Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL PAULISTA "JULIO DE MESQUITA FILHO"
FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS
CÂMPUS DE JABOTICABAL
AVALIAÇÃO QUANTI E QUALITATIVA DE OÓCITOS DE
FÊMEAS SUÍNAS NO ESTRO E DE ESTRUTURAS
EMBRIONÁRIAS 24 E 144 HORAS APÓS MONTA NATURAL
Eliana Cristina Gazoto
Orientador: Prof. Dr. Cesar Roberto Esper
Dissertação apresentada à Faculdade de CiênciasAgrárias e Veterinárias – UNESP, Câmpus deJaboticabal, como parte das exigências para aobtenção do título de Mestre em Medicina Veterinária(Reprodução Animal).
JABOTICABAL – SÃO PAULO – BRASIL
Junho – 2006

iii
DADOS CURRICULARES DO AUTOR
ELIANA CRISTINA GAZOTO – nascida em Jaboticabal – SP, aos 04 dias do
mês de maio de 1977; concluiu o ensino médio no Colégio Santo André, na cidade de
Jaboticabal, em 1995; ingressou no curso de graduação em Medicina Veterinária na
Universidade de Uberaba – UNIUBE, em 1998; concluiu o curso superior em Medicina
Veterinária em fevereiro de 2003; realizou curso de Especialização em Reprodução em
Bovinos, ao nível de Pós-Graduação “Lato Sensu”, no Centro Universitário de Espírito
Santo do Pinhal – UNIPINAL, no período de 20 de março de 2003 a 14 de maio de
2004; ingressou no curso de Mestrado, no Programa de Pós-Graduação em Medicina
Veterinária (Reprodução Animal), na Faculdade de Ciências Agrárias e Veterinárias,
Câmpus de Jaboticabal da Universidade Estadual Paulista – UNESP, em março de
2004.

iv
Você Aprende...
“Depois de algum tempo você aprende a diferença, a sutil diferença, entre dar a mão eacorrentar uma alma.E você aprende que amar não significa apoiar-se, e que companhia nem sempresignifica segurança.E começa a aceitar suas derrotas com a cabeça erguida e olhos adiante, com a graçade um adulto e não com a tristeza de uma criança.E aprende a construir todas as suas estradas no hoje, porque o terreno do amanhã éincerto demais para os planos, e o futuro tem costume de cair em meio ao vão.E o que importa não é o que você tem na vida, mas quem você tem na vida.Aprende que verdadeiras amizades continuam a crescer mesmo a longas distâncias.E que bons amigos são a família que nos permitiram escolher.Aprende que as circunstâncias e os ambientes têm influência sobre nós, mas nóssomos responsáveis por nós mesmos.Começa a aprender que não se deve comparar com os outros, mas com o melhor quepode ser.Descobre que se leva muito tempo para tornar a pessoa que quer ser, e que o tempo écurto.Aprende que nunca se deve dizer a uma criança que sonhos são bobagens, poucascoisas são tão humilhantes e seria uma tragédia se ela acreditasse nisso.Aprende que não importa aonde já chegou, mas onde está indo, mas se você não sabepara onde está indo qualquer lugar serve.Aprende que, ou você controla seus atos ou eles o controlarão, e que ser flexível nãosignifica ser fraco ou não ter personalidade, pois não importa quão delicada e frágil sejauma situação, sempre existem dois lados.Aprende que heróis são pessoas que fizeram o que era necessário fazer, enfrentandoas conseqüências.Aprende que ter paciência requer muita prática.Aprende que maturidade tem mais a ver com os tipos de experiência que se teve, e oque você aprendeu com elas do que com quantos aniversários você celebrou.Aprende que há mais dos seus pais em você do que você supunha.Aprende que nem sempre é suficiente ser perdoado por alguém, algumas vezes vocêtem que aprender a perdoar a si mesmo.Aprende que com a mesma severidade com que julga, você será em algum momentocondenado.Aprende que não importa em quantos pedaços seu coração foi partido, o mundo nãopára para que você o concerte.Aprende que o tempo não é algo que possa voltar atrás.E você aprende que realmente pode suportar... que realmente é forte, e que pode irmuito mais longe depois de pensar que não se pode mais.E que realmente a vida tem valor e que você tem valor diante da vida!”.
Shakespeare

v
DEDICATÓRIA
Primeiramente a Deus... pela força, coragem, busca constante, fé; por ensinar-me o verdadeiro sentido da paciência, da gratidão,
perceverança e por manter minha alma e o coração leves.Obrigada Senhor.
Aos meus pais, Oswaldo e Cida,...por acreditarem sempre em mim.
...apoio nos momentos mais difíceis desta trajetória....pelo amor incondicional.
...pela vida....por serem minha inspiração e referência de vida.
Meus irmãos, César e Edson,...pela torcida.
...amor....pela luta constante dos meus objetivos.
Meu muito Obrigada.
À minha família, a maior herança que recebi de Deus, eu dedico...

vi
A todos os Mestres,que têm me ensinado
o valor de aprender,a necessidade da busca,
a riqueza dos verbos,a simpatia do sorriso,
a força da perseverança,a importância das mudanças,a necessidade de recomeçar,
a tolerância e a humildade,a evolução da ciência,
a força do amore a beleza de Deus.

vii
AGRADECIMENTOS
A Deus, por guiar o meu caminho....
Ao professor orientador, Prof. Dr. Cesar Roberto Esper, grande mestre e cientista, pelo
idealismo, confiança, amizade, paciência, ensinamentos e sempre acreditar em meus sonhos.
Aos professores do Departamento de Reprodução Animal da FCAV/UNESP – Jaboticabal,
Prof. Dr. Cesar Roberto Esper, Prof. Dr. Joaquim Mansano Garcia, Profa. Dra. Vera Fernanda
Martins Hossepian de Lima, Prof. Dr. Wilter Ricardo Russiano Vicente, Prof. Dr. Paulo Henrique
Franceschini, Prof. Dr. Gilson Hélio Toniollo e Prof. Dr. Francisco Guilherme Leite, pela amizade,
interesse e conhecimentos transmitidos.
À Prof. Dra Rosangela Zacarias Machado, pelo incentivo à vida acadêmica, amizade, atenção e
afeto. Muito Obrigada.
Aos funcionários do DRA, Roberta Vantini, Ivo Luiz de Almeida Jr., Isabel Aparecida Penariol
Natarelli e Paulo Sérgio da Silva, pelos momentos alegres e pela presença em todos os momentos do
experimento.
Ao Prof. Dr. Antonio Sergio Ferraudo, pela disponibilidade e auxílio estatístico.
Aos Professores Dr. Paulo Henrique Francesquini e Dr. Joaquim Mansano Garcia, pelo
conselho de vida e contribuição na qualificação da dissertação.
A CAPES, pela concessão da bolsa do Mestrado.
À Médica Veterinária Viviane Purri, pela indispensável ajuda durante o experimento, por sua
competência e preocupação em sempre fazer o melhor.
Ao Frigorífico Ligeirinho, pelo auxílio e disponibilidade em me atender mesmos em dias
inesperados.
Pelos funcionários da Fazenda São João das Palmeiras, pela dedicação aos animais, auxílio
durante todo experimento, momentos de descontração mesmo nas horas difíceis, vocês são verdadeiros
Mestres...
Ao Amigo e companheiro Júnior, que muito me ajudou durante todo o experimento e pela sua
animação e incentivo até o término deste trabalho.
À Juliana do Departamento de Ciências Exatas, por me ajudar na correção desta dissertação.
À amiga Kellen, pela assistência informática, tarde e noite no departamento, pelo incentivo.
Aos amigos de Laboratório, Ana Paula Perini, Ana, Aracelle, Christina Ramires Ferreira,
Danilas Salinet de Melo, Eliandra, Eric Castro, Erlon Gomes de Oliveira, Felipe Perecin, Giovana

viii
D’andrea Pavão, Juliana Corrêa Borges, Kellen de Sousa Oliveira, Letícia Barretto, Lorivaldo Paz
Landim Junior, Maricy, Max Vitória Resende, Naiara Zoccal, Rúbia Bueno da Silva, Simone Cristina
Méo Niciura, Tatiane Almeida Drummond Tetzner, Viviane Castro, pela amizade, apoio,
companheirismo, troca de conhecimentos e muitas risadas. Agradeço a oportunidade de encontrar
pessoas maravilhosas como vocês...
A todos estagiários que passaram pelo DRA, pelo entusiasmo, interesse e auxílio.
Ao Deny Munari Trevisani,Que sempre esteve ao meu lado, como Mestre, Doutor e namorado.
...obrigada pela motivação,...acolhimento,...orientação,
...por se orgulhar dos meus erros, das minhas confusões, alegrias, conquistas....por me ensinar que é preciso continuar caminhando, mesmo quando muitos te dizem para voltar.
...paciência principalmente no final da elaboração deste trabalho.
Aos meus avós, Américo, Idalina, Albino e Geni (in memorian), por serem a minha referência,
através das suas experiências.....vocês sempre estarão eternamente no meu coração.
Ao meu sobrinho Enzo Leite Gazoto, que através do seu sorriso me ensinou como é importante
sonhar e acreditar nesses.
Às minhas cunhadas Juliana e Renata, pelo amor e amizade.
À minha sogrinha Clélia, por torcer e acreditar nos meus sonhos.
À Faculdade de Ciências Agrárias e Veterinárias, Câmpus de Jaboticabal – UNESP, em
especial, ao Departamento de Medicina Veterinária Preventiva e Reprodução Animal e à Pós-
graduação, pelas instalações, condições de trabalho, acolhimento e oportunidade.
Ao Instituto Raiz, minha grande família, através de todos eu aprendi a nunca desistir... Que
crescer é difícil, mas vale a pena e que chorar é muito importante, pois aproxima o coração da alma.
Vocês são pessoas abençoadas. Obrigada por existirem.
À Susana, que me ensinou o verdadeiro significado de ser Mestre.
Ao amigo Cadu, com sua alegria e coração sempre me compreendeu e me animou a continuar.
Ao amigo Cléber Barbosa de Oliveira, pela amizade tão especial, companheirismo, auxílio,
disponibilidade sempre.
Ao amigo Saul Borsari, pelo interesse, apoio e por levantar meu astral quando estava cansada.
Os médicos Veterinários Wagner e Marcelo, por me auxiliarem no decorrer da realização deste
trabalho.
Às amigas Carolina, Márcia, Andreza, Érika, Claudinha, Carla Braga , Eveline , obrigada pela
presença mesmo com a distância.

ix
APOIO FINANCEIRO
À CAPES, pela bolsa concedida, no período de março de 2004 a fevereiro de 2006.

x
SUMÁRIO
Página
LISTA DE TABELAS................................................................................................ xiLISTA DE FIGURAS................................................................................................. xiiABREVIATURAS...................................................................................................... xiiiRESUMO.................................................................................................................. xvSUMMARY................................................................................................................ xvi1. INTRODUÇÃO................................................................................................... 12. REVISÃO DE LITERATURA.............................................................................. 2
2.1. Considerações gerais.............................................................................. 22.2. Ciclo estral............................................................................................... 32.3. Puberdade............................................................................................... 52.4. Exposição ao macho............................................................................... 62.5. Técnicas para a colheita de oócitos em animais de abatedouro............ 82.6. Tipos de folículos ovarianos.................................................................... 92.7. Aspectos moleculares da maturação dos oócitos................................... 102.8. Fertilização in vitro (FIV).......................................................................... 132.9. Aspectos moleculares relacionados à ativação do oocitária................... 142.10. Descondensação da cromatina do espermatozóide............................... 152.11. Polispermia.............................................................................................. 162.12. Mecanismos moleculares relacionados à prevenção da polispermia..... 212.13. Morfologia e qualidade dos embriões...................................................... 232.14. Perdas embrionárias e fetais................................................................... 24
3. OBJETIVOS....................................................................................................... 284. MATERIAL E MÉTODOS................................................................................... 29
4.1. Animais.................................................................................................... 294.2. Período de adaptação das fêmeas.......................................................... 294.3. Manejo e alimentação............................................................................. 304.4. Planejamento experimental..................................................................... 314.5. Avaliação dos oócitos (“status” nuclear e grânulos corticais).................. 324.6. Análise estatística.................................................................................... 34
5. RESULTADOS................................................................................................... 355.1. Experimento 1......................................................................................... 355.2. Experimento 2......................................................................................... 375.3. Experimento 3......................................................................................... 385.4. Experimento 4......................................................................................... 39
6. DISCUSSÃO...................................................................................................... 417. CONCLUSÕES.................................................................................................. 458. REFERÊNCIAS.................................................................................................. 46

xi
LISTA DE TABELAS
PáginaTabela 1. Efeito do ciclo sexual sobre o desenvolvimento do trato genital....... 6
Tabela 2. Efeito da idade do macho na sua habilidade em estimular apuberdade em leitoas........................................................................ 7
Tabela 3. Composição do arraçoamento das marrãs destinadas à reproduçãoe utilizadas no experimento............................................................... 31
Tabela 4. Média e desvio padrão das estruturas isoladas e analisadas paracada experimento.............................................................................. 35
Tabela 5. Resultado do “status” nuclear e dos grânulos corticais de oócitosobtidos da aspiração de ovários de 8 marrãs após terem tido oestro detectado.................................................................................. 36
Tabela 6. Número e características das estruturas obtidas após lavagem dastubas uterinas de 8 marrãs após 24 horas da monta natural............ 37
Tabela 7. Número e características das estruturas obtidas após lavagem doscornos uterinos de 8 marrãs após 6 dias decorridos da montanatural................................................................................................ 38
Tabela 8. Número de marrãs, nascimentos, médio de leitões nascidos,desmamados, natimortos e mortalidade após a desmama nos anosde 2004 e 2005 da propriedade................................................ 39

xii
LISTA DE FIGURAS
PáginaFigura 1. Aspecto visual de ovários de marrãs colhidos em matadouro. (A)
ovário de fêmea pré-púbere (somente folículos); (B) ovários defêmeas púberes (presença de CLs).................................................. 10
Figura 2. Reação que ocorre quando um espermatozóide fertiliza um óvuloem mamíferos (Adaptado de Alberts et al. - Molecular biology of thecell. 4th ed. 2004)........................................................................ 15
Figura 3. Reação cortical após a entrada de espermatozóide através da zonapelúcida (Fonte: Adaptado de Alberts et al. - Molecular biology ofthe cell. 4th ed. 2004)........................................................ 22
Figura 4. A) Fêmeas (Large White) e B) Cachaço (Duroque), utilizados noexperimento....................................................................................... 29
Figura 5. A) Limpeza do prepúcio; B e C) Reflexo de tolerância ao homem; d)Monta natural (MN)........................................................................ 30
Figura 6. Aspiração folicular de oócito suíno.................................................... 32
Figura 7. Fotomicrografias de oócitos aspirados de ovários (experimento 1)e examinados sob microscopia de fluorescência. A e B) Oócitosimaturos com GV (Hoechst 33342); C) Oócito imaturo ilustrandoCG (em "clusters") no citoplasma (Lens culinaris aglutininaconjugado a isotiocianato de fluorescesceína); D) Oócito imaturoiniciando migração dos CG. Aumento de 400 X................................ 36
Figura 8. Micrografias digitalizadas de estruturas embrionárias obtidos detubas uterinas de marrãs após 24 horas da monta natural. A)Embrião de 2 células; B) Embrião de 4 células; C) Embrião de 2células com a presença do 2º corpúsculo polar (como indica aseta). 80 X......................................................................................... 38
Figura 9. Estruturas encontradas após lavagem dos cornos uterinos. A e Bmórulas compactas (asterisco) e blastocistos (seta)......................... 39
Figura 10. Leitegada na maternidade................................................................. 40
Figura 11. Número de estruturas (oócitos e embriões) por animal e porexperimento (1, 2 e 3) com o modelo ajustado................................. 40

xiii
LISTA DE ABREVIATURAS
°C Graus celsius
ATP Adenosina trifosfato
BSA Albumina sérica bovina
CAMP AMP Cíclico
CCO’S Complexo-cumulus-oócito
CL Corpo Lúteo
DNA Ácido desoxirribonucléico
DNP Dias não produtivos
EGF Fator de crescimento epidérmico
FITC-LCA Lens culinaris aglutinina conjugado a isotiocianato de fluoresceína.
FIV Fertilização in vitro
FPP Peptídeo-promotor da fertilização
GC’s Grânulos corticais
GJC Comunicações via Gap Junctions
GSH Glutationa
GV Vesícula germinativa
GVBD Rompimento da vesícula germinativa
h Horas
IA Inseminação Artificial
Kg Kilo
LF Líquido folicular
LH Hormônio luteinizante
M Metro
MAP-quinase Mitogen-activated-protein-kinase
mI Microlitro
MII Metáfase II
Min minuto

xiv
MIV Maturação in vitro
Ml Metáfase I
Mm Milímetro
MN Monta Natural
MPF Maturation-promoting-factor
mTaLP Meio de tyrodes modificado
mTBM Meio tris-tamponado modificado
NaCl Cloreto de Sódio
NCSU 23 North Carolina State University 23 Medium
nm Nanômetro
O2 Oxigênio
PBS Solução Salina Fosfatada
PIV Produção in vitro
PKC Proteína quinase C
PNm Pronúcleo masculino
pOSP Glicoproteínas secretórias-específicas do oviduto suíno
PVS Espaço perivitelino
SFB Soro fetal bovino
SIPS Sistema intensivo de produção de suínos
TCM-199- Tissue culture medium
ZP Zona pelúcida

xv
AVALIAÇÃO QUANTI E QUALITATIVA DE OÓCITOS DE FÊMEASSUÍNAS NO ESTRO E DE ESTRUTURAS EMBRIONÁRIAS
24 E 144 HORAS APÓS MONTA NATURAL
RESUMO – Esta pesquisa teve por objetivo verificar o número e as características morfológicas de oócitos
e estruturas de fêmeas suínas no primeiro estro. Para tanto, as fêmeas (24) foram divididas em quatro grupos: no
primeiro logo após a detecção do primeiro cio foram sacrificadas e seus ovários transladados para o laboratório e
tiveram seus folículos aspirados (Experimento 1); no segundo, após a detecção do primeiro cio, foram submetidas à
monta natural (MN) e, após 24 horas, foram sacrificadas e seus sistemas genitais removidos e suas tubas uterinas
lavadas e as estruturas encontradas avaliadas quanto ao número e características (Experimento 2); no terceiro, após
detecção do primeiro estro e submetidas à MN, foram cobertas e, após 144 horas (6 dias), abatidas e seus genitais
removidos para análise das estruturas embrionárias encontradas (Experimento 3) e, finalmente, um quarto grupo foi
usado como referencial quanto ao número de nascimentos a partir de marrãs da granja nos anos de 2004 e 2005. Dos
animais do Experimento 1 foram colhidas 269 oócitos, tendo 134 sido corados com Hoechst 33342 (estádio nuclear)
e lecitina (Lens culinaris) para detecção dos grânulos corticais. Todos os oócitos do experimento 1 apresentaram-se
com núcleo sob a forma de vesícula germinativa e com os grânulos corticais sob a forma de “clusters”. No
Experimento 2, foram colhidas 56 estruturas (10 oócitos, e 46 embriões, sendo 41 de duas, 4 de três e 1 de quatro
células); no experimento 3 foram obtidas 15 estruturas classificadas como mórulas (5), mórulas compactas (4),
blastocistos iniciais (2), blastocistos propriamente ditos (1) e degenerados (3). No Experimento 4, de um total de 46
primíparas, resultaram 500 nascimentos, tendo 473 leitões sido desmamados. Os dados foram submetidos a um
ajuste e ao modelo teórico y = a + b log (t), onde t era o número de horas, y o número de oócitos e a e b o
parâmetros do modelo. O ajuste foi feito no software Statistica versão 7. Concluiu-se que os oócitos ovulados a partir
da aceitação da monta, passam por uma série de transformações de ordem estrutural e endócrina, antes de serem
fertilizados, resultando em embriões e prole viável.
Palavras-chave: Suíno, estro, ovulação, grânulos corticais, fertilização.

xvi
QUALI AND QUANTITATIVE EVALUATION OF SWINE OOCYTES DURING ESTRUS, 24 AND 144
HOURS EMBRYOS AFTER BREED
SUMMARY – This research has the objective to verify the number and morphological characteristics of
swine oocytes and embryos structures at the first estrus. For that, 24 females were equally divided in 4 groups:
(Experiment 1) after the estrus detection the females were sacrificed and their ovaries transported to the laboratory
for oocytes aspiration. (Experiment 2) After the first estrus detection the females were bred and after 24 hours they
were sacrificed, and their genitals were removed. Their uterine tubes washed to count and to evaluate the
characteristics of embryos. (Experiment 3) After the first estrus detection and breeding, the females were sacrificed
after 144 hours (6 days), their uterine tubes washed in order, to count and evaluate the characteristics of embryos;
(Experiment 4) referential group based on birth number in 2004 and 2005. From at the first experiment, 269 oocytes
were collected, which 134 were stained with Hoechst 33342 and lectine (Lens culinaris), to check nuclear
development and cortical granulus. All of the 134 structures presented as germinative vesicle with cortical granulus
as "clusters". At the second experiment, 56 structures were collected, 10 oocytes and 46 embryos (41 with two cells,
4 with three cells and 01 embryo with four cells); in the third experiment 15 structures were obtained, five morulae,
four compacted morulae, two initial blastocysts, one blastocyt, and one degenerated. In the fourth experiment, we
considered 46 primiparas, 500 birth and 473 weaned pigs. The data were adjusted by Statistica v.7 and submitted to a
theory model y = a + b log (t), where t was the number of hours, y the number of oocytes, a and b model parameters.
We concluded that oocytes ovulated after breeder pass to some structural and endocrine transformations, before their
fertilization, resulting in viable embryos and offspring.
Key words: Swine, estrus, ovulation, cortical granulus, fertilization.

