Embed Size (px)
Citation preview

i
VIVIANE PANEGASSI PERIM
"EFEITOS DO ÔMEGA-3 EM MARCADORES DE
ESTRESSE OXIDATIVO EM MÚSCULOS
DISTRÓFICOS DO CAMUNDONGO MDX"
Campinas, 2015

ii

iii

iv

v

vi

vii
RESUMO
A Distrofia Muscular de Duchenne (DMD) é uma doença causada pela ausência da proteína
distrofina e se caracteriza por degeneração muscular progressiva. A deficiência da distrofina
na DMD e nos camundongos mdx, modelo experimental da DMD, promove mionecrose. O
processo inflamatório que se instala exacerba a mionecrose e aumenta o estresse oxidativo. O
estresse oxidativo tem sido proposto como um fator importante para a progressão da doença.
As espécies reativas de oxigênio promovem danos nas fibras distróficas, comprometendo
proteínas e lipídios da membrana, gerando grandes quantidades de lipofuscina e 4-HNE,
indicadores de estresse oxidativo. Este trabalho tem como objetivo verificar se o ômega-3
diminui o estresse oxidativo em diferentes músculos distróficos (bíceps, diafragma e
quadríceps) do camundongo mdx jovem. Observamos que o ômega-3 diminuiu a lipofuscina
e o 4-HNE nos músculos estudados. A atividade das enzimas antioxidantes SOD, GPx e GR
estava aparentemente diminuída no diafragma distrófico, quando comparado ao diafragma
normal. No diafragma, o ômega-3 promoveu aumento discreto da atividade da SOD e da GPx.
O músculo quadríceps não apresentou alterações significativas destas enzimas, tanto no mdx
controle, quanto no mdx tratado com ômega-3. O ômega-3 promoveu melhora da
distrofinopatia (redução da CK, diminuição da inflamação e aumento da regeneração
muscular). Concluímos que o ômega-3 diminui o estresse oxidativo (reduz a lipofuscina e o
4-HNE), principalmente no músculo diafragma, provavelmente por este ser mais afetado que
o quadríceps. A atividade aparentemente reduzida das enzimas antioxidantes no diafragma do
mdx controle pode sugerir menor capacidade de tamponamento das espécies reativas de
oxigênio neste músculo. Isto pode contribuir, pelo menos em parte, para o maior estresse
oxidativo e maior acometimento do diafragma em relação ao quadríceps. Estes dados sugerem
que o ômega-3 possa ter uma ação positiva em melhorar o estresse oxidativo em diferentes
músculos distróficos, tornando-o potencialmente útil para a terapia da DMD.

viii

ix
ABSTRACT
Duchenne muscle dystrophy (DMD) is a disease caused by the absence of dystrophin
characterized by progressive muscle degeneration. In DMD and in the mdx mice model of
DMD, the inflammatory process exacerbates myonecrosis and increases oxidative stress.
Oxidative stress has been proposed as an important factor in disease progression. Reactive
oxygen species promote damage in dystrophic fibers, affecting proteins and lipids of the
membrane, generating large amounts of lipofuscin and 4-HNE, markers of oxidative stress.
This study aims to determine whether ômega-3 therapy reduces oxidative stress in different
dystrophic muscles (biceps, diaphragm and quadriceps) of mdx mouse, at earlier stages of the
disease. Ômega-3 decreased lipofuscin and 4-HNE in all the muscles studied. The activity of
the antioxidant enzymes SOD, GPx and GR was apparently reduced in mdx diaphragm control
as compared to normal mice. In the diaphragm, ômega-3 promoted a slight increase in the
activity of SOD and GPx. The quadriceps muscle showed no significant changes in the activity
of these enzymes in mdx control, and ômega-3 did not change this profile. Ômega-3 also
ammeliorated dystrophinopathy (reduced CK, decreased inflammation and increased muscle
regeneration). We conclude that ômega-3 reduces oxidative stress (by decreasing lipofuscin
and 4-HNE), mainly in the dystrophic diaphragm, possibly because this muscle is more
affected than the quadriceps. The apparent decreased activity of antioxidant enzymes in the
mdx diaphragm may suggest a poor capacity of this muscle to buffer the reactive oxygen
species. This may contribute, at least in part, to the increased oxidative stress in the diaphragm
and to the fact that this muscle will turn out to be the more affected muscle in the mdx. The
present results suggest that ômega-3 may have a positive effect in improving oxidative stress
in different dystrophic muscles, making it potentially useful for DMD therapy.

x

xi
SUMÁRIO
Resumo..................................................................................................................................vii
Abstract..................................................................................................................................ix
Abreviaturas......................................................................................................................xxiii
1. Introdução.........................................................................................................................01
1.1 Distrofia Muscular de Duchenne......................................................................................01
1.2 Camundongo mdx.............................................................................................................02
1.3 Degeneração muscular......................................................................................................03
1.4 Regeneração muscular......................................................................................................04
1.5 Ômega-3...........................................................................................................................06
1.6 Espécies Reativas de Oxigênio e Estresse Oxidativo.......................................................09
1.7 Sistema antioxidante.........................................................................................................12
1.8 Espécies Reativas de Oxigênio e a Distrofia Muscular de Duchenne..............................14
1.9 Lipofuscina.......................................................................................................................16
2.Objetivo..............................................................................................................................20
2.1 Objetivos específicos........................................................................................................20
3. Materiais e Métodos.........................................................................................................20
3.1 Animais.............................................................................................................................20
3.2 Protocolo experimental.....................................................................................................21
3.3 Procedimento cirúrgico para obtenção dos músculos.......................................................21
3.4 Medida da força................................................................................................................22
3.5 Análise de Creatina Quinase (CK) no plasma sanguíneo.................................................22
3.6 Preparo e análise dos músculos para a técnica de Hematoxilina e Eosina.......................23
3.7 Preparo e análise dos músculos para análise da lipofuscina.............................................23
3.8 Western Blotting...............................................................................................................24
3.9 Determinação da atividade enzimática da GPx, GR e SOD.............................................25
4. Análise Estatística...............................................................................................................26

xii
5.Resultados...........................................................................................................................27
5.1 Massa corporal..................................................................................................................27
5.2 Medida de força.................................................................................................................27
5.3 Análise da CK (U/L).........................................................................................................28
5.4 Análise morfológica..........................................................................................................29
5.4.1 Análise qualitativa das fibras musculares (HE).................................................29
5.4.2 Análise quantitativa das fibras musculares (HE):..............................................33
5.5 Análise qualitativa e quantitativa dos grânulos de lipofuscina.........................................34
5.6 Níveis de 4-HNE (Western Blot)......................................................................................39
5.7 Atividade Enzimática........................................................................................................41
5.7.1 Atividade Enzimática da SOD...........................................................................41
5.7.2 Atividade Enzimática da GPx e da GR..............................................................41
6. Discussão............................................................................................................................43
6.1 Peso corporal.....................................................................................................................44
6.2 Creatina Quinase (CK) e Força de Tração........................................................................44
6.3 Inflamação e Regeneração nos músculos BB, DIA e QDR..............................................45
6.4 Sobre a análise da Lipofuscina..........................................................................................46
6.5 Sobre a análise do 4-HNE.................................................................................................48
6.6 Sobre a atividade enzimática da SOD, GPx e GR............................................................49
7. Conclusões.........................................................................................................................53
8. Referências Bibliográficas................................................................................................54
9. Comissão de ética: certificado..........................................................................................65
10. Anexo................................................................................................................................66

xiii
Dedico...
Ao Danilo, meu amor, pelo privilégio de ter você ao meu lado. Agradeço a Deus por
tê-lo colocado em minha vida. Muito obrigada pelo seu amor, respeito, confiança e apoio
incondicional em todas as alegrias e tristezas do meu caminho. Te amo!!!

xiv

xv
Agradeço...
Aos meus pais José Carlos e Cristina e a minha irmã Vanessa, pelo amor e pelo
exemplo de honestidade e dignidade, que uma família deve ser. Muito obrigada pela honra
de fazer parte desta família! Amo vocês!
À família Armellini, levo no meu coração todo carinho, dedicação, amor e orações
de vocês, que são essenciais no meu desenvolvimento pessoal e profissional. Muito obrigada!
À Deus, por tudo que acontece em minha vida. Nunca sei o que ele tem para dar,
mas Ele conhece meu coração, meus medos e minhas necessidades.
“Você pode fazer muito pouco com a fé. Mas você não pode fazer nada sem ela.”
(Samuel Butler)

xvi

xvii
Agradecimento especial
À Professora Dra. Maria Júlia Marques, exemplo de dedicação e inteligência, por
ter aberto as portas desta Universidade, pela compreensão e apoio em um dos momentos mais
difíceis da minha vida e por ter acreditado neste trabalho. Muito obrigada!

xviii

xix
AGRADECIMENTOS
Ao Programa de Pós-graduação em Biologia Celular e Estrutural pela contribuição no
desenvolvimento profissional.
Ao Prof. Dr. Humberto Santo Neto pela colaboração durante a realização deste trabalho.
À Profa. Dra. Elaine Minatel pelos ensinamentos e sugestões compartilhados como
coorientadora e por disponibilizar seu laboratório para a realização de muitos experimentos
essenciais para este trabalho;
Aos Professores Doutores Edson Rosa Pimentel, Ana Cristina Prado Veiga-Menoncello
e Heidi Dolder pelas valiosas considerações no exame de qualificação.
Aos Professores Doutores Marcondes Cavalcante Franca Junior, Leonardo dos Reis e
Marcelo Cunha pelas importantes considerações na pré-banca de mestrado.
À Sra. Liliam Alves Senne Panagio pela atenção, auxílio e prontidão durante o mestrado.
Aos funcionários do Departamento de Anatomia, Sr. Norivaldo Celestino, Sr. Marco
Aurélio Ribeiro de Paula, Sr. Paulo Afonso Bernardes, Sr. Paulo Francisco dos Santos,
Sr. Toni Donizeti dos Santos, Sra. Marlene Lima Francisco, Sr. Walter, Srta. Stella
Maris Fick de Ferraz e Srta. Érika da Silva Campos pela prestatividade ao longo desses
dois anos.
Aos amigos do laboratório Adriana Fogagnolo Maurício, Luana Marcela, Túlio Hermes,
Juliano Alves Pereira, Samara Carvalho, Drielen de Oliveira e à Dra. Cíntia
Matsumura, pela amizade e contribuição para a realização dos experimentos e da elaboração
deste trabalho.
Ao Dr. Felipe Meira de Faria por ensinar e compartilhar informações para a realização de
técnicas utilizadas neste trabalho.

xx
Ao CNPq, CAPES, CAPES/PROEX, CAPES/DS e FAPESP pelo apoio financeiro à
pesquisa.
À todos os amigos, colegas docentes e funcionários que contribuíram de alguma forma para
a realização deste trabalho.

xxi
“Leve na sua memória para o resto de sua vida as coisas boas que surgiram no meio das
dificuldades. Elas serão uma prova de sua capacidade em vencer as provas e lhe darão
confiança na presença divina, que nos auxilia em qualquer situação, em qualquer
tempo, diante de qualquer obstáculo.”
(Chico Xavier)

xxii

xxiii
ABREVIATURAS
AA – Ácido Araquidônico
AG – Ácidos graxos
AGE – Ácidos graxos essenciais
BB – Músculo bíceps braquial
CAT – Enzima catalase
COX – Cicloxigenase
CDG – Complexo distrofina-glicoproteínas
CK- Creatina Quinase
ClC3 – Canal-3 de cloreto
DHA – Ácido Docosahexaenóico
DIA – Músculo diafragma
DMD – Distrofia Muscular de Duchenne
DNA – Ácido desoxirribonucléico
EPA – Ácido Eicosapentaenóico
GPx – Glutationa peroxidase
GR – Glutationa redutase
GSH – Glutationa reduzida
GSSH – Glutationa oxidada
4-HNE – 4-hidroxinonenal
HE – Hematoxilina-eosina
IL-1 – Interleucina 1
INFL/REG – Inflamação e Regeneração
LOX – Lipoxigenase

xxiv
mdx – Murine dystrophin X-liked
NADPH – Nicotinamida Adenina-dinocleotídeo Fosfato
NO – Óxido nítrico
nNOS – Óxido nítrico sintase neural
NF-kB – Fator Nuclear kappa B
NC – Núcleo central
NP - Núcleo periférico
PBS – Tampão fosfato salina
PUFA – Ácido graxo poli-insaturado
PLA2 – Fosfolipase A2
QDR – Músculo quadríceps femoral
RL – Radicais livres
ROS – Reactive oxigen species
SAC – stretch-activated calcium chanels
SOD – Superóxido dismutase
SOD1 – Superóxido dismutase citosólica
SOD2 – Superóxido dismutase mitocondrial
TNF-α – Fator de Necrose Tumoral alfa

1
1. Introdução
1.1 Distrofia Muscular de Duchenne
As distrofias musculares constituem um grupo de doenças genéticas e hereditárias que se
caracterizam pela degeneração muscular progressiva.
Entre os diversos tipos de distrofinopatias destaca-se a Distrofia Muscular de Duchenne
(DMD), considerada a mais comum e devastadora forma de distrofia muscular (ENGEL et al.,
1994; FAIRCLOUGHF et al, 2011). Afeta cerca de um a cada 3.500 meninos nascidos vivos
e está associada à fraqueza muscular esquelética e cardíaca (BIGGAR et al., 2006). A Distrofia
Muscular de Duchenne é caracterizada por degeneração muscular progressiva, causada por
mutação no cromossomo X, no gene que expressa a proteína distrofina (HOFFMAN et al.,
1987).
A distrofina é uma proteína do citoesqueleto, cuja função é manter a estabilidade da
membrana da célula muscular ou sarcolema (DECONINCK & DAN, 2006). Com a ausência
da distrofina, a fibra muscular perde suas características morfológicas e funcionais e ocorre
saída de enzimas do músculo para o plasma sanguíneo, como por exemplo, a creatina-quinase
(CK) (STRAUB et al., 1997). No paciente distrófico, a quantificação desta enzima no plasma,
sendo elevada, auxilia no diagnóstico da DMD.
Os sinais clínicos aparecem em torno dos três aos seis anos de idade, caracterizados pela
fraqueza muscular (BIGGAR et al., 2006). Os meninos começam a cair com facilidade, têm
dificuldade em subir as escadas, correr, levantar-se do chão (ENGEL et al.,1994). Cansam-se
rapidamente, passam a caminhar na ponta dos pés e se levantam com a ajuda das mãos. Entre
os 7-12 anos de idade ocorre comprometimento da ambulação e o paciente passa a utilizar
cadeira de rodas (CHAKKALAKAL et al., 2005). Com o déficit muscular cada vez maior, o
paciente passa a ter, também, comprometimento da musculatura cardíaca e respiratória,
levando à morte dos pacientes entre a segunda e terceira década de vida por falência
cardiorrespiratória (ENGEL et al.,1994; BOGDANOVICK et al., 2004).
Para o tratamento são utilizados corticosteróides, como a prednisona e o deflazacort. Por
um lado, o uso dessas substâncias traz melhora significativa no estado clínico do paciente, ao
preservar a função cardíaca e pulmonar e retardar o aparecimento da escoliose (BIGGAR et.
al., 2012). Porém, por outro lado, com o tempo, os efeitos colaterais se sobressaem, como a

2
baixa estatura, osteoporose, aumento do peso, problemas comportamentais e endócrinos, que
afetam negativamente a qualidade de vida do paciente (ENGEL et al.,1994).
Portanto, os estudos têm se voltado na busca de fármacos que possam substituir, ou pelo
menos ser uma opção alternativa aos corticoides, fármacos estes que retardem a evolução da
doença e que permitam uso de doses menores de corticoides proporcionando diminuição dos
seus efeitos indesejados.
1.2 Camundongos mdx
Como modelo experimental para os estudos das distrofinopatias utiliza-se o camundongo
mdx (sigla em inglês para distrofia muscular com mutação no cromossomo X) (GROUNDS
et al., 2008). Este apresenta mutação gênica espontânea para a distrofina, a qual é ausente no
músculo, com elevado nível plasmático de creatina-quinase (CK) e degeneração muscular
(BULFIELD et al., 1984). Características que também ocorrem nos pacientes distróficos. Por
outro lado, os camundongos mdx apresentam ciclos sucessivos de regeneração muscular,
diferentemente dos pacientes com DMD. Acredita-se que isto seja devido ao fato destes
animais possuírem capacidade de regeneração eficiente, compensando os ciclos de
degeneração. Como resultado, a distrofia dessa linhagem de camundongos é menos severa se
comparada a DMD (GROUNDS et al., 2008).
A partir dos vinte e um dias de vida pós-natal, os músculos estriados esqueléticos do mdx
começam a degenerar, sendo observados ciclos de degeneração e regeneração. O pico de
degeneração muscular ocorre entre o vigésimo primeiro e o trigésimo dia de vida. No músculo
quadríceps, observa-se com 30 dias, até 60% das fibras em processo de regeneração ou
regeneradas (MARQUES et al., 2008). Em torno de seis meses de idade, alguns músculos,
tais como o tibial anterior e o sóleo, apresentam-se com poucas áreas em degeneração e
próximos da completa regeneração, caracterizados pela presença de 80-90% de suas fibras
com núcleo central (PASTORET e SEBILLE, 1995). Ao redor dos nove meses de idade,
alguns músculos, como o diafragma e coração, perdem a capacidade regenerativa, levando ao
aparecimento de fibrose. No mdx idoso, a partir de 01 ano de idade ocorre decréscimo
significativo da capacidade de regeneração muscular e, consequentemente, a degeneração e a
fibrose tornam-se predominantes. Nesta fase, a patologia no mdx se aproxima daquela
observada na distrofia humana (LEFAUCHEUR et al., 1995).

