Embed Size (px)
Citation preview

- 1 -
CLASSIFICAÇÃO E ORDENAÇÃO DA COMUNIDADE ARBÓREA DA
FLORESTA OMBRÓFILA MISTA DA FLONA DE SÃO FRANCISCO
DE PAULA, RS
________________________________
Sylviane Beck Ribeiro

- 2 -
UFSM
Tese de Doutorado
CLASSIFICAÇÃO E ORDENAÇÃO DA COMUNIDADE
ARBÓREA DA FLORESTA OMBRÓFILA MISTA DA
FLONA DE SÃO FRANCISCO DE PAULA, RS
________________________________
Sylviane Beck Ribeiro
PPGEF
Santa Maria, RS, Brasil
2004

- 3 -
Universidade Federal de Santa Maria
Centro de Ciência Rurais
CLASSIFICAÇÃO E ORDENAÇÃO DA COMUNIDADE
ARBÓREA DA FLORESTA OMBRÓFILA MISTA DA
FLONA DE SÃO FRANCISCO DE PAULA, RS
________________________________
por
Sylviane Beck Ribeiro
Tese apresentada ao Programa de Pós-Graduação em
Engenharia Florestal, Área de Concentração em Silvicultura,
da Universidade Federal de Santa Maria (UFSM, RS), como
requisito parcial para o grau de
Doutor em Engenharia Florestal
PPGEF
Santa Maria, RS, Brasil
2004

- 4 -
Programa de Pós-Graduação em Engenharia Florestal
A Comissão Examinadora, abaixo assinada,
aprova a Tese de Doutorado
CLASSIFICAÇÃO E ORDENAÇÃO DA COMUNIDADE ARBÓREA DA
FLORESTA OMBRÓFILA MISTA DA FLONA DE SÃO FRANCISCO
DE PAULA, RS
elaborada por
Sylviane Beck Ribeiro
como requisito parcial para obtenção do grau de
Doutor em Engenharia Florestal
COMISSÃO EXAMINADORA:
______________________________________
Prof. Dr. Solon Jonas Longhi (Presidente/Orientador)
_____________________________________
Profa. Dra. Sônia Maria Eisinger
______________________________________ Prof. Dr. Sylvio Péllico Netto
______________________________________
Prof. Dr. Miguel Antão Durlo
_______________________________________ Prof. Dr. Doádi Antônio Brena
Santa Maria, 26 de julho de 2004

- 5 -
Dedico aos meus pais Demetrio Ribeiro e Dormecy Beck Ribeiro (in memorian), e às minhas irmãs Rosa Emilia Beck Ribeiro e Rosamari Beck Ribeiro que me apoiaram em
todos os momentos de minha vida.

- 6 -
AGRADECIMENTOS
Não é fácil agradecer todas as pessoas que participam no desenvolvimento de uma tese de doutorado, logo, peço desculpas a qualquer pessoa que esquecer o nome.
Agradeço ao meu orientador Prof. Solon Jonas Longhi, pelas inúmeras vezes que
me apoiou; e pela tranqüilidade e paz que me transmitiu para trabalhar. Ao meu co-orientador Doádi Antônio Brena pelo apoio e disponibilidade da logística
que foi necessária para o desenvolvimento do trabalho. Aos professores do Curso de Pós-Graduação em Engenharia Florestal da UFSM. Aos amigos Andréa Magalhães Freitas, Maristela Machado Araujo, Juliana
Fernandes Gomes, Igor da Silva Narvaes, André Terra Nascimento, Tales Eduardo Rodrigues, Angelise Vieira Mendes, Luis Fernando Alberti.
Ao Programa de Pós-Graduação em Engenharia Florestal, secretárias Lílian e Tita. Ao Projeto de Longa Duração PELD-CNPQ, pelo aporte financeiro e logístico, ao
longo de toda a coleta de dados.

- 7 -
SUMÁRIO
Lista de Tabelas x
Lista de Figuras xiv
Resumo xv
Abstract xvii
1 INTRODUÇÂO 1
2 REVISÃO DA LITERATURA 5
2.1 Caracterização geral da Região 5
2.1.1 Geologia 5
2.1.2 Relevo 5
2.1.3 Solo 6
2.1.4 Clima 7
2.1.5 Vegetação primitiva 8
2.2 Fatores Ecológicos e de concorrência 12
2.2.1 Umidade do solo 14
2.2.2 Inclinação do solo 16
2.2.3 Profundidade do solo 17
2.2.4 Cor do solo 18
2.2.5 Exposição à luz 19
2.2.6 Quantidade de pedra 21
2.2.7 Densidade do sub-bosque 22
2.3 Parâmetros Fitossociológicos 23
2.4 Diversidade 27
2.5 Análise de Similaridade 29
2.6. Classificação e Ordenação 31
2.6.1 Técnicas de Classificação 32
2.6.1.1 Análise de Agrupamento (Cluster) 33
2.6.1.2 Análise de Discriminante 38
2.6.2 Técnicas de Ordenação 40
2.6.2.1 Componentes Principais 40
2.6.2.2 Análise Fatorial 42

- 8 -
2.6.2.3 Análise de Correspondência Canônica (CCA) 43
2.7 Regressão Logística 44
3 MATERIAL E MÉTODOS 47
3.1 Caracterização geral da FLONA 47
3.2 Histórico da FLONA 49
3.3 Métodos de levantamentos 51
3.3.1 Método de amostragem 51
3.3.2 Coleta de dados 52
3.3.2.1 Inclinação e exposição do terreno 53
3.3.2.2 Área basal, distribuição de freqüência e densidade do sub-
bosque 54
3.3.2.3 Fatores físicos do solo 54
3.4 Métodos de Análise 57
3.4.1 Análises Fitossociológica e Biométrica 59
3.4.2 Análise de Agrupamento (Cluster) 62
3.4.3 Análise do Discriminante 63
3.4.4 Análise de Similaridade 64
3.4.5 Análise de Correspondência Canônica 65
3.4.6 Análise de Regressão Logística 66
4 RESULTADOS E DISCUSSÃO 71
4.1 Caracterização geral da floresta 71
4.1.1 Composição florística 71
4.1.2 Estrutura fitossociológica 76
4.1.3 Parâmetros dendrométricos 78
4.2 Análise de agrupamento (Cluster - TWINSPAN) 80
4.3 Significância dos agrupamentos (Análise
Discriminante) 83
4.4 Similaridade entre os grupos florísticos obtidos 97
4.5 Caracterização geral dos grupos florísticos obtidos 100
4.5.1 Grupo 1 – Formação Podocarpus 100
4.5.1.1 Composição florística 100
4.5.1.2 Estrutura fitossociológica 101

- 9 -
4.5.1.3 Parâmetros dendrométricos 103
4.5.2 Grupo 2 – Formação Sebastiania 104
4.5.2.1 Composição florística 104
4.5.2.2 Estrutura fitossociológica 105
4.5.2.3 Parâmetros dendrométricos 108
4.5.3 Grupo 3 – Formação Araucaria 109
4.5.3.1 Composição florística 109
4.5.3.2 Estrutura fitossociológica 110
4.5.3.3 Parâmetros dendrométricos 113
4.6 Análise de gradientes ambientais (CCA) 114
4.6.1 Grupo 1 – Formação Podocarpus 114
4.6.2 Grupo 2 – Formação Sebastiania 118
4.6.3 Grupo 3 – Formação Araucaria 121
4.7 Regressão Logística 125
4.7.1 Podocarpus lambertii (Pinheiro-bravo) 126
4.7.2 Araucaria angustifolia (Pinheiro - brasileiro) 129
4.7.3 Cryptocarya aschersoniana (Canela-vick) 132
4.7.4 Sebastiania commersoniana (Branquilho-comum) 135
4.7.5 Sebatiania brasiliensis (Branquilho-leiteiro) 138
4.8 Contribuição silvicultural 142
5 CONCLUSÕES E RECOMENDAÇÕES 143
5.1 Conclusões 143
5.2 Recomendações 147
6 REFERÊNCIAS BIBLIOGRÁFICAS 149
ANEXOS 162

- 10 -
LISTA DE TABELAS
TABELA 1 - Classificação topográfica do terreno de acordo com
os graus de declividade do solo 17
TABELA 2 - Classes e valores utilizados para a avaliação da
profundidade do solo 17
TABELA 3 - Classificação da pedregosidade na superfície do
terreno 22
TABELA 4 - Tabela de contingência entre duas amostras 1 e 2 29
TABELA 5 - Modelo da matriz de dados com a densidade das
espécies para análise de agrupamento pelo
programa TWINSPAN 62
TABELA 6 - Modelo da primeira matriz de dados para Análise
Canônica pelo programa TWINSPAN (densidade
das espécies x parcelas) 65
TABELA 7 - Modelo da segunda matriz de dados para Análise de
Correspondência Canônica (fatores ecológicos x
parcelas) 66
TABELA 8 - Modelo de matriz de dados com a
presença/ausência das espécies e fatores
ecológicos e de concorrência por parcela para
Análise de Regressão Logística 67
TABELA 9 - Parâmetros estimados para o modelo de regressão
logística 68
TABELA 10 - Espécies encontradas na área da FLONA de São
Francisco de Paula, RS, com as respectivas famílias,
nomes científicos e populares 72
TABELA 11 - Parâmetros fitossociológicos para as espécies
arbóreas e arbustivas, amostra com CAP ≥ 30 cm,
na FLONA de São Francisco de Paula, RS 77
TABELA 12 Parâmetros dendrométricos da Floresta Ombrófila
Mista da FLONA de São Francisco de Paula, RS e
do Estado 79

- 11 -
TABELA 13 - Estatística do teste de seleção de espécies
(variáveis) discriminantes para a matriz de dados
brutos.
84
TABELA 14 - Estatística do teste de seleção de variáveis
discriminantes, em ordem de entrada, após 41
Steps. 88
TABELA 15 - Estatística do teste de seleção das funções
discriminantes: autovalores, variância explicada e
correlação canônica. 89
TABELA 16 - Estatística do teste de seleção das funções
discriminantes: Lambda de Wilks, Chi-quadrado,
grau de liberdade e significância. 89
TABELA 17 - Coeficientes padronizados das funções
discriminantes canônicas, para a matriz de dados
brutos. 91
TABELA 18 - Coeficientes das funções discriminantes para cada
um dos grupos florísticos obtidos. 93
TABELA 19 - Centróides das funções discriminantes canônicas
avaliados para os três grupos florísticos
encontrados. 94
TABELA 20 - Número de casos e percentagens de classificações
das parcelas nos três grupos florísticos
determinados. 96
TABELA 21 - Diversidade e Similaridade entre os grupos florísticos
na Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, RS 97
TABELA 22 - Espécies exclusivas dos grupos florísticos da
Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, RS 98
TABELA 23 - Estrutura horizontal do Grupo 1, da FLONA de São
Francisco de Paula, RS 102

- 12 -
TABELA 24 - Parâmetros dendrométricos dos grupos florísticos da
Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, RS 104
TABELA 25 - Estrutura horizontal do Grupo 2, da FLONA de São
Francisco de Paula, RS 107
TABELA 26 - Estrutura horizontal do Grupo 3, da FLONA de São
Francisco de Paula, RS 111
TABELA 27 - Coeficiente de correlação entre as variáveis
ambientais e os dois primeiros eixos de ordenação
da análise de correspondência canônica do Grupo 1.
FLONA de São Francisco de Paula, RS 115
TABELA 28 - Coeficiente de correlação entre as variáveis
ambientais e os dois primeiros eixos de ordenação
da análise de correspondência canônica do Grupo 2.
FLONA de São Francisco de Paula, RS 118
TABELA 29 - Coeficiente de correlação entre as variáveis
ambientais e os dois primeiros eixos de ordenação
da análise de correspondência canônica do Grupo 3.
FLONA de São Francisco de Paula, RS 122
TABELA 30 - Tabela de classificação da regressão logística para a
variável dependente Podocarpus lambertii 127
TABELA 31 - Variáveis independentes selecionadas pela
regressão logística pelo método Forward Stepwise,
para Podocarpus lambertii 127
TABELA 32 - Tabela de classificação da regressão logística para a
variável dependente Araucaria angustifolia 130
TABELA 33 - Variáveis independentes selecionadas pela
regressão logística pelo método Forward Stepwise,
para Araucaria angustifolia 130
TABELA 34 - Tabela de classificação da regressão logística para a
variável dependente Cryptocarya aschersoniana 133

- 13 -
TABELA 35 - Variáveis independentes selecionadas pela
regressão logística pelo método Forward Stepwise,
para Cryptocaria aschersoniana 134
TABELA 36- Tabela de classificação da regressão logística para a
variável dependente Sebastiania commersoniana 136
TABELA 37 - Variáveis independentes selecionadas pela
regressão logística pelo método Forward Stepwise,
para Sebatiania commersoniana 137
TABELA 38 - Tabela de classificação da regressão logística para a
variável dependente Sebatiania brasiliensis 140
TABELA 39 - Variáveis independentes selecionadas pela
regressão logística pelo método Forward Stepwise
para Seatiania brasiliensis 140
TABELA 40 - Características dos grupos florísticos obtidos na
FLONA de São Francisco de Paula, RS 143

- 14 -
LISTA DE FIGURAS
FIGURA 1 - Estado do Rio Grande do Sul, com a localização da
Cidade de São Francisco de Paula e Área da FLONA 48
FIGURA 2 - Conglomerado (parcela) quadrado de 10.000 m2 de
superfície, subdividido em 100 subunidades de 100m2 51
FIGURA 3 - Fluxograma do procedimento de análise 58
FIGURA 4 - Classificação das unidades amostrais avaliadas na
FLONA de São Francisco de Paula, em três grupos
florísticos 81
FIGURA 5 - Mapa territorial dos grupos florísticos determinados,
com a localização de seus respectivos centróides. 95
FIGURA 6 - Diagrama de ordenação produzidos pela análise de
correspondência canônica, baseado nos dados das
espécies mais abundantes do Grupo 1. FLONA de
São Francisco de Paula, RS 116
FIGURA 7 - Diagrama de ordenação produzidos pela análise de
correspondência canônica, baseado nos dados das
espécies mais abundantes do Grupo 2. FLONA de
São Francisco de Paula, RS 119
FIGURA 8 - Diagrama de ordenação produzidos pela análise de
correspondência canônica, baseado nos dados das
espécies mais abundantes do Grupo 3 da FLONA de
São Francisco de Paula, RS 123

- 15 -
RESUMO Tese de Doutorado
Programa de Pós-Graduação em Engenharia Florestal Universidade Federal de Santa Maria, RS, Brasil
CLASSIFICAÇÃO E ORDENAÇÃO DA COMUNIDADE ARBÓREA DA FLORESTA OMBRÓFILA MISTA DA FLONA DE SÃO FRANCISCO DE PAULA, RS
Autora: Sylviane Beck Ribeiro Orientador: Solon Jonas Longhi
Local e Data da Defesa: Santa Maria, 26 de julho de 2004.
A Floresta Ombrófila Mista é um ecossistema bastante complexo, que se encontra atualmente bastante fragmentado com escassos remanescentes, que precisam ser manejados e, para isso, necessitam de estudos que visam o entendimento de sua estrutura e dinâmica. A área onde foi realizado o estudo pertence à Floresta Nacional de São Francisco de Paula (FLONA), com área de 1.606,70 ha. Localiza-se a 930 m de altitude, no município de São Francisco de Paula a 27 Km da cidade, entre as coordenadas geográficas 29023’ e 29027' de latitude sul e 50023' e 50025’ de longitude oeste, na serra gaúcha, nordeste do Estado do Rio Grande do Sul, Brasil, na microrregião dos Campos de Cima da Serra. O trabalho teve como objetivos: (i) identificar as espécies que constituem a Floresta Ombrófila Mista de São Francisco de Paula; (ii) encontrar os fatores ambientais que delimitam as comunidades que compõem a floresta e como estes atuam sobre ela; (iii) determinar agrupamentos florísticos distintos e assim; (iv) fornecer subsídios básicos, acerca do comportamento destas espécies na Floresta Ombrófila Mista, podendo tais informações contribuírem para o seu estudo dentro da Ciência Florestal, mais precisamente nas áreas de Manejo Florestal, Silvicultura e Ecologia. A amostragem utilizada para a coleta de dados consistiu-se de 10 conglomerados permanentes de 100 m x 100 m (10.000 m2), os quais foram divididos em 10 faixas de 10 m x 100 m (1.000 m2) e estas subdivididas em 10 subunidades de 10 m x 10 m (100 m2), totalizando 100 subunidades amostrais em cada conglomerado. A Análise de Cluster, utilizando o programa TWINSPAN, foi utilizada para determinar os agrupamentos florísticos da vegetação arbórea da FLONA e a Análise de Correspondência Canônica (CCA) através do programa CANOCO para a determinação dos fatores ecológicos limitantes para a ocorrência das espécies indicadoras dentro dos grupos. As análises delinearam três grandes grupos. O primeiro grupo teve como espécie indicadora Podocarpus lambertii, Myrciaria tenella e Eugenia uruguayensis e os fatores limitantes do grupo foram: cor do solo (50 – 75 e > 75 de profundidade), profundidade do solo e quantidade de pedra. Obteve uma baixa diversidade (H’=1,7986) e uma alta similaridade com os outros grupos florísticos. O segundo grupo apresentou como espécies indicadoras Sebastiania commersoniana, Sebastiania brasiliensis e Cryptocarya aschersoniana e, como fatores ecológicos, umidade, quantidade de pedra, cor do solo e profundidade do solo. Teve, também, baixa diversidade florística (H’ = 1,6192) e alta similaridade com os outros grupos. A Araucaria angustifolia e Ilex paraguaiensis foram as espécies indicadoras do terceiro grupo, que apresentou como fatores limitantes inclinação, umidade do solo e cor do solo 0-50. Apresentou uma diversidade baixa de (H’= 1,4653) e também

- 16 -
uma alta similaridade com os outros grupos florísticos. Conclui-se que os grupos florísticos encontrados não diferem muito entre si, devido à baixa diversidade e alta similaridade florística, porém, as espécies formadoras dos grupos são exigentes a diferentes tipos de fatores ecológicos para sua ocorrência. A Regressão Logística, aplicada às espécies preferencias dos grupos, veio corroborar com os resultados encontrados pela CCA, isto é, foram encontrados os mesmos fatores ambientais relevantes para a presença das espécies indicadoras dos grupos.
Palavras-chave: Classificação, Ordenação, Floresta Ombrófila Mista

- 17 -
ABSTRACT Tese de Doutorado
Programa de Pós-Graduação em Engenharia Florestal Universidade Federal de Santa Maria, RS, Brasil
CLASSIFICATION AND ORDINATION OF THE ARBOREAL COMMUNITY OF THE MIXED OMBROPHYLOUS FOREST OF SÃO FRANCISCO DE PAULA’S FLONA.
Authora: Sylviane Beck Ribeiro Adviser: Solon Jonas Longhi
Place and Date of defence: Santa Maria, july, 26, 2004.
At present, the mixed ombrophylous forest is a very complex kind of phytogeographycal ecosystem, highly fragmented, with rare remanescents, and about which there are few studies. The area where this study was realized is located in the municipality of São Francisco de Paula – Rio Grande do Sul State, Brazil – 27 km far from the city, between 29°23’ and 29°27’S, and 50°23’ and 50°25’W., in the “serra gaúcha”, northeastern Rio Grande do Sul, in the micro region of the “Campos de Cima da Serra”. São Francisco de Paula’s FLONA’s total area is 1,606.70 ha and altitude of 930 m. This work aims to: (i) identify which species constitute São Francisco de Paula’s mixed ombrophylous forest; (ii) find which environmental factors delimit the communities that compound the forest and how such factors act on it; (iii) determine different floristic groups and then; (iv) provide basic subsidies about these species’ behavior in the mixed ombrophylous forest, since such information may contribute for its study in Forest Sciences, mainly in the areas of Forest Handling, Silviculture and Ecology. As sampling unities, we applied ten 100 m X 100 m lasting conglomerates (10,000 m² surface), divided into 10 lines of 10 m X 100 m (1,000 m²) and these ones were subdivided into 10 subunities of 10 m X 10 m (100 m²), in a total of 100 subunities. Cluster’s Analysis (TWINSPAN program) was used to determine the floristic groups that occurred in the FLONA. And Canonic Correspondence Analysis (CANOCO program, version 3.1) showed the limiting ecological factors to the occurrence of the indicatory species inside the groups. The analyses outlined three big groups. The first group obtained low diversity (H’=1,7986) and consequently a high similarity to the other floristic groups. It had Podocarpus lambertii , Myrciaria tenella and Eugenia uruguayensis, as its indicatory species, and its limiting factors were: soil color (50-75 e > 75 depth), soil depth and quantity of stone. The second group showed as indicatory species Sebastiania commersoniana, Sebastiania brasiliensis and Cryptocarya aschersoniana, and as ecological factors inclination, humidity, quantity of stone, soil color, and soil depth. Diversity, calculated by Shannon’s Index, was low (H’=1,6192) and similarity, calculated by Sorensen’s Index, was high among the groups. Araucaria angustifolia and Ilex paraguariensis was the indicatory species of the third group, which showed low diversity (H’= 1,4653), else a high similarity to other floristic groups. The conclusion is that the groups are not different one each other, because of low diversity and high floristic similarity, however the groups’ forming species demand different kinds of ecological factors for their occurrence. Logistic regression corroborates the results found

- 18 -
by CCA, that is, the groups’ indicatory species are determined by the same relevant environmental factors. Keywords: Classification, ordination; community, Mixed Ombrophylous forest

- 19 -
1 INTRODUÇÃO
A destruição das florestas é uma das maiores preocupações de nossa época. A
cada ano, segundo Perlin (1989), o mundo perde cerca de 15 milhões de hectares de
floresta. De acordo com as estimativas das Nações Unidas, entre 1950 e 1980, foram
destruídas quase 40% das florestas da América Central.
Entre os problemas relacionados com o desmatamento estão o esgotamento dos
estoques de madeira, que ainda é fonte primária de energia de três quartos da população
dos países em desenvolvimento, graves inundações, destruição acelerada do solo,
desertificação gradativa e diminuição da produtividade da terra. Se a taxa atual de
desmatamento não diminuir, esses problemas alcançarão enormes conseqüências, como
por exemplo, o desaparecimento da maioria das florestas tropicais ainda existentes e,
com elas, muitas espécies de plantas e animais da terra.
O Brasil apresenta uma expressiva diversidade de ecossistemas florestais devido à
sua grande área física e aos diversos tipos de clima e solo existentes em seu território.
Porém, os seus ecossistemas vêm sendo explorados de maneira predatória provocando
uma redução drástica de diversas comunidades vegetais com características especiais.
A Floresta Ombrófila Mista é um destes ecossistemas que se encontram
atualmente bastante fragmentados com escassos remanescentes. Existem poucos
estudos sobre a dinâmica e estrutura deste tipo fitogeográfico, bastante complexo,
constituído de diversos tipos de comunidades, variáveis ao longo de sua área de
distribuição natural. Tais estudos são importantes e necessários como subsídios para o
estabelecimento de uma política de conservação eficiente desses ecossistemas
(Nascimento, 2000).
Segundo Braun-Blanquet (1979) o conhecimento seguro do comportamento das
espécies em uma comunidade é de vital importância para o entendimento dos aspectos
fitossociológicos e dinâmicos das mesmas. Estudos florísticos da vegetação sem um
conhecimento suficiente das espécies são cientificamente inúteis. Os objetivos destes
estudos são reconhecer a importância das espécies e suas formas de vida na
comunidade e, assim, determinar as leis que regulam as relações dos organismos na
mesma. Portanto, não é possível alcançar uma definição precisa das unidades

- 20 -
fitossociológicas de uma comunidade, sem um conhecimento da composição florística e
do habitat (conjunto de fatores externos que atuam sobre a comunidade) das mesmas.
Muitos desses conhecimentos podem ser obtidos pela Fitossociologia, que
segundo Braun-Blunquet (1979), é a ciência que estuda os agrupamentos de plantas,
suas inter-relações e a dependência frente ao meio ambiente vivo e inanimado.
Estudos desta natureza foram utilizados para avaliar a composição, estrutura e
inter-relações ambientais do remanescente de Floresta Ombrófila Mista localizada na
Floresta Nacional (FLONA) de São Francisco de Paula - RS.
A FLONA de São Francisco de Paula possui área de 1.606 ha, das quais 901,2 ha
estão cobertas por florestas nativas e as restantes 704,8 ha por reflorestamentos
homogêneos com Araucaria angustifolia (Bertol.) Kuntze, Pinus spp. e Eucalyptus spp.,
açudes, barragens, estradas e outros. Tem uma produção florestal estimada, pelo plano
de manejo para comercialização de madeira, de 7.000 m3/ano. Possui uma fauna
abundante e bastante diversificada, sendo local propício para estudos de ecologia de
espécies, algumas ameaçadas de extinção. Há quantidade de água de boa qualidade,
com inúmeras nascentes que possibilitam a criação de peixes e aves aquáticas.
A FLONA possui como principais atividades a exploração de produtos e sub-
produtos florestais, como a madeira, a erva-mate e as sementes de Araucária; a
educação ambiental; a pesquisa e a investigação; a conservação e a preservação do
Patrimônio Natural, mantendo sempre o contato permanente com unidades e instituições
de pesquisa para o desenvolvimento de projetos.
O presente trabalho está vinculado a um "projeto-mãe", projeto PELD (Pesquisas
Ecológicas de Longa Duração), denominado "Conservação e Manejo Sustentável de
Ecossistemas Florestais", com previsão de estudos para até 2009. O "projeto-mãe"
assumiu características de transdisciplinaridade, pois envolve atividades conjuntas de três
Universidades brasileiras, a Universidade Federal do Paraná, a Universidade Federal de
Santa Maria e a Pontifícia Universidade Católica do Paraná. Terão a cooperação de
pesquisadores de outras três Universidades do exterior, Universidade de Freiburg -
Alemanha, Universidade de Ehime - Japão e Universidade de Michigan - USA.
Inicialmente o projeto tinha como enfoque caracterizar detalhada e integradamente
os processos, os mecanismos e os fatores relevantes em ecossistemas florestais,
principalmente no que diz respeito ao comportamento dinâmico das florestas e de sua

- 21 -
capacidade de regeneração natural, para assim propor indicativos concretos sobre a
dinâmica de funcionamento destes ecossistemas.
Após a coalizão das três Universidades estrangeiras, ao "projeto-mãe", seu objetivo
principal passou a ser:
"Quantificar e qualificar a longo prazo o grau de
alteração na produtividade (biomassa) e biodiversidade
de diversos ecossistemas florestais brasileiros em
função da magnitude e da intensidade das atividades
antrópicas."
Entre as metas, no campo científico, que o "projeto-mãe" pretende atingir, estão: (i)
montagem de uma coletânea do material de todas as espécies vegetais ocorrentes nos
ecossistemas abordados, que permitirá a correta identificação das espécies encontradas,
cujas exsicatas serão incorporadas aos herbários dos cursos de Engenharia Florestal; (ii)
caracterização da estrutura horizontal e vertical dos ecossistemas abordados, bem como
avaliar o comportamento das comunidades arbóreas ocorrentes, para oferecer subsídios
básicos para a proposta de Conservação e Manejo destes recursos florestais e; (iii)
caracterização dos ecossistemas abordados nas diferentes unidades experimentais
incluídas no projeto, para assim conhecer mais profundamente o nível de evolução dos
estágios sucessionais nos diferentes ecossistemas abordados para melhor formalizar a
proposta de sua conservação e manejo sustentável.
O presente trabalho pretende fornecer subsídios ao projeto PELD “Conservação e
Manejo Sustentável de Ecossistemas Florestais”, entre eles da FLONA de São Francisco
de Paula. Tem como objetivos:
a) identificar quais as espécies que constituem a Floresta Ombrófila Mista de São
Francisco de Paula;
b) encontrar quais fatores ambientais que delimitam as comunidades que compõem
a floresta e como estes atuam sobre ela;
c) determinar agrupamentos florísticos distintos e, assim;
d) fornecer subsídios básicos acerca do comportamento destas espécies na Floresta
Ombrófila Mista, podendo tais informações contribuir para o seu estudo dentro da
Ciência Florestal, mais precisamente nas áreas de Manejo Florestal, Silvicultura e
Ecologia.

- 22 -
2 REVISÃO DE LITERATURA
2.1 Caracterização geral da Região
2.1.1 Geologia
Na Região Sul o vulcanismo continental está representado por espessos e
extensos derrames de lavas, bem como por diques e soleiras, com pequenos e eventuais
corpos de rochas, sedimentares associados. Tal conjunto de litologias constitui a
Formação Serra Geral, dividida em duas porções – A Seqüência Básica e a Seqüência
Ácida (Kaul, 1990).
A Seqüência Básica da Formação da Serra Geral, compreende derrames de
basalto, andesito e basalto com vidro, além de brechas vulcânicas e sedimentares, diques
e soleiras de diabásico e corpos de arenito interderrames. Cada derrame basáltico pode
aparecer diferenciado, estruturalmente, em quatro zonas: zona basal, de disjunção
horizontal; zona central, de disjunção colunar, normalmente ampla; zona superior, de
disjunção horizontal; e zona vesicular, no topo.
A Seqüência Ácida da Formação da Serra Geral, que corresponde a áreas de
relevo menos dissecado e menos arrasado, compreende derrames de dacitos pórfiros,
dacitos felsíticos, riolitos felsíficos, riocitos felsíficos, basaltos pórfiricos e fenobasaltos
vítreos. Os dois últimos tipos de rochas têm características petrográficas de vulcânicas
básicas; entretanto, quimicamente são vulcânicas ácidas. A seqüência é, essencialmente,
produto de contaminação de magma básico, gerado no Manto Superior, com rochas
siálicas da Crosta Inferior (Kaul, 1990). Autores, indicam que até o momento, que a
formação originou-se em tempos juracretácicos.
2.1.2 Relevo
Na unidade de Relevo Planalto das Araucárias, na sua parte leste, ocorrem cotas
altimétricas que ultrapassam 1200 m próximo à escarpa conhecida como Serra Geral. Em
alguns pontos registram-se cotas altimétricas superiores a 1500 m, com a presença de

- 23 -
relevos residuais acimade 1800 m, como o Morro da Igreja, com 1822 m de altitude,
localizado próximo a São Joaquim (SC) (Hermann & Rosa, 1990).
A costa da Serra Geral, apresenta um desnível médio de 400 m, aparecendo em
quase toda a borda oriental do planalto. Já, na porção oeste, as cotas decaem
gradativamente em direção à parte central da Bacia Sedimentar do Paraná, atingindo, no
máximo, 300 m. A mudança do desnível está relacionado ao mergulho das camadas da
bacia sedimentar, caracterizando o relevo da unidade como um planalto monoclinal.
O relevo tem essa variação altimétrica em função do aprofundamento da drenagem
dos rios principais, como o Iguaçu e o Uruguai. Esses rios apresentam vários trechos com
desníveis entre as partes interfluviais e o fundo do vale acentuados em função da
potência e do gradiente do rio. Outro fator importante para compreender a topografia da
área são os desníveis existentes nos contatos entre as áreas conservadas e as áreas
dissecadas que, em muitos setores, é marcado por forte ruptura do declive (Hermann e
Rosa, 1990).
2.1.3 Solo
Com base no Sistema Brasileiro de Classificação de Solos – SBCS (Embrapa,
1999), são encontrados na FLONA, CAMBISSOLO HÚMICO Alumínico, CHERNOSSOLO
ARGILÚVICO Férrico e NEOSSOLO LITÓLICO Eutrófico (Streck et al., 2002).
Os Cambissolos são solos em processo incipientes de formação, variam de rasos a
profundos, a drenagem altera de bem drenados a imperfeitamente drenados, dependendo
da posição que ocupa na paisagem. No perfil dos cambissolos ocorre a presença de
fragmentos de rochas, assim, pouca intemperização do material (Streck et al., 2002).
O Cambissolo Húmico alumínico é o um solo com média quantidade de matéria
orgânica, com Al trocável ≥ 4 cmol/kg; saturação por Al ≥ 50%), logo, extremamente
ácido. Este tipo de solo ocorre em ambientes onde a alta pluviosidade e as baixas
temperaturas favorecem a acumulação de matéria orgânica, na Região dos Campos de
Cima da Serra, em relevo ondulado a forte ondulado (Streck et al., 2002).
Os Chernossolos são solos escuros com alta fertilidade química, variam de rasos a
profundos. Se caracterizam por apresentar razoáveis teores de material orgânico, o que
lhe confere cores escuras ao horizonte superficial que é do tipo A chernozêmico e tem

- 24 -
alta fertilidade química (saturação por bases ≥ 65%) e alta CTC (Capacidade de Troca
Catiônica) em todo o perfil. Um elevado teor de ferro (≥18%) identifica os Chernossolos
Argilúvicos férricos, os quais ocupam áreas de pequena extensão, em relevo ondulado a
fortemente ondulado, exigem práticas conservacionistas intensivas e oferecem condições
para o uso com culturas anuais, fruticulturas e reflorestamento (Streck et al., 2002).
Os Neossolos são solos novos, isso é, pouco desenvolvidos; são rasos ou
profundos encontrados nas mais diversas condições de relevo e drenagem. Os Neossolos
Litólicos, têm pequena espessura, ocorrem em regiões de relevo forte ondulado e
montanhoso, geralmente com pedregosidade e afloramento de rochas, têm baixa
capacidade de infiltração e armazenamento de água no solo e alta suscetibilidade à
erosão hídrica, impossibilitando o seu uso para culturas anuais. Aréas de Neossolos
Litólicos com declividade superior a 30% devem ser mantidas com cobertura vegetal
natural, constituindo-se área de preservação permanente (Streck et al., 2002).
2.1.4 Clima
O clima da área de estudo, de acordo com a classificação de Köppen, é do tipo
“Cfb”, mesotérmico médio. Este clima domina as cotas altimétricas entre 1.100 e 1.000m
no norte do Rio Grande do Sul. Compreende apenas 0,7% do Estado, estando
relacionado às cotas altimétricas mais elevadas da região, com sua ocorrência esparsa
em numerosos e pequenos locais montanhosos, geralmente pouco povoados. Entretanto,
duas áreas de ocorrência deste tipo climático merecem destaque pela sua extensão
territorial e pelos povoamentos nelas existentes. Trata-se da superfície da chapada de
Palmas e da chapada de Vacaria-Lages-São Joaquim, estando a área de estudo
compreendida nesta última (Nimer, 1990).
Nesta região, de clima característico, há pelo menos um mês com temperatura
média inferior a 10°C, possuindo inverno acentuado, cujo frio é uma constante dia e noite,
e verão onde calor é praticamente ausente, pelo efeito da altitude .
Resulta daí que, na área em estudo, a amplitude térmica é menos importante do
que as verificadas nas superfícies baixas da Região Sul, o que determina uma
temperatura média anual muito baixa, situando-se entre 12 e 14°C na superfície de
Vacaria-Lages-São Joaquim. Mesmo assim, enquanto o verão se caracteriza por

- 25 -
temperaturas muito amenas, o inverno é caracterizado por constante e acentuado frio.
Como efeito, a média do mês mais quente (janeiro) oscila em torno de 20°C; a média das
máximas diárias, em torno de 26°C, e a máxima absoluta tem pouca possibilidade de
ultrapassar 34°C. No inverno, entretanto, a temperatura cai diariamente a níveis próximos
de 0°C. Por este motivo, a média das mínimas diárias mantém-se abaixo de 6°C durante
todo o inverno e o número de noites frias varia de 15 a 20 dias por ano. A nevada, embora
não seja um fenômeno muito comum, não constitui uma raridade, como acontece nas
áreas de Clima Mesotérmico Brando. Decorre daí que o inverno, nestas áreas de Clima
Mesotérmico Médio “Cfb”, possui média térmica inferior a 10°C, pelo menos em julho, seu
mês mais frio (Nimer, 1990).
2.1.5 Vegetação primitiva
Rambo (1956) divide o Estado do Rio Grande do Sul em duas formações vegetais,
a do campo e a da floresta. Da área total do Estado, cerca de 131.896 km² (46,26%) são
campos, 98,327 km² (34,47%) matas e o restante, atribuído à vegetação litorânea,
banhados inundáveis e outras formações.
Para o mesmo autor, as variações climáticas das diversas partes do Estado não
são suficientes para explicar a presença dessas duas formações, uma vez que a
formação climática conveniente no Estado do Rio Grande do Sul é a da Floresta Alta
Subtropical. Os campos são formações climáticas e edáficas na sua origem e relitos
históricos ou manchas no tempo atual.
Segundo Jarenkow (1994), a distribuição atual das formações vegetais do sul do
Brasil resulta de um processo histórico, cujo entendimento remete a abordagens
multidisciplinares, em diferentes momentos de sua evolução, principalmente aqueles
ocorridos a partir do final do Terciário.
Para o autor, outras formações ou sub-formações florestais, de maior ou menor
importância, podem ainda ocorrer no Estado, no interior das diferentes regiões
fitogeográficas, como Matas de Restinga, Matas Insulares, Matas de Galeria, entre outras.
Segundo Leite & Klein (1990) a vegetação do Estado do Rio Grande do Sul
compreende nove regiões fitoecológicas ou fitogeográficas: Região da Floresta Ombrófila
Densa (Floresta Atlântica); Região da Floresta Ombrófila Mista (Floresta de Araucária);

- 26 -
Região da Floresta Estacional Semidecidual (Floresta Subcaducifólia); Região da Floresta
Estacional Decidual (Floresta Caducifólia); Região da Savana (Cerrado e Campo); Região
da Estepe (Campanha Gaúcha); Região da Savana Estépica (Campanha Gaúcha); Áreas
das Formações Pioneiras de Influência Marinha (Restingas e Dunas); e Área de Tensão
Ecológica (contatos).
Na FLONA de São Francisco de Paula o tipo fitogeográfico predominante é a
Floresta Ombrófila Mista ou Floresta de Araucária ou Floresta de Pinheiro. Segundo
Rambo (1956), o pinheiro ocorre em toda a borda superior livre do planalto, a começar do
norte de Santa Maria até o extremo nordeste; nos vales superiores e nas cabeceiras dos
Rios Caí, Taquari, das Antas, Jacuí e Pelotas; em grupos isolados ou densos sociedades,
nos capões disseminados por todo o planalto; em indivíduos solitários em pleno campo,
como se observa a leste de Cruz Alta; em mistura com a floresta virgem do Alto Uruguai,
ao norte de Passo Fundo e Lagoa Vermelha.
O mesmo autor afirma que o pinheiro é exclusivo do planalto, ocorrendo em
altitudes entre 500 m a Oeste e 1000 m ao Leste. Nunca desce, a não ser em manchas
ocasionais. Diferencia-se três núcleos principais do pinheiral: na aba do setor meridional
da escarpa, entre o Rio Taquari e o Rio dos Sinos; na borda dos Aparados entre o Rio
Maquiné e o Rio das Antas; e em pleno planalto central, no curso superior do Rio Jacuí,
ao sul de Passo Fundo.
Segundo Leite & Klein (1990) a concepção de Floresta Ombrófila Mista procede da
ocorrência da mistura de floras de diferentes origens, definindo padrões fitofisionômicos
típicos em zona climática pluvial. A área onde a coexistência de representantes da flora
tropical (afro-brasileira) e temperada (austro-brasileira), com marcada relevância
fisionômica de elementos Coniferales e Laurales, é denominada Planalto Meridional
Brasileiro, área de dispersão natural do pinheiro-brasileiro ou do pinheiro-do-paraná, a
Araucaria angustifolia ou “curiirama” dos indígenas, espécie gregária de alto valor
econômico e paisagístico.
Segundo Klein apud Leite & Klein (1990) a araucária e outros elementos de origem
temperada, em face às suas características heliófilas, encontram-se hoje desfavorecidos,
não só pela intervenção destruidora do homem, mas pela incompatibilidade com o clima
atual.

- 27 -
Os terrenos entre aproximadamente, os 500 e os 800 m de altitude estão
enquadrados na formação montana (Leite e Sohn, apud Leite & Klein, 1990) e
caracterizam-se por um clima sem época seca, com período frio (Temperatura média Tm
≤ 15º C) curto ou ausente e período quente longo (Tm ≥ 20º C). A área mais típica e
representativa da Floresta Ombrófila Mista é aquela das altitudes superiores aos 800 m,
principalmente dos terrenos altomontanos. Seu clima é o mais frio da região e com
maiores índices de geadas noturnas. Caracteriza-se pela ausência de período seco e
ocorrência de longo período frio (Tm ≤ 15º C). O período quente anual (Tm ≥ 20º C) é
geralmente curto ou ausente.
Sob estas condições climáticas e de acordo com a diversificação de outros
parâmetros ambientais, poder-se-ia determinar, na área típica da Floresta Ombrófila
Mista, dois grupos distintos de comunidades com araucária e lauráceas: um, onde o
pinheiro se distribuía de forma esparsa por sobre bosque contínuo, no qual 70 a 90% das
árvores pertenciam às espécies: imbuia (Ocotea porosa), espécie mais representativa,
canela-amarela (Nectandra lanceolata), canela-preta (Nectandra magapotamica), canela-
fogo ou canela-pururuca (Cryptocarya aschersoniana) acompanhadas da sapopema
(Sloanea monosperma), por vezes bastante freqüente, da guabirobeira (Campomanesia
xanthocarpa) e erva-mate (Ilex paraguariensis); outro grupo, onde a araucária formava um
estrato de 60 a 80% de folhosas, principalmente das espécies: canela-lageana (Ocotea
pulchella), espécie dominante, canela-amarela (Nectandra lanceolata), canela-guaicá
(Ocotea puberula), canela-fedida (Nectandra grandiflora), comboatá-vermelho (Cupania
vernalis) e comboatá-branco (Matayba elaeagnoides), acompanhadas de casca-d’anta
(Drimys brasiliensis), pimenteira (Capsicodendron dinisii), guabirobeira (Campomanesia
xanthocarpa) e diversas mirtáceas e aquifoliáceas.
Acompanhando planícies sedimentares recentes dispersas em diferentes altitudes
e latitudes e sujeitas a periódicas inundações, ocorre um tipo de formação definida como
Aluvial. Nela o pinheiro-do-paraná geralmente consorcia-se com branquilho (Sebastiania
commersoniana), jerivá (Syagrus romanzoffiana), murta (Blepharocalyx salicifolius),
corticeira-do-brejo (Erythrina cristagalli), tarumã (Vitex megapotamica), açoita-cavalo
(Luehea divaricata), salgueiro (Salix humboldtiana), além de aroeira-vermelha (Schinus
terebinthifolius) e diversas espécies de mirtáceas (Leite & Klein, 1990).

- 28 -
Na região de Floresta Ombrófila Mista é comum a ocorrência de campos. Nestes
se verifica grande número de capões e bosques, muitas vezes com a presença de
Araucaria angustifolia, denotando o lento processo de invasão das florestas nas áreas de
campo. Muito comum nestes campos é a ocorrência de araucária isoladas junto aos
capões (SUDESUL, 1978).
Segundo Rambo (1956), os capões também são típicos na porção leste do Planalto
(São Leopoldo), principalmente nos mananciais de água. Em sua composição destacam-
se as espécies arborescentes do parque campestre, que são as aroeiras (Schinus spp.),
assim como exemplares da mata virgem como cedro (Cedrela fissilis), cangerana
(Cabralea canjerana), louro (Cordia trichotoma), figueira (Ficus luschnathiana), jerivá
(Syagrus romanzoffiana), chá-de-bugre (Casearia sylvestris), mamica-de-cadela
(Zanthoxylum sp.) e camboatá (Matayba elaeagnoides). No interior cresce vegetação
lenhosa baixa, constituída de laranjeira-do-mato (Actinostemum concolor = Gymnanthes
concolor), cincho (Sorocea bonplandii), rubiáceas e monimiáceas arbustivas,
salsaparrilhas (Smylax sp.), entrelaçados por cipós. É uma mata virgem em escala
reduzida.
2.2 Fatores Ecológicos e de Concorrência
A palavra “ecologia” deriva do grego oikos que significa “casa” e logos que significa
“estudo”, logo, ecologia é o estudo do ambiente da casa, incluindo todos os organismos
contidos nela e todos os processos funcionais que a tornam habitável. Deste modo,
ecologia é o estudo do “lugar onde se vive” (Odum, 1988).
Uma definição concisa da ecologia moderna é dada por Edelbauer (1988): “Ciência
da inter-relação e das interinfluências das diferentes formas de vida e seu ambiente”.
Em épocas remotas os homens primitivos, quando escolhiam as plantas para a
alimentação e para curar determinadas doenças, além de locais adequados para a caça e
pesca, demonstravam o domínio do saber ecológico (Laroca, 1995).
As primeiras obras que faziam referências à ecologia foram as de Hipócrates e
Aristóteles (384-322 aC). Este último, filósofo e naturalista grego, organizou o
conhecimento sobre animais existentes na época, sendo considerado, por certos autores,

- 29 -
como um dos precursores da ecologia. No Brasil, o interesse pela ecologia moderna teve
início desde a publicação de Ecologia Vegetal (“Plantesamfund”), pelo dinamarquês
EUGENE WARMING, em 1895, e da classificação das formas de vida dos vegetais, por
RAUNKIAER, em 1903. Assim, a ciência que estuda as inter-relações das plantas, entre si e
com o meio, é relativamente nova, pois teve um desenvolvimento gradativo durante a
história e muito ainda deve ser estudada e desenvolvida (Ferri, 1963; Laroca, 1995;
Souza et. al., 1998).
Segundo Laroca (1995), a ecologia pode ser dividida em duas grandes áreas: a
auto-ecologia e a sinecologia. A AUTO-ECOLOGIA trata da ecologia do indivíduo e das
populações e, nesta área, pode-se incluir a genética ecológica, comportamento animal
(etologia), ecologia fisiológica, entre outras. A SINECOLOGIA envolve sistemas de muitas
espécies: a comunidade, como um todo, ou parte das mesmas, ou ecossistemas
(terrestres e aquáticos) e, portanto, nesta grande área pode-se incluir a fitossociologia.
Os primeiros trabalhos em fitossociologia no Brasil foram realizados por Davis e
Veloso em 1945 apud Souza et al. (1998). Antes disso não existiam dados sobre a
vegetação brasileira. Somente a partir da década de 70, surgiram trabalhos nos quais
foram avaliados a identificação taxonômica e os parâmetros fitossociológicos quantitativos
absolutos e relativos.
Atualmente fala-se muito sobre ecologia, fatores ecológicos, meio ambiente e
manejo sustentado dos recursos naturais renováveis (Rio+10, encontro em Johanesburgo
– África do Sul, 2002), mas apenas uma pequena parte da população mundial possui
conhecimento para entender a dinâmica e as inter-relações dos ecossistemas.
Em um ecossistema natural, cada componente é dependente do outro e, portanto,
devem ser estudados conjuntamente, para assim determinar suas influências sobre os
seres vivos. Existem fluxos de energia e de matéria em cada ecossistema de forma a
manter um equilíbrio ecológico. Com a modificação do equilíbrio ecológico, algumas
espécies sucumbem ao passo que outras se multiplicam maciçamente. Assim, torna-se
importante à realização de estudos sobre influência dos fatores ecológicos sobre o
desenvolvimento das espécies vegetais (Odum, 1988).
Durante a sua evolução o homem, muitas vezes, modificou os ecossistemas
naturais de maneira a satisfazer suas necessidades. As intervenções humanas
geralmente foram feitas sobre a vegetação, sistema hídrico e solo. Em função do

- 30 -
desenvolvimento, os ecossistemas naturais, através da retirada da vegetação nativa e do
cultivo, se tornaram ecossistemas modificados (Odum, 1988). Estes ecossistemas
modificados acabam forçando determinadas espécies a adaptarem-se às novas
intensidades que os fatores ambientais ou fatores ecológicos atuam sobre elas.
Segundo o autor, os fatores ecológicos são divididos em dois grandes grupos: (i)
ABIÓTICOS (fatores físicos e químicos) e; (ii) BIÓTICOS (patógenos, parasitas, predadores,
simbiontes, entre outros). Os componentes biótico e abiótico estão intimamente
associados no solo, o qual, por definição, consiste na camada intemperizada da crosta
terrestre com os organismos vivos e os produtos da sua decomposição intermisturados.
2.2.1 Umidade do solo
A água é uma necessidade fisiológica para todo o ser vivo e, em relação ao ponto
de vista ecológico, um fator limitante, principalmente em ambientes terrestres (Odum,
1988).
De acordo com o autor, o ritmo diário da umidade da natureza (alta à noite, baixa
durante o dia) afeta a umidade do solo juntamente com a temperatura e a luminosidade,
ajudam a regular as atividades dos organismos e a limitar sua distribuição.
A importância da água para o solo é evidente, pelo simples fato de ser um dos mais
ativos agentes de destruição e desagregação das rochas e minerais. A água é um dos
componentes do protoplasma das plantas, sendo obrigatória nas reações metabólicas,
além de funcionar como transportadora de elementos nutritivos e dos componentes que
se formam durante o metabolismo da planta e entre a planta e o solo (Vieira et al., 1988).
Para os mesmos autores a capacidade do solo de reter água no estado líquido é
resultante da ação conjunta e complexa de duas forças principais que são: (i) força de
adesão: a atração que as partículas do solo exercem sobre as moléculas de água e; (ii)
força de coesão: a atração que as moléculas de água exercem entre si. Não se deve
esquecer que a importância da cada um desses fatores depende do estado de umidade
do solo.
Segundo Silva Júnior (1998), os solos associados a comunidades com SOLOS
ÚMIDOS (glay húmico) apresentam teores mais altos de MO, Al, H + Al, P, Fe, Cu, Zn, e
saturação de Al e os maiores conteúdos de areia fina e grossa. As comunidades com

- 31 -
SOLOS SECOS podem ter níveis de pH, Ca, Mg, K, Mn e saturação de bases. Já os solos de
comunidades INTERMEDIÁRIAS apresentam os mais altos teores de argila e silte.
O abastecimento de água é muito importante para o crescimento da floresta, mas a
significação deste fator ecológico é naturalmente muito maior e não se limita só às
árvores.
A floresta regula o lançamento da água local e regional em grande escala, e este
sim, é um fator significante para a cultura da terra. Todas as águas de precipitação
retornam pela evaporação à atmosfera. Tanto dos continentes, como dos oceanos, sobe
vapor à atmosfera sem cessar, sob influência do calor da radiação solar e dos ventos.
Neste processo predominam os mares, por sua enorme extensão, como fornecedor da
umidade (Silva Júnior, 1998).
As circunstâncias de umidade de uma região climática serão determinadas, não
somente pelas precipitações, mas também através da temperatura e distribuição de
precipitação, ou seja, estes fatores determinarão em conjunto, se o clima será árido ou
úmido (Silva Júnior, 1998).
Logo, um fator relevante na manutenção da água no solo, é o relevo do lugar, pois
sobre um terreno plano a quantidade de água será igual a da chuva. Já em terrenos com
declividade, a água da precipitação, terá distribuição diferente, devido a sua lixiviação
(Nagy & Cunha,1986).
A floresta tem uma grande influência na quantidade de água da precipitação que
chega ao solo. O dossel das copas forma uma superfície, que representa uma superfície-
limite. Para cada chuva, uma certa parte da água consegue chegar ao solo da floresta
sem obstáculos e o que vai influenciar nesta quantidade de chuva será: (i) as espécies de
árvores da floresta; (ii) a densidade do povoamento; (iii) a idade; e (iv) o fechamento do
dossel. Portanto, quanto mais fechado for o povoamento, menos água de precipitação
chega ao solo (Nagy & Cunha, 1986).
Para os autores, as características do solo causam diferenças consideráveis na
umidade do solo de um sítio, porém, as maiores diferenças e oscilações da sua umidade
serão provocadas pela vegetação do sítio.

