Embed Size (px)
Citation preview

UNIVERSIDADE DE BRASÍLIA FACULDADE DE CEILÂNDIA
CURSO DE FARMÁCIA
LUSIANE DA CONCEIÇÃO GOMES LIMA
ANÁLISE DA PRODUÇÃO HEPÁTICA DE ESPÉCIES REATIVAS DE OXIGÊNIO DURANTE A FEBRE EM RATOS.
BRASÍLIA - DF 2016

LUSIANE DA CONCEIÇÃO GOMES LIMA
ANÁLISE DA PRODUÇÃO HEPÁTICA DE ESPÉCIES REATIVAS DE OXIGÊNIO DURANTE A FEBRE EM RATOS.
Trabalho de Conclusão de Curso apresentado ao Curso de Farmácia da Universidade de Brasília, Faculdade de Ceilândia, como requisito parcial para obtenção do grau de Farmacêutico. Orientadora: Profa. Dra. Fabiane Hiratsuka Veiga de Souza
BRASÍLIA - DF 2016

LIMA, Lusiane da Conceição Gomes. Análise Da Produção Hepática De Espécies Reativas De Oxigênio Durante A Febre Em Ratos/Lusiane da Conceição Gomes Lima. Brasília. UnB/ Faculdade de Ceilândia, Distrito Federal - Brasília, 2016. 42p. Trabalho de Conclusão de Curso apresentado ao Curso de Farmácia como requisito parcial para obtenção do grau de Farmacêutico. Universidade de Brasília, Faculdade de Ceilândia, Distrito Federal – Brasília – DF.

LUSIANE DA CONCEIÇÃO GOMES LIMA
ANÁLISE DA PRODUÇÃO HEPÁTICA DE ESPÉCIES REATIVAS DE OXIGÊNIO DURANTE A FEBRE EM RATOS.
BANCA EXAMINADORA
__________________________________________________ Orientadora: Profa. Dra. Fabiane Hiratsuka Veiga de Souza
(FCE/ Universidade de Brasília)
___________________________________________________ Prof. Dr. Paulo Gustavo Barboni Dantas do Nascimento
(FCE/ Universidade de Brasília)
___________________________________________________ Profa. Dra. Vivian da Silva Santos (FCE/ Universidade de Brasília)
BRASÍLIA - DF 2016

AGRADECIMENTOS
Agradeço primeiramente а Deus, o centro de tudo, o meu guia. A luta não foi
fácil, mas em momento algum pensei em desistir ou fraquejar, pois o Teu amor, e a fé
que em Ti deposito me levantaram quando as forças pareciam querer me abandonar,
obrigada por tudo meu Pai! A Ti dedico essa conquista, na certeza de que continuarás
a conduzir os meus passos em direção a Sua vontade.
À Santíssima Virgem Maria, que me acolheu e me sustentou nos momentos
mais difíceis. Agradeço-te, Mãezinha, por me ser uma fortaleza em minha vida, por
ser um exemplo pra mim e por não me deixar esquecer que Deus é a força que conduz
a minha vida. Agradeço imensamente aos meus pais Eleni e José por me transmitirem
valores fundamentais. Dedico essa, bem como as minhas demais conquistas a vocês,
muito obrigada por estarem sempre ao meu lado e lutarem para a realização desse
sonho. Mãe eu te agradeço também pela dedicação diária, pelas orações, por todo o
incentivo, por não me deixar desanimar e por me ensinar sempre a esperar o melhor
de Deus para mim, você é o meu maior exemplo.
Aos meus irmãos Cristiane e Francisco por acompanharem de perto essa
jornada e por serem fonte de incentivo e referências para mim. Agradeço também ao
meu precioso sobrinho Ryan Lucas que chegou logo no início dessa trajetória e trouxe
uma felicidade enorme para a minha vida. Essa conquista eu compartilho com vocês
com muita alegria, pois vocês são parte dessa vitória!
Ao meu namorado e melhor amigo, Jean, por todo amor, carinho e
compreensão dedicados a mim ao longo desses meses. Muito obrigada por me ajudar
e me acalmar sempre, o seu apoio constante fez com que eu me sentisse forte e
confiante. Serei eternamente grata a você por tudo que fez e faz por mim.
À minha orientadora Profa. Drª Fabiane, pelo empenho e dedicação, a admiro
muito, os seus ensinamentos contribuíram imensamente para o desenvolvimento
deste trabalho.
E a todos qυе direta оυ indiretamente fizeram parte dessa caminhada, о mеυ
muito obrigada.

RESUMO
A febre é o sinal clínico mais importante da resposta de fase aguda frente à uma infecção. Estudos sugerem que as espécies reativas de oxigênio (ERO’s) são liberadas por macrófagos como uma defesa contra a invasão bacteriana, apresentando um papel importante no controle de muitos processos fisiológicos. Sabendo que o fígado representa um componente importante do sistema imune, sendo altamente povoado por diversas células imunes durante processos infecciosos e possuindo componentes necessários para a defesa de primeira linha, é de suma importância avaliar a produção de ERO’s por esse órgão durante a resposta febril. Neste estudo avaliou-se a produção de ERO’s no fígado de ratos durante a febre induzida por LPS. Para isso, foram utilizados ratos Wistar machos e fêmeas. Os animais receberam injeção endovenosa de LPS (5 μg/kg) ou do seu veículo (salina 0,9%, 2 mL/Kg). A partir de então, cada animal teve sua temperatura corporal aferida a cada 15 minutos, durante 6 h. Em seguida os animais foram anestesiados e eutanasiados por decapitação para a retirada do fígado. A quantificação de ERO’s foi feita por ressonância paramagnética eletrônica (RPE) utilizando o marcador CMH. Os resultados do presente estudo demonstram que os animais tratados com LPS apresentam maior concentração de ERO’s no fígado, esse aumento pode ser decorrente de uma maior produção de ERO’s e/ou da redução dos sistemas antioxidantes. Palavras-chave: Febre, ERO’s, Lipopolissacarídeo, LPS, Fígado.

ABSTRACT Fever is the most important clinical sign in an acute phase caused by an infection. Studies suggest that reactive oxygen species (ROS) are released by macrophages as a defense against bacterial invasion, having an important role controlling many physiological processes. Knowing that the liver is an important component of the immune system, being highly populated by immune cells during infection processes and having components necessary for first-line defense it is indubitably important to evaluate the ROS role by this organ during a fever response. In this study was been evaluated the ROS production in rat liver during fever induced by LPS. For this, were used Wistar rats, males and females. The animals received an intravenous injection of LPS (5 mg / kg) or its vehicle (0.9% saline, 2 ml / kg). From then on, each animal had its body temperature measured every 15 minutes for 6 h. Then, the animals were euthanized and the liver was removed. ROS quantification was made by electron paramagnetic resonance using standard CMH. The results of this study demonstrate that animals treated with LPS had a higher concentration of ROS in the liver. This increase may be due to greater production of ROS and/or the reduction of antioxidant agents. Keywords: Fever.ROS. Lipopolysaccharide. LPS. Liver.

LISTA DE FIGURAS
Figura 1: Mecanismo pelo qual a IL-1 (e possivelmente TNF e IL-6) age sobre o “organum vasculosum laminae terminalis” (OVLT) para produzir a febre..................14
Figura 2: Mecanismo da febre de origem infecciosa.................................................15
Figura 3: Compartimentação subcelular de ERO’s................................................... 19
Figura 4: Participação de ERO’s mitocondrial na regulação de diversos processos celulares.....................................................................................................................20
Figura 5: Efeito da administração de LPS sobre a temperatura corporal..................30
Figura 6: Amplitude do sinal de RPE em função da [CP•] em padrões de calibração de [CP•] dissolvidos em tampão Krebs-Hepes (círculos)...........................................31
Figura 7: Espectro típico de CM• a partir de amostras do fígado de animais após a administração de LPS ou salina..................................................................................32
Figura 8: Aumento na [ERO’S] no fígado de ratos após a administração endovenosa de LPS........................................................................................................................32

LISTA DE SÍMBOLOS E ABREVIATURAS
AA - ácido araquidônico
AINE - antiinflamatório não esteroidal
APOHA - área pré-óptica do hipotálamo anterior
AMPc - adenosina monofosfato cíclico
ATP - adenosina trifosfato
°C - graus Celsius
C - cascata de complemento
Cm - centímetros
cK - células de Kupffer
COX - ciclooxigenase
C5a - componente 5a do complemento
DNA - ácido desoxirribonucleico
EP - receptor de prostaglandina
ERO’s - espécies reativas de oxigênio
g - gramas
GHz - gigahertz
h - horas
H2O2 - peróxido de hidrogênio
icv - intracerebroventicular
IL - interleucina
IL-1β - interleucina 1beta
i.p - intraperitoneal
i.v - intravenosa
Kg - quilogramas
LPS - lipopolissacarídeo
mg - miligramas
min - minutos
mL - mililitros
mM - milimolar
mW - megawatt
n - número de animais
NADH - nicotinamida-adenina-dinucleotídio

