Embed Size (px)
Citation preview

Prof. Didier SalmonMSc Cristiane S. Lessa
GLICÓLISE
Dezembro 2015
Bioquímica para Enfermagem
16/12/15

• Oxidação da glicose a piruvato
O que acontece com a glicose que ingerimos?
Matriz extracelular e polissacarídeos da
parede celular
Glicogênio, amido, sacarose
PiruvatoRibose-5-fosfato
GLICOSE
Armazenamento
Oxidação por glicólise
Síntese de polímeros estruturais
Oxidação pela via da pentose-fosfato

• Do grego antigo glykýs, adocicado e lýsis, quebra, degradação;
• É a sequência metabólica de oxidação da glicose;
• Possui 10 reações;
• Rota de produção de ATP;
• Presente em todos os tipos de tecido.
O que é a glicólise?
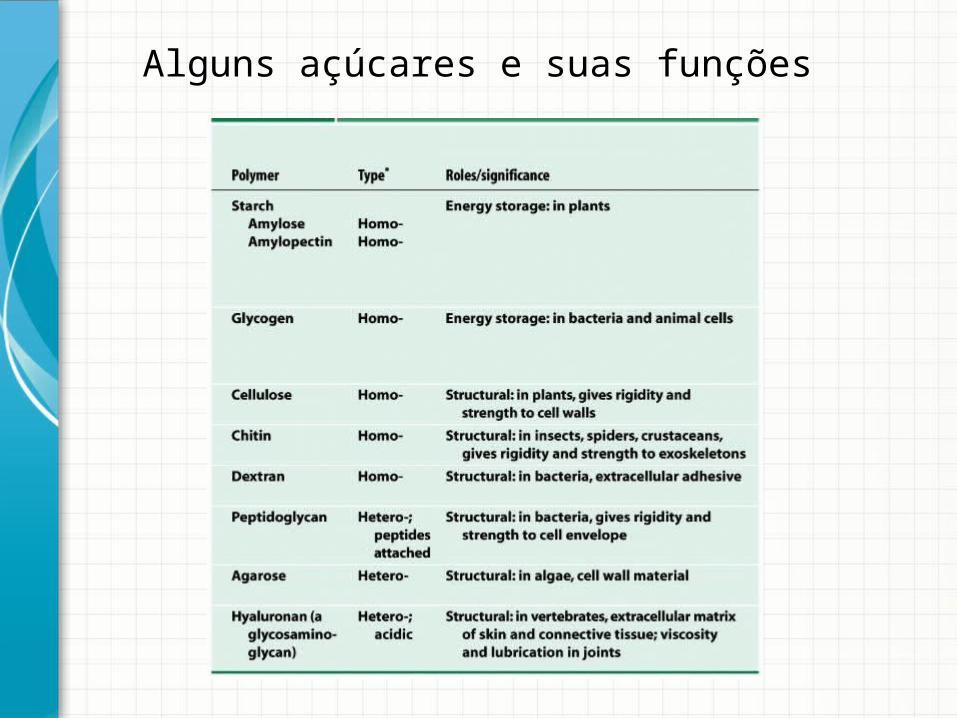
Alguns açúcares e suas funções

GlicóliseQuebra do açúcar (‘glykys’
‘lysis’)
Via fundamental para a oxidação da molécula de glicose em duas
moléculas de piruvato, conservando energia na forma
de ATP e NADH
Possui duas fases: preparatória e pagamento

hexoquinaseG0’ = - 16,7 kJ/mol
triose fosfato isomeraseG0’ = + 7,5 kJ/mol
aldolaseG0’ = + 23,8 kJ/mol
fosfofrutoquinase-1G0’ = - 14,2 kJ/mol
fosfoglicose isomeraseG0’ = + 1,7 kJ/mol
fosfoglicerato mutaseG0’ = + 4,4 kJ/mol
fosfoglicerato quinaseG0’ = - 18,8 kJ/mol
gliceraldeído 3-fosfato desidrogenase
G0’ = + 6,3 kJ/mol
enolase
G0’ = + 7,5 kJ/molpiruvato quinase
G0’ = - 31,4 kJ/mol
Fase preparatória: fosforilação da
glicose e conversão em gliceraldeído-3-
fosfato
Fase de pagamento:
conversão oxidativa da
gliceraldeído-3-fosfato em piruvato e
formação acoplada de ATP e NADH

hexocinase
Fosfofrutocinase
Piruvato-cinase
• As 3 reações irreversíveis tem grande ΔG negativa;• As outras reações tem próximo de 0 (equilíbrio);• Contorno das reações irreversíveis na gliconeogênese.

1ª etapa – fase preparatória: dupla fosforilação da glicose à custa de 2 ATP

Hexocinase
A glicose sofre fosforilação no
carbono 6, formando glicose-6-fosfato,
sendo ATP o doador do grupo fosfato.
Reação 1
O Mg2+ protege as cargas negativas do grupo fosforil do ATP, tornando o átomo de fósforo terminal um alvo mais fácil para o ataque nucleofílico por um grupo –OH da glicose

Fosfoglicose Isomerase
A enzima catalisa a isomerização
reversível da glicose-6-fosfato em frutose-
6-fosfato
Reação 2
Ligação na enzima e abertura do anel
Catálise ácida e formação da frutose-6-fosfato
Dissociação e fechamento do anel
Isomerização de uma aldose (glicose) em cetose (frutose)

Fosfofrutocinase-1 (PFK-1)
Catálise da transferência do grupo fosforil do ATP para a
frutose-6-fosfato
Reação 3
Etapa “comprometida” da via
Direcionamento para a glicólise com a formação da frutose-1,6-bifosfato
Essa é a primeira etapa “comprometida” da via glicolítica; a glicose-6-fosfato e a frutose-6-fosfato tem outros destinos possíveis, mas a frutose-1,6-bifosfato é direcionada para a glicólise

2ª etapa: clivagem da hexose produzindo 2 trioses fosforiladas, que são
interconvertíveis
Nas concentrações pequenasde reagentes a reação é reversível

AldolaseClivagem da hexose produzindo 2 trioses fosforiladas, que são
interconvertíveis
Catálise de condensação aldólica
A clivagem da frutose-1,6-bifosfato gera duas trioses-
fosfato: gliceraldeído-3-fosfato e di-
hidroxiacetona-fosfato.