- 32 -
2.2.2 Inclinação do terreno
Segundo Silva Júnior (1998) a inclinação (topografia), que condiciona o regime de
água nos solos, tem sido indicada como determinante de algumas características
ambientais e, conseqüentemente, da distribuição da vegetação. Enfatiza, também, o
papel direto da topografia no processo de um regime diferenciado da água.
Furley e Oliveira-Filho, citados por Silva Júnior (1998), relatam que a topografia
direciona a drenagem para os vales, resultando na presença do lençol freático próximo à
superfície, durante quase todo o ano. Ressaltam, então, que a topografia é considerada
um determinante primário da vegetação e pode definir a fisionomia, a composição
florística, a riqueza em espécies e a densidade de árvores.
Vieira et al. (1988) classificam as condições topográficas do relevo de uma região
segundo os graus de declividade em: plana, suave ondulada, ondulada, forte ondulada e
montanhosa (Tabela 1).
TABELA 1 – Classificação topográfica de acordo com os graus de declividade do solo.
Especificação Porcentagem de declividade Plana < 30 Suave ondulada 3 – 80 Ondulada 8 – 200 Forte Ondulada 20 – 450
Montonhosa > 45 0
Fonte: Vieira et al. (1988)
2.2.3 Profundidade do solo
Esta característica está relacionada com a espessura máxima do solo em que o
sistema radicular das plantas não encontra dificuldade ou barreira física para penetrar
livremente, facilitando a sua fixação e servindo de meio para a absorção de água e de
nutrientes (Lepsch,1993; Sanchez,1981).
A profundidade do solo tem influência sobre a fertilidade natural, especialmente se
um ou mais nutrientes tem sua disponibilidade limitada, bem como sobre as práticas de
manejo a serem utilizadas. Também tem influência no sistema ar-água, o qual, por sua
vez, influencia a disponibilidade de nutrientes e sistema radicular das árvores (Vieira et
al., 1988).

- 33 -
Os autores avaliam a profundidade dos solos através de 4 classes: raso, pouco
profundo, profundo e muito profundo (Tabela 2).
TABELA 2 – Classes e valores utilizados para a avaliação da profundidade do solo.
Especificação Profundidade do solo (cm)
Raso ≤ 50
Pouco profundo 50 –100
Profundo 100-200
Muito profundo >200
Fonte: Vieira et al. (1988)
2.2.4 Cor do solo
A cor do solo é uma das características que mais chama a atenção ao se estudar o
solo. As várias tonalidades de coloração ajudam a separação dos horizontes e também a
evidenciar condições de extrema importância, tais como: teores de matéria orgânica;
quantidade de argila, silte e os componentes de ferro, permitem também a avaliação de
propriedades relacionadas com a composição, aeração e drenagem do solo (Streck et al.,
2002).
Existem várias cores de solos, como: escuros, negros, cinzentos, vermelhos,
amarelos e ocasionalmente brancos, nas mais variadas intensidades (Streck et al., 2002).
Segundo Vieira et al. (1988) e Streck et al. (2002) , a matéria orgânica nos
horizontes O, H e A e em alguns horizontes B, é responsável pelas cores escuras, porém,
a coloração dos horizontes pode, à medida que aumenta o teor de matéria orgânica,
variar do branco ao negro. Já as cores brancas são proporcionadas, principalmente, pelo
conteúdo de sílica na forma de quartzo, e pela ausência de materiais pigmentantes
(matéria orgânica e óxidos de ferros) .
As cores vermelhas dependem do conteúdo de sesquióxidos e óxidos de Fe não-
hidratados, enquanto as cores amarelas e cinza-amareladas, dos óxidos hidratados. O
solo é tanto mais vermelho quanto menos hidratados forem os compostos de ferro. A cor
avermelhada, apresentada pelos compostos de ferro, pode, também, dar seguras
indicações sobre o grau de drenagem dos solos e, em geral, indica boa oxidação, boa
drenagem e boa aeração, enquanto que as cores cinzentas ou cinza-azuladas indicam

- 34 -
condições redutoras e drenagem pobre. Já a cor amarelada (brunada) também pode
significar um ambiente mais úmido, onde tempo de residência da água no solo é maior
(Vieira et al., 1988 e Streck et al., 2002).
Segundo Streck et al. (2002), as cores acinzentadas, deve-se à redução e remoção
de óxidos de ferro, podem estar relacionadas com ambientes de solo saturados com água
durante períodos prolongados.
Portanto, a cor do solo é um fator ambiental de significativa importância para as
propriedades do solo, assim como o conteúdo de matéria orgânica, o grau de hidratação
dos sesquióxidos e as condições de drenagem. Podem, ainda, dar uma idéia do seu grau
de aquecimento, pois solos de cores escuras absorvem calor mais rapidamente e o
refletem com menor intensidade (Vieira et al.,1988).
Segundo os mesmos autores, as cores são grandemente influenciadas pelo teor de
umidade e como conseqüência diminuem de intensidade à medida que aumenta o teor de
umidade.
2.2.5 Exposição à luz
Os organismos sobre a superfície terrestre ou perto dela recebem constantemente
radiação solar e fluxo de radiação térmica de grande comprimento de onda. Os dois tipos
de comprimento de onda contribuem para o ambiente climático (temperatura, evaporação
da água, movimento de ar e água), porém apenas uma pequena fração da radiação solar
pode ser convertida pela fotossíntese em energia para os componentes do ecossistema
(Odum, 1988).
A luz solar que atinge a biosfera é de 2 gcal/cm2/min, mas esta é atenuada
enquanto passa pela atmosfera para 1,34 gcal/cm2/mim ou 67% (Gates apud Odum,
1988).
Tendo que atravessar as nuvens, as partículas da água e a vegetação, a radiação
solar é atenuada ainda mais e a distribuição espectral da sua energia é grandemente
alterada. A entrada diária de luz solar na camada autotrófica de um ecossistema varia
principalmente entre 100 e 800 gcal/cm2. O fluxo de energia diária recebida por um
organismo exposto pode ser várias vezes maior ou consideravelmente menor do que a
radiação solar que entra (Odum, 1988).

- 35 -
Ainda, segundo Odum (1988), nos diversos estratos de um ecossistema florestal a
variação no fluxo de energia total de radiação é grande, variando entre as estações ou
locais da superfície terrestre. Isto afeta a distribuição dos organismos no ecossistema.
Segundo Cunha & Peichl (1986), na freqüência onde os olhos humanos estão
sensíveis à luz fica, também, a parte da radiação que tem importância decisiva para o
processo de fotossíntese e, com isso, para a produção de matéria das plantas.
Para os autores, a luz tem também outros efeitos importantes dentro das plantas,
tais como, o desenvolvimento e a diferenciação dos tecidos vegetais, especialmente a
formação das folhas umbrófilas e das folhas heliófilas (composição da luz e do
abastecimento com água) (Cunha & Peichl, 1986).
Em povoamentos idosos, fechados, uniformes e simples, existe uma correlação
estreita entre a intensidade da exposição e a densidade de copas no dossel. A
intensidade da exposição das espécies florestais, as quais formam o dossel das copas,
pode ser quantificada pelo número de árvores por hectare. O crescimento em altura
geralmente não é afetado pelo sombreamento, o que não ocorre com o crescimento em
diâmetro.
Geralmente, as diferentes espécies de árvores têm, nos primeiros anos de idade e
nos sítios com abundância de nutrientes e umidade, uma tolerância elevada à sombra,
comparando às árvores idosas e aos sítios com baixo teor de nutrientes. Logo, a
necessidade de luz não varia somente com a espécie, mas também com a idade do
indivíduo, sendo as exigências no estado juvenil geralmente inferior aquelas das idades
superiores (Cunha & Peichl, 1986).
Os autores colocam, como regra geral, que é possível constatar que a diminuição
da intensidade da luz causa uma redução da matéria orgânica das plantas. Esta redução
faz-se notar, principalmente, nas raízes, isto é, causam prejuízos às raízes. Isto significa
para a planta, menos absorção dos nutrientes e da água e, assim, diminuição da força de
competição com as árvores vizinhas.
A redução da luz no povoamento influencia na quantidade de luz disponível para
cada árvore na parte mais baixa da copa. Isto limita a fotossíntese nesta parte da copa.
Portanto, a intensidade da luz causa muitos efeitos sobre a morfologia das plantas.
Quando uma espécie possui uma exigência elevada de luz, há uma influência muito

- 36 -
grande sobre a formação do tronco e copa e, no caso das florestas nativas, nota-se
claramente esta influência na tortuosidade ou forma das árvores.
Pinto (1997) diz que disponibilidade de energia está relacionada com a localização
dos continentes e dos oceanos em relação ao equador e também com os movimentos da
terra em relação ao sol. Estes fatores dão origem às variações sazonais bem demarcadas
na disponibilidade de energia radiante com reflexos diretos no comprimento dos dias e na
temperatura, além de inúmeros fenômenos climáticos, tais como a evaporação da água e
a movimentação das massas de ar. Estas complexas interações resultam nas formações
ecológicas (florestais) bem definidas.
2.2.6 Quantidade de pedra
Os elementos químicos disponíveis às comunidades da biosfera são aqueles que,
pela sua natureza geoquímica, tendem a estar contidos nos tipos de rochas que afloram à
superfície. Os elementos abundantes no manto da Terra estão escassos na superfície. O
fósforo é um dos elementos cuja escassez na superfície da Terra muitas vezes limita o
crescimento vegetal (Odum,1988).
Segundo Vieira et al. (1988), a pedregosidade superficial, ou a quantidade de
pedras na superfície de solo, é uma característica ambiental, importante de ser avaliada,
pois está relacionada diretamente à retenção de água e aos componentes químicos que
compõem o solo. O autor classifica os afloramentos rochosos na superfície do terreno e 4
classes, conforme mostra a Tabela 3.
TABELA 3 – Classificação da pedregosidade na superfície do terreno.
Especificação Pedregosidade (%)
Pedregosidade menor < 0,1% da superfície do terreno
Pedregosidade entre 0,1 – 3%
Pedregosidade entre 3 – 15%
Pedregosidade superior > 15%
Fonte: Vieira et al. (1988)

- 37 -
2.2.7 Densidade do sub-bosque
Segundo Laroca (1995), os efeitos que o ambiente exerce sobre os seres vivos,
recaem sobre duas categorias: (i) efeitos dependentes da densidade; e (ii) independentes
da densidade.
Os efeitos dependentes da densidade modificam a taxa de nascimento ou de
mortalidade assim como a função do tamanho ou densidade populacional (N). As
competições intra1 e interespecífica são fatores que têm efeitos dependentes da
densidade, pois estas têm influências diretas ou indiretas, isto é, alteram o ambiente
através de secreções. Algumas vezes os efeitos são positivos, mas geralmente tendem a
ser negativos, diminuindo a taxa de nascimento e/ou aumento da taxa de mortalidade. No
entanto não se pode esquecer que estes efeitos variam de espécie para espécie (Laroca,
1995).
De acordo com Pizzato (1999), a dinâmica de uma floresta inicia-se com a
formação de clareiras, que provocam mudanças nas características edafoclimáticas,
ocasionando o processo de sucessão florestal. A abertura de clareiras é o principal fator
para que diversas espécies existam na floresta, sendo renovadas e sustentadas pela
dinâmica de perda de indivíduos mais velhos permitindo a existência de outros indivíduos.
Odum (1988) diz que a dinâmica de uma floresta, relacionada a fisiologia, estrutura
e funcionamento desta, envolve diversos processos de organização como: sucessão,
mortalidade, ingresso e crescimentos.
Para Pizzato (1999) na dinâmica das florestas, durante o curso de sucessão, os
indivíduos que morrem são substituídos pelo crescimento dos indivíduos vizinhos ou pelo
ingresso de novos indivíduos da mesma ou de diferentes espécies.
Barros (1986) relata que o ingresso, crescimento e mortalidade são resultado final
do processo da dinâmica das florestas. O contínuo aparecimento de novos indivíduos no
estrato inferior da floresta que limita o espaço de crescimento das árvores novas, devido
ao aumento da densidade, propiciam a eliminação de elementos menos capazes de
competir. As aberturas ocorridas no estrato superior da floresta, em decorrência da morte
de grandes árvores, são ocupadas pelas árvores que se encontram no estrato
1 A competição intra-específica se dá quando organismos da mesma espécie utilizam recursos limitados. Se os recursos não são limitados a competição ocorre quando os organismos em busca de recursos, manifestam comportamentos agressivos uns com outros.

- 38 -
imediatamente inferior que, por sua vez, proporcionam o estabelecimento de novos
indivíduos, tornando heterogênea a distribuição de idades das árvores na floresta.
Para o mesmo autor a densidade do sub-bosque influencia nos processos da
dinâmica florestal, sendo responsável tanto pela mudança da comunidade como pela
modificação do espaço desta. Tais processos se manifestam através da extinção e
imigração local de populações, assim como flutuação na abundância relativa de
populações dentro da comunidade.
2.3 Parâmetros Fitossociológicos
Montoya-Maquin & Matos apud Longhi (1997) afirmam que a vegetação natural é
complexa e está relacionada com os diversos fatores do meio, como climáticos,
pedológicos e biológicos. Estes fatores podem ser quantificados por vários parâmetros,
como os métodos baseados no estudo dos diversos elementos da vegetação, que são os
métodos florísticos ou taxonômicos e os baseados na estrutura e na fisionomia.
De acordo com Förster (1973) a análise estrutural da vegetação deve ser baseada
no levantamento e na interpretação de critérios de conteúdo mensuráveis. Análise dessa
natureza permite comparações entre diferentes tipos de florestas.
Segundo Souza et al. (1998), as análises florísticas (parâmetros florísticos)
contemplam as listas de espécies arbóreas ocorrentes no local de estudo, ou seja,
determinam a população de uma determinada área.
Para análise da estrutura horizontal das comunidades vegetais utiliza-se os
parâmetros de densidade (abundância), freqüência, dominância, valor de importância e
valor de cobertura, que revelam informações sobre a distribuição espacial das populações
e sua participação no contexto do ecossistema. A estrutura vertical ou o arranjo dos
indivíduos dentro do espaço vertical deve basear-se na distribuição dos indivíduos em
estratos (Guapyassü apud Longhi, 1997).
(i) Densidade: é o número de indivíduos, quer de uma espécie, quer de todas as
outras espécies em conjunto, por unidade de área. A Densidade Absoluta (DA) trata do
número de indivíduos da espécie por unidade de área considerada e a Densidade

- 39 -
Relativa (DR) é a proporção do número de indivíduos de uma espécie em relação ao
número total de indivíduos amostrados, em percentagem (Martins, 1991).
Já o termo Abundância, também utilizado por alguns pesquisadores, refere-se às
estimativas visuais da densidade das espécies, agrupando-as, assim, em classes de
abundância (muito rara, rara, ocasional, abundante, muito abundante). Porém, densidade
refere-se às contagens efetivas de indivíduos, em um espaço contínuo (Finol, 1971,
Förster, 1973; Longhi, 1980).
(ii) Freqüência: freqüência absoluta (FA) é a porcentagem de unidades de
amostragem com ocorrência da espécie, em relação ao número total de unidades de
amostragem. Freqüência Relativa (FR) é a proporção da freqüência absoluta de uma
espécie em relação à soma das freqüências absolutas de todas as espécies, em
porcentagem (Martins, 1991). Para melhor entendimento, freqüência indica a
uniformidade de distribuição de uma espécie sobre uma determinada área, isto é, a sua
dispersão média. Já para Souza et al. (1998), é a percentagem de ocorrência de uma
espécie em um número de áreas de igual tamanho, dentro de uma comunidade.
Segundo Longhi (1997), para determinar a freqüência, deve-se controlar a
presença/ausência da espécie, em uma série de amostras de tamanho uniforme,
independente do número de indivíduos. Portanto, se uma espécie aparece em todas as
unidades amostrais, tem uma freqüência de 100%. Pode-se dizer, portanto, que
freqüência refere-se à probabilidade de encontrar uma espécie na área estudada.
(iii) Dominância: expressa a proporção de tamanho, volume ou cobertura de cada
espécie, em relação ao espaço ou volume da fitocenose (Martins, 1991).
Pesquisadores como Förster (1973), Font-Quer (1975) e Longhi (1997), dizem que
dominância é a medida da projeção total do corpo das plantas, logo, a dominância de uma
espécie representa a soma de todas as projeções horizontais dos indivíduos pertencentes
à espécie.
Porém, em florestas muito densas, é praticamente impossível determinar os
valores de projeção horizontal das copas das árvores, pois a existência de estratos
inferiores forma uma estrutura vertical e horizontal muito complexa. Em função disso, Cain

- 40 -
et al. apud Longhi (1997) propuseram o uso da área basal como substituição à projeções
das copas, pelo fato de existir correlação entre ambas.
A Dominância Absoluta (DoA) de uma espécie consiste, portanto, na soma da área
basal de todos os indivíduos da espécie, presentes na amostragem. Dominância Relativa
(DoR) é a relação percentual entre a área basal total da espécie e a área basal por
hectare (Martins, 1991).
(iv) Valor Importância (VI): Os valores relativos de densidade, freqüência e
dominância revelam aspectos essenciais na composição florística das florestas, com
enfoques parciais, os quais isolados, não podem informar sobre a estrutura florística de
uma vegetação em conjunto (Martins, 1991).
Os parâmetros quantitativos de Densidade, Dominância e Freqüência (relativos) de
cada espécie, podem ser interpretados como de importância na fitocenose, isto vai
depender do que o pesquisador considera relevante (Longhi, 1997).
Segundo Förster (1973), para análise da vegetação, é importante encontrar um
valor que permita uma visão ou caracterização da importância de cada espécie, no
conglomerado total da floresta. Este valor pode ser dado pelo Valor de Importância (VI),
representado pela soma dos valores relativos de densidade, freqüência e dominância.
Martins (1991), diz que o Valor de Importância tem-se revelado muito útil, tanto para
separar tipos diferentes de florestas, como para relacioná-los a fatores ambientais ou para
relacionar a distribuição de espécies a fatores abióticos.
(v) Valor de cobertura (VC): refere-se ao número de árvores e a valores de superfície
coberta por elas. Obtêm-se esses valores através da soma da densidade e dominância
relativa da espécie (Blaun-Blanquet, 1979).
Förster (1973) diz que o Valor de Importância tem uma importância relativa, por
isso dever ser tratado de forma breve, o autor considera que uma espécie torna-se
importante na floresta pelo seu número de árvores e suas dimensões, isto é, pela sua
densidade e dominância, pois estas determinam o espaço dentro da biocenose, não
importando muito a freqüência da espécie.

- 41 -
Sendo assim, o autor caracteriza as espécies pelo seu Valor de Cobertura, isto é,
pela sua densidade relativa somada à dominância relativa, método citado por Braun-
Blanquet (1979), o que indica a potência da espécie na comunidade.
2.4 Diversidade
Segundo Magurran (1989) é muito difícil definir diversidade, já que se compõe de
dois elementos: (i) variação e (ii) abundância de espécies. Logo, pode-se medir a
diversidade, registrando o número de espécies e avaliando sua abundância relativa ou
usando uma medida que combine os dois componentes.
O conceito de diversidade de espécies possui dois componentes: (i) riqueza,
também chamada de densidade de espécies, baseada no número total de espécies
presentes e, (ii) uniformidade, baseada na abundância relativa de espécies e no grau da
sua dominância ou falta desta (Odum, 1988).
Para o autor, a diversidade de espécies tende a aumentar quanto maior o tamanho
da área. Já, em comunidades bióticas que sofrem estresse, que foram reduzidas pela
competição em comunidades antigas e ambientes físicos estáveis, a diversidade tende a
diminuir.
De acordo com Magurran (1989) as medidas de diversidade devem realizar-se no
contexto de duas áreas de aplicação. Em uma delas, a da supervisão ambiental, as
medidas de diversidade são usadas inteligentemente e têm-se aplicado com profusão. Na
outra, da conservação da natureza, obtém-se grandes êxitos, protegendo a diversidade,
que na maioria dos casos, se define como riqueza de espécies. A supervisão ambiental
demonstra que as medidas de diversidade podem ser, empiricamente muito úteis.
Para Cox (1976), diversidade de espécie é uma medida biológica, característica
única em nível de comunidade de organizações ecológicas. Além do mais, é uma
característica que reflete padrões organizacionais importantes no funcionamento da
comunidade. A composição em diversidade de espécie está relacionada ao grau de
estabilidade da comunidade, desde que uma comunidade com maior riqueza ou
diversidade de espécies, possuir uma rede trófica mais complexa ao longo da qual
mecanismos de controle de densidade possam operar.

- 42 -
Whittaker apud Odum (1988), descreve três diferentes maneiras de classificar a
diversidade. A diversidade dentro do habitat ou tipo de comunidade não pode ser
confundida com a diversidade de uma paisagem ou de uma região que contém uma
mistura de habitats. Portanto, a diversidade pode ser classificada em: (i) diversidade alfa,
para diversidade dentro de um habitat ou dentro de uma comunidade; (ii) diversidade
beta, para diversidade entre habitats e; (iii) diversidade gama, para diversidade de uma
grande área regional, bioma, continente e ilha, entre outros.
A análise florística da vegetação pode ser feita através dos índices de Diversidade
de espécies, entre os quais os Índices de Shannon e de Simpson, que julgam a afinidade
entre comunidades (Pielou, 1975).
O Índice de diversidade de Shannon é calculado pela fórmula:
onde: Pi é a relação entre o número de indivíduos da espécie (ni) e o número total de
indivíduos (N) ou seja Pi = ni/N.
Segundo Pielou (1975) e Martins (1991) seus valores variam de 1,5 a 3,5,
raramente passando de 4,5.
Já o Índice de diversidade de Simpson, é determinado pela equação:
Dn n
N N
i i=−
−
∑
( )
( )
1
1
Onde ni é o número de indivíduos na i-ésima espécie e N é o número total de indivíduos.
À medida que D se incrementa, a diversidade decresce e o índice de Simpson é,
portanto, expresso normalmente como 1-D.
2.5 Análise de Similaridade
Orlóci (1978) utiliza o termo semelhança como uma propriedade de objetos
mensuráveis, ou grupos de objetos e como uma função característica que os objetos
possuem. Como objetos, entendem-se os indivíduos das espécies, a comunidade, além
∑−= PiPiH ln.'

- 43 -
de outras qualidades.
Nas comparações numéricas das comunidades usam-se técnicas estatísticas que,
partindo das tabelas brutas ou matrizes primárias de atributos/amostras e, mediante uma
série de tratamentos matemáticos, permitem obter matrizes secundárias de semelhança
ou similaridades. A matriz de semelhança substitui os conjuntos de atributos presentes
por índices que medem a similaridade das amostras ou comunidades, em função da
coincidência de presença e de abundância dos atributos do par de comunidades a
comparar ou, de semelhança entre espécimes, segundo o número de amostras em que
aparecem juntas ou separadas. Estas matrizes secundárias constituem a entrada de
quase todos os sistemas numéricos e de alguns dos sistemas informais de classificação e
ordenação da vegetação (Matteucci & Colma, 1982).
As funções de semelhança podem ser dadas a partir de variáveis binárias ou
qualitativas (presença/ausência), ou de dados quantitativos (Matteucci & Colma, 1982 e
Johnson & Wichern, 1982). Trata-se de comparações entre espécies ou entre amostras
ou entre comunidades e calculam-se a partir de tabelas de contingência de 2 x 2 (Tabela
4).
TABELA 4: Tabela de contingência entre duas amostras 1 e 2
Amostra 1
+ -
Total
+
Amostra 2
-
a b
c d
a+b
c+d
Total a+c b+d p=a+b+c+d
onde:
a = número de espécies comuns nas amostras 1 e 2 (número de igualdade (1,1))
b = número de espécies exclusivas da amostra 2 (número de desigualdade (1,0))
c = número de espécies exclusivas da amostra 1 (número de desigualdade (0,1))
d = número de espécies ausentes nas duas amostras (número de igualdade (0,0))
p = número total de espécies.
Os índices de similaridade são expressões matemáticas utilizadas para comparar a

- 44 -
flora de duas ou mais comunidades vegetais. Indicam o grau de semelhança entre elas
(Mueller-Dombois & Ellenberg, 1974).
Saiz (1980), Matteucci & Colma (1982) e Johnson & Wichern (1982) citam um
grande número de Índices de Similaridade ou Coeficientes de Similaridade, entre os quais
os mais freqüentemente utilizados são:
A) Para dados qualitativos (presença/ausência):
a = elementos exclusivos da amostra 1
b = elementos exclusivos da amosta 2
c = elementos comuns por presença nas amostras 1 e 2
d = elementos comuns por ausência nas amostras 1 e 2
n = total de elementos considerados em 1 e 2 = a + b + c + d
A = número total de elementos da amostra 1
B = número total de elementos da amostra 2
1) Índice de Jaccard ou Coeficiente de Comunidade de Jaccard (SJ):
Sc
a b cJ = + +
=+ −c
A B c , 0 ≤ SJ ≤ 1
Desconsidera a igualdade (0,0) no numerador e denominador.
2) Índice de Sörensen ou Coeficiente de Comunidade de Sörensen (SS):
Sc
a b c
c
A BS = + +
=+
2
2
2 , 0 ≤ SS ≤ 1
Desconsidera a igualdade (0,0) no numerador e denominador e dá peso em dobro para a igualdade (1,1).
2.6 Classificação e Ordenação
Classificação e ordenação são duas técnicas multivariadas eficientes de
sumarização ou simplificação de dados numéricos, para comparações estatísticas. As
técnicas de classificação baseiam-se no agrupamento de amostras ou de espécies que
tenham propriedades em comum, enquanto que as técnicas de ordenação dispõem as

- 45 -
amostras ou as espécies ao longo de eixos de variação contínua (Mateucci & Colma,
1982).
Greig-Smith et al. (1967) consideram que o elevado número de espécies e a
ausência de predominância de uma ou de determinadas delas impediram o sucesso da
aplicação de procedimentos tradicionais de classificação em regiões tropicais.
Webb et al. (1967b) mencionam que o elevado número de espécies nas florestas
tropicais, torna-se uma grande dificuldade para as análises numéricas, porque demanda
um tempo de processamento de dados muito alto, podendo inviabilizar o processo. Além
disso, qualquer método que requer a extração dos autovalores a autovetores de uma
matriz de similaridade interespecífica é impraticável. Como cada espécie contém
pouquíssima informação, os autovalores de tal matriz decrescem muito lentamente.
De acordo com Greig-Smith et al. (1967), a ordenação é capaz de evidenciar,
satisfatoriamente, as principais relações da composição da vegetação de uma forma mais
clara, onde as informações podem ser apresentadas graficamente.
Em florestas naturais, heterogêneas, Webb et al. (1967a,b) recomendam que a
classificação seja conduzida primeiro e a ordenação seja reservada àqueles casos em
que os métodos classificatórios provarem ser ineficientes. Laven (1982) sugere que a
ordenação deve suceder a análise de agrupamentos para auxiliar na interpretação dos
resultados.
Segundo Orlóci (1978), o problema de escolha entre métodos de classificação e
ordenação não deve estar associado a pressuposições rígidas. Ambas devem ser
encaradas como técnicas de análise de dados, que podem auxiliar o usuário a alcançar
seus objetivos.
Do ponto de vista prático, Goodall (1982) observa que o tipo de análise a escolher
dependerá da preferência do investigador e, mais ainda, do objetivo de estudo, que da
natureza da vegetação. Quando o objetivo de estudo é descrever a vegetação, a
classificação é mais adequada. Se o objetivo é determinar relações entre a vegetação e o
ambiente, a ordenação simplifica as interpretações.
2.6.1 Técnicas de classificação
A classificação consiste em agrupar as amostras ou as espécies, segundo suas

- 46 -
características. Consiste em dividir o sistema multidimensional em compartimentos ou
células, em cada um dos quais se reúnem os pontos que apresentam maior similaridade
entre si (Matteucci & Colma, 1982). Para Gauch & Whittaker (1972 e 1981) este sistema
multidimensional é uma matriz primária espécies-parcelas que representa o espaço
espécie, no qual as espécies são os eixos do espaço multidimensional e as parcelas são
os pontos locados por seus escores, para cada espécie.
Sokal & Sneath (1963), Orlóci (1978), Gauch & Whittaker (1981) e Matteucci &
Colma (1982) observam que, quando se consideram as características (propriedades que
descrevem um indivíduo e que podem assumir um valor) que intervêm na formação dos
agrupamentos, as técnicas de classificação podem ser monotéticas e politéticas. As
técnicas monotéticas, de acordo com Lambert & Dale (1964) e Lambert & Williams (1962),
empregam uma só característica em cada fusão ou subdivisão, enquanto as técnicas
politéticas empregam uma função de semelhança ou função objetiva baseada num
conjunto de características.
Para Sokal & Sneath (1963), em ecologia, o termo monotético tem sido usado para
uma classificação, na qual os grupos são formados, baseando-se na presença ou
ausência de uma dada espécie.
Quando fusões ou subdivisões são condicionadas sobre uma forte discriminação
de duas ou mais espécies, em qualquer passo, no processo de agrupamento, o algoritmo
é dito ser politético (Orlóci, 1967).
Lambert & Williams (1962), Lambert & Dale (1964) e Matteucci & Colma (1982)
consideram dois tipos de técnicas de classificação, as que alocam indivíduos a classes
previamente estabelecidas (Análise Discriminante) e aquelas que criam as classes ou
agrupamentos, a partir das informações contidas nos dados (Análise de Agrupamento).
2.6.1.1 Análise de Agrupamento (Cluster)
Entende-se por Análise de Cluster um método de classificação numérica, onde se
buscam e caracterizam grupos, com diferentes graus de similaridade em uma quantidade
dada de dados (Bockor, 1975).
Segundo Sneath & Sokal (1973), Orlóci (1978) e Johnson & Wichern (1982), o
critério de agrupamento empregado, na maioria dos algoritmos, é uma medida de

- 47 -
similaridade ou de distância estatística entre os elementos de uma matriz X.
Essas medidas constituem na entrada (Input) para os vários algoritmos e definem
uma função dos valores dos vetores representativos dos elementos de X, para os quais
se calcula uma medida de similaridade ou distância, comumente denominada,
respectivamente, Coeficiente de Similaridade ou Medida de Distância (Orlóci, 1978).
Para Johnson & Wichern (1982), a distância Euclidiana é a métrica de maior
emprego nas análises de agrupamentos e a que apresenta maior facilidade de cálculo.
Contudo, quando os objetos não podem ser representados por medidas p-dimensionais,
os pares de objetos são comparados com base na presença ou ausência de certas
características.
As técnicas de agrupamento são utilizadas em muitas áreas de conhecimento,
como Medicina, Filosofia, Política, Geologia, entre outras. Contudo, pouco se sabe de sua
utilização em dados de vegetação. A esse respeito, Orlóci (1978) comenta que, ao aceitar
uma vegetação como uma população multiespécie, implica em um agregado de unidades
naturais. Se estas unidades existem, certamente necessitam de uma forte base natural
para identificação ou, pelo menos, são somente reconhecíveis com ambigüidade, por ser
de composição complexa, cujos limites não são facilmente definidos.
A hipótese de que a vegetação é um agregado de unidades naturais reconhecíveis
é defendida por muitos fitossociólogos das principais Escolas Européias como Greig-
Smith (1964), Kershaw (1973), Orlóci (1978), Braun-Blanquet (1979), Havel (1980a,b),
Whittaker (1975 e 1982) e Holdridge (1982). Eles afirmam que além de serem distintas, as
unidades tendem a formar tipos naturais (associações).
Dentre os procedimentos de uma classificação, dois enfoques têm sido abordados:
os procedimentos informais (descritivos) e os formais. Os informais têm sido amplamente
utilizados em estudos de vegetação, por detectarem classes naturais (tipos, associações,
etc.) com razoável consistência. Já os procedimentos formais possibilitam classificar
conjunto de dados visualmente heterogêneos, muito complexos e muito volumosos para
serem eficientemente manipulados por outros métodos (Whittaker, 1962).
A importância do emprego dos métodos formais de agrupamento em análise de
vegetação é mostrada por diversos autores, entre os quais, Williams & Dale (1965),
Anderberg (1973), Matteucci & Colma (1982).
Matteucci & Colma (1982) consideram que, os métodos formais de classificação

- 48 -
partem da matriz de similaridade e identificam as classes mediante uma série de cálculos.
Segundo Johnson & Wichern (1982) e Matteucci & Colma (1982), os métodos de
Agrupamento de Cluster podem ser de dois tipos: Hierárquico e Não-hierárquico.
As técnicas de agrupamento hierárquico baseaiam-se em uma série de fusões
sucessivas ou uma série de divisões sucessivas (Johnson & Wichern, 1982). Para
Matteucci & Colma (1982) este é o método mais utilizado em levantamentos com fatores
ecológicos, pois, os níveis de similaridade e dissimilaridade podem ser visualizados em
dendrogramas.
De acordo com o procedimento utilizado na formação das classes, as técnicas
podem ser divisivas ou aglomerativas. Para Matteucci & Colma (1982), as técnicas
divisivas começam com a população completa e subdivisões sucessivas vão formando
grupos cada vez menores. Em cada etapa da subdivisão buscam-se as diferenças dentro
dos grupos para separar subgrupos que diferem entre si. Por outro lado, as técnicas
aglomerativas, conforme Sneath & Sokal (1973), Orlóci (1978), Gauch & Whittaker (1981)
e Matteucci & Colma (1982), começam pela fusão sucessiva dos indivíduos, que se
combinam por suas semelhanças, repetindo o procedimento até esgotar as possibilidades
de combinação ou até que não fiquem indivíduos isolados.
Assim, as técnicas aglomerativas buscam a similaridade entre indivíduos, enquanto
as técnicas divisivas buscam a dissimilaridade (Orlóci, 1978 e Matteucci & Colma, 1982).
Segundo Crisci & Armengol (1983) em relação à quantidade de características que
intervêm na formação de classes, as técnicas de classificação são Monotéticas ou
Politéticas. As técnicas monotéticas, só podem ser divisivas (as técnicas monotéticas
aglomerativas resultam em dados não confiáveis), e utilizam só uma característica em
cada subdivisão. Por outro lado, as técnicas politétitas empregam uma função de
semelhança baseada em um conjunto de características.
Para esses autores o grupo monotético é um grupo constituído por rígidas e
sucessivas divisões lógicas, no qual todos os membros compartilham um conjunto de
atributos que são condições necessárias e suficientes para converter-se em um integrante
do grupo. Já, o grupo politético é um grupo onde seus membros compartilham um grande
número de atributos, porém nenhum desses atributos é compartilhado por todos os
membros, com a exceção do atributo de pertencer ao grupo. Cada membro possui alguns
atributos do conjunto. Portanto, nenhum atributo é condição necessária o suficiente para

- 49 -
converter um objeto em um integrante do grupo.
Orlóci (1978) cita que os métodos de agrupamento hierárquico, na sua grande
maioria, podem ser utilizados para classificar tanto variáveis (espécies), quanto unidades
amostrais, utilizando qualquer medida de associação (similaridade) com exceção de uns
poucos que utilizam o quadrado da distância Euclidiana, para as amostras, quando
utilizadas como as entidades a serem agrupadas.
Johnson & Wichern (1982) apontam que no método aglomerativo hierárquico existe
no início, tantos grupos, quanto objetos. Muitos objetos semelhantes são agrupados
primeiro e, estes grupos iniciais são fundidos de acordo com suas similaridades.
Relaxando no critério de similaridade, todos os grupos são fundidos dentro de um único
grupo. O método divisivo hierárquico faz o trabalho em direção oposta. O grupo inicial de
objetos é dividido em dois grupos, de modo que os objetos em um grupo são distantes
dos objetos do outro. Estes subgrupos são, então, melhor divididos em outros subgrupos
não semelhantes e o processo continua até que existam tantos subgrupos quanto objetos.
De acordo com os autores o agrupamento dos objetos ou indivíduos é feito por
ligações. Estas podem ser simples (método da mínima distância ou do vizinho mais
próximo), completa (método da máxima distância ou do vizinho mais longe) e média
(distância média). Para Orlóci (1967) e Matteucci & Colma (1982) um outro método de
ligação, muito usado em matrizes muito grandes, é conhecido por método da mínima
variância.
Descrições mais detalhadas desses métodos podem ser vistas em Anderberg
(1973), Hartigan (1975), Orlóci (1967, 1978), Mardia; Kent & Bibby (1979), Johnson &
Wichern (1982) e Matteucci & Colma (1982).
O agrupamento não-hierárquico é uma técnica usada, quando se deseja formar k-
grupos de itens ou objetos. O número de grupos, k, pode, igualmente, ser especificado
com antecipação ou determinado a partir do processo de agrupamento. Como a matriz de
distância (ou similaridade) não pode ser determinada e os dados básicos não podem ser
armazenados durante a computação, o método não-hierárquico pode ser aplicado para
muito mais dados que técnicas hierárquicas (Johnson & Wichern, 1982).
Para os autores, o método de agrupamento mais usado é o das k-médias, sugerido
por Macqueen em 1967, para descrever algoritmo que designa todos os itens do grupo,
tendo o centróide (média) mais próximo. Nessa mais simples versão, o processo é

- 50 -
composto por três passos:
a) partição dos itens em k-grupos iniciais;
b) prossegue com a lista de itens, onde cada um é colocado no grupo, cuja média
(centróide) esteja mais próxima. Usualmente calcula-se a distância Euclidiana com
observações padronizadas ou não. Recalcula-se o centróide para os grupos envolvidos
na troca do novo item e para as perdas de grupo.
c) repete-se o segundo passo, até que não restem recolocações a serem feitas.
O programa de computador TWINSPAN (TWo-way INdicator SPecies ANalyis),
utiliza um processo multivariado, ou seja, é um método hierárquico, divisivos e politético
(Hill, 1979).
TWINSPAN faz uma classificação das espécies em função das parcelas em que
estas ocorrem e das parcelas em função das espécies que nelas ocorrem. Esta análise
multivariada permite o descobrimento de padrões de dados, através da ordenação e
classificação. Permitem organizar os dados de comunidades baseadas na densidade de
espécies, não considerando os fatores ambientais. A ordenação arranja as espécies e
parcelas num espaço dimensional restrito, isto é, as parcelas que ficam próximas são
semelhantes e as que têm uma certa distância ficam distantes. Já a classificação
determina o local onde se encontram as espécies e parcelas (Felfili et al., 2001a).
Segundo Felfili et al. (2001b), o programa TWINSPAN, constrói uma tabela
bidimensional pela identificação de espécies preferenciais. As parcelas são classificadas
primeiro por dicotimização sucessivas e as espécies pelo mesmo processo. Para obter a
dicotimização as ordenações são divididas pela metade. Este método faz três
ordenações: (i) na ordenação primária, a direção de variação é determinada nos dados;
(ii) na segunda ordenação, a dicotomia preliminar é obtida pela divisão da ordenação pela
metade e ocorre a identificação de espécies preferenciais (são espécies com as mesmas
preferências ecológicas e, desta forma sua presença pode ser utilizada para identificar
condições ambientais particulares) em um ou outro lado da dicotomia e; (iii) a terceira
ordenação, é a ordenação por indicador, que é baseada num pequeno número de
espécies fortemente preferenciais (Hill, 1979).
Em cada divisão é calculado um autovalor (eigenvalue), que indica o quanto da
variação dos dados da comunidade foi explicada no eixo de ordenação (McCune &
Mefford apud Araujo, 2002). Normalmente, o autovalor em torno de 0,30 é utilizado para

- 51 -
explicar as variações ocorrentes na natureza, logo, quando a divisão atingir um autovalor
menor que 0,30, esta pode ser descartada (Araujo, 2002).
O DENDROGRAMA ou DIAGRAMA EM ÁRVORE é a forma visual, mais utilizada,
mundialmente, para representar a seqüência de agrupamentos. No dendrograma, cada
ligação corresponde a um valor numérico (um índice, que é a escala ou nível de
agregação) que representa o nível, no qual têm lugar os agrupamentos. Quanto maior o
índice, mais heterogêneo são os indivíduos agrupados. Uma vez formado o dendrograma,
a separação entre os grupos é realizada. Pode ser feito um corte em um determinado
nível, que é geralmente na metade da maior distância, ou segundo um critério subjetivo e
pessoal (Longhi, 1997).
2.6.1.2 Análise Discriminante
Segundo Anderson (1971), se medidas são realizadas em amostras aleatórias
pertencentes a diversas categorias ou grupos, então cada grupo será visualizado como
um conjunto de pontos num espaço multidimensional. A análise discriminante reduz o
número de medidas realizadas para um número menor de parâmetros que são funções
discriminantes linearmente dependentes das medidas originais. Desta forma, os grupos
(conjuntos de pontos) poderão ser visualizados num espaço multidimensional menor que
o anterior e, os coeficientes das funções discriminantes indicarão a contribuição relativa
das medidas originais, para cada função discriminante, que segundo Rao apud Batista
(1990), é aproximadamente um teste de significância de separação dos grupos.
Souza (1989) observa que a análise discriminante e a classificação têm como
objetivos descrever algébrica e/ou graficamente, num espaço reduzido, as características
diferenciais dos objetos de vários agrupamentos conhecidos e ordenar os objetos dentro
de classes pré-definidas, dando ênfase à regra de alocação ótima de novos objetos aos
agrupamentos.
De acordo com Marriott (1974), a análise discriminante consiste em investigar
como e quando é possível fazer distinções entre os membros de g agrupamentos, com
base nas observações feitas sobre eles. Tem como objetivo testar diferenças estatísticas
significantes a um dado nível de probabilidade entre g agrupamento; determinar o número
de funções discriminantes; construir regras de alocação para identificar um indivíduo

- 52 -
como membro de um dos g agrupamentos; e estimar as probabilidades de classificações
corretas.
Para Johnson & Wichern (1982), a técnica multivariada conhecida como Análise
Discriminante e Classificação trata dos problemas relacionados em separar conjuntos
distintos de objetos (observações) e em alocar novos objetos (observações) em grupos
previamente definidos. A terminologia discriminar e classificar foi introduzida por FISHER,
em 1938, no primeiro tratamento moderno dos problemas de separação.
Segundo os mesmos autores, a idéia básica do método de Fisher consiste em
separar duas classes de objetos ou fixar um novo objeto, em uma das duas classes. É
conveniente denominar as classes (populações) de π1 e π2. Os objetos são
ordinariamente separados ou classificados com base nas medidas das p-variáveis
aleatórias associadas [ ]′ =X X X X p1 2, , ,L .
A idéia de Fisher foi transformar as observações multivariadas X, nas observações
univariadas Y, tal que os Y’s derivados das populações π1 e π2 sejam separados tanto
quanto possível. Fisher sugeriu o uso da combinação linear das variáveis aleatórias de X
para a obtenção dos Y’s, porque são funções simples e fáceis de obter matematicamente.
A função discriminante linear de Fisher é dada pela combinação linear:
y c X X= ′ = − ′ −( )µ µ1 2
1Σ
2.6.2 Técnicas de Ordenação
Com os métodos de ordenação, obtêm-se seqüências ou gradientes, ao dispor os
indivíduos (amostras ou atributos), ao longo de eixos de variação contínua. Reduz-se o
número de dimensões, expressando a variação da vegetação em poucos eixos, nos quais
recupera-se a máxima quantidade de informação possível (Matteucci & Colma, 1982).
Segundo Morrison (1976), Orlóci (1978), Matteucci & Colma (1982) e Johnson &
Wichern (1982), existe um número bastante grande de técnicas que tratam da ordenação
de dados, conhecidos como Análise Multivariada. A aplicação destas técnicas, em
estudos da vegetação, está diretamente relacionada com o tipo de dados a serem
analisados, com a estrutura dos mesmos e, particularmente, com o objetivo da análise.
Três são as principais técnicas de ordenação: a Análise dos Componentes

- 53 -
Principais, a Análise Fatorial e a Análise de Correlação Canônica (Análise de
Correspondência Canônica) que, segundo Johnson & Wichern (1982), são conhecidas por
técnicas de Análise da Estrutura de Covariância.
2.6.2.1 Componentes Principais
Segundo Mallo (1985), a Análise de Componentes Principais foi descrita
inicialmente por Karl Pearson, em 1901 e posteriormente desenvolvido por Hotelling, em
1933. Sua finalidade principal consiste em condensar o essencial da informação dada, por
uma série de variáveis interdependentes, observadas diretamente sobre um conjunto de
indivíduos, em um número mais restrito de variáveis fundamentais independentes. Estas
novas variáveis originais possuem as seguintes características:
a) redução dimensional - são escolhidas em número menor que as originais;
b) não são correlacionadas;
c) significatividade, devem explicar a maior proporção possível da variabilidade
total, com poucas componentes.
Para Foguet & Arias (1989), a finalidade da análise de componentes principais é
construir novas variáveis latentes, as componentes, que recolham a variação do sistema
sem redundâncias, reduzindo a dimensionalidade do espaço original, com distorções
mínimas da informação.
Asensio (1989) observa que a análise de componentes principais tem como
objetivo facilitar o estudo das relações existentes entre as variáveis e, facilitar a análise da
dispersão das observações, detectando as variáveis responsáveis.
Segundo Johnson & Wichern (1982), a Análise dos Componentes Principais
procura explicar a estrutura de variância-covariância, mediante combinações lineares, não
correlacionadas, das variáveis originais. Em geral, tem como objetivo reduzir os dados e
facilitar a interpretação.
Com referência aos componentes principais, Pla (1986) observa que este é um dos
métodos de análise multivariada mais difundidos, permitindo a estruturação de um
conjunto de dados multivariados obtidos de uma população, cuja distribuição de
probabilidades não necessita ser conhecida, tratando-se de uma técnica matemática que
não requer um modelo estatístico para explicar a estrutura probabilística dos erros.

- 54 -
2.6.2.2 Análise Fatorial
A análise fatorial é uma técnica, onde muitas variáveis podem ser representadas
por uma função linear, com um número menor de variáveis não observáveis, chamadas
fatores (Morrison, 1976).
De acordo com Johnson & Wichern (1982), a análise fatorial tem provocado muita
controvérsia ao longo da história. O princípio moderno encontra-se no início do século XX,
no esforço de Karl Pearson, Charles Spearman e outros, em definir medidas de
inteligência. A dificuldade nos cálculos impediu um desenvolvimento da técnica, o que só
ocorreu com o advento dos computadores.
Segundo os autores, a proposta inicial da análise fatorial é descrever, se possível,
a covariância dos relacionamentos entre muitas variáveis, em termos de poucas variáveis
fundamentais, não observáveis, chamadas fatores. Supõe-se que variáveis possam ser
agrupadas por suas correlações. Isto é, todas as variáveis dentro de um grupo particular
são altamente correlacionadas entre si, mas com correlações relativamente baixas ou
negligenciáveis com as variáveis de um grupo diferente. É admissível que cada grupo de
variáveis represente um fator, que é responsável pelas correlações observadas.
De acordo com Batista (1990), a escolha do número de fatores pode ser feito a
partir de diversos critérios:
a) a estrutura física das variáveis pode sugerir um número de fatores
intrínsecos;
b) quando se usa a matriz de correlação, é recomendável que se tome tantos
fatores, quanto forem os autovalores maiores que 1;
c) pode-se, ainda, escolher um número de fatores que expliquem uma porção
específica da variância total.
Para o autor, a interpretação dos fatores é mais precisa, se cada variável tem peso
alto em, no máximo, um fator e, se todos os pesos dos fatores são grandes e positivos ou
próximos de zero. Assim, as variáveis são agrupadas em conjuntos distintos, cada um dos
quais associados a um fator. Às vezes é necessário fazer uma rotação dos fatores para
melhor interpretação.

