Embed Size (px)
Citation preview

UNIVERSIDADE DE MOGI DAS CRUZES
DANIELA GARCIA SGAI
GERAÇÃO E CARACTERIZAÇÃO DO CAMUNDONGO
DUPLO NOCAUTE PARA OS GENES DA LEPTINA
E RECEPTOR B1 DE CININAS
Mogi das Cruzes, SP
2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

UNIVERSIDADE DE MOGI DAS CRUZES
DANIELA GARCIA SGAI
GERAÇÃO E CARACTERIZAÇÃO DO CAMUNDONGO
DUPLO NOCAUTE PARA OS GENES DA LEPTINA
E RECEPTOR B1 DE CININAS
Prof. Orientador: Dr. Ronaldo de Carvalho Araújo
Mogi das Cruzes, SP
2007
Dissertação de mestrado apresentada ao programa de pós-graduação da Universidade de Mogi das Cruzes como parte dos requisitos para obtenção do título de Mestre em Biotecnologia.

FINANCIAMENTO
CAPES, FAEP
FICHA CATALOGRÁFICA Universidade de Mogi das Cruzes - Biblioteca Central
Sgai, Daniela Garcia
Geração e caracterização do camundongo duplo nocaute para os genes da leptina e receptor B1 de cininas / Daniela Garcia Sgai. -- 2007.
77 f.
Dissertação (Mestrado em biotecnologia) Universidade de Mogi das Cruzes, 2007
Área de concentração: Ciências biológicas Orientador: Prof. Dr. Ronaldo de Carvalho Araújo
1. Obesidade 2. Diabetes 3. Leptina 4. Receptor B1 5. Medicina experimental I. Título II. Araújo, Ronaldo de Carvalho
CDD 619.93


DEDICATÓRIA
Dedico este trabalho primeiramente a Deus.
À minha família, e especialmente aos meus pais, Wagner e Ivete, por todo o amor,
estrutura emocional e esforços imensuráveis despendidos a mim desde a infância, na
realização dos meus sonhos.
Aos meus avós, Walter e Florentina, José e Conchietta, por toda afeição e carinho com
que participam de minha vida.
Ao novo integrante da família, meu marido e meu grande amor, Evandro, pela
dedicação incondicional, paciência e apoio emocional tão precioso durante todo o
desenvolvimento deste trabalho.
Aos meus amigos que colaboraram direta ou indiretamente para a realização de mais
um sonho.
Meu amor e admiração por todos crescem a cada dia...

"Eu aprendi que todos querem viver no topo da montanha,
mas toda felicidade e crescimento ocorrem quando
você a está escalando.”
(William Shakespeare)

AGRADECIMENTOS
A todos que colaboraram e participaram da realização deste trabalho, seja pelo apoio
técnico, emocional e até mesmo financeiro. Sem isto, nada seria possível...
À minha família e amigos pela dedicação, paciência e apoio incondicionais.
Ao meu orientador, Prof. Dr. Ronaldo de Carvalho Araújo, pelos ensinamentos e
amizade, me orientando da melhor forma possível.
A todos os professores doutores que participaram da minha formação e da realização
deste trabalho, compartilhando conhecimentos valiosos que contribuíram para a sua
conclusão. À Lúcia Garcez do Carmo, João Lúcio de Azevedo, Jorge Luiz Pesquero,
João Bosco Pesquero, Michael Bader, Regina Lúcia Batista da Costa de Oliveira,
Renata Guimarães Moreira, Suma Imura Shimuta, Rui Cury e Ubiratan Machado.
Ao prezado amigo e técnico João Daivison da Silva Ramalho, pela prestatividade,
amizade e suporte nos experimentos.
A todos os colegas do laboratório, minha segunda família durante todo esse período,
pela colaboração e apoio.

RESUMO
A obesidade atualmente é apontada pela Organização Mundial de Saúde (OMS) como epidemia mundial. Sabe-se que é fator de risco para o desenvolvimento de muitas patologias, como síndrome metabólica, distúrbios inflamatórios, doenças cardiovasculares e diabetes mellitus tipo II. A hiperglicemia crônica observada no diabetes é considerada importante no desenvolvimento de retinopatias, doenças cardiovasculares e complicações renais. O sistema calicreína-cininas está envolvido em muitos processos biológicos, como resposta inflamatória, controle da pressão arterial e contratilidade gastrintestinal. Atua na captação de glicose por células musculares e adipócitos, através da translocação do GLUT4. Os animais nocaute para o receptor B1 de cininas foram gerados e mostraram-se resistentes à obesidade induzida por dieta hiperlipídica e mais sensíveis à leptina exógena. Ressaltou-se o envolvimento do receptor B1 na homeostase de glicose e funcionalidade das ilhotas pancreáticas. A leptina é importante na gênese e manutenção da obesidade, atua no processo de estocagem e controle energético, sendo também um eficiente sinalizador nutricional. É anorexígena em nível central e induz lipólise. Para investigação do papel da leptina no organismo, o camundongo ob/ob se torna uma importante ferramenta, por produzir uma leptina truncada. Isto confere ao animal uma obesidade hereditária severa, diabetes mellitus tipo II, número e tamanho de adipócitos elevado, infertilidade, entre outros. Levando em consideração resultados glicêmicos e relacionados à inflamação nos B1 nocaute, além do papel da leptina no apetite e lipólise, o objetivo deste trabalho foi gerar um camundongo deficiente para a leptina e receptor B1 de cininas (ob/ob;B1KO), visando investigar o papel deste na gênese e manutenção da obesidade e do diabetes. Após sucessivos cruzamentos, este foi gerado e caracterizado através da análise do DNA genômico. Observou-se ausência de atividade contrátil do estômago nos camundongos ob/ob;B1KO em banho de contração com o agonista do receptor B1. Como alternativa na obtenção dos animais, foi realizado o transplante de tecido adiposo branco, e os camundongos ob/ob tiveram sua fertilidade restabelecida dois meses após a cirurgia. Aos quatro meses de vida, os animais ob/ob;B1KO apresentaram menor massa corporal, comparados aos ob/ob selvagens (ob/ob;WT). Não foram observadas diferenças entre a ingestão dos grupos. Após eutanásia, vários órgãos foram retirados. Foi observada maior massa relativa de tecido adiposo inguinal e menor massa dos rins, coração e ventrículo esquerdo. A análise histológica mostrou menor diâmetro das fibras musculares do ventrículo esquerdo, indicando uma proteção parcial contra a hipertrofia cardíaca comumente observada nos animais ob/ob. No teste de tolerância à glicose, exibiram maior captação de glicose, não havendo diferença no teste de sensibilidade à insulina. Mostraram-se também mais sensíveis à leptina exógena, provavelmente por mecanismo central. Apesar do receptor B1 de cininas não ter se mostrado relacionado à gênese da obesidade mórbida com deficiência de leptina, nossos resultados sugerem seu envolvimento na patogênese de doenças associadas, como o diabetes tipo II e distúrbios cardiovasculares, além de um papel na sensibilidade à leptina. Palavras-chave: Sistema calicreína-cininas, leptina, obesidade, diabetes.

ABSTRACT
Obesity is classified by World Health Organization (WHO) as a worldwide epidemic problem. It is known that is a risk factor to various pathologies, as metabolic syndrome, inflammatory dysfunction, cardiovascular diseases and type 2 diabetes mellitus. The cronic hyperglicemia observed in diabetes is considered important to development of retinopathies, cardiovascular diseases and renal complications. The kallikrein-kinin system is involved in many biological processes, as inflammatory response, blood pressure control and gastrintestinal motility. It works on glucose uptake in muscle cells and adipocytes by GLUT4 translocation. Knockout animals for kinin B1 receptor (B1KO) has been generated. They were resistants to hyperlipidic diet-induced obesity and showed more sensibility to exogenous leptin. It has been demonstrated that B1 receptor is involved in glucose homeostasis and pancreatic islets function. Leptin is important in genesis and manteinance of obesity, acts on storage process and energetic balance, being also an efficient nutritional signaller. It is anorexigenic centrally and induces lipolisis. In order to investigate the role of leptin in the organism, the ob/ob mouse becomes an important tool because it sintetizes a defective leptin. It conferes to the animal an hereditary morbid obesity, type 2 diabetes mellitus, elevated number and size of adipocytes, infertility, and another ones. Considering glycemic results and the ones related to inflammation in B1KO mice, and even the role of leptin in apetite and lipolisis, the objective of this study was to generate a leptin and B1 receptor deficient (ob/ob;B1KO) mouse, aiming to investigate the role of this receptor on the genesis and manteinance of obesity and diabetes. After successive crosses, the ob/ob;B1KO mouse has been generated and characterized by genomic DNA analyses. It was observed no contractile activity in stomach of the ob/ob;B1KO mice when incubated with B1 receptor agonist in contraction bath. It has been done a transplant of white adipose tissue as an alternative method of obtaining the animals and, after two months of surgery, the ob/ob animals have got their fertility restored. The ob/ob;B1KO animals presented lower body mass, compared to the the wild ob/ob (ob/ob;B1WT), four months after birth. It was not observed difference between the ingestion of the two groups. After euthanasia, many organs were extracted. It was observed higher relative mass of inguinal adipose tissue, and a lower mass of kidneys, heart and left ventricle. The hystologic analysis showed muscle fibers in left ventricle with lower diameter in the ob/ob;B1KO ones, indicating a parcial protection against the cardiac hypertrophy, which is normally observed in the ob/ob animals. In glucose tolerance test, the ob/ob;B1KO showed an improvement in glucose uptake, and there were no differences in insulin tolerance test. They also presented a higher sensibility to exogenous leptin, probably because of a central mechanism. Despite the kinin B1 receptor has not been shown to be related to the genesis of morbid obesity with leptin deficiency, our results suggest its envolvement with pathogenesis of associated diseases, as type 2 diabetes and cardiovascular dysfunctions, and also a role in the leptin sensibility.
Keywords: Kallikrein-kinin system, leptin, obesity, diabetes.

LISTA DE ILUSTRAÇÕES
Figura 1 Vias de sinalização da insulina . ....................................................18 Figura 2 Representação esquemática da relação entre excesso de adiposidade visceral e desenvolvimento de doenças cardiovasculares e diabetes mellitus tipo II....................................20 Figura 3 Representação esquemática do sistema calicreína-cininas. ..........26 Figura 4 Eletroforese do fragmento do gene OB amplificado por PCR ........45 Figura 5 Representação dos sítios de reconhecimento da enzima de
restrição DdeI, com a presença ou ausência da mutação no fragmento do gene OB amplificado por PCR................................45
Figura 6 Eletroforese do fragmento do gene OB amplificado por PCR e submetido à digestão com a enzima de restrição DdeI ...............47 Figura 7 Eletroforese dos fragmentos do gene para o receptor B1 de cininas (A) e do cassete de neomicina (B), amplificados por PCR ..............................................................................................48 Figura 8 Registros de resposta contrátil de fundus de estômago de camundongos ob/ob;B1KO e ob/ob;B1WT (in vitro), frente aos agonistas dos receptores B1 e B2 de cininas..........................49 Figura 9 Gráfico de resposta contrátil de fundus de estômago de camundongos ob/ob;B1KO e ob/ob;B1KO (in vitro), frente aos agonistas dos receptores B1 e B2 de cininas..........................50 Figura 10 Foto de uma fêmea ob/ob;B1KO submetida ao transplante de tecido adiposo aos dois meses de vida. (A) Antes ; e (B) dois meses após a intervenção cirúrgica .................................51 Figura 11 Gráfico de acompanhamento da massa corporal de camundongos ob/ob;B1KO e ob/ob;B1WT, do primeiro ao quarto mês de vida........................................................................52 Figura 12 Gráfico de acompanhamento de ingestão relativa diária
média (g /g de peso vivo) dos camundongos ob/ob;B1KO e ob/ob;B1WT ..................................................................................52

Figura 13 Gráfico de glicemia em jejum dos camundongos ob/ob;B1KO e ob/ob;B1WT aos 2 meses de vida .........................53 Figura 14 Teste de tolerância à glicose nos camundongos ob/ob;B1KO e ob/ob;B1WT aos 3 meses de vida .........................53 Figura 15 Teste de sensibilidade à insulina nos camundongos ob/ob;B1KO e ob/ob;B1WT aos 3 meses e meio de vida .............54 Figura 16 Gráfico da concentração sérica de insulina basal dos camundongos ob/ob;B1KO e ob/ob;B1WT aos quatro meses e meio de vida....................................................................54 Figura 17 Teste de sensibilidade à leptina exógena. Gráfico de acompanhamento diário da ingestão alimentar média (A) e massa corporal média (B) dos camundongos ob/ob;B1KO e ob/ob;B1WT aos quatro meses de vida, durante cinco dias consecutivos à administração da leptina ................................55 Figura 18 Gráfico de massa relativa de órgãos extraídos dos camundongos ob/ob;B1KO e ob/ob;B1WT. (A) Tecido adiposo inguinal ; (B) rins ; (C) coração ; e (D) ventrículo esquerdo.......................................................................................56 Figura 19 Microscopia (400X) de lâminas histológicas de ventrículo esquerdo dos camundongos ob/ob;B1KO e ob/ob;B1WT............57

LISTA DE ABREVIATURAS E SIGLAS
% Porcentagem
ηg/µL Nanograma(s) por microlitro
°C Graus centígrados
µg Micrograma(s)
µL Microlitro(s)
B1+/- Camundongo heterozigoto para o receptor B1 de cininas
B1KO Camundongo nocaute para o gene do receptor B1 de cininas
B1WT Camundongo selvagem para o gene do receptor B1 de cininas
BK Bradicinina
CO2 Dióxido de carbono
DBK Des-Arg9-bradicinina
DdeI Enzima de restrição isolada de Desulfovibrio desulfuricans
DMID Diabetes mellitus insulino-dependente ou diabetes mellitus tipo I
DMNID Diabetes mellitus não insulino-dependente ou diabetes mellitus tipo II
DNA Ácido desoxiribonucléico
HDL Lipoproteína de alta densidade
i.p. intra-peritoneal
IL-1β Interleucina - 1 beta
IL-6 Interleucina - 6
IMC Índice de massa corporal
IRS Substrato do receptor de insulina

