Embed Size (px)
Citation preview

1
DEFICIÊNCIA DA GLICOSE-6-FOSFATO DESIDROGENASE EM INDIVÍDUOS RESIDENTES EM ÁREAS ENDÊMICAS DE MALÁRIA DA CIDADE DE MANAUS,
AMAZONAS
MARLI STELA SANTANA MACIEL
MANAUS 2012
UNIVERSIDADE DO ESTADO DO AMAZONAS
FUNDAÇÃO DE MEDICINA TROPICAL DR. HEITOR VIEIRA DOURADO
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL
DOUTORADO EM DOENÇAS TROPICAIS E INFECCIOSAS
...
...
..

2
MARLI STELA SANTANA MACIEL
DEFICIÊNCIA DA GLICOSE-6-FOSFATO DESIDROGENASE EM INDIVÍDUOS RESIDENTES EM ÁREAS ENDÊMICAS DE MALÁRIA DA CIDADE DE MANAUS,
AMAZONAS
Tese apresentada ao Programa de Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas em Convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, para obtenção do título de Doutor em Doenças Tropicais e Infecciosas.
Orientadora: Profª. Drª. MARIA DAS GRAÇAS COSTA ALECRIM
Co-Orientador: Prof. Dr. MARCUS VINÍCIUS GUIMARÃES DE LACERDA
MANAUS 2012
.i.
...
...

3
Ficha catalográfica elaborada pela Biblioteca Central da Universidade do Estado do Amazonas
M152d Maciel, Marli Stela Santana
Deficiência da glicose-6-fosfato desidrogenase em indivíduos residentes em áreas endêmicas de malária da cidade de Manaus,
Amazonas / Marli Stela Santana Maciel. – Manaus, AM : [s.n], 2012. 117 f. : il., 29 cm. Orientador: Maria das Graças Costa Alecrim ; Co-Orientador: Marcus Vinicius Guimarães de Lacerda. Tese (Doutorado), Universidade do Estado do Amazonas, 2012.
1- Malária – Manaus 2- Glicose-6-fosfato desidrogenase - deficiência. 3- Diabetes. I- Alecrim, Maria das Graças Costa (Orient.). II- Lacerda, Marcus Vinicius Guimarães (Co- Orient.). III- Universidade do Estado do Amazonas, Programa de Pós-Graduação em Medicina Tropical, Doutorado em Doenças Tropicais e infecciosas. IV- Título.
CDU 1997 -- 616.936(043)
.i.
...
...
...
...
..

4
FOLHA DE JULGAMENTO
DEFICIÊNCIA DA GLICOSE-6-FOSFATO DESIDROGENASE EM INDIVÍDUOS
RESIDENTES EM ÁREAS ENDÊMICAS DE MALÁRIA DA CIDADE DE MANAUS,
AMAZONAS
MARLI STELA SANTANA MACIEL
“Esta tese foi julgada adequada para obtenção do Título de Doutor em
Doenças Tropicais e Infecciosas, aprovada em sua forma final pelo Programa
de Pós-Graduação em Medicina Tropical da Universidade do Estado do
Amazonas em convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira
Dourado”.
Banca Julgadora:
__________________________________________ Profa. Maria das Graças Costa Alecrim, Dra
Presidente
__________________________________________ Profa. Flor Ernestina Martinez Espinosa, Dra
Membro
__________________________________________ Prof. Marcelo Cordeiro dos Santos, Dr
Membro
__________________________________________ Profa. Leny Nascimento da Motta Passos, Dra
Membro
__________________________________________ Prof. Sergio Roberto Lopes Albuquerque, Dr
Membro
ii

5
Dedico este trabalho aos meus pais, Maria Estela e José Cândido de Santana (in memoriam), fontes de inspiração, exemplos de esforço e dedicação.
.iii.
.....
.

6
AGRADECIMENTOS
Ao concluir mais esta etapa da minha formação acadêmica, desejo aqui registrar
minha gratidão:
Quero agradecer, em primeiro lugar, a Deus, pela força, iluminação e coragem durante
toda esta longa caminhada.
À minha família, na figura de minha mãe, Maria Estela Santana, pelo exemplo de força
e dignidade.
Ao meu companheiro, José Alves Maciel Júnior, por incentivar meu crescimento
pessoal. Por suportar minhas ausências: amor eterno.
À minha mãe-sogra, Belkiss Alves Maciel, quem muito me apoiou de várias maneiras.
Aos orientadores, a Professora Dra. Maria das Graças Costa Alecrim, e Professor Dr.
Marcus Vinícius Guimarães de Lacerda, pela confiança que depositaram em mim.
À Universidade do Estado do Amazonas, através da coordenação do Programa de
Pós-graduação em Medicina Tropical e seu corpo docente, os quais foram muito
importantes em minha formação acadêmica.
À Dra. Maria das Graças Vale Barbosa, pelos bons conselhos, e pelo exemplo de
excelência profissional.
Aos secretários do Programa de Pós-Graduação em Medicina Tropical da
Universidade do Estado do Amazonas, em especial à Sra. Maria da Conceição dos
Santos Tufic, sempre paciente e gentil.
À Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, por apoiar-me no
exercício de “fazer ciência” desde a iniciação científica.
iv

7
À Fundação de Amparo à Pesquisa do Amazonas e ao Programa de Pesquisa para o
Sistema Único de Saúde, pelo suporte financeiro.
À Dra. Monica Regina Farias Costa, pelo apoio logístico na execução deste trabalho.
Aos acadêmicos do curso de Farmácia do Centro Universitário do Norte, e do curso
de Enfermagem da UEA, em especial a Gloria Lima, Elizabeth Santos Galusso,
Evaulino Ferreira Itapirema, pelo grande apoio na coleta de amostras e auxílio no
laboratório.
À Fundação de Vigilância em Saúde, através do Sr. Vanderson Sampaio, dos técnicos
e agentes de saúde pelo grande apoio no trabalho de campo.
Ao Felicien Gonçalves Vásquez, pela colaboração na análise estatística dos
resultados.
Ao amigo Wuelton Marcelo Monteiro pelas excelentes ideias na confecção dos artigos
da tese.
Às queridas amigas do Ministério da Mulher pelo apoio através de suas orações.
A todos os que embora não nomeados, têm parte neste trabalho, a certeza da minha
gratidão.
v

8
Das Utopias Se as coisas são inatingíveis... ora! não é motivo para não querê-las. Que tristes os caminhos, se não fora a mágica presença das estrelas!
Mário Quintana
vi

9
RESUMO
P. vivax é responsável pela maioria das infecções maláricas no território brasileiro, e a cura radical da doença é possível somente pelo uso da primaquina, uma droga cujas reações adversas não podem ser negligenciadas em razão de seu grande efeito oxidativo às células do hospedeiro, principalmente em indivíduos geneticamente deficientes da glicose-6-fosfato desidrogenase (G6PD). Cerca de 400 milhões de pessoas sofrem dessa enzimopatia. Este estudo teve como objetivo estudar os aspectos moleculares e bioquímicos da deficiência da glicose-6-fosfato desidrogenase, em pacientes susceptíveis à malária, de Manaus, Amazonas. Foram incluídos no estudo 1.478 indivíduos do sexo masculino, residentes nas áreas endêmicas de malária da cidade de Manaus, situadas nas Zonas Leste, Oeste e Norte da cidade. Todos foram triados para a deficiência da G6PD, pelo método quantitativo, e uma vez confirmada a deficiência, foram realizados testes moleculares para identificação das variantes da G6PD e suas mutações, com prevalência de 4.5%. Cinquenta e seis deficientes G6PD (3,8%) apresentaram a mutação compatível com a variante G6PD A-, e 10 indivíduos (0,7%) gravemente deficientes foram genotipados como portadores da variante G6PD Mediterrânea. Indivíduos portadores das variantes deficientes da G6PD foram associados com menos de três episódios de malária ao longo de suas vidas (p<0,001) e associação com a necessidade de transfusão de sangue provocado por episódios anteriores de malária (p<0,001). As análises bioquímicas mostraram que indivíduos deficientes da G6PD são mais propensos a hiperglicemia e diabetes, com uma forte associação estatística, com risco 5 vezes maior que indivíduos com atividade da G6PD normal (p<0,001). Ambas as variantes deficientes da G6PD também evidenciaram frequências significativamente maiores de diabetes do que os não-deficientes da G6PD (p<0,001). Estudar a deficiência da G6PD sob as bases bioquímicas e moleculares oferece uma melhor compreensão sobre os riscos envolvidos na deficiência da G6PD, principalmente o da hemólise, constituída como grave problema de saúde pública, principalmente em áreas endêmicas de malária por P. vivax, onde o uso da primaquina se faz necessário. Palavras-chaves: Malária. Deficiência da G6PD. Caracterização molecular. Diabetes.
vii

10
ABSTRACT P. vivax is responsible for most malarial infection in the Brazilian territory, and the radical cure of the disease is possible only by the use of primaquine, a drug whose side effects cannot be neglected because of its large oxidative effect on host cells, especially in those with genetically impaired glucose-6-phosphate dehydrogenase (G6PD). About 400 million people worldwide suffer from this enzymopathy. This study aimed to study the molecular and biochemical aspects of glucose-6-phosphate dehydrogenase in patients susceptible to malaria, Manaus, Amazonas. The study included 1478 male subjects, living in malaria-endemic areas of the city of Manaus, located in the East, West and North of the city. All were screened for G6PD deficiency by quantitative method, and once confirmed the deficiency, tests were performed to identify the molecular variants of G6PD and its mutations, with a prevalence of 4.5%. Fifty-six G6PD deficient (3.8%) had mutations consistent with the variant G6PD A-, and 10 subjects (0.7%) severely disabled were genotyped as having the G6PD Mediterranean variant. Individuals with the G6PD deficient variants were associated with less than three episodes of malaria throughout their lives (p<0.001) and association with the need for blood transfusions caused by previous episodes of malaria (p<0.001). Biochemical analyzes showed that G6PD deficient are more prone to impaired fasting glucose and diabetes, with a strong statistical association, a risk five times greater than individuals with normal G6PD activity (p<0.001). Both G6PD deficient variants also showed significantly higher frequencies of diabetes than non-deficient G6PD (p <0.001). Study of G6PD deficiency in the biochemical and molecular basis provides a better understanding of the risks involved in G6PD deficiency, especially of hemolysis, formed as a serious public health problem, especially in endemic areas of malaria by P. vivax, where the use of primaquine is necessary. Key-words: Malaria. G6PD deficiency. Molecular characterization. Diabetes.
viii

11
LISTA DE FIGURAS E QUADROS Figura 1. Áreas de risco de transmissão de malária............................................ 2 Figura 2. Ciclo biológico da malária..................................................................... 6 Figura 3. A via das pentoses-fosfato................................................................... 10 Figura 4. Atuação da G6PD na remoção de peróxidos........................................ 11 Figura 5. Corpos de Heinz em hemácias deficientes da G6PD........................... 13 Figura 6. Distribuição da deficiência da G6PD no Brasil...................................... 23 Figura 7. Distribuição mundial da deficiência da G6PD....................................... 29 Figura 8. Fluxograma da execução do estudo.....................................................
Quadro 1. Drogas e substâncias que a serem evitadas por deficientes da G6PD.....................................................................................................................
34 14
ix

12
LISTA DE TABELAS
Tabela 1. Primers utilizados na detecção das mutações na deficiência da G6PD................................................................................................................... Tabela 2. Enzimas de restrição usadas na RFLP para detecção das variantes da deficiência da G6PD.......................................................................................
38
39
x

13
LISTA DE ABREVIATURAS E SIGLAS AHNEC Anemia hemolítica não-esferocítica crônica ATP Nucleotídeo de adenosina tri-fosfato BIND Disfunção neurológica induzida pela bilirrubina
(Bilirubin-induced neurologic dysfunction) DM Diabetes mellitus DMSO Dimetilsulfóxido DNA Ácido desoxirribonucleico dNTP Desoxinucleosídeo trifosfato EDTA Ácido etilenodiaminotetracético ELISA Teste imunoenzimático (Enzyme Linked Immuno Sorbent Assay) FAPEAM Fundação de Amparo à Pesquisa do Amazonas FMT-HVD Fundação de Medicina Tropical Dr. Heitor Vieira Dourado FVS Fundação de Vigilância em Saúde GE Gota espessa GSH Glutatião reduzido GSSG Glutatião oxidado G6P Glicose-6-fosfato G6PD Glicose6-fosfato-desidrogenase G6PD Gene da glicose 6-fosfato-desidrogenase IFI Imunofluorescência indireta NADPH Dinucleotídeo fosfatado de adenina nicotinamida reduzido NADP Dinucleotídeo fosfatado de adenina PCR-PFLP Reação em cadeia de polimerase polimorfismo de tamanho do
fragmento de restrição PPSUS Programa de Pesquisa para o SUS RAVREDA Rede Amazônica de Vigilância da Resistência às Drogas Antimaláricas RNAm Ácido ribonucleico mensageiro RNAsi RNA interferente ROS Oxigênios reativos SDS Lauril sulfato de sódio SSCP Polimorfismo Conformacional de Cadeia Simples
(Single Strand Conformation Polymorphism) SUFRAMA Superintendência da Zona Franca de Manaus UV Ultravioleta
xi

14
LISTA DE SÍMBOLOS E UNIDADES DE MEDIDA
°C Grau Celsius HCl Ácido clorídrico H2O2 Peróxido de hidrogênio kb Quilobases KCl Cloreto de potássio kDa Quilodálton M Molar MgCl2 Cloreto de magnésio min mg/dL
Minuto Miligrama por decilitro
mL mM
Mililitro Milimolar
NaCl Cloreto de sódio ng Nanograma PM Picomol
U/g Hb Unidades em gramas de hemoglobina rpm Rotação por minuto μg/ml Micrograma por mililitro µL Microlitro
xii

15
SUMÁRIO
1 INTRODUÇÃO................................................................................................... 1
1.1 Epidemiologia da Malária................................................................................ 2 1.2 Transmissão da Malária.................................................................................. 4 1.3 Ciclo Biológico da Malária................................................................................ 5 1.4 Diagnóstico da Malária.................................................................................... 1.5 Tratamento da Malária....................................................................................
7 7
1.6 Bioquímica Celular.......................................................................................... 8 1.7 A Deficiência da Glicose-6-Fosfato Desidrogenase......................................... 10 1.8 Manifestações Clínicas da Deficiência da Glicose-6-Fosfato Desidrogenase......................................................................................................
12
1.9 Anemia Hemolítica Aguda............................................................................... 12 1.9.1Hemólise por Drogas..................................................................................... 1.9.2 Hemólise por Infecção.................................................................................. 1.9.3 Hemólise por Acidose Diabética...................................................................
14 14 15
1.9.4 Favismo........................................................................................................ 15 1.9.5 Anemia Hemolítica Não-Esferocítica Crônica............................................... 1.9.6 Icterícia Neonatal......................................................................................... 1.10 Deficiência da Glicose- 6-Fosfato Desidrogenase e o Diabetes..................... 1.11 Deficiência da Glicose 6-Fosfato Desidrogenase e a Malária........................ 1.12 Epidemiologia da Deficiência da Glicose-6-Fosfato Desidrogenase.............. 1.13 Aspectos Genéticos da Deficiência da Glicose-6-Fosfato Desidrogenase......................................................................................................
16 16 17 20 23
24
1.14 Polimorfismos no Gene Glicose-6-Fosfato Desidrogenase em Populações Humanas...............................................................................................................
24
1.15 Polimorfismos no Gene Glicose-6-Fosfato Desidrogenase no Brasil..................................................................................................................... 1.16 Aspectos Étnicos da População de Manaus..................................................
26 27
2 OBJETIVO S...................................................................................................... 31 2.1 Geral............................................................................................................... 32 2.2 Específicos...................................................................................................... 32 3 MATERIAL E MÉTODOS................................................................................. 33 3.1 Tipo de Estudo................................................................................................ 34 3.2 População e Área de Estudo.......................................................................... 34 3.3 Análises Laboratoriais..................................................................................... 35 3.3.1 Amostras de Sangue................................................................................... 35 3.3.2 Determinação Quantitativa da G6PD............................................................ 3.3.3 Dosagem de Glicose....................................................................................
35 36
3.3.4 Análise do DNA............................................................................................ 37 3.3.5 Diagnóstico da Malária................................................................................. 39 3.4 Tamanho da Amostra...................................................................................... 39 3.5 Seleção dos Sujeitos da Pesquisa................................................................... 39 3.6 Critérios de Elegibilidade................................................................................. 3.7 Aspectos Éticos............................................................................................... 3.8 Análise Estatística...........................................................................................
40 41 41
xiii

16
3.9 Financiamento................................................................................................. 41
4 RESULTADOS .................................................................................................. 42 4.1 ARTIGO 1........................................................................................................ 4.2 ARTIGO 2........................................................................................................ 4.3 ARTIGO 3........................................................................................................
43 56 72
5 DISCUSSÃO................................................................................................... 5.1 Prevalência da deficiência da G6PD e seus polimorfismos na população da Amazônia Ocidental.............................................................................................. 5.2 Proteção das variantes da G6PD contra a malária........................................... 5.3 Complicações decorrentes da deficiência da G6PD........................................ 5.4 Deficiência da G6PD e diabetes......................................................................
80
81 84 84 85
6 CONCLUSÕES............................................................................................... 7 RECOMENDAÇÕES.........................................................................................
88
90
8 REFERÊNCIAS BIBLIOGRÁFICAS.................................................................. 92 9 ANEXOS............................................................................................................ 9.1 Mapa da cidade de Manaus (AM), com a localização das zonas de coleta de amostras............................................................................................................... 9.2 Termo de Consentimento Livre e Esclarecido................................................. 9.2.1 Termo de Consentimento Livre e Esclarecido (pais ou responsável)............ 9.3 Ficha Cadastral/Dados Epidemiológicos........................................................ 9.4 Aprovação junto ao Comitê de Ética em Pesquisa da FMT-HVD....................
111
112 113 114 115 116
xiv

1
1 INTRODUÇÃO

2
1.1 Epidemiologia da Malária
A malária é a doença parasitária mais importante do mundo que acomete seres
humanos. Apesar de ser uma doença infecciosa prevenível e tratável, continua
causando grande impacto na saúde pública das áreas tropicais e subtropicais do
globo, devido ao seu alto grau de morbidade e mortalidade, onde as condições sócio-
econômicas são precárias (Figura 1). Cerca de 3,3 bilhões de pessoas viveram em
áreas de risco de malária no ano de 2010, e 91% das mortes ocorreram entre crianças
abaixo de cinco anos de idade e mulheres grávidas, habitantes do continente africano
(WHO, 2011).
Figura 1. Áreas de risco de transmissão de malária (WHO, 2011)
De acordo com a Organização Panamericana de Saúde, cerca de 36% da
população do continente americano vive em áreas de risco de transmissão de malária.
No Brasil, foram registrados no ano de 2009 mais de 300.000 casos da doença

3
(PAHO, 2010), sendo que 99,5% dos casos de malária ocorrem na região Amazônica,
que compreende os estados do Acre, Amazonas, Roraima, Amapá, Rondônia, Pará,
Mato Grosso, Tocantins e porção ocidental do Maranhão, em áreas onde existem
condições precárias de habitação e trabalho, as quais se encontram próximas a
florestas e a coleções de água (Gonçalves e Alecrim, 2004; Brasil, 2005).
No ano de 2011, a região Amazônica apresentou 275.031 casos positivos de
malária e, dos nove estados que compõem essa região, o Amazonas notificou 59.096
casos da doença, o que representa 21.5% do total de casos. Em Manaus, apesar do
melhoramento das condições socioeconômicas e urbanização da periferia, neste
mesmo ano foram registrados 16.709 casos, sendo 14.485 (86.7 %) causadas pelo
Plasmodium vivax, 212 (1.3%) pelo Plasmodium falciparum e 12 (0.07%) por malária
mista (P. vivax + P. falciparum) (SIVEP, 2011). Em comparação com os dados do ano
de 2010, houve um declínio dos casos de malária no estado do Amazonas (menos
14.996 casos) e no município de Manaus (menos 1.318 casos), cabendo ressaltar a
importância das ações das entidades governamentais de saúde, que têm contribuído
para a diminuição da incidência da doença na região.
A maioria dos casos de malária no Brasil relacionam-se ao P. vivax (WHO, 2011).
No entanto, apesar dessa forma de malária ter sido negligenciada por muito tempo,
por ser considerada benigna, tem-se observado, que algumas infecções pelo P. vivax
podem evoluir para casos graves, muito semelhantes aos relacionados com o P.
falciparum. Um estudo realizado em mulheres grávidas e infectadas pelo P. vivax,
demonstrou a presença de pigmento malárico na placenta (McGready et al., 2004).
Outros estudos clínicos têm apresentado achados de síndrome respiratória aguda,
malária cerebral e trombocitopenia em pacientes com malária vivax (Kochar et al.,
2005; Lomar et al., 2005; Lacerda et al., 2008). Resistência do P. vivax à cloroquina
(Alecrim et al., 1999; Swmawinata et al., 2003) ou limitação do tratamento por
deficiência da glicose-6-fosfato desidrogenase (G6PD) também são aspectos
desafiantes no estabelecimento da cura parasitológica dessa espécie de malária.

4
1.2 Transmissão da Malária
A malária humana pode ser causada por parasitos da Ordem Coccidiida, Sub-
Ordem Haemosporidiidea, Família Plasmodiidae, Gênero Plasmodium. Atualmente
são conhecidas cinco espécies do gênero Plasmodium capazes de infectar o homem
– Plasmodium falciparum, Plasmodium vivax, Plasmodium ovale, Plasmodium
malariae e Plasmodium knowlesi, dentre as quais se destaca o P. falciparum devido
ao seu alto grau de mortalidade, sendo espécie predominante no continente africano.
A malária vivax, considerada uma forma branda da doença, consiste na forma mais
amplamente distribuída em regiões extra-africanas. O P. malariae encontra-se restrito
a pequenas áreas das regiões tropicais e temperadas. O P. ovale está restrito a
algumas áreas da África e encontrado ocasionalmente na Ásia, não ocorrendo casos
autóctones no Brasil. O P. knowlesi, pode ser encontrado principalmente em regiões
do Sudeste Asiático (Singh et al., 2004; White, 2008a).
Todos os seres humanos são passíveis de contrair malária, mesmo aqueles poli-
infectados, uma vez que a imunidade não confere uma proteção total, ocorrendo
apenas uma sintomatologia branda, muito frequente em áreas endêmicas da doença,
denominada comumente de “malária assintomática”. No entanto, para que haja um
processo infeccioso bem-sucedido, há necessidade de ocorrer perfeita sincronia entre
as células do hospedeiro e o Plasmodium. Existem algumas condições relacionadas
às características das hemácias que podem conferir uma resistência natural à doença,
as quais representam formas de proteção apenas parciais, porém suficientes para
evitar situações de gravidade. Podemos incluir entre estes fatores algumas
hemoglobinopatias, as talassemias, o antígeno Duffy, o sistema ABO, a deficiência de
glicose-6- fosfato desidrogenase (G6PD) e, a deficiência de piruvato kinase (Rowe et
al., 2007; Albuquerque et al., 2010; Berghout et al., 2012; Millimono et al., 2012).
Um fator importante e determinante para a infecção malárica está na presença
do vetor da doença, um culicídeo do gênero Anopheles, cujos sub-gêneros são:
Anopheles Meigen, 1918, Cellia Theobald, 1902, Nyssorhyncus Blanchard, 1902 e
Kertezia Theobald, 1905. Há cerca de 400 espécies de mosquitos do gênero
Anopheles no mundo e, em torno de 60 delas são vetoras sob condições naturais e,
destas, por volta de 30 se apresentam com importância epidemiológica (Sallum et al.,

5
2000). Segundo Tadei e Dutary-Thatcher (2000), em estudo dos fatores determinantes
da transmissão da malária, observaram que das 33 espécies de Anopheles que
ocorrem na Região Amazônica, apenas oito estavam infectadas pelo Plasmodium,
destacando o Anopheles darlingi como o principal vetor, considerado o mais
antropofílico e eficiente da Amazônia Brasileira. Através da manipulação genética dos
vetores há possibilidade de controle da malária, impedindo – os de serem infectados
pelo Plasmodium, permitindo que se procriem e transmitam esta característica
antimalárica às populações posteriores de mosquitos (Windbichler et al., 2011).
O surgimento da resistência à quimioterapia também é um dos grandes
desafios no controle da transmissão da malária, e deve-se ao processo da evolução
das espécies, onde a presença de fármacos age como parte do mecanismo de
seleção natural dos parasitas resistentes (Target et al., 2001; Nosten e Brasseur,
2002; Hastings e Donnelly, 2005). Elevadas taxas da doença, aliadas ao uso
inadequado dos medicamentos, contribuem para o surgimento de resistência (Guerin
et al., 2002; Daily, 2006). A capacidade de adaptação dos parasitos da malária
ocorreu principalmente com o P. falciparum, que desenvolveu resistência a
praticamente todos os antimaláricos sintéticos desenvolvidos a partir da década de
40 (Hildebrando e Oliveira, 2002; Cowman e Crabb, 2006). A fim de monitorar a
resistência aos fármacos de toda a Região Amazônica, no ano de 2001 foi criada a
Rede Amazônica de Vigilância da Resistência às Drogas Antimaláricas (RAVREDA)
utilizando protocolos padronizados para avaliar a suscetibilidade dos parasitas aos
medicamentos, com suporte técnico da Organização Panamericana de Saúde. Fazem
parte da rede amazônica os países: Bolívia, Brasil, Equador, Colômbia, Guiana, Peru,
Suriname e Venezuela (PAHO, 2001).
1.3 Ciclo Biológico da Malária
O ciclo biológico da malária é basicamente o mesmo para todas as espécies
do gênero Plasmodium, com duas fases reprodutivas: uma fase em que o parasito se
reproduz assexuadamente e que ocorre no homem (hospedeiro intermediário), e outra
fase denominada sexuada que ocorre no mosquito (hospedeiro definitivo) (Figura 2).
Ao realizar o repasto sanguíneo, a fêmea do mosquito do gênero Anopheles, inocula
através da derme e vasos sanguíneos, as formas denominadas esporozoítos, os quais

6
migram através da corrente sanguínea para células do parênquima hepático, onde se
multiplicam assexuadamente (esquizogonia), produzindo os esquizontes teciduais
primários. Estas formas se rompem, liberando os merozoítos, inicialmente em
vesículas (merossomos), e em seguida atingem as células sanguíneas, fixando-se nas
hemácias, transformando-se aí em trofozoítos jovens, podendo ainda alcançar a forma
de esquizonte. Os merozoítos diferenciam-se nas formas sexuadas do parasito,
denominadas gametócitos, e quando ingeridos pelo mosquito, dão origem ao ciclo de
vida do parasito no invertebrado. Nas infecções causadas por P. vivax e P.ovale,
alguns esporozoítos permanecem nos hepatócitos, formando hipnozoítos,
responsáveis pelas recaídas da doença (Thibergue et al., 2007). No tubo digestivo do
mosquito, os gametócitos masculinos sofrem gametogênese e fertilizam o gametócito
feminino. O zigoto desenvolve-se na parede do tubo digestivo sob a forma de oocisto,
originando os esporozoítos que migram para as glândulas salivares do mosquito, onde
poderão ser novamente transmitidos ao homem.
Figura 2. Ciclo biológico da malária. (Winzeler, 2008)

7
1.4 Diagnóstico da Malária
A utilização de dados epidemiológicos e clínicos são critérios importantes no
processo de suspeita da infecção malárica, sendo a indicação de exame laboratorial
utilizada na confirmação da doença. Dentre os métodos diagnósticos da malária mais
conhecidos podemos citar o método do esfregaço sanguíneo, o imunocromatográfico,
o da Imunofluorescência Indireta (IFI), o ELISA (Imunoenzimático) e da Reação em
Cadeia da Polimerase (PCR), este último de grande importância em pesquisa, porém,
não usado rotineiramente para diagnóstico laboratorial. No entanto, o método
diagnóstico laboratorial amplamente empregado é o da gota espessa, considerado
como o “padrão ouro”, tanto na operacionalização do controle, quanto da eliminação
da doença. O mesmo é feito por meio de punção digital e distribuição adequada da
gota de sangue em lâmina de vidro, realizando a coloração e leitura ao microscópio.
A técnica permite a identificação do parasito de acordo com a espécie, estágio de
desenvolvimento, assim como quantificação da parasitemia (Power, 1982; Ministério
da Saúde, 2005; O'Meara et al., 2005; WHO, 2009; The malERA, 2011). Todavia, a
qualidade do diagnóstico pode ser inadequada e limitada por fatores relacionados ao
preparo da lâmina, devido a utilização de equipamentos de baixa qualidade e ainda
imperícia do técnico.
1.5 Tratamento da Malária
Drogas antimaláricas são utilizadas tanto na prevenção da infecção, quanto no
controle da transmissão da doença e ainda na eliminação das formas dormentes do
fígado. Baseando-se na descrição da experiência fármaco-clínica ao longo das últimas
décadas, sabe-se que as drogas que agem contra os estágios do P. falciparum
geralmente serão efetivas contra o P. malariae, e que as drogas que agem contra o
P. vivax serão eficazes contra o P. ovale e P. knowlesi. Apesar dos avanços no
tratamento da malária, há muito a ser feito no contexto da erradicação da doença,
especialmente nas áreas endêmicas, destacando-se como uma das prioridades
dessas ações, o estabelecimento de pesquisas para o desenvolvimento de novas
drogas, que possam substituir as utilizadas atualmente com perfil de resistência.
Essas prioridades também incluem a produção de novas drogas de primeira linha para
tratar a malária falciparum e vivax, em suas formas brandas e graves, drogas