3
1.3 Degeneração muscular
O sarcolema possui canais de cálcio, os quais são essenciais para o processo de contração
e relaxamento muscular. Na distrofia muscular, devido à ausência da distrofina, o sarcolema
torna-se instável. A instabilidade mecânica, associada a alterações no funcionamento dos
canais de cálcio, contribui para a degeneração do músculo distrófico (WHITEHEAD et al.,
2006).
A distrofina é uma proteína encontrada na face citoplasmática do sarcolema ligada aos
miofilamentos e às glicoproteínas de membrana, formando o complexo distrofina-
glicoproteínas (CDG - Figura1), que conecta o citoesqueleto da fibra à matriz extracelular
(DECONINCK & DAN, 2006; PALMA et al., 2014). Esta proteína é fundamental para a
integridade do CDG e do sarcolema durante o processo de transferência da força contrátil
gerada pelas proteínas contráteis para o meio extracelular (RANDO, 2001; PALMA et al.,
2014). A ausência da distrofina leva à alteração da interação do complexo com a matriz e com
o citoesqueleto, tornando o sarcolema instável (DECONINCK & DAN, 2006). Além disso, o
sarcolema sofre pequenas rupturas, decorrentes da força contrátil do sarcômero no processo
de contração e relaxamento (GROUNDS et al., 2005). A ruptura da membrana, somada à sua
instabilidade, faz com que haja perda de conteúdo intracelular da fibra muscular para o meio
extracelular caracterizando a degeneração do músculo (Figura 2).
Figura 1: Esquema do complexo distrofina-glicoproteínas representando a α e β distroglicanas; α, β, γ, δ
sarcoglicanas; sintrofinas, distrobrevina e a distrofina. A distrofina exerce papel importante na
estabilização do sarcolema conectando a F-actina citoesquelética à cadeia da laminina, localizada na
lâmina basal via complexo de glicoproteínas.

4
O CDG também participa da manutenção dos canais de cálcio da membrana, sendo que
com a ausência da distrofina ocorre funcionamento anormal dos mesmos, provocando influxo
aumentado de cálcio na fibra muscular (WHITEHEAD et al., 2006). A entrada de cálcio se dá
através de diversos canais: voltagem-dependente; canais ativados por alongamento (stretch-
activated calcium channels – SAC) e do tipo leak (WHITEHEAD et al., 2006).
O aumento do influxo de cálcio gera uma série de reações, tais como ativação de proteases
endógenas (exemplo: calpaína) (VERCHERAT et al., 2009), ativação de enzimas
(fosfolipases A2) responsáveis pela digestão dos fosfolípidios da membrana e a produção das
espécies reativas de oxigênio (ROS) na mitocôndria (WHITEHEAD et al., 2006). Tais
processos influem na inflamação e na mionecrose das fibras musculares. No paciente
distrófico, com o tempo, as fibras musculares são substituídas por gordura e tecido conjuntivo
e o músculo perde sua função contrátil.
1.4 Regeneração muscular
Na regeneração ocorre formação de novas fibras musculares - processo conhecido como
miogênese. Neste processo, células satélites, que são células mononucleadas e indiferenciadas
presentes entre a lâmina basal e o sarcolema das fibras musculares (FOSCHINI et al., 2004),
proliferam e se diferenciam em mioblastos, os quais expressam miogenina, originando os
miotubos. Os miotubos, devido ao aumento da expressão e da ação da miogenina, se fundem
e se diferenciam, originando as fibras musculares. Tais células são ditas “dormentes”, pois são
acionadas somente quando o músculo sofre algum tipo de dano (BASSOLI, 2003).
Quando o tecido muscular é lesionado, ocorre morte das fibras musculares e em seguida
infiltração de células inflamatórias como neutrófilos, monócitos e macrófagos. Na inflamação,
tais células fagocitam as fibras musculares necróticas, com a finalidade de reparação (PRISK
& HUARD, 2003).
A inflamação é característica marcante nos músculos distróficos e contribui para o avanço
da doença. Em pacientes com DMD e no camundongo mdx, as fibras em degeneração liberam
várias substâncias, dentre as quais citocinas pró-inflamatórias e fatores quimiostáticos
(TIDBALL, 2005). Juntos, eles estimulam a migração de células inflamatórias,
principalmente macrófagos e neutrófilos, para o local da lesão. Macrófagos e neutrófilos
liberam proteases que degradam os fragmentos celulares e os fagocitam. Estas proteases

5
podem provocar danos ao tecido íntegro que circunda a lesão e aumentar a área lesada,
aumentando a resposta inflamatória local (SPENCER & TIDBALL, 2001; TIDBALL, 2005).
Como exemplo de mediador químico pode-se citar o NF-kB (Fator Nuclear kappa B) e a
citocina TNF-α (Fator de Necrose Tumoral alfa). O TNF-α é uma citocina pró-inflamatória
que se encontra em grande concentração no sangue dos pacientes distróficos. Estudos têm
demonstrado que altas concentrações de TNF-α induzem a necrose celular e a apoptose
(MAGGE et al, 2008). A fase proliferativa é caracterizada pela proliferação dos fibroblastos
e das células satélites. Os primeiros têm a finalidade de depositar uma nova matriz extracelular
e as células satélites têm a finalidade de gerar novas fibras musculares. Macrófagos da fase
inflamatória estimulam as células satélites a se dividirem. Após se proliferarem, estas se
diferenciam em células precursoras miogênicas, os mioblastos. Em seguida, os mioblastos
pela expressão da miogenina, originam os miotubos que se alinham na lâmina basal para
reestruturar a fibra muscular. (FAGANELLO, 2003; FERRARI, 2005; PINTO & CASTILLO,
1999). Por fim, se diferenciam, originando as miofibras multinucleadas (DAWHAN &
RANDO, 2005; LE GRAND & RUDNICKI, 2007).
Nos pacientes portadores da DMD, ainda no início da infância, a capacidade de
regeneração muscular pelas células satélites diminui e com o tempo, a degeneração das fibras
Figura 2: Esquema representando hipoteticamente o mecanismo que resulta em inflamação e necrose da
fibra muscular distrófica.

6
torna-se predominante e o músculo passa a ser substituído pelo tecido fibroadiposo (ENGEL,
1994). Já nos camundongos mdx, a capacidade de regeneração é menor quando ficam idosos,
pois o número de células satélites diminui e, consequentemente, há diminuição da capacidade
regenerativa do músculo (PASTORET & SEBILLE, 1995).
A distrofia tem como causa primária a ausência da distrofina. Entretanto, deve-se ressaltar
que a inflamação e o comprometimento da regeneração são fatores que agravam e contribuem
para a progressão da doença (ABDEL-SALAM et al., 2009; PALMA et al., 2014).
1.5 Ômega-3
Os ácidos graxos (AG) são classificados de acordo com o tamanho da cadeia carbônica
(quantidade de átomos de Carbono), o grau de saturação (número de duplas ligações) e a
posição da primeira dupla ligação de Carbonos (CALDER, 2012). Os AG poli-insaturados
(PUFAs) possuem duas ou mais duplas ligações em suas moléculas e são representados pelos
ácidos graxos essenciais (AG linoléico e linolênico), o ácido eicosapentaenóico (EPA) e
docosahexaenóico (DHA). Os ácidos graxos essenciais (AGE) não são produzidos pela
espécie humana e portanto devem ser adquiridos por fontes dietéticas. Dos ácidos graxos
essenciais, o AG linoléico é conhecido como ômega-6 e o AG linolênico, como ômega-3
(CALDER, 2008).

7
Os PUFAs do tipo ômega-3 e ômega-6 são classificados como de cadeia longa. O ômega-
3 possui 14 a 22 átomos de carbono, mais de uma dupla ligação e recebe a denominação
ômega-3 por conter a primeira dupla ligação no carbono 3 a partir do radical metil (CALDER,
2012). O ômega-6 possui dezoito átomos de carbonos na cadeia e tem duas duplas ligações,
sendo a primeira dupla ligação localizada no sexto carbono a partir do radical metil
(CALDER, 2002) (Figura 3).
Os AGE são encontrados nos vegetais (CALDER, 2012). O linoléico é encontrado, por
exemplo, no milho, no girassol e também na carne bovina e produtos lácteos, enquanto o
linolênico pode ser encontrado na soja e na semente de linhaça. EPA e DHA são encontrados
em maior quantidade em óleos de peixes marinhos, como por exemplo, na cavala, na sardinha
e no salmão (CALDER, 2012).
Os PUFAs participam da composição das membranas celulares e a incorporação de AGE
pode determinar alterações estruturais (ação direta na superfície de membrana ou nos
receptores intracelulares dos ácidos graxos) e funcionais (correto funcionamento das proteínas
de membrana, fluidez e cascatas de sinalização intracelulares) da membrana fosfolipídica
(CALDER, 2012) e assim influenciar processos biológicos importantes, como a síntese de
mediadores e reguladores inflamatórios que incluem os eicosanoides (LO et al., 1999;
CALDER, 2012; CALDER, 2013)
Com exceção dos eritrócitos, todas as células dos mamíferos produzem os eicosanoides,
os quais tem como característica funcionarem no mesmo local onde são produzidos e de
causarem profundos efeitos fisiológicos em pequenas concentrações. AG ômega-3 e ômega-
6 são precursores dos eicosanoides reguladores das funções inflamatória e imune (CALDER,
2013). A diferença está no fato de que, os eicosanoides provenientes do metabolismo dos
PUFA ômega-6 são potentes mediadores inflamatórios e os do PUFA ômega-3 são potentes
anti-inflamatórios (CALDER, 2002). A via de síntese dos eicosanoides é realizada pelas
enzimas cicloxigenase (COX) e lipoxigenase (LOX), as quais produzem respectivamente,
prostanóides (tromboxanos, prostaglandinas), leucotrienos e lipoxinas (Figura 4)
(SIMOPOULOS, 2002). Os eicosanóides provenientes do metabolismo do PUFA ômega-6,
particularmente o ácido araquidônico, são da série par, ou seja, a quantidade de duplas ligações
presentes em cada molécula é número par. Já o ácido graxo alfa-linolênico (ômega-3) pode
ser convertido em ácido eicosapentaenóico (EPA) e docosahexaenóico (DHA) (CALDER,

8
2012). Estes competem com o AA pelas vias enzimáticas da COX e da LOX e também
formam eicosanóides, porém da série ímpar (quantidade de duplas ligações presentes em cada
molécula é número ímpar) (SIMOPOULOS, 2002; CALDER, 2002). Os eicosanóides da
classe ímpar, produzidos pelos PUFA ômega-3 têm menor poder inflamatório que os da classe
par produzido pelos PUFA ômega-6.
Além da capacidade do ácido graxo poli-insaturado ômega-3 em antagonizar a produção
de eicosanóides derivados do ácido graxo poli-insaturado ômega-6 refletindo seu poder anti-
inflamatório, o PUFA ômega-3 também pode modular a produção de citocinas (CALDER,
2002; CALDER, 2013).
A ingestão excessiva de ácidos graxos ômega-6, além de aumentar os eicosanóides,
principalmente as prostaglandinas pró-inflamatórias, também aumenta a produção de
citocinas (CALDER, 2013). As citocinas são secretadas pelos monócitos ou macrófagos e
estimulam outros tipos de células inflamatórias (linfócitos) e teciduais (músculo esquelético
e tecido adiposo) (CALDER, 2002). Dentre suas funções, a que se destaca para este trabalho
é o fato das citocinas serem mediadoras de respostas inflamatórias. A interleucina 1 (IL-1)
inclui-se no grupo das citocinas, cuja principal atividade é justamente ser mediadora da
inflamação. Além disso, ela está associada ao Fator de Necrose Tumoral Alfa (TNF-α), que,
juntos, são os principais mediadores da inflamação (SIMOPOULOS, 2002).
Vários estudos têm demonstrado que o uso de ácidos graxos ômega-3 conseguiram inibir
ou reduzir a produção de fator de necrose tumoral alfa (TNF-α) e de interleucinas, como por
exemplo, da interleucina 1 (IL-1) após estimulação de monócitos, macrófagos e linfócitos
(ENDRES et al., 1989; SIMOPOULOS, 2002). No caso do TNF-α, o PUFA ômega-3 parece
inibir o deslocamento do NF-kB, presente no citoplasma, para o núcleo da célula e, dessa
forma, também inibe a síntese do TNF-α (BABCOCK et al, 2000; XI et al., 2001; CALDER,
2013). Portanto, os ácidos graxos ômega-3 no óleo de peixe interferem não só na via
metabólica dos eicosanoides, mas também na produção das citocinas (CALDER, 2002;
CALDER, 2013). Essa ação conjunta acarreta na diminuição da inflamação. Desta forma,
pacientes que tomaram suplementos de ácidos graxos ômega-3 tiveram seus sintomas
inflamatórios amenizados (SIMOPOULOS, 2002).
Por fim, estudos têm demonstrado que a ingestão de ácidos graxos pode alterar a tensão
da membrana fosfolipídica e, consequentemente, modificar a atividade dos canais iônicos

9
(JUDÉ et al., 2005). Logo, os PUFAs ômega-3 (EPA e DHA) interagem diretamente com os
canais iônicos, bloqueando-os e alterando a entrada de cálcio (JUDÉ et al., 2005).
1.6 Espécies Reativas de Oxigênio e Estresse Oxidativo
Radicais livres (RL) constituem um grupo de substâncias químicas que possuem um ou
mais elétrons desemparelhados em uma de suas órbitas externas (DROGE, 2002;
HALLIWELL, 1991; CLARKSON & THOMPSON, 2000). Tal fato gera instabilidade
química e reatividade aos RL, lhes permitindo captar (oxidantes) ou ceder (redutores) elétrons
e/ou átomos de hidrogênio para outras moléculas que os circundam (GOMEZ-CABRERA et
al., 2005; CLARKSON & THOMPSON, 2000). Deste modo, elevadas concentrações de RL
podem induzir alterações graves na estrutura de moléculas que são fundamentais na
manutenção da homeostasia celular, o que pode resultar em uma possível perda de
funcionalidade ou viabilidade da célula (HALLIWELL, 1991). Atualmente, os RL têm sido
muito estudados em situações de patologia que afetam os músculos esqueléticos, já que se
encontram em concentrações elevadas nessas situações (HALLIWELL, 1991).
A designação “espécies reativas” engloba os RL e moléculas que, apesar de não
possuírem átomos com elétrons, são potencialmente geradoras desses radicais. As espécies
Figura 4: Esquema hipotético da formação dos eicosanoides da série par e ímpar pela ação das
enzimas COX e LOX. A seta vermelha mostra o mecanismo de competição do ômega-3. Quando o
EPA compete com o AA, este deixa de ser esterificado pela membrana e EPA consegue se ligar à ela
levando à diminuição da inflamação e do influxo de cálcio.