Reação da Aldolase (classe I)
(delocalização eletrônica +clivagem da ligação C-C)
Resíduos de lisina e tirosina da enzima se ligam covalentemente ao substrato, estabilizando o estado intermediário
Formação da base de Schiff ocorre quando há ligação dupla carbono-nitrogênio

Interconversão de trioses
Triose-fosfato-isomerase
Reação 5
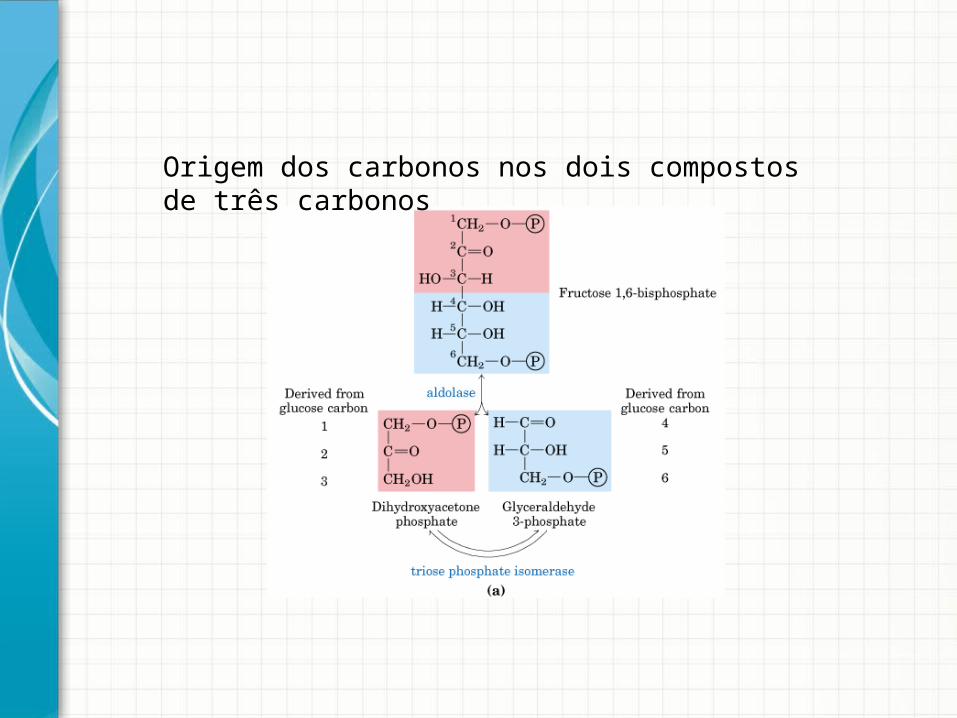
Origem dos carbonos nos dois compostos de três carbonos

3ª Etapa: Oxidação e nova fosforilação das trioses fosfato (por Pi), formando 2 moléculas de 1 intermediário com 2 grupos fosfato
Fase de pagamento
Energia conservada na forma de ATP e NADH

Gliceraldeído-3-fosfato desidrogenase
O grupo aldeído do gliceraldeído-3-fosfato
é oxidado em um anidrido de ácido
carboxílico com ácido fosfórico, chamado
acil-fosfato.
Reação 6

A oxidação do carbono torna a entrada do Fosfato inorgânico (Pi) mais favorável
1. Oxidação do aldeído (gliceraldeído 3-fosfato) a ácido carboxílico, com redução de NAD. Reação termodinamicamente favorável.
2. Ligação do Ác. Carboxílico com Pi, formando anidrido carboxílico-fosfórico, que é endergônica.
As reações ocorrem acopladas por um intermediário rico em energia.
Reação pode ser inibida pelo arseniato que compete com o fosfato

Oxidação pelo NAD+
fosforólise
Inibição de GAP desidrogenase
Reação da gliceraldeído-3-fosfato-desidrogenase
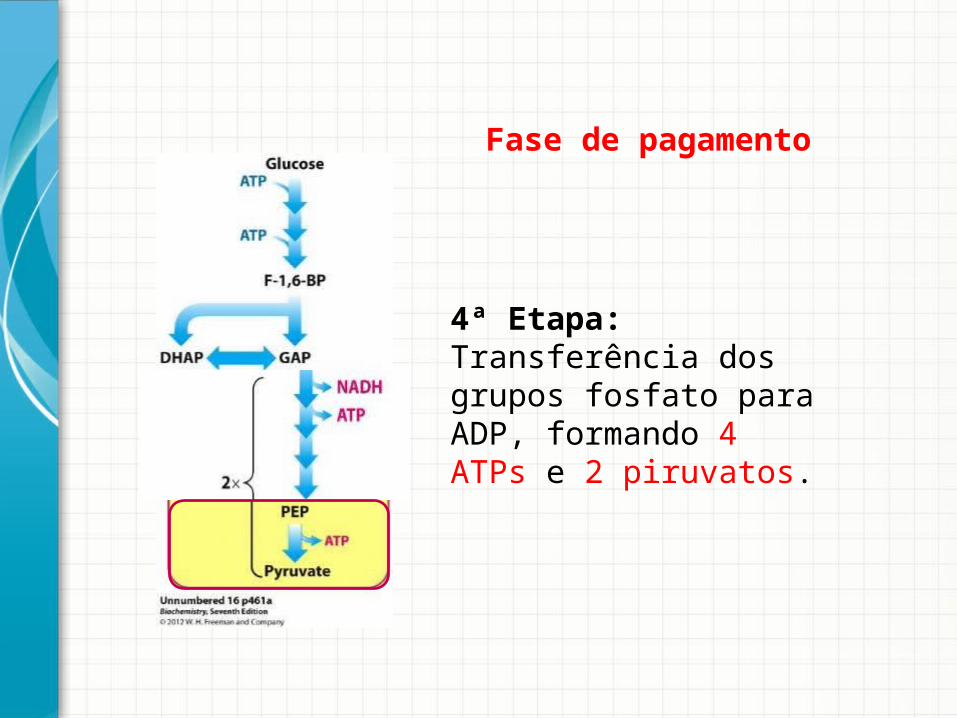
4ª Etapa: Transferência dos grupos fosfato para ADP, formando 4 ATPs e 2 piruvatos.
Fase de pagamento

