Embed Size (px)
Citation preview

RESPOSTA IMUNE CONTRA ARTRÓPODES PARASITOS
Cleber Oliveira Soares¹
Carlos Luiz Massard²
Cesar A. Mora Hernandez³
Adivaldo Henrique da Fonseca4
¹MsC, Doutorando do Curso de Pós-graduação em Medicina Veterinária - Parasitologia Veterinária, Universidade Federal
Rural do Rio de Janeiro (UFRRJ), Km 47, Rod. Rio-São Paulo, Seropédica-RJ, 23890-000, e-mail: [email protected]; ²PhD, Professor Titular, Depto. de Parasitologia Animal, UFRRJ; ³PhD, Professor Adjunto, Universidade Centroamericana,
Manágua, Nicarágua; 4PhD, Professor Titular, Depto. de Epidemiologia e Saúde Pública, UFRRJ.
Página
INTRODUÇÃO ....................................................................................................................................................................... 1
HIPERSENSIBILIDADE ....................................................................................................................................................... 3
IMUNIDADE ADQUIRIDA ................................................................................................................................................... 3
IMUNIDADE INDUZIDA ...................................................................................................................................................... 6
VACINAS CONTRA ARTRÓPODES .................................................................................................................................. 8
NÍVEIS DE ANTICORPOS CONTRA CARRAPATOS .............................................................................................................. 10
EFEITO DOS IMUNÓGENOS SOBRE OS CARRAPATOS ........................................................................................... 13
EFEITO SOBRE A BIOLOGIA DO CARRAPATO .................................................................................................................... 13 ALTERAÇÕES HISTOLÓGICAS DO TRATO INTESTINAL ..................................................................................................... 15
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................................................................. 17
INTRODUÇÃO
As respostas imunes contra os artrópodes são designadas, essencialmente, contra os antígenos
presentes na saliva, os quais são inoculados no hospedeiro durante o repasto alimentar. Estas respostas
podem ser de três tipos:
1 – Alguns antígenos salivares com baixo peso molecular (haptenos) se associam às proteínas
da pele do hospedeiro para estimular uma resposta imune, de base celular. Numa exposição
subseqüente estes haptenos estimulam uma reação de hipersensibilidade tardia. A dermatite alérgica a
picadas de pulgas, no cão, é caracterizada por este tipo de resposta;

2 2 – Os antígenos salivares podem se ligarem às células de Langerhans presentes na epiderme e
induzirem uma hipersensibilidade cutânea do tipo basofílica, associada à produção de imunoglobulina
da classe G (IgG) e, com a infiltração basofílica;
3 – Os antígenos salivares estimulam a produção de IgE, desencadeando uma reação de
hipersensibilidade do tipo I. Esta resposta induz a uma severa inflamação na pele, ocorrendo o prurido
e a dor.
Estes mecanismos imunológicos podem modificar a pele do hospedeiro, de maneira que o
repasto do artrópode é prejudicado. Entretanto, os artrópodes são hábeis em evadir o sistema imune dos
hospedeiros. As respostas imunes contra antígenos salivares são pouco eficientes, pois não determinam
danos consideráveis no artrópode, são de curta duração e a saliva dos artrópodes possui efeito
imunossupressor (Ribeiro et al., 1985; Tizard, 1995; Wikel, 1996). Os mecanismos de resposta imune
aos artrópodes são similares entre os diferentes grupos destes parasitas.
Os carrapatos são artrópodes ectoparasitas de diferentes grupos de animais. Estes possuem
grande importância por determinarem injúrias aos hospedeiros, pelo hematofagismo, inoculação de
toxinas, determinação de lesões e são responsáveis por transmitirem biológica e/ou mecanicamente a
maior diversidade de agentes patogênicos. Consequentemente, este parasito tem sido alvo de trabalhos
elucidativos para a compreensão da interrelação hospedeiro/ectoparasita.
A expressão da resposta imune a estes parasitas é reconhecida desde o início do século, com
estudos sobre a imunidade adquirida ao Boophilus microplus (Canestrini, 1887), segundo Johnston &
Bancroft (1918). A resistência imunológica adquirida aos carrapatos é expressa, principalmente, sobre
o estágio larval e se faz, especialmente, pela participação de células da inflamação e substâncias que
produzem. Na resistência induzida, a expressão ocorre sobre o estágio adulto do ixodídeo, sendo este
tipo de imunidade determinada por inoculação de antígenos brutos ou particulados (Willadsen, 1980).
Uma grande variedade de imunógenos foram testados objetivando o controle de carrapatos e
outros artrópodes: desde extratos brutos derivados de órgãos do ixodídeo; antígenos "ocultos"
recombinantes, a partir de proteínas de células digestivas; até peptídeos sintetizados. Na resistência
adquirida pouco se conhece sobre o mecanismo que determina a interrupção do repasto sangüineo do
carrapato. Entretanto, na imunidade induzida, mais eficiente na atualidade, o principal mecanismo
decorre da atuação dos anticorpos específicos, bloqueando a atividade endocítica das células digestivas
(Kemp et al., 1986; Wikel, 1996).
Baseado nos conhecimentos de expressão da imunidade dos animais aos ixodídeos, duas
alternativas no controle imunológico foram propostas atualmente: uma através da seleção de indivíduos

3imunocompetentes e outra pelo uso de antígenos purificados como indutor da resistência através da
imunização direta.
HIPERSENSIBILIDADE
A resposta imune humoral dos vertebrados às repetidas infestações por artrópodes é constatada
com a elevação dos níveis de IgM, IgG e IgE, sendo estas duas últimas envolvidas na reação de
hipersensibilidade resultante das infestações. Durante as infestações há uma dinâmica celular, na pele, e
a relação entre linfócitos T CD4 para T CD8 aumenta após suscessivas infestações (Mbow et al.,
1994).
A hipersensibilidade tipo I é um mecanismo associado à resistência ao carrapato, pois observa-
se aumento de mastócitos degranulados e imunoglobulinas da classe E; aumento de infiltrado
eosinofílico na pele e aumento da concentração de IgM e IgG. Ainda na hipersensibilidade, os
antígenos salivares ativam a fração C5 do complemento, que tem função de anafilatoxina e
quimiotática, importantes na reação inflamatória (Gordon & Allen, 1991).
IMUNIDADE ADQUIRIDA
O fenômeno de resistência imune adquirida a carrapatos tem como manifestação primária o
decréscimo do número da população infestante que completa o ciclo após novos desafios (Roberts,
1968). O estágio de larva é o mais afetado neste tipo de imunidade (Willadsen, 1987). Os carrapatos
encontram dificuldades ao realizarem o repasto em bovinos previamente infestados; apresentando estes
artrópodes peso corporal inferior em relação àqueles alimentados em animais sem contato prévio
(Kemp et al., 1976). A fixação dos carrapatos nos hospedeiros previamente infestados induz uma
marcada reação de hipersensibilidade local, fenômeno importante na mediação da resistência
(Willadsen, 1987) (Tabela 2).
Nos sítios de alimentação dos carrapatos, em animais resistentes, desenvolvem-se reações
cutâneas constituídas principalmente de basófilos e eosinófilos (Allen, 1973). Os basófilos possuem um
papel importante na limitação do repasto sangüineo. A expressão da resistência adquirida pode ser
reduzida pela administração de anticorpos anti-basófilos (Brown et al., 1982). A degranulação de
basófilos ocorre durante repetidas infestações na presença de antígenos salivares, e outros tipos
celulares degranulam durante a exposição primária e subseqüente, quando da formação do sítio
alimentar (Brossard & Fivaz, 1982). Enzimas salivares do carrapato podem hidrolisar a membrana