10
reativas de oxigênio (ROS – Reactive oxigen species) são constantemente geradas nas células
através das vias metabólicas. Aumento na produção de ROS ou diminuição de sua remoção
causam seu acúmulo na célula, caracterizando o estresse oxidativo (VERCHERAT et al.,
2009), ou seja, ocorre um desequilíbrio entre a ação dos agentes oxidantes e dos antioxidantes,
a favor dos primeiros (MOREL & BAROUKI, 1999). Como consequência, há um
desequilíbrio biológico caracterizando diversas doenças, tais como neurodegeneração,
diabetes e Distrofia Muscular de Duchenne (VERCHERAT et al., 2009).
Dentre as espécies reativas de oxigênio pode-se citar o radical superóxido (O₂⁻), o radical
hidroxil (OH⁻) e o peróxido de hidrogênio (H₂O₂). Este, por não conter um elétron
desemparelhado na sua órbita, não é considerado um verdadeiro radical (HALLIWELL, 1991;
CLARKSON & THOMPSON, 2000). Porém, esta molécula é uma potencial geradora de
radicais livres, por interagir com metais de transição e, portanto, a sua consequência orgânica,
fisiológica ou tóxica para a célula deve ser levada em consideração (HALLIWELL, 1991).
O metabolismo celular, mesmo em situações basais, promove continuamente a formação
de ROS, pelo fato de consumirem e reduzirem a molécula de oxigênio (MOREL &
BAROUKI, 1999; BECKMAN & AMES, 1998; IDE et al., 1999). A célula pode produzir
ROS em vários locais, tais como no citosol, em mitocôndrias, no retículo endoplasmático, nos
lisossomos e peroxissomos (MOREL & BAROUKI, 1999; AMES et al., 1993; BECKMAN
& AMES, 1998). No músculo esquelético, a mitocôndria parece ser a principal fonte de
produção das espécies reativas de oxigênio (AMES et al., 1993; BECKMAN & AMES, 1998).
A mitocôndria é a organela responsável pela fosforilação oxidativa celular e portanto, a
principal fonte de energia das células eucarióticas. Neste processo, a energia utilizada por esta
organela é proveniente da transferência de elétrons entre os complexos da cadeia
transportadora de elétrons com a finalidade de bombear prótons da matriz para o espaço
intermembranas (AMES et al., 1993). Este processo gera um gradiente eletroquímico
transmembrana com produção de água (AMES et al., 1993). Porém, pode ocorrer que uma
pequena porção desses elétrons reaja diretamente com o oxigênio molecular e
consequentemente forme as ROS (MOREL & BAROUKI, 1999; HALLIWELL, 1991, IDE
et al., 1999).
O oxigênio apresenta em sua órbita externa dois elétrons desemparelhados com spins
paralelos, tornando-o uma molécula potencialmente oxidativa pois ele é propenso a ganhar

11
elétrons (BECKMAN & AMES, 1998; MOREL & BAROUKI, 1999; CLARKSON &
THOMPSON, 2000). Assim, a redução da molécula de oxigênio com um, dois ou três
elétrons, origina respectivamente, o O₂⁻, o H₂O₂ e o OH⁻ (MOREL & BAROUKI, 1999)
(Figura 5A). Essa sequência de reações contribui para a peroxidação lipídica, danos no DNA
e degradação protéica durante os eventos de estresse. No processo de peroxidação lipídica
ocorre destruição dos lipídeos de membrana da célula resultando na formação de peróxidos
lipídicos e aldeídos, como por exemplo do malondialdeído (MDA) e do 4-hidroxinonenal
(4HNE) que são utilizados como biomarcadores do estresse oxidativo (ESTERBAUER &
CHEESEMAN, 1990).
Acredita-se que com o aumento da contração muscular e com o consequente aumento da
taxa de consumo de oxigênio, a síntese de ROS também aumenta agravando a oxidação
muscular. ROS são bastante reativas e desta forma são capazes de modificar moléculas
biológicas (DNA e proteínas) e destruir membranas, já que reage com os ácidos graxos poli-
insaturados, levando à lesão celular (IDE et al., 1999) (Figura 5B).
As células têm capacidade de reestabelecer o equilíbrio intracelular se os efeitos do
estresse oxidativo forem menores. Porém, níveis elevados de ROS podem levar à morte
celular, apoptose e necrose. O DNA oxidado é reparado por uma série de glicosilases que são
específicas para pares de bases peroxidadas. Proteínas oxidadas são degradadas por proteases.
Hidroxiperóxidos de lipídios são destruídas pela Glutationa Peroxidase. Quase todas essas
defesas parecem ser induzidas em resposta ao dano celular (AMES et al., 1993).
Desta forma, a redução univalente do oxigênio produz uma série de radicais livres e
espécies reativas de oxigênio que interagem com lipídios, DNA e proteínas. Esta interação
resulta na degradação protéica e promove quebra na molécula de DNA. Estas espécies reativas
afetam os lipídios, comprometendo a integridade dos ácidos graxos poli-insaturados e
consequentemente afeta a homeostasia celular (CLARKSON & THOMPSON, 2000).

12
1.7 Sistema antioxidante
Antioxidante pode ser definido como qualquer substância que, presente em menores
concentrações que as do substrato oxidável, é capaz de retardar ou inibir a oxidação deste de
maneira eficaz (DROGE, 2002; HALLIWELL, 1991). Tais ações podem se dar sobre os
radicais livres ou espécies não-radicais, diretamente neutralizando-os ou indiretamente,
participando dos sistemas enzimáticos com esta capacidade (HALIWELL et al., 2004). O
sistema antioxidante pode ser dividido como enzimático e não-enzimático.
O sistema não-enzimático tem como representantes os compostos antioxidantes de
origem dietética, tais como vitaminas, minerais e compostos fenólicos. O sistema enzimático
inclui as enzimas Superóxido Dismutase (SOD), Catalase (CAT) e Glutationa Peroxidase
(GPx). Essas enzimas atuam por meio de mecanismos de prevenção, impedindo e/ou
controlando a formação de radicais livres e espécies não-radicais, envolvidos com a iniciação
das reações em cadeia que resultam em propagação e amplificação do processo e,
consequentemente, com a ocorrência de danos oxidativos (FERREIRA & MATSUBARA,
1997; SCHENEIDER & OLIVEIRA, 2004).
A SOD tem como função catalisar a dismutação do superóxido (O₂¯) em peróxido de
hidrogênio (H₂O₂) e O₂ (MOREL & BAROUKI, 1999; HALLIWELL, 1991). A SOD pode
ser encontrada no citosol e na mitocôndria. No citosol, SOD é dependente de cobre e de zinco
Figura 5: A. Redução da molécula de oxigênio à molécula de água, com o conjunto de reações
intermediárias justificando a formação de espécies reativas de oxigênio na cadeia transportadora de
elétrons. B. Interação das espécies reativas de oxigênio com diferentes componentes da célula, que
induzem a lesão celular.

13
como co-fatores (Cu, Zn-SOD ou SOD1) e na mitocôndria de manganês (Mn-SOD ou SOD2)
(POWERS & LENNON, 1999). Tais co-fatores são importantes na atividade enzimática
efetiva da enzima. No músculo esquelético, 15-35% da atividade total da SOD está localizada
nas mitocôndrias e os restantes 65-85% no citosol (LEEUWENBURGH & JI, 1996).
As enzimas CAT e GPx impedem o acúmulo de peróxido de hidrogênio, convertendo-o
em H₂O e O₂. A GPx dos mamíferos apresenta maior afinidade pelo H₂O₂ do que a CAT. Isto
indica que, em baixas concentrações de H₂O₂, sua remoção celular é muito mais ativada pela
GPx (MOREL & BAROUKI, 1999). A CAT tem como co-fator o Fe³⁺ e pode ser encontrada
nos peroxissomos e no citosol (POWERS & LENNON, 1999). A GPx distribui-se pela
mitocôndria e pelo citosol. No músculo esquelético, em torno de 45% da atividade da GPx é
encontrada no citosol e os restantes 55% na mitocôndria (JI, 1999). Estas duas localizações,
mitocondrial e citoplasmática, permite a proximidade da GPx às diversas fontes de formação
de hidroperóxidos, fazendo desta enzima um dos principais neutralizadores de hidroperóxidos
provenientes das diferentes fontes celulares.
A ação da GPx na redução do peróxido é dependente do ciclo de redução-oxidação da
glutationa, que se apresenta como glutationa reduzida (GSH) e glutationa oxidada (GSSG)
(FERREIRA & MATSUBARA, 1997; SCHENEIDER & OLIVEIRA, 2004; ROVER et al.,
2001). A GPx reduz o H₂O₂ à água. Porém, o faz à custa da conversão da glutationa reduzida
em oxidada. No sentido de reestabelecer a integridade do ciclo redox da glutationa, a
Glutationa Redutase (GR) converte a GSSG em GSH. Essa reação utiliza como co-fator a
Nicotinamida Adenina-dinucleotídeo Fosfato (NADPH) na sua forma reduzida (DROGE,
2002; HALLIWELL, 1991). Em condições normais, a GR mantém mais de 98% da GSH
intracelular reduzida e consequentemente o meio intracelular no seu estado reduzido.
Portanto, a GPx e a GR atuando em conjunto no citosol e na mitocôndria são muito
importantes para o funcionamento normal do sistema antioxidante e da manutenção da
homeostasia celular, regulando quantidades adequadas da GSH dentro da célula (Figura 6).

14
1.8 Espécies Reativas de Oxigênio e a Distrofia Muscular de Duchenne
Espécies reativas de oxigênio têm sido propostas, ao longo dos anos, como um possível
mediador da degeneração do músculo distrófico (VERCHERAT et al., 2009; RENJINI et al.,
2012). Muitos trabalhos têm evidenciado que os músculos dos camundongos mdx são mais
suscetíveis à degeneração induzida pelos ROS e que o aumento na produção dos ROS causa
a peroxidação lipídica em períodos que precedem a necrose muscular dos camundongos mdx
(WHITEHEAD et al., 2006). A oxidação de proteínas e lipídios, características de lesão
oxidativa, é evidente nos músculos distróficos (VERCHERAT et al., 2009). Além disso, a
ausência da distrofina nos músculos torna os danos oxidativos suscetíveis de ocorrer
(VERCHERAT et al., 2009).
Níveis elevados de estresse oxidativo podem promover a invasão de células inflamatórias,
exarcebar os danos e interferir na sinalização celular responsável em promover a reparação
(TIDBALL & WEHLING-HENRICKS, 2007). Nos pacientes com DMD, a distrofia muscular
também pode ser mais suscetível ao estresse oxidativo (LAWLER, 2011) quando associado a
perturbações no complexo distrofina-glicoproteína (DGC) (RANDO et al., 1998).
Em relação à presença de ROS no músculo distrófico, acredita-se que isto seja devido à
captação anormal de cálcio pela mitocôndria, já que estudos demonstraram que a fosforilação
Figura 6: Ação integrada dos diferentes mecanismos antioxidantes enzimáticos (GPx – Glutationa
Peroxidase; SOD – Superóxido Dismutase; CAT - Catalase; GR – Glutationa Redutase; GSH e GSSG –
Formas reduzida e oxidada da Glutationa; DNA – Ácido Desoxirribonucléico; RNA – Ácido
Ribonucléico).

15
oxidativa mitocondrial mostrou-se prejudicada nos camundongos mdx e isto foi postulado
como sendo decorrente da sobrecarga de cálcio na mitocôndria (WHITEHEAD et al., 2006).
Experimentos mostram que a elevação intracelular de cálcio nos músculos esqueléticos
distróficos conduz ao aumento da permeabilidade de membrana. ROS tem a capacidade de
gerar danos musculares, já que ele afeta as proteínas da fibra muscular assim como a ativação
do processo inflamatório (WHITEHEAD et al., 2006).
Há também trabalhos que mostram que a fosfolipase A2 (PLA2), que é dependente de
cálcio, tem concentração significativamente maior nos músculos dos pacientes DMD
comparado com músculos normais. A fosfolipase A2 (PLA2) pode constituir uma fonte de
produção de ROS (LEEUWENBURGH & HEINECKE, 2001). Esta enzima está localizada
no sarcolema, na membrana de diferentes organelas, no sarcoplasma e no interior dos
lisossomos. Ela utiliza os fosfolipídios das membranas para sintetizar Ácido Araquidônico
(AA), o qual é substrato para a ação da cicloxigenase e da lipoxigenase resultando, na
formação de prostaglandinas, leucotrienos, tromboxanos e também de ROS
(LEEUWENBURGH & HEINECKE, 2001). O AA age nas membranas celulares diminuindo
a permeabilidade seletiva do sarcolema, o que favorece a perda da homeostasia celular ao íon
cálcio e da difusão de compostos sarcoplasmáticos para o meio intracelular (DROGE, 2002).
As espécies reativas de oxigênio também têm sido relacionadas com o NF-κB (fator de
transcrição que participa da regulação de citocinas pró-inflamatórias, tais como TNF-α e IL-
1β) (WHITEHEAD et al., 2006). O NF-κB transloca-se para o núcleo, onde se liga ao DNA
(ativação) e aumenta a produção de ROS e de proteínas e peptídeos pró-inflamatórios
(KUMAR et al., 2004). As citocinas pró-inflamatórias levam à inflamação do músculo e à
necrose, pois uma vez produzidas, atraem as células inflamatórias para o local da lesão
muscular que, junto do ROS, causam a degradação das proteínas e, por conseguinte, à necrose.
O aumento na concentração de ROS aumenta a atividade de NF-κB, a qual, por sua vez, ativa
TNF-α e IL-1β. A citocina TNF-α pode estimular a produção de ROS pela mitocôndria,
gerando um feedback positivo (WHITEHEAD et al., 2006).
As células inflamatórias, tais como neutrófilos e macrófagos, os quais são atraídos no
local da lesão muscular, também podem produzir ROS. Estes, porém, podem ser removidos
através da regulação do oxido nítrico (NO) (WHITEHEAD et al., 2006). Na miofibra normal,
a sintase neuronal do óxido nítrico (nNOS) catalisa a produção de óxido nítrico, presente no

16
sarcolema e sarcoplasma da célula. Porém, na DMD e no camundongo mdx, esta síntese é
ausente (NAKAE et al., 2004). Acredita-se que a diferença na expressão e na localização das
NOS entre as miofibras deficientes de distrofina e das miofibras normais pode ser responsável
pelos danos patológicos dos pacientes DMD e dos músculos dos mdx e, portanto, serem mais
suscetíveis ao estresse oxidativo (NAKAE et al., 2004). A diminuição de NO no músculo
distrófico agrava a inflamação e os danos na membrana destes músculos (WEHLING et al.,
2001).
Estudos tem relatado o papel da NAD(P)H oxidase como uma potencial fonte de estresse
oxidativo e na contribuição da patologia dos músculos esqueléticos na DMD (WHITEHEAD
et al., 2008). Esta oxidase se localiza na membrana celular e nas células inflamatórias
(NGUYEN & TIDBALL, 2003). NAD(P)H oxidase lança o ânion superóxido (O₂⁻) no espaço
intersticial, onde consegue entrar na célula facilmente via canal-3 de cloreto (ClC3) (FISHER,
2009). Já o peróxido de hidrogênio (H₂O₂), seguido de dismutação, têm sua difusão pela
membrana celular facilitada pelos canais de água (FISHER, 2009). Como já relatado, o
aumento desses radicais na célula leva à lesão celular, uma vez que reage com os diferentes
componentes de membranas.
O estresse oxidativo aumenta nos músculos com DMD e nos músculos dos camundongos
mdx (NAKAE et al., 2004). Este fato implica que os danos oxidativos das biomoléculas
celulares aceleram a progressão da doença nos músculos deficientes de distrofina (NAKAE et
al., 2004; TIDBALL & WEHLING-HENRICKS, 2007; LAWER, 2011).
1.9 Lipofuscina
A lipofuscina é um pigmento que indica a idade celular, considerada um marcador
biológico de envelhecimento (BRUNK & TERMAN, 2002). Quanto maior a concentração de
lipofuscina na célula, mais envelhecida ela está (YIN, 1996; NAKAE et al., 2001).
Muitos estudos têm demonstrado a presença da lipofuscina em músculos esqueléticos e
sua relação com a DMD (NAKAE et al., 2004). Acredita-se que o aumento da concentração
de radicais livres, produtos do estresse oxidativo celular, somados a ausência da distrofina,
resulta em aumento de lipofuscina nos músculos (NAKAE et al., 2004). No caso de pacientes
DMD e nos músculos dos camundongos mdx, estudos mostram que a lipofuscina começa a se
acumular muito cedo nos músculos esqueléticos. Este fato é observado nos pacientes DMD

17
com aproximadamente dois anos de idade e nos camundongos mdx com quatro semanas de
vida, quando comparados a um músculo normal, em que a lipofuscina é detectada em torno
de 20 semanas de vida (NAKAE et al., 2004).
A lipofuscina aparece como grânulos intracitoplasmáticos, de coloração pardo-
amarelada. Tais grânulos são constituídos principalmente de proteínas (30-70%) e lipídios
(20%-50%), sob a forma de polímeros não degradáveis, resultantes da modificação oxidativa
das proteínas e da degradação lipídica (BRUNK & TERMAN, 2004). Os resíduos de proteína
mostram um conteúdo variável de aminoácidos e os componentes lipídicos são principalmente
representados por triglicerídeos, ácidos graxos livres, colesterol e fosfolipídios (BRUNK &
TERMAN, 2004).
Várias técnicas de histoquímica são capazes de detectar a lipofuscina, tais como:
coloração de lipídios e carboidratos; microscopia eletrônica para detectar os grânulos de
lipofuscina na forma de corpos perinucleares através da membrana lisossomal que os rodeia
(BRUNK & TERMAN, 2004); imunocitoquímica através de microscopia de luz e de elétrons,
para revelar enzimas lisossomais associadas com lipofuscina (BRUNK & ERICSSON, 1972);
técnica de autofluorescência de largo espectro para a quantificação da lipofuscina, que é uma
de suas propriedades principais (BRUNK & TERMAN, 2004).
Acredita-se que a lipofuscina representa um material polimérico intralisossomal que não
pode ser degradado pelas hidrolases lisossomais e que se origina de componentes celulares
autofagocitados que se tornaram oxidados fora ou dentro do compartimento lisossomal
(BRUNK & TERMAN, 2004). Os materiais presentes no interior dos lisossomos entram
através dos processos de macroautofagia, microautofagia e autofagia mediada por chaperonas
e são decompostas em moléculas, tais como, aminoácidos e ácidos graxos. Porém, os
lisossomos não conseguem degradar todas essas substâncias que foram auto ou
heterofagocitadas e lentamente, conforme vão se acumulando, passam a ser chamados de
grânulos de lipofuscina. (BRUNK & TERMAN, 2004). Quanto mais velho o indivíduo for,
mais grânulos de lipofuscina se acumulam e menos funcional se torna a célula.
No interior do autofagolisossomo, o ambiente é ácido (pH em torno de 4,5) pois as
enzimas hidrolíticas presentes no seu interior também são ácidas e necessitam de pH baixo
para exercerem atividade ótima. Além disso, metaloproteínas são degradadas liberando íons
ferro, os quais reagem com o peróxido de hidrogênio produzindo o radical hidroxil (OH⁻)