Fosfoglicerato-cinase
A enzima catalisa a transferência do grupo fosforil de alta energia do grupo carboxil do 1,3-bifosfoglicerato para o ADP, formando ATP e 3-
fosfoglicerato
Reação 7
Acoplamento das reações GAPDH e PGK (reações 6 e 7):GAP + Pi + NAD+ → 1,3-BPG + NADH G 0’ = +6.3 kJ/mol 1,3-BPG + ADP → 3PG + ATP G 0’ = -18.5 kJ/mol
G 0’ = -12.2 kJ/mol
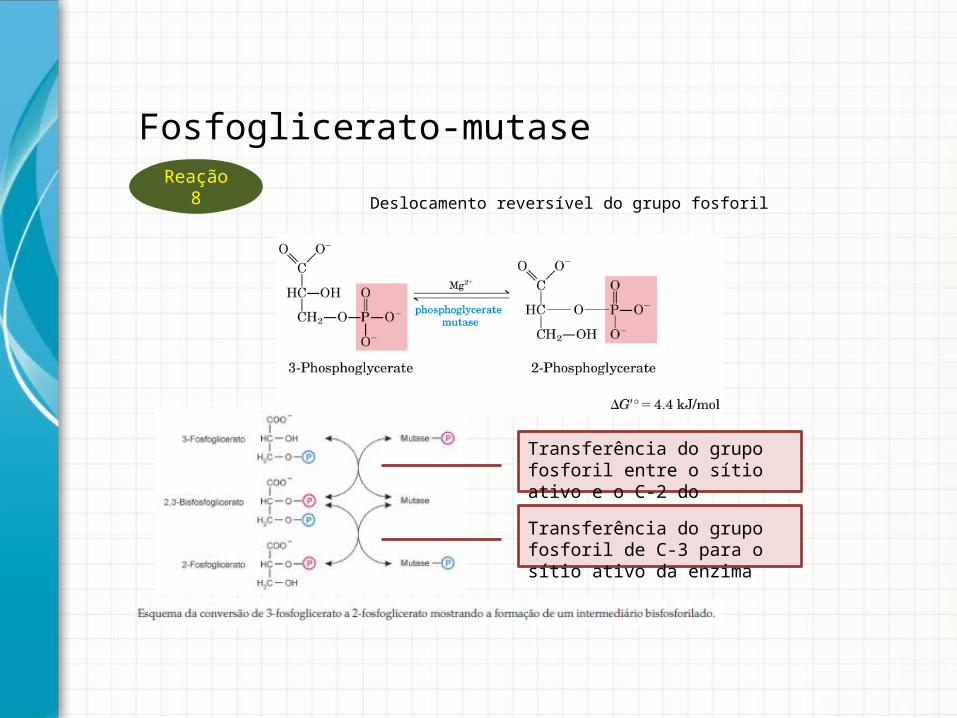
Deslocamento reversível do grupo fosforil
Transferência do grupo fosforil entre o sítio ativo e o C-2 do substrato
Transferência do grupo fosforil de C-3 para o sítio ativo da enzima
Fosfoglicerato-mutaseReação 8
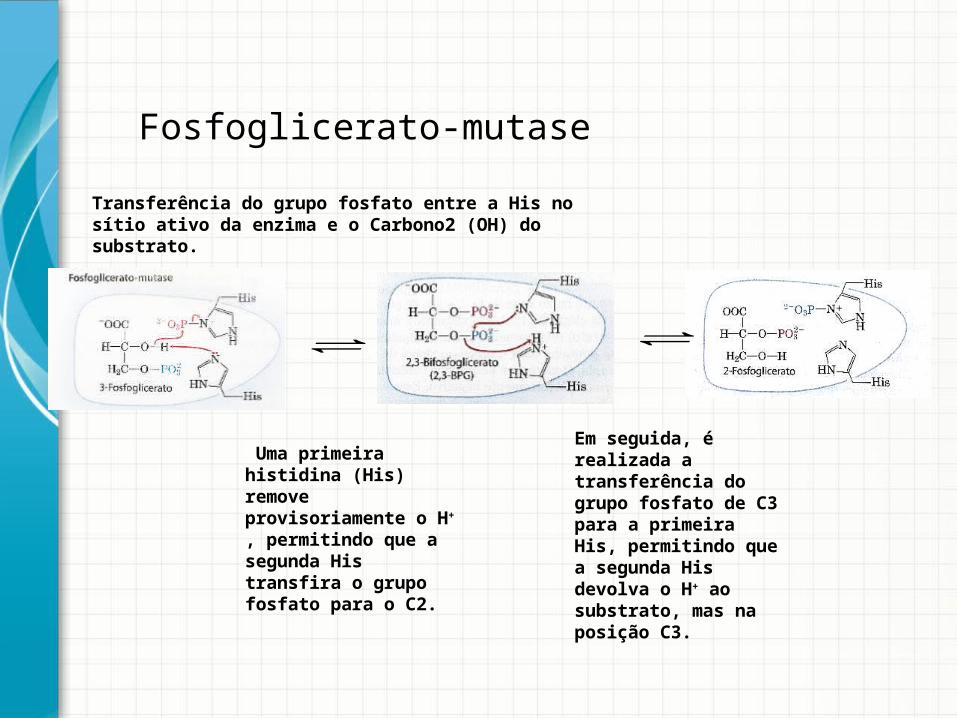
Fosfoglicerato-mutase
⇌ ⇌
Transferência do grupo fosfato entre a His no sítio ativo da enzima e o Carbono2 (OH) do substrato.
Uma primeira histidina (His) remove provisoriamente o H+ , permitindo que a segunda His transfira o grupo fosfato para o C2.
Em seguida, é realizada a transferência do grupo fosfato de C3 para a primeira His, permitindo que a segunda His devolva o H+ ao substrato, mas na posição C3.
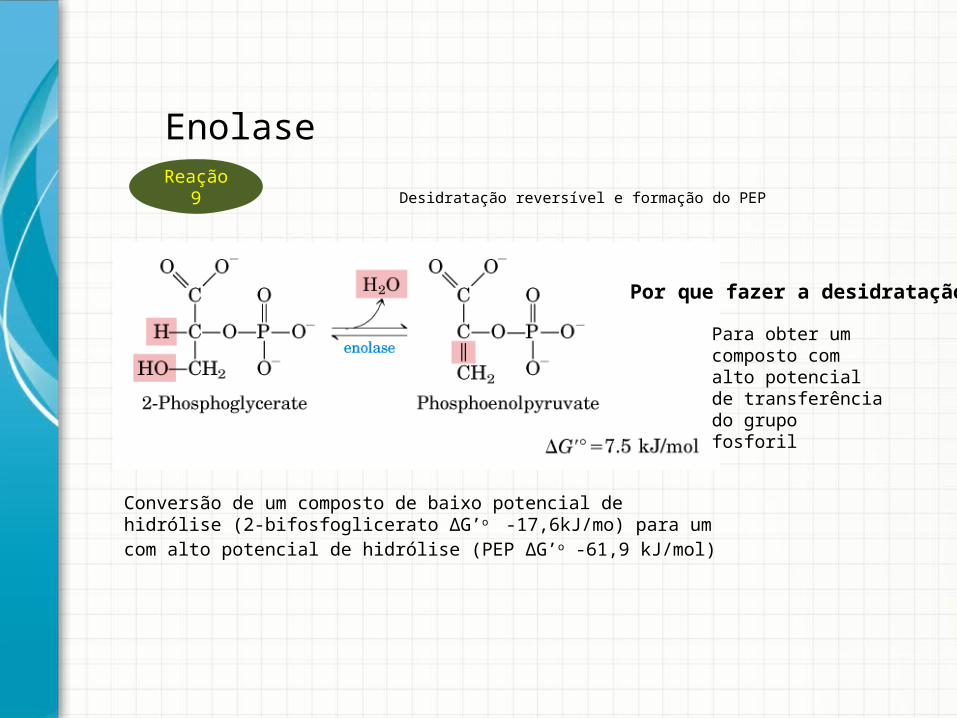
Desidratação reversível e formação do PEP
Conversão de um composto de baixo potencial de hidrólise (2-bifosfoglicerato ΔG’o
-17,6kJ/mo) para um com alto potencial de hidrólise (PEP ΔG’o -61,9 kJ/mol)
EnolaseReação 9
Por que fazer a desidratação?
Para obter um composto com alto potencial de transferência do grupo fosforil

