Embed Size (px)
Citation preview

Bioinorgânica do Níquel e do Cobalto Prof. Fernando R. Xavier
UDESC 2018

2
Características gerais
Ao examinarmos as reações químicas catalisadas por enzimas de níquel e por cobalto
ao longo do processo evolutivo, nota-se que ambos estiveram presentes e atuantes
na era pré oxigênio (atmosfera redutora). Tal fato reflete os altos níveis destes metais
em um grande número de bactérias anaeróbicas.
Em contrapartida, os níveis destes metais em
mamíferos é cerca de 100 vezes menor quando
comparado aos índices de zinco, ferro e cobre.
Enquanto o cobalto é importante para as enzimas
dependentes da vitamina B12, em seres
superiores, o níquel é encontrado na apenas
urease de plantas é praticamente inexistente em
células eucariontes. Staphylococcus aureus a 50,000x

3
Considerando a química ácido-base biológica, íons
níquel e cobalto podem substituir facilmente
íons zinco.
Estes dois metais se tornaram obsoletos na era pós-fotossíntese.
Já para química redox da era pós-oxigênio estes
podem ser substituídos por manganês, ferro e
cobre, que são muito mais abundantes.
Ambos níquel e cobalto, assim como o ferro possuem a características de serem ricos
em elétrons. Quando em baixos estados de oxidação alguns de seus elétrons 3d
tendem a ficar em orbitais mais expostos podendo atuar na transferência de 1
elétron aos moldes de radicais livres.

4
Enzimas de Níquel
Sete de oito enzimas de níquel conhecidas estão envolvidas na produção de gases
(CO, CO2, CH4, H2, NH3 e O2) os quais todos possuem papel importante nos ciclos
globais de carbono, nitrogênio e oxigênio.
ARD – Aciredutona dioxigenase; MCR – metil-CoM redutase; CODH – CO
deshidrogenase; ACS – Acetil-CoA sintetase.

5
Uma antiga enzima de níquel, a glioxilase (GlxI) catalisa a reação do metilglioxal (tóxico
pois pode formar adutos com o DNA) em lactato. Neste caso os íons níquel atuam
apenas como um ácido de Lewis, ou seja, não há química redox presente.
Nos seres humanos este metal foi substituído por zinco, sendo então mais uma
evidência de que a substituição de metais em enzimas é mais comum do que se pode
imaginar.

6
Enzimas de níquel apresentam uma grande adaptabilidade tanto em termos de
ambiente de coordenação quando em sua química redox. A Ni-SOD, por exemplo, deve
possuir uma faixa ampla de trabalho com potenciais variando desde + 890 até -160 mV.
Redutases de níquel tais como a MCR e a CODH podem apresentar potenciais de
redução de até -600 mV. Desta forma nota-se que enzimas de níquel podem atuar em
reações redox em um intervalo de até 1,5 Volts!
Uma vez que a concentração de íons níquel(2+) é muito baixa em condições ambientais
típicas, sistemas que possuem alta afinidade por estes íons foram desenvolvidos pela
natureza (metalochaperonas).

7

8
Urease
Historicamente, a primeira enzima contendo níquel descoberta foi a Urease. Esta
enzima foi extraída de feijões “jack beans” e cristalizada por James Sumner1 em 1926.
Somente 50 anos depois foi possível comprovar a presença de níquel em seu sítio ativo.
Esta enzima catalisa a hidrólise da ureia em amônia e carbamato que, espontaneamente
se decompõe em CO2, água e amônia.

9
A urease tem um papel chave no metabolismo de microrganismos e plantas pois
geram amônia, sendo então uma fonte de nitrogênio para a mesma. Ela possui um sitio
ativo binuclear de níquel onde estes estão a 3,5 Å um do outro ponteados por um grupo
OH.

10
Além da ponte μ-OH exógena, há uma ponte constituída por uma lisina carbamilada.
Estudos ureases submetidas a mutações sítio dirigidas apontam que na ausência desta
ponte endógena, esta enzima perde sua capacidade catalítica.
Um grupo hidróxido coordenado ao átomo de
níquel irá então atacar o substrato dando
sequência a reação química.
A alta barreira energética para a hidrólise
da ureia pode ser vencida pela
coordenação da carbonila da mesma a um
dos centros metálicos que irá ativá-la.

