Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO ESTADO DE SÃO PAULO
MARIA CAROLINA LEAL ALMEIDA
AVALIAÇÃO DE ALTERAÇÕES DA MATRIZ
EXTRACELULAR EM CULTURA DE CÉLULAS APÓS
LASERTERAPIA
SÃO PAULO
2013

MARIA CAROLINA LEAL ALMEIDA
AVALIAÇÃO DE ALTERAÇÕES DA MATRIZ
EXTRACELULAR EM CULTURA DE CÉLULAS APÓS
LASERTERAPIA
SÃO PAULO
2013
Dissertação apresentada ao Departamento de Bioquímica, Disciplina de Biologia Molecular da Universidade Federal de São Paulo, para obtenção do título de Mestre em Ciências. Orientador: Profa. Dra. Maria Aparecida da Silva Pinhal

Almeida, Maria Carolina Leal Avaliação de alterações da matriz extracelular em cultura de células após Laserterapia / Maria Carolina Leal Almeida - São Paulo, 2013. – 68 f. Dissertação de Mestrado – Universidade Federal de São Paulo. Campus São Paulo. Programa de Pós graduação em Ciências Biológicas (Biologia Molecular). Orientador: Maria Aparecida da Silva Pinhal 1. Laserterapia 2. Matriz extracelular 3. Glicosaminoglicanos

Aos meus pais, Vitor e Telma por sempre
apoiarem e respeitarem as minhas escolhas,
mesmo diante das dificuldades, sempre
servindo de exemplo e inspiração para que eu
pudesse me tornar alguém melhor e que
lutasse pelos meus sonhos. Amo vocês!!!

A toda minha família, em especial a
minha madrinha Maria, minha avó
Hilda, minha irmã Ana, minha
sobrinha Isadora e meu primo
Vinicius, pelos momentos de alegria,
descontração, carinho, amizade,
preocupação e paciência. Amo vocês!!!

E por fim e não menos importante dedico
este trabalho a Professora Cida, pelo
exemplo de paixão e dedicação à
pesquisa, pela honestidade, ética e
sinceridade.
Obrigada pelo voto de confiança,
amizade, conselhos, puxões de orelha e
por toda a atenção dispensada para que
este projeto fosse realizado.
Muito obrigada!!!!

AGRADECIMENTOS
Vamos lá no decorrer desses quatro anos de faculdade conheci muita gente boa, pessoas que
indiretamente ou diretamente fizerem parte dessa história:
Ao nosso grupo de trabalho orientado pela professora Cida, Teka, Juan, Dr. Leandro, Dr.
Luciano, Cintia, Eloah, Carina e Bethania, não tenho palavras para descrever o quanto sou
grata por ter conhecido vocês e pelo convívio diário no laboratório, regado de muitas risadas
e conversas, possibilitando uma experiência única, Obrigada!!!
As amigas Eliete, Risa, Thama, Teka, Carina, Bethania, Tati, Eloah, e a Maria Alice por
sempre estarem comigo nos momentos bons, mas principalmente nos momentos difíceis,
quando minha vontade era desistir.
Aos professores do departamento, profa. Giselle Zenker Justo por me ceder às células
HaCaT, prof. Hugo Pequeno Monteiro por ceder às células A431, prof. João Roberto Martins
por ceder as enzimas condroitinases AC e ABC, prof. Ivarne Tersariol, profa. Yara Michellaci
e profa. Leny Toma.
Aos meus colegas da bio mol do Laboratório da Faculdade de Medicina do ABC e da Unifesp
à galera do quinto andar em especial à Tatiana (Tati), sempre com seus conselhos sábios,
aos técnicos do laboratório pela ajuda prestada, a todos que direta ou indiretamente
contribuíram para a realização deste trabalho e pela convivência diária, pelas pipocas,
lanchinhos da tarde, festas de aniversários e risadas e lágrimas!
Aos meus colegas de plantão que eu adotei como parte da minha família Sandra, Beth, Maria
Alice (mais uma vez), ao pessoal da enfermagem. Muito obrigada por fazerem parte de mais
2 anos da minha vida!!!
A Lilian pela amizade, pela paciência e por toda a ajuda que você me deu, me ensinando a
cultivar células e por fazer praticamente todos os experimentos deste projeto comigo! Muito
obrigada!

Aos meus amigos da Gakkai pelo apoio, pelas palavras de incentivo e por entenderem a
minha dificuldade de participar das atividades.
Á Profa. Dra. Helena Nader pela oportunidade de fazer parte da equipe de biologia
molecular.
À Universidade Federal de São Paulo e Faculdade de Medicina do ABC.
Agradeço o apoio das agências de fomento que possibilitou o desenvolvimento deste
trabalho, CNPq – Conselho Nacional de Desenvolvimento Científico e Tecnológico, CAPES –
Coordenação de aperfeiçoamento de Pessoal de Nível Superior, pela bolsa de mestrado,
FAPESP – Fundação de Amparo à Pesquisa de Estado de São Paulo, pelo apoio financeiro
concedido ao laboratório na forma de Auxílios à Pesquisa.
Muitíssimo obrigada a todos,
Carol

“A causa da derrota não se encontra no obstáculo ou no rigor das circunstâncias;
está no retrocesso na determinação e na desistência da própria pessoa. Se falasse
em dificuldades, tudo realmente era difícil. Se falasse em impossibilidades, tudo
realmente era impossível. Quando o ser humano regride em sua decisão os
problemas que se erguem em sua frente acabam parecendo maiores e confundem-
no como uma realidade imutável. A derrota encontra-se exatamente nisso...
Somente o conhecimento não é suficiente. Somente quando o conhecimento alia-se
a sabedoria é que uma pessoa pode atingir a vitória na vida.“
Daisaku Ikeda

Resumo
A pele corresponde ao órgão de maior exposição a radiação por diferentes tipos de luz.
Alguns estudos demonstram o efeito de terapias de luz laser de baixa intensidade em
determinadas afecções da pele como manchas, processos inflamatórios e em diferentes
tipos de tumores. O laser de baixa potência tem sido utilizado na prática clínica há
aproximadamente 20 anos. A laserterapia quando utilizada em tecidos e células provoca
efeitos fotoquímicos, fotofísicos e/ou fotobiológicos, sem a produção de calor, estimulando a
proliferação celular, ativação de linfócitos e mastócitos, promove aumento na geração ATP
entre outros efeitos. Poucos artigos relacionam a laserterapia com as alterações de
componentes da matriz extracelular em cultura celulares. Portanto, o presente estudo tem
por objetivo investigar as possíveis alterações de componentes da matriz extracelular em
células HaCaT (queratinócitos humanos) e A431 (carcinoma epidermóide humano), que
representam dois tipos distintos de linhagens celulares da pele. Dentre os componentes da
matriz extracelular foram analisados glicosaminoglicanos sulfatados (GAG), ácido
hialurônico, enzimas que participam da degradação de heparam sulfato (heparanase) e
enzimas envolvidas na degradação e biossíntese de ácido hialurônico (hialuronidases e
ácido hialurônico sintases), respectivamente. As alterações dos componentes da matriz
extracelular foram avaliados comparando-se células submetidas ao tratamento com
laserterapia de baixa potência e células não tratadas (controle). A viabilidade celular de
células HaCaT e A431 também foi avaliada. Os GAG sulfatados foram analisados por
marcação metabólica utilizando sulfato radioativo, degradação enzimática e eletroforese. O
ácido hialurônico foi quantificado por método de ELISA indireto. Ensaios de PCR em tempo
real foram realizados para análise da expressão das enzimas HAS (ácido hialurônico
sintases) HYAL (Hialuronidases) e HPSE-1 (Heparanase). Os resultados obtidos no
demonstraram diminuição significativa na viabilidade celular em ambas as linhagens (HaCaT
e A431), após laserterapia. Houve alteração no perfil de GAG após tratamento com luz no
comprimento de onda associado (470 e 660 nm), sendo que as células HaCaT
apresentaram aumento significativo de heparam sulfato e condroitim sulfato. O ácido
hialurônico aumentou na fração celular, após laserterapia em células HaCaT, enquanto, não
foi observado alteração nas células A431. A analise por qPCR demonstrou diminuição da
expressão em células HaCaT e das células A431. Os resultados obtidos em nosso estudo
demonstraram que o tratamento com laserterapia de baixa intensidade altera componentes
da matriz extracelular e tais alterações diferem com relação ao tipo celular (células não
neoplásicas ou células de carcinoma epidermóide humano), e o comprimento de onda
utilizado.

LISTA DE ABREVIATURAS, SIGLAS E SÍMBOLOS
A431 – Linhagem estabelecida de carcinoma epidermóide humano
AH – Ácido hialurônico
ATP – Adenosina Trifosfato
AU - Ácido Urônico
BSA – Albumina sérica bovina
C - Carbono
CBC – Carcinoma basocelular
CEC – Carcinoma espinocelular
CO2 - Gás Carbônico
CS – Condroitim sulfato
Da – Dalton
DMEM – Meio Eagle Modificado por Dulbecco
DMSO - dimetilsulfóxido
DS - Dermatam sulfato
EBSS – Solução salina balanceada de Earl sem cálcio e sem magnésio
EGF – Fator de crescimento epidérmico
F-12 – Meio de cultura contendo mistura de nutrientes com L-glutamina e sem
bicarbonato de sódio
FGF – Fator de crescimento de fibroblastos
FGF-2 - Fator de crescimento básico de fibroblastos
FGF-7 - Fator de crescimento de queratinócitos
GAG – Glicosaminoglicano
Gal – Galactose
GlcNac:N-acetilglucosamina
GlcUA: ácido glucurônico
GPI - glicosilfosfatidilinositol
GTP - guanosina trifosfato
HaCaT – Linhagem estabelecida de queratinócitos humano
HAS - Ácido Hialurônico Sintase
HCl – Ácido Clorídrico
HEP – Heparina
HPSE - Heparanase

HS – Heparam sulfato
Hx – Hexosamina
HYAL - Hialuronidase
J - Joule
KDa – Kilodalton
KS – Queratam sulfato
LASER – Amplificação da luz por emissão estimulada de radiação
MEC - Matriz Extracelular
MTT - 3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazólio
NaCl – Cloreto de sódio
nm – Nanômetro
PBS – Tampão Fosfato-Salino
PCR - polymerase chain reaction
PDA – 1,3-Diaminopropano-acetato
PDT – Terapia Fotodinâmica
PG - Proteoglicanos
PGHS – Proteoglicanos de heparam sulfato
PGKS – Proteoglicanos de queratam sulfato
RNA – Ácido ribonucléico
ROS – Espécies reativas de oxigênio
RT-PCR - reverse transcriptase polymerase chain reaction
SFB – Soro Fetal Bovino
TGF-β - Fator de Crescimento Transformante Beta

ÍNDICE
1. INTRODUÇÃO ...................................................................................................................................... 1
1.1. LASERTERAPIA OU TERAPIA COM LUZ LASER DE BAIXA INTENSIDADE .............................................. 1
1.1.1. Efeitos da laserterapia nos tecidos biológicos ....................................................................... 2
1.2. PELE .................................................................................................................................................. 4
1.3. MATRIZ EXTRACELULAR .................................................................................................................... 8
1.3.1. Proteoglicanos ............................................................................................................................ 8
1.4. GLICOSAMINOGLICANOS.................................................................................................................... 9
1.4.1. Ácido Hialurônico ...................................................................................................................... 10
1.4.2. Condroitim Sulfato, Dermatam Sulfato e Queratam Sulfato .............................................. 13
1.4.3. Heparam Sulfato ....................................................................................................................... 14
2. OBJETIVOS .........................................................................................................................................17
3. MATERIAIS ..........................................................................................................................................18
3.1. LINHAGEM CELULAR ........................................................................................................................ 18
3.2. CULTURA DE CÉLULAS ..................................................................................................................... 18
3.3. ENZIMAS .......................................................................................................................................... 18
3.4. LÍQUIDO DE CINTILAÇÃO .................................................................................................................. 19
3.5. RADIOISÓTOPO ................................................................................................................................ 19
3.6. REAGENTES GERAIS E OUTROS MATERIAIS .................................................................................... 19
3.7. EQUIPAMENTOS ............................................................................................................................... 20
4. MÉTODOS ............................................................................................................................................22
4.1. CULTURA DE CÉLULAS ..................................................................................................................... 22
4.1.1. Descongelamento de células .................................................................................................. 22
4.1.2. Manutenção das células .......................................................................................................... 22
4.1.3. Subcultivo das células ............................................................................................................. 22
4.1.4. Contagem das células em Câmara de Neubauer ............................................................... 23
4.2. TRATAMENTO DA CULTURA DE CÉLULA COM RADIAÇÃO A LASER ................................................... 23
4.3. QUANTIFICAÇÃO DE GLICOSAMINOGLICANOS SULFATADOS SINTETIZADOS POR CÉLULAS. ........... 25
4.3.1. Marcação metabólica ............................................................................................................... 25
4.3.2. Extração de GAG marcados metabolicamente .................................................................... 25
4.3.3. Eletroforese em gel de agarose com tampão PDA ............................................................. 26
4.3.4. Análise e quantificação dos GAG marcados metabolicamente ........................................ 26
4.3.5. Dosagem de proteínas por método de Coomassie Blue .................................................... 27
4.4. DEGRADAÇÃO ENZIMÁTICA .............................................................................................................. 27
4.5. ENSAIO DE VIABILIDADE CELULAR ................................................................................................... 28
4.6. DOSAGEM DE ÁCIDO HIALURÔNICO ................................................................................................. 28
4.7. EXTRAÇÃO DE RNA TOTAL ............................................................................................................. 30
4.7.1. QUANTIFICAÇÃO DO RNA TOTAL .................................................................................................... 31
4.7.2. OBTENÇÃO CDNA A PARTIR DO RNA TOTAL ................................................................................. 31
4.8. ANÁLISE DA EXPRESSÃO POR PCR EM TEMPO REAL (QPCR) ....................................................... 31
4.9. ANÁLISE ESTATÍSTICA ..................................................................................................................... 32
5. RESULTADOS E DISCUSSÃO ..........................................................................................................33
5.1. VIABILIDADE CELULAR EM CÉLULAS HACAT E A431 ............................................................................... 33
5.4. DOSAGEM DE ÁCIDO HIALURÔNICO ................................................................................................ 42
5.5. EXPRESSÃO GÊNICA POR PCR EM TEMPO REAL DAS ENZIMAS DE BIOSSÍNTESE E DEGRADAÇÃO
DO ÁCIDO HIALURÔNICO ............................................................................................................................... 44

ÍNDICE
6. CONCLUSÕES ....................................................................................................................................47
7. REFERÊNCIA BIBLIOGRÁFICA .......................................................................................................48

ÍNDICE
ÍNDICE DE FIGURAS
Figura 1: Esquema que ilustra o poder de penetração dos diferentes tipos de luz em função do
comprimento de onda ---------------------------------------------------------------------------------------------------------- 3
Figura 2: Espectro de absorção dos cromóforos da pele. ----------------------------------------------------------- 4
Figura 3. Representação esquemática das camadas estruturais da pele ----------------------------------- 5
Figura 4: Processo de diferenciação de queratinócitos -------------------------------------------------------------- 6 Figura 5. Sitio de clivagem da heparanase e mecanismos de ação dos oligossacarídeos
gerados na MEC ------------------------------------------------------------------------------------------------------------------15 Figura 6. Representação esquemática do ensaio fluorimétrico para quantificação de ácido
hialurônico --------------------------------------------------------------------------------------------------------------------------30
Figura 8. Perfil de GAG sulfados sintetizados por HaCaT e A431 ----------------------------------------------36
Figura 11. Expressão relativa de Heparanse-1 (HPSE-1). -----------------------------------------------------------41
Figura 12. Quantificação de Ácido Hialurônico --------------------------------------------------------------------------43 Figura 13. Expressão relativa de Ácido Hialurônico Sintases (HAS1-3) e Hialuronidases
(HYAL1-2) ----------------------------------------------------------------------------------------------------------------------------45

ÍNDICE
ÍNDICE DE TABELAS
Tabela 1. Proteoglicanos presentes no tecido epitelial. ......................................................................10
Tabela 2. Características estruturais dos glicosaminoglicanos ........................................................11
Tabela 3. Parâmetros de Irradiação .........................................................................................................24
Tabela 4. Sequências de Oligonucleotídeos iniciadores ....................................................................32