8
específicas para mulheres grávidas e crianças, para profilaxia e com ação
hipnozoiticida para o P. vivax (The malERA, 2011).
A primaquina é uma droga antimalárica que tem recebido atenção especial dos
programas de erradicação da doença (Greenwood, 2008; White, 2008b). Também se
constitui na única droga capaz de estabelecer cura radical sobre as formas
parasitárias do fígado, e eliminação de formas sexuadas de todas as espécies de
Plasmodium (Shekalaghe et al., 2007; Leslie et al., 2008). Um estudo recente sugeriu
um modelo matemático com a combinação de drogas contendo a primaquina, a qual
resultou eficazmente na interrupção da transmissão da malária (Lawpoolsri et al.,
2009). No entanto, seu uso implica em manifestações clínicas desagradáveis, devido
a seu efeito hemolítico (Taylor e White, 2004; WHO, 2006a), presente em indivíduos
recebendo doses terapêuticas do fármaco, (Alving et al, 1960; Taylor e White, 2004),
durante tratamento prolongado (Clyde, 1981), e hemólise clinicamente significativa,
em indivíduos deficientes da enzima glicose-6-fosfato desidrogenase (Beutler et al.,
1989a; WHO, 2006a; White, 2008b), uma alteração genética muito comum em áreas
endêmicas de malária (Ruwende et al., 1995). Nos indivíduos deficientes da G6PD,
uma única dose de primaquina pode resultar numa significativa diminuição da taxa de
hemoglobina (Shekalaghe et al., 2007), cursando em anemia grave (Pannacciulli et
al., 1969; Reeve et al., 1992), hemoglobinúria (Sardinha, 2006) e metemoglobinemia
(Santana et al., 2007).
1.6 Bioquímica Celular
Considerando que a farmacodinâmica da primaquina pode afetar as hemácias
do hospedeiro da malária, torna-se relevante notar que a vida destas células depende
de uma série de eventos complexos e inúmeras reações químicas catalizadas por
enzimas necessárias à degradação e formação de moléculas importantes,
constituindo assim, o metabolismo celular. Devido à ausência de núcleo e
mitocôndrias, as hemácias maduras dispõem de duas vias metabólicas para a
metabolização da glicose: a via glicolítica de Embden-Meyerhof e a das pentoses-
fosfato. A via glicolítica de Embden-Meyerhof transforma a glicose em adenosina-
trifosfato (ATP). A via das pentoses-fosfato metaboliza a glicose em nicotinamida-

9
adenina-dinucleotídio fosfato reduzido (NADPH), sendo a única via utilizada pela
hemácia para produzir NADPH (Luzzatto, 2006; Beutler, 2008).
Anormalidades enzimáticas das hemácias podem desenvolver condições
clínicas diversas, incluindo anemia hemolítica, metemoglobinemia, policitemia, e
alterações não hematológicas como falhas do desenvolvimento e neurológicas. A
deficiência da G6PD é uma das mais prevalentes enzimopenias, apresentando
inúmeras mutações. Apesar disso, a maioria das variantes genéticas da G6PD com
baixa atividade não oferece graus elevados de morbi-mortalidade (Mason et al., 2007).
A G6PD é uma enzima que, em sua forma inativa, é um monômero de 515
aminoácidos, com massa molecular de 59 Quilodalton (kDa) (Rattazzi, 1968). Para
ativá-la, é necessária uma forte ligação a um substrato (NADP), e pela formação de
um dímero ou tetrâmero (Wrigley et al., 1972). Encontra-se presente no citoplasma de
todas as células, realizando papel essencial à integridade da hemácia contra danos
oxidativos. Na via das pentoses-fosfato, a G6PD age oxidando Glicose-6-fosfato
(G6P) em 6-fosfoglucolactona pela retirada dos hidrogênios (Figura 3). Estes
hidrogênios são transferidos a nicotinamida-adenina-dinucleotídeo-fosfato (NADP),
que pela ação da G6PD é reduzida a adenina-dinucleotídio-fosfato (NADPH). Os
produtos finais desta via são a pentose D-ribose-5-fosfato, utilizada na síntese de
nucleotídeos, e a coenzima NADPH, principal doadora de hidrogênio em diversas
reações enzimáticas que mantêm o status reduzido da célula (Luzzatto e Battistuzzi,
1985; Coelho et al., 2010).
O NADPH possui a função de doar hidrogênio ao glutatião oxidado
(GSSG), a fim de convertê-lo em glutatião reduzido (GSH). O GSH doa seus átomos
de hidrogênio à hemoglobina, reduzindo-a, tornando-se oxidado (GSSG) neste
processo. A conversão de GSSG a GSH é catalizada pela enzima glutatião redutase.
O GSH é responsável pela regeneração dos grupos sulfidrilas da cadeia β da
hemoglobina que se oxidaram durante o transporte de oxigênio. O NADPH também
atua na remoção dos peróxidos (H2O2) produzidos por agentes oxidantes, como certas
drogas, capazes de causar injúria na célula (Figura 4) (Luzzatto, 2006; Coelho et al.,
2010).

10
Figura 3. A via das pentoses-fosfato (Berg et al., 2011)
A atividade enzimática da G6PD gera NADPH que é utilizado para a redução
da glutationa. A glutationa reduzida restaura então a hemoglobina para a forma
solúvel. Assim, a manutenção de altas concentrações de glutationa reduzida
representa a principal defesa contra danos oxidativos à hemoglobina (Prchal e Gregg,
2005; Beutler e Duparc, 2007).
1.7 A Deficiência da Glicose-6-Fosfato Desidrogenase
A deficiência da G6PD foi descoberta durante uma investigação da ocorrência
de anemia hemolítica aguda em indivíduos com malária, tratados com drogas 6-
methoxy-8-aminoquinolinas (Cordes, 1926). No entanto, somente 30 anos depois o
mecanismo da hemólise poderia ser compreendido, ao se estudar a primaquina como

11
droga responsável por desencadear resposta hemolítica em indivíduos com hemácias
sensíveis ao fármaco, devido à deficiência da G6PD (Alving et al., 1956).
Figura 4. Atuação da G6PD na remoção de peróxidos (Beutler e Duparc, 2007)
Apenas uma parcela de indivíduos enzimopênicos apresenta manifestações
clínicas (Beutler et al., 1989a; Luzzatto e Notaro, 2001). Assim, a deficiência da G6PD
pode ser caracterizada como uma alteração genética que leva à diminuição da
atividade e estabilidade da enzima.

12
1.8 Manifestações Clínicas da Deficiência da Glicose-6-Fosfato Desidrogenase
Hemácias deficientes da G6PD, ao serem expostas a agentes oxidantes como
o peróxido de hidrogênio (H2O2) ou radicais livres, levam à formação de um complexo
hemoglobínico de conformação estrutural modificada, com oxidação dos grupos
sulfidrilas internos e consequente desnaturação da globina. O H2O2 também
desencadeia a desestruturação da membrana celular, principalmente das cadeias de
fosfolipídeos, ocasionando perda do conteúdo celular, especialmente a hemoglobina
(Srivastava e Beutler, 1970; Mashino et al., 1983; Silva et al., 2000). Por sua vez, a
globina desnaturada forma compostos insolúveis denominados corpos de Heinz,
(Figura 5) que se ligam às hemácias, tornando-as deformadas, sendo reconhecidas
pelo organismo como estranhas, e removidas da circulação pelos macrófagos, o que
leva à hemólise extravascular (Jacob e Winterhalter, 1970).
Indivíduos deficientes da G6PD são normalmente assintomáticos. No entanto,
uma vez que as hemácias sejam expostas a agentes oxidantes, ocorre desnaturação
da hemoglobina e rompimento da membrana celular. As principais consequências
clínicas da deficiência incluem anemia hemolítica aguda, anemia hemolítica crônica,
icterícia neonatal, podendo, em alguns casos, ser fatal (Beutler, 1996). A anemia
hemolítica ocorre principalmente pelo uso de algumas drogas, incluindo antimaláricos
(Beutler e Duparc, 2007). Além disso, pode ocorrer hemólise devido a outros fatores
como processos infecciosos, por ingestão de alguns alimentos (ex.: feijão fava), por
acidose diabética e diante da condição de metemoglobinemia (Beutler, 1994; Haroun,
2005; Santana et al., 2007; Cappellini e Fiorelli, 2008).
1.9 Anemia Hemolítica Aguda
Anemia hemolítica aguda é a manifestação clínica mais frequentemente associada
à deficiência da G6PD (Dhaliwal et al., 2004). Ocorre normalmente de 24 a 72 horas
após exposição ao agente estressor, com resolução da condição hemolítica de quatro
a sete dias (Edwards, 2002). Uma vez que as hemácias adultas deficientes da G6PD

13
Figura 5. Corpos de Heinz em hemácias deficientes da G6PD (Beutler, 2008)
sofram lise primeiro, os tecidos hematopoiéticos buscam compensar a perda lançando
na circulação hemácias jovens, denominadas reticulócitos, os quais possuem altos
níveis de atividade da G6PD, e são capazes de ajudar a minimizar os danos oxidativos
(Prchal e Gregg, 2005). Clinicamente, a hemólise aguda causa náusea, vômito, dor
abdominal, hemoglobinúria, insuficiência renal, esplenomegalia (Burgoine et al.,
2010), icterícia (surge quando mais da metade da população de hemácias tenham
sido hemolizadas) (Edwards, 2002). Estudos têm sugerido, ainda, que indivíduos
deficientes da G6PD são predispostos ao desenvolvimento de sepse e complicações
diante de processos hemolíticos graves, pois assim como a G6PD possui papel
importante no mecanismo anti-oxidativo, também pode ter um impacto importante na
produção e sinalização celular por meio de citocinas (Spolarics et al., 2001; Wilmanski
et al., 2005).

14
1.9.1 Hemólise por Drogas
Pessoas deficientes da G6PD possuem susceptibilidade à hemólise
intravascular aguda, quando tratados com drogas com efeito oxidante (Beutler, 1994).
Segundo a Organização Mundial da Saúde (2006a), o tratamento da malária pode
trazer grandes danos a indivíduos enzimopênicos, especialmente a primaquina, droga
utilizada na eliminação dos reservatórios de hipnozoítos do P. vivax e P. ovale
instituídos nos hepatócitos humanos e responsáveis pelas recaídas da doença. A
primaquina pode causar graves eventos hemolíticos, havendo uma sugestão de que
antes de iniciar o tratamento com este antimalárico, ou qualquer agente (Quadro 1)
comprovadamente capaz de causar injúria celular, seja realizado um teste para
deficiência da G6PD, a fim de prevenir anemia hemolítica (Beutler et al., 1989a;
Fanello et al., 2008). Nem mesmo mulheres grávidas ou que amamentem, com
atividade da G6PD normal, podem utilizar drogas oxidantes, pois os fetos e crianças
de até um ano de idade são deficientes fisiológicos da G6PD e são, portanto,
vulneráveis à hemólise (Corchia et al., 1995; Beutler, 2008).
Quadro 1. Drogas e Substâncias a serem evitadas por deficientes da G6PD
Acetanilida Glibenclamida Fenazopiridina Sulfanilamida
Cloranfenicol Azul de metileno Fenilhidrazina Sulfapiridina
Ciprofloxacino Naftaleno Primaquina Sulfasalazina
Dapsona Ácido Nalidixico Quinacrina Tiazolesulfona
Doxorubicina Niridazol Sulfacetamida Trinitrotolueno
Furazolidona Nitrofurantoína Sulfadimidina Urato oxidase
(Beutler, 1994; Vinzio et al., 2004; Mason et al., 2007)
1.9.2 Hemólise por Infecção
Um estudo realizado com 73 pacientes deficientes da G6PD durante episódio
de hemólise sugere que a infecção é o fator mais comum desencadeante de hemólise,
superando até mesmo a exposição a drogas oxidantes (Burka et al., 1966). Há, no
entanto, pouco esclarecimento quanto ao mecanismo envolvido nesse processo.

15
Sabe-se que, durante uma infecção, os leucócitos produzem substâncias oxidantes
durante a fagocitose, a fim de causar injúria ao antígeno, mas que também causam
stress nas hemácias. Todavia, tal explanação ainda é insuficiente para explicar o
mecanismo causador da hemólise, levando-se em conta a diversidade de agentes
infecciosos causadores de hemólise em deficientes da G6PD, incluindo Escherichia
coli, Salmonella, Streptococus beta-hemolítico, Ricketsia, Vírus da Hepatite e
Influenza (Frank, 2005).
1.9.3 Hemólise por Acidose Diabética
Em indivíduos deficientes da G6PD, a hemólise pode ocorrer entre pacientes
diabéticos tipo I e tipo II (Vanelli et al., 1996; Galtrey e Pathansali, 2008), como
resultado da hipoglicemia em insulino-dependentes (Shalev et al., 1985), cetoacidose
em deficientes da G6PD com variante Africana (Le Pommelet et al., 2006)
descartando, no entanto, a variante Mediterrânea (Shalev et al., 1984).
A hipótese que envolve o mecanismo da hemólise por acidose supõe que
alterações em um ou em múltiplos genes envolvidos, que atuam tanto na regulação
da secreção de insulina como na expressão da G6PD pelas células-beta, poderiam
predispor indivíduos diabéticos à cetose (Sobngwi et al., 2005). Embora pacientes
diabéticos desenvolvam hemólise durante o tratamento de cetoacidose diabética,
raramente necessitam de transfusão sanguínea (Messina et al., 2004), visto que
hemácias jovens irão surgir, com atividade da G6PD normal, e, portanto capazes de
gerar quantidade suficiente de NADPH, a fim de preservar a célula de danos oxidativos
(Errico et al., 2009).
1.9.4 Favismo
O feijão fava (Vicia faba) tem sido reconhecido ao longo da história como
agente indutor de hemólise. O matemático e filósofo Pitágoras, que viveu na Grécia
antiga, aconselhou seus seguidores a não fazerem ingestão da leguminosa,
descrevendo o “favismo”, como uma patologia desenvolvida por alguém quando
ingerisse o feijão, ou até mesmo inalasse o seu pólen (Luzzatto e Mehta, 1995;
Bhutani et al., 2004). Todos os indivíduos que apresentam “favismo”, ou seja, quadro

16
clínico de hemólise após ingestão desta qualidade de feijão é deficiente da G6PD.
Tem-se sugerido que substâncias presentes no feijão, identificados como glicosídeos
sejam responsáveis pelo desenvolvimento da hemólise (McMillan et al., 2001). No
entanto, nem todas as pessoas que são enzimopênicas e ingerem o feijão fava irão
apresentar episódio hemolítico, inferindo diretamente em indivíduos portadores de
variantes G6PD de Classe II, como é o caso da variante Mediterrânea, raramente
ocorrendo em indivíduos com variantes da classe III (ex. variante Africana) (Beutler,
1994; Mason, 1996).
1.9.5 Anemia Hemolítica Não-Esferocítica Crônica
Uma das manifestações clínicas da deficiência da G6PD é conhecida por
anemia hemolítica não-esferocítica crônica (AHNEC). Descrita primeiramente como
síndrome por Crosby (1950), mas foi através de Newton e Bass (1958) que a doença
foi relacionada à deficiência da G6PD. A AHNEC é determinada pela mutação de um
gene, propiciando hemólise, mesmo sob as condições metabólicas normais da
hemácia (Beutler, 1994; Mason, 1996). Embora na ausência de agentes estressores,
mas sob a condição de certas variantes da G6PD da classe I (ex. variante Campinas),
a hemólise, pontuada por anemia e icterícia persistentes, ocorre de modo constante
desde os primeiros anos de vida, havendo muitas vezes a necessidade de transfusão
sanguínea, principalmente diante de crises hemolíticas intensificadas por agentes
oxidantes (Fiorelli et al., 2000).
1.9.6 Icterícia Neonatal
De todas as consequências clínicas relacionadas à deficiência da G6PD,
nenhuma pode causar tantos danos fisiológicos permanentes quanto à icterícia
neonatal, que ocorre de modo frequente e grave em recém-nascidos, relacionada
principalmente à variante Mediterrânea, não descartando, entretanto, a associação
com outras variantes (Kaplan e Hammermann, 2004). Tal condição pode levar ao
kernicterus, termo utilizado para designar a disfunção neurológica induzida pela
bilirrubina, uma vez que o fígado do infante não é capaz de conjugar a bilirrubina.
Grande parte da bilirrubina indireta livre atravessa a barreira hemato-encefálica e se
deposita no cérebro, causando danos irreversíveis às células, podendo causar

17
distúrbios cognitivos, auditivos e visuais, e até mesmo levar à morte (Bhutani et al.,
2004). Assim, a deficiência da G6PD necessita ser considerada em recém-nascidos
que apresentem quadro de icterícia e hiperbilirrubinemia nas primeiras horas de vida,
pois dados da literatura indicam que 71% dos recém-nascidos com icterícia, são
deficientes da G6PD (Kaplan e Hammermann, 2004), a fim de recorrer a intervenções
capazes de minimizar as consequências devastadoras impostas por tal condição.
1.10 Deficiência da Glicose-6-Fosfato Desidrogenase e o Diabetes
O Diabetes Mellitus (DM) é uma desordem metabólica clinicamente e
geneticamente heterogênea, cujos mecanismos afetam o metabolismo dos
carboidratos, lipídios e proteínas (Conget, 2002). Considerado uma epidemia global,
atinge aproximadamente 171 milhões de pessoas no mundo, estimando-se um
aumento para 366 milhões no ano de 2030 (Wild et al., 2004). No Brasil, cerca de
7,6% da população, na faixa etária de 30 a 69 anos, são portadoras dessa
enfermidade sistêmica (Malerbi e Franco, 1992). O diabetes representa um grave
problema de saúde pública, em razão de sua elevada prevalência, acentuada
morbidade e mortalidade, além de suas repercussões econômicas e sociais
decorrentes do impacto dessas complicações (Wild et al., 2004). Caracteriza-se pela
elevação dos níveis de glicose sanguínea, causada pela deficiência total ou parcial da
secreção de insulina ou pela resistência dos receptores celulares à sua ação, o que
ao longo de sua evolução crônica gera danos a órgãos vitais, incluindo coração, rins,
olhos, fígado e sistema vascular (AAD, 2012).
A Associação Americana de Diabetes (2012) planificou a classificação atual do
diabetes mellitus, baseada na fisiopatologia de cada forma da doença. As duas
principais formas da doença são o diabetes mellitus tipo I e o diabetes mellitus tipo II,
compreendendo as duas principais formas de diabetes mellitus em importância clínica
e prevalência. O diabetes melittus tipo 1, também denominado insulino-dependente,
é uma doença auto-imune que pode desenvolver-se em qualquer faixa etária, sendo
mais comum em crianças ou adultos, antes dos 30 anos de idade (Lernmark, 1999).
É caracterizada pela destruição das células beta produtoras de insulina no pâncreas,
levando a uma perda total ou parcial da atividade secretora de insulina (Mathis et al.,
2001; Kawasaki et al., 2004). Por outro lado, o diabetes mellitus tipo II, também

18
chamado de não- insulino dependente, é caracterizado pela redução da sensibilidade
das células-alvo ao efeito da insulina, pela incapacidade da célula beta em manter
uma adequada secreção de insulina (Mahler e Adler, 1999), e ainda pelo aumento da
produção hepática de glicose, sendo mais frequente a partir dos 40 anos de idade, e
relacionada a fatores de risco como obesidade, tabagismo, stress e sedentarismo
(Lyra et al., 2006). Um agravante do diabetes tipo II é que cerca de 50% de seus
portadores desconhecem o diagnóstico, pois a hiperglicemia geralmente se
desenvolve de forma gradual e nos estágios iniciais raramente evidenciam-se os
sintomas clássicos da doença (Malerbi e Franco, 1992).
Tanto no diabetes mellitus tipo I quanto no diabetes mellitus tipo II ocorre perda
de células beta, de maneiras distintas. No diabetes tipo I, as células betas são
destruídas por um processo autoimune, enquanto no diabetes tipo II, a atividade
fisiológica das células beta diminui gradualmente. A exposição constante à glicose
sanguínea contribui grandemente para o decréscimo das células beta, que também
são sensíveis a agentes oxidantes, principalmente oxigênios reativos, capazes de
promover a falência destas células. Ao contrário disso, estudos têm demonstrado que
a minimização da ação do stress oxidativo promove melhora da função das células
beta (Robertson et al., 2005; Wajchenberg, 2007).
A associação da deficiência da G6PD com o diabetes tem sido alvo de estudos
ao longo dos anos (Saha, 1979; Niazi, 1991; Zhang et al., 2000; Gaskin et al., 2001;
Sobngwi et al., 2005; Xu et al., 2005; Nkhoma et al., 2009; Zhang et al., 2010; Lee et
al., 2011). Outros estudos têm ainda demonstrado significância estatística em
elevadas prevalências de diabetes melittus entre indivíduos deficientes da G6PD
(12.4% a 19.6%), principalmente em indivíduos do gênero masculino, independente
da idade, sugerindo desta maneira, que todos os pacientes com diabetes sejam
triados para a enzimopatia, a fim de evitar o uso de drogas ou agentes hemolíticos
(Saeed et al., 1985; Niazi, 1991). Uma das hipóteses em torno dessa associação
demonstra que as alterações nos genes que controlam tanto a secreção de insulina
quanto a ação da G6PD regulada por agentes antioxidantes, como o NADPH,
contribuem para uma predisposição ao surgimento do diabetes (West, 2002).

19
A despeito de todos os elementos do sistema antioxidante serem importantes
para a manutenção da célula, a enzima G6PD é ímpar para a produção de NADPH,
que é o principal redutor intracelular a promover ação antioxidante às peroxidases
(Pandolfi et al., 1995; Martini e Ursini, 1996). A G6PD é a enzima que atua na via das
pentoses-fosfato, produzindo ao final da via ribose-5-fosfato e NADPH (Salvemini et
al., 1999; Matsui et al., 2006). O NADPH é usado pelos sistemas glutatião e
tiorredoxina a fim de regenerar formas reduzidas que serão utilizadas como
antioxidantes. A catalase, por exemplo, que converte peróxido de hidrogênio em água
e oxigênio, não usa NADPH diretamente, mas um sítio alostérico essencial do NADPH
mantém a catalase em sua conformação tetramérica mais ativa e protegida contra a
toxicidade do peróxido de hidrogênio (H2O2) (Kirkman et al., 1999). Portanto, a
redução da atividade da G6PD e, como resultado, diminuição dos níveis de NADPH,
irá prejudicar o sistema antioxidante completamente, levando à destruição de células
beta pancreáticas.
A hiperglicemia também está associada a distúrbios que afetam o potencial
redutor das células, em especial o NADPH e níveis de glutatião reduzido, que são
diretamente dependentes da atividade da G6PD. Um estudo realizado por Díaz-Flores
et al., (2006) demonstrou que a G6PD apresentou sensibilidade diante das diferentes
condições de hiperglicemia e ainda mostrou que o NADPH/NADP é regulado pela
G6PD tanto no fígado, quanto nas células beta-pancreáticas, onde os níveis de
glutatião reduzido são dependentes do NADPH.
A fim de melhor elucidar o mecanismo envolvido na hiperglicemia e inibição da
atividade da G6PD, Zhang et al., (2010) analisaram as ilhotas de Langherans de ratos
e humanos, as células MIN6, que são células pertencentes a uma linha de insulinoma,
incluindo a avaliação de ratos deficientes da G6PD, mostrando que altos níveis de
glicose inibem tanto a expressão quanto a atividade da G6PD nas ilhotas. A inibição
da G6PD pelo siRNA (pequeno fragmento de RNA envolvido no controle da expressão
gênica) levou a um aumento de oxigênios reativos (ROS) e apoptose, com proliferação
diminuída, prejudicando a secreção de insulina. A glicose diminuiu a secreção de
insulina, o que foi melhorado pela superexpressão da G6PD. A expressão da G6PD
nas ilhotas de Langherans foi baixa, comparada à de outras células pancreáticas, no
entanto, o estudo mostrou que as ilhotas de ratos deficientes da G6PD, apresentaram-

20
se menores e com intolerância à glicose, em comparação com os ratos-controle, o
que sugere que a deficiência de G6PD, por si só leva à disfunção e morte das células
beta. Assim, este estudo buscou mostrar a interdependência entre a deficiência da
G6PD e o diabetes: a deficiência da G6PD pode levar ao diabetes, pela destruição
das células beta; e o diabetes, diante da hiperglicemia inibe a expressão da G6PD,
cursando em um decréscimo da atividade enzimática.
1.11 Deficiência da Glicose-6-Fosfato Desidrogenase e a Malária
O conhecimento de que algumas drogas antimaláricas podem induzir hemólise
diante da deficiência da G6PD (Alving et al., 1956) fez sobressair a observação sobre
extensas áreas geográficas endêmicas de malária onde coincidentemente existem
altas prevalências da deficiência da G6PD (Motulsky, 1960; Peters e Noorden, 2009).
Daí o investimento em pesquisas que buscam avaliar a relação entre a malária e a
deficiência da G6PD, as quais buscam sustentar a hipótese de proteção à malária
diante da deficiência da enzima.
Uma vez que a deficiência da G6PD encontra-se amplamente distribuída em
áreas endêmicas de malária, principalmente na África, Ásia, Europa Mediterrânea e
algumas regiões das Américas, estudos têm sugerido a deficiência como forma de
proteção contra infecções maláricas pelo P. falciparum, assim como as formas graves
da doença (Greene, 1999; Tishkoff et al., 2001; Guindo et al., 2007; Nkhoma et al.,
2009).
Estudos in vitro têm comparado o crescimento dos parasitas da malária em
hemácias deficientes da G6PD com células normais, mostrando que há significativo
decréscimo da densidade parasitária em células deficientes (Roth et al., 1983; Roth e
Schulman, 1988). Beutler (1973) relatou que a deficiência da G6PD protegeu soldados
afro-americanos no Vietnã, os quais não haviam tido nenhum contato prévio com a
doença, e cujos níveis de parasitemia, tanto na malária vivax quanto na falciparum,
foram significativamente maiores em indivíduos normais do que em deficientes da
G6PD.

21
A distribuição geográfica da deficiência da G6PD, e de outros fatores
sanguineos como a talassemia α, hemoglobina S, ovalocitose, e do grupo sanguíneo
Duffy-negativo são alguns exemplos de que a genética das diferentes populações
evoluiu para se proteger contra a malária (Kwiatkowski, 2005). A teoria de que a
resistência à malária grave esteja baseada na alta frequência da deficiência da G6PD
em áreas endêmicas de malária e que hemizigotos e heterozigotos encontram-se em
situação de vantagem foi estabelecida por Rwende et al., (1995), através de dois
estudos de caso-controle realizados com mais de 2.000 crianças africanas. Concluiu-
se que a variante enzimática da deficiência comum na África (G6PD A-) estava
associada a 46 a 58% da redução do risco de malária grave, tanto em heterozigotos
do gênero feminino, quanto em hemizigotos masculinos.
O possível mecanismo envolvido na resistência natural de indivíduos
deficientes da G6PD se deve ao fato do metabolismo essencial à sobrevivência do
parasito da malária encontrar-se comprometido, uma vez que o mesmo invade as
hemácias a fim de evadir-se do sistema imune. Um dos produtos da oxidação
hemoglobínica é o ferro oxidado, altamente tóxico ao parasita, devido à diminuição da
atividade da G6PD dentro das hemácias, que é o sistema capaz de providenciar
glutatião reduzido à célula, impedindo sua oxidação (Tripathy e Reddy, 2007). A
diminuição da atividade do sistema antioxidante também pode facilitar a ocorrência de
danos à membrana celular, seguida de fagocitose das hemácias parasitadas
(Cappadoro et al., 1998).
No entanto, enquanto estudos buscam evidências clínicas para dar suporte à
hipótese que a deficiência da G6PD confere diminuição do risco de infecção malárica
grave em homens hemizigotos (Guindo et al., 2007) e mulheres heterozigotas para a
deficiência da G6PD (Parikh et al., 2004), ainda há muitas controvérsias em ambos os
casos (Bienzle et al., 1972; Rwende et al., 1995; Guindo et al., 2007; Clark et al.,
2008), tanto em mostrar que não há efeito algum sobre a ocorrência de malária não
complicada em homens hemizigotos ou mulheres heterozigotas (Enevold et al., 2005),
devido à diversidade nos modelos dos estudos, nos vários métodos diagnósticos
empregados na detecção da deficiência da G6PD ou ainda da variabilidade de
fenótipos enzimáticos.