- 55 -
2.6.2.3 Análise de Correspondência Canônica (CCA)
A Análise de Correspondência Canônica (CCA), é uma técnica desenvolvida por
Ter Braak (1986). A CCA é diferente de todos os métodos de ordenação já discutidos. A
CCA é um método que apresenta a relação entre a distribuição das espécies e a
distribuição dos fatores ambientais, associados a gradientes (Kent & Coker, 1992). A CCA
permite ainda testar a probalidade de acerto nas relações encontradas por meio do teste
de permutação de “Monte Carlo” (Aubert e Oliveira-Filho, 1994).
Todos os métodos de ordenação como ordenação polar, análise de componentes
principais e análise fatorial têm este objetivo, porém, são indiretos em cada análise,
executando nos primeiros somente para espécies, e quando a interpretação a ambiental é
feita por acréscimo dos dados ambientais sobre as parcelas de ordenação (Aubert e
Oliveira-Filho, 1994).
Algumas análises podem ir até onde a correlação e regressão do eixo da parcela
marcam com fatores ambientais, mas, por várias razões, isto não é sempre satisfatório.
A CCA difere das outras técnicas clássicas indiretas, porque nesta análise é
incorporado a correlação à regressão, entre os dados florísticos e fatores ambientais
dentro da própria análise de ordenação (Botrel et al., 2002).
A entrada de dados consiste na criação de duas matrizes de dados. A primeira
matriz é uma que tem a relação entre espécies (densidade maior que 5) x parcelas, e a
segunda matriz é a relação entre os fatores ambientais x parcelas. Esta aproximação das
espécies e de dados ambientais no processo de ordenação atual é conhecida como uma
forma de Análise Canônica (Aubert e Oliveira-Filho, 1994).
O diagrama de ordenação resultante expressa, não só, padrões de variação da
composição florística, mas, também demonstra as relações principais entre as espécies e
cada uma das variáveis ambientais (Martins et al., 2003).
Os pontos, apresentados no diagrama, representam as espécies individuais e as
setas representam cada variável ambiental plotada no diagrama. O comprimento da seta
é proporcional à magnitude de mudança naquela direção, e para propósito de
interpretação cada seta pode também ser estendida no sentido contrário da origem
central. Aqueles fatores ambientais que na seta longa estão mais correlatos na ordenação
do que aqueles com seta curta, e são mais importantes na influência da variação da

- 56 -
comunidade (Kent & Coker, 1992). A ordem em que se apresentam os pontos projetados
sobre a seta, da origem à extremidade, indicam a influência do fator ambiental em relação
à espécie, isto é, quanto mais próximo da extremidade da seta, maior influência do fator
ambiental sobre a espécie.
2.7 Regressão Logística
Segundo Hosmer & Lemeshow (1989), os métodos de regressão têm se tornado um
componente integral de qualquer análise de dados relacionados com a descrição da
relação entre uma variável resposta (variável dependente) e uma ou mais variáveis
explanatórias (variável independente).
De acordo com os autores a regressão logística estima diretamente a probabilidade
de um evento dicotômico ocorrer, isto é, a presença de uma determinada espécie em uma
área pode ser considerada com um fenômeno dicotômico. Existem duas hipóteses: ou a
espécie ocorre ou não ocorre (Hosmer & Lemeshow, 1989).
Para estimar os parâmetros na regressão logística, é utilizado o método da máxima
verossimilhança, isto é, os coeficientes que fazem os resultados observados mais
prováveis são selecionados através de um processo interativo. Existem alguns
procedimentos para a seleção de modelo na regressão logística, por exemplo, para
adicionar ou retirar variáveis no modelo pode-se usar o método Forward Stepwise e
Backward Stepwise e para adicionar ou retirar propositadamente variáveis no modelo
pode-se usar o método ENTER.
Conforme Hasenauer apud Caldato (1998), a regressão logística produz valores de
probabilidade que variam de 0 (zero) e 1 (um), possibilitando, a simulação da ocorrência
de um determinado evento, ao se comparar o valor resultante da função em cada caso
específico com valores de números randômicos de distribuição uniforme ou de Poisson,
gerados no mesmo intervalo (0 – 1).
Para Hosmer & Lemeshow (1989), o modelo geral da regressão logística é:
Probabilidade (evento) = Ze−+1
1

- 57 -
Onde Z é a combinação linear ou não linear:
pp XBXBXBBZ +++++= ...22110
sendo B0, B1, B2 e Bp, coeficientes estimados dos dados; X1, X2 e Xp são as variáveis
independentes; e é a base do logaritmo natural, aproximadamente 2,718.
A interação de qualquer modelo requer que se tirem inferências práticas dos
coeficientes estimados no modelo. Os coeficientes estimados para as variáveis
independentes representam a inclinação ou a taxa de mudança da variável dependente,
em função da modificação de uma unidade da variável independente. A formulação lógica
de modelo geral, ou seja, a seleção das variáveis independentes é, pois, a base para a
carreta utilização da regressão logística (Hosmer & Lemeschow, 1989).
Segundo Caldato (1998), uma das maneiras de avaliar o quanto o modelo é
apropriado é pela comparação da precisão do modelo com dados reais, através de uma
tabela de classificação, entretanto, não revela a distribuição das probabilidades estimadas
para um fato em dois grupos. Para cada grupo estimado, todas as tabelas mostram-se a
probabilidade estimada é maior ou menor que 50%.
Para Hosmer & Lemeshow (1989) uma outra maneira para verificar o modelo
adequado, é pela observação da semelhança entre os dados amostrais e os previstos
pelo modelo, através da estátistica -2LL (2 Logarithm of the Likelihood = -2 vezes o
logaritmo da verossimilhança). Um bom modelo é o que tem uma alta semelhança entre
os dados e o resultado do modelo, isto é, o que leva um pequeno valor de -2LL, quando o
valor da verossimilhança observada for 1 e o -2LL é zero, o modelo descreve
perfeitamente os dados (Caldato, 1998).
Ainda, segundo os autores, existe outra estatística que pode ser usada para testar
o ajuste do modelo que é a Qualidade do ajuste (Goodness of fit), que compara as
probabilidades observadas com aquelas estimadas pelo modelo, tal que:
onde o resíduo é a diferença entre o valor observado (Yi), e o valor estimado (Pi).
)1(
Re 22
PiPi
siduoZ i
−=∑

2 REVISÃO DE LITERATURA
2.1 Caracterização geral da Região
2.1.1 Geologia
Na Região Sul o vulcanismo continental está representado por espessos e
extensos derrames de lavas, bem como por diques e soleiras, com pequenos e eventuais
corpos de rochas, sedimentares associados. Tal conjunto de litologias constitui a
Formação Serra Geral, dividida em duas porções – A Seqüência Básica e a Seqüência
Ácida (Kaul, 1990).
A Seqüência Básica da Formação da Serra Geral, compreende derrames de
basalto, andesito e basalto com vidro, além de brechas vulcânicas e sedimentares, diques
e soleiras de diabásico e corpos de arenito interderrames. Cada derrame basáltico pode
aparecer diferenciado, estruturalmente, em quatro zonas: zona basal, de disjunção
horizontal; zona central, de disjunção colunar, normalmente ampla; zona superior, de
disjunção horizontal; e zona vesicular, no topo.
A Seqüência Ácida da Formação da Serra Geral, que corresponde a áreas de
relevo menos dissecado e menos arrasado, compreende derrames de dacitos pórfiros,
dacitos felsíticos, riolitos felsíficos, riocitos felsíficos, basaltos pórfiricos e fenobasaltos
vítreos. Os dois últimos tipos de rochas têm características petrográficas de vulcânicas
básicas; entretanto, quimicamente são vulcânicas ácidas. A seqüência é, essencialmente,
produto de contaminação de magma básico, gerado no Manto Superior, com rochas
siálicas da Crosta Inferior (Kaul, 1990). Autores, indicam que até o momento, que a
formação originou-se em tempos juracretácicos.
2.1.3 Relevo
Na unidade de Relevo Planalto das Araucárias, na sua parte leste, ocorrem cotas
altimétricas que ultrapassam 1200 m próximo à escarpa conhecida como Serra Geral. Em
alguns pontos registram-se cotas altimétricas superiores a 1500 m, com a presença de

95
relevos residuais acimade 1800 m, como o Morro da Igreja, com 1822 m de altitude,
localizado próximo a São Joaquim (SC) (Hermann & Rosa, 1990).
A costa da Serra Geral, apresenta um desnível médio de 400 m, aparecendo em
quase toda a borda oriental do planalto. Já, na porção oeste, as cotas decaem
gradativamente em direção à parte central da Bacia Sedimentar do Paraná, atingindo, no
máximo, 300 m. A mudança do desnível está relacionado ao mergulho das camadas da
bacia sedimentar, caracterizando o relevo da unidade como um planalto monoclinal.
O relevo tem essa variação altimétrica em função do aprofundamento da drenagem
dos rios principais, como o Iguaçu e o Uruguai. Esses rios apresentam vários trechos com
desníveis entre as partes interfluviais e o fundo do vale acentuados em função da
potência e do gradiente do rio. Outro fator importante para compreender a topografia da
área são os desníveis existentes nos contatos entre as áreas conservadas e as áreas
dissecadas que, em muitos setores, é marcado por forte ruptura do declive (Hermann e
Rosa, 1990).
2.2.3 Solo
Com base no Sistema Brasileiro de Classificação de Solos – SBCS (Embrapa,
1999), são encontrados na FLONA, CAMBISSOLO HÚMICO Alumínico, CHERNOSSOLO
ARGILÚVICO Férrico e NEOSSOLO LITÓLICO Eutrófico (Streck et al., 2002).
Os Cambissolos são solos em processo incipientes de formação, variam de rasos a
profundos, a drenagem altera de bem drenados a imperfeitamente drenados, dependendo
da posição que ocupa na paisagem. No perfil dos cambissolos ocorre a presença de
fragmentos de rochas, assim, pouca intemperização do material (Streck et al., 2002).
O Cambissolo Húmico alumínico é o um solo com média quantidade de matéria
orgânica, com Al trocável ≥ 4 cmol/kg; saturação por Al ≥ 50%), logo, extremamente
ácido. Este tipo de solo ocorre em ambientes onde a alta pluviosidade e as baixas
temperaturas favorecem a acumulação de matéria orgânica, na Região dos Campos de
Cima da Serra, em relevo ondulado a forte ondulado (Streck et al., 2002).
Os Chernossolos são solos escuros com alta fertilidade química, variam de rasos a
profundos. Se caracterizam por apresentar razoáveis teores de material orgânico, o que
lhe confere cores escuras ao horizonte superficial que é do tipo A chernozêmico e tem

96
alta fertilidade química (saturação por bases ≥ 65%) e alta CTC (Capacidade de Troca
Catiônica) em todo o perfil. Um elevado teor de ferro (≥18%) identifica os Chernossolos
Argilúvicos férricos, os quais ocupam áreas de pequena extensão, em relevo ondulado a
fortemente ondulado, exigem práticas conservacionistas intensivas e oferecem condições
para o uso com culturas anuais, fruticulturas e reflorestamento (Streck et al., 2002).
Os Neossolos são solos novos, isso é, pouco desenvolvidos; são rasos ou
profundos encontrados nas mais diversas condições de relevo e drenagem. Os Neossolos
Litólicos, têm pequena espessura, ocorrem em regiões de relevo forte ondulado e
montanhoso, geralmente com pedregosidade e afloramento de rochas, têm baixa
capacidade de infiltração e armazenamento de água no solo e alta suscetibilidade à
erosão hídrica, impossibilitando o seu uso para culturas anuais. Aréas de Neossolos
Litólicos com declividade superior a 30% devem ser mantidas com cobertura vegetal
natural, constituindo-se área de preservação permanente (Streck et al., 2002).
2.1.4 Clima
O clima da área de estudo, de acordo com a classificação de Köppen, é do tipo
“Cfb”, mesotérmico médio. Este clima domina as cotas altimétricas entre 1.100 e 1.000m
no norte do Rio Grande do Sul. Compreende apenas 0,7% do Estado, estando
relacionado às cotas altimétricas mais elevadas da região, com sua ocorrência esparsa
em numerosos e pequenos locais montanhosos, geralmente pouco povoados. Entretanto,
duas áreas de ocorrência deste tipo climático merecem destaque pela sua extensão
territorial e pelos povoamentos nelas existentes. Trata-se da superfície da chapada de
Palmas e da chapada de Vacaria-Lages-São Joaquim, estando a área de estudo
compreendida nesta última (Nimer, 1990).
Nesta região, de clima característico, há pelo menos um mês com temperatura
média inferior a 10°C, possuindo inverno acentuado, cujo frio é uma constante dia e noite,
e verão onde calor é praticamente ausente, pelo efeito da altitude .
Resulta daí que, na área em estudo, a amplitude térmica é menos importante do
que as verificadas nas superfícies baixas da Região Sul, o que determina uma
temperatura média anual muito baixa, situando-se entre 12 e 14°C na superfície de
Vacaria-Lages-São Joaquim. Mesmo assim, enquanto o verão se caracteriza por

97
temperaturas muito amenas, o inverno é caracterizado por constante e acentuado frio.
Como efeito, a média do mês mais quente (janeiro) oscila em torno de 20°C; a média das
máximas diárias, em torno de 26°C, e a máxima absoluta tem pouca possibilidade de
ultrapassar 34°C. No inverno, entretanto, a temperatura cai diariamente a níveis próximos
de 0°C. Por este motivo, a média das mínimas diárias mantém-se abaixo de 6°C durante
todo o inverno e o número de noites frias varia de 15 a 20 dias por ano. A nevada, embora
não seja um fenômeno muito comum, não constitui uma raridade, como acontece nas
áreas de Clima Mesotérmico Brando. Decorre daí que o inverno, nestas áreas de Clima
Mesotérmico Médio “Cfb”, possui média térmica inferior a 10°C, pelo menos em julho, seu
mês mais frio (Nimer, 1990).
2.1.5 Vegetação primitiva
Rambo (1956) divide o Estado do Rio Grande do Sul em duas formações vegetais,
a do campo e a da floresta. Da área total do Estado, cerca de 131.896 km² (46,26%) são
campos, 98,327 km² (34,47%) matas e o restante, atribuído à vegetação litorânea,
banhados inundáveis e outras formações.
Para o mesmo autor, as variações climáticas das diversas partes do Estado não
são suficientes para explicar a presença dessas duas formações, uma vez que a
formação climática conveniente no Estado do Rio Grande do Sul é a da Floresta Alta
Subtropical. Os campos são formações climáticas e edáficas na sua origem e relitos
históricos ou manchas no tempo atual.
Segundo Jarenkow (1994), a distribuição atual das formações vegetais do sul do
Brasil resulta de um processo histórico, cujo entendimento remete a abordagens
multidisciplinares, em diferentes momentos de sua evolução, principalmente aqueles
ocorridos a partir do final do Terciário.
Para o autor, outras formações ou sub-formações florestais, de maior ou menor
importância, podem ainda ocorrer no Estado, no interior das diferentes regiões
fitogeográficas, como Matas de Restinga, Matas Insulares, Matas de Galeria, entre outras.
Segundo Leite & Klein (1990) a vegetação do Estado do Rio Grande do Sul
compreende nove regiões fitoecológicas ou fitogeográficas: Região da Floresta Ombrófila
Densa (Floresta Atlântica); Região da Floresta Ombrófila Mista (Floresta de Araucária);

98
Região da Floresta Estacional Semidecidual (Floresta Subcaducifólia); Região da Floresta
Estacional Decidual (Floresta Caducifólia); Região da Savana (Cerrado e Campo); Região
da Estepe (Campanha Gaúcha); Região da Savana Estépica (Campanha Gaúcha); Áreas
das Formações Pioneiras de Influência Marinha (Restingas e Dunas); e Área de Tensão
Ecológica (contatos).
Na FLONA de São Francisco de Paula o tipo fitogeográfico predominante é a
Floresta Ombrófila Mista ou Floresta de Araucária ou Floresta de Pinheiro. Segundo
Rambo (1956), o pinheiro ocorre em toda a borda superior livre do planalto, a começar do
norte de Santa Maria até o extremo nordeste; nos vales superiores e nas cabeceiras dos
Rios Caí, Taquari, das Antas, Jacuí e Pelotas; em grupos isolados ou densos sociedades,
nos capões disseminados por todo o planalto; em indivíduos solitários em pleno campo,
como se observa a leste de Cruz Alta; em mistura com a floresta virgem do Alto Uruguai,
ao norte de Passo Fundo e Lagoa Vermelha.
O mesmo autor afirma que o pinheiro é exclusivo do planalto, ocorrendo em
altitudes entre 500 m a Oeste e 1000 m ao Leste. Nunca desce, a não ser em manchas
ocasionais. Diferencia-se três núcleos principais do pinheiral: na aba do setor meridional
da escarpa, entre o Rio Taquari e o Rio dos Sinos; na borda dos Aparados entre o Rio
Maquiné e o Rio das Antas; e em pleno planalto central, no curso superior do Rio Jacuí,
ao sul de Passo Fundo.
Segundo Leite & Klein (1990) a concepção de Floresta Ombrófila Mista procede da
ocorrência da mistura de floras de diferentes origens, definindo padrões fitofisionômicos
típicos em zona climática pluvial. A área onde a coexistência de representantes da flora
tropical (afro-brasileira) e temperada (austro-brasileira), com marcada relevância
fisionômica de elementos Coniferales e Laurales, é denominada Planalto Meridional
Brasileiro, área de dispersão natural do pinheiro-brasileiro ou do pinheiro-do-paraná, a
Araucaria angustifolia ou “curiirama” dos indígenas, espécie gregária de alto valor
econômico e paisagístico.
Segundo Klein apud Leite & Klein (1990) a araucária e outros elementos de origem
temperada, em face às suas características heliófilas, encontram-se hoje desfavorecidos,
não só pela intervenção destruidora do homem, mas pela incompatibilidade com o clima
atual.

99
Os terrenos entre aproximadamente, os 500 e os 800 m de altitude estão
enquadrados na formação montana (Leite e Sohn, apud Leite & Klein, 1990) e
caracterizam-se por um clima sem época seca, com período frio (Temperatura média Tm
≤ 15º C) curto ou ausente e período quente longo (Tm ≥ 20º C). A área mais típica e
representativa da Floresta Ombrófila Mista é aquela das altitudes superiores aos 800 m,
principalmente dos terrenos altomontanos. Seu clima é o mais frio da região e com
maiores índices de geadas noturnas. Caracteriza-se pela ausência de período seco e
ocorrência de longo período frio (Tm ≤ 15º C). O período quente anual (Tm ≥ 20º C) é
geralmente curto ou ausente.
Sob estas condições climáticas e de acordo com a diversificação de outros
parâmetros ambientais, poder-se-ia determinar, na área típica da Floresta Ombrófila
Mista, dois grupos distintos de comunidades com araucária e lauráceas: um, onde o
pinheiro se distribuía de forma esparsa por sobre bosque contínuo, no qual 70 a 90% das
árvores pertenciam às espécies: imbuia (Ocotea porosa), espécie mais representativa,
canela-amarela (Nectandra lanceolata), canela-preta (Nectandra magapotamica), canela-
fogo ou canela-pururuca (Cryptocarya aschersoniana) acompanhadas da sapopema
(Sloanea monosperma), por vezes bastante freqüente, da guabirobeira (Campomanesia
xanthocarpa) e erva-mate (Ilex paraguariensis); outro grupo, onde a araucária formava um
estrato de 60 a 80% de folhosas, principalmente das espécies: canela-lageana (Ocotea
pulchella), espécie dominante, canela-amarela (Nectandra lanceolata), canela-guaicá
(Ocotea puberula), canela-fedida (Nectandra grandiflora), comboatá-vermelho (Cupania
vernalis) e comboatá-branco (Matayba elaeagnoides), acompanhadas de casca-d’anta
(Drimys brasiliensis), pimenteira (Capsicodendron dinisii), guabirobeira (Campomanesia
xanthocarpa) e diversas mirtáceas e aquifoliáceas.
Acompanhando planícies sedimentares recentes dispersas em diferentes altitudes
e latitudes e sujeitas a periódicas inundações, ocorre um tipo de formação definida como
Aluvial. Nela o pinheiro-do-paraná geralmente consorcia-se com branquilho (Sebastiania
commersoniana), jerivá (Syagrus romanzoffiana), murta (Blepharocalyx salicifolius),
corticeira-do-brejo (Erythrina cristagalli), tarumã (Vitex megapotamica), açoita-cavalo
(Luehea divaricata), salgueiro (Salix humboldtiana), além de aroeira-vermelha (Schinus
terebinthifolius) e diversas espécies de mirtáceas (Leite & Klein, 1990).

100
Na região de Floresta Ombrófila Mista é comum a ocorrência de campos. Nestes
se verifica grande número de capões e bosques, muitas vezes com a presença de
Araucaria angustifolia, denotando o lento processo de invasão das florestas nas áreas de
campo. Muito comum nestes campos é a ocorrência de araucária isoladas junto aos
capões (SUDESUL, 1978).
Segundo Rambo (1956), os capões também são típicos na porção leste do Planalto
(São Leopoldo), principalmente nos mananciais de água. Em sua composição destacam-
se as espécies arborescentes do parque campestre, que são as aroeiras (Schinus spp.),
assim como exemplares da mata virgem como cedro (Cedrela fissilis), cangerana
(Cabralea canjerana), louro (Cordia trichotoma), figueira (Ficus luschnathiana), jerivá
(Syagrus romanzoffiana), chá-de-bugre (Casearia sylvestris), mamica-de-cadela
(Zanthoxylum sp.) e camboatá (Matayba elaeagnoides). No interior cresce vegetação
lenhosa baixa, constituída de laranjeira-do-mato (Actinostemum concolor = Gymnanthes
concolor), cincho (Sorocea bonplandii), rubiáceas e monimiáceas arbustivas,
salsaparrilhas (Smylax sp.), entrelaçados por cipós. É uma mata virgem em escala
reduzida.
2.3 Fatores Ecológicos e de Concorrência
A palavra “ecologia” deriva do grego oikos que significa “casa” e logos que significa
“estudo”, logo, ecologia é o estudo do ambiente da casa, incluindo todos os organismos
contidos nela e todos os processos funcionais que a tornam habitável. Deste modo,
ecologia é o estudo do “lugar onde se vive” (Odum, 1988).
Uma definição concisa da ecologia moderna é dada por Edelbauer (1988): “Ciência
da inter-relação e das interinfluências das diferentes formas de vida e seu ambiente”.
Em épocas remotas os homens primitivos, quando escolhiam as plantas para a
alimentação e para curar determinadas doenças, além de locais adequados para a caça e
pesca, demonstravam o domínio do saber ecológico (Laroca, 1995).
As primeiras obras que faziam referências à ecologia foram as de Hipócrates e
Aristóteles (384-322 aC). Este último, filósofo e naturalista grego, organizou o
conhecimento sobre animais existentes na época, sendo considerado, por certos autores,

101
como um dos precursores da ecologia. No Brasil, o interesse pela ecologia moderna teve
início desde a publicação de Ecologia Vegetal (“Plantesamfund”), pelo dinamarquês
EUGENE WARMING, em 1895, e da classificação das formas de vida dos vegetais, por
RAUNKIAER, em 1903. Assim, a ciência que estuda as inter-relações das plantas, entre si e
com o meio, é relativamente nova, pois teve um desenvolvimento gradativo durante a
história e muito ainda deve ser estudada e desenvolvida (Ferri, 1963; Laroca, 1995;
Souza et. al., 1998).
Segundo Laroca (1995), a ecologia pode ser dividida em duas grandes áreas: a
auto-ecologia e a sinecologia. A AUTO-ECOLOGIA trata da ecologia do indivíduo e das
populações e, nesta área, pode-se incluir a genética ecológica, comportamento animal
(etologia), ecologia fisiológica, entre outras. A SINECOLOGIA envolve sistemas de muitas
espécies: a comunidade, como um todo, ou parte das mesmas, ou ecossistemas
(terrestres e aquáticos) e, portanto, nesta grande área pode-se incluir a fitossociologia.
Os primeiros trabalhos em fitossociologia no Brasil foram realizados por Davis e
Veloso em 1945 apud Souza et al. (1998). Antes disso não existiam dados sobre a
vegetação brasileira. Somente a partir da década de 70, surgiram trabalhos nos quais
foram avaliados a identificação taxonômica e os parâmetros fitossociológicos quantitativos
absolutos e relativos.
Atualmente fala-se muito sobre ecologia, fatores ecológicos, meio ambiente e
manejo sustentado dos recursos naturais renováveis (Rio+10, encontro em Johanesburgo
– África do Sul, 2002), mas apenas uma pequena parte da população mundial possui
conhecimento para entender a dinâmica e as inter-relações dos ecossistemas.
Em um ecossistema natural, cada componente é dependente do outro e, portanto,
devem ser estudados conjuntamente, para assim determinar suas influências sobre os
seres vivos. Existem fluxos de energia e de matéria em cada ecossistema de forma a
manter um equilíbrio ecológico. Com a modificação do equilíbrio ecológico, algumas
espécies sucumbem ao passo que outras se multiplicam maciçamente. Assim, torna-se
importante à realização de estudos sobre influência dos fatores ecológicos sobre o
desenvolvimento das espécies vegetais (Odum, 1988).
Durante a sua evolução o homem, muitas vezes, modificou os ecossistemas
naturais de maneira a satisfazer suas necessidades. As intervenções humanas
geralmente foram feitas sobre a vegetação, sistema hídrico e solo. Em função do

102
desenvolvimento, os ecossistemas naturais, através da retirada da vegetação nativa e do
cultivo, se tornaram ecossistemas modificados (Odum, 1988). Estes ecossistemas
modificados acabam forçando determinadas espécies a adaptarem-se às novas
intensidades que os fatores ambientais ou fatores ecológicos atuam sobre elas.
Segundo o autor, os fatores ecológicos são divididos em dois grandes grupos: (i)
ABIÓTICOS (fatores físicos e químicos) e; (ii) BIÓTICOS (patógenos, parasitas, predadores,
simbiontes, entre outros). Os componentes biótico e abiótico estão intimamente
associados no solo, o qual, por definição, consiste na camada intemperizada da crosta
terrestre com os organismos vivos e os produtos da sua decomposição intermisturados.
2.2.1 Umidade do solo
A água é uma necessidade fisiológica para todo o ser vivo e, em relação ao ponto
de vista ecológico, um fator limitante, principalmente em ambientes terrestres (Odum,
1988).
De acordo com o autor, o ritmo diário da umidade da natureza (alta à noite, baixa
durante o dia) afeta a umidade do solo juntamente com a temperatura e a luminosidade,
ajudam a regular as atividades dos organismos e a limitar sua distribuição.
A importância da água para o solo é evidente, pelo simples fato de ser um dos mais
ativos agentes de destruição e desagregação das rochas e minerais. A água é um dos
componentes do protoplasma das plantas, sendo obrigatória nas reações metabólicas,
além de funcionar como transportadora de elementos nutritivos e dos componentes que
se formam durante o metabolismo da planta e entre a planta e o solo (Vieira et al., 1988).
Para os mesmos autores a capacidade do solo de reter água no estado líquido é
resultante da ação conjunta e complexa de duas forças principais que são: (i) força de
adesão: a atração que as partículas do solo exercem sobre as moléculas de água e; (ii)
força de coesão: a atração que as moléculas de água exercem entre si. Não se deve
esquecer que a importância da cada um desses fatores depende do estado de umidade
do solo.
Segundo Silva Júnior (1998), os solos associados a comunidades com SOLOS
ÚMIDOS (glay húmico) apresentam teores mais altos de MO, Al, H + Al, P, Fe, Cu, Zn, e
saturação de Al e os maiores conteúdos de areia fina e grossa. As comunidades com

103
SOLOS SECOS podem ter níveis de pH, Ca, Mg, K, Mn e saturação de bases. Já os solos de
comunidades INTERMEDIÁRIAS apresentam os mais altos teores de argila e silte.
O abastecimento de água é muito importante para o crescimento da floresta, mas a
significação deste fator ecológico é naturalmente muito maior e não se limita só às
árvores.
A floresta regula o lançamento da água local e regional em grande escala, e este
sim, é um fator significante para a cultura da terra. Todas as águas de precipitação
retornam pela evaporação à atmosfera. Tanto dos continentes, como dos oceanos, sobe
vapor à atmosfera sem cessar, sob influência do calor da radiação solar e dos ventos.
Neste processo predominam os mares, por sua enorme extensão, como fornecedor da
umidade (Silva Júnior, 1998).
As circunstâncias de umidade de uma região climática serão determinadas, não
somente pelas precipitações, mas também através da temperatura e distribuição de
precipitação, ou seja, estes fatores determinarão em conjunto, se o clima será árido ou
úmido (Silva Júnior, 1998).
Logo, um fator relevante na manutenção da água no solo, é o relevo do lugar, pois
sobre um terreno plano a quantidade de água será igual a da chuva. Já em terrenos com
declividade, a água da precipitação, terá distribuição diferente, devido a sua lixiviação
(Nagy & Cunha,1986).
A floresta tem uma grande influência na quantidade de água da precipitação que
chega ao solo. O dossel das copas forma uma superfície, que representa uma superfície-
limite. Para cada chuva, uma certa parte da água consegue chegar ao solo da floresta
sem obstáculos e o que vai influenciar nesta quantidade de chuva será: (i) as espécies de
árvores da floresta; (ii) a densidade do povoamento; (iii) a idade; e (iv) o fechamento do
dossel. Portanto, quanto mais fechado for o povoamento, menos água de precipitação
chega ao solo (Nagy & Cunha, 1986).
Para os autores, as características do solo causam diferenças consideráveis na
umidade do solo de um sítio, porém, as maiores diferenças e oscilações da sua umidade
serão provocadas pela vegetação do sítio.

104
2.2.2 Inclinação do terreno
Segundo Silva Júnior (1998) a inclinação (topografia), que condiciona o regime de
água nos solos, tem sido indicada como determinante de algumas características
ambientais e, conseqüentemente, da distribuição da vegetação. Enfatiza, também, o
papel direto da topografia no processo de um regime diferenciado da água.
Furley e Oliveira-Filho, citados por Silva Júnior (1998), relatam que a topografia
direciona a drenagem para os vales, resultando na presença do lençol freático próximo à
superfície, durante quase todo o ano. Ressaltam, então, que a topografia é considerada
um determinante primário da vegetação e pode definir a fisionomia, a composição
florística, a riqueza em espécies e a densidade de árvores.
Vieira et al. (1988) classificam as condições topográficas do relevo de uma região
segundo os graus de declividade em: plana, suave ondulada, ondulada, forte ondulada e
montanhosa (Tabela 1).
TABELA 1 – Classificação topográfica de acordo com os graus de declividade do solo.
Especificação Porcentagem de declividade Plana < 30 Suave ondulada 3 – 80 Ondulada 8 – 200 Forte Ondulada 20 – 450
Montonhosa > 45 0
Fonte: Vieira et al. (1988)
2.2.3 Profundidade do solo
Esta característica está relacionada com a espessura máxima do solo em que o
sistema radicular das plantas não encontra dificuldade ou barreira física para penetrar
livremente, facilitando a sua fixação e servindo de meio para a absorção de água e de
nutrientes (Lepsch,1993; Sanchez,1981).
A profundidade do solo tem influência sobre a fertilidade natural, especialmente se
um ou mais nutrientes tem sua disponibilidade limitada, bem como sobre as práticas de
manejo a serem utilizadas. Também tem influência no sistema ar-água, o qual, por sua

105
vez, influencia a disponibilidade de nutrientes e sistema radicular das árvores (Vieira et
al., 1988).
Os autores avaliam a profundidade dos solos através de 4 classes: raso, pouco
profundo, profundo e muito profundo (Tabela 2).
TABELA 2 – Classes e valores utilizados para a avaliação da profundidade do solo.
Especificação Profundidade do solo (cm)
Raso ≤ 50
Pouco profundo 50 –100
Profundo 100-200
Muito profundo >200
Fonte: Vieira et al. (1988)
2.2.4 Cor do solo
A cor do solo é uma das características que mais chama a atenção ao se estudar o
solo. As várias tonalidades de coloração ajudam a separação dos horizontes e também a
evidenciar condições de extrema importância, tais como: teores de matéria orgânica;
quantidade de argila, silte e os componentes de ferro, permitem também a avaliação de
propriedades relacionadas com a composição, aeração e drenagem do solo (Streck et al.,
2002).
Existem várias cores de solos, como: escuros, negros, cinzentos, vermelhos,
amarelos e ocasionalmente brancos, nas mais variadas intensidades (Streck et al., 2002).
Segundo Vieira et al. (1988) e Streck et al. (2002) , a matéria orgânica nos
horizontes O, H e A e em alguns horizontes B, é responsável pelas cores escuras, porém,
a coloração dos horizontes pode, à medida que aumenta o teor de matéria orgânica,
variar do branco ao negro. Já as cores brancas são proporcionadas, principalmente, pelo
conteúdo de sílica na forma de quartzo, e pela ausência de materiais pigmentantes
(matéria orgânica e óxidos de ferros) .
As cores vermelhas dependem do conteúdo de sesquióxidos e óxidos de Fe não-
hidratados, enquanto as cores amarelas e cinza-amareladas, dos óxidos hidratados. O
solo é tanto mais vermelho quanto menos hidratados forem os compostos de ferro. A cor

106
avermelhada, apresentada pelos compostos de ferro, pode, também, dar seguras
indicações sobre o grau de drenagem dos solos e, em geral, indica boa oxidação, boa
drenagem e boa aeração, enquanto que as cores cinzentas ou cinza-azuladas indicam
condições redutoras e drenagem pobre. Já a cor amarelada (brunada) também pode
significar um ambiente mais úmido, onde tempo de residência da água no solo é maior
(Vieira et al., 1988 e Streck et al., 2002).
Segundo Streck et al. (2002), as cores acinzentadas, deve-se à redução e remoção
de óxidos de ferro, podem estar relacionadas com ambientes de solo saturados com água
durante períodos prolongados.
Portanto, a cor do solo é um fator ambiental de significativa importância para as
propriedades do solo, assim como o conteúdo de matéria orgânica, o grau de hidratação
dos sesquióxidos e as condições de drenagem. Podem, ainda, dar uma idéia do seu grau
de aquecimento, pois solos de cores escuras absorvem calor mais rapidamente e o
refletem com menor intensidade (Vieira et al.,1988).
Segundo os mesmos autores, as cores são grandemente influenciadas pelo teor de
umidade e como conseqüência diminuem de intensidade à medida que aumenta o teor de
umidade.
2.2.6 Exposição à luz
Os organismos sobre a superfície terrestre ou perto dela recebem constantemente
radiação solar e fluxo de radiação térmica de grande comprimento de onda. Os dois tipos
de comprimento de onda contribuem para o ambiente climático (temperatura, evaporação
da água, movimento de ar e água), porém apenas uma pequena fração da radiação solar
pode ser convertida pela fotossíntese em energia para os componentes do ecossistema
(Odum, 1988).
A luz solar que atinge a biosfera é de 2 gcal/cm2/min, mas esta é atenuada
enquanto passa pela atmosfera para 1,34 gcal/cm2/mim ou 67% (Gates apud Odum,
1988).
Tendo que atravessar as nuvens, as partículas da água e a vegetação, a radiação
solar é atenuada ainda mais e a distribuição espectral da sua energia é grandemente
alterada. A entrada diária de luz solar na camada autotrófica de um ecossistema varia

107
principalmente entre 100 e 800 gcal/cm2. O fluxo de energia diária recebida por um
organismo exposto pode ser várias vezes maior ou consideravelmente menor do que a
radiação solar que entra (Odum, 1988).
Ainda, segundo Odum (1988), nos diversos estratos de um ecossistema florestal a
variação no fluxo de energia total de radiação é grande, variando entre as estações ou
locais da superfície terrestre. Isto afeta a distribuição dos organismos no ecossistema.
Segundo Cunha & Peichl (1986), na freqüência onde os olhos humanos estão
sensíveis à luz fica, também, a parte da radiação que tem importância decisiva para o
processo de fotossíntese e, com isso, para a produção de matéria das plantas.
Para os autores, a luz tem também outros efeitos importantes dentro das plantas,
tais como, o desenvolvimento e a diferenciação dos tecidos vegetais, especialmente a
formação das folhas umbrófilas e das folhas heliófilas (composição da luz e do
abastecimento com água) (Cunha & Peichl, 1986).
Em povoamentos idosos, fechados, uniformes e simples, existe uma correlação
estreita entre a intensidade da exposição e a densidade de copas no dossel. A
intensidade da exposição das espécies florestais, as quais formam o dossel das copas,
pode ser quantificada pelo número de árvores por hectare. O crescimento em altura
geralmente não é afetado pelo sombreamento, o que não ocorre com o crescimento em
diâmetro.
Geralmente, as diferentes espécies de árvores têm, nos primeiros anos de idade e
nos sítios com abundância de nutrientes e umidade, uma tolerância elevada à sombra,
comparando às árvores idosas e aos sítios com baixo teor de nutrientes. Logo, a
necessidade de luz não varia somente com a espécie, mas também com a idade do
indivíduo, sendo as exigências no estado juvenil geralmente inferior aquelas das idades
superiores (Cunha & Peichl, 1986).
Os autores colocam, como regra geral, que é possível constatar que a diminuição
da intensidade da luz causa uma redução da matéria orgânica das plantas. Esta redução
faz-se notar, principalmente, nas raízes, isto é, causam prejuízos às raízes. Isto significa
para a planta, menos absorção dos nutrientes e da água e, assim, diminuição da força de
competição com as árvores vizinhas.
A redução da luz no povoamento influencia na quantidade de luz disponível para
cada árvore na parte mais baixa da copa. Isto limita a fotossíntese nesta parte da copa.

108
Portanto, a intensidade da luz causa muitos efeitos sobre a morfologia das plantas.
Quando uma espécie possui uma exigência elevada de luz, há uma influência muito
grande sobre a formação do tronco e copa e, no caso das florestas nativas, nota-se
claramente esta influência na tortuosidade ou forma das árvores.
Pinto (1997) diz que disponibilidade de energia está relacionada com a localização
dos continentes e dos oceanos em relação ao equador e também com os movimentos da
terra em relação ao sol. Estes fatores dão origem às variações sazonais bem demarcadas
na disponibilidade de energia radiante com reflexos diretos no comprimento dos dias e na
temperatura, além de inúmeros fenômenos climáticos, tais como a evaporação da água e
a movimentação das massas de ar. Estas complexas interações resultam nas formações
ecológicas (florestais) bem definidas.
2.2.6 Quantidade de pedra
Os elementos químicos disponíveis às comunidades da biosfera são aqueles que,
pela sua natureza geoquímica, tendem a estar contidos nos tipos de rochas que afloram à
superfície. Os elementos abundantes no manto da Terra estão escassos na superfície. O
fósforo é um dos elementos cuja escassez na superfície da Terra muitas vezes limita o
crescimento vegetal (Odum,1988).
Segundo Vieira et al. (1988), a pedregosidade superficial, ou a quantidade de
pedras na superfície de solo, é uma característica ambiental, importante de ser avaliada,
pois está relacionada diretamente à retenção de água e aos componentes químicos que
compõem o solo. O autor classifica os afloramentos rochosos na superfície do terreno e 4
classes, conforme mostra a Tabela 3.
TABELA 3 – Classificação da pedregosidade na superfície do terreno.
Especificação Pedregosidade (%)
Pedregosidade menor < 0,1% da superfície do terreno
Pedregosidade entre 0,1 – 3%
Pedregosidade entre 3 – 15%
Pedregosidade superior > 15%
Fonte: Vieira et al. (1988)

109
2.2.7 Densidade do sub-bosque
Segundo Laroca (1995), os efeitos que o ambiente exerce sobre os seres vivos,
recaem sobre duas categorias: (i) efeitos dependentes da densidade; e (ii) independentes
da densidade.
Os efeitos dependentes da densidade modificam a taxa de nascimento ou de
mortalidade assim como a função do tamanho ou densidade populacional (N). As
competições intra2 e interespecífica são fatores que têm efeitos dependentes da
densidade, pois estas têm influências diretas ou indiretas, isto é, alteram o ambiente
através de secreções. Algumas vezes os efeitos são positivos, mas geralmente tendem a
ser negativos, diminuindo a taxa de nascimento e/ou aumento da taxa de mortalidade. No
entanto não se pode esquecer que estes efeitos variam de espécie para espécie (Laroca,
1995).
De acordo com Pizzato (1999), a dinâmica de uma floresta inicia-se com a
formação de clareiras, que provocam mudanças nas características edafoclimáticas,
ocasionando o processo de sucessão florestal. A abertura de clareiras é o principal fator
para que diversas espécies existam na floresta, sendo renovadas e sustentadas pela
dinâmica de perda de indivíduos mais velhos permitindo a existência de outros indivíduos.
Odum (1988) diz que a dinâmica de uma floresta, relacionada a fisiologia, estrutura
e funcionamento desta, envolve diversos processos de organização como: sucessão,
mortalidade, ingresso e crescimentos.
Para Pizzato (1999) na dinâmica das florestas, durante o curso de sucessão, os
indivíduos que morrem são substituídos pelo crescimento dos indivíduos vizinhos ou pelo
ingresso de novos indivíduos da mesma ou de diferentes espécies.
Barros (1986) relata que o ingresso, crescimento e mortalidade são resultado final
do processo da dinâmica das florestas. O contínuo aparecimento de novos indivíduos no
estrato inferior da floresta que limita o espaço de crescimento das árvores novas, devido
ao aumento da densidade, propiciam a eliminação de elementos menos capazes de
competir. As aberturas ocorridas no estrato superior da floresta, em decorrência da morte
de grandes árvores, são ocupadas pelas árvores que se encontram no estrato
2 A competição intra-específica se dá quando organismos da mesma espécie utilizam recursos limitados. Se os recursos não são limitados a competição ocorre quando os organismos em busca de recursos, manifestam comportamentos agressivos uns com outros.

110
imediatamente inferior que, por sua vez, proporcionam o estabelecimento de novos
indivíduos, tornando heterogênea a distribuição de idades das árvores na floresta.
Para o mesmo autor a densidade do sub-bosque influencia nos processos da
dinâmica florestal, sendo responsável tanto pela mudança da comunidade como pela
modificação do espaço desta. Tais processos se manifestam através da extinção e
imigração local de populações, assim como flutuação na abundância relativa de
populações dentro da comunidade.
2.3 Parâmetros Fitossociológicos
Montoya-Maquin & Matos apud Longhi (1997) afirmam que a vegetação natural é
complexa e está relacionada com os diversos fatores do meio, como climáticos,
pedológicos e biológicos. Estes fatores podem ser quantificados por vários parâmetros,
como os métodos baseados no estudo dos diversos elementos da vegetação, que são os
métodos florísticos ou taxonômicos e os baseados na estrutura e na fisionomia.
De acordo com Förster (1973) a análise estrutural da vegetação deve ser baseada
no levantamento e na interpretação de critérios de conteúdo mensuráveis. Análise dessa
natureza permite comparações entre diferentes tipos de florestas.
Segundo Souza et al. (1998), as análises florísticas (parâmetros florísticos)
contemplam as listas de espécies arbóreas ocorrentes no local de estudo, ou seja,
determinam a população de uma determinada área.
Para análise da estrutura horizontal das comunidades vegetais utiliza-se os
parâmetros de densidade (abundância), freqüência, dominância, valor de importância e
valor de cobertura, que revelam informações sobre a distribuição espacial das populações
e sua participação no contexto do ecossistema. A estrutura vertical ou o arranjo dos
indivíduos dentro do espaço vertical deve basear-se na distribuição dos indivíduos em
estratos (Guapyassü apud Longhi, 1997).
(vi) Densidade: é o número de indivíduos, quer de uma espécie, quer de todas as
outras espécies em conjunto, por unidade de área. A Densidade Absoluta (DA) trata do
número de indivíduos da espécie por unidade de área considerada e a Densidade

111
Relativa (DR) é a proporção do número de indivíduos de uma espécie em relação ao
número total de indivíduos amostrados, em percentagem (Martins, 1991).
Já o termo Abundância, também utilizado por alguns pesquisadores, refere-se às
estimativas visuais da densidade das espécies, agrupando-as, assim, em classes de
abundância (muito rara, rara, ocasional, abundante, muito abundante). Porém, densidade
refere-se às contagens efetivas de indivíduos, em um espaço contínuo (Finol, 1971,
Förster, 1973; Longhi, 1980).
(vii) Freqüência: freqüência absoluta (FA) é a porcentagem de unidades de
amostragem com ocorrência da espécie, em relação ao número total de unidades de
amostragem. Freqüência Relativa (FR) é a proporção da freqüência absoluta de uma
espécie em relação à soma das freqüências absolutas de todas as espécies, em
porcentagem (Martins, 1991). Para melhor entendimento, freqüência indica a
uniformidade de distribuição de uma espécie sobre uma determinada área, isto é, a sua
dispersão média. Já para Souza et al. (1998), é a percentagem de ocorrência de uma
espécie em um número de áreas de igual tamanho, dentro de uma comunidade.
Segundo Longhi (1997), para determinar a freqüência, deve-se controlar a
presença/ausência da espécie, em uma série de amostras de tamanho uniforme,
independente do número de indivíduos. Portanto, se uma espécie aparece em todas as
unidades amostrais, tem uma freqüência de 100%. Pode-se dizer, portanto, que
freqüência refere-se à probabilidade de encontrar uma espécie na área estudada.
(viii) Dominância: expressa a proporção de tamanho, volume ou cobertura de cada
espécie, em relação ao espaço ou volume da fitocenose (Martins, 1991).
Pesquisadores como Förster (1973), Font-Quer (1975) e Longhi (1997), dizem que
dominância é a medida da projeção total do corpo das plantas, logo, a dominância de uma
espécie representa a soma de todas as projeções horizontais dos indivíduos pertencentes
à espécie.
Porém, em florestas muito densas, é praticamente impossível determinar os
valores de projeção horizontal das copas das árvores, pois a existência de estratos
inferiores forma uma estrutura vertical e horizontal muito complexa. Em função disso, Cain

112
et al. apud Longhi (1997) propuseram o uso da área basal como substituição à projeções
das copas, pelo fato de existir correlação entre ambas.
A Dominância Absoluta (DoA) de uma espécie consiste, portanto, na soma da área
basal de todos os indivíduos da espécie, presentes na amostragem. Dominância Relativa
(DoR) é a relação percentual entre a área basal total da espécie e a área basal por
hectare (Martins, 1991).
(ix) Valor Importância (VI): Os valores relativos de densidade, freqüência e
dominância revelam aspectos essenciais na composição florística das florestas, com
enfoques parciais, os quais isolados, não podem informar sobre a estrutura florística de
uma vegetação em conjunto (Martins, 1991).
Os parâmetros quantitativos de Densidade, Dominância e Freqüência (relativos) de
cada espécie, podem ser interpretados como de importância na fitocenose, isto vai
depender do que o pesquisador considera relevante (Longhi, 1997).
Segundo Förster (1973), para análise da vegetação, é importante encontrar um
valor que permita uma visão ou caracterização da importância de cada espécie, no
conglomerado total da floresta. Este valor pode ser dado pelo Valor de Importância (VI),
representado pela soma dos valores relativos de densidade, freqüência e dominância.
Martins (1991), diz que o Valor de Importância tem-se revelado muito útil, tanto para
separar tipos diferentes de florestas, como para relacioná-los a fatores ambientais ou para
relacionar a distribuição de espécies a fatores abióticos.
(x) Valor de cobertura (VC): refere-se ao número de árvores e a valores de superfície
coberta por elas. Obtêm-se esses valores através da soma da densidade e dominância
relativa da espécie (Blaun-Blanquet, 1979).
Förster (1973) diz que o Valor de Importância tem uma importância relativa, por
isso dever ser tratado de forma breve, o autor considera que uma espécie torna-se
importante na floresta pelo seu número de árvores e suas dimensões, isto é, pela sua
densidade e dominância, pois estas determinam o espaço dentro da biocenose, não
importando muito a freqüência da espécie.

113
Sendo assim, o autor caracteriza as espécies pelo seu Valor de Cobertura, isto é,
pela sua densidade relativa somada à dominância relativa, método citado por Braun-
Blanquet (1979), o que indica a potência da espécie na comunidade.
2.4 Diversidade
Segundo Magurran (1989) é muito difícil definir diversidade, já que se compõe de
dois elementos: (i) variação e (ii) abundância de espécies. Logo, pode-se medir a
diversidade, registrando o número de espécies e avaliando sua abundância relativa ou
usando uma medida que combine os dois componentes.
O conceito de diversidade de espécies possui dois componentes: (i) riqueza,
também chamada de densidade de espécies, baseada no número total de espécies
presentes e, (ii) uniformidade, baseada na abundância relativa de espécies e no grau da
sua dominância ou falta desta (Odum, 1988).
Para o autor, a diversidade de espécies tende a aumentar quanto maior o tamanho
da área. Já, em comunidades bióticas que sofrem estresse, que foram reduzidas pela
competição em comunidades antigas e ambientes físicos estáveis, a diversidade tende a
diminuir.
De acordo com Magurran (1989) as medidas de diversidade devem realizar-se no
contexto de duas áreas de aplicação. Em uma delas, a da supervisão ambiental, as
medidas de diversidade são usadas inteligentemente e têm-se aplicado com profusão. Na
outra, da conservação da natureza, obtém-se grandes êxitos, protegendo a diversidade,
que na maioria dos casos, se define como riqueza de espécies. A supervisão ambiental
demonstra que as medidas de diversidade podem ser, empiricamente muito úteis.
Para Cox (1976), diversidade de espécie é uma medida biológica, característica
única em nível de comunidade de organizações ecológicas. Além do mais, é uma
característica que reflete padrões organizacionais importantes no funcionamento da
comunidade. A composição em diversidade de espécie está relacionada ao grau de
estabilidade da comunidade, desde que uma comunidade com maior riqueza ou
diversidade de espécies, possuir uma rede trófica mais complexa ao longo da qual
mecanismos de controle de densidade possam operar.

114
Whittaker apud Odum (1988), descreve três diferentes maneiras de classificar a
diversidade. A diversidade dentro do habitat ou tipo de comunidade não pode ser
confundida com a diversidade de uma paisagem ou de uma região que contém uma
mistura de habitats. Portanto, a diversidade pode ser classificada em: (i) diversidade alfa,
para diversidade dentro de um habitat ou dentro de uma comunidade; (ii) diversidade
beta, para diversidade entre habitats e; (iii) diversidade gama, para diversidade de uma
grande área regional, bioma, continente e ilha, entre outros.
A análise florística da vegetação pode ser feita através dos índices de Diversidade
de espécies, entre os quais os Índices de Shannon e de Simpson, que julgam a afinidade
entre comunidades (Pielou, 1975).
O Índice de diversidade de Shannon é calculado pela fórmula:
onde: Pi é a relação entre o número de indivíduos da espécie (ni) e o número total de
indivíduos (N) ou seja Pi = ni/N.
Segundo Pielou (1975) e Martins (1991) seus valores variam de 1,5 a 3,5,
raramente passando de 4,5.
Já o Índice de diversidade de Simpson, é determinado pela equação:
Dn n
N N
i i=−
−
∑
( )
( )
1
1
Onde ni é o número de indivíduos na i-ésima espécie e N é o número total de indivíduos.
À medida que D se incrementa, a diversidade decresce e o índice de Simpson é,
portanto, expresso normalmente como 1-D.
2.5 Análise de Similaridade
Orlóci (1978) utiliza o termo semelhança como uma propriedade de objetos
mensuráveis, ou grupos de objetos e como uma função característica que os objetos
possuem. Como objetos, entendem-se os indivíduos das espécies, a comunidade, além
∑−= PiPiH ln.'