INTRODUÇÃO
1
1. INTRODUÇÃO
1.1. Laserterapia ou Terapia com luz laser de baixa intensidade
A laserterapia de baixa potência é investigada e utilizada na prática clínica há
aproximadamente 20 anos, Theodore H. Maiman construiu o primeiro emissor de
laser de rubi em 1960 e, em 1965, Sinclair e Knoll adaptaram esta radiação à prática
terapêutica. Um dos pioneiros na aplicação do laser de baixa intensidade nas áreas
biomédicas foi Endre Mester no início da década de 70, na Europa (FUKUDA T Y,
2008; LIN et al., 2010; MESTER; TOTA, 1967).
A palavra laser advém da abreviação de seu próprio significado (Light
Amplification by Stimulated Emisson of Radiation), ou seja, amplificação da luz por
emissão estimulada de radiação. Assim definimos laser como onda eletromagnética
não ionizante com características especiais (KARU, 1987; LIN et al., 2010; TANZI;
LUPTON; ALSTER, 2003). As características especiais do laser diferenciam
totalmente da luz natural e é definida como luz coerente, monocromática e paralela
(TANZI; LUPTON; ALSTER, 2003).
Os lasers são classificados em duas famílias, conforme sua potência e
capacidade de interação com os tecidos. O tratamento com laser de baixa
intensidade de energia (LILT ou Low Intensity Level Treatment) causa efeitos como
bioestimulação, analgesia, apresentam ação anti-inflamatória e podem diminuir o
edema. Por outro lado, o tratamento com laser de alta intensidade de energia (HILT,
Hight Intensity Laser Treatment) pode causar desnaturação de proteínas, são
utilizados para acelerar processos de coagulação sanguínea e causam vaporização
devido à produção de calor. Portanto, a literatura sugere que lasers de baixa
intensidade de energia são estimulatórios, enquanto os de alta intensidade de
energia possuem efeito inibitório e grande parte de sua utilização são para fins
terapêuticos (HARRIS, 1988; LIN et al., 2010).
Outro fator importante diz respeito aos conceitos físicos de irradiância, fluência
e energia depositada, pois como pode ser verificadas em diversos estudos as doses
utilizadas no tratamento deve levar em consideração estas propriedades físicas a fim
de padronizar e obter resultados satisfatórios do tratamento, visto que, a irradiância
influência no efeito biológico. Defini-se Irradiância como sinônimo de densidade de

INTRODUÇÃO
2
potência (DP) e esta define a potência óptica útil do laser e expressa em watts (W),
dividida pela área irradiada, expressa em centímetros quadrado (cm2). Então temos:
Irradiância = potência óptica laser (W) / Área irradiada (cm2). Enquanto a fluência
define a taxa de energia aplicada no tecido biológico, é expressa por: Irradiância (W
/ cm2) x tempo de exposição = densidade de energia (fluência) ou dose de energia
(DE) expressa em joule (J / cm2). Estes dados refletem de forma direta o efeito da
irradiação em tecidos biológicos, pois a irradiância vai determinar se há dano térmico
ou não. (LOPES, [S.d.]).
1.1.1. Efeitos da laserterapia nos tecidos biológicos
Há um crescente interesse tem sido demonstrado na utilização do laser.
Entretanto, ainda existe muita divergência com relação aos resultados obtidos de
diferentes estudos que descrevem os efeitos da utilização do laser. Os dispositivos
da bioestimulação estão sendo usados sem o conhecimento suficiente dos
mecanismos de ação do tipo de luz em cada tecido. Ainda, existe também
deficiência nos métodos de padronização para utilização da laserterapia, como por
exemplo intensidade da luz, tempo de aplicação e distância. Existem inúmeros
relatos sobre a eficácia da laserterapia no efeito analgésico, antiinflamatório e de
regeneração tecidual (HARRIS, 1988; LIN et al., 2010; TANZI; LUPTON; ALSTER,
2003), porém, muitos dos relatos são controversos por diferenças significativas na
metodologia aplicada.
Na área dermatológica os lasers mais utilizados vão de 308 (UV-B) até 10,600
(infravermelho) (CARROLL; HUMPHREYS, 2006), o protocolo de tratamento de pele
típico envolve a irradiação do local afetado com a luz laser de baixa intensidade e o
efeito da luz aparentemente cumulativo é repetido em intervalos de dias, podendo
ser absorvida, refletida, transmitida ou espalhada (HARRIS, 1988; TANZI; LUPTON;
ALSTER, 2003). A Figura 1 ilustra o poder de penetração das diferentes camadas
que compõem a pele, utilizando diferentes tipos de luz de acordo com o seu
comprimento de onda.
Quando a luz interage com as células e tecidos, é necessário observar que
lasers no espectro visível possuem pouca penetração nos tecidos, sendo de grande
importância a utilização de uma dose adequada, onde certas funções celulares

INTRODUÇÃO
3
podem ser estimuladas, tais como ativação de linfócitos, ativação de mastócitos,
aumento na produção de ATP mitocondrial, proliferação celular e apoptose. Também
foi observado efeito da luz sobre componentes da matriz extracelular como estimulo
da proliferação de fibroblastos, estímulo da síntese de fibras elásticas e colágenas,
aumento da migração celular de queratinócitos, estímulo da liberação do fator de
crescimento de fibroblastos (FGF) e estímulo no processo de cicatrização de feridas
(BASSO et al., 2012; GAO; XING, 2009; HOPKINS et al., 2004; LIN et al., 2010;
LOPES, [S.d.]; TANZI; LUPTON; ALSTER, 2003).
Figura 1: Esquema que ilustra o poder de penetração da radiação em função do comprimento
de onda. (LOPES, [S.d.])
A utilização de lasers no comprimento de onda de 470 nm (luz azul), apresenta
ação importante no processo bactericida e antifúngico. A utilização da luz azul pode
ser realizada em associação com fotossensibilizadores, causando fotoexcitação de
porfirinas endógenas de bactérias fotossensíveis e posterior produção de espécies
reativas de oxigênio (LUCCA; WILLIAMS; BHATNAGAR, 2012)
A laserterapia promove estresse nos tecidos por promover a formação de
espécies reativas de oxigênio e também induz diversas respostas como proliferação
celular e processos de cicatrização (GROSSMAN et al., 1998).

INTRODUÇÃO
4
A formação de espécies reativas de oxigênio (ROS) causado pela laserterapia
induz proliferação, produção de moléculas inflamatórias e aumento na produção de
ATP (CHEN; HUNG; HSU, 2008; FUJIMOTO et al., 2012; GAO; XING, 2009).
Lasers de baixa potência são frequentemente empregados no tratamento de
patologias, sendo associados a diferentes técnicas, como por exemplo, Terapia
Fotodinâmica (PDT), onde se utiliza fotossensibilizador capaz de absorver a luz em
comprimento de onda específico (CASTRO et al., 2007; WAGNER et al., 2012),
sendo assim a laserterapia difere da PDT por utilizar cromóforos endógenos. Na a
pele, os principais cromóforos endógenos são a riboflavina, bilirrubina, melanina,
protoporfirina e hemoglobina, como demonstrado na Figura 2 (MAHMOUD et al.,
2008).
Figura 2: Espectro de absorção dos cromóforos da pele. Adaptado de (MAHMOUD et al., 2008)
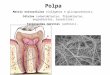
1.2. Pele
A pele é o maior órgão do corpo humano e representa 16% do peso corpóreo,
está dividida em camadas denominadas epiderme, derme e hipoderme, como
representado na Figura 3.

INTRODUÇÃO
5
Figura 3. Representação esquemática das camadas estruturais da pele. Adaptado de (“Cancer
Information,” [S.d.]).
A epiderme representa a camada mais superficial da pele e é composta de
camadas estratificadas denominadas estrato basal, estrato espinhoso, estrato
granuloso e estrato córneo (Figura 4).
A estrutura e função da pele dependem de um equilíbrio entre a proliferação e
diferenciação de queratinócitos, que sofrem alterações morfológicas e bíoquímicas
importantes, conferindo rigidez e força ao tecido epitelial, bem como proteção contra
a invasão de patógenos, agressões físicas e químícas. A integridade e resistência
da pele se deve à presença de uma extensa camada de filamentos de actina no
citoesqueleto dos queratinócitos e interações célula-célula (desmossomos) e célula-
matriz (hemidesmossomos) (SIMPSON; PATEL; GREEN, 2012).

INTRODUÇÃO
6
Figura 4: Processo de diferenciação de queratinócitos. Adaptado de PROKSCH; BRANDNER;
JENSEN, 2008.
Os queratinócitos representam cerca de 90% da epiderme e são as células
responsáveis pela produção da queratina, num processo denominado queratinização
(KANITAKIS, 2002). O estrato basal representa a porção mais interna da epiderme
em contato com a membrana basal. Neste estrato estão presentes queratinócitos
com aspecto cubóide, melanócitos responsáveis pela síntese de melanina e células
de Merkel, responsáveis pela sensação do tato. A camada basal germinativa garante
a renovação dos queratinócitos e renovação da pele, que ocorre em até 21 dias.
As camadas suprabasais da epiderme subdividem-se em estrato córneo, que
compreende a camada mais externa, constituída por células anucleadas, achatadas
e queratina, responsável por formar uma barreira de proteção que surge a partir da

INTRODUÇÃO
7
diferenciação específica dos queratinócitos da camada espinhosa e granulosa. No
estrato granuloso, além de queratinócitos com formato mais achatado encontram-se
células de Langerhans, que participam das reações imunológicas da epiderme.
Em diferentes situações patológicas, como lesão da epiderme, queratose
actínica, hiperpigmentação e sarda (lentigo), inicia-se um programa complexo de
reestruturação que promove reepitelização, migração e proliferação intensa de
queratinócitos. Tais processos são coordenados pela interação de fatores de
crescimento, como fator de crescimento epidérmico (EGF), fatores de crescimento
de fibroblastos (FGF) e fator de crescimento transformante β (TGF-β) (IRIYAMA et
al., 2011; NASHAN; MEISS; MÜLLER, 2013; SADOWSKI et al., 2003; SEPPÄNEN-
PASONEN et al., 2003).
A pele está sujeita a diferentes tipos de estresse, como, por exemplo, a
frequente exposição à luz UV, e sua alteração pode facilmente progredir ao
aparecimento de tumores classificados como melanomas malignos e tumores não-
melanomas, este último inclui o carcinoma espinocelular (CEC) e carcinoma
basocelular (CBC), que representam respectivamente 25% e 70% dos casos de
câncer de pele não-melanoma (“INCA,” [S.d.]; LUKIC D et al., 2012; PROKSCH;
BRANDNER; JENSEN, 2008; WU et al., 2012).
O carcinoma espinocelular (CEC) é uma lesão das células epiteliais originada a
partir da transformação de queratinócitos, e o segundo câncer não-melanoma mais
frequente na população branca. Sua principal origem é a presença de queratose
actínica causada pela exposição solar, sendo classificado de acordo com grau o
diferenciação dos queratinócitos e a invasão epidérmica (KIM; ARMSTRONG, 2012).
O crescimento do tumor é controlado pela interação das células tumorais com o
microambiente, envolvendo ativação de fibroblastos, células endoteliais e
inflamatórias e interações com diversos componentes da matriz extracelular,
promovendo proliferação celular, angiogênese, inibição de apoptose e invasão
tumoral (RODUST; FECKER; STOCKFLETH, 2012; WILLHAUCK et al., 2007).
O estudo com células HaCaT demonstra um grande potencial na investigação
da fisiologia e patologias da pele, visto que as células HaCaT possuem a
capacidade de se diferenciar in vitro, reproduzindo todas as camadas constituintes
da epiderme. Cumpre ser observado que trabalhos recentes demonstram
claramente que o meio de cultura é importante para o processo de diferenciação das
diversas camadas da epiderme. Dentre as diferentes variáveis do meio de cultura

INTRODUÇÃO
8
celular, a concentração ideal de cálcio e tempo de cultura permitem a presença de
todas as camadas epidérmicas em cultura de células HaCaT, em média 13 dias para
chegar ao estágio final de diferenciação (BORANIC et al., 1999; DEYRIEUX;
WILSON, 2007).
A linhagem não neoplásica de queratinócitos humanos imortalizada (HaCaT),
bem como células de carcinoma epidermóide humano (A431), são frequentemente
utilizadas como modelo in vitro de estudos (DEYRIEUX; WILSON, 2007; HSIEH; YU;
LYU, 2010).
1.3. Matriz Extracelular
1.3.1. Proteoglicanos
A matriz extracelular (MEC) é constituída por uma rede complexa de
macromoléculas composta por colágenos, glicoproteínas, fibras elásticas,
proteoglicanos (PG) e ácido hialurônico (AH) (KRESSE; SCHÖNHERR, 2001;
SCHONHERR; HAUSSER, 2000). A MEC desempenha diversas funções como
participação em mecanismos de sinalização celular, reservatório para fatores de
crescimento, citocinas e modulação de processos como proliferação, adesão e
migração celular (BAUER; JACKSON; JIANG, 2009; KRESSE; SCHÖNHERR,
2001; SCHONHERR; HAUSSER, 2000).
Os proteoglicanos são macromoléculas constituídas por um esqueleto protéico
ligado a cadeias de glicosaminoglicanos e encontram-se presentes na superfície
celular, em grânulos intracelulares e na MEC. Os proteoglicanos modulam a ação de
citocinas, fatores angiogênicos e fatores de crescimento. Os efeitos dos
proteoglicanos em vários mecanismos celulares em geral são modulados por
interações com as cadeias de glicosaminoglicanos (GAG) ou por interações entre o
esqueleto protéico dos proteoglicanos. Os proteoglicanos também possuem
importante papel na organização de fibras de colágeno e participam de fenômenos
biológicos como diferenciação, manutenção e organização da MEC (SCHONHERR;
HAUSSER, 2000).
No tecido epitelial são frequentemente encontrados proteoglicanos de
superfície celular, como PG da família dos sindecans, glipicans, família do CD44 e
proteoglicanos de MEC (versicam) e lâmina basal (perlecam). Os PG de baixo peso
molecular, ricos em resíduos de leucina como decorim, biglicam e fibromodulina

INTRODUÇÃO
9
também são característicos da MEC da pele. A Tabela 1 evidencia os diferentes
tipos de PG encontrados na pele.
1.4. Glicosaminoglicanos
Os glicosaminoglicanos (GAG) são heteropolissacarídeos lineares, formados
por unidades dissacarídicas repetitivas, constituídos por um resíduo de hexosamina
(glucosamina ou galactosamina), ligada a um resíduo de açúcar não aminado, que
pode ser ácido urônico (D-ácido glucurônico ou L-ácido idurônico) ou D-galactose. A
massa molecular dos glicosaminoglicanos pode variar entre 10-100 kDa. Com
exceção do ácido hialurônico, todos os GAG encontram-se ligados a um esqueleto
protéico formando os proteoglicanos, já mencionados (GANDHI; MANCERA, 2008;
RADEMACHER; PAREKH; DWEK, 1988).
Os GAG podem interagir com proteínas distintas, incluindo quimiocinas,
citocinas, fatores de crescimento, enzimas e moléculas de adesão, promovendo a
regulação de diversas funções biológicas (GALLO, 2000; GANDHI; MANCERA,
2008)
Os GAG participam de mecanismos de sinalização em vários processos
fisiológicos e patológicos como angiogênese, crescimento axonal, progressão de
tumores, metástases tumorais e mecanismos de coagulação.
Os GAG sulfatados incluem o condroitim sulfato (CS), dermatam sulfato (DS),
queratam sulfato (KS), heparina (HEP) e o heparam sulfato (HS). Enquanto o ácido
hialurônico (AH) representa a classe de GAG não sulfatado, a Tabela 1 representa a
nomenclatura proposta por Jeanloz e unidades dissacarídicas constituintes
(JEANLOZ, 1960).
A pele apresenta grande quantidade de ácido hialurônico, dermatam sulfato,
heparam sulfato e queratam sulfato, que modulam processos de adesão, migração e
proliferação celular (GALLO, 2000; MALMSTRÖM et al., 2012; ZHAO et al., 2013).

INTRODUÇÃO
10
Tabela 1. Proteoglicanos presentes no tecido epitelial*.
Proteoglicano GAGProteína
(KDa)Tecido Atividade
Sindecam CS 31
HS 82-88
Glipicam HS 62
Perlecam CS/HS 400
Versicam CS 265
Decorim CS/DS 36
Colágeno XVIII HS 150
Adesão celular
Fibromodulina KS 42 ConjuntivoAdesão celular,
fibrilogênese
CD44
CS
facultativo 32-38
Epitélio e
fibroblasto
Membrana basal
Fibroblasto
Conjuntivo
Membrana basal,
epitélio
Epitélio e
linfócitos
Morfogênese, adesão
celular
Endocitose
Estrutural
Adesão celular,
fibrilogênese
Interação célula-célula
Suporte mecânico,
migração celular
Epitélio e
fibroblasto
* Adaptada de (ESKO, 1991; SARRAZIN; LAMANNA; ESKO, 2011)
1.4.1. Ácido Hialurônico
O ácido hialurônico (AH) é um GAG não sulfatado, formado por unidades
repetidas de β-D-1,3-N-acetilglicosamina e β-1,4-ácido D-glucurônico, com peso
molecular que varia entre 105 a 107 Da (BRIMACOMBRE; WEBER, 1964; MEYER,
1958).
Na MEC o ácido hialurônico encontra-se associado à proteoglicanos como o
agrecam e versicam, formando agregados unidos por domínios de ligação,
conhecidos como proteína de ligação ou link protein, proporcionando estabilidade à
molécula e dificultando a degradação do AH (RODRIGUEZ; ROUGHLEY, 2006;
SEYFRIED et al., 2005).