Kg/m² Quilograma(s) por metro quadrado
LBK Calidina ou lisil-bradicinina
mg/dL Miligrama(s) por decilitro
mg/mL Miligrama(s) por mililitro
MgCl2 Cloreto de magnésio
min Minuto(s)
mL Mililitro(s)
mm Milímetro(s)
mM milimolar
n Número de camundongos utilizados por grupo
NaCl Cloreto de sódio
NO Óxido Nítrico
O2 Oxigênio gasoso
OB Gene da leptina
OB/OB Camundongo magro, selvagem para a mutação do gene da leptina
ob/ob Camundongo obeso, homozigoto para a mutação do gene da leptina
OB/ob Camundongo magro, heterozigoto para a mutação do gene da leptina
ob/ob;B1KO Camundongo duplo nocaute para os genes da leptina e receptor B1
de cininas
ob/ob;B1WT Camundongo nocaute para o gene da leptina e selvagem para o
gene do receptor B1 de cininas, utilizado como controle do animal
duplo nocaute
OMS Organização Mundial da Saúde
pb Pares de bases nitrogenadas

PCR Reação de polimerase em cadeia
pH Potencial hidrogeniônico
RNA Ácido ribonucléico
rpm Rotações por minuto
s Segundo(s)
SCC Sistema calicreína-cininas
TNF-α Fator de necrose tumoral - alfa
TTG Teste de tolerância à glicose
U/µL Unidade(s) arbitrária(s) por microlitro

SUMÁRIO
1 INTRODUÇÃO .................................................................................. 15
1.1 Obesidade ..............................................................................................15
1.2 Diabetes Mellitus ...................................................................................17
1.2.1 Diabetes Mellitus Tipo I ...............................................................19
1.2.2 Diabetes Mellitus Tipo II ..............................................................19
1.3 Leptina ...................................................................................................22
1.3.1 Camundongo ob/ob.....................................................................23
1.4 Sistema Calicreína-cininas ...................................................................24
1.4.1 Camundongos Nocaute para o Receptor B2 de Cininas (B2KO).........................................................................................29
1.4.2 Camundongos Nocaute para os Receptores B1 e B2 de Cininas (B1B2KO) .......................................................................30
1.4.3 Camundongos Nocaute para o Receptor B1 de Cininas (B1KO)........................................................................................30
2 OBJETIVOS...................................................................................... 32
3 MÉTODO........................................................................................... 33
3.1 Geração do Camundongo ob/ob;B1KO................................................33
3.2 Genotipagem .........................................................................................34
3.2.1 Extração do DNA Genômico.......................................................34
3.2.2 Amplificação do Gene OB...........................................................35
3.2.3 Amplificação do Gene do Receptor B1 de Cininas e Cassete de Neomicina................................................................37
3.3 Análise Funcional do Receptor B1 de Cininas ....................................38

3.4 Transplante de Tecido Adiposo Branco...............................................39
3.5 Controle de Massa Corporal .................................................................39
3.6 Controle de Ingestão Alimentar............................................................40
3.7 Glicemia em Jejum ................................................................................40
3.8 Teste de Tolerância à Glicose...............................................................40
3.9 Teste de Sensibilidade à Insulina.........................................................41
3.10 Teste de Sensibilidade à Leptina..........................................................41
3.11 Testes Bioquímicos...............................................................................42
3.12 Eutanásia dos Animais .........................................................................42
3.12.1 Análise Histológica .....................................................................43
4 RESULTADOS.................................................................................. 44
5 DISCUSSÃO ..................................................................................... 58
6 CONCLUSÕES E SUGESTÕES....................................................... 64
REFERÊNCIAS..................................................................................... 65
ANEXO A .............................................................................................. 76

15
1 INTRODUÇÃO
1.1 Obesidade
Atualmente, a obesidade apresenta-se como um grave problema de saúde
pública, sendo inclusive apontada pela Organização Mundial da Saúde (OMS) como
uma epidemia mundial (YORK et al., 2004; POIRIER et al., 2006). Sua prevalência
na população vem crescendo acentuadamente nas últimas décadas, inclusive nos
países em desenvolvimento (ECKEL et al., 2004).
É uma doença multifatorial que já acomete quase um terço da população
mundial. Além do fator hereditário, seu desenvolvimento está intimamente ligado a
fatores comportamentais, como sedentarismo e hábitos alimentares desregrados
(NOBRE et al., 2006).
Basicamente, quando a ingestão calórica supera o gasto energético, ocorre
um desbalanço energético e ganho de peso, e conseqüentemente, todas as
complicações associadas à obesidade são desencadeadas (SCHUTZ, 1995;
WEYER et al., 2000).
Uma das metodologias mais difundidas na avaliação do grau de obesidade é
o cálculo do índice de massa corporal (IMC) do indivíduo, obtido pela fração do peso
(em quilogramas) pela altura (em metros) ao quadrado (MULLIS et al., 2004).
Considera-se ideal um IMC entre 20 e 24,9Kg/m². O sobrebeso é caracterizado entre
25 e 29,9Kg/m² e a obesidade mórbida, a partir de 40Kg/m² (WILSON et al., 2007).
Inúmeros estudos demonstram que a obesidade com acúmulo de tecido
adiposo visceral, caracterizada pela deposição predominante de tecido adiposo na
região do abdômen, é a que representa maior fator de risco para doenças

16
cardiovasculares, estando também envolvida no desenvolvimento da síndrome
metabólica (EMPANA et al., 2004; BERGMAN et al., 2007).
A síndrome metabólica é uma associação de fatores de risco importante na
predisposição do indivíduo à morbidade ou mortalidade de origem cardiovascular. As
principais alterações observadas são: intolerância à glicose, obesidade, aumento da
pressão arterial, elevação do triglicérides e baixos níveis séricos de HDL (DAY,
2007).
Dentre os distúrbios mais freqüentes observados na associação deste tipo de
obesidade e esta síndrome, estão as dislipidemias, aumento dos níveis plasmáticos
de ácidos graxos livres, hipertensão, resistência à insulina, hiperinsulinemia,
diabetes mellitus tipo II, inflamações e trombose (POIRIER et al., 2006; BAHIA et al.,
2006; BERGMAN et al., 2007; WASSINK et al., 2007).
O acúmulo de adiposidade visceral promove um aumento de ácidos graxos,
triglicérides e colesterol circulantes; e de citocinas inflamatórias, como interleucina -
1β (IL-1β), proteína C reativa e fator de necrose tumoral - α (TNF-α). Promove
também hiperglicemia, hiperinsulinemia e aumento de resistina plasmática. Além
disso, pode levar à diminuição de adiponectina, responsável pela proteção das
paredes arteriais, e do tipo de colesterol HDL, uma lipoproteína de alta densidade
envolvida na manutenção da luz vascular (WASSINK et al., 2007). Essa cascata
desencadeada pode levar ao desenvolvimento da aterosclerose e outras doenças
cardiovasculares, além da resistência à insulina e diabetes mellitus tipo II
(CATERSON et al., 2004; WASSINK et al., 2007).

17
1.2 Diabetes Mellitus
O diabetes é uma doença que acomete mais de 170 milhões de pessoas no
mundo (FOX et al., 2006).
A hiperglicemia, característica desta patologia, ocorre devido a uma
deficiência na produção de insulina ou resistência à mesma (WITHERS et al., 1998).
A insulina é secretada pelas células β-pancreáticas e desenvolve um papel
importante na homeostase de glicose, modulando o metabolismo de carboidratos,
lipídios e proteínas por intermédio de suas vias de sinalização intracelular.
O receptor de insulina pertence à família dos receptores de membrana com
atividade da tirosina quinase. Uma vez ativado pela sua ligação específica com a
molécula de insulina, desencadeia a fosforilação de proteínas intracelulares, como
os IRSs e Src, além de Shc ou ainda Gab-1.
Conseqüentemente, vários efetores protéicos intracelulares são recrutados e
promovem a ativação de vias de sinalização intracelular, como as vias de
fosfatidilinositol 3-quinase (PI3-K) e MAP quinase, levando a uma variedade de
efeitos biológicos. Dentre eles, estão o transporte de glicose para o meio intracelular
via recrutamento da proteína transportadora GLUT4, síntese de glicogênio,
lipogênese ou glicólise, síntese de proteínas, bem como atuação no crescimento
celular (CARVALHEIRA et al., 2002). (Figura 1).
A principal ação metabólica deste receptor ocorre em músculos, fígado e
tecido adiposo (BAUDRY et al., 2002; CARVALHEIRA et al., 2002; RHODES e
WHITE, 2002; BARROSO et al., 2003).
A hiperglicemia crônica é considerada o principal fator no desenvolvimento de
retinopatias, doenças cardiovasculares e complicações renais associadas (BAUDRY

18
et al., 2002). Estes dados contribuem para o diabetes ser uma das doenças mais
pesquisadas atualmente no mundo.
Vários estudos têm sido realizados no intuito de determinar suas bases
fisiopatológicas, almejando desvendar um tratamento mais eficaz e até mesmo uma
possível cura.
Figura 1: Vias de sinalização da insulina (CARVALHEIRA et al., 2002).
É classificada basicamente em dois subtipos: diabetes mellitus insulino-
dependente (DMID) ou tipo I, e diabetes mellitus não insulino-dependente (DMNID)
ou tipo II.

19
1.2.1 Diabetes Mellitus Tipo I
O DMID é responsável por aproximadamente 10% dos casos de diabetes,
sendo de manifestação precoce. Ocorre principalmente em crianças e jovens.
Fatores de risco determinantes nesses casos são mães com diabetes mellitus tipo I,
com idade mais avançada ou mesmo que desenvolveram pré-eclampsia durante a
gravidez (MCKINNEY et al., 1999). Pode decorrer também de ação medicamentosa
(VANTYGHEM e PRESS, 2006). É o tipo de diabetes mais agressivo, visto que a
disfunção na produção de insulina pelas células β-pancreáticas ocorre devido à sua
destruição pelo próprio sistema imune. Esta deficiência na produção da insulina
torna o indivíduo dependente de insulina exógena.
1.2.2 Diabetes Mellitus Tipo II
O DMNID é uma doença poligênica complexa, caracterizada principalmente
por resistência à insulina e disfunção das células β-pancreáticas (DE MEYTS, 1993;
KAHN, 1994; SACKS et al., 1996; BARROSO et al., 2003).
Observada em aproximadamente 90% dos pacientes com diabetes, está
ligada a padrões hereditários complexos, sendo manifestada tardiamente. Fatores
comportamentais, como sedentarismo e hábitos alimentares pouco saudáveis,
também podem contribuir com o seu desenvolvimento (BAUDRY et al., 2002). Na
maioria dos pacientes, vem acompanhada de obesidade com acúmulo de
adiposidade visceral, compartilhando os fatores de risco (CUEVAS-ALVAREZ et al.,

20
2006; WILSON et al., 2007). Associada à obesidade, sua incidência tem aumentado
progressivamente, a exemplo dos Estados Unidos da América (FOX et al., 2006).
A obesidade e a resistência à insulina estão associadas aos fatores de risco
de doenças cardiovasculares, assim como se relacionam com os níveis alterados de
marcadores inflamatórios e disfunção endotelial. (WEYER et al., 2002)
Muitos estudos sobre obesidade têm ressaltado o papel do tecido adiposo
como um órgão endócrino, responsável pela secreção de inúmeros mediadores de
complicações vasculares e metabólicas observadas (BAHIA et al., 2006). (Figura 2)
Figura 2: Representação esquemática da relação entre excesso de adiposidade visceral e desenvolvimento de doenças cardiovasculares e diabetes mellitus tipo II .
Dois hormônios secretados por adipócitos foram recentemente identificados:
adiponectina e resistina. Estão envolvidos na resistência à insulina e risco
cardiovascular ligados à obesidade (STEPPAN et al., 2001; WEYER et al., 2002;
SHETTY et al., 2004).

21
A adiponectina é encontrada em altas concentrações na circulação periférica
(TSAO et al., 2002), e seus níveis circulantes estão extremamente diminuídos na
obesidade, diabetes tipo II e resistência à insulina (ARITA et al., 1999; WEYER et al.,
2001). Está associada a marcadores inflamatórios, como a proteína C reativa e o
fator de necrose tumoral α (TNF-α) (KRAKOFF et al., 2003; OUCHI et al., 2003),
atuando na supressão de respostas inflamatórias (YOKOTA et al., 2000).
Correlaciona-se inversamente com inflamação e disfunção endotelial (KRAKOFF et
al., 2003).
Pode estar envolvida na proteção vascular, sendo que altos níveis séricos
estão relacionados a um menor risco coronariano em homens com ou sem diabetes
(PISCHON et al., 2004; SCHULZE et al., 2005)
Desempenha também importante papel na sensibilidade à insulina (KUBOTA
et al., 2002; LINDSAY et al., 2002), aterogênese (OUCHI et al., 1999; OUCHI et al.,
2001), metabolismo de lipídeos e hiperlipidemia (KUBOTA et al., 2002;
MATSUBARA et al., 2002).
Já a resistina foi recentemente apontada como atuante na resistência à
insulina associada à obesidade (STEPPAN et al., 2001). Possui estrutura similar às
citocinas pró-inflamatórias, como a interleucina-6 (IL-6) e TNF-α, ambas envolvidas
em eventos de disfunções cardiovasculares relacionadas à obesidade. No entanto, a
associação da resistina com os marcadores inflamatórios parece ser independente
do IMC, sugerindo que tenha um papel pró-inflamatório direto ou que module seus
efeitos por mecanismos ainda desconhecidos (VGONTZAS et al., 2000).
A leptina é um outro hormônio que parece ter envolvimento importante com a
síndrome metabólica observada no DMNID. Múltiplos efeitos periféricos deste
hormônio têm sido descritos, incluindo a modulação da secreção de insulina pelas

22
células β-pancreáticas, modulação da sua ação e controle do metabolismo
energético em células adiposas e musculares esqueléticas (HOUSEKNECHT et al.,
1998).
O estágio inicial da doença se caracteriza por hiperinsulinemia decorrente da
resistência apresentada, não havendo dependência de insulina exógena. Com sua
evolução, pode ocorrer uma deficiência da produção de insulina pelas células β-
pancreáticas, iniciando um processo de desenvolvimento do DMID (BAUDRY et al.,
2002).
1.3 Leptina
A leptina é um hormônio expresso principalmente em tecido adiposo e, em
níveis mais baixos, em epitélio gástrico e placenta (ZHANG et al., 1994; MASUZAKI
et al., 1997).
Juntamente ao seu receptor, é muito importante no processo de estocagem e
manutenção energética, sendo também um importante sinalizador nutricional para
muitos outros sistemas fisiológicos, modulando suas funções. Em nível central, é
anorexígena e induz aumento de lipólise.
Seus níveis circulantes estão ligados à quantidade de tecido adiposo presente
no organismo (FRIEDMAN et al., 1998), porém, podem estar relacionados também à
resistência à leptina (MAFFEI et al., 1995; HAMILTON et al., 1995). Normalmente, os
casos de obesidade mórbida observados em humanos são decorrentes dessa
resistência central à leptina, mais especificamente por uma dificuldade de passagem
pela barreira hematoencefálica (CONSIDINE et al., 1995; CARLSSON et al., 1997).

