Embed Size (px)
Citation preview

Instituto Nacional do Câncer Programa de Pós-Graduação Strictu-Sensu em Oncologia
Março de 2009
CAROLINA LAZZAROTTO-SILVA
Estudos Moleculares de
Células Tronco Mesenquimais
Cultivadas In Vitro

CAROLINA LAZZAROTTO-SILVA
Estudos Moleculares de
Células Tronco Mesenquimais
Cultivadas In Vitro
Tese submetida ao Programa de Pós-Graduação em Oncologia Strictu-Sensu para
obtenção do grau de Doutor em Oncologia
Orientador: Eliana Saul Furquim Werneck Abdelhay
Rio de Janeiro
2009

L432e Lazzarotto-Silva, Carolina.
Estudos moleculares de células tronco mesenquimais cultivadas in vitro.
/ Carolina Lazzarotto-Silva. - Rio de Janeiro: INCA, 2009.
159 f. il.
Tese (Doutorado) - Instituto Nacional de Câncer. Programa de Pós-
graduação Stricto Sensu em Oncologia do Instituto Nacional de Câncer
(INCA-RJ), 2009. Orientadora: Eliana Saul Furquim Werneck Abdelhay.
1. Células Tronco Mesenquimais. 2. In Vitro. 3. Medula Óssea. 4.
Proteoma. 5. Expressão Gênica. I. Abdelhay, Eliana Saul Furquim
Werneck. II. Título.
CDD 616.02774

Carolina Lazzarotto-Silva
Estudos Moleculares de Células Tronco Mesenquimais
Cultivadas In Vitro
Tese de Doutorado
Instituto Nacional do Câncer
Pós-Graduação Strictu-Sensu em Oncologia
Data de Aprovação: _____/_____/_____
Dra. Nance Beyer Nardi ______________________________________
Dr. Alex Balduino de Souza ____________________________________
Dr. Hector Seuánez Abreu ____________________________________
Dr. Fábio Passetti __________________________________________
Dra. Teresa de Souza Fernandes _______________________________
Dra. Maria Eugênia Leite Duarte ________________________________

Agradecimentos
AGRADECIMENTOS
A todos que, de alguma maneira, contribuíram para o desenvolvimento
deste trabalho.
Mais importante ainda, agradeço a todos que ajudaram no meu
desenvolvimento científico e pessoal durante esses 4 anos, pois uma tese de
Doutorado é, acima de tudo, uma experiência de vida. E nessa minha
experiência, contei com a ajuda de muitos, cada um colocando um pedacinho
para construir a pessoa que sou agora.
Agradeço à minha orientadora Eliana, que exerce essa função desde
1996, quase 13 anos. Agradeço por toda sua dedicação, ensinamentos e
paciência, e pela oportunidade de partilhar sua companhia e experiência na
ciência.
Agradeço às minhas queridas amigas Luciana e Renata. Luciana, por ser
nossa mentora no mundo do proteoma, pela fundamental ajuda nas análises,
pelas relevantes discussões sobre nossos dados, e por ser a mãe mais linda
do ano. Renata, por ser praticamente meu braço direito e esquerdo nesse
trabalho, mas principalmente pelo carinho e companheirismo em todos os
momentos. E por ser uma mulher e amiga maravilhosa.
À Júlia e Bárbara, que contribuíram de forma essencial nos experimentos,
quando ainda eram alunas de IC, e agora que já trilham seus próprios projetos.
E pelo apoio nos momentos bons e nos difíceis.

Agradecimentos
Ao André, peça fundamental nas últimas análises desse trabalho, que
além de ser nosso mentor no PCR em tempo real, tem a paciência para
aguentar mais de 10 mulheres com seus problemas existenciais.
Aos queridos companheiros do Laboratório de Célula Tronco, incluindo
aqueles que já passaram e os que ainda estão (por favor, sintam-se todos
incluídos), pela amizade, companheirismo, parcerias científicas e festivas, e por
terem proporcionado momentos tão bons nesses anos. E à Dani pequena, por
não ter feito nada em especial, mas também por não ter atrapalhado.
À Teresa Fernandes, minha primeira orientadora, e que agora volta a me
ajudar nas análises citogenéticas.
A todos da Divisão de Laboratórios do CEMO, pela parceria no nosso
espaço.
Aos meus queridos pais e ao Juca, que sempre acreditaram e apostaram
em mim e na minha vitória, me proporcionando as melhores condições para eu
ser quem eu sou hoje, sobretudo amor e exemplo. Vocês são os maiores
homenageados hoje.
Ao meu amor Antônio, que é a pessoa mais maravilhosa que eu já
conheci, e que tem me ajudado a entender melhor o mundo, e a mim mesmo.
Enfim, agradeço a todos, e principalmente a Deus, por ter passado por
esse período de sacrifícios e vitórias da melhor forma possível, e poder
defender essa tese.

Abstract
ABSTRACT
Mesenchymal Stem Cells (MSC) can be expanded ex vivo and are able to
differentiate along multiple lineages, including chondrocytes, osteoblasts and
adipocytes. They secrete a number of cytokines and regulatory molecules
implicated in different aspects of hematopoiesis, and seem to modulate the
immune system. Because of these characteristics, MSC represent a promissing
tool for cellular therapies, especially that related to bone marrow
transplantation. The aim of this work was to improve the molecular
characterization of MSC cultures using genomic and proteomic tools. Protein
expression profile was analyzed by bidimensional electrophoresis and in
tandem mass spectrometry. In a window of observation (pH 4-7, MW 10-220
KDa), 84 distinct proteins were identified in MSC cultures. Our analyses
demonstrated a very similar proteomic profile of MSC cultures of the first
passage derived from different donors, suggesting that these cells have the
same expression pattern. Additionally, cells derived from different donors were
able to equally inhibit lymphocyte proliferation. Comparative analyses of
genomic and proteomic pattern of MSC under different culture passages
showed that the in vitro cultivation procedure is able to standardize the cells, in
terms of their molecular profile. Although MSC maintain their in vitro
immunomodulatory potential, after passage 5 they developed karyotypic
alterations, indicating genetic instability of cultured cells. Proteomic analyses of
MSC submitted to adipogenic and osteogenic differentiation suggest that these
processes strongly modify their molecular profile, affecting the expression of
various proteins, such as those involved in actin skeleton of cells.

Resumo
RESUMO
Células Tronco Mesenquimais (CTM) podem ser expandidas ex vivo e são
capazes de se diferenciar em diversas linhagens, incluindo condrócitos,
osteoblastos e adipócitos. Elas secretam uma série de citocinas e moléculas
regulatórias implicadas em diferentes aspectos da hematopoese, e parecem
modular o sistema imune. Devido a essas características, as CTM representam
promissoras ferramentas para a terapia celular, sobretudo aquelas associadas
ao transplante de medula óssea. O objetivo deste trabalho foi melhorar a
caracterização molecular de culturas de CTM, através da utilização de
ferramentas genômicas e proteômicas. O perfil de expressão proteica de
culturas de CTM foi analisado através de ensaios de eletroforese bidimensional
e espectrometria de massas in tandem. Na janela de análise utilizada (pH 4-7,
PM 10-220 KDa), foram identificadas 84 proteínas distintas em culturas de
CTM. Nossas análises mostraram um padrão proteômico muito similar entre
culturas de CTM derivadas de diferentes doadores já na primeira passagem,
sugerindo que as células apresentam o mesmo padrão de expressão protéica.
Além disso, culturas derivadas de diferentes doadores são igualmente capazes
de inibir a proliferação linfocitária in vitro. A análise comparativa dos padrões
genômicos e proteômicos de CTM submetidas a diferentes passagens em
cultura, mostrou que o procedimento de cultivo in vitro de células estromais é
capaz de uniformizar a população em cultura, pelo menos no que diz respeito a
seu perfil molecular. Apesar de manterem o potencial imunomodulatório in vitro,
as células adquiriram, depois da passagem 5, alterações no seu cariótipo,
indicando que elas apresentam instabilidade genética em cultura. Análises

Resumo
proteômicas de CTM submetidas à diferenciação adipogênica e osteogênica
sugerem ainda que a diferenciação in vitro das CTM altera de forma importante
o perfil molecular das culturas, afetando a expressão de diversas proteínas,
como as envolvidas na estrutura do esqueleto de actina das células.

Siglas e Abreviaturas
SIGLAS E ABREVIATURAS
µµµµg - micrograma
µµµµL – microLitro
µµµµM – microMolar
ACN - ACetoNitrila
ACTB - actina citoplasmática 1 (β-actina)
ACTBL2 - proteína 2 tipo ββββ-actina (κ-actina)
Ang-1 - Angiopoetina-1
ASB-14 – detergente Zwitteriônico
ββββ2M - ββββ-2 Microglobulina
BLAST - Basic Local Alignment Search Tool (ferramenta de procura por
alinhamento de sequências)
BSA – Bovine Serum Albumin, (Albumina Bovina Sérica)
CapG - proteína de capeamento de macrófagos
CD – Cluster of Differentiation (grupo de diferenciação)
cDNA – complementary DesoxyriboNucleic Acid (ácido desoxiribonucleico
complementar)
CEMO – Centro de Transplante de Medula Óssea
CFSE - CarboxyFluorescein Succinimidyl Ester
cGy – centiGray
cm – centímetro
CML - Culturas Mistas de Linfócitos
CMN - Células MonoNucleares
CMNe – Células MonoNucleares estimuladoras
CMNr – Células MonoNucleares respondedoras
CPH - Células Progenitoras Hematopoéticos
CTH – Célula Tronco Hematopoética
CTM – Célula Tronco Mesenquimal
dATP - deoxyAdenosine TriPhosphate (trifosfato de desoxiadenosina)
dCTP – deoxyCytidine TriPhosphate (trifosfato de desoxicitidina)
DEPC - DiEtil PiroCarbonato

Siglas e Abreviaturas
dGTP – deoxyGuanosine TriPhosphate (trifosfato de desoxiguanosina)
DMEM - Dulbecco’s Modified Eagle’s Medium (meio de Eagle modificado por
Dulbecco)
dNTP - deoxyNucleotIde TriPhosphate (trifosfato de desoxinucleotídeo)
DTT - DiTiTreitol
dTTP – deoxyThymidine TriPhosphate (trifosfato de deoxitimidina)
EDTA - Ethylene Dyamin Tetracetic Acid (Ácido etileno diamintetracético)
EGTA - Ethylene Glycol Tetracetic Acid (ácido etileno glicoltetraacético)
eIF5A - isoforma 2 do fator eucariótico de iniciação da tradução 5A-1
EUA – Estados Unidos da América
FGF-2 - Fibroblast Growth Factor 2 (fator de crescimento de fibroblastos 2 )
FGF-7 - Fibroblast Growth Factor 7 (fator de crescimento de fibroblastos 7)
FITC - Fluorescein IsoThioCyanate (isotiocianato de fluoresceína)
Flt3 - FMS-like tyrosine kinase 3 (tirosina quinase 3 tipo FMS)
g – grama
Gal-1 – Galectina 1
G-CSF – Granulocyte-Colony Stimulating Factor (fator estimulador de colônias
granulocíticas)
GM-CSF – Granulocyte/Macrophage-Colony Stimulating Factor (fator
estimulador de colônias granulocítica-macrofágicas)
IDT - Integrated DNA Technologies (tecnologias integradas de DNA)
IEF - IsoEletroFocalização
INCa – Instituto Nacional de Câncer
IPG - Immobilized pH Gradients (gradients imobilizados de pH)
IPI - International Protein Index (índice internacional de proteínas)
L – Litro
LIF - Leukemia-Inhibiting Factor (fator inibitório de leukemia)
LPL - LipoProteína Lipase
M – Molar
MALDI-TOF-TOF – Matrix Assisted Laser Desorption/Ionization – Time Of
Flight - Time Of Flight
MCAM/CD146 - Melanoma-associated Cell Adhesion Molecule (molécula de
adesão celular associada a melanoma)

Siglas e Abreviaturas
M-CSF – Macrophage-Colony Stimulating Factor (fator estimulador de colônias
macrofágicas)
mg –miligrama
mL – mililitro
mM – milimolar
MS – Mass Spectrometry (espectrometria de massas)
NaCl - cloreto de sódio
NaF - fluoreto de sódio
ng – nanograma
NGF - Neural Growth Factor (fator de crescimento neural)
NH4HCO3 - bicarbonato de amônio
NK - Natural Killer
nM – nanolitro
nM – nanomolar
Pat - Plasminogen activator inhibitor (ativador do plasminogênio tecidual)
PBS – Phosphate Buffer Saline (salina de tampão fosfato)
PCR - Polymerase Chain Reaction (reação em cadeia da polimerase)
PDGF - Platelet-Derived Growth Factor (fator de crescimento derivado de
plaquetas)
PE - PhycoEritrin (ficoeritrina)
PFA – ParaFormAldeído
pg - picograma
pH – potencial Hidrogeniônico
pI - ponto Isoelétrico
pM – picoLitro
pM – picoMolar
PPARγ - Peroxisome Proliferator-Activated Receptor γ
RNAm - messenger RiboNucleic Acid
RNAmi – micro RNA
SBF - Soro Bovino Fetal
Sca-1 - Stem Cells Antigen-1 (antígeno 1 de células tronco)
SCF - Stem Cell Factor (fator de células tronco)
SDS-PAGE - Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
(eletroforese em gel de poliacrilamida com sulfato de dodecilsódico)

Siglas e Abreviaturas
αSMA – alfa Smooth Muscle Actin (αactina de músculo liso)
TCTH – Transplante de Células Tronco Hematopoéticas
TFA - TriFluoroacetic Acid, (ácido trifluoroacético)
TMO - Transplante de Medula Óssea
TNF-αααα - Tumor Necrosis Factor-alpha (fator alfa de necrose tumoral)
TUB - TUBulina
TUBA4B - αααα-TUBulina 4B
TUBA1A – cadeia αααα-1A da TUBulina
TUBB – cadeia ββββ-1ββββ-2 da TUBulina
TUBB2A – cadeia ββββ-2A da TUBulina
TUBB3 – cadeia ββββ-3 da TUBulina
UCHL - Ubiquitin carboxyl-terminal hydrolase isozyme L1
UFC-F – Unidades Formadoras de Colônia de Fibroblastos
VCL - isoforma 1 da vinculina
VEGF – Vascular Endothelial Growth Factor (fator de crescimento endotelial
vascular)
Wdr1 - Isoforma 2 da proteína 1 contendo repetição WD

Índice
ÍNDICE
Página
INTRODUÇÃO 1
Diferenciação das CTM 8
Caracterização Antigênica 11
Identidade In Vivo 14
Migração em Humanos e Modelos Animais 17
Envelhecimento e Efeito da Expansão in vitro na Biologia das CTM 18
Caracterização Molecular 24
Modulação do Sistema Imune 26
Potencial em terapias celulares 32
Terapias Celulares associadas ao Transplante de Medula Óssea 39
OBJETIVOS 43
OBJETIVO GERAL 43
OBJETIVOS ESPECÍFICOS 43
METODOLOGIA 44
1. Isolamento e Cultivo de CTM 44
2. Caracterização das Culturas de CTMh 46
2.1 Análise ao Microscópio Ótico 46
2.2 Análise de Marcadores de Superfície 46
2.3 Ensaios de diferenciação in vitro 46
2.3.1 Diferenciação Osteogênica 47
2.3.2 Diferenciação Adipogênica 47
3 Caracterização Molecular 47

Índice
3.1 Análise da Expressão Gênica 47
3.1.1 Extração de mRNA (messenger RiboNucleotidic Acid) 47
3.1.2 Elaboração dos iniciadores 48
3.1.3 Síntese de cDNA 49
3.1.4 Amplificação do cDNA por PCR em Tempo Real 50
3.2 Análise Proteômica 51
3.2.1 Preparação de extratos protéicos totais 51
3.2.2 Quantificação dos extratos protéicos 52
3.2.3 Análise da integridade das proteínas no extrato 52
3.2.4 Eletroforese Bidimensional 52
IsoEletroFocalização IEF (PRIMEIRA DIMENSÃO) 52
Eletroforese SDS-PAGE (SEGUNDA DIMENSÃO) 53
3.2.5 Análise dos Padrões Proteômicos 54
3.2.6 Espectrometria de Massas 54
4 Análise do potencial imunossupressor in vitro 56
5 Análise Citogenética 57
RESULTADOS 59
1 Definição do Protocolo de Estabelecimento das Culturas de CTM 59
2. Caracterização Molecular das Culturas
2.1 Análise Protéica Global
67
2.2 Comparação entre culturas derivadas de diferentes doadores
numa mesma passagem
74
2.2.1 Análise do Perfil Proteômico 74

Índice
2.2.2 Inibição In vitro de Células T Mediada por CTM de um terceiro
indivíduo (Third-Party)
79
2.3 Comparação entre Culturas de um Mesmo Doador em Diferentes
Passagens
81
2.3.1 Análise do Perfil de Expressão de RNA 83
2.3.2 Análise do Perfil Proteômico 85
2.3.3 Análise do Perfil Citogenético 91
2.3.4 Análise do Potencial de Inibição In Vitro de Células T 94
2.4 Comparação entre culturas submetidas à diferenciação e culturas
não diferenciadas
96
DISCUSSÃO 100
CONCLUSÕES 115
REFERÊNCIAS BIBLIOGRÁFICAS 117
ANEXOS 145
Anexo 1 – Artigo publicado na Revista Cytotherapy 145

Introdução
INTRODUÇÃO
Na medula óssea, a manutenção da Célula Tronco Hematopoética (CTH)
e a regulação de sua auto-renovação e diferenciação in vivo dependem do
microambiente específico ao qual está submetida, conhecido historicamente
como “nicho da célula tronco hematopoética” (Wilson & Trumpp 2006).
O nicho hematopoético da medula óssea adulta é formado por um grupo
heterogêneo de células não hematopoéticas, derivadas do mesênquima
(mesenquimais), que inclui fibroblastos, adipócitos, precursores osteogênicos e
células reticulares.
Dentro deste grupo, acredita-se na existência de um conjunto de células
conhecido como células tronco ou progenitoras mesenquimais (Anjos-Afonso &
Bonnet 2007), ou células tronco esqueléticas (Bianco et al. 2006).
O conceito de células tronco mesenquimais (CTM) vem de trabalhos do
grupo de Alexander Friedenstein há quase 4 décadas atrás (Friedenstein et al.
1974). Eles notaram que, quando células da medula óssea eram submetidas à
aderência ao plástico, uma rara população se desenvolvia em Unidades
Formadoras de Colônia semelhantes a Fibroblastos (UFC-F). Após expansão in
vitro, culturas clonais derivadas de UFC-F individuais puderam ser introduzidas
em modelos experimentais, onde a formação de osso, cartilagem e elementos
estromais foi observada (Castro-Malaspina et al. 1980). A freqüência dessas
UFC-F era extremamente baixa, cerca de 1/10.000 a 1/100.000 das células
mononucleares da medula óssea (Castro-Malaspina et al. 1980).
Sua origem a partir das células estromais da medula óssea, e sua
possível natureza de célula tronco, levaram à adoção do termo, por Owen e
Friedenstein (1988), de “células tronco estromais da medula óssea”. O termo

Introdução
“Células Tronco Mesenquimais” (CTM, como tais células serão denominadas
nesse trabalho) foi proposto posteriormente para evidenciar a natureza não-
hematopoética das células tronco derivadas da medula óssea, muito embora
tais células não compartilhem todas as propriedades das células do
mesênquima embrionário, que se refere a um tipo celular embrionário com
capacidade de gerar não apenas tecido conectivo, mas também tecido
hematopoético e vascular (Bianco et al. 2006).
As CTM podem ser expandidas ex vivo e induzidas in vitro a se diferenciar
em diferentes tipos celulares esqueléticos encontrados em vários estágios do
desenvolvimento, assim como em sítios anatômicos específicos. Tais tipos
celulares incluem osteoblastos (osso), condrócitos (cartilagem), adipócitos
(estroma medular), fibroblastos (periósteo), e células reticulares (estroma
medular) (Bianco et al. 2008). Além disso, tem sido demonstrado que elas
podem ser induzidas experimentalmente a sofrer diferenciação não
convencional tanto in vitro como in vivo, formando células do tipo neural,
endodermal, e cardiomiócitos, derivadas, portanto, de folhetos embrionários
diferentes do mesenquimal. O tema da diferenciação não convencional é,
porém, bastante controverso, e muitos autores questionam a veracidade
dessas evidências (Bianco et al. 2006).
De maneira geral, ainda não é possível suprir em laboratório todos os
fatores extrínsecos gerados no nicho fisiológico de uma célula tronco (da Silva
Meirelles et al. 2008). Uma célula tronco fora do seu ambiente original não se
comporta da mesma maneira que em seu habitat natural. Assim, podemos
distinguir duas entidades celulares distintas, a CTM isolada e cultivada em
laboratório (CTM in vitro) e a CTM encontrada no organismo (CTM in vivo).

Introdução
Devido à ausência de marcadores específicos para isolar as CTM, ainda não
existe prova direta de que as células isoladas em cultura desempenham um
papel fisiológico in vivo.
Além disso, ainda não foi demonstrado que as CTM sejam realmente
células tronco. Uma célula tronco é definida por sua capacidade de auto-
renovação e sua habilidade em gerar pelo menos um ou mais progenitores
diferenciados. Até o momento, não foi possível demonstrar que a CTM seja
capaz de regenerar ou manter um compartimento tecidual ao nível de uma
única célula in vivo. As CTM não puderam ser transplantadas, isoladas e
serialmente transplantadas (Deans & Moseley 2000, Javazon et al. 2004).
Como ainda não é possível se isolar a CTM e caracterizá-la em um estado não
manipulado, sua definição está baseada em uma combinação de propriedades
morfológicas, fenotípicas e funcionais. Tais propriedades variam de acordo com
o método de isolamento/expansão empregado para o seu cultivo, e não existe
uma combinação específica de marcadores para a identificação dessas células
in vivo ou in vitro (Anjos-Afonso & Bonnet 2007). A ausência de marcadores
específicos torna a comparação de resultados entre os diversos grupos de
pesquisa difícil e retarda o progresso nesse campo.
Na tentativa de diminuir essa variação e uniformizar a terminologia
utilizada para essas células, o Comitê de Células Tronco Mesenquimais e
Teciduais da Sociedade Internacional de Terapia Celular (SITC) propôs
critérios mínimos para definir as CTM humanas. Primeiro, as CTM devem ser
“aderentes ao plástico” (aderentes ao substrato), quando mantidas em
condições padrão de cultivo in vitro. Segundo, as CTM devem expressar as
moléculas de superfície CD105, CD73 e CD90, e não expressar as moléculas

Introdução
de superfície CD45, CD34, CD14 ou CD11b, CD79α ou CD19, e HLA-DR.
Terceiro, as CTM devem se diferenciar in vitro em osteoblastos, adipócitos e
condrócitos. Esses critérios ainda precisam ser revistos, conforme o
conhecimento da biologia das CTM for aprofundado, porém são essenciais
para uniformizar a caracterização das CTM e facilitar a troca de dados entre os
diferentes grupos de pesquisa (Dominici et al. 2006).
CTM já foram isoladas em diversas espécies como humanos (Pittenger et
al. 1999), camundongos (Pereira et al. 1995, da Silva Meirelles & Nardi 2003),
ratos (Wakitani et al. 1995), cachorros (Kadiyala et al. 1997), gatos (Martin et
al. 2002) e babuínos (Devine et al. 2003). O estroma da medula óssea é a fonte
mais freqüentemente utilizada para o isolamento de progenitores
mesenquimais. Em humanos, as CTM podem ser isoladas a partir de alíquotas
de medula óssea de doadores normais obtidas por aspiração para fins de
transplante alogenêico. Após o plaqueamento das células mononucleares de
baixa densidade em um meio basal suplementado com lotes selecionados de
soro bovino fetal, a população resultante de células aderentes é considerada a
fonte primária ex vivo de CTM (Prockop 1997, Conget & Minguell 1999).
Além da medula óssea, já foi demonstrado que as CTM podem ser
encontradas em vários órgãos e no sangue periférico de fetos jovens
(Campagnoli et al. 2001), no endotélio e subendotélio da veia do cordão
umbilical (Covas et al. 2003), no sangue de cordão umbilical (Lee OK et al.
2004), na placenta (In’t Anker et al. 2004) e no fluido amniótico no segundo
trimestre de gestação (Tsai et al. 2004), de onde já puderam ser isoladas e
expandidas em cultura. CTM isoladas do fluido amniótico expressam, além dos
marcadores usuais de progenitores mesenquimais, a proteína Oct-4, fator de

Introdução
transcrição expresso em células tronco embrionárias e células germinativas,
sugerindo um alto potencial proliferativo e de diferenciação para os
progenitores embrionários. CTM foram também isoladas de vários tecidos
adultos, como tecido adiposo, músculo e derme (Zuk et al. 2001, Young et al.
2001). Leonardi e colaboradores (2006) derivaram culturas de CTM a partir da
medula óssea femoral obtida durante cirurgias de artroplastia total de quadril.
Culturas de CTM humanas (CTMh) contêm uma população de células
com morfologia de fibroblastos bipolares e algumas células adipocíticas,
poligonais e arredondadas, que desaparecem à medida em que as culturas são
replaqueadas. As células fibroblastóides apresentam ciclo celular de cerca de
33 horas e possuem um potencial de expansão in vitro grande, porém variável
(Minguell et al. 2000). Enquanto algumas preparações de CTMh podem ser
expandidas mais de 15 vezes, outras páram de replicar após 4 subculturas.
Essa variabilidade pode estar relacionada à baixa freqüência de CTM na
medula (2-5 CTM por 106 células mononucleares) (Koc et al. 1999), à idade e à
condição física do doador da medula óssea de onde foi preparada a cultura.
CTM derivadas de doadores mais velhos apresentam menor taxa de
proliferação, menor freqüência na medula óssea morfologia mais larga e
achatada, além de alterações no seu potencial de diferenciação (Roobrouck et
al. 2008). Em um trabalho recente, Stolzing e colaboradores (2008) realizaram
ensaios de UFC-F associados à análise de marcadores de envelhecimento
celular, de dano oxidativo e de senescência, e mostraram que, além do número
de CTMh obtidas da medula óssea diminuir conforme a idade do doador, o
“fitness” das CTM também é reduzido (termo atribuído pelos autores para a

Introdução
menor propensão das células a sofrer senescência), o que pode ter
conseqüências práticas em sua utilização como fonte para terapias celulares.
Além disso, o potencial de proliferação das CTM aparece de forma
bastante variável na literatura, devido às diferentes metodologias empregadas
na derivação das culturas de CTM e às diferentes técnicas utilizadas para
avaliação de sua capacidade de proliferação. A densidade inicial de células é
também um fator crucial no potencial de expansão de uma cultura de CTM:
diversos trabalhos têm demonstrado que as culturas têm capacidade de
expansão maior quando plaqueadas em baixa densidade, provavelmente
porque, em alta densidade, o contato célula-célula ou fatores secretados no
meio levam a taxas de crescimento mais baixas (Colter et al. 2000, 2001,
Sekiya et al. 2002a).
A habilidade de células individuais de gerar colônias dentro de culturas de
células estromais, quando plaqueadas em baixa densidade, reflete diretamente
sua competência para o crescimento “independente da densidade” (Bianco et
al. 2006). Em outras palavras, apenas quando cultivados em baixa densidade,
progenitores mesenquimais mais primitivos, capazes de se diferenciar em
adipócitos, osteoblastos e condrócitos, e capazes de proliferar independente de
estímulos parácrinos, podem se sobressair na cultura, não concorrendo com
progenitores que proliferam mais rapidamente, mas que já estão em estágios
posteriores de diferenciação. A formação de colônias e a clonogenicidade
coincidem entre si apenas quando as colônias são formadas em densidade
clonal (baixas densidades). Colônias também aparecem quando suspensões
celulares são plaqueadas em densidades não clonais. Nessas condições as

Introdução
colônias não necessariamente são clonais, e a sua formação não denota um
crescimento do tipo “independente da densidade” (Bianco et al. 2006).
Análises clonais mostram que a população em cultura é uma mistura de
progenitores multi e bipotentes, precursores linhagem-restritos, e uma pequena
quantidade de fibroblastos sem capacidade de diferenciação nas linhagens do
tecido conectivo (Phinney et al 1999, DiGirolamo et al 1999, Muraglia et al
2000, Baksh et al. 2004). Após períodos prolongados de exposição ao soro
bovino fetal, os progenitores mais primitivos dão origem a precursores mais
comprometidos que crescem rapidamente e se diferenciam terminalmente.
Vários tipos de progenitores comprometidos estão presentes em culturas de
CTM derivadas da medula óssea: progenitores quadri-linhagem (capazes de
seguir a diferenciação osteo/condro/adipogênica/estromal), tri-linhagem
(osteo/condro/adipogênica), bi-linhagem (osteo/condrogênica) e progenitores
osteogênicos, o que parece refletir a hierarquia de diferenciação a partir da
célula tronco mesenquimal não comprometida (Muraglia et al. 2000).
Mets & Verdonk (1981) descreveram pela primeira vez 2 tipos de células
morfologicamente distintas nas culturas de CTM: as células tipo I, que são
fusiformes e crescem rapidamente, e as células tipo II, que são achatadas e
crescem lentamente. Posteriormente baseando-se no tamanho, morfologia e
granulosidade, Colter et al. (2001) demonstraram que as culturas contêm
também células muito pequenas com grande capacidade proliferativa (células
RS), que parecem ser os progenitores mais primitivos e com maior potencial de
diferenciação multilinhagem. Tais progenitores diferem das células mais
maduras numa mesma cultura pela expressão de marcadores protéicos
específicos (Colter et al. 2001).

Introdução
Estudos in vitro têm demonstrado que culturas de células estromais
expressam baixos níveis de fatores adipogênicos, como C/EBPα e PPARγ
(Kuznetsov et al. 2001), fatores osteogênicos, como Runx2 e Osx (Satomura et
al. 2002, Nakashima et al. 2002), e fatores condrogênicos, como Sox9
(Akiyama et al. 2002). Duas possibilidades podem explicar tal achado: ou os
fatores específicos de uma linhagem reprimem a expressão dos fatores das
outras linhagens, gerando um estado de repressão mútua; ou a presença de
diferentes reguladores linhagem-específicos nas culturas acontece, pois estas
representam uma população heterogênea de células (Anjos-Afonso & Bonnet
2007), composta por diferentes tipos de progenitores comprometidos.
Devido à inabilidade de serem identificadas em sua localização natural no
organismo, a caracterização atualmente disponível das CTM foi feita apenas
com as células em cultura e, portanto, a biologia das CTM é uma biologia “fora
de contexto” (Javazon et al. 2004).
DIFERENCIAÇÃO DAS CTM
Quando submetidas a estímulos apropriados em cultura, as CTM podem
se diferenciar terminalmente em diversos tipos de células de origem
mesenquimal. O potencial de diferenciação in vitro das CTM tem sido testado
em diversos estudos, onde as condições ideais de cultura, os estímulos
apropriados e os métodos para identificação do fenótipo diferenciado vêm
sendo determinados.
In vitro já foi possível estabelecer a diferenciação das CTM em células
ósseas, células adiposas, células estromais da medula, condrócitos e
cardiomiócitos (Phinney et al. 1999, Johnstone et al. 1998, Wakitani et al. 1995,

Introdução
Caplan 1994, Xu et al. 2004). A diferenciação adipogênica, por exemplo, pode
ser induzida pelo tratamento com 1-metil-3-isobutilxantina, dexametasona e
indometacina. A indução pode ser evidenciada pela acúmulo de vacúolos ricos
em lipídeos dentro das células. Para promover a diferenciação condrogênica,
CTM são removidas da monocamada e distribuídas em estruturas
tridimensionais, na ausência de soro e na presença de fatores de crescimento
da superfamília TGF (transforming growth factor), como TGF-β1, TGF-β3, BMP
(bone morphogenetic protein)-2, BMP-6 e BMP-9. As culturas desenvolvem
uma morfologia multicamada rica em matriz e com presença maciça de
marcadores condrogênicos. Sob a influência de dexametasona, β-glicerol
fosfato e ácido ascórbico, na presença de soro fetal, células mesenquimais
isoladas formam agregados e aumentam sua expressão de fosfatase alcalina
(diferenciação osteogênica) (Pittenger et al. 1999). Células diferenciadas a
partir das CTM podem suportar a hematopoese in vitro, através do
estabelecimento de culturas de longa-duração (diferenciação estromal)
(Majumdar et al. 1998).
A capacidade de diferenciação das CTM em osteoblastos, adipócitos e
condrócitos reflete sua contribuição para o nicho hematopoético da medula
óssea, fornecendo o microambiente adequado ao controle da maturação,
diferenciação e sobrevivência das células e progenitores hematopoéticos. Além
disso, populações CTM multipotentes também fazem parte do nicho medular.
Neste contexto, o contato direto célula a célula, através de proteínas de adesão
específicas e da matriz extracelular, tais como fibronectina 1, caderina 11,
molécula de adesão vascular-1, conexina 43, integrina-β1 e TGFβ1, tem
mostrado importante papel na manutenção do estado indiferenciado dos

Introdução
progenitores hematopoéticos. Wagner et al. (2007a) demostraram que
monocamadas de CTM fornecem adequado nicho para células progenitoras
hematopoéticas (CPH) humanas. Posteriormente, o mesmo grupo mostrou
que várias moléculas de adesão estão envolvidas na manutenção das CPH
primitivas (Wagner et al. 2007), em particular a glicoproteína CD44 (Wagner et
al. 2008a).
Alguns trabalhos mostraram a diferenciação in vitro de CTM para células
com morfologia de neurônio e glia e que expressavam marcadores neuronais e
gliais após estimulação com ácido retinóico, NGF, FGFb, ácido valpróico,
forscolina e GGF (Sanchez-Ramos et al. 2000, Tohill et al. 2004, Qian &
Saltzman 2004, Safford et al. 2004), demonstrando o potencial de trans-
diferenciação dos progenitores mesenquimais primitivos a células derivadas de
outros folhetos embrionários (no caso, neuroectoderma). Kopen e
colaboradores (1999) mostraram que CTM podem adotar fenótipos neurais
quando expostas ao microambiente do cérebro, após sua implantação em
cérebros de camundongos neo-natos. Rose e colaboradores (2008), porém,
mostraram que CTM de camundongos, quando co-cultivadas em
cardiomiócitos de ratos, passam a expressar marcadores cardíacos (como fator
natriuretico atrial, Nkx2.5 e α-actina cardíaca), mas que as células resultantes
não são capazes de gerar potenciais de ação ou correntes iônicas típicas de
cardiomiócitos, sugerindo que as células retêm o fenótipo estromal. A
diferenciação in vitro das CTM em tipos celulares endodérmicos (Sato et al.
2005) e ectodérmicos (Kopen et al. 1999) poderia ser uma conseqüência do
próprio cultivo celular, mais do que refletir um potencial intrínseco de
diferenciação in vivo (da Silva Meirelles et al. 2008).