INTRODUÇÃO
11
O AH encontra-se altamente expresso em células epiteliais, desempenha
importante papel no remodelamento, reparo e regeneração do tecido, participando
também em processos patológicos como inflamação e desenvolvimento tumoral
(FRENKEL, 2012; KASTNER; THOMAS; JENKINS, 2007; NEUMAN et al., 2011;
PAIVA et al., 2005).
Tabela 2. Características estruturais dos glicosaminoglicanos
Açúcar não Ligação Hexosamina Ligação *Sulfato (s)
nitrogenado glicosídica (Hx) glicosídica
Ácido β-D- β (1-3) β-D-N- β (1-4) -
Glucurônico Acetilglucosamina
Ácido β-D- β (1-3) β-D-N- β (1-4) C4 ou C6
Glucurônico Acetilgalactosamina (Hx)
α-L-idurônico β (1-3) β-D-N- β (1-4) C4
Ácido β-D-Glucurônico α (1-3) Acetilgalactosamina (Hx)
β-D- β (1-4) β-D-N- β (1-3) C6 (Hx)
Galactose Acetilglucosamina C6 (Gal)
Ácido β-D- β (1-4) α-D-N- α (1-4) C6 (Hx)
Glucurônico Acetilglucosamina
α-D-Glucosamina
N-Sulfato
α-L- α (1-4) α-D-Glucosamina α (1-4) C6 (Hx)
idurônico N-Sulfato C2 (AU)
GAG
Ácido
Hialurônico
Condroitim
sulfato
Dermatam
sulfato
Queratam
sulfato
Heparam
sulfato
Heparina
*Posições de sulfatação: GAG, glicosaminoglicanos; Hx, Hexosamina; AU, Ácido urônico; N,
grupamento amino; C, carbono; Gal, galactose.
O AH apresenta-se distribuído no estrato basal e espinhoso da epiderme,
enquanto, os estratos granular, córneo, bem como a membrana basal são negativos
para a marcação do AH (TAMMI et al., 1988).
A associação do AH com diferentes tipos de receptores, como por exemplo na
superfície de células tronco e em células neoplásicas apresenta correlação com a
progressão de tumores. Dentre os receptores de AH encontramos CD44, RHAMM e
ICAM (BOURGUIGNON et al., 2012).
Três isoformas de enzimas participam da biossíntese do AH, denominadas
ácido hialurônico sintases (HAS-1, HAS-2 e HAS-3). As ácido hialurônico sintases
encontram-se ancoradas na membrana plasmática, onde o AH é sintetizado e
liberado para o espaço extracelular, em geral são pouco expressas, mas produzem

INTRODUÇÃO
12
uma grande quantidade de AH (KOSUNEN et al., 2004). As HAS-1 e HAS-2
produzem AH de alto peso molecular, entre 2-4 x106 Da, enquanto, a HAS-3 produz
fragmentos de AH de baixo peso molecular, entre 0,4-2,5 x 105 Da (AVERBECK;
GEBHARDT; VOIGT, 2006).
Os genes que codificam as diferentes ácido hialurônico sintases humanas
estão presentes em diferentes cromossomos, porém, possuem homologia entre si. O
gene da enzima HAS-1 está localizado no cromossomo 19q13.3-q13.4. O gene da
HAS-2 encontra-se no cromossomo 8 e o gene da HAS-3 encontra-se no
cromossomo 16q22.1 (NOBLE; LIANG; JIANG, 2011; NYKOPP et al., 2009).
Estudos mostram que a inibição de HAS-2 na fase embrionária causa
letalidade e/ou anormalidades no desenvolvimento embrionário (CAMENISCH et al.,
2000). Já as enzimas HAS-2 e HAS-3 são importantes no processo de diferenciação
e proliferação da epiderme (SEPPÄNEN-PASONEN et al., 2003).
As enzimas que participam da degradação do AH são denominadas
hialuronidases (HYAL). Existem seis isoformas conhecidas, HYAL1-4, PH 20/SPAM1
e HYALPI, sendo que as isoformas HYAL1 e HYAL2 são mais expressas em células
somáticas. A HYAL1 apresenta atividade em pH ácido e pode degradar AH de alto
ou baixo peso molecular, gerando tetra- e hexassacarídeos. A HYAL2 cliva
moléculas de AH de alto peso molecular, sendo encontrada em baixas
concentrações (CSOKA; FROST; STERN, 2001; GEORGE; STERN, 2004). A
HAYL3 apresenta-se expressa na medula óssea e testículo, enquanto a HAYL4 é
encontrada no músculo esquelético e placenta (CSOKA, ANTONEI BENJAMIN
SCHERER STEPHEN; STERN, 1999). A enzima PH 20/SPAM1 localiza-se no
cromossomo 7q31 e encontra-se expressa nos testículos (JONES et al., 1995;
PRIMAKOFF; HYATT; MYLES, 1985). A presença e ação das HAYL apresenta um
importante papel tanto na fisiologia quanto em processos patológicos e conforme
descrito na literatura sua presença é tecido-dependente.
O AH também pode ser degradado de forma não enzimática por mecanismo
que envolve radicais livres, em presença de agentes redutores, como ácido
ascórbico, tióis e íons cobre (STERN; MAIBACH, 2008).

INTRODUÇÃO
13
1.4.2. Condroitim Sulfato, Dermatam Sulfato e Queratam Sulfato
O condroitim sulfato (CS) e dermatam sulfato (DS) pertencem à classe de
galactosaminoglicanos, pois a hexosamina constituinte é a galactosamina. O CS é
composto por unidades dissacarídicas repetitivas de ácido D-glucurônico e N-
acetilgalactosamina unidas por ligações β1-4 e β1-3. O CS pode ser sulfatado na
posição C6 ou C4 dos resíduos de N-acetilgalactosamina. O DS difere do CS por
apresentar resíduos de ácido L-idurônico, além de ácido D-glucurônico
(BRIMACOMBRE; WEBER, 1964; MALMSTRÖM et al., 2012).
O padrão de sulfatação, desacetilação e epimerização cria uma diversidade de
famílias de CS com funções específicas de acordo com sua estrutura. Em
mamíferos, a unidade mais frequente de galactosamina é a forma monosulfatada, na
posição C4, também denominada CS-A ou CS-C quando apresenta sulfatação em
C6. Outras formas de CS têm sido descritos, tais como CS-D e CS-E, ambos
dessulfatados (MALMSTRÖM et al., 2012).
O condroitim ocorre em processos de crescimento, ossificação e manutenção
da cartilagem normal (MICHELACCI et al., 1979; MOURÃO et al., 1976).
Estudos em diversos tecidos neoplásicos mostram um aumento de CS na
matriz extracelular, principalmente do condroitim-6-sulfato (DIETRICH et al., 1980).
O DS é o GAG mais abundante na pele e talvez seja o principal GAG
associado ao colágeno, estando inclusive envolvido com a organização e
fibrilogênese. O DS também participa de processos de cicatrização, através da
interação com o cofator II da heparina e ativação de fatores de crescimento, como
FGF-2 e fator de crescimento de queratinócitos (FGF-7) (TAYLOR; RUDISILL;
GALLO, 2005; TOLLEFSEN, 1994).
O queratam sulfato (KS) é composto por D-galactose, ligada a N-
acetilglucosamina por ligação do tipo β1-4 e β1-3 (BRIMACOMBRE; WEBER, 1964).
O KS é um componente importante de vários tecidos ricos em colágeno e
representa um dos principais GAG presentes na córnea (HAYASHIDA et al., 2006).
Os proteoglicanos de queratam sulfato (PGKS), como lumicam, keratocam e
osteoglicina, apresentam um papel regulador na manutenção da estrutura córnea
em um grande número de espécies. Acredita-se que os PGKS formam a arquitetura
da córnea através de interações com as fibras colágenas (BLOCHBERGER et al.,
1992; CORPUZ et al., 1996; FUNDERBURGH et al., 1997).

INTRODUÇÃO
14
Na pele, o lumicam desempenha um papel central no colágeno da pele
fibrilogênese e cicatrização de feridas (YEH et al., 2010), e sua presença foi
detectada em células de melanoma humano, mas não em células de melanócitos
normais. (BRÉZILLON et al., 2007; SIFAKI et al., 2006).
1.4.3. Heparam Sulfato
O heparam sulfato (HS) foi descoberto há 50 anos por Jorpes e Gardell, no
entanto, suas funções biológicas até hoje são alvo de extensiva investigação. O HS
é amplamente distribuído no reino animal, possuindo as mais variadas funções e
interações na MEC (DIETRICH et al., 1998).
O HS faz parte dos GAG sulfatados e ocorre geralmente associado a um
esqueleto protéico formando proteoglicanos (PGHS). Os PGHS ligam-se
especificamente a uma variedade de proteínas (IOZZO, 2001).
A estrutura do HS é composta por resíduos de ácido L-idurônico e ácido D-
glucurônico ligados a N-acetilglicosamina (ligação β1-4) (NADER et al., 1987, 1989).
O padrão de sulfatação do HS influencia a interação e consequente modulação e
ativação de diferentes proteínas. Algumas funções biológicas do HS envolvem o
reconhecimento célula-célula, interação com componentes da MEC, tais como
laminina, fibronectina e colágeno, adesão e migração celular, além da ativação de
enzimas (IRIE et al., 2008; PARISH, 2006; PORCIONATTO; NADER; DIETRICH,
1999).
O HS participa do processo de divisão celular de duas maneiras diferentes:
como modulador positivo ou negativo, ou como responsável por estímulos
mitogênicos. Sua ação como modulador positivo da proliferação celular deve-se à
capacidade de ligar-se e agir como um co-receptor para fatores de crescimento
(PORCIONATTO; NADER; DIETRICH, 1999). As células não podem responder ao
fator de crescimento básico de fibroblastos (FGF-2) na ausência de HS. Somente a
presença de um receptor de fator de crescimento de fibroblasto (FGF) funcional na
célula não basta, o complexo só se torna ativo quando o HS está presente (ORNITZ;
WILEY, 2000).
Observou-se que a epiderme e membrana basal apresentam respectivamente
sindecam-1, sindecam-4 e perlecam (BEHRENS et al., 2012; JUNG et al., 2012). O

INTRODUÇÃO
15
HS parece ser essencial na constituição do complexo de ancoragem entre derme e
epiderme (IRIYAMA et al., 2011).
O heparam sulfato é degradado endogenamente pela enzima heparanase
(HPSE-1), uma endo-beta-glucuronidase que cliva cadeias intrassacarídicas do
heparam sulfato, propiciando a formação de oligossacarídeos que apresentam alta
afinidade por fatores de crescimento, citocinas e fatores angiogênicos. Os
oligossacarídeos gerados por ação da heparanase participam, portanto, de
processos biológicos como proliferação celular, inflamação, vascularização,
angiogênese e invasão tumoral, como mostra o esquema da Figura 5 (VLODAVSKY
et al., 1999; ZCHARIA et al., 2005).
Figura 5. Sitio de clivagem da heparanase e mecanismos de ação dos oligossacarídeos
gerados na MEC. Adaptado de (VLODAVSKY; FRIEDMANN, 2001).
Estudos recentes evidenciam o importante papel da heparanase na fisiologia
da pele e demonstra que a ativação desta enzima induz hiperplasia da epiderme,
angiogênese e rugas (IRIYAMA; MATSUNAGA; AMANO, 2010). A hiperpigmentação
da pele em humanos causada por exposição solar é induzida por heparanase, sendo
tal fato decorrência da perda de heparam sulfato essencial para a junção derme-
epiderme (IRIYAMA et al., 2011).
Foi evidenciado que a exposição à luz UV promove aumento na expressão e
atividade da enzima heparanase (KURDYKOWSKI et al., 2012). Pacientes com

INTRODUÇÃO
16
melanoma, bem como cultura de células de melanoma, demonstraram aumento na
expressão da heparanase (CHEN et al., 2012; WAGNER et al., 2012).
É evidente o papel dos glicosaminoglicanos e da enzima heparanase
(HSPE1) para a integridade e manutenção das funções da pele. Sabemos que a
laserterapia ou aplicação do laser de baixa frequência é bastante utilizada em
tratamentos de pele com a finalidade de remover manchas em geral e acne. Diante
do exposto resolvemos investigar possíveis alterações do perfil de
glicosaminoglicanos e da expressão da heparanase causadas pela irradiação com
laser de baixa freqüência (laserterapia) em modelo experimental in vitro. Em tais
estudos decidimos utilizar a linhagem celular característica de queratinócitos
humanos (HaCaT) e comparar os possíveis efeitos da laserterapia com células
neoplásicas de carcinoma epidermóide humano (A431). Este estudo poderá
elucidar o efeito da laserterapia em células não neoplásicas e células tumorais da
pele, auxiliando o entendimento dos efeitos e alterações moleculares
proporcionados por este tipo de terapia.

OBJETIVOS
17
2. OBJETIVOS
Avaliar o efeito da irradiação de baixa frequência (laserterapia) na
viabilidade celular em células não neoplásicas (HaCaT),
comparativamente com linhagem tumoral (A431).
Analisar possíveis alterações do perfil de GAG em queratinócitos humanos
(HaCaT) e em células de carcinoma epidermóide humano (A431) após
irradiação de baixa frequência (laserterapia).
Investigar possíveis alterações da expressão gênica da heparanase, ácido
hialurônico sintases e hialuronidases em células HaCaT e A431 após a
utilização de irradiação de baixa frequência (laserterapia).

MATERIAL E MÉTODOS
18
3. MATERIAIS
3.1. Linhagem celular
Para a realização deste trabalho foram utilizadas linhagens de células
estabelecidas de queratinócitos humanos (HaCaT), gentilmente cedida pela Profa.
Dra. Giselle Zenker Justo e de Carcinoma epidermóide humano (A431), gentilmente
cedida pelo Prof. Dr. Hugo Pequeno Monteiro.
3.2. Cultura de células
- Meio Eagle Modificado por Dulbecco (DMEM) baixa glicose da Nutricell,
Campinas SP, Brasil;
- Meio Eagle Modificado por Dulbecco (DMEM) alta glicose da Nutricell,
Campinas SP, Brasil;
- PBS (tampão fosfato salino) 0,1M, contendo NaCl 137 mM, Na2HPO4 4,3 mM,
KH2PO4 1,8 mM e KCL 2,7 mM em água destilada e filtrada em sistema Milli-QTM
(Millipore®, Billerica, MA, EUA);
- Placas para cultura de poliestireno P35 (35x10 mm) e P60 (60x10 mm)
falcon® (Lincon Park, New Jersey, EUA);
- Solução EBSS (solução salina balanceada de Earl sem cálcio e sem
magnésio), NaCl 116 mM; KCl 5 mM; Na2HPO4 8 mM; KH2PO4 1,46 mM; NaH2HPO4.
1H2O 1 mM; NaHCO3 8 mM (Merck®, Darmstadt, Alemanha); D-glucose 1% e
vermelho de fenol 0,01% (Sigma Chemical Co.®, St. Louis, MO, EUA) em um litro de
água destilada e deionizada em sistema Milli-QTM (Millipore®, Billerica, MA, EUA);
- Soro fetal bovino (SFB) da Cultilab® (Campinas, SP, Brasil);
- Tampão para proteólise, Tris-HCl 50 mM, contendo 3,5 M uréia, pH 7,4;
- Antibióticos: Estreptomicina e Penicilina.
3.3. Enzimas
- Maxatase (protease alcalina P126) foi adquirida da Biocon do Brasil Indústria
Ltda.® (Rio de Janeiro, RJ, Brasil);
- Tripsina 2,5% da Cultilab® (Campinas, SP, Brasil);