23
Considerada também uma citocina, é estimulada por processos inflamatórios
e tem importante função na hematopoiese (FAGGIONI et al., 2001).
Perifericamente, tem papel importante no metabolismo ósseo, inibindo o
crescimento desse tecido (DUCY et al., 2000). Esta propriedade tem sido
demonstrada por vários autores, como Elefteriou et al. (2004), que concluíram que
baixos níveis de leptina aumentam a densidade óssea e vice-versa; e Hamrick et al.
(2004), que utilizaram um modelo animal obeso e naturalmente nocaute para a
leptina, o camundongo ob/ob, verificando aumento da composição mineral óssea
normal nas vértebras e diminuição no fêmur.
1.3.1 Camundongo ob/ob
Para melhor investigação do papel da leptina no desenvolvimento de
obesidade e manutenção energética, o camundongo ob/ob se torna uma importante
ferramenta, visto que sintetiza uma leptina truncada, sem atividade biológica. Isso
decorre de uma mutação autossômica recessiva espontânea do gene OB,
cromossomo 6, na qual ocorre a substituição do códon para o aminoácido arginina
por um códon de parada, interrompendo a síntese da proteína (ZHANG et al., 1994).
A deficiência da leptina confere ao animal uma obesidade hereditária severa,
diabetes mellitus com resistência à insulina, número e tamanho de adipócitos
superior em relação aos camundongos selvagens (JOHNSON, 1972), e outras
anormalidades, como infertilidade, termorregulação alterada, hipercortisolemia
(COLEMAN et al., 1978), hiperglicemia e hiperinsulinemia, além de uma imaturidade

24
do eixo hipotalâmico-pituitário (SWERDLOFF et al., 1976) e de grande incidência de
hipertrofia cardíaca (BAROUCH et al., 2003).
Para a surpresa de vários pesquisadores o camundongo ob/ob apresentou
uma pressão arterial menor do que seus irmãos magros (MARK et al., 1999). Os
mecanismos envolvidos neste fenômeno, como o aumento de substâncias
vasodilatadoras ou mesmo a diminuição de peptídeos vasoconstritores, ainda não
foram investigados e podem abrir caminhos para o entendimento da hipertensão
associada à obesidade.
Abe et al. (2007) demonstraram também que os animais ob/ob apresentam
modificações de expressão do receptor B1 e B2 em vários tecidos relacionados com
a ingestão alimentar e gasto energético.
Em experimento recente, o transplante de tecido adiposo normal nesses
animais parece tê-los protegido contra a obesidade e restabelecido a sensibilidade à
insulina e sua fertilidade (KLEBANOV et al., 2005). Apesar de ser uma mutação rara
em humanos, os quais apresentam resistência à leptina na barreira
hematoencefálica (CONSIDINE et al., 1995; CARLSSON et al., 1997), este modelo é
muito utilizado nos estudos experimentais, visto que reproduz muitos distúrbios
observados em humanos com resistência à leptina (MAFFEI et al., 1995,
CONSIDINE et al., 1996).
1.4 Sistema Calicreína-cininas
O sistema calicreína-cininas (SCC) está envolvido em muitos processos
biológicos, dentre os quais os mecanismos de resposta inflamatória (BHOOLA et al.,

25
1992), nocicepção e transmissão de dor (DRAY e PERKINS, 1993), controle da
pressão arterial (SCHÖLKENS, 1996), hipertrofia cardíaca, fluxo local de sangue,
transporte de eletrólitos e glicose, e proliferação celular (BHOOLA et al., 1992),
atuando também no trato gastrintestinal (ALLOGHO et al., 1998). Além disso, está
presente no processo de captação de glicose por células musculares e adipócitos,
por meio da translocação do seu principal carreador de glicose (GLUT4), para a
membrana citoplasmática, o que seria facilitado pela ação da bradicinina sobre os
receptores B2 de cininas (KISHI et al., 1998).
De um modo geral, esse sistema é composto por: a) substratos glicoprotéicos
precursores (cininogênios), envolvidos na cascata de coagulação e inflamação; b)
enzimas proteolíticas da família das serino-proteases (calicreínas tecidual e
plasmática), presentes em células glandulares, neutrófilos e fluidos biológicos; c)
peptídeos vasoativos (cininas), provenientes da hidrólise dos cininogênios pelas
calicreínas; d) dois receptores transmembrânicos específicos (B1 e B2), acoplados à
proteína G (REGOLI, 1990; BHOOLA et al., 1992); e) enzimas que promovem
rapidamente o catabolismo das cininas (cininases), de 10 a 50 segundos após sua
formação, liberando os agonistas do receptor B1 ou peptídeos inativos. Este sistema
está representado de forma esquemática na figura 3.
A ação das calicreínas tecidual e plasmática em humanos libera os peptídeos
vasoativos calidina (LBK) e bradicinina (BK), respectivamente. O primeiro, gerado
pela ação sobre o cininogênio de baixo peso molecular; e o segundo, sobre o
cininogênio de alto peso molecular (BHOOLA et al, 1992).
As carboxipeptidases N e M, em especial, têm grande importância no SCC
por clivarem a arginina C-terminal da LBK e BK (agonistas do receptor B2) e
gerarem os agonistas do receptor B1, des-Arg10-calidina e des-Arg9-BK,

26
respectivamente. O metabolismo destas cininas geradas é diferente das primeiras
nos seguintes aspectos: as des-Arg-cininas não sofrem a ação das arginino-
carboxipeptidases, a ECA tem afinidade muito menor por elas, e a posição Phe8 na
porção C-terminal parece protegê-las da degradação por neprilisina (MARCEAU et
al., 1998).
Figura 3: Representação esquemática do sistema calicreína-cininas.
As cininas agem basicamente por ativação dos receptores B1 e B2.
O subtipo B2 é constitutivamente expresso, sendo predominante em células
endoteliais, e mostrando-se envolvido principalmente no controle da pressão arterial,
Cininogênio
Calicreína
Bradicinina Receptor B2
Cininases
ECA Arginino-carboxipeptidases
Des-Arg-Cininas Peptídeos
Inativos
Peptídeos
Inativos
Receptor B1

27
mediando resposta vasodilatadora perifericamente, e vasoconstritora em ventrículos
cerebrais (LINDSEY et al., 1988), também regulando a excreção de sais e água do
organismo (MUKAI et al., 1996). Destaca-se também na captação de glicose
independente de insulina (KISHI et al., 1998).
Recentemente, Christopher et al. (2001), demonstraram o aumento da
expressão do receptor B2 de bradicinina nas células musculares lisas vasculares em
hiperglicemia diabética. A glicose induz a ativação de proteína quinase C, que seria
mediadora da regulação positiva desses receptores, talvez podendo explicar as
alterações vasculares típicas em diabetes.
O receptor B1, descrito em musculatura lisa e fibroblastos de várias espécies,
apresenta-se pouco expresso em condições normais, porém sua expressão é
ativada em respostas inflamatórias (BHOOLA et al., 1992). É expresso
constitutivamente apenas em alguns poucos vasos sangüíneos (BURCH e KYLE,
1992).
Os mecanismos de segundos mensageiros envolvidos na ativação dos
receptores B1 e B2 de cininas vêm sendo também amplamente estudados, devido a
sua possível ligação com muitos dos efeitos metabólicos mediados por eles.
Sabe-se que ambos os receptores têm a capacidade de induzir aumento de
cálcio intracelular em músculo liso (MATHIS et al., 1996), podendo estar envolvidos
na liberação de óxido nítrico (NO) a partir do aminoácido L-arginina via cálcio-
dependente (MARCEAU, 1995). Destaca-se o papel do NO em muitos processos,
como no controle da contratilidade vascular (THIEMERMANN, 1997), hipertensão e
contratilidade cardíaca (MONCADA et al., 1991), reações imunológicas e agregação
de plaquetas (THIEMERMANN, 1997), processos infecciosos, e regulação da

28
ingestão alimentar e esvaziamento gástrico em modelo animal de obesidade
(MORLEY e FLOOD, 1991).
A recente clonagem do receptor B1 de cininas de coelho (MACNEIL et al.,
1995) e de camundongo (PESQUERO et al., 1996) permitiu a realização de uma
série de estudos acerca do papel deste receptor no metabolismo. Experimentos
executados no laboratório do Dr. João Bosco Pesquero, UNIFESP, utilizando-se
cultura de pré-adipócitos (linhagem de células 3T3L1) tratados com agonistas do
receptor B1, mostram que as cininas devem ter papel primordial na diferenciação
das células adiposas. Estas células se diferenciaram mais rapidamente do que
aquelas não tratadas.
Tanto Pesquero et al. (2000) como Araújo et al. (2001) demonstraram o
envolvimento do receptor B1 de cininas em respostas inflamatórias.
Além disso, em 2006, Araújo et al. observaram a participação deste na
homeostase de glicose e funcionalidade das ilhotas pancreáticas, assim como uma
maior sensibilidade à leptina exógena pelos animais nocaute para este receptor.
Tendo-se em vista a grande abrangência do SCC quanto aos processos
fisiológicos já descritos em que está envolvido, animais nocaute para os receptores
de cininas têm sido gerados no intuito de investigar e estabelecer sua participação
no metabolismo e em variados processos patológicos. Esta nova metodologia
desenvolvida permitiu a avaliação do papel dos receptores de cininas sem a
necessidade de intervenção farmacológica para supressão dos seus genes
(PESQUERO e BADER, 2006).

29
1.4.1 Camundongos Nocaute para o Receptor B2 de Cininas (B2KO)
O camundongo deficiente para o receptor B2 de cininas foi gerado por
Borkowski et al., em 1995. Este animal não apresentou comprometimento da
fertilidade.
O camundongo deficiente para o receptor B2 de cininas é hiperglicêmico e
apresenta resistência à insulina, comparado aos animais selvagens (DUKA et al.,
2001). Esses dados condizem com Bhoola et al. (1992), que demonstraram a
capacidade do receptor B2 de induzir a captação de glicose.
Madeddu et al. (1997) indicaram um quadro hipertensivo nesses animais.
Entretanto, Milia et al. (2001) mostraram que não há diferença significante na
pressão arterial. Todavia, os animais B2KO apresentaram um aumento da pressão
arterial quando submetidos à alta ingestão de sal (ALFIE et al., 1997).
Apresenta hipertrofia cardíaca, especialmente do ventrículo esquerdo,
sugerindo o envolvimento do receptor B2 de cininas no crescimento e remodelação
cardíaca (EMANUELI et al., 1999).
Resultados do nosso grupo mostram que estes animais são resistentes à
dieta hiperlipídica, e utilizam preferencialmente gorduras. Ainda foi observado um
quadro hiperglicêmico, além de uma menor captação de glicose. Apresentaram
maior teor de proteína e menor quantidade de tecido adiposo. Esse fato deve-se,
provavelmente, à maior massa muscular e à hipertrofia muscular observadas.
Sabe-se que o receptor B2 está envolvido no controle da pressão arterial,
homeostase da glicose, crescimento, remodelação cardíaca, além de ser importante
na excreção de sal (BHOOLA et al, 1992). Contudo, o seu papel na patogenia da
obesidade mórbida e na síndrome metabólica ainda não foi descrito.

30
1.4.2 Camundongos Nocaute para os Receptores B1 e B2 de
Cininas (B1B2KO)
Em 2006, o grupo do Dr. Michael Bader, do MDC da Alemanha, gerou o
camundongo duplo nocaute para os genes dos receptores B1 e B2 de cininas. Os
animais não apresentaram anomalias óbvias, hipertensão ou infertilidade.
Entretanto, assim como os animais B1KO, são resistentes à hipotensão induzida
pelo LPS. (Artigo submetido ao Journal of Biological Chemistry)
1.4.3 Camundongos Nocaute para o Receptor B1 de Cininas (B1KO)
Em 2000, Pesquero et al. geraram o camundongo transgênico nocaute para o
receptor B1 de cininas (B1KO). Esses animais não apresentaram nenhuma
disfunção reprodutiva, sendo sadios e férteis. Não foram observadas disfunções
cardiovasculares, sendo os animais normotensos e sem lesões cardíacas.
Mostraram-se ainda resistentes à hipotensão mediada pelo lipopolissacarídeo de
bactérias gram-negativas (LPS).
Visando verificar o papel desse receptor em processos inflamatórios, um outro
estudo foi realizado com estes camundongos. Para tanto, uma peritonite foi induzida,
sendo observada nesses animais menor migração de neutrófilos para os focos de
inflamação e menor índice de apoptose espontânea dos neutrófilos (ARAÚJO et al.,
2001).
Foi demonstrado também que a deficiência do receptor B1 de cininas nos
vasos pancreáticos pode ser responsável pela menor produção de insulina

31
observada no pâncreas, e conseqüentemente, pela hiperglicemia constatada
(ARAÚJO et al., 2006).
Em nosso laboratório, resultados anteriores mostraram que esses animais
apresentam valores muito baixos de leptina circulante, bem como uma maior
sensibilidade à leptina exógena (dados não publicados).
Além disso, foi realizado um tratamento crônico com dieta hiperlipídica nos
animais B1KO e B1 selvagens (B1WT). Comparando-se camundongos B1KO com
B1WT, tanto no controle quanto no tratamento hiperlipídico, a dosagem de insulina
mostrou-se inalterada, apesar de ambos os modelos tratados apresentarem níveis
mais elevados em relação aos modelos com ração controle. Os animais B1KO
também apresentaram menor elevação dos níveis séricos de leptina e maior
tolerância à glicose após o tratamento.
Observando-se que esses e outros experimentos realizados em nosso
laboratório obtiveram resultados que sugeriram importante ligação dos receptores de
cininas ao controle do metabolismo energético, pretendemos estabelecer a relação
do receptor B1 de cininas e outros fatores associados com o desenvolvimento da
obesidade.
Levando em consideração os resultados glicêmicos e os relacionados à
inflamação nos camundongos nocaute para o receptor B1, além do papel da leptina
no apetite e lipólise, um animal duplo nocaute para os genes da leptina e receptor
B1 de cininas foi gerado para evidenciar o papel do receptor B1 na obesidade e
doenças correlacionadas, como o diabetes tipo II.

