Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS- GRADUAÇÃO EM CIÊNCIAS FARMACÊUTICAS ÁREA DE CONCENTRAÇÃO: BIOANÁLISES
IgA SÉRICA, SECRETORA, IgE TOTAL E ESTADO NUTRICIONAL EM CRIANÇAS COM INFECÇÕES POR ENTEROPARASITAS
VALÉRIA CRISTINA RIBEIRO DANTAS
Orientadora: Profa. Dra. VALÉRIA SORAYA DE FARIAS SALES
Natal/RN2003

VALÉRIA CRISTINA RIBEIRO DANTAS
IgA SÉRICA, SECRETORA, IgE TOTAL E ESTADO NUTRICIONAL EM CRIANÇAS COM INFECÇÕES POR ENTEROPARASITAS
Dissertação submetida ao Programa de Pós-Graduação em Ciências Farmacêuticas para obtenção do Grau de Mestre em Ciências Farmacêuticas – Área de Concentração: Bioanálises.
Orientadora: PROFa. Dra. VALÉRIA SORAYA DE FARIAS SALES
Natal/ 2003

Aquela que representa amor, força, coragem e que com palavras certas na hora
exata me reergueram durante toda minha vida, e que sem sua educação e esforço
jamais poderia ter chegado até aqui.
Dedico este trabalho a minha grande e querida Mãe
Meu maior orgulho!

Ao meu companheiro no amor e no trabalho
Que me ensina todos os dias, sentimentos importantes da vida
Pelo seu amor, sua compreensão e paciência!
Eu consegui chegar ao meu objetivo
Ralfo, à você dedico este trabalho, que com sua ajuda
Pode ser Concluído, obrigado por ser tudo isso!!!!

Aos meus irmãos, André e Ana Conceição e a Cláudio.
Que representam orgulho e sabedoria, exemplos de perseverança.
Constante apoio e equilíbrio
Á Thiago que nos encheu de Alegrias e Orgulho!
Obrigado pelos ensinamentos de vida!
Dedico à vocês, este trabalho!

A Todas as crianças que vivem e sobrevivem a tantas dificuldades
Verdadeira demonstração de vitória e esperança
Deus certamente está com vocês!
Meu obrigado pela ajuda inocente e voluntária na
Execução deste trabalho na procura de um
Mundo melhor.

AGRADECIMENTOS
“A Deus, luz e força em minha vida!”
A minha Mãe, pelas constantes orações, preocupações, apóio, carinho e
compreensão.
Ao meu esposo Ralfo, pelo seu companheirismo, amor, dedicação, compreensão,
paciência e orientações durante todo este tempo, além de sua ajuda profissional,
muito importante para o desenvolvimento e término deste trabalho.
A Cláudio, Ana Conceição, Thiago e André, pelas correções, orientações, carinho,
momentos de alegrias e constante apóio no desenvolvimento deste trabalho.
A Professora Dra. Valéria Soraya de Farias Sales, pela orientação e inteligência,
pelos ensinamentos compartilhados, e dedicados neste trabalho, como também
sua amizade, credibilidade e incentivos a ciência. O meu obrigado!
A Professora Dra. Maria das Graças Almeída Thornton, que durante este tempo
colaborou com o incentivo, apoio, palavras e empenho indispensáveis à realização
deste trabalho e ao Laboratório LIATEC, pela contribuição nas análises
hematológicas além de todo o apóio, dos colegas Sérgio e Jhon Kenedy.
A Professora Dra.Tereza Maria Dantas de Medeiros, que esteve junto comigo
durante esta longa caminhada, participante efetivamente de etapas importantes
que constituíram momentos de alegrias e dificuldades. Pelo seu jeito humano,
sensível, amizade e carinho comigo, muito obrigado!!
A amiga de todos os dias e todas as horas, Ana Célia Souza Lucena, que sempre
esteve nos momentos mais difíceis e nas horas mais importantes, passando
sempre muita força e confiança. Obrigado pelas constantes preucupações e força,
minha amiga!

A Nutricionista Ana Kalina Dantas dos Santos pela colaboração e ajuda na
avaliação nutricional das crianças.
A Professora Dra. Lúcia de Fátima Campos Pedroza Schxarzschild, pela
colaboração, receptividade e ajuda na avaliação nutricional das crianças e no
enriquecimento de dados nutricionais ao trabalho.
Aos Professores da Universidade Potiguar do curso de Farmácia e Bioquímica,
Alexandre Augusto, Íris Cristina, Carla Elenuska e José Queiroz Filho, pelo
constante apoio e ajuda que me deram durante todo este período, em particular
aos Professores Alexandre Augusto e Íris Cristina pela grande ajuda na execução
de exames parasitológicos e análises bioquímicas.
Ao Laboratório de Alergia e Biologia Molecular dos Departamentos de Pediatria e
Biologia Molecular da Faculdade de Medicina de Ribeirão Preto – USP, em
especial a Dra. Luiza Carla de Paula Arruda, pela colaboração e presteza
dispensadas a este trabalho.
A amiga Rita de Cássia Oliveira, pela ajuda em todas as coletas realizadas nas
crianças, com tanto empenho e dedicação e pela sua amizade, carinho e
preocupações dispensadas à minha pessoa. O meu muito obrigado!
A Creche Nossa Senhora de Lourdes, em especial a professora Kátia que com os
seus esforços e dedicação pelas crianças, foi possível obter êxito no cumprimento
das amostras tão necessários para a concretização deste trabalho.
A Estatística Francielle do Departamento de Estatística da Universidade Federal
do Rio Grande do Norte, pela presteza e paciência, com que auxiliou na analises
estatísticas deste trabalho.

A Nutricionista Clélia de Oliveira Lyra, pela ajuda e orientação concedida no
trabalho, além de sua amizade, atenção e ajuda nas análises estatísticas, deste
trabalho. Meu obrigado!!
A todos os Professores do Departamento de Análises Clínicas e Toxicológicas da
UFRN, em especial, aos Professores dos Laboratórios de Parasitologia Clínica e
Bioquímica Clínica.
Aos Professores e amigos do Departamento de Análises Clínicas e Toxicológicas
Sandra Rezende, Julia Fernandes, Edna Marques, Ivanaldo Amâncio e Maíza, que
sempre estiveram ao meu lado, incentivando e apoiando durante todas as fases
de realização deste trabalho.
As Professoras e amigas Tereza Neuma, Zélia Maria, Polliana, Adriana Rezende e
Maria Goretti, pelo incentivo e constante preocupação na caminhada desta
jornada, além da contribuição imprescindível nas análises das Eletroforeses de
Proteínas pela professora Zelinha.
Ao Professor Eider de Araújo Carvalho – Diretor do NUPLAN pela gentileza na
doação dos medicamentos para o tratamento das crianças.
Aos colegas de curso: Alexandre, Janaína, Mabel, Cristiane, Valéria Morgiana,
Alaíze, Jucimare, Karine, Cristina, Edilson e Severina Carla, pela convivência
harmoniosa e cumplicidade compartilhada no decorrer deste período de trabalho.
A todos os funcionários do Laboratório de Análises Clínicas da Universidade
Potiguar – LACT, em especial à Fernanda Carla Lopes Mendes, pelo apóio na
obtenção de amostras de medicamentos para o tratamento das crianças.
A Eliane da Silva Rodrigues e Cristiane Alves de Moura, pelo carinho e ajuda na
coleta de material deste trabalho.

A professora Dra. Fernanda Nervo Raffim, pelas orientações e credibilidade
depositadas em mim, como também aos demais professores que compõem o
corpo Docente do Curso de Pós-Graduação em ciências Farmacêuticas pelos
ensinamentos e incentivos dedicados a ciência, pesquisa e empenho na
implantação deste programa.
As funcionárias Hilda Maria Rodrigues do Laboratório de Imunologia Clínica e
Maria da Luz do Laboratório de Parasitologia Clínica do Departamento de Análises
Clínicas e Toxicológicas da UFRN, pelo apoio e ajuda concedidos neste trabalho.
As amigas e colegas profissionais Adriana Amaral, Luciana Emília, Ednalva Lobo,
Jacira e Ana Cristina, pelo constante apoio e compreensão comigo, nestes últimos
meses.
A todos que direta ou indiretamente contribuíram para o êxito e término deste
trabalho, o meu muito Obrigado!!
A Deus que me deu forças, inteligência e saúde para sobreviver a tantas dificuldades e adversidades!
Obrigado a todos vocês!

Vivemos com o que recebemos,
mas marcamos a vida com o que damos.
(Winston Churchill)

RESUMO
Infecções por parasitas intestinais são uma das principais causas de
morbidade em humanos e, suas relações com níveis sócio-econômicos e
condições de higiene em países em desenvolvimento já são bem estabelecidas.
Muitos trabalhos, porém, estão sendo realizados para elucidar as complexas
interações entre nutrição, estas infecções e resposta imunológica, pois é visto que
desnutrição compromete a imunidade aumentando a susceptibilidade para
doenças infecciosas e estas por sua vez podem prejudicar o estado nutricional
humano.
Sabe-se que parasitas helmínticos estimulam síntese de IgE tanto policlonal
como específica para antígeno dos mesmos e que IgA secretora, principal
imunoglobulina de defesa das mucosas, pode atuar contra protozoários como a
Giardia lamblia e contra helmintos como Trichuris tichiura e Strongyloides
stercorales. Alguns estudos mostram que a desnutrição energético protéica
influencia na produção destas respostas, mas outros autores mostram resultados
divergentes.
Neste trabalho avaliou-se os níveis de IgE total, IgA sérica e secretora,
contagem de eosinófilos sanguíneos, níveis de proteínas séricas e estado
nutricional, em 103 crianças de baixo nível sócio-econômico, para se averiguar
uma correlação entre esses e infecção por enteroparasitas.
Participaram do estudo crianças de ambos os sexos, com idade de 3 a 6
anos, freqüentadoras da mesma creche e residentes em um bairro com precárias
condições de higiene e saneamento básico, na cidade do Natal.
Os resultados obtidos mostraram que as deficientes condições ambientais e
sócio-econômicas favoreceram a uma alta freqüência de infecção por
enteroparasitas, principalmente Trichuris trichiura e Ascaris lumbricoides entre os
helmintos e Endolimax nana e Giárdia lamblia entre os protozoários. Desnutrição
leve sem déficit protéico foi observada em 30% das crianças, as quais também

não apresentaram deficiências significativas de IgA sérica e secretora. As crianças
parasitadas apresentaram eosinofilia sanguínea e níveis séricos de IgE total
elevados confirmando a importante participação das mesmas na resposta imune
contra helmintos. Pode-se, portanto, sugerir que as crianças apesar de
poliparasitadas não estavam com sua resposta imune de mucosa contra parasitas,
prejudicada, provavelmente por ainda não estarem intensamente infectados, como
observado na contagem de ovos por grama de fezes e também por não terem seu
estado nutricional gravemente comprometido.

ABSTRACT
Infections for intestinal parasites are one of the main morbidade causes in
humans and, its relationships with socioeconomic levels and hygiene conditions in
countries in development are already very established. Many works, even so, they
are being accomplished to elucidate the complex interactions among nutrition,
these infections and answer imunológica, because it is seen that malnutrition
commits the immunity increasing the susceptibilidade for infectious diseases and
these for its time can harm the state human nutricional.
It is known that sponge helmínticos they stimulate synthesis of IgE so
much policlonal as specific for the same ones and that IgA secretora, main
imunoglobulina of defense of the mucous ones, can act against protozoa as the
Giardia lamblia and against helmintos as Trichuris tichiura and Strongyloides
stercorales. Some studies show that the malnutrition energy protéica influences in
the production of these answers, but some authors show results divergentes.
In this work it was evaluated the levels of total IgE, IgA sérica and secretora,
contagem of sanguine eosinófilos, levels of proteins séricas and state nutricional,
in 103 children of low socioeconomic level, to discover a correlation between those
and infection for enteroparasitas.
They participated in the study children of both sexes, with age of 3 to 6
years, visitors of the same creche and residents in a neighborhood with precarious
hygiene conditions and basic saneamento, in the city of Christmas.
The obtained results showed that the faulty environmental and
socioeconomic conditions favored to a high infection frequency for enteroparasitas,
mainly Trichuris trichiura and Ascaris lumbricoides between the helmintos and
Endolimax sleep and Giárdia lamblia among the protozoa. Light malnutrition
without deficit protéico was observed in 30% of the children, which didn't also
present significant deficiencies of IgA sérica and secretora. The sponged children

presented sanguine eosinofilia and levels séricos of high total IgE confirming the
important participation of the same ones in the immune answer against helmintos.
It cannot him, therefore, to suggest that the children in spite of poliparasitadas
were not with its immune answer of mucous harmed, probably for they be not still
intensely infected, as observed in the contagem of eggs by gram of feces and also
for they have not its state nutricional seriously committed.

LISTA DE FIGURAS
FIGURA 1 Níveis de proteínas totais e estado nutricional de 101
crianças de baixo nível socioeconômico, freqüentadoras da
Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
56
FIGURA 2 Concentração de albumina sérica e estado nutricional de 101
crianças baixo nível socioeconômico, freqüentadoras da
Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
57
FIGURA 3 Concentrações de globulinas séricas e estado nutricional de
101 crianças de baixo nível socioeconômico, freqüentadoras
da Creche Nossa Senhora de Lourdes, na cidade do Natal-
RN.
58
FIGURA 4 Quantificação do número de ovos de A. lumbricoides, por
grama de fezes, e estado nutricional de 101 crianças de
baixo nível socioeconômico.
62
FIGURA 5 Quantificação do número de ovos de T. trichiura, por grama
de fezes e estado nutricional de 101 crianças de baixo nível
socioeconômico.
63
FIGURA 6 Número absoluto de eosinófilos no sangue periférico de
acordo com a idade em 103 crianças, freqüentadoras da
Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
64

FIGURA 7 Número absoluto de eosinófilos no sangue periférico de
acordo com o sexo de 103 crianças de baixo nível
socioeconômico, freqüentadoras da Creche Nossa Senhora
de Lourdes, na cidade do Natal-RN.
65
FIGURA 8 Número absoluto de eosinófilos no sangue periférico de
acordo com o estado nutricional de 101 crianças de baixo
nível socioeconômico, freqüentadoras da Creche Nossa
Senhora de Lourdes, na cidade do Natal-RN.
66
FIGURA 9 Correlação entre o número absoluto de eosinófilos,
quantificação do número de ovos de A. lumbricoides e T.
trichiura por grama de fezes.
67
FIGURA 10 Correlação entre IgA sérica, IgA secretora e quantidade de
ovos de Ascaris lumbricoides por grama de fezes.
73
FIGURA 11 Correlação entre IgA sérica, IgA secretora e quantidade de
ovos de Trichuris trichiura por grama de fezes.
74
FIGURA 12 Correlação entre níveis de IgA sérica e IgA secretora de 103
crianças de baixo nível socioeconômico, freqüentadoras da
Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
75

FIGURA 13 Correlação entre as concentrações de IgA sérica, IgA
secretora e IgE total.
76
FIGURA 14 Correlação entre o número absoluto de eosinófilos, IgA
secretora e IgA sérica.
77
FIGURA 15 Correlação entre a concentração sérica de IgE total e a
quantidade de ovos de A. lumbricoides e T. trichiura por
grama de fezes.
81
FIGURA 16 Correlação entre níveis de IgE total e número absoluto de
eosinófilos de 103 crianças de baixo nível socioeconômico,
freqüentadoras da Creche Nossa Senhora de Lourdes, na
cidade do Natal-RN.
82
FIGURA 17 Concentração de IgE total e estado nutricional de 101
crianças de baixo nível socioeconômico, freqüentadoras da
Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
83

LISTA DE TABELAS
TABELA 1 Distribuição por sexo e idade de 103 crianças,
freqüentadoras, da Creche Nossa Senhora de Lourdes, na
cidade do Natal-RN.
52
TABELA 2 Distribuição segundo os indicadores sociais mínimos das
famílias de 103 crianças, freqüentadoras da Creche Nossa
Senhora de Lourdes, na cidade do Natal-RN.
54
TABELA 3 Avaliação do estado nutricional de 101 crianças de baixo
nível socioeconômico, freqüentadoras da Creche Nossa
Senhora de Lourdes, na cidade do Natal-RN.
55
TABELA 4 Parasitismo, segundo o número de espécies infectantes em
103 crianças de baixo nível socioeconômico, freqüentadoras
da Creche Nossa Senhora de Lourdes, na cidade do Natal-
RN.
59
TABELA 5 Freqüência de parasitas intestinais em 103 crianças de baixo
nível socioeconômico, freqüentadoras da Creche Nossa
Senhora de Lourdes, na cidade do Natal-RN.
60
TABELA 6 Freqüência de parasitas intestinais presentes nas 103
crianças de baixo nível socioeconômico, freqüentadoras da
Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
61

TABELA 7 Mediana dos níveis de IgA sérica e IgA secretora de 103
crianças de baixo nível socioeconômico, freqüentadoras da
Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
68
TABELA 8 Mediana das concentrações de IgA sérica e IgA secretora,
segundo o estado nutricional de 101 crianças de baixo nível
socioeconômico, freqüentadoras da Creche Nossa Senhora
de Lourdes, na cidade do Natal-RN.
69
TABELA 9 Parasitismo segundo o número de espécies infectantes e
relação com as concentrações de IgA sérica em 103 crianças
baixo nível socioeconômico.
70
TABELA 10 Parasitismo segundo o número de espécies infectantes e
relação com as concentrações de IgA secretora em 103
crianças de baixo nível socioeconômico.
71
TABELA 11 Mediana das concentrações de IgE total de 103 crianças de
baixo nível socioeconômico, freqüentadoras da Creche
Nossa Senhora de Lourdes, na cidade do Natal-RN.
78
TABELA 12 Parasitismo segundo o número de espécies infectantes e
relação com as concentrações de IgE sérica em 103 crianças
de baixo nível socioeconômico.
79

ÍNDICE
DEDICATÓRIA 3
AGRADECIMENTOS 7
LISTA DE FIGURAS 14
LISTA DE TABELAS 17
RESUMO 19
1 INTRODUÇÃO 21
2 REVISÃO DE LITERATURA 23
2.1 Sistema Imune das Mucosas 23
2.2 Imunoglobulina A (IgA) 26
2.3 Imunoglobulina E (IgE) 31
2.4 Eosinófilos e Infecções Parasitárias 34
2.5 Infecção Parasitária 36
2.6 Imunidade e Nutrição 38
3 OBJETIVOS 41
3.1 Objetivo Geral 41 3.2 Objetivos Específicos 41
4 CASUÍSTICA, MATERIAL E MÉTODOS 42
4.1 População de Estudo 42
4.2 Materiais 43

4.2.1 Amostras Biológicas 43
4.3 Métodos 44
4.3.1 Dosagens de IgA sérica, IgA secretora e IgE total 44
4.3.1.1 Determinação de IgA sérica 44
4.3.1.2 Determinação de IgA secretora 45
4.3.1.3 Determinação de IgE total 45
4.3.2 Determinação de Proteínas Totais 46
4.3.2.1 Determinação de Albumina sérica 46
4.3.2.2 Determinação das Globulinas 47
4.3.3 Eletroforese de Proteínas 47
4.3.4 Contagem Absoluta de Eosinófilos 47
4.3.5 Contagem Relativa de Eosinófilos 48
4.3.6 Exame Parasitológico de Fezes (EPF) 48
4.3.6.1 Método de Hoffman, Pons & Janer 48
4.3.6.2 Método de Faust et al. 49
4.3.6.3 Método de Baermann-Moraes 49
4.3.6.4 Método de Kato-Katz 49
4.3.7 Avaliação Nutricional 50
4.3.8 Análise Estatística dos Resultados 51
5 RESULTADOS 52
5.1 Características da Amostra 52
5.2 Fatores socioeconômicos 53
5.3 Avaliação Nutricional 55

5.3.1 Proteínas Totais e Frações 56
5.4 Infecção Parasitária 59
5.4.1 Carga Parasitária 62
5.5 Contagem Absoluta de Eosinófilos 64
5.6 Dosagem de Imunoglobulinas 68
5.6.1 Dosagens de IgA sérica e IgA secretora 68
5.6.2 Dosagem de IgE total 78
6 DISCUSSÃO 84
7 CONCLUSÕES 93
ABSTRACT 95
8 REFERÊNCIAS BIBLIOGRÁFICAS 97
ANEXOS 113

1 INTRODUÇÃO
O sistema imune das mucosas é formado pelos tecidos linfóides que estão
associados às superfícies mucosas do trato gastrintestinal, das vias respiratórias e
do trato urogenital. Sua principal função consiste em propiciar defesa ao hospedeiro
nas superfícies mucosas que são permanentemente provocadas por antígenos e
devem desenvolver uma resposta efetiva para limitar eventual invasão por agentes
infecciosos, mas também preservar a integridade anatômica e funcional das
estruturas que as superfícies mucosas protegem (HIROI & KIYONO, 2000; SILVA,
2001).
A imunoglobulina A (IgA) exibe várias propriedades que permitem a sua
atuação eficiente no ambiente das mucosas. Foram descritos receptores para IgA
em células inflamatórias que podem promover a destruição de patógenos celulares,
como parasitas, através do mecanismo de citotoxicidade celular dependente de
anticorpo (ADCC). Pode também exercer importante função quando é formado o
complexo chamado de IgA secretório que apresenta propriedades especiais as quais
explicam sua predominância nas mucosas, como a resistência à proteólise e a
inibição da aderência e colonização de vírus e bactérias na superfície dos
enterócitos, contribuindo assim para proteger as superfícies epiteliais contra reações
inflamatórias (RENEGAR et al., 1998; PEEBLES et al., 2001).
Sabe-se que a imunoglobulina E (IgE) está relacionada a processos alérgicos
e também constitui importante defesa nas infecções parasitárias principalmente
contra helmintos. As infecções helmínticas quando ocorrem em crianças de baixo
nível socioeconômico sugerem que o índice de exposição a agentes parasitários
pode estar relacionado ao desenvolvimento do estado nutricional. Fatores como a
freqüência na exposição e a intensidade da carga parasitária podem contribuir para
a obtenção de uma resposta imunológica inadequada (ROSÁRIO FILHO et al., 1982;
SUTTON, 1993).
Deficiências nutricionais podem influenciar no estado de infecção parasitária
por meio da modulação da resposta imunológica. A má nutrição energético proteíca
pode comprometer a produção de IgA que tem um importante papel na imunidade
da mucosa gastrintestinal. Por outro lado, a produção de IgE também pode ser

22
influenciada pelo estado nutricional (CASTILHO DE ARIZA et al., 1983; CHANDRA,
1984).
Embora se reconheça que as células imunocompetentes da mucosa intestinal
estejam implicadas na patogenia das doenças parasitárias, o relevante papel destas
células e a modulação da produção de anticorpos no intrigante complexo da relação
parasita-hospedeiro ainda carecem de elucidações. Diante destes fatores,
pretendemos estudar a relação entre IgA sérica, IgA secretora, IgE total, estado
nutricional e enteroparasitoses em crianças carentes que freqüentam creche na
cidade do Natal.

23
2 REVISÃO DE LITERATURA
2.1 Sistema Imune das Mucosas
As superfícies das mucosas contam com uma proteção natural onde atuam
mecanismos inespecíficos como movimento peristáltico, transporte muco-ciliar,
enzimas e mecanismos adaptativos, como: o sistema imune de mucosas, que
compreende o MALT (“Mucosal associated lymphoid tissue”), o qual nas vias
respiratórias é denominado BALT (“Bronchial associated lymphoid tissue”) e nas vias
digestivas, GALT (“Gut associated lymphoid tissue”) (STROBER et al., 1981;
KAGNOFF, 1993).
Os MALTs se distribuem anatomicamente pelas mucosas, e seus
componentes individuais incluem: a) um anel de estruturas linfóides circundando a
faringe (anel de Waldeyer), formado principalmente pelas amígdalas nasofaríngeas
(NALT – tecido linfóide associado ao nariz) e orofaríngeas, que podem ainda incluir
tonsilas linguais e tubárias em algumas espécies; b) tecido linfóide associado à
laringe (LALT); c) placas de Peyer intestinais; d) apêndice vermiforme; e) nódulos
linfóides agregados ao cólon; f) nódulos linfóides isolados dispersos em toda a
mucosa intestinal, do esôfago ao ânus; g) tecido linfóide associado ao brônquio
(BALT); h) tecido linfóide associado à conjuntiva (CALT). Esses tecidos estão
concentrados em locais de grande proliferação microbiana e de estimulação
antigênica. As respostas imunes das mucosas ocorrem tanto nos MALTs como em
seus linfonodos de drenagem. O “MALT” serve para amplificar as respostas de
memória através da atividade de localização de antígenos (TEODÓSIO et al., 1993;
CUTULO & LAMEGO, 1994).
Os MALTs são constituídos basicamente de uma rede de reticulina
preenchida por linfócitos e macrófagos. Os folículos linfóides são povoados
principalmente de linfócitos B que expressam proporções variáveis de
imunoglobulinas, principalmente IgA, constituindo 60% das células da mucosa
brônquica e até 85% da mucosa intestinal, de acordo com a exposição antigênica e
tipo de antígeno. A área parafolicular lateral ao folículo é uma área com linfócitos T-
dependentes, consistindo cerca de 20% do número total de células do MALT (MADI
et al., 2001).

24
O tecido linfóide nas superfícies das mucosas é constituído de linfócitos
distribuídos frouxamente ou de linfócitos organizados em folículos e agregados.
Existe uma compartimentalização dessas células, com uma população linfóide muito
distinta encontrada no epitélio, os linfócitos intra-epiteliais, compostos principalmente
de linfócitos T CD8+ com função supressora e citotóxica. Tais linfócitos intra-
epiteliais exercem papel importante na regulação do epitélio. Considerando-se as
observações de MILLER & NAWA (1979), os linfócitos T podem regular o número de
células caliciformes no epitélio e sua atividade secretora. Células T apresentando
receptor do tipo gama-delta também são encontradas em grande número no epitélio
das mucosas (STROBER et al., 1981; KAGNOFF, 1993).
Os MALTs são recobertos por um epitélio especializado, o linfoepitélio, que
captura ativamente macromoléculas e microorganismos. Os folículos linfóides
recobertos pelo linfoepitélio estão envolvidos internamente na lâmina própria da
mucosa. O linfoepitélio forma um ambiente favorável para o contato entre antígenos,
linfócitos intra-epiteliais e células apresentadoras de antígeno. Os sítios de captação
de antígeno são o linfoepitélio e outras áreas da superfície epitelial, incluindo as
zonas de extrusão vilosa do intestino. Assim, o potencial para interação entre
antígeno e células linfóides se espalha por toda a superfície mucosa (STROBER et
al., 1981). A capacidade do linfoepitélio em captar antígenos concentra as respostas
imunes nos MALTs e seus linfonodos de drenagem (KAGNOFF, 1993; MADI et al.,
2001).
As mucosas são permanentemente estimuladas por antígenos e precisam
desenvolver um sistema imune efetivo para limitar eventual invasão por agentes
infecciosos, mas também preservar a integridade anatômica e funcional das frágeis
estruturas que elas protegem. Uma quebra desse equilíbrio pode resultar em dano
permanente e disseminado da mucosa. Mecanismos imunes locais interagem com
respostas que ocorrem em outros sítios, sejam em mucosas ou não. Anticorpos
produzidos nas mucosas, assim como reações mediadas por células, devem induzir
resistência contra bactérias, vírus, parasitas, toxinas, alérgenos e etc. (ABBAS et al.,
2000; ROITT et al., 2003).
Essas respostas imunes são iniciadas após estimulação antigênica, e o
linfoepitélio especializado desempenha um papel maior em captar estes antígenos e
transferi-los por células apresentadoras de antígenos no MALT. Nestes locais são
apresentados para linfócitos T, iniciando assim a cascata de eventos que resultam

25
na resposta imune. Os produtos dos linfócitos B locais são anticorpos específicos
que potencializam a fagocitose, promovem a eliminação de bactérias e neutralizam
toxinas. Os linfócitos T efetores podem ser citotóxicos, ou podem agir através da
produção de citocinas, que medeiam respostas inflamatórias subagudas ou crônicas
(CUTULO & LAMEGO, 1994; SOUZA, 2001).
Uma vez que um contato antigênico tenha iniciado, o MALT tem duas
importantes funções: a de reconhecer patógenos e organizar uma resposta imune
adequada, que envolve principalmente IgA, ou a de suprimir as respostas locais ou
periféricas contra substâncias inócuas, através do mecanismo de tolerância. A
maioria dos contatos com antígenos estranhos se dá nas superfícies das mucosas, e
o mais freqüente é um estado de hiporresponsividade ou tolerância (CARVALHO et
al., 1998; PAINTLIA et al., 2002). Tais como os folículos linfóides do baço e dos
linfonodos, as regiões centrais de folículos mucosos são áreas ricas em células B,
que freqüentemente contêm centros germinativos (ABBAS et al., 2000).
O GALT é constituído de células imunocompetentes infiltrantes das mucosas
e de estruturas estrategicamente distribuídas por toda superfície mucosa, como as
placas de Peyer e os linfonodos mesentéricos, povoados por células de diversas
subpopulações com as mais variadas funções, inclusive a de produzir anticorpos
secretores, com predominância absoluta de IgA (ABBAS et al., 2000; ELIA et al.,
2001; ROITT et al., 2003).
As placas de Peyer encerram também um pequeno número de células T
CD4+, principalmente nas regiões interfoliculares. Algumas das células epiteliais
abaixo das placas de Peyer são células membranosas (células M), especializadas,
que não tem vilosidades, são pinocíticas ativas e transportam macromoléculas do
lúmem intestinal para os tecidos subepiteliais. São admitidas como exercendo um
importante papel na liberação de antígenos às placas de Payer. Folículos que são
morfologicamente e funcionalmente semelhantes são abundantes no apêndice e são
encontrados em menor número em grande parte dos tratos gastrintestinal e
respiratório (ABBAS et al., 2000; JANEWAY et al., 2000; ELIA et al., 2001; ROITT et
al., 2003).
As células M estão localizadas nos sítios indutores, pequenas regiões da
membrana da mucosa que estão situadas sobre os folículos linfóides. Os antígenos
transportados através da membrana da mucosa pelas células M podem ativar as
células B destes folículos linfóides. Estas células B ativadas se diferenciam em

26
células plasmáticas, que deixam os folículos e secretam IgA, que são transportados
através das células epiteliais e são liberados para o lúmem, com a IgA secretora,
onde podem interagir com antígenos (GOLDSBY et al., 2002).
A lâmina própria intestinal, que se localiza sob a camada epitelial, contém
uma população mista de células. Essa população inclui linfócitos T, a maioria dos
quais são CD4+ e têm o fenótipo das células ativadas. Há probabilidade de que as
células T inicialmente reconheçam e respondam aos antígenos dos linfonodos
mesentéricos regionais e remigrem para o intestino, para povoar a lâmina própria,
que contém também grande número de linfócitos B ativados e de plasmócitos, bem
como macrófagos, células dendríticas, eosinófilos e mastócitos (SEIDEL et al.,
2001).
O meio mucóide favorece e influencia o desenvolvimento de resposta do tipo
2 (Th2), com produção de IL-4, IL-5, IL-10 e de outras citocinas antiinflamatórias,
como o TGF-β, que também tem capacidade para induzir a produção preferencial de
IgA (GREWAL et al., 2000). Desta maneira, a produção de citocinas antiinflamatórias
pode favorecer a supressão de respostas imunes no local do antígeno (PAINTLIA et
al., 2002).
A grande maioria dos anticorpos é encontrada em líquidos corporais e ativam
as células efetoras somente após a ligação a antígenos específicos. A superfície da
mucosa intestinal, que representa uma das vias mais importantes de entrada de
patógenos no hospedeiro, é constituída de células epiteliais que desempenham um
importante papel no transporte de fluídos e secreções, na função de barreira
mecânica frente às agressões do meio externo e na interação dos componentes
luminares com as células imunológicas da lâmina própria (SCROFERNEKER &
POHLMANN, 1998; STITTES et al., 2000).
2.2 Imunoglobulina A (IgA) Um dos aspectos que distingue o sistema imunológico das mucosas, são as
respostas humorais induzidas nos folículos das mucosas que resultam
predominantemente na produção de anticorpos do tipo IgA (PAINTLIA et al., 2002).