1. INTRODUÇÃO
Nos últimos 20 anos foram conduzidos numerosos estudos sobre a produção
in vitro (PIV) de animais de interesse zootécnico, impulsionados tanto pela pesquisa
fundamental, como instrumento para estudar fenômenos fisiológicos relacionados aos
gametas masculino e feminino e os fatores relacionados à embriogênese, como pelo
interesse em se implementar a produção animal através de uma melhoria na eficiência
reprodutiva. Adicionalmente, a PIV tem respaldado o desenvolvimento de inúmeras
biotécnicas como clonagem por transferência nuclear, sexagem de espermatozóides e
embriões e das técnicas de preservação de oócitos e embriões (WILMUT et al., 1997).
A espécie suína dentro deste contexto vem despertando um lugar de destaque
devido à sua fisiologia ser bastante similar com a humana, tornando o suíno um doador
em potencial de tecidos e órgãos para xenotransplantes, assim como para a produção
de proteínas específicas (OKTAY et al., 2000).
Ao mesmo tempo, os sistemas de criação suína para a produção de carne
continuam a sofrer grandes transformações, tornando-os bastante receptivos à
implementação de novas tecnologias reprodutivas como forma de reduzir os custos de
produção, bem como aumentar a eficiência produtiva (NUNES, 2001).
Portanto, a eficiência da reprodução animal é registro fundamental para a
continuidade da espécie, bem como para o aproveitamento industrial (BONNEAU,
1998). O suinocultor objetiva produzir o maior número de leitões terminados por porca
por ano e para isso é necessário garantir a performance reprodutiva do plantel. Assim,
a reprodução é considerada, como um dos fatores mais importantes associados à
rentabilidade e sustentabilidade da atividade suinícula (HOLANDA et al., 2000).
O objetivo deste trabalho foi verificar o número e as características morfológicas
de oócitos obtidos de fêmeas suínas no primeiro estro, bem como dos embriões às 24 e
144 horas (6 dias) após monta natural, relacionando-os com o número de animais
nascidos numa granja suinícola.

2
2. REVISÃO DE LITERATURA
2.1. Considerações gerais
A suinocultura nos dias atuais tem merecido bastante atenção por parte de
produtores como uma forma alternativa de produção que possibilita a obtenção de
retornos financeiros num espaço de tempo menor, em relação a outras atividades rurais
(HOLANDA et al., 2000).
Segundo NUNES (2001), a crescente industrialização da suinocultura abre novas
oportunidades para agregar maior valor aos produtos derivados, gerando novos
empregos e uma maior atividade econômica. O mesmo autor acrescenta ainda, que no
Brasil, o consumo de produtos industrializados representa cerca de 70% da produção
de suínos do país.
A importância da suinocultura, no contexto nacional, reside não só no grande
contingente de produtores envolvidos, como também no volume de empregos diretos e
indiretos gerados (2,5 milhões nos Estados do Sul, São Paulo e Minas Gerais) e pela
capacidade de produzir grande quantidade de proteína de alta qualidade em reduzido
espaço físico e curto espaço de tempo, quando comparada a outras espécies animais
de médio e grande porte (GOMES et al., 1992).
Segundo GOMES et al. (1992), cerca de 80% dos suínos são produzidos em
áreas de até 100 hectares, constituindo-se em uma das mais importantes fontes de
receita para 46,5% das 5,8 milhões de propriedades no país, empregando mão-de-obra
tipicamente familiar e por isso constitui, também, um importante fator de estabilidade
social.
A suinocultura brasileira caracteriza-se por apresentar uma grande variabilidade
no que diz respeito ao tipo de criadores, propriedades, produtividade e tecnologia
empregada. As regiões sul e sudeste se destacam em importância econômica nesta
atividade. Porém, na região sul predominam propriedades e criações pequenas. Já na
região sudeste, as unidades são de maior porte, com características empresariais. Na
área econômica são freqüentes as queixas dos produtores quanto à instabilidade
econômica para manter uma atividade suinícola (TALAMINI, 1996).
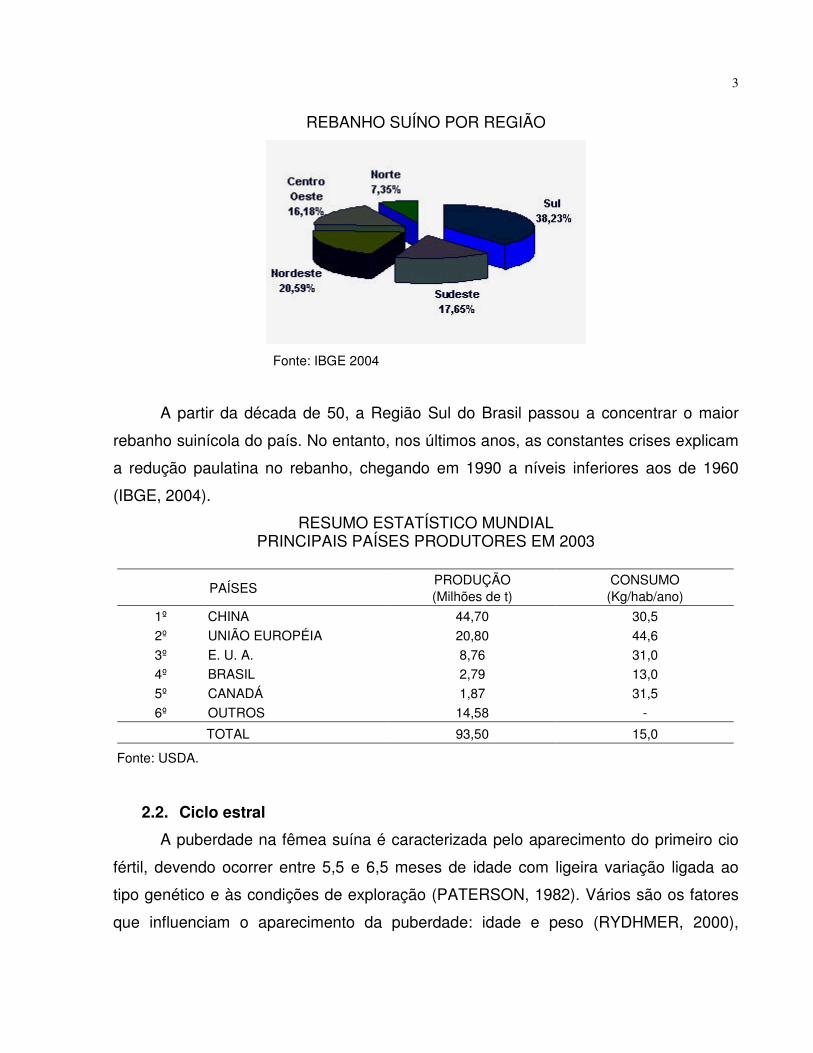
3
REBANHO SUÍNO POR REGIÃO
Fonte: IBGE 2004
A partir da década de 50, a Região Sul do Brasil passou a concentrar o maior
rebanho suinícola do país. No entanto, nos últimos anos, as constantes crises explicam
a redução paulatina no rebanho, chegando em 1990 a níveis inferiores aos de 1960
(IBGE, 2004).
RESUMO ESTATÍSTICO MUNDIALPRINCIPAIS PAÍSES PRODUTORES EM 2003
PAÍSESPRODUÇÃO(Milhões de t)
CONSUMO(Kg/hab/ano)
1º CHINA 44,70 30,52º UNIÃO EUROPÉIA 20,80 44,63º E. U. A. 8,76 31,04º BRASIL 2,79 13,05º CANADÁ 1,87 31,56º OUTROS 14,58 -
TOTAL 93,50 15,0
Fonte: USDA.
2.2. Ciclo estral
A puberdade na fêmea suína é caracterizada pelo aparecimento do primeiro cio
fértil, devendo ocorrer entre 5,5 e 6,5 meses de idade com ligeira variação ligada ao
tipo genético e às condições de exploração (PATERSON, 1982). Vários são os fatores
que influenciam o aparecimento da puberdade: idade e peso (RYDHMER, 2000),

4
genótipo (HASLEY e LEE, 1993), nutrição (BELTRANENA et al., 1991), alojamento,
transporte, sazonalidade (LOVE et al., 1993) e efeito do macho (BROOKS, 1999).
Um fator relevante é o nutricional, uma vez que a restrição alimentar pode
retardar o aparecimento da puberdade em mais de uma semana, comparativamente à
alimentação oferecida ad libitum (BELTRANENA et al., 1991). No entanto, MUNIZ et al.
(2002) reduziram a alimentação em 20%, não encontrando diferenças quanto à idade e
puberdade.
Os efeitos de longo prazo no manejo nutricional durante o período de
crescimento da leitoa e sua influência na produtividade futura ainda não foram
claramente definidos. No estudo de BELTRANENA et al. (1991) a idade mínima para a
puberdade ficou ao redor dos 160 dias e pode ser alcançada com uma taxa média de
crescimento de cerca de 600 g diárias, desde o nascimento até a puberdade,
encontrando-se em taxas mais elevadas de crescimento um efeito quadrático, explicado
pelo retardamento da idade à puberdade. Os animais melhorados geneticamente e com
alimentação ad libitum, portanto, terão taxas de crescimento mais elevadas. A
conseqüência da alimentação deste tipo de manejo alimentar seria a de provocar um
descompasso entre desenvolvimento genital e corporal, apesar das leitoas mostrarem-
se mais pesadas no momento da cobertura. Assim, é possível que nos animais com
rápido crescimento corporal, a maturação do eixo reprodutivo não acompanhe a
velocidade de crescimento e/ou exista a necessidade de os animais atingirem um limiar
de porcentagem de gordura corporal para que os eventos fisiológicos indutores da
puberdade sejam deflagrados (BELTRANENA et al., 1991).
Condições climáticas relacionadas à época do ano, também de semelhante
importância podem influenciar o desenvolvimento sexual de leitoas, pois o percentual
de leitoas que atingem a puberdade no verão é menor que no inverno. O fotoperíodo,
que influencia muitos aspectos ligados à reprodução suína, também afeta a idade à
puberdade (LOVE et al., 1993). Os mesmos autores relataram uma diminuição da idade
à puberdade após a exposição de leitoas a dias de luz crescente.
O manejo com o macho adulto (acima de 11 meses de idade) junto a fêmeas
imaturas estimula, sabidamente, à puberdade. O efeito da presença do macho ocorre

5
devido a uma ação sinérgica de vários componentes estimulatórios do cachaço. Essa
ação não estaria restrita apenas aos estímulos olfatórios de feromônios, mas também a
outros odores corporais, estímulos táteis e possivelmente visuais e auditivos (BROOKS,
1999). O autor sugeriu que o mecanismo da atuação dos estímulos do macho deveria
ocorrer por intermédio do aumento dos níveis de LH na leitoa, com o desenvolvimento
folicular e aumento dos níveis plasmáticos de estrógeno.
A idade da primeira cobertura da leitoa pode ter importantes implicações na
eficiência total de sua vida reprodutiva. Economicamente, justifica-se a cobrição ou a IA
o mais cedo possível, uma vez que reduz o seu período improdutivo no rebanho.
Entretanto, o número médio de ovulações aumenta do primeiro ao terceiro cio, de tal
forma que, realizando a primeira cobrição, por ocasião do terceiro cio, melhora-se a
prolificidade da fêmea. Isso parece ser mais evidente no caso de leitoas submetidas à
restrição alimentar se comparado com leitoas com ração ad libitum (ROZEBOOM et al.,
1996). FOXCROFT & AHERNE (2000) ressaltaram a importância de se atingir a
puberdade o mais precocemente, por volta dos 120 a 140 dias de idade, pois a
estimulação precoce da marrã daria a oportunidade a esta de experimentar maior
número de ciclos estrais antes da cobrição, melhorando o tamanho da primeira
leitegada.
2.3. Puberdade
A puberdade representa o momento em que as leitoas estão aptas para a
atividade reprodutiva, sendo também chamada de maturidade sexual. Essa
competência reprodutiva é evidenciada através dos sintomas do primeiro estro ou cio,
resultando de um processo interno de maturação dos órgãos reprodutivos e de
funcionamento de todo um sistema hormonal (eixo hipotalâmico-hipofisário).
(DETTWEILER, 1924)
Diversos fatores podem influenciar a idade na qual as leitoas apresentam sua
puberdade. Entre outros, podemos citar a genética, a idade, o peso, a reserva de
gordura corporal, a temperatura, o fotoperíodo e o programa nutricional. (HUGHES,
1982).

6
Dentro de um ótimo manejo reprodutivo de leitoas, o primeiro ponto a ser
seguido é a adoção de práticas que possibilitem a antecipação da idade à puberdade, o
que normalmente não é observado em condições de campo. (LAMBERSON et al.,
1991). Estes autores indicaram os seguintes procedimentos para se atingir tal objetivo:
a) identificar mais prontamente o histórico de ciclo estral de cada leitoa, facilitando os
manejos de sincronização do estro das mesmas; b) permitir que fêmeas tardias e/ou
que estarão em anestro sejam mais rapidamente descartadas do plantel, reduzindo o
desperdício e os DNP (dias não produtivos); c) antecipar o primeiro estro, aumentarmos
o percentual de leitoas cobertas já ao terceiro estro (ou mais) sem para isso estarem
em idade excessiva, permitindo maior desenvolvimento do trato genital (Tabela 1) e
chances de melhores desempenhos reprodutivos; d) identificar precocemente as leitoas
que apresentam cios muito curtos (<24 hs) e associarmos à ficha das mesmas a
necessidade de atenção e diferenciação no manejo de inseminação artificial; e) evitar
que muitas leitoas atinjam o peso de mercado (100 Kg) antes de chegarem à
puberdade. Com a evolução genética, é comum trabalharmos com fêmeas que
cheguem a esse peso antes de atingirem a puberdade, principalmente se esta não for
bem estimulada ou induzida.
Tabela 1. Efeito do ciclo sexual sobre o desenvolvimento do trato genital.
Leitoa Pré-Pubere 1º Cio 2º Cio 3º CioIdade (dias) 150+/7,2 196+/-8,1 223+/-7,9 249+/-11,6Nº de fêmeas 20 20 20 20Tamanho do útero (cm) 38,1+/-5,9 54,1+/-3,5 61+/-4,3 75+/-6,8
Fonte: Rillo, S.M (1999).
2.4. Exposição ao macho
A exposição ao macho representa a forma mais comum e efetiva de indução à
puberdade na espécie suína. Em algumas situações práticas, observa-se uma falha na
indução da puberdade através do contato com o cachaço. Nesses casos, o que
7
normalmente ocorre é a falha em alguma das etapas do processo de estímulo.
(BORTOLOZZO, 1999).
Sabe-se que o chamado “efeito macho” é reflexo da junção de componentes
olfativos, táteis, visuais e auditivos. O componente olfativo é comprovadamente o efeito
primário, sem o qual os demais componentes não agem de forma eficaz na estimulação
da fêmea ao estro. O mecanismo pelo qual o estímulo olfativo interfere no “status”
endócrino da fêmea passa pelos ferormônios encontrados, principalmente, na saliva
dos machos, embora uma quantidade menor também seja encontrada na urina. Os dois
principais ferormônios reconhecidos, a 5α-androsterona e o 3α-androsterol, são
produzidos principalmente pelas glândulas salivares sub-maxilares em resposta à
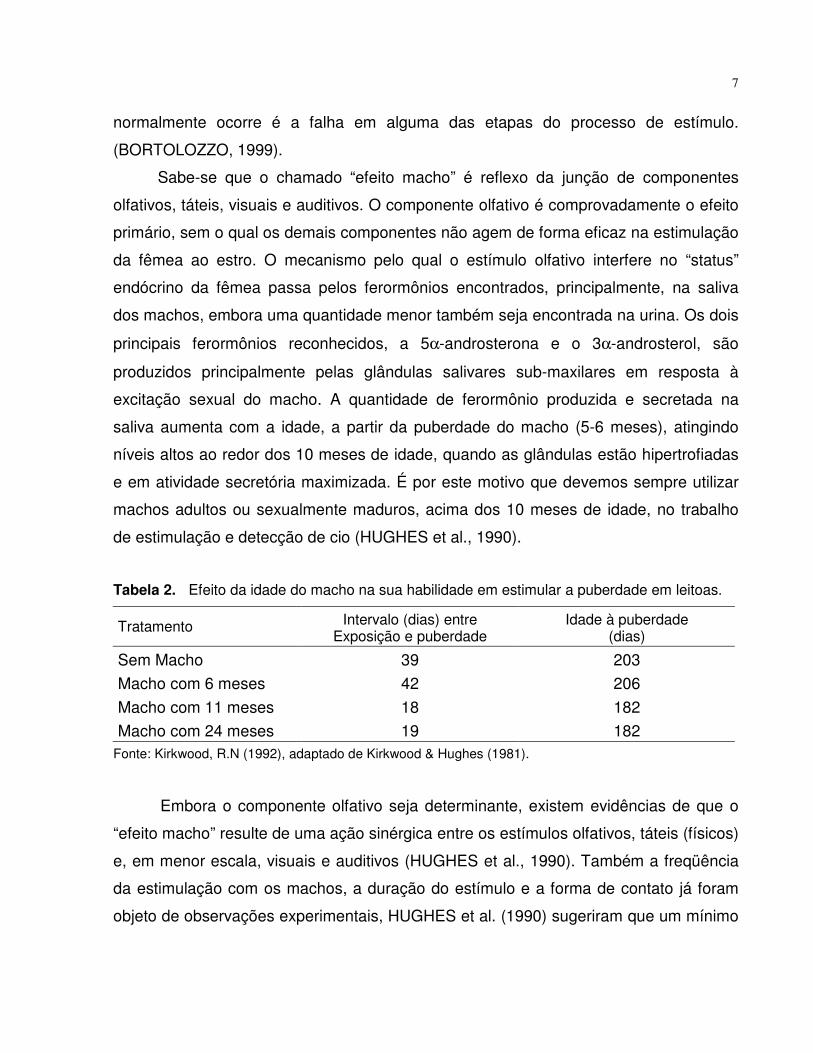
excitação sexual do macho. A quantidade de ferormônio produzida e secretada na
saliva aumenta com a idade, a partir da puberdade do macho (5-6 meses), atingindo
níveis altos ao redor dos 10 meses de idade, quando as glândulas estão hipertrofiadas
e em atividade secretória maximizada. É por este motivo que devemos sempre utilizar
machos adultos ou sexualmente maduros, acima dos 10 meses de idade, no trabalho
de estimulação e detecção de cio (HUGHES et al., 1990).
Tabela 2. Efeito da idade do macho na sua habilidade em estimular a puberdade em leitoas.
Tratamento Intervalo (dias) entreExposição e puberdade
Idade à puberdade(dias)
Sem Macho 39 203Macho com 6 meses 42 206Macho com 11 meses 18 182Macho com 24 meses 19 182
Fonte: Kirkwood, R.N (1992), adaptado de Kirkwood & Hughes (1981).
Embora o componente olfativo seja determinante, existem evidências de que o
“efeito macho” resulte de uma ação sinérgica entre os estímulos olfativos, táteis (físicos)
e, em menor escala, visuais e auditivos (HUGHES et al., 1990). Também a freqüência
da estimulação com os machos, a duração do estímulo e a forma de contato já foram
objeto de observações experimentais, HUGHES et al. (1990) sugeriram que um mínimo

