Embed Size (px)
Citation preview

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA UNIVERSIDADE FEDERAL DO AMAZONAS – UFAM
DIVERSIDADE CARIOTÍPICA ENTRE DEZ ESPÉCIES DO
GÊNERO ANCISTRUS (SILURIFORMES, LORICARIIDAE)
DA BACIA AMAZÔNICA: ESTRUTURA E MECANISMOS
DE EVOLUÇÃO CROMOSSÔMICA
Renildo Ribeiro de Oliveira
Dissertação apresentada ao Programa de Pós-
Graduação em Biologia Tropical e Recursos
Naturais do convênio INPA/UFAM, como parte
dos requisitos para a obtenção do título de mestre
em Ciências Biológicas, área de concentração em
Biologia de Água Doce e Pesca Interior.
MANAUS – AM
2006

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA UNIVERSIDADE FEDERAL DO AMAZONAS – UFAM
DIVERSIDADE CARIOTÍPICA ENTRE DEZ ESPÉCIES DO
GÊNERO ANCISTRUS (SILURIFORMES, LORICARIIDAE)
DA BACIA AMAZÔNICA: ESTRUTURA E MECANISMOS
DE EVOLUÇÃO CROMOSSÔMICA
Renildo Ribeiro de Oliveira
Dissertação apresentada ao Programa de Pós-
Graduação em Biologia Tropical e Recursos
Naturais do convênio INPA/UFAM, como parte
dos requisitos para a obtenção do título de mestre
em Ciências Biológicas, área de concentração em
Biologia de Água Doce e Pesca Interior.
Orientadora: Eliana Feldberg, Dra.
Co-orientador: Jansen Alfredo Sampaio Zuanon, Dr.
MANAUS – AM
2006

FICHA CATALOGRÁFICA
FICHA CATALOGRÁFICA
Oliveira, Renildo Ribeiro de.
Diversidade cariotípica entre dez espécies do gênero Ancistrus
(Siluriformes, Loricariidae) da Bacia Amazônica: estrutura e mecanismos
de evolução cromossômica / Renildo Ribeiro de Oliveira. Manaus: 2006.
81 f.: il. color.
Dissertação (mestrado). INPA/UFAM, 2006.
1. Amazônia; 2. Peixes; 3. Hypostominae; 4. Citogenética; 5. Cromossomos
sexuais.
CDD 19 ed. 537.5
Sinopse Com intuito de verificar quais os mecanismos de evolução cromossômica existentes
nas espécies amazônicas pertencentes ao gênero Ancistrus (Hypostominae,
Loricariidae), foram estudadas dez espécies provenientes de diversos pontos da
Bacia Amazônica. As análises revelaram uma grande diversidade cariotípica, tanto
na macroestrutura (número e forma cromossômica) como também nos padrões de
regiões organizadoras de nucléolos e de heterocromatina constitutiva, entre as
espécies desse gênero. A ocorrência de baixos números diplóides sugere que fusões
cêntricas tenham sido eventos importantes na evolução cromossômica deste grupo.
Em contraste com a família Loricariidae, há a presença de sistemas de cromossomos
sexuais em muitas das espécies de Ancistrus. Características biológicas e etológicas
tais como, baixa vagilidade, territorialismo e cuidado parental, podem estar
associadas à grande diversidade cariotípica encontrada nesses peixes.
Palavras–chave: Amazônia; Peixes; Hypostominae; Citogenética; Cromossomos
sexuais.

Dedico essa dissertação às pessoas
que mais amo nessa vida, os meus
pais: João e Santilia, pelo incentivo e
amor nos momentos em que mais
precisei.

A REALIZAÇÃO DESTE TRABALHO SÓ FOI POSSÍVEL GRAÇAS:
Ao programa de Pós-Graduação em Biologia de Água Doce e Pesca Interior do
Instituto Nacional de Pesquisas da Amazônia / Universidade Federal do Amazonas.
Ao Laboratório de Genética de Peixes, onde este trabalho foi desenvolvido,
financiado pelos Projetos de Pesquisas Institucionais: PPI: 2-3750, PRONEX/FINEP e
PNOPG/CNPq.
WWF; PDBFF–INPA/Instituto Smithsonian, Fundação O Boticário de Proteção
à Natureza, pelo suporte financeiro às coletas.
Ao Conselho Nacional de desenvolvimento Cientifico e Tecnológico-CNPq,
pela concessão da minha bolsa de mestrado sem a qual este trabalho não teria sido
realizado.

AGRADECIMENTOS
A Deus, por ter me dado força para superar os obstáculos e dificuldades
encontradas no transcorrer deste trabalho.
À Dra. Eliana Feldberg, por me acolher como um filho, pela cuidadosa
orientação, pelas críticas e sugestões e pela confiança em mim depositada.
Ao Dr. Jansen Zuanon, pela co-orientação, pela amizade e confiança, pelas
valiosas sugestões e críticas, que muito contribuíram para a realização deste trabalho, e
pela realização de algumas coletas.
Aos professores do curso BADPI, pelos ensinamentos que muito contribuiu para
o meu crescimento como Biólogo.
Aos amigos do Laboratório de Citogenética de peixes do INPA: Maria Claudia,
Carlos, Aldaléia, Celeste, Maria Leandra, Eduardo, Denise, Germano, Maria José,
Arlindo e Jorge Porto, pelas discussões surgidas no decorrer deste período, as quais
contribuíram muito para a realização deste trabalho.
À Maria Claudia Gross, pela amizade e pela ajuda na preparação e interpretação
dos resultados da meiose.
À Maria do Carmo Arruda (Carminha) secretária do curso BADPI, pelo carinho
e atenção com todos os alunos do curso.
À Dra. Ângela Varella, pela atenção, paciência e por nos emprestar sua máquina
fotográfica sem a qual esta dissertação não teria sido entregue no prazo previsto.
Aos professores doutores que constituíram a banca avaliadora deste trabalho:
Orlando Moreira Filho (UFSCar), Roberto Ferreira Artoni (UEPG), Paulo César Venere
(UFMT), Cleusa Yoshiko Nagamachi (UFPA) e Lucia Helena Rapp Py-Daniel (INPA),
pelas contribuições.
Aos meus colegas de turma: Cilene, Luiza, Marcos Valério (Valdir), Lian,
Fábio, Michel, Márcio, Marcelo, Leonardo, Rodrigo, Maria Claudia, Fabiola, Gelson,
Daniela (Dani Tucuxi), Daniela Magalhães (Dani Boto) e Daniel Previateli.
À Rafaela, Liana, Gina, e Silvia, por terem me acolhido em sua casa quando
cheguei a Manaus.
À Ana Maria Dias, pela amizade, pelo carinho e por ser esta pessoa maravilhosa
que sempre me ajudou quando precisei.

À Rosa Maria (Dona Rosinha) e Christianny Oliveira (Chris), pela amizade,
carinho, por terem me acolhido como um integrante da família e pelas comidas
maravilhosas preparadas sempre com muito amor pela Dona Rosinha.
À toda minha família, pela confiança e carinho que foram muito importantes
para suportar a saudade de casa.
À Janaína Brito (Chaveirinho), minha “irmãzinha”, pela amizade verdadeira,
carinho e pela troca de conhecimento e por me agüentar desde a época de graduação.
Aos meus cunhados Marcos, Roberto, Sebastião e Ricardo, por terem se tornado
os irmãos que eu não tive.
Aos meus queridos sobrinhos, Diogo, Thalma, Pamela, Maikon, Rangel,
Rodrigo, Geovana, Milena e Guilherme, por fazer minha vida mais feliz.
Às minhas irmãzinhas, Raimunda, Cristina, Maria Madalena e Carla, pelo amor,
carinho, atenção e pela união que é a melhor característica da nossa família. Amo muito
vocês.
À Maeda, pela ajuda nos momentos que precisei, por ser esta pessoa doce e
carinhosa e principalmente pelo amor que têm dedicado a mim.
Enfim, a todas as pessoas que contribuíram direta ou indiretamente para a
realização deste trabalho.

i
SUMÁRIO
LISTA DE FIGURAS.................................................................................................... iii
LISTA DE TABELAS................................................................................................... vi
RESUMO....................................................................................................................... vii
ABSTRACT ................................................................................................................. viii
1. INTRODUÇÃO .......................................................................................................... 1
1.1. CONSIDERAÇÕES GERAIS ........................................................................................ 1
1.2. O GÊNERO ANCISTRUS KNER, 1854 ......................................................................... 3
1.3. CITOGENÉTICA DE LORICARIIDAE........................................................................... 4
2. MATERIAL E MÉTODOS ..................................................................................... 10
2.1.ORIGEM E CARACTERÍSTICAS MORFOLÓGICAS DAS ESPÉCIES ESTUDADAS ............. 10
2.2. MÉTODOS ............................................................................................................. 18
2.2.1. Indução de mitoses ....................................................................................... 18
2.2.2. Obtenção de cromossomos mitóticos ........................................................... 18
2.2.3. Obtenção de cromossomos meióticos de machos......................................... 18
2.2.4. Detecção das Regiões Organizadoras de Nucléolos (RONs)....................... 19
2.2.5. Detecção da heterocromatina constitutiva (HC).......................................... 20
2.2.6. Análise cariotípica........................................................................................ 20
3. RESULTADOS ......................................................................................................... 22
3.1. ANCISTRUS RANUNCULUS ....................................................................................... 22
3.2. ANCISTRUS SP.1 “PURUS” ...................................................................................... 25
3.3. ANCISTRUS SP.2 “CATALÃO” ................................................................................. 28
3.4. ANCISTRUS SP.3 “BARCELOS”................................................................................ 31
3.5. ANCISTRUS SP.4 “VERMELHO”............................................................................... 34
3.6. ANCISTRUS SP.5 “RIO BRANCO” ............................................................................ 37
3.7. ANCISTRUS SP.6 “PIAGAÇÚ” .................................................................................. 41
3.8. ANCISTRUS SP.7 “DIMONA” ................................................................................... 44
3.9. ANCISTRUS SP.8 “BALBINA” .................................................................................. 47
3.10. ANCISTRUS SP.9 “TROMBETAS” ........................................................................... 50
4. DISCUSSÃO ............................................................................................................. 53

ii
4.1. DIVERSIDADE CARIOTÍPICA E MECANISMOS DE EVOLUÇÃO CROMOSSÔMICA NO
GÊNERO ANCISTRUS...................................................................................................... 53
4.1.1. MECANISMOS DE CROMOSSOMOS SEXUAIS ................................................... 59
4.2. RELAÇÃO ENTRE A DIVERSIDADE CARIOTÍPICA E CARACTERÍSTICAS DA BIOLOGIA E
ETOLOGIA DOS PEIXES DO GÊNERO ANCISTRUS ............................................................. 65
5. CONSIDERAÇÃO FINAIS..................................................................................... 68
6. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................... 70

iii
LISTA DE FIGURAS
Figura 1. Espécies estudadas (nome e valores de comprimento total). A) Ancistrus
ranunculus (11 cm), B) Ancistrus sp.1 “Purus” (9 cm), C) Ancistrus sp.2 “Catalão” (13
cm), D) Ancistrus sp.3 “Barcelos” (11 cm), E) Ancistrus sp.4 “Vermelho” (14 cm), F)
ancistrus sp.5 “Rio Branco” (17 cm), G) Ancistrus sp.6 “Piagaçu” (7 cm), H) Ancistrus
sp.7 “Dimona” (10,5 cm), I) Ancistrus sp.8 “Balbina” (9,5 cm), J) Ancistrus sp.9
“Trombetas” (5,3 cm) ..................................................................................................... 16
Figura 2. Mapa da Amazônia brasileira, em destaque os locais de coleta .................... 17
Figura 3. Cariótipo de macho e fêmea de Ancistrus ranunculus, com coloração
convencional em Giemsa, evidenciado (caixas) os cromossomos sexuais e os portadores
de RONs impregnadas com nitrato de prata................................................................... 23
Figura 4. A e B- Metáfases de Ancistrus ranunculus preparadas para observação de
bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva nesta
espécie ............................................................................................................................ 24
Figura 5. Cariótipo de macho e fêmea de Ancistrus sp.1 “Purus”, com coloração
convencional em Giemsa, evidenciado (caixas) os cromossomos sexuais e os portadores
de RONs impregnadas com nitrato de prata................................................................... 26
Figura 6. A e B- Metáfases de Ancistrus sp.1 “Purus”preparadas para observação de
bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva nesta
espécie ............................................................................................................................ 27
Figura 7. Cariótipo de macho e fêmea de Ancistrus sp.2 “Catalão”, com coloração
convencional em Giemsa, evidenciado (caixas) os cromossomos sexuais e os portadores
de RONs impregnadas com nitrato de prata................................................................... 29
Figura 8. A e B- Metáfases de Ancistrus sp.2 “Catalão” preparadas para observação de
bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva nesta
espécie ............................................................................................................................ 30
Figura 9. Cariótipo de macho e fêmea de Ancistrus sp.3 “Barcelos”, com coloração
convencional em Giemsa, evidenciado (caixas) cromossomos sexuais e os portadores de
RONs impregnadas com nitrato de prata........................................................................ 32

iv
Figura 10. A e B- Metáfases de Ancistrus sp.3 “Barcelos” preparadas para observação
de bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva
nesta espécie ................................................................................................................... 33
Figura 11. Cariótipo de macho de Ancistrus sp.4 “Vermelho”, com coloração
convencional em Giemsa, evidenciado (caixas) os cromossomos portadores de RONs
impregnadas com nitrato de prata................................................................................... 35
Figura 12. A e B- Metáfases de Ancistrus sp.4 “Vermelho” preparadas para observação
de bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva
nesta espécie. .................................................................................................................. 36
Figura 13. Cariótipo de macho e fêmea de Ancistrus sp.5 “Rio Branco”, com coloração
convencional em Giemsa, evidenciado (caixa) os cromossomos sexuais e os portadores
de RONs impregnadas com nitrato de prata................................................................... 38
Figura 14. A e B- Metáfases de Ancistrus sp.5 “Rio Branco” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição da heterocromatina
constitutiva nesta espécie ............................................................................................... 39
Figura 15. Células meióticas de Ancistrus sp.5 “Rio Branco” observadas em coloração
convencional: A- paquíteno evidenciando cromossomos sexuais com heteropicnose
positiva (seta); B e C- célula diplotênica evidenciando 22 bivalentes autossômicos e o
bivalente sexual com heteropicnose negativa (seta); D- diacinese 22 bivalentes +
sexuais em heteropicnose negativa (seta); E- metáfase I evidenciando cromossomos
dispostos na placa equatorial e sexual com heteropicnose negativa (seta)..................... 40
Figura 16. Cariótipo de macho e fêmea de Ancistrus sp.6 “Piagaçu”, com coloração
convencional em Giemsa, evidenciado (caixa) os cromossomos sexuais e os portadores
de RONs impregnadas com nitrato de prata................................................................... 42
Figura 17. A e B- Metáfases de Ancistrus sp.6 “Piagaçu” preparadas para observação
de bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva
nesta espécie ................................................................................................................... 43
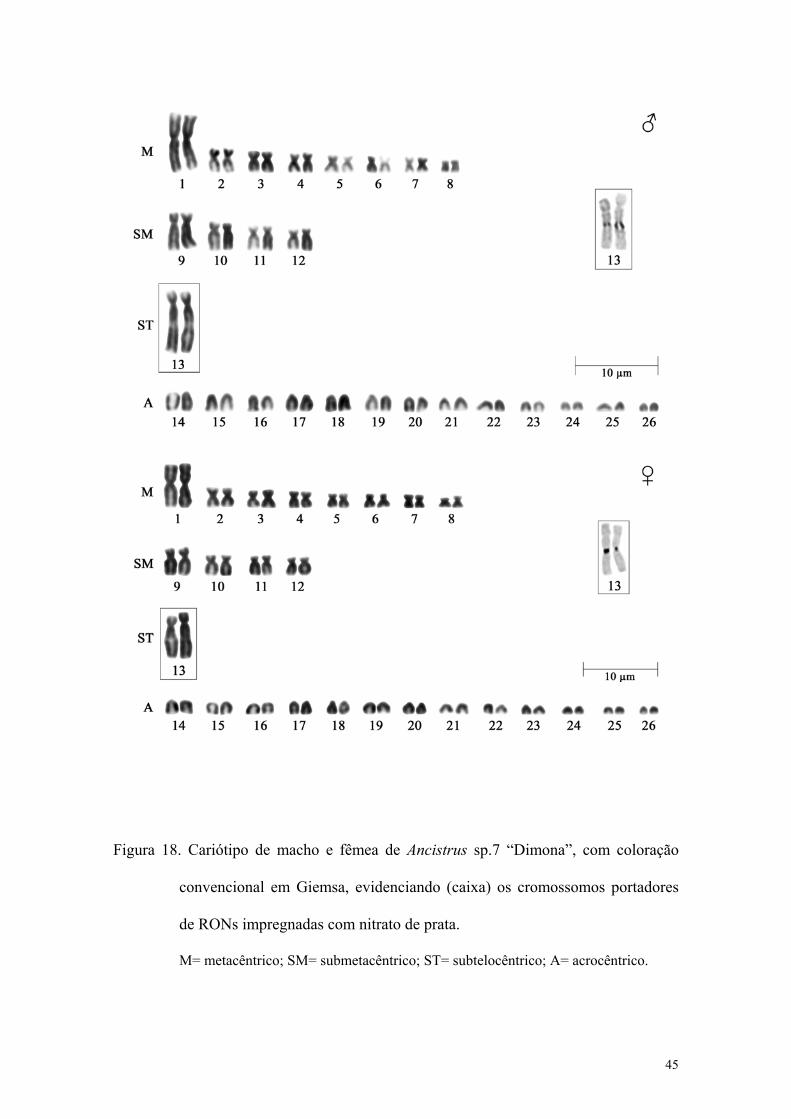
Figura 18. Cariótipo de macho e fêmea de Ancistrus sp.7 “Dimona”, com coloração
convencional em Giemsa, evidenciado (caixa) os cromossomos portadores de RONs
impregnadas com nitrato de prata................................................................................... 45