32
2 OBJETIVOS
A partir do conhecimento prévio das características fenotípicas e de alguns
parâmetros bioquímicos e hormonais dos camundongos nocaute para o receptor B1
de cininas e dos nocautes naturais para a leptina (ob/ob), objetivamos gerar um
camundongo ob/ob;B1KO, nocaute tanto para o receptor B1 de cininas quanto para
a leptina, visando investigar e estabelecer a correlação funcional entre a sinalização
metabólica do receptor B1, mediadores, segundos mensageiros e a leptina; e assim,
os possíveis mecanismos envolvidos no processo de metabolismo energético e
desenvolvimento de obesidade e diabetes.

33
3 MÉTODO
3.1 Geração do Camundongo ob/ob;B1KO
Os camundongos nocaute machos para o receptor B1 de cininas (B1KO) e as
fêmeas heterozigotas OB/ob foram obtidos no CEDEME, UNIFESP/EPM, com a
colaboração do Dr. João Bosco Pesquero.
O processo de geração do camundongo B1KO foi realizado baseando-se na
recombinação homóloga, na qual o éxon codificador do receptor B1 de cininas foi
substituído por uma seqüência denominada “cassete de neomicina”. Este cassete
consistia da seqüência do gene da aminoglicosídeo fosfotransferase, flanqueada pela
porção 3’ e 5’ terminais do gene do receptor B1 de cininas. Foram utilizadas células
totipotentes transformadas por eletroporação, sendo estas injetadas em infundíbulo de
fêmeas receptoras após seleção positiva (PESQUERO et al., 2000).
O primeiro cruzamento foi realizado entre três fêmeas OB/ob;B1WT e três
machos OB/OB;B1KO, em três gaiolas diferentes.
Os filhotes obtidos eram desmamados aos 21 dias de vida e então uma pequena
biópsia de extremidade de cauda (~7mm) era realizada, prosseguindo-se com os
passos de genotipagem para o gene OB, cassete de neomicina e para o gene do
receptor B1 de cininas.
Após seleção genética, os animais de interesse eram submetidos ao
cruzamento, e sua ninhada era avaliada da mesma maneira.
Foram realizados sucessivos cruzamentos e genotipagens, até que fosse
possível estabelecer parentais de genótipos OB/ob;B1KO e OB/ob;B1WT em

34
quantidade suficiente para gerar um número satisfatório dos animais de interesse
ob/ob;B1KO e ob/ob;B1KO.
A partir de um heredograma pudemos analisar as probabilidades de obtermos
nosso animal a cada cruzamento.
3.2 Genotipagem
3.2.1 Extração do DNA Genômico
Os animais da primeira ninhada (F1) foram genotipados somente para a
presença do gene mutado ob, diferentemente dos posteriores (a partir de F2), que
também foram genotipados para o receptor B1 e o cassete de neomicina.
Primeiramente, para a extração do DNA genômico, realizava-se uma pequena
biópsia de extremidade de cauda (~7mm). Para a digestão das amostras, adicionavam-
se 700µL de tail buffer e 35µL de proteinase K (10mg/mL) a cada uma, submetendo a
preparação à agitação constante de 850 rpm e temperatura de 55°C no agitador
térmico Thermomixer, por aproximadamente 15 horas. A extração do DNA genômico
iniciava-se da amostra totalmente digerida. Ao ser retirada do Thermomixer, esta
permanecia no gelo por 10 minutos. Adicionavam-se 300µL de solução de NaCl
saturado e homogeneizava-se manualmente, seguindo-se a centrifugação a 4°C,
14000 rpm, por 20min. O precipitado era desprezado, e o sobrenadante, transferido
para um tubo de 2mL e incubado com 5µL de RNase (4mg/mL) a 37°C, por 15 minutos.

35
Adicionava-se 1mL de isopropanol 100% e homogeneizava-se lentamente por
inversão, submetendo a amostra à centrifugação a 4°C, 14000 rpm, por 30min. O
sobrenadante era rejeitado e ao precipitado adicionavam-se 500µL de etanol 70%,
homogeneizando-se a amostra lentamente por inversão. Novamente o material era
submetido à centrifugação a 4°C, 14000 rpm, por 5min, o sobrenadante era rejeitado e
o tubo permanecia invertido e aberto sobre um papel absorvente por aproximadamente
1 hora, até secagem completa. O precipitado era ressuspendido em 30µL de solução
Tris/EDTA, pH 8,0 (T.E).
As amostras tinham sua concentração dosada em espectrofotômetro, e eram
acondicionadas a uma temperatura de –20°C.
Seguiram-se os passos de genotipagem para o gene mutado ob, receptor B1 e
cassete de neomicina.
3.2.2 Amplificação do Gene OB
Para identificação do gene ob, inicialmente foi realizada uma reação de
polimerase em cadeia (PCR), com primers específicos para o gene OB (OBS e OBAS),
mutado ou não, no intuito de amplificá-lo para os próximos procedimentos. A seqüência
do primer OBS (sense) é: 5’ CCC TGC TCC AGC AGC TGC 3’ ; e a do OBAS
(antisense) é: 5’ CAT GAT TCT TGG GAG CCT GG 3’. O protocolo utilizado foi o
seguinte:

36
PCR
1 µg do DNA extraído
5,0 µL de Tampão de PCR (10x)
2,5 µL de MgCl2 (50 mM)
2,0 µL de dNTPs (10 mM)
1,0 µL de primer OBS (50 ηg/µL)
1,0 µL de primer OBAS (50 ηg/µL)
0,5 µL de Taq DNA polimerase (5 U/µL)
X µL de H2O milli-Q autoclavada
50 µL total
O fragmento amplificado pela reação era analisado por eletroforese em gel de
agarose 3% contendo brometo de etídeo (0,5µg/mL), e a banda esperada era de 355
pares de bases nitrogenadas.
Após a verificação da integridade do DNA e amplificação do gene por PCR, as
amostras eram submetidas à diálise sobre pequenas membranas próprias de DNA
posicionadas sobre água milli-Q autoclavada, visando eliminar parte dos reagentes
utilizados na reação.
Após a diálise, promovia-se uma digestão pela enzima de restrição DdeI, a 37°C
por 12 horas, sendo utilizados ~3,5µg da amostra dialisada, 2,0µL de tampão REACT 3
(10X), 1,0µL de DdeI (3,5U/µL) e água milli-Q autoclavada, para um volume final de
20µL.
Novamente, o resultado era analisado por eletroforese, nas mesmas condições
anteriores.
Programa para OB
1. 94°C por 4 min 2. 80°C por 5 s 3. 94°C por 50 s 4. 56°C por 50 s 5. 72°C por 1min e 20 s 6. Voltar ao passo 3 - 40 vezes 7. 72°C por 7 min

37
Cassete de Neomicina
3.2.3 Amplificação do Gene do Receptor B1 de Cininas e Cassete de
Neomicina
A genotipagem para o receptor B1 e cassete de neomicina, realizou-se por
reação de polimerase em cadeia (PCR) com primers específicos para cada um, sendo
eles:
Oligonucleotídeos Utilizados para Genotipagem
Primer1:BKES2 (sense): 5’GGA AGC CAT CAC TCA ACA TCC 3’
Primer2:NEO1L (antisense): 5’CCT GCG TGC AAT CCA TCT TGT TCA ATG 3’
Primer1:BK135(7) (sense): 5´CGG AAG CCT GGG ATC TGC TG 3´
Primer2:BK135(3) (antisense): 5´CCA GCA ACC TGT AGC GGT CC 3´
PCR
0,5 µg do DNA extraído
2,5 µL de Tampão de PCR (10x)
1,25 µL de MgCl2 (50 mM)
1,25 µL de dNTPs (10 mM)
0,5 µL de primer 1 (50 ηg/µL)
0,5 µL de primer 2 (50 ηg/µL)
0,5 µL de Taq DNA polimerase (5 U/µL)
X µL de H2O milli-Q autoclavada
25 µL total
Receptor B1

38
Os fragmentos amplificados pela reação eram analisados por eletroforese em
gel de agarose 1,5% contendo brometo de etídeo (0,5µg/mL). A banda esperada para o
receptor B1 era de aproximadamente 320 pb, e para o cassete de neomicina contendo
uma seqüência da região 5’ do receptor B1 e da região codificadora, de
aproximadamente 1300 pb.
3.3 Análise Funcional do Receptor B1 de Cininas
Para confirmação funcional dos animais B1KO, foi realizada análise funcional do
receptor B1 nos animais obesos (ob/ob) de genótipos B1KO (n=2) e B1WT como
controle positivo (n=2), e nos animais magros (OB/OB) também de genótipos B1KO
(n=2) e B1WT como controle positivo (n=2). Para tanto, os animais foram sacrificados
por deslocamento cervical após jejum de 12 horas. O estômago foi retirado e lavado
com uma solução de Krebs, pH 7,4. O fundus do estômago foi isolado e montado em
tiras longitudinais ligadas a um transdutor e um fisiógrafo. O preparado ficou imerso em
banho com 5mL de solução de Krebs a 37ºC, com 95% de O2 e 5% de CO2. Foi
aplicada tensão de 0,5g ao preparado, equilibrando-se em aproximadamente 2 horas.
Programa para receptor B1 de cininas
1. 94°C por 4 min 2. 80°C por 5 s 3. 94°C por 50 s 4. 56°C por 50 s 5. 72°C por 1min e 20 s 6. Voltar ao passo 3 - 40 vezes 7. 72°C por 7 min
Programa para cassete de Neomicina
1. 94°C por 5 min 2. 94°C por 45 s 3. 56°C por 45 s 4. 72°C por 1 min e 20 s 5. Voltar ao passo 2 - 35 vezes 6. 72°C por 7 min

39
O registro da força contrátil foi obtido administrando-se os agonistas carbacol,
bradicinina e des-Arg9-bradicinina.
3.4 Transplante de Tecido Adiposo Branco
Visando desenvolver uma metodologia alternativa na obtenção dos animais
duplo nocaute (e seu controle) em maior número, foi realizado transplante de tecido
adiposo branco inguinal. Utilizaram-se doadores normais para fêmeas (n=10) e
machos (n=10) obesos (ob/ob), de genótipos B1KO e B1WT, com três meses de idade
e com um mês e meio de idade. Os doadores eram irmãos de mesma ninhada e do
mesmo sexo dos obesos receptores, sendo sacrificados por deslocamento cervical.
Aos receptores foi administrada anestesia geral dissociativa por via intra-peritoneal.
Foram implantados 160mg de tecido, distribuídos em quatro micro-incisões dorsais.
Após cirurgia, foram administrados antibiótico e anti-inflamatório via intra-peritoneal,
para evitar infecção pós-cirúrgica. Os animais foram monitorados diariamente e o
acompanhamento quanto à massa corporal foi realizado semanalmente. Após dois
meses de acompanhamento, os animais foram utilizados para cruzamento.
3.5 Controle de Massa Corporal
O desmame dos animais era realizado aos 30 dias de idade, quando ocorria a
primeira pesagem em todos da ninhada. As pesagens subseqüentes eram realizadas
aos dois meses de idade em todos, e aos três, três e meio, e quatro meses somente

40
nos animais que seriam submetidos a todos os testes subseqüentes, os obesos (ob/ob)
de genótipos B1KO (n=10) e B1WT (n=10).
3.6 Controle de Ingestão Alimentar
Foi realizado o controle de ingestão alimentar utilizando-se ração padrão para
roedores. Os animais foram separados em gaiolas individuais e monitorados a partir
dos três meses de idade. No primeiro mês de acompanhamento, o controle foi
realizado diariamente, passando a ser semanal após esse período.
3.7 Glicemia em Jejum
Aos dois meses de idade, todos os animais foram submetidos à dosagem de
glicemia após jejum de 6 horas, com água ad libitum. As amostras de sangue foram
obtidas de extremidade de cauda, e a dosagem realizada com auxílio do glicosímetro
comercial Accu-Chek Advantage.
3.8 Teste de Tolerância à Glicose
Aos três meses de idade, foi realizado o teste de tolerância à glicose nos
animais. Após jejum de 20 horas, com água ad libitum, foi administrada glicose (1mg/g

41
de peso vivo, i.p.). As dosagens de glicemia foram realizadas utilizando-se sangue de
extremidade de cauda, nos tempos zero, 15, 30, 60, 120 e 180 minutos, com auxílio do
glicosímetro comercial Accu-Chek Advantage.
3.9 Teste de Sensibilidade à Insulina
Aos três meses e meio de idade, foi realizado o teste de sensibilidade à insulina.
Após jejum de 20 horas, com água ad libitum, foi administrada insulina humana
recombinante monocomponente de curta duração (1U/Kg de peso vivo, i.p.- Biohulin
N/Biobrás).
As dosagens de glicemia foram realizadas utilizando-se sangue de extremidade
de cauda, nos tempos zero, 15, 30, 60, 120 e 180 minutos, com auxílio do glicosímetro
comercial Accu-Chek Advantage.
3.10 Teste de Sensibilidade à Leptina
Aos quatro meses de idade, foi realizado o teste de sensibilidade à leptina.
Administrou-se leptina recombinante de camundongo (1,8µg/g de peso vivo, i.p.)
durante cinco dias consecutivos. Neste período, a massa corporal e a ingestão
alimentar de ração padrão para roedores foram monitoradas diariamente.
A ingestão alimentar diária média durante os quinze dias anteriores ao teste foi
considerada como consumo basal.

