, membros do
grupo estão presentes em ambientes nutricionalmente ricos como vegetais, leites, carnes, e
alguns compõem a microbiota normal como cavidade oral, trato intestinal e vaginal de
mamíferos 23; 24
.
Uma importante diferença dentre os subgrupos das BAL são os produtos formados
durante a fermentação dos carboidratos. Um grupo, denominado de homofermentativo,
produz ácido láctico como único ou principal produto, enquanto que o outro, denominado
heterofermentativo, produz ácido lático, etanol e/ou ácido acético e CO2. Os gêneros
Leuconostoc, Oenococcus, Weissella e um subgrupo de lactobacilos são heterofermentativos
(Quadro 2), enquanto que todas as outras BAL e um subgrupo de lactobacilos são
homofermentativos 23
.
Quadro 2 - Diferenciação dos principais gêneros de BAL
Gênero Formato e arranjo Fermentação Produto metabólico
Streptococcus cocos em cadeias homofermentativo ácido lático L (+),
acetaldeído, diacetil
Leuconostoca cocos em cadeias heterofermentativo ácido lático D(-), diacetil
Pediococcus cocos em tétrades homofermentativo ácido lático DL
Lactobacillus bacilos geralmente em
cadeias
bhomofermentativo
cheterofermentativo
ácido lático DL
Enterococcus cocos em cadeias homofermentativo ácido lático L(+)
Lactococcusd cocos em cadeias homofermentativo ácido lático L(+)
aLeuconostoc mesenteroides subsp. cremoris também produz etanol e CO2
bLactobacillus acidophilus, L. gasserie,L. helveticus,L. johnsonii e L. kefiranofaciens
cLactobacillus casei,L. reuteri,L. plantarum,L. fermentum,L. rhamnosus e L. kefiri produzem ácido lático L (+)
dLactococcus lactis subsp. biovar diacetylactis produz diacetil e CO2
Fonte: Madigan, Martinko et al. 25
; Tamime 2

24
Ácido lático L (+) e D (-) são isômeros ópticos do ácido 2-hidroxipropanóico. Os dois
isômeros ópticos podem ocorrer na forma pura ou na forma racêmica D,L - ácido lático,
contendo uma mistura de ambas as formas 26
.
Historicamente, o gênero Bifidobacterium também é considerado pertencente ao grupo
de BAL. Embora espécies de Bifidobacterium se enquadrem nas descrições acima, estão mais
relacionadas filogeneticamente ao grupo Actinomycetaceae de bactérias Gram-positivas.
Além disso, têm uma via metabólica única para fermentação do açúcar, própria do gênero,
que claramente as separa dos outros membros do grupo de BAL.
As BAL há muitos séculos, vem sendo utilizadas na fermentação de vários produtos e
seu papel conservante e benéfico para saúde são reconhecidos, pois possuem ação antagônica
contra numerosos patógenos responsáveis por toxinfecções intestinais 27
, bem como fungos e
micotoxinas 28
. Algumas BAL podem prevenir a aderência, o estabelecimento, e a invasão de
patógenos ou a produção de toxina pelos mesmos 29; 30
. Além disso, podem inibir o
crescimento de bactérias patogênicas pela redução do pH, resultado da produção de ácidos
orgânicos como o acético, o propiônico e o ácido láctico 27; 31; 32; 33
ou pela produção de
peróxido de hidrogênio 34
, dióxido de carbono e diacetil 35
. As BAL também podem competir
por nutrientes ou sítios de adesão do patógeno e, além disso, podem secretar peptídeos
antimicrobianos conhecidos como bacteriocinas 31; 34; 36
. Sua aplicação em produtos
fermentados deve também contribuir para melhoria da qualidade sensorial do produto 37
.
2.1.1.1.1 Gênero Lactobacillus
Lactobacillus spp. é o gênero mais amplo do grupo de BAL, incluindo cerca de 80
espécies; muito heterogêneo, abrangendo espécies com grande variedade de propriedades
fenotípicas, bioquímicas e fisiológicas. A heterogeneidade é refletida pelo taxa de
porcentagem do conteúdo de Guanina e Citosina (GC) do DNA das espécies incluídas neste
gênero. Três subgrupos de classificação dos lactobacilos foram criados baseando-se nas
características fisiológicas de diferentes espécies, relacionadas com a presença ou ausência de
enzimas essenciais no metabolismo homo ou heterofermentativo do açúcar. Sendo assim,
podem ser classificadas como homofermentativos obrigatórios, heterofermentativos
obrigatórios ou facultativos 23
.
Este gênero exerce papel crucial na produção de alimentos fermentados, como
vegetais, carnes e produtos lácteos. Nas últimas décadas vem sendo amplamente aplicados na

25
indústria de alimentos, como cultura iniciadora (starter) e cultura adjunta (incluindo
probióticos), aumentando o interesse no estudo de suas funções e metabolismo. Geralmente
são os mais ácido-tolerantes do grupo das BAL e, portanto, estão ativos no final de muitos
processos fermentativos espontâneos, como fermentação de silagens e vegetais 23; 24
.
2.1.1.1.2 Gênero Lactococcus
Lactococcus spp. são cocos Gram-positivos que ocorrem em pares ou em cadeias, são
imóveis e não produzem a enzima catalase, e pertencem ao grupo sorológico N de Lancefield,
anteriormente classificados como Streptococcus. Não são β-hemolíticos e raramente α-
hemolíticos 38
.
Lactococcus spp. é comumente encontrado em superfícies de plantas, peles de animas
e em leite. As duas subespécies mais utilizadas como cultura starter na indústria de produtos
lácteos, principalmente na fabricação de queijos, são L. lactis subsp. lactis e L. lactis subsp.
cremoris. 38; 39
.
Os microrganismos pertencentes a este gênero são homofermentativos, com exceção
da subespécie lactis que utiliza o citrato (Quadro 2), recentemente reconhecida como L. lactis
subsp. lactis biovar diacetylactis2.
Nos últimos anos, estudos mostraram que esse microrganismo foi capaz de melhorar o
equilíbrio da microbiota intestinal de hospedeiro animal, sendo utilizado como microrganismo
probiótico 39
. O principal papel do lactococos na fermentação de alimentos é a acidificação,
principalmente pela produção de ácido lático L(+). Este gênero também contribui para o
desenvolvimento da textura, devido à produção de exopolissacarídeos, ou para o flavor devido
à produção de compostos aromáticos (álcools, cetonas e aldeídos) ou pelos metabólitos
gerados pela utilização do citrato, aminoácidos e lipídeos. Também podem ser utilizados para
conservação, devido a sua habilidade em produzir ácidos orgânicos e bacteriocinas, sendo a
nisina a mais caracterizada e reconhecida dentre estas bacteriocinas38
.
2.1.1.1.3 Gênero Leuconostoc
Membros do gênero Leuconostoc são cocos Gram-positivos (em pares ou cadeias),
anaeróbios facultativos, que não produzem a enzima catalase. Morfologicamente são similares
aos Streptococcus, entretanto são heterofermentativos. A seleção desse gênero pode ser

26
facilitada pelo uso de vancomicina no meio de cultura, uma vez que todas as espécies de
Leuconostoc são intrinsicamente resistentes a este antibiótico. Esta resistência intrínseca é
consequência da ausência do dipeptídeo D-alanina-D-alanina de um pentapeptídeo, que nestes
microrganismos é substituído naturalmente por D-alanina-D-lactato 40
.
Estes microrganismos eram geralmente encontrados em plantas e a partir deste habitat
natural, disseminaram-se para vários nichos, incluindo leite cru e produtos lácteos
refrigerados, exercendo papel importante na indústria e processos de fermentação de
alimentos como linguiças, vegetais, cereais e produtos lácteos.
Na tecnologia de obtenção de produtos lácteos estão naturalmente presentes em uma
variedade de queijos, bebidas fermentadas (kefir) e amplamente utilizados como starters 41
.
Além disso, também possuem papel essencial na formação da textura e aroma dos produtos
lácteos. Em particular, espécies L. mesenteroides e L. lactis metabolizam o citrato, produzindo
os compostos aromáticos diacetil e acetoína. Algumas linhagens produzem dextrana quando
cultivados em sacarose 40; 41
, característica que pode ser aplicada na produção de
oligossacáridos, que influenciam na textura, além da aplicação funcional como prebiótico 41
.
Alguns estudos já mostraram que este gênero é capaz de produzir bacteriocinas em produtos
lácteos, mas a aplicação industrial destas bacteriocinas, ainda não foi desenvolvida 40
.
2.1.1.2 Bactérias Ácido Acéticas (BAA)
As BAA apresentam-se em forma elíptica ou em forma de bastonetes, isoladas, em
pares ou em cadeias, flageladas ou não, Gram-negativas ou Gram-variáveis, aeróbicas, não
formadoras de esporos, produtoras da enzima catalase, mas não da oxidase. O pH ótimo para
crescimento está entre 5,0 e 6,5, porém podem crescer em pH mais baixo, entre 3,0 e 4,0 42; 43
.
As BAA foram classificadas em dois gêneros principais: Acetobacter e
Gluconobacter, mas atualmente doze gêneros já são reconhecidos e enquadrados na família
Acetobacteriaceae 42
. Estas bactérias são bem conhecidas por sua habilidade em oxidar
açúcares e álcoois, resultando na acumulação de ácidos orgânicos como produto final. As
BAA estão envolvidas em alguns processos industriais importantes, como na indústria do
vinagre, pois podem produzir altas concentrações de ácido acético a partir do etanol 42
.
Também exercem importante papel no produto lácteo kefir, pois além da contribuir para as
propriedades sensoriais, são capazes de produzir vitamina B12 e estimular o crescimento de
outros microrganismos 2; 44
.

27
2.1.1.3 Leveduras
Leveduras são fungos unicelulares, pertencentes a dois filos dentro do reino fungi:
ascomicetos e basidiomicetos. A classe de leveduras hemiascomicetos é constituída por
fungos unicelulares, frequentemente associados às indústrias de processamento de alimentos e
bebidas 45
. As células de leveduras são maiores que as bacterianas, variando em tamanho de 4
a 8 µm de largura, por 7 a 12 µm de comprimento. As células são esféricas, ovais ou
cilíndricas, e a divisão celular geralmente ocorre por brotamento. Embora a maioria das
leveduras reproduzam-se apenas como células individuais, algumas podem formar filamentos,
sob certas condições. Os fungos podem ter duas formas, uma sexual (teleomorfo), que é
considerada perfeita, e uma assexual (anamorfo), que é considerada imperfeita. Em alguns
casos, apenas a forma imperfeita é conhecida 45
. A espécie Saccharomyces cerevisiae tem
sido estudada, há muitos anos, como um organismo modelo em estudos de genética e biologia
molecular, sendo o primeiro eucarioto a ter seu genoma completamente sequenciado 25; 45
.
Na maioria das vezes, o crescimento de leveduras é indesejável em produtos lácteos,
pois geralmente estão associados a processos de deterioração. Como são capazes de crescer
em uma ampla faixa de pH, os produtos lácteos com baixo pH, como o leite fermentado, o
creme de leite, o cream cheese e as bebidas lácteas são alvos de deterioração por leveduras.
Em manteiga, geralmente causam deterioração por lipólise e proteólise. Estas leveduras
contaminam o leite durante a ordenha ou após o tratamento térmico, ocorrendo uma
recontaminação do produto. Entretanto, também são utilizadas na fabricação de alguns
produtos lácteos, sendo importantes no desenvolvimento da superfície maturada de queijos
macios e semi-duros devido a sua relação sinérgica com BAL, Brevibacterium linens,
Microbacterium lacticum e micrococos. Produtos lácteos fermentados tradicionais, como o
kefir e kumys, sofrem fermentação ácido lática e também alcóolica, que é realizada pelas
leveduras 46
.
2.2 ALIMENTOS FUNCIONAIS
O termo “funcional” implica que o alimento possui algum valor benéfico para a saúde,
incluindo a redução do risco de doença para quem o esteja consumindo 10
. Substâncias
biologicamente ativas encontradas nos alimentos funcionais podem ser classificadas em

28
grupos, tais como: probióticos e prebióticos; alimentos sulfurados e nitrogenados; pigmentos
e vitaminas; compostos fenólicos, ácidos graxos poli-insaturados e fibras 11
.
2.2.1 Probióticos
O termo probiótico foi proposto por Vergio 47
, quando comparou os efeitos adversos
(antibiotika) que os antibióticos exerciam sobre a microbiota intestinal com as ações
benéficas (probiotika) exercidas por outros fatores não determinados na época. Atualmente,
probiótico foi definido por uma comissão da Fao/Who 12
como microrganismos vivos, que
quando ingeridos em quantidades adequadas, conferem benefícios à saúde do hospedeiro. A
influência benéfica dos probióticos sobre a microbiota intestinal humana inclui fatores como
efeitos antagônicos, competição e efeitos imunológicos, resultando em um aumento da
resistência contra patógenos (Quadro 3). Assim, a utilização de culturas bacterianas
probióticas estimula a multiplicação de bactérias benéficas, em detrimento à proliferação de
bactérias potencialmente prejudiciais, reforçando os mecanismos naturais de defesa do
hospedeiro 48
.
Entre os microrganismos mais empregados e selecionados com fins probióticos,
destacam-se as BAL, particularmente as do gênero Lactobacillus 24
e Bifidobacterium spp. 5;
49; 50. Duas leveduras também são utilizadas: a Saccharomyces boulardii, na medicina humana
e a Saccharomyces cerevisiae, na medicina veterinária 6; 51
.
A principal vantagem no uso de leveduras como probiótico é que não são afetadas pela
administração de antibióticos 51
. A levedura S. boulardii é um dos poucos microrganismos
utilizados como probióticos que não são de origem humana. Esta levedura foi isolada
inicialmente da fruta lichia (Litchi chinensis) em 1950 52
, e possui a seu favor o grande
número de ensaios laboratoriais e clínicos, comprovando seu potencial probiótico 51
.
2.2.2 Efeitos benéficos dos probióticos
O uso de probióticos está associado com um grande número de efeitos benéficos,
dentre os quais já foram citados: melhora da intolerância a lactose 53
; modulação do sistema
imunológico 54; 55
; redução da hipercolesterolemia 56; 57
e hipertensão 58; 59
; proteção contra
enfermidades infecciosas, como diarreia crônica causada por Clostridium difficile 60
e infecção
por Helicobacter pylori 61; 62; 63
; redução da colonização da mucosa nasal por S. aureus, S.

29
pneumoniae e Haemophilus influenzae 64
; redução do risco de alergias e atopias 65
; e
tratamento e prevenção de certos tipos de câncer e doenças inflamatórias crônicas 66; 67; 68
. A
administração oral de alguns probióticos também poderia exercer um efeito analgésico no
intestino similar ao efeito da morfina 69
, além de exercer um papel benéfico para pessoas
diabéticas 57; 70
.
Entretanto, o conhecimento sobre os mecanismos de ação continua sendo uma das
grandes lacunas nesse campo (Quadro 3). Nesse sentido, não são conhecidos os mecanismos
exatos pelos quais certos probióticos modulam a composição microbiana das mucosas, ainda
que tradicionalmente, sejam citados fenômenos de competição por nutrientes, por receptores,
ou a produção de substâncias antimicrobianas. Além disso, as propriedades benéficas não
podem ser consideradas para todas as bactérias láticas e bifidobactérias, ou até mesmo para
cepas de uma mesma espécie 6. As mesmas são incomparáveis e podem possuir áreas de
aderência distintas, efeitos imunológicos específicos e mecanismos de ação distintos, sobre a
mucosa saudável e a inflamada 37
. O efeito benéfico de uma estirpe depende das condições de
seu emprego e, muito particularmente, da dose consumida 6.
Quadro 3– Potenciais efeitos benéficos de alimentos funcionais contendo probiótico.
Efeito benéfico Possíveis causas e mecanismos
Aumento da digestibilidade Degradação parcial de proteínas, lipídeos e hidratos de
carbono.
Aumento do valor nutritivo Níveis elevados das vitaminas do complexo B e de alguns
aminoácidos, p. ex. metionina, lisina e triptofano.
Melhor utilização da lactose Níveis reduzidos de lactose no produto e maior
disponibilidade de β-galactosidase.
Ação antagônica contra
agentes patogênicos entéricos
Distúrbios tais como diarréia, colite mucosa, colite ulcerosa,
diverticulite e colite antibiótica controlada pela acidez,
inibidores microbianos (H2O2, bacteriocinas e sais biliares) e
inibição da adesão e ativação dos patógenos (consumo de
ferro).
Colonização do intestino Sobrevivência no ácido gástrico, resistência à lisozima e a
tensão superficial baixa do intestino, adesão ao epitélio
intestinal, multiplicação no trato gastrointestinal, modulação
imunitária.
Ação anticarcinogênica Conversão de potencial pré-carcinogênicos em compostos
menos danosos;
Ação inibitória perante alguns tipos de câncer, p.ex. câncer do
trato gastrointestinal por degradação dos pré-carcinogênicos,
redução das enzimas promotoras de carcinogênicos e
estimulação do sistema imunitário.
Ação hipocolesterolêmica Produção de inibidores da síntese do colesterol
Utilização do colesterol por assimilação e precipitação com
sais biliares desconjugados.
Modulação imunitária Melhor produção de macrófagos, estimulação da produção de
células supressoras e γ-interferon Fonte: Holzapfel e Schillinger
71; Kalantzopoulos
72.

30
Adicionalmente, é possível que os determinantes dos efeitos benéficos de produtos
lácteos contendo probióticos sejam multifatoriais, não apenas pela presença da bactéria viva,
mas também por outros componentes do leite fermentado, como cálcio altamente absorvível,
ácidos linoleicos conjugados, ácido butírico e peptídeos bioativos. Benefícios fisiológicos têm
sido atribuídos também a microrganismos mortos, pois certos mecanismos de ação, como
liberação de algumas enzimas, não requerem células vivas. Entretanto, independentemente da
funcionalidade, microrganismos mortos não são probióticos 73
.
Resultados de estudos in vitro e in vivo indicam que, efetivamente, existem certas
estirpes capazes de proporcionar efeitos benéficos para a saúde do consumidor. Em qualquer
caso, é necessário realizar estudos bem planejados e aprofundar o desenvolvimento de
marcadores biológicos validados, que permitam a comprovação interlaboratorial dos efeitos
probióticos produzidos in vivo 6; 50; 73; 74; 75
. Atualmente, os probióticos estão sendo
rigorosamente avaliados em diversos hospitais e centros médicos da Europa quanto aos seus
efeitos benéficos adjuntos no tratamento e prevenção de enfermidades como disbiose,
síndrome do intestino irritável, doença diverticular, entre outras, sendo seus resultados
promissores 66
.
2.2.3 Critérios de seleção de bactérias probióticas
Nas últimas décadas, bactérias probióticas são adicionadas em leites fermentados e
iogurtes, devido aos seus reconhecidos efeitos benéficos 76
. Contudo, a introdução no mercado
de alimentos probióticos não lácteos, como produtos cárneos fermentados, sucos ou outros
alimentos de origem vegetal, é crescente. Há uma grande variedade de bactérias
potencialmente probióticas, e, provavelmente, nos próximos anos, este número continuará
aumentando, devido ao interesse, por parte das instituições de pesquisa e indústrias, pelo
desenvolvimento de novas estirpes potencialmente probióticas 6.
A seleção de novas estirpes com fins probióticos não é um trabalho simples, pois não
existem testes in vitro, que sirvam para predizer a atividade probiótica de uma determinada
estirpe. Por isso, não existe uma uniformidade entre os critérios propostos para a seleção
destas bactérias 71
.
No processo de seleção de um microrganismo probiótico vários aspectos, incluindo
características de segurança, funcionais e tecnológicas devem ser levados em consideração 6;
49; 73; 76. Tradicionalmente, recomenda-se que cepas probióticas sejam isoladas de amostras da

31
mesma espécie na qual será aplicada, baseando-se na hipótese de que as estirpes de origem
humana se adaptariam e colonizariam o epitélio gastrointestinal humano com maior facilidade
do que aquelas isoladas de outras fontes 71
. No entanto, outros estudos já demonstraram que a
capacidade de cepas probióticas para aderir ao epitélio do hospedeiro não depende de sua
origem 6. Nem todas as cepas destinadas ao uso humano, disponíveis atualmente no mercado,
são de origem humana e, além disso, é praticamente impossível conhecer a origem de uma
cepa, uma vez que a mesma pode advir originalmente de um alimento ou, ainda, do animal ou
vegetal que proporcionou a matéria prima para elaborá-lo 6; 12
.
O comitê criado pela Fao/Who 12
sugere que a seleção de um microrganismo
probiótico deve estar baseada, independente de sua origem, em quatro critérios fundamentais:
(1) correta identificação, em nível de estirpe por meios de técnicas moleculares; (2)
capacidade de chegar viável, e em número suficiente ao seu sítio de ação; (3) segurança; e (4)
efetividade real, determinada mediante testes clínicos in vivo planejados, de acordo com
padrões bem aceitos.
2.2.3.1 Aspectos funcionais
Tradicionalmente, em relação aos aspectos funcionais, a seleção das bactérias
probióticas tem como base os seguintes critérios: apresentar tolerância à passagem pelo Trato
Gastrointestinal (TGI); capacidade de aderir à mucosa intestinal e de colonizar, ao menos
temporariamente, o TGI humano; e possuir propriedades antagonistas de imunomodulação e
anticancerígenas 6; 12; 48; 49; 73; 77
.
Para que as cepas possam exercer seus efeitos benéficos, devem ser capazes de resistir
às condições ambientais (suco gástrico, pancreático e sais biliares) existentes durante o
trânsito pelo aparelho digestivo e colonizar o TGI, ou ainda, ser consumido em um alimento
veículo que permita a sobrevivência das estirpes à passagem pelo estômago e exposição aos
sais biliares 12
. Para tanto, são comumente utilizados testes in vitro, como acidificação e/ou
adição de sais biliares ao meio de cultura, o emprego de secreções gastrointestinais obtidas a
partir de indivíduos saudáveis ou, ainda, o uso de modelos dinâmicos mais sofisticados que
simulam as condições gastrointestinais in vivo 6.
A sobrevivência dos probióticos ao intestino pode ser estudada in vivo, utilizando-se
técnicas de intubação intestinal e biópsias do cólon ou, ainda, analisando-se sua presença nas
fezes de pessoas ou animais que os tenham ingerido. Em geral, estima-se que a concentração

32
dos probióticos viáveis que deve chegar ao colón para obter um efeito benéfico deve ser de
aproximadamente ≥ 108
Unidades Formadoras de Colônias (UFC)/g. Porém, no intestino
grosso existe uma população bacteriana de 109-10
12 UFC/g e, ao se estudar a cepa probiótica
no TGI, técnicas moleculares devem ser estabelecidas para distinguir a cepa probiótica
ingerida de outros milhares de microrganismos que compõem o ecossistema do TGI 6; 48; 73
Quando os microrganismos alcançam o intestino, são capazes de permanecerem ali
durante um curto período de tempo em estado não proliferativo. A adesão à mucosa intestinal
é o primeiro passo para a colonização, e a maioria dos autores a considera como um pré-
requisito para exercer efeitos benéficos no hospedeiro, tal como a exclusão competitiva de
bactérias enteropatogênicas ou imunomodulação 30; 54; 78
. Geralmente, o probiótico é detectado
nas fezes durante os dias e/ou semanas seguintes a sua ingestão, desaparecendo gradualmente,
o que indica uma colonização transitória na ausência de sua ingestão continuada 6; 48; 79
.
Devido às dificuldades do estudo de aderência das bactérias in vivo, métodos in vitro
foram desenvolvidos para estimar a capacidade da cepa em colonizar o intestino. Os modelos
mais empregados utilizam linhagem celular derivada de adenocarcinoma de cólon humano,
como a Caco-2 e HT-29. As células da linhagem Caco-2 se diferenciam e se polarizam
espontaneamente quando confluem, mostrando uma membrana apical funcional com as
microvilosidades completamente desenvolvidas; por isto é constantemente utilizada em
estudos in vitro da organização e função de células intestinais humanas. Da mesma forma, a
linhagem HT-29-MTX secreta muco, podendo simular o ambiente mucoso do intestino
humano. Vários trabalhos já demonstraram a utilidade de ambas as linhagens no estudo da
aderência de lactobacilos 78
, obtendo-se uma boa correlação entre a capacidade adesiva das
cepas probióticas e os resultados de estudos in vivo 6; 30; 50; 80; 81; 82
, quando da utilização desses
modelos.
2.2.3.2 Aspectos de segurança
Os aspectos de segurança incluem especificações quanto a sua origem (TGI humano
saudável), ao histórico de não patogenicidade e à ausência de genes determinantes de
resistência a antibióticos 48; 49
. Portanto, é indispensável avaliar a segurança e eficácia do
produto probiótico final, realizando, inicialmente, testes laboratoriais, posteriormente, em
modelo animal e finalmente, estudos com humanos 5.

33
Segundo Martín e colaboradores 6 há quatros tipos de efeitos adversos que as bactérias
podem causar na população em geral ou em certos grupos específicos, como os
imunodeprimidos: (1) atividades metabólicas indesejáveis; (2) infecções sistêmicas; (3)
transferência de genes (como de resistência a antibióticos); e (4) imunomodulação excessiva.
Alguns autores 83; 84
propõem uma série de testes para avaliar a segurança das estirpes,
ainda que alguns possam parecer obsoletos 6, como produção de D-lactato, ou, ainda, difíceis
de justificar diante do comitê de ética de experimentação animal, como o emprego de cobaias
irradiadas. Dentre as atividades metabólicas indesejáveis, destacam-se: produção de D(-)
ácido lático, produção de aminas biogênicas, atividade de enzimas produtoras de sais biliares
secundários (atividade 7α-deshidroxilase), produção de enzimas que degradam a mucina,
atividades azoredutase e/ou nitroredutase, β-glucoronidase e β-glucosidase 68
. Entretanto, a
maior parte destas atividades foi estudada de forma isolada, o que torna difícil obter
conclusões sobre seus efeitos in vivo 6.
A produção do isômero D(-) do ácido lático é objeto de grande controversia6. Há
alguns anos atrás, um comitê sobre aditivos alimentares, criado pela Fao/Who 85
relatou que
havia evidências de que neonatos apresentavam dificuldade em utilizar o ácido lático D(-),
restringindo o uso deste isômero ou da forma racêmica como aditivo em alimentos infantis,
principalmente naqueles destinados às crianças com menos de três meses de idade. Em 1981,
o padrão recomendado pelo Codex Alimentarius 86
para fórmulas infantis permitia o uso
somente de culturas microbianas produtoras de ácido lático L(+). A maior preocupação estava
relacionada com o uso de bactérias produtoras do lactato D(-) como ingrediente probiótico em
fórmulas infantis que poderiam ser prejudiciais por um possível aumento dos níveis
plasmáticos de ácido lático D(-), podendo levar a uma acidose metabólica87; 88
. Porém,
segundo Connolly e Lönnerdal 89
não há estudos disponíveis na literatura científica que
comprovem que indivíduos saudáveis de qualquer idade poderiam ser afetados em função da
suplementação da dieta com estirpes produtoras deste isômero. A literatura médica descreve
exemplos raros de acidose metabólica causada por ácido lático D(-) em humanos, quase que
exclusivamente estudados em pacientes com síndrome do intestino curto 6; 89; 90
.
As aminas biogênicas são produzidas em certos alimentos como consequência da
atividade descarboxilase de alguns microrganismos, como alguns Lactobacillus spp., e podem
causar problemas toxicológicos nos consumidores quando encontram-se em concentrações
elevadas. A histamina e tiramina possuem propriedades vasoativas e psicoativas, enquanto

34
que as diaminas putrescina e cadaverina além de potencializar a ação tóxica das primeiras, são
precursoras de nitrosaminas carcinogênicas.
As bactérias intestinais também podem influenciar nos processos de biotransformação
de sais biliares, formando sais biliares secundários. Estes ácidos podem se acumular na
circulação enterohepática e participar de patologias como câncer de cólon, formação de
cálculos biliares e outras enfermidades gastrointestinais 6; 83
.
Várias enzimas bacterianas estão envolvidas na degradação da mucina no intestino
grosso. Embora possa ser considerado uma atividade desejável da microbiota colônica, a
integridade do muco gástrico e do intestino delgado é uma importante barreira para infecção
por patógenos e danos ao epitélio por meios químicos e físicos.
Alguns autores acreditam também que o efeito carcinogênico de alguns compostos
tóxicos endógenos poderia ser influenciado pela atividade das enzimas azoredutase,
nitroredutase, β-glucoronidase e β-glicosidase 6; 68; 83; 84
. Em geral, a presença deste tipo de
atividade não é frequente em BAL, diferentemente do que ocorre com os clostrídios,
bacteróides e enterobactérias. De fato, observa-se que a administração de BAL e
bifidobactérias costuma reduzir estas atividades no intestino humano 6. Curiosamente, estudos
recentes indicam que um aumento da atividade β-glicosidase poderia supor um benefício para
a saúde, mediante a liberação de flavonóides bioativos 91; 92; 93
.
A maior parte das infecções humanas já relatadas parece ser causada por estirpes de
origem endógena em pacientes com algum fator predisponente, particularmente alterações em
válvulas cardíacas, presença de cateteres ou estado de imunodepressão, que costuma resultar
em endocardites subagudas, septicemias ou abscessos hepáticos 24; 94
. Pacientes
imunodeprimidos são geralmente mais vulneráveis à infecção por patógenos reconhecidos e a
infecções oportunistas. Entre as espécies associadas a tais casos destacam-se L. rhammnosus,
L. casei, L. paracasei e L. plantarum 24
. Os poucos casos descritos de infecções relacionadas
com o emprego de lactobacilos probióticos comerciais, tem como protagonista a espécie L.
rhammnosus GG. De fato, esta espécie é a única do gênero, que contém cepas que podem ser
consideradas patógenos oportunistas, uma vez que são capazes de afetar não só pessoas
imunodeprimidas, mas também imunocompetentes, com histórico de substituição de válvulas
cardíacas ou que padeçam de endocartite reumática 94
.
Com relação à resistência a antimicrobianos, a informação genética, especialmente a
codificada em plasmídeos e outros elementos móveis, pode ser transferida de um
microrganismo para outro. Este fenômeno biológico levanta a questão de que é possível que

35
ocorra transferência genética entre os microrganismos presentes nos alimentos e naqueles que
possam existir na microbiota intestinal, incluindo os patógenos 95; 96; 97
. A troca de material
genético dentro de uma comunidade microbiana pode realizar-se mediante transdução,
conjugação ou transformação, sendo as duas últimas as que oferecem maior potencial de
intercâmbio de DNA entre bactérias pertencentes a distintas espécies 6; 96
. Em qualquer caso,
ao comparar perfis de resistência a antibiótico, é importante se realizar uma diferenciação
entre a resistência intrínseca e a adquirida, mediada por elementos genéticos específicos.
Resistência intrínseca é inerente a espécies de bactérias e é típica de todas as estirpes daquela
espécie. Em contraste, quando uma cepa de uma espécie tipicamente susceptível é resistente a
um dado fármaco antimicrobiano, isto é considerado resistência adquirida. A resistência
adquirida pode ocorrer devido a genes adicionados (genes adquiridos pela bactéria via DNA
exógeno) ou por mutação de genes autóctones 97
.
Muitas espécies de Lactobacillus, Pediococcus e Leuconostoc são intrinsicamente
resistentes à vancomicina devido à estrutura que adotam os precursores dos peptidoglicanos
de suas paredes celulares. Em consequência, este fenótipo não pode ser transferido a nenhum
outro microrganismo presente nos alimentos ou no intestino 6; 98
.
2.2.3.3 Aspectos tecnológicos
Para a utilização de culturas probióticas na tecnologia de fabricação de produtos
alimentícios, seu desempenho tecnológico também deve ser considerado. Culturas probióticas
com boas propriedades tecnológicas devem apresentar boa multiplicação no leite, promover
propriedades sensoriais adequadas no produto e ser estáveis, viáveis e funcionais durante o
armazenamento 6; 77; 99
.
Ao introduzir um probiótico no mercado deve-se assegurar que o produto é seguro,
estável e funcional, e que a rotulagem está de acordo com a legislação vigente do país 6; 11
. No
Brasil, são permitidas as alegações que fazem referência à manutenção geral da saúde, ao
papel fisiológico dos nutrientes e não nutrientes, e à redução de risco de doenças. Porém, não
são permitidas alegações de saúde que façam referência à cura ou prevenção da doença 100; 101
.
No quadro a seguir (Quadro 4) consta a alegação de propriedade funcional e de saúde revisada
e aprovada pela Comissão de Assessoramento Técnico-Científico em Alimentos Funcionais e
Novos Alimentos (CTCAF), criada pela Agencia Nacional de Vigilância Sanitária (ANVISA)
para produtos probióticos devidamente registrado pelo órgão 7.

36
Inicialmente, pesquisas sobre probióticos ocorrem em escala laboratorial, onde os
volumes dos cultivos são medidos em mililitros ou litros, enquanto que em escala industrial,
mede-se em metros cúbicos. Em muitas situações, o processo em larga escala não é
presumível, uma vez que à medida que o volume aumenta, podem ocorrer variações notáveis
em fatores importantes, como a transferência de oxigênio e calor, a pressão ou a densidade
celular 6. Portanto se faz necessário considerar todas as características dos probióticos, que
podem influenciar na sua produção industrial, para que assegure a viabilidade das cepas em
número suficiente e a manutenção de suas propriedades funcionais durante o processo
produtivo, o armazenamento e a distribuição das cepas e/ou alimento probiótico. Porém, sua
presença não deve afetar negativamente as propriedades sensoriais do alimento final 48; 73
.
Quadro 4 - Alegação de propriedade funcional e de saúde aprovada pela ANVISA para produtos probióticos
Microrganismo probiótico
Lactobacillus acidophilus, Lactobacillus casei shirota, Lactobacillus casei variedade rhamnosus,
Lactobacillus casei variedade defensis, Lactobacillus paracasei, Lactococcus lactis, Bifidobacterium
bifidum, Bifidobacterium animallis (incluindo a subespécie B. lactis), Bifidobacterium longum,
Enterococcus faecium
Alegação
“O (indicar a espécie do microrganismo) (probiótico) contribui para o equilíbrio da microbiota
intestinal. Seu consumo deve estar associado a uma alimentação equilibrada e hábitos de vida
saudáveis”.
Requisitos específicos
A quantidade mínima viável para os probióticos deve estar situada na faixa de 108 a 10
9 UFC na
recomendação diária do produto pronto para o consumo, conforme indicação do fabricante. Valores
menores podem ser aceitos, desde que a empresa comprove sua eficácia.
A documentação referente à comprovação de eficácia, deve incluir:
- Laudo de análise do produto que comprove a quantidade mínima viável do microrganismo até o final
do prazo de validade.
- Teste de resistência da cultura utilizada no produto à acidez gástrica e aos sais biliares.
A quantidade do probiótico em UFC, contida na recomendação diária do produto pronto para
consumo, deve ser declarada no rótulo, próximo à alegação. Os microorganismos Lactobacillus
delbrueckii (subespécie bulgaricus) e Streptococcus salivarius (subespécie thermophillus) foram
retirados da lista tendo em vista que, além de serem espécies necessárias para produção de iogurte, não
possuem efeito probiótico cientificamente comprovado. Fonte: Brasil
7

37
2.3 KEFIR
2.3.1 Definição
No RTIQ de Leites Fermentados 20
, publicado pelo Ministério da Agricultura Pecuária e
Abastecimento (MAPA) kefir é definido como “produto resultante da fermentação do leite
pasteurizado ou esterilizado, por cultivos ácido-láticos elaborados com grãos de kefir,
Lactobacillus kefir, espécies do gênero Leuconostoc, Lactococcus e Acetobacter com
produção de ácido láctico, etanol e dióxido de carbono”. E os grãos de kefir devem ainda ser
“constituídos por leveduras fermentadoras de lactose (Kluyveromyeces marxianus) e
leveduras não fermentadoras de lactose (Saccharomyces omnisporus e Saccharomyces
cerevisiae e Saccharomyces exiguus), Lactobacillus casei, Bifidobacterium spp. e
Streptococcus salivarius subsp. thermophilus”, devendo apresentar os requisitos físico-
químicos e microbiologicos constantes no quadro 5.
Quadro 5- Requisitos físico-químicos e microbiológicos do kefir
Kefir
Físico-químico
Acidez (g de ácido láctico/100g) <1,0
Etanol (% v/m) 0,5 a 1,5
Matéria gorda láctea (g/100g)
Creme Mín 6,0
Integral 3,0 a 5,9
Parcialmente desnatado 0,6 a 2,9
Desnatado Max. 0,5
Proteínas lácteas (g/100g) Mín. 2,9
Microbiológico
Contagem de BAL totais (UFC/g) Mín 107
Contagem de leveduras
específicas (UFC/g)
Mín 104
Fonte: Brasil 20
2.3.2 Aspectos gerais
O kefir, também conhecido como kefyr, kephir, kefer, kiaphur, quefir, knapon, kepi e
kippi 102; 103; 104
, é uma das bebidas fermentadas mais antigas do mundo, e a tecnologia de
produção e o uso da cultura starter específica vem sendo aplicada ao longo dos anos, em
vários países 2. Tradicionalmente, a fermentação do leite era realizada em bolsas
confeccionadas com peles de animais e potes de barro, nos quais leites de vaca, cabra, ovelha,
camelos e búfalos eram usados como substrato para fermentação de forma contínua, que, após
o consumo do kefir adicionava-se uma nova quantidade de leite fresco. Os microrganismos
formavam uma fina camada e posteriormente aglomeravam-se sobre as superfícies dos

38
recipientes. Acredita-se que este processo de formação do filme microbiano foi potencializado
pelas condições climáticas do Cáucaso 2.
Kefir é uma bebida láctea revigorante, refrescante, viscosa, levemente carbonatada,
ácida e com baixo teor alcóolico 18
. A principal diferença entre esta bebida e os outros leites
fermentados tradicionais, deve-se ao fato de que o kefir é produzido utilizando grãos de kefir
como cultura iniciadora e, ao fim do processo de fermentação, esses grãos podem ser
recuperados e reutilizados em um novo processo 16; 44
. O nome kefir é derivado da palavra
turca keyif, que significa “sentir-se bem” 16; 18
.
Historicamente, grãos de kefir eram considerados um presente de Alá para os povos
muçulmanos, do norte das montanhas do Cáucaso, e eram passados de geração a geração. As
tribos acreditavam que os grãos eram abençoados e possuíam propriedades excepcionais para
promoção da saúde. Por esse motivo, esses povos mantinham sigilo sobre a existência dos
grãos e o método de fermentação da bebida, até que a família russa Blandov, proprietária de
uma fábrica de latícinios, foi designada a obter os grãos e desvendar o mistério da produção
do kefir. Então, Irina Sakharova foi enviada à corte do príncipe Bek-Mirza Barchorov, na
tentativa de obter uma pequena quantidade dos grãos. O príncipe ficou encantado com Irina,
mas recusou-se a doá-los ou vendê-los. Ao retornar para Moscou, Irina foi sequestrada pelos
súditos do príncipe e obrigada a casar-se com ele. Porém, os irmãos Blandov conseguiram
resgatá-la antes do casamento, e o príncipe foi obrigado a recompensá-la dando-lhe uma
grande quantidade de grãos de kefir. Em 1908, as primeiras garrafas de kefir produzidas pelos
irmãos Blandov chegaram a Moscou e foram usadas primeiramente por médicos e, logo
depois, já era a bebida láctea fermentada mais popular da Rússia. Em 1973, Irina Sakharova
recebeu uma carta do ministro da Indústria de Alimentos da antiga União Soviética,
agradecendo-a por seu papel heróico no fornecimento do kefir para população russa 105
.
Atualmente, o autêntico kefir tradicional pode ser preparado pela adição dos grãos de
kefir em leite fresco ou pasteurizado por famílias de todo o mundo. Há anos, é muito popular
na antiga União Soviética, Hungria e Polônia. Também é conhecido na Suécia, Noruega,
Finlândia e Alemanha e, mais recentemente, na Grécia, Áustria, Brasil e Israel. Nos Estados
Unidos, é considerada como uma bebida étnica e está disponível comercialmente há mais de
20 anos e, no Japão sua popularidade vem aumentando 106
.

39
2.3.3 Grãos de kefir
Tradicionais do Cáucaso, os grãos surgiram a partir de uma fermentação natural
realizada em bolsas confeccionadas com pele de animais, que ficavam dentro das casas
durante o inverno e fora, durante o verão. O leite fermentado era removido para consumo,
adicionando-se leite fresco aos resíduos remanescentes na bolsa. Ao longo do tempo, os
microrganismos se acumularam e foram incorporados a um material composto por proteínas e
polissacarídeos, formando o grão 44
.
Os grãos podem ter surgido de forma independente em locais diferentes, e podem estar
associados a populações microbianas específicas responsáveis pela produção de bebidas com
propriedades sensoriais distintas 107
. Portanto, ainda não está claro se todos os grãos são
provenientes de uma única cultura starter, ou não, já que análises microbiológicas de grãos,
provenientes de diferentes regiões, apresentam diferenças em suas respectivas populações
microbianas 18
.
Figura 1 – Grãos de Kefir
Fonte: Carneiro 8
Grãos de kefir assemelham-se as flores da couve-flor, sua cor varia do branco ao
branco amarelado, têm uma textura viscosa e firme, multi-lobular irregular, unida por uma
seção central única com um tamanho que varia de 0,3 a 3,0 cm de diâmetro (Figura 1). Estes
grãos são compostos por proteína e polissacarídeos onde a microbiota fica revestida por estas
macromoléculas 18; 44; 102; 108; 109
. A composição química dos grãos compreende 890-900 g/kg
água, 2 g/kg lipídio, 30 g/kg de proteína, 60 g/kg de açúcar e 7 g/kg de cinza 102; 110
. A
microbiota dos grãos é descrita como uma associação simbiótica, notavelmente estável,
acoplada a uma matriz composta por proteína (caseína) e polissacarídeos. O principal
polissacarídeo produzido pelos microrganismos é denominado kefiran 108
.

40
Embora algumas tentativas para produção de grãos já tenham sido realizadas, a partir
de culturas puras ou mistas comumente presentes nos grãos, nenhum resultado positivo foi
relatado 111
. Muitos autores acreditam que novos grãos de kefir só podem ser obtidos pela
propagação dos grãos já existentes 16; 102; 110
. Entretanto, Motaghi e colaboradores 112
sugeriram que novos grãos poderiam ser produzidos utilizando o mesmo método rústico
descrito na história. Para isso, adicionaram leite pasteurizado e microbiota intestinal de ovelha
à bolsa confeccionada com pele de cabra, incubando-se a mistura a 24-26°C por 48 horas,
agitando-se a cada hora. Após a coagulação do leite, 75% do leite fermentado era trocado por
leite fresco, durante 12 semanas. Ao final do processo, observaram que uma camada de
polissacarídeo foi formada na superfície. Esta camada foi removida e propagada em leite de
vaca. Dessa forma, os autores relataram que obtiveram grãos semelhantes aos grãos
tradicionais (0,5 – 3,2 cm em diâmetro), e alguns microrganismos isolados foram similares
aos frequentemente encontrados no kefir.
Provavelmente, diferentes fatores exercem influência no aumento da biomassa dos
grãos de kefir, dentre os quais se destacam: a renovação de leite em intervalos regulares, a
temperatura de cultivo, a lavagem dos grãos e a presença de nutrientes essenciais na
concentração correta no meio de crescimento 113
. Os grãos se mantêm viáveis por sua
transferência diária em leite fresco e, ao crescerem por aproximadamente 20 horas, podem
aumentar cerca de 25% de sua biomassa 18
. Quando estocados diretamente à temperatura
ambiente ou a 4°C, apresenta um perfil microbiológico diferente dos grãos frescos 114
.
Garrote, Abraham e De Antoni 115
demonstraram que ao estocá-los ressuspensos em leite à -
80 ou -20°C por 120 dias, os grãos mantiveram suas propriedades de fermentação.
No Brasil, o kefir ainda não é fabricado industrialmente, mas existe como um produto
de fabricação doméstica. Esta bebida difere dos outros produtos lácteos porque não é
resultado da atividade metabólica de uma única ou poucas espécies, o que confere ao produto
final (kefir) propriedade sensorial singular 102
.
2.3.4 Aspectos microbiológicos dos grãos e do kefir
Embora variações sazonais na microbiota tenham sido reportadas, a população
microbiana que compõe os grãos parece ser relativamente constante ao longo do tempo 102; 116
.
Ainda que os microrganismos presentes em diferentes fermentações possam diferir uns dos
outros, algumas espécies estão sempre presentes 16; 102
, entre as quais, bactérias dos gêneros

41
lactobacilos, lactococos e leuconostoc. A presença do gênero acetobacter, embora considerada
por muitos anos como contaminante, atualmente é aceita como membro natural da microbiota
dos grãos e kefir 102; 109
, além das leveduras que também fazem parte deste consórcio.
A proporção dos organismos presentes no kefir geralmente é de 80% de Lactococcus
spp. e Leuconostoc spp., 10-15% de leveduras e 5-10% de Lactobacillus spp., enquanto que
os grãos de kefir contém 65-80% de Lactobacillus spp., 10-15% de leveduras e 5-25% de
Lactococcus spp. e Leuconostoc spp. 2; 18; 104
. Normalmente, as contagens presuntivas de
microrganismos, presentes nos grãos estão na ordem de 108-10
9 UFC/g de lactobacilos, 10
8-
109 UFC/g de lactococos e leuconostoc, 10
5-10
8 UFC/g de BAA e 10
6-10
8 UFC/g de
leveduras; enquanto que para o produto fermentado são um pouco mais baixas, estimadas em
106-10
8 UFC/mL de lactobacilos, 10
8-10
9 UFC/mL de lactococos e leuconostoc, 10
4-10
5
UFC/mL de BAA e 104-10
5 UFC/mL de leveduras
44; 102; 104; 108; 109; 117; 118. O perfil microbiano
do produto final (kefir) não é igual ao perfil dos grãos, devido a várias condições do processo
de fermentação 104
, como o tempo de fermentação, a temperatura, o grau de agitação, o tipo de
leite, a proporção inóculo grão/leite, entre outros 2; 104; 119
.
A distribuição dos microrganismos no grão também pode influenciar o perfil
microbiológico do kefir. Em vários estudos foram utilizados Microscopia Eletrônica de
Varredura (MEV) para avaliar a estrutura e distribuição dos microrganismos em grãos de
kefir, e os resultados obtidos são controversos44; 109; 117; 120; 121; 122
. Apesar de L. lactis ser um
dos microrganismos predominantemente isolado da bebida kefir, vários pesquisadores 109; 117;
121 não detectaram cocos nas micrografias obtidas com os grãos, provavelmente devido à fraca
aderência dos lactococos aos grãos durante o crescimento, o que facilitaria a liberação dos
mesmos no processo de lavagem 44; 109; 117; 121
. Segundo Rea et al. 44
que observaram a rara
presença de cocos na porção externa de grãos, esta fraca aderência dos cocos à superfície dos
grãos justificaria a maior contagem de lactococos frequentemente encontrada na bebida kefir,
diferentemente do perfil dos grãos 102
. Além disso, também é provável que o interior dos
grãos possua um pH baixo, devido à produção de ácidos por lactobacilos, o que dificulta o
crescimento dos lactococos neste microambiente 44
.
Os microrganismos de superfície são aqueles que provavelmente causam um maior
impacto no processo de fermentação da produção do kefir, fazendo com que as diferenças
observadas entre as porções internas e externas do grão sejam relevantes 102
. Lin, Chen e Liu
120 concluíram que o local de origem dos grãos por si só pode explicar as diferenças que vêm
sendo reportadas em vários estudos de microscopia eletrônica.

42
Figura 2 - Micrografia eletrônica de grão de kefir mostrando bactérias e leveduras na matriz de polissacarídeo e
proteína. (foto do autor)
Diferentes espécies de bactérias e leveduras já foram isoladas e identificadas a partir
dos grãos de kefir e de sua respectiva bebida. Porém, devido à associação simbiótica dos
microrganismos presentes nos grãos (Figura 2), o crescimento e a sobrevivência de estirpes
individuais são dependentes da presença de outras. Alguns autores relataram dificuldade em
estudar e monitorar os grãos por métodos microbiológicos convencionais, pois quando os
microrganismos são separados como culturas puras não crescem bem em leite ou apresentam
redução de sua atividade bioquímica. Sendo assim, o isolamento e a identificação dos
microrganismos presentes no grão e no kefir depende da escolha de meios de cultura
adequados (Quadros 6 e 7) para o crescimento da maior parte das populações que compõem a
microbiota 102
.
A maioria das bactérias está intimamente relacionada entre si 23
, dificultando sua
identificação, uma vez que várias de suas características fenotípicas e bioquímicas são
semelhantes, gerando identificações errôneas 123; 124
. Atualmente, métodos moleculares de
identificação são considerados confiáveis e definitivos na classificação e identificação dos
microrganismos e, por este motivo, estão sendo amplamente aplicados no estudo do kefir e de
seus grãos 107; 125; 126
, fazendo com que a identificação de vários microrganismos descrita
anteriormente esteja sendo revisada.
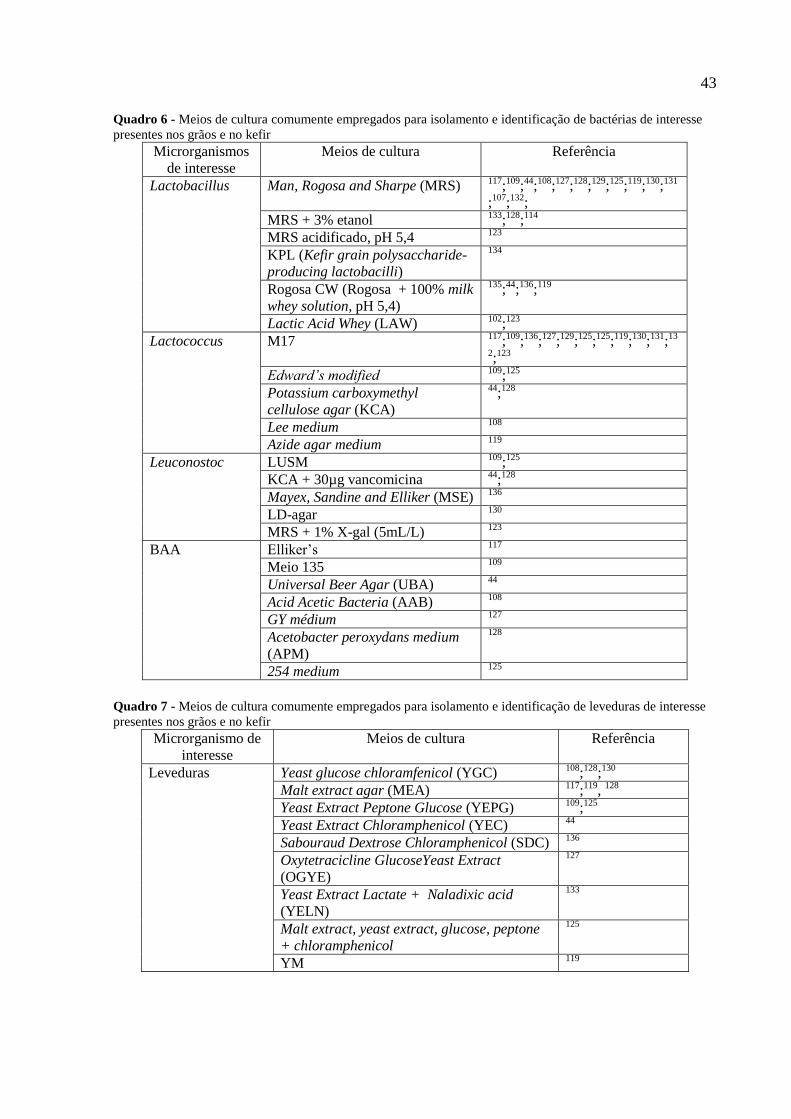
43
Quadro 6 - Meios de cultura comumente empregados para isolamento e identificação de bactérias de interesse
presentes nos grãos e no kefir
Microrganismos
de interesse
Meios de cultura Referência
Lactobacillus Man, Rogosa and Sharpe (MRS) 117
;109
;44
;108
;127
;128
;129
;125
;119
;130
;131
;107
;132
;
MRS + 3% etanol 133
;128
;114
MRS acidificado, pH 5,4 123
KPL (Kefir grain polysaccharide-
producing lactobacilli)
134
Rogosa CW (Rogosa + 100% milk
whey solution, pH 5,4)
135;44
;136
;119
Lactic Acid Whey (LAW) 102
;123
Lactococcus M17 117
;109
;136
;127
;129
;125
;125
;119
;130
;131
;13
2;123
Edward’s modified 109
;125
Potassium carboxymethyl
cellulose agar (KCA)
44;128
Lee medium 108
Azide agar medium 119
Leuconostoc LUSM 109
;125
KCA + 30µg vancomicina 44
;128
Mayex, Sandine and Elliker (MSE) 136
LD-agar 130
MRS + 1% X-gal (5mL/L) 123
BAA Elliker’s 117
Meio 135 109
Universal Beer Agar (UBA) 44
Acid Acetic Bacteria (AAB) 108
GY médium 127
Acetobacter peroxydans medium
(APM)
128
254 medium 125
Quadro 7 - Meios de cultura comumente empregados para isolamento e identificação de leveduras de interesse
presentes nos grãos e no kefir
Microrganismo de
interesse
Meios de cultura Referência
Leveduras Yeast glucose chloramfenicol (YGC) 108
;128
;130
Malt extract agar (MEA) 117
;119
, 128
Yeast Extract Peptone Glucose (YEPG) 109
;125
Yeast Extract Chloramphenicol (YEC) 44
Sabouraud Dextrose Chloramphenicol (SDC) 136
Oxytetracicline GlucoseYeast Extract
(OGYE)
127
Yeast Extract Lactate + Naladixic acid
(YELN)
133
Malt extract, yeast extract, glucose, peptone
+ chloramphenicol
125
YM 119

44
Em alguns casos, a nomenclatura atribuída a várias espécies bacterianas foi
modificada, e alguns autores especulam que, nos últimos anos, a composição da comunidade
microbiana dos grãos e do kefir pode ter sido superestimada 102; 123
. Além disso, métodos
independentes de cultivo, como as técnicas genéticas de fingerprints estão sendo aplicados
para a análise do complexo consórcio microbiano existente nos grãos e no kefir, a fim de
complementar os estudos realizados com métodos de microbiologia clássica dependentes de
cultivo 107
.
2.3.4.1 Bactérias
A caracterização da diversidade bacteriana presente nos grãos de kefir tem sido
avaliada utilizando-se tanto os métodos tradicionais dependentes de cultivo, quanto
independentes de cultivo. Várias espécies de Lactobacillus já foram isoladas tanto da bebida
kefir quanto dos grãos, incluindo L. kefiri, L. kefiranofaciens, L. kefirgranum e L. parakefiri
compondo a população dominante 44; 108; 119; 133
. Além desses, L. casei, L. parabuchneri, L.
paracasei e L. plantarum também foram identificados 107; 121; 123; 132; 137; 138
. Lactococcus lactis
subsp. lactis e Lactococcus lactis subsp. cremoris são comumente reportados 121
,
predominando no produto fermentado 119; 126; 131; 132
. Leuconostoc spp. e outras espécies de
Lactobacillus são isoladas em menores números 119; 121; 123
.
O L. kefiranofaciens, isolado frequentemente do kefir, é de particular interesse, devido
à produção do polissacarídeo kefiran. Já foi relatado que a produção de kefiran é estimulada
quando o L. kefiranofaciens cresce com S. cerevisiae, mimetizando a simbiose presente nos
grãos de kefir 139
.
As espécies BAA têm sido isoladas e identificadas tanto dos grãos quanto do kefir.
Entre as quais, já foram citadas: Acetobacter aceti, A. lovaniensis, A. rasens, A. syzygii e
Gluconobacter japonicus 44; 107; 109
. No entanto, em alguns países, a presença destas espécies é
considerada indesejável 2; 102; 104
. Estes microrganismos têm recebido menor atenção, apesar
de seu papel essencial tanto no consórcio microbiano 110
quanto nas características sensoriais
do produto final.

45
2.3.4.2 Leveduras
As leveduras são menos estudadas do que as bactérias, embora seja evidente que as
primeiras promovam um ambiente favorável ao crescimento das últimas, produzindo
metabólitos que contribuem para o flavor e sabor típico do kefir 119
.
Isoladas do kefir, as leveduras podem ser classificadas tanto como fermentadoras de
lactose, quanto como não fermentadoras, sendo as últimas dominantes 104
. Recentemente,
métodos dependentes e independentes de cultivo foram utilizados para estudo e identificação
da diversidade de leveduras, destacando-se: Kluyveromyces marxianus, Torulaspora
delbrueckii, Saccharomyces cerevisiae, Candida kefyr, Saccharomyces unisporus, Pichia
fermentans, Kazachastania aerobia, Lachancea meyersii, Yarrowia lipolytica e Kazachstania
unispora 109; 119; 121; 140; 141
.
A fonte de carbono para a propagação de culturas de leveduras consiste usualmente de
açúcares facilmente assimiláveis, como glicose, frutose, manose, galactose, maltose, lactose
hidrolisada, entre outros. As leveduras produzem, predominantemente, as vitaminas do
complexo B, constituindo assim uma excelente fonte destas vitaminas para a nutrição humana
e animal 43
.
As leveduras são as principais responsáveis pela produção do etanol e CO2 no kefir.
Segundo Simova e colaboradores 119
, uma contagem mínima de 105
UFC/g de leveduras deve
estar presente nos grãos, para que as propriedades sensoriais desejadas no produto sejam
atingidas. No entanto, alguns lactobacilos heterofermentativos como L. kefiri, também são
capazes de produzir moderadas quantidades de etanol e CO2 104
.
2.3.4.3 Interações entre microrganismos no kefir
As características de um produto lácteo fermentado espontaneamente são resultantes
da combinação do metabolismo de vários microrganismos 16
. As possíveis interações entre
leveduras e bactérias em leites fermentados já foram descritas por alguns autores, mas ainda
não foram completamente elucidadas 139; 142
.
Leveduras e bactérias sobrevivem e se propagam, compartilhando seus bioprodutos
como fontes de energia ou estimulantes de crescimento. Leveduras exercem um importante
papel no processo de fermentação do kefir, produzindo fatores essenciais de crescimento,
como aminoácidos, vitaminas, alteração do pH, secreção de etanol e produção CO2 16
. A

46
interação entre leveduras e bactérias láticas pode ser estimulada ou inibida pelo crescimento
de uma, ou ambas, em co-culturas. Estes microrganismos podem competir por nutrientes para
crescimento, ou podem produzir metabólitos que inibam ou estimulem umas as outras 16
.
Os produtos da fermentação do kefir são, predominantemente, oriundos das
fermentações láticas por BAL. A tendência é que os lactococos cresçam mais rápido no leite
do que leveduras, produzindo ácido lático a partir da fermentação da lactose e forneçam um
ambiente adequado para o crescimento das leveduras. As BAL diminuem rapidamente o pH
com acúmulo de ácido lático 2, até que essa produção seja inibida e os componentes que
originam o flavor sejam produzidos (acetaldeído, diacetil, etc) juntamente com aroma de leite
fermentado 16
.
Algumas espécies possuem atividade proteolítica e lipolítica, aumentando a
concentração de aminoácidos livres e ácidos graxos durante o seu crescimento, como por
exemplo, Yarrowia lipolytica. Leveduras e Acetobacter spp. também podem produzir
vitaminas que estimulam o crescimento de BAL.
Várias espécies de leveduras, como Debaromyces hansenni e Y. lipolytica, podem
assimilar ácido lático e em co-cultura com BAL, podem gerar um aumento no pH, o que
permite o crescimento e o metabolismo adicional da lactose por bactérias láticas, aumentando,
assim, a produção de ácido lático e desencadeando o início de todo o ciclo metabólico 16
.
2.3.5 Características do kefir
As características químicas, microbiológicas e sensoriais do kefir são influenciadas
pelos microrganismos presentes nos grãos e origem dos mesmos, atributos químicos do leite
empregado, condições de estocagem, manipulação e tecnologia de processamento, como
agitação e temperatura de incubação 110; 127
. Algumas dessas características foram
selecionadas, e estão descritas nas próximas seções.
2.3.5.1 Químicas
A composição química do kefir 106
varia de acordo com o tipo de leite empregado, sua
origem e teor de gordura, composição microbiológica dos grãos ou culturas iniciadoras e
processo tecnológico empregado 129
.

47
Os principais produtos finais da fermentação do kefir são ácido láctico, CO2 e etanol
110; 143. Compostos aromáticos tais como acetaldeído, acetoína, diacetil
110; 130; 143; 144; 145 ácido
hipúrico, pirúvico, acético, propiônico e butírico 118
também estão presentes no kefir. O
lactato L(+) é o ácido orgânico presente em mais alta concentração após a fermentação, da
lactose original do leite 18
.
O kefir tradicional geralmente apresenta pH entre 4.2 a 4.6 102; 106
, cerca de 0,6 a 1%
(p/p) de ácido láctico, álcool entre 0,01 a 2,0% (m/v); 1,0 – 1,33 g/L de CO2 e teor de gordura,
dependente do leite utilizado 19; 102; 110
. Segundo Zourari e Anifantakis 110
, uma proporção
ótima de 3:1 entre diacetil e acetaldeído define o aroma típico do kefir. Propionaldeíno,
isobutanol, n-propanol, álcool isoamílico e ácido acético são compostos igualmente
importantes para contribuição do flavor, mas seus teores variam consideravelmente durante a
maturação. As características típicas do kefir são seu sabor ácido, pungente e levemente
efervescente, resultando em uma bebida refrescante 19; 143; 144
.
Considerando o Codex Alimentarius 21
, a composição físico-química do kefir deve ser
constituída por menos de 10% gordura láctea (% p/p), mínimo de 2,7% de proteína láctea (%
p/p) e mínimo de 0,6% de acidez titulável expressa como % de ácido lático (% p/p). No RTIQ
de Leites Fermentados, publicado pelo MAPA também constam os requisitos da composição
físico-química do kefir produzido no Brasil (Quadro 5).
2.3.5.2 Nutricionais
As quantidades de proteína, gordura e minerais do kefir dependem daquelas presentes
no leite em que foi produzido possuindo, portanto, um valor nutricional relacionado ao
mesmo. A bebida é palatável e tem alta digestibilidade, pois no processo de fermentação
ocorre desnaturação das proteínas do leite e hidrólise de algumas, resultando em estruturas
menores que são mais susceptíveis à digestão pelos sucos gástrico e intestinal 102
. Vitaminas,
minerais e aminoácidos essenciais também estão presentes no kefir, auxiliando no tratamento
e manutenção das funções normais do corpo 106
.
O kefir é rico em vitamina B1, B12, B6, cálcio, aminoácidos, ácido fólico, vitamina K 18;
106; 118 e biotina, uma vitamina do complexo B, que auxilia a assimilação de outras vitaminas,
como ácido fólico, pantotênico e B12. O teor de vitaminas presentes na bebida é influenciado
pela fonte do leite empregado e pela microbiota dos grãos de kefir 102
. Um aumento no teor de
piridoxina (B6) foi observado no kefir produzido com leite de ovelha, cabra e égua, enquanto

48
que o ácido fólico (B9) teve sua concentração aumentada com o uso de leite de ovelha, vaca e
cabra. Entretanto, outros autores observaram uma redução no teor de vitamina B12 (34-37%)
devido à inclusão de Propionibacterium peterssoni e Propionibacterium pituitosum 118.
Com relação ao teor de minerais, foi relatada a presença de macro-elementos como
potássio, cálcio, magnésio, fosforo, e micro-elementos como cobre, zinco, ferro, manganês,
cobalto e molibdênio 106
.
As proteínas do kefir são parcialmente digeridas, facilitando a sua utilização pelo
organismo. Guzel-Seydim, Seydim e Greene 146 relataram que mudanças no perfil de
aminoácidos são resultantes do crescimento microbiano que ocorre durante a fermentação do leite.
Portanto, observaram que o perfil de aminoácidos presentes no kefir apresentou maior quantidade
de treonina, serina, alanina, lisina e amônia do que no leite ou iogurte. Outro pesquisador reportou
a presença de valina, isoleucina, metionina, lisina, treonina, fenilalanina e triptofano 118.
Além disso, este leite fermentado contém uma variedade de microrganismos que tem
potencial para auxiliar na digestão de lactose através da atividade da enzima β-galactosidase.
Sendo assim, é um alimento indicado na dieta de indivíduos intolerantes a lactose, que são
incapazes de digerir quantidades significantes deste açúcar predominante no leite 102; 106
.
2.3.5.3 Compostos bioativos no kefir
Na área de alimentos funcionais, o interesse por compostos bioativos é crescente, uma
vez que o mesmos conferem benefícios à saúde e resistência a doenças. Uma subclasse dos
alimentos funcionais são os alimentos probióticos, a partir dos quais há várias fontes de
componentes bioativos. Há vários compostos no kefir que podem ter propriedades bioativas:
os próprios microrganismos (vivos ou mortos), metabólitos produzidos por microrganismos
durante a fermentação (polissacarídeos, bacteriocinas), ou a quebra de produtos da matriz
alimentar (peptídeos). Todos estes compostos podem ser responsáveis por efeitos benéficos e
podem agir independentemente, ou em conjunto, para produzir tais efeitos ao longo do
consumo de kefir 18
.
Alguns exopolissacarídeos com estrutura e composição diferentes são produzidos por
uma variedade de BAL, incluindo os gêneros Lactobacillus, Streptococcus, Lactococcus e
Leuconostoc 147; 148
. Estes carboidratos conferem propriedades protetoras e adaptativas para as
bactérias produtoras e são secretados para o meio extracelular 148
. Nos alimentos, os
exopolissacarídeos contribuem para características sensoriais e estabilidade do produto 18; 139
.

49
Um polissacarídeo solúvel em água, denominado kefiran é produzido por L. kefiranofaciens
sob condições anaeróbias. A proporção de kefiran nos grãos é de aproximadamente 45%,
sendo composto por D-glicose e D-galactose em uma proporção de 1:1. A principal função do
kefiran nos grãos é descrita como uma proteção dos microrganismos contra dessecação 116; 139;
147. O potencial das propriedades benéficas do kefiran à saúde tem sido reportado por vários
grupos 18; 102; 149
.
Durante a produção tradicional do kefir, os grãos não são manipulados assepticamente
e, apesar disso, nenhuma contaminação significativa com microrganismos indesejáveis é
reportada. A alta concentração de ácidos orgânicos e presença de outras substâncias
antimicrobianas, tais como bacteriocinas, poderiam explicar a ausência de patógenos nos
grãos manipulados por centenas de anos 150
.
Muitos isolados bacterianos de grãos de kefir possuem alta atividade proteolítica, o
que aumenta a possibilidade de que peptídeos bioativos estejam presentes no kefir. 18
, fato que
pode auxiliar no crescimento de microrganismos. Além disso, estes compostos liberados por
bactérias e acumulados no leite afetam o potencial nutricional e o valor biológico do leite
fermentado 151
.
Os peptídeos bioativos são definidos como fragmentos de proteínas específicas e
exercem um impacto positivo nas funções ou condições corporais, como por exemplo, os
peptídeos bioativos isolados de produtos lácteos fermentados, incluindo o kefir 152
, que
possuem atividade inibitória à enzima conversora de angiotensina-I (ECA), responsável pela
hidrólise de substratos envolvidos na regulação da pressão arterial 152
.
2.3.5.4 Terapêuticas
Dentre os vários efeitos benéficos relacionados com o consumo de kefir, já foram
citados a redução dos efeitos de intolerância a lactose 153
, a imunomodulação 154; 155
, a
proteção contra microrganismos patogênicos 156; 157; 158
, o balanço da microbiota intestinal 159
,
a atividade anticarcinogênica 160
, a regeneração hepática 161
e a atividade anti-inflamatória 162
.

50
2.4 MÉTODOS DEPENDENTES E INDEPENDENTES DE CULTIVO EMPREGADOS
NO ESTUDO DE COMUNIDADES MICROBIANAS
2.4.1 Métodos tradicionais
Considerados como “padrão ouro” para análises de alimentos, métodos convencionais
empregados para detecção, identificação e classificação de microrganismos envolvem o
cultivo destes em meios seletivos, análise da morfologia e propriedades bioquímicas e/ou
imunológicas, sendo muitas vezes insuficientes para alguns grupos bacterianos.
Métodos de cultivo do microrganismo em meios seletivos e diferenciais são,
aparentemente, os mais simples. Entretanto, estes métodos são mais demorados e podem não
ser representativos, principalmente em análises de microrganismos mais relacionados entre si,
onde a identificação baseada em características fenotípicas e bioquímicas, pode ser ambígua 6;
24; 123.
Os métodos anteriormente descritos não são altamente confiáveis em estudos de
ecologia microbiana, devido à existência de bactérias não cultiváveis, às exigências de certos
microrganismos por ambientes anaeróbios, e a outros fatores que podem subestimar a
diversidade microbiana estudada, como por exemplo, a adequação nutricional e seletividade
do meio de cultura, temperatura e tempo de incubação empregados 6; 163
. Dessa forma, o
emprego de métodos independentes de cultivo tornou-se uma necessidade metodológica no
campo da ecologia microbiana de alimentos, principalmente em estudos de comunidades
complexas como o kefir.
2.4.2 Métodos moleculares
Uma das áreas da microbiologia molecular aplicada está envolvida no
desenvolvimento de métodos moleculares para identificação e monitoramento de
microrganismos em ecossistemas naturais. Nos últimos anos, técnicas moleculares baseadas
no estudo do gene rrs, codificador do RNA ribossomal 16S, fragmento alvo de amplificação
pela reação em cadeia da polimerase ou PCR (Polymerase Chain Reaction), tem se tornado
uma ferramenta importante no estudo taxonômico e de comunidades bacterianas 6; 164
. Essas
técnicas facilitam a caracterização e o monitoramento de uma enorme variedade de grupos
microbianos, incluindo populações não cultiváveis, possibilitando inclusive, identificação dos
componentes individuais presentes em diversos grupos e seu estado fisiológico de atividade 6.