v
Figura 19. A e B- Metáfases de Ancistrus sp.7 “Dimona” preparadas para observação
de bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva
nesta espécie ................................................................................................................... 46
Figura 20. Cariótipo de macho e fêmea de Ancistrus sp.8 “Balbina”, com coloração
convencional em Giemsa, evidenciado (caixa) os cromossomos sexuais e os portadores
de RONs impregnadas com nitrato de prata................................................................... 48
Figura 21. A e B- Metáfases de Ancistrus sp.8 “Balbina” preparadas para observação
de bandas C; C- Idiograma mostrando a distribuição da heterocromatina constitutiva
nesta espécie ................................................................................................................... 49
Figura 22. Cariótipo de macho de Ancistrus sp.9 “Trombetas”, com coloração
convencional em Giemsa, evidenciado (caixa) os cromossomos portadores de rons
impregnadas com nitrato de prata................................................................................... 51

vi
LISTA DE TABELAS
Tabela 1. Informações citogenéticas disponíveis para espécies de loricariídeos da Tribo
Ancistrini (sensu Armbruster, 2004) ................................................................................ 8
Tabela 2. Alguns dados morfométricos das espécies de Ancistrus do presente estudo. 14
Tabela 3. Síntese das características cariotípicas das dez espécies de Ancistrus
analisadas no presente estudo......................................................................................... 52

vii
RESUMO
A tribo Ancistrini pertencente à subfamília Hypostominae (Siluriformes: Loricariidae),
compõe um grupo de peixes cuja sistemática é ainda pouco conhecida e estudos
filogenéticos são escassos. Ancistrus, gênero tipo da tribo Ancistrini, engloba 53
espécies válidas e pelo menos seis espécies amazônicas ainda não descritas. Com o
objetivo de verificar os mecanismos de evolução cromossômica existentes em espécies
pertencentes ao gênero Ancistrus, foram estudadas dez espécies provenientes de
diversos pontos da bacia amazônica. As análises revelaram uma grande diversidade
cariotípica, tanto na macroestrutura (número e fórmula cromossômica) como também
nos padrões de regiões organizadoras de nucléolos e de heterocromatina constitutiva. O
número diplóide variou de 34 a 52 cromossomos e o número de braços de 68 a 82. A
RON variou tanto na localização no cromossomo (terminal, intersticial, braço longo e
braço curto) como na posição no cariótipo. Poucos cromossomos apresentaram
heterocromatina constitutiva, as quais foram distribuídas em blocos discretos dispostos
em posição centromérica, terminal ou intersticial. A ocorrência de números diplóides
baixos (menor que 52) sugere a presença de fusões cêntricas na evolução cromossômica
deste grupo. Além da diversidade cariotípica, sistemas de cromossomos sexuais foram
registrados para sete das 10 espécies analisadas sendo: duas ZZ/ZW, três XX/XY e dois
sistemas múltiplos XX/XY1Y2 e Z1Z1Z2Z2/Z1Z2W1W2. Peixes do gênero Ancistrus
exibem baixa vagilidade, territorialismo e cuidado parental. Estas características
etológicas podem criar uma barreira ao fluxo gênico entre populações, permitindo
especiações micro-alopátricas e resultando na grande diversidade cariotípica encontrada
nestas espécies.

viii
ABSTRACT
The tribe Ancistrini belongs to the subfamily Hypostominae (Siluriformes:
Loricariidae). Phylogenetic studies of this group of fishes are few, leaving their
systematics relatively unknown. Ancistrus, the type genus of the Ancistrini tribe,
contains 53 valid species and at least six Amazonian species that have yet to be
described. In order to explore mechanisms of chromosome evolution in the genus
Ancistrus, we studied 10 species collected from locations across the Amazon basin. We
found a large karyotypic diversity among the ten species, in both macrostructure
(chromosome number and formula) and pattern of nucleolar organizer regions and
constitutive heterochromatin. The diploid number varied between 34 and 52
chromosomes and the number of arms varied between 68 and 82. The RON varied in
both chromosome location (terminal, interstitial, long arm, short arm) and karyotype
position. Few chromosomes presented constitutive heterochromatin; when present, this
heterochromatin was distributed in discrete blocks in the centromeric position, terminal
or interstitial. The occurrence of low diploid numbers suggests the presence of centric
fusions in the chromosomal evolution of this group. Besides documenting karyotypic
diversity, we also studied the sex chromosome systems of seven of the 10 species. We
found two cases of ZZ/ZW, three cases of XX/XY, and two multiple systems:
XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2. Fish of the genus Ancistrus exhibit low vagility,
territoriality and parental care. These ethological characteristics may create a gene flow
barrier among populations, allowing microallopatric speciation and resulting in the large
karyotypic diversity found in these species.

1
1. INTRODUÇÃO
1.1. Considerações gerais
A bacia amazônica abriga a maior e mais diversa ictiofauna do mundo,
com estimativas que variam de 1500 a 6000 espécies (Goulding, 1980; Reis et
al., 2003). A família Loricariidae, a maior dentre os Siluriformes com
aproximadamente 683 espécies válidas (Reis et al., 2003; Armbruster, 2004)
constitui um grupo muito abundante e diversificado nesse ecossistema (Fisch-
Muller et al., 2001), ocupando uma grande diversidade de habitats, que vão
desde o leito dos grandes rios, áreas de várzeas e de igapós, até igarapés de
terra-firme (Junk et al., 1983; Sabino & Zuanon, 1998; Anjos, 2005).
Os loricariídeos representam um importante recurso sócio-econômico
na região amazônica, tanto para a pesca ornamental (Ferraz, 1999; Zuanon,
1999), como também para a pesca artesanal e comercial, onde compõem uma
parte significativa dos desembarques (Petrere Jr., 1978; Goulding, 1980;
Batista & Petrere Jr., 2003). Além disso, têm grande participação na pesca de
subsistência, sendo um recurso alimentar muito utilizado por moradores
ribeirinhos (Batista et al., 1998).
Na Amazônia, os loricariídeos são popularmente conhecidos como
acaris ou bodós. Caracterizam-se por apresentar o corpo totalmente recoberto
por placas ósseas, dispostas em várias séries; boca situada em posição ventral
e em forma de ventosa; dentes alongados e geralmente bífidos, ou em forma de
colher, dispostos em uma única série em cada maxila. Possuem intestino
enovelado e extremamente longo, reflexo de seus hábitos alimentares,

2
predominantemente, detritívoros-iliófagos, (Ferreira et al., 1998; Power,
1990). Muitas espécies apresentam respiração acessória ou aérea, realizada
através do estômago, o que lhes permite sobreviver em ambientes aquáticos
extremamente pobres em oxigênio (Val & Almeida-Val, 1995; Lowe-
McConnell, 1999).
A morfologia altamente diferenciada faz dos loricariídeos um dos
grupos mais bem caracterizados entre os Siluriformes (de Pinna, 1998;
Zuanon, 1999). Segundo Armbruster (2004), cinco subfamílias são
reconhecidas entre os Loricariidae: Loricariinae (~209 espécies),
Hypoptopomatinae (~79 espécies), Neoplecostominae (~7 espécies),
Upsilodinae (~5 espécies) e Hypostominae (~386 espécies) (o número de
espécies baseado em Reis et al., 2003).
Apesar desta recente classificação, os loricariídeos representam um
grupo ainda relativamente confuso no que se refere aos aspectos taxonômicos e
sistemáticos. Muitas classificações divergentes foram feitas para essa família
(e.g., Kner, 1854; Bleeker, 1862; Eigenmann & Eigenmann, 1890; Regan,
1904; Gosline, 1945; Isbrücker, 1980; Howes, 1983; Schaefer, 1987). Tal
situação reflete a carência de estudos filogenéticos para esse grupo; além
disso, muitas espécies novas de loricariídeos continuam sendo coletadas em
áreas pobremente amostradas de diferentes sistemas hidrográficos,
especialmente na Amazônia (Armbruster, 2003).
Pertencente à subfamília Hypostominae, a tribo Ancistrini (~217
espécies) compõe um grupo de peixes cuja sistemática é ainda pouco

3
conhecida, não sendo muito numerosos os estudos filogenéticos feitos sobre
essa tribo. Com base em estudos morfológicos, esta já foi considerada uma
subfamília (Isbrücker, 1980), já comportou gêneros que não formavam um
grupo natural, sendo portanto, um grupo não monofilético, incluindo
representantes de Hypostominae (Howes, 1983), como também um grupo
monofilético bem definido (Schaefer, 1987) de composição mais restrita. Com
base em análises filogenéticas, utilizando seqüências parciais de rDNA
mitocondrial de 58 espécies de loricariídeos, foi considerada um grupo
parafilético, incluindo Loricariinae e parte de Hypostominae (Montoya-Burgos
et al., 1998).
1.2. O gênero Ancistrus Kner, 1854
Ancistrus, gênero tipo da tribo Ancistrini, é composto por um grupo de
espécies amplamente distribuídas no Neotrópico e especialmente diversificado
no sistema do rio Amazonas. Engloba 53 espécies válidas, e pelo menos seis
espécies amazônicas ainda não descritas (Fisch-Muller et al., 2001; Fisch-
Muller, 2003).
As espécies do gênero Ancistrus apresentam o corpo largo e deprimido,
cuja largura pode ser cinco a seis vezes maior que a sua altura; cabeça grande;
focinho largo com borda nua (sem placas), sendo a borda nua mais estreita nas
fêmeas e mais larga nos machos. Um número variável de tentáculos cutâneos
apresenta-se dispostos sobre o focinho, sendo pequenos nas fêmeas e maiores e

4
bifurcados nos machos. As espécies são geralmente de pequeno porte, podendo
atingir 15 cm de comprimento (Burgess, 1989).
Espécies de Ancistrus, bem como loricariídeos de uma maneira geral,
possuem pouca vagilidade e permanecem escondidos em troncos, fendas de
pedras e outros abrigos, durante o dia. Via de regra, possuem hábitos noturnos,
são territorialistas e apresentam cuidado parental, construindo ninhos e
protegendo seus ovos e filhotes (Burgess, 1989; Zuanon, 1999).
1.3. Citogenética de Loricariidae
A família Loricariidae pode ser considerada um grupo de peixes ainda
pouco estudado em nível cromossômico. Segundo Kavalco et al. (2005), os
dados citogenéticos disponíveis para esse grupo referem-se a aproximadamente
70 espécies, o que representa cerca de 10% do total estimado de espécies,
segundo Reis et al. (2003). Estudos têm mostrado que esta família apresenta
uma grande diversidade cariotípica, com o número diplóide variando de 36 em
Rineloricaria latirostris (Loricariinae) (Giuliano-Caetano, 1998) a 96 em
Upsilodus sp. (Upsilodinae) (Kavalco et al., 2005).
A maioria dos estudos citogenéticos, nessa família, foi conduzida com
espécies da subfamília Hypostominae, na qual o número diplóide varia de 38 a
80 cromossomos (Artoni, 1996; Artoni & Bertollo, 1996; Alves et al., 2003),
sendo que na tribo Ancistrini o número diplóide varia de 38 a 52 cromossomos
(Tabela 1); em Loricariinae, este número varia de 2n=36 a 2n=74 (Artoni,
1996; Giuliano-Caetano, 1998; Alves et al., 2003); nas subfamílias

5
Hypoptopomatinae e Neoplecostominae o número diplóide de 54 cromossomos
parece ser o mais comum entre as espécies estudadas (Andreata et al., 1992);
e, em Upsilodinae, há um único registro de 2n=96 cromossomos em Upsilodus
sp. (Kavalco et al., 2005).
As alterações mais comumente envolvidas na evolução cromossômica
dos peixes são os rearranjos Robertsonianos (fusões e fissões cêntricas) e os
rearranjos não Robertsonianos (inversões, deleções, duplicações e
translocações) (Denton, 1973). Em Ancistrini, as inversões pericêntricas e
paracêntricas são provavelmente os principais rearranjos cromossômicos
envolvidos no processo de evolução cariotípica (Alves et al., 2003). Tais
formas de rearranjo mantêm o número modal 2n=52 cromossomos,
predominantemente dos tipos metacêntrico e submetacêntrico, na maioria das
espécies estudadas. No entanto, esses mesmos autores sugerem que fusão
cêntrica seja o mecanismo responsável pela redução do número diplóide
encontrado em algumas espécies desta tribo (Lara, 1998; Alves et al., 2003;
Mariotto, 2003; Souza, 2003) (Tabela 1).
A maioria das espécies de peixes Neotropicais estudada
citogeneticamente não apresenta cromossomos sexuais diferenciados (Moreira-
Filho et al., 1993; Centofante et al., 2002). As espécies da família Loricariidae
até agora analisadas, também apresentam, em geral, um homomorfismo em
relação à presença destes tipos de cromossomos (Artoni & Bertollo, 2001;
Alves et al., 2005; Kavalco et al., 2005). Entretanto, alguns casos de
cromossomos sexuais em Loricariidae foram descritos e são dos tipos XX/XY

6
e ZZ/ZW. Entre espécies de Hypostominae, mecanismos do tipo XX/XY foram
observados em Hypostomus macrops (Michelle et al., 1977) e em uma
população de Ancistrus cf. dubius (Mariotto, 2003) e do tipo ZZ/ZW em
Hypostomus sp.G (Artoni et al., 1998), em uma população de Ancistrus cf.
dubius (Mariotto et al., 2004) e em Hemiancistrus spilomma (de Oliveira et
al., no prelo). Em Hypoptopomatinae, observou-se o tipo XX/XY em
Pseudocinclus tietensis (Andreata et al., 1992) e o tipo ZZ/ZW em duas
populações de Microlepidogaster leucofrenatus (Andreata et al., 1993; 1994).
Já entre os Loricariinae, o tipo ZZ/ZW foi observado em Loricariichthys
platymetopom (Scavone & Júlio Jr., 1995).
Quanto às regiões organizadoras de nucléolos (RONs), a ocorrência de
sistemas simples, com sítios na posição terminal, é a mais comum para os
loricariídeos (Artoni & Bertollo, 1996; Kavalco et al., 2005). Contudo, há
registros de sistemas múltiplos, com esses sítios localizados em posição
intersticial para algumas espécies (Artoni, 1996; Artoni & Bertollo, 1996;
Artoni & Bertollo, 2001; Souza, 2003; de Oliveira et al., no prelo.).
Sabe-se que a diversidade cromossômica pode fornecer informações
valiosas para o entendimento das relações filogenéticas entre os peixes. Dessa
forma, informações cariotípicas como a posição do centrômero, a distribuição
da heterocromatina, a posição, a localização e o número das regiões
organizadoras de nucléolos, podem ser utilizadas para auxiliar na elucidação
de problemas taxonômicos e filogenéticos (Pieczarka, 1995; Almeida-Toledo,
1998). Feldberg et al. (2003), usando informações citogenéticas disponíveis

7
para a família Cichlidae e comparando-as com propostas filogenéticas
existentes para esse grupo, puderam inferir sobre os principais rearranjos
cromossômicos e caminhos que provavelmente ocorreram no processo
evolutivo deste grupo.
1.4. Objetivo
Diante disso, o presente trabalho teve como objetivo caracterizar,
citogeneticamente, espécies do gênero Ancistrus, de várias localidades da
bacia amazônica e inferir sobre os possíveis mecanismos de evolução
cromossômica entre as espécies deste gênero.

