Embed Size (px)
Citation preview

PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA ANIMAL
FABRÍCIO BARRETO TERESA
Diversidade funcional de comunidades de peixes de riachos
Tese apresentada para obtenção do título de Doutor em Biologia Animal junto ao Programa de Pós-Graduação em Biologia Animal do Instituto de Biociências, Letras e Ciências Exatas da Universidade Estadual Paulista “Júlio de Mesquita Filho”, Campus de São José do Rio Preto. Orientador: Profª. Drª. Lilian Casatti
São José do Rio Preto - SP
2012

Teresa, Fabrício Barreto. Diversidade funcional de comunidades de peixes de riachos / Fabrício Barreto Teresa. - São José do Rio Preto : [s.n.], 2012. 101 f. : il. ; 30 cm. Orientador: Lilian Casatti Tese (doutorado) – Universidade Estadual Paulista, Instituto de Biociências, Letras e Ciências Exatas 1. Ecologia animal. 2. Ictiofauna. 3. Peixe de riacho. 4. Peixe – Estrutura de comunidades. 5. Diversidade biológica. I. Casatti, Lilian. II. Universidade Estadual Paulista, Instituto de Biociências, Letras e Ciências Exatas. III. Título.
CDU – 591.5
Ficha catalográfica elaborada pela Biblioteca do IBILCE
Campus de São José do Rio Preto - UNESP

Este trabalho foi realizado no Laboratório de Ictiologia, do Departamento de
Zoologia e Botânica do Instituto de Biociências, Letras e Ciências Exatas, UNESP de
São José do Rio Preto, com auxílio financeiro da FAPESP na forma de bolsa de
doutorado
(processo 2008/03583-9)

Fabrício Barreto Teresa
Diversidade funcional de comunidades de peixes de riachos Tese apresentada para obtenção do título de Doutor em Biologia Animal junto ao Programa de Pós-Graduação em Biologia Animal do Instituto de Biociências, Letras e Ciências Exatas da Universidade Estadual Paulista “Júlio de Mesquita Filho”, Campus de São José do Rio Preto.
Banca Examinadora
Profª. Drª. Lilian Casatti UNESP – São José do Rio Preto Orientadora Prof. Dr. Maurício Cetra Universidade Federal de São Carlos (UFSCar) Prof. Dr. Marcus Vinícius Cianciaruso Universidade Federal de Goiás (UFG) Profª. Drª. Katharina Eichbaum Esteves Instituto de Pesca Profª. Drª. Denise de Cerqueira e Rossa Feres UNESP – São José do Rio Preto
São José do Rio Preto
08/fevereiro/ 2012

“Dedico este trabalho a minha família”

AGRADECIMENTOS .
A defesa de uma tese marca o término de um ciclo que começou muito tempo antes do
ingresso na pós-graduação. Por isso, sou grato a todas as pessoas que contribuíram de
alguma forma para a minha formação pessoal e profissional.
A minha orientadora, Profa. Dra. Lilian Casatti pela confiança, amizade e por tudo que
me ensinou. Durante minha carreira espero um dia conseguir chegar ao menos próximo
da sua dedicação, seriedade, dinamismo e competência.
A Profa. Dra. Eliane Gonçalves-de-Freitas, que além de acompanhar o
desenvolvimento do meu projeto de doutorado, orientou-me durante a graduação e o
mestrado, ensinando-me a prática científica e despertando-me para o gosto pelo ensino.
A Profa Dra Maria Stela M.C. Noll e ao Dr. Mateus F. Ferrareze pelas críticas e
sugestões proferidas durante a qualificação.
A todos os professores da graduação e pós-graduação, em especial a Profa Dra Denise
de Cerqueira e Rossa Feres e ao Prof Dr. Francisco Langeani pela amizade, incentivo
e ensinamentos ao longo de toda a minha vida acadêmica.
Ao Prof Dr. Marcus Vinícius Cianciaruso por gentilmente ter me recebido em seu
laboratório na UFG e pela solicitude no ensino de metodologias de análise de
diversidade funcional.
Aos novos e antigos amigos do laboratório de Ictiologia André, Breno, Diego, Flávio,
Jaquelini, Mariela, Márcio, Paulo, Mateus, Lucas, em especial Ângelo, Camilo,
Cristiane, Gustavo, Fernanda, Fernando, Mônica, Renato e Roselene pela amizade,
momentos agradáveis que compartilhamos e pela ajuda em diferentes etapas da tese.
A André L.S. Navarro, Angélica Amaya, Angelo R. Manzoti, Bruno Luiz R. Silva,
Camilo Roa Fuentes, Cristiane P. Ferreira, Danilo G.H. Silva, Dayane Boracini, Diogo
B. Provete, Edis Belini, Felipe Nuvuoli, Fernanda Martins, Fernando R. Silva, Getúlio
M. Tanaka, Gustavo Baviera, José Luis V. Junior, Larissa, Leonardo Gedraite, Luciana

S. Ondei, Manuel A.M. Cabajal, Mariana Garcia, Mônica C. Bastos, Paulinho
(motorista), Renato M. Romero, Roselene S. C. Ferreira, Samuel V. Vieira e Thiago
Galego pela ajuda nos trabalhos de campo.
Ao Bruno Luiz Rodrigues da Silva, amigo de longa data pela inestimável ajuda nas
diferentes etapas deste trabalho, desde as viagens de reconhecimento até a última coleta,
sempre disposto e bem-humorado. Também sou grato a sua família pela amizade e por
acolher-me em sua casa no período final do doutorado
Ao Renato de Mei Romero, especialista em proporcionar apoio moral, pela amizade,
companheirismo e pela ajuda em diversas etapas da tese, assim como em outros
trabalhos.
Aos funcionários do IBILCE pela convivência e amizade nos últimos 12 anos, em
especial Silvia E. Kazama, Vitor B. Thomazine e Rosemar R. C. Brena da seção de
Pós-Graduação pela solicitude e paciência com os pedidos fora de prazo.
A Jane Dilvana Lima, pela amizade, incentivo, paciência e disponibilidade no
atendimento das minhas frequentes solicitações para revisão dos capítulos dessa tese;
ao Dr. Robert Vadas-Jr e Dr. Fábio Cop Ferreira pela avaliação crítica do capítulo I;
ao Diogo B. Provete e Thiago Gonçalves de Souza pela leitura crítica do capítulo III.
A todos os amigos futebolistas do IBILCE pelos agradáveis momentos compartilhados
nas “peladas” semanais que contribuíram para manter minha salubridade mental e
física.
A toda minha família em especial aos meus pais Maria Luiza Ribeiro Barreto e José
Teresa pelo amor, carinho e valores que alicerçam minha vida. Às minhas irmãs
Amanda e Débora, sobrinhas Anelise e Heloisa e cunhado Alexandre pelo carinho e
companheirismo, tornando adoráveis os momentos que passamos juntos.
À minha esposa Luciana de Souza Ondei pelo amor, paciência, companheirismo,
compreensão e pelos momentos felizes que compartilhamos. Como dizem nossos
amigos Celso e Lídia de Goiânia, sou um dependente fisiológico e comportamental de

você. A toda a família Mendes Ondei, pelo carinho, atenção e por terem me acolhido
como a um filho.

"O verdadeiro mestre é aquele que
acredita que a sua palavra sempre fará
diferença"
Rubem Alves

RESUMO .
Neste estudo avaliaram-se os efeitos do desmatamento das zonas ripárias sobre a
composição e diversidade funcional de comunidades de peixes de riachos. Para isso, foram
elaborados três capítulos a partir de amostragens da ictiofauna em 126 segmentos de cinco
metros de extensão em seis riachos, sendo três florestados e três desmatados, localizados
na bacia do rio São José dos Dourados, sistema do Alto rio Paraná, Noroeste do Estado de
São Paulo, Brasil. No CAPÍTULO I duas características que descrevem a distribuição
longitudinal das espécies, a preferência por fluxo e por profundidade, foram avaliadas. As
espécies apresentaram diferentes padrões de preferência, mas as respostas ao fluxo foram
mais consistentes entre riachos florestados e desmatados do que em relação à
profundidade. No CAPÍTULO II os padrões de diversidade taxonômica e funcional foram
comparados entre os riachos florestados e desmatados, considerando a heterogeneidade
estrutural em uma escala de meso-hábitat e no CAPÍTULO III foi avaliado se as alterações
provocadas pelo desmatamento representam filtros ambientais que modulam a composição
funcional das comunidades. Os padrões de diversidade e composição funcional diferiram
em função das alterações ambientais nos riachos associadas ao desmatamento. As
características funcionais associadas com o uso de hábitat, ecologia trófica, tolerância à
hipóxia e tamanho influenciaram a distribuição das espécies dentro e entre riachos. Os
resultados sugerem que as mudanças na disponibilidade de hábitat e recursos alimentares,
assim como nas condições físico-químicas representam filtros ambientais importantes na
organização das comunidades. Ao relacionar o desmatamento com alterações em outros
componentes da diversidade que não somente taxonômico, os resultados obtidos
possibilitam uma avaliação mais completa sobre os impactos biológicos desse tipo de
alteração.
Palavras-chave: ictiofauna, desmatamento, zona ripária, filtros ambientais

ABSTRACT .
The effects of riparian deforestation on the functional composition and diversity of stream
fish communities were evaluated. This study was divided into three chapters whose data
were obtained by sampling fish fauna in 126 five meters long mesohabitats in forested (n =
3) and deforested (n = 3) streams, located in the São José dos Dourados river Basin, upper
Paraná river system, northwestern São Paulo State, Brazil. In CHAPTER I two traits that
describe the longitudinal distribution of the species, the preference for velocity and depth
were evaluated. The species showed different patterns of preference, but their responses
were more consistent between forested and deforested streams for velocity than for depth.
In CHAPTER II the patterns of taxonomic and functional diversity were compared
between forested and deforested streams in a mesohabitat scale and in CHAPTER III it
was evaluated if the environmental modifications caused by deforestation represent
environmental filters that modulate the functional composition of communities. Patterns of
functional diversity and composition differed according to the environmental changes in
streams related to deforestation. The distribution of species within and between streams
was influenced by traits such as habitat use, trophic ecology, tolerance to hypoxia and size.
The results suggest that changes in the habitat and food availability and physicochemical
conditions represent environmental filters acting in the organization of communities. By
linking deforestation to changes in other components of diversity rather than only
taxonomic, the results obtained here provide a more complete knowledge about the
biological impacts of the loss of riparian forest.
Keywords: ichthyofauna, deforestation, riparian zone, environmental filters

SUMÁRIO
INTRODUÇÃO GERAL................................................................................................ 14
CAPÍTULO ...................................................................................................................... 18
Abstract........................................................................................................................... 19
Introduction..................................................................................................................... 20
Methods.......................................................................................................................... 22
Study area.................................................................................................................. 22
Fish sampling and environmental descriptors .......................................................... 22
Development of habitat suitability criteria and test for transferability….………… 23
Results............................................................................................................................ 24
Discussion....................................................................................................................... 27
References....................................................................................................................... 31
CAPÍTULO 2................................................................................................................... 35
Abstract........................................................................................................................... 36
Introduction..................................................................................................................... 37
Material and Methods..................................................................................................... 39
Study area and general sampling design………………………………………….. 39
Data sampling............................................................................................................ 40
Trait and trait categories............................................................................................ 42
Functional diversity metrics...................................................................................... 44
Data analysis........................................................................................................... 45
Results......................................................................................................................... 45
Discussion.................................................................................................................... 48
References................................................................................................................... 52
Supporting information............................................................................................... 59
CAPÍTULO 3................................................................................................................. 64
Resumo........................................................................................................................ 65
Introdução.................................................................................................................... 66
Material e Métodos...................................................................................................... 68
Área de estudo........................................................................................................ 68
Amostragem da ictiofauna...................................................................................... 69
Caracterização das unidades amostrais…………………………………............ 70

Características funcionais……………………………………………………… 71
Análise dos dados................................................................................................... 73
Resultados.................................................................................................................... 75
Discussion.................................................................................................................... 81
Referências Bibliográficas........................................................................................... 85
Apêndice...................................................................................................................... 90
DISCUSSÃO GERAL................................................................................................... 93
CONCLUSÕES.............................................................................................................. 96
REFERÊNCIAS GERAIS............................................................................................ 97

14
INTRODUÇÃO GERAL .
A conservação de ecossistemas aquáticos de água doce é um desafio limitado
pela falta de conhecimento sobre a sua estrutura e funcionamento, bem como sobre as
formas pelas quais esses sistemas são afetados pelas interferências antrópicas
(BARLETTA et al., 2010). Dentre as atividades humanas que exercem impactos
negativos sobre os rios e riachos destacam os represamentos, substituição de vegetação
nativa por culturas agrícolas ou pecuária, introdução de espécies exóticas e poluição
(ALLAN; FLECKER, 1993). A supressão da vegetação nativa dentro das bacias de
drenagem e, especialmente nas margens dos rios (zonas ripárias), são uma das
alterações cujos efeitos ainda não são completamente compreendidos, principalmente
em ecossistemas tropicais (BOJSEN; BARRIGA, 2002; LORION; KENNEDY, 2009).
No Brasil, a importância do desmatamento como fonte de impactos aos rios e
riachos varia regionalmente (BARLETTA et al., 2010). Em regiões populosas e
industrializadas esse tipo de alteração é uma das principais ameaças. Um exemplo
marcante é a região Noroeste do Estado de São Paulo, uma das mais desmatadas do
país, apresentando apenas 4% de vegetação nativa remanescente (SMA/IF, 2005). A
perda da vegetação nativa atinge 75% das zonas ripárias dos rios e riachos desta região
(SILVA et al., 2007) que por sua vez apresentam baixa integridade física e biológica
(CASATTI et al., 2006, 2009a).
A perda da vegetação ripária pode interferir na biota aquática diretamente,
através das modificações nas condições abióticas como temperatura e oxigênio
dissolvido, afetando o desempenho dos organismos em diferentes etapas do seu ciclo de
vida (LOWE-MCCONNELL, 1999), assim como no fornecimento de material alóctone
(galhos, frutos, sementes, folhas e troncos) para dentro do canal, que servem como
alimento, sítio reprodutivo e refúgio para diversas espécies (FAUSCH; NORTHCOTE,
1992; SCHNEIDER; WINEMILLER, 2008). Os efeitos indiretos incluem mudanças na
disponibilidade de alimento (e.g., algas e detrito) em função do aumento da
produtividade primária devido à perda do sombreamento (BOJSEN; BARRIGA, 2002;
LORION; KENNEDY, 2009) e a redução da retenção de sedimentos e outras
substâncias provenientes do ambiente terrestre adjacente (GREGORY et al., 1991),
influenciando a qualidade da água e a composição do substrato (RABENI; SMALE,
1995). A perda da vegetação ripária também proporciona a proliferação de gramíneas

15
nos barrancos (PUSEY; ARTHINGTON, 2003, GROWNS et al., 2003, ROCHA et al.,
2009), que por sua vez constituem micro-hábitats favoráveis para a ocorrência de
algumas espécies oportunistas (ROCHA et al., 2009).
Os efeitos do desmatamento das zonas ripárias sobre os organismos e
populações traduzem-se também em nível de comunidade e são representadas por
mudanças na composição de espécies e nos padrões de diversidade (e.g., Pusey &
Arthington, 2003; CASATTI et al., 2009b). Bojsen; Barriga (2002) evidenciaram
substituição de espécies e aumento da abundância de peixes em riachos submetidos a
desmatamento das suas zonas ripárias na Amazônia equatoriana. Além desses efeitos,
Lorion; Kennedy (2009) observaram também maior riqueza de espécies em riachos
desprovidos de matas ripárias na Costa Rica, resultados corroborados por Teresa;
Casatti (2010) no sudeste do Brasil.
Além dos impactos sobre os padrões de composição e diversidade dentro de
uma perspectiva taxonômica, existem evidências de que o declínio e/ou favorecimento
das espécies em riachos submetidos ao desmatamento não é aleatório e dependem de
algumas características dos organismos que influenciam o seu desempenho nesses
ecossistemas (características funcionais). Por exemplo, Casatti et al. (2009b)
compararam a fauna de peixes entre riachos mais afetados pela Interferências antrópica,
com ausência de vegetação ripária arbórea e riachos submetidos a menor pressão
antrópica, com zona ripária arborizada. Esses dois grupos de riachos abrigaram
comunidades muito distintas, com as espécies tolerantes e generalistas dominando os
riachos do primeiro grupo e as espécies reofílicas sendo mais associadas ao segundo
grupo (CASATTI et al., 2009b). De forma congruente, Teresa; Casatti (2010)
evidenciaram que o aumento da riqueza de espécies nos riachos desmatados é
representado pelo aumento no número de espécies tolerantes à degradação física do
hábitat.
Se as características funcionais são importantes para a definição da persistência
ou declínio das espécies em ambientes antropizados, como sugerido anteriormente, a
utilização de métricas que incorporem esse componente biológico (i.e., diversidade
funcional) pode fornecer informações úteis para a compreensão da estrutura e
organização das comunidades ao longo de gradientes de degradação antrópica. A
diversidade funcional pode ser definida como a variabilidade de características
biológicas relacionadas com a função que as espécies desempenham no ecossistema

16
(DIAZ; CABIDO, 2001). A importância de se considerar esse componente da
biodiversidade é reforçada por estudos que têm revelado que a diversidade funcional
tem maior poder preditivo sobre os processos ecossistêmicos do que a diversidade
taxonômica (TILMAN et al., 1997; DIAZ; CABIDO, 2001; MOUILLOT et al., 2011).
A explicação para isso reside no fato de que a abordagem taxonômica assume que as
espécies são diferentes sem a ponderação sobre a equivalência ou complementaridade
do papel que elas desempenham no ambiente (DIAZ; CABIDO, 2001). Nesse contexto,
a avaliação dos padrões de diversidade funcional das comunidades em ambientes
submetidos às interferências antrópicas pode contribuir também para a predição de
mudanças em processos ecológicos. No caso dos riachos submetidos ao desmatamento,
os padrões de diversidade funcional poderiam indicar o grau de modificação de
processos (e.g., produtividade, sedimentação, dinâmica trófica) que são
conhecidamente influenciados por esse tipo de alteração (MURPHY; HALL, 1981;
SWEENEY et al., 2004).
Alguns estudos têm utilizados métricas de diversidade funcional dentro de uma
perspectiva aplicada com o intuito de verificar os efeitos biológicos das interferências
antrópicas (e.g., ERNST et al., 2006; EROS et al., 2009; FLYNN et al., 2009;
VILLÉGER et al., 2010; BARRAGÁN et al., 2011). Por exemplo, Ernst et al. (2006)
observaram redução da diversidade funcional de comunidades de anuros em fragmentos
florestais degradados por desmatamento, embora não tenham diagnosticado mudanças
na diversidade taxonômica. Nesse caso, a alteração ambiental resultou na substituição
de espécies funcionalmente distintas por espécies redundantes. Dentro dessa abordagem
aplicada, é evidente que a incorporação de métricas de diversidade funcional fornece
subsídios para uma avaliação mais completa sobre os impactos das alterações humanas
sobre a biodiversidade. Com relação aos riachos, embora os efeitos das alterações
antrópicas sobre a estrutura e composição taxonômica das comunidades sejam
relativamente bem compreendidos (BOJSEN; BARRIGA, 2002; PUSEY;
ARTHINGTON, 2003; LORION; KENNEDY, 2009), o mesmo não acontece com
relação à perspectiva funcional.
Assim, o objetivo geral deste estudo foi avaliar os efeitos do desmatamento das
zonas ripárias sobre a composição e diversidade funcional de comunidades de peixes de
riachos. Baseado no conhecimento de que o desmatamento influencia as comunidades
de peixes (BOJSEN; BARRIGA, 2002; LORION; KENNEDY, 2009) e nas evidências

17
de que tais Efeitos são dependentes das características funcionais exibidas pelas
espécies (CASATTI et al., 2009b; TERESA; CASATTI, 2010), hipotetizamos que as
modificações ambientais causadas pelo desmatamento nos riachos refletem em
mudanças na composição e diversidade funcional das comunidades. Para o
cumprimento do objetivo geral, este estudo foi dividido em três capítulos que
correspondem a manuscritos, cujos dados foram obtidos a partir das amostragens da
ictiofauna em seis riachos, sendo três florestados e três desmatados, localizados na
bacia do rio São José dos Dourados, sistema do Alto rio Paraná, Noroeste do Estado de
São Paulo, Brasil.
Uma das dificuldades para a utilização da abordagem funcional para alguns
táxons é o conhecimento limitado sobre as características funcionais exibida pelas
espécies (PETCHEY; GASTON, 2006), como é o caso dos peixes de riachos. Assim,
inicialmente (Capítulo I) foi avaliada a preferência por fluxo e profundidade, duas
características associadas com a distribuição das espécies ao longo do gradiente
longitudinal. Posteriormente (Capítulo II), os padrões de diversidade taxonômica e
funcional foram comparados entre os riachos florestados e desmatados, considerando a
heterogeneidade estrutural em escala de meso-hábitat. Finalmente (Capítulo III), foi
avaliado se as alterações provocadas pelo desmatamento representam filtros ambientais
e para isto foram comparados padrões de diferenciação funcional entre as comunidades
dos riachos florestados e desmatados com padrões obtidos aleatoriamente.

18
Manuscrito submetido para Ecohydrology
- CAPÍTULO I -
Development of habitat suitability criteria for some Neotropical stream
fishes and an assessment of their transferability to streams with
different conservation status

19
ABSTRACT
We assessed the preference of 10 fish species for depth and velocity conditions in
forested streams from southeastern Brazil using habitat suitability criteria (HSC curves).
We also tested whether preference patterns observed in forested streams can be
transferred to deforested streams. We used data from fish sampled in 62 five-meter
quadrats in three forested streams to construct preference curves. Astyanax altiparanae,
A. fasciatus, Knodus moenkhausii, and Piabina argentea showed a preference for deep
slow habitats, whereas Aspidoras fuscoguttatus, Characidium zebra, Cetopsorhamdia
iheringi, Pseudopimelodus pulcher, and Hypostomus nigromaculatus showed an
opposite pattern: preference for shallow fast habitats. Hypostomus ancistroides showed
a multimodal pattern of preference for depth and velocity. To evaluate whether patterns
observed in forested streams may be transferred to deforested streams, we sampled 64
five-meters quadrats in three deforested streams using the same methodology. The
preference for velocity was more consistent than for depth, as success in the
transferability criterion was 86% and 29% of species, respectively. This indicates that
velocity is a good predictor of species abundance in streams, regardless of their
condition.
Keywords: habitat preference; velocity; depth; riparian forest; fish assemblages

20
INTRODUCTION
The recognition that species are distributed non-randomly in habitats,
preferentially occupying suitable sites for feeding, reproduction, and survival (Grinnell,
1917; Hutchinson, 1957), constitutes the basis for developing predictive models of
species distribution along environmental gradients (e.g., Ahmadi-Nedushan et al.,
2006). This approach has a broad application for natural resources management, as
since it allows prediction of changes in biological communities from changes in
environmental conditions caused by human activities. However, the predictive power of
these models depends on knowledge of the response of species in relation to the
gradient of environmental conditions. This information can be obtained by developing
habitat suitability criteria (HSC; Bovee, 1982, 1986), a component for conducting
physical habitat simulation (PHABSIM), a tool used to assess the consequences of
changes in river flows on habitat availability (Bovee, 1982).
The lack of knowledge about traits exhibited by species has limited the use of
the functional approach in studies focusing in diversity patterns. In this way, the
definition of HSC also may be useful, since the preference of the species for habitat
features may be used as traits in functional diversity studies. The HSC can be assessed
by curves defining the degree of preference by a given species in relation to habitat
variables alone, such as depth, velocity, substrate and cover. Efforts to develop HSC for
multiple species have increased, owing to the degradation of water resources and
growing concern over biodiversity loss (e.g., Lamouroux et al., 1999; Vadas & Orth,
2001; Strakosh et al., 2003). However, this approach has not been applied in
Neotropical environments, limiting the use of predictive models and the ability to
quantitatively predict the biological impacts from changes in hydraulic variables of
aquatic environments. This is especially alarming because changes in the natural flow
regime are among the major anthropogenic threats for fish conservation in Neotropical
lotic environments (Barletta et al., 2010).
Several factors, such as environmental heterogeneity, food availability,
predation, competition, and habitat availability influence the patterns of species
occupancy in riverine patches (Power, 1984; Thomas & Bovee, 1993; Jackson et al.,
2001; Leal et al., 2011). The differential effects of these factors may limit the
applicability of HSC between different environments (Groshens & Orth, 1993; Freeman
et al., 1997; Vadas & Orth, 2001). In these cases, the development of specific criteria

21
for each type of stream are recommended (Bovee, 1986), which brings high costs and
operational difficulties. However, other studies have found consistency in preference
patterns across streams (Thomas & Bovee, 1993; Strakosh et al., 2003). This highlights
the importance of testing HSC validity to the site of interest (transferability) before
developing new criteria.
Our present goal is to evaluate the preference of different fish species for depth
and velocity conditions in forested streams from southeastern Brazil, based on the
development of HSC curves. In our study region, the clearing of native vegetation
affects about 75% of riparian zones (Silva et al., 2007), and most streams have some
degradation of their physical integrity that is directly or indirectly related to
deforestation (Casatti et al., 2006). Here, we also evaluate the consistency of habitat
preference patterns of species in forested streams and test HSC transferability to
deforested streams.