18
(BRUNK & TERMAN, 2004). O peróxido de hidrogênio (H₂O₂) é formado como produto de
oxidação celular, mas principalmente da respiração mitocondrial, onde é formado pela
dismutação do O₂⁻ (BRUNK & TERMAN, 2002). Uma vez formado, o H₂O₂ é reduzido em
água rapidamente, pela ação da catalase ou da glutationa peroxidase. Porém, uma pequena
quantidade pode se difundir facilmente pelo autofagolisossomo e reagir com os íons ferro ali
presentes. Os radicais hidroxil induzem a peroxidação de macromoléculas auto e
heterofagocitadas, e isso resulta na formação de material não degradado (lipofuscina)
(BRUNK & TERMAN, 2004).
Dando suporte a esses dados, têm-se demonstrado que o efeito do estresse oxidativo sobre
a formação da lipofuscina (lipofuscinogênese) é aumentada quando combinada com a inibição
das proteases lisossomais, o que atrasa a degradação dos materiais autofagocitados e permite
mais tempo para sua oxidação (BRUNK & TERMAN, 2002). Vários sistemas de modelos
experimentais têm comprovado que as células tornam-se carregadas com lipofuscina em
função do estresse oxidativo, ou seja, forma-se lipofuscina muito mais lentamente quando as
células são mantidas em concentração de oxigênio a 8% do que em uma concentração de 40%
(TERMAN & BRUNK, 1998). Assim, tanto a formação da lipofuscina quanto a lesão da
célula seriam processos decorrentes da ação dos radicais livres (O₂⁻, OH⁻, H₂O₂) produzidos
pelo metabolismo normal da célula a partir do oxigênio molecular.
A autofagocitose da mitocôndria também parece ser um grande contribuinte para a
formação da lipofuscina. A mitocôndria é o principal local de formação de ROS e também o
principal alvo para seus ataques. Uma vez danificadas pela oxidação, esta organela pode
conter ainda algumas macromoléculas peroxidadas que não foram degradadas, íons ferro e
O₂⁻. No lisossomo, essa produção de O₂⁻ e a catalisação pelos íons ferro na oxidação de
componentes mitocondriais ainda pode continuar por um tempo (BRUNK & TERMAN,
2002) (Figura 7). O envolvimento dos constituintes mitocondriais com a formação da
lipofuscina é suportado pelo fato de que pelo menos uma proteína integrante da mitocôndria
– ATP sintase - é um componente predominante da lipofuscina de neurônios envelhecidos
(ELLEDER et al., 1997).
Resumindo, a formação e o acúmulo do pigmento lipofuscina decorrem dos seguintes
fatores: 1. desequilíbrio no processo contínuo de autofagocitose de componentes celulares
oxidados dentro ou fora do compartimento lisossomal; 2. diminuição da produção e da

19
atividade das enzimas lisossômicas; 3. incapacidade da célula em eliminar os resíduos da
autodigestão.
Desta forma, a lipofuscina passa a ser um material polímero intralisossomal, o qual não
pode ser degradado pelas hidrolases lisossomais e também não pode ser exocitado (BRUNK
& TERMAN, 2004; NAKAE et al., 2004).
A lipofuscina se acumula no interior dos lisossomos das células pós-mitóticas, tais como
miócitos cardíacos e células do músculo esquelético (NAKAE et al., 2001). Acredita-se que
altas concentrações desse pigmento levem à disfunção celular progressiva, isto é, as células
perdem sua funcionalidade até morrerem (BRUNK & TERMAN, 2004).
Em estudos anteriores realizados pelo nosso grupo, foi observado que tanto o EPA
purificado quanto as cápsulas de ômega-3 protegeram a distrofinopatia nos músculos
esqueléticos dos camundongos jovens (MACHADO et al., 2011; MAURÍCIO et al., 2012).
No presente trabalho, vamos complementar, estudando os efeitos das cápsulas de ômega-3 no
estresse oxidativo e desta forma fornecer mais dados sobre o seu uso como terapia adicional
para DMD.
Figura 7: Esquema ilustrando um mecanismo básico da lipofuscinogênese. Peróxido de hidrogênio, um
produto do metabolismo normal da célula é eliminado pela catalase e pela glutationa peroxidase. Porém,
uma parte se difunde através da membrana lisossomal e na presença dos íons ferro, reagem produzindo
radicais hidroxil OH⁻. Estes atacam macromoléculas (partes de mitocôndria e outros componentes
celulares), oxidando-as, resultando na formação de material não degradado, a lipofuscina.

20
2. Objetivo
Verificar se o óleo da cápsula de ômega-3, contendo uma mistura de EPA + DHA, diminui
o estresse oxidativo em diferentes músculos distróficos (bíceps, diafragma e quadríceps) do
camundongo mdx, nas fases iniciais da doença.
2.1 Objetivos específicos:
1. Análise do estresse oxidativo: -Verificar se o ômega-3 diminui a concentração dos grânulos
de lipofuscina nos músculos bíceps, diafragma e quadríceps dos camundongos distróficos.
-Verificar se o ômega-3 diminui o 4 HNE nos camundongos distróficos.
-Verificar o efeito do ômega-3 na atividade das seguintes enzimas antioxidantes: glutationa
peroxidase, glutationa redutase e superóxido dismutase.
2. Análise morfológica: verificar se o ômega-3 protege, nos músculos bíceps, diafragma e
quadríceps, a área de inflamação/regeneração em camundongos distróficos.
3. Verificar o efeito do ômega-3 sobre a massa corporal, a força muscular e os níveis de CK
em camundongos distróficos.
3.Materiais e métodos
3.1 Animais
Neste trabalho foram utilizados camundongos de 14 dias de ambos os sexos, da linhagem
mdx e da linhagem controle C57BL/10. Os animais foram obtidos do Biotério Central da
UNICAMP e foram mantidos no Biotério do Departamento de Anatomia do Instituto de
Biologia, UNICAMP. Durante o experimento, os animais permaneceram em caixas padrão
em condições controladas (12 horas de ciclo claro/escuro) com ração e água ad libitum.
Os animais foram divididos em três grupos experimentais: camundongos C57BL/10
controle (CTRL; n=28); camundongos mdx tratados com nujol (mdx-nujol; n=28) e
camundongos mdx tratados com ômega-3 (mdx-ômega3; n=28).

21
3.2 Protocolo experimental
Grupo Controle
Foram utilizados animais C57BL/10 de 30 dias de idade. Estes não receberam tratamento.
Grupo mdx-nujol
Os camundongos distróficos foram tratados com 300mg/Kg/dia de óleo mineral (Nujol; 100%
óleo mineral-Mantecorp Indústria Química e Farmacêutica Ltda.) via gavagem. O tratamento
teve início quando os animais completaram 14 dias de idade e foi realizado durante 16 dias.
Grupo mdx-ômega3
Os camundongos distróficos foram tratados com 300mg/Kg/dia com óleo da cápsula ômega-
3 EPA 1000 mg, (Fabricado por FDC vitamins, Inc., Miami, Flórida, EUA. Importado e
distribuído exclusivamente por Fedco Ind. Ltda.) contendo 0.4 g de EPA, 0.2 g de DHA, 2 mg
de vitamina E, 0.9 de proteína, 2.0 g de gordura total, 0.4 g de gordura saturada, 0.0 g de
gordura trans, 0.0 g de gordura monossaturada e 1.0 g de gordura poli-insaturada, via
gavagem, recebendo a mesma dosagem que o grupo anterior no mesmo período de tratamento.
Todos os experimentos foram realizados de acordo com as diretrizes para experimentação
animal de nossa Instituição, sob o processo 3068-1 a Comissão de Ética no Uso de Animais
(CEUA-IB-UNICAMP) e esteve de acordo com os princípios éticos adotados pelo Colégio
Brasileiro de Experimentação Animal (COBEA).
3.3 Procedimento cirúrgico para obtenção dos músculos
Após 30 dias de vida, os animais foram sacrificados com dose letal de cloridrato de
xilazina (6,8 mg/kg, 2% Virbaxyl, Virbac, São Paulo, Brasil) e cloridrato de cetamina (130
mg/kg, Francotar,Virbac, São Paulo, Brasil), via injeção intraperitoneal. Em seguida foram
retirados os músculos diafragma (DIA), bíceps braquial (BB) e quadríceps (QDR) dos animais
de cada grupo. Todos os músculos foram fixados em suportes de madeira com tragacanthgum,
imersos em n-hexano a 90ºC por um minuto e imediatamente colocados em nitrogênio líquido
a -159ºC. Após estes procedimentos, os músculos foram retirados do nitrogênio e
armazenados a -80ºC em biofreezer (BioFreezer SoLow V85-13, EUA, Departamento de
Anatomia, IB). Para a obtenção dos cortes, os músculos foram descongelados por

22
aproximadamente 30 minutos até atingirem a temperatura de -23ºC, quando foram
seccionados transversalmente na espessura de 8μm utilizando-se criostato (Leica CM1860,
Alemanha, Departamento de Anatomia, IB). Os cortes foram utilizados para a análise
histopatológica em hematoxilina-eosina (HE) e para a quantificação da lipofuscina.
3.4 Medida da força
Os camundongos de cada grupo tiveram a força obtida no início e no fim do tratamento.
A força foi quantificada no aparelho de medida de força horizontal (Newprimer®). Os animais
foram estabilizados pela cauda e estes, com as patas dianteiras, seguraram uma tela de nylon
ligada a um transdutor, que transmite a força de contração dos músculos para o aparelho. O
pico de força de cinco tentativas, em quilogramas, mostradas no diplay do aparelho, foram
colhidas e utilizadas para composição da média de cada animal, sendo o resultado final
expresso em Kg/força (MESSINA et al, 2009; TANIGUTI, 2011).
Figura 8: Equipamento para medida de força dos membros anteriores. O animal segura a argola puxando-
a para trás, exercendo assim força muscular que é registrada no visor digital, em quilogramas.
3.5 Análise de Creatina Quinase (CK) no plasma sanguíneo
A quantificação de CK no plasma sanguíneo é um indicador do processo de necrose
muscular. O sangue dos animais foi coletado sob anestesia, conforme descrito anteriormente.
Após a toracotomia e exposição do coração, uma seringa heparinizada foi introduzida no
ventrículo direito do animal e um volume de sangue coletado e transferido para um ependorf
de 1,5 ml. Em seguida, o sangue foi centrifugado (centrifuga refrigerada Sigma® 3-18k) nas
seguintes condições: 12000 rpm, 4°C por 15 minutos, sendo o sobrenadante utilizado para

23
análise. Para quantificação da CK foi utilizado o kit CK da Bioclin®. As absorbâncias das
amostras foram lidas a 25°C utilizando-se espectrofotômetro U.V (ThermoElectron
Corporation®Spectrophotometer Genesys 20) com comprimento de onda de 340 nm e cubetas
de quartzo de 1cm de caminho óptico. Os valores foram expressos em U/L.
3.6 Preparo e análise dos músculos para a técnica de Hematoxilina e Eosina
Para a coloração com hematoxilina-eosina (HE), os cortes congelados foram lavados com
etanol 70%. Em seguida, as lâminas foram lavadas em água corrente por dez minutos e os
cortes corados com hematoxilina de Harris e depois com eosina, respectivamente.
Posteriormente, os cortes foram desidratados em séries de etanol, diafanizados em xilol e as
lâminas montadas em Entelan (Sigma) para observação em microscópio de luz.
As lâminas coradas com HE foram observadas com auxílio de objetiva de 10X no
microscópio (Carl Zeiss). A quantificação foi feita na forma de áreas com fibras de núcleo
periférico, áreas com fibras de núcleo central, áreas de inflamação e áreas em regeneração,
expressas em porcentagem em relação à área total do corte. As imagens foram capturadas pelo
fotomicroscópio (Nikon Eclipse E-400) com objetiva de 10X acoplada a um computador com
software (Image Pro-Express Version 4). Para os três músculos, BB, QDR e DIA, e a
quantificação foi expressa em porcentagem em relação a área total do corte.
3.7 Preparo e análise dos músculos para análise da lipofuscina
Os cortes obtidos pelo procedimento descrito anteriormente foram coletados em lâmina e
montados em meio de montagem DABCO, fotografadas em objetiva de 40X em microscópio
invertido de fluorescência (Nikon, Eclipse TS100/ TS100F). As imagens foram capturadas
por uma vídeo câmera (Nikon® Express Series) acoplada ao microscópio de luz e conectada
a um microcomputador com o software Image ProExpress®.
Foram utilizados 15 animais (5 C57BL/10; 5 mdx-nujol; 5 mdx-ômega3) e para cada
músculo foram escolhidos cinco cortes. Os grânulos de lipofuscina foram contados de campo
em campo, até varrer o corte inteiro. Foi feito a média de cada animal e somados os resultados
dos músculos direito e esquerdo. Completada a análise de todos os animais, a média geral foi
obtida somando os valores de cada animal.

24
A análise da lipofuscina se baseou nos seguintes critérios: 1. Forma: estruturas granulares
arredondadas ou ovaladas; 2. Local: grânulos associados ou não ao sarcolema e no interstício
celular; 3. Autofluorescência nos comprimentos de onda 488 nm e 546 nm, simultaneamente.
3.8 Western Blotting
Para a quantificação do 4 HNE foram utilizados 45 camundongos jovens (15 C57BL/10;
15 mdx-nujol; 15 mdx-ômega). Os camundongos foram anestesiados com cloridrato de
cetamina e cloridrato de xilazina e perfundidos com PBS. Os músculos diafragma, bíceps e
quadríceps foram retirados e homogeneizados em tampão para homogeneização (Tris-HCl
100 mM, pH 7,4 contendo Triton X-100 1%, pirofosfato de sódio 100 mM, fluoreto de sódio
100 mM, EDTA 10 mM, ortovanadato de sódio 10 mM, PMSF 2 mM e 0,1 mg/ml de
aprotinina) a 4 ºC com homogeneizador do tipo Polytron PTA 20S (PT 10/35; Kinematica
Ag) operado em velocidade máxima por 30 s. Os extratos foram centrifugados a 11000rpm
a 4ºC por 20 minutos e o sobrenadante foi utilizado para análise do extrato total. A
determinação das proteínas foi realizada pelo método de Bradford e colaboradores (1976).
As amostras do extrato proteico foram tratadas com tampão Laemmli (azul de
bromofenol 0,1% e fosfato de sódio 1 M pH 7,0, glicerol 50% e SDS 10%), acrescido de
ditiotreitol 100mM, aquecidas em água fervente por 5 minutos e centrifugadas por 1 minuto.
Em seguida, 30 μg de proteína foram aplicadas em gel SDS-poliacrilamida 8-15% em
aparelho para eletroforese da Bio-Rad (mini-Protean, Bio-Rad Laboratories, Richmond, CA,
EUA). A eletrotransferência do gel para a membrana de nitrocelulose foi realizada em 90
minutos a 120 V em aparelho de transferência da Bio-Rad.
As membranas para marcação do 4 HNE foram incubadas com solução basal (Trisma 10
mM, cloreto de sódio 150 mM e Tween-20 0,02%) contendo 3% e 5% de leite desnatado,
por uma hora, em temperatura ambiente, para reduzir a ligação inespecífica de proteínas.
Posteriormente, foram incubadas com anticorpo primário (4 HNE: Goat polyclonal antibody;
Santa Cruz Biotechnology, Santa Cruz, CA, USA) diluídos em 10 ml de solução basal
contendo 3% de leite desnatado a 4ºC durante a noite. No dia seguinte, as membranas foram
lavadas por 30 minutos com solução basal e depois, incubadas com anticorpo secundário
(IgG goat conjugado à peroxidase correspondente (H + L) (KPL, Gaithersburg, MD, USA)
diluídos em 10 ml em solução basal contendo 3% de leite desnatado por duas horas, em

25
temperatura ambiente. Posteriormente, as membranas foram lavadas por 30 minutos com
solução basal. Em seguida, as membranas foram imersas em solução de quimiluminescência
(Super Signal West Pico Chemiluminescente, Pierce Biotechnology, Rockford, Illinois,
USA) por 5 minutos seguido de exposição a um filme Kodak XAR (Eastman KodaK,
Rochester, N.Y, USA). As densidades das bandas e das amostras sobre o filme foram
‘escaneadas’ e ‘salvas’ em discos de computador para quantificação da densitometria ótica,
usando um scanner e o programa ImageJ (Image Processing and Analysis in Java).
Para a normalização dos dados obtidos foi realizado o controle interno através da
incubação das amostras com o anticorpo GAPDH (FL-335, rabbit polyclonal antibody, Santa
Cruz Biotechnology). O protocolo consiste na reutilização das membranas utilizadas para
marcação de 4 HNE. Para tanto, as referidas membranas foram lavadas com TBS-T (solução
basal) 3 vezes, por 10 minutos. Em seguida, foram incubadas com 10 mL de Stripping
Buffer (10mM Tris-HCl pH 7.5; β-Mercaptoethanol 0.1M; Uréia 8M) durante 1 hora, à 60°C.
Após esse período, as membranas foram reequilibradas e lavadas com TBS-T. A seguir foram
incubadas com 10μg do anticorpo primário GAPDH e 2,5 μg do anticorpo secundário,
seguindo o protocolo acima descrito.
3.9 Determinação da atividade enzimática da GPx, GR e SOD
A atividade enzimática foi realizada apenas nos músculos diafragma e quadríceps, por
permitirem maior quantidade de material para análise. O músculo BB, por ser de dimensões
menores, não oferecia material suficiente para execução da técnica de atividade enzimática.
Para o diafragma foram utilizados 4 pools de 2 animais, totalizando 24 animais (8 C57BL/10;
8 mdx-nujol; 8 mdx-ômega3). Para o quadríceps foram utilizados 24 animais individuais (8
C57BL/10; 8 mdx-nujol; 8 mdx-ômega3).
Atividade enzimática da GPx, GR e SOD
As amostras foram homogeneizadas em homogeneizador de tecidos com 1 ml de solução
tampão fosfato tamponado (PBS), imediatamente imersas em nitrogênio líquido e
armazenadas em biofreezer (-70 ºC). Posteriormente foram utilizadas para quantificação da
atividade enzimática.