42
3.11 Testes Bioquímicos
Aos quatro meses e meio de idade, os animais foram anestesiados para
possibilitar punção retrorbital com auxílio de pipetas Pasteur. Foram coletadas
amostras de sangue para posterior análise bioquímica de insulina pelo método de
imunoensaio enzimático (ELISA).
As amostras foram centrifugadas a 1200 rpm durante 10 minutos, e o soro foi
coletado e armazenado a -80ºC para realização dos testes com os respectivos kits
diagnósticos.
3.12 Eutanásia dos Animais
Após a punção, os animais foram sacrificados por deslocamento cervical.
Imediatamente o coração foi retirado, limpo em solução fisiológica (NaCl a 0,9%) e sua
massa foi mensurada. As câmaras foram separadas em átrios, ventrículo direito e
ventrículo esquerdo, sendo suas massas mensuradas separadamente.
Foram retirados e tiveram sua massa mensurada também o tecido adiposo
inguinal e perirenal, fígado, pâncreas, baço e rins.
Todos os órgãos e câmaras do coração foram dissecados, sendo metade
imediatamente congelada em gelo seco e armazenada a -80ºC, para posterior extração
de RNA.
A outra metade foi imersa em solução de formol a 10%, para conservação dos
tecidos e posterior análise histológica.

43
3.12.1 Análise Histológica
Amostras de tecido conservadas a uma temperatura de 4°C em solução de
formol a 10% foram submetidas isoladamente a um banho de parafina. Após
resfriamento do preparado, os órgãos se localizaram inseridos em blocos de parafina.
Um micrótomo foi utilizado para realizar cortes laminares de espessura
micrométrica, possibilitando a obtenção de amostras de tecido em vários níveis do
órgão.
As amostras laminares eram afixadas em lâminas de vidro, coradas pelo método
hematoxilina-eosina (HE), e recobertas por uma lamínula de vidro.
As lâminas de ventículo esquerdo dos dois grupos foram preparadas por corte
sagital para permitir melhor análise das fibras, e avaliadas por comparação em
microscopia de aumento 400X.

44
4 RESULTADOS
Nos primeiros cruzamentos, o genótipo de interesse nas ninhadas era OB/ob,
visto que somente animais OB/OB e OB/ob poderiam ser obtidos. Obtivemos somente
fêmeas do genótipo-alvo. Estas foram cruzadas com seus pais e as ninhadas
resultantes foram submetidas à genotipagem para o gene OB, até que se
conseguissem heterozigotos OB/ob de ambos os sexos.
Em um segundo momento, os casais iniciais deram origem a três ninhadas,
dentre as quais obtivemos machos e fêmeas OB/ob;B1+/-, possibilitando o cruzamento
entre eles. A cada ninhada subseqüente, os animais eram genotipados e selecionados
para posterior cruzamento, sendo os heterozigotos OB/ob separados em B1KO e
B1WT.
Todos os resultados dos testes foram expressos por comparação entre os
camundongos obesos (ob/ob) dos genótipos B1KO (nocautes para o receptor B1 de
cininas) e B1WT (selvagens para este receptor, utilizados como controle). Ambos os
grupos foram gerados após sucessivos cruzamentos e genotipagens.
Os animais ob/ob, nos genótipos B1KO e B1WT, apresentavam cerca do dobro
da massa corporal dos animais selvagens OB, já aos 3 meses de idade.
A seleção genética foi realizada durante todo o trabalho (figuras 4 a 7) e nos
permitiu analisar a viabilidade de cada cruzamento para a obtenção desses animais.
A digestão do fragmento amplificado do gene OB pela enzima de restrição DdeI
nos permitiu a identificação da presença da mutação no gene. Trata-se de uma
mutação pontual no códon 105, na qual a base nitrogenada citosina é substituída pela
base nitrogenada timina.

45
603 pb 310 pb
Figura 4: Eletroforese do fragmento do gene OB, de aproximadamente 355 pares de bases, amplificado pela reação de polimerase em cadeia (PCR) em gel de agarose 3%. Marcadores de massa molecular: Φλ (esquerda) e 100 pb (direita).
Em animais portadores da mutação no gene OB, gera-se um novo sítio de
reconhecimento para esta enzima, além dos sítios convencionais. Observa-se, nesses
casos, um padrão de fragmentação diferente do animal selvagem para OB, com
fragmentos adicionais. (Figura 5)
Figura 5: Gene OB, de 355 pb, e os possíveis sítios de ligação da enzima DdeI: convencionais (azul) e no sítio mutado (vermelho).

46
As bandas esperadas, em pares de bases nitrogenadas, para cada genótipo
eram:
OB/OB - animal selvagem, magro
OB/ob - animal heterozigoto, magro
ob/ob - animal homozigoto, obeso
Para identificação da presença do gene mutado em um ou em ambos os alelos,
em gel de agarose, a banda de 103pb é ideal, visto que a de 48pb (que também
poderia ser usada) não permite identificação positiva confiável, por ser próxima ao
tamanho da banda de 59pb. (Figura 6)
Já para diferenciação entre os animais heterozigotos e ob/ob, utilizamos o
fenótipo dos mesmos, visto que o animal homozigoto apresenta obesidade severa
(cerca do dobro da massa corporal do selvagem), e o heterozigoto apresenta o mesmo
fenótipo do animal sem a mutação (magro).
Os resultados da genotipagem para o gene do receptor B1 de cininas e para o
cassete de neomicina foram avaliados em conjunto na determinação dos genótipos
B1KO e B1WT. Os animais selvagens (B1WT) apresentavam somente a banda
específica do receptor B1; os nocautes (B1KO), somente a banda do cassete de
neomicina; e os heterozigotos (B1+/-), apresentavam ambas as bandas. (Figura 7)
OB/OB OB/ob ob/ob
59 59 59
145 145 145
151 151 /
/ 103 103
/ 48 48

47
+ + +
Figura 6: Eletroforese das amostras provenientes da digestão do fragmento do gene OB pela enzima DdeI em gel de agarose 3%. Nota-se a primeira banda, mais forte, de 145 pb ou 151 pb, e a última banda de 48 pb ou 59 pb em todos os animais. Os animais indicados com “+” possuem uma banda adicional de 103 pb, indicando a presença da mutação em um ou nos dois alelos (OB/ob ou ob/ob, respectivamente). O marcador de massa molecular utilizado foi o 100 pb.
Além da confirmação genotípica, os animais nocautes (B1KO) foram submetidos
à confirmação funcional no teste de verificação de força contrátil em músculo liso.
Esses experimentos foram delineados para a confirmação da ausência de resposta
mediada por agonista do receptor B1. A análise foi realizada em relação à resposta
obtida pela aplicação do agonista carbacol no banho. Em relação à bradicinina (BK),
ambos os grupos responderam de maneira semelhante. Já na aplicação de des-Arg9-
bradicinina (DBK), agonista do receptor B1, somente o grupo dos animais selvagens
respondeu, sendo essa reposta significativamente maior em relação aos animais
nocautes (p<0,01). (Figuras 8 e 9)
145/151 pb
103 pb 48/59 pb

48
A +/- -/- +/+ +/+ +/-
B
Figura 7: Eletroforese dos fragmentos do gene do receptor B1 de cininas (A) e do cassete de neomicina (B) obtidos por reação de polimerase em cadeia (PCR), em gel de agarose 1,5%. As colunas identificadas com o símbolo +/+ representam animais B1WT, -/- representa o B1KO, e +/- representa os camundongos heterozigotos para B1. Marcador de massa molecular: Φλ
Os transplantes realizados de tecido adiposo normal, apesar de serem pilotos,
apresentaram interessantes resultados fenotípicos. Em seis semanas, houve uma
redução de aproximadamente 40% da massa corporal dos ob/ob transplantados aos
três meses de idade (de 40,1g ± 0,55 para 24,2g ± 0,37), tanto nos machos quanto nas
603 pb 310 pb
1.353pb

49
fêmeas, os quais passaram a apresentar um fenótipo magro característico dos
camundongos normais (OB/OB). A fertilidade desses animais foi restabelecida
parcialmente dois meses após a cirurgia, permitindo cruzamentos bem sucedidos entre
eles, com a obtenção de 100% da ninhada composta de obesos (ob/ob). Isso facilitou
muito todos os experimentos finais devido a uma probabilidade maior de obtenção dos
animais específicos de cada grupo. (Figura 10)
Figura 8: Registros de fisiógrafo para força contrátil de fundus de estômago dos camundongos obesos (ob/ob), de genótipos B1KO e B1WT, em resposta a diferentes drogas. A força de contração registrada em resposta ao agonista carbacol (10-6M) foi utilizada como referência para os outros registros. Ao aplicar-se bradicinina (10-6M), os tecidos dos B1KO e dos B1WT apresentaram força contrátil semelhante. A resposta à des-Arg9-bradicinina (10-6M) foi nula nos animais B1KO e significativamente maior nos B1WT.
Bradicinina (BK)
(10-6M)
Des-Arg9-Bradicinina (DBK)
(10-6M)
Carbacol
(10-6M)
ob/ob;B1WT
ob/ob;B1KO

50
O desenvolvimento corporal dos camundongos não se apresentou
significativamente diferente entre os grupos até os três meses e meio de idade. Aos
quatro meses de idade, os animais B1KO já apresentavam menor massa corporal em
relação aos animais controle B1WT (p<0,05). (Figura 11)
A ingestão alimentar não apresentou diferença entre os grupos de obesos
durante os quatro meses de acompanhamento. Não foi constatada alteração na
ingestão relativa diária média entre os camundongos B1KO e B1WT (Figura 12)
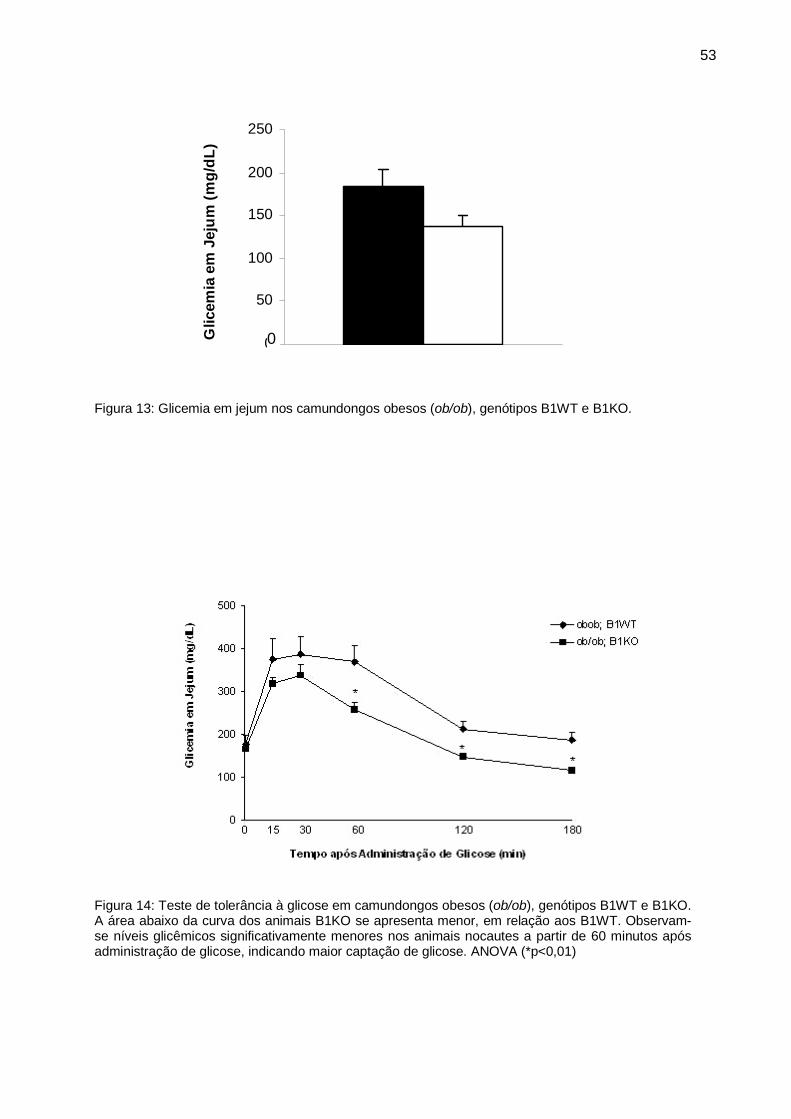
Em relação à glicemia em jejum, não houve diferença entre os animais (figura
13). Posteriormente, no teste de tolerância à glicose (TTG), os animais ob/ob;B1KO
apresentaram uma maior captação de glicose durante sua realização. Foi observada
diferença significativa a partir de 60 minutos de teste (p<0,01). (Figura 14)
Figura 9: Força contrátil de fundus de estômago na presença de bradicinina (BK) e des-Arg9-bradicinina (DBK) em relação à contração promovida pelo carbacol (%). Na presença de BK, os camundongos obesos (ob/ob) B1KO apresentaram uma tendência de menor força contrátil em relação aos ob/ob B1WT. Na presença da DBK (agonista do receptor B1) os B1KO não apresentaram resposta, enquanto os controles B1WT apresentaram resposta de 70%. ANOVA (** p<0,01)
0
10
20
30
40
50
60
70
80
90
100
Drogas Aplicadas
Res
po
sta
Co
ntr
átil
(%)
B1WT
B1KO
**
BK DBK

51
A
B
Figura 10: Foto da fêmea obesa (ob/ob) B1KO submetida ao transplante de tecido adiposo branco proveniente de irmã normal (OB/OB) da mesma ninhada. Notam-se os sinais de obesidade severa e acúmulo de tecido adiposo antes do transplante (A). Após 40 dias da intervenção cirúrgica, a mesma fêmea apresenta uma reversão da obesidade (B), exibindo um fenótipo semelhante ao das irmãs normais da mesma ninhada.