27
A IgA foi identificada pela primeira vez por HEREMANS et al.(1959), e é
considerada a principal imunoglobulina presente em secreções como saliva, muco
intestinal, secreções brônquicas, líquido prostático, suor, suco gástrico e lágrima,
além de estar presente também no colostro e no leite, considerado no neonato a
principal fonte de proteção contra patógenos intestinais (SILVA, 2001; BEYER et al.,
2002).
A IgA é importante na defesa imunológica primária contra infecções locais em
áreas como o trato respiratório e gastrintestinal. Seu efeito protetor deve-se a
capacidade de impedir que um agente invasor se ligue e penetre na superfície do
epitélio. A predominância de IgA nas secreções mucosas se explica pela
concentração seletiva de plasmócitos produtores de IgA nas mucosas. Essas células
podem derivar de respostas locais ou à distância (CARVALHO et al., 1998).
A molécula de IgA é constituída de duas cadeias leves (L) do tipo kappa (K)
ou lambda (λ) e duas cadeias pesadas (H) do tipo alfa (α). A IgA pode apresentar-se
na forma monomérica, que tem peso molecular de aproximadamente 165.000 Da, e
na forma dimérica, apresentando um peso molecular de 400.000 Da, e é constituída
de glicoproteínas produzidas por plasmócitos. A IgA representa 15 a 20% das
imunoglobulinas e se divide em duas subclasses: IgA1 (93%) e IgA2 (7%)
(SHELDRAKE et al., 1984).
A IgA1 predomina no soro com uma meia-vida de 5,5 dias, onde é encontrada
sob a forma de monômeros e pode sofrer a ação de proteases de IgA, que são
produzidas por algumas bactérias patogênicas que agem sobre seqüências de
aminoácidos na região de dobradiça da IgA1 (RÚPULO et al., 1998). A IgA2 não é
sensível à ações de enzimas, como as proteases de IgA, por ter uma deleção em
seqüências de aminoácidos e também por conter o componente secretor, o que
favorece sua adaptação, especialmente, em sítios intensamente colonizados, como
o cólon e a faringe. A IgA2 é encontrada nas secreções brônquicas sob a forma
dimérica ou ainda como multímeros e é mais freqüente nas secreções urogenitais e
partes distais do trato gastrintestinal, embora as duas classes possam ser
encontradas em proporções equilibradas, variando de um sítio para outro
(RENEGAR et al., 1998).
A maioria dos plasmócitos intestinais secreta IgA e, preferencialmente, a
subclasse IgA2, ao contrário do que é observado no sangue na forma circulante. Os
mecanismos que regulam a produção e a especificidade das subclasses de

28
imunoglobulinas registram a possibilidade do envolvimento das populações de
linfócitos T helper e supressor, a expressão de receptores específicos e a produção
de citocinas como IL-4, IL-5 e IL-6 (LEBMAN & COFFMAN, 1988).
Nas células B estimuladas por antígenos, a troca de isótipos da IgA é
estimulada pelo fator β de crescimento e transformação (TGF-β), que pode também
ser induzidas por células T e por células estromais não linfóides e IL-5. A razão pela
qual são produzidas maiores quantidades de IgA no sistema imune das mucosas do
que nos outros tecidos é, em parte, a troca de isótipos para a IgA, que ocorre mais
eficientemente no tecido linfóide da mucosa, e em parte porque as células T
auxiliares produtoras de IL-5 são mais numerosas nas mucosas do que nos outros
tecidos linfóides. As células B produtoras de IgA podem também ter uma especial
propensão para residirem nos tecidos mucosos (ISLAM et al., 1991; HIROI et al.,
1998; VAN GINKE et al., 1999; CHUNG et al., 2002; ROITT et al., 2003).
Depois de secretada pelos plasmócitos na lâmina própria, a IgA é
transportada seletivamente através do epitélio da mucosa para o lúmen do órgão.
Esta molécula secretada liga-se pela região Fc ao componente secretor, um
polipeptídio de 90 kDa produzido pelas células epiteliais das membranas das
mucosas. A formação de polímeros de IgA e a ligação ao componente secretor são
essenciais para o transporte através do epitélio (MELLANDER et al., 1985; STOLL et
al., 1993).
A IgA secretora é um dímero com quatro cadeias leves e quatro cadeias
pesadas unidas pelo polipeptídeo denominado de cadeia J (de “joining”) e o
componente secretor. O anticorpo da classe A originado na lâmina própria intestinal,
se liga a um receptor poli-Ig na membrana basolateral de células epiteliais, e é
internalizado por endocitose do complexo IgA e componente secretor, mediado pelo
receptor em vesículas e a migração destas vesículas ocorre para a superfície apical
da célula. O receptor de poli-Ig, que é constituído de glicoproteínas produzidas pelo
epitélio intestinal, encontra-se nas porções basolaterais das células epiteliais. Após
um processo de clivagem proteolítica na região apical do epitélio, o componente
secretor permanece ligado à membrana celular e os dímeros de IgA são liberados na
luz intestinal (STITES et al., 2000; GOLDSBY et al., 2002).
O componente secretor mascara os sítios suscetíveis à clivagem proteolítica
na região flexível da IgA secretora, permitindo à molécula polimérica uma existência
mais prolongada no meio da mucosa que é rica em proteases. A capacidade da IgA

29
dimérica de ligar-se ao componente secretor aumenta a sua capacidade de atuar na
luz intestinal, e o aspecto mais importante é que esta ligação constitui a etapa inicial
básica no mecanismo de transporte de IgA, permitindo ao sistema imunológico das
mucosas fornecer a IgA nos sítios da mucosa (SHELDRAKE et al., 1984;
TEODÓSIO et al., 1993).
A extremidade Fc e o componente secretor da IgA são hidrofílicos e
mucofílicos, e a ligação da IgA a um microorganismo retarda a sua fixação às células
epiteliais, aprisionado-o na camada de muco. Em resumo, as propriedades da região
Fc/componente secretor da IgA permitem a IgA impedir a colonização de patógenos,
sem induzir inflamação. Naturalmente, trata-se de uma propriedade importante numa
região do organismo repleta de substâncias que têm o potencial de induzir respostas
inflamatórias (NAGAO et al., 1995).
Há algumas evidências de que a IgA pode mediar a ADCC através da ligação
ao receptor para Fc. Além disso, a IgA interage através de sua região Fc com a
lactoferrina e lactoperoxidase e, desta maneira, aumenta a função destes elementos
inespecíficos na defesa do hospedeiro. Foram descritos receptores para IgA em
células inflamatórias, que podem promover a destruição de patógenos celulares,
como parasitas, através do mecanismo de ADCC (CARVALHO et al., 1998; SEIDEL
et al., 2001; ROITT et al., 2003).
A IgA secretora dificulta a absorção de partículas antigênicas pelo epitélio
intestinal e impede a aderência e colonização de bactérias, vírus e parasitas na
superfície dos enterócitos (BUENO et al., 2000; GREWAL et al., 2000). Sendo
portanto juntamente com a IgE importante fator nos mecanismos antiparasitários do
sistema imunológico (PEEBLES et al., 2001).
Uma das principais funções dos anticorpos secretores é promover a exclusão
imune, que consiste em um mecanismo não inflamatório para manter fora do
organismo toxinas, microorganismos e outros materiais antigênicos (BRANDTZAEG,
1995).
ELIA (2001a), descreveu que existem muitos indivíduos com deficiência de
IgA, embora sejam saudáveis. A ausência desta imunoglobulina é freqüentemente
associada com infecções do trato respiratório, algumas doenças auto-imunes, asma,
e outras alergias. IgA secretora representa uma barreira local importante para a
destruição de antígenos, pois manifestações gastrintestinais são descritas em
indivíduos com deficiência desta imunoglobulina. Pode ocorrer ocasionalmente uma

30
compensação parcial pelo aumento na síntese de IgM na mucosa e nas secreções,
mas que apresenta uma ação ineficiente. A subclasse mais comprometida na
deficiência de IgA é a IgA1, sendo assim, a preservação relativa de IgA2, predominante nas secreções gastrintestinais, pode manter a função protetora
inalterada (RUPULO et al.,1998).
De acordo com SIRISINHA et al. (1975), diversos estudos experimentais
sugerem que a desnutrição altera os mecanismos de defesa imunológica das
mucosas e compromete a morfologia e a função da barreira intestinal, contra a
invasão de microorganismos e agentes patogênicos em geral. É possível que em
crianças com desnutrição energético protéica exista uma falha na síntese de IgA, ou
que elas apresentem algum bloqueio no transporte de IgA para a superfície das
mucosas (CHANDRA, 1989).
Trabalho realizado por REDDY et al. (1976), demonstrou que o estado
nutricional severo pode comprometer a resposta mediada por IgA. Ao investigar um
grupo de crianças com desnutrição que apresentavam Kwashiorkor e Marasmo, a
concentração de IgA em fluído duodenal, salivar, secreções nasais e lágrimas
estavam significativamente reduzidas quando foram admitidas, e voltaram ao normal
quatro semanas após tratamento. Porém, a concentração de IgA secretora em
crianças com desnutrição energético protéica era semelhante à concentração em
crianças normais, sugerindo que a deficiência de IgA secretora pode ser um
importante fator, promovendo o desenvolvimento de infecções bacterianas,
correspondendo a elevada incidência de infecções severas na mucosa de crianças
com desnutrição protéico calórica na forma grave (FOGAÇA, 2001).
Existem poucas referências na literatura sobre a relação das infecções por
enteroparasitas e níveis de IgA sérica e secretora. ORTIZ et al. (2000) estudando
dois grupos de crianças de duas comunidades diferentes em estágios de
desnutrição, descreveu que deficiências nutricionais em crianças infectadas por
Ascaris lumbricoides e Trichuris trichiura podem interferir e estar diretamente
relacionadas com as concentrações de IgA secretora, como também IgE total. Os
resultados mostram que a medida que aumenta a intensidade da carga parasitária e
a desnutrição, se observa que há uma tendência a diminuir a IgA secretora, com
aumento de IgE total, sugerindo que a desnutrição altera o mecanismo de defesa
das mucosas e compromete a barreira intestinal contra a invasão de patógenos

31
intestinais, embora estes resultados apresentem significado quando comparam-se
as duas populações de crianças entre si (ELIA, 2001b).
2.3 Imunoglobulina E (IgE)
A IgE é uma imunoglobulina monomérica e tem peso molecular de 190.000
Da, constituindo apenas cerca de 0,004% das imunoglobulinas totais do soro
(BACHARIER & GEHA, 2000).
A IgE está presente no cordão umbilical, mucosas, colostro, sangue e tecidos,
nos quais encontra-se fixada aos mastócitos. Os plasmócitos produtores de IgE
distribuem-se quase que completamente nos tecidos linfáticos adjacentes ao trato
respiratório e gastrintestinal (GEHA, 1984; BELL, 1996; RODRIGUES, 2001).
A ação da IgE é ampliada dependendo das atividades dos receptores aos
quais ela está ligada (SUTTON & GOULD, 1993). A IgE se liga aos receptores Fc
nas membranas dos basófilos sangüíneos e dos mastócitos tissulares (RIVERA et
al., 1998). A ligação cruzada das moléculas de IgE, aos receptores pelo antígeno,
induz a degranulação dos basófilos e dos mastócitos, como resultado, há liberação
de vários mediadores farmacologicamente ativos presentes nos grânulos, causando
manifestações alérgicas. A degranulação dos mastócitos e basófilos induzida pela
IgE pode também liberar mediadores que participam e ajudam no recrutamento de
várias células, como macrófagos, necessárias para a defesa antiparasitária
(SUTTON & GOULD, 1993; ATTA et al., 1999; GOLDSBY et al., 2002). A liberação
de mediadores inflamatórios lipídicos, citocinas e quimiocinas nos locais das reações
desencadeadas pela IgE, resulta no recrutamento de eosinófilos e basófilos,
aumentando a resposta de hipersensibilidade do tipo Ι ou hipersensibilidade
imediata (SUTTON & GOULD, 1993; BONNEFOY et al., 1994).
A IgE pode encontrar-se ligada a mastócitos e basófilos pela fração constante
(Fc) à qual liga-se a receptores presentes nessas células. O Fc∈RI e Fc∈RII são
receptores específicos para a IgE (ATTA et al., 1993).

32
Indivíduos com doenças alérgicas ou infecções parasitárias que tem elevados
níveis de IgE, apresentam marcante aumento da expressão de receptores Fc∈RI na
superfície dos mastócitos, com aumento da sensibilidade dessas células à ativação
por baixas concentrações de antígeno específico e à liberação de mediadores e
citocinas dependentes de IgE. O segundo receptor de IgE, o Fc∈RII, é uma molécula
que se liga a IgE com baixa afinidade e é encontrado em muitos tipos celulares
diferentes, incluindo as células B, células T ativadas, monócitos, eosinófilos,
plaquetas, células dendríticas foliculares e algumas células epiteliais tímicas,
apresentando importante função nas células apresentadoras de antígeno, na captura
de antígenos através da IgE específica (CAPRON & CAPRON, 1994; JANEWAY et
al., 2000; TAKAMOTO et al., 2001).
Em infecções por helmintos, e os antígenos liberados por estes estimulam
macrófagos e linfócitos T a interagirem com linfócitos B, os quais produzem
anticorpos, principalmente da classe IgE, que podem se elevar consideravelmente
nos portadores destes parasitas (HAGEL et al., 1993c; JAIN, 1994).
Valores elevados de IgE total e a indução de IgE específica são
características típicas do parasitismo por helmintos. A principal causa para a
elevação dos níveis de IgE, em populações de países em desenvolvimento, é a
síntese policlonal de IgE, a qual é induzida pelas helmintíases intestinais e
dependentes de IL-4 (ELSE & FINKELMAN, 1998; LYNCH et al., 1998; CORRY &
KHERADMAND, 1999).
A produção de grande quantidade de IgE apresenta um aspecto característico
da resposta imune causada por helmintos. Somente parte desta IgE é específica
para o parasita, e o restante representa IgE policlonal não específica (HAGEL et al.,
1995).
As células T auxiliares Th, de acordo com seu perfil de produção de citocinas,
são classificadas em dois tipos: Th1 e Th2. Nos processos em que estas células
secretam IL-4, IL-5 e IL-10 são classificadas como Th2. A IL-4 é uma das principais
citocinas que estimulam linfócitos B a produzirem anticorpos IgE, enquanto o INF-γ é
capaz de retardar tal síntese (SELL, 1996; BACHARIER & GEHA, 2000).
Os anticorpos IgE são produzidos devido a antígenos liberados de parasitas e
apresentados em associação com moléculas de MHC classe II por células
apresentadoras fagocíticas que tendem a ativar células TCD4+ do tipo Th2, as quais

33
secretam IL-4, induzindo a produção de anticorpos IgE pelas células B. A
interleucina 13 também é produzida pela subpopulação Th2 e tem importância na
resposta imune contra parasitas, assim como a IL-4 pode atuar nos linfócitos B,
induzindo produção de IgE, e também podendo contribuir para contração da
musculatura intestinal, auxiliando no processo de expulsão de alguns parasitas
intestinais (MATTHAEI et al., 1997; MEEUSEN & BALIC, 2000).
A elevação de IgE em pacientes parasitados tem sido estudada na Etiópia e
está relacionada principalmente com infestações por Ascaris. CASTILHO DE ARIZA
et al. (1983) demonstrou que a elevação de IgE tem correlação com a presença de
parasitas intestinais, não somente Ascaris, mas também Trichuris (SALDIVA et al.,
1993; MACDONALD et al., 1994).
HAGEL et al. (1993b) em estudo realizado na Venezuela, relacionou a
resposta mediada por IgE em crianças infectadas com A. lumbricoides e o grau de
pobreza. Os resultados demonstraram que a elevada prevalência de helmintíases
nessas crianças estavam relacionadas com o aumento dos níveis de IgE total e
eosinófilos no sangue circulante, relacionando estas variáveis com o baixo nível
socioeconômico da população e as precárias condições de higiene. Esta relação não
foi observada para resposta específica a esses parasitas, pois a positividade para o
teste cutâneo para antígenos de A. lumbricoides e os níveis de IgE específica anti-
Ascaris foram baixos nas crianças mais pobres, que tinham as mais elevadas
incidências de helmintos e os mais altos níveis de IgE total. Isto sugere que baixos
níveis socioeconômicos e a falta de condições sanitárias podem, por influência da
resposta por IgE, comprometer a resistência dessas crianças às infecções
parasitárias.
Porém, LYNCH et al. (1993) avaliando níveis séricos de IgE em crianças de
diferentes idades, de áreas pobres de Caracas, onde a presença de helmintos
intestinais era endêmica, observou que as crianças acima de 5 anos apresentavam
IgE alta e tinham níveis de IL-4 detectáveis no soro, enquanto que as crianças com
menos de 5 anos apresentaram níveis baixos de IgE e tinham IL-4 indetectáveis,
sugerindo que as helmintíases nas crianças com menos de 5 anos não induzem
estimulação policlonal da síntese de IgE, possivelmente devido a ausência de
produção de IL-4 pelo sistema imune imaturo.
Em trabalho realizado por RODRIGUES (2001), na cidade do Natal, em área
de elevados índices de helmintíases intestinais e precárias condições de higiene em

34
crianças de baixo nível socioeconômico com idades de 3 aos 6 anos, os resultados
revelaram altos níveis de IgE total, inclusive com produção de IgE específica para A.
lumbricoides, sugerindo que crianças menores de 5 anos podem desenvolver, após
estímulo, uma resposta imune mediada por IgE específica e policlonal, revelando
resultados diferentes dos apresentados por HAGEL et al. (1993a) e LYNCH et al.
(1993).
2.4 Eosinófilos e Infecções Parasitárias Os eosinófilos são células leucócitos granulocíticos que apresentam vida
média de aproximadamente treze dias, sendo seis dias em desenvolvimento na
medula óssea, um dia na circulação e seis dias nos tecidos. São derivados da
linhagem progenitora mielóide na medula óssea, por estímulo das citocinas IL-3, IL-5
e GM-CSF, e assim chamados por apresentarem grânulos que contêm proteínas
básicas ricas em arginina, que se coram de laranja brilhante pelo corante ácido
eosina (MENDES et al., 1998).
Eosinófilos são células importantes na defesa contra infecções parasitárias.
Foi observado que em presença de antígenos parasitários, estas células apresentam
um tempo de geração medular menor e emergem da medula após 18 h, em vez de
ficarem armazenados alguns dias. Depois disto, ficam em circulação de 6 a 12 h
antes de migrarem para a pele, pulmões e, principalmente ao trato gastrintestinal.
Normalmente não retornam à circulação (McEWEN, 1992; ABBAS et al., 2000;
ROITT et al., 2003).
Dos grânulos encontrados no interior dos eosinófilos destacam-se dois tipos,
que são os pequenos grânulos primários, os quais contêm: arilsulfatase, peroxidase
eosinofílica, fosfatase ácida e os grânulos cristalóides que contêm quatro proteínas
principais: proteína básica maior (PBM), proteína catiônica eosinofílica (PCE),
peroxidase eosinofílica (POE) e neurotoxina derivada de eosinófilo (NDE). Os
grânulos cristalóides consistem de um núcleo de PBM circundado por uma matriz
que contêm PCE, POE e NDE (SPRY, 1985). Entre as substâncias liberadas pelos
eosinófilos para destruição dos parasitas, destaca-se a proteína básica principal

35
(PBP), que é mais efetiva na eliminação destes parasitas (MENDES et al., 1998;
PAWANKAR, 2001).
Assim como os mastócitos, os eosinófilos também atuam nas respostas de
hipersensibilidade e de defesa imune contra parasitas. Estas células liberam
proteínas altamente tóxicas e radicais livres dos seus grânulos, que podem destruir
microrganismos e parasitas, como também podem produzir lesão tecidual
significativa nas reações alérgicas (HAGEL et al., 1993a; ORTIZ et al., 2000).
Os eosinófilos produzem algumas moléculas, incluindo as prostaglandinas,
leucotrienos e citocinas, que amplificam a resposta inflamatória, recrutando e
ativando mais eosinófilos e outros leucócitos (CAPRON & DESREUMAUX, 1997). O
aumento do número de eosinófilos no sangue periférico ocorre em resposta a vários
estímulos específicos e inespecíficos (McEWEN, 1992). Os mastócitos quando
ativados também produzem substâncias, como as prostaglandinas, leucotrienos e
citocinas, que amplificam a resposta inflamatória recrutando e ativando mais
eosinófilos (CAPRON & DESREUMAUX, 1997; CORRY & KHERADMAND, 1999).
As infecções parasitárias constituem causa comum de eosinofilia, em
particular, as infecções por helmintos que invadem os tecidos durante a fase larvária
de desenvolvimento (CARDOSO, 1997; TUNON DE LARA et al., 2000).
Os eosinófilos combatem os parasitas de duas formas: através da liberação
de substâncias que atuam na superfície dos parasitas, e através da ADCC, na qual
estes anticorpos, preferencialmente da classe IgE, promovem uma maior
proximidade das células com o parasita, facilitando o processo de degranulação
destas (ALBUQUERQUE et al., 1990).
Por ocasião de infecções parasitárias, os eosinófilos apresentam um maior
número de receptores para fração Fc de IgG, IgE e IgA, e também para algumas
frações do complemento (C3b e C4), o que evidencia a influência do parasita na
maturação celular (ABU-GHAZALEH et al., 1989; MOTEGI & KITA, 1998; DE LA
RUE, 2001).
A destruição de parasitas deve-se a liberação do conteúdo de grânulos dos
eosinófilos no meio circundante. Normalmente, os eosinófilos degranulam e liberam
o conteúdo granular sobre a cutícula do verme (JARRETT & MILLER, 1982; LYNCH
et al., 1998; DE LA RUE, 2001).
Este conteúdo granular inclui os produtos da explosão respiratória, tais como
o superóxido, peróxido de hidrogênio e outros radicais livres gerados pela

36
peroxidase e pelas enzimas líticas eosinofílicas, como lisofosfolipase e a fosfolipase
D (SPRY, 1985). A tendência destas células em descarregar sua peroxidase
extracelularmente, sugere que seu papel inicial básico seja de defesa contra
parasitas invasores teciduais, e que o mecanismo de resposta anti-helmíntica
mediada por eosinófilos IgE-dependente deva ser o mais importante mecanismo de
resistência a helmintos (SPRY, 1985; WELLER, 1991; ATTA et al., 2002).
DE LA RUE (2001) refere que a relação entre eosinofilia e protozoários é
pouco analisada na literatura científica, provavelmente devido a sua menor
importância, entretanto, está bem descrita a presença de eosinófilos nos tecidos em
processos patológicos causados por protozoários, mas sua presença aumentada no
sangue circulante não tem sido normalmente observada.
MEEUSEN & BALIC (2000), relatam em trabalhos, que em relação aos
helmintos, é importante considerar que as formas que estão na luz intestinal
produzem eosinofilia expressiva, ao contrário das formas teciduais, onde esta se
manifesta significativamente. Entre os nematóides intestinais, Ancylostomatidae e
Strongyloides são os que produzem eosinofilia mais elevada. A. lumbricoides,
quando encontrado na forma adulta, pode originar discreta elevação destas células
em um terço dos pacientes, como também com Enterobius vermicularis onde
metade dos pacientes apresentam discreta eosinofilia (HASWELL-ELKINS, 1992).
2.5 Infecção Parasitária
As infecções por helmintos intestinais são as de maior prevalência entre as
infecções humanas, afetando aproximadamente um quarto da população mundial,
principalmente nos países em desenvolvimento, onde a maioria dos indivíduos vive
em condições precárias de higiene e saneamento básico. A disposição na forma de
transmissão tem sido vinculada a variações das características do meio em que se
encontram, imunidade do hospedeiro e a resistência das formas parasitárias ao
ambiente. Indivíduos que apresentam elevadas cargas parasitárias representam
uma grande importância na dinâmica de transmissão destas infecções. A severidade
dessas enteroparasitoses pode estar relacionada com o nível de higiene da

37
população, localização e o equilíbrio parasita – hospedeiro (FORRESTER et al.,
1988; MORALES et al., 1999).
O ambiente exerce um importante papel na transmissão das parasitoses
intestinais, já que ovos de helmintos, alguns embrionados como os ovos de A.
lumbricoides, quando eliminados no solo pelas fezes do hospedeiro, não possuem
capacidade para infecção. Essa capacidade só é adquirida após um processo
evolutivo que dura cerca de três ou quatro semanas, necessitando para isso de
lugares úmidos, quentes e sombreados, podendo levar à contaminação da água e
alimentos (MACEDO et al., 1999; MORALES et al., 1999; CAMPOS et al., 2002;
CARNEIRO et al., 2002).
A maioria das infecções parasitárias passa por repetidos ciclos vitais
complexos. Uma das características fundamentais destas infecções é sua
cronicidade, na qual existem muitas possibilidades para que ela ocorra, incluindo a
diminuída imunidade inata e a capacidade dos parasitas de se evadirem ou
apresentarem resistência à eliminação pela resposta imune específica. Indivíduos
que residem em áreas endêmicas exigem repetidos ciclos de quimioterápicos em
razão da constante exposição aos parasitas. A principal resposta imune inata aos
protozoários é dada principalmente pelos fagócitos, porém, muitos desses parasitas
são resistentes aos mecanismos de fagocitose e podem se multiplicar. Os fagócitos
também apresentam mecanismos contra helmintos e secretam substâncias
microbicidas para destruir organismos que são muito grandes para serem
fagocitados. Muitos helmintos apresentam tegumento espesso que os tornam
resistentes (CARDOSO, 1997; KIGHTLINGER et al., 1998; ABBAS et al., 2000).
As parasitoses intestinais são importantes causas de alterações imunológicas
em indivíduos de países em desenvolvimento e a gravidade dessas alterações pode
ser correlacionada ao grau de parasitismo. Além disso, estas infecções estão
freqüentemente associadas à desnutrição, que contribuem para o estado de
imunodeficiência, e esta presença de desnutrição pode modular a resposta
imunológica, alterando os mecanismos de produção dos anticorpos, principalmente
IgA e IgE (CASTILHO DE ARIZA et al., 1983; CARVALHO et al., 1998).
Estudos realizados por GUIMARÃES & SOGAYAR (2002) avaliaram
anticorpos séricos anti-Giardia lamblia em crianças de creches em área endêmica e
obtiveram um total de 63,3% de crianças com a presença de G. lamblia, sendo que
destas 68% apresentaram anticorpos anti-Giardia lamblia, salientando a importância