51
A taxonomia molecular baseada no estudo das moléculas de DNA e RNA pode levar à
inferência de relações evolutivas sobre grupos de microrganismos, complementando as
características fenotípicas e resolvendo muitos problemas polêmicos de identificação e
classificação de diversos grupos taxonômicos complexos 165
. O aumento na velocidade e
acuidade da técnica de sequenciamento de ácidos nucléicos associado ao método de PCR é o
mais importante avanço nos estudos de sistemática microbiana 166
.
A mólecula do RNAr é amplamente aplicada como marcador filogenético, em estudos
de taxonomia e ecologia microbiana, pois está presente em todas as células, sendo detectada
com facilidade. Esta mólecula possui numerosas regiões variáveis e um elevado número de
sequências depositadas nas bases de dados, correspondentes a um amplo espectro de
microrganismos 6. Uma particularidade do gene RNAr 16S é a presença de regiões altamente
conservadas, as quais permitem o desenho de iniciadores de PCR e regiões altamente
variadas, as quais são utilizadas para inferir relações filogenéticas entre os microrganismos,
representadas em forma de árvores filogenéticas 6.
De maneira geral, os métodos moleculares podem ser divididos em dois grupos:
métodos que analisam a estrutura de uma comunidade a partir de um perfil de amplificação de
genes que codificam o RNAr 16S via PCR, conhecido como fingerprints; e métodos que
envolvem o sequenciamento do gene codificador do RNAr 16S dos membros da comunidade
e posterior identificação destas sequências em bancos de dados 167
.
2.4.2.1 Técnicas de fingerprinting de comunidades microbianas
As técnicas de fingerprinting fornecem uma visão global da estrutura genética da
comunidade microbiana. De uma maneira simplificada, a diversidade das sequências de DNA
obtidas a partir da amostra estudada é estimada através da migração eletroforética diferencial
em géis de agarose ou acrilamida, em função do tamanho ou composição destas sequências. O
fingerprint genético fornece um perfil de bandas, que reflete a complexidade da comunidade
microbiana analisada. Entretanto, estes perfis não podem ser diretamente interpretados em
termos de riqueza e abundância, uma vez que uma mesma banda pode ter se originado de
diferentes espécies, e/ou uma célula pode ser representada por várias bandas.
Assim, as técnicas de fingerprinting são especialmente adequadas para a comparação
de comunidades, uma vez que os perfis de bandas podem ser analisados em termos de
similaridades e as relações podem ser representadas por um dendrograma de similaridade.

52
Dessa maneira, correlações entre alterações na estrutura genética de comunidades microbianas
e variações temporais no ecossistema podem ser estabelecidas 166
.
A técnica de PCR, juntamente com eletroforese em gel com gradiente desnaturante
(Denaturing Gradient Gel Electrophoresis- DGGE) descrita por Muyzer, De waal e
Uitterlinden 164
, é uma ferramenta altamente utilizada para estudo de ecossistemas
microbianos complexos, pois é capaz de produzir um fingerprint da comunidade microbiana
em amostras de alimentos, após a extração do DNA da comunidade e amplificação de
sequências do gene codificador do RNAr 16S 168
.
A técnica de DGGE é um método de eletroforese capaz de separar os produtos de PCR
de mesmo tamanho, mas com sequências diferentes, com base na composição de seus
nucleotídeos. Isto ocorre à medida que os fragmentos de DNA migram em um gel de
poliacrilamida contendo gradiente desnaturante composto por uréia e formamida, no qual os
fragmentos são separados baseados em seus diferentes perfis de desnaturação. Além disso, a
adição de uma sequência rica em GC (entre 30 e 50 pb), conhecida como grampo na
extremidade 5’ de um dos iniciadores utilizados durante a amplificação, permite a
incorporação de uma zona de fixação ou ancoragem em um dos extremos de todos os
produtos de PCR, impedindo a separação total da dupla fita de DNA. A eletroforese é
comumente realizada à uma temperatura de 60ºC e a identificação das espécies pode ser
alcançada pela excisão, purificação, reamplificação e sequenciamento das bandas obtidas com
o DGGE 6.
A tipagem de microrganismos, isto é, a capacidade de identificação em nível da
espécie e de discriminação entre indivíduos da mesma espécie, apresentou grandes avanços
nos últimos anos, tendo sido apoiada por uma série de novos métodos que fazem uso da
variação encontrada no DNA destes microrganismos 166
.
Embora o método de análise de restrição do DNA ribossomal amplificado (Amplified
Ribosomal DNA Restriction Analysis – ARDRA) tenha sido frequentemente utilizado para
caracterização de isolados de bactérias, na teoria, este método também pode ser usado para
análise de populações bacterianas, caso estas apresentem baixa diversidade de grupos
taxonômicos. A técnica de ARDRA envolve o uso de um par de iniciadores universais para a
amplificação do gene codificador do RNAr 16S ou da região espaçadora entre o RNAr 16S e
o RNAr 23S, seguida da digestão com enzimas de restrição e da análise dos perfis obtidos
com os fragmentos resultantes da digestão. Delfederico et al. 124
, ao estudarem lactobacilos
isolados de grãos de kefir, relataram que esta técnica foi capaz discriminar taxonomicamente

53
os microrganismos, em nível de espécie, enquanto que os perfis obtidos com a amplificação
aleatória do DNA polimórfico (Random Amplification of Polymorphic DNA – RAPD)
permitiram a diferenciação em nível de estirpe.
A técnica de RAPD-PCR consiste na amplificação do DNA utilizando um iniciador
randômico geralmente composto de 10 nucleotídeos à baixa temperatura de anelamento
(36°C). Este método não requer o conhecimento prévio da sequência a ser amplificada e pode
ser aplicado para qualquer espécie a partir do DNA purificado. Os produtos são diretamente
analisados em um gel de agarose, e o perfil eletroforético gerado pode servir para diferenciar
e caracterizar microrganismos 6; 124; 137
. Porém, a reação é muito sensível às condições de
amplificação, qualidade e concentração de DNA, os quais influem na reprodutibilidade do
método. Sendo assim, a avaliação com mais de um iniciador para caracterizar as estirpes deve
ser sempre utilizada 6; 169
.
Outro método empregado para gerar fingerprints, sem o uso de enzimas de restrição é
o rep-PCR (repetitive element sequence based PCR), que envolve o uso de iniciadores
baseados em sequências curtas, repetidas ao longo do genoma dos procariotos. Quando um
destes elementos repetitivos é detectado durante a reação de PCR, produtos amplificados de
tamanhos característicos são obtidos, gerando-se um padrão de fingerprint em gel de agarose
170 específicos e discriminativos para bactérias, em nível de cepas ou isolados
169; 170.
2.4.2.2 Técnicas de sequenciamento dos membros da comunidade microbiana
Há pouco tempo atrás, os estudos de ecologia microbiana por sequenciamento
baseava-se na combinação da PCR do gene rrs, com a clonagem de fragmentos resultantes e
posterior sequenciamento dos mesmos, o que permitia acesso direto à diversidade filogenética
dos microrganismos existentes em comunidades complexas. A biblioteca de clones incluía o
maior número possível de clones únicos da comunidade, os quais se podiam sequenciar e
submeter a uma análise filogenética 6. Entretanto, além de ser uma técnica demorada e
laboriosa, os altos custos da construção das bibliotecas, manutenção dos clones e o
sequenciamento baseado em capilaridade do DNA (métodos de Sanger) frequentemente
limitavam o número de sequências obtidas nas pesquisas 167; 171
.
Técnicas mais recentes de sequenciamento massivo de moléculas de DNA, conhecidas
também como pirosequenciamento, começaram a ser aplicadas, pois permitem analisar
centenas de milhares de sequências diferentes em apenas algumas horas, sem a necessidade da

54
exaustiva etapa de clonagem 172; 173; 174
. Estas plataformas de sequenciamento de nova
geração, como a 454 Life Sciences (Roche Applied Sciences), são utilizadas para explorar a
diversidade microbiana nos mais variados ambientes, inclusive na área de alimentos 173; 175
.
A técnica foi descrita por Ronaghi e colaboradores 176
que demonstraram que a
molécula de pirofosfato (PPi) produzida durante a reação de polimerização do DNA, poderia
ser utilizada para detectar a incorporação de um nucleotídeo específico. Este método foi
denominado sequenciamento pela síntese, pois a sequência é determinada à medida que a fita
complementar é sintetizada. Para a reação de PCR é necessário o preparo de uma solução
contendo o tampão da reação, os iniciadores, o DNA molde, o conjunto de enzimas e seus
substratos. Os desoxinucleotídeos utilizados na síntese da fita complementar são inseridos na
reação por etapas. Quando um dos nucleotídeos adicionados é complementar à fita molde,
este é incorporado pela DNA polimerase, liberando uma mólecula de PPi como subproduto da
reação (Figura 3). Neste momento, a enzima ATP sulfurilase vai agir, convertendo o PPi em
ATP (pelo substrato APS), e então a enzima luciferase utiliza o ATP gerado para oxidar a
luciferina, produzindo um sinal luminoso, de intensidade proporcional à quantidade de
nucleotídeos incorporados, que é capturado por uma câmera CCD (charge-coupled device)
acoplada ao sistema. Uma vez que o nucleotídeo adicionado é conhecido, a sequência pode
ser determinada. O ciclo, então, é finalizado com a atividade da enzima aspirase, que degrada
os nucleotídeos e moléculas de ATP que sobram da reação 167; 171; 172
.
A preparação da amostra começa com a fragmentação do DNA ou amplificação do
gene de interesse, seguida da ancoragem de sequências chamadas de adaptadores-454 ao final
dos fragmentos de DNA e por uma sequência de “código de barras” ou barcodes, específico
para cada amostra analisada. Esses adaptadores são complementares às sequências de
microesferas magnéticas e permitem que os fragmentos gerados (por fragmentação ou PCR)
sejam capturados individualmente pelas microesferas, fazendo com que as etapas posteriores
fiquem restritas a uma única fita simples por microesfera. As microesferas são
compartimentalizadas dentro de gotículas de uma emulsão, contendo os reagentes da reação
de PCR, onde os fragmentos únicos de fita simples são amplificados individualmente 174
.
Após a PCR em emulsão, as microesferas ligadas a milhares de cópias idênticas de
fragmentos de fita simples são depositadas individualmente em poços distintos, presentes em
uma placa, onde os reagentes para o sequenciamento são distribuídos, ocorrendo reações de
sequenciamento em cada poço, para um único tipo de fragmento ligado à microsfera 171; 174
.

55
Figura 3 – Esquema ilustrativo do sequenciamento na plataforma 454.
Fonte: Carvalho e Silva 171
; Margulies et al. 174
A ideia de se utilizar “códigos de barras” nas sequências produzidas foi introduzida
por Binladen e colaboradores 177
e permitiu reduzir o custo de uma única corrida. O princípio
é bastante simples e consiste em incluir nos iniciadores uma sequência de nucleotídeos curta
que permite a separação das sequências geradas por amostras ao fim do processo.

56
3 JUSTIFICATIVA
O hábito de consumo de kefir é crescente em vários países, em função do seu alto
valor nutricional e de suas propriedades funcionais benéficas à saúde. Entretanto, o método
tradicional de obtenção do kefir, utilizando-se os grãos como cultura starter, gera bebidas
fermentadas não padronizadas, o que dificulta a implantação industrial dessa tecnologia. Esta
dificuldade tem sido atribuída à complexidade e à alteração dinâmica da microbiota do grão
de kefir ao longo de sucessivas fermentações, prejudicando a obtenção de um produto
uniforme e estável.
O kefir se diferencia de outros produtos probióticos comerciais por ser mais complexo,
apresentando uma população microbiana mista de bactérias e leveduras, cuja composição
relativa pode variar em função de diversos fatores. Entretanto, devido à intensa simbiose entre
leveduras e bactérias, a população microbiana resultante exibe características muito similares,
tornando difícil o isolamento e a identificação de espécies individuais. Sendo assim, a
aplicação de técnicas moleculares em estudos de ecologia microbiana do kefir parece ser
necessária para identificação e monitoramento de microrganismos neste ecossistema.
Estas ferramentas moleculares permitirão estudar a ecologia microbiana desse produto,
a fim de avaliar a importância de cada microrganismo envolvido, tanto no processo de
fermentação e estocagem, quanto nos benefícios à saúde relatados ao longo de seu histórico
de consumo. Dessa forma, as técnicas de biologia molecular serão associadas aos métodos
convencionais, para auxiliar no esclarecimento de questões relacionadas à microbiota presente
nos grãos e nas etapas de produção e estocagem do produto final, viabilizando o melhor
entendimento das bases tecnológicas que podem ser empregadas em escala de produção
industrial.
Devido às suas características, os grãos de kefir podem servir como fonte de cepas
probióticas para aplicação na fabricação de novos produtos ou ainda de uma bebida comercial
“tipo kefir”. Desta forma, o uso destas técnicas poderá ser útil para a caracterização das cepas
isoladas com potencial probiótico.

57
4 OBJETIVOS
4.1 OBJETIVO GERAL
Estudar a ecologia da comunidade microbiana presente dos grãos do kefir, caracterizar
a bebida kefir durante a fermentação e estocagem, do ponto de vista microbiológico e
químico; além de avaliar o potencial probiótico das bactérias isoladas.
4.2 OBJETIVOS ESPECÍFICOS
caracterizar a diversidade microbiana presente em diferentes grãos de kefir, utilizando
as técnicas independentes de cultivo: PCR-DGGE e pirosequenciamento;
comparar a comunidade microbiana e monitorar possíveis mudanças durante
fermentação e estocagem da bebida kefir por técnicas dependentes e independentes de
cultivo;
determinar as modificações da composição de ácidos orgânicos, etanol e carboidratos
durante a fermentação e estocagem da bebida kefir;
avaliar o potencial probiótico de BAL isoladas dos grãos de kefir.

58
5 ESTRATÉGIA EXPERIMENTAL
Neste estudo, três grãos de kefir (AD, AV e AF) foram obtidos de pesquisadores de
Universidades Brasileiras (UFLA, AFV, UNIFAL, respectivamente), sendo o AF cultivado
em açúcar mascavo; e um quarto grão (AR) doado por uma família brasileira, que o cultivava
artesanalmente em leite. Devido ao baixo teor de gordura, e consequente minimização dos
efeitos inibitórios nas reações, moleculares, o leite desnatado foi eleito como o mais adequado
para ser utilizado neste estudo. Os grãos foram ativados em leite UHT desnatado (10% p/p)
por 20h a 20°C, três vezes consecutivas.
Grãos cultivados em leite (AD, AV e AR) foram submetidos à análise da comunidade
de bactérias e leveduras por PCR-DGGE e a diversidade bacteriana dos grãos, foi
caracterizada por pirosequenciamento (Cap. II). Em paralelo, o grão AR foi selecionado
devido à sua maior capacidade de aumento da biomassa em relação aos demais, sendo assim
utilizado como cultura starter para fermentação de leite UHT desnatado a 25°C por 24h.
Durante diferentes tempos da fermentação (0, 6, 12, 18 e 24 h) e estocagem (2, 7, 14, 28 dias),
alíquotas do kefir AR foram coletadas para a caracterização dos microrganismos cultiváveis
que foram enumerados e identificados em meios de cultura específicos, enquanto que a
comunidade microbiana foi avaliada por PCR-DGGE e RT-PCR-DGGE. A composição
química da bebida foi caracterizada quanto à concentração de ácidos orgânicos, carboidratos e
etanol durante a fermentação e estocagem, por CLAE (Cap. III). Além disso, as BAL isoladas
de todos os grãos foram avaliadas quanto ao seu potencial probiótico (Cap. IV) (Fig. 4).
Figura 4– Desenho experimental

59
6 CAPÍTULOS
O presente estudo culminou na produção de quatro artigos científicos que foram
traduzidos e expostos nos capítulos a seguir. O primeiro capítulo engloba uma revisão da
literatura sobre os aspectos terapêuticos, microbiológicos e tecnológicos da bebida kefir. O
segundo refere-se à caracterização da microbiota de grãos de kefir, no terceiro reporta-se a
análise da comunidade microbiana e a determinação dos metabólitos produzidos durante a
fermentação e estocagem da bebida kefir, e no quarto relata-se a avaliação do potencial
probiótico das estirpes de bactérias láticas isoladas dos grãos de kefir.
6.1 CAPÍTULO I - KEFIR: UMA BEBIDA PROBIÓTICA NATURAL1
RESUMO
Kefir é um leite fermentado produzido pela ação de bactérias e leveduras que existem em
associação simbiótica e compõem os grãos de kefir. A produção artesanal da bebida kefir
fundamenta-se na tradição dos povos do Cáucaso, difundida para outras partes do mundo, a
partir no final do século passado, integrando suas indicações nutricionais e terapêuticas ao
cotidiano alimentar atual. O grande número de microrganismos presentes no kefir, a
variabilidade de possíveis compostos bioativos e os vários benefícios associados ao seu
consumo fazem com que esta bebida seja considerada um probiótico natural, designado como
o iogurte do século XXI. Vários estudos têm demonstrado que o kefir e seus constituintes
possuem atividade antimicrobiana, antitumoral, anticarcinogênica, imunomoduladora,
melhora da digestão da lactose, entre outras. Esta revisão inclui dados sobre composição
microbiológica do kefir, seus aspectos tecnológicos e principais efeitos benéficos para saúde
humana.
INTRODUÇÃO
A palavra kefir é derivada do turco keyif, que significa “sentir-se bem” após sua
ingestão 2; 16
. A bebida kefir é originária das montanhas do Cáucaso, e é um produto
tradicional e altamente consumido na Europa Oriental, Rússia e sudoeste Asiático 2.
Atualmente, relata-se um aumento de seu consumo em vários países, devido a suas
propriedades sensoriais únicas e sua longa história associada aos efeitos benéficos à saúde
humana 2; 18; 106
. No Brasil, o conhecimento desta bebida, bem como os benefícios da inclusão
1 Apêndice A

60
deste alimento probiótico na dieta, não são muito difundidos, e sua fabricação é
exclusivamente artesanal 125
.
O kefir é caracterizado por seu flavor distinto, típico de levedura e um efeito
efervescente sentido na boca 16; 104
. Os principais produtos da fermentação no kefir são ácido
lático, etanol e CO2, que conferem viscosidade, acidez e um baixo teor alcóolico. Compostos
minoritários também podem ser encontrados, incluindo diacetil, acetaldeído, aminoácidos
livres e acetato, contribuindo para a composição do flavor 104
. Esta bebida difere de outros
produtos lácteos fermentados porque não é resultado da atividade metabólica de uma única
espécie 102
.
GRÃOS DE KEFIR
Os grãos de kefir desempenham papel de cultura starter durante a produção do kefir e
são recuperados após o processo de fermentação por coagem do leite 104
. Estes grãos são
compostos por uma microbiota imobilizada em uma matriz de polissacarídeo e proteína, onde
várias espécies de bactérias e leveduras coexistem em associação simbiótica 102; 150
. Neste
ecossistema, há uma população de microrganismos relativamente estável, que interage e
influencia outros membros da comunidade. Esta população determina a síntese de metabólitos
bioativos, que são essenciais para o crescimento dos grãos e inibição de microrganismos,
como patógenos e contaminantes do alimento 150
.
Grãos de kefir (Figura 5) variam em tamanho de 0,3 a 3,0 cm de diâmetro, são
caracterizados por uma superfície irregular, multi-lobular, unida por uma seção central única,
e sua cor varia do branco ao branco amarelado. Os grãos são elásticos, e possuem textura
viscosa e firme 44; 102; 109
.
Figura 5 – Estrutura macroscópica de grãos de kefir
Embora a bebida kefir possa ser encontrada em muitos países, no Brasil os grãos não
estão comercialmente disponíveis, e culturalmente são doados de pessoa para pessoa.

61
ASPECTOS MICROBIOLÓGICOS
No kefir, Bactérias Ácido Láticas (BAL) são os principais responsáveis pela conversão
da lactose presente no leite em ácido lático, que resulta na queda do pH e na preservação do
leite. Outros constituintes microbianos do kefir incluem leveduras, que podem fermentar a
lactose produzindo etanol e CO2. Leveduras não fermentadoras de lactose e Bactérias Ácido
Acéticas (BAA) também participam do processo 104; 109
. Após a fermentação, os grãos
aumentam cerca de 5-7% da sua biomassa. Durante o crescimento em leite, as proporções de
microrganismos presentes nos grãos diferem daqueles presentes no produto final 2; 104
. Esta
diferença está associada às condições do processo de fermentação, como: tempo de
fermentação, temperatura, grau de agitação, tipo de leite, proporção inóculo grão/leite,
distribuição dos microrganismos nos grãos, entre outros 2; 104; 119
.
Tradicionalmente, métodos microbiológicos clássicos são utilizados para estudar a
microbiota do kefir 44; 119
. Embora estes métodos sejam úteis, em alguns casos, não são
discriminativos o suficiente para identificar espécies intimamente relacionadas entre si ou
novas espécies. Devido à associação simbiótica dos microrganismos presentes nos grãos, o
crescimento e sobrevivência de cepas individuais são dependentes da presença uma das
outras. Frequentemente, microrganismos quando isolados dos grãos, não crescem bem em
leite e/ou apresentam redução de sua atividade bioquímica 102
. Portanto, métodos
independentes de cultivo têm sido utilizados de forma complementar aos métodos
convencionais no estudo da microbiota dos grãos de kefir. A técnica de reação em cadeia da
polimerase, aliada à eletroforese em gel com gradiente desnaturante (PCR-DGGE) tem se
mostrado adequada para analisar consórcios microbianos complexos 121; 125; 140; 178
, enquanto
que o sequenciamento parcial do gene codificante para o RNAr 16S tem sido utilizado para
identificação das espécies 104; 123
. Entretanto, alguns estudos mostram que a técnica de PCR-
DGGE não permite a detecção de mudanças significativas durante a fermentação do kefir 125
,
provavelmente devido à relativa estabilidade da população dominante presente na
comunidade.
A complexidade desta comunidade pode ser mais bem entendida a partir da introdução
de técnicas de sequenciamento massivo, como pirosequenciamento. Dois estudos recentes, um
deles realizado por nosso grupo de pesquisa 126; 178
utilizaram esta técnica para avaliar a
diversidade microbiana nos grãos e kefir e, detectaram a presença de microrganismos
pertencentes a grupos minoritários, não reportados anteriormente por técnicas dependentes e
independentes de cultivo. Além disso, Leite e colaboradores 178
comparando três grãos de

62
kefir brasileiros relataram que, ainda que de origens distintas, certos grupos de
microrganismos, como espécies do gênero Lactobacillus estavam presentes em todos os
grãos. Os autores sugerem que a população minoritária e específica de cada grão pode
contribuir para as características sensoriais particulares apresentadas nas bebidas fermentadas
178.
Algumas espécies raramente isoladas dos grãos e do kefir não são microrganismos
chave e, geralmente são considerados contaminantes. Recentemente, os fungos Dipodascus
capitus e Trichosporon coremiiforme, considerados patógenos, foram identificados por
sequenciamento em grãos de kefir104
. Alguns contaminantes bacterianos também já foram
descritos, como Pseudomonas spp. 121; 126; 178
e membros da família Enterobacteriaceae 126
e
Clostridiaceae 126
, quando a microbiota dos grãos foi caracterizada pela técnica de
pirosequenciamento e PCR-DGGE.
Distribuição dos microrganismos em grãos de kefir
A distribuição dos microrganismos dentro dos grãos tem sido estudada, e os resultados
obtidos são controversos. Um grupo de pesquisadores defende a hipótese de que geralmente
as leveduras são encontradas no interior e zona intermediária, predominando na área
superficial bastonetes e raros lactococos 120; 122
. Entretanto, outros pesquisadores 44; 109; 117; 121;
140 descrevem que as leveduras estão distribuídas tanto na parte externa quanto na interna,
sendo que a principal diferença encontrada entre as duas áreas está relacionada com a
morfologia dos bastonetes. Contrariando resultados reportados anteriormente 44; 122
, Guzel-
Seydim e colaboradores 117
observaram um predomínio de bastonetes, sem qualquer evidência
da presença de leveduras na porção interna. Além disso, alguns autores109; 121
relataram uma
menor quantidade de células observada na porção interna, quando comparada com a porção
externa. O material fibrilar correlacionado com a presença do polissacarídeo kefiran, também
foi observado por todo grão109; 117; 125; 140
.
Nosso grupo de pesquisa também avaliou a distribuição dos microrganismos de três
grãos de kefir brasileiros (Figura 6) e observou uma relativa variação na distribuição dos
mesmos em função da origem dos grãos. De forma geral, bastonetes foram observados tanto
na porção interna (Figura 6 B,D,F), quanto na externa (Figura 6 A,C,E), enquanto que as
leveduras foram mais frequentes na porção externa (Figura 6 A,C,E). Em um dos grãos, cocos
foram observados na porção externa. Provavelmente, o material granulado observado
correspondia às partículas de leite coagulado aderidas à superfície do grão e o material fibrilar
ao polissacarídeo kefiran, distribuídos por todas as partes dos grãos 44; 109; 117
.

63
Apesar de L. lactis ser um dos microrganismos predominantemente isolado na bebida
kefir, vários pesquisadores 109; 117; 121
não observaram cocos nas micrografias, provavelmente
devido à fraca aderência dos lactococos aos grãos durante o crescimento, o que facilitaria a
liberação dos mesmos no processo de lavagem 44; 109; 117; 121
. Além disso, também é provável
que o pH ácido do interior dos grãos, dificulte o crescimento deste microrganismo neste
microambiente 44
.
Provavelmente os microrganismos de superfície causam um maior impacto no
processo de fermentação do kefir 102
. Contudo, trabalhos anteriores demonstraram que uma
grande variação na população microbiana ocorre entre diferentes grãos e dentro do mesmo
grão. Lin, Chen e Liu 120
concluíram que o local de origem dos grãos por si só pode explicar
as diferenças que vêm sendo relatadas em vários estudos de microscopia eletrônica.
Bactérias do kefir
BAL homofermentativas, incluindo espécies de Lactobacillus, como L. delbrueckii
subsp. bulgaricus, L. helveticus, L. kefiranofaciens subsp. kefiranofaciens, L. kefiranofaciens
subsp. kefirgranum e L. acidophilus; Lactococcus spp. como L. lactis subsp. lactis e L. lactis
Figura 6 - Microscopia eletrônica de varredura da microbiota de grãos de kefir brasileiros. A, C, E : porções
externas; B,D,F: porções internas. Setas: micrografia A: cocos; micrografia D: material fibrilar – polissacarídeo
kefiran; micrografia E: seta 1- material granular – proteína coagulada, seta 2 – diferentes leveduras

64
subsp. cremoris; e Streptococcus thermophilus já foram identificados nos grãos de kefir e na
bebida fermentada, assim como BAL heterofermentativas, incluindo L. kefiri, L. parakefiri, L.
fermentum e L. brevis 104; 178
; e estirpes citrato-positivas de L. lactis (L. lactis subsp. lactis
biovar diacetylactis), Leuconostoc mesenteroides subsp. cremoris, e Leuconostoc
mesenteroides subsp. mesenteroides 16; 104; 178
. A utilização de citrato pelas estirpes citrato-
positivas resulta na produção de compostos chave para o flavor típico do kefir 104
.
O kefiran produzido por L. kefiranofaciens, é um polissacarídeo ramificado, solúvel
em água e contém quantidades iguais de D-glicose e D-galactose. A produção deste
polissacarídeo é estimulada quando o L. kefiranofaciens cresce em co-cultura com S.
cerevisiae 139
.
Espécies BAA têm sido isoladas e identificadas, tanto dos grãos quanto do kefir. No
entanto, em alguns países, a presença destas espécies é considerada indesejável 2; 102; 104
e tem
recebido menor atenção, apesar de seu papel essencial, tanto no consórcio microbiano, quanto
nas características sensoriais do produto final 2; 44
.
Leveduras do kefir
Embora produzam metabólitos que contribuem para as propriedades sensoriais
desejáveis e típicas do kefir 104; 119
, as leveduras são menos estudadas que as bactérias do
kefir. As principais capazes de fermentar a lactose encontradas no kefir e nos grãos são
Kluyveromyces marxianus/Candida kefyr, Kluyveromyces lactis var. lactis, Debaryomyces
hansenii e Dekkera anomala, enquanto que as não fermentadoras de lactose incluem
Saccharomyces cerevisiae, Torulaspora delbrueckii, Pichia fermentans, Kazachstania
unispora, Saccharomyces turicensis, Issatchenkia orientalis, e Debaryomyces occidentalis .
Interações entre os microrganismos do kefir
As complexas interações entre leveduras e bactérias no grão de kefir e sua
interdependência não são completamente entendidas. Mas, quando as bactérias dos grãos são
separadas das leveduras, não crescem tão eficientemente 102; 104; 139
.
Devido a sua grande capacidade de metabolizar a lactose 44
, o gênero Lactococcus
tende a crescer mais rápido do que as leveduras no leite 2; 44
e hidrolisam a lactose,
produzindo ácido lático e ambiente adequado ao crescimento de leveduras 2. Além disso,
leveduras sintetizam vitaminas do complexo B, hidrolisam proteínas do leite, utilizam
oxigênio para produzir CO2 e álcool 2; 16
. A interação entre leveduras e bactérias láticas pode
ser estimulada ou inibida, pelo crescimento de uma ou ambas, em co-culturas. Estes
microrganismos podem competir por nutrientes para o crescimento, ou podem produzir

65
metabólitos, que inibam ou estimulem umas às outras 16
. Algumas espécies de leveduras são
proteolíticas ou lipolíticas, disponibilizando aminoácidos e ácidos graxos 104
. Espécies como
Debaryomyces hansenii e Yarrowia lipolytica assimilam o ácido lático formado por BAL,
elevando o pH e estimulando o crescimento de bactérias. A produção de vitamina B por
Acetobacter spp. também favorece o crescimento de outros microrganismos presentes nos
grãos 16; 44
.
ASPECTOS TECNOLÓGICOS
Durante a fermentação, os grãos aumentam em tamanho e número, e geralmente são
recuperados do leite fermentado e reutilizados 150
. Se cuidadosamente preservados podem
reter sua atividade por anos 16; 104
. O principal marcador para avaliar a relação simbiótica entre
os diferentes microrganismos é o aumento da biomassa durante a fermentação 150
.
Preservação dos grãos
Grãos de kefir podem ser preservados liofilizados, secos ou úmidos 150
, mas a
constante lavagem reduz sua viabilidade 102
. No entanto, Pintado e colaboradores 114
observaram que grãos estocados nestas condições, apresentam um perfil microbiológico
diferente dos grãos frescos. Grãos desidratados mantém sua atividade por 12-18 meses,
enquanto que grãos úmidos mantém atividade por 8 a 10 dias 150
. Diferentes métodos de
conservação foram testados 115
, sendo o congelamento considerado o melhor. O processo de
liofilização dos grãos também testado resultou na redução da metabolização da lactose, além
da modificação do perfil bacteriano, distinto ao dos grãos originais 102
.
Produção do kefir
Há três maneiras principais de produção do kefir: (1) artesanal, (2) comercial pelo
método russo e (3) comercial com culturas puras 18; 104; 106
. Outros substratos também podem
ser utilizados como leites de outras espécies animais, leite de coco, leite de soja, sucos de
frutas, solução de açúcar e melaços 104; 125; 129
.
A produção artesanal tradicional envolve a inoculação do leite com uma quantidade
variável de grãos, e a fermentação por um período entre 18-24h a 20 - 25°C. Ao final da
fermentação, os grãos são peneirados (Figura 7) e podem ser utilizados para uma nova
fermentação, ou conservados (1-7 dias) em leite fresco, enquanto que o kefir é estocado a
4°C, pronto para o consumo 19; 102; 106
.

66
Figura 7 - Produção artesanal do kefir. (1) Grãos de kefir são adicionados ao leite (2), e mantidos a temperatura
ambiente por 18-24 h (3); Após a fermentação, são coados (4) e o leite fermentado (kefir) está pronto para
consumo (5) e os grãos (1), podem ser reutilizados
A concentração do inóculo inicial de grãos (proporção grão/leite) afeta o pH,
viscosidade, concentração final de lactose e perfil microbiológico do produto final 119; 179
. A
agitação durante a fermentação também influencia a composição microbiana do kefir,
favorecendo o desenvolvimento de lactococos homofermentativos e leveduras 2; 102; 104
.
Temperaturas de incubação superiores a 30°C favorecem o crescimento de BAL termofílicas,
ao passo que desfavorecem o crescimento de leveduras e BAL mesofílicas 104
.
O segundo método, conhecido como “método russo” (Figura 8A), permite a produção
do kefir em maior escala, e utiliza fermentações em série, a partir do percolado resultante da
primeira fermentação com os grãos (fermentado sem os grãos ou cultura-mãe) 102; 104
.
No processo industrial do kefir, diferentes métodos podem ser utilizados, mas
baseiam-se no mesmo princípio (Figura 8B). O leite é inoculado com culturas puras isoladas
de grãos de kefir ou culturas comerciais 2; 19; 104
.

67
Figura 8 - Fluxograma de produção da bebida kefir. (A) Método russo; (B) Método com culturas puras (bebida
“tipo kefir”). Adaptado Otles e Cagindi 106
A etapa de maturação pode ser realizada ou não, e consiste em manter o kefir a 8-10°C
por até 24h 19; 104
, de forma a permitir o crescimento dos microrganismos, principalmente das
leveduras que contribuem para sabor específico do produto 19
. A omissão desta etapa está
associada a desenvolvimento de flavor atípico no kefir 19; 104
.
Durante o armazenamento, a produção de CO2 por leveduras ou BAL
heterofermentativas pode causar um estufamento na embalagem do produto, fato este que
deve ser considerado na escolha da embalagem 102; 180
.
Embora a bebida comercial esteja disponível em muitos países, nem sempre todas as
propriedades do kefir tradicional estão presentes 16; 102
. Assadi, Pourahmad e Moazami 181
testaram várias relações de culturas iniciadoras isoladas dos grãos (BAL, levedura e BAA), e
concluíram que o kefir tradicional produzido com os grãos, foi mais bem aceito, que o obtido
com a cultura iniciadora. Rossi e Gobbetti 182
produziram uma bebida “tipo-kefir”, que
apresentou menor viscosidade e ausência de alguns dos componentes voláteis, frequentemente
encontrados no kefir tradicional. Já Carneiro 8 desenvolveu uma cultura iniciadora, a partir de
microrganismos isolados de grãos de kefir, e o produto obtido foi mais bem aceito que o kefir
tradicional. Beshkova e colaboradores 19
sugeriram dois métodos de fermentação do kefir:
método por fermentação simultânea e por fermentação sucessiva. Para tal, utilizaram uma
cultura iniciadora que consistia de bactérias e leveduras isoladas de grãos, combinada a duas

68
estirpes comumente utilizadas na fabricação de iogurte. Leveduras foram adicionadas à
cultura iniciadora com sacarose, tanto no início (fermentação simultânea), quanto após a etapa
de fermentação ácido lática (fermentação sucessiva). Os dois processos de fermentação
produziram kefir com alto número de lactococos e lactobacilos viáveis, com propriedades
sensoriais similares ao kefir tradicional.
O uso de culturas comerciais pode padronizar a produção comercial do kefir, se a
seleção de espécies e estirpes de leveduras e bactérias for realizada de forma rigorosa e
cuidadosa, possibilitando produção de uma bebida “tipo kefir”, com flavor aceitável e boas
propriedades de conservação 8; 19
. A bebida comercial pode ter validade comercial de até 28
dias, enquanto que recomenda-se que o kefir produzido com os grãos seja consumido entre 3
a 12 dias. No entanto, o produto “tipo de kefir” pode não apresentar as mesmas propriedades
terapêuticas e probióticas presentes no kefir tradicional 104
.
O desenvolvimento de uma bebida “tipo-kefir” não é a única aplicação industrial
investigada. Grãos de kefir também já foram estudados quanto à produção de células ricas em
proteínas (Single Cell Protein – SCP) na bioconversão de soro de queijo e aplicação destas
proteínas na indústria de alimentos, a fim de melhorar as características sensoriais dos
produtos 183
.
As principais falhas na fabricação do kefir podem ser atribuídas ao sabor e aroma
desagradáveis, típico de levedura 2. O último pode ser causado pela rápida multiplicação de S.
cerevisiae, acompanhado por um aroma típico de vinagre 2. A produção excessiva de ácido
acético pode influenciar o aroma do kefir, e ocorre devido ao intenso crescimento de
Acetobacter spp. ou presença de Dekkera spp. nos grãos. Um sabor amargo no kefir pode ser
causado por fungos (por exemplo. Geotrichum candidum) e/ou atividade de algumas
leveduras atípicas que podem estar presentes no produto 2.
ASPECTOS TERAPEUTICOS
Historicamente o kefir tem sido recomendado para tratamento de vários quadros
clínicos como problemas gastrointestinais, hipertensão, alergia e doença cardíaca isquêmica
102; 104. Porém, a variabilidade inerente às condições utilizadas na produção do kefir em
diferentes ensaios, dificulta a comparação entre os resultados científicos relatados 18; 102; 104
.
Fermentações a partir de vários substratos com grãos têm sido avaliadas 18; 125; 129
, e
uma grande variedade de compostos bioativos tem sido encontrada como ácidos orgânicos,
CO2, H2O2, etanol, peptídeos bioativos, exopolissacarídeos (kefiran), e bacteriocinas. Estes

69
compostos podem agir independentemente ou em conjunto para produzir os vários benefícios
à saúde atribuídos ao consumo de kefir 104; 150
.
Atividade antimicrobiana
Santos e colaboradores 157
observaram o comportamento antagônico dos isolados de
lactobacilos de grãos de kefir contra E. coli, L. monocytogenes, Salmonella Typhimurium, S.
Enteritidis, Shigella flexneri e Y. enterocolitica. Silva e colaboradores 158
observaram inibição
de Candida albicans, Salmonella Typhi, Shigella sonnei, Staphylococcus aureus e E. coli por
kefir cultivado em açúcar mascavo. Já Chifiriuc, Cioaca e Lazar 156
, observaram que todos os
leites fermentados com grãos de kefir apresentaram atividade antimicrobiana contra Bacillus
subtilis, S. aureus, E. coli, E. faecalis e S. Enteritidis, mas não inibiram P. aeruginosa e C.
albicans. Em todos estes estudos os pesquisadores indicaram que a atividade antimicrobiana
do kefir está associada à produção de ácidos orgânicos, peptídeos (bacteriocinas), dióxido de
carbono, peróxido de hidrogênio, etanol e diacetil. Estes compostos podem exercer efeitos
benéficos, não só na redução de patógenos alimentares e bactérias deteriorantes, durante a
produção do alimento e estocagem, mas também no tratamento e prevenção de gastroenterites
e infecções vaginais 18; 118
. Além disso, a atividade antimicrobiana do polissacarídeo kefiran,
também foi comprovada contra bactérias e C. albicans 149
.
Atividade anti-inflamatória e cicatrizante
A atividade cicatrizante do kefir e kefiran em camundongos foi evidenciada após sete
dias de tratamento com gel de kefir 149
assim como a atividade anti-inflamatória 162
sobre
modelo de indução de tecido granulomatoso e contorções abdominais induzidas por ácido
acético, em camundongos. Huseini et al. 184
comprovaram a atividade cicatrizante em
queimaduras infectadas com Pseudomonas aeruginosa, em camundongos.
Impacto no Trato Gastrintestinal (TGI)
O efeito provocado pelo consumo de kefir na composição da microbiota intestinal
pode ser devido a uma combinação de fatores, como: inibição direta de patógenos pela
produção de ácidos e bacteriocinas, além da exclusão competitiva de patógenos na mucosa
intestinal 104
. Segundo Marquina e colaboradores 185
o consumo de kefir aumentou
significativamente as contagens de BAL na mucosa intestinal e reduziu a população de
enterobactérias e clostrídios. Além disso, preveniu a colonização de Campylobacter jejuni no
ceco de pintos 186
, e foi eficaz no tratamento pós-operatório e em pacientes com desordem
gastrintestinal 118
. Na Rússia, o kefir tem sido utilizado por pesquisadores no tratamento de
pacientes com úlcera péptica no estômago e duodeno 102
.

70
Efeitos anticarcinogênicos
O papel anticarcinogênico dos produtos lácteos fermentados pode ser atribuído, de
maneira geral, à prevenção de câncer e supressão de tumores iniciais, pelo retardo das
atividades enzimáticas, que convertem compostos pro-carcinogênicos a carcinogênicos, ou
pela ativação do sistema imune 118
. Kubo et al. 160
reportaram a inibição da proliferação do
tumor ascítico de Ehrlich, transplantado subcutaneamente, em camundongos. Liu e
colaboradores 187
observaram inibição do crescimento tumoral em camundongos, indução da
lise celular por apoptose do tumor e aumento significativo dos níveis de IgA, sugerindo
potencial antitumoral da bebida e promotor da resistência de mucosa à infecções intestinais.
Guven e Gulmez 188
relataram que camundongos tratados com kefir, apresentaram maior
efeito protetor contra os danos induzidos pelo tetracloreto de carbono, indicando que o kefir
pode agir como antioxidante.
Estimulação do sistema imune
Durante o processo de fermentação ou digestão, a formação de peptídeos bioativos,
tem mostrado uma variedade de atividades fisiológicas, incluindo a estimulação do sistema
imune, em modelos animais 18
. Thoreux e Schmucker 189
após alimentar camundongos com
kefir, observaram um aumento da resposta imune específica de mucosa (IgA) contra toxina da
cólera. A estimulação do sistema imune também pode ocorrer devido à ação de
exopolissacarídeos encontrados nos grãos de kefir 18; 190
. Medrano e colaboradores 191
observaram que o kefiran foi capaz de modificar o equilíbrio das células imunes na mucosa
intestinal.
Vinderola e colaboradores 192
demonstraram a capacidade de imunomodulação de
kefir, na resposta imune de mucosa intestinal de camundongos. A administração de kefir
também induziu a resposta na mucosa intestinal, sugerindo que componentes do kefir podem
estimular células do sistema imunológico inato, suprimindo resposta imune do fenótipo Th2
ou promovendo respostas imunes mediadas por célula, contra tumores e infecções de
patógenos intracelulares 155
. Recentemente, Hong e colaboradores 154
mostraram in vitro, a
capacidade imunomoduladora de BAL isoladas de grãos de kefir, sugerindo a influência na
secreção de citocinas pro-inflamatórias IL-6 e TNF-α através de TLR-2.
Efeito hipocolesterolêmico
Os possíveis mecanismos propostos para a atividade hipocolesterolêmica de BAL
podem envolver a inibição de absorção de colesterol exógeno no intestino delgado, pela
ligação e incorporação do colesterol em células bacterianas, assimilação do colesterol, bem

71
como uma supressão da reabsorção dos ácidos biliares, pela desconjugação enzimática dos
sais biliares, promovida pela enzima bacteriana Bile Salt Hydrolase (BSH) 193
. Wang e
colaboradores 193
observaram uma redução significativa nos níveis séricos de colesterol total,
lipoproteínas de baixa densidade (LDL-C) e triacilgliceróis, enquanto que não houve mudança
no nível de lipoproteínas de alta densidade (HDL-C), em camundongos alimentados com dieta
rica em colesterol, suplementada com Lactobacillus plantarum MA2 isolado de Kefir do
Tibet. Além disso, o colesterol total e triacilgliceróis do fígado também foram reduzidos. Já o
colesterol e triacilgliceróis das fezes dos animais aumentaram significativamente. Em outro
estudo 194
também foi observado uma redução dos níveis séricos de triacilglicerol e colesterol,
principalmente da fração não-HDL-C. Um resultado contraditório foi relatado por St-Onge e
colaboradores 195
, onde o consumo de kefir não reduziu os níveis de colesterol total, LDL-C,
HDL-C ou triglicerídeos, mas aumentou as concentrações de ácidos isobutíricos, isovaléricos
e propiônicos, assim como, a quantidade total de ácidos graxos de cadeia curta nas fezes.
Kefir e a intolerância a lactose
A capacidade de redução na concentração de lactose e presença de atividade β-
galactosidase, em produtos lácteos fermentados, os tornam adequados para consumo por
pessoas classificadas como intolerantes a lactose 102; 118
. Já foi demonstrado que alguns grãos
de kefir possuem atividade da enzima β-galactosidase, que se mantem ativa quando
consumida e que o kefir contém menos lactose que o leite 18; 118
. Um kefir comercial 153
foi tão
efetivo quanto o iogurte, na redução de hidrogênio expirado e flatulência em adultos
intolerantes a lactose, quando comparado à ingestão de leite. De Vrese et al. 196
mostraram
que porcos alimentados com kefir, apresentaram um aumento significativo nas concentrações
plasmáticas de galactose, sugerindo melhora da hidrólise intestinal da lactose pela enzima β-
galactosidase microbiana.
CONSIDERAÇÕES FINAIS
O kefir tradicional é um exemplo da coexistência de bactérias e leveduras e a
importância desta relação simbiótica parece clara, uma vez que é necessária para produção de
compostos benéficos para saúde e para obtenção de um produto final de boa qualidade.
Embora as evidências não possam ser conclusivas e mais estudos devam ser conduzidos, os
estudos já realizados comprovam os benefícios à saúde relatados empiricamente pelo seu
consumo histórico. Atualmente, a aplicação de probióticos na indústria de alimentos está em
franca expansão e o entendimento das relações simbióticas entre diferentes microrganismos

72
presentes neste alimento, assim como, suas interações poderiam auxiliar na melhoria dos
processos tecnólogicos.
A qualidade do kefir tradicional é principalmente influenciada por microrganismos
presentes nos grãos e condições de processamento. Embora cientistas e indústrias alimentícias
tentem desenvolver uma bebida “tipo-kefir” elaborada por diferentes culturas comerciais de
BAL mesofílicas e termofiílicas, ou até mesmo culturas puras isoladas de grãos de kefir, seu
sucesso quando comparado ao kefir tradicional é limitado. Porém, do ponto de vista
industrial, estes desenvolvimentos são bem-vindos, face à dificuldade de padronização na
produção e comercialização do kefir tradicional.
AGRADECIMENTOS
Os autores agradecem a ajuda do prof. Dr. Sérgio Seabra, do técnico Eliandro Lima da
Universidade Estadual da Zona Oeste (UEZO) e ao MSc Laidson Paes Gomes pelo apoio
técnico no processamento das amostras e micrografias obtidas por MEV.

73
6.2 CAPÍTULO II - AVALIAÇÃO DA DIVERSIDADE MICROBIANA DE GRÃOS DE
KEFIR2
6.2.1 Material e Métodos
6.2.1.1 Amostras de grãos de kefir
Os três grãos de kefir estudados foram obtidos de diferentes cidades do Sudeste do
Brasil, com condições climáticas similares e distância média de 400 km (AR, Niterói, Rio de
Janeiro; AV, Viçosa, Minas Gerais; e AD, Lavras, Minas Gerais). Grãos AV e AD foram
gentilmente doados por pesquisadores de Universidades brasileiras (UFLA e UFV), enquanto
que o terceiro grão (AR) pertencia a uma família que tradicionalmente cultivava o grão
artesanalmente, para consumo próprio. No laboratório, grãos foram ativados em leite
reconstituído (10% w/v) a 25ºC por 24 h, coados em uma peneira para remoção do leite
coagulado, e lavados com água estéril. Esta etapa de ativação foi repetida três vezes.
6.2.1.2 Isolamento do DNA microbiano total
Para extração do DNA genômico microbiano, os grãos de kefir ativados foram
homegeneizados em citrato de sódio 2%, com auxílio de gral e pistilo. Alíquotas de 2 mL de
cada homogeneizado foi centrifugado por 10 min a 10,000 g e o DNA total foi extraído e
purificado usando o kit FastDNA Spin®
(QBIOgene), de acordo com as instruções do
fabricante. O DNA obtido foi quantificado usando o fluorímetro Qubit (Invitrogen Detection
Technologies).
6.2.1.3 Análise de DGGE dos grãos de kefir
6.2.1.3.1 Amplificação por PCR de fragmentos do gene codificador do RNAr 16S e RNAr 26S
O DNA genômico foi utilizado como molde na reação de amplificação da região V3
do gene codificador do RNAr 16S bacteriano, utilizando o par de iniciador universal F357-
GC (5’–TACGGGAGGCAGCAG–3’ e R518 (5’–ATTACCGCGGCTGCTGG–3’), como
reportado por Muyzer, De Waal e Uitterlinden 164
. Iniciadores grupo-específico para detecção
de BAL também foram usados, incluindo o par Lac1 (5’–AGCAGTAGGGAATCTTCCA–3’)

74
e Lac2-GC (5’–GATTYCACCGCTACACATG–3’) para deteção de membros dos gêneros
Lactobacillus, Pediococcus, Leuconostoc e Weissella 197, e o par Lac3 (5’–
AGCAGTAGGGAATCTTCGG–3’) e Lac2-GC para detecção de membros dos gêneros
Lactococcus, Streptococcus, Enterococcus, Tetragenococcus and Vagococcus 198
. O domínio
D1, do gene codificador do RNAr 26S para fungos, foi amplificado utilizando o par de
iniciador NL1-GC (5’–GCCATATCAATAAGCGGAGGAAAG–3’) e LS2 (5’–
ATTCCCAAACAACTCGACTC–3’), como descrito por Cocolin e colaboradores 199
. Todos
os iniciadores continham uma sequência grampo-GC de 39 pb acoplada à extremidade 5’ de
um dos iniciadores, para prevenir a completa desnaturação dos fragmentos amplificados. A
reação de PCR foi realizada em um volume final de 50 µL, contendo Taq-DNA polimerase
master mix (Ampliqon) utilizando como molde 100 ng de cada amostra de DNA e 0.2 mM
de cada oligonucleotídeo iniciador.
6.2.1.3.2 Condições de eletroforese e identificação das bandas
O DGGE foi realizado utilizando equipamento DCode (Bio-Rad) a 60°C, em géis de
poliacrilamida (8%) contendo gradiente desnaturante de 40-60% para bactérias totais, 40-50%
para BAL grupo específico e 30-50% para fungos. A eletroforese foi conduzida a 75 V por 16
h e 130 V por 4.5 h para produtos de amplificações bacterianas e fungicas, respectivamente.
Os géis de DGGE foram corados com brometo de etídio (0,5 µg/mL) e as bandas visualizadas
sob luz UV e fotodocumentadas no aparelho Gel Doc 1000 (BioRad).
As bandas detectadas nos géis foram excisadas, e o DNA foi eluído em 50 µL de água
estéril a 4ºC por 20 h. O DNA foi reamplificado com os mesmos iniciadores sem sequência
grampo-GC e sequenciado no ABI 373 DNA (Applied Biosystems). A identidade das
sequências foi determinada pelo algoritmo BLASTN no banco de dados GenBank
(http://www.ncbi.nlm.nih.gov/BLAST/).
6.2.1.4 Análise de pirosequenciamento dos grãos de kefir
6.2.1.4.1 Iniciadores e condições de amplificação do gene rrs
Os iniciadores universais Y1 (5’–TGGCTCAGGACGAACGCTGGCGGC–3’)
(posição 20-43 relativa a sequencia de Escherichia coli do gene rrs) e Y2 (5’–

75
CCTACTGCTGCCTCCCGTAGGAGT–3’) (posições 361-338)200
, foram utilizados na
reação de PCR para amplificação de um fragmento de 348 pb, relativo as regiões variáveis V1
e V2 do gene rrs. Adaptadores-454 foram incluídos em ambos os primers senso (5’–
CGTATCGCCTCCCTCGCGCCATCAG–3’) e anti-senso (5’–
CTATGCGCCTTGCCAGCCCGCTCAG–3’), seguido por uma sequência barcode de 10-pb
amostra específica. As reações de amplificação foram conduzidas como descrito
anteriormente, nas seguintes condições: 95°C por 5 minutos, 25 ciclos de 94°C por 30
segundos, 52°C por 40 segundos e 72°C por 30 segundos, e uma etapa de extensão final a
72°C por 10 minutos.
Os produtos de PCR foram purificados com o kit GenEluteTM
PCR Clean-Up columns
(Sigma-Aldrich), e a concentração e qualidade do DNA foi mensurada em um
espectofotômetro de micro volume Epoch (BioTek Instruments). Quantidades iguais das três
amostras foram agrupadas, para uma concentração total de 100 ng. O DNA de todas as
amostras foi reunido e amplificado em uma reação de PCR em emulsão e, então,
sequenciados na plataforma 454 (Roche). As sequências obtidas foram depositadas no arquivo
de sequências do NCBI (SRA) sob número de acesso SRA045648.2, SRR340042.2,
SRR340043.1 and SRR340041.1.
6.2.1.4.2 Tratamento das sequências e análise de bioinformática
As sequências iniciais obtidas do pirosequenciamento foram processadas na
plataforma do Ribosomal Database Project (RDP) para pirosequencimento
(http://wildpigeon.cme.msu.edu/pyro/index.jsp). Aquelas que apresentavam tamanho do
fragmento menor que 300 pb ou ausência de um dos iniciadores foram consideradas de baixa
qualidade e excluídas das análises posteriores. As sequências parcias de alta qualidade do
gene rrs foram classificadas utilizando a ferramenta “RDP-II classifier” utilizando um limite
de confiança de 80%, para obter o alinhamento taxonômico e a abundância relativa dos
diferentes grupos bacterianos 201
. Um alinhamento múltiplo das sequências de cada amostra
foi realizado usando a ferramenta Aligner (configurações padrão) no website do RDP. Os
alinhamentos obtidos serviram como entrada para construção da matriz de distância e o
agrupamento das sequências em unidades taxonômicas operacionais (OTUs), usando o
software MOTHUR v. 1.14.0 202
. Os clusters foram construídos a 3% de dissimilaridade
como valor de corte e serviram como OTUs para gerar modelos de rarefação preditivos e,

76
determinar os índices de riqueza (ACE e Chao 1) 203
e o índice de diversidade de Shannon 204
.
O programa MOTHUR também foi utilizado para realizar o teste Fast UniFrac, que foi
empregado na comparação da estrutura filogenética das bibliotecas obtidas e construção do
diagrama de Venn. Uma árvore filogenética foi construída com as sequências representativas
de cada OTU selecionada no MOTHUR. Essas sequências foram comparadas com as
sequências do banco de dados do RDP, utilizando a opção Seqmatch, a fim de selecionar as
espécies mais relacionadas entre si. Todas as sequências foram alinhadas utilizando o
software MEGA 5.0 205
e o modelo de Jukes-Cantor foi aplicado. A sequência equivalente a
archea Halococcus saccharolyticus (AB004876) foi utilizada como grupo externo na
construção da árvore.
6.2.2 Resultados
6.2.2.1 Análise por PCR-DGGE das comunidades bacterianas e de leveduras
Os fingerprints obtidos no DGGE das comunidades microbianas dos três grãos de
kefir brasileiros foram simples, com um perfil variando de um a cinco bandas (Fig. 9, A - D) e
a maioria das bandas foi compartilhada por todas as amostras. O perfil das espécies de
bactérias totais amplificadas com iniciadores universais foi composto por até cinco bandas
correspondentes a três espécies diferentes (Fig. 9 A). Bandas correspondentes a Lb.
kefiranofaciens (bandas 1, 2 e 5) e a Lb. kefiri (banda 4) foram detectadas em todas as
amostras. Uma banda adicional presente na amostra AV (banda 3) foi identificada como Lc.
lactis. Essas três espécies também foram detectadas utilizando os iniciadores grupo-
específicos para lactobacilos e lactococos (Figura 9, C e D, respectivamente). Os fingerprints
obtidos no DGGE para comunidade de leveduras também foram simples e similares para os
três grãos. Uma banda de alta intensidade presente em todas as amostras foi identificada como
S. cerevisiae (banda 6, Fig. 9 B). No grão de kefir AD, uma banda de baixa intensidade foi
detectada e identificada como Kazachstania unispora (banda 7, Fig. 9 B).
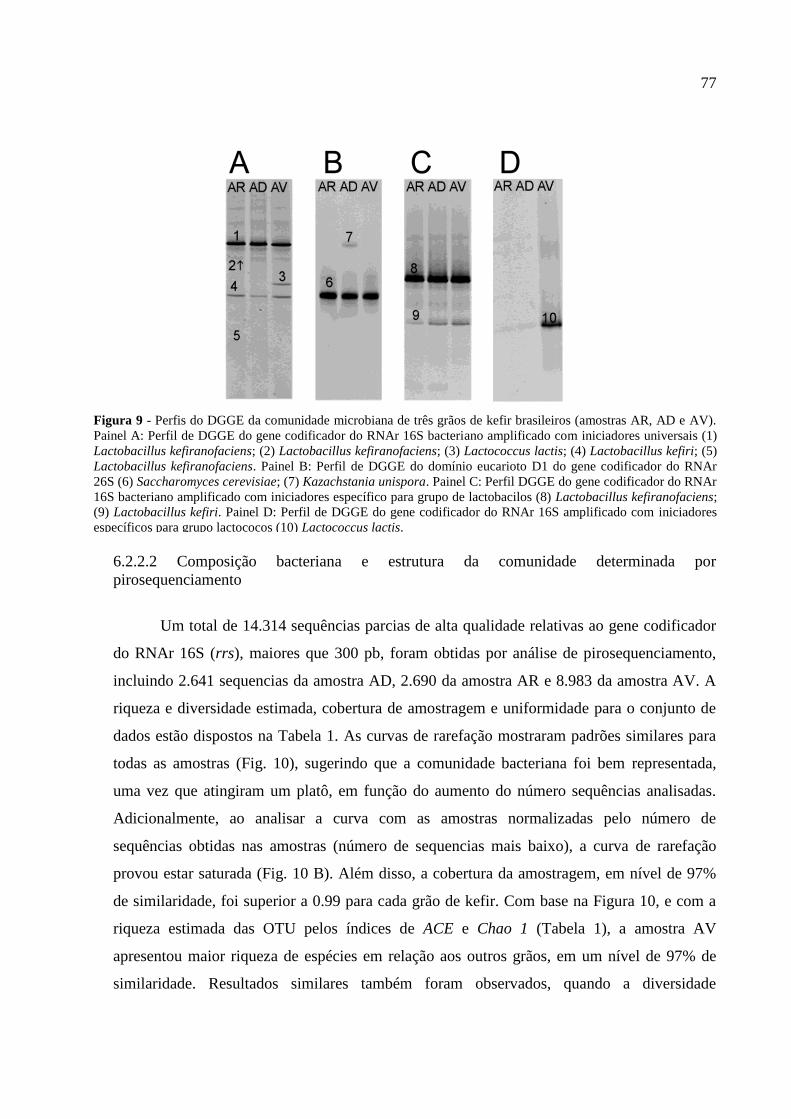
77
6.2.2.2 Composição bacteriana e estrutura da comunidade determinada por
pirosequenciamento
Um total de 14.314 sequências parcias de alta qualidade relativas ao gene codificador
do RNAr 16S (rrs), maiores que 300 pb, foram obtidas por análise de pirosequenciamento,
incluindo 2.641 sequencias da amostra AD, 2.690 da amostra AR e 8.983 da amostra AV. A
riqueza e diversidade estimada, cobertura de amostragem e uniformidade para o conjunto de
dados estão dispostos na Tabela 1. As curvas de rarefação mostraram padrões similares para
todas as amostras (Fig. 10), sugerindo que a comunidade bacteriana foi bem representada,
uma vez que atingiram um platô, em função do aumento do número sequências analisadas.
Adicionalmente, ao analisar a curva com as amostras normalizadas pelo número de
sequências obtidas nas amostras (número de sequencias mais baixo), a curva de rarefação
provou estar saturada (Fig. 10 B). Além disso, a cobertura da amostragem, em nível de 97%
de similaridade, foi superior a 0.99 para cada grão de kefir. Com base na Figura 10, e com a
riqueza estimada das OTU pelos índices de ACE e Chao 1 (Tabela 1), a amostra AV
apresentou maior riqueza de espécies em relação aos outros grãos, em um nível de 97% de
similaridade. Resultados similares também foram observados, quando a diversidade
Figura 9 - Perfis do DGGE da comunidade microbiana de três grãos de kefir brasileiros (amostras AR, AD e AV).
Painel A: Perfil de DGGE do gene codificador do RNAr 16S bacteriano amplificado com iniciadores universais (1)
Lactobacillus kefiranofaciens; (2) Lactobacillus kefiranofaciens; (3) Lactococcus lactis; (4) Lactobacillus kefiri; (5)
Lactobacillus kefiranofaciens. Painel B: Perfil de DGGE do domínio eucarioto D1 do gene codificador do RNAr
26S (6) Saccharomyces cerevisiae; (7) Kazachstania unispora. Painel C: Perfil DGGE do gene codificador do RNAr
16S bacteriano amplificado com iniciadores específico para grupo de lactobacilos (8) Lactobacillus kefiranofaciens;
(9) Lactobacillus kefiri. Painel D: Perfil de DGGE do gene codificador do RNAr 16S amplificado com iniciadores
específicos para grupo lactococos (10) Lactococcus lactis.
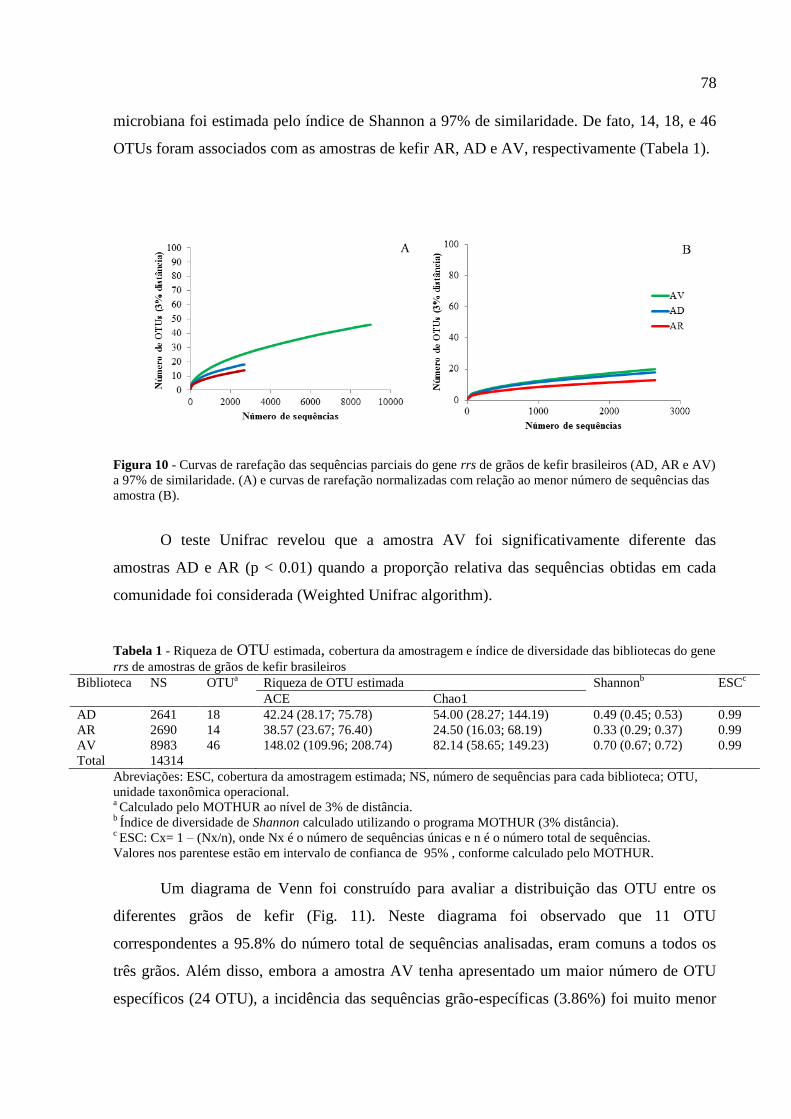
78
microbiana foi estimada pelo índice de Shannon a 97% de similaridade. De fato, 14, 18, e 46
OTUs foram associados com as amostras de kefir AR, AD e AV, respectivamente (Tabela 1).
Figura 10 - Curvas de rarefação das sequências parciais do gene rrs de grãos de kefir brasileiros (AD, AR e AV)
a 97% de similaridade. (A) e curvas de rarefação normalizadas com relação ao menor número de sequências das
amostra (B).
O teste Unifrac revelou que a amostra AV foi significativamente diferente das
amostras AD e AR (p < 0.01) quando a proporção relativa das sequências obtidas em cada
comunidade foi considerada (Weighted Unifrac algorithm).
Tabela 1 - Riqueza de OTU estimada, cobertura da amostragem e índice de diversidade das bibliotecas do gene
rrs de amostras de grãos de kefir brasileiros Biblioteca NS OTU
a Riqueza de OTU estimada Shannon
b ESC
c
ACE Chao1
AD 2641 18 42.24 (28.17; 75.78) 54.00 (28.27; 144.19) 0.49 (0.45; 0.53) 0.99
AR 2690 14 38.57 (23.67; 76.40) 24.50 (16.03; 68.19) 0.33 (0.29; 0.37) 0.99
AV 8983 46 148.02 (109.96; 208.74) 82.14 (58.65; 149.23) 0.70 (0.67; 0.72) 0.99
Total 14314
Abreviações: ESC, cobertura da amostragem estimada; NS, número de sequências para cada biblioteca; OTU,
unidade taxonômica operacional. a Calculado pelo MOTHUR ao nível de 3% de distância.
b Índice de diversidade de Shannon calculado utilizando o programa MOTHUR (3% distância).
c ESC: Cx= 1 – (Nx/n), onde Nx é o número de sequências únicas e n é o número total de sequências.
Valores nos parentese estão em intervalo de confianca de 95% , conforme calculado pelo MOTHUR.
Um diagrama de Venn foi construído para avaliar a distribuição das OTU entre os
diferentes grãos de kefir (Fig. 11). Neste diagrama foi observado que 11 OTU
correspondentes a 95.8% do número total de sequências analisadas, eram comuns a todos os
três grãos. Além disso, embora a amostra AV tenha apresentado um maior número de OTU
específicos (24 OTU), a incidência das sequências grão-específicas (3.86%) foi muito menor

79
do que aquelas compartilhadas por todas as amostras (95.8%). Da mesma forma, as OTU
específicas das outras duas amostras (AR e AD) foram representadas por um baixo percentual
das sequências.
Figura 11- Diagrama de Venn com disposição das OTU comuns e específicas em grãos de kefir brasileiros AD,
AR e AV, e o percentural de ocorrência das sequências totais (entre parênteses).
As leituras das sequências bacterianas foram agrupadas em três filos diferentes:
Firmicutes, Actinobacteria, e Proteobacteria. Destes, Firmicutes foi o filo mais abundante e
dominado por membros da classe Bacilli, pertencente à ordem Lactobacillales. Dentre as
sequências pertencentes a esta ordem, as famílias Leuconostocaceae, Streptococcaceae e
Lactobacillaceae foram detectadas. A família Lactobacillaceae foi predominante em todas as
amostras, e representada por somente um gênero, Lactobacillus, que contribuiu para 99.7,
93,9, e 99,6% das leituras dos grãos AR, AV e AD, respectivamente (Fig. 12). Na família
Streptococcaceae, o genêro Streptococcus correspondeu a somente 0,01% e 0,04% de todas as
sequências identificadas nos grãos AV e AD respectivamente, enquanto que o gênero
Lactococcus foi detectado somente no grão de kefir AV (4,87% das leituras). O gênero
Leuconostoc também ocorreu em baixo nível nas amostras AV (0,12%) e AD (0,23%).
Poucas sequências foram alinhadas ao filo Proteobacteria, que contribuiu para 0.3% do total
das sequências alinhadas para o grão AR, 1% para AV e 0,04% para AD. As sequências deste
filo pertenciam ao gênero Acetobacter na amostra AR (0,26%) e AD (0,04%), e ao gênero
Pseudomonas (0,99%) na amostra AV. O filo Actinobacteria foi representado por leituras
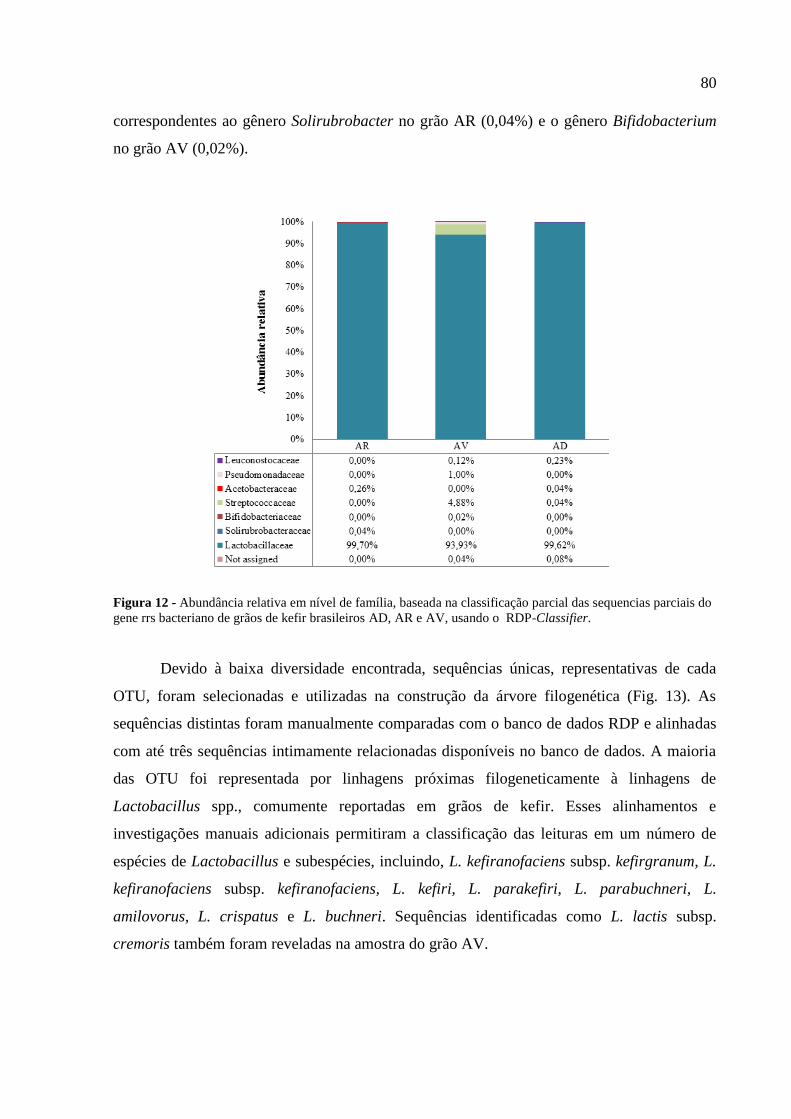
80
correspondentes ao gênero Solirubrobacter no grão AR (0,04%) e o gênero Bifidobacterium
no grão AV (0,02%).
Figura 12 - Abundância relativa em nível de família, baseada na classificação parcial das sequencias parciais do
gene rrs bacteriano de grãos de kefir brasileiros AD, AR e AV, usando o RDP-Classifier.
Devido à baixa diversidade encontrada, sequências únicas, representativas de cada
OTU, foram selecionadas e utilizadas na construção da árvore filogenética (Fig. 13). As
sequências distintas foram manualmente comparadas com o banco de dados RDP e alinhadas
com até três sequências intimamente relacionadas disponíveis no banco de dados. A maioria
das OTU foi representada por linhagens próximas filogeneticamente à linhagens de
Lactobacillus spp., comumente reportadas em grãos de kefir. Esses alinhamentos e
investigações manuais adicionais permitiram a classificação das leituras em um número de
espécies de Lactobacillus e subespécies, incluindo, L. kefiranofaciens subsp. kefirgranum, L.
kefiranofaciens subsp. kefiranofaciens, L. kefiri, L. parakefiri, L. parabuchneri, L.
amilovorus, L. crispatus e L. buchneri. Sequências identificadas como L. lactis subsp.
cremoris também foram reveladas na amostra do grão AV.
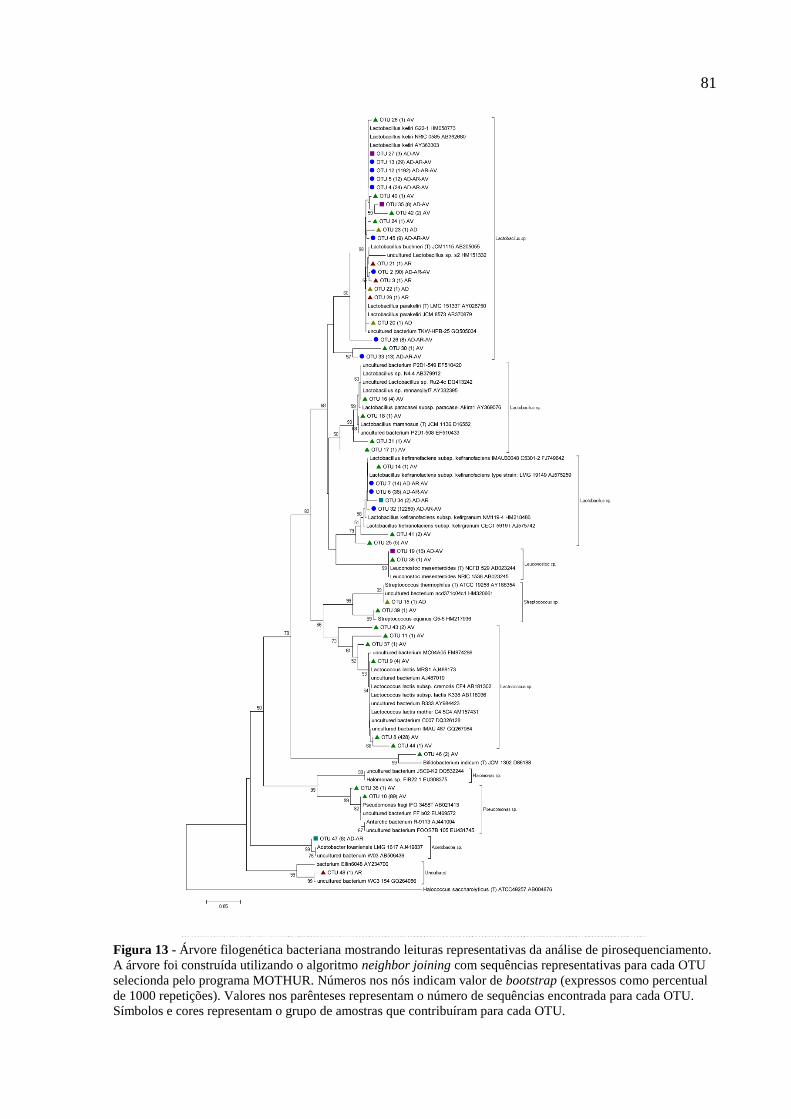
81
Figura 13 - Árvore filogenética bacteriana mostrando leituras representativas da análise de pirosequenciamento.
A árvore foi construída utilizando o algoritmo neighbor joining com sequências representativas para cada OTU
selecionda pelo programa MOTHUR. Números nos nós indicam valor de bootstrap (expressos como percentual
de 1000 repetições). Valores nos parênteses representam o número de sequências encontrada para cada OTU.
Símbolos e cores representam o grupo de amostras que contribuíram para cada OTU.