NO - óxido nítrico
O2 - oxigênio
OH - hidroxila
OVLT – organum vasculosum laminae terminalis
PG - prostaglandina
PGE2 - prostaglandina E
PGG2 - prostaglandina G2
PGH2 - prostaglandina H2
RNAm - ácido ribonucleico mensageiro
RPE - ressonância paramagnética eletrônica
SEC - células endoteliais sinusoidais
SC - células estreladas
TLR-4 - toll-like receptor 4
TNF- α - fator de necrose tumoral α
TXA2 - tromboxano A2
μg - microgramas
μL - microlitros
VDAC - canal de ânion dependente de voltagem
v/v - volume/ volume

SUMÁRIO
1. INTRODUÇÃO ................................................................................................... 12
1.1. Termorregulação ........................................................................................ 12
1.2. Resposta Febril .......................................................................................... 12
1.3. Pirógenos .................................................................................................... 16
1.4. Espécies Reativas de Oxigênio- ERO’s .................................................... 17
1.5. Fígado ......................................................................................................... 21
2. JUSTIFICATIVA ................................................................................................. 24
3. OBJETIVOS ....................................................................................................... 25
3.1. Objetivo Geral ............................................................................................. 25
3.2. Objetivo Específico .................................................................................... 25
4. MATERIAIS E MÉTODOS .................................................................................. 26
4.1. Drogas ......................................................................................................... 26
4.2. Animais ....................................................................................................... 26
4.3. Cirurgia para Implante de Transmissores de Temperatura na Cavidade Peritoneal .............................................................................................................. 27
4.4. Mensuração Da Temperatura Corporal Dos Ratos.................................. 27
4.5. Procedimentos Experimentais .................................................................. 28
4.6. Ressonância Paramagnética Eletrônica (RPE) ........................................ 28
4.7. Estatística ................................................................................................... 29
5. RESULTADOS ................................................................................................... 30
5.1. Efeito da Administração de LPS sobre a Temperatura Corporal ........... 30
5.2. Efeito da Administração de LPS sobre a Produção de ERO’s ............... 31
6. DISCUSSÃO ...................................................................................................... 34
7. CONCLUSÃO ..................................................................................................... 37
8. REFERÊNCIAS BIBLIOGRÁFICAS .................................................................. 38
9. ANEXOS ............................................................................................................. 42
9.1. Anexo 1 ....................................................................................................... 42

12
1. INTRODUÇÃO
1.1. Termorregulação
A termorregulação consiste na manutenção da temperatura corporal central
relativamente constante. A temperatura corporal normal em humanos é de
aproximadamente 37°C, podendo variar entre 33,2°C e 38,2°C. Tais oscilações
podem ocorrer ao longo do dia (ritmo circadiano), ao longo de um mês (ciclo
menstrual) e ao longo de toda a vida (envelhecimento) (TANSEY & JOHNSON, 2015).
Os mecanismos termorreguladores autonômicos têm grande importância na
homeostase fisiológica. Para garantir a função fisiológica ideal e a sobrevivência, os
seres humanos devem ser capazes de preservar a temperatura corporal frente aos
desafios da temperatura ambiente. Neste contexto, vários mecanismos
termorreguladores visam à manutenção da temperatura corporal normal, tais como os
mecanismos de conservação e produção de calor (vasoconstrição, piloereção) e os
mecanismos de dissipação de calor (sudorese, vasodilatação cutânea) (TANSEY
& JOHNSON, 2015).
É importante destacar que a termorregulação ocorre por meio de um sistema
complexo, sendo que o centro termorregulatório está localizado na área pré-óptica do
hipotálamo anterior (APOHA). Os neurônios termossensíveis dessa região integram
sinais aferentes sobre a temperatura corporal central e periférica e provocam várias
respostas comportamentais e fisiológicas, controlando a produção ou a dissipação do
calor (BOULANT et al., 1997).
A frequência de disparo dos neurônios dessa região é afetada tanto por
variações na temperatura sanguínea da área adjacente, como por influência de
conexões diretas com termorreceptores distribuídos na pele e nos músculos
(BOULANT, 2000).
Vale ressaltar que desvios anormais da temperatura desafiam a
termorregulação do organismo. Sendo assim, variações da temperatura fora do
intervalo normal podem ser fatais (TANSEY & JOHNSON, 2015).
1.2. Resposta Febril
Desde os tempos antigos a resposta febril tem sido reconhecida como um
importante sintoma de doença, sendo tratada durante muitos anos como um
fenômeno que ameaçava a vida. Porém, atualmente a febre tem sido considerada
parte importante dos mecanismos de defesa do organismo (HARDEN et al., 2015).

13
A febre é o sinal clínico mais importante da resposta de fase aguda frente a
uma infecção bacteriana, de modo que estudos realizados no século passado, e nas
últimas décadas, têm contribuído muito para a compreensão deste processo complexo
e dos mecanismos responsáveis por sua indução (SOARES et al., 2012; NETEA;
KULLBERG; VAN DER MEER, 2000).
Kluger (1991) define a febre como uma resposta biológica complexa, altamente
integrada, caracterizada pela elevação controlada da temperatura corporal, resultante
da atividade neuronal hipotalâmica. Diante desse conceito, é importante salientar que
apesar das infecções serem a causa mais comum da febre, outras várias doenças não
infecciosas também podem ter a febre como sua apresentação clínica primária
(DINARELLO, 1996; CURY, 1994).
Como foi mencionado anteriormente, a temperatura corporal normal é regulada
por um centro no hipotálamo que controla o equilíbrio entre a perda e a produção de
calor. Desse modo, a febre ocorre quando há um desequilíbrio do termostato
hipotalâmico, o que provoca a elevação do ponto de ajuste da temperatura corporal
(RANG et al., 2007).
A febre ocorre pela ação de algumas citocinas (pirógenos endógenos) sobre os
centros termorregulares do hipotálamo, elevando o limiar térmico e desencadeando
respostas metabólicas de produção e conservação de calor (tremores, vasoconstrição
periférica, aumento do metabolismo basal) (VOLTARELLI, 2011).
Estudos demonstram que a prostaglandina E2 (PGE2) tem papel chave nesse
processo, visto que ela promove o aumento de AMPc e faz com que o hipotálamo
promova a elevação da temperatura corporal, levando ao aumento da geração de
calor e a redução da sua perda (HARDMAN & LIMBIRD, 2006).
Quando um microrganismo invade um hospedeiro e entra em sua corrente
sanguínea, a estimulação de leucócitos e de outros tipos de células promove a síntese
e liberação de um grupo de moléculas que podem induzir a resposta febril (NETEA;
KULLBERG; VAN DER MEER, 2000).
Dentre essas substâncias, o fator de necrose tumoral (TNF-α), a interleucina-
1β (IL-1β) e a interleucina-6 (IL-6) são consideradas as principais responsáveis pela
mediação da resposta pirogênica durante a infecção (DINARELLO & BUNN, 1997).
Ao alcançarem a circulação essas substâncias estimulam a produção de
prostaglandina E2 por várias células (endoteliais, macrófagos e até neurônios) na
vizinhança dos centros termorreguladores hipotalâmicos, sendo que as maiores

14
concentrações de PGE2 estão próximas dos órgãos circunventriculares (organum
vasculosum laminae terminalis, OVLT) (Figura 1) (VOLTARELLI, 2011). A PGE2
produzida por essas células é considerada um importante mediador pirogênico da
febre (EVANS; REPASKY; FISHER, 2015).
Figura 1: Mecanismo pelo qual a IL-1 (e possivelmente TNF e IL-6) age sobre o
“organum vasculosum laminae terminalis” (OVLT) para produzir febre. A IL-1
proveniente da circulação sistêmica estimula neurônios, células endoteliais e
macrófagos do OVLT a produzirem PGE2, que se difunde para o centro
termorregulador (POM: núcleo pré-óptico medial), onde vai inibir a ação de neurônios
sensíveis ao calor e elevar o limiar térmico. CA: comissura anterior. POME: núcleo
pré-óptico mediano. Adaptado de: VOLTARELLI, 1994.
A prostaglandina E2 (PGE2) é derivada de eicosanóides, formada a partir da
ciclooxigenase, uma enzima responsável pela ciclização e oxigenação do ácido
araquidônico. A PGE2 atua sobre os receptores EP3 na APOHA para desencadear a
resposta febril, ao se ligar a esses receptores ocorre a diminuição dos níveis

15
intracelulares de adenosina monofosfato cíclico (AMPc), com consequente redução
da atividade neuronal (FOSTER et al., 2015; OISHI et al., 2015).
Como resultado há a inibição dos neurônios sensíveis ao calor e excitação dos
neurônios termossensíveis, elevando assim o limiar térmico. Estas respostas
involuntárias aumentam a temperatura corporal central em diferentes graus,
dependendo da magnitude da infecção, constituindo uma resposta imunológica crucial
(FOSTER et al., 2015; OISHI et al., 2015).
A Figura 2 descreve o mecanismo da febre, em que a estimulação de leucócitos
por produtos bacterianos induz a síntese de citocinas, com uma indução subsequente
da síntese de PGE2 na APOHA, desencadeando a resposta febril.
Figura 2: Mecanismo da febre de origem infecciosa. A invasão microbiana
periférica aciona a resposta imunológica não específica, ativando leucócitos e fazendo
com que os mesmos liberem substâncias pró-inflamatórias, tais como as citocinas
pirogênicas IL-6, IL-1β e TNF-α. Estas, por sua vez, estimulam a produção de PGE2
dentro da APOHA. Os neurônios situados nessa região orquestram a resposta febril
após a PGE2 se ligar aos seus receptores. Adaptado de: ARONOFF & NEILSON,
2011.