11
Propostas mecanísticas

12
Proteínas Níquel-Ferro-Enxofre
Três das oito enzimas contendo níquel são sistemas Ni-Fe-S. Entre elas estão as
hidrogenases, as CO-deshidrogenases (CODH) e a acetil-CoA sintetase (ACS).
Hidrogenases tem um papel crucial no metabolismo de microrganismos por
catalisarem a oxidação reversível do hidrogênio.
Três tipos de hidrogenases são conhecidos: As [NiFe], [FeFe] e Fe-hidrogenases. Em
ambientes de baixa concentração de níquel, Fe-hidrogenases são predominantes.

13
14
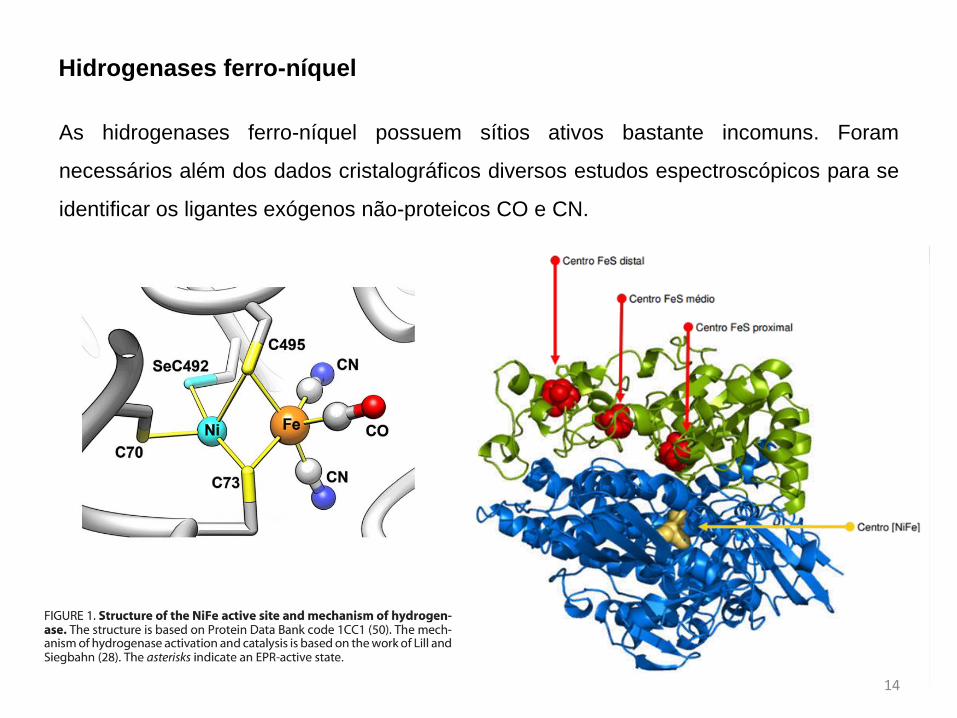
Hidrogenases ferro-níquel
As hidrogenases ferro-níquel possuem sítios ativos bastante incomuns. Foram
necessários além dos dados cristalográficos diversos estudos espectroscópicos para se
identificar os ligantes exógenos não-proteicos CO e CN.