Duas partes: - ADP ataca a fosforila do PEP formando ATP e enolpiruvato - tautomerização do PEP a piruvato
Formação de ATP por transferência do grupo fosforil
Piruvato-cinaseReação 10
Acoplamento das reações: -61.9 kJ/mol (hidrólise de PEP
Metade da energia está armazenada durante a formação do ATP (∆G= -30.5 kJ/mol) e o restante (∆G= -31,4 kJ/mol) serve para impulsionar a reação.
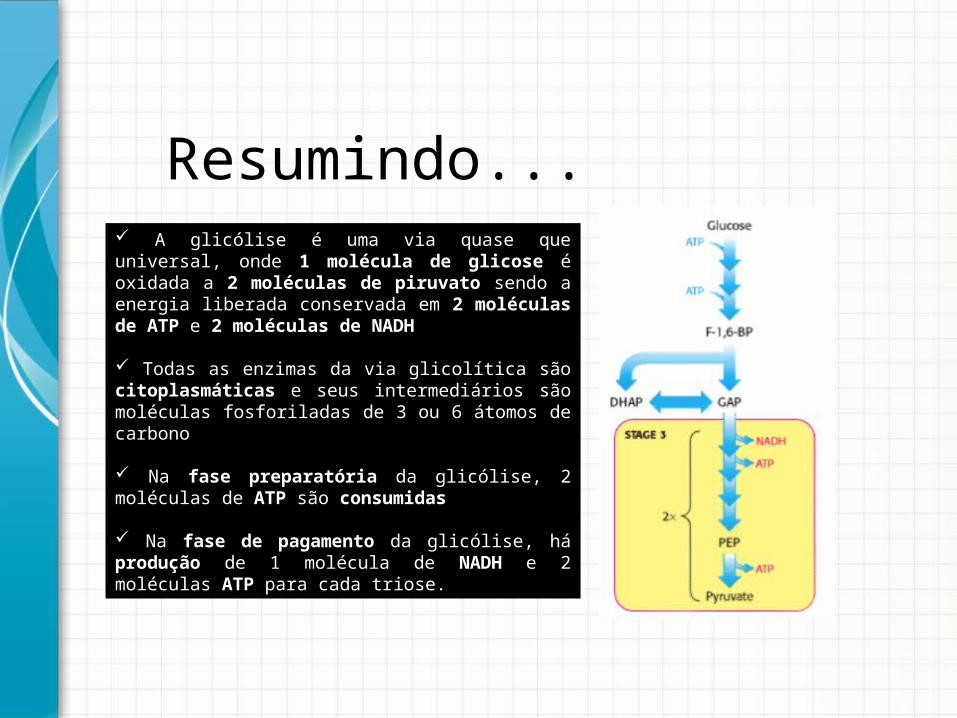
A glicólise é uma via quase que universal, onde 1 molécula de glicose é oxidada a 2 moléculas de piruvato sendo a energia liberada conservada em 2 moléculas de ATP e 2 moléculas de NADH
Todas as enzimas da via glicolítica são citoplasmáticas e seus intermediários são moléculas fosforiladas de 3 ou 6 átomos de carbono
Na fase preparatória da glicólise, 2 moléculas de ATP são consumidas
Na fase de pagamento da glicólise, há produção de 1 molécula de NADH e 2 moléculas ATP para cada triose.
Resumindo...

Equação geral da glicólise
Glicose + 2ATP + 2NAD+ + 4ADP + 2Pi 2Piruvatos + 2ADP + 2NADH + H+ + 4ATP + H2O
2 2
Glicose + 2NAD+ + 2ADP + 2Pi 2Piruvatos + 2NADH + H+ + 2ATP + H2O
Saldo final

De onde vem a glicose circulante?
Gliconeogênese
Glicogênio7% do peso líquido

Digestão de Carboidratos• Inicia-se na boca• a-amilase salivar: rompimento das ligações a(1→4)• Amido = amilose a(1→4) e amilopectina • Amilopectina e glicogênio possuem ligações a(1→6), e sendo
assim, o produto da digestão da a-amilase contém uma mistura de moléculas de oligossacarídeos menores e ramificados.

Digestão de Carboidratos
• A digestão dos carboidratos cessa temporariamente no estômago, devido ao pH que inativa a a-amilase salivar
• O conteúdo gástrico é neutralizado pelo bicarbonato secretado pelo pâncreas, e a a-amilase pancreática continua o processo digestivo no intestino
Amilase pancreática gera maltose e dextrinas, que são degradadas até glicose por enzimas do epitélio intestinal com borda em escova

Digestão de Carboidratos• A digestão final ocorre pela ação de enzimas da mucosa intestinal –dissacaridases e
oligossacaridases• Há a absorção de monossacarídeos pelas células da mucosa intestinal
Mono GlicoseDi MaltoseTri Maltotriose

Como os açúcares que ingerimos na alimentação entram na via glicolítica?

Frutose• A frutose livre presente em frutas ou formada pela hidrolise da sacarose é fosforilada
pela hexoquinase. Esta é a principal via pela qual a frutose entra na via glicolítica.
• Frutose + ATP → frutose-6-fosfato + ADP
• No fígado a frutose entra na via glicolítica através da frutoquinase que catalisa a fosforilação do C1 da frutose.
• Frutose + ATP → frutose-1-fosfato + ADP
• A frutose-1-fosfato é clivada em gliceraldeído e diidroxicetona-fosfato pela frutose 1-fosfato aldolase. A diidroxicetona-fosfato é convertida em gliceraldeído-3-fosfato e o gliceraldeído é fosforilado. Portanto os produtos frutose-1-fosfato entram na via glicolítca como gliceraldeído-3-fosfato.