16
De acordo com Voltarelli (2011) a febre vai atuar no aumento da velocidade da
quimiotaxia de neutrófilos e da secreção de substâncias antibacterianas (peróxidos,
superóxidos, lisozima e lactoferrina), no aumento da produção de interferons, na
estimulação das fases de reconhecimento e sensibilização da resposta imunológica,
promovendo uma interação mais eficiente entre macrófago e linfócito T, e na redução
da disponibilidade de ferro, a qual limita a proliferação bacteriana. Através destas
alterações fisiológicas, a febre apresenta um papel importante na sobrevivência do
hospedeiro durante episódios infecciosos (EVANS; REPASKY; FISHER, 2015).
É importante ressaltar que embora existam dados consideráveis indicando que
febres moderadas são benéficas para o hospedeiro infectado, a elevação da
temperatura corporal acima de certa temperatura (por exemplo, 42°C) pode ser muito
prejudicial ao organismo (TANSEY & JOHNSON, 2015).
1.3. Pirógenos
O termo pirógeno é utilizado para descrever qualquer substância que provoque
febre. Os pirógenos exógenos são derivados de fora do hospedeiro, sendo a grande
maioria produtos microbianos, toxinas ou micróbios, que induzem a produção e
liberação de proteínas imunorreguladoras denominadas citocinas, essas substâncias
atuam como mediadores endógenos da febre (DINARELLO, 1996; ROTH & SOUZA,
2001).
As citocinas são pequenas proteínas, produzidas principalmente por células
imunocompetentes, que têm como principal função a regulamentação e a
coordenação das respostas imunitárias. Elas podem ser classificadas de acordo com
a sua função em citocinas pró-inflamatórias e citocinas antiinflamatórias. As citocinas
pró-inflamatórias incluem IL-1β, TNF, IL-6, IL-12 e IL-18, enquanto que as citocinas
antiinflamatórias incluem IL-4, IL-10 e IL 13 (CONTI et al., 2004; DINARELLO, 1996).
Existe uma enorme variedade de pirógenos exógenos, mas um exemplo
clássico é o lipopolissacarideo (LPS) de bactérias gram-negativas. A injeção de LPS
é um modelo experimental de inflamação sistêmica comumente utilizada, sendo um
importante agente causador da patogênese do choque séptico (VOLTARELLI, 2011;
LEON et al., 1999).
O LPS é um componente da parede celular de bactérias gram-negativas que
possui 3 partes: O-polissacarideo, R-core e lipídeo A. Dependendo da dose, da

17
temperatura ambiente e da via de administração, a administração de
lipopolissacarideo causa uma febre estereotipada e reprodutível que consiste em
várias fases e dura de 6 a 8 horas, sendo que a destruição do lipídeo A reduz ou
elimina os efeitos pirogénicos (DINARELLO & WOLFF, 1982; RIETSCHIEL & BRADE,
1992; SOARES et al., 2012).
É amplamente aceito que o LPS induz febre por meio da ativação de receptores
identificados como toll-like receptors (TLR)-4 que são expressos por populações de
células imunitárias inatas, incluindo macrófagos, neutrófilos e células dendríticas
(EVANS; REPASKY; FISHER, 2015).
A estimulação dessas células, especialmente monócitos/macrófagos e
neutrófilos, leva à síntese de mediadores pró-inflamatórios que atua como pirógenos
endógenos para estimular diretamente o centro termorregulador do hipotálamo e
causar febre (NETEA; KULLBERG; VAN DER MEER, 2000; OGAWA & KANOH,
1984).
Segundo Blatteis (2007), o início das respostas febris, após a administração de
LPS, está correlacionado com o aparecimento do LPS em células de Kupffer do fígado
(cK), a sua chegada ativa imediatamente a cascata do complemento (C) e a
consequente produção do componente 5a do complemento (C5a). O C5a é ativado,
estimulando as células de Kupffer a liberar prostaglandina E2 (PGE2) pela ação das
enzimas ciclooxigenases COX-1 e -2.
A febre induzida por LPS ocorre por meio de mecanismos autonômicos
acionados pela PGE2, que se liga a receptores de prostaglandinas EP3 expressos por
neurônios de termorregulação no núcleo da área pré-óptica medial do hipotálamo. A
elevação da temperatura corporal se dá por meio da liberação de noradrenalina, que
aumenta a termogênese no tecido adiposo marrom e induz vasoconstrição nas
extremidades para reduzir a perda de calor passivo. A PGE2 é considerada um
mediador pirogênico crucial da febre (EVANS; REPASKY; FISHER, 2015).
1.4. Espécies Reativas de Oxigênio - ERO’s
Na maioria das vezes, os termos ''radicais livres'' e ''espécies reativas de
oxigênio (ERO’s)'' são usados alternadamente (LUSHCHAK, 2014).
Entretanto, o termo espécies reativas de oxigênio (ERO’s) inclui as espécies
radicais livres e outras, que embora não contenham elétrons desemparelhados são
altamente reativas em decorrência de sua instabilidade (RIBEIRO et al., 2005).

18
Os radicais livres são moléculas ou fragmentos de moléculas que contém um
ou mais elétrons não emparelhados (CORDOVA & NAVAS, 2000).
De acordo com Manente et al., (2011) os radicais livres são espécies altamente
reativas geradas nos organismos vivos com a finalidade de proteção, entretanto, em
algumas circunstâncias, estes são responsáveis pela ocorrência ou o agravo de danos
teciduais, a sua ativação pode causar processos traumáticos nos tecidos pelo
desencadeamento de diversas cadeias de reações (CORDOVA & NAVAS, 2000).
Já as espécies reativas de oxigênio (ERO’s) são moléculas geradas por meio
da redução parcial do oxigênio molecular (O2) (HERNÁNDEZ-GARCÍA et al., 2010).
O oxigênio é utilizado como aceptor final de elétrons pelos organismos
aeróbicos, pois permite uma elevada produção de energia na respiração, em
consequência do seu alto potencial eletroquímico. Entretanto, devido a sua
configuração eletrônica o oxigênio pode sofrer reduções parciais e levar a formação
de radicais livres (RIBEIRO et al., 2005).
As ERO’s possuem elétrons desemparelhados em um átomo de oxigênio, estas
propriedades lhes conferem meia vida curta e elevada reatividade, visto que elas
reagem rapidamente com outras moléculas químicas, a fim de emparelhar seus
elétrons. Estas reações muitas vezes levam a formação de grandes quantidades de
outros radicais, gerando uma cadeia de reações que irá ativar várias vias de
sinalização (GUTTERIDGE & HALLIWELL, 2000).
As principais espécies reativas são o ânion superóxido (O2-), o radical hidroxila
(OH-) e o peróxido de hidrogênio (H2O2) (ARAUZ; RAMOS-TOVAR; MURIEL, 2016).
Quanto ao potencial de reatividade, pode-se afirmar que os radicais hidroxila
são os mais reativos, causando degradações subletais ou letais (toxicidade), enquanto
que o superóxido e o peróxido de hidrogénio têm uma reatividade mais baixa, mas
podem causar danos locais ou ativação da cascata de sinalização, quando presentes
em maiores concentrações (TAFANI et al, 2015).
Cinco compartimentos principais contêm ERO’s: mitocôndria, citoplasma,
organelas individuais ligadas à membrana (peroxissomos, endossomos e
fagossomos), exossomos e líquidos extracelulares, incluindo o plasma. As ERO’s são
produzidas pela ação de diferentes enzimas, sendo que a mitocôndria é o principal
compartimento celular responsável pela produção de espécies reativas de oxigênio.
As ERO’s produzidas na mitocôndria podem ser desintoxicadas ou podem deixar a
19
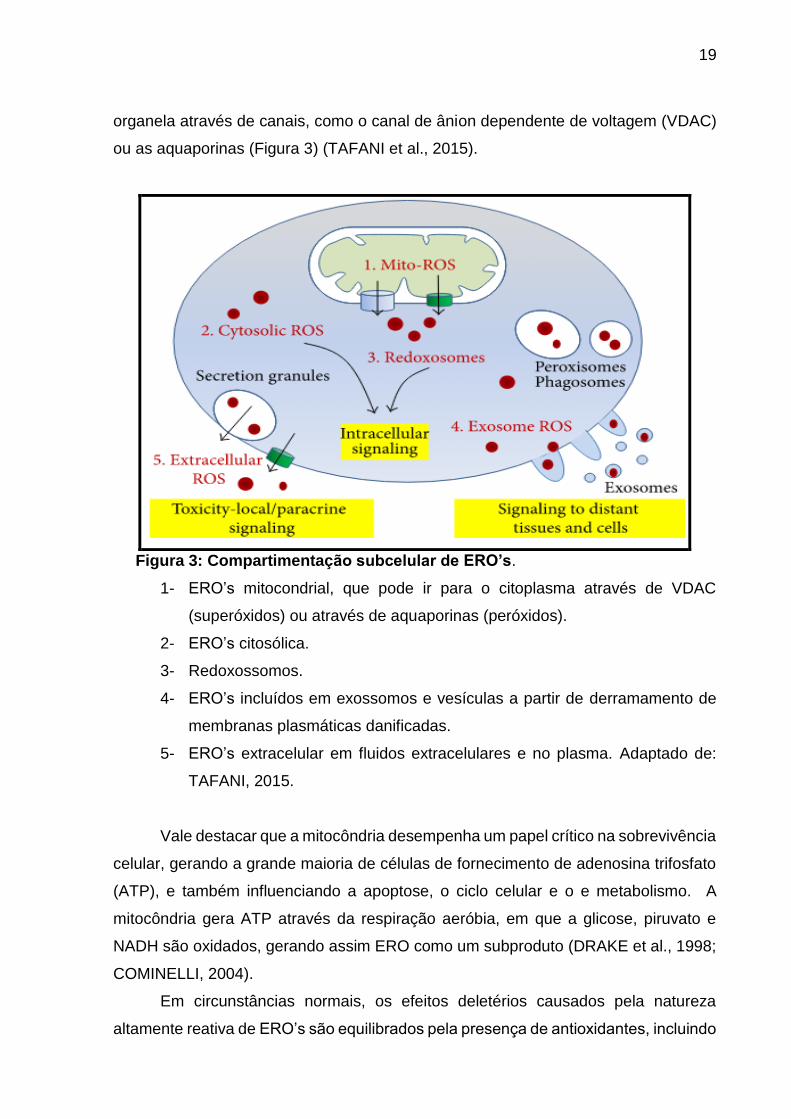
organela através de canais, como o canal de ânion dependente de voltagem (VDAC)
ou as aquaporinas (Figura 3) (TAFANI et al., 2015).
Figura 3: Compartimentação subcelular de ERO’s.
1- ERO’s mitocondrial, que pode ir para o citoplasma através de VDAC
(superóxidos) ou através de aquaporinas (peróxidos).
2- ERO’s citosólica.
3- Redoxossomos.
4- ERO’s incluídos em exossomos e vesículas a partir de derramamento de
membranas plasmáticas danificadas.
5- ERO’s extracelular em fluidos extracelulares e no plasma. Adaptado de:
TAFANI, 2015.
Vale destacar que a mitocôndria desempenha um papel crítico na sobrevivência
celular, gerando a grande maioria de células de fornecimento de adenosina trifosfato
(ATP), e também influenciando a apoptose, o ciclo celular e o e metabolismo. A
mitocôndria gera ATP através da respiração aeróbia, em que a glicose, piruvato e
NADH são oxidados, gerando assim ERO como um subproduto (DRAKE et al., 1998;
COMINELLI, 2004).
Em circunstâncias normais, os efeitos deletérios causados pela natureza
altamente reativa de ERO’s são equilibrados pela presença de antioxidantes, incluindo