8
Tabela 1. Informações citogenéticas disponíveis para espécies de loricariídeos da Tribo Ancistrini (sensu Armbruster, 2004). (2n=número diplóide; M=metacêntrico; SM=submetacêntrico; ST=subtelocêntrico; A=acrocêntrico; C. sex= cromossomos sexuais; RONs=regiões organizadoras de nucléolos; p=braço curto; q=braço longo).
Espécie Rio/Estado Brasileiro 2n Fórmula cariotípica C. sex Localização das RONs Ref.
Ancistrus n. sp.1 São Francisco/Acre 38 30M/SM+8ST (M) 5p, intersticial 1
Ancistrus cf. dubius Pantanal/ Mato Grosso 42 24M+10SM+8ST (SM) 16p, intersticial 2
Ancistrus cf. dubius Pantanal/ Mato Grosso 42 24M+10SM+8ST (XX/XY) (SM) 16p, intersticial 2
Ancistrus cf. dubius Pantanal/ Mato Grosso 44 18M+10SM+8ST+8A (ZZ/ZW) (SM) 13p, intersticial 3
Ancistrus sp. Iguaçu/Paraná 48 18M+14SM+12ST+4A (ST) p, terminal 4
Ancistrus ranunculus Xingu/Pará 48 8M+18SM+6ST+16A (ST) 15, proximal 5
Ancistrus n. sp.2 Betari/São Paulo 52 32M/SM+20ST/A (ST) 24p, terminal 1
Ancistrus multispinnis Itapocu/Santa Catarina 52 28M/SM+24ST/A (ST) 17p, terminal 1
Hemiancistrus sp. Araguaia/Mato Grosso 52 20M+20SM+8ST+4A (SM) 9p, terminal 6
Hemiancistrus spinosissimus Araguaia/Mato Grosso 52 26M+22SM+4ST (SM) 17p, terminal 8
Hemiancistrus spilomma Araguaia/Mato Grosso 52 ♀25M+21SM+6ST ♂24M+22SM+6ST
(ZZ/ZW) (M) 29p, terminal; (SM) 8q, terminal; (M) 3p, terminal
8
Megalancistrus aculeatus Paraná/Paraná 52 26M+26SM (SM) p, Intersticial 4
Panaque cf. nigrolineatus Araguaia/Mato Grosso 52 26M+20SM+6ST (A) p, terminal 6
Baryancistrus cf. niveatus Xingu/Pará 52 16M+32SM+4ST (M) 3p, intersticial 7
Baryancistrus sp.1 Jarí/Pará 52 8M+34SM+10ST (SM) 18p, intersticial 5
Baryancistrus sp.2 Xingu/Pará 52 18M+30SM+4ST (M) 6p, intersticial 5
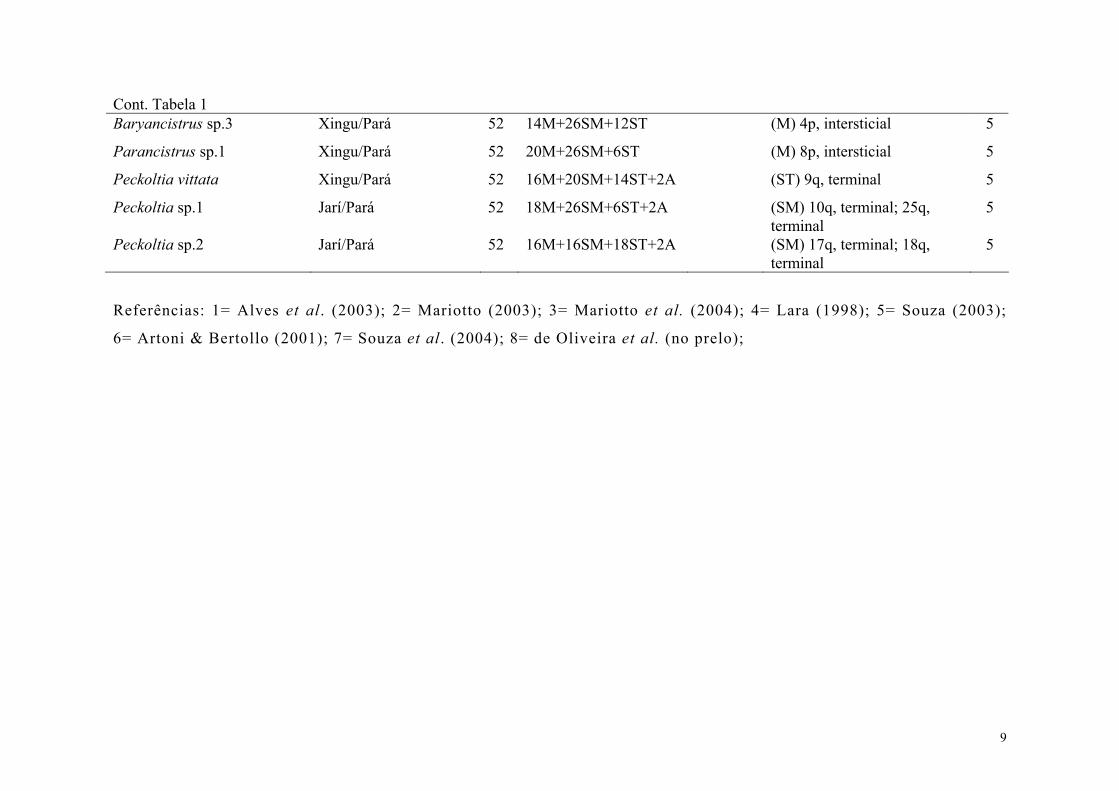
9
Cont. Tabela 1 Baryancistrus sp.3 Xingu/Pará 52 14M+26SM+12ST (M) 4p, intersticial 5
Parancistrus sp.1 Xingu/Pará 52 20M+26SM+6ST (M) 8p, intersticial 5
Peckoltia vittata Xingu/Pará 52 16M+20SM+14ST+2A (ST) 9q, terminal 5
Peckoltia sp.1 Jarí/Pará 52 18M+26SM+6ST+2A (SM) 10q, terminal; 25q, terminal
5
Peckoltia sp.2 Jarí/Pará 52 16M+16SM+18ST+2A (SM) 17q, terminal; 18q, terminal
5
Referências: 1= Alves et al. (2003); 2= Mariotto (2003); 3= Mariotto et al. (2004); 4= Lara (1998); 5= Souza (2003);
6= Artoni & Bertollo (2001); 7= Souza et al. (2004); 8= de Oliveira et al. (no prelo);

10
2. MATERIAL E MÉTODOS
2.1. Origem e características morfológicas das espécies estudadas
Foi analisada, no presente estudo, a estrutura cariotípica de dez
espécies de peixes pertencentes ao gênero Ancistrus, as quais foram
coletadas em vários pontos da bacia amazônica. Algumas coletas foram
realizadas aproveitando excursões de outros projetos com objetivo de obter
um número maior de espécies de diferentes localidades. A princípio, cada
lote de exemplares, de cada localidade, foi considerado como uma entidade
biológica distinta, para posterior análise e discussão dos resultados obtidos
(Figura 1 e 2). Para uma melhor caracterização das espécies foi feita uma
breve descrição morfológica, baseada em algumas proporções corporais e
nos padrões de colorido (material fresco), de modo a facilitar a
identificação das espécies em ocasiões futuras (Segundo S. Fisch-Muller
comunicação pessoal, a maioria dessas espécies ainda não foram descritas).
Lotes de exemplares-testemunho foram depositados na coleção de peixes do
INPA (Tabela 2).
Ancistrus ranunculus: cinco exemplares (3 ♂ e 2 ♀) foram
adquiridos junto a exportadores de peixes ornamentais (Turkys Aquarium),
originários do rio Xingu, região de Altamira, Pará. Esta espécie apresenta
dorsalmente, coloração de fundo escura e/ou preta uniforme e não
apresentando pintas pelo corpo; ventralmente, coloração cinza uniforme; a
nadadeira dorsal possui coloração preta uniforme (Figura 1A).
Ancistrus sp.1 “Purus”: 12 exemplares (5 ♂ e 7 ♀) foram adquiridos
junto a exportadores de peixes ornamentais (K2), originários do rio Purus

11
na região de (Santa Maria). Esta espécie apresenta dorsalmente, coloração
de fundo marrom com pintas amareladas maiores que a pupila distribuídas
pelo corpo; ventralmente, coloração marrom com manchas difusas; a
nadadeira dorsal possui coloração de fundo preta com listras horizontais
claras (Figura 1B).
Ancistrus sp.2 “Catalão”: 12 exemplares (5 ♂ e 7 ♀) foram
coletados no Lago Catalão, localizado na região de confluência dos rios
Solimões e Negro (S03º10’45.0”, W059º54’25.4”). Esta espécie apresenta
dorsalmente, coloração de fundo preta com pintas amareladas menores que
a pupila distribuídas pelo corpo; ventralmente, coloração escura, com
manchas pequenas difusas; a nadadeira dorsal possui coloração de fundo
preta com listras horizontais claras (Figura 1C).
Ancistrus sp.3 “Barcelos”: 11 exemplares (7 ♂ e 4 ♀) foram
coletados no rio Demeni, médio rio Negro. Esta espécie apresenta
dorsalmente, coloração de fundo preta com pintas brancas muito pequenas
(pontilhado fino); ventralmente, coloração escura com pintas claras bem
definidas maiores que as do corpo; a nadadeira dorsal possui coloração
preta com pontilhado branco e borda branca (Figura 1D).
Ancistrus sp.4 “Vermelho”: cinco exemplares (5 ♂) foram coletados
no rio Demeni, médio rio Negro. Esta espécie apresenta dorsalmente,
coloração de fundo cinza/amarronzado sem pintas; ventralmente, coloração
amarronzada com manchas claras bem definidas e muito pequenas;
coloração da nadadeira dorsal cinza uniforme (Figura 1E).

12
Ancistrus sp.5 “Rio Branco”: 12 exemplares (6 ♂ e 6 ♀) foram
coletados no igarapé Macoari, afluente do rio Branco, no estado de Roraima
(S01º10’30,9”, W061º51’05,3”). Esta espécie apresenta dorsalmente,
coloração de fundo marrom escuro com pintas vermelhas menores que a
pupila; ventralmente, coloração escura com pequenas manchas difusas; a
nadadeira dorsal possui coloração escura com pintas vermelhas (Figura 1F).
Ancistrus sp.6 “Piagaçu”: sete exemplares (3 ♂ e 4 ♀) foram
coletados no Lago Aiapuá, localizado na margem esquerda do rio Purus, na
Reserva de Desenvolvimento Sustentável Piagaçú-Purus (S04º27’26”,
W062º11’56”). Esta espécie possui dorsalmente, coloração de fundo cinza
escura com pintas brancas minúsculas; ventralmente, coloração cinza
amarelado com pontilhado bem definido, que nos exemplares menores,
assemelha a ocelos; coloração da nadadeira dorsal cinza escura com
pontilhado minúsculo (Figura 1G).
Ancistrus sp.7 “Dimona”: oito exemplares (6 ♂ e 2 ♀) foram
coletados em um igarapé localizado a 80 km ao norte de Manaus, na
Fazenda Dimona (S02º19’45,5”,W60°04’39,0”) que é uma das áreas de
estudo do projeto Dinâmica Biológica de Fragmentos Florestais
(INPA/Smithsonian Institute). Esta espécie apresenta dorsalmente,
coloração de fundo preta com pontilhado branco esverdeado muito pequeno;
ventralmente, coloração escura com pontilhado bem definido; possui
nadadeira dorsal transparente (membranas hialinas) com pontilhados sobre
os raios (Figura 1H).

13
Ancistrus. sp.8 “Balbina”: 12 exemplares (6 ♂ e 6 ♀) foram
coletados no igarapé Barretinho, município de Balbina à 180 km ao norte de
Manaus, (S01º58’20”, W059º29’48”). Esta espécie possui dorsalmente,
coloração de fundo marrom com pintas grandes, maiores que a pupila, de
coloração olivácea; ventralmente, coloração escura com pintas grandes e
bem definidas; a nadadeira dorsal possui coloração com faixas pretas e
brancas alternadas (Figura 1I).
Ancistrus. sp.9 “Trombetas”: 1 exemplar (1 ♂) foi coletado no rio
Trombetas estado do Pará. Esta espécie apresenta dorsalmente, coloração de
fundo cinza escuro com pintas brancas menores que a pupila; ventralmente,
coloração escura com pontilhado bem definido coloração da nadadeira
dorsal marrom/acinzentada (Figura 1J).
Algumas medidas morfométricas podem ser observadas na tabela 2.

14
Tabela 2. Alguns dados morfométricos das espécies de Ancistrus do presente estudo. Nº. Reg.= número do registro no Laboratório de Genética de Peixes do INPA; N.R.D.= número de raios da nadadeira dorsal; N.P.L.= número de placas da linha lateral; D.O/I.O.= diâmetro do olho/interorbital; C.C.= comprimento da cabeça; C.P.= comprimento padrão (mm).
Espécie Nº. Reg. N.R.D. N.P.L. D.O/I.O. C.C. C.P. Número INPA
Ancistrus ranunculus 7236 6744 6748
I+7 26 0,23 0,34 0,40
38,7 19,8 17,4
90,8 50,7 45,5
25624
Ancistrus sp.1 “Purus” 6649 6662 6658
I+7 24 0,38 0,39 0,37
19,2 19,9 18,8
59,9 64
60,9 25625
Ancistrus sp.2 “Catalão” 6940 6922 6937
I+7 23-25 0,31 0,35 0,34
27,2 31,6 26,9
89 95,2 88,8
25626
Ancistrus sp.3 “Barcelos” 7254 7248 7247
I+8 a I+9 22-23
0,35 0,47 0,48
35 26,9 25,4
101 82,5 81,2
25627
Ancistrus sp.4 “Vermelho” 7241 7265 7266
I+7 23 0,36 0,39 0,41
35,9 33,8 28,7
111,3 107 87
25628
Ancistrus sp.5 “Rio Branco” 7407 7262 7409
I+8 23-24 0,36 0,34 0,38
41,4 44,9 33
135,2 134,7 105,5
25629

15
Cont. tabela 2
Ancistrus sp.6 “Piagaçu” 6745 6737 6738
I+8 23 0,35 0,39 0,49
44,7 26,7 17,9
125,5 80
53,2 25630
Ancistrus sp.7 “Dimona” 7373 7374 7376
I+7 24 0,40 0,40 0,40
23 17,3 14,3
82,6 64,7 57,8
25631
Ancistrus sp.8 “Balbina” 7380 7448 7449
I+7 23-24 0,39 0,41 0,44
27,2 25,9 19,9
90,3 86,9 71,4
25632
Ancistrus sp.10 “Trombetas” 7406 I+7 23 0,46 13,46 37,12 25633

16
Figura 1. Espécies estudadas (Nome e valores de comprimento total). A)
Ancistrus ranunculus (11 cm), B) Ancistrus sp.1 “Purus” (9 cm), C) Ancistrus sp.2 “Catalão” (13 cm), D) Ancistrus sp.3 “Barcelos” (11 cm), E) Ancistrus sp.4 “Vermelho” (14 cm), F) Ancistrus sp.5 “Rio branco”(17 cm), G) Ancistrus sp.6 “Piagaçu”(7 cm), H) Ancistrus sp.7 “Dimona” (10,5 cm), I) Ancistrus sp.8 “Balbina” (9,5 cm), J) Ancistrus sp.9 “Trombetas” (5,3 cm).

17
Figura 2: Mapa da Amazônia brasileira, em destaque os locais de coleta.
1= Rio Xingu; 2= Rio Purus; 3= Área do Catalão; 4= Rio Demeni; 5= Igarapé
Macoari; 6= Lago Aiapuá; 7= Igarapé Dimona; 8= Igarapé Barretinho; 9= Rio
Trombetas.