115
de outras qualidades.
Nas comparações numéricas das comunidades usam-se técnicas estatísticas que,
partindo das tabelas brutas ou matrizes primárias de atributos/amostras e, mediante uma
série de tratamentos matemáticos, permitem obter matrizes secundárias de semelhança
ou similaridades. A matriz de semelhança substitui os conjuntos de atributos presentes
por índices que medem a similaridade das amostras ou comunidades, em função da
coincidência de presença e de abundância dos atributos do par de comunidades a
comparar ou, de semelhança entre espécimes, segundo o número de amostras em que
aparecem juntas ou separadas. Estas matrizes secundárias constituem a entrada de
quase todos os sistemas numéricos e de alguns dos sistemas informais de classificação e
ordenação da vegetação (Matteucci & Colma, 1982).
As funções de semelhança podem ser dadas a partir de variáveis binárias ou
qualitativas (presença/ausência), ou de dados quantitativos (Matteucci & Colma, 1982 e
Johnson & Wichern, 1982). Trata-se de comparações entre espécies ou entre amostras
ou entre comunidades e calculam-se a partir de tabelas de contingência de 2 x 2 (Tabela
4).
TABELA 4: Tabela de contingência entre duas amostras 1 e 2
Amostra 1
+ -
Total
+
Amostra 2
-
a b
c d
a+b
c+d
Total a+c b+d p=a+b+c+d
onde:
a = número de espécies comuns nas amostras 1 e 2 (número de igualdade (1,1))
b = número de espécies exclusivas da amostra 2 (número de desigualdade (1,0))
c = número de espécies exclusivas da amostra 1 (número de desigualdade (0,1))
d = número de espécies ausentes nas duas amostras (número de igualdade (0,0))
p = número total de espécies.
Os índices de similaridade são expressões matemáticas utilizadas para comparar a

116
flora de duas ou mais comunidades vegetais. Indicam o grau de semelhança entre elas
(Mueller-Dombois & Ellenberg, 1974).
Saiz (1980), Matteucci & Colma (1982) e Johnson & Wichern (1982) citam um
grande número de Índices de Similaridade ou Coeficientes de Similaridade, entre os quais
os mais freqüentemente utilizados são:
A) Para dados qualitativos (presença/ausência):
a = elementos exclusivos da amostra 1
b = elementos exclusivos da amosta 2
c = elementos comuns por presença nas amostras 1 e 2
d = elementos comuns por ausência nas amostras 1 e 2
n = total de elementos considerados em 1 e 2 = a + b + c + d
A = número total de elementos da amostra 1
B = número total de elementos da amostra 2
1) Índice de Jaccard ou Coeficiente de Comunidade de Jaccard (SJ):
Sc
a b cJ = + +
=+ −c
A B c , 0 ≤ SJ ≤ 1
Desconsidera a igualdade (0,0) no numerador e denominador.
2) Índice de Sörensen ou Coeficiente de Comunidade de Sörensen (SS):
Sc
a b c
c
A BS = + +
=+
2
2
2 , 0 ≤ SS ≤ 1
Desconsidera a igualdade (0,0) no numerador e denominador e dá peso em dobro
para a igualdade (1,1).
2.6 Classificação e Ordenação
Classificação e ordenação são duas técnicas multivariadas eficientes de
sumarização ou simplificação de dados numéricos, para comparações estatísticas. As
técnicas de classificação baseiam-se no agrupamento de amostras ou de espécies que
tenham propriedades em comum, enquanto que as técnicas de ordenação dispõem as

117
amostras ou as espécies ao longo de eixos de variação contínua (Mateucci & Colma,
1982).
Greig-Smith et al. (1967) consideram que o elevado número de espécies e a
ausência de predominância de uma ou de determinadas delas impediram o sucesso da
aplicação de procedimentos tradicionais de classificação em regiões tropicais.
Webb et al. (1967b) mencionam que o elevado número de espécies nas florestas
tropicais, torna-se uma grande dificuldade para as análises numéricas, porque demanda
um tempo de processamento de dados muito alto, podendo inviabilizar o processo. Além
disso, qualquer método que requer a extração dos autovalores a autovetores de uma
matriz de similaridade interespecífica é impraticável. Como cada espécie contém
pouquíssima informação, os autovalores de tal matriz decrescem muito lentamente.
De acordo com Greig-Smith et al. (1967), a ordenação é capaz de evidenciar,
satisfatoriamente, as principais relações da composição da vegetação de uma forma mais
clara, onde as informações podem ser apresentadas graficamente.
Em florestas naturais, heterogêneas, Webb et al. (1967a,b) recomendam que a
classificação seja conduzida primeiro e a ordenação seja reservada àqueles casos em
que os métodos classificatórios provarem ser ineficientes. Laven (1982) sugere que a
ordenação deve suceder a análise de agrupamentos para auxiliar na interpretação dos
resultados.
Segundo Orlóci (1978), o problema de escolha entre métodos de classificação e
ordenação não deve estar associado a pressuposições rígidas. Ambas devem ser
encaradas como técnicas de análise de dados, que podem auxiliar o usuário a alcançar
seus objetivos.
Do ponto de vista prático, Goodall (1982) observa que o tipo de análise a escolher
dependerá da preferência do investigador e, mais ainda, do objetivo de estudo, que da
natureza da vegetação. Quando o objetivo de estudo é descrever a vegetação, a
classificação é mais adequada. Se o objetivo é determinar relações entre a vegetação e o
ambiente, a ordenação simplifica as interpretações.
2.6.1 Técnicas de classificação
A classificação consiste em agrupar as amostras ou as espécies, segundo suas

118
características. Consiste em dividir o sistema multidimensional em compartimentos ou
células, em cada um dos quais se reúnem os pontos que apresentam maior similaridade
entre si (Matteucci & Colma, 1982). Para Gauch & Whittaker (1972 e 1981) este sistema
multidimensional é uma matriz primária espécies-parcelas que representa o espaço
espécie, no qual as espécies são os eixos do espaço multidimensional e as parcelas são
os pontos locados por seus escores, para cada espécie.
Sokal & Sneath (1963), Orlóci (1978), Gauch & Whittaker (1981) e Matteucci &
Colma (1982) observam que, quando se consideram as características (propriedades que
descrevem um indivíduo e que podem assumir um valor) que intervêm na formação dos
agrupamentos, as técnicas de classificação podem ser monotéticas e politéticas. As
técnicas monotéticas, de acordo com Lambert & Dale (1964) e Lambert & Williams (1962),
empregam uma só característica em cada fusão ou subdivisão, enquanto as técnicas
politéticas empregam uma função de semelhança ou função objetiva baseada num
conjunto de características.
Para Sokal & Sneath (1963), em ecologia, o termo monotético tem sido usado para
uma classificação, na qual os grupos são formados, baseando-se na presença ou
ausência de uma dada espécie.
Quando fusões ou subdivisões são condicionadas sobre uma forte discriminação
de duas ou mais espécies, em qualquer passo, no processo de agrupamento, o algoritmo
é dito ser politético (Orlóci, 1967).
Lambert & Williams (1962), Lambert & Dale (1964) e Matteucci & Colma (1982)
consideram dois tipos de técnicas de classificação, as que alocam indivíduos a classes
previamente estabelecidas (Análise Discriminante) e aquelas que criam as classes ou
agrupamentos, a partir das informações contidas nos dados (Análise de Agrupamento).
2.6.1.1 Análise de Agrupamento (Cluster)
Entende-se por Análise de Cluster um método de classificação numérica, onde se
buscam e caracterizam grupos, com diferentes graus de similaridade em uma quantidade
dada de dados (Bockor, 1975).
Segundo Sneath & Sokal (1973), Orlóci (1978) e Johnson & Wichern (1982), o
critério de agrupamento empregado, na maioria dos algoritmos, é uma medida de

119
similaridade ou de distância estatística entre os elementos de uma matriz X.
Essas medidas constituem na entrada (Input) para os vários algoritmos e definem
uma função dos valores dos vetores representativos dos elementos de X, para os quais
se calcula uma medida de similaridade ou distância, comumente denominada,
respectivamente, Coeficiente de Similaridade ou Medida de Distância (Orlóci, 1978).
Para Johnson & Wichern (1982), a distância Euclidiana é a métrica de maior
emprego nas análises de agrupamentos e a que apresenta maior facilidade de cálculo.
Contudo, quando os objetos não podem ser representados por medidas p-dimensionais,
os pares de objetos são comparados com base na presença ou ausência de certas
características.
As técnicas de agrupamento são utilizadas em muitas áreas de conhecimento,
como Medicina, Filosofia, Política, Geologia, entre outras. Contudo, pouco se sabe de sua
utilização em dados de vegetação. A esse respeito, Orlóci (1978) comenta que, ao aceitar
uma vegetação como uma população multiespécie, implica em um agregado de unidades
naturais. Se estas unidades existem, certamente necessitam de uma forte base natural
para identificação ou, pelo menos, são somente reconhecíveis com ambigüidade, por ser
de composição complexa, cujos limites não são facilmente definidos.
A hipótese de que a vegetação é um agregado de unidades naturais reconhecíveis
é defendida por muitos fitossociólogos das principais Escolas Européias como Greig-
Smith (1964), Kershaw (1973), Orlóci (1978), Braun-Blanquet (1979), Havel (1980a,b),
Whittaker (1975 e 1982) e Holdridge (1982). Eles afirmam que além de serem distintas, as
unidades tendem a formar tipos naturais (associações).
Dentre os procedimentos de uma classificação, dois enfoques têm sido abordados:
os procedimentos informais (descritivos) e os formais. Os informais têm sido amplamente
utilizados em estudos de vegetação, por detectarem classes naturais (tipos, associações,
etc.) com razoável consistência. Já os procedimentos formais possibilitam classificar
conjunto de dados visualmente heterogêneos, muito complexos e muito volumosos para
serem eficientemente manipulados por outros métodos (Whittaker, 1962).
A importância do emprego dos métodos formais de agrupamento em análise de
vegetação é mostrada por diversos autores, entre os quais, Williams & Dale (1965),
Anderberg (1973), Matteucci & Colma (1982).
Matteucci & Colma (1982) consideram que, os métodos formais de classificação

120
partem da matriz de similaridade e identificam as classes mediante uma série de cálculos.
Segundo Johnson & Wichern (1982) e Matteucci & Colma (1982), os métodos de
Agrupamento de Cluster podem ser de dois tipos: Hierárquico e Não-hierárquico.
As técnicas de agrupamento hierárquico baseaiam-se em uma série de fusões
sucessivas ou uma série de divisões sucessivas (Johnson & Wichern, 1982). Para
Matteucci & Colma (1982) este é o método mais utilizado em levantamentos com fatores
ecológicos, pois, os níveis de similaridade e dissimilaridade podem ser visualizados em
dendrogramas.
De acordo com o procedimento utilizado na formação das classes, as técnicas
podem ser divisivas ou aglomerativas. Para Matteucci & Colma (1982), as técnicas
divisivas começam com a população completa e subdivisões sucessivas vão formando
grupos cada vez menores. Em cada etapa da subdivisão buscam-se as diferenças dentro
dos grupos para separar subgrupos que diferem entre si. Por outro lado, as técnicas
aglomerativas, conforme Sneath & Sokal (1973), Orlóci (1978), Gauch & Whittaker (1981)
e Matteucci & Colma (1982), começam pela fusão sucessiva dos indivíduos, que se
combinam por suas semelhanças, repetindo o procedimento até esgotar as possibilidades
de combinação ou até que não fiquem indivíduos isolados.
Assim, as técnicas aglomerativas buscam a similaridade entre indivíduos, enquanto
as técnicas divisivas buscam a dissimilaridade (Orlóci, 1978 e Matteucci & Colma, 1982).
Segundo Crisci & Armengol (1983) em relação à quantidade de características que
intervêm na formação de classes, as técnicas de classificação são Monotéticas ou
Politéticas. As técnicas monotéticas, só podem ser divisivas (as técnicas monotéticas
aglomerativas resultam em dados não confiáveis), e utilizam só uma característica em
cada subdivisão. Por outro lado, as técnicas politétitas empregam uma função de
semelhança baseada em um conjunto de características.
Para esses autores o grupo monotético é um grupo constituído por rígidas e
sucessivas divisões lógicas, no qual todos os membros compartilham um conjunto de
atributos que são condições necessárias e suficientes para converter-se em um integrante
do grupo. Já, o grupo politético é um grupo onde seus membros compartilham um grande
número de atributos, porém nenhum desses atributos é compartilhado por todos os
membros, com a exceção do atributo de pertencer ao grupo. Cada membro possui alguns
atributos do conjunto. Portanto, nenhum atributo é condição necessária o suficiente para

121
converter um objeto em um integrante do grupo.
Orlóci (1978) cita que os métodos de agrupamento hierárquico, na sua grande
maioria, podem ser utilizados para classificar tanto variáveis (espécies), quanto unidades
amostrais, utilizando qualquer medida de associação (similaridade) com exceção de uns
poucos que utilizam o quadrado da distância Euclidiana, para as amostras, quando
utilizadas como as entidades a serem agrupadas.
Johnson & Wichern (1982) apontam que no método aglomerativo hierárquico existe
no início, tantos grupos, quanto objetos. Muitos objetos semelhantes são agrupados
primeiro e, estes grupos iniciais são fundidos de acordo com suas similaridades.
Relaxando no critério de similaridade, todos os grupos são fundidos dentro de um único
grupo. O método divisivo hierárquico faz o trabalho em direção oposta. O grupo inicial de
objetos é dividido em dois grupos, de modo que os objetos em um grupo são distantes
dos objetos do outro. Estes subgrupos são, então, melhor divididos em outros subgrupos
não semelhantes e o processo continua até que existam tantos subgrupos quanto objetos.
De acordo com os autores o agrupamento dos objetos ou indivíduos é feito por
ligações. Estas podem ser simples (método da mínima distância ou do vizinho mais
próximo), completa (método da máxima distância ou do vizinho mais longe) e média
(distância média). Para Orlóci (1967) e Matteucci & Colma (1982) um outro método de
ligação, muito usado em matrizes muito grandes, é conhecido por método da mínima
variância.
Descrições mais detalhadas desses métodos podem ser vistas em Anderberg
(1973), Hartigan (1975), Orlóci (1967, 1978), Mardia; Kent & Bibby (1979), Johnson &
Wichern (1982) e Matteucci & Colma (1982).
O agrupamento não-hierárquico é uma técnica usada, quando se deseja formar k-
grupos de itens ou objetos. O número de grupos, k, pode, igualmente, ser especificado
com antecipação ou determinado a partir do processo de agrupamento. Como a matriz de
distância (ou similaridade) não pode ser determinada e os dados básicos não podem ser
armazenados durante a computação, o método não-hierárquico pode ser aplicado para
muito mais dados que técnicas hierárquicas (Johnson & Wichern, 1982).
Para os autores, o método de agrupamento mais usado é o das k-médias, sugerido
por Macqueen em 1967, para descrever algoritmo que designa todos os itens do grupo,
tendo o centróide (média) mais próximo. Nessa mais simples versão, o processo é

122
composto por três passos:
a) partição dos itens em k-grupos iniciais;
b) prossegue com a lista de itens, onde cada um é colocado no grupo, cuja média
(centróide) esteja mais próxima. Usualmente calcula-se a distância Euclidiana com
observações padronizadas ou não. Recalcula-se o centróide para os grupos envolvidos
na troca do novo item e para as perdas de grupo.
c) repete-se o segundo passo, até que não restem recolocações a serem feitas.
O programa de computador TWINSPAN (TWo-way INdicator SPecies ANalyis),
utiliza um processo multivariado, ou seja, é um método hierárquico, divisivos e politético
(Hill, 1979).
TWINSPAN faz uma classificação das espécies em função das parcelas em que
estas ocorrem e das parcelas em função das espécies que nelas ocorrem. Esta análise
multivariada permite o descobrimento de padrões de dados, através da ordenação e
classificação. Permitem organizar os dados de comunidades baseadas na densidade de
espécies, não considerando os fatores ambientais. A ordenação arranja as espécies e
parcelas num espaço dimensional restrito, isto é, as parcelas que ficam próximas são
semelhantes e as que têm uma certa distância ficam distantes. Já a classificação
determina o local onde se encontram as espécies e parcelas (Felfili et al., 2001a).
Segundo Felfili et al. (2001b), o programa TWINSPAN, constrói uma tabela
bidimensional pela identificação de espécies preferenciais. As parcelas são classificadas
primeiro por dicotimização sucessivas e as espécies pelo mesmo processo. Para obter a
dicotimização as ordenações são divididas pela metade. Este método faz três
ordenações: (i) na ordenação primária, a direção de variação é determinada nos dados;
(ii) na segunda ordenação, a dicotomia preliminar é obtida pela divisão da ordenação pela
metade e ocorre a identificação de espécies preferenciais (são espécies com as mesmas
preferências ecológicas e, desta forma sua presença pode ser utilizada para identificar
condições ambientais particulares) em um ou outro lado da dicotomia e; (iii) a terceira
ordenação, é a ordenação por indicador, que é baseada num pequeno número de
espécies fortemente preferenciais (Hill, 1979).
Em cada divisão é calculado um autovalor (eigenvalue), que indica o quanto da
variação dos dados da comunidade foi explicada no eixo de ordenação (McCune &
Mefford apud Araujo, 2002). Normalmente, o autovalor em torno de 0,30 é utilizado para

123
explicar as variações ocorrentes na natureza, logo, quando a divisão atingir um autovalor
menor que 0,30, esta pode ser descartada (Araujo, 2002).
O DENDROGRAMA ou DIAGRAMA EM ÁRVORE é a forma visual, mais utilizada,
mundialmente, para representar a seqüência de agrupamentos. No dendrograma, cada
ligação corresponde a um valor numérico (um índice, que é a escala ou nível de
agregação) que representa o nível, no qual têm lugar os agrupamentos. Quanto maior o
índice, mais heterogêneo são os indivíduos agrupados. Uma vez formado o dendrograma,
a separação entre os grupos é realizada. Pode ser feito um corte em um determinado
nível, que é geralmente na metade da maior distância, ou segundo um critério subjetivo e
pessoal (Longhi, 1997).
2.6.1.2 Análise Discriminante
Segundo Anderson (1971), se medidas são realizadas em amostras aleatórias
pertencentes a diversas categorias ou grupos, então cada grupo será visualizado como
um conjunto de pontos num espaço multidimensional. A análise discriminante reduz o
número de medidas realizadas para um número menor de parâmetros que são funções
discriminantes linearmente dependentes das medidas originais. Desta forma, os grupos
(conjuntos de pontos) poderão ser visualizados num espaço multidimensional menor que
o anterior e, os coeficientes das funções discriminantes indicarão a contribuição relativa
das medidas originais, para cada função discriminante, que segundo Rao apud Batista
(1990), é aproximadamente um teste de significância de separação dos grupos.
Souza (1989) observa que a análise discriminante e a classificação têm como
objetivos descrever algébrica e/ou graficamente, num espaço reduzido, as características
diferenciais dos objetos de vários agrupamentos conhecidos e ordenar os objetos dentro
de classes pré-definidas, dando ênfase à regra de alocação ótima de novos objetos aos
agrupamentos.
De acordo com Marriott (1974), a análise discriminante consiste em investigar
como e quando é possível fazer distinções entre os membros de g agrupamentos, com
base nas observações feitas sobre eles. Tem como objetivo testar diferenças estatísticas
significantes a um dado nível de probabilidade entre g agrupamento; determinar o número
de funções discriminantes; construir regras de alocação para identificar um indivíduo

124
como membro de um dos g agrupamentos; e estimar as probabilidades de classificações
corretas.
Para Johnson & Wichern (1982), a técnica multivariada conhecida como Análise
Discriminante e Classificação trata dos problemas relacionados em separar conjuntos
distintos de objetos (observações) e em alocar novos objetos (observações) em grupos
previamente definidos. A terminologia discriminar e classificar foi introduzida por FISHER,
em 1938, no primeiro tratamento moderno dos problemas de separação.
Segundo os mesmos autores, a idéia básica do método de Fisher consiste em
separar duas classes de objetos ou fixar um novo objeto, em uma das duas classes. É
conveniente denominar as classes (populações) de π1 e π2. Os objetos são
ordinariamente separados ou classificados com base nas medidas das p-variáveis
aleatórias associadas [ ]′ =X X X X p1 2, , ,L .
A idéia de Fisher foi transformar as observações multivariadas X, nas observações
univariadas Y, tal que os Y’s derivados das populações π1 e π2 sejam separados tanto
quanto possível. Fisher sugeriu o uso da combinação linear das variáveis aleatórias de X
para a obtenção dos Y’s, porque são funções simples e fáceis de obter matematicamente.
A função discriminante linear de Fisher é dada pela combinação linear:
y c X X= ′ = − ′ −( )µ µ1 2
1Σ
2.6.2 Técnicas de Ordenação
Com os métodos de ordenação, obtêm-se seqüências ou gradientes, ao dispor os
indivíduos (amostras ou atributos), ao longo de eixos de variação contínua. Reduz-se o
número de dimensões, expressando a variação da vegetação em poucos eixos, nos quais
recupera-se a máxima quantidade de informação possível (Matteucci & Colma, 1982).
Segundo Morrison (1976), Orlóci (1978), Matteucci & Colma (1982) e Johnson &
Wichern (1982), existe um número bastante grande de técnicas que tratam da ordenação
de dados, conhecidos como Análise Multivariada. A aplicação destas técnicas, em
estudos da vegetação, está diretamente relacionada com o tipo de dados a serem
analisados, com a estrutura dos mesmos e, particularmente, com o objetivo da análise.
Três são as principais técnicas de ordenação: a Análise dos Componentes

125
Principais, a Análise Fatorial e a Análise de Correlação Canônica (Análise de
Correspondência Canônica) que, segundo Johnson & Wichern (1982), são conhecidas por
técnicas de Análise da Estrutura de Covariância.
2.6.2.1 Componentes Principais
Segundo Mallo (1985), a Análise de Componentes Principais foi descrita
inicialmente por Karl Pearson, em 1901 e posteriormente desenvolvido por Hotelling, em
1933. Sua finalidade principal consiste em condensar o essencial da informação dada, por
uma série de variáveis interdependentes, observadas diretamente sobre um conjunto de
indivíduos, em um número mais restrito de variáveis fundamentais independentes. Estas
novas variáveis originais possuem as seguintes características:
a) redução dimensional - são escolhidas em número menor que as originais;
b) não são correlacionadas;
c) significatividade, devem explicar a maior proporção possível da variabilidade
total, com poucas componentes.
Para Foguet & Arias (1989), a finalidade da análise de componentes principais é
construir novas variáveis latentes, as componentes, que recolham a variação do sistema
sem redundâncias, reduzindo a dimensionalidade do espaço original, com distorções
mínimas da informação.
Asensio (1989) observa que a análise de componentes principais tem como
objetivo facilitar o estudo das relações existentes entre as variáveis e, facilitar a análise da
dispersão das observações, detectando as variáveis responsáveis.
Segundo Johnson & Wichern (1982), a Análise dos Componentes Principais
procura explicar a estrutura de variância-covariância, mediante combinações lineares, não
correlacionadas, das variáveis originais. Em geral, tem como objetivo reduzir os dados e
facilitar a interpretação.
Com referência aos componentes principais, Pla (1986) observa que este é um dos
métodos de análise multivariada mais difundidos, permitindo a estruturação de um
conjunto de dados multivariados obtidos de uma população, cuja distribuição de
probabilidades não necessita ser conhecida, tratando-se de uma técnica matemática que
não requer um modelo estatístico para explicar a estrutura probabilística dos erros.

126
2.6.2.3 Análise Fatorial
A análise fatorial é uma técnica, onde muitas variáveis podem ser representadas
por uma função linear, com um número menor de variáveis não observáveis, chamadas
fatores (Morrison, 1976).
De acordo com Johnson & Wichern (1982), a análise fatorial tem provocado muita
controvérsia ao longo da história. O princípio moderno encontra-se no início do século XX,
no esforço de Karl Pearson, Charles Spearman e outros, em definir medidas de
inteligência. A dificuldade nos cálculos impediu um desenvolvimento da técnica, o que só
ocorreu com o advento dos computadores.
Segundo os autores, a proposta inicial da análise fatorial é descrever, se possível,
a covariância dos relacionamentos entre muitas variáveis, em termos de poucas variáveis
fundamentais, não observáveis, chamadas fatores. Supõe-se que variáveis possam ser
agrupadas por suas correlações. Isto é, todas as variáveis dentro de um grupo particular
são altamente correlacionadas entre si, mas com correlações relativamente baixas ou
negligenciáveis com as variáveis de um grupo diferente. É admissível que cada grupo de
variáveis represente um fator, que é responsável pelas correlações observadas.
De acordo com Batista (1990), a escolha do número de fatores pode ser feito a
partir de diversos critérios:
a) a estrutura física das variáveis pode sugerir um número de fatores
intrínsecos;
b) quando se usa a matriz de correlação, é recomendável que se tome tantos
fatores, quanto forem os autovalores maiores que 1;
c) pode-se, ainda, escolher um número de fatores que expliquem uma porção
específica da variância total.
Para o autor, a interpretação dos fatores é mais precisa, se cada variável tem peso
alto em, no máximo, um fator e, se todos os pesos dos fatores são grandes e positivos ou
próximos de zero. Assim, as variáveis são agrupadas em conjuntos distintos, cada um dos
quais associados a um fator. Às vezes é necessário fazer uma rotação dos fatores para
melhor interpretação.

127
2.6.2.3 Análise de Correspondência Canônica (CCA)
A Análise de Correspondência Canônica (CCA), é uma técnica desenvolvida por
Ter Braak (1986). A CCA é diferente de todos os métodos de ordenação já discutidos. A
CCA é um método que apresenta a relação entre a distribuição das espécies e a
distribuição dos fatores ambientais, associados a gradientes (Kent & Coker, 1992). A CCA
permite ainda testar a probalidade de acerto nas relações encontradas por meio do teste
de permutação de “Monte Carlo” (Aubert e Oliveira-Filho, 1994).
Todos os métodos de ordenação como ordenação polar, análise de componentes
principais e análise fatorial têm este objetivo, porém, são indiretos em cada análise,
executando nos primeiros somente para espécies, e quando a interpretação a ambiental é
feita por acréscimo dos dados ambientais sobre as parcelas de ordenação (Aubert e
Oliveira-Filho, 1994).
Algumas análises podem ir até onde a correlação e regressão do eixo da parcela
marcam com fatores ambientais, mas, por várias razões, isto não é sempre satisfatório.
A CCA difere das outras técnicas clássicas indiretas, porque nesta análise é
incorporado a correlação à regressão, entre os dados florísticos e fatores ambientais
dentro da própria análise de ordenação (Botrel et al., 2002).
A entrada de dados consiste na criação de duas matrizes de dados. A primeira
matriz é uma que tem a relação entre espécies (densidade maior que 5) x parcelas, e a
segunda matriz é a relação entre os fatores ambientais x parcelas. Esta aproximação das
espécies e de dados ambientais no processo de ordenação atual é conhecida como uma
forma de Análise Canônica (Aubert e Oliveira-Filho, 1994).
O diagrama de ordenação resultante expressa, não só, padrões de variação da
composição florística, mas, também demonstra as relações principais entre as espécies e
cada uma das variáveis ambientais (Martins et al., 2003).
Os pontos, apresentados no diagrama, representam as espécies individuais e as
setas representam cada variável ambiental plotada no diagrama. O comprimento da seta
é proporcional à magnitude de mudança naquela direção, e para propósito de
interpretação cada seta pode também ser estendida no sentido contrário da origem
central. Aqueles fatores ambientais que na seta longa estão mais correlatos na ordenação
do que aqueles com seta curta, e são mais importantes na influência da variação da

128
comunidade (Kent & Coker, 1992). A ordem em que se apresentam os pontos projetados
sobre a seta, da origem à extremidade, indicam a influência do fator ambiental em relação
à espécie, isto é, quanto mais próximo da extremidade da seta, maior influência do fator
ambiental sobre a espécie.
2.7 Regressão Logística
Segundo Hosmer & Lemeshow (1989), os métodos de regressão têm se tornado um
componente integral de qualquer análise de dados relacionados com a descrição da
relação entre uma variável resposta (variável dependente) e uma ou mais variáveis
explanatórias (variável independente).
De acordo com os autores a regressão logística estima diretamente a probabilidade
de um evento dicotômico ocorrer, isto é, a presença de uma determinada espécie em uma
área pode ser considerada com um fenômeno dicotômico. Existem duas hipóteses: ou a
espécie ocorre ou não ocorre (Hosmer & Lemeshow, 1989).
Para estimar os parâmetros na regressão logística, é utilizado o método da máxima
verossimilhança, isto é, os coeficientes que fazem os resultados observados mais
prováveis são selecionados através de um processo interativo. Existem alguns
procedimentos para a seleção de modelo na regressão logística, por exemplo, para
adicionar ou retirar variáveis no modelo pode-se usar o método Forward Stepwise e
Backward Stepwise e para adicionar ou retirar propositadamente variáveis no modelo
pode-se usar o método ENTER.
Conforme Hasenauer apud Caldato (1998), a regressão logística produz valores de
probabilidade que variam de 0 (zero) e 1 (um), possibilitando, a simulação da ocorrência
de um determinado evento, ao se comparar o valor resultante da função em cada caso
específico com valores de números randômicos de distribuição uniforme ou de Poisson,
gerados no mesmo intervalo (0 – 1).
Para Hosmer & Lemeshow (1989), o modelo geral da regressão logística é:
Probabilidade (evento) = Ze−+1
1

129
Onde Z é a combinação linear ou não linear:
pp XBXBXBBZ +++++= ...22110
sendo B0, B1, B2 e Bp, coeficientes estimados dos dados; X1, X2 e Xp são as variáveis
independentes; e é a base do logaritmo natural, aproximadamente 2,718.
A interação de qualquer modelo requer que se tirem inferências práticas dos
coeficientes estimados no modelo. Os coeficientes estimados para as variáveis
independentes representam a inclinação ou a taxa de mudança da variável dependente,
em função da modificação de uma unidade da variável independente. A formulação lógica
de modelo geral, ou seja, a seleção das variáveis independentes é, pois, a base para a
carreta utilização da regressão logística (Hosmer & Lemeschow, 1989).
Segundo Caldato (1998), uma das maneiras de avaliar o quanto o modelo é
apropriado é pela comparação da precisão do modelo com dados reais, através de uma
tabela de classificação, entretanto, não revela a distribuição das probabilidades estimadas
para um fato em dois grupos. Para cada grupo estimado, todas as tabelas mostram-se a
probabilidade estimada é maior ou menor que 50%.
Para Hosmer & Lemeshow (1989) uma outra maneira para verificar o modelo
adequado, é pela observação da semelhança entre os dados amostrais e os previstos
pelo modelo, através da estátistica -2LL (2 Logarithm of the Likelihood = -2 vezes o
logaritmo da verossimilhança). Um bom modelo é o que tem uma alta semelhança entre
os dados e o resultado do modelo, isto é, o que leva um pequeno valor de -2LL, quando o
valor da verossimilhança observada for 1 e o -2LL é zero, o modelo descreve
perfeitamente os dados (Caldato, 1998).
Ainda, segundo os autores, existe outra estatística que pode ser usada para testar
o ajuste do modelo que é a Qualidade do ajuste (Goodness of fit), que compara as
probabilidades observadas com aquelas estimadas pelo modelo, tal que:
onde o resíduo é a diferença entre o valor observado (Yi), e o valor estimado (Pi).
)1(
Re 22
PiPi
siduoZ i
−=∑

130
4 RESULTADOS E DISCUSSÃO
4.1 Caracterização geral da floresta
4.1.1 Composição florística
A vegetação da FLONA de São Francisco de Paula pertence à Floresta Ombrófila
Mista, tipo fitogeográfico bastante expressivo do sul do Brasil, que hoje, segundo Leite e
Sohn apud Leite & Klein (1990), está reduzida a pouco mais de 10% de sua área original.
Nas 10 parcelas (10 ha) estudadas foram encontradas 130 espécies arbóreas
pertencentes a 79 gêneros e 45 famílias botânicas, entre os indivíduos com CAP ≥ 30 cm,
além de árvores não identificadas, mortas, cipós e uma exótica (Citrus sp.) (Tabela 10).
As famílias Myrtaceae, com 11 gêneros e 27 espécies e Lauraceae, com 4 gêneros
e 9 espécies foram as mais representativas da floresta, seguidas de Flacourtiaceae, com
3 gêneros e 7 espécies; Solanaceae, com 2 gêneros e 7 espécies; Asteraceae e
Fabaceae, com 4 gêneros e 5 espécies; Rutaceae, com 3 gêneros e 5 espécies;
Aquifoliaceae, com 1 gênero e 5 espécies; Euphorbiaceae, com 3 gêneros e 4 espécies;
Myrsinaceae, com 1 gênero e 4 espécies; Mimosaceae, com 2 gêneros e 4 espécies;
Sapindaceae, com 3 gêneros e 3 espécies; Monimiaceae, com 2 gêneros e 3 espécies;
Annonaceae e Symplocaceae, com 1 gênero e 3 espécies; Cunoniaceae, Meliaceae,
Rhamnaceae, Rubiaceae e Verbenaceae, com 2 gêneros e 2 espécies; e Celestraceae e
Icacinaceae, com 1 gênero e 2 espécies cada. As 23 famílias restantes, apresentaram 1
única espécie.
Jarenkow (1985) também encontrou, em uma Floresta Ombrófila Mista no
município de Esmeralda, Rio Grande do Sul, a família Myrtaceae como a mais
característica e representativa do ponto de vista florístico.

131
TABELA 10 – Espécies encontradas na FLONA de São Francisco de Paula, RS, com
respectivas famílias, nomes científicos e populares.
FAMÍLIA / NOME CIENTÍFICO NOME POPULAR ANACARDIACEAE
Lithraea brasiliensis Marchand Aroeira-bugre ANNONACEAE
Rollinia rugulosa Schltdl. Araticum-quaresma Rollinia salicifolia Schltdl. Araticum-salso
AQUIFOLIACEAE Ilex brevicuspis Reissek Caúna-da-serra Ilex dumosa Reissek Congonha
Ilex microdonta Reissek Caúna Ilex paraguariensis A. St.-Hil. Erva-mate
Ilex sp. Caúna ARALIACEAE
Oreapanax fulvus Marchal Tamanqueira ARAUCARIACEAE
Araucaria angustifolia (Bertol.) Kuntze Pinheiro-brasileiro ARECACEAE
Syagrus romanzoffiana (Cham.) Glassman Jerivá ASTERACEAE
Dasyphyllum spinescens (Less.) Cabrera Açucara Dasyphyllum tomentosum (Spreng.) Cabrera Açucará-piloso
Gochnatia polymorpha (Less.) Cabrera Cambará Piptocharpha tomentosa Baker Pau-toucinho Vernonia discolor (Spreng.) Less. Vassourão-preto
BIGNONIACEAE Tabebuia umbellata (Sond.) Sandwith Ipê-amarelo
CARICACEAE Carica quercifolia (A. St.-Hil.) Hieron Mamoeiro-do-mato
CELASTRACEAE Maytenus aquifolia Mart. Concorosa-arbórea
Maytenus evonimoides Reissek Coração de bugre CLETHRACEAE
Clethra scabra Sleumer Caujuja-de-ule CUNONIACEAE
Lamanonia ternata Vell. Guaperê Weinmania paulliniifolia Pohl ex Ser. Gramimunha
CYATHEACEAE Alsophila sp. Xaxim-de-espinho
DICKSONIACEAE Dicksonia sellowiana Hook. Xaxim
EUPHORBIACEAE Sapium glanutum (Vell.) Pax. Pau-leiteiro
Sebastiania brasiliensis Spreng. Branquilho-leiteiro Sebastiania commersoniania (Baill.) L. B. Sm. et Downs Branquilho
Stillingia oppositifolia Baill. ex Müll. Arg. Leiterinho FABACEAE
Dalbergia frutenscens (Vell.) Britton Rabo-de-bugio Erythrina cristagalli L. Corticeira-do-
banhado Lonchocarpus campestris Mart. ex Benth. Pau-canzil Lonchocarpus nitidus (Vogel) Benth. Farinha-seca

132
Continua TABELA 10 – Continuação
FAMÍLIA / NOME CIENTÍFICO NOME POPULAR Machaerium paraguariense Hassl. Canela-do-brejo
GRISELINIACEAE Griselinia ruscifolia (Clos) Taub. Griselina
FLACOURTIACEAE Banara parviflora (A. Gray) Benth. Guaçatunga-preta
Banara tomentosa Clos Guaçatunga-branca Casearia decandra Jacq. Guaçatunga Casearia obliqua Spreng. Casearia sylvestris Sw. Chá-de-bugre
Xylosma pseudosalzmannii Sleumer Espinho-de-judeu Xylosma tweedianum (Clos) Eichler Sucará
ICACINACEAE Citronella gongonha (Mart.) Howard Gongonha-espinhosa Citronella paniculata (Mart.) Howard Gongonha
LAMIACEAE Cunila insisa Benth. Poejo
LAURACEAE Cinnamomum amoenum (Nees) Kosterm. Canela Cinnamomum glaziovii (Mez) Kosterm. Canela
Cryptocarya aschersoniania Mez Canela-vick Cryptocarya moschata Nees Canela-cega
Nectandra lanceolata Nees et Mart. ex Nees Canela-amarela Nectandra megapotamica (Spreng.) Nez Canela-preta
Ocotea indecora (Schott) Mez Canela Ocotea puberula (A. Rich.) Nees Canela-guaicá
Ocotea pulchella Mart. Canela-langeana LOGANIACEAE
Strychnos brasiliensis (Spreng.) Mart. Anzol-de-lontra MELIACEAE
Cabralea canjerana (Vell.) Mart. Canjerana Cedrela fissilis Vell. Cedro
MIMOSACEAE Acacia bonariensis Gill. ex Hook. et Arn. Unha-de-gato
Inga marginata Willd. Ingá-feijão Inga sessilis (Vell.) Mart. Ingá
/cut /cut Willd. Ingá-banana MONIMIACEAE
Hennecartia omphalandra J. Poiss. Canema Mollinedia elegans Tul. Pimentinha Mollinedia sp. Capixim MYRSINACEAE
Myrsine coriacea (Sw.) R. Br. Capororoquinha Myrsine lorentziana (Mez) Arecav. Capororoca-d’Água
Myrsine umbelata Mart. Capororocão Myrsine sp. Capororoca
MYRTACEAE Acca sellowiana (O. Berg) Burret Goiaba-serrana Blepharocalyx salicifolius (Kunth) O. Berg Murta Calyptranthes concinna DC. Guamirim-de-facho Campomanesia rhombea O. Berg Guabiroba-miúda Campomanesia xanthocarpa O. Berg Guabiroba

133
Continua TABELA 10 – Continuação
FAMÍLIA / NOME CIENTÍFICO NOME POPULAR Eugenia involucrata DC. Cerejeira-do-mato Eugenia pluriflora DC. Jaboticaca-do-campo Eugenia psidiiflora O. Berg Guamirim Eugemia uruguayensis Cambess. Myrceugenia cucullata D. Legrand Guamirim-quebradiço Myrceugenia foveolata (O. Berg) Sobral Guamirim Myrceugenia glaucescens (Cambess.) D. Legrand et Kausel Conserva-branca Myrceugenia miersiana (Gardner) D. Legrand et Kausel Guamirim-piloso
Myrceugenia myrcioides (Cambess.) O. Berg Guamirim Myrceugenia oxysepala (Berret) D. Legrand et Kausel Guamirim Myrcia arborescens O. Berg Guamirim-ferro Myrcia bombycina (O. Berg) Nied. Guamirim-do-campo Myrcia lajeana D. Legrand Myrcia oligantha O. Berg Guamirim Myrcia palustris DC. Guamirinzinho Myrcianthes gigantea (D. Legrand) D. Legrand lcuti-do-mato Myrcianthes pungens (O. Berg) D. Legrand Guabijú Myrciaria delicatula (DC.) O. Berg Camboim Myrciaria floribunda (West ex Willd.) O. Berg Camboim Myrciaria tenella (DC.) O. Berg Camboinzinho Myrrhinium atropurpureum Schott Murtilho Siphoneugena reitzii D. Legrand Camboim-de-reitz
NYCTAGINACEAE Guapira opposita (Vell.) Teitz Pau-cebola
PHYTOLACCACEAE Seguieria aculeata L. Cipó-umbú PODOCARPACEAE
Podocarpus lambertii Klotzsch ex Endl. Pinheiro-bravo PROTEACEAE
Roupala brasiliensis Klotzsch Carvalho-brasileiro RHAMNACEAE
Rhamnus sphaerosperma Sw. Pau-cangica Scutia buxifolia Reissek Coronilha
ROSACEAE Prunus myrtifolia (L.) Urb. Pessegueiro-do-mato
RUBIACEAE Coussarea contracta (Walp.) Müll. Arg. Pimenteira Coutarea hexandra (Jacq.) K. Schum. Quina
RUTACEAE Citrus sp. Laranjeira
Policarpus pennatifolius Lem. Cutia Zanthoxylum fagara (L.) Sarg. Coentrilho
Zanthoxylum kleinii (Cowan) P. G. Waterman Juvevê Zanthoxylum rhoifolium L. Mamica-de-cadela
SAPINDACEAE Allophylus edulis (A. St.-Hil. Et al.) Radlk. Chal-chal
Cupania vernalis Cambess. Camboáta-vermelho Matayba elaeagnoides Radlk. Camboáta-branco
SIMAROUBACEAE Picramnia parvifolia Engl. Pau-amargo

134
Continua TABELA 10 – Continuação
FAMÍLIA / NOME CIENTÍFICO NOME POPULAR SOLANACEAE Cestrum sp. Coerana
Solanum mauritianum Scop. Fumo-bravo Solanum pabstii L. B. Sm. & Downs Canema
Solanum pseudocapsicum L. Tomatinho Solanum pseudoquina A. St.-Hil. Peloteira Solanum sanctae-catharinae Dunal Joá-manso
Solanum sp. Fuminho STYRACACEAE
Styrax leprosus Hook. Et Arn. Carne-de-vaca SYMPLOCACEAE
Symplocos pentandra Occhioni Sete-sangrias Symplocos tetrandra (Mart.) Miq. Sete-sangrias Symplocos uniflora (Pohl) Benth. Sete-sangrias
THEACEAE Gordonia /cutifólia (Wawra) H. Keng Santa-rita
TILIACEAE Luehea divaricata Mart. Et Zucc. Açoita-cavalo
ULMACEAE Celtis iguanaea (Jacq.) Sargent Esporão-de-galo
VERBENACEAE Citharexylum solonaceum Cham. Tarumã
Duranta vestita Cham. Duranta Cipós Mortas
Não identificada
Klein apud Nascimento (2000) cita 96 espécies da família Myrtaceae para o Estado
do Rio Grande do Sul e destas, 41 espécies são encontradas na Floresta Ombrófila Mista.
A família desempenha, portanto, um papel importante na composição florística deste tipo
fitogeográfico do Estado.
Observa-se, também, que 51% das famílias encontradas na área da FLONA estão
representadas por uma única espécie. Das famílias restantes, 34% estão representadas
por 2 a 4 espécies e apenas 15% por mais de 5 espécies.
O Índice de Diversidade de Shannon encontrado (H’=3,194) comprova que a
floresta da FLONA apresenta alta diversidade florística. Valor semelhante (H’=3,651) foi
encontrado por Guapyassú (1994) em Floresta Ombrófila Mista no Paraná. Segundo
Magurran (1989) o índice de Shannon usualmente situa-se entre 1,5 e 3,5 e em
raríssimos casos, ultrapassa 5. Este índice estima a probabilidade de se identificar
corretamente um indivíduo escolhido ao acaso em uma população (Pielou, 1977).

135
4.1.2 Estrutura Fitossociológica
Na Tabela 11 estão relacionados os parâmetros das principais espécies em ordem
do Valor de Importância, que são as mais características e importantes da área estudada.
Estas espécies são as mais abundantes, dominantes e freqüentes da floresta.
Araucaria angustifolia é a espécie mais importante da comunidade estudada,
representando 15,56% do Valor de Importância e 22,49% do Valor de Cobertura.
Destaca-se na fisionomia da floresta por apresentar alta densidade e dominância.
A predominância de Araucaria é fator característico de qualquer formação de
Floresta Ombrófila Mista, conforme pode ser observado em trabalhos realizados neste
tipo fitogeográfico (Longhi, 1980 e 1997; Jarenkow, 1985; Nascimento, 2000; Rio Grande
do Sul, 2002).
São também importantes as espécies Blepharocalyx salicifolius, Ilex brevicuspis,
Chryptocarya aschersoniana, Sebastiania commersoniana, Ilex paraguariensis,
Sebastiania brasiliensis, Ocotea pulchella e Nectandra megapotamica, com 39% do Valor
de Importância e 52% do Valor de Cobertura total das espécies da floresta.
Estas oito espécies, mais a Araucaria angustifolia, representam 42,07% do Valor
de Importância e 55,36% do Valor de Cobertura total das espécies.
.

136
TABELA 11 – Parâmetros fitossociológicos para as espécies arbóreas e arbustivos, amostras com CAP ≥ 30 cm, na FLONA de São
Francisco de Paula, RS.
Espécie
DR FR DoR VI (%)
VI (%) Acum.
VC (%) VC(%) Acum
Araucaria angustifolia 11,67 1,70 33,31 15,56 15,56 22,49 22,49 Blepharocalyx salicifolius 5,08 1,70 6,98 4,57 20,13 6,00 28,49 Ilex brevicuspis 4,39 1,70 6,06 4,05 24,18 5,22 33,71 Cryptocarya aschersoniana 3,43 1,70 5,76 3,63 27,81 4,60 38,31 Morta 4,65 0.85 2,82 2,77 30,58 3,73 42,04 Sebatiania commersoniana 3,83 0,51 3,84 2,73 33,31 3,83 45,87 Ilex paraguariensis 3,96 1,53 2,03 2,51 35,82 2,99 48,86 Ocotea pulchella 2,05 1,36 2,64 2,02 37,84 2,35 51,21 Sebatiania brasiliensis 3,31 1,02 1,00 1,97 39,81 2,45 53,66 Nectadra megapotamica 2,09 1,53 2,14 1,92 41,73 2,11 55,77 Dicksonia sellowiana 1,83 1,70 1,45 1,66 43,39 1,64 57,41 Podocarpus lambertii 1,63 1,36 1,93 1,64 45,03 1,78 59,19 Matayba elaeagnoides 1,74 1,53 1,26 1,51 46,54 1,49 60,68 Sapium glandulatum 2,14 0,85 1,31 1,43 47,97 1,72 62,40 Banara parviflora 2,25 1,36 0,66 1,42 49,39 1,46 63,86 Lamonia ternata 1,46 0,51 1,91 1,29 50,68 1,68 65,54 Campomanesia rhombea 1,61 1,53 0,72 1,27 51,95 1,16 66,70 Campomanesia xanthocarpa 1,44 1,53 0,83 1,26 53,21 1,14 67,84 Vernonia discolor 0,73 1,53 1,09 1,11 54,32 0,91 68,75 Casearia decandra 2,21 0,17 0,85 1,07 55,39 1,52 70,27 Myrciaria floribunda 1,80 0,51 0,89 1,06 56,45 1,34 71,61
Sub-total 63,30 27,20 78,59 56,45 71,61 Restantes 36,70 73,80 21,41 43,55 28,39 Total 100,00 100,00 100,00 100,00 100,00

137
As 19 espécies listadas na Tabela 11 mais as árvores mortas, que representam
16% do total de espécies amostradas, somam 63,30% da Densidade relativa (número de
indivíduos), 27,20% da Freqüência relativa, 78,59% da Dominância relativa (área basal),
56,45% do Valor de Importância e 71,61% do Valor de Cobertura total das espécies
encontradas. São, portanto, as espécies mais características e importantes da Floresta
Ombrófila Mista da FLONA de São Francisco de Paula.
As espécies restantes, incluindo uma não identificadas e cipós, representaram
36,70 % da Densidade relativa; 73,80% da Freqüência relativa; 21,41% da Dominância
relativa; 43,55% do Valor de Importância e 28,39% do Valor de Cobertura do total
encontrado.
As árvores mortas aparecem em quinto lugar na ordem de importância das espécies,
com 2,77% do Valor de Importância das espécies. A presença significativa de árvores
mortas é comum em todos os tipos fitogeográficos e em todos os remanescentes das
florestas nativas do Estado, conforme observado no Inventário Florestal Contínuo do Rio
Grande do Sul (Rio Grande do Sul, 2002) e em outros trabalhos de fitossociologia.
Portanto, as árvores mortas são significativas na composição das comunidades e
constituiem um fenômeno natural de substituição dos indivíduos na dinâmica sucessional
das florestas.
4.1.3 Parâmetros Dendrométricos
Os parâmetros dendrométricos da Floresta Ombrófila Mista da FLONA de São
Francisco de Paula podem ser observados na Tabela 12, juntamente com os parâmetros
determinados no Inventário Florestal Contínuo do Rio Grande do Sul (Rio Grande do Sul,
2002) para este tipo fitogeográfico de todo o Estado.
Os parâmetros dendrométricos das árvores amostradas na FLONA de São
Francisco de Paula, indicam um diâmetro médio de 21,95 cm, maior que o encontrado
para o Estado, de 19,42 cm. O diâmetro máximo encontrado foi de 143,63 cm pertencente
a um indivíduo de Araucaria angustifolia (Pinheiro-brasileiro). O coeficiente de variação
médio dos diâmetros foi de 65,31%, superior ao determindo para este tipo fitogeográfico
do Estado, indicando maiores variações de diâmetros das espécies na floresta.