38
da avaliação imunológica de crianças em idade escolar e habitante de áreas
contaminadas.
Biópsias de cólon de crianças com síndrome diarréica por T. trichiura,
mostraram aumento do número de células contendo TNF-alfa, quando comparado
ao grupo controle. É possível que uma inflamação mediada por macrófagos e por
uma produção local de citocinas produzam função reguladora da inflamação
intestinal na infecção por T. trichiura (LILLYWHITE et al., 1991; MACDONALD et al.,
1994).
Desnutrição e infecções parasitárias continuam sendo as principais causas de
morbidade e mortalidade infantil nos países em desenvolvimento, sendo assim,
quando associadas constituem a principal causa de óbito, principalmente em
crianças menores de dois anos. É necessário conhecer as alterações imunológicas
determinadas pela má nutrição para melhor combater as infecções parasitárias
(GROSS et al., 1989; CARDOSO,1997; NASCIMENTO et al., 1999).
2.6 Imunidade e Nutrição
A presença de desnutrição energético protéica e as deficiências de múltiplos
nutrientes com enfermidades associadas aumentam a morbidade e a mortalidade, e
na maioria das vezes contribuem para o aumento da susceptibilidade às infecções
(CHANDRA, 1972).
Na desnutrição crônica, observa-se que todos os órgãos linfóides, inclusive o
timo, estão atrofiados e depletados de linfócitos. A desnutrição grave pode
determinar disfunções no sistema imunológico e alterações importantes nas
principais respostas imunoefetoras, caracterizadas por alterações da imunidade
celular e humoral, com diminuição da produção de anticorpos direcionados a
antígenos T-dependentes, diminuição da afinidade dos anticorpos, alterações nas
funções fagocíticas dos leucócitos, no sistema complemento, diminuição da
atividade das células NK (Natural Killer) e síntese de citocinas (REDDY et al., 1976).
Ocorre, na desnutrição indução significativamente reduzida da concentração
de IgA secretora, presente na mucosa intestinal e em suas secreções, conseqüente

39
à diminuição da formação de células precursoras de linfócitos B e da diferenciação
para células B-IgA+ dependentes de linfócitos T CD4+ nas placas de Peyer. Da
mesma forma, a regulação dos plasmócitos produtores de IgA também depende das
linfocinas produzidas por linfócitos T CD4+ na mucosa intestinal, por isso, é
necessária a presença de células T CD4+ funcionantes no intestino para que haja o
desenvolvimento de todo o mecanismo de defesa dependente da resposta de
plasmócitos locais (REDDY et al.,1976; SUSKIND et al.,1976).
HAGEL et al. (1995), relataram que crianças mal nutridas podem ter uma
potencialização da síntese policlonal de IgE induzida por helmintos, pois, ao
comparar crianças bem nutridas com mal nutridas, observou-se que estas últimas
tinham níveis de IL-4 e de IgE total elevado, mesmo após tratamento com anti-
helmínticos, o que não ocorria com os níveis de IgE específica para A. lumbricoides,
que já eram baixos, e assim, continuavam mesmo após tratamento. Foi então
sugerido que a desnutrição, por afetar a resposta mediada por IgE, pode diminuir a
resistência das crianças às infecções helmínticas, e que, nesta situação, os
mecanismos protetores não são reativados, mesmo após tratamento anti-helmíntico.
Em trabalho realizado por CHANDRA (1983), estudando o sangue periférico
de crianças que apresentavam desnutrição energético protéica, evidenciou que na
ausência de infecção, o número de linfócitos foi menor que nos pacientes controles,
com uma diminuição de subpopulações de células T CD3+, CD4+ e CD8+, mas com
alterações mais significativas na subpopulação CD4+. Ainda mostrou que apesar do
número de linfócitos B estarem discretamente diminuídos em relação aos controles,
em presença de desnutrição, estes apresentavam alterações funcionais que
desapareciam após a repleção nutricional.
SIRISINHA et al. (1975), demonstrou que crianças com distúrbios nutricionais
podem apresentar alterações morfoestruturais na mucosa intestinal. A mesma
constatou que crianças com desnutrição protéica energética crônica e grave,
conhecida como Kwashiorkor, apresentavam níveis diminuídos de IgA secretora e
que após a reabilitação nutricional, estes níveis se normalizavam.
As condições nutricionais da população brasileira melhoraram nas últimas três
décadas, mesmo assim, é pouco conhecido o papel da assistência oferecida por
instituições públicas, que atendem crianças pré-escolares de baixa renda e de maior
risco nutricional. Nos períodos de 1974 a 1996, houve, em território nacional, uma
diminuição importante na prevalência de retardo de crescimento, mantendo-se

40
diferenças regionais desfavoráveis para as regiões Norte e Nordeste, que em
menores de cinco anos, apresentaram déficit de altura de 17% e 13%,
respectivamente (CORREA et al., 1999).

41
3 OBJETIVOS
3.1 Objetivo Geral
Avaliar níveis de IgA sérica, IgA secretora salivar, IgE total e estado
nutricional de uma população de crianças de baixo nível socioeconômico,
freqüentadoras de creche, e correlacionar com infecções parasitárias.
3.2 Objetivos Específicos
1. Correlacionar as infecção por enteroparasitas com os níveis de IgA sérica e
secretora;
2. Estudar a relação entre o estado nutricional das crianças com os níveis das
imunoglobulinas dosadas (IgA sérica, IgA secretora e IgE total);
3. Avaliar o número de eosinófilos no sangue destas crianças;
4. Analisar o estado nutricional e correlacionar com níveis de proteínas totais,
albumina e globulina;
5. Avaliar a correlação entre infecção por enteroparasitas e estado nutricional das
crianças.

42
4 CASUÍSTICA, MATERIAL E MÉTODOS 4.1 População de estudo
A população estudada foi representada por 103 crianças de ambos os sexos,
sendo 54 do sexo masculino e 49 do sexo feminino, com idades variando dos 3 aos
6 anos, todas freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do
Natal. O trabalho teve início após ter sido aprovado pelo Comitê de Ética em
Pesquisa da Universidade Federal do Rio Grande do Norte (CEP-UFRN). A coleta de dados teve início após a assinatura do “Termo de
Consentimento Consciente”, segundo a resolução 196/96 do Conselho Nacional
de Saúde (Anexo 1), pelos pais ou responsável, os quais responderam a um
protocolo de pesquisa (Anexo 2), onde foi avaliada a história pessoal e familiar das
crianças.
As amostras de sangue, fezes e saliva, foram obtidas durante o período de
Março a Junho de 2002, juntamente com a avaliação antropométrica, a qual foi
realizada por nutricionista da Universidade Federal do Rio Grande do Norte – UFRN.
Das 103 crianças, 02 não foram submetidas a avaliação nutricional.
As crianças eram todas moradoras do mesmo bairro, e segundo o protocolo
respondido pelos pais, todas encontravam-se com as vacinas básicas em dia e
viviam em condições socioeconômicas similares, sem saneamento básico e com
precárias condições de higiene. A renda média observada era de um salário mínimo,
segundo a classificação dos indicadores mínimos (IBGE, 1998).
Foi utilizado como critério de exclusão: crianças com idades inferiores a três
anos e superiores a 6, que faziam uso de alguma medicação, e as que
apresentavam quadros febris ou alguma patologia diagnosticada no momento da
entrevista com os responsáveis.

43
4.2 Materiais 4.2.1 Amostras Biológicas
• Sangue
As amostras de sangue periférico foram coletadas às 7 horas da
manhã através de punção venosa, com seringas de 5 mL e agulhas
descartáveis, transferidos para tubos contendo etilenodiamino tetracético
tripotássico – EDTA-K3 (VACUTEINER) para realização do hemograma e
para tubos sem anticoagulantes (VACUTEINER), para obtenção de soro a
partir da coagulação do sangue total em temperatura ambiente, seguido de
centrifugação por 15 min, a 5.000 Xg e 26-27oC. O soro foi utilizado para a
realização das dosagens de proteínas totais e frações, eletroforese de
proteínas, IgA e IgE total.
• Fezes
As fezes foram colhidas em três (03) amostras seriadas, em coletores
descartáveis (coletores universal), e encaminhados para análise sem anti-
sépticos ou preservativos.
• Saliva
A saliva foi colhida pela manhã, com a criança sem ingestão prévia de
alimentos sólidos para evitar que restos de alimentos se misturassem à saliva.
A amostra foi coletada em coletores estéreis e descartáveis (coletores
universal), sem anti-sépticos ou preservativos, e conservadas à –20oC até a
sua análise.

44
4.3 Métodos 4.3.1 Dosagens de IgA sérica, IgA secretora e IgE total
As dosagens de IgA sérica foram realizadas por turbidimetria,
utilizando-se o sistema automatizado TURBIQUANT (Dade Behring), no
Laboratório Antônio Prudente, Natal/RN, enquanto as dosagens de IgE total
foram realizadas pelo método de fluorimetria, através do sistema UniCAP 100
da Pharmacia & UpJonh, no Laboratório de Alergia e Biologia Molecular da
Faculdade de Medicina de Ribeirão Preto/SP. A IgA secretora foi determinada
por nefelometria em automação, no sistema Beckman Array 360, no Instituto
de Patologia Clínica Hermes Pardini, em Belo Horizonte/MG.
4.3.1.1 Determinação de IgA Sérica
A determinação dos níveis séricos de IgA foi realizada pela técnica de
turbidimetria, utilizando kit comercial (IgA TURBIQUANT- Dade Behring),
seguindo as especificações do fabricante. Em resumo, foi pipetado 500 µL de
tampão contendo anticorpos monoclonais de coelho anti-IgA humana para 50
µL da amostra diluída de 1:21 em solução fisiológica, e então colocadas em
uma cubeta dentro do equipamento. A turbidez formada é proporcional a
concentração de IgA na amostra, formando um precipitado resultante do
complexo antígeno-anticorpo.
As concentrações presentes foram determinadas quantitativamente
pela medição turbidimétrica da velocidade máxima da reação, através do
método da taxa de pico (peack-rate). Os valores de referência utilizados foram
de 0,7 a 4,0 g/L.

45
4.3.1.2 Determinação de IgA Secretora A determinação dos níveis de IgA secretora foi realizada pela técnica
de nefelometria, utilizando kit comercial (Beckman IgA – Immunoglobulin A
quantificação), seguindo todas as especificações do fabricante. A técnica
consiste, em resumo, em depositar 150µL da amostra diluída em uma razão
de 1:36 em microtubos colocados no equipamento, juntamente com o
reagente (IGA) que é formado por uma solução com anticorpos monoclonais
anti-IgA, peça secretora, para a formação de um precipitado no qual foi obtido
leitura através de nefelômetro. Após a análise, o resultado foi calculado
baseado na diluição utilizada.
As amostras foram enviadas seguindo todas as orientações e
recomendações de coleta fornecidas pelo laboratório. Os valores de
referência utilizados de acordo com os padrões de IgA secretora foram de 3,5
a 36,8 mg/L. 4.3.1.3 Determinação de IgE Total
A determinação dos níveis séricos de IgE total foi realizada pela técnica
de fluorimetria (Enzyme Linked Immuno sorbent Assay), utilizando kit UniCAP
(Pharmacia & Upjonh), seguindo as especificações do fabricante. A técnica
consiste na captura de antígenos (IgE) utilizando anticorpos monoclonais anti-
IgE ligados em fase sólida. Em uma primeira fase, foram incubados 40 µL de
soro durante 30 min, seguido de lavagem após o período de incubação. Na
segunda fase, foram adicionados 50 µL do conjugado (enzima β-
galactosidase conjugada a anti-IgE de rato) e incubados durante 30 min à
37°C. Logo após a segunda etapa de incubação, foram realizadas lavagens
para remover o conjugado que não se ligou ao imunocomplexo, e adicionado
50 µL da substância cromogênica (4-Metilumbeliferil-β-D-galactosídeo a
0,01%). Após o período de 10 min de incubação ao abrigo da luz, a reação foi
bloqueada com 600 µL de carbonato de sódio a 4%, e a leitura da
fluorescência realizada em seguida. A curva de calibração foi traçada com
calibradores de IgE total nas concentrações de 2, 10, 50, 500, 1000 e 5000

46
KU/I, padronizadas segundo as especificações do 2nd International Reference
Preparation 75/502 of Human Serum Imunoglobulin E.
As amostras foram enviadas seguindo todas as orientações e
recomendações de coleta fornecidas pelo laboratório. Os valores de
referência utilizados de IgE total por idade foram: 3 anos (até 32 KU/L), 4
anos (até 40 KU/L), 5 anos (até 48 KU/L) e 6 anos (até 56 KU/L).
4.3.2 Determinação de Proteínas Totais A determinação dos níveis de proteínas totais foi realizada pela técnica
de colorimetria, utilizando kit comercial (Proteínas-Kit bioMérieux), em
analisador automatizado (ALIZÉ, Inc. France), seguindo as especificações do
fabricante. Nesta análise, foram colocados nas cubetas 150 µL de soro e
incubados por 3 min com 300 µL de biureto, constituído de uma solução de
sulfato de cobre, citrato trissódico, carbonato de sódio e hidróxido de sódio.
Após a incubação, o complexo formado de coloração violeta foi lido em
espectrofotômetro a 545 nm (KACHMAR, 1970).
Os valores de referência no soro para indivíduos normais variam de 6,2
a 8,0 g/dL, de acordo com os padrões de comparação do fabricante 4.3.3 Determinação de Albumina Sérica
A determinação dos níveis de albumina sérica foi realizada pela técnica
colorimétrica, utilizando kit comercial (Albumina - Kit bioMérieux), em
Analisador automatizado (ALIZÉ, Inc. France), seguindo as especificações do
fabricante. Nesta análise, 180 µL de soro foram incubados com 250 µL de
verde de bromocresol em pH alcalino por 4 min em cubetas, no interior do
equipamento, o que resultou em um composto verde, que foi proporcional à
concentração de albumina da amostra. A leitura foi obtida por
espectrofotômetro a 628 nm. (KACHMAR, 1970).
De acordo com o fabricante, os valores de referência para indivíduos normais
variam de 3,8 a 5,4 g/L.

47
4.3.3.1. Determinação das Globulinas
A determinação das globulinas séricas foi feita através da diferença
(subtração) entre as proteínas totais e a albumina sérica.
Os valores de referência utilizados em indivíduos normais, variam de 1,0
a 3,0 g/dL (KACHMAR, 1970).
4.3.4 Eletroforese de Proteínas
As frações protéicas (albumina, α-1 globulina, α-2 globulina, β-globulina e
γ-globulina) no soro foram separadas por corrida eletroforética em acetato de
celulose (2,5 x 14 cm – marca CELLOGEL), sob a diferença de potencial de
200 volts, durante 25 min, utilizando tampão Tris-Glicina-Pirofosfato de Sódio,
pH 9,0.
A amostra foi depositada sobre o suporte com a ajuda do micro aplicador
(0,5µL de soro), sendo submetida a ação de campo elétrico em cuba
eletroforética durante 35 min (tensão de 200 V). Após separação, as frações
foram coradas pelo Ponceau S (Ácido tricloroacético 5%) durante 8 min e
posteriormente descorada em solução de ácido acético a 5% e submetidas a
um processo de transparentização em solução de metanol: ácido acético:
glicerina (21,25: 3,5 :0,5 mL), seguido de secagem por 5 min em estufa a
60˚C, sendo quantificadas em densitômetro BTS 235 da Biosystems
(NOGUEIRA et al., 1990).
4.3.5 Contagem Absoluta de Eosinófilos
A contagem absoluta dos eosinófilos foi realizada em analisador
automático ADVIA 60 (BAYER-DIAGNOSTICS), utilizando 30 µL de sangue
total, previamente homogeneizado, analisado pelo método de impedância. Os
valores de referência variam até 650 eosinófilos por mm3 de sangue, de
acordo com a faixa etária das crianças estudadas (SPRY, 1988).

48
4.3.6 Contagem Relativa de Eosinófilos
O número relativo de eosinófilos no sangue foi determinado em distenção
sanguínea, obtida de sangue periférico, como descrito no item (4.2.1). A
lâmina foi coberta por corante de Leishman durante 7 min e 30 seg, seguida
de lavagem em água destilada. Após secagem da lâmina, foram contados 200
leucócitos em microscópio óptico em objetiva de imersão (100 x). O critério de
classificação para eosinofilia foi o proposto por Naveira: Normal (1 a 4%),
Grau I (5 a 10%), Grau II (11 a 20%), Grau III (21 a 50%) e Grau IV (acima de
50%) (NAVEIRA, 1960).
4.3.7 Exame Parasitológico de Fezes (EPF)
Foram avaliadas 03 amostras seriadas de fezes de cada criança para a
detecção de ovos, cistos e larvas de parasitas, empregando-se os métodos
de sedimentação espontânea, descrito como método de HOFFMAN, PONS &
JANER (HOFFMAN et al., 1934), o método de FAUST et al. (FAUST et al.,
1938), que se baseia na centrifugo-flutuação das formas parasitárias em
sulfato de zinco a 33%, e o terceiro e último método foi o de BAERMANN–
MORAES (BAERMANN-MORAES, 1948), baseada no hidro e termotropismo
das larvas. O método de KATO-KATZ (Kato-Katz apud NETO, 1991) foi
empregado quando necessário, para a quantificação de ovos de helmintos.
4.3.7.1 Método de HOFFMAN, PONS & JANER
Neste método, 5 g de fezes foram emulsionadas em água destilada,
filtrada através de gaze (dobrada quatro vezes) em cálice de sedimentação e
o volume final foi completado com água destilada. Após repouso de 2 a 24 h
foi decantado o sobrenadante e examinado o sedimento em microscópio
óptico em lâmina com uma gota de lugol (40x) (HOFFMAN, et al., 1934).

49
4.3.7.2 Método de FAUST et al. Para execução deste método foi preparada uma suspensão
homogeneizada de fezes, com aproximadamente 2 g de fezes para 20 mL de
água destilada, utilizando um bastão de vidro. Esta suspensão foi filtrada
através de gaze (dobrada quatro vezes) para tubos de vidro (13 x 100 mm) e
centrifugado por 1 min, a 5000 Xg, em temperatura ambiente. O sobrenadante
foi decantado e o sedimento ressuspenso com 3 mL de água destilada e foi
centrifugado novamente por 1 min, a 5.000 Xg, em temperatura ambiente. A
operação foi repetida até que o sobrenadante ficasse transparente, sendo
então decantado, e o sedimento ressuspenso em 3 mL de solução de sulfato
de zinco a 33% e centrifugado a 5.000 Xg durante 1 min. A película formada
na superfície do líquido foi retirada com alça bacteriológica de platina dobrada
em um ângulo de 90º, colocada sobre lâmina com uma gota de lugol e coberta
com lamínula. A leitura foi feita em microscópio óptico comum (40 x) (FAUST
et al., 1938).
4.3.7.3 Método de BAERMANN – MORAES Neste método, 8 a 10 g de fezes foram espalhadas em gaze (dobrada
quatro vezes) sobre tela de nylon encaixada em cálice de sedimentação. Em
seguida, foi adicionada água destilada aquecida a 45oC, até que as fezes
ficassem parcialmente submersas. Após repouso de 60 min, foram retirados 4
mL do sedimento com pipeta de Pasteur e depositado em vidro de relógio por
5 min, para ser examinado em microscópio entomológico (4x) (BAERMANN-
MORAES, 1948).
4.3.7.4 Método de KATO-KATZ Neste método, foi determinado o número de ovos de helmintos (método
de quantificação para ovos). As fezes foram colocadas sobre papel
absorvente, e sobre a mesma comprimiu-se uma tela de nylon para que uma

50
pequena parte da amostra passasse através das malhas. Logo após, a parte
da amostra que passou sobre a tela, foi colocada em um orifício de uma placa
perfurada com ajuda de uma pequena espátula plástica descartável que ficou
sobre a lâmina.
Depois de retirada a placa perfurada que estava sobre a lâmina, um
cilindro de fezes foi formado e coberto por lamínula de papel celofane tratada
previamente com solução de verde malaquita em glicerina. A lamínula foi
pressionada com o polegar para que o material se espalhasse
uniformemente. Após repouso por 60 min a preparação foi analisada em
microscópio óptico para a contagem dos ovos de helmintos, utilizando objetiva
de 40x (Kato-Katz apud NETO, 1991).
4.3.8 Avaliação Nutricional
O estado nutricional das crianças foi determinado pela avaliação
antropométrica a partir da obtenção de medidas de peso e altura. O peso foi
aferido com auxílio de balança digital (Filizola®), com capacidade para 150
kg, precisão de gramas e considerando uma casa decimal para a sua leitura.
As crianças foram pesadas de pé, descalças e com o mínimo de roupa.
A altura foi medida com o auxílio de fita métrica, graduada em
centímetros, com precisão de milímetros, considerando para a sua leitura o
0,5 cm mais próximo. As crianças estavam descalças, em posição ereta, de
costas para o marcador, com os pés unidos, calcanhares e a parte posterior
da cabeça apoiada à parede (ANJOS, 1989).
Para a avaliação antropométrica foram utilizados os indicadores
Peso/Idade (P/I), Peso/Altura (P/A) e Altura/Idade (A/I), os quais foram
comparados utilizando-se os ESCORES DE DESVIO PADÃO (ESCORES-Z),
derivados dos dados de referência do National Center for Health Statistics
(NCHS), segundo recomendações da World Health Organization (WHO) (1983).
As crianças foram classificadas nos seguintes estados nutricionais:
nutridas (eutróficas) e desnutridas (desnutrição leve e moderada). O estado
nutricional foi definido pela presença do mesmo parâmetro (desnutridas ou

51
nutridas), apresentado em no mínimo dois indicadores antropométricos,
dando prioridade, ao indicador da altura.
4.3.9 Análise Estatística dos Resultados
Os resultados obtidos nos protocolos descritos neste projeto foram
divididos em duas partes. A primeira parte foi à análise descritiva para os
tipos de parasitas e uma outra parte foi à análise inferencial com a aplicação
do teste não-paramétrico (Teste da Mediana) para os grupos estudados
(idade, sexo e estado nutricional) e aplicação da correlação de Pearson entre
as variáveis estudadas, no qual o valor mínimo de significância utilizado foi de
5%.
Os dados foram analisados no software STATISTICA versão 5
(StatSoft, inc.2000).

52
5 RESULTADOS 5.1 Características da Amostra Na amostra estudada, verificou-se uma distribuição homogênea em relação
ao sexo das crianças e as idades de 3 e 4 anos encontraram-se uniformemente
distribuídas (77,6%), enquanto que as crianças de 5 e 6 anos apresentaram
números menores 21,4% e 1,0%, respectivamente, como podemos observar na
Tabela 1.
TABELA 1: Distribuição por sexo e idade de 103 crianças, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
POPULAÇÃO
NÚMERO ABSOLUTO PERCENTUAL (%)
SEXO
Masculino
Feminino IDADE 3 Anos 4 Anos 5 Anos 6 Anos
54
49
40
40
22
01
52,4
47,6
38,8
38,8
21,4
1,0

53
5.2 Fatores Socioeconômicos O perfil sócio-econômico encontrado nas famílias das crianças estudadas, foi
de renda familiar de até um salário mínimo, os pais com ocupação de baixa
remuneração ou mesmo desempregados, com moradia sem esgoto ou fossa e a
água de consumo sendo originada de torneira ou filtro. A Tabela 2 mostra a
distribuição das famílias das crianças segundo os indicadores sociais mínimos
(IBGE,1998).
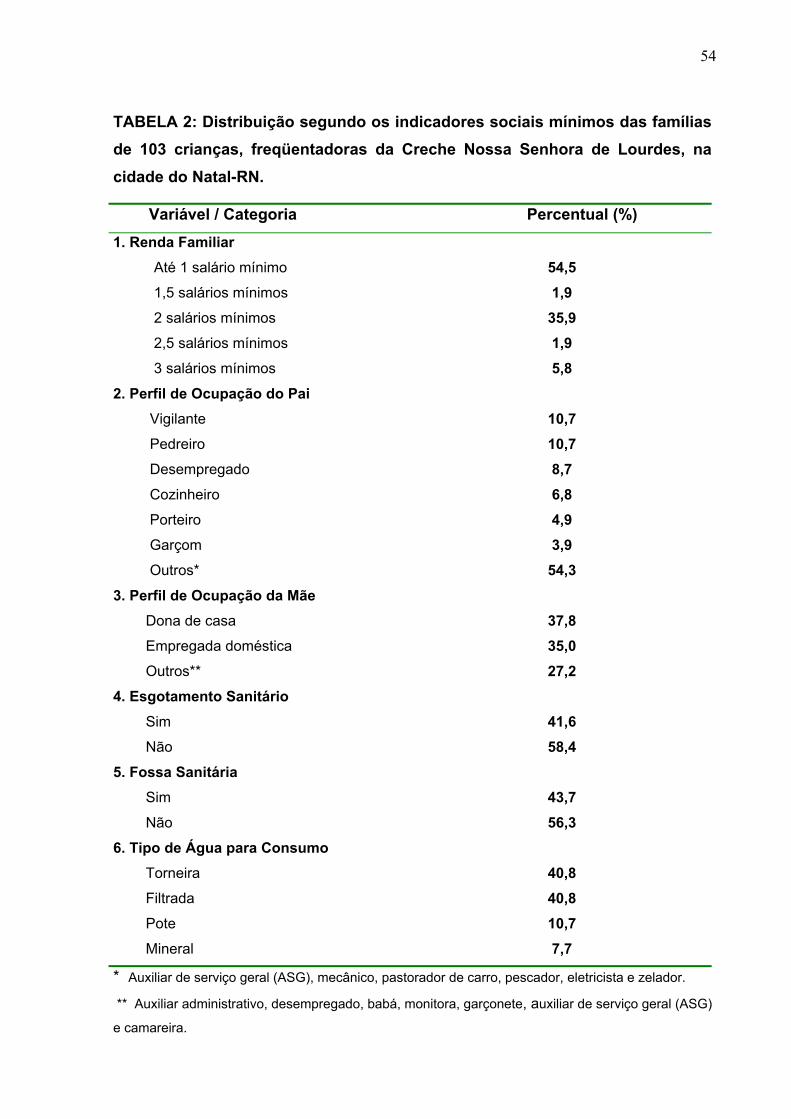
54
TABELA 2: Distribuição segundo os indicadores sociais mínimos das famílias de 103 crianças, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
Variável / Categoria Percentual (%) 1. Renda Familiar Até 1 salário mínimo
1,5 salários mínimos
2 salários mínimos
2,5 salários mínimos
3 salários mínimos
2. Perfil de Ocupação do Pai Vigilante
Pedreiro
Desempregado
Cozinheiro
Porteiro
Garçom
Outros*
3. Perfil de Ocupação da Mãe Dona de casa
Empregada doméstica
Outros**
4. Esgotamento Sanitário Sim
Não
5. Fossa Sanitária Sim
Não
6. Tipo de Água para Consumo Torneira
Filtrada
Pote
Mineral
54,5 1,9
35,9 1,9 5,8
10,7 10,7 8,7 6,8 4,9 3,9
54,3
37,8 35,0 27,2
41,6 58,4
43,7 56,3
40,8 40,8 10,7 7,7
* Auxiliar de serviço geral (ASG), mecânico, pastorador de carro, pescador, eletricista e zelador.
** Auxiliar administrativo, desempregado, babá, monitora, garçonete, auxiliar de serviço geral (ASG)
e camareira.