52
Figura 11: Acompanhamento de massa corporal (g) dos camundongos obesos (ob/ob), de genótipos B1KO e B1WT. Nota-se um menor ganho de massa dos animais ob/ob;B1KO aos quatro meses de vida. ANOVA (*p<0,01)
Figura 12: Acompanhamento de ingestão relativa diária média (g /g de peso vivo) dos camundongos obesos (ob/ob), de genótipos B1KO e B1WT.
53
Figura 13: Glicemia em jejum nos camundongos obesos (ob/ob), genótipos B1WT e B1KO.
Figura 14: Teste de tolerância à glicose em camundongos obesos (ob/ob), genótipos B1WT e B1KO. A área abaixo da curva dos animais B1KO se apresenta menor, em relação aos B1WT. Observam-se níveis glicêmicos significativamente menores nos animais nocautes a partir de 60 minutos após administração de glicose, indicando maior captação de glicose. ANOVA (*p<0,01)
0
50
100
150
200
250
Glic
emia
em
Jej
um
(m
g/d
L)
0

54
No teste de sensibilidade à insulina, não houve diferença significativa nos níveis
glicêmicos obtidos (figura 15), revelando que a melhor captação de glicose observada
no teste de tolerância à glicose não foi proveniente de maior sensibilidade à insulina.
Figura 15: Teste de sensibilidade à insulina em camundongos obesos (ob/ob), genótipos B1WT e B1KO.
A dosagem bioquímica de insulina nesses animais sugeriu maior concentração
sérica deste hormônio nos animais ob/ob;B1KO, condizendo com a hipótese da sua
ligação com a melhor captação de glicose apresentada. (Figura 16)
Figura 16: Insulina sérica (ηg/mL) em camundongos obesos (ob/ob), genótipos B1WT e B1KO (direita). Os camundongos indicados como OB/OB são utilizados como referência na reação. ANOVA (*p<0,05)

55
Quanto ao teste de sensibilidade à leptina, os resultados sugerem maior
sensibilidade dos animais ob/ob;B1KO à leptina exógena, apresentando maior inibição
na ingestão diária, e diminuição na massa corporal mais acentuada (figura 17). Foi
observada uma diferença significativa maior entre os grupos no segundo dia, tendendo
a desaparecer a partir do quinto e sexto dias de experimento. Considerou-se inibição
de ingestão a diminuição significativa do consumo basal diário médio de cada grupo.
Figura 17: Teste de sensibilidade à leptina. Os animais ob/ob;B1KO apresentaram maior inibição da
ingestão alimentar (A) e maior perda de massa corporal (B) em relação ao controle. ANOVA (*p<0,05)
B
A
0 1 2 3 4 5
0 1 2 3 4 5

56
Na extração dos órgãos dos animais, uma observação macroscópica inicial
permitiu constatar grande acúmulo de gordura envolta no pericárdio de ambos os
grupos, além de uma esteatose hepática mais acentuada nos animais B1KO. Quanto à
relação da massa dos tecidos com a massa corporal, esta se apresentou
significativamente elevada em tecido adiposo inguinal (p<0,001), e significativamente
mais baixa em rins (p<0,01), coração (p<0,05) e ventrículo esquerdo isolado (p<0,05)
dos B1KO em relação ao controle. (Figura 18)
Figura 18: Massa relativa dos órgãos (mg/g de peso vivo) em camundongos obesos (ob/ob), genótipos B1WT (controle) e B1KO. Os animais B1KO apresentaram maior quantidade de tecido adiposo branco inguinal em relação aos B1WT (A), e menor massa nos rins (B), coração (C), e ventrículo esquerdo (D). ANOVA (* p<0,05; ** p<0,01; *** p<0,001)
C
0
10
20
30
40
50
60
70
Rel
ação
Tec
ido
Ad
ipo
so B
ran
co
Ing
uin
al /
Mas
sa C
orp
ora
l (m
g/g
) ob/ob; B1WT
ob/ob; B1KO
***
A
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
9,0R
elaç
ão R
ins
/ Mas
sa
Co
rpo
ral (
mg
/g)
ob/ob; B1WT
ob/ob; B1KO **
B
0,0
0,5
1,0
1,5
2,0
2,5
Rel
ação
Ven
tríc
ulo
Esq
uer
do
/ M
assa
Co
rpo
ral (
mg
/g)
ob/ob; B1WT
ob/ob; B1KO
*
D
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
Rel
ação
Co
raçã
o /
Mas
sa
Co
rpo
ral (
mg
/g)
ob/ob; B1WT
ob/ob; B1KO
*

57
Para investigar se a menor massa observada em coração e ventrículo esquerdo
isolado dos camundongos ob/ob;B1KO estava relacionada com uma menor hipertrofia
cardíaca nesses animais, foi realizada a análise histológica do ventrículo esquerdo.
Uma análise inicial permitiu verificar que os nossos animais apresentavam
cardiomiócitos de menor diâmetro. (Figura 19)
Figura 19: Análise histológica de ventrículo esquerdo de camundongos obesos (ob/ob), genótipos B1WT e B1KO (400X). Os animais B1WT (A) apresentaram cardiomiócitos de maior diâmetro em relação aos B1KO (B), como indicado pelas setas amarelas, caracterizando uma hipertrofia de maior grau.
A B

58
5 DISCUSSÃO
Inicialmente, este trabalho apresentou certa dificuldade de desenvolvimento
em relação à geração do animal obeso (ob/ob) e nocaute para o receptor B1 de
cininas. Devido à infertilidade característica dos camundongos ob/ob, foi necessário
selecionar por genotipagem apenas os heterozigotos OB/ob para os cruzamentos.
Estes heterozigotos eram selvagens para o receptor B1 de cininas. Em
contrapartida, os animais nocautes para o receptor B1 eram selvagens para ob
(OB/OB).
A partir de um heredograma pudemos analisar as probabilidades de obtermos
nosso animal a cada cruzamento. No caso da primeira ninhada, todos poderiam
apresentar somente o genótipo B1+/-. Quanto ao gene da leptina, as probabilidades
eram de 50% para OB/OB e 50% para OB/ob, mas obtivemos uma média de 37% de
OB/ob.
Na terceira geração, proveniente do cruzamento das fêmeas de duas
ninhadas (OB/ob;B1+/-) com seus pais (OB/OB;B1KO), esperávamos 50% de
genótipo B1+/- e 50% de genótipo B1KO. Em relação à leptina, esperava-se 50% de
animais OB/OB e 50% de animais OB/ob, mas não obtivemos o genótipo de
interesse. O genótipo de escolha seria OB/ob;B1KO, porém, foi necessária a
utilização dos animais OB/ob;B1+/-, para aumentarmos nosso número de
cruzamentos, já que a probabilidade de obtermos machos e fêmeas OB/ob;B1KO a
cada ninhada era muito baixa.
Na quarta geração, obtida pelo cruzamento entre irmãos OB/ob;B1+/-, as
probabilidades seriam melhores: 25% de B1WT e de B1KO, e 50% de B1+/-; 25% de
OB/OB e de ob/ob, e 50% de OB/ob. Os animais mais adequados para cruzamentos

59
posteriores seriam os de genótipo OB/ob;B1KO (já que os ob/ob têm problemas de
fertilidade), e a probabilidade de os obtermos seria de 12%, sendo que deveriam ser
machos e fêmeas desse genótipo; os de interesse para os experimentos eram os
ob/ob;B1KO, porém a probabilidade era muito baixa (6%).
Foram necessárias várias gerações até conseguirmos separar parentais de
heterozigotos para o gene da leptina OB (OB/ob), dos genótipos B1KO e B1WT, e
para obtermos animais obesos B1KO (ob/ob;B1KO) e seus controles (ob/ob;B1WT)
em número suficiente para os experimentos. Sendo assim, o processo de geração
dos animais foi constante durante todo o decurso dos experimentos com os animais.
Além da confirmação genotípica dos animais gerados, a análise funcional do
receptor B1 permitiu a confirmação funcional. Os animais obesos B1KO e B1WT
responderam ao agonista do receptor B2 (bradicinina), havendo maior força contrátil
nos ob/ob;B1WT. Pode ter ocorrido uma metabolização de parte da bradicinina em
des-Arg9-bradicinina, que também estaria atuando, no receptor B1 dos animais
B1WT (ALLOGHO et al., 1998). A resposta ao agonista do receptor B1 (des-Arg9-
bradicinina) foi nula nos animais ob/ob;B1KO, e significativamente maior nos
controles (ob/ob;B1WT).
Os transplantes de tecido adiposo foram realizados, e propiciaram o
restabelecimento do fenótipo normal dos animais. O tecido adiposo implantado
parece ter atuado no metabolismo por meio da secreção de leptina e outros
hormônios endócrinos (KLEBANOV et al., 2005).
Pode ser que estes animais transplantados tenham maior sensibilidade à
leptina, visto sua deficiência nesse hormônio. Foi possível restabelecer também a
fertilidade dos camundongos ob/ob, sendo constatada por cruzamentos bem
sucedidos entre eles. Esse fato foi muito importante para a finalização do nosso

60
trabalho, pois começamos a obter 100% de filhotes obesos (ob/ob), provenientes
dos parentais magros, férteis e de genótipo ob/ob.
Essa metodologia foi utilizada como alternativa na obtenção do animal em
maior quantidade, visto que a metodologia de seleção de heterozigotos OB/ob por
genotipagem era mais lenta e produzia um número limitado de animais obesos por
ninhada.
Após a geração e determinação dos grupos, seguiram-se os testes nos
animais.
A hipótese da ingestão alimentar estar contribuindo para que os animais
ob/ob;B1WT apresentassem maior massa corporal foi testada. Porém, não houve
diferença entre os dois grupos, indicando que o receptor B1 de cininas não está
relacionado com a hiperfagia dos camundongos ob/ob.
A glicemia dos animais duplo nocaute não apresentou diferença significativa
dos animais controle. O teste de tolerância à glicose indicou maior tolerância à
sobrecarga glicídica nesses camundongos, observada por uma menor área abaixo
da curva glicêmica. Já a resistência à insulina não parece ter sido atenuada nesses
animais, como esperado. A maior captação de glicose deve estar ligada a outros
fatores endócrinos, como uma maior liberação de insulina durante o teste. O
resultado das dosagens séricas de insulina basal sugerem que os animais
ob/ob;B1KO tenham maior concentração basal deste hormônio. No entanto, ainda é
necessária a realização de testes com maior número de animais.
A bradicinina, agonista do receptor B2 de cininas, é capaz de induzir a
translocação do GLUT4 e promover a captação de glicose via insulino-independente,
principalmente por células musculares (KISHI et al., 1998). Além disso, a deficiência
de leptina pode alterar a expressão dos receptores de cininas (ABE et al., 2007).

61
Observando esses dados, uma outra possibilidade para justificar nosso resultado é a
superexpressão do receptor B2 de cininas, inclusive como mecanismo
compensatório à ausência do receptor B1 de cininas, promovendo uma melhor
captação da glicose via insulino-independente.
Sabe-se que a insulina atua na modulação da produção e secreção de leptina
pelos adipócitos (LEVY et al., 2001). Em nosso laboratório, resultados anteriores
mostraram que a deficiência do receptor B1 em animais magros induz um aumento
de sensibilidade à leptina exógena (dados não publicados). Assim como nos animais
ob/ob selvagens (COLEMAN, 1973), nossos animais obesos B1KO e B1WT
mostraram-se sensíveis à leptina exógena, apesar de possuírem deficiência na
produção deste hormônio, revelando que não havia comprometimento de seus
receptores. Além disso, os camundongos obesos do grupo ob/ob;B1KO também
apresentaram uma resposta central mais acentuada em relação ao grupo controle,
sugerindo maior sensibilidade desses animais à leptina. Sabe-se que o cortisol e a
insulina são potentes estimuladores da expressão e secreção da leptina
(HOUSEKNECHT et al., 1998). A hiperinsulinemia observada no grupo pode estar
contribuindo para uma maior sensibilidade dos receptores de leptina, já que a
produção e secreção deste hormônio estão prejudicadas nesses animais. Ainda em
1998, Houseknecht et al. atribuíram a potencialização dos efeitos obtidos pela
administração de leptina a uma taxa metabólica elevada nos animais tratados.
Nossos animais poderiam estar apresentando uma taxa metabólica mais elevada em
relação ao controle, indicando resultados mais expressivos quanto à sensibilidade à
leptina exógena. A falta do receptor B1 também pode estar relacionada a este
fenômeno a nível central, por meio de mecanismos ainda não determinados e que
demandam uma investigação mais complexa. Entretanto, demonstrou-se que o

62
peptídeo opióide endógeno dinorfina A induz o influxo de cálcio por canais de cálcio
voltagem-dependentes em neurônios sensoriais, via ativação dos receptores de
cininas pela atuação como agonista sobre eles (LAI et al., 2006). Portanto, outra
hipótese plausível seria a superexpressão do receptor B2 de cininas, facilitando a
atuação deste opióide na potencialização dos efeitos observados pelo tratamento
com leptina exógena.
Com relação à massa relativa dos rins, esta foi menor nos animais
ob/ob;B1KO. A ausência do receptor B1 parece ter atenuado os efeitos do diabetes
sobre eles, os quais normalmente apresentam insuficiência, necrose e até falência
renal (TSUCHIDA et al., 1999). Entretanto, nossos dados não suportam ainda que a
ausência do receptor B1 possa proteger de afecções renais. Outros experimentos
são necessários para testar esta hipótese.
Avaliando-se o coração, visto que a hipertrofia cardíaca é freqüentemente
observada nos animais ob/ob (BAROUCH et al., 2003), o ventrículo esquerdo
apresentou menor massa relativa e cardiomiócitos de menor diâmetro nos
camundongos duplo nocaute, caracterizando uma menor hipertrofia cardíaca nesses
animais. Os mesmos obtiveram uma proteção parcial ou total do órgão, levando-nos
a pensar em uma possível superexpressão do receptor B2 de cininas como
mecanismo compensatório à falta do receptor B1 de cininas (XU et al., 2005;
LAGNEUX et al., 2002). Para melhor julgamento desses dados, devemos verificar os
níveis de expressão dos receptores B1 e B2 em todos os tecidos desse modelo de
obesidade mórbida.
Uma recente publicação mostra que a expressão dos receptores B1 e B2 de
cininas está modificada em vários tecidos importantes para o metabolismo
energético dos camundongos ob/ob, os quais apresentaram baixa expressão de

63
RNA mensageiro para o receptor B2 de cininas em importantes sítios de modulação
neuroendócrina (ABE et al, 2007). Esses resultados sugeriram a ligação do SCC e
obesidade, salientando o possível envolvimento desse sistema na etiopatogênese
da síndrome metabólica, freqüentemente ligada ao diabetes tipo II e obesidade.

64
6 CONCLUSÕES E SUGESTÕES
De acordo com os nossos resultados obtidos com o camundongo duplo nocaute
para os genes da leptina e receptor B1 de cininas, gerado ao início do trabalho,
pudemos concluir que o receptor B1 de cininas está relacionado a este modelo de
obesidade severa, sugerindo seu envolvimento na patogênese do diabetes e
doenças associadas.
No decorrer do trabalho, a partir da obtenção de parentais fixos, aumentamos
nosso número de camundongos, possibilitando a realização de testes
complementares conclusivos que serão realizados em estudos posteriores.
Realizaremos uma análise histológica mais detalhada de ventrículo esquerdo,
rins, pâncreas e fígado, os quais estão ligados à obesidade e diabetes.
Será verificada também a expressão do receptor B2 de cininas no ventrículo
esquerdo, e de citocinas no tecido adiposo branco, bem como na musculatura
esquelética desses camundongos.
Além disso, avaliaremos o consumo de oxigênio, visando estabelecer a
relevância do gasto energético em nossos resultados, e a liberação de insulina
durante o teste de tolerância à glicose, almejando determinar seu papel na maior
tolerância à sobrecarga glicídica observada nos animais duplo nocaute gerados.
Posteriormente, pretendemos aplicar também aos ob/ob transplantados todos
os testes apresentados neste trabalho, visando verificar o restabelecimento da
sensibilidade à insulina e do perfil metabólico considerado normal.

