Embed Size (px)
Citation preview

Membrana Plasmática, Citoplasma e Ribossomo.
Introdução
O que é uma célula? Bem, em certo nível, é uma bolsa de gosma. A membrana
plasmática - o limite externo da célula - é a bolsa, e o citoplasma é a gosma.
Naturalmente, uma célula é muito mais do que apenas uma bolsa de gosma. É uma
unidade complexa, altamente organizada, a unidade estrutural básica de todas as coisas
vivas. E a membrana plasmática e o citoplasma são, de fato, muito sofisticados.
A membrana é uma estrutura delicada, de duas camadas de lipídios e proteínas, e
controla o que pode entrar e sair da célula. Similarmente, o citoplasma de uma célula
eucariótica consiste não somente de citosol - uma substância similar a um gel composta de
água, íons e macromoléculas - mas também de organelas e de proteínas estruturais que
compõem o citoesqueleto, ou "esqueleto da célula".
A membrana plasmática
As células procarióticas e eucarióticas têm uma membrana plasmática, uma dupla
camada de lipídios que separa o interior da célula do ambiente exterior. Essa dupla camada
consiste em grande parte de lipídios especializados chamados de fosfolipídios.
Um fosfolipídio é feito de uma cabeça de fosfato hidrofílica, atraída pela água, com
duas caudas de ácidos graxos hidrofóbicas, que repelem água. Os fosfolipídios
espontaneamente se organizam em uma estrutura de dupla camada, com suas caudas
hidrofóbicas apontando para dentro e suas cabeças hidrofílicas viradas para fora. Essa
estrutura de duas camadas, energeticamente favorável, chamada de bicamada fosfolipídica,
é encontrada em muitas membranas biológicas.

Como mostrado abaixo, as proteínas também são importantes componentes da
membrana plasmática. Algumas delas atravessam toda a membrana, servindo como canais ou
receptores de sinais, enquanto outras são apenas um anexo da borda. Tipos diferentes de
lipídios, como colesterol, podem ser encontrados na membrana celular e afetar sua fluidez.
Crédito de imagem: modificada de OpenStax Biology
Uma imagem da membrana plasmática mostra a bicamada lipídica, proteínas
inseridas, e moléculas de colesterol. A membrana separa o espaço extracelular, fora da célula,
do citosol dentro da célula.

A membrana plasmática é a fronteira entre o interior e o exterior de uma célula.
Assim, controla a passagem de várias moléculas - incluindo açúcares, aminoácidos, íons e
água - para dentro e para fora da célula. A facilidade das moléculas cruzarem a membrana
depende de seu tamanho e polaridade. Algumas moléculas pequenas, apolares, como
oxigênio, podem passar diretamente pela porção fosfolipídica da membrana. Moléculas
maiores e mais polares, hidrofílicas, como aminoácidos, devem cruzar a membrana por
canais proteicos, um processo que geralmente é regulado pela célula.
A área de superfície da membrana plasmática limita a troca de materiais entre a célula
e seu ambiente. Algumas células são especializadas na troca de resíduos ou nutrientes e têm
modificações para aumentar a área da membrana plasmática. Por exemplo, as membranas de
algumas células de absorção de nutrientes são dobradas em projeções parecidas com dedos
chamadas de microvilosidades. Células com microvilosidades cobrem a superfície interior
do instestino delgado, o órgão que absorve nutrientes de alimentos digeridos. As
microvilosidades ajudam as células intestinais a maximizar sua absorção de nutrientes da
comida por aumentar a área de superfície da membrana plasmática.
Crédito de imagem: OpenStax Biology. Micrografia é uma modificação do trabalho de Louisa
Howard.
Diagrama e micrografia de células instestinais, mostrando os "dedos" protuberantes da
membrana plasmática - chamados de microvilosidades - que entram em contato com o líquido
dentro do intestino delgado.

O citoplasma
A parte da célula chamada de citoplasma é um pouco diferente em eucariontes e
procariontes. Em células eucarióticas, que têm um núcleo, o citoplasma é tudo entre a
membrana plasmática e o envelope nuclear. Em procariontes, que não têm núcleo, o
citoplasma significa simplesmente tudo que é encontrado dentro da membrana plasmática.
Um componente principal do citoplasma tanto em procariontes quanto em eucariontes
é o citosol gelatinoso, uma solução à base de água que contém íons, pequenas moléculas, e
macromoléculas. Nos eucariontes, o citoplasma inclui organelas envoltas por membranas,
suspensas no citosol. O citoesqueleto, uma rede de fibras que sustentam a célula e dão sua
forma, também é parte do citoplasma e ajuda a organizar os componentes da célula.
Apesar de o citosol ser em grande parte água, tem uma consistência quase sólida,
como gel, por conta das várias proteínas nele suspensas. O citosol contém um rico caldo de
macromoléculas e pequenas moléculas orgânicas, incluindo glicose e outros açúcares
simples, polissacarídeos, aminoácidos, ácidos nucleicos e ácidos graxos. Íons de sódio,
potássio, cálcio, e outros elementos também são encontrados no citosol. Muitas reações
metabólicas, incluindo a síntese proteica, acontecem nesta parte da célula.
Ribossomos
Como mencionado acima, os ribossomos são as máquinas moleculares
responsáveis pela síntese proteica. Um ribossomo é feito de RNA e proteínas, e cada
ribossomo consiste de dois conjuntos de RNA-proteína distintos, conhecidos como as
subunidades menor e maior. A subunidade maior fica em cima da subunidade menor, com
uma molécula de RNA mensageiro entre elas, formando um "sanduíche". (Um ribossomo se
parece um pouco com um hambúrguer, com um pão arredondado em cima e um "recheio" de
RNA espalhado.)
Nos eucariontes, os ribossomos recebem suas instruções do núcleo para a síntese
proteica, onde pedaços de DNA (genes) são transcritos para fazer RNAs mensageiros
(RNAms). Um RNAm viaja até o ribossomo, que utiliza a informação que ele contém para
construir uma proteína com uma sequência específica de aminoácidos. Este processo é
chamado de tradução. Os procariontes não possuem núcleo, assim seus RNAms são
transcritos no citoplasma e podem ser traduzidos pelos ribossomos imediatamente.