Introdução
CARACTERIZAÇÃO ANTIGÊNICA
Embora anticorpos para vários antígenos de superfície possam ser
usados para reconhecer as CTM, não existem marcadores específicos para
identificá-las. Estudos realizados em culturas de CTM não clonais mostram
certo perfil característico: CD34-, CD45-, CD14-, CD13+, CD29+, CD44+,
CD49a+, CD63+, CD90+, CD105+, CD106+, CD146+, CD166+ (Barry et al. 1999,
Deschaseaux & Charbord 2000, Filshie et al. 1998, Gronthos et al. 1999,
Stewart et al. 2003, Vogel et al. 2003, Zannettino et al. 2003, revisado em
Bianco et al. 2006). Esse fenótipo pode ser modulado em cultura, e não reflete
o fenótipo in vivo. Grande parte desses marcadores é expresso em diferentes
níveis em linhagens de fibroblastos, que não se comportam como as células
estromais da medula óssea tanto in vivo quanto in vitro (Bianco et al. 2006).
Na busca de marcadores para as CTM, Jones e colaboradores (Jones et
al. 2002, 2006) identificaram uma rara população celular na medula óssea,
distinta das células hematopoéticas por sua baixa expressão de CD45 e por
seu maior tamanho. O marcador Stro-1 estava uniformemente expresso nessa
população, mas a reação cruzada deste marcador em outras populações da
medula óssea limitou sua utilização na identificação da CTM. O mesmo grupo
identificou outros marcadores muito expressos na população CTM, mas pouco
expressos nas outras populações da medula óssea. O receptor para o fator de
crescimento neural de baixa afinidade (Low-affinity Nerve Growth Factor
Receptor, LNGFR), também conhecido como CD271, foi identificado como o
marcador mais diferencialmente expresso nessa população (Jones et al. 2002,
2006). Outra molécula neural, o GangliosíDeo 2 (GD2), teve sua expressão
recentemente demonstrada em populações in vivo de CTM e após seu cultivo

Introdução
in vitro. Estudos recentes mostraram que SSEA-1 e SSEA-4, antígenos
embrionários estágio-específicos, são marcadores para células mesenquimais
primitivas na medula óssea humana e murina (Anjos-Afonso & Bonnet 2007).
Os progenitores mesenquimais compartilham fenótipo antigênico com
células endoteliais, epiteliais e musculares, e não expressam marcadores
hematopoéticos típicos, como CD45, CD34 e CD14. O padrão de expressão de
citocinas e receptores para fatores de crescimento das CTM é dinâmico e
complexo, e sugere que os progenitores mesenquimais na medula óssea
contribuem para a formação e função do microambiente estromal. Dentre esses
fatores estão moléculas de matriz extracelular, incluindo fibronectina, laminina,
colágeno e proteoglicanos (Prockop 1997, Chichester et al. 1993, Pittenger et
al. 1999), e receptores de matriz e de interações célula-célula (Conget &
Minguell 1999, Pittenger et al. 1999, Gronthos & Simmons 1995, Satomura et
al. 1998). De particular relevância é a forte expressão de CD44 (Conget &
Minguell 1999, Pittenger et al. 1999), receptor para vários ligantes importantes
na organização da matriz extracelular na medula e no osso, além da expressão
das proteínas SH-2, SH-3 e SH-4. As CTM secretam uma série de citocinas e
fatores de crescimento com atividades parácrinas e autócrinas, tais como Il-6,
Il-7, Il-8, Il-11, Il-12, Il-14, Il-15, fator estimulador de colônias macrofágicas (M-
CSF), ligante de Flt-3 e Stem Cell Factor (SCF), e são induzidas por Il-1α a
produzir Il-1α, leukemia-inhibiting factor (LIF), fator estimulador de colônias
granulocíticas (G-CSF) e fator estimulador de colônias granulocítica-
macrofágicas (GM-CSF) (Majumdar et al. 1998, Kim et al. 2005). Elas também
produzem reguladores negativos da hematopoese, como Il-8/CXCL8, proteína
inflamatória de macrófagos (MIP-1α/CCL3), TGF-β, e citocinas que induzem a

Introdução
síntese de outras citocinas por células do sistema imune (Majumdar et al.
1998). Os fatores secretados produzem uma série de respostas no
microambiente medular e no sistema imune local, além de potencializar a
angiogênese, e induzir a proliferação e diferenciação de células tronco
teciduais (Caplan & Dennis 2006). As células estromais expressam também
uma série de fatores pro-angiogênicos, como a Angiopoetina-1 (Ang-1), o fator
de crescimento endotelial vascular (VEGF), fator de crescimento derivado de
plaquetas (PDGF), fator de crescimento de fibroblastos 2 (FGF-2), FGF-7,
citocinas (Il-6 e TNF-α) e o ativador do plasminogênio tecidual (PAt). Além de
promover a angiogênese, as CTM induzem a expressão de proteínas de junção
e um aumento na integridade microvascular (Zacharek et al. 2007).
A molécula de adesão celular associada a melanoma (melanoma-
associated cell adhesion molecule, MCAM/CD146) foi descrita recentemente
como possível marcador da população de progenitores estromais na medula
óssea humana, capaz de regenerar osso e estroma, e reestabelecer o
microambiente medular in vivo (Sacchetti et al. 2007). A alta expressão de
MCAM/CD146 nesses progenitores é compartilhada com células subendoteliais
de sinusóides da medula óssea, mas não por células osteoblásticas capazes
de formar apenas osso heterotrópico (Sacchetti et al. 2007). Os progenitores
CD146+ descritos pelo grupo de Sacchetti são capazes de se auto-renovar in
vivo, expressam Ang-1 e, portanto, parecem participar ativamente dos eventos
angiogênicos para o estabelecimento de sinusóides anterior à hematopoese
(2007). Sorrentino e colaboradores (2008) descreveram uma metodologia
eficiente para isolamento e cultivo de células estromais mesenquimais
multipotentes CD146+, população com maior potencial de proliferação e

Introdução
manutenção em cultura, capaz de se diferenciar em osteoblastos, condrócitos,
adipócitos e de fornecer suporte para a hematopoese em ensaios de cultura a
longo termo. Essa população pode representar um modelo in vitro para estudos
do nicho hematopoético (Sorrentino et al. 2008).
IDENTIDADE IN VIVO
Embora as CTM sejam estudadas há mais de três décadas, a natureza
exata do progenitor presente in vivo está apenas começando a ser
caracterizada. Sua localização e identidade in vivo são questões ainda pouco
conhecidas. Algumas hipóteses podem ser levantadas no que se refere à sua
localização. As CTM poderiam estar localizadas em algum(s) sítio(s)/órgão(s)
específico(s) do organismo e serem mobilizadas, através da circulação, para
outros sítios/órgãos que necessitassem de reposição de suas populações
celulares (da Silva Meirelles et al. 2008). A aparente impossibilidade de se
isolar culturas de CTM do sangue periférico em condições fisiológicas é um
forte argumento contra essa hipótese (Lazarus et al. 1997, Wexler et al. 2003).
Em camundongos, porém, UFC-F circulam no sangue e representam uma
população de células estromais que podem migrar em órgãos hematopoéticos
(Piersma et al. 1983). Em humanos, células com características de
progenitores mesenquimais foram detectadas no sangue periférico de
pacientes com câncer de mama após mobilização com fatores de crescimento,
mas não no sangue de doadores (Fernandez et al. 1997).
Uma segunda possibilidade baseia-se no fato de que culturas
semelhantes às de CTM já foram isoladas de vários tecidos pós-natais, além
da medula óssea, como tecido adiposo (Zuk et al. 2001), tendão

Introdução
(Salingcarnboriboon et al. 2003), ligamento peridontal (Seo et al. 2004),
músculo (Lee et al. 2000, Kuroda et al. 2006), membrana sinovial (De Bari et al.
2001), pele (Toma et al. 2001), e pulmões (Sabatini et al. 2005). Em
camundongos, culturas semelhantes morfológica, fenotípica e funcionalmente
às de CTM puderam ser isoladas de vários tecidos e órgãos, mesmo quando
os animais tinham o sangue de suas veias previamente retirado por perfusão
(da Silva Meirelles et al. 2006), impedindo a circulação de progenitores.
Entretanto, a potência dos progenitores isolados de tecidos não-
hematopoéticos e da medula óssea não foi sistematicamente comparada por
ensaios in vivo (Bianco et al. 2008), e é possível que eles tenham
características diferentes. Por exemplo, UFC-F derivadas da polpa dentária
cultivadas e transplantadas em condições idênticas às utilizadas para UFC-F
da medula óssea, formam dentina e não osso (Gronthos et al. 2002).
Uma terceira hipótese, defendida por diversos autores (Shi et al. 2003,
Farrington-Rock et al. 2004, Bianco et al. 2001, da Silva Meirelles et al. 2006,
2008, Covas et al. 2008), considera a relação direta das CTM com células
perivasculares, ou seja, os pericitos seriam a identidade in vivo das CTM. Essa
possibilidade poderia explicar por que as CTM podem ser isoladas de vários
tecidos, uma vez que sua associação com a vasculatura aumentaria sua
disponibilidade para a substituição celular. Neste cenário, lesões em quaisquer
tecidos liberariam as CTM do seu sítio perivascular (Da Silva Meirelles et al.
2008), podendo assim secretar fatores tróficos e imunorregulatórios
necessários para a reconstituição/regeneração local. Evidências que favorecem
essa hipótese incluem o estabelecimento de culturas de CTM de vasos
sanguíneos isolados, incluindo o glomérulo encapsulado (da Silva Meirelles et

Introdução
al. 2006). Além disso, o possível marcador MCAM/CD146 (Sacchetti et al.
2007) é também expresso por pericitos (Li et al. 2003).
Crisan e colaboradores (2008) identificaram células perivasculares em
múltiplos órgãos humanos, como músculo esquelético, pâncreas, tecido
adiposo e placenta, com expressão nativa de CD146, αSMA, NG2, fosfatase
alcalina e PDGF-Rβ, e ausência de marcadores hematopoéticos, endoteliais e
miogênicos. Essas células, quando cultivadas por longos períodos, mantêm a
expressão dos marcadores perivasculares, apresentando também expressão
de antígenos típicos de CTM, tais como CD10, CD13, CD44, CD73, CD90 e
CD105. Em contraste, as células em cultura não expressam CD56, CD133,
CD324, CD326, CD344 e HLA-DR, antígenos tipicamente não expressos em
CTM. Além disso, células perivasculares derivadas de tecidos musculares e
não musculares, tais como pâncreas, gordura, medula óssea e placenta,
apresentam grande potencial miogênico. Esses resultados reforçam a hipótese
de que as paredes dos vasos sanguíneos abrigam um reservatório de células
progenitoras, que podem ser a origem das CTM e de outras células tronco
adultas.
MIGRAÇÃO EM HUMANOS E MODELOS ANIMAIS
Devido ao seu grande tamanho, e pelo fato de expressarem diversas
moléculas de adesão, tem sido demonstrado que CTM injetadas por via
intravenosa ficam inicialmente retidas nos pulmões (Fox et al. 2007).
Apesar disso, várias evidências mostram que CTM injetadas por via
intravenosa são capazes de migrar em modelos animais e se distribuir por
vários tecidos, como tecidos gastro-intestinais, fígado, rins, timo, pele e medula

Introdução
óssea (Pereira et al. 1995, Ferrari et al. 1998, Awad et al. 1999, Devine et al.
2003). A migração para tecidos que sofreram injúria é particularmente
importante após injeção sistêmica. CTM de ratos e humanos foram detectadas
em sítios de injúria no cérebro de ratos após isquemia e injeção por via
intravenosa (Mahmood et al. 2003). Ratos que sofreram rejeição de
aloenxertos de coração foram tratados com infusão intravenosa de CTM, e foi
observado que estas migraram para o sítio de rejeição e melhoraram a enxertia
(Wu et al. 2003).
Pouco se sabe, no entanto, a respeito dos mecanismos da migração
sistêmica transendotelial de CTM e das moléculas de adesão envolvidas. É
possível que o tecido lesado expresse receptores ou ligantes específicos que
facilitem o tráfego, adesão e infiltração ao sítio de injúria, de forma similar ao
recrutamento de leucócitos aos sítios de inflamação. Várias moléculas
envolvidas na adesão e transmigração de leucócitos da corrente sanguínea
para os tecidos são expressas nas CTM, incluindo integrinas, selectinas e
receptores de quimiocinas (Krampera et al. 2006), o que reforça essa hipótese
(Fox et al. 2007).
ENVELHECIMENTO E EFEITO DA EXPANSÃO IN VITRO NA BIOLOGIA DAS CTM
O envelhecimento é um processo complexo e a identificação dos
mecanismos que governam o envelhecimento de populações celulares requer
que estas possam ser isoladas e caracterizadas com relativa homogeneidade.
A maioria das células adultas sofre no máximo 50 a 60 divisões antes de se
tornar senescente. Essa limitada capacidade replicativa é conhecida como o
“limite de Hayflick”, termo utilizado por Leonardo Hayflick (1965) para descrever

Introdução
o processo resultante do progressivo encurtamento dos telômeros. A
transformação celular é o processo in vitro pelo qual células escapam desse
limite, através da aquisição de mutações em genes específicos (Roobrouck et
al. 2008).
Como o isolamento prospectivo da CTM ainda não foi possível, sua
utilização com propósitos terapêuticos depende do isolamento e expansão em
cultura. No entanto, os métodos de isolamento e as condições de cultivo para
selecionar e expandir as CTM são muito variáveis. Além disso, sua definição
em cultura se baseia em parâmetros físicos, fenotípicos e funcionais, e não em
marcadores definidos e únicos. A qualidade das preparações realizadas em
diferentes laboratórios varia enormemente.
CTM humanas apresentam morfologia fibroblastóide e usualmente param
de proliferar antes de 40 duplicações populacionais (population doublings,
PDs), quando se tornam maiores e mais achatadas (Bruder et al. 1997), o que
está possivelmente associado ao fenômeno da senescência in vitro. Um dos
mecanismos que levam à senescência celular é o encurtamento dos telômeros
sofrido durante o cultivo in vitro. Diversos autores têm estudado o tamanho dos
telômeros das CTM, bem como o encurtamento sofrido durante o cultivo in
vitro. Como esperado, os telômeros de CTM derivadas de doadores jovens são
maiores que os de doadores velhos (Baxter et al. 2004, Guillot et al. 2007,
Mareschi et al. 2006). Esses estudos concordam que ocorre uma perda no
tamanho dos telômeros a cada passagem in vitro, em torno de 1,5-2 kb,
embora o tamanho original dos telômeros varie em cada um.
As células tronco adultas apresentam baixos níveis de atividade da
telomerase, e as CTM não são uma exceção à regra (Roobrouck et al. 2008).

Introdução
Assim, uma estratégia adotada por diversos grupos para estender o tempo de
vida em cultura das CTM é a expressão forçada de hTERT, subunidade
catalítica da enzima telomerase humana, que é capaz de aumentar a
capacidade replicativa in vitro das CTM em mais de 80 (Simonsen et al. 2002)
e 260 (Shi et al. 2002) duplicações populacionais. As CTM transduzidas com
hTERT retêm o potencial de diferenciação in vitro para as linhagens osteo,
condro e adipogênica, e de diferenciação osteogênica in vivo (Simonsen et al.
2002, Shi et al. 2002).
Mesmo sendo expressa em baixos níveis nas CTM, e não tendo influência
direta na replicação celular, a telomerase parece ter um papel na diferenciação
in vitro dessas células, uma vez que CTM isoladas do camundongo knockout
para telomerase não são capazes de se diferenciar em adipócitos e
condrócitos, mesmo nas primeiras passagens (Liu et al. 2004).
Utilizando um modelo de envelhecimento acelerado que combina a
mutação Wrn (da síndrome de Werner) e o encurtamento acelerado dos
telômeros pela ausência da telomerase (camundongo duplo mutante Wrn-/- e
Terc-/-), Pignolo e colaboradores (2008) mostraram que CTM isoladas desse
camundongo apresentam diminuição no tempo de vida em cultura e
senescência prematura, além de redução do potencial de diferenciação
osteogênica in vitro. O duplo mutante apresenta diminuição na massa óssea,
resultando em osteoporose precoce, o que parece estar relacionado à
deficiência na diferenciação osteoblástica, uma vez que a diferenciação
osteoclástica se mantém intacta. Os dados desse modelo sugerem que o
envelhecimento replicativo de precursores osteoblásticos in vivo é um
importante mecanismo na osteoporose senil.

Introdução
Além do encurtamento dos telômeros, outros fatores como a inativação da
expressão de p16INK4A, um dos reguladores do ciclo celular, estão implicados
na indução da senescência (Roobrouck et al. 2008). Shibata e colaboradores
(2007) mostraram que a expressão de p16INK4A está fortemente associada à
senescência in vitro, e que o cultivo prolongado de CTM humanas pode induzir
a inativação do gene de p16, levando a aberrações cromossômicas.
Outra questão importante na biologia das CTM in vitro está relacionada à
idade do doador. CTM derivadas de doadores mais velhos apresentam menor
taxa de proliferação, mesmo em passagens iniciais (Banfi et al. 2000, Baxter et
al. 2004, Stolzing et al. 2008, Zhou et al. 2008), menor freqüência na medula
óssea (Baxter et al. 2004, Stolzing et al. 2008), medida pelo ensaio de UFC-F,
morfologia mais larga e achatada (Roobrouck et al. 2008), mesmo nas
passagens iniciais, além de alterações no seu potencial de diferenciação
(D’Ippolito et al. 1999, Mendes et al 2002, Baxter et al. 2004, Zhou et al. 2008).
Em relação a alterações na capacidade de diferenciação das CTM de
acordo com a idade do doador, a maioria dos trabalhos descreve uma redução
na geração de células positivas para fosfatase alcalina em UFC-F, diretamente
relacionada ao aumento na idade do doador (D’Ippolito et al. 1999, Baxter et al.
2004, Zhou et al. 2008). O grupo de Stenderup, no entanto, não observou
diferenças com a idade do doador no potencial de diferenciação ostogênica e
adipogênica in vitro, nem de diferenciação ostogênica in vivo, determinada
após implante das células associadas a matrizes de fosfato de cálcio em
camundongos NOD/SCID (Stenderup et al. 2003). Mendes e colaboradores
mostraram que CTM derivadas tanto de doadores jovens como de velhos eram
capazes de formar osso in vivo, embora a formação de osso diminuísse

Introdução
significativamente com a idade (Mendes et al. 2002). Para estudar a influência
da idade do doador no potencial condrogênico das CTM, Zheng e
colaboradores compararam a produção de matriz extracelular cartilaginosa
entre células derivadas de ratos de 1 semana, 12 semanas e 1 ano (Zheng et
al 2007). Somente as CTM derivadas de ratos de 1 semana produziram matriz,
sugerindo que o potencial condrogênico também diminui com a idade.
O trabalho do grupo de Muraglia, mostrando que os progenitores
mesenquimais da medula óssea seguem um modelo hierárquico de
diferenciação in vitro, sugere também que o potencial de diferenciação in vitro
das CTM diminui com a idade (Muraglia et al. 2000). Progenitores tripotentes e
bipotentes apresentam frequência absoluta maior em doadores jovens. Clones
de CTM cultivados in vitro perdem primeiro seu potencial adipogênico, o que
vai de encontro à observação in vivo de que a adipogênese na medula óssea
aumenta com a idade, enquanto que a osteogênese diminui (Roobrouck et al.
2008).
Para investigar a suscetibilidade de transformação de CTM em cultura e
determinar se as propriedades biológicas das células permanecem apropriadas
para terapia celular, Bernardo e colaboradores cultivaram CTM de doadores
humanos saudáveis até atingirem senescência, ou chegarem à passagem 25, e
realizaram a caracterização genética das culturas ao longo de várias
passagens (Bernardo et al. 2007). Uma grande variabilidade em termos do
potencial de proliferação e de vida útil da cultura foi observada entre diferentes
doadores. Não foram detectadas anormalidades cromossômicas nas culturas
analisadas, assim como não foram observadas atividade da enzima telomerase
e expressão de transcritos de hTERT, mostrando que os telômeros sofreram

Introdução
encurtamento durante o período de cultura (Bernardo et al. 2007). Esses
resultados sugerem que as CTM derivadas da medula óssea podem ser
seguramente expandidas in vitro, que elas não são suscetíveis à transformação
maligna, o que as torna adequadas para utilização em terapias celulares.
Para caracterizar o efeito da expansão in vitro na biologia da CTM,
Izadpanah e colaboradores compararam culturas derivadas da medula óssea e
do tecido adiposo de humanos e de macacos Rhesus (Izadpanah et al. 2008).
Análises cariotípicas revelaram o desenvolvimento de alterações citogenéticas
somente em células derivadas de macacos em passagens tardias (a partir da
passagem 20), mas não em células humanas. Não foram observadas
alterações cromossômicas em CTM humanas cultivadas por longos períodos,
enquanto que uma alta freqüência de aneuploidias foi detectada em culturas de
CTM de macacos Rhesus. Poucas alterações na expressão gênica, no entanto,
foram observadas em uma mesma amostra ao longo de diferentes passagens
(Izadpanah et al. 2008).
Com o objetivo de estudar o impacto da senescência replicativa em
preparações de CTM humanas, Wagner e colaboradores realizaram análises
morfológicas, imunofenotípicas e de diferenciação de CTM submetidas à
expansão in vitro, além de analisar como o padrão de expressão de RNAm e
RNAmi muda durante o cultivo e propagação in vitro (Wagner et al 2008b). Os
autores pretendiam estudar os efeitos moleculares da senescência replicativa
em passagens tardias, o que poderia ter implicações na qualidade de
preparações de CTM usadas em aplicações terapêuticas. Diversas alterações
foram observadas em passagens mais tardias (entre 7 e 12), tais como
anormalidades morfológicas, alterações na expressão de marcadores

Introdução
específicos, diminuição da proliferação, do potencial adipogênico e aumento do
potencial osteogênico. Além disso, foi observado que mudanças no padrão de
expressão gênica global não estão restritas a passagens tardias, mas são
adquiridas ao longo do subcultivo das CTM, o que sugere que a senescência
replicativa das preparações de CTM é um processo contínuo, que se inicia logo
nas primeiras passagens, e que precisa ser considerada para sua aplicação
terapêutica (Wagner et al 2008b).
A diminuição na capacidade proliferativa e a aquisição de defeitos na
diferenciação de populações em cultura são responsáveis pelo fenômeno do
envelhecimento, que tem implicação direta em terapias celulares. Diversos
estudos têm demonstrado o declínio na capacidade de diferenciação e
expansão em cultura das CTM, de acordo com o envelhecimento do organismo
(Roobrouck et al. 2008). A extensão dessa redução, no entanto, ainda não está
clara, sobretudo devido à ausência de um consenso na caracterização
fenotípica e condições de isolamento e cultivo das CTM. O melhor
entendimento nos efeitos da expansão in vitro sobre a biologia das CTM, e os
efeitos relacionados ao envelhecimento celular, tem grande impacto em seu
uso clínico como terapia celular.
CARACTERIZAÇÃO MOLECULAR
Devido à ausência atual de marcadores prospectivos para o isolamento e
caracterização das CTM, sua definição se baseia em propriedades físicas,
fenotípicas e funcionais (Dominici et al. 2006). Entretanto, a variabilidade de
protocolos empregados para estabelecer culturas de CTM, assim como a
diversidade das fontes iniciais, como a medula óssea, o tecido adiposo, o

Introdução
sangue e segmento de cordão umbilical e o sangue periférico, torna necessário
o desenvolvimento de estudos mais aprofundados de caracterização molecular
das preparações celulares.
A análise da expressão gênica e protéica surgiu, no campo de estudo das
CTM, como importante ferramenta para a caracterização molecular das
preparações celulares. Na definição do perfil de expressão gênica das CTM,
diversos estudos de análise em larga escala têm se focalizado na sua
diferenciação in vitro (Doi et al. 2002, Sekiya et al. 2002b, Qi et al. 2003,
Nakamura et al. 2003, Sekiya et al. 2004), na comparação direta entre
diferentes preparações de CTM (Majumdar et al. 1998, Jiang et al. 2002, Lee et
al. 2002), na estimativa absoluta da expressão de RNAm na CTM (Tremain et
al. 2001, Jiang et al. 2002, Wieczorek et al. 2003, Silva et al. 2003, Lee RH et
al. 2004), e na comparação entre CTM isoladas de diferentes fontes, como a
medula óssea, tecido adiposo e o cordão umbilical, e linhagens de fibroblastos
terminalmente diferenciados (Panepucci et al. 2004, Wagner et al. 2005).
Enquanto as propriedades biológicas únicas codificadas no genoma das
CTM podem ser analisadas ao nível do transcriptoma da célula, o fenótipo e
função celular são determinados em última instância pelas proteínas e enzimas
traduzidas. A análise proteômica tem facilitado a caracterização molecular e
permitido a identificação de proteínas essenciais para o papel das CTM no
microambiente hematopoético da medula óssea e na modulação das respostas
imunes.
Um dos primeiros trabalhos a determinar um mapa proteômico
bidimensional de referência de CTM foi realizado em 2006 por Wagner e
colaboradores. Os autores analisaram o padrão proteômico e transcriptômico

Introdução
diferencial de preparações de CTM da medula óssea humana, isoladas e
cultivadas em diferentes meios de cultura (10% ou 2% de soro bovino fetal).
Cento e trinta e quatro spots protéicos foram identificados por MADI-TOF-MS e
subsequente análise de bioinformática, e correspondiam a 81 proteínas
diferentes, a maior parte pertencente à categoria de proteínas do citoesqueleto.
Os spots protéicos encontrados nos géis das células cultivadas em diferentes
meios foram selecionados para análise por microarray (63 proteínas
representadas por 73 spots protéicos, que estavam representados por 105
clones de cDNA). Os resultados demonstraram uma intensidade de sinal
significativamente maior nos clones de cDNA que correspondiam às proteínas
mais expressas identificadas por proteoma. Entretanto, conforme demonstrado
por estudos anteriores (Griffin et al. 2002, Anderson & Seilhamer 1997), a
relação entre a quantidade absoluta de proteínas em uma célula não está
diretamente correlacionada à quantidade de RNAm, provavelmente devido às
modificações pós-transcricionais e pós-traducionais sofridas pelo RNAm e pela
proteína (Wagner et al. 2006).
Posteriormente, outros grupos realizaram análises do perfil proteômico de
preparações de CTM, nos mais diversos contextos. Park e colaboradores, em
2007, revisaram as análises proteômicas já publicadas, descrevendo cerca de
264 proteínas distintas presentes em culturas de CTM humanas (Park et al.
2007). Várias tecnologias foram empregadas nessas análises, tais como
Eletroforese Bidimensional (EB), Cromatografia Líquida (CL), Espectrometria
de Massas (EM), bioinformática e arrays de proteínas. Esses e outros estudos
de caracterização em larga escala têm ajudado a elucidar mecanismos de ação

Introdução
dessa população celular, contribuindo cada vez mais para a utilização das CTM
como importante ferramenta em terapias nas mais diversas áreas da Medicina.
MODULAÇÃO DO SISTEMA IMUNE
Diversos estudos têm demonstrado dois pontos cruciais no que se refere
à relação das CTM com o sistema imune: 1) elas são consideradas
imunoprivilegiadas (Deans & Moseley 2000, DiNicola et al. 2002, Krampera et
al. 2003).; e 2) elas parecem modular o sistema imune (Aggarwal & Pittenger
2005, Rasmusson 2006, Uccelli et al. 2007), tendo ação direta sobre células T
(Bartholomew et al. 2002, DiNicola et al. 2002, Le Blanc et al. 2003, Tse et al.
2003, Krampera et al. 2006, Benvenuto et al. 2007, Nasef et al. 2007, Sato et
al. 2007), células B (Corcione et al. 2006, Rasmusson et al. 2007), células
Natural Killer (NK) (Spaggiari et al. 2006) e células dendríticas (Ramasamy et
al. 2007, Djouad et al. 2007).
As primeiras evidências do papel das CTM sobre o sistema imune vêm do
trabalho de Bartholomew e colaboradores, que mostraram que a administração
intravenosa de CTM em babuínos foi capaz de prolongar a sobrevivência de
enxertos alogenêicos de pele, de forma similar a potentes drogas
imunossupressoras utilizadas rotineiramente na clínica (Bartholomew et al.
2002).
Posteriormente seu papel na regulação do sistema imune começou a ser
estudado in vitro, inicialmente na função das células T. Krampera e
colaboradores mostraram que as CTM diminuem a resposta de células T naive
e de células T de memória a seus antígenos específicos, um efeito que parece
ser independente de células dendríticas (Krampera et al. 2002). Foi também

Introdução
demonstrado que as CTM são capazes de atenuar a produção de Il-2 por
células T, o que resulta na diminuição da formação de células T CD8
citotóxicas (Rasmusson et al. 2003). Essa inibição afeta a proliferação de
células T induzida por aloantígenos, mitógenos, assim como por anticorpos
para CD3 e CD28 (Nauta & Fibbe 2007).
Uma questão controversa a respeito do efeito inibitório das CTM sobre as
células T é se o contato entre as duas células é necessário, ou se tal efeito é
exercido por fatores solúveis. A maioria dos estudos concorda que fatores
solúveis estejam envolvidos, muito embora outros sugiram que o contato
célula-a-célula seja essencial para esse efeito (Nauta & Fibbe 2007).
Sobrenadantes de culturas de CTM humanas e murinas não apresentaram
efeito inibitório, exceto quando as CTM haviam sido co-cultivadas com células
mononucleares (Potian et al. 2003, Djouad et al. 2003, Le Blanc et al. 2004a),
sugerindo que o fator supressor não seja secretado constitutivamente pelas
CTM, mas que seja necessária a interação dinâmica entre a CTM e a célula T
(Nauta & Fibbe 2007). Diversos fatores são candidatos para mediar o efeito
supressor das CTM sobre as células T, como TGF-β e HGF (Di Nicola et al.
2002), PGE2 (Tse et al. 2003), e InDOleamina 2,3-dioxigenase (IDO) (Meisel et
al. 2004).
A indução de anergia das células T tem sido proposta como potencial
mecanismo para o efeito imunossupressor das CTM. Os baixos níveis de
expressão de moléculas co-estimulatórias, como CD40, CD80 e CD86, tem
sido relacionado ao potencial estado de anergia das células T induzido pelas
CTM (Nauta & Fibbe 2007).

Introdução
O efeito de modulação in vitro sobre as células T depende da
concentração de CTM. Alguns trabalhos mostram que quando adicionadas a
culturas mistas linfocitárias (CML) e a linfócitos ativados por mitógenos, as
CTM são capazes de induzir inibição em altas doses, enquanto que um
aumento na proliferação de células T foi observado quando as CTM estavam
em baixas doses. Além disso, esse efeito parece ser independente do HLA (Le
Blanc et al. 2003).
Fang e colaboradores, estudando a aloreatividade de células T na
presença de CTM, levantaram mais uma hipótese relacionada ao
imunoprivilégio das CTM, e que poderia explicar as controvésias encontradas
nessa área. Eles mostraram que as CTM são capazes de induzir a proliferação
de células T e secreção de Il-2 em níveis mensuráveis apenas em baixas
concentrações (taxa CTM/célula T ≥0, 1:1) (Fang et al. 2006). Tal achado
sugere que o estado imunoprivilegiado da CTM existe apenas quando sua
capacidade de inibição do sistema imune, através da produção de fatores
imunomodulatórios, se sobrepõe ao seu próprio potencial aloestimulatório (por
se tratar de uma célula estranha ao sistema imune receptor). Portanto, o
imunoprivilégio das CTM representa um balanço de seus efeitos supressores e
estimuladores (Fang et al. 2006).
Sua atuação direta em células B foi pouco estudada até o momento.
Corcione e colaboradores mostraram inibição da proliferação de células B, em
co-culturas com CTM humanas, devido à parada do ciclo celular na fase
G0/G1. Nesse estudo, foi demonstrado também que as CTM parecem afetar as
propriedades quimiotáticas das células B, embora não afetassem sua produção
de citocinas (Corcione et al. 2006). Recentemente, porém, Rasmusson e

Introdução
colaboradores mostraram que as CTM estimulam a secreção de anticorpos por
células B in vitro. Dependendo do nível de estimulação das células B, as CTM
podem induzir ou inibir a secreção de IgG pelas células B (Rasmusson et al.
2007).
A relação das CTM com as células NK também tem sido investigada.
Spaggiari e colaboradores mostraram que células NK ativadas são capazes de
matar as CTM, enquanto que as CTM inibem fortemente a proliferação de
células NK induzidas por Il-2, suas atividades citotóxicas e de produção de
citocinas (Spaggiari et al. 2006).
As CTM atuam de forma importante sobre a diferenciação, maturação e
função das células dendríticas (English et al. 2008). As CTM são capazes de
bloquear a diferenciação de monócitos e precursores da medula óssea em
células dendríticas e inibir o aumento da expressão de CD1A, CD40, CD80
CD86 e HLA-DR durante a maturação das células dendríticas, mantendo-as em
um estado imaturo. Mais recentemente, foi demonstrado que as CTM induzem
a expressão de Il-10, uma citocina “tolerogênica”, pelas células apresentadoras
de antígeno (Gur-Wahnon et al. 2007), o que sugere que as CTM poderiam
induzir um fenótipo supressor ou inibitório das células dendríticas. Em
camundongos, esse achado foi visto como um fator protetor para o
desenvolvimento da Doença do Enxerto-Contra-Hospedeiro (DECH) (Sato et al.
2003).
O fenótipo imunogênico de CTM tanto humanas como de animais parece
explicar o conceito de imunoprivilégio atribuído às CTM, e seu papel ativo na
modulação das respostas imunes. Elas expressam baixos níveis de antígenos

Introdução
co-estimulatórios, como B7-1, B7-2, CD40 e CD40L, e de HLA classe II (Deans
& Moseley 2000, DiNicola et al. 2002, Krampera et al. 2003).
Evidências mais recentes, no entanto, contestam o imunoprivilégio das
CTM e mostram que, pelo menos em modelos murinos, as CTM são capazes
de induzir respostas de células T de memória após a injeção em hospedeiros
imunocompetentes (Eliopoulos et al. 2005, Nauta et al. 2006). Dando suporte a
esse conceito, a injeção de CTM humanas no miocárdio de ratos infartados
mostrou a “pega” das células xenogenêicas associada, porém, com significante
infiltração imune local (Grinnemo et al. 2004). Infiltrados imunes foram
observados em hospedeiros alogeneicos injetados com CTM murinas
modificadas geneticamente para expressar BMP-2, apesar de as células
injetadas terem se diferenciado em tecido ósseo e cartilaginoso (Noel et al.
2003). Além disso, o potencial imunomodulatório in vivo das CTM tem sido
questionado por alguns grupos, que obtiveram resultados que vão de encontro
à hipótese de que elas modulam o sistema imune. Min e colaboradores
mostraram que a injeção precoce de CTM após o transplante de medula óssea
não foi capaz de atenuar a severidade da DECH aguda, exceto quando as
CTM expressavam constitutivamente a citocina anti-inflamatória IL-10 (Min et al
2007), enquanto Sudres e colaboradores mostraram que células capazes de
suprimir a proliferação de linfócitos in vitro falharam na prevenção da DECH em
camundongos submetidos ao TCTH (Sudres et al 2006).
Como ainda não é possível se isolar as CTM do seu nicho in vivo, todo
conhecimento atual de sua biologia deriva de estudos realizados com as
células expandidas ex vivo. O conceito da imunomodulação exercida pelas
CTM está embasado principalmente em estudos in vitro e utilizando-se o que

Introdução
hoje denominamos de CTM. O potencial imunossupressor das CTM também
tem sido avaliado in vivo. No modelo experimental murino da esclerose múltipla
humana, a chamada encefalomielite auto-imune experimental, a administração
intravenosa de CTM antes e no início da doença, mas não após sua
estabilização, parece prevenir/melhorar seu desenvolvimento (Zappia et al.
2005). Esses estudos utilizaram as células manipuladas ex vivo, e que podem
não corresponder totalmente à identidade da CTM in vivo.
Algumas evidências sinalizam para um papel fisiológico da CTM na
manutenção da homeostase imune (Nauta & Fibbe 2007). CTM derivadas da
medula óssea de pacientes com anemia aplástica severa não foram capazes
de suprimir a proliferação de linfócitos in vitro (Bacigalupo et al. 2005), embora
a relevância desses defeitos na patogênese da anemia aplástica ainda seja
desconhecida. Progenitores endoteliais e estromais em pacientes com
esclerose sistêmica também foram descritos como funcionalmente defeituosos,
apresentando capacidade de proliferação e diferenciação reduzidas. O defeito
no microambiente da medula óssea poderia ser importante no prejuízo à
vasculogênese observado nesses pacientes (Del Papa et al. 2006). Assim, é
possível formular a hipótese de que a capacidade imunossupressora das CTM
poderia ser importante na criação de um sítio imunoprivilegiado dentro da
medula óssea que protegesse as CTH primitivas contra respostas imunes
locais (Nauta & Fibbe 2007).
Além disso, a observação de que as CTM humanas podem ser isoladas
da decídua (In’t Anker et al. 2004), do fluido amniótico (In’t Anker et al. 2003b),
do sangue fetal (In’t Anker et al. 2003a) e do sangue de cordão umbilical
(Bieback et al. 2004) são possíveis indicativos de um papel das CTM in vivo na

Introdução
tolerância fetal. Dados atuais sobre o mecanismo do imunoprivilégio no útero
de mulheres grávidas mostram similaridade aos efeitos imunossupressores já
descritos para as CTM, incluindo a produção de IDO (Barry et al. 2005).
Uma questão importante relacionada ao papel imunomodulatório das CTM
envolve sua utilização na terapia coadjuvante ao transplante de medula óssea,
uma vez que o efeito inibitório das CTM atingiria as células T do doador que
reagem contra o receptor, um efeito protetor para o desenvolvimento da DECH.
Entretanto, esse efeito inibitório poderia atingir também as células T do doador
que reagem contra a leucemia, diminuindo o efeito do Enxerto Contra
Leucemia (ECL). Nenhum estudo foi realizado até momento para avaliar esse
paradigma. No entanto, Djouad e colaboradores mostraram que a injeção de
CTM em conjunto com linhagens de melanoma em camundongos
imunocompetentes favoreceu o desenvolvimento de tumor, em comparação
com camundongos injetados somente com a linhagem tumoral, que não
desenvolveram tumor, indicando que a imunossupressão global pode favorecer
o crescimento do tumor (Djouad et al. 2003).
POTENCIAL EM TERAPIAS CELULARES
A multipotencialidade e a facilidade de isolamento e expansão em cultura
das CTM, suas propriedades imunomodulatórias e sua capacidade de migrar
para sítios de inflamação quando injetadas por via intravenosa (Chapel et al.
2003, Ortiz et al. 2003), têm gerado um grande interesse em sua utilização em
terapias celulares para o tratamento de diversas doenças.
Até recentemente, o conceito de tratamento para doenças do
coração,através da geração de novo tecido miocárdico funcional, era

Introdução
considerada fisiologicamente impossível. Esse conceito vem sendo
questionado, e células derivadas da medula óssea têm sido utilizadas para
testar a possibilidade de regeneração deste tecido. Diversos autores têm
demonstrado que a injeção intracoronariana de populações de células da
medula óssea pode representar uma estratégia simples e promissora para o
tratamento de doenças do coração (Orlic et al. 2001, Ohnishi & Nagaya 2007).
No entanto, ainda não está claro se a população responsável por esse efeito é
a de CTH, CTM ou de progenitores diferenciados.
Após a injeção direta de CTM no miocárdio infartado de um modelo de
porco, foi observada a diferenciação das células injetadas para a linhagem
miogênica, expressando proteínas musculares específicas, e as células
injetadas foram capazes de atenuar a disfunção cardíaca dos animais (Shake
et al. 2002). Além da diferenciação em células cardiomiogênicas, tem sido
proposto que as CTM exerçam seus efeitos na regeneração cardíaca através
de efeitos parácrinos, pela secreção de fatores angiogênicos, anti-apoptóticos e
mitogênicos, como VEGF, HGF, adrenomedulina (AM), e IGF-1 (Kinnaird et al.
2004, Nagaya et al. 2005). A injeção de CTM em um modelo de rato de
cardiomiopatia dilatada melhorou a função cardíaca dos animais, aumentando
a densidade capilar (Nagaya et al. 2005). Em ratos com miocardite aguda, a
administração intravenosa de CTM melhorou as alterações inflamatórias e a
função cardíaca (Ohnishi et al. 2007).
Com a experiência acumulada nos modelos animais, alguns grupos têm
testado a utilização de CTM e células da medula óssea em ensaios clínicos
para o tratamento de disfunções do coração. A administração intracoronariana
de células mononucleares autólogas de medula óssea foi capaz de induzir a

Introdução
melhora de alguns parâmetros cardíacos de pacientes com infarto agudo do
miocárdio (Wollert et al. 2004), enquanto que um resultado mais promissor foi
obtido quando a administração foi feita com populações de CTM derivadas da
medula óssea (Chen et al. 2004). Além disso, o mesmo ensaio realizado em
pacientes portadores de cardiomiopatias isquêmicas crônicas mostrou
resultados semelhantes na melhora da função cardíaca dos pacientes (Chen et
al. 2006). Katritsis e colaboradores mostraram que o transplante
intracoronariano de CTM contribui para a regeneração regional do tecido
miocárdico, mostrando melhora na contratilidade cardíaca de um ou mais
segmentos do miocárdio no grupo que recebeu as CTM, mas não no controle
(Katritsis et al. 2005). Trabalhos de outros grupos mostraram resultados
similares (Assmus et al. 2002, Katritsis et al. 2007), o que sugere que a terapia
celular com populações de células da medula óssea é possível, segura e pode
contribuir para regeneração do tecido do miocárdio lesado. Além disso, o maior
número de células multipotentes presente na população de células cultivadas
ex vivo, quando comparado com a população de células mononucleares da
medula óssea, é necessário para um efeito mais promissor da terapia.
O principal fator na homeostase óssea é o balanço entre a função das
células produtoras de osso (osteoblastos, derivadas das CTM) e a função das
células que reabsorvem osso (osteoclastos, derivadas das CTH). A CTM
influencia diretamente a homeostase óssea, pela taxa de geração de
osteoblastos e pela produção de fatores que afetam o balanço
osteoblasto/osteócito. A substituição óssea, necessária no tratamento de
tecidos lesados por trauma, doenças congênitas e degenerativas, e câncer, é
um dos maiores desafios atuais na pesquisa em ortopedia e traumatologia.