26
GPx (glutationa peroxidase)
O homogenato de tecido muscular foi diluído em tampão fosfato (1: 10). Em 100 μL desta
solução foi adicionado 50 μL H2O2 (0.25 mM), 20 μL de glutationa reduzida (GSH) (10 mM),
20 μL NADPH (4 mM), 10 μL (1 U) de enzima glutationa redutase em PBS, pH 7,8. A
absorbância foi lida em espectrofotômetro a 365 nm. Os resultados foram expressos em
mM/min por mg de proteína.
GR (glutationa redutase)
A atividade dessa enzima foi determinada seguindo-se espectrofotometricamente a
oxidação de NADPH a 340 nm. A reação enzimática foi constituída de 100 μL do homogenato
previamente diluído em tampão fosfato (1:10), 50 μL de tampão fosfato (1M), pH=7, 10 μL
de EDTA (0,2 mM, 20 μL de GSSG (1 mM) e 20 μL de NADPH (0,1 mM). O consumo de
NADPH foi determinado pela diminuição da absorbância à 340 nm. A atividade da enzima
foi definida como nM NADPH consumido/min/mg de proteína.
SOD (superóxido dismutase)
A atividade da SOD foi determinada pela inibição da redução do NBT (nitro blue
tetrazolium) pelo radical superóxido gerado através do sistema hipoxantina/ xantina oxidase
(XO) à 37º C. O homogenato de tecido foi diluído em tampão fosfato (1:10). Em 100 μL desta
solução, foram adicionados 40 μL de tampão fosfato 0.1 M (pH: 7.4), xantina 0.07 U/ml, 20
μL de hipoxantina (100 μM) e 20 μL de NBT. A absorbância foi lida em espectrofotômetro a
560 nm. Os resultados foram expressos em U/mg de proteína.
4. Análise Estatística
A análise estatística foi realizada no software Bioestat 5.0. Os valores foram expressos
em média ± desvio padrão. A análise de variância, ANOVA, foi utilizada para a comparação
entre os grupos experimentais e a significância foi de p≤0,05.
27
5. Resultados
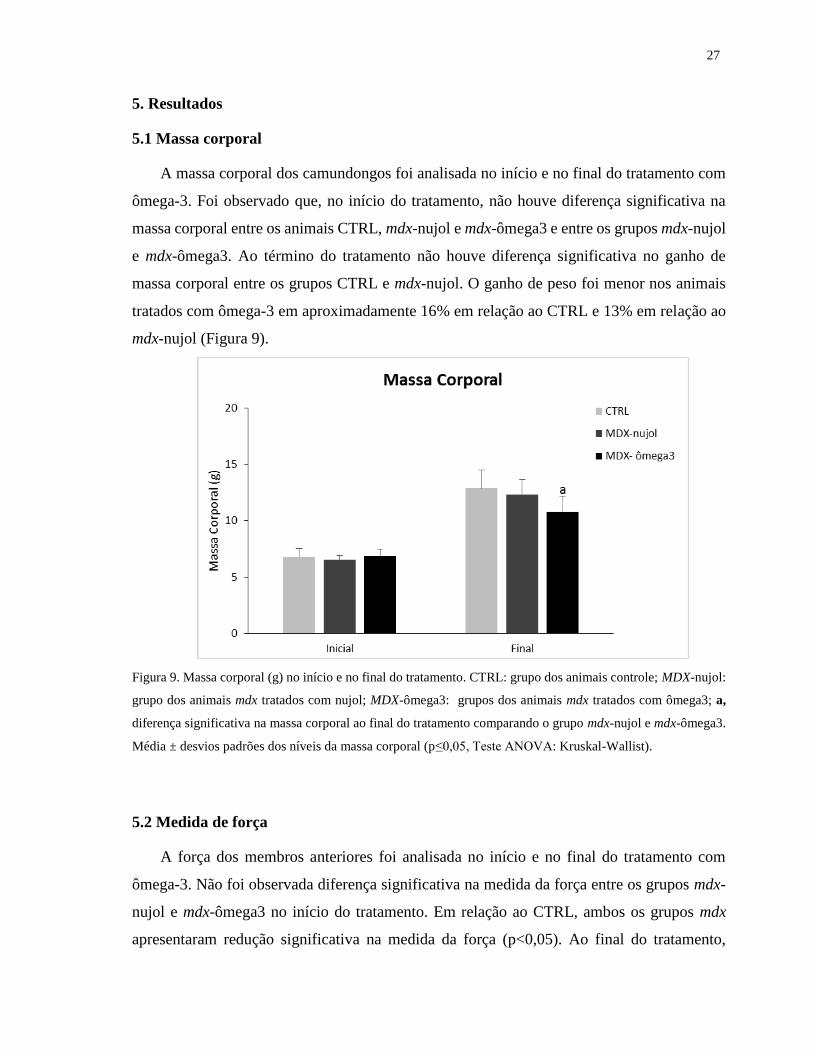
5.1 Massa corporal
A massa corporal dos camundongos foi analisada no início e no final do tratamento com
ômega-3. Foi observado que, no início do tratamento, não houve diferença significativa na
massa corporal entre os animais CTRL, mdx-nujol e mdx-ômega3 e entre os grupos mdx-nujol
e mdx-ômega3. Ao término do tratamento não houve diferença significativa no ganho de
massa corporal entre os grupos CTRL e mdx-nujol. O ganho de peso foi menor nos animais
tratados com ômega-3 em aproximadamente 16% em relação ao CTRL e 13% em relação ao
mdx-nujol (Figura 9).
Figura 9. Massa corporal (g) no início e no final do tratamento. CTRL: grupo dos animais controle; MDX-nujol:
grupo dos animais mdx tratados com nujol; MDX-ômega3: grupos dos animais mdx tratados com ômega3; a,
diferença significativa na massa corporal ao final do tratamento comparando o grupo mdx-nujol e mdx-ômega3.
Média ± desvios padrões dos níveis da massa corporal (p≤0,05, Teste ANOVA: Kruskal-Wallist).
5.2 Medida de força
A força dos membros anteriores foi analisada no início e no final do tratamento com
ômega-3. Não foi observada diferença significativa na medida da força entre os grupos mdx-
nujol e mdx-ômega3 no início do tratamento. Em relação ao CTRL, ambos os grupos mdx
apresentaram redução significativa na medida da força (p<0,05). Ao final do tratamento,

28
observou-se redução significativa da força dos grupos mdx-nujol e mdx-ômega3 em relação
ao CTRL (p<0,05). Em relação aos grupos distróficos, o mdx-ômega3 apresentou aumento
significativo da força em relação ao mdx-nujol (p<0,05) (Figura 10).
Figura 10. Medida de força no início e no final do tratamento. CTRL: grupo dos animais controle; MDX-nujol:
grupo dos animais mdx tratados com nujol; MDX-ômega3: grupos dos animais mdx tratados com ômega-3; a,
diferença significativa na força no início e no final do tratamento comparando o grupo CTRL com os grupos
mdx-nujol e mdx-ômega3. b, diferença significativa na força ao final do tratamento comparando o grupo mdx-
nujol com mdx-ômega-3. Média ± desvios padrões da medida de força (p≤0,05, Teste ANOVA:Kruskal-Wallist).
5.3 Análise da CK (U/L)
Os animais mdx-nujol apresentaram cerca de 78% de aumento nos níveis de CK em
relação ao animal controle. O ômega-3 diminui significativamente os níveis de CK em cerca
de 22% em relação ao mdx-nujol (Figura 11).

29
Figura 11. Níveis da Creatina Quinase (CK), expressos em U/L. a, diferença significativa entre os grupos CTRL
e mdx-nujol; b, diferença significativa entre mdx-nujol com mdx ômega3. Média ± desvios padrões dos níveis da
creatina quinase (p≤0,05, Teste ANOVA:Kruskal-Wallist).
5.4 Análise morfológica
5.4.1 Análise qualitativa das fibras musculares (HE)
Os músculos esqueléticos dos grupos estudados (CTRL, mdx-nujol e mdx- ômega3)
apresentaram fibras musculares com características histológicas diferentes. Foram observadas
fibras com núcleo periférico, fibras em regeneração, fibras completamente regeneradas e áreas
de inflamação em todos os músculos do grupo mdx (Figura 12 A, B e C). As fibras com núcleo
periférico são indicativas de fibras que não sofreram processo de degeneração-regeneração;
apresentam formato poligonal e diâmetro pouco variável (Fibra com núcleo periférico, Figura
12 A, B e C; seta). Fibras regeneradas (Figura 12 A, B e C; cabeça de seta) foram
caracterizadas pelo núcleo centralizado, condensado, com citoplasma eosinófilo e diâmetro
da fibra muscular próximo ao de uma fibra muscular esquelética com núcleo periférico. As
fibras musculares em regeneração foram caracterizadas por pequenos miócitos com núcleo
central, entremeados por infiltrado inflamatório, em áreas de grande celularidade (Área em
regeneração, Figura 12 A, B e C; asterisco).

30
Figura 12A. Seção transversal do músculo Bíceps (BB) corados com Hematoxilina & Eosina (HE). CTRL: grupo
dos animais controle não tratados; Mdx: grupo dos animais mdx tratados com nujol; Ômega-3: grupo dos animais
mdx tratados com ômega-3. Núcleo periférico (seta); núcleo central (cabeça de seta); área em regeneração
(asterisco) Barra: 100µm

31
Figura 12B. Seção transversal do músculo Diafragma (DIA) corados com Hematoxilina & Eosina (HE). CTRL:
grupo dos animais controle não tratados; Mdx: grupo dos animais mdx tratados com nujol; Ômega-3: grupo dos
animais mdx tratados com ômega-3. Núcleo periférico (seta); núcleo central (cabeça de seta); área em
regeneração (asterisco) Barra: 100µm

32
Figura 12C. Seção transversal do músculo Quadríceps (QDR) corados com Hematoxilina & Eosina (HE). CTRL:
grupo dos animais controle não tratados; Mdx: grupo dos animais mdx tratados com nujol; Ômega-3: grupo dos
animais mdx tratados com ômega-3. Núcleo periférico (seta); núcleo central (cabeça de seta); área em
regeneração (asterisco) Barra: 100µm

33
5.4.2 Análise quantitativa das fibras musculares (HE)
Nos músculos BB e DIA, a porcentagem da área de fibras com NP dos animais tratados
com ômega-3 estava aumentada, respectivamente, em 5% e 21%. No DIA, a porcentagem da
área de fibras com NC diminuiu significativamente (64%) com o tratamento. O ômega-3
protegeu os dois músculos da distrofinopatia, promovendo diminuição significativa da área
de inflamação/regeneração em 85% no BB e 46% no DIA (Figura 13- A e B).
Em relação ao QDR, no grupo mdx-ômega3, comparado com o grupo mdx- nujol houve
diminuição da área das fibras com núcleo periférico (28%) e aumento da área de fibras com
núcleo central (34%). Nos animais tratados com ômega-3 a área de inflamação/regeneração
foi maior (98%) que nos mdx tratados com nujol (Figura 13-C).

34
5.5 Análise qualitativa e quantitativa dos grânulos de lipofuscina
Grânulos de lipofuscina possuem formato aproximadamente arredondado, distribuídos no
interior da fibra muscular e/ou no meio extracelular (Figura 14, seta). Tipicamente, grânulos
de lipofuscina fluorescem ao serem incididos com luz nos comprimentos de onda de 488nm e
568nm. Assim, grânulos fluorescentes em apenas um ou outro comprimento de luz não foram
considerados como sendo de lipofuscina, não sendo computados para quantificação.
A Figura 14 A, B e C mostra cortes transversais dos músculos BB, DIA e QDR dos
animais CTRL e mdx observados ao microscópio de fluorescência. Os cortes em verde (à
esquerda no painel) foram excitados com luz de 488nm; os cortes em vermelho (à direita no
painel) foram excitados com luz de 568 nm. Em todos os músculos estudados dos animais
mdx-nujol e mdx-ômega3 foram observados grânulos de lipofuscina na idade estudada.
O tratamento com ômega-3 diminuiu a concentração dos grânulos de lipofuscina em
61.5% no BB, 23% no DIA e de 73% no QDR (Figura 15).
Figura 13: Comparação entre a porcentagem de fibras com núcleo periférico (NP), porcentagem de fibras com
núcleo central (NC) e porcentagem da área de inflamação/regeneração (INFL/REG) dos músculos: A) Bíceps
braquial; B) Diafragma e C) Quadríceps em animais CTRL, mdx-nujol e animais mdx- ômega3. a, diferença
significativa entre os grupos CTRL e mdx-nujol; b, diferença significativa entre mdx-nujol com mdx ômega3
(p≤0,05, Teste ANOVA:Kruskal-Wallist).

35
Figura 14A. Grânulos de lipofuscina ao microscópio de fluorescência no músculo: Bíceps. Seta: grânulos
fluorescentes, nos dois nos comprimentos de onda de 488nm (painéis em verde) e 568nm (painéis em vermelho),
respectivamente. Barra: 100µm.

36
Figura 14B. Grânulos de lipofuscina ao microscópio de fluorescência no músculo: Diafragma. Seta: grânulos
fluorescentes, nos dois nos comprimentos de onda de 488nm (painéis em verde) e 568nm (painéis em vermelho),
respectivamente. Barra: 100µm.

37
Figura 14C. Grânulos de lipofuscina ao microscópio de fluorescência no músculo: Quadríceps. Seta: grânulos
fluorescentes, nos dois nos comprimentos de onda de 488nm (painéis em verde) e 568nm (painéis em vermelho),
respectivamente. Barra: 100µm.
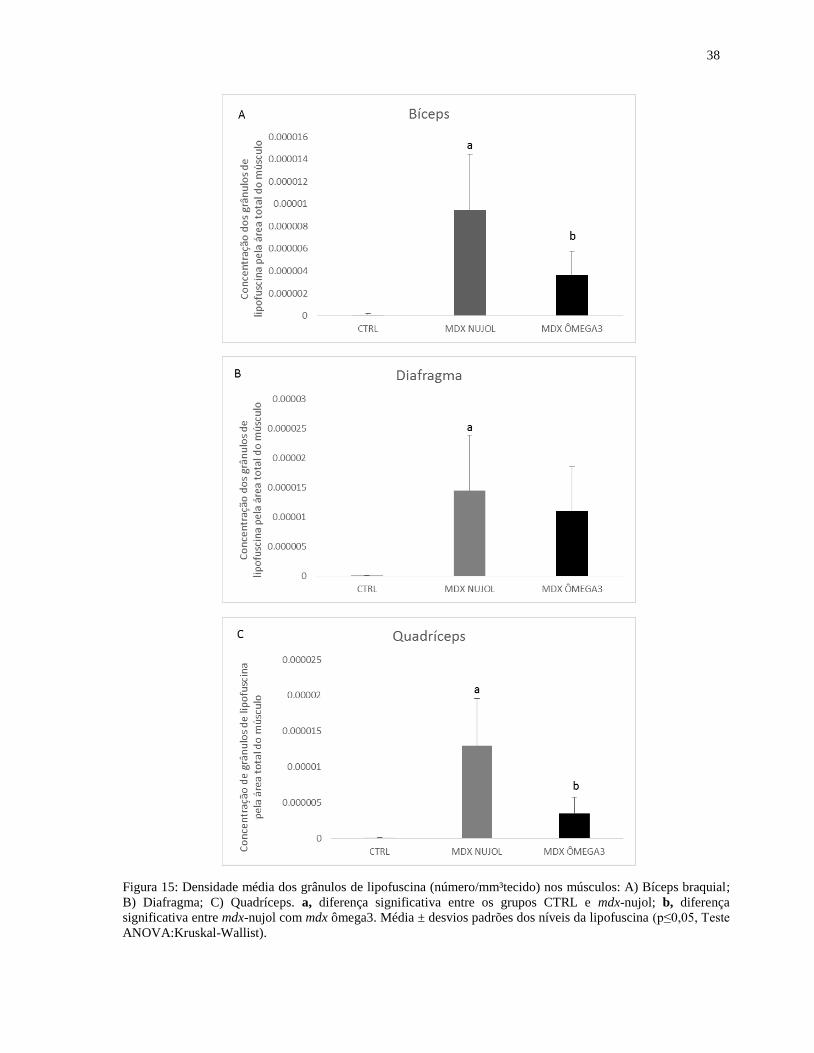
38
Figura 15: Densidade média dos grânulos de lipofuscina (número/mm³tecido) nos músculos: A) Bíceps braquial;
B) Diafragma; C) Quadríceps. a, diferença significativa entre os grupos CTRL e mdx-nujol; b, diferença
significativa entre mdx-nujol com mdx ômega3. Média ± desvios padrões dos níveis da lipofuscina (p≤0,05, Teste
ANOVA:Kruskal-Wallist).