8
de 15 minutos diários de contato direto seria recomendável para se obter os efeitos
desejados. Ainda os mesmos autores comprovaram a diferença na resposta das leitoas
quando se comparava o contato direto com um contato naso - nasal através de uma
divisória.
Uma melhor resposta observada pelo contato direto fica evidenciada pela
importância do contato físico (tátil), provavelmente ligado à liberação intensa de cortisol
após esse contato, de forma semelhante ao que ocorre em situações de estresse de
transporte e reagrupamento. Essa liberação de cortisol pode estar ligada a uma maior
secreção de LH e, conseqüentemente, contribuição com o processo de ciclicidade
ovariana (PEARCE et al., 1988, citado por HUGHES et al., 1990).
2.5. Técnicas para a colheita de oócitos em animais de abatedouro
A obtenção dos oócitos é etapa fundamental para a realização da biotécnica de
PIV em animais domésticos e estes podem ser colhidos in vitro de ovários obtidos em
abatedouros que têm sido a fonte de oócitos, de menor custo e mais abundante para a
produção de embriões em larga escala (MARTINO et al., 1994; WANI et al., 2002), por
ovariectomia, ou colhidos in vivo por aspiração de folículos utilizando-se métodos de
laparotomia, laparoscopia ou via transvaginal guiada por ultra-som (ARMSTRONG et
al., 1997).
Os ovários são transportados ao laboratório em solução salina a 0,9% (NaCl) ou
PBS (solução salina fosfatada) suplementado com antibióticos. A temperatura e o
tempo entre a coleta dos ovários e o início da aspiração folicular podem influenciar a
qualidade dos oócitos. Por isso, a manutenção da viabilidade dos oócitos durante esse
período é considerada um ponto crítico para que se alcance o sucesso na maturação
dos oócitos (ABEYDEERA, 2002). Oócitos e embriões suínos possuem uma maior
sensibilidade a baixas temperaturas quando comparados a outras espécies de
mamíferos (POLLARD et al., 1994). Esta sensibilidade está relacionada com o fato dos
embriões suínos possuírem um maior conteúdo lipídico que as demais espécies
(NAGASHIMA et al., 1994). WILMUT (1972) já havia demonstrado que embriões
suínos, desde a clivagem até o estágio de blastocisto, são bastante sensíveis a

9
temperaturas inferiores a 15°C. WALTERS & GRAVES (1998) examinaram o efeito de
várias temperaturas de estocagem de ovários (25, 30 e 37°C), por diferentes tempos (2,
6, 10, 14 e 26h) sobre a maturação dos oócitos provenientes destes ovários. Em todas
as temperaturas testadas, a taxa de maturação foi significativamente baixa quando o
intervalo de estocagem foi prolongado. Mais de 75% dos oócitos completaram a
maturação quando estocados por até 5 horas a 25°C. A maturação dos oócitos foi
comprometida quando os ovários foram mantidos por mais de 5 horas em temperaturas
superiores ou inferiores a 25°C. Portanto, é importante que o período entre a colheita
dos ovários e o início da maturação dos oócitos não seja superior a 5h para evitar que
os oócitos percam a habilidade de maturação e conseqüente potencial para fertilização
(ABEYDEERA, 2002).
2.6. Tipos de folículos ovarianos
A qualidade do oócito recuperado é determinada, entre outras características
pela presença ou ausência de células do cumulus. O papel destas células no
desenvolvimento completo da competência dos oócitos tem sido investigado
(LEIBFRIED & FIRST, 1979). Os estudos mostram que não há maturação ou esta
ocorre em menor escala, quando as células do cumulus são removidas antes dos
oócitos serem maturados in vitro (GONÇALVES et al., 2002).
Ovários suínos contêm folículos de diferentes diâmetros, sendo classificados de
acordo com o seu tamanho, em pequenos (<3 mm), médios (3 a 6 mm), e grandes (>6
mm). Os oócitos utilizados para rotinas de MIV são normalmente recuperados de
folículos de tamanho médio (ABEYDEERA, 2002).
Em suínos, o antrum folicular é totalmente diferenciado em folículos de 0.5mm,
com o oócito correspondente chegando a apenas três-quartos (¾) de seu tamanho final
(MOTLIK et al., 1984). Estes oócitos têm uma capacidade muito limitada de iniciar a
maturação nuclear (MOTLIK et al., 1986).
Muitos oócitos destes folículos iniciam a maturação meiótica, porém, não
ultrapassam o estádio de metáfase I (MI). A habilidade dos oócitos em continuar e
completar a meiose não é atingida de uma só vez, em um determinado ponto do seu

10
desenvolvimento, mas é adquirida de maneira gradativa. Dos oócitos competentes,
presentes nos folículos antrais, apenas aqueles que tiverem alcançado um adequado
estágio de desenvolvimento irão responder ao estímulo para maturação, e iniciar as
mudanças morfológicas e funcionais, incluindo o reinício da meiose (ABEYDEERA,
2002).
YOON et al. (2000) relataram que um maior número de oócitos de grandes
folículos (3 a 8 mm) alcançou o estágio de MII (91% vs 58%), formaram o pronúcleo (90
vs 81%) e desenvolveram-se até o estádio de blastocisto (10% vs 2%), comparados
com aqueles provenientes de folículos pequenos (<3 mm). Estes resultados sugeriram
que o completo potencial para completar a meiose e a subseqüente competência para
continuar o seu desenvolvimento é adquirido com o crescimento folicular. Entretanto, a
heterogenicidade dos oócitos isolados de folículos de diferentes tamanhos, podem
também influenciar na progressão da meiose, na maturação citoplasmática e na
competência para suportar desenvolvimento.
Figura 1. Aspecto visual de ovários de marrãs colhidos em matadouro. (A) ovário de fêmeapré-púbere (somente folículos); (B) ovários de fêmeas púberes (presença de CLs).
2.7. Aspectos moleculares da maturação dos oócitos
A maturação de oócitos compreende dois aspectos: a maturação nuclear e a
maturação citoplasmática. Geralmente um oócito é reconhecido como maturado quando
ocorre a extrusão do primeiro corpúsculo polar (maturação nuclear), e o oócito alcança
o estágio de MII da meiose. Oócitos que completaram sua maturação nuclear podem
ser fertilizados, contudo, o potencial de seu desenvolvimento pode ser incompleto, em

11
razão de serem deficientes em certos fatores citoplasmáticos. Portanto, o completo
potencial de desenvolvimento, requer uma síncrona maturação nuclear e citoplasmática
(SUN et al., 2003).
Maturação nuclear (in vitro)
Como foi descrito anteriormente, oócitos suínos em crescimento não são
capazes de completar a meiose in vitro. Esta incapacidade dos oócitos em crescimento
para completarem a meiose não é resultado da falta do fator intracelular promotor da
metáfase, MPF (maturation promoting factor), e/ou da MAP quinase (mitogen-activated-
protein-kinase), dois importantes sinalizadores moleculares que controlam a meiose
(SUN et al., 2003). A quantidade das subunidades do MPF, p34cdc2
(proteína de 34kD) e
ciclina B (proteína de 45kD), nestes oócitos em crescimento, são comparável aos níveis
encontrados em oócitos que completaram seu crescimento, entretanto, os oócitos em
crescimento possuem a p24, duas isoformas da MAP-quinase, que oócitos com
crescimento completo, porém, a MAP-quinase não é ativada durante o cultivo destes
oócitos (SUN et al., 2001b). KANAYAMA et al. (2002), concluíram que a competência
para os oócitos completar a meiose com sucesso, está relacionada com a sua
habilidade em ativar a MPF e a MAP-quinase.
Maturação citoplasmática (in vitro)
Muitos oócitos maturados in vitro parecem ser deficientes em alguns fatores
citoplasmáticos, pois, estes não possuem competência plena para se desenvolver.
Embora fatores citoplasmáticos não tenham sido plenamente estudados o movimento
incompleto das mitocôndrias para o interior do citoplasma pode, portanto, afetar a
maturação citoplasmática (SUN et al., 2001d). Além disso, a adição de cisteína,
cisteamina, glutamina, gonadotropinas, fator de crescimento epidérmico (EGF), ß-
mercaptoethanol, líquido folicular, ou a presença de células do cumulus podem auxiliar
a maturação citoplasmática dos oócitos tornando-os mais aptos para continuar o
desenvolvimento (HUNTER, 1991; DING & FOXCROFT, 1994; GRUPEN et al., 1995;

12
WANG & NIWA, 1995; SINGH et al., 1997; ABEYDEERA et al., 1998; BING, et al.,
2001; JEONG & YANG, 2001).
A maturação in vitro de oócitos suínos pode ser realizada em vários tipos de
meios de cultura, contendo soro fetal bovino (SFB) ou líquido folicular (LF),
gonadotrofinas e fatores de crescimento (COSKUN et al., 1994; DING et al., 1994;
PROCHAZKA et al., 2000; MCCAULEY et al., 2001). Entretanto, a suplementação com
SFB ou LF introduz vários fatores desconhecidos e dificulta a identificação dos fatores
chave na regulação da maturação normal dos oócitos (BECKMANN et al., 1993).
O desenvolvimento de um meio livre do soro irá auxiliar a identificação das
condições ótimas de cultura, e também irá reduzir a variabilidade entre os resultados
obtidos em diferentes laboratórios. Em geral, os oócitos selecionados para rotinas de
MIV (maturação in vitro) contêm mais de três camadas de células do cumulus (células
que envolvem os oócitos). Durante o processo de MIV, as células do cumulus podem
apresentar variáveis graus de expansão, e podem estar funcionalmente relacionadas à
maturação nuclear ou citoplasmática do oócito. O grau de expansão das células do
cumulus pode servir como indicador de sucesso na maturação nuclear ou
citoplasmática. Na maioria dos mamíferos, no momento da ovulação, os oócitos estão
inclusos em uma massa expandida de células do cumulus, rica em mucina
(ABEYDEERA, 2002).
A presença de células do cumulus durante a maturação influencia
significativamente a maturação meiótica até o estágio de metáfase II (MII), o nível de
GSH (glutadiona) intracelular, a taxa de penetração, a formação do pronúcleo
masculino, e a atividade da Histona H1 quinase (YAMAUCHI et al., 1999). Além disso,
as células do cumulus estabilizam a distribuição dos grânulos corticais (GC’s), evitando
uma migração prematura e exocitose parcial dos mesmos. Deve-se levar em conta que
a completa maturação nuclear não resulta necessariamente em uma maturação
citoplasmática normal. A presença das células do cumulus parece desempenhar um
importante papel na estimulação deste processo. (ABEYDEERA, 2002).
Embora diversos meios de MIV tenham resultado em uma taxa satisfatória de
maturação até o estágio de MII, SALUSTRI et al. (1989) estudaram eventos nucleares

13
relacionados à maturação, tais como o rompimento da vesícula germinativa e a
extrusão do primeiro corpúsculo polar parecerem ter ocorrido normalmente, o
citoplasma do oócito geralmente não promovia a formação do pronúcleo masculino
após a penetração do esperma. Concluiu-se então que, a maturação citoplasmática era
inadequada em certas condições da MIV. Esta falha talvez esteja relacionada com
deficiências inerentes aos próprios oócitos, ou por condições sub-ótimas dos meios de
cultura. Em geral, a aparência do oócito antes da maturação e do estágio de meiose
após a maturação in vitro era usada para predizer a qualidade do oócito. Contudo, está
ficando bastante claro que estas medidas são amplamente inadequadas (ABEYDEERA,
2002).
2.8. Fertilização in vitro (FIV)
A penetração de oócitos maturados in vitro tem sido realizada com sucesso em
diversos meios de fertilização utilizando sêmen resfriado ou congelado. Devido às
dificuldades em se encontrar uma técnica eficiente de criopreservação, o sêmen
resfriado continua sendo a fonte mais comum de espermatozóides nas rotinas de FIV
(ABEYDEERA, 2002). Além disso, grandes variações têm sido observadas entre
reprodutores suínos, bem como entre frações do mesmo ejaculado, no que diz respeito
a penetração nos oócitos e a taxa de polispermia (XU et al., 1996).
Os dois principais entraves na produção de embriões suínos através das
técnicas de MIV – FIV são a pequena taxa de pequena formação do pronúcleo
masculino e alta taxa de polispermia (ABEYDEERA, 2001).
Vários melhoramentos aplicados a MIV e técnicas de cultivo, como adição de
líquido folicular, co-cultura junto a células somática limitar a exposição à gonadotrofinas
e suplementação com EGF (fator de crescimento epidérmico) ou cisteína, trouxeram
significativos avanços em termos de maturação citoplasmática, evidenciada por uma
maior formação de pronúcleos masculinos após a penetração dos espermatozóides
(ABEYDEERA, 1997; DAY et al., 2000). O aumento na formação de pronúcleos
masculinos induzida pela suplementação de cisteína é correlacionada com altas
concentrações de GSH intracelular em oócitos maturados (YOSHIDA et al., 1993).

14
Um efeito semelhante nos níveis intracelulares de GSH também foi observado
quando foi procedida a co-cultura junto a células somáticas (ABEYDEERA et al., 1998).
A síntese de GSH durante a maturação dos oócitos é conhecida como pré-
requisito para a descondensação da cromatina nos espermatozóides dos hamsters e
para a boa formação do pronúcleo masculino (PERREAULT et al., 1998). Também foi
constatado que a formação de prónucleos masculinos em oócitos com baixos índices
de GSH intracelular é comprometida. A habilidade do citoplasma em transformar um
espermatozóide que tenha penetrado em um pronúcleo é mais eficiente em oócitos que
tenham sidos maturados e fertilizados na presença de células do cumulus (MATTIOLI et
al., 1998; KIKUCHI et al., 1993; KA et al., 1997). O nível intracelular de GSH parece ser
dependente da presença de células do cumulus durante a cultura (FUNAHASHI et al.,
1995).
Uma das rotas utilizadas pelas células do cumulus para transportar fatores para
dentro dos oócitos é através das GJC (comunicações via Gap Junctions) (COSKUN et
al., 1994). MORI et al. (2000), sugeriram que as GJC representam um importante papel
na regulação do transporte do GSH produzido das células do cumulus para dentro dos
oócitos, e está correlacionado com a formação dos pró-núcleos masculinos. Esta
evidência reforça a importância do papel desempenhado pelas células do cumulus na
maturação citoplasmática.
2.9. Aspectos moleculares relacionados à ativação do oocitária
Durante a FIV em suínos, a penetração espermática inicia 3h após a
inseminação. A rápida penetração dos espermatozóides induz o reinício da meiose e a
reação cortical que determinará o bloqueio fisiológico da polispermia, conforme pode
ser visto na Figura 2.

15
Figura 2. Reação que ocorre quando um espermatozóide fertiliza um óvulo em mamíferos(Adaptado de Alberts et al. - Molecular biology of the cell. 4th ed. 2004).
Após 5h pode-se observar descondensação da cromatina dos espermatozóides
e a metade dos óvulos em anáfase II. Passadas 8h, os pronúcleos masculino e
feminino estarão formados (DING et al., 1992).
2.10.Descondensação da cromatina do espermatozóide
A fertilização é o primeiro passo para o desenvolvimento embrionário. Antes da
fertilização, o DNA no núcleo do espermatozóide está altamente condensado e possui
proteínas básicas de ligação ao DNA, denominadas proteaminas, ou proteínas
específicas do núcleo do espermatozóide. Para que seja possível completar a
fertilização, quando a cabeça do espermatozóide penetra o oócito, iniciam-se uma série
de mudanças citológicas e bioquímicas (NAKAZAWA et al., 2002).
No momento da fertilização, a cromatina do espermatozóide que se encontra
altamente condensada e incapaz de realizar a transcrição, inicia uma remodelação em

16
uma cromatina descondensada e competente para realizar a transcrição. Inicialmente, a
cromatina torna-se dispersa e então volta a se condensar em uma pequena massa logo
após adentrar o ooplasma. Esta mudança morfológica coincide e parece ser
dependente da transição das proteaminas específicas dos espermatozóides em
histonas e da organização da cromatina em nucleossomos. Então, a cromatina volta a
se descondensar resultando na formação do pronúcleo masculino, e então, adquire
muitas das proteínas que estão associadas a cromatina materna (McLAY & CLARKE,
2003.).
SHIMADA et al. (2000) demonstraram que a associação das proteaminas ocorre
em oócitos suínos fertilizados logo após a entrada do espermatozóide, porém, antes da
descondensação do núcleo do espermatozóide. Esta observação indica que a
remodelação das proteaminas em histonas inicia-se logo após a penetração do
espermatozóide e se completa durante a descondensação da cromatina do
espermatozóide, antes da síntese do DNA em pronúcleo masculino.
Estudos comprovaram que a presença de células foliculares no meio de
maturação aumenta a formação do pronúcleo masculino (PNm) (NAGAI et al., 1993).
Da mesma forma, a presença de células do cumulus durante a fertilização aumenta a
taxa de formação de pronúcleos se comparado aos oócitos em que as células do
cumulus foram removidas (KA et al., 1997).
Outros estudos indicam que o aumento do conteúdo intracelular de GSH
promove a formação de PNm logo após a fertilização (YOSHIDA et al., 1993; KA et al.,
1997; NAGAI, 2001). Entretanto, o exato mecanismo através do qual células somáticas
afetam a formação do PNm é desconhecido.
Portanto, é bastante possível que o GSH medie a descondensação do
acrossoma do espermatozóide e, portanto, incremente a formação do PNm, assim
como, em outras espécies (SUN et al., 2001a).
2.11.Polispermia
Entende-se como polispermia a penetração dos oócitos por mais de um
espermatozóide.