15
Mecanismo de ação

16
CO-Deshidrogenase (CODH)
De uma forma conceitual, a maneira mais simples de preparar um composto orgânico é
construir este um carbono de cada vez. Este é justamente o caminho percorrido pela
enzima CO-deshidrogenase durante síntese da Acetil-CoA atuando na redução do
CO2.
17
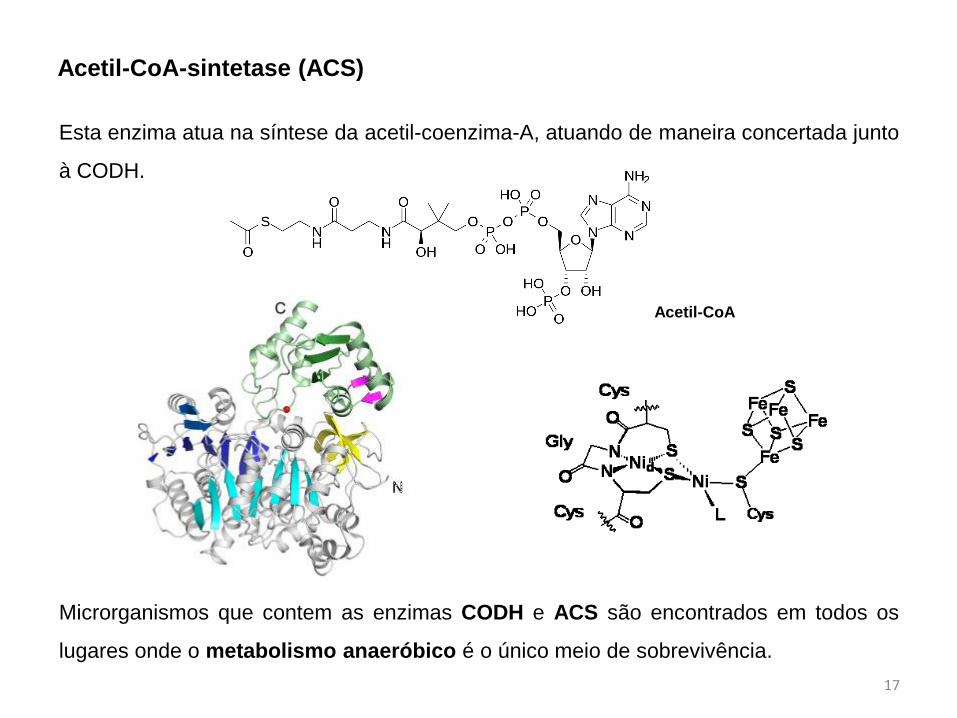
Acetil-CoA-sintetase (ACS)
Esta enzima atua na síntese da acetil-coenzima-A, atuando de maneira concertada junto
à CODH.
Acetil-CoA
Microrganismos que contem as enzimas CODH e ACS são encontrados em todos os
lugares onde o metabolismo anaeróbico é o único meio de sobrevivência.

18

19

20
Metil-coenzima-M redutase (MCR)
Atualmente estima-se que cerca de 109 toneladas de CH4 são geradas por bactérias
anaeróbicas metanogênicas. O metano gerado deste processo é fruto da atividade da
MCR.
Cofator F430

21
Estrutura cristalina, sítio ativo e mecanismo reacional

22
Cobalaminas e proteínas de cobalto
A vitamina B12 foi identificada em 1925 como imprescindível para evitar a anemia
perniciosa. Sua estrutura consiste em um macrociclo corrínico com um átomo de
cobalto(1+) ao centro, um quinto ligante: O 5,6-dimetilbenzimidazol nucleotídeo (Dmb).
O sexto ligante pode variar, gerando
então diferentes compostos:
• R = CN: Vitamina B12;
• R = 5’-deoxadenosina: coenzima
B12;
• R = CH3: alquil cobalamina;