MATERIAL E MÉTODOS
19
- Condroitinase AC e ABC da Sigma Chemical Co.® (St. Louis, MO, EUA);
- Transcriptase Reverse - kit ImProm-IITM Reverse Transcription System
(Promega, Madison, Wisconsin);
- Oligo (dT)12-18 da Invitrogen® (Life Technologies™ Corporation).
3.4. Líquido de cintilação
- Ultima Gold™ (PerkinElmer Life And Analytical Sciences®, Inc.; Wellesley,
MA, USA).
3.5. Radioisótopo
- Sulfato de sódio (Na2[35S]O4) do Instituto de Pesquisas Energéticas e
Nucleares (IPEN, São Paulo, SP, Brasil);
3.6. Reagentes gerais e outros materiais
Todos os reagentes foram utilizados de acordo com as normas técnicas:
- Agarose da Bio Laboratories (Richmond, CA, EUA);
- Albumina sérica bovina (BSA) da Sigma Chemical Co.® (St. Louis, MO, EUA);
- Azul de toluidina da Fisher Scientific Co.® (Fair Lawn, NY, EUA);
- Solução de Coomassie Brilliant Blue G, contendo corante Coomassie Brilliant
Blue G 117μM (Sigma Chemical Co.®, St. Louis, MO, EUA), etanol 4,8% e ácido
fosfórico 8,5% (Merck®, Darmstadt, Alemanha).
- Cetavlon (brometo de cetiltrimetilamônia da Merck®, Darmstadt, Alemanha);
- Cloreto de Sódio (Merck®, Darmstadt, Alemanha);
- DMSO (dimetilsulfóxido) da Aldrich Chemical Company, Inc. (Millwaukee, WI,
EUA);
- Garrafas de cultura, 75, 100 ou 150cm2 da Corning Laboratory Sciences Co.®
(Corning, NY, EUA);
- Placas “multiwell” para cultura com 96 poços da Falcon® (Lincon Park, New
Jersey);
- Tris da Gibco® (Grand Island, NY, USA);
- Uréia ultra pura da Sigma Chemical Co.® (St. Louis, MO, EUA);

MATERIAL E MÉTODOS
20
New Jersey, EUA);
- TRIzol da Invitrogen® (Life Technologies Corporation, Carlsbad, CA);
- Maxima ® SYBR Green qPCR Master Mix (2x) (Fermentas®, Life Sciences)
3.7. Equipamentos
- Agitador Vortex-Genie 2 modelo G250, Scientific Industries® (Bohemia, NY,
EUA);
- Autoclave vertical e horizontal da FABBE® (São Paulo, SP, Brasil);
- Banhos de temperatura constante FANEM Ltda.® (São Paulo, SP, Brasil);
- Câmaras para eletroforese em gel de agarose, modelo desenvolvido por
Jaques e col. (1968), manufaturado pela Técnica Permatron Ltda. (São Paulo, SP,
Brasil)
- Contador de cintilação Micro Beta Jet® da Perkin Elmer, Inc. (Fremont, CA,
EUA);
- Cyclone™ Sistema de estoque de fósforo, da Packard Instrument Company
(Meriden, CT, EUA), com software OptiQuant™, para análise de imagem;
- Espectrofotômetro modelo Spectronic 2000, da Bauch&Lomb® (Rochester,
NY, EUA);
- Filmes radiossensíveis Multipurpose (12,5 x 19,2; 12,5 x 25,2 e 12,5 x 43,0)
da Packard Instruments Co.® (Meriden, CT, EUA);
- Fluxo Laminar vertical da Vecco® (Campinas, SP, Brasil);
- Incubadora de CO2, modelo 315, acoplada a torpedos de CO2 pelo Gás Guard
modelo 3030 da Forma Scientific® (Marietta, OH, EUA);
- Leitor de Elisa modelo ELX 800 (Universal microplate reader) da Bio-tek
instruments Inc. (Highland Park, WI, EUA);
- Microcentrifuga 14.000 rpm modelo 5415, Eppendorf® (Hamburg, Alemanha);
- Microscópio invertido de campo claro e contraste de fase, modelo TE-100,
com câmera fotográfica digital acoplada, modelo Coolpix 990, ambos da Nikon,
Nippon Kogakuk® (Tóquio, Japão);
- Microplacas de 96 poços MaxiSorp/FluoroNunc™.
- NanoDrop® 1000 Spectrophotometer da Thermo Fischer Scientific INC
- pHmetro modelo 420ª da Orion Research Incorporated® (Boston, MA, EUA);

MATERIAL E MÉTODOS
21
- Purificador de água Milli-Q™ ZD2- 115 84 da Millipore Corporation® (Bedford,
MA, EUA);
- Termociclador ABI Prism 7500 da Applied Biosystems (Life Technologies
Corporation, Carlsbad, CA)

MATERIAL E MÉTODOS
22
4. MÉTODOS
4.1. Cultura de células
4.1.1. Descongelamento de células
Para o descongelamento das células, o fluxo laminar vertical foi previamente
limpo com álcool 70% e tratado com luz ultravioleta por 20 minutos seguido por
equilíbrio da ventilação por 10 minutos. As células, armazenadas em nitrogênio
líquido, foram descongeladas em banho-maria a 37ºC, ressuspensas em 5 mL de
meio de cultura estéril e centrifugadas para eliminação dos eventuais resíduos dos
reagentes de congelamento. O sobrenadante desta centrifugação foi descartado e
as células foram ressuspensas em 1mL de meio DMEM baixa glicose com 10% de
SFB para a linhagem estabelecida HaCaT e DMEM alta glicose com 10% de SFB
para a linhagem estabelecida A431 e plaqueadas em garrafas plásticas específicas
para cultura.
4.1.2. Manutenção das células
A linhagem estabelecida de queratinócito (HaCaT) e Carcinoma epidermóide
(A431) foram cultivadas em garrafas/placas de cultura, contendo meio de cultura
estéril DMEM. Este meio foi preparado acrescentando-se 10% de SFB e antibióticos
(100 μg/mL de estreptomicina e 100 Ul/mL de penicilina).
A cada dois dias o meio de cultura foi trocado, por aspiração com o auxilio de
uma pipeta Pasteur acoplada à bomba de vácuo. O meio de cultura contém o
indicador de pH vermelho de fenol que apresenta coloração vermelha em pH neutro
e amarela em pH ácido. Devido ao metabolismo celular o pH tende a acidificar,
tornando o meio de cultura amarelo, indicando, portanto, a necessidade de trocar o
meio. O pH ideal para o cultivo das células é neutro, próximo a 7,4.
4.1.3. Subcultivo das células
Quando as células apresentaram semi-confluência, o meio de cultura foi aspirado
e a placa foi lavada com tampão EBSS para remover qualquer resíduo de SFB, para

MATERIAL E MÉTODOS
23
que este não inative a enzima proteolítica utilizada na remoção das células na placa.
Para a remoção das células utilizou-se da enzima tripsina. A tripsina diluída 1:10 foi
adicionada a cada placa, mantendo-se a placa na estufa a 37ºC, por
aproximadamente 10 minutos, para que as células aderidas se soltassem da
garrafa/placa de cultura. Após essa etapa as células foram removidas da placa e
colocadas em tubo de plástico de 15mL contendo 3 mL de meio DMEM com 10%
SFB. O SFB apresenta inibidores de proteases e foi usado para inativar a tripsina, e
assim, evitar lise celular. Este tubo foi centrifugado a uma velocidade de 2.500 rpm
por 5 minutos e o sobrenadante descartado. As células foram ressuspensas em 5
mL de meio de cultura para serem plaqueadas novamente.
4.1.4. Contagem das células em Câmara de Neubauer
A contagem foi realizada para determinar a quantidade de células a ser
distribuída em cada placa (2,0x105 células/placa). A contagem foi feita em câmara
de Neubauer, a partir das células ressuspensas e homogeneizadas em 5 mL de
DMEM com 10% de SFB, colocou-se 10 μl do volume total de células na câmara. A
câmara de Neubauer é composta de quatro quadrantes laterais maiores, que são
subdivididos em dezesseis quadrantes cada. A contagem foi feita com o auxílio do
microscópio óptico invertido. Foram contados os quatro quadrantes e foi calculada a
média de células por quadrante. Este resultado foi multiplicado por 1,0x104 e o
resultado final correspondeu ao número de células por mililitro de suspensão de
células. As células foram incubadas a 37ºC, contendo 5% de CO2 até tornarem-se
70% confluentes.
4.2. Tratamento da cultura de célula com radiação a laser
O equipamento utilizado para a irradiação foi o Quasar Esthétique (DMC
equipamentos Ltda., Brasil), utilizado comumente no tratamento de acne, manchas
na pele, olheiras e marcas de expressão. Este equipamento utilizado para irradiar
nas diferentes células em cultura apresenta as seguintes especificações: Tensão de
Operação 90V ~ 240V; Emissor Visível comprimento de Onda 460 – 480 nm; Diodo
Laser Vermelho 660 – 685 nm; Potência Útil 70 mW.

MATERIAL E MÉTODOS
24
O equipamento possui um feixe de LED no comprimento de onda de 470 nm,
com 180 mW de potência, área do feixe de 78,5 cm2, sendo a aplicação da luz de
470 nm em cultura de células, foi realizada a 10 cm de distância da cultura celular,
por 3 minutos, numa densidade de energia de 0,417 J/cm2.
O feixe de diodo no comprimento de onda de 660 nm, foi aplicado nas
culturas celulares com 112 mW de potência, área do feixe de 78,5 cm2, aplicação a
10 cm de distância da cultura celular, por 1 minuto numa densidade de energia de
0,0856 J/cm2.
Para o tratamento associado, utilizando ambos os comprimentos de onda
(470 nm por 3 minutos e 660 nm por 1 minuto), os parâmetros utilizados foram os
mesmos descritos anteriormente, numa densidade de energia de 0,4983 J/cm2 . As
células foram submetidas a 2 aplicações diárias, seguindo as condições de
irradiação para cada comprimento de onda descritas na Tabela 3, considerando um
intervalo de 5 horas entre cada aplicação, totalizando 6 aplicações no período de 3
dias. Após a irradiação as células foram submetidas aos testes e incubadas por 18
horas a 37ºC, em estufa umidificada contendo 5% de CO2. A Tabela 3 esquematiza
os parâmetros de irradiação utilizados para o tratamento das diferentes linhagens
celulares (HaCaT e A431) em cultura.
Doses de 0,05-10 J/cm2 ocasionam estímulo, no entanto densidades acima 10
J/cm2 disto exercem efeito oposto, e a potência abaixo de 400 W/cm2. (HARRIS,
1988), ajudam a identificar os efeitos não-térmico da laserterapia.
Tabela 3. Parâmetros de Irradiação
PARÂMETROS DE IRRADIAÇÃO
Comprimento de onda 470 nm 660 nm 470 + 660 nm
Potência 180 mW 112 mW 180 + 112 mW
Densidade de energia total 2,4762 J/cm2 0,5136 J/cm
2 2.9898 J/cm
2
Área do feixe 78,5 cm2 78,5 cm
2 78,5 cm
2
Distância da cultura 10 cm 10 cm 10 cm
Tempo Total 18 minutos 6 minutos 18 min + 6 min

MATERIAL E MÉTODOS
25
4.3. Quantificação de glicosaminoglicanos sulfatados sintetizados por
células.
4.3.1. Marcação metabólica
As culturas de células de queratinócitos (HaCaT) e Carcinoma epidermóide
(A431) foram submetidas ao crescimento até atingirem aproximadamente 70% de
confluência e foram subcultivadas em placa de 6 poços (35 mm), em meio DMEM
com 10% de soro fetal bovino. As células foram incubadas por 18 horas a 37ºC em
estufa umidificada contendo 5% CO2. As células foram submetidas à irradiação com
laser a 470 nm, 660 nm e 470 nm + 660 nm, conforme descrito na metodologia (item
4.2). Após o terceiro dia de tratamento das culturas celulares nas diferentes
condições, foram adicionados 200 µCi/mL de [35S]-sulfato ao meio de cultura em
meio F12 na ausência de soro fetal bovino (SFB). Este ensaio permitiu a marcação
das moléculas de GAG sintetizadas e secretadas pelas células. A incubação foi
realizada durante 18 horas, 37ºC, em estufa sob atmosfera de 5% de CO2.
4.3.2. Extração de GAG marcados metabolicamente
Após o período de incubação das células irradiadas, contendo sulfato radioativo,
o meio de cultura foi removido e uma alíquota de 500 μl foi submetida à proteólise e
outra alíquota de 500 μl foi armazenada para posterior análise de ácido hialurônico.
Às células aderidas foram adicionados 500 μl do tampão Tris HCl 25 mM,
contendo uréia 7 M, pH 7,5, e com auxílio de um “scrapper”, esfregou-se o fundo da
placa para remoção das células e matriz extracelular. A fração celular (células +
matriz extracelular), foi homogeneizada e dividida em dois tubos contendo 250 μl
cada.
A proteólise da fração celular e meio de cultura foi realizada com adição de 125
μl de Maxatase (1 mg/mL) em presença de Tris-HCl 0,05 M e NaCl 0,15 M pH 8,0
(alíquota do meio de cultura e fração celular) e incubando-se o material a 60ºC por
24 horas em banho-maria, seguindo eletroforese em gel de agarose em tampão
PDA.

MATERIAL E MÉTODOS
26
4.3.3. Eletroforese em gel de agarose com tampão PDA
Finalizada a proteólise, uma alíquota de 5 µL do meio de cultura e da fração
celular, foi submetida à eletroforese em gel de agarose 0,55% em tampão 1,3-
diaminopropano acetato (PDA) 0,05 M, pH 9,0. Em seguida, o gel foi submetido à
eletroforese (100 V), durante 1 hora em câmara refrigerada a 4°C. Após a corrida
eletroforética, os GAG foram precipitados por imersão do gel em solução de cetil-
trimetilamônio (Cetavlon®) 0,1% por 2 horas. Em seguida, o gel foi totalmente
desidratado sob calor e ventilação. Após a secagem, o gel foi corado com azul de
toluidina 0,1% em ácido acético 1% e etanol 50%, por 15 minutos. O excesso de
corante foi removido com uma solução descorante contendo ácido acético 1% e
etanol 50%.
4.3.4. Análise e quantificação dos GAG marcados metabolicamente
A lâmina de eletroforese marcada radioativamente foi posicionada para
impressão em filme radiossensível (Multipurpose, Packard Instruments Co).
A revelação do filme radiosensível foi realizada em câmara escura, no
equipamento Cyclone® (Storage Phosphor System, Packard Instr.). As bandas
correspondentes à migração dos GAG tiveram como referência a migração de GAG
padrão. Os dados foram analisados utilizando-se o software OptiQuanti.
As bandas referentes aos GAG foram recortadas do gel desidratado, com o
auxílio de uma pinça e quantificadas em 5 mL de solução de líquido de cintilação,
Ultima Gold™. Esta solução intensifica a radiação emitida pelo sulfato radioativo e
permite a quantificação dos GAG expressa em cpm (contagem por minuto). A
quantificação de cada GAG foi corrigida por proteína total, dosada em cada
experimento antes da proteólise pelo método convencional de Coomassie Blue.
Assim, foi obtido a quantificação dos GAG sintetizados pelas linhagens estabelecida
de queratinócitos e de carcinoma epidermóide mantidas em cultura, em valores
expressos em cpm/µg de proteínas totais.