18
2.2. Métodos
2.2.1. Indução de mitoses
Para indução à formação de um número de metáfases satisfatório por
indivíduo preparado, foi aplicada uma solução de fermento biológico na
proporção de 1ml para cada 100g de peso vivo, conforme metodologia
descrita por Oliveira (1987).
2.2.2. Obtenção de cromossomos mitóticos Os exemplares foram sacrificados e retirou-se o rim, que é o órgão
hematopoiético em peixes. Para obtenção de cromossomos mitóticos,
aplicou-se a técnica de suspensão celular por “air drying”, adaptada para
peixes por Bertollo et al. (1978), com as seguintes modificações:
Injetou-se, na região intraperitonial, colchicina a 0,0125% na
proporção de 1ml para cada 100g de peso vivo. Após isso, o peixe foi
deixado em um aquário bem aerado por um período de 45 a 50 minutos.
Para a análise do material, pingou-se duas a três gotas da suspensão
obtida em uma lâmina limpa, aquecida a 60ºC, em banho-maria. A mesma
foi corada com Giemsa a 5% em solução tampão fosfato (pH=6,8), durante
10 minutos. Em seguida, lavou-se a lâmina em água corrente, deixando-se
secar ao ar.
2.2.3. Obtenção de cromossomos meióticos de machos Para o estudo dos cromossomos meióticos foram utilizadas células
gonadais masculinas, empregando-se a técnica descrita por Kligerman &

19
Bloom (1977) adaptada para peixes por Bertollo et al. (1978), com as
seguintes modificações:
O modo de aplicação da colchicina e o intervalo para o sacrifício do peixe são
os mesmos da técnica usada para obtenção de cromossomos mitóticos. Após o sacrifício
do peixe, os testículos foram seccionados em fragmentos bem pequenos e colocados em
solução hipotônica de KCl a 0,075M por 30 minutos. Logo após, o material foi
transferido para uma cubeta contendo fixador Carnoy (metanol 3: 1 ácido acético)
recém-preparado, por 20 minutos. O processo de fixação foi repetido por mais duas
vezes. Em seguida, o material foi guardado em refrigerador a 4º C, em tubos tipo
“eppendorf”, com fixador Carnoy. Para continuação da técnica, os fragmentos foram
retirados do fixador e secos em papel de filtro. Em seguida, este foi colocado sobre uma
lâmina limpa e macerado em 1ml de ácido acético 50%, com o auxílio de um bastão de
vidro. A lâmina foi levada a uma placa aquecedora a 40º C, até secar. Posteriormente,
esta foi corada com Giemsa 5% em tampão fosfato 0,06M e pH 6,8 por 10 minutos,
sendo em seguida lavada em água corrente e seca diretamente ao ar.
Para interpretação da meiose foram consideradas as seguintes fases:
leptóteno/zigóteno, paquíteno, diplóteno, diacinese, metáfase I e metáfase
II, dando maior ênfase ao diplóteno, diacinese, metáfase I e metáfase II, as
quais são, em geral, mais elucidativas na determinação de rearranjos
cromossômicos (John, 1990).
2.2.4. Detecção das Regiões Organizadoras de Nucléolos (RONs) Para a observação das RONs foi utilizada a metodologia de
impregnação com nitrato de prata (AgNO3) em metáfases fixadas, descrita
por Howell & Black (1980), com as seguintes adaptações:

20
Foi suprimido o tratamento da lâmina em HCl a 1 N a 60°C em
banho-maria, por 4 minutos; e após pingar sobre uma lâmina, preparada
conforme a técnica adotada para cromossomos mitóticos, três gotas de
solução aqüosa de gelatina a 2%, acrescida de ácido fórmico (1:100ml) e
adicionar três gotas de solução aqüosa de nitrato de prata a 50%, foram
acrescentadas três gotas de água destilada.
2.2.5. Detecção da heterocromatina constitutiva (HC) Para análise da heterocromatina constitutiva foi utilizada a técnica
descrita por Sumner (1972), com as seguintes adaptações:
A lâmina foi tratada com ácido clorídrico (HCl) 0,2N por 2 minutos,
em banho-maria a 47ºC. Em seguida, incubou-se a lâmina em solução
2XSSC (Cloreto de sódio 0,3M e citrato trisódico 0,03M, pH 7,0), a 60ºC,
por 10 minutos. Posteriormente, tratou-se a lâmina em solução de hidróxido
de bário (5%), recém preparada e filtrada, a 47ºC, durante 25 segundos, e
lavou-se a lâmina, rapidamente, em HCl (0,2N).
Para finalizar, incubou-se a lâmina novamente em solução 2XSSC a
60ºC, durante 20 minutos e corou-se com solução de Giemsa a 5% em
solução tampão fosfato pH 6,8 durante 10 minutos. É importante salientar
que, após cada tratamento, a lâmina foi lavada em água destilada e deixada
ao ar livre, até secar totalmente.
2.2.6. Análise cariotípica As preparações cromossômicas foram analisadas em microscópio
óptico, e, após a contagem dos cromossomos em cerca de 30 metáfases por
indivíduo, foi estabelecido o valor modal do número diplóide para cada

21
espécie. Para saber a localização dos blocos de heterocromatina constitutiva
foi feito banda C seqüencial em todas as espécies.
As melhores metáfases, isto é, aquelas que apresentaram um bom
espalhamento e morfologia mais nítida dos cromossomos, foram
fotografadas em microscópio Olympus BH-2 com adaptação do sistema PM-
6 para fotografia e com objetiva de imersão. Os negativos foram obtidos em
filmes KODAK IMAGELINK – HQ e AGFA, regulados para asa 25, com
revelação processada em revelador Dektol Kodak. Os negativos
selecionados foram copiados em papel Kodabrome Print RC F3.
Os cromossomos foram recortados, medidos utilizando-se o software
ImageJ (Rasband, 1997), e em seguida, emparelhados. Para a classificação
dos cromossomos foram medidas ambas as cromátides e determinado o
comprimento médio do braço maior (BM), do braço menor (Bm) e o
comprimento médio total (Ct) de cada cromossomo. A relação de braços
(RB) foi calculada dividindo-se o comprimento médio do braço maior pelo
comprimento médio do braço menor (RB=BM/Bm).
Os cromossomos homólogos foram emparelhados em ordem
decrescente de tamanho e separados em grupos de acordo com a
nomenclatura proposta por Levan et al. (1964) em metacêntricos (RB de 1,0
a 1,70), submetacêntricos (RB de 1,71 a 3,0), subtelocêntricos (RB de 3,01
a 7,0) e acrocêntricos (RB > 7,0). Para se determinar o número fundamental
(NF), que é o número de braços cromossômicos do cariótipo, foram
considerados os cromossomos metacêntricos, submetacêntricos e
subtelocêntricos como tendo dois braços e acrocêntricos como tendo um só
braço.

22
3. RESULTADOS 3.1. Ancistrus ranunculus
Esta espécie apresentou número modal de 2n=48 cromossomos e
número fundamental igual a 82, para ambos os sexos. Entretanto, a fórmula
cariotípica foi divergente entre machos e fêmeas, sendo
20M+8SM+6ST+14A e 19M+9SM+6ST+14A, respectivamente. Com base
no heteromorfismo do cariótipo feminino, sugere-se um mecanismo de
cromossomos sexuais do tipo ZZ/ZW, onde o cromossomo Z é representado
por um pequeno metacêntrico (10° par do complemento) e o W por um
submetacêntrico médio (Figura 3).
As RONs estão localizadas na região proximal dos braços longos do
16° par, sendo coincidentes com blocos de heterocromatina e portanto,
banda C positivas. Foi observado heteromorfismo no tamanho das RONs
entre os homólogos (Figura 3).
Foram observados blocos de heterocromatina conspícuos na região
distal dos pares 18°, 20°, 21° e 24°, na região proximal do 16° par e um
pequeno bloco pericentromérico no 9° par. Um padrão interessante foi
detectado no cromossomo W, que além de possuir pequenos blocos
pericentroméricos, também observados em Z, apresentou uma considerável
porção de heterocromatina no braço longo (Figuras 4A e B).

23
Figura 3. Cariótipo de macho e fêmea de Ancistrus ranunculus, com coloração
convencional em Giemsa, evidenciando (caixas) os cromossomos sexuais e
os portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.

24
Figura 4. A e B- Metáfases de Ancistrus ranunculus preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição da
heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positiva e os sexuais.

25
3.2. Ancistrus sp.1 “Purus”
Esta espécie apresentou número modal de 2n=34 cromossomos e
número fundamental igual a 68, para ambos os sexos. Entretanto, a fórmula
cariotípica foi divergente entre machos e fêmeas, sendo 21M+11SM+2ST e
20M+12SM+2ST, respectivamente. Com base no heteromorfismo do
cariótipo masculino, sugere-se um mecanismo de cromossomos sexuais do
tipo XX/XY, onde o cromossomo X é representado por um submetacêntrico
grande (11° par do complemento) e o Y por um metacêntrico pequeno
(Figura 5).
As RONs estão localizadas na região distal dos braços curtos do 4°
par (Figura 5), sendo coincidentes com blocos de heterocromatina e,
portanto, banda C positivas.
Detectaram-se blocos heterocromáticos pericentroméricos nos pares
1, 2 e 3, blocos distais nos braços curtos dos pares 4 e 12, um bloco
proximal nos braços curtos do par 8, e um pequeno bloco intersticial nos
braços curtos do 2° par (Figuras 6 A, B e C).

26
Figura 5. Cariótipo de macho e fêmea de Ancistrus sp.1 “Purus”, com coloração
convencional em Giemsa, evidenciando (caixas) os cromossomos sexuais e os
portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico.

27
Figura 6. A e B- Metáfases de Ancistrus sp.1 “Purus” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positivas.

28
3.3. Ancistrus sp.2 “Catalão”
Esta espécie apresentou número modal de 2n=34 cromossomos e
número fundamental igual a 68 para ambos os sexos. A fórmula cariotípica
foi 22M+8SM+4ST para machos e fêmeas. Contudo, foi detectado um
heteromorfismo no tamanho entre os homólogos do 5° par do complemento
no cariótipo masculino, evidenciando um mecanismo de cromossomos
sexuais do tipo XX/XY, sendo o cromossomo X representado por um
metacêntrico de tamanho médio (5° par do complemento) e o Y por um
pequeno metacêntrico (Figura 7).
As RONs estão localizadas na região distal dos braços curtos do 4°
par (Figura 7), sendo coincidentes com blocos de heterocromatina e,
portanto, banda C positivas.
Foram observados pequenos blocos pericentroméricos de
heterocromatina nos pares 1 e 3, um bloco proximal no par 9, blocos
intersticiais pequenos nos braços curtos dos pares 2, 6 e 7 e um bloco mais
conspícuo na região distal dos braços curtos do 4º par (Figuras 8A, B e C).

29
Figura 7. Cariótipo de macho e fêmea de Ancistrus sp.2 “Catalão”, com coloração
convencional em Giemsa, evidenciando (caixas) os cromossomos sexuais e os
portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico.

30
Figura 8. A e B- Metáfases de Ancistrus sp.2 “Catalão” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positivas.

31
3.4. Ancistrus sp.3 “Barcelos”
Esta espécie apresentou número modal de 2n=52 cromossomos e
número fundamental igual a 78 para machos e 77 para as fêmeas. A fórmula
cariotípica foi 12M+12SM+4ST+24A para machos e 11M+12SM+4ST+25A
para fêmeas. Com base no heteromorfismo do cariótipo feminino, sugere-se
um mecanismo de cromossomos sexuais do tipo Z1Z1Z2Z2/Z1Z2W1W2. O
cromossomo Z1 é um pequeno metacêntrico (5° par do complemento), o
cromossomo Z2 é o maior submetacêntrico do complemento (7° par), o W1 é
um acrocêntrico pequeno e o W2 é um submetacêntrico médio (Figura 9).
As RONs foram localizadas nos braços curtos do 23° par (Figura 9),
sendo coincidentes com blocos de heterocromatina e, portanto, banda C
positivas.
Pequenos blocos pericentroméricos de heterocromatina foram
observados nos pares 15, 16 e 17, blocos conspícuos foram localizados nos
braços curtos dos pares 14 e 23, e em posição intersticial nos braços longos
dos pares 1 e 11 e nos braços curtos do 2º par. Não foram encontrados
blocos de heterocromatina nos cromossomos sexuais. (Figuras 10A, B e C).

32
Figura 9. Cariótipo de macho e fêmea de Ancistrus sp.3 “Barcelos”, com coloração
convencional em Giemsa, evidenciando (caixas) cromossomos sexuais e os
portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.
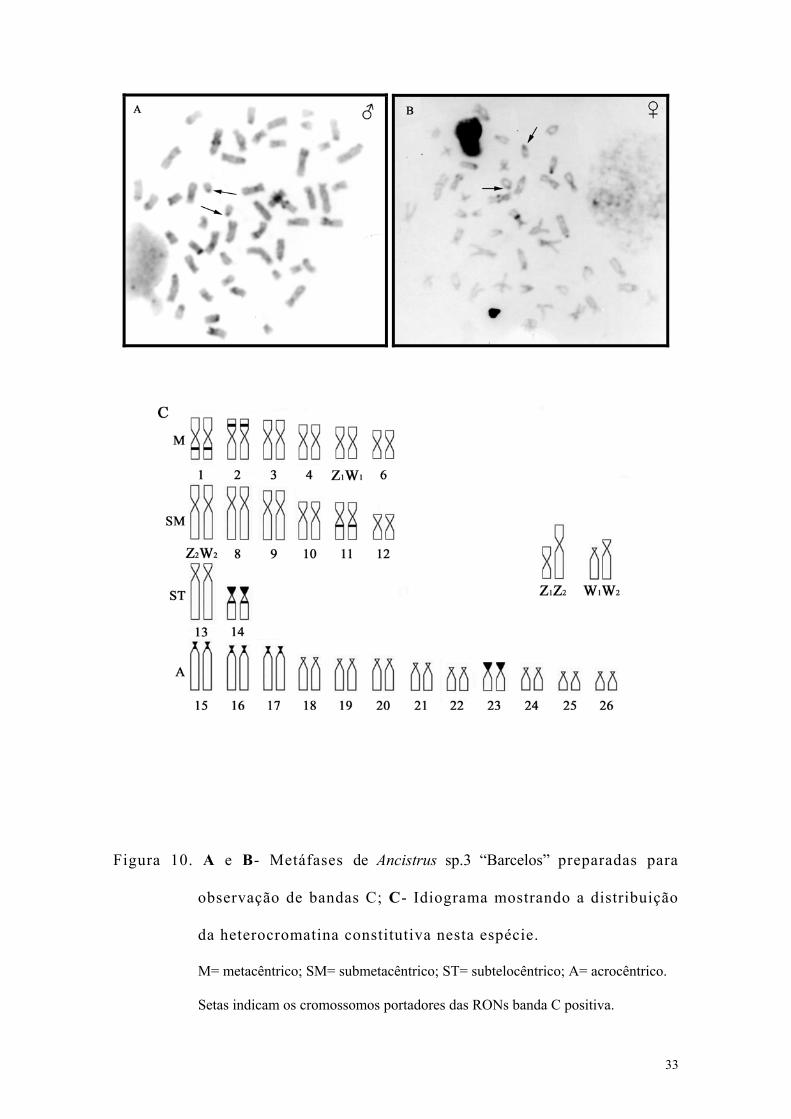
33
Figura 10. A e B- Metáfases de Ancistrus sp.3 “Barcelos” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positiva.

34
3.5. Ancistrus sp.4 “Vermelho”
Desta espécie foram analisados apenas machos e o número modal
observado foi de 2n=42 cromossomos, número fundamental igual a 78 e
fórmula cariotípica 26M+6SM+4ST+6A (Figura 11).
As RONs foram localizadas na região terminal dos braços longos do
20° par (Figura 11), sendo coincidentes com blocos de heterocromatina e,
portanto, banda C positivas.
Detectou-se blocos intersticiais de heterocromatina nos braços
curtos dos pares 2, 3, 6, 14 e 15 e nos braços longos dos pares 3, 4, 5 e 8.
Blocos distais foram localizados nos braços curtos do par 7 e nos braços
longos dos pares 14 e 20. Foram observados dois blocos proximais nos
pares 17 e 19 e dois pequenos blocos pericentroméricos nos pares 4 e 15
(Figuras 12A e B).

35
Figura 11. Cariótipo de macho de Ancistrus sp.4 “Vermelho”, com coloração
convencional em Giemsa, evidenciando (caixas) os cromossomos
portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.

36
Figura 12. A e B- Metáfases de Ancistrus sp.4 “Vermelho” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positivas.