Introdução
Enxertos ósseos de vários tipos (autólogos, homólogos, próteses) podem ser
utilizados para a reconstrução de defeitos esqueléticos, cada um deles
apresentando vantagens e desvantagens (Niemeyer et al. 2006). Entretanto, as
propriedades osteocondutoras, osteoindutoras e osteogênicas estão presentes
apenas pela combinação de biomateriais artificiais com propriedades
osteocondutoras com células osteogênicas viáveis, como osteócitos e
progenitores mesenquimais. Neste sentido, CTM autólogas têm sido utilizadas
em conjunto com matrizes reabsorvíveis para a reconstituição de lesões
ósseas. Em modelos caninos, de coelhos e de ovelhas, a regeneração de
defeitos ósseos foi muito melhor nos grupos que receberam a matriz
colonizada com CTM, do que nos grupos que receberam somente a matriz
(Bruder et al. 1998a, Bruder et al. 1998b, Perka et al. 2000, Quarto et al. 2001).
A origem do novo osso formado nesses modelos é uma questão controversa:
enquanto alguns grupos sugerem que as CTM se diferenciam diretamente em
precursores osteogênicos, que formam posteriormente osteócitos maduros que
induzem a formação de novo osso (Pittenger et al. 1999), Bianco e Robey
sugerem que, em resposta a estímulos locais e sistêmicos (por exemplo,
fraturas), CTM se diferenciam em osteoprogenitores que formam matriz
extracelular (Bianco & Robey 2000). Tal matriz fornece as condições
ambientais osteogênicas ideais para que osteoblastos locais migrem e formem
osso novo. Independentemente do mecanismo biológico, o fato é que
resultados em modelos animais de diferentes grupos apontam para melhor
cicatrização óssea na presença das CTM (Niemeyer et al. 2006). Embora os
resultados desses estudos sejam bem promissores, ainda não existem relatos

Introdução
clínicos confiáveis desse tipo de tratamento na reconstrução óssea em
pacientes humanos (Petite et al. 2000).
Comparada ao osso, a cartilagem possui uma menor capacidade de
regeneração, devido à avascularidade e à acelularidade do tecido. Defeitos da
cartilagem articular podem levar à osteoartrtite e à perda da função da
articulação. Enquanto o transplante de condrócitos autólogos pode predispor a
articulação à osteoartrite, devido à retirada de tecido sadio da região, a
utilização de CTM autólogas derivadas de biópsias da medula óssea surge
como uma promissora opção terapêutica. Defeitos da cartilagem do côndilo
femoral de um modelo canino foram tratados com sucesso pelo transplante de
CTM autólogas em gel de colágeno (Wakitani et al. 1994). Em um modelo
caprino de osteoartrite, Murphy e colaboradores (2003) mostraram que a
injeção direta de CTM autólogas na região intra-articular potencializou a
regeneração do menisco medial. A terapia com CTM alogeneicas foi bem-
sucedida na prevenção do dano tecidual em um modelo de artrite induzida por
colágeno (Augello et al. 2007). Em humanos, o potencial do transplante de
CTM autólogas para o tratamento de lesões de cartilagem foi testado com
sucesso. Pacientes com defeitos no côndilo femoral medial receberam CTM
autólogas, com melhores resultados clínicos que o grupo controle (Wakitani et
al. 2002). Em um relato de caso recente, Centeno e colaboradores mostraram
que o transplante de CTM autólogas resultou no aumento do volume da
cartilagem do menisco (Centeno et al. 2008). Embora esses resultados sejam
bastante promissores, diversos problemas técnicos e biológicos têm sido
observados, como a formação de condrócitos hipertróficos a partir das CTM e a
migração das CTM no osso subcondral, e mais estudos ainda são necessários

Introdução
para desenvolver métodos e biomateriais funcionais para o controle da
diferenciação condrogênica e de sua incorporação nas camadas mais
profundas que margeiam o osso.
O grupo liderado por Horwitz fez avanços significativos na utilização de
progenitores mesenquimais para o tratamento da osteogenesis imperfecta (OI),
desordem genética das células mesenquimais caracterizada pela expressão de
uma forma defeituosa do colágeno tipo I. Os pacientes com OI severa
apresentam numerosas fraturas, deformidades esqueléticas progressivas e
crescimento ósseo retardado. Primeiro, o grupo demonstrou o potencial
terapêutico do transplante alogenêico de medula óssea convencional em
crianças com osteogenesis imperfecta (Horwitz et al. 1999, 2001). Embora em
curto prazo este procedimento tenha resultado em significativa melhora no
quadro dos pacientes transplantados, como formação de nova massa óssea,
aumento na taxa de crescimento e redução na freqüência de fraturas, em longo
prazo esses parâmetros voltaram aos níveis de antes do transplante.
Posteriormente, o mesmo grupo infundiu CTM isoladas da medula óssea de
doadores alogenêicos, em crianças com OI severa que haviam sido
submetidas anteriormente a transplante alogenêico de medula óssea (Horwitz
et al. 2002). O principal achado desse trabalho foi a demonstração da presença
de células diferenciadas derivadas das CTM infundidas no osso, no estroma
medular e na pele, uma vez que elas haviam sido marcadas por transdução de
retrovírus contendo o gene de resistência a Neomicina. Esses resultados
indicam que CTM alogenêicas administradas em crianças com OI severa
podem ser incorporadas em ossos geneticamente defeituosos e se diferenciar

Introdução
em osteoblastos capazes de estender os benefícios clínicos gerados pelo
transplante de medula óssea. Apesar dos resultados promissores, o
desenvolvimento de protocolos clínicos efetivos para o tratamento da OI ainda
não é uma realidade.
Devido à não existência de modalidades terapêuticas efetivas para o
tratamento de lesões do sistema nervoso central, causadas por trauma ou por
doenças neurodegenerativas, a utilização de terapias celulares surge como
uma alternativa para essas patologias. Vários estudos demonstram que o
transplante de CTM resulta em melhora clínica em modelos animais de
isquemia cerebral e de lesões da medula espinhal (Li et al. 2005), embora os
mecanismos envolvidos ainda não estejam definidos. Substituição celular,
liberação de fatores tróficos, e imunomodulação estão entre os possíveis
mecanismos (Kan et al. 2007). Estudos pré-clínicos em um modelo murino de
esclerose lateral amiotrófica, o camundongo SOD1-G93A (Zhao et al. 2007,
Vercelli et al. 2008) mostraram aumento na sobrevida e retardo no
desenvolvimento da doença nos animais que receberam injeção intravenosa de
CTM. Mazzini e colaboradores conduziram um estudo piloto com a
administração intra-espinhal de CTM derivadas da medula óssea em um
pequeno grupo de pacientes portadores dessa doença, que foram
acompanhados por 4 anos (Mazzini et al.2008). Os resultados foram
promissores, mas estão sendo repetidos em um estudo clínico de fase 1 com
maior número de pacientes.

Introdução
TERAPIAS CELULARES ASSOCIADAS AO TRANSPLANTE DE MEDULA ÓSSEA
Tem sido demonstrado na literatura que as CTM facilitam o crescimento
de células hematopoéticas em sistemas in vitro (Mourcin et al. 2005, Li et al.
2007) e em modelos animais. O cotransplante de CTM e CTH humanas
(células CD34+) em fetos de ovelha (Almeida-Porada et al. 1999, 2000) e em
camundongos NOD/SCID (Angelopoulou et al. 2003, in’t Anker et al. 2003,
Bensidhoum et al. 2004) foi capaz de induzir uma “pega” mais eficiente do que
o transplante isolado de células CD34+ humanas, ou seja, um menor número de
células CD34+ foi necessário para um transplante eficiente quando CTM eram
também injetadas. Esses estudos forneceram o racional para a utilização das
CTM em ensaios clínicos de terapias celulares associadas ao transplante de
CTH, com o objetivo de melhorar a reconstituição hematológica dos pacientes.
Após demonstrar, em um estudo piloto (Lazarus et al. 1995), a segurança da
infusão em pacientes de CTM autólogas expandidas in vitro, Koç e
colaboradores (2000) mostraram, num grupo de 28 pacientes com câncer de
mama submetidos à quimioterapia e transplante de células progenitoras do
sangue periférico que a infusão de CTM é factível e parece ter um impacto
positivo na hematopoese, com ausência de efeitos adversos significativos. Em
um ensaio subseqüente com 46 pacientes, a infusão de CTM foi bem tolerada
e o tratamento resultou na melhora da recuperação hematopoética na maioria
dos pacientes (Lazarus et al. 2005).
O transplante de CTH evoluiu nos últimos 30 anos em uma terapia efetiva
para doenças malignas hematológicas e desordens hereditárias ou adquiridas
do sistema hematológico/imune. Após submeterem-se a um regime de
condicionamento mieloablativo para combater a doença hematológica, os

Introdução
pacientes dessas desordens recebem uma infusão de células da medula
óssea, ou de CTH mobilizadas do sangue periférico provenientes do doador
(transplante de CTH), com o objetivo de reconstituir seu sistema
hematopoético. Em desordens malignas, a reconstituição da imunidade de
células T (provenientes do doador) tem um potente efeito antitumoral (ECL),
mas resulta também em um ataque aos tecidos do receptor, levando ao
desenvolvimento da DECH. A eficácia do transplante depende do balanço
desses dois efeitos, o que é bastante complicado, e a DECH atualmente é a
maior causa de morbidade em pacientes submetidos ao transplante de medula
óssea (Bolaños-Meade 2008).
As propriedades imunomodulatórias observadas in vitro geraram um
grande interesse na aplicação das CTM como uma nova terapia
imunossupressora para a DECH. Em modelos animais de TCTH alogeneico,
porém, não há consenso quanto ao potencial imunossupressor das CTM.
Alguns trabalhos demonstraram que a infusão de CTM foi eficaz na prevenção
da DECH, levando a um aumento na sobrevivência de camundongos que
receberam transplante de doador totalmente incompatível (Chung et al. 2004).
Ainda, a injeção subcutânea de células de melanoma B16 levou ao
crescimento de tumor em animais apenas quando CTM foram co-injetadas
(Djouad et al. 2003), o que poderia ser atribuído à diminuição do efeito ECL
devido à imunossupressão exercida pelas CTM. Em outros trabalhos, os
resultados obtidos foram opostos, o que tem levado à controvérsia a respeito
do potencial imunomodulatório das CTM em modelos animais. Eliopoulos et al.
(2005) e Nauta et al. (2006) mostraram que CTM alogeneicas são
imunogênicas e capazes de sofrer rejeição em camundongos

Introdução
imunocompetentes. Usando um modelo animal de DECH, Min et al. (2007)
mostraram que CTM foram capazes de atenuar a severidade da doença
apenas quando as células haviam sido transduzidas para expressar
constitutivamente a citocina antiinflamatória Il-10. Sudres et al. (2006)
mostraram que células capazes de suprimir a proliferação de linfócitos in vitro
falharam na prevenção da DECH em camundongos submetidos ao TCTH,
enquanto Tisato et al. (2007) mostraram ainda que CTM derivadas de sangue
do cordão umbilical podem previnir a GVHD, mas não são capazes de tratar a
doença.
A despeito da ausência de dados pré-clínicos consistentes, a utilização de
CTM na clínica tem evoluído rapidamente como um novo tratamento para a
DECH, em particular para a DECH refratária a esteróides. Neste contexto, o
primeiro trabalho a mostrar o uso clínico de CTM para o tratamento da DECH
foi desenvolvido pelo grupo de Le Blanc, em 2004, e tornou-se um marco para
os grupos que trabalham com aplicação clínica das CTM. Após infusão
intravenosa de CTM provenientes de sua mãe, um paciente com leucemia
linfoblástica aguda, que havia sido submetido a transplante alogenêico de CTH,
apresentou significativa melhora clínica e foi o único a sobreviver (1 ano após o
transplante) de um grupo de 25 pacientes que desenvolveram DECH (Le Blanc
et al. 2004b). Em 2006, Ringden e colaboradores mostraram a melhora na
DECH aguda refratária a esteróides na maioria dos pacientes submetidos ao
transplante após receber infusão de CTM (Ringden et al. 2006). Um ensaio
clínico de fase II foi desenhado para observar a toxicidade e segurança do
tratamento com CTM para DECH aguda refratária, e recentemente publicado
(Le Blanc et al. 2008). Trinta e nove de 55 pacientes com DECH aguda severa

Introdução
e resistente a esteróides responderam ao tratamento com CTM, com maiores
taxas de sobrevivência e menores de mortalidade relacionada ao transplante.
Conforme demonstrado nesses e em outros estudos, o grande potencial
das CTM como terapia celular para o tratamento de várias doenças, em
particular nas terapias associadas ao transplante de medula óssea, é atribuído
em grande parte às suas propriedades in vitro, tais como potencial
imunomodulatório, efeitos parácrinos sobre células e progenitores
indiferenciados, seu próprio potencial de diferenciação, e possível efeito
angiogênico.
O potencial terapêutico das CTM começou a ser utilizado há
aproximadamente duas décadas, após as descobertas de Friedenstein e
colaboradores, quando empresas de biotecnologia começaram a explorar este
potencial (Bianco et al. 2006). Apesar de controvérsias nos estudos em
modelos animais, e do pouco entendimento acerca dos mecanismos pelos
quais elas exercem seus efeitos, as CTM têm sido empregadas em grandes
ensaios clínicos no mundo inteiro, com alguns resultados bastante
promissores. Tal paradoxo assemelha-se ao que ocorreu com o medicamento
aspirina (ácido acetilsalicílico), que é utilizado há mais de 100 anos como uma
das mais potentes drogas antiinflamatórias, antipiréticas e analgésicas do
mercado, apesar de seu mecanismo de ação só ter sido desvendado 70 anos
depois que a droga foi lançada no mercado (Giordano et al. 2007).
Nesse contexto, estudos de caracterização molecular, em particular os de
larga escala, podem fornecer pistas importantes sobre seus mecanismos de

Introdução
ação terapêutica, possíveis efeitos adversos, além da influência de seu
isolamento e cultivo ex vivo sobre suas características essenciais.

Objetivos
OBJETIVOS
Tendo em vista a variação de resultados obtidos na literatura em relação
à biologia básica e às propriedades in vitro e in vivo das CTM, estudos de
caracterização molecular, em particular os de larga escala, podem fornecer
pistas importantes sobre seus mecanismos de ação terapêutica, possíveis
efeitos adversos, além da influência de seu isolamento e cultivo ex vivo sobre
suas características essenciais. Assim, os objetivos dessa tese estão descritos
a seguir.
OBJETIVO GERAL
� Caracterizar molecularmente culturas de CTM humanas estabelecidas a
partir de amostras da medula óssea de doadores de CTH no CEntro de
Transplante de Medula Óssea (CEMO) / Instituto Nacional de Câncer
(INCa).
OBJETIVOS ESPECÍFICOS
� Estabelecer protocolo para o isolamento e cultivo de CTM a partir de
amostras da medula óssea de doadores de CTH;
� Caracterizar as culturas como CTM;
� Identificar características moleculares importantes, através de estudos
proteômicos, para sua utilização em terapias celulares associadas ao
Transplante de Medula Óssea (TMO).

Metodologia
METODOLOGIA
1. ISOLAMENTO E CULTIVO DE CTM
Esse projeto foi aprovado pelo Comitê de Ética e Pesquisa do INCa
(processo CEP INCa n°034/06) e todo material foi utilizado após assinatura por
parte dos doadores de medula óssea de um Termo de Consentimento Livre e
Esclarecido.
Amostras de medula óssea de doadores do CEMO, obtidas por aspiração
da crista ilíaca posterior em Cloreto de Sódio 0,9% com 10U/mL de heparina,
foram processadas para o cultivo in vitro de Células MonoNucleares (CMN)
aderentes: 5-10mL do aspirado foram primeiro diluídos em igual volume de
soro fisiológico (Cloreto de Sódio 0,9%); essa suspensão foi adicionada
cuidadosamente sobre cerca de 5-10mL de Ficoll-PaqueTM Plus, 1,077g/L
(Amersham Biosciences, Suécia); após centrifugação a 500g por 30 minutos a
10oC, as CMN foram removidas da interface do gradiente e lavadas 2 vezes em
soro fisiológico.
Para determinação da densidade inicial de plaqueamento das CMN da
medula óssea, elas foram distribuídas em poços de 9cm2 em sucessivas
densidades (2x103 células/cm2, 2x104 células/cm2, 2x105 células/cm2 e 2x106
células/cm2), em meio DMEM com baixa glicose (Dulbecco’s Modified Eagle’s
Medium, Invitrogen, EUA), 100U/mL de penicilina, 100µg/mL de estreptomicina,
2mM de glutamina e 15% de soro bovino fetal - SBF (meio CTM), e incubadas
em estufa a 37oC e 5% de CO2. Após vinte e quatro horas, as culturas foram
lavadas 2 vezes em PBS 1x (Phosphate Buffer Saline, KCl 2,7mM, K3PO3
1,8mM, NaCl 137mM, Na3PO3 10,1mM) para remoção das células não

Metodologia
aderentes. Após 13 dias de cultivo, as células foram lavadas em PBS 1x,
fixadas em metanol, e coradas em Giemsa (KaryoMAX Giemsa Stain improved
R66 solution “Gurr”, GibcoBRL/Invitrogen, EUA). O número de colônias obtidas
foi contado a olho nu, e comparado entre as diversas concentrações iniciais de
células. Este número foi correlacionado ao número de UFC-F presentes na
amostra de medula óssea.
Determinada a densidade inicial de plaqueamento para o estabelecimento
das culturas de CTM, as amostras de células da medula óssea passaram a ser
distribuídas em garrafas de 25cm2, na concentração de 2 x 105 células/cm2.
Quando atingiam 80% da confluência, as culturas eram submetidas a uma
solução de tripsina 0,05%-ácido etileno diamino tetraacético 0.53mM (Ethylene
Dyamin Tetracetic Acid, EDTA) (InVitrogen, EUA), para que as células
aderentes se soltassem da placa, e fossem distribuídas em novas
placas/garrafas, na proporção de 500-1000 células/cm2 (passagem 1).
2. CARACTERIZAÇÃO DAS CULTURAS DE CTM
2.1 Análise ao Microscópio Ótico
As culturas foram rotineiramente observadas em microscópio invertido de
contraste de fase (Zeiss, Alemanha). Para uma análise morfológica mais
detalhada, as células foram coradas em Giemsa após lavagem em PBS 1x e
fixação em metanol 100%.
2.2 Análise de Marcadores de Superfície
A caracterização dos marcadores de superfície das CTM foi realizada
através de ensaios de citometria de fluxo. A uma suspensão de 1 x 106 células,

Metodologia
obtidas após tripsinização de culturas de CTM, foram adicionados anticorpos
conjugados aos fluorocromos FITC, PE e Cy5-FITC. As células foram
analisadas em um citômetro de fluxo Fluorescence Activated Cell Analyser
(FACScan, Becton Dickson, EUA) e o software CELLQuestTM (Becton
Dickson, EUA). Células positivas foram identificadas por comparação com
controles isotípicos. Os anticorpos utilizados reconhecem os seguintes
marcadores de superfície: CD45, CD34 e HLA-DR.
2.3 Ensaios de diferenciação in vitro
O potencial de diferenciação das CTM foi testado em várias passagens
das culturas, começando pela passagem 1. Foram realizados ensaios para a
diferenciação adipogênica e osteogênica, utilizando-se protocolos específicos
para cada caso.
2.3.1 Diferenciação Osteogênica
Cerca de 104 células foram cultivadas em poços de 9cm2 em meio
osteogênico (DMEM, 10% SBF, 0,01µM dexametasona, 10mM β-glicerol
fosfato, 0,2mM ácido ascórbico), por no mínimo 3 semanas, com troca de meio
2 vezes por semana.
A diferenciação das células mesenquimais foi detectada pela observação
de depósitos de Cálcio nas culturas, através da coloração com Vermelho de
Alizarina pH4,2 por 5 minutos a temperatura ambiente.
2.3.2 Diferenciação Adipogênica

Metodologia
Cerca de 104 células foram cultivadas em poços de 9cm2 em meio
adipogênico (DMEM em 10%SBF, 0,01µM dexametasona, 5µg/mL insulina) de
2 a 3 semanas, com troca de meio 2 vezes por semana.
O fenótipo adipogênico das culturas foi confirmado pela presença de
gotículas de gordura, após fixação das células em PFA (ParaFormAldeído) 4%
por 1 hora e coloração com corante Oil Red O em isopropanol por 5 minutos.
3. CARACTERIZAÇÃO MOLECULAR
3.1 Análise da Expressão Gênica
3.1.1 Extração de mRNA (messenger RiboNucleotidic Acid)
Culturas de CTM foram submetidas a uma solução de tripsina 0,05%-
EDTA, para individualização das células, que foram posteriormente lavadas em
PBS 1X por 2 vezes e adicionadas a 1mL de Trizol (Invitrogen, EUA), na
proporção de 106 células/mL de Trizol. Após homogeneização, as células foram
centrifugadas por 10 minutos a 12.000g a 4°C, e o sobrenadante foi recuperado
em um tubo novo. Ao sobrenadante foram adicionados 200µL de clorofórmio
para cada 1mL de Trizol, e em seguida a solução foi misturada vigorosamente
por 15 segundos e incubada por 2 a 3 minutos a temperatura ambiente. O tubo
foi então centrifugado por 15 minutos a 12.000g a 4°C, e a fase aquosa do
sobrenadante foi recuperada em um tubo novo. Foram adicionados então
500µL de isopropanol para cada 1mL de Trizol, e a mistura incubada por 10
minutos a temperatura ambiente. Após a incubação, o material foi centrifugado
por 10 minutos a 12.000g a 4°C e o sobrenadante descartado. O sedimento foi
lavado com 1mL de etanol 75% e submetido à agitação vigorosa em vórtex GV
Lab (Gilson, Inglaterra), sendo centrifugado por 5 minutos a 7.500g a 4°C, e o

Metodologia
sobrenadante descartado. O sedimento foi ressuspendido em 20µL de água
tratada com DiEtil PiroCarbonato (DEPC) e guardado a -70°C. Para a
quantificação da massa de RNA extraída das culturas, foi realizada análise em
espectrofotômetro Nanodrop ND-100 (Thermo Scientific, EUA) a 260nm. A
avaliação da qualidade do RNA foi realizada através de eletroforese em gel de
agarose 0,8%.
3.1.2 Desenho dos iniciadores
Os iniciadores utilizados para análise da expressão gênica por PCR
(Polymerase Chain Reaction) foram desenhados de acordo com instruções
contidas no manual do Power SYBR® Green PCR Master Mix and RT-PCR
(Applied Biosystems, EUA), com auxílio da ferramenta disponível on-line na
página da IDT (Integrated DNA Technologies - www.idt.org). As seqüências dos
RNAm foram obtidas no banco de dados do genoma humano
(www.ncbi.nlm.nih.gov/nucleotide). A especificidade dos oligonucleotídeos
foram determinadas previamente in silico diante de sua similaridade aos dados
de RNAm humanos através da ferramenta BLAST (Basic Local Alignment
Search Tool) / HumanGenome (www.ncbi.nlm.nih.gov/Blast.cgi). Na Tabela 1,
está o resumo dos iniciadores usados.
Tabela 1: Oligonucleotídeos (iniciadores) utilizados para amplificação por PCR Nome Sequência No acesso GenBank
BMPR1A 5’-CGAAGTGCTGGACGAAAGCCT
5’-TTGACACACACAACCTCACGCA
NM_004329+2
CD44 5’-AGACCCACTCAGCCAAATCT
5’-CTTTCACAGGAGAGAACAGGTG
NM_000610+3

Metodologia
SCF 5’-ATGCAAATGGTTGCCTGGTCCC
5’TGCCAATGTCCTTCTTGGCAGA
NM_003994+4
Runx2 5’-CCTTGACCATAACCGTCTTCA
5’-ATGCGCCCTAAATCACTGHAG
NM_004348+4
LPL 5’-GGGCATGTTGACATTTACCC
5’-TGGAACTGCACCTGTAGGC
NM_000237+2
3.1.3 Síntese de cDNA (complementary DeoxyNucleotidic Acid, transcrição
reversa)
Para a síntese do cDNA foram utilizados 1µg de RNA, 250ng de Oligo
DT primer (Invitrogen, EUA), 10mM de dNTPs (10mM de dATP, 10mM de
dCTP, 10mM de dTTP e 10mM de dGTP, Invitrogen, EUA) e água MiliQ para
completar um volume final de 12µL. A reação foi incubada por 5 minutos a
65°C e mantida no gelo. Posteriormente, foram adicionados 4µL de tampão 5x,
2 µL de DTT (DiThioThreitol) e 1µL de inibidor de ribonuclease RNAguard (GE
Healthcare, Suécia), completando um volume final de 19µL. A reação foi
misturada gentilmente, e incubada por 2 minutos a 42oC. Após a incubação,
foram adicionadas 200 unidades da enzima Superscript II RT (Invitrogen, EUA),
completando o volume final para 20µL, e a reação foi incubada por 50 minutos
a 42 °C, e por 15 minutos a 70°C, e estocada a -20 °C.
3.1.4 Amplificação do cDNA por PCR em Tempo Real
Inicialmente, foram realizadas reações testes para a determinação das
condições ideais do ensaio (curva padrão e curva de dissociação), onde foram
utilizadas quantidades sucessivas de cDNA (50ng, 20ng, 5ng, 500pg), 2µM de

Metodologia
cada oligo específicos (senso e anti-senso) e 1X Power SYBR Green PCR
Master Mix (Applied Biosystems, EUA) em 20µL de reação. As reações para
análise por PCR em tempo real foram submetidas à incubação inicial no
termociclador (RotorGene RG-3000, Corbett, Austrália) de 95°C por 10min, e
por 50 ciclos de 95°C por 20s e 60°C por 1min. Como normalizador da
amplificação, foram utilizados oligonucleotídeos específicos para o gene da ββββ-2
Microglobulina (ββββ2M) humana (3 e 0,5 uM de cada iniciador). As alterações nos
níveis de MRNA foram calculadas pelo método baseado na curva padrão de
amplificação dos genes em questão. A quantidade de MRNA dos pontos
experimentais foi calculado com auxilio da equação da reta obtida da curva
padrão. Em seguida, os resultados foram normalizados pelos níveis de MRNA
de β2M. A diferença dos níveis de MRNA entre as condições experimentais foi
calculada em relação a uma amostra calibradora, estabelecida de acordo com
cada questão experimental.
3.2 Análise Proteômica
3.2.1 Preparação de extratos protéicos totais
Culturas de CTM foram tripsinizadas e lavadas três vezes em PBS (1X)
Após serem ressuspensas em Tampão de Extração de Proteínas a 4oC [50mM
de Tris pH7.5, 5mM EDTA, 10mM ácido etileno glicol tetraacético (Ethylene
Glycol Tetracetic Acid, EGTA), 50mM fluoreto de sódio (NaF) 20mM β-
Glicerolfosfato, 250mM cloreto de sódio (NaCl), 0.1% de Triton X-100, 20Mm
ortovanadato de sódio, e mix de inibidores de protease (2,4µg/mL 4-(2-
aminoetil) benzenosulfonil fluoreto hidrocloreto, 0,1µg/mL bestatina,
0,1µg/mLpepstatina, 0,1 µg/mL leupeptina, 1,8µg/mL E-64, Calbiochem, EUA],

Metodologia
as células foram incubadas durante trinta minutos a 4°C e centrifugadas a
12.000g por 30 minutos. O sobrenadante foi retirado e utilizado como extrato
total de proteínas. Após a clarificação dos extratos, estes foram quantificados e
armazenados a -70°C.
Para a análise proteômica das células após diferenciação, 103
células/cm2 na passagem 3 foram cultivadas em meio osteogênico (DMEM,
10% SBF, 0,01µM dexametasona, 10mM β-glicerol fosfato, 0,2mM ácido
ascórbico), ou adipogênico (DMEM em 10%SBF, 0,01µM dexametasona,
5µg/mL insulina) durante 3 semanas, sendo então tripsinizadas e submetidas
ao mesmo procedimento descrito acima. Concomitantemente, amostras das
mesmas células eram submetidas à diferenciação em poços de 9cm2
(conforme descrito no item 2.2.1), e o fenótipo diferenciado era evidenciado
como anteriormente.
3.2.2 Quantificação dos extratos protéicos
Os extratos protéicos foram quantificados segundo o método de Bradford
(1976), que se baseia na interação entre o corante Coomassie Brilliant G-250 e
macromoléculas de proteínas que contém aminoácidos com cadeias laterais
básicas ou aromáticas, o que altera a coloração da amostra. Após a construção
da curva padrão de proteínas, obtida pela medida da densidade ótica de
soluções de concentrações conhecidas de Albumina Sérica Bovina (Bovine
Serum Albumin, BSA) e reagente de Bradford (0,01% Coomassie Brilliant G-
250, 4,75% Etanol e 8,5% Ácido Fosfórico), a concentração dos extratos
protéicos foi determinada.

Metodologia
3.2.3 Análise da integridade das proteínas no extrato
Para análise da integridade e qualidade dos extratos protéicos, 15 µg de
cada extrato foram submetidos à eletroforese unidimensional em gel de
poliacrilamida SDS-PAGE (Sodium Dodecyl Sulfate Polyacrylamide Gel
Electrophoresis) 10%, submerso em tampão de corrida (250 mM Tris-base,
192mM Glicina). Os géis foram corados em 0,1% Coomassie Blue R-250, 30%
Etanol e 0,5% Ácido Acético, descorados por 15 minutos em solução
descorante (25% Metanol e 5% Ácido Acético) e secos em solução de secagem
(30% Metanol e 3% Glicerol) por 15 minutos.
3.2.4 Eletroforese Bidimensional
IsoEletroFocalização IEF (PRIMEIRA DIMENSÃO)
Os extratos foram inicialmente precipitados com o kit 2D Clean up (GE
Healthcare, Suécia) e ressuspendidos em 200µL de tampão de reidratação. As
análises iniciais foram realizadas com tampão de reidratação 1 [8M uréia, 15mM
DiTiTreitol (DTT), 2% NP40, 0,5% tampão IPG pH 3-10, GE Healthcare, Suécia,
e traços de azul de bromofenol]. Outras análises foram realizadas com tampão
de reidratação 2 [6M uréia, 2M tiouréia, 15mM DTT, 2% ASB14, 0,5% tampão
IPG pH 3-10, e traços de azul de bromofenol]. Após dissolução no tampão, as
proteínas foram reidratadas a 40V, em tiras cilíndricas de poliacrilamida com
gradientes lineares de pH 4 a 7 com 11cm de comprimento, Immobiline DryStrip
(GE Healthcare, Suécia). A focalização foi realizada, no sistema Ettan IPGphor
II (GE Healthcare, Suécia), em um total de 30550VhT.