55
5.3 Avaliação Nutricional Os resultados da avaliação nutricional de 101 crianças estudadas foram
obtidos através de análise antropométrica.
Na Tabela 3 pode-se verificar que, 69,3% das crianças analisadas foram
classificadas como nutridas (eutróficas) e 30,7% desnutridas (desnutrição leve ou
moderada). TABELA 3: Avaliação do estado nutricional de 101 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
ESTADO NUTRICIONAL*
NÚMERO ABSOLUTO
PERCENTAGEM (%)
Nutridas
Desnutrido*
70
31
69,3
30,7
* Desnutrido: Desnutrição leve (N=29) e moderada (N=2).
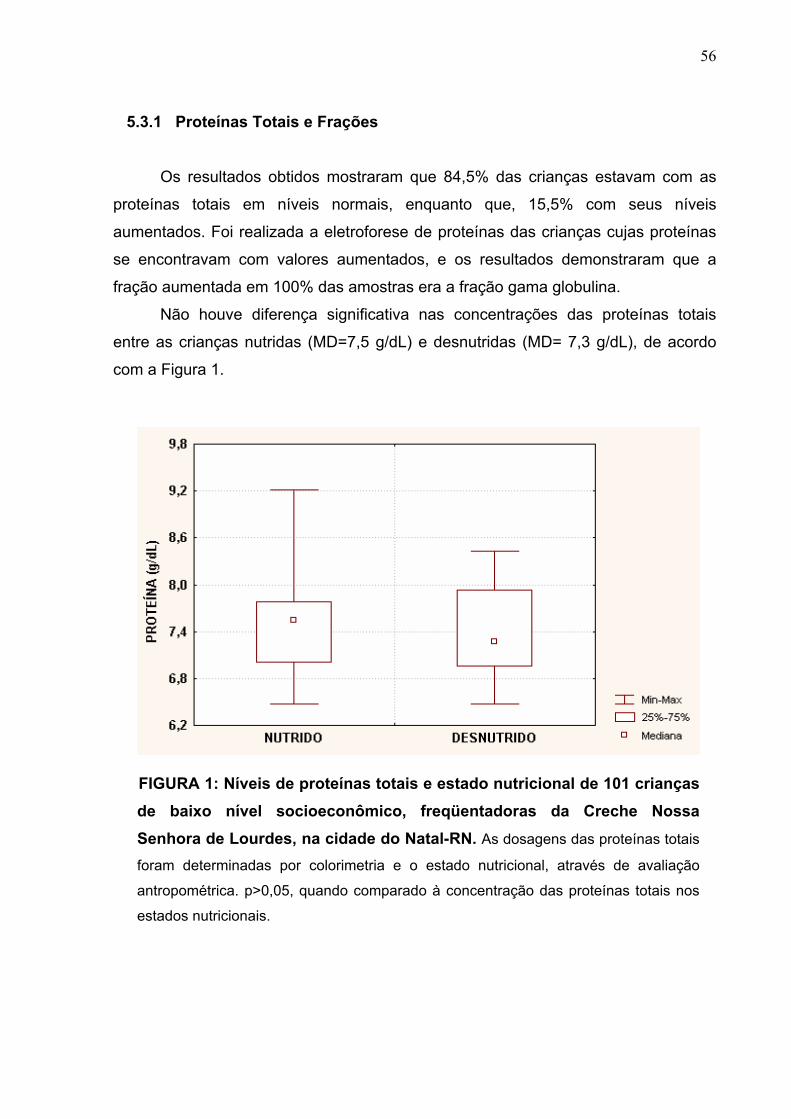
56
5.3.1 Proteínas Totais e Frações Os resultados obtidos mostraram que 84,5% das crianças estavam com as
proteínas totais em níveis normais, enquanto que, 15,5% com seus níveis
aumentados. Foi realizada a eletroforese de proteínas das crianças cujas proteínas
se encontravam com valores aumentados, e os resultados demonstraram que a
fração aumentada em 100% das amostras era a fração gama globulina.
Não houve diferença significativa nas concentrações das proteínas totais
entre as crianças nutridas (MD=7,5 g/dL) e desnutridas (MD= 7,3 g/dL), de acordo
com a Figura 1.
FIGURA 1: Níveis de proteínas totais e estado nutricional de 101 crianças
de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. As dosagens das proteínas totais
foram determinadas por colorimetria e o estado nutricional, através de avaliação
antropométrica. p>0,05, quando comparado à concentração das proteínas totais nos
estados nutricionais.

57
As Figuras 2 e 3 mostram as medianas das concentrações de albumina
e globulina, obtidas entre as crianças nutridas (MD=4,74 g/dL) e (MD=2,77 g/dL) e
desnutridas (MD=4,67 g/dL) e (MD=2,36 g/dL) respectivamente, nas quais, observa-
se que não houve diferença significativa entre estes dois grupos relacionados.
FIGURA 2: Concentração de albumina sérica e estado nutricional de 101 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. A dosagem de Albumina foi determinada
por colorimetria e o estado nutricional através de avaliação antropométrica. p>0,05, quando
comparado à concentração de albumina nos estados nutricionais.
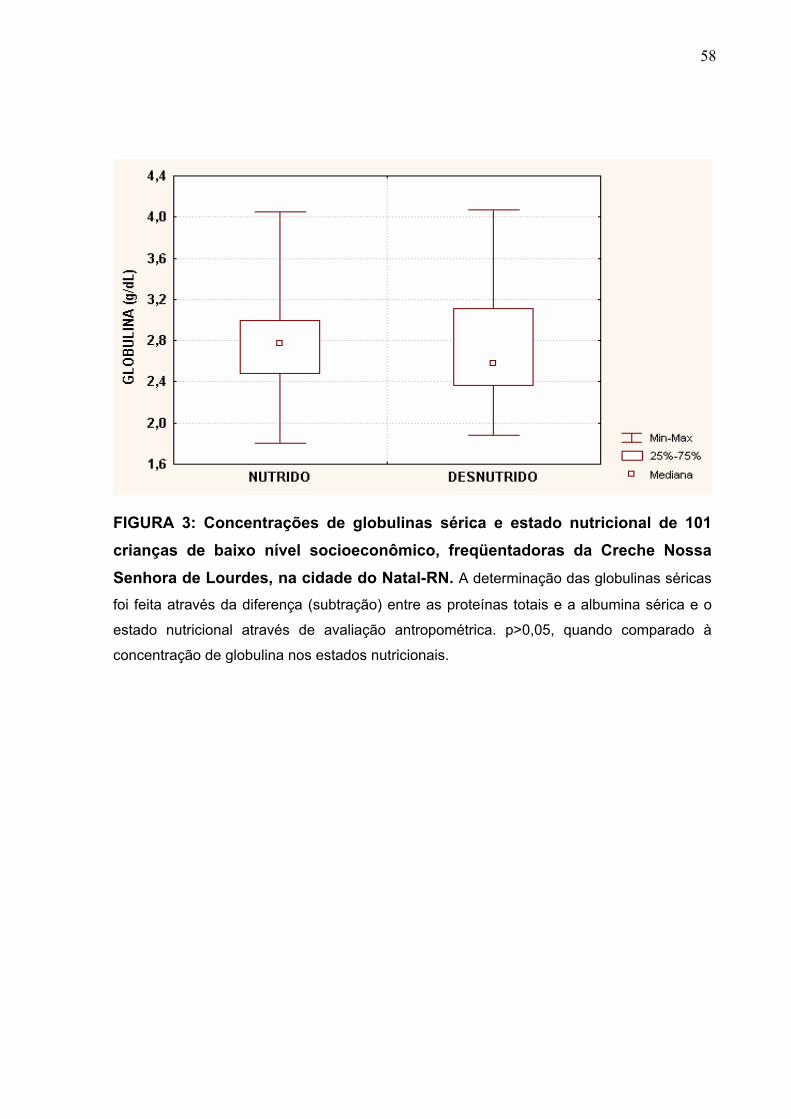
58
FIGURA 3: Concentrações de globulinas sérica e estado nutricional de 101 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. A determinação das globulinas séricas
foi feita através da diferença (subtração) entre as proteínas totais e a albumina sérica e o
estado nutricional através de avaliação antropométrica. p>0,05, quando comparado à
concentração de globulina nos estados nutricionais.

59
5.4 Infecção Parasitária Para a pesquisa de helmintos e protozoários intestinais nas crianças
freqüentadoras da Creche Nossa Senhora de Lourdes, os dados obtidos (Tabela 4),
apresentaram uma alta prevalência de enteroparasitoses (94,2%), estando a grande
maioria das crianças (64,1%) infectadas com três ou mais espécies de parasita.
Apenas 6 amostras (5,8%) não apresentaram ovos e larvas de helmintos e/ou cistos
de protozoários. Não houve diferença significativa quando se relacionou a
distribuição dos parasitas intestinais com a idade e sexo das crianças.
TABELA 4: Parasitismo, segundo o número de espécies infectantes em 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
PARASITISMO*
NÚMERO ABSOLUTO
PERCENTAGEM (%)
Ausência de Parasitas Monoparasitado Biparasitado Poliparasitado
06
13
18
66
5,8
12,6
17,5
64,1
TOTAL
103
100,0
* As amostras de fezes foram processadas e analisadas pelos métodos de Hoffman, Pons
& Janer; Baermann-Morais e Faust et al.

60
Na Tabela 5 pode-se observar que de 97 crianças com presença de parasitas
intestinais, a grande maioria apresentou associação entre helmintos e protozoários
(64,1%). Apenas 8 crianças estavam infectadas por helmintos e 23 com
protozoários. Conforme se observa na Tabela 6, T. trichiura (52,4%) e A.
lumbricoides (49,5%) foram os hemintos mais freqüentes, enquanto que E. nana
(68,0%) e G. lamblia (49,5%) foram os protozoários mais comumente presentes nas
amostras analisadas.
TABELA 5: Freqüência de parasitas intestinais em 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
PARASITAS INTESTINAIS*
NÚMERO ABSOLUTO
PERCENTAGEM (%)
Ausente Presente Helmintos Protozoários Helmintos e Protozoários
06
97
08
23
66
5,8
94,2
7,8
22,3
64,1
* As amostras de fezes foram processadas e analisadas pelos métodos de Hoffman,Pons &
Janer; Baermann-Morais e Faust et al.

61
TABELA 6: Parasitas intestinais presentes em 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
PARASITAS INTESTINAIS*
NÚMERO ABSOLUTO
PERCENTAGEM (%)
HELMINTOS
Trichuris trichiura Ascaris lumbricoides Hymenolepis nana Strongyloides stercoralis Enterobius vermicularis Ancylostoma duodenale PROTOZOÁRIOS Endolimax nana Giardia lamblia Entamoeba coli Entamoeba histolytica Iodamoeba butschlii
54
51
12
07
04
04
70
51
34
16
07
52,4
49,5
11,7
6,8
3,9
3,9
68,0
49,5
33,0
15,5
6,8
* As amostras de fezes foram processadas e analisadas pelos métodos de Hoffman, Pons &
Janer; Baermann-Morais e Faust et al.
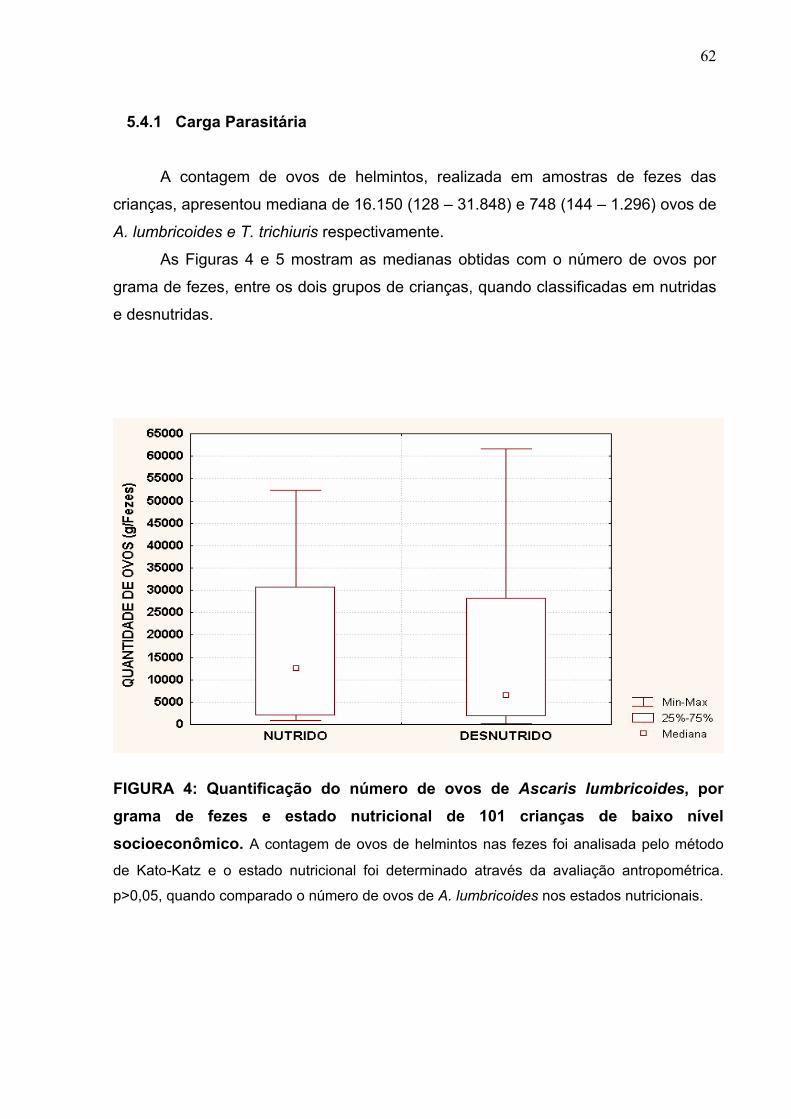
62
5.4.1 Carga Parasitária A contagem de ovos de helmintos, realizada em amostras de fezes das
crianças, apresentou mediana de 16.150 (128 – 31.848) e 748 (144 – 1.296) ovos de
A. lumbricoides e T. trichiuris respectivamente.
As Figuras 4 e 5 mostram as medianas obtidas com o número de ovos por
grama de fezes, entre os dois grupos de crianças, quando classificadas em nutridas
e desnutridas.
FIGURA 4: Quantificação do número de ovos de Ascaris lumbricoides, por grama de fezes e estado nutricional de 101 crianças de baixo nível socioeconômico. A contagem de ovos de helmintos nas fezes foi analisada pelo método
de Kato-Katz e o estado nutricional foi determinado através da avaliação antropométrica.
p>0,05, quando comparado o número de ovos de A. lumbricoides nos estados nutricionais.
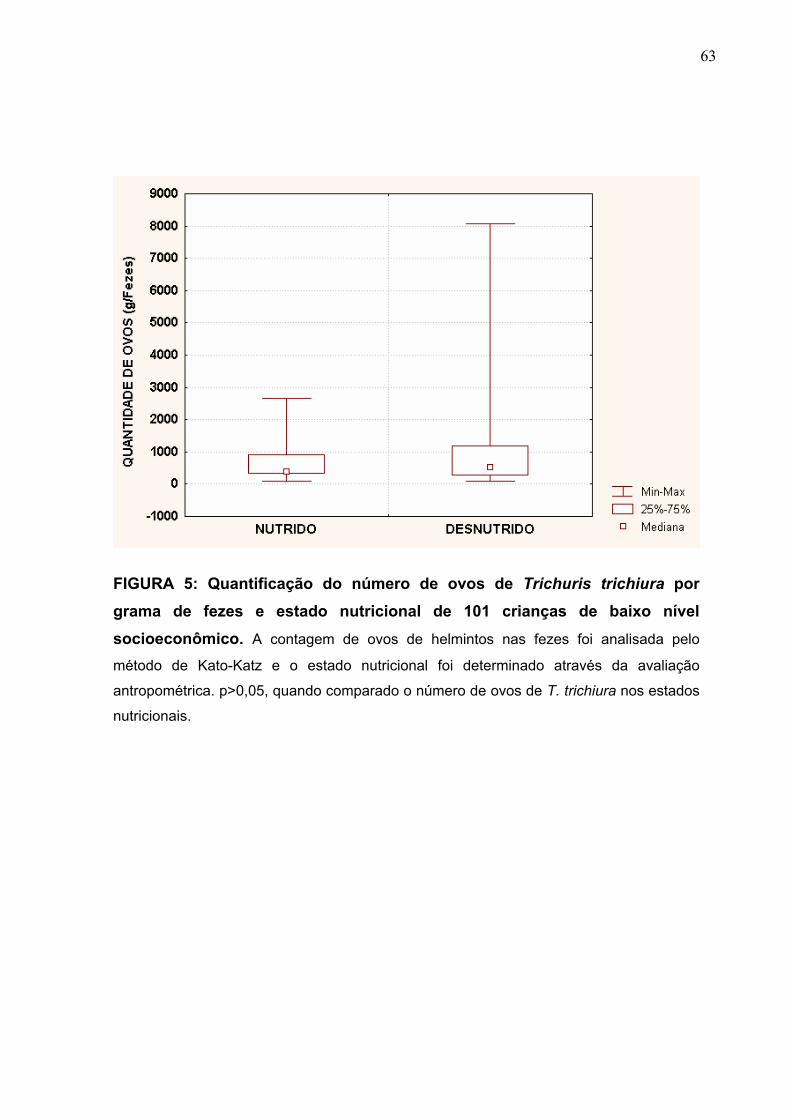
63
FIGURA 5: Quantificação do número de ovos de Trichuris trichiura por grama de fezes e estado nutricional de 101 crianças de baixo nível socioeconômico. A contagem de ovos de helmintos nas fezes foi analisada pelo
método de Kato-Katz e o estado nutricional foi determinado através da avaliação
antropométrica. p>0,05, quando comparado o número de ovos de T. trichiura nos estados
nutricionais.
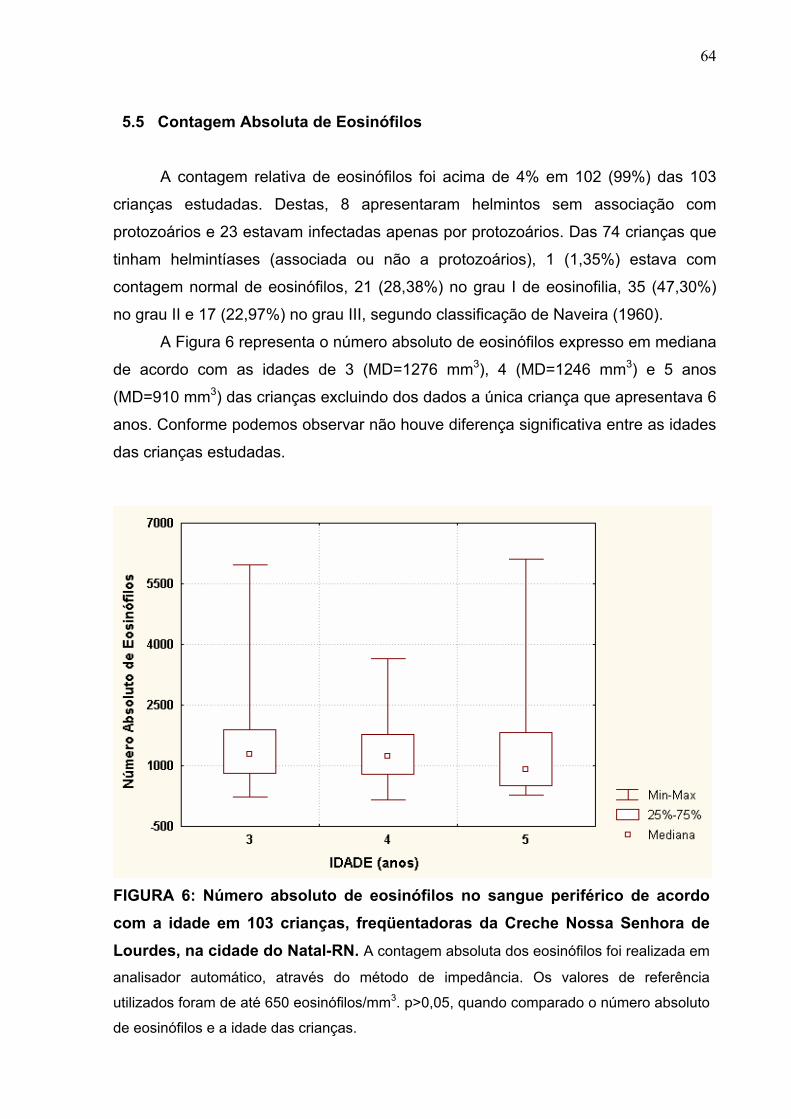
64
5.5 Contagem Absoluta de Eosinófilos A contagem relativa de eosinófilos foi acima de 4% em 102 (99%) das 103
crianças estudadas. Destas, 8 apresentaram helmintos sem associação com
protozoários e 23 estavam infectadas apenas por protozoários. Das 74 crianças que
tinham helmintíases (associada ou não a protozoários), 1 (1,35%) estava com
contagem normal de eosinófilos, 21 (28,38%) no grau I de eosinofilia, 35 (47,30%)
no grau II e 17 (22,97%) no grau III, segundo classificação de Naveira (1960).
A Figura 6 representa o número absoluto de eosinófilos expresso em mediana
de acordo com as idades de 3 (MD=1276 mm3), 4 (MD=1246 mm3) e 5 anos
(MD=910 mm3) das crianças excluindo dos dados a única criança que apresentava 6
anos. Conforme podemos observar não houve diferença significativa entre as idades
das crianças estudadas.
FIGURA 6: Número absoluto de eosinófilos no sangue periférico de acordo com a idade em 103 crianças, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. A contagem absoluta dos eosinófilos foi realizada em
analisador automático, através do método de impedância. Os valores de referência
utilizados foram de até 650 eosinófilos/mm3. p>0,05, quando comparado o número absoluto
de eosinófilos e a idade das crianças.
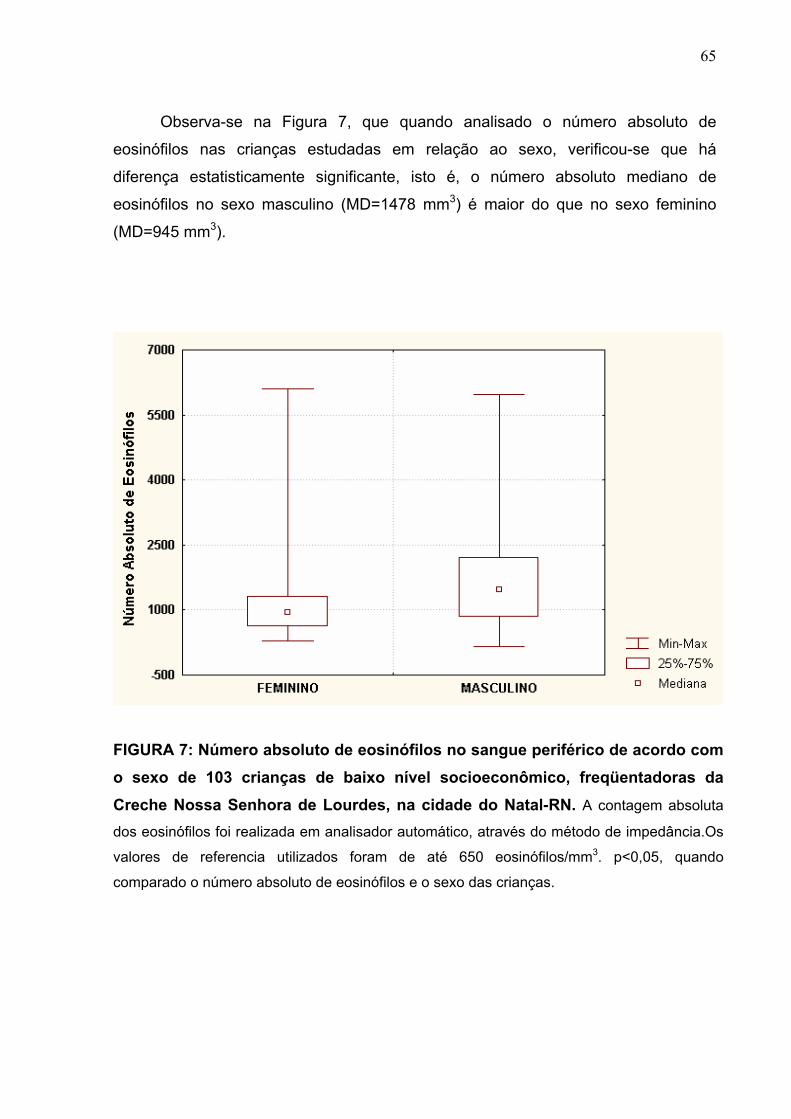
65
Observa-se na Figura 7, que quando analisado o número absoluto de
eosinófilos nas crianças estudadas em relação ao sexo, verificou-se que há
diferença estatisticamente significante, isto é, o número absoluto mediano de
eosinófilos no sexo masculino (MD=1478 mm3) é maior do que no sexo feminino
(MD=945 mm3).
FIGURA 7: Número absoluto de eosinófilos no sangue periférico de acordo com o sexo de 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. A contagem absoluta
dos eosinófilos foi realizada em analisador automático, através do método de impedância.Os
valores de referencia utilizados foram de até 650 eosinófilos/mm3. p<0,05, quando
comparado o número absoluto de eosinófilos e o sexo das crianças.
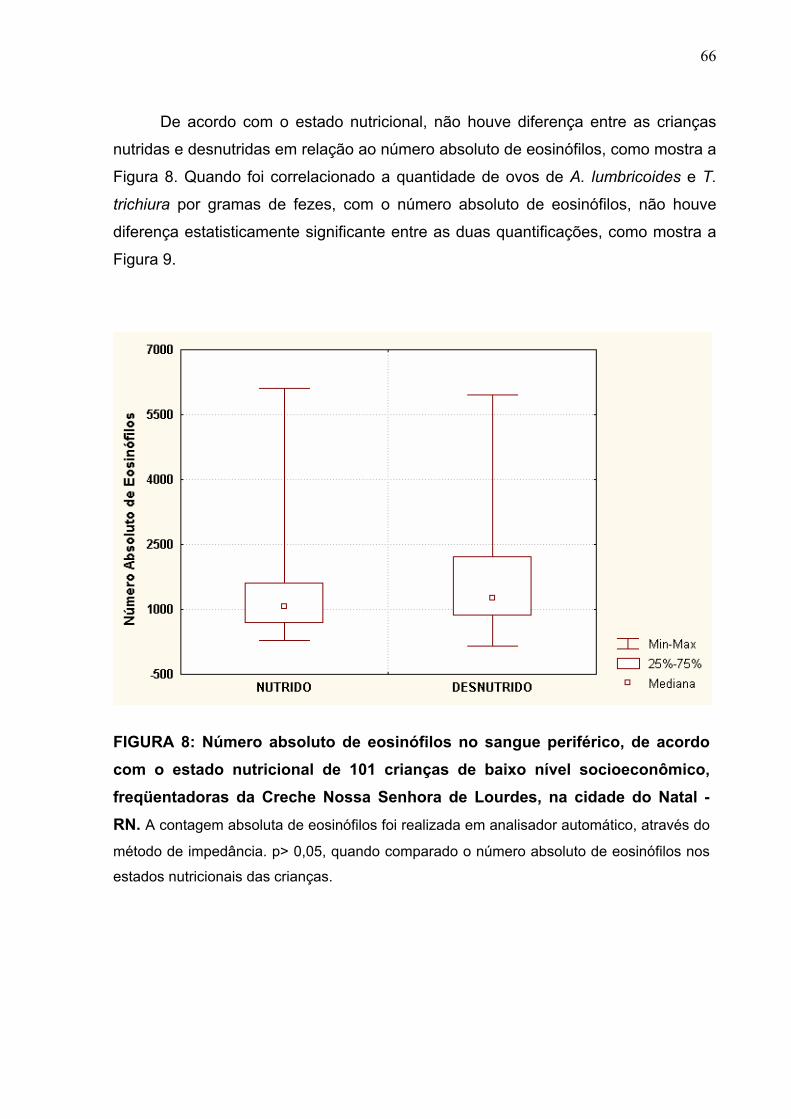
66
De acordo com o estado nutricional, não houve diferença entre as crianças
nutridas e desnutridas em relação ao número absoluto de eosinófilos, como mostra a
Figura 8. Quando foi correlacionado a quantidade de ovos de A. lumbricoides e T.
trichiura por gramas de fezes, com o número absoluto de eosinófilos, não houve
diferença estatisticamente significante entre as duas quantificações, como mostra a
Figura 9.
FIGURA 8: Número absoluto de eosinófilos no sangue periférico, de acordo com o estado nutricional de 101 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal - RN. A contagem absoluta de eosinófilos foi realizada em analisador automático, através do
método de impedância. p> 0,05, quando comparado o número absoluto de eosinófilos nos
estados nutricionais das crianças.
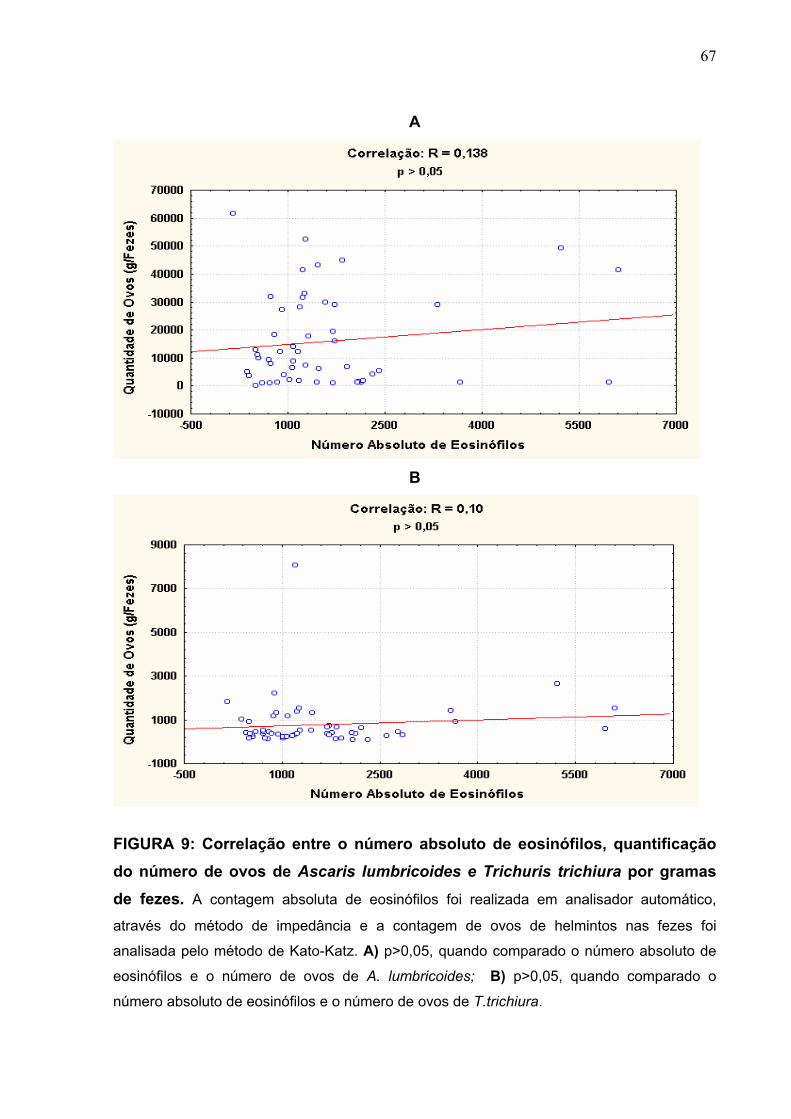
67
A
B
FIGURA 9: Correlação entre o número absoluto de eosinófilos, quantificação do número de ovos de Ascaris lumbricoides e Trichuris trichiura por gramas de fezes. A contagem absoluta de eosinófilos foi realizada em analisador automático,
através do método de impedância e a contagem de ovos de helmintos nas fezes foi
analisada pelo método de Kato-Katz. A) p>0,05, quando comparado o número absoluto de
eosinófilos e o número de ovos de A. lumbricoides; B) p>0,05, quando comparado o
número absoluto de eosinófilos e o número de ovos de T.trichiura.