22
A análise do gene G6PD também revela que os alelos da deficiência
demonstram alguns sinais de seleção quanto à malária (Tripathy e Reddy, 2007). Um
exemplo é a distribuição geográfica das variantes da G6PD (Motulski, 1960), onde o
alelo G6PD* A-, com atividade enzimática de 10% a 50%, ou seja, variantes que
predispõe o indivíduo à hemólise somente mediante exposição a agentes oxidantes
como a primaquina, do contrário permanecendo assintomáticos (Beutler et al., 1989b;
Rwende et al., 1995). Uma das variantes da deficiência da G6PD que tem sido alvo
de estudo é a G6PD A-, muito frequente em regiões endêmicas de malária da África,
sugerindo que esta variante limita a apresentação do P. falciparum ao sistema imune
mediado por células, levando a uma fraca resposta de anticorpos específicos (Courtin
et al., 2011).
Quanto às espécies de malária, a maioria dos estudos tem sugerido que a
deficiência da G6PD possui efeito protetor à malária falciparum (Roth et al., 1983;
Guindo et al., 2007), havendo escassos estudos relacionados ao P. vivax. Ao contrário
das vastas regiões malaríngeas da África, a malária por P. vivax representa cerca de
85% dos casos da doença na Amazônia Brasileira, consistindo em um grave problema
de saúde pública, que necessita ser considerado.
Um estudo conduzido na Região da Amazônia Ocidental sugeriu uma proteção
significativa contra episódios de malária vivax, em homens deficientes da G6PD,
independente da faixa etária (Santana et al., 2009). Assim, estudos baseados nos
aspectos clínicos e epidemiológicos da malária por P. vivax têm sido realizados nesta
região, a fim de buscar melhor esclarecimento quanto aos principais aspectos
envolvidos na repercussão dessa espécie de malária, tais como tratamento,
patogênese e mecanismo de resistência às drogas (Ramos Júnior et al., 2010; Gama
et al., 2011; Lacerda et al., 2012).

23
1.12 Epidemiologia da Deficiência da Glicose-6-Fosfato Desidrogenase
Cerca de 400 milhões de pessoas da população mundial são afetadas pela
deficiência da enzima G6PD (Beutler et al., 1989a; Nkhoma et al. 2009). As
frequências dessa enzimopenia na população mundial podem alcançar Índices de
70%, como é o caso dos Judeus Kurdos (Nkhoma et al., 2009) e até sua total ausência
nas populações indígenas do Brasil (Weimer et al., 1993).
Estudos desenvolvidos em diferentes regiões do Brasil demonstraram
prevalências dessa enzimopenia em torno de 3.0 % e 8.9%, principalmente estudos
realizados entre indivíduos do gênero masculino ((Marques e Campos 1975; Kuhn et
al., 1983; Hamel et al., 2002; Katsuragawa et al., 2004; Castro et al., 2006; Oliveira et
al., 2009; Santana et al., 2009; Maia et al., 2010; Cardoso et al., 2012) (Figura 6).
Figura 6. Distribuição da deficiência da G6PD no Brasil (Marques e Campos 1975;
Kuhn et al., 1983; Hamel et al., 2002; Katsuragawa et al., 2004; Castro et al., 2006;
Oliveira et al., 2009; Santana et al., 2009; Maia et al., 2010; Cardoso et al., 2012).

24
No entanto, estudos realizados na Amazônia Brasileira confirmaram a ausência ou
baixa frequência da deficiência da G6PD, assim como a de outras doenças
hematológicas, entre populações indígenas (Neel et al., 1964; Saldanha et al., 1976;
Callegari-Jacques et al., 1996).
1.13 Aspectos Genéticos da Deficiência da Glicose-6-Fosfato Desidrogenase
A clonagem do gene G6PD e a descrição de sua estrutura molecular trouxeram o
conhecimento de que o gene que codifica a G6PD está localizado no braço longo do
cromossomo X (locus q28), contendo 13 exons e 12 introns, distribuídos em uma
sequência com cerca de 21 kilobases (kb) de comprimento (Martini et al., 1986;
Takizaua et al., 1986; Chen et al., 1991; Toniolo et al., 1991; Vulliamy et al., 2007). A
região do éxon 1 e parte do éxon 2 corresponde à porção não traduzida do mRNA.
Desta maneira, os outros 12 éxons formam o mRNA e serão traduzidos na síntese
enzimática (Martini et al., 1986).
A enzimopenia possui um padrão de herança ligada ao sexo (X recessivo) e define
as representantes do gênero feminino como deficientes homozigotas ou
heterozigotas, enquanto os representantes do gênero masculino deficientes em
hemizigotos (Beutler, 1996). A heterozigoze feminina ocorre devido a certo grau de
lyonização (Lyon, 1961), ou seja, inativação aleatória do cromossomo X,
apresentando uma população mista de hemácias, uma parte deficiente e outra com
função normal da G6PD (Davidson et al., 1963), o que dificulta muitas vezes o
diagnóstico da enzimopenia feminina por métodos bioquímicos. O mesmo pode ser
feito com segurança através de técnicas moleculares.
1.14 Polimorfismos no Gene Glicose-6-Fosfato Desidrogenase em Populações
Humanas
A deficiência da G6PD também pode ser avaliada do ponto de vista de suas
mutações (Hirono e Beutler, 1988; Beutler et al., 1989b), cujo gene exibe notável
polimorfismo, com 185 mutações descritas pela literatura (Minucci et al., 2012). Em
sua maioria, as mutações do gene G6PD são pontuais, com pequenas deleções
determinantes dos defeitos estruturais, grande parte caracterizada por substituições

25
de nucleotídeos (Beutler e Vulliamy, 2002), e instabilidade da enzima, esta última
causada pela diminuição da afinidade da enzima G6PD a seus substratos (NADP ou
G6P) (Luzzatto, 2006). Além do mais, podem-se observar várias mutações
relacionadas à mesma variante, como é o caso da variante A-, que representa um
grupo de variantes que tem a mesma mutação, 202 G-A, a mais usual; ou ao contrário
disso, a mesma mutação, em termos de sequência de DNA, presente em variantes
distintas, condição relacionada, por exemplo, às variantes Matera e Betica (Beutler et
al., 1989b).
O alelo G6PD* A (ou G6PD A- ) o mais frequente em populações humanas, e
contém duas mutações, G-A no nucleotídeo 376 e G-A no nucleotídeo 202 (Takizawa
e Yoshida, 1987). A mutação 376 ocorre no alelo G6PD* A, responsável pela variante
G6PD A – ou Africana, mas somente associada à mutação 202 G-A é capaz de causar
deficiência (Hirono e Beutler, 1988). Tal associação pode ser ainda encontrada junto
a outras mutações, originando tres conjuntos de mutações diferentes responsáveis
distintamente pelo fenótipo G6PD A-, cada qual decorrente de dois pontos de mutação
em diferentes nucleotídeos do gene: 376G/202A, 376G/968C e 376G/680T (Beutler et
al., 1989b; Beutler e Kuhl, 1990). Esses três conjuntos de genótipos podem
apresentar mudanças em vários aminoácidos (Hirono e Beutler, 1988; Vulliamy et al.,
1992), mas duas das mutações encontradas em cada um dos três conjuntos de alelos
A- são necessárias para produzir o fenótipo deficiente (Town et al.,1992).
Os polimorfismos da G6PD variam imensamente nas diferentes populações do
mundo (Figura 7), sendo que a variante africana (G6PD A-) constitui a de maior
abrangência, muito frequente entre os afrodescendentes e populações do Sul da Itália,
Portugal, Espanha e Península Arábica, e apresenta prevalências de 3.6 a 28%. A
variante G6PD Mediterrânea apresenta a forma mais grave da deficiência, prevalente
em populações originárias da região do Mar Mediterrâneo, especialmente em regiões
da Itália como Sicília e Sardenha, árabes, persas e judeus orientais, com frequências
de 2 a 20% (Beutler, 1996). As variantes da G6PD possuem estrutura e estabilidade
diversificadas, e são determinantes no grau de hemólise - moderada, intermediária e
grave - podendo ser agrupadas em quatro classes principais, de acordo com a
atividade da enzima e manifestações clínicas (Beutler et al., 1989a; Beutler, 1994;
Minucci et al., 2012):

26
Classe I: graves mutações, que ocorrem provavelmente na região da enzima onde
se liga o NADP1 ou G6P, geralmente associadas à anemia hemolítica não-
esferocítica crônica (<10% de atividade). Ex. variante Campinas. Com significado
clínico (Baronciani et al., 1993).
Classe II: deficiência grave (<10% de atividade). Ex. variante Mediterrânea
(Vulliamy et al., 1988) e Santamaria (Beutler et al., 1991; Cittadella et al., 1997).
Com significado clínico, mas assintomática. Diante de stress oxidativo, apresenta
hemólise não-limitada, ou seja, tanto hemácias jovens, quanto adultas, e a
hemólise cessa somente após remoção dos agentes estressores (Beutler et al.,
1954; Dern et al., 1954).
Classe III: deficiência moderada (10 a 60%). Ex. variante Africana (Takizawa e
Yoshida, 1987). Com significado clínico, porém assintomática. Variantes com as
maiores prevalências, apresenta hemólise auto-limitada, capaz de afetar somente
hemácias adultas (Beutler e Duparc, 2007).
Classe IV: assintomática, de atividade normal ou levemente diminuída (60 a
100%). Ex. variante São Borja (Weimer et al., 1993). Sem significado clínico e
assintomática.
1.15 Polimorfismos no Gene Glicose-6-Fosfato Desidrogenase no Brasil
A heterogeneidade do gene G6PD encontra-se representada na população
brasileira, onde foi relatada a presença de variantes já descritas na literatura, com
predominância das variantes Africana/G6PD A- (202 G-A; 376 A-G) e G6PD
Mediterrânea (563 C-T) (Saad et al., 1997; Compri et al., 2000; Castro et al., 2007;
Oliveira et al., 2009; Cardoso et al., 2012). Outras variantes já conhecidas por estudos
genéticos anteriores também foram identificadas, tais com G6PD Seatlle (844 G-C)
(Weimer et al., 1998; Hamel et al., 2002; Mezzacappa et al., 2010), G6PD Chatam
(Saad et al., 1997), G6PD Santamaria (542 A-T, 376 A-G e G6PD Tokyo (1246 G-A)
(Hamel et al., 2002), assim como a identificação de novas variantes: G6PD Campinas
(1463G-T) (Baronciani et al., 1993); G6PD Sumaré (1272 T-G) (Arruda et al., 1997);
Lages (40G--A), Farroupilha (977C-A) (Weimer et al., 1998); G6PD Belém (409 C-T),

27
G6PD Ananindeua (376 A-G, 871 G-A), G6PD Crispim (375 G-T, 379 G-T, 383 T-C, e
384 C-T) e G6PD Amazonia (185 C-A) (Hamel et al., 2002).
1.16 Aspectos Étnicos da População de Manaus
Compreender a deficiência da G6PD como elemento útil no estudo da
composição genética da população, pode trazer à tona riqueza social e histórica da
ocupação do território brasileiro a partir do ano de 1500, assim como da região
Amazônica, que teve seu início no século XVII, com a colonização portuguesa (Santos
e Guerreiro, 1995). Neste período da história os indígenas eram maioria, numa
estimativa de cerca de 6.8 milhões de nativos (Cunha, 1992) espalhados ao longo do
espaço nacional. Todavia, foram desaparecendo aos poucos do ambiente selvático,
devido aos conflitos de posse territorial, epidemias de doenças infecciosas, inclusive
a malária, e ainda pela incorporação do elemento indígena ao contexto urbano do
homem branco, por meio de casamentos mestiços politicamente incentivados por
favorecimentos com armas, terras e dinheiro, estendidos principalmente aos soldados
portugueses (Cruz, 1973).
A chegada dos negros se deu a partir de 1538, com o início do tráfico de negros
para o Brasil, de modo que em 1798 somente 39% da população brasileira era
constituída por famílias europeias, ou seja, indígenas e negros eram a maioria da
população. Esse quadro foi mudando à medida que o governo português passou a
incentivar a imigração europeia compreendida por espanhóis, italianos e alemães. A
chegada dos negros na Amazônia se intensificou a partir da criação, da Companhia
Geral de Comércio do Grão-Pará e Maranhão, em 1755. Pretendia-se substituir a sua
mão-de-obra pela negra, no trabalho agrícola. O Amazonas, no período colonial, não
recebeu grandes contingentes de escravos negros (Reis, 1989). Documentos ainda
indicam que o número de escravos, que a província registrava em 1860, era de 1026
escravos (Ferreira, 2007).
Os princípios da recolonização brasileira ocorreram de fato, após a abolição da
escravatura, quando a mão-de-obra escrava deu vez ao trabalho dos imigrantes que
chegaram para trabalhar principalmente nas lavouras de café, a maioria deles
italianos, oportunizando um momento político para a chegada de imigrantes de outras

28
origens como portugueses, espanhóis, japoneses e alemães (Seyferth, 2002). Judeus
e árabes vindos, a maioria, do Marrocos e Oriente Médio, iniciaram sua chegada à
Amazônia, a partir do ano de 1810, encontrando no Brasil condições políticas
favoráveis, muitos imigraram para o Amazonas, em busca de sobrevivência e
liberdade religiosa (Safady, 1973; Benchimol, 2008).
Considerando que a deficiência da G6PD encontra-se relacionada à etnia,
constitui elemento útil para estudar sua ocorrência nas comunidades brasileiras, que
foram incorporando ao longo de sua história, povos de várias origens e com diversos
graus de miscigenação, como é o caso de Manaus, capital do Estado do Amazonas,
uma cidade marcada pelos traços étnicos herdados dos ameríndios, que iniciaram a
ocupação humana na Amazônia.
A deficiência da G6PD encontra-se frequente em cerca de 3.0% da população
masculina de Manaus (Santana et al., 2009). No entanto, sua presença nesta
população não pode ser atribuída ao ancestral ameríndio, uma vez que estudos sobre
a deficiência da G6PD entre indígenas da Amazônia demonstram que a enzimopenia
entre os mesmos é praticamente inexistente (Neel et al., 1964; Saldanha et al., 1976).
Também não parece estar ligada ao elemento negro, pois segundo o censo de 2000,
realizado pelo Instituto Nacional de Geografia e Estatística (IBGE), mais de 10 milhões
de brasileiros foram classificados como “pretos”, o que corresponde a 6,21% da
população total. Destes, mais de 87 mil estão no Amazonas e pouco mais de 34 mil,
em Manaus, o que representa somente 2.43% da raça (IBGE, 2000), sendo que a
contribuição do elemento negro na constituição da população de Manaus, do ponto
de vista genético, não pode ser considerada significante, quando comparada a de
outras linhagens genéticas originalmente Africanas (Carvalho et al., 2010).

29
Fig
ura
7.
Dis
trib
uiç
ão
mu
nd
ial da
deficiê
ncia
da
G6
PD
. W
HO
, 19
89a
, A
da
pta
do
po
r P
ete
rs e
No
ord
en
, 2
00
9;
Be
utle
r e
Du
pa
rc.
G6
PD
De
ficie
ncy W
ork
ing
Gro
up
, 2
00
7, A
dap
tado
.

30
Estudar as correntes imigratórias para o Brasil, em especial para a região
Amazônica, é importante, pois evidências epidemiológicas demonstram que a
deficiência da G6PD ocorre com frequência nas populações que imigraram para o
Amazonas provenientes da África, Ásia, Oriente Médio, Europa Mediterrânea (Frank,
2005), podendo assim trazer maior esclarecimento diante principalmente das
possíveis variantes da deficiência da G6PD que venham surgir a partir de estudos
genéticos.
A cidade de Manaus também possui em sua geografia áreas endêmicas de
malária, formadas pelas zonas leste, norte e oeste da cidade, com infecções causadas
em sua maioria pelo P. vivax, o que constitui fator de risco a essas populações,
considerando que indivíduos afetados por essa condição genética podem sofrer
hemólise por uso da primaquina, droga usada prioritariamente na cura desta espécie
de malária. Todavia, a literatura relacionada à caracterização molecular da deficiência
da G6PD nessas áreas malaríngeas ainda é escassa, o que representa uma
oportunidade de através deste estudo buscar identificar as possíveis alterações
genéticas e bioquímicas que dizem respeito a essa enzimopenia.

31
2 OBJETIVOS

32
2.1 Objetivo Geral
Estudar a deficiência da G6PD em indivíduos residentes em áreas endêmicas
de malária da cidade de Manaus, Amazonas.
2.2 Objetivos Específicos
2.2.1 Estimar a frequência da deficiência da G6PD.
2.2.2 Quantificar a atividade da G6PD.
2.2.3 Identificar os polimorfismos relacionados à deficiência da G6PD.
2.2.4 Estudar a relação da deficiência da G6PD e a malária.
2.2.5 Estudar a relação entre a deficiência da G6PD e o diabetes.

33
3 MATERIAL E MÉTODOS

34
3.1 Tipo de Estudo
Tratou-se de um estudo transversal e descritivo, para estudo do perfil molecular
e bioquímico da deficiência da G6PD. O estudo foi conduzido de acordo com esquema
da Figura 8, desde a seleção dos sujeitos até a caracterização molecular das
amostras.
Figura 8. Fluxograma da execução do estudo
3.2 População e Área de Estudo
A população de estudo foi constituída por indivíduos que concordaram em
participar do estudo, residentes nas zonas Leste, Oeste e Norte do município de
Manaus, Amazonas (ANEXO 9.1), cujas áreas apresentam os maiores IPAs (Índice
Parasitário Anual) do município: zona Leste: 3.5; zona Oeste 0.5; zona Norte: 103.0,
com suas respectivas populações estimadas em número de habitantes: 554.723,
555.649, 218.485 (IBGE, 2011; SIVEP, 2011).

35
Quanto às características da cidade de realização do estudo, Manaus é a
capital do estado do Amazonas, localizada sob as coordenadas uma
-03° 06' 07'' latitude sul e -60° 01' 30'' longitude oeste, na região norte do Brasil,
constituída por uma população de 1.832.423 habitantes e densidade populacional de
160,72 habitante/km2 (IBGE, 2011).
O clima da cidade de Manaus é considerado equatorial úmido tipo Af (Peel et
al., 2007) com temperatura média anual de 33 °C, tendo uma umidade relativa elevada
durante o ano, com médias mensais entre 76 e 89%, e precipitação média anual de
2.194,9 mm, sendo o inverno Amazônico marcado por elevado índice pluviométrico
que chega a 310,2 mm. O clima, a temperatura e o índice de chuvas influenciam
diretamente a densidade vetorial e o ciclo da malária (Souza-Santos, 2002).
3.3 Análises Laboratoriais
As determinações laboratoriais foram conduzidas nos Laboratórios de Malária,
de Virologia e de Análises Clínicas da Fundação de Medicina Tropical Dr. Heitor Vieira
Dourado (FMT-HVD) e no laboratório de Genética Belo Horizonte.
3.3.1 Amostras de Sangue
Foram coletadas amostras de sangue de cada indivíduo por punção venosa 10
mL em tubo com anticoagulante EDTA (ácido etilenodiaminotetracético) para a
dosagem quantitativa da G6PD e para identificação de variantes da G6PD por PCR-
PFLP. Para a dosagem da glicose foram coletados 5.0 mL de sangue em tubo a vácuo
sem anticoagulante, para a dosagem da glicose, com o devido preparo em jejum de 8
a 12 horas (AAD, 2012).
3.3.2 Determinação Quantitativa da G6PD
Foram coletados 10 ml de amostra de sangue total de cada indivíduo, em tubos
à vácuo contendo anticoagulante EDTA, armazenados à temperatura de 2° a 8°C e
processadas dentro de 48 horas na realização do rastreamento da deficiência da
G6PD e até 72 horas nas análises moleculares. A atividade enzimática foi mensurada

36
utilizando-se kit Neolisa® G6PD (Interscientífica Corporation, USA), utilizando-se 5µL
de sangue total, transferidos para uma placa de microtitulação com fundo em “U”,
contendo 96 poços. Adicionou-se 75µL de reagente de eluição. Colocou-se a placa
sobre um agitador orbital por 20 minutos, à temperatura ambiente. Transferiu-se 15µL
do eluato para a placa de fundo chato, adicionando-se a seguir 75µL do reagente de
trabalho. Realizou-se, imediatamente, a leitura em 405nm (DO) de ponto final, sob a
leitora digital da marca Expert Plus ASYS HITECH GMBH, para mensurar a
quantidade de hemoglobina (Hb) da amostra. Incubou-se (temperatura ambiente) a
placa no agitador por 10 minutos, e após isso, adicionou-se o reagente de cor.
Incubou-se a placa novamente por 10 minutos, à temperatura ambiente, realizando
finalmente a segunda leitura em 570nm em modo cinético, por 10 minutos, com
intervalo de 60 segundos entre as leituras, para medida do NADPH produzido. As
amostras-controle foram cedidas pelo fabricante do kit, designados em níveis de
atividade da enzima G6PD: Atividade normal= 15.8 U/g Hb; Atividade intermediária=
4.7 U/g Hb e Atividade deficiente= 1.3 U/g Hb. Os resultados foram gerados a partir
das fórmulas incorporadas em uma planilha do Excel, expressos em unidades por
grama de hemoglobina (U/g Hb). Os valores de referência foram considerados de
acordo com orientações do fabricante do kit Neolisa® G6PD Intercientífica para
mensuração da atividade da G6PD:
<2.5 U/g Hb – deficiente
2.6 a 7.8 U/g Hb – parcialmente deficiente
>7.9 U/g Hb – normal
3.3.3 Dosagem da Glicose
Os pacientes que tiveram as amostras positivas para a deficiência da G6PD
foram contatados para agendamento de uma segunda coleta, para dosagem da
glicose sérica, com preparo em jejum de 8 a 12 horas, por método enzimático, com
utilização de equipamento automatizado ISE 900/P 800 Hitachi – Roche. Trata-se
de um exame auxiliar útil no diagnóstico e monitoração do diabetes mellitus e outros
distúrbios relacionados ao metabolismo dos carboidratos, em situações de
desidratação, acidoses metabólicas, hipoglicemia e disfunção na secreção de
insulina. Utilizando-se como valores de referência (WHO 2006b):
- ≤ 109 mg/dL - normal.

37
- 110 a 125 mg/dL - glicemia de jejum aumentada.
- ≥ 126 mg/dL - diabetes mellitus.
3.3.4 Análise do DNA
As amostras positivas para a deficiência da G6PD foram analisadas
molecularmente. O DNA genômico foi extraído pelo método inorgânico com sal,
conforme técnica descrita por Miller et al., 1988, com modificações: Após
centrifugação de 5mL de sangue periférico coletado com EDTA por 3 minutos em
5.000 rpm, o pellet de leucócitos foi recolhido em tubo plástico de 1.5 mL. Foi
adicionado às células 1.0 mL de tampão de lise I*. A mistura resultante foi centrifugada
por 5 minutos, 5.000 rpm, e então ressuspensa em 250 μL de tampão de lise II** e 5
μL de SDS 20%. Após homogeinização em vortex, adicionou-se 100 μL de NaCl.
Homogeinizou-se a mistura em vortex e centrifugou-se por 3 minutos, 5.000 rpm. O
sobrenadante resultante foi transferido para um em tubo de polipropileno,
adicionando-se 300 μL de isopropanol absoluto. O DNA foi centrifugado por 5 minutos,
5.000 rpm e lavado duas vezes com solução de etanol 75%. Após repouso de 12 h a
temperatura ambiente, o DNA foi hidratado com 50 μL de água milli-Q ultra pura
autoclavada. *Tampão de Lise I: Sacarose 0.3M, Tris-HCl 10mM (pH = 7,5), MgCl
5mM, Triton x-100 1% - conservado sob refrigeração de 40C.
**Tampão de Lise II: NaCl 0.075M, Na-EDTA 0.024M (pH=8.0) – conservado a
temperatura ambiente.
Após a extração, o DNA foi amplificado, utilizando a técnica de Reação em
Cadeia de Polimerase (PCR), nas condições apropriadas para os iniciadores (primers,
listados na Tabela 1), para análise das principais mutações encontradas na população
brasileira: G6PD A-, com as mutações 202 G-A / 376 A-G e G6PD Mediterrânea, com
a mutação 563 C-T.

38
Tabela 1. Primers utilizados detecção das mutações na deficiência da G6PD
Mutação: 202 G-A
Sense primer 5’-GTGGCTGTTCCGGGATGGCCTTCTG-‘3
Antisense primer 5’-GTTTGTCTCACTCGGGAAGAAGTTC-‘3
Temperatura de anelamento 560
Mutação: 376 A-G
Sense primer 5’-CAGTACGATGATGGCAGC-‘3
Antisense primer 5‘-TGGCGGAAGAAGATGGAC-‘3
Temperatura de anelamento 560
Mutação: 563 C-T
Sense primer 5'GGT GAG GCT CCT GAG TAC CA 3'
Antisense primer 5'AGC TGT GAT CCT CAC TCC CC 3'
Temperatura de anelamento 620
Para o ensaio da PCR foram utilizados foram utilizados 100 ng DNA, num
volume final de 25µL amplificados por um termociclador (AppliedBioSystems®, USA),
com adição dos seguintes reagentes (Invitrogen®): 1 X 500 mM de tampão KCl (pH
8.4), 2mM de dNTP, 5mM MgCl2, 20 pM de cada primer e 1U de Taq DNA polimerase.
Para aumentar a atividade da DNA polimerase, foram adicionados em todas as PCRs,
10% de DMSO (Sigma). A amplificação da PCR ocorreu em 34 ciclos, com as
temperaturas 950 (30’’) - 61 a 620 (30’’) -720 (45’’). 10µL do produto final da PCR foram
digeridos a 370 através do Polimorfismo de Tamanho do Fragmento de Restrição
(RFLP), usando as enzimas de restrição NlaIII, FokI e MboII, para detecção das duas
variantes mais frequentes na população brasileira: G6PD A- e a G6PD Mediterrânea
(Tabela 2), seguido de eletroforese em gel de agarose a 1.5% corado com brometo
de etídeo (0.25 μg/ml), e analisada por transiluminação UV. As reações da PCR-
RFLP foram realizadas conforme técnicas descritas por Bouanga et al., (1998) e Kurdi-
Haidar et al., (1990), com modificações.