82
6.2.3 Discussão
A diversidade microbiana de grãos de kefir de diferentes origens vem sendo analisada
tanto por métodos dependentes de cultivo 107; 119; 128; 132; 141
quanto por métodos independentes
de cultivo 107; 126; 132; 206
. Neste trabalho, a diversidade e a estrutura microbiana de grãos de
kefir (AV, AR e AD), provenientes de três diferentes regiões do Brasil, foram avaliadas por
dois métodos independentes de cultivo. As populações dominantes foram identificadas pela
técnica de PCR-DGGE, enquanto que com o pirosequenciamento foi possível obter uma visão
mais completa da comunidade presente nos grãos.
Os fingerprints obtidos na PCR-DGGE originaram um pequeno número de bandas,
assim como já demonstrado anteriormente 121; 132; 138; 140
. Estas bandas corresponderam a
espécies de Lactobacillus que tem sido reportadas como prevalentes. Em estudos anteriores,
L. kefiranofaciens 121; 132; 140
e L. kefiri 107
também foram representados pelas bandas mais
intensas, detectadas por esta metodologia e como dominantes no perfil microbiológico de
diferentes grãos, por técnicas de cultivo 107; 123; 132
. A comunidade de leveduras apresentou um
perfil menos complexo, como já observado em outros grãos de kefir. Entretanto, neste estudo,
S. cerevisiae foi a espécie predominante, enquanto que em trabalhos prévios, além de
Saccharomyces spp., outros gêneros como Kluyveromyces lactis, Kazachstania spp. e
Candida spp. haviam sido reportados como predominantes 119; 121; 138; 141; 207
.
A técnica de pirosequenciamento reflete o estado da arte, quando empregada para
análise de populações microbianas de diferentes ecossistemas. Atualmente, vem sendo
utilizada para estudar diversos tipos de alimentos fermentados 175; 208; 209
. A técnica de
pirosequenciamento já havia sido utilizada na análise da comunidade bacteriana de um grão
de kefir irlandês e da bebida fermentada 126
.
No presente estudo, na análise por pirosequenciamento foi revelado que o filo
Firmicutes foi dominante em todos os grãos analisados, representando mais do que 99% das
sequencias. Este filo é composto de um grupo de bactérias Gram-positivas, com baixo
conteúdo de GC, que incluem as BAL. Firmicutes também foram dominantes no kefir
irlandês, no qual tanto o interior quanto o exterior dos grãos foram analisados 126
. Tais autores
também demonstraram que todos os outros filos detectados (Actinobacteria, Proteobacteria e
Bacteriodetes) eram componentes minoritários da comunidade, no interior dos grãos. Neste
estudo, Pseudomonas spp., pertencente ao filo Proteobacteria foi identificada no grão AV, a
qual também foi detectada anteriormente e considerada oriunda de uma contaminação

83
ambiental 126
. O gênero Acetobacter (subgrupo Proteobacteria) foi detectado somente em
dois grãos (AR e AD). Embora frequentemente, BAA seja mencionada como um dos
principais componentes da população bacteriana de grãos de kefir 44; 107; 108
, estas de fato, são
microrganismos detectados ocasionalmente 107; 121; 126; 132; 138
.
De forma geral, as duas técnicas independentes de cultivo utilizadas neste estudo para
avaliar a diversidade bacteriana de grãos de kefir de origens distintas, foram consistentes com
relação a detecção das bactérias predominantes. Entretanto, algumas bactérias identificadas
por pirosequenciamento não foram detectadas pela análise de DGGE, provavelmente porque
faziam parte da população minoritária, presentes em menor número nos grãos. Esta limitação
do método de PCR-DGGE foi relatada previamente por Ercolini 210
, que reportou que grupos
bacterianos minoritários, em comunidades complexas podem não estar representados nos
perfis de DGGE. Os resultados obtidos neste estudo mostraram que o pirosequenciamento
permite a detecção destes grupos, raramente associados à comunidade deste ecossistema e que
não fazem parte da comunidade dominante, como por exemplo, a detecção de bifidobacteria,
também reportada por pirosequenciamento por Dobson e colaboradores 126
. Esses
microrganismos embora minoritários parecem contribuir para características sensoriais,
particulares de cada bebida fermentada por seus respectivos grãos, via produção de
metabólitos, como ácidos orgânicos, etanol e compostos aromáticos, o que torna esta bebida
única.
A baixa diversidade bacteriana encontrada, em função da predominância do gênero
lactobacilos, leva a suposição de que embora os grãos tivessem origens distintas, a população
predominante na comunidade foi mantida. Farnworth e Mainville 102
, levantaram a hipótese
de que as bactérias e leveduras presentes nos grãos não devem variar significativamente de
uma parte do mundo para outra se condições de cultivos similares forem empregadas,
cuidados na manutenção e condições sanitárias mínimas forem mantidas. Entretanto, ao longo
do tempo e sob diferentes condições de crescimento, grãos de kefir podem mudar sua
composição microbiana e propriedades de fermentação. Neste estudo, mesmo considerando
que os três grãos tenham sido cultivados em diferentes regiões ao longo dos anos, a
microbiota dominante foi similar, variando somente a comunidade minoritária. Estas
pequenas diferenças entre os grupos microbianos específicos presentes em cada grão pode
estar associada com o perfil sensorial distinto que cada grão pode proporcionar no produto
final 44; 114; 119
, como já mencionado.

84
6.3 CAPÍTULO III - CARACTERÍSTICAS MICROBIOLÓGICAS E QUÍMICAS DA
BEBIDA KEFIR DURANTE A FERMENTAÇÃO E ESTOCAGEM2
6.3.1 Material e Métodos
6.3.1.1 Obtenção do grão de kefir
O Grão de kefir (AR) foi obtido de uma família de Niterói/RJ, que consumia
tradicionalmente a bebida kefir, cultivada em leite. Os grãos foram ativados em leite (1:10)
em pó desnatado reconstituído (10% p/v) a 25°C por 24 h, coados e lavados com água estéril.
A etapa de ativação foi repetida três vezes consecutivas 132
.
6.3.1.2 Produção do kefir
Os grãos ativados (30 g - peso úmido) foram inoculados em leite UHT (1000 mL) e
incubados estaticamente a 25°C por 24 h. Alíquotas de 1 mL da bebida foram coletadas
assepticamente a cada 6 h e as fermentações foram realizadas em triplicata.
6.3.1.3 Análises microbiológicas
A contagem dos microrganismos foi realizada pela técnica de semeadura em
superfície, a partir de diluições apropriadas (alíquotas de 0,1 mL) do leite fermentado em
diferentes meios de cultura, em triplicata. Uma solução de citrato de sódio a 2% foi utilizada
no preparo das diluições para as análises microbiológicas. As bactérias ácido láticas (BAL)
foram enumeradas em ágar MRS (Difco) e ágar M17 suplementado com 0,5% de glicose
(Difco), enquanto que as bactérias ácido acéticas (BAA) foram enumeradas em ágar AAB 211
.
Todos os meios para enumeração bacteriana foram suplementados com 200 ppm de
cicloheximide (Sigma-Aldrich) para inibir o crescimento de fungos filamentosos e leveduras.
As leveduras foram enumeradas em ágar YGC (Difco). As placas foram incubadas a 30°C por
5 dias para crescimento bacteriano, sob anaerobiose, usando sistema de anaerobiose Gaspak
EZ (Difco) e sob aerobiose a 25°C por 5 dias para leveduras. Após a contagem, médias e
desvios-padrões foram calculados.
2 Apêndice C

85
Alguns isolados foram aleatoriamente selecionados das placas para identificação, a
fim de confirmar a identidade das colônias enumeradas. O esfregaço pelo método de Gram e
teste de catalase foram utilizados na seleção inicial das estirpes pertencentes ao grupo de BAL
e BAA, que foram posteriormente identificadas por métodos moleculares. Todas as estirpes
isoladas foram estocadas em caldo MRS com 20% de glicerol a -80°C.
6.3.1.4 Análise de restrição do DNA ribossomal amplificado (ARDRA)
A extração do DNA das culturas puras de bactérias e leveduras isoladas foi realizada
de acordo com método descrito por Randazzo e colaboradores 212
e Del Aguila e
colaboradores 213
, respectivamente.
Os oligonucleotídeos iniciadores (primers) 27f (5’–AGAGTTTGATCCTGGCTCAG–
3’) e 1512r (5’– CGGCTACCTTGTTACGACT – 3’) foram utilizados na reação de
amplificação do gene codificador do RNAr 16S, enquanto que o par de iniciador ITS1 (5’-
TCCGTAGGTGAACCTGCGG-3’) e ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) foi
utilizado para amplificar a região do espaçador não-codificante interno (ITS), conforme
descrito por Wang e colaboradores 214
e Naumova, Ivannikova e Naumov 215
para bactérias e
fungos, respectivamente. Posteriormente, produtos da reação de PCR foram separados por
eletroforese em gel de agarose a 1%. Os produtos amplificados foram purificadas utilizando o
kit Illustra GFX PCR DNA and Gel Band Purification (GE Healthcare Life Sciences), de
acordo com as instruções do fabricante. Posteriormente, 10 µL dos produtos purificados
foram digeridos com enzimas de restrição FastDigest® HaeIII e HhaI para BAL, FastDigest
®
AluI e ScrF1 para BAA e FastDigest® HhaI e DdeI para leveduras conforme recomendado
pelo fabricante (Fermentas Inc.). Os produtos de PCR digeridos foram então separados por
eletroforese em gel de agarose a 1,5 %, por 1 h e 45 min a 75V. Os géis foram corados com
GelRed® 1X (Uniscience) e visualizados sob luz ultra violeta (dados não mostrados).
Os perfis representativos de cada espécie observado no ARDRA foram selecionados
para sequenciamento e, então, a análise das sequências parciais do gene codificador do RNAr
16S e da região espaçadora não-codificante interna ITS foi realizada em um sequenciador
3130 (Applied Biosystems) e utilizada para identificação, em nível de espécie, dos isolados
representativos de bactéria e levedura. As sequências obtidas foram analisadas e comparadas
por similaridade com as depositadas na biblioteca do GenBank, verificando as que revelaram
maior similaridade com as obtidas.

86
6.3.1.5 Extração de DNA e RNA da bebida kefir e grãos
Amostras de kefir coletadas durante a fermentação e estocagem foram congeladas e
estocadas a -80°C. Uma amostra do grão (1g) utilizado como inóculo também foi coletada e
homogeneizada em solução de citrato de sódio a 2%. Um mililitro desse homogeneizado e das
amostras de kefir foram centrifugadas por 10 min a 10,000 g.
O DNA genômico foi extraído e purificado dos pellets utilizando o kit FastDNA Spin®
(QBIOgene) de acordo com as instruções do fabricante.
O RNA total foi extraído dos pellets em tubos eppendorfs contendo 0,3 g de pérolas de
vidro e 1 mL do reagente TRIzol (Invitrogen). As amostras foram lisadas em FastPrep® (Bio
101) com dois ciclos de 40 segundos à velocidade 6,0, e intervalos de 1 min em banho de
gelo. Posteriormente, os tubos foram centrifugados a 12000 g a 4°C por 10 min, e 200 µL de
clorofórmio foram adicionados ao sobrenadante. Os tubos foram centrifugados novamente, a
12000 g a 4°C por 10 min, e a fase aquosa precipitada com 1 mL de etanol absoluto
refrigerado.
O RNA foi coletado a 14000 g/4°C por 10 min, e os pellets foram lavados com etanol
75% e secos a temperatura ambiente. Uma alíquota de 50 µL de água DEPC estéril foi
adicionada e mantida por um período de 30 minutos a 45°C para facilitar a solubilização do
ácido nucléico. Posteriormente, 1 µL de enzima DNase, livre de Rnase (Roche Diagnostics)
foi adicionada para digestão do DNA a 37°C por 1 h. A solução de RNA foi examinada
quanto presença de quantidades residuais de DNA, através de uma reação de amplificação.
Caso os produtos de amplificação fossem obtidos após a reação de PCR, o tratamento com
DNAse era repetido para eliminar qualquer DNA residual.
6.3.1.6 Protocolo do PCR e RT-PCR
O DNA bacteriano obtido foi utilizado como molde para amplificação da região V3 do
gene codificador do RNAr 16S, utilizando os iniciadores universais F338-GC (5’–
TACGGGAGGCAGCAG–3’ e R518 (5’–ATTACCGCGGCTGCTGG–3’), como reportado
por Muyzer, De Waal e Uitterlinden 164
. A amplificação do domínio D1 do gene codificador
do RNAr 26S de fungo foi realizada utilizando os primers NL1-GC (5’-

87
GCCATATCAATAAGCGGAGGAAAG–3’) e LS2 (5’–ATTCCCAAACAACTCGACTC–
3’), conforme reportado por Cocolin e colaboradores 199
.
A reação de PCR (volume final 50 μL) continha tampão da enzima 1 X concentrado,
0.2 mM de cada dNTP (Fermentas Inc.), 1.5 mM MgCl2, 25 pmol of cada primer
(Invitrogen), 50 ng of cada DNA molde e 1.5 U de Taq platinum (Invitrogen). Reação de PCR
(30 ciclos) foi realizada no termociclador Mycycler (Bio-Rad) nas seguintes condições:
desnaturação a 94ºC por 30 seg, anelamento a 55°C para a reação para bactéria e 52°C para a
reação para levedura, por 30 seg, e elongação a 72°C por 1 min, seguido de uma etapa de
elongação final a 72°C por 7 min.
O RT-PCR foi realizado utilizando o kit Ready-To-Go RT-PCR beads (GE Healthcare
Life Sciences), em que 1 µg do RNA total foi adicionado a 25 pmol de cada iniciador (338f-
518r para bactéria e NL1-LS2 para levedura), e o volume final corrigido para 50 µL com água
estéril livre de RNAse (Invitrogen). A reação de RT-PCR foi realizada em um termociclador e
incubada a 42°C por 20 min, e após a etapa a 95°C por 10 min, os ciclos de amplificação
foram realizados, conforme descrito acima. Os produtos da amplificação (5 µL) foram
analisados por eletroforese em gel de agarose a 1,2% corados com GelRed® 1X.
6.3.1.7 Análise do kefir por DGGE
O DGGE foi realizado utilizando equipamento DCode (Bio-Rad) a 60°C, com gel de
poliacrilamida 8% contendo um gradiente desnaturante de 30-55% para bacterias e 30-50%
para leveduras, em tampão 1X TAE a 50V por 10 min, seguido por 150V por 4 h 30 min.
Após a eletroforese, os geis foram corados com SYBR Green (1:10.000 diluição; Invitrogen)
por 60 min e o padrão de bandas do DGGE foi visualizado sob luz ultravioleta com o
transiluminador MiniBisPro (BioAmerica Inc.). O perfil de bandas obtido no DGGE foi
normalizado, comparado e agrupado utilizando o software Bionumerics 6.5 (Applied Maths).
As bandas foram automaticamente detectadas e marcadas, e posteriormente, correções
manuais foram realizadas. Dendrogramas de similaridade foram gerados com auxílio do
software Bionumerics, utilizando o coeficiente de Dice e, para agrupamento, o algoritmo
UPGMA.
Além disso, bandas dos géis de PCR-DGGE e RT-PCR-DGGE foram identificadas
pela análise da sequencia dos nucleotídeos. Para isto, as mesmas foram excisadas do gel de
acrilamida e purificadas com o kit Qiaex II DNA extraction (Qiagen), de acordo com as

88
recomendações do fabricante. O DNA eluído de cada banda foi reamplificado com o mesmo
par de iniciador, sem o grampo GC. Posteriormente, uma eletroforese em gel de agarose dos
produtos da segunda reação PCR foi realizada, as bandas foram excisadas e o DNA foi
purificado com o kit Illustra GFX PCR DNA and GEL Band Purification (GE Healthcare Life
Sciences), de acordo com as instruções do fabricante. Produtos de PCR purificados foram
ligados diretamente ao vetor pGEM-T® Easy Vector (Promega) seguindo as instruções do
fabricante. Os produtos da ligação foram transformados em células competentes de
Escherichia coli DH5-α. e os clones positivos (colônias brancas) cresceram em ágar LB
contendo ampicilina (100 µg/mL), IPTG (isopropyl-β-D-1-thiogalactopyranoside; 0.5 mM) e
X-gal (5-bromo-4-chloro-3-indolyl-β-D-galacto-pyranoside; 80 µg/mL). Posteriormente, os
plasmídeos foram extraídos utilizando o método de lise alcalina miniprep 216
e o
sequenciamento do inserto foi realizado utilizando o sistema Big Dye Terminator e um
sequenciador automático em capilar ABI 3130 (Applied Biosystems). A identificação das
sequências foi determinada por comparação com as sequências depositadas no banco de dados
do GenBank (http://www.ncbi.nlm.nih.gov/BLAST/).
6.3.1.8 Análise dos compostos químicos por CLAE
A extração dos ácidos orgânicos, etanol e carboidratos da bebida kefir foi realizada
como descrito por González De Llano, Rodriguez e Cuesta 217
. Uma solução de H2SO4 a 45
mmol/L (25 mL) foi adicionada a 5 mL do leite UHT ou 5 mL do kefir, e após 1 h sob
agitação constante a 250 rpm, o sobrenadante foi separado por centrifugação a 6000 g e
filtrado em membranas de 0.45 µm (Millipore).
Amostras filtradas foram injetadas (50 µL), em triplicata, em uma sistema de
Cromatografia Líquida de Alta Eficiência (CLAE) (Shimadzu) equipado com uma coluna
HPX-87H (Aminex Monitoring Fermentation column - 150 X 7.8 mm) (Bio-Rad) e protegida
por uma pré-coluna (Microguard cartridge - 30 X 4.6 mm) (Bio-Rad) mantida a 65°C. Os
ácidos orgânicos (ácidos lático, acético, cítrico, butírico e propionico) foram quantificados
utilizando um detector por arranjo de diodos modelo SPD-M20A (Shimadzu, Tokyo, Japan),
e a absorbância monitorada a 210 nm. Lactose, galactose, glicose e etanol foram quantificados
utilizando um detector de índice de refração, modelo RID-10A (Shimadzu). A fase móvel
(vazão de fluxo isocrático a 0.7 mL/min) utilizada foi H2SO4 a 3 mM. Os cromatogramas e a
quantificação dos compostos foram obtidos utilizando o software LC Solution (Shimadzu).

89
Curvas padrão, baseadas na área do pico, foram calculadas para os ácidos orgânicos,
carboidratos e etanol abrangendo uma ampla faixa de concentração, pela comparação com as
soluções-padrão. Padrões (Sigma-Aldrich) foram preparados em água deionizada e filtradas
em membrana de 0.45 µm (Millipore).
6.3.1.9 Análises estatísticas
Análises de variância (ANOVA) das médias foram utilizadas para identificar
diferenças nas enumerações microbianas, concentrações de ácidos orgânicos, carboidratos e
etanol ao longo do tempo de fermentação e estocagem do kefir. Quando o valor de F
significativo foi observado, um teste adicional de Tukey (post hoc test) foi realizado com
nível de significância 0,05 (p<0,05). Todas as análises foram feitas com o software Statistica
versão 7.0 (Statsoft, Inc).
6.3.2 Resultados
6.3.2.1 Enumeração microbiana durante a fermentação e estocagem da bebida kefir
Microrganismos presentes na bebida kefir foram enumerados durante 24 h de
fermentação do leite (3% grãos de kefir) a 25°C e estocagem do kefir a 4°C (Fig 14) por 28
dias (Fig 14). A contagem do número de células presuntivas do grupo de BAL (ágar M17)
aumentou consideravelmente (4 unidades logarítimicas de UFC/mL) com 12 h de fermentação
(p < 0.05), enquanto que contagens no ágar MRS, no mesmo ponto de amostragem, mostrou
um aumento de aproximadamente 2 log10 UFC/mL, atingindo seu valor máximo (10 log10
UFC/mL) com 18 h de fermentação. Na contagem presuntiva de BAA houve um aumento
significativo em 12 h atingindo valor máximo em 24 h (7,8 log10 UFC/mL) de fermentação.
Até 12 h de fermentação, foi observado um aumento na contagem de leveduras, mantendo-se
constante na faixa de 6 log10 UFC/mL.
Os microrganismos também foram enumerados durante a estocagem do kefir a 4°C
por 672 h (28 dias) (Fig 14). Neste período, contagens de leveduras (aproximadamente 6 log10
UFC/mL) e contagem do grupo de BAL (aproximadamente 10 log10 UFC/mL) mantiveram-se
constantes até o final do período de estocagem (p > 0.05), enquanto que a contagem
presuntiva de BAA foi reduzida para 7,2 log10 UFC/mL em 672 h de estocagem.
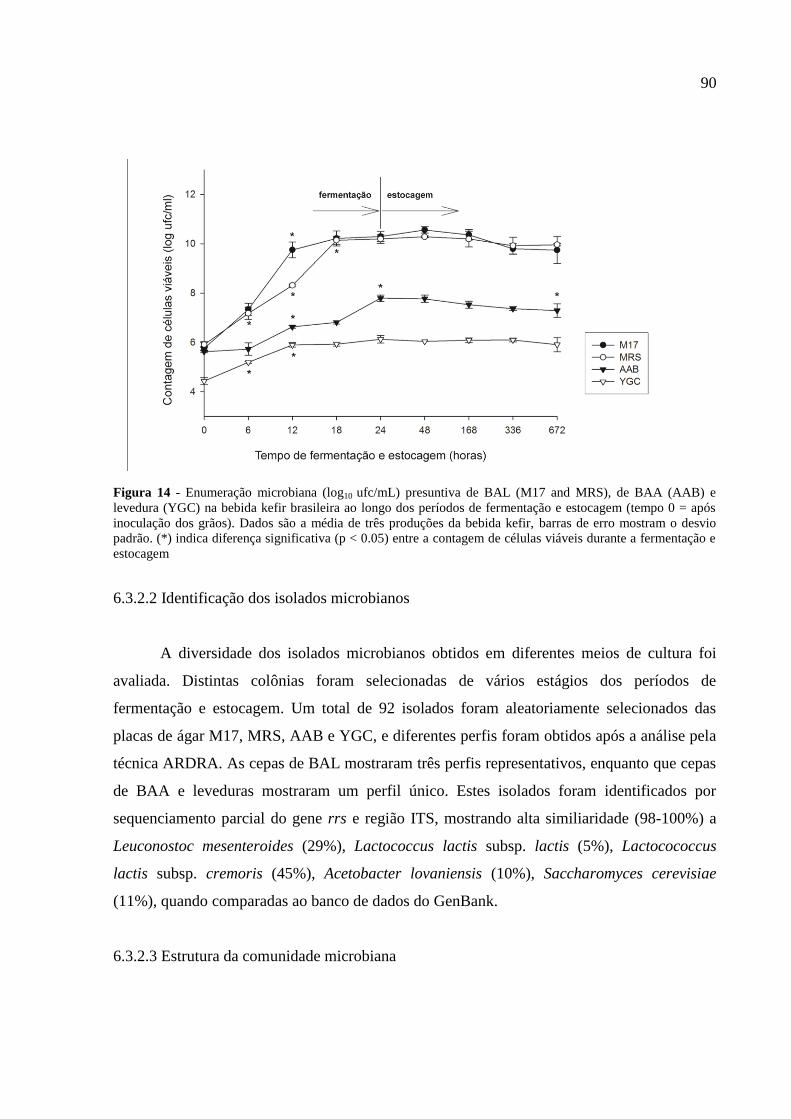
90
Figura 14 - Enumeração microbiana (log10 ufc/mL) presuntiva de BAL (M17 and MRS), de BAA (AAB) e
levedura (YGC) na bebida kefir brasileira ao longo dos períodos de fermentação e estocagem (tempo 0 = após
inoculação dos grãos). Dados são a média de três produções da bebida kefir, barras de erro mostram o desvio
padrão. (*) indica diferença significativa (p < 0.05) entre a contagem de células viáveis durante a fermentação e
estocagem
6.3.2.2 Identificação dos isolados microbianos
A diversidade dos isolados microbianos obtidos em diferentes meios de cultura foi
avaliada. Distintas colônias foram selecionadas de vários estágios dos períodos de
fermentação e estocagem. Um total de 92 isolados foram aleatoriamente selecionados das
placas de ágar M17, MRS, AAB e YGC, e diferentes perfis foram obtidos após a análise pela
técnica ARDRA. As cepas de BAL mostraram três perfis representativos, enquanto que cepas
de BAA e leveduras mostraram um perfil único. Estes isolados foram identificados por
sequenciamento parcial do gene rrs e região ITS, mostrando alta similiaridade (98-100%) a
Leuconostoc mesenteroides (29%), Lactococcus lactis subsp. lactis (5%), Lactocococcus
lactis subsp. cremoris (45%), Acetobacter lovaniensis (10%), Saccharomyces cerevisiae
(11%), quando comparadas ao banco de dados do GenBank.
6.3.2.3 Estrutura da comunidade microbiana

91
A ecologia do kefir brasileiro foi avaliada por DGGE utilizando os pares de
iniciadores 338-518 e NL1-LS2 para amplificação das sequências de DNA e RNA de
bactérias e leveduras, respectivamente. Bandas selecionadas foram excisadas (Fig 15 e Fig
16) e sequenciadas após a reamplificação, seguidas por clonagem e identificação pelo
alinhamento com as sequências disponíveis no GenBank (Tabela 2).
Tabela 2 – Identificação dos microrganismos baseados na comparação com as sequências disponíveis no banco
de dados do Genbank, após clonagem e sequenciamento das bandas selecionadas do PCR e RT-PCR-DGGE,
utilizando iniciadores universais para bactéria (RNAr 16S) e leveduras (RNAr 26S).
Banda Sequência identica % Identidade n° de acesso GenBank
Bacteria
a Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
b Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
c Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
d Lactobacillus kefiri 99 AB429371.1/ FJ749410.1
e Lactobacillus parakefiri 99 AB370879.1/ NR_029039.1
f Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
i Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
J Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
k Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
l Lactobacillus kefiri 99 AB429371.1/ FJ749410.1
m Lactobacillus kefiri 99 AB429371.1/ FJ749410.1
n Lactobacillus parakefiri 100 AB370879.1/ NR_029039.1
o Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
Levedura
g Saccharomyces cerevisiae 99 EU441887.1/ BK006945.2
h Saccharomyces cerevisiae 100 EU441887.1/ BK006945.2
p Saccharomyces cerevisiae 100 EU441887.1/ BK006945.2
q Saccharomyces cerevisiae 100 EU441887.1/ BK006945.2
A análise dos dendrogramas de similaridade obtidos com o pefil da comunidade de
bactérias e leveduras permitiu a identificação de dois grupos principais, correspondentes aos
estágios iniciais de fermentação e estocagem, com um coeficiente de similaridade entre eles
de aproximadamente 69% e 60% para bactéria e levedura, respectivamente.
O perfil de bandas obtido com análise do DNA da comunidade de bactérias das
amostras coletadas nos períodos iniciais de fermentação (0-18 h) foi agrupado em um cluster
(Fig 15 – A.1) com um coeficiente de similaridade de 75%, enquanto que um segundo cluster
(90% de similaridade) correspondeu às amostras do período final da fermentação (24 h) e
àquelas analisadas durante as 672 h de estocagem a 4°C. O leite UHT desnatado e os grãos
de kefir não foram agrupados nestes clusters principais e mostraram um coeficiente de
similaridade de 46% e 59%, respectivamente (Fig 15).
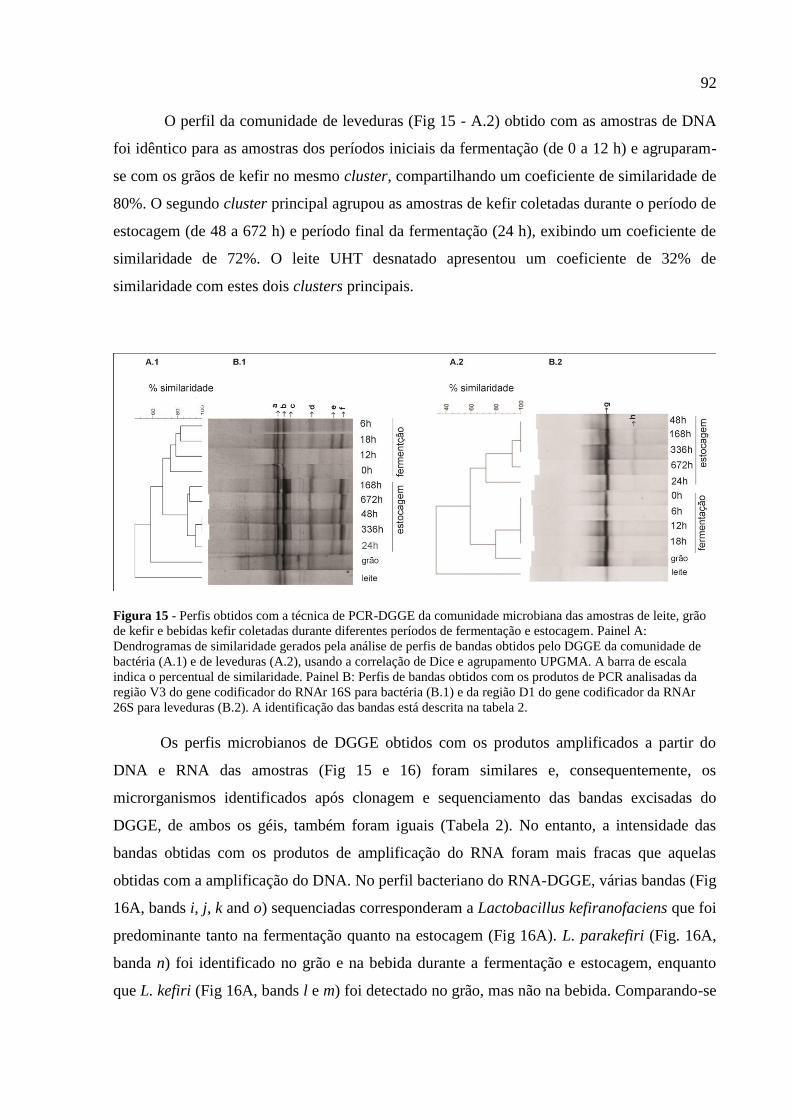
92
O perfil da comunidade de leveduras (Fig 15 - A.2) obtido com as amostras de DNA
foi idêntico para as amostras dos períodos iniciais da fermentação (de 0 a 12 h) e agruparam-
se com os grãos de kefir no mesmo cluster, compartilhando um coeficiente de similaridade de
80%. O segundo cluster principal agrupou as amostras de kefir coletadas durante o período de
estocagem (de 48 a 672 h) e período final da fermentação (24 h), exibindo um coeficiente de
similaridade de 72%. O leite UHT desnatado apresentou um coeficiente de 32% de
similaridade com estes dois clusters principais.
Figura 15 - Perfis obtidos com a técnica de PCR-DGGE da comunidade microbiana das amostras de leite, grão
de kefir e bebidas kefir coletadas durante diferentes períodos de fermentação e estocagem. Painel A:
Dendrogramas de similaridade gerados pela análise de perfis de bandas obtidos pelo DGGE da comunidade de
bactéria (A.1) e de leveduras (A.2), usando a correlação de Dice e agrupamento UPGMA. A barra de escala
indica o percentual de similaridade. Painel B: Perfis de bandas obtidos com os produtos de PCR analisadas da
região V3 do gene codificador do RNAr 16S para bactéria (B.1) e da região D1 do gene codificador da RNAr
26S para leveduras (B.2). A identificação das bandas está descrita na tabela 2.
Os perfis microbianos de DGGE obtidos com os produtos amplificados a partir do
DNA e RNA das amostras (Fig 15 e 16) foram similares e, consequentemente, os
microrganismos identificados após clonagem e sequenciamento das bandas excisadas do
DGGE, de ambos os géis, também foram iguais (Tabela 2). No entanto, a intensidade das
bandas obtidas com os produtos de amplificação do RNA foram mais fracas que aquelas
obtidas com a amplificação do DNA. No perfil bacteriano do RNA-DGGE, várias bandas (Fig
16A, bands i, j, k and o) sequenciadas corresponderam a Lactobacillus kefiranofaciens que foi
predominante tanto na fermentação quanto na estocagem (Fig 16A). L. parakefiri (Fig. 16A,
banda n) foi identificado no grão e na bebida durante a fermentação e estocagem, enquanto
que L. kefiri (Fig 16A, bands l e m) foi detectado no grão, mas não na bebida. Comparando-se
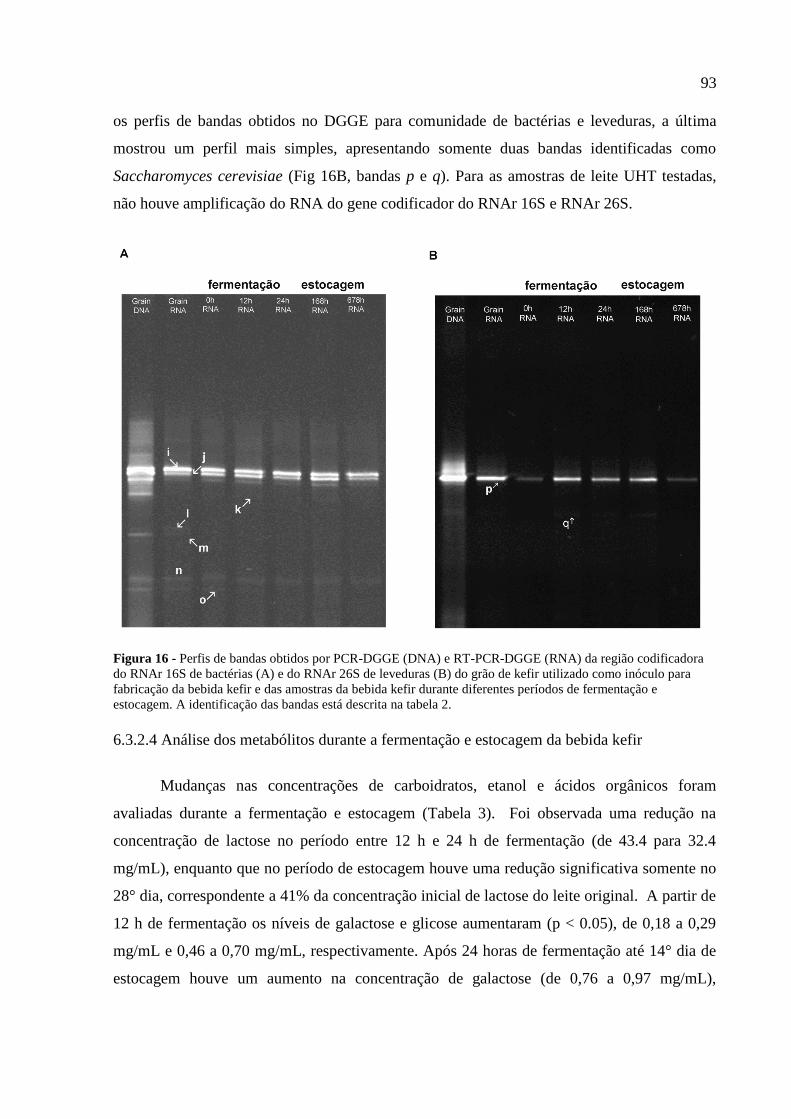
93
os perfis de bandas obtidos no DGGE para comunidade de bactérias e leveduras, a última
mostrou um perfil mais simples, apresentando somente duas bandas identificadas como
Saccharomyces cerevisiae (Fig 16B, bandas p e q). Para as amostras de leite UHT testadas,
não houve amplificação do RNA do gene codificador do RNAr 16S e RNAr 26S.
Figura 16 - Perfis de bandas obtidos por PCR-DGGE (DNA) e RT-PCR-DGGE (RNA) da região codificadora
do RNAr 16S de bactérias (A) e do RNAr 26S de leveduras (B) do grão de kefir utilizado como inóculo para
fabricação da bebida kefir e das amostras da bebida kefir durante diferentes períodos de fermentação e
estocagem. A identificação das bandas está descrita na tabela 2.
6.3.2.4 Análise dos metabólitos durante a fermentação e estocagem da bebida kefir
Mudanças nas concentrações de carboidratos, etanol e ácidos orgânicos foram
avaliadas durante a fermentação e estocagem (Tabela 3). Foi observada uma redução na
concentração de lactose no período entre 12 h e 24 h de fermentação (de 43.4 para 32.4
mg/mL), enquanto que no período de estocagem houve uma redução significativa somente no
28° dia, correspondente a 41% da concentração inicial de lactose do leite original. A partir de
12 h de fermentação os níveis de galactose e glicose aumentaram (p < 0.05), de 0,18 a 0,29
mg/mL e 0,46 a 0,70 mg/mL, respectivamente. Após 24 horas de fermentação até 14° dia de
estocagem houve um aumento na concentração de galactose (de 0,76 a 0,97 mg/mL),

94
enquanto que a glicose permaneceu constante ao longo da fermentação, aumentando no 2° dia
de estocagem (de 0,29 mg/mL a 0,36 mg/mL).
Durante os períodos de fermentação e estocagem, os valores de pH (Tabela 3)
reduziram progressivamente (p < 0.05) variando de 6,55 a 4,31, como consequência da
produção de ácidos orgânicos. A concentração de ácido lático aumentou rápida e
significativamente (p < 0.05) durante as 24 h de fermentação, atingindo 7,38 mg/mL e
aumentando para 9,54 mg/mL no final do período de estocagem (28◦
dia). Ácido acético
também foi produzido a partir de 12 h, atingindo valores máximos de 0,93 e 1,16 mg/mL após
fermentação (24 h) e estocagem (672h), respectivamente (Tabela 3).
O etanol (Tabela 3) foi produzido após 18 h de fermentação (0,14 mg/mL) até 24 h,
quando atingiu a concentração de 0,32 mg/mL. Durante a estocagem, a concentração de
etanol aumentou para 0,45 mg/mL, atingindo valor máximo (1,36 mg/mL) em 28 dias de
estocagem.
Mais de 35% do citrato (Tabela 3) presente no leite original foi consumido após 12 h
de fermentação (de 4,29 para 2,73 mg/mL), mantendo-se constante até o final do período de
estocagem, enquanto os ácidos propiônico e butírico aumentaram com 6 e 12 horas de
fermentação, respectivamente. Foi observado também um aumento significativo na produção
de ácido butírico durante a fermentação (24 h) e estocagem (2, 7, e 28 dias), enquanto que o
ácido propiônico aumentou com 12 h de fermentação (de 0,80 a 1,09 mg/mL), mantendo-se
constante por todo período, exceto por uma redução significativa em 28 dias (para 0,96
mg/mL, p < 0.05) de estocagem (Tabela 3).
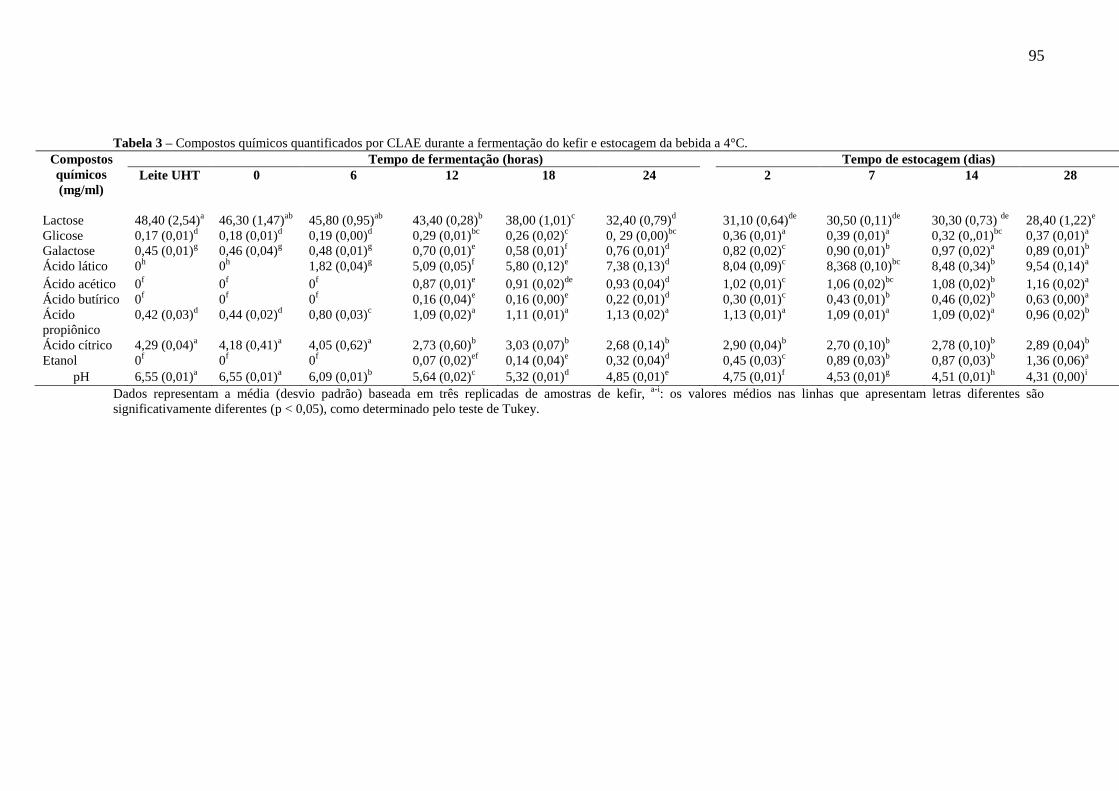
95
Tabela 3 – Compostos químicos quantificados por CLAE durante a fermentação do kefir e estocagem da bebida a 4°C. Compostos
químicos
(mg/ml)
Tempo de fermentação (horas) Tempo de estocagem (dias)
Leite UHT 0 6 12 18 24 2 7 14 28
Lactose 48,40 (2,54)a
46,30 (1,47)ab
45,80 (0,95)ab
43,40 (0,28)b 38,00 (1,01)
c 32,40 (0,79)
d 31,10 (0,64)
de 30,50 (0,11)
de 30,30 (0,73)
de 28,40 (1,22)
e
Glicose 0,17 (0,01)d
0,18 (0,01)d 0,19 (0,00)
d 0,29 (0,01)
bc 0,26 (0,02)
c 0, 29 (0,00)
bc 0,36 (0,01)
a 0,39 (0,01)
a 0,32 (0,,01)
bc 0,37 (0,01)
a
Galactose 0,45 (0,01)g
0,46 (0,04)g 0,48 (0,01)
g 0,70 (0,01)
e 0,58 (0,01)
f 0,76 (0,01)
d 0,82 (0,02)
c 0,90 (0,01)
b 0,97 (0,02)
a 0,89 (0,01)
b
Ácido lático 0h 0
h 1,82 (0,04)
g 5,09 (0,05)
f 5,80 (0,12)
e 7,38 (0,13)
d 8,04 (0,09)
c 8,368 (0,10)
bc 8,48 (0,34)
b 9,54 (0,14)
a
Ácido acético 0f 0
f 0
f 0,87 (0,01)
e 0,91 (0,02)
de 0,93 (0,04)
d 1,02 (0,01)
c 1,06 (0,02)
bc 1,08 (0,02)
b 1,16 (0,02)
a
Ácido butírico 0f 0
f 0
f 0,16 (0,04)
e 0,16 (0,00)
e 0,22 (0,01)
d 0,30 (0,01)
c 0,43 (0,01)
b 0,46 (0,02)
b 0,63 (0,00)
a
Ácido
propiônico
0,42 (0,03)d
0,44 (0,02)d 0,80 (0,03)
c 1,09 (0,02)
a 1,11 (0,01)
a 1,13 (0,02)
a 1,13 (0,01)
a 1,09 (0,01)
a 1,09 (0,02)
a 0,96 (0,02)
b
Ácido cítrico 4,29 (0,04)a
4,18 (0,41)a 4,05 (0,62)
a 2,73 (0,60)
b 3,03 (0,07)
b 2,68 (0,14)
b 2,90 (0,04)
b 2,70 (0,10)
b 2,78 (0,10)
b 2,89 (0,04)
b
Etanol 0f 0
f 0
f 0,07 (0,02)
ef 0,14 (0,04)
e 0,32 (0,04)
d 0,45 (0,03)
c 0,89 (0,03)
b 0,87 (0,03)
b 1,36 (0,06)
a
pH 6,55 (0,01)a 6,55 (0,01)
a 6,09 (0,01)
b 5,64 (0,02)
c 5,32 (0,01)
d 4,85 (0,01)
e 4,75 (0,01)
f 4,53 (0,01)
g 4,51 (0,01)
h 4,31 (0,00)
i
Dados representam a média (desvio padrão) baseada em três replicadas de amostras de kefir, a-i
: os valores médios nas linhas que apresentam letras diferentes são
significativamente diferentes (p < 0,05), como determinado pelo teste de Tukey.

96
6.3.3 Discussão
Além do estudo de diferentes grãos de kefir, também foi abordada a caracterização da
bebida kefir produzida a partir dos grãos AR, ao longo da fermentação e estocagem do
produto, reproduzindo as condições comumente utilizadas por famílias que tradicionalmente,
consomem o leite fermentado. Desta forma, técnicas independentes de cultivo e dependentes
de cultivo foram realizadas para avaliar o comportamento das populações no produto final,
além da quantificação de ácidos orgânicos, etanol e carboidratos.
Na análise de DGGE, não foi observada nenhuma mudança significativa na estrutura
das comunidades microbianas (bactérias e leveduras), durante as fases da fermentação e
estocagem estudadas. Tal comportamento já tinha sido observado em outros estudos 125; 140
.
Este resultado pode ser devido à limitação da técnica de DGGE, capaz de detectar somente
populações dominantes, à semelhança dos resultados obtidos com os grãos, aliado ao intenso
consórcio microbiano, dominado por certas populações. Sendo assim, uma reação de PCR por
transcriptase reversa aliada a técnica de DGGE (RT-PCR-DGGE) foi realizada a fim de
avaliar através dos RNAm presentes, quais populações predominantes, estariam ativas na
comunidade.
Alguns autores realizaram a reação da transcriptase reversa para avaliar a viabilidade
de populações, em comunidades microbianas 212; 218; 219; 220
. Randazzo e colaboradores 212;
Cocolin e colaboradores 221
foram capazes de diferenciar componentes ativos (derivados do
RNAr) da diversidade total (derivado do DNAr) da comunidade, combinando técnicas de RT-
PCR-DGGE e PCR-DGGE. Entretanto, estudos prévios de dinâmicas de populações baseado
na quantificação de diferentes moléculas alvos, sugeriram que outros genes podem ser mais
adequados para avaliar a viabilidade 222; 223; 224
. De acordo com Falentin e colaboradores 219
a
molécula RNAr 16S é estável e a presença de seu transcrito na comunidade, não prova que as
células ainda estejam metabolicamente ativas. Por contraste, outros pesquisadores mostraram
que o RNAr 16S despareceu relativamente rápido após tratamento letal extremo 225; 226
,
dependendo da escolha do fragmento amplificado, do tratamento de inativação e condições do
pós-tratamento 227
.
Na análise de DGGE dos transcritos de cDNA, referente ao gene rrs, foi possível notar
que todos os microrganismos estavam ativos, durante a fermentação e estocagem, com
exceção do L. kefiri presente somente nos grãos, sugerindo que este organismo desempenha
um papel simbiótico e ecológico nos mesmos. Entretanto, Magalhães e colaboradores 109
isolaram este microrganismo do kefir durante a fermentação. Talvez a presença de tal espécie

97
no leite fermentado dependa das condições de processamento aplicadas na produção e da
origem dos grãos. Bandas correspondentes a L. kefiranofaciens foram as mais intensas ao
longo da fermentação e estocagem, tanto no fingerprint obtido com RNA, quanto com DNA
das amostras, reforçando o papel dominante desta espécie, conforme já reportado neste
trabalho (cap. II) e por outros autores 121; 132; 140
.
Poucos pesquisadores têm analisado a microbiota resultante na bebida kefir, produzida
tradicionalmente com grãos de kefir. Magalhães e colaboradores 140
avaliaram a estrutura da
comunidade microbiana de kefir, produzido com diferentes substratos lácteos, por PCR-
DGGE, e os microrganismos detectados foram similares aos reportados neste trabalho,
utilizando a mesma técnica. Posteriormente, o mesmo grupo 109
isolou várias espécies de
Lactobacillus spp., L. lactis, A. lovaniensis e leveduras não fermentadoras e fermentadoras de
lactose, em meios de cultura específicos, durante a fermentação de kefir cultivado em leite.
Neste estudo, os microrganismos ativos detectados na bebida fermentada kefir pela
técnica RT-PCR-DGGE, foram identificados como Lactobacillus kefiranofaciens,
Lactobacillus parakefiri. Outrossim, as colônias isoladas nos meios de cultura foram
identificadas como Lactococcus lactis subsp. lactis, L. lactis subsp. cremoris, Leuconostoc
mesenteroides, Acetobacter lovaniensis. Ao comparar os resultados obtidos pelos métodos
dependentes e independentes de cultivo, não houve correlação entre o perfil de bactérias
analisado por cada um dos métodos. Já em relação às leveduras, Saccharomyces cerevisiae foi
detectada por ambas metodologias. Bandas correspondentes a S. cerevisiae foram dominantes
no perfil de RT-PCR-DGGE, e também identificadas no isolamento com ágar YGC,
confirmando sua viabilidade e predominância na comunidade.
Lactobacillus kefiranofaciens foi predominante na análise de DGGE, mas não foi
isolado no meio seletivo. Embora o número de isolados (92 no total) tenha sido relativamente
pequeno para avaliar a diversidade microbiana cultivável do kefir, a escolha do meio de
cultivo (MRS) pode ter sido uma das causas do não isolamento de tal microrganismo. Toba e
colaboradores 228
desenvolveram o ágar KPL (Kefir Grain Polysaccharide-producing
Lactobacilli), para isolamento de lactobacilos homofermentativos do kefir, produtores de
polissacarídeo. Posteriormente, Fujisawa e colaboradores 134
classificaram estes isolados
homofermentativos como L. kefiranofaciens, e observaram que os mesmos não foram capazes
de crescer nos meios BL e MRS, sendo recuperados somente em ágar KPL a 30°C. Kojima e
colaboradores 135
obtiveram melhores resultados ao utilizar o meio Rogosa-CW (meio Rogosa
modificado com 100% de solução de soro de leite, pH 5,4), para isolar e cultivar lactobacilos
de grãos de kefir. Recentemente, Farnworth e Mainville 102
compararam os meios MRS, KPL

98
e Rogosa-CW com o meio LAW (Lactic Acid Whey), e concluíram que o meio LAW
apresentou maiores taxas de recuperação e crescimento para L. kefiranofaciens subsp.
kefirgranum. Portanto, o não isolamento e recuperação de L. kefiranofaciens no presente
estudo, pode ter ocorrido por falta de suplementos ou nutrientes específicos para o
crescimento da espécie, no meio de cultura utilizado (MRS).
Além disso, alguns meios utilizados não foram seletivos. Por exemplo, Leuconostoc
mesenteroides foi isolado no ágar M17 e MRS, assim como, Lactococcus spp. foi isolado no
ágar MRS. Esta falta de seletividade já foi discutida anteriormente 132; 133; 212
, e por este
motivo, alguns autores usam estratégias para ajustar os meios ou condições de cultivo, em
favor do crescimento de certas estirpes, que sem um conhecimento prévio de sua presença na
comunidade, poderiam ser negligenciadas. Portanto, métodos de cultivo podem sobrestimar
ou subestimar a diversidade microbiana, e nem sempre são ferramentas representativas de
comunidades complexas 132; 133; 212
.
A técnica de PCR-DGGE elimina a necessidade de isolamento e evita erros inerentes
ao método tradicional de cultivo, uma vez que algumas espécies, não são capazes de crescer
sob condições experimentais específicas, como já discutido anteriormente. No entanto, vários
autores 131; 132; 212; 229
já notaram que técnicas independentes de cultivo regularmente falham
para identificar espécies isoladas com métodos dependentes de cultivo. Estes dois tipos de
métodos revelam diferentes imagens da mesma comunidade 132; 212; 229
. Neste estudo, as
estirpes de BAL identificadas nos meios de cultura foram diferentes daquelas detectadas no
PCR-DGGE. Pode ser que o número de células de certas espécies de BAL estava abaixo do
limite de detecção da técnica PCR-DGGE 221
; ou que a grande quantidade de DNA molde
proveniente das espécies mais abundantes, competiram pelos iniciadores durante a reação de
amplificação, resultando na formação de produtos de PCR, somente das populações
dominantes 168; 230
. Desta forma, a aplicação das duas técnicas combinadas, dependentes e
independentes de cultivo, pode ser a maneira mais adequada para se obter uma visão geral e
real da estrutura da comunidade microbiana 131; 132; 220; 229
.
Em geral, os resultados obtidos neste estudo com o plaqueamento tradicional
corroboram com estudos anteriores 109; 117; 136
, que relataram um aumento crescente na
contagem microbiana durante as primeiras 24 h de fermentação do kefir. Durante o período de
estocagem, Öner, Karahan e Çakmakçi 129
também observaram uma estabilidade na contagem
dos microrganismos na bebida fermentada, produzida com leite de vaca. Entretanto, Irigoyen
e colaboradores 127
e Grønnevik, Falstad e Narvhus 130
reportaram uma redução na unidade
logarítimica (log) das contagens presuntivas de lactobacilos e lactococos, ao longo de quatro

99
semanas de estocagem, enquanto que as contagens de levedura e BAA mantiveram-se
constantes. Grønnevik, Falstad e Narvhus 130
demonstraram um aumento significativo nas
contagens de leveduras, após três semanas de estocagem da bebida kefir comercial
norueguesa. Os valores de unidades log reportados neste estudo foram, de maneira geral,
distintos dos encontrados por outros autores 127; 129; 130
, provavelmente devido às condições de
produção do kefir.
A Organização das Nações Unidas 21
e a legislação brasileira 20
preconizam que a
bebida kefir contenha, no mínimo, até o final da estocagem 107
UFC/g e 104
UFC/g, como
contagem total mínima de bactérias e leveduras, respectivamente. Sendo assim, as
quantidades encontradas no kefir estudado estão de acordo com estas especificações. A
estabilidade e relação da composição dos microrganismos no kefir entre a produção e
consumo são importantes para aqueles consumidores com interesse nas propriedades
probióticas, promovidas pela microbiota do kefir 102
. Segundo a regulamentação de
probióticos no Brasil 7, a quantidade mínima viável para probióticos deve estar situada na
faixa de 108 a 10
9 UFC, considerando a recomendação diária do produto para o consumo.
Desta forma, o kefir produzido enquadra-se nos limites estipulados até o período final de
estocagem, se considerado o consumo de uma porção diária de 200 mL do produto 231
.
A análise dos metabólitos produzidos e substratos utilizados, durante a fermentação e
estocagem do kefir também foi avaliada, por cromatografia líquida de alta eficiência (CLAE).
A redução da lactose já era esperada, uma vez que é hidrolisada a glicose e galactose pelos
lactococos e lactobacilos 130; 143; 232
. As bactérias identificadas no kefir estudado foram tanto
homofermentativas, quanto heterofermentativas, Ambas as vias são empregadas pelas
bactérias e o processo fermentativo é caracterizado pela acumulação de ácidos orgânicos e
redução do valor do pH. Na via homofermentativa, 2 moles de ácido lático são formados por
cada molécula de glicose. No entanto, a via heterofermentativa rende teoricamente 1 mol de
ácido lático, 1 mol de etanol e 1 mol de CO2 143
.
A concentração de ácido lático produzida, durante a fermentação (16%) e estocagem
(23%), foi maior do que a observada em estudos anteriores 143; 233;
, enquanto que a
concentração de ácido acético foi cerca de 66% mais baixa que a encontrada por Magalhães e
colaboradores 109
. Ácido acético é um intermediário do metabolismo do citrato, assim como,
um dos produtos finais do metabolismo heterofermentativo da lactose, e também pode ser
produzido por BAA pela oxidação do etanol 130; 142
. Citrato também foi reduzido,
provavelmente por BAL, uma vez que é considerado o substrato preferencial para formação
de ácido acético, acetoína e diacetil por estas bactérias. González De Llano, Rodriguez e

100
Cuesta 217
à semelhança deste estudo, também encontraram ácidos butírico e propiônico,
avaliando a atividade de culturas de Lactococcus spp. e Leuconostoc spp. na fermentação do
leite e atribuíram o aumento de tais ácidos, a atividades esterases inespecíficas por estirpes de
lactococos 234
.
A produção de etanol aumentou ao longo do processo de fermentação e estocagem e,
em 24h de fermentação, o valor encontrado foi 40% mais baixos que aquele quantificado
previamente em kefir brasileiro por CLAE 109
. A produção de etanol é derivada do
metabolismo de leveduras, como Saccharomyces cerevisiae, identificada neste trabalho pelas
técnicas dependentes e independentes de cultivo. Entretanto, bacterias heterofermetnativas,
como Lactobacillus kefiri e Leuconostoc spp. também são capazes de produzir etanol 143; 232;
235, uma vez que possuem atividade álcool desidrogenase, enzima capaz de converter o
acetaldeído a etanol. Parte do conteúdo de etanol pode ter sido convertido a ácido acético
pelas BAA, como o gênero Acetobacter 125
. Os produtos finais da fermentação de levedura,
como etanol e CO2, são fundamentais para o flavor refrescante exótico e aroma de levedura
típico do kefir 143; 233
.
Diferenças nas concentrações de ácidos orgânicos, encontradas por outros autores,
durante a fermentação e estocagem do kefir 109; 140; 143; 233
podem estar associadas a variações
empregadas na produção da bebida (proporção de grão inoculado, condições de cultivo, etc),
assim como, a composição da microbiota dos diferentes grãos de kefir 127; 143
.

101
6.4 CAPÍTULO IV - POTENCIAL PROBIÓTICO DAS ESTIRPES BACTERIANAS
ISOLADAS DE GRÃOS DE KEFIR 3
6.4.1 Material e métodos
6.4.1.1 Isolamento de bactérias
BAL foram isoladas de quatro grãos de kefir coletados de diferentes cidades do Brasil,
utilizando o método de diluição em ágar. Uma alíquota de 10 g de cada amostra foi
homogeneizada em 90 mL de citrato de sódio (2%). A partir deste homogeneizado, diluições
decimais seriadas foram obtidas e uma alíquota de 0.1 mL dessas diluições foi inoculada em
ágar MRS e M17 (Difco) suplementado com 200 ppm cicloheximide (Sigma-Aldrich) e,
incubado em condições de aerobiose e anaerobiose com auxílio do sistema de anaerobiose
Gaspak EZ (Difco) a 30°C por 72 h. Colônias significativas foram aleatoriamente coletadas e
purificadas por subcultivo no mesmo meio. Seleção presuntiva de BAL foi realizada por
coloração de esfregaço pelo método de Gram e teste da catalase. Culturas puras foram
estocadas em caldo MRS contendo 20% de glicerol a −80 °C.
Para todos os testes realizados, cepas de BAL foram ativadas no caldo MRS a 30°C
por 18-24, e subcultivado em ágar MRS a 30°C por 18-24h.
6.4.1.2 Sobrevivência dos isolados em pH ácido
A habilidade das estirpes crescerem em pH ácido foi avaliada em caldo MRS-THIO
(MRS suplementado com 0.2% tioglicolato de sódio) acidificado com HCl a pH 3,0, como
descrito por Nishida e colaboradores 236
. O crescimento das culturas incubadas a 30°C
overnight foi estimado por densidade ótica a 600 nm (DO600) para fase logarítimica (DO600 =
0.5 – 1.0). As células foram coletadas por centrifugação e o pellet foi ressuspendido em buffer
salina fosfato (PBS, pH 6.5) a fim de obter uma DO600 = 0.5. As suspensões celulares foram
diluídas 10 vezes em meio MRS acidificado (pH 3.0) e incubadas a 37°C por 3 h. Após este
período, a tolerância das culturas ao pH ácido foi determinada por enumeração de células
viáveis em ágar MRS incubado a 37°C por 48 h. Tubos contendo MRS convencional foram
submetidos as mesmas condições e utilizados como controle para cada amostra. Nos casos em
3 Apêndice D

102
que o número de UFC/mL enumeradas no ágar MRS foi maior do que as contagens obtidas no
ágar MRS acidificado o crescimento foi considerado negativo.
6.4.1.3 Tolerância à bile das culturas
A tolerância à bile bovina (Oxgall - Difco) foi avaliada pelo crescimento dos isolados
em ágar e caldo seguindo o procedimento descrito por Dunne e colaboradores 81
e Guo e
colaboradores 237
, respectivamente. O crescimento das culturas foi estimado em caldo MRS-
THIO com (teste) e sem (controle) 0.3% de sais biliares Oxgall (Difco). Culturas overnight
(1%) incubadas a 37 °C por 18h foram adicionados ao meio, cultivadas a 37°C em banho-
maria, e seu crescimento monitorado por 9h em espectofotômetro (DO620) ou até que uma
diferença de 0,3 unidades na absorbância fosse atingida entre o teste e controle. O efeito foi
mensurado em base do tempo requerido para aumentar a absorbância a 620 nm (DO620) por
0,3 unidades em caldo MRS-THIO com e sem 0,3% de sais biliares. Essa diferença no tempo
(horas) de crescimento das culturas presentes nos tubos teste e controle foi considerada como
fase lag (LT) como descrito por Guo e colaboradores 237
e Usman e Hosono 238
.
Adicionalmente, colônias individuais foram selecionadas do ágar MRS e suspensas em 2-5
mL de solução salina estéril a 0.85% a uma densidade correspondente a escala 1 de
McFarland. Alíquotas dessa suspensão (10 µl) foram inoculadas em ágar MRS contendo bile
(0.3%, 0.5%, 1% e 2%) 239
e as placas foram incubadas a 37°C, sob condições aneróbicas e, o
crescimento foi observado após 24-48h. Uma placa sem bile foi utilizada como controle
positivo.
6.4.1.4 Identificação de BAL
Os padrões de fermentação de carboidratos e testes bioquímicos foram realizados para
as estirpes selecionadas no teste anterior, utilizando o sistema API 50-CHL e API 20 Strep
(bioMérieux), respectivamente de acordo com as instruções do fabricante. Esses ensaios
foram utilizados como identificação preliminar dos gêneros de BAL, sem atingir nível de
espécie.
Finalmente, a identificação das culturas selecionadas foi confirmada por análises
moleculares. As estirpes selecionadas foram recuperadas no meio correspondente e seus DNA
genômicos foram extraídos e purificados utilizando o kit GenElute Bacterial Genomic DNA
kit (Sigma–Aldrich Inc.), seguindo as recomendações do fabricante. Os DNA obtidos foram

103
utilizados como molde na amplificação do fragmento correspondente ao gene codificador do
RNAr 16S pela técnica de PCR conforme descrito por Wang e colaboradores 214
utilizando os
iniciadores universais S-D-Bact-0008-a-S-20 (27F) (5’-AGAGTTTGATCCTGGCTCAG-3’)
e S-*-Univ-1492R-b-A-21 (1492R) (5’-GGTTACCTTGTTACGACTT-3’).
Os produtos da amplificação foram digeridos com as enzimas de restrição HaeIII e
HhaI (Invitrogen Ltd.) e a análise de restrição do DNA ribossomal e o sequenciamento parcial
dos fragmentos amplificados foram realizados conforme descrito anteriormente na seção
6.3.1.4. As sequências que apresentaram um percentual de identidade maior ou igual a 98%
foram classificadas como pertencentes à mesma espécie comparada 240
.
6.4.1.5 Análise de tipificação molecular
Os isolados de lactobacilos foram analisados e agrupados pela combinação das
técnicas de tipificação rep-PCR e RAPD-PCR, a fim de excluir possíveis clones entre as
estirpes selecionadas. Para isso foram utilizados os iniciadores BOXA1R (5’–
CTACGGCAAGGCGACGCTGACG–3’) e BOXA2R (5’-
ACGTGGTTTGAAGAGATTTTCG-3’), como reportado por Koeuth, Versalovic e Lupski 170
para técnica de rep-PCR, e o iniciador M13 (5’-GAGGGTGGCGGTTCT-3’), conforme
reportado por Rossetti e Giraffa 241
, para técnica de RAPD-PCR.
Dendrogramas de similaridade foram construídos a partir da média dos padrões de
bandas obtidos com cada iniciador, utilizando o coeficiente de Dice e para o agrupamento, o
algoritmo UPGMA, por meio do software Bionumerics 6.5 (Applied Maths).
6.4.1.6 Propriedades antimicrobianas
6.4.1.6.1 Inibição de patógenos
A capacidade das cepas em inibir um grupo de patógenos alimentares foi determinada
utilizando o teste de spot em ágar como descrito por Annuk e colaboradores 242
e Ripamonti e
colaboradores 243
. As culturas testadas foram inoculadas (2 µL) na superficie do agar MRS
(sem citrato de amônio e acetato de sódio) e incubadas anaerobicamente (GasPak system) por
24 h, a 30°C para estirpes de lactococos e leuconostoc, e a 37°C para lactobacilos. Após o
crescimento, as colônias foram inativadas utilizando o gás clorofórmio por 30 minutos. As
estirpes dos patógenos indicadores utilizados foram Escherichia coli ATCC 25922,

104
Salmonella enterica var. Enteritidis ATCC 13076, Staphylococcus aureus ATCC 25923,
Listeria monocytogenes ATCC 15313. Alíquotas de 100 µL do crescimento overnight das
bactérias indicadoras foram adicionadas em 10 mL de agar BHI semi-sólido (0.7%), e vertidas
sobre as placas de MRS com as colônias de BAL inativadas, e incubadas aerobicamente a
37°C. Após 24 h de incubação, a leitura dos halos de inibição foi realizada. Quando os halos
de inibição observados eram claros, nítidos e possuíam mais do que 1 mm ao redor do spot
eram considerados positivos. A estirpe de L.acidophilus ATCC 4356 foi utilizada como
controle negativo 243
e cada teste foi realizado em duplicata.
6.4.1.6.2 Produção de bacteriocina
A capacidade de produção de substâncias com atividade antimicrobiana foi avaliada
utilizando o teste de spot em ágar e posteriormente o teste de difusão em poços. As estirpes
indicadoras (descritas como susceptíveis a bacteriocinas) utilizadas foram Lactobacillus sakei
CECT 906, Lactococcus lactis IL 1403 31; 244; 245
e Listeria monocytogenes ATCC 15313. O
cultivo overnight das BAL isoladas foi inoculado na superfície do ágar M17 e MRS (0,2%
glucose), e as placas foram incubadas por 24h a 30°C para estirpes de lactococos e
leuconostoc e a 37°C para lactobacilos, respectivamente. Após esse período, as placas
contendo os spots foram submetidas a vapor de gás clorofórmio 246
por 30 min para inativação
do crescimento das culturas. Posteriormente, essas placas foram recobertas com 10 mL de
agar semi-sólido (0,75%), inoculado previamente com a bactéria indicadora e incubada sob as
condições exigidas por cada indicador,
As estirpes que mostraram resultados postivos no teste de spot em ágar foram
avaliadas pelo teste de difusão em ágar, como descrito por Schillinger e Lucke 33
. O meio BHI
semi-sólido a 45°C (20 mL) foi homogeneizado com 200 µL do cultivo overnight da estirpe
indicadora e vertido sobre placas de petri. Os sobrenadantes do cultivo overnight das estirpes
de BAL testadas foram neutralizados a pH 6,5 - 7,0 com NaOH 0,1 M, centrifugados a 12,000
g por 5 min, e filtrados em membrana de 0,22 µm (Millipore). Alíquotas de 50 µL de cada
sobrenadante foram depositadas em poços feitos no BHI semi-sólido e antes da incubação, as
placas foram mantidas a 4°C por 1 h. A inibição do crescimento das estirpes indicadoras foi
avaliada após 24 h de incubação sob condições de cultivo apropriadas para cada indicador.
Os sobrenadantes que mostraram atividade inibitória a qualquer um dos indicadores
testados foram investigados quanto a sua natureza protéica. Para isso, poços adicionais foram
confeccionados no ágar próximo àqueles contendo os sobrenadantes neutralizados (50 µL)

105
com atividade inibitória. Esses novos poços foram preenchidos com 50 µL de solução
contendo proteinase K ou pronase (cada uma a 20 mg/mL) e após a incubação, as placas
foram avaliadas quanto à sensibilidade da substância inibitória à proteólise 33; 247
. A estirpe
Lactococcus lactis 1A6, produtora de nisina 31
foi utilizada como controle positivo. Os testes
foram realizados em duplicata.
6.4.1.7 Ensaio de adesão
A linhagem de células de epitélio intestinal Caco-2 foi utilizada para avaliar a
habilidade de adesão de quatro estirpes de lactobacilos selecionadas. O cultivo e a
manutenção das linhagens foram realizados de acordo com procedimentos descritos
anteriormente 248
, utilizando o meio DMEM suplementado com 10% de soro fetal bovino
(Sigma-Aldrich) e dois antibióticos (50 µg/mL gentamicina e 1,25 µg/mL anfotericina B)
(Sigma-Aldrich). A linhagem celular foi utilizada após atingir estágio de confluência
diferenciado (13 ± 1 dias). A estirpe de Lactobacillus rhamnosus LMG 18243 (também
nomeada como GG) foi utilizada como controle positivo de BAL devido a sua capacidade de
adesão bem reconhecida 249
.
Quatro estirpes de lactobacilos, crescidas overnight em caldo MRS (10 mL) a 37°C,
foram coletadas por centrifugação, lavadas duas vezes em solução tampão PBS Dulbecco
(Sigma-Aldrich) e suspensas no meio DMEM ou McCoy sem antibiótico, a uma concentração
de cerca de 108 UFC/mL. Monocamadas da linhagem celular também foram lavadas duas
vezes em tampão PBS Dulbecco, para remoção de antibióticos. As suspensões bacterianas
(108 UFC/ml) foram, então, adicionadas na proporção de 10:1 de célula bacteriana : eucariota.
As placas foram incubadas por 1h a 37°C, 5% CO2 e, posteriormente, os poços foram
gentilmente lavados (3 vezes) com tampão PBS Dulbecco para remoção das bactérias não
aderidas. As monocamadas tratadas com solução tripsina-EDTA (Sigma-Aldrich) e as
contagens bacterianas foram realizadas em ágar MRS. Os resultados do teste de adesão foram
expressos como percentual de bactéria aderida em relação à quantidade de bactéria adicionada
(% UFC de bactéria aderida /UFC bactéria adicionada). Dois experimentos em replicata foram
conduzidos para cada isolado utilizando duas microplacas independentes (cada isolado foi
testado em duplicata em cada placa).