65
REFERÊNCIAS
ABE, K.C.; DA SILVA MORI, M.A.; PESQUERO, J.B. Leptin deficiency leads to the regulation of kinin receptors expression in mice. Regul Pept., 138(2-3): 56-8, 2007.
ALFIE, M.E.; SIGMON, D.H.; POMPOSIELLO, S.I.; CARRETERO, O.A. Effect of high salt intake in mutant mice lacking bradykinin-B2 receptors. Hypertension, 29(1-Pt 2): 483-7, 1997.
ALLOGHO, S.; GOBEIL, F.; PERRON, S.I.; HESS, J.F.; REGOLI, D. Effects of kinins on isolated stomachs of control and transgenic knockout B2 receptor mice. Naunyn Schmiedebergs Arch Pharmacol., 357(3): 191-196, 1998.
ARAUJO, R.C.; KETTRITZ, R.; FICHTNER, I.; PAIVA, A.C.; PESQUERO, J.B.; BADER, M. Altered neutrophil homeostasis in kinin B1 receptor-deficient mice. Biol Chem., 382(1): 91-5, 2001.
ARAÚJO, R.C.; MORI, M.A.; MERINO, V.F.; BASCANDS, J.L.; SCHANSTRA, J.P.; ZOLLNER, R.L.; VILLELA, C.A.; NAKAIE, C.R.; PAIVA, A.C.M.; PESQUERO, J.L.; BADER, M.; PESQUERO, J.B. Role of the kinin B1 receptor in insulin homeostasis and pancreatic islet function. Biol. Chem., 387: 431-436, 2006.
ARITA, Y.; KIHARA, S.; OUCHI, N.; TAKAHASHI, M.; MAEDA, K.; MIYAGAWA, J.; HOTTA, K.; SHIMOMURA, I.; NAKAMURA, T.; MIYAOKA, K.; KURIYAMA, H.; NISHIDA, M.; YAMASHITA, S.; OKUBO, K.; MATSUBARA, K.; MURAGUCHI, M.; OHMOTO, Y.; FUNAHASHI, T.; MATSUZAWA, Y. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun., 257(1): 79-83, 1999.
BAHIA, L.; AGUIAR, L.G.; VILLELA, N.; BOTTINO, D.; GODOY-MATOS, A.F.; GELONEZE, B.; TAMBASCIA, M.; BOUSKELA, E. Relationship between adipokines, inflammation, and vascular reactivity in lean controls and obese subjects with metabolic syndrome. Clinics, 61(5): 433-40, 2006.
BAROUCH, L.A.; BERKOWITZ, D.E.; HARRISON, R.W.; O’DONNELL, C.P.; HARE, J.M. Disruption of leptin signaling contributes to cardiac hypertrophy independently of body weight in mice. Circulation, 108: 754-759, 2003.

66
BARROSO, I.; LUAN, J.; MIDDELBERG, R.P.; HARDING, A.H.; FRANKS, P.W.; JAKES, R.W.; CLAYTON, D.; SCHAFER, A.J.; O'RAHILLY, S.; WAREHAM, N.J. Candidate gene association study in type 2 diabetes indicates a role for genes involved in beta-cell function as well as insulin action. PLoS Biol., 1(1): E20, 2003.
BAUDRY, A.; LEROUX, L.; JACKEROTT, M.; JOSHI, R.L. Genetic manipulation of insulin signaling, action and secretion in mice: insights into glucose homeostasis and pathogenesis of type 2 diabetes. Review. EMBO Rep., 3(4): 323-8, 2002.
BERGMAN, R.N.; KIM, S.P.; HSU, I.R.; CATALANO, K.J.; CHIU, J.D.; KABIR, M.; RICHEY, J.M.; ADER, M. Abdominal obesity: role in the pathophysiology of metabolic disease and cardiovascular risk. Am J Med., 120(2-Suppl 1): S3-8, 2007.
BHOOLA, K.D.; FIGUEROA, C.D.; WORTHY, K. Bioregulation of kinins: kallikreins, kininogens and kininases. Pharmacol. Rev., 44: 1-79, 1992.
BORKOWSKI, J.A.; RANSOM, R.W.; SEABROOK, G.R.; TRUMBAUER, M.; CHEN, H.; HILL, R.G.; STRADER, C.D.; HESS JF. Targeted disruption of a B2 bradykinin receptor gene in mice eliminates bradykinin action in smooth muscle and neurons. J Biol Chem., 270(23): 13706-10, 1995.
BURCH, R.M.; KYLE, D.J. Recent developments in the understanding of bradykinin receptors. Life Sci., 50(12): 829-838, 1992.
CARLSSON, B.; LINDELL, K.; GABRIELSSON, B.; KARLSSON, C.; BJARNASON, R.; WESTPHAL, O. Obese (ob) gene defects are rare in human obesity. Obes Res, 5(1): 30-5, 1997.
CATERSON, I.D.; HUBBARD, V.; BRAY, G.A.; GRUNSTEIN, R.; HANSEN, B.C.; HONG, Y.; LABARTHE, D.; SEIDELL, J.C.; SMITH JR, S.C. Obesity, a worldwide epidemic related to heart disease and stroke: worldwide comorbidities of obesity. Circulation, 110(18): e476-83, 2004.
CARVALHEIRA, J.B.C.; ZECCHIN, H.G.; SAAD, M.J.A. Vias de sinalização da insulina. Atualização. Arq Bras Endocrinol Metab., 46(4): 419-425, 2002.
CHRISTOPHER, J.; VELARDE, V.; ZHANG, D.; MAYFIELD, R.K.; JAFFA, A.A. Regulation of B2-kinin receptors by glucose in vascular smooth muscle cells. Am. J. Phisiol. Heart Circ. Physiol., 280: H1537-1546, 2001.

67
COLEMAN, D.L. Effects of parabiosis of obese with diabetes and normal mice. Diabetologia, 9(4): 294-8, 1973.
COLEMAN, D.L. Obese and diabetes: two mutant genes causing diabetes-obesity syndromes in mice. Diabetologia, 14: 141-148, 1978.
CONSIDINE, R.V.; CONSIDINE, E.L.; WILLIAMS, C.J.; NYCE, M.R.; MAGOSIN, S.A.; BAUER, T.L. Evidence against either a premature stop codon or the absence of obese gene mRNA in human obesity. J Clin Invest, 95(6): 2986-8, 1995.
CONSIDINE, R.V.; SINHA, M.K.; HEIMAN, M.L.; KRIAUCIUNAS, A.; STEPHENS, T.W.; NYCE, M.R. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med, 334(5): 292-5, 1996.
CUEVAS-ALVAREZ, N.A.; VELA-OTERO, Y.; CARRADA-BRAV, T. Identification of risk factors in relatives of type-2 diabetics. Rev Med Inst Mex Seguro Soc., 44(4): 313-20, 2006.
DAY, C. Metabolic syndrome, or What you will: definitions and epidemiology. Diab Vasc Dis Res., 4(1): 32-8, 2007.
DE MEYTS, P. The diabetogenes concept of NIDDM. Review. Adv Exp Med Biol., 334: 89-100, 1993.
DRAY, A.; PERKINS, M. Bradykinin and inflammatory pain. TINS., 16(3): 99-104, 1993.
DUCY, P.; AMLING, M.; TAKEDA, S.; PRIEMEL, M.; SCHILLING, A.F.; BEIL, F.T.; SHEN, J.; VINSON, C.; RUEGER, J.M.; KARSENTY, G. Leptin inhibits bone formation through a hypothalamic relay: a central control of bone mass. Cell, 100: 197-207, 2000.
DUKA, I.; SHENOUDA, S.; JOHNS, C.; KINTSURASHVILI, E.; GAVRAS, I.; GAVRAS, H. Role of the B(2) receptor of bradykinin in insulin sensitivity. Hypertension, 38(6): 1355-60, 2001.
ECKEL, R.H.; YORK, D.A.; ROSSNER, S.; HUBBARD, V.; CATERSON, I.; ST JEOR, S.T.; HAYMAN, L.L.; MULLIS, R.M.; BLAIR, S.N. Obesity, a worldwide epidemic related to heart disease and stroke: executive summary. Circulation, 110(18): 2968-75, 2004.

68
ELEFTERIOU, F.; TAKEDA, S.; EBIHARA K.; MAGRE, J.; PATANO, N.; KIM, C.A.; OGAWA, Y.; LIU, X.; WARE, S.M.; CRAIGEN, W.J.; ROBERT, J.J.; VINSON, C.; NAKAO, K.; CAPEAU, J.; KARSENTY, G. Serum leptin level is a regulator of bone mass. Proc. Natl. Acad. Sci. USA, 101: 3258-3263, 2004.
EMANUELI, C.; MAESTRI, R.; CORRADI, D.; MARCHIONE, R.; MINASI, A.; TOZZI, M.G.; SALIS, M.B.; STRAINO, S.; CAPOGROSSI, M.C.; OLIVETTI, G.; MADEDDU P. Dilated and failing cardiomyopathy in bradykinin B(2) receptor knockout mice. Circulation, 100(23): 2359-65, 1999.
EMPANA, J.P.; DUCIMETIERE, P.; CHARLES, M.A.; JOUVEN, X. Sagittal abdominal diameter and risk of sudden death in asymptomatic middle-aged men: the Paris Prospective Study I. Circulation, 110(18): 2781-5, 2004.
FAGGIONI, R.; FEINGOLD, K.R.; GRUNFELD, C. Leptin regulation of the immune response and the immunodeficiency of malnutrition. FASEB J., 15(14): 2565-71, 2001.
FOX, C.S.; PENCINA, M.J.; MEIGS, J.B.; VASAN, R.S.; LEVITZKY, Y.S.; AGOSTINO, R.B. Trends in the incidence of type 2 diabetes mellitus from the 1970s to the 1990s: the Framingham Heart Study. Circulation, 113(25): 2914-8, 2006.
FRIEDMAN, J.M.; HALAAS, J.L. Leptin and the regulation of body weight in mammals. Nature, 395: 763-770, 1998.
HAMILTON, B.S.; PAGLIA, D.; KWAN, A.Y.M.; DEITEL, M. Increased obese mRNA expression in omental fat cells from massively obese humans. Nat. Med., 9: 953-956, 1995.
HAMRICK, M.W.; PENNINGTON, C.; NEWTON, D.; XIE, D.; ISALES, C. Leptin deficiency produces contrasting phenotypes in bones of the limb and spine. Bone, 34: 376-383, 2004.
HOUSEKNECHT, K.L.; BAILE, C.A.; MATTERI, R.L.; SPURLOCK, M.E. The Biology of Leptin: A Review. J. Anim. Sci., 76: 1405–1420, 1998.
JOHNSON, P.R.; HIRSCH, J. Cellularity of adipose depots in six strains of genetically obese mice. J. Lipid Res., 13: 2-11, 1972.

69
KAHN, C.R. Banting Lecture. Insulin action, diabetogenes, and the cause of type II diabetes. Review. Diabetes, 43(8): 1066-84, 1994.
KISHI, K.; MUROMOTO, N.; NAKAYA, Y.; MYATA, I.; HAGI, A.; HAYASHI, H.; EBINA, Y. Bradykinin directly triggers GLUT4 translocation via an insulin-independent pathway. Diabetes, 47: 550-558, 1998.
KLEBANOV, S.; ASTLE, C.M.; DESIMONE, O.; ABLAMUNITS, V.; HARRISON, D.E. Adipose tissue transplantation protects ob/ob mice from obesity, normalizes insulin sensitivity and restores fertility. J Endocrinol., 186(1): 203-11, 2005.
KRAKOFF, J.; FUNAHASHI, T.; STEHOUWER, C.D.; SCHALKWIJK, C.G.; TANAKA, S.; MATSUZAWA, Y.; KOBES, S.; TATARANNI, P.A.; HANSON, R.L.; KNOWLER, W.C.; LINDSAY, R.S. Inflammatory markers, adiponectin, and risk of type 2 diabetes in the pima indian. Diabetes Care, 26(6): 1745-51, 2003.
KUBOTA, N.; TERAUCHI, Y.; YAMAUCHI, T.; KUBOTA, T.; MOROI, M.; MATSUI, J.; ETO, K.; YAMASHITA, T.; KAMON, J.; SATOH, H.; YANO, W.; FROGUEL, P.; NAGAI, R.; KIMURA, S.; KADOWAKI, T.; NODA, T. Disruption of adiponectin causes insulin resistance and neointimal formation. J Biol Chem., 277(29): 25863-6, 2002.
LAGNEUX, C.; BADER, M.; PESQUERO, J.B.; DEMENGE, P.; RIBUOT, C. Detrimental implication of B1 receptors in myocardial ischemia: evidence from pharmacological blockade and gene knockout mice. Int Immunopharmacol., 2(6): 815-22, 2002.
LAI, J.; LUO, M.C.; CHEN, Q.; MA, S.; GARDELL, L.R.; OSSIPOV, M.H.; PORRECA, F. Dynorphin A activates bradykinin receptors to maintain neuropathic pain. Nat Neurosci., 9(12): 1534-40, 2006.
LEVY, J.R.; STEVENS, W. The effects of insulin, glucose, and pyruvate on the kinetics of leptin secretion. Endocrinology, 142: 3558-3562, 2001.
LINDSAY, R.S.; FUNAHASHI, T.; HANSON, R.L.; MATSUZAWA, Y.; TANAKA, S.; TATARANNI, P.A.; KNOWLER, W.C.; KRAKOFF, J. Adiponectin and development of type 2 diabetes in the Pima Indian population. Lancet., 360(9326): 57-8, 2002.
LINDSEY, C.J.; FUJITA, K.; MARTIN, T.O. The central effect of bradykinin in normotensive and hypertensive rats. Hypertension, 11(2): I126-I129, 1998.