20
a glutationa, carotenóides e enzimas antioxidantes, tais como a catalase e a glutationa
peroxidase (REUTER et al., 2010). Essas substâncias são capazes de inativar
moléculas reativas de oxigênio, restabelecendo o equilíbrio e evitando o estresse
oxidativo, no entanto, um desequilíbrio entre a estabilidade das reações de oxiredução
e a quantidade de ERO’s é altamente prejudicial (JESUS et al., 2015).
Altas concentrações de espécies reativas de oxigênio provocam danos
celulares. Os alvos biológicos mais vulneráveis são proteínas, lipídeos e DNA,
resultando numa condição denominada estresse oxidativo, a qual pode estar
relacionada ao surgimento de condições patológicas no organismo, incluindo doenças
neurodegenerativas, diabetes, câncer e envelhecimento prematuro (MRAKIC-
SPOSTA, et al., 2012; SENA & CHANDEL, 2012).
Apesar dos efeitos danosos das ERO's, estudos indicam que sua formação
nem sempre é deletéria. As espécies reativas de oxigênio apresentam um papel muito
importante no controle de muitos processos fisiológicos, como na resposta
inflamatória aguda e na defesa contra processos infecciosos (REUTER et al., 2010).
Figura 4: Participação de ERO’s mitocondrial na regulação de diversos processos celulares. Adaptado de: SENA & CHANDEL, 2012.
As ERO’s desempenham função fundamental na destruição de
microrganismos, constituindo um fator essencial no combate a infecções, além disso,
atuam na regulação de processos celulares variados, incluindo a diferenciação,

21
apoptose, adaptação metabólica, autofagia e regulação da longevidade (Figura 4)
(SENA & CHANDEL, 2012; ZHU et al., 2016).
Nas últimas décadas, a elucidação dos papeis fisiológicos de ERO’s,
juntamente com os seus efeitos danosos, têm ampliado os conceitos de estresse
oxidativo e de sinalização redox (ZHU et al., 2016).
1.5. Fígado
O fígado é a maior víscera do corpo humano, desempenhando um grande
número de funções vitais à saúde do organismo. Com base nisso a compreensão da
fisiologia hepática é fundamental para a análise dos processos patológicos que
acometem o órgão (SCHINONI, 2006).
O fígado exerce o trabalho de manter a homeostasia metabólica do corpo. Isso
inclui o processamento de aminoácidos, carboidratos, lipídios e vitaminas da dieta, a
síntese de muitas proteínas plasmáticas e a detoxificação e excreção para a bile de
produtos de eliminação endógenos e xenobióticos. Dessa forma, o fígado está
vulnerável a ampla variedade de insultos metabólicos, tóxicos, microbianos e
circulatórios (KUMAR; ABBAS; ASTER, 2013).
A alta atividade metabólica deste órgão é muito importante para a geração de
radicais livres. Vários radicais livres são induzidos por enzimas no fígado, incluindo a
diamina oxidase, a aldeído desidrogenase, a triptofano dupla oxidase, a
desidrogenase hepática e o complexo enzimático citocromo P450 (ARAUZ; RAMOS-
TOVAR; MURIEL, 2016).
As ERO’s são liberadas por macrófagos hepáticos após a ativação de citocinas,
LPS e prostaglandinas, como uma defesa contra a invasão bacteriana (ARTHUR;
KOWALSKI-SANDERS; WRIGHT, 1988).
As mitocôndrias e citocromo P450 nos hepatócitos, as células de Kupffer e os
neutrófilos são responsáveis pela produção de ERO’s, as quais podem agir positiva
ou negativamente dependendo do funcionamento da célula, da intensidade e da
duração da tensão oxidativa produzida na célula (ARAUZ; RAMOS-TOVAR; MURIEL,
2016).
O fígado é um importante componente do sistema imune, embora não seja
classificado como um órgão de função primariamente imunológica. Componentes da
resposta imune inata e adaptativa estão presentes ou são sintetizados no fígado
(SCHINONI, 2006).

22
Por ser um órgão altamente vascularizado, o fígado é altamente povoado por
diversas células imunes durante processos infecciosos e inflamatórios sistêmicos,
suprindo os metabólitos essenciais para a resposta de estresse e os componentes
necessários para a defesa de primeira linha no sítio de inflamação, restringindo os
limites da lesão tecidual, clareando agentes agressores e auxiliando no reparo celular
(VOLTARELLI, 2011; SCHINONI, 2006).
O revestimento sinusoidal do fígado contém as populações de células não
parenquimatosas, que consistem em células de Kupffer (cK), células endoteliais
sinusoidais (SEC) e células estreladas (SC). Todos os três tipos de células parecem
desempenhar um papel crucial na homeostase do fígado (SMEDSROD et al., 1994).
A maioria das bactérias que entram na corrente sanguínea são absorvidas e
eliminadas no fígado. Os mecanismos específicos que estão subjacentes à função do
fígado na resolução de infecções bacterianas sistêmicas continuam a ser
indeterminados, no entanto a grande maioria dos estudos realizados até o momento
tem destacado a função dos macrófagos teciduais residentes (células de Kupffer) que
revestem os sinusóides hepáticos (GREGORY & WING, 2002).
O fígado é rico em células de Kupffer, que estão em contato direto com a
circulação (DINARELLO,1982) essas células são ativadas por vários estímulos
bacterianos, incluindo o lipopolissacarídeo bacteriano (LPS) (SEKI et al., 2000).
A principal função das células de Kupffer é a remoção, por fagocitose, de corpos
estranhos e materiais particulados, bem como a captação e a detoxicação de
endotoxinas (SOARES, 2001). Sendo assim, essas células parecem exercer um papel
fundamental nas respostas imunes inatas e na defesa do hospedeiro através da
expressão e secreção de vários mediadores inflamatórios, incluindo radicais livres e
citocinas (WINWOOD & ARTHUR, 1993; MIRANDA et al., 2004).
Além disso, a eliminação rotineira de organismos estranhos a partir do fígado
parece depender em parte da infiltração de neutrófilos. Estudos sugerem que os
neutrófilos que se infiltram no fígado após uma infecção exercem um amplo efeito
sobre as defesas do hospedeiro por meio da modulação da produção de citocinas e a
síntese de óxido nítrico (NO) por células de Kupffer (GREGORY & WING, 2002).
Os neutrófilos ativados, isto é, aqueles que expressam proteínas de adesão,
fixam-se e migram através do endotélio, causando destruição de tecidos pela
liberação de radicais livres, enzimas proteolíticas e peroxidase (MIRANDA et al.,
2004).