17
Apesar dos significativos avanços obtidos na definição de melhores métodos
para a fecundação in vitro nos suínos, uma alta taxa de polispermia ainda persiste. Sob
condições in vivo a fertilização ocorre dentro de poucas horas após a ovulação, e na
maioria das vezes, ocorre uma penetração monoespérmica (ABEYDEERA, 2002).
Em comparação com o que ocorre em condições in vivo, oócitos maturados in
vitro são expostos a um excessivo número de espermatozóides por um período mais
longo de tempo. Estas condições podem predispor os oócitos à penetração por mais de
um espermatozóide, caracterizando a polispermia (ABEYDEERA, 2002). O ideal de um
sistema de FIV (fecundação in vitro) seria proporcionar uma alta taxa de penetração
(>80%), com uma baixa incidência de polispermia (<10%). Entretanto, uma alta
correlação foi relatada entre o número absoluto de espermatozóides por oócito na
fertilização e o grau de polispermia (RATH, 1992). Teoricamente, o problema da
polispermia poderia ser superado através da redução do número de espermatozóides
nas gotas de FIV. Entretanto, na maioria dos casos, estes ajustes levariam uma baixa
taxa de penetração dos espermatozóides nos oócitos (ABEYDEERA, 2002).
Em mamíferos a penetração do espermatozóide provoca a ativação dos oócitos
e a subseqüente exocitose dos GC’s (grânulos corticais). Os GC’s entram em exocitose
em resposta a elevação intracelular de cálcio provocada pela penetração dos
espermatozóides (HOODBHOY et al., 1994). O conteúdo liberado pelos GC’s no
espaço perivitelino (PVS) são responsáveis pelas modificações na ZP (zona pelúcida) e
por estabelecer o bloqueio da polispermia. Durante a maturação, ocorre uma extensa
redistribuição das organelas intracelulares (mitocôndrias e grânulos corticais). Portanto,
uma melhor distribuição destas organelas durante a maturação in vitro pode
desempenhar um importante papel na prevenção da polispermia (CRAN et al., 1986).
Entretanto, a observação de oócitos maturados in vitro, no microscópio confocal
6 horas após FIV revelou que não há diferenças na liberação dos GC’s, quando
comparados com oócitos ovulados, porém, a incidência de polispermia foi maior nos
oócitos maturados in vitro (WANG et al., 1998). É possível que esta diferença ocorra em
razão do atraso na exocitose dos GC’s nos momentos seguintes a penetração dos
espermatozóides (ABEYDEERA et al., 1997, WANG et al., 1997a). Paralelamente, apesar

18
dos GC’s serem liberados imediatamente após a penetração dos espermatozóides, o
menor espaço perivitelínico (PVS) em oócitos maturados in vitro pode interferir com
uma adequada distribuição do conteúdo dos GC’s, e pode atrasar o bloqueio da zona.
Em ambas as situações, espermatozóides acessórios podem conseguir entrar antes do
estabelecimento do bloqueio da zona (ABEYDEERA et al., 1997).
Oócitos suínos maturados no meio de WHITTEN, contendo baixos níveis de
NaCl (cloreto de sódio), resultam em oócitos com um PVS mais amplo, propiciando uma
diminuição da polispermia (FUNAHASHI et al., 1994). Portanto, o desenvolvimento de
um meio de FIV que resulte em oócitos maturados com um PVS mais amplo, poderia
pelo menos em parte, resolver o problema da polispermia (ABEYDEERA, 2002).
Como mencionado anteriormente, oócitos ovulados são significativamente
menos polispérmicos que aqueles maturados e fertilizados in vitro (WANG et al., 1998).
Segundo ABEYDEERA (2002), quando comparados com os maturados in vivo, oócitos
maturados in vitro são similares àqueles presentes nos folículos pré-ovulatórios.
DAY et al. (2000), compararam os parâmetros morfológicos, físicos e referentes
à fertilização em óocitos maturados in vitro, ovulados e pré-ovulatórios. Em contraste
com os oócitos que atingiram a ovulação, os óocitos maturados in vitro e os pré-
ovulatórios foram similares na maioria dos parâmetros analisados. Ficou claro que,
após a ovulação, importantes mudanças ocorrem nos oócitos durante a passagem pelo
oviduto que são importantes para a prevenção da polispermia.
O mesmo estudo demonstrou ainda que, após transferência cirúrgica de oócitos
maturados in vitro para ovidutos de fêmeas receptoras em estro, por um período de 4h,
resultaram em mudanças similares em termos morfológicos e físicos, daqueles
observados nos oócitos ovulados. Além disso, os oócitos maturados in vitro e expostos
ao ambiente do oviduto tiveram taxa de penetração semelhante (83 vs. 87%), porém,
foram significantemente menos polispérmicos (67 vs. 26%) que os oócitos controle.
Ao menos nos suínos, o desenvolvimento de todo o mecanismo para prevenir a
polispermia parece somente se completar após a passagem dos oócitos pelo oviduto.
Diversos estudos demonstraram os efeitos benéficos das células do oviduto e/ou meios
condicionados na redução da polispermia. Nestes estudos foram utilizadas células do

19
oviduto tanto de fêmeas pré-púberes como de animais apresentando sinais de estro.
(NAGAI & MOOR, 1990; KANO et al., 1994; BUREAU et al., 2000; ROMAR et al., 2000).
VATZIAS & HAGEN (1999) observaram que a suplementação do meio de
fertilização in vitro com um meio derivado da cultura de células retiradas de ovidutos de
animais que estavam próximos a período da ovulação, reduziu a incidência de
polispermia, comparado a meios condicionados com células retiradas de ovidutos no
meio da fase luteal.
Estas informações, tomadas em conjunto, reforçam a hipótese que as secreções
do oviduto contêm alguns fatores que interagem com os oócitos e/ou com os
espermatozóides agindo na prevenção ou diminuindo a entrada de múltiplos
espermatozóides.
O oviduto, em resposta aos hormônios ovarianos, sintetiza e secreta múltiplas
proteínas. Isso cria um ambiente capaz de possibilitar os eventos da fertilização e o
desenvolvimento dos embriões. Uma grande variedade de proteínas sintetizadas e
secretadas pelo oviduto suíno já foi identificada (ABEYDEERA, 2002). A maior parte é
composta por glicoproteínas estrógeno-dependentes, identificadas como glicoproteínas-
secretórias-específicas do oviduto suíno (pOSP) (BUHI et al., 2000).
A exposição dos oócitos a pOSP semi-purificadas, por um período de 4h antes
da FIV e durante a fertilização proporcionou uma redução na polispermia (61 vs. 29%)
sem comprometer a taxa de penetração dos espermatozóides (74 vs. 63%) (KOUBA et
al., 2000). Além disso, a exposição dos oócitos a pOSP também reduziu a ligação dos
espermatozóides a ZP. Não foi totalmente esclarecido se a redução da polispermia por
parte da exposição às pOSP, está relacionado a sua interação com os oócitos e/ou
espermatozóides, em razão dos oócitos terem sidos expostos as proteínas antes e
durante a fertilização. Porém, em recente estudo, pode-se comprovar que a exposição
dos oócitos as pOSP, foi suficiente para reduzir a polispermia (McCAULEY et al., 2001).
Somado ao seu papel desempenhado na expansão das células do cumulus, o
uso do ácido hialorurônico durante a pré-incubação dos espermatozóides, ou na co-
incubação do complexo esperma-oócito pode reduzir a polispermia sem qualquer efeito
adverso na taxa de penetração (SUZUKI et al., 2000). O ácido hialorônico está presente

20
nos momentos peri e pós-ovulatórios no fluído do oviduto dos suínos, e tem efeito
estimulante na capacitação in vitro de espermatozóides sem induzir reação do
acrossoma (TIENTHAI et al., 2000). Portanto, aliado as glicoproteínas do oviduto, o
ácido hialurônico pode desempenhar um importante papel no controle da penetração
espermática e redução da polispermia (RODRIGUEZ et al., 1997).
A maioria dos meios usados para FIV em suínos são suplementados com
cafeína, um inibidor das fosfodiesterases, que possui a função de aumentar o cAMP
(AMP ciclíco). Em recente estudo foram examinados o efeito da cafeína, do peptídeo-
promotor da fertilização (FPP), e da adenosina em parâmetros de fecundação de
oócitos maturados in vitro. Na comparação com FPP (75%) e adenosina (71%), uma
porcentagem significativamente maior de oócitos foram penetrados quando o meio de
fertilização continha cafeína (98%). Entretanto, a maior parte dos oócitos penetrados na
presença de cafeína apresentou polispermia (87%), quando comparados com FPP
(25%) ou adenosina (21%). A menor incidência de polispermia foi atribuída a habilidade
do FPP e da adenosina em estimular a capacitação e inibir as reações espontâneas do
acrossoma (FUNAHASHI et al., 2000). Portanto, a suplementação do meio de fertilização
com adenosina, ao invés da cafeína parece uma alternativa razoável para minimizar os
problemas da polispermia.
Além dos componentes adicionados durante a fertilização, o tipo de meio de
cultura utilizado no cultivo dos oócitos, também pode afetar a incidência da polispermia
(ABEYDEERA, 2002).
MARTINEZ-MADRID et al. (2001) compararam dois meios de maturação, o Tris-
tamponado modificado (mTBM) e o meio Tyrodes modificado (mTaLP), quanto as taxas
de penetração espermática e polispermia. A FIV no meio mTAPL com uma concentração
espermática de 0.5 a 1x 106/ml resultou numa maior taxa de penetração (92 a 94% vs.
61 a 77%), comparado ao meio mTBM. Entretanto, a maior parte dos oócitos fertilizados
com o mTALP mostraram-se polispérmicos.
KIDSON et al. (2001), em estudo semelhante, utilizando uma baixa concentração
espermática (4 x 105/ml), tanto a penetração (54 vs. 32%) quanto à polispermia (40 x 105)
foram mais altas quando no meio mTALP comparado ao mTBM. Entretanto, duplicando

21
a concentração espermática, a taxa de penetração aumentava de maneira proporcional
(82 vs. 79%), porém, a taxa de polispermia era significantemente mais baixa no meio
mTBM (76 vs. 26%). Com base nesses estudos, parece que a seleção de um meio
apropriado para FIV também é um fator importante para reduzir o problema da
polispermia.
2.12.Mecanismos moleculares relacionados à prevenção da polispermia
A polispermia é geralmente considerada um fenômeno patológico nos mamíferos.
Entretanto, uma incidência especialmente alta tem sido encontrada na fertilização de
suínos. Em condições in vivo a incidência da polispermia em suínos pode alcançar
30-40%, enquanto em condições in vitro pode alcançar índices superiores a 65% (XIA
et al., 2001). A polispermia tem sido considerada o principal entrave a ser superado
para melhorar a eficiência da fertilização in vitro nos suínos. Os mecanismos
envolvidos na alta incidência de polispermia em oócitos suínos não estão bem
esclarecidos, e relativamente pouco é conhecido a respeito dos mecanismos para
preveni-la (ABEYDEERA, 2002).
Como em outras espécies, oócitos suínos contêm GC’s que são ricos em
enzimas (YOSHIDA et al., 1993). Durante a maturação dos oócitos os GC’s são
submetidos a uma migração centrífuga para formar uma única camada junto a
membrana plasmática (WANG et al., 1998).
Após a penetração de um espermatozóide, no momento da fecundação acontece uma
reação cortical que libera esse conteúdo cortical para o espaço perivitelino (PVS),
prevenindo a penetração de mais espermatozóides. Este bloqueio ocorre em razão do
espessamento da ZP. A reação da ZP é, portanto um mecanismo de defesa contra a
polispermia na espécie suína, visto que através da indução da reação cortical pode-se
prevenir a penetração de espermatozóides em oócitos com a ZP intacta, porém não,
em oócitos sem ZP (WANG et al., 1998). Entretanto, espermatozóides no PVS são
comumente observados, o que sugere existir um bloqueio secundário na superfície
vitelina (HUNTER, 2000).

22
Figura 3. Reação cortical após a entrada de espermatozóide através da zona pelúcida.(Fonte: Adaptado de Alberts et al. - Molecular biology of the cell. 4th ed. 2004).
Tanto os oócitos maturados in vitro ou in vivo possuem habilidade para liberar os
GC’s após a penetração dos espermatozóides (WANG et al., 1997b), portanto, acredita-
se que a alta incidência de polispermia nos oócitos inseminados in vitro possa não estar
relacionada com um atraso, ou uma incompleta liberação dos GC’s, e sim, por um
atraso na reação da ZP e/ou por penetração simultânea de mais de um espermatozóide
(SUN et al., 1992).
A respeito dos aspectos moleculares da reação cortical, estudos preliminares
demonstraram que a proteína-G, o aumento do íon Ca2+ e a proteína quinase C (PKC)
participam no processo de exocitose dos GC’s. A fertilização nos suínos induz
oscilações do íon Ca2+, e um dos primeiros eventos que pode ser considerado cálcio-
dependente é a exocitose dos GC’s. Estimulantes da PKC induzem 90% dos oócitos
suínos a liberarem parcialmente, ou totalmente seus GC’s, e este efeito pode ser
suprimido quando feita uma inibição a PKC (SUN et al., 2001c). Foi demonstrado que o
co-cultivo de espermatozóides e células do oviduto pode reduzir a incidência de
polispermia cerca de 40 a 50% (NAGAI & MOOR, 1990).

23
Portanto, é possível que as secreções oriundas do oviduto contenham fatores
que podem interagir com os oócitos e/ou espermatozóides prevenirem a entrada de
múltiplos espermatozóides (ABEYDEERA, 2002).
2.13. Morfologia e qualidade dos embriões
Apesar dos avanços na produção in vitro (PIV) de embriões, diferenças
morfológicas têm sido observadas entre embriões produzidos in vitro e in vivo, incluído
o estágio de blastocisto. Embriões com blastômeros bem definidos nos estágios iniciais
do desenvolvimento e uma massa celular interna proeminente no estágio de blastocisto
são evidentes em embriões coletados in vivo (WANG et al., 1998).
Segundo PAPAIONNOU & EBERT (1998), o número de células nos blastocistos
produzidos in vitro é menor que nos produzidos in vivo. De acordo com WANG et al.,
(1998), a clivagem anormal e o baixo número de células nos balstocistos produzidos in
vitro estão relacionados à deficiências nos filamentos de actina do citoplasma. Uma
maturação citoplasmática inadequada dos oócitos maturados in vitro e/ou condições
sub-ótimas podem ser responsáveis pela baixa qualidade embrionária. A transferência
de zigotos provenientes de MIV/FIV para o oviduto de fêmeas receptoras (FUNAHASHI
et al., 1994) e a cultura de zigotos originados in vivo (MACHATY et al., 1998), sugerem
que tanto as condições de cultivo, quanto o processo de maturação por si só, são
responsáveis pela baixa qualidade dos embriões, no que tange ao número de células.
Evidências mais recentes mostraram que o desenvolvimento até o estágio
blastocistos e o número de células aumentaram quando os embriões produzidos in vitro
foram cultivados no meio NSCU 23 (North Carolina State University 23 Medium), sob
baixa tensão de O2 (5%) (MACHATY et al., 2001).
O mesmo estudo relatou que a inibição parcial da fosforilação oxidativa no
estágio de mórula tem efeitos benéficos no desenvolvimento dos blastocistos e no
incremento do número de células. Entretanto, sob baixas tensões de O2, a produção de
ATP no estágio de mórula pode favorecer a glicólise sobre a fosforilação oxidativa.

24
2.14.Perdas embrionárias e fetais
Em Sistemas Intensivos de Produção de Suínos (SIPS) a rentabilidade do
sistema se eleva com o aumento do número de leitões desmamados por porca por ano.
Considerando o fato de que para cada leitão desmamado, em média, mais de dois
oócitos foram inicialmente fertilizados, fica claro que a mortalidade do concepto
durante a prenhez (VONNAHME et al., 2002), do feto no parto e do leitão na lactação
(ABRAHÃO et al., 2003) são essenciais para a maximização da produção.
O tamanho de leitegada ao nascimento e o número de nascidos vivos são
índices importantes nos SIPS, no entanto, suas herdabilidades são muito baixas (em
torno de 0,10 ou menos), (ROTHSCHILD, 1996). Portanto, quaisquer tentativas de
seleção direta para estas características apresentam resultados insatisfatórios. Muitos
experimentos têm sido conduzidos com o objetivo de se aumentar o tamanho de
leitegada, sendo evidenciado o alto grau de variabilidade da característica decorrente
da influência de fatores ambientais, genéticos, manejo e nutricional, determinando o
insucesso do processo de seleção (BENNETT & LEYMASTER, 1990; LAMBERSOM et
al., 1991).
O número de ovulações, a taxa de fertilização e a mortalidade embrionária e fetal
são os principais pontos de controle das perdas até o parto.
O número de ovulações é uma característica que apresenta um alto grau de
variabilidade, podendo representar as perdas embrionárias quando se avaliam os
ovários no matadouro de fêmeas abatidas logo após a cobrição ou mais tarde, durante
a gestação. HALEY & LEE (1993) relataram um declínio da sobrevivência pré-natal de
2,5% para cada oócito ovulado. Existem maneiras de se elevar o seu número, ou
utilizando-se gonadotrofinas em doses de superovulação (HAZELEGER et al., 2000) ou
através de melhoramento genético, visto que a herdabilidade do número de ovulações é
médio-alta, em torno de 0,39 (ROTHSCHILD, 1996). A superovulação não é utilizada
comercialmente com a finalidade de aumentar o tamanho da leitegada devido ao alto
número de ovulações estar aliada à alta taxa de mortalidade embrionária e fetal (WU
et al., 1987), como já descrito anteriormente.

25
A taxa de fertilização (número de oócitos que foram fecundados por um
espermatozóide/total de ovulações) geralmente é muito elevada, cerca de 95-100%, em
condições normais (HUNTER, 1972). A Inseminação Artificial (IA) torna-se numa
ferramenta importante em um SIPS, mas na dependência de sua prática poderá
interferir negativamente sobre a fertilização dos oócitos, caso seja feita de forma
inadequada (KEMP & SOEDE, 1997).
Muitos estudos têm sido realizados a fim de determinar o momento exato da
ovulação e realização da IA. Atualmente sabe-se que o cio em porcas pode durar de 24
a 108 h, com média de 55 h (SOEDE et al., 1994), sendo que a ovulação acontece
quando decorridos 70% do cio, ou seja, 20 a 50 h após o início do cio. Estudos
realizados utilizando a ultra-sonografia para detecção do momento da ovulação
mostraram que a realização de apenas uma IA nas 24 horas em porcas e 12 horas em
marrãs, prévias à ovulação, apresentaram altos índices de fertilização (SOEDE &
KEMP, 1997). A grande variação na duração do cio e, conseqüentemente no momento
da ovulação, faz com que utilizemos mais de uma dose nos esquemas de IA, a fim de
garantir que pelo menos uma dose coincida com os intervalos referidos.
Define-se a perda embrionária como a mortalidade antes dos 35 dias de
prenhez. Está bem estabelecido que a taxa média de mortalidade embrionária em
suínos, submetidos às condições normais, é de 20-30% (PERRY & ROWLANDS, 1962;
VONNAHME et al., 2002). Ao contrário do que ocorre no número de ovulações, a
mortalidade embrionária varia muito entre raças e linhagens. Por exemplo, a raça
Meishan, raça hiperprolífica Chinesa, apresenta número de ovulações dentro do
considerado normal (±20 ovulações), porém apresenta dois a três leitões nascidos a
mais que as raças normais utilizadas comercialmente. Isto se deve a uma maior
sobrevivência embrionária apresentada por esta raça (VONNAHME et al., 2002).
A literatura aponta que a maior parte da mortalidade embrionária se concentra
até 17 dias de gestação (POPE et al., 1985; VAN DER LENDE et al., 1990), mas são
ainda muito discutidos os fatores que causam a mortalidade.
A fase da gestação na fêmea que corresponde ao embrião trata-se de um
período muito sensível, em que fatores externos influenciam significativamente. A

26
manutenção de um ambiente calmo, sem qualquer tipo de estresse (evitar bater nas
porcas, brigas entre as fêmeas, etc.) é primordial para haver menor perda. Além disso,
qualquer elevação de temperatura corpórea (estresse, doenças, temperatura ambiente
elevada, etc.) ou situações estressantes, principalmente no período antes da
implantação, pode levar a uma maior mortalidade embrionária (OMTVEDT et al., 1971;
WETTEMAN et al., 1985). Logo após a cobrição da fêmea é recomendado fornecer
uma ração com baixa quantidade de energia, pois pode interferir na sobrevivência
embrionária.
A taxa média de perda fetal em suínos é relativamente baixa e varia de 5-15%
(POPE et al., 1985). A mortalidade fetal é em média maior em populações com altas
taxas de ovulação (LONGENECKER & DAY, 1968). Segundo KNIGHT et al. (1977), o
período de 40-70 dias após a fertilização é tido como período crítico onde ocorre a
maioria das mortes fetais. Uma evidência apontou, neste estudo, que a placenta seria
um fator limitante do desenvolvimento fetal, pois um feto com uma placenta pouco
desenvolvida terá uma taxa de crescimento baixo, diminuindo a viabilidade do feto
devido a um aporte insuficiente de nutrientes e uma sub-ótima remoção dos resíduos
fetais. Um outro fator limitante é a “Capacidade uterina”, ou seja, a capacidade do útero
em gestar embriões e fetos (tamanho e peso dos cornos uterinos, espaço para a
implantação dos embriões e desenvolvimento dos fetos, produção e secreção do “leite
uterino”, etc.). FENTON et al. (1972) e BAZER et al. (1969) estudaram o período em
que o tamanho uterino passa a exercer efeito negativo sobre a sobrevivência pré-natal,
estabelecendo que após 25 dias de gestação começa a ocorrer uma competição para o
estabelecimento de uma área de superfície adequada para trocas de nutrientes entre a
circulação materno-fetal, resultando em um comprometimento do tamanho de leitegada.
Como medida de prevenção da mortalidade fetal, a capacidade uterina deve ser
aumentada, por exemplo, pela seleção genética (CHRISTENSON, 1993) ou pela
seleção de leitoas que apresentam uma maior eficiência placentária (FORD, 1997;
WILSON et al., 1999).
O desenvolvimento placentário também é um fator limitante para o
desenvolvimento fetal, pois a placenta é um órgão que transporta os gases

27
respiratórios, nutrientes e resíduos entre o sistema materno-fetal. Portanto, a sua
principal função é a de suprir os subtratos metabólicos necessários para o suporte do
crescimento fetal (REYNOLDS & REDMER, 1995). Nesta mesma revisão os autores
relataram a influência de outros fatores sobre o crescimento fetal, como o genótipo
materno, número de fetos, má alimentação materna e estresse ambiental. Os pesos
fetais e placentários também são reduzidos quando a área de superfície uterina é
reduzida experimentalmente (KNIGHT et al., 1977). A área de contato da placenta com
o endométrio é também uma variável importante no desenvolvimento fetal, porque a
placenta do suíno é do tipo epitélio-corial, apresentando estruturas absortivas
especializadas por toda a placenta (auréolas), responsáveis pelas trocas de nutrientes
entre a mãe e o feto.
Existem algumas doenças que podem levar a abortamentos e mumificações
(morte fetal, sem contaminação bacteriana, em qualquer estágio de seu
desenvolvimento, apresentando absorção de fluído corpóreo). Atualmente a Brucelose,
Leptospirose, Doença de Aujezsky, Parvovirose são as principais doenças que levam a
perdas reprodutivas (VANROOSE et al., 2000).