138
TABELA 12 – Parâmetros dendrométricos da Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, RS e do Estado.
PARÂMETROS DENDROMÉTRICOS FLONA ESTADO DO RS Diâmetro médio (cm) 21,95 19,42 CV dos diâmetros (%) 65,31 51,33 Altura total média (m) 13,12 11,65 CV da altura total (%) 32,97 31,46 Altura comercial média (m) 7,79 6,28 CV da altura comercial (%) 46,55 44,10 Número de indivíduos por ha 852 831 Área basal média (m2/ha) 45,69 31,79 Volume comercial médio (m3/ha) 337,72 205,59 Índice de diversidade de Shennon (H’) 3,1939 2,5778
A altura total média dos indivíduos da floresta da FLONA foi estimada em 13,12 m
e a altura comercial média foi de 7,79 m, também superiores aos valores encontrados
para o tipo fitogeográfico do Estado, que foram de 11,65 m e 6,28 m, respectivamente
para altura total e altura comercial médias. A altura total mínima medida foi de 1,50 m e a
altura total máxima amostrada foi de 33,50 m, cujos coeficientes de variação médios
foram de 32,97% para altura total e 46,55% para altura comercial.
O número médio de árvores da floresta da FLONA, considerando todos os
indivíduos com CAP ≥ 30 cm, foi estimado em 852 árvores/ha, também superior ao
encontrado para o Estado, que foi de 831 indivíduos.
Conseqüentemente, a área basal média e o volume comercial da floresta da
FLONA foram superiores aos valores encontrados para o Estado. Na área de estudo
encontrou-se 45,69 m2/ha de área basal e 337,72 m3/ha de volume comercial. No Estado
os valores foram de 31,79 m2/ha e 205,59 m3/ha, respectivamente.
Desta forma, pode-se afirmar que a Floresta Ombrófila Mista da FLONA de São
Francisco de Paula apresenta maior estoque de madeira por hectare do que a média das
outras formações de Araucaria do Estado. Isso se deve ao estado de conservação das
florestas, pouco exploradas, importante para a manutenção da diversidade genética.
Além disso as florestas da FLONA apresentaram maior índice de diversidade, que
foi de 3,1939 contra 2,5778 para o Estado, comprovando o melhor estado de conservação
das mesmas.

139
4.2 Análise de Agrupamento (Cluster - TWINSPAN)
A Análise de Cluster (TWINSPAN) definiu três grupos florísticos distintos entre as
parcelas da FLONA (Figura 4), os quais diferenciaram-se em relação a suas
características ecológicas, que foram determinadas através da densidade das espécies
(Crisci & Armengol, 1983). Observa-se que a análise relaciona as espécies indicadoras de
cada grupo, que são espécies com preferências ecológicas que podem identificar as
condições ambientais das subunidades (Hill et al., 1975).
A primeira divisão das subunidades amostrais formou dois grandes grupos. O
AUTOVALOR de 0,3659 (>0,30) mostrou que houve diferenciação entre eles. Essa primeira
divisão apontou que existe similaridade florística entre as parcelas relacionadas na
esquerda da Figura 5 que, por sua vez, são diferentes das relacionadas na direita, que
também são similares entre si. A maioria das espécies não se mostraram exclusivas,
porém ocorrem em maior abundância em determinado grupo, de acordo com suas
preferências ecológicas.
Sebastiania commersoniana, Cryptocarya aschersoniana, Eugenia psidiiflora e
Sebastiania brasiliensis foram às espécies indicadoras do primeiro grande agrupamento e
a inclusão de Ocotea pulchella como espécie preferencial.

140
FIGURA 4 – Classificação das unidades amostrais avaliadas na FLONA de São Francisco de Paula, RS; em três grupos florísticos
Dendrograma das parcelas da FLONA/Twinspan Autovalor da 10 divisão 0,3659
Parcelas: 1, 2, 3, 4, 5, 6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 30, 31, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 60, 61, 62, 64, 65, 66, 67, 68, 70, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97 , 98, 99, 100, 101, 102, 103, 105 , 106 , 107 , 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148 , 149, 150, 151, 152, 153, 154, 155, 156, 158, 161, 162, 163, 164, 165, 166, 167, 168, 170, 171, 172, 174, 175, 176, 177, 178, 179, 180, 181, 183, 184, 185, 188, 189, 190, 191, 193, 195, 196, 197, 198, 200, 203, 205, 213, 218, 220, 221, 222, 231, 233, 238, 252, 256, 262, 266, 293, 296, 299, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 371, 372,
373, 374, 375, 376, 377, 378, 380, 383, 385, 386, 387, 389, 390, 395, 396, 397, 398, 399, 400, 401, 403, 404, 406, 407, 409, 412, 413, 414, 415, 416, 417, 420, 423, 424, 426, 427, 428, 436, 441, 446, 448, 449, 451, 452, 458, 459, 462, 467, 468, 471, 472, 477, 478, 481, 483, 486, 492, 494, 495, 496, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 513, 514, 515, 516, 517, 518, 519, 520, 521, 522, 523, 524, 525, 526, 527, 528, 529, 530, 531, 532, 533, 534, 535, 536, 537, 538, 539, 540, 541, 542, 543, 544, 545, 546, 547, 548, 549, 550, 551, 552, 553, 554, 555, 556, 557, 558, 559, 560, 561, 562, 563, 564, 565, 566, 567, 568, 569, 570, 571, 572, 573, 574, 575, 576, 577, 578, 579, 580, 581, 582, 584, 585, 586, 587, 588, 589, 590, 591, 592, 593, 594, 595, 596, 597, 598, 599, 600, 635, 701, 702, 703, 705, 706, 707, 708, 710, 712, 713, 714, 715, 716, 717, 719, 720, 722, 723, 724, 726, 729, 730, 732, 733, 734, 735, 736, 737, 738, 739, 740, 741, 742, 743, 744, 745, 747, 748, 749, 750, 751, 752, 753, 754, 755, 756, 757, 758, 759, 760, 761, 762, 763, 764, 766, 768 769, 771, 772, 773, 774, 775, 776, 777, 778, 780, 781, 782, 783, 784, 785, 787, 789, 791, 792, 793, 795, 796, 797, 798, 799, 801, 804, 811, 815,
816, 823, 828, 831, 856, 867, 884, 885, 890, 894, 895, 907, 949, 981
Espécies indicadoras: Sebastiania commersoniana; Cryptocarya aschersoniana, Eugenia psidiiflora e Sebaiania brasiliensis
Espécies preferenciais: Cryptocarya aschersoniana 1(160, 28) Ocotea pulchela 1(116, 44) Sebastiania commersoniania 1(138, 22) Eugenia psidiiflora 1(200, 8)
Parcelas: 10, 29, 32, 52, 63, 69, 71, 104, 157, 159, 160, 169, 170, 171, 172, 173, 182, 186, 187, 192, 199, 200, 201, 202, 204, 206, 207, 208, 209, 210, 211, 212, 215, 216, 217, 219, 223, 224, 225, 226, 227, 229, 230, 232, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 250, 251, 252, 253, 254, 255, 257, 259, 260, 261, 263, 264, 265, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 292, 294, 295, 297, 298, 300, 341, 369, 370, 379,
380, 381, 382, 384, 388, 391, 392, 393, 394, 402, 405, 408, 410, 411, 418, 419, 421, 422, 425, 429, 430, 431, 432, 433, 435, 437, 438, 439, 440, 442, 443, 444, 445, 447, 450, 453, 454, 455, 456, 457, 460, 461, 463, 464, 465, 466, 469, 470, 473, 474, 475, 476, 478, 479, 480, 481, 482, 484, 485, 487, 488, 489, 490, 491, 493, 495, 496, 497, 498, 499, 500, 583, 601, 602, 603, 604, 605, 606, 607, 608, 609, 610, 611, 612, 613, 614, 615, 616, 617, 618, 619, 620, 621, 622, 623, 624, 625, 626, 627, 628, 629, 630, 631, 632, 633, 634, 636,
637, 638, 639, 640, 641, 642, 643, 644, 645, 646, 647, 648, 649, 650, 651, 652, 653, 654, 655, 656, 657, 658, 659, 660, 661, 662, 663, 664, 665, 666, 667, 668, 669, 670, 671, 672, 673, 674, 675, 676, 677, 678, 679, 680, 681, 682, 683, 684, 685, 686, 687, 688, 689,
690, 691, 692, 693, 694, 695, 696, 697, 698, 700, 704, 709, 711, 718, 721, 723, 724, 725, 727, 728, 731, 746, 765, 770, 779, 786, 788, 790, 794, 800, 802, 803, 805, 806, 807, 808, 809, 810, 812, 813, 814, 817, 818, 819, 820, 821, 822, 824, 825, 826, 827, 829, 830, 832, 833, 834, 835, 836, 837, 838, 839, 840, 841, 842, 843, 844, 845, 846, 847, 848, 849, 850, 851, 852, 853, 854, 855, 857, 858, 859,
860, 861, 862, 863, 864, 865, 866, 868, 869, 870, 871, 872, 873, 874, 875, 876, 877, 878, 879, 880, 881, 882, 883, 886, 887, 888, 889, 891, 892, 893, 896, 897, 898, 899, 900, 901, 902, 903, 904, 906, 908, 909, 910, 911, 912, 913, 914, 915, 916, 917, 918, 919, 920,
921, 922, 923, 924, 925, 926, 927, 928, 929, 930, 931, 932, 933, 934, 935, 936, 937, 938, 939, 940, 941, 942, 943, 944, 945, 946, 947, 948, 950, 951, 952, 953, 954, 955, 956, 957, 958, 959, 960, 961, 962, 963, 964, 965, 966, 967, 968, 969, 970, 971, 972, 973, 974, 975,
976, 977, 978, 979, 980, 982, 983, 984, 985, 986, 987, 988, 989, 990, 991, 992, 993, 994, 995, 996, p997, 998
Espécies indicadoras Araucaria angustifólia, Ilex paraguariensis Espécies preferenciais: Araucária angustifólia 1(148,140), Banara parvifolia (49,
110), Ilex paraguariensis (88, 158), Araucária angustifólia 2 (10,223)
Autovalor da 20 divisão: 0,4155
Parcelas: 39, 100, 101, 103, 105, 106, 107, 108, 110, 111, 112, 113, 114, 115, 116, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130,
Espécies indicadoras: Podocarpus lambertii, Myrciaria
tenela e Eugenia uruguariensis
Espécies preferenciais: Lamonia Ternata 1 (31,34); Lithraea
brasiliensis 1 (20, 4); Myrciaria tenela 1 (36, 5); Vernonia discolor 1(30, 5); Eugenia
uruguayensis 1 (41,30) e Myrciaria floribunda 1 (34, 31)
Grupo Grupo 2
Grupo 3
Parcelas: 1, 2, 3, 4, 5, 6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 30, 31, 33, 34, 35, 36, 37, 38, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 60, 61, 62, 64, 65, 66, 67, 68, 70, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97 , 98, 99, 103, 109, 117, 118, 148, 163, 196, 200, 203, 205, 213, 218, 220, 221, 222, 231, 233, 238, 252, 256, 262, 266, 293, 296, 299, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360,
361, 362, 363, 364, 365, 366, 367, 368, 371, 373, 374, 375, 376, 377, 378, 380, 383, 385, 386, 387, 389, 390, 395, 396, 397, 399, 400, 401, 403, 404, 406, 407, 409, 412, 413, 415, 416, 417, 420, 423, 424, 426, 427, 428, 436, 441, 446, 448, 449, 451, 452, 458, 459, 462, 467, 468, 471, 472, 477, 478, 481, 483, 486, 492, 494, 495, 496, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 514, 515, 516, 517, 518, 519, 520, 521, 523, 524, 526, 527, 528, 529, 530, 531, 532, 533, 534, 535, 536, 537, 538, 539, 540, 541, 543, 544, 545, 546, 547, 548, 549, 550, 551, 552, 553, 554, 555, 556, 557, 558, 559, 560, 561, 562, 563, 564, 565, 566, 567, 568, 569, 570, 571, 572, 573, 574, 575, 576, 577, 578, 579, 580, 581, 582, 584, 585, 586, 587, 588, 589, 590, 591, 592, 593, 594, 595, 596, 597, 598, 599, 600, 635, 701, 702, 703, 705, 706, 707, 708, 710, 712, 713, 714, 715, 716, 717, 719, 720, 722, 723, 724, 726, 729, 730, 732, 733, 734, 735, 736, 737, 738, 739, 740, 741, 742, 743, 744, 745, 747, 748, 749, 750, 751, 752, 753, 754, 755, 756, 757, 758, 759, 760, 761, 762, 763, 764, 766, 768, 769, 771, 772, 773, 774, 775, 776, 777, 778, 780, 782, 783, 784, 785, 787, 789, 791, 792, 793, 795, 796, 797, 798, 801, 804, 811, 815, 816, 823, 828, 856, 867, 884, 890, 894, 895, 907, 981
Espécies indicadoras: Sebastiana commersoniania, Cryptocarya aschersoniana e Sebastiana brasiliensis Espécies preferenciais: Cryptocarya aschersoniana 1 (14, 146); Blepharocalyx salicifolius 1 (8, 140), Sebatiania brasiliensis 1 (1, 115), Sebastiana commersoniania (90, 138)

141
O primeiro agrupamento apresentou como espécies indicadoras Eugenia psidiiflora,
Cryptocarya aschersoniana e Sebastiania commersoniana e, a inclusão de Ocotea
pulchella, como espécies preferenciais. Pelas exigências das espécies, constitui o grupo
das partes mais baixas da área, com solos úmidos. As duas espécies do gênero
Sebastiania são, segundo Smith et al. (1988), seletivas higrófilas e adaptadas à luz difusa.
O segundo agrupamento apresentou Araucaria angustifolia e Ilex paraguariensis
como espécies indicadoras. È o agrupamento constituído pela maioria das parcelas
amostradas, tendo a inclusão da Banara parviflora como espécie preferencial. Pelas
exigências das espécies constitui o grupo das partes mais altas da área com melhor
drenagem do solo.
Na segunda divisão, com um AUTOVALOR 0, 4155 (>0,30), separou o primeiro
agrupamento em dois grupos. Um grupo menor tendo Podocarpus lambertii, Myrciaria
tenella e Eugenia uruguayensis como espécies indicadoras e Lamanonia ternata, Lithraea
brasiliensis, Myrciaria tenella, Vernonia discolor e Myrciaria floribunda como espéies
preferenciais e um grupo maior tendo Sebatiania commersoniania, Sebastiania
brasiliensis e Cryptocarya aschersoniania, como espécie indicadora e Blepharocalyx
salicifolius como espécie preferencial.
Na terceira divisão, com AUTOVALOR 0,3214, separou o segundo agrupamento em
dois grupos, considerado similares do ponto de vista das exigências das espécies.
Embora o autovalor da divisão tenha sido >0,30, considerou-se como um grupo distinto.
No dendrograma ou tabela bidimensional (Figura 4), pode-se visualizar as divisões
realizadas.
Os agrupamentos formados geraram novas divisões, porém, pelas observações de
campo e pelos objetivos desejados no estudo, considerou-se a diferenciação de três
grupos florísticos distintos, que demonstraram claramente as características ecológicas da
área. Foram, de acordo com as espécies indicadoras, assim denominadas:
Grupo 1 - Formação Podocarpus
Grupo 2 - Formação Sebastiania;
Grupo 3 - Formação Araucaria.
Segundo Hill (1979) fica a critério do pesquisador a escolha do número de grupos,
caso as divisões ainda continuarem atingindo autovalores maiores que 0,30.

142
Conforme a Figura 4, o número após a espécie refere-se à pseudoespécies (nível
de corte), enquanto que o número entre parênteses refere-se às parcelas onde estão
contidas as pseudoespécies, em ambos os lados da divisão.
Analisando os dados complementares, presentes nas matrizes de fatores
ecológicos e os relatórios de campo, foi possível estabelecer tais diferenças presentes,
sobretudo em relação à profundidade do solo e teor de umidade em que se encontram as
unidades amostrais. Mais detalhes são apresentados no item 4.6, na análise dos
gradientes ambientais.
4.3 Significância dos agrupamentos (Análise Discriminante)
Após obtidos os grupos, realizou-se uma análise discriminante, visando determinar
funções que permitam classificar novas comunidades, dentro dos grupos para as quais
apresentam maior probabilidade de pertencerem, bem como verificar a precisão da
classificação e indicar as parcelas mal classificadas.
Da matriz de dados brutos com a densidade das espécies obteve-se, pela análise
Discriminante, a matriz de Correlação entre as variáveis, pelo Método STEPWISE.
Desta matriz determinou-se os valores de Lambda de Wilks (Λ*), F e a Significância
obtida para cada variável (espécie), que permitem interpretar a real capacidade de
discriminação das mesmas, na formação dos grupos. Tais valores podem ser observados
na Tabela 13.

143
TABELA 13 - Estatística do teste de seleção de espécies (variáveis) discriminantes para a matriz de dados brutos. VARIÁVEL LAMBDA DE
WILKS (ΛΛΛΛ*) F TOLERÂNCIA
Acacia bonariensis 0,999 0,288 1,000 Acca selloviana 0,991 4,418 1,000 Allophylus edulis 0,996 2,020 1,000 Alsophila sp. 0,998 0,897 1,000 Araucaria angustifolia 0,726 187,869 1,000 Banara parviflora 0,965 17,788 1,000 Banara tomentosa 0,999 0,313 1,000 Blepharocalyx salicifolius 0,983 8,735 1,000 Cabralea cangerana 0,999 0,559 1,000 Calyptranthes concinna 0,929 38,058 1,000 Campomanesia rhombea 0,992 3,915 1,000 Campomanesia xanthocarpa 0,991 4,553 1,000 Carica quercifolia 0,999 0,559 1,000 Casearia decandra 0,993 3,567 1,000 Casearia obliqua 0,993 3,619 1,000 Casearia sylvestris 0,999 0,668 1,000 Cedrela fissilis 1,000 0,228 1,000 Celtis iguanaea 0,998 1,198 1,000 Cestrum sp. 0,999 0,454 1,000 Cinnamomum amoenum 0,954 23,838 1,000 Cinnamomum glaziovii 0,997 1,656 1,000 Cipós 0,986 7,143 1,000 Citharexylum solanaceum 0,998 0,833 1,000 Citronella gongonha 0,993 3,273 1,000 Citronella paniculata 0,999 0,559 1,000 Citrus sp. 1,000 0,114 1,000
Clethra uleana 0,991 4,522 1,000 Caussarea contracta 0,998 1,008 1,000 Coutarea hexandra 0,998 1,120 1,000 Cryptocarya aschersoniana 0,907 50,893 1,000 Cryptocarya moschata 1,000 0,203 1,000 Cunila incisa 0,998 0,944 1,000 Cupania vernalis 0,992 4,155 1,000 Dalbergia frutescens 1,000 0,114 1,000 Dasyphyllum spinescens 0,970 15,526 1,000 Dasyphyllum tomentosum 0,999 0,649 1,000 Dicksonia sellowiana 0,979 10,909 1,000 Duranta vestita 0,997 1,254 1,000 Erithryna cristagalli 0,999 0,559 1,000 Eugenia involucrata 0,991 4,604 1,000 Eugenia pluriflora 0,988 6,129 1,000 Eugenia psidiifolia 0,802 122,674 1,000 Eugenia uruguayensis 0,896 57,439 1,000
Continua

144
TABELA 13 – Continuação
VARIÁVEL LAMBDA DE WILKS (ΛΛΛΛ*)
F TOLERÂNCIA
Gochnatia polymorpha 0,999 0,668 1,000 Gordonia acutifolia 0,977 11,838 1,000 Griselina ruscifolia 0,999 0,668 1,000 Guapira opposita 0,997 1,684 1,000 Hennecartia omphalandra 0,996 2,007 1,000 Ilex brevicuspis 0,996 2,197 1,000 Ilex dumosa 0,991 4,365 1,000 Ilex microdonta 0,985 7,387 1,000 Ilex paraguariensis 0,968 16,300 1,000 Ilex sp. 0,998 1,008 1,000 Inga marginata 0,998 1,120 1,000 Inga Vera 0,989 5,306 1,000 Lamanonia ternata 0,935 34,730 1,000 Lithraea brasiliensis 0,905 52,501 1,000 Lonchocarpus campestris 0,995 2,719 1,000 Lonchocarpus nitidus 0,996 2,211 1,000 Luehea divaricata 0,962 19,918 1,000 Machaerium paraguariense 0,993 3,280 1,000 Matayba elaeagnoides 0,964 18,373 1,000 Maytenus aquifolia 0,984 8,308 1,000 Maytenus evonymoides 0,972 14,358 1,000 Mollinedia elegans 0,996 2,031 1,000 Mollinedia sp. 0,999 0,559 1,000 Morta 0,993 3,747 1,000 Myrceugenia cucullata 0,988 5,969 1,000 Myrceugenia foveolata 0,999 0,668 1,000 Myrceugenia glaucescens 0,999 0,559 1,000
Myrceugenia miersiana 0,989 5,497 1,000 Myrceugenia myrcioides 0,997 1,459 1,000 Myrceugenia oxysepala 0,993 3,376 1,000 Myrcia arborescens 0,998 1,008 1,000 Myrcia bombycina 0,998 1,131 1,000 Myrcia lajeana 0,982 9,136 1,000 Myrcia oligantha 0,973 13,573 1,000 Myrcia palustris 0,999 0,597 1,000 Myrcianthes gigantea 0,964 18,537 1,000 Myrcianthes pungens 0,999 0,654 1,000 Myrciaria delicatula 0,977 11,956 1,000 Myrciaria floribunda 0,871 73,538 1,000 Myrciaria tenella 0,760 157,407 1,000 Myrrhinium atropurpureum 0,948 27,405 1,000 Myrsine coriacea 0,997 1,448 1,000 Myrsine lorentziana 0,957 22,618 1,000 Myrsine sp. 0,997 1,365 1,000
Myrsine umbellata 0,934 35,373 1,000 Continua

145
TABELA 13 - Continuação
VARIÁVEL LAMBDA DE WILKS (ΛΛΛΛ*)
F TOLERÂNCIA
Nectandra lanceolata 0,999 0,457 1,000 Nectandra megapotamica 0,996 2,098 1,000 NI 0,971 14,854 1,000 Ocotea indecora 0,994 3,212 1,000 Ocotea puberula 0,968 16,203 1,000 Ocotea pulchella 0,972 14,158 1,000 Oreopanax fulvus 0,999 0,302 1,000 Picramnia parvifolia 0,998 0,808 1,000 Pilocarpus pennatifolius 0,998 1,027 1,000 Piptocarpha tomentosa 0,999 0,559 1,000 Podocarpus lambertii 0,729 184,986 1,000 Prunus myrtifolia 0,996 2,063 1,000 Rhamnus sphaerosperma 1,000 0,228 1,000 Rollinia rugulosa 0,997 1,361 1,000 Rolinia salicifolia 0,999 0,716 1,000 Rollinia sylvatica 1,000 0,126 1,000 Roupala brasiliensis 0,932 36,330 1,000 Sapium glandulatum 0,971 14,713 1,000 Scutia buxifolia 0,993 3,256 1,000 Sebastiania brasiliensis 0,976 12,425 1,000 Sebastiania commersoniana 0,893 59,598 1,000 Seguieria aculeata 1,000 0,228 1,000 Sihoneugena reitzii 0,939 32,420 1,000 Solanum mauritianum 0,996 2,126 1,000 Solanum pabstii 0,992 3,825 1,000 Solanum pseudocapsicum 0,999 0,559 1,000 Solanum pseudoquina 0,999 0,624 1,000 Solanum sanctae-catharinae 0,992 4,001 1,000 Solanum sp. 0,992 3,957 1,000 Stillingia oppositifolia 0,999 0,734 1,000 Strychnos brasiliensis 1,000 0,114 1,000 Styrax leprosus 0,988 6,069 1,000 Syagrus romanzoffiana 0,999 0,559 1,000 Symplocos pentandra 0,993 3,576 1,000 Symplocos tetandra 0,881 67,424 1,000 Symplocos uniflora 0,947 27,839 1,000 Tabebuia umbellata 0,999 0,559 1,000 Vernonia discolor 0,828 103,580 1,000 Weinmania paullinifolia 0,996 1,827 1,000 Xylosma pseudosalzmannii 0,978 11,265 1,000 Xylosma tweedianum 0,991 4,765 1,000 Zanthoxylum fagara 0,998 0,897 1,000 Zanthoxylum kleinii 0,999 0,559 1,000 Zanthoxylum rhoifolium 0,984 8,032 1,000

146
Nota-se, pela tabela, que Araucaria angustifolia é a espécie que apresenta maior
valor de F e, conseqüentemente, menor valor de Λ*. Foi a primeira variável selecionada
para entrar na análise. Tem o mais alto valor do critério de seleção. Esta variável é
pareada com as demais, uma de cada vez, para selecionar a segunda melhor variável
discriminante, e assim sucessivamente. Após 38 Steps (passos), foram selecionadas 38
variáveis (espécies) com real poder de discriminação (Tabela 14).
A estatística Λ* mostrou, ao nível de 0,01 % de significância, que as espécies
relacionadas na Tabela 14, têm alto poder discriminante.
As dezesseis primeiras espécies selecionadas estão entre espécies indicadoras e
preferenciais obtidas pela análise de agrupamento (Figura 4). Observa-se, também, que
estas 38 espécies estão entre as mais características e importantes da vegetação
analisada.
As demais espécies, com valores de F menores que 42,174, após os 38 Steps,
foram eliminadas da análise. Apresentam importância limitada na separação dos grupos,
isto é, possuem baixo poder de discriminação.
Analisando o valor do teste F, da Tabela 13, nota-se que Myrsine umbellata,
Cinnamomum amoenum, Myrcianthes gigantea, Maytenus evonymoides, Sebastiania
brasiliensis, Myrciaria delicatula, Gordonia acutifolia, Myrcia lajeana e Blepharocalyx
salicifolius deveriam ser selecionadas na análise (F> 8,000). No entanto, após os 38
Steps elas foram substituídas por Styrax leprosus, Cupania vernalis, Solanum sanctae-
catharinae, Casearia decandra, Citronella gongonha e Solanum mauritianum, que
apresentaram valor de F baixo na análise (< 8,000). Isso ocorre, em função de que, à
medida que uma variável é selecionada e retirada da matriz, nova combinação é
realizada, alterando os valores de F e, conseqüentemente, a ordem de seleção.
Após a definição das variáveis discriminantes, procedeu-se a determinação das
funções discriminantes, importantes na análise das contribuições dessas variáveis. Estes
coeficientes foram computados, de tal forma que seus escores acham-se padronizados,
com média zero e variância unitária.

147
TABELA 14 - Estatística do teste de seleção de variáveis discriminantes, em ordem de
entrada, após 38 Steps.
STEPS VARIÁVEL LAMBDA WILKS (ΛΛΛΛ*)
F SIG.
1 Araucaria angustifolia 0,726 187,869 0,000 2 Podocarpus lambertii 0,530 185,530 0,000 3 Myrciaria tenella 0,442 166,683 0,000 4 Eugenia psidiifolia 0,377 155,513 0,000 5 Myrciaria floribunda 0,340 141,513 0,000 6 Sebastiania commersoniana 0,312 130,352 0,000 7 Vernonia discolor 0,288 121,957 0,000 8 Cryptocaria aschersoniana 0,270 114,044 0,000 9 Eugenia uruguayensis 0,254 108,057 0,000 10 Lamanonia ternata 0,240 102,733 0,000 11 Siphoneugena reitzii 0,230 97,026 0,000 12 Matayba elaeagnoides 0,222 92,134 0,000 13 Banara parviflora 0,214 87,889 0,000 14 Ilex paraguariensis 0,208 83,731 0,000 15 Lihtraea brasiliensis 0,202 80,055 0,000 16 Calyptranthes concinna 0,197 76,713 0,000 17 Roupala brasiliensis 0,192 73,728 0,000 18 Sapium glandulatum 0,188 71,079 0,000 19 Ocotea puberula 0,183 68,681 0,000 20 Dicksonia sellowiana 0,179 66,467 0,000 21 Symplocos uniflora 0,176 64,348 0,000 22 Dasyphyllum spinescens 0,172 62,392 0,000 23 Luehea divaricata 0,169 60,610 0,000 24 Ocotea pulchella 0,166 58,858 0,000 25 NI 0,164 57,180 0,000 26 Solanum mauritianum 0,161 55,593 0,000 27 Styrax leprosus 0,159 54,150 0,000 28 Myrrhinium atropurpureum 0,157 52,786 0,000 29 Zanthoxylum rhoifolium 0,154 51,504 0,000 30 Xylosma pseudosalzmannii 0,152 50,341 0,000 31 Myrsine lorentziana 0,150 49,245 0,000 32 Myrcia oligantha 0,148 48,147 0,000 33 Symplocos tetranda 0,146 47,082 0,000 34 Cupania vernalis 0,145 46,026 0,000 35 Casearia decandra 0,144 45,006 0,000 36 Citronella gongonha 0,142 44,035 0,000 37 Solanum sanctae-catharinae 0,141 43,083 0,000 38 Maytenus aquifolia 0,140 42,174 0,000
Os testes estatísticos concernentes ao procedimento de seleção do número de
discriminantes acham-se nas Tabelas 15 e 16. Pela observação dos autovalores, nota-se
que não houve nenhuma predominância de uma função sobre a outra, pois a proporção

148
da variância total explicada para cada função foi semelhante. Tal comportamento foi
confirmado pelos elevados valores dos coeficientes de correlação canônica, que mostram
um alto grau de relacionamento entre as duas funções discriminantes obtidas e o grupo
de variáveis.
Nota-se, também, que foram necessárias 2 funções discriminantes, para
representar 100% da variância total explicada. A primeira função discriminante
representa aproximadamente 62,6 % dessa variância, considerado baixo em termos
estatísticos (Tabela 15).
TABELA 15 - Estatística do teste de seleção das funções discriminantes: autovalores, variância explicada e correlação canônica. Função Autovalor % da variância % da variância
acumulada Correlação canônica
1 2,139 62,6 62,6 0,825 2 1,276 37,4 100,0 0,749
TABELA 16 - Estatística do teste de seleção das funções discriminantes: Lambda de Wilks, Chi-quadrado, grau de liberdade e significância. Teste da função ΛΛΛΛ* X2 GL Sig.
1 de 2 0,140 1920,111 76 0,000 2 0,439 803,087 37 0,000
O teste de significância da estatística Qui-quadrado (X2) e o valor Lambda de Wilks
(Λ*) mostram como as informações nas sucessivas funções discriminantes são retiradas.
Antes de nenhuma função ser retirada, o valor Λ*, de 0,140, muito baixo, indica que a
série de variáveis que estão sendo usadas possuem um considerável poder de
discriminação. Depois de uma parte dessa capacidade de discriminação ser alocada na
Primeira Função Discriminante, o Λ* aumenta pouco e o valor alto do teste X2 assegura
que, estatisticamente, uma quantia significativa de informações discriminantes ainda
exista. Na segunda função discriminante o valor de Λ*, de 0,439, ainda baixo, indica que
as variáveis continuam contendo poder de discriminação. Isso pode ser comprovado pelo
alto valor do teste X2 (803,087) e alta probabilidade (> 99,9 %).
Conforme como cada função discriminante foi gerada, segundo a magnitude do seu

149
autovalor associado, a estatística Λ* e o teste X2 avaliam a quantidade de informação
discriminante não incorporada pelas funções precedentes. Assim, ao nível de significância
de 0,01, o teste X2 indicou que nenhuma função discriminante deveria ser eliminada da
análise. As duas funções discriminantes são necessárias para descrever a série de
pontos definidos pelos agrupamentos.
Analisando os coeficientes padronizados das funções discriminantes (Tabela 17),
nota-se que não houve predominância total de uma ou de determinadas espécies nas
funções discriminantes. Isto se deve ao fato de que estes coeficientes, bem como as
percentagens dos autovalores associados (Tabela 15), com suas funções, apresentarem-
se muito baixos.
Estudos feitos por Webb at al. (1967a,b) e Souza (1989) comprovam que o elevado
número de espécies, a não predominância de uma ou de determinadas espécies sobre as
demais e a grande incidência de espécies semelhantes ecologicamente, analisadas em
matrizes de dados muito grandes, contribuem para que seus autovalores e autovetores
decresçam muito lentamente. Isso dificulta a extração de informações sobre a influência
das espécies mais importantes, em cada função discriminante.

150
TABELA 17 - Coeficientes padronizados das funções discriminantes canônicas, para a matriz de dados brutos.
VARIÁVEIS Função 1 Função 2 Araucaria angustifolia -0,381 0,467 Banara parviflora -0,141 0,130 Calyptranthes concinna 0,188 0,019 Casearia decandra 0,108 -0,061 Citronella gongonha -0,103 0,074 Cryptocaria aschersoniana 0,061 -0,344 Cupania vernalis 0,006 0,144 Dasyphyllum spinescens -0,113 0,148 Dicksonia sellowiana 0,007 -0,216 Eugenia psidiifolia 0,029 -0,403 Eugenia uruguayensis 0,229 0,119 Ilex paraguariensis -0,150 0,123 Lamanonia ternata 0,257 0,094 Lihtraea brasiliensis 0,163 0,031 Luehea divaricata -0,082 0,163 Matayba elaeagnoides -0,166 0,167 Maytenus aquifolia -0,082 0,081 Myrrhinium atropurpureum 0,088 0,132 Myrciaria floribunda 0,409 0,090 Myrsine lorentziana 0,153 0,052 Myrcia oligantha 0,003 -0,148 Myrciaria tenella 0,361 0,156 NI -0,018 -0,153 Ocotea puberula -0,159 0,115 Ocotea pulchella 0,058 -0,158 Podocarpus lambertii 0,331 0,149 Roupala brasiliensis 0,012 -0,181 Sapium glandulatum 0,174 -0,087 Sebastiania commersoniana 0,031 -0,369 Siphoneugena reitzii 0,246 0,071 Solanum mauritianum 0,139 0,013 Solanum sanctae-catharinae -0,074 0,096 Styrax leprosus 0,167 -0,006 Symplocos tetranda 0,160 -0,002 Symplocos uniflora 0,104 0,117 Vernonia discolor 0,331 0,128 Xylosma pseudosalzmannii -0,122 0,092 Zanthoxylum rhoifolium 0,138 0,082
Mesmo assim, é possível afirmar que na primeira função discriminante padronizada
destacam-se Myrciaria floribunda, Myrciaria tenella, Podocarpus lambertii, Vernonia

151
discolor, Lamanonia ternata, Siphoneugena reitzii, Eugenia uruguayensis, Calyptranthes
concinna, Sapium glandulatum, Styrax leprosus, Lithraea brasiliensis e Symplocos
tetandra e na segunda, Araucaria angustifolia, Eugenia psidiifolia, Sebastiania
commersoniana, Cryptocaria aschersoniana; Dicksonia sellowiana, Roupala brasiliensis,
Matayba elaeagnoides, Luehea divaricata, Dasyphyllum spinescens, Myrcia oligantha e
Cupania vernalis.
As demais espécies listadas na Tabela 17, também foram importantes na
discriminação dos grupos, mas pouco contribuíram nas funções discriminantes, onde
apresentaram escores baixos.
Nota-se que Podocarpus lambertii, Myrciaria floribunda, Myrciaria tenella, Vernonia
discolor, Lamanonia ternata, Eugenia uruguayensis e Lithraea brasiliensis, com maiores
pesos na segunda função discriminante, são espécies indicadoras e preferenciais do
Grupo 1; Araucaria angustifolia e Ilex paraguariensis, a primeira de maior peso na
segunda função discriminante é a espécies indicadora que, juntamente com a segunda
caracterizam e definem o Grupo 3; Eugenia psidiifolia, Sebastiania commersoniana,
Cryptocaria aschersoniana; Roupala brasiliensis e Ocotea pulchella com maiores pesos
na segunda função discriminante, caracterizam e definem o Grupo 2.
Na Tabela 18 encontram-se os coeficientes das funções discriminantes para as 38
variáveis selecionadas na análise em cada um dos três grupos obtidos pela análise de
Twinspan.
Nota-se, pela referida tabela, que Styrax leprosus, Solanum mauritianum, Vernonia
discolor, Myrciaria tenella, Myrsine lorentziana, Siphoneugena reitzii, Symplocos uniflora,
Podocarpus lambertii, Lamanonia ternata, Symplocos tetandra, Zanthoxylum rhoifolium,
Myrrhinium atropurpureum, Calyptranthes concinna, Myrciaria floribunda, Cupania
vernalis, Eugenia uruguayensis, Sapium glandulatum e Casearia decandra, foram as
espécies mais importantes do grupo 1. No grupo 2 são características as espécies
Myrcia oligantha, Styrax leprosus, Ocotea pulchella, Eugenia psidiiflora, Cryptocaria
aschersoniana, Dicksonia sellowiana, Sebastiania commersoniana e Roupala brasiliensis.

152
TABELA 18 - Coeficientes das funções discriminantes para cada um dos grupos florísticos
obtidos.
GRUPOS VARIÁVEIS (Espécies)
1 2 3 Araucaria angustifolia 0,120 0,276 1,321 Banara parviflora 0,203 0,628 1,616 Calyptranthes concinna 3,323 0,795 0,168 Casearia decandra 1,344 0,879 0,385 Citronella gongonha -1,653 -0,020 2,346 Cryptocaria aschersoniana 0,398 1,322 0,188 Cupania vernalis 2,760 0,571 2,243 Dasyphyllum spinescens 0,209 0,322 1,677 Dicksonia sellowiana 0,343 1,219 0,446 Eugenia psidiifolia 0,233 1,357 0,280 Eugenia uruguayensis 2,918 0,326 0,282 Ilex paraguariensis 0,305 0,679 1,356 Lamanonia ternata 3,455 0,478 0,211 Lihtraea brasiliensis 3,920 0,447 -0,210 Luehea divaricata 0,220 0,023 0,803 Matayba elaeagnoides 0,092 0,460 1,554 Maytenus aquifolia -0,278 0,155 1,380 Myrrhinium atropurpureum 3,390 -0,360 0,626 Myrciaria floribunda 3,274 0,616 0,165 Myrsine lorentziana 5,710 1,085 0,606 Myrcia oligantha 0,548 2,297 0,797 Myrciaria tenella 6,274 0,507 0,181 NI -0,324 1,300 0,222 Ocotea puberula -0,320 0,452 1,593 Ocotea pulchella 1,002 1,391 0,569 Podocarpus lambertii 4,165 0,215 0,023 Roupala brasiliensis -0,121 1,071 -0,055 Sapium glandulatum 1,976 1,239 0,553 Sebastiania commersoniana 0,287 1,228 0,302 Siphoneugena reitzii 4,756 0,782 0,260 Solanum mauritianum 7,620 0,323 -0,515 Solanum sanctae-catharinae 0,227 0,453 2,704 Styrax leprosus 7,925 1,942 -0,150 Symplocos tetranda 3,527 0,732 -0,199 Symplocos uniflora 4,684 -0,132 0,739 Vernonia discolor 6,472 0,608 0,141 Xylosma pseudosalzmannii -0,015 0,763 2,026 Zanthoxylum rhoifolium 3,510 0,669 0,703
(Constante) -13,501 -3,736 -3,914

153
Já no grupo 3 destacaram-se as espécies Solanum sanctae-catharinae, Citronella
gongonha, Cupania vernalis, Xylosma pseudosalzmannii, Dasyphyllum spinescens,
Ocotea puberula, , Matayba elaeagnoides, Ilex paraguariensis, Banara parvifolia e
Araucaria angustifolia. A maioria destas espécies são as indicadoras e preferenciais
determinadas na análise de agrupamento (Figura 4).
Na Tabela 19 encontram-se os centróides ou pontos médios de cada grupo, nas
duas funções discriminantes, demarcados no Mapa Territorial (Figura 5), considerando as
Funções Discriminantes Canônicas 1 e 2.
TABELA 19 - Centróides das funções discriminantes canônicas avaliados para os
três grupos florísticos encontrados.
GRUPOS Função 1 Função 2 1 4,015 1,345 2 0,196 -1,295 3 -1,030 0,889
Observa-se pela Figura 5, que houve uma nítida separação entre os três grupos
obtidos, onde foram localizados os centróides dos grupos, comprovando a precisão dos
agrupamentos.
Na Tabela 20, encontram-se os percentuais de classificações corretas e incorretas
das unidades amostrais. Observa-se que 83 % das parcelas foram classificadas
corretamente no grupo 1, 92,3 % no grupo 2 e 92,8 % no grupo 3. Por outro lado, no
grupo 1, 2,6 % das parcelas foram classificados erradas no grupo 2 e 0,6 % no grupo 3;
no grupo 2, 12,0 % das parcelas foram classificadas erradas no grupo 1 e 6,6 % no grupo
3; enquanto que no grupo 3, 5,0 % das parcelas foram classificadas erradas no grupo 1 e
6,1 % no grupo 2.
No geral, houve 91,2 % de classificações corretas, indicando precisão da técnica
de agrupamento. Desta forma, pode-se considerar que as duas funções discriminantes
podem ser usadas para classificar novos indivíduos, dentro dos grupos estabelecidos.

154
FIGURA 5 - Mapa territorial dos grupos florísticos determinados, com a localização
de seus respectivos centróides.

155
TABELA 20 - Número de casos e percentagens de classificações das parcelas nos três
grupos florísticos determinados.
GRUPOS CLASSIFICAÇÃO EM CADA GRUPO TOTAL
1 2 3
Contagem 1
2
3
83
11
3
12
390
31
5
26
437
100
427
471
% 1
2
3
83,0
2,6
0,6
12,0
91,3
6,6
5,0
6,1
92,8
100,0
100,0
100,0
TOTAL DE CASOS CLASSIFICADOS CORRETAMENTE: 91,2 %
Para alocação de uma nova comunidade em um dos grupos, calcula-se o valor de
cada discriminante para essa comunidade (Y1, Y2), através da Tabela 17 e as distâncias
Euclidianas do ponto calculado, aos centróides de cada grupo (d1, d2, d3) da Tabela 19,
através da equação:
22
2
21
1)()(),( XYXYXYd −+−=
A menor distância Euclidiana obtida indica o grupo em que a nova comunidade
deve ser alocada.
A técnica de Análise Discriminante mostrou-se bastante eficiente como
complemento na análise de agrupamentos florísticos de comunidades vegetais, pois
permitiu classificar e reclassificar de maneira precisa, populações diferenciadas pela
Análise de Cluster (método de Twinspan), além de permitir alocação de novas populações
em um dos grupos obtidos.

156
0≤≤≤≤ Ss ≤≤≤≤ 1
4.4 Similaridade entre os grupos florísticos obtidos
A similaridade entre comunidades pode ser analisada através de comparações
qualitativas, baseadas na presença e na ausência das espécies e quantitativas, baseadas
na densidade das espécies (Kent & Coker, 1992).
Para comparar os três grupos obtidos quanto à similaridade florística, foi construída
uma matriz de presença e ausência das espécies nos grupos. A partir dessa matriz foi
calculada a similaridade florística através do Índice de Sorensen (Tabela 21). Os valores
variam de 0-1 (Kent & Coker ,1992).
cba
cSs
2
2
++=
onde:
a = número de espécies exclusivas do grupo 1;
b = número de espécies exclusivas do grupo 2;
c = número de espécies comuns aos dois grupos.
TABELA 21 – Diversidade e Similaridade entre os grupos florísticos na Floresta Ombrófila Mista
da FLONA de São Francisco de Paula, RS
Similaridade
Grupos (Ss) Grupo Diversidade
(H’) 1 2 3
1 1,799 0,667
2 1,619 0,758
3 1,465 0,691
Pelos resultados da Tabela 21 pode-se deduzir que existe similaridade florística
entre o três grupos obtidos, o que é lógico por pertencerem à formação de Floresta
Ombrófila Mista. Maior similaridade existe entre os grupos 1 (Formação Podocarpus) e 2
(Formação Sebastiania) cujo índice de similaridade foi de 0,758. Observando o índice de

157
diversidade nota-se que ambos os grupos (1 e 2) apresentam semelhança,
respectivamente com valores de 1,799 e 1,691. Por sua vez o grupo 3 é o que apresenta
menor similaridade com os demais e é o que apresenta maior diversidade florística
(H’=1,799).
Os índices altos de similaridade encontrados indicam coincidência entre a maioria
das espécies amostradas nos grupos, isto é, existe um grande número de espécies
comuns entre os três grupos (Anexo 1). São as espécies adaptadas a todas situações
ecológicas, embora a densidade das mesmas seja bastante variável.
Felfili et al. (2001a) encontraram resultados semelhantes em um estudo
comparando a similaridade florística de comunidades do Cerrado sensu stricto da
Chapada do Espigão Mestre do São Francisco, isto é, encontraram grande número de
espécies comuns nas diferentes comunidades estudadas.
Assim mesmo existem espécies exclusivas para cada grupo florístico determinado
(Tabela 22). São as espécies limitadas a certas características do habitat.
TABELA 22 – Espécies exclusivas dos grupos florísticos da Floresta Ombrófila Mista da
FLONA de São Francisco de Paula, RS.
Grupos Espécie
1 2 3 Alsophila sp. X
Cabralea canjerana X Carica quercifolia X Casearia obliqua X
Cestrum sp. X Citronella paniculata X Clethra uleana X
Coussarea contracta X
Coutarea hexandra X
Drimys brasiliensis X Continua

158
TABELA 22 – Continuação
Grupos Espécie
1 2 3 Erythrina cristagalli X
Eugenia pluriflora X
Griselinia ruscifolia X
Guapira opposita X
Hennecartia omphalandra X Ilex sp. X
Inga marginata X Inga sessilis X Lonchocarpus nitidus X Mollinedia sp. X
Myrceugenia foveolata X
Myrceugenia glaucescens X
Myrcia arborescens X
Myrcia lajeana X
Myrcia palustris X
Nectandra lanceolata X
Ocotea indecora X
Piptocarpha tomentosa X
Rhamnus sphaerosperma X
Rollinia sylvatica X
Solanum mauritianum X
Solanum pabstii X
Solanum pseudocapsicum X
Syagrus romanzoffiana X
Tabebuia umbellata X
Zanthoxylum kleinii X
TOTAL 3 8 25
No grupo 1 (Formação Podocarpus) ocorreu um pequeno número de expécies
exclusivas (%), Eugenia pluriflora, Drimys brasiliensis e Myrcia lajeana, todas seletivas
higrófitas, esciófitas ou de luz difusa e até heliófitas, tornando-se não raro ou até
localmente muito freqüente.

159
No grupo 2 (Formação Sebastiania), de áreas mais úmidas, ocorreram espécies
exclusivas, tais como: Alsophila sp., Clethra uleana, Erythrina cristagalli, Griselinia
ruscifolia, Myrceugenia foveolata, Nectandra lanceolata, Ocotea indecora e Zanthoxylum
kleinii.
No grupo 3 (Formação Araucaria) ocorreu um grande número de espécies
exclusivas, pelo fato de ser o grupo típico da Floresta Ombrófila Mista de locais altos com
solos mais profundos e permeáveis. Entre estas espécies destacam-se Cabralea
canjerana, Casearia obliqua, Citronella paniculata, Coussarea contracta, Coutarea
hexandra, Guapira opposita, Hennecartia omphalandra, Inga sessilis, Lonchocarpus
nitidus, Mollinedia sp., Myrceugenia glaucescens, Myrcia arborescens, Myrcia palustris,
Piptocarpha tomentosa, Rhamnus sphaerosperma, Rollinia sylvatica, Solanum mauritianum,
Solanum pabstii, solanum peseudocapsicum, Syagrus romanzoffiana, Tabebuia umbellata, entre
outras.
4.5 Caracterização geral dos grupos florísticos obtidos
4.5.1 Grupo 1 – Formação Podocarpus
4.5.1.1 Composição florística
O resultados da composição florística do grupo 1, determinado na Floresta
Ombrófila Mista da FLONA de São Francisco de Paula estão relacionadas no Anexo 2.
Foram encontradas 75 espécies arbóreas pertencentes a 51 gêneros e 29 famílias
botânicas, entre os indivíduos com CAP ≥ 30 cm, além de algumas árvores de não
identificados, mortas e cipós.
As famílias Myrtaceae, com 11 gêneros e 16 espécies, Lauraceae, com 4 gêneros
e 6 espécies; Flacourtiaceae, com 3 gêneros e 5 espécies; Aquifoliaceae e Myrsinaceae,
com 1 gênero e 4 espécies e as famílias Asteraceae e Sapindaceae, com 3 gêneros e 3
espécies e Euphorbiaceae, com 2 gêneros e 3 espécies foram as mais representativas
da floresta. Destacaram-se também as famílias Cunoniaceae e Verbenaceae, com com 2
gêneros e 2 espécies; Annonaceae, Solanaceae e Symplocaceae, com 1 gênero e 2

160
espécies. As 16 famílias restantes estavam representadas por 1 única espécie.
A predominância de Myrtaceae é comum nas florestas do Rio Grande do Sul e,
principalmente neste tipo Fitogeográfico, independente das comunidades ecológicas que
ocorrem associadas. Tal afirmação pode ser comprovada em trabalhos de Jarenkow
(1994), Klein apud Nascimento (2000) e Rio Grande do Sul (2002).
4.5.1.2 Estrutura Fitossociológica
Os parâmetros fitossociológicos das espécies encontradas no grupo 1 podem ser
observados no Anexo 2. Na Tabela 23 estão relacionadas as principais espécies do
grupo, em ordem do valor de importância. São as espécies mais características e
importantes dessa formação, pelo fato de apresentarem maiores valores de densidade,
dominância, freqüência, valor de importância e valor de cobertura.
Observou-se que Podocarpus lambertii e Myrciaria floribunda foram as espécies
mais importantes do grupo, somando respectivamente 7,92% e 5,32% do valor de
importância e 11,33% e 7,82% do valor de cobertura. Nota-se que, pelo valor de
cobertura, Podocarpus lambertii fica como a principal espécie, cujo destaque deve-se ao
elevado valor de dominância (grandes diâmetros) e baixo valor da freqüência.
Pela análise de agrupanto, Podocarpus lambertii é a espécie indicadora do grupo 1,
ou seja, ela caracteriza o grupo. Segundo Marchiori (1996), a espécie é encontrada na
composição do tipo florestal estudado e ocorre em morros e ravinas, tais características
são similares às observadas nas parcelas que caraterizam o grupo. Desenvolve-se tanto
em solos férteis como também em solos pobres, rasos ou rochosos, como é o caso na
Serra Sudeste. Portanto, é uma espécie esciófila e heliófita, isto é, ocorre tanto em áreas
sombrias, quanto expostas ao sol, e é difundida pelos pássaros que se alimentam do arilo
das sementes (Reitz et al., 1988).
As nove primeiras espécies, juntamente com Myrciaria floribunda e Podocarpus
lambertii representam 43,04% do valor de importância e 52,57% do valor de cobertura
total das espécies do grupo.