23
Quando o cofator está em sua forma livre, este pode coexistir em três formas básicas,
conforme indicado abaixo:
A forma ligada do Dmb é predominante em pH fisiológico. Em algumas enzimas B12
dependentes, o grupo Dmb pode ser substituído por um resíduo de histidina.
24
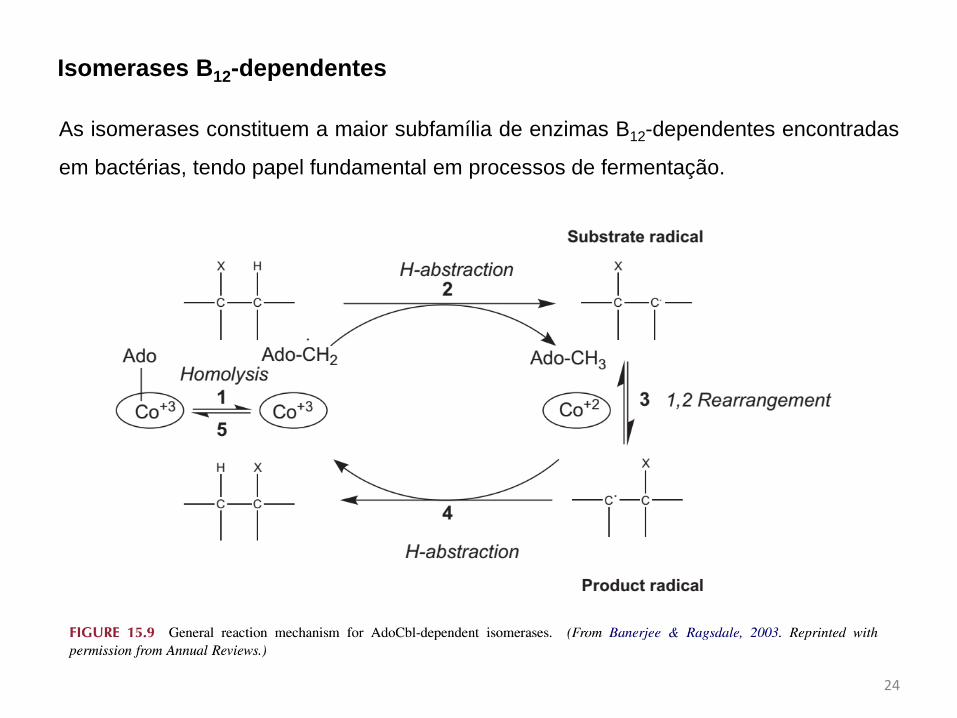
Isomerases B12-dependentes
As isomerases constituem a maior subfamília de enzimas B12-dependentes encontradas
em bactérias, tendo papel fundamental em processos de fermentação.

25
1. Cisão homolítica e formação do radical 5’-deoxiadenosil;
Etapas:
2. Abstração de um próton para formar o substrato radical;
3. Processo de rearranjo do tipo 1,2;
4. Uma nova abstração de próton leva a formação do radical 5’-deoxiadenosil;
5. Reconstituição da coenzima B12.
Como já visto em outras enzimas tais como nas ribonucleotídeo redutases de ferro, a
formação do radical tilil é feita pela coenzimas B12.

26
Ribonucleotídio redutase (RNR)

27
Metiltransferases B12-dependentes
Estas enzimas estão envolvidas no metabolismo de microrganismos anaeróbicos que
fixam CO2. Estas macromoléculas atuam ainda no metabolismo de aminoácidos em
muitos organismos, incluindo o humano.
A proteína que contém o grupo B12 tua como carreador “intermediário” do grupamento
metila.

28
As metiltransferases envolvem 03 componentes proteicos, cada um localizados em um
domínio da proteína.
O domínio MT1 se liga ao grupo doador de metila e a transfere para o complexo B12; O
domínio MT2 catalisa a transferência da metila ligada ao complexo B12 para o destino
final.

29
30
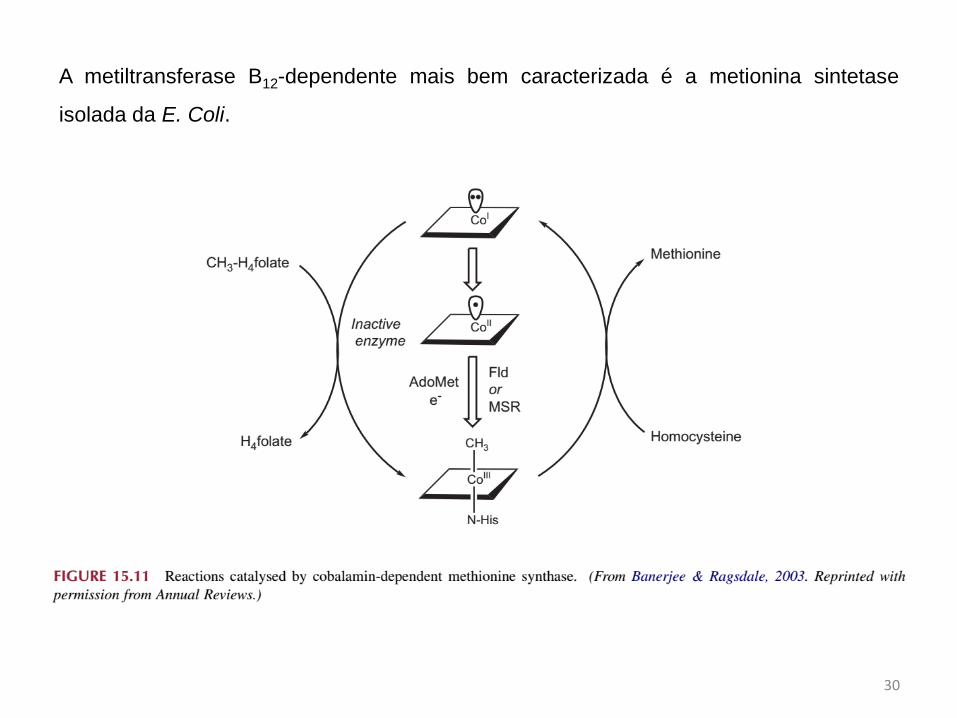
A metiltransferase B12-dependente mais bem caracterizada é a metionina sintetase
isolada da E. Coli.