Metodologia
Eletroforese SDS-PAGE (SEGUNDA DIMENSÃO)
Após a primeira dimensão, as tiras foram equilibradas por 15 minutos
em tampão de equilíbrio (6M uréia, 30% glicerol, 2% SDS em tampão 0,05M
Tris-HCl pH 8,8, e traços de azul de bromofenol) contendo 100mg de DTT, e
incubadas por mais 15 minutos em tampão de equilíbrio contendo 250mg de
iodocetamida. As tiras foram submetidas a uma eletroforese desnaturante em
gel de gradiente de 8 a 18% de poliacrilamida (ExcelGel SDS 8-18%, GE
Healthcare, Suécia), utilizando-se o sistema Multiphor II (GE Healthcare,
Suécia), durante 1hora e 50 minutos, contendo 10µL de padrão de peso
molecular Benchmark (Invitrogen, EUA). As proteínas separadas na primeira
dimensão (por unidade de pH, que corresponde ao seu ponto Isoelétrico, pI)
foram então separadas por peso molecular, podendo ser identificadas em um
eixo cartesiano. Os géis preparativos foram corados em Comassie Blue
colloidal (2% Comassie Blue G-250, Sigma-Aldrich, EUA) por 72 horas e
descorados em água destilada.
3.2.5 Análise dos Padrões Proteômicos
Após a eletroforese bidimensional os géis foram escaneados utilizando
o software LabScan, analisados e comparados no software Image Master 2D
Platinum 6.0 (GE Healthcare, Suécia). Os pI foram determinados utilizando-se
uma distribuição linear de 4-7, e o peso molecular foi determinado baseando-se
no padrão Benchmark (Invitrogen, EUA), o que gerou uma curva logarítmica. Os
spots visualizados em cada gel foram comparados e tiveram seus volumes
normalizados. A intensidade de cada spot foi quantificada pelo cálculo do
volume do spot após normalização da imagem utilizando-se o método de

Metodologia
normalização do volume total do spot multiplicado pela área total de todos os
spots. Foram então geradas comparações qualitativas e quantitativas entre as
diferentes amostras. Para garantir a confiabilidade dos dados, apenas spots
apresentando diferenças maiores que 2x foram considerados diferencialmente
expressos.
Após esta análise, os spots proteicos de cada amostra foram retirados
para identificação, sendo transferidos individualmente para microtubos
siliconizados de 1500µL, e lavados em 50% de ACetoNitrila (ACN) / 25mM de
bicarbonato de amônio (NH4HCO3) pH 8.0 durante 30 minutos, até ficarem
completamente descorados. Os spots foram então desidratados em ACN 100%
por 10 minutos, e completamente secos a temperatura ambiente.
3.2.6 Espectrometria de Massas
Os spots selecionados para identificação foram digeridos pelo método
InGel Digest. Após a desidratação e secagem, os spots foram reidratados a 4oC
em tampão de digestão (25 mM NH4HCO3 pH 8.0) contendo 15 ng/µL de
tripsina (Sequence Grade Modified Trypsin, Porcine – Promega, EUA), e
incubados por 12 horas a 37°C. Os peptídeos gerados foram então extraídos do
gel com 50% de ACN/5% de Ácido Trifluoroacético (TriFluoroacetic Acid, TFA).
Os extratos digeridos foram concentrados em um concentrador Hetovac
(HetoLab Equipment, Dinamarca), e posteriormente redissolvidos em 3µL de
50% de ACN/1% de TFA.
Para as análises por espectrometria de massas, 0,5µL de cada amostra
foram misturados a 0,5 µL de matriz α-ciano recristalizada (10mg/mL em 50%
ACN e 0,1% TFA, Sigma-Aldrich, EUA), e aplicados em uma placa de MALDI,
juntamente com os calibrantes do kit Sequazyme Peptide Mass Standards

Metodologia
(Perseptive Biosystems, EUA). Os espectros foram adquiridos em um
espectrômetro MALDI-TOF-TOF (Matrix Assisted Laser Desorption/Ionization –
Time Of Flight - Time Of Flight) 4700 (Applied Biosystems, EUA), no modo
refletor de íons positivos. Os espectros de massas foram analisados por
fingerprinting das massas dos peptídeos gerados (MS) e confirmados pelo
perfil de fragmentação dos peptídeos (MS/MS) (Figura 1). Foram analisados
peptídeos de 800 a 4000Da. Os espectros adquiridos permitiram a identificação
das proteínas através do programa Mascot Daemon searching engine (Matrix
Science, EUA) contra a base de dados de sequências proteicas International
Protein Index (IPI, versão 3.12). Os parâmetros de busca para MS e MS/MS
foram 0,2Da, e os critérios para identificação positiva de proteínas foram um
mínimo de 2 peptídeos com score peptídico no Mascot ≥ 20.
Figura 1: Esquema de funcionamento do espectrômetro MALDI-TOF-TOF. O extrato de peptídeos gerado pela digestão in gel de cada proteína (1), após ser misturado com matriz de ácido forte (α-ciano), foi ionizado com laser (2). Os íons voam em um tubo a vácuo e o tempo de chegada de cada íon à placa detectora gera um espectro. Os espectros foram analisados por fingerprinting das massas dos peptídeos gerados (MS) e confirmados pela sequenciamento dos peptídeos (MS/MS) (3) (Modificado de Aebersold & Mann, 2003).

Metodologia
4. ANÁLISE DO POTENCIAL IMUNOSSUPRESSOR IN VITRO
Para testar a capacidade de culturas de CTM de inibir a proliferação de
linfócitos em resposta à estimulação por aloantígenos, foram realizadas
Culturas Mistas de Linfócitos (CML), que consistiram na estimulação de CMN
com CMN alogeneicas, na presença ou não de CTM.
Após serem ressuspendidas em meio RPMI completo (RPMI 1640
suplementado com 10% SBF, 2mM L-glutamina, 50U/mL penicilina e 50 µg/ml
estreptomicina), CMN derivadas de sangue periférico de dois doadores
distintos foram denominadas de CMN estimuladoras (CMNe) ou CMN
respondedoras (CMNr). As CMNe foram irradiadas a 2500 cGy e as CMNr
foram marcadas com CFSE (CarboxyFluorescein Succinimidyl Ester) 2,5 µM
por 5 minutos à temperatura ambiente, e a marcação foi interrompida
adicionando-se 5%SBF gelado. Ambas as células, CMNe e CMNr, foram
lavadas em PBS 1x, centrifugadas a 300g por 5 minutos, e ressuspendidas na
concentração de 5x106 células/mL em meio RPMI completo. As culturas foram
feitas em placas de 24 poços, adicionando-se 5x105 CMNr e 5x105 CMNe na
presença ou não de 5x104 CTM (10% CTM), num volume final de 1000µL. As
células foram cultivadas a 37°C em atmosfera de 5% de CO2, durante 7 dias,
sendo posteriormente analisadas por citometria de fluxo. Os dados foram
adquiridos no citômetro de fluxo FACScan (Becton Dickson, EUA) e analisados
com o software CELLQuest (Becton Dickson, EUA) e com teste T-student,
onde valores de P<0,05 foram considerados estatisticamente significativos. As
análises estatísticas e representações gráficas foram realizados através do
software GraphPad PrismTM (GraphPad, EUA).

Metodologia
5. ANÁLISE CITOGENÉTICA
Culturas de CTM em diversas passagens foram cultivadas em garrafas de
25cm2 até 70-80% da confluência. As culturas foram incubadas com 0.05µg/mL
colchicina (Sigma-Aldrich, EUA) por 2 horas a 37oC e 5% CO2., sendo então
tripsinizadas, lavadas em PBS 1X, e o precipitado obtido foi submetido ao
choque hipotônico, utilizando-se solução de 0.075M cloreto de potássio. O
material foi mantido a 37oC por 15 minutos. Em seguida, foi feita fixação com
Solução Carnoy 3:1 (metanol:ácido acético) por 20 min. O material foi
ressuspendido em um pequeno volume de fixador para preparo das lâminas.
Esse procedimento foi feito pingando-se uma gota da suspensão em lâmina e,
em seguida, o material foi corado em corante Giemsa (Merck, EUA) diluído em
tampão fosfato (14,08g/L NaH2PO4 + 13,91g/L Na2HPO4). Uma das lâminas foi
observada em microscopia ótica para verificação do índice mitótico e da
qualidade das metáfases. As outras foram submetidas à técnica de
bandeamento GTG (Seabright 1971). As lâminas foram envelhecidas de 1 a 2
dias. Em seguida, foram incubadas em tripsina 0,1% diluída em solução
Dulbeco pH 7,8 (8g/L NaCl, 200mg/L KCl, 200mg/L KH2PO4, 1,5g/L NaH2PO4)
aquecida a 37°C, em tempos que variaram de um segundo a um minuto.
Posteriormente, as lâminas foram lavadas em solução de soro fisiológico e
coradas em corante Giemsa 2% (Merck, EUA) por 15 minutos.
Os cromossomos foram identificados e classificados de acordo com o
Sistema Internacional de Nomenclatura de Citogenética Humana de 2005
(ISCN, 2005). A análise cromossômica foi realizada em microscopia óptica
utilizando-se no mínimo 20 metáfases por cultura. Foram adquiridas 5

Metodologia
metáfases para a montagem dos cariótipos de cada passagem da cultura,
através do Sistema de Cariotipagem Cytovision (Applied Image, EUA).

Resultados
RESULTADOS
1. Definição do Protocolo de Estabelecimento das Culturas de CTM e sua
Caracterização
A primeira etapa deste trabalho consistiu na determinação das condições
ideais para o cultivo e estabelecimento das culturas de CTM a partir das células
de medula óssea de doadores. Para isso, foram testados alguns protocolos, e o
sucesso no estabelecimento das culturas de CTM foi obtido quando utilizamos
o anel de células mononucleares, DMEM de baixa glicose (1g/L) e 15% de soro
bovino fetal. Aspirados de medula óssea foram obtidos de 41 doadores do sexo
masculino (média de idade de 40,3 anos, variando entre 30 e 59 anos) e 18
doadores do sexo feminino (média de idade de 40,4 anos, variando entre 32 e
47 anos), em um total de 59 amostras. As amostras foram processadas e
colocadas em cultura para obtenção de culturas de CTM. Não foi possível o
obter sucesso no estabelecimento de culturas de CTM de 9 amostras de
medula óssea. Consideramos sucesso no estabelecimento quando as culturas
puderam ser expandidas por mais de 4 passagens, ainda mostrando a mesma
aparência morfológica e potencial proliferativo das passagens iniciais (0 e 1).
Os motivos que levaram ao não sucesso de algumas culturas incluíram
problemas técnicos do laboratório, e somente de 2 amostras, as células na
passagem 0 não proliferaram, o que podemos atribuir à própria quantidade de
CTM presentes na amostra de medula óssea coletada, ou a algum problema no
cultivo que não pudemos determinar.

Resultados
Nesse trabalho, foi adotada a seguinte terminologia: para cada cultura de
CTM (derivada de um doador de medula óssea) foi atribuída uma sigla, CTM
XX, com XX sendo o número da amostra, seguido de PY, com Y
correspondendo à passagem da cultura, por exemplo, CTM14 P1 é a cultura
estabelecida a partir da amostra de medula óssea de número 14, na primeira
passagem, ou seja, após o primeiro repique.
A melhor concentração de células mononucleares para o cultivo primário
foi determinada através da análise de UFC-F obtidas após o plaqueamento de
amostras de células mononucleares em diversas concentrações. Foram
distribuídas concentrações sucessivas de células mononucleares em poços de
9cm2, cultivadas por 13 dias, e o número de colônias obtidas foi avaliado e
comparado. Podemos observar na Figura 2 que as melhores concentrações
variaram de 105 a 106 células/cm2, e a concentração escolhida para o
estabelecimento das culturas foi a de 2 x 105 células/cm2, uma vez que era
suficiente para obtenção não clonal de culturas de CTM (Bianco et al 2006),
que atingiam a confluência após 7 a 10 dias de cultivo.
Figura 2: Determinação da melhor concentração de células mononucleares para o plaqueamento inicial da cultura de CTM.
0
5
10
15
20
25
2 x 10E3 2 x 10E4 2 x 10E5 2 x 10E6 2 x 10E7
CMN plaqueadas / cm2
UF
C-F

Resultados
Outra etapa importante para o cultivo das CTM foi a determinação da
concentração ideal de células para o plaqueamento secundário das culturas,
isto é, qual a quantidade mais adequada para a distribuição em novas placas
de células obtidas da tripsinização de culturas primárias. Pela análise da
literatura e experiência no cultivo das células, estabelecemos o número de 500
a 1.000 células/cm2 como ideal para o plaqueamento secundário das CTM.
Logo após o primeiro repique, a morfologia das culturas passou a ser mais
homogênea, apesar de ainda serem observados alguns tipos celulares
diferentes entre si: células fibroblastóides retas e compridas, células
fibroblastóides com mais de 2 prolongamentos, células com corpo celular mais
espraiado, células arredondadas (Figura 3).
As culturas foram observadas diariamente ao microscópio ótico e
fotografadas (Figura 4). Podemos observar que, na passagem zero (culturas
primárias), havia células de diversos tipos e origens, uma vez que se tratavam
de todas as células mononucleares presentes na medula óssea, tais como
células hematopoéticas, tanto células mais diferenciadas quanto progenitores e
precursores, células de gordura, fibroblastos do estroma, entre outros
elementos presentes no ambiente da medula óssea.
Os resultados iniciais deste trabalho levaram ao desenvolvimento de um
protocolo de estabelecimento de culturas de CTM a partir de células da medula
óssea (Quadro 1), o que constitui uma etapa fundamental para o
desenvolvimento de futuros protocolos terapêuticos utilizando tais células.

Figura 3: Análise ao microscópio ótico de culturas de CTM coradas em May-Grünwald/Giemsa; (A), (B), (C) e (D) CTM 26 P1; (E) e (F) CTM24 P5 (aumento de 100X).
A C B
F E D

Figura 4: Análise ao microscópio ótico de culturas de células mononucleares da medula óssea para o estabelecimento de CTM (CTM16 P0);(A) e (C) 5 dias de cultivo, (B) 8 dias de cultivo; (A e B em aumento de 100X, C em aumento de 200X).

Resultados
Protocolo de Estabelecimento de CTM a partir da medula óssea
� Separar as células mononucleares da amostra de medula óssea, por
extração em Ficoll-HypaqueTM;
� Lavar as células mononucleares em PBS 1x ;
� Distribuir em placas de cultura 2 x 106 células/cm2 em meio DMEM
com baixa glicose (1g/L), 15% de SBF, 2mM de glutamina, 100U/mL de
penicilina e 100µg/mL de estreptomicina (passagem 0); incubar em
estufa a 37oC em 5% de CO2;
� Observar diariamente as culturas, para a presença de células
aderentes;
� Após 12 horas, lavar vigorosamente as culturas em PBS 1x, a fim de
retirar as células não aderidas, e adicionar meio novo;
� Quando as culturas atingirem de 80-90% da confluência (cerca de 7-10
dias após o plaqueamento inicial), submeter as culturas à tripsinização
com solução de tripsina 0,05%-EDTA;
� Distribuir cerca de 500-1000 células/cm2 por placa de cultura
(passagem 1);
� Observar as culturas diariamente, e submetê-las à expansão, ou outros
ensaios específicos, quando a confluência for alcançada. O tempo
médio para o subcultivo é de 7-10 dias. Em passagens posteriores (a
partir da 5 ou 6), o subcultivo deve ser feito utilizando-se acima de
1.000 células/cm2, a fim de otimizar o tempo para confluência, uma vez
que as células parecem ter sua capacidade de proliferação diminuída à
medida em que são subcultivadas (observações feitas durante o
cultivo).
Quadro 1: Protocolo para o estabelecimento e cultivo de CTM a partir de amostras de medula óssea de doadores saudáveis.

Resultados
Devido à ausência de marcadores específicos e ensaios de repopulação
in vivo, a capacidade de uma população em se diferenciar em células da
linhagem adipogênica, osteogênica e condrogênica tem sido utilizada como
ensaio para identificação das CTMs (ensaios funcionais). Neste trabalho,
desenvolvemos protocolos de diferenciação para corroborar a identificação das
células estabelecidas a partir da medula óssea para as linhagens osteogênicas
e adipogênicas, e garantir que nosso protocolo de estabelecimento é confiável.
Na Figura 5, observamos ensaios de diferenciação adipogênica e
osteogênica realizados com células na passagem 1. Em 5A, as células foram
cultivadas em meio indutor adipogênico, suplementado com insulina e
dexametasona, e coradas em Oil Red O, corante que evidencia gotículas de
gordura, acumuladas em células adiposas. Em 5B, pode ser observada a
diferenciação osteogênica das células, após o cultivo em meio suplementado
com dexametasona, β-glicerol fosfato e ácido ascórbico, e demonstração dos
depósitos de Cálcio pela coloração com corante Vermelho de Alizarina. Em 5C,
observamos células cultivadas em meio DMEM + 15% SBF, na ausência de
indutores de diferenciação.
As culturas foram também analisadas por citometria de fluxo para os
marcadores CD34, CD45 e HLADR, que são conhecidos marcadores negativos
de culturas de CTM. Na Figura 6, podemos observar a ausência dos
marcadores hematopoéticos CD34 e CD45, e a baixa expressão de HLA-DR,
em uma das culturas estabelecidas na passagem 3, e esse padrão estava
presente em todas as outras culturas. Esse padrão está de acordo com a
literatura.

Resultados
Figura 5: Diferenciação in vitro de culturas de CTM na passagem 1. A e C) Células cultivadas em meio DMEM baixa glicose + 15% SBF; B) Células cultivadas em meio indutor adipogênico, e coradas em corante Oil Red, evidenciando o acúmulo de gotículas lipidícas; e D) Células cultivadas em meio indutor osteogênico, e coradas em Vermelho de Alizarina, que mostra os depósitos de Cálcio nas células.
Figura 6: Perfil imunofenotípico de uma cultura na passagem 3 (CTM50 P3). Os histogramas mostram a expressão das moléculas CD34, CD45 e HLA-DR (pico vermelho), comparados ao controle isotípico (pico verde). Como podemos observar, as células analisadas não expressam os marcadores negativos CD34 e CD45, e expressam baixos níveis de HLA-DR.
CD34 HLA-DR CD45
M1 M2
A
D C
B

Resultados
2. Caracterização Molecular das Culturas
2.1 Análise Protéica Global
Para a utilização de CTM em protocolos terapêuticos, alguns parâmetros
precisam ser estabelecidos, como por exemplo se células de diferentes
doadores apresentam as mesmas características e podem ser empregadas em
protocolos terapêuticos independentemente da compatibilidade imunológica, ou
se existe uma variabilidade importante entre as células derivadas de diferentes
passagens. Essas definições são necessárias para avaliarmos a equivalência
entre células derivadas de diferentes doadores e até qual passagem uma
cultura poderia ser utilizada em protocolos clínicos, ou se mais de uma
passagem poderia ser utilizada. Nesse trabalho, foi realizado um estudo do
perfil de expressão protéica e de RNA de culturas de CTM, utilizando-se o
proteoma e o PCR em tempo real como ferramentas de análise.
Extratos proteicos totais foram obtidos a partir das células em cultura, e a
quantidade obtida de proteínas foi determinada pelo método de Bradford
(1976). Na figura 7, podemos observar um exemplo de curva padrão (A), e a
análise da integridade das proteínas em gel de poliacrilamida SDS-PAGE 10%
de 2 amostras (B).
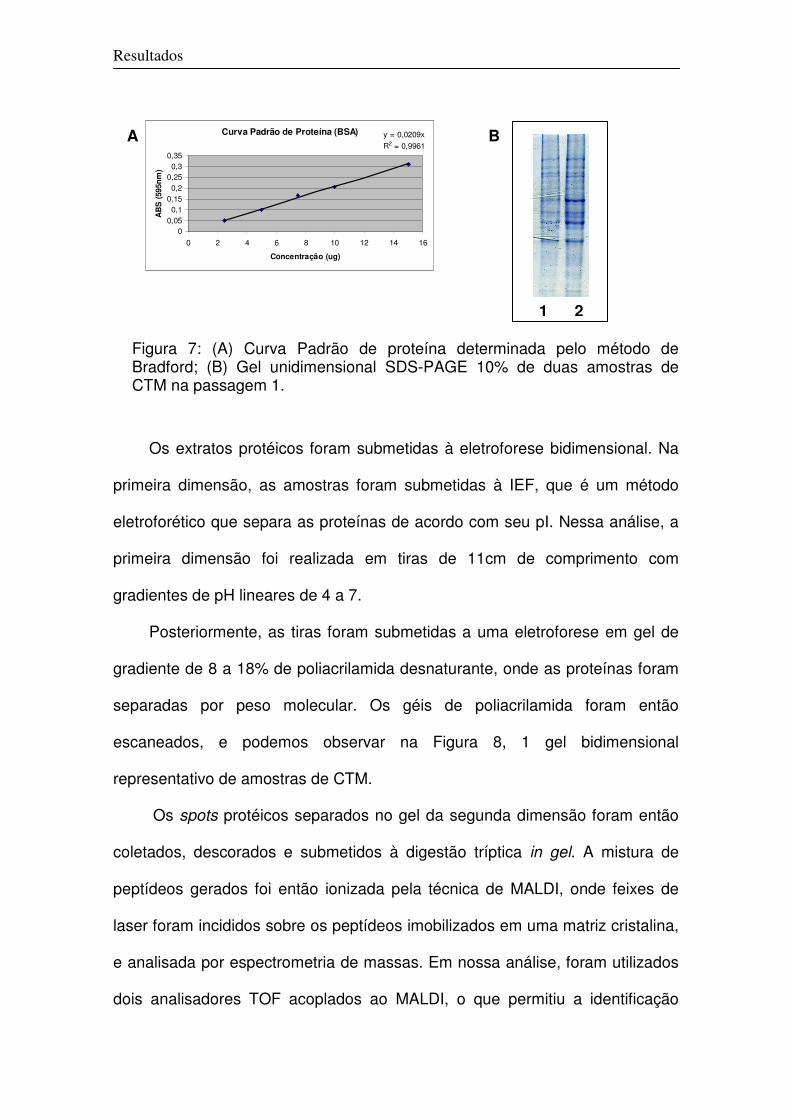
Resultados
Figura 7: (A) Curva Padrão de proteína determinada pelo método de Bradford; (B) Gel unidimensional SDS-PAGE 10% de duas amostras de CTM na passagem 1.
Os extratos protéicos foram submetidas à eletroforese bidimensional. Na
primeira dimensão, as amostras foram submetidas à IEF, que é um método
eletroforético que separa as proteínas de acordo com seu pI. Nessa análise, a
primeira dimensão foi realizada em tiras de 11cm de comprimento com
gradientes de pH lineares de 4 a 7.
Posteriormente, as tiras foram submetidas a uma eletroforese em gel de
gradiente de 8 a 18% de poliacrilamida desnaturante, onde as proteínas foram
separadas por peso molecular. Os géis de poliacrilamida foram então
escaneados, e podemos observar na Figura 8, 1 gel bidimensional
representativo de amostras de CTM.
Os spots protéicos separados no gel da segunda dimensão foram então
coletados, descorados e submetidos à digestão tríptica in gel. A mistura de
peptídeos gerados foi então ionizada pela técnica de MALDI, onde feixes de
laser foram incididos sobre os peptídeos imobilizados em uma matriz cristalina,
e analisada por espectrometria de massas. Em nossa análise, foram utilizados
dois analisadores TOF acoplados ao MALDI, o que permitiu a identificação
Curva Padrão de Proteína (BSA) y = 0,0209x
R2 = 0,9961
00,050,1
0,150,2
0,250,3
0,35
0 2 4 6 8 10 12 14 16
Concentração (ug)
AB
S (
595n
m)
A B
1 2

Resultados
mais específica das proteínas analisadas. As proteínas foram identificadas pelo
mapeamento das massas dos peptídeos gerados (peptide-mass fingerprinting),
e pelo sequenciamento posterior desses peptídeos. Os espectros adquiridos
permitiram a identificação das proteínas através do programa Mascot Daemon
contra a base de dados de sequências protéicas IPI.
Figura 8: Gel bidimensional de proteínas totais extraídas de culturas de CTM na passagem 1. A primeira dimensão foi realizada em tiras de 11cm de comprimento com gradientes de pH lineares de 4 a 7, que foram então submetidas a uma eletroforese em gel de gradiente de 8 a 18% de poliacrilamida desnaturante
Dezoito amostras protéicas foram submetidas à análise proteômica na
primeira etapa deste trabalho. Dos géis bidimensionais, foram obtidos em
média 181±±±±10 spots protéicos por gel, dos quais 132 puderam ser
identificados, e correspondiam a 84 proteínas distintas (vários spots
representam isoformas da mesma proteína). Na Tabela 2, estão listadas as
proteínas identificadas com seus respectivos números de identificação, o
220 KDa
10 KDa
pH 7.0 pH 4.0

Resultados
número no IPI, a razão, que corresponde à média dos volumes normalizados
de cada spot, o peso molecular e o pI teóricos, o score no programa Mascot,
que descreve a significância do resultado na análise, e a porcentagem de
cobertura da sequência identificada em relação à seqüência total da proteína
(Cob).
A identificação dessa lista revelou 40 novas proteínas nunca antes
descritas em análises proteômicas de CTM, permitindo a expansão do perfil
proteômico descrito de CTM (Park et al 2007). Dentre elas, a proteína
Galectina-1, que nunca havia sido descrita em análises proteômicas de CTM
em larga escala, e que tem sido relacionada à prevenção da DECH em
modelos animais (Baum et al 2003).
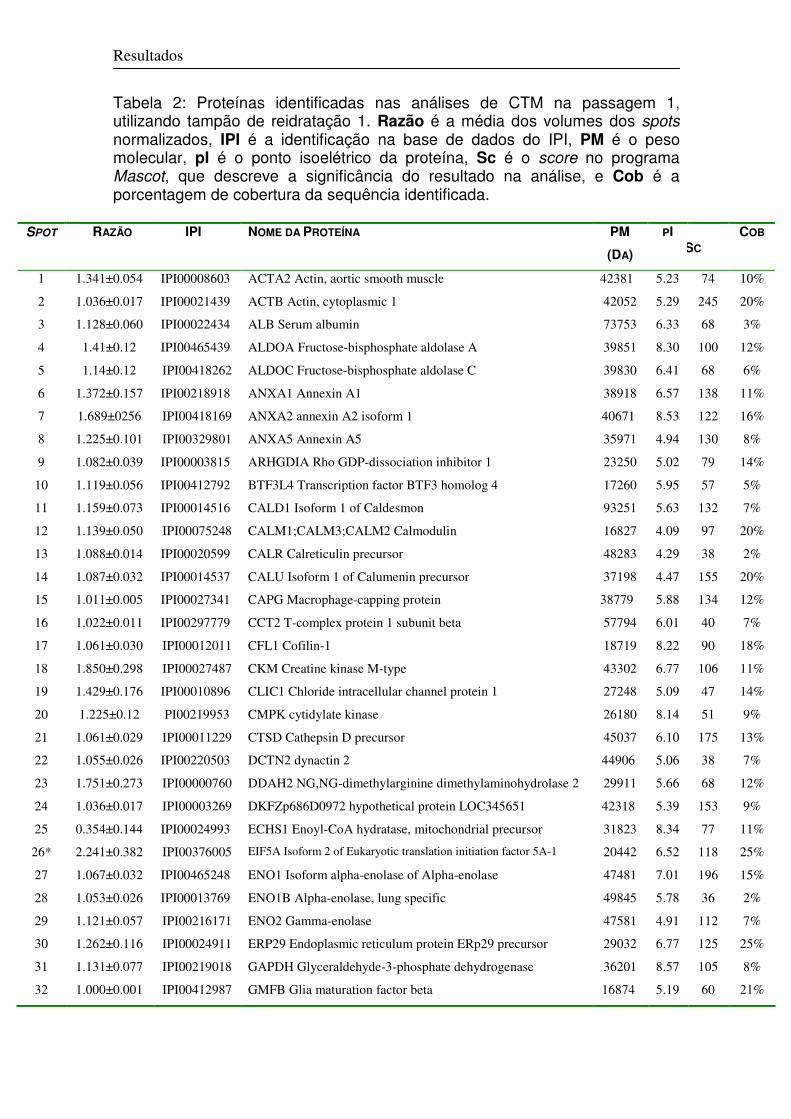
Resultados
Tabela 2: Proteínas identificadas nas análises de CTM na passagem 1, utilizando tampão de reidratação 1. Razão é a média dos volumes dos spots normalizados, IPI é a identificação na base de dados do IPI, PM é o peso molecular, pI é o ponto isoelétrico da proteína, Sc é o score no programa Mascot, que descreve a significância do resultado na análise, e Cob é a porcentagem de cobertura da sequência identificada.
SPOT RAZÃO IPI NOME DA PROTEÍNA PM
(DA)
PI SC
COB
1 1.341±0.054 IPI00008603 ACTA2 Actin, aortic smooth muscle 42381 5.23 74 10%
2 1.036±0.017 IPI00021439 ACTB Actin, cytoplasmic 1 42052 5.29 245 20%
3 1.128±0.060 IPI00022434 ALB Serum albumin 73753 6.33 68 3%
4 1.41±0.12 IPI00465439 ALDOA Fructose-bisphosphate aldolase A 39851 8.30 100 12%
5 1.14±0.12 IPI00418262 ALDOC Fructose-bisphosphate aldolase C 39830 6.41 68 6%
6 1.372±0.157 IPI00218918 ANXA1 Annexin A1 38918 6.57 138 11%
7 1.689±0256 IPI00418169 ANXA2 annexin A2 isoform 1 40671 8.53 122 16%
8 1.225±0.101 IPI00329801 ANXA5 Annexin A5 35971 4.94 130 8%
9 1.082±0.039 IPI00003815 ARHGDIA Rho GDP-dissociation inhibitor 1 23250 5.02 79 14%
10 1.119±0.056 IPI00412792 BTF3L4 Transcription factor BTF3 homolog 4 17260 5.95 57 5%
11 1.159±0.073 IPI00014516 CALD1 Isoform 1 of Caldesmon 93251 5.63 132 7%
12 1.139±0.050 IPI00075248 CALM1;CALM3;CALM2 Calmodulin 16827 4.09 97 20%
13 1.088±0.014 IPI00020599 CALR Calreticulin precursor 48283 4.29 38 2%
14 1.087±0.032 IPI00014537 CALU Isoform 1 of Calumenin precursor 37198 4.47 155 20%
15 1.011±0.005 IPI00027341 CAPG Macrophage-capping protein 38779 5.88 134 12%
16 1.022±0.011 IPI00297779 CCT2 T-complex protein 1 subunit beta 57794 6.01 40 7%
17 1.061±0.030 IPI00012011 CFL1 Cofilin-1 18719 8.22 90 18%
18 1.850±0.298 IPI00027487 CKM Creatine kinase M-type 43302 6.77 106 11%
19 1.429±0.176 IPI00010896 CLIC1 Chloride intracellular channel protein 1 27248 5.09 47 14%
20 1.225±0.12 PI00219953 CMPK cytidylate kinase 26180 8.14 51 9%
21 1.061±0.029 IPI00011229 CTSD Cathepsin D precursor 45037 6.10 175 13%
22 1.055±0.026 IPI00220503 DCTN2 dynactin 2 44906 5.06 38 7%
23 1.751±0.273 IPI00000760 DDAH2 NG,NG-dimethylarginine dimethylaminohydrolase 2 29911 5.66 68 12%
24 1.036±0.017 IPI00003269 DKFZp686D0972 hypothetical protein LOC345651 42318 5.39 153 9%
25 0.354±0.144 IPI00024993 ECHS1 Enoyl-CoA hydratase, mitochondrial precursor 31823 8.34 77 11%
26* 2.241±0.382 IPI00376005 EIF5A Isoform 2 of Eukaryotic translation initiation factor 5A-1 20442 6.52 118 25%
27 1.067±0.032 IPI00465248 ENO1 Isoform alpha-enolase of Alpha-enolase 47481 7.01 196 15%
28 1.053±0.026 IPI00013769 ENO1B Alpha-enolase, lung specific 49845 5.78 36 2%
29 1.121±0.057 IPI00216171 ENO2 Gamma-enolase 47581 4.91 112 7%
30 1.262±0.116 IPI00024911 ERP29 Endoplasmic reticulum protein ERp29 precursor 29032 6.77 125 25%
31 1.131±0.077 IPI00219018 GAPDH Glyceraldehyde-3-phosphate dehydrogenase 36201 8.57 105 8%
32 1.000±0.001 IPI00412987 GMFB Glia maturation factor beta 16874 5.19 60 21%

Resultados
33 1.225±0.120 IPI00219757 GSTP1 Glutathione S-transferase P 23569 5.43 82 12%
34 1.482±0.194 IPI00148063 HEBP1 Heme-binding protein 1 21198 5.71 58 8%
35 1.412±0.170 IPI00013881 HNRPH1 Heterogeneous nuclear ribonucleoprotein H 49484 5.89 136 15%
36 1.032±0.015 IPI00003362 HSPA5 HSPA5 protein 72492 5.07 221 11%
37 1.088±0.042 IPI00003865 HSPA8 Isoform 1 of Heat shock cognate 71 kDa protein 71082 5.37 88 6%
38 1.109±0.052 IPI00007765 HSPA9 Stress-70 protein, mitochondrial precursor 73920 5.87 67 7%
39 1.289±0.126 IPI00025512 HSPB1 Heat shock protein beta-1 22826 5.98 256 20%
40 1.326±0.140 IPI00022433 HSPB6 Heat-shock protein beta-6 18886 7.90 92 21%
41 1.463±0.187 IPI00472102 HSPD1 61 kDa protein 61346 5.70 230 15%
42 1.651±0.245 IPI00219219 LGALS1 Galectin-1 15048 5.34 286 37%
43 1.055±0.026 IPI00303882 M6PRBP1 Iso BofMannose-6-phosphate receptor-binding protein1 47189 5.30 87 7%
44 1.188±0.086 IPI00219301 MARCKS Myristoylated alanine-rich C-kinase substrate 31707 4.47 36 15%
45 1.072±0.034 IPI00179589 MTPN Myotrophin 13058 5.27 154 32%
46 1.619±0.196 IPI00335168 MYL6 Isoform Non-muscle of Myosin light polypeptide 6 17090 4.56 92 25%
47 1.072±0.035 IPI00012048 NME1 Nucleoside diphosphate kinase A 17309 5.83 108 31%
48 1.474±0.191 IPI00171664 NUP43 48 kDa protein 49082 5.75 34 2%
49 1.194±0.088 IPI00298547 PARK7 Protein DJ-1 20050 6.33 141 26%
50 1.648±0.244 IPI00025252 PDIA3 Protein disulfide-isomerase A3 precursor 57146 5.98 292 14%
51 1.332±0.142 IPI00014850 PEA15 Astrocytic phosphoprotein PEA-15 15088 4.93 83 22%
52 1.486±0.195 IPI00000051 PFDN1 Prefoldin subunit 1 14202 6.32 45 9%
53 1.738±0.269 IPI00006052 PFDN2 Prefoldin subunit 2 16695 6.20 72 9%
54 1.044±0.021 IPI00015361 PFDN5 Prefoldin subunit 5 17374 5.93 181 33%
55 1.502±0.160 IPI00216691 PFN1 Profilin-1 15216 8.44 38 20%
56 1.069±0.033 IPI00029997 PGLS 6-phosphogluconolactonase 27815 5.70 91 22%
57 1.738±0.269 IPI00419585 PPIA;PPIAL3;LOC654188 Peptidyl-prolyl cis-trans isomerase A 18229 7.68 66 20%
58 1.136±0.063 IPI00027350 PRDX2 Peroxiredoxin-2 22049 5.66 192 14%
59 1.101±0.047 IPI00023788 PRSS7 Enteropeptidase precursor 114905 4.90 35 1%
60 1.306±0.132 IPI00016832 PSMA1 Isoform Short of Proteasome subunit alpha type 1 29822 6.15 63 16%
61 1.850±0.298 IPI00008223 RAD23B UV excision repair protein RAD23 homolog B 43202 4.79 43 5%
62 1.468±0.189 IPI00015842 RCN1 Reticulocalbin-1 precursor 38866 4.86 119 16%
63* 4.495±0.636 IPI00101037 RCN3 Reticulocalbin-3 precursor 37470 4.74 139 9%
64 1.709±0.261 IPI00335280 RPE Isoform 1 of Ribulose-phosphate 3-epimerase 25139 5.33 44 8%
65 0.094±0.366 IPI00013895 S100A11 Protein S100-A11 11847 6.56 100 23%
66 1.293±0.128 IPI00025318 SH3BGRL SH3 domain-binding glutamic acid-rich-like protein 12766 5.22 72 8%
67 1.853±0.299 IPI00010402 SH3BGRL3 Hypothetical protein 24108 9.27 125 12%
68 1.926±0.316 IPI00745465 Similar to Checkpoint protein HUS1 15655 8.96 65 54%
69 1.026±0.013 IPI00218733 SOD1 16 kDa protein 16340 5.87 86 8%
70 1.438±0.179 IPI00479946 STIP1 STIP1 protein 68687 7.81 38 3%
71 1.009±0.004 IPI00479997 STMN1 Stathmin 17292 5.76 90 23%
72 1.215±0.097 IPI00217236 TBCA Tubulin-specific chaperone A 12904 5.25 93 10%
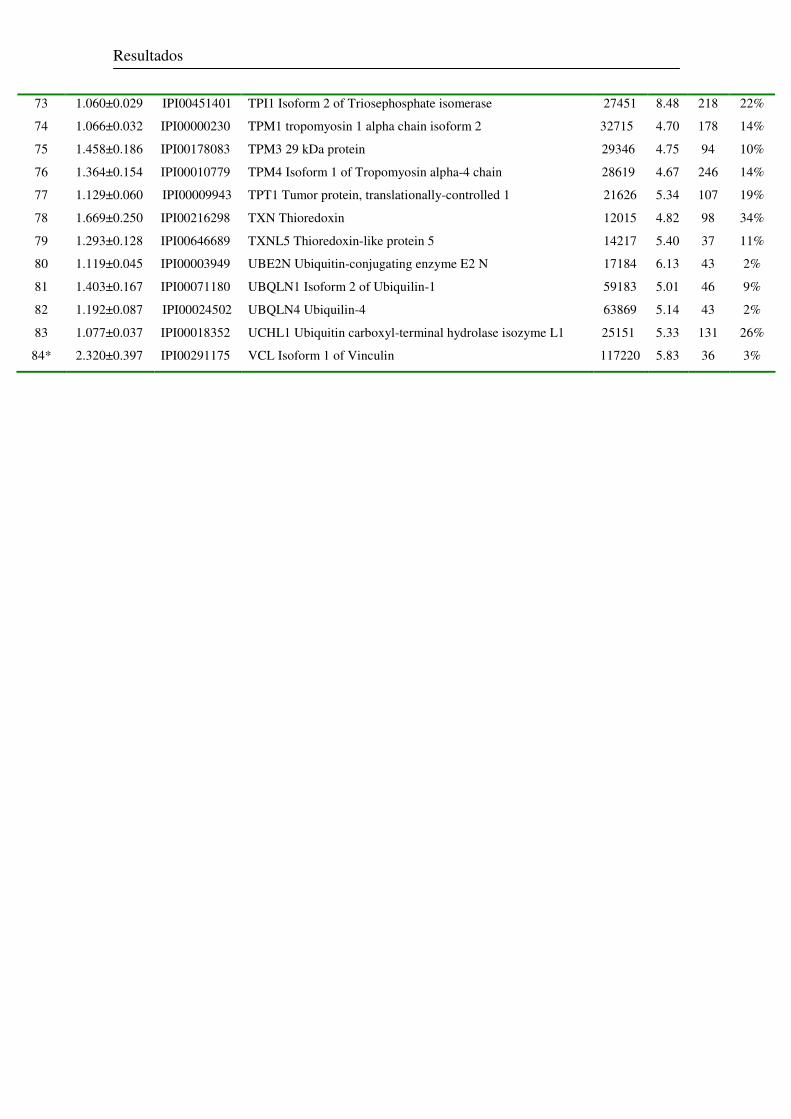
Resultados
73 1.060±0.029 IPI00451401 TPI1 Isoform 2 of Triosephosphate isomerase 27451 8.48 218 22%
74 1.066±0.032 IPI00000230 TPM1 tropomyosin 1 alpha chain isoform 2 32715 4.70 178 14%
75 1.458±0.186 IPI00178083 TPM3 29 kDa protein 29346 4.75 94 10%
76 1.364±0.154 IPI00010779 TPM4 Isoform 1 of Tropomyosin alpha-4 chain 28619 4.67 246 14%
77 1.129±0.060 IPI00009943 TPT1 Tumor protein, translationally-controlled 1 21626 5.34 107 19%
78 1.669±0.250 IPI00216298 TXN Thioredoxin 12015 4.82 98 34%
79 1.293±0.128 IPI00646689 TXNL5 Thioredoxin-like protein 5 14217 5.40 37 11%
80 1.119±0.045 IPI00003949 UBE2N Ubiquitin-conjugating enzyme E2 N 17184 6.13 43 2%
81 1.403±0.167 IPI00071180 UBQLN1 Isoform 2 of Ubiquilin-1 59183 5.01 46 9%
82 1.192±0.087 IPI00024502 UBQLN4 Ubiquilin-4 63869 5.14 43 2%
83 1.077±0.037 IPI00018352 UCHL1 Ubiquitin carboxyl-terminal hydrolase isozyme L1 25151 5.33 131 26%
84* 2.320±0.397 IPI00291175 VCL Isoform 1 of Vinculin 117220 5.83 36 3%

Resultados
2.2 Comparação entre culturas derivadas de diferentes doadores numa
mesma passagem
2.2.1 Análise do Perfil Proteômico
A análise visual preliminar de géis bidimensionais sugeriu que o perfil
protéico global é muito semelhante entre amostras de CTM na passagem 1
derivadas de diferentes doadores (Figura 9). A comparação visual mais
minuciosa foi realizada entre pequenos pedaços do gel, como pode ser
observado nas Figuras 10 e 11, mostrando mais uma vez o alto grau de
semelhança entre géis de diferentes culturas na passagem 1. Nessas figuras,
está mostrada também a identificação das bandas evidenciadas. A análise
comparativa dos géis foi também realizada no software Image Master 2D
Platinium 6.0 (GE Healthcare, Suécia), após normalização dos spots. Como
observado no scatter plotter (Figura 12), a comparação entre os diferentes
doadores revelou um alto grau de similaridade (de 94,4%) e de
reprodutibilidade, com diferenças quantitativas maiores que 2x em apenas três
proteínas (Tabela 2). Esses resultados sugerem que o perfil proteômico de
culturas de CTM humanas derivadas de diferentes doadores é similar nesta
janela de análise, mesmo em uma passagem tão precoce (passagem 1). No
entanto, a similaridade do perfil proteômico isoladamente não representa uma
informação importante do ponto de vista da utilização clínica das CTM. Para
isso, ensaios independentes de imunomodulação in vitro foram realizados
utilizando-se culturas de CTM na passagem 1 derivadas de diferentes
doadores.