68
5.6 Dosagem de Imunoglobulinas 5.6.1 Dosagens de IgA sérica e IgA secretora Apesar de não haver diferença estatisticamente significante entre as
concentrações de IgA sérica e IgA secretora das 103 crianças estudadas e os
valores de referência utilizados para indivíduos normais, 24,3% das crianças
apresentaram valores abaixo do normal para IgA sérica e 47,1% para IgA secretora.
A Tabela 7 representa as medianas (MD) das concentrações séricas de IgA sérica e
IgA secretora.
TABELA 7: Mediana dos níveis de IgA sérica e IgA secretora de 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
IMUNOGLOBULINAS (Igs)
1°
QUARTIL (25%)
MEDIANA (50%)
3°
QUARTIL (75%)
IgA Sérica
IgA Secretora
0,702
2,6
0,938
(0,0-2,50)
3,7 (1,1-22,8)
1,2
5,7
As dosagens de IgA sérica e IgA secretora foram determinadas por turbidimetria e
Nefelometria respectivamente e as concentrações foram obtidas através de curva de
calibração de IgA sérica e IgA secretora. Os valores de referência utilizados para indivíduos
normais são de 0,7 a 4,0 g/L para IgA sérica e 3, 5 a 36,5 mg/L para IgA secretora.

69
Não houve diferença estatisticamente significante quando correlacionou-se o
estado nutricional e as concentrações de IgA sérica e IgA secretora. Porém
podemos observar que as crianças desnutridas apresentaram medianas menores
para IgA secretora e maiores para IgA sérica, do que as crianças nutridas, como
mostra a Tabela 8.
TABELA 8: Mediana das concentrações de IgA sérica e IgA secretora, segundo o estado nutricional de 101 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN.
Ig(s)
1° Quartil (25%)
Nutrido Mediana
(MD)
3° Quartil (75%)
1° Quartil (25%)
Desnutrido Mediana (MD)
3° Quartil (75%)
IgA*
(Sérica)
IgA** (Secretora)
0,75
2,3
0,91
3,9
1,2
5,7
0,54
2,3
0,96
3,3
1,3 4,6
As dosagens de IgA sérica e IgA secretora foram determinadas por turbidimetria e
Nefelometria respectivamente e as concentrações foram obtidas através de curva de
calibração de IgA sérica e IgA secretora. Os valores de referência utilizados para indivíduos
normais são de 0,7 a 4,0 g/L para IgA sérica e 3, 5 a 36,5 mg/L para IgA secretora. p>0,05,
quando correlacionado a IgA sérica com os estados nutricionais e p>0,05, quando
correlacionado a IgA secretora e os estados nutricionais.
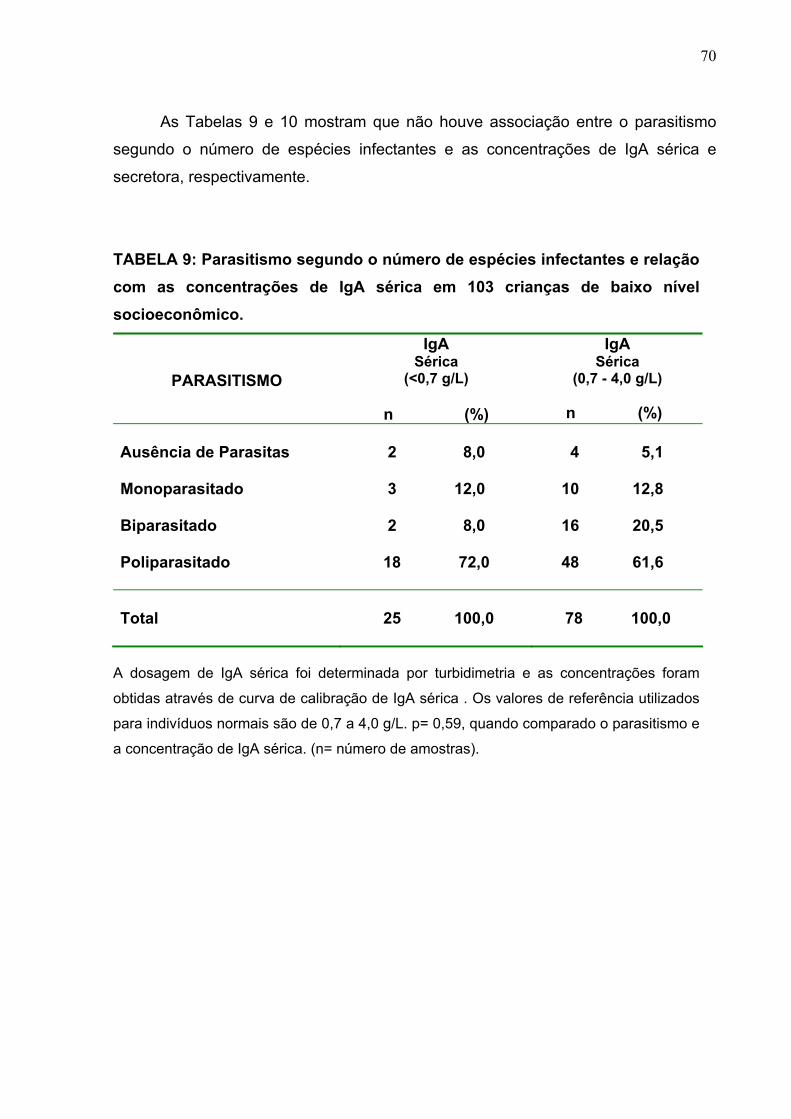
70
As Tabelas 9 e 10 mostram que não houve associação entre o parasitismo
segundo o número de espécies infectantes e as concentrações de IgA sérica e
secretora, respectivamente.
TABELA 9: Parasitismo segundo o número de espécies infectantes e relação com as concentrações de IgA sérica em 103 crianças de baixo nível socioeconômico.
PARASITISMO
IgA Sérica
(<0,7 g/L)
n (%)
IgA Sérica
(0,7 - 4,0 g/L)
n (%) Ausência de Parasitas Monoparasitado Biparasitado Poliparasitado
2 8,0 3 12,0 2 8,0 18 72,0
4 5,1 10 12,8 16 20,5 48 61,6
Total
25 100,0
78 100,0
A dosagem de IgA sérica foi determinada por turbidimetria e as concentrações foram
obtidas através de curva de calibração de IgA sérica . Os valores de referência utilizados
para indivíduos normais são de 0,7 a 4,0 g/L. p= 0,59, quando comparado o parasitismo e
a concentração de IgA sérica. (n= número de amostras).
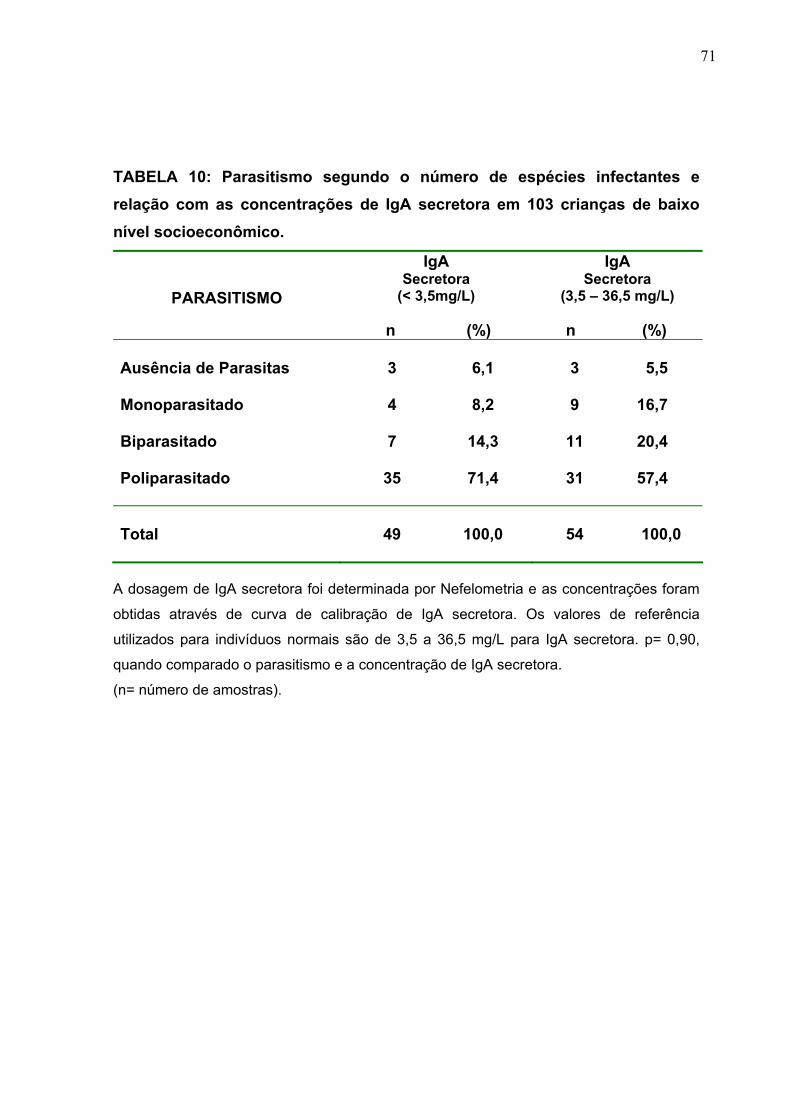
71
TABELA 10: Parasitismo segundo o número de espécies infectantes e relação com as concentrações de IgA secretora em 103 crianças de baixo nível socioeconômico.
PARASITISMO
IgA Secretora
(< 3,5mg/L)
n (%)
IgA Secretora
(3,5 – 36,5 mg/L)
n (%) Ausência de Parasitas Monoparasitado Biparasitado Poliparasitado
3 6,1 4 8,2 7 14,3 35 71,4
3 5,5 9 16,7 11 20,4 31 57,4
Total
49 100,0
54 100,0
A dosagem de IgA secretora foi determinada por Nefelometria e as concentrações foram
obtidas através de curva de calibração de IgA secretora. Os valores de referência
utilizados para indivíduos normais são de 3,5 a 36,5 mg/L para IgA secretora. p= 0,90,
quando comparado o parasitismo e a concentração de IgA secretora.
(n= número de amostras).

72
• Correlação das concentrações de IgA sérica, IgA secretora e o número de ovos de A. lumbricoides e T. trichiura por grama de fezes
A Figura 10 mostra que as concentrações de IgA sérica e secretora das
crianças que estavam infectadas por A. lumbricoides, não apresentaram correlação
estatisticamente significante com o número de ovos deste helminto nas fezes
analisadas.
Quando analisado as concentrações de IgA sérica e IgA secretora das
crianças que estavam infectadas por T. trichiura, com o número de ovos por grama
de fezes deste helminto, esta correlação não apresentou positividade significante
de acordo com a Figura 11.
• Correlação entre as concentrações de IgA sérica, IgA secretora e IgE total sérica
Não houve correlação estatisticamente significante entre as concentrações de
IgA sérica e secretora como mostra a Figura 12.
Na Figura 13 observa-se que não houve uma correlação positiva entre os
níveis séricos de IgE total com as concentrações de IgA sérica e secretora.
• Correlação entre as concentrações de IgA sérica, IgA secretora e número absoluto de eosinófilos
Na Figura 14 observa-se que as concentrações de IgA sérica e secretora não
apresentaram correlação estatisticamente significante com o número absoluto de
eosinófilos nas 103 crianças estudadas.
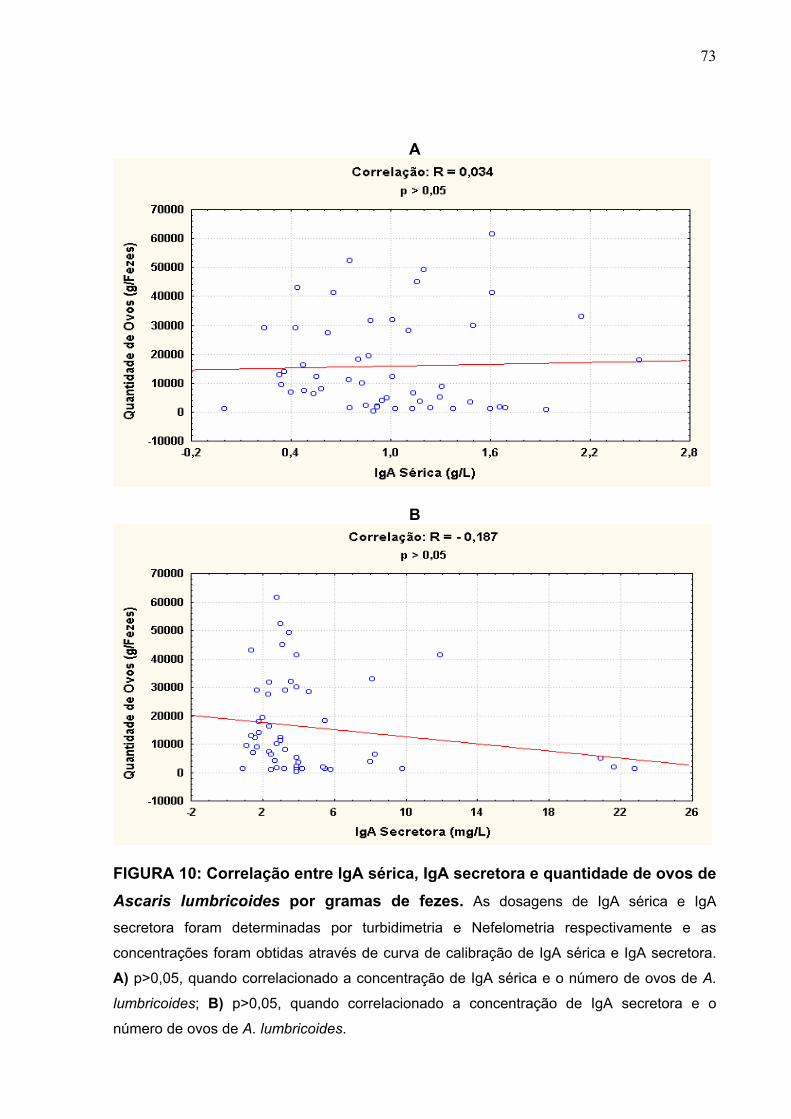
73
A
B
FIGURA 10: Correlação entre IgA sérica, IgA secretora e quantidade de ovos de Ascaris lumbricoides por gramas de fezes. As dosagens de IgA sérica e IgA
secretora foram determinadas por turbidimetria e Nefelometria respectivamente e as
concentrações foram obtidas através de curva de calibração de IgA sérica e IgA secretora.
A) p>0,05, quando correlacionado a concentração de IgA sérica e o número de ovos de A.
lumbricoides; B) p>0,05, quando correlacionado a concentração de IgA secretora e o
número de ovos de A. lumbricoides.
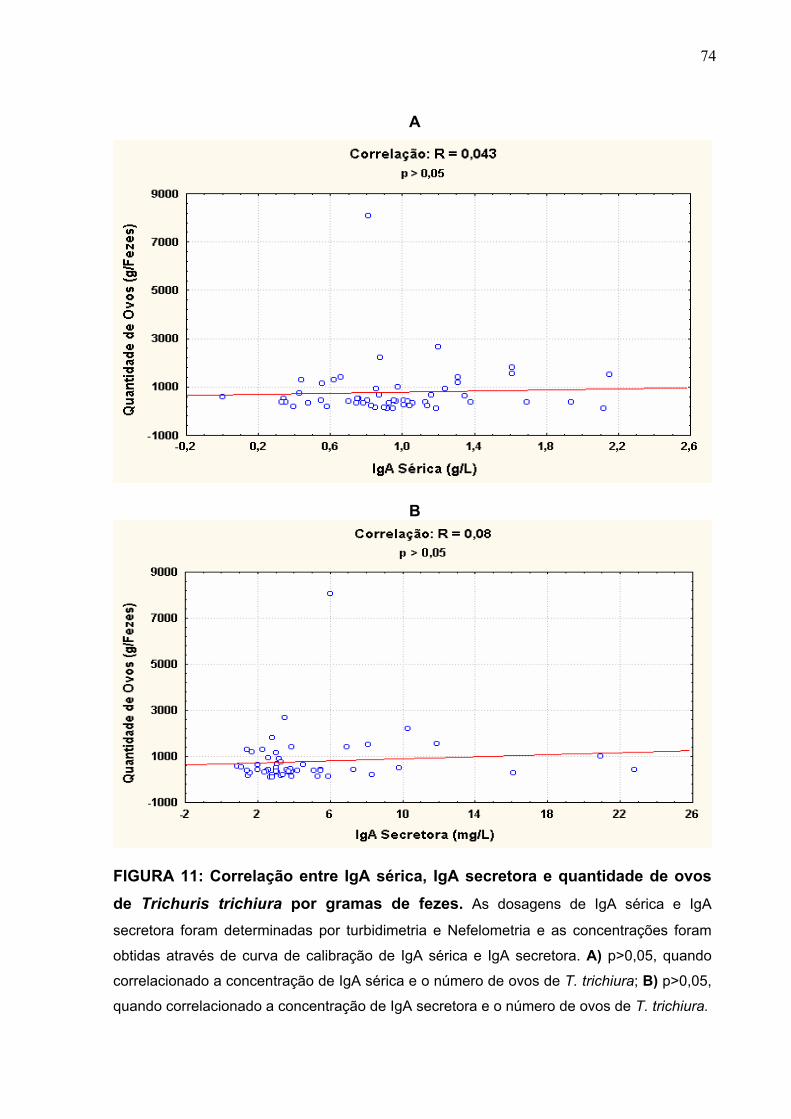
74
A
B
FIGURA 11: Correlação entre IgA sérica, IgA secretora e quantidade de ovos de Trichuris trichiura por gramas de fezes. As dosagens de IgA sérica e IgA
secretora foram determinadas por turbidimetria e Nefelometria e as concentrações foram
obtidas através de curva de calibração de IgA sérica e IgA secretora. A) p>0,05, quando
correlacionado a concentração de IgA sérica e o número de ovos de T. trichiura; B) p>0,05,
quando correlacionado a concentração de IgA secretora e o número de ovos de T. trichiura.
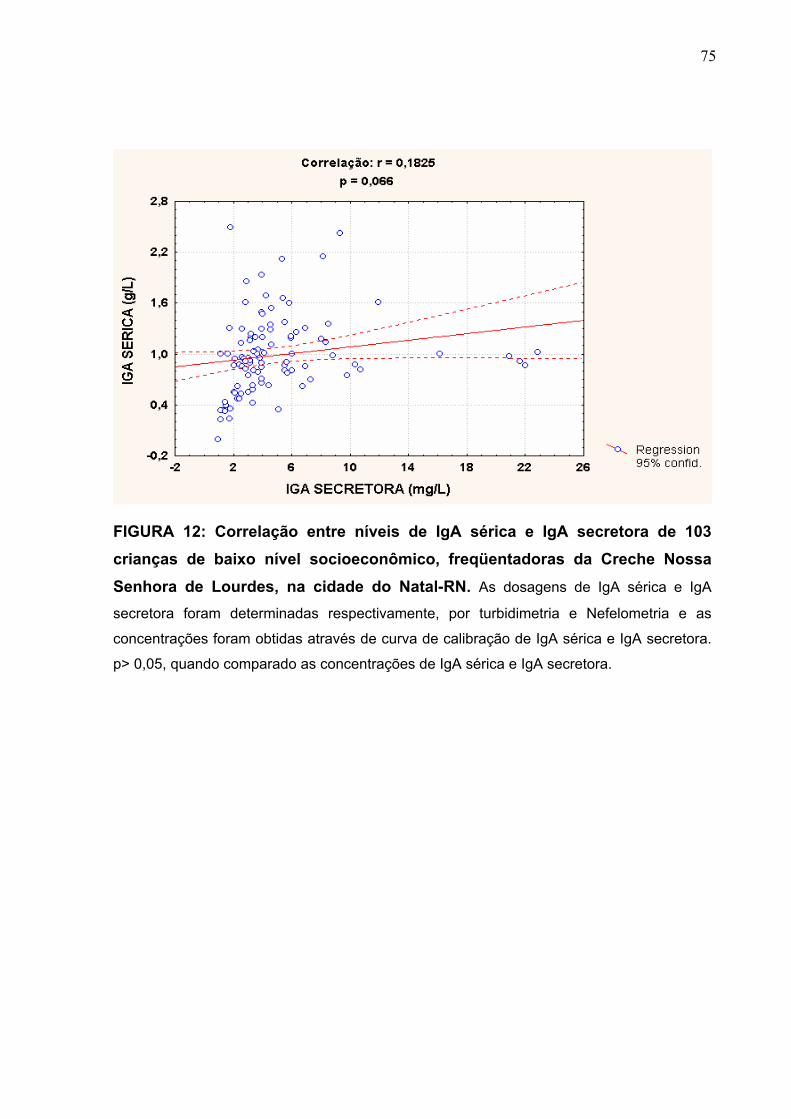
75
FIGURA 12: Correlação entre níveis de IgA sérica e IgA secretora de 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. As dosagens de IgA sérica e IgA
secretora foram determinadas respectivamente, por turbidimetria e Nefelometria e as
concentrações foram obtidas através de curva de calibração de IgA sérica e IgA secretora.
p> 0,05, quando comparado as concentrações de IgA sérica e IgA secretora.
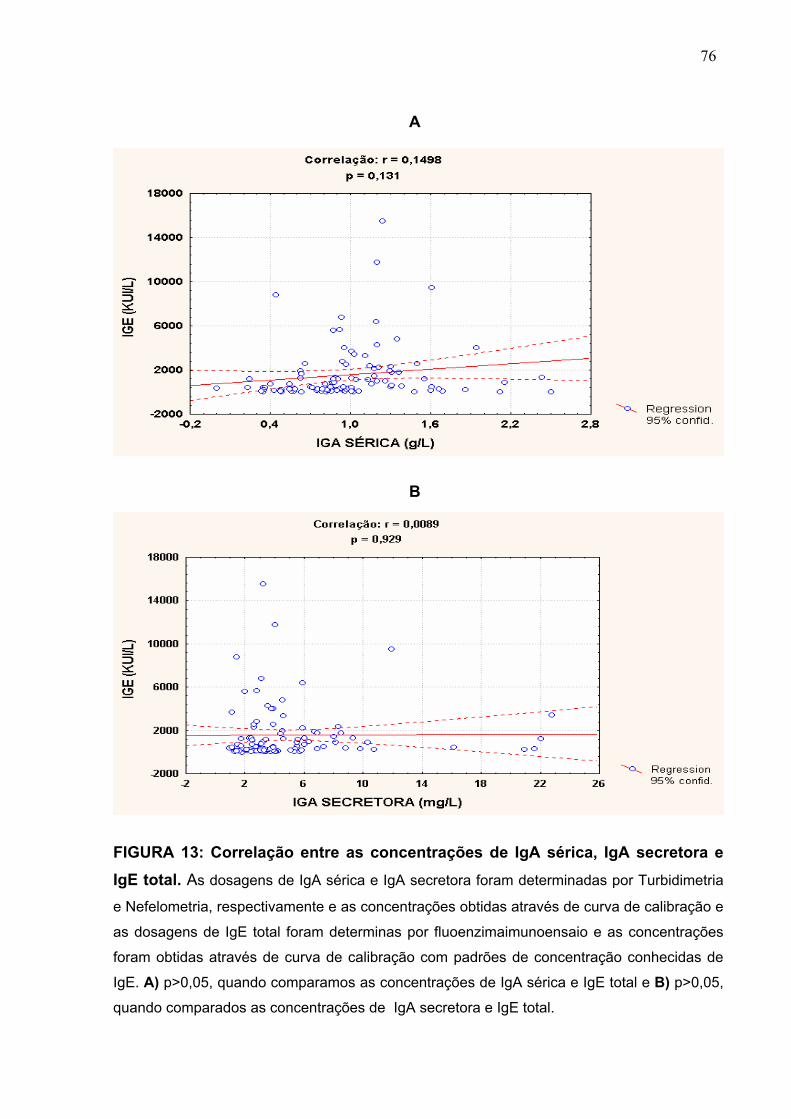
76
A
B
FIGURA 13: Correlação entre as concentrações de IgA sérica, IgA secretora e IgE total. As dosagens de IgA sérica e IgA secretora foram determinadas por Turbidimetria
e Nefelometria, respectivamente e as concentrações obtidas através de curva de calibração e
as dosagens de IgE total foram determinas por fluoenzimaimunoensaio e as concentrações
foram obtidas através de curva de calibração com padrões de concentração conhecidas de
IgE. A) p>0,05, quando comparamos as concentrações de IgA sérica e IgE total e B) p>0,05,
quando comparados as concentrações de IgA secretora e IgE total.
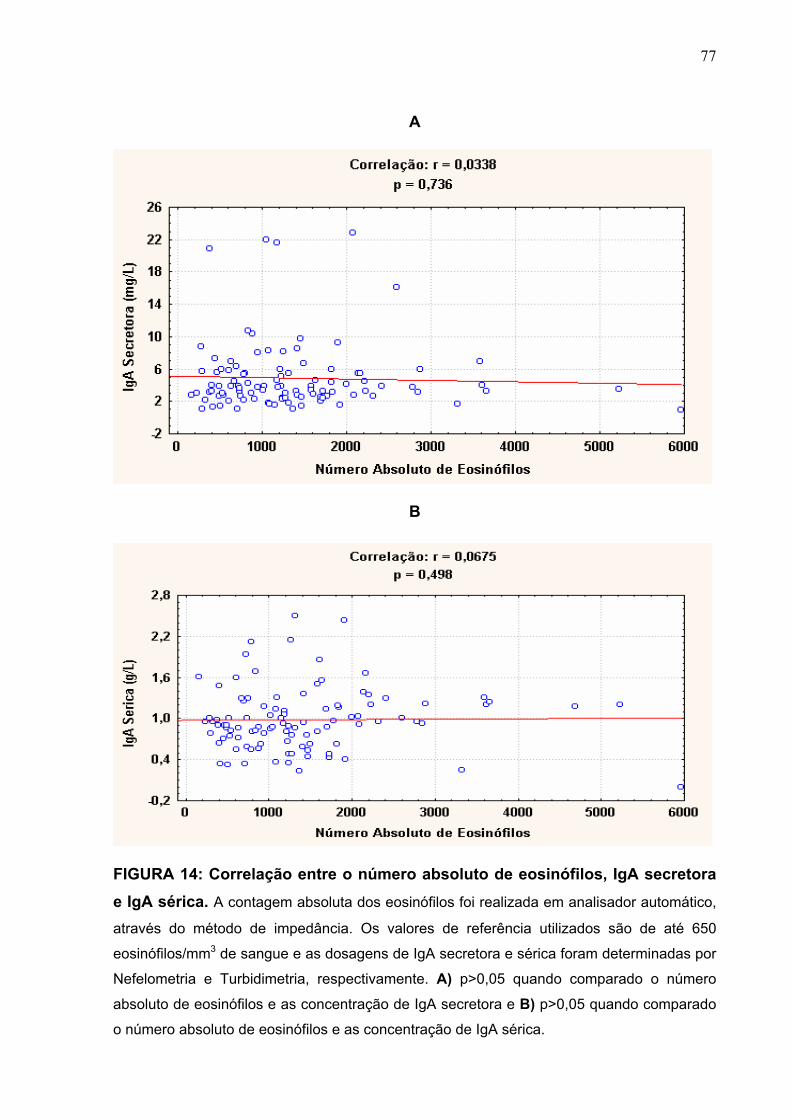
77
A
B
FIGURA 14: Correlação entre o número absoluto de eosinófilos, IgA secretora e IgA sérica. A contagem absoluta dos eosinófilos foi realizada em analisador automático,
através do método de impedância. Os valores de referência utilizados são de até 650
eosinófilos/mm3 de sangue e as dosagens de IgA secretora e sérica foram determinadas por
Nefelometria e Turbidimetria, respectivamente. A) p>0,05 quando comparado o número
absoluto de eosinófilos e as concentração de IgA secretora e B) p>0,05 quando comparado
o número absoluto de eosinófilos e as concentração de IgA sérica.