161
TABELA 23 – Estrutura horizontal do grupo 1, da FLONA de São Francisco de Paula, RS.
ESPÉCIE
DENSIDADE FREQÜÊNCIA DOMINÂNCIA VALOR DE IMPORTÂNCIA
VALOR DE COBERTURA
DA DR FA FR DoA DoR VI (%) VI ac. VC (%) VC ac. Podocarpus lambertii 93,93 9,01 8,08 1,12 5,69 13,64 7,92 7,92 11,33 5,31 Myrciaria floribunda 94,95 9,21 2,02 0,28 2,69 6,45 5,32 13,24 7,82 19,15 Eugenia uruguariensis 55,56 5,39 37,37 5,16 1,60 3,84 4,80 18,04 4,62 23,77 Vernonia discolor 45,45 4,41 3,03 0,42 3,97 9,53 4,78 22,82 6,97 30,74 Araucária angustifolia 27,27 2,64 22,22 3,07 5,57 8,56 4,75 27,58 5,61 36,35 Lamonia ternata 44,44 4,31 21,21 2,93 2,37 5,68 4,30 31,89 4,99 41,34 Myrciaria tenela 57,58 5,58 16,16 2,23 1,45 3,49 3,77 35,65 4,53 45,87 Ilex brevicuspis 52,53 5,09 7,07 0,98 2,12 5,08 3,72 39,36 5,08 50,95 Ocotea pulchella 20,20 1,96 56,57 7,81 0,53 1,28 3,68 43,04 1,62 52,57 Morta 61,62 5,97 1,01 0,14 1,90 4,56 3,55 46,56 5,26 57,83 Casearia decandra 32,32 3,13 20,20 2,79 0,66 1,58 2,50 49,06 3,85 61,68 Symplocos tetrandra 24,24 2,35 2,02 0,28 1,97 4,72 2,45 51,51 3,54 65,22 Calyptranthes concinna 29,29 2,84 21,21 2,93 0,38 0,90 2,22 53,73 1,87 67,09 Sapium glandulatum 33,33 3,23 2,02 0,28 1,13 2,71 2,07 55,80 2,97 70,06 Lithraea brasiliensis 24,24 2,35 1,01 0,14 1,48 3,55 2,01 57,81 2,95 73,01 Xylosma tweedianum 1,01 0,10 42,42 5,86 0,02 0,04 1,99 59,80 0,07 73,08 Myrsine lorentziana 12,12 1,18 21,21 2,93 0,71 1,71 1,93 61,74 1,44 74,52 Myrsine umbellata 31,31 3,04 6,06 0,84 0,79 1,90 1,93 63,67 2,47 76,99 Myrcianthes pungens 1,01 0,10 37,37 5,16 0,02 0,05 1,77 65,44 2,66 79,65 Siphoneigena reitzii 25,25 2,45 4,04 0,56 0,69 1,65 1,55 66,99 2,05 81,70
Sub-total 768,04 70,03 332,30 45,91 35,98 80,92 66,99 81,70 Restantes 261,25 29,97 391,94 54,09 5,66 19,08 33,01 18,30 Total 1029,29 100,00 724,24 100,00 41,64 100,00 100,00 100,00
DA = Densidade absoluta; DR = Densidade relativa; FA = Freqüência absoluta; FR = Freqüência relativa; DoA = Dominância absoluta; DoR = Dominância relativa; VI (%) = Valor de importância em %; VI ac. = Valor de importância acumulado; VC (%) = Valor de cobertura em %; VC ac. = Valor de cobertura acumulado.

162
As dez espécies restantes listadas na Tabela 23 contribuem com mais 23,43% do
valor de importância e 23,87 % do valor de cobertura.
As árvores mortas aparecem em decimo lugar na ordem de importância das
espécies. Portanto, são significativas na composição da comunidade e constitui um
fenômeno natural de substituição dos indivíduos na dinâmica da área estudada. A
presença destacada de árvores mortas é comum nas florestas nativas do Estado,
comprovada pelo Inventário Florestal Contínuo do Estado do Rio Grande do Sul (Rio
Grande do Sul, 2002).
As 19 principais espécies listadas na Tabela 23, juntamente com as árvores mortas
representam aproximadamente 70% da densidade, 46% da freqüência, 81% da
dominância, 67% do valor de importância e 82% do valor de cobertura total das espécies
da floresta.
As demais espécies representam apenas 30% da densidade, 54% da freqüência,
19% da dominância, 33% do valor de importância e 18% do valor de cobertura total das
espécies.
4.5.1.3 Parâmetros Dendrométricos
Os parâmetros dendrométricos das parcelas amostradas no grupo 1, indicam um
diâmetro médio de 20,46 cm, cujo coeficiente de variação médio dos diâmetros foi de
47,43%.
A altura total média do grupo foi estimada em 11,97 m e o coeficiente de variação
médio da altura total foi de 24,20%.
A altura comercial média foi estimada em 6,50 m e o coeficiente de variação médio
das alturas comerciais foi de 39,69%.
Observa-se pela Tabela 24, que o grupo 1 foi o grupo que apresentou menores
valores médios para diâmetro, altura total e comercial entre os grupos encontrados na
Floresta Ombrófila Mista de São Francisco de Paula.
Por outro lado, apresentou maior número médio de indivíduos por hectare (1031,31
indivíduos), indicando que o grupo apresenta indivíduos de menor porte, mas em maior
quantidade. Apresentou, também, maior diversidade florística, com índice de Shannon de
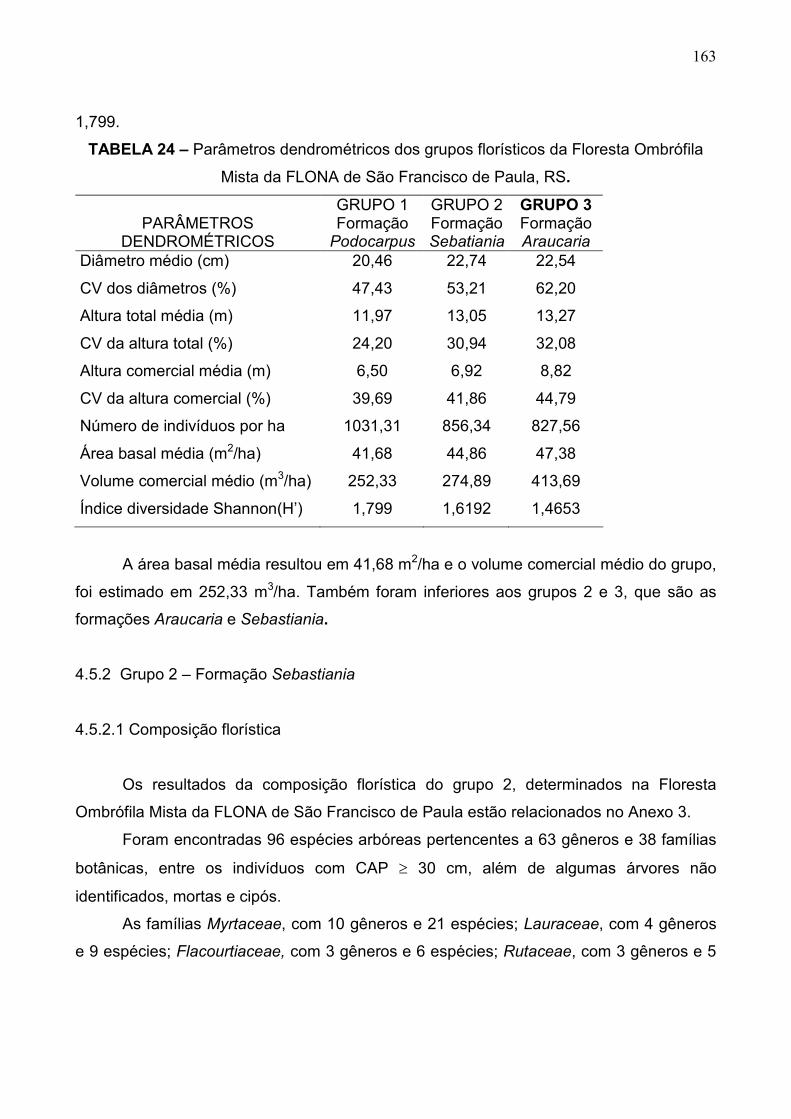
163
1,799.
TABELA 24 – Parâmetros dendrométricos dos grupos florísticos da Floresta Ombrófila
Mista da FLONA de São Francisco de Paula, RS.
PARÂMETROS
DENDROMÉTRICOS
GRUPO 1 Formação Podocarpus
GRUPO 2 Formação Sebatiania
GRUPO 3 Formação Araucaria
Diâmetro médio (cm) 20,46 22,74 22,54
CV dos diâmetros (%) 47,43 53,21 62,20
Altura total média (m) 11,97 13,05 13,27
CV da altura total (%) 24,20 30,94 32,08
Altura comercial média (m) 6,50 6,92 8,82
CV da altura comercial (%) 39,69 41,86 44,79
Número de indivíduos por ha 1031,31 856,34 827,56
Área basal média (m2/ha) 41,68 44,86 47,38
Volume comercial médio (m3/ha) 252,33 274,89 413,69
Índice diversidade Shannon(H’) 1,799 1,6192 1,4653
A área basal média resultou em 41,68 m2/ha e o volume comercial médio do grupo,
foi estimado em 252,33 m3/ha. Também foram inferiores aos grupos 2 e 3, que são as
formações Araucaria e Sebastiania.
4.5.2 Grupo 2 – Formação Sebastiania
4.5.2.1 Composição florística
Os resultados da composição florística do grupo 2, determinados na Floresta
Ombrófila Mista da FLONA de São Francisco de Paula estão relacionados no Anexo 3.
Foram encontradas 96 espécies arbóreas pertencentes a 63 gêneros e 38 famílias
botânicas, entre os indivíduos com CAP ≥ 30 cm, além de algumas árvores não
identificados, mortas e cipós.
As famílias Myrtaceae, com 10 gêneros e 21 espécies; Lauraceae, com 4 gêneros
e 9 espécies; Flacourtiaceae, com 3 gêneros e 6 espécies; Rutaceae, com 3 gêneros e 5

164
espécies; Fabaceae, com 4 gêneros e 4 espécies; Euphorbiaceae, com 3 gêneros e 4
espécies; Aquifoliaceae e Myrsinaceae, com 1 gênero e 4 espécies; Sapindaceae, com 3
gêneros e 3 espécies e Solanaceae, com 1 gênero e 3 espécies. Destacaram-se também
as famílias Mimosaceae, com 2 gêneros e 2 espécies; Celastraceae e Symplocaceae,
com 1 gênero e 2 espécies. As 24 famílias restantes apresentaram apenas 1 única
espécie.
4.5.2.2 Estrutura Fitossociológica
Os parâmetros fitossociológicos das espécies encontradas no grupo 2 podem ser
observados no Anexo 3. Na Tabela 25 estão relacionadas as principais espécies do grupo
em ordem do valor de importância. São as espécies mais caracteristicas e importantes
dessa formação, pelo fato de apresentarem maiores valores de densidade, dominância,
freqüência, valor de importância e valor de cobertura.
Observou-se que Cryptocarya aschersoniana, Eugenia psidiiflora, Sebastiania
commersoniana, Blepharocalyx salicifolius, Ilex brevicuspis foram as espécies mais
importantes do grupo, somando, em conjunto, 36,88% do valor de importância e 38,34%
do valor de cobertura. São as espécies mais características dessa formação de ambientes
úmidos da Floresta Ombrófila Mista de São Francisco de Paula.
As espécies Sebatiania commersoniania e Sebastiania brasiliensis, esta também
importante, segundo Araujo (2002), ocorrem em locais com flutuação do lençol freático ou
solos preferencialmente úmidos. São, portanto, adaptadas a estas condições.
Já a Cryptocarya aschersoniania é uma árvore de ampla dispersão, espécie
higrófita, freqüente ao longo dos rios, planícies aluviais, inícios de encostas, solos úmidos
e mais profundos (Vattimo, 1979).
As árvores mortas aparecem em sexto lugar na ordem de importância das
espécies. Portanto, são significativas na composição da comunidade e constituem um
fenômeno natural de substituição dos indivíduos na dinâmica da área estudada. A
presença destacada de árvores mortas é comum nas florestas nativas do Estado,
comprovado pelo Inventário Florestal Contínuo do Estado do Rio Grande do Sul (Rio
Grande do Sul, 2002).

165
TABELA 25 – Estrutura horizontal do grupo 2, da FLONA de São Francisco de Paula, RS
ESPÉCIE DENSIDADE FREQÜÊNCIA DOMINÂNCIA VALOR DE
IMPORTÂNCIA VALOR DE COBERTURA
DA DR FA FR DoA DoR VI (%) VI ac. VC (%) VC ac. Cryptocarya aschersoniana 56,34 6,58 33,80 5,67 5,52 12,30 8,18 8,18 9,44 9,44 Blepharocalyx salicifolius 44,84 5,24 31,69 5,31 5,60 12,48 7,68 15,85 8,86 18,30 Eugenia psidiiflora 89,91 10,50 46,95 7,87 1,94 4,33 7,57 23,42 7,41 25,71 Sebastiania commersoniana 69,25 8,09 31,69 5,31 3,93 8,75 7,38 30,80 5,60 31,31 Ilex brevicuspis 39,67 4,63 24,88 4,17 4,23 9,44 6,08 36,88 7,03 38,34 Morta 36,62 4,28 30,99 5,20 1,54 3,43 4,30 41,18 3,85 42,19 Ocotea pulchella 28,12 3,29 23,24 3,90 2,47 5,51 4,23 45,41 4,40 46,59 Sebastiania brasiliensis 42,02 4,91 26,29 4,41 0,60 1,33 5,32 50,73 3,12 49,71 Araucaria angustifolia 18,54 2,17 12,91 2,17 2,32 5,18 3,17 53,90 3,67 53,38 Sapium glandulatum 26,29 3,07 18,08 3,03 0,88 1,96 2,68 56,58 2,51 55,89 Nectandra megapotamica 18,08 2,11 14,79 2,48 1,33 2,98 2,52 59,10 2,54 58,43 Ilex paraguariensis 23,47 2,74 17,37 2,91 0,79 1,76 2,47 61,57 2,25 60,68 Dicksonia sellowiana 25,35 2,96 12,44 2,09 1,02 2,27 2,44 64,01 2,61 63,29 Myrceugenia cucullata 24,28 2,82 17,14 2,87 0,60 1,33 2,34 68,69 2,08 65,37 Roupala brasiliensis 21,13 2,47 16,20 2,72 0,66 1,47 2,22 70,91 1,97 67,34 Casearia decandra 19,01 2,22 16,20 2,72 0,34 0,75 1,89 72,80 1,48 68,82 Campomanesia xanthocarpa 15,73 1,85 12,21 2,05 0,56 1,25 1,71 74,51 1,54 70,36 Campomanesia rhombea 15,02 1,75 13,15 2,20 0,45 1,01 1,65 76,16 1,38 71,74 Lamanonia ternata 9,15 1,07 7,75 1,30 1,10 2,45 1,60 77,76 1,76 73,50 Eugenia involucrata 12,44 1,45 10,56 1,77 0,38 0,84 1,35 79,11 1,14 74,64
Sub-total 663,43 74,20 402,13 66,25 36,26 76,82 79,11 74,64 Restantes 192,91 25,80 194,11 33,75 8,60 23,18 20,89 25,36 Total 856,34 100,00 596,24 100,00 44,86 100,00 100,00 100,00
DA = Densidade absoluta; DR = Densidade relativa; FA = Freqüência absoluta; FR = Freqüência relativa; DoA = Dominância absoluta; DoR = Dominância relativa; VI (%) = Valor de importância em %; VI ac. = Valor de importância acumulado; VC (%) = Valor de cobertura em %; VC ac. = Valor de cobertura acumulado.

166
As demais espécies relacionadas na Tabela 25 representam 40% do valor de importância
e 36,30% do valor de cobertura total da floresta do grupo.
As 19 principais espécies listadas na Tabela 25, juntamente com as árvores mortas e não
identificadas representam 74,20% da densidade, 66,25% da freqüência, 76,82% da dominância,
79,11% do valor de importância e 74,64% do valor de cobertura total das espécies da floresta.
As demais espécies representam apenas 25,80% da densidade, 33,75% da
freqüência, 23,28% da dominância, 20,89% do valor de importância e 25,36% do valor de
cobertura total das espécies.
4.5.2.3 Parâmetros Dendrométricos
Os parâmetros dendrométricos das parcelas amostradas no grupo 2, indicam um
diâmetro médio de 22,74 cm, cujo coeficiente de variação médio dos diâmetros foi de
53,21%.
A altura total média do grupo foi estimada em 13 m e o coeficiente de variação
médio da altura total foi de 30,94%.
A altura comercial média foi estimada em 6,92 m e o coeficiente de variação médio
das alturas comerciais foi de 41,86%.
Observa-se pela Tabela 24, que o grupo 3 foi o grupo que apresentou valor médios
para a maioria dos parâmetros dendrométricos, isto é, para altura, área basal, volume e
índice de diversidade, exceto para diâmetro. Para este parâmetro foi o grupo que
apresentou maiores valores, indicando que, embora localiza-se em locais úmidos,
apresenta indivíduos de grandes diâmetros.
O número médio de árvores do grupo, considerando todos os indivíduos com CAP
≥ 30 cm, foi de aproximadamente 856 indivíduos/ha.
A área basal média resultou em 44,86 m2/ha, o volume comercial médio do grupo,
foi estimado em 275 m3/ha.
4.5.3 Grupo 3 – Formação Araucaria
4.5.3.1 Composição florística

167
Os resultados da composição florística do grupo 3, determinados na Floresta Ombrófila Mista da FLONA de São Francisco de Paula, estão relacionadas no Anexo 4.
Foram encontradas 110 espécies arbóreas pertencentes a 69 gêneros e 39 famílias botânicas, entre os indivíduos com CAP ≥ 30 cm, além de algumas árvores de não identificados, mortas e cipós.
As famílias Myrtaceae, com 10 gêneros e 22 espécies; Solanaceae, com 2 gêneros
e 7 espécies; Lauraceae, com 4 gêneros e 6 espécies; Flacourtiaceae, com 3 gêneros e 6
espécies; Aquifoliaceae, com 1 gênero e 5 espécies; Asteraceae, Fabaceae e Rutaceae,
com 3 gêneros e 4 espécies; Mimosaceae, com 2 gêneros e 4 espécies; Myrsinaceae, 1
gênero e 4 espécies; Sapindaceae, 3 gêneros e 3 espécies; Euphorbiaceae e
Monimiaceae, com 2 gêneros e 3 espécies; Annonaceae, com 1 gênero e 3 espécies;
Cunoniaceae, Meliaceae, Rhamnaceae e Rubiaceae, com 2 gêneros e 2 espécies foram
as mais representantes da floresta. Destacaram-se também as famílias, Celastraceae,
Icacinaceae e Symplocaceae, com 1 gênero e 2 espécies. As18 famílias restantes
apresentaram apenas1 única espécie.
A predominância de Myrtaceae continua ocorrendo neste grupo, que é a formação
de Floresta Ombófila Mista típica. Esta família domina as formações florestais do Estado,
conforme pode ser observado pelos diversos trabalhos realizados (Jarenkow, 1985; Klein
apud Nascimento, 2000; Rio Grande do Sul, 2002).
4.5.3.2 Estrutura Fitossociológica
Os parâmetros fitossociológicos das espécies encontradas no grupo 2 podem ser
observados no Anexo 4. Na Tabela 26 estão relacionadas as principais espécies do
grupo, em ordem do valor de importância. São as espécies mais características e
importantes dessa formação, pelo fato de apresentarem maiores valores de densidade,
dominância, dreqüência, valor de importância e valor de cobertura.
Observou-se que Araucaria angustifolia foi a espécie mais importante do grupo,
somando respectivamente 33,44% do valor de importância e 42,86% do valor de
cobertura. Representou, também, 23,60% da densidade, 14,60% da freqüência e 62,12%
da dominância total da floresta do grupo.
É, portanto, a principal espécie do grupo e foi, pela análise de agrupamento a
espécie indicadora e a que mais caracteriza o grupo 3.

168
A Araucaria angustifolia geralmente é encontrada formando agrupamentos densos
na Floresta Ombrófila Mista típica, principalmente na parte leste e central do planalto
meridional do Brasil. É encontrada desde uma altitude média de 500 metros, em terrenos
medianamente ondulados, enquanto nos terrenos fortemente ondulados está sendo
substituídas pela floresta latifoliada da fralda da Serra Geral (Reitz et al., 1988).

169
TABELA 26 – Estrutura horizontal do Grupo 3, da FLONA de São Francisco de Paula, RS
ESPÉCIE
DENSIDADE FREQÜÊNCIA DOMINÂNCIA VALOR DE IMPORTÂNCIA
VALOR DE COBERTURA
DA DR FA FR DoA DoR VI (%) VI ac. VC (%) VI ac. Araucaria angustifolia 195,30 23,60 77,14 14,60 29,43 62,12 33,44 33,44 42,86 42,86 Blepharocalyx salicifolius 49,57 5,99 30,56 5,78 1,60 3,37 5,05 38,49 4,68 47,54 Ilex paraguariensis 45,73 5,53 33,33 6,31 1,04 2,20 4,68 43,17 3,86 51,40 Morta 39,10 4,73 31,20 5,90 1,03 2,17 4,26 43,43 3,45 54,85 Ilex brevicuspis 33,97 4,11 20,50 3,88 1,61 3,40 3,79 51,22 3,75 58,60 Sebastiania brasiliensis 32,26 3,90 21,37 4,04 0,42 0,89 2,94 54,16 2,40 61,00 Banara parviflora 28,63 3,46 23,29 4,41 0,45 0,96 2,94 57,01 2,21 63,21 Matayba elaeagnoides 24,79 3,00 15,81 2,99 0,78 1,64 2,54 59,64 1,54 64,75 Nectandra megapotamica 19,66 2,38 16,03 3,03 0,75 1,58 2,33 61,97 1,98 66,73 Ocotea puberula 17,09 2,07 12,18 2,30 1,21 2,55 2,31 64,28 2,31 69,04 Luehea divaricata 24,36 2,94 12,18 2,30 0,76 1,60 2,28 66,56 2,27 71,31 Casearia decandra 16,88 2,04 11,75 2,22 0,37 0,78 1,68 68,24 1,41 72,72 Dasyphyllum spinescens 14,74 1,78 10,04 1,90 0,43 0,90 1,53 69,77 1,34 74,06 Campomanesia rhombea 15,81 1,91 9,40 1,78 0,30 0,64 1,44 71,21 1,27 75,33 Ocotea pulchella 11,97 1,45 10,04 1,90 0,40 0,85 1,40 72,61 1,45 76,78 Myrceugenia cucullata 14,32 1,73 10,47 1,98 0,22 0,46 1,39 74,00 1,09 77,87 Campomanesia xanthocarpa 11,54 1,39 8,12 1,54 0,29 0,61 1,18 76,18 1,00 78,87 Dicksonia sellowiana 9,83 1,19 7,26 1,37 0,47 0,98 1,18 77,36 1,08 79,95 Lamonia ternata 8,97 1,08 7,26 1,37 0,36 0,76 1,07 78,43 0,93 80,88 Cryptocarya aschersoniana 8,33 1,01 6,20 1,17 0,46 0,98 1,05 79,48 0,99 81,87
Sub-total 622,85 73,25 351,48 64,05 41,87 89,44 79,48 81,87 Restantes 204,72 26,75 177,00 35,95 6,00 10,56 20,52 18,13 Total 827,57 100,00 528,42 100,00 47,38 100,00 100,00 100,00
DA = Densidade absoluta; DR = Densidade relativa; FA = Freqüência absoluta; FR = Freqüência relativa; DoA = Dominância absoluta; DoR = Dominância relativa; VI (%) = Valor de importância em %; VI ac. = Valor de importância acumulado; VC (%) = Valor de cobertura em %; VC ac. = Valor de cobertura acumulado.

170
Destacaram-se também as espécies Blepharocalyx salicifolius, com
5,05% do VI e 4,68% do VC e Ilex paraguariensis, com 4,68% do VI e
3,86% do VC. O destaque dessas duas espécies deve-se principalmente
à densidade de indivíduos.
Estas duas espécies, juntamente com a Araucária, somam 43,17%
do valor de importância e 51,40% do valor de cobertura total das espécies
do grupo.
Blepharocalyx salicifolius, distribui-se praticamente por todo o Rio
Grande do Sul. Freqüentes em matas de galerias, onde, por vezes, se
torna uma das espécies dominantes; também freqüentes nas submatas
dos pinhais situados em solos úmidos e compactos, solos de aclive suave
e de drenagem bastante lenta, como nas depressões. Segundo
pesquisadores, é uma espécie higrófita e heliófita, que se desenvolve nos
mais variados ambientes da vegetação, desde a vegetação baixa e
esparsa até os subbosques bastante desenvolvidos, ou seja, possui
grande potencialidade de adaptação (Reitz et al., 1988).
Segundo o mesmo autor Ilex paraguariensis é uma espécie
vastamente difundida por toda região dos pinhais. Espécie esciófita e
seletiva higrófita que cresce preferencialmente nas associações mais
evoluídas dos pinhais, onde predomina a Ocotea pulchella. Tem
preferência por terrenos compactos e pouco íngremes, onde forma
agrupamentos bastante densos.
Ilex brevicuspis, Banara parviflora, Ocotea puberula, Sebastiania
commersoniana, Sebastiania brasiliensis, Nectandra megapotamica,
Mollinedia sp., Campomanesia rhombea, Ocotea pulchella, Matayba
elaeagnoides, Cryptocarya aschersoniana, Luehea divaricata,
Campomanesia xanthocarpa, Casearia decandra e Dasyphyllum
spinescens são também importantes da formação Araucaria. Estas 15
espécies representam 36% do valor de importância e 30,47% do valor de
cobertura total das espécies.

171
As árvores mortas aparecem em quarto lugar na ordem de
importância das espécies. São significativas na composição da
comunidade e comprovam sua importância na dinâmica das florestas.
As principais espécies listadas na Tabela 26, juntamente com as
árvores mortas representam 73,25% da densidade, 64,05% da
freqüência, 89,44% da dominância, 79,48% do valor de importância e
81,87% do valor de cobertura total das espécies da floresta.
As demais especies representam apenas 26,75% da densidade,
35,95% da freqüência, 10,56% da dominância, 20,52% do valor de
importância e 18,13% do valor de cobertura total das espécies.
4.5.3.3 Parâmetros Dendrométricos
Os parâmetros dendrométricos das parcelas amostradas no grupo
2, indicam um diâmetro médio de 22,53 cm, cujo coeficiente de variação
médio dos diâmetros foi de 62,20%.
A altura total média do grupo foi estimada em 13,27 m e o
coeficiente de variação médio da altura total foi de 32,08%.
A altura comercial média foi estimada em 8,82 m e o coeficiente de
variação médio das alturas comerciais foi de 44,79%.
Observa-se que o grupo 2 foi o grupo que apresentou maiores
valores médios para altura total e comercial comparando com os demais
grupos (formação Podocarpus e formação Sebastiania) e um diâmetro
médio aproximadamente igual ao grupo 2 (formação Sebastiania).
Por outro lado, apresentou menor número médio de indivíduos por
hectare (827 indivíduos).
Conforme pode-se observar pela tabela 24, o grupo apresentou
elevado coeficiente da variação dos diâmetros (62,20%), indicando a
existência de indivíduos com diâmetros aos extremos, onde os maiores
corresponderam as Araucárias, resultando maiores valores médios para
área basal (47,38 m2/ha) e para volume comercial (413,69 m3/ha).

172
O grupo apresentou valores mais baixos para a diversidade
florística, em comparação com os demais grupos (H’= 1,4653).
4.6 Análise de gradientes ambientais (CCA)
A Análise de Correspondência Canônica (CCA) é um dos métodos
de análise multivariada que tem se mostrado mais indicado para estudos
que visam compreender as correlações existentes entre a vegetação e
fatores ambientais.
Basicamente esta técnica analisa e sintetiza os dados da vegetação em
um diagrama de ordenação multidimensional, formado por eixos
perpendiculares, os quais expressam as variações na densidade das
espécies em função das mudanças nas condições ambientais, além de
mostrar, concomitantemente, quais os fatores ambientais são
responsáveis pela máxima variação entre as características da vegetação
(Martin et al., 2003).
4.6.1 Grupo 1 – Formação Podocarpus
A partir das matrizes de espécies e variáveis ambientais, o programa
CANOCO produziu uma ordenação com dois eixos, onde a distribuição
das espécies foi representada por pontos, enquanto que as variáveis
ambientais foram representadas através de setas indicando a direção do
gradiente máximo desta, sendo o comprimento da seta proporcional à
correlação da variável com os eixos.
Os resultados da análise de correspondência canônica estão

173
apresentados na Tabela 27 e Figura 6.
Os autovalores para os dois primeiros eixos de ordenação canônica foram
0,158 (eixo 1), 0,094 (eixo 2) e 0,071 (eixo 3), estes valores são
considerados baixos, ou seja, as espécies se distribuem igualmente por
todo o grupo, com algumas delas variando apenas na abundância relativa
(Botrel et al., 2002).
TABELA 27 – Coeficiente de correlação entre as variáveis ambientais e os dois
primeiros eixos de ordenação da análise de correspondência canônica do Grupo 1.
FLONA de São Francisco de Paula, RS.
Variáveis ambientais Eixo 1 Eixo 2
Eixo 3
Umidade -0,040 0,152 -0,254 Inclinação -0,177 -0,143 -0,157 Profundidade do solo 0,439 -0,114 -0,227 Cor do solo (0-50) 0,022 0,023 -0,044 Cor do solo (50-75) 0,215 -0,012 -0,068 Cor do solo (> 75) 0,263 -0,146 -0,135 Exposição -0,225 -0,182 -0,117 Quantidade de pedra 0,028 -0,017 -0,161 Densidade do sub-bosque -0,025 -0,333 0,399 Área Basal -0,150 0,066 0,026 Freqüência -0,180 -0,371 -0,203
Os dois primeiros eixos apresentaram uma variância global dos dados de
6,8%, que segundo Botrel et al. (2002) é muito baixa e indica não ocorrer
variância do remanescente, sendo o primeiro eixo explicando 3,3%, o segundo
2,0%, e o terceiro 1,5% da variância total dos dados. Apesar disso, a significância
das relações espécie-ambiente não foi prejudicada, pois o teste de permutação de
Monte Carlo indicou altas correlações espécie-ambiente nos dois primeiros eixos:

174
0,652 (eixo 1), 0,675 (eixo 2) e 0,626 (eixo 3) (Martins et al., 2003). O teste
mostrou que as correlações entre a abundância das espécies nas parcelas e as
variáveis ambientais foram significativas ao nível de 5% de probabilidade.
Observa-se, de acordo com a Tabela 27, que as variáveis mais fortemente
correlacionadas com o primeiro eixo foram: profundidade do solo, cor do solo nas
profundidades de 50-75 cm e maior que 75 cm, exposição, inclinação do terreno,
freqüência e área basal. Para o segundo eixo destacam-se umidade, densidade do
sub-bosque, freqüência exposição e exposição, e já para o terceiro eixo foram
densidade do sub-bosque, umidade, inclinação, profundidade do solo e área basal.
Aru ang
Ban par
Ble salCal com
Cas dec
Cry asc
Cup ver
Ile bre
Ile par
Lam ter
Lit bra
M yr mie
Myr gigM yr tem
Myr at r
M yr lor
Myr umb
Nec lan
Nec meg
Oco pul
Pod lam
Pru myr
Sap gla
Sip rei
Sym uni
Ver disZan rho
Mor
Eug uru
Myr f loGor acu
Cin amo
Sym tet
May evo
resltado1
Axis 1
Axis 2
Fr
I
C>75

175
FIGURA 6 – Diagrama de ordenação produzido pela análise de correspondência
canônica, baseado nos dados das espécies mais abundantes do Grupo 1. FLONA
de São Francisco de Paula, RS
Pela análise dos valores positivos e negativos das correlações deduz-se
que este grupo, que tem Podocarpus lambertii como espécie preferencial
ocorre em locais altos, inclinados, expostos, com solos profundos e bem
drenados.
Pode-se observar, também, pela Figura 6, que outras espécies como
Ocotea pulchella, Blepharocalyx salicifolius, Araucaria angustifolia,
Lithraea brasiliensis, Nectandra megapotamica, Siphoneugena reitzii,
Myrsine lorentziana e Myrciaria floribunda apresentam as mesmas
exigências a estes tipos de habitats.
Segundo Kent & Coker (1992) o comprimento da seta é proporcional à
magnitude de mudança naquela direção. Aqueles fatores ambientais onde
a seta é longa estão mais correlatos na ordenação do que aquelas com
seta curta e são mais importantes na influência da variação da
comunidade. A ordem em que se apresentam os pontos projetados sobre
a seta, da origem à extremidade, indicam a influência do fator ambiental
em relação à espécie, isto é, quanto mais próximo da extremidade da
seta, maior influência do fator ambiental sobre a espécie.
Através da Análise Cluster (TWINSPAN) determinou-se que as
espécies indicadoras do grupo 1 foram Podocarpus lambertii e Myrciaria
floribunda. Segundo a Análise de Correspondência Canônica os fatores
ambientais que têm influência na ocorrência das espécies são: cor do solo
na profundidade > 75 cm influência negativa; inclinação, profundidade do
solo de forma positiva e freqüência.
Observa-se que este grupo ocorreu em solos profundos, cujo solo
na profundidade entre 50-75 cm e > 75 cm apresentaram cores
avermelhadas. Segundo Vieira et al. (1988), esse tipo de solo apresenta

176
compostos de ferro e são bem drenados. Apresentam baixa quantidade
de pedra e, conseqüentemente, baixa retenção de água. São, portanto,
solos bem drenados (Lima, 1989).
A profundidade do solo está relacionada com a espessura máxima
do solo em que o sistema radicular das plantas não encontra dificuldade
ou barreira física para penetrar livremente, facilitando a sua fixação e
servindo de meio para a absorção de água e de nutrientes (Lepsch,1993;
Sanchez,1981, Liberman et al., 1985).
Pela Análise de Correspondência Canônica pode-se conhecer os
fatores ambientais que mais influenciaram para a ocorrência das espécies
no grupo 1. Deduz-se então que as espécies encontradas neste grupo
têm como fatores limitantes para sua ocorrência solos ricos em ferro, bem
drenados e profundos.
4.6.2 Grupo 2 – Formação Sebastiania
Os resultados da análise de correspondência canônica estão
apresentados na Tabela 28 e Figura 7.
TABELA 28 – Coeficiente de correlação entre as variáveis ambientais e
os dois primeiros eixos de ordenação da análise de correspondência
canônica do Grupo 2. FLONA de São Francisco de Paula, RS
Variáveis ambientais Eixo 1 Eixo 2 Eixo 3
Umidade 0,130 -0,513 - 0,100 Inclinação -0,057 0,058 - 0,495 Profundidade do solo -0,492 -0,027 -0, 075 Cor do solo (0-50) 0,062 0,261 0,015 Cor do solo (50-75) -0,132 0,127 0,027 Cor do solo (> 75) -0,250 -0,174 - 0, 093 Exposição -0,188 0,215 - 0,147 Quantidade de pedra 0,592 -0,074 0,120 Densidade do sub-bosque 0,090 0,195 - 0,249

177
Área Basal -0,011 0,197 0,065 Freqüência -0,038 -0,074 0,240
Os autovalores para os eixos de ordenação canônica foram 0,177 (eixo
1), 0,111 (eixo 2) e 0,101 (eixo 3). São considerados baixos, ou seja, as
espécies se distribuem igualmente por todo o grupo, com algumas delas
variando apenas na abundância relativa (Botrel et al., 2002).
Os eixos apresentaram uma variância global dos dados de 7,7%
que, segundo os autores, é muito baixa e indica não ocorrer variância do
remanescente, sendo o primeiro eixo explicando 1,6%, o segundo 2,6% e
o terceiro 3,5% da variância total dos dados. Apesar disso, a significância
das relações espécie-ambiente não foi prejudicada, pois, o teste de
permutação de Monte Carlo indicou altas correlações espécie-ambiente
nos eixos foram: 0,683 (eixo 1), 0,627 (eixo 2) e 0,574 (eixo 3) (Martins et
al., 2003). O teste mostrou que as correlações entre a abundância das
espécies nas parcelas e as variáveis ambientais foram significativas ao
nível de 5% de probabilidade.
A ll edu
A ru ang
Asp par
Ban par
B le sal
Cal com
Com rom
Cam xanCas dec
Cel iguCry asc
Cup ver
Das sp i
Das t om
Eug inv
Ile b re
Ile par
Lam ter
Lon cam
M at ela
M yr cucM yr mie
M yr pug
M yr at r Nec meg Oco pub
Oco pul
Pil pen
Pod lam
Pru myr
Ro l sal
Rou b ra
Sap g la
St i opp
Ver d isWei pauM or
Oco lanCun inc
M yr delM yr o li
Eug uru
M yr oxy
Cipos
Gor acu
Oco ind
Sym tet
Eug psi
Cry mos
P.SQ.P
0 10 20 30 40
20
40
Axis 1
Axis 2
All edu
Aru ang
Asp par
Ban par
Ble sal
Cal com
Com rom
Cam xanCas dec
Cel iguCry asc
Cup ver
Das spi
Eug inv
Ile bre
Ile par
Lam ter
Lon cam
Mat ela
Myr pug
Myr atr Nec meg Oco pub
Oco pul
Pil pen
Pod lam
Pru myr
Rou bra
Sap gla
Sti opp
Oco lanCun inc
Myr delMyr oli
Eug uru
Myr oxy
Cipos
Gor acu
Oco ind
Sym tet
Eug psi
Cry mos
P.S 20
40
Axis 1
Axis 2

178
FIGURA 7 – Diagrama de ordenação produzido pela análise de correspondência
canônica, baseado nos dados das espécies mais abundantes do Grupo 2. FLONA
de São Francisco de Paula, RS
A análise do Twinspan apresentou Sebastiania commersoniana,
Sebastiania brasiliensis e Cryptocarya aschersoniania como as espécies
indicadoras do grupo. A primeira espécie, Sebastiania commersoniana,
está restrita a solos rochosos úmidos e onde a vegetação é mais esparsa,
depressões de terrenos, nascentes e outros locais muito úmidos (Smith et
al., 1988).
Segundo os autores a espécie é mais freqüente nos sub-bosques
dos pinhais e nos capões, devido haver mais luz no seu interior, tendo a
sua maior densidade nas matas de galeria do planalto, onde os rios senis
apresentam planícies aluviais largas, extremamente úmidas e que durante
as enchentes ficam inundados.
Sebastiania brasiliensis é uma espécie de luz difusa higrófita, desenvolve-
se preferencialmente no interior dos capões e dos sub-bosques dos
pinhais situados em solos úmidos, beira dos rios e regatos, na restinga

179
arbustiva do litoral, localizado, em pequenas depressões (Smith et al.,
1988).
Já a Cryptocarya aschersoniana é uma espécie seletiva higrófita,
freqüente ao longo dos rios, planícies aluviais, início de encostas, solos
úmidos e mais profundos (Vattimo, 1979). Encontra-se nos sub-bosques
dos pinhais, sobretudo em solos bastante fertéis e em terrenos
medianamente ondulados (Smith et al., 1988).
As variáveis mais fortemente correlacionadas com o primeiro eixo foram:
quantidade de pedra, profundidade do solo, cor do solo na profundidade
maior que 75 cm, exposição, cor do solo na profundidade entre 50-75 cm
e umidade. O segundo eixo apresentou as variáveis umidade, cor do solo
em todas as profundidades, exposição, área basal e densidade do sub-
bosque. Já o terceiro eixo, apresentou as variáveis: umidade, inclinação,
exposição, densidade do sub-bosque e freqüência.
Pela análise dos valores positivos e negativos das correlações deduz-se
que este grupo, que tem as espécies de Sebastiania como preferenciais
ocorre em locais planos, expostos, com solos bastante úmidos, às vezes
rasos e pedregosos (onde ocorre Sebastiania brasiliensis) e às vezes
profundos (onde ocorre Sebastiania commersoniana).
Pode-se observar, também, pela Figura 7, que outras espécies como
Myrsine lorentziana, Cryptocarya aschersoniana, Vernonia discolor,
Myrciaria floribunda, Myrceugenia cucullata, Ilex brevicuspis, Casearia
decandra, Ilex paraguariensis, Dicksonia sellowiana, Roupala brasiliensis,
entre outras, apresentam as mesmas exigências a estes tipos de habitats.
Foi observada alta umidade nas parcelas do grupo que, segundo
Silva Júnior (1998) e Pinto (1997) correspondem a solos úmidos com
teores mais altos de MO, Al, H +, P, Fe, Cu, Zn, CEC e saturação de Al e
os maiores conteúdos de areia fina e grossa. Foi também observada uma
grande quantidade de pedra nestes locais. Segundo Vieira et al. (1988)
quanto maior a quantidade de pedra maior será a retenção de água no
solo. Solos profundos facilitam o sistema radicular das espécies penetrar

180
livremente e a absorver água e nutrientes. Bazzaz & Pickett (1979), dizem
que qualquer tipo de vegetação com solos profundos possuem uma maior
concentração de água a partir de 50 cm de profundidade.
Este grupo representa habitats caracterizados por apresentarem
solos úmidos profundos e com alta quantidade de pedras. São locais
especiais, limitantes para o desenvolvimento de um grande número de
espécies.
4.6.3 Grupo 3 – Formação Araucaria
A análise do Twinspan apresentou a Araucaria angustifolia como a
espécie indicadora do grupo. Segundo Reitz et al. (1988) a espécie é
encontrada desde uma altitude média de 500 metros, em terrenos
medianamente ondulados. O pinheiro ocupa preferencialmente as
depressões dos campos, curso d’água, onde iniciam os capões e matas
de galeria, que se estendem por quase todos os campos e encontram-se
nas mais variadas formas de aglomeração e desenvolvimento, desde os
capões de poucos exemplares, até as pujantes matas de galeria ao longo
dos rios maiores.
Segundo os autores, o pinheiro é uma espécie pioneira, muito apropriada
para o reflorestamento em campo aberto, porém, devem-se fazer mais
estudos em relação ao seu “habitat”, principalmente em relação à
qualidade e profundidade dos solos, ao clima, entre outros. Daí, a
importância do presente estudo que visa a determinar as condições
propícias de sítios para a espécie.
Os resultados da análise de correspondência canônica para o grupo 3
(Formação Araucaria) estão apresentados na Tabela 29 e Figura 8.

181
TABELA 29 – Coeficiente de correlação entre as variáveis ambientais e os dois
primeiros eixos de ordenação da análise de correspondência canônica do Grupo
3. FLONA de São Francisco de Paula, RS
Variáveis ambientais Eixo 1 Eixo 2
Eixo3
Umidade 0,198 - 0,034 0,283 Inclinação 0,040 0,343 - 0,327 Profundidade do solo - 0,018 - 0,001 - 0,017 Cor do solo (0-50) - 0,053 - 0,367 - 0,324 Cor do solo (50-75) - 0,083 0,205 0,193 Cor do solo (> 75) 0,007 0,169 0,193 Exposição 0,061 - 0,052 - 0,071 Quantidade de pedra 0,090 0,232 - 0,010 Densidade do sub-bosque 0,099 0,223 - 0,213 Área Basal 0,125 - 0,181 0,011 Freqüência 0,551 - 0,075 0,058
Os autovalores para os dois primeiros eixos de ordenação canônica foram
0,157 (eixo 1), 0,112 (eixo 2) e 0,110 (eixo 3), valores considerados
baixos, ou seja, as espécies se distribuem igualmente por todo o grupo,
com algumas delas variando apenas na abundância relativa (Botrel et al.,
2002).

182
FIGURA 8 – Diagrama de ordenação produzido pela análise de correspondência
canônica, baseado nos dados das espécies mais abundantes do Grupo 3. FLONA
de São Francisco de Paula, RS
Os eixos apresentaram uma variância global dos dados de 3,1%
que, segundo os autores, é muito baixa e indica não ocorrer variância do
Aru ang
Asp par
Ban par
Ban tomBle sal
Com rom
Cam xan
Cas dec
Cit gon
Cry asc
Cup ver
Das spi
Das tom
Dic sel
Eug inv
Acc sel
Hen omp
Ile bre
Ile dum
Ile par
Ing ver
Lam ter
Lon cam
Lue div
Mac par
Mat ela
May aqu
Myr cucMyr mie
Myr gig
Myr pug
Myr atr
Myr corMyr umbNec meg
Oco pub
Oco pulPil pen
Pod lam
Pru myr
Rol rug
Rol sal
Sap gla Scu bux
Seb braSeb com
Sip rei
Sol sp
Sol pse
Sol san
Sti opp
Ver dis
Xyl pse
Zan rho
Mor
Mac gla
Oco lan
NI
Pic par
Eug uru
Myr flo
Cipos
Cin gla
Eug psi
Cry mos
May evo
Sol pab
Cit myr
Cas obl
U.I.
C 0_50
0
0
40 80
20
40
60
80
Axis 1
Axis 2
FqDs

183
remanescente, sendo o primeiro eixo explicando 1,3%, o segundo 0,9% e
o terceiro 0,9% da variância total dos dados. Apesar disso, a significância
das relações espécie-ambiente não foi prejudicada, pois o teste de
permutação de Monte Carlo indicou altas correlações entre as espécies e
as variáveis ambientais nos eixos: 0,596 (eixo 1), 0,568 (eixo 2) e 0,580
(eixo 3). O teste mostrou que a correlação entre a abundância das
espécies nas parcelas e as variáveis ambientais foram significativas ao
nível de 5% de probabilidade.
As variáveis mais fortemente correlacionadas com o primeiro eixo foram:
freqüência, umidade do solo e área basal. Para o segundo eixo, destaca-
se a inclinação, cor do solo na profundidade ente 0-50 cm, cor do solo nas
profundidades de 50-75 cm e maior que 75 cm, densidade do sub-
bosque, quantidade de pedra e área basal. E para o segundo eixo foram
umidade, inclinação, cor do solo na profundidade ente 0-50 cm, cor do
solo nas profundidades de 50-75 cm e maior que 75 cm e densidade do
sub-bosque.
Pela análise dos valores positivos e negativos das correlações deduz-se
que este grupo, que tem Araucaria angustifolia como espécie preferencial
ocorre em locais inclinados com solos pouco profundos, bem drenados e
com alta concorrência.
Pode-se observar, também, pela Figura 8, que outras espécies como
Luehea divaricata, Dasyphyllum tomentosum, Blepharocalyx salicifolius,
Ilex paraguariensis, Banara tomentosa, Banara parviflora, Lamanonia
ternata, Dicksonia sellowiana, Sebastiania brasiliensis, entre outras,
apresentam as mesmas exigências a estes tipos de habitats.
Este grupo, denominado Formação Araucaria apresentou, como
fatores ambientais limitantes a umidade, inclinação, cor do solo na
profundidade entre 0-50 cm, freqüência e densidade do sub-bosque.
A cor do solo na profundidade entre 0-50 cm, nas parcelas que
caracterizam o grupo, é vermelho-amarelo, ou seja, tem uma hidratação
média dos compostos de ferro (Vieira et al., 1988). A inclinação varia de

184
ondulada a altamente ondulada, isto é, inclinação entre 200 – 450 que,
segundo Botrel et al. (2002), determina uma boa drenagem e estão
relacionadas com o transporte de material (Fe, P, K, Ca e Mg) das partes
mais altas para as partes mais baixas das parcelas, tornando os solos,
nas baixadas, com uma maior condição de fertilidade. Péllico Netto et al.
(2002), em estudos de serapilheira em Floresta Ombrofila Mista,
detectaram variações estacionais ao longo do ano nos teores desses
elementos.
O grupo ocorre em solos com umidade média. Segundo Van Der
Berg (1995) são solos com teores altos de argila e silte. Zahner (1967)
afirma que nestes solos a água é removida mais prontamente da região
de maior concentração de raízes uma vez que esta concentração é
inversamente proporcional à profundidade do solo, sendo maior na
superfície onde há uma melhor aeração e maior concentração de
nutrientes.
4.7 Regressão Logística
Para ratificar os resultados da análise de correspondência canônica, em relação aos fatores ecológicos que determinaram a ocorrência das espécies nas parcelas, realizou-se para cada espécie indicadora dos grupos uma análise de regressão logística. Por esta análise foi possível determinar quais fatores são determinantes na ocorrência das espécies.
O modelo geral testado para explicar a ocorrência das espécies
indicadoras dos grupos foi:
.)..........( )75()7550()500(1
1lFreqkGSjDPiQhExpSgCSfCSeCSdPcInclbUa
eP +++++++++++− >−−+=
O modelo foi testado pelo método FORWARD STEPWISE, tendo
como variável dependente a presença ou ausência das espécies e como

185
variáveis independentes a umidade, inclinação do terreno, profundidade
do solo, cor do solo na profundidade de 0-50 cm, cor do solo na
profundidade de 50-75 cm, cor do solo na profundidade maior que 75 cm,
exposição do terreno, quantidade de pedras, densidade do sub-bosque,
área basal e distribuição de freqüência.
4.7.1 Podocarpus lambertii (Pinheiro-bravo)
Para a ocorrência de Podocarpus lambertii foram significativas as
variáveis: freqüência, quantidade de pedras e cor do solo na profundidade
maior que 75 cm.
Desta forma, o modelo específico para estimativa da probabilidade
de ocorrência de Podocarpus lambertii, na Floresta Ombrófila Mista da
FLONA de São Francisco de Paula, ficou assim definido:
).(.. )75(1
1>+++−+
=dCordosolocQPbFreqalambPod
eP
Na tabela de classificação da regressão logística para a variável
dependente “presença de Podocarpus lambertii” (Tabela 30), pode-se
verificar que as 903 parcelas sem a presença de Podocarpus lambertii
(valor 0) foi corretamente estimada pelo modelo como não tendo a
espécie. Similarmente, 3 parcelas com presença de Podocarpus lambertii
(valor 1) foi corretamente estimada como tendo a espécie. A diagonal
secundária da tabela mostra quantas parcelas foram incorretamente
classificadas, ou seja, (88+6=94). Das parcelas sem a presença da
espécie, 99,34% foram corretamente classificadas e das parcelas com
pelo menos uma árvore de Podocarpus lambertii, 3,30% foram
corretamente classificadas. No total, 90,60% das parcelas foram
corretamente classificadas.