MATERIAL E MÉTODOS
27
4.3.5. Dosagem de proteínas por método de Coomassie Blue
Este método utiliza um reagente que apresenta afinidade por proteína e possui
diferente intensidade de coloração dependendo da quantidade de proteínas
presentes. O reagente contém 117 μM de Coomassie Blue G250, 4,8% de etanol e
8,5% de ácido fosfórico preparado em nosso laboratório.
A dosagem foi realizada diluindo-se 5 e 10 μl de cada amostra da fração celular
sem proteólise das células irradiadas, em 100 μl do tampão Tris-HCl 25 mM com
uréia 3,5M, pH 7,5 e adicionou-se 2,9 mL do reagente Coomassie Blue. Cada
mistura foi homogeneizada e a leitura foi efetuada em espectrofotômetro com
comprimento de onda ajustado a 595 nm. A leitura da absorbância é proporcional a
quantidade de proteínas presente na fração celular. A quantificação relativa à leitura
das amostras foi determinada por equação da reta de uma curva padrão de
albumina.
A dosagem de proteínas da fração celular de cada placa foi realizada com o
intuito de corrigir a quantidade de proteínas produzidas pela célula, assim o
resultado apresentado demonstra apenas a influência da irradiação na síntese e
secreção dos GAG.
4.4. Degradação enzimática
O ensaio de degradação enzimática foi realizado para certificar-se que as
bandas marcadas no pulso de sulfato correspondiam aos GAG condroitim sulfato e
dermatam sulfato.
A condroitinase AC é uma enzima que atua degradando condroitim 4,6-
sulfatado, produzindo dissacarídeos insaturados 4- e 6- sulfatados (ΔU-GalNAc,4S e
ΔU-GalNAc,6S) e dissacarídeo insaturado não sulfatado (ΔDi-0S), uma vez que esta
enzima quebra, especificamente, ligações glicosídicas do tipo β(1-4), entre resíduos
de N-acetil-hexosamina que podem ou não apresentar sulfatação em C-4 ou C-6 e
resíduos de ácido glucurônico (GlcUA). A condroitinase B é uma enzima que
degrada ligações glicosídicas do tipo β(1-4), envolvendo N-acetil-hexosamina e
ácido-L-idurônico degradando dermatam sulfato, gerando dissacarídeos insaturados
4- e 6- sulfatados (ΔU-GalNAc,4S e ΔU-GalNAc,6S) (MICHELACCI; HORTON e
POBLACIÓN, 1987). A condroitinase ABC é uma liase que atua por meio de uma

MATERIAL E MÉTODOS
28
reação de β-eliminação, sobre sulfato de condroitim-4-sulfato (C4S), condroitim-6-
sulfato (C6S) e dermatam sulfato (DS) (ROBAT et al., 2012).
O teste foi realizado com as amostras de proteólise do pulso de sulfato. Em
diferentes tubos colocou-se 20 μl da enzima condroitinase AC e condroitinase ABC e
acrescentou-se 5 μl de cada amostra. Incubou-se as amostras em banho Maria a 37ºC
por 24 horas. Após esse período foi feita eletroforese em gel de agarose e impressão do
gel em filme conforme já descrito.
4.5. Ensaio de viabilidade celular
O ensaio de viabilidade celular foi realizado utilizando-se o “kit Vybrant MTT Cell
Proliferation Assay”, de acordo com o protocolo sugerido pelo fabricante. O ensaio
de redução do MTT (brometo de 3-(4,5)-dimetiltialzolil-2,5-difeniltetrazólio) é um
ensaio colorimétrico para medir a atividade da redutase mitocondrial em células
vivas. Esta metodologia baseia-se na redução do sal de tetrazólio em cristais de
formazan por células metabolicamente ativas.
As células foram subcultivadas em uma microplaca de 96 poços, sendo que cada
amostra foi semeada em nove poços e incubada por 24 horas, a 37ºC, em estufa
umidificada contendo 5% de CO2. Após a incubação as células foram submetidas ao
tratamento com irradiação a laser com luz de 470 nm, 660 nm ou a associação (470
+ 660 nm), sendo utilizado como controle do ensaio células não tratadas com
laserterapia. O sobrenadante foi removido e adicionado 100 μL de MTT. A
microplaca foi protegida por papel alumínio e incubada por 3 horas, em estufa a
37°C, e 5% de CO2. Em seguida, a solução foi removida e foram adicionado 50 μL
de DMSO em cada poço. Após incubação de 10 minutos, a leitura foi realizada em
espectrofotômetro no comprimento de onda de 540 nm. O ensaio de viabilidade
celular foi determinado pela média da absorbância, seguida de desvio padrão.
4.6. Dosagem de ácido hialurônico
Utilizamos um método fluorimétrico para a determinação do AH, que se utiliza de
sondas de ligação do AH. A sonda corresponde à proteína de ligação, isolada de
cartilagem nasal bovina. A proteína de ligação compreende à porção globular da
proteína que promove a ligação do agrecam (proteoglicano de queratam sulfato e

MATERIAL E MÉTODOS
29
condroitim sulfato) ao AH. A proteína de ligação é imobilizada em microplacas de 96
poços MaxiSorp/FluoroNunc™.
Em cada poço da placa contendo a sonda adsorvida foram adicionados 100 μl de
solução de diferentes concentrações de AH padrão (0 a 500 μg/L), diluídas no
tampão de ensaio Tris-HCl 0,05 M, pH 7,75 e BSA 1% (albumina bovina sérica), ou
alíquotas de meio de cultura celular e fração celular, diluídas 1:100 no mesmo
tampão. Cada ensaio foi realizado em triplicata. A incubação foi realizada a 4°C, por
12 horas, seguindo três lavagem consecutivas com tampão Tris-HCl 0,05 M, pH
7,75. Em seguida, à cada poço foram adicionados 100 μl da proteína de ligação
(1mg/ml) conjugada com biotina. Após a adição da proteína de ligação conjugada
com biotina foi realizada agitação por 2 horas, seguindo lavagens exaustivas com
tampão de lavagem. Após lavagem, foram adicionados 100 μl de estreptavidina
marcada com európio (diluição 1:10000) em tampão de ensaio durante 30 minutos.
Como a estreptavidina tem afinidade pela biotina conjugada à proteína de ligação
forma-se um complexo (sanduíche). Após várias lavagens foi adicionado uma
solução de “enhancement” (280 μl em cada poço) e, após agitação por cinco
minutos, o európio foi deslocado do complexo com estreptoavidina. A fluorescência
emitida pelo európio livre foi quantificada em um fluorímetro. A Figura 6 representa o
esquema das etapas descritas. A quantificação de ácido hialurônico de cada
amostra foi determinada em valores expressos por ng de AH / ug de proteína total.

MATERIAL E MÉTODOS
30
Figura 6. Representação esquemática do ensaio fluorimétrico para quantificação de ácido
hialurônico. A - Placas 96 poços MaxiSorp/FluoroNunc™ foram revestidas com a proteína de ligação
(proteína ligante de proteoglicanos ao ácido hialurônico); B - As amostras da fração celular e meio de
cultura de células HaCaT e A431 ligada a proteína de ligação; C – Adição de biotina conjugada com a
proteína de ligação; D – Revelação com estreptavidina marcada com európio E - Após a liberação do
európio com solução "enhancement" a fluorescência final foi quantificada em fluorímetro. Este método
foi padronizado por Martins e colaboradores (MARTINS et al., 2003).
4.7. Extração de RNA total
As células foram cultivadas e plaqueadas em placas de 60 mm e tratadas
conforme metodologia já descrita . Após incubação de 18 horas, as células foram
removidas com tripsina. O sobrenadante foi aspirado utilizando a bomba a vácuo e
ao precipitado foi adicionado 1 mL de TRIZOL® e homogeneizado por 5 minutos em
temperatura ambiente. A amostra foi transferida para um eppendorf de 1,5 mL e foi
adicionado 0,2 mL de clorofórmio, misturando gentilmente por 3 minutos em
temperatura ambiente. O clorofórmio promove a extração do RNA na fase aquosa.
Em seguida, as amostras foram centrifugadas a 10.000 x g, 4ºC, durante 15 minutos.
A solução contendo RNA foi transferida para outro tubo e acrescentou-se 0,5 mL de
isopropanol para a precipitação do RNA. As amostras foram incubadas por mais 10
A
D C
B
E

MATERIAL E MÉTODOS
31
minutos, temperatura ambiente e, em seguida, centrifugadas a 4ºC, a 10.000 x g, por
10 minutos. O precipitado obtido foi lavado com 1 mL de etanol 75%,
homogeneizado e centrifugado a 7.500 x g, 4ºC, por 5 minutos. O etanol foi
desprezado e as amostras submetidas a banho seco (55°C por 3 minutos). As
amostras de RNA foram solubilizadas em 25 µL de água milliQ tratada com DPEC
(livre de RNAse). Finalmente, o RNA total foi quantificado por espectrofotometria.
4.7.1. Quantificação do RNA Total
A dosagem foi realizada por espectrofotometria, em aparelho NanoDrop,
previamente calibrado. Os valores obtidos foram expressos em ng / μl.
4.7.2. Obtenção cDNA a partir do RNA Total
A obtenção do cDNA foi realizada utilizando-se o kit de RT Improm II®. Na
primeira etapa da reação foi adicionado de 1 μg de RNA total e 10 μM de Oligo dT
ou oligonucleotídeo iniciador (incubação 70°C, por 5 minutos e 4°C por 5 minutos).
Em seguida, foi adicionado ao tubo 10 μM de dNTPs (desoxirribonucleotídeos
trifosfatados), 2,4 μl de MgCl2 (25 mM), 4 μl do tampão de reação 5X e 1 μl da
enzima transcriptase reversa. A reação de transcrição reversa ocorreu a 42°C, por
60 minutos. As reações foram armazenadas à -20°C para posterior amplificação do
cDNA com oligonucleotídeos iniciadores específicos.
4.8. Análise da expressão por PCR em tempo real (qPCR)
A expressão do RNAm da heparanase (HPSE), ácido hialurônico sintases (HAS1,
HAS2 e HAS3) e hialuronidases (HYAL1 e HYAL2) foram realizadas em amostras
coletadas das culturas de células HaCaT e A431, após o tratamento com irradiação
a laser, comparando-se com o controle (células não submetidas ao tratamento). A
partir do cDNA obtido pela reação de transcrição reversa, anteriormente descrito,
aproximadamente 1μg/μL das amostras de cDNA foram incubadas com
oligonucleotídeos iniciadores sense e antisense (1,5 μM), 6 μl da enzima DNA
polimerase SYBR Green® e 0,025 μl de Rox (normalizador de fluorescência para
reação de qRT-PCR). As amostras foram homogeneizados em tubos cônicos de 0,2

MATERIAL E MÉTODOS
32
mL específicos para qPCR. As amostras foram submetidas à reação de qPCR numa
ciclagem de 95ºC por 10 minutos iniciais e 40 ciclos (95ºC, 15 minutos; 60ºC, 60
segundos). Todos os oligonucleotídeos iniciadores utilizados apresentavam
temperatura de desnaturação (Tm), entre 60ºC a 95ºC. Os genes endógenos
utilizados como controle foram da enzima gliceraldeído-3-fosfato-desidrogenase
(GAPDH) e da subunidade 60S da proteína ribossomal L13A (RPL13A). Os
oligonucleotídeos iniciadores utilizados no presente estudo estão mostrados na
Tabela 4.
Os resultados obtidos da reação de qRT-PCR representam valores relativos
obtidos por média geométrica considerando a expressão de ambos os genes
endógenos constitutivos, GAPDH e RPL13A, (-ΔCt). Os valores foram expressos em
2-ΔΔCt.
Tabela 4. Sequências de Oligonucleotídeos iniciadores
mRNA Foward Reverse
HPSE1 5' TGG CAA GAA GGT CTG GTT AGG AGA 3' 5' GCA AAG GTG TCG GAT AGC AAG GG 3'
HAS1 5' CAA GGC GCT CGG AGA TTC 3’ 5’ CCA ACC TTG TGT CCG AGT CA 3’
HAS2 5’ CAG ACA GGC TGA GGA GGA CTT TAT 3’ 5’ GGA TAC ATA GAA ACC TCT CAC AAT GC 3’
HAS3 5’ GGC GAT TCG GTG GAC TAC AT 3’ 5’ CGA TGG TGC AGG CTG GAR 3’
HAYL1 5’ TAA CCC TGC CAG TTT CTC CAT CCA 3’ 5’ ACA GCC ATC TGT GCC TGA TCT TCA 3’
HAYL2 5’ AGG ACA AAG ATG TGT ATC GCC GGT 3’ 5’ GAA CTG CTG TGC TGC GAA CTC AAA 3’
RPL13a 5´ TTG AGG ACC TCT GTG TAT TTG TCA A 3´ 5´ CCT GGA GGA GAA GAG GAA AGA GA 3´
GAPDH 5´ TCG ACA GTC AGC CGC ATC TTC TTT 3´ 5´ GCC CAA TAC GAC CAA ATC CGT TGA 3´
4.9. Análise Estatística
A analise estatística foi realizada utilizando-se o programa GraphPad Prism 5.0 (La
Jolla, CA, USA). Para verificar a ocorrência de diferenças significativas entre os
grupos estudados foi utilizado o teste não paramétrico Kruskal-Wallis. Em todas as
foram considerados valores estastiticamente significantes p < 0,05).

RESULTADOS E DISCUSSÃO
33
5. RESULTADOS E DISCUSSÃO
5.1. Viabilidade Celular em células HaCaT e A431
O uso da laserterapia está frequentemente associado ao aumento da
proliferação celular e resposta inflamatória (GAO; XING, 2009). No presente estudo
decidimos investigar o efeito da irradiação em diferentes comprimentos de onda
sobre a proliferação de células epiteliais normais (HaCaT) e células tumorais (A431).
A escolha da dose e quantidade de aplicações foi determinada conforme o protocolo
utilizado em ambulatório médico.
Observamos na Figura 7 diminuição da viabilidade celular, após laserterapia
utilizando os comprimentos de onda 470 nm, 660 nm, bem como a associação de
ambos os tipos de luz. Em células HaCaT e A431 foi observado diminuição da
viabilidade celular de aproximadamente 32, 67 e 55 % (células HaCaT) e 35, 47 e 28
% (Células A431), respectivamente, apresentando relevância estatística nos dois
tipos celulares (* < 0,05). A viabilidade celular foi analisada utilizando o teste MTT
descrito na metodologia. O resultado encontra-se expresso em porcentagem de
células viáveis.
Resultados semelhantes ao encontrado em nosso estudo, demonstraram
diminuição da viabilidade de células HaCaT e A431, quando utilizada a luz UVB 280-
315 nm (JUAN et al., 2012). O tratamento das células HaCaT e A431 com luz UVB
evidenciou alteração do ciclo celular, aumentando o número de células em G1-S e
diminuição da viabilidade celular dose dependente (JUAN et al., 2012). Ainda, tais
efeitos sobre o ciclo celular e viabilidade celular parecem ser mediados pelo fator de
transcrição nuclear kappa-B (NF-κB), quinase dependente de ciclina 6 (CDK6) e
proteínas da família BCL-2 (JUAN et al., 2012).
A utilização de luz UVC (200-290nm), em células A431, também demonstrou
inibição da proliferação celular (TAI et al., 2012).
Outros estudos utilizando cultura primária de células epiteliais humanas,
demonstraram aumento da proliferação celular, após irradação com laser de 660 nm
e 780 nm (EDUARDO et al., 2007). Este dado contrário ao obtido no presente
estudo pode ser explicado pelo fato de que a densidade de energia utilizada em tais
estudos foi menor àquela utilizada em nossos experimentos (3 e 5 J/cm2).

RESULTADOS E DISCUSSÃO
34
Viabilidade Celular
CTR
470
nm
660
nm
470+
660
nmCTR
470
nm
660
nm
470+
660
nm
0
20
40
60
80
100
120
*
*
*
*
*
HaCaT A431
% d
e c
élu
las
viá
veis
Figura 7. Viabilidade Celular após laserterapia. As células HaCaT e A431 foram submetidas a seis
irradiações com diferentes comprimentos de onda, como indicado em métodos. Os valores obtidos
expressam a média e desvio padrão de ensaios realizados em quintuplicata para células HaCaT e
nonoplicatas para células A431. * representa p < 0,05 utilizando-se o teste não paramétrico Kruskal-
Wallis.
Estudos realizados com outros tipos celulares como células endoteliais
HUVEC (CHEN; HUNG; HSU, 2008), cultura primária de fibroblastos humano
(ESMAEELINEJAD; BAYAT, 2013; SEKHEJANE; HOURELD; ABRAHAMSE, 2011)
e pré-osteoblastos (PAGIN et al., 2012), evidenciaram grande diversidade nos
protocolos experimentais que utilizam laserterapia, variando a intensidade de
energia, frequência de aplicações, distância, área, potência e tempo, gerando
diferentes efeitos biológicos, que dificilmente podem ser comparados.
Vale salientar que a utilização do laser de 400-500 nm apresenta atividade
bactericida, evidenciando diminuição da viabilidade em bactérias E. coli. Efeito
oposto pôde ser observado em estudos com laser de 400-500 nm em bactérias S.
aureus (LIPOVSKY et al., 2010), mostrando também aumento na geração de
espécies reativas de oxigênio (ROS), que explica o efeito bactericida. Entretanto, em
estudos com S. aureus e utilização de laser de 500-800 nm foi observado menor

RESULTADOS E DISCUSSÃO
35
produção de ROS, sugerindo que o efeito bactericida é mais eficiente em
comprimentos de onda menor (LIPOVSKY et al., 2010).
5.2. Efeito da laserterapia na síntese de glicosaminoglicanos sulfatados em
células HaCaT e A431
Os glicosaminoglicanos sulfatados exercem um importante papel na
sinalização celular e no remodelamento de matriz extracelular. De acordo com o tipo
celular existe grande variabilidade estrutural dos glicosaminoglicanos sulfatados. Tal
diversidade estrutural das cadeias de glicosaminoglicanos está diretamente
relacionada com a função que desempenham (DIETRICH et al., 1998;
PORCIONATTO; NADER; DIETRICH, 1999). Alterações dos glicosaminoglicanos
sulfatados estão relacionadas com o desenvolvimento de várias doenças como
câncer, doenças inflamatórias, doenças degenerativas e em processos de
cicatrização (KOZLOWSKI; PAVÃO; BORSIG, 2011; PECCHI et al., 2012; TAYLOR;
RUDISILL; GALLO, 2005; TEIXEIRA et al., 2012).
Ainda, o estudo de componentes da matriz extracelular, como proteoglicanos,
proteínas fibrosas, glicoproteínas, metaloproteases e glicosidases, podem elucidar
alterações moleculares diretamente relacionadas com o desenvolvimento de tais
doenças (MOSCATELLO et al., 1998; PECCHI et al., 2012; THEODORO et al.,
2007).
Atualmente, são conhecidos os benefícios da laserterapia no tratamento de
processos inflamatórios e cicatrização de feridas, porém, pouco se conhece sobre os
efeitos da utilização da terapia de laser de baixa potência na matriz extracelular.
Diante de tais observações é interessante investigar se existe alteração do
perfil de glicosaminoglicanos em cultura celular, após o tratamento laserterapia.
Observamos que as células HaCaT e A431, apresentaram CS/DS e HS por
análise eletroforética (Figura 8). A confirmação dos galactosaminoglicanos
sulfatados (CS e DS), foi determinada após degradação enzimática com
condroitinase AC e condroitinase ABC (Figura 9). A análise desses resultados
demonstrou a presença de DS sintetizado e secretado para o meio de cultura de
ambas linhagens celulares, HaCaT e A431 (Figura 9).