39
Tabela 2. Enzimas de restrição usadas na RFLP para detecção das variantes da deficiência da G6PD
Variante da G6PD
Mutação Enzima de restrição
Exon Amplificado
G6PD A- 202 G-A NlaIII 4
376 A-G FokI 5
Mediterrânea 563 C-T MboII 6
3.3.5 Diagnóstico da Malária
A confecção das lâminas, para diagnóstico da malária, foi realizada pela técnica
da gota espessa (GE), bem como a leitura ao microscópio óptico foram efetuadas por
agentes da Fundação de Vigilância em Saúde do Amazonas (FVS/AM), de acordo
com padronização do Ministério da Saúde (2005).
3.4 Tamanho da Amostra
A amostra de 1.340 indivíduos foi calculada em programa estatístico EPI
INFO®, versão 2004, com base na população de Manaus, de 1.680.000 habitantes,
com precisão de 2%; prevalência de 3% e intervalo de confiança de 95%.
3.5 Seleção dos Sujeitos da Pesquisa
O método de captação de sujeitos da pesquisa consistiu de uma abordagem
inicial do colaborador, através de explanação dos objetivos da pesquisa, destacando
a importância da participação destes num estudo que beneficiará a produção de
conhecimento relacionado à deficiência da enzima G6PD. A amostra de indivíduos foi
dividida entre três zonas da cidade de Manaus: Oeste, Norte e Leste, sendo
selecionado, segundo amostragem aleatória simples, um indivíduo por residência
visitada, e por conveniência, aproveitando a ocasião da visita de agentes da Fundação
de Vigilância e Saúde do Amazonas (FVS/AM) às residências, através de busca ativa
por casos de malária.
Foram também analisadas amostras de 15 indivíduos com diagnóstico clínico
de hemólise por primaquina atendidos na demanda espontânea do serviço de

40
urgência, enfermaria ou ambulatório da Fundação de Medicina Tropical Dr. Heitor
Vieira Dourado (FMT-HVD), integrantes do projeto “Fenótipo e Genótipo da
Deficiência da Glicose-6-Fosfato Desidrogenase”, aprovado pelo Comitê de Ética em
Pesquisa da Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (FMT-HVD),
sob o registro No. 0774-2007. Tais sujeitos da pesquisa foram triados para a
deficiência da G6PD, cujas amostras de sangue também foram analisadas
molecularmente, a fim de identificar os polimorfismos do gene G6PD envolvidos no
processo hemolítico e colaborar com a compreensão dos resultados do presente
estudo (dados não publicados).
3.6 Critérios de Elegibilidade
Foram convidados a participar do estudo indivíduos do gênero masculino pelo
fato da G6PD ser uma enzima cujo loco inativado encontra-se no cromossomo
X, frequentemente encontrada em indivíduos do gênero masculino, eliminando-
se a análise de fenótipos heterozigotos que ocorrem com infrequência na
população feminina, em virtude da inativação casual da mesma região do par
de cromossomos X (Lyon, 1961; Beutler, 1996).
Outro critério de inclusão relacionou-se com idade, somente para indivíduos
acima de 1 ano (porque menores de 1 ano são fisiologicamente deficientes da
G6PD (WHO, 1990), com limite de 65 anos (devido ao decréscimo da atividade
enzimática na fase senil) (Benzi et al., 1989; Nohl, 1993).
Procurou-se incluir indivíduos não consanguíneos para que houvesse uma
melhor representatividade da deficiência da enzima.
Também contou como critério de inclusão os indivíduos serem residentes nas
áreas de estudo que compreederam as Zonas Oeste, Norte e Leste do
município de Manaus.

41
3.7 Aspectos Éticos
Após explanação acerca dos objetivos do projeto, houve assinatura de um
Termo de Consentimento Livre e Esclarecido (ANEXO 9.2) e entrevista para obtenção
de dados epidemiológicos (ANEXO 9.3). O estudo foi aprovado pelo Comitê de Ética
em Pesquisa da Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (FMT-
HVD), sob o registro No. 2399-2006 (ANEXO 9.4). Houve concordância da parte da
Fundação de Vigilância em Saúde do Amazonas (FVS/AM) no desenvolvimento de
parceria na busca dos sujeitos da pesquisa.
3.8 Análise Estatística
Os dados foram analizados utilizando o programa STATA (versão 11.1) e Epi-
Info versão 3.5.3 para Windows. A distribuição normal dos dados foi avaliada o teste
de Kolmogorov-Smirnov. Para testar as diferenças nas proporções os dados foram
analizados com o teste Chi-quadrado e teste exato de Fisher, quando necessário. O
nível de significância fixado para os testes foi de 5% e Intervalo de Confiança de 95%.
3.9 Financiamento
O estudo teve o apoio financeiro da Superintendência da Zona Franca de
Manaus (SUFRAMA), da Fundação de Amparo à Pesquisa (FAPEAM) and Programa
de Pesquisa para o SUS (PPSUS), através do repasse de verbas para o projeto
“Caracterização Molecular da Deficiência da Glicose-6-Fosfato Desidrogenase em
Indivíduos Residentes em Áreas Endêmicas de Malária da Cidade de Manaus,
Amazonas, Brasil”, sob a responsabilidade técnica da MSc. Monica Regina Farias
Costa.

42
4 RESULTADOS

43
4.1 ARTIGO 1 Santana MS, de Lacerda MVG, Barbosa MdGV, Alecrim WD, Alecrim MdGC, 2009 Glucose-6-Phosphate Dehydrogenase Deficiency in an Endemic Area for Malaria in Manaus: A Cross-Sectional Survey in the Brazilian Amazon. PLoS ONE 4(4): e5259. Publicado em 16 de abril de 2009.

44
Glucose-6-Phosphate Dehydrogenase Deficiency in an Endemic Area for Malaria in Manaus: A Cross-Sectional Survey in the Brazilian Amazon
Marli Stela Santana1*, Marcus Vinícius Guimarães de Lacerda1,2,3*, Maria das Graças Vale Barbosa1,2,3, Wilson Duarte Alecrim 2,3, Maria das Graças Costa Alecrim1,3
1University of the State of Amazonas, Manaus, Amazonas, Brazil, 2Tropical Medicine Foundation of Amazonas, Manaus, Amazonas, Brazil, 3Nilton Lins Universitary Center, Manaus, Amazonas, Brazil * Corresponding author: [email protected] (MS); [email protected]
(ML)

45
Abstract
Background: There is a paucity of information regarding glucose-6-phosphate
dehydrogenase (G6PD) deficiency in endemic areas for malaria in Latin America.
Methodology/Principal Findings: This study determined the prevalence of the G6PD
deficiency in 200 male nonconsanguineous individuals residing in the Ismail Aziz
Community, on the outskirts of Manaus (Brazilian Amazon). Six individuals (3%) were
deficient using the qualitative Brewer’s test. Gel electrophoresis showed that five of
these patients were G6PD A-. The deficiency was not associated with the ethnic origin
(P = 0.571). In a multivariate logistic regression analysis, G6PD deficiency protected
against three or more episodes of malaria (P = 0.049), independently of the age, and
was associated with a history of jaundice (P = 0.020) and need of blood transfusion (P
= 0.045) during previous treatment for malarial infection, independently of the age and
the previous malarial exposure.
Conclusions/Significance: The frequency of G6PD deficiency was similar to other
studies performed in Brazil and the finding of a predominant G6PD A- variant will help
the clinical management of patients with drug-induced haemolysis. The history of
jaundice and blood transfusion during previous malarial infection may trigger the
screening of patients for G6PD deficiency. The apparent protection against multiple
malarial infections in an area primarily endemic for Plasmodium vivax needs further
investigation.
Received July 30, 2008; Accepted March 12, 2009; Published April 16, 2009.
Funding: The study was supported by Superintendência da Zona Franca de Manaus
(SUFRAMA). The funders had no role in study design, data collection and analysis,
decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.

46
Introduction
Malaria remains the most important tropical disease in tropical and sub-tropical areas
of the world [1]. In Brazil, approximately 99.5% of malaria cases occur within the
Amazon Region, with 544,615 in 2006. The State of Amazonas reported 181,973
cases, representing 33.4% of the total. In Manaus, the capital of the state, 51,228
cases were reported in that same year, and 40,679 (79.4%) of these were caused by
Plasmodium vivax [2]. The continued high incidence of malaria in the municipality of
Manaus is related to environmental and socio-economic factors that include high
temperatures, humidity, vector density, unplanned expansion of the urban peripheral
areas and resistance to antimalarials [3,4].
To obtain the radical cure of vivax malaria, primaquine is prescribed for all patients [5].
Despite a low fatality rate, P. vivax infection may lead to severe clinical complications
related to the side effects of the drugs, such as haemolysis induced by primaquine in
individuals genetically deficient for G6PD. The worldwide distribution of this deficiency
is a vital consideration in the development of new antimalarials that may also have the
potential of causing haemolysis [6]. G6PD is an enzyme present in the cytoplasm of all
cells, acting specifically in the maintenance of the integrity of the erythrocytes,
preventing the oxidation of hemoglobin and other cellular proteins [7].
G6PD deficiency is X-linked and predisposes to hemolysis and to a lesser extent to
methemoglobinemia in those persons in use of a substance with oxidative properties
[8]. The degree of drug induced hemolysis may be determined by the G6PD variants
involved, grouped in five classes, identified according to the activity of the enzyme by
electrophoretic and molecular characteristics [9,10]. G6PD deficiency occurs most
frequently in individuals of African descent, with a frequency ranging from 3.6 to 28.0%
[11,12]. In Asia the deficiency prevalence ranges from 6.0 to 15.8% [13,14], in India it
is 10.5% [15], and in the Middle East the prevalence varies from 3 to 29% [16,17]. In
Brazil, a few studies have found prevalences between 1.7 and 6.0% with a
predominance of the African variant (A2), with new mutations already described only
in non-endemic areas for malaria [18–21]. To date, there is no community-based
estimate of the frequency of G6PD deficiency in highly-endemic areas for malaria in
Latin America, such as Manaus, and the major variants predominating in this

47
population are unknown. Therefore, this study aimed to estimate the prevalence of
G6PD in an endemic area for malaria in Manaus, in the Brazilian Amazon.
Results
Of the 200 analyzed samples, six (3%; CI95% 0.97–5.03%) were deficient for G6PD
according to the first screening using the phenotypic Brewer’s test. The deficiency was
confirmed in all the samples by electrophoresis in agarose gel. Five samples with
G6PD deficiency showed the type A2 standard on electrophoresis. One sample
disclosed an indeterminate band in the gel, which did not permit a valid interpretation
of the type of deficiency. All the patients were afebrile at the time of the thick blood
smear collection, but three samples were positive for Plasmodium sp. (two for P. vivax
and one for P. falciparum), disclosing an infection prevalence of 1.5% (CI95% 0.49–
2.51). In Table 1, it is shown that G6PD was not associated with the ethnic origin, but
was associated with less than three episodes of malaria throughout their lives and
strongly associated with jaundice and need of blood transfusion during previous
malarial infections. Table 2 shows that the same results were confirmed after a
multivariate analysis (adjusting for age and previous malarial infections).
Discussion
Since G6PD deficiency is a recessive trait linked to the X chromosome, studying the
population frequency of the disorder exclusively in men is an optimal approach [6].
Measuring the deficiency in hospital-based studies may underestimate the true
magnitude of the problem, because patients in a haemolytic crisis may increase their
number of peripheral reticulocytes with normal G6PD values, thereby leading to a
false-negative screening for G6PD deficiency. The qualitative Brewer’s test for the
reduction of methemoglobin is a simple method that uses low-cost reagents. Although
only yielding qualitative results, it is a good screening test and it has been widely
utilized in various studies of selected populations [20,22].
The biochemical characterization of the isoenzymes demonstrated a high frequency of
the phenotype A- G6PD deficiency. To a certain extent this confirms findings in other
Brazilian studies with different study designs [22,23]. Since one sample in our study

48
could not be characterized through the biochemical approach, molecular studies are
needed to confirm new possible variants in further studies. The common African variant
G6PD A- is usually a mild/moderate deficiency (10–15% of normal activity in
hemizygous males). In this case, G6PD activity is greatest in younger cells, so as the
red cell population recovers, G6PD activity increases and hemolysis is controlled. This
is what makes drug induced G6PD deficiency-related hemolytic anemia in G6PD A-
subjects a self-limiting problem in most cases [6]. Therefore, our study supports
conservative management of hemolytic crisis for patients from Manaus, based on their
predominant genotype.
Among the six individuals with the G6PD A- variant, five were non-white (but not black)
and one was white. This illustrates the impact of the racial mixing which occurred in
the recent history of Brazil, especially in the Amazon region. A significant immigration
from various parts of Brazil and abroad took place in the nineteenth century due to
rubber extraction activities and during the twentieth century because of the creation of
a tax-free zone and a growing industrial base in Manaus. The large number of
immigrants from Portugal and Lebanon in Manaus probably explains this predominant
variant found in the studied population, since G6PD A2 is frequently seen in these
groups [7,17,18].
Epidemiologic studies have suggested that G6PD deficiency protects against malaria
and severe malaria by P. falciparum [24–26]. The precise mechanism of protection
remains unknown. We observed a significant protection against more than three
episodes of malaria in the G6PD deficient men enrolled in this study, independently of
their age, suggesting some degree of protection confered by the deficiency against P.
vivax malaria, which is the major species in the area of the study. This potential
protection conferred by G6PD deficiency needs further investigation. The impact of
G6PD deficiency in this population affected mostly by P. vivax infection is reflected in
their history of significantly more jaundice and more blood transfusions. This
information could be useful as a first screening for G6PD deficient patients, to
determine who should have a Brewer’s test performed. This test is not routinely offered
to the general population with a diagnosis of P. vivax malaria in Brazil even though
they will receive a standard 7-day treatment regimen with primaquine (0.5 mg/kg/day)
for radical cure of the parasite. G6PD deficiency therefore increases the morbidity of

49
P. vivax infection itself and the side effects of primaquine. In both cases the costs of
treatment and hospitalization are increased. These additional expenses threaten
financial resources and difficulty achieving a radical cure complicates the control of P.
vivax malaria.
Since only a few studies have been published estimating the prevalence of G6PD
deficiency in malaria endemic areas of Latin America, this study adds information to
the global mapping of the problem, despite a low external validity, as most of the
literature on this issue. Understanding the burden of G6PD deficiency will guide local
antimalarial treatment policies and enable the proper design of clinical trials for P. vivax
radical cure with new antimalarials having the potential of hemolysis in this population,
such as tafenoquine [27]. Despite the small number of patients enrolled in this
population based survey, we detected a 3% prevalence of G6PD deficiency in an
endemic area for malaria in Manaus. Accurate information about this deficiency and its
relation to acute hemolysis with the use of primaquine in the Brazilian Amazon is
scarce. G6PD A- was confirmed through electrophoretic isoenzyme detection in 5 of 6
patients after the deficiency was initially detected via screening with the qualitative
Brewer’s test. G6PD A- was not associated with a black ethnicity, probably because of
the genetically mixed population of Manaus. The history of jaundice and blood
transfusions associated with prior malaria infections was associated with G6PD
deficiency and may be used as a simple clinical marker for this entity.
Materials and Methods
The study was carried out in the municipality of Manaus (03u089 S/60u009 W), in the
Ismail Aziz Community, with an estimated population of 1,500 inhabitants (750 male),
including natives and immigrants (Figure 1). Census data were used to estimate the
population size. The community was chosen due to a high annual parasite index in
2006 (234/1,000 persons) [2]. The sample size was calculated for a population of 750
nonconsaguineous men (in order to avoid the consanguinity bias) with an expected
prevalence of G6PD deficiency of 3%, power of 80% and significance level below 5%.
Therefore, 200 nonconsanguineous men between 1 and 65 years-old (children under
one year are physiologically G6PD deficient and over sixty-five, due to the decrease
of enzyme activity during senile) were randomly selected (selection of the fourth man

50
in the visit to all the houses of the community, except those who were consanguineous
relatives of other selected subjects) and enrolled in a cross-sectional survey of G6PD
deficiency, after informed consent has been obtained. Ethnic origin, previous malaria
infection and previous clinical complications triggered by malarial infection were
acquired through an individual interview, and all the data were recorded in a standard
questionnaire.
Laboratorial techniques
Ten milliliters of venous blood were collected from each person using EDTA tubes (BD
Vacutainer H). A qualitative G6PD screening test by the methemoglobin reduction
method was performed [28]. In all those initially found to be G6PD deficient, after the
whole blood samples were hemolysed, the material was submitted to electrophoresis
[29] for the confirmation of the deficiency by isoenzyme analysis. The diagnosis of the
malaria was based in the method of the thick blood smear by Walker and examinated
directly to the optic microscope, according to norms of the World Health Organization,
was carried through in all the individuals [30]. Statistics The statistical analyses were
performed in Epi Info 3.3 2004 (CDC/Atlanta). 95% CIs (CI95%) were estimated for all
the observed frequencies of the study. To assess the extent to which ethnic origin,
previous malaria exposure and previous clinical complications triggered by malarial
infection were associated with G6PD deficiency, odds ratios (OR) with 95% CIs
(CI95%) were estimated by univariate logistic regression analysis (Table 1, Table 2).
Qualitative data were analyzed with the x2-test or the Fisher exact-test when indicated.
Adjustment for multiple variables was performed by adding the covariates in a set of
multiple logistic-regression models. Test for trend was conducted through the exact
test for trend (modified Wilcoxon test for trend). A two tailed value of P,0.05 was
considered statistically significant.
Ethical clearance
The study was approved by the Ethics Committee Board of the Tropical Medicine
Foundation of Amazonas. Informed written consent was obtained from all participants.

51
Acknowledgments
We are grateful for Pricila Santos, Suzi Silva, Walber Brandao, Sabrina Silva, Carmem
Oliva, Gloria Silva, Jason Brune, Ericilda Araujo, Raimunda Silva, Marli Marques,
Monica Costa, Jose Alves Jr., Ricardo Faria, Marilia Fernandes, Rubenita Costa, and
Leıla Coelho who helped in the field and laboratory activities. Special thanks to Donald
Skillman, for the assistance with the English version of the manuscript.
Author Contributions
Conceived and designed the experiments: MSS MVL MGB WDA MGA. Performed the
experiments: MSS. Analyzed the data: MSS MVL MGB WDA MGA. Contributed
reagents/materials/analysis tools: MSS. Wrote the paper: MSS MVL MGB WDA MGA.
References 1. World Health Organization (2008) World Health Report. Available: http://www.who.int/whr/en/index.html. Accessed 2008 Dec 10. 2. Brazilian Ministry of Health (2006) Epidemiological Bulletin of Malaria (SIVEP). Available: http://www.saude.gov.br_sivep. Accessed 2007 Jun 20. 3. Alecrim MGC, Alecrim WD, Macedo V (1999) Plasmodium vivax resistance to chloroquine (R2) and mefloquine (R3) in Brazilian Amazon region. Rev Soc Bras Med Trop 32: 67–68. 4. Gonçalves MJF, Alecrim WD (2004) Non-planed urbanization as a contributing factor for malaria incidence in Manaus-Amazonas, Brazil. Rev Salud Publica (Bogota) 6: 156–166. 5. FUNASA (2001) Malaria Therapeutics Guidelines. Brası´lia: National Foundation of Health, Brazilian Ministry of Health. 6. Beutler E, Duparc S (2007) G6PD Deficiency Working Group. Glucose-6-phosphate dehydrogenase deficiency and antimalarial drug development. Am J Trop Med Hyg 77: 779–789. 7. Luzzatto L, Mehta A, Meloni T (1995) Haemoglobinuria and haptoglobin in G6PD deficiency. Br J Haematol 91: 511–512. 8. Santana MS, Arcanjo ARL, Rocha MAF, Sardinha JFJ, Alecrim WD, et al. (2007) Association of methemoglobinemia and glucose-6-phosphate dehydrogenase deficiency in malaria patients treated with primaquine. Rev Soc Bras Med Trop 40: 533–536.

52
9. Beutler E, Vulliamy TJ (2002) Hematologically important mutations: glucose-6-phosphate dehydrogenase. Blood Cells Mol Dis 28: 93–103. 10. Yoshida A, Beutler E, Motulsky AG (1971) Human glucose-6-phosphate dehydrogenase variants. Bull World Health Organ 45: 243–253. 11. Badens C, Martinez di Montemuros F, Thuret I, Michel G, Mattei JF, et al. (2000) Molecular basis of haemoglobinopathies and G6PD deficiency in the Comorian population. Hematol J 1: 264–268. 12. De Araujo C, Migot-Nabias F, Guitard J, Pelleau S, Vulliamy T, et al. (2006) The role of the G6PD AEth376G/968C allele in glucose-6-phosphate dehydrogenase deficiency in the seerer population of Senegal. Haematologica 91: 262–263. 13. Iwai K, Hirono A, Matsuoka H, Kawamoto F, Horie T, et al. (2001) Distribution of glucose-6-phosphate dehydrogenase mutations in Southeast Asia. Hum Genet 108: 445–449. 14. Louicharoen C, Nuchprayoon I (2005) G6PD Viangchan (871G.A) is the most common G6PD-deficient variant in the Cambodian population. J Hum Genet 50: 448–452. 15. Sukumar S, Mukherjee MB, Colah RB, Mohanty D (2004) Molecular basis of G6PD deficiency in India. Blood Cells Mol Dis 33: 141–145. 16. Al-Riyami A, Ebrahim GJ (2003) Genetic Blood Disorders Survey in the Sultanate of Oman. J Trop Pediatr 49: 1–20. 17. Usanga EA, Ameen R (2000) Glucose-6-phosphate dehydrogenase deficiency in Kuwait, Syria, Egypt, Iran, Jordan and Lebanon. Hum Hered 50: 158–61. 18. Weimer TA, Salzano FM, Westwood B, Beutler E (1993) Molecular characterization of glucose-6-phosphate dehydrogenase variants from Brazil. Hum Biol 65: 41–47. 19. Saad ST, Salles TS, Carvalho MH, Costa FF (1997) Molecular characterization of glucose-6-phosphate dehydrogenase deficiency in Brazil. Hum Hered 47: 17–21. 20. Compri MB, Saad ST, Ramalho AS (2000) Genetic-epidemiological and molecular investigation of G-6-PD deficiency in a Brazilian community. Cad Sau´de Pu´blica 16: 335–342. 21. Castro S, Weber R, Dadalt V, Tavares V, Giugliani R (2006) Prevalence of G6PD deficiency in newborns in the south of Brazil. J Med Screen 13: 85–86. 22. Hamel AR, Cabral IR, Sales TS, Costa FF, Saad STO (2002) Molecular heterogeneity of G6PD deficiency in an Amazonian population and description of four new variants. Blood Cells Mol Dis 28: 399–406. 23. Silva MC, Santos EB, Costal EG, Filho MG, Guerreiro JF, et al. (2004) Clinical and laboratorial alterations in Plasmodium vivax malaria patients and glucose-6-phosphate

53
dehydrogenase deficiency treated with primaquine at 0.50 mg/kg/day. Rev Soc Bras Med Trop 37: 215–217. 24. Greene LS, McMahon L, DiIorio J (1993) Co-evolution of glucose-6-phosphate dehydrogenase deficiency and quinine taste sensitivity. Ann Hum Biol 20: 497–500. 25. Guindo A, Fairhurst RM, Doumbo OK, Wellems TE, Diallo DA (2007) XLinked G6PD deficiency protects hemizygous males but not heterozygous females against severe malaria. PLoS Med 4: e66. doi:10.1371/journal.pmed.0040066. 26. Roth EF Jr, Raventos-Suarez C, Rinaldi A, Nagel RL (1983) Glucose-6-phosphate dehydrogenase deficiency inhibits in vitro growth of Plasmodium falciparum. Proc Natl Acad Sci USA 80: 298–299. 27. Lacerda MVG, Zackiewicz C, Alecrim WD, Alecrim MGC (2007) The neglected Plasmodium vivax: are researchers from endemic areas really concerned about new treatment options? Rev Soc Bras Med Trop 40: 489–90. 28. Brewer GJ, Tarlov AR, Alving AS (1962) The methaemoglobin reduction test for primaquine-type sensitivity of erythrocytes. A simplified procedure for detecting a specific hypersusceptibility to drug haemolysis. JAMA 180: 386–388. 29. Forbes J, Steytler JG, van Heerden R (1991) Agarose gel electrophoresis of glucose-6-phosphate-dehydrogenase isoenzymes. Clin Chim Acta 199: 279–282. 30. World Health Organization (1975) Microscopic Diagnosis of Malaria, 4th edition.

54
Table 1. Crude odds ratios and respective CI95% in the univariate logistic regression
analysis of the association between G6PD deficiency and ethnic origin, previous
malaria exposure and previous clinical complications triggered by malarial infection.
Table 2. Adjusted odds ratios and respective CI95% for the variables associated with
G6PD deficiency in multiple logistic-regression models.

55
Figure 1. Geographic localization of the Ismail Aziz Community in an endemic area for
malaria in Manaus, Amazonas State, Brazil.

56
4.2 ARTIGO 2
Santana MS, Monteiro WM, Siqueira AM, Costa MRF, Sampaio V, Lacerda MVG,
Alecrim MGC, 2012. Glucose-6-phosphate dehydrogenase deficient variants are
associated with reduced susceptibility to malaria in the Brazilian Amazon. Transactions
of the Royal Society of Tropical Medicine and Hygiene. Publicado em 10 de março de
2013.

57
Glucose-6-phosphate dehydrogenase deficient variants are associated with reduced susceptibility to malaria in the Brazilian Amazon
Marli Stela Santana*1,2, Wuelton Marcelo Monteiro1,2,3,
André Machado Siqueira1,2, Mônica Regina Farias Costa2, Vanderson Sampaio,
Marcus Vinícius Guimarães Lacerda*1,3,4, Maria Graças Costa Alecrim1,2
1 Universidade do Estado do Amazonas, Manaus, Amazonas, Brazil
2 Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, Manaus, Amazonas,
Brazil
3 Department of Public Health, Universidade Federal do Amazonas, Manaus,
Amazonas, Brazil
4 Universidade Nilton Lins, Manaus, Amazonas, Brazil *Corresponding authors: Tel: + 55 92 2127-3443; E-mail: [email protected];
Address: Gerência de Malária/FMT-HVD – Av. Pedro Teixeira 25, Dom Pedro,
69040-000, Manaus, Amazonas, Brazil.
Adresses each afiliation:
1 , 2 Av. Pedro Teixeira , 25, D. Pedro - CEP 69040-000, Manaus – Amazonas- Brasil; 3 Av. General Rodrigo Otávio, 3000, Coroado- CEP 69077-000, Manaus – Amazonas-Brasil; 4 Av. Prof. Nilton Lins, 3.259, Pq. das Laranjeiras, CEP 69058-040, Manaus-Amazonas-Brasil.

58
ABSTRACT
Background: Glucose-6-phosphate dehydrogenase deficiency (G6PDd) has been
shown to protect against malaria infection and severe manifestations in African and
Asia, but there is a scarcity of studies in the Americas. This study aimed to study the
prevalence of G6PDd and its association with malaria occurrence in the Brazilian
Amazon.
Methods: A cross-sectional study was conducted in the male population to estimate
the prevalence of G6PDd and malaria infection. G6PD deficient samples were
genotyped to identify the deficient variant. Number of previous malaria episodes and
need for blood transfusion during malaria episodes were recorded by applying
a standardized questionary.
Results: From a sample of 1478 male individuals, 66 were detected as G6PD
deficient, resulting in a prevalence of of 4.5% (95% CI = 3.44–5.56%). Fifty six G6PD
deficient individuals (3.8%; 95% CI = 2.82–4.77) presented the G6PD A-variant
mutation, while 10 individuals (0.7%; 95% CI = 0.42–0.97) severely deficient were
genotyped as carriers of the G6PD Mediterranean variant. After adjusting for age,
G6PD deficient individuals were less likely to report the occurrence of malaria
episodes, and the protective effect was related to the enzyme activity, with carriers of
the GG6PD A-variant presenting a 88% reduction (AOR: 0.119; 95% CI = 0.057–0.252;
p<0.001) and carriers of the Meditarrenean variant presenting 99% lower risk (AOR:
0.010; 95% CI = 0.002–0.252; p<0.001) when compared to non-deficient individuals.
On the other hand, G6PD deficient subjects reported higher need of transfusion during
malaria episodes (p<0.001).
Conclusion: G6PD enzyme activity was directly related to susceptibility to malaria in
the Brazilian Amazon, where P. vivax predominates. Severe G6PDd was associated
with considerable higher risk of malaria-related transfusions.