28
3. OBJETIVOS
♣ Verificar o número e características dos oócitos recuperados após aspiração
folicular de marrãs no primeiro estro.
♣ Verificar o número e as características morfológicas de embriões após 24 e
144 horas da primeira monta natural.
♣ Identificar e caracterizar o "status" nuclear de oócitos suínos a partir da
coloração com Hoechst 33342.
♣ Verificar pela coloração com Lens culinaris, a distribuição e localização dos
grânulos corticais no citplasma de oócitos de suínos.

29
4. MATERIAL E MÉTODOS
4.1. Animais
Para a realização dos experimentos foram selecionados 24 (vinte e quatro)
fêmeas da espécie suína, provenientes do cruzamento entre as raças Landrace x Large
White (F1), com idades entre 5½ a 6 meses de idade, e um macho da raça Duroc (1½
ano de idade) de 160 Kg para a realização da Monta Natural, pertencentes à Fazenda
São João das Palmeiras em Jaboticabal - São Paulo, como mostra a Figura 4.
Figura 4. A) Fêmeas (Large White) e B) Cachaço (Duroc), utilizados no experimento.
4.2. Período de adaptação das fêmeas.
Durante o período de adaptação, as fêmeas pré-púberes (3-4 meses) foram
separadas em baias e numeradas para controle dos experimentos onde começaram a
receber ração balanceada para fase de gestação. Nesse período, um cachaço, com 8
meses, foi colocado em contato com as marrãs duas vezes ao dia (manhã e tarde) por
30 minutos em cada período, visando estimulação das marrãs à antecipação da
ciclicidade.
Esse procedimento foi o mesmo utilizado para detecção do cio por reflexo de
tolerância ao macho e ao homem. Uma fêmea, foi considerada no estro, quando
permitia a cópula obedecendo ao manejo reprodutivo acima citado de acordo como
ilustrado na Figura 5.
A B

30
Figura 5. A) Limpeza do prepúcio; B e C) Reflexo de tolerância ao homem; D) Monta natural(MN).
4.3. Manejo e alimentação
As fêmeas (marrãs) destinadas ao experimento, bem como o reprodutor, foram
submetidas ao arraçoamento convencional da granja de acordo com a sua categoria,
como apresentado na Tabela 3.
As fêmeas suínas púberes, com idades de 180 dias, foram mantidas em baias de
6m x 7m com uma lotação de 8 animais por baia em sistema intensivo de criação
durante todo o experimento, ou seja, ração balanceada no cocho oferecida de acordo
com o manejo já utilizado na granja (média de 2.5 Kg/animal/dia, distribuídas em duas
etapas - manhã e tarde). O cachaço, da raça Duroc, com peso de 160 Kg e 8 meses de
idade, recebia 2 Kg/animal/dia de ração (em duas etapas). Todos os animais tinham
acesso à água ad libitum.
A B
C D

31
Tabela 3. Composição do arraçoamento das marrãs destinadas à reprodução e utilizadas noexperimento.
Ingredientes Peso (Kg) Milho 60,6 Farinha de carne 5,0 Farelo de soja 18,0 Farelo de trigo 15,0 Calcário 0,5 Sal comum 0,5 Suicrescimento 0,2 Minermix 0,2 Energia digestível de suínos 4,12 Fibra bruta 4,3 Gordura 3,49 Proteína bruta 15,04 Estrato Etéreo 3,5 Minerais 5,7 Lisina 0,67
Fonte: composição alimentar adotada na propriedade
4.4. Planejamento experimental
A parte experimental foi composta de quatro experimentos com objetivo de
avaliar o número de estruturas colhidas, sua classificação (SHIMADA & TERADA, 2001)
“status” nuclear (MEO, 2002) e dos grânulos corticais (CHERR et al., 1988). Para tanto,
em cada experimento, por repetição, foram constituídos grupos com diferentes horários
após abate e diferentes estruturas colhidas.
As 24 fêmeas, com idades de 180 dias (ao início do experimento), foram
divididas aleatoriamente nos seguintes grupos:
Experimento1- Oito animais foram sacrificados em abatedouro logo após a
detecção do primeiro cio e seus ovários removidos e transladados ao laboratório.
Experimento 2- Oito animais, após detecção do primeiro cio foram submetidos à
monta natural, abatidos 24 horas depois e seus sistemas genitais removidos e
transferidos ao laboratório.

32
Experimento 3- Oito animais, após a detecção do primeiro cio foram submetidos
à monta natural e sacrificados seis dias após a primeira cobertura e, como no item
anterior, tiveram seus genitais removidos e transferidos ao laboratório.
Experimento 4- Um quarto grupo denominado “controle”, foi utilizado como
referencial quanto ao número de nascimentos oriundos de marrãs no cio da granja nos
anos de 2004 e 2005 (Tabela 8).
Experimento 1
Após detecção do estro, 8 fêmeas foram abatidas em Frigorífico, seus ovários
removidos e conduzidos em solução salina (0,9% NaCl) a 30-35ºC em recipiente
isotérmico até o Laboratório de Fecundação In Vitro. Em seguida, os ovários foram
lavados em solução salina e seus folículos antrais, (de 3 a 8 mm de diâmetro) foram
aspirados com agulha 18-G acoplada a uma seringa descartável de 20mL (Figura 6). O
fluido folicular aspirado foi transferido para um tubo de poliestireno de 50mL onde
permaneciam por 15 min para decantação. Posteriormente, o sedimento formado foi
transferido para placas de poliestireno de 60 mm de diâmetro, quando se procedia a
seleção dos oócitos sob microscópio estereoscópico (IKEDA & TAKAHASHI, 2003).
Figura 6. Aspiração folicular de oócito suíno.
4.5. Avaliação dos oócitos (“status” nuclear e grânulos corticais)
Oócitos suínos foram obtidos a partir de aspiração folicular de ovários
proveniente de fêmeas abatidas, como descrito anteriormente.

33
Foram selecionados oócitos com aspecto homogêneo de citoplasma e
compactação das células do cumulus, com pelo menos três a quatro camadas de
células de revestimento. Os oócitos selecionados foram lavados duas vezes em meio
TCM-199 contendo tampão Hepes, suplementado com 10% de SFB, 0,20mM de
piruvato e 83,4ì g/mL de amicacina (meio de lavagem) e então, foram desnudados,
incubando-se os complexos-cumulus-oócitos em hialuronidase (2mg/mL) a 38,5ºC por
5 minutos procedendo-se, em seguida, pipetação durante 3 a 5 minutos.
Posteriormente, os oócitos foram submetidos à fixação e coloração de cromatina e
grânulos corticais de acordo com a metodologia descrita por CHERR et al. (1988).
Oócitos tiveram inicialmente, a zona pelúcida removida com a incubação em pronase a
0,5% em PBS, por aproximadamente 5 minutos. Em seguida, foram fixados em 3% de
formaldeído em PBS por 30 minutos, à temperatura ambiente e incubados a 4ºC
durante a noite em solução de bloqueio – SB (PBS com 1mg/mL de BSA, 100mM de
glicina e 0,2% de azida de sódio). Para a permeabilização, os oócitos foram tratados
por 5 minutos a 38ºC em SB com a adição de 0,1% de Triton X-100. Os oócitos foram
então incubados com 10ì g/mL de Lens culinaris aglutinina conjugado a isotiocianato
de fluoresceína (FITC-LCA) em SB por 15 minutos e, com 10ì g/mL de Hoechst 33342
diluído em meio TCM 199 Hepes durante 10 minutos. Em seguida, foram lavados em
SB três vezes (5 minutos cada vez), montados em lâmina com 90% de glicerol em PBS
e visualizados em microscópio de epifluorescência (Olympus – IX-FLA-70, Tokyo,
Japão) sob dois filtros de excitação, sendo BP 330-385nm com barreira de 420nm para
avaliação do estádio nuclear e BP 335nm com barreira de 515nm para avaliação da
distribuição de grânulos corticais no interior dos oócitos.
Experimento 2
Neste experimento, 8 fêmeas (marrãs), após a detecção do primeiro estro, foram
submetidas à monta natural e, após 24 horas, foram levadas ao frigorífico, abatidas e
suas tubas uterinas removidas e levadas para o laboratório. As tubas foram lavadas
com PBS com auxílio de uma seringa e agulha e todo seu conteúdo depositado em

34
uma placa de poliestireno de 60mm. Em seguida, as estruturas (oócitos e/ou embriões)
foram recuperadas, contadas e classificadas sob microscópio estereoscópico (80 X).
As estruturas não fecundadas (eventualmente presentes) foram coradas quanto
aos grânulos corticais como descrito anteriormente.
Experimento 3
Oito fêmeas (marrãs) como no experimento 2, tiveram seu primeiro estro
detectado, submetidas à monta natural e, após um interstício de 6 dias, foram abatidas
e seus genitais removidos e levados para o laboratório. Após os procedimentos quanto
à antissepsia, como descrito anteriormente, os cornos uterinos (isolados) foram lavados
com PBS e todo conteúdo foi depositado em placa de poliestireno de 60mm.
Posteriormente, procedia-se o isolamento, contagem e classificação das estruturas
encontradas.
Experimento 4
Objetivando verificar a leitegada média de marrãs (1ª parição) da granja, foram
obtidos dados relativos aos nascimentos de leitões durante os anos de 2004 e 2005.
Este grupo foi utilizado como “controle” para fins de análise comparativa e as montas
naturais foram realizadas no terceiro cio com intervalos aproximados de 12 horas a
partir da detecção do estro.
4.6. Análise estatística
Visando explicar ao longo do tempo o número de oócitos e/ou estruturas obtidos
nos momentos 1 (Experimento 1), 2 (Experimento 2) e 3 (Experimento 3), os dados
foram submetidos a um ajuste e, o modelo teórico que melhor se adaptou foi y = a + b
log(t), onde t é o número de horas, y é o número de oócitos e a e b são os parâmetros
do modelo. O ajuste foi feito no software Statistica versão 7, desenvolvido pela Statsoft
(2004), tendo como resultados o modelo y = 32,14 – 15,24 log(t) sendo o coeficiente de
determinação de ajuste R2 = 70,6%, ou seja, 70,6% da variabilidade pode ser atribuída
ao modelo. O instante inicial foi considerado t=1 pelo fato da definição de logaritmo
exigir t>0. A derivada da função dy/dt = - 6,6/t, expressa a perda de oócitos por hora.

35
5. RESULTADOS
A Tabela 4 apresenta Média e Desvio Padrão das estruturas isoladas e analisadas
para cada experimento.
Tabela 4. Média e desvio padrão das estruturas isoladas e analisadas para cada experimento.
Animal nº Experimento 1 Experimento 2 Experimento 31 55 12 32 50 10 23 42 0 44 24 6 25 26 15 36 19 13 17 20 0 08 33 0 0
Média 33,625 7 1,875Desvio Padrão 13,866 6,347103 1,457738
5.1. Experimento 1
Os oócitos de 8 marrãs foram avaliadas quanto à configuração da cromatina e
grânulos corticais e os resultados estão expostos na Tabela 5 e Figura 7.
Das 8 marrãs, ou seja, dos 16 ovários aspirados no experimento 1, foram
colhidas 269 estruturas, classificadas como oócitos imaturos apresentando vesícula
germinativa (GV), identificados pela coloração com Hoechst (Figura 7 A e B)
Foram identificadas algumas variações quanto à distribuição dos grânulos
corticais (CG) no ooplasma. A maioria apresentava os CG sob a forma de "clusters", ou
seja, distribuídos de forma aglomerada pelo ooplasma.

36
Tabela 5. Resultado do “status” nuclear e dos grânulos corticais de oócitos obtidos da aspiraçãode ovários de 8 marrãs após terem tido o estro detectado.
Grupo 1(8 marrãs)
Quantidade deoócitos após
seleção
Quantidade deoócitos após
coloração
Coloraçãop/Hoechst(GVBK)
Coloração dosgrânulos corticais
(“clusters”)1 55 18 + +2 50 22 + +3 42 20 + +4 24 10 + +5 26 18 + +6 19 10 + +7 20 17 + +8 33 19 + +
Total 269 134
Figura 7. Fotomicrografias de oócitos aspirados de ovários (experimento 1) e examinados sobmicroscopia de fluorescência. A e B) Oócitos imaturos com GV (Hoechst 33342);C) Oócito imaturo ilustrando CG (em "clusters") no citoplasma (Lens culinarisaglutinina conjugado a isotiocianato de fluorescesceína); D) Oócito imaturo iniciandomigração dos CG. Aumento de 400 X.
A
B
DC