22
METHODS
Study area
All streams studied (second and third order) are located in the drainage of the
São José dos Dourados, Upper Paraná River basin, northwestern region of the state of
São Paulo, Brazil (Figure 1). The climate of the region is influenced by equatorial and
tropical masses, resulting in a tropical climate with dry and rainy periods, with higher
rainfall and higher temperatures from December to February and lower rainfall and
milder temperatures between June and August (IPT, 1999). The forested streams are
located in forest fragments, representing what is best preserved in the region, whereas
the deforested streams are located away from forest fragments where the riparian zone
is dominated by pasture.
Figure 1. Location of the study sites in the São José dos Dourados basin, São Paulo
State, Brazil (A and B). Forested sites (dark circles) are located within the greatest forest
fragments and deforested (gray circles) streams lack arboreal vegetation in their riparian
zone and are located in areas dominated by pasture.
Fish sampling and environmental descriptors
We collectively sampled 62 five-meter quadrats in three forested streams to
identify the preference of fish species for depth and velocity and thus define habitat
suitability criteria. In each stream we selected enough 19-24 quadrats to maximize
structural variation between quadrats to better represent hydrological variability while

23
reducing variation within each quadrat. The fish samplings were carried out during the
dry season (September to November 2009 and April to July 2010) using electrofishing
preceded by the up- and downstream isolation of each quadrat. The electrofishing
sampling consisted of successive passes along the segment. The sample ended when no
new individual was captured after one passage.
After the fish sampling, we characterized the segments in relation to depth and
velocity. To accomplish this, we divided each quadrat into four equidistant, transverse
transects, where were done measurements at five equidistant points from one margin to
another. We used a mechanical flowmeter model 2030 (General Oceanics®) to measure
water velocity at ~ 0.6 of total depth. Data from transects were lumped together to
obtain the average values for the whole quadrat.
Development of habitat suitability criteria and test for transferability
Only species that occurred in at least 10 samples in forested streams were
included in the habitat suitability criteria for depth and velocity. We adopted this
procedure because infrequent species may have been under-sampled, resulting in higher
estimated errors of preference. We classified the depth gradient into five classes (D1 = Ø
to 0.2 m; D2 = 0.21 to 0.30 m; D3 = 0.31 to 0.40 m; D4 = 0.41 to 0.50 m; D5 > 0.51 m)
and velocity into four classes (V1 = 0-0.20 m/s; V2 = 0.21 to 0.40 m/s; V3 = 0.41-0.60
m/s; V4 > 0.61 m/s). The limits for the classes were defined to represent discrete units of
the hydrological gradient while ensuring a comparable number of samples in each class.
We built HSC curves (type III, sensu Bovee, 1986) from preference values that were
calculated the abundance ratio of species in each class of depth and velocity, divided by
the proportionate number of habitat in the class in relation to all habitats sampled
(Freeman et al., 1997). With this procedure, we considered both the relative abundance
of species in the habitats and habitat availability. Posteriorly, we standardized the
preference values to vary from 0 to 1, thereby obtaining the suitability index (SI). For
the class with the highest preference value, we assigned an index of 1 and proportionally
smaller values to other classes.
We tested if the HSC developed for forested streams could be transferred to
deforested streams. To do this, we sampled fish and environmental descriptors in 64
five-meter quadrats in three deforested streams (between 16 and 24 segments in each
stream) during the same period and using the same methodology applied to forested
streams. We considered optimal those SI values for depth and velocity conditions

24
greater than 0.7, according to the criteria developed for forested streams. We choose 0.7
as a reference value because it included most of the raw values of habitat preference
higher than 1, i.e., whose relative abundance is higher than the proportional availability
of habitats.
To test the transferability of HSC from forested to deforested streams, we
considered only species that occurred in at least 10 quadrats in the deforested streams.
We classified the quadrats according to the optimum/non-optimum threshold defined
from the HSC curves from forested streams. For the criterion to be considered
transferable, the abundance of a species should be higher in “optimal” samples of
deforested streams, as defined for forested streams. Thus, we conducted a comparison
of the mean species abundance between optimum and non-optimum quadrats using a
one-tailed t-test.
RESULTS
We recorded 31 species in forested streams, of which 10 occurred in at least 10
samples (Table I). The evaluation of the HSC curves for these species for depth and
velocity demonstrated three main patterns (Figure 2). Astyanax altiparanae, A.
fasciatus, Knodus moenkhausii, and Piabina argentea had high SI values in slow-
flowing and deep segments. Another four species showed an opposite pattern, with
higher SI values associated with shallow and fast-flowing habitats. This was the case of
Characidium zebra, Cetopsorhamdia iheringi, Pseudopimelodus pulcher, and
Hypostomus nigromaculatus. Aspidoras fuscoguttatus and Hypostomus ancistroides
showed distinct patterns from the above species and had the highest SI values in
multiple classes of depth and in habitats with high and low water velocity, respectively
(Figure 2).
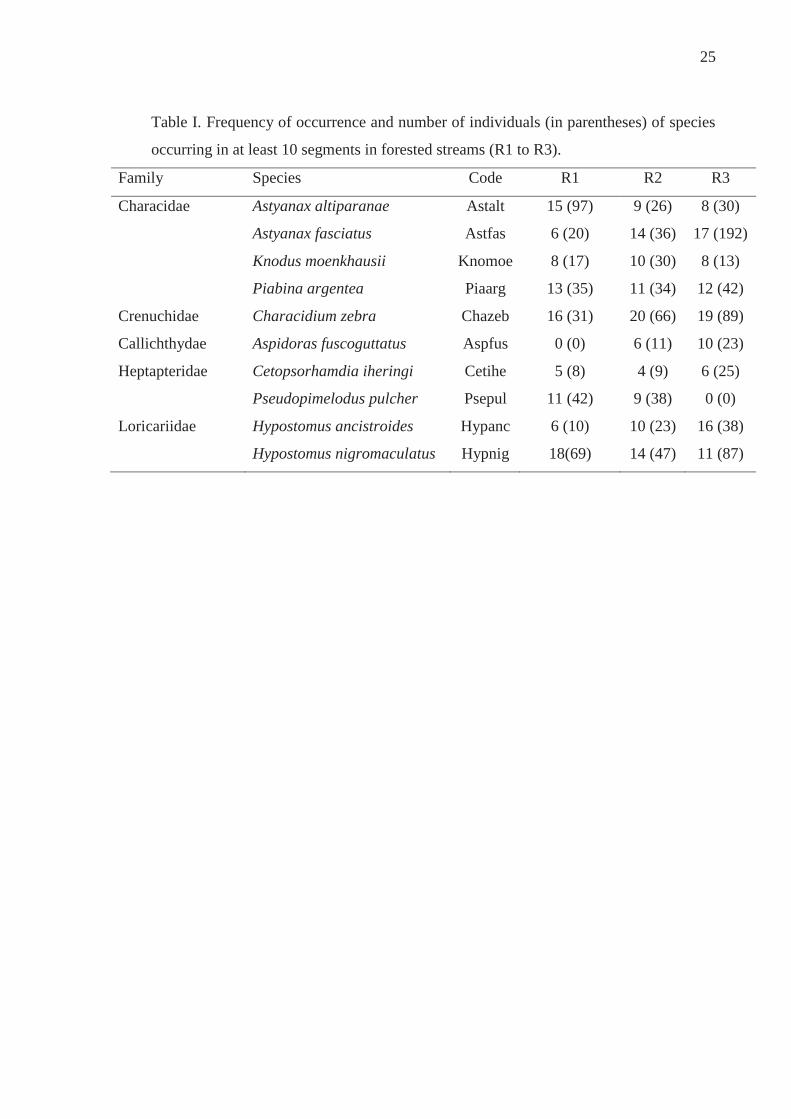
25
Table I. Frequency of occurrence and number of individuals (in parentheses) of species
occurring in at least 10 segments in forested streams (R1 to R3).
Family Species Code R1 R2 R3
Characidae Astyanax altiparanae Astalt 15 (97) 9 (26) 8 (30)
Astyanax fasciatus Astfas 6 (20) 14 (36) 17 (192)
Knodus moenkhausii Knomoe 8 (17) 10 (30) 8 (13)
Piabina argentea Piaarg 13 (35) 11 (34) 12 (42)
Crenuchidae Characidium zebra Chazeb 16 (31) 20 (66) 19 (89)
Callichthydae Aspidoras fuscoguttatus Aspfus 0 (0) 6 (11) 10 (23)
Heptapteridae Cetopsorhamdia iheringi Cetihe 5 (8) 4 (9) 6 (25)
Pseudopimelodus pulcher Psepul 11 (42) 9 (38) 0 (0)
Loricariidae Hypostomus ancistroides Hypanc 6 (10) 10 (23) 16 (38)
Hypostomus nigromaculatus Hypnig 18(69) 14 (47) 11 (87)
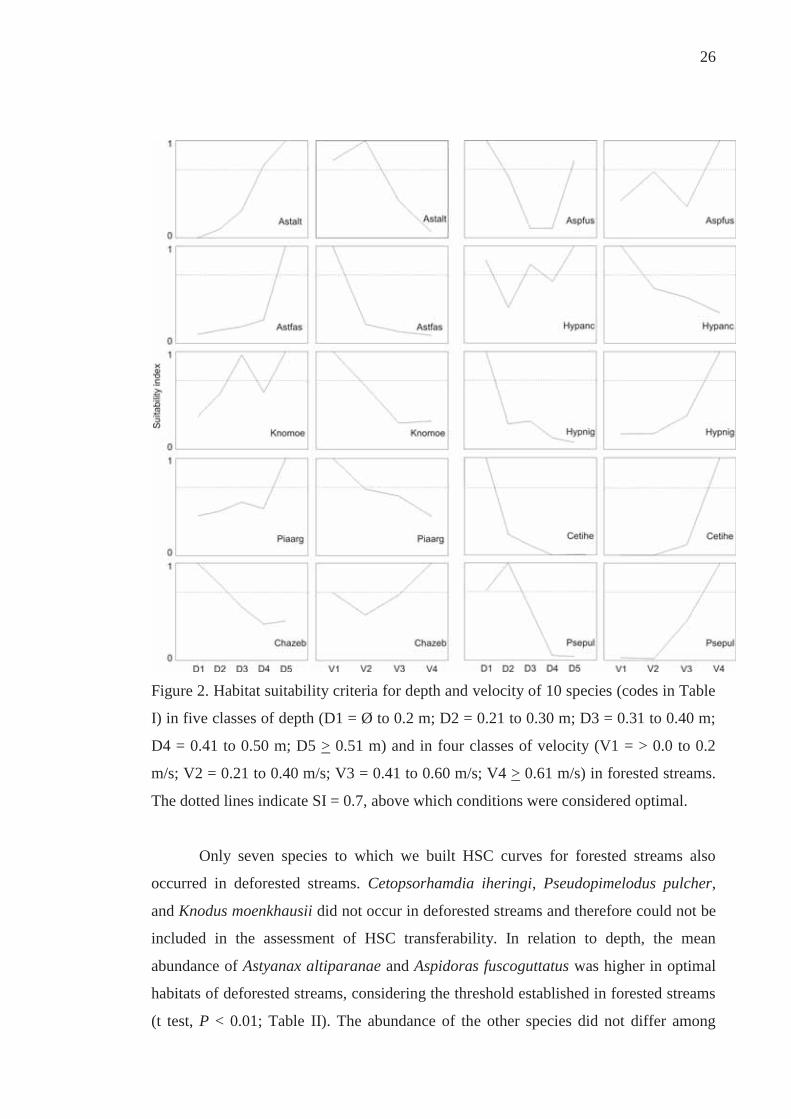
26
Figure 2. Habitat suitability criteria for depth and velocity of 10 species (codes in Table
I) in five classes of depth (D1 = Ø to 0.2 m; D2 = 0.21 to 0.30 m; D3 = 0.31 to 0.40 m;
D4 = 0.41 to 0.50 m; D5 > 0.51 m) and in four classes of velocity (V1 = > 0.0 to 0.2
m/s; V2 = 0.21 to 0.40 m/s; V3 = 0.41 to 0.60 m/s; V4 > 0.61 m/s) in forested streams.
The dotted lines indicate SI = 0.7, above which conditions were considered optimal.
Only seven species to which we built HSC curves for forested streams also
occurred in deforested streams. Cetopsorhamdia iheringi, Pseudopimelodus pulcher,
and Knodus moenkhausii did not occur in deforested streams and therefore could not be
included in the assessment of HSC transferability. In relation to depth, the mean
abundance of Astyanax altiparanae and Aspidoras fuscoguttatus was higher in optimal
habitats of deforested streams, considering the threshold established in forested streams
(t test, P < 0.01; Table II). The abundance of the other species did not differ among

27
optimal and non-optimal habitats in deforested streams (t test, P > 0.15; Table II). In
relation to water velocity, all species had higher abundances in optimal habitats of
deforested streams, except for Hypostomus ancistroides, according to the criteria
established for forested streams (t test, P < 0.02; Table II).
Table II. Transferability test of HSC from forested to deforested streams. The + sign
indicates that the HSC developed for forested streams was able to predict the abundance
of a given species in deforested streams, and the - sign indicates the opposite.
Species
Abundance
Depth Velocity
Astyanax altiparanae + +
Astyanax fasciatus - +
Piabina argentea - +
Characidium zebra - +
Aspidoras fuscoguttatus + +
Hypostomus ancistroides - -
Hypostomus nigromaculatus - +
DISCUSSION
In this study we established the preference of ten species of stream fishes for
depth and velocity conditions. In general, species showed well-defined preference
patterns for depth and velocity, reinforcing the importance of hydraulic variables as
predictors of fish distribution in lotic environments (Lamouroux et al., 1999; Vadas &
Orth, 2001; Schwartz & Herricks 2008; Leal et al., 2011). One of the patterns was the
association of Astyanax altiparanae, A. fasciatus, Knodus moenkhausii, and Piabina
argentea in deep, slow-flowing habitats. Indeed, the morphology of these species is
compatible with this habitat preference, as their compressed bodies, well-developed
caudal and pectoral fins, and lateral eyes represent morphological adaptations to live in
lentic habitats (Gatz, 1979; Oliveira et al., 2010). Behavioral patterns displayed by these
species also favor the occupation of lentic and deep habitats, as they are mid water
swimmers that feed on drift items in the water column (Casatti & Castro, 1998; Casatti
et al., 2001; Ceneviva-Bastos et al., 2010). Probably they are also favored in deep
habitats with a higher habitat volume (i.e., water column) and where slow velocity

28
requires less energy expenditure for movement and feeding (Casatti & Castro, 1998;
Bürhnheim, 2002).
An opposite pattern was observed for Characidium zebra, Cetopsorhamdia
iheringi, Pseudopimelodus pulcher, and Hypostomus nigromaculatus that showed
preference for shallow and fast-flowing habitats. These fish have morphological and
behavioral adaptations congruent with life in these conditions. For example, these
species share depressed bodies and expanded pectoral fins (Casatti et al. 2005; Casatti
& Castro, 2006), which allow short movements in fast-flowing environments (Watson
& Balon, 1984). In addition, the diet composed of periphyton (H. nigromaculatus) and
aquatic insects (remaining species) is consistent with the high availability of these items
in shallow, fast-flowing environments (Angermeier & Karr, 1983).
The conditions considered non-optimal in this study (SI < 0.7) are not
necessarily uninhabitable. Within this range, one could still discriminate between a
range of habitable or marginal conditions and another as uninhabitable (Thomas &
Bovee, 1993), which would represent the conditions in which a given species could not
live. Most species that preferred shallow, fast-flowing habitats were less plastic, with a
wider range of conditions that may be considered uninhabitable (SI values close to zero
in two classes of depth and velocity), compared with species that preferred deep, slow-
flowing habitats. The only exception to this pattern was Characidium zebra, whose non-
optimal, but occupied conditions could be considered habitable, as the SI values were
higher than 0.37. Species that preferred deep, slow-flowing habitats showed SI values
close to zero in a maximum of one class of depth and velocity. This indicates that
species that preferred shallow, fast-flowing habitats are more specialized and probably
more sensitive to changes in hydraulic conditions. In fact, the loss of rheophilic species
in streams from our study region has been attributed to changes in hydraulic conditions
and simplification of habitats (Casatti et al., 2006; Casatti et al., 2009a; Teresa and
Casatti, 2010). For example, Cetopsorhamdia iheringi and Pseudopimelodus pulcher
did not occur in the degraded streams studied by Casatti et al. (2009a), a fact that they
attributed to the loss of riffles from siltation and habitat simplification.
The habitat suitability criteria obtained for Hypostomus ancistroides and
Aspidoras fuscoguttatus were distinct from other species with respect to depth.
Normally, the HSC results in unimodal patterns (Thomas and Bovee, 1993; Vadas and
Orth, 2001; Strakosh et al., 2003), although different patterns can also be observed
(Freeman et al., 1997). The bimodal pattern observed for A. fuscoguttatus might be an

29
artifact of small sample sizes. Because this species occurred in only two out of three
forested streams, we did not consider the samples from the stream in which it was
absent to calculate the habitat availability. Consequently, only five samples fell into the
deepest depth class. Because the calculation of SI involves the relative abundance ratio
of the proportional availability of habitat, the occurrence of a few individuals (seven) in
two samples resulted in high SI values. One of the procedures that can be adopted in
this case is either group together two adjacent classes to increase sample size or not take
into account the SI value obtained (see Freeman et al., 1997). With the adoption of
these procedures, the preference pattern of A. fuscoguttatus would be similar to species
with a preference for shallow habitats, and therefore consistent with the pattern of
habitat use reported for this species (Araújo and Garutti, 2003). Although the criterium
developed here is consistent in deforested streams, this was due to the preference of this
fish for shallowest depth class, as no individual was recorded in the deepest depth class.
This strengthens our suggestion that the second peak of the HSC curve for this species
has no biological basis. The habitat use criterion for velocity indicated a preference for
fast-flowing habitats, but similar to Characidium zebra, other velocity conditions were
considered habitable (SI values > 0.32). This is consistent with the plasticity of this
species, which occurs in streams with different hydrological characteristics (Casatti et
al., 2009a,b).
The multimodal pattern of the depth preference curve of Hypostomus
ancistroides suggests that this factor has little influence in determining its abundance
patterns. On the other hand, velocity seems to be a good predictor of its abundance in
forested streams, via a more uniform relationship, indicating a preference for lentic
habitats, as also found by Uieda et al. (1997) and Casatti et al. (2005). Interestingly, this
pattern is uncommon for species of Hypostomus, which are usually associated with fast-
flowing habitats, like H. nigromaculatus in this study.
The analysis of HSC transferability from forested to deforested streams revealed
that the preference of fish species is partially consistent across environments of different
conservation status. Depth HSCs could be transferred to only two out of seven species
(29%), whereas velocity HSCs could be transferred to six out of seven species (86%).
These results indicate that the need to develop specific criteria for each condition is
dependent on the variable considered, with velocity having higher transferability than
depth. The failure to transfer HSCs may be due to different factors, such as (i) the
preferred conditions are not available, (ii) interaction with other factors in deforested

30
streams and (iii) failure of the criterion to describe optimal habitats. The first
explanation may, at least partially, justify the failure in transferring the criteria for
depth, especially for species whose optimum condition was assigned to the last depth
class (> 0.51 m) like Astyanax fasciatus. In deforested streams, only four segments (~
6%), three of which in the same stream, fell into the deepest depth class. According to
Bovee (1986), the successful transfer of HSC is dependent on the availability of
conditions in the site to which one wants to transfer the criterion.
The second explanation (interaction with other factors) may also apply if
environmental factors influenced fish differently in forested as deforested streams. For
example, the high abundance of marginal submerged vegetation in deforested streams,
as a result of the proliferation of exotic grasses, provide favorable microhabitats for
some species (Casatti et al., 2009) and may influence their response to other variables.
Such grasses can attenuate water flow along stream banks, providing slow-flowing
microhabitats even in stretches with high velocities. As a result, species with
preferences for slow-flowing habitats would be favored, notably Hypostomus
ancistroides in my study. In fact, individuals of this species were observed in the
vegetation on the banks of deforested streams, even in stretches with high water flow.
On the other hand, this species was restricted to the mid-channel in forested streams,
where submerged grasses are lacking and supposedly the influence of velocity is higher.
This could explain the failure to transfer the criterion for flow to H. ancistroides.
The failure in transferring HSCs also may be related to the fact that they do not
always describe a habitat preferred by species. For example, in this study, depth and
velocity co-varied in both forested and deforested streams (Pearson’s product-moment
correlation performed a posteriori, r = 0.57, P < 0.01; r = 0.29, P < 0.01, respectively),
so that the preference patterns observed for a variable (e.g., depth) can be influenced by
a second variable (velocity). If the pattern for depth preference of species is an artifact
of their association with velocity, then one would expect a distinct preference pattern
for this variable (i.e., failure to transfer HSCs) only if the velocity-depth relationship
was different between forested and deforested streams. In fact this happened, as a
higher proportion of segments classified into the fourth flow class (> 0.6 m/s) were
grouped into classes with higher depth (D2, D3, and D4) in the deforested streams, as
shown by chi-square test performed a posteriori (M-L Chi-square, P = 0.03). This could
explain the failure to transfer depth HSCs among streams for species that showed

31
preference for fast-flowing habitats, like Hypostomus nigromaculatus and Characidium
zebra.
The results indicated a high consistency in the response of species for velocity,
suggesting that this variable is a good predictor of species abundance and that its HSC
has great potential to be transferred to streams of different conservation status. The
scale of this study is relevant, because the transferability of criteria is usually tested
between streams (Thomas and Bovee, 1993) or between regional models built for
individual streams (Lamouroux et al., 1999). The data presented in this study should
represent part of an effort to develop criteria for Neotropic species, encompassing a
broader spectrum of conditions, as well as other variables known to be important for
fish, like substrate and cover (Vadas and Orth, 2001; Wright and Flecker, 2004; Leal et
al., 2011).
REFERENCES
Ahmadi-Nedushan B, St-Hilaire A, Bérubé M, Robichaud E, Thiémonge N, Bobée B.
2006. A review of statistical methods for the evaluation of aquatic habitat suitability
for instream flow assessment. River Research and Applications 22: 503–523.
Angermeier PL, Karr JR. 1983. Fish communities along environmental gradients in a
system of tropical streams. Environmental Biology of Fishes 9: 117–135.
Araujo RB, Garutti V. 2003. Ecology of a stream from upper Paraná river basin
inhabited by Aspidoras fuscoguttatus Nijssen & Isbrücker, 1976 (Siluriformes,
Callichthyidae). Brazilian Journal of Biology 63: 363–372.
Barletta M, Jaureguizar AJ, Baigun C, Fontoura NF, Agostinho AA, Almeida-Val
VMF, Val AL, Torres RA, Jimenes-Segura LF, Giarrizzo T, Fabré NN, Batista VS,
Lasso C, Taphorn DC, Costa MF, Chaves PT, Vieira JP, Corrêa MFM. 2010. Fish
and aquatic habitat conservation in South America: a continental overview with
emphasis on Neotropical systems. Journal of Fish Biology 76: 2118–2176.
Bovee KD. 1982. A guide to stream habitat analysis using the instream flow
incremental methodology. Instream Flow Information Paper 12. U.S. Fish and
Wildlife Service FWS?OBS-82/26: Washington, D. C.
Bovee KD. 1986. Development and evaluation of habitat suitability criteria for use in
the instream flow incremental methodology. Instream Flow Information Paper 21.
U.S. Fish and Wildlife Service Biological Report 86 (7): Washington, D. C.

32
Bürhnheim CM. 2002. Heterogeneidade de habitats: rasos x fundos em assembléias de
peixes de igarapés de terra firme na Amazônia Central. Revista Brasileira de
Zoologia 19: 889–905.
Casatti L, Castro RMC. 1998. A fish community of the São Francisco river headwater
riffles, southeastern Brazil. Ichthyological Exploration of Freshwaters 9: 229-242.
Casatti L, Castro RMC. 2006. Testing the ecomorphological hypothesis in a headwater
riffles fish assemblage of the rio São Francisco, southeastern Brazil. Neotropical
Ichthyology 4: 203–214.
Casatti L, Langeani F, Castro RMC. 2001. Peixes de riacho do Parque Estadual Morro
do Diabo, bacia do alto rio Paraná, SP. Biota Neotropica 1: 1–15.
Casatti L, Rocha FC, Pereira DC. 2005. Habitat use by two species of Hypostomus
(Pisces, Loricariidae) in southeastern brazilian streams. Biota Neotropica 5: 157–
165.
Casatti L, Langeani F, Silva AM, Castro RMC. 2006. Stream fish, water and habitat
quality in a pasture dominated basin, southeastern Brazil. Brazilian Journal of
Biology 66: 681–696.
Casatti L, Ferreira CP, Carvalho FR. 2009a. Grass-dominated stream sites exhibit low
fish species diversity and dominance by guppies: an assessment of two tropical
pasture river basins. Hydrobiologia 623: 173–189.
Casatti L, Veronezi-Júnior JL, Ferreira CP. 2009b. Dieta do cascudo Aspidoras
fuscoguttatus (Ostariophysi, Callichthyidae) em riachos com diferentes
características limnológicas e estruturais. Biota Neotropica 9: 1–9.
Ceneviva-Bastos M, Casatti L, Rossa-Feres DC. 2010. Meso and microhabitat analysis
and feeding habits of small nektonic characins (Teleostei: Characiformes) in
Neotropical streams. Zoologia 27: 191–200.
Freeman MC, Bowen ZH, Crance JH. 1997. Transferability of habitat suitability criteria
for fishes in warmwater streams. North American Journal of Fisheries Management
17: 20–31.
Gatz AJ Jr. 1979. Ecological morphology of freshwater stream fishes. Tulane Studies in
Zoology and Botany 21: 91–124.
Grinnell J. 1917. The niche-relationship of the California thrasher. Auk 34:427–33.
Groshens TP, Orth DJ. 1993. An assessment of the transferability of habitat suitability
criteria for smallmouth bass. Virginia Water Resources Research Center Bulletin
180.