4celular causando a liberação de substâncias vasoativas (Sauer et al., 1995). Antígenos salivares,
imunoglobulinas e complemento foram localizados na junção dermo-epidermal de animais resistentes
(Allen et al., 1979), e a salivação contínua do carrapato permite a formação de complexos antígeno-
anticorpo circulantes que se depositam no sítio alimentar.
Moléculas biorreativas oriundas de células da inflamação influenciam a biologia dos carrapatos,
causando-lhes injúrias e alterações comportamentais. Histamina, leucotrienos, prostaglandinas,
proteínas eosinofílicas, enzimas, fixação de complemento e outros mediadores da resposta imune e
inflamatória contribuem na formação de lesão nos sítios de fixação, afetando o ingurgitamento (Wikel,
1996). Por outro lado, elevados níveis de substâncias vasoativas, induzidas pela salivação, aumentam a
permeabilidade vascular, além de outras alterações endoteliais.
As células de Langerhans são importantes na resistência adquirida dos animais aos carrapatos,
pois estas decrescem de número durante a infestação primária e aumentam na fase inicial da infestação
secundária. Atuam, ainda, como células acessórias apresentadoras de antígeno na expressão da
resistência adquirida (Nithivthai & Allen, 1984). Anticorpos contra antígenos salivares não garantem
imunidade sólida; visto que determinam proliferação de linfócitos e degranulação celular, além de
serem glicoproteínas que possuem reações cruzadas interespecíficas (Wheeler et al., 1991) (Tabela 1).

5Tabela 1: Principais formas de expressão de imunidade adquirida dos hospedeiros aos carrapatos.
CARRAPATO HOSPEDEIRO CARRAPATO
Ação: Efeito: Ação:
Fixação Hipersensibilidade local N° de larvas
Sítio de alimentação Basófilos e eosinófilos Limitação do repasto sanguíneo
Antígenos salivares Degranulaçao de basófilos Limitação do repasto sanguíneo
Enzimas salivares Liberação de substâncias vasoativas Limitação do repasto sanguíneo
Salivação contínua imunocomplexos no sítio alimentar Ingurgitamento
Saliva Moléculas biorreativas Injúrias e alterações comportamentais
Saliva Permeabilidade vascular, alterações
endoteliais
---
Bos taurus infestados com adultos de Dermacentor andersoni, apresentaram linfócitos
sangüíneos periféricos reativos in vitro com extrato de glândula salivar da mesma espécie de carrapatos
(Wikel & Osburn, 1982). Enquanto que B. indicus infestados com Amblyomma americanum
desenvolveram linfócitos menos responsivos, in vitro, ao extrato salivar deste carrapato (George et
al.,1985). A resistência adquirida pelo bovino ao B. microplus, até certo grau, é hereditária, o que
justifica a seleção de indivíduos menos susceptíveis (Hewetson, 1978).
O uso de anticorpos anti-saliva para detectar a imunidade à carrapatos, quanto à resistência
adquirida, é um erro potencial, pois estes apenas contribuem na expressão da resistência e muitos não
são reativos com moléculas envolvidas nos processos biológicos do artrópode (Wikel, 1996). A saliva
dos carrapatos possui substâncias farmacologicamente ativas com propriedades antiinflamatória,
antihemostática e imunossupressiva o que reduz a defesa do hospedeiro (Ribeiro et al., 1985). A
imunossupressão mediada por carrapatos contribui ao sucesso no estabelecimento de infecção por
hematozoários transmitidos por estes ixodídeos (Urioste et al., 1994).
A intensidade de fixação das larvas de B. microplus sobre animais resistentes está relacionada à
hipersensibilidade e a prolongada irritação; além da existência da imunidade humoral, o que foi
comprovado pela transfusão sérica de animais resistentes à animais susceptíveis (Oberen, 1984).
Willadsen & Williams (1976) identificaram na saliva de B. microplus as enzimas esterase hidrolítica,
enzima proteolítica e, as de inibição da coagulação sangüínea e de fixação do complemento. Todas
estas enzimas contribuem na reação de hipersensibilidade imediata em diferentes níveis, de acordo a
resistência individual (Willadsen, 1980). Pouco se conhece sobre o mecanismo da interrupção no

6repasto sangüíneo do carrapato, prejudicando a produção de ovos e a viabilidade destes. As substâncias
biologicamente ativas liberadas por basófilos, eosinófilos e outros tipos celulares são apenas fatores
que podem contribuir no fenômeno da resistência.
IMUNIDADE INDUZIDA
Os primeiros avanços no conhecimento da imunidade induzida ao carrapatos foram reportados
por Trager (1939), imunizando cobaios e coelhos com extratos brutos de D. variabilis. Este autor
observou que larvas e ninfas submetidas aos desafios apresentavam menores índices de
engurgitamento. A partir da década de 40 muitos estudos foram desenvolvidos relativos à resistência
induzida aos ixodídeos. A redução no número de adultos de Hyalomma anatolicum excavatum foi
observada em coelhos após inoculação com extrato de glândula salivar (Wikel, 1996). Garin &
Grabarev (1972) obtiveram sucesso na imunização de coelhos e cobaios com extrato de glândula
salivar de Rhipicephalus sanguineus. Em teste com cobaios imunizados com 1 - 4 mg de extrato de
larva de Ixodes holocyclus, verificou-se, após três semanas, o percentual de rejeição para larvas de 29 -
68 % (Bagnall, 1978).
O extrato de glândula salivar de B. microplus inoculado em bovinos é capaz de induzir
resistência (Brossard, 1976). A purificação do alérgeno 2 (inibidor da enzima proteolítica e da
coagulação sangüínea), a partir de larvas de B. microplus, quando inoculado em bovinos, demonstrou
sucesso parcial na resposta imune, caracterizada pela reação de hiperssensibilidade. Assim, Willadsen
et al. (1978) sugeriram a necessidade do encontro de antígenos internos que induzissem resposta imune
satisfatória. Bovinos mestiços (B. indicus x B. taurus) foram imunizados com a combinação de
antígenos solúveis e particulados, preparados de B. microplus, onde observou-se uma redução
altamente significativa na carga parasitária (Johnston et al., 1986).
Os melhores resultados na imunização contra os ixodídeos têm sido obtidos ao inocular-se nos
animais antígenos derivados de porções dos carrapatos que não estão diretamente expostas no momento
do repasto sangüíneo. Para estes antígenos foi proposta a denominação de "antígenos ocultos" ou
"antígenos escondidos" (Ackerman et al., 1980). Diferente da imunidade adquirida à carrapato, a
imunidade induzida por antígenos particulados, principalmente os “ocultos”, não apresenta reações de
hipersensibilidade; atua principalmente sobre o estágio adulto do carrapato; os carrapatos sobreviventes
são pequenos e apresentam sinais de lesões internas, com cor avermelhada e, ao exame histológico,