23
Dessa forma, o fígado constitui um importante sítio de regulação do sistema
imune o que, de certa forma o torna mais susceptível e vulnerável à resposta imune,
como em processos sépticos, por exemplo (SCHINONI, 2006).

24
2. JUSTIFICATIVA
A febre é uma elevação regulada na temperatura corporal central, resultante de
uma alteração no termostato endógeno, sendo considerada parte essencial dos
mecanismos de defesa do organismo contra agentes infecciosos. Contudo, os
mecanismos envolvidos nesse processo não estão completamente elucidados
(KLUGER, 1991).
Estudos sugerem que a estimulação das células do sistema imune por diversos
pirógenos exógenos, tais como o LPS, leva à síntese de mediadores pró-inflamatórios,
incluindo espécies reativas de oxigênio, essas substâncias parecem desempenhar
uma importante função na destruição de microrganismos, constituindo um fator
significativo no combate a infecções.
Considerando a complexidade das vias moleculares que coordenam a resposta
febril e em vista da necessidade de se conhecer melhor as propriedades e atividades
específicas das ERO’s durante a febre induzida por LPS, percebe-se a importância de
mais pesquisas acerca do assunto.
Deste modo, caracterizar e quantificar as espécies reativas de oxigênio durante
a febre pode revelar vias de sinalização importantes para este processo, que poderão
ser extremamente úteis para uma maior compreensão dos mecanismos envolvidos na
resposta febril e para o desenvolvimento de novas terapias antitérmicas.

25
3. OBJETIVOS
3.1. Objetivo Geral
Avaliar a produção de espécies reativas de oxigênio (ERO’s) durante a febre
em ratos.
3.2. Objetivo Específico
- Analisar a produção de ERO’s no fígado de ratos controles e febris.

26
4. MATERIAIS E MÉTODOS
4.1. Drogas
As seguintes drogas foram usadas nos experimentos:
Cloridrato de oxitetraciclina (Terramicina®, Pfizer, São Paulo, Brasil);
Dexametasona (Azium®, Mantecorp Ind. Quím. e Farm. Ltda., Rio de
Janeiro, Brasil);
Cloridrato de S (+) cetamina (Ketamin®, Cristália, São Paulo, Brasil);
Xilazina 2% (Calmiun®, Agener União, São Paulo, Brasil);
Cloridratos de lidocaína e fenilefrina (Novocol 100®, S. S. White Artigos
Dentários Ltda, Rio de Janeiro, Brasil);
LPS (endotoxina de E. coli0111: B4, Sigma ChemCo., St. Louis, EUA).
CP
CMH
4.2. Animais
Os experimentos foram conduzidos utilizando ratos (Rattus norvegicus),
variedade Wistar, fêmeas e machos, pesando entre 180 e 200 g. Foram utilizados 8
ratos, divididos em dois grupo: LPS e controle. Os animais foram provenientes e
alojados no Biotério do Instituto de Biologia (IB) da Universidade de Brasília (UnB) em
ambiente de temperatura de 24º ±1ºC, sob um ciclo claro-escuro de 12 horas, com
água e alimentos fornecidos ad libitum. Os procedimentos cirúrgicos e os
experimentos farmacológicos foram realizados no Laboratório de Bioquímica e
Química de Proteínas do IB/UnB, sempre no período das 08 às 18 h. O projeto foi
aprovado pelo Comitê de Ética no Uso Animal (CEUA) do Instituto de Biologia da
Universidade de Brasília, UnB Doc. 147474/2015 (Anexo 1).
4.3. Cirurgia para Implante de Transmissores de Temperatura na Cavidade
Peritoneal
A temperatura corporal dos animais foi medida por meio de transmissores de
temperatura (Data Loggers, Subcue, Calgary Canada) que foram implantados na
cavidade peritoneal dos ratos. Antes das cirurgias, os transmissores foram
desinfectados em solução de álcool 70% (v/v; imersão por 30 min). Os animais foram
anestesiados com o uso de uma mistura de ketamina e xilazina (60 mg/Kg e 10 mg/Kg,

27
respectivamente, i.p.). Após tricotomia e antissepsia da pele, foi feita uma incisão de
aproximadamente 2 cm na pele e músculos peritoneais. O transmissor foi lavado com
solução salina estéril e então inserido na cavidade peritoneal e, em seguida, o músculo
e a pele foram suturados separadamente. Esse procedimento foi realizado uma
semana antes dos experimentos.
4.4. Mensuração da Temperatura Corporal dos Ratos
A mensuração da temperatura corporal dos ratos durante os experimentos foi
realizada por transmissores implantados cirurgicamente na cavidade abdominal,
descritos anteriormente. O procedimento experimental foi realizado dentro da zona
termoneutra para ratos, ou seja, a temperatura da sala experimental foi mantida a
27°±1°C. Os animais permaneceram nesse ambiente por pelo menos uma hora antes
do início dos experimentos, para a ambientação dos mesmos. Para o processo de
leitura da temperatura corporal por telemetria, os transmissores implantados na
cavidade peritoneal foram programados para serem acionados no dia anterior ao
experimento. As temperaturas basais foram determinadas pela média das 4 medições
anteriores a qualquer tratamento.
4.5. Procedimentos Experimentais
Cada animal foi utilizado apenas uma vez. Os animais foram transportados
para a sala de experimentação, onde permaneceram em repouso por uma hora. Após
esse período suas temperaturas basais foram determinadas por 4 vezes, a intervalos
de 15 minutos.
Os animais foram alocados em grupos experimentais e receberam injeção
endovenosa de LPS (5 μg/kg) ou do seu veículo (salina 0,9%, 1 mL/Kg). A dose de
LPS usada foi padronizada para os experimentos em nosso laboratório. A partir de
então, cada animal teve sua temperatura corporal aferida a cada 15 minutos, durante
6 h. Em seguida os animais foram anestesiados e eutanasiados por decapitação. Um
pedaço do lobo esquerdo do fígado foi retirado, cortado em fatias e imerso em 147 µL
de tampão Krebs-Hepes e 3 µL de CMH (10 mM), atingindo concentração final de 200
µM de CMH. Em seguida essa solução foi incubada a 37C durante 60 min. Após esse
período o sobrenadante foi coletado, transferido para microtubos, imediatamente
congelado em nitrogênio líquido e armazenado a -80ºC até a realização da análise da
produção de ERO’s total por ressonância paramagnética eletrônica (RPE). Sabe-se

28
que a oxidação de CMH leva a formação de CM•(3-metoxi-carbonilproxilnitróxido)
paramagnético. Os espectros de RPE de CM• e CP• são idênticos a 150 K, por esta
razão as amostras para a curva de calibração foram obtidas a partir de uma solução
estoque do padrão 3-carboxi-proxil (CP•) (Noxygen, Alemanha) preparado em tampão
Krebs-Hepes e diluído nas concentrações de 10, 25, 50, 100 e 200 µM.
4.6. Ressonância Paramagnética Eletrônica (RPE)
Devido a sua alta reatividade e meia-vida extremamente curta, os radicais livres
são difíceis de detectar. A RPE é uma técnica de detecção baseada no
comportamento de radicais livres submetidos a um campo magnético. Os radicais
livres contém elétrons desemparelhados que podem atuar como paramagnéticos e,
portanto se alinharem no campo magnético gerado pelo espectrômetro de RPE.
Quando uma fonte de energia externa sob a forma de microondas é aplicada a
amostra, esses elétrons livres mudam de um estado de baixa para um estado de alta
energia. Esta absorção de energia pode ser medida, e correlaciona-se diretamente
com a quantidade de radicais livres presentes na amostra.
Para a detecção de ERO’s por RPE é necessário o uso de spin traps e/ou spin
probes para estabilizar as ERO’s e aumentar sua meia-vida para que o sinal possa
ser detectado por RPE. Para a detecção de ERO’s total foi utilizado o spin probe 1-
Hidroxi-3-metoxicarbonil-2, 2, 5,5-tetrametilpirrolidina (CMH) (Noxygen, Alemanha),
ERO’s total representa todas as espécies reativas de oxigênio, entretanto as principais
espécies marcadas pelo spin probe usado são ânions superóxidos, peróxido de
hidrogênio e radicais hidroxila.
As medidas de RPE foram realizadas em um espectrômetro Bruker EMX500,
utilizando a banda X (9,35 GHz), potência 20 mW e campo de modulação de 1 Gauss.
Todas as medidas foram feitas a 150 K.
Padrões de calibração de 10, 25, 50, 100 e 200 µM de CP•, dissolvidos em
tampão Krebs-Hepes, foram utilizados para fazer a curva de calibração. A relação
entre a amplitude do sinal de RPE (l) e a concentração de radicais [ERO’s] (µM) pode
ser calculada pelo teste t de Student. A inclinação da curva de calibração foi indicada
como s (µM-1) e a intersecção no eixo y foi indicada como l0. A concentração de
radicais nas amostras foi calculada da seguinte forma: [ERO’s] = (l-l0/s).