37
5.2. Experimento 2
Avaliação e contagem das estruturas encontradas a partir da lavagem da tuba
uterina de marrãs abatidas 24 horas após monta natural. Os resultados estão
expressos na Tabela 6 e Figura 8.
No Experimento 2 foram colhidas 56 estruturas classificadas como oócitos,
embriões de 2, 3 e 4 células (blastômeros). Como pode ser visto, o número de
estruturas não correspondeu ao número esperado e/ou desejado. Nota-se pela análise
da figura 6 que houve uma queda real de estruturas do momento 1 (Experimento 1) da
ordem de 79,18%. Quanto ao aspecto morfológico os embriões apresentaram-se com
blastômeros distintos após a primeira clivagem, ou seja, com 2 células bem
evidenciadas. Quanto ao aspecto, os blastômeros mostravam-se escuros e com espaço
perivitelíneo bem delimitado.
Saliente-se o fato do animal n0 2 ter apresentado somente oócitos não
fecundados, o que destoa dos outros 7 que produziram embriões de 2 a 4 células.
Tabela 6. Número e características das estruturas obtidas após lavagem das tubas uterinas de8 marrãs após 24 horas da monta natural.
Grupo 2Nº de
estruturasencontradas
OócitosEstruturascom duas
células
Estruturascom trêscélulas
Estruturascom quatro
células
1 12 0 11 0 1
2 10 10 0 0 0
3 0 0 0 0 0
4 6 0 5 1 0
5 15 0 14 1 0
6 13 0 11 2 0
7 0 0 0 0 0
8 0 0 0 0 0
Total 56 10 41 4 1
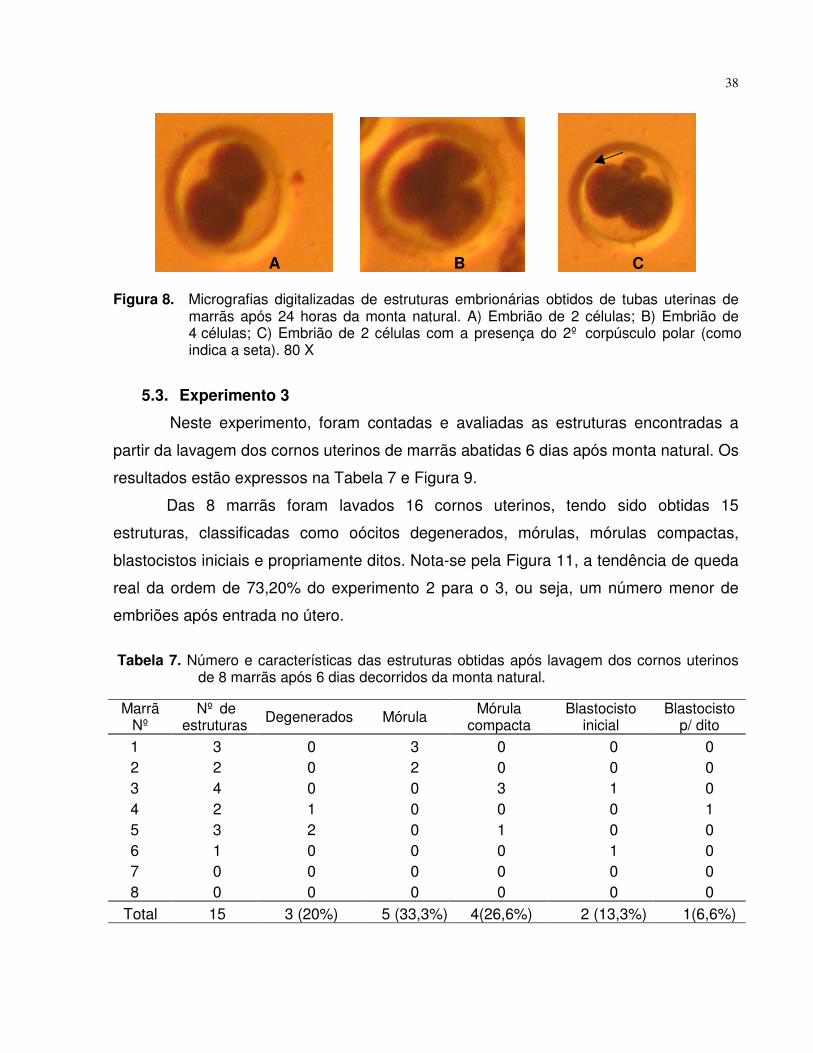
38
Figura 8. Micrografias digitalizadas de estruturas embrionárias obtidos de tubas uterinas demarrãs após 24 horas da monta natural. A) Embrião de 2 células; B) Embrião de4 células; C) Embrião de 2 células com a presença do 2º corpúsculo polar (comoindica a seta). 80 X
5.3. Experimento 3
Neste experimento, foram contadas e avaliadas as estruturas encontradas a
partir da lavagem dos cornos uterinos de marrãs abatidas 6 dias após monta natural. Os
resultados estão expressos na Tabela 7 e Figura 9.
Das 8 marrãs foram lavados 16 cornos uterinos, tendo sido obtidas 15
estruturas, classificadas como oócitos degenerados, mórulas, mórulas compactas,
blastocistos iniciais e propriamente ditos. Nota-se pela Figura 11, a tendência de queda
real da ordem de 73,20% do experimento 2 para o 3, ou seja, um número menor de
embriões após entrada no útero.
Tabela 7. Número e características das estruturas obtidas após lavagem dos cornos uterinosde 8 marrãs após 6 dias decorridos da monta natural.
MarrãNº
Nº deestruturas Degenerados Mórula Mórula
compactaBlastocisto
inicialBlastocisto
p/ dito 1 3 0 3 0 0 0 2 2 0 2 0 0 0 3 4 0 0 3 1 0 4 2 1 0 0 0 1 5 3 2 0 1 0 0 6 1 0 0 0 1 0 7 0 0 0 0 0 0 8 0 0 0 0 0 0
Total 15 3 (20%) 5 (33,3%) 4(26,6%) 2 (13,3%) 1(6,6%)
A B C
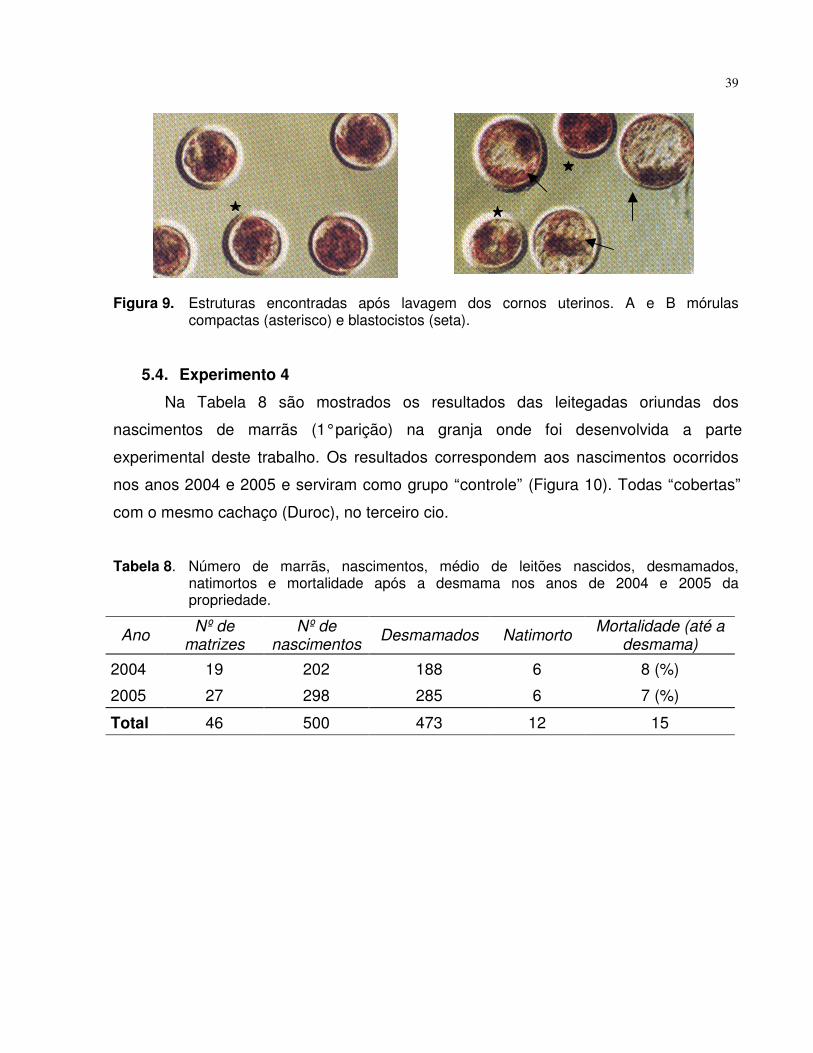
39
Figura 9. Estruturas encontradas após lavagem dos cornos uterinos. A e B mórulascompactas (asterisco) e blastocistos (seta).
5.4. Experimento 4
Na Tabela 8 são mostrados os resultados das leitegadas oriundas dos
nascimentos de marrãs (1°parição) na granja onde foi desenvolvida a parte
experimental deste trabalho. Os resultados correspondem aos nascimentos ocorridos
nos anos 2004 e 2005 e serviram como grupo “controle” (Figura 10). Todas “cobertas”
com o mesmo cachaço (Duroc), no terceiro cio.
Tabela 8. Número de marrãs, nascimentos, médio de leitões nascidos, desmamados,natimortos e mortalidade após a desmama nos anos de 2004 e 2005 dapropriedade.
Ano Nº dematrizes
Nº denascimentos Desmamados Natimorto Mortalidade (até a
desmama)
2004 19 202 188 6 8 (%)
2005 27 298 285 6 7 (%)
Total 46 500 473 12 15
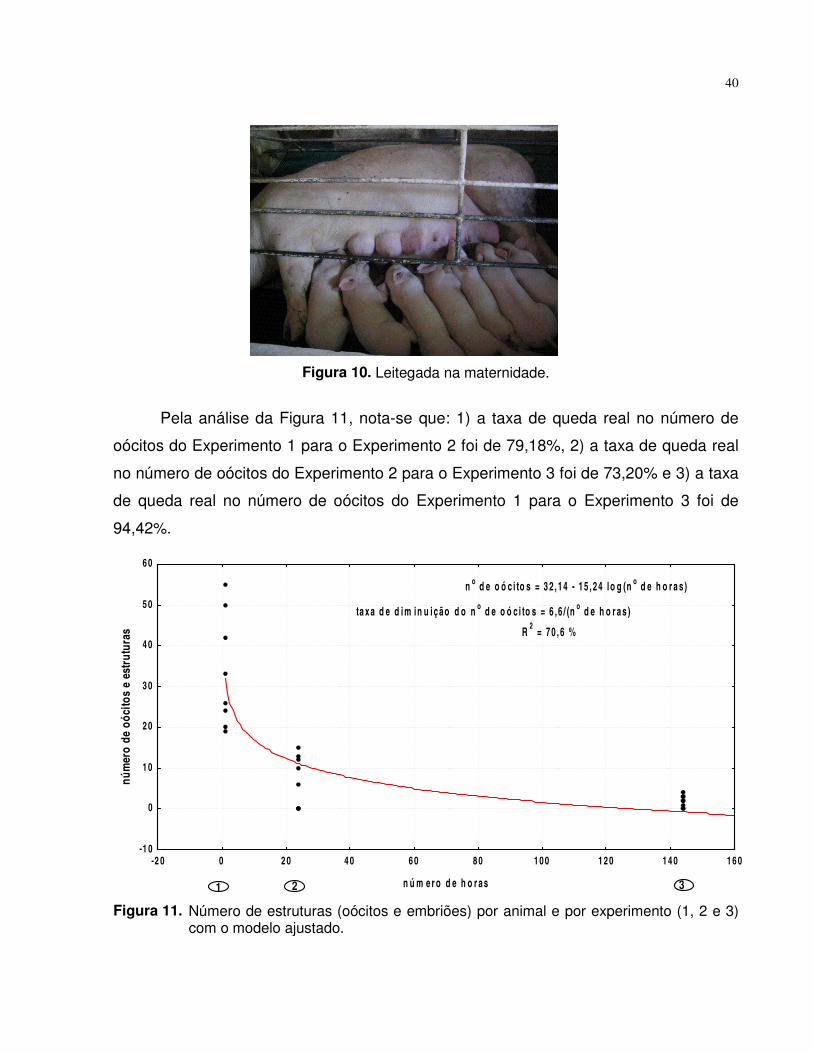
40
-2 0 0 2 0 4 0 6 0 8 0 1 0 0 1 2 0 1 4 0 1 6 0
n ú m e r o d e h o r a s
-1 0
0
1 0
2 0
3 0
4 0
5 0
6 0
núm
ero
de o
ócito
s e
estru
tura
s
n o d e o ó c i to s = 3 2 ,1 4 - 1 5 , 2 4 lo g (n o d e h o r a s )
ta x a d e d i m in u i ç ã o d o n o d e o ó c i to s = 6 ,6 / (n o d e h o r a s )
R 2 = 7 0 , 6 %
1 2 3
Figura 10. Leitegada na maternidade.
Pela análise da Figura 11, nota-se que: 1) a taxa de queda real no número de
oócitos do Experimento 1 para o Experimento 2 foi de 79,18%, 2) a taxa de queda real
no número de oócitos do Experimento 2 para o Experimento 3 foi de 73,20% e 3) a taxa
de queda real no número de oócitos do Experimento 1 para o Experimento 3 foi de
94,42%.
Figura 11. Número de estruturas (oócitos e embriões) por animal e por experimento (1, 2 e 3)com o modelo ajustado.

41
6. DISCUSSÃO
O inicio da puberdade em suínos não é um evento distinto, constituindo-se numa
complexa interação de fatores associados, muitos deles ainda por serem esclarecidos.
Além da diversidade genética, fatores como “status” endócrino e neuroendócrino,
manejo, plano nutricional e, em particular, a presença de um cachaço maturo, podem
influenciar a idade à puberdade (CUPPS, 1991). Raças Chinesas como a Meishan
desenvolvem a puberdade antes dos 100 dias de idade, possuem placenta menor e
uma grande leitegada quando comparada a raças Inglesas continentais (SUN, et al.,
2002). Neste trabalho, foram usadas fêmeas (marrãs) sintéticas (Landrace x Large
White) com idades de 5½ a 6 meses quando atingiam a puberdade. Normalmente, o
ciclo estral de marrãs manifesta-se a cada 21 dias (DZIUK, 1987) e a duração da
ovulação (intervalo da ovulação do primeiro ao último folículo durante o estro) varia
entre 1 e 4 horas (SOEDE et al., 1992 e SOEDE & KEMP, 1993). Não foi objeto desta
pesquisa, observar o número de corpos lúteos e sua relação com o número de oócitos
aspirados e/ou ovulados. No Experimento 1, foi obtida uma média de 33,6 oócitos
aspirados de ovários de fêmeas no estro apresentando folículos de 3 a 8 mm de
diâmetro, especificamente 1 hora após o aceite da monta (HALEY & LEE, 1993). Sabe-
se que o número de ovulações e o comprimento do útero determinam, respectivamente,
o potencial de embriões e sobrevivência dos fetos (WU & DZIUK, 1995). Embora estas
duas características sejam herdáveis, a correlação entre elas é baixa (CHEN & DZIUK,
1993). Por outro lado, WU & DZIUK (1995) não encontraram correlação entre fetos
vivos e número de corpos lúteos e que, em alguns casos, a perda pré-natal era
desproporcionalmente alta ao grande número de corpos lúteos, concluindo que o
comprimento de úteros pré-púberes dava uma indicação do comprimento pós-
puberdade e o potencial do tamanho da leitegada em suínos.
As estruturas (oócitos) encontradas no Experimento 1, caracterizavam-se por
apresentar de uma a três camadas de cumulus oophorus, zona pelúcida espessa,
citoplasma homogêneo, apresentando grânulos corticais em “clusters” e,
invariavelmente, com núcleo no estádio de vesícula germinativa. A redução média de

42
33 oócitos para 7 estruturas de 2 a 4 células (Tabela 4), indica que as ovulações não
foram simultâneas, o que era de se esperar pelo tempo de ovulação variável de 1 a 4
horas, como enfatizado por SOEDE et al. (1992) e SOEDE & KEMP (1993). Partindo-se
da premissa que a duração do estro pode variar de 24 a 96 horas entre porcas e que, o
momento da ovulação após o início do estro, pode variar de 10 a 85 horas,
conseqüentemente, este último não é um bom indicador para o tempo de ovulação
(SOEDE & KEMP, 1997). Por outro lado, esta diminuição de estruturas do
Experimento 2, poderia ser justificada pelo fato de raças sintéticas apresentarem
menor taxa de ovulação quando comparadas com a prolífica chinesa Meishan
(CHRISTENSON, 1993). Folículos pré-ovulatórios das fêmeas Meishan apresentam
estágios mais avançados da meiose, bem como maior concentração de estradiol no
fluido folicular do que as híbridas, sugerindo que a maturação mais avançada das
primeiras, antes da ovulação, pode ser importante para garantir a maior prolificidade
desta raça (FAILLACE & HUNTER, 1994).
Outros fatores também podem estar envolvidos ao se considerar esta
discrepância em relação aos Experimentos 1 e 2. Desde que a duração do estro pode
variar de 24 a 96 horas e o momento da ovulação de 10 a 85 horas de seu início entre
porcas, torna-se difícil detectar a hora da ovulação (SOEDE & KEMP, 1997). O pico de
LH (30 ± 3 h) e elevação da concentração de progesterona (13 ± 3 h) em porcas
multíparas são bons indicadores do momento da ovulação (SOEDE et al., 1994).
Considerando que, o tempo da inseminação ou monta natural, normalmente se baseia
no início do estro (“standing”) (SOEDE et al., 1994), a aspiração e colheita de oócitos
no Experimento 1 foi realizada no máximo 1 hora após as marrãs terem aceitado a
detecção do estro desta maneira. Isto provavelmente poderia ser outra possível
explicação pela quais todos os oócitos do Experimento 1 tenham se apresentado, na
sua totalidade, com núcleo sob a forma de vesícula germinativa. Pelas observações de
IKEDA & TAKAHASHI (2003) a baixa competência para desenvolvimento de oócitos de
fêmeas pré-púberes podem ser conseqüência de sua inabilidade de maturação
citoplasmática e que isto não se devia às alterações na cinética de maturação nuclear,
nem da atividade kinase p34cdc2. Acrescentaram também que oócitos rotineiramente

43
colhidos de folículos ovarianos de marrãs pré-púberes apresentavam citoplasma em
crescimento, imaturos e com diâmetro menor.
Outro aspecto relevante, concomitante à presença dos núcleos do experimento 1
no estágio de vesícula germinativa, foi forma de distribuição dos grânulos corticais.
Todos os oócitos examinados (Tabela 5) apresentaram os grânulos corticais (CG) sob a
forma de “clusters”. Desde que os grânulos corticais em oócitos de mamíferos migram
para a periferia da membrana do ooplasma após a liberação de LH e a meiose II é
completada (YANAGIMACHI, 1994), deduz-se que oócitos aspirados de fêmeas no
momento da aceitação da monta (cachaço ou homem), não estariam aptos para
fertilização. Mas, WANG et al., (1998), ao compararem morfologicamente oócitos
suínos, naturalmente ovulados e colhidos de ovidutos, com maturados in vitro,
observaram que os primeiros apresentavam áreas mais claras no córtex citoplasmático,
mais grânulos corticais “agregados”, um largo espaço perivitelíneo e uma espessa e
dura zona pelúcida. A taxa de polispermia foi significantemente mais alta nos oócitos
maturados in vitro (65%) do que nos in vivo (28%).
No Experimento 2 (Tabela 6) era de se esperar que, devido à diminuição do
número de estruturas encontradas em relação ao Experimento 1, seriam detectados
oócitos e/ou estruturas fecundadas apresentando polispermia, caracterizando outro
fator limitante para um número maior de prole. Mas, como observado por FUNAHASHI
(2003), a fertilização polispérmica in vivo é rara em porcas cobertas e/ou inseminadas
artificialmente no inicio do estro e antes da ovulação. Isto está coerente com as
observações de GRUPEN et al. (1997) de que a retomada da meiose é desencadeada
in vivo pelo aparecimento do hormônio luteinizante (LH), sendo a mesma completada
40 horas mais tarde. Portanto, no Experimento 2, sendo a monta realizada quando da
detecção do estro e a maturação completada após 40 horas, deve ter ocorrido
assincronia em relação à capacitação espermática e conseqüente fertilização,
resultando em baixo número de estruturas. Por outro lado, a não detecção de
polispermia pode ser explicada pela monta “precoce” em relação às possíveis
ovulações tardias, diminuindo a viabilidade do gameta masculino. A fertilização
polispérmica em suínos tem sua incidência aumentada com a “idade” dos oócitos pós-

44
ovulação in vivo (HUNTER, 1991). Taxas de concepção e tamanho de leitegadas
baixas são observadas quando IAs eram feitas no início do estro, sendo o tempo da IA,
portanto, de suma importância para uma fertilização eficiente (DZIUK, 1970).
No Experimento 3 (Tabela 7) foram encontrados 12 embriões viáveis após a
entrada no útero aos 6 dias pós-monta. Este baixo número de embriões vem
novamente comprovar as multifacetadas causas entre o número de ovulações e prole
final. Os resultados dos Experimentos 2 e 3 também são dependentes do gameta
masculino. O ejaculado do cachaço pode conter de 10 a 100 x 109 spt em 50-400 ml de
fluido, a cópula pode se estender por vários minutos e, dentro de poucas horas, o útero
está relativamente livre de sêmen e um “reservatório” de spt é estabelecido na junção
útero-tubárica (DZIUK, 1991). Observações recentes (TIENTHAI et al., 2004) têm
mostrado que capacitação massiva de sptz não ocorre in vivo no “reservatório” no
período pré e perí-ovulatório sob condições espontâneas de estro, mas aumentam
significativamente pós-ovulação.
A Figura 11 teve como objetivo demonstrar a significativa queda da proporção de
potenciais folículos a serem ovulados (Experimento 1), após fertilização (Experimento 2)
e quando da entrada no útero (Experimento 3). As razões implicadas nesta “redução”
do potencial de possíveis ovulações e conseqüentes leitegadas (experimento 4) estão
relacionadas a múltiplos fatores envolvendo maturação do oócito, capacitação
espermática, fertilização propriamente dita e aqueles inerentes à gestação
(manutenção, migração e espaço para os embriões). Mas, parece que, como
concluíram XIE et al. (1990), as interações entre o útero e embrião no dia 12, estão
predispostas às diferenças nas seqüências dos folículos ovulados no estro. Ou seja,
possíveis mecanismos devem existir dando preferência à sobrevivência para embriões
resultantes de oócitos inicialmente ovulados do que aqueles ovulados mais tarde.

45
7. CONCLUSÕES
♣ Oócitos aspirados de ovários de marrãs sintéticas (Landrace X Large White)
em primeiro estro e no momento de aceitação de monta, apresentam núcleos
em fase de vesícula germinativa e grânulos corticais sob a forma de “clusters”.
♣ O número de estruturas embrionárias colhidas 24 e 144 horas após a
realização da monta natural estão diminuídas em relação ao potencial de
oócitos passíveis de serem ovulados de marrãs sintéticas (Landrace X Large
White).
♣ A leitegada resultante da realização de três montas naturais viabiliza um maior
número de nascimentos quando comparadas ao potencial resultante da
aceitação de uma monta no momento do estro.

Normas da ABNT-NR 6023
46
5. REFERÊNCIAS BIBLIOGRÁFICAS
ABEYDEERA, L.R. In vitro fertilization and embryo development in the pigs. In:
INTERNATIONAL CONFERENCE OF PIG REPRODUCTION, 16., 2001, Columbia.
Preceedings… p. 159-173.
ABEYDEERA, L.R. In vitro production of embryos in swine. Theriogenology, Los
Altos, v. 57, n. 1, p. 257-273, 2002.
ABEYDEERA, L. R.; DAY, B.N. Fertilization and subsequent development in vitro of pig
oocytes inseminated in a modified Tris-buffered medium with frozen-thawed ejaculated
spermatozoa. Biology of Reproduction, Champaign, v. 57, n. 4, p. 729-734, 1997.
ABEYDEERA, L. R. et al. Presence of beta-mercaptoethanol can increase gluthatione
content in pig oocytes matured in vitro and the rate of blastocist development after in
vitro fertilization. Theriogenology, Los Altos, v. 50, n. 5, p. 747-756, 1998.
ABRAHÃO, A.A.F.; VIANNA, W.L.; MORETTI, A.S. Estudo das causas de mortalidade
em leitões neonatos de um sistema intensivo de criação de suínos. In: CONGRESSO
BRASILEIRO DE VETERINÁRIOS ESPECIALISTAS EM SUÍNOS, 11., 2003, Goiânia.
Anais… Goiânia: Associação Brasileira de Veterinários Especializados em Suínos,
2003. p. 425-426.
ANEL, L. et al. Repeated laparoscopic follicular aspiration in lambs. Theriogenology,
Los Altos, v. 47, n. 1, p. 152, 1997.
ARMSTRONG, D.T. Advances in production of embryos in vitro from juvenile and
prepubertal oocytes from the calf and lamb. Reproduction, Fertility and Development,
East Melbourn, v. 9, n. 3, p. 333-339, 1997.
ARMSTRONG, D.T.; EVANS, G. Factors influencing success of embryo transfer in
goats. Theriogenology, Los Altos, v. 19, n. 1, p. 31-42, 1983.