78
5.6.2 Dosagem de IgE total Das 103 crianças estudadas 98 (95,1%) apresentaram níveis séricos de IgE
total acima dos valores de referência, com mediana bastante elevada, de 516 KU/L
(21,6 KU/L - 15.520 KU/L). A Tabela 11 representa a mediana das concentrações de
IgE de acordo com as idades das crianças de 3, 4, e 5 anos, excluindo-se a única
criança de 6 anos. Verificamos que não houve diferença estatisticamente significante
entre as idades estudadas.
TABELA 11: Mediana das concentrações de IgE total de 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN
IDADE
IgE Total KU/L (MD)
3 Anos
4 Anos
5 Anos
365,85
509,50
979,00
As dosagens de IgE total foram determinadas por fluorenzimaimunoensaio e as
concentrações foram obtidas através de curva de calibração com padrões de concentrações
conhecidas de IgE total. Os valores de referência de IgE total por idade utilizados foram: 3
anos até 32K U/L, 4 anos até 40 KU/L, 5 anos até 48 KU/L e 6 anos até 56 KU/L.
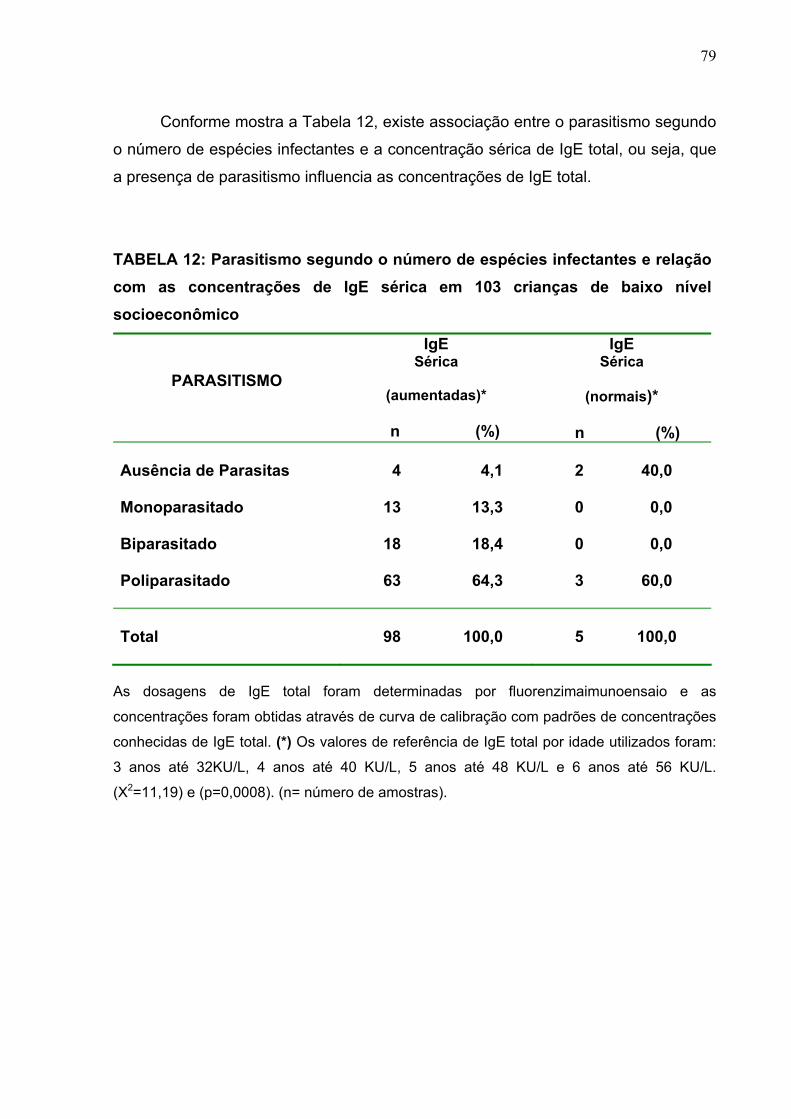
79
Conforme mostra a Tabela 12, existe associação entre o parasitismo segundo
o número de espécies infectantes e a concentração sérica de IgE total, ou seja, que
a presença de parasitismo influencia as concentrações de IgE total.
TABELA 12: Parasitismo segundo o número de espécies infectantes e relação com as concentrações de IgE sérica em 103 crianças de baixo nível socioeconômico
PARASITISMO
IgE Sérica
(aumentadas)*
n (%)
IgE Sérica
(normais)*
n (%)
Ausência de Parasitas Monoparasitado Biparasitado Poliparasitado
4 4,1 13 13,3 18 18,4 63 64,3
2 40,0 0 0,0 0 0,0 3 60,0
Total
98 100,0
5 100,0
As dosagens de IgE total foram determinadas por fluorenzimaimunoensaio e as
concentrações foram obtidas através de curva de calibração com padrões de concentrações
conhecidas de IgE total. (*) Os valores de referência de IgE total por idade utilizados foram:
3 anos até 32KU/L, 4 anos até 40 KU/L, 5 anos até 48 KU/L e 6 anos até 56 KU/L.
(X2=11,19) e (p=0,0008). (n= número de amostras).

80
• Correlação das concentrações de IgE sérica total e o número de ovos de A. lumbricoides e T. trichiura por grama de fezes
Verificou-se na Figura 15 que as concentrações de IgE total das crianças que
estavam infectadas por A. lumbricoides, não apresentaram correlação
estatisticamente significante com o número de ovos deste helminto nas fezes
analisadas.
Quando foi analisado as concentrações de IgE total das crianças que estavam
infectadas por T. trichiura, com o número de ovos por grama de fezes deste
helminto, esta correlação não apresentou positividade significante de acordo com a
Figura 15.
• Correlação entre a concentração de IgE total e o número absoluto de eosinófilos
Na Figura 16 pode-se observar que uma correlação positiva foi encontrada
entre as concentrações de IgE total e o número absoluto de eosinófilos nas 103
crianças estudadas.
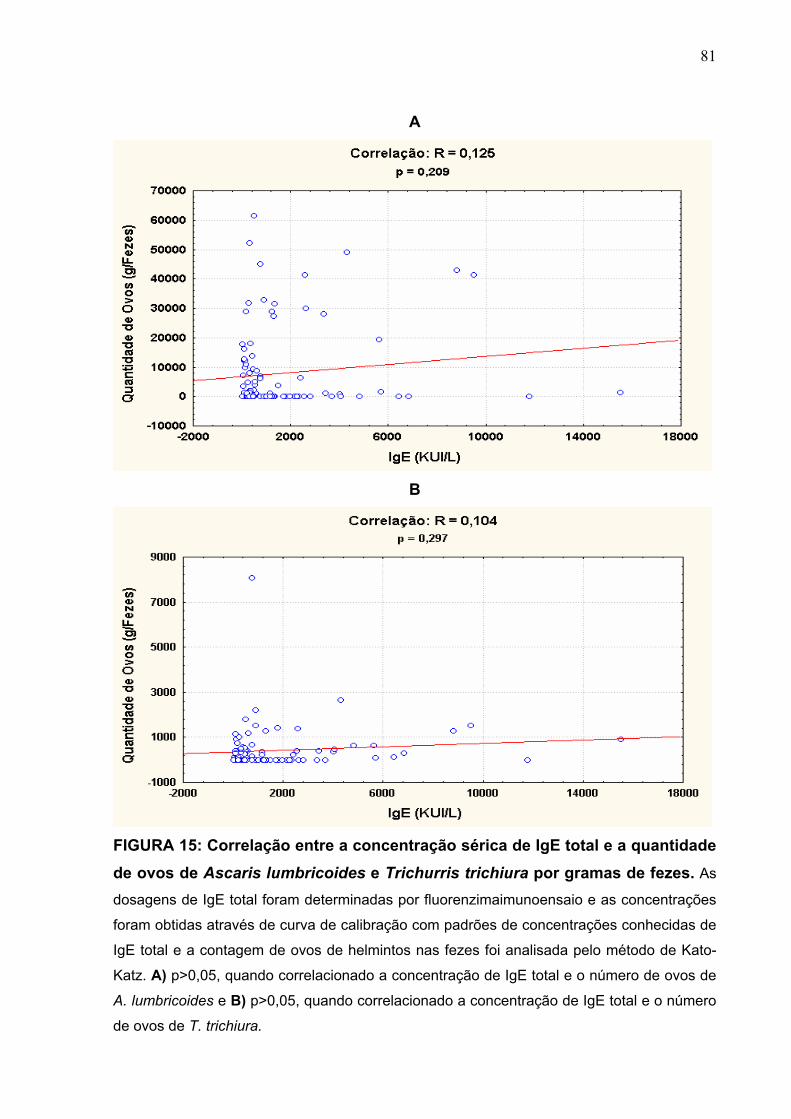
81
A
B
FIGURA 15: Correlação entre a concentração sérica de IgE total e a quantidade de ovos de Ascaris lumbricoides e Trichurris trichiura por gramas de fezes. As
dosagens de IgE total foram determinadas por fluorenzimaimunoensaio e as concentrações
foram obtidas através de curva de calibração com padrões de concentrações conhecidas de
IgE total e a contagem de ovos de helmintos nas fezes foi analisada pelo método de Kato-
Katz. A) p>0,05, quando correlacionado a concentração de IgE total e o número de ovos de
A. lumbricoides e B) p>0,05, quando correlacionado a concentração de IgE total e o número
de ovos de T. trichiura.
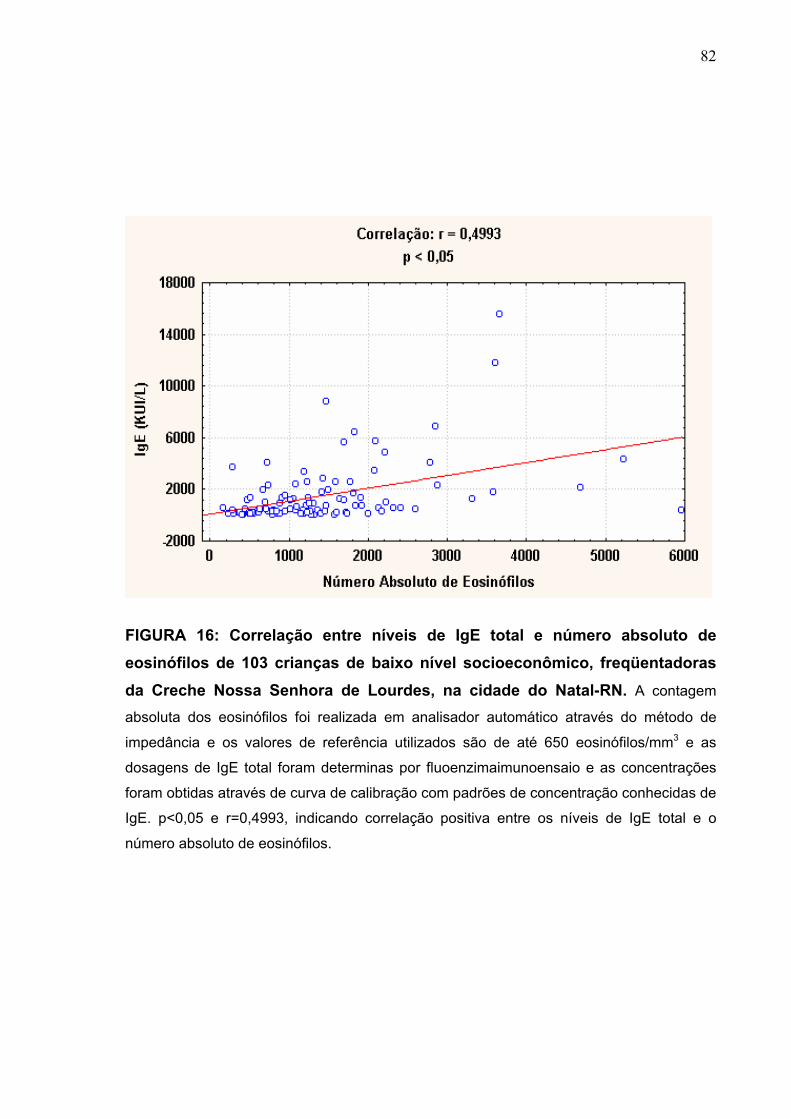
82
FIGURA 16: Correlação entre níveis de IgE total e número absoluto de eosinófilos de 103 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. A contagem
absoluta dos eosinófilos foi realizada em analisador automático através do método de
impedância e os valores de referência utilizados são de até 650 eosinófilos/mm3 e as
dosagens de IgE total foram determinas por fluoenzimaimunoensaio e as concentrações
foram obtidas através de curva de calibração com padrões de concentração conhecidas de
IgE. p<0,05 e r=0,4993, indicando correlação positiva entre os níveis de IgE total e o
número absoluto de eosinófilos.
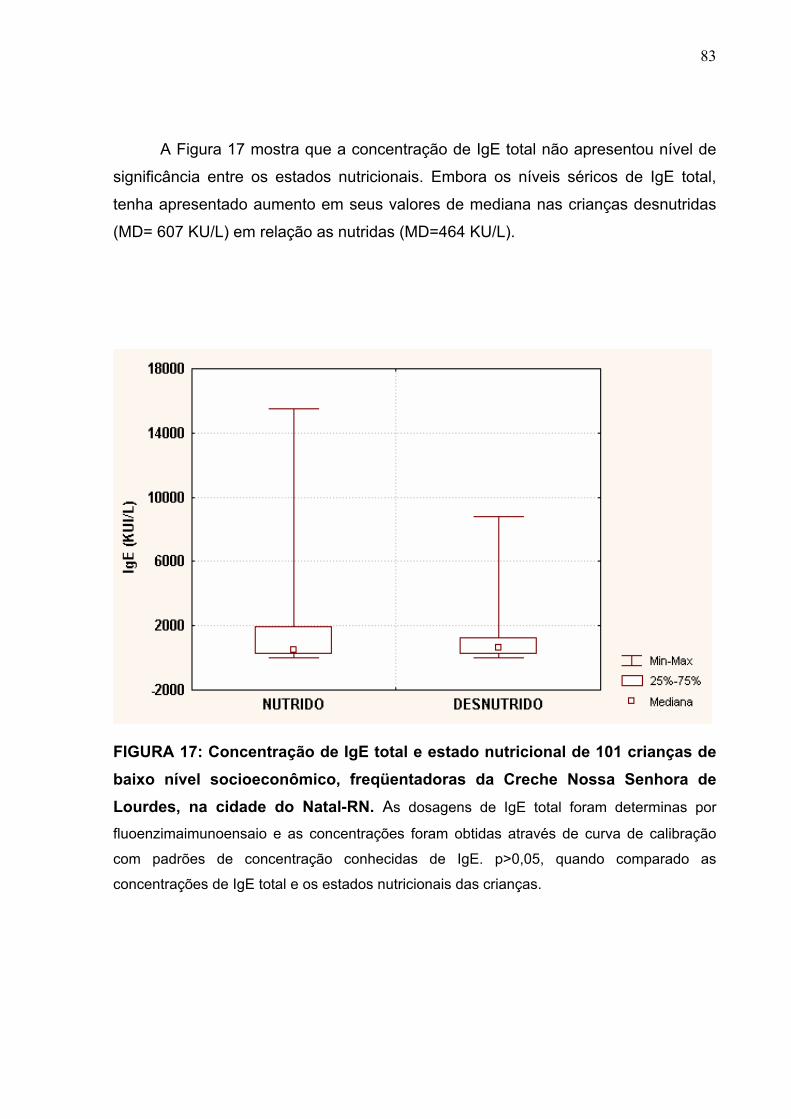
83
A Figura 17 mostra que a concentração de IgE total não apresentou nível de
significância entre os estados nutricionais. Embora os níveis séricos de IgE total,
tenha apresentado aumento em seus valores de mediana nas crianças desnutridas
(MD= 607 KU/L) em relação as nutridas (MD=464 KU/L).
FIGURA 17: Concentração de IgE total e estado nutricional de 101 crianças de baixo nível socioeconômico, freqüentadoras da Creche Nossa Senhora de Lourdes, na cidade do Natal-RN. As dosagens de IgE total foram determinas por
fluoenzimaimunoensaio e as concentrações foram obtidas através de curva de calibração
com padrões de concentração conhecidas de IgE. p>0,05, quando comparado as
concentrações de IgE total e os estados nutricionais das crianças.

84
6 DISCUSSÃO
As infecções intestinais por parasitas estão entre as mais prevalentes das
infecções humanas, principalmente em países em desenvolvimento.
Classicamente, os estudos epidemiológicos têm procurado explicar as
desigualdades na saúde da população segundo fatores sociais e econômicos tais
como renda, ocupação, educação, habitação, ambiente ou de uma maneira geral, as
assim denominadas condições de vida. É vasta a literatura que aborda os
diferenciais na saúde como reflexo das desigualdades de uma sociedade. Um
estudo internacionalmente conhecido, realizado na Grã-Bretanha, (“The Black
Report”), revelou grandes disparidades na situação de saúde, demonstrando que
aqueles situados no limite inferior da escala social têm condições bem piores de
saúde do que aqueles pertencentes aos estratos mais favorecidos (SZWARCWALD,
et al., 1999).
LYNCH et al. (1983), demonstrou a existência de uma relação direta entre as
condições sócio-econômicas da população, inexistência de educação sanitária e
maior exposição às infecções causadas por parasitas em indivíduos menos
favorecidos, isto é, pertencentes a classes sociais mais baixas.
O ecossistema e esses fatores socioeconômicos que permitem aos parasitas
sobreviverem em nosso ambiente também promovem o desenvolvimento de
desnutrição, a qual compromete a imunidade aumentando a susceptibilidade para
doenças infecciosas, e estas por sua vez, podem prejudicar o estado nutricional
humano.
Com base em estudo anteriormente realizado no nosso laboratório, o qual
mostrou que crianças de baixo nível socioeconômico, residentes em área com
precárias condições de higiene e saneamento básico na cidade do Natal,
apresentavam-se poliparasitadas, com eosinofilia sanguínea e elevadas
concentrações de IgE total e específica para A. lumbricoides (RODRIGUES, 2001),
resolveu-se pesquisar as possíveis correlações entre estes, estado nutricional e IgA
sérica e secretora, a qual é a principal imunoglobulina protetora de mucosas.
No nosso estudo, trabalhamos com uma população de crianças de ambos os
sexos, na faixa etária de 3 a 6 anos (Tabela 1), residentes no mesmo bairro e

85
freqüentadoras da mesma creche. A grande maioria vivia em precárias condições de
higiene, sem saneamento básico e com baixo nível socioeconômico, segundo os
indicadores sociais mínimos do IBGE (1999) (Tabela 2). Estes índices apresentados
revelaram um perfil nada diferente de outros centros urbanos do país (GROSS et al.,
1989; BECKER et al., 2002) que bem retratam as condições precárias de
saneamento básico e o baixo nível sócio econômico das populações com alta
incidência de parasitoses intestinais (SILVA, 1998).
No contexto dos grandes problemas de saúde pública as infecções por
parasitas intestinais figuram com um dos maiores índices de prevalência na grande
massa populacional. Fatores como péssimas condições de moradia, má
alimentação, falta de água tratada, esgoto e principalmente educação e
conscientização da necessidade, desde o primeiro ano escolar, de criar no âmbito
familiar a cultura de hábitos de higiene e limpeza levam sempre a prejuízos trazidos
pelas parasitoses intestinais, principalmente as de curso crônico e assintomático que
têm como conseqüência o déficit de desenvolvimento físico e mental além do
comprometimento imunológico (HASWELL-ELKINS et al., 1992; CROMPTON, 1992;
MORALES et al., 1999; BECKER et al., 2002; CARVALHO et al., 2002).
Situação também caracterizada em nosso trabalho, pois houve uma grande
freqüência de parasitoses (Tabela 5), possivelmente devido aos fatores
socioeconômicos, e às precárias condições em que as crianças e suas famílias
vivem. Constatou-se uma elevada prevalência das enteroparasitoses, sendo o A.
lumbricoides e T. trichiura, os helmintos mais freqüentes (Tabela 6). A maioria das
crianças estudadas estava infectada por no mínimo dois tipos diferentes de parasitas
intestinais, sendo mais comum à associação entre estes helmintos com protozoários,
como a E. nana e G. lamblia (Tabelas 4, 5 e 6).
Os resultados encontrados concordaram com estudos realizados em várias
regiões de países subdesenvolvidos (HAGEL et al., 1993; LYNCH et al., 1998;
MORALES et al., 1999; ANDRADE et al., 2001) e com outros realizados em regiões
brasileiras, inclusive em diferentes locais da nossa cidade (ROSÁRIO FILHO et al.,
1982; MACEDO et al., 1998; NASCIMENTO et al., 1999; BECKER et al., 2002).
A presença de poliparasitismo, detectada em nosso estudo, agrava a doença
parasitária com quadros de maior patogenia, o que parece ser mais comum
principalmente em crianças. Não houve interferência na freqüência das parasitoses
intestinais com a idade das crianças, possivelmente por estas praticarem os mesmos

86
hábitos desfavoráveis de higiene, encontrarem-se em uma faixa etária próxima e em
mesmas condições sociais. Esta elevada freqüência de parasitismo encontra-se
compatível com as condições socioeconômicas, nas quais crianças que geralmente
nascem em áreas endêmicas para enteroparasitoses, podem albergá-los por um
longo período, devido ao número grande de exposições repetidas a estes parasitas
como também, a inabilidade de poder desenvolver uma imunidade protetora
(SALDIVA et al., 1993; PRATA, 1994; MACEDO et al., 1999).
Como notado no presente estudo há um predomínio de A. lumbricoides, T.
trichiura e G. lamblia, parasitas que traduzem a falta de rede de esgoto na região
bem como a presença de córrego a céu aberto, contribuindo sobremaneira para a
disseminação de muitas doenças em especial destas enteroparasitoses.
KIGHTLINGER et al. (1998), relacionou a intensidade da carga parasitaria em
infecções por A. lumbricoides em crianças com determinantes em relação ao
ambiente e ao nível socioeconômico, e os achados mostraram que a carga
parasitária aumentou à medida que diminuiu a higiene alimentar, o nível cultural e
econômico da região estudada. Isto é quanto maior a pobreza, maior o número de A.
lumbricoides albergados nas crianças (CARNEIRO et al., 2002).
A intensidade das infecções helmínticas foi classificada baseada nos critérios
recomendados pela OMS (1987), segundo os quais a infecção é atribuída como
leve, quando a contagem de ovos por grama de fezes for menor que 5.000,
moderada quando a contagem estiver entre 5.000 e 50.000, e maciça quando
ultrapassar 50.000 ovos por grama de fezes, isto para A. lumbricoides, e quando for
para T. trichiura a infecção é atribuída como leve se o número de ovos for inferior a
1.000, moderada quando estiver compreendida entre 1.000 a 10.000 e maciça se a
contagem for maior que 10.000 ovos por grama de fezes.
Baseados nesta classificação os resultados da quantificação do número de
ovos por grama de fezes, mostraram que as infecções por A. lumbricoides e T.
trichiura, encontravam-se como infecções parasitárias de leve e moderada
intensidade. Quando se correlacionou as cagas parasitárias com as concentrações
de IgA sérica, IgA secretora e IgE total (Figuras 10, 11 e 15), observou-se que não
houve diferença estatisticamente significante, achados parcialmente semelhantes foi
encontrado por ROSÁRIO FILHO et al. (1982) no estado do Paraná, quando
relacionou o número de ovos com a concentração sérica de IgE.