RESULTADOS E DISCUSSÃO
36
Figura 8. Perfil de GAG sulfados sintetizados por HaCaT e A431. Este emsaio representa a
radioautografia da eletroforese realizada para identificação e quantificação dos GAG sulfatados. As
células HaCaT e A431 foram marcadas com [35
S]-sulfato e analisadas após 6 aplicações de
laserterapia, exceto o grupo controle (como indicado em métodos). Painel A e B análise da linhagem
HaCaT; Painel C e D análise da linhagem A431; os números 1 e 2, representam amostras não sem
tratamento (controles); os números 3 e 4, indicam amostras submetidas ao tratamento 470 nm; os
números 5 e 6, indicam amostras submetidas ao tratamento 660 nm; os números 7 e 8, representam
amostras submetidas ao tratamento 470+660 nm.
É importante ser observado que até o presente, não existem trabalhos
na literatura que caracterizem os glicosaminoglicanos sulfatados de células HaCaT e
A431. Portanto, os resultados aqui apresentados são importantes para o melhor
entendimento das alterações de tais componentes após laserterapia.
Após a eletroforese em gel de agarose, os glicosaminoglicanos sulfatados
foram quantificados e os valores obtidos estão representados na Figura 10.
É importante observar que a irradiação induz alterações cutâneas, como
fibrose, que envolvem o acúmulo de componentes da MEC, como proteoglicanos de
heparam sulfato, que sabidamente são requeridos para os mecanismos de reparo
HaCaT
A431
A B
C D

RESULTADOS E DISCUSSÃO
37
tecidual, visto que tais compostos interagem e modulam a função de uma variedade
de outras moléculas como exemplo fatores de crescimento.
Figura 9. Radioautografia da eletroforese após degradação enzimática dos GAG sulfados
sintetizados por HaCaT e A431. As células HaCaT e A431 foram marcadas com [35
S]-sulfato e
submetidas a seis aplicações de laserterapia, como descrito em métodos. O extrato obtido da fração
celular e meio condicionado foram submetidos à degradação com condroitinases AC e ABC, como
descrito detalhadamente em métodos. Painel A e B, Células; Painel C e D, Céçulas A431; os
números 1,4,7 e 10, indicam amostras quenão foram submetidas à degradação enzimática
(controle); os números 2, 5, 8 e 11 representam as amostras digeridas com condroitinase AC e os
números 3,6,9 e 12 representam as amostras digeridas com condroitinase ABC.
A avaliação do perfil de glicosaminoglicanos sulfatados em células HaCaT
após o tratamento com a associação de 470 nm e 660 nm, sugere um aumento do
heparam sulfato de aproximadamente três vezes, como mostra a Figura 10.
Entretanto, análises estatísticas não confirmam que tal diferença seja significativa.
Nas células tumorais A431, não foi observado alteração expressiva do heparam
sulfato após aplicação com os diferentes comprimentos de onda (Figura 10).
Considerando a linhagem tumoral A431, observamos claramente que houve
diminuição significativa do HS na fração celular em relação ao controle e o DS não
apresentou alterações. No meio de cultura não houve alterações significativas na
secreção de HS e DS após o tratamento (Figura 10).
A431
HaCaT
A B
C D

RESULTADOS E DISCUSSÃO
38
Considerando a quantificação do DS na linhagem HaCaT, após aplicação dos
diferentes comprimentos de onda, observamos que a associação (470 nm e 660
nm), desencadeou um aumento de aproximadamente duas e três vezes,
repectivamente para a fração celualr e meio de cultura. Novamente, análises
estatísticas não comprovam significância para este aumento (Figura 10).
Figura 10. Quantificação dos glicosaminoglicanos sintetizados por células HaCaT e A431. HS,
heparam sulfato; DS, Dermatam sulfato; Fração celular, quantificação de glicosaminoglicanos
sintetizados por células e matriz extracelular. Meio, glicosaminoglicanos secretados para o meio de
cultura; CTR, controle (células sem tratamento com laser); 470 nm, após seis aplicações do laser com
470 nm; 660 nm, após seis aplicações com laser de 660 nm; 470 + 660 nm, após seis aplicações
com associação do laser de 470 + 660 nm. Os ensaios foram realizados em triplicatas e os valores
expressam a média e desvio padão.
A avaliação da biossíntese do heparam sulfato em células HaCaT foi
investigada em estudos de transfecção que promovem a superexpresão de enzimas
A431 DS Fração Celular DS Meio
CTR
470
nm
660
nm
470+
660
nmCTR
470
nm
660
nm
470
+ 66
0 nm
0
10000
20000
30000
40000
cp
m/
g p
rote
ína
HaCaT DS Fração Celular DS Meio
CTR
470
nm
660
nm
470
+ 66
0 nm
CTR
470
nm
660
nm
470
+ 66
0 nm
0
500
1000
1500
2000
25005000
10000
15000
20000
25000
cp
m/
g p
rote
ína
A431 HS Fração Celular HS Meio
CTR
470
nm
660
nm
470
+ 66
0 nm
CTR
470
nm
660
nm
470
+ 66
0 nm
0
50000
100000
150000
200000
cp
m/
g p
rote
ína
HaCaT HS Fração Celular HS Meio
CTR
470
nm
660
nm
470
+ 660
nm
CTR
470
nm
660
nm
470
+ 660
nm
0
2000
4000
6000
8000
20000
40000
60000
80000
100000
cp
m/
g p
rote
ína

RESULTADOS E DISCUSSÃO
39
anti-oxidantes. Em tais estudos, foi observado que houve um aumento significativo
no nível de expressão de cinco enzimas envolvidas na biossíntese do heparam
sulfato, xilosiltransferase II, glicosiltransferase-2 (EXTL2), D-glucuronosil-C5-
epimerase, iduronosil-2-O-sulfotransferase e 6-O-sulfo transferase (NAKAYAMA et
al., 2008).
Deve ser lembrado que heparina e heparam sulfato compartilham a mesma
via de biossíntese e também apresentam semelhança estrutural. Portanto, a
diminuição do heparam sulfato presente na fração celular em células A431, após o
tratamento com laserterapia, pode explicar a diminuição acentuada na viabilidade de
células A431 (Figura 7). Um dado que corrobora com esta hipótese foi descrito na
literatura demonstrando um efeito anti-proliferativo em células A431 mediado por
heparina (VANNUCCHI et al., 1991).
O perlecam é um proteoglicano de HS de membrana basal conhecido por
impedir a difusão descontrolada de diversos fatores da epiderme para a derme e sua
degradação pode afetar a junção dermo-epidérmica, promovendo o aumento da
ligação de fatores de crescimento, como fator de crescimento de hepatócitos (HGF),
fator de crescimento de queratinócitos (KGF) e fator de crescimento de fibroblastos
(FGF-2), transferindo e estimulando a melanogênese (processo bioquímico de
formação do pigmento de melanina) (IRIYAMA et al., 2011).
Foi observado em estudo de biópsias de pele humana que ocorrem
alterações na síntese de HS e perlecam até 72 horas após a exposição com luz UV,
evidenciando diminuição da expressão de perlecam (JUNG et al., 2012). A análise
de carcinoma espinocelular oral bem diferenciado, demonstrou baixa intensidade de
marcação de heparam sulfato (MORI et al., 1998).
É importante observar que além das alterações na quantidade de heparam
sulfato, alterações da estrutura de tal composto também pode estar relacionada com
o desenvolvimento de tumores. Foi demonstrado em carcinoma epidermóide de
pulmão que as cadeias de heparam sulfato apresentam diferenças estruturais no
padrão de dissacarídeos, comparativamente com tecido não tumoral (NACKAERTS
et al., 1997). Mais uma vez fica evidente a importância do heparam sulfato na
organização da estrutura da pele e no potencial metastático após sua clivagem.
O presente estudo claramente mostra aumento do HS com o tratamento de
laserterapia, quando se associa comprimentos de onda (470 e 660 nm). O aumento
da síntese de heparam sulfato, após laserterapia pode favorecer a adesão estrutural

RESULTADOS E DISCUSSÃO
40
dermo-epidérmica e ainda, podemos sugerir que o aumento do heparam sulfato,
após laserterapia poderia estar relacionado com menor probabilidade de
desenvolvimento de carcinoma epitelial.
Estudos sobre a importância do DS associado ao FGF-10 (Fator de
crescimento fibroblástico-10), demonstram que a presença de 10 ou 20
dissacarídeos, contendo ácido idurônico, estimulam o receptor de FGF-10,
promovendo aumento da migração e proliferação de queratinócitos e aceleram o
processo de reparo de feridas (RADEK; TAYLOR; GALLO, 2009).
No entanto, outros estudos demonstraram em carcinoma de células
escamosas de esôfago que a alta expressão de CS/DS e das enzimas epimerase-1
(DS-epi1), 6-O- e 4-O-sulfotransferases estavam significativamente aumentadas
nestes tumores. Com a finalidade de investigar melhor o papel do ácido idurônico
em células tumorais, este grupo estudou células deficientes em dissacarídeos de DS
que continham ácido idurônico em sua estrutura, e foi observado que houve uma
diminuição da capacidade de migração e invasão tumoral in vitro, associada à
redução da ligação com fatores de crescimento de hepatócitos (HGF) (THELIN et al.,
2012).
Os dados obtidos em nosso estudo demonstram estímulo da síntese de DS
em queratinócitos normais (HaCaT), somente após o tratamento associado (470 nm
+ 660 nm), o que pode sugerir que a laserterapia pode efetivamente desempenhar
um papel importante na diferenciação de tais células, devido ao estímulo da
quantidade de dermatam sulfato.
5.3. Expressão gênica por PCR em Tempo Real da HPSE-1
Após o tratamento à laser, foi observado diminuição significativa da expressão
de HPSE-1 em células HaCaT, comparativamente com o controle de células HaCaT
não tratadas. Porém, tal diminuição não foi estatisticamente significante quando foi
avaliada a expressão de HPSE-1 em células tumorais A431, comparativamente com
o controle de células A431 não tratadas (Figura 11).
Podemos observar na que a expressão relativa de HPSE-1 é maior nas
células A431, comparativamente às células HaCaT, visto que os ensaios foram
realizados nas mesmas condições (mesma quantidade de RNA total e cDNA nas
reações de amplificação, como descrito em métodos). Estes resultados corroboram

RESULTADOS E DISCUSSÃO
41
com dados da literatura, que evidenciam aumento da HPSE-1 em células tumorais,
comparativamente com células não neoplásicas (BRUN et al., 2009; VLODAVSKY et
al., 2012).
qPCR - HaCaT e A431
CTR
470
nm
660
nm
470+
660
nm
0.0
0.1
0.2
0.3
0.8
1.0
1.2 HaCaT
A431
*
HPSE-1
Ex
pre
ss
ão
Re
lati
va
**
Figura 91. Expressão relativa de Heparanase-1 (HPSE-1). CTR, Controle, células HaCaT e A431
sem tratamento; Barras brancas indicam a expressão do RNA mensageiro da HPSE-1 em células
HaCaT e Barras pretas indicam a expressão de RNA mensageiro em células A431, tratadas com seis
aplicações do laser de baixa potência como descrito em métodos. Os valores expressos são
referentes a média e desvio padrão, obtidos em ensaios realizados em triplicatas. * p < 0,05.
É evidente a menor quantidade de heparam sulfato, da fração celular em
células A431, comparativamente com células HaCaT (Figuras 8 e 9).
Consequentemente, este dado poderia ser explicado por maior expressão de
HPSE-1 na linhagem tumoral A431 (Figura 10), após laserterapia.
Sabe-se que a HPSE-1 está frequentemente aumentada em tumores, e
confere um caráter invasivo e metastático a essas células. Sabendo-se que a
irradiação com luz UVB é prejudicial à pele e sua ação pode desencadear o
aparecimento de câncer de pele, demonstrou-se aumento da expressão e atividade
da HPSE-1 em cultura de queratinócitos (KURDYKOWSKI et al., 2012). Outro
estudo que utilizou aplicação tópica de heparanase recombinante, demonstrou que a
HPSE-1 acelerou significativamente a cicatrização de feridas, devido à epitelização e
maturação dos vasos sanguíneos, aumentando a angiogênese da ferida,

RESULTADOS E DISCUSSÃO
42
possivelmente por gerar oligossacarídeos de heparam sulfato que apresentam alta
afinidade por fatores angiogênicos e intensificam sua ação. Consequentemente, a
maior intensidade de angiogênese no processo de reparação do tecido sugere um
efeito direto mediado por ação da HPSE-1 (ZCHARIA et al., 2005).
Ainda, a ativação da HPSE-1 na pele induz hiperplasia da epiderme, aumento
dos vasos linfáticos e formação de rugas, sendo que tais mecanismos são mediados
pela degradação das cadeias de heparam sulfato presentes na pele (IRIYAMA;
MATSUNAGA; AMANO, 2010).
5.4. Dosagem de Ácido Hialurônico
A análise da dosagem de ácido hialurônico (AH) em células HaCaT
demonstrou aumento na fração celular e na secreção deste composto, após os
diferentes tratamentos com laserterapia (Figura 12A). É importante ser observado
que a administração concomitante dos comprimentos de onda 470 e 660 nm
promoveu um aumento de aproximadamente 3 vezes na secreção de ácido
hialurônico em células HaCaT, comparativamente ao controle (sem tratamento).
As células A431 não apresentaram alterações na concentração de AH
presente na fração celular e secretado para o meio de cultura, independente do
comprimento de onda utilizado (Figura 12B).
Um resultado bastante interessante de ser observado foi o fato de que células
não neoplásicas (HaCaT), sintetizam e secretam quantidades maiores de AH,
comparativamente com células tumorais A431 (Figura 12).
Foi descrito na literatura que a redução do AH em amostras de biópsias de
pacientes com carcinoma oral espinocelular está diretamente relacionada a um
prognóstico desfavorável. A diminuição da imunomarcação de AH mostrou uma
correlação direta com menor sobrevida dos pacientes (KOSUNEN et al., 2004).
Portanto, a diminuição da quantidade de AH em células A431,
comparativamente com células não neoplásicas HaCaT, corrobora com a hipótese
de que células menos diferenciadas apresentam menor quantidade de AH.