Frutose-1-fosfato gliceraldeído + di-hidroxicetona-fosfatoFrutose-1-fosfato-aldolase
gliceraldeído-3-fosfato
Triose quinase
Frutose-1-fosfato-aldolase
VIA GLICOLÍTICA
Frutose
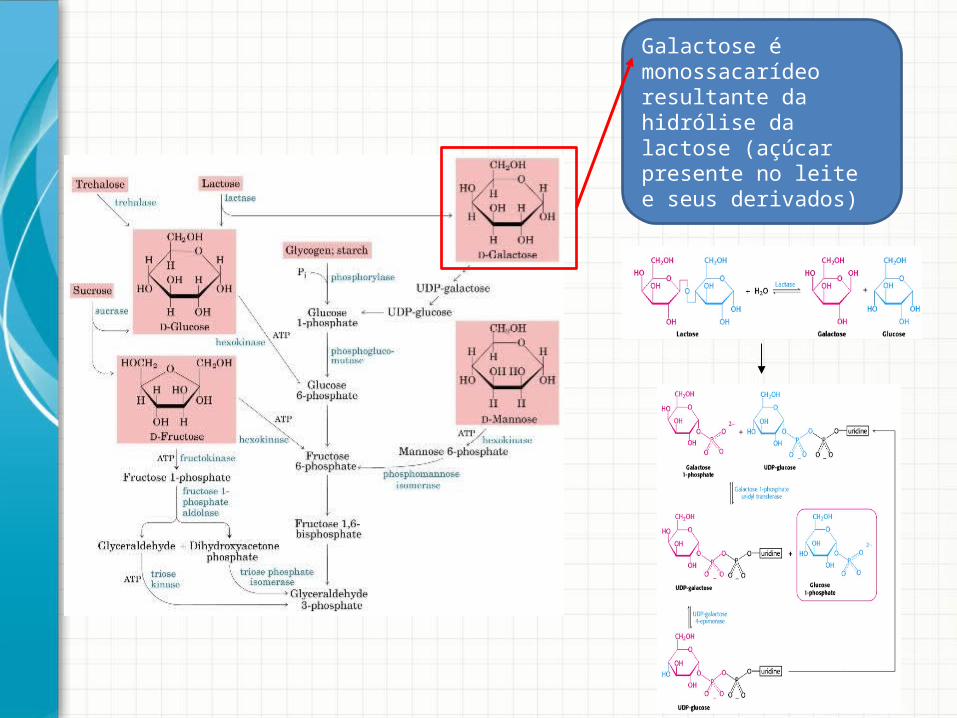
Galactose é monossacarídeo resultante da hidrólise da lactose (açúcar presente no leite e seus derivados)

Galactose
• Galactose é um monossacarídeo resultante da hidrolise da lactose (açúcar presente no leite e seus derivados)
Galactoquinase
Difosfato de uridina (UDP)Coenzima transportadora de
grupos hexoses
Deficiência de enzimas?
Gera acúmulo de galactose, sendo convertido a galactiol (açúcar alcoólico)
Galactiol leva à formação de catarata

Galactosemia• Ausência das enzimas de conversão da galactose;• Altas concentrações de galactose no sangue e urina;• Conversão de galactose em galactitol;• Acumulo de galactitol no cristalino causa catarata na infância.
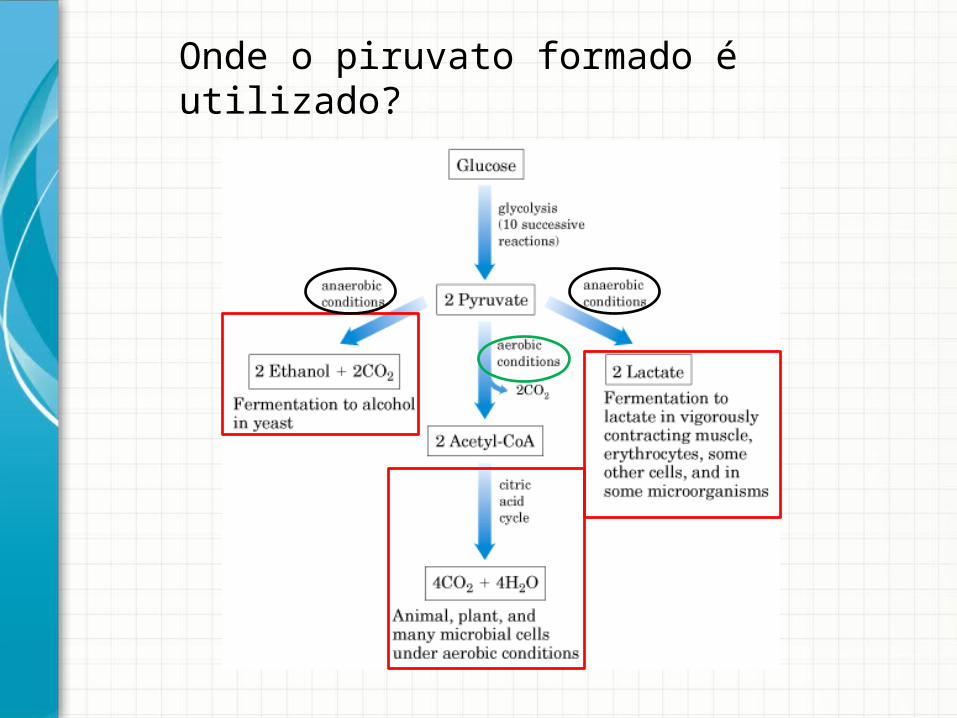
Onde o piruvato formado é utilizado?

Destinos do piruvato em anaerobiose• Fermentação
alcoólica• Fermentação lática

Um pouco de história...• Louis Pasteur
– 1861: crescimento de leveduras, por grama de glicose, maior na presença do que na ausência de ar. “EFEITO PASTEUR”: Glicose consumida mais lentamente na presença de ar do que na ausência.
– Teoria vitalista – fermentação era catalisada por uma ‘força vital’ dentro da célula (fermentos).
• Eduard Buchner– 1907 – Prêmio Nobel– Derruba a Teoria vitalista – a fermentação ocorre sem vida organizada –
Zimases (enzima do levedo de cerveja, provocando degradação da glicose em álcool e CO2)
• Harden e Young– 1909: isolamento do primeiro intermediário da via glicolítica– 1929: Arthur Harden - Prêmio Nobel– Descoberta de um procedimento para acelerar a fermentação: adição de
Pi ao meio.A zimase podia ser separada em 2 extratos: um contendo moléculas grandes e sensíveis ao calor (as enzimas) e uma fração de moléculas menores e pouco sensíveis ao calor (as coenzimas), e que estes só fermentavam o açúcar quando juntos.