29
4.7. Estatística
Os dados são apresentados como média ± EPM (erro padrão da média). As
comparações estatísticas entre os grupos SAL e LPS foram efetuadas por test t de
Student, com a utilização do programa estatístico Prism 5.0. O nível de significância
considerado foi de 5% (p < 0,05).

30
5. RESULTADOS
5.1. Efeito da Administração de LPS sobre a Temperatura Corporal
Os animais foram alocados em grupos experimentais e receberam injeção
endovenosa de LPS (5 μg/kg) ou do seu veículo (salina 0,9%, 2 mL/Kg). Logo após
as injeções de LPS observou-se a elevação da temperatura dos animais tratados com
LPS e dos que receberam o controle (Figura 05). Esse aumento ocorreu devido à
manipulação dos animais e segue o padrão de resposta febril induzida por esse
estímulo em ratos.
Os ratos tratados com LPS (5 μg/kg, iv) e o seu veículo apresentaram um
aumento significativo na temperatura corporal, com perfil bifásico iniciando-se após
1,5 h, atingindo o primeiro pico em cerca de 2,5 h e o segundo em cerca de 5 h após
a injeção, sendo a temperatura corporal mantida elevada durante as 6 h de
observação (Figura 05).
O animais do grupo salina não apresentaram variações significativas da
temperatura corporal durante todo o período de observação, exceto na primeira hora
após a administração de salina, em decorrência da manipulação para a realização das
injeções endovenosas.
0 1 2 3 4 5 636.0
36.5
37.0
37.5
38.0
38.5
39.0
39.5
40.0
Veículo/Salina
Veículo/LPS
________________________***
Tempo (h)
Tem
pera
tura
(C
)
Figura 5: Efeito da administração de LPS sobre a temperatura corporal. Os
animais receberam LPS (símbolos cheios) por via intravenosa na dose de 5 μg/Kg/mL,
ou o mesmo volume de solução salina 0,9% (símbolos vazios). Os pontos

31
representam a média ± EPM da temperatura corporal (em °C) dos animais, medida
por telemetria.
5.2. Efeito da Administração de LPS sobre a Produção de ERO’s
Os padrões de calibração de 10, 25, 50, 100 e 200 µM de CP·, dissolvidos em
tampão Krebs-Hepes foram utilizados para fazer a curva de calibração (Figura 6).
Nesse caso, a inclinação da curva foi de s= 0,0779 µM-1 e a intersecção no eixo y foi
l0=-0,22.
Figura 6: Amplitude do sinal de RPE em função de [CP•] em padrões de calibração de [CP•] dissolvidos em tampão Krebs-Hepes (círculos). Curva de calibração calculada pelo teste t de Student.
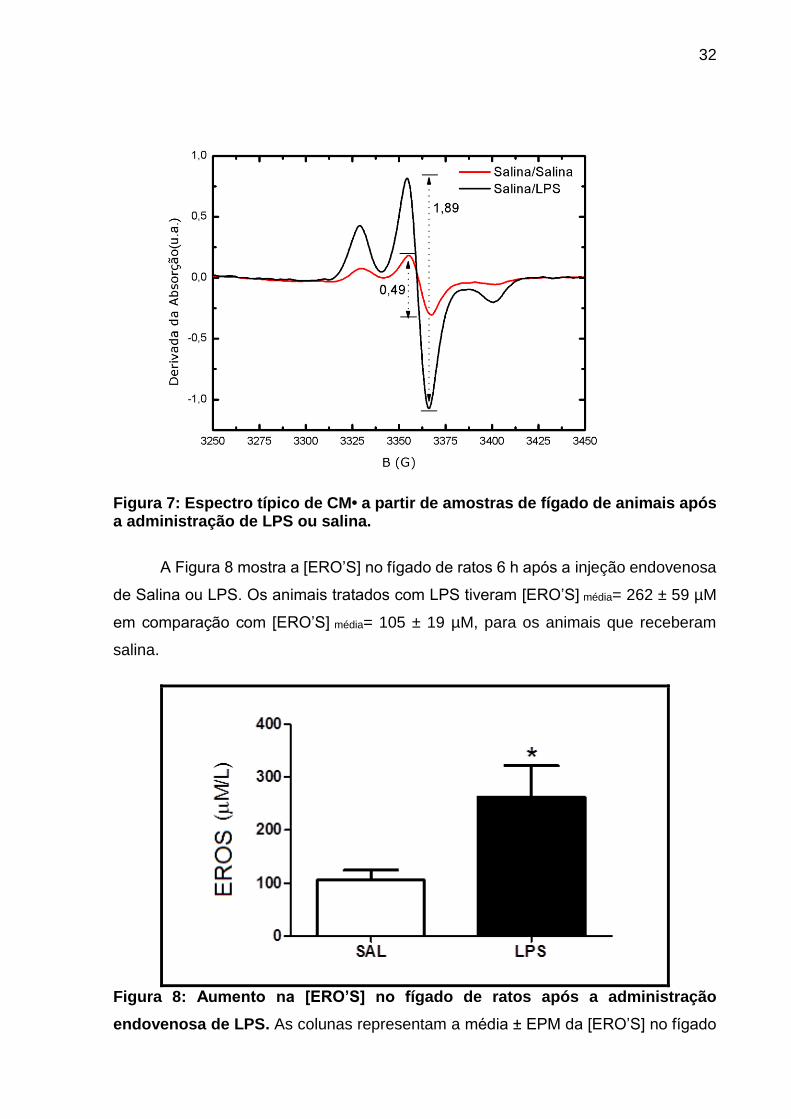
A figura 7 apresenta o espectro de RPE típico de CM•, obtido a partir de
amostras de fígado de ratos tratados com LPS ou Salina. A amplitude do sinal foi
definida como os valores de pico a pico, nesse caso demonstra-se o valor de 0,49
para o um animal do grupo Salina/Salina e de 1,89 para um animal do grupo
Salina/LPS
32
Figura 7: Espectro típico de CM• a partir de amostras de fígado de animais após a administração de LPS ou salina.
A Figura 8 mostra a [ERO’S] no fígado de ratos 6 h após a injeção endovenosa
de Salina ou LPS. Os animais tratados com LPS tiveram [ERO’S] média= 262 ± 59 µM
em comparação com [ERO’S] média= 105 ± 19 µM, para os animais que receberam
salina.
Figura 8: Aumento na [ERO’S] no fígado de ratos após a administração
endovenosa de LPS. As colunas representam a média ± EPM da [ERO’S] no fígado

33
de animais tratados com solução salina 0,9% (SAL) ou LPS 5μg/Kg/mL.

34
6. DISCUSSÃO
Há algumas décadas, as espécies reativas de oxigênio foram destacadas por
causarem dano oxidativo em biomoléculas, contribuindo para o desenvolvimento de
uma variedade de doenças. No entanto, estudos mais recentes sugeriram que ERO’s
intracelulares representam um componente importante da cascata de sinalização
intracelular (ZHU et al., 2016). Deste modo, este trabalho buscou consolidar antigas e
novas ideias sobre o papel fisiológico e patológico de ERO’s para uma melhor
compreensão das suas propriedades e atividades específicas no fígado durante a
febre induzida por LPS.
Grande parte dos conhecimentos atuais acerca dos mecanismos moleculares
envolvidos na febre provém de estudos em animais tratados com lipopolissacarídeo
(LPS). O LPS é uma molécula constituinte da parede de bactérias gram-negativas que
após se ligar a receptores (TLR)-4 induz a ativação de diversas vias de sinalização as
quais aumentam a produção e secreção de moléculas pró-inflamatórias, tais como o
fator de necrose tumoral (TNF) e a interleucina-6 (IL- 6). Essas citocinas são liberadas
por macrófagos do fígado (isto é, células de Kupffer), bem como por macrófagos de
outros órgãos. Neste modelo, a prostaglandina E2 (PGE2) produzida por células
endoteliais vasculares cerebrais é considerada um mediador importante da resposta
pirogênica (ROTH & BLATTEIS, 2014; ZHU et al., 2016).
A dose de LPS usada neste estudo (5 μg/Kg, iv) induziu resposta febril
moderada e contínua. Nos primeiros momentos ocorreu um aumento típico da
temperatura corporal decorrente da manipulação dos animais, em seguida, foram
observados picos de temperatura cerca de 2,5h e 5h após a injeção de LPS. Sabe-se
que administração de LPS, dependendo da dose, temperatura ambiente e via de
administração induz uma febre estereotipada e reprodutível que consiste em várias
fases e dura de 6 a 8 horas, de modo que a elevação da temperatura observada
durante as 6 horas é semelhante ao descrito na literatura para a resposta típica a
doses moderadas de LPS. (SOARES et al., 2012).
Neste trabalho, investigou-se a hipótese de que a administração endovenosa
de LPS poderia induzir o aumento na concentração de ERO’s no fígado de ratos e
buscou-se avaliar a importância dessas espécies reativas no controle da resposta
febril.
Primeiramente é importante destacar o papel dual do oxigênio. O oxigênio é um
elemento não-metálico altamente reativo que forma compostos com a maioria dos