106
6.4.1.8 Avaliação das propriedades de segurança
6.4.1.8.1 Produção de hemolisinas
A produção de hemolisinas foi avaliada em placas de ágar Columbia contendo 5% de
sangue de carneiro (bioMérieux). A presença de hemólise é indicada pela formação de zonas
claras ao redor das colônias.
6.4.1.8.2 Susceptibilidade a antimicrobianos
A concentração minima inibitória (MIC) de antibióticos capaz de inibir o crescimento
dos quatro isolados de lactobacilos foi determinada, por microdiluição em placas de VetMIC®
para BAL (SVA, National Veterinary Institute of Sweden), seguindo as recomendações do
fabricante. Testes de microdiluição VetMICTM
Lact-1 e Lact-2 foram utilizados para
determinar a MIC para gentamicina (faixa de concentração testada: 0,5-256 µg/mL),
kanamicina (2-1024 µg/ml), estreptomicina (0,5-256 μg/ml), neomicina (0,5–256 μg/ml),
tetraciclina (0,12–64 μg/ml), etritromicina (0,016–8 μg/ml), clindamicina (0,03–16 μg/ml),
cloranfenicol (0,12–64 μg/ml), ampicilina (0,03-16 μg/ml), penicilina (0,03-16 μg/ml),
vancomicina (0,25-128 μg/ml), virginiamicina (0,016-8 μg/ml), linezolida (0,03-16 μg/ml),
trimetoprim (0,12-64 μg/ml), ciprofloxacina (0,25-128 μg/ml) e rifampicina (0,12-64 μg/ml).
Colônias crescidas em ágar LSM 250
foram suspensas em 5 mL de solução salina
estéril a 0,9% para obter uma densidade correspondente a escala 1 de McFarland. Logo, as
suspensões foram diluídas 1:1000 em LSM e 100 µL do inóculo foi adicionado em cada poço
da placa VetMICTM
e as placas foram incubadas a 28 °C por 48 h. As MIC foram definidas
como a menor concentração de antibiótico onde nenhum crescimento bacteriano foi
observado visualmente.
6.4.1.9 Atividades enzimáticas
As atividades enzimáticas dos isolados foram mensuradas pelo teste comercial semi-
quantitativo API-ZYM (bioMérieux), seguindo as recomendações do fabricante. Colônias
crescidas overnight foram suspensas em solução salina estéril a 0,9% e uma alíquota de 65
µL, correspondente a escala 5 de McFarland foi inoculada dentro de cada poço presente nas
fitas API-ZYM. As atividades enzimáticas foram avaliadas após 4h de incubação anaeróbia a

107
37°C e registradas utilizando uma escala de 0 a 5, baseada em uma escala de cores, fornecida
pelo fabricante do kit API ZYM. O número aproximado de nmol liberados de substrato
hidrolisado foi estimado pela cartela de cores: 0, nenhuma atividade; 1, liberação de 5 nmols;
2, 10 nmols; 3, 20 nmols; 4, 30 nmols; 5, ≥ 40 nmols.
6.4.2 Resultados
6.4.2.1 Tolerância ao pH e sais biliares
Dentre 150 colônias de BAL testadas, 37 foram capazes de sobreviver a 3 h de
incubação a pH 3,0, mas nenhuma delas foi capaz de crescer neste pH (Tabela 4). Dentre as
quais, 33 (85%) mostraram resistência aos ácidos biliares e foram capazes de crescer a 0,3%
(Tabela 4), embora um atraso de 1 até 4 h tenha sido observado para o crescimento celular,
quando comparado com o controle (MRS sem bile). No teste em placa, cerca de 65% das
estirpes de BAL foram capazes de crescer em 0,5% de sais biliares, 35% em 1% de de sais
biliares (Tabela 4), enquanto que somente 19% foram capazes de crescer em placas de MRS
contendo 2% de sais biliares.
6.4.2.2 Identificação de BAL
Dentre as 33 estirpes de BAL selecionadas, resistentes ao pH e sais biliares, quatro
perfis representativos de cada espécie foram observados na análise de ARDRA e selecionados
para o sequenciamento parcial do gene rrs (dados não mostrados). A identificação desses 33
isolados foi obtida e suas espécies e respectivas origens estão dispostas na tabela 5.
Sequências parciais do DNA dos microrganismos representativos mostraram alta homologia
(98-100%) com Lactobacillus casei/paracasei (15%), Lactococcus lactis subsp. lactis (9%),
Lactococcus lactis subsp. cremoris (24%) e Leuconostoc mesenteroides (52%). Em relação ao
gênero Lactobacillus não foi possível obter uma diferenciação entre as espécies casei e
paracasei e, assim, os cinco isolados (MR1, MRN4, MRS59, M1743, MRS55) foram
descritos neste trabalho como L. casei/paracasei.

108
6.4.2.3 Propriedades antimicrobianas
6.4.2.3.1 Atividade antimicrobiana frente a patógenos
No teste de spot em ágar (Tabela 4), todas as 33 estirpes testadas inibiram a maioria
dos patógenos. Um total de 23 e 21 estirpes, incluindo aquelas pertencentes ao gênero
Lactobacillus, inibiram E. coli e S. Enteritidis, respectivamente; enquanto que 24 estirpes
apresentaram atividade inibitória contra L. monocytogenes, e somente oito inibiram S. aureus.
As estirpes positivas no teste de spot em ágar foram submetidas ao teste de difusão em ágar
(Tabela 4) e após a neutralização dos respectivos sobrenadantes livre de células, nenhuma
atividade inibitória foi observada.
6.4.2.3.2 Produção de substância semelhante à bacteriocina
No teste de spot em ágar, o indicador L. sakei CECT 906 foi inibido por seis estirpes,
L. lactis IL 1403 por 14 estirpes e L. monocytogenes por 24 estirpes. Os isolados que
apresentaram atividade antibacteriana contra qualquer um dos indicadores foram submetidos
ao teste de difusão em ágar. Nenhuma estirpe de Leuconostoc mostrou inibição contra os
indicadores testados neste segundo teste, enquanto que L. lactis IL 1403 foi inibido por três
estirpes de lactococos (MRS26, M171 and MRS52), L. sakei CECT 906 foi inibido por
estirpes de lactococos e lactobacilos (MRS26, M171, MRS 52, MRS 20, MRS 55) e L.
monocytogenes ATCC 15313 por quatro estirpes (MRS 26, M171, MRS 52, MRS 55) Os
sobrenadantes testados, considerados como positivos, exibiram um halo pequeno (cerca de 3
mm) mas com efeitos inibitórios claros contra os indicadores testados (Tabela 4). Após o
tratamento com proteinase, as atividades inibitórias desapareceram, confirmando a natureza
protéica da substância antibacteriana produzida pelas cinco estirpes (Fig. 17).
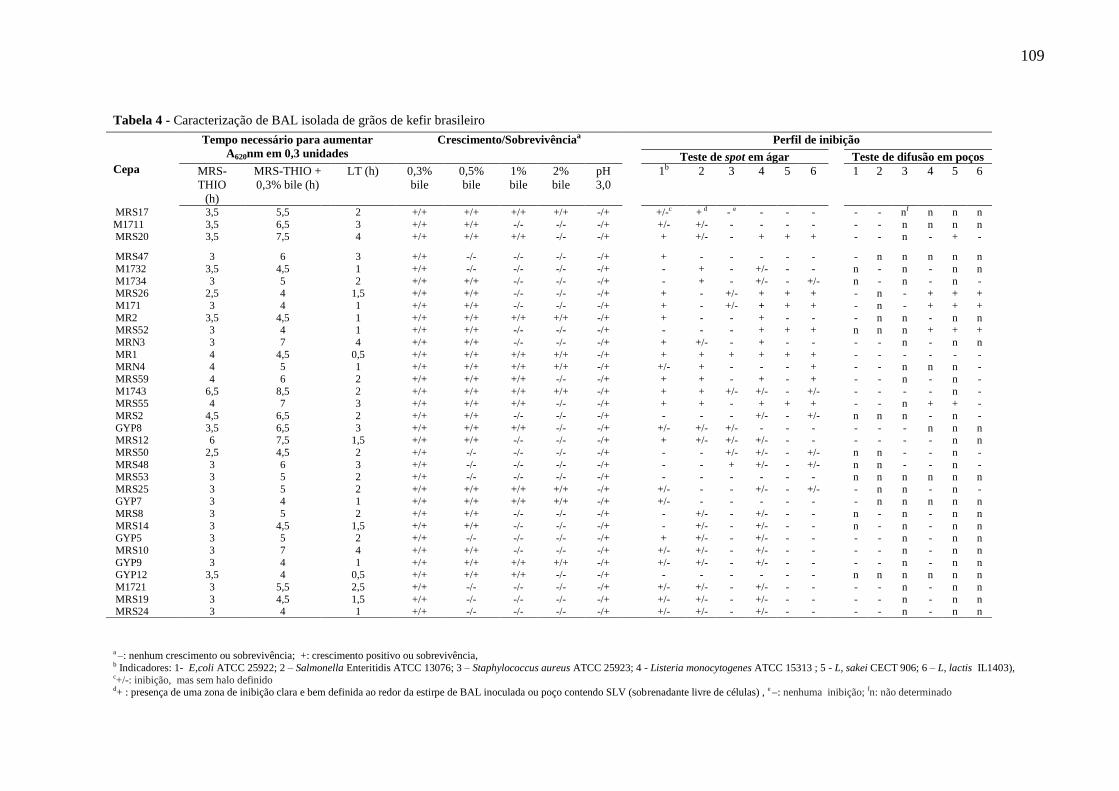
109
Tabela 4 - Caracterização de BAL isolada de grãos de kefir brasileiro
a –: nenhum crescimento ou sobrevivência; +: crescimento positivo ou sobrevivência, b Indicadores: 1- E,coli ATCC 25922; 2 – Salmonella Enteritidis ATCC 13076; 3 – Staphylococcus aureus ATCC 25923; 4 - Listeria monocytogenes ATCC 15313 ; 5 - L, sakei CECT 906; 6 – L, lactis IL1403), c+/-: inibição, mas sem halo definido d+ : presença de uma zona de inibição clara e bem definida ao redor da estirpe de BAL inoculada ou poço contendo SLV (sobrenadante livre de células) , e –: nenhuma inibição; fn: não determinado
Cepa
Tempo necessário para aumentar
A620nm em 0,3 unidades
Crescimento/Sobrevivênciaa Perfil de inibição
Teste de spot em ágar Teste de difusão em poços
MRS-
THIO
(h)
MRS-THIO +
0,3% bile (h)
LT (h) 0,3%
bile
0,5%
bile
1%
bile
2%
bile
pH
3,0
1b 2 3 4 5 6 1 2 3 4 5 6
MRS17 3,5 5,5 2 +/+ +/+ +/+ +/+ -/+ +/-c + d - e - - - - - nf n n n
M1711 3,5 6,5 3 +/+ +/+ -/- -/- -/+ +/- +/- - - - - - - n n n n MRS20 3,5 7,5 4 +/+ +/+ +/+ -/- -/+ + +/- - + + + - - n - + -
MRS47 3 6 3 +/+ -/- -/- -/- -/+ + - - - - - - n n n n n
M1732 3,5 4,5 1 +/+ -/- -/- -/- -/+ - + - +/- - - n - n - n n
M1734 3 5 2 +/+ +/+ -/- -/- -/+ - + - +/- - +/- n - n - n - MRS26 2,5 4 1,5 +/+ +/+ -/- -/- -/+ + - +/- + + + - n - + + +
M171 3 4 1 +/+ +/+ -/- -/- -/+ + - +/- + + + - n - + + +
MR2 3,5 4,5 1 +/+ +/+ +/+ +/+ -/+ + - - + - - - n n - n n MRS52 3 4 1 +/+ +/+ -/- -/- -/+ - - - + + + n n n + + +
MRN3 3 7 4 +/+ +/+ -/- -/- -/+ + +/- - + - - - - n - n n
MR1 4 4,5 0,5 +/+ +/+ +/+ +/+ -/+ + + + + + + - - - - - -
MRN4 4 5 1 +/+ +/+ +/+ +/+ -/+ +/- + - - - + - - n n n -
MRS59 4 6 2 +/+ +/+ +/+ -/- -/+ + + - + - + - - n - n -
M1743 6,5 8,5 2 +/+ +/+ +/+ +/+ -/+ + + +/- +/- - +/- - - - - n - MRS55 4 7 3 +/+ +/+ +/+ -/- -/+ + + - + + + - - n + + -
MRS2 4,5 6,5 2 +/+ +/+ -/- -/- -/+ - - - +/- - +/- n n n - n -
GYP8 3,5 6,5 3 +/+ +/+ +/+ -/- -/+ +/- +/- +/- - - - - - - n n n MRS12 6 7,5 1,5 +/+ +/+ -/- -/- -/+ + +/- +/- +/- - - - - - - n n
MRS50 2,5 4,5 2 +/+ -/- -/- -/- -/+ - - +/- +/- - +/- n n - - n -
MRS48 3 6 3 +/+ -/- -/- -/- -/+ - - + +/- - +/- n n - - n - MRS53 3 5 2 +/+ -/- -/- -/- -/+ - - - - - - n n n n n n
MRS25 3 5 2 +/+ +/+ +/+ +/+ -/+ +/- - - +/- - +/- - n n - n - GYP7 3 4 1 +/+ +/+ +/+ +/+ -/+ +/- - - - - - - n n n n n
MRS8 3 5 2 +/+ +/+ -/- -/- -/+ - +/- - +/- - - n - n - n n
MRS14 3 4,5 1,5 +/+ +/+ -/- -/- -/+ - +/- - +/- - - n - n - n n GYP5 3 5 2 +/+ -/- -/- -/- -/+ + +/- - +/- - - - - n - n n
MRS10 3 7 4 +/+ +/+ -/- -/- -/+ +/- +/- - +/- - - - - n - n n
GYP9 3 4 1 +/+ +/+ +/+ +/+ -/+ +/- +/- - +/- - - - - n - n n GYP12 3,5 4 0,5 +/+ +/+ +/+ -/- -/+ - - - - - - n n n n n n
M1721 3 5,5 2,5 +/+ -/- -/- -/- -/+ +/- +/- - +/- - - - - n - n n
MRS19 3 4,5 1,5 +/+ -/- -/- -/- -/+ +/- +/- - +/- - - - - n - n n MRS24 3 4 1 +/+ -/- -/- -/- -/+ +/- +/- - +/- - - - - n - n n

110
Tabela 5 - Espécies e origem das cepas de BAL isoladas de grãos de kefir brasileiros
Figura 17 - Atividade antagônica e sensibilidade à proteólise de estirpes de L. lactis isoladas de grãos de kefir.
Painel A: Inibição de E. coli ATCC 25922 pelas estirpes L. lactis subsp. cremoris MRS26 (1), MRS20 (2) e
M171 (3). Painel B: Atividade antimicrobiana da estirpe MRS26 contra o indicador Lb. sakei CECT 906 e sua
sensibilidade à proteólise. Poços: 1 – sobrenadante livre de células neutralizado da estirpe L. lactis subsp.
cremoris MRS26 ; PK - proteinase K (20mg/ml) e PN – pronase (20 mg/ml).
Espécies Cepas
isoladas
Região geográfica
L. lactis cremoris MRS17 Niterói, RJa
M1711 Niterói, RJ
MRS20 Niterói, RJ
MRS47 Lavras, MGb
M1732 Niterói, RJ
M1734 Niterói, RJ
MRS26 Niterói, RJ
M171 Niterói, RJ
L. lactis lactis MR2 Viçosa, MG
MRS52 Lavras, MG
MRN3 Viçosa, MG
L. casei/paracasei MR1 Viçosa, MG
MRN4 Niterói, RJ
MRS59 Alfenas, MG
M1743 Alfenas, MG
MRS55 Alfenas, MG
Leuconostoc mesenteroides MRS2 Niterói, RJ
GYP8 Niterói, RJ
MRS12 Niterói, RJ
MRS50 Lavras, MG
MRS48 Alfenas, MG
MRS53 Alfenas, MG
MRS25 Niterói, RJ
GYP7 Niterói, RJ
MRS8 Niterói, RJ
MRS14 Niterói, RJ
GYP5 Niterói, RJ
MRS10 Niterói, RJ
GYP9 Niterói, RJ
GYP12 Niterói, RJ
M1721 Niterói, RJ
MRS19 Niterói, RJ
MRS24 Niterói, RJ

111
6.4.2.4 Tipificação das estirpes de Lactobacillus casei/paracasei
A tipificação dos isolados de Lactobacillus casei/paracasei foi realizada por
diferenciação molecular, através da análise dos perfis de bandas obtidos com as técnicas de
rep-PCR e RAPD-PCR. Um dendrograma de similaridade (Fig.18) foi gerado pela
combinação dos perfis obtidos com os iniciadores BOXA1R, BOXA2R e M13. O valor de
92% de similaridade foi utilizado como limite de discriminação entre as estirpes e, então,
quatro perfis distintos (MRN4, MRS55, MRS59 e M1743) foram obtidos e selecionados para
caracterização adicional do seu potencial probiótico.
Figura 18 - Dendrograma de similaridade construído a partir da combinação dos perfis obtidos com as técnicas
rep-PCR e RAPD-PCR, usando os iniciadores BOXA1R, BOXA2R e M13.
6.4.2.5 Atividades enzimáticas
L. casei/paracasei MRS 55 e MRN4 foram capazes de hidrolisar 15 substratos,
enquanto que as outras estirpes hidrolisaram cerca de 13 a 14 substratos (Tabela 6). A estirpe
MRS55 apresentou altos níveis de atividade enzimática para maior parte das enzimas
analisadas. Todos os isolados mostraram atividade para fosfatase alcalina, esterase (C4),
esterase-lipase (C8), lipase, leucina arilamidase, valina arilamidase, cisteína arilamidase, α–
quimotripsina, fosfatase ácida, naftol-AS-BI-fosfohidrolase, β–galactosidase, α–glicosidase e
β-glicosidase (Tabela 6). As estirpes de Lactobacillus mostraram atividades máximas para
leucina arilamidase, valina arilamidase, enquanto, níveis mais baixos foram detectados para
cisteína arilamidase. Os quatro isolados também apresentaram moderada atividade para
esterase (C4), esterase-lipase (C8) e baixa atividade para lipase (C14), enquanto que
atividades para N-acetil-β-glicosaminidase e tripsina foram observadas para dois isolados e a
atividade de α-fucosidase foi observada somente para uma estirpe. Atividades de α-
galactosidase, β-glucuronidase e α-mannosidase não foram observadas para nenhum isolado.
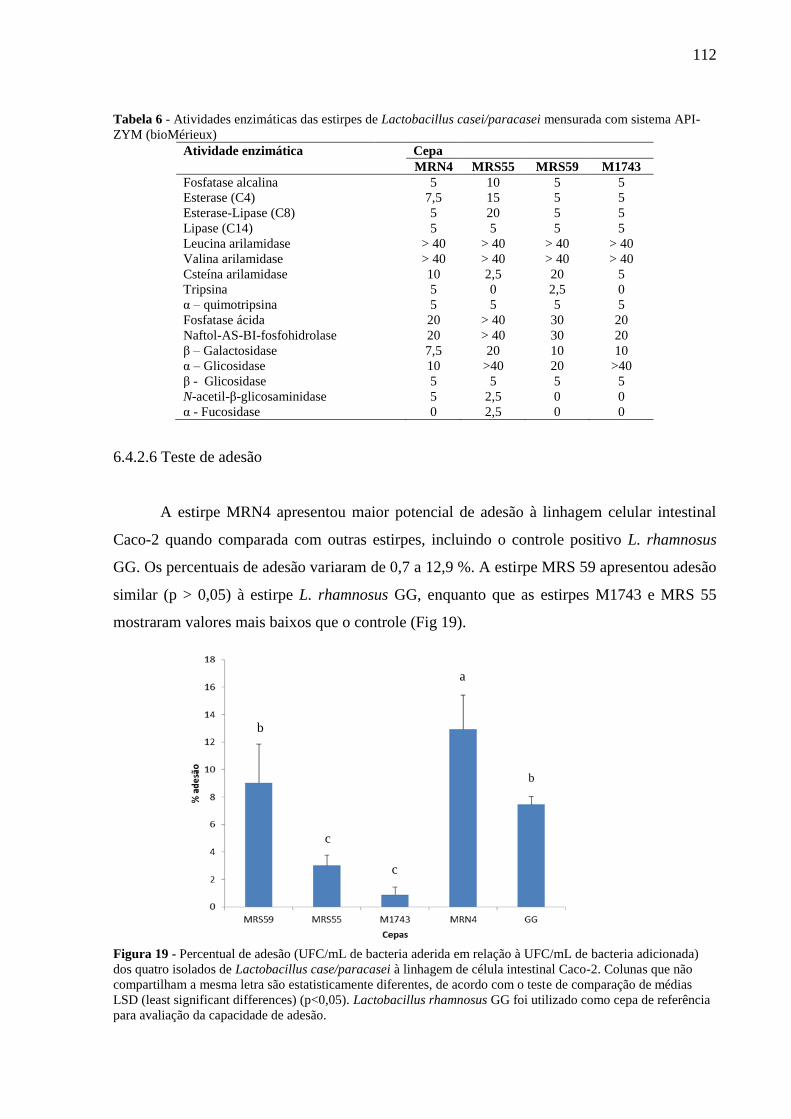
112
Tabela 6 - Atividades enzimáticas das estirpes de Lactobacillus casei/paracasei mensurada com sistema API-
ZYM (bioMérieux)
Atividade enzimática Cepa
MRN4 MRS55 MRS59 M1743
Fosfatase alcalina 5 10 5 5
Esterase (C4) 7,5 15 5 5
Esterase-Lipase (C8) 5 20 5 5
Lipase (C14) 5 5 5 5
Leucina arilamidase > 40 > 40 > 40 > 40
Valina arilamidase > 40 > 40 > 40 > 40
Csteína arilamidase 10 2,5 20 5
Tripsina 5 0 2,5 0
α – quimotripsina 5 5 5 5
Fosfatase ácida 20 > 40 30 20
Naftol-AS-BI-fosfohidrolase 20 > 40 30 20
β – Galactosidase 7,5 20 10 10
α – Glicosidase 10 >40 20 >40
β - Glicosidase 5 5 5 5
N-acetil-β-glicosaminidase 5 2,5 0 0
α - Fucosidase 0 2,5 0 0
6.4.2.6 Teste de adesão
A estirpe MRN4 apresentou maior potencial de adesão à linhagem celular intestinal
Caco-2 quando comparada com outras estirpes, incluindo o controle positivo L. rhamnosus
GG. Os percentuais de adesão variaram de 0,7 a 12,9 %. A estirpe MRS 59 apresentou adesão
similar (p > 0,05) à estirpe L. rhamnosus GG, enquanto que as estirpes M1743 e MRS 55
mostraram valores mais baixos que o controle (Fig 19).
Figura 19 - Percentual de adesão (UFC/mL de bacteria aderida em relação à UFC/mL de bacteria adicionada)
dos quatro isolados de Lactobacillus case/paracasei à linhagem de célula intestinal Caco-2. Colunas que não
compartilham a mesma letra são estatisticamente diferentes, de acordo com o teste de comparação de médias
LSD (least significant differences) (p<0,05). Lactobacillus rhamnosus GG foi utilizado como cepa de referência
para avaliação da capacidade de adesão.
b
b
b
b
c
c
c
c
c
c
c
a
b
b
b
b
c
c
c
c
c
c
c
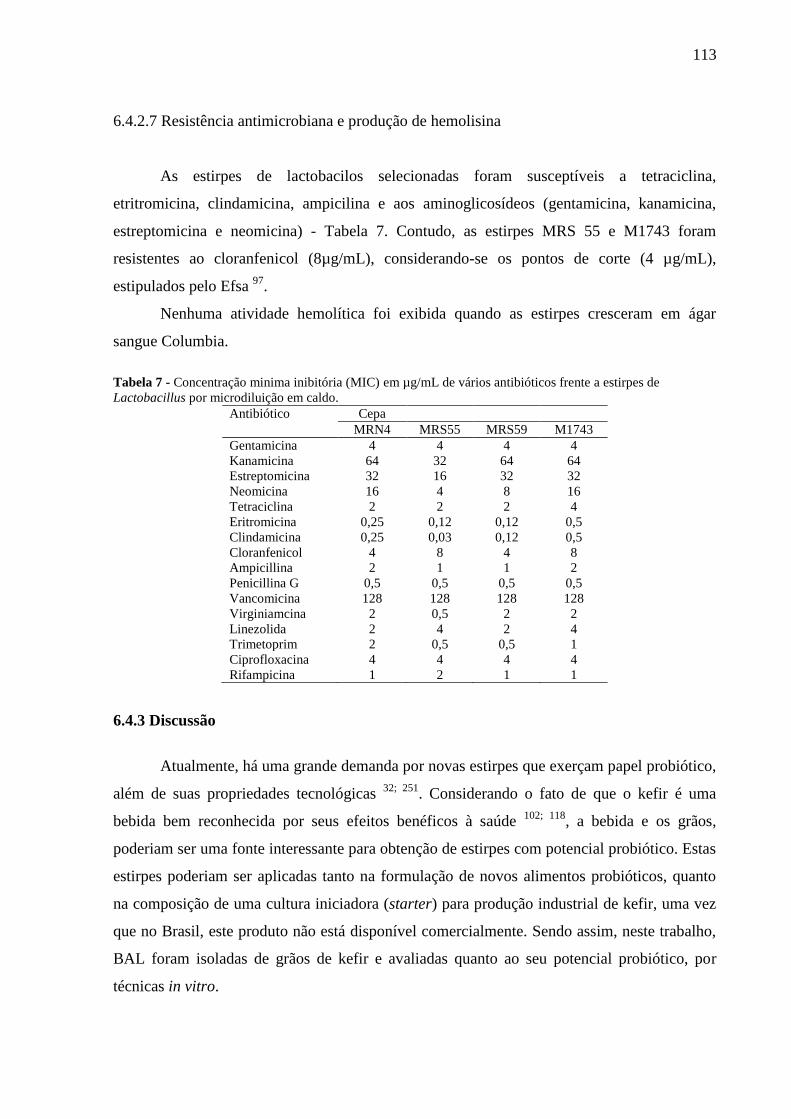
113
6.4.2.7 Resistência antimicrobiana e produção de hemolisina
As estirpes de lactobacilos selecionadas foram susceptíveis a tetraciclina,
etritromicina, clindamicina, ampicilina e aos aminoglicosídeos (gentamicina, kanamicina,
estreptomicina e neomicina) - Tabela 7. Contudo, as estirpes MRS 55 e M1743 foram
resistentes ao cloranfenicol (8µg/mL), considerando-se os pontos de corte (4 µg/mL),
estipulados pelo Efsa 97
.
Nenhuma atividade hemolítica foi exibida quando as estirpes cresceram em ágar
sangue Columbia.
Tabela 7 - Concentração minima inibitória (MIC) em µg/mL de vários antibióticos frente a estirpes de
Lactobacillus por microdiluição em caldo.
Antibiótico Cepa
MRN4 MRS55 MRS59 M1743
Gentamicina 4 4 4 4
Kanamicina 64 32 64 64
Estreptomicina 32 16 32 32
Neomicina 16 4 8 16
Tetraciclina 2 2 2 4
Eritromicina 0,25 0,12 0,12 0,5
Clindamicina 0,25 0,03 0,12 0,5
Cloranfenicol 4 8 4 8
Ampicillina 2 1 1 2
Penicillina G 0,5 0,5 0,5 0,5
Vancomicina 128 128 128 128
Virginiamcina 2 0,5 2 2
Linezolida 2 4 2 4
Trimetoprim 2 0,5 0,5 1
Ciprofloxacina 4 4 4 4
Rifampicina 1 2 1 1
6.4.3 Discussão
Atualmente, há uma grande demanda por novas estirpes que exerçam papel probiótico,
além de suas propriedades tecnológicas 32; 251
. Considerando o fato de que o kefir é uma
bebida bem reconhecida por seus efeitos benéficos à saúde 102; 118
, a bebida e os grãos,
poderiam ser uma fonte interessante para obtenção de estirpes com potencial probiótico. Estas
estirpes poderiam ser aplicadas tanto na formulação de novos alimentos probióticos, quanto
na composição de uma cultura iniciadora (starter) para produção industrial de kefir, uma vez
que no Brasil, este produto não está disponível comercialmente. Sendo assim, neste trabalho,
BAL foram isoladas de grãos de kefir e avaliadas quanto ao seu potencial probiótico, por
técnicas in vitro.

114
Primeiramente, as cepas isoladas foram testadas quanto à tolerância à acidez e à bile.
A capacidade de resistir à secreção de ácido gástrico e a passagem pelo estômago constituem
os mecanismos de defesa primários, que todos os microrganismos ingeridos devem superar
para manter a viabilidade, incluindo aqueles com potencial probiótico 81
. No estômago
humano, o pH varia de 1 a 4,5, e a digestão do alimento pode levar até 3h 32; 252
. Ainda que as
estirpes de lactobacilos isolados neste estudo não tenham crescido quando expostas a pH 3,0
por 3h, foram resistentes e sobreviveram ao ambiente ácido. Estes resultados estão de acordo
com o de estudos anteriores 157; 236; 239; 253
em que a maior parte das estirpes de lactobacilos foi
incapaz de crescer, mas capaz de sobreviver em meios onde o pH varia de 2,5-3,0. Além
disso, é importante considerar que bactérias probióticas são consumidas principalmente em
produtos lácteos, na presença de proteínas do leite. As proteínas do leite podem exercer uma
ação protetora adicional nas culturas starters, auxiliando a sobrevivência bacteriana no
ambiente ácido do estômago 252; 254
.
Em contraste, a maioria das cepas testadas cresceu na presença dos sais biliares
(0,3%), simulando o ambiente do intestino delgado 99
. A bile é sintetizada no fígado, estocada
na vesicular biliar, e secretada no duodeno na forma conjugada, após a ingestão do alimento
81; 255. Esta secreção digestiva exerce um importante papel na emulsificação e solubilização
dos lipídeos 256
. Porém, esta é tóxica para microrganismos, uma vez que afeta a estrutura das
membranas bacterianas 237
, e desta forma, a tolerância aos sais biliares é uma propriedade
essencial para a sobrevivência de BAL e subsequente colonização do trato gastrintestinal 12
.
Em muitos estudos foi notado que a maioria das estirpes de BAL sobrevive a esta condição, o
que ocasiona uma possível recuperação dos níveis iniciais do número de células, durante a
passagem pelo intestino delgado 237; 239; 253; 254; 257; 258
. A resistência à bile varia entre espécies
e cepas de Lactobacillus e o mecanismo ainda não é conhecido. Os resultados obtidos
mostraram que 100% e 60% dos lactobacilos isolados e testados foram capazes de resistir e
crescer em 1% e 2% de bile, respectivamente. Outros autores também já reportaram alta
resistência à bile entre estirpes de lactobacilos 32; 81; 239
.
As estirpes selecionadas foram então avaliadas quanto a sua atividade antagonista
contra patógenos e produção de substância semelhante à bacteriocina. Estudos anteriores 32;
157; 243; 253; 259 tem ressaltado que os lactobacilos são capazes de inibir bactérias patogênicas,
pela produção de metabólitos como ácidos orgânicos, ácidos graxos, peróxido de hidrogênio,
etanol e/ou bacteriocinas 31; 36; 260
. Estes relatos estão de acordo com os resultados obtidos
neste estudo, já que no ensaio de inibição de patógenos, o efeito antagônico mostrado pelas
estirpes selecionadas pode estar relacionado à produção de ácidos orgânicos ou peróxido de

115
hidrogênio, enquanto que no ensaio de produção de bacteriocina por cinco isolados, incluindo
um lactobacilo e quatro lactococos, produziram compostos semelhantes à bacteriocina. A
atividade antimicrobiana exibida pelas cepas de BAL isoladas de grãos de kefir poderia
sugerir a atividade da microbiota no TGI, exercendo um importante papel na manutenção do
ecossistema intestinal 27; 99
. Ryan e colaboradores 245
isolaram uma estirpe de L. lactis (DPC
3147) de grãos de kefir da Irlanda, capaz de produzir uma bacteriocina (lactococina 3147),
que possui amplo espectro de inibição. Em outro estudo, o pesquisador demonstrou que
Lactobacillus plantarum ST8KF isolado de kefir, produziu uma bacteriocina de 3,5 kDa, ativa
contra Lactobacillus casei, Lactobacillus salivarius, Lactobacillus curvatus e Listeria
innocua 261
.
Membros do gênero Lactobacillus são um dos mais comumente utilizados no
desenvolvimento de alimentos probióticos 12; 50; 81; 258
. Neste trabalho, a análise das sequências
parciais do gene rrs falhou na diferenciação entre as espécies L. casei e L. paracasei
pertencentes ao gênero Lactobacillus, como já descrito previamente 262; 263
. Além disso, um
método de tipagem molecular foi realizado para agrupar os isolados deste gênero baseado na
heterogeneidade exibida pelo perfil de bandas obtido com cada uma das estirpes 99; 137; 169; 264;
265, o que permitiu selecionar quatro diferentes estirpes de lactobacilos para caracterização
adicional de suas propriedades probióticas.
A atividade enzimática é uma característica importante a ser avaliada em estirpes com
intenção de uso como culturas probióticas ou starters, em função de suas habilidades
funcionais e/ou tecnológicas 258; 266; 267
. Todas as estirpes produziram β-galactosidase, uma
enzima benéfica para ambos os aspectos, probióticos e tecnológicos, auxiliando na redução da
intolerância a lactose, assim como na acidificação do leite 267
. Esta enzima também pode
contribuir para a formação de galacto-oligossacarídeos (GOS). GOS são oligossacarídeos não
digeríveis que agem como substrato promotor de crescimento para bifidobactéria no intestino
humano e suprime bactérias potencialmente nocivas, devido ao ambiente ácido criado pelas
mesmas 268; 269
. Em relação às atividades enzimáticas relacionadas com o catabolismo de
carboidrato, todos os isolados apresentaram atividade de α-glicosidase e β-glicosidase, que
podem contribuir para digestão de polissacarídeos 266; 270
. No entanto, corroborando com
Monteagudo-Mera e colaboradores 267
, α-glicosidase catalisa o etapa final da digestão de
carboidratos, e sua presença pode ser indesejável na dieta de pacientes obesos e/ou diabéticos.
Apesar da β-glicosidase já ter sido descrita como uma enzima nociva para saúde, devido a sua
possível atividade procarcinogênica 68; 93
, estudos recentes indicaram que a mesma é desejável
para saúde, por sua ação na absorção das isoflavonas, promovendo a hidrólise das formas

116
glicosiladas às formas agliconas, biologicamente ativas, e que estão frequentemente
associadas com a prevenção do câncer, desordens cardiovasculares, osteoporose, síndromes
pós-menopausa, hipercolesterolemia e efeitos antioxidantes 91; 92; 93; 271
. A atividade α–
fucosidase que pode ser responsável pela colonização seletiva do intestino por bactéria 267
, foi
detectada somente para estirpe MRS55.
Além disso, foi observado que todas as estirpes testadas mostraram atividades de
aminopeptidases e lipolíticas. Atividade aminopeptidase é uma característica tecnológica
importante para cepas de BAL, porque contribuem para o desenvolvimento de flavors
desejáveis em produtos lácteos 267
, assim como esterases e lipases podem estar associadas à
degradação de gorduras em produtos lácteos 264
.
Nenhuma das estirpes de lactobacilos estudadas produziu a enzima β-glucoronidase,
considerada uma enzima com potencial carcinogênico. Esta enzima pode converter
substâncias precarcinogênicas em carcinogênicas, estimulando o desenvolvimento de câncer
de cólon humano 267
.
A capacidade de adesão à mucosa intestinal é outra propriedade importante para cepas
candidatas a probiótico, já que devem colonizar o intestino do hospedeiro proporcionando
efeitos benéficos à saúde 21
. Geralmente, a linhagem celular de adenocarcinoma de cólon
humano Caco-2 é utilizada porque exibe in vitro, as características de enterócitos maduros do
intestino delgado 30; 272
. Comparando os valores de percentual de adesão obtidos com a
bactéria probiótica Lb. rhamnosus GG, utilizada como controle positivo neste ensaio, duas
cepas isoladas neste estudo (MRN4 e MRS59) mostraram capacidade de adesão às
monocamadas de Caco-2, igual ou superior a do controle, sugerindo que estas duas estirpes
seriam capazes de aderir-se nas células epiteliais. Porém, testes in vivo são necessários para
confirmar essa capacidade de adesão 21; 82
.
A espécie L. casei/ paracasei isolada neste estudo tem uma longa história de uso
seguro em produtos lácteos, e é encontrada em muitos produtos probióticos comerciais 21; 94;
262. Está incluída na lista proposta pela Efsa
94, com status QPS (Qualified Presumption
Safety) qualificado como seguro, baseado nos dados atuais disponíveis, sobre segurança para
consumo humano. Entretanto, antes de sua aplicação tecnológica e funcional, as propriedades
de segurança devem ser investigadas para cada estirpe destinada para uso em alimentos,
especialmente com relação à susceptibilidade a antibióticos 12; 24
. Neste estudo, duas estirpes
apresentaram concentração mínima inibitória (Minimun Inhibitory Concentration - MIC) para
cloranfenicol maior que o ponto de corte, estipulado pela EFSA (4 µg/mL). No entanto, se o
MIC fosse comparado com o valor (≥ 32 µg/mL) proposto pela Clsi 273
, nenhuma das cepas

117
poderia ser considerada resistente. Pesquisas prévias 252; 274; 275; 276
também encontraram cepas
de L. casei/paracasei com padrões de MIC similares para cloranfenicol, e alguns destes
autores 275; 276
ainda sugeriram que o ponto de corte (MIC > 8 µg/mL) fosse mais alto para
este antibiótico, para estas espécies destinadas ao uso probiótico ou nutricional. Outros
estudos também confirmaram a baixa resistência de L. casei/paracasei ao cloranfenicol 257; 277;
278; 279. Desta forma, pode ser sugerido que estas duas estirpes não possuem mecanismo de
resistência adquirida, mas pesquisas adicionais devem ser conduzidas para assegurar seu uso
como probiótico.
Embora, culturas starter em produtos lácteos não representem uma importante fonte
de propagação de genes de resistência a agentes antimicrobianos, cuidados devem ser
tomados com perfis de resistência a antibióticos de novas estirpes utilizadas como starters ou
probióticos em alimentos. É importante distinguir entre resistência intrínseca e adquirida a um
antimicrobiano, uma vez que somente a resistência adquirida é considerada uma ameaça à
saúde, devido a grandes chances de transferência 97
. Resistência intrínseca ocorre quando é
inerente à determinada espécie bacteriana e típica a todas as estirpes daquela espécie. Um
exemplo bem reconhecido é a resistência intrínseca à vancomicina, de lactobacilos
heterofermentativos facultativos e obrigatórios. Dados do presente estudo confirmam esta
afirmação, devido a altos valores de MIC para vancomicina em todas as cepas testadas. A
resistência intrínseca desta espécie à vancomicina ocorre pela presença de D-Ala-D-Lac ao
invés do dipeptídeo habitual D-Ala-D-Ala em seu peptidoglicano 96; 280
.
Já a resistência adquirida, pode ocorrer devido à inserção de genes (DNA exógeno) ou
pela mutação de genes cromossomiais. O último mecanismo tem baixo risco de disseminação
horizontal e é considerada aceitável pelo comitê FEEDAP, enquanto que a inserção de DNA
exógeno apresenta um alto risco de disseminação horizontal de genes de resistência 97
.

118
7 CONCLUSÕES FINAIS
As técnicas independentes de cultivo foram úteis para avaliar a diversidade microbiana de
diferentes grãos de kefir, evidenciando os microrganismos dominantes nas comunidades e
os componentes bacterianos minoritários, raramente associados ao ecossistema dos grãos;
A bebida kefir foi melhor caracterizada a partir da combinação de técnicas dependentes e
independentes de cultivo, proporcionando uma visão mais completa e real da comunidade
microbiana do kefir;
Duas cepas de L.casei/paracasei (MRS59 e MRN4) mostraram-se adequadas para uso
como probiótico, embora estudos adicionais sejam necessários para assegurar suas
propriedades benéficas à saúde e de segurança, incluindo ensaios in vivo.
Quatro cepas de L. lactis (MRS 52, MRS 26, MRS 20 e M171) produziram substância
semelhante à bacteriocina, que deve ser melhor caracterizada e suas propriedades
tecnológicas avaliadas a fim de serem utilizadas como cultura starter ou adjunta na
indústria láctea.
O conhecimento de microrganismos envolvidos, assim como, seus metabólitos
produzidos ao longo da fermentação e estocagem do produto final podem servir como
base para uma futura produção do kefir no Brasil em escala industrial.

119
REFERÊNCIAS
1 PRAJAPATI, J. B.; NAIR, B. M. The History of Fermented Foods. In:
FARNWORTH, E. R. (Ed.). Handbook of fermented functional foods. 2. Boca
Raton: CRC Press, 2008. cap. 1, p.1-25.
2 TAMIME, A. Y. Production of Kefir, Koumiss and Other Related Products. In:
TAMIME, A. Y. (Ed.). Fermented Milk. Oxford, UK: Blackwell Science Ltd, 2006.
cap. 8, p.174-216.
3 VASILJEVIC, T.; SHAH, N. P. Probiotics—From Metchnikoff to bioactives.
International Dairy Journal, v. 18, n. 7, p. 714-728, 2008.
4 STANTON, C. et al. Fermented functional foods based on probiotics and their
biogenic metabolites. Current Opinion in Biotechnology, v. 16, n. 2, p. 198-203,
2005.
5 DEL PIANO, M. et al. Probiotics: from research to consumer. Digestive and Liver
Disease, v. 38, p. S248-S255, 2006.
6 MARTÍN, R. et al. Probióticos: concepto, criterios de selección y seguridad. In:
RODRÍGUEZ, J. M. G. (Ed.). La leche humana, un alimento vivo: bactérias
probióticas en leche humana. Granada: Puleva Food, S.L., 2010. cap. 104-121,
p.234.
7 BRASIL. Alimentos com Alegações de Propriedades Funcionais e ou de Saúde,
Novos Alimentos/Ingredientes, Substâncias Bioativas e Probióticos. Disponível
em:http://www.anvisa.gov.br/alimentos/comissoes/tecno_lista_alega.htm#,2008.
Acesso em: 04/10/2012.
8 CARNEIRO, R. P. Desenvolvimento de uma cultura iniciadora para produção de
kefir. 2010. 143 Mestrado (Dissertação (Mestre)). Faculdade de Farmácia,
Universidade Federal de Minas Gerais, Belo Horizonte.
9 SANDERS, M. E. Overview of Functional Foods: Emphasis on Probiotic Bacteria.
International Dairy Journal, v. 8, n. 5–6, p. 341-347, 1998.
10 STRINGHETA, P. C. et al. Alimentos Funcionais - Conceitos, contextualização e
regulamentação. Juiz de Fora: Templo, 2007. 246p.
11 MORAES, F. P.; COLLA, L. M. Alimentos funcionais e nutracêuticos: definições,
legislação e benefícios à saúde. Revista Eletrônica de Farmácia, v. 3, n. 2, p. 109-
122, 2006.

120
12 FAO/WHO. Probiotic in foods. Health and nutritional properties and guidelines
for evaluation. Rome: FAO Food and Nutrition Paper. 85 2006.
13 BRANDÃO, S. C. C. Novas gerações de produtos lácteos funcionais. . São Paulo,
p.64-66. 2002
14 SMOLYANSKY, M. About Kefir. Disponível em:
http://www.lifeway.net/LifewayWorld/OurStory/AboutKefir.aspx. Acesso em:
13/04/2012.
15 MANS, J. Inside Lifeway Foods: from Russia with kefir. FindArticles.com.
http://findarticles.com/p/articles/mi_m3301/is_n2_v95/ai_15237058/. 1994
16 LOPITZ-OTSOA, F. et al. Kefir: a symbiotic yeasts-bacteria community with alleged
healthy capabilities. Revista Iberoamericana de Micología, v. 23, n. 2, p. 67-74,
2006.
17 GORSKI, D. Kefir: 21st century yogurt? Dairy Foods, v. 95, p. 49, 1994.
18 FARNWORTH, E. R. Kefir - a complex probiotic. Food Science and Techology
Bulletin: Functional Foods, v. 2, p. 1-17, 2005.
19 BESHKOVA, D. et al. Pure cultures for making kefir. Food Microbiology, v. 19, p.
537-544, 2002.
20 BRASIL. Aprova Regulamento Técnico de Identidade e Qualidade de Leites
Fermentados Instrução Normativa. Brasília: Diário Oficial da República Federativa
do Brasil. 46 2007.
21 FAO/WHO. CODEX Standard for Fermented Milks. 243-2003. STAN, C.: 1-11 p.
2003.
22 JAY, J. M. Fermentação e Produtos Lácteos Fermentados. In: JAY, J. M. (Ed.).
Microbiologia de Alimentos. 6. Porto Alegre: Artmed, 2005. cap. 7, p.131-147.
23 AXELSSON, L. Lactic Acid Bacteria: Classification and Physiology. In:
SALMINEN, S.;VON WRIGHT, A., et al (Ed.). Lactic Acid Bacteria:
Microbiological and Functional Aspects. 3. New York: Marcel Dekker, Inc., 2004.
24 BERNARDEAU, M. et al. Safety assessment of dairy microorganisms: the
Lactobacillus genus. International Journal of Food Microbiology, v. 126, n. 3, p.
278-85, 2008.

121
25 MADIGAN, M. T.; MARTINKO, J. M.; PARKER, J. Microbiologia de Brock. 10.
São Paulo: Prentice Hall, 2004. 608p.
26 GAWEHN, K. D-(-)-Lactate. In: BERMEYER, H. U. (Ed.). Methods of Enzymatic
Analysis. 3. Weinheim: Verlag Chemie, 1984. p.588-592.
27 SERVIN, A. L. Antagonistic activities of lactobacilli and bifidobacteria against
microbial pathogens. FEMS Microbiology Reviews, v. 28, n. 4, p. 405-40, Oct 2004.
28 DALIÉ, D. K. D.; DESCHAMPS, A. M.; RICHARD-FORGET, F. Lactic acid
bacteria – Potential for control of mould growth and mycotoxins: A review. Food
Control, v. 21, n. 4, p. 370-380, 2010.
29 CARASI, P. et al. Surface proteins from Lactobacillus kefir antagonize in vitro
cytotoxic effect of Clostridium difficile toxins. Anaerobe, v. 18, n. 1, p. 135-42, Feb
2012.
30 COCONNIER, M. H. et al. Antagonistic activity of Lactobacillus acidophilus LB
against intracellular Salmonella enterica serovar Typhimurium infecting human
enterocyte-like Caco-2/TC-7 cells. Applied and Environmental Microbiology, v. 66,
n. 3, p. 1152-7, 2000.
31 ALEGRIA, A. et al. Bacteriocins produced by wild Lactococcus lactis strains isolated
from traditional, starter-free cheeses made of raw milk. International Journal of
Food Microbiology, v. 143, n. 1-2, p. 61-6, 2010.
32 JAMALY, N.; BENJOUAD, A.; BOUKSAIM, M. Probiotic Potential of Lactobacillus
strains Isolated from Known Popular Traditional Moroccan Dairy Products. British
Microbiology Research Journal v. 1, n. 4, p. 79-94, 2011.
33 SCHILLINGER, U.; LUCKE, F. K. Antibacterial activity of Lactobacillus sake
isolated from meat. Applied and Environmental Microbiology, v. 55, n. 8, p. 1901-
6, 1989.
34 DE VUYST, L.; LEROY, F. Bacteriocins from lactic acid bacteria: production,
purification, and food applications. Journal of Microbiology Methods, v. 13, n. 4, p.
194-9, 2007.
35 LIU, S.-N.; HAN, Y.; ZHOU, Z.-J. Lactic acid bacteria in traditional fermented
Chinese foods. Food Research International, v. 44, n. 3, p. 643-651, 2011.
36 GALVEZ, A. et al. Application of bacteriocins in the control of foodborne pathogenic
and spoilage bacteria. Critical Reviews in Biotechnology, v. 28, n. 2, p. 125-52,
2008.

122
37 ISOLAURI, E.; SALMINEN, S.; OUWEHAND, A. C. Probiotics. Best Practice
& Research Clinical Gastroenterology, v. 18, n. 2, p. 299-313, 2004.
38 CASALTA, E.; MONTEL, M.-C. Safety assessment of dairy microorganisms: The
Lactococcus genus. International Journal of Food Microbiology, v. 126, n. 3, p.
271-273, 2008.
39 NOMURA, M. et al. Phenotypic and molecular characterization of Lactococcus lactis
from milk and plants. Journal of Applied Microbiology, v. 101, n. 2, p. 396-405,
2006.
40 OGIER, J. C. et al. Safety assessment of dairy microorganisms: The Leuconostoc
genus. International Journal of Food Microbiology, v. 126, n. 3, p. 286-290, 2008.
41 HOLLAND, R.; LIU, S. Q. Lactic Acid Bacteria | Leuconostoc spp. In: Fukay, J.W.
(Ed.). Encyclopedia of Dairy Sciences (Second Edition). San Diego: Academic
Press, 2011. p.138-142.
42 SENGUN, I. Y.; KARABIYIKLI, S. Importance of acetic acid bacteria in food
industry. Food Control, v. 22, n. 5, p. 647-656, 2011.
43 AQUARONE, E. et al. Biotecnologia Industrial. São Paulo: Blücher, 2001. 523
44 REA, M. C. et al. Irish kefir-like grains : their structure, microbial composition and
fermentation kinetics. Journal of Applied Bacteriology, v. 81, p. 83-94, 1996.
45 JACQUES, N.; CASAREGOLA, S. Safety assessment of dairy microorganisms: The
hemiascomycetous yeasts. International Journal of Food Microbiology, v. 126, n. 3,
p. 321-326, 2008.
46 BÜCHL, N. R.; SEILER, H. YEASTS AND MOLDS | Yeasts in Milk and Dairy
Products. In: EDITOR-IN-CHIEF: JOHN, W. F. (Ed.). Encyclopedia of Dairy
Sciences (Second Edition). San Diego: Academic Press, 2011. p.744-753.
47 VERGIO, F. Anti- und Probiotika. Hippokrates, v. 25, p. 16-119, 1954.
48 PUUPPONEN-PIMIÄ, R. et al. Development of functional ingredients for gut health.
Trends in Food Science & Technology, v. 13, n. 1, p. 3-11, 2002.
49 SAARELA, M. et al. Probiotic bacteria: safety, functional and technological
properties. Journal of Biotechnology, v. 84, n. 3, p. 197-215, 2000. ISS
50 GUEIMONDE, M.; SALMINEN, S. New methods for selecting and evaluating
probiotics. Digestive and Liver Disease, v. 38, p. S242-S247, 2006.

123
51 MARTINS, F. S. et al. Utilização de leveduras como probióticos. Revista de
Biologia e Ciências da Terra, v. 5, n. 2, 2005.
52 HATTINGH-LOURENS, A.; VILJOEN, B. C. Growth and survival of a probiotic
yeast in dairy products. Food Research International, v. 34, p. 791-796, 2001.
53 GUARNER, F. et al. Should yoghurt cultures be considered probiotic? Britsh
Journal of Nutrition, v. 93, n. 6, p. 783-6, 2005.
54 CANDELA, M. et al. Interaction of probiotic Lactobacillus and Bifidobacterium
strains with human intestinal epithelial cells: Adhesion properties, competition against
enteropathogens and modulation of IL-8 production. International Journal of Food
Microbiology, v. 125, n. 3, p. 286-292, 2008.
55 CONNOLLY, E. et al. In situ colonisation and immunomodulation by the probiotic
lactobacillus reuteri (ATCC 55730) in the human gastrointestinal tract. Clinical
Nutrition, v. 22, Supplement 1, n. 0, p. S57, 2003.
56 WANG, J. et al. Selection of potential probiotic lactobacilli for cholesterol-lowering
properties and their effect on cholesterol metabolism in rats fed a high-lipid diet.
Journal of Dairy Science, v. 95, n. 4, p. 1645-1654, 2012.
57 EJTAHED, H. S. et al. Effect of probiotic yogurt containing Lactobacillus
acidophilus and Bifidobacterium lactis on lipid profile in individuals with type 2
diabetes mellitus. Journal of Dairy Science, v. 94, n. 7, p. 3288-3294, 2011.
58 MINERVINI, F. et al. Fermented goats’ milk produced with selected multiple starters
as a potentially functional food. Food Microbiology, v. 26, n. 6, p. 559-564, 2009.
59 GOBBETTI, M.; MINERVINI, F.; RIZZELLO, C. G. Angiotensin I-converting-
enzyme-inhibitory and antimicrobial bioactive peptides. International Journal of
Dairy Technology, v. 57, n. 2-3, p. 173-188, 2004.
60 MCFARLAND, L. V. Evidence-based review of probiotics for antibiotic-associated
diarrhea and Clostridium difficile infections. Anaerobe, v. 15, n. 6, p. 274-280, 2009.
ISSN 1075-9964.
61 MYLLYLUOMA, E. et al. Effects of anti-Helicobacter pylori treatment and probiotic
supplementation on intestinal microbiota. International Journal of Antimicrobial
Agents, v. 29, n. 1, p. 66-72, 2007.

124
62 MYLLYLUOMA, E. et al. Probiotic intervention decreases serum gastrin-17 in
Helicobacter pylori infection. Digestive and Liver Disease, v. 39, n. 6, p. 516-523,
2007.
63 GARCIA, A. C. et al. Probiotic properties of Lactobacillus spp. isolated from gastric
biopsies of Helicobacter pylori infected and non-infected individuals. Revista médica
de Chile v. 137, p. 369-376, 2009.
64 GLUCK, U.; GEBBERS, J. O. Ingested probiotics reduce nasal colonization with
pathogenic bacteria (Staphylococcus aureus, Streptococcus pneumoniae, and beta-
hemolytic streptococci). The American Journal of Clinical Nutrition, v. 77, n. 2, p.
517-20, 2003.
65 VILJANEN, M. et al. Induction of inflammation as a possible mechanism of probiotic
effect in atopic eczema–dermatitis syndrome. Journal of Allergy and Clinical
Immunology, v. 115, n. 6, p. 1254-1259, 2005.
66 IANNITTI, T.; PALMIERI, B. Therapeutical use of probiotic formulations in clinical
practice. Clinical Nutrition, v. 29, n. 6, p. 701-725, 2010.
67 JIMÉNEZ, M. B. Treatment of irritable bowel syndrome with probiotics. Revista
Española de Enfermidades Digestivas, v. 101, n. 8, p. 553-564, 2009.
68 WOLLOWSKI, I.; RECHKEMMER, G.; POOL-ZOBEL, B. L. Protective role of
probiotics and prebiotics in colon cancer. The American Journal of Clinical
Nutrition, v. 73, n. 2 Suppl, p. 451S-455S, 2001.
69 ROUSSEAUX, C. et al. Lactobacillus acidophilus modulates intestinal pain and
induces opioid and cannabinoid receptors. Nature Medicine, v. 13, n. 1, p. 35-37,
2007.
70 EJTAHED, H. S. et al. Probiotic yogurt improves antioxidant status in type 2 diabetic
patients. Nutrition, v. 28, n. 5, p. 539-543, 2012.
71 HOLZAPFEL, W. H.; SCHILLINGER, U. Introduction to pre- and probiotics. Food
Research International, v. 35, n. 2–3, p. 109-116, 2002.
72 KALANTZOPOULOS, G. Fermented Products with Probiotic Qualities. Anaerobe,
v. 3, n. 2–3, p. 185-190, 1997. ISSN 1075-9964.
73 SANDERS, M. E. Probiotics: definition, sources, selection, and uses. Clinical
Infectious Diseases, v. 46 Suppl 2, p. S58-61; discussion S144-51, 2008.

125
74 HIRAYAMA, K.; RAFTER, J. The role of probiotic bacteria in cancer prevention.
Microbes and Infection, v. 2, n. 6, p. 681-686, 2000.
75 SALMINEN, S. Probiotics: Scientific Support for Use. Food Technology. 53: 66 p.
1999.
76 PITHVA, S. et al. Potential of Probiotic Lactobacillus Strains as Food Additivies.
Food Additive. EL-SAMRAGY, Y.: InTech: 175-190 p. 2012.
77 SAAD, S. M. I. Probióticos e prebióticos: o estado da arte. Revista Brasileira de
Ciências Farmacêuticas, v. 42, n. 1, p. 2-16, 2006.
78 BERNET, M. F. et al. Lactobacillus acidophilus LA 1 binds to cultured human
intestinal cell lines and inhibits cell attachment and cell invasion by enterovirulent
bacteria. Gut, v. 35, n. 4, p. 483-9, 1994.
79 LINK-AMSTER, H. et al. Modulation of a specific humoral immune response and
changes in intestinal flora mediated through fermented milk intake. FEMS
Immunology and Medical Microbiology, v. 10, n. 1, p. 55-63, 1994.
80 AYENI, F. A. et al. Evaluation of the functional potential of Weissella and
Lactobacillus isolates obtained from Nigerian traditional fermented foods and cow's
intestine. International Journal of Food Microbiology, v. 147, n. 2, p. 97-104, May
27 2011.
81 DUNNE, C. et al. Probiotics: from myth to reality. Demonstration of functionality in
animal models of disease and in human clinical trials. Antonie Van Leeuwenhoek, v.
76, n. 1-4, p. 279-92, Jul-Nov 1999.
82 CENCIC, A.; LANGERHOLC, T. Functional cell models of the gut and their
applications in food microbiology--a review. International Journal of Food
Microbiology, v. 141 Suppl 1, p. S4-14, 2010.
83 O'BRIEN, J. et al. Safety evaluation of probiotics. Trends in Food Science &
Technology, v. 10, n. 12, p. 418-424, 1999.
84 SALMINEN, S. et al. Functional food science and gastrointestinal physiology and
function. Br J Nutr, v. 80 Suppl 1, p. S147-71, 1998.
85 FAO/WHO. Toxicological evaluation of certain food additivies 5. SERIES, W. F.
A. Geneva: WHO/FAO. 53: 40 p. 1974.
86 ______. Standard for Infant Formula and Formulas for Special Medical
Purposes Intended for Infats. 72. STAN, C.: 21 p. 1981.

126
87 EWASCHUK, J. B.; NAYLOR, J. M.; ZELLO, G. A. D-lactate in human and
ruminant metabolism. J Nutr, v. 135, n. 7, p. 1619-25, 2005.
88 HOVE, H. Lactate and short chain fatty acid production in the human colon:
implications for D-lactic acidosis, short-bowel syndrome, antibiotic-associated
diarrhoea, colonic cancer, and inflammatory bowel disease. Dan Med Bull, v. 45, n.
1, p. 15-33, 1998.
89 CONNOLLY, E.; LÖNNERDAL, B. D(-)-Lactid acid-producing bacteria: Safe to use
in infant formulas. Nutrafoods Research, v. 3, n. 3, p. 37-49, 2004.
90 CONNOLLY, E.; ABRAHAMSSON, T.; BJÖRKSTÉN, B. Safety of D(-)-Lactic
Acid Producing Bacteria in the Human Infant. Journal of Pediatric
Gastroenterology and Nutrition, v. 41, n. 4, p. 489-492, 2005.
91 OTIENO, D. O.; ASHTON, J. F.; SHAH, N. P. Role of microbial strain and storage
temperatures in the degradation of isoflavone phytoestrogens in fermented soymilk
with selected β-glucosidase producing Lactobacillus casei strains. Food Research
International, v. 40, n. 3, p. 371-380, 2007.
92 YANG, S. et al. Hydrolysis of soybean isoflavone glycosides by a thermostable β-
glucosidase from Paecilomyces thermophila. Food Chemistry, v. 115, n. 4, p. 1247-
1252, 2009.
93 BHATIA, Y.; MISHRA, S.; BISARIA, V. S. Microbial beta-glucosidases: cloning,
properties, and applications. Critical Reviews in Biotechnology, v. 22, n. 4, p. 375-
407, 2002.
94 EFSA. EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the
maintenance of the list of QPS biological agents intentionally added to food and
feed (2011 update), v. 9, n. 12, p. 82, 2011.
95 AMMOR, M. S.; FLOREZ, A. B.; MAYO, B. Antibiotic resistance in non-
enterococcal lactic acid bacteria and bifidobacteria. Food Microbiology, v. 24, n. 6, p.
559-70, 2007.
96 MATHUR, S.; SINGH, R. Antibiotic resistance in food lactic acid bacteria--a review.
International Journal of Food Microbiology, v. 105, n. 3, p. 281-95, 2005.
97 EFSA. Technical guidance prepared by the Panel on Additives and Products or
Substances used in Animal Feed (FEEDAP) on the update of the criteria used in the
assessment of bacterial resistance to antibiotics of human or veterinary importance., v.
732, p. 1-15, 2008.

127
98 BILLOT-KLEIN, D. et al. Modification of peptidoglycan precursors is a common
feature of the low-level vancomycin-resistant VANB-type Enterococcus D366 and of
the naturally glycopeptide-resistant species Lactobacillus casei, Pediococcus
pentosaceus, Leuconostoc mesenteroides, and Enterococcus gallinarum. Journal of
Bacteriology, v. 176, n. 8, p. 2398-405, 1994.
99 VINDEROLA, G. et al. Usefulness of a set of simple in vitro tests for the screening
and identification of probiotic candidate strains for dairy use. LWT - Food Science
and Technology, v. 41, n. 9, p. 1678-1688, 2008.
100 BRASIL. Aprova o Regulamento Técnico que Estabelece as Diretrizes Básicas
para Análise e Comprovação de Propriedades Funcionais e ou de Saúde Alegadas
em Rotulagem de Alimentos Resolução. Brasília: Diário Oficial da União Federativa
do Brasil. Resolução nº18 1999.
101 ______. Aprova o Regulamento Técnico de Procedimentos para Registro de
Alimento com Alegaçao de Propriedades Funcionais ou de Saúde em sua
Rotulagem. Resolução. Brasilia: Diário Oficial da União Federativa do Brasil. 19
1999.
102 FARNWORTH, E. R.; MAINVILLE, I. Kefir - A Fermented Milk Product. In:
FARNWORTH, E. R. (Ed.). Handbook of Fermented Functional Foods. 2. Boca
Raton, London, New York: CRC Press Taylor & Francis Group 2008. cap. 4, p.89-127
103 KEMP, N. Kefir, the champagne of cultured dairy products. Cultured Dairy
Products Journal, v. 19, p. 29-30, 1984.
104 RATTRAY, F. P.; O’CONNELL, M. J. Fermented Milks | Kefir. In: Fukay, J. W.
(Ed.). Encyclopedia of Dairy Sciences (Second Edition). San Diego: Academic
Press, 2011. p.518-524.
105 SHAVIT, E. Renewed interest in kefir, the ancient elixir of longevity. Fungi. 1: 14-
18 p. 2008.
106 OTLES, S.; CAGINDI, O. Kefir: A probiotic dairy-composition, nutritional and
therapeutic aspects Pakistan Journal of Nutrition, v. 2, n. 2, p. 54-59, 2003.
107 MIGUEL, M. G. D. C. P. et al. Diversity of bacteria present in milk kefir grains using
culture-dependent and culture-independent methods. Food Research International,
v. 43, n. 5, p. 1523-1528, 2010.
108 GARROTE, G. L.; ABRAHAM, A. G.; DE ANTONI, G. L. Chemical and
microbiological characterisation of kefir grains. Journal of Dairy Research, v. 68, n.
04, p. 639-652, 2001.

128
109 MAGALHÃES, K. T. et al. Brazilian Kefir: Structure, Microbial Communities and
Chemical Composition. Brazilian Journal of Microbiology, v. 42, p. 693-702, 2011.
110 ZOURARI, A.; ANIFANTAKIS, E. M. Le kéfir caractères physico-chimiques,
microbiologiques et nutritionnels. Technologie de production. Une revue. Le Lait, v.
68, n. 4, p. 373-392, 1988.
111 LIBUDZISZ, Z.; PIATKIEWICZ, A. Kefir production in Poland. Dairy Industries
International v. 55, n. 7, p. 31-33, 1990.
112 MOTAGHI, M. et al. Kefir production in Iran. World Journal of Microbiology and
Biotechnology, v. 13, n. 5, p. 579-581, 1997.
113 CHEN, T. H. et al. Microbiological and chemical properties of kefir manufactured by
entrapped microorganisms isolated from kefir grains. Journal of Dairy Science, v.
92, n. 7, p. 3002-3013, 2009.
114 PINTADO, M. E. et al. Microbiological and rheological studies on Portuguese kefir
grains. International Journal of Food Science & Technology, v. 31, n. 1, p. 15-26,
1996.
115 GARROTE, G. L.; ABRAHAM, A. G.; DE ANTONI, G. Preservation of Kefir
Grains, a Comparative Study. Lebensmittel-Wissenschaft und-Technologie, v. 30, p.
77-84, 1997.
116 LA RIVIÈRE, J. W. M.; KOOMAN, P. Kefiran, a novel polysaccharide produced in
the kefir grain by Lactobacillus brevis. Archiv für Mikrobiologie, v. 59, p. 269-279,
1967.
117 GUZEL-SEYDIM, Z. et al. Turkish kefir and kefir grains: microbial enumeration and
electron microscobic observation†. International Journal of Dairy Technology, v.
58, n. 1, p. 25-29, 2005.
118
SARKAR, S. Potential of kefir as a dietetic beverage – a review. British Food
Journal, v. 109, n. 4, p. 280-290, 2007.
119 SIMOVA, E. et al. Lactic acid bacteria and yeasts in kefir grains and kefir made from
them. Journal of Industrial Microbiology & Biotechnology v. 28, p. 1-6, 2002.
120 LIN, C. W.; CHEN, H. L.; LIU, J. R. Identification and characterization of lactic acid
bacteria and yeasts isolated from kefir grains in Taiwann. Australian Journal of
Dairy Technology, v. 54, p. 14-18, 1999.