87
As infecções gastrintestinais podem também contribuir para o
desenvolvimento da desnutrição infantil pela redução da ingestão de alimentos,
prejudicando a absorção intestinal e alterando o metabolismo dos nutrientes e têm
maior probabilidade de ocorrer em crianças com déficit nutricional pré-existente.
Episódios de diarréia podem ter duração mais longa, apresentar maior gravidade e
levar à morte crianças desnutridas (RIKIMARU et al., 1998; SILVA & STURION,
1998; CHANDRA, 1989; BERN, 1992; BERKMAN et al., 2002).
A carga parasitária também foi verificada com a finalidade de avaliar se as
crianças desnutridas apresentavam maiores cargas, mas não houve diferença
significativa (Figuras 4 e 5). As relações foram feitas com os helmintos A.
lumbricoides e T. trichiura individualmente, como as crianças são poliparasitadas
talvez possa existir uma relação positiva quando considerar-se cargas de infecções
concomitantes.
O nexo nutrição-infecção-imunidade é um conjunto de interações complexas.
A imunodepressão na desnutrição protéico energética é hoje aceita como um
componente de risco de morbidade e mortalidade por infecção. Estudos têm
mostrado que infecção por A. lumbricoides é realmente um fator na etiologia e
persistência de desnutrição infantil (GUPTA et al 1977; WILLETT et al 1979) e em
alguns casos de infecções concomitantes de A. lumbricoides e T. trichiura a
desnutrição infantil pode exacerbar-se (COOPER & BUNDY, 1988; FORRESTER,
1988).
Tanto a resposta imune celular como a humoral são importantes em limitar a
invasão dos parasitas e em eliminá-los. Desnutrição prejudica estas respostas
imunes e é importante determinante no resultado final da interação parasita-
hospedeiro (STIEHM, 1980; CHANDRA, 1983). Os efeitos da desnutrição energético
protéica na imunidade do hospedeiro têm sido averiguados, porém muitos dos
estudos clínicos têm sido realizados em crianças com desnutrição severa, muitas
vezes hospitalizadas, fazendo-se necessário então, mais estudos em crianças
desnutridas de intensidades leve e moderada, pois estas representam mais de 90%
da população infantil com problemas de desnutrição (OLIVEIRA & THÉBAUD-
MONY, 1998; JINABHAI et al., 2001).
Os dados deste trabalho referentes ao estado nutricional mostram que 30,7%
das crianças apresentaram desnutrição (Tabela 3), destas 93,5% foram classificadas

88
em desnutrição leve e 6,5% como moderada, não havendo assim nenhum caso de
desnutrição grave.
Apesar das crianças apresentarem um poliparasitismo a grande maioria delas
encontravam-se nos estados de desnutrição leve. Isto pode ser relacionado ao fato
das crianças freqüentarem um ambiente de creche, que mesmo com déficit de
alimentos consegue suprir as carências básicas de vitaminas e proteínas, no
momento em que estas crianças permanecem o dia, recebendo as três refeições
básicas.
CORREA et al. (1999) mostrou em trabalho desenvolvido visando avaliar o
impacto nutricional de um programa municipal de assistência alimentar, no Brasil
durante um ano, que já nos 3 primeiros meses de fornecimento dos alimentos houve
uma melhora no status nutricional em 32% das crianças, permitindo assim concluir
que em crianças o aporte de alimentos, através de programas específicos de
suplementação teve conseqüências benéficas, não só sobre o crescimento, mas
também sobre a atividade, cognição e ainda compensação de energia destacando a
modificação do efeito negativo da diarréia sobre o crescimento (SILVA & STURION,
1998).
É importante reconhecer que a criança desnutrida em decorrência de ingestão
inadequada ou de infecção recorrente tem déficit nas reservas de proteínas, energia,
vitaminas e minerais (SUSKIND et al., 1976). Quando dosamos as proteínas totais
observamos que 84,5% das crianças encontravam-se com seus níveis normais,
enquanto que 15,5% estavam com seus níveis aumentados (Figura 1). Para ser
observada em maiores detalhes esta elevação optou-se em realizar a eletroforese
de proteínas no soro das crianças, e os resultados mostraram que a fração gama
globulina estava aumentada em 100% das crianças que estavam com as proteínas
totais aumentadas. Esta elevação pode ser devido a um aumento de outras
imunoglobulinas não estudadas, como as IgM e IgG, cujas subclasses IgG1, IgG2 e
IgG4 são importantes nas infecções, principalmente, causadas por T. trichiura
(BUNDY et al., 1991), que foi o helminto mais freqüente em nosso trabalho, ou
também devido o estado de infecção crônica destas crianças se refletir em um
estímulo de produção das proteínas de fase aguda.
Os resultados da alta prevalência parasitária também se refletiram nos
elevados números de leucócitos sanguíneos quando determinado o número de
eosinófilos no sangue dessas crianças estudadas. Os dados da contagem relativa

89
do número de eosinófilos demonstraram que 99% das crianças estudadas
apresentam eosinofilia no sangue periférico (NAVEIRA, 1960)
ROSÁRIO FILHO et al. (1982), mostrou que indivíduos infectados com T.
trichiura apresentavam contagens significativamente maiores do que o grupo
controle utilizado por ele. Esta eosinofilia encontrada em nosso trabalho reflete
provavelmente a presença de helmintos intestinais como A. lumbricoides e T.
trichiura, como resultado de um possível aumento na resposta imune contra os
mesmos.
A mediana do número de eosinófilos, não mostrou diferença com relação à
idade das crianças (Figura 6), porém em relação ao sexo foi observada uma
diferença estatisticamente significante maior no sexo masculino (Figura 7),
concordando com trabalho de MENDES et al. (1998).
Observamos, portanto que a eosinofilia encontrada, pode ser devido as
parasitoses, pois é visto na literatura que a resposta imune a parasitas é mediada
com a participação dos eosinófilos como células efetoras na destruição destes, e
esta ação é dada pela liberação de grânulos citoplasmáticos tóxicos (MENDES et al.,
1998; YING et al., 2002), sendo a mesma importante para destruir ou danificar
estruturas não fagocitáveis, como por exemplo, os helmintos em fase de migração
tecidual (SMITH, 2001).
Nas helmintíases que provocam eosinofilia é importante destacar alguns
fatores, como a espécie do parasita, a duração da infecção, a quantidade de
parasitas e sua localização (LILLYWHITE et al., 1991; MACDONALD et al., 1994).
As helmintíases que freqüentemente cursam com eosinofilia acentuada, são as que
evoluem com invasão nos tecidos, porém a eosinofilia é mais bem destacada
durante a fase aguda de invasão, seguida do desenvolvimento larvário e migração
pelos tecidos (NEEDHAM & LILLYWHITH, 1994).
Quando se relaciona a eosinofilia em algumas crianças não parasitadas,
destacamos que, apesar de ter sido realizado a pesquisa em três amostras de fezes,
dependendo do estágio de vida do ciclo evolutivo de alguns parasitas intestinais e da
localização anatômica em que eles se encontram no organismo do hospedeiro,
torna-se difícil sua detecção através de técnicas coprológicas de rotina utilizadas
neste estudo (CHAVES et al., 1979; CARNEIRO et al., 2002).
É sugerido também a hipótese de que estas crianças poderiam estar
parasitadas por Enterobius vermicularis, já que não foi realizada pesquisa específica

90
para este agente e a presença de outras causas, tais como: escabiose, outros
processos infecciosos e atopia, uma vez que não foi pesquisado no presente
trabalho, mas é sabido que esta se correlaciona com a presença de eosinofilia
(HAGEL et al., 1993; LYNCH et al., 1998; TUNON DE LARA et al., 2000).
Na criança desnutrida, o número de células B é normal ou aumentado bem
como os níveis de imunoglobulinas circulantes. Ocasionalmente algumas crianças
podem ter baixas concentrações de IgA e IgG séricas (CHANDRA,1992).
Postulou-se que os níveis normais ou aumentados são secundários a uma
diminuição da população de células T supressoras ou a uma elevada exposição a
vários antígenos. Enquanto existem níveis normais ou aumentados de
imunoglobulinas e células B circulantes não se pode presumir resposta competente
do sistema imune humoral aos antígenos estranhos. A produção de anticorpos em
resposta aos antígenos parece seletiva nas crianças desnutridas, comprometida
contra alguns e adequada em relação a outros (CHANDRA, 1972; ORTIZ et al.,
2000).
Enquanto a resposta de anticorpo sérica em desnutridos é freqüentemente
normal, IgA secretora é reduzida. É possível que exista uma falha na síntese desta
imunoglobulina ou bloqueio no seu transporte para as superfícies mucosas. As IgA
produzidas localmente na mucosa são importantes na imunidade para infecção e isto
pode ter implicações clínicas em termos de uma lenta recuperação incompleta e
propagação sistêmica de infecção. No caso de parasitas, tem-se observado que,
desnutrição facilita infecção causada por E. histolytica, G. lamblia e S. stercoralis
(CHANDRA 1984; WEKMAN, 1987).
Quando foram analisadas as 103 crianças verificou-se que apesar de não
haver significância estatística entre os valores de IgA sérica, IgA secretora e os
valores de referência, uma percentagem de 24% e 47% das crianças, apresentaram
os valores abaixo dos de referência para IgA sérica e IgA secretora, respectivamente
(Tabela 7).
Vários trabalhos (FOGAÇA, 1990; BUNDY et al., 1991; LILLYWHITE, 1991;
SALDIVA et al., 1983; NEEDHAM & LILLYWHITE, 1994) mostram que as infecções
parasitárias apresentam uma tendência a diminuir os níveis de IgA sérica e/ou
secretora.
Trabalho desenvolvido em crianças de duas comunidades carentes da
Venezuela demonstrou que quanto maior foi à intensidade da carga parasitária por

91
A. lumbricoides e T trichiura, menores foram os valores da IgA secretora quando
correlacionou-se uma comunidade com a outra. Observou-se também neste trabalho
que a comunidade mais desnutrida era a mais parasitada, e a que apresentou
menores valores para a IgA secretora (ORTZ et al., 2000).
Nossos resultados demonstram que as crianças desnutridas apresentaram os
valores das medianas para IgA secretora menores do que o grupo nutrido, enquanto
que a IgA sérica apresentou uma mediana maior, mais quase desapercebida nas
crianças desnutridas (Tabela 8). Esta tendência de diminuição da IgA secretora, que
foi observado em nosso trabalho, não foi estatisticamente significante,
provavelmente pelo fato destas crianças encontrarem-se a maioria delas, no grau de
desnutrição leve.
Também não foi encontrada nenhuma associação entre parasitismo e
intensidade de infecção com os níveis de IgA sérica e secretora (Tabelas 9 e 10).
A IgE desempenha uma função importante nos mecanismos de proteção
contra as infecções helmínticas e essa resposta é influenciada pelo estado
nutricional. Tem-se determinado que os helmintos não só estimulam respostas para
IgE, como também provocam uma síntese policlonal de IgE (ROSÁRIO FILHO,
1982; ORTIZ et al.,2000).
Os níveis séricos de IgE estavam elevados em 95,1% das 103 crianças,
estudadas, não apresentado nenhuma diferença entre as idades (Tabela 11), sendo,
porém, estatisticamente significante no parasitismo, segundo o número de espécies
infectantes. (Tabela 12). Já está bem estabelecido na literatura o aumento de IgE em
crianças parasitadas e este reproduz a capacidade que os antígenos dos helmintos
têm de induzir a produção de IL-4, com conseqüente síntese de IgE (LYNCH et al.,
1998; HAGEL et al., 1983c; ORTIZ at al., 2000). Esta resposta da IgE relacionada a
policlonalidade e a produção de IgE específica para A. lumbricoides foi observado
em estudos já realizados por nosso laboratório na UFRN.
Nossos resultados mostraram níveis elevados de IgE total em crianças
menores de 5 anos, concordantes com trabalho desenvolvido na região
(RODRIGUES, 2001), os quais mostraram que crianças menores de 5 anos também
apresentaram valores de IgE elevado em 97,3% delas. Já trabalhos realizados por
LYNCH et al. (1983) diferem quando constataram, na região de Caracas, onde
helmintos intestinais eram endêmicos, que crianças maiores de 5 anos tinham IgE

92
total em altos níveis, porém as menores de 5 anos apresentaram níveis baixos de
IgE e IL-4 indetectável.
Quando se avaliou a intensidade de infecção através da análise da
quantidade de ovos por grama de fezes, verificou-se que as crianças tinham
infecções de intensidade leve a moderada para A. lumbricoides e T. trichiura,
portanto o que provavelmente levou a ausência de correlação com o número de
eosinófilos, e a concentrações de IgA sérica, IgA secretora e IgE total.
A presença de correlação entre IgE e eosinófilos (Figura 16), enfatiza a
importância destas células e desta imunoglobulina na defesa contra parasitas
intestinais. Entre os mecanismos antiparasitários do sistema imunológico podem-se
destacar fatores importantes nos quais podemos relacionar a resposta da IgA e a
IgE. A ação protetora desses anticorpos, principalmente a IgA, ainda carece de
maiores elucidações visto que os trabalhos relatam com maior destaque a resposta
mediada pela IgE (PEEBLES et al., 2001; PAINTLIA et al., 2002).
Com base nos dados obtidos neste estudo pode-se verificar a grande
complexidade da relação existente entre parasitoses, nutrição e resposta imune em
crianças poliparasitadas cronicamente por vários tipos de parasitas. Não foi possível
esclarecer através de dados concretos com a IgA sérica e IgA secretora, porém
verificou-se que não houve interferência da produção de IgE sobre a IgA. Os poucos
estudos com relação à desnutrição leve e moderada associados a questões
deixadas por este trabalho, nos motiva a continuar esta linha de pesquisa.

93
7 CONCLUSÕES
- Este estudo mostrou que crianças pertencentes a famílias de níveis
socioeconômico baixos e residente em áreas com precárias condições de higiene,
apresentaram alta freqüência de infecções por enteroparasitas, sendo os helmintos
mais freqüentes Trichuris trichiura e Ascaris lumbricoides e os protozoários
Endolimax nana e Giardia lamblia.
- Apesar do poliparasitismo, a maioria das crianças, encontrava-se nutrida e as
demais com desnutrição em grau leve sem déficit protéico. Não houve diferença na
intensidade de infecção por Trichuris trichiura e Ascaris lumbricoides entre crianças
nutridas e desnutridas.
- A maioria das crianças apresentou eosinofilia no sangue periférico, não havendo
diferença estatisticamente significante entre a mesma e idades estudadas, estado
nutricional, número de ovos de Trichuris trichiura e Ascaris lumbricoides e
concentração de IgA sérica e secretora. Porém, existiu correlação positiva entre
eosinofilia e concentração de IgE total.
- A intensidade da carga parasitária encontra para ovos de Ascaris lumbricoides e
Trichuris trichiura por grama de fezes, foi uma carga leve e modera respectivamente,
e não apresentou correlação com as concentrações de IgA sérica, IgA secretora ,
IgE total e o número de eosinófilos.

94
- Existiu uma correlação positiva entre o parasitismo, segundo o número de espécies
infectantes e as concentrações de IgE total, o que não ocorreu entre as
concentrações de IgA sérica e IgA secretora.
- As concentrações de IgA sérica e IgA secretora, não apresentaram diferenças
estatisticamente significantes, quando correlacionadas com os valores de
referência utilizados, porém observou-se valores abaixo do normal em 24,3% para
IgA sérica e 47,1% para IgA secretora, das crianças parasitadas.
- Nossos resultados deixam claro o envolvimento da IgE na resposta a infecções
causadas por parasitas helmínticos e sugerem que apesar do grande aumento de
sua síntese, por antígenos destes parasitas, não houve desvio na produção de IgA.
- Ainda são necessários muitos estudos envolvendo as infecções causadas por
parasitas intestinais, visando-se a resposta imune específica, para se ter uma
definição do perfil de resposta de cada um deles e elucidar os padrões de defesa
imunológica.
- São necessários muitas investigações sobre o que diz respeito a desnutrição leve e
moderada e o comprometimento do sistema imunológico, pois o maior
conhecimento que se tem é a respeito da desnutrição grave.

95
ABSTRACT
Infections for intestinal parasites are one of the main morbidade causes in
humans and, its relationships with socioeconomic levels and hygiene conditions in
countries in development are already very established. Many works, even so, they
are being accomplished to elucidate the complex interactions among nutrition, these
infections and answer imunológica, because it is seen that malnutrition commits the
immunity increasing the susceptibilidade for infectious diseases and these for its time
can harm the state human nutricional.
It is known that sponge helmínticos they stimulate synthesis of IgE so
much policlonal as specific for the same ones and that IgA secretora, main
imunoglobulina of defense of the mucous ones, can act against protozoa as the
Giardia lamblia and against helmintos as Trichuris tichiura and Strongyloides
stercorales. Some studies show that the malnutrition energy protéica influences in
the production of these answers, but some authors show results divergentes.
In this work it was evaluated the levels of total IgE, IgA sérica and secretora,
contagem of sanguine eosinófilos, levels of proteins séricas and state nutricional, in
103 children of low socioeconomic level, to discover a correlation between those and
infection for enteroparasitas.
They participated in the study children of both sexes, with age of 3 to 6 years,
visitors of the same creche and residents in a neighborhood with precarious hygiene
conditions and basic saneamento, in the city of Christmas.
The obtained results showed that the faulty environmental and socioeconomic
conditions favored to a high infection frequency for enteroparasitas, mainly Trichuris
trichiura and Ascaris lumbricoides between the helmintos and Endolimax sleep and
Giárdia lamblia among the protozoa. Light malnutrition without deficit protéico was
observed in 30% of the children, which didn't also present significant deficiencies of
IgA sérica and secretora. The sponged children presented sanguine eosinofilia and
levels séricos of high total IgE confirming the important participation of the same
ones in the immune answer against helmintos. It cannot him, therefore, to suggest
that the children in spite of poliparasitadas were not with its immune answer of

96
mucous harmed, probably for they be not still intensely infected, as observed in the
contagem of eggs by gram of feces and also for they have not its state nutricional
seriously committed.

97
8 REFERÊNCIAS BIBLIOGRÁFICAS
ABBAS, A. K.; LICHTMAN, A. H.; POBER, J. S. Cellular and Molecular Immunology., 4 ed., Philadelphia: W. B. Sauders Company, 2000. Cap. 2, p.31-32,
cap. 13, p. 332-333.
ABU-GHAZALEH, R. I.; FUJISAWA, T.; MESTECKY, J.; KYLE, R. A.; GLEICH, G. J.
IgA-Induced eosinophil degranulation. J. Immunol., v. 142, n. 7, p. 2393-2400, 1989.
ALBUQUERQUE, L. M. M.; JUNIOR, G. B. C.; RIBEIRO, M. D. Eosinofilia sanguínea
em crianças com helmintíases intestinais. Rev. Bras. Anál. Clín., v. 22, n. 2, p. 26-
30, 1990.
ANDRADE, C.; ALAVA, T.; DE PALÁCIO, I. A.; DEL POGGIO, P.; JAMOLETTI, C.;
GULLETTA, M.; MONTRESOR, A. Prevalence and intensity of Soil-transmitted
Helminthiasis in the City of Portoviejo (Ecuador). Mem. Inst. Oswaldo Cruz, v. 96, n.
8, p. 1075-1079, 2001.
ANJOS, L. A. Índices antroprométicos e estado nutricional de escolares de baixa
renda de um município do estado do Rio de Janeiro (Brasil): Um estudo piloto. Rev. Saúde Publ. v. 23, n. 3, p. 221-229, 1989.
ATTA, M. L. B. S.; ARAUJO, M. I.; JÚNIOR, A. D.; JESUS, A. R.; ALMEIDA, R. P.;
ATTA, A. M.; CARVALHO, E. M. Detection of specific IgE antibodies in parasite
diseases. Braz. J. Med. Res., v. 32, p. 1101-1105, 1999.
ATTA, A. M.; ATTA, M. L. B. S.; D’OLIVEIRA, A.; ALMEIDA, R. P.; ARAÚJO, M. I.;
CARVALHO, E. M. IgG Anti-IgE Autoantibodies in Visceral Leishmaniasis. Mem. Inst. Oswaldo Cruz, v. 97, n. 1, p. 101-103, 2002.
BAERMANN, A.; MORAES, R. G. Contribuição parao estudo do Strogyloides
stercorale e da estrongiloidíase no Brasil. Rev. Serv. Esp. Saúde Publ., v.1, p. 507-
524, 1948.

98
BACHARIER, L. B.; GEHA, R. S. Molecular mechanisms of IgE regulation. J. Allergy Clin. Immunol., v. 105, p. 547-558, 2000.
BECKER, A. A.; IOSCHPE, R.; DELWING, D.; DELWING, D.; CANALI, J. Incidence
of intestinal parasitosis in students from public school in Novo Hamburgo-RS. Rev. Bras. Anal. Clin., v. 34, n. 2, p. 85-87, 2002.
BELL, R. G. IgE, allergies and helminth parasites: A new perspective on an old
conundrum. Immunol. Cell Biol., v. 74, p. 337-345, 1996.
BERKMAN, D. S.; LESCANO, A. G.; GILMAN, R. H.; LOPEZ, S. L.; BLACK, M. M.
Effects of stunting, diarrhoeal disease, and parasitic infection during infancy on
cognition in late childhood: a follow-up study. Lancet, v. 359, n. 9306, p. 564-571,
2002.
BERN, C.; MARTINES, J.; DE ZOYSA, I. The magnitude of the global problem of
diarrhoeal disease: a ten-year update. WHO, v. 70, p. 705-714, 1992.
BEYER, K.; CASTRO, R.; BIRNBAUM, A.; BENKOV, K.; PITTMAN, N.; SAMPSON,
H. A. Human milk-specific mucosal lymphocytes of the gastrointestinal tract display a
Th 2 cytokine profile. J. Allergy Clin. Immunol., v. 109, n. 4, p. 707-713, 2002.
BONNEFOY, J. Y.; AURBRY, J. P.; GAUCHAT, J. F.; GRABER, P.; LIFE, P.;
FLORES ROMO, L.; MAZZEI, G. Receptor for IgE. Curr. Opin. Immunol., v. 5, p.
944-949, 1993.
BRANDTZAEG, P. Molecular and cellular aspects of the secretory immunoglobulin
system. APMIS., v.103, p. 1-19, 1995.
BUENO, E. C.; VAZ, A. J.; MACHADO, L. R.; LIVRAMENTO, J. A. Detection of IgG,
IgA and IgE antibodies in cerebrospinal fluid, serum and saliva samples by Elisa with
Taenia solium and Taenia crassiceps antigens. Arq. Neuropsiquiatr., v. 58, n.1, p.
18-24, 2000.

99
BUNDY, D. A. P.; LILLYWHITE, J. E.; DIDIER, J. M.; SIMMONS, I.; BIANCO, A. E.
Age-dependency of infection status and serum antibody levels in human whipworm
(Trichuris trichiura) infection. Parasitol. Immunol., v. 13, n 6, p. 629-39, 1991.
CAMPOS, M. R.; VALENCIA, L. I. O.; FORTES, B. P. M. D.; BRAGA, R. C. C.;
MEDRONHO, R. A. Spatial distribution of Ascaris lumbricoides infection. Rev. Saúde Pública, v. 36, n.1, p. 64-74, 2002.
CAPRON, M.; CAPRON, A. Immunoglobulin E and Effector Cells in Schistosomiasis.
Science, v. 264, p. 1876-1877, 1994.
CAPRON, M., DESREUMAUX, P. Immunobiology of eosinophils in allergy and
inflammation. Rev. Immunol., v. 148, p. 29-33, 1997.
CARDOSO, R. R. A. O papel protetor das infestações por helmintos. Rev. Bras. Alerg. Imunopatol., v. 20, p.105-106, 1997.
CARNEIRO, F. F.; CIFUENTES, E.; TELLEZ-ROJO, M. M.; ROMIEU, I. The risk of
Ascaris lumbricoides infection in children as an environmental health indicator to
guide preventive activities in Caparaó and Alto Caparaó, Brazil. Wor. Heal. Organ., v. 80, p. 40-46, 2002.
CARVALHO, B. T. C.; NUDELMAN, V.; SAMPAIO, M. M. S. C. Mecanismos de
defesa contra infecções. J. Pediatr., v. 74, n.1, 1998.
CASTILHO DE ARIZA, M.; ALMONTE, P. R.; RICHARDO, J. S.; GUILLEARD, J. C.
Estudo comparativo de IgE em ninos desnutridos parasitados y no parasitados.
Arch. Domin. Pediat., v. 19, p. 93-96, 1983.
CHANDRA, R. K. Immunocompetence in undernutrition. J. Pediatr., v. 81, n. 6, p.
1194-1200, 1972.
CHANDRA, R. K. Nutrition, immunity and infection: present knowledge and future
directions. Lancet. v. 3, p. 688-691, 1983.

100
CHANDRA, R. K. Parasitic infection, nutrition and immune response. Fed. Proc., v.
43, p. 251-255, 1984.
CHANDRA, R. K. Nutritional Regulation of Immunity and Risk of Illness. Indian J. Pediatr., v. 56, p. 607-611, 1989.
CHANDRA, R. K. Nutritional and Immunoregulation. Significance for Host Resistance
to Tumors and Infectious Diseases in Humans and Rodents. Am. J. Nutr., v. 122, p.
754-757, 1992.
CHAVES, A.; ALCANTARA, O. S.; CARVALHO, O. S.; SANTOS, J. S. Estudo
comparativo dos métodos coprológicos de LUTZ, KATO-KATZ e FAUST modificado.
Rev. Saúde Pública, v. 13, p. 348-352, 1979.
CHUNG, H. L.; HWANG, J. B.; PARK, J. J.; KIM, S. G. Expression of transforming
growth factor β1, transforming growth factor type I and III receptors, and TNF-α in the
mucosa of the small intestine in infants with food protein-induced enterocolitis
syndrome. J. Allergy Clin. Immunol., v. 109, n. 1, p. 150-154, 2002.
COOPER, E. S. ; BUNDY, D. A. P. Trichuris is not trivial. Parasitol. Today., v. 4, p.
301-306, 1988.
CORRÊA, A. M. S.; GONÇALVES, N. N. S.; GONÇALVES, A.; LEITE, G. P. R.;
PADOVANI, C. R. Evolução da relação entre peso e altura e peso e idade em
crianças de 3 meses a 6 anos assistidas em creche, Sorocaba (SP), Brasil. Rev. Panam. Salud. Publ., v. 6, n. 1, p. 26-30, 1999.
CORRY, D. B.; KHERADMAND, F. Induction and regulation of the IgE response.
Nature, v. 402, p. 18-23, 1999.
CROMPTON, D. W. T. Ascariasis and childhood malnutrition. Trans. R. Soc. Trop. Med. Hyg, v.86, p. 577-579, 1992.

101
CUTULO, L. R. A.; LAMEGO, A. C. Fatores Imunológicos do Aleitamento Materno.
Arq. Catar. Med., v. 23, n.3, p. 191-198, 1994.
DE LA RUE, M. L. Eosinofilia devida a parasitas. Rev. Bras. Anal. Clín., v. 33, n.4,
p. 221-223, 2001.
ELIA, C. C. S.; PIMENTEL, M. L.; CARNEIRO, A. J. V. ; CARVALHO, A. T. P.
Estrutura e Componentes Celulares do Sistema Imunitário da Mucosa Intestinal. In:
ELIA, C. C. S.; SOUZA, H. S. P. Imunologia da mucosa intestinal: da bancada ao leito, 1 ed. Atheneu, 2001. Cap. 3, p. 41-57.
ELIA, C. C. S. Deficiência de IgA. In: ELIA, C. C. S.; SOUZA, H. S. P. Imunologia da mucosa intestinal: da bancada ao leito, 1 ed. Atheneu, 2001. Cap. 7.2, p. 129 -
133 a .
ELIA, C. C. S. Parasitoses Intestinais. In: ELIA, C. C. S.; SOUZA, H. S. P.
Imunologia da mucosa intestinal: da bancada ao leito, 1 ed. Atheneu, 2001. Cap.
7.5, p. 137-140 b.
ELSE, K. J.; FINKELMAN, F. D. Intestinal nematode parasites, cytokines and effector
mechanisms. J. Parasitol., v. 28, p. 1145-1158, 1998.
FAUST, E. C.; D´ANTONI, J. S.; ODON, V.; MILLER, M. J.; PEREZ, C.; SAWITZ, W.;
THOMEN, L. F.; TOBLE, J.; WALKER, J. H. A critical study of clinical laboratory
technics of the diagnosis of protozoan cysts and helmints eggs in feces. I –
Preliminary communication. Am. J. Trop. Med., v. 18, p. 169, 1938.
FOGAÇA, H. S.; ELIA, C. C. S.; MADI, K.; OLIVEIRA, A. Estudo das imunoglobulinas
intestinais na estrongiloidíase. F. Méd., v. 100, n. 4, p. 229-235, 1990.
FOGAÇA, H. S. Desnutrição. In: ELIA, C. C. S.; SOUZA, H. S. P. Imunologia da mucosa intestinal: da bancada ao leito, 1 ed. Atheneu, 2001. Cap. 7.8, p. 159-
163.

102
FORRESTER, J. E.; SCOTT, M. E.; BUNDY, D. A. P.; GOLDEN, M. H. N. Clustering
of Ascaris lumbricoides and Trichuris trichiura infections within households. Trans. R. Soc. Trop. Med. Hyg., v. 82, n. 2, p. 282-288, 1988.
GEHA, R. S. Human IgE. Allergy. Clin. Immunol., v. 74, p. 109-120, 1984.
GOLDSBY, R. A.; KINDT, T. J.; OSBORNE, B. A. KUBY IMUNOLOGIA. 4 ed. Rio de
Janeiro: REVINTER, 2002, Cap. 2, p. 54-57, cap. 4, p. 100-101.
GREWAL, H. M. S.; KARLSEN, T. H.; VETVIK, H.; AHRÉN, C.; GJESSING, H. K.;
SOMMERFELT, H.; HANEBERG, B. Measurement of specific IgA in faecal extracts
and intestinal lavage fluid for monitoring of mucosal immune responses. Science, v.
239, p. 53-62, 2000.
GROSS, R.; SCHELL, B.; MOLINA, M. C. B.; LEÃO, M. A. C.; STRACK, U. The
impact of improvement of water supply and sanitation facilities on diarrhea and
intestinal parasites: A Brazilian experience with children in two low-income urban
communites. Rev. Saúde Pública, v. 23, n. 3, p. 214-220, 1989.
GUPTA, M. C. ARORA, K. L.; MITHAL, S.; TANDON, B. N. Effect of periodic
deworming on nutritional status of Ascaris-infested preschool children receiving
supplementary food. Lancet., n. 2, p. 108-110, 1977.
GUIMARÃES, S.; SORGAYAR, M. I. L. Detection of anti-Giardia lamblia serum
antibody among children of day care centers. Rev. Saúde Pública, v. 36, n.1, p. 63-
68, 2002.
HAGEL, I.; LYNCH, N. R.; PEREZ, M.; DI PRISCO, M. C.; LOPEZ, R.; ROJAS, E.
Modulation of allergic reactivity of slum children by helminthic infection. Parasitol. Immunol., v. 15, n.6, p. 311-315, 1993a.
HAGEL, I.; LYNCH, N. R.; DI PRISCO, M. C.; LÓPEZ, R.; GARCIA, N. M. Allergic
Reactivity of Children of Different Socioeconomic Levels in Tropical Populations. Int. Arch. Allergy Immunol., v. 101, p. 209-214, 1993b.