Figura: OpenStax Biology.
Imagem de um ribossomo com as subunidades maior e menor, com um RNAm ligado
e uma cadeia polipeptídica sendo formada. Uma molécula de RNAt é mostrada fazendo
ligação com o RNAm e trazendo o aminoácido adequado para a posição certa para que ele
seja acrescentado à cadeia.
Os ribossomos eucarióticos podem estar livres, flutuando no citoplasma, ou ligados ao
retículo endoplasmático ou ao exterior do envelope nuclear. Como a síntese proteica é uma
função essencial de todas as células, os ribossomos são encontrados em praticamente
todos os tipos de células dos organismos multicelulares, assim como nos procariontes, tais
como as bactérias. No entanto, as células eucarióticas que se especializam em produzir
proteínas têm um número particularmente grande de ribossomos. Por exemplo, o pâncreas é
responsável pela produção e secreção de grandes quantidades de enzimas digestivas, assim as
células pancreáticas que fazem estas enzimas têm um número de ribossomos
descomunalmente elevado.
Para terminar, um fato curioso: como um testemunho da importância do ribossomo, o
Prêmio Nobel de Química de 2009 foi entregue a três pesquisadores que mapearam sua
estrutura e movimentos até o nível atômico, usando uma técnica conhecida como
cristalografia de raios X.

O citoesqueleto
Introdução
O que aconteceria se alguém entrasse sorrateiramente durante a noite e roubasse o seu
esqueleto? Só para deixar claro, não é provável que isso aconteça, biologicamente falando.
Porém, se isso realmente acontecer, a ausência do seu esqueleto poderia fazer com que seu
corpo perdesse grande parte de sua estrutura. Sua forma externa mudaria, alguns de seus
órgãos internos se deslocariam e, muito provavelmente, seria bem difícil andar, falar ou se
mover.
Curiosamente, o mesmo aconteceria com uma célula. Nós frequentemente pensamos
sobre as células como geleias macias e não-estruturadas. Mas na verdade, elas são altamente
estruturadas em grande parte do mesmo jeito que nossos corpos. Elas têm uma rede de
filamentos conhecida como citoesqueleto (literalmente, "esqueleto da célula"), que não
apenas suporta a membrana plasmática e dá a célula uma forma geral, mas também que
ajuda no correto posicionamento das organelas, proporciona caminhos para o
transporte de vesículas e (em muitos tipos de células) permitem que a célula se mova.
Nos eucariontes, existem três tipos de fibras proteicas no citoesqueleto: os
microfilamentos, filamentos intermediários e microtúbulos. Aqui, examinaremos cada
tipo de filamento, bem como algumas estruturas especializadas relacionadas com o
citoesqueleto.
Microfilamentos
Dos três tipos de fibras proteicas no citoesqueleto, os microfilamentos são os mais
finos. Eles têm um diâmetro de cerca de 7 nm e são feitos de muitos monômeros ligados de
uma proteína chamada de actina, combinados numa estrutura que se assemelha a uma
hélice dupla. Pelo fato de serem feitos de microfilamentos de monômeros de actina, os
microfilamentos também são conhecidos como filamentos de actina. Os filamentos de actina
têm direcionamento, ou seja, eles têm dois terminais estruturalmente diferentes.

Imagem adaptada de OpenStax Biology.
Filamento de actina, uma estrutura helicoidal composta de monômeros de actina
(subunidades).
Os filamentos de actina têm vários papéis importantes na célula. Por exemplo, eles
servem como caminhos para o movimento de uma proteína motora chamada de miosina, que
também pode formar filamentos. Por causa de sua relação com a miosina, a actina está
envolvida em muitos eventos celulares que envolvem movimento.
Por exemplo, na divisão celular animal, um anel feito de actina e miosina atuam para
dividir uma célula gerando duas células filhas. A actina e miosina também são abundantes
nas células musculares, onde elas formam estruturas organizadas de sobreposição de
filamentos chamados de sarcômeros. Quando os filamentos de actina e miosina de um
sarcômero deslizam um sobre o outro, seus músculos se contraem.
Os filamentos de actina também podem servir como rodovias para o interior das
células para o transporte de cargas, incluindo vesículas contendo proteínas e ainda organelas.
Estas cargas são transportadas pelos motores individuais de miosina, que "andam" ao longo
de pacotes de filamento de actina.