7observa-se severas lesões no trato digestivo (Agbede & Kemp, 1986; Mora Hernandez et al.,1997)
acompanhadas de redução no peso e postura das fêmeas que ingurgitam (Kemp et al., 1986; Massard et
al., 1995). Assim, esta imunidade expressa seus efeitos principalmente nos sistemas digestivos e
reprodutivos do carrapato (Tabela 2).
Os antígenos do intestino médio de B. microplus são os que induziram melhores níveis de
imunidade a infestação por estes carrapatos. Opdebeeck et al. (1988 a) observaram uma redução de
87% da carga parasitária de B. microplus em bezerros Hereford submetidos ao desafio com antígenos
intestinais. Posteriormente, o intestino do B. microplus foi processado e fracionado em uma parte
insolúvel (membranas) e outra solúvel e observou-se que a parte insolúvel conferiu 91% de proteção e
a mistura de ambas 82%. A fração solúvel não induziu proteção significativa (Opdebeeck et al., 1988
b).
Tabela 2: Comparação entre a imunidade adquirida e a imunidade induzida de bovinos ao carrapato
Boophilus microplus (Canestrini, 1887).
EFEITOS TIPO DE IMUNIDADE
ADQUIRIDA INDUZIDA
- Estágio mais afetado Larva Adultos
- Efeitos da imunidade:
1. Número de carrapatos ingurgitados Variável, a maioria Usualmente grande redução
2. Peso dos carrapatos ingurgitados Pequeno efeito Grande efeito
3. Peso da postura Não afeta Grande efeito
4. Viabilidade da postura Não afeta Algum efeito
- Expressão da imunidade Rejeição das larvas pouco afetadas Severos danos e morte dos adultos
- Mecanismo imunológico Primordialmente por hipersensibilidade imediata Principalmente por anticorpos
Adaptado de: Willadsen, P. (1987). Inter. J. Parasitol., 17(2): 671-677.
Alguns fatores podem interferir na resposta do hospedeiro à resistência ao carrapato, e dentre
eles destacam-se:
1 - fatores externos que induzem ao estresse: de origem nutricional e ambiental;
2 - condição reprodutiva do hospedeiro e imunossupressão por drogas;
3 – infecção por hemoparasitos como Babesia sp., Anaplasma sp., e Trypanosoma sp., que
podem aumentar a susceptibilidade aos carrapatos (Heller-Haupt et al., 1983; Oberen,
1984).

8Os estudos na imunidade induzida passaram pela utilização de extratos brutos dos diferentes
estágios dos carrapatos, extratos de glândulas salivares, extratos purificados, extratos de ovos
embrionados e antígenos particulados a partir de diferentes tecidos, sendo o derivado do trato digestivo
aquele que induziu melhor proteção (Wikel, 1996; Toro-Ortiz et al., 1997). Duas alternativas no
controle imunológico de carrapatos foram resumidas e propostas por Willadsen (1987), sendo uma
através da seleção de indivíduos imunocompetentes e outra pelo uso de antígenos purificados como
indutor da resistência pela imunização direta.
VACINAS CONTRA ARTRÓPODES
Várias tentativas vêm sendo realizadas para a obtenção de vacinas contra artrópodes. O uso de
antígenos particulados de Stomoxys calcitrans para induzir imunidade reduziu o grau de ingurgitamento
deste muscídeo (Webster et al., 1992). A redução da fecundidade de Aedes aegypti foi observada
quando estes insetos ingeriram sangue de animais imunizados com o extrato bruto deste culicídeo
(Sutherland & Ewen, 1974). Pesquisas com Glossina morsitans (Otieno et al.,1984) e Hypoderma sp
(Baron & Weintraub, 1986) foram desenvolvidas imunizando-se coelhos e bovinos, respectivamente,
contra estes artrópodes. Utilizando extratos de membrana peritrófica de Lucilia cuprina, para imunizar
ovinos, foi observado a inibição do crescimento da larva deste díptero no hospedeiro (East et al., 1993;
Eisemann & Binnington, 1994). Mumcuoglu et al. (1996) localizaram antígenos no trato digestivo de
Pediculus humanus que conferem imunidade ao homem.
Os efeitos desses imunógenos são variados sobre os artrópodes, sendo observado interferência
na fecundidade, oviposição, sobrevida, índices nutricionais e reprodutivos, ruptura de órgãos, paralisia
e morte (Opdebeeck, 1994). Os antígenos vacinais são derivados de diferentes órgãos, tecidos ou
células; como por exemplo para Ctenocephalides felis, onde a membrana intestinal está sendo utilizada
como antígeno (Opdebeeck & Slacek, 1993). Diferentes tipos de antígenos “ocultos” de artrópodes têm
sido utitlizados na tentativa de produção de imunógenos (Tabela 3). Para os imunógenos contra os
artrópodes, os melhores resultados foram encontrados contra carrapatos (Opdebeeck, 1994); este fato
pode estar relacionado, possivelmente, à digestão intracelular dos ixodídeos, enquanto nos insetos a
digestão se processa na luz intestinal dentro da membrana peritrófica.

9Tabela 3: Tentativas atuais de produção de vacinas contra insetos de importância Médico Veterinária.
ARTRÓPODE TIPO DE ANTÍGENO
Stomoxys calcitrans Antígenos particulados
Aedes aegypti Extratos bruto
Glossina morsitans Extrato intestinal
Hypoderma sp Extrato bruto
Lucilia cuprina Membrana peritrófica (extrato)
Pediculus humanus Antígenos intestinais
Ctenocephalides felis Membrana intestinal
O uso de antígenos a partir de glândula salivar de carrapatos foi pouco expressivo como
imunógenos (Wikel, 1988; 1996). Os estudos tenderam ao encontro de antígenos “ocultos” em órgãos
internos dos ixodídeos que não estão em contato direto com o hospedeiro. Assim, foram identificados e
quantificados os componentes séricos do hospedeiro, entre eles imunoglobulinas, haptoglobulinas,
albumina, transferrinas e anticorpos específicos, na hemolinfa do ixodídeo; fato que contribuiu ao
desenvolvimento de imunógenos (Ackerman et al., 1981; Tracey-Patte et al., 1987; Ben-Yakir, 1989).
Fracionando antígeno bruto total de porções intestinais de B. microplus, a atividade protetora foi
obtida com frações contendo proteínas entre 79 - 205 Kilodaltons (Kd) e com ponto isoelétrico de 5,1 -
5,6. A imunidade induzida por estas frações foi superior àquela obtida com o material bruto (Willadsen
et al., 1988). Genes, responsáveis por estes antígenos, foram identificados, conseguindo-se purificar e
caracterizar uma glicoproteína da superfície de membrana das células digestivas no intestino médio de
B. microplus, a qual possui peso molecular de 89Kd com ponto isoelétrico entre 5,1 - 5,6,
posteriormente denominada Bm 86 (Willadsen et al., 1989). A glicoproteína Bm 86 encontra-se em
grande quantidade na superfície externa da membrana de células do “tipo digestivas” do trato intestinal
do B. microplus (Figura 1).
Figura 1a: Corte transversal do intestino médio de fêmea de Boophilus microplus: visão do conjunto de células digestivas, predomínio de células do “tipo digestivas”. Tricrômico de Masson, 250x.
Figura 1b: Corte transversal do intestino médio de fêmea de Boophilus microplus: visão do conjunto de células digestivas: células secretoras S1, S2, células basofílicas, células basais e células do “tipo digestivas”. Hematoxilina-Eosina, 160x.