Resultados
Figura 9: Géis representativos do perfil proteômico de amostras de CTM, derivadas de 2 doadores diferentes, na passagem 1, mostrando um padrão bastante semelhante entre as culturas. Os eixos horizontais representam o valor de pI, e os eixos verticais representam o peso molecular.

Figura 10: Comparação visual entre pequenos pedaços dos géis de 4 diferentes doadores, com a respectiva identificação das bandas envolvidas, mostrando a grande semelhança entre os perfis proteômicos.

Figura 11: Comparação visual entre pequenos pedaços dos géis de 4 diferentes doadores, com a respectiva identificação das bandas envolvidas, mostrando a grande semelhança entre os perfis proteômicos.

Resultados
Figura 12: Comparação entre 2 géis representativos de amostras de CTM na passagem 1 derivadas de 2 doadores diferentes, realizada no software Image Master 2D Platinium 6.0 (GE Healthcare, Suécia), após normalização dos spots. A análise revelou o alto grau de similaridade entre os géis (94,4%). mostrando a grande semelhança entre os perfis proteômicos de CTM derivadas de diferentes doadores.
2.2.2 Inibição In vitro de Células T Mediada por CTM de um terceiro
indivíduo (Third-Party)
Para testar a habilidade de culturas de CTM derivadas de diferentes
doadores de inibir a proliferação de linfócitos dirigida por aloantígenos, foram
realizados ensaios de CML na presença e ausência de CTM. CTM foram
adicionadas a CML no dia 0, e as culturas foram analisadas após 7 dias. Como
demonstrado na Figura 13A, CTM thirdy-party na passagem 1, derivadas de 5
doadores diferentes, foram capazes de inibir a proliferação de linfócitos em
resposta a aloantígenos em 12 experimentos independentes (p<0,0001). O
resultado de um ensaio representativo pode ser observado na Figura 13B. A
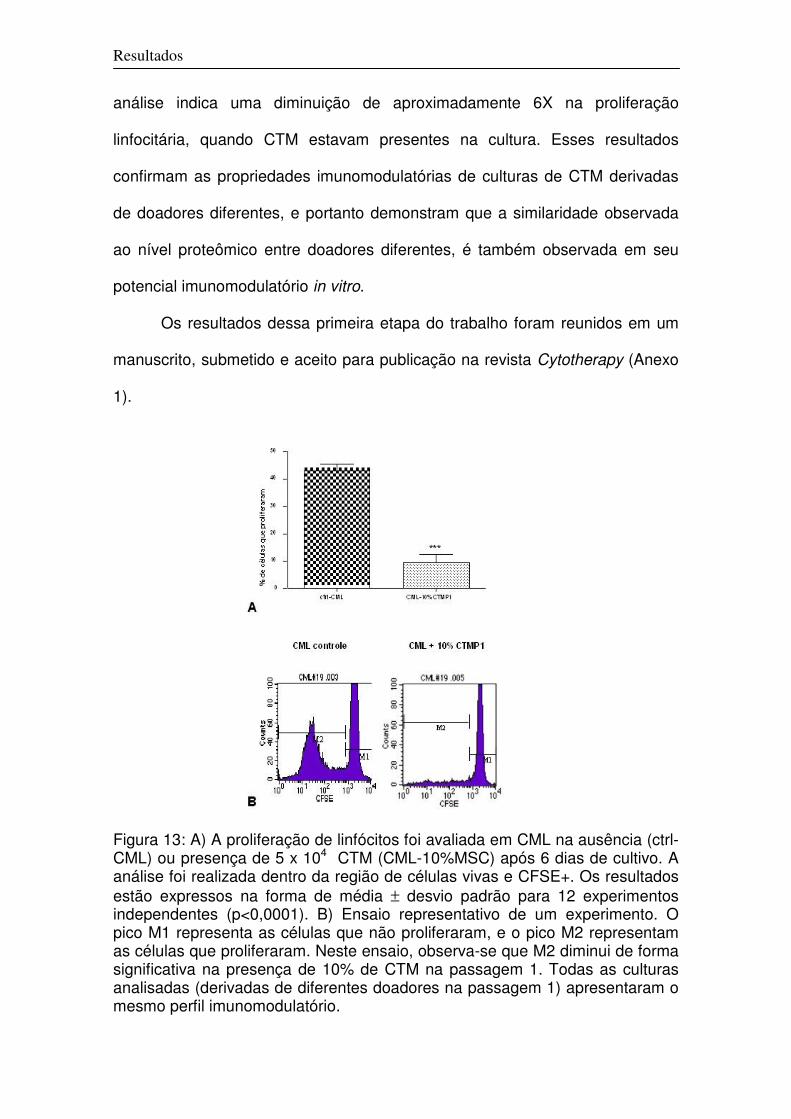
Resultados
análise indica uma diminuição de aproximadamente 6X na proliferação
linfocitária, quando CTM estavam presentes na cultura. Esses resultados
confirmam as propriedades imunomodulatórias de culturas de CTM derivadas
de doadores diferentes, e portanto demonstram que a similaridade observada
ao nível proteômico entre doadores diferentes, é também observada em seu
potencial imunomodulatório in vitro.
Os resultados dessa primeira etapa do trabalho foram reunidos em um
manuscrito, submetido e aceito para publicação na revista Cytotherapy (Anexo
1).
Figura 13: A) A proliferação de linfócitos foi avaliada em CML na ausência (ctrl-CML) ou presença de 5 x 104 CTM (CML-10%MSC) após 6 dias de cultivo. A análise foi realizada dentro da região de células vivas e CFSE+. Os resultados estão expressos na forma de média ± desvio padrão para 12 experimentos independentes (p<0,0001). B) Ensaio representativo de um experimento. O pico M1 representa as células que não proliferaram, e o pico M2 representam as células que proliferaram. Neste ensaio, observa-se que M2 diminui de forma significativa na presença de 10% de CTM na passagem 1. Todas as culturas analisadas (derivadas de diferentes doadores na passagem 1) apresentaram o mesmo perfil imunomodulatório.

Resultados
2.3 Comparação entre Culturas de um Mesmo Doador em Diferentes
Passagens
Foi realizada também análise do perfil proteômico e transcriptômico de
culturas derivadas de um mesmo doador em diferentes passagens, com o
objetivo de se verificar se CTM submetidas a mais passagens in vitro
preservam as características das CTM de passagens mais precoces. Esse tipo
de análise é de fundamental importância para a utilização clínica das CTM,
devido à necessidade de seu cultivo ex vivo antes de sua aplicação
terapêutica. Assim, mais conhecimento a respeito da influência do cultivo in
vitro sobre a biologia básica das CTM, e sobre seu potencial terapêutico, é
ainda imprescindível.
Culturas de CTM derivadas de um mesmo doador foram estabelecidas e,
em cada passagem (da passagem 1 até 10), foi realizada extração de
proteínas e de RNA das células para análise proteômica e transcriptômica.
Células em diferentes passagens foram também submetidas à análise
citogenética, com o objetivo de verificar se o cultivo in vitro é capaz de induzir a
aquisição de anomalias cromossômicas. Foi também realizada análise do
potencial imunomodulatório das CTM em diversas passagens, através da
realização de CML na ausência ou presença de CTM.
Durante o cultivo, sinais de senescência nas células, tais como aumento
no tamanho celular, achatamento da célula e aumento no número de vacúolos
intracelulares, começaram a ser observados a partir da passagem 9 (Figura
14), quando seu potencial de proliferação diminuiu em relação ao de
passagens mais precoces (dados não mostrados).

Resultados
Figura 14: Análise ao microscópio ótico de uma cultura de CTM na passagem 9, mostrando sinais morfológicos de senescência, tais como aumento no tamanho celular e no número de vacúolos intracelulares.
2.3.1 Análise do Perfil de Expressão de RNA
Em cada passagem da cultura, as células tiveram seu conteúdo de RNA
extraído, quantificado, e submetido à análise por PCR em tempo real, para
determinação da variação da expressão de alguns genes marcadores de
indiferenciação das CTM, segundo Park e colaboradores (Park et al 2007).
Na Figura 15, observamos a análise por PCR em tempo real da
expressão dos genes CD44, SCF e BMPR1, de culturas derivadas de um
doador da passagem 1 até a passagem 5. Essa análise nos mostrou que o
perfil de expressão para esses genes se manteve constante da passagem 1

Resultados
até a passagem 5, sugerindo que as células encontravam-se em um estado
indiferenciado. A partir da passagem 6, algumas análises indicam um perfil não
uniforme para a expressão destes genes .
Figura 15: Análise por PCR em tempo real da expressão dos genes CD44 (A), SCF (B) e BMPR-I (C) em culturas de CTM da passagem 1 até a passagem 5. Os gráficos mostram que o perfil de expressão dos genes se mantém constante ao longo do subcultivo in vitro.
CD44
0,5
0,6
0,7
0,8
0,9
1
1,1
P1 P2 P3 P4 P5
PASSAGENS
SCF
0,5
0,6
0,7
0,8
0,9
1
1,1
P1 P2 P3 P4 P5
PASSAGENS
BMPRI
0,5
0,6
0,7
0,8
0,9
1
P1 P2 P3 P4 P5
PASSAGENS

Resultados
2.3.2 Análise do Perfil Proteômico
Nessa segunda parte do trabalho, os extratos protéicos foram dissolvidos
no tampão de reidratação 2. Essa mudança permitiu a visualização de um
maior número de spots protéicos visualizados no gel 2D (de 181±±±±10 spots
obtidos com o tampão 1 para 271±±±±8 spots obtidos com o tampão 2), o que
pode ser atribuído à maior solubilização das proteínas pelo tampão 2, devido à
sua composição específica, como a presença do sal caotrópico tiouréia, e a
substituição do detergente zwitteriônico de NP40 por ASB14 (Hebert 1999).
Assim, foi gerada uma nova lista de proteínas identificadas em culturas de
CTM. Foram identificadas 149 proteínas, das quais 107 não haviam sido
identificadas em nossas análises anteriores. Na Tabela 3, estão relacionadas
as 107 novas proteínas identificadas nessa etapa do trabalho, e dessas 79
nunca haviam sido descritas em análises proteômicas de culturas de CTM
(Park et al 2007). A análise comparativa (visual e por scatter plotter) entre géis
bidimensionais de diferentes doadores na passagem 1 mostrou que a
similaridade entre doadores se manteve com a utilização do tampão de
reidratação 2
A comparação visual entre os géis das diversas passagens mostrou que
as culturas mantêm um perfil proteômico bastante constante ao longo do
subcultivo in vitro, com poucas diferenças quantitativas. A partir da passagem
9, porém, observa-se uma mudança importante nesse padrão, com uma
redução geral na expressão protéica, conforme pode ser observado na figura
16, onde estão os géis de culturas de CTM de um mesmo doador na passagem
1 e culturas na passagem 9. Vale lembrar que a quantidade de proteína
aplicada nos dois géis é a mesma. Algumas proteínas diferencialmente

Resultados
expressas entre as duas populações foram identificadas, como a proteína
Eif5A (isoforma 2 do fator eucariótico de iniciação da tradução 5A-1), Wdr1
(Isoforma 2 da proteína 1 contendo repetição WD), CapG (proteína macrófago-
capping), UCHL (Ubiquitin carboxyl-terminal hydrolase isozyme L1), ACTB
(actina citoplasmática 1 ou β-actina), ACTBL2 (proteína 2 tipo β-actina ou κ-
actina) e VCL (isoforma 1 da vinculina), todas com expressão bastante
diminuída em P9. Outras proteínas diferencialmente expressas ainda
necessitam ser identificadas.

Resultados
Tabela 3: Proteínas identificadas nas análises de CTM na passagem 1, utilizando tampão de reidratação 2. As proteínas listadas não foram identificadas nas análises com o tampão 1 (Tabela 2). IPI é a identificação na base de dados do IPI, PM é o peso molecular, pI é o ponto isoelétrico da proteína, Sc é o score no programa Mascot, que descreve a significância do resultado na análise, e Cob é a porcentagem de cobertura da sequência analisada.
SPOT IPI NOME DA PROTEÍNA PM
(Da) pI Sc Cob
1 IPI00063827 ABHD14B Isoform 1 of Abhydrolase domain-containing protein 14B 22446 5.94 83 11%
2 IPI00003269 ACTBL2 Beta-actin-like protein 2
42318 5.39 173 9% 3 IPI00013808 ACTN4 Alpha-actinin-4 105245 5.27 172 12% 4 IPI00029468 ACTR1A Alpha-centractin
42701 6.19 85 14% 5 IPI00028091 ACTR3 Actin-related protein 3 47797 5.61 95 15% 6 IPI00216773 ALB protein
46442 5.77 38 3% 7 IPI00103467 ALDH1B1 Aldehyde dehydrogenase X, mitochondrial precursor 57658 6.36 68 9% 8 IPI00002459 ANXA6 annexin VI isoform 2 75571 5.46 207 14% 9 IPI00002460 ANXA7 Isoform 1 of Annexin A7 53078 5.52 36 2%
10 IPI00005161 ARPC2 Actin-related protein 2/3 complex subunit 2 34426 6.84 234 23% 11 IPI00303476 ATP5B ATP synthase subunit beta, mitochondrial precursor 56525 5.26 69 14% 12 IPI00007812 ATP6V1B2 Vacuolar ATP synthase subunit B, brain isoform 56807 5.57 38 4% 13 IPI00301434 BOLA2;BOLA2B BolA-like protein 2 17150 8.42 63 16% 14 IPI00410226 BOLA2;BOLA2B Isoform 2 of BolA-like protein 2 7008 5.65 58 43% 15 IPI00056357 C19orf10 UPF0556 protein C19orf10 precursor 18897 6.20 34 13%
16 IPI00014230 C1QBP Complement component 1 Q subcomponent-binding protein, mitochondrial 31742 4.74 49 4%
17 IPI00218694 CALD1 Isoform 2 of Caldesmon 65725 6.37 179 13% 18 IPI00789155 CALU Calumenin precursor 38141 4.47 308 23% 19 IPI00025084 CAPNS1 Calpain small subunit 1 28469 5.05 76 21% 20 IPI00026185 CAPZB Isoform 1 of F-actin-capping protein subunit beta 31616 5.36 91 8% 21 IPI00010720 CCT5 T-complex protein 1 subunit epsilon 60089 5.45 41 4% 22 IPI00027626 CCT6A T-complex protein 1 subunit zeta 58444 6.23 51 4% 23 IPI00001560 CDKN2A Cyclin-dependent kinase inhibitor 17986 6.37 33 24% 24 IPI00001960 CLIC4 Chloride intracellular channel protein 4 28982 5.45 140 33% 25 IPI00216393 CLTA Isoform Non-brain of Clathrin light chain A 23704 4.45 44 13% 26 IPI00291136 COL6A1 Collagen alpha-1(VI) chain precursor 109602 5.26 50 3% 27 IPI00017704 COTL1 Coactosin-like protein 16049 5.54 126 38% 28 IPI00295741 CTSB Cathepsin B precursor 38766 5.88 75 5% 29 IPI00010182 DBI Isoform a 1 of Acyl-CoA-binding protein 10038 6.12 46 12% 30 IPI00473014 DSTN Destrin 18950 8.06 141 16% 31 IPI00412497 DYNLRB1 Dynein, light chain, roadblock-type 1 16527 6.58 100 18%

Resultados
32 IPI00023048 EEF1D Elongation factor 1-delta 31217 4.90 84 16% 33 IPI00186290 EEF2 Elongation factor 2 96246 6.41 99 6% 34 IPI00328328 EIF4A2 Isoform 1 of Eukaryotic initiation factor 4A-II 46601 5.33 155 14% 35 IPI00003766 ETHE1 protein, mitochondrial precursor 28368 6.35 44 6% 36 IPI00219684 FABP3 Fatty acid-binding protein, heart 14906 6.29 183 40% 37 IPI00007797 FABP5;FABP5L7 Fatty acid-binding protein, epidermal 15497 6.60 84 47% 38 IPI00303300 FKBP10 FK506-binding protein 10 precursor 64717 5.36 35 4% 39 IPI00182126 FKBP9 FK506-binding protein 9 precursor 63500 4.91 59 4% 40 IPI00382931 FTH1 Ferritin 19649 8.78 38 13% 41 IPI00554521 FTH1 Ferritin heavy chain 21383 5.30 41 18% 42 IPI00375676 FTL Ferritin light polypeptide variant 21340 5.70 60 8% 43 IPI00026314 GSN Isoform 1 of Gelsolin precursor 86043 5.90 35 1% 44 IPI00018246 HK1 Isoform 1 of Hexokinase-1 103561 6.36 33 3% 45 IPI00219344 HPCAL1 Hippocalcin-like protein 1 22413 5.21 44 12%
46 IPI00382470 HSP90AA1 heat shock protein 90kDa alpha (cytosolic), class A member 1 isofo 98670 5.07 193 7%
47 IPI00414676 HSP90AB1 Heat shock protein HSP 90-beta 83554 4.97 229 11% 48 IPI00027230 HSP90B1 Endoplasmin precursor 92696 4.76 149 10% 49 IPI00556538 HSP90B2P Heat shock protein 94b 46343 5.14 48 7% 50 IPI00784154 HSPD1 60 kDa heat shock protein, mitochondrial precursor 61187 5.70 175 12%
51 IPI00030702 IDH3A Isoform 1 of Isocitrate dehydrogenase [NAD] subunit alpha, mitochondrial 40022 6.47 40 2%
52 IPI00021405 LMNA Isoform A of Lamin-A/C 74380 6.57 72 10% 53 IPI00886929 LOC100131813 similar to hCG2041218 26501 8.48 67 5% 54 IPI00869284 LOC171220 Pseudogene candidate 16866 8.78 95 11% 55 IPI00017596 MAPRE1 Microtubule-associated protein RP/EB family member 1 30151 5.02 34 13% 56 IPI00033494 MRLC2 Myosin regulatory light chain 19824 4.71 92 29% 57 IPI00219365 MSN Moesin 67892 6.08 85 3% 58 IPI00000105 MVP Major vault protein 99551 5.34 134 8% 59 IPI00019502 MYH9 Myosin-9 227646 5.50 59 3% 60 IPI00027681 NNMT Nicotinamide N-methyltransferase 30011 5.56 75 3%
61 IPI00017672 NP CDNA FLJ25678 fis, clone TST04067, highly similar to PURINE NUCLEOSIDE P 32758 6.71 115 20%
62 IPI00220740 NPM1 Isoform 2 of Nucleophosmin 29617 4.47 104 20% 63 IPI00010796 P4HB Protein disulfide-isomerase precursor 57480 4.76 384 18% 64 IPI00246058 PDCD6IP PDCD6IP protein 97385 6.13 36 3% 65 IPI00893541 PDIA3 14 kDa protein 13739 6.78 78 21% 66 IPI00299571 PDIA6 Isoform 2 of Protein disulfide-isomerase A6 precursor 54380 5.17 115 7% 67 IPI00107555 PFN2 Isoform IIb of Profilin-2 15421 5.78 51 7% 68 IPI00220644 PKM2 Isoform M1 of Pyruvate kinase isozymes M1/M2 58538 7.60 50 4%
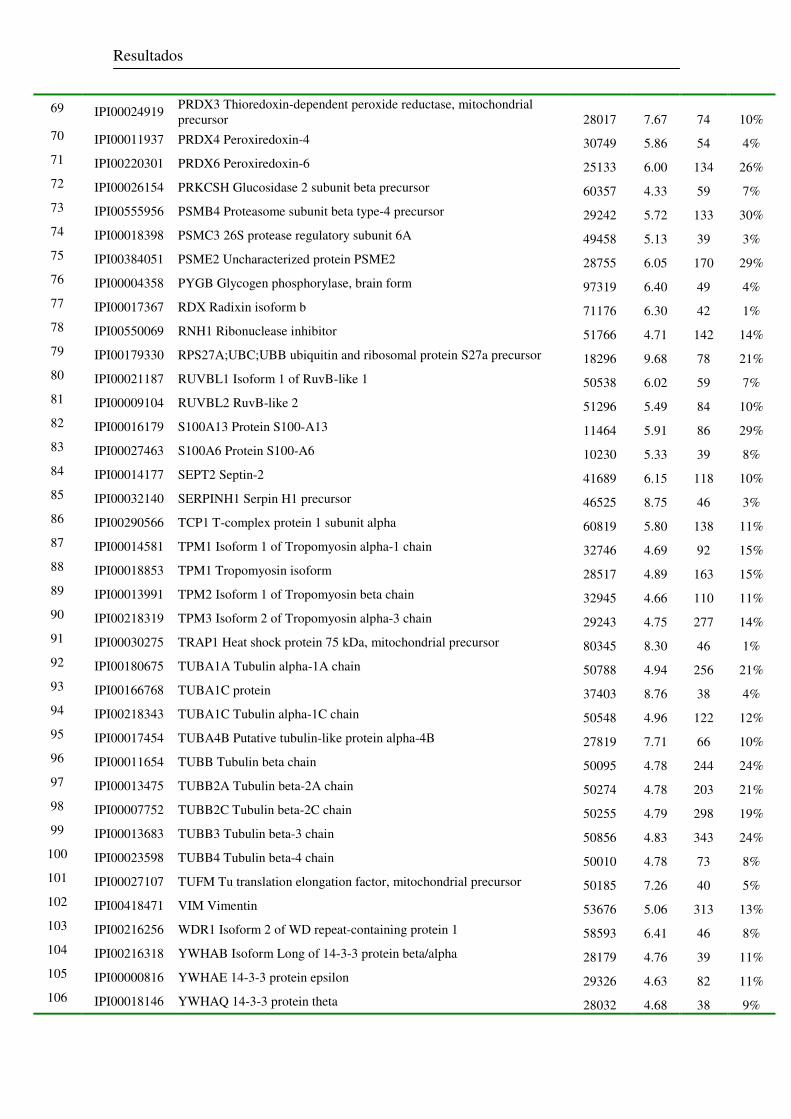
Resultados
69 IPI00024919 PRDX3 Thioredoxin-dependent peroxide reductase, mitochondrial precursor 28017 7.67 74 10%
70 IPI00011937 PRDX4 Peroxiredoxin-4 30749 5.86 54 4% 71 IPI00220301 PRDX6 Peroxiredoxin-6 25133 6.00 134 26% 72 IPI00026154 PRKCSH Glucosidase 2 subunit beta precursor 60357 4.33 59 7% 73 IPI00555956 PSMB4 Proteasome subunit beta type-4 precursor 29242 5.72 133 30% 74 IPI00018398 PSMC3 26S protease regulatory subunit 6A 49458 5.13 39 3% 75 IPI00384051 PSME2 Uncharacterized protein PSME2 28755 6.05 170 29% 76 IPI00004358 PYGB Glycogen phosphorylase, brain form 97319 6.40 49 4% 77 IPI00017367 RDX Radixin isoform b 71176 6.30 42 1% 78 IPI00550069 RNH1 Ribonuclease inhibitor 51766 4.71 142 14% 79 IPI00179330 RPS27A;UBC;UBB ubiquitin and ribosomal protein S27a precursor 18296 9.68 78 21% 80 IPI00021187 RUVBL1 Isoform 1 of RuvB-like 1 50538 6.02 59 7% 81 IPI00009104 RUVBL2 RuvB-like 2 51296 5.49 84 10% 82 IPI00016179 S100A13 Protein S100-A13 11464 5.91 86 29% 83 IPI00027463 S100A6 Protein S100-A6 10230 5.33 39 8% 84 IPI00014177 SEPT2 Septin-2 41689 6.15 118 10% 85 IPI00032140 SERPINH1 Serpin H1 precursor 46525 8.75 46 3% 86 IPI00290566 TCP1 T-complex protein 1 subunit alpha 60819 5.80 138 11% 87 IPI00014581 TPM1 Isoform 1 of Tropomyosin alpha-1 chain 32746 4.69 92 15% 88 IPI00018853 TPM1 Tropomyosin isoform 28517 4.89 163 15% 89 IPI00013991 TPM2 Isoform 1 of Tropomyosin beta chain 32945 4.66 110 11% 90 IPI00218319 TPM3 Isoform 2 of Tropomyosin alpha-3 chain 29243 4.75 277 14% 91 IPI00030275 TRAP1 Heat shock protein 75 kDa, mitochondrial precursor 80345 8.30 46 1% 92 IPI00180675 TUBA1A Tubulin alpha-1A chain 50788 4.94 256 21% 93 IPI00166768 TUBA1C protein 37403 8.76 38 4% 94 IPI00218343 TUBA1C Tubulin alpha-1C chain 50548 4.96 122 12% 95 IPI00017454 TUBA4B Putative tubulin-like protein alpha-4B 27819 7.71 66 10% 96 IPI00011654 TUBB Tubulin beta chain 50095 4.78 244 24% 97 IPI00013475 TUBB2A Tubulin beta-2A chain 50274 4.78 203 21% 98 IPI00007752 TUBB2C Tubulin beta-2C chain 50255 4.79 298 19% 99 IPI00013683 TUBB3 Tubulin beta-3 chain 50856 4.83 343 24%
100 IPI00023598 TUBB4 Tubulin beta-4 chain 50010 4.78 73 8% 101 IPI00027107 TUFM Tu translation elongation factor, mitochondrial precursor 50185 7.26 40 5% 102 IPI00418471 VIM Vimentin 53676 5.06 313 13% 103 IPI00216256 WDR1 Isoform 2 of WD repeat-containing protein 1 58593 6.41 46 8% 104 IPI00216318 YWHAB Isoform Long of 14-3-3 protein beta/alpha 28179 4.76 39 11% 105 IPI00000816 YWHAE 14-3-3 protein epsilon 29326 4.63 82 11% 106 IPI00018146 YWHAQ 14-3-3 protein theta 28032 4.68 38 9%
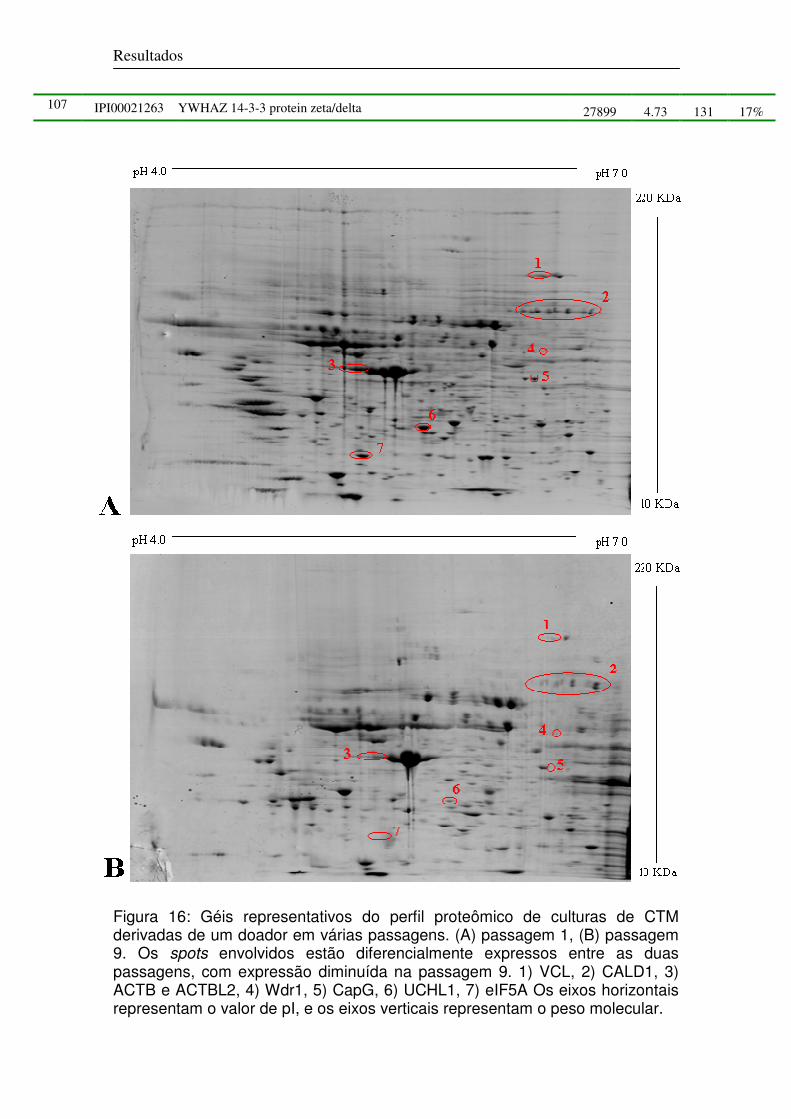
Resultados
107 IPI00021263 YWHAZ 14-3-3 protein zeta/delta 27899 4.73 131 17%
Figura 16: Géis representativos do perfil proteômico de culturas de CTM derivadas de um doador em várias passagens. (A) passagem 1, (B) passagem 9. Os spots envolvidos estão diferencialmente expressos entre as duas passagens, com expressão diminuída na passagem 9. 1) VCL, 2) CALD1, 3) ACTB e ACTBL2, 4) Wdr1, 5) CapG, 6) UCHL1, 7) eIF5A Os eixos horizontais representam o valor de pI, e os eixos verticais representam o peso molecular.