Os filamentos de actina podem montar-se e desmontar-se rapidamente, e esta
propriedade permite que eles tenham um papel importante na motilidade (movimentação)
celular, tal como na movimentação de uma célula branca sanguínea no seu sistema
imunológico.
Finalmente, os filamentos de actina têm um papel estrutural chave na célula. Na
maioria das células animais, a rede de filamentos de actina é encontrada na região do
citoplasma no limite da célula. Esta rede, que está ligada à membrana plasmática por
conectores proteicos especiais, dá à célula a sua forma e estrutura.
Filamentos intermediários
Os filamentos intermediários são um tipo de elemento do citoesqueleto composto de
múltiplos filamentos de proteínas fibrosas juntas. Como seu nome sugere, os filamentos
intermediários têm um diâmetro médio entre 8 e 10 nm, entre os diâmetros dos
microfilamentos e microtúbulos (discutidos abaixo).
Crédito de imagem: "The cytoplasm and cellular organelles," por OpenStax College (CC BY 3.0).

Filamentos intermediários em uma célula, formando uma rede que mantém o núcleo e
outras organelas no lugar.
Os filamentos intermediários aparecem em diferentes variedades, cada uma composta
de um tipo diferente de proteína. Uma proteína que forma os filamentos intermediários é a
queratina, uma proteína fibrosa encontrada nos cabelos, unhas e pele. Por exemplo, você
pode ter visto um anúncio de xampu que diz suavizar a queratina de seus cabelos!
Diferentemente dos filamentos de actina, que podem crescer e desmontar-se
rapidamente, os filamentos intermediários são mais permanentes e atuam essencialmente
na função estrutural da célula. Eles são especializados em suportar tensão, e sua função
inclui manter a forma da célula e ancorar o núcleo e outras organelas em seus lugares.
Microtúbulos
Apesar do "micro" em seu nome, os microtúbulos são os maiores entre os três tipos
de fibras do citoesqueleto, com um diâmetro de cerca de 25 nm. Um microtúbulo é composto
de proteínas tubulinas organizadas na forma de um tubo oco, como um canudo, e cada
proteína tubulina consiste de duas subunidades, α-tubulina e β-tubulina.
Os microtúbulos, como os filamentos de actina, são estruturas dinâmicas: elas
podem esticar e encolher rapidamente pela adição ou remoção de proteínas tubulina.
Também igualmente aos filamentos de actina, os microtúbulos têm direcionalidade, isto é,
eles têm dois terminais que são estruturalmente diferentes um do outro. Numa célula, os
microtúbulos têm um papel estrutural chave, ajudando a célula a resistir às forças de
compressão.

Figura: OpenStax Biology.
Esquerda: modelo 3D de um microtúbulo, mostrando que é um cilindro oco de
proteínas. Direita: diagrama de um microtúbulo, mostrando que é composto de dois tipos
diferentes de subunidades (alfa e beta). As subunidades formam dímeros, e os dímeros estão
conectados na forma de espiral para formar o tubo oco do microtúbulo.
Além de fornecer um suporte estrutural, os microtúbulos atuam numa variedade de
funções mais especializadas numa célula. Por exemplo, eles fornecem rotas para as
proteínas motoras chamadas de cinesinas e dineínas, que transportam vesículas e outras
cargas pelo interior da célula. Durante a divisão celular, os microtúbulos se montam numa
estrutura chamada de fuso mitótico, que separa os cromossomos.
Flagelos, cílios e centrossomos
Os microtúbulos são também componentes chaves de três estruturas celulares
especializadas de eucariontes: flagelos, cílios e centrossomos. Você pode se lembrar que
nossos amigos, os procariontes também têm estruturas com flagelos, que eles usam para se
mover. Não fique confuso - os flagelos dos eucariontes que vamos discutir têm praticamente
a mesma função, mas uma estrutura muito diferente.
Os flagelos (do latim flagellum) são estruturas longas que parecem cabelos que se
estendem da superfície da célula e são usados para mover a célula inteira, tal como num
espermatozoide. Se uma célula tem qualquer flagelo, ela normalmente tem um ou uns
poucos. Cílios motores (do latim cilium) são similares, mas são mais curtos e normalmente
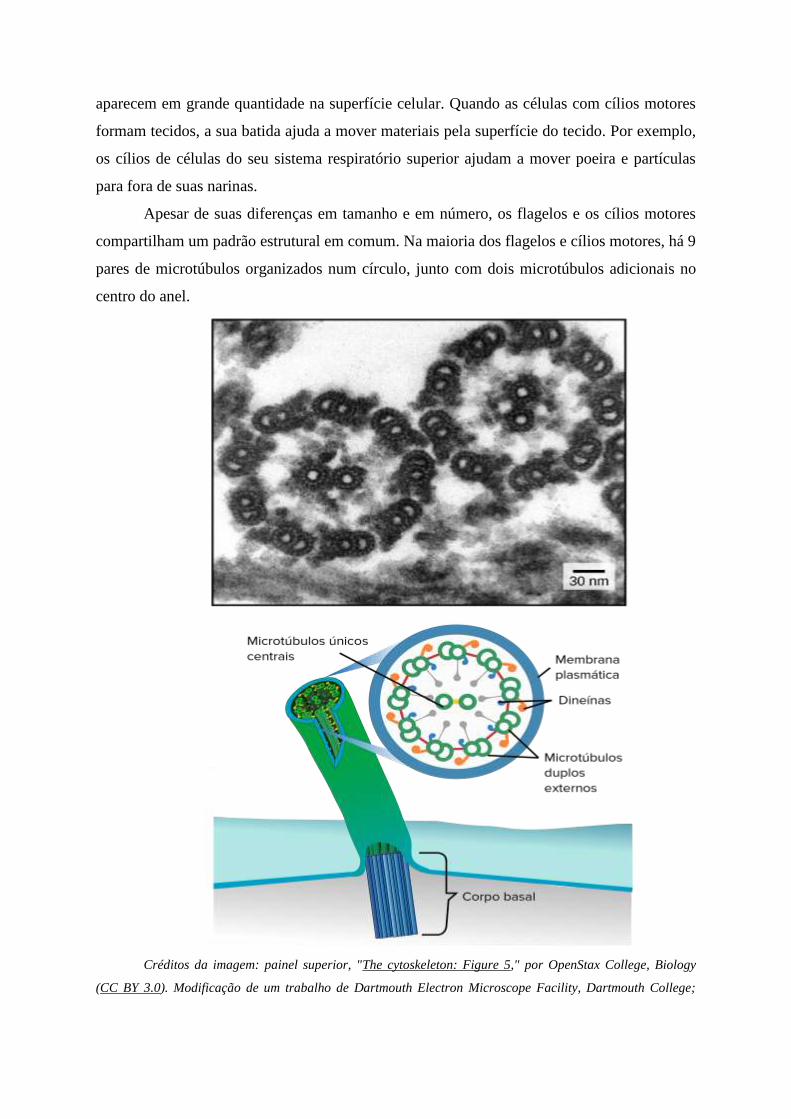
aparecem em grande quantidade na superfície celular. Quando as células com cílios motores
formam tecidos, a sua batida ajuda a mover materiais pela superfície do tecido. Por exemplo,
os cílios de células do seu sistema respiratório superior ajudam a mover poeira e partículas
para fora de suas narinas.
Apesar de suas diferenças em tamanho e em número, os flagelos e os cílios motores
compartilham um padrão estrutural em comum. Na maioria dos flagelos e cílios motores, há 9
pares de microtúbulos organizados num círculo, junto com dois microtúbulos adicionais no
centro do anel.
Créditos da imagem: painel superior, "The cytoskeleton: Figure 5," por OpenStax College, Biology
(CC BY 3.0). Modificação de um trabalho de Dartmouth Electron Microscope Facility, Dartmouth College;