Normas da ABNT-NR 6023
47
ARMSTRONG, D.T. et al. Gonadotropin stimulation regimens for follicular aspiration
and in vitro embryo production from calf oocytes. Theriogenology, Los Altos, v. 42,
n. 7, p. 1227-1236, 1994.
BALDASSARRE, H. et al. Technique for efficient recovery of sheep oocytes by
laparoscopic folliculocentesis. Animal Reproduction Science, Amsterdam, v. 35, n. 1-
2, p. 145-150, 1994.
BALDASSARRE, H. et al. In vitro production of sheep embryos using laparoscopic
folliculocentesis: alternative gonadotrophin treatments for stimulation of oocytes donors.
Theriogenology, Los Altos, v. 45, n. 3, p. 707-717, 1996.
BALDASSARRE, H. et al. Advances in the production and propagation of transgenic
goats using laparoscopic ovum pick-up and in vitro embryo production technologies.
Theriogenology, Los Altos, v. 57, n. 1, p. 275-284, 2002.
BARIL, G.; BREBION, P.; CHESNÉ, P. Manuel de formation pratique pour la
transplantation embryonnaire chez la brebis et la chévre. Rome: FAO, 1993.
183 p.
BATT, P.A.; KILLEEN, I. D.; CAMERON, A.W.N. Use of single or multiple injections of
FSH in embryo collection programmes in goats. Reproduction, Fertility and
Development, East Melbourne, v. 5, n. 1, p. 49-56, 1993.
BAZER, F.W. et al. Uterine capacity at two stages of gestation in gilts following embryo
superinduction. Journal of Animal Science, Champaign, v. 29, n. 1, p. 30-34, 1969.
BECKMANN, L.S.; DAY, B.N. Effects of media NaCl concentration and osmolarity on
the culture of early-stage porcine embryos and the viability of embryos cultured in a
selected superior medium. Theriogenology, Los Altos, v. 39, n. 3, p. 611-622, 1993.
BELTRANENA, E. et al. Effects of pre- and postpubertal feeding on production traits at
first and second estrus in gilts. Journal of Animal Science, Champaign, v. 69, n. 3,
p. 886-893, 1991.

Normas da ABNT-NR 6023
48
BENNETT, G.L.; LEYMASTER, K.A. Genetic implications of a simulation model of litter
size in swine based on ovulation rate, potential embryonic viability and uterine capacity:
II. Simulated selection. Journal of Animal Science, Champaign, v. 68, n. 4, p. 980-
986, 1990.
BEZERRA, M.B. et al. Aspectos quantitativos e qualitativos da foliculogênese pré-natal
na espécie caprina. Ciência Animal, Fortaleza, v. 8, n. 2, p. 47-56, 1998.
BONNEAU, M. The cost of building and maintaining an isolation unit. In: ALLEND.
LEMAN SWINE CONFERENCE, 25., 1998, Minnesota. Anais…Minnesota: Veterinary
Outreach Programs, 1998. p. 103-111.
BING, Y.Z.; RODRIGUEZ-MARTINEZ, H.; NAGAI, T. Effects of cysteamine, FSH, and
estradiol -17ß on in vitro maturation of porcine oocytes. Theriogenology, Stoneham,
v. 55, n. 4, p. 867-876, 2001.
BORTOLOZZO, F.P.; WENTZ, I. Manejo reprodutivo da fêmea suína de reposição. A
Hora Veterinária, Porto Alegre, v. 19, n.110, p. 47-54, 1999.
BROADBENT, P. J. et al. Effect of frequency of follicle aspiration on oocyte yield and
subsequent superovulatory response in cattle. Theriogenology, Los Altos, v. 47, n. 5,
p. 1027-1040, 1997.
BROOKS, P. H. Management of the replacement gilt: the “male effect” thirty years on.
The Pig Journal, Malmesbury, v. 43, n. 1, p. 92-109, 1999.
BUHI, W.C.; ALVAREZ, I.M.; KOUBA, A.J. Secreted proteins of the oviduct. Cells
Tissues Organs, Basel, v. 166, n. 2, p. 165-179, 2000.
BUREAU, M.; BAILEY, J.L.; SIRARD, M.A. Influence of oviduct cells and conditioned
media on porcine gametes. Zygote, Cambridge, v. 8, n. 2, p. 139-144, 2000.
CHERR, G.N.; DROBNIS, E.Z.; KATZ, D.F. Localizacion of cortical granule constituents
before and after exocytosis in the hamster egg. Journal of Experimental Zoology,
New Cork, v. 246, n. 1, p. 81-93, 1988.

Normas da ABNT-NR 6023
49
CHRISTENSON, R.K. Ovulation rate and Embryonic Survivel in Chinese Meishan
White Crossbred Pigs. Journal Animal Science, Champaign, v. 71, n. 11, p. 3060-
3066, 1993.
CLEN, Z.Y; DZIUK, P.J. Influence of initial length of uterus per embryo and gestation
stage on prenatal survivel, development and sex ratio in the pig. Journal Animal
Science, Champaign, v. 71, n. 7, p. 1895-1901, 1993.
COGNIÉ, Y. State of the art in sheep-goat embryo transfer. Theriogenology, Los
Altos, v. 51, n. 1, p. 105-116, 1999.
CORDEIRO, M.F. Produção e criopreservação de embriões ovinos da raça Santa
Inês. 2001. 61 f. Dissertação (Mestrado em Ciências Veterinárias) - Universidade
Estadual do Ceará, Fortaleza, 2001.
COSKUN, S.; LIN, Y.C. Effect of transforming growth factor and activin-A on in vitro
porcine oocyte maturation. Molecular Reproduction and Development, New York,
v. 38, n. 2, p. 153-159, 1994.
CRAN, D.G.; CHENG, W.T.K. The cortical reaction of pig oocytes during in vivo and in
vitro fertilization. Gamete Research, New York, v. 13, n. 3, p. 241-251, 1986.
CUPPS, P.T. Reproduction in Domestic Animals. 4.ed. Department of Animal Science
University of California, Davis, 1991.
DAY, B.N.; ABEYDEERA, L.R.; PRATHER, R.S. Recent progress in pig embryo
production through in vitro maturation and fertilization techniques. In: JOHNSON, L.A.;
GUTHRIE, H.D. (Ed.). Boars Semen preservation IV. Kansas: Allen Press, 2000.
p. 81-92.
DETTWEILER, F. Die zucht dês schweines. In: DETTWEILER, F.; MULLER, K. (Ed.).
Lehrbuch der schweinezucht. Berlin: Editora, 1924. p. 339-344.
DING, J.; FOXCROFT, G.R. Epidermal growth enhances oocyte maturation in pigs.
Molecular Reproduction and Development, New York, v. 39, n. 1, p. 30-40, 1994.

Normas da ABNT-NR 6023
50
DING, J. et al. Protein and nuclear changes in pigs at fertilization. Molecular
Reproduction and Development, New York, v. 31, n. 4, p. 287-296, 1992.
DZIUK, P.J. Estimation of optimum time for insemination of gilts and ewes by double-
mating at certain times relative to ovulation. Journal of Reproduction and Fertility,
Cambridge, v. 22, n. 2, p. 277-282, 1970.
DIZIUK, P. Reproduction in the pig. In: CUPPS, P.T. Reproduction in domestic
animals. 4. ed., California: Academic Press, 1991. chap., 14, p.471-489.
FAILLACE, L.S; HUNTER, M.G. Follicle development and oocyte maturation during the
immediate preovulatory period in Meishan and white hybrid gilts. Journal of
Reproduction and Fertility, Nottingham, v. 101, n. 3, p. 571-576, 1994.
FENTON, F.R. et al. Effect of quantity of uterus on uterine capacity in gilts. Journal of
Animal Science, Champaign, v. 31, n. 3, p. 104-106, 1972.
FORD, S.P. Embryonic and fetal development in different genotypes in pigs. Journal
of Reproduction and Fertility, Cambridge, v. 52, p. 165-176, 1997.
FOXCROFT, G.; AHERNE, F. Manejo da marrã de reposição e da porca de primeiro
parto: Parte II. Selecionando marrãs para uma melhor vida útil reprodutiva. In:
SIMPÓSIO INTERNACIONAL DE REPRODUÇÃO E INSEMINAÇÃO ARTIFICIAL EM
SUÍNOS, 7., 2000, Foz do Iguaçu. Anais... Foz do Iguaçu: Associação Internacional de
Reprodução e Inseminação Artificial em Suínos, 2000. p. 91-98.
FREITAS, V.J.F.; SIMPLÍCIO, A.A. Transferência de embriões em caprinos. In:
GONÇALVES, P. B. D.; FIGUEIREDO, J. R.; FREITAS, V. J. F. Biotécnicas aplicadas
à reprodução animal. São Paulo: Livraria Varela, 2002. p. 179-194.
FRY, R. C. et al. The collection of oocytes from bovine ovaries. Theriogenology, Los
Altos, v. 7, n. 5, p. 977-987, 1997.
FUNAHASHI, H. Polyspermic penetration in porcine IVM-IVF systems. Reproduction,
Fertility and Development, Tokio, v. 15, p. 167-177, 2003.

Normas da ABNT-NR 6023
51
FUNAHASHI, H.; DAY, B.N. Effects of cumulus cells on glutathione content of porcine
oocytes during in vitro maturation. Journal of Animal Science, Champaign, v. 73,
supl. 1, p. 90, 1995.
FUNAHASHI, H. et al. Both fertilization promoting peptide and adenosine stimulate
capacitation but inhibit spontaneous acrosome loss in ejaculated boar spermatozoa in
vitro. Molecular Reproduction Development, New York, v. 55, n. 1, p. 117-124, 2000.
FUNAHASHI, H. et al. Use of low-salt culture medium for in vitro maturation of porcine
oocytes is associated with elevated oocyte glutathione levels and enhanced male
pronuclear formation after in vitro fertilization. Biology of Reproduction, Champaign,
v. 51, n. 4, p. 633-639, 1994.
GALLI, C. et al. Embryo production by ovum pick-up from live donors. Theriogenology,
Los Altos, v. 55, n. 6, p. 1341-1357, 2001.
GOMES, M.F.M. et al. Análise prospectiva do complexo agroindustrial de suínos
no Brasil. Concórdia : EMBRAPA-CNPSA, 1992. 108 p.
GONÇALVES, P.B.D.; FIGUEIREDO, J.R.; FREITAS, V.J.F. Biotécnicas aplicadas à
reprodução animal. São Paulo: Livraria Varela, 2002. p.198-199.
GRAFF, K.J. et al. Transvaginal ultrasound-guided oocyte retrieval following FSH
simulation of domestic goats. Theriogenology, Los Altos, v. 51, n. 6, p. 1099-1119,
1999.
GRUPEN, C. G.; NAGASHIMA; NOTTLE, M. B. Asynchronous meiotic progression in
porcine oocytes matured in vitro: a cause of polyspermic fertilization?. Reproduction ,
Fertility and development, East Melbourne, v. 9, n. 1, p. 187-91, 1997.
GRUPEN, C.G.; NAGASHIMA, H.; NOTTLE, M.B. Cysteamine enhances in vitro
development of porcine oocytes matured and fertilized in vitro. Biology of
Reproduction, Madison, v. 53, n. 1, p. 173-178, 1995.

Normas da ABNT-NR 6023
52
HASLEY, C.S.; LEE, G.J. Genetic basis of prolificacy in Meishan pigs. Journal of
Reproduction and Fertility, Cambridge, v. 48, n. 3, p. 247-259, 1993.
HAZELEGER, W. et al. Effect of superovulation induction on embryonic development
on day 5 and subsequent development and survival after nonsurgical embryo transfer in
pigs. Theriogenology, Sotoneham, v. 53, n. 5, p. 1063-1070, 2000.
HOLANDA, M.C.R. et al. Natimortalidade e Mortalidade até 21 dias de idade em leitões
da raça Large White. Revista Brasileira de Zootecnia, Viçosa, v. 6, n. 29, p. 2276-
2282, 2000.
HOODBHOY, T.; TALBOT, P. Mammalian cortical granules: contents, fate, and
function. Molecular Reproduction and Development, New York, v. 39, n. 4, p. 439-
448, 1994.
HUGHES, P.E. Factors affecting the natural attainment of puberty in the gilt. In: COLE,
D.J.A.; FOXCROFT, G.R. (Ed.). Control of pig reproduction. London: Butterworths,
1982. p. 117-138.
HUGHES, P.E.; PEARCE, G.P.; PATERSON, A.M. Mechanisms mediating the
stimulatory effects of the boar on gilt reproduction. Journal of Reproduction and
Fertility, Cambridge, suppl. 40, p. 323-341, 1990.
HUNTER, M.G. Oocyte maturation and ovum quality in pigs. Reviews of
Reproduction, Cambridge, v. 5, n. 2, p. 122-130, 2000.
HUNTER, R.H. Oviduct function in pigs, with particular reference to the pathological
condictin of the polyspermy. Molecular Reproduction and Development, New York,
v. 29, n. 4, p. 385-391, 1991.
HUNTER, R.H.F. Fertilization in the pig: sequence of nuclear and cytoplasmic events.
Journal of Reproduction and Fertility, Cambridge, v. 29, n. 3, p. 395-406, 1972.

Normas da ABNT-NR 6023
53
IKEDA, K.; TAKAHASHI, Y. Comparison of maturational and development parameters
of oocytes recovered from prepubertal and adult pigs. Reproduction, Fertility and
Development, East Melbourne, v. 15, n. 4, p. 215-221, 2003.
ISHWAR, A.K.; MEMON, M.A. Embryo transfer in sheep and goats: a review. Small
Ruminant Research, Amsterdam, v. 19, n. 1, p. 35-43, 1996.
JEONG, B.S.; YANG, X. Cysteine, glutathione, and percoll treatments improve porcine
oocyte maturation and fertilization in vitro. Molecular Reproduction Development,
New York, v. 59, n. 3, p. 330-335, 2001.
KA, H.H. et al. Amino acids in maturation medium and presence of cumulus cells at
fertilization promote male pronuclear formation in porcine oocytes matured and
penetrated in vitro. Biology of Reproduction, Champaign, v. 57, n. 6, p. 1478-1483,
1997.
KANAYAMA, N.; MIYANO, I.; LEE, J. Aquisition of meiotic competence in growing pigs
oocytes correlates with their ability to activate Cdc2 kinase and MAP kinase. Zygote,
Cambridge, v. 10, n. 3, p. 261-270, 2002.
KANO, K.; MIYANO, T.; KATO, S. Effect of oviduct epithelial cells on fertilization of pig
oocytes in vitro. Theriogenology, Los Altos, v. 42, n. 6, p. 1061-1068, 1994.
KEMP, B.; SOEDE, N.M. Consequences of variation in interval from insemination to
ovulation on fertilization in pigs. Journal of Reproduction and Fertility, Cambridge,
v. 52, n.1, p. 79-89, 1997.
KIDSON, A. et al. Polyspermia in the pig is dependent on both IVF medium and sperm
dose during fertilization in vitro. In: INTERNATIONAL CONFERENCE OF PIG
REPRODUCTION, 6., 2001, Columbia. Proceedings…p. 75.
KIKUCHI, K. et al. Effect of follicle cells on in vitro fertilization of pig follicular oocytes.
Theriogenology, Los Altos, v. 39, n. 3, p. 593-599, 1993.

Normas da ABNT-NR 6023
54
KIRKWOOD, R.N.; THACKER, P.A. Management of replacement breeding animals.
Veterinary Clinics of North America: Food Animal Practice, Swine Reproduction,
Philadelphia, v. 8, n. 3, p. 575-587, 1992.
KNIGHT, J.W. et al. Conceptus development in intact and unilaterally hysterecttomized-
ovariectomized gilts: interralations among hormonal status, placental development, fetal
fluids and fetal growth. Journal of Animal Science, Champaign, v. 44, n. 4, p. 620-
637, 1977.
KOUBA, A.J. et al. Effects of the porcine oviduct-specific glycoprotein on fertilization,
polyspermy and embryonic development in vitro. Biology of Reproduction,
Champaign, v. 63, n. 1, p. 242-250, 2000.
KÜHHOLZER, B. et al. Repeated endoscopic ovum pick-up in hormonally untreated
ewes: a new technique. Theriogenology, Los Altos, v. 48, n. 4, p. 545-550, 1997.
LAMBERSON, W.R. et al. Direct responses to selection for increased litter size,
decreased age at puberty, or random selection following selection for ovulation rate in
swine. Journal Animal Science. Champaign, v. 69, n. 8, p. 3129-3143, 1991.
LEIBFRIED, L.; FIRST, N.L. Characterization of bovine follicular oocytes and their
ability to mature in vitro. Journal Animal Science, Savoy, v. 48, n. 1, p. 76-86, 1979.
LENZ, S.; LAURITSEN, J.G. Ultrasonically guided percutaneous aspiration of human
follicles under local anaesthesia: a new method of collecting oocytes for in vitro
fertilization. Fertility and Sterility, Birmingham, v. 38, n. 6, p. 673-677, 1982.
LONGENECKER, D.E.; DAY, B.N. Fertility level of sows superovulated at post weaning
estrus. Journal of Animal Science, Champaign, v.27, n. 3, p. 709-711, 1968.
LOVE, R.J.; EVANS, G.; KLUPIEC, C. Seasonal effects on fertility in gilts and sows.
Journal of Reproduction and Fertility, Cambridge, v. 48, n. 1, p. 191-206, 1993.
MACHATY, Z.; DAY, B.N.; PRATHER, R.S. Development of early porcine embryos in
vitro and in vivo. Biology of Reproduction, Champaign, v. 59, n. 2, p. 451-455, 1998.

Normas da ABNT-NR 6023
55
MACHATY, Z. et al. Inhibitors of mitochondrial ATP production at the time of
compaction improve development of in vitro produced porcine embryos. Molecular
Reproduction and Development, New York, v. 58, n. 1, p. 39-44, 2001.
MARCHAL, R. et al. Effect of follicular size on meiotic and developmental competence
of porcine oocytes. Theriogenology, Los Altos, v. 57, n. 5, p. 1523-1532, 2002.
MARTINEZ-MADRID, B. et al. Effect of IVF medium and sperm concentration on
fertilization parameters. In: INTERNATIONAL OF CONFERENCE OF PIG
REPRODUCTION, 6., 2001, Colombia. Proceedings…p. 74.
MARTINO, A. et al. Influence of the collection technique of prepubertal goat oocytes on
in vitro maturation and fertilization. Theriogenology, Los Altos, v. 42, n. 5, p. 859-873,
1994.
MATTIOLI, M.; GALEATI, G.; SEREN, E. Effect of follicle somatic cells during pig
oocyte maturation on egg penetrability and male pronucleus formation. Gamete
Research, New York , v. 20, n. 2, p. 177-183, 1998.
McCAULEY, T.C. Exposure of oocytes to porcine oviduct-specific glycoprotein reduces
the incidence of polyspermic penetration in vitro. In: INTERNATIONAL CONFERENCE
OF PIG REPRODUCTION, 6., 2001, Columbia. Proceedings… p. 47.
MCLAY, D.W.; CLARKE, H.J. Remodelling the paternal cromatin at fertilization in
mammals. Journal of Reproduction and Fertility, Cambridge, v. 125, n. 5, p. 625-
633, 2003.
MÉO, S.C. Ativação partenogenética e desenvolvimento embrionário inicial de oócitos
bovinos tratados com estrôncio, ionomicina e 6-dimetilaminopurina. 2002. 86f.
Dissertação (Mestrado em Reprodução Animal) – Faculdade de Ciências Agrárias e
Veterinárias “Júlio de Mesquita Filho”, Universidade Estadual Paulista, Jaboticabal,
2002.