129
121 JIANZHONG, Z. et al. Analysis of the microflora in Tibetan kefir grains using
denaturing gradient gel electrophoresis. Food Microbiology, v. 26, n. 8, p. 770-775,
2009.
122 BOTTAZZI, V.; BIANCHI, F. A Note on Scanning Electron Microscopy of Micro-
organisms associated with the Kefir Granule. Journal of Applied Microbiology, v.
48, n. 2, p. 265-268, 1980.
123 MAINVILLE, I. et al. Polyphasic characterization of the lactic acid bacteria in kefir.
Systematic and Applied Microbiology, v. 29, n. 1, p. 59-68, Jan 2006.
124 DELFEDERICO, L. et al. Molecular identification and typing of lactobacilli isolated
from kefir grains. Journal of Dairy Research, v. 73, n. 1, p. 20-7, Feb 2006.
125 MAGALHÃES, K. T. et al. Microbial communities and chemical changes during
fermentation of sugary Brazilian kefir. World Journal of Microbiology and
Biotechnology, v. 26, n. 7, p. 1241-1250, 2010.
126 DOBSON, A. et al. High-throughput sequence-based analysis of the bacterial
composition of kefir and an associated kefir grain. FEMS Microbiology Letters, v.
320, n. 1, p. 56-62, 2011.
127 IRIGOYEN, A. et al. Microbiological, physicochemical, and sensory characteristics
of kefir during storage. Food Chemistry, v. 90, n. 4, p. 613-620, 2005.
128 WITTHUHN, R. C.; SCHOEMAN, T.; BRITZ, T. J. Isolation and characterization of
the microbial population of different South African kefir grains. International
Journal of Dairy Technology, v. 57, n. 1, p. 33-37, 2004.
129 ÖNER, Z.; KARAHAN, A. G.; ÇAKMAKÇI, M. L. E. Effects of different milk types
and starter cultures on kefir. Gida, v. 35, p. 177-182, 2010.
130 GRØNNEVIK, H.; FALSTAD, M.; NARVHUS, J. A. Microbiological and chemical
properties of Norwegian kefir during storage. International Dairy Journal, v. 21, n.
9, p. 601-606, 2011.
131 KESMEN, Z.; KACMAZ, N. Determination of Lactic Microflora of Kefir Grains and
Kefir Beverage by Using Culture-Dependent and Culture-Independent Methods.
Journal of Food Science, v. 76, n. 5, p. M276-M283, 2011.
132 CHEN, H.-C.; WANG, S.-Y.; CHEN, M.-J. Microbiological study of lactic acid
bacteria in kefir grains by culture-dependent and culture-independent methods. Food
Microbiology, v. 25, n. 3, p. 492-501, 2008.

130
133 WITTHUHN, R. C.; SCHOEMAN, T.; BRITZ, T. J. Characterisation of the microbial
population at different stages of Kefir production and Kefir grain mass cultivation.
International Dairy Journal, v. 15, n. 4, p. 383-389, 2005.
134 FUJISAWA, T. et al. Lactobacillus kefiranofaciens sp. nov. isolated from kefir
grains. International Journal of Systematic Bacteriology, v. 38, n. 1, p. 12-14,
1988.
135 KOJIMA, S. et al. An improved medium for the isolation of lactobacilli from kefir
grains. Bioscience, Biotechnology and Biochemistry, v. 57, n. 1, p. 119-120, 1993.
136 GARCÍA FONTÁN, M. C. et al. Microbiological and chemical changes during the
manufacture of Kefir made from cows’ milk, using a commercial starter culture.
International Dairy Journal, v. 16, n. 7, p. 762-767, 2006.
137 GOLOWCZYC, M. A. et al. Characterization of homofermentative lactobacilli
isolated from kefir grains: potential use as probiotic. Journal of Dairy Research, v.
75, n. 2, p. 211-7, 2008.
138 GARBERS, I. M.; BRITZ, T. J.; WITTHUHN, R. C. PCR-based Denaturing Gradient
Gel Electrophoretic typification and identification of the microbial consortium present
in Kefir grains. World Journal of Microbiology and Biotechnology, v. 20, n. 7, p.
687-693, 2004.
139 CHEIRSILP, B.; SHIMIZU, H.; SHIOYA, S. Enhanced kefiran production by mixed
culture of Lactobacillus kefiranofaciens and Saccharomyces cerevisiae. Journal of
Biotechnology, v. 100, n. 1, p. 43-53, 2003.
140 MAGALHÃES, K. T. et al. Production of fermented cheese whey-based beverage
using kefir grains as starter culture: Evaluation of morphological and microbial
variations. Bioresource Technology, v. 101, n. 22, p. 8843-8850, 2010.
141 WANG, Y. S. et al. Identification of Yeasts and Evaluation of their Distribution in
Taiwanese Kefir and Viili Starters. Journal of Dairy Science, v. 91, p. 3798-3805,
2008.
142 PARAMITHIOTIS, S. et al. Interactions between Saccharomyces cerevisiae and
lactic acid bacteria in sourdough. Process Biochemistry, v. 41, n. 12, p. 2429-2433,
2006. ISSN 13595113.
143 GÜZEL-SEYDIM, Z. B. et al. Determination of Organic Acids and Volatile Flavor
Substances in Kefir during Fermentation. Journal of Food Composition and
Analysis, v. 13, p. 35-43, 2000.

131
144 MAGALHÃES, K. T. et al. Comparative study of the biochemical changes and
volatile compound formations during the production of novel whey-based kefir
beverages and traditional milk kefir. Food Chemistry, v. 126, n. 1, p. 249-253, 2011.
145 BESHKOVA, D. M. et al. Production of volatile aroma compounds by kefir starter
cultures. International Dairy Journal, v. 13, n. 7, p. 529-535, 2003.
146 GUZEL-SEYDIM, Z. B.; SEYDIM, A. C.; GREENE, A. K. Comparison of amino
acid profiles of milk, yogurt and Turkish kefir. Milchwissenschaft, v. 58, n. 3-4, p.
158-160, 2003.
147 BADEL, S.; BERNARDI, T.; MICHAUD, P. New perspectives for Lactobacilli
exopolysaccharides. Biotechnology Advances, v. 29, n. 1, p. 54-66, Jan-Feb 2011.
148 RUAS-MADIEDO, P.; DE LOS REYES-GAVILÁN, C. G. Invited Review: Methods
for the Screening, Isolation, and Characterization of Exopolysaccharides Produced by
Lactic Acid Bacteria. Journal of Dairy Science, v. 88, n. 3, p. 843-856, 2005.
149 RODRIGUES, K. L. et al. Antimicrobial and healing activity of kefir and kefiran
extract. International Journal of Antimicrobial Agents, v. 25, n. 5, p. 404-8, 2005.
150 GARROTE, G. L.; ABRAHAM, A. G.; DE ANTONI, G. Microbial Interactions in
Kefir: A Natural Probiotic Drink. In: MOZZI, F.;RAYA, R. R., et al (Ed.).
Biotechnology of Lactic Acid Bacteria - Novel Applications. Iowa: Blackwell
Publishing, 2010. cap. 18, p.327-340.
152 SIMOVA, E. et al. Amino acid profiles of lactic acid bacteria, isolated from kefir
grains and kefir starter made from them. International Journal of Food
Microbiology, v. 107, n. 2, p. 112-23, 2006.
152 QUIRÓS, A. et al. Angiotensin-converting enzyme inhibitory activity of peptides
derived from caprine kefir. Journal of Dairy Science, v. 88, p. 3480-3487, 2005.
153 HERTZLER, S. R.; CLANCY, S. M. Kefir improves lactose digestion and tolerance
in adults with lactose maldigestion. Journal of the American Dietetic Association, v.
103, n. 5, p. 582-7, 2003.
154 HONG, W.-S. et al. Effects of kefir supernatant and lactic acid bacteria isolated from
kefir grain on cytokine production by macrophage. International Dairy Journal, v.
19, n. 4, p. 244-251, 2009.
155 VINDEROLA, G. et al. Effects of kefir fractions on innate immunity.
Immunobiology, v. 211, n. 3, p. 149-56, 2006.

132
156 CHIFIRIUC, M. C.; CIOACA, A. B.; LAZAR, V. In vitro assay of the antimicrobial
activity of kephir against bacterial and fungal strains. Anaerobe, v. 17, n. 6, p. 433-5,
2011.
157 SANTOS, A. et al. The Antimicrobial Properties of Different Strains of Lactobacillus
spp. Isolated from Kefir. Systematic and Applied Microbiology, v. 26, n. 3, p. 434-
437, 2003.
158 SILVA, K. R. et al. Antimicrobial activity of broth fermented with kefir grains.
Applied Biochemistry and Biotechnology, v. 152, n. 2, p. 316-25, 2009.
159 URDANETA, E. et al. Intestinal beneficial effects of kefir-supplemented diet in rats.
Nutrition Research, v. 27, n. 10, p. 653-658, 2007.
160 KUBO, M. et al. Pharmacological study on kefir--a fermented milk product in
Caucasus. I. On antitumor activity Yakugaku Zasshi, v. 112, n. 7, p. 489-95, 1992.
161 SCHMIDT, P.; VASS, A.; SZAKALY, S. Effect of fermented milk diets on
regeneration of the rat liver. Acta Med Hung, v. 41, n. 2-3, p. 163-9, 1984.
162 DINIZ, R. O. et al. Atividade antiinflamatória de quefir, um probiótico da medicina
popular. Revista Brasileira de Farmacognosia, v. 13, p. 19-21, 2003.
163 JAY, J. M. Microbiologia de Alimentos 6.Porto Alegre: Artmed, 2005. 711
164 MUYZER, G.; DE WAAL, E. C.; UITTERLINDEN, A. G. Profiling of complex
microbial populations by denaturing gradient gel electrophoresis analysis of
polymerase chain reaction-amplified genes coding for 16S rRNA. Applied and
Environmental Microbiology, v. 59, n. 3, p. 695-700, 1993.
165 WOESE, C. R. Bacterial evolution. Microbiological Reviews, v. 51, n. 2, p. 221-71,
1987.
166 OLIVEIRA, V. M. Uso da técnica denaturing gradient gel electrophoresis
(DGGE) para caracterização da diversidade microbiana. UFRJ. Rio de Janeiro
2006.
167 DUARTE, R. T. D. Micro-organimos em ambientes criogênicos: gelo glacial, solos
expostos por recuo de geleiras, e permafrost polares. 2010. Tese (Doutorado em
Biotecnologia). Instituto Butantan, Universidade de São Paulo, São Paulo, 2010.
168 JANY, J. L.; BARBIER, G. Culture-independent methods for identifying microbial
communities in cheese. Food Microbiology, v. 25, n. 7, p. 839-48, 2008.

133
169 URRAZA, P. J. et al. DNA fingerprinting of thermophilic lactic acid bacteria using
repetitive sequence-based polymerase chain reaction. Journal of Dairy Research, v.
67, p. 381-392, 2000.
170 KOEUTH, T.; VERSALOVIC, J.; LUPSKI, J. R. Differential subsequence
conservation of interspersed repetitive Streptococcus pneumoniae BOX elements in
diverse bacteria. Genome Research, v. 5, n. 4, p. 408-18, 1995.
171 CARVALHO, M. C. C. G.; SILVA, D. C. G. Sequenciamento de DNA de nova
geração e suas aplicações na genômica de plantas. Ciência Rural, v. 40, n. 3, p. 735-
744, 2010.
172 RONAGHI, M. Pyrosequencing Sheds Light on DNA Sequencing. Genome
Research, v. 11, p. 3-11, 2001.
173 HALL, N. Advanced sequencing technologies and their wider impact in microbiology.
The Journal of Experimental Biology, v. 210, n. Pt 9, p. 1518-25, 2007.
174 MARGULIES, M. et al. Genome sequencing in microfabricated high-density picolitre
reactors. Nature, v. 437, n. 7057, p. 376-80, 2005
175 HUMBLOT, C.; GUYOT, J. P. Pyrosequencing of tagged 16S rRNA gene amplicons
for rapid deciphering of the microbiomes of fermented foods such as pearl millet
slurries. Applied and Environmental Microbiology, v. 75, n. 13, p. 4354-61, 2009.
176 RONAGHI, M. et al. Real-Time DNA Sequencing Using Detection of Pyrophosphate
Release. Analytical Biochemistry, v. 242, n. 1, p. 84-89, 1996.
177 BINLADEN, J. et al. The Use of Coded PCR Primers Enables High-Throughput
Sequencing of Multiple Homolog Amplification Products by 454 Parallel Sequencing.
PLoS ONE, v. 2, n. 2, p. e197, 2007.
178
LEITE, A. M. O. et al. Assessment of the microbial diversity of Brazilian kefir grains
by PCR-DGGE and pyrosequencing analysis. Food Microbiology, v. 31, n. 2, p. 215-
221, 2012.
179 GARROTE, G. L.; ABRAHAM, A. G.; DE ANTONI, G. Characteristics of kefir
prepared with different grain:milk ratios. Journal of Dairy Research, v. 65, p. 149-
154, 1998.
180 SARKAR, S. Biotechnological innovations in kefir production: a review. British
Food Journal, v. 110, n. 3, p. 283-295, 2008.

134
181 ASSADI, M. M.; POURAHMAD, R.; MOAZAMI, N. Use of isolated kefir starter
cultures in kefir production. World Journal of Microbiology and Biotechnology, v.
16, n. 6, p. 541-543, 2000.
182 ROSSI, J.; GOBBETTI, M. Impiego di un multistarter per la produzione in
continuo di kefir. Milano, ITALIE: Dipartimento di scienze e tecnologie alimentari e
microbiologiche dell' Universitá degli studi di Milano, 1991.
183 PARASKEVOPOULOU, A. et al. Functional properties of single cell protein
produced by kefir microflora. Food Research International, v. 36, n. 5, p. 431-438,
2003.
184 HUSEINI, H. F. et al. Evaluation of wound healing activities of kefir products.
Burns, v. 38, n. 5, p. 719-723, 2012.
185 MARQUINA, D. et al. Dietary influence of kefir on microbial activities in the mouse
bowel. Letters in Applied Microbiology, v. 35, n. 2, p. 136-40, 2002.
186 ZACCONI, C. et al. Competitive exclusion of Campyloacter jejuni by kefir
fermented milk Annals of Microbiology, v. 53, n. 2, p. 179-187, 2003.
187 LIU, J. R. et al. Antitumor activity of milk kefir and soy milk kefir in tumor-bearing
mice. Nutrition Cancer, v. 44, n. 2, p. 183-7, 2002.
188 GUVEN, A.; GULMEZ, M. The effect of kefir on the activities of GSH-Px, GST,
CAT, GSH and LPO levels in carbon tetrachloride-induced mice tissues. Journal of
veterinary medicine. B, Infectious diseases and veterinary public health, v. 50, n.
8, p. 412-6, 2003.
189 THOREUX, K.; SCHMUCKER, D. L. Kefir milk enhances intestinal immunity in
young but not old rats. The Journal of Nutrition, v. 131, n. 3, p. 807-12, 2001.
190 FURUKAWA, N. et al. The effect of oral administration of water soluble fraction
from kefir grain on antibody production in mice. Animal Science and Technology, v.
63, n. 4, p. 428-436, 1992.
191 MEDRANO, M. et al. Oral Administration of Kefiran Induces Changes in the
Balance of Immune Cells in a Murine Model. Journal of Agricultural and Food
Chemistry, v. 59, n. 10, p. 5299-5304, 2011.
192 VINDEROLA, C. G. et al. Immunomodulating capacity of kefir. Journal of Dairy
Research, v. 72, n. 2, p. 195-202, May 2005.

135
193 WANG, Y. et al. Effects of Lactobacillus plantarum MA2 isolated from Tibet kefir
on lipid metabolism and intestinal microflora of rats fed on high-cholesterol diet.
Applied Microbiology and Biotechnology, v. 84, n. 2, p. 341-347, 2009.
194 LIU, J. R. et al. Hypocholesterolaemic effects of milk-kefir and soyamilk-kefir in
cholesterol-fed hamsters. Britsh Journal of Nutrition, v. 95, n. 5, p. 939-46, 2006.
195 ST-ONGE, M.-P. et al. Kefir consumption does not alter plasma lipid levels or
cholesterol fractional synthesis rates relative to milk in hyperlipidemic men: a
randomized controlled trial. BMC Complementary and Alternative Medicine, v. 2,
n. 1, p. 1, 2002.
196 DE VRESE, M.; KELLER, B.; BARTH, C. A. Enhancement of intestinal hydrolysis
of lactose by microbial beta-galactosidase (EC 3.2.1.23) of kefir. Britsh Journal of
Nutrition, v. 67, n. 1, p. 67-75, 1992.
197 WALTER, J. et al. Detection of Lactobacillus, Pediococcus, Leuconostoc, and
Weissella species in human feces by using group-specific PCR primers and denaturing
gradient gel electrophoresis. Applied and Environmental Microbiology, v. 67, n. 6,
p. 2578-85, Jun 2001.
198 ENDO, A.; OKADA, S. Monitoring the lactic acid bacterial diversity during shochu
fermentation by PCR-denaturing gradient gel electrophoresis. Journal of Bioscience
and Bioengeneering, v. 99, n. 3, p. 216-21, Mar 2005.
199 COCOLIN, L. et al. An application of PCR-DGGE analysis to profile the yeast
populations in raw milk. International Dairy Journal, v. 12, n. 5, p. 407-411, 2002.
200 YOUNG, J. P. W.; DOWNER, H. L.; EARDLY, B. D. Phylogeny of the phototrophic
Rhizobium strain BTAil by polymerase chain reaction-based sequencing of a 16S
rRNA gene segment. Journal of Bacteriology, v. 173, n. 2271-2277, 1991.
201 WANG, Q. et al. Naive Bayesian classifier for rapid assignment of rRNA sequences
into the new bacterial taxonomy. Applied and Environmental Microbiology, v. 73,
n. 16, p. 5261-7, 2007.
202 SCHLOSS, P. D. et al. Introducing mothur: open-source, platform-independent,
community-supported software for describing and comparing microbial communities.
Applied and Environmental Microbiology, v. 75, n. 23, p. 7537-41, 2009.
203 CHAO, A.; BUNGE, J. Estimating the Number of Species in a Stochastic Abundance
Model. Biometrics, v. 58, n. 3, p. 531-539, 2002.
204 SHANNON, C. E.; WEAVER, W. The mathematical theory of information AT & T
Technical Journal, v. 27, p. 359-423, 1949.

136
205 TAMURA, K. et al. MEGA5: Molecular Evolutionary Genetics Analysis Using
Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods.
Molecular Biology and Evolution, v. 28, n. 10, p. 2731-2739, October 1, 2011 2011.
206 NINANE, V.; MUKANDAYAMBAJE, R.; BERBEN, G. Identification of lactic acid
bacteria within the consortium of a kefir grain by sequencing 16S rDNA variable
regions. Journal of AOAC International, v. 90, n. 4, p. 1111-7, Jul- 2007.
207 LATORRE-GARCÍA, L.; DEL CASTILLO-AGUDO, L.; POLAINA, J. Taxonomical
classification of yeasts isolated from kefir based on the sequence of their ribosomal
RNA genes. World Journal of Microbiology and Biotechnology, v. 23, n. 6, p. 785-
791, 2007.
208 ROH, S. W. et al. Investigation of archaeal and bacterial diversity in fermented
seafood using barcoded pyrosequencing. ISME Journal, v. 4, n. 1, p. 1-16, 2010.
209 JUNG, J. Y. et al. Metagenomic analysis of kimchi, a traditional Korean fermented
food. Applied and Environmental Microbiology, v. 77, n. 7, p. 2264-74, 2011.
210 ERCOLINI, D. PCR-DGGE fingerprinting: novel strategies for detection of microbes
in food. Journal of Microbiology Methods, v. 56, n. 3, p. 297-314, 2004.
211 CARR, J. G.; PASSMORE, S. M. Methods for identifying acetic acid bacteria. In:
SHINNER, F. A. e LOVELOCK, D. W. (Ed.). Indentification methods for
microbiologist. London: Academic Press, 1979.
212 RANDAZZO, C. L. et al. Diversity, Dynamics, and Activity of Bacterial
Communities during Production of an Artisanal Sicilian Cheese as Evaluated by 16S
rRNA Analysis. Applied and Environmental Microbiology, v. 68, n. 4, p. 1882-
1892, 2002.
213 AGUILA, E. M.; SILVA, J. T.; PASCHOALIN, V. N. M. F. Expression of the yeast
calcineurin subunitsCNA1andCNA2during growth and hyper-osmotic stress. FEMS
Microbiology Letters, v. 221, n. 2, p. 197-202, 2003.
214 WANG, X. et al. Diversity of a stable enrichment culture which is useful for silage
inoculant and its succession in alfalfa silage. FEMS Microbiology Ecology v. 57, n.
1, p. 106-15, 2006.
215 NAUMOVA, E.; IVANNIKOVA, Y.; NAUMOV, G. Genetic Differentiation of the
Sherry Yeasts Saccharomyces cerevisiae. Applied Biochemistry and Microbiology,
v. 41, n. 6, p. 578-582, 2005.

137
216 NOT, F. et al. New Insights into the Diversity of Marine Picoeukaryotes. PLoS ONE,
v. 4, n. 9, p. e7143, 2009.
217 GONZÁLEZ DE LLANO, D.; RODRIGUEZ, A.; CUESTA, P. Effect of lactic starter
cultures on the organic acid composition of milk and cheese during ripening—analysis
by HPLC. Journal of Applied Microbiology, v. 80, n. 5, p. 570-276, 1996.
218 DOLCI, P. et al. Microbial diversity, dynamics and activity throughout manufacturing
and ripening of Castelmagno PDO cheese. International Journal of Food
Microbiology, v. 143, n. 1-2, p. 71-5, 2010.
219 FALENTIN, H. et al. Reverse transcription quantitative PCR revealed persistency of
thermophilic lactic acid bacteria metabolic activity until the end of the ripening of
Emmental cheese. Food Microbiology, v. 29, n. 1, p. 132-40, 2012.
220
RANTSIOU, K. et al. Microflora of Feta cheese from four Greek manufacturers.
International Journal of Food Microbiology, v. 126, n. 1–2, p. 36-42, 2008.
221 COCOLIN, L. et al. Study of the Ecology of Fresh Sausages and Characterization of
Populations of Lactic Acid Bacteria by Molecular Methods. Applied and
Environmental Microbiology, v. 70, n. 4, p. 1883-1894, 2004.
222 ANDERSON, I. C.; PARKIN, P. I. Detection of active soil fungi by RT-PCR
amplification of precursor rRNA molecules. Journal of Microbiology Methods, v.
68, n. 2, p. 248-53, 2007.
223 CENCIARINI, C. et al. Influence of Long Time Storage in Mineral Water on RNA
Stability of Pseudomonas aeruginosa and Escherichia coli after Heat Inactivation.
PLoS ONE, v. 3, n. 10, p. e3443, 2008.
224 FALENTIN, H. et al. Specific metabolic activity of ripening bacteria quantified by
real-time reverse transcription PCR throughout Emmental cheese manufacture.
International Journal of Food Microbiology, v. 144, n. 1, p. 10-9, 2010.
225 BENTSINK, L. et al. Amplification of RNA by NASBA allows direct detection of
viable cells of Ralstonia solanacearum in potato. Journal of Applied Microbiology,
v. 93, n. 4, p. 647-55, 2002.
226 MCKILLIP, J. L.; JAYKUS, L. A.; DRAKE, M. rRNA stability in heat-killed and
UV-irradiated enterotoxigenic Staphylococcus aureus and Escherichia coli O157:H7.
Applied and Environmental Microbiology, v. 64, n. 11, p. 4264-8, 1998.
227 CHURRUCA, E. et al. Detection of Campylobacter jejuni and Campylobacter coli in
chicken meat samples by real-time nucleic acid sequence-based amplification with

138
molecular beacons. International Journal of Food Microbiology, v. 117, n. 1, p. 85-
90, 2007.
228 TOBA, T. et al. A medium for isolation of capsular bacteria from kefir grains.
Agricultural and Biological Chemistry, v. 50, n. 10, p. 2673-2674, 1986.
229 FLÓREZ, A. B.; MAYO, B. PCR–DGGE as a tool for characterizing dominant
microbial populations in the Spanish blue-veined Cabrales cheese. International
Dairy Journal, v. 16, n. 10, p. 1205-1210, 2006.
230 PRAKITCHAIWATTANA, C. J.; FLEET, G. H.; HEARD, G. M. Application and
evaluation of denaturing gradient gel electrophoresis to analyse the yeast ecology of
wine grapes. FEMS Yeast Research, v. 4, n. 8, p. 865-77, 2004.
231 BRASIL. Aprova Regulamento Técnico de Porções de Alimentos Embalados para
Fins de Rotulagem Nutricional. Resolução RDC 359. SANITÁRIA., A. N. D. V.
Diário Oficial da República Federativa do Brasil 2003.
232 MAGALHÃES, K. T. et al. Chemical composition and sensory analysis of cheese
whey-based beverages using kefir grains as starter culture. International Journal of
Food Science & Technology, v. 46, n. 4, p. 871-878, 2011.
233 GÜZEL-SEYDIM, Z. B. et al. Determination of Organic Acids and Volatile Flavor
Substances in Kefir during Fermentation. Journal of Food Composition and
Analysis, v. 13, n. 1, p. 35-43, 2000.
234 HOLLAND, R. et al. Esterases of lactic acid bacteria and cheese flavour: Milk fat
hydrolysis, alcoholysis and esterification. International Dairy Journal, v. 15, n. 6-9,
p. 711-718, 2005.
235 KEENAN, T. W. Production of acetic acid and other volatile compounds by
Leuconostoc citrovorum and Leuconostoc dextranicum. Applied Microbiology, v. 16,
n. 12, p. 1881-5, 1968.
236 NISHIDA, S. et al. Evaluation of the probiotic potential of Lactobacillus paracasei
KW3110 based on in vitro tests and oral administration tests in healthy adults. The
Journal of General and Applied Microbiology, v. 54, n. 5, p. 267-76, 2008.
237 GUO, Z. et al. In vitro comparison of probiotic properties of Lactobacillus casei
Zhang, a potential new probiotic, with selected probiotic strains. LWT - Food Science
and Technology, v. 42, n. 10, p. 1640-1646, 2009.
238 USMAN; HOSONO, A. Bile Tolerance, Taurocholate Deconjugation, and Binding of
Cholesterol by Lactobacillus gasseri Strains. Journal of Dairy Science, v. 82, n. 2, p.
243-248, 1999.

139
239 DELGADO, S. et al. Subtractive screening for probiotic properties of lactobacillus
species from the human gastrointestinal tract in the search for new probiotics. Journal
of Food Science, v. 72, n. 8, p. M310-5, 2007.
240 PALYS, T.; NAKAMURA, L. K.; COHAN, F. M. Discovery and classification of
ecological diversity in the bacterial world: the role of DNA sequence data.
International Journal of Systematic Bacteriology, v. 47, n. 4, p. 1145-56, 1997.
241 ROSSETTI, L.; GIRAFFA, G. Rapid identification of dairy lactic acid bacteria by
M13-generated, RAPD-PCR fingerprint databases. Journal of Microbiology
Methods, v. 63, n. 2, p. 135-144, 2005.
242 ANNUK, H. et al. Characterization of intestinal lactobacilli as putative probiotic
candidates. Journal of Applied Microbiology, v. 94, p. 403-412, 2003.
243 RIPAMONTI, B. et al. Screening of species-specific lactic acid bacteria for veal
calves multi-strain probiotic adjuncts. Anaerobe, v. 17, n. 3, p. 97-105, 2011.
244 GONZALEZ, B. et al. Detection, purification, and partial characterization of
plantaricin C, a bacteriocin produced by a Lactobacillus plantarum strain of dairy
origin. Applied and Environmental Microbiology, v. 60, n. 6, p. 2158-63, 1994.
245 RYAN, M. P. et al. An application in Cheedar Cheese Manufacture for a Strain of
Lactococcus lactis producing a novel broad-spectrum bacteriocin, Lacticin 3147.
Applied and Environmental Microbiology, v. 62, n. 2, p. 612-619, 1996.
246 MIGUEL, M. A.L.; DIAS, A. C. C.; LEITE, S. F. G. Inhibition of vancomycin and
high-level aminoglycoside-resistant enterococci strains and Listeria monocytogenes by
bacteriocin-like substance produced by Enterococcus faecium E86. Current
Microbiology, v. 57, n. 5, p. 429-36, 2008.
247 VAN BELKUM, M. J. et al. Cloning of two bacteriocin genes from a lactococcal
bacteriocin plasmid. Applied and Environmental Microbiology, v. 55, n. 5, p. 1187-
91, 1989.
248 SÁNCHEZ, B. et al. Technological and probiotic selection criteria of a bile-adapted
Bifidobacterium animalis subsp. lactis strain. International Dairy Journal, v. 20, n.
11, p. 800-805, 2010.
249 TUOMOLA, E. M.; SALMINEN, S. J. Adhesion of some probiotic and dairy
Lactobacillus strains to Caco-2 cell cultures. International Journal of Food
Microbiology, v. 41, n. 1, p. 45-51, 1998.

140
250 KLARE, I. et al. Evaluation of new broth media for microdilution antibiotic
susceptibility testing of Lactobacilli, Pediococci, Lactococci, and Bifidobacteria.
Applied and Environmental Microbiology, v. 71, n. 12, p. 8982-6, 2005.
251 PLESSAS, S. et al. Microbial ecology and quality assurance in food fermentation
systems. The case of kefir grains application. Anaerobe, v. 17, n. 6, p. 483-5, 2011.
252 MARAGKOUDAKIS, P. A. et al. Probiotic potential of Lactobacillus strains isolated
from dairy products. International Dairy Journal, v. 16, n. 3, p. 189-199, 2006.
253 GU, R. X. et al. Probiotic properties of lactic acid bacteria isolated from stool samples
of longevous people in regions of Hotan, Xinjiang and Bama, Guangxi, China.
Anaerobe, v. 14, n. 6, p. 313-7, 2008.
254 CHARTERIS, W. P. et al. Development and application of an in vitro methodology to
determine the transit tolerance of potentially probiotic Lactobacillus and
Bifidobacterium species in the upper human gastrointestinal tract. Journal of Applied
Microbiology, v. 84, n. 5, p. 759-68, 1998.
255 BEGLEY, M.; HILL, C.; GAHAN, C. G. Bile salt hydrolase activity in probiotics.
Applied and Environmental Microbiology, v. 72, n. 3, p. 1729-38, 2006.
256 BEGLEY, M.; GAHAN, C. G.; HILL, C. The interaction between bacteria and bile.
FEMS Microbiology Reviews, v. 29, n. 4, p. 625-51, 2005.
257 MATHARA, J. M. et al. Functional characteristics of Lactobacillus spp. from
traditional Maasai fermented milk products in Kenya. Internantional Journal of
Food Microbiology, v. 126, n. 1-2, p. 57-64, 2008.
258 ZAGO, M. et al. Characterization and probiotic potential of Lactobacillus plantarum
strains isolated from cheeses. Food Microbiology, v. 28, n. 5, p. 1033-1040, 2011.
259 BATDORJ, B. et al. Isolation, taxonomic identification and hydrogen peroxide
production by Lactobacillus delbrueckii subsp. lactis T31, isolated from Mongolian
yoghurt: inhibitory activity on food-borne pathogens. Journal of Applied
Microbiology, v. 103, n. 3, p. 584-93, 2007.
260 BELLEI, B. et al. Purification of a bacteriocin produced by Enteroccus faecium and
its effectiveness for preservation of fresh-cut lettuce. Journal of Microbiology and
Antimicrobials, v. 3, n. 5, p. 119-125, 2011.
261 POWELL, J. E. et al. Characterization of bacteriocin ST8KF produced by a kefir
isolate Lactobacillus plantarum ST8KF. International Dairy Journal, v. 17, n. 3, p.
190-198, 2007.

141
262 GUEIMONDE, M. et al. Viability and diversity of probiotic Lactobacillus and
Bifidobacterium populations included in commercial fermented milks. Food
Research International, v. 37, n. 9, p. 839-850, 2004.
263 MORI, K. et al. Comparative sequence analyses of the genes coding for 16S rRNA of
Lactobacillus casei-related taxa. International Journal of Systematic Bacteriology,
v. 47, n. 1, p. 54-7, 1997.
264 FERNÁNDEZ, E. et al. Phenotypic, genetic and technological characterization of
Lactococcus garvieae strains isolated from a raw milk cheese. International Dairy
Journal, v. 20, n. 3, p. 142-148, 2010.
265 ADIGUZEL, G. C.; ATASEVER, M. Phenotypic and Genotypic Characterization of
lactic acid bacteria isolated from Turkish dry fermented sausage. Romanian
Biotechological Letters, v. 14, n. 1, p. 4130-4138, 2009.
266 PAPAMANOLI, E. et al. Characterization of lactic acid bacteria isolated from a
Greek dry-fermented sausage in respect of their technological and probiotic properties.
Meat Science, v. 65, n. 2, p. 859-67, 2003.
267 MONTEAGUDO-MERA, A. et al. Characterization of certain bacterial strains for
potential use as starter or probiotic cultures in dairy products. Journal of Food
Protection, v. 74, n. 8, p. 1379-86, 2011.
268 PARK, A. R.; OH, D. K. Galacto-oligosaccharide production using microbial beta-
galactosidase: current state and perspectives. Applied Microbiology and
Biotechnology, v. 85, n. 5, p. 1279-86, 2010.
269 NERI, D. F. M. et al. Galactooligosaccharides production by β-galactosidase
immobilized onto magnetic polysiloxane–polyaniline particles. Reactive and
Functional Polymers, v. 69, n. 4, p. 246-251, 2009.
270 LU, Z. H. et al. Isolation, characterization and identification of lactic acid bacteria and
yeasts from sour Mifen, a traditional fermented rice noodle from China. Journal of
Applied Microbiology, v. 105, n. 3, p. 893-903, 2008.
271 OTIENO, D. O.; ASHTON, J. F.; SHAH, N. P. Evaluation of enzymic potential for
biotransformation of isoflavone phytoestrogen in soymilk by Bifidobacterium
animalis, Lactobacillus acidophilus and Lactobacillus casei. Food Research
International, v. 39, n. 4, p. 394-407, 2006.
272 PINTO, M. et al. Enterocyte-like differentiation and polarization of the human colon
carcinoma cell line Caco-2 in culture. Biology of the Cell, v. 47, p. 323-330, 1983.

142
273 CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-
First Informational Supplement. CLSI document M 100-S21. INSTITUTE, C. A. L.
S. Wayne, PA: Clinical and Laboratory Institute 2011.
274 D'AIMMO, M. R.; MODESTO, M.; BIAVATI, B. Antibiotic resistance of lactic acid
bacteria and Bifidobacterium spp. isolated from dairy and pharmaceutical products.
International Journal of Food Microbiology, v. 115, n. 1, p. 35-42, 2007.
275 FLOREZ, A. B.; DELGADO, S.; MAYO, B. Antimicrobial susceptibility of lactic
acid bacteria isolated from a cheese environment. Canandian Journal of
Microbiology, v. 51, n. 1, p. 51-8, 2005.
276 KLARE, I. et al. Antimicrobial susceptibilities of Lactobacillus, Pediococcus and
Lactococcus human isolates and cultures intended for probiotic or nutritional use.
Journal of Antimicrobial Chemother, v. 59, n. 5, p. 900-12, 2007.
277 TEMMERMAN, R. et al. Identification and antibiotic susceptibility of bacterial
isolates from probiotic products. International Journal of Food Microbiology, v. 81,
n. 1, p. 1-10, 2003.
278 FLOREZ, A. B. et al. Antimicrobial susceptibility profiles of 32 type strains of
Lactobacillus, Bifidobacterium, Lactococcus and Streptococcus spp. International
Journal of Antimicrobial Agents, v. 31, n. 5, p. 484-6, 2008.
279 DELGADO, S.; FLOREZ, A. B.; MAYO, B. Antibiotic susceptibility of Lactobacillus
and Bifidobacterium species from the human gastrointestinal tract. Current
Microbiology, v. 50, n. 4, p. 202-7, 2005.
280 KLEIN, G. et al. Exclusion of vanA, vanB and vanC type glycopeptide resistance in
strains of Lactobacillus reuteri and Lactobacillus rhamnosus used as probiotics by
polymerase chain reaction and hybridization methods. Journal of Applied
Microbiology, v. 89, n. 5, p. 815-24, 2000.

143
APÊNDICES

144
APÊNDICE A – ARTIGO I 1
2
Microbiological, technological and therapeutic properties of kefir: a natural probiotic 3
beverage 4
Analy Machado de Oliveira Leite1*
; Marco Antonio Lemos Miguel2; Raquel Silva Peixoto
2; 5
Alexandre Soares Rosado 2
; Joab Trajano Silva1; Vania Margaret Flosi Paschoalin
1 6
1Instituto de Química, Universidade Federal do Rio de Janeiro, Avenida Athos da Silveira 7
Ramos, 149-Bloco A, Sala 545, 21941-909-Cidade Universitária Rio de Janeiro, RJ, Brazil; 8
2Instituto de Microbiologia, Universidade Federal do Rio de Janeiro, Avenida Carlos Chagas 9
Filho, 373, 21941-904-Cidade Universitária, Rio de Janeiro, RJ, Brazil. 10
*Corresponding author. Mailing address: Instituto de Química, Universidade Federal do 11
Rio de Janeiro, Avenida Athos da Silveira Ramos, 149-Bloco A, Sala 545, 21941-909-Cidade 12
Universitária Rio de Janeiro, RJ, Brazil, Phone:+55 21 25627362, Fax: +55 21 25627266, 13
email: [email protected] 14
Running title: Kefir: a natural probiotic beverage 15
Abstract 16
Kefir is a fermented milk beverage produced by the action of bacteria and yeasts that exist in 17
symbiotic association in kefir grains. The artisanal production of the kefir is based on the 18
tradition of the peoples of Caucasus, which has spread to other parts of the world, from the 19
late 19th
century, and nowadays integrates its nutritional and therapeutic indications to the 20
everyday food choices of several populations. The large number of microorganisms present in 21
kefir and their microbial interactions, the possible bioactive compounds resulting of microbial 22
metabolism, and the benefits associated with the use this beverage confers kefir the status of a 23
natural probiotic, designated as the 21th
century yoghurt. Several studies have shown that kefir 24
and its constituents have antimicrobial, antitumor, anticarcinogenic and immunomodulatory 25

145
activity and also improve lactose digestion, among others. This review includes data on the 26
technological aspects, the main beneficial effects on human health of kefir and its 27
microbiological composition. Generally, kefir grains contain a relatively stable and specific 28
microbiota enclosed in a matrix of polysaccharides and proteins. Microbial interactions in 29
kefir are complex due to the composition of kefir grains, which seems to differ among 30
different studies, although some predominant Lactobacillus species are always present. 31
Besides, the specific populations of individual grains seem to contribute to the particular 32
sensory characteristics present in fermented beverages. This review also includes new 33
electron microscopy data on the distribution of microorganisms within different Brazilian 34
kefir grains, which showed a relative change in its distribution according to grain origin. 35
36
Keywords: probiotic, microbial diversity, kefir grains, fermented milk, kefir 37
38
Introduction 39
The word kefir is derived from the Turkish word keyif, which means "feeling good" 40
after its ingestion (27, 48). The kefir beverage is originally from the Caucasus Mountains, a 41
traditional product highly consumed in Eastern Europe, Russia and Southwest Asia (48). 42
Currently, an increase in kefir consumption in many countries has been reported, due to its 43
unique sensory properties and long history associated with beneficial effects on human health 44
(10, 35, 48). In Brazil, the knowledge of this beverage, as well as the benefits of the inclusion 45
of probiotic food in a regular diet, are not very widespread, and kefir manufacturing is 46
exclusively artisanal (4, 29). 47
Kefir is characterized by its distinct flavour, typical of yeast, and an effervescent effect 48
felt in the mouth (27, 38). The main products of kefir fermentation are lactic acid, ethanol and 49
CO2, which confer this beverage viscosity, acidity and low alcohol content. Minor 50

146
components can also be found, including diacetyl, acetaldehyde, ethyl and amino acids 51
contributing to the flavour composition (38). This drink differs from other fermented dairy 52
products because it is not the result of the metabolic activity of a single or a few microbial 53
species (11). 54
Kefir grains 55
Kefir grains play a natural starter culture role during the production of kefir and are 56
recovered after the fermentation process by milk straining (38). These grains are composed of 57
microorganisms immobilized on a polysaccharide and protein matrix, where several species 58
of bacteria and yeast coexist in symbiotic association (11, 15). In this ecosystem there is a 59
relatively stable microorganism population, which interacts with and influences other 60
members of the community. This population provides the synthesis of bioactive metabolites, 61
which are essential for grain growth and microorganism inhibition, such as food pathogens 62
and contaminants (15). 63
Kefir grains vary in size, from 0.3 to 3.0 cm in diameter (Fig 1), are characterized by 64
an irregular, multi-lobular surface, united by a single central section, and their color varies 65
from white to yellowish white. The grains are elastic and have a viscous and firm texture (11, 66
28, 39). 67
Figure 1 68
Although the kefir drink can be found in many countries, in Brazil the grains are not 69
available commercially, and are culturally donated from person to person. 70
Microbiological aspects 71
In kefir, lactic acid bacteria (LAB) are primarily responsible for the conversion of the 72
lactose present in milk into lactic acid, which results in a pH decrease and milk preservation. 73
Other kefir microbial constituents include lactose-fermenting yeasts that produce ethanol and 74
CO2. Non-lactose fermenting yeast and acetic acid bacteria (AAB) also participate in the 75

147
process (28, 38). After fermentation the grains increase in about 5-7% of their biomass. 76
During their growth in milk, the microorganism proportions in the grains differ from those 77
present in the final product (38, 48). This difference is associated with the fermentation 78
process conditions such as fermentation time, temperature, degree of agitation, type of milk, 79
grain/milk inoculum ratio and microorganism distribution, among others (38, 46, 48). 80
Traditionally, classical microbiological methods are used to study kefir microbiota 81
(39, 46). While these methods are useful, in some cases they are not discriminating enough to 82
identify closely related or new species. Because of the microbial symbiotic association present 83
in the grains, the growth and survival of individual strains are dependent on the presence of 84
each other. Often, when microorganisms are isolated from the grains, they do not grow well in 85
milk and/or show reduced biochemical activity (11). Therefore, independent cultivation 86
methods have been used as a complement to conventional methods in the study of kefir grain 87
microbiota. The polymerase chain reaction technique, coupled to electrophoresis in 88
denaturing gradient gel (PCR-DGGE) has proved appropriate for analyzing complex 89
microbial consortia (21, 23, 29, 30), while the partial sequencing of the gene coding for 16S 90
rRNA has been used for species identification (31, 38). However, some studies show that the 91
PCR-DGGE technique does not allow the detection of significant changes during kefir 92
fermentation (29), probably due to the relative stability of the dominant population in this 93
community. 94
The complexity of this community can be better understood by the introduction of 95
massive sequencing techniques, such as pyrosequencing. Two recent studies, one conducted 96
by our research group (9, 23) used this technique to evaluate kefir microbial diversity and 97
detected the presence of microorganisms belonging to minority groups, not previously 98
reported, by dependent or independent culture techniques. Furthermore, Leite et al. (23), 99
comparing three Brazilian kefir grains, reported that, although of different origins of grains, 100

148
certain microorganism groups such as Lactobacillus species were always present in all grains. 101
The authors suggest that the minority and specific populations of individual grains can 102
contribute to the particular sensory characteristics present in fermented beverages (23). 103
Some species rarely isolated from kefir grains and the kefir beverage are not key 104
species, and are usually considered contaminants. Recently, the fungi Dipodascus capitus and 105
Trichosporon coremiiforme, considered pathogens, were identified by gene sequencing in 106
kefir grains (38). Some bacterial contaminants have also been described, such as 107
Pseudomonas spp. (9, 21, 23) and members of the Enterobacteriaceae (9) and Clostridiaceae 108
(9) families, when kefir grain microbiota was characterized by pyrosequencing and PCR-109
DGGE. The presence of these microorganisms may be associated with contamination during 110
handling of kefir grains or improper practices adopted during the preparation of the kefir 111
beverage. 112
Microorganism distribution in kefir grains 113
The distribution of microorganisms inside kefir grains has been studied, and results are 114
controversial. A group of researchers supports the hypothesis that yeasts are generally found 115
in the inner and intermediate grain zone, with rod-shaped bacteria and rare lactococci 116
predominantly on the surface area (3, 24). Conversely, other researchers (17, 21, 28, 39) 117
describe that yeasts are distributed in both the outer and the inner grain areas, with the main 118
difference between the two areas being related to rod-shaped bacteria morphology. Contrary 119
to previously reported results (3, 39), Guzel-Seydim et al. (17) observed a predominance of 120
rod-shaped bacteria, without any evidence of yeast in the inner grain area. Furthermore, some 121
authors (21, 28) have reported a lower number of cells observed in the inner grain portion as 122
compared with the outer portion. Some authors have postulated that fibrillar material observed 123
in the grain may be in fact the polysaccharide kefiran present throughout the grain (17, 28, 29, 124
30). 125

149
Our research group also evaluated the distribution of microorganisms from three 126
Brazilian kefir grains (Figure 2) and observed a relative variation in their distribution 127
according to grain origin. In general, rod-shaped bacteria were observed both in the inner 128
(Figure 2 b, d, f) and the outer grain portions (Figure 2 a, c, e), whereas yeasts were most 129
frequent in the outer portion (Figure 2 a, c, e). Cocci were observed in the outer portion of one 130
of the grains. In all probability, the granular material observed corresponded to particles of 131
coagulated milk adhered to the surface of the grain and the fibrillar material to the 132
polysaccharide kefiran, spread over all parts of the grains (17, 28, 39). 133
Figure 2 134
Although L. lactis is one of the predominant microorganisms isolated from the kefir 135
drink, several investigators (17, 21, 28) have not observed cocci in micrographs, probably due 136
to poor lactococcal adhesion to the grains during growth, which would facilitate lactococcal 137
release in the washing process (17, 21, 28, 39). Furthermore, it is likely that the acidic pH in 138
the grain interior would interfere in the growth of this microorganism in this particular 139
microenvironment (39). 140
It is probable that surface microorganisms cause a greater impact on the kefir 141
fermentation process (11). However, studies have shown that a wide variation in microbial 142
population occurs between different grains and within the same grain. Lin et al. (24) 143
concluded that grain origin site alone can explain the differences that have been reported in 144
several studies by electron microscopy. 145
Kefir bacteria 146
Homofermentative LAB, including Lactobacillus species, such as L. delbrueckii 147
subsp. bulgaricus, L. helveticus, L. kefiranofaciens subsp. kefiranofaciens, L. kefiranofaciens 148
subsp. kefirgranum and L. acidophilus; Lactococcus spp. Such as L. lactis subsp. lactis and L. 149
lactis subsp. cremoris and Streptococcus thermophilus have been identified in kefir grains and 150

150
in the fermented beverage, as well as heterofermentative LAB, including L. kefiri, L. 151
parakefiri, L. fermentum and L. brevis (23, 38), and citrate-positive strains of L. lactis (L. 152
lactis subsp. lactis biovar diacetylactis), Leuconostoc mesenteroides subsp. cremoris, and 153
Leuconostoc mesenteroides subsp. mesenteroides (23, 27, 38). The use of citrate by citrate-154
positive strains results in the production of key compounds that contribute to typical kefir 155
flavour (38). 156
Kefiran produced by L. kefiranofaciens is a branched, water-soluble polysaccharide, 157
containing equal amounts of D-glucose and D-galactose. The production of this 158
polysaccharide is stimulated when L. kefiranofaciens grows in co-culture with S. cerevisiae 159
(5). 160
AAB species have been isolated and identified in both kefir grain and the kefir 161
beverage. However, in some countries, the presence of these species is considered undesirable 162
(11, 38, 48) and has received less attention, even though they play an essential role in both the 163
microbial consortium and the sensory characteristics of the final product (39, 48). 164
Kefir yeast 165
Although they produce metabolites that contribute to the desirable and typical kefir 166
sensory properties (38, 46), kefir yeast are less studied than kefir bacteria. The main yeast 167
capable of fermenting lactose found in kefir and kefir grains are Kluyveromyces 168
marxianus/Candida kefyr, Kluyveromyces lactis var. lactis, Debaryomyces hansenii e 169
Dekkera anomala, while the non-lactose fermenters include Saccharomyces cerevisiae, 170
Torulaspora delbrueckii, Pichia fermentans, Kazachstania unispora, Saccharomyces 171
turicensis, Issatchenkia orientalis and Debaryomyces occidentalis . 172
Interactions between kefir microorganisms 173
The complex interactions between yeast and bacteria and their interdependence in 174
kefir grains are not completely understood. However, when the bacteria are separated from 175

151
the grain, yeast will not grow as efficiently (5, 11, 38). 176
Due to its high capacity to metabolize lactose (39), the genus Lactococcus tends to 177
grow faster than yeast in milk (39, 48). This genus hydrolyzes lactose, producing lactic acid 178
and a suitable environment for yeast growth (48). Moreover, yeasts synthesize complex B 179
vitamins and hydrolyze milk proteins, using oxygen to produce CO2 and ethanol (27, 48). The 180
interaction between yeast and lactic acid bacteria can be stimulated or inhibited by the growth 181
of one or both, in co-cultures. These microorganisms can compete for nutrients for growth, or 182
may produce metabolites that inhibit or stimulate one another (27). Some yeast species are 183
proteolytic or lipolytic, providing amino acids and fatty acids (38). Species such as 184
Debaryomyces hansenii and Yarrowia lipolytica assimilate the lactic acid formed by LAB, 185
raising the pH and stimulating bacteria growth. The production of vitamin B by Acetobacter 186
spp. also favors the growth of other microorganisms present in kefir grains (27, 39). 187
Technological aspects 188
During fermentation, the grains increase in size and number, and are usually recovered 189
from the fermented milk and reutilized (15). If carefully preserved, they may retain their 190
activity for years (27, 38). The main marker to assess the symbiotic relationship between the 191
different microorganisms is increased biomass during fermentation (15). 192
Grain preservation 193
Kefir grains can be preserved lyophilized, dry or wet (15), but constant washing 194
reduces their viability (11). However, Pintado, et al. (37) observed that grain stored in these 195
conditions present different microbiological profiles than fresh grain. Dried grains maintain 196
their activity for 12-18 months while wet grains maintain activity for 8-10 days (15). Different 197
preservation methods have been tested (13), with freezing being considered the best method. 198
Grain lyophilization has also been tested, but resulted in reduced lactose metabolism, as well 199
as modifications in the bacterial profile, which was different from the original grain profile 200

152
(11). 201
Kefir production 202
There are three main ways of producing kefir (I) the artisanal process, (ii) the 203
commercial process by the Russian method and (iii) the commercial process using pure 204
cultures (10, 35, 38). Other substrates may also be used, such as milk from other animal 205
species, coconut milk, soybean milk, fruit juices and/or sugar and molasses solutions (29, 34, 206
38). 207
The traditional artisanal production involves milk inoculation with a variable amount 208
of grains and fermentation for a period between 18-24h at 20-25 °C. At the end of the 209
fermentation process the grains are sieved and can be used for a new fermentation or kept (1-7 210
days) in fresh milk, while the kefir beverage is stored at 4 °C, ready for consumption (2, 11, 211
35). 212
The initial inoculum concentration of the grains (grain/milk proportion) affects the pH, 213
viscosity, final lactose concentration and the microbiological profile of the final product (14, 214
46). Agitation during fermentation also influences kefir microbial composition, favoring the 215
development of homofermentative lactococci and yeast (11 38, 48). Incubation at 216
temperatures above 30 °C stimulates the growth of thermophilic LAB, while being a 217
disadvantage for yeast growth and mesophilic LAB (38). 218
The second method, known as the "Russian method", allows for the production of 219
kefir on a larger scale, and uses a process of fermentation in series, from the percolate 220
resulting from the first fermentation of the grains (fermented without the grains or mother 221
culture) (11, 38). 222
Different methods can be used in the industrial process of kefir production, but all 223
based on the same principle. The milk is inoculated with pure cultures isolated from kefir 224
grains and commercial cultures (2, 38, 48). The maturation phase can be performed or not, 225

153
consisting of maintaining the kefir at 8-10 °C for up to 24h (2, 38), to allow microorganism, 226
primarily yeast, growth, contributing to the specific flavour of the product (2). Omission of 227
this step is associated with development of atypical flavour in kefir (2, 38). 228
During storage, the CO2 production by yeast or heterofermentative LAB can cause 229
bloating in the product package, a fact that should be considered in the choice of packaging 230
(11, 44). 231
Although the commercial beverage is available in many countries, not all the 232
properties of traditional kefir are always present (11, 27). Assadi et al. (1) tested several ratios 233
of starter cultures isolated from the grains (LAB, yeast, AAB), and found that traditional kefir 234
produced with kefir grains has been better accepted than kefir obtained by using the starter 235
culture. Rossi and Gobbetti (41) produced a "kefir-type" beverage, with lower viscosity and 236
the absence of some of the volatile components often found in traditional kefir. Carneiro (4) 237
on the other hand, developed a starter culture from microorganisms isolated from Kefir grains 238
and the product was more accepted that the traditional kefir. Beshkova et al. (2) proposed two 239
methods of kefir fermentation: one by simultaneous fermentation and one by successive 240
fermentation. Therefore, they used a starter culture consisting of bacteria and yeast isolated 241
from kefir grains, combined to two strains commonly used in yogurt manufacturing. Yeasts 242
were added to the starter culture with sucrose, both at the beginning (simultaneous 243
fermentation) and after the lactic acid fermentation step (successive fermentation). The two 244
fermentation processes produced kefir with high number of viable lactococci and lactobacilli, 245
with sensory properties similar to traditional kefir. 246
The use of commercial cultures can standardize the commercial production of kefir, if 247
the selection of species and strains of yeasts and bacteria is carried out accurately and 248
carefully, thus allowing for the production of a "kefir-type" beverage with acceptable flavour 249
and good conservation properties (2, 4). The commercial beverage may have a commercial 250

154
life period of up to 28 days, while it is recommended that kefir produced with grains be 251
consumed between 3-12 days. However, the "kefir-type" beverage may not present the same 252
therapeutic and probiotic properties present in traditional kefir (38). 253
The development of a "kefir-type" beverage is not the only industrial application that 254
has been investigated. Kefir grains have also been studied regarding the production of single 255
cell protein (SCP) in the bioconversion of cheese whey and their application in the food 256
industry, to improve the sensory characteristics of certain products (36). 257
The main deficiencies in kefir manufacturing can be attributed to unpleasant taste and 258
aroma typical of yeast (48). The latter can be caused by rapid growth of S. cerevisiae, 259
accompanied by a typical vinegar aroma (48). The excessive production of acetic acid can 260
also influence kefir aroma, and occurs due to the intense growth of Acetobacter spp. or the 261
presence of Dekkera spp. in the grains. A bitter taste may be caused by fungi (for example 262
Geotrichum candidum) and/or the activity of some atypical yeasts which may be present in 263
the product (48). 264
Therapeutic aspects 265
Historically, kefir has been recommended for the treatment of several clinical 266
conditions such as gastrointestinal problems, hypertension, allergies, and ischemic heart 267
disease (11, 38). However, the variability inherent in kefir production conditions in different 268
assays makes it difficult to conduct comparisons between reported scientific results (10, 11, 269
38). 270
Kefir grain fermentations from various substrates have been evaluated (10, 29, 34) and 271
a wide variety of bioactive compounds have been observed, such as organic acids, CO2, H2O2, 272
ethanol, bioactive peptides, exopolysaccharides (kefiran), and bacteriocins. These compounds 273
may act independently or together to produce the various health benefits attributed to kefir 274
consumption (15, 38). 275

155
Antimicrobial activity 276
Santos et al. (42) observed the antagonistic behavior of lactobacilli isolated from kefir 277
grains against E. coli, L. monocytogenes, Salmonella Typhimurium, S. Enteritidis, Shigella 278
flexneri and Y. enterocolitica. Silva et al. (45) observed the inhibition of Candida albicans, 279
Salmonella typhi, Shigella sonnei, Staphylococcus aureus and E. coli by kefir cultured in 280
brown sugar. On the other hand, Chifiriuc et al. (5), observed that all milk fermented with 281
kefir grains had antimicrobial activity against Bacillus subtilis, S. aureus, E. coli, E. faecalis 282
and S. Enteritidis, but did not inhibit P. aeruginosa and C. albicans. All these studies indicate 283
that kefir antimicrobial activity is associated with the production of organic acids, peptides 284
(bacteriocins), carbon dioxide, hydrogen peroxide, ethanol and diacetyl. These compounds 285
may have beneficial effects not only in the reduction of food borne pathogens and 286
deteriorating bacteria during beverage production and storage, but also in the treatment and 287
prevention of gastroenteritis and vaginal infections (10, 43). In addition, the antimicrobial 288
activity of the polysaccharide kefiran was also demonstrated against bacteria and C. albicans 289
(40). 290
Anti-inflammatory and healing activity 291
Healing and anti-inflammatory activities of kefir and kefiran in mice were observed 292
after a seven-day treatment with kefir gel (40), as well as anti-inflammatory activitiy (8) on a 293
model in which granulomatous tissue and writhing is induced by acetic acid in mice. Husseini 294
et al. (20) corroborated the healing activity in burns infected with Pseudomonas aeruginosa in 295
mice. 296
Impact on the gastrointestinal tract (GIT) 297
The effect caused by kefir consumption in the intestinal microbiota composition may 298
be due to a combination of factors, such as direct pathogen inhibition by acids and bacteriocin 299
production, besides competitive pathogen exclusion in the intestinal mucosa (38). According 300

156
to Marquina et al. (32) kefir consumption significantly increased LAB counts in the intestinal 301
mucosa and reduced enterobacteria and clostridia populations. Furthermore, kefir 302
consumption also prevented C. jejuni colonization in chick ceca (53), and was effective in 303
postoperative treatments and in patients with gastrointestinal disorders (43). In Russia, kefir 304
has been used by researchers in the treatment of peptic ulcers in the stomach and duodenum 305
of human patients (11). 306
Anticarcinogenic effects 307
The anticarcinogenic role of fermented dairy products can be attributed, in general, to 308
cancer prevention and the suppression of early-stage tumors, by the delaying of enzyme 309
activities that convert pro-carcinogenic compounds to carcinogens, or by the activation of the 310
immune system (43). Kubo et al. (22) reported the inhibition of the proliferation of 311
subcutaneously transplanted Ehrlich ascites tumors in mice. Liu et al. (26) observed the 312
inhibition of tumor growth, the induction of apoptotic cell lysis in tumors and significant 313
increases in IgA levels in mice, suggesting that kefir potentially has anti-tumoral properties 314
and promotes resistance of the mucosa to intestinal infections. Guven and Gulmez (16) 315
reported that mice treated with kefir had a higher protective effect against damage induced by 316
carbon tetrachloride, indicating that kefir can also act as an antioxidant. 317
Stimulation of the immune system 318
The formation of bioactive peptides during fermentation or digestion processes has 319
shown a variety of physiological activities, including stimulation of the immune system in 320
animal models (10). Thoreux and Schmucker (49), after feeding mice with kefir, observed an 321
increase in the specific mucosal immune response (IgA) against the cholera toxin. Stimulation 322
of the immune system may also occur due to the action of exopolysaccharides found in kefir 323
grains (10, 12). Medrano et al. (33) observed that kefiran was able to modify the balance of 324
the immune cells in the intestinal mucosa. Vinderola et al. (50) demonstrated the 325

157
immunomodulation ability of kefir in the immune response of the intestinal mucosa of mice. 326
The administration of kefir also induced a response in the intestinal mucosa, suggesting that 327
components of kefir can stimulate cells of the innate immune system, suppressing the immune 328
response of the Th2 phenotype or promoting cell-mediated immune responses against tumors 329
and intracellular pathogen infections (26). Recently, Hong et al. (19) demonstrated, in vitro, 330
the immunomodulating capacity of LAB isolated from kefir grains, suggesting their influence 331
on the secretion of the proinflammatory cytokines IL-6 and TNF-α by TLR-2. 332
Hypocholesterolemic effect 333
Possible mechanisms proposed for the hypocholesterolemic activity in LAB may 334
involve the inhibition of the exogenous cholesterol absorption in the small intestine, by the 335
binding and incorporation of cholesterol to bacterial cells and cholesterol uptake, as well as 336
suppression of bile acid reabsorption by enzymatic deconjugation of bile salts, promoted by 337
the bacterial bile salt enzyme, hydrolase (BSH) (52). Wang et al. (52) observed a significant 338
reduction in serum levels of total cholesterol, low density lipoproteins (LDL) and 339
triglycerides, while there was no change in high density lipoprotein (HDL-C) levels in mice 340
fed cholesterol-rich a diet supplemented with Lactobacillus plantarum MA2. Moreover, total 341
cholesterol and triglycerides in the liver were also reduced. Cholesterol and triglycerides in 342
animal feces, on the other hand, increased significantly. Another study (25) also observed a 343
reduction of serum triglyceride and cholesterol levels, especially the non-HDL-C fraction. A 344
contradictory result was reported by St-Onge et al. (47), where kefir consumption did not 345
reduce total cholesterol, LDL-C, HDL-C and triglycerides levels, but increased the 346
concentrations of isobutiric, propionic and isovaleric acids, as well as the total amount of 347
short chained fatty acids in faeces. 348
Kefir and lactose intolerance 349
The ability to decrease lactose concentrations and the presence of β-galactosidase 350

158
activity in fermented milk products make them suitable for consumption by people classified 351
as lactose intolerant (11, 43). It has been demonstrated that some kefir grains show β-352
galactosidase enzyme activity, which stays active when consumed, and that kefir contains less 353
lactose than milk (10, 43). A commercial kefir (18) proved to be as effective as yoghurt in 354
reducing expired hydrogen and flatulence in lactose intolerant adults when compared to the 355
ingestion of milk. de Vrese et al. (7) demonstrated that pigs fed with kefir showed a 356
significant increase in galactose plasma concentrations, suggesting improvement of intestinal 357
lactose hydrolysis by the microbial enzyme β-galactosidase. 358
Final considerations 359
Kefir is a traditional example of the co-existence of bacteria and yeasts and the 360
importance of this symbiotic relationship seems clear, since it is necessary to produce 361
compounds beneficial to health. Although the evidence is not conclusive and further studies 362
should be conducted, the existing scientific studies demonstrate the health benefits reported 363
empirically by historical kefir consumption. Currently, the application of probiotics in the 364
food industry is in expansion and understanding the symbiotic relationships between different 365
microorganisms present in food, as well as their interactions, could assist in the improvement 366
of technological processes. 367
The quality of traditional kefir is mainly influenced by the microorganisms present in 368
kefir grains and kefir processing conditions. Although scientists and food companies 369
attempted to develop a commercial "kefir-type" beverage produced by different cultures and 370
mesophilic and thermophilic LAB, or even pure cultures isolated from kefir grains, their 371
success when compared to traditional kefir is limited. We suggest that this limitation is due to 372
the microbial diversity present in kefir grains and their interactions, which can determine the 373
probiotic and therapeutic properties of final product, as well as peculiarities conferred by 374
certain minority groups present in different grains. However, from an industrial point of view, 375

159
these developments are welcome, given the lack of standardization in the production and 376
marketing of traditional kefir. 377
Acknowledgements 378
The authors acknowledge the help of Prof. Dr. Sergio Seabra and lab technician Eliandro 379
Lima State of the Universidade Estadual da Zona Oeste (UEZO) and M.Sc. Laidson Paes 380
Gomes for technical assistance in the sample processing and SEM micrographs. We also 381
thank to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação 382
de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) and Coordenação de 383
Aperfeiçoamento de Pessoal de Nível Superior (CAPES). 384
References 385
1. Assadi, M. M., Pourahmad, R., & Moazami, N. (2000). Use of isolated kefir starter 386
cultures in kefir production. World J. Microbiol. Biotechnol., 16(6), 541-543. 387
2. Beshkova, D., Simova, E. D., Simov, Z. I., Frengova, G. I., & Spasov, Z. N. (2002). Pure 388
cultures for making kefir. Food Microbiol., 19, 537-544. 389
3. Bottazzi, V., & Bianchi, F. (1980). A Note on Scanning Electron Microscopy of Micro-390
organisms associated with the Kefir Granule. J. Appl. Microbiol., 48(2), 265-268. 391
4. Carneiro, R. P. (2010). Desenvolvimento de uma cultura iniciadora para produção de 392
kefir. Belo Horizonte, Brasil, 142p. (M.Sc. Dissertation. Faculdade de Farmácia. UFMG). 393
5. Cheirsilp, B., Shimizu, H., & Shioya, S. (2003). Enhanced kefiran production by mixed 394
culture of Lactobacillus kefiranofaciens and Saccharomyces cerevisiae. J. Biotechnol., 395
100(1), 43-53. 396
6. Chifiriuc, M. C., Cioaca, A. B., & Lazar, V. (2011). In vitro assay of the antimicrobial 397
activity of kephir against bacterial and fungal strains. Anaerobe, 17(6), 433-435. 398
7. de Vrese, M., Keller, B., & Barth, C. A. (1992). Enhancement of intestinal hydrolysis of 399
lactose by microbial beta-galactosidase (EC 3.2.1.23) of kefir. Br. J. Nutr., 67(1), 67-75. 400

160
8. Diniz, R. O., Perazzo, F. F., Carvalho, J. C. T., & Schneenedorf, J. M. (2003). Atividade 401
antiinflamatória de quefir, um probiótico da medicina popular. Rev. Bras .Farmacogn., 402
13, 19-21. 403
9. Dobson, A., O'Sullivan, O., Cotter, P. D., Ross, P., & Hill, C. (2011). High-throughput 404
sequence-based analysis of the bacterial composition of kefir and an associated kefir 405
grain. FEMS Microbiol. Lett., 320(1), 56-62. 406
10. Farnworth, E. R. (2005). Kefir - a complex probiotic. Food Sci Technol Bull: Functional 407
Foods, 2, 1-17. 408
11. Farnworth, E. R., & Mainville, I. (2008). Kefir - A Fermented Milk Product. In: 409
Farnworth, E. R. (2th
ed.), Handbook of Fermented Functional Foods (2 ed ). CRC Press 410
Taylor & Francis Group, Boca Raton, London, New York, p. 89-127. 411
12. Furukawa, N., Liyama, R., Takahashi, T., & Yamanka, Y. (1992). The effect of oral 412
administration of water soluble fraction from kefir grain on antibody production in mice. 413
Anim. Sci. Technol., 63(4), 428-436. 414
13. Garrote, G. L., Abraham, A. G., & De Antoni, G. (1997). Preservation of Kefir Grains, a 415
Comparative Study. Lebensm-Wiss u-Technol, 30, 77-84. 416
14. Garrote, G. L., Abraham, A. G., & De Antoni, G. (1998). Characteristics of kefir 417
prepared with different grain:milk ratios. J. Dairy Res., 65, 149-154. 418
15. Garrote, G. L., Abraham, A. G., & De Antoni, G. (2010). Microbial Interactions in Kefir: 419
A Natural Probiotic Drink. In F. Mozzi, R. R. Raya & G. M. Vignolo (Eds.), 420
Biotechnology of Lactic Acid Bacteria - Novel Applications (pp. 327-340). Iowa: 421
Blackwell Publishing. 422
16. Guven, A., & Gulmez, M. (2003). The effect of kefir on the activities of GSH-Px, GST, 423
CAT, GSH and LPO levels in carbon tetrachloride-induced mice tissues. J. Vet. Med. B 424
Infect. Dis. Vet. Public Health, 50(8), 412-416. 425

161
17. Guzel-Seydim, Z., Wyffels, J. T., Seydim, A. C., & Greene, A. K. (2005). Turkish kefir 426
and kefir grains: microbial enumeration and electron microscobic observation†. Int. 427
J. Dairy Technol., 58(1), 25-29. 428
18. Hertzler, S. R., & Clancy, S. M. (2003). Kefir improves lactose digestion and tolerance in 429
adults with lactose maldigestion. J Am Diet Assoc 103(5), 582-587. 430
19. Hong, W.-S., Chen, H.-C., Chen, Y.-P., & Chen, M.-J. (2009). Effects of kefir 431
supernatant and lactic acid bacteria isolated from kefir grain on cytokine production by 432
macrophage. Int. Dairy J., 19(4), 244-251. 433
20. Huseini, H. F., Rahimzadeh, G., Fazeli, M. R., Mehrazma, M., & Salehi, M. (2012). 434
Evaluation of wound healing activities of kefir products. Burns, 38 (5), 719-723. 435
21. Jianzhong, Z., Liu, X., Jiang, H., & Dong, M. (2009). Analysis of the microflora in 436
Tibetan kefir grains using denaturing gradient gel electrophoresis. Food Microbiol., 437
26(8), 770-775. 438
22. Kubo, M., Odani, T., Nakamura, S., Tokumaru, S., & Matsuda, H. (1992). 439
Pharmacological study on kefir--a fermented milk product in Caucasus. I. On antitumor 440
activity. Yakugaku Zasshi, 112(7), 489-495. 441
23. Leite, A. M. O., Mayo, B., Rachid, C. T. C. C., Peixoto, R. S., Silva, J. T., Paschoalin, V. 442
M. F., & Delgado, S. (2012). Assessment of the microbial diversity of Brazilian kefir 443
grains by PCR-DGGE and pyrosequencing analysis. Food Microbiol., 31(2), 215-221. 444
24. Lin, C. W., Chen, H. L., & Liu, J. R. (1999). Identification and characterization of lactic 445
acid bacteria and yeasts isolated from kefir grains in Taiwann. Aust. Journal of Dairy 446
Tech., 54, 14-18. 447
25. Liu, J. R., Wang, S. Y., Chen, M. J., Chen, H. L., Yueh, P. Y., & Lin, C. W. (2006). 448
Hypocholesterolaemic effects of milk-kefir and soyamilk-kefir in cholesterol-fed 449
hamsters. Br. J. Nutr., 95(5), 939-946. 450