33
Hutchinson MF. 1957. Concluding remarks. Cold Spring Harbour Symposium on
Quantitative Biology 22:415–427
IPT (Instituto de Pesquisas Tecnológicas do Estado de São Paulo). 1999. Diagnóstico
da situação atual dos Recursos Hídricos e estabelecimento de diretrizes técnicas
para a elaboração do Plano da Bacia Hidrográfica do Turvo/Grande. Relatório nº
40.515. Secretaria de Recursos Hídricos, Saneamento e Obras, São Paulo.
Jackson DA, Peres-Neto PR Olden JD. 2001. What controls who is where in freshwater
fish communities – the roles of biotic, abiotic and spatial factors. Canadian Journal
of Fisheries and Aquatic Sciences 58: 157–170.
Lamouroux N, Capra H, Pouilly M, Souchon Y. 1999. Fish habitat preferences in large
streams of southern France. Freshwater Biology 42: 673–687.
Leal CG, Junqueira NT, Pompeu PS. 2011. Morphology and habitat use by fishes of the
Rio das Velhas basin in southeastern Brazil. Environmental Biology of Fishes
90:143–157.
Oliveira EF, Goulart E, Breda L, Minte-Vera CV, Paiva LRS, Vismara MR. 2010.
Ecomorphological patterns of the fish assemblage in a tropical floodplain: effects of
trophic, spatial and phylogenetic structures. Neotropical Ichthyology 8: 569–586.
Power ME. 1984. Depth distribution of armored catfish: predator-induced resource
avoidance? Ecology 65: 523–528.
São Paulo. 2006. Relatório da qualidade ambiental do Estado de São Paulo 2006.
Secretaria do Estado do Meio Ambiente – MMA, São Paulo.
Schwartz JS, Herricks EE. 2008. Fish use of ecohydraulic-based mesohabitat units in a
low-gradient Illinois stream: implications for stream restoration. Aquatic
Conservation: Marine and Freshwater Ecosystems 18: 852–866.
Silva AM, Nalon MA, Kronka FJAN, Álvares CA, Camargo PB, Martinelli LA. 2007.
Historical land-cover/use in different slope and riparian buffer zones in watershed of
the State of São Paulo, Brazil. Scientia Agricola 64: 325–335.
Strakosh TR, Neumann RM, Jacobson RA. 2003. Development and assessment of
habitat suitability criteria for adult brown trout in southern New England rivers.
Ecology of Freshwater Fish 12: 265–274.
Teresa FB, Casatti L. 2010. Importância da vegetação ripária em região intensamente
desmatada no sudeste do Brasil: um estudo com peixes de riacho. Pan-American
Journal of Aquatic Sciences 5: 444–453.

34
Thomas JA, Bovee KD. 1993. Application and testing of a procedure to evaluate
transferability of habitat suitability criteria. Regulated Rivers: Research and
Management 8: 285–294.
Uieda VS, Buzzato P, Kikuchi RM. 1997. Partilha de recursos alimentares em peixes
em um riacho de serra no sudeste do Brasil. Anais da Academia Brasileira de
Ciências 69: 243–252.
Vadas RL Jr, Orth DJ. 2001. Formulation of habitat suitability models for stream fish
guilds: do the standard methods work? Transactions of the American Fisheries
Society 130: 217–235.
Watson DJ, Balon EK. 1984. Ecomorphological analysis of fish taxocenes in rainforest
streams of northern Borneo. Journal of Fish Biology 25: 371–384.
Wright JP, Flecker AS. 2004. Deforesting the riverscape: the effects of wood on fish
diversity in a Venezuelan piedmont stream. Biological Conservation 120: 439–447.

35
Manuscrito aceito para publicação na Ecology of Freshwater Fish
- CAPÍTULO II -
Influence of forest cover and mesohabitats types on functional and
taxonomic diversity of fish communities in Neotropical lowland
streams

36
ABSTRACT
In this study, we investigated how taxonomic and functional diversity of fish
communities is influenced by forest cover and mesohabitat types in Neotropical
lowland streams. We sampled fish fauna of 126 five meter-long mesohabitats using an
electrofishing unit in forested (n = 3) and deforested (n = 3) streams in the upper Paraná
River basin of southeastern Brazil. According to velocity depth, three mesohabitat types
have been considered: riffles (shallow and fast-flowing habitat), pools (deep and slow-
flowing habitat) and run (intermediate depth and velocity). Seven functional traits and
27 trait categories related to ecological, behavioral and life history aspects of fish were
considered. Our results indicate that forest cover and mesohabitat type influence
differently functional patterns of communities. While deforestation affects communities
primarily through changes in diversity (functional and taxonomic), mesohabitat types
determine changes in the functional composition. The increased diversity in deforested
mesohabitats is driven by a decrease of species turnover among habitat patches within
streams. This can be attributed to new feeding opportunities and microhabitat
availabilities in deforested streams and favors the occurrence species with a particular
set of traits, indicating a strong habitat-trait relationship.
Keywords: ichthyofauna, functional traits, riparian forest, guilds, environmental
impact.

37
INTRODUCTION
Deforestation of the riparian zone is one of the most important threats to the
conservation of freshwater fish species (Naiman & Décamps 1997; Sweeney et al.
2004). Deforestation primarily affects streams, whose physical and biological integrity
depends on native forests in riparian zones (Pinto et al. 2006). Changes resulting from
deforestation include modifications to aquatic communities and ecosystem services
(Sweeney et al. 2004). The consequences of deforestation on fish communities have
been primarily examined under a taxonomic perspective (Bojsen & Barriga 2002;
Casatti et al. 2010). These studies have shown changes in species composition and
indicators of species diversity such as the richness, diversity, and dominance of species
(Bojsen & Barriga 2002; Wright & Flecker 2004; Teresa & Casatti 2010). However,
these metrics represent a limited set of traits of biological communities because they do
not take accont the relative functional differences between species and often fail to
detect changes caused by anthropogenic interference (Rabeni & Smale 1995; Ernst et al.
2006).
Recently, there has been remarkable growth in the number of studies
incorporating aspects of the functional diversity of communities (Petchey & Gaston
2002; Hoeinghaus et al. 2007; Poff et al. 2010). This approach takes into account both
the functional traits of species and the ecological roles they play in the ecosystem, and it
provides additional information otherwise not obtainable through a taxonomic approach
(Eros et al. 2009). In addition, the functional structure of communities is considered a
better predictor of ecosystem functioning than species diversity itself (Tilman et al.
1997; Mokany et al. 2008; Moulliot et al. 2011). The study of the anthropogenic
impacts on the functional structure of communities is fundamental because it may
predict not only communities patterns but also processes operating in ecosystems
(Mayfield et al. 2010).
Studies examining the organization of fish communities based on their
functional composition have shown that habitat use is influenced by the functional
traits of a species (Poff & Allan 1995; Jones III et al. 1999; Irz et al. 2007; Eros et al.
2009; Montaña & Winemiller 2010). Thus, anthropogenic impacts affecting the
physical structure of habitats have the potential to influence functional aspects of fish
communities. In support of this idea, the addition and/or loss of species in deforested
streams appears to be non-random (Jones III et al. 1999; Goldstein & Meador 2005),

38
suggesting that deforestation limits or favors certain species according to their traits.
For example, Rabeni & Smale (1995) demonstrated that species that rely on hard
substrates for reproduction tend to disappear from environments subject to siltation.
In addition to functional composition, functional diversity is another way in
which community structure can be described. Indices that have been developed recently
deal with functional traits within a multivariate framework (e.g., Petchey & Gaston
2006; Villéger et al. 2008). This approach has also been useful in identifying functional
changes along gradients of deforestation (Ernst et al. 2006; Barragán et al. 2011), and
its use in streams is promising because little is known about how different components
of biodiversity in these environments respond to human impacts.
Streams are heterogeneous systems composed of a mosaic of patches recognized
at different scales (Frissel et al. 1986). At the mesohabitat scale, three physiognomies
are easily recognized according to their geomorphological and hydrological features:
riffles represent shallow, fast waters; pools are deep, slow waters that usually have fine
substrate; runs are intermediate between riffles and pools. These mesohabitats influence
the composition and structure of fish fauna (Bührnheim 2002; Langeani et al. 2005;
Schwartz & Herricks 2008; Rezende et al. 2010), as well as some characteristics of fish
species (e.g., diet, reproductive strategies and preference for substrate) (Berkman &
Rabeni 1987; Casatti & Castro 1998; Goldstein & Meador 2004). For example, fish
with less diversified diets predominate in riffles, while the increased variety of food
items available in pools would support communities composed of more generalist
species (Angermeier & Karr 1983; Berkman & Rabeni 1987). Thus, it is expected that
different mesohabitats and their associated communities respond differently to
environmental disturbances.
To our knowledge, no studies have investigated how deforestation influences
the functional diversity and composition of Neotropical stream fish assemblages in
different mesohabitat types. Therefore, in this study, we investigated whether
functional diversity metrics vary between forested and deforested mesohabitats and/or
among mesohabitats types. The study was conducted in forested and deforested streams
of southeastern Brazil, one of the most developed regions of the country, where
deforestation affects more than 75% of the riparian zone of streams (Silva et al. 2007).
To characterize the fish fauna, we used multiple traits including behavioral and
ecological (e.g., life-history, feeding tactic, current preference) aspects.

39
MATERIAL AND METHODS
Study area and general sampling design
All streams studied are located in the drainage of the São José dos Dourados
River in the upper Paraná River watershed of southeastern Brazil (Figure 1). The
forested streams (n = 3) are located within the greatest forest fragments (seasonal
semideciduous forest > 1.600 ha) of the basin and the deforested streams (n = 3) lack
arboreal vegetation in their riparian zone and are located in areas dominated by pasture
(Figure 1). There are no representative forest remnants associated with deforested
streams upstream or downstream of the study sites. These streams were selected from a
group of 80 previously visited streams in the region. They were chosen because they are
representative of forested and deforested streams, present mesohabitat variability,
represent second or third order streams with similar volume of water, have no evidence
of chemical pollution, and differ only in respect to structural features resulting from
differences in forest cover, as described below. Due to the scarcity of large forest
fragments and the difficulty of finding comparable deforested areas, sites were not
randomly distributed in the landscape but rather were clustered in specific areas.
However, all these streams were considered to be influenced by the same regional
species pool (e.g., Casatti et al. 2006; 2009), as the distances between watercourses do
not explain the variation in the species composition among streams distributed within
the study area (Casatti et al. 2012).
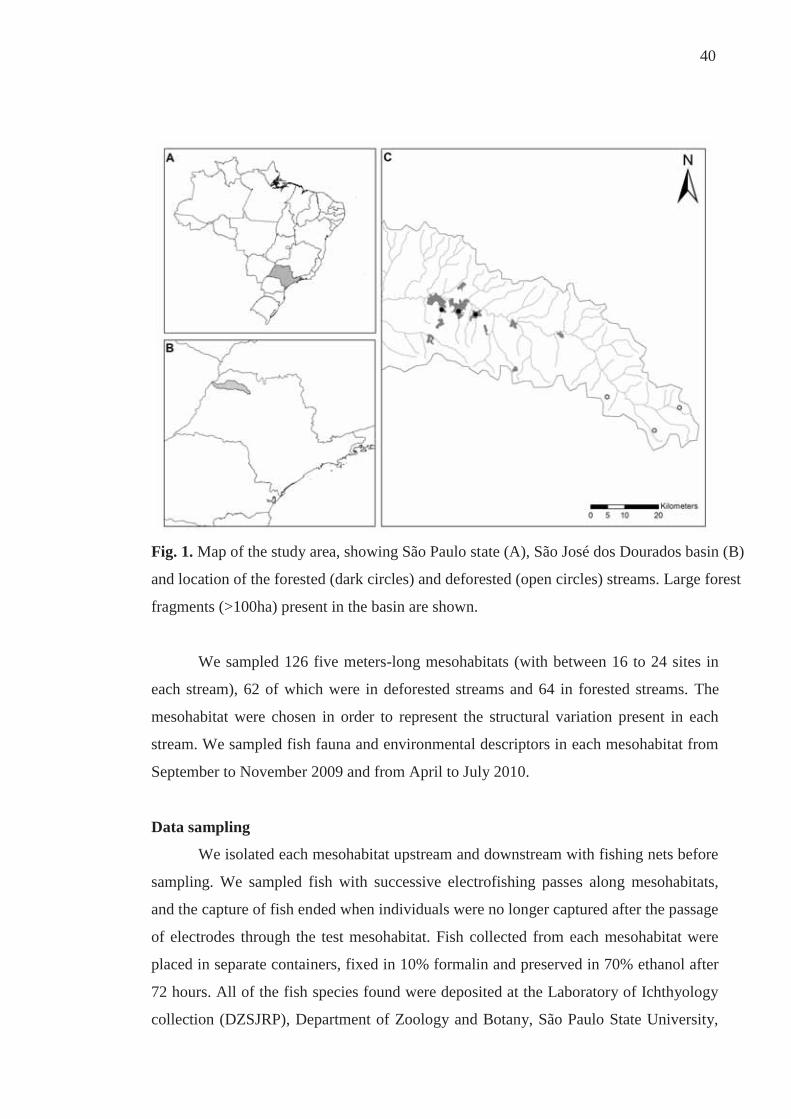
40
Fig. 1. Map of the study area, showing São Paulo state (A), São José dos Dourados basin (B)
and location of the forested (dark circles) and deforested (open circles) streams. Large forest
fragments (>100ha) present in the basin are shown.
We sampled 126 five meters-long mesohabitats (with between 16 to 24 sites in
each stream), 62 of which were in deforested streams and 64 in forested streams. The
mesohabitat were chosen in order to represent the structural variation present in each
stream. We sampled fish fauna and environmental descriptors in each mesohabitat from
September to November 2009 and from April to July 2010.
Data sampling
We isolated each mesohabitat upstream and downstream with fishing nets before
sampling. We sampled fish with successive electrofishing passes along mesohabitats,
and the capture of fish ended when individuals were no longer captured after the passage
of electrodes through the test mesohabitat. Fish collected from each mesohabitat were
placed in separate containers, fixed in 10% formalin and preserved in 70% ethanol after
72 hours. All of the fish species found were deposited at the Laboratory of Ichthyology
collection (DZSJRP), Department of Zoology and Botany, São Paulo State University,

41
São José do Rio Preto, Brazil (see Table A1, Supporting Information, for a species list
of each stream).
We collected data on the physical structure of habitat after fish sampling. To do
this, we sub-divided each mesohabitat into four transversal, equidistant transects where
we recorded the following data: width, measured at the surface and at the middle of the
water column; depth, measured at five equidistant points from one margin to another;
velocity (with a mechanical flowmeter), measured at the middle of the water column;
streambed substrate composition (i.e., the relative proportion of each substrate
component) visually estimated according to Cummins (1962) and the volume occupied
by the submerged vegetation (roots, leaves and stems from the submerged terrestrial
vegetation) on the banks of each margin, based on the measures of the height and the
width of vegetation on stream banks of each transect. We classified the streambed
substrate as either stable, which allows colonization by benthic fauna (in the
consolidated clay, gravel, pebble and rock) or unstable, with sand and unconsolidated
clay. The values of each mesohabitat unit were calculated by the mean of the values
obtained in the transects.
The mesohabitat units were classified according to their depth and velocity
characteristics. These two variables were chosen because they represent important
predictors of fish community structure in streams (Lamouroux et al. 1999; Bürhnheim
2002). Moreover, both forested and deforested streams had similar mesohabitats
considering these two variables. After performing a hierarchical agglomerative cluster
analysis with Euclidean distance by using UPGMA method, mesohabitats were
classified in three types: (i) shallow, fast-flowing habitat; (ii) deep, slow-flowing
habitat; (iii) intermediate conditions in relation to these two descriptors. These three
groups of mesohabitats were named riffles, pools, and runs, respectively. We then
pooled and averaged the structural descriptors data to represent the mesohabitats in each
stream.
A Principal Component Analysis (PCA) was used to describe the variation of
data in the five descriptors of habitat structure measured (depth, width, velocity,
percentage of stable substrates and volume of submerged marginal vegetation). The
variables were first standardized so that their means were zero and their standard
deviation equal to one. We also carried out a Multivariate Analysis of Variance
(MANOVA) to test the significance of the structural patterns exhibited by mesohabitat
types and streams with different forest cover.

42
Traits and trait categories
We adopted a multidimensional approach to characterize fish species traits. We
considered seven traits and 27 trait categories, including behavioral and ecological
aspects (Table 1). We defined traits and their categories based on the species
information recorded in literature and in this study. The species characterization
followed a binary approach, with each species being associated to one trait category for
each trait. We adopted this practice because there is little information on the variability
of traits for many species. This approach was conservative, but it was sufficient for an
analysis of the communities’ general patterns (Poff et al. 2006). The most common trait
categories expressed by each species were obtained from the literature, from data
collected in this study and based on the author’s experience (Table A2 – Supporting
Information).

43
Table 1. Description of seven functional traits and 27 trait categories used to
characterize fish species with their respective codes, including aspects of ecology and
feeding behavior, habitat use, life history and tolerance.
Traits Trait categories
Ecology and feeding behavior
Diet1 Omnivore, periphytivore, insectivore, carnivore or
detritivore
Feeding tactic,3 Water column feeding, substrate speculation, stealth
predation, grazing, nocturnal predation or digging
Habitat use2,4
Velocity preference Slow, intermediate or fast
Stable substrate preference High, moderate or low
Habitat use
Surface, nektonic, nectobenthic, margins or benthic
Life history5
Size Small (< 50 mm), intermediate (50-150 mm) or large
(> 150 mm)
Tolerance6
Tolerance to hypoxia Tolerant or intolerant 1 Diet analysis made by us 2 Underwater observations made by us 3 Keenleyside (1979), Sazima (1986), Sabino & Zuanon (1998) 4 Casatti & Castro (1998), Casatti et al. (2001), Casatti (2002) and personal observation 5 Biometry made by us 6 Kramer & Mehegan (1981), Araujo & Garutti (2003), Bozzetti & Schulz (2004) and
Casatti et al. (2006)
For additional information on habitat use and feeding tactics, we made
observations during 1,200 minutes while snorkeling along 100-meter stretches in the six
studied streams. We also performed a diet analysis by examining the stomach contents
of 10 specimens of each species (except for those with less than 10 individuals)
representative of the streams sampled. Only species with more than five individuals
were analyzed. We identified and classified food items into broad categories (detritus,
insects, other invertebrates, periphyton, fragments of higher plants and algae). When

44
assigning a trophic group to a species, we considered the most important items in their
diet. Species with less than five individuals had their diet characterized based on data
available in literature.
We evaluated the independence or redundancy of traits prior to the calculation of
functional diversity metrics. This allowed the identification of non-informative and
autocorrelated traits that should be removed from the analysis to avoid under or
overestimation of functional diversity (Petchey & Gaston 2006). We performed a matrix
correlation analysis (2Stage analysis) using the software PRIMER 6.0 (Clarke & Gorley
2006) from binary matrices for each trait (species as samples and trait categories as
variables). The results of this analysis indicated that the seven traits had low
autocorrelation (< 0.53), meaning that the previously selected traits represent different
aspects of species. Thus, we considered all seven traits in additional analyses.
Functional diversity metrics
We used functional richness, functional evenness (Villéger et al. 2008), and
functional dispersion (Laliberte & Legendre 2010) to represent functional diversity.
These indices measure different aspects of species composition in the multidimensional
functional space, and they are independent of species richness (Villéger et al. 2008) and
very flexible when using distinct trait types (Laliberté & Legendre 2010). The
functional richness represents the volume of the convex hull generated by positioning
species distribution in the n-dimensional species trait space (Villéger et al. 2008). The
functional evenness measures the regularity with which the functional space is occupied
by species weighted by their abundance (Villéger et al. 2008). The functional evenness
is high when abundance is evenly distributed among species or when the functional
distance between species is regular (Villéger et al. 2008). The functional dispersion was
first developed as a measure of beta diversity (Anderson et al. 2006), but it was recently
incorporated as a metric of functional diversity. In contrast to functional richness,
functional dispersion may account for the abundance of species within communities,
and it is not highly affected by outliers (Laliberté & Legendre 2010). This index
consists of measuring the mean distance of the multidimensional functional space from
each species to the centroid of all species, which in turn is dislocated towards the most
abundant species (Laliberté & Legendre 2010). We used the function dbFD in the
package FD of the R software (R Development Core Team 2011) to calculate the three
indices of functional diversity. This package incorporates changes that make the indices

45
proposed by Villéger et al. (2008) more flexible, making possible the use of exclusively
qualitative traits (Laliberté & Legendre 2010), as is the case in our study.
Data analysis
We used two-way ANOVA to investigate how the number of species within each
trait category varied depending on forest cover and mesohabitats types in the R software
(R Development Core Team 2011). Because part of the data did not fit assumptions of
normality and/or homogeneity of variance, we used a more conservative significance
level (p ≤ 0.01) to minimize type I error. We evaluated the effects of forest cover and
mesohabitats on functional diversity metrics and species richness with the same
analytical strategy. In order to evaluate the contribution of species exclusive to forested
or deforested streams to the general patterns of diversity, we also conducted an analysis
considering only the species shared between these groups of streams. In these cases, data
had normal distribution and homogeneous variances; therefore, we used p ≤ 0.05 to
attribute significance. We considered different mesohabitats types and streams as
independent samples. Prior to hypothesis testing, we pooled data by mesohabitat type
within streams, producing a matrix with 18 samples corresponding to the three types of
mesohabitats sampled in the six streams.
RESULTS
The first two axes of PCA accounted for 79.5% of the variation in the physical
habitat structure data, with axis 1 accounting for 54.7% of the variation and
representing water velocity, percentage of stable substrate, and depth (eigenvectors = -
0.536, - 0.492, 0.485, respectively; Figure 2). The second axis accounted for 24.8% of
the variation and represented the width and the amount of submerged marginal
vegetation (eigenvectors = 0.681, -0.569, respectively; Figure 2). The first axis
distinguished the mesohabitats with fast water velocity, less depth and larger proportion
of stable substrates associated with riffles than those with slow water velocity, high
depth and a low proportion of stable substrates associated with pools; intermediate
values along this gradient were associated with runs (Figure 2). Axis 2 differentiated
forested from deforested streams. Accordingly, deforested streams were associated to a
higher amount of submerged marginal vegetation, whereas forested streams were
associated to greater width (Figure 2). The separation of the samples in accordance to
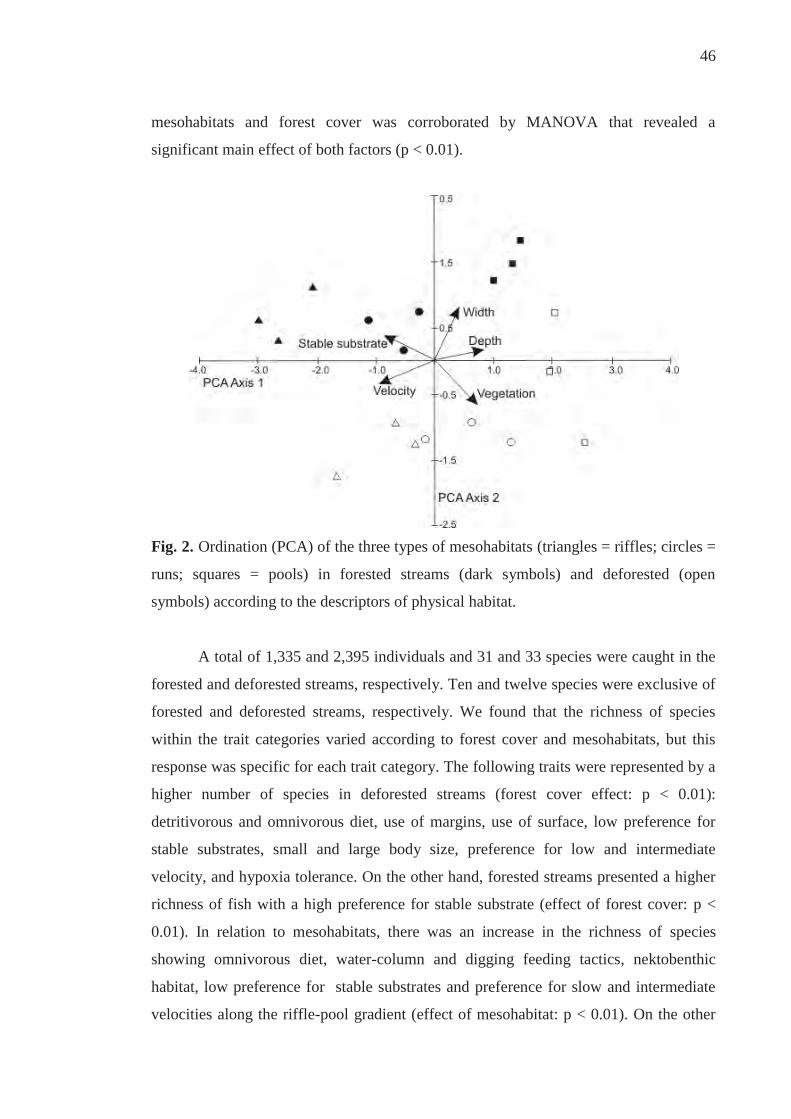
46
mesohabitats and forest cover was corroborated by MANOVA that revealed a
significant main effect of both factors (p < 0.01).
Fig. 2. Ordination (PCA) of the three types of mesohabitats (triangles = riffles; circles =
runs; squares = pools) in forested streams (dark symbols) and deforested (open
symbols) according to the descriptors of physical habitat.
A total of 1,335 and 2,395 individuals and 31 and 33 species were caught in the
forested and deforested streams, respectively. Ten and twelve species were exclusive of
forested and deforested streams, respectively. We found that the richness of species
within the trait categories varied according to forest cover and mesohabitats, but this
response was specific for each trait category. The following traits were represented by a
higher number of species in deforested streams (forest cover effect: p < 0.01):
detritivorous and omnivorous diet, use of margins, use of surface, low preference for
stable substrates, small and large body size, preference for low and intermediate
velocity, and hypoxia tolerance. On the other hand, forested streams presented a higher
richness of fish with a high preference for stable substrate (effect of forest cover: p <
0.01). In relation to mesohabitats, there was an increase in the richness of species
showing omnivorous diet, water-column and digging feeding tactics, nektobenthic
habitat, low preference for stable substrates and preference for slow and intermediate
velocities along the riffle-pool gradient (effect of mesohabitat: p < 0.01). On the other