dados de escala-bar de Matt Russell. Painel inferior, modificação de "Eukaryotic cilium diagram," por
Mariana Ruiz Villareal (domínio público)._
Superior: Corte transversal do flagelo em microscopia eletrônica de transmissão,
mostrando a organização da estrutura de microtúbulos 9+2. Inferior: Diagrama de um cílio
motor, mostrando os microtúbulos únicos no centro, os microtúbulos duplos organizados num
círculo em volta do microtúbulo único e as dineínas conectadas aos microtúbulos duplos.
Toda a estrutura é cercada pela membrana plasmática. Na base do cílio fica um corpo basal,
que é também feito de microtúbulos.
Nos flagelos e cílios motores, proteínas motoras chamadas dineínas se movem ao
longo dos microtúbulos, gerando uma força que faz o flagelo ou cílio bater. As conexões
estruturais entre os pares de microtúbulos e a coordenação do movimento das dineínas
permitem a atividade dos motores para produzir um padrão regular de batida.
Você pode notar uma outra característica no diagrama acima: o cílio ou flagelo tem
um corpo basal localizado em sua base. O corpo basal é composto de microtúbulos e atua em
um papel importante na montagem do cílio ou flagelo. Uma vez que a estrutura está montada,
ele também regula quais proteínas podem entrar ou sair.
O corpo basal é na verdade um centríolo modificado. Um centríolo é um cilindro de
nove trios de microtúbulos, agrupados por proteínas de sustentação. Os centríolos são mais
conhecidos por seu papel nos centrossomos, estruturas que atuam como centros
organizadores de microtúbulos nas células animais. Um centrossomo consiste de dois
centríolos orientados em ângulos retos entre si, rodeado por uma massa de "material
pericentriolar", que fornece campos ancoragem para os microtúbulos.

Crédito da imagem: adaptação de "Centriole," de Kelvinsong (CC BY 3.0)
Imagem de um centrossomo. O centrossomo contém dois centríolos posicionados
perpendiculares um ao outro.
O centrossomo é duplicado antes que uma célula se divida e o centrossomo
emparelhado parece atuar como organizador de microtúbulos que separa os cromossomos
durante a divisão celular. Entretanto, a exata função dos centríolos neste processo ainda não é
clara. As células com seus centrossomos removidos ainda podem se dividir, e em células de
plantas, que não possuem centrossomos, a divisão simplesmente acontece.
Sistema de endomembranas: Organelas
Introdução
Imagine que você seja uma célula pancreática. Seu trabalho é secretar enzimas
digestivas que são transportadas para o intestino delgado e ajudam a digerir os nutrientes do
alimento. Para realizar este trabalho, você, de alguma forma, tem que conseguir que essas
enzimas sejam enviadas de seu local de síntese - dentro da célula - para seu local de ação -
fora da célula.