35
outros elementos, ele possui dois elétrons desemparelhados o que o torna
paramagnético. O paramagnetismo refere-se ao estado magnético de uma espécie
química com um ou mais elétrons não emparelhados. A característica paramagnética
permite a detecção e quantificação de O² em sistemas biológicos utilizando
espectrometria de ressonância paramagnética eletrônica (EPR), técnica específica
para a detecção de radicais que foi utilizada neste estudo (ZHU et al., 2016).
O oxigênio é uma molécula essencial para a vida aeróbia, contudo a sua
utilização resulta na formação de espécies de oxigênio reativas (ERO’s), que em altas
concentrações podem causar dano oxidativo aos constituintes celulares, o que
representa a base molecular da toxicidade do oxigênio (ZHU et al., 2016).
Embora os efeitos nocivos do oxigênio tenham sido relatados, estudos recentes
sugerem que ERO’s intracelulares são um componente importante da cascata de
sinalização intracelular, possuindo um papel duplo na fisiologia. Por um lado, as
ERO’s derivadas de O² contribuem para a morte de microorganismos patogênicos por
células fagocíticas, promovendo assim a sobrevivência do hospedeiro. Por outro lado,
ERO’s atuam como segundos mensageiros para ativar a transdução de sinal celular,
levando a respostas fisiológicas desejadas (WEIDINGER & KOZLOV, 2015; ZHU et
al., 2016). Os resultados do presente estudo indicam que os animais tratados com o
LPS apresentaram uma maior concentração de ERO’s no fígado.
ERO’s estão relacionados a uma ampla variedade de doenças humanas.
SZUSTER-CIESIELSKA e colaboradores (2004) investigaram a produção de espécies
reativas de oxigênio no sangue de pacientes com carcinoma de laringe e de pacientes
saudáveis e observaram uma maior concentração de ERO’s nos pacientes com o
carcinoma.
O estresse oxidativo foi documentado também em tecido adiposo de
camundongos, o principal objetivo deste estudo foi verificar se a presença ou ausência
de proteínas do tipo UCPs afeta a geração de ERO’s (SHABALINA et al., 2014).
Além disso, o aumento na produção de EROS no cérebro após o tratamento
com LPS já foi avaliado em roedores (PINHEIRO, 2015), porém, até o momento não
se conhece estudos que tenham demonstrado o aumento dessas espécies no fígado
de animais durante a febre.
Sabendo que o fígado representa um componente importante do sistema
imune, sendo altamente povoado por diversas células imunes durante processos
infecciosos e inflamatórios sistêmicos e possuindo componentes necessários para a

36
defesa de primeira linha, é de suma importância avaliar o papel de ERO’s sobre esse
órgão durante a resposta febril (SCHINONI, 2006).
Sabe-se que as células de Kupffer que revestem os sinusóides hepáticos
exercem possivelmente um papel essencial nas respostas imunes inatas e na defesa
do hospedeiro através da expressão e secreção de vários mediadores inflamatórios,
incluindo espécies reativas de oxigênio e citocinas, constituindo um componente
importante do sistema imune. As células de Kupffer (cK) são ativadas por vários
estímulos bacterianos, incluindo o lipopolissacarídeo e liberam essas substâncias
como uma defesa contra a invasão bacteriana (MIRANDA et al., 2004; SEKI et al.,
2000).
Os dados obtidos revelaram um aumento de espécies reativas de oxigênio em
amostras de fígados de ratos tratados com LPS quando comparados com o grupo
controle. É importante ressaltar que o aumento na concentração de ERO’s pode ser
decorrente de um aumento na sua produção e/ou da redução dos sistemas
antioxidantes.
As defesas antioxidantes enzimáticas são responsáveis pela proteção do
organismo contra a ação oxidativa das ERO’s e incluem a atividade da superóxido
dismutase, catalase, glutationa peroxidase, glutationa redutase e glutationa S-
transferase (HALIWELL, 2012).
Estudos recentes sugerem que quando a produção de espécies reativas de
oxigênio está sob controle homeostático, elas desempenham funções fisiológicas
importantes, assim o aumento das espécies reativas de oxigênio pode representar um
importante evento no controle da resposta febril (ZHU et al., 2016).
Ainda há muito a entender sobre os papéis de ERO’s produzidas no fígado
durante a resposta febril, é possível que elas contribuam para o aumento da resposta
febril ou que sejam uma consequência do aumento de temperatura desencadeado
durante a febre. Mais estudos estão sendo realizados em nosso laboratório na
tentativa de elucidar essas questões.

37
7. CONCLUSÃO
As espécies reativas de oxigênio são temas de pesquisa nos últimos anos,
devido à sua importância vários ensaios clínicos buscam avaliar o seu papel em
processos celulares. Os dados obtidos neste estudo revelam um aumento na
concentração de espécies reativas de oxigênio no fígado de ratos durante a resposta
febril induzida por LPS. Dada a relevância dessas espécies em processos fisiológicos,
esse aumento pode representar um fator importante no controle da resposta febril. No
entanto, é necessário mais estudos para que essa hipótese seja confirmada.

38
8. REFERÊNCIAS BIBLIOGRÁFICAS
ARAUZ, J.; RAMOS-TOVAR, E. R.; MURIEL, P. Redox state and methods to evaluate oxidative stress in liver damage: From bench to bedside, Annals of Hepatology, v. 15, n. 2, p. 160-173. 2016.
ARONOFF, D. M.; NEILSON, E. G. Antipyretics: Mechanisms of Action and Clinical Use in Fever Suppression, Am. J. Med, v. 111, p. 304-315. 2001.
ARTHUR, M. J.; KOWALSKI-SANDERS, P.;WRIGHT, R. Effect of endotoxin on release of reactive oxygen intermediates by rat hepatic macrophages, Gastroenterology, v. 95, n. 6, p.1588-1594. 1988.
BLATTEIS, C. M. The onset of fever: new insights into its mechanism, Prog Brain Res, v. 162, p. 3-14. 2007.
BOULANT, J. A.; CHOW, A. R.; GRIFFIN, J. D. Determinants of hypothalamic neuronal thermosensitivity, Ann N Y Acad Sci, p.133-138. 1997.
BROWN, G. C. Mechanisms of inflammatory neurodegeneration: iNOS and NADPH oxidase, Biochem SocTrans, v. 35, p. 1119–1921. 2007.
COMINELLI, F. Cytokine-based therapies for Crohn’s disease- new paradigms, N. Engl. J. Med, v. 351, p. 2045-2048. 2004.
CONTI, B.; TABAREAN, I.; ANDREI, C.; BARTFAI, T. Cytokines and fever, Front Biosci, v. 9, p. 433-449. 2004.
CORDOVA, A.; NAVAS, F. J. Os radicais livres e o dano muscular produzido pelo exercício: papel dos antioxidantes, Bras Med Esporte, v. 6, n.5, p. 204-208. 2000.
CURY, C. P. L.; SAMARA, A. M. A febre nas doenças reumáticas, Revista de Ciências Médicas- PUCCAMP, v. 3, n.3, p. 69-75. 1994.
DINARELLO, C. A. Thermoregulation and the pathogenesis of fever, Infectious Disease Clinics of North America, v.10, n. 2, p. 433-449. 1996.
DINARELLO, C. A. Infection, fever, and exogenous and endogenous pyrogens: some concepts have changed, Journal of Endotoxin Research, v. 10, n. 4, p. 201-222. 2004.
DINARELLO, C. A.; BUNN, P. A. Fever, Semin. Oncol, v. 24, p. 288-298. 1997.
DINARELLO, C. A.; WOLFF, S. M. Exogeneous pyrogens, In: Milton, A. S., eds. Hand book of experimental pharmacology, Springer-Vergan, v. 60, p. 73–112. 1982.
DRAKE, I. M.; MAPSTONE, N. P.; SCHORAH, C. J.; WHITE, K. L.; CHALMERS, D. M.; DIXON, M. F.; AXON, A.T. R. Reactive oxygen species activity and lipid peroxidation in Helicobacter pylori associated gastritis: relation to gastric mucosal ascorbic acid concentrations and effect of H pylori eradication, Gut, v. 42, n.6, p.768–771. 1998.
EVANS, S. S.; REPASKY, E. A.; FISHER, D. T. Fever and the thermal regulation of immunity: the immune system feels the heat, Nat Rev Immunol, v. 15, n. 6. p. 335-349. 2015.
FERREIRA, T. R.; LOPES, L. C. Analysis of analgesic, antipyretic, and non steroidal anti-inflammatory drug use in pediatric prescriptions, J pediatr, v. 92, n. 1, p. 81-87. 2016.