Um pouco de história...O efeito Pasteur é um efeito inibidor de oxigénio sobre o processo de fermentação
Enquanto a concentração de oxigénio é baixo, o produto da glicólise, é transformado em etanol e dióxido de carbono, e a eficiência de produção de energia é baixo
Se a concentração de oxigénio aumenta, o piruvato é convertido em acetil CoA, que pode ser usado no ciclo do ácido cítrico Para cada molécula de glicose, terei 30 a 32 moléculas de ATP formadas
Sob condições anaeróbicas, a taxa de metabolismo da glicose é mais rápido, mas a quantidade de ATP produzido é menor. Quando exposto a condições aeróbias, os aumentos de produção de ATP e a taxa de glicólise diminui, porque o ATP produzido atua como um inibidor alostérico para fosfofrutoquinase 1, a terceira enzima da via da glicólise.
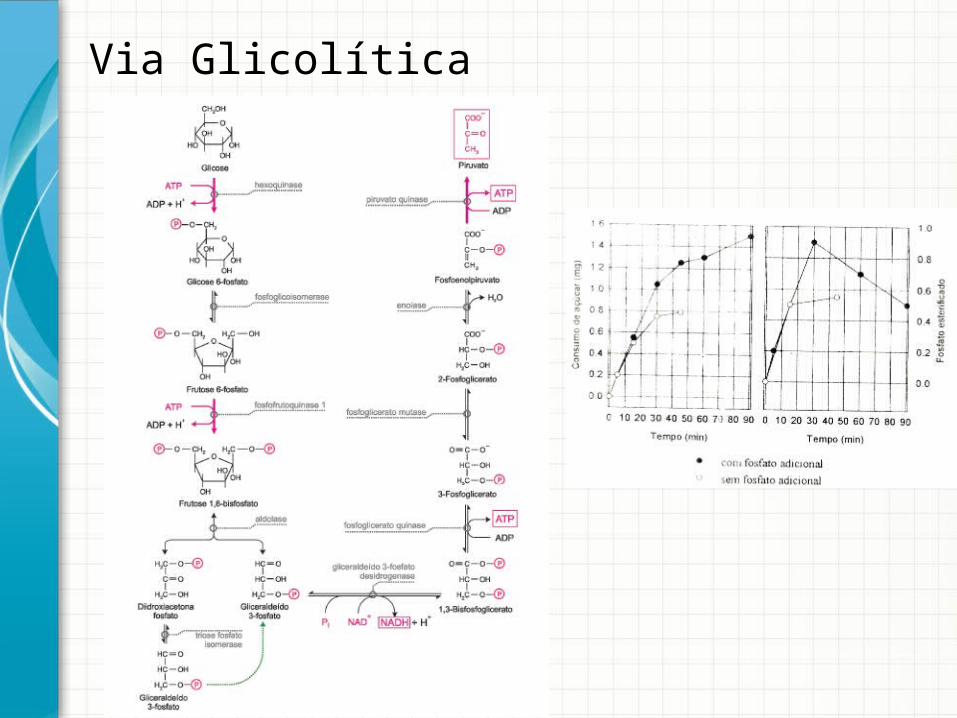
Via Glicolítica

Otto Meyerhoff
1922: Prêmio Nobel - Descoberta da correlação entre o consumo de oxigênio e o metabolismo do ácido lático nos músculos de coelho.
Ativador: obtido por autólise de levedura. O ativador perde a atividade se aquecido por 1 minuto a 50 ºC e conserva-se bem em gelo.
Você pode imaginar a natureza desse ativador?
Para você é espantoso que se obtenha um ativador de músculo de coelho a partir de levedura?

Metabolismo do Etanol no fígado
Álcool desidrogenase
Acetaldeído desidrogenase
ALDH
ADHÁlcool produzido pela digestão e quebra de carboidratos (pouca quantidade)
Por bactérias em nosso intestino (grande quantidade)
Ingestão de bebida alcoolica= aumento dos níveis de acetaldeído
Taquicardia, sudorese, náusea...

Sensibilidade ao álcoolConsumo de álcool segundo diferentes padrões levou a uma evolução divergente.
Existem várias enzimas ADH no homem: dímeros (5 genes). ADH são essenciais pois quebram e metabolizam as moléculas de álcool (tóxico) que é absorvida para o sangue.
População do Sudeste Asiático: maior intolerância ao álcool – acúmulo de acetaldeído – rubor alcoólico (“Asian flush”)
Alcoolismo (tolerância ao álcool)-Populações europeias: alelos ADH2 e ADH3 menos ativas metabolizam lentamente o etanol
- Sudeste asíatico: ~ 50 % pop. possui o alelo mutante ALDH2*2 (8% da atividade do gene wt)

Regulação da Glicólise

1. Número de enzimas (por controles transcricionais/traducionais; Ocorre em minutos ou até horas).
2. Mudança da atividade enzimática (ocorre em segundos) por: Modificação covalente ou ligação a proteína reguladora Regulação alostérica Seqüestro da enzima ou do substrato em compartimentos diferentes
Fluxo de metabólitos de uma via metabólica pode ser modulado por:
Regulação da via glicolítica

Via glicolítica
• Reações irreversíveis– Hexoquinase– Fosfofrutoquinase-1– Piruvato quinase
Gº muito negativo
G0’ = - 16,7 kJ/mol
G0’ = - 14,2 kJ/mol
G0’ = - 31,4 kJ/mol
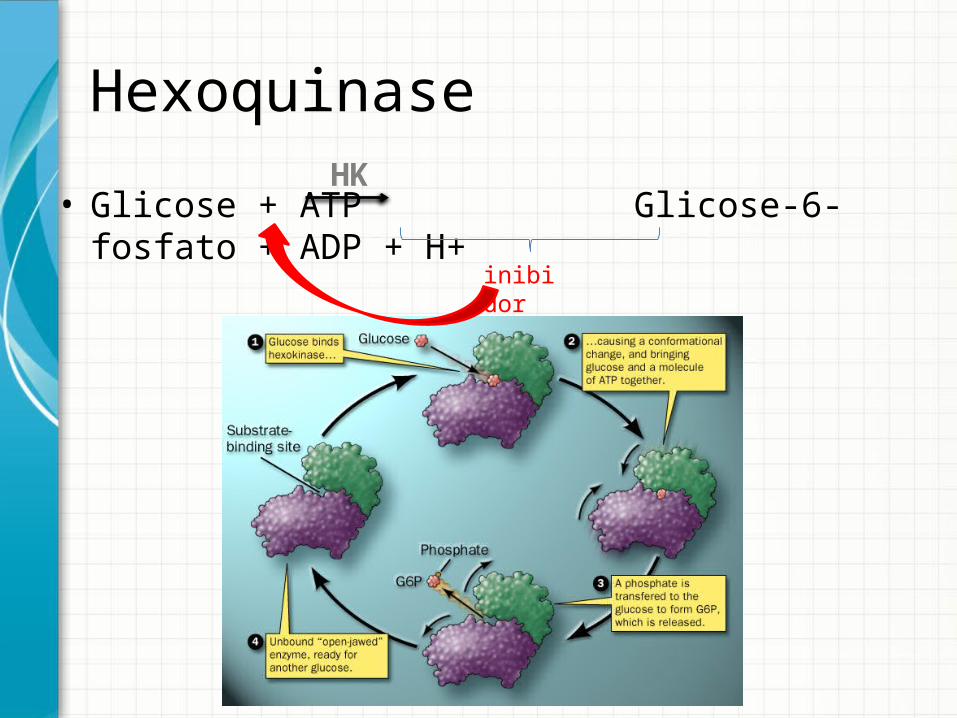
Hexoquinase
• Glicose + ATP Glicose-6-fosfato + ADP + H+HK
inibidor