Como você faria isto? Num momento de desespero você poderia até pensar em
despachar pelos Correios, mas pensando mais friamente você se lembraria: tenho o sistema
endomembranar!
O que é sistema endomembranar?
O sistema endomembranar (endo- = "dentro") é um grupo de membranas e
organelas das células eucarióticas que trabalham em conjunto para modificar, empacotar e
transportar lipídios e proteínas. Ele inclui uma variedade de organelas, tais como o
envoltório nuclear e lisossomos, que talvez você já conheça, e o retículo endoplasmático e
complexo de Golgi, que abordaremos em breve.
Embora não esteja tecnicamente dentro da célula, a membrana plasmática também faz
parte do sistema endomembranar. Como veremos, a membrana plasmática interage com
outras organelas endomembranares, e é por ela que as proteínas secretadas (como as enzimas
pancreáticas da introdução) são exportadas. Nota importante: o sistema endomembranar
não inclui as mitocôndrias, cloroplastos ou peroxissomos.
Vamos conhecer as diferentes partes do sistema endomembranar e como elas
funcionam no transporte de proteínas e lipídios.
Retículo endoplasmático
O retículo endoplasmático (RE) tem um papel chave na modificação de proteínas e
na síntese de lipídios. Ele consiste de uma rede de túbulos membranosos e bolsas achatadas.
Os discos e túbulos do RE são ocos, e o espaço em seu interior é chamado de lume ou luz.
RE rugoso
O retículo endoplasmático rugoso (RER) recebe seu nome dos inúmeros
ribossomos aderidos à sua superfície citoplasmática. Conforme estes ribossomos sintetizam
proteínas, eles entregam as novas cadeias de proteínas em formação ao lume. Algumas são
completamente transferidas ao RE e flutuam em seu interior, enquanto outras são ancoradas à
membrana.
No interior do RE, as proteínas passam por dobramentos e modificações, como a
adição de cadeias laterais de carboidratos. Estas proteínas modificadas serão incorporadas nas
membranas celulares—na membrana do RE ou naquelas de outras organelas—ou secretadas
para fora da célula.

Se as proteínas modificadas não estiverem destinadas a permanecer no RE, elas serão
empacotadas em vesículas, pequenas esferas de membrana que são usadas para transporte, e
enviadas ao complexo de Golgi. O RER também produz fosfolipídios para outras membranas
celulares, também transportados na forma de vesículas.
Crédito da imagem: esquerda, "Sistema endomembranar e proteínas: figura 2" da Faculdade
OpenStax, Biologia (CC BY 3.0), adaptação do trabalho por Lousia Howard; direita, adaptação de "Estrutura
celular animal" por Mariana Ruiz, domínio público
Micrografia e diagrama do retículo endoplasmático. A micrografia mostra o RE
rugoso como uma série de dobras da membrana ao redor do núcleo. O diagrama fornece uma
representação 3D de RE rugoso e RE liso, juntamente com o núcleo celular.
Como o RER ajuda a modificar as proteínas que serão secretadas pelas células, as
células que secretam grandes quantidades de enzimas ou outras proteínas, como as células do
fígado, possuem muitos RER.
RE liso
O retículo endoplasmático liso (REL) é contínuo com o RER, mas possui poucos ou
nenhum ribossomos em sua superfície citoplasmática. As funções do REL incluem:
Síntese de carboidratos, lipídios e hormônios esteroides (hormônios).
Desintoxicação de medicamentos e venenos
Armazenamento de íons cálcio

Nas células musculares, um tipo especial de REL chamado retículo sarcoplasmático é
responsável pelo armazenamento de íons cálcio que são necessários para a ativação das
contrações coordenadas das células musculares.
Há também pequenos trechos "lisos" de retículo endoplasmático encontrados dentro
do RER. Esses trechos funcionam como sítios de saída para as vesículas que germinam do
RER e são chamados REs de transição.
O complexo de Golgi
Quando as vesículas irrompem do RE, para onde elas vão? Antes de alcançar seu
destino final, os lipídios e proteínas nas vesículas de transporte precisam ser organizados,
empacotados e etiquetados para que eles terminem em seus devidos lugares. Esta
organização, empacotamento e distribuição ocorrem no complexo de Golgi (ou aparelho de
Golgi), organela feita por discos achatados de membrana.
Crédito da imagem: "Sistema endomembranar e proteínas: figura 3" da Faculdade OpenStax, Biologia
(CC BY 3.0), adaptado por Lousia Howard
Micrografia do aparelho de Golgi mostrando uma série de discos de membrana
achatados em corte transversal
O lado receptor do complexo de Golgi é chamado de face cis e o lado oposto é
chamado de face trans. As vesículas transportadoras do RE viajam até a face cis, fundem-se a
ela, e esvaziam seus conteúdos no lume do complexo de Golgi.