70
MACNEIL, T.; BIERILO, K.K.; MENKE, J.G.; HESS, J.F. Cloning and pharmacological characterization of a rabbit bradykinin B1 receptor. Biochim Biophys Acta., 1264(2): 223-8, 1995.
MADEDDU, P.; VARONI, M.V.; PALOMBA, D.; EMANUELI, C.; DEMONTIS, M.P.; GLORIOSO, N.; DESSI-FULGHERI, P.; SARZANI, R.; ANANIA, V. Cardiovascular phenotype of a mouse strain with disruption of bradykinin B2-receptor gene. Circulation, 96(10): 3570-8, 1997.
MAFFEI, M.; HALAAS, J.; RAVUSSIN, E.; PRATLEY, R.E.; LEE, G.H.; ZHANG, Y.; FEI, H.; KIM, S.; LALLONE, R.; RANGANATHAN, S.; KERN, P.A.; FRIEDMAN, J.M. Leptin levels in human and rodent: measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nature Med., 1: 1155-1161, 1995.
MARCEAU, F. Kinin B1 receptors: a review. Immunopharmacology, 30: 1-26, 1995.
MARCEAU, F.; HESS, J.F.; BACHVAROV, D.R. The B1 receptors for kinins. Pharmacol. Rev., 50(3): 357-386, 1998.
MARK, A.L.; SHAFFER, R.A.; CORREIA, M.L.; MORGAN, D.A.; SIGMUND, C.D.; HAYNES, W.G. Contrasting blood pressure effects of obesity in leptin-deficient ob/ob mice and agouti yellow obese mice. J Hypertens., 17(12 - Pt 2): 1949-53, 1999.
MASUZAKI, H.; OGAWA, Y.; SAGAWA, N.; HOSODA, K.; MATSUMOTO, T.; MISE, H.; NISHIMURA, H.; YOSHIMASA, Y.; TANAKA, I.; MORI, T.; NAKAO, K. Nonadipose tissue production of leptin: leptin as a novel placenta-derived hormone in humans. Nature Med., 3: 1029-1033, 1997.
MATHIS, A.S.; CRISCIMAGNA, N.L.; LEEB-LUNDEBERG, L.M. B1 and B2 kinin receptors mediate distinct patterns of intracellular Ca2+ signaling in single cultured vascular muscle cells. Molecular Pharmacology, 50: 128-139, 1996.
MATSUBARA, M.; MARUOKA, S.; KATAYOSE, S. Inverse relationship between plasma adiponectin and leptin concentrations in normal-weight and obese women. Eur J Endocrinol., 147(2): 173-80, 2002.
MCKINNEY, P.A.; PARSLOW, R.; GURNEY, K.A.; LAW, G.R.; BODANSKY, H.J.; WILLIAMS, R. Perinatal and neonatal determinants of childhood type 1 diabetes. A case-control study in Yorkshire, U.K. Diabetes Care, 22(6): 928-32, 1999.

71
MILIA, A.F.; GROSS, V.; PLEHM, R.; DE SILVA, J.A.JR.; BADER, M.; LUFT, F.C. Normal blood pressure and renal function in mice lacking the bradykinin B(2) receptor. Hypertension, 37(6): 1473-9, 2001.
MONCADA, S.; PALMER, R.M.J.; HIGGS, E.A. Nitric oxide: physiology, patophysiology and pharmacology. Pharmacologycal Reviews, 43(2): 109-142, 1991.
MORLEY, J.E.; FLOOD, J.F. Evidence that nitric oxide modulates food intake in mice. Life Science, 49: 707-711, 1991.
MUKAI, H.; FITZGIBBON, W.R.; BOZEMAN, G.; MARGOLIUS, H.S.; PLOTH, D.W. Bradykinin B2 receptor antagonist increases chloride and water absorption in rat medullary collecting duct. Am J Physiol., 271(2Pt2): R352-R360, 1996.
MULLIS, R.M.; BLAIR, S.N.; ARONNE, L.J.; BIER, D.M.; DENKE, M.A.; DIETZ, W.; DONATO, K.A.; DREWNOWSKI, A.; FRENCH, S.A.; HOWARD, B.V.; ROBINSON, T.N.; SWINBURN, B.; WECHSLER, H. Obesity, a worldwide epidemic related to heart disease and stroke: prevention/treatment. Circulation, 110(18): e484-8, 2004.
NOBRE, M.R.; DOMINGUES, R.Z.; DA SILVA, A.R.; COLUGNATI, F.A.; TADDEI, J.A. Prevalence of overweight, obesity and life style associated with cardiovascular risk among middle school students. Rev Assoc Med Bras., 52(2): 118-24, 2006.
OUCHI, N.; KIHARA, S.; ARITA, Y.; MAEDA, K.; KURIYAMA, H.; OKAMOTO, Y.; HOTTA, K.; NISHIDA, M.; TAKAHASHI, M.; NAKAMURA, T.; YAMASHITA, S.; FUNAHASHI, T.; MATSUZAWA, Y. Novel modulator for endothelial adhesion molecules: adipocyte-derived plasma protein adiponectin. Circulation, 100(25): 2473-6, 1999.
OUCHI, N.; KIHARA, S.; ARITA, Y.; NISHIDA, M.; MATSUYAMA, A.; OKAMOTO, Y.; ISHIGAMI, M.; KURIYAMA, H.; KISHIDA, K.; NISHIZAWA, H.; HOTTA, K.; MURAGUCHI, M.; OHMOTO, Y.; YAMASHITA, S.; FUNAHASHI, T.; MATSUZAWA, Y. Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation, 103(8): 1057-63, 2001.

72
OUCHI, N.; KIHARA, S.; FUNAHASHI, T.; NAKAMURA, T.; NISHIDA, M.; KUMADA, M.; OKAMOTO, Y.; OHASHI, K.; NAGARETANI, H.; KISHIDA, K.; NISHIZAWA, H.; MAEDA, N.; KOBAYASHI, H.; HIRAOKA, H.; MATSUZAWA, Y. Reciprocal association of C-reactive protein with adiponectin in blood stream and adipose tissue. Circulation, 107(5): 671-4, 2003.
PESQUERO, J.B.; PESQUERO, J.L.; OLIVEIRA, S.M.; ROSCHER, A.A.; METZGER, R.; GANTEN, D.; BADER, M. Molecular cloning e functional characterization of a mouse bradykinin B1 receptor gene. Biochem Biophys Res Commun., 224(1): 281, 1996.
PESQUERO, J.B.; ARAUJO, R.C.; HEPPENSTALL, P.A.; STUCKY, C.L.; SILVA, J.A.JR.; WALTHER, T.; OLIVEIRA, S.M.; PESQUERO, J.L.; PAIVA, A.C.; CALIXTO, J.B.; LEWIN, G.R.; BADER, M. Hypoalgesia and altered inflammatory responses in mice lacking kinin B1 receptors. Proc Natl Acad Sci USA, 97(14): 8140-5, 2000.
PESQUERO, J.B.; BADER, M. Genetically altered animal models in the kallikrein-kinin system. Review. Biol Chem., 387(2): 119-26, 2006.
PISCHON, T.; GIRMAN, C.J.; HOTAMISLIGIL, G.S.; RIFAI, N.; HU, F.B.; RIMM, E.B. Plasma adiponectin levels and risk of myocardial infarction in men. JAMA., 291(14): 1730-7, 2004.
POIRIER, P.; GILES, T.D.; BRAY, G.A.; HONG, Y.; STERN, J.S.; PI-SUNYER, F.X.; ECKEL, R.H. Obesity and cardiovascular disease: pathophysiology, evaluation, and effect of weight loss. Arterioscler Thromb Vasc Biol., 26(5): 968-76, 2006.
POIRIER, P.; GILES, T.D.; BRAY, G.A.; HONG, Y.; STERN, J.S.; PI-SUNYER, F.X.; ECKEL, R.H. Obesity and cardiovascular disease: pathophysiology, evaluation, and effect of weight loss: an update of the 1997 American Heart Association Scientific Statement on Obesity and Heart Disease from the Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism. Review. Circulation, 113(6): 898-918, 2006.
REGOLI, D.; DRAPEAU, G.; ROVERO, P.; DION, S.; RHALEB, N.E. Kinin receptor subtypes. J. Cardiovasc. Pharmacol., 15(S6): 30-38, 1990.
RHODES, C.J.; WHITE, M.F. Molecular insights into insulin action and secretion. Eur J Clin Invest., 32(Suppl 3): 3-13, 2002.

73
SACKS, D.B.; MCDONALD, J.M. The pathogenesis of type II diabetes mellitus. A polygenic disease. Am J Clin Pathol., 105(2): 149-56, 1996.
SCHÖLKENS, B. Kinin in the cardiovascular system. Immunopharmacology, 33: 209-216, 1996.
SCHULZE, M.B.; SHAI, I.; RIMM, E.B.; LI, T.; RIFAI, N.; HU, F.B. Adiponectin and future coronary heart disease events among men with type 2 diabetes. Diabetes, 54(2): 534-9, 2005.
SCHUTZ, Y. Abnormalities of fuel utilization as predisposing to the development of obesity in humans. Obes Res., 3 (Suppl 2): 173S-178S, 1995.
SHETTY, G.K.; ECONOMIDES, P.A.; HORTON, E.S.; MANTZOROS, C.S.; VEVES, A. Circulating adiponectin and resistin levels in relation to metabolic factors, inflammatory markers, and vascular reactivity in diabetic patients and subjects at risk for diabetes. Diabetes Care, 27(10): 2450-7, 2004.
STEPPAN, C.M.; BAILEY, S.T.; BHAT, S.; BROWN, E.J.; BANERJEE, R.R.; WRIGHT, C.M.; PATEL, H.R.; AHIMA, R.S.; LAZAR, M.A. The hormone resistin links obesity to diabetes. Nature, 409(6818): 307-12, 2001.
SWERDLOFF, R.S.; BATT, R.A.; BRAY, G.A. Reproductive hormonal function in the genetically obese (ob/ob) mouse. Endocrinology, 98: 1359-1364, 1976.
THIEMERMANN, C. Nitric oxide and septic shock. Gen. Pharmac., 29(2): 159-166, 1997.
TSAO, T.S.; LODISH, H.F.; FRUEBIS, J. ACRP30, a new hormone controlling fat and glucose metabolism. Eur J Pharmacol., 440(2-3): 213-21, 2002.
TSUCHIDA, S.; MIYAZAKI, Y.; MATSUSAKA, T.; HUNLEY, T.E.; INAGAMI, T.; FOGO, A.; ICHIKAWA, I. Potent antihypertrophic effect of the bradykinin B2 receptor system on the renal vasculature. Kidney Int., 56(2): 509-16, 1999.
VANTYGHEM, M.C.; PRESS, M. Management strategies for brittle diabetes. Ann Endocrinol. (Paris), 67(4): 287-96, 2006.

74
VGONTZAS, A.N.; BIXLER, E.O.; PAPANICOLAOU, D.A.; CHROUSOS, G.P. Chronic systemic inflammation in overweight and obese adults. JAMA., 283(17): 2235-2236, 2000.
WASSINK, A.M.; OLIJHOEK, J.K.; VISSEREN, F.L. The metabolic syndrome: metabolic changes with vascular consequences. Eur J Clin Invest., 37(1): 8-17, 2007.
WEYER, C.; PRATLEY, R.E.; SALBE, A.D.; BOGARDUS, C.; RAVUSSIN, E.; TATARANNI, P.A. Energy expenditure, fat oxidation, and body weight regulation: a study of metabolic adaptation to long-term weight change. J Clin Endocrinol Metab., 85(3): 1087-94, 2000.
WEYER, C.; FUNAHASHI, T.; TANAKA, S.; HOTTA, K.; MATSUZAWA, Y.; PRATLEY, R.E.; TATARANNI, P.A. Hypoadiponectinemia in obesity and type 2 diabetes: close association with insulin resistance and hyperinsulinemia. J Clin Endocrinol Metab., 86(5): 1930-5, 2001.
WEYER, C.; YUDKIN, J.S.; STEHOUWER, C.D.; SCHALKWIJK, C.G.; PRATLEY, R.E.; TATARANNI, P.A. Humoral markers of inflammation and endothelial dysfunction in relation to adiposity and in vivo insulin action in Pima Indians. Atherosclerosis, 161(1): 233-42, 2002.
WILSON, C.; GILLILAND, S.; MOORE, K.; ACTON, K. The epidemic of extreme obesity among American Indian and Alaska Native adults with diabetes. Prev Chronic Dis., 4(1): A06, 2007.
WITHERS, D.J.; GUTIERREZ, J.S.; TOWERY, H.; BURKS, D.J.; REN, J.M.; PREVIS, S.; ZHANG, Y.; BERNAL, D.; PONS, S.; SHULMAN, G.I.; BONNER-WEIR, S.; WHITE, M.F. Disruption of IRS-2 causes type 2 diabetes in mice. Nature, 391(6670): 900-4, 1998.
XU, J.; CARRETERO, O.A.; SUN, Y.; SHESELY, E.G.; RHALEB, N.E.; LIU, Y.H.; LIAO, T.D.; YANG, J.J.; BADER, M.; YANG, X.P. Role of the B1 kinin receptor in the regulation of cardiac function and remodeling after myocardial infarction. Hypertension., 45(4): 747-53, 2005.
YOKOTA, T.; ORITANI, K.; TAKAHASHI, I.; ISHIKAWA, J.; MATSUYAMA, A.; OUCHI, N.; KIHARA, S.; FUNAHASHI, T.; TENNER, A.J.; TOMIYAMA, Y.; MATSUZAWA, Y. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood, 96(5): 1723-32, 2000.

75
YORK, D.A.; ROSSNER, S.; CATERSON, I.; CHEN, C.M.; JAMES, W.P.; KUMANYIKA, S.; MARTORELL, R.; VORSTER, H.H. Obesity, a worldwide epidemic related to heart disease and stroke: worldwide demographics of obesity. Circulation, 110(18): e463-70, 2004.
ZHANG, Y.; PROENCA, R.; MAFFEI, M.; BARONE, M.; LEOPOLD, L.; FRIEDMAN, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature, 372: 425-432, 1994.

ANEXO A - PARECER DA COMISSÃO DE ÉTICA EM
MANIPULAÇÃO E EXPERIMENTAÇÃO ANIMAL
(CEMEA/UMC)


Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