162
26. Liu, J. R., Wang, S. Y., Lin, Y. Y., & Lin, C. W. (2002). Antitumor activity of milk kefir 451
and soy milk kefir in tumor-bearing mice. Nutr. Cancer, 44(2), 183-187. 452
27. Lopitz-Otsoa, F., Rementeria, A., Elguezabal, N., & Garaizar, J. (2006). Kefir: a 453
symbiotic yeasts-bacteria community with alleged healthy capabilities. Rev. Iberoam. 454
Micol., 23(2), 67-74. 455
28. Magalhães, K. T., Pereira, G. V. M., Campos, C. R., Dragone, G., & Schwan, R. F. 456
(2011). Brazilian Kefir: Structure, Microbial Communities and Chemical Composition. 457
Braz. J. Microbiol., 42, 693-702. 458
29. Magalhães, K. T., Pereira, G. V. M., Dias, D. R., & Schwan, R. F. (2010). Microbial 459
communities and chemical changes during fermentation of sugary Brazilian kefir. World 460
J. Microbiol. Biotechnol., 26(7), 1241-1250. 461
30. Magalhães, K. T., Pereira, M. A., Nicolau, A., Dragone, G., Domingues, L., Teixeira, J. 462
A., Silva, J.B.A., Schwan, R.F. (2010). Production of fermented cheese whey-based 463
beverage using kefir grains as starter culture: Evaluation of morphological and microbial 464
variations. Bioresour. Technol., 101(22), 8843-8850. 465
31. Mainville, I., Robert, N., Lee, B., & Farnworth, E. R. (2006). Polyphasic characterization 466
of the lactic acid bacteria in kefir. Syst. Appl. Microbiol., 29(1), 59-68. 467
32. Marquina, D., Santos, A., Corpas, I., Munoz, J., Zazo, J., & Peinado, J. M. (2002). 468
Dietary influence of kefir on microbial activities in the mouse bowel. Lett. Appl. 469
Microbiol., 35(2), 136-140. 470
33. Medrano, M., Racedo, S. M., Rolny, I. S., Abraham, A. a. G., Pérez, P. F. (2011). Oral 471
Administration of Kefiran Induces Changes in the Balance of Immune Cells in a Murine 472
Model. J. Agric. Food. Chem., 59(10), 5299-5304. 473
34. Öner, Z., Karahan, A. G., & Çakmakçi, M. L. E. (2010). Effects of different milk types 474
and starter cultures on kefir. Gida, 35, 177-182. 475

163
35. Otles, S., & Cagindi, O. (2003). Kefir: A probiotic dairy-composition, nutritional and 476
therapeutic aspects. Pakistan J Nutr, 2(2), 54-59. 477
36. Paraskevopoulou, A., Athanasiadis, I., Kanellaki, M., Bekatorou, A., Blekas, G., & 478
Kiosseoglou, V. (2003). Functional properties of single cell protein produced by kefir 479
microflora. Food Res. Int., 36(5), 431-438. 480
37. Pintado, M. E., Da Silva, J. A. L., Fernandes, P. B., Malcata, F. X., & Hogg, T. A. 481
(1996). Microbiological and rheological studies on Portuguese kefir grains. Int 482
J. Food Sci. Technol., 31(1), 15-26. 483
38. Rattray, F. P., & O'Connell, M. J. (2011). Fermented Milks | Kefir. In: Fukay, J. W. (ed.), 484
Encyclopedia of Dairy Sciences (2th
ed). Academic Press , San Diego, USA, p. 518-524. 485
39. Rea, M. C., Lennartsson, T., Dillon, P., Drina, F. D., Reville, W. J., Heapes, M., & 486
Cogan, T. M. (1996). Irish kefir-like grains : their structure, microbial composition and 487
fermentation kinetics. J. Appl. Microbiol., 81, 83-94. 488
40. Rodrigues, K. L., Caputo, L. R., Carvalho, J. C., Evangelista, J., & Schneedorf, J. M. 489
(2005). Antimicrobial and healing activity of kefir and kefiran extract. Int. J. Antimicrob. 490
Agents, 25(5), 404-408. 491
41. Rossi, J., & Gobbetti, M. (1991). Impiego di un multistarter per la produzione in 492
continuo di kefir (Vol. 41). Milano, ITALIE: Dipartimento di scienze e tecnologie 493
alimentari e microbiologiche dell' Universitá degli studi di Milano. 494
42. Santos, A., San Mauro, M., Sanchez, A., Torres, J. M., & Marquina, D. (2003). The 495
Antimicrobial Properties of Different Strains of Lactobacillus spp. Isolated from Kefir. 496
Syst. Appl. Microbiol., 26(3), 434-437. 497
43. Sarkar, S. (2007). Potential of kefir as a dietetic beverage – a review. Br. Food J., 109(4), 498
280-290. 499

164
44. Sarkar, S. (2008). Biotechnological innovations in kefir production: a review. Br. Food 500
J., 110(3), 283-295. 501
45. Silva, K. R., Rodrigues, S. A., Filho, L. X., & Lima, A. S. (2009). Antimicrobial activity 502
of broth fermented with kefir grains. Appl. Biochem. Biotechnol., 152(2), 316-325. 503
46. Simova, E., Beshkova, D., Angelov, A., Hristozova, T., Frengova, G., & Spasov, Z. 504
(2002). Lactic acid bacteria and yeasts in kefir grains and kefir made from them. J. Ind. 505
Microbiol. Biotechnol. 28, 1-6. 506
47. St-Onge, M.-P., Farnworth, E., Savard, T., Chabot, D., Mafu, A., & Jones, P. (2002). 507
Kefir consumption does not alter plasma lipid levels or cholesterol fractional synthesis 508
rates relative to milk in hyperlipidemic men: a randomized controlled trial. BMC 509
Complement Altern Med, 2(1), 1. 510
48. Tamime, A. Y. (2006). Production of Kefir, Koumiss and Other Related Products. In: 511
Tamime, A. Y. (ed.), Fermented Milk Blackwell Science Ltd , Oxford, UK, p. 174-216. 512
49. Thoreux, K., & Schmucker, D. L. (2001). Kefir milk enhances intestinal immunity in 513
young but not old rats. J Nutr, 131(3), 807-812. 514
50. Vinderola, C. G., Duarte, J., Thangavel, D., Perdigon, G., Farnworth, E., & Matar, C. 515
(2005). Immunomodulating capacity of kefir. J Dairy Res, 72(2), 195-202. 516
51. Vinderola, G., Perdigon, G., Duarte, J., Thangavel, D., Farnworth, E., & Matar, C. 517
(2006). Effects of kefir fractions on innate immunity. Immunobiology, 211(3), 149-156. 518
52. Wang, Y., Xu, N., Xi, A., Ahmed, Z., Zhang, B., & Bai, X. (2009). Effects of 519
Lactobacillus plantarum MA2 isolated from Tibet kefir on lipid metabolism and 520
intestinal microflora of rats fed on high-cholesterol diet. Appl Microbiol Biotechnol, 521
84(2), 341-347. 522
53. Zacconi, C., Scolari, G., Vescovo, M., & Sarra, P. G. (2003). Competitive exclusion of 523
Campyloacter jejuni by kefir fermented milk. Ann Microbiol, 53(2), 179-187. 524

165
Legends 525
Figure 1 - Macroscopic structure of kefir grains 526
Figure 2 - Scanning electron microscopy of Brazilian kefir grain microbiota. A, C, E: outer 527
grain portions, B, D, F: inner grain portions. Arrows – Micrography A: cocci; Micrography D: 528
fibrillar material – kefiran polysaccharide; Micrography E: arrow 1 - granular material - 529
coagulated protein, arrow 2 - different yeast species. 530
531
532

166
Figure 1 533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
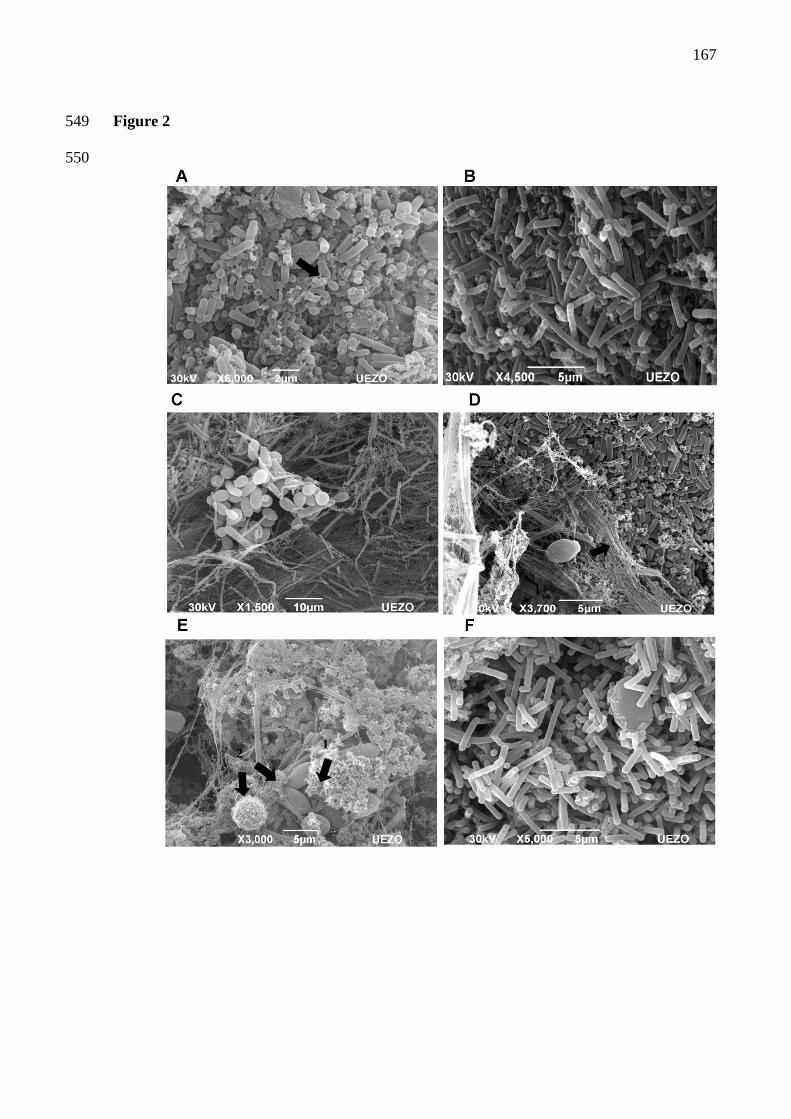
167
Figure 2 549
550
1 2

168
APÊNDICE B – ARTIGO II

at SciVerse ScienceDirect
Food Microbiology 31 (2012) 215e221
Contents lists available
Food Microbiology
journal homepage: www.elsevier .com/locate/ fm
Assessment of the microbial diversity of Brazilian kefir grains by PCR-DGGEand pyrosequencing analysis
A.M.O. Leite a,b, B. Mayo a, C.T.C.C. Rachid c, R.S. Peixoto c, J.T. Silva b, V.M.F. Paschoalin b, S. Delgado a,*
aDepartamento de Microbiología y Bioquímica, Instituto de Productos Lácteos de Asturias (IPLA-CSIC), Carretera de Infiesto s/n, 33300 Villaviciosa, Asturias, SpainbDepartamento de Bioquímica, Instituto de Química, Universidade Federal do Rio de Janeiro, Avenida Athos da Silveira Ramos, 149-Bloco A, Sala 545, 21941-909 Cidade Universitária,Rio de Janeiro, RJ, BrazilcDepartamento de Microbiologia, Instituto de Microbiologia Geral, Universidade Federal do Rio de Janeiro, Avenida Carlos Chagas Filho, 373, 21941-904 Cidade Universitária,Rio de Janeiro, RJ, Brazil
a r t i c l e i n f o
Article history:Received 25 November 2011Received in revised form29 February 2012Accepted 19 March 2012Available online 1 April 2012
Keywords:Kefir grainsCulture-independent microbiologyPCR-DGGEPyrosequencingLactobacillus
* Corresponding author. Tel.: þ34 985 892131; fax:E-mail address: [email protected] (S. Delgado)
0740-0020/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.fm.2012.03.011
a b s t r a c t
The microbial diversity and community structure of three different kefir grains from different parts ofBrazil were examined via the combination of two culture-independent methods: PCR-denaturinggradient gel electrophoresis (PCR-DGGE) and pyrosequencing. PCR-DGGE showed Lactobacillus kefir-anofaciens and Lactobacillus kefiri to be the major bacterial populations in all three grains. The yeastcommunity was dominated by Saccharomyces cerevisiae. Pyrosequencing produced a total of 14,314partial 16S rDNA sequence reads from the three grains. Sequence analysis grouped the reads into threephyla, of which Firmicutes was dominant. Members of the genus Lactobacillus were the most abundantoperational taxonomic units (OTUs) in all samples, accounting for up to 96% of the sequences. OTUsbelonging to other lactic and acetic acid bacteria genera, such as Lactococcus, Leuconostoc, Streptococcusand Acetobacter, were also identified at low levels. Two of the grains showed identical DGGE profiles anda similar number of OTUs, while the third sample showed the highest diversity by both techniques.Pyrosequencing allowed the identification of bacteria that were present in small numbers and rarelyassociated with the microbial community of this complex ecosystem.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Kefir is a viscous, acidic, and mildly alcoholic milk beverageproduced via the fermentation of milk using kefir grain as a starterculture (FAO/WHO, 2003). Thought to be native to the Caucasus andMiddle East, production and consumption of kefir has now spreadthroughout the world, spurred by its long history of beneficialhealth effects (Urdaneta et al., 2007; Farnworth and Mainville,2008; Chifiriuc et al., 2011). Kefir grains are white to yellowish-white, cauliflower-like “florets” composed of an inert poly-saccharide/protein matrix containing a stable and specific micro-bial community of different lactic acid bacteria (LAB), acetic acidbacteria (AAB) and yeast species in a complex symbiotic relation-ship (Farnworth, 2005; Magalhães et al., 2010, 2011). Kefir grainsare supposed to have developed spontaneously in milk stored incontainers made from animal skins, intestines or bladders. Grainsmay have arisen independently in different locations and may be
þ34 985 892233..
All rights reserved.
associated with specific microbial populations that producebeverages with distinct sensory properties (Rea et al., 1996; Miguelet al., 2010).
The microbial diversity of kefir has traditionally been assessedby culturing methods by which different LAB species have beenidentified. Awide variety of Lactobacillus species have been isolatedfrom both kefir beverages and grains, including Lactobacillus kefiri,Lactobacillus kefiranofaciens, Lactobacillus kefirgranum and Lacto-bacillus parakefiri, which constitute dominant populations (Reaet al., 1996; Kuo and Lin, 1999; Garrote et al., 2001; Simova et al.,2002). Lactococcus lactis subsp. lactis and Lactococcus lactis subsp.cremoris are also commonly reported; these bacteria are thought tobe loosely associated with the grains and responsible for acidifi-cation (Magalhães et al., 2011). Not surprisingly, Lc. lactis has beenidentified as dominant in the fermented product by both culturingand culture-independent techniques (Simova et al., 2002; Chenet al., 2008; Dobson et al., 2011). Leuconostoc and other Lactoba-cillus species have also been isolated in small numbers (Simovaet al., 2002; Mainville et al., 2006). AAB have received less atten-tion, although they are presumed to be essential in both themicrobial consortium and the organoleptic characteristics of the

A.M.O. Leite et al. / Food Microbiology 31 (2012) 215e221216
final product (Rea et al., 1996; Magalhães et al., 2011). Among theyeasts, Kluyveromyces marxianus, Torulaspora delbrueckii, Saccha-romyces cerevisiae, Candida kefir, Saccharomyces unisporus, Pichiafermentans, Kazachastania aerobia, Lachancea meyersii, Yarrowialipolytica and Kazachstania unispora have all been detected (Simovaet al., 2002; Wang et al., 2008; Magalhães et al., 2010, 2011).
Culturing methods have proven to be unreliable for thecomplete characterization of microbial ecosystems, including thoseof food fermentation (Giraffa and Neviani, 2001; Jany and Barbier,2008). Culture-independent microbial techniques, such as dena-turing gradient gel electrophoresis (DGGE) and the constructionand analysis of libraries of conserved genes such as the 16S rRNAgene are therefore now used, and indeed have already beenemployed to study the microbiology of kefir grains (Wang et al.,2006; Ninane et al., 2007; Chen et al., 2008). These techniqueshave detected all of the commonly cultured species, as well as somepreviously undetected microorganisms. However, in spite of thisextensive knowledge, the inventory of the microbial species asso-ciated with the kefir grains is thought to be far from complete.
Pyrosequencing, an automated high-throughput sequencingtechnique that involves the synthesis of single-stranded deoxy-ribonucleic acid and the detection of the light generated by pyro-phosphate released in a coupled reactionwith luciferase (Margulieset al., 2005), has recently made its debut in the study of foodfermentation (Humblot and Guyot, 2009; Roh et al., 2010; Junget al., 2011). This technique allows the rapid and accuratesequencing of nucleotide sequences that can then be used toanalyze the population structure, gene content and metabolicpotential of the microbial communities in an ecosystem. Pyrose-quencing has recently been used to study the diversity anddynamics of the bacterial populations of an Irish kefir grain and itscorresponding fermented product (Dobson et al., 2011).
The aim of the present work was to characterize the microbialdiversity of three different kefir grains collected in different regionsof Brazil. Several studies of Brazilian kefir have already beenundertaken (Magalhães et al., 2010, 2011; Miguel et al., 2010) butmost of these focused on the microbial composition of the kefirbeverage during fermentation (Magalhães et al., 2010, 2011). Thepresent work catalogues the microbial species identified in threekefir grains using two culture-independent microbial methods e
PCR-DGGE and barcode pyrosequencing e and compares theresults obtained with those reported in the literature.
2. Material and methods
2.1. Kefir grain samples
The three kefir grains used in this study were collected fromdifferent cities with similar climates and an average distance of400 km in southeastern Brazil (AR, Niterói, Rio de Janeiro; AV,Viçosa, Minas Gerais; and AD, Lavras, Minas Gerais). Grains AV andAD were kindly provided by researchers from Brazilian universities(Universidade Federal de Lavras e UFLA and Universidade Federalde Viçosa e UFV), meanwhile the third grain (AR) belonged toa family that traditionally cultivated the kefir grain in privatehousehold for self consumption. At the laboratory, grains wereactivated in sterile reconstituted skim milk (10% w/v) at 25 �C for24 h, filtered through a sieve to remove the clotted milk, and rinsedwith sterile water. This activation step was repeated three times.
2.2. Isolation of total microbial DNA
For microbial genomic DNA extraction, activated kefir grainswere first homogenized in 2% sodium citrate, and 2 ml of eachhomogenatewas centrifuged for 10min at 10,000 g. Total DNA from
the pellets was extracted and purified using the FastDNA Spin kit(QBIOgene, Carlsbad, CA, USA) according to the manufacturer’sinstructions. The DNA obtained was quantified using a Qubitflourometer apparatus (Invitrogen Detection Technologies, Eugene,OR, USA).
2.3. DGGE analysis of kefir grains
2.3.1. PCR amplification of 16S and 26S rDNA sequencesGenomic DNAwas used as a template in PCR amplifications of the
V3 region of the bacterial 16S rRNA gene, using the universal primersF357-GC (50eTACGGGAGGCAGCAGe30) and R518 (50eATTACCGCGGCTGCTGGe30), as reported by Muyzer et al. (1993). Group-specific primers for the detection of LAB were also used. Thesewere the pair Lac1 (50eAGCAGTAGGGAATCTTCCAe30) and Lac2-GC(50eGATTYCACCGCTACACATGe30) to detect members of the generaLactobacillus, Pediococcus, Leuconostoc and Weissella (Walter et al.,2001), and Lac3 (50eAGCAGTAGGGAATCTTCGGe30) and Lac2-GC todetect members of the genera Lactococcus, Streptococcus, Entero-coccus, Tetragenococcus and Vagococcus (Endo andOkada, 2005). TheD1 domain of the 26S rRNA gene of fungi was amplified using theprimers NL1-GC (50eGCCATATCAATAAGCGGAGGAAAGe30) and LS2(50eATTCCCAAACAACTCGACTCe30), as reported by Cocolin et al.(2002). All GC primers contained a 39 bp GC-clamp sequence attheir 50 end to prevent the complete denaturation of amplicons. PCRwas performed in 50 ml reaction volumes using a Taq-DNA poly-merase master mix (Ampliqon, Skovlunde, Denmark) withw100 ngof each DNA sample as a template and 0.2 mM of each primer.
2.3.2. Electrophoretic conditions and identification of bandsDGGE was performed by using a DCode apparatus (Bio-Rad,
Richmond, CA, USA) at 60 �C and employing 8% polyacrylamide gelswith a denaturing range of 40e60% for total bacteria, 40e50% forgroup-specific LAB and 30e50% for fungi. Electrophoresis wasperformed at 75 V for 16 h and 130 V for 4.5 h for bacterial andfungal amplifications respectively. Bands were visualized under UVlight after staining with ethidium bromide (0.5 mg ml�1) andphotographed.
Bands in the gels were identified by sequencing. Bands wereexcised from the gels and DNA eluted overnight in 50 ml of sterilewater at 4 �C. The DNAwas re-amplified with the same primer pairwithout the GC-clamp and sequenced by cycle extension in an ABI373 DNA sequencer (Applied Biosystems, Foster City, CA, USA). Theidentity of the sequences was determined by the BLASTN algorithmin the GenBank database (http://www.ncbi.nlm.nih.gov/BLAST/).
2.4. Pyrosequencing analysis of kefir grains
2.4.1. Primers and 16S rRNA gene amplification conditionsTwo universal primers, Y1 (50eTGGCTCAGGACGAACGCTGG
CGGCe30) (position 20e43 on 16S rRNA gene, Escherichia colinumbering) and Y2 (50eCCTACTGCTGCCTCCCGTAGGAGTe30)(positions 361e338) (Young et al., 1991), were used to PCR-amplifya 348-bp stretch of DNA embracing the V1 and V2 variable regionsof theprokaryotic 16S rDNA. 454-adaptorswere included in both theforward (50eCGTATCGCCTCCCTCGCGCCATCAGe30) and reverse(50eCTATGCGCCTTGCCAGCCCGCTCAGe30) primers, followed bya 10-bp sample-specific barcode sequence. Amplifications wereperformed as described above using the following PCR conditions:95 �C for 5min, 25 cycles of 94 �C for 30 s, 52 �C for 40 s and 72 �C for30 s, and a final extension step at 72 �C for 10 min.
Amplicons were purified through GenElute� PCR Clean-Upcolumns (SigmaeAldrich, St. Louis, MO, USA), and the DNAconcentration and quality was measured using an Epoch micro-volume spectrophotometer (BioTek Instruments, Winooski, VT,

Fig. 1. DGGE profiles of the microbial community from three Brazilian kefir grains(samples AR, AD and AV). Panel A: DGGE profile of the eubacterial 16S rRNA geneobtained with universal primers (1) Lactobacillus kefiranofaciens; (2) Lactobacilluskefiranofaciens; (3) Lactococcus lactis; (4) Lactobacillus kefiri; (5) Lactobacillus kefir-anofaciens. Panel B: DGGE profile of the eukaryotic domain D1 of the 26S rRNA gene (6)Saccharomyces cerevisiae; (7) Kazachstania unispora. Panel C: DGGE profile of the 16SrRNA gene obtained with specific primers for the lactobacilli group (8) Lactobacilluskefiranofaciens; (9) Lactobacillus kefiri. Panel D: DGGE profile of the 16S rRNA geneobtained with specific primers for the lactococcus group (10) Lactococcus lactis.
A.M.O. Leite et al. / Food Microbiology 31 (2012) 215e221 217
USA). Equal amounts of the three samples were pooled, for a totalamount of 100 ng. Pooled DNAwas then amplified in PCR-mixture-oil emulsions and sequenced in different lanes of a PicoTiterPlate ona 454 Genome Sequencer 20 system (Roche, Basel, Switzerland).The sequences obtained were uploaded at the NCBI SequenceRead Archive (SRA) under accession numbers SRA045648.2,SRR340042.2, SRR340043.1 and SRR340041.1.
2.4.2. Sequence treatment and bioinformatics analysisRaw sequenceswere processed through the Ribosomal Database
Project (RDP) pyrosequencing pipeline (http://wildpigeon.cme.msu.edu/pyro/index.jsp). Sequences were excluded from the anal-ysis if they were of low quality, if the read length was less than300 bp, or if one of the primer sequences was missing. The high-quality partial 16S rDNA sequences were submitted to the RDP-IIclassifier using an 80% confidence threshold to obtain the taxo-nomic assignment and the relative abundance of the differentbacterial groups (Wang et al., 2007). Multiple sequence alignmentsfor each sample were made using the Aligner tool (default settings)on the RDP website. These alignments served as inputs for con-structing the distance matrix and for clustering the sequences intooperational taxonomic units (OTUs) using MOTHUR v. 1.14.0 soft-ware (Schloss et al., 2009). The clusters were constructed at a 3%
Table 1Estimated OTU richness, sample coverage and diversity index of 16S rDNA libraries of ke
Library NS OTUsa Estimated OTU richness
ACE
AD 2641 18 42.24 (28.17; 75.78)AR 2690 14 38.57 (23.67; 76.40)AV 8983 46 148.02 (109.96; 208.74)Total 14,314
Abbreviations: ESC, estimated sample coverage; NS, number of sequences for each libraValues in brackets are 95% confidence intervals as calculated by MOTHUR.
a Calculated by MOTHUR at the 3% distance level.b Shannon diversity index calculated using MOTHUR (3% distance).c ESC: Cx ¼ 1 � (Nx/n), where Nx is the number of unique sequences and n is the tot
dissimilarity cut-off and served as OTUs for generating predictiverarefaction models, and for determining the ACE and Chao1 rich-ness (Chao and Bunge, 2002) and Shannon diversity indices(Shannon and Weaver, 1949). The MOTHUR programwas also usedto perform the Fast UniFrac test, which was employed to comparethe phylogenetic structure of the libraries and to generate the Venndiagrams. A neighbour-joining tree was constructed with repre-sentative sequences of each OTU selected by MOTHUR. Thesesequences were compared against the RDP database using theSeqmatch option to select for the nearest neighbours. All sequenceswere then aligned using MEGA 5.0 software (Tamura et al., 2011)and the JukeseCantor model. The equivalent sequence of thearchaea Halococcus saccharolyticus (AB004876) was used as anoutgroup to root the tree.
3. Results
3.1. PCR-DGGE analysis of bacterial and yeast communities
DGGE fingerprints of the microbial communities in the threeBrazilian kefir grains were rather simple, containing from one tofive different bands (Fig. 1, panels A through D). Most bands wereshared by all three samples. The species profile of the total bacteriaamplified with the universal primers involved up to five bandsreflecting three different species (Fig. 1, panel A). Bands corre-sponding to Lb. kefiranofaciens (bands 1, 2 and 5) and to Lb. kefiri(band 4) were found in all samples. An additional band present insample AV (band 3) was identified as Lc. lactis. The same threespecies were also found using the group-specific primers for lac-tobacilli and lactococci (Fig. 1, panels C and D, respectively). TheDGGE fingerprints of the yeast community were also simple andsimilar for the three grains. A high-intensity band was present in allsamples, andwas identified as S. cerevisiae (band 6, Fig.1 panel B). Alow-intensity band corresponding to K. unispora was revealed inkefir grain AD (band 7, Fig. 1 panel B).
3.2. Bacterial composition and community structure determined bypyrosequencing
A total of 14,314 high-quality partial 16S rDNA sequences, longerthan 300 bp, were obtained by pyrosequencing analysis, including2641 from sample AD, 2690 from sample AR and 8983 from sampleAV. Diversity richness, coverage, and evenness estimations calcu-lated for each data set are presented in Table 1. Rarefaction curvesshowed similar patterns for all samples (Fig. 2), and suggested thatthe bacterial community was well represented since they becameflatter while the number of sequences analyzed increased. Addi-tionally, when re-sampling analyses were performed, normalizingby sample size, the rarefaction curves proved to be saturated (Fig. 2panel B). Moreover, the coverage at the 97% similarity level wasabove 0.99 for each of the kefir grains. According to Fig. 2, and the
fir grain samples.
Shannonb ESCc
Chao1
54.00 (28.27; 144.19) 0.49 (0.45; 0.53) 0.9924.50 (16.03; 68.19) 0.33 (0.29; 0.37) 0.9982.14 (58.65; 149.23) 0.70 (0.67; 0.72) 0.99
ry; OTU, operational taxonomic unit.
al number of sequences.
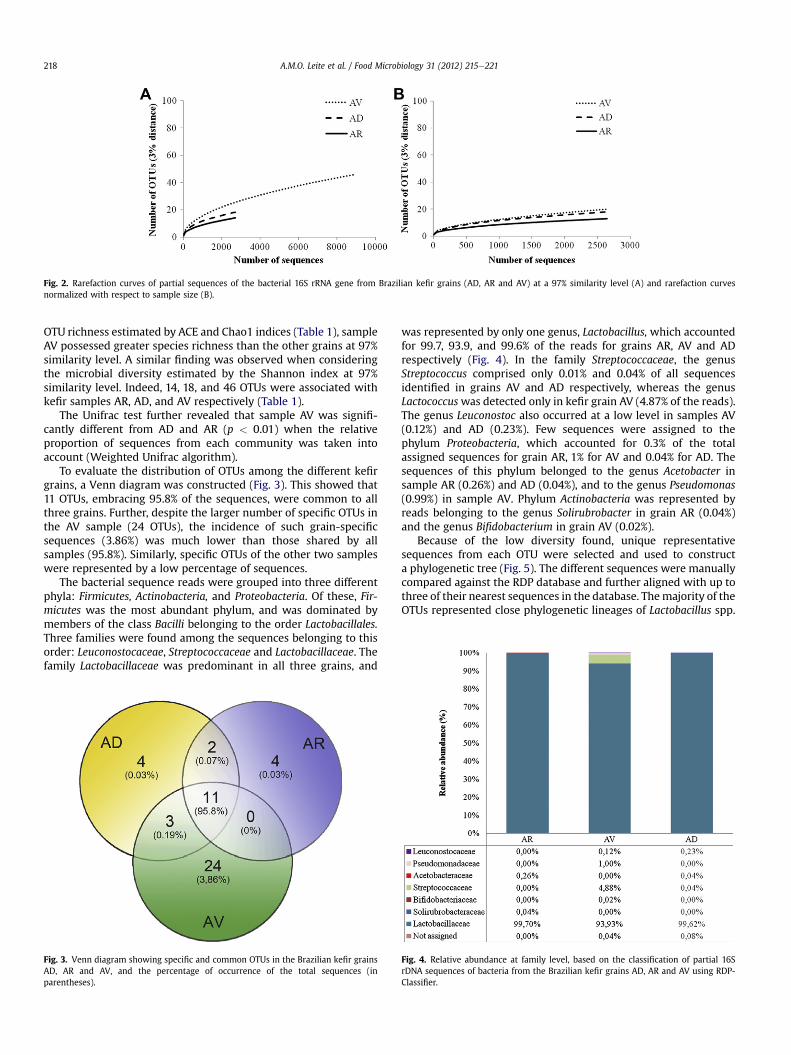
Fig. 2. Rarefaction curves of partial sequences of the bacterial 16S rRNA gene from Brazilian kefir grains (AD, AR and AV) at a 97% similarity level (A) and rarefaction curvesnormalized with respect to sample size (B).
A.M.O. Leite et al. / Food Microbiology 31 (2012) 215e221218
OTU richness estimated by ACE and Chao1 indices (Table 1), sampleAV possessed greater species richness than the other grains at 97%similarity level. A similar finding was observed when consideringthe microbial diversity estimated by the Shannon index at 97%similarity level. Indeed, 14, 18, and 46 OTUs were associated withkefir samples AR, AD, and AV respectively (Table 1).
The Unifrac test further revealed that sample AV was signifi-cantly different from AD and AR (p < 0.01) when the relativeproportion of sequences from each community was taken intoaccount (Weighted Unifrac algorithm).
To evaluate the distribution of OTUs among the different kefirgrains, a Venn diagram was constructed (Fig. 3). This showed that11 OTUs, embracing 95.8% of the sequences, were common to allthree grains. Further, despite the larger number of specific OTUs inthe AV sample (24 OTUs), the incidence of such grain-specificsequences (3.86%) was much lower than those shared by allsamples (95.8%). Similarly, specific OTUs of the other two sampleswere represented by a low percentage of sequences.
The bacterial sequence reads were grouped into three differentphyla: Firmicutes, Actinobacteria, and Proteobacteria. Of these, Fir-micutes was the most abundant phylum, and was dominated bymembers of the class Bacilli belonging to the order Lactobacillales.Three families were found among the sequences belonging to thisorder: Leuconostocaceae, Streptococcaceae and Lactobacillaceae. Thefamily Lactobacillaceae was predominant in all three grains, and
Fig. 3. Venn diagram showing specific and common OTUs in the Brazilian kefir grainsAD, AR and AV, and the percentage of occurrence of the total sequences (inparentheses).
was represented by only one genus, Lactobacillus, which accountedfor 99.7, 93.9, and 99.6% of the reads for grains AR, AV and ADrespectively (Fig. 4). In the family Streptococcaceae, the genusStreptococcus comprised only 0.01% and 0.04% of all sequencesidentified in grains AV and AD respectively, whereas the genusLactococcuswas detected only in kefir grain AV (4.87% of the reads).The genus Leuconostoc also occurred at a low level in samples AV(0.12%) and AD (0.23%). Few sequences were assigned to thephylum Proteobacteria, which accounted for 0.3% of the totalassigned sequences for grain AR, 1% for AV and 0.04% for AD. Thesequences of this phylum belonged to the genus Acetobacter insample AR (0.26%) and AD (0.04%), and to the genus Pseudomonas(0.99%) in sample AV. Phylum Actinobacteria was represented byreads belonging to the genus Solirubrobacter in grain AR (0.04%)and the genus Bifidobacterium in grain AV (0.02%).
Because of the low diversity found, unique representativesequences from each OTU were selected and used to constructa phylogenetic tree (Fig. 5). The different sequences were manuallycompared against the RDP database and further aligned with up tothree of their nearest sequences in the database. Themajority of theOTUs represented close phylogenetic lineages of Lactobacillus spp.
Fig. 4. Relative abundance at family level, based on the classification of partial 16SrDNA sequences of bacteria from the Brazilian kefir grains AD, AR and AV using RDP-Classifier.
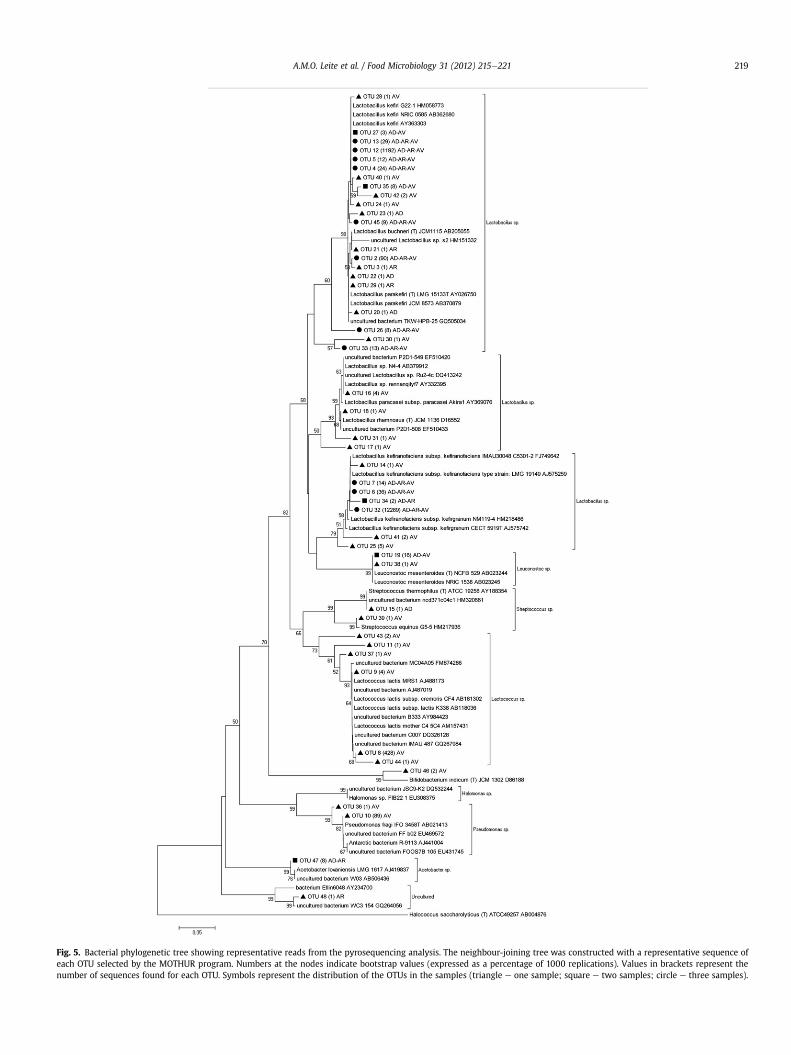
Fig. 5. Bacterial phylogenetic tree showing representative reads from the pyrosequencing analysis. The neighbour-joining tree was constructed with a representative sequence ofeach OTU selected by the MOTHUR program. Numbers at the nodes indicate bootstrap values (expressed as a percentage of 1000 replications). Values in brackets represent thenumber of sequences found for each OTU. Symbols represent the distribution of the OTUs in the samples (triangle e one sample; square e two samples; circle e three samples).
A.M.O. Leite et al. / Food Microbiology 31 (2012) 215e221 219

A.M.O. Leite et al. / Food Microbiology 31 (2012) 215e221220
commonly reported in kefir grains. These alignments and manualinvestigations further allowed the classification of the reads ina number of Lactobacillus species and subspecies, including amongothers Lb. kefiranofaciens subsp. kefirgranum, Lb. kefiranofacienssubsp. kefiranofaciens, Lb. kefiri, Lb. parakefiri, Lactobacillus para-buchneri, Lactobacillus amilovorus, Lactobacillus crispatus andLactobacillus buchneri. Sequences identified as Lc. lactis subsp. cre-moris were also revealed in kefir sample AV.
4. Discussion
The microbial diversity of kefir grains of different origins hasbeen analyzed using both culturing (Simova et al., 2002; Witthuhnet al., 2005; Mainville et al., 2006; Chen et al., 2008; Wang et al.,2008; Miguel et al., 2010) and culture-independent techniques(Garbers et al., 2004; Wang et al., 2006, 2008; Ninane et al., 2007;Chen et al., 2008; Miguel et al., 2010; Dobson et al., 2011). In thepresent study, two culture-independent techniques were used toevaluate the microbial diversity and community structure of threedifferent kefir grains from different locations in Brazil. The domi-nant populations were identified using PCR-DGGE, while the next-generation sequencing technology of pyrosequencing alloweda more complete view of the grain communities’ overallcomposition.
As in previous studies (Garbers et al., 2004; Chen et al., 2008;Jianzhong et al., 2009; Miguel et al., 2010; Magalhães et al.,2010), the bacterial PCR-DGGE profiles were shown to becomposed of a small number of bands. These corresponded toseveral Lactobacillus species that have always been reported asprevalent. Lb. kefiranofaciens (Chen et al., 2008; Jianzhong et al.,2009; Magalhães et al., 2010) and Lb. kefiri (Miguel et al., 2010)have been described as accounting for the more intense DGGEbands. These two bacterial species have also been reported asdominant by culturing in different kefir grains (Mainville et al.,2006; Chen et al., 2008; Miguel et al., 2010). The small numberof DGGE bands seen for the yeast profile has also been reported formany other kefir grains (Garbers et al., 2004; Wang et al., 2008;Jianzhong et al., 2009; Magalhães et al., 2010). The dominantyeasts found in these earlier investigations were Saccharomycesspp., Kluyveromyces lactis, Kazachtania spp. and Candida spp.(Garbers et al., 2004; Wang et al., 2008; Jianzhong et al., 2009).S. cerevisiae was the main yeast species detected in the presentwork. This and other related species have also been identified asmajority via culturing (Simova et al., 2002; Latorre-García et al.,2007).
Pyrosequencing is becoming the state-of-the-art technique forthe analysis of microbial populations from different ecosystems. Ithas been used to study several types of food fermentation(Humblot and Guyot, 2009; Roh et al., 2010; Jung et al., 2011).Indeed, one report exists in which a kefir grain and its fermentedmilk were analyzed by this technique (Dobson et al., 2011). Thepresent pyrosequencing analysis revealed the phylum Firmicutes tobe strongly dominant in the examined grains, accounting for morethan 99% of the sequences. This phylum is composed of a group oflow-GC-content Gram-positive bacteria, which includes LAB. Fir-micutes was also found dominant in the study of the Irish kefir, inwhich both the interior and exterior of the grain were analyzed(Dobson et al., 2011). These authors also showed that all other phyladetected (Actinobacteria, Proteobacteria and Bacteriodetes) wereminor components of the kefir community in the interior of thegrain. Within the phylum Proteobacteria, Pseudomonas spp. wasidentified in the grain AV, which has been suggested to be anenvironmental contamination (Dobson et al., 2011). The genusAcetobacter (Proteobacteria subgroup) was found in only two of theBrazilian grains (AR and AD). Although AAB have often been
mentioned as one of the main components of the bacterial pop-ulation of kefir grains (Rea et al., 1996; Garrote et al., 2001; Miguelet al., 2010), they have in fact only occasionally been detected(Garbers et al., 2004; Chen et al., 2008; Jianzhong et al., 2009;Miguel et al., 2010; Dobson et al., 2011).
In general, the two techniques used in this study were consis-tent with respect to the detection of the predominant bacteria.However, somemicroorganisms identified by pyrosequencing werenot detected by DGGE analysis, probably because they were part ofminority populations in the grains. This limitation of the PCR-DGGEmethodwas previously noted by Ercolini (2004), who reported thatminor bacterial groups in complex communities may not be rep-resented in the DGGE profiles. The present results show thatpyrosequencing allows the detection of microorganisms that arenot part of the dominant community such as bifidobacteria. Theseminor microorganisms may contribute to the particular sensorycharacteristics of the kefir beverage fermented by each grain (fiz-ziness, acidic taste, and refreshing flavour), via the production ofmetabolites such as organic acids, ethanol and aromaticcompounds.
Traditional culturing and molecular techniques have indicatedthat a few specific microbial genera and species may be constantlypresent in kefir grains, whereas others may or may not occur(Simova et al., 2002; Witthuhn et al., 2005; Mainville et al., 2006;Wang et al., 2006, 2008; Ninane et al., 2007; Chen et al., 2008;Miguel et al., 2010; Dobson et al., 2011). Further, as Farnworthand Mainville (2008) have noted, the list of bacteria and yeasts inkefir grains should not vary significantly from one part of the worldto another if good care, similar growth conditions, and propersanitary conditions are maintained. However, over time and underdifferent growing conditions, kefir grains may change theirmicrobial make up and fermentation properties. In the presentwork, even considering that the three kefir grains may have beengrown in different locations over the years, the dominant micro-biota was similar; only the minority communities varied. Thesesmall microbial differences may be associated with distinctivegrain-specific sensory profiles (Pintado et al., 1996; Rea et al., 1996;Simova et al., 2002).
5. Conclusions
Two culture-independent methods were used to evaluate themicrobial diversity of three Brazilian kefir grains: PCR-DGGE andpyrosequencing. Both techniques showed Lb. kefiranofaciens to bedominant, while DGGE showed S. cerevisiae to be the maineukaryotic microorganism. Pyrosequencing analysis also allowedthe identification of minor bacterial components. For the completedescription of the microbial communities of the kefir grains,pyrosequencing analysis using specific primers for eukaryotic andarchaea organisms should also be performed.
Acknowledgements
The study was supported by the CAPES Foundation (processnumber PDEE 5019109) and a project from the Spanish Ministry ofScience and Innovation (MICINN) (reference AGL2007-61869-ALI).S. Delgado was supported by a research contract from MICINNunder the Juan de la Cierva Program (reference JCI-2008-02391).
References
Chao, A., Bunge, J., 2002. Estimating the number of species in a stochastic abun-dance model. Biometrics 58, 531e539.
Chen, H.C., Wang, S.Y., Chen, M.-J., 2008. Microbiological study of lactic acid bacteriain kefir grains by culture-dependent and culture-independent methods. FoodMicrobiology 25, 492e501.

A.M.O. Leite et al. / Food Microbiology 31 (2012) 215e221 221
Chifiriuc, M.C., Cioaca, A.B., Lazar, V., 2011. In vitro assay of the antimicrobial activityof kephir against bacterial and fungal strains. Anaerobe 17, 433e435.
Cocolin, L., Aggio, D., Manzano, M., Cantoni, C., Comi, G., 2002. An application ofPCR-DGGE analysis to profile the yeast populations in raw milk. InternationalDairy Journal 12, 407e411.
Dobson, A., O’sullivan, O., Cotter, P.D., Ross, P., Hill, C., 2011. High-throughputsequence-based analysis of the bacterial composition of kefir and an associatedkefir grain. FEMS Microbiology Letters 320, 56e62.
Endo, A., Okada, S., 2005. Monitoring the lactic acid bacterial diversity duringshochu fermentation by PCR-denaturing gradient gel electrophoresis. Journal ofBioscience and Bioengineering 99, 216e221.
Ercolini, D., 2004. PCR-DGGE fingerprinting: novel strategies for detection ofmicrobes in food. Journal of Microbiological Methods 56, 297e314.
FAO/WHO, 2003. CODEX Standard for Fermented Milks. Codex Stan 243-2003.Reviewed 2010, second ed. URL: http://www.codexalimentarius.net/download/standards/400/CXS_243e.pdf.
Farnworth, E.R., 2005. Kefir e a complex probiotic. Food Science and TechnologyBulletin: Functional Foods 2, 1e17.
Farnworth, E.R., Mainville, I., 2008. Kefir e a fermented milk product. In: Farn-worth, E.R. (Ed.), Handbook of Fermented Functional Foods, second ed. CRCPress Taylor & Francis Group, Boca Raton, FL, pp. 89e127.
Garbers, I.-M., Britz, T.J., Witthuhn, R.C., 2004. PCR-based denaturing gradient gelelectrophoretic typification and identification of the microbial consortiumpresent in kefir grains. World Journal of Microbiology and Biotechnology 20,687e693.
Garrote, G.L., Abraham, A.G., de Antoni, G.L., 2001. Chemical and microbiologicalcharacterisation of kefir grains. Journal of Dairy Research 68, 639e652.
Giraffa, G., Neviani, E., 2001. DNA-based, culture-independent strategies for eval-uating microbial communities in food-associated ecosystems. InternationalJournal of Food Microbiology 67, 19e34.
Humblot, C., Guyot, J.P., 2009. Pyrosequencing of tagged 16S rRNA gene ampliconsfor rapid deciphering of the microbiomes of fermented foods such as pearlmillet slurries. Applied and Environmental Microbiology 75, 4354e4361.
Jany, J.L., Barbier, G., 2008. Culture-independent methods for identifying microbialcommunities in cheese. Food Microbiology 25, 839e848.
Jianzhong, Z., Xiaoli, L., Hanhu, J., Mingsheng, D., 2009. Analysis of the microflora inTibetan kefir grains using denaturing gradient gel electrophoresis. FoodMicrobiology 26, 770e775.
Jung, J.Y., Lee, S.H., Kim, J.M., Park, M.S., Bae, J.W., Hahn, Y., Madsen, E.L., Jeon, C.O.,2011. Metagenomic analysis of kimchi, a traditional Korean fermented food.Applied and Environmental Microbiology 77, 2264e2274.
Kuo, C.-Y., Lin, C.W., 1999. Taiwanese kefir grains: their growth, microbial andchemical composition of fermented milk. Australian Journal of Dairy Tech-nology 54, 19e23.
Latorre-García, L., Castillo-Agudo, L., Polaina, J., 2007. Taxonomical classification ofyeasts isolated from kefir based on the sequence of their ribosomal RNA genes.World Journal of Microbiology and Biotechnology 23, 785e791.
Mainville, I., Robert, N., Lee, B., Farnworth, E.R., 2006. Polyphasic characterization ofthe lactic acid bacteria in kefir. Systematic and Applied Microbiology 29, 59e68.
Magalhães, K.T., Pereira, M.A., Nicolau, A., Dragone, G., Domingues, L., Teixeira, J.A.,Silva, J.B.A., Schwan, R.F., 2010. Production of fermented cheese whey-basedbeverage using kefir grains as starter culture: evaluation of morphologicaland microbial variations. Bioresource Technology 101, 8843e8850.
Magalhães, K.T., Pereira, G.V.M., Campos, C.R., Dragone, G., Schwan, R.F., 2011.Brazilian kefir: structure, microbial communities and chemical composition.Brazilian Journal of Microbiology 42, 693e702.
Margulies, M., Egholm, M., Altman, W.E., Attiya, S., Bader, J.S., Bemben, L.A., Berka, J.,Braverman, M.S., Chen, Y.J., Chen, Z., Dewell, S.B., Du, L., Fierro, J.M., Gomes, X.V.,Godwin, B.C., He, W., Helgesen, S., Ho, C.H., Irzyk, G.P., Jando, S.C., Alenquer, M.L.,Jarvie, T.P., Jirage, K.B., Kim, J.B., Knight, J.R., Lanza, J.R., Leamon, J.H.,Lefkowitz, S.M., Lei, M., Li, J., Lohman, K.L., Lu, H., Makhijani, V.B., McDade, K.E.,
McKenna, M.P., Myers, E.W., Nickerson, E., Nobile, J.R., Plant, R., Puc, B.P.,Ronan, M.T., Roth, G.T., Sarkis, G.J., Simons, J.F., Simpson, J.W., Srinivasan, M.,Tartaro, K.R., Tomasz, A., Vogt, K.A., Volkmer, G.A., Wang, S.H., Wang, Y.,Weiner, M.P., Yu, P., Begley, R.F., Rothberg, J.M., 2005. Genome sequencing inmicrofabricated high-density picolitre reactors. Nature 437, 376e380.
Miguel, M.G.C.P., Cardoso, P.G., Lago, L.A., Schwan, R.F., 2010. Diversity of bacteriapresent in milk kefir grains using culture-dependent and culture-independentmethods. Food Research International 42, 1523e1528.
Muyzer, G., De Waal, E.C., Uitterlinden, A.G., 1993. Profiling of complex microbialpopulations by denaturing gradient gel electrophoresis analysis of polymerasechain reaction-amplified genes coding for 16S rRNA. Applied and Environ-mental Microbiology 59, 695e700.
Ninane, V., Mukandayambaje, R., Berben, G., 2007. Identification of lactic acidbacteria within the consortium of a kefir grain by sequencing of 16S rDNAvariable regions. Journal of AOAC International 90, 1111e1117.
Pintado, M.E., Lopes Da Silva, J.A., Fernandes, P.B., Malcata, F.X., Hogg, T.A., 1996.Microbiological and rheological studies on Portuguese kefir grains. Interna-tional Journal of Food Science and Technology 31, 15e26.
Rea, M.C., Lennartsson, T., Dillon, P., Drinan, F.D., Reville, W.J., Heapes, M.,Cogan, T.M., 1996. Irish kefir-like grains: their structure, microbial compositionand fermentation kinetics. Journal of Applied Bacteriology 81, 83e94.
Roh, S.W., Kim, K.H., Nam, Y.D., Chang, H.W., Park, E.J., Bae, J.W., 2010. Investigationof archaeal and bacterial diversity in fermented seafood using barcoded pyro-sequencing. The ISME Journal 4, 1e16.
Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R., Hartmann, M., Hollister, E.B.,Lesniewski, R.A., Oakley, B.B., Parks, D.H., Robinson, C.J., Sahl, J.W., Stres, B.,Thallinger, G.G., Van Horn, D.J., Weber, C.F., 2009. Introducing mothur: open-source, platform-independent, community-supported software for describingand comparing microbial communities. Applied and Environmental Microbi-ology 75, 7537e7541.
Shannon, C.E., Weaver, W., 1949. The mathematical theory of information. AT&TTechnical Journal 27, 359e423.
Simova, E., Beshkova, D., Angelov, A., Hristozova, T., Frengova, G., Spasov, Z., 2002.Lactic acid bacteria and yeasts in kefir grains and kefir made from them. Journalof Industrial Microbiology and Biotechnology 28, 1e6.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolu-tionary distance, and maximum parsimony methods. Molecular Biology andEvolution. doi:10.1093/molbev/msr121.
Urdaneta, E., Barrenetxe, J., Aranguren, P., Irigoyen, A., Marzo, F., Ibáñez, F.C., 2007.Intestinal beneficial effects of kefir-supplemented diet in rats. NutritionResearch 27, 653e658.
Walter, J., Hertel, C., Tannock, G.W., Lis, C.M., Munro, K., Hammes, W.P., 2001.Detection of Lactobacillus, Pediococcus, Leuconostoc, and Weissella species inhuman feces by using group-specific PCR primers and denaturing gradient gelelectrophoresis. Applied and Environmental Microbiology 67, 2578e2585.
Wang, Y.Y., Li, H.R., Jia, S.F., Wu, Z.F., Guo, B.H., 2006. Analysis of bacterial diversity ofkefir grains by denaturing gradient gel electrophoresis and 16S rDNAsequencing. Wei Sheng Wu Xue Bao 46, 310e313.
Wang, Q., Garrity, G.M., Tiedje, J.M., Cole, J.R., 2007. Naïve Bayesian classifier forrapid assignment of rRNA sequences into the new bacterial taxonomy. Appliedand Environmental Microbiology 73, 5261e5267.
Wang, S.Y., Chen, H.C., Liu, J.R., Lin, Y.C., Chen, M.J., 2008. Identification of yeasts andevaluation of their distribution in Taiwanese kefir and viili starters. Journal ofDairy Science 91, 3798e3805.
Witthuhn, R.C., Schoeman, T., Britz, T.J., 2005. Characterisation of the microbialpopulation at different stages of kefir production and kefir grain mass culti-vation. International Dairy Journal 15, 383e389.
Young, J.P.W., Downer, H.L., Eardly, B.D., 1991. Phylogeny of the phototrophicRhizobium strain BTAil by polymerase chain reaction-based sequencing of a 16SrRNA gene segment. Journal of Bacteriology 173, 2271e2277.

176
APÊNDICE C – ARTIGO III 1
2
Microbiological and chemical characteristics of Brazilian kefir during fermentation and 3
storage processes 4
Leite, A.M.O.1*
; Leite, D. C. A.2; Del Aguila, E.M.
1; Alvares, T.S.
1; Peixoto, R. S.
2; Miguel, M. 5
A. L.2; Silva, J. T.
1; Paschoalin, V.M.F.
1 6
1Departamento de Bioquímica, Instituto de Química, Universidade Federal do Rio de Janeiro, 7
Avenida Athos da Silveira Ramos, 149-Bloco A, Sala 545, 21941-909-Cidade Universitária Rio 8
de Janeiro, RJ, Brazil 9
2Departamento de Microbiologia, Instituto de Microbiologia Geral, Universidade Federal do Rio 10
de Janeiro, Avenida Carlos Chagas Filho, 373, 21941-904-Cidade Universitária, Rio de Janeiro, 11
RJ, Brazil. 12
13
14
Keywords: Brazilian kefir, PCR-DGGE, RT-PCR-DGGE, culture-dependent and –independent 15
methods, microbial community, HPLC. 16
Short title: Ecology of Brazilian kefir during fermentation and storage 17
*Correspondence to: Analy Machado de Oliveira Leite, Universidade Federal do Rio de Janeiro, 18
Instituto de Química, Departamento de Bioquímica, Avenida Athos da Silveira Ramos, 149- CT, 19
Bloco A, Sala 545. CEP: 21941-909-Cidade Universitária, Rio de Janeiro, RJ, Brazil. Phone: 20
+55 21 25627362. Fax number: +55 21 25627266 21
e-mail: [email protected] 22
23

177
Abstract 24
The microbial community composition and chemical characteristics of milk Brazilian kefir 25
produced during its manufacturing and refrigerated storage were investigated by culture-26
dependent and –independent methods and high performance liquid chromatography (HPLC). 27
Lactococcus lactis subsp. cremoris and subsp. lactis, Leuconostoc mesenteroides, Acetobacter 28
lovaniensis and Saccharomyces cerevisiae were isolated, whereas the detected DGGE bands 29
corresponded to Lactobacillus kefiranofaciens, L. kefiri, L. parakefiri and S. cerevisiae. After 30
fermentation, lactic acid bacteria (LAB) were present at levels of 10 log units, whereas acetic 31
acid bacteria (AAB) and yeast were present at levels of 7.8 and 6 log units, respectively. The 32
LAB and yeast counts remained constant, while AAB counts decreased to 7.2 log units during 33
storage. From fermentation to final storage, the pH, lactose content and citric acid of the kefir 34
beverage decreased, followed by an increase in the concentrations of glucose, galactose, ethanol 35
and lactic, acetic, butyric and propionic acids. 36
1 Introduction 37
Kefir is a traditional dairy beverage originally from Caucasian and Eastern Europe 38
regions, produced by the direct addition of kefir grains to milk. The bacteria and yeast that 39
compose kefir grains are enclosed in a polysaccharide and protein matrix. These microorganisms 40
are responsible for the acid-alcoholic fermentation of the milk, producing a fermented milk 41
product with unique properties. This product also has a long history of beneficial health effects, 42
which has contributed significantly to the increase in the consumption of kefir and interest in 43
this product in several Western countries (Farnworth & Mainville, 2008; Tamime, Wszolek, 44
Božanić, & Özer, 2011). 45
The complex symbiotic relationship that exists between the microorganisms that 46
compose the kefir grains makes them difficult to study, and also promotes a relative stability and 47
specificity of the kefir microbial community, which is generally composed of different lactic 48
acid bacteria (LAB), acetic acid bacteria (AAB) and yeast species (Farnworth, 2005). However, 49

178
some factors, such as the grain to milk ratio, incubation time and temperature, agitation and 50
storage conditions can affect the microbiological and chemical characteristics of the final kefir 51
product (Farnworth, 2005; Garrote, Abraham, & De Antoni, 2001; Irigoyen, Arana, Castiella, 52
Torre, & Ibáñez, 2005; Öner, Karahan, & Çakmakçi, 2010). 53
After milk fermentation with grains or commercial starter cultures, lactic acid, acetic 54
acid, CO2, ethanol and aromatic compounds are formed. Kefir’s exotic aroma and flavor, a 55
refreshing feature and slightly acidic taste are the result of the coexistence of yeast and lactic 56
acid bacteria in a symbiotic association, and depends on the diversity of the microbiota of each 57
kefir grain (Güzel-Seydim, Seydim, Greene, & Bodine, 2000b; Leite, et al., 2012; Magalhães, 58
Pereira, Campos, Dragone, & Schwan, 2011; Öner, et al., 2010). 59
Although previous information exists regarding the microbiological diversity of kefir 60
grains from different origins (Chen, Wang, & Chen, 2008; Garrote, et al., 2001; Jianzhong, Liu, 61
Jiang, & Dong, 2009; Miguel, Cardoso, Lago, & Schwan, 2010; Pintado, Da Silva, Fernandes, 62
Malcata, & Hogg, 1996) and of the fermented milk (final product) produced by these grains 63
(Magalhães, Pereira, Nicolau, et al., 2010; Mainville, Robert, Lee, & Farnworth, 2006; Simova, 64
et al., 2002) using dependent- and independent-culture methods, few studies have been 65
published with regard to microorganism enumeration and microbial characterization of the 66
different microorganism groups present during milk fermentation by the traditional kefir grain 67
(Guzel-Seydim, Wyffels, Seydim, & Greene, 2005; Magalhães, Pereira, et al., 2011) and during 68
the kefir beverage storage (Irigoyen, et al., 2005; Öner, et al., 2010). 69
Traditional methods based on cultivation are not able to detect viable but not cultivable 70
bacteria, which can considerably underestimate microbial diversity. Although culturing methods 71
have the advantage of not detecting dead microorganisms (Cenciarini, Courtois, Raoult, & La 72
Scola, 2008), they may not be sufficiently selective, depending on the complexity of the studied 73
ecosystem and, furthermore, are labor intensive and time-consuming for monitoring population 74
dynamics (Cocolin, et al., 2004; Randazzo, Torriani, Akkermans, de Vos, & Vaughan, 2002). 75

179
Molecular approaches, especially those based on the use of 16S rRNA gene sequences, 76
have provided new insights into the ecology of complex communities (Jany & Barbier, 2008; 77
Randazzo, et al., 2002). Today, the analysis of PCR products generated with universal primers 78
by using of denaturant gradient gel electrophoresis (DGGE) is one of the most widely used 79
techniques to study microbial communities in dairy products. This technique allows for the 80
separation of PCR amplicons of the same size with different sequences and, thus, has the 81
potential to provide information concerning variations in target genes in a microbial population 82
(Jany & Barbier, 2008). 83
Although, molecular methods have been routinely applied to directly detect DNA in 84
microbial ecosystems, the direct targeting of 16S rRNA transcripts may be a strategy to provide 85
information about the activity of the microbial community (Dolci, Alessandria, Rantsiou, 86
Bertolino, & Cocolin, 2010; Falentin, et al., 2010; Randazzo, et al., 2002). 87
The commercial product “kefir” is not available in Brazil and, therefore, kefir grains are 88
traditionally used as starters in the production of homemade kefir for personal consumption. 89
Recently, some reports concerning the microbial and chemical characterization of Brazilian kefir 90
milk beverages have been described, although only during the fermentation process (Magalhães, 91
Dias, et al., 2011; Magalhães, Pereira, et al., 2011), not evaluating the product during storage. 92
The aim of this study was to evaluate the microbiological and chemical changes that occur not 93
only in the different fermentation stages in kefir manufacturing but also during storage. For this 94
purpose, samples were analyzed by traditional plating, isolation of microorganisms and direct 95
DNA and RNA extraction, followed by amplification (PCR and RT-PCR) of bacterial 16S 96
rRNA and yeast 26S rRNA genes and DGGE analysis. Organic acid, ethanol production, and 97
carbohydrate concentration at these different stages were also determined. Additionally, strains 98
of LAB, AAB and yeast were isolated and subsequently identified by DNA sequencing. 99
2 Material and Methods 100
2.1 Kefir grain 101

180
Brazilian kefir grains were obtained from a private household in Niterói city, which is 102
located in the state of Rio de Janeiro, Brazil. Grains were activated in sterile reconstituted 103
skimmed milk (10% w/v) at 25 °C for 24 h, filtered to remove the clotted milk and rinsed with 104
sterile water. The activation step was repeated three times (Chen, et al., 2008). 105
2.2 Milk kefir production 106
The activated grains (30 g) were inoculated in commercial ultra-high-temperature (UHT) 107
skimmed milk (1000 mL) and statically incubated at 25°C for 24 h. Samples (1 mL) of the 108
beverage were aseptically taken every 6 h. Triplicate fermentations were performed. 109
2.3 Microbiological analyses 110
Microorganism counting was carried out by the surface spreading technique of 111
appropriate sodium citrate (2%) dilutions (0.1 mL aliquots) of the fermented milk, in different 112
culture media, in triplicate. Lactic acid bacteria (LAB) were enumerated on Lactobacilli deMan, 113
Rogosa and Sharpe (MRS) agar (Difco - BD Diagnostics, Sparks, MD, USA) and on M17 agar 114
(Difco - BD Diagnostics, Sparks, MD, USA) supplemented with 0.5% glucose (Difco - BD 115
Diagnostics, Sparks, MD, USA). Acetic acid bacteria (AAB) were enumerated on AAB medium 116
(Carr & Passmore, 1979). All media for bacterial enumeration were supplemented with 200 ppm 117
cycloheximide (Sigma-Aldrich, St. Louis, MO, USA) to inhibit yeast and fungi growth. Yeast 118
were enumerated on yeast extract-glucose-chloramphenicol agar (YGC) (Difco - BD 119
Diagnostics, Sparks, MD, USA). Plates were incubated at 30 °C for 5 days for bacteria growth, 120
under anaerobic conditions using a Gaspak EZ Anaerobic System (Difco) and for yeast growth, 121
under aerobic conditions at 25 °C for 5 days. After counting, means and standard deviations 122
were calculated. 123
In order to confirm the identity of the enumerated colonies, some isolates were taken at 124
random from the agar plates for identification. Gram staining and catalase testing were used to 125
screen the isolate strains as belonging to the LAB and AAB group and the samples were then 126

181
analyzed by molecular methods. All isolated strains were stored in MRS broth with 20% 127
glycerol at -80°C, until subsequent analyses. 128
2.4 Amplified ribosomal DNA restriction analysis (ARDRA) 129
DNA template preparation from the isolated pure bacteria or yeast cultures was 130
performed by the methods described by Randazzo, et al. (2002) and Del Aguila, Silva, and 131
Paschoalin (2003), respectively. 132
PCR primers 27f (5’–AGAGTTTGATCCTGGCTCAG–3’) and 1512r (5’– 133
CGGCTACCTTGTTACGACT – 3’) were used to amplify the 16S rRNA gene sequence, while 134
primers ITS1 (5’- TCCGTAGGTGAACCTGCGG-3’) and ITS4 (5’-135
TCCTCCGCTTATTGATATGC-3’) were used to amplify internal transcribed spacer (ITS) 136
regions, as described by Wang, et al. (2006) and Naumova, Ivannikova, and Naumov (2005) for 137
bacteria and fungi, respectively. PCR products were then separated by electrophoresis on 1% 138
agarose gels. Amplicon bands were extracted and purified using the Illustra GFX PCR DNA and 139
Gel Band Purification Kit (GE Healthcare Life Sciences,Uppsala, Sweden), according to the 140
manufacturer’s instructions. Ten microliters of purified amplicons were digested with the 141
following restriction enzymes: FastDigest® HaeIII and HhaI for LAB, FastDigest® AluI and 142
ScrF1 for AAB and FastDigest® HhaI and DdeI for yeast as recommended by the manufacturer 143
(Fermentas Inc., Glen Burnie, MD, USA). Electrophoresis runs were then conducted in agarose 144
gels (1.5 %) for 1 hour and 45 min at 75 V. Gels were stained with GelRed (Biotium, Hayward, 145
CA) diluted at 1:10,000 and visualized under UV light (data not shown). 146
Representative profiles of each species observed in the ARDRA analysis were selected 147
for sequencing. The sequencing analysis of partial 16S rRNA gene and ITS region was 148
accomplished with a 3130 sequencer (Applied Biosystems, Carlsbad, CA, USA) and 149
subsequently used for identification to the species level of the bacteria and yeast isolates. The 150
identity of the sequences was determined by using the BLASTN algorithm at the GenBank 151
database (GenBank ID: BA123456). 152

182
2.5 DNA and RNA extraction of kefir beverage and kefir grains 153
Kefir samples collected during fermentation and storage were stored at -80°C. Grain 154
samples (1 g) used as inoculums were also collected and homogenized in 2% sodium citrate. 1 155
mL of each homogenate and 1 mL of kefir samples were then centrifuged for 10 min at 10,000 156
g. 157
Genomic DNA was extracted and purified from the pellets using the FastDNA Spin kit 158
(QBIOgene, Carlsbad, CA, USA) according to the manufacturer’s instructions. 159
Total RNA was extracted from the pellets in screw-cap tubes containing 0.3 g of glass 160
beads and 1 mL of TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Subsequently, two 40-161
second treatments at the maximum speed, with an interval of 1 min in an ice bath, were 162
performed in a FastPrep® Instrument (Bio 101, Vista, CA, USA.). Tubes were then centrifuged 163
at 12,000 g at 4°C for 10 min, and two hundred microliters of chloroform were added to 164
supernatant. The tubes were centrifuged again at 12,000 g at 4°C for 10 min and the aqueous 165
phase was precipitated with 1 mL of ice-cold absolute ethanol. 166
RNA was obtained after centrifugation at 14,000 g at 4 °C for 10 min, where the pellets 167
were washed with 75% ethanol and dried at room temperature. Fifty microliters of sterile water 168
were added, and a 30-min period at 45 °C was used to facilitate nucleic acid solubilization. One 169
microliter of DNase, Rnase-free (Roche Diagnostics, Indianapolis, IN, USA) was added to 170
digest the DNA, at 37 °C for 1 h. The RNA solution was checked for the presence of residual 171
amounts of DNA by performing PCR amplification. When positive signals were detected, the 172
DNase treatment was repeated until the remaining DNA was eliminated. 173
2.6 PCR and RT-PCR protocol 174
Bacterial DNA was used as template for amplifications of the V3 region of the 16S 175
rRNA gene using the universal primers F338-GC (5’–TACGGGAGGCAGCAG–3’ and R518 176
(5’–ATTACCGCGGCTGCTGG–3’), as reported by Muyzer, de Waal and Uitterlinden (1993). 177
The amplification of the D1 domain of the 26S rRNA gene of fungi was accomplished by using 178

183
the primers NL1-GC (5’-GCCATATCAATAAGCGGAGGAAAG–3’) and LS2 (5’–179
ATTCCCAAACAACTCGACTC–3’), as reported by Cocolin, Aggio, Manzano, Cantoni and 180
Comi (2002). All GC primers contained a 39 bp GC clamp sequence at their 5’ end, to prevent 181
the complete denaturation of amplicons. 182
RT-PCR was performed using the kit Ready-To-Go RT-PCR beads (GE Healthcare Life 183
Sciences, Uppsala, Sweden), in which one microgram of total RNA was mixed with 25 pmol of 184
each primer (338f-518r for bacteria and NL1-LS2 for yeasts), and the volume was brought up to 185
50 µL with RNase-free (Invitrogen, Carlsbad, CA, USA) sterile water. The reaction mixtures 186
were incubated at 42 °C for 20 min, and after a 95 °C incubation for 10 min, the PCR conditions 187
described by Muyzer, et al. (1993) and Cocolin, et al. (2002) were carried out for bacteria and 188
yeast, respectively. 189
2.7 DGGE analysis of kefir 190
DGGE was performed using a DCode apparatus (Bio-Rad, Hercules, CA, USA) at 60 °C 191
and employing 8 % polyacrylamide gels with a denaturing range of 30-55 % for bacteria and 30-192
50 % for fungi in 1X Tris-acetate-EDTA buffer at a constant voltage of 50V for 10 min, 193
followed by 150V for 4 h and 30 min. After electrophoresis, the gels were stained with SYBR 194
Green nucleic acid gel stain (1:10.000 dilution; Invitrogen, Carlsbad, CA, USA) for 60 min and 195
the DGGE band patterns were visualized under UV with the MiniBisPro (BioAmerica Inc., 196
Miami, FL, USA). 197
In addition, bands from the PCR-DGGE and RT-PCR-DGGE gels were identified by 198
nucleotide sequence analyses. For this purpose, the bands were excised from the acrylamide gels 199
and processed by the Qiaex II DNA extraction kit (Qiagen, Hilden, Germany) according to the 200
manufacturer’s recommendations. The eluted DNA of each band was re-amplified with the same 201
primer pair without the GC-clamp. Agarose gel electrophoresis of the PCR products was 202
performed prior to purification and DNA was purified using the Illustra GFX PCR DNA and 203
GEL Band Purification Kit (GE Healthcare Life Sciences, Uppsala, Sweden) according to the 204