Conforme as proteínas e lipídeos viajam pelo complexo, eles passam por
modificações adicionais. Cadeias curtas de moléculas de açúcares podem ser adicionadas ou
removidas, ou grupos fosfato incluídos como etiquetas. O diagrama mostra o processamento
dos carboidratos, conforme o grupo carboidrato (em roxo), ligado à proteína, ganha ou perde
ramificações.
Imagem adaptada de "Sistema endomembranar e proteínas: figura 1" da Faculdade OpenStax,
Biologia (CC BY 3.0), adaptado por Magnus Manske
A imagem mostra o transporte de uma proteína de membrana do retículo
endoplasmático rugoso através do complexo de Golgi para a membrana plasmática. A
proteína é inicialmente modificada pela adição de cadeias ramificadas de carboidratos no RE
rugoso; estas cadeias são então aparadas e repostas com outras cadeias ramificadas no

complexo de Golgi. A proteína, com sua composição final de cadeias de carboidratos, é então
transportada à membrana plasmática por uma vesícula de transporte. A vesícula se funde à
membrana plasmática e suas cargas de lipídios e proteínas se tornam parte dela.
Finalmente, as proteínas modificadas são organizadas (com base em marcadores tais
como sequências de aminoácidos e etiquetas químicas) e empacotadas em vesículas que
irrompem da face trans do complexo de Golgi. Algumas destas vesículas despejam seus
conteúdos em outras partes da célula, como nos lisossomos ou vacúolos, onde serão usados.
Outras se fundem à membrana plasmática, disponibilizando as proteínas úteis para a
membrana e secretando outras proteínas para o meio extracelular.
As células que secretam muitas proteínas como células das glândulas salivares
secretoras de enzimas digestivas ou células do sistema imunológico secretoras de
anticorpos—possuem muitas cisternas de Golgi. Nas células vegetais o complexo de Golgi
também produz polissacarídeos (carboidratos de cadeia longa), sendo que alguns são
incorporados na parede celular.
Lisossomos
O lisossomo é uma organela que contém enzimas digestivas e atua como usina de
reciclagem das organelas na célula animal. Ele degrada as estruturas obsoletas para
reutilizar suas moléculas. Os lisossomos são parte do sistema endomembranar, e algumas
vesículas que deixam o Golgi são direcionadas para o lisossomo.
Os lisossomos também podem digerir proteínas externas que são trazidas para o
interior da célula. Por exemplo, vamos considerar o macrófago, um tipo de glóbulo branco do
sangue que é parte do sistema imunológico humano. Em um processo conhecido
como fagocitose, uma parte da membrana plasmática do macrófago invagina-se - dobra-se
para dentro - para engolfar um patógeno, como mostrado abaixo.

Crédito da imagem: adaptada de "Sistema endomembranar e proteínas: figura 4" da Faculdade
OpenStax, Biologia (CC BY 3.0)
Diagrama de fagocitose, no qual o fagossomo gerado pelo englobamento de uma
partícula se funde com um lisossomo, permitindo a digestão desta.
A seção invaginada com o patógeno em seu interior tem origem na membrana
plasmática e forma uma estrutura chamada fagossomo. O fagossomo então se funde a um
lisossomo, formando um compartimento combinado onde enzimas digestivas destroem o
patógeno.
Vacúolos
As células vegetais não possuem lisossomos. Em vez disso, elas possuem um outro
tipo de organela chamada vacúolo. O grande vacúolo central armazena água e resíduos, isola
materiais deletérios e possui enzimas que podem degradar macromoléculas e componentes
celulares, como ocorre no lisossomo. Os vacúolos vegetais também atuam no equilíbrio
hídrico e podem ser usados para armazenar compostos como toxinas e pigmentos (partículas
coloridas).

Lisossomos x peroxissomos
Um ponto que pode ser confuso é a diferença entre lisossomos e peroxissomos.
Ambos os tipos de organelas estão envolvidos na degradação de moléculas e neutralização
dos perigos à célula. Ainda, ambos se apresentam como pequenas bolhas arredondadas nos
diagramas.
No entanto, o peroxissomo é uma organela diferente, com propriedades e funções
singulares na célula. Ele abriga enzimas envolvidas nas reações de oxidação, que produzem
peróxido de hidrogênio como subproduto. As enzimas degradam ácidos graxos e
aminoácidos e elas também desintoxicam algumas substâncias que entram no corpo. Por
exemplo, o álcool é desintoxicado pelos peroxissomos encontrados nas células hepáticas.
Observe que os peroxissomos—diferentemente dos lisossomos—não são parte do
sistema endomembranar. Isto significa que eles não recebem vesículas do complexo de Golgi.
Mitocôndrias e Cloroplastos
Pontos Principais:
Mitocôndrias são as "usinas de energia" da célula, quebrando moléculas de
combustível e capturando energia na respiração celular.
Cloroplastos são encontrados em plantas e algas. Eles são responsáveis pela captura
de energia luminosa para fabricar açúcares na fotossíntese.
Mitocôndria e cloroplastos provavelmente começaram como bactérias que foram
englobadas por células maiores (a teoria endossimbiótica).
Introdução
Você deve saber que o seu corpo é feito de células (trilhões delas). Você também deve
saber que a razão pela qual você precisa de comida — como vegetais — é para que você
tenha energia para fazer coisas como praticar esportes, estudar, andar e até mesmo respirar.
Mas o que acontece exatamente no seu corpo para transformar a energia armazenada
nos brócolis de forma que o seu corpo consiga usá-la? E como a energia acaba armazenada
nos brócolis?