37
3.6. Ancistrus sp.5 “Rio Branco”
Esta espécie apresentou número modal de 2n=46 cromossomos e
número fundamental igual 81 para machos e 82 para fêmeas. A fórmula
cariotípica foi 18M+11SM+6ST+11A para machos e 18M+12SM+6ST+10A
para fêmeas. Com base no heteromorfismo do cariótipo masculino, sugere-
se um mecanismo de cromossomos sexuais do tipo XX/XY. Os
cromossomos X são pequenos submetacêntricos (15° par do complemento) e
o cromossomo Y é um acrocêntrico médio (Figura 13).
As análises da meiose evidenciaram no paquíteno a presença de
cromossomos sexuais com heteropicnose positiva. No diplóteno e diacinese
foram visualizados 22 bivalentes autossômicos e o bivalente sexual com
heteropicnose negativa. Na metáfase I os bivalentes mostraram-se dispostos
no equador da célula, com o bivalente sexual com heteropicnose negativa
(Figura 15A-E).
As RONs foram localizadas na região distal dos braços curtos do
19° par (Figura 13), sendo coincidentes com blocos de heterocromatina e,
portanto, banda C positivas.
Foram observados blocos intersticiais de heterocromatina nos
braços longos dos pares 2, 19 e 21. Blocos distais conspícuos localizaram-
se nos braços curtos dos pares 17 e 19 e nos braços longos do 20º par, e um
pequeno bloco distal foi localizado nos braços curtos do 2º par. Um bloco
pericentromérico foi observado no 20º par (Figuras 14A-C).

38
Figura 13. Cariótipo de macho e fêmea de Ancistrus sp.5 “Rio Branco”, com coloração
convencional em Giemsa, evidenciando (caixa) os cromossomos sexuais e os
portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.

39
Figura 14. A e B- Metáfases de Ancistrus sp.5 “Rio Branco” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positivas.

40
Figura 15. Células meióticas de Ancistrus sp.5 “Rio Branco” observadas em
coloração convencional: A- paquíteno evidenciando cromossomos
sexuais com heteropicnose positiva (seta); B e C- célula
diplotênica evidenciando 22 bivalentes autossômicos e o
bivalente sexual com heteropicnose negativa (seta); D- diacinese
22 bivalentes + sexuais em heteropicnose negativa (seta); E-
metáfase I evidenciando os bivalentes dispostos na placa
equatorial e o sexual com heteropicnose negativa (seta).

41
3.7. Ancistrus sp. 6 “Piagaçú”
Esta espécie apresentou número modal de 2n= 52 cromossomos e número
fundamental igual a 78 para os machos e 79 para as fêmeas. A fórmula cariotípica foi
16M+8SM+2ST+26A e 16M+9SM+2ST+25A, para machos e fêmeas, respectivamente.
Com base no heteromorfismo do cariótipo feminino, sugere-se um mecanismo de
cromossomos sexuais do tipo ZZ/ZW, sendo os cromossomos Z representados por
acrocêntricos grandes (16° par do complemento) e o W por um submetacêntrico
pequeno (Figura 16).
As RONs foram localizadas nos braços curtos do 26° par (Figura 16), sendo
coincidentes com grandes blocos de heterocromatina e, portanto, banda C positivas.
Foram observados pequenos blocos pericentroméricos de heterocromatina nos
pares 1, 9, 14, 15, 17, 22, 23, 24 e 25, blocos intersticiais nos braços longos dos pares 2
e 4 e nos braços curtos do par 2, um bloco distal pequeno nos braços curtos do 2° par e
um bloco distal bem conspícuo nos braços curtos do 26° par (Figuras 17A-C).

42
Figura 16. Cariótipo de macho e fêmea de Ancistrus sp.6 “Piagaçu”, com coloração
convencional em Giemsa, evidenciando (caixa) os cromossomos sexuais e os
portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.

43
Figura 17. A e B- Metáfases de Ancistrus sp.6 “Piagaçu” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positiva.

44
3.8. Ancistrus sp.7 “Dimona”
Esta espécie apresentou número modal de 2n=52 cromossomos e número
fundamental igual a 78. A fórmula cariotípica foi 16M+8SM+2ST+26A, para machos e
fêmeas. Não foram encontradas diferenças cromossômicas relacionadas ao sexo. (Figura
18).
As RONs foram localizadas na região intersticial dos braços longos do 13° par
(Figura 18), sendo coincidentes com blocos de heterocromatina e, portanto, banda C
positivas.
Foram observados blocos de heterocromatina intersticial nos braços longos dos
pares 1 e 13 e blocos pericentroméricos nos pares 2, 4 e 21 e em um dos homólogos dos
pares 1 e 13 (Figuras 19A-C).
45
Figura 18. Cariótipo de macho e fêmea de Ancistrus sp.7 “Dimona”, com coloração
convencional em Giemsa, evidenciando (caixa) os cromossomos portadores
de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.

46
Figura 19. A e B- Metáfases de Ancistrus sp.7 “Dimona” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positiva.

47
3.9. Ancistrus sp.8 “Balbina”
Esta espécie apresentou número modal de 2n=39 cromossomos, número
fundamental igual a 78 e fórmula cariotípica 27M+10SM+2ST para machos e número
modal de 2n=38 cromossomos, número fundamental igual a 76 e fórmula cariotípica
26M+10SM+2ST para fêmeas. Com base na diferença numérica encontrada no
cariótipo masculino, em relação ao feminino, sugere-se um mecanismo de cromossomos
sexuais do tipo XX/XY1Y2, sendo os cromossomos X representados por metacêntricos
grandes (2° par do complemento) e os cromossomos Y por pequenos metacêntricos (14°
par do complemento masculino) (Figura 20).
As RONs foram localizadas na região terminal dos braços longos do 12° par
(Figura 20), sendo coincidentes com blocos de heterocromatina e, portanto, banda C
positivas.
Foram observados blocos intersticiais de heterocromatina nos braços curtos
dos pares 3, 4 e 14 e nos braços longos do par 6, blocos pericentroméricos nos pares 8 e
9, blocos distais nos braços longos do par 12 e blocos proximais nos braços longos do
19° par (Figuras 21A-C).

48
Figura 20. Cariótipo de macho e fêmea de Ancistrus sp.8 “Balbina”, com coloração
convencional em Giemsa, evidenciando (caixa) os cromossomos sexuais e os
portadores de RONs impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico.

49
Figura 21. A e B- Metáfases de Ancistrus sp.8 “Balbina” preparadas para
observação de bandas C; C- Idiograma mostrando a distribuição
da heterocromatina constitutiva nesta espécie.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico.
Setas indicam os cromossomos portadores das RONs banda C positiva.

50
3.10. Ancistrus sp.9 “Trombetas”
Desta espécie foi analisado apenas um macho e o número modal observado foi
de 2n=38 cromossomos, número fundamental igual a 73 e fórmula cariotípica
22M+8SM+5ST+3A. Não foi possível emparelhar dois cromossomos do complemento,
o que sugere um mecanismo de cromossomos sexuais do tipo XX/XY. No entanto, este
heteromorfismo só poderá ser confirmado quando for analisado o cariótipo feminino
(Figura 22).
As RONs foram localizadas na região pericentromérica do 5° par (Figura 22).

51
Figura 22. Cariótipo de macho de Ancistrus sp.9 “Trombetas”, com coloração
convencional em Giemsa, evidenciando (caixa) os portadores de RONs
impregnadas com nitrato de prata.
M= metacêntrico; SM= submetacêntrico; ST= subtelocêntrico; A= acrocêntrico.

52
Tabela 3. Síntese das características cariotípicas das dez espécies de Ancistrus analisadas no presente estudo. (2n=número diplóide; NF=número fundamental; M=metacêntrico; SM=submetacêntrico; ST=subtelocêntrico; A=acrocêntrico; RON=regiões organizadora de nucléolo; p=braço curto; q=braço longo; ?=provável XX/XY).
Espécie Nº de individuos 2n NF Localização da
RON Formula Cariotípica Mecanismo Par sexual Local de coleta
♂ ♀
A. ranunculus 3 2 48 82 16ºSTq, intersticial
♂ 20M+8SM+6ST+14A ♀ 19M+9SM+6ST+14A ZZ/ZW 10ºM Rio Xingu/ PA
Ancistrus sp 1 5 7 34 68 4ºMp, terminal ♂ 21M+11SM+2ST ♂ 20M+12SM+2ST XX/XY 11ºSM Rio Purus/ AM
Ancistrus sp 2 6 6 34 68 4ºMp, terminal ♂ 21M+9SM+4ST ♀ 22M+8SM+4ST XX/XY 5ºM Lago Catalão/ AM
Ancistrus sp3 7 4 52 ♂ 78♀ 77 23ºAp, terminal ♂ 12M+12SM+2ST+26A
♀ 11M+11SM+3ST+27A Z1Z1Z2Z2/ Z1Z2W1W2
5ºM, 7ºSM Rio Demeni/ AM
Ancistrus sp 4 5 - 42 ♂ 78 20ºAq, terminal ♂ 26M+6SM+4ST+6A -------- ---------- Rio Demeni/ AM
Ancistrus n. sp 5 5 6 46 ♂ 81♀ 82 19ºAp, terminal ♂ 18M+11SM+6ST+11A
♀ 18M+12SM+6ST+10A XX/XY 15ºSM Rio Branco/ RR
Ancistrus sp 6 3 4 52 ♂ 78♀ 79 26ºAp, terminal ♂ 14M+10SM+2ST+26A
♀ 14M+11SM+2ST+25A ZZ/ZW 16ºA Lago Aiapuá/ AM
Ancistrus sp 7 6 2 52 78 13ºSTq, intersticial
♂ 16M+8SM+2ST+26A ♀ 16M+8SM+2ST+26A -------- ---------- Igarapé Dimona/ AM
Ancistrus sp 8 6 6 ♂ 39♀ 38
♂ 78♀ 76 12ºMq, terminal ♂ 27M+10SM+2ST
♀ 26M+10SM+2ST XX/XY1Y2 2ºM Igarapé Barretinho/ AM
Ancistrus sp 9 1 - 38 ♂ 73 5°M, pericentromérica ♂ 22M+8SM+5ST+3A ? 17°ST Rio Trombetas/ PA

53
4. DISCUSSÃO
4.1. Diversidade cariotípica e mecanismos de evolução cromossômica no gênero
Ancistrus
Estudos citogenéticos conduzidos na família Loricariidae indicam que o
número diplóide de 54 cromossomos, predominantemente dos tipos meta e
submetacêntrico, representa a condição plesiomórfica para este grupo de peixes (Artoni,
1996; Artoni & Bertollo, 2001; Kavalco et al., 2005). Em Hypoptopomatinae, salvo
Otocinclus aff. vestitus (Andreata et al., 1994) com 2n= 72, todas as espécies estudadas
apresentam 2n=54 cromossomos. Em Loricariinae, o número diplóide basal é
encontrado nas espécies do gênero Loricariichthys (Scavone & Julio Jr., 1995). Em
Neoplecostominae, todas as espécies estudadas possuem número diplóide de 54
cromossomos (Alves et al., 2005; Kavalco et al., 2005). Em Hypostominae, esta
característica também foi observada em Hypostomus plecostomus (Muramoto et al.,
1968), Rhinelepis aspera e Pogonopoma wertheimeri (Artoni & Bertollo, 2001). A
hipótese de que o número diplóide de 54 cromossomos representa uma condição basal
entre os loricariídeos é corroborada pela ocorrência deste número diplóide em grupos
próximos a Loricariidae, tais como, Tricomycteridae (Torres et al., 1998; Sato et al.,
2004) e Callichthyidae (Porto & Feldberg, 1993).
Embora os dados citogenéticos disponíveis para Ancistrini ainda sejam
escassos em vista da quantidade de espécies que compõe esta tribo, há indícios de que
este grupo apresente como número diplóide ancestral 52 cromossomos (Artoni &
Bertollo, 2001; Alves et al., 2003), principalmente dos tipos meta e submetacêntricos
(Tabela 1), uma sinapomorfia em relação aos outros grupos de Loricariidae. Dentre os
gêneros pertencentes à Ancistrini com número diplóide conhecido, somente Ancistrus

54
apresenta registros de números diplóides diferentes do considerado ancestral, possuindo
uma grande diversidade cariotípica, onde o número diplóide varia de 34 a 52
cromossomos (incluindo dados do presente estudo). Isto demonstra que intensos
processos de rearranjos cromossômicos podem estar associados à evolução cariotípica
de Ancistrus, os quais indicam uma tendência evolutiva para a redução do número
diplóide neste táxon.
Alves et al. (2003) acreditam que rearranjos cromossômicos do tipo fusões
cêntricas tenha predominado na evolução cariotípica de Ancistrus. Deste modo, a
presença de 2n=34 cromossomos em Ancistrus sp.1 “Purus” e Ancistrus sp.2 “Catalão”,
menor número diplóide já registrado para Ancistrini e para Loricariidae, corroboram
esta hipótese. Contudo, inversões pericêntricas e translocações também devem ter
ocorrido na diversificação cariotípica deste gênero. Tais mecanismos explicariam a
presença de cromossomos acrocêntricos, principalmente nas espécies que apresentam
número diplóide igual a 52 cromossomos, onde a maior parte do complemento é
constituída por cromossomos deste tipo. Além disso, tais mecanismos também
explicariam a grande diversidade no número fundamental, que variou de 68 em
Ancistrus sp.1 “Purus” e Ancistrus sp.2 “Catalão a 82 em Ancistrus ranunculus e
Ancistrus sp.5 ”Rio Branco” (Tabelas 1 e 3).
Eventos de fusões cêntricas não são ocorrências comuns na evolução
cariotípica de Loricariidae, sendo, até o momento, registrados apenas na subfamília
Loricariinae, em populações de Rineloricaria latirostris com 2n variando de 36 a 48
cromossomos (Giuliano-Caetano, 1998). Para outros loricariídeos (e.g.,
Hypoptopomatinae e Neoplecostominae), a evolução por meio de inversões
pericêntricas e paracêntricas parece ser a explicação mais aceitável para a manutenção
do número diplóide constante. Em Upsilodinae e em alguns grupos de Hypostominae,

55
(principalmente no gênero Hypostomus), o aumento do número diplóide e de
cromossomos do tipo acrocêntrico indica que eventos de fissões cêntricas foram os
mecanismos mais importantes na evolução cromossômica destes grupos (Artoni &
Bertollo, 1996; Kavalco et al., 2005). Em espécies de Ancistrini, a manutenção do
número diplóide de 52 cromossomos parece ter ocorrido por meio de rearranjos não
Robertsonianos, principalmente inversões pericêntricas e paracêntricas. Assim, pode-se
sugerir que as espécies do gênero Ancistrus apresentam uma evolução cariotípica
divergente, e que contrasta tanto com a evolução conservativa das demais espécies de
Ancistrini, quanto com os outros mecanismos encontrados nos demais grupos de
Loricariidae.
Artoni (1996) propôs que sítios ribossomais em posição terminal no braço
longo de um par de cromossomos metacêntricos grandes seja o fenótipo de regiões
organizadoras de nucléolo ancestral para Loricariidae. Entretanto, fenótipos variados de
RONs são encontrados com freqüência entre as espécies desta família, sendo essa
divergência observada tanto no tamanho dos cromossomos nucleolares, como na
posição destes sítios (Artoni & Bertollo, 1996; Mariotto et al., 2004; Alves et al., 2005;
Kavalco et al., 2005).
As RONs em Ancistrus apresentaram uma grande diversidade, tanto em relação
à sua localização no cromossomo quanto à sua posição no cariótipo. Entretanto, uma
grande similaridade foi observada entre Ancistrus sp.1 “Purus” e Ancistrus sp.2
“Catalão”, que apresentaram sítios ribossomais na região distal do braço curto do 4º par
do complemento, e entre Ancistrus sp.9 “Trombetas” e Ancistrus n. sp.1 (Alves et al.,
2003), que apresentaram as RONs em posições semelhantes no 5º par do complemento.
É possível que estas similaridades sejam homologias e que ocorram devido à

56
conservação da macroestrutura cariotípica numérica e estrutural, em espécies com
fenótipos de RONs semelhantes.
Similaridades menores foram observadas entre as espécies Ancistrus sp.3
“Barcelos”, Ancistrus sp.5 “Rio Branco” e Ancistrus sp.6 “Piagaçu”, onde as RONs
ocupam posição terminal no braço curto de cromossomos acrocêntricos coincidentes
com as constrições secundárias; contudo, os cromossomos portadores dos sítios
ribossomais são muito diferentes quanto ao tamanho. Ancistrus ranunculus e Ancistrus
sp.7 “Dimona” também possuem os sítios ribossomais em posições semelhantes em
cromossomos do tipo subtelocêntrico. Contudo, estas semelhanças não devem
representar a conservação desse caráter, mas sim eventos ocasionais gerados pelos
intensos rearranjos cromossômicos que ocorreram na evolução cariotípica destas
espécies, envolvendo também os organizadores nucleolares. Entretanto, seria necessário
comparar tais resultados com uma proposta de arranjo filogenético para as espécies do
gênero, de forma a testar estas hipóteses.
Segundo Almeida-Toledo (1998) e Feldberg et al. (1999; 2003), o número, a
posição e a morfologia das RONs permite fazer inferências sobre suas tendências
evolutivas para alguns grupos de peixes. Além de ser uma importante ferramenta para
distinguir espécies. Contudo, pode-se dizer que, embora permita identificar cada espécie
a RON não é um bom marcador para ser usado na construção de inferências
filogenéticas para espécies de Ancistrus, que mostram grande diversidade em relação a
esse caráter, tornando difícil a detecção de um padrão e consequentemente a sua
polarização.
Embora não observado em todas as células analisadas, um heteromorfismo no
tamanho das RONs foi registrado entre os homólogos de quase todas as espécies
analisadas no presente estudo, com exceção de Ancistrus sp.1 “Purus”, Ancistrus sp.6