59
Introduction
The 2011 WHO malaria report shows a worldwide progress on reducing the
transmission of the disease, with decline in the estimated number of malaria cases and
deaths.1 Mapping studies have demonstrated that Plasmodium vivax is more widely
distributed than Plasmodium falciparum and that this species is a potential cause of
morbidity and mortality amongst the 2.85 billion people living at risk of infection, most
of them in Central and South East Asia and Latin America (LA).2 In the last continent,
from 2000 to 2007, 7 554 993 cases of malaria were reported, of which 72.9% were
caused by P. vivax.3 Since the mid-1990s, P. vivax has increasingly become the
predominant malária species in Brazil,4 where Manaus, located in the Western
Brazilian Amazon, with around two million inhabitants, is one of the cities with higher
numbers of malaria episodes in LA.5 One of the reasons why P. vivax malaria is a more
difficult parasite to be controlled when compared to P. falciparum is the existence in
the former one’s cycle of the hypnozoites in the liver, which can result in relapses and
provide an infectious reservoir.6 Glucose-6-phosphate dehydrogenase deficiency
(G6PDd) is one of the most common X-linked genetic mutations, which explains a
higher prevalence of the functional deficiency in males.7 Red blood cells with deficient
enzyme activity become more susceptible to oxidative stress triggered specially by
oxidant drugs or its metabolytes.8 This issue is especially relevant on the treatment of
P. vivax infection, as radical cure requires the use of primaquine, which can lead to
acute intravascular hemolysis in G6PD deficient individuals, resulting in severe anemia
and acute renal failure.9,10 The public health consequences of this condition deserve
special attention on the impossibility of using primaquine in regions where there is a
high prevalence of this deficiency, further hampering transmission control efforts for
this parasite.11 Conversely, the G6PD deficient status has been associated with
protection against P. falciparum malaria.12,13 The mechanism of falciparum malaria
resistance in deficient subjects is likely related to an impaired antioxidante defense in
deficient ring-stage parasitized red cell evolving to membrane damage, which triggers
an increasing removal by phagocytosis before parasite maturation to trophozoite-stage
parasitized red cell and to schizonts.14 More recently, this has also been shown to be
protective against malaria episodes caused by P. vivax.15,16 It has been estimated that
G6PDd affects around 353 million people worldwide, with 96 million in the malaria
endemic countries.17 Indeed, G6PDd prevalence in a given region has been proposed

60
as an important marker of the selective pressure exerted by malaria.14 In Latin
America, where malaria was probably only introduced after European colonization,18,19
possibly the first work concerning G6PDd in areas of P. vivax circulation was that of
Best,20 who found no G6PD deficiente individuals amongst 228 Peruvian Indians living
in a malarial zone. Studies from different areas of the Brazilian Amazon confirmed an
absence or a very low frequency of G6PDd in Amerindian populations.21–25 However,
previous studies with non-Amerindians in malarial areas from Amazonian countries
showed prevalence of G6PDd ranging from 3.0–12.0%,15,26–28 which probably reflects
the intense European and African migration in the past. In Manaus the prevalence of
G6PDd among males from malaria endemic areas was estimated to vary from 3.0–
8.7%.15,28 Despite the clinical and epidemiological significance of the interaction
between G6PDd and malaria, the extent of its occurrence and consequences has not
been properly measured in this distinct population with high admixture rates. In Brazil,
where primaquine is systematically prescribed with chloroquine, to all patients with
microscopical diagnosis of vivax malaria, in the dosage of 0.5mg/kg/day for 7 days, the
risk of adverse events in G6PD deficient patients can be even more relevant. Recent
evidence of protection against P. vivax infections conferred by G6PDd indicates the
need for further investigation on the impact of different variants of G6PD on the burden
of malária in P. vivax predominant areas.15,16 In this work, we investigated the
prevalence of G6PD deficient variants amongst males and its association with self-
reported malaria episodes and need for blood transfusion in an area of unstable
transmission with predominance of P. vivax in the Brazilian Amazon.
Materials and methods
Study population
Considering that a previous study found a G6PDd prevalence of 3% in the area,15 the
sample size calculated to detect a variant with prevalence of 1%, with 95% CI ranging
from 0.5–1.5% and design effect of 1, was of 1478 individuals. The study sample was
calculated based on the population of the city of Manaus and distributed evenly
between the three áreas of the city where malaria transmission occurs throughout the
year (Eastern Zone n= 489; Western Zone n= 497; Northern Zone n= 492). Annual
parasite incidences for these three areas in 2009 were, respectively, 0.5, 103.0 and
3.5 cases/1000 people. The relative incidence of P. falciparum and P. vivax in this area
was around 50% each in 1990 but in the recent years P. vivax was responsible for

61
more than 90% of the cases.29 Male individuals from 1–65 years old were selected
from the periodical census performed by the Amazonas Foundation for Health
Surveillance (FVS) personnel. From March 2009 to March 2010, the cross-sectional
study conducted by health agents of the FVS was performed to estimate the
prevalence of the G6PDd together with active case detection. During the agents’ visit
to the residences, the study and procedures were explained, and the eldest male
individual of each family was invited to participate in the study. This study was
restricted to the male population because the mutations leading G6PDd are X-linked
and the presumable higher prevalence of the deficiency in this group.7 If written
consent by the patient or a guardian was provided, a standardized questionnaire
(Supplementary File) containing demographic, and clinico-epidemiological data
(number of previous malaria episodes, need for blood transfusion during malária
episodes) was applied by four trained and calibrated interviewers. Malaria treatment is
only available in public health units and only provided after confirmed microscopic
diagnosis by thick blood smear, providing considerable reliability to the self-report
information.
Laboratory procedures
During the visits, a finger prick was performed for malaria diagnosis and venous blood
was drawn for G6PD phenotyping and genotyping studies. Thick blood smears for
malaria diagnosis were collected and read by a local microscopist as recommended
by the Brazilian Ministry of Health.30 Afterwards, the slides were sent to a central
laboratory and reviewed by an experienced microscopist, who confirmed the diagnosis.
Ten milliliters of venous blood were collected from each person using BD Vacutainer
EDTA tubes (Becton, Dickinson and Company, Franklin Lakes, NJ USA) for G6PD
activity assays. The material was stored at 2–88C and processed within 48 hours for
screening of G6PDd and 72 hours for molecular analyses. Hemoglobin and enzymatic
activity (EA) was measured using Interscientific Neolisa G6PD Kit (Interscientific
Corporation, Hollywood, FL, USA), according the the manufacturer’s protocol. The
results were expressed as international units per gram of haemoglobin (IU/gHb). The
G6PD status was categorized according to G6PD activity: 6.0 IU/gHb as normal;
between 2.0 and 6.0 IU/ gHb as intermediate deficiency; and 2.0 IU/gHb as severe
deficiency. Negative and positive controls were included for each reaction. For
molecular characterization of G6PD deficient and partially deficient samples, DNA was

62
obtained from phenol-chlorophorm extractions. Genotyping was made by RFLP-PCR
analysis using oligonucleotide primers artificially creating restriction sites and following
protocols previously described to detect the G6PD A-, (African variant: mutations 202
G-A and 376 A-G)31 and the G6PD Med (Mediterranean variant: mutation 563 C-T),32
with some modifications. In summary, the amplification reaction mixture (25 mL)
contained 100 ng DNA, 1x buffer KCl 500 mM (pH 8.4), 20 pM of each primer, 2 mM
dNTP, 5mM MgCl2 and 1 U of Taq DNA Polymerase from Invitrogen (Life
Technologies Corporation, Carlsbad, CA, USA). It was added 10% DMSO (Sigma-
Aldrich, St Louis, MO, USA) in each reaction. The reactions were performed on a DNA
thermal cycler Applied BioSystems (Life Technologies). After 5 min incubation at 94°C,
a total of 34 amplification cycles were performed under the following conditions:
denaturation at 94°C for 10 s, annealing at 568C for 20 s and elongation at 728C for
12 s. These steps were followed by final elongation at 72°C for 7 min. Fragments were
digested overnight with appropriate restriction enzymes (New England Biolabs,
Ipswich, MA, USA) according to the manufacturer’s specifications: FokI to recognize
the 376 A-G mutation, NlaIII for the 202 G-A mutation and MboII for the 563 C-T
mutation. Products were analyzed by agarose gel (1.5%) electrophoresis and
visualized under ultraviolet light after ethidium bromide staining.
Statistical analysis
The prevalence of G6PDd and variants was calculated as proportions with 95% CIs.
Clinical and demographical characteristics were compared between the groups using
ANOVA for continuous variables (age) and X2 and Fisher’s exact test, when
appropriate, for categorical and binary variables. The association between G6PDd and
Plasmodium sp. infection was assessed with Fisher’s test. The association between
G6PD variant and history of malaria was analysed through logistic regression adjusted
for age. As more than 97% of individuals referred only having lived in Amazon rural
areas, age was the only covariate added to the logistic regression models being treated
linearly in years. The association between malaria-related transfusion and G6PD
status was assessed only between individuals with positive history of malaria using
Fisher’s exact test. The data was analysed using Stata version 11.1 (STATA Corp,
College Station, TX, USA). Ethical approval Informed written consent was obtained
from all participants. For those under 18 years old, parents were instructed about the
objectives of the study and signed together with the participant an informed consent.

63
All patients tested positive in the thick blood smear during the cross-sectional study
were treated according to the anti-malarial treatment guidelines from the Brazilian
Ministry of Health.
Results
A total of 1478 individuals participated in the study, of whom 66 were diagnosed with
G6PDd in the enzymatic screening, resulting in a prevalence of 4.5% (95% CI 3.44–
5.56%). Out of these, 56 (85%) individuals presented moderate deficiency, while 10
(15%) were classified with severe deficiency. All the G6PD deficiente individuals
presented mutations at the molecular analysis as revealed by DNA electrophoretic
profiles, with the 56 carriers of intermediate deficiency having the mutation compatible
with the G6PD A- variant (202 G-A and 376 A-G), an estimated prevalence of 3.8%
(95% CI 2.82–4.77%), and the 10 individuals with severe deficiency genotyped as
carriers of the G6PD Mediterranean variant (563 C-T), a prevalence of 0.7% (95% CI
0.42–0.97%) for this variant (Table 1). In the active search for asymptomatic carriers
of Plasmodium sp., 54 (3.7%) individuals were found infected by blood smear reading.
Among these, 52 (96%) were infected with P. vivax and two (4%) with P. falciparum.
Only one (2%) G6PD deficiente individual was diagnosed with malaria, compared to
53 (3.8%) non-deficient subjects, but this difference was not significant (Table 2).
G6PD deficient variants were associated with a protective effect against self-reported
history of malaria, with carriers of the African variant having 87.7% lower risk of malaria
history (AOR: 0.123; 95% CI 0.058–0.259) and carriers of the Mediterranean variant,
98.9% lower risk (AOR: 0.011; 95% CI 0.002–0.043) when compared to non-deficient
individuals, therefore demonstrating an inverse relationship between enzyme activity
and susceptibility to malaria (Table 2). Regarding the need for blood transfusion related
to malária episodes there was a strong association between genotype and transfusion
(p<0.001), with all three carriers of the Mediterranean variant who experienced malaria
reporting receiving blood transfusion, compared to 40% of G6PD A-carriers, and only
0.15% of non-deficient individuals.
Discussion
Plasmodium vivax is the most widespread parasite causing human malaria, with an
estimated 2.85 billion living at risk of infection worldwide.2 High prevalence of G6PDd
in many malaria-endemic countries adds considerable difficulty in malaria erradication

64
efforts.23 Testing individuals for G6PD status in field conditions is currently unrealistic
due to costs and logistic aspects, and, therefore, most countries opt not to administrate
primaquine to avoid drug-related haemolysis. This highlights the need for
comprehensive estimates of G6PDd in malaria endemic regions and of its clinical
epidemiological consequences. In this study we reported 4.5% of G6PDd in a malaria-
exposed population in the periurban area of Manaus, which is comparable with a
previous study of 200 men residing in a distinct area of this city, which had found a
prevalence of 3.0%, but no genotyping was performed at the time.15 Among the 66
G6PD deficiente subjects, 56 (85%) showed deficiency compatible with the G6PD A-
variant and 10 (15%) individuals with severe deficiency were genotyped as carriers of
the G6PD Mediterranean variant. The predominance of the African variant agrees with
data from other molecular studies in other regions of Brazil, where this variant appears
to be the most prevalent.33,34 The occurrence of G6PD Mediterranean variant has also
been described in Southern Brazil,35 but this is the first time it has been identified in
Northern Brazilian populations. In the Brazilian Amazon, the prevalence of G6PD
deficiente variants was previously investigated in the city of Belém, where 82.1% of
196 asymptomatic and unrelated male G6PD deficiente blood donors were identified
as carriers of G6PD A-, with the additional description of eight minor variants in lower
frequencies.33
In Manaus, gel electrophoresis from six deficient individuals with enzyme deficiency
showed that five were G6PD A-.15 In a review of G6PDd populational-based surveys
conducted in South America that included 4558 European descendants, 2484 admixed
Black/Indian/White or Black/White, and 10 298 Amerindian subjects, the G6PD
Mediterranean variant was only found in 20 cases (0.1%), all of them living in Southern
Brazil.35 The presence of G6PDd is almost inexistent in Amerindians,21–25 and the
presence of the Mediterranean variant in Manaus can be explained by the presence of
Arab and Jewish migrants, who arrived mainly from the Middle East and Morocco.36
The geographic distribution of G6PDd suggests that some polymorphisms confer
resistance to falciparum malaria.17 This phenomenon has been investigated mainly for
the African variant (G6PD A-), showing that it also confers protection against lethal
falciparum malaria.37 The higher prevalence of G6PDd in malaria endemic countries is
indicative that malaria infection has exterted a strong selective pressure in many
human populations.14,38 In P. falciparum infection it has been demonstrated that shorter

65
half-life and rapid clearance of red cells of G6PDd individuals make them less
susceptible to malária attacks from these parasites,39 and it is likely that a similar
pathophysiological mechanism could occur in P. vivax infected cells. There is,
however, the need to further explore and comprehend the mechanisms by which
individuals with G6PDd become less susceptible to infection. P. vivax invades
reticulocytes preferentially, but P. falciparum can invade erythrocytes of all ages. This
is a significant phenomenon since G6PD activity is markedly reduced in older
erythrocytes. Thus, P. vivax infected red cells probably are less vulnerable to the
oxidant stress produced by parasitization. The precise mechanism by which G6PD
deficiency promotes less susceptibility to vivax malaria remains to be established. In
this study, it was possible to detect that G6PD deficiente subjects presented a
significant lower number of self-reported malaria episodes throughout their lives than
non-deficients in an area where P. vivax is the predominant species. Although malaria
history was obtained retrospectively, and thus subject to recall bias, the fact that in
Brazil all the febrile cases in the endemic area are submitted to a thick blood smear
and positive cases reported to a unified surveillance system4 provides considerable
reliability to the information. These findings contribute to the understanding that both
Mediterranean and African variants reduce susceptibility against vivax malaria and that
this reduction is inversely related to the degree of enzyme activity. In a case-control
study performed in Afghan refugees, the G6PD Mediterranean variant was associated
with protection in hemizygous deficient males and in females carrying the deficiency
gene as heterozygotes or homozygotes.16 Due to the low number of subjects with
Plasmodium infection during the cross-sectional, it was not possible to properly assess
the association between G6PD variants and malarial infection. As this study was
restricted to males, the extent to which the susceptibility to malarial infection is
influenced in heterozygote and hemizygote female carriers of G6PD in Latin America
is yet to be determined. Among subjects who reported the need for blood transfusion
related to malarial infection (and supposedly after the use of primaquine, which is
systematically prescribed without prior G6PDd testing in all patients with vivax malaria
in Brazil), more than 90% (21/23) had G6PDd. Both African and Mediterranean
variants were associated with a higher frequency of need for blood transfusion.
Interestingly, we observed that all the three carriers of G6PD Mediterranean variant
who reported history of malaria referred having received blood tranfusion previously,
which was considerably higher than the proportion of malaria-related transfusion

66
reported by G6PD A- carriers. The G6PD Mediterranean variant in certain ethnic
groups originating in the Mediterranean and West Asia has minimal G6PD activity,
which was confirmed in our data, making them more vulnerable to severe life
threatening hemolysis with primaquine.40 Thus, these data suggest that African and
Mediaterranean G6PD variants in the Brazilian Amazon populations could be
responsible for clinical complications in patients with vivax malaria, such as severe
anemia and renal failure, triggered specifically by the use of primaquine in the higher
daily dosage (0.5mg/kg/day).10 Data on clinical complications in vivax malaria patients
during treatment are scarce. A previous report from Manaus describes the
development of hemolysis in 18 G6PD deficiente patients treated with chloroquine and
primaquine, with hospitalizations accompanied by severe hemolysis and the need of
red blood cells transfusion and acute renal failure, highlighting the impact upon the
public health system in the tropics.10 Indeed, an autopsy series of P. vivax-infected
individuals in Manaus, also describes two deaths related to primaquine-induced
haemolysis, demonstrating the lethal potential of this condition.41 Likewise,
methaemoglobin levels were also found to be higher in G6PD deficient patients
following primaquine therapy.28
Conclusion
For the first time the G6PD Mediterranean variant has been described amongst males
living in malaria endemic areas in the Brazilian Northern region. Carriers of the G6PD
A- and G6PD Mediterranean variants had lower rates of reporting the occurrence of
malaria episodes during their lifetime in the Brazilian Amazon region, where P. vivax
predominates. Furthermore, the lower susceptibility to presenting febrile cases due to
malária appeared to be inversely related to the degree of enzyme activity. Although
the G6PDd prevalence amongst males in the region was 4.5%, they presented a
significant higher need for blood transfusion during malaria episodes. These findings
suggest that G6PDd may contribute to a considerable proportion of malaria-related
complications, which has yet to be further investigated in studies that measure the real
extent to which it is a burden to health systems. We hope this results will highlight the
need for the study of better strategies to diagnose and manage G6PDd in malaria field
conditions and to the Malaria Control Programs in Latin America not to neglect this
condition in their national guidelines.

67
Supplementary data
Supplementary data are available at Transactions Online
(http://trstmh.oxfordjournals.org/).
Authors’ contributions: MSS, MFC, VS, MVL and MGA conceived the study; MSS,
MVL and MGA designed the study; MSS, WMM and AMS analyzed the data; MSS,
WMM, AMS and MVL interpreted the data; MSS, WMM, AMS and MVL drafted the
manuscript; AMS, MFC, VS, MVL and MGA critically revised the manuscript for
intellectual content. All authors read and approved the final version. MVL and MGA are
guarantors of the paper.
Acknowledgements: We are grateful to all the technical support from health agents
from the Foundation of Health Surveillance. Special thanks to Gloria Lima, Elizabeth
Santos Galusso, Evaulino Ferreira Itapirema, to all the undergraduate students from
UNINORTE and UEA, and to José Alves Maciel Jr., who helped in the field and
laboratory activities. We thank the reviewers for considerably contributing to the
improvement of the manuscript. Funding: The study was supported by Fundação de
Amparo à Pesquisa do Amazonas (FAPEAM)/Programa de Pesquisa para o SUS
(PPSUS) (grant number 193-8767348/2006). The funders had no role in study design,
data collection and analysis, decision to publish or preparation of the manuscript.
Competing interests: None declared.
Ethical approval: The study was approved by the Ethics Committee Board of the
Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (approval number
2399/2006).
References 1 WHO. World Malaria Report 2011. Geneva: World Health Organization; 2011. 2 Guerra CA, Howes RE, Patil AP et al. The international limits and population at risk
of Plasmodium vivax transmission in 2009. PLoS Negl Trop Dis 2010;4:e774.
3 PAHO. Report on the Situation of Malaria in the Americas. Washington: Pan American Health Organization; 2008.

68
4 Oliveira-Ferreira J, Lacerda MV, Brasil P et al. Malaria in Brazil: an overview. Malar J 2010;9:115.
5 Saraiva MG, Amorim RD, Moura MA et al. Urban expansion and spatial distribution
of malaria in the municipality of Manaus, State of Amazonas. Rev Soc Bras Med Trop 2009;42:515–22.
6 Cogswell FB. The hypnozoite and relapse in primate malaria. Clin Microbiol Rev
1992;5:26–35.
7 Cappellini MD, Fiorelli G. Glucose-6-phosphate dehydrogenase deficiency. Lancet 2008;371:64–74.
8 Ganesan S, Chaurasiya ND, Sahu R et al. Understanding the mechanisms for
metabolism-linked hemolytic toxicity of primaquine against glucose 6-phosphate dehydrogenase deficient human erythrocytes: evaluation of eryptotic pathway. Toxicology 2012; 294:54–60.
9 Beutler E, Duparc S. Glucose-6-phosphate dehydrogenase deficiency and
antimalarial drug development. Am J Trop Med Hyg 2007; 77:779–89.
10 Ramos-Junior WM, Sardinha JF, Costa MR et al. Clinical aspects of hemolysis in patients with P. vivax malaria treated with primaquine, in the Brazilian Amazon. Braz J Infect Dis 2010;14:410–2.
11 White NJ. The role of anti-malarial drugs in eliminating malaria. Malar J 2008;7:29.
12 Mockenhaupt FP, Mandelkow J, Till H et al. Reduced prevalence of Plasmodium falciparum infection and of concomitant anaemia in pregnant women with heterozygous G6PD deficiency. Trop Med Int Health 2003;8:118–24.
13 Clark TG, Fry AE, Auburn S et al. Allelic heterogeneity of G6PD deficiency in West
Africa and severe malaria susceptibility. Eur J Hum Genet 2009;17:1080–5.
14 Ruwende C, Khoo SC, Snow RW et al. Natural selection of hemi- and heterozygotes for G6PD deficiency in Africa by resistance to severe malaria. Nature 1995;376:246–9.
15 Santana MS, Lacerda MV, Barbosa MG et al. Glucose-6-phosphate dehydrogenase deficiency in an endemic area for malaria in Manaus: a cross-sectional survey in the Brazilian Amazon. PloS One 2009;4:e5259.
16 Leslie T, Briceno M, Mayan I et al. The impact of phenotypic and genotypic G6PD
deficiency on risk of Plasmodium vivax infection: a case-control study amongst Afghan refugees in Pakistan. PLoS Med 2010;7:e1000283.
17 Howes RE, Piel FB, Patil AP et al. G6PD deficiency prevalence and estimates of
affected populations in malaria endemic countries: a geostatistical model-based map. PLoS Med 2012;9:e1001339.

69
18 Li J, Collins WE, Wirtz RA et al. Geographic subdivision of the range of themalaria parasite Plasmodiumvivax. Emerg Infect Dis 2001;7:35–42.
19 Yalcindag E, Elguero E, Arnathau C et al. Multiple independente introductions of
Plasmodium falciparum in South America. Proc Natl Acad Sci USA 2012;109:511–6.
20 Best WR. Absence of erythrocyte glucose-6-phosphate dehydrogenase deficiency
in certain Peruvian Indians. J Lab Clin Med 1959;54:791.
21 Neel JV, Salzano FM, Junqueira PC et al. Studies on the Xavante Indians of the Brazilian Mato Grosso. Am J Hum Genet 1964;16:52–140.
22 Saldanha PH, Lebensztajn B, Itskan SB. Activity of glucose-6-phosphate
dehydrogenase among Indians living in a malarial region of Mato Grosso and its implication to the Indian-mixed populations in Brazil. Hum Hered 1976;26:241–51.
23 Black FL, Salzano FM, Layrisse Z et al. Restriction and persistence of
polymorphisms of HLA and other genetic traits in the Parakanã Indians of Brazil. Am J Phys Anthropol 1980;52:119–132.
24 Neel JV, Mohrenweiser HW, Rothman ED et al. Revised indirect estimate of
mutation rates in Amerindians. Am J Hum Genet 1986;38:649–666.
25 Callegari-Jacques SM, Salzano FM, Weimer TA et al. The Wai Wai Indians of South America: history and genetics. Ann Hum Biol 1996;23:189–201.
26 Moyano M, Mendez F. Erythrocyte defects and parasitemia density in patients with
Plasmodium falciparum malaria in Buenaventura, Colombia. Rev Panam Salud Publica 2005;18:25–32.
27 Katsuragawa TH, Cunha RP, Souza DC et al. Malaria and hematological aspects
among residents to be impacted by reservoirs for the Santo Antonio and Jirau Hydroelectric Power Stations, Rondonia State, Brazil. Cad Saude Publica 2009;25:1486–92.
28 Santana MS, Rocha MA, Arcanjo AR et al. Association of methemoglobinemia and
glucose-6-phosphate dehydrogenase deficiency in malaria patients treated with primaquine. Rev Soc Bras Med Trop 2007;40:533–6.
29 Brazilian Ministry of Health. Epidemiological Bulletin of Malaria. Brasilia: Brazilian
Ministry of Health; 2012.
30 Brazilian Ministry of Health. Guideline for Laboratorial Diagnosis of Malaria in Brazil. Brasilia: Brazilian Ministry of Health; 2005.
31 Bouanga JC, Mouele R, Prehu C et al. Glucose-6-phosphate dehydrogenase
deficiency and homozygous sickle cell disease in Congo. Hum Hered 1998;48:192–7.

70
32 Kurdi-Haidar B, Mason PJ, Berrebi A et al. Origin and spread of the glucose-6-phosphate dehydrogenase variant (G6PD-Mediterranean) in the Middle East. Am J Hum Genet 1990;47:1013–9.
33 Hamel AR, Cabral IR, Sales TS et al. Molecular heterogeneity of G6PD deficiency
in an Amazonian population and description of four new variants. Blood Cell Mol Dis 2002;28:399–406.
34 Oliveira RA, Oshiro M, Hirata MH et al. A novel point mutation in a class IV glucose-
6-phosphate dehydrogenase variant (G6PD Sao Paulo) and polymorphic G6PD variants in Sao Paulo State, Brazil. Genet Mol Biol 2009;32:251–4.
35 Weimer TA, Salzano FM,Westwood B et al. G6PD variants in three South American
ethnic groups: population distribution and description of two new mutations. Hum Hered 1998;48:92–6.
36 Benchimol S. Amazonia: Social and Cultural Formation [in Portuguese]. Manaus:
Editora Valer; 1999.
37 Guindo A, Fairhurst RM, Doumbo OK et al. X-linked G6PD deficiency protects hemizygous males but not heterozygous females against severe malaria. PLoS Med 2007;4:e66.
38 Louicharoen C, Patin E, Paul R et al. Positively selected G6PD-Mahidol mutation
reduces Plasmodium vivax density in Southeast Asians. Science 2009;326:1546–9.
39 Cappadoro M, Giribaldi G, O’Brien E et al. Early phagocytosis of glucose-6-
phosphate dehydrogenase (G6PD)-deficient erythrocytes parasitized by Plasmodium falciparum may explain malária protection in G6PD deficiency. Blood 1998;92:2527–34.
40 Baird JK, Surjadjaja C. Consideration of ethics in primaquine therapy against
malaria transmission. Trends Parasitol 2011; 27:11–16.
41 Lacerda MV, Fragoso SC, Alecrim MG et al. Postmortemcharacterization of extent does this parasite kill? Clin Infect Dis 2012: 55: e67-74.

71

72
4.3 ARTIGO 3
Santana MS, Monteiro WM, Siqueira AM, Costa MRF, Sampaio V, Lacerda MVG,
Alecrim MGC, 2012. High frequency of diabetes in patients with glucose-6-phosphate
dehydrogenase deficiency, Western Brazilian Amazon. Artigo submetido em 17 de
janeiro de 2013 para a revista The American Journal of Tropical Medicine and Hygiene.

73
Short Communication
Can concurrent diabetes and glucose-6-phosphate dehydrogenase deficiency increase the risk of haemolysis in vivax malaria patients treated with
primaquine?
Marli Stela Santana*1,2, Wuelton Marcelo Monteiro1,2,3, Monica Regina Farias Costa2,
Vanderson de Souza Sampaio, Marcus Vinícius Guimarães de Lacerda*1,3,4, Maria das
Graças Costa Alecrim1,2
1 Universidade do Estado do Amazonas, Manaus, Amazonas, Brazil
2 Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, Manaus, Amazonas,
Brazil
3 Department of Public Health, Universidade Federal do Amazonas, Manaus,
Amazonas, Brazil
4 Universidade Nilton Lins, Manaus, Amazonas, Brazil
*Corresponding authors: Tel: + 55 92 2127-3443; E-mail: [email protected];
Address: Gerência de Malária/FMT-HVD – Av. Pedro Teixeira 25, Dom Pedro,
69040-000, Manaus, Amazonas, Brazil.