Hexoquinase• Isoformas
– I, II e III – cinética michaelliana• Km < 0,1 mM – alta afinidade
• [plasmática] de glicose = 4 a 5 mM– Ou seja, isoformas I, II e III funcionam sempre na Vmáx

Glicoquinase• Hexoquinase (músculo): I, II, e III • Glicoquinase ou Hexoquinase IV – presente no fígado: menor
afinidade pela glicose.• Ligada a uma proteína reguladora forma um complexo inativo.

Glicoquinase• Não é inibida por glicose-6-fosfato• Maior Km pela glicose: 10 mM
– Regulada pela [glicose]plasmática – Regulação por sequestro no núcleo celular
Estado Alimentado
> 10 mM
Hepatócito
Glicose não é desperdiçada quando estiver abundante, síntese de glicogênio e ácidos graxos

Glicoquinase• Não é inibida por glicose-6-fosfato• Maior Km pela glicose: 10 mM
– Regulada pela [glicose]plasmática
– Regulação por sequestro no núcleo celular
Jejum
< 10 mM
Hepatócito
Fígado não compete com demais órgãos pela glicose escassa. Prioridade cérebro e músculo

Fosfofrutoquinase-1 (PFK-1)• Fru 6-F → Fru 1,6-bF
– A partir desse ponto o açúcar está comprometido com a via glicolítica
• Reação altamente exergônica e irreversível– G0’ = - 14,2 kJ/mol
• Além do sítio ativo essa enzima possui diversos sítios onde inibidores e ativadores alostéricos se ligam.

Reguladores Alostéricos da PFK-1• Negativos: ATP (retroinibição) e Citrato (CK)
• Positivos: AMP (músculo esquelético) e frutose 2,6-bifosfato
Negativo= diminuição da velocidade de reação
Positivo= aumento da velocidade de reação

Reguladores Alostéricos da PFK-1• Negativos: ATP (retroinibição) e Citrato (CK)
• Positivos: AMP (músculo esquelético) e frutose 2,6-bifosfato
Regulação positiva – aumento da velocidade de reação
Regulação negativa – diminuição da velocidade de reação

Reguladores Alostéricos da PFK-1• Negativos: ATP (retroinibição) e Citrato (CK)
• Positivos: AMP (músc. Esq.) e frutose 2,6-bifosfato Quando a glicose é abundante, há um excesso de frutose-6-fosfato, que passa a ser fosforilada também pela PFK2, gerando frutose-2,6-bifosfato.
A frutose2,6fosfato é um modulador alostérico da PFK-1, aumentando a atividade dessa enzima

Frutose-2,6-Bifosfato• O ativador mais potente da via glicolítica• Ativa PFK-1• Aumenta a velocidade da via

Enzima bifuncional (PFK2/F2,6BPase)• Atividades 6-fosfofruto-2-quinase e frutose-2,6-bifosfatase
• Regulação Alostérica
PFK2
Alta concentração de precursores biossintéticos
Citrato

Regulação por Controle Covalente

Piruvato quinase• Último passo da via glicolítica• Fosfoenolpiruvato + ADP → ATP + piruvato• Tetrâmero apresentando diferentes isoformas
– L (fígado) e M (músculo)
• Regulação Alostérica
Ativação anterógrada
ATP /
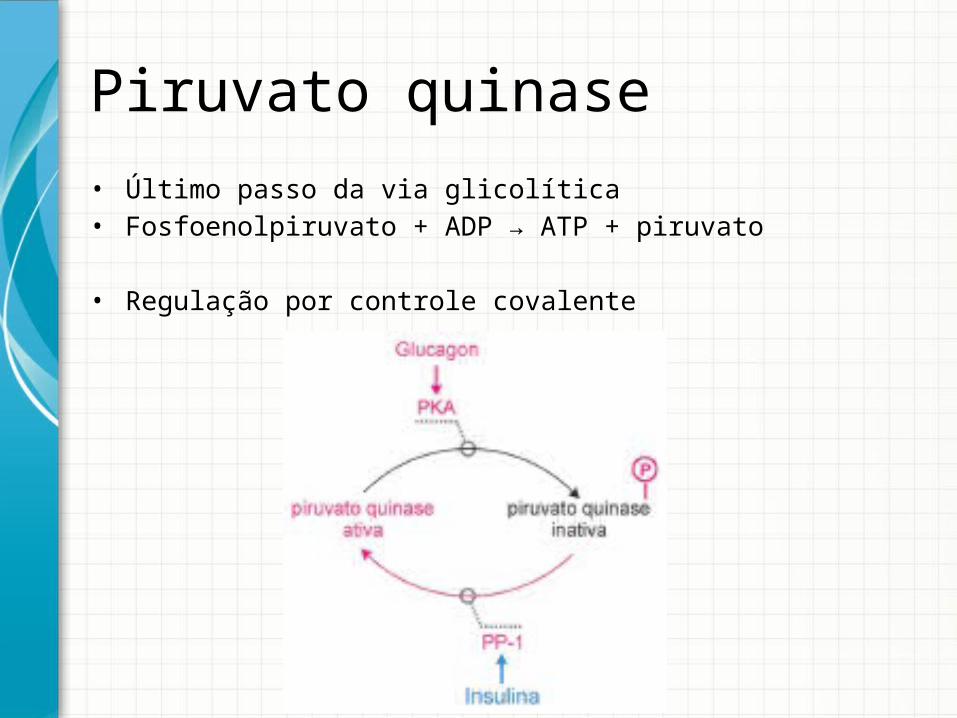
Piruvato quinase• Último passo da via glicolítica• Fosfoenolpiruvato + ADP → ATP + piruvato
• Regulação por controle covalente

Piruvato quinase• Também é um tetrâmero apresentando diferentes isoformas em
diferentes tecidos.• Isoforma L (fígado) e isoforma M (músculo).• Diferença: regulação por ligação covalente (fosforilação)
Fígado deixa de fazer glicólise quando a [glicose] no sangue cai