Resultados
2.3.3 Análise do Perfil Citogenético
Devido à necessidade de expansão in vitro das CTM, é importante
examinar a estabilidade citogenética antes de seu uso terapêutico. O cariótipo
é um indicador confiável para avaliar a estabilidade genética e transformação
de células cultivadas in vitro (Zhang et al 2007), como é o caso das CTM.
Nesse trabalho, culturas de CTM derivadas de um mesmo doador foram
analisadas citogeneticamente ao longo de várias passagens in vitro, com o
objetivo de se avaliar até que passagem as culturas mantêm o perfil
citogenético inicial. Na Tabela 3, observamos o padrão cromossômico em
várias passagens de uma dessas culturas. Notamos que a partir da passagem
6 foi detectada uma alteração cromossômica, a pseudodiploidia, levando à
trissomia do 6 e à perda do cromossomo sexual X. Foi observado também, em
algumas células, perda cromossômica. Essas alterações cromossômicas se
mantiveram até a passagem 8. Na passagem 10, o índice mitótico foi
extremamente baixo, não apresentando condições para definição do padrão
cromossômico. Na Figura 17, podem ser observados cariótipos representativos
de culturas na passagem 3 (46, XX) e na passagem 6 (46,X-X+16).
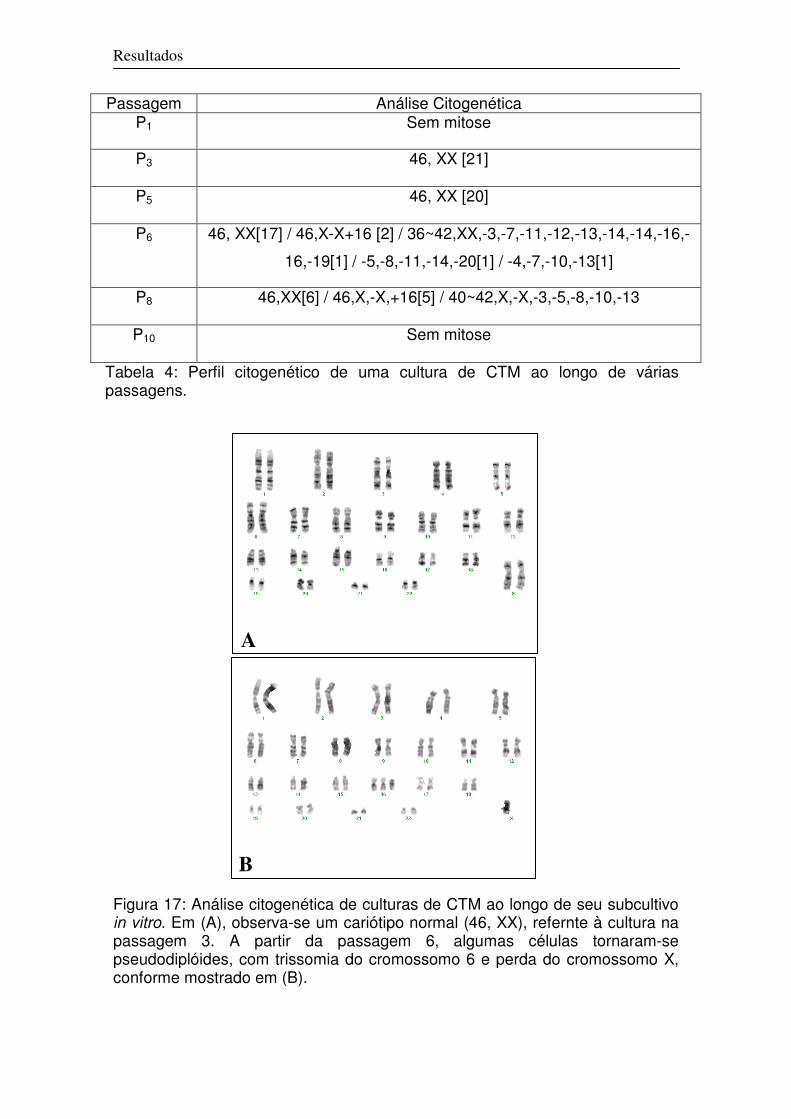
Resultados
Passagem Análise Citogenética P1 Sem mitose
P3 46, XX [21]
P5 46, XX [20]
P6 46, XX[17] / 46,X-X+16 [2] / 36~42,XX,-3,-7,-11,-12,-13,-14,-14,-16,-
16,-19[1] / -5,-8,-11,-14,-20[1] / -4,-7,-10,-13[1]
P8 46,XX[6] / 46,X,-X,+16[5] / 40~42,X,-X,-3,-5,-8,-10,-13
P10 Sem mitose
Tabela 4: Perfil citogenético de uma cultura de CTM ao longo de várias passagens.
Figura 17: Análise citogenética de culturas de CTM ao longo de seu subcultivo in vitro. Em (A), observa-se um cariótipo normal (46, XX), refernte à cultura na passagem 3. A partir da passagem 6, algumas células tornaram-se pseudodiplóides, com trissomia do cromossomo 6 e perda do cromossomo X, conforme mostrado em (B).
A
B
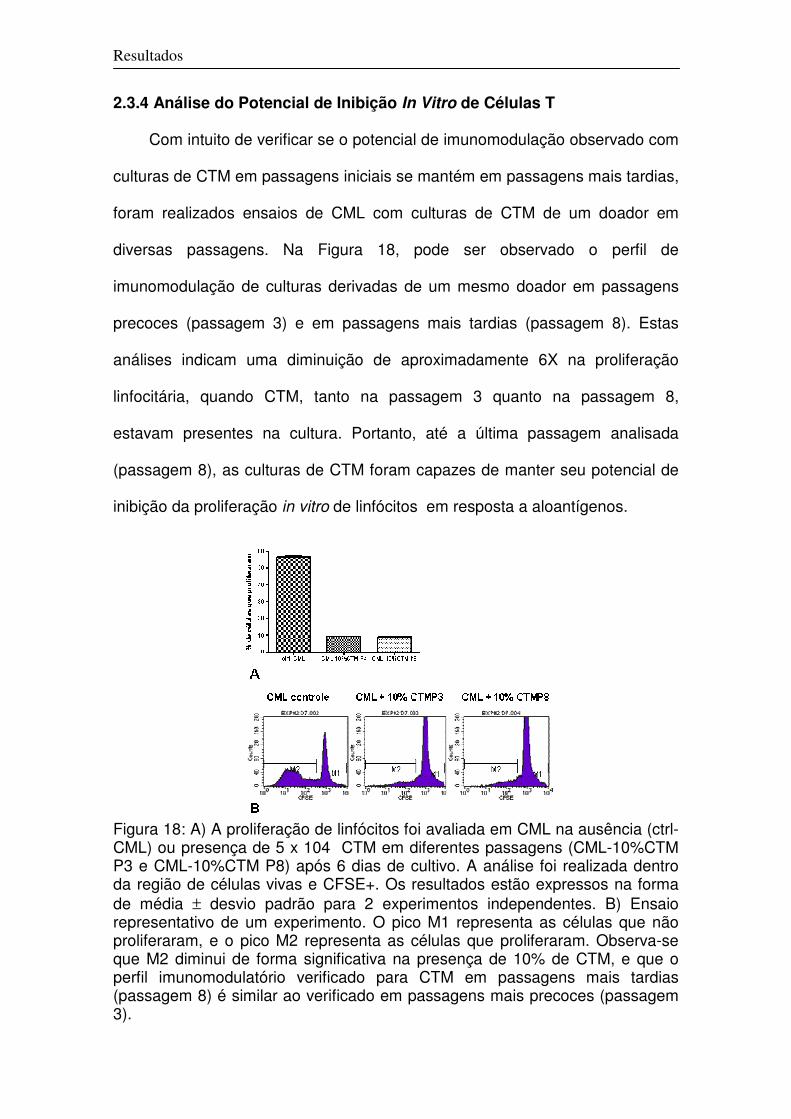
Resultados
2.3.4 Análise do Potencial de Inibição In Vitro de Células T
Com intuito de verificar se o potencial de imunomodulação observado com
culturas de CTM em passagens iniciais se mantém em passagens mais tardias,
foram realizados ensaios de CML com culturas de CTM de um doador em
diversas passagens. Na Figura 18, pode ser observado o perfil de
imunomodulação de culturas derivadas de um mesmo doador em passagens
precoces (passagem 3) e em passagens mais tardias (passagem 8). Estas
análises indicam uma diminuição de aproximadamente 6X na proliferação
linfocitária, quando CTM, tanto na passagem 3 quanto na passagem 8,
estavam presentes na cultura. Portanto, até a última passagem analisada
(passagem 8), as culturas de CTM foram capazes de manter seu potencial de
inibição da proliferação in vitro de linfócitos em resposta a aloantígenos.
Figura 18: A) A proliferação de linfócitos foi avaliada em CML na ausência (ctrl-CML) ou presença de 5 x 104 CTM em diferentes passagens (CML-10%CTM P3 e CML-10%CTM P8) após 6 dias de cultivo. A análise foi realizada dentro da região de células vivas e CFSE+. Os resultados estão expressos na forma de média ± desvio padrão para 2 experimentos independentes. B) Ensaio representativo de um experimento. O pico M1 representa as células que não proliferaram, e o pico M2 representa as células que proliferaram. Observa-se que M2 diminui de forma significativa na presença de 10% de CTM, e que o perfil imunomodulatório verificado para CTM em passagens mais tardias (passagem 8) é similar ao verificado em passagens mais precoces (passagem 3).

Resultados
2.4 Comparação entre culturas submetidas à diferenciação e culturas não
diferenciadas
Com o objetivo de estudar o padrão proteômico das CTM após sua
diferenciação in vitro, e identificar proteínas diferencialmente expressas durante
esse processo, foi realizada análise proteômica das culturas após sua
diferenciação para as linhagens osteogênica e adipogênica.
Para a diferenciação osteogênica, CTM na passagem 3 foram cultivadas
em meio de diferenciação osteogênico, que continha 10% SBF, 0,01µM
dexametasona, 10mM β-glicerol fosfato e 0,2mM ácido ascórbico, em DMEM
de alta glicose, durante 3 semanas. As células foram então tripsinizadas, sendo
preparados seus extratos protéicos totais. Os extratos foram quantificados e
submetidos à eletroforese bidimensional, para análise do perfil proteômico das
células diferenciadas.
Para a diferenciação adipogênica, CTM na passagem 3 foram cultivadas
em meio de diferenciação adipogênico, que continha 10%SBF, 0,01µM
dexametasona, 5µg/mL insulina em DMEM de alta glicose, durante 3 semanas.
Após esse período, as culturas foram tripsinizadas, e foi preparado extrato
protéico total delas. Após quantificação pelo método de Bradford, os extratos
foram submetidos à eletroforese bidimensional para análise do perfil
proteômico das células após diferenciação.
Os ensaios de diferenciação realizados em paralelo confirmaram o
fenótipo diferenciado das células, tanto para a linhagem osteogênica, quanto
para a linhagem adipogênica.

Resultados
Foi também realizada, através de ensaios de PCR em tempo real, a
análise da expressão de genes marcadores de diferenciação das CTM, tais
como LPL (LipoProteína Lipase) para a diferenciação adipogênica, e Runx2
para a diferenciação osteogênica. Na figura 19, pode ser observado o perfil de
expressão dos genes em células diferenciadas para o fenótipo adipogênico e
em células diferenciadas para o fenótipo osteogênico, mostrando aumento
considerável dos genes marcadores de diferenciação (mais de 7.000 vezes
para o RNAm de LPL após a diferenciação adipogênica, e quase 5 vezes para
o RNAm de Runx2 após a diferenciação osteogênica). Essa análise confirmou
que as culturas de CTM sofreram diferenciação no meio indutor específico.
DIFERENCIAÇÃO OSTEOGÊNICA
1,10
5,42
0,00
1,00
2,00
3,00
4,00
5,00
6,00
CTRL OSTEO DIF OSTEO
ng
DE
mR
NA
DE
RU
NX
2/B
2M
Figura 19: Análise por PCR em tempo real dos genes Runx2 e LPL em culturas de CTM submetidas à diferenciação in vitro para as linhagens osteogênica e adipogênica, respectivamente.
DIFERENCIAÇÃO ADIPOGÊNICA
5,1E-04
9,0E+00
0,0E+00
1,0E+00
2,0E+00
3,0E+00
4,0E+00
5,0E+00
6,0E+00
7,0E+00
8,0E+00
9,0E+00
1,0E+01
CTRL ADIPO DIF ADIPO
ng
DE
mR
NA
DE
LP
L/B
2M

Resultados
Na figura 20, podemos observar o padrão proteômico de CTM não
diferenciadas, de células após a diferenciação osteogênica e após a
diferenciação adipogênica.
As imagens dos géis foram comparadas visualmente, e os spots
expressos exclusivamente em amostras diferenciadas, ou nas não
diferenciadas, foram retirados, processados e estão sendo identificados por
espectrometria de massas. Conforme demonstrado na figura, podemos
observar a modificação do perfil de expressão protéica das culturas após a
diferenciação in vitro. Algumas proteínas encontravam-se diferencialmente
expressas, como por exemplo, a proteína Caldesmon (CALD1), que aparece
no gel em diversos spots, que representam suas diversas isoformas,
decorrentes de modificações pós-traducionais. Esse conjunto de proteínas
desaparece nos géis das culturas após a diferenciação. As proteínas TUBA4B
(α-TUBulina 4B), TUBA1A (cadeia α-1A da TUBulina), TUBB (cadeia β-1β-2
da TUBulina), TUBB2A (cadeia β-2A da TUBulina), TUBB3 (cadeia β-3 da
TUBulina) também estão ausentes nas culturas após a diferenciação (tanto
osteo como adipo). Outras proteínas diferencialmente expressas ainda estão
sendo identificadas por espectrometria de massas.

Figura 20: Géis representativos do perfil proteômico de culturas de CTM, na passagem 3, após diferenciação adipogênica (A) e osteogênica (B). Em C, o perfil proteômico de uma cultura de CTM não diferenciada na também na passagem 3. Os spots envolvidos estão diferencialmente expressos entre as culturas diferenciadas e as não diferenciadas. 1) Várias isoformas da proteína CALD1, 2) TUBA4B, TUBA1A, TUBB, TUBB2A, TUBB3. Os spots envolvidos, mas sem numeração, ainda não foram identificados, mas apresentam variações importantes de expressão entre as células diferenciadas e não diferenciadas. Os eixos horizontais representam o valor de pI, e os eixos verticais representam o peso molecular.

Discussão
DISCUSSÃO
CTM são definidas como progenitores multipotentes descritos in vitro como
populacões heterogêneas de células estromais. Devido ao seu potencial de originar
diferentes linhagens mesodermais, como osteócitos, condrócitos e adipócitos, sua
função imunomodulatória in vitro e in vivo, e sua relativa facilidade de expansão e
manipulação ex vivo, tais células representam ferramentas promissoras para
terapia celular. O uso clínico das CTM é um campo em ascensão para o tratamento
de diversas doenças, incluindo defeitos ósseos e fraturas, artrite inflamatória,
desordens cardiovasculares (Abdallah & Kassem 2009), DECH (Ringden et al
2006), e na melhora da reconstituição hematopoética após o transplante de medula
óssea (Giordano et al 2007). Como as CTM se popularizaram em cima de sua
promissora aplicação terapêutica, existe ainda pouco conhecimento acerca de
marcadores moleculares específicos para essa população, e do impacto dos
métodos utilizados para sua expansão em cultura (Wagner et al 2008b).
Desde a descoberta de seu potencial para terapias celulares, algumas
questões relacionadas à natureza dessas células surgiram como possíveis
problemas em sua aplicação terapêutica. As CTM são derivadas ex vivo por
adesão à placa de cultura, e sua definição está baseada em ensaios funcionais,
especialmente porque essas células nunca foram isoladas de seu nicho natural.
Culturas de CTM são uma mistura de progenitores com diferentes potenciais
(Muraglia et al 2000). A amplificação ex vivo, necessária para obtenção de número
suficiente de células para terapia, leva inevitavelmente à senescência celular,
acumulando risco de alterações genéticas e epigenéticas. Todas essas questões
poderiam ser melhor entendidas se as características moleculares envolvidas em

Discussão sua função terapêutica e em sua biologia básica em cultura pudessem ser
definidas.
O rápido desenvolvimento das tecnologias de análise molecular em larga
escala nas últimas décadas, em particular a tecnologia proteômica, permitiu a
comparação entre o perfil molecular de diferentes tipos de células tronco. Em
relação ao estudo das CTM, abordagens proteômicas têm sido aplicadas na
identificação de moléculas regulatórias e potenciais biomarcadores. Devido à
ausência de marcadores prospectivos para se isolar e identificar as CTM, parte da
pesquisa nesse campo tem se concentrado no estabelecimento de um painel de
características moleculares para delinear os padrões para o controle de qualidade
das preparações de CTM (Wagner et al. 2006). Ferramentas proteômicas são
também importantes para a elucidação dos mecanismos moleculares envolvidos na
diferenciação das CTM. Diversos estudos têm identificado o proteoma de CTM,
conforme revisado por Park e colaboradores (2007). No entanto, nenhuma das
proteínas identificadas parece ser específica para essa população, sugerindo
novamente a ausência de marcadores únicos para as CTM. Os estudos
proteômicos realizados nessa área compararam CTM derivadas de diferentes
tecidos humanos (Li et al. 2009, Noël et al. 2008, Feldmann et al. 2005), CTM
submetidas a diferentes condições de cultivo (Wagner et al. 2006, Wang et al.
2004), CTM induzidas à diferenciação in vitro (Lee et al. 2006, Sun et al. 2006),
mas em nenhum deles foram analisadas as diferenças entre culturas derivadas de
diferentes doadores de medula óssea, ou entre células de diversas passagens em
cultura.
Nesse estudo, estabelecemos inicialmente um perfil de proteínas expressas
em culturas de CTM na passagem 1, numa janela de análise que cobre proteínas

Discussão com pI de 4 a 7, e com peso molecular de 10 a 220kDa. Quarenta novas proteínas
foram identificadas além das proteínas já descritas em estudos proteômicos de
CTM (Park et al. 2007), provavelmente porque os parâmetros experimentais
utilizados para esta análise por eletroforese bidimensional foram diferentes dos
utilizados anteriormente. Dentre elas, identificamos a proteína Galectina-1 (Gal-1),
membro de uma família evolutivamente conservada de lecitinas com afinidade por
β-galactosídeos. A lecitina está envolvida em uma variedade de processos normais
e patológicos, como adesão celular, regulação do crescimento celular,
imunomodulação, inflamação, apoptose, embriogênese e progressão do câncer
(Vas et al. 2005). A expressão de Gal-1 foi identificada em órgãos linfóides e em
sítios imunoprivilegiados, como placenta e córnea, sugerindo um papel importante
na geração e manutenção da tolerância imune (Kiss et al. 2007). O papel de Gal-1
como regulador efetivo das respostas imunológicas foi demonstrado em modelos
animais de doenças auto-imunes, tais como ratos Lewis suscetíveis à
encefalomielite (Offner et al. 1990), coelhos com miastenia autoimune (Levi et al.
1983), e modelos experimentais de artrite reumatóide (Rabinovich et al. 2000). Em
um modelo animal, o tratamento com Gal-1 foi capaz de prevenir eficientemente o
desenvolvimento de DECH (Baum et al. 2003). Kiss e colaboradores demonstraram
ainda que Gal-1 afeta a mobilização de Células Progenitoras Hematopoéticas
(CPH) induzida por drogas como a ciclofosfamida, e citocinas como G-CSF
(Granulocyte-Colony Stimulating Factor). A expressão de Gal-1 em CTM foi
recentemente detectada e implicada na diferenciação das células da medula óssea
(Panepucci et al. 2004, Kadri et al. 2005). Devido às suas propriedades
imunomodulatórias, seu possível envolvimento nas funções imunológicas das CTM
pode ser sugerido.

Discussão
Este é o primeiro estudo proteômico em larga escala a identificar Gal-1 em
CTM, embora ela já tenha sido anteriormente associada às CTM (Kadri et al.
2005). Essa proteína, assim como a grande maioria das proteínas identificadas,
estava expressa em níveis quase idênticos em todas as amostras de CTM na
passagem 1 analisadas, o que demonstra que CTM derivadas de diferentes
doadores têm um padrão de expressão protéica muito similar.
A semelhança observada no perfil proteômico de culturas de CTM em
passagens tão precoces é de fato muito interessante, uma vez que tem sido
demonstrada uma grande variabilidade no perfil proteômico de células
mononucleares da medula óssea, do sangue periférico, ou do sangue de cordão
umbilical (Zenzmaier et al. 2003, 2005), derivadas de diferentes doadores. Nossas
análises sugerem que o cultivo in vitro, mesmo por apenas uma passagem, é
capaz de uniformizar o perfil proteômico das células entre diferentes indivíduos, e
que sua capacidade de inibir a proliferação de linfócitos in vitro é similar entre eles,
confirmando a similaridade do potencial imunomodulatório entre os indivíduos.
Diversos estudos têm demonstrado que as CTM apresentam propriedades
imunossupressoras in vitro e in vivo (Rasmusson 2006), produzem citocinas que
dão suporte à hematopoese e são capazes de potencialmente melhorar a
recuperação medular após quimio ou radioterapia (Ball et al. 2007, Le Blanc et al.
2004, 2007, Le Blanc & Pittenger 2005, Lee et al. 2002, Koç et al. 2000). Assim, as
CTM têm sido utilizadas em associação ao transplante alogeneico de medula óssea
com o objetivo de facilitar a “pega” da CTH e diminuir o desenvolvimento da DECH
(Le Blanc et al. 2004, 2007, Lee et al. 2002, Koç et al. 2000).
Todos os estudos mencionados anteriormente avaliaram o uso de infusões
de CTM derivadas de um terceiro doador, diferente do doador de medula óssea.

Discussão Além disso, Le Blanc e colaboradores (2003) demonstraram, através de ensaios in
vitro, que o efeito supressor sobre a proliferação de linfócitos, exercido pelas CTM,
independe da compatibilidade de HLA. Uma maior atividade supressora foi
demonstrada por Maccario e colaboradores (2005), quando CTM eram de um
terceiro doador, mais do que quando as células eram autólogas. Em um estudo
piloto utilizando CTM para tratar pacientes com DECH severa após TCTH,
Ringdén e colaboradores (2006) demonstraram que a infusão de CTM alogeneicas
derivadas tanto de doadores gêmeos idênticos, de doadores haploidênticos
relacionados, ou mesmo de doadores totalmente incompatíveis, tem efeito positivo
na imunossupressão da DECH, e na melhora do epitélio intestinal. Essas
evidências sugerem que a fonte das CTM de medula óssea não é um fator crucial
para seu uso em terapias celulares. Assim, é importante estabelecer
molecularmente as diferenças relevantes entre culturas de CTM derivadas de
diferentes indivíduos, corroborando os dados in vitro e in vivo, e demonstrar que,
mesmo CTM derivadas de um terceiro indivíduo, são capazes de diminuir
respostas imunológicas. Isto encoraja o uso dessas células como doadores
universais, independentemente do complexo de histocompatibilidade principal.
Esse é o primeiro estudo a demonstrar a similaridade do perfil proteômico de
CTM derivadas de diferentes indivíduos, mesmo em células da primeira passagem
em cultura. Esses resultados encorajam o uso de CTM third-party em terapias
celulares. Além disso, nossas análises in vitro mostram que CTM third-party na
primeira passagem são capazes de inibir a proliferação de linfócitos de forma
similar. Nossa hipótese, porém, ainda precisa ser corroborada por análises
proteômicas adicionais realizadas com proteínas de membrana, e proteínas

Discussão secretadas, que são os verdadeiros efetores das respostas imunes disparadas por
células no organismo hospedeiro.
Outras análises do perfil proteômico foram realizadas nesse trabalho,
utilizando-se um tampão de reidratação para as amostras protéicas diferente do
utilizado nas análises anteriores (tampão 2), sendo assim obtido um maior número
de spots protéicos no gel bidimensional. Mais proteínas foram identificadas nas
novas análises, provavelmente devido à melhor capacidade de solubilização
proteica pelo tampão 2, que contém o detergente ASB14 e o sal caotrópico
tiouréia. Nesta nova análise, 107 novas proteínas foram identificadas em culturas
de CTM na passagem 1, e dessas 79 nunca haviam sido descritas em análises
proteômicas de culturas de CTM (Park et al. 2007), expandindo portanto o perfil de
proteínas descritas no proteoma de CTM.
Em nossa análise do perfil molecular de culturas de CTM ao longo de suas
passagens, pôde-se observar que o padrão proteômico determinado para células
na passagem 1 se manteve até a passagem 8. Numa análise visual minuciosa,
observou-se que os géis bidimensionais de passagens posteriores apresentaram
diferenças qualitativas e quantitativas em relação às passagens anteriores. A partir
da passagem 9, sinais de senescência foram observados nas células, com
diminuição de seu potencial de proliferação, sugerindo que a mudança do perfil
proteômico das passagens iniciais acompanhou a progressão das células para um
estado de senescência. Wagner e colaboradores (Wagner et al 2008b) mostraram,
através da análise da expressão gênica global, que mudanças nesse padrão não
estão restritas a passagens tardias, mas são adquiridas ao longo do subcultivo das
CTM, sugerindo que a senescência replicativa das preparações de CTM é um

Discussão processo contínuo, que se inicia logo nas primeiras passagens. Nossos métodos
de análise, porém, não foram capazes de detectar o processo descrito pelo grupo
do Wagner, provavelmente porque alterações nos níveis de RNA ocorrem
anteriormente às dos níveis de proteínas. Além disso, nossa análise in vitro do
perfil imunomodulatório das CTM mostrou que mesmo células em passagens
tardias preservam seu potencial de inibição da proliferação de linfócitos, de
maneira semelhante ao observado em nossas análises anteriores com células na
passagem 1.
A partir da passagem 9, foi observada uma redução global da expressão
protéica, que se manteve na passagem 10. A comparação visual dos géis permitiu
a observação de algumas proteínas com diferenças de expressão entre as
populações de diferentes passagens. As proteínas identificadas (eIF5A, Wdr1,
CapG, UCHL, ACTB, CALD1 e VCL) apresentavam expressão diminuída em
relação às passagens anteriores. Essas proteínas apresentam papéis relacionados
principalmente à motilidade e forma celular, à proliferação e controle do ciclo celular
e à síntese protéica, processos que provavelmente sofrem redução com o cultivo
celular prolongado.
A proteína eIF5A foi originalmente identificada como um fator de tradução
altamente conservado de arqueobactérias a mamíferos. O fator é a única proteína
celular que sofre uma modificação pós-traducional chamada hiposinação, que está
envolvida no controle do ciclo celular e da proliferação em vários organismos
(Zanelli et al 2007). eIF5A foi recentemente relacionada à diferenciação do músculo
esquelético, uma vez que concentrações aumentadas da proteína foram
encontradas em células satélites em diferenciação (Luchessi et al 2009). Seu papel

Discussão como mediador das ações neurotróficas e neuroprotetoras de NGF (Neural Growth
Factor) foi também demonstrado (Huang et al 2007).
A proteína Wdr1, também conhecida como Aip1 (proteína 1 de interação com
actina), é uma proteína com repetições WD altamente conservada em eucariotos, e
que promove a desmontagem dos filamentos de actina mediada por cofilina. A
proteína parece ter um papel fundamental na conclusão da citocinese, por
promover a desmontagem dos filamentos de actina na telófase, além de funcionar
na migração celular direcionada restringindo as protusões de membrana em uma
direção via cofilina (Kato et al 2008). Fujibuchi e colaboradores mostraram que a
supressão de AIP1/Wdr1 induz o achatamento de células mitóticas, o que sugere
que a atividade de desmontagem dos filamentos de actina seja necessária para
induzir alterações morfológicas das células, especialmente seu arredondamento
durante a mitose (Fujibuchi et al 2005). Um dos sinais de senescência observados
em células em cultura é seu achatamento. A função de Wdr1em células
hematopoéticas, especificamente em megacariócitos e neutrófilos, foi demonstrada
pelo grupo de Kile. O grupo mostrou que enquanto a perda de função do locus
Wdr1 causa letalidade embrionária e macrotrombocitopenia (resultado de defeitos
na maturação de megacariócitos), camundongos com alelos hipomórficos
desenvolvem uma doença autoinflamatória, caracterizada por infiltração massiva de
neutrófilos (Kile et al 2007). Esses achados indicam que a dinâmica dos filamento
de actina, mediada por cofilina e Wdr1, é crítica no desenvolvimento e função de
ambas as células.
CapG (proteína reguladora de actina) é uma proteína sensível ao Cálcio, da
família da gelsolina, que bloqueia reversivelmente as terminações barbed de
filamentos de actina. O grupo de Hubert propôs um papel para a proteína como

Discussão mediador da interação entre o citoesqueleto de actina e organelas ligadas aos
microtúbulos que regulam a divisão celular (Hubert et al 2009). Foi demonstrada a
expressão aumentada de CapG em câncer de mama, especialmente durante a
metástase, e o gene que codifica CapG tem sido considerado como um putativo
oncogene (Renz et al 2008). Além disso, o aumento na expressão de CapG induz
a motilidade celular (Renz et al 2008).
A enzima UCHL (Ubiquitin carboxyl-terminal hydrolase isozyme L1) é uma tiol
protease que reconhece e hidroliza a ligação peptídica no C-terminal da glicina da
ubiquitina, estando envolvida tanto no processamento de precursores de ubiquitina,
quanto de proteínas ubiquitiniladas. A proteína UCHL é responsável por desmontar
cadeias de poliubiquitina, para aumentar a disponibilidade de ubiquitinas
monoméricas livres para o sistema proteassomo/ubiquitina, favorecendo a
degradação protéica. Mutações no gene UCHL-1 têm sido associadas à doença de
Parkinson (Barrachina et al 2006).
As actinas são proteínas altamente conservadas que estão envolvidas em
vários tipos de mobilidade celular e são expressas ubiquitariamente em todas as
células eucarióticas. A proteína ACTBL2, ou κ-actina, foi encontrada em níveis
aumentados em carcinoma hepatocelular, possivelmente substituindo as β-actinas
do tecido como maiores constituintes do citoesqueleto celular (Chang et al 2006).
A proteína VCL (vinculina), também conhecida como metavinculina, está
presente em complexos de adesão focal, que conectam a matriz extracelular ao
citoesqueleto de actina da célula. Esses complexos são essenciais para a
morfologia e locomoção celular (Schäfer et al 2008).

Discussão
A proteína CALD1 (Caldesmon 1) é uma proteína regulatória envolvida na
contração actina-miosina e na estabilidade dos filamento de actina (Mayanagi et al
2008).
Todas essas proteínas apresentam funções relacionadas principalmente à
motilidade e forma celular, proliferação e ciclo celular, processos cujo controle
diminui em células senescentes. Nesta passagem em cultura, a quantidade de
células senescentes é provavelmente bem alta, contribuindo para a mudança no
perfil de expressão protéica observado.
Neste trabalho, foi observada a aquisição de alterações citogenéticas nas
culturas de CTM a partir da passagem 6, que se mantiveram até a última
passagem analisada (P10). O desenvolvimento de alterações cromossômicas em
culturas de CTM foi observado por alguns grupos, mas não por outros,
provavelmente devido a diferenças nas metodologias de cultivo das células. O
grupo de Bochkov mostrou uma incidência aumentada de alterações citogenéticas
em culturas de CTM humanas quando comparadas com culturas de linfócitos de
sangue periférico. O papel dessas alterações não foi esclarecido, mas os autores
sugerem que seja pouco provável que elas não sejam importantes para atestar a
qualidade das células para terapias celulares, mostrando a necessidade de um
controle citogenético das células para tal fim (Bochkov et al. 2007). Já o grupo de
Izadpanah mostrou que CTM humanas cultivadas in vitro não desenvolvem
alterações cromossômicas, apesar de ter sido detectada uma alta frequência de
aneuploidias cromossômicas em CTM de macacos Rhesus em passagens mais
avançadas (Izadpanah et al. 2008). O grupo de Bernardo também mostrou que
CTM humanas cultivadas in vitro não desenvolvem anormalidades cromossômicas

Discussão (Bernardo et al. 2007). CTM de camundongos, porém, apresentam uma alta taxa
de instabilidade cromossômica, conforme demonstrado pelo desenvolvimento de
aberrações estruturais e numéricas mesmo nas primeiras passagens em cultura
(Miura et al. 2006).
Alguns genes marcadores, embora não exclusivos de CTM, foram escolhidos
para a análise molecular das células ao longo das passagens em cultura. O RNAm
da citocina SCF é constitutivamente expresso por CTM indiferenciadas, e seus
níveis de expressão são mantidos após diferenciação em meio osteogênico. Tanto
CTM pré-cultivadas em meio controle quanto em meio osteogênico apresentam
capacidade similar de dar suporte a culturas de longa duração (Majumdar et al.
2000). Vacanti e colaboradores mostraram em culturas de CTM porcinas que a
expressão de SCF, assim como de outras citocinas hematopoéticas como G-CSF
e LIF, diminui em células submetidas a mais passagens em cultura (Vacanti et al.
2005). CD44 é uma glicoproteína de adesão envolvida no contato de HPC com
CTM humanas, conforme demonstrado por Wagner e colaboradores, e que é
expressa em culturas indiferenciadas de CTM (Wagner et al. 2008a). As proteínas
BMP 2/4 desempenham papéis fundamentais na embriogênese e no
desenvolvimento esquelético. Elas agem em suas células alvo através de dois
tipos de receptores serina/treonina quinase, BMPR-I e BMPR-II (Watanabe et al.
2007). Nossas análises para a expressão dos três genes, SCF, CD44 e BMPR-IA,
em culturas de CTM ao longo de suas passagens mostram que as células mantêm
um perfil constante até a passagem 5. Tais genes são conhecidamente expressos
em células indiferenciadas, e nossos resultados sugerem que, até a passagem 5,
as culturas apresentam células em estado indiferenciado.

Discussão
Nossos resultados da segunda parte do trabalho sugerem que o procedimento
de cultivo in vitro de células estromais é capaz de uniformizar a população em
cultura, pelo menos no que diz respeito a seu perfil molecular, tanto em relação ao
padrão de expressão de genes de indiferenciação, quanto em relação ao padrão
proteômico. Apesar de o cultivo in vitro induzir a senescência de células em cultura,
nossos dados mostram que o padrão molecular observado em passagens iniciais
se mantém até passagens mais avançadas. As alterações observadas no perfil
proteômico a partir da passagem 9 sugerem ainda modificações em proteínas
relacionadas principalmente às funções do citoesqueleto de actina. No entanto,
alterações cromossômicas foram detectadas mesmo em passagens em que o perfil
molecular e de modulação do sistema imune das culturas de CTM se manteve,
indicando que a análise citogenética é a metodologia mais adequada para o
controle de qualidade das preparações de CTM, e que a utilização dessas
preparações em protocolos de terapia celular deve ser realizada com cautela e
associada a um rigoroso controle citogenético.
Neste trabalho, foi realizado também o estudo proteômico de CTM
submetidas à diferenciação in vitro para as linhagens osteogênica e adipogênica. A
identificação de proteínas diferencialmente expressas entre CTM não diferenciadas
e células submetidas à diferenciação in vitro através de abordagens proteômicas já
foi realizada por alguns grupos nesta área. Foster e colaboradores (2005) utilizaram
a cromatografia líquida associada à espectrometria de massas tandem (LC-MS/MS)
para identificar variações de proteínas de membrana após a indução da
diferenciação osteogênica de CTM estabelecidas em cultura pela superexpressão
da telomerase humana. Neste trabalho, foi possível a identificação de 463
proteínas, incluindo os marcadores conhecidos de CTM, como SH3 (CD71), SH2

Discussão (CD105), CD166, CD44, Thy1, CD29 e CD63 (HOP26). Dentre essas proteínas,
estão também incluídas proteínas integrais, ancoradas ou associadas à membrana,
integrinas e moléculas de adesão celular, receptores e GTPases relacionadas a
Ras. Após a diferenciação osteogênica, proteínas como fosfatase alcalina (ALP),
proteína versican core e tenascina tiveram um aumento de expressão de pelo
menos 2 vezes, enquanto outras diminuíram sua expressão na proporção inversa.
Entretanto, esse estudo tem certas limitações, como a utilização de uma célula que
superexpressa hTERT, o que poderia afetar as funções fisiológicas da célula; o
método para diferenciação das CTM foi o tratamento a curto prazo com calcitriol (4
dias), que induziu a expressão de vários RNA marcadores de diferenciação
osteoblástica, mas que a coloração para fosfatase alcalina revelou
heterogeneidade, sugerindo que nem todas as células estavam sincronizadas no
fenótipo diferenciado.
Outro trabalho que estudou os perfis de expressão protéica de CTM
submetidas à diferenciação osteogênica foi realizado por Zhang e colaboradores
(2007), utilizando eletroforese bidimensional e MALDI-TOF-MS. Mapas proteômicos
foram gerados de células não diferenciadas e de células induzidas em meio
osteogênico após 3 e 7 dias de cultivo. Cinqüenta e duas proteínas foram
identificadas como diferencialmente expressas entre as células não diferenciadas e
as diferenciadas, a maioria envolvida no metabolismo celular, como ciclo de Krebs,
metabolismo de aminoácidos, glicólise e biossíntese de proteínas. Apesar de terem
sido identificadas proteínas de conhecido papel na osteogênese, como Anexina A1,
Anexina A2, Anexina V e OSPBL7, o período de indução osteogênica das CTM foi
curto, de apenas 7 dias, e não foi realizado nenhum ensaio para demonstrar a
diferenciação das células no momento da extração de proteínas. As proteínas

Discussão identificadas como diferencialmente expressas provavelmente estão envolvidas no
início do comprometimento osteogênico das CTM.
O proteoma de membrana de CTM submetidas à diferenciação adipogênica foi
analisado por Jeong e colaboradores (2007), utilizando cromatografia líquida (LC) e
eletroforese bidimensional associada à espectrometria de massas. Setecentos e
sete proteínas foram identificadas nas culturas não diferenciadas pela metodologia
de cromatografia líquida, das quais metade estava associada direta ou
indiretamente ao sistema de membrana celular. A análise de proteínas de
membrana contendo ectodomínio revelou uma série de novos marcadores de
membrana para as CTM da medula óssea, adequados para o desenvolvimento de
anticorpos para isolamento e identificação celular. Análises por cromatografia
líquida mostraram grandes diferenças na constituição da membrana celular entre a
população de células diferenciadas e não diferenciadas, tais como a emergência de
várias enzimas do metabolismo lipídico.
Em nossa análise, identificamos dois grupos de proteínas cuja expressão
aparece bastante reduzida nos géis bidimensionais de culturas de CTM após a
diferenciação: as várias isoformas das cadeias da proteína Tubulina (TUB) e da
proteína Caldesmon (CALD1). TUB é o maior constituinte dos microtúbulos. Liga-se
à actina e à miosina, e está implicada na regulação da interação entre as duas
proteínas, em células musculares e não musculares, enquanto que CALD1 é uma
proteína regulatória envolvida na contração actina-miosina e na estabilidade dos
filamento de actina (Mayanagi et al 2008). Nossos dados sugerem que a
diferenciação das CTM afeta a estrutura do esqueleto de actina das células, e que
este seja um evento importante durante o processo.

Discussão
Mais análises ainda precisam ser realizadas para identificar novas proteínas
envolvidas na diferenciação in vitro das CTM. No entanto, a metodologia
proteômica se mostrou uma ferramenta bastante promissora neste sentido,
revelando novos mecanismos, e permitindo o estudo de proteínas envolvidas nesse
processo.