10 Outras proteínas foram isoladas e caracterizadas, de B. microplus, como a Bm 91, porém não
conferiram imunidade satisfatória quando usada isoladamente (Wikel, 1996). Antígenos obtidos a partir
do cultivo de células de A. americanum induziram significativo nível de imunidade às infestações de
adultos de A. americanum e D. variabilis (Wikel, 1985), no entanto, pouco se sabe sobre estes tipos
antigênicos.
Clones de cDNA da glicoproteína Bm 86 foram isolados, purificados, homegeneizados, sendo
determinado a seqüência de fragmentos peptídicos. A recombinação gênica foi expressa inicialmente
em Escherichia coli (Rand et al, 1989). Posteriormente, a Bm 86 foi também recombinada aos fungos
Aspergillus nidulans e A. niger (Turnbull et al., 1990) e em baculovírus (Richardson et al., 1993). Este
antígeno recombinante induziu proteção ao mesmo nível que a rBm 86 em E. coli (Opdebeeck, 1994).
Rodriguez et al. (1994) isolaram e amplificaram o gene que codifica a Bm 86, expressando-o em vários
sistemas, incluindo a levedura metilotrófica Pichia pastoris. Esta expressão induziu um aumento no
potencial imunogênico por ser secretada na forma glicosilada, originando partículas de 20 - 36 nm.
Estas recombinações gênicas possuem a vantagem de apresentarem cadeias regulares de polipeptídeos
do antígeno, as quais se dobram homogeneamente.
Quanto a importância dos antígenos recombinantes, estes potencialmente diferem dos nativos
tanto pela configuração, como pelos padrões no processo de glicosilação (Willadsen & Mckenna,
1991). O principal mecanismo de atuação dos anticorpos contra os antígenos recombinantes rBm 86 se
dá pelo bloqueio da atividade endocítica das células digestivas do carrapato, associada por conseguinte
às lesões (Kemp et al., 1986; Mora Hernandez et al.,1997). Os antígenos recombinantes da Bm 86 têm
sido utilizados como imunógenos em alguns países da América Latina como Cuba, Brasil, Argentina,
Colômbia, México e Nicarágua; e na Austrália, apresentando resultados satisfatórios (De La Fuente,
1995; Massard et al., 1995; Willadsen et al., 1995). Estes antígenos foram demonstrados e
reconhecidos em diversas cepas de B. microplus (Penichet et al., 1994).
Outro modelo de imunógenos, considerados eficazes contra carrapatos, são os peptídeos
sintéticos, inicialmente testados por Sharp et al. (1990). Estes foram sintetizados, purificando
oligopeptídeos a partir de cálculos preditivos da seqüência de aminoácidos da glicoproteína Bm 86. Ao
avaliar os peptídeos em coelhos e bovinos, os anticorpos produzidos reconheceram as proteínas Bm 86
e rBm 86 pelo ensaio Imunoblotting.
Níveis de Anticorpos Contra Carrapatos