103
HAGEL, I.; LYNCH, N. R.; PÉREZ, M.; DI PRISCO, M. C.; LÓPEZ, R.; ROJAS, E.
Relationship between the degree of poverty and the IgE response to Ascaris
infection in slum children. Trans. R. Soc. Trop. Med. Hyg., v. 87, n. 1, p. 16-18,
1993c.
HAGEL, I.; LYNCH, N. R.; DI PRISCO, M. C.; SANCHEZ, J.; PÉREZ, M. Nutritional
status and the IgE response against Ascaris lumbricoides in children from a tropical
slum. Trans. R. Soc. Trop. Med. Hyg., v. 5, n. 89, p. 562-565, 1995.
HASWELL-ELKINS, M. R.; LEONARD, H.; KENNEDY, M. W.; ELKINS, D. B.;
MAIZELS, R. M. Immunoepidemiology of Ascaris lumbricoides: relationships between
antibody specificities, exposure and infection in a human community. Parasitol., v.
104, p. 1:153-1:159, 1992.
HEREMANS, J. F.; HEREMANS, M. T. ; SCHULTZE, H. E. Isolation and description
of a few properties of the A Globulin of human serum. Clin. Chim. Acta., p. 4-96,
1959.
HIROI, T.; YANAGITA, M.; TAKATSU, K.; KIYONO, H. IL-5R+, B-1 cells: Are they
CMIS independent B cells for mucosal IgA plasma cells? Muc. Immunol. Update, p.
20-22, 1998.
HIROI, T.; KIYONO, H. Role of Interleukin 15 and IL-15R for Common Mucosal
Immune System-Independent IgA B-1 Cells. Muc. Immunol. Update, v. 6, n. 4, p.
14-15, 2000.
HOFFMAN, W. A. ; PONS, J. A.; JANER, J. L. The sedimentation concentration
method in schistosomiasis mansoni. J. Publ. Health. Trop. Med., v. 9, p. 238-298,
1934.
IBGE (Instituto Brasileiro de Geografia e Estatística). Pesquisa sobre padrões de vida - 1996- 1997. Rio de Janeiro: Departamento de População e Indicadores
Sociais, 1998.

104
ISLAM, K. B.; NILSSON, L.; SIDERAS, P.; HAMMARSTEROM, L.; SMITH, C. I.
TGF-beta-1 induces germ-line transcripts of both IgA suclasses in human B
lynphocytes. International Immunol., v. 3, p. 1099-1106, 1991.
JAIN, N. C. A briel review of the pathophysiology of eosinophils. Comp. Cont. Educ. Pract. Vet., v. 16, p. 1212-1218, 1994.
JANEWAY, C. A.; TRAVERS, P.; WALPORT, M.; CAPRA, J. D. Imunobiologia: O Sistema Imunológico na Saúde e na Doença, 4 ed. Porto Alegre. Artes Médicas
Sul, 2000. Cap. 3, p. 79-113, cap. 12, p. 461-488.
JARRETT, E. E. E.; MILLER, H. R. P. Production and Activities of IgE in Helminth
Infection. Prog. Allergy, v. 31, p. 178-233, 1982.
JINABHAI, C. C.; TAYLOR, M.; COUTSOUDIS, A.; COOVADIA, H. M.; TOMKINS, A.
M.; SULLIVAN, K. R. A health and nutritional profile of rural school children in
KwaZulu-Natal, South Africa. An. Trop. Pediatr., v. 21, p. 50-58, 2001.
JOKIPII, L.; MIETTINEN, A.; JOKIPII, A. M. M. Antibodies to Cysts of Giardia lamblia
in Primary Giardiasis and in the Absence of giardiasis. J. Clin. Microbiol., v. 26, n. 1,
p. 121-125, 1988.
KAGNOFF, M. Immunology of the Intestinal Tract. Gastroent., v. 105, p. 1275-1280,
1993.
KACHMAR, J. F. Proteins and amino acids. In: TIETZ, N. W. Fundamentals of Clinical Chemistry, Philadelphia Saunders Company, 1970. Cap. 5, p. 177-262.
KIGHTLINGER, L. K.; SEED, J. R.; KIGHTLINGER, M. B. Ascaris lumbricoides
intensity in relation to environmental, socioeconomic, and behavioral determinants of
exposure to infection in children from southeast Madagascar. J. Parasitol., v. 84, n.
3, p. 480-484, 1998.

105
LEBMAN, D. A.; COFFMAN, R. L. The effects of IL-4 and IL-5 on the IgA response
by murine peyer’s patch B cell subpopulations. J. Immunol., v. 141, n. 6, p. 1050-
1056, 1988.
LILLYWHITE, J. E.; BUNDY, D. A. P.; DIDIER, J. M.; COOPER, E. S.; BIANCO, A.
E. Humoral immune response in human infection with the whipworm Trichuris
trichura. Parasitol. Immunol., v. 13, n. 5, p. 491-507, 1991.
LYNCH, N. R.; HAGEL, I. A.; PÉREZ, M.; DI PRISCO M. C.; LOPEZ, R.; ALVAREZ,
N. Effect of anthelmintic treatment on the allergic reativity of children in a tropical
slum. J. Allergy Clin. Immunol., v. 92, p. 404-411, 1993.
LYNCH, N. R.; HAGEL, I. A.; PALENQUE, M. E.; DI PRISCO, M. C.; ESCUREDO, J.
E.; CORAO, L. A.; SANDIA, J. A.; FERREIRA, L. J.; BOTTO, C.; PEREZ, M.; Lê
SOUEF, P. N. Relationship between helminthic infection and IgE response in atopic
and nonatopic children in a tropical environment. J. Allergy Clin. Immunol., v. 101,
n. 2, p. 217-221, 1998.
MACDONALD, T. T.; SPENCER, J.; MURCH, S. H.; CHOY, M. Y.; VENUGOPAL, S.;
BUNDY, D. A. P.; COOPER, E. S. Immunoepidemiology of intestinal helminthic
infections. Trans. R. Soc. Trop. Méd. Hyg., v. 88, p. 265-268, 1994.
MACEDO, L. M. C.; COSTA, M. C. E.; ALMEIDA, L. M. Parasitismo por Ascaris
lumbricoides em crianças menores de dois anos: Estudo populacional em
comunidade do Estado do Rio de Janeiro. Cad. Saúde Pública, v. 15, n. 1, p. 173-
178, 1999.
MACEDO, L. M. C.; SILVA, J. R. M.; SILVA, R. R.; OLIVEIRA, L. M.; VIANNA, M. S.
R. Intestinal parasitoses in low-income children under five years of age in Rio de
Janeiro, Brazil. Cad. Saúde Publica, v. 14, n. 4, p. 851-855, 1998.
MADI, K.; ZATMAN, C.; TAKIYA, C. M. Arquitetura da Mucosa Intestinal e sua
Plasticidade. In: ELIA, C. C. S.; SOUZA, H. S. P. Imunologia da mucosa intestinal: da bancada ao leito, 1 ed. Atheneu, 2001. Cap. 2, p. 11-39.

106
MATTHAEI, K. I.; FOSTER, P. S.; YOUNG, I. G. The role of interleukin-5 (IL-5) in
vivo: studies with IL-5 deficient mice. Mem. Inst. Oswaldo Cruz., v. 92, suppl. 2, p.
63-68, 1997.
Mc EWEN, B. J. Eosinophils: A review. Vet. Res. Communic., v. 16, p. 11-42, 1992.
MEEUSEN, E. N. T.; BALIC, A. Do Eosinophils have a Role in the Killing of Helminto
Parasites? Parasitol. Today, v. 16, v. 3, p. 95-101, 2000.
MELLANDER, L.; CARLSSON, B.; JALIL, F.; SODERSTROM, T.; HANSON, L. A.
Secretory IgA antibody response against Escherichia coli antigens in infants in
relation to exposure. J. Pediatr., v. 107, n. 3, p. 430-432, 1985.
MENDES, D. M.; CAMARGO, M. F.; AUN, V. V.; FERNANDES, M. F. M.; AUN, W.
T.; MELLO, J. F. Eosinofilia. Rev. Bras. Alerg. Imunop. v. 23, p. 84-91, 1998.
MILLER, H. R. P.; NAWA Y. Nippostrongylus brasiliensis: Intestinal goblet cell
response in adoptively immunized rats. Exp. Parasitol., v. 47, p. 81-90, 1979.
MORALES, G.; LOAIZA, L.; PINO, L. A. Marcadores de riesgo para indivíduos com
altas cargas de Ascaris lumbricoides em uma comunidad rural Del Estado Cojedes,
Venezuela. Bol. Chil. Parasitol., v. 54, p. 3-4, 1999.
MOTEGI, Y.; KITA, H. Interaction with Secretory Component Stimulates Effector
Functions of Human Eosinophils But Not of Neutrophils. Am. Assoc. Immunol., v.
161, p. 4340-4346, 1998.
NAGAO, A. T.; PILAGALLO, M.; PEREIRA, A. B.; SAMPAIO, M. M. S. C.; HANSON,
L. A. Quantification of salivary, urinary and fecal secretory IgA, as well as in saliva
titers and avidities of IgA antibodies in children living at different levels of antigenic
exposure and undernutrition. Adv. Mucos. Immunol., p. 507-511, 1995.

107
NASCIMENTO, E. G.; FRANÇA, J. F.; ARAÚJO E. M. Prevalência de parasitoses
intestinais em crianças da favela do Maruim-Natal/RN. Rev. Saúde, v.13, p. 15-20,
1999.
NAVEIRA, J. B. Aspectos da biologia dos eosinófilos. Rev. Bras. Maraliolog. Doenças Trop., v. 12, p. 103, 1960.
NEEDHAM, C. S.; LILLYWHITH, J. E. Immunoepidemiology of intestinal helminthic
infections. Immunological correlates with patterns of Trichuris Infection. Trans. R. Soc. Trop. Med. Hyg., v. 88, n. 3, p. 262-264, 1994.
NETO V. A.; CORRÊA, L. L. Exames parasitológicos de fezes, 5 ed. São Paulo.
Savier ,1991.
NOGUEIRA, D. M.; STRUFALDI, B.; HIRATA, M. H.; ABDALLA, D. S. P.; HIRATA,
R. D. C. Métodos de bioquímica clínica – técnica e interpretação, São Paulo:
Pancast Editorial, 1990, Cap. 3, p. 41-50.
OLIVEIRA, S. P.; THÉBAUD-MONY, A. Hábitos e práticas alimentares em três
localidades da cidade de São Paulo (BRASIL). Rev. Nutr., v. 11, n. 1, p. 37-50,
1998.
ORTIZ, D.; AFONSO C.; HAGEL, I.; RODRIGUEZ, O.; ORTIZ, C.; PALENQUE, M.;
LYNCH, N. R. Influencia de lãs infecciones helmínticas y estado nutricional em la
respuesta immunitaria de ninos venezoelanos. Rev. Panam. Salud Pública, v. 8, p.
3-8, 2000.
PAINTLIA, M. K.; KAUR, S.; GUPTA, I.; GANGULY, N. K.; MAHAJAN, R. C.; MALLA,
N. Specific IgA response, T-cell subtype and cytokine profile in experimental
intravaginal trichomoniasis. Parasitol. Res., v. 88, p. 338-343, 2002.
PAWANKAR, R. Anti-IgE treatment in Allergic Disease. Allergy Clin. Immunol., v.
56, p.13-21, 2001

108
PEEBLES, J. R. S.; HAMILTON, R. G.; LICHTENSTEIN, L. M.; SCHLOSBERG, M.;
LIU, M. C.; PROUD, D.; TOGIAS, A. Antigen-specific IgE and IgA antibodies in
bronchoalveolar lavage fluid are associated with stronger antigen-induced late phase
reactions. Clin. Exper. Allergy., v. 31, p. 239-244, 2001.
PRATA, P. R. Desenvolvimento econômico, desigualdade e saúde. Cad. Saúde Publ., v. 10, n. 3, p. 387-391, 1994.
REDDY, V.; RAGHURAMULU, N.; BHASKARAM, C. Secretory IgA in protein-calorie
malnutrition. Arc. Disease Childh., v. 51, p. 871-874, 1976.
RENEGAR, K. B.; JACKSON, G. D. F.; MESTECKY, J. In vitro comparison of the
biologic activities of monoclonal monomeric IgA, polymeric IgA, and secretory IgA. J. Immunol., v. 160, p. 1219-1223, 1998.
RIKIMARU, T.; TANIGUCHI, K.; YARTEY, J. E.; KENNEDY, D. O.; NKRUMAH, F. K.
Humoral and cell-mediated immunity in malnourished children in Ghana. Europ. J. Clin. Nutrit., v. 52, p. 344-350, 1998.
RIVERA, J.; GANZALEZ-ESPINOSA, C.; KOVAROVA, M.; PARRAVICINI, V. The
architecture of IgE-Dependent Mast Cell Signaling. ACI Intern., v. 14, p. 25-36, 2002.
RODRIGUES, C. E. F. B. Correlação entre níveis séricos de IgE e proteína catiônica eosinofílica, eosinofilia, testes cutâneo de hipersensibilidade imediata e chiado em crianças enteroparasitadas. 2001. 99 f. Dissertação
(mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Ciências da
Saúde. Programa de Pós-graduação em Ciências Farmacêuticas.
ROSÁRIO FILHO, N. A.; CARNEIRO FILHO, M.; FERREIRA, E.; BARANSKI, M. C.;
CAT, I. Níveis de IgE total no Soro e Contagens de Eosinófilos em Crianças com
Enteroparasitoses: Efeito do Tratamento Anti-Helmíntico. J. Pediatr., n. 52, n. 4, p.
209-215, 1982.

109
ROITT, I.; BROSTOFF J.; MALE, D. Imunologia, 6 ed. São Paulo, 2003. Cap. 2, p.
40-44, cap. 16, p. 261-275.
RÚPULO, B. S.; MIRA, J. G. S.; JÚNIOR, O. K. Deficiência de IgA J. Pediatr., v. 74,
p. 6-9, 1998.
SALDIVA, S. R.; SILVEIRA, A. S.; PHILIPPI, S. T.; TORRES, D. M.; MANGINI, A. C.;
DIAS, R. M. S.; SILVA, R. M.; BURATINI, M. N.; MASSAD, E. Ascaris-Trichuris
association and malnutrition in Brazilian children. Pediatr. Perin. Epidemiol., v. 13,
p. 89-98, 1993.
SCROFERNEKER, M. L.; POHLMANN P. R. Imunologia Básica e Aplicada, 1 ed.
Porto Alegre, Sagra Luzzatto, 1998, cap. 1, p. 24-25.
SEIDEL, B. M.; SCHUBERT, S.; SCHULZE, B.; BORTE, M. Secretory IgA, free
secretory component and IgD in saliva of newborn infants. Science, v. 62, n. 2, p.
159-164, 2001.
SELL, S. Immunology – Immunopathology and Immunity, 5 ed. Stamford, 1996,
Cap. 14, p. 385-429.
SHELDRAKE, R. F.; HUSBAND, A. J.; WATSON, D. L.; CRIPPS, A. W. Selective
transport of serum derived IgA into mucosal secretions. J. Immunol., v. 132, n. 1, p.
363-368, 1984.
SIRISINHA, S.; SUSKIND, R.; EDELMAN, R.; ASVAPAKA, C.; OLSON, R. E.
Secretory and Serum IgA in Children With Protein-Calorie Malnutrition. Pediatr., v.
55, n. 2, p. 166-169, 1975.
SILVA, M. V.; STURION, G. L. Attendance at day-care centers and other conditioning
factors in child nutricional status. Rev. Nutr., v. 11, n. 1, p. 58-68, 1998.

110
SILVA, J. R. L. Sistema Imunitário das Mucosas: Introdução ao tema. In: ELIA, C. C.
S.; SOUZA, H. S. P. Imunologia da mucosa intestinal: da bancada ao leito, 1 ed.
Atheneu, 2001. Cap. 1, p. 1-9.
SMITH, H. M.; DEKAMINSKY, R. G.; NIWAS, S.; SOTO, R. J.; JOLLY, P. E.
Prevalence and Intensity of Ascaris lumbricoides and Trichuris trichiura and
Associated Socio-demographic Variables in Four Rural Honduran Communities.
Mem. Inst. Oswaldo Cruz, v. 96, n. 3, p. 303-314, 2001.
SOLIMAN, M. M.; TAGHI-KILANI, R.; ABOU-SHADY, A. F. A.; EL-MAGEID, S. A. A.;
HANDOUSA, A. A.; HEGAZI, M. M.; BELOSEVIC, M. Comparasion of serum
antibody responses to Giardia lamblia of symptomatic and asymptomatic patients.
Am. J. Trop. Med. Hyg., v. 58, n. 2, p. 232-239, 1998.
SOUZA, H. S. P. Doenças Inflamatórias Intestinais. In: ELIA, C. C. S.; SOUZA, H. S.
P. Imunologia da mucosa intestinal: da bancada ao leito, 1 ed. Atheneu, 2001.
Cap. 7.1, p. 109-127.
SPRY, C. F. J. Synthesis and secretion of eosinophil granule substances. Immunol Today, v. 6, p. 332-335, 1985.
SPRY, C. F. J. Eosinophils: a comprehensive review and guide to the scientific and medical literatute. Oxford. Oxford University Press, 1988.
STIEHM, E. R. Humoral immunity in malnutrition. Fed. Proc., v. 39, n. 13, p. 3093-
3097, 1980.
STITTES, D. P.; TERR, A I.; PARSLOW, T. G. Imunologia Médica, 9 ed. Rio de
Janeiro, Guanabara Koogan, 2000, cap. 13, p. 152-161.
STOLL, B.; LEE, F. K.; HALE, E.; SCHWARTZ, D.; HOLMES, R.; ASHBY, R.;
CZERKINSKY, C.; NAHMIAS, A. J. Immunoglobulin secretion by the normal and the
infected newborn infant. J. Pediatr., v. 22, n. 5, p. 780-786, 1993.

111
STROBER, W.; RICHMAN, L. K.; ELSON, C. O. The regulation of gastrointestinal
immune response. Immunol. Today, v. 8, p. 156-162, 1981.
SUSKIND, R.; SIRISINHA, S.; VITHAYASAI, V.; EDELMAN, R.; DAMRONGASAK,
D.; CHARUPATANA, C.; OLSON, R. E. Immunoglobulins and antibody response in
children with protein-calorie malnutrition. Am. J. Clin. Nutr., v. 29, p. 836-841, 1976.
SUTTON, B. J.; GOULD, H. J. The human IgE network. Nature, v. 366, p. 421-428,
1993.
SZWARCWALD, C. L.; BASTOS, F. I.; ESTEVES, M. A. P.; ANDRADE, C. L. T.;
PAEZ, M. S.; MÉDICI, E. V.; DERRIÇO, M. Income inaquality and health: the case of
Rio de Janeiro. Cad. Saúde Publica, v. 15, n. 1, p. 15-28, 1999.
TAKAMOTO, M.; WANG, Z.; WATANABE, N.; SUGANE, K. The measurement of
parasite antigen-specific IgE levels using anti-IgE monoclonal antibodies and
biotinylated antigens. Parasitol. Res., v. 87, p. 919-923, 2001.
TEODÓSIO, M. R.; PEREIRA, A. B.; NAGAO, A.; MAGALHÃES, M. F. IgA secretória
na urina de crianças portadoras de infecção do trato urinário (ITU) e diferentes
estados nutricionais: seguimento de 12 meses. J. Pediatr., v. 69, n. 3, p. 180-185,
1993.
TEODÓSIO, M. R.; OLIVEIRA, E. C. M. Urinary secretory IgA after nutritional
rehabilitation. Braz. J. Méd. Biol. Res., v. 32, n. 4, p. 421-426, 1999.
TRAJMAN, A.; MACDONALD, T. T.; ELIA, C. C. S. Intestinal immune cells in
Strongyloides stercoralis infection. J. Clin. Pathol., v. 50, p. 991-995, 1997.
TUNON DE LARA, J. M.; MAGNAN, A.; HUMBERT, M. La résponse à IgE. Rev. Mal. Respir., v. 17, p. 183-193, 2000.
VAN GINKE, F. W.; WAHL, S. M.; KEARNEY, J. F.; KWEON, M.; FUJIHASHI, K.;
BURROWS, P. D.; KIYONO, H.; MCGHEE, J. R. Partial IgA-Deficiency with

112
Increased Th 2-Type Cytokines in TGF-β1 Knockout Mice. Am. Assoc. Immunol., v.
163, p. 1951-1957, 1999.
YING, S.; MENG, Q.; SMITH, S. J.; LARCHÉ, M.; ROBINSON, D. S.; KAY, A. B.
Methods for Identifying Human Eosinophils in Blood and Tissue. ACI Intern., v. 14, p.
64-71, 2002.
WELLER, P. F. The imunobiology of eosinophils. N. Engl. J. Med., v. 324, p. 110-
118, 1991.
WENMAN, W. M. Human Immune Response to Giardia lamblia Infection. J. Infect. Dis., v. 155, n. 1, p. 137-140, 1987.
WHO (World Health Organization), Physical Status: The Use and Interpretation of
Anthropometry: Report of a WHO Expert Committee, WHO Technical Report Series
854. Switzerland: WHO, 1983.
WHO (World Health Organization), Public health significance of intestinal parasitic infections. Bulletin of the Wold Health Organization. v. 65, p.575-588,
1987.
WILLETT, W. C.; KILAMA, W. L.; KIHAMIA, C. M. Ascaris and growth rates: a
randomized trial of treatment. Am. J. Publ. Health., n. 69, p. 987-991, 1979.

113
ANEXO 01
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS FARMACÊUTICAS ÁREA DE CONCENTRAÇÃO: BIOANÁLISES
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO
Título do Projeto: IgA SÉRICA, SECRETORA, IGE TOTAL E ESTADO NUTRICIONAL EM CRIANÇAS COM INFECÇÕES POR ENTEROPARASITAS. Pesquisadores: Professora Dra. Valéria Soraya de Farias Sales (Orientador) Valéria Cristina Ribeiro Dantas (Aluno)
OBJETIVO O Objetivo desta pesquisa é avaliar o estado parasitário das crianças e correlacionar com os níveis séricos de IgA sérica, secretora e IgE total das crianças. PROCEDIMENTO Procedimentos a serem realizados com pesquisadores (Itens 2, 3 e 4) e responsáveis (Item 1) que concordarem em participar do projeto:
1. Responder a uma entrevista; 2. Realizar coleta de fezes em 03 amostras em dias alternados das crianças; 3. Realizar coleta de sangue com material descartável; 4. Realizar coleta de saliva em recipiente estéril e descartável.
RISCOS Os riscos possíveis associados à participação neste estudo são aqueles referentes ao exame de sangue: sangramento, infecções e desmaios. Riscos estes considerados mínimos. BENEFICIOS Os benefícios em particular deste estudo, são: contribuir com uma melhor condição de saúde das crianças, tendo em vista, a detecção de parasitoses, como também, obter dados que contribuirão para o entendimento da resposta imune a parasitas.

114
CONFIDENCIAL DO ESTUDO O registro da participação neste estudo será mantido confidencial, até o limite permitido por lei. No entanto, o Comitê de Ética da Universidade Federal do Rio Grande do Norte pode inspecionar e copiar registros pertinentes à pesquisa, e estes podem conter informações identificadoras. Nós guardamos os registros de cada indivíduo, e somente os pesquisadores que estão trabalhando na pesquisa terão acesso a estas informações. Cada indivíduo receberá um número para ser utilizado no laboratório. Se qualquer relatório ou publicação resultar deste trabalho, a identificação do paciente não será revelada. Os resultados serão relatados de forma sumarizada e o indivíduo não será identificado. DANO ADVINDO DA PESQUISA Se algum dano resultar diretamente desta pesquisa nenhum ressarcimento financeiro estará disponível advindos da UFRN, a não ser que esta injúria seja resultante diretamente da negligência do pesquisador. PARTICIPAÇÃO VOLUNTÁRIA Não há penalidade alguma em desistir, em qualquer época, da participação voluntária desta pesquisa. O responsável pelo paciente pode vir a desistir sem resultar em qualquer dano para o indivíduo ou seu responsável. CONSENTIMENTO PARA PARTICIPAÇÃO Estou de acordo com o estudo descrito acima, submetendo e autorizando a participação do paciente. Fui devidamente esclarecido quanto aos objetivos da pesquisa, aos procedimentos aos quais o paciente será submetido, e dos possíveis riscos que possam advir de tal participação. Foram garantidos esclarecimentos sobre quaisquer questões que venham a surgir durante o curso da pesquisa e o direito de desistir da participação em qualquer momento, sem que nossa desistência implique em qualquer prejuízo à minha pessoa ou de minha guarda. A nossa participação na pesquisa não implica em custos ou prejuízos adicionais, sejam estes de caráter econômico, social, psicológico ou moral. Nos foi garantido o anonimato e o sigilo dos dados referentes à nossa identificação. NOME DA CRIANÇA: _________________________________________________ RESPONSÁVEL: _____________________________________________________ TESTEMUNHA: ______________________________________________________

115
COMPROMISSO DO INVESTIGADOR Eu discuti as questões acima apresentadas com os responsáveis dos pacientes e participantes. É de minha opinião que o indivíduo entenda os riscos, benefícios e obrigações relacionadas a este projeto
Assinatura do investigador (Orientador) Valéria Soraya de Farias Sales
Assinatura do investigador (Aluno) Valéria Cristina Ribeiro Dantas

116
ANEXO 02
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS FARMACÊUTICAS ÁREA DE CONCENTRAÇÃO: BIOANÁLISES
FICHA DE ANAMNESE
FICHA N_ْ_______
HISTÓRIA DA CRIANÇA:
Nome:.............................................................................................. Idade:................... Data Nascimento:........................................Parto:....................................................... Filiação: Pai:................................................................................................................. Mãe:............................................................................................................................... Naturalidade:................................................................................................................. Endereço:...................................................................................................................... Renda familiar:......................Profissão da mãe:............................Pai:....................... Grau de escolaridade da Mãe:........................................................Pai:...................... Observações:........................................................................................................................................................................................................................................................
Cirurgias: Não ( ); Sim ( ),Quais?...............................................................................
Já esteve internada?...................Porque?..................................................................
Estado de saúde atual:................................................................................................ Vacinação:..................................................................................................................... Mamou no peito?...........................Quanto tempo?....................................................
HISTÓRIA FAMILIAR:
Algum tipo de alergia na família? Não ( ); Sim ( ), Quais?..........................................
Outras doenças na família? Quais?..............................................................................

117
POSSUIDOR DE ALGUMA ALERGIA (ANTECEDENTES ALÉRGICOS): OLHOS: ( ) Prurido / conjuntivite ( ) Lacrimejamento
NARIZ: ( ) Espirros ( ) Coriza ( ) Obstrução ( ) Secreção
OUVIDO: ( )Secreção ( ) Prurido
OROFARINGE:( ) Prurido ( ) Ronco ( ) Salivação /baba ( ) Amígdala ( ) Adenóide
PULMÃO: ( ) Sibilo /chiado ( ) Tosse ( ) Dispnéia /falta de ar ( ) Bronquite
PELE: ( ) Picada de inseto ( ) Prurido ( ) Dermatite ( ) Alergia ( ) Urticária
REAÇÃO À DROGA: ( ) Urticária ( ) Edema ( ) Olhos inchados
REAÇÃO À ALIMENTOS: ( ) Não; ( ) Sim. Quais?...................................................
REAÇÃO À LÃ? ( ) Sim ( )Não.
FATORES QUE AGRAVAM OU MELHORAM O ESTADO DE SAÚDE/CRIANÇA:
FATORES CLIMÁTICOS: ( )Verão ( )Inverno ( )Outros fatores ambientais
REALIZAÇÃO DE EXAMES (nos últimos 6 meses):
( )Sangue ( )Urina ( )Fezes ( )Outros. Quais?.....................................................
........................................................................................................................................
JÁ TEVE ALGUMA PARASITOSE? ( )Não ( )Sim. Quais?....................................
........................................................................................................................................
REALIZADO TRATAMENTO? ( )Não ( )Sim.
MEDICAMENTOS EM USO:.......................................................................................... ........................................................................................................................................
SINTOMAS OU QUEIXAS GÁSTRICAS:
Diarréia: ( ) Sim ( ) Não. Freqüência:........................................................................
Vômito: ( ) Sim ( ) Não. Freqüência:...........................................................................
Náusea: ( ) Sim ( ) Não. Freqüência:..........................................................................
Dor abdominal: ( ) Sim ( ) Não. Freqüência:...............................................................
Constipação: ( ) Sim ( ) Não. Freqüência:..................................................................

118
CONDIÇÕES DE MORADIA DA CRIANÇA:
Tipo de casa: ( )Alvenaria ( )Taipa ( )Outros. Quais?...........................................
Presença de: ( )Tapetes ( )Cortina ( )Livros ( ) Bicho de pelúcia ( )Planta.
Quais?.......................................... ( )Animais. Quais?.................................................
Dorme de: ( )Cama ( )Rede ( )Outros.
N.º de cômodos:............. N.º de pessoas:................................................................... Existe barata em casa? ( )Sim ( )Não.
Freqüência da faxina:................................................................................................... Usa água encanada?....................................................................................................
Bebe água: ( )Filtrada ( )Mineral ( ) Pote ( ) Outro Qual?.........................................
A Casa possui: ( )Esgoto ( )Fossa ( )Outro Qual?....................................................
Costuma lavar as mãos antes das refeições? ( )Sim ( )Não
Costuma lavar as frutas e verduras? ( )Sim ( )Não
AVALIAÇÃO NUTRICIONAL:.......................................................................................
• Peso:...................................................................................................................
• Altura:.................................................................................................................
Natal, ___/___/___.





