39
5.6 Níveis de 4-HNE (Western Blot)
Os níveis de 4-HNE do músculo BB dos camundongos mdx-nujol e mdx- ômega3 foram
54% e 21% maiores em relação aos animais CTRL. O tratamento com ômega-3 reduziu os
níveis de 4-HNE em 21% (Figura 16A).
Os níveis de 4-HNE do músculo DIA dos animais mdx-nujol e mdx-ômega3 foram de
29% e 2.4% maiores comparados ao CTRL. O tratamento reduziu os níveis de 4-HNE em
21% (Figura 16B).
No músculo QDR, os grupos mdx apresentaram níveis semelhantes de 4-HNE. Ambos
apresentaram uma redução de 17% (mdx-nujol) e 19% (mdx-ômega3) dos níveis de 4-HNE
em relação ao grupo CTRL (Figura 16C).
170kd
43kd
35kd
4-HNE
GAPDH

40
Figura 16: Western blot dos músculos bíceps braquial (A), diafragma (B) e quadríceps (C) de animais CTRL, mdx-
nujol e mdx-ômega3. Bandas imunorreativas do 4HNE e gráfico representando os níveis de 4HNE expressos em
pixels. a, diferença significativa entre os grupos CTRL e mdx-nujol; b, diferença significativa entre mdx-nujol com
mdx ômega-3. Média ± desvios padrões dos níveis do 4HNE (p≤0,05, Teste ANOVA:Kruskal-Wallist).
GAPDH
4-HNE 43kd
25kd
4-HNE
GAPDH
170kd
23kd 25kd
43kd 35kd
130kd

41
5.7 Atividade Enzimática
5.7.1 Atividade Enzimática da SOD
A atividade da SOD no músculo DIA dos animais mdx-nujol foi 43% menor em relação
ao CTRL. O ômega-3 aumentou os níveis da SOD em 41% comparado ao mdx-nujol (Figura
17).
No músculo QDR, a atividade da SOD estava cerca de 24.5% aumentada em relação ao
grupo CTRL. O tratamento com ômega-3 elevou em 7.6% a SOD (Figura 17).
Figura 17: Atividade enzimática da SOD nos músculos diafragma e quadríceps de animais CTRL, mdx- nujol e
mdx tratado com ômega-3. Média ± desvios padrões dos níveis da SOD (p≤0,05, Teste ANOVA:Kruskal-Wallist).
5.7.2 Atividade Enzimática da GPx e da GR
A atividade da GPx no músculo DIA dos animais mdx foi 37.8% menor comparado ao
CTRL. O ômega-3 aumentou a atividade da GPx em 20% (Figura 18A). Em relação ao CTRL,
o grupo mdx-nujol apresentou atividade semelhante da GR e no grupo mdx-ômega3 ocorreu
aumento de 16% da atividade desta enzima. O ômega-3 levou a um aumento de 21.8% na
atividade da GR, comparado ao mdx-nujol (Figura 18A).
No músculo QDR, a atividade da GPx no mdx-nujol foi semelhante a observada no animal
CTRL. O tratamento com ômega-3, quando comparado ao mdx-nujol, promoveu um aumento
de 8.72% na atividade da GPx (Figura 18B). A atividade da GR estava maior (6.3%) no mdx

42
em relação ao grupo CTRL. Os grupos mdx-nujol e mdx-ômega3 apresentaram atividades
semelhantes da GR (Figura 18B).
Figura 18: Atividade enzimática da GPx e da GR nos músculos: A) diafragma e B) quadríceps de animais CTRL,
mdx-nujol e mdx tratado com ômega-3. Média ± desvios padrões dos níveis da GPx e da GR (p≤0,05, Teste
ANOVA:Kruskal-Wallist).

43
6. Discussão
A DMD destaca-se dentre as distrofias musculares por ser a mais comum e devastadora,
com intensa degeneração e fraqueza muscular progressiva (ENGEL et. al., 1994). Como
modelo experimental para os estudos das distrofinopatias utiliza-se o camundongo mdx
(GROUNDS et.al, 2008). Na DMD e no camundongo mdx a proteína distrofina está ausente
ou não funcional na fibra muscular e como consequência o CDG se desorganiza tornando o
músculo mais frágil durante os ciclos de contração/relaxamento, aumentando o influxo de
cálcio na fibra muscular e a produção de radicais livres levando a necrose celular (GROUNDS
et. al., 2008; WHITEHEAD et al., 2006).
Nos animais mdx, a degeneração se inicia por volta dos 21 dias de vida pós-natal, com
ciclos de degeneração e regeneração e picos de inflamação. Além disso, há evidências de que
os músculos dos camundongos mdx são mais suscetíveis à degeneração induzida pelos ROS
(WHITEHEAD et al., 2006). Os radicais livres têm a capacidade de gerar danos musculares,
ativação do processo inflamatório (WHITEHEAD et al., 2006) e lesão oxidativa dos
componentes intracelulares resultando na produção e acúmulo dos grânulos de lipofuscina
(NAKAE et al., 2001).
Anti-inflamatórios esteroides como a prednisolona e o deflazacort são amplamente
utilizados como terapia para a Distrofia Muscular de Duchenne (DMD), pois comprovaram
ser eficientes na redução dos prejuízos funcionais da DMD. Porém, o seu uso prolongado
provoca efeitos colaterais significativos e prejudiciais ao organismo (MALIK et. al., 2012).
O ômega-3 é um ácido graxo poli-insaturado com ação anti-inflamatória encontrado em
peixes marinhos ou no óleo de peixe (CALDER, 2012). A ingestão de ômega-3 tem mostrado
ser eficaz na proteção de algumas situações fisiológicas, tais como em desordens inflamatórias
e doenças cardiovasculares (SIMOPOULOS, 2002) através da ação em promover mudanças
na produção das citocinas pró-inflamatórias e em antagonizar a produção de eicosanóides
derivados do ácido graxo poli-insaturado ômega-6 (BABCOCK et. al., 2000). O AA é
convertido em eicosanóides (prostaglandinas, prostaciclinas, tromboxanos e leucotrienos)
através da ação das enzimas cicloxigenase (COX) e lipoxigenase (LOX). Sugere-se que o EPA
iniba, competitivamente, a atividade da cicloxigenase e lipoxigenase, diminuindo a produção
dos eicosanoides da série par e, consequentemente, diminuindo a inflamação (CALDER,
2002).

44
No presente trabalho, avaliamos se as cápsulas de ômega-3 também diminuem o estresse
oxidativo nos músculos esqueléticos bíceps, diafragma e quadríceps de camundongos mdx, na
fase inicial da distrofinopatia.
6.1 Peso corporal
Observamos que o grupo mdx-ômega3 teve menor ganho de peso em relação ao CTRL e
mdx-nujol. Este fato não está de acordo com nossos estudos anteriores, em que não
observamos diferenças no ganho de peso em animais tratados com EPA purificado ou cápsulas
de ômega-3 (MAURÍCIO, 2012; MACHADO, 2011). Uma possiblidade para explicar este
achado estaria relacionada ao tamanho das ninhadas. A ninhada dos animais tratados com
ômega 3 era composta, em média, por 10 animais. Já as ninhadas dos animais CTRL e mdx-
nujol continham cerca de 8-9 animais. É sabido que ninhadas com número superior a 8 filhotes
podem favorecer a perda de peso entre os filhotes (BRAGGIO et al., 2003). Assim,
acreditamos que o menor ganho de peso observado nos animais tratados com ômega-3 seja
devido ao tamanho da ninhada e não a algum efeito colateral da droga.
6.2 Creatina Quinase (CK) e Força de Tração
A análise quantitativa da enzima creatina quinase (CK) expressa o quanto ela é liberada
pelos músculos esqueléticos e proporciona dados relacionados ao processo de degeneração
muscular. Embora seja um dos parâmetros mais indicados para o diagnóstico de lesões
musculares (GROUNDS et al., 2008), o nível sérico da CK pode apresentar considerável
variação individual. Assim, os valores de CK devem servir apenas como uma análise
complementar para o diagnóstico e acompanhamento da evolução das doenças
neuromusculares.
Em camundongos mdx, os níveis de CK se mostram altos ao longo da vida dos animais
(BULFIELD et al., 1984) e na DMD, os níveis estão cerva de 50 a 100 vezes acima dos limites
dos valores de referência (ENGEL et al., 1994). Neste trabalho, a creatina-quinase estava
aumentada significativamente nos grupos mdx comparados ao CTRL. O tratamento com
ômega-3 diminuiu os níveis séricos da CK em relação ao mdx-nujol. Acredita-se que esta
diminuição esteja relacionada a proteção contra a mionecrose, melhorando o quadro da
distrofinopatia. Estes dados condizem com resultados anteriores que relatam proteção contra

45
a mionecrose em camundongos tratados com EPA (MACHADO et al., 2011) e com as
cápsulas de ômega-3 (MAURÍCIO et al., 2012).
A força muscular foi medida no início e no final do tratamento. Houve aumento da força
no grupo mdx-ômega3 comparado ao mdx-nujol. Acredita-se que este aumento esteja
relacionado com a proteção contra o processo inflamatório. Os camundongos distróficos
apresentam degeneração e inflamação a partir dos vinte e um dias de vida pós-natal. Um
estudo anterior em nosso laboratório demonstrou que o tratamento com EPA reduziu os níveis
de TNF-α (citocina pró-inflamatória) e protegeu as fibras musculares dos camundongos
distróficos da degeneração (MACHADO et al., 2011).
Concluindo, estes dados mostram que o ômega-3 foi eficaz em reduzir a mionecrose
(diminuição da CK) e melhorar a força muscular de camundongos distróficos, sugerindo que
seu uso possa ser uma alternativa para a terapia das distrofinopatias.
6.3 Inflamação e Regeneração nos músculos BB, DIA e QDR
Houve diminuição na porcentagem da área de inflamação e de regeneração nos músculos
bíceps (BB) e diafragma (DIA) tratados com ômega-3. Os ciclos de degeneração/regeneração
também parecem ter diminuído, pois houve aumento da porcentagem de núcleos periféricos,
que são indicativos de fibras que não sofreram o processo de degeneração. Estes fatos são
condizentes com dados observados em trabalhos anteriores que utilizaram como tratamento o
EPA purificado (MACHADO et al., 2011) e as cápsulas de ômega-3 (MAURÍCIO et al., 2012)
e que mostram que a diminuição da mionecrose ocorre junto à preservação de maior número
de fibras com núcleo periférico.
No QDR dos animais mdx, o achado mais expressivo foi a regeneração muscular, que foi
estimulada pelo ômega-3. Estudos anteriores em nosso laboratório já haviam observado a
grande capacidade regenerativa do quadríceps em comparação ao diafragma e bíceps e os
efeitos positivos do ômega-3 sobre a regeneração. Possivelmente, a diminuição do NF-kb pelo
ômega-3 (APOLINÁRIO, 2012) possa colaborar para este achado, uma vez que o NF-kb
parece estar envolvido na diminuição da regeneração muscular (MESSINA et al., 2011).
Possivelmente, o quadríceps possua uma linhagem de células satélites diferenciadas em
relação ao DIA e BB, com maior capacidade regenerativa, mas estudos futuros serão
necessários para melhor entendimento destes resultados.

46
Assim, o tratamento com ômega-3 apresentou-se eficaz quanto a sua capacidade anti-
inflamatória nos músculos BB e DIA de camundongos mdx jovens.
6.4 Sobre a análise da Lipofuscina
A lipofuscina é considerada um marcador biológico de envelhecimento (NAKAE et al.,
2004). Muitos estudos têm demonstrado a presença da lipofuscina em músculos esqueléticos
e sua relação com a DMD. Acredita-se que o aumento da concentração de radicais livres,
produtos do estresse oxidativo celular, somados a ausência da distrofina, resulta em aumento
de lipofuscina nos músculos (NAKAE et al., 2004). Os grânulos autofluorescentes de
lipofuscina estão presentes em miofibras, células satélites e células intersticiais, incluindo
macrófagos (NAKAE et al., 2001). Em músculos normais jovens, não se observam grânulos
de lipofuscina em grandes quantidades. Porém, estudos tem demonstrado que em pacientes
DMD e camundongos mdx, a lipofuscina começa a se acumular muito cedo nos músculos
esqueléticos (NAKAE et al., 2004). Nos pacientes DMD com aproximadamente dois anos de
idade e nos camundongos mdx com quatro semanas de vida, quando comparados a um
músculo normal, em que a lipofuscina é detectada em torno de 20 semanas de vida (NAKAE
et al., 2004). No envelhecimento, estes grânulos aumentam em consequência de lesão
oxidativa de macromoléculas celulares (YIN, 1996; TERMAN & BRUNK, 1998).
As análises qualitativa e quantitativa mostraram a presença de um número considerável
de grânulos de lipofuscina em todos os músculos estudados, BB, DIA e QDR, nos grupos mdx
jovens. O acúmulo destes grânulos foi maior nos músculos mdx comparados ao grupo controle
na mesma idade, no qual quase nenhum grânulo foi observado. A análise qualitativa mostrou
que o acúmulo dos grânulos de lipofuscina nos músculos distróficos estavam condizentes com
os critérios pré-estabelecidos para este trabalho. Primeiro, os grânulos se apresentaram, em
geral, de forma arredondada ou ovalada. Segundo, os grânulos se apresentaram
autofluorescentes nos dois comprimentos de onda, 488 nm e 546 nm. Terceiro, a localização
dos grânulos de lipofuscina, estavam na maioria das vezes, associados ao sarcolema e
eventualmente, eles estavam presentes no sarcoplasma e no meio extracelular, provavelmente
associados as células inflamatórias. Estes dados são condizentes com os estudos realizados
por Nakae et. al., 2001 e 2004, os quais relatam a presença dos grânulos de lipofuscina nos
mesmos locais observados, no músculo diafragma dos grupos mdx e controle jovens.

47
No presente trabalho, observamos que a lipofuscina começa a se acumular muito cedo
nos músculos esqueléticos dos animais distróficos e que o tratamento com as cápsulas de
ômega-3 diminuiu o número desses grânulos em 61.5% no BB, 23% no DIA e de 73% no
QDR. Estes dados sugerem que a presença da lipofuscina em animais distróficos reflete o
aumento do estresse oxidativo e a debilitação da degradação lisossomal, assim como já
relatado em doenças degenerativas. Os radicais livres são produtos da lesão oxidativa e
Disatnik et al. (1998) verificou que a expressão de enzimas antioxidantes e a oxidação lipídica
estavam aumentadas nos músculos dos membros posteriores nos mdx de 15 a 20 dias de idade
e sugeriram que o nível de espécies reativas de oxigênio aumentou nos músculos na fase pré-
necrótica. Estas descobertas estão condizentes com Nakae et. al. (2004) que fornece a primeira
constatação de que o estresse oxidativo aumenta de maneira anormal nos músculos dos mdx
nos estágios iniciais de vida e isto implica que os danos oxidativos das biomoléculas celulares
aceleram a desordem nos músculos deficientes de distrofina.
Sabe-se que células tornam-se carregadas de lipofuscina em função do estresse oxidativo,
o qual gera inibição das proteases lisossomais, atrasando a degradação dos materiais
autofagocitados e favorecendo a oxidação dos mesmos (BRUNK & TERMAN, 2002).
O mecanismo pelo qual o ômega-3 diminuiu o número de grânulos de lipofuscina nos
músculos distróficos ainda não foi esclarecido. O DHA é o ácido graxo ômega-3 presente em
maior quantidade na membrana celular e estudos tem demonstrado que em diversos tecidos,
como no esquelético, o conteúdo de ácidos graxos ômega-3 aumenta em resposta a uma dieta
com DHA e EPA (ARTERBURN et al., 2006). Sabe-se também que a concentração do DHA
na membrana é significativamente menos abundante nos camundongos mdx (TUAZZON &
HENDERSON, 2012). Os ácidos graxos poli-insaturados ômega-3 influenciam algumas
propriedades fisiológicas da membrana, tais como fluidez, espessura e forma (ARTERBURN
et al., 2006). Nos animais mdx, a falta da distrofina no sarcolema da fibra muscular gera
instabilidade no CDG, alterando o funcionamento dos canais iônicos, o que gera um aumento
no influxo de cálcio para o meio intracelular (JUDÈ et al., 2006). No músculo distrófico, o
aumento da concentração do cálcio intracelular ativa proteases e fosfolipases que levam à
mionecrose (WHITEHEAD et al., 2006). Sabe-se também que a captação anormal de cálcio
pela mitocôndria pode resultar no aumento da produção de ROS, os quais reagem com os
componentes da membrana celular (lipídios e proteínas), degradando-os. Tais reações