57
“Piagaçu” e Ancistrus sp.8 “Balbina”. Esta diferença pode ser resultado da duplicação
gênica “em tandem” das seqüências de DNAr em um dos homólogos (Gold, 1984).
Outra explicação é que a presença de heterocromatina constitutiva entre os genes
ribossomais pode possibilitar permutas desiguais entre as cromátides, causando este
polimorfismo em relação ao tamanho (Sola et al., 1988), tal situação pode ter ocorrido
entre as espécies analisadas no presente trabalho, já que todas apresentaram RONs
positivas para banda C.
A relação entre a heterocromatina constitutiva e a RON é muito discutida e
muitas vezes gerou opiniões controversas. Alguns pesquisadores sugerem que a
heterocromatina teria a função de proteger o DNAr, não permitindo que mutações ou
permutas alterassem os cístrons ribossomais 18S e 28S (King, 1980; Schmid, 1982). Por
outro lado, há os que admitem que a presença da heterocromatina adjacente ou
coincidente com as RONs facilite a ocorrência de rearranjos envolvendo essas regiões,
propiciando uma diversificação cariotípica deste caráter (Moreira-Filho et al., 1984).
Análises da heterocromatina constitutiva são ainda escassas entre os
Siluriformes, apesar do fato de sua distribuição e composição serem importantes para a
compreensão da evolução cariotípica deste grupo (Kavalco et al., 2004). Segundo Gold
et al. (1990), uma pequena quantidade de heterocromatina constituindo o genoma é o
padrão observado para muitos teleósteos, incluindo os Siluriformes. Nas espécies de
Ancistrus nem todos os cromossomos apresentaram heterocromatina constitutiva, a qual
foi distribuída em blocos discretos dispostos em posições centromérica, telomérica ou
intersticial e que muitas vezes apresentaram-se pálidos. Os blocos mais conspícuos
foram aqueles em associação com as RONs.
No gênero Hypostomus, Artoni & Bertollo (1999) observaram dois padrões de
distribuição da heterocromatina constitutiva. Um deles apresenta pouca heterocromatina

58
localizada nas regiões teloméricas e/ou centroméricas; enquanto o outro tem um número
maior dessas regiões, distribuídas também em posição intersticial de diversos
cromossomos acrocêntricos. Esses autores associaram tais padrões com o número
diplóide das espécies, sendo o primeiro padrão presente em espécies com número
diplóide baixo, e o segundo naquelas com número diplóide alto. Assim, sugeriu-se a
existência de um relacionamento entre número diplóide e conteúdo de heterocromatina,
e que mudanças na heterocromatina poderiam ocorrer paralelamente às fissões
cêntricas, provavelmente devido ao aumento do DNA repetitivo.
O gênero Ancistrus apresentou uma grande diversidade nos padrões de
heterocromatina, tanto na distribuição como no tamanho dos blocos, sendo, portanto,
distinto para cada espécie, e constituindo um importante marcador citogenético para
este grupo. Uma possível homologia foi a presença de um bloco intersticial no braço
curto de um par de cromossomos metacêntricos de tamanho médio, que pode estar
localizado do segundo ao quarto par do complemento, em seis das 10 espécies
estudadas (Figuras 6,8,10,12,17 e 21). Entretanto, como ocorre com o marcador RON, o
padrão de heterocromatina também não permite inferir sobre as tendências evolutivas
neste grupo.
A espécie Ancistrus sp.7 “Dimona” manteve o padrão heterocromático
observado nas demais espécies analisadas no presente estudo, com poucos blocos de
heterocromatina distribuídos no complemento cromossômico. No entanto, foi verificada
a presença de um bloco pericentromérico de tamanho médio, localizado em somente um
dos homólogos do primeiro par metacêntrico e primeiro par subtelocêntrico (Figura 19).
A explicação para este fato é que tenha ocorrido um rearranjo do tipo translocação entre
estes cromossomos, com a migração do bloco de heterocromatina do homólogo

59
metacêntrico para o subtelocêntrico, uma vez que blocos pericentroméricos no primeiro
par metacêntrico foram observados em algumas espécies deste trabalho.
No gênero Pimelodus (Siluriformes: Pimelodidae), as espécies analisadas
apresentam diferentes números diplóides, mas com cariótipos bem semelhantes,
constituídos de um mesmo número de cromossomos metacêntricos de tamanhos
similares e RONs localizadas em posição terminal no braço longo do 20º par do
complemento cromossômico (Garcia & Moreira-Filho, 2005). Tal conservação não foi
observada entre as espécies que compõem o gênero Ancistrus. Apesar de terem sido
detectados sistemas de RONs simples em todas as espécies, não foi observado fenótipo
semelhante ao considerado ancestral. A grande diversidade cariotípica dificultou a
determinação de padrões gerais na evolução da localização das RONs e distribuição da
heterocromatina constitutiva.
Entretanto, é importante ressaltar que ainda se faz necessário ampliar a
investigação citogenética para mais espécies de Ancistrus, associada a investigações
morfológica, ecológica e molecular bem como a comparação com propostas
filogenéticas para esse grupo, para que se possa elucidar os caminhos evolutivos
trilhados por esses peixes.
4.1.1. Mecanismos de cromossomos sexuais
De acordo com Centofante et al. (2002), 55 ocorrências de cromossomos
sexuais heteromórficos haviam sido registradas para a ictiofauna Neotropical, das quais
64% referiam-se a machos heterogaméticos e 36% a fêmeas heterogaméticas. Embora o
número de espécies de peixes Neotropicais com registro citológico de cromossomos
sexuais diferenciados seja pequeno, vários casos de diferenciação têm sido observados
desde as primeiras descrições em peixes Characiformes dos gêneros Hoplias (Bertollo

60
et al., 1978), Apareiodon (Moreira-Filho et al., 1980) e Leporinus (Galetti Jr. et al.,
1981) e em Gymnotiformes do gênero Eigenmannia (Almeida-Toledo, 1978).
Segundo Moreira-Filho et al. (1993), cinco sistemas de cromossomos sexuais
já foram descritos em peixes Neotropicais: ZZ/ZW, XX/XY, X1X1X2X2/X1X2Y,
XX/XY1Y2 e ZZ/ZW1W2. Em 2001, Valentim observou mais um sistema, sendo este do
tipo XX/XO em Potamotrygon sp. (Chondrichthyes: Rajiformes: Potamotrygonidae).
Na família Loricariidae, a presença de sistemas de cromossomos sexuais tem
sido considerada uma situação atípica, visto que a grande maioria das espécies não
possui tais mecanismos (Artoni & Bertollo, 2001; Alves et al., 2003; Alves et al., 2005;
Kavalco et al., 2005). Entretanto, diferentes sistemas já foram encontrados em algumas
espécies. Estes são mecanismos de cromossomos sexuais simples dos tipos ZZ/ZW e
XX/XY, observados em Hypostomus macrops (Michelle et al., 1977), Hypostomus sp.
G (Artoni et al, 1998), Pseudocinclus tietensis (Andreata et al., 1992),
Microlepidogaster leucofrenatus (Andreata et al., 1993; 1994), Loricariichthys
platymetopom (Scavone & Júlio Jr., 1995) e Hemiancistrus spilomma (de Oliveira et al.,
no prelo). No presente estudo, sete das 10 espécies apresentaram cromossomos sexuais
que somado às duas ocorrências de heteromorfismo sexual já descritas para este grupo,
faz um total de 56% das espécies analisadas, um número alto se comparado com as
demais subfamílias de Loricariidae.
No gênero Ancistrus, sistema simples de determinação do sexo do tipo ZZ/ZW
foi observado em três espécies. Em Ancistrus cf. dubius do Pantanal de Mato Grosso
(Mariotto et al., 2004), onde o cromossomo Z é facilmente identificado por um grande
bloco de heterocromatina no braço longo, o W é o menor cromossomo acrocêntrico do
complemento e não possui blocos heterocromáticos. Em Ancistrus ranunculus (presente
trabalho) o cromossomo Z é um metacêntrico pequeno e possui um pequeno bloco de

61
heterocromatina em posição centromérica, enquanto o W é um submetacêntrico médio,
quase todo heterocromático (Figura 3 e 4). Entretanto, Souza (2003) não detectou
cromossomos sexuais nesta espécie (Tabela 1). Já, em Ancistrus sp.6 “Piagaçu”
(presente trabalho) o cromossomo W é um submetacêntrico pequeno, enquanto o
cromossomo Z é um acrocêntrico grande. No entanto, nesta espécie não foi observado
nenhum bloco de heterocromatina nos cromossomos sexuais (Figura 16 e 17).
Mecanismo do tipo XX/XY havia sido descrito para uma população de
Ancistrus cf. dubius do pantanal de Mato Grosso (Mariotto, 2003). No presente
trabalho, três novos registros são apresentados: Ancistrus sp.1 “Purus”, Ancistrus sp.2
“Catalão” e Ancistrus sp.5 “Rio Branco”. Nestas espécies, os cromossomos sexuais são
bem caracterizados morfologicamente, não deixando dúvidas quanto ao
heteromorfismo, embora não apresentem nenhum bloco de heterocromatina (Figuras 6,
8 e 14, respectivamente).
No mecanismo XX/XY, como no ZZ/ZW, também foi observado uma variação
no tamanho dos cromossomos sexuais: em Ancistrus sp.1 “Purus” e Ancistrus sp.2
“Catalão” o cromossomo X é muito maior do que o Y, em Ancistrus sp.5 “Rio Branco”
o cromossomo X é um submetacêntrico pequeno, ao passo que o Y é um acrocêntrico de
tamanho médio e maior que o X. Segundo (Galetti & Foresti, 1986), no gênero
Leporinus os cromossomos Z e W apresentam forma e tamanho similar em todas as
espécies em que ocorrem, o que pode indicar que o sistema ZZ/ZW provavelmente
envolva o mesmo par cromossômico nas diferentes espécies. Por outro lado, no gênero
Ancistrus, a evolução dos cromossomos sexuais deve ter ocorrido como eventos
independentes envolvendo diferentes pares de cromossomos.
Dois caminhos básicos têm sido postulados para a origem e desenvolvimento
dos cromossomos sexuais em vertebrados. No primeiro, a diferenciação destes

62
cromossomos estaria relacionada a rearranjos estruturais, resultando em um processo de
prevenção das recombinações meióticas (Beçak & Beçak, 1969). No segundo caso, a
diferenciação entre os cromossomos sexuais seria a partir de um processo de
heterocromatinização (Singh et al., 1980).
Em peixes Neotropicais, a hipótese mais aceita para explicar a diferenciação de
cromossomos sexuais é o acúmulo de heterocromatina constitutiva, seguido de outros
rearranjos estruturais (e.g., Galetti & Foresti, 1986; Almeida-Toledo & Foresti, 2001;
Centofante et al., 2002; Venere et al., 2004). Contudo, no gênero Ancistrus o processo
de heterocromatinização está envolvido somente na diferenciação sexual de A.
ranunculus (presente trabalho) e de Ancistrus cf. dubius (Mariotto et al., 2004). Assim,
pode-se sugerir que rearranjos estruturais tenha sido a primeira etapa na diferenciação
dos cromossomos sexuais de várias espécies deste gênero. Esta hipótese é corroborada
pela ausência de heterocromatina constitutiva nos cromossomos sexuais de seis das oito
espécies que apresentam mecanismos de diferenciação, bem como pela ocorrência de
numerosos rearranjos cromossômicos na história evolutiva deste grupo. Cromossomos
sexuais originados a partir de rearranjos estruturais em peixes já foram registrados em
Eigenmania sp. (Almeida-Toledo et al., 1984) e em algumas populações de Hoplias
malabaricus (Bertollo et al., 1997) e é fortemente postulado como principal evento na
formação de sistemas múltiplos (Moreira-Filho et al., 1980)
Segundo Ohno (1967), o primeiro passo para a diferenciação dos cromossomos
sexuais é o aparecimento de um mecanismo que impeça o crossing-over entre os
homólogos. Contudo, a evolução de cromossomos sexuais heteromórficos, a partir de
um par originalmente homomórfico, não seria o resultado de uma única mudança
estrutural, mas do desenvolvimento de vários passos seqüenciais. Desta forma, apesar

63
da heterocromatinização dos cromossomos sexuais ser um evento raro em Ancistrus,
isso não descarta a possibilidade de que tenha ocorrido nas espécies deste gênero.
No presente trabalho, também foram evidenciados dois tipos de sistemas
múltiplos de cromossomos sexuais. Em Ancistrus sp.8 “Balbina” foi observado o
mecanismo do tipo XX/XY1Y2, (Figura 20) já descrito em Hoplias malabaricus
(Bertollo & Mestriner, 1998). Moreira-Filho et al. (1980) observaram em Apareiodon
affinis (Characiformes: Parodontidae) um mecanismo do tipo ZZ/ZW1W2. Embora,
neste sistema as fêmeas é que constituem o sexo heterogamético, a explicação para a
origem deste sistema, seria análoga para o mecanismo observado em Ancistrus sp.8
“Balbina”. Acredita-se que um processo de fissão cêntrica, envolvendo um dos
cromossomos metacêntricos do segundo par do cariótipo masculino, tenha originado um
par de acrocêntricos, que após sofrerem outros rearranjos cromossômicos (como por
exemplo, inversões pericêntricas), originaram um par de pequenos metacêntricos, o que
explicaria a ocorrência de um cromossomo adicional nos machos desta espécie.
O segundo mecanismo de cromossomos sexuais múltiplos proposto é do tipo
Z1Z1Z2Z2/Z1Z2W1W2 observado em Ancistrus sp.3 “Barcelos” e constitui o primeiro
registro de um sistema deste tipo em peixes (Figura 9). Mecanismo semelhante somente
havia sido registrado para uma espécie de pulga (Nosopsyllus fasciatus; Aphaniptera),
contudo o sexo heterogamético era o masculino (Bayreuther, 1969).
De acordo com Guerra (1988), os sistemas de cromossomos sexuais múltiplos
se originam a partir de sistemas simples pré-existentes no grupo. Tais sistemas
múltiplos surgem, principalmente, como resultado de rearranjos do tipo translocação
recíproca (troca de segmentos cromossômicos entre cromossomos não-homólogos)
envolvendo cromossomos sexuais e autossomos. A ausência do processo de

64
heterocromatinização tanto na diferenciação dos sistemas simples como nos sistemas
múltiplos nas espécies de Ancistrus, parece corroborar esta afirmativa.
Em Ancistrus sp.5 “Rio Branco”, a presença dos cromossomos sexuais também
foi evidenciada pelas análises meióticas onde os cromossomos sexuais apresentaram-se
heteropicnóticos (Figura 15). Essa condensação precoce do par sexual evita que durante
o crossing-over, este faça trocas genéticas com os pares heteromórficos (John, 1990). A
análise meiótica pode ser uma prática importante para detecção de cromossomos
sexuais em Ancistrus, uma vez que a ausência de heterocromatina no par sexual, na
maioria das espécies, impossibilita o uso da técnica de bandamento C como ferramenta
para melhor caracterização desses mecanismos.
Cromossomos sexuais diferenciados apresentam uma distribuição muito
peculiar entre os peixes. O mesmo sistema pode ser encontrado em todas as espécies de
um grupo, como ZZ/ZW em Triportheus (Falcão, 1988; Bertollo & Cavallaro, 1992) ou
em grande parte de um grupo, como ZZ/ZW em Leporinus (Galetti & Foresti, 1986;
Venere et al., 2004). Entretanto, um mesmo sistema pode ser encontrado entre famílias
e gêneros diferentes, geralmente sem relações filogenéticas próximas. Sempre que há
espécies com sistemas de cromossomos sexuais diferenciados, frequentemente há outras
proximamente relacionadas com cromossomos homomórficos. Assim, o surgimento de
cromossomos sexuais parece ocorrer diversas vezes e de forma independente entre os
peixes (Solari, 1994). Diante disto, a presença ou ausência de mecanismo de
cromossomos sexuais em Ancistrus não pode ser interpretada, a priori, como uma
proximidade filogenética entre as espécies.
Segundo Almeida-Toledo & Foresti (2001), grupos de peixes com diferentes
sistemas de cromossomos sexuais, tais como: machos e fêmeas heterogaméticos,
sistemas múltiplos e simples, em espécies do mesmo gênero ou diferentes populações