11 Os níveis de anticorpos contra a glicoproteína Bm 86, recombinada em P. pastoris, foram
avaliados em bovinos, através de um ensaio imunoenzimático (ELISA) (Rodriguez et al., 1994). Estes
pesquisadores procederam o controle do B. microplus em populações de bovinos, a campo em Cuba,
vacinados com a rBm 86, acompanhando sorologicamente durante 33 semanas. Observaram que os
anticorpos surgem entre a primeira e segunda semana após a vacinação e atingem pico máximo por
volta da décima semana, permanecendo detectáveis com títulos consideráveis até a 32ª semana
(Rodriguez et al. 1995a). O mesmo modelo foi utilizado no Brasil, ao avaliar a imunidade de bovinos
B. taurus puros e mestiços com B. indicus, frente ao antígeno recombinante. Os animais apresentaram
perfil de anticorpos similar ao observado em Cuba (Rodriguez et al. 1995b).
Na Argentina, Lamberti et al. (1995) observaram, através do ELISA, em bovinos mestiços
imunizados com a rBm 86 uma curva de anticorpos próxima aos relatos de Rodriguez et al. (1995 a, b).
Anticorpos monoclonais contra o B. microplus foram analisados quanto aos seus efeitos na eficiência
reprodutiva do carrapato, através de ELISA e Imunoblotting (Toro-Ortiz et al., 1997). Modelos
matemáticos por computador foram propostos e utilizados para avaliação da dinâmica populacional de
B. microplus ao nível de campo, empregando acaricidas e imunógenos. Os dados determinados
sorologicamente foram utilizados para a determinação das épocas ideais de vacinação (Floyd et al.,
1995; Lodos et al., 1995).
Uma metodologia prática foi desenvolvida por Machado & Santizo (1995), para determinar os
níveis de anticorpos em bovinos imunizados com a proteína recombinante rBm 86, a campo. Esta
técnica utiliza placas de poliestireno branco com escavações circulares sensibilizadas com o antígeno
rBm 86. A análise sorológica está baseada em um imunoensaio indireto em fase sólida, que emprega
sondas de ouro coloidal amplificadas com reveladores de prata. O conjugado adicionado contém
proteína A (recombinante) marcada com ouro coloidal, para a detecção de imunocomplexos formados.
A reação é amplificada pela adição de reveladores físicos, à base de íons de prata, produzindo reações
in situ insolúveis, de cor marrom, cuja intensidade é proporcional à concentração de anticorpos
presentes na amostra.
Pesquisando anticorpos em bovinos no Brasil, imunizados com a rBm 86, Mora Hernandez et
al. (1998), utilizaram o ensaio desenvolvido por Machado & Santizo (1995). Neste estudo,
acompanhou-se o perfil de anticorpos durante 65 semanas, observando uma variação do negativo à
títulos de 1:10240. Após o segundo inóculo (semana 4), verificou-se uma resposta imune crescente, que
se manteve elevada durante as 65 semanas. O declínio dos níveis de anticorpos ocorreu próximo às
revacinações, entretanto, estes níveis foram superiores à linha de corte adotada, não sendo relevante o
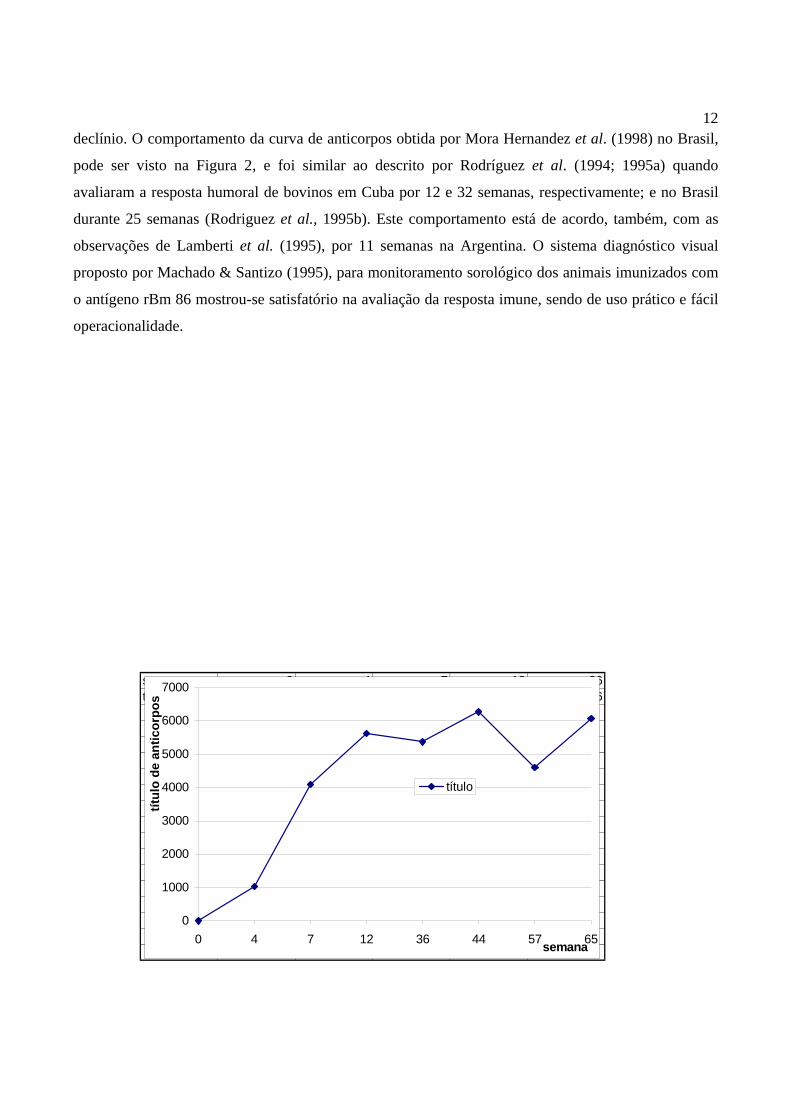
12declínio. O comportamento da curva de anticorpos obtida por Mora Hernandez et al. (1998) no Brasil,
pode ser visto na Figura 2, e foi similar ao descrito por Rodríguez et al. (1994; 1995a) quando
avaliaram a resposta humoral de bovinos em Cuba por 12 e 32 semanas, respectivamente; e no Brasil
durante 25 semanas (Rodriguez et al., 1995b). Este comportamento está de acordo, também, com as
observações de Lamberti et al. (1995), por 11 semanas na Argentina. O sistema diagnóstico visual
proposto por Machado & Santizo (1995), para monitoramento sorológico dos animais imunizados com
o antígeno rBm 86 mostrou-se satisfatório na avaliação da resposta imune, sendo de uso prático e fácil
operacionalidade.
semana 0 4 7 12 36título 0 1024 4096 5632 5376
0
1000
2000
3000
4000
5000
6000
7000
0 4 7 12 36 44 57 65semana
títu
lo d
e an
tico
rpo
s
título

13Figura 2. Curva dos níveis de anticorpos, média aritmética dos títulos, em bovinos mestiços leiteiros
imunizados com o antígeno recombinante rBm 86, no Brasil. Avaliação durante 65 semanas.
EFEITO DOS IMUNÓGENOS SOBRE OS CARRAPATOS
Na relação hospedeiro-parasito hematófago, em especial nos carrapatos, é conhecido que o
mecanismo que leva ao desenvolvimento de resistência adquirida está acompanhado de reações de
hipersensibilidade (Koudstaal et al., 1978). Um fato importante da diferenciação entre a imunidade
adquirida e a induzida é que, embora ambas atuem para elevar os níveis de imunidade frente aos
carrapatos, a primeira age principalmente contra estágio larvar, enquanto a segunda sobre o estágio
adulto (Tabela 2).
O antígeno recombinante rBm 86 induz a formação de altos níveis de anticorpos e de outros
componentes séricos que mediam a resposta imune dos bovinos, como o complemento. Os anticorpos
específicos se fixam ao antígeno, in situ e, neste caso, à superfície de membrana das células digestivas
de B. microplus, ocasionando severos danos morfofisiológicos, quando o sangue dos animais vacinados
é ingerido pelos carrapatos adultos (Agbede & Kemp, 1986; Kemp et al., 1986). Estes danos
manifestam-se principalmente das seguintes formas:
1 - na redução do número e tamanho das fêmeas alimentadas;
2 - na redução da ovoposição;
3 - na redução da fertilidade dos ovos;
4 - na diminuição do potencial reprodutivo dos carrapatos em gerações sucessivas, o que leva ao
decréscimo das populações nas pastagens.
Efeito Sobre a Biologia do Carrapato
Pouco são os estudos referentes a influência de anticorpos anti-rBm 86 nos parâmetros
biológicos dos carrapatos. Na fase parasitária tem-se observado, em diversas pesquisas, que as fêmeas
na sua maioria encontram-se semi-ingurgitadas, mortas, aderidas à pele do hospedeiro, apresentando

14aspecto "seco". As fêmeas sobreviventes possuem colorações anormais que variam do róseo ao azul
escuro (Willadsen, 1987; Massard et al., 1995; Rodriguez et al., 1995b).
Para a fase não parasitária, as avaliações sobre o efeito dos anticorpos anti-rBm 86 podem ser
determinadas pelo período de pré postura, ovoposição, produção e incubação dos ovos, viabilidade da
postura, peso das teleóginas, percentual de fêmeas que ovopositaram, índices nutricionais e
reprodutivos, e sobrevivência e viabilidade das larvas. Os resultados obtidos, para estas análises por
vários pesquisadores, são variados, porém todos concordam em buscar a eficiência do imunógeno,
baseando-se na redução desses índices e parâmetros.
Pouco são, no entanto, os registros sobre a sobrevivência e longevidade das larvas provenientes
de fêmeas de carrapatos alimentadas em bovinos imunizados com diferentes antígenos de B. microplus,
com a Bm 86 ou a rBm 86. Observações originais foram realizadas no Brasil, no que se refere à menor
sobrevivência e longevidade das larvas, oriundas de fêmeas de B. microplus sobreviventes de animais
imunizados (Mora Hernandez, 1996) (Figura 3).
Hemoglobina
Heme (Fe) + Globina
5 - 10% do Heme ingerido
Oócitos (desenvolvimento) Ovos
Heme + Proteína (do carrapato)
Hemixodovina Hemolinfa
Ovos, Embriões (proteína de reserva)
Larvas (intestino como reserva energética)
Proteína + Hematina (pela ação da digestão)
Pigmento violeta-púrpuro