47
hand, some trait categories became more representative along the pool-riffle gradient,
such as an insectivorous diet, benthic habitat, substrate speculation, high preference for
stable substrates, and a preference for fast velocity (effect of mesohabitat: p < 0.01). A
carnivorous diet was exclusive of fish from deforested streams and found with great
frequency in pools (interaction “forest cover” vs. “mesohabitats”; p < 0.01). Forest
cover or mesohabitats did not influence (p > 0.04) the following trait categories:
periphytivorous diet, nektonic habitat, grazing, stealth predation, nocturnal predation,
moderate preference for stable substrate, and preference for intermediate velocity.
The taxonomic richness and functional richness was higher in deforested than in
forested mesohabitats (effect of forest cover: p = 0.01), but did not differ among
mesohabitats (effect of mesohabitat: p = 0.34, Figure 3). Neither forest cover nor
mesohabitat had an effect on functional evenness (p > 0.42, Figure 3). The functional
dispersion was higher in deforested streams, but this effect was mesohabitat-dependent
(interaction “forest cover” vs. mesohabitats: p = 0.01), being significant only in riffles
(Tukey test, p < 0.01; Figure 3). The qualitative results were the same for the three
indices when only those species shared between forested and deforested streams were
considered.
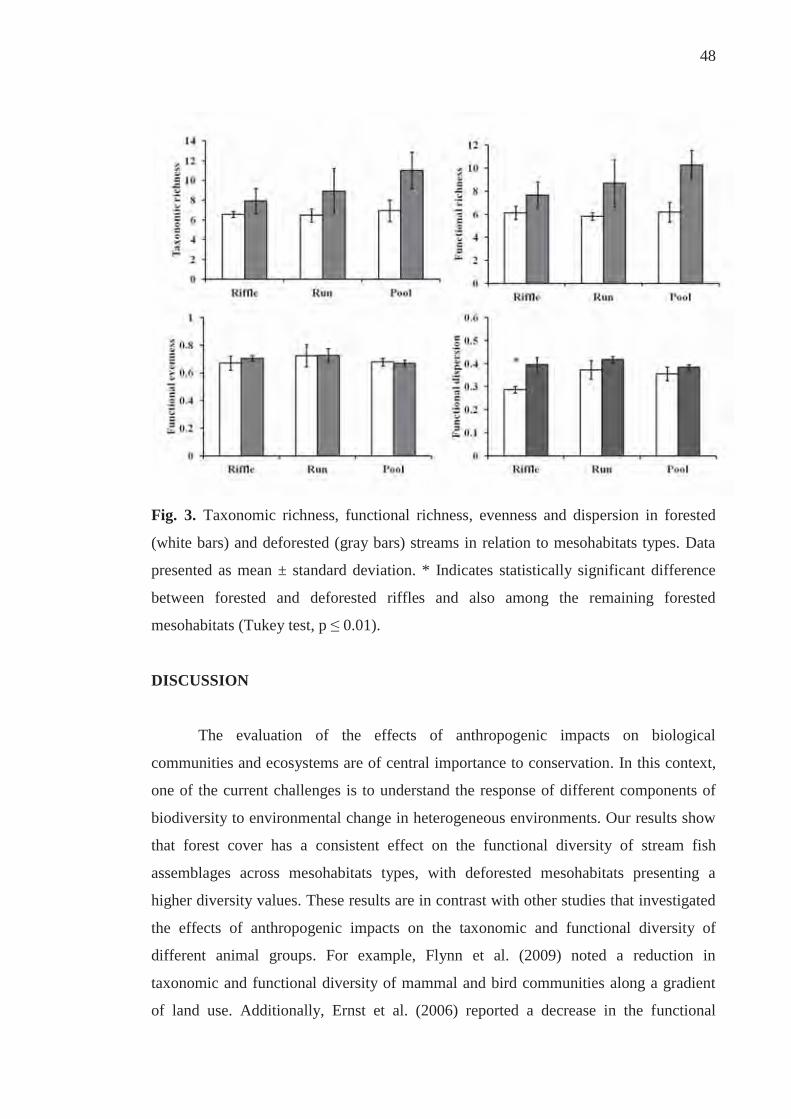
48
Fig. 3. Taxonomic richness, functional richness, evenness and dispersion in forested
(white bars) and deforested (gray bars) streams in relation to mesohabitats types. Data
presented as mean ± standard deviation. * Indicates statistically significant difference
between forested and deforested riffles and also among the remaining forested
mesohabitats (Tukey test, p ≤ 0.01).
DISCUSSION
The evaluation of the effects of anthropogenic impacts on biological
communities and ecosystems are of central importance to conservation. In this context,
one of the current challenges is to understand the response of different components of
biodiversity to environmental change in heterogeneous environments. Our results show
that forest cover has a consistent effect on the functional diversity of stream fish
assemblages across mesohabitats types, with deforested mesohabitats presenting a
higher diversity values. These results are in contrast with other studies that investigated
the effects of anthropogenic impacts on the taxonomic and functional diversity of
different animal groups. For example, Flynn et al. (2009) noted a reduction in
taxonomic and functional diversity of mammal and bird communities along a gradient
of land use. Additionally, Ernst et al. (2006) reported a decrease in the functional

49
diversity in amphibian communities from different deforestation regimes, but no effect
on taxonomic richness along the gradient of degradation. By contrast, taxonomic
richness increased as functional diversity decreased in estuarine fish communities of a
degraded lake associated with the Gulf of Mexico (Villéger et al. 2010). These patterns
are consistent with habitat loss and simplification in degraded ecosystems, which in turn
result in the loss of species and functional traits (Flynn et al. 2009) or in the
replacement of species with unique traits by functionally redundant species (Ernst et al.
2006; Villéger et al. 2010).
The increased diversity in deforested mesohabitats would be due to the addition
of functionally distinct species in these streams. However, while species were added to
the deforested streams, an equivalent number of species occurred exclusively in the
forested streams, corroborating other studies and evidencing a pattern of species
substitution along the gradient of deforestation (Bojsen & Barriga 2002; Casatti et al.
2009). When considering all of the species and excluding species absent in either
deforested or forested streams, the congruence of the patterns of functional diversity
suggest that the exclusive species influenced the functional diversity of both group of
streams in a similar way. For example, while a carnivorous diet (Hoplias malabaricus
and Synbranchus marmoratus) and the combination of a periphytivorous diet and the
use of the margins (Hisonotus francirochai) are unique traits of species occurring
exclusively in deforested streams, an insectivorous diet with speculation tactics
(Cetopsorhamdia iheringi and Pseudopimelodus pulcher) and with a large body size
(Pimelodus maculatus) are unique combinations represented by species exclusive to
forested streams. Thus, although the exclusive species may be important to increasing
the functional diversity within each group of streams, they were not responsible for the
differences between them. The increase of taxonomic and functional diversity in
deforested mesohabitats was mainly due to higher evenness in the distribution of
species within these streams. This conclusion is based on the fact that deforested
streams presented a higher mean richness per mesohabitat, but a similar number of
species to forested ones when considering the streams as a whole (31 and 33 species,
respectively). Furthermore, even considering only the species shared between both
groups of streams, deforested mesohabitats show a higher taxonomic and functional
richness. This indicates that deforested mesohabitats support a higher taxonomic and
functional diversity mainly by providing conditions that increase the frequency of
species that were formerly present.

50
The functional evenness in both groups of streams was relatively high (> 0.6)
regardless of mesohabitat, indicating that the functional space was evenly occupied.
These results also suggest that the species added in deforested mesohabitats filled
unoccupied niches, leading to a more even use of resources. In fact, when the number of
species in a community increases, local assemblages can accommodate new species by
increasing the volume of niche space (Barragán et al. 2011). The increase of the niche
space in deforested streams may be represented by rare or formerly absent opportunities
that become available, favoring the occurrence of opportunistic and/or invasive species
(Lorion & Kennedy 2009). Some of the new opportunities in deforested streams can be
represented by the greater availability of microhabitats (Casatti et al. 2009; Teresa &
Casatti 2010) and food items, such as detritus (Rocha et al. 2009), algae and periphyton
(Rounick et al. 1982; Bojsen & Barriga 2002; Lorion & Kennedy 2009), and this would
favor, for example, the occurrence of omnivorous species. One the most prominent
habitat alterations recorded was the greater abundance of marginal herbaceous
vegetation in deforested streams (see Figure 2). This vegetation is composed mainly of
exotic grasses (Brachiaria spp.) that tend to invade stream banks of deforested areas
(Casatti et al. 2009) and may provide substrate for shelter and reproduction (Growns et
al. 2003; Teresa & Romero, 2010). This would favor the occurrence of species usually
associated with submerged vegetation, like Crenicichla britskii, Gymnotus carapo,
Hoplias malabaricus, and Hysonotus francirochai. The high abundance of marginal
submerged vegetation may have contributed to the higher richness of species with a
preference for slow or intermediate velocities, as they can attenuate water velocity
along stream banks and provide slow-flowing microhabitats even in areas with fast
velocities. The vegetation can also trap detritus in their leaves and roots (Rocha et al.
2009), which are favored by detritivores (Casatti et al. 2009) like Corydoras aeneus,
Geophagus brasiliensis, Hypostomus ancistroides and Poecilia reticulata, all of which
became more frequent in the deforested streams.
Some of the species favored by submerged marginal vegetation (e.g., G. carapo,
G. brasiliensis and H. malabaricus) present also large body, which may also face a
lesser risk of predation by birds and mammals when in vegetated environments.
Vegetated habitats may also provide camouflage and cover to conceal prey (Padial et al.
2009), which could explain the higher richness of this trait in deforested mesohabitats.
However, it is important to note that the association between trait categories represented
by a few species with the factors investigated here should be taken with caution, as they

51
are more likely to autocorrelate with other trait categories. Hoplias malabaricus and
Synbranchus marmoratus can be cited as examples of carnivores that have been
associated with degraded environments (Casatti et al. 2006; Casatti et al. 2009, this
study), but other traits (e.g., hypoxia tolerance or margin habitat) rather than the diet
may be important in determining their association with deforested streams.
Furthermore, the higher richness of carnivorous fish in pools can also be associated with
the use of these sites as refuge during periods of inactivity (Power 1984).
Traits like tolerance to hypoxia, small body size and the use of surface may
favor the colonization of shallow, warm, vegetated microhabitats and are shared by
opportunistic species like Phalloceros harpagos, Serrapinnus notomelas and the exotic
Poecilia reticulata, which are dominant in the grass-dominated banks of degraded
streams (Casatti et al. 2009). The increased richness of fish tolerant to hypoxia in
deforested streams is probably related to rising temperatures and reduced oxygen levels,
conditions commonly found in silted deforested streams (Heartsill-Scalley & Aide
2003; Casatti et al. 2006). The higher number of species with a low preference for
stable substrates reflects the physical alterations commonly reported for structurally
degraded streams (Gregory et al. 1991; Walser & Bart 1999; Casatti et al. 2006) that
experienced higher fine-sediment deposition, as shown in the PCA plot (see Figure 2).
Despite the fact that mesohabitat types did not influence the diversity patterns of
the communities, they were important in determining their functional composition. The
dominance of fish species with an omnivorous diet, nektonics, and water column
feeders showing low preference for stable substrates and a preference for slow and
intermediated water velocity in pools is compatible with the low current velocity and
often rich food resources present in this mesohabitat (Bürhnheim 2002; Angermeier &
Karr 1983; Langeani et al. 2005; this study). These traits are shared by a group of
species, mostly small characins, which represent a significant proportion of the fish
communities in Neotropical streams (Castro 1999). The nektobenthic habitat and
digging tactic are traits found with greater frequency in pools and are shared by
detritivorous/omnivorous species that feed in soft bottom sites. In contrast to the
patterns observed in the pools, a higher number of species with a high preference for
stable substrates and fast velocity habitats were associated with riffles, which is
compatible with the hydraulic conditions and substrate composition present in these
mesohabitats (Casatti & Castro 1998; Schwartz & Herricks 2008). The relationship
between traits and conditions present in the riffles was more pronounced in forested

52
streams, as forested riffles had a lower functional dispersion than deforested riffles and
other mesohabitats. This result indicates that, on average, species are functionally more
similar to each other on these sites. In fact, two groups of species predominated in
forested riffles: benthic, grazers and periphytivorous fish (e.g., Hypostomus
nigromaculatus) and benthic, speculators and insectivorous fish (e.g., Pseudopimelodus
pulcher and Cetopsorhamdia iheringi). The fast water velocity and limited availability
of food items in the riffles (Angermeier & Schlosser 1989) might represent more
restrictive environmental filters, limiting the occurrence of a great diversity of traits and
resulting in highly similar assemblages. The increased functional dispersion in
deforested streams might indicate a relaxation of these filters. For example, the
constraint imposed by the fast velocity of water would be minimized in these streams by
an abundance of submerged marginal vegetation (Pusey & Arthington 2003).
In summary, our results indicate that forest cover and mesohabitat types
influence different aspects related to the functional patterns of communities. While
deforestation affects communities primarily through changes in diversity (both
functional and taxonomic), mesohabitat types mainly determine changes in the
functional composition. The increased diversity in deforested mesohabitats is driven by
a decrease in species turnover among habitat patches within streams. The new
opportunities available in deforested streams favor species with a particular set of traits
(e.g., detritivores, species inhabiting stream margins, tolerant to hypoxia), indicating a
strong habitat-trait relationship. Knowledge of how functional patterns and processes
operate on aquatic communities over a wider range of degradation contexts, and
whether the patterns observed herein are consistent with other scales, are promising
topics for future research.
REFERENCES
Anderson, M.J.,Ellingsen, K.E. & McArdle, B.H. 2006. Multivariate dispersion as a
measure of beta diversity. Ecology Letters. 9: 683-693.
Angermeier, P.L. & Karr, J.R. 1983. Fish communities along environmental gradients
in a system of tropical streams. Environmental Biology of Fishes 9: 117–135.
Angermeier, P. & Schlosser, I.J. 1989. Species-area relationship for stream fishes.
Ecology 70: 1450–1462.

53
Araujo, R.B. & Garutti, V. 2003. Ecology of a stream from upper Paraná river basin
inhabited by Aspidoras fuscoguttatus Nijssen and Isbrücker, 1976 (Siluriformes,
Callichthyidae). Brazilian Journal of Biology 63: 363–372.
Barragán, F., Moreno, C.E., Escobar, F., Halffter, G. & Navarrete, D. 2011. Negative
impacts of human land use on dung beetle functional diversity. PLoS One 6:
e17976.
Berkman, H.E. & Rabeni, C.F. 1987. Effect of siltation on stream fish communities.
Experimental Biology of Fishes 18: 285–294.
Bojsen, B.H. & Barriga, R. 2002. Effects of deforestation on fish community structure
in Ecuadorian Amazon streams. Freshwater Biology 47: 2246–2260.
Bozzetti, M. & Schulz, U.H. 2004. An index of biotic integrity based on fish
assemblages for subtropical streams in southern Brazil. Hydrobiologia 529: 133–
144.
Bürhnheim, C.M. 2002. Heterogeneidade de habitats: rasos x fundos em assembléias de
peixes de igarapés de terra firme na Amazônia Central. Revista Brasileira de
Zoologia 19: 889–905.
Casatti, L. 2002. Alimentação dos peixes em um riacho do Parque Estadual Morro do
Diabo, bacia do Alto Rio Paraná, sudeste do Brasil. Biota Neotropica 2: 1-15.
Casatti, L. 2010. Alterações no Código Florestal Brasileiro: impactos potenciais sobre a
ictiofauna. Biota Neotropica 10: 31–34.
Casatti, L. & Castro, R.M.C. 1998. A fish community of the São Francisco river
headwater riffles, southeastern Brazil. Ichthyological Exploration of Freshwaters 9:
229–242.
Casatti, L., Langeani, F. & Castro, R.M.C. 2001. Peixes de riacho do Parque Estadual
Morro do Diabo, bacia do alto rio Paraná, SP. Biota Neotropica 1: 1–15.
Casatti, L., F. Langeani, A.M. Silva & Castro, R.M.C. 2006. Stream fish, water and
habitat quality in a pasture dominated basin, southeastern Brazil. Brazilian Journal
of Biology 66: 681–696.
Casatti, L., Ferreira, C.P. & Carvalho, F.R. 2009. Grass-dominated stream sites exhibit
low fish species diversity and dominance by guppies: an assessment of two tropical
pasture river basins. Hydrobiologia 632: 273–283.
Casatti, L., Teresa, F.B., Gonçalves-Souza, T., Bessa, E., Manzotti, A.R., Gonçalves,
C.S. & Zeni, J.O. 2012. From forests to cattail: how does the riparian zone
influence stream fish? Neotropical Ichthyology. In press.

54
Castro, R.M.C. 1999. Evolução da ictiofauna de riachos sul-americanos: padrões gerais
e possíveis processos causais. In: Caramaschi, E.P., Mazzoni, R., Bizerril, C.R.S.F.,
Peres-Neto, P.R., eds. Ecologia de Peixes de Riachos: Estado Atual e Perspectivas.
Rio de Janeiro: Oecologia Brasiliensis, v. VI, pp. 139–155.
Clarke, K.R. & Gorley, R.N. 2006. PRIMER v6: User Manual/Tutorial. Plymouth:
PRIMER-E. 192 pp
Cummins, K.H. 1962. An evaluation of some techniques for the collection and analysis
of benthic samples with special emphasis on lotic waters. The American Midland
Naturalist 67: 477–504.
Ernst, R., Linsenmair, K.E. & Rödel, M.O. 2006. Diversity erosion beyond the species
level: dramatic loss of functional diversity after selective logging in two tropical
amphibian communities. Biological Conservation 133: 143–155.
Eros, T., Heino, J., Schmera, D.& Rask, M. 2009. Characterising functional trait
diversity and trait–environment relationships in fish assemblages of boreal lakes.
Freshwater Biology 54: 1788–1803.
Flynn, D.F.B., Gogol-Prokurat, M., Nogeire, T., Molinari, N., Richers, B.T., Lin, B.B.,
Simpson, N., Mayfield, M.M. & DeClerck, F. 2009. Loss of functional diversity
under land use intensification across multiple taxa. Ecology Letters 12: 22–33.
Goldstein, R.M. & Meador, M.R. 2004. Comparisons of fish species traits from small
streams to large rivers. Transactions of the American Fisheries Society 133: 971–
983.
Goldstein, R.M. & Meador, M.R. 2005. Multilevel assessment of fish species traits to
evaluate habitat degradation in streams of the Upper Midwest. North American
Journal of Fisheries Management 25: 180–194.
Gregory, S.V., Swanson, F.J., McKee, W.A. & Cummins, K.W. 1991. An ecosystem
perspective of riparian zones. BioScience 41: 540–551.
Growns, I., Gerke, P.C., Astles, K.L. & Pollard, D.A. 2003. A comparison of fish
assemblages associated with different riparian vegetation types in the Hawkesbury–
Nepean river system. Fisheries Management & Ecology 10: 209–220.
Heartsill-Scalley, T. & Aide, T. M. 2003. Riparian vegetation and stream condition in a
tropical agriculture-secondary forest mosaic. Ecological Applications 13: 225–234.
Hoeinghaus, D.J., Winemiller, K.O. & Birnbaum, J.S. 2007. Local and regional
determinants of stream fish assemblage structure: inferences based on taxonomic vs.
functional groups. Journal of Biogeography 34: 324–338.

55
Irz, P., Michonneau, F., Oberdorff, T., Whittier T.R., Lamoroux, N., Mouillot, D. &
Argillier, C. 2007. Fish community comparisons along environmental gradients in
lakes of France and north-east USA. Global Ecology and Biogeography 16: 350–
366.
Jones III, E.B.D., Helfman, G.S., Harper, J.O. & Bolstad, P.V. 1999. Effects of riparian
forest removal on fish assemblages in southern Appalachian streams. Conservation
Biology 13: 1454–1465.
Keenleyside, M.H.A. 1979. Diversity and adaptation in fish behaviour. Berlin: Springer.
208 pp
Kramer, D.L. & Mehegan, J.P. 1981. Aquatic surface respiration, an adaptative
response to hypoxia in the guppy, Poecilia reticulata (Pisces, Poeciliidae).
Environmental Biology of Fishes 6: 299–313.
Laliberté, E. & Legendre, P. 2010. A distance-based framework for measuring
functional diversity from multiple traits. Ecology 91: 299–305.
Lamouroux, N., Capra, H. Pouilly, M. & Souchon, Y. 1999. Fish habitat preferences in
large streams of southern France. Freshwater Biology 42: 673–687.
Langeani, F., L. Casatti, H.S. Gameiro, A.B. Carmo & Rossa-Feres, D.C. 2005. Riffle
and pool fish communities in a large stream of southeastern Brazil. Neotropical
Ichthyology 3: 305–311.
Lorion, C. M. & Kennedy, B. P. 2009. Riparian forest buffers mitigate the effects of
deforestation on fish assemblages in tropical headwater streams. Ecological
Applications 19: 468–479.
Mayfield, M.M., Bonser, S.P., Morgan, J.W. Aubin, I., McNamara, S. & Vesk, P.A.
2010. What does species richness tell us about functional trait diversity?
Predictions and evidence for responses of species and functional trait diversity to
land-use change. Global Ecology and Biogeography 19: 423–431.
Mokany, K., Ash, J. & Roxburgh, S. 2008. Functional identity is more important than
diversity in influencing ecosystem processes in a temperate native grassland.
Journal of Ecology 96: 884–893.
Montaña, C.G. & Winemiller, K.O. 2010. Local-scale habitat influences morphological
diversity of species assemblages of cichlid fishes in a tropical floodplain river.
Ecology of Freshwater Fish 19: 216–227.

56
Mouillot, D., Villéger, S., Scherer-Lorenzen, M. & Mason, N.W.H. 2011. Functional
structure of biological communities predicts ecosystem multifunctionality. PLoS
One 6: e17476.
Naiman, R.J. & Décamps, H. 1997. The ecology of interfaces: riparian zones. Annual
Review of Ecology and Systematics 28: 621–658.
Padial, A.A., Thomaz, S.M. & Agostinho, A.A. 2009. Effects of structural
heterogeneity provided by the floating macrophyte Eichhornia azurea on the
predation efficiency and habitat use of the small Neotropical fish Moenkhausia
sanctaefilomenae. Hydrobiologia 624: 161–170.
Petchey, O.L. & Gaston, K.J. 2002. Functional diversity (FD), species richness and
community composition. Ecology Letters 5: 402–411.
Petchey, O.L. & Gaston K.J. 2006. Functional diversity: back to basics and looking
forward. Ecology Letters 9: 741–758.
Pinto, B.C.T., Araujo, F.G. & Hughes, R.M. 2006. Effects of landscape and riparian
condition on a fish index of biotic integrity in a large southeastern Brazil river.
Hydrobiologia 556: 69–83.
Poff, N.L. & Allan, J.D. 1995. Functional organization of stream fish assemblages in
relation to hydrologic variability. Ecology 76: 606–627.
Poff, N.L., Olden, J.D., Vieira, N.K.M., Finn, D.S., Simmons, M.P. & Kondratieff, B.C.
2006. Functional trait niches of North American lotic insects: traits-based
ecological applications in light of phylogenetic relationships. Journal of the North
American Benthological Society 25: 730–755.
Poff, N.L., Pyne, M.I., Bledsoe, B.P., Cuhaciyan, C.C. & Carlisle, D.M. 2010.
Developing linkages between species traits and multiscaled environmental variation
to explore vulnerability of stream benthic communities to climate change. Journal
of the North American Benthological Society 29: 1441–1458.
Power, M.E. 1984. Depth distribution of armored catfish: predator-induced resource
avoidance? Ecology 65: 523–528.
Pusey, B.J.& Arthington, A.H. 2003. Importance of the riparian zone to the
conservation and management of freshwater fish: a review. Marine and Freshwater
Research 54: 1–16.
R Development Core Team. 2005. R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria.