184
manufacturer’s instructions. Purified amplicons were directly ligated into the pGEM-T Easy 205
Vector (Promega, Madison, WI, USA) following the manufacturer’s instructions. The ligation 206
products were transformed into DH5-α Escherichia coli competent cell. Positive clones (white 207
colonies) were grown in LB agar containing ampicilin (100 µg mL-1
), IPTG (isopropyl-β-D-1-208
thiogalactopyranoside; 0.5 mM) and X-gal (5-bromo-4-chloro-3-indolyl-β-D-galacto-209
pyranoside; 80 µg mL-1
). Plasmids were extracted using a miniprep alkaline lyses method (Not, 210
del Campo, Balagué, de Vargas, & Massana, 2009). Sequencing of the insert was performed 211
using a Big Dye Terminator system and an ABI 3730 automatic capillary sequencer (Applied 212
Biosystems, Carlsbad, CA, USA). The identity of sequences was determined by using the 213
BLASTN algorithm at the GenBank database (GenBank ID: BA123456). 214
2.8 HPLC analysis 215
Extraction of organic acids, ethanol and carbohydrates from the kefir beverage was 216
carried out as described by González de Llano, Rodriguez and Cuesta (1996). Briefly, 25 mL of 217
H2SO4 (45 mmol L-1
) were added to 5 mL of the kefir beverage and homogenized for 1 h in a 218
rotatory shaker at 250 rpm. The supernatant fluid was then separated by centrifugation at 6,000 g 219
and filtered through 0.45 µm filters (Millipore, Billerica, MA, USA). 220
Filtered samples were injected (50 µL) in triplicate into a high performance liquid 221
chromatography (HPLC) system (Shimadzu) equipped with an HPX-87H Aminex fermentation 222
monitoring column (150 X 7.8 mm, I.D.) and protected by a cation H+ microguard cartridge (30 223
X 4.6 mm, I.D.)(Bio-Rad Laboratories INC., Hercules, CA, USA) maintained at 65 °C. Organic 224
acids (lactic, acetic, citric, butyric and propionic acids) were quantified by using a diode array 225
detector model SPD-M20A (Shimadzu, Tokyo, Japan) monitoring the absorbance at 210 nm. 226
Carbohydrates and ethanol were quantified by using a refractive index detector model RID-10A 227
(Shimadzu, Tokyo, Japan). The mobile phase (isocratic flow rate at 0.7 mL min -1
) used was 3 228
mM H2SO4. HPLC chromatograms and compound quantification were obtained using the LC 229
Solution software (Shimadzu, Tokyo, Japan). Standard curves based on peak area were 230

185
calculated for the individual organic acids, carbohydrates and ethanol, covering a broad range of 231
concentrations, by the comparison with standard solutions. Standards (Sigma-Aldrich, St. Louis, 232
MO, USA) were prepared in deionized water filtered through 0.45 µm filters (Millipore, 233
Billerica, MA, USA). 234
2.9 Statistical analyses 235
A one-way analysis of variance with repeated measures was used to identify differences 236
between microbial enumerations, organic acids, carbohydrates and ethanol content over the kefir 237
beverage’s fermentation and storage periods. When a significant F was found, additional post 238
hoc tests with Tukey’s adjustment were performed. The statistical significance was set at a 0.05 239
level of confidence. All analyses were performed using a commercially available statistical 240
package (Statsoft, Inc. Version 7.0 Tulsa, Oklahoma). 241
DGGE patterns were normalized, compared and clustered using the Bionumerics 6.5 242
software (Applied Maths, Sint-Martens-Latem, Belgium). Bands were automatically detected 243
and matched, and further corrections were applied manually. Dendrograms relating band pattern 244
similarities were automatically calculated with the Dice coefficient and UPGMA (unweighted 245
pair group method with arithmetic mean). 246
3 Results 247
3.1 Microbial enumeration during kefir beverage fermentation and storage 248
Conventional microorganism enumeration was carried out during the 24 h milk 249
fermentation (3 % kefir grain) at 25 °C. Counts of presumptive LAB groups (M17 agar) 250
increased 4 log units at 12 h of fermentation (P < 0.05), whereas counts on MRS agar at this 251
same sampling point showed an increase of approximately 2 log units, reaching maximum 252
values (10 log units) within 18 h of fermentation. Presumptive AAB counts also increased 253
significantly at 12 h, reaching maximum values (7.8 log units) at 24 h. Yeast counts increased 254
until 12 h, remaining constant at around 6 log units over the fermentation period. 255

186
Microorganisms were also enumerated during kefir storage at 4 °C for 672 h (28 days) 256
(Fig 1). During this period, yeast (approximately 6 log units) and LAB group counts 257
(approximately 10 log units) remained constant until the end of the storage period (P > 0.05), 258
while the count of presumptive AAB decreased to 7.2 log units at 672 h of the storage period. 259
3.2 Identification of microbial isolates 260
In order to evaluate the diversity of the microbial isolates from the different growth 261
media, distinct colonies were randomly selected at various stages within the fermentation and 262
storage periods. Ninety-two isolates were randomly selected from agar plates (M17, MRS, AAB 263
and YGC) and grouped into representative profiles obtained after ARDRA analysis. LAB strains 264
showed three different representative profiles, while AAB and yeast strains showed only one. 265
These isolates were identified by the sequence analysis of partial 16S rRNA gene and ITS 266
region, as the following: Leuconostoc mesenteroides (29%), Lactococcus lactis subsp. lactis 267
(5%), Lactococcus lactis subsp. cremoris (45%), Acetobacter lovaniensis (10%), Saccharomyces 268
cerevisiae (11 %), based on homology identity (98-100 %) searches at the GenBank database 269
(GenBank ID: BA123456). 270
3.3 Microbial community structure 271
The microbial communities of Brazilian kefir were evaluated by DGGE using primer 272
pairs 338-518 and NL1-LS2 for the amplification of bacterial and yeast DNA and RNA 273
sequences. Selected bands were excised (Fig 2 and Fig 3) and sequenced after reamplification, 274
followed by cloning and identification by sequence alignment according to the GenBank 275
database (GenBank ID: BA123456) (Table 1). 276
The cluster analysis for bacteria and yeast profiles allowed the identification of two main 277
clusters, corresponding to the initial stages of fermentation and storage, with a coefficient of 278
similarity between them of approximately 69% and 60% for bacteria and yeast, respectively. 279
DNA bacterial profiles from samples taken during the initial periods of fermentation (0-280
18 h) were grouped in one cluster (Fig 2 – A.1) with a coefficient of similarity of 75 %, whereas 281

187
the second cluster (90 % similarity) was comprised of samples taken during the final 282
fermentation period (24 h) and those analyzed during 672 h storage at 4 °C. The UHT skimmed 283
milk and grain kefir samples were not grouped into these main clusters and showed a coefficient 284
of similarity of 46% and 59%, respectively. 285
The fingerprints of the yeast community (Fig 2 - A.2), at the initial fermentation periods 286
(from 0 to 12 h) were shown to be identical and grouped in the same cluster as kefir grains, 287
sharing a coefficient of similarity of 80 %. The second main cluster was comprised of kefir 288
samples taken during the storage period (from 48 to 672 h) and at the end of the fermentation 289
period (24 h), exhibiting a coefficient of similarity of 72 %. The UHT skimmed milk showed 290
only 32 % of similarity with these two main clusters. 291
The DGGE profiles were similar when comparing DNA and RNA amplicons (Fig 2 and 292
3), and, consequently, the microorganisms identified after cloning and sequencing of the DGGE 293
bands were also similar (Table 1). However, RNA band intensities were weaker than the ones 294
obtained from DNA amplification. Concerning the RNA DGGE bacterial profiles (Fig 3), 295
Lactobacillus kefiranofaciens was the predominant microorganism in both stages, fermentation 296
and storage, and was identified by sequencing of bands i, j, k and o (Fig 3A). L. parakefiri (Fig. 297
3A, band n) was identified in both the grain and kefir beverage during fermentation and storage, 298
whereas L. kefiri (Fig 3A, bands l and m) was detected in the kefir grain, but not in the beverage. 299
Moreover, the yeast profile (Fig 3B) was much simpler than the bacterial DGGE 300
fingerprints, since the yeast community presented only two bands, identified as Saccharomyces 301
cerevisiae (Fig 3B, bands p and q). The UHT milk sample showed no amplification at the RNA 302
level targeting 16S and 26S rRNA genes. 303
3.4 HPLC analysis of metabolites during kefir beverage fermentation and storage 304
When tracking the changes in carbohydrates, ethanol and organic acids during 305
fermentation and storage (Table 2), it was demonstrated that lactose content decreased after 12 h 306
and remained low until 24 h fermentation (from 43.4 to 32.4 mg mL-1
), while during the storage 307

188
period there was only a significant reduction at 28 days, corresponding to 41 % of the original 308
milk content. Galactose and glucose levels showed a significant (P < 0.05) increase at 12 h 309
fermentation, from 0.18 to 0.29 mg mL-1
and 0.46 to 0.70 mg mL-1
, respectively. After 310
fermentation for 24 hours until 14 days of storage, there was an increase in galactose content 311
(from 0.76 to 0.97 mg mL-1
), while glucose content remained constant throughout the 312
fermentation period, increasing at 2 days of storage (from 0.29 mg mL-1
to 0.36 mg mL-1
). 313
During the fermentation and storage periods, pH values (Table 2) progressively 314
decreased (P < 0.05), ranging from 6.55 to 4.31, as a consequence of organic acid production. 315
Lactic acid increased rapidly and significantly (P < 0.05) over the 24 h fermentation period, to 316
7.38 mg mL-1
and increasing to 9.54 mg mL-1
at the final storage period (28 days). Acetic acid 317
was also produced at 12 h of the kefir fermentation process, reaching maximum values of 0.93 318
and 1.16 mg mL-1
after 24 h fermentation and storage periods, respectively (Table 2). 319
Ethanol (Table 2) was produced after 18 h of fermentation (0.14 mg mL-1
) until 24 h, 320
when levels reached 0.32 mg mL -1
. During storage, ethanol concentrations increased to 0.45 mg 321
mL-1
, reaching maximum values (1.36 mg mL-1
) at the 28th
day of storage. 322
The citrate (Table 2) present in the original milk decreased by more than 35 % after 12 h 323
of fermentation (from 4.29 to 2.73 mg mL-1
), remaining constant until the end of the storage 324
period, whereas propionic and butyric acids increased at 6 h and 12 h of fermentation, 325
respectively. A significant increase of the production of butyric acid during kefir fermentation 326
(24 h) and storage (2, 7 and 28 days) was observed, while propionic acid increased at 12 h of 327
fermentation (from 0.80 to 1.09 mg mL-1
), remaining constant throughout the whole storage 328
period, except for a significant decrease (to 0.96 mg mL-1
, P < 0.05) at the 28th
storage-day 329
(Table 2). 330
4 Discussion 331
Monitoring changes in the microbial and chemical composition of milk kefir during its 332
manufacture and storage is required so that the product may be fully understood according to a 333

189
scientific point of view. A polyphasic approach, using culture-dependent and -independent 334
methods, was performed in order to investigate the dynamics of the microbial populations from 335
Brazilian kefir beverage. 336
In general, the results of traditional plating are in accordance with previous studies 337
(García Fontán, Martínez, Franco, & Carballo, 2006; Guzel-Seydim, et al., 2005; Magalhães, 338
Pereira, et al., 2011), which reported a great increase in the microbial count during the first 24 h 339
of kefir production. However, in the present study, the values at 24 h of fermentation were not 340
similar to those published by other authors, most likely due to the kefir production conditions 341
(García Fontán, et al., 2006; Guzel-Seydim, et al., 2005; Magalhães, Pereira, et al., 2011) and 342
the particular behavior of each grain. In the storage period, the results described in the present 343
study corroborated those reported by Öner, et al. (2010) for kefir produced from cow milk, 344
where the microorganism counts remained stable during the first 15 days of storage. However, 345
Irigoyen, et al. (2005) and Grønnevik, Falstad, and Narvhus (2011) reported a significant 346
decrease in the counts of presumptive lactobacilli and lactococci throughout 4 weeks of storage, 347
whereas yeast and AAB counts remained constant in the findings described by Irigoyen et al. 348
(2005). On the other hand, Grønnevik, et al. (2011), showed a significant increase in yeast 349
counts from Norwegian commercial kefir after 3 weeks of storage. 350
The microorganisms enumerated in the Brazilian kefir beverage are in agreement with 351
the specifications of FAO/WHO (2003), that recommend that kefir beverages should contain up 352
107
colony forming unit/g (cfu g-1
) and 104
cfu g-1
as minimum total bacteria and yeast counts, 353
respectively, at the end of the storage period. 354
The analyses of bacteria and yeast communities by DGGE fingerprints showed no 355
significant changes in the community structures during kefir fermentation or storage, probably 356
due the existence of a dominant microbial consortium, corroborating the data reported by 357
Magalhães, Pereira, Nicolau, et al. (2010). For this reason, the RT-PCR-DGGE analysis was 358

190
performed on certain sampling points, in order to evaluate the most predominant 359
microorganisms in the active population at different stages of kefir fermentation and storage. 360
Several studies have performed analyses on reverse-transcribed RNA to reveal 361
population viability, within the microbial community (Dolci, et al., 2010; Falentin, et al., 2012; 362
Randazzo, et al., 2002; Rantsiou, Urso, Dolci, Comi, & Cocolin, 2008), and some authors 363
(Cocolin, et al., 2004; Randazzo, et al., 2002) have already been able to differentiate the active 364
components (rRNA-derived) from the total diversity (rDNA-derived) of the community, by 365
combining RT-PCR-DGGE and PCR-DGGE. However, other studies on populations dynamics 366
based on the quantification of different targets suggest that other genes may be better markers of 367
cell viability than rRNA (Cenciarini, et al., 2008; Falentin, et al., 2010). 368
In this study, the RT-PCR-DGGE analysis demonstrated that all microorganisms were 369
active during fermentation and storage, with the exception of L. kefiri, only present in the kefir 370
grain. It seems that L. kefiri was present but not active in the kefir beverage, participating in the 371
composition of the grain with the likely function of balancing the microflora and ecology of the 372
grains. Meanwhile, L. kefiranofaciens was shown to be the most predominant and active 373
microorganism in the kefir beverage, a result also reported by Magalhães, Pereira, Nicolau, et al. 374
(2010). 375
L. kefiranofaciens, L. parakefiri and S. cerevisiae were shown to be active 376
microorganisms in Brazilian kefir beverage. Additionaly, L. lactis subsp. lactis, L. lactis subsp. 377
cremoris, L. mesenteroides, A. lovaniensis and S. cerevisiae also make up the cutivable 378
microbiota. Several researchers have studied the composition of the microbiota in kefir grain, by 379
culture-dependent (Garrote, et al., 2001; Guzel-Seydim, et al., 2005; Miguel, et al., 2010; 380
Simova, et al., 2002; Witthuhn, Schoeman, & Britz, 2005) and -independent techniques (Chen, 381
et al., 2008; Jianzhong, et al., 2009; Kesmen & Kacmaz, 2011; Leite, et al., 2012; Magalhães, 382
Pereira, Nicolau, et al., 2010; Miguel, et al., 2010), but little is known about microbiota of kefir 383
beverage. Magalhães, Pereira, et al. (2011) isolated a variety of Lactobacillus species in 384

191
appropriate culture media, as well as A. lovaniensis and non-lactose- and lactose-fermenting 385
yeast during the fermentation of Brazilian milk kefir. On the other hand, the microorganisms 386
found in this study by the PCR-DGGE analysis were similar to those reported by Magalhães, 387
Pereira, Nicolau, et al. (2010) in Brazilian kefir grain and kefir manufactured with different 388
dairy substrates by independent-culture methods. 389
Comparing the results obtained in this study by both culture-dependent and –independent 390
methods, we observed correlations for the yeast community, whereas no correlation was found 391
for the bacterial community. With regard to the yeast community, the plate counts and 392
identification of isolated strains correlated well with the PCR-DGGE and RT-PCR-DGGE 393
profiles. Predominant S. cerevisiae bands were obtained in both DGGE fingerprints and isolated 394
by the culturing method throughout the analyzed period. RT-PCR-DGGE profiles confirm the 395
viability and predominance of this non-lactose-fermenting yeast in the community during the 396
studied kefir beverage fermentation and storage. 397
On the other hand, no specific DGGE signals for Lactococcus spp., Leuconostoc spp. and 398
AAB were found at the DNA or RNA levels, while there was a presumptive count for these 399
microorganism groups and, in fact, they were isolated during culture. L. kefiranofaciens was the 400
dominant organism in the kefir samples by PCR-DGGE analysis, but was not isolated on the 401
selective media. This most likely occurred due to the relatively small number of strains 402
evaluated (92 total), which could not reveal an overview of the community and/or the culture 403
media used in the study was not suitable, since perhaps the lack of supplements and nutrients 404
necessary for the growth of these species in the culture media may have influenced these results 405
(Kesmen & Kacmaz, 2011). 406
These results have already been noted by other authors (Chen, et al., 2008; Flórez & 407
Mayo, 2006; Kesmen & Kacmaz, 2011; Randazzo, et al., 2002) using polyphasic approaches in 408
studies regarding food microbial ecology. Some possibilities include, perhaps, the fact that the 409
cell numbers of some LAB species were lower than the detection limit of PCR-DGGE (Cocolin, 410

192
et al., 2004); or species from more abundant populations in the mixture might give greater 411
amounts of template DNA that compete for primers during the amplification reaction, resulting 412
in the formation of PCR products only for the dominant species (Jany & Barbier, 2008; 413
Prakitchaiwattana, Fleet, & Heard, 2004). 414
Moreover, we also observed that some media were not selective. For example, 415
Leuconostoc mesenteroides was isolated on M17 and MRS media and Lactococcus spp. was 416
isolated on MRS agar. This non-specificity of culture media has already been discussed (Chen, 417
et al., 2008; Randazzo, et al., 2002; Witthuhn, et al., 2005). Thus, some authors have used 418
strategies to adjust the media or conditions for cultivation in order to favor the growth of these 419
strains. Without this, their presence would be overlooked. Therefore, cultivation methods may 420
over- or underestimate the microbial diversity, not always being representative of this complex 421
ecosystem (Chen, et al., 2008; Randazzo, et al., 2002; Witthuhn, et al., 2005). 422
Thus, the application of a polyphasic approach, as performed in the present study, using 423
culture-dependent and -independent methods, may be worthwhile in order to obtain a more 424
accurate view of the structure of the microbial community (Chen, et al., 2008; Flórez & Mayo, 425
2006; Kesmen & Kacmaz, 2011; Rantsiou, et al., 2008). 426
Considering the chemical composition of the kefir beverage, the lactose reduction was 427
expected, because lactose is readily hydrolyzed to glucose and galactose by Group N 428
streptococci, Lactobacillus and by some strains of Kluyveromces (Grønnevik, et al., 2011; 429
Güzel-Seydim, et al., 2000b; Magalhães, Dias, et al., 2011). The most frequently isolated 430
bacteria from kefir in previous reports, and also found in the present study, were both 431
homofermentative and heterofermentative bacteria, such as Lactobacillus, Lactococcus, 432
Leuconostoc and Acetobacter (Grønnevik, et al., 2011; Güzel-Seydim, et al., 2000b; Magalhães, 433
Pereira, et al., 2011; Rea, et al., 1996). Both pathways are employed by these bacteria and the 434
fermentative process is characterized by the accumulation of organic acids and drops in pH 435
values. In the homofermentative pathway, glucose is further metabolized by the Embden-436

193
Meyerhoff-Parnas (EMP) pathway to pyruvate and 2 mol of lactic acid are formed per glucose 437
molecule. However, the heterofermentative pathway yields, theoretically, 1 mol each of acid 438
lactic, ethanol and CO2 (Güzel-Seydim, et al., 2000b). The produced organic acids contribute not 439
only to the flavor and aroma of fermented dairy products but also to their preservation (Güzel-440
Seydim, et al., 2000b; Magalhães, Dias, et al., 2011). 441
Acetic acid is an intermediary in citrate metabolism. It can be produced by the oxidation 442
of ethanol and it is another end product of the heterofermentative lactose metabolism 443
(Grønnevik, et al., 2011; Paramithiotis, Gioulatos, Tsakalidou, & Kalantzopoulos, 2006), formed 444
probably by heterofermentative lactic acid and acetic acid bacteria. Citrate was also reduced in 445
the studied kefir beverage, probably due to citrate metabolism by LAB, since this is considered 446
the preferred substrate for acetic acid, acetoin and diacetyl formation by some lactic acid 447
bacteria (Güzel-Seydim et al., 2000). González de Llano, et al. (1996) also found butyric and 448
propionic acids when evaluating the activity of Lactococcus spp. and Leuconostoc spp. cultures 449
in milk fermentation and attributed the increase of the butyric and propionic acids to unspecific 450
esterase activities by lactococcal strains. 451
Ethanol production increased during the fermentation and storage processes, and the 452
values found were lower than those quantified previously in Brazilian kefir by HPLC 453
(Magalhães, Dias, et al., 2011; Magalhães, Pereira, et al., 2011). Ethanol production is typically 454
derived from yeast metabolism, such as S. cerevisiae, identified in this study. However, 455
heterofermentative bacteria such as Lactobacillus kefiri and Leuconostoc spp. are also capable of 456
producing ethanol (Güzel-Seydim, et al., 2000b; Keenan, 1968; Magalhães, Dias, et al., 2011), 457
since they show alcohol-dehydrogenase activity, which is an enzyme capable of converting 458
acetaldehyde to ethanol. Part of the ethanol content may also be converted to acetic acid by 459
heterofermentative bacteria, such as bacteria belonging to the genus Acetobacter (Magalhães, 460
Pereira, Dias, & Schwan, 2010). The end products of yeast fermentation, ethanol and CO2, are 461

194
critical in producing the exotic refreshing flavor and yeasty aroma of authentic kefir (Güzel-462
Seydim, Seydim, Greene, & Bodine, 2000a; Güzel-Seydim, et al., 2000b). 463
Differences between the organic acid contents found in the present study and other 464
reports involving kefir fermentation and storage (Grønnevik, et al., 2011; Güzel-Seydim, et al., 465
2000a, 2000b; Magalhães, Pereira, Nicolau, et al., 2010; Magalhães, Pereira, et al., 2011) might 466
be due to the variations in the ratio of inoculated kefir grains and the microorganism species 467
present in the different kefir grains (Güzel-Seydim, et al., 2000b; Irigoyen, et al., 2005). 468
5 Conclusions 469
Culture-dependent and-independent techniques were combined to obtain a more 470
complete view of the microbial ecosystem present in kefir. The identification of microorganisms 471
involved in the fermentation and present during the storage period of Brazilian kefir, as well as 472
their generated metabolites exert a strong influence in the unique characteristics of the end 473
product. This may serve as a basis for the future industrial production of this beverage in Brazil. 474
Acknowledgements 475
The authors acknowledge financial support for this project from FAPERJ, CNPq and 476
CAPES. 477
References 478
479
Del Aguila, E. M., Silva, J. T., & Paschoalin, V. M. F. (2003). Expression of the yeast 480
calcineurin subunits CNA1 and CNA2 during growth and hyper-osmotic stress. FEMS 481
Microbiology Letters, 221, 197-202. 482
Carr, J. G., & Passmore, S. M. (1979). Methods for identifying acetic acid bacteria. In F. A. 483
Shinner & D. W. Lovelock (Eds.), Indentification methods for microbiologist. London: 484
Academic Press. 485
Cenciarini, C., Courtois, S., Raoult, D., & La Scola, B. (2008). Influence of Long Time Storage 486
in Mineral Water on RNA Stability of Pseudomonas aeruginosa and Escherichia coli 487
after Heat Inactivation. PLoS ONE, 3, e3443. 488
Chen, H.-C., Wang, S.-Y., & Chen, M.-J. (2008). Microbiological study of lactic acid bacteria in 489
kefir grains by culture-dependent and culture-independent methods. Food Microbiology, 490
25, 492-501. 491
Cocolin, L., Aggio, D., Manzano, M., Cantoni, C., & Comi, G. (2002). An application of PCR-492
DGGE analysis to profile the yeast populations in raw milk. International Dairy Journal, 493
12, 407-411. 494

195
Cocolin, L., Rantsiou, K., Iacumin, L., Urso, R., Cantoni, C., & Comi, G. (2004). Study of the 495
Ecology of Fresh Sausages and Characterization of Populations of Lactic Acid Bacteria 496
by Molecular Methods. Applied and Environmental Microbiology, 70, 1883-1894. 497
Dolci, P., Alessandria, V., Rantsiou, K., Bertolino, M., & Cocolin, L. (2010). Microbial 498
diversity, dynamics and activity throughout manufacturing and ripening of Castelmagno 499
PDO cheese. International Journal of Food Microbiology, 143, 71-75. 500
Falentin, H., Postollec, F., Parayre, S., Henaff, N., Le Bivic, P., Richoux, R., Thierry, A., & 501
Sohier, D. (2010). Specific metabolic activity of ripening bacteria quantified by real-time 502
reverse transcription PCR throughout Emmental cheese manufacture. International 503
Journal of Food Microbiology, 144, 10-19. 504
Falentin, H., Henaff, N., Le Bivic, P., Deutsch, S. M., Parayre, S., Richoux, R., Sohier, D., 505
Thierry, A., Lortal, S., & Postollec, F. (2012). Reverse transcription quantitative PCR 506
revealed persistency of thermophilic lactic acid bacteria metabolic activity until the end 507
of the ripening of Emmental cheese. Food Microbiology, 29, 132-140. 508
FAO/WHO. (2003). CODEX Standard for Fermented Milks. In C. Stan (Ed.), 243-2003 (2nd 509
ed., pp. 1-11). 510
Farnworth, E. R. (2005). Kefir - a complex probiotic. Food Science and Technology Bulletin: 511
Functional Foods, 2, 1-17. 512
Farnworth, E. R., & Mainville, I. (2008). Kefir - A Fermented Milk Product. In E. R. Farnworth 513
(Ed.), Handbook of Fermented Functional Foods (2 ed., pp. 89-127 ). Boca Raton, 514
London, New York: CRC Press Taylor & Francis Group 515
Flórez, A. B., & Mayo, B. (2006). PCR–DGGE as a tool for characterizing dominant microbial 516
populations in the Spanish blue-veined Cabrales cheese. International Dairy Journal, 16, 517
1205-1210. 518
García Fontán, M. C., Martínez, S., Franco, I., & Carballo, J. (2006). Microbiological and 519
chemical changes during the manufacture of Kefir made from cows’ milk, using a 520
commercial starter culture. International Dairy Journal, 16, 762-767. 521
Garrote, G. L., Abraham, A. G., & De Antoni, G. L. (2001). Chemical and microbiological 522
characterisation of kefir grains. Journal of Dairy Research, 68, 639-652. 523
González de Llano, D., Rodriguez, A., & Cuesta, P. (1996). Effect of lactic starter cultures on 524
the organic acid composition of milk and cheese during ripening—analysis by HPLC. 525
Journal of Applied Microbiology, 80, 570-276. 526
Grønnevik, H., Falstad, M., & Narvhus, J. A. (2011). Microbiological and chemical properties of 527
Norwegian kefir during storage. International Dairy Journal, 21, 601-606. 528
Guzel-Seydim, Z., Wyffels, J. T., Seydim, A. C., & Greene, A. K. (2005). Turkish kefir and 529
kefir grains: microbial enumeration and electron microscopic observation†. International 530
Journal of Dairy Technology, 58, 25-29. 531
Güzel-Seydim, Z. B., Seydim, A. C., Greene, A. K., & Bodine, A. B. (2000a). Determination of 532
Organic Acids and Volatile Flavor Substances in Kefir during Fermentation. Journal of 533
Food Composition and Analysis, 13, 35-43. 534
Güzel-Seydim, Z. B., Seydim, A. C., Greene, A. K., & Bodine, A. B. (2000b). Determination of 535
Organic Acids and Volatile Flavor Substances in Kefir during Fermentation. Journal of 536
Food Composition and Analysis, 13, 35-43. 537
Irigoyen, A., Arana, I., Castiella, M., Torre, P., & Ibáñez, F. C. (2005). Microbiological, 538
physicochemical, and sensory characteristics of kefir during storage. Food Chemistry, 539
90, 613-620. 540
Jany, J. L., & Barbier, G. (2008). Culture-independent methods for identifying microbial 541
communities in cheese. Food Microbiology, 25, 839-848. 542
Jianzhong, Z., Liu, X., Jiang, H., & Dong, M. (2009). Analysis of the microflora in Tibetan kefir 543
grains using denaturing gradient gel electrophoresis. Food Microbiology, 26, 770-775. 544

196
Keenan, T. W. (1968). Production of acetic acid and other volatile compounds by Leuconostoc 545
citrovorum and Leuconostoc dextranicum. Applied Microbiology, 16, 1881-1885. 546
Kesmen, Z., & Kacmaz, N. (2011). Determination of Lactic Microflora of Kefir Grains and 547
Kefir Beverage by Using Culture-Dependent and Culture-Independent Methods. Journal 548
of Food Science, 76, M276-M283. 549
Leite, A. M. O., Mayo, B., Rachid, C. T. C. C., Peixoto, R. S., Silva, J. T., Paschoalin, V. M. F., 550
& Delgado, S. (2012). Assessment of the microbial diversity of Brazilian kefir grains by 551
PCR-DGGE and pyrosequencing analysis. Food Microbiology, 31, 215-221. 552
Magalhães, K. T., Pereira, G. V. M., Dias, D. R., & Schwan, R. F. (2010). Microbial 553
communities and chemical changes during fermentation of sugary Brazilian kefir. World 554
Journal of Microbiology and Biotechnology, 26, 1241-1250. 555
Magalhães, K. T., Pereira, M. A., Nicolau, A., Dragone, G., Domingues, L., Teixeira, J. A., 556
Silva, J. B. A., & Schwan, R. F. (2010). Production of fermented cheese whey-based 557
beverage using kefir grains as starter culture: Evaluation of morphological and microbial 558
variations. Bioresource Technology, 101, 8843-8850. 559
Magalhães, K. T., Dias, D. R., Pereira, G. V. M., Oliveira, J. M., Domingues, L., Teixeira, J. A., 560
Silva, J. B. A., & Schwan, R. F. (2011). Chemical composition and sensory analysis of 561
cheese whey-based beverages using kefir grains as starter culture. International Journal 562
of Food Science & Technology, 46, 871-878. 563
Magalhães, K. T., Pereira, G. V. M., Campos, C. R., Dragone, G., & Schwan, R. F. (2011). 564
Brazilian Kefir: Structure, Microbial Communities and Chemical Composition. Brazilian 565
Journal of Microbiology, 42, 693-702. 566
Mainville, I., Robert, N., Lee, B., & Farnworth, E. R. (2006). Polyphasic characterization of the 567
lactic acid bacteria in kefir. Systematic and Applied Microbiology, 29, 59-68. 568
Miguel, M. G. d. C. P., Cardoso, P. G., Lago, L. d. A., & Schwan, R. F. (2010). Diversity of 569
bacteria present in milk kefir grains using culture-dependent and culture-independent 570
methods. Food Research International, 43, 1523-1528. 571
Muyzer, G., de Waal, E. C., & Uitterlinden, A. G. (1993). Profiling of complex microbial 572
populations by denaturing gradient gel electrophoresis analysis of polymerase chain 573
reaction-amplified genes coding for 16S rRNA. Applied and Environmental 574
Microbiology, 59, 695-700. 575
Naumova, E., Ivannikova, Y., & Naumov, G. (2005). Genetic Differentiation of the Sherry 576
Yeasts Saccharomyces cerevisiae. Applied Biochemistry and Microbiology, 41, 578-582. 577
Not, F., del Campo, J., Balagué, V., de Vargas, C., & Massana, R. (2009). New Insights into the 578
Diversity of Marine Picoeukaryotes. PLoS ONE, 4, e7143. 579
Öner, Z., Karahan, A. G., & Çakmakçi, M. L. E. (2010). Effects of different milk types and 580
starter cultures on kefir. Gida, 35, 177-182. 581
Paramithiotis, S., Gioulatos, S., Tsakalidou, E., & Kalantzopoulos, G. (2006). Interactions 582
between Saccharomyces cerevisiae and lactic acid bacteria in sourdough. Process 583
Biochemistry, 41, 2429-2433. 584
Pintado, M. E., Da Silva, J. A. L., Fernandes, P. B., Malcata, F. X., & Hogg, T. A. (1996). 585
Microbiological and rheological studies on Portuguese kefir grains. International 586
Journal of Food Science & Technology, 31, 15-26. 587
Prakitchaiwattana, C. J., Fleet, G. H., & Heard, G. M. (2004). Application and evaluation of 588
denaturing gradient gel electrophoresis to analyse the yeast ecology of wine grapes. 589
FEMS Yeast Research, 4, 865-877. 590
Randazzo, C. L., Torriani, S., Akkermans, A. D. L., de Vos, W. M., & Vaughan, E. E. (2002). 591
Diversity, Dynamics, and Activity of Bacterial Communities during Production of an 592
Artisanal Sicilian Cheese as Evaluated by 16S rRNA Analysis. Applied and 593
Environmental Microbiology, 68, 1882-1892. 594

197
Rantsiou, K., Urso, R., Dolci, P., Comi, G., & Cocolin, L. (2008). Microflora of Feta cheese 595
from four Greek manufacturers. International Journal of Food Microbiology, 126, 36-596
42. 597
Rea, M. C., Lennartsson, T., Dillon, P., Drina, F. D., Reville, W. J., Heapes, M., & Cogan, T. M. 598
(1996). Irish kefir-like grains: their structure, microbial composition and fermentation 599
kinetics. Journal of Applied Bacteriology, 81, 83-94. 600
Simova, E., Beshkova, D., Angelov, A., Hristozova, T., Frengova, G., & Spasov, Z. (2002). 601
Lactic acid bacteria and yeasts in kefir grains and kefir made from them. Journal of 602
Industrial Microbiology & Biotechnology, 28, 1-6. 603
Tamime, A. Y., Wszolek, M., Božanić, R., & Özer, B. (2011). Popular ovine and caprine 604
fermented milks. Small Ruminant Research, 101, 2-16. 605
Wang, X., Haruta, S., Wang, P., Ishii, M., Igarashi, Y., & Cui, Z. (2006). Diversity of a stable 606
enrichment culture which is useful for silage inoculant and its succession in alfalfa 607
silage. FEMS Microbiology Ecology, 57, 106-115. 608
Witthuhn, R. C., Schoeman, T., & Britz, T. J. (2005). Characterization of the microbial 609
population at different stages of Kefir production and Kefir grain mass cultivation. International 610
Dairy Journal, 15, 383-389. 611
612
613
Legends 614 Figure 1 – Microbial enumeration (log10 cfu ml
-1) of presumptive LAB (M17 and MRS), 615
presumptive acid acetic bacteria (AAB) and yeast (YGC) in Brazilian kefir beverage over the 616
fermentation and storage periods (time 0 = after grain inoculation). Data are the means of three 617
of kefir beverage production; error bars show standard deviation. (*) indicates significant 618
difference (P < 0.05) 619
Figure 2 – PCR-DGGE profiles of the microbial community from milk, kefir grain and kefir 620
samples taken during Brazilian kefir manufacture and storage periods. Panel A: Dendrograms 621
generated by UPGMA clustering (Dice correlation coefficient) from bacteria DGGE (A.1) and 622
yeast DGGE (A.2) profiles. The scale bar indicates the percentage of similarity. Panel B: DGGE 623
profiles of the PCR products obtained from V3 region of the 16S rRNA gene for bacteria (B.1) 624
and D1 region of the 26S rRNA gene for yeasts (B.2). The identification of microorganisms was 625
reported in Table 1. 626
Figure 3 – PCR-DGGE (DNA) and RT-PCR-DGGE (RNA) profiles targeting bacteria 16S 627
rRNA gene (A) and yeast 26S rRNA gene (B) of kefir grain inoculum and samples of kefir 628
beverage taken during fermentation and storage. The identification of microorganisms was 629
reported in Table 1. 630
631
632
633
634
635
636
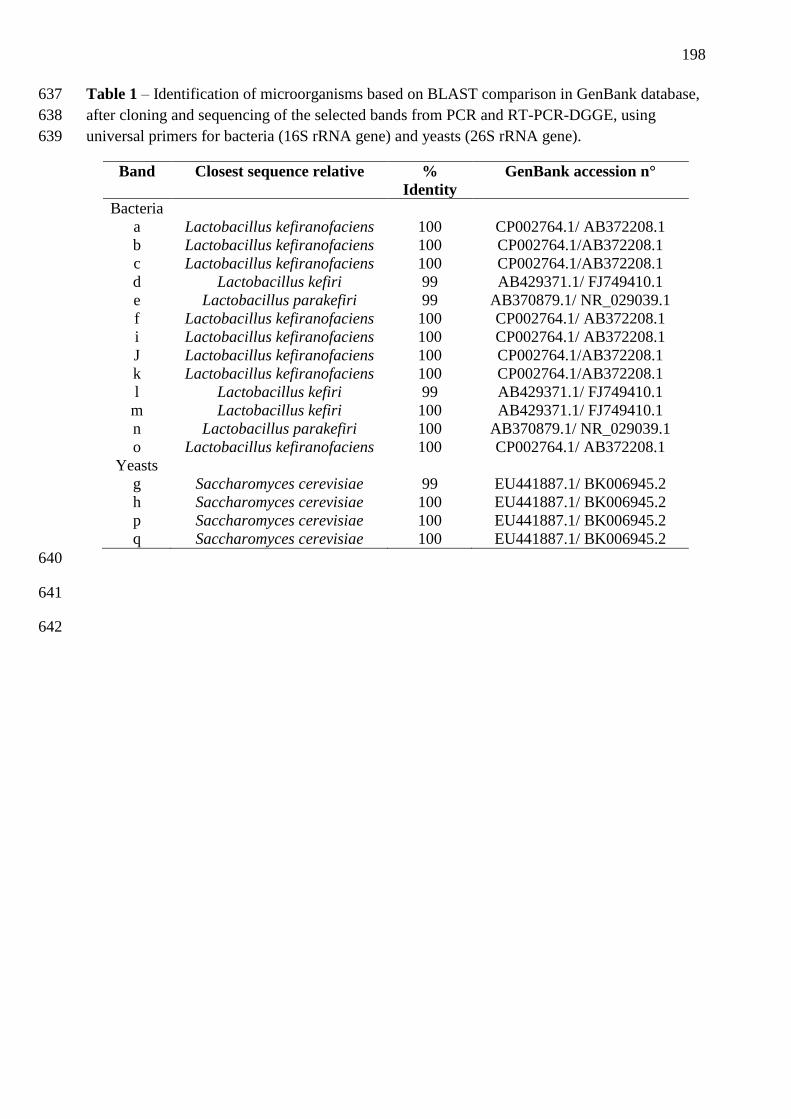
198
Table 1 – Identification of microorganisms based on BLAST comparison in GenBank database, 637
after cloning and sequencing of the selected bands from PCR and RT-PCR-DGGE, using 638
universal primers for bacteria (16S rRNA gene) and yeasts (26S rRNA gene). 639
Band Closest sequence relative %
Identity
GenBank accession n°
Bacteria
a Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
b Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
c Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
d Lactobacillus kefiri 99 AB429371.1/ FJ749410.1
e Lactobacillus parakefiri 99 AB370879.1/ NR_029039.1
f Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
i Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
J Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
k Lactobacillus kefiranofaciens 100 CP002764.1/AB372208.1
l Lactobacillus kefiri 99 AB429371.1/ FJ749410.1
m Lactobacillus kefiri 100 AB429371.1/ FJ749410.1
n Lactobacillus parakefiri 100 AB370879.1/ NR_029039.1
o Lactobacillus kefiranofaciens 100 CP002764.1/ AB372208.1
Yeasts
g Saccharomyces cerevisiae 99 EU441887.1/ BK006945.2
h Saccharomyces cerevisiae 100 EU441887.1/ BK006945.2
p Saccharomyces cerevisiae 100 EU441887.1/ BK006945.2
q Saccharomyces cerevisiae 100 EU441887.1/ BK006945.2
640
641
642
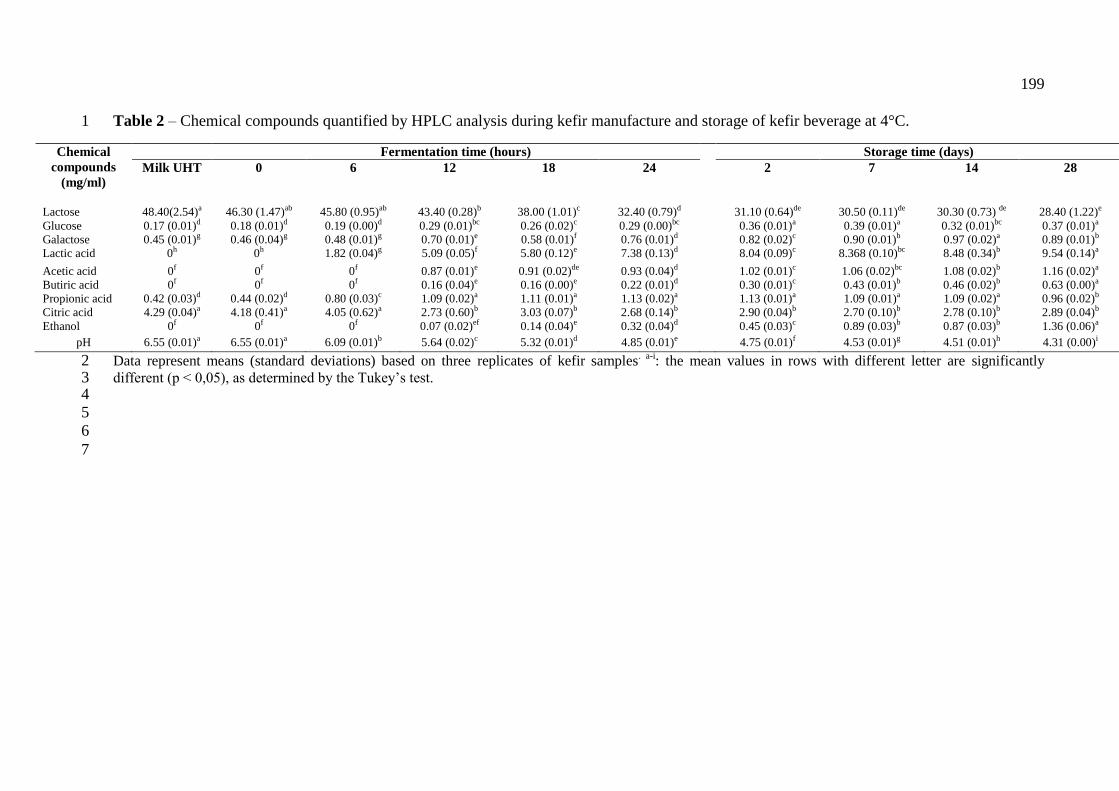
199
Table 2 – Chemical compounds quantified by HPLC analysis during kefir manufacture and storage of kefir beverage at 4°C. 1
Chemical
compounds
(mg/ml)
Fermentation time (hours) Storage time (days)
Milk UHT 0 6 12 18 24 2 7 14 28
Lactose 48.40(2.54)a 46.30 (1.47)ab 45.80 (0.95)ab 43.40 (0.28)b 38.00 (1.01)c 32.40 (0.79)d 31.10 (0.64)de 30.50 (0.11)de 30.30 (0.73) de 28.40 (1.22)e
Glucose 0.17 (0.01)d 0.18 (0.01)d 0.19 (0.00)d 0.29 (0.01)bc 0.26 (0.02)c 0.29 (0.00)bc 0.36 (0.01)a 0.39 (0.01)a 0.32 (0.01)bc 0.37 (0.01)a
Galactose 0.45 (0.01)g 0.46 (0.04)g 0.48 (0.01)g 0.70 (0.01)e 0.58 (0.01)f 0.76 (0.01)d 0.82 (0.02)c 0.90 (0.01)b 0.97 (0.02)a 0.89 (0.01)b
Lactic acid 0h 0h 1.82 (0.04)g 5.09 (0.05)f 5.80 (0.12)e 7.38 (0.13)d 8.04 (0.09)c 8.368 (0.10)bc 8.48 (0.34)b 9.54 (0.14)a
Acetic acid 0f 0f 0f 0.87 (0.01)e 0.91 (0.02)de 0.93 (0.04)d 1.02 (0.01)c 1.06 (0.02)bc 1.08 (0.02)b 1.16 (0.02)a
Butiric acid 0f 0f 0f 0.16 (0.04)e 0.16 (0.00)e 0.22 (0.01)d 0.30 (0.01)c 0.43 (0.01)b 0.46 (0.02)b 0.63 (0.00)a
Propionic acid 0.42 (0.03)d 0.44 (0.02)d 0.80 (0.03)c 1.09 (0.02)a 1.11 (0.01)a 1.13 (0.02)a 1.13 (0.01)a 1.09 (0.01)a 1.09 (0.02)a 0.96 (0.02)b
Citric acid 4.29 (0.04)a 4.18 (0.41)a 4.05 (0.62)a 2.73 (0.60)b 3.03 (0.07)b 2.68 (0.14)b 2.90 (0.04)b 2.70 (0.10)b 2.78 (0.10)b 2.89 (0.04)b
Ethanol 0f 0f 0f 0.07 (0.02)ef 0.14 (0.04)e 0.32 (0.04)d 0.45 (0.03)c 0.89 (0.03)b 0.87 (0.03)b 1.36 (0.06)a
pH 6.55 (0.01)a 6.55 (0.01)a 6.09 (0.01)b 5.64 (0.02)c 5.32 (0.01)d 4.85 (0.01)e 4.75 (0.01)f 4.53 (0.01)g 4.51 (0.01)h 4.31 (0.00)i
Data represent means (standard deviations) based on three replicates of kefir samples. a-i
: the mean values in rows with different letter are significantly 2 different (p < 0,05), as determined by the Tukey’s test. 3 4
5
6
7
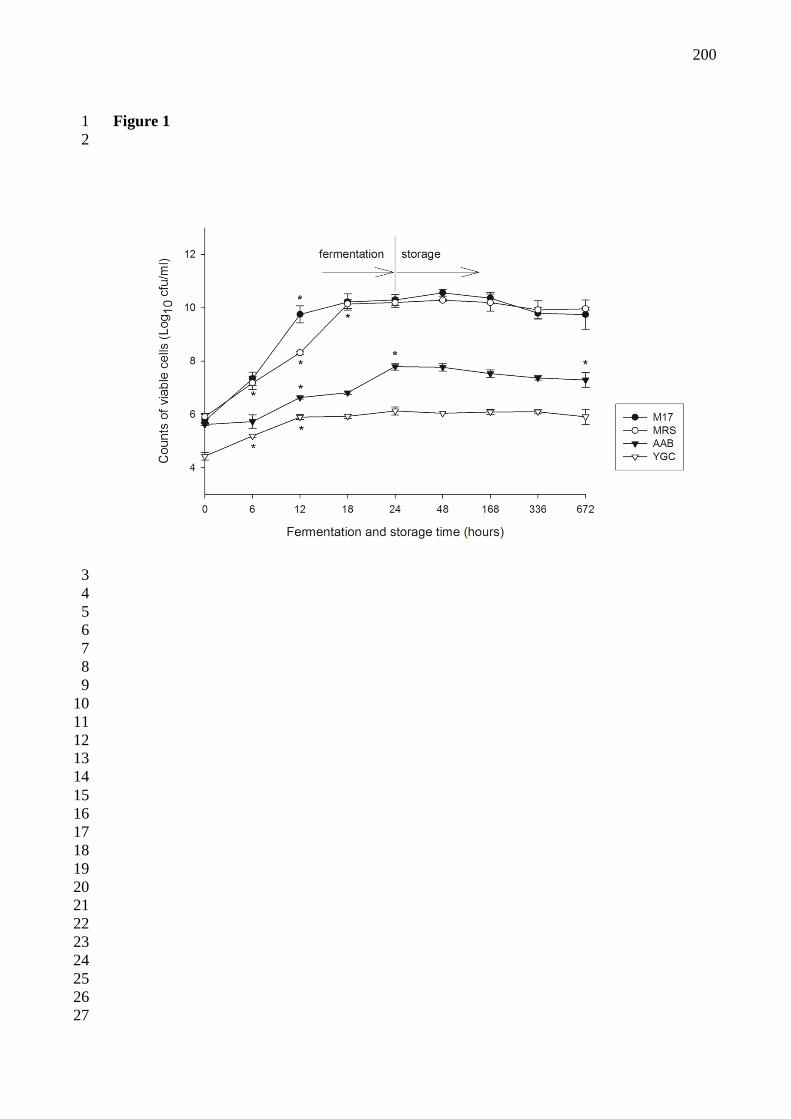
200
Figure 1 1
2
3
4
5
6
7
8
9
10
11
12 13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
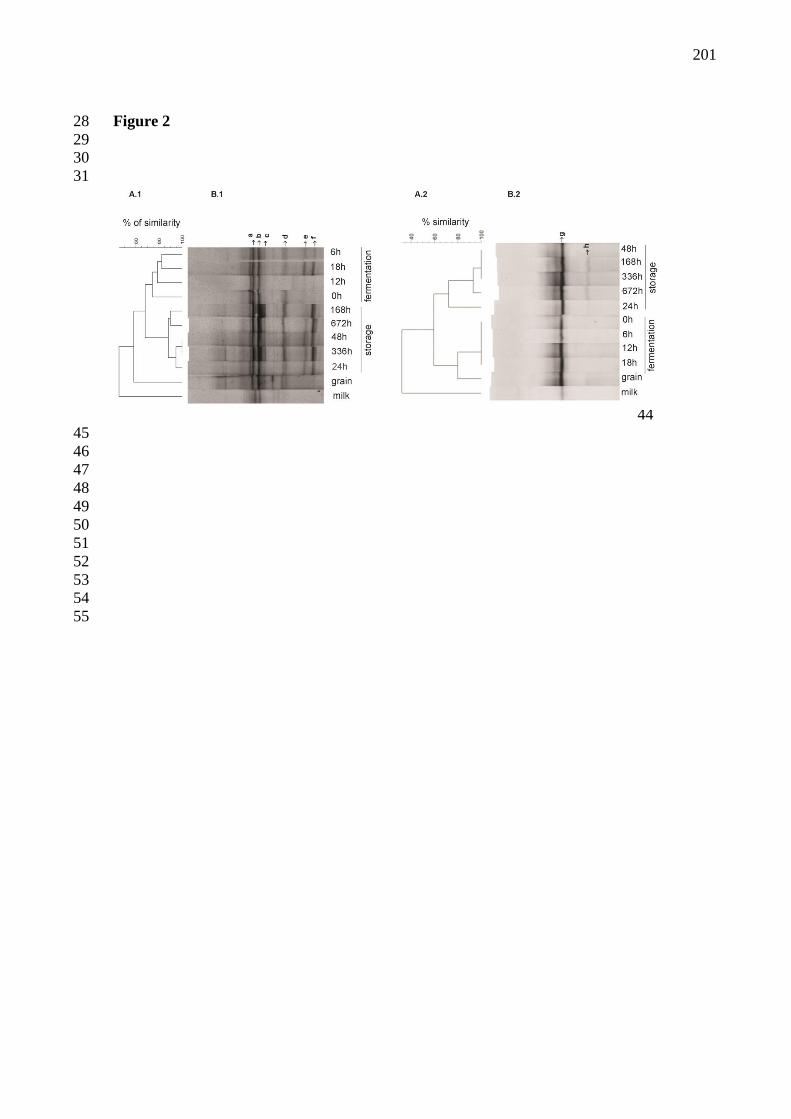
201
Figure 2 28
29 30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55

202
1
APÊNDICE C – ARTIGO IV 2
3
Identification of cultivable lactic acid bacteria from Brazilian kefir grains and probiotic 4
potential of lactobacilli strains 5
Leite, A.M.O.a,c
; Miguel; M.A.L.b; Peixoto, R.S.
b;Ruas-Madiedo,P.
a,; Silva, J.T.
c Paschoalin, 6
V.M.F.c; Delgado, S.
a; Mayo, B.
a* 7
8
Instituto de Productos Lácteos de Asturias (IPLA-CSIC), Carretera de Infiesto s/n, 33300-9
Villaviciosa, Asturias, Spaina, 10
Instituto de Microbiologia Paulo de Goes, Universidade Federal do Rio de Janeiro, Avenida 11
Carlos Chagas Filho, 373, 21941-904-Cidade Universitária, Rio de Janeiro, RJ, Brazilb, 12
Instituto de Química, Universidade Federal do Rio de Janeiro, Avenida Athos da Silveira 13
Ramos, 149-Bloco A, Sala 545, 21941-909-Cidade Universitária Rio de Janeiro, RJ, Brazilc. 14
15
Keywords: probiotic properties; Lactobacillus; kefir grains 16
17
Short title: Probiotic properties of lactobacilli from kefir grains 18
19
*Correspondence to: Baltasar Mayo, Instituto de Productos Lácteos de Asturias (IPLA-CSIC), 20
Departamento de Microbiología y Bioquímica, Carretera de Infiesto s/n, 33300-Villaviciosa, 21
Asturias, Spain, Phone/fax number: +34 985 892131/+34 985 892233 22
23
24

203
Abstract 25
A total of 33 lactic acid bacteria (LAB) obtained from Brazilian kefir grains were 26
selected from 150 initial isolates using their ability to survive acidic pH and to grow on 0.3% 27
bile salts, as a criterion to select strains for potential use as probiotics. Genotypic 28
characterization of those 33 isolates was performed by amplified ribosomal DNA restriction 29
analysis (ARDRA) followed by partial sequencing of 16S rRNA gene identified them as 30
Lactobacillus casei/paracasei (15%), Lactococcus lactis subsp. lactis (9%), L. lactis subsp. 31
cremoris (24%) and Leuconostoc mesenteroides (52%). All strains showed some inhibition 32
against food pathogens tested. Among them, five strains belonging to genera Lactobacillus or 33
Lactococcus produced bacteriocin-like inhibitory substances against indicator organisms 34
assayed. None of the selected lactobacilli showed undesirable enzymatic or haemolytic 35
activities. All picked lactobacilli showed susceptibility towards most of the antibiotics and 36
intrinsic resistance to vancomycin. Adhesion of two lactobacilli strains to human Caco-2 37
epithelial cells was comparable or higher than observed for L. rhamnnosus GG. These results 38
suggest that two L. casei/paracasei strains (MRN4 and MRS59) could be good candidate to 39
probiotic use, although further research, including in vivo analyses, should be performed 40
concerning their health benefit properties and safety properties. 41
42

204
Introduction 43
Kefir is fermented milk originated in the Northern Caucasus. The name kefir is 44
derived from the Turkish language and the word “keyif” means “good feeling” for the sense 45
experienced after drinking it (Lopitz-Otsoa, et al., 2006). The fermented beverage is acidic, 46
viscous, slightly carbonated and presents small amounts of alcohol (Magalhães, et al., 2011). 47
In Eastern Europe, kefir has a long tradition of offering health benefits and due to its 48
functional characteristics is becoming popular around the world. There are several compounds 49
in kefir that may have bioactive properties, such as exopolysaccharides and bioactive peptides 50
(Santos, et al., 2003, Farnworth, 2005). 51
The kefir beverage is considered a probiotic food (Sarkar, 2007, Farnworth & 52
Mainville, 2008) “defined as live microorganisms which when administered in adequate 53
amounts confer several benefits to human health” (Reid, et al., 2003, FAO/WHO, 2006, 54
Sanders, 2008). It is associated generally with welfare (Otles & Cagindi, 2003), including 55
reducing effects of lactose intolerance (Hertzler & Clancy, 2003), immunomodulation 56
(Vinderola, et al., 2006, Hong, et al., 2009), antimicrobial activity against pathogenic 57
microorganisms (Ota, 1999, Chifiriuc, et al., 2011), balance of intestinal microbiota (Lopitz-58
Otsoa, et al., 2006, Urdaneta, et al., 2007), anticarcinogenic effects (Shiomi, et al., 1982, 59
Kubo, et al., 1992, Otles & Cagindi, 2003), liver regeneration (Schmidt, et al., 1984), anti-60
inflammatory and cicatrizing activity (Rodrigues, et al., 2005, Huseini, et al., 2012) and 61
positive effects on cholesterol metabolism (Liu, et al., 2006, Farnworth & Mainville, 2008). 62
Many of these beneficial effects has been attributed to kefiran (Rodrigues, et al., 2005, 63
Moreira, et al., 2008), such as, antibacterial, antimycotic, antitumour activity (Santos, et al., 64
2003, Moreira, et al., 2008), anti-inflammatory and cicatrizing activities (Rodrigues, et al., 65
2005). 66

205
Traditional kefir is made only from kefir grains, which are the starter culture 67
composed by a complex microbiota (Magalhães, et al., 2011). Kefir grains are lobed 68
resembling a cauliflower, measures 0.3 – 3.5 cm in diameter, white to yellowish-white, and 69
have a slimy but firm texture. The grains are composed of lactic acid bacteria (Lactobacillus, 70
Lactococcus, Leuconostoc), acetic acid bacteria and yeast enclosed by a protein and 71
polysaccharide matrix (Garrote, et al., 2001, Farnworth & Mainville, 2008, Miguel, et al., 72
2010). The principal polysaccharide is a water-soluble substance known as kefiran. The 73
constituent microorganisms of kefir grain coexist in a complex symbiotic relationship, 74
meaning that they survive or propagate by sharing their bioproducts as an energy source or 75
growth stimulating source (Lopitz-Otsoa, et al., 2006, Tamime, 2006), which can make the 76
identification and study of their microbiota even more difficult (Kesmen & Kacmaz, 2011). 77
Most of selected probiotic bacteria belong to the lactic acid bacteria (LAB) group as 78
Lactobacillus spp. and Bifidobacterium sp. (Del Piano, et al., 2006, Gueimonde & Salminen, 79
2006, Bernardeau, et al., 2008), and the efficacy and safety of probiotics is strain and product-80
specific (FAO/WHO, 2006, Gueimonde & Salminen, 2006). 81
When selecting a probiotic strain, the microorganism must fulfill different 82
technological, functional and safe requisites, for human and animal consumption (Saarela, et 83
al., 2000, Pithva, et al., 2012). Probiotic strains must overcome biological and chemical 84
barriers such as acid and bile in the gastrointestinal (GI) tract. Alternatively, they can be 85
consumed in a food vehicle that allows them to survive passage through the stomach and 86
exposure to bile, and reach their site of action. They also must be able to proliferate, act and 87
persist in the gut environment providing health benefit effects (FAO/WHO, 2006, Gueimonde 88
& Salminen, 2006, Sanders, 2008). Furthermore, it is required that the strain retains viability 89
and maintenance of desirable characteristics during manufacturing and storage, as well as 90
provides good sensorial properties for the product (Sanders, 2008, Vinderola, et al., 2008). 91

206
Even though many authors advocate the importance of human origin as a selection criteria for 92
probiotic strains, fairly recent an expert panel proposed by FAO/WHO (2006) suggested that 93
the specificity of probiotic action is more important than the source of microorganism. Thus, 94
many Lactobacillus spp. have been isolated from dairy products with a intended use in 95
commercial probiotic products (Golowczyc, et al., 2008, Ayeni, et al., 2011, Jamaly, et al., 96
2011, Zago, et al., 2011). 97
Although there is a plausible number of well characterized probiotic strains available 98
commercially around the world (Vasiljevic & Shah, 2008) , the screening for novel strains is 99
still of great interest from both health and industrial points of view (Vinderola, et al., 2008, 100
Ayeni, et al., 2011, Plessas, et al., 2011). Due to long history of benefits effects associated to 101
consumption of kefir , this traditional product can be an interesting source of strains of lactic 102
acid bacteria well adapted in the microenvironment and with specific functional properties, 103
which might be used for development of new functional products for industrial dairy 104
application (Santos, et al., 2003, Golowczyc, et al., 2008, Plessas, et al., 2011). 105
The aim of this study was to characterize lactic acid bacteria strains isolated from 106
traditional kefir grains, selecting the most suitable ones according to their probiotic potential 107
for a possible use in dairy industry. 108
Material and Methods 109
Bacterial strains isolation 110
Lactic acid bacteria were isolated from four kefir grains collected from different 111
regions of Brazil, using the dilution agar method. Briefly, 10 g of each sample were 112
homogenized in 90 ml of sodium citrate (2%). Serial decimal dilutions were obtained and a 113
volume of 0.1 ml of the dilutions was plated on MRS and M17 agar media (Difco - BD 114
Diagnostics, Sparks, MD, USA) supplemented with 200 ppm cycloheximide (Sigma-Aldrich, 115
St. Louis, MO, USA), and incubated in aerobic and anaerobic conditions using Gaspak EZ 116

207
anaerobic system (Difco - BD Diagnostics, Sparks, MD, USA) at 30°C for 72 h. Significative 117
colonies were taken randomly and purified by subculturing on the same media. Presumptive 118
selection of LAB was performed by Gram staining and catalase testing. Pure cultures were 119
stored at −80 °C until use. 120
For all assays performed, LAB strains were activated in MRS broth at 30°C for 18-24, 121
and subcultured in MRS agar at 30°C for 18-24h. 122
Survival of the isolates at low pH 123
The ability of the isolates to grow at low pH was checked in MRS-THIO broth (MRS 124
supplemented with 0.2% sodium thioglycolate) acidified with HCl to a final pH ranging 3.0, 125
as described by Nishida, et al. (2008). Overnight cultures growth were estimated by optical 126
density at 600 nm (OD600) to logarithmic phase (OD600 = 0.5 – 1.0).Cells were harvested by 127
centrifugation and the pellet was suspended on phosphate-saline-buffer (PBS, pH 6.5) to 128
obtain an OD600 = 0.5. Cell suspension was diluted 10 times with acidic MRS medium, 129
adjusted to pH 3.0 and was incubated at 37°C for 3 h. pH tolerance of the cells was 130
determined by enumerating the viable cells on MRS agar plates after 48 h of incubation. A 131
culture in conventional MRS was used as control and if the number of colonies counted on 132
MRS agar was higher than in acidic MRS, the growth was considered “negative”. 133
Bile tolerance of cultures 134
The tolerance to bovine bile (Oxgall - Difco) was assayed by growing the isolates in 135
agar plate and media broth following the procedure of Dunne, et al. (1999) and Guo, et al. 136
(2009), respectively. Cultures growth was estimated in MRS-THIO broth with (test) and 137
without (control) 0.3% Oxgall bile salts (Difco). Overnight cultures (1%) incubated at 37 °C 138
for 18h were added into medium, cultured at 37°C in water bath, and monitored for 9h or until 139
a 0.3 unit difference in absorbance, between test and control was reached. The effect was 140
measured on basis of time required to increase the absorbance at 620 nm (OD620) by 0.3 units 141

208
in MRS-THIO broth with and without 0.3% bile salts. The difference in time (hours) between 142
the culture media was considered as the lag time (LT) as described by Guo, et al. (2009) and 143
Usman and Hosono (1999). Additionally, individual colonies picked on MRS agar plates were 144
suspended in 2-5 ml of sterile saline 0.85% at a density corresponding to McFarland standard 145
1. Aliquots of the suspensions (10 µl) were spotted onto bile-containing (0.3%, 0.5%, 1% and 146
2%) agar plates (Delgado, et al., 2007). These plates were incubated at 37°C under anaerobic 147
conditions and growth was recorded after 24-48h. A plate without bile was used as positive 148
control. 149
LAB identification 150
Carbohydrate fermentation patterns and biochemical tests were recorded for the 151
selected strains in the previous assay, using the API 50-CHL and API 20 Strep systems 152
(bioMérieux, Montalieu-Vercieu, France), respectively, according to the manufacturer’s 153
instructions. These assays were useful only to give preliminary identification of the LAB 154
genera without precision about species. 155
Finally, the identification of the selected cultures was confirmed by molecular 156
analysis. Selected strains were recovered in the corresponding media and total genomic DNA 157
was purified using the GenElute Bacterial Genomic DNA kit (Sigma–Aldrich Inc., St. Louis, 158
MO, USA), following the supplier’s recommendations, and was used as DNA templates to 159
amplify a segment of the 16S rRNA gene by the polymerase chain reaction (PCR) technique 160
as described by Wang, et al. (2006) using the universal prokaryotic primers S-D-Bact-0008-a-161
S-20 (27F) (5’-AGAGTTTGATCCTGGCTCAG-3’) and S-*-Univ-1492R-b-A-21 (1492R) 162
(5’-GGTTACCTTGTTACGACTT-3’). Amplicons were purified through GenEluteTM
PCR 163
Clean-Up columns (Sigma–Aldrich), digested with HaeIII and HhaI restriction enzymes 164
(Invitrogen Ltd., Pasley, UK) and electrophoresed in agarose gels. Gels were stained with 165
ethidium bromide (0.5 mg/ml) and documented under UV light. Representatives profiles of 166

209
each species of microorganisms observed in amplified ribosomal DNA restriction analysis 167
(ARDRA) were selected for sequencing analysis. Sequence analysis of partial 16S rRNA was 168
accomplished by ABI 373 DNA sequencer (Applied Biosystems, Carlsbad, CA, USA) and 169
used for identification of representative bacteria to species level. An average of 850 bp were 170
obtained per sequence and were compared with those available in the GenBank database 171
using the BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/) to determine the closest 172
known relatives of the partial ribosomal DNA sequences obtained. Sequences with a 173
percentage of identity of 98% or higher were allocated to the same species (Palys, et al., 174
1997). 175
Molecular typing analysis 176
In order to exclude possible clones, lactobacilli isolates were grouped by combined 177
technique of repetitive sequence-based PCR (rep-PCR) typing using primer BOXA1R (5’–178
CTACGGCAAGGCGACGCTGACG–3’) and BOXA2R (5’-179
ACGTGGTTTGAAGAGATTTTCG-3’) as reported by Koeuth, et al. (1995), and random 180
amplification of polymorphic RAPD-PCR typing with primer M13 (5’-181
GAGGGTGGCGGTTCT-3’), as reported by Rossetti and Giraffa (2005). 182
Banding patterns were examined for a band matching using Dice coefficient and 183
Bionumerics 6.5 software (Applied Maths, Belgium). Cluster analysis of composite data 184
obtained with rep-PCR and RAPD was achieved using the unweighted pairgroup method with 185
arithmetic averages (UPGMA). 186
Antimicrobial properties 187
Inhibition of pathogens 188
The capacity of the strains to inhibit a group of food pathogens was determined using 189
agar spot test described by Annuk, et al. (2003) and (Ripamonti, et al., 2011). The test 190
cultures were spotted (2 µl) on the surface of modified MRS agar (without ammonium citrate 191

210
and sodium acetate) and incubated anaerobically (GasPak system) for 24 h at 30°C to develop 192
the spots and thereafter inactivated using chloroform gas for 30 min. The indicator strains 193
included Escherichia coli ATCC 25922, Salmonella enterica var. Enteritidis ATCC 13076, 194
Staphylococcus aureus ATCC 25923, Listeria monocytogenes ATCC 15313. A 100 µl 195
volume of an overnight culture of the indicator bacteria was mixed with 10 ml of brain heart 196
infusion (BHI) soft agar (0.7%), and poured over the MRS plates which were incubated 197
aerobically at 37°C. After 24 h of incubation, inhibition zones were read. A clear zone of 198
more than 1 mm around a spot was scored as positive. L.acidophilus ATCC 4356 was used as 199
negative control (Ripamonti, et al., 2011). Each test was performed twice. 200
Bacteriocin production 201
Production of substances with antimicrobial activity was successively examined using 202
the agar spot test and well-diffusion assay. The indicator strains used (described as suceptible 203
to bacteriocins) were Lactobacillus sakei CECT 906, Lactococcus lactis IL 1403 (Gonzalez, 204
et al., 1994, Ryan, et al., 1996, Alegria, et al., 2010) and Listeria monocytogenes ATCC 205
15313. Overnight cultures of the isolates were spotted on the surface of M17 and MRS agar 206
plates (0.2% glucose), incubated for 24h at 30°C for lactocococci, leuconostoc strains and at 207
37°C for lactobacilli, respectively, and then inactivated by chloroform gas (Lemos Miguel, et 208
al., 2008). Spots were then covered with 10 ml of soft agar (0.75%), inoculated with the 209
indicator bacteria and incubated under the conditions required by each indicator strain. 210
Positive cultures were tested by a well-diffusion assay as described by Schillinger and 211
Lucke (1989). Twenty milliliters of agar medium at 45°C was vigorously mixed with 200 µl 212
of an overnight culture of the indicator strain and poured into Petri dishes. Supernatants from 213
overnight cultures of the producing strains were neutralized to pH 6.5-7.0 with NaOH 0.1 M, 214
centrifuged at 12,000 g for 5 min, and filter-sterilized through a 0.20 µm pore membrane. 215
(Millipore, Bedford, MA, USA). Aliquots of 50 µl of each supernatant were placed in a well-216