48
favorecem o aumento da lesão celular e da inflamação, aumentando a produção de citocinas e
eicosanoides pró-inflamatórios. Muitos estudos já demonstraram que o ácido graxo poli-
insaturado ômega-3 (EPA + DHA) é um potencial anti-inflamatório e tem ação eficaz na
diminuição da mionecrose (MAURÍCIO et al., 2012). Seu uso diminui a produção de
eicosanóides derivados do ácido graxo poli-insaturado ômega-6, por competir pelas mesmas
vias enzimáticas da COX e da LOX, diminui a produção de citocinas pró-inflamatórias, tais
como a TNF-α (CALDER, 2002; CALDER, 2013) e consequentemente diminui a mionecrose
muscular.
A partir dessas informações, nossos resultados sugerem que a incorporação do ômega-3
(EPA + DHA) pode promover a estabilização do sarcolema, possivelmente pela incorporação
do DHA na membrana, diminuindo o influxo de cálcio e em consequência, a mionecrose. O
consumo de ômega-3 também pode ter melhorado a distrofinopatia ao modular as espécies
reativas de oxigênio (XIE et al., 2011; KAJAKAWIA et al., 2010). Estes dados juntos podem
explicar a diminuição dos grânulos de lipofuscina com o tratamento, já que com a diminuição
na entrada de cálcio na fibra muscular, pode diminuir a captação de cálcio pela mitocôndria e
consequentemente, a produção de ROS diminui. Juntos, tais processos possivelmente
diminuem a peroxidação e a oxidação dos componentes celulares que serão
auto/heterofagocitados pelos lisossomos reduzindo assim a produção de lipofuscina.
6.5 Sobre a análise do 4-HNE
O 4-hidroxinonenal (4-HNE) é considerado um marcador da peroxidação lipídica
(ESTERBAUER et al., 1991). Este processo causa a destruição de lipídios presentes na
membrana celular, resultando na formação de peróxidos lipídicos e aldeídos, tais como 4-
HNE e o malondialdeído (MDA) (ESTERBAUER & CHEESEMAN, 1990). Assim, o 4-HNE
é um produto final da peroxidação lipídica e tem sido aceito como um possível indutor do
estresse oxidativo, estando envolvido na patogênese de várias doenças degenerativas como o
Alzheimer e a aterosclerose (UCHIDA, 2003; ESTERBAUER et al., 1991). Estudos que
realizaram experimentos utilizando camundongos mdx observaram que, a peroxidação lipídica
da membrana celular causada pelo aumento da produção de ROS aconteceu no período que
antecede o início da degeneração das fibras musculares (DISATNIK et al., 1998). Assim,

49
nossos experimentos utilizaram o 4-HNE como marcador para analisar a peroxidação lipídica
decorrente da degeneração do músculo distrófico.
Nossos resultados demonstraram que o tratamento com as cápsulas de ômega-3 reduziu
o 4-HNE nos músculos BB e DIA. Concomitante a isso, as áreas de fibras com núcleo
periférico em ambos os músculos estavam aumentadas, indicativo de fibras integras, que não
sofreram processo de degeneração. De maneira geral, estes dados indicam que as cápsulas de
ômega-3 foram eficazes na redução do processo de peroxidação lipídica, provavelmente por
gerar uma estabilização no sarcolema e reduzir a degradação dos ácidos graxos poli-
insaturados pelas ROS e assim, diminuir a degeneração muscular nos músculos distróficos.
Estes fatos são condizentes com o trabalho de Maurício et al., 2012 que também utilizou como
tratamento as cápsulas de ômega-3 nos músculos dos camundongos mdx e cujos resultados
mostraram aumento da integridade do sarcolema (redução da degeneração), diminuição da
mionecrose e também do 4-HNE, protegendo o tecido muscular.
6.6 Sobre a atividade enzimática da SOD, GPx e GR.
O estresse oxidativo é resultado do desequilíbrio entre a ação dos agentes oxidantes e dos
antioxidantes, a favor dos primeiros (MOREL & BAROUKI, 1999). O estresse oxidativo
aumenta de maneira anormal nos músculos com DMD e nos músculos dos camundongos mdx
(NAKAE et al., 2004). Este fato implica que os danos oxidativos aceleram a lesão dos
músculos deficientes de distrofina (NAKAE et al., 2004; TIDBALL & WEHLING-
HENRICKS, 2007; LAWER, 2011).
Em relação à presença de ROS no músculo distrófico, várias hipóteses têm sido
levantadas. Acredita-se que com o influxo aumentado de cálcio para o espaço intracelular,
decorrente da instabilidade da membrana, gera uma captação anormal de cálcio pela
mitocôndria, o que prejudica a fosforilação oxidativa mitocondrial com consequente aumento
na produção de ROS (WHITEHEAD et al., 2006). Há também trabalhos sobre o papel da
fosfolipase A2 (PLA2), que é dependente de cálcio e presente no sarcolema, como fonte de
produção de ROS (LEEUWENBURGH & HEINECKE, 2001). Ela libera o ácido
araquidônico (AA), o qual é substrato para a ação da cicloxigenase e da lipoxigenase,
resultando na formação de prostaglandinas, leucotrienos, tromboxanos e também de ROS
(LEEUWENBURGH & HEINECKE, 2001).

50
As espécies reativas de oxigênio também têm sido relacionadas com o NF-κB (fator de
transcrição que participa da regulação de citocinas pró-inflamatórias, tais como TNF-α e IL-
1β; WHITEHEAD et al., 2006). O NF-κB transloca-se para o núcleo, onde se liga ao DNA
(ativação) e aumenta a produção de ROS e de proteínas e peptídeos pró-inflamatórios
(KUMAR et al., 2004). As células inflamatórias, tais como neutrófilos e macrófagos, os quais
são atraídos no local da lesão muscular, também podem produzir ROS. Estudos tem relatado
o papel da NAD(P)H oxidase como uma potencial fonte de estresse oxidativo e na
contribuição da patologia dos músculos esqueléticos na DMD (WHITEHEAD et al., 2008),
já que, uma vez presente na membrana celular, ela facilita a entrada do ânion superóxido (O₂⁻)
no espaço intersticial (FISHER, 2009) e do peróxido de hidrogênio (H₂O₂), os quais levam à
lesão celular, uma vez que reage com os diferentes componentes de membranas.
Antioxidante é qualquer substância que é capaz de retardar ou inibir a oxidação de
substratos oxidáveis de maneira eficaz (DROGE, 2002; HALLIWELL, 1991). Tais ações
podem agir sobre os radicais livres ou espécies não-radicais, diretamente neutralizando-os ou
indiretamente, participando dos sistemas enzimáticos com esta capacidade (HALIWELL et
al., 2004).
O sistema não-enzimático é representado pelos compostos antioxidantes de origem
dietética, tais como vitaminas, minerais e compostos fenólicos. O sistema enzimático inclui
as enzimas Superóxido Dismutase (SOD), Catalase (CAT) e Glutationa Peroxidase (GPx).
Essas enzimas atuam por meio de mecanismos de prevenção, impedindo e/ou controlando a
formação de radicais livres e espécies não-radicais, envolvidos com a iniciação das reações
em cadeia que resultam em propagação e amplificação do processo e, consequentemente, com
a ocorrência de danos oxidativos (FERREIRA & MATSUBARA, 1997; SCHENEIDER &
OLIVEIRA, 2004).
O presente trabalho mostra que a atividade enzimática da SOD, GPx e GR apresentou-se
diferenciada, dependendo do músculo estudado. Considerando-se o musculo quadríceps,
observamos que o mdx-nujol não apresentou alteração na atividade das enzimas antioxidantes,
em relação ao animal controle. Adicionalmente, o ômega-3 não levou a nenhuma alteração,
apenas uma tendência a aumentar a atividade da SOD e da GPx. Assim, o quadríceps distrófico
se assemelha ao quadríceps normal, sugerindo que o quadríceps distrófico tem capacidade
normal para tamponar as espécies reativas de oxigênio. Entretanto, é importante ressaltar que

51
nesta fase o quadríceps apresentava um fenótipo com pouca inflamação e intensa regeneração,
o que pode justificar a atividade normal destas enzimas, pelo menos nesta etapa da doença.
Muito embora a lipofuscina estivesse significativamente aumentada no quadríceps distrófico,
os níveis de 4-HNE, produto de peroxidação lipídica (ESTERBAUER & CHEESEMAN,
1990), estavam semelhantes ao do músculo controle. A lipofuscina está relacionada a
degeneração muscular. A regeneração do quadríceps sugere que, em estágios anteriores, este
tenha sofrido degeneração, o que pode explicar os altos níveis de lipofuscina. Em conjunto,
estes dados podem sugerir que os níveis de estresse oxidativo no quadríceps sejam menores
do que os observados no diafragma, o que pode justificar a atividade normal das enzimas
antioxidantes.
Considerando o diafragma distrófico, observamos altos níveis de lipofuscina e 4-HNE,
indicando que o estresse oxidativo estava em andamento, mesmo nesta fase inicial da doença.
De fato, a inflamação também foi uma característica deste músculo. Entretanto, a atividade
das enzimas antioxidantes não estava aumentada no diafragma do mdx-nujol (comparado ao
controle), o que seria esperado, como uma tentativa de tamponar as espécies reativas de
oxigênio que provavelmente estão sendo produzidas em maior quantidade. Por outro lado,
podemos entender que a diminuição da atividade destas enzimas no mdx explique, pelo menos
em parte, sua baixa capacidade para modular o estresse oxidativo, o qual é aumentado nos
músculos distróficos (NAKAE et al., 2004).
Trabalhos prévios demonstraram que no músculo distrófico, tanto em pacientes com
DMD, como no camundongo mdx, essas enzimas estão aumentadas (TIDBALL e WHELING-
HENRICKS, 2007; DISATNIK et al., 1998). Entretanto, os dados parecem ser discrepantes.
Assim, em alguns estudos (Ragusa et al.,1996) não foi demonstrada, em camundongos mdx,
uma correlação entre a atividade enzimática da SOD e da GPx e a severidade da
distrofinopatia, em diferentes músculos. Outro estudo mostrou que a avaliação destas enzimas
após a fase pré-necrótica pode gerar dados distintos, pois a degeneração muscular, assim como
o processo secundário de infiltração das células inflamatórias que ocorre em resposta a necrose
celular, podem estar associados com o aumento do estresse oxidativo (HALLIWELL &
GUTTERIDGE, 1989). Todas as medidas bioquímicas de lesão oxidativa obtidas durante a
fase ativa da doença podem ser uma consequência, e não uma causa, do processo degenerativo.
Desta forma, as discrepâncias entre os resultados de diferentes estudos podem ser devidas a

52
variações na intensidade da necrose, da substituição do tecido muscular por tecido adiposo e
conjuntivo, na quantidade de regeneração e na extensão da resposta inflamatória (DISATNIK
et al., 1998).
Observamos uma tendência do ômega-3 em aumentar a atividade das enzimas
antioxidantes. Cabe ressaltar que a metodologia empregada no presente estudo requisita
grande quantidade de material para ser analisado, o que nos levou a utilizar pools de músculos,
de vários animais e mesmo inviabilizou o uso do bíceps braquial. Este fato pode explicar o
desvio padrão elevado e a não significância entre as médias. É sabido haver grande
variabilidade entre camundongos mdx, em vários parâmetros de avaliação da progressão da
doença (GROUNDS et al., 2008), o que pode ter afetado o fato de usarmos pools de músculos
em uma técnica bioquímica sensível que mede um fenômeno complexo. Não obstante,
acreditamos que o ômega-3 possa ter um efeito também sobre a atividade destas enzimas, uma
vez que houve diminuição da lipofuscina e do 4-HNE, marcadores do estresse oxidativo.
Seria importante estudarmos os efeitos do ômega-3 em outras vias relacionadas ao
estresse oxidativo, tais como as relacionadas a NADPH-oxidase (GROTE et al., 2003). Este
sistema enzimático situa-se nas membranas celulares e possivelmente pode ter sua função
alterada pela falta da distrofina, promovendo o aumento da produção de ROS nos músculos
distróficos. Tal sistema enzimático poderia ser afetado diretamente pelo ômega-3, pois
acredita-se que este ácido graxo seja incorporado na membrana (ARTERBURN et al., 2006),
o que poderia promover a estabilidade molecular da mesma.

53
7. CONCLUSÕES
- O ômega-3 diminuiu de forma significativa os marcadores do estresse oxidativo (lipofuscina
e 4-HNE) em diferentes músculos distróficos, nos estágios iniciais da doença.
Adicionalmente, houve tendência do ômega-3 em melhorar a atividade de enzimas
antioxidantes. Isto sugere a ação antioxidante deste ácido graxo poli-insaturado, nos estágios
iniciais da distrofinopatia.
- Estes dados sugerem que o ômega-3, além de seus efeitos na mionecrose, inflamação e
regeneração muscular, também apresenta efeitos positivos sobre o estresse oxidativo,
tornando-o potencialmente útil para a terapia da DMD. Entretanto, deve-se salientar a
importância de estudos complementares sobre as doses adequadas para uso humano e os
possíveis efeitos colaterais que o uso contínuo do ômega-3 possa trazer para os pacientes com
DMD.

54
8. REFERÊNCIAS BIBLIOGRÁFICAS
ABDEL-SALAM E.; ABDEL-MEGUID I.; KORRAA S.S. Markers of degeneration and
regeneration in Duchenne muscular dystrophy. Acta mycological: myopathies and
cardiomyopathies: official journal of the Mediterranean society of myology.; 23(3): 94-
100, 2009.
AMES B. N., SHIGENAGA M; HAGEN T. Oxidants, antioxidants, and the degenerative
diseases of aging. Proc. Natl. Acad. Sci.; 90(17): 7915-7922, 1993.
APOLINÁRIO, M.L. “Ácido eicosapentaenoico x deflazacort: Mecanismos de ação e
comparação de efeitos no tratamento de camundongos distróficos da linhagem mdx.”, 2012.
Tese (Mestrado em Biologia Celular e Estrutural) -Instituto de Biologia, Universidade
Estadual de Campinas, SP. 2012.
ARTERBURN L.M.; HALL E.B.; OKEN H. Distribution, interconversion, and dose response
of n3 fatty acids in humans. Am. J. Clin. Nutr.; 83:1467S-1476S, 2006.
BABCOCK T.; HELTON W. S.; and JOSEPH E. N. Eicosapentaenoic Acid (EPA): An
Antiinflammatory Omega-3 Fat with Potential Clinical Applications. Nutrition.; 16:1116-
1118, 2000.
BASSOLI, D. A. A importância das células satélites na regeneração de músculos esqueléticos.
In: CLARETIANO - Revista do Centro Universitário.; 1(3): 166-174, 2003.
BECKMAN K, AMES B. The free radical theory of aging matures. Physiol. Rev.; 78(2): 547-
581, 1998.

55
BIGGAR, W.D.; HARRIS, V.A.; ELIASOPH, L.; ALMAN B. Long-term benefits of
deflazacort treatment for boys with Duchenne Muscular Dystrophy in their second decade.
Neuromuscular Disorders.; 16:249–255, 2006.
BOGDANOVICH, S.; PERKINS, K.J.; KRAG, T.O.B.; KHURANA, T.S. Therapeutics for
Duchenne muscular dystrophy: current approaches and future directions. Journal of
Molecular Medicine.; 82(2): 102-115, 2004.
BULFIELD, G.; SILLER, W.G.; WIGHT, P.A.L.; MOORE, K. X-chromosome-linked
muscular dystrophy (mdx) in the mouse. Processings of the National Academy of Sciences
of United States of America.; 81(4): 1189-1192, 1984.
BRAGGIO, M.M.; MARTINS, A.R.S.; VALEROS, V.B. Influência do manejo na
produtividade e no desenvolvimento de camundongos (Mus Musculus). Arq. Inst. Biol.;
70(2): 149-153, 2003.
BRUNK, U.; TERMAN A. Lipofuscin: mechanisms of age-related accumulation and
influence on cell function. Free Radical Biology & Medicine.; 33(5): 611–619, 2002.
BRUNK, U.; ERICSSON, J.L.E. Electron microscopical studies of ratbrain neurons.
Localization of acid phosphatase and mode offormation of lipofuscin bodies. J. Ultrastruct.
Res.; 38:1–15; 1972.
CALDER, P.C. Dietary modification of inflammation with lipids. Nutrition Society.;
61:345–358, 2002.
CALDER P.C. Polyunsaturated fatty acids, inflammatory processes and inflammatory bowel
diseases. Molecular Nutrition and Food research.; 52:885-897, 2008.
CALDER, P.C. Long-chain fatty acids and inflammation. Nutrition Society.; 71: 284–289,
2012.