57
Rabeni, C.F. & Smale, M.A. 1995. Effects on siltation on stream fishes and the
potential mitigating role of the buffering riparian zone. Hydrobiologia 303: 211–
219.
Rezende, C.F., Moraes, M., Manna, L.R., Leitão, R.P., Caramaschi, E.P. & Mazzoni, R.
2010. Mesohabitat indicator species in a coastal stream of the Atlantic rainforest,
Rio de Janeiro-Brazil. Revista de Biología Tropical 58: 1479–1487.
Rocha, F.C., Casatti, L., Carvalho, F.R. & Silva, A.M.S. 2009. Fish assemblages in
stream stretches occupied by cattail (Typhaceae, Angiospermae) stands in Southeast
Brazil. Neotropical Ichthyology 7: 241–250.
Rounick J.S., Winterbourn, M.J., & Lyon, G.L. 1982. Differential utilization of
allochthonous and autochthonous inputs by aquatic invertebrates in some New
Zealand streams: a stable carbon isotope study. Oikos 39: 191–198
Sabino, J. & Zuanon, J. 1998. A stream fish assemblage in Central Amazonia:
distribution, activity patterns and feeding behavior. Ichthyological Exploration of
Freshwaters 8: 201–210.
Sazima, I. 1986. Similarities in feeding behaviour between some marine and freshwater
fishes in two tropical communities. Journal of Fish Biology, 29: 53–65.
Schwartz, J.S. & Herricks, E.E. 2008. Fish use of ecohydraulic-based mesohabitat units
in a low-gradient Illinois stream: implications for stream restoration. Aquatic
Conservation: Marine and Freshwater Ecosystems 18: 852–866.
Silva, A.M., Nalon, M.A., Kronka, F.J.A.N., Álvares, C.A., Camargo, P.B. &
Martinelli, L.A. 2007. Historical land-cover/use in different slope and riparian
buffer zones in watershed of the State of São Paulo, Brazil. Scientia Agricola 64:
325–335.
Sweeney, B.W.; Bott, T.L.; Jackson, J.K.; Kaplan, L.A.; Newbold, J.D.; Standley, L.J.;
Hession, W.C.; Horwitz, R.J. & Colman, M.G. 2004. Riparian Deforestation,
Stream Narrowing, and Loss of Stream Ecosystem Services. Proceedings of the
National Academy of Sciences of the United States of America 101: 14132–14137.
Teresa, F.B. & Casatti, L. 2010. Importância da vegetação ripária em região
intensamente desmatada no sudeste do Brasil: um estudo com peixes de riacho. Pan-
American Journal of Aquatic Sciences 5: 444–453.
Teresa, F.B. & Romero, R.M. 2010. Influence of the riparian zone phytophysiognomies
on the longitudinal distribution of fishes: evidence from a Brazilian savanna stream.
Neotropical Ichthyology 8: 163–170.

58
Tilman, D., Lehman, C.L. & Thomson, K.T. 1997. Plant diversity and ecosystem
productivity: theoretical considerations. Proceedings of Natural Academy of
Sciences of the United States of America 94: 1857–1861.
Villéger, S., Mason, N.W.H. & Mouillot, D. 2008. New multidimensional functional
diversity indices for a multifaceted framework in functional ecology. Ecology 89:
2290–2301.
Villéger, S., Miranda, J.R., Hernáandez, D.F. & Mouillot, D. 2010. Contrasting changes
in taxonomic vs. functional diversity of tropical fish communities after habitat
degradation. Ecological Applications 20: 1512–1522.
Walser, C. A. & Bart, H.L. 1999. Influence of agriculture on instream habitat and fish
assemblage structure in Piedmont watersheds of the Chattahoochee River system.
Ecology of Freshwater Fish 8: 237–246.
Wright, J.P. & Flecker, A.S. 2004. Deforesting the riverscape: the effects of wood on
fish diversity in a Venezuelan piedmont stream. Biological Conservation 120: 439–
447.
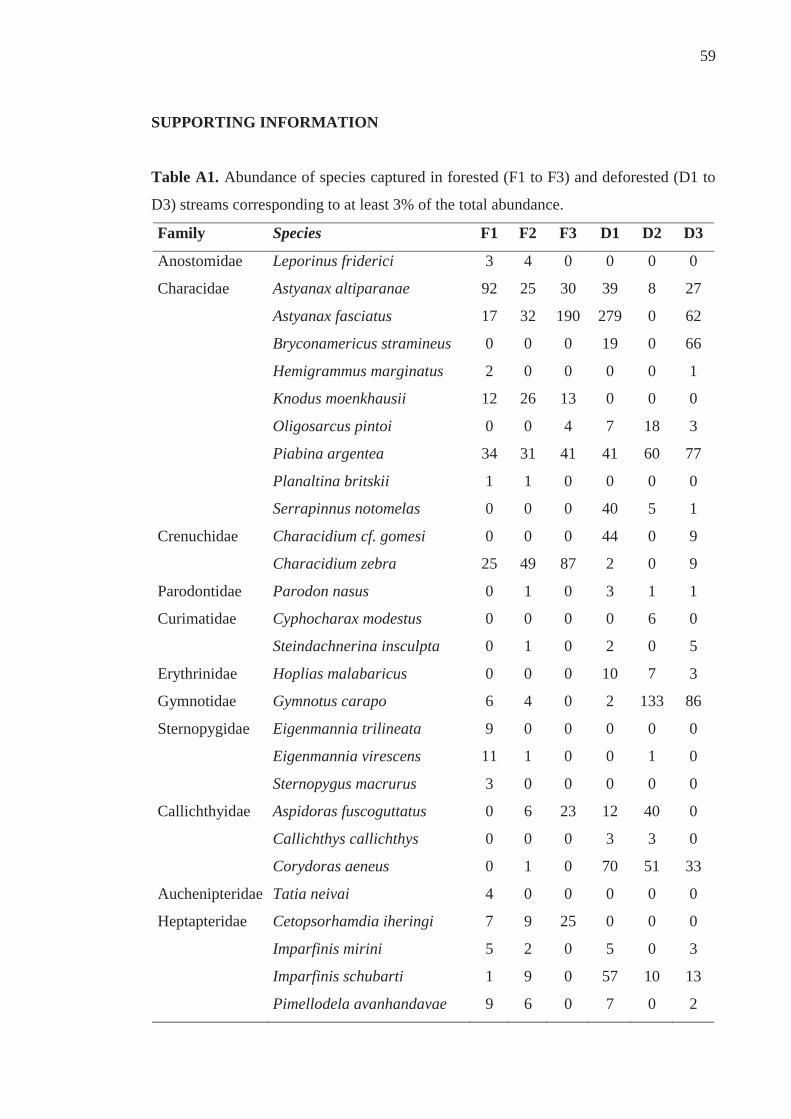
59
SUPPORTING INFORMATION
Table A1. Abundance of species captured in forested (F1 to F3) and deforested (D1 to
D3) streams corresponding to at least 3% of the total abundance.
Family Species F1 F2 F3 D1 D2 D3
Anostomidae Leporinus friderici 3 4 0 0 0 0
Characidae Astyanax altiparanae 92 25 30 39 8 27
Astyanax fasciatus 17 32 190 279 0 62
Bryconamericus stramineus 0 0 0 19 0 66
Hemigrammus marginatus 2 0 0 0 0 1
Knodus moenkhausii 12 26 13 0 0 0
Oligosarcus pintoi 0 0 4 7 18 3
Piabina argentea 34 31 41 41 60 77
Planaltina britskii 1 1 0 0 0 0
Serrapinnus notomelas 0 0 0 40 5 1
Crenuchidae Characidium cf. gomesi 0 0 0 44 0 9
Characidium zebra 25 49 87 2 0 9
Parodontidae Parodon nasus 0 1 0 3 1 1
Curimatidae Cyphocharax modestus 0 0 0 0 6 0
Steindachnerina insculpta 0 1 0 2 0 5
Erythrinidae Hoplias malabaricus 0 0 0 10 7 3
Gymnotidae Gymnotus carapo 6 4 0 2 133 86
Sternopygidae Eigenmannia trilineata 9 0 0 0 0 0
Eigenmannia virescens 11 1 0 0 1 0
Sternopygus macrurus 3 0 0 0 0 0
Callichthyidae Aspidoras fuscoguttatus 0 6 23 12 40 0
Callichthys callichthys 0 0 0 3 3 0
Corydoras aeneus 0 1 0 70 51 33
Auchenipteridae Tatia neivai 4 0 0 0 0 0
Heptapteridae Cetopsorhamdia iheringi 7 9 25 0 0 0
Imparfinis mirini 5 2 0 5 0 3
Imparfinis schubarti 1 9 0 57 10 13
Pimellodela avanhandavae 9 6 0 7 0 2

60
Pimelodus maculatus 1 0 0 0 0 0
Pseudopimelodus pulcher 38 34 0 0 0 0
Rhamdia quelen 1 10 19 29 10 1
Loricariidae Hisonotus francirochai 0 0 0 88 19 6
Hypostomus ancistroides 9 17 38 172 105 118
Hypostomus nigromaculatus 61 41 83 39 1 2
Hypostomus sp1 1 0 0 0 0 0
Hypostomus sp2 0 0 0 1 1 1
Poeciliidae Phalloceros harpagos 0 0 0 2 20 5
Poecilia reticulata 0 4 0 102 90 48
Cichlidae Cichlasoma paranaense 1 0 0 0 3 0
Crenicichla britskii 2 2 0 8 30 10
Geophagus brasiliensis 0 0 0 41 0 31
Oreochromis niloticus 0 0 0 0 0 1
Synbranchidae Synbranchus marmoratus 0 0 0 2 2 2
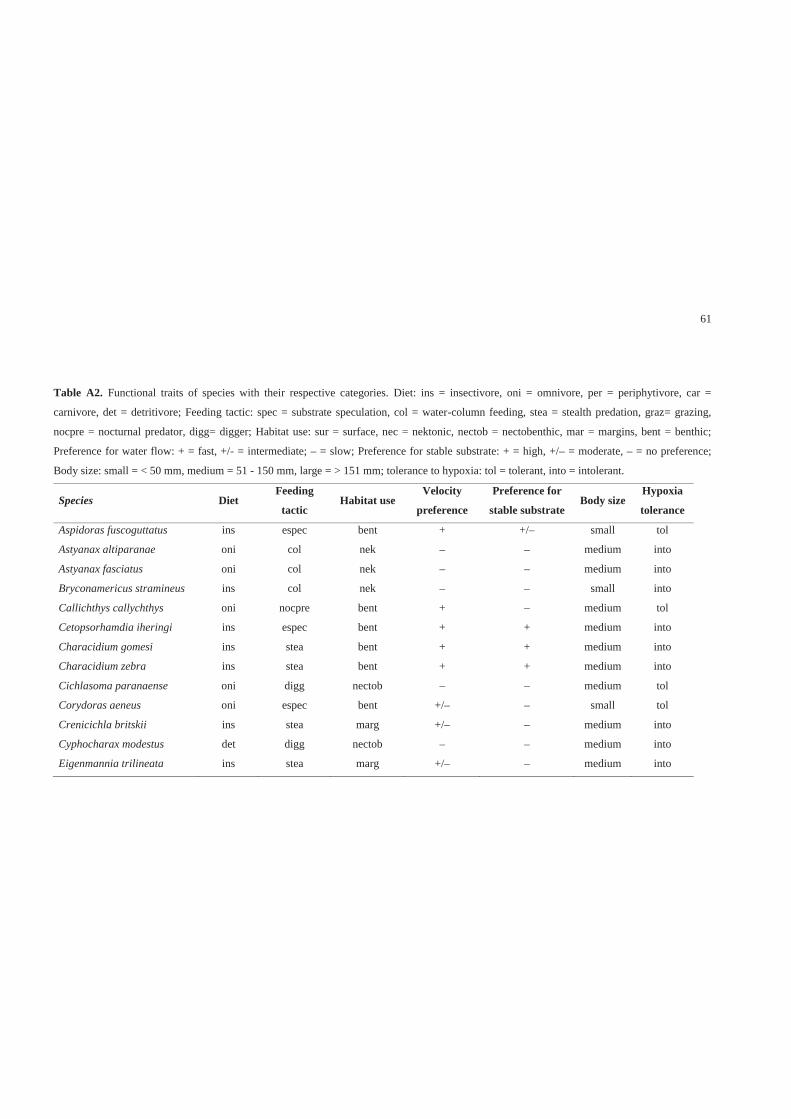
61
Table A2. Functional traits of species with their respective categories. Diet: ins = insectivore, oni = omnivore, per = periphytivore, car =
carnivore, det = detritivore; Feeding tactic: spec = substrate speculation, col = water-column feeding, stea = stealth predation, graz= grazing,
nocpre = nocturnal predator, digg= digger; Habitat use: sur = surface, nec = nektonic, nectob = nectobenthic, mar = margins, bent = benthic;
Preference for water flow: + = fast, +/- = intermediate; – = slow; Preference for stable substrate: + = high, +/– = moderate, – = no preference;
Body size: small = < 50 mm, medium = 51 - 150 mm, large = > 151 mm; tolerance to hypoxia: tol = tolerant, into = intolerant.
Species Diet Feeding
tactic Habitat use
Velocity
preference
Preference for
stable substrate Body size
Hypoxia
tolerance
Aspidoras fuscoguttatus ins espec bent + +/– small tol
Astyanax altiparanae oni col nek – – medium into
Astyanax fasciatus oni col nek – – medium into
Bryconamericus stramineus ins col nek – – small into
Callichthys callychthys oni nocpre bent + – medium tol
Cetopsorhamdia iheringi ins espec bent + + medium into
Characidium gomesi ins stea bent + + medium into
Characidium zebra ins stea bent + + medium into
Cichlasoma paranaense oni digg nectob – – medium tol
Corydoras aeneus oni espec bent +/– – small tol
Crenicichla britskii ins stea marg +/– – medium into
Cyphocharax modestus det digg nectob – – medium into
Eigenmannia trilineata ins stea marg +/– – medium into
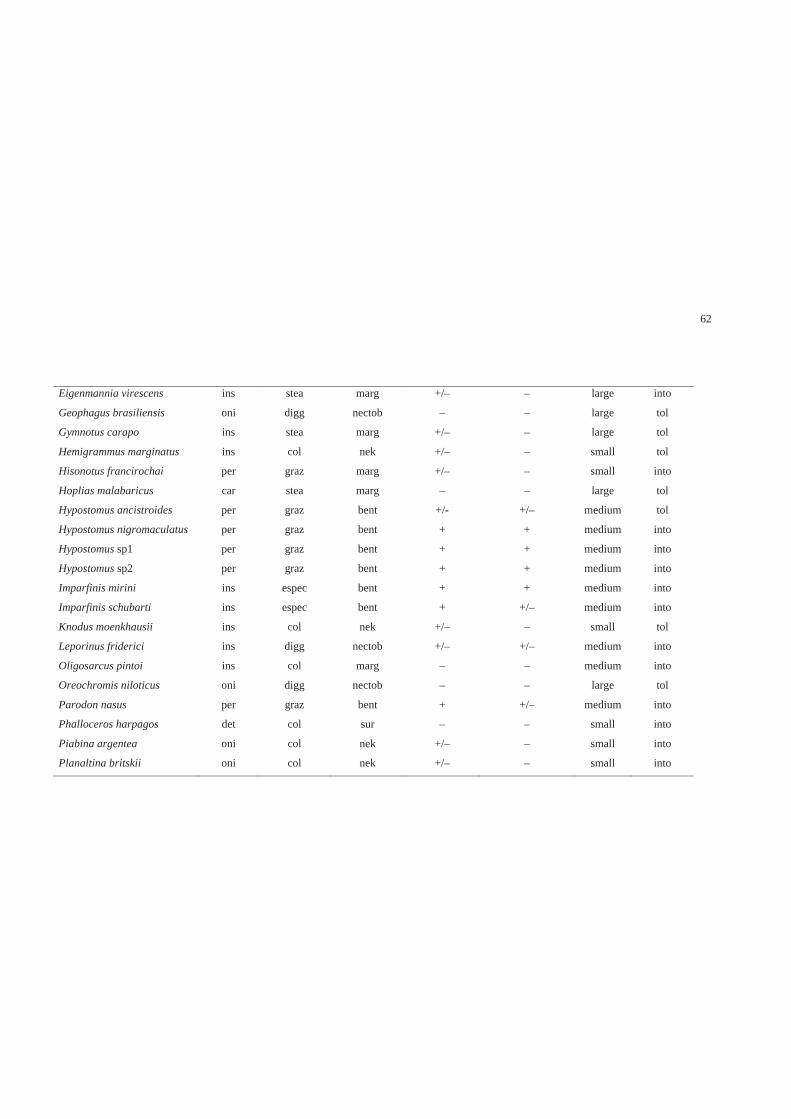
62
Eigenmannia virescens ins stea marg +/– – large into
Geophagus brasiliensis oni digg nectob – – large tol
Gymnotus carapo ins stea marg +/– – large tol
Hemigrammus marginatus ins col nek +/– – small tol
Hisonotus francirochai per graz marg +/– – small into
Hoplias malabaricus car stea marg – – large tol
Hypostomus ancistroides per graz bent +/- +/– medium tol
Hypostomus nigromaculatus per graz bent + + medium into
Hypostomus sp1 per graz bent + + medium into
Hypostomus sp2 per graz bent + + medium into
Imparfinis mirini ins espec bent + + medium into
Imparfinis schubarti ins espec bent + +/– medium into
Knodus moenkhausii ins col nek +/– – small tol
Leporinus friderici ins digg nectob +/– +/– medium into
Oligosarcus pintoi ins col marg – – medium into
Oreochromis niloticus oni digg nectob – – large tol
Parodon nasus per graz bent + +/– medium into
Phalloceros harpagos det col sur – – small into
Piabina argentea oni col nek +/– – small into
Planaltina britskii oni col nek +/– – small into
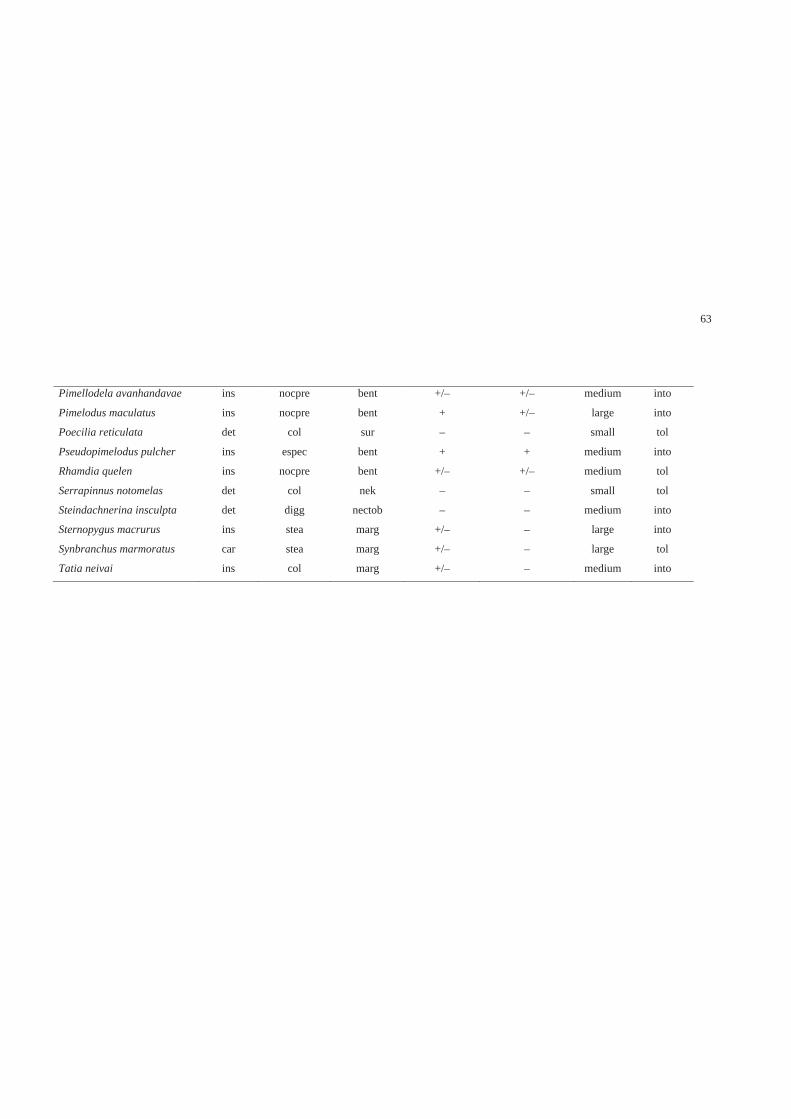
63
Pimellodela avanhandavae ins nocpre bent +/– +/– medium into
Pimelodus maculatus ins nocpre bent + +/– large into
Poecilia reticulata det col sur – – small tol
Pseudopimelodus pulcher ins espec bent + + medium into
Rhamdia quelen ins nocpre bent +/– +/– medium tol
Serrapinnus notomelas det col nek – – small tol
Steindachnerina insculpta det digg nectob – – medium into
Sternopygus macrurus ins stea marg +/– – large into
Synbranchus marmoratus car stea marg +/– – large tol
Tatia neivai ins col marg +/– – medium into

64
Manuscrito será submetido para Journal of Tropical Ecology
- CAPÍTULO III -
Diferenciação funcional de comunidades de peixes em riachos com
diferentes status de conservação

65
RESUMO
As alterações ambientais causadas pelo desmatamento nos riachos podem
representar filtros ambientais, restringindo a ocorrência de espécies a partir das
características funcionais que elas exibem. Testamos essa hipótese em riachos do
sudeste do Brasil, avaliando se riachos em contextos contrastantes com relação à
cobertura florestal podem ser diferenciados a partir da contribuição não aleatória de
grupos funcionais específicos. As amostragens foram realizadas em seis riachos, sendo
três florestados e três desmatados. Foram consideradas cinco características funcionais:
dieta, uso de hábitat, preferência por fluxo, tamanho e tolerância à hipóxia. A
diferenciação dos riachos, em especial entre os riachos florestados e desmatados, foi
maior do que o esperado ao acaso e correspondeu à contribuição diferencial de três
grupos funcionais. Nos riachos desmatados predominaram espécies tolerantes à hipóxia,
que apresentaram combinação variável das outras características. Nos riachos
florestados predominaram espécies de hábitos especializados, bentônicas, com
preferência por alto fluxo de água e dieta insetívora. As alterações nos riachos
provocadas pelo desmatamento relacionadas com a disponibilidade de hábitat, recursos
alimentares e condições físico-químicas (e.g., hipóxia) representam filtros ambientais
que restringem a ocorrência de grupos de espécies especializadas em detrimento do
beneficiamento de espécies tolerantes e oportunistas. Dessa forma, é possível predizer
que a intensificação desse tipo de alteração poderá resultar em modificação do pool
funcional regionalmente.
Palavras-chave: ictiofauna, características funcionais, filtros ambientais, partição da
diversidade

66
INTRODUÇÃO
De acordo com modelos baseados no nicho, a abundância e distribuição das
espécies dependem das suas características funcionais, que por sua vez, determinam a
forma pela qual as espécies são influenciadas pelo ambiente (Townsend & Hildrew,
1994; Goldstein & Meador, 2004; Blanc et al., 2007). Assim, no tempo ecológico, a
composição de espécies em um local é o resultado da seleção não aleatória de espécies a
partir de um pool regional, refletindo a ação de processos baseados na limitação da
similaridade e/ou filtros ambientais (Leibold, 1998; Cornwell et al., 2006).
Comunidades estruturadas por filtros ambientais apresentam espécies com
características mais similares do que esperado ao acaso, resultado da seleção de um
conjunto de espécies com características compatíveis com as condições ambientais
locais (Cornwell et al., 2006). Por outro lado, comunidades estruturadas por processos
de limitação da similaridade apresentam maior dissimilaridade interespecífica do que o
esperado ao acaso, como resultado da exclusão competitiva de espécies similares
(Weiher & Keddy, 1995).
A estruturação de comunidades de riachos é o resultado da ação preponderante
dos filtros ambientais (Jackson et al., 2001; Peres-Neto, 2004). Isso é suportado por
estudos que demonstram a associação das variáveis abióticas com a composição de
espécies (p.ex., Poff, 1997; Goldstein & Meador et al., 2004; 2005). Nesse contexto, as
restrições ambientais manifestam-se em diferentes escalas (Poff, 1997) e, podem ser
geradas também por interferências antrópicas. Por exemplo, Casatti et al. (2009)
compararam a fauna de peixes de dois grupos de riachos em estado de conservação
contrastantes com relação à estrutura física do hábitat e a cobertura florestal, em uma
região submetida à forte interferência antrópica no sudeste do Brasil. Esses dois grupos
de riachos abrigaram comunidades muito distintas, com as espécies tolerantes e
generalistas dominando os riachos degradados e as espécies especialistas e sensíveis
associadas aos riachos em melhor estado de conservação (Casatti et al., 2009). Esses
resultados fornecem evidência de que as restrições na ocorrência das espécies em
riachos variam em função do seu estado de conservação, sugerindo que modificações
antrópicas possam atuar como filtros ambientais (Mayfield et al., 2005).
Se, de fato, as interferências antrópicas modificam as condições nos riachos,
representando novos filtros ambientais dentro de um contexto regional, seria esperado
encontrar convergência funcional, ou seja, comunidades funcionalmente mais similares

67
entre riachos degradados do que o esperado a partir de uma seleção aleatória de espécies
em um pool regional (Cornwell et al., 2006). Nesse sentido, a identificação de grupos de
espécies que compartilham características funcionais responsáveis pela diferenciação
entre riachos em estado de conservação contrastantes seria uma estratégia para a
identificação dos filtros ambientais específicos atuando na organização das
comunidades. Essa abordagem seria especialmente útil para a compreensão dos
processos responsáveis pela estruturação das comunidades em regiões submetidas
impactos antrópicos.
Neste estudo, avaliamos se riachos com diferentes graus de cobertura florestal
na zona ripária podem ser diferenciados a partir da composição funcional das
comunidades de peixes. Realizamos amostragens da ictiofauna em riachos florestados e
desmatados do sudeste do Brasil, em uma região onde a degradação da zona ripária é
uma das principais ameaças à conservação da ictiofauna (Casatti et al., 2006a). O
desmatamento da zona ripária influencia diversos aspectos importantes para peixes em
riachos, tais como disponibilidade de micro-hábitats (Casatti et al., 2009), recursos
alimentares (Ferreira et al., 2011), condições estruturais e fisico-químicas (Murphy &
Hall, 1981). Assim, hipotetizamos que as mudanças causadas pelo desmatamento nos
riachos representam filtros ambientais, resultando na seleção não aleatória de espécies
que compartilham características funcionais associadas à tolerância e à capacidade de
explorar as oportunidades que se tornam disponíveis. Desta forma, predizemos que a
diferenciação funcional entre as comunidades dos riachos florestados e desmatados será
maior do que o padrão obtido aleatoriamente em função da contribuição diferencial de
grupos funcionais específicos.
A estratégia analítica utilizada neste estudo foi adaptada de Pavoine et al.
(2009a), que propuseram a partição da diversidade filogenética ao longo dos ramos da
filogenia com a finalidade de avaliar a importância das linhagens filogenéticas para a
diferenciação das comunidades. De acordo com os próprios autores, essa metodologia
pode ser adaptada para uma abordagem funcional, através da substituição da filogenia
por árvores funcionais. Nesse caso, é possível avaliar se grupos de espécies que
compartilham características funcionais são importantes para a diferenciação de
comunidades de ambientes em diferentes estados de conservação, como é o caso neste
estudo.