Conclusões
CONCLUSÕES
� Esse trabalho mostrou que culturas de CTM podem ser obtidas a partir de
amostras de medula óssea de doadores saudáveis através de protocolos
padrão de processamento, cultivo e caracterização in vitro.
� A comparação entre o perfil proteômico e de inibição in vitro da proliferação
de linfócitos de culturas de CTM derivadas de diferentes doadores mostrou
a uniformização da expressão de proteínas majoritárias, mesmo após
somente uma passagem em cultura. Essa similaridade encoraja o uso de
CTM em protocolos terapêuticos de forma independente da compatibilidade
de seu HLA com o do receptor.
� Nossas análises do perfil citogenético sugerem que as células primárias em
cultura apresentam instabilidade genética, levando a alterações
citogenéticas após várias passagens. Mesmo apesar das anormalidades
cromossômicas adquiridas, as culturas continuam a apresentar o potencial
in vitro de inibição da proliferação de linfócitos. A mudança no perfil
proteômico das culturas ocorreu também posteriormente à aquisição da
anomalia cromossômica, indicando que tal ferramenta ainda não está
adequada para o controle de qualidade das preparações de CTM para
utilização em protocolos clínicos. Os dados desse trabalho sugerem que
culturas de CTM devem ser submetidas a poucos subcultivos ex vivo, e
devem passar por um rigoroso controle citogenético prévio à infusão em
pacientes.
� Nossas análises do perfil proteômico e molecular de culturas não
diferenciadas e culturas submetidas à diferenciação osteogênica e

Conclusões
adipogênica mostram que este perfil se altera de forma importante com a
diferenciação das células. No entanto mais análises são ainda necessárias
para melhor definir essas alterações, e estudar seu papel neste processo.
� Esse trabalho mostrou também que mesmo metodologias proteômicas
pouco sensíveis, como a eletroforese bidimensional associada à
espectrometria de massas in tandem, são importantes para o estudo de
células tronco, e capazes de fornecer pistas para a elucidação dos
mecanismos pelos quais as CTM exercem seus efeitos biológicos.

Referência Bibliográficas
REFERÊNCIAS BIBLIOGRÁFICAS
ABDALLAH BM, KASSEM M. The use of mesenchymal (skeletal) stem cells for
treatment of degenerative diseases: current status and future perspectives. J
Cell Physiol 2009;218:9-12.
AGGARWAL S, PITTENGER MF. Human mesenchymal stem cells modulate
allogeneic immune cell responses. Blood 2005; 105:1815-1822.
AKIYAMA H, CHABOISSIER MC, MARTIN JF, SCHEDL A, DE
CROMBRUGGHE B. The transcription factor Sox9 has essential roles in
successive steps of the chondrocyte differentiation pathway and is required for
expression of Sox5 and Sox6. Genes Dev 2002;16:2813-2828.
ALMEIDA-PORADA G, PORADA CD, TRAN N, ZANJANI ED. Cotransplantation
of human stromal cell progenitors into preimmune fetal sheep results in early
appearance of human donor cells in circulation and boosts cell levels in bone
marrow at later time points after transplantation. Blood 2000;95:3620-3627.
ALMEIDA-PORADA G, FLAKE AW, GLIMP HA, ZANJANI ED. Cotransplantation
of stroma results in enhancement of engraftment and early expression of donor
hematopoietic stem cells in utero. Exp Hematol 1999;27:1569-1575.
ANDERSON L, SEILHAMER J. A comparison of selected mRNA and protein
abundances in human liver. Electrophoresis 1997;18:533-537.
ANGELOPOULOU M, NOVELLI E, GROVE JE, RINDER HM, CIVIN C, CHENG
L, KRAUSE DS. Cotransplantation of human mesenchymal stem cells enhances
human myelopoiesis and megakaryocytopoiesis in NOD/SCID mice, Exp
Hematol 2003;31:413-420.
ANJOS-AFONSO F, BONNET D. Flexible and dynamic organization of bone
marrow stromal compartment. Br J Haematol 2007;139:373-384.
ASSMUS B, SCHÄCHINGER V, TEUPE C, BRITTEN M, LEHMANN R,
DÖBERT N, GRÜNWALD F, AICHER A, URBICH C, MARTIN H, HOELZER D,
DIMMELER S, ZEIHER AM. Transplantation of Progenitor Cells and
Regeneration Enhancement in Acute Myocardial Infarction (TOPCARE-AMI).
Circulation 2002;106:3009-3017.

Referência Bibliográficas
AUGELLO A, TASSO R, NEGRINI SM, CANCEDDA R, PENNESI G. Cell
therapy using allogeneic bone marrow mesenchymal stem cells prevents tissue
damage in collagen-induced arthritis. Arthritis Rheum 2007;56:1175-1186.
AWAD HA, BUTLER DL, BOIVIN GP, SMITH FN, MALAVIYA P, HUIBREGTSE
B, CAPLAN AI, Autologous mesenchymal stem cell-mediated repair of tendon,
Tissue Eng 1999;5:267-277.
BACIGALUPO A, VALLE M, PODESTÀ M, PITTO A, ZOCCHI E, DE FLORA A,
POZZI S, LUCHETTI S, FRASSONI F, VAN LINT MT, PIAGGIO G. T-cell
suppression mediated by mesenchymal stem cells is deficient in patients with
severe aplastic anemia. Exp Hematol 2005;33:819-827.
BAKSH D, SONG L, TSUAN RS. Adult mesenchymal stem cells:
characterization, differentiation, and application in cell and gene therapy, J Cell
Mol Med 2004;8:301-316.
BANFI A, MURAGLIA A, DOZIN B, MASTROGIACOMO M, CANCEDDA R,
QUARTO R. Proliferation kinetics and differentiation potential of ex vivo
expanded human bone marrow stromal cells: Implications for their use in cell
therapy. Exp Hematol 2000;28:707-715.
BARRY FP, MURPHY JM, ENGLISH K, MAHON BP. Immunogenicity of adult
mesenchymal stem cells: lessons from the fetal allograft. Stem Cells Dev
2005;14:252-265.
BARRY FP, BOYNTON RE, HAYNESWORTH S, MURPHY JM, ZAIA J. The
monoclonal antibody SH-2, raised against human mesenchymal stem cells,
recognizes an epitope on endoglin (CD105). Biochem Biophys Res Commun
1999;265:134-139.
BARTHOLOMEW A, STURGEON C, SIATSKAS M, FERRER K, MCINTOSH K,
PATIL S, HARDY W, DEVINE S, UCKER D, DEANS R, MOSELEY A,
HOFFMAN R. Mesenchymal stem cells supress lymphocyte proliferation in vitro
and prolong skin graft survival in vivo, Exp Hematol 2002;30:42-48.
BAUM LG, BLACKALL DP, ARIAS-MAGALLANO S, NANIGIAN D, UH SY,
BROWNE JM, HOFFMANN D, EMMANOUILIDES CE, TERRITO MC,
BALDWIN GC. Amelioration of graft versus host disease by galectin-1. Clin
Immunol. 2003;109:295-307.

Referência Bibliográficas
BAXTER MA, WYNN RF, JOWITT SN, WRAITH JE, FAIRBAIRN LJ,
BELLANTUONO I. Study of telomere length reveals rapid aging of human
marrow stromal cells following in vitro expansion. Stem Cells 2004;22:675-682.
BENSIDHOUM M, CHAPEL A, FRANCOIS S, DEMARQUAY C, MAZURIER C,
FOUILLARD L, BOUCHET S, BERTHO JM, GOURMELON P, AIGUEPERSE J,
CHARBORD P, GORIN NC, THIERRY D, LOPEZ M. Homing of in vitro
expanded Stro-1- or Stro-1+ human mesenchymal stem cells into the NOD/SCID
mouse and their role in supporting human CD34 cell engraftment. Blood
2004;103:3313-3319.
BENVENUTO F, FERRARI S, GERDONI E, GUALANDI F, FRASSONI F,
PISTOIA V, MANCARDI G, UCCELLI A. Human Mesenchymal Stem Cells
Promote Survival of T Cells in a Quiescent State. Stem Cells 2007;25:1753-
1760.
BERNARDO ME, ZAFFARONI N, NOVARA F, COMETA AM, AVANZINI MA,
MORETTA A, MONTAGNA D, MACCARIO R, VILLA R, DAIDONE MG,
ZUFFARDI O, LOCATELLI F. Human bone marrow derived mesenchymal stem
cells do not undergo transformation after long-term in vitro culture and do not
exhibit telomere maintenance mechanisms. Cancer Res 2007;67:9142-9149.
BIANCO P, ROBEY PG, SIMMONS PJ. Mesenchymal stem cells: revisiting
history, concepts, and assays.Cell Stem Cell 2008;2:313-319.
BIANCO P, KUZNETSOV SA, RIMUNICCI M, ROBEY PG. Postnatal Skeletal
Stem Cells. Methods in Enzimol 2006;419:117-148.
BIANCO P, RIMINUCCI M, GRONTHOS S, ROBEY PG. Bone marrow stromal
stem cells: nature, biology, and potential applications. Stem Cells 2001;19:180-
192.
BIANCO P, GEHRON ROBEY P. Marrow stromal stem cells. J Clin Invest
2000;105:1663-1668.
BOCHKOV NP, VORONINA ES, KOSYAKOVA NV, LIEHR T, RZHANINOVA
AA, KATOSOVA LD, PLATONOVA VI, GOL'DSHTEIN DV. Chromosome
variability of human multipotent mesenchymal stromal cells. Bull Exp Biol Med
2007;143:122-126.
BOLAÑOS-MEADE J, VOGELSANG GB.Chronic graft-versus-host disease.
Curr Pharm Des 2008;14:1974-1986.

Referência Bibliográficas
BRADFORD MM. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding, Anal Biochem
1976;72:248-254.
BRUDER SP, JAISWAL N, HAYNESWORTH SE. Growth kinetics, self-renewal,
and the osteogenic potential of purified human mesenchymal stem cells during
extensive subcultivation and following cryopreservation. J Cell Biochem
1997;64::278-294.
BRUDER SP, KRAUS KH, GOLDBERG VM, KADIYALA S. The effect of
implants loaded with autologous mesenchymal stem cells on the healing of
canine segmental bone defects. J Bone Joint Surg Am 1998a;80:985-996.
BRUDER SP, KURTH AA, SHEA M, HAYES WC, JAISWAL N, KADIYALA S.
Bone regeneration by implantation of purified, culture-expanded human
mesenchymal stem cells. J Orthop Res 1998b;16:155-162.
CAMPAGNOLI C, ROBERTS IA, KUMAR S, BENNETT PR, BELLANTUONO I,
FISK NM. Identification of mesenchymal stem/progenitor cells in human first-
trimester fetal blood, liver, and bone marrow, Blood 2001;98:2396-2402.
CAPLAN AI, DENNIS JE. Mesenchymal stem cells as trophic mediators. J Cell
Biochem 2006;98:1076-1084.
CAPLAN AI. The mesengenic process, Clin Plastic Surg 1994;21:429-435.
CASTRO-MALASPINA H, GAY RE, RESNICK G, KAPOOR N, MEYERS P,
CHIARIERI D, MCKENZIE S, BROXMEYER HE, MOORE MA. Characterization
of human bone marrow fibroblast colony-forming cells (CFU-F) and their
progeny. Blood 1980;56:289-301.
CENTENO CJ, BUSSE D, KISIDAY J, KEOHAN C, FREEMAN M, KARLI D.
Regeneration of meniscus cartilage in a knee treated with percutaneously
implanted autologous mesenchymal stem cells. Med Hypotheses 2008;71:900-
908.
CHANG KW, YANG PY, LAI HY, YEH TS, CHEN TC, YEH CT. Identification of a
novel actin isoform in hepatocellular carcinoma. Hepatol Res 2006;36:33-39.
CHAPEL A, BERTHO JM, BENSIDHOUM M, FOUILLARD L, YOUNG RG,
FRICK J, DEMARQUAY C, CUVELIER F, MATHIEU E, TROMPIER F,
DUDOIGNON N, GERMAIN C, MAZURIER C, AIGUEPERSE J, BORNEMAN J,
GORIN NC, GOURMELON P, THIERRY D. Mesenchymal stem cells home to

Referência Bibliográficas
injured tissues when co-infused with hematopoietic cells to treat a radiation-
induced multi-organ failure syndrome. J Gene Med 2003;5:1028-1038.
CHEN S, LIU Z, TIAN N, ZHANG J, YEI F, DUAN B, ZHU Z, LIN S, KWAN TW.
Intracoronary transplantation of autologous bone marrow mesenchymal stem
cells for ischemic cardiomyopathy due to isolated chronic occluded left anterior
descending artery. J Invasive Cardiol 2006;18: 552–556.
CHEN SL, FANG WW, YE F, LIU YH, QIAN J, SHAN SJ, ZHANG JJ,
CHUNHUA RZ, LIAO LM, LIN S, SUN JP. Effect on left ventricular function of
intracoronary transplantation of autologous bone marrow mesenchymal stem cell
in patients with acute myocardial infarction. Am J Cardiol 2004;94: 92–95.
CHICHESTER CO, FERNANDEZ M, MINGUELL JJ. Extracellular matrix gene
expression by human bone marrow stroma and by marrow fibroblasts, Cell
Adhes Commun 1993;1:93-99.
CHUNG NG, JEONG DC, PARK SJ, CHOI BO, CHO B, KIM HK, CHUN CS,
WON JH, HAN CW. Cotransplantation of marrow stromal cells may prevent
lethal graft-versus-host disease in major histocompatibility complex mismatched
murine hematopoietic stem cell transplantation. Int J Hematol 2004;80:370-376.
COLTER DC, SEKIYA I, PROCKOP DJ. Identification of a subpopulation of
rapidly self-renewing and multipotential adult stem cells in colonies of human
marrow stromal cells, Proc Natl Acad Sci USA 2001;98:7841-7845.
COLTER DC, CLASS R, DIGIROLAMO CM, PROCKOP DJ. Rapid expansion of
recycling stem cells in cultures of plastic-adherent cells from human bone
marrow, Proc Natl Acad Sci USA 2000;97:3213-3218.
CONGET PA, MINGUELL JJ. Phenotypical and functional properties of human
bone marrow mesenchymal progenitor cells, J Cell Physiol 1999;181:67-73.
CORCIONE A, BENVENUTO F, FERRETTI E, GIUNTI D, CAPPIELLO V,
CAZZANTI F. Human mesenchymal stem cells modulate B-cell functions. Blood
2006; 107: 367-372.
COVAS DT, PANEPUCCI RA, FONTES AM, SILVA WA JR, ORELLANA MD,
FREITAS MC, NEDER L, SANTOS AR, PERES LC, JAMUR MC, ZAGO MA.
Multipotent mesenchymal stromal cells obtained from diverse human tissues
share functional properties and gene-expression profile with CD146+
perivascular cells and fibroblasts. Exp Hematol 2008;36:642-654.

Referência Bibliográficas
COVAS DT, SIUFI JIC, SILVA ARL, ORELLANA MD. Isolation and culture of
umbilical vein mesenchymal stem cells, Braz J Med Biol Res 2003;36:1179-
1183.
CRISAN M, YAP S, CASTEILLA L, CHEN CW, CORSELLI M, PARK TS,
ANDRIOLO G, SUN B, ZHENG B, ZHANG L, NOROTTE C, TENG PN, TRAAS
J, SCHUGAR R, DEASY BM, BADYLAK S, BUHRING HJ, GIACOBINO JP,
LAZZARI L, HUARD J, PÉAULT B. A perivascular origin for mesenchymal stem
cells in multiple human organs. Cell Stem Cell 2008;3:301-313.
DA SILVA MEIRELLES L, CAPLAN AI, NARDI NB. In search of the in vivo
identity of mesenchymal stem cells. Stem Cells 2008;26:2287-2299.
DA SILVA MEIRELLES L, CHAGASTELLES PC, NARDI NB. Mesenchymal
stem cells reside in virtually all post-natal organs and tissues. J Cell Sci
2006;119:2204-2213.
DA SILVA MEIRELLES L, NARDI NB. Murine marrow-derived mesenchymal
stem cell: isoaltion in vitro expansion and characterization, Brit J Haemat
2003;123: 702-711.
DE BARI C, DELL'ACCIO F, TYLZANOWSKI P, LUYTEN FP. Multipotent
mesenchymal stem cells from adult human synovial membrane.Arthritis Rheum
2001;44:1928-1942.
DEANS RJ, MOSELEY AB. Mesenchymal stem cells: biology and potential
clinical uses, Exp Hematol 2000;28:875-884.
DEL PAPA N, QUIRICI N, SOLIGO D, SCAVULLO C, CORTIANA M,
BORSOTTI C, MAGLIONE W, COMINA DP, VITALI C, FRATICELLI P,
GABRIELLI A, CORTELEZZI A, LAMBERTENGHI-DELILIERS G. Bone marrow
endothelial progenitors are defective in systemic sclerosis. Arthritis Rheum
2006;54:2605-2615.
DESCHASEAUX F, CHARBORD P. Human marrow stromal precursors are
alpha 1 integrin subunit-positive. J Cell Physiol 2000;184:319-325.
DEVINE SM, COBBS C, JENNINGS M, BARTHOLOMEW A, HOFFMAN R.
Mesenchymal stem cells distribute to a wide range of tissues following systemic
infusion into nonhuman primates, Blood 2003;101:2999-3001.
DI NICOLA M, CARLO-STELLA C, MAGNI M, MILANESI M, LONGONI PD,
MATTEUCCI P, GRISANTI S, GIANNI AM. Human bone marrow stromal cells

Referência Bibliográficas
suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic
stimuli. Blood 2002;99:3838-3843.
DIGIROLAMO CM, STOKES D, COLTER D, PHINNEY DG, CLASS R,
PROCKOP DJ. Propagation and senescence of human marrow stromal cells in
culture: a simple colony-forming assay identifies samples with the greatest
potential to propagate and differentiate. Br J Haematol 1999;107:275-281.
D'IPPOLITO G, SCHILLER PC, RICORDI C, ROOS BA, HOWARD GA. Age-
related osteogenic potential of mesenchymal stromal stem cells from human
vertebral bone marrow. J Bone Miner Res 1999;14:1115-1122.
DJOUAD F, CHARBONNIER LM, BOUFFI C, LOUIS-PLENCE P, BONY C,
APPARAILLY F, CANTOS C, JORGENSEN C, NOËL D. Mesenchymal Stem
Cells Inhibit the Differentiation of Dendritic Cells Through an Interleukin-6-
Dependent Mechanism. Stem Cells 2007;25:2025–2032.
DJOUAD F, PLENCE P, BONY C, TROPEL P, APPARAILLY F, SANY J, NOËL
D, JORGENSEN C. Immunosuppressive effect of mesenchymal stem cells
favors tumor growth in allogeneic animals. Blood. 2003;102:3837-44.
DOI M, NAGANO A, NAKAMURA Y. Genome-wide screening by cDNA
microarray of genes associated with matrix mineralization by human
mesenchymal stem cells in vitro, Biochem Biophys Res Commun 2002;290:
381-390.
DOMINICI M, LE BLANC K, MUELLER I, SLAPER-CORTENBACH I, MARINI F,
KRAUSE D, DEANS R, KEATING A, PROCKOP D, HORWITZ E. Minimal
criteria for defining multipotent mesenchymal stromal cells: The International
Society for Cellular Therapy position statement, Cytotherapy 2006;8:315-317.
ELIOPOULOS N, STAGG J, LEJEUNE L, POMMEY S, GALIPEAU J. Allogeneic
marrow stromal cells are immune rejected by MHC class I– and class II–
mismatched recipient mice. Blood 2005;106:4057-4065.
ENGLISH K, BARRY FP, MAHON BP. Murine mesenchymal stem cells
suppress dendritic cell migration, maturation and antigen presentation. Immunol
Lett 2008;115:50-58.
FANG L, LANGE C, ENGEL M, ZANDER AR, FEHSE B. Sensitive balance of
suppressing and activating effects of mesenchymal stem cells on T-cell
proliferation. Transplantation 2006;82:1370-1373.

Referência Bibliográficas
FARRINGTON-ROCK C, CROFTS NJ, DOHERTY MJ, ASHTON BA, GRIFFIN-
JONES C, CANFIELD AE. Chondrogenic and adipogenic potential of
microvascular pericytes. Circulation 2004 12;110:2226-2232.
FELDMANN RE JR, BIEBACK K, MAURER MH, KALENKA A, BÜRGERS HF,
GROSS B, HUNZINGER C, KLÜTER H, KUSCHINSKY W, EICHLER H. Stem
cell proteomes: a profile of human mesenchymal stem cells derived from
umbilical cord blood. Electrophoresis 2005;26:2749-2758.
FERNANDEZ M, SIMON V, HERRERA G, CAO C, DEL FAVERO H,
MINGUELL JJ. Detection of stromal cells in peripheral blood progenitor cell
collections from breast cancer patients, Bone Marrow Transplant 1997;20:265-
271.
FERRARI G, ANGELIS GC, COLETTA M, PAOLUCCI E, STORNAIUOLO A,
COSSA G, MAVILIO F. Muscle regenaration by bone marrow-derived myogenic
progenitors, Science 1998;279:1528-1530.
FILSHIE RJ, ZANNETTINO AC, MAKRYNIKOLA V, GRONTHOS S, HENNIKER
AJ, BENDALL LJ, GOTTLIEB DJ, SIMMONS PJ, BRADSTOCK KF. MUC18, a
member of the immunoglobulin superfamily, is expressed on bone marrow
fibroblasts and a subset of hematological malignancies. Leukemia 1998;12:414-
421.
FOX JM, CHAMBERLAIN G, ASHTON BA, MIDDLETON J. Recent advances
into the understanding of mesenchymal stem cell trafficking. Br J Haematol
2007;137:491-502.
FRIEDENSTEIN AJ, GORSKAJA JF, KULAGINA NN. Fibroblast precursors in
normal and irradiated mouse hematopoietic organs, Exp Hematol 1976;4:267-
274.
FRIEDENSTEIN AJ, CHAILAKHYAN RK, LATSINIK NV, PANASYUK AF,
KEILISS-BOROK IV. Stromal cells responsible for transferring the
microenvironment of the hemopoietic tissues. Cloning in vitro and
retransplantation in vivo. Transplantation. 1974;17:331-340.
FUJIBUCHI T, ABE Y, TAKEUCHI T, IMAI Y, KAMEI Y, MURASE R, UEDA N,
SHIGEMOTO K, YAMAMOTO H, KITO K. AIP1/WDR1 supports mitotic cell
rounding. Biochem Biophys Res Commun 2005;327:268-275.

Referência Bibliográficas
GIORDANO A, GALDERISI U, MARINO IR. From the laboratory bench to the
patient's bedside: an update on clinical trials with mesenchymal stem cells. J
Cell Physiol 2007;211:27-35.
GRIFFIN TJ, GYGI SP, IDEKER T, RIST B, ENG J, HOOD L, AEBERSOLD R.
Complementary profiling of gene expression at the transcriptome and proteome
levels in Saccharomyces cerevisiae. Mol Cell Proteomics 2002;1:323-333.
GRINNEMO KH, MANSSON A, DELLGREN G. Xenoreactivity and engraftment
of human mesenchymal stem cells transplanted into infarcted rat myocardium. J
Thorac Cardiovasc Surg 2004;127:1293-1300.
GRONTHOS S, BRAHIM J, LI W, FISHER LW, CHERMAN N, BOYDE A,
DENBESTEN P, ROBEY PG, SHI S. Stem cell properties of human dental pulp
stem cells. J Dent Res 2002;81:531-535.
GRONTHOS S, ZANNETTINO AC, GRAVES SE, OHTA S, HAY SJ, SIMMONS
PJ. Differential cell surface expression of the STRO-1 and alkaline phosphatase
antigens on discrete developmental stages in primary cultures of human bone
cells. J Bone Miner Res 1999;14:47-56.
GRONTHOS S, SIMMONS PJ. The growth factor requirements of STRO-1-
positive human bone marrow stromal precursors under serum-deprived
conditions in vitro, Blood 1995;85:929-940.
GUILLOT PV, GOTHERSTROM C, CHAN J, KURATA H, FISK NM. Human first-
trimester fetal MSC express pluripotency markers and grow faster and have
longer telomeres than adult MSC. Stem Cells 2007;25:646-654.
GUR-WAHNON D, BOROVSKY Z, BEYTH S, LIEBERGALL M,
RACHMILEWITZ J. Contact-dependent induction of regulatory antigen-
presenting cells by human mesenchymal stem cells is mediated via STAT3
signaling. Exp Hematol 2007;35:426-433.
HAYFLICK L. The limited in vitro lifetime of human diploid cell strains. Exp Cell
Res 1965;37:614-636.
HEBERT, B. Advances in protein solubilisation for two-dimensional
eletrophoresis. Electrophoresis 1999;20:660-663.
HORWITZ EM, GORDON PL, KOO WKK, MARX JC, NELL MD, MCNAIL RY,
MUL L, HOFMANN T. Isolated allogenic bone marrow-derived mesenchymal
cells engraft and stimulate growth in children with osteogenesis imperfecta:

Referência Bibliográficas
Implications for cell therapy of bone, Proc Natl Acad Sci USA 2002;99: 8932-
8937.
HORWITZ EM, PROCKOP DJ, FITZPATRICK LA, KOO WW, GORDON PL,
NEEL M, SUSSMAN M, ORCHARD P, MARX JC, PYERITZ RE, BRENNER
MK. Clinical responses to bone marrow transplantation in children with severe
osteogenesis imperfecta. Blood 2001;97:1227-1231.
HORWITZ EM, PROCKOP DJ, FITZPATRICK LA, KOO WW, GORDON PL,
NEEL M, SUSSMAN M, ORCHARD P, MARX JC, PYERITZ RE, BRENNER
MK. Transplantability and therapeutic effects of bone marrow-derived
mesenchymal cells in children with osteogenesis imperfecta, Nat Med
1999;5:309-313.
HUANG Y, HIGGINSON DS, HESTER L, PARK MH, SNYDER SH. Neuronal
growth and survival mediated by eIF5A, a polyamine-modified translation
initiation factor. Proc Natl Acad Sci U S A 2007;104:4194-4199.
HUBERT T, VAN IMPE K, VANDEKERCKHOVE J, GETTEMANS J. The actin-
capping protein CapG localizes to microtubule-dependent organelles during the
cell cycle. Biochem Biophys Res Commun 2009;380:166-170.
IN 'T ANKER PS, SCHERJON SA, KLEIJBURG-VAN DER KEUR C, DE
GROOT-SWINGS GM, CLAAS FH, FIBBE WE, KANHAI HH. Isolation of
mesenchymal stem cells of fetal or maternal origin from human placenta. Stem
Cells 2004;22:1338-1345.
IN 'T ANKER PS, NOORT WA, KRUISSELBRINK AB, SCHERJON SA,
BEEKHUIZEN W, WILLEMZE R, KANHAI HH, FIBBE WE. Nonexpanded
primary lung and bone marrow-derived mesenchymal cells promote the
engraftment of umbilical cord blood-derived CD34(+) cells in NOD/SCID mice,
Exp Hematol 2003a; 31:881-889.
IN 'T ANKER PS, SCHERJON SA, KLEIJBURG-VAN DER KEUR C, NOORT
WA, CLAAS FH, WILLEMZE R, FIBBE WE, KANHAI HH. Amniotic fluid as a
novel source of mesenchymal stem cells for therapeutic transplantation. Blood
2003b;102:1548-1549.
IN 'T ANKER PS, NOORT WA, SCHERJON SA, KLEIJBURG-VAN DER KEUR
C, KRUISSELBRINK AB, VAN BEZOOIJEN RL, BEEKHUIZEN W, WILLEMZE
R, KANHAI HH, FIBBE WE. Mesenchymal stem cells in human second-trimester

Referência Bibliográficas
bone marrow, liver, lung, and spleen exhibit a similar immunophenotype but a
heterogeneous multilineage differentiation potential. Haematologica.
2003;88:845-852.
ISCN 2005: An international system for human cytogenetic nomenclature. In
Shaffe LG, Tommerup N, editors. Basel: S. Karger, 2005.
IZADPANAH R, KAUSHAL D, KRIEDT C, TSIEN F, PATEL B, DUFOUR J,
BUNNELL BA. Long-term in vitro expansion alters the biology of adult
mesenchymal stem cells. Cancer Res 2008;68:4229-4238.
JAVAZON EH, BEGGS KJ, FLAKE AW. Mesenchymal stem cells: paradoxes of
passaging. Exp Hematol 2004;32:414-25.
JEONG JA, KO KM, PARK HS, LEE J, JANG C, JEON CJ, KOH GY, KIM H.
Membrane proteomic analysis of human mesenchymal stromal cells during
adipogenesis. Proteomics 2007;7:4181-4191.
JIANG Y, VAESSEN B, LENVIK T, BLACKSTAD M, REYES M, VERFAILLIE
CM. Multipotent progenitor cells can be isolated from postnatal murine bone
marrow, muscle, and brain, Exp Hematol 2002;30:896-904.
JOHNSTONE B, YOO JU, BARRY F. In vitro chondrogenesis of bone marrow-
derived mesenchymal cells, Exp Cell Res 1998;238:265-2
JONES EA, ENGLISH A, KINSEY SE, STRASZYNSKI L, EMERY P, PONCHEL
F, MCGONAGLE D. Optimization of a flow cytometry-based protocol for
detection and phenotypic characterization of multipotent mesenchymal stromal
cells from human bone marrow. Cytometry B Clin Cytom 2006;70:391-399.
JONES EA, KINSEY SE, ENGLISH A, JONES RA, STRASZYNSKI L,
MEREDITH DM, MARKHAM AF, JACK A, EMERY P, MCGONAGLE D. Isolation
and characterization of bone marrow multipotential Mesenchymal progenitor
cells. Arthritis Rheum 2002;46:3349-3360.
KADIYALA S, YOUNG RG, THIEDE MA, BRUDER SP. Culture expanded
canine mesenchymal stem cells possess osteochondrogenic potential in vivo
and in vitro, Cell Transplant 1997;6:125-134.
KADRI T, LATAILLADE JJ, DOUCET C, MARIE A, ERNOU I, BOURIN P,
JOUBERT-CARON R, CARON M, LUTOMSKI D. Proteomic study of Galectin-1
expression in human mesenchymal stem cells. Stem Cells Dev 2005;14:204-
212.

Referência Bibliográficas
KAN I, MELAMED E, OFFEN D. Autotransplantation of bone marrow-derived
stem cells as a therapy for neurodegenerative diseases. Handb Exp Pharmacol
2007;(180):219-242.
KATO A, KURITA S, HAYASHI A, KAJI N, OHASHI K, MIZUNO K. Critical roles
of actin-interacting protein 1 in cytokinesis and chemotactic migration of
mammalian cells. Biochem J 2008;414:261-270.
KATRITSIS DG, SOTIROPOULOU P, GIAZITZOGLOU E, KARVOUNI E,
PAPAMICHAIL M. Electrophysiological effects of intracoronary transplantation of
autologous mesenchymal and endothelial progenitor cells. Europace
2007;9:167–171.
KATRITSIS DG, SOTIROPOULOU PA, KARVOUNI E, KARABINOS I,
KOROVESIS S, PEREZ SA, VORIDIS EM, PAPAMICHAIL M. Transcoronary
transplantation of autologous mesenchymal stem cells and endothelial
progenitors into infracted human myocardium. Catheter Cardiovasc Interv
2005;65:321–329.
KIM DH, YOO KH, CHOI KS, CHOI J, CHOI SY, YANG SE, YANG YS, IM HJ,
KIM KH, JUNG HL, SUNG KW, KOO HH. Gene expression profile of cytokine
and growth factor during differentiation of bone marrow-derived mesenchymal
stem cell, Cytokine 2005;31:119-26.
KINNAIRD T, STABILE E, BURNETT MS, LEE CW, BARR S, FUCHS S,
EPSTEIN SE. Marrow-derived stromal cells express genes encoding a broad
spectrum of arteriogenic cytokines and promote in vitro and in vivo
arteriogenesis through paracrine mechanisms. Circ Res 2004;94:678–685.
KISS J, KUNSTÁR A, FAJKA-BOJA R, DUDICS V, TÓVÁRI J, LÉGRÁDI A,
MONOSTORI E, UHER F. A novel anti-inflammatory function of human galectin-
1: inhibition of hematopoietic progenitor cell mobilization. Exp Hematol
2007;35:305-313.
KOC ON, GERSON SL, COOPER BW, DYHOUSE SM, HAYNESWORTH SE,
CAPLAN AI, LAZARUS HM. Rapid hematopoietic recovery after coinfusion of
autologous-blood stem cells and culture-expanded marrow mesenchymal stem
cells in advanced breast cancer patients receiving high-dose chemotherapy, J
Clin Oncol 2000;18:307-316.

Referência Bibliográficas
KOC ON, PETERS C, AUBOURG P, RAGHAVAN S, DYHOUSE S,
DEGASPERI R, KOLODNY EH, YOSEPH YB, GERSON SL, LAZARUS HM,
CAPLAN AI, WATKINS PA, KRIVIT W. Bone marrow-derived mesenchymal
stem cells remain host-derived despite successful hematopoietic engraftment
after allogeneic transplantation in patients with lysosomal and peroxisomal
storage diseases, Exp Hematol 1999;27:1675-1681.
KOPEN GC, PROCKOP DJ, PHINNEY DG. Marrow stromal cells migrate
throughout forebrain and cerebellum, and they differentiate into astrocytes after
injection into neonatal mouse brains. Proc Natl Acad Sci U S A 1999;96:10711-
10716.
KRAMPERA M, COSMI L, ANGELI R, PASINI A, LIOTTA F, ANDREINI A,
SANTARLASCI V, MAZZINGHI B, PIZZOLO G, VINANTE F, ROMAGNANI P,
MAGGI E, ROMAGNANI S, ANNUNZIATO F. Role for Interferon-γ in the
Immunomodulatory Activity of Human Bone Marrow Mesenchymal Stem Cells.
Stem Cells 2006;24:386–398.
KRAMPERA M, GLENNIE S, DYSON J, SCOTT D, LAYLOR R, SIMPSON E,
DAZZI F. Bone marrow mesenchymal stem cells inhibit the response of naïve
and memory antigen-specific T cells to their cognate peptide, Blood 2002;101:
3722-3729.
KURODA R, USAS A, KUBO S CORSI K, PENG H, ROSE T, CUMMINS J, FU
FH, HUARD J. Cartilage repair using bone morphogenetic protein 4 and muscle-
derived stem cells. Arthritis Rheum 2006;54:433– 442.
KUZNETSOV SA, MANKANI MH, GRONTHOS S, SATOMURA K, BIANCO P,
ROBEY PG. Circulating skeletal stem cells. J Cell Biol 2001;153:1133-1140.
LAZARUS HM, KOC ON, DEVINE SM, CURTIN P, MAZIARZ RT, HOLLAND
HK, SHPALL EJ, MCCARTHY P, ATKINSON K, COOPER BW, GERSON SL,
LAUGHLIN MJ, LOBERIZA FR JR, MOSELEY AB, BACIGALUPO A.
Cotransplantation of HLA-Identical Sibling Culture-Expanded Mesenchymal
Stem Cells and Hematopoietic Stem Cells in Hematologic Malignancy Patients.
Biol Blood Marrow Transplant 2005;11: 389-398.
LAZARUS HM, HAYNESWORTH SE, GERSON SL, CAPLAN AI. Human bone
marrow-derived mesenchymal (stromal) progenitor cells (MPCs) cannot be

Referência Bibliográficas
recovered from peripheral blood progenitor cell collections. J Hematother
1997;6:447-455.
LAZARUS HM, HAYNESWORTH SE, GERSON SL, ROSENTHAL NS,
CAPLAN AI. Ex vivo expansion and subsequent infusion of human bone
marrow-derived stromal progenitor cells (mesenchymal progenitor cells):
implications for therapeutic use, Bone Marrow Transplant 1995;16:557-564.
LE BLANC K, FRASSONI F, BALL L, LOCATELLI F, ROELOFS H, LEWIS I,
LANINO E, SUNDBERG B, BERNARDO ME, REMBERGER M, DINI G,
EGELER RM, BACIGALUPO A, FIBBE W, RINGDÉN O. Developmental
Committee of the European Group for Blood and Marrow Transplantation.
Mesenchymal stem cells for treatment of steroid-resistant, severe, acute graft-
versus-host disease: a phase II study. Lancet 2008;371:1579-1586.
LE BLANC K, SAMUELSSON H, GUSTAFSSON B, REMBERGER M,
SUNDBERG B, ARVIDSON J, LJUNGMAN P, LÖNNIES H, NAVA S, RINGDÉN
O. Transplantation of mesenchymal stem cells to enhance engraftment of
hematopoietic stem cells. Leukemia 2007;21:1733–1738.
LE BLANC K, RASMUSSON I, GÖTHERSTRÖM C, SEIDEL C, SUNDBERG B,
SUNDIN M, ROSENDAHL K, TAMMIK C, RINGDÉN O. Mesenchymal stem
cells inhibit the expression of CD25 (interleukin-2 receptor) and CD38 on
phytohaemagglutinin-activated lymphocytes. Scand J Immunol 2004a; 60:307-
315.
LE BLANC K, RASMUSSON I, SUNDBERG B, GOTHERSTROM C, HASSAN
M, UZUNEL M, RINGDEN O. Treatment of severe acute graft-versus-host
disease with third party haploidentical mesenchymal stem cells, Lancet
2004b;363:1439-1441.
LE BLANC K, TAMMIK L, SUNDBERG B, HAYNESWORTH SE, RINGDÉN O.
Mesenchymal stem cells inhibit and stimulate mixed lymphocyte cultures and
mitogenic responses independently of the major histocompatibility complex.
Scand J immunol 2003;57:11-20.
LEE HK, LEE BH, PARK SA, KIM CW. The proteomic analysis of an adipocyte
differentiated from human mesenchymal stem cells using two-dimensional gel
electrophoresis. Proteomics 2006;6:1223-1229.