65
de uma mesma espécie, tal como, em Hoplias malabaricus e Eigenmannia virescens,
caracterizam-se por estágios recentes ou por estratégias particulares de diferenciação
dos cromossomos sexuais. A presença de diferentes sistemas de cromossomos sexuais,
inclusive em populações de uma mesma espécie (Ancistrus cf. dubius, Mariotto, 2003),
pode estar indicando que Ancistrus também represente um grupo cuja diferenciação dos
cromossomos sexuais é ainda evolutivamente recente.
4.2. Relação entre a diversidade cariotípica e características da biologia e etologia
dos peixes do gênero Ancistrus
A enorme diversidade de espécies observadas entre os peixes indica que este
grupo provavelmente experimenta diferentes caminhos de evolução cromossômica. Há
grupos que mantiveram uma macroestrutura cariotípica muito similar, como no caso dos
Curimatidae (Venere & Galetti Jr., 1989; Feldbeg et al., 1992; 1993), Anostomidae
(Galetti Jr. & Foresti, 1986; 1987; Venere et al., 2004), Parodontidae (Moreira-Filho et
al., 1980; 1985; 1993; Centofante et al., 2002), ao passo que outros grupos apresentam
uma extensa diversidade cromossômica, tais como: Erythrinidae (Bertollo et al., 1978;
1997; 2004 ) Characidae (Falcão & Bertollo, 1985; Nakayama et al., 2002; Moreira-
Filho & Bertollo, 1991) Callichthyidae (Oliveira et al., 1993; Porto & Feldberg, 1993).
De acordo com King (1985), a conservação do cariótipo, isto é, a ausência de
uma associação entre mudanças cromossômicas e especiação em alguns organismos,
pode estar relacionada à impossibilidade de se formar populações isoladas, como
espécies migratórias ou altamente móveis. Em contraste, grupos com distribuição mais
limitada e exposta à grande heterogeneidade ambiental, podem apresentar diversidade
cromossômica mais extensa, incluindo diferenciação interpopulacional, ou seja,

66
pequenas populações facilitariam uma intensificação dos efeitos das trocas gênicas e a
fixação de rearranjos (Galleti Jr. et al., 2000).
A grande diversidade cariotípica encontrada entre as espécies do gênero
Ancistrus, como observado no presente estudo, pode estar relacionada às características
da biologia e etologia desses peixes, os quais apresentam preferências quanto ao uso de
microhábitats, permanecendo escondidos em troncos, fendas de pedras e outros abrigos,
onde estabelecem territórios e possuem pouca vagilidade (Power, 1984; 1990; Buck &
Sazima, 1995). Em relação à reprodução, possuem características muito peculiares: os
machos constroem ninhos dentro de abrigos e protegem seus ovos e filhotes de
predadores e machos rivais, mantendo o cuidado parental até 10 dias após a eclosão dos
ovos (Sabaj et al., 1999). Tais características poderiam ter favorecido o surgimento de
mecanismos de isolamento ou de barreiras ao fluxo gênico (Futuyma, 1997),
possibilitando a ocorrência de fixação de rearranjos cromossômicos e contribuindo para
o desenvolvimento de processos de especiação micro-alopátrica nesse gênero, assim
como nos ciclídeos dos grandes lagos Africanos (Lewis,1982; Lowe-McConnell, 1999).
Sabaj et al. (1999) acreditam que a presença de tentáculos cutâneos
muito desenvolvidos no focinho de machos de Ancistrus esteja relacionada
a uma estratégia reprodutiva de mimetismo de larvas. Estes autores
hipotetizam que as fêmeas de Ancistrus teriam preferência em acasalar-se
com machos que possuem larvas em seu ninho, pois isso demonstraria um
melhor cuidado parental despendido com a prole e maior sucesso
reprodutivo. Assim, os machos usariam seus tentáculos para simular a
presença de larvas e com isso atraírem mais fêmeas. Diante disso, machos
com mais tentáculos cutâneos e mais desenvolvidos teriam maiores chances

67
de reproduzir com mais fêmeas e, conseqüentemente, passarem seus genes
para um número maior de descendentes.
Foi observado em aquário, que um macho de Ancistrus pode
acasalar repetidamente com diversas fêmeas e cuidar de todos os ovos e
larvas simultaneamente (Sabaj et al., 1999). Comportamento similar foi
registrado durante as coletas de espécimes de Ancistrus sp.7 “Dimona” e
Ancistrus sp.8 “Balbina”, onde estes peixes apresentam distribuição
aparentemente agregada, formando pequenos grupos que contêm várias
fêmeas, alguns juvenis e geralmente, apenas um macho maduro. Embora
isso não tenha sido investigado para as demais espécies estudadas, é
provável que este comportamento se repita (J. Zuanon comunicação
pessoal). Assim, é possível que mutações cromossômicas ocorridas em
machos dominantes sejam fixadas mais rapidamente nas populações do que
em grupos de espécies que não apresentam este tipo de comportamento. Se
a diversidade cariotípica encontrada nas espécies de Ancistrus estiver
realmente associada às características mencionadas acima, estudos em
outros grupos que apresentem comportamentos similares tais como,
Cichlidae (principalmente em Apistogramma), Callichthyidae
(especialmente no gênero Corydoras), Trichomycteridae, Crenuchidae (em
Characidiinae) e Poeciliidae, poderiam ser especialmente elucidativos.
Além de contribuir para um melhor entendimento dos mecanismos de
especiação e evolução cromossômica em peixes.

68
5. CONSIDERAÇÕES FINAIS
1) As espécies do gênero Ancistrus apresentaram grande diversidade cariotípica, com
número diplóide variando de 34 a 52 cromossomos. Isto demonstra que intensos
processos de rearranjos cromossômicos, principalmente fusões cêntricas, estão
associados à evolução cariotípica deste gênero, os quais indicam uma tendência
evolutiva para a redução do número diplóide neste táxon.
2) Inversões pericêntricas e translocações também ocorreram na diversificação
cariotípica deste gênero. Tais mecanismos explicariam a presença do grande número de
cromossomos acrocêntricos nas espécies que apresentaram número diplóide igual a 52
cromossomos. Além disso, tais mecanismos também explicam a grande diversidade no
número fundamental, que variou de 68 a 82.
3) Um par de cromossomos portadores de RONs e positivos para banda C, foi
observado em todas as espécies, porém este caráter apresentou uma grande diversidade,
tanto em relação à sua localização no cromossomo quanto à sua posição no cariótipo.
Portanto, não pôde ser evidenciado nenhum padrão geral quanto à localização deste
marcador nas espécies desse gênero, uma vez que este caráter foi espécie-específico.
4) Poucos cromossomos apresentaram heterocromatina constitutiva, a qual distribuiu-se
em blocos discretos dispostos em posição centromérica, terminal, intersticial ou
proximal, que muitas vezes apresentaram-se pálidos. Blocos mais conspícuos foram
evidenciados em associação com as RONs, em todas as espécies.
5) Sete das 10 espécies apresentaram cromossomos sexuais heteromórficos. Destas,
duas apresentaram mecanismos do tipo ZZ/ZW, três do tipo XX/XY e dois sistemas

69
múltiplos, XX/XY1Y2 e Z1Z1Z2Z2/Z1Z2W1W2, ainda não descritos para Loricariidae,
sendo que o último ainda não havia sido descrito para peixes.
6) A grande diversidade cariotípica encontrada entre as espécies de Ancistrus pode ter
relação com as características da biologia e etologia desses peixes. Tais características
poderiam ter favorecido o surgimento de mecanismos de isolamento ou de barreiras ao
fluxo gênico, possibilitando a ocorrência e fixação de rearranjos cromossômicos,
contribuindo assim para o desenvolvimento de processos de especiação micro-
alopátrica nesse gênero.

70
6. REFERÊNCIAS BIBLIOGRÁFICAS
Almeida-Toledo, L.F. 1978. Contribuição à citogenética dos Gymnotoidei (Pisces,
Ostariophisi). Ph.D. thesis, instituto de biociências da Universidade de São Paulo,
São Paulo. 128p.
Almeida-Toledo, L.F. 1998. Cytogenetic Markers in Neotropical Freshwater Fishes. In:
Malabaraba, L.R.; Reis, R.E.; Vari, R.P.; Lucena, Z.M.S; Lucena; C.A.S. (Eds.).
Phylogeny and Classification of Neotropical Fishes. Edipucrs, Porto Alegre.
p.279-330.
Almeida-Toledo, L.F.; Foresti, F. 2001. Morphologically differentiated sex
chromosomes in neotropical freshwater fish. Genetica, 111: 91-100
Almeida-Toledo, L.F.; Foresti, F.; Toledo-Filho, S. de A. 1984. Complex sex
chromosome system in Eigenmannia sp. (Pisces, Gymnotiformes). Genetica, 64:
165-169
Alves, A.L.; Oliveira, C.; Foresti, F. 2003. Karyotype variability in eight species of the
subfamilies Loricariinae e Ancistrinae (Teleostei, Siluriformes, Loricariidae).
Caryologia, 56(1): 57-63
Alves, A.L.; Oliveira, C.; Foresti, F. 2005. Comparative cytogenetic analysis of eleven
species of subfamilies Neoplecostominae e Hypostominae (Siluriformes:
Loricariidae). Genetica, 128: 127-136
Andreata, A.A.; Almeida-Toledo, L.F.; Oliveira, C.; Toledo-Filho, S.A. 1992.
Chromosome studies in Hypoptopomatinae (Pisces, Siluriformes, Loricariidae): I.
XX/XY sex chromosomes heteromorphism in Pseudotocinclus tietensis.
Cytologia, 57: 369-372
Andreata, A.A.; Almeida-Toledo, L.F. de; Oliveira, C.; Toledo-Filho, S. de A. 1993.
Chromosome studies in Hypoptopomatinae (Pisces, Siluriformes, Loricariidae).
II. ZZ/ZW sex chromosome system, B chromosomes, and constitutive
heterochromatin differentiation in Microlepidogaster leucofrenatus. Cytogenetics
and Cell Genetics, 63(4): 215-220

71
Andreata, A.A.; Almeida-Toledo, L.F. de; Oliveira, C.; Toledo-Filho, S. de A. 1994.
Cytogenetic studies on the subfamily Hypoptopomatinae (Pisces, Siluriformes,
Loricariidae): III. Analysis of 7 species. Caryologia, 47(1): 27-37
Anjos, M.B. 2005. Estrutura de comunidades de peixes de igarapés de terra firme na
Amazônia Central: composição, distribuição e características tróficas.
Dissertação de Mestrado. Universidade Federal do Amazonas/Instituto
Nacional de Pesquisas da Amazônia – INPA. Manaus, Amazonas. 68p.
Armbruster, J.W. 2003. Peckoltia sabaji, a new species from the Guyana Shield
(Siluriformes: Loricariidae). Zootaxa, 344: 1-12
Armbruster, J.W. 2004. Phylogenetic relationships of the suckermouth armoured
catfishes (Loricariidae) with emphasis on the Hypostominae and the Ancistrinae.
Zoological Journal of the Society, 141: 1-80
Artoni, R.F. 1996. Estudos citogenéticos na família Loricariidae, com
ênfase no gênero Hypostomus Lacépede (1803) (Pisces, Loricariidae,
Hypostominae). Dissertação de Mestrado. Universidade Federal de São
Carlos, São Paulo. 162p.
Artoni, R.F.; Bertollo L.A.C. 1996. Cytogenetic studies on Hypostominae (Pisces,
Siluriformes, Loricariidae). Considerations on karyotype evolution in the genus
Hypostomus. Caryologia, 49: 81-90
Artoni, R.F.; Venere, P.C.; Bertollo L.A.C. 1998. A heteromorphic ZZ/ZW sex
chromosome system in fish, genus Hypostomus (Loricariidae). The Japan Mendel
Society, 63: 421-425
Artoni, R.F.; Bertollo L.A.C. 1999. Nature and distribution of constitutive
heterochromatin in fishes, genus Hypostomus (Loricariidae). Genetica, 106: 209-
214
Artoni, R.F.; Bertollo L.A.C. 2001. Trends in the karyotype evolution of Loricariidae
fish (Siluriformes). Hereditas, 134: 201-210

72
Batista, V.S.; Inhamuns, A.J.; Freitas, C.E.C.; Freire-Brasil, D. 1998. Characterization
of the fishery in river communities in the low-Solimões/high-Amazon region.
Fisheries Management and Ecology, 5: 419-435
Batista, V.S.; Petrere Jr., M. 2003. Characterization of the commercial fish production
landed at Manaus, Amazonas state, Brazil. Acta Amazonica, 33(1): 53-66
Bayreuther, K.1969. Die cytogenetik zweier norddeutscher populationen von
Nosopsyllus fasciatus bosc.(Aphaniptera). Chromosoma, 27: 20-46
Beçak, W.; Beçak, M.L. 1969. Cytotaxonomy and chromasomal evolution in serpentes.
Cytogenetics, 8: 247-262
Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho, O. 1978. Cytotaxonomic
considerations on Hoplias lacerdae (Pisces, Erythrinidae). Brasilian Journal
Genetics, 1(2): 103-120
Bertollo, L.A.C.; Cavalaro, Z.I.1992. A highly differentiated ZZ/ZW sex chromosome
system in a Characidae fish, Triportheus guentheri. Cytogenetics and Cell
Genetics, 60: 60-63
Bertollo, L.A.C.; Fontes, M.S.; Fenocchio, A.S.; Cano, J. 1997. The X1X2Y sex system
in the fish Hoplias malabaricus. I. G-, C- and chromosome replication banding.
Chromosome Research, 5: 493-499
Bertollo, L.A.C.; Mestriner, C. A. 1998. The X1X2Y sex chromosome system in the
fish Hoplias malabaricus. II. Meiotic analyses. Chromosome Research, 6: 141-
147
Bertollo, L.A.C.; Oliveira, C.; Molina, W.F.; Margarido, V.P.; Fontes, M.S.; Pastori,
M.C.; Falcão, J.das N.; Fenocchio, A.S. 2004. Chromosome evolution in the
erythrinid fish, Erythrinus erythrinus (Teleostei: Characiformes). Heredity, 93:
228-233
Bleeker, P. 1862. Atlas ichthyologique des Indes orientales neerlandaises. Siluroides,
Characoides et Heterobranchoides. 2:1-122.