15Figura 3. Seqüência do metabolismo da hemoglobina em Boophilus microplus.
Aproximadamente, 5-10% da partícula heme ingerida pelas fêmeas do carrapato passa para o
desenvolvimento dos oócitos. O complexo heme dos ovos é uma proteína diferente da hemoglobina do
hospedeiro. Após a quebra da hemoglobina pela digestão, algumas partes da molécula de ferro e do
heme passam à hemolinfa, ligando-se a proteínas específicas do carrapato (Kitaoka, 1961). Este novo
tipo de composto, ferro-proteína, é denominado hemixodovina (Bremner, 1959). A hemixodovina tem
a função de reserva protéica, durante a embriogênese, e de reserva energética nas larvas em jejum
(Balashov, 1972).
Outra possibilidade reportada por Mora Hernandez (1996) reside no fato de que, após o
desprendimento das fêmeas ingurgitadas, o retículo endoplasmático rugoso das células intestinais
começa a reorganizar-se, sintetizando grânulos, o que corrobora ao efeito dos componentes séricos
sobre as células digestivas das teleóginas e o conseqüente efeito sobre a ovoposição (Coons et al.,
1982; Agbede e Kemp, 1986). O material secretado por estas células de B. microplus, se faz nos
espaços extracelulares das dobras basais, as quais estão em contato com a hemolinfa (Agbede e Kemp,
1987). Desta forma, pode ser relacionado o período de sobrevivência das larvas, descendentes de
fêmeas que realizaram o repasto sangüineo em bovinos imunizados com o antígeno rBm 86.
A redução na população de carrapato é decorrente da alteração dos processos biológicos,
especialmente os processos metabólicos da digestão, afetando as gerações que se sucedem.
Conseqüentemente, a reprodução torna-se menos eficiente e a capacidade de sobrevivência e o poder
de fixação das larvas diminui substancialmente. As alterações nos parâmetros biológicos de B.
microplus contribuem para a redução das populações deste carrapato nas pastagens.
Alterações Histológicas do Trato Intestinal
A estrutura histológica do intestino dos carrapatos está composta por uma membrana basal
coberta de fibras musculares longitudinais e circulares, formando várias camadas (Balashov, 1972).
Três tipos celulares para o epitélio intestinal de Hyalomma asiaticum foram descritos por Balashov
(1972), os quais foram denominados como células de reserva, digestivas e secretoras, sugerindo que
estas originam-se de uma única célula fonte. Entretanto, Tatchell (1964) e Chinery (1964) afirmaram
que a parede do intestino destes artrópodes é constituida, essencialmente, por células epiteliais e suas

16estruturas e funções variam de acordo com o nível de ingurgitamento e com as fases do ciclo biológico
dos carrapatos.
Agbede & Kemp (1985) concordaram com o critério de Balashov (1972) quanto a origem dos
tipos celulares, acrescentando, no entanto, que as células basais encontram-se em todos os ínstares.
Estes autores classificaram as células intestinais de B. microplus em cinco tipos celulares: célula basal,
digestiva, secretora (S1), secretora (S2) e célula basofílica (Figura 1). Esta última persiste até a morte
das fêmeas.
O mecanismo de ação dos anticorpos anti-rBm 86 se dá pela fixação destes à membrana das
células digestivas (Kemp et al., 1986). Os anticorpos, junto ao complemento e outros componentes
séricos, provocam severos danos na ausência de leucócitos (Agbede & Kemp, 1986; Willadsen et al.,
1989). O complemento pode ou não ser essencial, uma vez que foi demonstrado que o soro de bovinos
imunizados reage com a superfície das células intestinais e que a intervenção de anticorpos contra as
células digestivas resulta em uma marcante inibição de suas atividades endocíticas, precedendo lesões
celulares detectáveis (Willadsen & Kemp, 1988). Severas alterações morfológicas no intestino de
carrapatos alimentados, in vitro e in vivo, foram registradas três semanas após a terceira dose do
imunógeno Bm 86; quando os níveis de anticorpos circulantes encontravam-se no pico máximo. Em B.
microplus coletados 12 horas após iniciada a alimentação, verificou-se que ocorre um bloqueio das
atividades endocíticas nas células digestivas (Agbede & Kemp, 1986; 1987).
No Brasil, estudos histológicos foram realizados no trato digestivo de B. microplus alimentados
em bovinos imunizados com a rBm 86, de fêmeas coletadas 30, 40, 50, 60 e 70 dias pós vacinação. Foi
observado a destruição de células digestivas, das secretoras (S1 e S2) e, em casos mais severos, das
células basofílicas, permanecendo apenas a lâmina basal. Ocorreu ainda erosão e ruptura da parede
intestinal e extravasamento do conteúdo digestivo para a cavidade celomática (Mora Hernandez et al.,
1997). Estas lesões incompatibilizam o processo de oogênese neste carrapato e interrompe seu ciclo
vital (Figura 4).
Figura 4a: Lesões no intestino médio de fêmea de Boophilus microplus pela ação de anticorpos anti-rBm 86: células digestivas com alterações citoplasmáticas e nucleares, células secretoras S1 e S2 em número reduzido com vacuolização citoplasmática, ausência de células do “tipo digestivas”. Hematoxilina-Eosina, 400x.
Figura 4b: Lesões no intestino médio de fêmea de Boophilus microplus pela ação de anticorpos anti-rBm 86: acentuada destruição de células do “tipo digestivas”, células secretoras S1 e S2, células basofílicas e da lâmina basal. Hematoxilina-Eosina, 400x.