As respostas para estas perguntas têm muito a ver com duas importantes organelas:
mitocôndrias e cloroplastos.
Cloroplastos são organelas encontradas nas células dos brócolis, assim como
nas células de outras plantas e algas. Elas capturam a energia da luz e a armazenam como
moléculas de combustível nos tecidos das plantas.
Mitocôndrias são encontradas dentro de suas células, e também nas células de
plantas. Elas convertem a energia armazenada nas moléculas dos brócolis (ou outras
moléculas combustíveis) em uma forma que a célula consegue utilizar.
Vamos estudar com mais detalhes estas duas organelas muito importantes.
Cloroplastos
Cloroplastos são encontrados somente em plantas e algas fotossintetizantes.
(Humanos e outros animais não possuem cloroplastos.) O papel do cloroplasto é realizar um
processo chamado fotossíntese.
Na fotossíntese, a energia luminosa é coletada e usada para construir açúcares a partir
do dióxido de carbono. Os açúcares produzidos na fotossíntese podem ser usados pela célula
da planta, ou podem ser consumidos por animais que comem plantas, como os humanos. A
energia contida nestes açúcares é colhida através de um processo chamado respiração celular,
o qual acontece dentro das mitocôndrias das células das plantas e dos animais.
Cloroplastos são organelas em forma de disco encontradas no citosol da célula. Eles
possuem membranas externa e interna com um espaço intermembranar entre elas. Se você
atravessasse as duas camadas de membrana e alcançasse o espaço no centro, você descobriria
que ele contém discos de membrana conhecidos como tilacóides, dispostos em pilhas
interconectadas chamadas de grana (singular, granum).

Imagem modificada de "Chloroplast mini," de Kelvin Ma (CC BY 3.0).
Diagrama de um cloroplasto, mostrando membrana externa, membrana interna,
espaço intermembranar, estroma e tilacoides em pilhas, chamadas grana.
A membrana de um disco tilacóide contém complexos de coleta de luz que incluem
a clorofila, um pigmento que dá às plantas a sua cor verde. Discos tilacóides são ocos e o
espaço dentro do disco é chamado de espaço tilacóide ou lúmen, enquanto o espaço
contendo fluido envolvendo os tilacóides é chamado de estroma.
Mitocôndrias
Mitocôndrias (singular, mitocôndria) são geralmente chamadas de usinas de energia
ou fábricas de energia da célula. A função delas é produzir um suprimento constante de
adenosina trifosfato (ATP), a principal molécula carregadora de energia da célula. O
processo de fabricar ATP usando energia química de combustíveis, tais como os açúcares, é
chamado de respiração celular, e muitos desses passos acontecem dentro da mitocôndria.
As mitocôndrias são suspensas no citosol gelatinoso da célula. Elas possuem formato
oval e possuem duas membranas: uma externa, envolvendo toda a organela, e uma interna,
com muitas saliências internas chamadas cristas que aumentam a área de superfície.
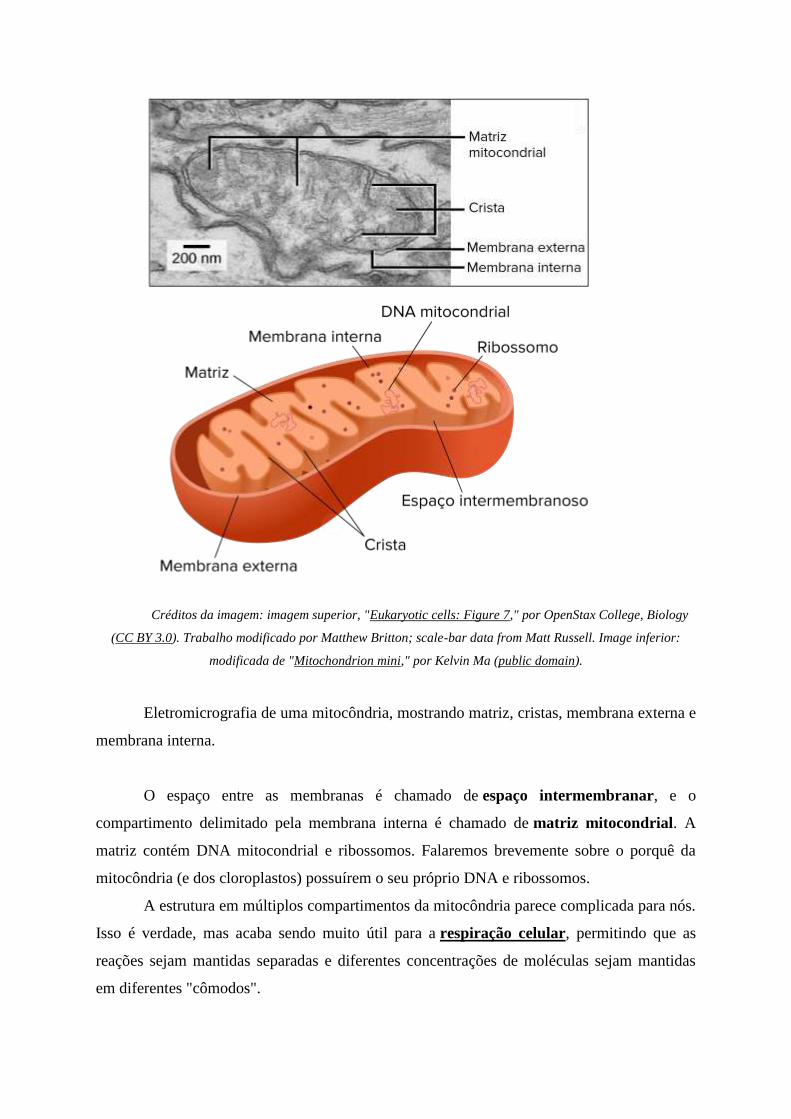
Créditos da imagem: imagem superior, "Eukaryotic cells: Figure 7," por OpenStax College, Biology
(CC BY 3.0). Trabalho modificado por Matthew Britton; scale-bar data from Matt Russell. Image inferior:
modificada de "Mitochondrion mini," por Kelvin Ma (public domain).
Eletromicrografia de uma mitocôndria, mostrando matriz, cristas, membrana externa e
membrana interna.
O espaço entre as membranas é chamado de espaço intermembranar, e o
compartimento delimitado pela membrana interna é chamado de matriz mitocondrial. A
matriz contém DNA mitocondrial e ribossomos. Falaremos brevemente sobre o porquê da
mitocôndria (e dos cloroplastos) possuírem o seu próprio DNA e ribossomos.
A estrutura em múltiplos compartimentos da mitocôndria parece complicada para nós.
Isso é verdade, mas acaba sendo muito útil para a respiração celular, permitindo que as
reações sejam mantidas separadas e diferentes concentrações de moléculas sejam mantidas
em diferentes "cômodos".