68
MATERIAL E MÉTODOS
Área de estudo
O estudo foi realizado na bacia do rio São José dos Dourados, sistema do alto rio
Paraná, sudeste do Brasil (Figura 1). Realizamos amostragens da ictiofauna em trechos
de aproximadamente 300 metros em riachos florestados e desmatados. Os riachos
florestados (n = 3) estão localizados dentro dos maiores fragmentos florestais presentes
na bacia (floresta estacional semi-decidual > 1.600 ha) e os riachos desmatados (n = 3)
não apresentaram vegetação arbórea na zona ripária, estando localizados em uma região
cuja pastagem é a matriz predominante (Figura 1). Não há fragmentos florestais à
montante e jusante dos trechos desmatados. Estes riachos foram selecionados porque
apresentaram características representativas de condições extremas dentro de um
gradiente de cobertura florestal e ao mesmo tempo são similares quanto ao volume de
água e disponibilidade de meso-hábitats (ver capítulo II). Em função da dificuldade de
se encontrar riachos comparáveis aos florestados, os pontos amostrais foram
distribuídos de forma não aleatória na paisagem, com os riachos desmatados
apresentando maior proximidade entre si do que entre os florestados. Entretanto, ambos
os grupos de riachos foram considerados pertencentes ao mesmo pool de espécies
regional (p.ex., Casatti et al. 2006b), já que a distância não explica a variação na
composição de espécies dentro desta bacia (Casatti et al., 2012).

69
Figura. 1. Mapa da área de estudo, destacando o estado de São Paulo (A), a bacia do rio São
José dos Dourados (B), onde estão localizados os riachos florestados (círculos escuros),
inseridos dentro dos maiores fragmentos florestais da bacia (manchas cinza) e os riachos
desmatados (círculos claros) (C).
Amostragem da ictiofauna
Amostramos a ictiofauna em 126 segmentos de cinco metros de extensão cada
(entre 16 e 24 segmentos por riacho), os quais foram distribuídos ao longo de trechos de
cerca de 300 metros em cada riacho. Os segmentos foram escolhidos visando
contemplar a variabilidade de meso-hábitats disponíveis em cada riacho. Antes do início
das coletas, isolamos cada segmento à montante e à jusante com redes. Para as coletas
utilizamos equipamento de pesca elétrica (gerador de corrente alternada, 220 V, 50-
60Hz, 3,4-4,1 A, 1.000 W), passando sucessivamente os eletrodos e puçás ao longo do
segmento. A captura em cada segmento foi finalizada quando nenhum indivíduo foi
capturado após uma passagem. Os animais capturados foram fixados no campo com
formaldeído (10%) e, posteriormente, transferidos para frascos com solução de etanol
(70%) para conservação. Todos os animais encontram-se depositados na coleção
ictiológica (DZSJRP) do Departamento de Zoologia e Botânica da Universidade
Estadual Paulista, São José do Rio Preto, SP, Brasil.

70
Caracterização das unidades amostrais
Após a coleta dos peixes, realizamos a caracterização dos riachos quanto a sua
estrutura física e condições físico-químicas. Essa caracterização foi realizada em cada
segmento (exceto para os parâmetros físico-químicos que foram medidos no ponto
médio de cada trecho), cujos valores foram agregados pela média para cada riacho.
Com relação à estrutura física, medimos a largura superficial, largura interna,
profundidade medida em cinco pontos eqüidistantes de uma margem a outra, fluxo
(medido com um fluxômetro mecânico General Oceanics , modelo 2030) no meio na
coluna d’água, composição do substrato (proporção relativa de cada um dos seus
componentes) (Cummins, 1962) e composição representatividade de estruturas
presentes nos barrancos em cada uma das margens. O substrato foi classificado em
estável, que inclui os tipos de substrato consolidados que permitem a colonização pela
fauna bentônica (argila consolidada, cascalho, seixo e rocha) e instável que inclui areia
e argila inconsolidada, elementos cuja dominância nos leitos dos riachos é indicador de
baixa integridade física (Casatti et al., 2006b). Com relação à vegetação marginal,
foram consideradas qualquer estrutura da vegetação ripária tais como raízes, folhas e
caules oriundos da vegetação terrestre submersos no hábitat interno. Nesse caso, o
volume ocupado pela vegetação submersa foi calculado a partir de medidas da altura e
da largura dos bancos de vegetação em cada transecto. Os parâmetros físico-químicos
avaliados foram oxigênio dissolvido com um oxímetro (LT Lutron, modelo DO 5510),
condutividade, pH, turbidez e temperatura com Horiba, modelo U-10.
Verificamos que os riachos florestados e desmatados diferiram com relação à
proporção de substrato estável e volume de vegetação marginal em contato com a água.
Os riachos florestados apresentaram maior proporção de substrato estável (t = 3,7, p <
0,01), enquanto os riachos desmatados apresentaram maior volume da vegetação
marginal em contato com a água (t = -5,9, p = 0,02). Além disso, os riachos florestados
apresentaram maiores valores de oxigênio dissolvido do que riachos desmatados
(Tabela 1).
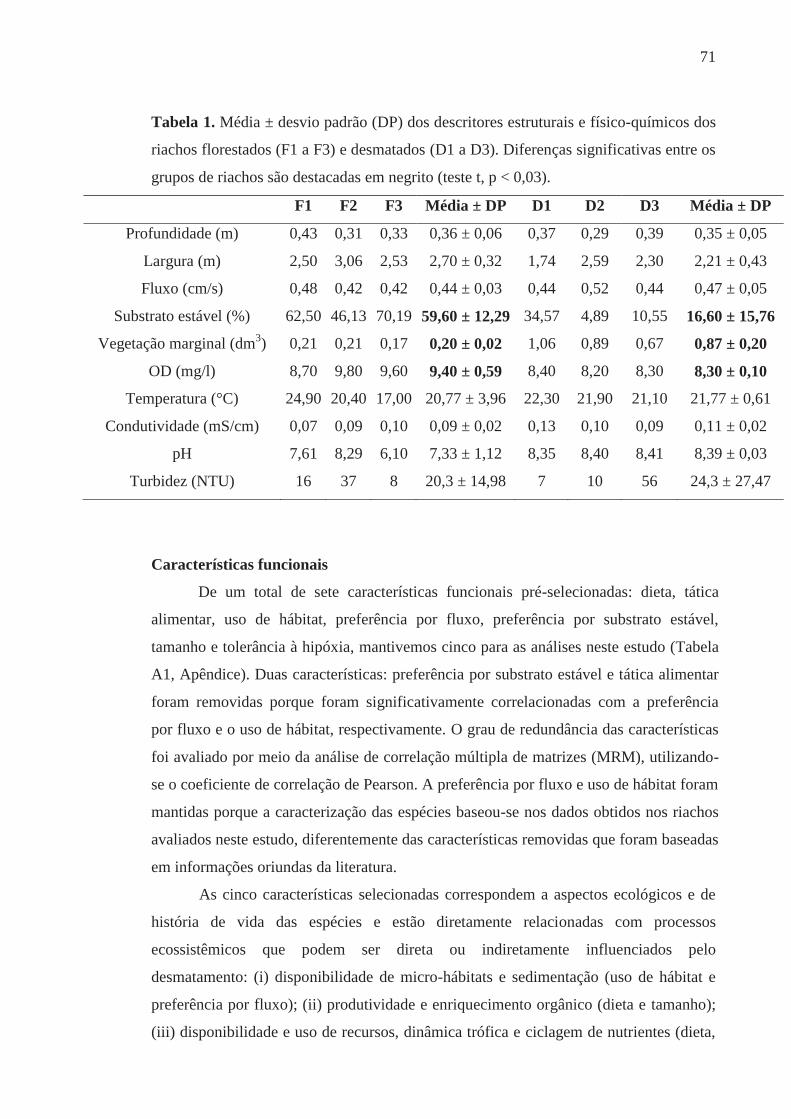
71
Tabela 1. Média ± desvio padrão (DP) dos descritores estruturais e físico-químicos dos
riachos florestados (F1 a F3) e desmatados (D1 a D3). Diferenças significativas entre os
grupos de riachos são destacadas em negrito (teste t, p < 0,03).
F1 F2 F3 Média ± DP D1 D2 D3 Média ± DP
Profundidade (m) 0,43 0,31 0,33 0,36 ± 0,06 0,37 0,29 0,39 0,35 ± 0,05
Largura (m) 2,50 3,06 2,53 2,70 ± 0,32 1,74 2,59 2,30 2,21 ± 0,43
Fluxo (cm/s) 0,48 0,42 0,42 0,44 ± 0,03 0,44 0,52 0,44 0,47 ± 0,05
Substrato estável (%) 62,50 46,13 70,19 59,60 ± 12,29 34,57 4,89 10,55 16,60 ± 15,76
Vegetação marginal (dm3) 0,21 0,21 0,17 0,20 ± 0,02 1,06 0,89 0,67 0,87 ± 0,20
OD (mg/l) 8,70 9,80 9,60 9,40 ± 0,59 8,40 8,20 8,30 8,30 ± 0,10
Temperatura (°C) 24,90 20,40 17,00 20,77 ± 3,96 22,30 21,90 21,10 21,77 ± 0,61
Condutividade (mS/cm) 0,07 0,09 0,10 0,09 ± 0,02 0,13 0,10 0,09 0,11 ± 0,02
pH 7,61 8,29 6,10 7,33 ± 1,12 8,35 8,40 8,41 8,39 ± 0,03
Turbidez (NTU) 16 37 8 20,3 ± 14,98 7 10 56 24,3 ± 27,47
Características funcionais
De um total de sete características funcionais pré-selecionadas: dieta, tática
alimentar, uso de hábitat, preferência por fluxo, preferência por substrato estável,
tamanho e tolerância à hipóxia, mantivemos cinco para as análises neste estudo (Tabela
A1, Apêndice). Duas características: preferência por substrato estável e tática alimentar
foram removidas porque foram significativamente correlacionadas com a preferência
por fluxo e o uso de hábitat, respectivamente. O grau de redundância das características
foi avaliado por meio da análise de correlação múltipla de matrizes (MRM), utilizando-
se o coeficiente de correlação de Pearson. A preferência por fluxo e uso de hábitat foram
mantidas porque a caracterização das espécies baseou-se nos dados obtidos nos riachos
avaliados neste estudo, diferentemente das características removidas que foram baseadas
em informações oriundas da literatura.
As cinco características selecionadas correspondem a aspectos ecológicos e de
história de vida das espécies e estão diretamente relacionadas com processos
ecossistêmicos que podem ser direta ou indiretamente influenciados pelo
desmatamento: (i) disponibilidade de micro-hábitats e sedimentação (uso de hábitat e
preferência por fluxo); (ii) produtividade e enriquecimento orgânico (dieta e tamanho);
(iii) disponibilidade e uso de recursos, dinâmica trófica e ciclagem de nutrientes (dieta,

72
uso de hábitat e tamanho); (iv) qualidade da água (tolerância à hipóxia) (Murphy &
Hall, 1981; Smale & Rabeni, 1995; Holmlund & Halmer, 1999; Bunn et al., 1999;
Esteves et al., 2008; Lorion & Kennedy, 2009).
Dieta
A dieta foi avaliada a partir da análise do conteúdo estomacal das espécies
coletadas. Os estômagos de 10 exemplares de cada espécie (exceto para aquelas que
apresentaram abundância inferior a 10 indivíduos) foram removidos e abertos para
inspeção e identificação do conteúdo estomacal. Os itens alimentares foram
identificados e classificados em grandes categorias (detrito, insetos, peixes, perifíton,
protista, fragmentos de vegetais superiores e algas). A composição porcentual de cada
item em cada estômago foi estimada visualmente para a identificação do item
dominante cuja freqüência foi combinada com a porcentagem de ocorrência do item nos
estômagos de cada espécie para a obtenção do índice de importância alimentar (IAi,
Bennemann et al., 2006):
ni
iOD
1
Onde Oi é o número proporcional de estômagos em que o item i ocorreu, e D a
proporção de estômagos em que o item i foi dominante. Os valores presentes na matriz
de dieta (variável fuzzy) correspondem aos valores de IAi.
Uso de hábitat
As espécies foram classificadas quanto ao uso do espaço (bentônico,
nectobentônico, nectônico, superfície e margem), atribuindo-se o valor “1” aos locais
que são eventualmente ocupados pela espécie e “2” para os locais onde a espécie ocorre
preferencialmente (variável fuzzy). As informações sobre o uso de hábitat foram obtidas
a partir de observações subaquáticas realizadas nos seis riachos deste estudo em um
período que antecedeu a coleta da ictiofauna, totalizando 1.200 horas de observações
subaquáticas. Complementarmente, foram utilizadas informações da literatura (Casatti
& Castro, 1998; Casatti et al., 2001; Casatti et al.,2005; Ceneviva-Bastos et al., 2010).
Preferência por fluxo

73
A preferência por fluxo (variável fuzzy) foi medida a partir do cálculo do índice
de preferência (IP) calculado para cada espécie em cada uma das quatro classes de fluxo
(F1 = 0,0 a 0,25 cm/s; F2 = 0,26 a 0,45 cm/s; F3 = 0,46 a 0,65 cm/s; F4 > 0,66 cm/s)
baseado na metodologia proposta por Freeman et al. (1997). O valor final de IP foi
padronizado para variar de 0 a 1 dividindo-se os valores de IP de cada espécie em cada
classe pelo valor máximo de IP encontrado para a espécie:
Onde, nsi é o número de indivíduos da espécie na classe i, N é o número total de
indivíduos da espécie coletados, nhi é o número de unidades de hábitats amostrados na
classe i, H é o número total de unidades de hábitats e max refere-se ao valor máximo.
Como a preferência por condições de fluxo são consistentes entre riachos florestados e
desmatados (Capítulo I) o valor final de IP para as espécies que ocorreram em ambos os
grupos de riachos foi representado pela média dos valores obtidos em cada grupo.
Tamanho e tolerância à hipóxia
Os maiores indivíduos de cada espécie amostrada tiveram seu comprimento
padrão mensurados (variável quantitativa) com o auxílio de um paquímetro. A
caracterização das espécies quanto à tolerância à hipóxia (variável nominal) baseou-se
em informações da literatura. Foram consideradas tolerantes as espécies que são
comumente encontradas em trechos com baixos níveis de oxigênio (Kramer &
Mehegan, 1981; Araujo & Garutti, 2003; Bozzetti & Schulz, 2004; Casatti et al., 2006b;
Rocha et al., 2009; Casatti et al., 2012).
Análise dos dados
Utilizamos para as análises somente espécies que ocorreram em mais do que 3%
dos segmentos amostrados, visando reduzir a possível influência de estimativas
equivocadas de dieta e preferência por fluxo baseadas em poucos indivíduos. Como o
número de segmentos amostrados diferiu entre os riachos, a matriz de espécies final foi
composta pela abundância média das espécies considerando os segmentos amostrados
em cada riacho.
O dendrograma funcional foi construído a partir de uma matriz de distância
representando a similaridade entre as espécies baseada nas cinco características
funcionais selecionadas para este estudo. Essa matriz foi obtida a partir de uma

74
generalização da aplicação do coeficiente de Gower proposta por Pavoine et al. (2009b)
que possibilita a combinação de características cujos dados são de natureza estatística
diferentes (e.g., fuzzy, nominal e quantitativa). A distância entre cada par de espécies
representa a média da distância entre as duas espécies, considerando cada característica
separadamente, após a sua padronização (variação de 0 a 1). O dendrograma funcional
final foi construído utilizando o método das médias aritméticas não ponderadas
(UPGMA).
A diversidade funcional foi calculada a partir do índice de entropia quadrática de
Rao (QE). Este índice é baseado na soma das distâncias entre os pares de espécies de
uma comunidade considerando sua abundância (Botta-Dukát, 2005) Quando QE é
aplicado para medir diversidade funcional a partir de um dendrograma, a distância entre
duas espécies é igual à soma do comprimento do braço do dendrograma que conecta as
duas espécies a um ancestral comum (Pavoine et al. 2005). Adaptando a metodologia de
Pavoine et al. (2009a) para lidar com árvore funcional ao invés da árvore filogenética,
calculamos o valor de QE para cada partição do dendrograma. Nesse caso, os grupos
funcionais que se originam em cada partição equivalem aos ramos terminais e a
abundância corresponde à soma das abundâncias das espécies que compõem o grupo.
Para avaliar as diferenças funcionais entre as comunidades (diversidade funcional β) e a
contribuição dos grupos funcionais formados nas diferentes partições do dendrograma,
realizamos a partição de QE nos componentes γ, α e β, assumindo que a diversidade
funcional regional (γ) corresponde à média da diversidade de cada riacho (α) somado à
diversidade entre os riachos (β) (Lande, 1996). Os valores de β/γ (i.e., variabilidade
entre riachos) observados para cada partição do dendrograma foram comparados com
valores simulados a partir da aleatorização dos ramos terminais, de forma que a conexão
entre a estrutura funcional do dendrograma e a abundância dos ramos terminais fosse
permutada (999 permutações) (Pavoine et al., 2009a). Valores de β/γ maiores do que o
esperado ao acaso indicam agregação funcional, ou seja, maior semelhança funcional
dentro das comunidades do que entre comunidades, resultado esperado em comunidades
organizadas por filtros ambientais (Pavoine & Bonsall, 2011).
Complementarmente, aplicamos a análise de coordenadas principais dupla
(DPCoA) (Pavoine et al., 2004) para identificar os grupos funcionais que foram mais
importantes na diferenciação funcional entre os riachos florestados e desmatados. A
DPCoA possibilita a ordenação das espécies, cujas posições relativas são definidas
pelas suas dissimilaridades funcionais, assim como das amostras cujas posições

75
relativas refletem as suas diferenças baseadas no QE (Pavoine et al., 2004). Todas as
análises foram realizadas no software R (R Development Core Team 2009), utilizando
os pacotes “ade4” (Dray & Dufour, 2007) e as funções “aptree”, “abg.aptree” e
“rtest.aptree” fornecidas por Pavoine et al. (2009a).
RESULTADOS
Foram amostradas 43 espécies (Tabela 2) das quais oito foram removidas das
análises por apresentarem baixa ocorrência, resultando em uma matriz final composta
por 35 espécies, que representaram 99,5% da abundância total. Essas espécies foram
agrupadas de acordo com suas semelhanças funcionais em um dendrograma composto
por 34 partições (Figura 2). A diferenciação dos riachos (β/γ) baseado na composição
funcional das comunidades foi maior do que o esperado ao acaso nas partições #30 e
#33 (p < 0,02). Essas partições separaram as espécies em três e seis grupos funcionais,
respectivamente (Figura 3). O primeiro eixo da DPCoA resumiu a maior parte da
variação referente às diferenças na composição funcional das comunidades (72,6%) e
permitiu destacar os grupos funcionais foram mais importantes para a diferenciação dos
riachos florestados e desmatados (Figura 4). Os três grupos originados na partição #33
foram importantes para a dissimilaridade funcional entre os riachos florestados e
desmatados, sendo que os grupos 1 e 2 foram mais associados aos riachos desmatados,
enquanto as espécies do grupo 3 foram mais abundantes nos riachos florestados
(Figuras 4). As espécies dos grupos 1 e 2 compartilham a tolerância à hipóxia e
complementarmente, as espécies do grupo 1 podem ser caracterizadas ainda pelo grande
porte, dieta carnívora/insetívora e por utilizarem as margens como hábitat. As espécies
do grupo 2 podem ser caracterizadas pelo pequeno porte, uso da superfície com dieta
detritívora (Poecilia reticulata e Phalloceros harpagos) e pelo hábitat bentônico com
preferência por fluxo de água intermediário e dieta insetívora (demais espécies) (Figura
3). O grupo 3 foi composto por espécies intolerantes à hipóxia e com variações nas
outras características (Figura 3).