186
TABELA 30 - Tabela de classificação da regressão logística para a
variável dependente Podocarpus lambertii.
Estimada
0 1 Percentagem correta
Observada
0 903 6 99,34%
1 88 3 3,30%
Total 90,60%
Os coeficientes logísticos e demais estatísticas da seleção das
variáveis independentes do modelo, isto é, quando a freqüência (Freq.),
quantidade de pedra (QP) e cor do solo na profundidade for maior que 75
cm (Cor do solo( >75)) são incluídos no modelo e podem ser observados na
Tabela 31.
TABELA 31 - Variáveis independentes selecionadas pela regressão
logística pelo método Forward Stepwise, para Podocarpus lambertii.
Variáveis B E.P. WALD GL Sig. R Exp. (B)
Freqüência 0,2102 0,0328 40,9710 1 0,000 -0,1714 0,2564
Quantidade de pedra
-1,3490 0,2898 21,6631 1 0,000 -0,1796 0,2595
Cor do solo (>75) -1,3609 0,3049 19,9188 1 0,000 -0,1714 0,2564
Constante -2,2724 0,4751 22,8742 1 0,000 - -
Como os níveis de significância observada, dos coeficientes para

187
as variáveis no modelo, foram menores que 0,05, estas variáveis foram
significativas e se mantêm no modelo. Assim, o modelo definitivo pode ser
escrito como da seguinte maneira:
)3609,13490,1.2102,02724,2(.. )75(1
1>−−+−−+
=CSQPFreqlambPod
eP
Observa-se que, dos fatores que influenciaram a presença do
Podocarpus lambertii na Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, a distribuição da freqüência das espécies atuou de
forma positiva, ou seja, quando a freqüência é alta as chances de ter a
espécie aumentam. Entretanto, o baixo valor do coeficiente B para a
freqüência indica que sua influência para a presença do Podocarpus não
é muito significante. O resultado pode ser deduzido pela observação do
Exp (B). Com o aumento de uma unidade da freqüência, a chance de
ocorrência de Podocarpus aumenta apenas 0,25 vezes. Além disso, deve-
se considerar que quando a espécie (Podocarpus) está presente, a
freqüência é alta justamente porque a presença dela ocasiona este
aumento.
A quantidade de pedras e a cor do solo na profundidade maior que
75 cm determinam uma menor probabilidade de ocorrência de
Podocarpus lambertii na área. Isto pode ser deduzido por serem
significantes e pelos valores negativos do coeficiente B. Pela
interpretação do Exp (B) as chances de ocorrência de Podocarpus em
solo pedregoso e cor do solo na profundidade maior que 75 cm são de
aproximadamente 0,25 vezes, ou seja, 75% menor do que em solo sem
pedras ou com cores não avermelhadas.
Pode-se observar que a cor do solo>75 (vermelha) e a quantidade
de pedra, são os principais fatores ecológicos que determinam a
ocorrência da espécie nas parcelas do grupo 1, de forma negativa, isto é,
suas chances de ocorrerem em solos pedregosos e vermelhos, na

188
profundidade maior que 75 cm, são menores. A influência destes fatores
foi determinada tanto na Análise de Correspondência Canônica quanto na
Análise de Regressão Logística.
Por outro lado, pode-se afirmar que as variáveis que não foram
selecionadas no modelo, como umidade, inclinação, profundidade do
solo, cor do solo de 0-50 cm e 50-75 cm, exposição, densidade do sub-
bosque e área basal, provavelmente não estão ligadas ou têm pouca
influência na presença de Podocarpus lambertii na Floresta Ombrófila
Mista da FLONA de São Francisco de Paula estudada.
4.7.2 Araucaria angustifolia (Pinheiro-brasileiro)
O modelo de regressão logística para a espécie foi testada tendo
como variável dependente a presença ou ausência de Araucaria
angustifolia e como independente a umidade, inclinação, profundidade do
solo, cor do solo (0-50), cor do solo (50-75), cor do solo (>75), exposição,
quantidade de pedra, densidade do sub-bosque, área basal e freqüência.
Foram significativas as variáveis Área basal (G), cor do solo na
profundidade de 0-50 cm (CS(0-50), cor do solo na profundidade maior que
75 cm (CS(>75)), Umidade (U) e Freqüência (Freq). Desta forma, o modelo
específico para a estimativa da probabilidade de ocorrência do pinheiro-
brasileiro no local do estudo ficou resumido a:
.)...(.. )75()500(1
1fFreqeUSdCScCbGaangArau
eP +++++− >−+
=
Na tabela de classificação da regressão logística para a variável
dependente “presença de Araucaria angustifolia” (Tabela 32), pode-se
verificar que as 481 parcelas sem a presença de Araucaria angustifolia
(valor 0) foi corretamente estimada pelo modelo como não tendo a
espécie. Similarmente, 180 parcelas com presença de pinheiro (valor 1)

189
foi corretamente estimada como tendo a espécie. A diagonal secundária
da tabela mostra quantas parcelas foram incorretamente classificadas, ou
seja, (237+102=339). Das parcelas sem a presença da espécie, 83,50%
foram corretamente classificadas e das parcelas com pelo menos uma
árvore de Araucaria angustifolia, 43,17% foram corretamente
classificadas. No total, 66,10% das parcelas foram corretamente
classificadas.
TABELA 32 - Tabela de classificação da regressão logística para a
variável dependente Araucaria angustifolia.
Estimada
0 1 Percentagem correta
Observada
0 481 102 83,50%
1 237 180 43,17%
Total 66,10%
Os coeficientes logísticos e demais estatísticas da seleção das
variáveis independentes do modelo, isto é, quando a área basal, cor do
solo nas profundidades de 0-50 cm e maior que 75 cm, umidade e
freqüência do sub-bosque são incluídos no modelo, podem ser
observados na Tabela 33.
TABELA 33 - Variáveis independentes selecionadas pela regressão
logística pelo método Forward Stepwise, para Araucaria angustifolia.
Variáveis B E.P. WALD GL Sig. R Exp. (B)
Área basal 2,5420 0,3146 65,2915 1 0,000 0,2158 12,7048
Cor do solo (0-50) 0,6304 0,1030 37,4846 1 0,000 0,1616 1,8784
Cor do solo (>75) -0,4730 0,1041 20,6433 1 0,000 -0,1171 0,6231
Umidade 0,3371 0,1160 8,4429 1 0,0037 0,0689 1,4009
Freqüência -0,0427 0,0216 3,9145 1 0,0479 -0,0375 0,9582
Constante -2,2471 0,3295 46,5114 1 0,000 - -

190
Como os níveis de significância observada dos coeficientes para as
variáveis no modelo foram menores que 0,05, estas variáveis foram
significativas e se mantém no modelo. Assim, o modelo definitivo pode ser
escrito como segue:
.)0427,0.3371,0.4730,0..6304,05420,22471,2(..)75()500(1
1FreqUSCSCGangArau
eP
−+−++−− >−+=
Observa-se que dos fatores que influenciaram a presença da
Araucaria angustifolia na Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, a área basal das espécies atuou de forma positiva,
ou seja, quando a área basal é alta as chances de ter a espécie
aumentam. O valor relativamente alto do coeficiente B para a área basal
indica que sua influência para a presença da espécie é significante. O
resultado pode ser deduzido pela observação do Exp (B). Com o aumento
de uma unidade da freqüência, a chance de ocorrência de Araucaria
aumenta 12,70 vezes. Além disso, deve-se considerar que quando a
Araucaria angustifolia está presente, a área basal é alta justamente
porque a presença dela ocosiona este aumento, devido às características
dominantes (grandes diâmetros) da espécie.
A cor so solo na profundidade de 0-50 cm e a umidade também
atuaram de forma positiva na distribuição da Araucaria na área.
Entretanto os valores baixos dos coeficientes B indicam que sua
influência na ocorrência da espécie não é tão significante, comprovado
também pelo baixo Exp (B). Com o aumento de uma unidade na cor do
solo na profundidade de 0-50 cm e na umidade, a chance de ocorrência
da Araucaria aumenta 1,87 e 1,40 vezes, respectivamente.
A cor do solo na profundidade maior que 75 cm e a densidade do
sub-bosque são também importantes na ocorrência de Araucaria
angustifolia na área, embora em menor proporção. Isto pode ser deduzido
por serem significantes e pelos valores negativos do coeficiente B. Pela

191
interpretação do Exp (B) as chances de ocorrência de Araucaria em solos
de cor vermelha na profundidade maior que 75 cm são de
aproximadamente 0,60 vezes e de ocorrência em locais de freqüência alta
são de 0,95 vezes.
Pode-se observar que cor do solo na profundidade de 0-50 cm
(vermelho-amarelo) com média hidratação dos compostos de ferro e
umidade baixa determinada pelo solo bem drenado, são os principais
fatores ecológicos que determinam a ocorrência da Araucaria nas
parcelas do grupo 2. Também tiveram influência de forma negativa a cor
vermelha do solo na profundidade maior que 75 cm e a freqüência. A
influência destes fatores foi determinada tanto na Análise de
Correspondência Canônica quanto na Análise de Regressão logística,
como fatores limitantes para a ocorrência da espécie no grupo.
Por outro lado, pode-se afirmar que as variáveis que não foram
selecionadas no modelo (profundidade do solo, cor do solo na
profundidade de 50-75 cm, exposição, quantidade de pedra e densidade
do sub-bosque) provavelmente não estão ligadas ou tem pouca influência
na presença de Araucaria angustifolia na Floresta Ombrófila Mista da
FLONA de São Francisco de Paula estudada. A ocorrência de pinheiro-
brasileiro é indiferente a estas características nas condições em que foi
executado o estudo.
4.7.3 Cryptocarya aschersoniana (Canela-vick)
Para esta espécie foram significativas as variáveis: área basal (G),
exposição (Exp.), quantidade de pedra (QP) e cor do solo na
profundidade maior que 75 cm (CS(>75)). Portanto, o modelo específico
para a estimativa da probabilidade de ocorrência de Cryptocarya
aschersoniana, no local do estudo ficou assim:

192
)....(.. )75(1
1>++++−+
=SeCPdQcExpbGaaschCryp
eP
Na tabela de classificação da regressão logística para a variável
dependente “presença de Cryptocarya aschersoniana” (Tabela 34), pode-
se verificar que 813 parcelas sem a presença de Cryptocarya
aschersoniana (valor 0) foram corretamente estimadas pelo modelo como
não tendo Cryptocarya aschersoniana. Já nenhuma parcela com
presença de Cryptocarya aschersoniana (valor 1) foi corretamente
estimada com tendo a espécie. A diagonal secundária da Tabela 34
mostra quantas parcelas foram incorretamente classificadas, ou seja, 187.
Das parcelas sem a presença da espécie, 100% foram corretamente
classificadas e das parcelas com pelo menos uma Cryptocarya
aschersoniana 0% foram corretamente classificadas. No total, 81,30% das
parcelas foram corretamente classificadas.
TABELA 34 - Tabela de classificação da regressão logística para a
variável dependente Cryptocarya aschersoniana.
Estimada
0 1 Percentagem correta
Observada
0 813 0 100,00%
1 187 0 0,00%
Total 81,30%
Os coeficientes logísticos e demais estatísticas da seleção das
variáveis independentes do modelo, isto é, quando a área basal,
exposição, quantidade de pedra e cor do solo na profundidade maior que
75 cm são incluídos no modelo, podem ser observados na Tabela 35.

193
TABELA 35 - Variáveis independentes selecionadas pela regressão
logística pelo método Forward Stepwise, para a Cryptocarya
aschersoniana
Variáveis B E.P. WALD GL Sig. R Exp. (B)
Área basal 0,8716 0,2977 8,5718 1 0,0034 0,0826 2,3907
Exposição -0,1102 0,0427 6,6513 1 0,0099 -0,0695 0,8957
Quantidade de pedra
0,2784 0,1035 7,2288 1 0,0072 0,0737 1,3210
Cor do solo (>75) 0,2869 0,1120 6,5620 1 0,0104 0,0688 1,3322
Constante -2,0130 0,2986 45,4517 1 0,000 - -
Como os níveis de significância observada dos coeficientes para as
variáveis foram menores que 0,05 de probabilidade, estas variáveis foram
significativas e se mantém no modelo. Assim, o modelo definitivo pode ser
escrito como segue:
).2869,0.2784,0.1102,08716,00130,2(.. )75(1
1>++−+−−+
=SCPQExpGaschCryp
eP
Observa-se que dos fatores que influenciaram a presença de
Cryptocarya aschersoniana na Floresta Ombrófila Mista da FLONA de
São Francisco de Paula, a área basal das espécies atuou de forma
positiva, ou seja, quando a área basal é alta as chances de ter a espécie
aumentam. O valor médio do coeficiente B para a área basal indica que
sua influência para a presença da espécie tem relativa significância. O
resultado pode ser deduzido pela observação do Exp (B). Com o aumento
de uma unidade da freqüência, a chance de ocorrência da espécie
aumenta 2,39 vezes. A importância da área basal deve-se à própria
presença de Cryptocarya aschersoniana, com seus grandes indivíduos
(grandes diâmetros).
A quantidade de pedra e a cor do solo na profundidade maior que
75 cm também atuaram de forma positiva na distribuição de Cryptocarya

194
aschersoniana na área. Entretanto os valores baixos dos coeficientes B
indicam que sua influência na ocorrência da espécie não é tão
significante, comprovado também pelo baixo Exp (B). Com o aumento de
uma unidade na quantidade de pedra e na cor do solo na profundidade
maior que 75 cm, a chance de ocorrência da espécie aumenta 1,32 e 1,33
vezes, respectivamente.
A exposição é também importante na ocorrência de Cryptocarya
aschersoniana na área, embora em menor proporção. Isto pode ser
deduzido por ser significante e pelo valor negativo do coeficiente B. Pela
interpretação do Exp (B) as chances de ocorrência da espécie em locais
mais expostos são de, aproximadamente 0,89 vezes.
Pode-se observar que a quantidade de pedra e a cor avermelhada
do solo na profundidade maior que 75 cm foram os principais fatores
ecológicos que determinaram a ocorrência de Cryptocarya aschersoniana
nas parcelas do grupo 3, além da área basal, que é um fator de
concorrência. Também, teve influência de forma negativa a exposição,
indicando que a espécie não se adapta a terrenos muito expostos. A
influência da quantidade de pedras foi também determinada na Análise de
Correspondência Canônica, como fator limitante para a ocorrência da
espécie no grupo.
As outras variáveis que não foram selecionadas (umidade,
inclinação, profundidade do solo, cor do solo nas profundidades de 0-50
cm e 50-75 cm, densidade do sub-bosque e freqüência) pelo modelo,
provavelmente não estão ligadas ou têm pouca influência na presença de
Cryptocarya aschersoniana na Floresta Ombrófila Mista da FLONA de
São Francisco de Paula. A ocorrência da espécie é indiferente a estas
características nas condições em que foi executado o estudo.
4.7.4 Sebastiania commersoniana (Branquilho-comum)
Para esta espécie foram significativas as variáveis: profundidade

195
do solo (PS), densidade do sub-bosque (DS) e umidade (U). O modelo
específico para a estimativa da probabilidade de ocorrência de
Sebastiania commersoniana no local do estudo ficou assim definido:
)...(.. 1
1dUScDSbPacommSeb
eP +++−+
=
Na tabela de classificação da regressão logística para a variável
dependente “presença de Sebastiania commersoniana” (Tabela 36),
pode-se verificar que 848 parcelas sem a presença de Sebastiania
commersoniana (valor 0) foram corretamente estimadas pelo modelo
como não tendo branquilho-comum. Já, 9 parcelas com presença de
Sebastiania commersoniana (valor 1) foram corretamente estimadas com
tendo a espécie. A diagonal secundária da Tabela 36 mostra quantas
parcelas foram incorretamente classificadas, ou seja, (138+5=143). Das
parcelas sem a presença da espécie, 99,41% foram corretamente
classificadas e das parcelas com pelo menos um indivíduo de
Sebastiania commersoniana 6,12% foram corretamente classificadas. No
total, 85,70% das parcelas foram corretamente classificadas.
TABELA 36 - Tabela de classificação da regressão logística para a
variável dependente Sebastiania commersoniana.
Estimada

196
0 1 Percentagem correta
Observada
0 848 5 99,41%
1 138 9 6,12%
Total 85,70%
Os coeficientes logísticos e demais estatísticas da seleção das
variáveis independentes do modelo, isto é, quando a profundidade do
solo, densidade do sub-bosque e umidade são incluídos no modelo,
podem ser observados na Tabela 37.
TABELA 37 - Variáveis independentes selecionadas pela regressão
logística pelo método Forward Stepwise, para Sebastiania
commersoniana
Variáveis B E.P. WALD GL Sig. R
Exp.
(B)
Profundidade do solo
1,0341 0,1425 52,6650 1 0,000 0,2463 2,8125
Densidade do sub-bosque
-0,7279 0,1602 20,6380 1 0,000 -0,1494 0,4829
Umidade 0,1119 0,0262 18,1938 1 0,000 0,1393 1,1185
Constante -3,1423 0,6069 26,8127 1 0,000 - -
Como os níveis de significância observada dos coeficientes para as
variáveis foram menores que 0,05, estas variáveis foram significativas e
se mantêm no modelo. Assim, o modelo definitivo pode ser escrito como
segue:
)1119,0.7279,0..0341,11423,3(.. 1
1USDSPcommSeb
eP +−+−−+
=
Observa-se que dos fatores que influenciaram a presença de

197
Sebastiania commersoniana na Floresta Ombrófila Mista da FLONA de
São Francisco de Paula, a profundidade do solo atuou de forma positiva,
ou seja, quando a profundiade do solo é alta as chances de ter a espécie
aumentam. O valor relativamente alto do coeficiente B para a
profundidade do solo indica que sua influência, para a presença da
espécie é significante. O resultado pode ser deduzido pela observação do
Exp (B). Com o aumento de uma unidade da freqüência, a chance de
ocorrência da espécie aumenta 2,81 vezes
Também a umidade do solo atua de forma positiva na distribuição
de Sebastiania commersoniana na área. Os valores um pouco menores
do coeficientes B indica que sua influência na ocorrência da espécie é um
pouco menor, comprovado pelo Exp (B). Com o aumento de uma unidade
na quantidade na umidade do solo, a chance de ocorrência da espécie
aumenta 1,11 vezes.
A densidade do sub-bosque é também importante na ocorrência de
Sebastiania commersoniana na área, embora em menor proporção. Isto
pode ser deduzido por ser significante e pelo valor negativo do coeficiente
B. Pela interpretação do Exp (B) as chances de ocorrência da espécie em
locais mais expostos são de aproximadamente 0,48 vezes.
Pode-se observar que a profundidade e umidade do solo são os
principais fatores ecológicos que determinam a ocorrência de Sebastiania
commersoniana nas parcelas do grupo 3. Também teve influência de
forma negativa a densidade do sub-bosque, indicando que a espécie não
tolera locais de maior adensamento. A influência desses fatores foi
também determinada na Análise de Correspondência Canônica, como
fator limitante para a ocorrência da espécie no grupo.
As outras variáveis que não foram selecionadas pelo modelo
(freqüência, inclinação, cor do solo nas diversas profundidades,
exposição, quantidade de pedra e área basal), provavelmente não estão
ligadas ou têm pouca influência na presença de Sebastiania
commersoniana na Floresta Ombrófila Mista da FLONA de São Francisco

198
de Paula. A ocorrência da espécie é indiferente a estas características
nas condições em que foi executado o estudo.
4.7.5 Sebatiania brasiliensis (Branquilho-leiteiro)
As variáveis significativas determinadas pela regressão logística
para a Sebatiania brasiliensis foram: esposição (Exp.), cor do solo na
profundidade de 0-50 cm (CS(0-50)), Freqüência (Freq.), Inclinação do
terreno (Incl.), quantidade de pedra (QP) e umidade (U). O modelo
específico para a estimativa da probabilidade de ocorrência de
Sebastiania brasiliensis, no local do estudo, ficou assim definido:
)...(.. )500(1
1gUfQPeIncldFreqcCSbExpabrasSeb
eP ++++++− −+
=
Na tabela de classificação da regressão logística para a variável
dependente “presença de Sebastiania brasiliensis” (Tabela 38), pode-se
verificar que 771 parcelas sem a presença de Sebastiania brasiliensis
(valor 0) foram corretamente estimadas pelo modelo como não tendo a
espécie. Já 16 parcelas com presença de Sebastiania brasiliensis (valor
1) foram corretamente estimadas com tendo a espécie. A diagonal
secundária da Tabela mostra quantas parcelas foram incorretamente
classificadas, ou seja, (198+15=213). Das parcelas sem a presença da
espécie, 98,09% foram corretamente classificadas e das parcelas com
pelo menos uma Sebastiania brasiliensis 7,48% foram corretamente
classificadas. No total, 78,70% das parcelas foram corretamente
classificadas.
Os coeficientes logísticos e demais estatísticas da seleção das

199
variáveis independentes do modelo, isto é, quando a exposição, cor do
solo na profundidade de 0-50 cm, quantidade de pedra, inclinação e
umidade são incluídos no modelo, podem ser observados na Tabela 39.
TABELA 38 - Tabela de classificação da regressão logística para a
variável dependente Sebatiania brasiliensis.
Estimada
0 1 Percentagem correta
Observada
0 771 15 98,09%
1 198 16 7,48%
Total 78,70%
TABELA 39 - Variáveis independentes selecionadas pela regressão
logística pelo método Forward Stepwise, para Sebatiania brasiliensis
Variáveis B E.P. WALD GL Sig. R Exp. (B)
Exposição -0,2116 0,0442 22,8637 1 0,000 -0,1417 0,8093
Cor do solo (0-50) 0,4414 0,1022 18,6669 1 0,000 0,1267 1,5549
Quantidade de pedra
0,0919 0,0235 15,3320 1 0,000 0,1133 1,0962
Inclinação -0,8800 0,2276 14,9515 1 0,000 -0,1117 0,4148
Umidade 0,2795 0,1298 4,6350 1 0,031 0,0504 1,3224
Constante -1,8002 0,4712 14,5958 1 0,000 - -
Como os níveis de significância observada dos coeficientes para as
variáveis no modelo foram menores que 0,05, estas variáveis foram
significativas e se mantêm no modelo. Assim, o modelo definitivo pode ser

200
escrito como segue:
)279,0.880,0092,0441,0.212,080,1(.. )500(1
1UInclQPCSEbrasSeb
eP +−++−−− −+
=
Observa-se que dos fatores que influenciaram a presença de
Sebastiania brasiliensis na Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, a cor do solo (vermelho-amarelo) na profundidade de
0-50 cm, umidade do solo e quantidade de pedra atuaram de forma
positiva, ou seja, os aumentos desses teores aumentam as chances de
ter a espécie. Os valores médios dos coeficientes B para esses elementos
indicam que suas influências para a presença da espécie têm relativa
significância. O resultado pode ser deduzido pela observação do Exp (B).
Com o aumento de uma unidade da cor do solo na profundidade de 0-50
cm, umidade e quantidade de pedra, a chance de ocorrência da espécie
aumenta 1,55, 1,32 e 1,09 vezes.
A exposição e inclinação são também importantes na ocorrência de
Sebastiania brasiliensis na área, embora em menor proporção. Isto pode
ser deduzido por ser significante e pelo valor negativo do coeficiente B.
Pela interpretação do Exp (B) as chances de ocorrência da espécie em
locais mais expostos são de aproximadamente 0,80 e o,41 vezes,
respectivamente.
Pode-se observar que a umidade do solo e a quantidade de pedra,
além da cor vermelha amarelada do solo nas profundidades ente 0-50 cm
são os principais fatores ecológicos que determinam a ocorrência de
Sebastiania brasiliensis nas parcelas do grupo 3. Também, teve influência
de forma negativa a exposição e inclinação do terreno, indicando que a
espécie não tolera locais expostos e inclinados. A influência desses
fatores foi também determinada na Análise de Correspondência
Canônica, como fatores limitantes para a ocorrência da espécie no grupo.
Conforme já citados por Silva Júnior (1998) e Pinto (1997) solos

201
úmidos apresentam teores mais altos de matéria orgânica, Al, P, Fe, Cu,
Zn. A saturação de Al é, possivelmente, o fator mais limitante ao
crescimento da maioria das espécies. A maior quantidade de pedras nos
solos desse grupo indica, segundo Vieira et al. (1988), uma maior
retenção de água.
As outras variáveis que não foram selecionadas pelo modelo
(profundidade do solo, cor do solo nas profundidades de 50-75 cm e
maior que 75 cm, densidade do sub-bosque, freqüência e área basal),
provavelmente não estão ligadas ou têm pouca influência na presença de
Sebastiania brasiliensis na Floresta Ombrófila Mista da FLONA de São
Francisco de Paula. A ocorrência da espécie é indiferente a estas
características nas condições em que foi executado o estudo.
4.8 Contribuição silvicultural
O bioma Floresta Ombrófila Mista, distribui-se em uma superfície
de cerca de 200.000 km2, ocorrendo 25% no Estado do Rio Grande do
Sul (Péllico Netto et al., 2002).
Devido as intervenções humanas, a diversidade dos
remanescentes florestais tem diminuído e, apesar de todas as leis e
fiscalizações, preservar integralmente os recursos naturais nos biomas
florestais é um enorme desafio.
A Floresta Ombrófila Mista é formada por uma vasta diversidade de
espécies de plantas, onde as espécies de porte arbóreo são dominantes.
A Araucaria angustifolia é a principal representante do bioma. Associada
a ela existe grande número de espécies.
O conhecimento sobre a composição florística, a estrutura
fitossociológica e as exigências ecológicas das espécies é fundamental

202
para o fornecimento de sibsídios para a aplicação de técnicas adequadas
de manejo silvicultural, visando aumentar a produção ou mesmo auxiliar
na sua conservação integral.
Com os resultados do presente trabalho é possivel afirmar que
para proceder um manejo na FLONA de São Francisco de Paula deve-se
considerar:
1) Os grupos determinados, que por sua vez representam as
situações ecológicas e florísticas locais distintas;
2) A composição florística dos distintos grupos, que fornecem
opção de escolha das espécies potenciais de utilização nos
planos de manejo;
3) As exigências ecológicas das espécies, ajuda preciosa para
efetivação de plantios de enriquecimento.
A Tabela 40 apresenta informações essenciais da real situação da
floresta da FLONA, que devem ser levadas em consideração na decisão
sobre a aplicação de planos de manejo silvicultural.
TABELA 40 – Características dos grupos florísticos obtidos na
FLONA de São Francisco de Paula, RS.
GRUPO FATORES AMBIENTAIS
ESPÉCIES POTENCIAIS
1
Solos ricos em ferro, drenados, pedregosos e inclinados
Podocarpus lambertii, Ocotea pulchella, Nectandra megapotamica, Cinnamomum amoenum, Cryptocarya aschersoniana, Ilex brevicuspis, Lamanonia ternata, Blepharocalyx salicifolius, Prunus myrtifolia, Roupala brasiliensis.
Sebastiania commersoniana, Sebastiania brasiliensis, Cryptocarya aschersoniana, Cryptocarya moschata, Blepharocalyx salicifolius, Ocotea pulchella, Ocotea indecora, Ocotea

203
2
Solos planos e profundos, úmidos e com alta quantidade de pedras
lancifolia, Nectandra megapotamica, Nectandra lanceolata, Cinammomum glaziovii, Cinammomum amoenum, Ilex brevicuspis, Ilex paraguariensis, Ilex theezans, Roupala brasiliensis, Prunus myrtifolia, Cupania vernalis, Matayba elaeagnoides, Luehea divaricata, Capsicodendron dinisii, Podocarpus lambertii, Erythrina cristagalli.
3
Solos secos, profundos, bem drenados .
Araucaria angustifolia, Podocarpus lambertii, Ilex paraguariensis, Ilex brevicuspis, Roupala brasiliensis, Nectandra megapotamica, Ocotea pulchella, Cryptocarya aschersoniana, Cryptocarya moschata, Cinnamomum glaziovii, Ocotea lancifolia, Endlicheria paniculata, Cinnamomum amoenum, Cedrela fissilis, Blepharocalyx salicifolius, Luehea divaricata, Matayba elaeagnoides, Cupania vernalis, Machaerium paraguariense, Lamanonia ternata, Prunus myrtifolia, Tabebuia umbellata, Tabebuia chrysotricha, Aspidosperma parvifolium.
Os resultados da pesquisa sugerem que nos locais onde
atualmente são realizados práticas de manejo em sistema silvipastoril,
manejo de araucária, clareiras naturais e provocadas por incêndios, sejam
recuperadas com a utilização das espécies indicadoras e das espécies
comuns entre os grupos (Anexo 1). São espécies fundamentais na
conservação, enriquecimento e recuperação desses locais. As espécies
poderão ser utilizadas, não somente nas áreas da FLONA, mas, também
em locais que apresentem as mesmas características ambientais, por
todo o Estado do Rio Grande do Sul, ou outros Estados nos quais a
Floresta Ombrófia Mista seja encontrada.
Além disso, segundo Araujo (2002), projetos de recuperação,
enriquecimento e conservação devem considerar as características da
área a ser trabalhada, utilizado-se espécies adaptadas a cada ambiente.
Daí, a importância dos resultados apresentados.

204
5 CONCLUSÕES e RECOMENDAÇÕES
5.1 Conclusões
Com os resultados apresentados no trabalho pode-se concluir que:

205
1) Foram encontradas na Floresta Ombrófila Mista da FLONA de
São Francisco de Paula, 130 espécies arbóreas pertencentes a
84 gêneros e 45 famílias botânicas, além de árvores não
identificados, árvores mortas e cipós;
2) A floresta apresentou alta diversidade, cujo índice de Shannon
foi de 3,1939;
3) As famílias Myrtaceae, Lauraceae, Fabaceae, Rutaceae e
Flacourtiaceae foram as mais representativas da floresta;
4) Araucaria angustifolia, Blepharocalyx salicifolius, Ilex
brevicuspis, Cryptocarya aschersoniana, Sebastiania
commersoniana, Ilex paraguariensis, Ocotea pulchella,
Sebastiania brasiliensis, Nectandra megapotamica, Dicksonia
sellowiana e Podocarpus lambertii foram as espécies mais
características e importantes da floresta;
5) A análise de agrupamento utilizando o TWINSPAN formou três
grandes grupos florísticos, que correspondem a situações
ecológicas distintas:
a) Grupo 1, denominado de Formação Podocarpus, ocorre em
locais altos e inclinados, com solos drenados ricos em ferro
e pedregosos, tem Podocarpus lambertii, Myrciaria tenella e
Eugenia uruguayensis como espécies indicadoras e
Lamanonia ternata, Lithraea brasiliensis, Vernonia discolor e
Myrciaria floribunda como espécies preferenciais;
b) Grupo 2, denominado Formação Sebastiania, ocorre em
locais expostos, planos, com solos úmidos, tem Sebastiania
commersoniana, Cryptocarya aschersoniana e Sebastiania
brasiliensis como espécies indicadoras e Blepharocalyx
salicifolius como espécie preferencial;
c) Grupo 3, denominado Formação Araucaria, ocorre em locais
planos, com solos drenados e profundos, tem Araucaria

206
angustifolia e Ilex paraguariensis como espécies indicadoras
e Banara parviflora como espécie preferencial;
6) Foram necessárias duas funções discriminantes para
representar os grupos obtidos pela análise de agrupamento,
com 91,2 % de classificações corretas;
7) No grupo 1 foram encontradas 75 espécies arbóreas
pertencentes a 50 gêneros e 28 famílias botânicas, cujas
famílias Myrtaceae, Lauraceae, Flaucartiaceae e Aquifoliaceae
foram as mais representativas no grupo;
8) No grupo 2 foram encontradas 99 espécies arbóreas
pertencentes a 76 gêneros e 39 famílias botânicas, cujas
famílias Myrtaceae, Lauraceae, Flacourtiaceae, e Aquifoliaceae
foram as mais representativas no grupo;
9) No grupo 3 foram encontradas 114 espécies arbóreas
pertencentes a 84 gêneros e 44 famílias botânicas, cujas
famílias Myrtaceae, Lauraceae, Flaucourtiaceae, Aquifoliaceae,
Euphorbiaceae, Fabaceae, Rutaceae e Solanaceae foram as
mais representativas no grupo;
10) A Análise de Correspondência Canônica revelou quais os
fatores ecológicos são determinantes para a formação dos
grupos e da ocorrência das espécies indicadoras, indicando ser
uma técnica importante na análise das correlações das
variáveis ambientais com as espécies;
11) Encontrou-se como fatores ecológicos relevantes para o
desenvolvimento das espécies a umidade do solo, inclinação do
terreno, profundidade do solo, cor do solo (em diferentes
profundidades), quantidade de pedra e densidade do sub-
bosque;
12) Podocarpus lambertii, espécie indicadora do grupo 1, prefere
locais altos, inclinados, expostos, com solos pedregosos e bem
drenados;

207
13) Sebastiania commersoniana e Sebastiania brasiliensis,
espécies indicadoras do grupos 2, preferem locais planos,
úmidos e profundos e alta quantidade de pedra;
14) Araucaria angustifolia, espécie indicadora do grupos 3, prefere
locais planos, com solos de profundidade média a profundos e
bem drenados;
15) A determinação dos fatores ecológicos preferenciais das
espécies e dos grupos obtidos foi essencial como subsídio para
utilizar racionalmente os recursos naturais da floresta e também
para a aplicação de práticas adequadas de manejo sustentável.
5.2 Recomendações
Visando compreender os processos dinâmicos da floresta,
importante para estabelecimento de planos de manejo florestal
sustentável, recomenda-se:
a) Realizar outros estudos sobre a dinâmica das espécies
componentes da Floresta Ombrófila Mista da FLONA de São
Francisco de Paula e dos grupos florísticos obtidos;
b) Realizar estudos visando definir sub-grupos florísticos, em
função de outros parâmetros, que representam os estágios
sucessionais na floresta;
c) Realizar estudos dos mecanismos ou fases da regeneração:
fluxos ou dispersão de sementes, banco de sementes do solo e
banco de plântulas;
d) Realizar estudos sobre a fenologia das principais espécies ocorrentes na floresta.

208
6 REFERÊNCIAS BIBLIOGRAFICAS
ANDERBERG, M. R. Cluster Analysis for Applications. London:
Academic Press, 1973. 359 p.
ANDERSON, A. J. B. An introduction to multivariate statistical
analysis. 2 ed. New York: John Wiley & Sons, 1971. 415 p.
ARAUJO, M.M. Vegetação e mecanismos de regeneração em fragmento
de Floresta Estacional Decidual Ripária, Cachoeira do Sul, RS, Brasil.
Santa Maria: 2002, UFSM. 154p. Tese (Doutorado em Engenharia
Florestal) – Universidade Federal de Santa Maria, 2002.
ASENSIO, L. J. Técnicas de análisis de datos multidimensionales:
Bases teóricas y aplicaciones en agricultura. Madrid: Ministerio de
Agricultura Pesca y Alimentacion, 1989. 301 p.
AUBERT, E.; OLIVEIRA-FILHO, Análise multivariada da estrutura
fitossociológica do sub-bosque de plantios experimentais de Eucalyptus

209
spp. Em Lavras (MG). Revista Árvore, Viçosa, v.18, n.3, p.194-214,
1994.
BARROS, P. L. C. de. Estudo fitossociológico de uma floresta tropical
úmida no planalto de Curuá-Una, Amazônia Brasileira. Curitiba: UFPR,
1986. 146p. Tese (Doutorado em Manejo Florestal) __ Universidade
Federal do Paraná, 1986.
BATISTA, J. L. M. Análise multivariada no planejamento de extensão
florestal: subsídios para uma política de recuperação de áreas.
Curitiba: 1990. 137 p. Dissertação (Mestrado em Ciências Florestais) -
Setor de Ciências Agrárias, Universidade Federal do Paraná, 1990.
BAZZAZ, F. A.; PICKETT, S. T. A. Physiological ecology of tropical
succession: a comparative review. Ann. Rev. Ecol. Syst., v.11, p. 287-
310, 1980.
BOCKOR, I. Aplicación de un metodo de classificación numerica para
diferenciar tipos de bosques. Rev. For. Venez., Mérida, v.18, n.28, p.23-
37, 1975.
BOTREL, R.; OLIVEIRA-FILHO, A. T; RODRIGUES, L.A.; CURI, N.
Influência do solo e topografia sobre as variações da composição florística
e estrutura da comunidade arbórea-arbustiva de uma floresta estacional
semidecidual em Ingaí, MG. Revista Brasileira de Botânica, v.25, n.2, p.
195-213, jun. 2002.
BRASIL. Ministério da Agricultura. Instituto Brasileiro de
Desenvolvimento Florestal. Inventário Florestal Nacional - Florestas
Nativas do Rio Grande do Sul. Brasília: 1983. 345 p.
BRAUN-BLANQUET, J. Fitosociologia: bases para el estudio de las
comunidades vegetales. Madrid: H Blume Ediciones, 1979. 820 p.

210
CAIN, S. A.; CASTRO, G. M. O.; PIRES, J. M.; SILVA, N. T. Aplication of
some phytosociological tecniques to Brazilian rain forests. American
Journal of Botany, v. 43, n. 3, p.911-941, 1956.
CALDATO, S. L. Dinâmica Populacional da Ocotea Porosa
(Lauraceae) na Floresta Ombrófila Mista em Caçador, SC. Santa
Maria: UFSM, 1998. 89f. Dissertação (Mestrado em Engenharia Florestal)
- Universidade Federal de Santa Maria, 1998.
COX, G.W. Measurement of species diversity. In: Laboratory Manual of
General Ecology. Iowa: Brow Company Publishers, 1976. p.162-164.
CRISCI, J. V.; ARMENGOL, M. F. L.; Introduccion a la teoria y practica
de la taxonomia numerica. Washington: Secretaría General de la
Organización de los Estados Americanos, Programa Regional de
Desarrollo Científico y Tecnológico, 1983. 131p.
CUNHA, N. T. S.; PEICHL, B., Manual técnico florestal, apostilas do
Colégio Florestal de Irati. In: Ecologia Florestal. Campo Largo: Ingra S.A,
1986. p.422-427. Vol. 1, Cap.4.
DAUBENMIRE, R. Plant communities - a textbook of plant synecology.
New York: Harper & Row, 1968. 300 p.
EDELBAUER, G.; Evolução dos problemas ecológicos e a morte das
árvores na Alemanha. In: CONGRESSO FLORESTAL ESTADUAL, 6.,
1988, Nova Prata/RS. Anais... Porto alegre: Pallotti, 1988. p.1297 – 1320.
EMBRAPA . Centro Nacional de Pesquisa de Solos. Sistema brasileiro
de classificação de solos. Brasília: 1999. 412 p.
FELFILI, J. M.; SERVILHA, A. C. Distribuição espacial de parcelas e de
espécies em quatro áreas de cerrado sensu stricto na Chapada do
Espigão Mestre de São Francisco. In: FELFILI, J. M.; SILVA Jr., M. C. da

211
(org.). Biogeografia do bioma Cerrado: estudo fitofisionômico na
Chapada do Espigão Mestre do São Francisco. Brasília: UnB, 2001, p.61-
74
FELFILI, J. M.; SERVILHA, A. C.; SILVA Jr., M. C. da. Comparação entre
as unidades fisiográficas Chapada Pratinha, Veadeiros e Espigão Mestre
do São Francisco. In: FELFILI, J. M.; SILVA Jr., M. C. da (org.).
Biogeografia do bioma Cerrado: estudo fitofisionômico na Chapada do
Espigão Mestre do São Francisco. Brasília: UnB, 2001a. p. 80-94.
FELFILI, J. M.; SILVA Jr., M. C. da; REZENDE, A. V.; HARIDASAN, M.;
FIGUEIRAS, T. S.; MENDONÇA, R. C. da; WALTER, B. M. T.;
NOGUEIRA, P. E. O projeto Biogeografia do Bioma Cerrado: hipótese
e padronização da metodologia. In: GARAY, I.; DIAS, B. F. S.
Conservação da biodiversidade em ecossistemas tropicais. Petrópolis:
Vozes. 2001b, p. 157-173.
FINOL, H.. Nuevos parametros a considerarse en el analisis estructural de
las selvas virgenes tropicales. Ver. For. Venez., Mérida, v. 14, n.21, p.
29-42, 1971.
FOGUET, J. M. B.; ARIAS, M. del R. M. Analisis multivariante: Analisis
en Componentes Principales. Barcelona: Editorial Hispano Europea,
1989. 130 p.
FONT-QUER, P. Dicionario de botánica. Barcelona: Labor, 1975. 1244
p.
FÖRSTER, M.. Strukturanalyse aines tropishen Regenwalds in
Kolumbien. Allg. Forst.-u. J.-Stg., Wien, v.144, n.1, p.1-8, 1973.
GAUCH, H. G.; WHITTAKER, R. H. Comparison of ordination techniques.
Ecology, Ithaca, v.53, n.5, p.868-875, 1972.

212
GAUCH, H. G.; WHITTAKER, R. H. Hierarchical classification of
community data. Jour. Ecol., Oxford, v.69, n.2, p.573-597, 1981.
GOODALL, D. W. Sample similarity and species correlation. In:
WHITTAKER, R. H. Ordination of plant communities. The Hague: Dr.
W. Junk Publishers, 1982. p.99-149.
GREIG-SMITH, P. Quantitative plant ecology. 2.ed. London: Butter
Worths, 1964. 256 p.
GREIG-SMITH, P.; AUSTIN, M. P.; WHITMORE, T. C. The application of
quantitative methods to vegetation survey. I-Association analysis and
principal component ordination of rain forest. Jour. Ecol., Oxford, v.55,
n.2, p.483-503, 1967.
GUAPYASSÚ, M. dos S. Caracterização fitossociológica de três
fases sucessionais de uma Floresta Ombrófila Densa
Submontana, Morretes-Paraná. Curitiba: 1994. 165 p. Dissertação
(Mestrado em Ciências Florestais) - Setor de Ciências Agrárias,
Universidade Federal do Paraná, 1994.
HARTIGAN, J. A. Clustering algorithms. New York: John Wiley, 1975.
351 p.
HAVEL, J. J. Application of fundamental synecological knowledge to
practical problems in forest management. I. Theory and Methods. For.
Ecol. and Manag., Amsterdam, v.3, n.2, p.1-29, 1980a.
HAVEL, J. J. Application of fundamental synecological knowledge to
practical problems in forest management. II-Application. For. Ecol. and
Manag., Amsterdam, v.3, n.2, p.81-111, 1980b.
HERRMANN, M. L. de P.; ROSA, R. de O. Relevo. In: IBGE. Fundação
Instituto Brasileiro de Geografia e Estatística. Geografia do Brasil:

213
Região Sul. Rio de Janeiro: 1990. p.55-84.
HILL, M. O. TWINSPAN: a FORTRAN program fo arranging multivariate
data in an ordered two way table by classification of individual and
attributes. Ithaca, NY: Cornell University, 1979. 60p.
HILL, M. O.; BUNCE, R. G. H.; SHAW, M. W. Indicator species analysis, a
divisive polythetic method of classification, and its application to a survey
of native pinewoods in Scotland. The Journal of Ecology, v. 63, n.2,
p.597-613, 1975.
HOLDRIDGE, L. R. Ecología basada en zonas de vida. San Jose: IICA-
Instituto Interamericano de Cooperación para la Agricultura, 1982. 216 p.
HOSMER, D. W. Jr.; LEMESHOW, S. Applied logistic regression. New
York: Wiley, 1989.
JARENKOW, J. A. Composição florística e estrutura da mata com
Araucária na Estação Ecológica de Aracuri, Esmeralda, Rio Grande
do Sul. Porto Alegre: UFRGS, 1985. 82 p. Dissertação (Mestrado em
Ciências Biológicas) - Departamento de Botânica, Universidade Federal
do Rio Grande do Sul, 1985.
JARENKOW, S. A. Estudo fitossociológico comparativo entre duas
áreas com mata de encosta no Rio Grande do Sul. São Carlos: USC,
1994. 125f. Tese (Doutorado em Biologia) – Universidade de São Carlos,
1994.
JOHNSON, R. A.; WICHERN, D. W. Applied Multivariate Statistical
Analysis. Madison: Prentice Hall International, 1982. 607 p.
KAUL, P. F. T. Geologia. In: IBGE. Fundação Instituto Brasileiro de
Geografia e Estatística. Geografia do Brasil: Região Sul. Rio de Janeiro:
1990. p.29-54.

214
KELLMAN, N. C. Plant geography. London: Meethmen, 1975. 135 p.
KENT, M.; COKER, P. Vegetation description analyses. London:
Behaven Press, 1992. 363p.
KERSHAW, K. A. Quantitative and dynamic plant ecology. Hamilton:
Edward Arnold, 1973. 308 p.
LAMBERT, J. M.; DALE, M. B. The use of statistics in phytossociology.
Adv. Ecol. Res., v.2, p.59-99, 1964.
LAMBERT, J. M.; WILLIAMS, W. T. Multivariate methods in plant ecology.
IV-Nodal analysis. Jour. Ecol., Oxford, v.50, n.4-6, p.775-802, 1962
LAMPRECHT, H. Ensaio sobre unos metodos para el analisis estructural
de los bosques tropicales. Rev. For. Venez., v.13, n. 2, p. 57-65, 1962.
LAROCA, S. Ecologia: princípios & métodos. Petrópolis – RJ: Vozes,
1995. 197p.
LAVEN, R. D. Stablishing homogeneity in studies of forest sucession.
For. Ecol. and Manag., Amsterdam, v.4, n.2, p.161-177, 1982.
LEITE, P. F.; KLEIN, R. M. Vegetação. In: IBGE. Fundação Instituto
Brasileiro de Geografia e Estatística. Geografia do Brasil: Região Sul.
Rio de Janeiro, 1990. p.113-150.
LEPSCH, I. F.; Manual para levantamento utilitário do meio físico e
classificação de terras no sistema de capacidade de uso. Campinas:
Sociedade Brasileira de Ciências do Solo, 1993. 175p.
LIBERMAN, D.; LIBERMAN, M.; PERALTA, R.; HARTSHORN, G.S.
Mortality patterns and stands tournover rates in a wet tropical forest in
Costa Rica. Journal of Ecology, v.72, n.1, p.915-924, 1985.
LIMA, W. de P. Funções hidrológicas da Mata Ciliar. In: BARBOSA, L.

215
Simpósio sobre Mata Ciliar. 1989, Campinas. Anais... Campinas:
Fundação Cargil, 1989. p.25-42.
LOETSCH, F.; ZORHER, F.; HALLER, K. E. Forest inventary. 2.ed.
Munich: BLV Vellagsgesellschaft, 1973. 469 p.
LONGHI, S. J. A estrutura de uma floresta natural da Araucaria
anguatifolia (Berth). Kuntze, no sul do Brasil. Curitiba: UFPR, 1980.
198f. Dissertação (Mestrado em Engenharia Florestal) - Universidade
Federal do Paraná, 1980.
LONGHI, S. J. Agrupamento e análise fitossociológica de
comunidades florestais na sub-bacia hidrográfica do Rio Passo
Fundo- RS. Curitiba: UFPR, 1997. 198f. Tese (Tese de Doutorado) - .
Universidade Federal do Paraná, 1997.
MAGURRAN, A. E. Diversidad Ecológica y su Medición. Barcelona:
Vedra, 1989. 200 p.
MALLO, F. Analisis de Componentes Principales y tecnicas
factoriales relacionadas: Teoria, computacion y aplicaciones. Leon:
Universidad de Leon, 1985. 523 p.
MARCHIORI, J. N. C. Dendrologia das gimnospermas. Santa
Maria: Ed. da UFSM, 1996. 158p.
MARDIA, K. V.; KENT, J. T.; BIBBY, J. M. Multivariate analysis. London:
Academic Press, 1979. 518 p.
MARRIOTT, F. H. C. The interpretation of multivariate observation.
London: Academic Press, 1974. 117 p.
MARTINS, F. R. Estrutura de uma floresta mesófila. São Paulo: Editora
da UNICAMP, 1991. 246p.

216
MARTINS, S. V; SILVA, N. R. S; SOUZA, A. L; NETO, J. A. A. M.
Distribuição de espécies arbóreas em um gradiente topográfico de
Floresta Estacional Semidecidual em Viçosa, MG. Scientia Florestalis,
n.64, p. 172 – 181, Dez. 2003.
MATTEUCCI, S. C.; COLMA, A. Metodologia para el estudio de la
vegetacion. Washington: Secretaria General de la Organizción de los
Estados Americanos - Programa Regional de Desarrolo Científico y
Tecnológico, 1982. 169p.
MORENO, J. A. Clima do Rio Grande do Sul. Porto Alegre: Governo do
Estado do Rio Grande do Sul, Secretaria da Agricultura, Diretoria de
Terras e Colonização, 1961. 42p.
MORRISON, D. F. Multivariate Statistical Methods. 2.ed. New York:
McGraw-Hill, 1976. 415 p.
MUELLER-DOMBOIS, D.; ELLENBERG, H., Aims and methods of
vegetation ecology. Ney York: John Wiley & Sons, 1974. 547p.
MUNSELL SOIL COLOR CHARTS. Munsell Color Division. Baltimor,
Maryland: Editora Kollmorgen Corporation, 1971. 18p.
NAGY, J. L. & CUNHA, N. T. S, Manual técnico florestal, apostilas do
Colégio Florestal de Irati. In: ____. As condições ecológicas na
floresta. Campo Largo: Ingra S.A, 1986. P.23-32. Vol. 1, Cap.1.
NASCIMENTO, A. R.T.; Análise Estrutural e Padrões de Distribuição
Espacial de Uma Amostra de Floresta Ombrófila Mista. Santa maria:
UFSM, 2000. 90f. Dissertação (Mestrado em Engenharia Florestal) -
Universidade Federal de Santa Maria, 2000.
NETTO, S. N; SANQUETA, C. R; BRENA, D. A.; Os sites e o
programa brasileiro de pesquisas ecológicas de longa duração. In: ____.