Embora as mitocôndrias sejam encontradas na maioria dos tipos celulares de humanos
(assim como na maioria dos tipos celulares de outros animais e plantas), os seus números
variam dependendo da função da célula e de sua demanda de energia. Por exemplo, as células
musculares possuem tipicamente alta demanda de energia e grande número de mitocôndrias,
enquanto que as células vermelhas do sangue, que são altamente especializadas para
transporte de oxigênio, não possuem mitocôndrias.
De onde estas organelas se originaram?
Tanto as mitocôndrias quanto os cloroplastos contêm o seu próprio DNA e
ribossomos. Por que estas organelas precisariam de DNA e ribossomos, quando existe DNA
no núcleo e ribossomos no citosol?
Fortes evidências apontam para a endossimbiose como resposta para o enigma.
Simbiose é a relação no qual organismos de duas espécies separadas vivem em uma relação
próxima e dependente. Endossimbiose (endo- = “dentro”) é um tipo específico de simbiose
onde um organismo vive dentro do outro.
Imagem modificada de "Origem eucarionte: Figura 4," de OpenStax College, Biology, (CC BY 4.0).

1. O primeiro evento endossimbiótico ocorreu: O ancestral eucarionte consumiu
bactéria aeróbicas que evoluíram em mitocôndrias.
2. Em um segundo evento endossimbiótico, o eucarionte ancestral consumiu
bactérias fotossintetizantes que evoluíram em cloroplastos.
Bactérias, mitocôndrias e cloroplastos são similares no tamanho. Bactérias também
possuem DNA e ribossomos similares àqueles de mitocôndrias e cloroplastos. Baseado nesta
e outras evidências, cientistas pensam que células hospedeiras e bactérias formaram
relações endossimbióticas há muito tempo, quando células hospedeiras individuais
englobaram bactérias aeróbicas (que usam oxigênio) e fotossintetizantes, mas não as
destruíram. Através de milhões de anos de evolução, as bactérias aeróbicas se tornaram
mitocôndrias e as bactérias fotossintetizantes se tornaram cloroplastos.
.
A parede celular
Apesar das plantas não produzirem colágeno, elas têm seu próprio tipo de estrutura
extracelular de suporte: a parede celular. A parede celular é uma cobertura rígida que
envolve a célula, protegendo-a e dando a ela suporte e forma. Você já percebeu que quando
morde um vegetal cru, como aipo (salsão), ele é crocante? Uma boa parte disso é causada
pela rigidez das paredes celulares do aipo.
Fungos também têm paredes celulares, assim como alguns protistas (um grupo
formado principalmente por eucariontes unicelulares) e a maioria dos procariontes - embora
eu não recomende morder nenhum deles para ver se são crocantes!
A parede celular vegetal é constituída por moléculas secretadas pela célula. A
principal molécula orgânica da parede celular vegetal é a celulose, um polissacarídeo
composto de unidades de glicose. A celulose organiza-se em fibras chamadas microfibrilas,
como mostrado no diagrama abaixo.

Crédito da imagem: "Diagrama da parede celular das plantas" por Mariana Ruiz Villareal, dominio
público
Imagem da parede celular vegetal, mostrando a rede de microfibrilas de celulose e
pectinas (sendo que as pectinas são especialmente abundantes na lamela média).
A maioria das paredes celulares vegetais contém uma variedade de diferentes
polissacarídeos e proteínas. Além da celulose, outros polissacarídeos comumente encontrados
na parede celular das plantas incluem hemicelulose e pectina, mostrados no diagrama acima.
A lamela média, mostrada ao longo da parte superior do diagrama, é uma camada adesiva
que ajuda a manter unidas as paredes celulares das células adjacentes.
Referência: Este texto foi copiado na íntegra do site “khanacademy” com algumas
modificações. Disponível em: https://pt.khanacademy.org/science/biology/structure-of-a-cell








![aula antimicrobianos 190410.ppt [Modo de Compatibilidade] · Aminoglicosídeos Mecanismo: Inibição da síntese protéica e redução da fidelidade de tradução do RNAm no ribossomo](https://img.document.onl/doc/110x75/5bf4389d09d3f21b2a8cd8f9/aula-antimicrobianos-modo-de-compatibilidade-aminoglicosideos-mecanismo.jpg)



