Normas da ABNT-NR 6023
56
MORI, T.; AMANO, T.; SHIMIZU, H. Roles of gap junctional communication of cumulus
cells in cytoplasmic maturation of porcine oocytes cultured in vitro. Biology of
Reproduction, Champaign, v. 62, n. 2, p. 913-919, 2000.
MOTLIK, J.; FULKA, J. Factors affecting meiotic competence in pig oocytes.
Theriogenology, Los Altos, v. 25, n. 1, p. 87-96, 1986.
MOTLIK, J.; CROZET, N.; FULKA, J. Meiotic competence in vitro of oocytes isolated
from early antral follicles. Journal of Reproduction and Fertility, Cambridge, v. 72,
n. 2, p. 323-328, 1984.
MUNIZ, A.; MORETTI, A.S.; RODRIGUES, P.H.M. Efeito da presença do macho na
indução e duração do estro em marrãs, submetidas à alimentação, à vontade e restrita.
In: CONGRESSO LATINO AMERICANO DE SUINOCULTURA, 1., 2002, Foz do
Iguaçu. Anais... Foz do Iguaçu: Associação Latinoamericana de Suinocultura, 2002.
p. 157-158.
MUZII, L. et al. Laparoscopic excision of ovarian cysts: is the stripping technique a
tissue-sparing procedure?. Fertility and Sterility, Birmingham, v. 77, n. 3, p. 604-614,
2002.
NAGAI, T. The improvement of in vitro maturation systems for bovine and porcine
oocytes. Theriogenology, Los Altos, v. 55, n. 6, p. 1291-1301, 2001.
NAGAI, T.; MOOR, R.M. Effect of oviduct cells on the incidence of polyspermy in pig
eggs fertilized in vitro. Molecular Reproduction and Development, New York, v. 26,
n. 4, p. 377-382, 1990.
NAGAI, T.; DING, J.; MOOR, R.M. Effect of follicle cells and steriodogenesis on
maturation and fertilization in vitro of pig oocytes. Journal Experimental Zoology, New
York, v. 266, n. 2, p. 146-151, 1993.
NAGASHIMA, H. et al. Removal of cytoplasmic lipid enhances the tolerance of porcine
embryos to chilling. Biology of Reproduction, Champaign, v. 51, n. 4, p. 618-622,
1994.

Normas da ABNT-NR 6023
57
NAKAZAWA, Y. et al. Replacement of nuclear protein by histone in pig sperm nuclei
during in vitro fertilization. Journal of Reproduction and Fertility, Cambridge, v. 124,
n. 4, p. 565-572, 2002.
NUNES, C.G.V. Efeito de um sistema de acondicionamento térmico do ambiente e
da nutrição sobre o desempenho reprodutivo de porcas gestantes. Dissertação
(Mestrado em Medicina Veterinária) Universidade Federal de Viçosa, Viçosa, 2001.
OKTAY, K.; KARLIKAYA, G.G.; AYDIN, B.A. Ovarian cryopreservation and
transplantation basic aspects. Molecular and Celular Endocrinology, Amsterdam, v.
169, n. 1-2, p. 105-108, 2000.
OMTVEDT, I. T. et al. Influence of heat stress during early, mid and late pregnancy of
gilts. Journal of Animal Science, Champaign, v. 32, n. 2, p. 312-317, 1971.
PAPAIOANNOU, V.E.; EBERT, K.M. The preimplantation pig embryo: Cell number and
allocation to trophectoderm and inner cell mass of the blastocyst in vivo and in vitro.
Development, Cambridge, v. 102, n. 1, p. 793-803, 1998.
PATERSON, A.M. The controled induction of puberty. In: D.J.A.; G.R. (Ed.). Control
of pig reproduction. London: Butterworths, 1982. p. 139-159.
PERREAULT, S.D.; BARBEE, R.R.; SLOTT, V.I. Importance of glutathione in the
acquisition and maintenance of sperm nuclear decondensing activity in maturing
hamster oocytes. Development of Biology, Orlando, v. 125, n. 1, p. 181-186, 1998.
PERRY, J.S.; ROWLANDS, I. W. Early pregnancy in the pig. Journal of
Reproduction and Fertility, Cambridge, v. 4, n. 1, p. 175-188, 1962.
POLLARD, J.W.; LEIBO S.B. Chilling sensitivity of mammalian embryos.
Theriogenology, Los Altos, v. 41, n. 1, p. 101-106, 1994.
POPE, W.F.; FIRST, N.L. Factors affecting the survival of pig embryos.
Theriogenology, Stonehan, v. 23, n. 1, p. 91-105, 1985.

Normas da ABNT-NR 6023
58
PROCHAZKA, R. et al. Devepmental regulation of effect of epidermal growth factor on
porcine oocyte-cumulus cells complexes: nuclear maturation, expansion, and F-actin
remoldeling. Molecular Reproduction and Development, New York, v. 56, n. 1, p. 63-
73, 2000.
PTAK, G. et al. Offspring from one-mounth-old lambs: studies on the developmental
capability. Biology of Reproduction, Wisconsin, v. 61, n. 6, p. 1568-1574, 1999.
RATH, D. Experiments to improve in vitro fertilization techniques for in vivo-matured
porcine oocytes. Theriogenology, Los Altos, v. 37, n. 4, p. 885-896, 1992.
REYNOLDS, L.P.; REDMER, D.A. Utero-placental vascular development and placental
function. Journal of Animal Science, Champaign, v. 73, n. 1, p. 1839-1851, 1995.
RENOU, P. et al. The collection of human oocytes for in vitro fertilization. I. An
instrument for maximizing oocyte recovery rate. Fertility and Sterility, Birmingham,
v. 35, n. 4, p. 409-412, 1981.
RILLO, S.M. et al. Efecto del aparato genital de la primeriza sobre la productividad de la
cerda. In: SIMPOSIUM INTERNACIONAL DE REPRODUCCIÓN E INSEMINATION
ARTIFICIAL PORCINA, 6.,1999, Madrid. Anais... Madrid: Ministerio de Agricultura,
Pesca y Alimentacion, 1999. p. 63-72.
RODRIGUEZ-MARTNEZ, H. et al. Production of hyaluronic acid by porcine oocyte-
cumulus cell complexes during in vitro maturation. In: INTERNATIONAL CONFERENCE
OF PIG REPRODUCTION, 5., 1997, Kerkrade. Proceedings…p. 55.
ROMAR, R. et al. In vitro fertilization of porcine oocytes pre-cultured in the presence of
porcine oviductal epithelial cells. Theriogenology, Los Altos, v. 53, n. 1 , p. 98-104,
2000.
ROTHSCHILD, M.F. Genetics and reproduction in the pig. Animal Reproduction
Science, Amsterdam, v. 42, n. 1, p. 143-151, 1996.

Normas da ABNT-NR 6023
59
ROZEBOOM, D.W. et al. Influence of gilt age and body composition at first breeding on
sow reproductive performance and longevity. Journal of Animal Science, Champaign,
v. 74, n.1, p. 138-150, 1996.
RYDHMER, L. Genetics of sow reproduction, including puberty, oestrus, pregnancy,
farrowing and lactation. Livestock Production Science, Oxon, v. 66, n. 1, p. 1-12,
2000.
SALUSTRI, A.; YANAGISHITA, M.; HASCALL, V.C. Synthesis and accumulation of
hyaluronic acid and proteoglycans in the mouse cumulus cell-oocyte complex during
follicle-stimulating hormone-induced mucification. Journal of Biological Chemistry,
Baltimore, v. 264, n. 1, p. 13840-13847, 1989.
SHIMADA, M.; TERADA, T. Phosphatidylinosotol 3- kinase in cumulus cells and
oocytes is responsible for activation of oocyte mitogen-activated protein kinase during
meiotic progression beyond the meiosis I stage in pigs. Biology of Reproduction,
Madison, v. 64, n. 4, p. 1106-1114, 2001.
SHIMADA, M.; MAEDA, T.; TERADA, T. Dinamic changes of connexin-43, gap
junctional protein, in outer layer of cumulus cells are regulated by PKC and PI 3-kinase
during meiotic resumption in porcine oocytes. Biology Reproduction, Madison, v. 64,
n. 1, p. 124-131, 2001.
SINGH, B. et al. Effects of epidermal growth factor an growth factor and follicle
stimulating hormone during in vitro maturation on cytoplasmic maturation of porcine
oocytes. Molecular Reproduction Development, New York, v. 46, n. 3, p. 401-407,
1997.
SOEDE, M.N; KEMP, B. Expression of oestrus and timing of ovulation in pigs. Journal
of Reproduction ans Fertility, Cambridge, v.52, suppl., p. 91-103, 1997.
SOEDE, M.N; KEMP, B. In synchronized pigs, the duration of ovulation is not affected
by insemination and is not a determinant for early embryonic diversity.
Theriogenology, Stoneham, v. 39, n. 5, p. 1043-1053, 1993.

Normas da ABNT-NR 6023
60
SOEDE, M.N.; HELMOND, F.A.; KEMP, B. Periovulatory profiles oestradiol, LH and
progesterone in relation to oestrus and embryo mortality in multiparous sows using
transrectal ultrasonography to detect ovulation. Journal of Reproduction and Fertility,
Cambridge, v. 101, n. 3, p. 633-641, 1994.
SOEDE, N.M; NOORDHUIZEN, J.P.T.M.; KEMP, B. The duration of ovulation in pigs,
studied by transrectal ultrasonography, is not related to early embryonic diversity.
Theriogenology, Wageningen, v. 38, n. 4, p. 653-666, 1992.
STANGL, M. et al. Repeated endoscopic ovum pick-up in sheep. Theriogenology, Los
Altos, v. 52, n. 4, p. 709-716, 1999.
STATSOFT. Statistica: data analysis software system, version 7. Disponível em:
www.statsoft.com. Acesso em: 20 Janeiro 2006.
STUBBINGS, R.B.; WOSIK, C.; ARMSTRONG, D.T. Ovarian response in calves to
multiple versus a single subcutaneous injection of Folltropin. Theriogenology, Los
Altos, v. 39, n. 1, p. 321, 1993.
SUN, F. Z. et al. A comparison of intracellular changes in porcine eggs after fertilization
and electroactivation. Development, Cambridge, v. 115, n. 4, p. 947-956, 1992.
SUN, Q.Y.; NAGAI, T. Molecular mechanisms underlying pig oocyte maturation and
fertilization. Journal of Reproduction and Development, Tokio, v. 49, n. 5, p. 347-
359, 2003.
SUN, Q.Y. et al. Cytoplasmic chances in relation to nuclear maturation and early
embryo developmental potencial of porcine oocytes: effects of gonadotropins, cumulus
cells, follicular size and prothein synthesis inhibition. Molecular Reproduction and
Development, New York, v. 59, n. 1, p. 131-137, 2001a.
SUN, Q.Y. et al. Dynamic events are differently mediated by microfilaments,
microtubules, and mitogen-activated protein kinase during porcine oocyte maturation
and fertilization in vitro. Biology of Reproduction, Champaign, v. 64, n. 3, p. 879-889,
2001b.

Normas da ABNT-NR 6023
61
SUN, Q.Y. et al. Microtubule assembly after treatment of pig oocytes with taxol:
correlation with chromosomes, gamma-tubulin, and MAP kinase. Molecular
Reproduction and Development, New York, v. 60, n. 4, p. 481-490, 2001c.
SUN, Q.Y. et al. Traslocation of active mitochondria during pig oocyte maturation,
fertilization and early embryo development in vitro. Reproduction, Cambridge, v. 122,
n. 1, p. 155-163, 2001d.
SUN, H.S. et al. Neonatal Meishan pigs show POU1F1 genotype effects on plasma GH
and PRL concentration. Animal Reproduction Science, Amsterdam, v.69, n.3-4,
p. 223-237, 2002.
SUZUKI, K. et al. Effect of hyaluronan on monospermic penetration of porcine oocytes
fertilized in vitro. International Journal of Andrology,Oxford, v. 23, n. 1, p. 13-21,
2000.
TALAMINI, D.J.D. As questões dos preços na suinocultura. Concórdia: TERVIT, H.
R. Laparoscopy/laparotomy oocyte recovery and juvenile breeding. Animal
Reproduction Science, Amsterdam, v. 42, n. 1-4, p. 227-238, 1996.
TIENTHAI, P. et al. Localization and quantitation of hyaluronan and sulfated
glycosaminoglycans in the tissues and intraluminal fluid of the pig oviductl.
Reproduction, Fertility and Development, East Melbourne, v. 12, n. 3-4, p. 173-182,
2000.
TIENTHAI, P; JOHANNISSON, A.; RODRIGUEZ-MARTINEZ, H. Sperm capacitation in
the porcine oviduct. Animal Reproduction Science, Amsterdam, v. 80, n. 1, p. 131-
146, 2004.
VALLET, J.C. et al. Feasibility and repeatability of embryo recoveries from dairy goats
under laparoscopy. In: SCIENTIFIC MEETING OF EUROPEAN EMBRYO TRANSFER
ASSOCIATION, 3., 1987, Lyon. Proceedings… p. 60.

Normas da ABNT-NR 6023
62
VAN DER LENDE, T.; SCHOENMAKER, G.J.W. The relationship between ovulation
rate and litter size before and after Day 35 of pregnancy in gilts and sows: an analysis of
published data. Livestock Production Science, Oxon, v. 26, n. 3, p. 217-229, 1990.
VANROOSE, G.; DE KRUIF, A.; VAN SOOM, A. Embryonic mortality and embryo-
pathogen interactions. Animal Reproduction Science, Amsterdam, v. 60-61, n. 1, p.
131-143, 2000.
VATZIAS, G.; HAGEN, D.R. Effects of porcine follicular fluid and oviduct-conditioned
media on maturation and fertilization of porcine oocytes in vitro. Biology of
Reproduction, Champaign, v. 60, n. 1, p. 42-48, 1999.
VONNAHME, K.A. et al. Impacts on conceptus survival in a commercial swine heard.
Journal of Animal Science, Champaign, v. 80, n. 3, p. 553-559, 2002.
XIA, P. et al. Ultrastructural study of polyspermy during early embryo development in
pigs observed by scanning electron microscope and transmission electron microscope.
Cell and Tissue Research, Berlin, v. 303, n. 2, p. 271-275, 2001.
XIE, S; BROERMANN, K.P; GEISERT, R.D; POPE, W.F. Ovulation and early
embryogenesis in Swine. Biology of Reproduction, Ohio, v. 43, n. 1, p. 236-240, 1990.
XU, X. et al. In vitro fertilization of in vitro matured pig oocytes: Effects of boar and
ejaculate fraction. Theriogenology, Los Altos, v. 45, n. 4, p. 745-755, 1996.
XU, X. et al. Semen dilution for assessment of boar ejaculate quality in pig IVM and IVF
systems. Theriogenology, Los Altos, v. 46, n. 8, p. 1325-1337, 1996.
WALTERS, E.M.; GRAVES, C.N. Transportation and storage effects on porcine
ovaries. Journal of Animal Science, Champaign, v. 76, suppl. 2, p. 69, 1998. Abstract.
WANG, W.H.; NIWA, K. Synergetic effects of epidermal growth factor and
gonadotropins on the cytoplasmatic maturation of pig oocytes in a serum free medium.
Zygote, Winnepeg, v. 51, n. 3, p. 345-350, 1995.

Normas da ABNT-NR 6023
63
WANG, W.H.; ABEYDEERA, L.R.; PRATHER, R.S.; DAY, B.N. Morphologic comparison
of ovulated and in vitro-matured porcine oocytes, with particular reference to polyspermy
after in vitro fertilization. Molecular Reproduction Development, Columbia, v. 49, n. 3,
p. 308-316, 1998.
WANG, W.H.; HOSOE, M.; SHIOYA, Y. Induction of cortical granule exocytosis of pig
oocytes by spermatozoa during meiotic maturation. Journal of Reproduction and
Fertility, Cambridge, v. 109, n. 2, p. 247-255, 1997a.
WANG, W.H. et al. Morphologic comparison of ovulated and in vitro-matured porcine
oocytes, with particular reference to polyspermy after in vitro fertilization. Molecular
Reproduction and Development, New York, v. 49, n. 3, p. 308-316, 1998.
WANG, W.H. et al. Quantified analysis of cortical granule distribuition and exocytosis of
porcine oocytes during meiotic maturation and activation. Biology of Reproduction,
Champaign, v. 56, n. 6, p. 1376-1382, 1997b.
WANI, N.A. In vitro maturation and in vitro fertilization of sheep oocytes. Small
Ruminant Research, Amsterdam, v. 44, n. 2, p. 89-95, 2002.
WETTEMAN, R.P.; BAZER, F.W. Influence of environmental temperature on prolificacy
of pigs. Journal of Reproduction and Fertility, Cambridge, v. 33, n. 2, p. 199-208,
1985.
WILMUT, I. The low temperature preservation of mammalian embryos. Journal of
Reproduction and Fertility, Cambridge, v. 31, n. 4, p. 513-514, 1972.
WILMUT, I.; et al. Viable offspring derived from fetal and adult mammalian cell. Nature,
London, n. 385, p. 810-813, 1997.
WILSON, M.E.; BIENSEN, N.J.; FORD, S.P. Novel insight into the control of litter size in
pigs, using placental efficiency as a selection tool. Journal of Animal Science,
Champaign, v. 77, n. 6, p. 1654-1658, 1999.

Normas da ABNT-NR 6023
64
WU, M.C; DIZIUK, P.J. Relationship of length of uterus in prepubertal pigs and number
of corpora lútea and fetuses at 30 days of gestation. Animal Reproduction Science,
Illinois, v. 38, n. 4 , p. 327-336, 1995.
WU, M.C.; HENTZEL, M.D.; DIZIUK, P.J. Relationships between uterine length and
number of fetuses and prenatal mortality in pigs. Journal of Animal Science,
Champaingn, v. 65, n. 3, p. 762-770, 1987.
YAMAUCHI, N.; NAGAI, T. Male pronuclear formation in denuded porcine oocytes after
in vitro maturation in the presence of cysteamine. Biology of Reproduction,
Champaign, v. 61, n. 3, p. 828-833, 1999.
YANAGIMACHI, R. Mammalian fertization, Ch 5. In: KNOBIL, E.; NEILL, J. D. (Ed.).
The physiology of reproduction, 2.ed., New York: Raven Press, 1994.
YOON, K.W. et al. Development of porcines oocytes from preovulatory follicles from
different sizes after maturation in media supplemented with follicular fluids.
Reproduction, Fertility and Development, Tokio, v. 12, n. 3-4, p. 133-139, 2000.
YOSHIDA, M.; CRAN, D.G.; PURSEL, V.G. Confocal and fluorescence microscopic
study using lectins of the distribution of cortical granules during the maturation and
fertilization of pig oocytes. Molecular Reproduction and Development, New York, v.
36, n. 4, p. 462-468, 1993.
YOSHIDA, M. et al. Glutathione concentration during maturation and after fertilization in
pig oocytes: relevance to the ability of oocytes to form male pronucleus. Biology of
Reproduction, Champaign, v. 49, n. 1, p. 89-94, 1993.
YOUNG, L.D.; LEYMASTER, K.; LUSTRA, D.D. Genetic- Variation in testicular
development and its relationship to female reproductive traits in swine. Journal of
Animal Science, Champaign, v. 63, n. 1, p. 17-26, 1986.





