73
Buck, S.; Sazima, I. 1995. Na assemblage of mailed catfishes (Loricariidae)
in southeastern Brazil: distribuition, activity, and feeding. Ichthyological
Explorer Freshwaters, 6(4): 325-332
Burguess, W.E. 1989. An Atlas of Freshwater and Marine Catfishes, A
Preliminary Survey of the Siluriformes. Publications, T.F.H. 784p.
Centofante L., Bertollo, L.A.C.; Moreira-Filho, O. 2002. A ZZ/ZW sex
chromosome system in a new species of the genus Parodon (Pisces,
Parodontidae). Caryologia, 55: 139-150
Denton, T.E. 1973. Fish chromosome methodology. Charles C Thomas
Publisher, USA, 166p.
Eigenmann, C.H.; Eigenmann, R.S. 1890. A revision of the South American
nematognathi, or cat-fishes. Occas. Papers California Academy Science,
1: 1-508.
Falcão, J. das N.; Betollo, L.A.C. 1985. Chromosome characterization in
Acestrorhynchinae and Cynopotaminae (Pisces, Characidae). Journal of
Fish Biology, 27: 603-610
Falcão, J. das N. 1988. Caracterização cariotípica em peixes do gênero
Triportheus (Teleostei, Characiformes, Characidae). Tese de
Doutorado. Faculdade de Medicina de Ribeirão Preto da Universidade de São
Paulo, Ribeirão Preto, SP. 137p.
Feldberg, E.; Porto, J.I.R.; Bertollo, L.A.C. 1992. Karyotype evolution in
Curimatidae (Teleostei, Characiformes) of the Amazon region. I. studies
on the genera Curimata, Psectrogaster, Steindachnerina and
Curimatella. Brasilian Journal Genetics, 15(2): 369-363
Feldberg, E.; Porto, J.I.R.; Nakayama, C.M. 1993. Karyotype evolution in
Curimatidae (Teleostei, Characiformes) from the Amazon Region. II.
Centric fissions in the genus Potamorhina. Genome, 36: 372-376
Feldberg, E.; Porto, J.I.R.; Santos, E.B.P.; Valentim, F.C.S. 1999.
Cytogenetic studies of two freshwater scianids of the genus Plagioscion

74
(Perciformes, Sciaenidae) from the central Amazon. Genetics and
Molecular Biology, 22(3): 351-356
Feldberg, E.; Porto, J.I.R.; Bertollo, L.A.C. 2003. Chromosomal changes
and adaptation of Cichlid fishes during evolution. In: Val, Adalberto
Luís, Kapoor, B.G. Fish Adaptations. Ed. Science Publishers, New
Dehli & New York. USA, p.285-308.
Ferraz, E. 1999. Management and diseases of the ornamental fish exported
from the rio Negro basin. In: Val, A.L; Almeida-Val, V.M.F. (Eds)
Biology of Tropical Fishes. Ed. INPA, Manaus, Amazonas. p.99-111.
Ferreira, E.J.G.; Zuanon, J.A.S.; Santos, G.M. dos. 1998. Peixes comerciais
do médio Amazonas: região de Santarém, Pará. Brasília: Edição
IBAMA. 211p.
Fisch-Muller, S.; Mazzoni, R.; Weber, C. 2001. Genetic and morphological
evidences for two new sibling species of Ancistrus (Siluriformes:
Loricariidae) in upper rio Tocantins drainage, Brazil. Ichthyological
Explorer Freshwaters, 12(4): 289-304
Fisch-Muller, S. 2003. Subfamily Ancistrinae. (Armored catfishes). In:
Reis, R.E; Kullander, S.O.; Ferraris, C.J. Jr. (Eds). Check list of the
Freshwater Fishes of South and Central America. Edipucrs, Porto
Alegre. p.373-400.
Futuyma, D.J. 1997. Biologia evolutiva. Ed. Ribeirão Preto, SBG/CNPq.
646p.
Galetti Jr., P.M.; Foresti, F.; Betollo, L.A.C.; Moreira-filho, O. 1981.
Heteromorphic sex chromosome in three species of the Leporinus
(Pisces, Anostomidae). Cytogenetics and Cell Genetics, 49: 138-142
Galetti Jr., P.M.; Foresti, F. 1986. Evolution of the ZZ/ZW system in
Leporinus (Pisces, Anostomidae) role of constitutive heterochromatin.
Cytogenetics and Cell Genetics, 43: 43-46

75
Galetti Jr., P.M.; Foresti, F. 1987. Two new cases of ZZ/ZW heterogamety
in Leporinus (Pisces, Anostomidae) and their relationships in the
phylogeny of the group. Brasilian Journal Genetics, 1: 135-140
Galetti Jr., P.M.; Aguilar, C.T.; Molina, W.F. 2000. An overiew of marine
fish cytogenetics. Hydrobiologia, 420: 55-62
Garcia, C.; Moreira-Filho, O. 2005. Cytogenetical analyses in three fish
species of the genus Pimelodus (Siluriformes: Pimelodidae) from rio
São Francisco: considerations about the kayotypical evolution in the
genus. Neotropical Ichthyology, 3(2): 285-290
Giuliano-Caetano, L. 1998. Polimorfismo cromossômico Robertsoniano em
populações de Rineloricaria latirostris (Pisces, Loricariinae). Tese de
Doutorado. Universidade Federal de São Carlos, São Carlos, São Paulo.
78p.
Gold, J.R.; 1984. Silver-staining and heteromorphism of chromosomal
nucleolus organizer regions in North American cyprinid fishes. Copeia,
133-139
Gold, J.R.; Li, Y.C; Shipley, N.S.; Powers, P.K. 1990. Improved methods
for working with fish chromosomes with a review of metaphase
chromosome banding. Journal of Fish Biology, 37: 563-575
Gosline, W.A. 1945. Catálogo dos nematognathos de água-doce da América
do Sul e Central. Boletim doMuseu Nacional do Rio de Janeiro, Série
Zoologia, 33: 1-138
Goulding, M. 1980. The Fishes and the Forest. Exploration in Amazonian
Natural History. University of California Press. Berkeley, USA. 280p.
Guerra, M.S. 1988. Introdução à Citogenética Geral. Ed. Guanabara, Rio de
Janeiro, 142p.
Howell, W.M.; Black, D.A. 1980. Controlled silver-staining of nucleolus organizer
regions with a protective colloidal developer: as 1-step method. Experientia, 36:
1014-1015

76
Howes, G.J. 1983. The cranial muscle of Loricarioid catfishes, their homologies and
value as taxonomic characters (Teleostei: Siluroidei). Bulletin Britannic Museum
History. (Zoology), 45(6): 309-345
Isbrücker, I.J.H. 1980. Classification and Catalogue of the mailed Loricariidae (Pisces,
Siluriformes). Verslagen en technische Genevens, Universiteit van Amsterdam,
181p.
John, B. 1990. Meiosis. Cambridge University Press, New York, USA. 396p.
Junk, W.J.; Soares, G. M.; Carvalho, F. M. 1983. Distribution of the fish species in a
lake of the Amazon River floodplain near Manaus (Lago Camaleão), with special
reference to extreme oxygen conditions. Amazoniana, 7 (4): 397-431
Kavalco, K.F.; Pazza, R.; Bertollo, L.C.B.; Moreira-Filho, O. 2004. Heterochromatin
characterization of four fish species of the family Loricariidae (Siluriformes).
Hereditas, 141: 237-242
Kavalco, K.F.; Pazza, R.; Bertollo, L.C.B.; Moreira-Filho, O. 2005. Karyotypic
diversity and evolution of Loricariidae (Pisces, Siluriformes). Heredity, 94: 180-
186
King, M. 1980. C-banding studies on australian hylid frogs: secondary cnstriction
structure and the concept of euchromatin transformation. Chromosoma, 80: 191-
217
King, M. 1985. The canalization model of chromosomal evolution: A critique.
Systematic Zoology, 34: 69-75
Kligerman, A.D.; Bloom, S.E. 1977. Rapid chromosome preparations from solid tissues
of fishes. Journal Fisheries Research Board of Canada, 34: 266-269
Kner, R. 1854. Die Hypostomiden. Zweite Hauptgruppe der familie der panzerfische
(Loricata vel Goniodontes). Denks. Kaiser. Akad. Wissens. Wien, mathem.-
naturwissen. Classe AM XXI: 65-98.

77
Lara, M. C. S. 1998. Aspectos citogenéticos de quatro espécies de peixes da subfamília
Ancistrinae (Siluriformes, Loricariidae) da bacia do rio Paraná. Dissertação de
Mestrado. Universidade Estadual de Maringá, Paraná. 52p.
Levan, A.; Fredga, K.; Sandberg, A.A. 1964. Nomenclature for centromeric position on
chromosomes. Hereditas, 52: 201-220.
Lewis, D. S. C. 1982. Problems of species definition in Lake Malawi cichlid fishes
(Pisces: Cichlidae). J.L.B Smith Institute of Ichthyology, Special publication. 23: 1-
5
Lowe-McConnel, R.H. 1999. Estudos Ecológicos de Comunidades de Peixes Tropicais.
Editora da Universidade de São Paulo, São Paulo. 533p.
Mariotto S. 2003. Citogenética de Cinco populações de Ancistrus cf. dubius (Pisces,
Loricariidae, Ancistrinae), da bacia do Pantanal, Mato Grosso, Brasil.
Dissertação de Mestrado. Universidade Federal de Mato Grosso, Mato Grosso.
77p.
Mariotto S.; Artoni, R.F.; Miyazawa, C.S. 2004. Occurrence of sexual chromosome, of
the type ZZ/ZW in Ancistrus cf. dubius (Loricariidae, Ancistrinae) of the Paraguay
River Basin, Mato Grosso, Brazil. Caryologia, 57 (4): 327-331
Michelle, J.L.; Takahashi, C.S.; Ferrari, I. 1977. Karyotypic studies of some species of
the family Loricariidae (Pisces). Cytologia, 42: 539-546
Montoya-Burgos, J.I.; Fisch-Muller, S.; Weber, C.; Pawlowski, J. 1998.
Phylogenetic relationships of the Loricariidae (Siluriformes) basead on
mitocondrial rDNA gene sequences. In: Malabaraba, L.R.; Reis, R.E.; Vari, R.P.;
Lucena, Z.M.S; Lucena; C.A.S. (Eds.). Phylogeny and Classification of
Neotropical Fishes. Edipucrs, Porto Alegre. p.279-330.
Moreira-Filho, O.; Bertollo, L.A.C.; Galetti Jr., P.M. 1980. Evidences for a multiple
sex chromosome system with female heterogamety in Apareiodon affinis (pisces,
Parodontidae). Caryologia, 33:83-91

78
Moreira-Filho, O.; Bertollo, L.A.C.; Galetti Jr., P.M. 1984. Structure and variability of
Nucleolar Organizer Regions in Parodontidae fish. Canadian Journal of Genetics
and Cytology, 26: 564-568
Moreira-Filho, O.; Bertollo, L.A.C.; Galetti Jr., P.M. 1985. Karyotipic study of some
species of family Paradontidae (Pisces - Cypriniformes). Caryologia, 38(1): 47-55
Moreira-Filho, O.; Bertollo, L.A.C. 1991. Astyanax scabripinnis (Pisces, Characidae):
a species complex. Brasilian Journal Genetics, 14: 331-357
Moreira-Filho, O.; Bertollo, L.A.C.; Galetti Jr., P.M. 1993. Distribution of sex
chromosome mechanisms in Neotropical fish and description of a ZZ/ZW system
in Parodon hilarii (Parodontidae). Caryologia, 46: 115-125
Muramoto, J.; Ohno, S.; Atkin, N.B. 1968. On the diploid state of the fish order
Ostariophysi. Chromosoma, 24: 59-66
Nakayama, C.M.; Porto, J.I.R.; Feldberg, E. 2002. A Comparative cytogenetic study of
five piranha species (Serrasalmus, Serrasalminae) from the Amazon basin.
Genetica, 114: 231-236
Ohno, S. 1967. Sex chromosomes and sex-linked genes. Springer-Verlag. New York,
192p.
Oliveira, C. 1987. Estudos citogenéticos no gênero Corydoras (Pisces, Siluriformes,
Callichthyidae). Dissertação de Mestrado. Universidade Federal de São Paulo, São
Paulo. 154p.
Oliveira, C.; Almeida-Toledo, L.F.; Mori, L.; Toledo-Filho, S.A. 1993. Cytogenetic
and DNA content in six genera of the family Callichthyidae (Pisces, Siluriformes).
Caryologia, 46(3): 171-188
de Oliveira, R.R.; Souza, I.L.; Venere, P.C. (no prelo). Karyotype description of three
species of Loricariidae (Siluriformes) and occurrence of the ZZ/ZW sexual system
in Hemiancistrus spilomma Cardoso & Lucinda, 2003. Neotropical Ichthyology
Petrere Jr., M. 1978. Pesca e esforço de pesca no estado do Amazonas. Acta Amazonica,
8(3): 439-454

79
Pieczarka, J.C. 1995. Variability in constitutive heterochromatin in South American
primates. Neotropical Primates, 3(3): 86-88
de Pinna, M.C. 1998. Phylogenetic relationships of Neotropical Siluriformes (Teleostei:
Ostariophysi): Historical overview and synthesis of hypotheses. In: Malabaraba,
L.R.; Reis, R.E.; Vari, R.P.; Lucena, Z.M.S; Lucena; C.A.S. (Eds.). Phylogeny and
Classification of Neotropical Fishes. Edipucrs, Porto Alegre. p.279-330.
Porto, J.I.R.; Feldberg, E. 1993. Is Callichthys Linné (Siluriformes, Callichthyidae) a
monotypic genus. Acta Amazonica, 24(2): 311-314
Power, M.E. 1984. The importance of sediment in the grazing ecology and size class
interactions of an armored catfish, Ancistrus spinosus. Environmental Biology of
Fishes, 10(3): 173-181
Power, M.E. 1990. Resource enhancement by indirect effects of grazers: armored
catfish, algae, a sediment. Ecology, 71(3): 897-904
Rasband, W.S. 1997. Image J, U.S. National Institute of Health, Bethesda, Maryland,
USA ( http:// rsb.info.nih.gov/ij/). Acesso: 25/10/05
Regan, C.T. 1904. A monograph of the fishes of the family Loricariidae. Trans. Zool.
Soc. London. 17: 191-351
Reis, R.E; Kullander, S.O.; Ferraris Jr., C.J. 2003. Check list of the
Freshwater Fishes of South and Central America. Edipucrs, Porto
Alegre. 729p.
Sabaj, M.H.; Armbruster, J.W.; Paje, L.M. 1999. Spawning in Ancistrus (Siluriformes:
Loricariidae) with comments on the evolution of snout tentacles as a novel
reproductive strategy: larval mimicry. Ichthyological Explorer Freshwaters, 10(3):
217-229
Sabino, J.; Zuanon, J.A. 1998. A stream fish assemblage in Central Amazônia:
distribution, activity patterns and feeding behavior. Ichthyological Explorer
Freshwaters, 8(3): 201-210

80
Sato, L.R.; Oliveira, C.; Foresti, F. 2004. Karyotype description of five species of
Trichomycterus (Teleostei: Siluriformes: Tricomicteridae). Genetics and Molecular
Biology, 27(1): 45-50
Scavone, M.D.P. & Júlio Jr., H.F. 1995. Cytogenetics analysis and heterochromatin
distribution in ZZ/ZW sex chromosomes of the mailed catfish Loricariichthys
platymetopon (Loricariidae, Siluriformes). Brazilian Journal of Genetics, 18(1):
31-35
Schaefer, A.S.,1987. Osteology of Hypostomus plecostomus (Linnaeus), with a
phylogenetic analysis of the Loricariid subfamilies (Pisces: Siluroidei).
Contribution Science Natural History Museum Los Angeles, 394: 1-31
Schmid, M. 1982. Chromosome banding in amphibian. VII. Analysis of the structure
and variability of NORs in Anura. Chromosoma, 87: 327-344
Singh, L.; Purdom, I.F.; Jones, K.W. 1980. Sex chromosome associated satellite DNA:
evolution and Conservation. Chromosoma, 79: 137-157
Sola, L.; Natili, G.L.; Cataudella, S. 1988. Cytogenetical Characterization of
Odontesthes bonariensis (Pises, Atherinidae), an Argentine species introduced in
Italy. Genetica, 77: 217-224
Solari, A.J. 1994. Sex chromosomes and sex determination in vertebrates. CRC, florida,
308p.
Souza, A.C.P. 2003. Descrição cariotípica de peixes dos gêneros Baryancistrus,
Parancistrus, Peckoltia e Ancistrus (Ancistrinae, Loricariidae) da Bacia
Amazônica. Dissertação de Mestrado. Zoologia, Universidade Federal do
Pará/Museu Paraense Emílio Goeldi. 130p.
Souza, A.C.P.; Nascimento, A.L.; Carvalho Jr. J.R.; Barros, R.M.S.; Feldberg, E.;
Nagamachi, C.Y.; Pieczarka, J.C. 2004. Karyotypic analysis of Baryancistrus aff.
niveatus (Ancistrinae, Loricariidae) by C-banding, Ag-Nor, CMA3, DAPI and
FISH. Caryologia, 57(3): 219-223
Sumner, A.T. 1972. A Simple technique for demonstrating centromeric heterocromatin.
Experimental Cell Research, 75: 304-306

81
Torres, R.A.; Oliveira, C.; Foresti, F. 1998. Comparative cytogenetic studies in
alopatric populations of Trichomycterus paolence Eigenmann, 1918 from
Southeastern Brazil (Siluriformes, Tricomycteridae). Cytologia, 63: 107-114
Val, A.L.; Almeida-Val, V.M.F., 1995. Fishes of the Amazon and their environment.
Physiological and biochemical features. Heidelberg, Springer Verlag. 224p.
Valentim, F.C. de S. 2001. Caracterização cromossômica em peixes da família
Potamotrygonidae (Chondrichthyes, Rajiformes) do médio Rio Negro. Dissertação
de Mestrado. Universidade do Amazonas/ Universidade Federal de São Carlos,
São Paulo. 79p.
Venere, P.C.; Galetti Jr., P.M. 1989. Chromosome evolution and phylogenetic
relationships of some Neotropical Characiformes of the family Curimatidae.
Brasilian Journal Genetics, 12(1): 17-25
Venere, P.C.; Ferreira, I.A.; Martins, C.; Galetti jr., P.M. 2004. A novel ZZ/ZW sex
chromosome system for the genus Leporinus (Pisces, Anostomidae,
Characiformes). Genetica, 121: 75-80
Zuanon, J.A.S. 1999. História Natural da ictiofauna de corredeiras do rio Xingu, na
região de Altamira, Pará. Tese de Doutorado. Instituto de Biologia da
Universidade Estadual de Campinas, Campinas, São Paulo. 199p.