31
Um exemplo: Metilmalonil-CoA mutase.
32
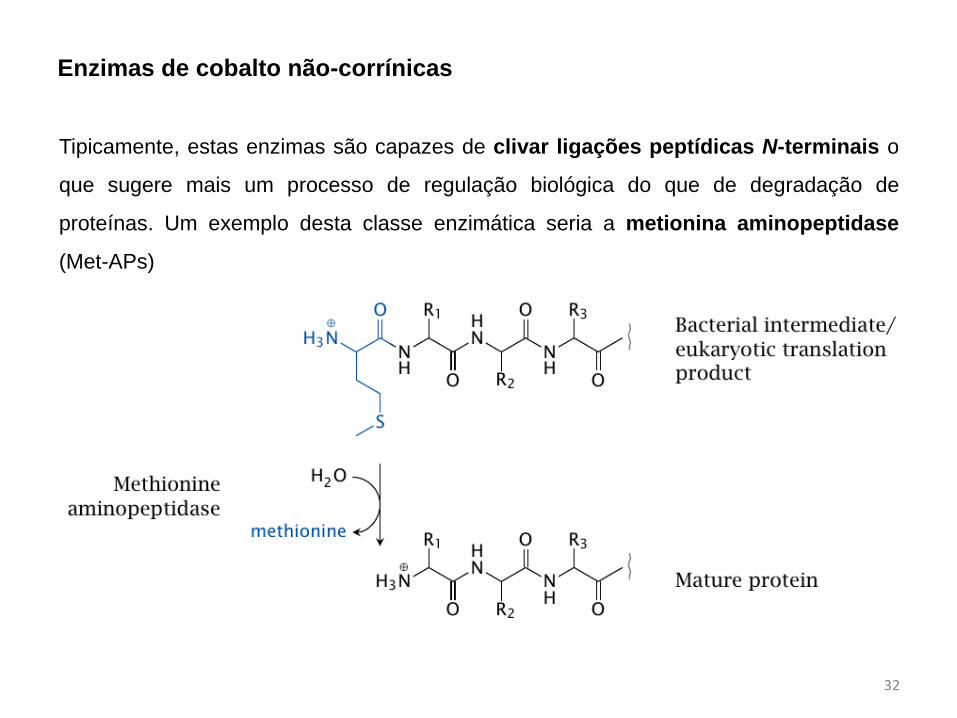
Enzimas de cobalto não-corrínicas
Tipicamente, estas enzimas são capazes de clivar ligações peptídicas N-terminais o
que sugere mais um processo de regulação biológica do que de degradação de
proteínas. Um exemplo desta classe enzimática seria a metionina aminopeptidase
(Met-APs)
33