211
made into the agar and before incubation; the plates were maintained at 4°C for 1 h. The 217
inhibition of indicator strain growth was evaluated after incubation 24 h under appropriate 218
culture conditions. 219
To investigate the proteinaceous nature of bacteriocin-like substances produced, an 220
additional well was made in the agar next to a well containing the supernatants (50 µl) with 221
inhibitory activity. The novel well was filled with 50 µl of a solution containing proteinase K 222
or pronase (each at concentration of 20 mg/ml). After incubation, plates were examined to 223
judge whether the inhibitory substance was sensitive to proteolysis (Schillinger & Lucke, 224
1989, van Belkum, et al., 1989). Lactococcus lactis 1A6 strain, nisin producer (Alegria, et al., 225
2010) was used as positive control. Each test was performed twice. 226
Adhesion assay 227
The epithelial intestinal cell line Caco-2 was used to assess the adhesion ability of the 228
4 selected lactobacilli. The culture and maintenance of the cell lines were carried out 229
following standard procedures (Sánchez, et al., 2010) using DMEM medium supplemented 230
with 10% fetal bovine serum (Sigma-Aldrich, St Louis, MO, USA) and a mixture of 231
antibiotics (50 µg/ml gentamicin and 1.25 µg/ml amphotericin B) (Sigma-Aldrich, St Louis, 232
MO, USA). The cell line was used after reaching the confluent differentiated stay (13 ± 1 233
days). The strain Lactobacillus rhamnosus LMG 18243 (also named GG) was used as positive 234
control of well-recognized adherent LAB (Tuomola & Salminen, 1998). 235
Four lactobacilli strains, grown overnight in 10 ml MRS under standard conditions, 236
were harvest by centrifugation, washed twice in Dulbecco’s PBS solution (Sigma-Aldrich, St. 237
Louis, MO, USA) and suspended in DMEM or McCoy’s media without antibiotics, at a 238
concentration of about 108 CFU/ml. Cell line monolayers were also washed twice in 239
Dulbecco’s PBS, to remove antibiotics, and bacterial suspensions (108 CFU/ml) were added at 240
a ratio of bacteria:eukaryotic cell of 10:1. Plates were incubated for 1h at 37°C, 5% CO2 and, 241

212
afterwards, wells were gently washed three times with Dulbecco’s PBS buffer to remove the 242
non-adhered bacteria. Then, monolayers were trypsinized with EDTA-trypsin solution 243
(Sigma-Aldrich, St. Louis, MO, USA) and bacterial counts were carried out in MRS agar. 244
Adhesion results were expressed as the percentage of bacteria adhered with respect to the 245
amount of bacteria added (% CFU bacteria adhered/CFU bacteria added). Two replicated 246
experiments were carried out for each isolate using two independent microplates (each 247
isolated tested by duplicated in each plate). 248
Safety assessment 249
Production of haemolysins 250
The production of haemolysins was analysed by assays in Columbia agar plates 251
containing 5% sheep blood (bioMérieux, Montalieu-Vercieu, France). The presence of 252
haemolysis is indicated by the formation of clear zones surrounding the colonies. 253
Antibiotic resistance 254
The minimum inhibitory concentration (MIC) against antibiotics was determined in 4 255
selected lactobacilli isolates by microdilution in VetMICTM
plates for lactic acid bacteria 256
(SVA, National Veterinary Institute of Sweden, Uppsala, Sweden), following the 257
manufacture’s recommendations. VetMICTM
Lact-1 and Lact-2 microdilution tests were used 258
to determine the MIC to gentamicin (concentration range tested: 0.5-256 µg/ml), kanamycin 259
(2-1024 µg/ml), streptomycin (0.5-256 μg/ml), neomycin (0.5–256 μg/ml), tetracycline (0.12–260
64 μg/ml), erythromycin (0.016–8 μg/ml), clindamycin (0.03–16 μg/ml), and chloramphenicol 261
(0.12–64 μg/ml), ampicillin (0.03-16 μg/ml), penicillin (0.03-16 μg/ml), vancomycin (0.25-262
128 μg/ml), virginiamycin (0.016-8 μg/ml), linezolid (0.03-16 μg/ml), trimethroprim (0.12-64 263
μg/ml), ciprofloxacin (0.25-128 μg/ml), rifampicin (0.12-64 μg/ml). 264
Colonies grown in LSM (Klare, et al., 2005) agar plates were suspended in 5 ml of 265
sterile saline solution 0.9% to obtain a density corresponding to McFarland standard 1. 266

213
Suspensions were further diluted 1:1000 in LSM. One hundred microliters of the inoculum 267
were added to each well of the VetMICTM
plate, and the plate was incubated at 28 °C for 48 h. 268
The MICs were defined as the lowest antibiotic concentration at which no visual growth was 269
observed. 270
Enzyme activities 271
Enzyme activities were measured by the commercial, semi quantitative API-ZYM 272
system (bioMérieux, Montalieu-Vercieu, France) following the manufacturer’s 273
recommendations. 65 µl corresponding to McFarland standard 5 were inoculated into each 274
well of the API-ZYM trips. Enzyme activities were evaluated after 4h of anaerobic incubation 275
at 37°C. The enzyme activity was recorded from 0 to 5 with API ZYM color reaction chart. 276
The approximate number of free nmol hydrolyzed substrate was estimated by the color chart: 277
0, no activity; 1, liberation of 5 nmols; 2, 10 nmols; 3, 20 nmols; 4, 30 nmols; 5, ≥ 40 nmols. 278
Results 279
pH and bile salt tolerance 280
37 from 150 LAB strains tested were able to survive to 3h incubation at pH 3.0, but 281
none of them were able to grow at this pH value (Table 2). Among them , 33 (85%) showed 282
resistance to bile salts and were able to grow at 0.3% (Table 1), although it has been observed 283
a delay from 1 h until 4 h in the growth cell comparing to the control (MRS without bile). In 284
the plate test, around 65% of the LAB strains were able to grow in 0.5% concentration, 35% 285
in 1% concentration (Table 1), whereas only 19% in MRS plates containing 2% of bile salts. 286
LAB Identification 287
Among 33 pH or bile salts-resistant LAB strains, four different representative profiles 288
of each species were screened in partial ARDRA analysis and selected for sequencing of 289
partial 16S rRNA gene (data not shown). Identification of the 33 isolates was obtained and 290
their species and origins were shown in table 2. DNA sequences of representative 291

214
microorganisms showed highest homology (98-100%) to Lactobacillus casei/paracsei (15%), 292
Lactococcus lactis subsp. lactis (9%), Lactococcus lactis subsp. cremoris (24%) and 293
Leuconostoc mesenteroides (52%). With regarding to the genus lactobacilli, the analysis of 294
the 16S rRNA gene sequences failed to differentiate the species L. casei and L. paracasei 295
(Table 2), as previously indicated by Mori, et al. (1997), Gueimonde, et al. (2004), so, the 296
five isolates (MR1, MRN4, MRS59, M1743, MRS55) were referred as L. casei/paracasei. 297
Antimicrobial properties 298
Antimicrobial activity against pathogens 299
In the agar spot test (Table 1), all the strains tested inhibited the most of the pathogens. 300
Twenty three and twenty one strains, including those belonging to genus lactobacilli, inhibited 301
E. coli and S. Enteritidis, respectively. Twenty four showed inhibitory properties against L. 302
monocytogenes, whereas only eight against S. aureus. For those positive strains in agar spot 303
test, the inhibitory ability was confirmed by agar well-diffusion assay (Table 1) and the 304
neutralization of cell-free supernatant affected the antibacterial activity of all strains (no 305
inhibition observed for any strain). 306
Bacteriocin-like substance production 307
In agar spot assay, L. sakei CECT 906 was inhibited by 06 strains, L. lactis IL 1403 by 308
14 strains and L. monocytogenes by 24 strains. The isolates with antibacterial activity against 309
any of the indicators were subsequently subjected to the well-diffusion assay. None 310
Leuconostoc strain showed inhibition against indicators in this second assay, meanwhile L. 311
lactis IL 1403 was inhibited by 03 lactococci strains (MRS26, M171 and MRS52), L. sakei 312
CECT 906 was inhibited by lactococci and lactobacilli strains (MRS26, M171, MRS 52, MRS 313
20, MRS 55) and L. monocytogenes ATCC 15313 by 04 strains (MRS 26, M171, MRS 52, 314
MRS 55), displaying a small halo (around 3 mm), but with clear inhibitory effects against all 315

215
indicators tested. The proteinaceous nature of the antibacterial substance produced by the 05 316
strains was confirmed by proteinase treatment, when their activities disappeared (Fig. 1). 317
Lactobacillus casei/paracasei typing analysis 318
Molecular differentiation was performed by typing Lactobacillus casei/paracasei 319
isolates with rep-PCR and RAPD analysis. The UPGMA dendrogram (Fig.2) was obtained by 320
combination of BOXA1R, BOXA2R and M13 profiles. Threshold 92% of similarity was 321
considered to discriminate between different strains. A total of 04 representative profiles 322
(MRN4, MRS55, MRS59 and M1743) of Lactobacillus casei/paracasei strains were chosen 323
for further characterization of their probiotic potential. 324
Enzymatic activities 325
L. casei/paracasei MRS 55 and MRN4 were able to hydrolyze 15 substrates, whereas 326
the other strains hydrolyzed between 14 and 13 substrates (Table 3). MRS55 showed higher 327
levels enzymatic activity to the most of enzymes assayed. All isolates displayed activities of 328
alkaline phosphatase, esterase (C4), esterase-lipase (C8), lipase, leucine arylamidase, valine 329
arylamidase, cysteine arylamidase, α–chymotrypsin, acid phosphatase, naphthol-AS-BI-330
phosphohydrolase, β–galactosidase, α–glucosidase and β-glucosidase (Table 3). Lactobacillus 331
strains showed maximum activities for leucine arylamidase, valine arylamidase, and lower 332
levels were detected for cysteine arylamidase. The four isolates also presented moderate 333
esterase (C4), esterase-lipase (C8) and low lipase (C14) activities, whereas N-acetyl-β-334
glucosaminidase and trypsin activities were observed by two isolates and α-fucosidase was 335
observed by only one strain. α-galactosidase, β-glucuronidase and α-mannosidase activities 336
were not observed. 337
Adhesion assay 338
MRN4 strain showed higher potential of adhesion in the intestinal cell line when 339
compared with other strains, including the positive control L. rhamnosus GG. Adhesion 340

216
percentages ranged from 0.7 to 12.9 %. MRS 59 showed similar adhesion (p > 0.05) to 341
control strain L. rhamnosus GG, whereas the M1743 and MRS 55 strains showed lower ones 342
(Fig 3). 343
Antibiotic resistance and haemolysin production 344
Selected strains were susceptible to tetracycline, erythromycin, clindamycin, 345
ampicillin and aminoglycosides (gentamicin, kanamycin, streptomycin and neomycin) - Table 346
4. On the other hand, MRS 55 and M1743 strains were resistant to chloramphenicol (8µg/ml), 347
considering the microbiological breakpoints (4 µg/ml), suggested by EFSA (2008). 348
None haemolytic activity was exhibited when strains were grown in Columbia blood 349
agar. 350
Discussion 351
Currently, there is an increasing demand for new strains that play a probiotic role, 352
beyond their technological function (Jamaly, et al., 2011, Plessas, et al., 2011). Kefir is a 353
beverage traditionally consumed in Eastern Europe and well known by its health effects 354
(Sarkar, 2007, Farnworth & Mainville, 2008). For this reason, it could be a source for 355
obtaining novel strains potentially probiotic, seeking for both future application in the 356
formulation of novel probiotic foods or even to render a culture starter for industrial 357
production of kefir, not yet commercialized in Brazil. 358
In this study LAB isolates from kefir grains were firstly tested for in vitro tolerance to 359
acidity and bile. The selected strains were assayed for their antagonistic activity against 360
pathogens and production of bacteriocin-like inhibitory substances. Additionally, important 361
features to enable the lactobacilli strains as potentially probiotic were tested by in vitro assays, 362
such as enzymatic activity profile, adherence to epithelial cells and safety properties. 363
Secretion of gastric acid and transit through stomach constitutes primary defense 364
mechanism that all the ingested microorganisms must overcome, including those potential 365

217
probiotics bacteria (Dunne, et al., 1999). pH in the human stomach ranges from 1 to 4.5, and 366
food digestion can take up to 3h (Maragkoudakis, et al., 2006, Jamaly, et al., 2011). Although 367
the lactobacilli strains isolated in this study did not grow when exposed to pH 3.0 for 3h, they 368
were resistant and survived at acidic environment. The results described here are in agreement 369
with similar ones (Santos, et al., 2003, Delgado, et al., 2007, Gu, et al., 2008, Nishida, et al., 370
2008) where most of Lactobacillus strains were unable to grow, but survived at pH of 2.5-3.0. 371
Furthermore, it is important to take into account that probiotic bacteria are mainly consumed 372
in the dairy product in the presence of milk proteins. Milk proteins could exert an additional 373
protective action on the starters, supporting bacterial survival in the acid environment of 374
stomach (Charteris, et al., 1998, Maragkoudakis, et al., 2006). In contrast, most strains tested 375
grew in the presence of bile salts (0.3%), simulating the small intestine environment 376
(Vinderola, et al., 2008). Most studies have reported that the majority of LAB strains survive 377
well under such conditions, suggesting a possible recuperation of the initial inoculum level 378
during passage through the small intestine (Charteris, et al., 1998, Delgado, et al., 2007, Gu, 379
et al., 2008, Mathara, et al., 2008, Guo, et al., 2009, Zago, et al., 2011). 380
Bile is synthesized in the liver, stored and concentrated in gallbladder, and secreted 381
into the duodenum in the conjugated form, after food intake (Dunne, et al., 1999, Begley, et 382
al., 2006). It is a digestive secretion that plays a major role in the emulsification and 383
solubilization of lipids (Begley, et al., 2005). Bile salts are toxic for microorganisms, since 384
they affect the bilayer structure of the bacterial membranes (Guo, et al., 2009), thus, bile salt 385
tolerance is an essential feature for LAB survival and subsequent colonization of the 386
gastrointestinal tract (FAO/WHO, 2006). The resistance to bile varies between Lactobacillus 387
species and strains and the mechanism is still unknown (Papamanoli, et al., 2003, Begley, et 388
al., 2005, Zago, et al., 2011). There are reports that the relevant physiological concentrations 389
of human bile ranges from 0.3% (Dunne, et al., 1999) to 0.5% (Vizoso Pinto, et al., 2006). 390

218
However, there is still no consensus about the precise concentration to which the potential 391
probiotic strain should be tolerant (Vizoso Pinto, et al., 2006, Zago, et al., 2011). In recent 392
studies (Vizoso Pinto, et al., 2006, Delgado, et al., 2007, Mathara, et al., 2008, Vinderola, et 393
al., 2008, Zago, et al., 2011) bile concentration has ranged from 0.3 to 2%, but in most of 394
them it has been used 0.3% Oxgall solution to substitute human bile salts (Del Piano, et al., 395
2006, Mathara, et al., 2008, Vinderola, et al., 2008, Ayeni, et al., 2011, Jamaly, et al., 2011, 396
Zago, et al., 2011). The resistance to bile varies between Lactobacillus species and strains and 397
the mechanism is still unknown (Papamanoli, et al., 2003, Begley, et al., 2005, Zago, et al., 398
2011). Our results showed that most of lactobacilli strains were able to resist and grow in 399
high bile concentrations (2%). Other authors have also reported high resistance to bile among 400
lactobacilli strains (Dunne, et al., 1999, Delgado, et al., 2007, Jamaly, et al., 2011). 401
Several studies (Santos, et al., 2003, Batdorj, et al., 2007, Gu, et al., 2008, Alegria, et 402
al., 2010, Jamaly, et al., 2011, Ripamonti, et al., 2011) demonstrated a desirable ability of 403
probiotic lactobacilli to inhibit pathogenic bacteria growth by the production of metabolites 404
such as organic acids, fatty acids, hydrogen peroxide and ethanol. In addition, LAB may 405
produce bacteriocins, which can inhibit the growth of closely related microbes or a wide range 406
of Gram positive bacteria, including foodborne pathogens and spoilage microorganisms such 407
as Bacillus cereus, Listeria monocytogenes, Staphylococcus aureus and Clostridium 408
tyrobutyricum (Galvez, et al., 2008, Alegria, et al., 2010, Bellei, et al., 2011). These reports 409
are in agreement with our findings, since in pathogens-inhibition assay, the antagonistic effect 410
displayed by the selected strains may be related to the production of organic acids or 411
hydrogen peroxide, whereas in the bacteriocin-production assays, five isolates, including 01 412
lactobacilli and 04 lactococci, produced bacteriocin-like compounds. The antimicrobial 413
properties exhibited by LAB strains isolated in this study could indicate that they could 414
protect the gastrointestinal tract GIT), playing an important role in the maintenance of the 415

219
intestinal ecosystem (Servin, 2004, Vinderola, et al., 2008). Ryan, et al. (1996) also isolated 416
from Irish kefir grain a bacteriocin (lacticin 3147) produced by Lactococcus lactis strain DPC 417
3147 that showed broad inhibition spectrum. The lacticin 3147-producing strain was 418
employed as starter culture in the manufacture of cheddar cheese and it was observed that the 419
starters provided useful in controlling of the proliferation of undesirable microorganisms 420
during cheese manufacture. The efficiency of the antimicrobial activity produced by the 421
isolated strains described here should be confirmed in food matrices. 422
Members of the Lactobacillus genus are one of the most commonly used in the 423
development of probiotic foods (Dunne, et al., 1999, FAO/WHO, 2006, Gueimonde & 424
Salminen, 2006, Zago, et al., 2011). In this study, a molecular typing method was performed 425
to group the lactobacilli isolates according to the heterogeneities exhibited in their band 426
profiles. The same technique has been used by several authors with similar purposes (Urraza, 427
et al., 2000, Golowczyc, et al., 2008, Vinderola, et al., 2008, Adiguzel & Atasever, 2009, 428
Fernández, et al., 2010) and as expected it allowed us to select 04 different lactobacilli strains 429
that were further characterized concerning to their probiotic properties. 430
Enzymatic activity is an important feature in strains envisaged as probiotics or starter 431
cultures, according to their functional and/or technological abilities required (Papamanoli, et 432
al., 2003, Monteagudo-Mera, et al., 2011, Zago, et al., 2011). None of isolated lactobacilli 433
strains showedβ-glucuronidase activity, which may have negative effects in the colon and has 434
been considered a carcinogenic enzyme, since it converts precarcinogeninto carcinogen-435
substances, which can stimulate the development of human colon cancer (Monteagudo-Mera, 436
et al., 2011). In contrast, all strains produced β-galactosidase, a beneficial enzyme considering 437
both probiotic and technological aspects, supporting the reduction of lactose intolerance as 438
well as acidification of milk (Monteagudo-Mera, et al., 2011). β-galactosidase also 439
contributes to the formation of galacto-oligossacharides (GOS), the non-digestible 440

220
oligossacharides that act as a growth promoting substrate for bifidobacteria in the human 441
intestine and suppress potentially harmful bacteria due to the acid environment created by 442
bifidobacteria (Neri, et al., 2009, Park & Oh, 2010). With regard to activities of the enzymes 443
correlated with carbohydrate catabolism, all isolates presented activities of α-glucosidase and 444
β-glucosidase that could contribute for the digestion of polysaccharides (Papamanoli, et al., 445
2003, Lu, et al., 2008). Nevertheless, according to Monteagudo-Mera, et al. (2011) α-446
glucosidase catalyzes the final step in the digestion of carbohydrates, and its presence can be 447
undesirable in the diet for diabetic and obese patients, since it can stimulate postprandial 448
hyperglycemia. Although β-glucosidase had been described previously as a harmful enzyme 449
to health due to its possible procarcinogenic activity (Wollowski, et al., 2001, Bhatia, et al., 450
2002), recent studies indicate that it may be desirable to break down the isoflavone 451
glucosides to the biologically active aglycones compounds that are often associated with 452
prevention of cancer, cardiovascular disorders, osteoporosis, postmenopausal syndromes, 453
hypercholesterolemia, and antioxidative effects (Bhatia, et al., 2002, Otieno, et al., 2006, 454
Otieno, et al., 2007, Yang, et al., 2009).α–fucosidase activity, which can be responsible for 455
the selective colonization of the intestine by bacteria (Monteagudo-Mera, et al., 2011), was 456
detected only in MRS55 strain . The presence of glycosidase activities in starter cultures 457
seems to have an impact on the sensory properties, since that flavour is often linked to sugars 458
utilization (Papamanoli, et al., 2003, Mesas, et al., 2011). Moreover, it was noteworthy that 459
all strains tested here showed aminopeptidases and lipolytic activities. Aminopeptidase 460
activity is an important technologic characteristic for LAB strains, because it contributes to 461
the development of desirable flavors in the dairy products (Monteagudo-Mera, et al., 2011), 462
as well as esterases and lipases activities can be associated to the degradation of lipids in the 463
dairy products (Fernández, et al., 2010). 464

221
Adhesion capacity to the intestinal mucosa is an important property for candidate 465
probiotic strain, since it should colonize the host gut enabling health benefit effects 466
(FAO/WHO, 2003). Generally, the human colon adenocarcinoma cell line Caco-2 is used 467
because it exhibits in vitro, the characteristics of a mature enterocyte of small intestine (Pinto, 468
et al., 1983, Coconnier, et al., 2000). It is widely accepted as a model for assessing the 469
adhesion ability of putative probiotic strains to epithelial intestinal cells (Gueimonde & 470
Salminen, 2006, Candela, et al., 2008, Ayeni, et al., 2011). Comparing the adhesion values 471
obtained with the probiotic bacteria, Lb. rhamnosus GG used as positive control in this assay, 472
with the two selected strains (MRN4 and MRS59), the isolates showed higher or similar 473
adhesion to Caco-2 monolayers. Although further in vivo tests are needed (FAO/WHO, 2003, 474
Cencic & Langerholc, 2010) to confirm the adhesion capability of MRN4 and MRS59 strains, 475
the results obtained point to promising potential of strains in adhering to the epithelial cells. 476
Although further in vivo tests are needed to confirm the adhesion capability of MRN4 and 477
MRS59 strains, the results obtained point to promising potential of strains in adhering to the 478
epithelial cells. 479
The L. casei/ paracasei isolated in this study has a long history of apparent safe use in 480
dairy foods, and it has been found in many commercial probiotic products (FAO/WHO, 2003, 481
Gueimonde, et al., 2004, EFSA, 2011). It was included in the list of taxonomic units proposed 482
by EFSA for Qualified Presumption Safety (QPS) status (EFSA, 2011) based on the current 483
available data about safety for human consumption. However, before its technological and 484
functional application, safety properties should be investigated for each strain intended to 485
have food use, especially regarding the susceptibility to antibiotics (FAO/WHO, 2006, 486
Bernardeau, et al., 2008). Here, we report two strains with MIC value higher than EFSA’s 487
breakpoints (4 µg/ml) for chloramphenicol. However, if MIC were compared to the clinical 488
breakpoint of this antibiotic (≥ 32 µg/ml) proposed by CLSI (2011), none of them could be 489

222
considered resistant. Previous studies (Florez, et al., 2005, Maragkoudakis, et al., 2006, 490
D'Aimmo, et al., 2007, Klare, et al., 2007) also found L. casei/paracasei strains with similar 491
MIC patterns to chloramphenicol, and some of these authors (Florez, et al., 2005, Klare, et 492
al., 2007) even suggested higher microbiological breakpoints values (8 µg/ml) of this 493
antibiotic for strains of L.casei/paracasei intended to have probiotic or nutritional use. Other 494
studies also confirmed the low resistance of this lactobacilli specie towards chloramphenicol 495
(Temmerman, et al., 2003, Delgado, et al., 2005, Florez, et al., 2008, Mathara, et al., 2008). 496
Thus, it can be suggested that these two strains have not acquired resistance mechanism, but 497
further searches for resistance genes should be conducted to ensure their potential use as 498
probiotic. 499
It is important to distinguish between intrinsic resistance and acquired resistance to an 500
antimicrobial, since that only acquired resistance is a threat to health, due to the serious 501
chance of it being transferred (EFSA, 2008). Analysis of MICs and their distribution helps 502
differentiate these two resistances mechanisms (Ammor, et al., 2007). Intrinsic resistance is 503
inherent to a bacterial species and it is typical for all strains from that specie. A well-known 504
example of intrinsic resistance is the resistance to vancomycin of heterofermentative and 505
facultative heterofermentative lactobacilli, confirmed by the high MIC found for all isolates 506
tested. The intrinsic resistance of this specie to vancomycin is due to the presence of D-Ala-507
D-Lac as the normal dipeptide in its peptidoglycan (Klein, et al., 2000, Mathur & Singh, 508
2005). Acquired resistance can to occur by the insertion of exogenous genes or due to a 509
mutation in the chromosomal genes. The latter mechanism presents a low risk of horizontal 510
dissemination and would generally be acceptable for the FEEDAP panel, whereas antibiotic 511
resistance by added genes presents the greatest risk for horizontal dissemination resistance 512
(EFSA, 2008). 513

223
In summary, in this study 33 isolates from kefir grains were identified and among 514
them 04 distinct Lactobacillus casei/paracasei strains were further selected to be evaluated in 515
vitro as potential probiotics. All lactobacilli strains showed good survival to conditions of 516
GIT, inhibition of intestinal pathogens and no harmful enzymatic activity, but regarding to 517
safety and adhesion properties, two L.casei/paracasei MRS59 and MRN4 strains were the 518
most suitable for probiotic use These results suggest that L.casei/paracasei MRS59 and 519
MRN4 isolated from kefir grains showed good potential for a possible application in 520
functional foods, although additional studies are needed concerning health benefit properties 521
and safety properties, as well as in vivo assays before the application of these isolates to the 522
formulation of functional probiotics products. Additionally, four L. lactis isolates produce 523
bacteriocin-like inhibitory substances that should be better characterized and their 524
technological properties evaluated, in order to be used as starter cultures or adjunct cultures in 525
the dairy industry. 526
Acknowledgements 527 The study was supported by FAPERJ, CAPES Foundation (process number PDEE 528
5019109) and a project from the Spanish Ministry of Science and Innovation (MICINN) 529
(reference AGL2007-61869-ALI). S. Delgado was supported by a research contract from 530
MICINN under the Juan de la Cierva Program (reference JCI-2008-02391). 531
532
References 533 Adiguzel GC & Atasever M (2009) Phenotypic and Genotypic Characterization of lactic acid 534
bacteria isolated from Turkish dry fermented sausage. Romanian Biotechological Letters 14: 535
4130-4138. 536
Alegria A, Delgado S, Roces C, Lopez B & Mayo B (2010) Bacteriocins produced by wild 537
Lactococcus lactis strains isolated from traditional, starter-free cheeses made of raw milk. 538
International Journal of Food Microbiology 143: 61-66. 539
Ammor MS, Florez AB & Mayo B (2007) Antibiotic resistance in non-enterococcal lactic 540
acid bacteria and bifidobacteria. Food Microbiology 24: 559-570. 541
Annuk H, Shchepetova J, Kulisaar T, Songisepp E, Zilmer M & Milkelsaar M (2003) 542
Characterization of intestinal lactobacilli as putative probiotic candidates. Journal of Applied 543
Microbiology 94: 403-412. 544
Ayeni FA, Sanchez B, Adeniyi BA, de Los Reyes-Gavilan CG, Margolles A & Ruas-545
Madiedo P (2011) Evaluation of the functional potential of Weissella and Lactobacillus 546
isolates obtained from Nigerian traditional fermented foods and cow's intestine. International 547
Journal of Food Microbiology 147: 97-104. 548
Batdorj B, Trinetta V, Dalgalarrondo M, et al. (2007) Isolation, taxonomic identification and 549
hydrogen peroxide production by Lactobacillus delbrueckii subsp. lactis T31, isolated from 550

224
Mongolian yoghurt: inhibitory activity on food-borne pathogens. Journal of Applied 551
Microbiology 103: 584-593. 552
Begley M, Gahan CG & Hill C (2005) The interaction between bacteria and bile. FEMS 553
Microbiology Reviews 29: 625-651. 554
Begley M, Hill C & Gahan CG (2006) Bile salt hydrolase activity in probiotics. Applied and 555
Environmental Microbiology 72: 1729-1738. 556
Bellei B, Miguel M, Del Aguila EM, Silva JT & Paschoalin VMF (2011) Purification of a 557
bacteriocin produced by Enteroccus faecium and its effectiveness for preservation of fresh-cut 558
lettuce. Journal of Microbiology and Antimicrobials 3: 119-125. 559
Bernardeau M, Vernoux JP, Henri-Dubernet S & Gueguen M (2008) Safety assessment of 560
dairy microorganisms: the Lactobacillus genus. International Journal of Food Microbiology 561
126: 278-285. 562
Bhatia Y, Mishra S & Bisaria VS (2002) Microbial beta-glucosidases: cloning, properties, and 563
applications. Critical Reviews in Biotechnology 22: 375-407. 564
Candela M, Perna F, Carnevali P, et al. (2008) Interaction of probiotic Lactobacillus and 565
Bifidobacterium strains with human intestinal epithelial cells: Adhesion properties, 566
competition against enteropathogens and modulation of IL-8 production. International 567
Journal of Food Microbiology 125: 286-292. 568
Cencic A & Langerholc T (2010) Functional cell models of the gut and their applications in 569
food microbiology--a review. International Journal of Food Microbiology 141 Suppl 1: S4-570
14. 571
Charteris WP, Kelly PM, Morelli L & Collins JK (1998) Development and application of an 572
in vitro methodology to determine the transit tolerance of potentially probiotic Lactobacillus 573
and Bifidobacterium species in the upper human gastrointestinal tract. Journal of Applied 574
Microbiology 84: 759-768. 575
Chifiriuc MC, Cioaca AB & Lazar V (2011) In vitro assay of the antimicrobial activity of 576
kephir against bacterial and fungal strains. Anaerobe 17: 433-435. 577
CLSI (2011) Performance Standards for Antimicrobial Susceptibility Testing; Twenty-First 578
Informational Supplement. (Institute CaLS, ed.^eds.), p.^pp. Clinical and Laboratory Institute, 579
Wayne, PA. 580
Coconnier MH, Lievin V, Lorrot M & Servin AL (2000) Antagonistic activity of 581
Lactobacillus acidophilus LB against intracellular Salmonella enterica serovar Typhimurium 582
infecting human enterocyte-like Caco-2/TC-7 cells. Applied and Environmental Microbiology 583
66: 1152-1157. 584
D'Aimmo MR, Modesto M & Biavati B (2007) Antibiotic resistance of lactic acid bacteria 585
and Bifidobacterium spp. isolated from dairy and pharmaceutical products. International 586
Journal of Food Microbiology 115: 35-42. 587
Del Piano M, Morelli L, Strozzi GP, et al. (2006) Probiotics: from research to consumer. 588
Digestive and Liver Disease 38: S248-S255. 589
Delgado S, Florez AB & Mayo B (2005) Antibiotic susceptibility of Lactobacillus and 590
Bifidobacterium species from the human gastrointestinal tract. Current Microbiology 50: 202-591
207. 592
Delgado S, O'Sullivan E, Fitzgerald G & Mayo B (2007) Subtractive screening for probiotic 593
properties of lactobacillus species from the human gastrointestinal tract in the search for new 594
probiotics. Journal of Food Science 72: M310-315. 595
Dunne C, Murphy L, Flynn S, et al. (1999) Probiotics: from myth to reality. Demonstration of 596
functionality in animal models of disease and in human clinical trials. Antonie Van 597
Leeuwenhoek 76: 279-292. 598

225
EFSA (2008) Technical guidance prepared by the Panel on Additives and Products or 599
Substances used in Animal Feed (FEEDAP) on the update of the criteria used in the 600
assessment of bacterial resistance to antibiotics of human or veterinary importance. 732: 1-15. 601
EFSA (2011) EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the 602
maintenance of the list of QPS biological agents intentionally added to food and feed (2011 603
update) 9: 82. 604
FAO/WHO (2003) CODEX Standard for Fermented Milks. (Stan C, ed.^eds.), p.^pp. 1-11. 605
FAO/WHO (2006) Probiotic in foods. Health and nutritional properties and guidelines for 606
evaluation. Vol. 85 ed.^eds.), p.^pp. FAO Food and Nutrition Paper, Rome. 607
Farnworth ER (2005) Kefir - a complex probiotic. Food Science and Techology Bulletin: 608
Functional Foods 2: 1-17. 609
Farnworth ER & Mainville I (2008) Kefir - A Fermented Milk Product. Handbook of 610
Fermented Functional Foods,(Farnworth ER, ed.^eds.), p.^pp. 89-127 CRC Press Taylor & 611
Francis Group Boca Raton, London, New York. 612
Fernández E, Alegría Á, Delgado S & Mayo B (2010) Phenotypic, genetic and technological 613
characterization of Lactococcus garvieae strains isolated from a raw milk cheese. 614
International Dairy Journal 20: 142-148. 615
Florez AB, Delgado S & Mayo B (2005) Antimicrobial susceptibility of lactic acid bacteria 616
isolated from a cheese environment. Canandian Journal of Microbiology 51: 51-58. 617
Florez AB, Ammor MS, Mayo B, van Hoek AH, Aarts HJ & Huys G (2008) Antimicrobial 618
susceptibility profiles of 32 type strains of Lactobacillus, Bifidobacterium, Lactococcus and 619
Streptococcus spp. International Journal of Antimicrobial Agents 31: 484-486. 620
Galvez A, Lopez RL, Abriouel H, Valdivia E & Omar NB (2008) Application of bacteriocins 621
in the control of foodborne pathogenic and spoilage bacteria. Critical Reviews in 622
Biotechnology 28: 125-152. 623
Garrote GL, Abraham AG & De Antoni GL (2001) Chemical and microbiological 624
characterisation of kefir grains. Journal of Dairy Research 68: 639-652. 625
Golowczyc MA, Gugliada MJ, Hollmann A, et al. (2008) Characterization of 626
homofermentative lactobacilli isolated from kefir grains: potential use as probiotic. Journal of 627
Dairy Research 75: 211-217. 628
Gonzalez B, Arca P, Mayo B & Suarez JE (1994) Detection, purification, and partial 629
characterization of plantaricin C, a bacteriocin produced by a Lactobacillus plantarum strain 630
of dairy origin. Applied and Environmental Microbiology 60: 2158-2163. 631
Gu RX, Yang ZQ, Li ZH, Chen SL & Luo ZL (2008) Probiotic properties of lactic acid 632
bacteria isolated from stool samples of longevous people in regions of Hotan, Xinjiang and 633
Bama, Guangxi, China. Anaerobe 14: 313-317. 634
Gueimonde M & Salminen S (2006) New methods for selecting and evaluating probiotics. 635
Digestive and Liver Disease 38: S242-S247. 636
Gueimonde M, Delgado S, Mayo B, Ruas-Madiedo P, Margolles A & de los Reyes-Gavilán 637
CG (2004) Viability and diversity of probiotic Lactobacillus and Bifidobacterium populations 638
included in commercial fermented milks. Food Research International 37: 839-850. 639
Guo Z, Wang J, Yan L, Chen W, Liu X-m & Zhang H-p (2009) In vitro comparison of 640
probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected 641
probiotic strains. LWT - Food Science and Technology 42: 1640-1646. 642
Hertzler SR & Clancy SM (2003) Kefir improves lactose digestion and tolerance in adults 643
with lactose maldigestion. Journal of the American Dietetic Association 103: 582-587. 644
Hong W-S, Chen H-C, Chen Y-P & Chen M-J (2009) Effects of kefir supernatant and lactic 645
acid bacteria isolated from kefir grain on cytokine production by macrophage. International 646
Dairy Journal 19: 244-251. 647

226
Huseini HF, Rahimzadeh G, Fazeli MR, Mehrazma M & Salehi M (2012) Evaluation of 648
wound healing activities of kefir products. Burns. 649
Jamaly N, Benjouad A & Bouksaim M (2011) Probiotic Potential of Lactobacillus strains 650
Isolated from Known Popular Traditional Moroccan Dairy Products. British Microbiology 651
Research Journal 1: 79-94. 652
Kesmen Z & Kacmaz N (2011) Determination of Lactic Microflora of Kefir Grains and Kefir 653
Beverage by Using Culture-Dependent and Culture-Independent Methods. Journal of Food 654
Science 76: M276-M283. 655
Klare I, Konstabel C, Muller-Bertling S, et al. (2005) Evaluation of new broth media for 656
microdilution antibiotic susceptibility testing of Lactobacilli, Pediococci, Lactococci, and 657
Bifidobacteria. Applied and Environmental Microbiology 71: 8982-8986. 658
Klare I, Konstabel C, Werner G, et al. (2007) Antimicrobial susceptibilities of Lactobacillus, 659
Pediococcus and Lactococcus human isolates and cultures intended for probiotic or nutritional 660
use. Journal of Antimicrobial Chemother 59: 900-912. 661
Klein G, Hallmann C, Casas IA, Abad J, Louwers J & Reuter G (2000) Exclusion of vanA, 662
vanB and vanC type glycopeptide resistance in strains of Lactobacillus reuteri and 663
Lactobacillus rhamnosus used as probiotics by polymerase chain reaction and hybridization 664
methods. Journal of Applied Microbiology 89: 815-824. 665
Koeuth T, Versalovic J & Lupski JR (1995) Differential subsequence conservation of 666
interspersed repetitive Streptococcus pneumoniae BOX elements in diverse bacteria. Genome 667
Research 5: 408-418. 668
Kubo M, Odani T, Nakamura S, Tokumaru S & Matsuda H (1992) Pharmacological study on 669
kefir--a fermented milk product in Caucasus. I. On antitumor activity Yakugaku Zasshi 112: 670
489-495. 671
Lemos Miguel MA, Dias de Castro AC & Ferreira Gomes Leite S (2008) Inhibition of 672
vancomycin and high-level aminoglycoside-resistant enterococci strains and Listeria 673
monocytogenes by bacteriocin-like substance produced by Enterococcus faecium E86. 674
Current Microbiology 57: 429-436. 675
Liu JR, Wang SY, Chen MJ, Chen HL, Yueh PY & Lin CW (2006) Hypocholesterolaemic 676
effects of milk-kefir and soyamilk-kefir in cholesterol-fed hamsters. Britsh Journal of 677
Nutrition 95: 939-946. 678
Lopitz-Otsoa F, Rementeria A, Elguezabal N & Garaizar J (2006) Kefir: a symbiotic yeasts-679
bacteria community with alleged healthy capabilities. Revista Iberoamericana de Micología 680
23: 67-74. 681
Lu ZH, Peng HH, Cao W, Tatsumi E & Li LT (2008) Isolation, characterization and 682
identification of lactic acid bacteria and yeasts from sour Mifen, a traditional fermented rice 683
noodle from China. Journal of Applied Microbiology 105: 893-903. 684
Magalhães KT, Dias DR, de Melo Pereira GV, et al. (2011) Chemical composition and 685
sensory analysis of cheese whey-based beverages using kefir grains as starter culture. 686
International Journal of Food Science & Technology 46: 871-878. 687
Maragkoudakis PA, Zoumpopoulou G, Miaris C, Kalantzopoulos G, Pot B & Tsakalidou E 688
(2006) Probiotic potential of Lactobacillus strains isolated from dairy products. International 689
Dairy Journal 16: 189-199. 690
Mathara JM, Schillinger U, Guigas C, et al. (2008) Functional characteristics of Lactobacillus 691
spp. from traditional Maasai fermented milk products in Kenya. Internantional Journal of 692
Food Microbiology 126: 57-64. 693
Mathur S & Singh R (2005) Antibiotic resistance in food lactic acid bacteria--a review. 694
International Journal of Food Microbiology 105: 281-295. 695

227
Mesas JM, Rodriguez MC & Alegre MT (2011) Characterization of lactic acid bacteria from 696
musts and wines of three consecutive vintages of Ribeira Sacra. Letters in Applied 697
Microbiology 52: 258-268. 698
Miguel MGdCP, Cardoso PG, Lago LdA & Schwan RF (2010) Diversity of bacteria present 699
in milk kefir grains using culture-dependent and culture-independent methods. Food Research 700
International 43: 1523-1528. 701
Monteagudo-Mera A, Caro I, Rodriguez-Aparicio LB, Rua J, Ferrero MA & Garcia-Armesto 702
MR (2011) Characterization of certain bacterial strains for potential use as starter or probiotic 703
cultures in dairy products. Journal of Food Protection 74: 1379-1386. 704
Moreira MEC, Santos MH, Pereira IO, Ferraz V, Barbosa LCA & Schneedorf JM (2008) 705
Atividade antiinflamatória de carboidrato produzido por fermentação aquosa de grãos de 706
quefir. Química Nova 31: 1738-1742. 707
Mori K, Yamazaki K, Ishiyama T, et al. (1997) Comparative sequence analyses of the genes 708
coding for 16S rRNA of Lactobacillus casei-related taxa. International Journal of Systematic 709
Bacteriology 47: 54-57. 710
Neri DFM, Balcão VM, Dourado FOQ, Oliveira JMB, Carvalho LB & Teixeira JA (2009) 711
Galactooligosaccharides production by β-galactosidase immobilized onto magnetic 712
polysiloxane–polyaniline particles. Reactive and Functional Polymers 69: 246-251. 713
Nishida S, Michinaka A, Nakashima K, Iino H & Fujii T (2008) Evaluation of the probiotic 714
potential of Lactobacillus paracasei KW3110 based on in vitro tests and oral administration 715
tests in healthy adults. The Journal of General and Applied Microbiology 54: 267-276. 716
Ota A (1999) Protection against an infectious disease by enterohaemorrhagic E. coli 0–157. 717
Medical hypotheses 53: 87-88. 718
Otieno DO, Ashton JF & Shah NP (2006) Evaluation of enzymic potential for 719
biotransformation of isoflavone phytoestrogen in soymilk by Bifidobacterium animalis, 720
Lactobacillus acidophilus and Lactobacillus casei. Food Research International 39: 394-407. 721
Otieno DO, Ashton JF & Shah NP (2007) Role of microbial strain and storage temperatures in 722
the degradation of isoflavone phytoestrogens in fermented soymilk with selected β-723
glucosidase producing Lactobacillus casei strains. Food Research International 40: 371-380. 724
Otles S & Cagindi O (2003) Kefir: A probiotic dairy-composition, nutritional and therapeutic 725
aspects Pakistan Journal of Nutrition 2: 54-59. 726
Palys T, Nakamura LK & Cohan FM (1997) Discovery and classification of ecological 727
diversity in the bacterial world: the role of DNA sequence data. International Journal of 728
Systematic Bacteriology 47: 1145-1156. 729
Papamanoli E, Tzanetakis N, Litopoulou-Tzanetaki E & Kotzekidou P (2003) 730
Characterization of lactic acid bacteria isolated from a Greek dry-fermented sausage in 731
respect of their technological and probiotic properties. Meat Science 65: 859-867. 732
Park AR & Oh DK (2010) Galacto-oligosaccharide production using microbial beta-733
galactosidase: current state and perspectives. Applied Microbiology and Biotechnology 85: 734
1279-1286. 735
Pinto M, Robine-Leon S, Appay MD, et al. (1983) Enterocyte-like differentiation and 736
polarization of the human colon carcinoma cell line Caco-2 in culture. Biology of the Cell 47: 737
323-330. 738
Pithva S, Ambalam P, Dave JM & Vvas BR (2012) Potential of Probiotic Lactobacillus 739
Strains as Food Additivies. (El-Samragy Y, ed.^eds.), p.^pp. 175-190. InTech. 740
Plessas S, Alexopoulos A, Voidarou C, Stavropoulou E & Bezirtzoglou E (2011) Microbial 741
ecology and quality assurance in food fermentation systems. The case of kefir grains 742
application. Anaerobe 17: 483-485. 743
Reid G, Sanders ME, Gaskins HR, et al. (2003) New scientific paradigms for probiotics and 744
prebiotics. Journal of Clinical Gastroenterology 37: 105-118. 745

228
Ripamonti B, Agazzi A, Bersani C, et al. (2011) Screening of species-specific lactic acid 746
bacteria for veal calves multi-strain probiotic adjuncts. Anaerobe 17: 97-105. 747
Rodrigues KL, Caputo LR, Carvalho JC, Evangelista J & Schneedorf JM (2005) 748
Antimicrobial and healing activity of kefir and kefiran extract. International Journal of 749
Antimicrobial Agents 25: 404-408. 750
Rossetti L & Giraffa G (2005) Rapid identification of dairy lactic acid bacteria by M13-751
generated, RAPD-PCR fingerprint databases. Journal of Microbiology Methods 63: 135-144. 752
Ryan MP, Rea MC, Hill C & Ross P (1996) An application in Cheedar Cheese Manufacture 753
for a Strain of Lactococcus lactis producing a novel broad-spectrum bacteriocin, Lacticin 754
3147. Applied and Environmental Microbiology 62: 612-619. 755
Saarela M, Mogensen G, Fondén R, Mättö J & Mattila-Sandholm T (2000) Probiotic bacteria: 756
safety, functional and technological properties. Journal of Biotechnology 84: 197-215. 757
Sánchez B, Fernández-García M, Margolles A, de los Reyes-Gavilán CG & Ruas-Madiedo P 758
(2010) Technological and probiotic selection criteria of a bile-adapted Bifidobacterium 759
animalis subsp. lactis strain. International Dairy Journal 20: 800-805. 760
Sanders ME (2008) Probiotics: definition, sources, selection, and uses. Clinical Infectious 761
Diseases 46 Suppl 2: S58-61; discussion S144-151. 762
Santos A, San Mauro M, Sanchez A, Torres JM & Marquina D (2003) The Antimicrobial 763
Properties of Different Strains of Lactobacillus spp. Isolated from Kefir. Systematic and 764
Applied Microbiology 26: 434-437. 765
Sarkar S (2007) Potential of kefir as a dietetic beverage – a review. British Food Journal 109: 766
280-290. 767
Schillinger U & Lucke FK (1989) Antibacterial activity of Lactobacillus sake isolated from 768
meat. Applied and Environmental Microbiology 55: 1901-1906. 769
Schmidt P, Vass A & Szakaly S (1984) Effect of fermented milk diets on regeneration of the 770
rat liver. Acta Med Hung 41: 163-169. 771
Servin AL (2004) Antagonistic activities of lactobacilli and bifidobacteria against microbial 772
pathogens. FEMS Microbiology Reviews 28: 405-440. 773
Shiomi M, Sasaki K, Murofushi M & Aibara K (1982) Antitumor activity in mice of orally 774
administered polysaccharide from Kefir grain. Japanese journal of medical science & biology 775
35: 75-80. 776
Tamime AY (2006) Production of Kefir, Koumiss and Other Related Products. Fermented 777
Milk,(Tamime AY, ed.^eds.), p.^pp. 174-216. Blackwell Science Ltd, Oxford, UK. 778
Temmerman R, Pot B, Huys G & Swings J (2003) Identification and antibiotic susceptibility 779
of bacterial isolates from probiotic products. International Journal of Food Microbiology 81: 780
1-10. 781
Tuomola EM & Salminen SJ (1998) Adhesion of some probiotic and dairy Lactobacillus 782
strains to Caco-2 cell cultures. International Journal of Food Microbiology 41: 45-51. 783
Urdaneta E, Barrenetxe J, Aranguren P, Irigoyen A, Marzo F & Ibáñez FC (2007) Intestinal 784
beneficial effects of kefir-supplemented diet in rats. Nutrition Research 27: 653-658. 785
Urraza PJ, Gómez-Zavaglia A, Lozano ME, Romanowski V & De Antoni G (2000) DNA 786
fingerprinting of thermophilic lactic acid bacteria using repetitive sequence-based polymerase 787
chain reaction. Journal of Dairy Research 67: 381-392. 788
Usman & Hosono A (1999) Bile Tolerance, Taurocholate Deconjugation, and Binding of 789
Cholesterol by Lactobacillus gasseri Strains. Journal of Dairy Science 82: 243-248. 790
van Belkum MJ, Hayema BJ, Geis A, Kok J & Venema G (1989) Cloning of two bacteriocin 791
genes from a lactococcal bacteriocin plasmid. Applied and Environmental Microbiology 55: 792
1187-1191. 793
Vasiljevic T & Shah NP (2008) Probiotics—From Metchnikoff to bioactives. International 794
Dairy Journal 18: 714-728. 795

229
Vinderola G, Perdigon G, Duarte J, Thangavel D, Farnworth E & Matar C (2006) Effects of 796
kefir fractions on innate immunity. Immunobiology 211: 149-156. 797
Vinderola G, Capellini B, Villarreal F, Suárez V, Quiberoni A & Reinheimer J (2008) 798
Usefulness of a set of simple in vitro tests for the screening and identification of probiotic 799
candidate strains for dairy use. LWT - Food Science and Technology 41: 1678-1688. 800
Vizoso Pinto MG, Franz CMAP, Schillinger U & Holzapfel WH (2006) Lactobacillus spp. 801
with in vitro probiotic properties from human faeces and traditional fermented products. 802
International Journal of Food Microbiology 109: 205-214. 803
Wang X, Haruta S, Wang P, Ishii M, Igarashi Y & Cui Z (2006) Diversity of a stable 804
enrichment culture which is useful for silage inoculant and its succession in alfalfa silage. 805
FEMS Microbiology Ecology 57: 106-115. 806
Wollowski I, Rechkemmer G & Pool-Zobel BL (2001) Protective role of probiotics and 807
prebiotics in colon cancer. The American Journal of Clinical Nutrition 73: 451S-455S. 808
Yang S, Wang L, Yan Q, Jiang Z & Li L (2009) Hydrolysis of soybean isoflavone glycosides 809
by a thermostable β-glucosidase from Paecilomyces thermophila. Food Chemistry 115: 1247-810
1252. 811
Zago M, Fornasari ME, Carminati D, et al. (2011) Characterization and probiotic potential of 812
Lactobacillus plantarum strains isolated from cheeses. Food Microbiology 28: 1033-1040. 813
814
815
816
817
818
819
820
821
822
823
824
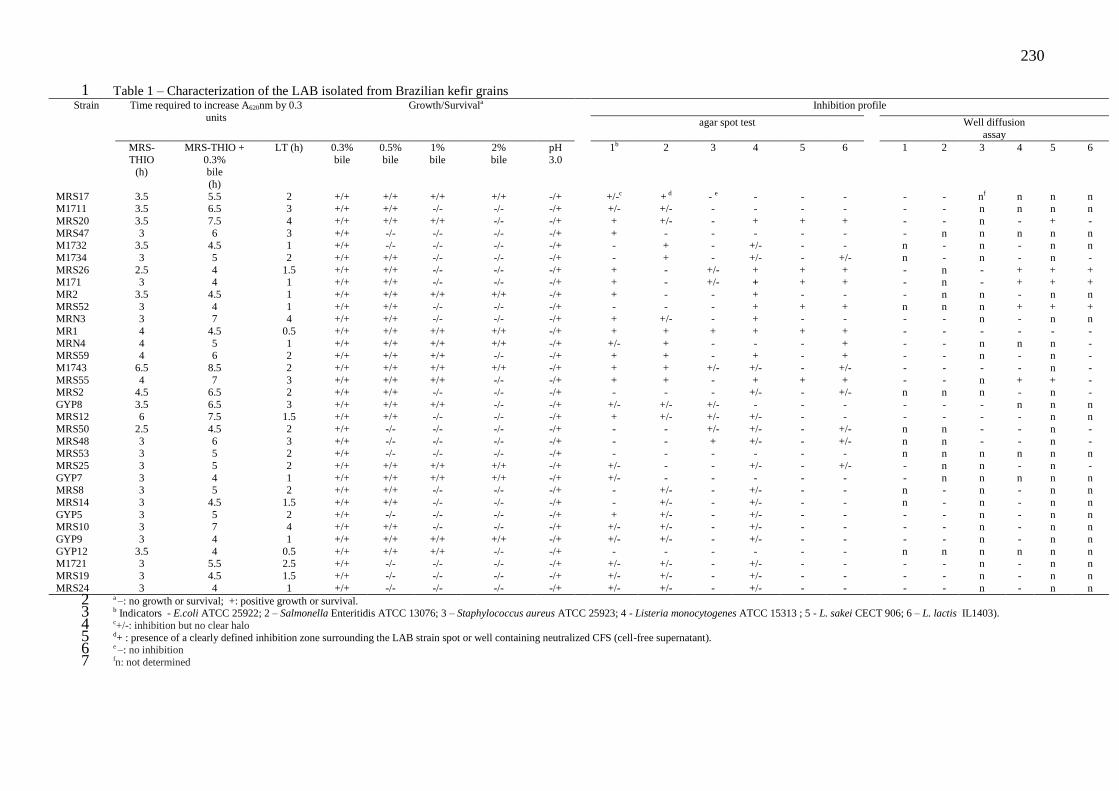
230
Table 1 – Characterization of the LAB isolated from Brazilian kefir grains 1
Strain Time required to increase A620nm by 0.3
units
Growth/Survivala Inhibition profile
agar spot test Well diffusion
assay
MRS-
THIO (h)
MRS-THIO +
0.3% bile
(h)
LT (h) 0.3%
bile
0.5%
bile
1%
bile
2%
bile
pH
3.0
1b 2 3 4 5 6 1 2 3 4 5 6
MRS17 3.5 5.5 2 +/+ +/+ +/+ +/+ -/+ +/-c + d - e - - - - - nf n n n M1711 3.5 6.5 3 +/+ +/+ -/- -/- -/+ +/- +/- - - - - - - n n n n
MRS20 3.5 7.5 4 +/+ +/+ +/+ -/- -/+ + +/- - + + + - - n - + -
MRS47 3 6 3 +/+ -/- -/- -/- -/+ + - - - - - - n n n n n M1732 3.5 4.5 1 +/+ -/- -/- -/- -/+ - + - +/- - - n - n - n n
M1734 3 5 2 +/+ +/+ -/- -/- -/+ - + - +/- - +/- n - n - n -
MRS26 2.5 4 1.5 +/+ +/+ -/- -/- -/+ + - +/- + + + - n - + + + M171 3 4 1 +/+ +/+ -/- -/- -/+ + - +/- + + + - n - + + +
MR2 3.5 4.5 1 +/+ +/+ +/+ +/+ -/+ + - - + - - - n n - n n
MRS52 3 4 1 +/+ +/+ -/- -/- -/+ - - - + + + n n n + + + MRN3 3 7 4 +/+ +/+ -/- -/- -/+ + +/- - + - - - - n - n n
MR1 4 4.5 0.5 +/+ +/+ +/+ +/+ -/+ + + + + + + - - - - - -
MRN4 4 5 1 +/+ +/+ +/+ +/+ -/+ +/- + - - - + - - n n n - MRS59 4 6 2 +/+ +/+ +/+ -/- -/+ + + - + - + - - n - n -
M1743 6.5 8.5 2 +/+ +/+ +/+ +/+ -/+ + + +/- +/- - +/- - - - - n -
MRS55 4 7 3 +/+ +/+ +/+ -/- -/+ + + - + + + - - n + + - MRS2 4.5 6.5 2 +/+ +/+ -/- -/- -/+ - - - +/- - +/- n n n - n -
GYP8 3.5 6.5 3 +/+ +/+ +/+ -/- -/+ +/- +/- +/- - - - - - - n n n MRS12 6 7.5 1.5 +/+ +/+ -/- -/- -/+ + +/- +/- +/- - - - - - - n n
MRS50 2.5 4.5 2 +/+ -/- -/- -/- -/+ - - +/- +/- - +/- n n - - n -
MRS48 3 6 3 +/+ -/- -/- -/- -/+ - - + +/- - +/- n n - - n - MRS53 3 5 2 +/+ -/- -/- -/- -/+ - - - - - - n n n n n n
MRS25 3 5 2 +/+ +/+ +/+ +/+ -/+ +/- - - +/- - +/- - n n - n -
GYP7 3 4 1 +/+ +/+ +/+ +/+ -/+ +/- - - - - - - n n n n n MRS8 3 5 2 +/+ +/+ -/- -/- -/+ - +/- - +/- - - n - n - n n
MRS14 3 4.5 1.5 +/+ +/+ -/- -/- -/+ - +/- - +/- - - n - n - n n
GYP5 3 5 2 +/+ -/- -/- -/- -/+ + +/- - +/- - - - - n - n n MRS10 3 7 4 +/+ +/+ -/- -/- -/+ +/- +/- - +/- - - - - n - n n
GYP9 3 4 1 +/+ +/+ +/+ +/+ -/+ +/- +/- - +/- - - - - n - n n
GYP12 3.5 4 0.5 +/+ +/+ +/+ -/- -/+ - - - - - - n n n n n n M1721 3 5.5 2.5 +/+ -/- -/- -/- -/+ +/- +/- - +/- - - - - n - n n
MRS19 3 4.5 1.5 +/+ -/- -/- -/- -/+ +/- +/- - +/- - - - - n - n n
MRS24 3 4 1 +/+ -/- -/- -/- -/+ +/- +/- - +/- - - - - n - n n a –: no growth or survival; +: positive growth or survival. 2 b Indicators - E.coli ATCC 25922; 2 – Salmonella Enteritidis ATCC 13076; 3 – Staphylococcus aureus ATCC 25923; 4 - Listeria monocytogenes ATCC 15313 ; 5 - L. sakei CECT 906; 6 – L. lactis IL1403). 3 c+/-: inhibition but no clear halo 4 d+ : presence of a clearly defined inhibition zone surrounding the LAB strain spot or well containing neutralized CFS (cell-free supernatant). 5 e –: no inhibition 6 fn: not determined7

231
1 2 Table 2 – Species and corresponding origin of LAB strains isolated from Brazilian kefir 3
grains 4 Species Isolated
strains
Geographic region
L. lactis cremoris MRS17 Niterói, RJa
M1711 Niterói, RJ
MRS20 Niterói, RJ
MRS47 Lavras, MGb
M1732 Niterói, RJ
M1734 Niterói, RJ
MRS26 Niterói, RJ
M171 Niterói, RJ
L. lactis lactis MR2 Viçosa, MG
MRS52 Lavras, MG
MRN3 Viçosa, MG
L. casei/paracasei MR1 Viçosa, MG
MRN4 Niterói, RJ
MRS59 Alfenas, MG
M1743 Alfenas, MG
MRS55 Alfenas, MG
Leuconostoc mesenteroides MRS2 Niterói, RJ
GYP8 Niterói, RJ
MRS12 Niterói, RJ
MRS50 Lavras, MG
MRS48 Alfenas, MG
MRS53 Alfenas, MG
MRS25 Niterói, RJ
GYP7 Niterói, RJ
MRS8 Niterói, RJ
MRS14 Niterói, RJ
GYP5 Niterói, RJ
MRS10 Niterói, RJ
GYP9 Niterói, RJ
GYP12 Niterói, RJ
M1721 Niterói, RJ
MRS19 Niterói, RJ
MRS24 Niterói, RJ aRio de Janeiro State;
bMinas Gerais State 5
6 7
8
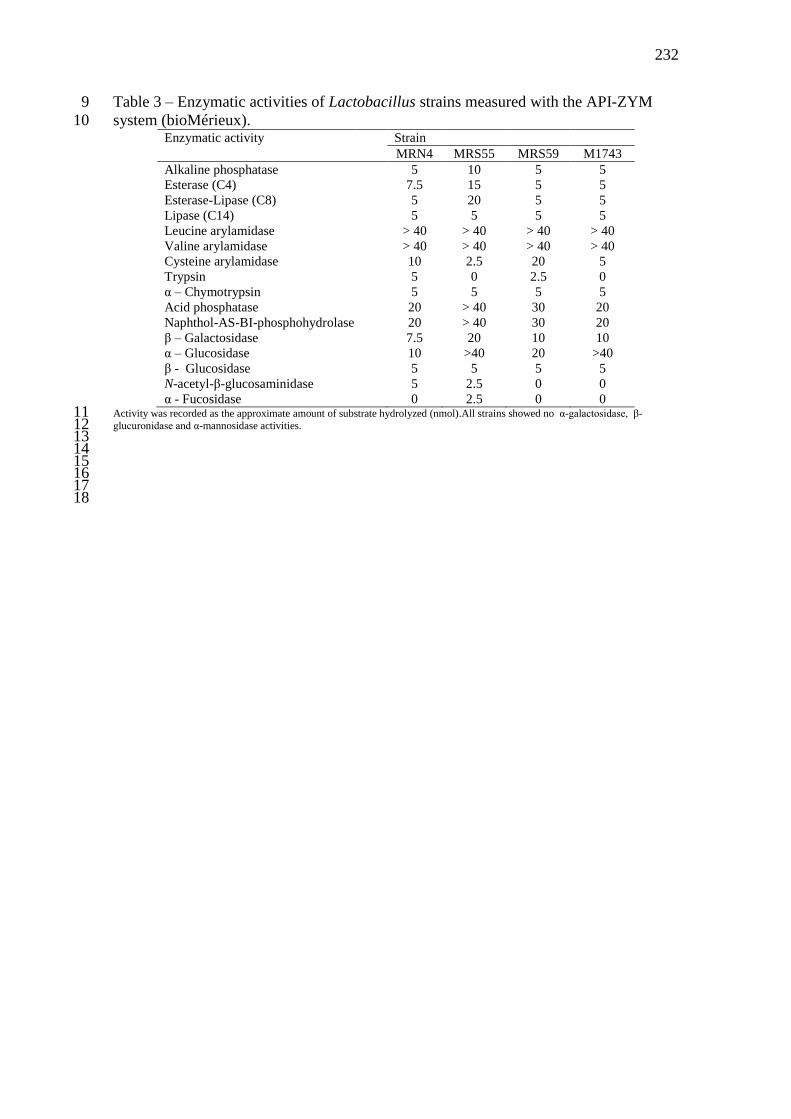
232
Table 3 – Enzymatic activities of Lactobacillus strains measured with the API-ZYM 9
system (bioMérieux). 10 Enzymatic activity Strain
MRN4 MRS55 MRS59 M1743
Alkaline phosphatase 5 10 5 5
Esterase (C4) 7.5 15 5 5
Esterase-Lipase (C8) 5 20 5 5
Lipase (C14) 5 5 5 5
Leucine arylamidase > 40 > 40 > 40 > 40
Valine arylamidase > 40 > 40 > 40 > 40
Cysteine arylamidase 10 2.5 20 5
Trypsin 5 0 2.5 0
α – Chymotrypsin 5 5 5 5
Acid phosphatase 20 > 40 30 20
Naphthol-AS-BI-phosphohydrolase 20 > 40 30 20
β – Galactosidase 7.5 20 10 10
α – Glucosidase 10 >40 20 >40
β - Glucosidase 5 5 5 5
N-acetyl-β-glucosaminidase 5 2.5 0 0
α - Fucosidase 0 2.5 0 0 Activity was recorded as the approximate amount of substrate hydrolyzed (nmol).All strains showed no α-galactosidase, β-11 glucuronidase and α-mannosidase activities. 12 13 14 15 16 17
18
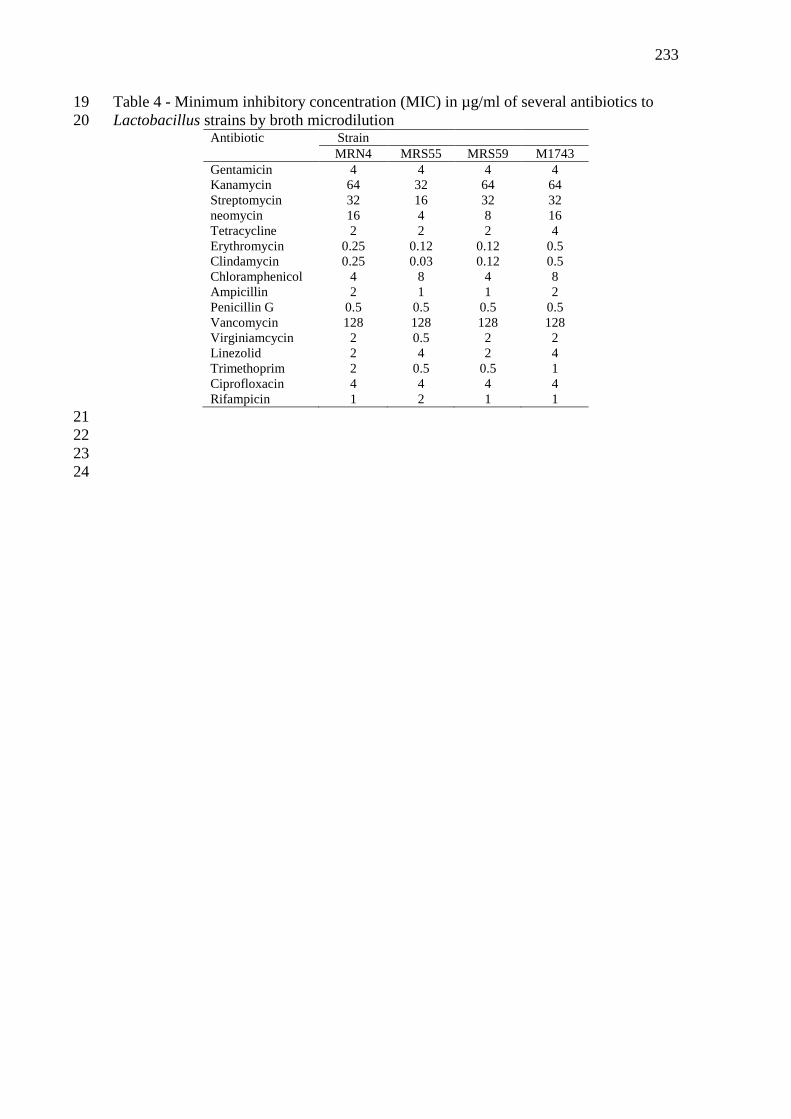
233
Table 4 - Minimum inhibitory concentration (MIC) in µg/ml of several antibiotics to 19
Lactobacillus strains by broth microdilution 20 Antibiotic Strain
MRN4 MRS55 MRS59 M1743
Gentamicin 4 4 4 4
Kanamycin 64 32 64 64
Streptomycin 32 16 32 32
neomycin 16 4 8 16
Tetracycline 2 2 2 4
Erythromycin 0.25 0.12 0.12 0.5
Clindamycin 0.25 0.03 0.12 0.5
Chloramphenicol 4 8 4 8
Ampicillin 2 1 1 2
Penicillin G 0.5 0.5 0.5 0.5
Vancomycin 128 128 128 128
Virginiamcycin 2 0.5 2 2
Linezolid 2 4 2 4
Trimethoprim 2 0.5 0.5 1
Ciprofloxacin 4 4 4 4
Rifampicin 1 2 1 1
21
22
23
24

234
25
26
27
28
29
30
31
32
33
34
35
36
37
38
Figure 1 – Antagonist activities and proteolysis sensitivity of L. lactis strains isolated 39
from kefir grains. A – Inhibition of E. coli ATCC 25922 by L. lactis subsp. cremoris 40
MRS26 (1), MRS20 (2) and M171 (3) strains. B - Antimicrobial activity of MRS26 41
strain against the indicator Lb. sakei CECT 906 and its sensitivity to proteolysis. Wells: 42
1 – neutralized cell- free supernatant of L. lactis subsp. cremoris MRS26; PK - 43
proteinase K (20mg/ml) and PN – pronase (20 mg/ml). 44
45

235
Figure 2 – Combined dendrogram obtained from rep-PCR and RAPD-PCR profiles, 46
using BOXA1R, BOXA2R and M13 primers. 47
48
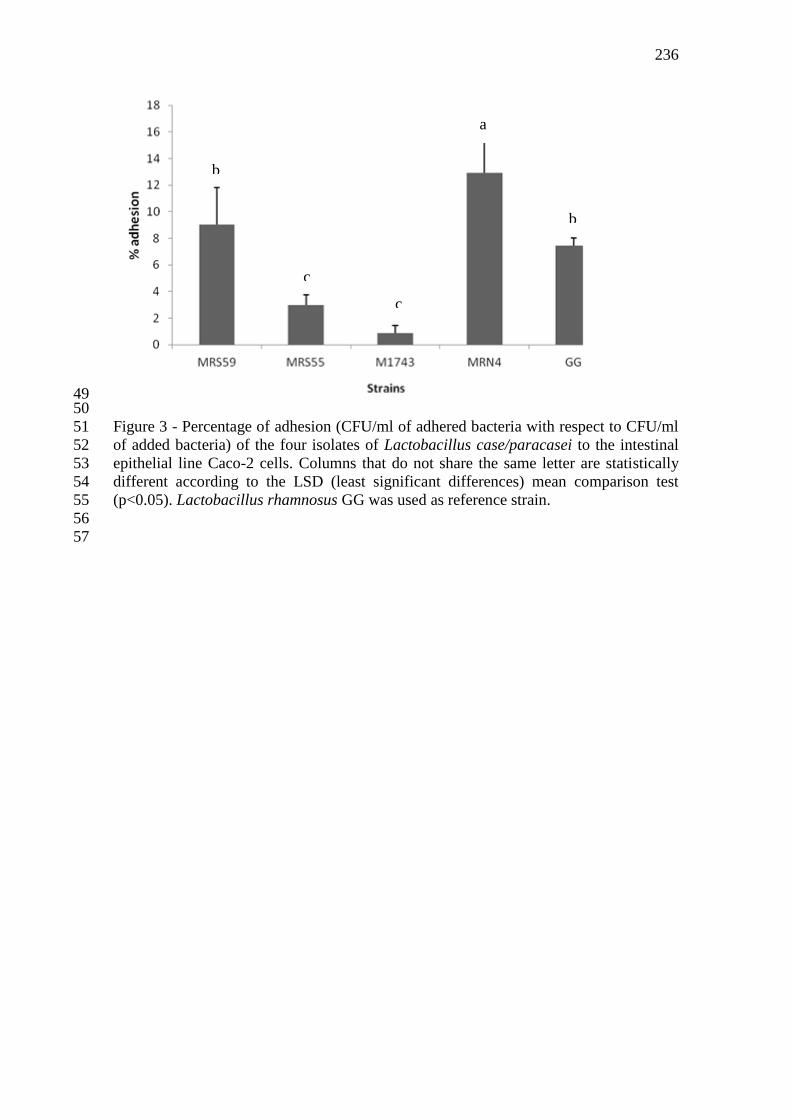
236
49 50
Figure 3 - Percentage of adhesion (CFU/ml of adhered bacteria with respect to CFU/ml 51
of added bacteria) of the four isolates of Lactobacillus case/paracasei to the intestinal 52
epithelial line Caco-2 cells. Columns that do not share the same letter are statistically 53
different according to the LSD (least significant differences) mean comparison test 54
(p<0.05). Lactobacillus rhamnosus GG was used as reference strain. 55
56
57
b
b
b
b b
b
b
b c
c
c
c
c
c
c
a
c
c
c
c
c
c
c