RESULTADOS E DISCUSSÃO
43
Figura 10. Quantificação de Ácido Hialurônico. A, células HaCaT; B, células A431. O ácido
hialurônico foi quantificado na Fração celular (células e matriz extarcelular), bem como no meio de
cultura. A dosagem de ácido hialurônico está expressa em ng/μg de proteína total. Os valores
representam a média e desvio padrão de ensaios realizado triplicata para células HaCaT e
nonoplicata para células A431.
O aumento na quantidade de AH, após laserterapia, observado somente nas
células não neoplásicas (HaCaT), pode sugerir que o tratamento favoreça
mecanismos de diferenciação celular e manutenção da homeostase celular,
associado ao aumento do ácido hialurônico nas células HaCaT, porém, tais
mecanismos celulares não são afetados em células tumorais.
A concentração de AH foi avaliada em ratos, após sofrerem injúria no fêmur.
Os resultados mostraram que após laserterapia, utilizando laser diodo de 660 nm
(100 mW, 240 J/cm-2), ocorre aumento de AH no tecido ósseo (DE SOUZA MERLI et
al., 2012). Estes resultados corroboram com a análise do nosso experimento em
células HaCaT, reforçando a teoria de que laserterapia poderia contribuir com
mecanismos de reparo e cicatrização da pele mediada pelo aumento de AH.
O ácido hialurônico é o componente mais abundante da pele e possui um
turnover de 1-2 dias no conteúdo epidermal (AVERBECK; GEBHARDT; VOIGT,
2006). A constante síntese e degradação do AH é mediada por ação de ácido
hialurônico sintases e hialuronidases.
Dosagem de Ácido Hialurônico em células A431 Fração Celular Meio de Cultura
CTR
470
nm
660
nm
470
+ 66
0 nm
CTR
470
nm
660
nm
470
+ 66
0 nm
0
20
40
60
80
100200
250
300
350
400
AH
ng
/g
pro
teín
a t
ota
l
Dosagem de Ácido Hialurônico em células HaCaT Fração Celular Meio de Cultura
CTR
470
nm
660
nm
470
+ 66
0 nm
CTR
470
nm
660
nm
470
+ 66
0 nm
0
200
400
600
8001000
1200
1400
1600
1800
2000
AH
ng
/ug
pro
teín
a t
ota
l
*
*
B A

RESULTADOS E DISCUSSÃO
44
Diante dos resultados obtidos até o momento decidimos investigar a
expressão das ácido hialurônico sintases e hialuronodases em células HaCaT e
A431.
5.5. Expressão gênica por PCR em Tempo Real das enzimas de biossíntese e degradação do ácido hialurônico
A análise por PCR em tempo real demonstrou diferenças na expressão de
ácido hialurônico sintases e hialuronidases entre as células HaCaT e A431, após
laserterapia. No entanto estatisticamente só pode ser observado significância nas
células HaCaT na expressão do gene HAS2 durante o tratamento de 660 nm e no
gene HAS3, nos tratamentos de 470 nm e associado.
Nas células HaCaT, foi observado diminuição da expressão de RNAm de
ácido hialurônico sintases e hialuronidases, independente do tipo de tratamento com
laserterapia (Figura 13A). O fato da quantidade de ácido hialurônico ter aumentado
em células HaCaT, após laserterapia, mesmo com diminuição da taxa de transcrição
das enzimas envolvidas com a síntese e degradação do AH, reflete que
possivelmente, em células não neoplásicas (HaCaT), a atividade das sintases
encontra-se significativamente aumentada. Esta hipótese pode justificar a
necessidade de altos níveis de ácido hialurônico em queratinócitos não neoplásicos
para manutenção da viscosidade, resiliência e permeabilidade, funções essenciais
na manutenção do epitélio.
Curiosamente o tratamento com laser promoveu aumento da expressão das
ácido hialurônico sintases em células A431, como mostra a Figura 13B.
Com relação às hialuronidases (HYAL), verificamos que a expressão da
HYAL-2 aumentou após os tratamentos com 470 nm e 660 nm, enquanto houve
diminuição da expressão da HYAL-1.

RESULTADOS E DISCUSSÃO
45
HaCaT
CTR
HAS 1
HAS 2
HAS 3
HYA
L 1
HYA
L 2
0.00
0.02
0.04
0.06
0.08
0.5
1.0
1.5
470 nm
660 nm470+660 nm
*
* *
AE
xp
res
sã
o R
ela
tiv
a
A431
CTR
HAS 1
HAS 2
HAS 3
HYAL 1
HYAL 2
0.00.20.40.60.81.01.52.02.53.03.54.04.04.55.05.56.0
470 nm
660 nm
470+660 nm
Ex
pre
ss
ão
Re
lati
va
B
Figura 113. Expressão relativa de Ácido Hialurônico Sintases (HAS1-3) e Hialuronidases
(HYAL1-2). Controle (CTR), sem tratamento; A. Células HaCaT tratada com seis aplicações do laser
de baixa potência. B. Células A431, tratada com seis aplicações do laser de baixa potência, como
descrito em métodos. Os valores expressos são referentes a média e desvio padrão, obtidos em
triplicata. * p < 0,05.
Sabendo que a HYAL-2 degrada especialmente o AH de alto peso molecular
(NOBLE; LIANG; JIANG, 2011), poderíamos sugerir que em células A431
preferencialmente existe tal composto. Ainda, é descrito na literatura que a HYAL-2
pode ser regulada por citocinas inflamatórias, enquanto a HYAL-1 não sofre tal
regulação (FLANNERY et al., 1998).
A diminuição da expressão das ácido hialurônico sintases e hialuronidases foi
observada em células HaCaT tratadas com radiação UVB (280 nm) (HAŠOVÁ et al.,
2011).

RESULTADOS E DISCUSSÃO
46
O estudo evidenciou alterações no perfil e expressão de moléculas da matriz
extracelular, sendo de grande relevância para o estudo dos efeitos do laser de baixa
freqüência em tratamentos dermatológicos, principalmente se ao utilizarmos a
laserterapia com doses inadequadas, pode-se causar alterações irreversíveis as
células epiteliais e facilitar uma progressão tumoral, entanto vale observar, que os
testes in vitro não representam em sua totalidade o tecido epitelial devido a ausência
de interação com outros moléculas, como por exemplo a melanina, e hemoglobina,
respectivamente constituintes de melanócitos e hemácias circulantes no tecido
epitelial, visto que a luz também exerce efeito sobre tais moléculas, e poderiam
modificar a resposta ao tratamento.

CONCLUSÃO
47
6. CONCLUSÕES
A utilização do laser de baixa frequência diminuiu a viabilidade celular de
ambas as linhagens celulares não neoplásicas (HaCaT) e tumoral (A431).
A utilização do laser de baixa frequência alterou o perfil de
glicosaminoglicanos sulfatados das células HaCaT e A431. Nas células
HaCaT observamos aumento do HS, quando utilizado o tratamento de
associação entre os comprimentos de onda 470 nm e 660 nm. Porém, na
linhagem tumoral A431, houve diminuição do HS na fração celular em todos
os tratamentos, após laserterapia.
Foram verificadas diferentes respostas com relação à quantidade de ácido
hialurônico nas diferentes linhagens estudadas. Após irradiação com laser de
baixa potência ocorreu aumento da quantidade de ácido hialurônico na
linhagem HaCaT, não ocorrendo nenhuma alteração para a linhagem tumoral
A431.
Foi observado diminuição da expressão do RNA mensageiro da enzima
HPSE-1, após laserterapia em ambas as linhagens celulares estudadas.
Porém, uma diminuição mais acentuada da expressão do RNA mensageiro
da HPSE-1 foi observada na linhagem HaCaT. Tal resultado possivelmente
corrobora com o aumento do HS observado para a linhagem HaCaT.
Na linhagem HaCaT foi demonstrado diminuição da expressão das ácido
hialurônico sintases e hialuronidases. Entretanto, a linhagem A431 o efeito foi
oposto, sendo observado aumento na expressão das ácido hialurônico
sintases e hialuronidase 2, mas não na expressão de hialuronidase 1.

REFERÊNCIA BIBLIOGRÁFICA
48
7. REFERÊNCIA BIBLIOGRÁFICA
AVERBECK, M.; GEBHARDT, C.; VOIGT, S. Differential regulation of hyaluronan metabolism in the epidermal and dermal compartments of human skin by UVB irradiation. The Society for Investigative Dermatology, p. 687-697, 2006.
BASSO, F. G. et al. In Vitro Wound Healing Improvement by Low-Level Laser Therapy Application in Cultured Gingival Fibroblasts. International Journal of Dentistry, p. 1-6, 2012.
BAUER, A. L.; JACKSON, T. L.; JIANG, Y. Topography of Extracellular Matrix Mediates Vascular Morphogenesis and Migration Speeds in Angiogenesis. Plos Computacional Biology, v. 5, n. 7, p. 1-18, 2009.
BEHRENS, D. T. et al. The Epidermal Basement Membrane Is a Composite of Separate Laminin- or Collagen IV-containing Networks Connected by Aggregated Perlecan , but Not by Nidogens. The Journal of biological chemistry, v. 287, n. 22, p. 18700-18709, 2012.
BLOCHBERGER, T. C. et al. cDNA to Chick Lumican ( Corneal Keratan Sulfate Proteoglycan ) Reveals Homology to the Small Interstitial Proteoglycan Gene Family and Expression in Muscle and Intestine. The Journal of biological chemistry, v. 267, n. 1, p. 347-352, 1992.
BORANIC, M. et al. Skin cell culture: utilization in plastic surgery and laboratory studies. Lijec Vjesn, v. 121, n. 4-5, p. 137-43, 1999.
BOURGUIGNON, L. Y. W. et al. Stem Cell Marker (Nanog) and Star-3 Signaling Promote MicroRNA-21 Expression and Chemoresistance in Hyaluronan/CD44-activated Head and Neck Squamous Cell Carcinoma Cells. Oncogene, v. 31, n. 2, p. 149-160, 2012.
BRIMACOMBRE, J. S.; WEBER, J. M. Mucopolysaccharides - chemical structure, distribuition and isolation. 6. ed. New York: Elsevier Publishing Company, 1964. p. 181
BRUN, R. et al. Heparanase expression by Barrett ’ s epithelium and during esophageal carcinoma progression. Modern Pathology, v. 22, n. 12, p. 1548-1554, 2009.
BRÉZILLON, S. et al. Expression of lumican , a small leucine-rich proteoglycan with antitumour activity , in human malignant melanoma. Experimental Dermatology, v. 32, p. 405-416, 2007.
CAMENISCH, T. D. et al. Disruption of hyaluronan synthase-2 abrogates normal cardiac morphogenesis and hyaluronan-mediated transformation of epithelium to mesenchyme. J. Clin. Invest, v. 106, n. 3, p. 349-360, 2000.
CARROLL, L.; HUMPHREYS, T. R. LASER-tissue interactions. Clinics in Dermatology, v. 24, p. 2-7, 2006.

REFERÊNCIA BIBLIOGRÁFICA
49
CASTRO, M. D. et al. Putative role of heparan sulfate proteoglycan expression and shedding on the proliferation and survival of cells after photodynamic therapy. The International Journal of Biochemistry & Cell Biology, v. 39, p. 1130-1141, 2007.
CHEN, C.; HUNG, H.; HSU, S. Low-Energy Laser Irradiation Increases Endothelial Cell Proliferation , Migration , and eNOS Gene Expression Possibly Via PI3K Signal Pathway. Lasers in Surgery and Medicine, v. 40, p. 46-54, 2008.
CHEN, Y. et al. Evaluation of heparanase and matrix metalloproteinase-9 in patients with cutaneous malignant melanoma. The Journal of Dermatology, v. 39, p. 339-343, 2012.
CORPUZ, L. M. et al. Molecular Cloning and Tissue Distribution of Keratocan. The Journal of biological chemistry, v. 271, n. 16, p. 9759-9763, 1996.
CSOKA, A. B.; FROST, G. I.; STERN, R. The six hyaluronidase-like genes in the human and mouse genomes. Matrix Biology, v. 20, p. 499-508, 2001.
CSOKA, ANTONEI BENJAMIN SCHERER STEPHEN, W.; STERN, R. Expression Analysis of Six Paralogous Human Hyaluronidase Genes Clustered on Chromosomes 3p21 and 7q31. Genomics, v. 60, p. 356–361, 1999.
Cancer Information. Disponível em: <www.ccsb.org/cancersummary/cdr0000552637>. Acesso em: 16 abr. 2013.
DE SOUZA MERLI, L. A. et al. The low level laser therapy effect on the remodeling of bone extracellular matrix. Photochemistry and photobiology, v. 88, n. 5, p. 1293-301, 2012.
DEYRIEUX, A. F.; WILSON, V. G. In vitro culture conditions to study keratinocyte differentiation using the HaCaT cell line. Cytotechnology, v. 54, p. 77-83, 2007.
DIETRICH, C. et al. Role of sulfated mucopolysaccharides in cell recognition and neoplastic transformation. An Acad Bras Cienc, v. 52, n. 1, p. 179-86, 1980.
DIETRICH, C. et al. Structure of heparan sulfate: identification of variable and constant oligosaccharide domains in eight heparan sulfates of different origins. Cell Mol Biol, v. 44, n. 3, p. 417-29, 1998.
EDUARDO, F. P. et al. Cultured Epithelial Cells Response to Phototherapy With Low Intensity Laser. Lasers in Surgery and Medicine, v. 39, p. 365-372, 2007.
ESKO, J. D. Genetic analysis of proteoglycan structure, function and metabolism. Curr Opin Cell Biol., v. 3, n. 5, p. 805-16, 1991.
ESMAEELINEJAD, M.; BAYAT, M. The effects of low-level laser irradiation on cellular viability and proliferation of human skin fibroblasts cultured in high glucose mediums. lasers med Sci, 2013.

REFERÊNCIA BIBLIOGRÁFICA
50
FLANNERY, C. R. et al. Expression and Activity of Articular Cartilage Hyaluronidases. Biochemical and biophsycal research comunications, v. 829, p. 824-829, 1998.
FRENKEL, J. S. The role of hyaluronan in wound healing. International Wound Journal, p. 1-7, 2012.
FUJIMOTO, T. et al. Induction of Different Reactive Oxygen Species in the Skin During Various Laser Therapies and Their Inhibition by Fullerene. Lasers in Surgery and Medicine, v. 44, p. 685-694, 2012.
FUKUDA T Y, M. C. A. Analysis of low-level laser therapy doses in Brazilian equipment. Revista Brasileira de Fisioterapia, v. 12, n. 1, p. 70-4, 2008.
FUNDERBURGH, J. L. et al. Mimecan , the 25-kDa Corneal Keratan Sulfate Proteoglycan , Is a Product of the Gene Producing Osteoglycin. The Journal of biological chemistry, v. 272, n. 44, p. 28089-28095, 1997.
GALLO, R. Proteoglycans and cutaneous vascular defense and repair. Journal of Investigative Dermatology Symposium …, p. 55-60, 2000.
GANDHI, N. S.; MANCERA, R. L. The Structure of Glycosaminoglycans and their Interactions with Proteins. Chem Biol Drug Des, v. 72, p. 455-482, 2008.
GAO, X.; XING, D. Molecular mechanisms of cell proliferation induced by low power laser irradiation. Journal of Biomedical science, v. 16, p. 31-37, 2009.
GEORGE, J.; STERN, R. Serum hyaluronan and hyaluronidase : very early markers of toxic liver injury. Clinica Chimica Acta, v. 348, p. 189-197, 2004.
GOMES, R. C. T. et al. Perfil de Glicosaminoglicanos Sulfatados no Útero de Camundongas Durante o Ciclo Estral. Rev Assoc Med Bras, v. 53, n. 3, p. 261-266, 2007.
GROSSMAN, N. et al. 780 nm Low Power Diode Laser Irradiation Stimulates Proliferation of Keratinocyte Cultures : Involvement of Reactive Oxygen Species. Lasers in Surgery and Medicine, v. 22, p. 212-218, 1998.
HAN, Z. et al. The Relationship between the Extracellular Matrix and the Angiogenesis and Metastasis of Laryngeal Carcinoma. ORL, v. 70, p. 352-358, 2008.
HARRIS, D. M. Laser Biostimulation: Review and Hypothesis. L.I.A. Laser Topics, v. 10, n. 3, p. 9-14, 1988.
HAŠOVÁ, M. et al. Hyaluronan minimizes effects of UV irradiation on human keratinocytes. Archives of dermatological research, v. 303, n. 4, p. 277-84, maio. 2011.