Regulação da via glicolítica

Via antagônica à glicólise: Gliconeogênese
• Síntese de glicose a partir de compostos que não são carboidratos: aminoácidos, lactato e glicerol
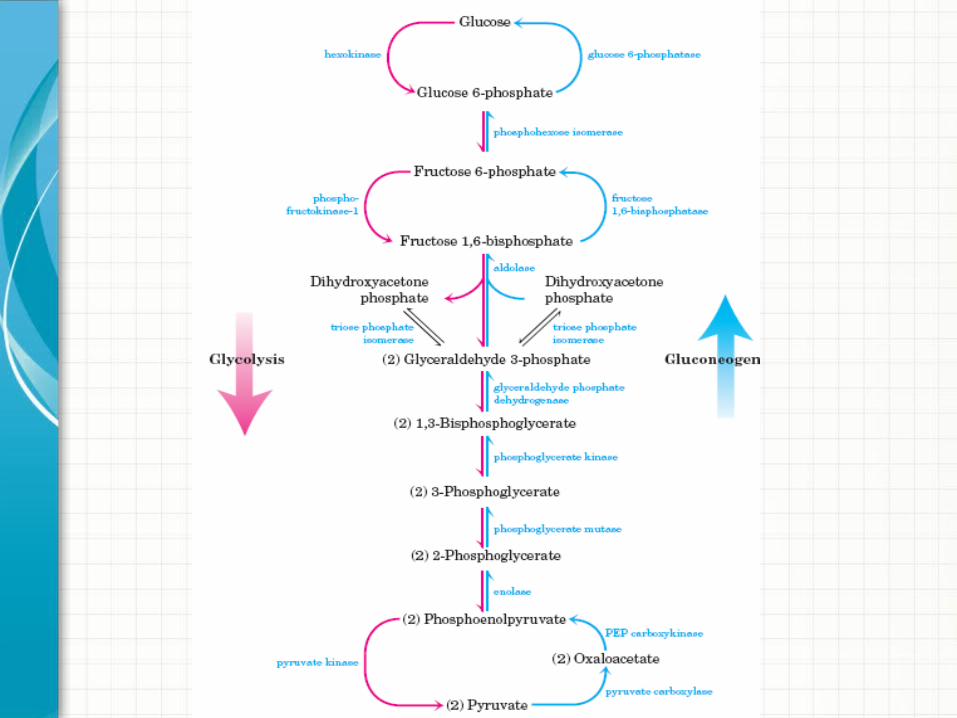

Reações de contorno Gliconeogênese
• Piruvato à fosfoenol piruvato:
Piruvato + bicarbonato +ATP oxaloacetato + ADP + Pi
Oxaloacetato + GTP Fosfoenolpiruvato + GDP + CO2
• A membrana mitocondrial não tem transportador de membrana para o oxaloacetato.
• Oxaloacetato convertido em malato para deixar a mitocôndria;• Malato novamente convertido em oxaloacetato.
Piruvato-carboxilase
PEP-carbocinase

Fosfofrutoquinase-1 e Frutose-1,6-Bifosfatase

Frutose-2,6-BifosfatoAtiva PFK-1
Ativa frutose-1,6-bifosfataseInibe PFK-1

Reações de contorno Gliconeogênese
• Glicose-6-fosfato à glicose:
Glicose-6-fosfato + H2O Glicose + Pi
• Ultima reação de contorno;• Enzima ativada por Mg2+ dos hepatócitos, células renais e
epiteliais do intestino delgado;• Fazem gliconeogênese o fígado e os rins;• Outros tecidos são incapazes de fazer gliconeogênese;
Glicose-6-fosfatase

Regulação Glicólise x Gliconeogênese

Aspectos clínicos

1) Isquemia (Infarto do miocárdio):
Isquemia: • Falta de suprimento sangüíneo para um tecido orgânico; • Necrose do tecido por isquemia
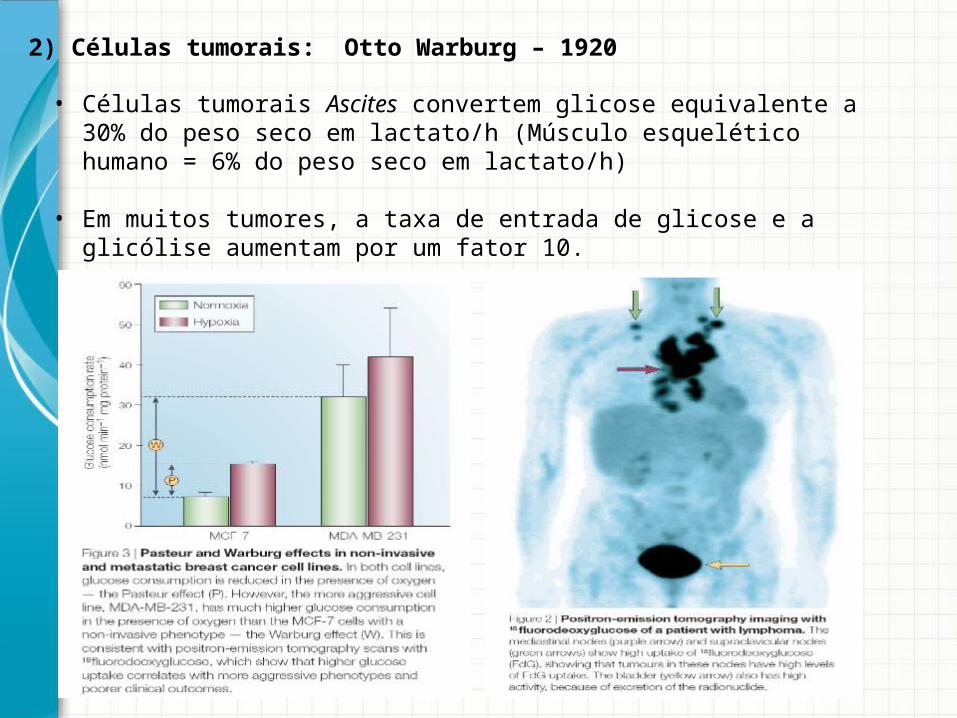
• Células tumorais Ascites convertem glicose equivalente a 30% do peso seco em lactato/h (Músculo esquelético humano = 6% do peso seco em lactato/h)
• Em muitos tumores, a taxa de entrada de glicose e a glicólise aumentam por um fator 10.
2) Células tumorais: Otto Warburg – 1920

Transformação de uma célula normal para tumoral:
• Mudança para um metabolismo glicolitico;• Tolerância a baixo pH;• Mais o tumor é agressivo maior é a sua taxa de fluxo glicolítico (superxpressão de enzimas glicolíticas e dos transportadores)





