74
ABSTRACT
Introduction: Glucose-6-phosphate dehydrogenase (G6PD) deficiency is one of the
most common human genetic abnormalities, with significant prevalence worldwide.
Evidence indicates that there may be association between diabetes and G6PD
deficiency. The purpose of this study was to estimate the frequency of diabetes among
G6PD deficient individuals in Manaus, in an area of unstable malaria transmission, in
the Western Brazilian Amazon. G6PD deficient males were more prone to impaired
fasting glucose and diabetes, with a strong statistical association (p <0.001).
Key-words Glucose-6-phosphate deficiency, diabetes, impaired fasting glucose.
The glucose-6-phosphate dehydrogenase (G6PD) deficiency is a genetic
disorder X-linked affecting about 400 million people living mainly in tropical regions,
which makes it the most common enzyme deficiency of the globe,1 and can be found
in the male population of Manaus, Amazon State.2 Moreover, diabetes mellitus is a
serious public health problem, due to its high prevalence, severe morbidity and
mortality and is widely distributed in the world population, being that 5.6% of the
Brazilian population has clinical diagnosis for diabetes.3 Epidemiological data suggest
that G6PD deficiency may be a risk factor for diabetes. Several mechanisms may be
involved around the association between diabetes and G6PD deficiency, especially
changes related to genes controlling both insulin secretion and action of G6PD
regulated by antioxidants, such as NADPH.4 In addition, it appears that the G6PD-
deficient diabetic patients do not respond to the disease as well as those who are
diabetic and not G6PD-deficient.5 The present study aimed to estimate the frequency
of diabetes mellitus among G6PD deficient individuals in Manaus, in the Western
Brazilian Amazon.
A group of 1.478 male subjects (aged 1-65 years) residents in areas of unstable
malaria transmission were screened for G6PD deficiency in the period March 2009 to
March 2010. We collected 10 ml blood sample from each individual in vacuum tubes
containing EDTA anticoagulant. Samples were stored at 2° to 8° C and processed
within 48 hours for enzyme activity measure (kit Neolisa G6PD Intercientífica®
Corporation, USA). Individuals presenting G6PD deficiency were contacted for the

75
preparation for a fasting of 8-12 hours, to perform collection of 3 ml blood sample in
vacuum tubes with gel separator used in the determination of glucose serum by the
enzymatic method. Deficient samples were submitted to molecular analysis through
Polymerase Chain Reaction Polymorphism and Restriction Fragment Size (PCR-
RFLP). For each G6PD deficient individual, two controls were selected among non-
G6PD deficient from the demand of patients in a tertiary healthcare center in tropical
diseases to serve as controls. The study was approved by the Ethics Committee Board
of the FMT-HVD (approval number 2399/2006). Informed written consent was obtained
from all participants. For boys, parents were instructed about the objectives of the study
and signed an informed consent. Individuals presenting glucose levels ≤ 109 were
classified as euglycemic, between 110 and 125 as impaired fasting glucose and ≥ 126
as diabetic.6 Patients diagnosed with impaired fasting glucose or diabetes were
referred to specialized treatment according to the guidelines of the Brazilian Ministry
of Health.
A total of 66 individuals were detected as G6PD deficients in the enzymatic
screening, resulting in an overall prevalence of G6PD deficiency of 4.5% (CI95% 3.44-
5.56). Of this group 56 (84.8%) individuals showed a moderate deficiency with the
carriage of the mutation compatible with the African variant (202 G→A) and 10 (15.2%)
were severely deficient and genotyped as carriers of the Mediterranean variant. G6PD
deficient variants were not associated with the frequency of positive thick blood smears
(p=0.614), although only one infected subject (1.5%) was found among G6PD
deficient. A total of 132 individuals were enrolled as controls. As seen in the Table 1,
G6PD deficient individuals are more prone to impaired fasting glucose and diabetes,
with a strong statistical association (a five times higher risk) (p<0.001). Both G6PD
deficient variants showed significantly higher frequencies of diabetes than non-G6PD
deficients (p<0.001). Similar frequencies of impaired fasting glucose and diabetes
were found among carriers of the African and Mediterranean variants (p=0.269).
Hemolytic crises were reported in 21 (31.8%) of the G6PD deficient individuals. No
report of hemolysis occurred in the non-G6PD deficients. Diabetes and impaired
fasting glucose were not associated with higher frequency of hemolysis in the general
population (p=0.183) or among the G6PD deficient individuals (p=0.427) (data not
shown).

76
In this work, G6PD deficient individuals are more prone to impaired fasting
glucose and diabetes, both for carriers of African and Mediterranean variants. This
result agrees with previous reports of hyperglycemia7 and higher risk of diabetes8 in
G6PD deficient individuals. Concurrence of these two entities may trigger great clinical
repercussions. There is an increased prevalence of proliferative retinopathy in patients
with type 1 diabetes who are deficient in glucose-6-phosphate dehydrogenase,9
suggesting that G6PD deficiency accelerates the microvascular complications
of diabetes. The same authors showed a trend for increased frequency of
microalbuminuria among G6PD-deficient patients with diabetes. A higher percentage
of the elevated glycated hemoglobin level was found among the G6PD deficient
diabetic patients than among the non-deficient diabetic patients.5 To our knowledge,
this is the first investigation focusing on the association of diabetes with the carriage
of specific G6PD variants and, interestingly, our result supports a strong association
irrespective the variant. However, contradictory data focusing this association have
been presented and it is difficult to judge the validity of the many claims that have been
made.10
In this work, the frequency of reported episodes of hemolysis was similar
between the group with G6PD deficiency associated with diabetes/impaired fasting
glucose and that with isolated G6PD deficiency. Diabetic acidosis can induce
hemolysis in G6PD deficient patients even caused by drugs used to treat diabetes type
2, such as glibenclamide.11 This study was not able to identify precisely which drugs
or clinical factors were involved in reports of hemolysis, but since hemolytic episodes
were recorded after a malaria attack we suggest that treatment with primaquine may
be contributing for this complication.
The occurrence of hemolysis in the context of diabetes crises has been reported
in carriers of the African variant,12 but not in that with the Mediterranean variant.13 The
relative high frequency of the Mediterranean variant in our sample would explain the
results presented here. Anyway, the impact of the concurrence of diabetes and G6PD
deficiency different variants needs to be clarified.
An interesting finding from our group is that G6PD deficiency promotes a
protection against malaria in a transmission area with predominance of Plasmodium

77
vivax in the Brazilian Amazon,2 since the diagnosis of malaria was positive in only one
of the individuals G6PD deficient in this study. In this context, one speculates that
malaria incidence and diabetes may have an inverse relatedness. Otherwise, G6PD
deficiency may be a cause of clinical complications such hemolysis with the need of
blood transfusion in patients presenting vivax malaria treated with primaquine.2 From
a practical standpoint, patients diagnosed with diabetes should be screened for G6PD
deficiency before primaquine is prescribed, when they present vivax malaria, in order
to prevent possible clinical complications.
In summary, we found a high prevalence of diabetics and impaired fasting
glucose in subjects with G6PD deficiency. Our data suggest that diabetic patients
attending health services are screened for G6PD deficiency after the diagnosis of vivax
malaria, even before starting treatment with primaquine.
Acknowledgements
We are grateful to all the technical support from health agents from the Foundation of
Health Surveillance (FVS). Special thanks to Gloria Lima, Elizabeth Santos Galusso,
Evaulino Ferreira Itapirema, to all the undergraduate students from UNINORTE and
UEA, and to José Alves Maciel Jr., who helped in the field and laboratory activities.
The study was supported by Fundação de Amparo à Pesquisa (FAPEAM) and
Programa de Pesquisa para o SUS (PPSUS). The funders had no role in study design,
data collection and analysis, decision to publish or preparation of the manuscript.
References
1. Nkhoma ET, Poole C, Vannappagari V, Hall SA, Beutler E, 2009. The global prevalence of glucose-6-phosphate dehydrogenase deficiency: a systematic review and meta-analysis. Blood Cells Mol Dis 42: 267-78. 2. Santana MS, De Lacerda MV, Barbosa MG, Alecrim WD, Alecrim MG, 2009. Glucose-6-phosphate dehydrogenase deficiency in an endemic area for malaria in Manaus: a cross-sectional survey in the Brazilian Amazon. PloS one 4: e5259. 3. Brasil. VIGITEL, 2011. Vigilância de fatores de risco e proteção para doenças crônicas por inquérito telefônico. Dados Sobre Diabetes. Brasília: Ministério da Saúde. Available: http://portalsaude.saude.gov.br/portalsaude/arquivos/pdf/2012/Mai/09/Vigitel_2011_d

78
iabetes_final.pdf [Accessed 26/06/2012]. 4. Zhang Z, Liew CW, Handy DE, Zhang Y, Leopold JA, Hu J, Guo L, Kulkarni RN, Loscalzo J, Stanton RC, 2010. High glucose inhibits glucose-6-phosphate dehydrogenase, leading to increased oxidative stress and beta-cell apoptosis. FASEB J 24: 1497-505. 5. Price MDJ, Cockburn B, 2001. Glucose-6-phosphate dehydrogenase activity in diabetes in Trinidad and Tobago. West Indian Med J 50: 34-35. 6. World Health Organization, 2006. Definition and Diagnosis of Diabetes Melittus and Intermediate Hyperglycaemia: 5-34. 7. Chanmugam D, Frumin AM, 1964. Abnormal Oral Glucose Tolerance Response in Erythrocyte Glucose-6-Phosphate Dehydrogenase Deficiency. N Engl J Med 271: 1202-4. 8. Niazi GA, 1991. Glucose-6-phosphate dehydrogenase deficiency and diabetes mellitus. Int J Hematol 54: 295-8. 9. Cappai G, Songini M, Doria A, Cavallerano JD, Lorenzi M, 2011. Increased prevalence of proliferative retinopathy in patients with type 1 diabetes who are deficient in glucose-6-phosphate dehydrogenase. Diabetologia 54: 1539-42. 10. Meloni T, Pacifico A, Forteleoni G, Meloni GF, 1992. G6PD deficiency and diabetes mellitus in northern Sardinian subjects. Haematologica 77: 94-5. 11. Vinzio S, Andres E, Perrin AE, Schlienger JL, Goichot B, 2004. Glibenclamide-induced acute haemolytic anaemia revealing a G6PD-deficiency. Diabetes Res Clin Pr 64: 181-3. 12. Le Pommelet C, Le Moullec N, Zunic P, 2006. Diabetic ketoacidosis revealing glucose-6-phosphate dehydrogenase deficiency: description of an adult case. Diabetes Metab 32: 636-7. 13. Shalev O, Wollner A, Menczel J, 1984. Diabetic ketoacidosis does not precipitate haemolysis in patients with the Mediterranean variant of glucose-6-phosphate dehydrogenase deficiency. BMJ 288: 179-80.

79
Table 1. Association between G6PD deficiency and impaired fasting glucose or diabetes.
Glicemia G6PD
deficient
Non-G6PD
deficient Total OR (IC95%) p*
Normal (≤ 110 mg/dL) 21 (16.5%) 106 (83.5%) 127
Impaired (110-125 mg/dL) 29 (65.9%) 15 (34.1%) 44 5.1 (2.5-11.2) <0.001
Diabetes (≥ 126 mg/dL) 16 (59.3%) 11 (40.7%) 27 5.3 (2.3-12.6) <0.001
* Adjusted by age.

80
5 DISCUSSÃO

81
5.1 Prevalência da deficiência da G6PD e seus polimorfismos na população
da Amazônia Ocidental
O primeiro aspecto a ser discutido diz respeito à realização de um estudo
preliminar para detecção da prevalência da deficiência da G6PD na população da
Amazônia Ocidental (Santana et al., 2009). A experiência inicial de um estudo de
base populacional serviu como parâmetro para a execução do presente trabalho, um
tanto mais abrangente e que buscou identificar os polimorfismos moleculares
relacionados à deficiência da G6PD, assim como suas implicações bioquímicas. Deste
modo, uma vez que a deficiência de G6PD é uma característica recessiva ligada ao
cromossomo X, ambos os estudos estiveram comprometidos em analisar somente
sujeitos da pesquisa do gênero masculino, encontrando nessa abordagem uma
estimativa real da enzimopatia (Beutler, 1996).
Do ponto de vista de triagem populacional, os 1.478 indivíduos mostraram-se
adequados para serem incluídos no estudo, de modo que encontrou-se 66 indivíduos
com deficiência da G6PD, enquanto 56 (84,8%) mostraram deficiência compatível
com a variante G6PD A- e 10 (15,2%) indivíduos deficientes foram genotipados como
portadores da variante G6PD Mediterrânea. O predomínio da variante Africana
concorda com dados de outros estudos moleculares em outras regiões do Brasil, onde
esta variante parece mais prevalente em relação às outras (Hamel et al., 2002; Oliveira
et al., 2009).
A variante G6PD A- é a mais frequente em alguns países árabes africanos
como Tunísia e Argélia, com taxas de 63,6% e 46% (Ben Daoud et al., 2008; Nafa et
al., 1994). Pode-se afirmar categoricamente que a variante africana é quase a única
mutação responsável pela deficiência de G6PD na África (Beutler, 1994) e é comum
em áreas onde etnias de origem africana estão presentes (Cappellini e Fiorelli, 2008).
Todavia, dados de estudos anteriores mostram que a variante G6PD A- não se
encontra restrita somente a indivíduos afrodescendentes, mas pode ser comumente
ser identificada em populações do sul da Itália, Portugal, Espanha e Península Arábica
(Weimer et al., 1993; Luzzato e Mehta, 1995).

82
No caso da população estudada, a presença da variante Africana pode ser
fracamente relacionada à presença do elemento negro em sua constituição genético-
epidemiológica, pois tal elemento encontra-se parcialmente representado dentro da
população de Manaus, em torno de 12% (Santos e Guerreiro, 1995) e também
segundo Carvalho et al., (2010) as linhagens afrodescendentes estão representadas,
em concentração insignificante dentro da avaliação de linhagens genéticas, numa
comparação com outros extratos humanos africanos. Portanto, a presença desta
variante neste estudo pode estar relacionada a outros grupos étnicos que formaram a
colonização do Amazonas, dentre eles portugueses, espanhóis, franceses e sírio-
libaneses (Benchimol, 1999), exceto o elemento indígena, uma vez que a enzimopenia
entre indígenas da Amazônia é praticamente inexistente (Neel et al., 1964; Saldanha
et al., 1976).
O fenótipo G6PD A- foi produzido somente pela mutação 202 G-A/376 A-G,
sendo um resultado compatível com o de outros estudos dos polimorfismos do gene
G6PD, os quais demonstraram ser a mutação 202 G-A/376 A-G, a mais prevalente
em populações brasileiras (Weimer et al., 1993; Saad et al., 1997; Weimer et al., 1998;
Hamel et al., 2002; Oliveira et al., 2009). Isoladamente, a mutação 376 G-A não seria
capaz de causar a deficiência (Hirono e Beutler, 1988), mas uma vez associada a uma
segunda mutação, que no caso da variante G6PD A- pode ocorrer de tres maneiras:
376G/202A, 376G/968C e 376G/680T (Hirono e Beutler, 1988; Vulliamy et al., 1992),
pode desta maneira produzir o fenótipo deficiente (Town et al.,1992). Um estudo
recente sugere que uma quarta mutação encontrada no Oeste africano, foi identificada
no nucleotídeo 542 e também pode estar relacionada ao fenótipo G6PD A- (Clark et
al., 2009), mas ainda não encontra-se oficialmente catalogada.
A outra variante identificada no estudo foi a G6PD Mediterrânea, conhecida
como fenótipo de grande significado clínico e epidemiológico caracteriza-se como a
mutação mais comum nos países árabes asiáticos e no Egito, com freqüências
variando de 53,6% na Jordânia a 91,2% em Bahrain (Al Fadhli et al., 2005; Al-Ali et
al., 2002 ; Al-Momen et al., 2004;. Arnaout et al., 2011; Bayoumi et al., 1996;
Karadsheh et al., 2005), embora seja a segunda variante mais comum em alguns
países africanos árabes, como a Argélia e a Tunísia, com frequências que variam
entre 23% e 11,4% (Ben Daoud et al., 2008; Nafa et al., 1994), também constitui a

83
mutação mais freqüente entre os iraquianos curdos (87,8%), na Turquia (80%), no Irã
(66,2% -91,2%), Sul da Europa (69-77%) e no subcontinente indiano (60,4-79,6 %).
Esta variante foi também descrita por outros autores, no caso brasileiros,
especialmente daqueles responsáveis por estudos realizados no Sudeste e Sul do
Brasil (Oliveira et al., 2009; Saad et al., 1997; Weimer et al., 1998; Compri et al., 2000;
Castro et al., 2007), mas de outra maneira, nunca antes identificada na população da
Amazônia, até mesmo pelo fato de poucos estudos moleculares envolvendo essa
deficiência enzimática foram realizados até o momento.
Na Amazônia brasileira, o trabalho mais detalhado sobre a prevalência de
variantes deficientes da G6PD foi realizado na cidade de Belém, onde 82,1% dos 196
homens assintomáticos e sem relação com doadores de sangue com deficiência da
G6PD foram diagnosticados como portadores da mais comum das variantes, a G6PD
A-, seguido por oito variantes menos encontradas (Hamel et al., 2002). Em Manaus,
a partir de seis indivíduos deficientes, a eletroforese em gel mostrou que cinco foram
G6PD A- (Santana et al., 2009). Em uma revisão realizada com inquéritos à população
G6PD realizados em 4.558 descendentes de europeus 2484 misturados
negro/índio/branco ou negro/ branco e 10.298 indivíduos descendentes de indígenas
que vivem na América do Sul, a variante G6PD mediterrânea foi encontrado em
apenas 20 casos (0,1%), porém todos provenientes da região Sul do Brasil (Weimer
et al., 1998).
A fim de explicar a identificação da variante Mediterrânea na população estudada,
buscou-se encontrar subsídio na formação étnica da população de Manaus, uma
cidade marcada pela presença de árabes e judeus, vindos principalmente do Oriente
Médio e Marrocos (Benchimol, 1999) o que poderia explicar a presença da variante
Mediterrânea entre os indivíduos do estudo (Al Fadhli et al., 2005; Al-Ali et al., 2002 ;
Al-Momen et al., 2004; Arnaout et al., 2011; Bayoumi et al., 1996; Karadsheh et al.,
2005; Ben Daoud et al., 2008), mas seria necessário um estudo genético de linhagem
ancestral a fim de avaliar a contribuição dessas ou de outras etnias na constituição do
substrato genético da população de Manaus.

84
5.2 Proteção das variantes da G6PD contra a malária
Na relação da deficiência da G6PD e a malária, poucos estudos têm sido
publicados a respeito desta associação na América Latina, todavia os resultados deste
estudo acrescentam informações para o mapeamento global do problema, apesar de
uma baixa validade externa, como a maioria da literatura sobre esta questão.
Compreender o ônus da deficiência de G6PD conduzirá as políticas locais de
tratamento contra a malária e permitirá a boa concepção dos ensaios clínicos para P.
vivax com novos antimaláricos para a cura radical da doença, tendo em consideração
o potencial de hemólise numa determinada população, como é o caso da tafenoquina
(Walsh et al., 2004; Lacerda et al., 2007).
No estudo piloto conduzido anteriormente a este se observou que a deficiência da
G6PD protegeu contra várias infecções maláricas em uma área endêmica
principalmente por P. vivax (Santana et al., 2009), havendo concordância com os
resultados atuais, onde verificou-se uma proteção significativa contra episódios de
malária vivax nos homens com deficiência de G6PD inscritos neste estudo. n-plos
Neste trabalho, tanto a variante G6PD A- quanto G6PD Mediterrânea foram
associadas à proteção contra a malária. Um estudo publicado recentemente mostrou
que a deficiência da G6PD foi menos comum entre os casos de malária vivax que os
controles. A análise genética mostrou que a variante G6PD Mediterrânea esteve
associada com proteção em homens deficientes e em mulheres portadoras do gene
da deficiência tanto na heterozigoze quanto na homozigoze (Usanga e Ameen, 2000).
No caso da variante Africana, até o momento há dados publicados que apontem
proteção somente contra infecções graves por P. falciparum (Guindo et al., 2007), e
não contra infecções por P. vivax, havendo necessidade de melhor investigação dessa
associção.
5.3 Complicações decorrentes da deficiência da G6PD
O impacto da deficiência da G6PD na população estudada em sua relação com as
infecções maláricas por P. vivax infecção se reflete em sua história de hemólise e a
necessidade de transfusões de sangue por uso de primaquina. Neste estudo, os
portadores de ambas variantes, Africana e Mediterrânea, foram associadas com uma

85
maior freqüência de necessidade de transfusão de sangue. Curiosamente, observou-
se que três pacientes identificados com a variante G6PD Mediterrânea, receberam
transfusão de sangue após malária. A variante G6PD Mediterrânea apresenta mínima
atividade da G6PD, tornando seus portadores vulneráveis à ameaça de crise
hemolítica (Santana et al., 2007). Assim, estes dados sugerem que tanto a variante
G6PD A- quanto a G6PD Mediterrânea poderiam ser responsáveis por complicações
clínicas em pacientes com malária vivax. No caso da variante Mediterrânea,
considerada um polimorfismo de grande repercussão clínica, o dano hemolítico
causado por exemplo pela primaquina, é verificado logo no primeiro contato com a
droga, e no caso da variante Africana, apesar de ser mais benigna do ponto de vista
clínico, na continuidade da exposição, a partir da terceira ou quarta dose (Ramos
Júnior et al., 2010), tudo isso determinado pelo mecanismo redox da hemácia (Beutler
e Duparc, 2007), condicionado pela capacidade funcional da enzima (Beutler et al.,
1989a; Beutler, 1994; Minucci et al., 2012).
Dados sobre complicações clínicas em pacientes com malária vivax durante
tratamento são escassos. Um relatório prévio de estudo conduzido em pacientes
atendidos em uma unidade terciária de Saúde de Manaus descreve o
desenvolvimento de hemólise em 18 pacientes com deficiência da G6PD tratados para
malária por P. vivax com cloroquina e primaquina. Os achados mais freqüentes que
acompanharam a hemólise foram febre, leucocitose, anemia que requereu transfusão
de sangue e insuficiência renal aguda. Esses autores concluem que a hemólise em
pacientes em uso de primaquina não é infreqüente e contribui para a morbidade da
infecção causada por P. vivax (Ramos Júnior et al., 2010). Santana et al., (2007)
mostraram que nesta região, os níveis de metemoglobina também parece ser maior
em pacientes com deficiência da G6PD, quando em um grupo de pacientes
desenvolveram metemoglobinemia após a terapia primaquina, 51% eram deficientes
da G6PD (deficiência da G6PD naqueles sem metahemoglobinemia foi de 8,7%).
5.4 Deficiência da G6PD e diabetes
O objetivo de identificar a presença do diabetes nos indivíduos estudados neste
trabalho mostrou que deficientes da G6PD foram significativamente mais propensos
a intolerância à glicose de jejum, e com risco maior ao diabetes, tanto para portadores

86
das variantes Africanas quanto para a Mediterrânea. Este resultado concorda com
relatos prévios de hiperglicemia (Chanmugam e Frumin 1964) e maior risco de
diabetes (Niazi, 1991) em deficientes da G6PD. A presença concomitante da
deficiência da G6PD e diabetes no diagnóstico clínico de um indivíduo pode
representar importantes repercussões clínicas, como um aumento da prevalência de
retinopatia proliferativa em pacientes com diabetes tipo 1, (Cappai et al., 2011),
sugerindo que a deficiência da G6PD acelera as complicações microvasculares do
diabetes. Os mesmos autores mostraram uma tendência de aumento da freqüência
de microalbuminúria entre pacientes deficientes da G6PD diabéticos. Price e
Cockburn (2001) encontraram a maior porcentagem do nível de hemoglobina glicada
elevada foi encontrada entre os pacientes diabéticos com deficiência da G6PD do que
entre os pacientes diabéticos não deficientes. Para nosso conhecimento, esta é a
primeira investigação enfocando a associação do diabetes com variantes específicas
da deficiência da G6PD e, curiosamente, nossos resultados demonstram uma forte
associação, independentemente da variante. No entanto, dados contraditórios
centrados nesta associação têm sido apresentadas e é difícil avaliar a validade das
afirmações que se têm sido feitas (Meloni et al., 1992).
Também foi encontrada semelhança na frequência dos episódios notificados
de hemólise entre o grupo com deficiência da G6PD associada com diabetes / glicose
de jejum aumentada e com a deficiência da G6PD isolada. A própria condição de
acidose diabética pode induzir hemólise em pacientes com deficiência da G6PD,
assim com a utilização de fármacos para tratar o diabetes tipo II, como a glibenclamida
(Vinzio et al., 2004). A ocorrência de hemólise, no contexto das crises de diabetes tem
sido relatada em portadores da variante Africana (Le Pommelet et al., 2006), mas não
com a variante Mediterrânea (Shalev et al., 1984). A freqüência relativamente alta da
variante Mediterrânea em nossa amostra poderia explicar os resultados aqui
apresentados, uma vez que se trata de uma variante cuja estrututa e atividade são
efetivamente prejudicadas (Beutler, 1994), o que influi nas manifestações clínicas
indesejadas ao hospedeiro. De qualquer forma, o impacto da ocorrência de diabetes
e deficiência da G6PD por G6PD Mediterrânea precisa ser melhor esclarecido.
Finalmente, os resultados encontrados nesse estudo mostram a importância da
triagem molecular de indivíduos portadores da deficiência da G6PD na população

87
estudada, contribuindo para um melhor conhecimento sobre esta condição genética e
suas possíveis manifestações clínicas, principalmente diante da suscetibilidade à
malária vivax, a despeito de seu efeito protetor contra a doença. Embora esses
indivíduos normalmente apresentem-se assintomáticos, podem manifestar hemólise
de graus variáveis, ao fazerem uso de drogas, especialmente pela primaquina, ou
ainda pelas relevantes implicações metabólicas envolvidas, como é o caso do
diabetes, que uma vez instalada, representa um fator determinante de morbidade ao
longo da vida dos seres humanos.

88
6. CONCLUSÕES

89
Na triagem inicial, o método por quantificação da atividade enzimática
demonstrou ser eficaz na detecção e estimativa da prevalência da enzimopenia
entre os indivíduos que participaram do estudo.
As variantes G6PD A- e Mediterrânea, encontradas nos indivíduos do estudo,
são atualmente o perfil genético da G6PD na região da Amazônia Ocidental.
Os dados são sugestivos de que os indivíduos enzimopênicos portadores das
variantes G6PD A- e Mediterrânea parecem apresentar-se menos susceptíveis
à infecção malárica, e num grau menor, com menores índices parasitários, do
que os indivíduos normais.
Conforme o relato clínico dos indivíduos, hemólise por uso de primaquina e a
necessidade de transfusões de sangue constituem o maior impacto da
deficiência da G6PD na população estudada em sua relação com as infecções
maláricas por P. vivax .
O estudo sugere que pode haver relação do surgimento do diabetes em
pacientes deficientes da G6PD. Daí a importância de diagnosticá-las, pois tal
associação pode representar danos extensivos à saúde de indivíduos que
moram em áreas endêmicas de malária, aumentando assim as chances de
ocorrer hemólise.
Tanto o grupo com deficiência da G6PD associada com diabetes / intolerância
à glicose quanto o que apresentou somente a deficiência da G6PD em si,
relataram ocorrência de hemólise, mostrando que o diabetes pode ser um fator
adicional a essa manifestação clínica, mas que a deficiência da G6PS sempre
será a determinante principal.

90
7 RECOMENDAÇÕES

91
De acordo com os resultados obtidos e considerando a importância do
diagnóstico da deficiência da G6PD, torna-se relevante a análise molecular da
deficiência da glicose-6-fosfato desidrogenase das populações das áreas
endêmicas de malária de Manaus de indivíduos deficientes em nossa
população, uma vez que expostos à ação do antimalárico primaquina, podem
manifestar hemólise.
Nossos dados sugerem que os pacientes diabéticos atendidos nos serviços de
saúde, sejam triados para a deficiência de G6PD ao serem diagnosticados com
malária vivax, antes mesmo de iniciar o tratamento com primaquina.

92
8 REFERÊNCIAS BIBLIOGRÁFICAS

93
1. Al-Ali AK, Al-Mustafa ZH, Al-Madan M, Qaw F, Al-Ateeq S. Molecular characterization of glucose-6-phosphate dehydrogenase deficiency in the eastern Province of Saudi Arabia. Clin Chem Lab Med 2002 Aug; 40(8): 814-6.
2. Albuquerque SR, Cavalcante Fde O, Sanguino EC, Tezza L, Chacon F, Castilho L, dos Santos MC. FY polymorphisms and vivax malaria in inhabitants of Amazonas State, Brazil. Parasitol Res. 2010 Apr; 106(5): 1049-53.
3. Alecrim MG, Alecrim WD, Macedo V. Plasmodium vivax resistance to
chloroquine (R2) and mefloquine (R3) in Brazilian Amazon region. Rev Soc Bras Med Trop 1999 Jan-Feb; 32(1): 67-8.
4. Al Fadhli S, Kaaba S, Elshafey A, Salim M, AlAwadi A, Bastaki L. Molecular characterization of glucose-6-phosphate dehydrogenase gene defect in the Kuwaiti population. Arch Pathol Lab Med 2005 Sep;129(9):1144-7.
5. Al-Momen N, Al Arrayed SS, Al Allawi A. Molecular homogeneity of GPD Deficiency. Bahrain Med Bull 2004 Dec; 26(4): 1–6.
6. Alving AS, Carson PE, Flanagan CL, Ickes CE. Enzymatic deficiency in primaquina sensitive erythrocytes. Science 1956 Sep 14; 124(3220): 484-5.
7. Alving AS, Johnson CF, Tarlov AR, Brewer GJ, Kellermeyer RW, Carson PE. Mitigation of the haemolytic effect of primaquine and enhancement of its action against exoerythrocytic forms of the Chesson strain of Plasmodium vivax by intermittent regimens of drug administration:a preliminary report. Bull World Health Organ 1960; 22: 621-31.
8. American Diabetes Association. Diagnosis and classification of Diabetes
Mellitus. Diabetes Care 2012 Jan; 35 Suppl 1: S64-71.
9. Arnaout NH, El-Gharbawy NM, Shaheen IA, Afifi RA, Abd El-Dayem OY. Incidence and Association of 563 C/T Mediterranean and the Silent 1311 C/T mutations in G6PD-deficient Egyptian Children. Lab Med 2011 June;42(6):355–360.
10. Arruda VR, Sonati MF, Saad ST, Salles TS, Costa FF. G6PD Sumare: a novel mutation in the G6PD gene (1292 T→G) associated with chronic nonspherocytic anemia. Hum Mutat 1997; 10(3): 245-7.
11. Baronciani L, Tricta F, Beutler E. G6PD "campinas:" a deficient enzyme with a
mutation at the far 3' end of the gene. Hum Muta. 1993; 2(1): 77-8.
12. Bayoumi RA, Nur-E-Kamal MS, Tadayyon M, Mohamed KK, Mahboob BH, Qureshi MM, Lakhani MS, Awaad MO, Kaeda J, Vulliamy TJ, Luzzatto L. Molecular characterization of erythrocyte glucose-6-phosphate dehydrogenase deficiency in Al-Ain District, United Arab Emirates. Hum Hered 1996 May-Jun; 46(3): 136-41.