17 As alterações no intestino de B. microplus, pela ação dos anticorpos anti-rBm 86, justificam as
observações macroscópicas encontradas em fêmeas mortas, ou parcialmente ingurgitadas aderidas à
pele do hospedeiro (Massard et al., 1995; Mora Hernandez, 1996; Mora Hernandez et al., 1997). As
lesões intestinais podem afetar a oviposição, por interrupção da digestão ou pelo bloqueio da síntese de
vitelogenina nas células intestinais (Agbede & Kemp, 1986). O motivo das larvas não serem afetadas
pelos anticorpos anti-rBm 86 deve-se ao fato de os componentes celulares do intestino deste ínstar
serem constituídos apenas por células basais (Balashov, 1972; Agbede & Kemp, 1985). Também é
conhecida a existência de enzimas inibidoras da proteólise, presentes, apenas, no estágio larval
(Willadsen et al., 1989).
A glicoproteína Bm 86 mantem-se conservada em diferentes cepas de B. microplus. Os
anticorpos de bovinos imunizados com a rBm 86 protegem e atuam contra distintas cepas de B.
microplus, mesmo as cepas resistentes a acaricidas como os organofosforados (Penichet et al., 1994).
REFERÊNCIAS BIBLIOGRÁFICAS
ACKERMAN, S.; FLOYD; M. & SONENSHINE, D. E. 1980. Artificial immunity to Dermacentor variabilis
(Acari: Ixodidae) vaccination using tick antigens. J. Medical Entomol., 17 (5):391-397.
ACKERMAN, S., BRIAN CLARE, F., MCGILL, T. W. & SONENSHINE, D. E. 1981. Passage of host serum
components, incluiding antibody, across the digestive tract of Dermacentor variabilis (Say). J. Parasitol.,
67 (5): 737-740.
AGBEDE, R. I. S. & KEMP, D. H. 1985. Digestion in the catle-tick Boophilus microplus: light microscope
study of the gut cells in nymphs and females. Intern. J. Parasitol., 15(2): 147-157.
AGBEDE, R. I. S. & KEMP, D. H. 1986. Immunization of cattle against Bophilus microplus using extracts
derived from adult female ticks: histopathology of ticks feeding on vaccinated cattle. Intern. J. Parasitol.,
16 (1): 35-41.
AGBEDE, R. I. S. & KEMP, D. H. 1987. Ultrastructure of secretory cells in the gut of the cattle-Tick Boophilus
microplus. Intern. J. Parasitol., 17 (6): 1089-1098.
ALLEN, J. R. 1973. Ticks resistence: basophils in skin reactions of resistant guinea pigs. Int. J. Parasitol., 3:
195-200.
ALLEN, J. R.; KHALIL, H. M. & GRAHAM, J. E. 1979. The location of tick salivary gland antigens,
complement and immunoglobulin in the skin of guinea pigs infested with Dermacentor andersoni larvae.
Immunology, 38: 467-472.

18BAGNALL, B. G. 1978. Cutaneous immunity to the tick Ixodes holocyclus. In: Tick-borne disease and their
vectors. WILDE, J. K. H. Ed. University of Edinburgh; p. 79-81.
BALASHOV, Y. S. 1972. Bloodsucking ticks (Ixodoidea) vector of diseases of man and animals. Miscellaneous
Publications Entomol. Soc. America, 8 (5): 276-296.
BARON, R. W. & WEINTRAUB, J. 1986. Immunizaton of cattle against hypodermatosis (Hypoderma lineatum
de Vill.) and H. bovis (L.) using H. lineatum antigens. Vet. Parasitol., 56(1-3) 225-238.
BEN-YAKIR, D. 1989. Quantitative studies of host immunoglobulin G in the Hemolymph of Ticks (Acari). J.
Med. Entomol., 26 (4) 243-246.
BREMNER, K. C. 1959. Studies on “haemoxidovin”, the pigments in the eggs of the cattle tick Boophilus
microplus (Acarina, Ixodidae). Aust. J. Biol. Sci., 12: 263-273.
BROSSARD, M. 1976. Relation immunologiques entre bovins et tiques, plus particuliérement entre bovins et
Boophilus microplus. Acta Tropica XXXIII, I: 15-36.
BROSSARD, M. & FIVAZ, V. 1982. Ixodes ricinus L.: Mast cells, basophils and eosinophils in the sequence of
cellur events in skin of infested or reinfested rabbits. Parasitology, 85: 583-592.
BROWN, S. J.; GALLI, S. J. GLEICG, G. J. & ASKENASE, P. W. 1982. Ablation of immunity to Amblyomma
americanum by anti-basophil serum: cooperation between basophils and eosinophils in expression of
immunity to ectoparasites (ticks) in guinea pigs. J. Immunol., 129: 790-796.
CHINERY, W. A. 1964. The midgut epitelium of the tick Haemaphysalis spinigera Neumann, 1897. Med.
Entomol., 1: 206-212.
COONS, L. B., TARNOWSKI, B. & QURTH, D. D. 1982. Rhipicephalus sanguineus: Localization of
vitelogenin synthesis by immunological methods and eletron microscpy. Experim. Parasitol., 54: 331-339.
DE LA FUENTE, J. 1995. Reconbinant vaccines for the control of cattle tick, Elfos Scientiae, Monograph,
Havana. Cuba. 241p.
EAST, I. J.; FITZGERALD, C. J.; PEARSON, R. D.; DONALDSON, R. A.; VUOCOLO, T.; CADOGAN, L.
C.; TELLAM, R. L. & EISEMANN, C. H. 1993. Lucilia cuprina: inhibition of growth induced by
immunization of host sheep with extracts of larval peritrophic membrane. Inter. J. Parasitol., 23 (4): 221-
229.
EISEMANN, C. H. & BINNINGTON, K. C. 1994. The Peritrophic membrane: its formation, struture, chemical
composition and permeability in relation to vaccination against ectoparasitic arthropods. . Inter. J.
Parasitol., 24(1): 15-26.
FLOYD, R. B., SUTHERST, R. W. & HUNGERFORD, J. 1995. Modelling the field efficacy of a genetically
engineered vaccine against the cattle tick, Boophilus microplus. Intern. J. Parasitol., 25: 285-291.

19GARIN, N. S. & GRABAREV, P. A. 1972. Protective reactions in rabbits and guinea pigs when repeatedly
exposed to Rhipicephalus sanguineus (Latr. 1806) ticks. Medit. Parazitolog. Parazitar. Bolez., 41: 274-
279.
GEORGE, J. E.; OSBURN, R. L. & WIKEL, S. K. 1985. Acquisition and expression of resistence by Bos
indicus and Bos indicus x Bos taurus calves to Amblyomma americanum infestation. J. Parasitol., 71:
174-182.
GORDON, J. R. & ALLEN, J. R.. 1991. Nonspecific activation of complement factor 5 by isolated Dermacentor
andersoni salivary antigens. J. Parasitol., 77(2): 296-301.
HELLER-HAUPT, A.; VARMA, M. C. R.; LANG, A. O. & ZETLIN, A. 1983. The effect of Trypanosoma
congolense infection on adcquired immunity to the tick Rhipicephalus appendiculatus. Ann. Trop. Med.
Parasitol., 77: 219-222.
HEWETSON, R. W. 1978. Selection of cattle for resistance against Boophilus microplus. In: WILDE, J. K. H.
(Ed.) Tick-Borne Diseases and Their Vectors. University of Edinburgh, pp 258-261.
JOHSNTON, T. H. & BANCROFT, M. J. 1918. A tick resistant condition in cattle. Proc. Roy. Soc. Qd., 41;
121-132.
JOHSTON, L. A. Y.; KEMP, D. H. & PEARSON, R. D. 1986. Immunization of cattle against Boophilus
microplus using extracts derived from adult female ticks: Effects of induced immunity on tick populations.
Intern. J. Parasitol., 16: 27-34.
KEMP, D. H.; KOUDSTAAL, D.; ROBERTS, J. A. & KERR, J. D. 1976. Boophilus microplus: The effect of
host resistance on larval attachment and growth. Parasitology, 73: 123-136.
KEMP, D. H.; AGBEDE, R. I. S.; JHONSTON, L. A. Y. & COUGH, J. M. 1986. Immunization of cattle against
Boophilus microplus using extracts derived from adult female ticks: Feeding and survival of the parasite
on vaccinated cattle. Intern. J. Parasitol., 16: 115-120.
KITAOKA, S. 1961. Physiological and ecological studies on somes ticks VI. Rate of digestion of blood-meal
and nitrogen, iron and, sterol economy in the tick. Bull. Naio. Inst. Ani. Hth., 1:105-112.
KOUDSTAAL, D.; KEMP, D. H. & KERR, J. D. 1978. Boophilus microplus: Rejection of Larvae from British
Breed Cattle. Parasitology., 76: 379-386.
LAMBERTI, J.; SIGNORINI, A.; MATTOS. C.; D’AGOSTINO, B.; CITRONI, D.; BACOS, E.;
RODRÍGUEZ, M. & DE LA FUENTE, J. 1995. Evaluation of the Recombinant Vaccine Against
Boophilus microplus in Grazing Cattle in Argentina. In: DE LA FUENTE, J. (Ed.). Recombinant Vaccines
for the Control of Cattle Tick, Elfos Scientiae, Monograph, Havana, 1995. 241p. p. 205-227.
LODOS, J.; OCHAGAVIA, M. E.; RODRIGUEZ, M. & DE LA FUENTE, J. 1995. Computer simulation of
Boophilus microplus cattle tick population dynamics emploing acaricide and vaccination control methods.
In: DE LA FUENTE, J. (Ed.). Recombinant Vaccines for the Control of Cattle Tick, Elfos Scientiae,
Monograph, Havana, 1995. 241p. p. 131-145.