76
Tabela 2. Abundância média das espécies (com seus respectivos códigos) que
ocorreram em mais do que 3% dos segmentos nos riachos florestados (F1 a F3) e
desmatados (D1 a D3), bacia do rio São José dos Dourados, Alto Paraná.
Família Espécies Códigos F1 F2 F3 D1 D2 D3
Anostomidae Leporinus friderici Lepfri 0.2 0.2 0.0 0.0 0.0 0.0
Characidae Astyanax altiparanae Astalt 4.0 1.2 1.6 1.6 0.4 1.7
Astyanax fasciatus Astfas 0.8 1.7 10.1 11.6 0.0 3.9
Bryconamericus stramineus Brystr 0.0 0.0 0.0 0.8 0.0 4.2
Knodus moenkhausii Knomoe 0.7 1.4 0.7 0.0 0.0 0.0
Oligosarcus pintoi Olipin 0.0 0.0 0.2 0.3 0.8 0.2
Piabina argentea Piaarg 1.5 1.6 2.2 1.7 2.7 4.9
Serrapinnus notomelas Sernot 0.0 0.0 0.0 1.7 0.2 0.1
Crenuchidae Characidium cf. gomesi Chagom 0.0 0.0 0.0 1.8 0.0 0.6
Characidium zebra Chazeb 1.3 3.1 4.7 0.1 0.0 0.6
Parodontidae Parodon nasus Parnas 0.0 0.0 0.0 0.1 0.0 0.1
Curimatidae Steindachnerina insculpta Steins 0.0 0.0 0.0 0.1 0.0 0.3
Erythrinidae Hoplias malabaricus Hopmal 0.0 0.0 0.0 0.4 0.3 0.2
Gymnotidae Gymnotus carapo Gymcar 0.3 0.2 0.0 0.1 6.1 5.4
Sternopygidae Eigenmannia trilineata Eigtri 0.4 0.0 0.0 0.0 0.0 0.0
Eigenmannia virescens Eigvir 0.5 0.0 0.0 0.0 0.0 0.0
Sternopygus macrurus Stemac 0.2 0.0 0.0 0.0 0.0 0.0
Callichthyidae Aspidoras fuscoguttatus Aspfus 0.0 0.5 1.2 0.5 1.9 0.0
Callichthys callichthys Calcal 0.0 0.0 0.0 0.1 0.1 0.0
Corydoras aeneus Coraen 0.0 0.0 0.0 2.9 2.3 2.1
Auchenipteridae Tatia neivai Tatnei 0.3 0.0 0.0 0.0 0.0 0.0
Heptapteridae Cetopsorhamdia iheringi Cetihe 0.3 0.4 1.3 0.0 0.0 0.0
Imparfinis mirini Impmir 0.2 0.1 0.0 0.2 0.0 0.2
Imparfinis schubarti Impsch 0.0 0.7 0.0 2.4 0.5 0.8
Pimellodela avanhandavae Pimava 0.5 0.3 0.0 0.3 0.0 0.1
Pseudopimelodus pulcher Psepul 1.8 1.8 0.0 0.0 0.0 0.0
Rhamdia quelen Rhaque 0.0 0.5 1.0 1.2 0.5 0.1
Loricariidae Hisonotus francirochai Hisfra 0.0 0.0 0.0 3.7 0.9 0.4
Hypostomus ancistroides Hypanc 0.4 1.1 2.0 7.2 5.0 7.7
77
Hypostomus nigromaculatus Hypnig 2.9 2.2 4.6 1.6 0.0 0.1
Poeciliidae Phalloceros harpagos Phahar 0.0 0.0 0.0 0.1 0.9 0.3
Poecilia reticulata Poeret 0.0 0.2 0.0 4.3 4.1 3.0
Cichlidae Crenicichla britskii Crebri 0.1 0.1 0.0 0.3 1.5 0.6
Geophagus brasiliensis Geobra 0.0 0.0 0.0 1.7 0.0 1.9
Synbranchidae Synbranchus marmoratus Synmar 0.0 0.0 0.0 0.1 0.1 0.1

78
Figura 2. Dendrograma de similaridade funcional das 35 espécies de peixes incluídas nas análises. A abundância média das espécies dentro dos
riachos florestados (F1-F3) e desmatados (D1-D3) são representadas pelo tamanho dos quadrados pretos. Os nomes das espécies encontram-se
abreviados de acordo com a tabela 2.

79
Três dos seis grupos funcionais que se originaram na partição #30 (grupos 1, 2 e
3 ) foram importantes na diferenciação funcional entre os riachos florestados e
desmatados (Figura 4). Os grupos 1 e 2 foram os mesmos originados na partição #33. O
grupo 3 foi composto por espécies de hábitos especializados, bentônicas, com
preferência por alto fluxo de água e dieta insetívora (Figura 3). Os grupos 1 e 2 foram
mais associados aos riachos desmatados e o grupo 3 concentrou-se nos riachos
florestados (Figura 4). O grupo 4, formado por espécies que vivem associadas às
margens de dieta insetívora, e o “grupo” 5, representado por uma única espécie
(Steindacherina insculpta), foram pouco abundantes e conseqüentemente tiveram
pequena contribuição para a diferenciação funcional dos riachos (Figura 2 e 4). As
espécies do grupo 6 que compartilham o hábito nectônico, dieta generalista/insetívora
com preferência por ambientes lênticos tiveram menor importância na diferenciação
funcional dos riachos por serem homogeneamente distribuídas entre as amostras (Figura
2 e 4).

80
Figura 3. Identificação dos grupos funcionais definidos pelas partições #30 e #33 com suas respectivas tendências de mudança na
representatividade em relação ao desmatamento. As características dominantes dos grupos funcionais que foram importantes para a diferenciação
dos riachos de acordo com a DPCoA (Figura 4) também são mostradas. Os nomes das espécies encontram-se abreviados de acordo com a tabela
2.

81
Figura 4. Representação gráfica da análise de coordenadas principais dupla,
descrevendo as diferenças funcionais entre os riachos florestados (F1-F3) e desmatados
(D1-D3). Os círculos pretos correspondem às espécies cujas posições relativas refletem
suas dissimilaridades funcionais. As espécies estão conectadas de acordo com os grupos
funcionais originados na partição #33 (a) e #30 (b). A identidade das espécies e suas
respectivas características funcionais são mostradas na figura 3.
DISCUSSÃO
Neste estudo, mostramos que os riachos submetidos ao desmatamento das suas
zonas ripárias diferem de riachos florestados com relação à composição de espécies
como resultado da restrição à ocorrência de grupos de espécies baseado nas suas
características funcionais. A adaptação da estratégia analítica desenvolvida por Pavoine
et al. (2009a) para lidar com uma árvore funcional em substituição à filogenia, permitiu
diferenciar os riachos desmatados e florestados a partir da estrutura funcional das
comunidades. A partir da decomposição da diversidade ao longo da árvore funcional
constatamos maiores valores de diversidade funcional beta do que o esperado ao acaso,
indicando que as comunidades dentro de cada riacho apresentam dominância de um ou
poucos grupos funcionais em relação ao esperado a partir do pool regional (alta β e
baixa α) (Pavoine et al., 2005). Os maiores valores de diversidade beta ocorreram nas
partições basais do dendrograma de similaridade funcional (#30 e #33) e foram
conseqüência da contribuição diferencial dos grupos funcionais 1, 2 e 3 entre os riachos,
sugerindo a ocorrência de convergência funcional dentro das comunidades, o que é
compatível com a influência de filtros ambientais (Cornwell et al., 2006; Mouillot et al.,

82
2007). As espécies de comunidades estruturadas por filtros ambientais tendem a ser
mais similares entre si devido às restrições do ambiente que favorece aquelas que
compartilham características compatíveis com as condições locais. A diferenciação
funcional foi mais acentuada entre os riachos florestados e desmatados, refletindo as
diferenças nas condições ambientais que, por sua vez, representam filtros ambientais
importantes para a estruturação das comunidades.
O beneficiamento de grupos funcionais em riachos desmatados, representado
neste estudo pelos grupos 1 e 2, também foi descrito por Bojsen & Barriga (2002) e
Lorion & Kennedy (2009). Estes autores mostraram um aumento da abundância e
biomassa de peixes herbívoros e detritívoros em riachos desmatados como
conseqüência da redução do sombreamento e aumento da produtividade primária. Por
outro lado, Casatti et al. (2006a,b) e Teresa & Casatti (2010) atribuíram o declínio de
espécies reofílicas à simplificação do hábitat e ao desmatamento, o que no presente
estudo corresponde ao declínio verificado no grupo 3 (#30). Esses padrões são
coerentes com a generalização de que a degradação ambiental resulta na ampliação da
distribuição de espécies oportunistas e generalistas em substituição às espécies sensíveis
e especialistas (Rahel, 2002; Devictor et al., 2008).
A associação dos grupos funcionais tolerantes à hipóxia (grupos 1 e 2) aos
riachos desmatados pode estar relacionada com as alterações em alguns processos
ecológicos. A perda da cobertura ripária e, consequentemente, o aumento da incidência
de radiação solar proporciona o aumento da produtividade primária em riachos (Murphy
& Hall, 1981; Gücker et al., 2009; Lorion & Kennedy, 2009). Isso foi evidenciado neste
estudo de maneira indireta, pela grande abundância de macrófitas e algas nos riachos
desmatados. O aumento da produtividade, associado à disponibilidade de nutrientes,
como é típico de áreas rurais (Brodie & Mitchell, 2005), pode resultar no aumento das
taxas de decomposição e, consequentemente redução da disponibilidade de oxigênio
dissolvido. Nas margens, esse efeito pode ser potencializado, uma vez que a vegetação
herbácea abundante nos barrancos desses riachos (Casatti et al., 2009; Rocha et al.,
2009) estende-se para o hábitat interno, contribuindo para a atenuação do fluxo de água
e interceptação de detritos, condições propícias para a aceleração dos processo de
decomposição (i.e., consumo de oxigênio) (Bunn et al., 1999). Assim, a suposta maior
disponibilidade de micro-hábitats com baixa disponibilidade de oxigênio nos riachos
desmatados pode estar associado com o favorecimento das espécies tolerantes à hipóxia
dos grupos 1 e 2 e declínio das espécies intolerantes do grupo 3 de ambas partições.

83
Além da tolerância à hipóxia, as condições encontradas nos riachos desmatados
podem filtrar outras características. Por exemplo, as espécies do grupo 1 caracterizam-
se pelo grande porte (> 180 mm) e inclui as únicas espécies piscívoras registradas neste
estudo (Hoplias malabaricus e Synbranchus marmoratus). O favorecimento de
predadores em riachos desmatados pode estar associado ao aumento da produtividade
primária que aumentaria a energia disponível no sistema, possibilitando a manutenção
de peixes maiores de níveis tróficos superiores (Brocksen et al., 1968; Murphy & Hall,
1981). Além disso, conforme discutido no capítulo II, a vegetação marginal abundante
pode conferir proteção para peixes de grande porte contra predadores terrestres.
O grupo funcional 2 inclui espécies que apresentam características variáveis
quanto ao uso de hábitat, preferência por fluxo e dieta e podem ser divididas em dois
subgrupos. Em um deles encontram-se Poecilia reticulata e Phalloceros harpagos que
apresentam em comum o uso da superfície, tamanho pequeno e detrito como um
importante item alimentar. Essas características favorecem a exploração de micro-
hábitats rasos e vegetados que são abundantes em riachos desmatados (Casatti et al.,
2009). As espécies pertencentes ao outro subgrupo dentro do grupo 2 são insetívoras,
bentônicas e generalistas quanto à preferência por fluxo. Essas espécies podem ser
consideradas equivalentes funcionais tolerantes à hipóxia do grupo 3 (#30), uma vez
que compartilham dieta, tamanho e uso de hábitat. Nesse sentido, o declínio do grupo 3
e favorecimento do grupo 2 nos riachos desmatados segue a tendência constatada para
peixes (Rahel, 2002; Casatti et al., 2009; Casatti et al., 2012) e outros grupos animais
(Warren et al., 2001; Devictor et al., 2008), de substituição de espécies especialistas por
generalistas tolerantes em ambientes antropicamente degradados. A redundância
funcional das espécies bentônicas do grupo 2 e as do grupo 3 (#30) quanto à dieta e uso
de hábitat e a sua complementaridade quanto à tolerância à hipóxia podem contribuir
para a manutenção de processos relacionados ao uso de recursos nos riachos
desmatados, mesmo com a mudança na composição de espécies. Isso acontece quando
as espécies compartilham características que exercem algum efeito sobre o ecossistema
(e.g., dieta baseada em insetos bentônicos), mas diferem na resposta às variações nas
condições do ambiente (e.g., tolerância à hipóxia) (Diaz & Cabido, 2001). Embora seja
plausível supor que esses grupos contribuem para processos como ciclagem de
nutrientes, dinâmica trófica e produtividade secundária (Taylor et al., 2006; Small et al.,
2011) nos dois grupos de riachos, não é possível inferir a magnitude e equivalência (ou
divergência) das suas contribuições sem a utilização de abordagens experimentais.

84
Três grupos funcionais não foram importantes para a diferenciação funcional
dos riachos florestados e desmatados, por conterem espécies raras e pouco abundantes
(grupos 4 e 5) e por não apresentarem ocorrência influenciada pela cobertura florestal
(grupo 6). As espécies do grupo 6 compartilham o hábito nectônico, dieta
insetívora/onívora e preferência por meso-hábitats com baixo fluxo de água (Capítulo I
e II) e são representadas, na sua maioria por pequenos caracídeos. Trata-se de um grupo
dominante em riachos Neotropicais (Castro, 1999), tendo sido bem representados nos
riachos deste estudo, exceto em D2, provavelmente em função de particularidades
presentes neste riacho como maior fluxo de água e menor disponibilidade de espaço na
coluna d’água livre de macrófitas no meio do canal. Essas espécies são conhecidas pela
sua plasticidade trófica e oportunismo, sendo capazes de explorar com sucesso os
recursos disponíveis mesmo em ambientes degradados (Melo, 2004; Ceneviva-Bastos et
al., 2010; Ferreira et al., 2012), o que pode explicar a sua baixa sensibilidade ao
desmatamento.
A abordagem utilizada neste estudo propiciou a identificação de combinações de
características necessárias para a formação de grupos de espécies que apresentam
respostas similares às variações no grau de cobertura florestal. Nesse sentido, os
resultados proporcionam suporte para a idéia de que a classificação de espécies em
grupos funcionais representa uma alternativa viável para a predição da integridade de
ambientes lóticos (Welcomme et al., 2006). Adicionalmente, a incorporação de aspectos
funcionais das comunidades torna essa abordagem útil também para a predição dos
efeitos das alterações antrópicas sobre processos e serviços ecossistêmicos (Dainly et
al., 2006).
Os resultados corroboram a hipótese e indicam que as variações na composição
funcional das comunidades dependem do estado de conservação dos riachos com
relação à cobertura florestal das suas zonas ripárias. Especificamente, a influência das
variações na cobertura florestal sobre a disponibilidade de hábitat e recursos
alimentares, assim como nas condições físico-químicas (e.g., hipóxia) parecem ser
importantes na organização das comunidades. Ao demonstrar que as modificações
ambientais causadas pelo desmatamento resultam na restrição de grupos de espécies que
compartilham características funcionais específicas, antecipamos que intensificação
dessa alteração poderá resultar além da homogeneização da fauna como sugerido por
Casatti et al. (2009), na limitação do pool funcional.

85
REFERÊNCIAS BIBLIOGRÁFICAS
Araujo, R.B. & Garutti, V. 2003. Ecology of a stream from upper Paraná river basin
inhabited by Aspidoras fuscoguttatus Nijssen and Isbrücker, 1976 (Siluriformes,
Callichthyidae). Brazilian Journal of Biology, 63: 363–372.
Bennemann, S.T., Casatti, L. & Oliveira, D.C. 2006. Alimentação de peixes: proposta
para análise de itens registrados em conteúdos gástricos. Biota Neotropica, 6(2): 1-
8.
Blanck, A., Tedesco, P.A. & Lamouroux, N. 2007. Relationships between life-history
strategies of European freshwater fish species and their habitat preferences
Freshwater Biology, 52: 843–859.
Botta-Dukát, Z. 2005. Rao’s quadratic entropy as a measure of functional diversity
based on multiple traits. Journal of Vegetation Science, 16: 533–540.
Bozzetti, M. & Schulz, U.H. 2004. An index of biotic integrity based on fish
assemblages for subtropical streams in southern Brazil. Hydrobiologia, 529: 133–
144.
Brocksen, R.W., Davis, G.E. & Warren, C.E. 1968. Competition, Food consumption,
and production of sculpins and trout in laboratory stream communities. The Journal
of Wildlife Management, 32(1): 51–75.
Brodie, J.E. & Mitchell, A.W. 2005. Nutrients in Australian tropical rivers: changes
with agricultural development and implications for receiving environments Marine
and Freshwater Research, 56(3): 279–302.
Bunn, S.E., Davies, P.M. & Mosisch, T.D. 1999. Ecosystem measures of river health
and their response to riparian and catchment degradation. Freshwater Biology, 41:
333–345.
Casatti, L. & Castro, R.M.C. 1998. A fish community of the São Francisco river
headwater riffles, southeastern Brazil. Ichthyological Exploration of Freshwaters,
9: 229–242.
Casatti, L., Langeani, F. & Castro, R.M.C. 2001. Peixes de riacho do Parque Estadual
Morro do Diabo, bacia do alto rio Paraná, SP. Biota Neotropica, 1: 1–15.
Casatti, L., Rocha, F.C. & Pereira, D.C. 2005. Habitat use by two species of
Hypostomus (Pisces, Loricariidae) in southeastern brazilian streams. Biota
Neotropica, 5: 157–165.

86
Casatti, L., Langeani, F. & Ferreira, C.P. 2006a. Effects of physical habitat degradation
on the stream fish assemblage structure in a Pasture Region. Environmental
Management, 38:974–982.
Casatti, L., Langeanim F., Silva, A.M. & Castro, R.M.C. 2006b. Stream fish, water and
habitat quality in a pasture dominated basin, southeastern Brazil. Brazilian Journal
of Biology, 66: 681–696.
Casatti, L., Ferreira, C.P. & Carvalho, F.R. 2009. Grass-dominated stream sites exhibit
low fish species diversity and dominance by guppies: an assessment of two tropical
pasture river basins. Hydrobiologia, 623: 173–189.
Casatti, L., Teresa, F.B., Gonçalves-Souza, T., Bessa, E., Manzotti, A.R., Gonçalves,
C.S. & Zeni, J.O. 2012. From forests to cattail: how does the riparian zone
influence stream fish? Neotropical Ichthyology. In press.
Castro, R.M.C. 1999. Evolução da ictiofauna de riachos sul-americanos: padrões gerais
e possíveis processos causais. In: Caramaschi, E.P., Mazzoni, R., Bizerril,
C.R.S.F., Peres-Neto, P.R., eds. Ecologia de Peixes de Riachos: Estado Atual e
Perspectivas. Rio de Janeiro: Oecologia Brasiliensis, v. VI, pp. 139–155.
Ceneviva-Bastos, M., Casatti, L. & Rossa-Feres, D.C. 2010. Meso and microhabitat
analysis and feeding habits of small nektonic characins (Teleostei: Characiformes)
in Neotropical streams. Zoologia, 27: 191–200.
Cornwell, W.K., Schwilk, D.W. & Ackerly, D.D. 2006. A trait-based test for habitat
filtering: convex hull volume. Ecology, 87(6): 1465–1471.
Costa, P.F. & Schulz, U.H. 2010. The fish community as an indicator of biotic integrity
of the streams in the Sinos River basin, Brazil. Brazilian Journal of Biology, 70(4):
1195–1205.
Cummins, K.H. 1962. An evaluation of some techniques for the collection and analysis
of benthic samples with special emphasis on lotic waters. The American Midland
Naturalist, 67: 477–504.
Daily, G., Alexander, S., Ehrlich, P.R., Gouler, L., Lubchenco, J., Matson, P.A.,
Mooney, H.A., Postel, S., Schneider, S.H., Tilman, D. & Woodwell, G.M.1997.
Ecosystem services: benefits supplied to human societies by natural ecosystems.
Issues in Ecology, 2: 1–18.Devictor, V., Julliard, R., Jiguet, J.C.F., Lee, A. &
Couvet, D. 2008. Functional biotic homogenization of bird communities in
disturbed landscapes. Global Ecology and Biogeography, 17: 252–261.

87
Diaz, S. & Cabido, M. 2001. Vive la différence: plant functional diversity matters to
ecosystem processes. Trends in Ecology and Evolution, 16(8): 646–655.
Dray, S. & Dufour, A.B. 2007. The ade4 package: implementing the duality diagram for
ecologists. Journal of Statistical Software, 22: 1–20.
Ernst, R., Linsenmair, K.E. & Rödel, M.O. 2006. Diversity erosion beyond the species
level: dramatic loss of functional diversity after selective logging in two tropical
amphibian communities. Biological Conservation, 133: 143–155.
Esteves, K.E., Lobo, A.V.P. & Faria, M.D.R. 2008. Trophic structure of a fish
community along environmental gradients of a subtropical river (Paraitinga River,
Upper Tietê River Basin, Brazil). Hydrobiologia, 598: 373–387.
Ferreira, A., Paula, F.R., Ferraz, S.F.B., Gerhard, P., Kashiwaqui, E.A.L., Cyrino, J.E.P.
& Martinelli, L.A. 2012. Riparian coverage affects diets of characids in neotropical
streams. Ecology of Freshwater Fish, 21(1): 12–22.
Freeman, M.C., Bowen, Z.H. & Crance, J.H. 1997. Transferability of habitat suitability
criteria for fishes in warmwater streams. North American Journal of Fisheries
Management, 17: 20–31.
Goldstein, R.M. & Meador, M.R. 2004. Comparisons of fish species traits from small
streams to large rivers. Transactions of the American Fisheries Society, 133: 971–
983.
Gücker, B., Boëchat, I.G. & Giani, A. 2009. Impacts of agricultural land use on
ecosystem structure and whole-stream metabolism of tropical Cerrado streams.
Freshwater Biology, 54: 2069–2085.
Holmlund, C.M. & Hammer, M. 1999. Ecosystem services generated by fish
populations. Ecological Economics, 29: 253–268.
Kawaguchi, Y. & Nakano, S. 2001. Contribution of terrestrial invertebrates to the
annual resource budget for salmonids in forest and grassland reaches of a
headwater stream. Freshwater Biology, 46: 303–316.
Kramer, D.L. & Mehegan, J.P. 1981. Aquatic surface respiration, an adaptative
response to hypoxia in the guppy, Poecilia reticulata (Pisces, Poeciliidae).
Environmental Biology of Fishes, 6: 299–313.
Lande, R. 1996. Statistics and partitioning of species diversity, and similarity among
multiple communities. Oikos, 76: 5–13.
Leibold, M. 1998. Similarity and local coexistence of species in regional biotas.
Evolution & Ecology, 12(1): 95–100.

88
Lorion, C.M. & Kennedy, B. P. 2009. Riparian forest buffers mitigate the effects of
deforestation on fish assemblages in tropical headwater streams. Ecological
Applications, 19: 468–479.
Melo, C.E., Machado, F.A. & Pinto-Silva, V. 2004. Feeding habits of fish from a stream
in the savanna of Central Brazil, Araguaia Basin. Neotropical Ichthyology, 2: 37–
44.
Mouillot, D., Dumay, O. & Tomasini, J.A. 2007. Limiting similarity, niche filtering and
functional diversity in coastal lagoon fish communities. Estuarine, Coastal and
Shelf Science, 71: 443–456.
Mouillot, D., Villéger, S., Scherer-Lorenzen, M. & Mason, N.W.H. 2011. Functional
structure of biological communities predicts ecosystem multifunctionality. PLoS
One, 6: e17476.
Murphy, M.L., & Hall, J.D. 1981. Varied effects of clear-cut logging on predators and
their habitat in small streams of the Cascade Mountains, Oregon. Canadian Journal
of Fisheries and Aquatic Sciences, 38: 137–145.
Pavoine, S. & Bonsall, M.B. 2011. Measuring biodiversity to explain community
assembly: a unified approach. Biological Reviews, 86: 702-812.
Pavoine, S., Dufour, A.B. & Chessel, D. 2004. From dissimilarities among species to
dissimilarities among communities: a double principal coordinate analysis. Journal
of Theoretical Biology, 228: 523–537.
Pavoine, S., Ollier, S. & Pontier, D. 2005. Measuring diversity from dissimilarities with
Rao’s quadratic entropy: are any dissimilarity indices suitable? Theoretical
Population Biology, 67: 231–239.
Pavoine, S., Love, M. & Bonsall, M. B. 2009a. Hierarchical partitioning of evolutionary
and ecological patterns in the organization of phylogenetically structured species
assemblages: application to rockfish (genus: Sebastes) in the Southern California
Bight. Ecology Letters, 12: 898–908.
Pavoine, S., Vallet, J., Dufour, A.-B., Gachet, S. & Daniel, H. 2009b. On the challenge
of treating various types of variables: application for improving the measurement
of functional diversity. Oikos, 118: 391–402.
Petchey, O.L. & Gaston K.J. 2006. Functional diversity: back to basics and looking
forward. Ecology Letters, 9: 741–758.