94
13. Benchimol S. Amazônia: Formação Social e Cultural. Manaus (AM): Editora Valer; 1999.
14. Benchimol S. Eretez Amazônia – os judeus na Amazônia. 3ª edição. Manaus (AM): Editora Valer; 2008.
15. Ben Daoud B, Mosbehi I, Prehu C, Chaouachi D, Hafsia R, Abbes S. Molecular characterization of erythrocyte glucose-6-phosphate dehydrogenase deficiency in Tunisia. Pathol Biol (Paris) 2008 Jul; 56(5): 260-7.
16. Benzi G, Pastoris O, Marzatico F, Villa RF. Age-related effect induced by oxidative stress on the cerebral glutathione system. Neurochem Res 1989 May; 14(5): 473-81.
17. Berg JM, Tymoczko JL, Stryer L. Bioquímica Fundamental. 6a edição. Rio de
janeiro: Guanabara Koogan; 2011.
18. Berghout J, Higgins S, Loucoubar C, Sakuntabhai A, Kain KC, Gros P. Genetic diversity in human erythrocyte pyruvate kinase. Genes Immun. 2012 Jan; 13(1): 98-102.
19. Beutler E. G-6-PD deficiency and malaria inblack Americans in Vietnam. Mil
Med. 1973 Mar;138(3):153-5.
20. Beutler E. G6PD deficiency. Blood [Review] 1994 Dec 1; 84(11): 3613-36
21. Beutler E. G6PD: population genetics and clinical manifestations. Blood Rev 1996 Mar; 10(1): 45-52.
22. Beutler E. Glucose-6-phosphate dehydrogenase deficiency: a historical
perspective. Blood 2008 Jan 1; 111(1): 16-24.
23. Beutler E, Dern RJ, Alving AS. The hemolytic effect of primaquine. IV. The relationship of cell age to hemolysis. J Lab Clin Med 1954 Sep; 44(3): 439-42.
24. Beutler E, Duparc S (G6PD Deficiency Working Group). Glucose-6-phosphate
dehydrogenase deficiency and antimalarial drug development. Am J Trop Med Hyg 2007 Oct; 77(4): 779-89.
25. Beutler E, Gaetani G, der Kaloustion V, Luzzatto l, Niwa S, Pannich V,
Sodeinde O, Belsey M, Kuliev AM, Modell B, Shah PM. Glucose-6-phosphate dehydrogenase deficiency. Bull World Health Organ 1989a; 67(6): 601-11.
26. Beutler E, Kuhl W. The NT 1311 polymorphism of G6PD: G6PD
Mediterranean mutation may have originated independently in Europe and Asia. Am J Hum Genet 1990 Dec; 47(6): 1008-12.

95
27. Beutler E, Kuhl W, Sáenz GF, Rodríguez W, Mutation analysis of glucose-6-phosphate dehydrogenase (G6PD) variants in Costa Rica, Hum Genet 1991 Aug;87(4):462-4.
28. Beutler E, Kuhl W, Vives-Corrons JL, Prchal JT. Molecular heterogeneity of glucose-6-phosphate dehydrogenase A-. Blood 1989b Nov 15; 74(7): 2550-5.
29. Beutler E, Vulliamy TJ. Hematologically important mutations: glucose-6-
phosphate dehydrogenase. Blood Cells Mol Dis 2002 Mar-Apr; 28(2): 93-103.
30. Bhutani VK, Johnson LH, Shapiro SM. Kernicterus in sick and preterm infants (1999-2002): a need for an effective preventive approach. Semin Perinatol 2004 Oct; 28(5): 319-25.
31. Bienzle U, Ayeni O, Lucas AO, Luzzatto L. Glucose-6-phosphate
dehydrogenase and malaria: greater resistance of female heterozygous for enzyme deficiency and of males with non-deficient variant. Lancet 1972 Jan 15; 1(7742): 107-10.
32. Bouanga JC, Mouele R, Prehu C, Wajcman H, Feingold J, Galacteros F.
Glucose-6-phosphate dehydrogenase deficiency and homozygous sickle cell disease in Congo. Hum Hered 1998 Jul-Aug; 48(4): 192-7.
33. Brasil. Situação Epidemiológica da Malária no Brasil – Ministério da Saúde;
2005. Disponível em: http://portal.saude.gov.br/portal/arquivos/pdf/be_malaria_01_2005.pdf.
34. Burgoine KL, Bancone G, Nosten F. The reality of using primaquine. Malar J
2010 Dec 27; 9: 376.
35. Burka ER, Weaver Z III, Marks PA. Clinical spectrum of hemolytic anemia associated with G 6 PD deficiency. Ann Intern Med 1966 April; 64: 817-25.
36. Callegari-Jacques SM, Salzano FM, Weimer TA, Franco MH, Mestriner MA,
Hutz MH, Schüler L. The Wai Wai Indians of South America: history and genetics. Ann Hum Biol 1996 May-Jun; 23(3): 189-201.
37. Cappadoro M, Giribaldi G, O'Brien E, Turrini F, Mannu F, Ulliers D, Simula G,
Luzzatto L, Arese P. Early phagocytosis of glucose-6-phosphate dehydrogenase (G6PD)-deficient erythrocytes parasitized by Plasmodium falciparum may explain malaria protection in G6PD deficiency. Blood 1998 Oct 1; 92(7): 2527-34.
38. Cappai G, Songini M, Doria A, Cavallerano JD, Lorenzi M. Increased
prevalence of proliferative retinopathy in patients with type 1 diabetes who are deficient in glucose-6-phosphate dehydrogenase. Diabetologia 2011 Jun; 54(6): 1539-42.
39. Cappellini MD, Fiorelli G. Glucose-6-phosphate dehydrogenase deficiency.
Lancet 2008 Jan 5; 371(9606): 64-74.

96
40. Cardoso MA, Scopel KK, Muniz PT, Villamor E, Ferreira MU. Underlying factors associated with anemia in Amazonian children: a population-based, cross-sectional study. PLoS One. 2012; 7(5): e36341.
41. Carvalho M, Brito P, Lopes V, Andrade L, Anjos MJ, Real FC, Gusmão L. Analysis of paternal lineages in Brazilian and African populations. Genet Mol Biol 2010 Jul; 33(3): 422-7.
42. Castro S, Weber R, Dadalt V, Tavares V, Giugliani R. Prevalence of G6PD
deficiency in newborns in the south of Brazil. J Med Screen 2006; 13(2): 85-6.
43. Castro S, Weber R, Matte U, Giugliani R. Molecular characterization of glucose-6-phosphate dehydrogenase deficiency in patients from the southern Brazilian city of Porto Alegre, RS. Genet Mol Biol 2008; 31(2): 423-26.
44. Chanmugam D, Frumin AM. Abnormal Oral Glucose Tolerance Response in
Erythrocyte Glucose-6-Phosphate Dehydrogenase Deficiency. N Engl J Med 1964 Dec 3; 271: 1202-4.
45. Chen EY, Cheng A, Lee A, Kuang WJ, Hillier L, Green P, Schlessinger D,
Ciccodicola A, D'Urso M. Sequence of human glucose-6-phosphate dehydrogenase cloned in plasmids and a yeast artificial chromosome. Genomics 1991 Jul; 10(3): 792-800.
46. Cittadella R, Civitelli D, Manna I, Azzia N, Di Cataldo A, Schilirò G, Brancati C.
Genetic heterogeneity of glucose-6-phosphate dehydrogenase deficiency in south-east Sicily. Ann Hum Genet 1997 May; 61(Pt 3): 229-34.
47. Clark TG, Fry AE, Auburn S, Campino S, Diakite M, Green A, Richardson A,
Teo YY, Small K, Wilson J, Jallow M, Sisay-Joof F, Pinder M, Sabeti P, Kwiatkowski DP, Rockett KA. Allelic heterogeneity of G6PD deficiency in West Africa and severe malaria susceptibility. Eur J Hum Genet 2009 Aug; 17(8): 1080-5.
48. Clark TD, Greenhouse B, Njama-Meya D, Nzarubara B, Maiteki-Sebuguzi C,
Staedke SG, Seto E, Kamya MR, Rosenthal PJ, Dorsey G. Factors determining the Heterogeneity of Malaria Incidence in Children in Kampala, Uganda. J Infect Dis 2008 Aug 1; 198(3): 393-400.
49. Clyde DF. Clinical problems associated with the use of primaquine as a tissue
schizontocidal and gametocytocidal drug. Bull World Health Organ 1981; 59(3): 391-5.
50. Coelho PM, Salvador A, Savageau MA. Relating mutant genotype to
phenotype via quantitative behavior of the NADPH redox cycle in human erythrocytes. PLoS One. 2010 Sep 28; 5(9). pii: e13031.
51. Compri MB, Saad STO, Ramalho AS. Investigação genético-epidemiológica e
molecular da deficiência de G-6-PD em uma comunidade brasileira. Cad Saude Publica 2000 Apr-Jun; 16(2): 335-42.

97
52. Conget L Diagnosis, classification and cathogenesis of diabetes mellitus. Rev Esp Cardiol 2002 May; 55(5): 528-35.
53. Corchia C, Balata A, Meloni GF, Meloni T. Favism in a female newborn infant whose mother ingested fava beans before delivery. J Pediatr 1995 Nov; 127(5): 807-8.
54. Cordes W. Experiences with plasmochin in malaria. In: Anonymous. 15th Annual Report of the United Fruit Company Medical Division. Boston, MA: United Fruit Co; 1926. p. 66-71.
55. Courtin D, Milet J, Bertin G, Vafa M, Sarr JB, Watier L, Deloron P, Troye-
Blomberg M, Garcia A, Migot-Nabias F. G6PD A-variant influences the antibody responses to Plasmodium falciparum MSP2. Infect Genet Evol. 2011 Aug; 11(6): 1287-92.
56. Cowman AF, Crabb BS. Invasion of red blood cells by malaria parasites. Cell
2006 Feb 24; 124(4): 755-66.
57. Crosby WH. Hereditary nonspherocytic hemolytic anemia. Blood 1950 Mar; 5(3): 233-53.
58. Cruz E. História do Pará. Governo do Estado do Pará, Belém; 1973. p. 148.
59. Cunha MC. Introdução a uma História Indígena. In: História dos Índios no Brasil. São Paulo: Companhia das Letras; 1992. p. 127.
60. Daily JP. Antimalarial drug therapy: The role of parasite biology and drug
resistance. J Clin Pharmacol 2006 Dec; 46(12): 1487-97.
61. Davidson RG, Nitowsky HM, Childs B. Demonstration of two populations of cells in the human female heterozygous for glucose-6-phosphate dehydrogenase variants. Proc Natl Acad Sci U S A 1963 Sep; 50: 481-5.
62. Dern RJ, Beutler E, Alving AS. The hemolytic effect of primaquine. II. The
natural course of the hemolytic anemia and the mechanism of its self-limited character. J Lab Clin Med 1954 Aug; 44(2): 171-6.
63. Dhaliwal G, Cornett PA, Tierney LM Jr. Hemolytic anemia. Am Fam Physician
2004 Jun 1; 69(11): 2599-606.
64. Díaz-Flores M, Ibáñez-Hernández MA, Galván RE, Gutiérrez M, Durán-Reyes G, Medina-Navarro R, Pascoe-Lira D, Ortega-Camarillo C, Vilar-Rojas C, Cruz M, Baiza-Gutman LA. Glucose-6-phosphate dehydrogenase activity and NADPH/NADP+ ratio in liver and pancreas are dependent on the severity of hyperglycemia in rat. Life Sci 2006 Apr 25; 78(22): 2601-7.
65. Edwards CQ. Anemia and the liver. Hepatobiliary manifestations of anemia.
Clin Liver Dis 2002 Nov; 6(4): 891-907, viii.

98
66. Enevold A, Vestergaard LS, Lusingu J, Drakeley CJ, Lemnge MM, Theander TG, Bygbjerg IC, Alifrangis M. Rapid screening for glucose-6-phosphate dehydrogenase deficiency and haemoglobin polymorphisms in Africa by a simple high-throughput SSOP-ELISA method. Malar J 2005 Dec 15; 4: 61.
67. Errico MK, Iovane B, Bernardini A, Gliati D, Scarabello C, Fainardi V, Chiari G,
Sav M, Vanelli M. Haemolysis during diabetic ketoacidosis treatment in two girls with incomplete glucose-6-phosphate dehydrogenase deficiency. Acta Biomed 2009 Apr; 80(1): 69-72.
68. Fanello CI, Karema C, Avellino P, Bancone G, Uwimana A, Lee SJ,
d'Alessandro U, Modiano D. High risk of severe anaemia after chlorproguanil-dapsone+artesunate antimalarial treatment in patients with G6PD (A-) deficiency. PLoS One 2008; 3(12): e4031.
69. Ferreira SMP. Federalismo, economia exportadora e representação política: o
Amazonas na República Velha: 1889-1914. Manaus (AM): Editora da Universidade Federal do Amazonas; 2007.
70. Fiorelli G, Martinez di Montemuros F, Cappellini MD. Chronic non-spherocytic
haemolytic disorders associated with glucose-6-phosphate dehydrogenase variants. Baillieres Best Pract Res Clin Haematol 2000 Mar; 13(1): 39-55.
71. Frank JE.Diagnosis and management of G6PD deficiency. Am Fam Physician
2005 Oct 1; 72(7): 1277-82.
72. Galtrey CM, Pathansali R. New onset diabetes complicated by haemolysis and rhabdomyolysis: a case report and review of the literature. J Med Case Rep 2008 May 16; 2: 159.
73. Gama BE, Lacerda MV, Daniel-Ribeiro CT, Ferreira-da-Cruz Mde F.
Chemoresistance of Plasmodium falciparum and Plasmodium vivax parasites in Brazil: consequences on disease morbidity and control. Mem Inst Oswaldo Cruz 2011 Aug; 106 Suppl 1: 159-66.
74. Gaskin RS, Estwick D, Peddi R. G6PD deficiency: its role in the high
prevalence of hypertension and diabetes mellitus. Ethn Dis 2001 Fall; 11(4): 749-54.
75. Gonçalves MJF, Alecrim WD. Non-planed urbanization as a contributing factor
for malaria incidence in Manaus-Amazonas, Brazil. Rev Salud Publica (Bogota) 2004 May-Aug; 6(2): 156-66.
76. Greene LS. Genetic and dietary adaptation to malaria in human populations.
Parassitologia 1999 Sep; 41(1-3): 185-92.
77. Greenwood BM. Control to elimination: implications for malaria research. Trends Parasitol 2008 Oct; 24(10): 449-54.

99
78. Guerin PJ, Olliaro P, Nosten F, Druilhe P, Laxminarayan R, Binka F, Kilama WL, Ford N, White NJ. Malaria: current status of control, diagnosis, treatment, and a proposed agenda for research and development. Lancet Infect Dis 2002 Sep; 2(9): 564-73.
79. Guindo A, Fairhurst RM, Doumbo OK, Wellems TE, Diallo DA. XLinked G6PD
deficiency protects hemizygous males but not heterozygous females against severe malaria. PLoS Med 2007 Mar; 4(3): e66.
80. Hamel AR, Cabral IR, Sales TS, Costa FF, Olalla Saad ST. Molecular
heterogeneity of G6PD deficiency in an Amazonian population and description of four new variants. Blood Cells Mol Dis 2002 May-Jun; 28(3): 399-406.
81. Haroun M. G6PD deficiency induced hemolysis in diabetic twins. Indian
Pediatr 2005 Jul; 42(7): 705-7.
82. Hastings IA, Donnelly MJ. Drug Resist Updat 2005 Feb-Apr; 8(1-2): 43-50.
83. Hildebrando L, Oliveira VEG. Ciência & Saúde Coletiva 2002; 7: 49.
84. Hirono A, Beutler E. Molecular cloning and nucleotide sequence of cDNA for human glucose-6-phosphate dehydrogenase deficiency. Proc Natl Acad Sci U S A 1988 Jun; 85(11): 3951-4.
85. Instituto Brasileiro de Geografia e Estatística (IBGE). Censo Demográfico
Brasileiro, 2000 [texto na Internet]. Rio de Janeiro; 2000. [Acesso em 06 de janeiro de 2012]. Disponível em: http://www.ibge.gov.br.
86. Instituto Brasileiro de Geografia e Estatística (IBGE). Censo Demográfico Brasileiro, 2010 [texto na Internet]. Rio de Janeiro; 2011. [Acesso em 10 de junho de 2012]. Disponível em: http://www.ibge.gov.br.
87. Jacob H, Winterhalter K. Unstable hemoglobins: the role of heme loss in Heinz
body formation. Proc Natl Acad Sci U S A 1970 Mar; 65(3): 697-701.
88. Kaplan M, Hammerman C. Glucose-6-phosphate dehydrogenase deficiency: a hidden risk for kernicterus. Semin Perinatol 2004 Oct; 28(5): 356-64.
89. Karadsheh NS, Moses L, Ismail SI, Devaney JM, Hoffman E. Molecular heterogeneity of glucose-6-phosphate dehydrogenase deficiency in Jordan. Haematologica 2005 Dec; 90(12): 1693-4.
90. Katsuragawa TH, Gil LHS, Stábile RG, Pires MG, Bonini-Domingos CR. Avaliação da incidência de glicose-6-fosfato desidrogenase (G6PD) e perfil hematológico em indivíduos de uma região de Rondônia. Rev Bras. Hematol Hemoter 2004; 26(4): 268-273.
91. Kawasaki E, Abiru N, Eguchi K. Pevention of type 1 diabetes: from the view
point of beta cell damage. Diabetes Res Clin Pract 2004 Dec; 66 Suppl 1: S27-32.

100
92. Kirkman H N, Rolfo M, Ferraris A M, Gaetani G F. Mechanisms of protection of catalase by NADPH. Kinetics and stoichiometry. J Biol Chem 1999 May 14; 274(20): 13908-14.
93. Kochar DK, Saxena V, Singh N, Kochar SK, Kumar SV, Das A 2005.
Plasmodium vivax malaria. Emerg Infect Dis 2005 Jan; 11(1): 132-4.
94. Kühn VL, Lisbôa V, de Cerqueira LP. Glucose-6-phosphate dehydrogenase deficiency in blood donors in a general hospital of Salvador, Bahia, Brazil. Rev Paul Med 1983 Sep-Oct; 101(5): 175-7.
95. Kurdi-Haidar B, Mason PJ, Berrebi A, Ankra-Badu G, al-Ali A, Oppenheim A,
Luzzatto L. Origin and spread of the glucose-6-phosphate dehydrogenase variant (G6PD-Mediterranean) in the Middle East. Am J Hum Genet 1990 Dec; 47(6): 1013-9.
96. Kwiatkowski DP. How malaria has affected the human genome and what human genetics can teach us about malaria. Am J Hum Genet 2005 Aug; 77(2): 171-92.
97. Lacerda MV, Hipolito JR, Passos LN. Chronic Plasmodium vivax infection in a patient with splenomegaly and severe thrombocytopenia. Rev Soc Bras Med Trop 2008 Sep-Oct; 41(5): 522-3.
98. Lacerda MV, Mourão MP, Alexandre MA, Siqueira AM, Magalhães BM,
Martinez-Espinosa FE, Filho FS, Brasil P, Ventura AM, Tada MS, Couto VS, Silva AR, Silva RS, Alecrim MG. Understanding the clinical spectrum of complicated Plasmodium vivax malaria: a systematic review on the contributions of the Brazilian literature. Malar J 2012 Jan 9; 11: 12.
99. Lacerda MV, Zackiewicz C, Alecrim WD, Alecrim MG. The neglected
Plasmodium vivax: are researchers from endemic areas really concerned about new treatment options? Rev Soc Bras Med Trop 2007 Jul-Aug; 40(4): 489-90.
100. Lawpoolsri S, Klein EY, Singhasivanon P, Yimsamran S, Thanyavanich N,
Maneeboonyang W. Optimally timing primaquine treatment to reduce Plasmodium falciparum transmission in low endemicity Thai-Myanmar border populations. Malar J 2009 Jul 15; 8: 159.
101. Lee JW, Choi AH, Ham M, Kim JW, Choe SS, Park J, Lee GY, Yoon KH, Kim
JB. G6PD up-regulation promotes pancreatic beta-cell dysfunction. Endocrinology 2011 Mar; 152(3): 793-803.
102. Lernmark A. Type I diabetes. Clin Chem 1999 Aug; 45(8 Pt 2): 1331-8.
103. Le Pommelet C, Le Moullec N, Zunic P. Diabetic ketoacidosis revealing
glucose-6-phosphate dehydrogenase deficiency: description of an adult case. Diabetes Metab 2006 Dec; 32(6): 636-7.

101
104. Leslie T, Mayan I, Mohammed N, Erasmus P, Kolaczinski J, Whitty CJ, Rowland M. A randomised trial of an eight-week, once weekly primaquine regimen to prevent relapse of plasmodium vivax in Northwest Frontier Province, Pakistan. PLoS One 2008 Aug 6; 3(8): e2861.
105. Lomar AV, Vidal JE, Lomar FP, Barbas CV, Matos GJ, Boulos M. Acute respiratory distress syndrome due to vivax malaria: case report and literature review. Braz J Infect Dis 2005 Oct; 9(5) :425-30.
106. Luzzatto L. Glucose 6-phosphate dehydrogenase deficiency: from genotype to
phenotype. Haematologica 2006 Oct; 91(10): 1303-6.
107. Luzzatto L, Battistuzzi G. Glucose-6-phosphate dehydrogenase. Adv Hum Genet 1985; 14: 217-329, 386-8.
108. Luzzatto L, Mehta A. Glucose-6-Phosphate Dehydrogenase Deficiency. In:
Seidman DS, Becktel J, Snow RW, Valle D, editors. The metabolic basis of inherited disease. 2a ed. New York: Mc Graw-Hill; 1995. p. 3367- 97.
109. Luzzatto L, Notaro R. Malaria. Protecting against bad air. Science 2001 Jul 20;
293(5529): 442-3.
110. Lyon MF. Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature 1961 Apr 22; 190: 372-3.
111. Lyra R, Oliveira M, Luís D, Cavalcanti N. Prevenção do Diabetes Mellitus Tipo
2. Arq Bras Endocrinol Metabol 2006 Apr; 50(2): 239-49. 112. Mahler RJ, Adler ML. Type 2 diabetes mellitus: update on diagnosis,
pathophysiology, and treatment. J Clin Endocrinol Metab 1999 Apr; 84(4): 1165-71.
113. Maia UM, Batista DCA, Pereira WO, Fernandes TAM. Prevalência da
deficiência da glicose-6-fosfato desidrogenase em doadores de sangue de Mossoró, Rio Grande do Norte. Rev Bras Hematol Hemoter 2010; 32(5): 422-23.
114. Malerbi DA, Franco LJ. Multicentric study of the prevalence of diabetes melito
and impaired tolerance in urban Brazilian population aged 30-69 yr. Diabetes Care 1992 Nov; 15(11): 1509-16.
115. Marques J, Campos JO. Incidência da deficiência de glicose-6-fosfato
desidrogenase em negros de Minas Gerais. Rev Assoc Med Bras 1975; 21:111-2.
116. Martini G, Toniolo D, Vulliamy T, Luzzatto L, Dono R, Viglietto G, Paonessa G,
D'Urso M, Persico MG. Structural analysis of the X-linked gene encoding human glucose 6-phosphate dehydrogenase. EMBO J 1986 Aug; 5(8): 1849-55.

102
117. Martini G, Ursini M V. A new lease of life for an old enzyme. Bioessays 1996 Aug; 18(8): 631-7.
118. Mashino K, Tanaka Y, Takahashi K, Inoue K, Nojima S. Hemolytic activities of
various phospholipids and their relation to the rate of transfer between membranes. J Biochem 1983 Sep; 94(3): 821-31.
119. Mason PJ. New insights into G6PD deficiency. Br J Haematol 1996 Sep; 94(4): 585-91.
120. Mason PJ, Bautista JM, Gilsanz F. G6PD deficiency: the genotype-phenotype
association. Blood Rev 2007 Sep; 21(5): 267-83.
121. Mathis D, Vence L, Benoist C. β- celldeath during progression to diabetes. Nature 2001 Dec 13; 414(6865): 792-8.
122. Matsui R, Xu S, Maitland KA, Mastroianni R, Leopold JA, Handy DE, Loscalzo
J, Cohen RA. Glucose-6-phosphate dehydrogenase deficiency decreases vascular superoxide and atherosclerotic lesions in apolipoprotein E(-/-) mice. Arterioscler Thromb Vasc Biol 2006 Apr; 26(4): 910-6.
123. McGready R, Davison BB, Stepniewska K, Cho T, Shee H, Brockman A,
Udomsangpetch R, Looareesuwan S, White NJ, Meshnick SR, Nosten F. The effects of Plasmodium falciparum and P. vivax infections on placental histopathology in an area of low malaria transmission. Am J Trop Med Hyg. 2004 Apr; 70(4): 398-407.
124. McMillan DC, Bolchoz LJ, Jollow DJ. Favism: effect of divicine on rat
erythrocyte sulfhydryl status, hexose monophosphate shunt activity, morphology, and membrane skeletal proteins. Toxicol Sci. 2001 Aug; 62(2): 353-9.
125. Meloni T, Pacifico A, Forteleoni G, Meloni GF. G6PD deficiency and diabetes
mellitus in northern Sardinian subjects. Haematologica 1992 Jan-Feb; 77(1): 94-5.
126. Messina MF, Lombardo F, Crisafulli G, Salzano G, Rossano M, Di Giorgio RM.
Hemolityc crisis in a non-ketotic and euglycemic child with Glucose-6-phosphate dehydrogenase deficiency and onset of type 1 diabetes mellitus. J Pediatr Endocrinol Metab 2004 Dec; 17(12): 1671-3.
127. Mezzacappa MA, Facchini FP, Pinto AC, Cassone AE, Souza DS, Bezerra
MA, Albuquerque DM, Saad ST, Costa FF. Clinical and genetic risk factors for moderate hyperbilirubinemia in Brazilian newborn infants. J Perinatol. 2010 Dec; 30(12): 819-26.
128. Miller SA, Dykes DD and Polesky HF. A simple salting out procedure for
extracting DNA from human nucleated cells. Nucleic Acids Res 1988 Feb 11; 16(3): 1215.