Referência Bibliográficas
LEE JY, QU-PETERSEN Z, CAO B KIMURA S, JANKOWSKI R, CUMMINS J,
USAS A, GATES C, ROBBINS P, WERNIG A, HUARD J. Clonal isolation of
muscle-derived cells capable of enhancing muscle regeneration and bone
healing. J Cell Biol 2000;150:1085–1100.
LEE OK, KUO TK, CHEN WM, LEE KD, HSIEH SL, CHEN TH. Isolation of
multipotent mesenchymal stem cells from umbilical cord blood, Blood
2004;103:1669-1675.
LEE RH, KIM B, CHOI I, KIM H, CHOI HS, SUH K, BAE YC, JUNG JS.
Characterization and expression analysis of mesenchymal stem cells from
human bone marrow and adipose tissue, Cell Physiol Biochem 2004;14:311-
324.
LEE ST, JANG JH, CHEONG JW, KIM JS, MAEMG HY, HAHN JS, KO YW, MIN
YH. Treatment of high-risk acute myelogenous leukaemia by myeloablative
chemoradiotherapy followed by co-infusion of T cell-depleted haematopoietic
stem cells and culture-expanded marrow mesenchymal stem cells from a related
donor with one fully mismatched human leucocyte antigen haplotype. Br J
Haematol 2002;118:1128-1131.
LEONARDI E, DEVESCOVI V, PERUT F, CIAPETTI G, GIUNTI A. Isolation,
characterisation and osteogenic potential of human bone marrow stromal cells
derived from the medullary cavity of the femur. Chir Organi Mov 2008;92:97-103.
LEVI G, TARRAB-HAZDAI R, TEICHBERG VI. Prevention and therapy with
electrolectin of experimental autoimmune myasthenia gravis in rabbits. Eur J
Immunol 1983;13:500-507.
LI G, ZHANG XA, WANG H, WANG X, MENG CL, CHAN CY, YEW DT, TSANG
KS, LI K, TSAI SN, NGAI SM, HAN ZC, LIN MC, HE ML, KUNG HF.
Comparative proteomic analysis of mesenchymal stem cells derived from human
bone marrow, umbilical cord, and placenta: implication in the migration.
Proteomics 2009;9:20-30.
LI N, FEUGIER P, SERRURRIER B, LATGER-CANNARD V, LESESVE JF,
STOLTZ JF, ELJAAFARI A. Human mesenchymal stem cells improve ex vivo
expansion of adult human CD34+ peripheral blood progenitor cells and decrease
their allostimulatory capacity. Exp Hematol 2007;35:507-515.

Referência Bibliográficas
LI Q, YU Y, BISCHOFF J, MULLIKEN JB, OLSEN BR. Differential expression of
CD146 in tissues and endothelial cells derived from infantile haemangioma and
normal human skin. J Pathol 2003;201:296-302.
LI Y, CHEN J, ZHANG CL, WANG L, LU D, KATAKOWSKI M, GAO Q, SHEN
LH, ZHANG J, LU M, CHOPP M. Gliosis and brain remodeling after treatment of
stroke in rats with marrow stromal cells. Glia 2005;49:407-417.
LIU L, DIGIROLAMO CM, NAVARRO PA, BLASCO MA, KEEFE DL.
Telomerase deficiency impairs differentiation of mesenchymal stem cells. Exp
Cell Res 2004;294:1-8.
LUCHESSI AD, CAMBIAGHI TD, HIRABARA SM, LAMBERTUCCI RH,
SILVEIRA LR, BAPTISTA IL, MORISCOT AS, COSTA-NETO CM, CURI R.
Involvement of eukaryotic translation initiation factor 5A (eIF5A) in skeletal
muscle stem cell differentiation. J Cell Physiol 2009;218:480-489.
MACCARIO R, PODESTÀ M, MORETTA A, COMETA A, COMOLI P,
MONTAGNA D, DAUDT L, IBATICI A, PIAGGIO G, POZZI S, FRASSONI F,
LOCATELLI F. Interaction of human mesenchymal stem cells with cells involved
in alloantigen-specific immune response favors the differentiation of CD4+ T-cell
subsets expressing a regulatory/suppressive phenotype. Haematologica
2005;90: 516–525.
MAHMOOD A, LU D, CHOPP M. Marrow stromal cell transplantation after
traumatic brain injury promotes cellular proliferation within the brain.
Neurosurgery 2004;55:1185–1193.
MAITRA B, SZEKELY E, GJINI K, LAUGHLIN MJ, DENNIS J, HAYNESWORTH
SE, KOÇ ON, Human mesenchymal stem cells support unrelated donor
hematopoietic stem cells and suppress T-cell activation, Bone Marrow
Transplant 2004;33: 597-604.
MAJUMDAR MK, THIEDE MA, HAYNESWORTH SE, BRUDER SP, GERSON
SL. Human marrow-derived mesenchymal stem cells (MSCs) express
hematopoietic cytokines and support long-term hematopoiesis when
differentiated toward stromal and osteogenic lineages. J Hematother Stem Cell
Res. 2000;9:841-848.
MAJUMDAR MK, THIEDE MA, MOSCA JD, MOORMAN M, GERSON SL.
Phenotypic and functional comparison of cultures of marrow-derived

Referência Bibliográficas
mesenchymal stem cells (MSCs) and stromal cells, J Cell Physiol 1998;76:57-
66.
MARESCHI K, FERRERO I, RUSTICHELLI D, ASCHERO S, GAMMAITONI L,
AGLIETTA M, MADON E, FAGIOLI F. Expansion of mesenchymal stem cells
isolated from pediatric and adult donor bone marrow. J Cell Biochem
2006;97:744-754.
MARTIN DR, COX NR, HATHCOCK TL, NIEMEYER GP, BAKER HJ. Isolation
and characterization of multipotential mesenchymal stem cells from feline bone
marrow, Exp Hematol 2002;30:879-886.
MAYANAGI T, MORITA T, HAYASHI K, FUKUMOTO K, SOBUE K.
Glucocorticoid receptor-mediated expression of caldesmon regulates cell
migration via the reorganization of the actin cytoskeleton. J Biol Chem
2008;283:31183-31196.
MAZZINI L, VERCELLI A, MARESCHI K, FERRERO I, TESTA L, FAGIOLI F.
Mesenchymal stem cells for ALS patients. Amyotroph Lateral Scler 2008:1-2.
MEISEL R, ZIBERT A, LARYEA M, GÖBEL U, DÄUBENER W, DILLOO D.
Human bone marrow stromal cells inhibit allogeneic T-cell responses by
indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood
2004;103:4619-4621.
MENDES SC, TIBBE JM, VEENHOF M, BAKKER K, BOTH S, PLATENBURG
PP, ONER FC, DE BRUIJN JD, VAN BLITTERSWIJK CA. Bone tissue-
engineered implants using human bone marrow stromal cells: effect of culture
conditions and donor age. Tissue Eng 2002;8:911-920.
METS T, VERDONK G. In vitro aging of human bone marrow derived stromal
cells. Mech Ageing Dev 1981;16:81-89.
MIN CK, KIM BG, PARK G, CHO B, OH IH. IL-10-transduced bone marrow
mesenchymal stem cells can attenuate the severity of acute graft-versus-host
disease after experimental allogeneic stem cell transplantation. Bone Marrow
Transplant 2007;39:637-645.
MINGUELL JJ, CONGET P, ERICES A. Biology and clinical utilization of
mesenchymal progenitor cells, Braz J Med Biol Res 2000;33:881-887.
MIURA M, MIURA Y, PADILLA-NASH HM, MOLINOLO AA, FU B, PATEL V,
SEO BM, SONOYAMA W, ZHENG JJ, BAKER CC, CHEN W, RIED T, SHI S.

Referência Bibliográficas
Accumulated chromosomal instability in murine bone marrow mesenchymal
stem cells leads to malignant transformation. Stem Cells 2006;24:1095-1103.
MOURCIN F, GRENIER N, MAYOL JF, LATAILLADE JJ, SOTTO JJ, HÉRODIN
F, DROUET M. Mesenchymal stem cells support expansion of in vitro irradiated
CD34(+) cells in the presence of SCF, FLT3 ligand, TPO and IL3: potential
application to autologous cell therapy in accidentally irradiated victims. Radiat
Res 2005;164:1-9.
MURAGLIA A, CANCEDDA R, QUARTO R. Clonal mesenchymal progenitors
from human bone marrow differentiate in vitro according to a hierarchical model,
J Cell Sci 2000;113:1161-1166.
MURPHY JM, FINK DJ, HUNZIKER EB, BARRY FP. Stem cell therapy in a
caprine model of osteoarthritis. Arthritis Rheum 2003;48:3464-3474.
NAGAYA N, KANGAWA K, ITOH T, IWASE T, MURAKAMI S, MIYAHARA Y,
FUJII T, UEMATSU M, OHGUSHI H, YAMAGISHI M, TOKUDOME T, MORI H,
MIYATAKE K, KITAMURA S. Transplantation of mesenchymal stem cells
improves cardiac function in a rat model of dilated cardiomyopathy. Circulation
2005;112: 1128–1135.
NAKAMURA T, SHIOJIMA S, HIRAI Y, IWAMA T, TSURUZOE N, HIRASAWA A,
KATSUMA S, TSUJIMOTO G. Temporal gene expression changes during
adipogenesis in human mesenchymal stem cells, Biochem Biophys Res
Commun 2003;303:306-312.
NAKASHIMA K, ZHOU X, KUNKEL G, ZHANG Z, DENG JM, BEHRINGER RR,
DE CROMBRUGGHE B. The novel zinc finger-containing transcription factor
osterix is required for osteoblast differentiation and bone formation. Cell
2002;108:17-29.
NASEF A, CHAPEL A, MAZURIER C, BOUCHET S, LOPEZ M, MATHIEU N,
SENSEBÉ L, ZHANG Y, GORIN NC, THIERRY D, FOUILLARD L. Identification
of IL-10 and TGF-beta transcripts involved in the inhibition of T-lymphocyte
proliferation during cell contact with human mesenchymal stem cells. Gene Expr
2007; 13: 217-226.
NAUTA AJ, FIBBE WE. Immunomodulatory properties of mesenchymal stromal
cells. Blood 2007;110:3499-3506.

Referência Bibliográficas
NAUTA AJ, WESTERHUIS G, KRUISSELBRINK AB, LURVINK EG, WILLEMZE
R, FIBBE WE. Donor-derived mesenchymal stem cells are immunogenic in an
allogeneic host and stimulate donor graft rejection in a nonmyeloablative setting.
Blood 2006;108:2114-2120.
NOËL D, CATON D, ROCHE S, BONY C, LEHMANN S, CASTEILLA L,
JORGENSEN C, COUSIN B. Cell specific differences between human adipose-
derived and mesenchymal-stromal cells despite similar differentiation potentials.
Exp Cell Res 2008;314:1575-1584.
NOEL D, GAZIT D, BOUQUET C. Short-term BMP-2 expression is sufficient for
in vivo osteochondral differentiation of mesenchymal stem cells. Stem Cells
2003;22:74-85.
NIEMEYER P, KRAUSE U, KASTEN P, KREUZ PC, HENLE P, SÜDKAM NP,
MEHLHORN A. Mesenchymal stem cell-based HLA-independent cell therapy for
tissue engineering of bone and cartilage. Curr Stem Cell Res Ther 2006;1:21-27.
OFFNER H, CELNIK B, BRINGMAN TS, CASENTINI-BOROCZ D, NEDWIN
GE, VANDENBARK AA. Recombinant human beta-galactoside binding lectin
suppresses clinical and histological signs of experimental autoimmune
encephalomyelitis. J Neuroimmunol 1990;28:177-184.
OHNISHI S, YANAGAWA B, TANAKA K, MIYAHARA Y, OBATA H, KATAOKA
M, KODAMA M, ISHIBASHI-UEDA H, KANGAWA K, KITAMURA S, NAGAYA N.
Transplantation of mesenchymal stem cells attenuates myocardial injury and
dysfunction in a rat model of acute myocarditis. J Mol Cell Cardiol 2007;42: 88–
97.
ORLIC D, KAJSTURA J, CHIMENTI S, JAKONIUK I, ANDERSON SM, LI B,
PICKEL J, MCKAY R, NADAL-GINARD B, BODINE DM, LERI A, ANVERSA P.
Bone marrow cells regenerate infarcted myocardium. Nature 2001;410:701-705.
ORTIZ LA, GAMBELLI F, MCBRIDE C, GAUPP D, BADDOO M, KAMINSKI N,
PHINNEY DG. Mesenchymal stem cell engraftment in lung is enhanced in
response to bleomycin exposure and ameliorates its fibrotic effects. Proc Natl
Acad Sci USA. 2003;100:8407-8411.
OWEN M, FRIEDENSTEIN AJ. Stromal stem cells: marrow-derived osteogenic
precursors. Ciba Found Symp 1988;136:42-60.

Referência Bibliográficas
PANEPUCCI RA, SIUFI JL, SILVA WA, PROTO-SIQUIERA R, NEDER L,
ORELLANA M, ROCHA V, COVAS DT, ZAGO MA. Comparison of gene
expression of umbilical cord vein and bone marrow-derived mesenchymal stem
cells, Stem Cells 2004;22:1263-1278.
PARK HW, SHIN JS, KIM CW. Proteome of mesenchymal stem cells.
Proteomics 2007;7:2881-2894.
PEREIRA RF, HALFORD KW, O’HARA MD, LEEPER DB, SOKOLOV BP,
POLLARD MD, BAGSARA O, PROCKOP DJ. Cultured adherent cells from
marrow can serve as long-lasting precursor cells for bone, cartilage and lung in
irradiated mice, Proc Natl Acad Sci USA 1995;92:4857-4861.
PERKA C, SCHULTZ O, SPITZER RS, LINDENHAYN K. The influence of
transforming growth factor beta1 on mesenchymal cell repair of full-thickness
cartilage defects. J Biomed Mater Res 2000;52:543-552.
PETITE H, VIATEAU V, BENSAÏD W, MEUNIER A, DE POLLAK C,
BOURGUIGNON M, OUDINA K, SEDEL L, GUILLEMIN G. Tissue-engineered
bone regeneration. Nat Biotechnol 2000;18:959-963.
PHINNEY DG, KOPEN G, RIGHTER W, WEBSTER S, TREMAIN N, PROCKOP
DJ. Donor variation in the growth properties and osteogenic potential of human
marrow stromal cells, J Cell Biochem 1999;75:424-436.
PIERSMA AH, PLOEMACHER RE, BROCKBANK KG. Transplantation of bone
marrow fibroblastoid stromal cells in mice via the intravenous route, Br J
Haematol 1983;54:285-290.
PIGNOLO RJ, SUDA RK, MCMILLAN EA, SHEN J, LEE SH, CHOI Y, WRIGHT
AC, JOHNSON FB. Defects in telomere maintenance molecules impair
osteoblast differentiation and promote osteoporosis. Aging Cell 2008;7::23-31.
PITTENGER MF, MACKAY AM, BECK SC, JAISWAL RK, DOUGLAS R,
MOSCA JD, MOORMAN MA, SIMONETTI DW, MARSHAK DR. Multilineage
potential of adult human mesenchymal stem cells, Science 1999;284:143-147.
POTIAN JA, AVIV H, PONZIO NM, HARRISON JS, RAMESHWAR P. Veto-like
activity of mesenchymal stem cells: functional discrimination between cellular
responses to alloantigens and recall antigens, J Immunol 2003;171:3426-3434.
PROCKOP DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues,
Science 1997;276:71-74.

Referência Bibliográficas
QI H, AGUIAR DJ, WILLIAMS SM, LA PEAN A, PAN W, VERFAILLIE CM.
Identification of genes responsible for osteoblast differentiation from human
mesodermal progenitor cells, Proc Nat Acad Sci USA 2003;100:3305-3310.
QIAN L, SALTZMAN WM. Improving the expansion and neuronal differentiation
of mesenchymal stem cells through culture surface modification, Biomat
2004;25:1331-1337.
QUARTO R, MASTROGIACOMO M, CANCEDDA R, KUTEPOV SM,
MUKHACHEV V, LAVROUKOV A, KON E, MARCACCI M. Repair of large bone
defects with the use of autologous bone marrow stromal cells. N Engl J Med
2001;344:385-386.
RABINOVICH GA, SOTOMAYOR CE, RIERA CM, BIANCO I, CORREA SG.
Evidence of a role for galectin-1 in acute inflammation. Eur J Immunol
2000;30:1331-1339.
RAMASAMY R, LAM EW, SOEIRO I, TISATO V, BONNET D, DAZZI F.
Mesenchymal stem cells inhibit proliferation and apoptosis of tumor cells: impact
on in vivo tumor growth. Leukemia 2007;21:304-310.
RASMUSSON I, LE BLANC K, SUNDBERG B, RINGDÉN O. Mesenchymal
stem cells stimulate antibody secretion in human B cells. Scand J Immunol
2007;65:336-343.
RASMUSSON I. Immune modulation by mesenchymal stem cells. Exp Cell Res
2006;312:2169-2179.
RASMUSSON I, RINGDÉN O, SUNDBERG B, LE BLANC K. Mesenchymal
stem cells inhibit the formation of cytotoxic T lymphocytes, but not activated
cytotoxic T lymphocytes or natural killer cells. Transplantation 2003;76:1208-
1213.
RENZ M, LANGOWSKI J. Dynamics of the CapG actin-binding protein in the cell
nucleus studied by FRAP and FCS. Chromosome Res 2008;16:427-437.
RINGDÉN O, UZUNEL M, RASMUSSON I, REMBERGER M, SUNDBERG B,
LÖNNIES H, MARSCHALL HU, DLUGOSZ A, SZAKOS A, HASSAN Z, OMAZIC
B, ASCHAN J, BARKHOLT L, LE BLANC K. Mesenchymal stem cells for
treatment of therapy-resistant graft-versus-host disease, Transplant
2006;81:1390-1397.

Referência Bibliográficas
ROOBROUCK VD, ULLOA-MONTOYA F, VERFAILLIE CM. Self-renewal and
differentiation capacity of young and aged stem cells. Exp Cell Res
2008;314:1937-1944.
ROSE R A, JIANG H, WANG X, HELKE S, TSOPORIS JN, GONG N, KEATING
SCJ, PARKER TG, BACKX PH, KEATING A. Bone Marrow-Derived
Mesenchymal Stromal Cells Express Cardiac-Specific Markers, Retain the
Stromal Phenotype and do not Become Functional Cardiomyocytes In Vitro.
Stem Cells 2008;26:2884-2892.
SABATINI F, PETECCHIA L, TAVIAN M, JODON DE VILLEROCHÉ V, ROSSI
GA, BROUTY-BOYÉ D. Human bronchial fibroblasts exhibit a mesenchymal
stem cell phenotype and multilineage differentiating potentialities. Lab Invest
2005;85:962-971.
SACCHETTI B, FUNARI A, MICHIENZI S, DI CESARE S, PIERSANTI S,
SAGGIO I, TAGLIAFICO E, FERRARI S, ROBEY PG, RIMINUCCI M, BIANCO
P. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a
hematopoietic microenvironment. Cell 2007;131:324-336.
SAFFORD KM, SAFFORD SD, GIMBLE JM, SHETTY AK, RICE HE.
Characterization of neuronal/glial differentiation of murine adipose-derived adult
stromal cells, Exp Neurol 2004;187:319-328.
SALINGCARNBORIBOON R, YOSHITAKE H, TSUJI K, OBINATA M,
AMAGASA T, NIFUJI A, NODA M. Establishment of tendon-derived cell lines
exhibiting pluripotent mesenchymal stem cell-like property. Exp Cell Res
2003;287:289-300.
SANCHEZ-RAMOS J, SONG S, CARDOZO-PELAEZ F, HAZZI C,
STEDEFORD T, WILLING A, FREEMAN TB, SAPORTA S, JANSSEN W,
PATEL N, COOPER DR, SANBERG PR. Adult bone marrow stromal cells
differentiate into neural cells in vitro, Exp Neurol 2000;164:247-256.
SATO K, OZAKI K, OH I, MEGURO A, HATANAKA K, NAGAI T, MUROI K,
OZAWA K. Nitric oxide plays a critical role in suppression of T-cell proliferation
by mesenchymal stem cells. Blood, 2007; 109: 228-234.
SATO K, YAMASHITA N, YAMASHITA N, BABA M, MATSUYAMA T.
Regulatory dendritic cells protect mice from murine acute graft-versus-host
disease and leukemia relapse. Immunity 2003;18:367-379.

Referência Bibliográficas
SATO Y, ARAKI H, KATO J, NAKAMURA K, KAWANO Y, KOBUNE M, SATO T,
MIYANISHI K, TAKAYAMA T, TAKAHASHI M, TAKIMOTO R, IYAMA S,
MATSUNAGA T, OHTANI S, MATSUURA A, HAMADA H, NIITSU Y. Human
mesenchymal stem cells xenografted directly to rat liver are differentiated into
human hepatocytes without fusion. Blood 2005;106:756-763.
SATOMURA K, DERUBOIS AR, FEDARKO NS, IBARAKI-O’CONNOR K,
KUZNETSOV SA, ROWE DW, YOUNG MF, GEHRON RP. Receptor tyrosine
kinase expression in human bone marrow stromal cells, J Cell Physiol
1998;177:426-438.
SCHÄFER C, BORM B, BORN S, MÖHL C, EIBL EM, HOFFMANN B. One step
ahead: Role of filopodia in adhesion formation during cell migration of
keratinocytes. Exp Cell Res 2008 Dec 3.
SEABRIGHT M. A rapid banding technique for human chromosomes. Lancet
1971;30;2:971-972.
SEKIYA I, LARSON BL, VUORISTO JT, CUI J-G, PROCKOP DJ. Adipogenic
differentiation of human adult stem cells from bone marrow stroma (MSCs), J
Bone Miner Res 2004;19:256-264.
SEKIYA I, LARSON BL, SMITH JR, POCHAMPALLY R, CUI J-G, PROCKOP
DJ. Expansion of human adult stem cells from bone marrow stroma: conditions
that maximize the yields of early progenitors and evaluate their quality, Stem
Cells 2002a;20:530-541.
SEKIYA I, VUORISTO JT, LARSON BL, PROCKOP DJ. In vitro cartilage
formation by human adult stem cells from bone marrow stroma defines the
sequence of cellular and molecular events during chondrogenesis. Proc Nat
Acad Sci USA 2002b;99:4397-4402.
SEO BM, MIURA M, GRONTHOS S, BARTOLD PM, BATOULI S, BRAHIM J,
YOUNG M, ROBEY PG, WANG CY, SHI S. Investigation of multipotent
postnatal stem cells from human periodontal ligament. Lancet 2004;364:149-
155.
SHAKE JG, GRUBER PJ, BAUMGARTNER WA, SENECHAL G, MEYERS J,
REDMOND JM, PITTENGER MF, MARTIN BJ. Mesenchymal stem cell
implantation in a swine myocardial infarct model: engraftment and functional
effects. Ann Thorac Surg 2002; 73:1919–1925.

Referência Bibliográficas
SHI S, GRONTHOS S. Perivascular niche of postnatal mesenchymal stem cells
in human bone marrow and dental pulp. J Bone Miner Res 2003;18:696-704.
SHI S, GRONTHOS S, CHEN S, REDDI A, COUNTER CM, ROBEY PG, WANG
CY. Bone formation by human postnatal bone marrow stromal stem cells is
enhanced by telomerase expression. Nat Biotechnol 2002;20:587-591.
SHIBATA KR, AOYAMA T, SHIMA Y, FUKIAGE K, OTSUKA S, FURU M,
KOHNO Y, ITO K, FUJIBAYASHI S, NEO M, NAKAYAMA T, NAKAMURA T,
TOGUCHIDA J. Expression of the p16INK4A gene is associated closely with
senescence of human mesenchymal stem cells and is potentially silenced by
DNA methylation during in vitro expansion. Stem Cells 2007;25:2371-2382.
SIMONSEN JL, ROSADA C, SERAKINCI N, JUSTESEN J, STENDERUP K,
RATTAN SI, JENSEN TG, KASSEM M. Telomerase expression extends the
proliferative life-span and maintains the osteogenic potential of human bone
marrow stromal cells. Nat Biotechnol 2002;20:592-596.
SORRENTINO A, FERRACIN M, CASTELLI G, BIFFONI M, TOMASELLI G,
BAIOCCHI M, FATICA A, NEGRINI M, PESCHLE C, VALTIERI M. Isolation and
characterization of CD146+ multipotent mesenchymal stromal cells. Exp
Hematol 2008;36:1035-1046.
SPAGGIARI GM, CAPOBIANCO A, BECCHETTI S, MINGARI MC, MORETTA
L. Mesenchymal stem cell-natural killer cell interactions: evidence that activated
NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2-induced
NK-cell proliferation. Blood 2006;107:1484-1490.
STENDERUP K, JUSTESEN J, CLAUSEN C, KASSEM M. Aging is associated
with decreased maximal life span and accelerated senescence of bone marrow
stromal cells. Bone 2003;33:919-926.
STEWART K, MONK P, WALSH S, JEFFERISS CM, LETCHFORD J,
BERESFORD JN. STRO-1, HOP-26 (CD63), CD49a and SB-10 (CD166) as
markers of primitive human marrow stromal cells and their more differentiated
progeny: a comparative investigation in vitro. Cell Tissue Res 2003;313:281-290.
STOLZING A, JONES E, MCGONAGLE D, SCUTT A. Age-related changes in
human bone marrow-derived mesenchymal stem cells: consequences for cell
therapies. Mech Ageing Dev 2008;129:163-173.

Referência Bibliográficas
SUDRES M, NOROL F, TRENADO A, GRÉGOIRE S, CHARLOTTE F,
LEVACHER B, LATAILLADE JJ, BOURIN P, HOLY X, VERNANT JP,
KLATZMANN D, COHEN JL. Bone Marrow Mesenchymal Stem Cells Suppress
Lymphocyte Proliferation In Vitro but Fail to Prevent Graft-versus-Host Disease
in Mice. J Immunol 2006; 176:7761-7767.
SUN HJ, BAHK YY, CHOI YR, SHIM JH, HAN SH, LEE JW. A proteomic
analysis during serial subculture and osteogenic differentiation of human
mesenchymal stem cell. J Orthop Res 2006;24:2059-2071.
TOHILL M, MANTOVANI C, WIBERG M, TERENGHI G. Rat bone marrow
mesenchymal stem cells express glial markers and stimulate nerve
regeneration, Neuroscience Let 2004;362:200-203.
TOMA JG, AKHAVAN M, FERNANDES KJ. Isolation of multipotent adult stem
cells from the dermis of mammalian skin. Nat Cell Biol 2001;3:778–784.
TREMAIN N, KORKKO J, IBBERSON D, KOPEN GC, DIGIROLAMO C,
PHINNEY DG. MicroSAGE analysis of 2,353 expressed genes in a single cell-
derived colony of undifferentiated human mesenchymal stem cells reveals
mRNAs of multiple cell lineages, Stem Cells 2001;19:408-418.
TISATO V, NARESH K, GIRDLESTONE J, NAVARRETE C, DAZZI F.
Mesenchymal stem cells of cord blood origin are effective at preventing but not
treating graft-versus-host disease. Leukemia 2007;21:1992-1999.
TSAI MS, LEE JL, CHANG YJ, HWANG SM. Isolation of human multipotent
mesenchymal stem cells from second-trimester amniotic fluid using a novel two-
stage culture protocol. Hum Reprod 2004 ;19:1450-1456.
TSE WT, PENDLETON JD, BEYER WM, EGALKA MC, GUINAN EC.
Suppression of allogeneic T-cell proliferation by human marrow stromal cells:
implications in transplantation. Transplantation 2003;75:389-397.
UCCELLI A, PISTOIA V, MORETTA L. Mesenchymal stem cells: a new strategy
for immunosuppression? Trends in Immunol, 2007;28: 219-226.
VACANTI V, KONG E, SUZUKI G, SATO K, CANTY JM, LEE T. Phenotypic
changes of adult porcine mesenchymal stem cells induced by prolonged
passaging in culture. J Cell Physiol 2005;205:194-201.

Referência Bibliográficas
VAS V, FAJKA-BOJA R, ION G, DUDICS V, MONOSTORI E, UHER F. Biphasic
effect of recombinant galectin-1 on the growth and death of early hematopoietic
cells. Stem Cells 2005;23:279-287.
VERCELLI A, MEREUTA OM, GARBOSSA D, MURACA G, MARESCHI K,
RUSTICHELLI D, FERRERO I, MAZZINI L, MADON E, FAGIOLI F. Human
mesenchymal stem cell transplantation extends survival, improves motor
performance and decreases neuroinflammation in mouse model of amyotrophic
lateral sclerosis. Neurobiol Dis 2008;31:395-405.
VOGEL W, GRÜNEBACH F, MESSAM CA, KANZ L, BRUGGER W, BÜHRING
HJ. Heterogeneity among human bone marrow-derived mesenchymal stem cells
and neural progenitor cells. Haematologica. 2003;88:126-133.
WAGNER W, WEIN F, RODERBURG C, SAFFRICH R, DIEHLMANN A,
ECKSTEIN V, HO AD. Adhesion of human hematopoietic progenitor cells to
mesenchymal stromal cells involves CD44. Cells Tissues Organs
2008a;188:160-169.
WAGNER W, HORN P, CASTOLDI M, DIEHLMANN A, BORK S, SAFFRICH R,
BENES V, BLAKE J, PFISTER S, ECKSTEIN V, HO AD. Replicative
senescence of mesenchymal stem cells: a continuous and organized process.
PLoS ONE 2008b;3:e2213.
WAGNER W, WEIN F, RODERBURG C, SAFFRICH R, FABER A, KRAUSE U,
SCHUBERT M, BENES V, ECKSTEIN V, MAUL H, HO AD. Adhesion of
hematopoietic progenitor cells to human mesenchymal stem cells as a model for
cell−cell interaction, Exp Hematol 2007;35:314-325.
WAGNER W, FELDMANN RE, WEIN F. The heterogeneity of human
mesenchymal stem cell preparations--evidence from simultaneous analysis of
proteomes and transcriptomes, Exp Hematol 2006;34:536-548.
WAKITANI S, IMOTO K, YAMAMOTO T, SAITO M, MURATA N, YONEDA M.
Human autologous culture expanded bone marrow mesenchymal cell
transplantation for repair of cartilage defects in osteoarthritic knees.
Osteoarthritis Cartilage 2002;10:199-206.
WAKITANI S, SAITO T, CAPLAN AI. Myogenic cells derived from rat bone
marrow mesenchymal stem cells exposed to 5-azactydine, Muscle Nerve
1995;18:57-66.

Referência Bibliográficas
WAKITANI S, GOTO T, PINEDA SJ, YOUNG RG, MANSOUR JM, CAPLAN AI,
GOLDBERG VM. Mesenchymal cell-based repair of large, full-thickness defects
of articular cartilage. J Bone Joint Surg Am 1994;76:579-592.
WANG D, PARK JS, CHU JS, KRAKOWSKI A, LUO K, CHEN DJ, LI S.
Proteomic profiling of bone marrow mesenchymal stem cells upon transforming
growth factor beta1 stimulation. J Biol Chem 2004;279:43725-43734.
WATANABE H, SHIONYU M, KIMURA T, KIMATA K, WATANABE H. Splicing
factor 3b subunit 4 binds BMPR-IA and inhibits osteochondral cell differentiation.
J Biol Chem. 2007;282:20728-20738.
WEXLER SA, DONALDSON C, DENNING-KENDALL P, RICE C, BRADLEY B,
HOWS JM. Adult bone marrow is a rich source of human mesenchymal 'stem'
cells but umbilical cord and mobilized adult blood are not. Br J Haematol
2003;121:368-374.
WIECZOREK G, STEINHOFF C, SCHULZ R, SCHELLER M, VINGRON M,
ROPERS HH, NUBER UA. Gene expression profile of mouse bone marrow
stromal cells determined by cDNA microarray analysis, Cell Tissue Res
2003;311:227-237.
WILSON A, TRUMPP A. Bone-marrow haematopoietic-stem-cell niches. Nat
Rev Immunol 2006;6:93-106.
WOLLERT KC, MEYER GP, LOTZ J, RINGES-LICHTENBERG S, LIPPOLT P,
BREIDENBACH C, FICHTNER S, KORTE T, HORNIG B, MESSINGER D,
ARSENIEV L, HERTENSTEIN B, GANSER A, DREXLER H. Intracoronary
autologous bone-marrow cell transfer after myocardial infarction: the BOOST
randomised controlled clinical trial. Lancet 2004;364:141-148.
WU GD, NOLTA JA, JIN YS, BARR ML, YU H, STARNES VA, CRAMER DV.
Migration of mesenchymal stem cells to heart allografts during chronic rejection.
Transplantation 2003;75:679-685.
XU M, WANI M, DAI YS, WANG J, YAN M, AYUB A, ASHRAF M. Differentiation
of bone marrow stromal cells into the cardiac phenotype requires intercellular
communication with myocytes, Circulation 2004;110:2658-2665.
YOUNG HE, STEELE TA, BRAY RA, HUDSON J, FLOYD JA, HAWKINS K,
THOMAS K, AUSTIN T, EDWARDS C, CUZZOURT J, DUENZL M, LUCAS PA,
BLACK AC JR. Human reserve pluripotent mesenchymal stem cells are present

Referência Bibliográficas
in the connective tissues of skeletal muscle and dermis derived from fetal, adult,
and geriatric donors, Anat Rec 2001;264:51-62.
ZACHAREK A, CHEN J, CUI X, LI A, LI Y, ROBERTS C, FENG Y, GAO Q,
CHOPP M. Angiopoietin1/Tie2 and VEGF/Flk1 induced by MSC treatment
amplifies angiogenesis and vascular stabilization after stroke. J Cereb Blood
Flow Metab 2007;27:1684-1691.
ZANELLI CF, VALENTINI SR. Is there a role for eIF5A in translation? Amino
Acids 2007;33:351-358.
ZANNETTINO AC, HARRISON K, JOYNER CJ, TRIFFITT JT, SIMMONS PJ.
Molecular cloning of the cell surface antigen identified by the osteoprogenitor-
specific monoclonal antibody, HOP-26. J Cell Biochem 2003;89:56-66.
ZAPPIA E, CASAZZA S, PEDEMONTE E, BENVENUTO F, BONANNI I,
GERDONI E, GIUNTI D, CERAVOLO A, CAZZANTI F, FRASSONI F,
MANCARDI G, UCCELLI A. Mesenchymal stem cells ameliorate experimental
autoimmune encephalomyelitis inducing T-cell anergy, Blood 2005;106:1755-
1761.
ZENZMAIER C, GESSLBAUER B, GROBUSCHEK N, JANDROSITZ A,
PREISEGGER KH, KUNGL AJ. Proteomic profiling of human stem cells derived
from umbilical cord blood. Biochem Biophys Res Commun 2005;328:968-972.
ZENZMAIER C, KOLLROSER M, GESSLBAUER B, JANDROSITZ A,
PREISEGGER KH, KUNGL AJ. Preliminary 2-D chromatographic investigation
of the human stem cell proteome. Biochem Biophys Res Commun
2003;310:483-490.
ZHAO CP, ZHANG C, ZHOU SN, XIE YM, WANG YH, HUANG H, SHANG YC,
LI WY, ZHOU C, YU MJ, FENG SW. Human mesenchymal stromal cells
ameliorate the phenotype of SOD1-G93A ALS mice. Cytotherapy 2007;9:414-
426.
ZHENG H, MARTIN JA, DUWAYRI Y, FALCON G, BUCKWALTER JA. Impact
of aging on rat bone marrow-derived stem cell chondrogenesis. J Gerontol A Biol
Sci Med Sci 2007;62:136-148.
ZHOU S, GREENBERGER JS, EPPERLY MW, GOFF JP, ADLER C, LEBOFF
MS, GLOWACKI J. Age-related intrinsic changes in human bone-marrow-

Referência Bibliográficas
derived mesenchymal stem cells and their differentiation to osteoblasts. Aging
Cell 2008;7:335-343.
ZUK PA, ZHU M, MIZUNO H, HUANG J, FUTRELL JW, KATZ AJ, BENHAIM P,
LORENZ HP, HEDRICK MH. Multilineage cells from human adipose tissue:
implications for cell-based therapies, Tissue Eng 2001;7:211-228.

Anexos
ANEXOS
Artigo publicado na revista Cytotherapy
Lazzarotto-Silva C, Binato R, Rocher BD, Costa JA, Pizzatti L, Bouzas LF, Abdelhay E. Similar proteomic profiles of human mesenchymal stromal cells from different donors. Cytotherapy. 2009;11(3):268-77.





