56
CALDER, P.C. n-3 Fatty acids, inflammation and immunity: new mechanisms to explain old
actions. Nutrition Society.; 72: 326–336, 2013.
CARVALHO, S.C. Populações de macrófagos em músculos esqueléticos de camundongos
mdx tratados com ácido eicosapentaenoico. 2012. 53 f. Tese (Mestrado em Biologia Celular
e Estrutural) - Instituto de Biologia, Universidade Estadual de Campinas, SP. 2012.
CHAKKALAKAL, J. et al. Molecular, cellular, and pharmacological therapies for
Duchenne/Becker muscular dystrophies. FASEB J.; 19:880-891, 2005.
CLARKSON P.; THOMPSON H. Antioxidants: what role do they play in physical activity
and health? Am. J. Clin. Nutr.; 72: 637S-646S, 2000.
DECONINCK N.; DAN B. Pathophysiology of Duchenne Muscular Dystrophy: Current
Hypotheses. Pediatric neurology.; 36(1): 1-7, 2006.
DHAWAN, J.; RANDO, T.A. Stem cells in postnatal myogenesis: molecular mechanisms of
satellite cell quiescence, activation and replenishment. TRENDS Cell. Biol.; 15(12): 666-
673, 2005.
DISATNIK M-H.; DHAWAN J.; YU Y.; BEAL M.F.; WHIRL M.M.; FRANCO A.A.;
RANDO T.A. Evidence of oxidative stress in mdx mouse muscle: Studies of the pre-necrotic
state. J. Neurol. Sci.; 161: 77–84, 1998.
DRÖGE, W. Free radicals in the physiological control of cell function. Physiol. Rev.; 82: 47-
95, 2002.
ELLEDER, M.; SOKOLOVA, J.; HREBICEK, M. Follow-up study of subunit c of
mitochondrial ATP synthase (SCMAS) in Batten disease and in unrelated lysosomal
disorders. Acta Neuropathol.; 93:379–390; 1997.

57
ENDRES S.; GHORBANI R.; KELLEY V.E.; GEORGILIS K.; LONNEMANN G.; VAN
DER MEER J.M.W.; CANNON J.G.; ROGERS T.S.; KLEMPNER M.S.; WEBER P.C.; et
al. The effect of dietary supplementation with n-3 polyunsaturated fatty acids on the synthesis
of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med.; 320:265–
71, 1989.
ENGEL, A.G.; YAMAMOTO, M.; FISCHBECK, K.H. Dystrophinopathies. In: Engel, AG,
Franzini-Armstrong, C, editors Myology. New York: McGraw-Hill.; 2: 1133-1187, 1994.
ESTERBAUER, H.; CHEESEMAN K. H. Determination of aldehydic lipid peroxidation
products: malonaldehyde and 4-hydroxynonenal. Methods Enzymol.; 186: 407-21, 1990.
ESTERBAUER, H.; SCHAUR R. J.; et al. Chemistry and biochemistry of 4-hydroxynonenal,
malonaldehyde and related aldehydes. Free Radic. Biol. Med.; 11(1): 81-128, 1991.
FAGANELLO, F.R. Ação do ultra-som terapêutico no processo de regeneração do músculo
esquelético. 2003. 86 f. Dissertação (Mestrado em Biodinâmica da Motricidade Humana) -
Instituto de Biociências, Universidade Estadual Paulista, Rio Claro, 2003.
FAIRCLOUGH, R.J.; BAREJA, A.; DAVIES, K.E. Progress in therapy for Duchenne
muscular dystrophy. Experimental Physiology.; 96(11): 1101-13, 2011.
FERRARI, R. J; et al. Processo de Regeneração na Lesão Muscular: uma revisão. In:Revista
Fisioterapia em Movimento.; 18(2): 63 -71, 2005.
FERREIRA, A.L.; MATSUBARA L.S. Free radicals: concepts, associated diseases, defense
system and oxidative stress. Rev. Assoc. Med. Bras.; 43(1): 61-8, 1997.
FISHER, A.B. Redox signaling across cell membranes. Antioxid. Redox Signal.; 11: 1349–
1356, 2009.

58
FOSCHINI, R.M.S.A.; RAMALHO, F.S.; BICAS H.E.A. Células satélites musculares. Arq.
Bras. Oftamol.; 67: 681-687, 2004.
GOMEZ-CABRERA M.; BORRÁS C.; PALLARDÓ F.; SASTRE J.; JI L.; VIÑA J.
Decreasing xanthine oxidase-mediated oxidative stress prevents useful cellular adaptations to
exercise in rats. J. Physiol.; 567(Pt 1): 113-120, 2005.
GROUNDS, M.D.; RADLEY, H.G.; LYNCH, G.S.; NAGARAJU, K.; DE LUCA, A.
Towards developing standard operating procedures for pre-clinical testing in the mdx mouse
model of Duchenne muscular dystrophy. Neurobiology of Disease.; 31: 119, 2008.
GROTE K.; FLACH I.; LUCHTEFELD M. et al. Mechanical stretch enhances mRNA
expression and proenzyme release of matrix metalloproteinase2 (MMP-2) via NAD(P)H
oxidase-derived reactive oxygen species. Circ. Res.; 92(11): E80–6, 2003.
HALLIWELL, B.; GUTTERIDGE J.M.C. Free Radicals in Biology and Medicine, 2nd ed.,
Oxford: Clarendon Press, 1989.
HALLIWELL, B. Reactive oxygen species in living systems: source, biochemistry, and role
in human disease. Am. J. Med.; 91(suppl 3C): 14S-22S, 1991.
HALLIWELL, B.;WHITEMAN M. Measuring reactive species and oxidative damage in vivo
and in cell culture: how should you do it and what do the results mean? Br. J. Pharmacol.;
142(2): 231-55, 2004.
HOFFMAN, E.P.; BROWN, R.H.; KUNKEL, L.M. Dystrophin: the protein of Duchenne
muscular dystrophy locus. Cell.; 51: 919-928, 1987.
IDE, T.; TSUTSUI, H.; KINUGAWA, S.; UTSUMI, H.; KANG, D.; HATTORI, N.;
UCHIDA, K.; ARIMURA, K.; EGASHIRA, K.; TAKESHITA, A. Mitochondrial electron

59
transport complex I is a potential source of oxygen free radicals in the failing myocardium.
Circul. Res.; 15: 357-363, 1999.
JI, L. Antioxidants and oxidative stress in exercise. Proc. Soc. Exp. Biol. Med.; 222: 283-
292, 1999.
JUDÈ, S.; ROGER, S.; MARTEL, E.; BESSON, P.; RICHARD, S.; BOUGNOUX, P.;
CHAMPEROUX, P.; LE GUENNEC, J.Y. Dietary long-chain omega-3 fatty acids of marine
origin: a comparison of their protective effects on coronary heart disease and breast cancers.
Progress in biophysics and molecular biology.; 90: 299-325, 2006.
KAJIKAWA, S.; IMADA, K.; TAKEUCHI, T.; SHIMIZU, Y.; KAWASHIMA, A.;
HARADA, T.; MIZUGUCHI, K. Eicosapentaenoic Acid Attenuates Progression oh Hepatic
Fibrosis with Inhibition of Reactive Oxygen Species Production in Rats Fed Methionine- and
Choline-Deficient Diet. Digestive diseases and sciences.; 56(4): 1065-74, 2010.
KUMAR, A.; TAKADA, Y.; BORIEK, A.M.; AGGARWAL, B.B. Nuclear factor-kB: its role
in health and disease. J. Mol. Med.; 82:434–448, 2004.
LAWLER, J. M. Exacerbation of pathology by oxidative stress in respiratory and locomotor
muscles with Duchenne muscular dystrophy. The Journal of Physiology.; 589(9):2161–
2170, 2011.
LE GRAND, F.; RUDNICKI, M.A. Skeletal muscle satellite cells and adult myiogenesis.
Curr. Opin. Cell Biol.; 19: 628-633, 2007.
LEEUWENBURGH, C.; JI, L. Alteration of glutathione and antioxidant status with exercise
in unfed and refed rats. J. Nutr.; 126(7): 1833-1843, 1996.
LEEUWENBURGH, C.; HEINECKE, J. Oxidative stress and antioxidants in exercise. Curr.
Med. Chem.; 8(7): 829-838, 2001.

60
LEFAUCHER, J.P.; PASTORET, C.; SEBILLE, A. Phenotype of distrophinophathy in old
mdx mice. The Anatomical record.; 242: 70 –76, 1995.
LO, C.H.; CHIU, K.C.; FU, M.; LO, R.; HELTON, S. Fish Oil Augments Macrophage
Cyclooxygenase II (COX-2) Gene Expression Induced by Endotoxin. Journal of Surirgical
Research.; 86: 103-107, 1999.
MACHADO, R.V.; MAURICIO, A.F.; TANIGUTI, A.P.T.; FERRETI, R.; SANTO NETO,
H.; MARQUES, MJ. Eicosapentaenoic acid decreases TNF-alpha and protects dystrophic
muscles of mdx mice from degeneration. Journal of Neuroimmunology.; 232: 145-50, 2011.
MAGEE, P.; PEARSON, S.; ALLEN, J. The omega-3 fatty acid, eicosapentaenoic acid
(EPA), prevents the damaging effects of tumor necrosis factor (TNF)-alpha during murine
skeletal muscle cell differentiation. Lipids in health and disease.; 7(24): 1-11, 2008.
MALIK, V.; RODINO-KLAPAC, L.; MENDELL, J.R. Emerging Drugs for Duchenne
Muscular Dystrophy. Expert Opin. Emerg. Drugs.; 17(2): 261–277, 2012.
MARQUES, M.J.; MACHADO, R.V.; MINATEL, E.; SANTO NETO, H. Disodium
cromoglycate protects dystrophin-deficient muscle fibers from leakiness. Muscle & Nerve.;
37(1): 61-67, 2008.
MAURÍCIO, A.F. Efeito do ômega-3 em músculos de animais distróficos da linhagem mdx.
2012. 53 f. Tese (Mestrado em Biologia Celular e Estrutural) - Instituto de Biologia,
Universidade Estadual de Campinas, SP. 2012.
MAURÍCIO, A.F., MINATEL, E., NETO, H.S., MARQUES, M.J. Effects of fish oil
containing eicosapentaenoic acid and docosahexaenoic acid on dystrophic mdx mice. Clinical
Nutrition, 2012.

61
MESSINA, S.; BITTO, A.; AGUENNOUZ M.; VITA, G.L.; POLITO, F.; IRRERA, N.;
ALTAVILLA, D.; MARINI, H.; MIGLIORATO, A.; SQUADRITO, F.; VITA, G. The soy
isoflavone genistein blunts nuclear factor kappa-B, MAPKs and TNF-a activation and
ameliorates muscle function and morphology in mdx mice. Neuromuscul. Disord.; 21: 579-
89, 2011.
MOREL, Y.; BAROUKI, R. Repression of gene expression by oxidative stress. Biochem J.;
342: 481-496, 1999.
NAKAE, Y.; STOWARD, P.J.; SHONO, M.; MATSUZAKI, T. Most apoptotic cells in mdx
diaphragm muscle contain accumulated lipofuscin. Histochem. Cell Biol.; 115:205-214,
2001.
NAKAE, Y.; STOWARD, P.J.; KASHIYAMA; SHONO, M.; AKAGI, A.; MATSUZAKI,
T.; NONAKA, I. Early onset of lipofuscin accumulation in dystrophin-deficient skeletal
muscles of DMD patients and mdx mice. Journal of Molecular Histology.; 35:489-499,
2004.
PALMA, C.; PERROTTA, C.; PELLEGRINO, P.; CLEMENTI, E.; CERVIA, D. Skeletal
muscle homeostasis in Duchenne muscular dystrophy: modulating autophagy as a promising
therapeutic strategy. Aging Neurosciense, 2014.
PASTORET, C.; SEBILLE, A. Mdx mice show progressive weakness and muscle
deterioration with age. Journal of Neurological Science.; 129: 97-105, 1995.
PINTO, S. S; CASTILLO, A. A. Lesão muscular: fisiopatologia e tratamento. Revista
Fisioterapia em Movimento.; 12(2): 23-36, out./98/mar./99 .
POWERS, S.; LENNON, S. Analysis of cellular responses to free radicals: focus on exercise
and skeletal muscle. Proc. Nutr. Soc.; 58(4): 1025-1033, 1999.
.

62
PRISK, V.; HUARD, J. Muscle injuries and repair: the role of prostaglandins and
inflammation. Histol. Histopathol.; 18(4):1243-56, 2003.
RAGUSA, R.J.; CHOW, C.K.; ST. CLAIR D.K., PORTER, J.D. Extraocular, limb, and
diaphragm muscle group-specific antioxidant enzyme activity patterns in control and mdx
mice. J. Neurol. Sci.; 139:180–6, 1996.
RANDO, T.A.; DISATNIK, M.H.; YU, Y.; FRANCO, A. Muscle cells from mdx mice have
an increased susceptibility to oxidative stress. Neuromuscul. Disord.; 8:14–21, 1998.
RANDO, T.A. Role of nitric oxide in the pathogenesis of muscular dystrophies: a “two hit”
hypothesis of the cause of muscle necrosis. Microscopy Research and Thechnique.; 55:
223-235, 2001.
RENJINI, R.; GAYATHRI, N.; NALINI, A.; SRINIVAS BHARATH, M. M. Oxidative
Damage in Muscular Dystrophy Correlates with the Severity of the Pathology: Role of
Glutathione Metabolism. Neurochem Res.; 37:885-898, 2012.
ROVER, JR. L.; HOEHR, N.F.; VELLASCO, A.P. Sistema antioxidante envolvendo o ciclo
metabólico da glutationa associado a métodos eletroanalíticos na avaliação do estresse
oxidativo. Química Nova.; 24(1): 112-9, 2001.
SIMOPOULOS, A.P. Ômega-3 fatty acids in inflammation and autoimmune diseases. J. Am.
Coll. Nutr.; 21:495-505, 2002.
SCHNEIDER, C. D.; OLIVEIRA, A. R. Radicais livres de oxigênio e exercício: mecanismos
de formação e adaptação ao treinamento físico. Rev. Bras. Med. Esporte.; v.10, n.4, 2004.
SPENCER, M.J; TIDBALL, J.G. Do immune cells promote the pathology of dystrophin
deficient myopathies? Neuromuscular Disorders.; 11(6-7): 556-564, 2001.

63
STRAUB, V.; CAMPBELL, K. Muscular dystrophies and the dystrophin-glycoprotein
complex. Curr. Opin. Neurol.; 10(6): 168-175, 1997.
TANIGUTI, AP. T. “ Fibrose muscular em camundongo mdx: efeitos do exercício físico e de
agente anti-fibrótico.”, 2011. Tese (Doutorado em Biologia Celular e Estrutural) -Instituto de
Biologia, Universidade Estadual de Campinas, SP. 2011.
TERMAN, A.; BRUNK, U.T. Lipofuscin: mechanisms of formation and increase with age.
APMIS.; 106:265–276; 1998.
TERMAN, A.; BRUNK, U.T. Molecules in focus Lipofuscin. The International Journal of
Biochemistry & Cell Biology.; 36:1400–1404, 2004.
TIDBALL, J.G. Inflammatory processes in muscle injury and repair. American Journal of
Physiology – Regulatory, Integrative and Comparative Physiology.; 288: 345-353, 2005.
TIDBALL J.G.; WEHLING-HENRICKS, M. The role of free radicals in the pathophysiology
of muscular dystrophy. J. Appl. Physiol.; 102(4): 1677-1686, 2007.
TUAZON, M.A.; HENDERSON, G.C. Fatty acid profile of skeletal muscle phospholipid is
altered in mdx mice and predictive of disease markers. Metabolism.; 61: 801-811, 2012.
UCHIDA, K. 4-Hydroxy-2-nonenal: a product and mediator of oxidative stress. Prog. Lipid
Res.; 42(4): 318-43, 2003.
XI, S.; COHEN, D.; BARVE, S.; CHEN, L.H. Fish oil suppressed cytokines and nuclear factor
kappaB induced by murine AIDS virus infection. Nutrition Research.; 21:865–878, 2001.
XIE, N.; ZHANG, W.; LI, J.; LIANG, H.; ZHOU, H.; DUAN, W.; XU, X.; YU, S.; ZHANG,
H.; YU, D. a-Linolenic acid intake attenuates myocardial ischemia/reperfusion injury through

64
anti-inflammatory and anti-oxidative stress effects in diabetic but not normal rats. Archives
of Medical Research.; 42: 171-181, 2011.
VERCHERAT, C.; CHUNG, T.K.; YALCIN, S.; GULBAGCI, N.; GOPINADHAN, S.;
GHAFFARI, S.; TANEJA, R. Stra13 regulates oxidative stress mediated skeletal muscle
degeneration. Human Molecular Genetics.; 18(22): 4304-4316, 2009.
YIN, D. Biochemical basis of lipofuscin, ceroid and age pigment-like fluorophores. Free
Radic. Biol. Med.; 21: 871 – 888, 1996.
WEHLING, M.; SPENCER, M.J.; TIDBALL, J.G. A nitric oxide synthase transgene
ameliorates muscular dystrophy in mdx mice. J. Cell Biol.; 155:123–131, 2001.
WHITEHEAD, N.P.; YEUNG, E.W.; ALLEN, D.G. Muscle damage in mdx (dystrophic)
mice: role of calcium and reactive oxygen species. Clinical and Experimental
Pharmacology & Physiology.; 33: 657-662, 2006.
WHITEHEAD, N.P.; PHAM, C.; GERVASIO, O.L.; ALLEN, D.G. N-Acetylcysteine
ameliorates skeletal muscle pathophysiology in mdx mice. J. Physiol.; 586(7): 2003–2014,
2008.

65

66
ANEXO





