217
A floresta de Araucária e transições – Site 9. Belo Horizonte: Fundação
Universidade Federal do Rio Grande do Sul – RS. Universidade Federal
de Minas Gerais. (2002).
NIMER, E. Clima. In: IBGE. Fundação Instituto Brasileiro de Geografia e
Estatística. Geografia do Brasil: Região Sul. Rio de Janeiro: 1990. p.151-
187.
ODUM, E. P. Ecologia. Rio de Janeiro: Guanabara Koogan, 1988. 434p.
OLIVEIRA-FILHO, A.T. e AUBERT, E., Análise multivariada da
estrutura fitossociológica do sub-bosque de plantios experimentais
de Eucalyptus spp. E Pinus spp. Em Lavras (MG)
ORLÓCI, L. An agglomerative method for classification of plant
communities. Jour. Ecol., Oxford, v.55, n.1, p.193-206, 1967.
ORLÓCI, L. Multivariate analysis in vegetation research. 2.ed. The
Hague: DR. W. Junk B. V. Publishers, 1978. 415 p.
PÉLLICO NETTO, S.; BRENA, D. A. Inventário Florestal. Curitiba:
Universidade Federal do Paraná / Universidade Federal de Santa Maria,
1997. 245 p.
PERLIN, J. História das Florestas. Rio de Janeiro: Ed. Imagro, 1989.
578p.
PIELOU, E. C. Ecological diversity. New York: John Wiley, 1975. 165 p.
PIELOU, E. C. Matematical ecology. New York: John Wiley & Sons,
1977. 385p.
PINTO, J. R. R. Levantamento florístico, estrutura da comunidade
arbóreo-arbustiva e suas correlações com variáveis ambientais em
uma floresta do Vale no Parque Nacional da Chapada dos

218
Guimarães, Mato Grosso. Lavras: UFL, 1997. 85f. Dissertação
(Mestrado em Engenharia Florestal) - Universidade Federal de Lavras,
1997.
PLA, L. E. Metodo de componentes principales. Washington:
Secretaria General de la Organización de los Estados Americanos, 1986.
89 p. (Monografia Científica, 27).
PRADO, H. Manual de classificação dos solos do Brasil. 20 edição.
FUNEP, Jaboticabal, 1995.
PRODAN, M. Holzmesslehre. Frankfurt: J. D. Sauerländer’s Verlag,
1965. 644p.
RAMBO, B. A fisionomia do Rio Grande do Sul. Porto Alegre: Livraria
Selbach, 1956. 456 p.
REITZ, R; KLEIN, R.M.; REIS, A. Projeto Madeira do Rio Grande
do Sul. Porto Alegre: Governo do Estado do Rio Grande do Sul,
Secretaria da Agricultura e Abastecimento / Herbário Barbosa Rodrigues,
1988. 525p.
RIO GRANDE DO SUL. Governo do Estado, Secretaria Especial do Meio
Ambiente. Inventário Florestal Contínuo do Rio Grande do Sul. Porto
Alegre: SEMA/FATEC, 2002. 706p. (Relatório Técnico).
SANCHEZ, P. A., Suelos del tropico: características y manejo. Costa
Rica: Instituto Interamericano de Cooperación para la Agricultura, 1981.
660p.
SAIZ, F.; Experiencias em el Uso de Criterios de Similitud en el Estudio de
Comunidades. Archos Biol. Med. Exp. , v.13, p. 387-402, 1980.
SCHNEIDER, P. R., Introdução ao Manejo Florestal. Santa Maria:
UFSM/CPEF/FATEC, 1993. 348p.

219
SILVA JÚNIOR, M. C. da.; Comunidades de árvores e sua relação com os
solos na Mata do Pitoco, Reserva Ecológica do IBGE, Brasília – DF.
Revista Árvore, Viçosa – Minas Gerais, v. 22, n. 1, p. 29-40, 1998.
SILVA, P. E. N. da. Florística, fitossociologia e nutrição mineral do
cerrado sentido restrito no Complexo Xavantina – MT. São Paulo:
USP, 2002. Tese (Doutorado em Ecologia) – Universidade de São Paulo,
2002.
SMITH, L. B.; DOWNS, R. J.; KLEIN, R. M. Euforbiaceas. In: Reitz, R.
Flora Ilustrada Catarinense. Itajaí: Herbário Barbosa Rodrigues, 1988.
p.308-313.
SNEATH, P. H.; SOKAL, R. R. Numerical Taxonomy. San Francisco:
Freeman, 1973. 573p.
SOKAL, R. R.; SNEATH, P. H. Principles of numerical taxonomy. San
Francisco: Freeman, 1963. 450p.
SOUZA, A. L. DE. Análise muitivariada para manejo de florestas
naturais: alternativas de produção sustentada de madeiras para serraria.
Curitiba: UFPR, 1989. 255p. Tese (Doutorado em Ciências Florestais) –
Universidade Federal do Paraná, 1989.
SOUZA, A. L.; NETO, J. A. A. M.; SCHETTINO, S. Avaliação florística,
fitossociológica e paramétrica de fragmento de florestas atlântica
secundária, município de Pedro Canário, Espirito Santo. Viçosa: SIF,
1998. 117p. (Documento SIF, 18).
STRECK, E. V. et al; Solos do Rio Grande do Sul. Porto Alegre:
EMATER/RS; UFRGS, 2002. 107p.
SUDESUL. Superintendência de Desenvolvimento da Região Sul. A
vegetação atual da Região Sul. Porto Alegre: 1978. 115p. (Série

220
Planejamento e Estudos Regionais, 2).
TER BRAAK, C. J. F. Canonical correspondence analysis: a new
eigenvector technique for multivariate direct analysis. Ecology, v.67,
p.1167-1179, 1986.
TER BRAAK, C. J. F. CANOCO – A FORTRAN program for canonical
community ordination by (Partial) (Detrended) (Canonical) correspondence
analysis, principal components analysis and redundancy analysis, version
2.1 (Technical report LWA-88-2, TNO). Wageningen: Institute of Applied
Computer Science, 1988. 95 p.
VAN DEN BERG, E. Estudo florístico e fitossociológico de uma
floresta ripária em Itutinga, MG, e análise das correlações entre
variáveis ambientais e a distribuição das espécies de porte arbóreo-
arbustivo. Lavras: UFL, 1995, 73f. Dissertação (Mestrado em Engenharia
Florestal) __ Universidade Federal de Lavras, 1995.
VATTIMO, I. Lauraceae. In: REITZ, R. Flora Ilustrada Catarinense. Itajaí:
Herbário Barbosa Rodrigues, 1979. p.14-19.
VIEIRA, L. S.; SANTOS, P. C. T.; VIEIRA, M. N.; Solos: propriedade,
classificação e manejo. Brasília: MEC/ABEAS, 1988. 154p.
WEBB, L. J.; TRACEY, J. G.; WILLIAMS, W. T.; LANCE, G. N. Studies in
the numerical analysis of complex rain-forest communities. I-A comparison
of methods applicable to site/species data. Jour. Ecol., Oxford, v.55, n.1,
p.171-191, 1967a.
WEBB, L. J.; TRACEY, J. G.; WILLIAMS, W. T.; LANCE, G. N. Studies in
the numerical analysis of complex rain-forest communities. II-The problem
of species-sampling. Jour. Ecol., Oxford, v.55, n.2, p.525-538, 1967b.
WHITTAKER, R. H. Classification of natural communities. Bot. Rev.,

221
v.28, n.4, p.1-239, 1962.
WHITTAKER, R. H. Communities and Ecosystems. 2.ed. New York:
MacMillan Publishing Co. Inc., 1975. 385 p.
WHITTAKER, R. H. Ordination of plant communities. The Hague: Dr.
W. Junk Publishers, 1982. 388 p.
WILLIAMS, W. T.; DALE, M. B. Fundamental problems in numerical
taxonomy. Adv. Bot. Res., n.2, p.35-68, 1965.
ZILLER, S. R. Análise fotossociológica de caxetais. Curitiba, UFPR,
1992. 101 f. Dissertação (Mestrado em Ciências Florestais) -
Universidade Federal do Paraná, 1992.

222
ANEXOS
223
ANEXO 1 – Relação das espécies em seus respectivos grupos florísticos encontrados na Floresta Ombrófila Mista de São Francisco de Paula, RS.
FAMÍLIA/ESPÉCIE NOME VULGAR GRUPO 1 2 3
ANACARDIACEAE
Lithraea brasiliensis Marchand Aroeira-bugre X X X
ANNONACEAE
Rollinia rugulosa Schltdl. Araticum-quaresma X X X
Rollinia salicifolia Schltdl. Araticum-salso X X
Rollinia sylvatica (A. St.-Hil.) Mart. Araticum-do-mato X
AQUIFOLIACEAE
Ilex brevicuspis Reissek Caúna-da-serra X X X
Ilex dumosa Reissek Congonha X X X
Ilex microdonta Reissek Caúna X X X
Ilex paraguariensis A. St.-Hil. Erva-mate X X X
Ilex sp. Caúna X
ARALIACEAE
Oreapanax fulvus Marchal Tamanqueira X X
ARAUCARIACEAE
Araucaria angustifolia (Bertol.) Kuntze Pinheiro-brasileiro X X X
ARECACEAE
Syagrus romanzoffiana (Cham.) Glassman Jerivá X
ASTERACEAE
Dasyphyllum spinescens (Less.) Cabrera Açucará X X
Dasyphyllum tomentosum (Spreng.) Cabrera Açucará-piloso X X X
Gochnatia polymorpha (Less.) Cabrera Cambará X
Piptocharpha tomentosa Baker Pau-toucinho X
Vernonia discolor (Spreng.) Less. Vassourão-preto X X X
BIGNONIACEAE
Tabebuia umbellata (Sond.) Sandwith Ipê-amarelo X
CARICACEAE
Carica quercifolia (A. St.-Hil.) Hieron Mamoeiro-do-mato X
CELASTRACEAE
Maytenus aquifolia Mart. Concorosa-arbórea X X
Maytenus evonymoides Reissek Coração-de-bugre X X X
CLETHRACEAE
Clethra uleana Sleumer Caujuja-de-ule X
CUNONIACEAE
Lamanonia ternata Vell. Guaperê X X
Weinmania paulliniifolia Pohl ex Ser. Gramimunha X X X
CYATHEACEAE
Alsophila sp. Xaxim-de-espinho X
Continua

224
ANEXO 1 – Continuação
FAMÍLIA/ESPÉCIE NOME VULGAR GRUPO 1 2 3
DICKSONIACEAE
Dicksonia sellowiana Hook. Xaxim X X X
EUPHORBIACEAE
Sapium gladulatum (Vell.) Pax. Pau-leiteiro X X
Sebastiania brasiliensis Spreng. Branquilho-leiteiro X X X
Sebastiania commersoniania (Baill.) L. B. Sm. et Downs Branquilho X X X
Stillingia oppositifolia Baill. ex Müll. Arg. Leiterinho X X
FABACEAE
Dalbergia frutenscens (Vell.) Britton Rabo-de-bugio X X
Erythrina cristagalli L. Corticeira-banhado X
Lonchocarpus campestris Mart. ex Benth. Pau-canzil X X
Lonchocarpus nitidus (Vogel) Benth. Farinha-seca X
Machaerium paraguariense Hassl. Canela-do-brejo X X X
GRISELINIACEAE
Griselinia ruscifolia (Clos) Taub. Griselina X
FLACOURTIACEAE
Banara parviflora (A. Gray) Benth. Guaçatunga-preta X X X
Banara tomentosa Clos Guaçatunga-branca X X
Casearia decandra Jacq. Guaçatunga X X X
Casearia obliqua Spreng. Estralador X
Casearia sylvestris Sw. Chá-de-bugre X X
Xylosma pseudosalzmannii Sleumer Espinho-de-judeu X X X
Xylosma tweedianum (Clos) Eichler Sucará X X X
ICACINACEAE
Citronella gongonha (Mart.) Howard Gongonha-espinho X X
Citronella paniculata (Mart.) Howard Gongonha X
LAMIACEAE
Cunila insisa Benth. Poejo X X X
LAURACEAE
Cinnamomum amoenum (Nees) Kosterm. Canela X X
Cinnamomum glaziovii (Mez) Kosterm. Canela X X
Cryptocarya aschersoniania Mez Canela-vick X X X
Cryptocarya moschata Nees Canela-cega X X X
Nectandra lanceolata Nees et Mart. ex Nees Canela-amarela X
Nectandra megapotamica (Spreng.) Nez Canela-preta X X X
Ocotea indecora (Schott) Mez Canela X
Ocotea puberula (A. Rich.) Nees Canela-guaicá X X X
Ocotea pulchella Mart. Canela-langeana X X X
LOGANIACEAE
Strychnos brasiliensis (Spreng.) Mart. Anzol-de-lontra X X
Continua

225
ANEXO 1 – Continuação
FAMÍLIA/ESPÉCIE NOME VULGAR GRUPO 1 2 3
MELIACEAE
Cabralea canjerana (Vell.) Mart. Canjerana X
Cedrela fissilis Vell. Cedro X X
MIMOSACEAE
Acacia bonariensis Gill. ex Hook. et Arn. Unha-de-gato X X
Inga marginata Willd. Ingá-feijão X
Inga vera Willd. Ingá-banana X X X
Inga sessilis (Vell.) Mart. X
MONIMIACEAE
Hennecartia omphalandra J. Poiss. Canema X
Mollinedia elegans Tul. Pimentinha X X
Mollinedia sp. Capixim X
MYRSINACEAE
Myrsine coriacea (Sw.) R. Br. Capororoquinha X X X
Myrsine lorentziana (Mez) Arecav. Capororoca-d’Água X X X
Myrsine umbelata Mart. Capororocão X X X
Myrsine sp. Capororoca X X X
MYRTACEAE
Acca sellowiana (O. Berg) Burret Goiaba-serrana X X
Blepharocalyx salicifolius (Kunth) O. Berg Murta X X X
Calyptranthes concinna DC. Guamirim-de-facho X X
Campomanesia rhombea O. Berg Guabiroba-miúda X X X
Campomanesia xanthocarpa O. Berg Guabiroba X X X
Eugenia _cutifólia_ DC. Cerejeira-do-mato X X X
Eugenia pluriflora DC. Jaboticaba-campo X
Eugenia psidiiflora O. Berg Guamirim X X X
Eugenia uruguayensis Cambess. Batinga-vermelha X X X
Myrceugenia cucullata D. Legrand Guamirim-quebrado X X X
Myrceugenia foveolata (O. Berg) Sobral Guamirim X
Myrceugenia glaucescens (Cambess.) D. Legrand et Kausel Conserva-branca X
Myrceugenia miersiana (Gardner) D. Legrand et Kausel Guamirim-piloso X X X
Myrceugenia myrcioides (Cambess.) O. Berg Guamirim X X
Myrceugenia oxysepala (Berret) D. Legrand et Kausel Guamirim X X
Myrcia arborescens O. Berg Guamirim-ferro X
Myrcia bombycina (O. Berg) Nied. Guamirim-do-campo X X X
Myrcia lajeana D. Legrand Cambuí X
Myrcia oligantha O. Berg Guamirim X X
Myrcia palustris DC. Guamirinzinho X
Myrcianthes _cutifóli (D. Legrand) D. Legrand Araça-do-mato X X X
Myrcianthes pungens (O. Berg) D. Legrand Guabijú X X X
Continua

226
ANEXO 1 – Continuação
FAMÍLIA/ESPÉCIE NOME VULGAR GRUPO 1 2 3
Myrciaria delicatula (DC.) O. Berg Camboim X X X
Myrciaria floribunda (West ex Willd.) O. Berg Camboim X X X
Myrciaria tenella (DC.) O. Berg Camboinzinho X X X
Myrrhinium atropurpureum Schott Murtilho X X X
Siphoneugena reitzii D. Legrand Camboim-de-reitz X X X
NYCTAGINACEAE
Guaipira _cutifól (Vell.) Reitz Pau-cebola X
PHYTOLACCACEAE
Seguieria _cutifól L. Cipó-umbú X X
PODOCARPACEAE
Podocarpus lambertii Klotzsch ex Endl. Pinheiro-bravo X X X
PROTEACEAE
Roupala brasiliensis Klotzsch Carvalho-brasileiro X X X
RHAMNACEAE
Rhamnus sphaerosperma Sw. Pau-cangica X
Scutia buxifolia Reissek Coronilha X X X
ROSACEAE
Prunus myrtifolia (L.) Urb. Pessegueiro-mato X X X
RUBIACEAE
Coussarea contracta (Walp.) Müll. Arg. Pimenteira X
Coutarea hexandra (Jacq.) K. Schum. Quina X
RUTACEAE
Citrus sp. Laranjeira X X
Policarpus pennatifolius Lem. Cutia X X
Zanthoxylum fagara (L.) Sarg. Coentrilho X X
Zanthoxylum kleinii (Cowan) P. G. Waterman Juvevê X
Zanthoxylum rhoifolium L. Mamica-de-cadela X X X
SAPINDACEAE
Allophylus edulis (A. St.-Hil. Et al.) Radlk. Chal-chal X X X
Cupania vernalis Cambess. Camboáta-vermelho X X X
Matayba elaeagnoides Radlk. Camboáta-branco X X X
SIMAROUBACEAE
Picramnia parvifolia Engl. Pau-amargo X X
SOLANACEAE
Cestrum sp. Coerana X
Solanum mauritianum Scop. Fumo-bravo X
Solanum pabstii L. B. Sm. & Downs Canema X
Solanum pseudocapsicum L. Tomatinho X
Solanum pseudoquina A. St.-Hil. Peloteira X X
Solanum sanctae-catharinae Dunal Joá-manso X X X
Continua

227
ANEXO 1 – Continuação
FAMÍLIA/ESPÉCIE NOME VULGAR GRUPO 1 2 3
Solanum sp. Fuminho X X X
STYRACACEAE
Styrax leprosus Hook. Et Arn. Carne-de-vaca X X
SYMPLOCACEAE
Symplocos pentandra Occhioni Sete-sangrias X X
Symplocos tetrandra (Mart.) Miq. Sete-sangrias X X
Symplocos uniflora (Pohl) Benth. Sete-sangrias X X
THEACEAE
Gordonia _cutifólia (Wawra) H. Keng Santa-rita X X
TILIACEAE
Luehea divaricata Mart. Et Zucc. Açoita-cavalo X X
ULMACEAE
Celtis iguanaea (Jacq.) Sargent Esporão-de-galo X X
VERBENACEAE
Citharexylum solanaceum Cham. Tarumã-de-espinho X X
Duranta vestita Cham. Duranta X X
WINTERACEAE
Drimys brasiliensis Miers Casca-d’anta X
Cipós X X X
Mortas X X X
Não identificada X X X
TOTAL 78 98 113

228
ANEXO 2 – Parâmetros fitossociológicos das espécies do grupo 1 (Formação Podocarpus) da Floresta Ombrófila Mista da FLONA de São Francisco de Paula, RS.
DENSIDADE FREQÜÊNCIA DOMINÂNCIA VALOR IMPORT.
VALOR COBERT.
ESPÉCIES DA DR FA FR DoA DoR VI VC Podocarpus lambertii 92,93 9,01 8,08 1,12 5,69 13,64 23,77 22,66 Myrciaria floribunda 94,95 9,21 2,02 0,28 2,69 6,45 15,93 15,65 Eugenia uruguayensis 55,56 5,39 37,37 5,16 1,60 3,84 14,39 9,23 Vernonia discolor 45,45 4,41 3,03 0,42 3,97 9,53 14,36 13,94 Araucaria angustifolia 27,27 2,64 22,22 3,07 3,57 8,56 14,27 11,21 Lamanonia ternata 44,44 4,31 21,21 2,93 2,37 5,68 12,92 9,99 Myrciaria tenella 57,58 5,58 16,16 2,23 1,45 3,49 11,30 9,07 Ilex brevicuspis 52,53 5,09 7,07 0,98 2,12 5,08 11,15 10,17 Ocotea pulchella 20,20 1,96 56,57 7,81 0,53 1,28 11,05 3,24 Morta 61,62 5,97 1,01 0,14 1,90 4,56 10,67 10,53 Casearia decandra 32,32 3,13 20,20 2,79 0,66 1,58 7,50 4,71 Symplocos tetrandra 24,24 2,35 2,02 0,28 1,97 4,72 7,35 7,08 Calyptranthes concinna 29,29 2,84 21,21 2,93 0,38 0,90 6,67 3,74 Sapium glandulatum 33,33 3,23 2,02 0,28 1,13 2,71 6,22 5,94 Lithraea brasiliensis 24,24 2,35 1,01 0,14 1,48 3,55 6,04 5,90 Xylosma tweedianum 1,01 0,10 42,42 5,86 0,02 0,04 5,99 0,14 Myrsine lorentziana 12,12 1,18 21,21 2,93 0,71 1,71 5,81 2,88 Myrsine umbellata 31,31 3,04 6,06 0,84 0,79 1,90 5,78 4,94 Não identificada 3,03 0,29 38,38 5,30 0,06 0,14 5,73 0,43 Myrcianthes pungens 1,01 0,10 37,37 5,16 0,02 0,05 5,31 0,15 Inga vera 2,02 0,20 33,33 4,60 0,04 0,10 4,90 0,30 Siphoneugena reitzii 25,25 2,45 4,04 0,56 0,69 1,65 4,66 4,10
Continua

229
ANEXO 2 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Gochnatia polymorpha 1,01 0,10 31,31 4,32 0,08 0,18 4,61 0,28 Maytenus evonymoides 18,18 1,76 2,02 0,28 0,98 2,36 4,40 4,12 Styrax leprosus 1,01 0,10 30,3 4,18 0,04 0,09 4,37 0,19 Blepharocalyx salicifolius 12,12 1,18 12,12 1,67 0,19 0,47 3,32 1,64 Ilex paraguariensis 17,17 1,67 2,02 0,28 0,48 1,16 3,10 2,83 Cryptocarya aschersoniana 12,12 1,18 1,01 0,14 0,63 1,52 2,83 2,69 Duranta vestita 1,01 0,10 19,19 2,65 0,01 0,02 2,77 0,12 Symplocos uniflora 15,15 1,47 1,01 0,14 0,45 1,09 2,69 2,55 Rollinia salicifolia 1,01 0,10 18,18 2,51 0,01 0,02 2,63 0,12 Myrrhinium atropurpureum 17,17 1,67 2,02 0,28 0,27 0,64 2,59 2,31 Gordonia acutifolia 8,08 0,78 7,07 0,98 0,30 0,72 2,48 1,50 Sebastiania brasiliensis 2,02 0,20 16,16 2,23 0,02 0,05 2,48 0,24 Cupania vernalis 5,05 0,49 12,12 1,67 0,13 0,30 2,47 0,79 Myrcianthes gigantea 15,15 1,47 1,01 0,14 0,33 0,79 2,39 2,25 Xylosma pseudosalzmannii 3,03 0,29 14,14 1,95 0,04 0,10 2,35 0,39 Solanum sanctae-catharinae 4,04 0,39 13,13 1,81 0,05 0,13 2,34 0,52 Zanthoxylum rhoifolium 16,16 1,57 1,01 0,14 0,24 0,57 2,28 2,14 Ocotea puberula 1,01 0,10 14,14 1,95 0,08 0,19 2,24 0,29 Ilex microdonta 2,02 0,20 13,13 1,81 0,09 0,21 2,22 0,40 Myrsine coriacea 2,02 0,20 12,12 1,67 0,12 0,30 2,17 0,49 Ilex dumosa 7,07 0,69 2,02 0,28 0,45 1,07 2,04 1,76 Myrcia bombycina 1,01 0,10 13,13 1,81 0,03 0,07 1,98 0,17 Banara parviflora 7,07 0,69 7,07 0,98 0,12 0,29 1,95 0,97
Continua

230
ANEXO 2 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Cryptocarya moschata 1,01 0,10 12,12 1,67 0,07 0,16 1,93 0,26 Cinnamomum amoenum 8,08 0,78 1,01 0,14 0,36 0,86 1,78 1,64 Prunus myrtifolia 8,08 0,78 1,01 0,14 0,13 0,31 1,24 1,10 Myrceugenia miersiana 9,09 0,88 1,01 0,14 0,09 0,21 1,23 1,09 Myrceugenia cucullata 3,03 0,29 6,06 0,84 0,03 0,07 1,20 0,36 Dasyphyllum tomentosum 1,01 0,10 5,05 0,70 0,16 0,39 1,19 0,49 Myrciaria delicatula 6,06 0,59 3,03 0,42 0,07 0,17 1,17 0,75 Myrsine sp. 2,02 0,20 6,06 0,84 0,04 0,10 1,13 0,30 Nectandra megapotamica 6,06 0,59 1,01 0,14 0,14 0,35 1,07 0,93 Eugenia psidiiflora 5,05 0,49 1,01 0,14 0,15 0,36 0,99 0,85 Cunila incisa 1,01 0,10 6,06 0,84 0,01 0,02 0,96 0,12 Machaerium paraguariense 2,02 0,20 1,01 0,14 0,25 0,60 0,93 0,80 Allophylus edulis 2,02 0,20 2,02 0,28 0,16 0,39 0,86 0,58 Weinmania paulliniifolia 1,01 0,10 1,01 0,14 0,25 0,60 0,84 0,70 Rollinia rugulosa 3,03 0,29 3,03 0,42 0,03 0,08 0,79 0,37 Casearia sylvestris 4,04 0,39 1,01 0,14 0,09 0,21 0,74 0,61 Roupala brasiliensis 3,03 0,29 1,01 0,14 0,13 0,30 0,74 0,60 Dicksonia sellowiana 2,02 0,20 1,01 0,14 0,12 0,28 0,62 0,48 Drimys brasiliensis 1,01 0,10 2,02 0,28 0,07 0,18 0,56 0,28 Eugenia pluriflora 2,02 0,20 2,02 0,28 0,04 0,08 0,56 0,28 Mollinedia elegans 1,01 0,10 3,03 0,42 0,01 0,04 0,55 0,13 Citharexylum solanaceum 1,01 0,10 3,03 0,42 0,01 0,03 0,55 0,13
Continua

231
ANEXO 2 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Myrcia lajeana 2,02 0,20 1,01 0,14 0,03 0,07 0,41 0,27 Myrceugenia oxysepala 2,02 0,20 1,01 0,14 0,02 0,05 0,39 0,25 Eugenia involucrata 2,02 0,20 1,01 0,14 0,02 0,04 0,37 0,23 Matayba elaeagnoides 1,01 0,10 1,01 0,14 0,04 0,10 0,34 0,20 Campomanesia xanthocarpa 1,01 0,10 1,01 0,14 0,03 0,08 0,32 0,18 Sebastiania commersoniana 1,01 0,10 1,01 0,14 0,03 0,06 0,30 0,16 Campomanesia rhombea 1,01 0,10 1,01 0,14 0,02 0,05 0,29 0,15 Solanum sp. 1,01 0,10 1,01 0,14 0,01 0,03 0,27 0,13 Acca sellowiana 1,01 0,10 1,01 0,14 0,01 0,02 0,26 0,12 Cipós 1,01 0,10 1,01 0,14 0,01 0,02 0,26 0,12 Scutia buxifolia 1,01 0,10 1,01 0,14 0,01 0,02 0,25 0,12 TOTAL (78) 1029,30 100 721,24 100 41,64 100 300 200

232
ANEXO 3 – Parâmetros fitossociológicos das espécies do grupo 2 (Formação Sebastiania) da Floresta Ombrófila Mista da FLONA de São Francisco de Paula, RS.
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Cryptocarya aschersoniana 56,34 6,58 33,80 5,67 5,52 12,30 24,55 18,88 Blepharocalyx salicifolius 44,84 5,24 31,69 5,31 5,60 12,48 23,03 17,72 Sebastiania commersoniana 69,25 8,09 31,69 5,31 3,93 8,75 22,16 16,84 Eugenia psidiiflora 89,91 10,50 46,95 7,87 1,94 4,33 22,15 14,82 Ilex brevicuspis 39,67 4,63 24,88 4,17 4,30 9,44 18,24 14,07 Morta 36,62 4,28 30,99 5,20 0,54 3,43 12,90 7,71 Ocotea pulchella 28,40 3,52 23,47 3,94 0,48 5,53 12,77 9,05 Sebastiania brasiliensis 42,02 4,91 26,29 4,41 0,60 1,33 10,65 6,24 Araucaria angustifolia 18,54 2,17 12,91 2,17 0,32 5,18 9,51 7,34 Sapium glandulatum 26,29 3,07 18,08 3,03 0,88 1,96 8,06 5,03 Nectandra megapotamica 18,08 2,11 14,79 2,48 0,33 2,98 7,57 5,09 Ilex paraguariensis 23,47 2,74 17,37 2,91 0,79 1,76 7,41 4,50 Dicksonia sellowiana 25,35 2,96 12,44 2,09 0,02 2,27 7,32 5,23 Myrceugenia cucullata 24,18 2,82 17,14 2,87 0,60 1,33 7,03 4,16 Roupala brasiliensis 21,13 2,47 16,20 2,72 0,66 1,47 6,65 3,94 Casearia decandra 19,01 2,22 16,20 2,72 0,34 0,75 5,69 2,97 Campomanesia xanthocarpa 15,73 1,84 12,21 2,05 0,56 1,25 5,14 3,09 Campomanesia rhombea 15,02 1,75 13,15 2,20 0,45 1,01 4,96 2,76 Eugenia uruguayensis 13,85 1,62 9,15 1,54 0,53 1,18 4,33 2,79 Eugenia involucrata 12,44 1,45 10,56 1,77 0,38 0,84 4,06 2,29 Não identificada 12,44 1,45 10,09 1,69 0,32 0,72 3,87 2,18 Banara parviflora 11,74 1,37 10,56 1,77 0,18 0,40 3,55 1,77
Continua

233
ANEXO 3 – Continuação.
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Myrciaria floribunda 12,21 1,43 8,69 1,46 0,27 0,60 3,48 2,02 Cinnamomum glaziovii 8,22 0,96 5,63 0,94 0,63 1,41 3,32 2,37 Prunus myrtifolia 8,69 1,01 7,04 1,18 0,33 0,73 2,93 1,75 Myrcia oligantha 9,62 1,12 8,22 1,38 0,17 0,38 2,89 1,51 Myrciaria delicatula 8,45 0,99 7,28 1,22 0,27 0,59 2,80 1,58 Cipós 9,39 1,10 7,98 1,34 0,13 0,29 2,72 1,38 Matayba elaeagnoides 6,34 0,74 5,87 0,98 0,44 0,98 2,70 1,72 Myrceugenia miersiana 9,39 1,10 7,51 1,26 0,14 0,31 2,67 1,41 Podocarpus lambertii 7,04 0,82 5,16 0,87 0,44 0,98 2,67 1,80 Ocotea puberula 4,69 0,55 3,99 0,67 0,37 0,82 2,04 1,37 Ilex microdonta 4,69 0,55 3,99 0,67 0,32 0,72 1,93 1,26 Calyptranthes concinna 6,81 0,79 4,93 0,83 0,12 0,26 1,88 1,05 Siphoneugena reitzii 5,40 0,63 4,69 0,79 0,15 0,33 1,75 0,96 Weinmania paulliniifolia 1,88 0,22 1,88 0,31 0,38 0,85 1,39 1,07 Dasyphyllum tomentosum 3,05 0,36 2,35 0,39 0,28 0,63 1,38 0,98 Inga vera 4,69 0,55 3,52 0,59 0,10 0,22 1,35 0,76 Xylosma tweedianum 3,29 0,38 3,05 0,51 0,14 0,32 1,22 0,70 Cryptocarya moschata 3,05 0,36 1,64 0,28 0,23 0,52 1,15 0,87 Myrcianthes gigantea 3,52 0,41 2,82 0,47 0,08 0,17 1,06 0,59 Ocotea indecora 2,11 0,25 1,41 0,24 0,22 0,49 0,98 0,74 Vernonia discolor 2,11 0,25 1,64 0,28 0,17 0,39 0,91 0,63 Pilocarpus pennatifolius 3,76 0,44 2,11 0,35 0,04 0,09 0,89 0,53 Cinnamomum amoenum 1,88 0,22 1,64 0,28 0,17 0,38 0,88 0,60
Continua
234
ANEXO 3 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Myrsine umbellata 2,58 0,30 2,35 0,39 0,07 0,15 0,85 0,45 Myrsine lorentziana 2,11 0,25 1,88 0,31 0,10 0,23 0,79 0,48 Xylosma pseudosalzmannii 2,35 0,27 2,11 0,35 0,05 0,12 0,75 0,39 Zanthoxylum rhoifolium 2,11 0,25 2,11 0,35 0,04 0,10 0,70 0,35 Stillingia oppositifolia 1,88 0,22 1,64 0,28 0,08 0,18 0,67 0,40 Dasyphyllum spinescens 1,41 0,16 1,41 0,24 0,11 0,24 0,64 0,41 Lonchocarpus campestris 1,41 0,16 1,17 0,20 0,11 0,26 0,62 0,42 Gordonia acutifolia 1,41 0,16 0,94 0,16 0,12 0,27 0,59 0,43 Scutia buxifolia 1,41 0,16 0,94 0,16 0,11 0,26 0,58 0,42 Allophylus edulis 1,88 0,22 1,64 0,28 0,03 0,07 0,57 0,29 Picramnia parvifolia 1,64 0,19 1,64 0,28 0,05 0,10 0,57 0,29 Myrciaria tenella 1,41 0,16 1,17 0,20 0,06 0,13 0,49 0,29 Myrsine coriacea 1,17 0,14 1,17 0,20 0,07 0,15 0,49 0,29 Cunila incisa 1,88 0,22 0,94 0,16 0,05 0,11 0,49 0,33 Myrceugenia oxysepala 1,64 0,19 1,41 0,24 0,02 0,05 0,48 0,25 Symplocos tetrandra 1,17 0,14 0,47 0,08 0,12 0,26 0,48 0,40 Solanum pseudoquina 1,41 0,16 1,41 0,24 0,01 0,03 0,43 0,19 Cupania vernalis 0,94 0,11 0,94 0,16 0,05 0,12 0,39 0,23 Lithraea brasiliensis 1,17 0,14 0,70 0,12 0,06 0,13 0,39 0,27 Myrrhinium atropurpureum 1,17 0,14 1,17 0,20 0,02 0,05 0,38 0,19 Rollinia rugulosa 1,17 0,14 1,17 0,20 0,01 0,03 0,37 0,17 Celtis iguanaea 1,41 0,16 0,94 0,16 0,02 0,03 0,36 0,20 Ilex dumosa 0,94 0,11 0,94 0,16 0,04 0,08 0,35 0,19
Continua

235
ANEXO 3 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Myrsine sp. 0,70 0,08 0,70 0,12 0,06 0,13 0,33 0,21 Maytenus aquifolia 0,94 0,11 0,70 0,12 0,04 0,09 0,32 0,20 Myrcia bombycina 0,94 0,11 0,94 0,16 0,01 0,03 0,30 0,14 Banara tomentosa 1,17 0,14 0,70 0,12 0,01 0,03 0,29 0,17 Maytenus evonymoides 0,70 0,08 0,47 0,08 0,06 0,13 0,29 0,21 Zanthoxylum fagara 0,70 0 ,08 0,70 0,12 0,02 0,04 0,24 0,12 Duranta vestita 0,70 0,08 0,47 0,08 0,03 0,07 0,23 0,15 Myrcianthes pungens 0,70 0,08 0,70 0,12 0,01 0,02 0,22 0,10 Alsophila sp. 0,70 0,08 0,70 0,12 0,01 0,02 0,22 0,10 Acacia bonariensis 0,94 0,11 0,47 0,08 0,01 0,02 0,21 0,13 Luehea divaricata 0,23 0,03 0,23 0,04 0,06 0,12 0,19 0,15 Cedrela fissilis 0,47 0,05 0,47 0,08 0,02 0,04 0,18 0,10 Citronella gongonha 0,47 0,05 0,47 0,08 0,02 0,05 0,18 0,10 Casearia sylvestris 0,47 0,05 0,23 0,04 0,03 0,07 0,17 0,13 Machaerium paraguariense 0,23 0,03 0,23 0,04 0,04 0,09 0,15 0,11 Seguieria aculeata 0,47 0,05 0,47 0,08 0,01 0,01 0,14 0,06 Myrceugenia myrcioides 0,47 0,05 0,47 0,08 0,01 0,01 0,14 0,07 Styrax leprosus 0,23 0,03 0,23 0,04 0,03 0,06 0,13 0,09 Griselinia ruscifolia 0,47 0,05 0,23 0,04 0,01 0,02 0,11 0,07 Solanum sp. 0,47 0,05 0,23 0,04 0,01 0,02 0 ,11 0,07 Clethra uleana 0,23 0,03 0,23 0,04 0,01 0,03 0,09 0,05 Nectandra lanceolata 0,23 0,03 0,23 0,04 0,01 0,01 0,08 0,04 Solanum sanctae-catharinae 0,23 0,03 0,23 0,04 0,01 0,01 0,08 0,04
Continua

236
ANEXO 3 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Citrus sp. 0,23 0,03 0,23 0,04 0,01 0,01 0,08 0,04 Dalbergia frutescens 0,23 0,03 0,23 0,04 0,01 0,01 0,07 0,03 Erythrina cristagalli 0,23 0,03 0,23 0,04 0,01 0,01 0,07 0,03 Strychnos brasiliensis 0,23 0,03 0,23 0,04 0,01 0,01 0,07 0,03 Zanthoxylum kleinii 0,23 0,03 0,23 0,04 0,01 0,01 0,07 0,03 Myrceugenia foveolata 0,23 0,03 0,23 0,04 0,01 0,01 0,07 0,03 Oreopanax fulvus 0,23 0,03 0,23 0,04 0,01 0,01 0,07 0,03 Symplocos pentandra 0,23 0,03 0,23 0,04 0,01 0,01 0,07 0,03 TOTAL (98) 856,34 100 596,24 100 4,86 100 300 200

237
ANEXO 4 – Parâmetros fitossociológicos das espécies do grupo 3 (Formação Araucaria) da Floresta Ombrófila Mista da FLONA de São Francisco de Paula, RS.
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Araucaria angustifolia 195,30 23,60 77,14 14,60 29,43 62,12 100,32 85,72 Blepharocalyx salicifolius 49,57 5,99 30,56 5,78 1,60 3,37 15,14 9,36 Ilex paraguariensis 45,73 5,53 33,33 6,31 1,04 2,20 14,03 7,73 Morta 39,10 4,73 31,20 5,90 1,03 2,17 12,80 6,90 Ilex brevicuspis 33,97 4,11 20,51 3,88 1,61 3,40 11,39 7,50 Banara parviflora 28,63 3,46 23,29 4,41 0,45 0,96 8,83 4,42 Sebastiania brasiliensis 32,26 3,90 21,37 4,04 0,42 0,89 8,83 4,79 Matayba elaeagnoides 24,79 3,00 15,81 2,99 0,78 1,64 7,62 4,63 Nectandra megapotamica 19,66 2,38 16,03 3,03 0,75 1,58 6,99 3,95 Ocotea puberula 17,09 2,07 12,18 2,30 1,21 2,55 6,92 4,62 Luehea divaricata 24,36 2,94 12,18 2,30 0,76 1,60 6,85 4,55 Casearia decandra 16,88 2,04 11,75 2,22 0,37 0,78 5,04 2,82 Dasyphyllum spinescens 14,74 1,78 10,04 1,90 0,43 0,90 4,58 2,68 Campomanesia rhombea 15,81 1,91 9,40 1,78 0,30 0,64 4,33 2,55 Ocotea pulchella 12,18 1,48 10,25 1,94 0,40 0,86 4,27 2,33 Myrceugenia cucullata 14,32 1,73 10,47 1,98 0,22 0,46 4,17 2,19 Xylosma pseudosalzmannii 10,47 1,27 8,76 1,66 0,30 0,62 3,55 1,89 Campomanesia xanthocarpa 11,54 1,39 8,12 1,54 0,29 0,61 3,54 2,01 Dicksonia sellowiana 9,83 1,19 7,26 1,37 0,47 0,98 3,54 2,17 Inga vera 11,54 1,39 7,91 1,50 0,28 0,59 3,48 1,98 Lonchocarpus campestris 13,46 1,63 4,49 0,85 0,36 0,76 3,24 2,39
Continua

238
ANEXO 4 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Lamanonia ternata 8,97 1,08 7,26 1,37 0,36 0,76 3,22 1,85 Cryptocarya aschersoniana 8,33 1,01 6,20 1,17 0,46 0,98 3,16 1,99 Maytenus evonymoides 9,40 1,14 6,62 1,25 0,24 0,51 2,90 1,65 Zanthoxylum rhoifolium 7,91 0,96 6,62 1,25 0,12 0,25 2,46 1,21 Maytenus aquifolia 6,84 0,83 5,13 0,97 0,27 0,58 2,37 1,40 Cinnamomum glaziovii 7,69 0,93 5,77 1,09 0,17 0,35 2,37 1,28 Eugenia involucrata 6,84 0,83 5,56 1,05 0,13 0,27 2,15 1,10 Sebastiania commersoniana 5,98 0,72 4,70 0,89 0,16 0,33 1,94 1,05 Myrsine umbellata 5,77 0,70 5,13 0,97 0,09 0,19 1,86 0,89 Prunus myrtifolia 5,34 0,65 5,13 0,97 0,11 0,23 1,84 0,87 Podocarpus lambertii 3,63 0,44 3,21 0,61 0,28 0,59 1,64 1,03 Cupania vernalis 4,70 0,57 4,27 0,81 0,06 0,13 1,51 0,70 Scutia buxifolia 4,70 0,57 3,42 0,65 0,11 0,24 1,45 0,81 Acca sellowiana 5,13 0,62 2,99 0,57 0,07 0,14 1,33 0,76 Eugenia uruguayensis 4,27 0,52 2,78 0,53 0,10 0,21 1,25 0,72 Citronella gongonha 2,99 0,36 2,78 0,53 0,11 0,23 1,11 0,59 Machaerium paraguariense 4,06 0,49 2,14 0,40 0,09 0,19 1,09 0,68 Myrrhinium atropurpureum 3,21 0,39 2,99 0,57 0,06 0,12 1,07 0,50 Solanum sp. 2,99 0,36 2,99 0,57 0,07 0,14 1,07 0,50 Eugenia psidiiflora 3,42 0,41 2,35 0,44 0,09 0,18 1,04 0,60 Rollinia salicifolia 3,21 0,39 2,99 0,57 0,03 0,07 1,03 0,46 Solanum sanctae-catharinae 2,78 0,34 2,56 0,49 0,06 0,13 0,95 0,47 Cipós 2,99 0,36 2,35 0,44 0,06 0,13 0,93 0,49
Continua

239
ANEXO 4 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Solanum pabstii 2,99 0,36 2,35 0,44 0,05 0,10 0,91 0,46 Rollinia rugulosa 2,56 0,31 2,56 0,49 0,03 0 ,06 0,86 0,37 Não identificada 2,35 0,28 2,14 0,40 0,05 0,11 0,80 0,40 Dasyphyllum tomentosum 1,92 0,23 1,71 0,32 0,10 0,21 0,77 0,44 Pilocarpus pennatifolius 2,78 0,34 1,71 0,32 0,03 0,07 0,73 0,40 Vernonia discolor 1,92 0,23 1,71 0,32 0,07 0,15 0,71 0,38 Lonchocarpus nitidus 2,56 0,31 0,85 0,16 0,08 0,16 0,63 0,47 Myrceugenia miersiana 2,35 0,28 1,28 0,24 0,03 0,06 0,58 0,34 Casearia obliqua 1,71 0,21 1,50 0,28 0,04 0,09 0,58 0,30 Ilex dumosa 2,14 0,26 1,50 0,28 0,02 0,03 0,57 0,29 Myrcianthes gigantea 1,28 0,15 1,28 0,24 0,06 0,12 0,52 0,27 Myrcianthes pungens 1,50 0,18 1,07 0,20 0,06 0,12 0,51 0,30 Banara tomentosa 1,50 0,18 1,28 0,24 0,02 0,04 0,46 0,22 Myrciaria floribunda 1,50 0,18 0,85 0,16 0,05 0,11 0,45 0,29 Picramnia parvifolia 1,28 0,15 1,28 0,24 0,01 0,03 0,43 0,18 Myrciaria delicatula 0,85 0,10 0,64 0,12 0,08 0,16 0,38 0,26 Lithraea brasiliensis 0,85 0,10 0,85 0,16 0,05 0,11 0,37 0,21 Cryptocarya moschata 1,50 0,18 0,64 0,12 0,03 0,07 0,37 0,25 Hennecartia omphalandra 1,07 0,13 0,85 0,16 0,03 0,07 0,36 0,20 Solanum mauritianum 1,06 0,13 1,06 0,20 0,01 0,04 0,36 0,16 Roupala brasiliensis 0,85 0,10 0,85 0,16 0,04 0,08 0,35 0,19 Solanum pseudoquina 1,07 0,13 0,85 0,16 0,01 0,02 0,31 0,15 Xylosma tweedianum 0,85 0,10 0,64 0,12 0,04 0,08 0,30 0,18
Continua

240
ANEXO 4 – Continuação
ESPÉCIES
DENSIDADE
FREQÜÊNCIA
DOMINÂNCIA
VALOR IMPORT.
VALOR COBERT.
DA DR FA FR DoA DoR VI VC Acacia bonariensis 0,85 0,10 0,85 0,16 0,01 0,02 0,29 0,12 Siphoneugena reitzii 0,85 0,10 0,64 0,12 0,01 0,03 0,25 0,13 Ilex sp. 0,85 0,10 0,64 0,12 0,01 0,02 0,25 0,13 Myrsine coriacea 0,64 0,08 0,64 0,12 0,02 0,05 0,24 0,12 Symplocos uniflora 0,64 0,08 0,43 0,08 0,04 0,08 0,24 0,16 Allophylus edulis 0,64 0 ,08 0,64 0,12 0,01 0,02 0,22 0,10 Stillingia oppositifolia 0,85 0,10 0,43 0,08 0,02 0,04 0,22 0,14 Oreopanax fulvus 0,64 0,08 0,64 0,12 0,01 0,02 0,21 0,09 Guapira opposita 0,85 0,11 0,42 0,08 0,01 0,02 0,20 0,12 Myrciaria tenella 0,43 0,05 0,43 0,08 0,03 0,06 0,19 0,11 Weinmania paulliniifolia 0,43 0,05 0,43 0,08 0,03 0,05 0,19 0,11 Myrcia oligantha 0,64 0,08 0,43 0,08 0,01 0,01 0,17 0,09 Myrcia arborescens 0,64 0,08 0,43 0,08 0,01 0,01 0,17 0,09 Coussarea contracta 0,64 0,08 0,43 0,08 0,01 0,01 0,17 0,09 Cunila incisa 0,43 0,05 0,43 0,08 0,01 0,02 0,16 0,08 Myrsine sp. 0,43 0,05 0,43 0,08 0,01 0,03 0,16 0,08 Cedrela fissilis 0,43 0,05 0,43 0,08 0,01 0,02 0,15 0,07 Cestrum sp. 0,43 0,05 0,43 0,08 0,01 0,02 0,15 0,07 Myrsine lorentziana 0,43 0,05 0,43 0,08 0,01 0,02 0,15 0,07 Seguieria aculeata 0,43 0,05 0,43 0,08 0,01 0,01 0,15 0,06 Zanthoxylum fagara 0,43 0,05 0,43 0,08 0,01 0,02 0,15 0,07 Symplocos pentandra 0,43 0,05 0,43 0,08 0,01 0,02 0,15 0,07 Inga marginata 0,43 0,05 0,43 0,08 0,01 0,01 0,14 0,06
Continua

241
ANEXO 4 – Continuação
DENSIDADE FREQÜÊNCIA DOMINÂNCIA VALOR IMPORT.
VALOR COBERT.
ESPÉCIES
DA DR FA FR DoA DoR VI VC Strychnos brasiliensis 0,43 0,05 0,43 0,08 0,01 0,01 0,14 0,06 Citronella paniculata 0,43 0,05 0,21 0,04 0,01 0,02 0,11 0,07 Tabebuia umbellata 0,43 0,05 0,21 0,04 0,01 0,02 0,11 0,07 Mollinedia sp. 0,21 0,03 0,21 0,04 0,01 0,02 0,09 0,04 Citharexylum solanaceum 0,21 0,03 0,21 0,04 0,01 0,01 0 ,08 0,04 Syagrus romanzoffiana 0,21 0,03 0,21 0,04 0,01 0,01 0,08 0,04 Cabralea canjerana 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Carica quercifolia 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Celtis iguanaea 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Dalbergia frutescens 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Ilex microdonta 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Inga sessilis 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Mollinedia elegans 0,21 0 ,03 0,21 0,04 0,01 0,01 0,07 0,03 Myrcia bombycina 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Rhamnus sphaerosperma 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Rollinia sylvatica 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Citrus sp. 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Myrceugenia myrcioides 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Myrceugenia glaucescens 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Myrcia palustris 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Coutarea hexandra 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Piptocarpha tomentosa 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 Solanum pseudocapsicum 0,21 0,03 0,21 0,04 0,01 0,01 0,07 0,03 TOTAL (113) 827,57 100 528,42 100 7,38 100 300 200





