103
129. Millimono TS, Loua KM, Rath SL, Relvas L, Bento C, Diakite M, Jarvis M, Daries N, Ribeiro LM, Manco L, Kaeda JS. High prevalence of hemoglobin disorders and glucose-6-phosphate dehydrogenase (G6PD) deficiency in the Republic of Guinea (West Africa). Hemoglobin. 2012; 36(1): 25-37.
130. Ministério da Saúde. Manual de diagnóstico laboratorial da malária; 2005. Acessado em 20 de dezembro de 2009. Disponível em http://portal.saude.gov.br/portal/arquivos/pdf/manual_diag_malaria.pdf.
131. Minucci A, Moradkhani K, Hwang MJ, Zuppi C, Giardina B, Capoluongo E.
Glucose-6-phosphate dehydrogenase (G6PD) mutations database: Review of the "old" and update of the new mutations. Blood Cells Mol Dis 2012 Mar 15; 48(3): 154-65.
132. Motulsky AG. Metabolic polymorphisms and the role of infectious diseases in
human evolution. Hum Biol 1960 Feb; 32: 28-62. 133. Nafa K, Reghis A, Osmani N, Baghli L, Aït-Abbes H, Benabadji M, Kaplan JC,
Vulliamy T, Luzzatto L. At least five polymorphic variants account for the prevalence of glucose 6-phosphate deficiency in Algeria. Hum Genet 1994 Nov; 94(5): 513-7.
134. Neel JV, Salzano FM, Junqueira PC, Keiter F, Maybury-Lewis D. Studies on
the Xavante Indians of the Brazilian Mato Grosso. Am J Hum Genet 1964 Mar; 16: 52-140.
135. Newton WA Jr, Bass JC. Glutathione sensitive chronic non-spherocytic
hemolytic anemia. Am J Dis Child. 1958 Oct; 96:501-2. 136. Niazi GA. Glucose-6-phosphate dehydrogenase deficiency and diabetes
mellitus. Int J Hematol 1991 Aug; 54(4): 295-8.
137. Nkhoma ET, Poole C, Vannappagari V, Hall SA, Beutler E. The global prevalence of glucose-6-phosphate dehydrogenase deficiency: a systematic review and meta-analysis. Blood Cells Mol Dis 2009 May-Jun; 42(3): 267-78.
138. Nohl H. Involvement of free radicals in ageing: a consequence or cause of
senescence. Br Med Bull 1993 Jul; 49(3): 653-67. 139. Nosten F, Brasseur P. Combination therapy for malaria: the way forward?
Drugs 2002; 62(9): 1315-29. 140. Oliveira RA, Oshiro M, Hirata MH, Hirata RD, Ribeiro GS, Medeiros TM, de O
Barretto OC. A novel point mutation in a class IV glucose-6-phosphate dehydrogenase variant (G6PD São Paulo) and polymorphic G6PD variants in São Paulo State, Brazil. Genet Mol Biol 2009 Apr; 32(2): 251-4.
141. O'Meara WP, McKenzie FE, Magill AJ, Forney JR, Permpanich B, Lucas C,
Gasser RA Jr, Wongsrichanalai C. Sources of variability in determining

104
malaria parasite density by microscopy. Am J Trop Med Hyg 2005 Sep; 73(3): 593-8.
142. Pan American HealthOrganization. Third Meeting of the Surveillance Network for Emerging Infectious Diseases in the Amazon Countries. Monitoring Resistance to Antimalarial Drugs: Treatment Policies; 2001. Disponível da URL: http://www.paho.org/english/ad/dpc/cd/reunion2001.htm. Acesso em 12 de janeiro de 2012.
143. Pan American HealthOrganization. Health Situation in The Americas: Basic
Indicators 2010. 144. Pandolfi P P, Sonati F, Rivi R, Mason P, Grosveld F, Luzzatto L. Targeted
disruption of the housekeeping gene encoding glucose 6-phosphate dehydrogenase (G6PD): G6PD is dispensable for pentose synthesis but essential for defense against oxidative stress. EMBO J 1995 Nov 1; 14(21): 5209-15.
145. Pannacciulli I, Salvidio E, Tizianello A, Parravidino G. Hemolytic effects of
standard single dosages of primaquine and chloroquine on G-6-PD-deficient Caucasians. J Lab Clin Med 1969 Oct; 74(4): 653-61.
146. Parikh S, Dorsey G, Rosenthal PJ. Host polymorphisms and the incidence of
malaria in Ugandan children. Am J Trop Med Hyg 2004 Dec; 71(6): 750-3. 147. Peel MC, Finlayson BL, McMahon TA. Updated world map of the Köppen-
Geiger climate classification. Hydrol Earth Syst Sci 2007; 11: 1633-44.
148. Peters AL, Van Noorden CJ. Glucose-6-phosphate dehydrogenase deficiency and malaria: cytochemical detection of heterozygous G6PD deficiency in women. J Histochem Cytochem. 2009 Nov; 57(11): 1003-11.
149. Power KT. The Romanowsky stains: A review. Am J Med Technol 1982 Jun;
48(6): 519-23. 150. Prchal JT, Gregg XT. Red Cell Enzymes. Hematology (Am Soc Hematol Educ
Program) [série online] 2005; (1):19. Acesso em: fev 2006. Disponível na URL: http://www.asheducationbook.org/cgi/content/full/2005/1/19.
151. Price MDJ, Cockburn B. Glucose-6-phosphate dehydrogenase activity in diabetes in Trinidad and Tobago. W INDIAN MED J 2001; 50: 34-5.
152. Ramos Júnior WM, Sardinha JF, Costa MR, Santana MS, Alecrim MG,
Lacerda MV. Clinical aspects of hemolysis in patients with P. vivax malaria treated with primaquine, in the Brazilian Amazon. Braz J Infect Dis 2010 Jul-Aug; 14(4): 410-2.
153. Rattazzi MC. Glucose 6-phosphate dehydrogenase from human erythrocytes:
molecular weight determination by gel filtration. Biochem Biophys Res Commun 1968 Apr 5; 31(1): 16-24.

105
154. Reeve PA, Toaliu H, Kaneko A, Hall JJ, Ganczakowski M. Acute intravascular haemolysis in Vanuatu following a single dose of primaquine in individuals with glucose-6-phosphate dehydrogenase deficiency. J Trop Med Hyg 1992 Oct; 95(5): 349-51.
155. Reis, ACF. História do Amazonas. In: Manaus: Superintendência Cultural do
Amazonas 2ª. ed. Belo Horizonte: Itatiaia; 1989. p. 232. 156. Robertson R P, Tanaka Y, Takahashi H, Tran P O, Harmon J S. Prevention of
oxidative stress by adenoviral overexpression of glutathione-related enzymes in pancreatic islets. Ann N Y Acad Sci 2005 Jun; 1043: 513-20.
157. Roth EF, Raventos-Suarez C, Rinaldi A, Nagel RL. Glucose-6-phosphate
Dehydrogenase deficiency inhibits in vitro growth of Plasmodium falciparum . Proc Natl Acad Sci U S A 1983 Jan; 80(1): 298-9.
158. Roth EF Jr, Schulman S. The adaptation of Plasmodium. falciparum to
oxidative stress in G6PD deficient human erythrocytes. Br J Haematol 1988 Nov; 70(3): 363-7.
159. Rowe JA, Handel IG, Thera MA, Deans AM, Lyke KE, Koné A, Diallo DA, Raza A, Kai O, Marsh K, Plowe CV, Doumbo OK, Moulds JM. Blood group O protects against severe Plasmodium falciparum malaria through the mechanism of reduced rosetting. Proc Natl Acad Sci U S A. 2007 Oct 30;104(44):17471-6.
160. Ruwende C, Khoo SC, Snow RW, Yates SN, Kwiatkowski D, Gupta S, Warn P, Allsopp CE, Gilbert SC, Peschu N. Natural selection of hemi- and heterozygotes for G6PD deficiency in Africa by resistance to severe malaria. Nature 1995 Jul 20; 376(6537): 246-9.
161. Saad STO, Salles TSI, Carvalho MH and Costa FF (1997) Molecular
characterization of glucose-6-phosphate dehydrogenase deficiency in blood donors from Brazil. Hum Hered 1997 Jan-Feb; 47(1): 17-21.
162. Saeed TK, Hamamy HA, Alwan AAS. Association of Glucose-6-phosphate dehydrogenase deficiency with diabetes mellitus. Diabet Med 1985 Mar; 2(2): 110-2.
163. Safady J. O Café e o Mascate. Vol. 2 in Obras Completas. São Paulo: Ed.
Comercial Safady Ltda; 1973.
164. Saha N. Association of glucose-6-phosphate dehydrogenase deficiency with diabetes mellitus in ethnic groups of Singapore. J Med Genet 1979 Dec; 16(6): 431-4.
165. Saldanha PH, Lebensztajn B, Itskan SB. Activity of glucose-6-phosphate
dehydrogenase among Indians living in a malarial region of Mato Grosso and its implication to the Indian-mixed populations in Brazil. Hum Hered 1976; 26(4): 241-51.

106
166. Sallum MAM, Schultz TR, Wilkerson RC. Phylogeny of Anophelinae (Diptera, Culicidae) based on morphological characters. Ann Entomol Soc Am 2000; 93: 745-75.
167. Salvemini F, Franze A, Iervolino A, Filosa S, Salzano S, Ursini M V. Enhanced
glutathione levels and oxidoresistance mediated by increased glucose-6-phosphate dehydrogenase expression. J Biol Chem 1999 Jan 29; 274(5): 2750-7.
168. Santana MS, de Lacerda MVG, Barbosa MdGV, Alecrim WD, Alecrim MdGC.
Glucose-6-Phosphate Dehydrogenase Deficiency in an Endemic Area for Malaria in Manaus: A Cross-Sectional Survey in the Brazilian Amazon. PLoS One 2009; 4(4): e5259.
169. Santana MS, da Rocha MA, Arcanjo AR, Sardinha JF, Alecrim WD, Alecrim
MG. Association of methemoglobinemia and glucose-6-phosphate dehydrogenase deficiency in malaria patients treated with primaquine. Rev Soc Bras Med Trop 2007 Sep-Oct; 40(5): 533-6.
170. Santos SEB, Guerreiro JF. The Indigenous Contribution to the Formation of the Population of the Brazilian Amazon Region. Rev Bras Genetica 1995; 18: 311-15.
171. Sardinha JFJ. Prevalência da deficiência de glicose-6-fosfato desidrogenase e metemoglobinemia em pacientes com malária tratados com primaquina [Dissertação]. Manaus (AM): Universidade do Estado do Amazonas; 2006.
172. Seyferth G. Colonização, imigração e a questão racial no Brasil. In REVISTA
USP, São Paulo, n.53; março/maio 2002. p. 117-49.
173. Shalev O, Eliakim R, Lugassy GZ, Menczel J. Hypoglycemia-induced hemolysis in glucose-6-phosphate dehydrogenase deficiency. Acta Haematol 1985; 74(4): 227-9.
174. Shalev O, Wollner A, Menczel J. Diabetic ketoacidosis does not precipitate
haemolysis in patients with the Mediterranean variant of glucose-6-phosphate dehydrogenase deficiency. Br Med J (Clin Res Ed) 1984 Jan 21; 288(6412): 179-80.
175. Shekalaghe S, Drakeley C, Gosling R, Ndaro A, van Meegeren M, Enevold A, Alifrangis M, Mosha F, Sauerwein R, Bousema T. Primaquine clears submicroscopic Plasmodium falciparum gametocytes that persist after treatment with sulphadoxine-pyrimethamine and artesunate. PLoS One 2007 Oct 10; 2(10): e1023.
176. Silva MMC, Madeira VMC, Almeida LM, Custódio JBA. Hemolysis of human
erythrocytes induced by tamoxifen is related to disruption of membrane structure. Biochim Biophys Acta 2000 Mar 15; 1464(1): 49-61.

107
177. Singh B, Kim Sung L, Matusop A, Radhakrishnan A, Shamsul SS, Cox-Singh J, Thomas A, Conway DJ. A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 2004 Mar 27; 363(9414): 1017-24.
178. Sistema de Vigilância Epidemiológica (SIVEP) - Boletim Epidemiológico de
Malária, Estado – 2011. Acesso em: 06 de janeiro de 2012. Disponível na URL: http://www.saude.gov.br.
179. Sobngwi E, Gautier JF, Kevorkian JP, Villette JM, Riveline JP, Zhang S,
Vexiau P, Leal SM, Vaisse C, Mauvais-Jarvis F. High prevalence of glucose-6-phosphate dehydrogenase deficiency without gene mutation suggests a novel genetic mechanism predisposing to ketosis prone diabetes J Clin Endocrinol Metab 2005 Aug; 90(8): 4446-51.
180. Souza-Santos R. Seasonal distribution of malaria vectors in Machadinho
d'Oeste, Rondônia State, Amazon Region, Brazil. Cad Saude Publica 2002 Nov-Dec; 18(6): 1813-8.
181. Spolarics Z, Siddiqi M, Siegel JH, Garcia ZC, Stein DS, Ong H, Livingston DH,
Denny T, Deitch EA. Increased incidence of sepsis and altered monocyte functions in severely injured type glucose-6-phosphate dehydrogenase-deficient African American trauma patients. Crit Care Med 2001 Apr; 29(4): 728-36.
182. Srivastava SK, Beutler E. Glutathione metabolism of the erythrocyte. The
enzymic cleavage of glutathione-haemoglobin preparations by glutathione reductase. Biochem J 1970 Sep; 119(3): 353-7.
183. Sumawinata IW, Bernadeta, Leksana B, Sutamihardja A, Purnomo, Subianto
B, Sekartuti, Fryauff DJ, Baird JK. Very high risk of therapeutic failure with chloroquine for uncomplicated Plasmodium falciparum and P. vivax malaria in Indonesian Papua. Am J Trop Med Hyg 2003 Apr; 68(4): 416-20.
184. Tadei WP, Dutary-Thatcher B. Malaria vector in the Brasilian Amazon:
Anopheles of the subgenus Nyssorhynchus. Rev Inst Med Trop Sao Paulo 2000 Mar-Apr; 42(2): 87-94.
185. Takizawa T, Huang IY, Ikuta T, Yoshida A. Human glucose-6-phosphate
dehydrogenase: primary structure and cDNA cloning. Proc Natl Acad Sci U S A 1986 Jun; 83(12): 4157-61.
186. Takizawa T, Yoshida A. Molecular abnormality of the common glucose-6-
phosphate dehydrogenase variant, G6PD A( +), and restriction-fragment-length polymorphism. Am J Hum Genet 1987; 41: A241.
187. Targett G, Drakeley C, Jawara M. J Infect Dis 2001 Apr 15; 183(8): 1254-9.
188. Taylor, WR, White NJ. Antimalarial drug toxicity: a review. Drug Saf 2004;
27(1): 25-61.

108
189. The malERA Consultative Group on Monitoring, Evaluation, and Surveillance. A research agenda for malaria eradication: Monitoring, evaluation, and surveillance. PLoS Med 2011 Jan 25; 8(1): e1000400.
190. Thiberge S, Blazquez S, Baldacci P, Renaud O, Shorte S, Ménard R, Amino
R. In vivo imaging of malaria parasites in the murine liver. Nat Protoc 2007; 2(7): 1811-8.
191. Tishkoff SA, Varkonyi R, Cahinhinan N, Abbes S, Argyropoulos G, Destro-
Bisol G, Drousiotou A, Dangerfield B, Lefranc G, Loiselet J, Piro A, Stoneking M, Tagarelli A, Tagarelli G, Touma EH, Williams SM, Clark AG. Haplotype diversity and linkage disequilibrium at human G6PD: recent origin of alleles that confer malarial resistance. Science 2001 Jul 20; 293(5529): 455-62.
192. Toniolo D, Filippi M, Dono R, Lettieri T, Martini G. The CpG island in the 5’
region of the G6PD gene of man and mouse. Gene 1991 Jun 30; 102(2): 197-203.
193. Town M, Bautista JM, Mason PJ, Luzzatto L. Both mutations in G6PD A- are necessary to produce the G6PD deficient phenotype. Hum Mol Genet 1992 Jun; 1(3): 171-4.
194. Tripathy V, Reddy BM. Present status of understanding on the G6PD
deficiency and natural selection. J Postgrad Med 2007 Jul-Sep; 53(3): 193-202.
195. Usanga EA, Ameen R. Glucose-6-phosphate dehydrogenase deficiency in
Kuwait, Syria, Egypt, Iran, Jordan and Lebanon. Hum Hered 2000 May-Jun; 50(3): 158-61.
196. Vanelli M, Lucentini L, Picco P, Galanello R, Chiari G, Adinolfi B, Street ME.
Blood glucose normalization induced haemolysis in three adolescents with type 1 diabetes mellitus at onset and unknown G6PD deficiency. J Pediatr Endocrinol Metab 1996 Mar-Apr; 9(2): 193-6.
197. Vinzio S, Andres E, Perrin AE, Schlienger JL, Goichot B. Glibenclamide-induced
acute haemolytic anaemia revealing a G6PD-deficiency. Diabetes Res Clin Pract 2004 Jun; 64(3): 181-3.
198. Vulliamy TJ, D'Urso M, Battistuzzi G, Estrada M, Foulkes NS, Martini G, Calabro V, Poggi V, Giordano R, Town M. Diverse point mutations in the human glucose-6-phosphate dehydrogenase gene cause enzyme deficiency and mild or severe hemolytic anemia. Proc Natl Acad Sci U S A 1988 Jul; 85(14): 5171-5.
199. Vulliamy T, Mason P, Luzzatto L. The molecular basis of glucose-6-phosphate
dehydrogenase deficiency. Review. Trends Genet 1992 Apr; 8(4): 138-43. 200. Vulliamy T, Mehta A, Luzzatto L: Chapter 179: In The Online Metabolic and
Molecular Bases of Inherited Disease Edited by Valle D, Beaudet A,

109
Vogelstein B, Kinzler K, Antonarakis S, Ballabio A; 2007. Glucose 6-Phosphate Dehydrogenase Deficiency. [Acessed in: http:/ / www.ommbid.com/ OMMBID/ the_online_metabolic_and_molecular_ bases_of_inherited_disease/ b/ fulltext/ part19/ ch179].
201. Wajchenberg BL. Beta-cell failure in diabetes and preservation by clinical
treatment. Endocr Rev 2007 Apr; 28(2): 187-218. 202. Walsh DS, Wilairatana P, Tang DB, Heppner DG Jr, Brewer TG, Krudsood S,
Silachamroon U, Phumratanaprapin W, Siriyanonda D, Looareesuwan S. Randomized trial of 3-dose regimens of tafenoquine (WR238605) versus low-dose primaquine for preventing Plasmodium vivax malaria relapse. Clin Infect Dis 2004 Oct 15; 39(8): 1095-103.
203. Weimer TA, Salzano FM, Westwood B, Beutler E. Molecular characterization
of glucose-6-phosphate dehydrogenase variants from Brazil. Hum Biol 1993 Feb; 65(1): 41-7.
204. Weimer TA, Salzano FM, Westwood B, Beutler E. G6PD variants in three
South American ethnic groups: population distribution and description of two new mutations, Hum Hered 1998 Mar-Apr; 48(2): 92-6.
205. West IC. Glucose-6-phosphate dehydrogenase: a candidate gene for
diabetes. Diabet Med 2002 Feb; 19(2): 172-4. 206. White NJ. Plasmodium knowlesi: The Fifth Human Malaria Parasite. Clin Infect
Dis 2008a Jan 15; 46(2): 172-3. 207. White NJ. The role of anti-malarial drugs in eliminating malaria. Malar J 2008b
Dec 11; 7 Suppl 1: S8. 208. Wild SH, Roglic G, Green A, Sicree R, King H. Global prevalence of diabetes
estimates for the year 2000 and projections for 2030. Diabetes Care 2004 May; 27(5): 1047-53.
209. Wilmanski J, Siddiqi M, Deitch EA, Spolarics Z. Augmented IL-10 production
and redox-dependent signaling pathways in glucose-6-phosphate dehydrogenase-deficient mouse peritoneal macrophages. J Leukoc Biol 2005 Jul; 78(1): 85-94.
210. Windbichler N, Menichelli M, Papathanos PA, Thyme SB, Li H, Ulge UY,
Hovde BT, Baker D, Monnat RJ Jr, Burt A, Crisanti A. A synthetic homing endonuclease-based gene drive system in the human malaria mosquito. Nature 2011 May 12; 473(7346): 212-5.
211. Winzeler EA. Malaria research in the post-genomic era. Nature 2008 Oct 9;
455(7214): 751-6. 212. Wrigley NG, Heather JV, Bonsignore A, De Flora FA. Human erythrocyte
glucose 6-phosphate dehydrogenase: electron microscope studies on

110
structure and interconversion of tetramers, dimers and monomers. J Mol Biol 1972 Jul 28; 68(3): 483-99.
213. World Health Organization. Practical Chemotherapy of Malaria. Geneva; 1990.
p. 34-5. 214. World Health Organization. Guidelines for the treatment of malaria.
WHO/HTM/MAL/2006.1108. World Health Organization, Geneva, Switzerland; 2006a.
215. World Health Organization. Definition and Diagnosis of Diabetes Melittus and
Intermediate Hyperglycaemia; 2006b. 216. World Health Organization. Malaria microscopy quality assurance manual -
version 1. Manila: World Health Organization, Regional Office for the Western Pacific; 2009.
217. World Health Organization. Roll Back Malaria. World Malaria Report. 2011.
Acesso em: 20 jan 2012. Disponível na URL: http://www.rbm.who.int/wrm. 218. Xu Y, Osborne BW, Stanton RC. Diabetes causes inhibition of glucose-6-
phosphate dehydrogenase via activation of PKA, which contributes to oxidative stress in rat kidney cortex. Am J Physiol Renal Physiol 2005 Nov; 289(5): F1040-7.
219. Zhang Z, Apse K, Pang J, Stanton RC. High glucose inhibits glucose-6-
phosphate dehydrogenase via cAMP in aortic endothelial cells. J Biol Chem 2000 Dec 22; 275(51): 40042-7.
220. Zhang Z, Liew CW, Handy DE, Zhang Y, Leopold JA, Hu J, Guo L, Kulkarni
RN, Loscalzo J, Stanton RC. High glucose inhibits glucose-6-phosphate dehydrogenase, leading to increased oxidative stress and beta-cell apoptosis. FASEB J 2010 May; 24(5): 1497-505.

111
9 ANEXOS

112
Anexo 9.1 Mapa da cidade de Manaus (AM), com a localização das zonas de coleta de amostras.
Zona Oeste
N=497 Zona Norte
N=492 Zona Leste
N=489

113
Anexo 9.2 Termo de Consentimento Livre e Esclarecido
CARACTERIZAÇÃO MOLECULAR DA DEFICIÊNCIA DA GLICOSE 6-FOSFATO-DESIDROGENASE EM INDIVÍDUOS DE ÁREAS ENDÊMICAS DE MALÁRIA DE
MANAUS, AMAZONAS
Patrocinador: Fundação de Amparo à Pesquisa (FAPEAM) e Programa de Pesquisa para o
SUS (PPSUS).
Equipe responsável:
Ma das Graças Costa Alecrim (Orientadora) Marli Stela Santana Maciel (Orientanda)
Descrição do projeto: Este é um estudo que estamos fazendo no Hospital Tropical (FMT-
HVD), através da Universidade do Estado do Amazonas (UEA), com o objetivo de estudar a
malária e a deficiência da glicose-6-fosfato desidrogenase em indivíduos moradores nas
zonas norte, leste e oeste da cidade de Manaus (AM). Para isso, é preciso que seja feita
retirada de amostra de sangue do paciente, sendo uma parte da amostra reservada a estudos
futuros. Riscos associados ao estudo: Depois de colher sangue do braço, pode ficar com
uma mancha roxa e doer um pouco. Em casos de danos/despesas que venham ocorrer
mediante a participação nesse estudo há garantia de indenização/ressarcimento de nossa
parte. Benefícios: Participando desse estudo não será recebido qualquer benefício adicional,
nem dinheiro, mas estará contribuindo para o conhecimento da deficiência da G6PD.
Confidencialidade: A participação nesse estudo será confidencial. Os resultados dos
exames serão mostrados às pessoas do Hospital Tropical que trabalham com malária ou
pesquisadores de outras cidades ou países, mas o nome da pessoa que participa nunca será
revelado. Direito à retirada do estudo: A pessoa que participa da pesquisa tem todo o direito
de fazer qualquer pergunta referente aos riscos potenciais ou conhecidos, ou de dizer que
não quer participar dela. Participação voluntária: A participação é voluntária. Se houver
qualquer recusa em participar neste estudo, não ocorrerá qualquer perda de benefícios. Em
caso de dúvida ou nova informação relacionada ao estudo, serei capaz de contatar a
Dra. Marli (Bioquímica) no número de telefone 8807-1243.
Eu,..............................................................................................................................., recebi a
devida informação sobre o conteúdo deste termo, livremente, sem qualquer pressão por
parte dos pesquisadores, expresso meu consentimento em participar desta pesquisa:
......................................................................... Data...../...../.....
Pesquisador que conversou com o paciente
.........................................................................
Assinatura do paciente

114
Anexo 9.2.1 Termo de Consentimento Livre e Esclarecido (pais ou responsável)
CARACTERIZAÇÃO MOLECULAR DA DEFICIÊNCIA DA GLICOSE 6-FOSFATO-DESIDROGENASE EM INDIVÍDUOS DE ÁREAS ENDÊMICAS DE MALÁRIA DE
MANAUS, AMAZONAS
Patrocinador: Fundação de Amparo à Pesquisa (FAPEAM) e Programa de Pesquisa para o
SUS (PPSUS).
Equipe responsável:
Ma das Graças Costa Alecrim (Orientadora) Marli Stela Santana Maciel (Orientanda)
Descrição do projeto: Este é um estudo que estamos fazendo no Hospital Tropical (FMT-
HVD), através da Universidade do Estado do Amazonas (UEA), com o objetivo de estudar a
malária e a deficiência da glicose-6-fosfato desidrogenase em indivíduos moradores nas
zonas norte, leste e oeste da cidade de Manaus (AM). Para isso, é preciso que seja feita
retirada de amostra de sangue dele, sendo uma parte da amostra reservada a estudos
futuros. Riscos associados ao estudo: Depois de colher sangue do braço, pode ficar com
uma mancha roxa e doer um pouco. Em casos de danos/despesas que venham ocorrer
mediante a participação nesse estudo há garantia de indenização/ressarcimento de nossa
parte. Benefícios: Participando desse estudo, ele não receberá qualquer benefício adicional,
nem dinheiro, mas estará contribuindo para o conhecimento da deficiência da G6PD.
Confidencialidade: A participação nesse estudo será confidencial. Os resultados dos
exames serão mostrados às pessoas que trabalham neste projeto ou pesquisadores de
outras cidades ou países, mas o nome de quem participa do estudo nunca será revelado.
Direito à retirada do estudo: Você tem o direito de fazer qualquer pergunta referente aos
riscos potenciais ou conhecidos, ou de dizer que não quer que ele participe dela.
Participação voluntária: A participação dele é voluntária. Se houver qualquer recusa em
participar neste estudo, não ocorrerá qualquer perda de benefícios. Em caso de dúvida ou
nova informação relacionada ao estudo, serei capaz de contatar a Dra. Marli
(Bioquímica) no número de telefone 8807-1243.
Eu,..................................................................................................recebi a devida informação
sobre o conteúdo deste termo, sem qualquer pressão por parte dos pesquisadores, expresso
meu consentimento que:...........................................................................................
........................................................................................................, participe desta
pesquisa.......................................................................... Data...../...../.....
Pesquisador que conversou com o responsável
.........................................................................
Assinatura do responsável

115
Anexo 9.3 Ficha Cadastral/Dados Epidemiológicos
CARACTERIZAÇÃO MOLECULAR DA DEFICIÊNCIA DA GLICOSE 6-FOSFATO-DESIDROGENASE EM INDIVÍDUOS DE ÁREAS ENDÊMICAS DE MALÁRIA DE MANAUS,
AMAZONAS
Data:....../....../...... nº........................
Nome:...............................................................................................................................
.........................................................................................................................................
Endereço:Rua............................................................................................nº..................
Bairro:.............................................................................................................................
Fone res.:.......................................cel:.........................................................................
Idade: ....................
Naturalidade: município: ................................................................................
Estado: AC ( ) AL ( ) AM( ) AP( ) BA( ) CE( ) DF ( ) ES( ) GO ( )
MA( ) MG( ) MS( ) MT( ) PA( ) PB( ) PE( ) PI( ) PR( ) RJ( )
RN( )RO( ) RR( ) RS( ) SC( ) SE( ) SP( ) TO( )
País:Brasil( ) Outros:......................................
Antecedente de malária: ( )0 ( )1X ( )2X ( )3X ou +
Febre SIM( ) NÃO( ) Cefaléia SIM( ) NÃO( ) Calafrio SIM( ) NÃO( )
Gota espessa: ( )P. vivax ( )P. falciparum ( )<+ ( )+ ( )++ ( )+++ ( )Fg
Já fez transfusão após malária? SIM( ) NÃO( )
G6PD UI/gHb............................................ Variante:.............................
Obs.:..........................................................................................................................
Diabético? SIM( ) NÃO( ) em tratamento? SIM( ) NÃO( )
Glicemia em jejum -g/dL............................
Preenchido por:.......................................................................................

116
Anexo 9.4 Aprovação junto ao Comitê de Ética em Pesquisa da FMT-HVD














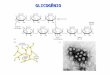

![Controle da [glicose] plasmática InsulinaInsulinaGlucagonGlucagon [glicose] estímulo liberação insulina](https://img.document.onl/doc/110x75/552fc12b497959413d8d0782/controle-da-glicose-plasmatica-insulinainsulinaglucagonglucagon-glicose-estimulo-liberacao-insulina.jpg)