89
Poff, N.L. 1997. Landscape filters and species traits: towards mechanistic understanding
and prediction in stream ecology. Journal of the North American Benthological
Society, 16(2): 391–409.
R Development Core Team. 2009. R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria.
Rocha, F.C., Casatti, L., Carvalho, F.R. & Silva, A.M.S. 2009. Fish assemblages in
stream stretches occupied by cattail (Typhaceae, Angiospermae) stands in
Southeast Brazil. Neotropical Ichthyology, 7: 241–250.
Tilman, D., Lehman, C.L. & Thomson, K.T. 1997. Plant diversity and ecosystem
productivity: theoretical considerations. Proceedings of Natural Academy of
Sciences of the United States of America, 94: 1857–1861.
Villéger, S., Miranda, J.R., Hernández, D.F. & Mouillot, D. 2010. Contrasting changes
in taxonomic vs. functional diversity of tropical fish communities after habitat
degradation. Ecological Applications, 20: 1512–1522.
Warren, M.S., Hill, J.K., Thomas, J.A., Asher, J., Fox, R., Huntley,B., Roy, D.B.,
Telfer, M. G., Jeffcoate, S., Harding, P., Jeffcoate, G., Willis, S.G., Greatorex-
Davies, J.N., Moss D. & Thomas, C.D. 2001 Rapid responses of British butterflies
to opposing forces of climate and habitat change. Nature, 414: 65–69.
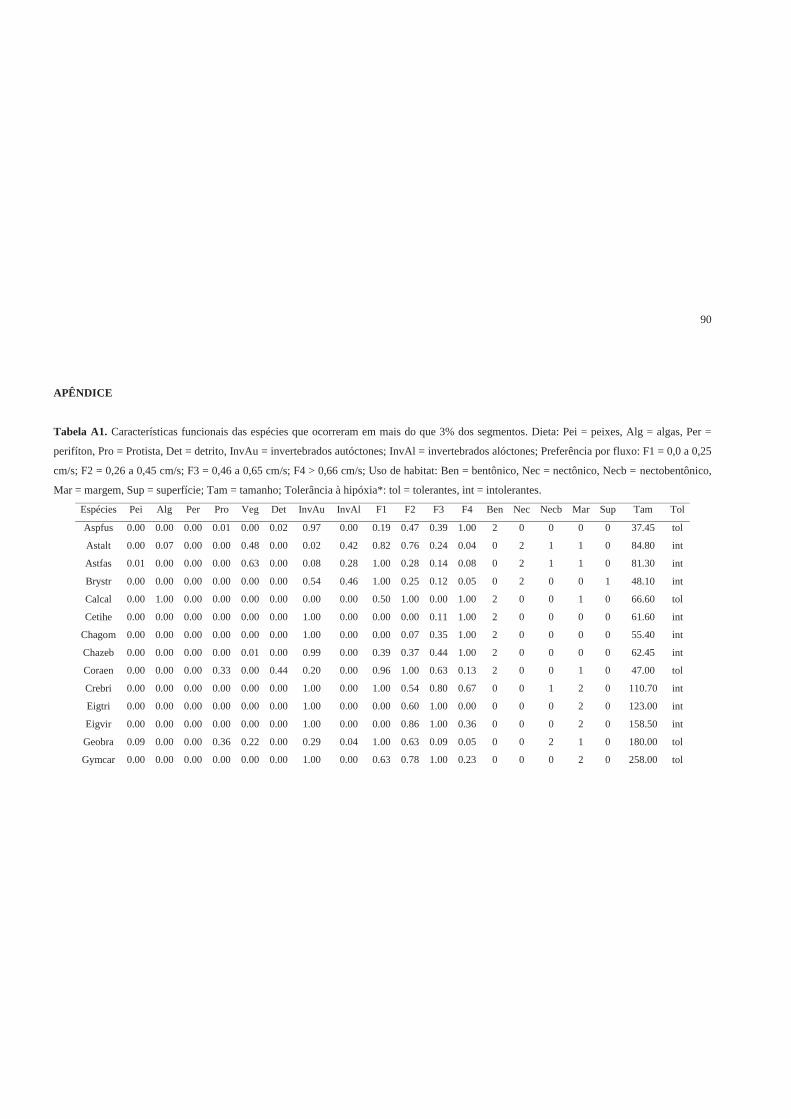
90
APÊNDICE
Tabela A1. Características funcionais das espécies que ocorreram em mais do que 3% dos segmentos. Dieta: Pei = peixes, Alg = algas, Per =
perifíton, Pro = Protista, Det = detrito, InvAu = invertebrados autóctones; InvAl = invertebrados alóctones; Preferência por fluxo: F1 = 0,0 a 0,25
cm/s; F2 = 0,26 a 0,45 cm/s; F3 = 0,46 a 0,65 cm/s; F4 > 0,66 cm/s; Uso de habitat: Ben = bentônico, Nec = nectônico, Necb = nectobentônico,
Mar = margem, Sup = superfície; Tam = tamanho; Tolerância à hipóxia*: tol = tolerantes, int = intolerantes.
Espécies Pei Alg Per Pro Veg Det InvAu InvAl F1 F2 F3 F4 Ben Nec Necb Mar Sup Tam Tol
Aspfus 0.00 0.00 0.00 0.01 0.00 0.02 0.97 0.00 0.19 0.47 0.39 1.00 2 0 0 0 0 37.45 tol
Astalt 0.00 0.07 0.00 0.00 0.48 0.00 0.02 0.42 0.82 0.76 0.24 0.04 0 2 1 1 0 84.80 int
Astfas 0.01 0.00 0.00 0.00 0.63 0.00 0.08 0.28 1.00 0.28 0.14 0.08 0 2 1 1 0 81.30 int
Brystr 0.00 0.00 0.00 0.00 0.00 0.00 0.54 0.46 1.00 0.25 0.12 0.05 0 2 0 0 1 48.10 int
Calcal 0.00 1.00 0.00 0.00 0.00 0.00 0.00 0.00 0.50 1.00 0.00 1.00 2 0 0 1 0 66.60 tol
Cetihe 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.00 0.00 0.11 1.00 2 0 0 0 0 61.60 int
Chagom 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.00 0.07 0.35 1.00 2 0 0 0 0 55.40 int
Chazeb 0.00 0.00 0.00 0.00 0.01 0.00 0.99 0.00 0.39 0.37 0.44 1.00 2 0 0 0 0 62.45 int
Coraen 0.00 0.00 0.00 0.33 0.00 0.44 0.20 0.00 0.96 1.00 0.63 0.13 2 0 0 1 0 47.00 tol
Crebri 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 1.00 0.54 0.80 0.67 0 0 1 2 0 110.70 int
Eigtri 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.00 0.60 1.00 0.00 0 0 0 2 0 123.00 int
Eigvir 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.00 0.86 1.00 0.36 0 0 0 2 0 158.50 int
Geobra 0.09 0.00 0.00 0.36 0.22 0.00 0.29 0.04 1.00 0.63 0.09 0.05 0 0 2 1 0 180.00 tol
Gymcar 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.63 0.78 1.00 0.23 0 0 0 2 0 258.00 tol

91
Hisfra 0.00 0.00 0.82 0.00 0.00 0.18 0.00 0.00 0.86 1.00 1.00 0.72 0 0 0 2 0 39.40 int
Hopmal 0.95 0.00 0.00 0.00 0.00 0.00 0.05 0.00 0.93 1.00 0.10 0.07 0 0 1 2 0 198.00 tol
Hypanc 0.00 0.00 0.77 0.00 0.15 0.08 0.00 0.00 0.95 0.71 0.65 0.69 2 0 0 1 0 91.10 tol
Hypnig 0.00 0.00 0.98 0.00 0.02 0.00 0.00 0.00 0.30 0.14 0.43 1.00 2 0 0 0 0 81.80 int
Impmir 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.13 0.27 0.33 1.00 2 0 0 1 0 83.90 int
Impsch 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.03 0.38 0.27 1.00 2 0 0 1 0 54.25 int
Knomoe 0.00 0.00 0.00 0.00 0.00 0.00 0.98 0.02 1.00 0.79 0.33 0.36 0 2 1 0 1 35.80 int
Lepfri 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 1.00 0.14 0.33 0.18 0 1 2 0 0 62.50 int
Olipin 0.00 0.00 0.00 0.00 0.00 0.00 0.97 0.00 1.00 0.49 0.30 0.00 0 1 0 2 0 52.00 int
Parnas 0.00 0.03 0.10 0.00 0.07 0.50 0.30 0.00 0.00 0.00 1.00 0.50 2 0 0 0 0 90.35 int
Phahar 0.00 0.00 0.00 0.00 0.00 0.56 0.44 0.00 0.22 1.00 0.80 0.36 0 0 0 1 2 32.10 tol
Piaarg 0.00 0.00 0.00 0.00 0.04 0.00 0.92 0.04 1.00 0.78 0.79 0.32 0 1 2 0 0 60.65 int
Pimava 0.02 0.00 0.00 0.00 0.00 0.00 0.91 0.00 0.90 0.31 0.74 0.15 2 0 0 0 0 90.400 int
Poeret 0.00 0.00 0.00 0.00 0.00 0.82 0.18 0.00 0.27 0.50 0.20 0.52 0 0 0 1 2 30.000 tol
Psepul 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.03 0.01 0.41 1.00 2 0 0 0 0 66.100 int
Rhaque 0.00 0.00 0.00 0.00 0.02 0.00 0.98 0.00 0.71 0.91 0.46 0.64 2 0 0 1 0 139.600 tol
Sernot 0.00 0.15 0.00 0.00 0.15 0.61 0.10 0.00 1.00 0.10 0.02 0.00 0 2 0 0 1 32.300 int
Steins 0.00 0.07 0.00 0.07 0.07 0.74 0.05 0.00 1.00 0.00 0.00 0.00 0 0 2 0 0 93.750 int
Stemac 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.00 1.00 0.00 0.00 0 0 0 2 0 202.000 int
Synmar 1.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.87 0.65 0 0 0 2 0 260.000 tol
Tatnei 0.00 0.00 0.00 0.00 0.00 0.00 0.00 1.00 1.00 0.75 0.33 0.33 0 0 0 1 2 57.600 int

92
* Segundo Kramer & Mehegan (1981), Araujo & Garutti (2003), Bozzetti & Schulz (2004), Casatti et al. (2006b), Rocha et al. (2009) e Casatti et
al. (2012).

93
DISCUSSÃO GERAL .
Existe certo consenso de que o desmatamento das zonas ripárias é uma das
principais ameaças à conservação da ictiofauna (NAIMAN; DÉCAMPS, 1997;
BARLETTA et al., 2010). Isso foi reforçado nos estudos apresentados aqui, onde foi
possível constatar que esse tipo de alteração afeta, além da diversidade taxonômica
(Capítulo II), outros componentes da biodiversidade, como a composição e diversidade
funcional (Capítulos II e III). As modificações que o desmatamento da zona ripária
provoca nos riachos representam filtros ambientais que limitam a ocorrência de grupos
de espécies especializadas, em especial as reofílicas (Capítulo III), enquanto favorece
outras espécies que são capazes de explorar as novas oportunidades que se tornam
disponíveis (Capítulo II e III).
As métricas utilizadas nos capítulos II e III capturam aspectos diferentes da
diversidade funcional. A dispersão funcional (FDis) é uma medida de diversidade
funcional que é ponderada pela abundância das espécies, sendo portanto, menos
sensível a presença ou ausência de características diferenciadas exibidas por espécies
raras do que outros índices, como por exemplo a riqueza funcional (FRic). FDis é um
índice que apresenta alta correlação com o índice de entropia quadrática de Rao
(LALIBERTÉ; LEGENDRE, 2010), o qual foi utilizado no capítulo III e, embora seja
calculado de forma distinta, também considera a abundância das espécies. Quando os
riachos florestados e desmatados foram comparados na escala de meso-hábitats
constatou-se que, apesar dos corredores e poços desmatados abrigarem maior número
de espécies funcionalmente distintas (maior FRic), a estrutura quantitativa (medida pelo
FDis) das suas comunidades foi similar à dos florestados. Isso, provavelmente, deve-se
a dominância de espécies nectônicas, limnofílicas e de dieta insetívora/generalista
(grupo 6 do capítulo III) nesses meso-hábitats, cuja ocorrência é pouco influenciada
pela cobertura florestal (Capítulo III).
Somente as corredeiras florestadas apresentaram estrutura quantitativa
diferenciada, com menores valores de FDis, indicando maior redundância funcional, o
que foi atribuído a maior restrição ambiental presente nesse meso-hábitat (Capítulo II),
representada pelo alto fluxo de água e a baixa variabilidade na disponibilidade de
alimento (ANGERMEIER; KARR, 1983). As espécies que ocupam as corredeiras
(grupo 3 da partição #30 do Capítulo III) apresentam especializações para lidar com
essas restrições (CASATTI; CASTRO, 1998), sendo altamente sensíveis à

94
descaracterização desses meso-hábitats. Embora as corredeiras estejam presentes nos
riachos desmatados (Capítulo II) sua distribuição é limitada, além de apresentarem
estrutura física com maior grau de alteração (e.g., sedimentação, atenuação do fluxo
pelas macrófitas marginais). Isso provavelmente explica o declínio desse grupo de
espécies quando os riachos desmatados foram considerados como um todo (Capítulo
III). Esses resultados sugerem que o comprometimento da estrutura física do hábitat
causado pelo desmatamento é uma importante fonte de alteração da estrutura funcional
das comunidades de riachos.
Os resultados obtidos no capítulo II, em especial o aumento da riqueza de
espécies e de FRic nos riachos desmatados reforçam outros estudos com um enfoque
taxonômico (LORION; KENNEDY, 2009; TERESA; CASATTI, 2010) e proporcionam
suporte à idéia de que maiores valores de diversidade não são necessariamente um
indicativo de integridade biológica. Uma explicação que é usualmente evocada quando
maiores valores de diversidade são encontrados em ambientes submetidos às
interferências antrópicas é a Hipótese do Distúrbio Intermediário (HDI) (CONNELL,
1978). Essa hipótese baseia-se na idéia de que comunidades submetidas a distúrbios
muito freqüentes são representadas por espécies tolerantes e oportunistas, enquanto que
em níveis baixos de distúrbio as espécies mais especializadas, com maior habilidade
competitiva prosperam sobre as primeiras. Em níveis intermediários de distúrbios as
comunidades são compostas por uma combinação de espécies de ambos os extremos e,
consequentemente, são mais diversas (CONNELL, 1978). Portanto, para a HDI ser uma
explicação para o padrão observado no capítulo II seria necessário assumir que os
riachos desmatados estudados representam um estágio intermediário de degradação e
que os maiores valores de diversidade devem-se à co-ocorrência de espécies de ambos
os extremos de degradação. De fato, apesar dos riachos desmatados apresentarem
elementos indicadores de degradação como menor proporção de substrato estável e
dominância de macrófitas no hábitat interno (CASATTI et al., 2009b), eles ainda
possuem variabilidade de meso-hábitats (poços, corredores e corredeiras) e substrato
estável em proporção significativa, que são elementos associados a ambientes em
melhor estado de conservação (CASATTI, 2010). Entretanto, para avaliar se a HDI
pode explicar os padrões observados seria necessário contemplar um espectro maior de
condições de desmatamento, incluindo também riachos em estado de degradação
extrema (sensu CASATTI, 2010). Dessa forma, seria possível avaliar se o padrão de
diversidade obtido no capítulo II representa um pico seguido por um declínio em

95
riachos mais degradados (compatível com a HDI), ou se representa uma tendência
linear de aumento da diversidade.
Os padrões de diversidade observados também podem ser interpretados dentro
de uma perspectiva de funcionamento dos ecossistemas. A retirada da vegetação ripária
e aumento da radiação solar, combinado à menor retenção de nutrientes provenientes do
ambiente terrestre (Brodie & Mitchell, 2005) podem criar condições propícias para o
aumento da produtividade (Bunn et al., 1999). Considerando que a diversidade
funcional correlaciona-se bem com o funcionamento dos ecossistemas (Tilman et al.,
1997), esse aumento no metabolismo geral dos riachos desmatados pode estar associado
ao aumento da diversidade observado nestes ambientes (e.g., FRic). Além da
produtividade, outros processos ecossistêmicos podem estar associados com as
mudanças na composição funcional provocadas pelo desmatamento. Por exemplo, a
retirada da vegetação ripária aumenta a sedimentação, modificando a composição do
substrato e reduzindo o fluxo (Rabeni & Smale, 1995; Casatti et al., 2006), o que por
sua vez pode resultar no favorecimento das espécies bentônicas menos especializadas
em detrimento de espécies reofílicas de hábitos mais especializados (Capítulo III).
A problemática envolvendo o desmatamento é de extrema relevância,
principalmente ao considerar que as eminentes mudanças na legislação ambiental
brasileira (revisão do Código Florestal: Projeto de Lei nº 1.876/1999) prevêem redução
da área florestada ao redor dos rios e riachos. Diversos estudos têm alertado para os
impactos desse tipo de alteração sobre diversos aspectos (Toledo et al., 2010; Tundisi &
Matsumura-Tundisi, 2010), inclusive sobre a conservação da ictiofauna (Casatti, 2010;
Magalhães et al., 2011). Os resultados obtidos nos estudos apresentados aqui permitem
antecipar que a intensificação do desmatamento, que em algumas regiões já atinge a
maioria dos cursos d’água (Casatti et al., 2006; Silva et al., 2007), pode resultar em
mudanças no pool funcional dentro de um contexto regional. Isso porque o
desmatamento provoca homogeneização funcional da ictiofauna dentro dos riachos
(Capítulo II) e supostamente entre riachos (Casatti et al., 2009b), por meio da limitação
da ocorrência de espécies funcionalmente distintas daquelas que são favorecidos neste
contexto de degradação (Capítulo III).

96
CONCLUSÕES .
Os resultados obtidos nos capítulos I, II e III complementam o conhecimento
acumulado a partir da utilização da abordagem taxonômica sobre os impactos do
desmatamento em comunidades de peixes tropicais e fornecem subsídios para uma
avaliação mais completa dos efeitos desse tipo de alteração sobre a biota aquática. Os
padrões de diversidade e composição funcional diferiram em função das alterações
provocadas nos riachos pelo desmatamento, corroborando a hipótese geral. As
características funcionais associadas com o uso de hábitat, ecologia trófica, tolerância à
hipóxia e tamanho influenciaram a distribuição das espécies dentro (entre meso-
hábitats) e entre riachos em contextos de degradação contrastantes. Nesse sentido, as
modificações na estrutura funcional das comunidades causadas pelo desmatamento
parecem ser mediadas pelos efeitos desse tipo de alteração sobre a disponibilidade de
hábitat e recursos alimentares, assim como nas condições físico-químicas (e.g.,
hipóxia), que por sua vez representam filtros ambientais importantes na organização das
comunidades.

97
REFERÊNCIAS GERAIS .
ALLAN, J. D.; FLECKER, A. S. Biodiversity conservation in running waters.
BioScience, v. 43, n. 1, p. 32–43, 1993.
ANGERMEIER, P. L.; KARR, J. R. Fish communities along environmental gradients
in a system of tropical streams. Environmental Biology of Fishes, v. 9, p. 117–135,
1983.
BARLETTA, M. et al. Fish and aquatic habitat conservation in South America: a
continental overview with emphasis on Neotropical systems. Journal of Fish Biology,
v. 76, p. 2118–2176, 2010.
Barragán, F. et al. Negative impacts of human land use on dung beetle functional
diversity. PLoS One, v. 6, p. e17976, 2011.
BOJSEN, B. H.; BARRIGA, R. Effects of deforestation on fish community structure in
Ecuadorian Amazon streams. Freshwater Biology, v. 47, p. 2246–2260, 2002.
BRODIE, J. E;& MITCHELL, A. W. Nutrients in Australian tropical rivers: changes
with agricultural development and implications for receiving environments. Marine
and Freshwater Research, v. 56, n. 3, p. 279–302, 2005.
BUNN, S. E.; DAVIES, P. M; MOSISCH, T. D. Ecosystem measures of river health
and their response to riparian and catchment degradation. Freshwater Biology, v. 41, p.
333–345, 1999.
CASATTI, L. Alterações no Código Florestal Brasileiro: impactos potenciais sobre a
ictiofauna. Biota Neotropica, v. 10, n. 4, p. 31-34, 2010.
CASATTI, L.; CASTRO, R. M. C. A fish community of the São Francisco river
headwater riffles, southeastern Brazil. Ichthyological Exploration of Freshwaters, v.
9, p. 229–242, 1998.
CASATTI, L. et al. Stream fish, water and habitat quality in a pasture dominated basin,
southeastern Brazil. Brazilian Journal of Biology, v. 66, p. 681–696, 2006.
CASATTI, L.; FERREIRA, C. P.; LANGEANI, F. A fish-based biotic integrity index
for assessment of lowland streams in southeastern Brazil. Hydrobiologia, v. 623, n. 1,
p. 173–189, 2009a.
CASATTI, L.; FERREIRA, C. P.; CARVALHO, F. R. Grass-dominated stream sites
exhibit low fish species diversity and dominance by guppies: an assessment of two
tropical pasture river basins. Hydrobiologia, v. 632, p. 273–283, 2009b.

98
DIAZ, S.; CABIDO, M. Vive la différence: plant functional diversity matters to
ecosystem processes. Trends in Ecology and Evolution, v. 16, n. 8, p. 646-655, 2001.
CONNELL, J. H. Diversity in tropical rain forest and coral reefs. Science, v. 199, p.
1302-1310, 1978.
ERNST, R.; LINSENMAIR, K. E.; RÖDEL, M. O. Diversity erosion beyond the
species level: dramatic loss of functional diversity after selective logging in two tropical
amphibian communities. Biological Conservation, v. 133, p. 143–155, 2006.
EROS, T. et al. Characterizing functional trait diversity and trait–environment
relationships in fish assemblages of boreal lakes. Freshwater Biology, v. 54, p. 1788–
1803, 2009.
FAUSCH, K. D.; NORTHCOTE, T. G. Large woody debris and salmonid habitat in a
small coastal British Columbia stream. Canadian Journal of Aquatic Sciences, v. 49,
n. 4, p. 682-693, 1992.
FLYNN, D. F. B. et al. Loss of functional diversity under land use intensification across
multiple taxa. Ecology Letters, v. 12, p. 22–33, 2009.
GREGORY, S. V. An ecosystem perspective of riparian zones. BioScience, v. 41, p.
540–551, 1991.
GROWNS, I. et al. A comparison of fish assemblages associated with different riparian
vegetation types in the Hawkesbury–Nepean river system. Fisheries Management &
Ecology, v. 10, p. 209–220, 2003.
LALIBERTÉ, E.; LEGENDRE, P. A distance-based framework for measuring
functional diversity from multiple traits. Ecology, v. 91, p. 299–305, 2010.
LORION, C. M. & KENNEDY, B. P. Riparian forest buffers mitigate the effects of
deforestation on fish assemblages in tropical headwater streams. Ecological
Applications, v. 19, p. 468–479, 2009.
LOWE-MCCONNELL, R. H. Estudos ecológicos de comunidades de peixes
tropicais. São Paulo: Edusp, 1999.
MAGALHÃES, A. L. B.; CASATTI, L.; VITULE, J. R. S. Alterações no código
florestal brasileiro favorecerão espécies não-nativas de peixes de água doce. Natureza
& Conservação, v. 9, n. 1, p. 121-124, 2011.
MOUILLOT, D. et al. Functional structure of biological communities predicts
ecosystem multifunctionality. PLoS One, v. 6, p. e17476, 2011.

99
MURPHY, M. L.; HALL, J. D. Varied effects of clear-cut logging on predators andtheir
habitat in small streams of the Cascade Mountains, Oregon. Canadian Journal of
Fisheries and Aquatic Sciences, v. 38, p. 137-145, 1981.
NAIMAN, R. J.; DÉCAMPS, H. The ecology of interfaces: riparian zones. Annual
Review of Ecology and Systematics, v. 28, p. 621–658, 1997.
PETCHEY, O. L.; GASTON, K. J. Functional diversity: back to basics and looking
forward. Ecology Letters, v. 9, p. 741–758, 2006.
PUSEY, B. J.; ARTHINGTON, A. H. Importance of the riparian zone to the
conservation and management of freshwater fish: a review. Marine and Freshwater
Research, v. 54, p 1–16, 2003.
RABENI, C. F.; SMALE, M. A. Effects on siltation on stream fishes and the potential
mitigating role of the buffering riparian zone. Hydrobiologia, v. 303, p. 211–219, 1995.
ROCHA, F. C. et al. Fish assemblages in stream stretches occupied by cattail
(Typhaceae, Angiospermae) stands in Southeast Brazil. Neotropical Ichthyology, v. 7
p. 241–250, 2009.
SCHNEIDER, K. N.; WINEMILLER, K. O. Structural complexity of woody debris
patches influences fish and macroinvertebrate species richness in a temperate
floodplain-river system. Hydrobiologia, v. 610, p. 235–244, 2008.
SILVA, A. M. Historical land-cover/use in different slope and riparian buffer zones in
watershed of the State of São Paulo, Brazil. Scientia Agricola, v. 64, p. 325–335, 2007.
SMA/IF (Secretaria do Meio Ambiente / Instituto Florestal). Inventário florestal da
vegetação natural do Estado de São Paulo. São Paulo: Imprensa Oficial do Estado de
São Paulo, 2005.
SWEENEY, B. W. et al. Riparian deforestation, stream narrowing, and loss of stream
ecosystem services. Proceedings of the National Academy of Sciences of the United
States of America, v. 101, p. 14132–14137, 2004.
TERESA, F. B.; CASATTI, L. Importância da vegetação ripária em região
intensamente desmatada no sudeste do Brasil: um estudo com peixes de riacho. Pan-
American Journal of Aquatic Sciences, v. 5, p. 444–453, 2010.
TILMAN, D.; LEHMAN, C. L.; THOMSON, K. T. Plant diversity and ecosystem
productivity: theoretical considerations. Proceedings of Natural Academy of Sciences
of the United States of America, v. 94, p. 1857–1861, 1997.
TOLEDO, L. F. et al. A revisão do Código Florestal Brasileiro: impactos negativos para
a conservação dos anfíbios. Biota Neotropica, v. 10, n. 4, p. 35–38, 2010.

100
TUNDISI, J. G.; MATSUMURA-TUNDISI, T.M. Impactos potenciais das alterações
do Código Florestal nos recursos hídricos. Biota Neotropica, v. 10, n. 4, p. 67–76,
2010.
VILLÉGER, S. et al. Contrasting changes in taxonomic vs. functional diversity of
tropical fish communities after habitat degradation. Ecological Applications, v. 20, p.
1512–1522, 2010.

101
Autorizo a reprodução xerográfica para fins de pesquisa.
São José do Rio Preto, 01/03/2012
Fabrício Barreto Teresa
![UNIVERSIDADE FEDERAL DE SANTA CATARINA Centro de ...Estrutura das comunidades de peixes de costão rochoso em ilhas costeiras do estado de santa Catarina [dissertação] / -Florianópolis,](https://img.document.onl/doc/110x75/611f9db7d88fcf7aa73ba2ff/universidade-federal-de-santa-catarina-centro-de-estrutura-das-comunidades-de.jpg)




