39
FOSTER, J.; MAUGER, A. R.; CHRISMAS, B. C. R.; THOMASSON, K.; TAYLOR, L. Is prostaglandin E2 (PGE2) involved in the thermogenic response to environmental cooling in healthy humans? Med Hypotheses, v.85, n.5. p. 60-611. 2015.
GREGORY, S. H.; WING, E. J. Neutrophil-Kupffer cell interaction: a critical component of host defenses to systemic bacterial infections, Journal of Leukocyte Biology, v. 72, n. 2, p. 239-248. 2002.
GUTTERIDGE, J. M.; HALLIWELL, B. Free radicals and antioxidants in theyear. A historical look to the future, Ann. N.Y. Acad. Sci, v. 899, p. 136–147. 2000.
HALIWELL, B. Free radicals and antioxidants: updating a personal view. Nutr Rev, v. 70, p. 257-265. 2012.
HARDEN, L. M.; KENT, S.; PITTMAN, Q. J.; ROTH, J. Fever and sickness behavior: Friend or foe? Brain, Behavior, and Immunity, v.50, p.322–333. 2015.
HARDMAN, J. G.; LIMBIRD, L. E. Goodman & Gilman As Bases Farmacológicas da Terapêutica, Mc Graw Hill, 11ª ed. 2006.
JESUS, P.R.; BAUERMANN, L. F.; GUEZ, C. G.; REGINATO, F. Z.; SILVA, A. R. H.;LOPES, G. H. H. Atividade antioxidante após tratamento subagudo com oliva em fígado e rim de ratos wistar, v. 7, n.2. 2015.
KLUGER, M.J. Fever: role of pyrogens and cryogens, Physiological Reviews, v.71, n.1, p.93-127. 1991.
KLUGER, M. J.; RINGLER, D. H.; ANVER, M. R. Fever and survival, Science, v. 188, p. 166–168. 1975.
KUMAR, V.; ABBAS, A. K.; ASTER, J. C. Robbins Patologia Básica. 9. Ed, Elsevier, 2013.
LEON, L. R.; KOZAK, W.; RUDOLPH, K.; KLUGER, M. J. An antipyretic role for interleukin-1 in LPS fever in mice, Am J Physiol, v. 276. 1999.
LUSHCHAK, V. I. Free radicals, reactive oxygen species, oxidative stress and its classification, Chemico-Biological Interactions, v. 224 p. 164–175. 2014.
MANENTE, F. A.; MELLO, L. R. D.A.; KHALIL, O. A. K.; CARVALHO, C. T. D.; BANNACH, G.; VELLOSA, J. C. R. Effect of metal complexation to anti-inflammatory over the action against oxidative and free radicals: ketoprofen action, Eclética Química, v. 36, n. 2, p.107-127. 2011.
MIRANDA, L. C. C.; VIARO, F.; CENEVIVA, R.; EVORA, P. R. B. As bases experimentais da lesão por isquemia e reperfusão do fígado, Acta Cirúrgica Brasileira, v. 19, n. 1, p. 3-11. 2004.
MRAKIC-SPOSTA, S.; GUSSONI, M.; MONTORSI, M.; PORCELLLI, S.; VEZZOLI, A. Assessment of a Standardized ROS Production Profile in Humans by Electron Paramagnetic Resonance, Oxidative Medicine and Cellular Longevity, p. 1-10. 2012.
NETEA, M. G.; KULLBERG, B. J.; VAN DER MEER, J. W. M. Circulating Cytokines as Mediators of Fever, Clinical Infectious Diseases, v. 31, p.178–84. 2000.
OGAWA, Y; KANOH, S. Involvement of Central Action of Lipopolysaccharide in Pyrogen Fever, Jpn J Pharmacol, v. 36, n. 3, p. 389-95. 1984.

40
OISHI, Y.; YOSHIDA, K.; SCAMMELL, T. E.; URADE, Y.; LAZARUS, M.; SAPER, C.B. The roles of prostaglandin E2 and D2 in lipopolysaccharide-mediated changes in sleep, Brain Behav Immun, v. 47, p. 172-177. 2015.
PINHEIRO, G. J. Análise da produção de oxigênio singleto durante a febre em ratos. 2015.
RANG, H. P.; DALE, M. M.; RITTER, J. M.; GARDNER, P. Farmacologia 6ª ed, Elsevier. 2007.
REUTER, S.; GUPTA, S. C.; CHATURVEDI, M. M.; AGGARWAL, B. B. Oxidative stress, inflammation, and cancer: how are they linked? Free Radic, Biol. Med, v. 49, p. 1603–1616. 2010.
RIBEIRO, S. M. R., QUEIROZ, J. H., PELUZO, M. C. G., COSTA, N. M. B., MATTA, S. L. P. A formação e os efeitos das espécies reativas de oxigênio no meio biológico, Biosci. J, v. 21, n. 3, p. 133-149. 2005.
RIETSCHIEL, E.T.; BRADE, H. Bacterial endotoxins, Sci. Am, v. 267, p. 26-33. 1992.
ROTH, J.; BLATTEIS, C. M. Mechanisms of Fever Production and Lysis: Lessons from Experimental LPS Fever, Comprehensive Physiology, v.4, n. 4, p.1553-1604. 2014.
ROTH, J.; SOUZA, G. E. P. Fever induction pathways: evidence from responses to systemic or local cytokine formation, Braz J Med Biol Res, v. 34, n. 3, p. 301-314. 2001.
SCHINONI, M. A.FisiologiaHepática, Gazméd, p. 5-9. 2006.
SEKI, S.; HABU, Y.; KAWAMURA, T.; TAKEDA, K.; DOBASHI, H.; OHKAWA, T.; HIRAIDE, H. The liver as a crucial organ in the first line of host defense: the roles of Kupffer cells, natural killer (NK) cells and NK1.1 Ag+ T cells in T helper 1 immune responses, Immunol Rev, v. 174, p. 35-46. 2000.
SENA, L. A.; CHANDEL, N. S. Physiological roles of mitochondrial reactive oxygen species. Mol Cell, v. 48, n.2, p. 158-167. 2012.
SHABALINA, I. G.; VRBACKY; PECINOVA, A.; KALINOVICH, A. V.; DRAHOTA, Z.; HOUSTEK, J.; MRACEK, T.; CANNON, B.; NEDERGAARD, J. ROS production in brown adipose tissue mitochondria: the question of UCP1-dependence, Biochim Biophys Acta, v. 12, p.2017-2030. 2014.
SHAMI, N. J. I.; MOREIRA, E. A. M. Licopeno como agente antioxidante, Rev. Nutr, v. 17, n. 2, p. 227-236. 2004.
SILVA, R. R.; OLIVEIRA, T. T; NAGEM, T.J.; LEÃO M. A. Efeito de flavonoides no metabolismo do ácido araquidônico, Medicina, Ribeirão Preto, v. 35, p.127-133. 2002.
SMEDSROD, B.; DE BLESER, P.J.; BRAET, F.; LOVISETTI, P.; VANDERKERKEN, K.; WISSE, E.; GEERTS, A. Cell biology of liver endothelial and Kupffer cells, Gut, v. 35, n. 1, p. 1509-1516. 1994.
SOARES, D. M.; FIGUEIREDO, M. J.; MARTINS, J. M.; MACHADO, R. R.; SORGI, C.; FACIOLLI, L. H.; ALVES-FILHO, J.C.; CUNHA, F. Q.; SOUZA, G. E. P.A crucial role for IL-6 in the CNS of rats during fever induced by the injection of live E. coli, Medical Microbiology and Immunology, v. 201, p. 47-60. 2012.

41
SOARES, E. C. Fígado e hormônios. In “Doenças do fígado e vias biliares”. Gayotto e Alves. 1ª. Edição, Savvier; p. 53-57. 2001.
SOUZA, G.E.P.; CARDOSO, R.A.; MELO, M.C.C.; FABRICIO, A.S.C.; SILVA, V.M.S.; LORA, M.; DE BRUM-FERNANDES, A.J.; FERREIRA, S.H.; ZAMPRONIO, A.R. A comparative study of antipyretic profiles of indomethacin and dipyrone in rats. Inflam. Res, v. 51, p. 24-32. 2002.
SZUSTER-CIESIELSKA, A; HRYCIUK-UMER, E; STEPULAK, A; KUPISZ, K, KANDEFER-SZERSZEN, M. Reactive oxygen species production by blood neutrophils of patients with laryngeal carcinoma and antioxidative enzyme activity in their blood, Acta Oncol, v. 43, n.3, p. 252-258. 2004.
TANSEY, E. A.; JOHNSON, C. D. Recent advances in thermoregulation, Adv Physiol Educ, v. 39, n. 3, p.139-148. 2015.
TAFANI, M.; SANSONE, L.; LIMANA, F.; ARCANGELI, T.; DE SANTIS, E.; POLESE, M.; FINI, M.; RUSSO, M. A. The Interplay of Reactive Oxygen Species, Hypoxia, Inflammation, and Sirtuins in Cancer Initiation and Progression, Oxidative Medicine and Cellular Longevity, p. 1-18. 2015.
VOLTARELLI J. C. Febre e inflamação, Medicina, Ribeirão Preto, v. 27, n.1/2, p. 7-48. 1994.
WINWOOD, P. J.; ARTHUR, M. J. Kupffer cells: their activation and role in animal models of liver injury and human liver disease, Semin Liver Dis, v.13, n.1. p. 50-59. 1993.
ZHU, H.; TRAORE, K.; SANTO, A.; TRUSH, M, A.; LI, Y. R. Oxygen and Oxyge Toxicity: The Birth of Concepts, Reactive Oxygen Species, v. 1, p. 1-8. 2016.

42
9. ANEXOS
9.1. Anexo 1





