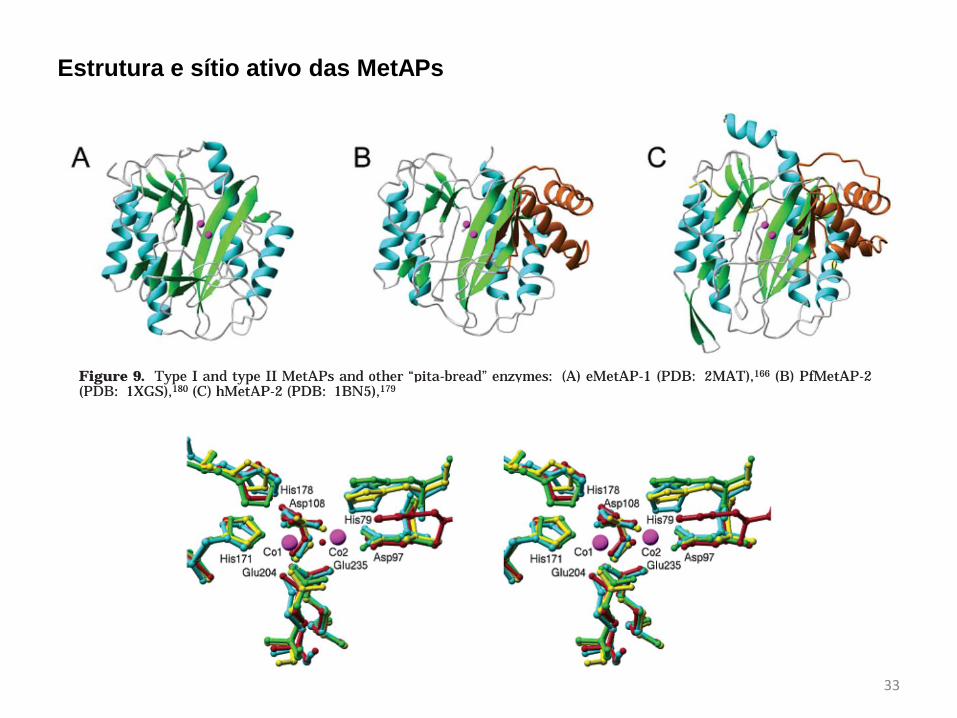
Estrutura e sítio ativo das MetAPs
34
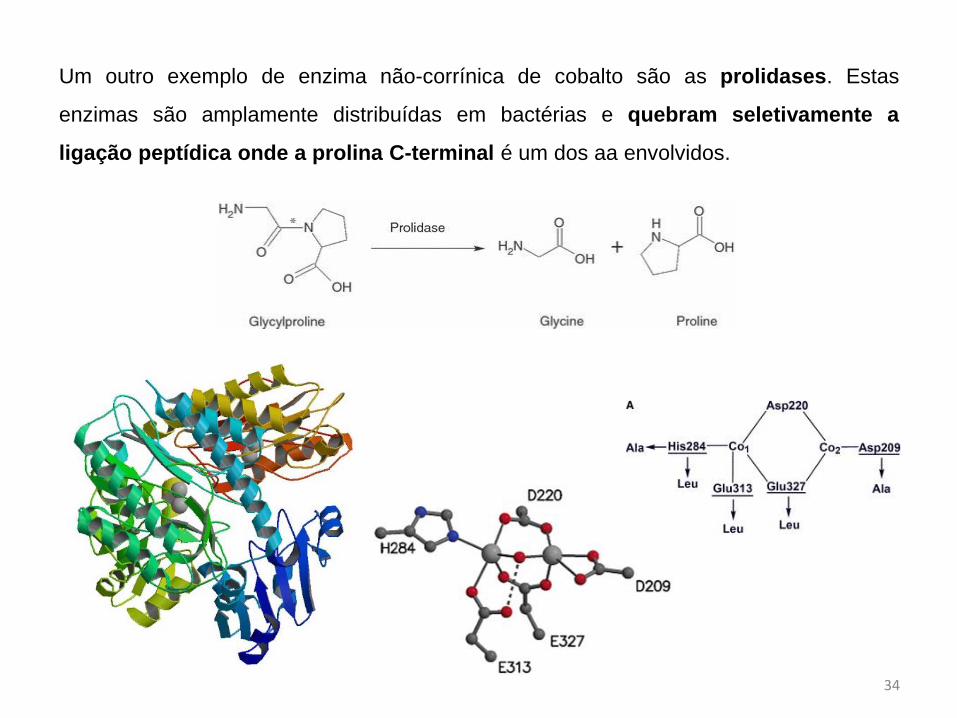
Um outro exemplo de enzima não-corrínica de cobalto são as prolidases. Estas
enzimas são amplamente distribuídas em bactérias e quebram seletivamente a
ligação peptídica onde a prolina C-terminal é um dos aa envolvidos.





