REFERÊNCIA BIBLIOGRÁFICA
51
HOPKINS, J. T. et al. Low-Level Laser Therapy Facilitates Superficial Wound Healing in Humans: A Triple-Blind, Sham-Controlled Study. Journal of Athletic Training, v. 39, n. 3, p. 223-229, 2004.
HSIEH, Y.; YU, J.; LYU, P. Characterization of photodynamic therapy responses elicited in A431 cells containing intracellular organelle-localized photofrin. Journal of cellular biochemistry, v. 111, p. 821-833, 2010.
INCA. Disponível em: <http://www2.inca.gov.br/wps/wcm/connect/tiposdecancer/site/home/pele_nao_melanoma>. Acesso em: 1 abr. 2013.
IOZZO, R. V. Heparan sulfate proteoglycans : intricate molecules with intriguing functions. The Journal of Clinical Investigation, v. 108, n. 2, p. 165-167, 2001.
IRIE, F. et al. Heparan sulfate regulates ephrin-A3 / EphA receptor signaling. PNAS, v. 105, n. 34, p. 12307-12312, 2008.
IRIYAMA, S. et al. Hyperpigmentation in human solar lentigo is promoted by heparanase-induced loss of heparan sulfate chains at the dermal–epidermal junction. Journal of Dermatological Science, v. 64, n. 3, p. 223–228, 2011.
IRIYAMA, S.; MATSUNAGA, Y.; AMANO, S. Heparanase activation induces epidermal hyperplasia , angiogenesis , lymphangiogenesis and wrinkles. Experimental Dermatology, v. 19, p. 965-972, 2010.
JEANLOZ, R. W. The Nomenclature of mucopolysaccharides. Arthritis Rheum, v. 3, p. 233-237, 1960.
JONES, M. H. et al. Expression analysis, genomic structure, and mapping to 7q31 of the human sperm adhesion molecule gene SPAM1. Genomics, v. 29, p. 796–800, 1995.
JUAN, M. Z. et al. Differential Responses to UVB Irradiation in Human Keratinocytes and Epidermoid Carcinoma Cells. Biomed Environ Sci, v. 25, n. 5, p. 583-589, 2012.
JUNG, J. et al. Acute UV Irradiation Increases Heparan Sulfate Proteoglycan Levels in Human Skin. Journal Korean Med Sci, v. 27, p. 300-306, 2012.
KANITAKIS, J. Anatomy, histology and immunohistochemistry of normal human skin. Eur J Dermatol, v. 12, n. 4, p. 390-401, 2002.
KARU, T. I. Photobiological Fundamentals of Low-Power Laser Therapy. IEEE Journal of Quantum Electronics, v. QE-23, p. 1703-1717, 1987.
KASTNER, S.; THOMAS, G.; JENKINS, R. Hyaluronan induces the selective accumulation of matrix-and cell-associated proteoglycans by mesangial cells. The american Journal of Pathology, v. 171, n. 6, p. 1811-1821, 2007.

REFERÊNCIA BIBLIOGRÁFICA
52
KIM, R. H.; ARMSTRONG, A. W. Nonmelanoma Skin Cancer. Dermatologic Clinics, v. 30, n. 1, p. 125-139, 2012.
KOSUNEN, A. et al. Reduced expression of hyaluronan is a strong indicator of poor survival in oral squamous cell carcinoma. Oral Oncology, v. 40, p. 257-263, 2004.
KOZLOWSKI, E.; PAVÃO, M.; BORSIG, L. Ascidian dermatan sulfates attenuate metastasis , inflammation and thrombosis by inhibition of P-selectin. journal of Thrombosis and Haemostasis, v. 9, p. 1807-1815, 2011.
KRESSE, H.; SCHÖNHERR, E. Proteoglycans of the extracellular matrix and growth control. Journal of cellular physiology, v. 274, n. July, p. 266-274, 2001.
KURDYKOWSKI, S. et al. Biology Ultraviolet-B irradiation induces epidermal up-regulation of heparanase expression and activity. Journal of Photochemistry & Photobiology, B: Biology, v. 106, p. 107-112, 2012.
LIN, F. et al. Lasers , stem cells , and COPD. Journal of Translational Medicine, v. 8, n. 16, p. 1-10, 2010.
LIPOVSKY, A. et al. Visible Light-Induced Killing of Bacteria as a Function of Wavelength : Implication for Wound Healing. Lasers in Surgery and Medicine, v. 42, p. 467-472, 2010.
LOPES, L. A. Laserterapia: conceitos e aplicações. Disponível em: <http://www.nupen.com.br/Revista_port/fund_fisicos4_2.php>. Acesso em: 23 jun. 2013a.
LOPES, L. A. Laserterapia Conceitos e aplicações. Disponível em: <http://www.nupen.com.br/Revista_port/introducao.php>. Acesso em: 14 abr. 2013b.
LUCCA, A. J. D.; WILLIAMS, K. A.; BHATNAGAR, D. Blue light ( 470 nm ) effectively inhibits bacterial and fungal growth. Letters Applied microbiology, v. 55, p. 460-466, 2012.
LUKIC D et al. The results of treatment of basoceltular carcinomas of the head skin. Med Arh, v. 66, n. 3, p. 169-72, 2012.
MAHMOUD, B. H. et al. Effects of Visible Light on the Skin. Photochemistry and Photobiology, v. 84, p. 450-462, 2008.
MALMSTRÖM, A. et al. Iduronic Acid in Chondroitin/Dermatan Sulfate: Biosynthesis and Biological Function. Journal of Histochemistry & Cytochemistry, v. 60, p. 916-925, 2012.
MARTINS, R. M. et al. Practical determination of hyaluronan by a new noncompetitive fluorescence-based assay on serum of normal and cirrhotic patients. Analytical Biochemistry, v. 319, p. 65-72, 2003.

REFERÊNCIA BIBLIOGRÁFICA
53
MESTER, E.; TOTA, J. Effect of laser on hair growth of mice. Kiserl Orvostud, v. 19, p. 628-631, 1967.
MEYER, K. Chemical structure of hyaluronic acid. Fed Proc, v. 17, n. 4, p. 1075-7, 1958.
MICHELACCI, Y. M. et al. Chondroitin sulfates and proteoglycans from normal and arthrosic human cartilage. Connect Tissue Res, v. 7, p. 29, 1979.
MORI, S. et al. A novel evaluation system of metastatic potential of oral squamous cell carcinoma according to the histopathological and histochemical grading. Oral Oncology, v. 34, p. 549-557, 1998.
MOSCATELLO, D. K. et al. Decorin Suppresses Tumor Cell Growth by Activating the Epidermal Growth Factor Receptor. J. Clin. Invest, v. 101, n. 2, p. 406-412, 1998.
MOURÃO, P. A. et al. The distribution of chondroitin sulfates in articular and growth cartilages of human bones. Biochim Biophys Acta, v. 428, p. 19, 1976.
NACKAERTS, K. et al. Heparan Sulfate Proteoglycan Expression in Human Lung-Cancer Cells. Int. J. Cancer, v. 74, p. 335–345, 1997.
NADER, H. et al. Heparin sequences in the heparan sulfate chains of an endothelial cell proteoglycan. Proc Nat Acad Sci USA, v. 84, p. 3565-9, 1987.
NADER, H. et al. Heparin stimulates the synthesis and modifies the sulfatation pattern of heparan sulfate proteoglycan from endothelial cells. J Cell Physiol, v. 140, p. 305-10, 1989.
NAKAYAMA, F. et al. Hidrogen Peroxide as a Potential Mediator of the Transcriptional Regulation of Heparan Sulphate Biosynthesis in Keratinocytes. Cellular & Molecular Biology Letters, v. 13, p. 475-492, 2008.
NASHAN, D.; MEISS, F.; MÜLLER, M. Therapeutic strategies for actinic keratoses - a systematic review. Eur J Dermatol, v. 23, n. 1, p. 14-32, 2013.
NEUMAN, M. G. et al. Damage in Skin Cells In vitro Anti-Inflammatory Effects of Hyaluronic Acid in Ethanol-Induced. J Pharm Pharmaceut Sci, v. 14, n. 3, p. 425-437, 2011.
NOBLE, W. P.; LIANG, J.; JIANG, D. Hyaluronan as an Immune Regulator in Human Diseases. Physiol Rev., v. 91, n. 1, p. 221-264, 2011.
NYKOPP, T. K. et al. ( HYAL1 – 2 ) in Serous Ovarian Carcinomas : Inverse Correlation between HYAL1 and Hyaluronan Content. BMC Cancer, v. 9, p. 1-9, 2009.
ORNITZ, D. M.; WILEY, J. FGFs , heparan sulfate and FGFRs : complex interactions essential for development. BioEssays, v. 22, p. 108-112, 2000.

REFERÊNCIA BIBLIOGRÁFICA
54
PAGIN, M. T. et al. Laser and light-emitting diode effects on pre-osteoblast growth and differentiation. lasers med Sci, 2012.
PAIVA, P. et al. Expression patterns of hyaluronan , hyaluronan synthases and hyaluronidases indicate a role for hyaluronan in the progression of endometrial cancer. Gynecologic Oncology, v. 98, p. 193-202, 2005.
PARISH, C. Te role of heparan sulphate in inflammation. Nat Rev Immunol, v. 6, p. 633-43., 2006.
PECCHI, E. et al. A potential role of chondroitin sulfate on bone in osteoarthritis: inhibition of prostaglandin E₂ and matrix metalloproteinases synthesis in interleukin-1β-stimulated osteoblasts. Osteoarthritis Cartilage, v. 20, n. 2, p. 127-35, 2012.
PORCIONATTO, M.; NADER, H.; DIETRICH, C. Heparan sulfate and cell division. Braz J Med Biol Res, v. 32, n. 5, p. 539-44, 1999.
PRIMAKOFF, P.; HYATT, H.; MYLES, D. G. A role for the migrating sperm surface antigen PH-20 in guinea pig sperm binding to the egg zona pellucida. Cell. Biol., v. 101, p. 2239–2244, 1985.
PROKSCH, E.; BRANDNER, J. M.; JENSEN, J. The skin : an indispensable barrier. Experimental Dermatology, v. 17, p. 1063-1072, 2008.
RADEK, K. A.; TAYLOR, K. R.; GALLO, R. L. FGF-10 and specific structural elements of dermatan sulfate size and sulfation promote maximal keratinocyte migration and cellular proliferation. Wound Repair and Regeneration, v. 17, p. 118-126, 2009.
RADEMACHER, T. .; PAREKH, R. .; DWEK, R. . Glycobiology. Annu Rev Biochem, v. 57, p. 785-838, 1988.
ROBAT, M. INAZARI- et al. Co-solvent mediated thermal stabilization of chondroitinase ABC I form Proteus vulgaris. International Journal of Biological Macromolecules, v. 50, p. 487–492, 2012.
RODRIGUEZ, E.; ROUGHLEY, P. Link protein can retard the degradation of hyaluronan in proteoglycan aggregates. Osteoarthritis and Cartilage, v. 14, n. 8, p. 823-829, 2006.
RODUST, P. M.; FECKER, L. F.; STOCKFLETH, E. Activation of mitochondrial apoptosis pathways in cutaneous squamous cell carcinoma cells by diclofenac / hyaluronic acid is related to upregulation of Bad as well as downregulation of Mcl-1 and Bcl-w. Experimental Dermatology, v. 221, p. 520-525, 2012.
RÖCK, K. et al. Estradiol Protects Dermal Hyaluronan / Versican Matrix during Photoaging by Release of Epidermal Growth Factor from. The Journal of biological Chemistry, v. 287, n. 24, p. 20056-20069, 2012.

REFERÊNCIA BIBLIOGRÁFICA
55
SADOWSKI, T. et al. Matrix Metalloproteinase 19 Regulates Insulin-like Growth Factor-mediated Proliferation , Migration , and Adhesion in Human Keratinocytes through Proteolysis of Insulin-like Growth Factor Binding Protein-3. Molecular Biology of the Cell, v. 14, n. November, p. 4569-4580, 2003.
SARRAZIN, S.; LAMANNA, W. C.; ESKO, J. D. Heparan sulfate proteoglycans. Cold Spring Harbor perspectives in biology, v. 3, n. 7, jul. 2011.
SCHONHERR, E.; HAUSSER, H. R. Extracellular Matrix and Cytokines: A Functional Unit. Developmental Immunology, v. 7, n. 2-4, p. 89-101, 2000.
SEKHEJANE, P. R.; HOURELD, N. N.; ABRAHAMSE, H. Irradiation at 636 nm Positively Affects Diabetic Wounded and Hypoxic Cells in Vitro. Photomedicine and Laser Surgery, v. 29, n. 8, p. 521-530, 2011.
SEPPÄNEN-PASONEN, S. et al. EGF Upregulates, Whereas TGF-b Downregulates, the Hyaluronan Synthases Has2 and Has3 in Organotypic Keratinocyte Cultures: Correlations with Epidermal Proliferation and Di¡erentiation. The Society for Investigative Dermatology, v. 120, p. 1038-1044, 2003.
SEYFRIED, N. T. et al. Expression and purification of functionally active hyaluronan-binding domains from human cartilage link protein, aggrecan and versican: formation of ternary complexes with defined hyaluronan oligosaccharides. The Journal of biological chemistry, v. 280, n. 7, p. 5435-48, 18 fev. 2005.
SIFAKI, M. et al. Lumican , a small Leucine-rich Proteoglycan substituted with Keratan Sulfate Chains is expressed and secreted by Human Melanoma Cells and not normal Melanocytes. IUBMB Life, v. 58, n. 10, p. 606-610, 2006.
SIMPSON, C. L.; PATEL, D. M.; GREEN, K. J. Deconstructing the skin: cytoarchitectural determinants of epidermal morphogenesis. Nat Rev Mol Cell Biol, v. 12, n. 9, p. 565-580, 2012.
STERN, R.; MAIBACH, H. I. Hyaluronan in skin: aspects of aging and its pharmacologic modulation. Clin Dermatol, v. 26, p. 106-22, 2008.
TAI, M. et al. Ultraviolet C Irradiation Induces Different Expression of Cyclooxygenase 2 in NIH 3T3 Cells and A431 Cells: The Roles of COX-2 Are Different in Various Cell. International Journal of Molecular Sciences, v. 13, p. 4351-4366, 2012.
TAMMI, R. et al. Localization of epidermal hyaluronic acid using the hyaluronate binding region of cartilage proteoglycan as a specific probe. j Invest dermatol., v. 90, p. 412-4, 1988.
TANZI, E. L.; LUPTON, J. R.; ALSTER, T. S. Lasers in dermatology : Four decades of progress. J Am Acad Dermatol, v. 49, n. 1, p. 1-34, 2003.

REFERÊNCIA BIBLIOGRÁFICA
56
TAYLOR, K. R.; RUDISILL, J. A; GALLO, R. L. Structural and sequence motifs in dermatan sulfate for promoting fibroblast growth factor-2 (FGF-2) and FGF-7 activity. The Journal of biological chemistry, v. 280, n. 7, p. 5300-6, 18 fev. 2005.
TEIXEIRA, L. et al. Heparanase expression and glycosaminoglycans profile in renal cell carcinoma. International Journal of Urology, v. 19, p. 1036-1040, 2012.
THELIN, M. et al. Dermatan sulfate is involved in the tumorigenic properties of esophagus squamous cell carcinoma. Cancer Res., v. 15, n. 8, p. 1943-52, 2012.
THEODORO, T. R. et al. Heparanase Expression in Circulating Lymphocytes of Breast Cancer Patients Depends on the Presence of the Primary Tumor and / or Systemic Metastasis 1. Neoplasia, v. 9, n. 6, p. 504-510, 2007.
TOLLEFSEN, D. The interaction of glycosaminoglycans with heparin cofactor II. Ann N Y Acad Sci., v. 714, p. 21-31, 1994.
VANNUCCHI, S. et al. Heparin inhibits A431 cell growth independently of serum and ECF mitogcnic signalling. FEBS, v. 281, p. 141-144, 1991.
VLODAVSKY, I. et al. Mammalian heparanase: gene cloning, expression and function in tumor progression and metastasis. Nature Med, v. 5, n. 7, p. 793-802, 1999.
VLODAVSKY, I. et al. Significance of Heparanase in Cancer and Inflammation. Cancer Microenvironment, v. 5, p. 115-132, 2012.
VLODAVSKY, I.; FRIEDMANN, Y. Molecular properties and involvement of heparanase in cancer metastasis and angiogenesis. The Journal of Clinical Investigation, v. 108, n. 3, p. 341-347, 2001.
WAGNER, M. et al. Methylene blue photodynamic therapy in malignant melanoma decreases expression of proliferating cell nuclear antigen and heparanases. Clinical and Experiemental Dermatology, v. 37, p. 527-533, 2012.
WILLHAUCK, M. J. et al. Reversion of tumor phenotype in surface transplants of skin SCC cells by scaffold-induced stroma modulation. carcinogenesis, v. 28, n. 3, p. 595-610, 2007.
WU, D. et al. Upregulation of TCTP expression in human skin squamous cell carcinoma increases tumor cell viability through anti-apoptotic action of the protein. Experimental and Therapeutic Medicine, v. 3, p. 437-442, 2012.
YEH, J. et al. Impaired skin wound healing in lumican-null mice. British Journal of Dermatology, v. 163, p. 1174-1180, 2010.
ZCHARIA, E. et al. Heparanase accelerates wound angiogenesis and wound healing in mouse and rat models. The FASEB Journal, v. 19, p. 211-221, 2005.

REFERÊNCIA BIBLIOGRÁFICA
57
ZHAO, X. et al. Sequence Analysis and Domain Motifs in the Porcine Skin Decorin Glycosaminoglycan Chain. Journal of Biological Chemistry, v. 288, n. 13, p. 9226-9237, 2013.