20MACHADO, H., SANTIZO, C. 1995. A rapid visual immunoassay for the quantification of bovine antibodies
against recombinant Boophilus microplus Bm 86 antigen. In: DE LA FUENTE, J. (Ed.). Recombinant
vaccines for the control of cattle tick, Elfos Scientiae, Monograph, Havana, 1995. 241p. p. 103-111.
MASSARD, C. L.; FONSECA, A. H.; BITTENCOURT,V. R.; OLIVEIRA, J. B. & SILVA, K. M. 1995.
Avaliação da eficácia da vacina recombinante Rbm 86- “GAVAC” contra o carrapato Boophilus
microplus no Brasil. Rev. Bras. Med. Vet., 17 (4): 167-173.
MBOW, M. L.; RUTTI, B.; BROSSARD, M. 1994. Infiltration of CD4 CD8 T cell, and expression of ICAM-1,
Ia antigens, IL1-alpha and TNF-alpha in the skin lesion of Balb/c mice undergoing repeated infestations
with nynphal Ixodes ricinus ticks. Immunology, 82(4): 596-602.
MORA HERNANDEZ, C. 1996. Avaliação a campo do imunógeno recombinante rBm 86 no controle do carrapato
Boophilus microplus (Canestrini, 1887) no Brasil. Tese de Doutorado, UFRRJ, Seropédica-RJ. 100p.
MORA HERNANDEZ, C.; MASSARD, C. L.; SOARES, C. O. & FONSECA, A. H. 1997. Alterações histológicas
do trato digestivo de Boophilus microplus pela ação de anticorpos anti-rBm 86. Rev. Bras. Parasitol. Vet.,
6(1): 33-37.
MORA HERNANDEZ, C.; MASSARD, C. L.; SOARES, C. O. & FONSECA, A. H. 1998. Resposta imune de
bovinos vacinados com o antígeno recombinante Bm 86. Rev. Bras. Med. Vet., 20(4): 187-189.
MUMCUOGLU, K. Y.; RAHAMIM, E.; BEN-YAKIR, D.; OCHANDA, J. O. & GALUN, R. 1996.
Localisation of immunogenic antigens on midgut of the human body louse Pediculus humanus humanus
(Anoplura: Pediculisdae). J. Med. Entomol., 33(1): 74-77.
NITHIVTHAI, S. & ALLEN, J. R. 1984. Signifiance changes in epitelial Langerhans cells of guinea pigs
infested with ticks (Dermacentor andersoni). Immunology, 51: 133-141.
OBEREN, P.T. 1984. The immunological basis of host resistance to ticks - A review. J. South African Vet. Ass.,
55 (4) 215-217.
OPDEBEECK, J. P.; WONG, J. Y .M.; JACKSON, L. A. & DOBSON, C., 1988a. Vaccine to protect hereford
cattle tick, Boophilis microplus. Immunology, 63: 363-367.
OPDEBEECK. J. P.; WONG, J. Y .M.; JACKSON, L. A. & DOBSON, C. 1988b. Hereford cattle immunized
and protected against Boophilus microplus with soluble and membrane associated antigens from the
midgut of ticks. Parasite Immunol., 10: 405-410.
OPDEBEECK. J. P. 1994. Vaccines against blood-sucking arthropods. Vet. Parasitol., 54: 205-222.
OPDEBEECK. J. P. & SLACEK, B. 1993. An attempt to protect cats against infestation with Ctenocephalides
felis felis using gut membrane antigens as a vaccine. Intern. J. Parasitol., 23(8): 1063-1067.
OTIENO, L. H.; VUNDLA, R. M. W. & MONGI, A. 1984. Observations on Glossina morsitans morsitans
maintained on rabbits immunized with crude tsetse midgut proteases. Insect Sci. Appl., 5: 297-302.
PENICHET, M.; RODRIGUEZ, M.; CASTELLANO, O.; MANDADO, S.; ROJAS, Y. RUBIERA, R.;
SANCHEZ, P.; LLEONART, R. & DE LA FUENTE, J. 1994. Detection of Bm 86 antigen in different

21strains of Boophilus microplus and effectiveness of immunization with recombinant Bm 86. Parasite
Immunol., 16 (9): 493-500.
RAND, K. N.; MOORE, T.; SRISKANTHA, A.; SPRING, K.; TELLAN, R.; WILLADSEN, P. & COBON,
G.S., 1989. Cloning and expression of a protective antigen from the cattle tick Boophilus microplus. Proc.
Natl, Acad. Sci. USA, 86 : 9657- 9661. Biochem.
RIBEIRO, J. M. C.; MAKOUL, G. T.; LEVINE, J. ROBINSON, D. R. & SPILMAN, A. 1985. Antihaemostatic,
antinflammatory and immunosuopressive properties of the saliva of a tick, Ixodes dammini. J. Exp. Med.,
161: 332-344.
RICHARDSON, M. A.; SMITH, D. R. J.; KEMP, D. H. & TELLAN, R. L. 1993. Native and baculovirus-
expressed forms of the immunoprotective protein Bm 86 from Boophilus microplus are anchored to the
cell membrane by a glycosyl-phosphatidyl inositol linkage. Insect Molecular Biol., 1: 1-9.
ROBERTS, J. A. 1968. Resistance of cattle to tick Boophilus microplus (Canestrini). I. Development of ticks on
Bos taurus. J. Parasitol., 54 (4): 663-666.
RODRÍGUEZ, M.; RUBIERA, R.; PENICHET, M.; MONTESINOS, R.; CREMATA, J.; FALCÓN, V. I.;
SÁNCHEZ, G.; BRINGAS, R.; CORDOVÉS, C.; VALDÉS, M.; LLEONART, R.; HERRERA, L.& DE
LA FUENTE, J. 1994. High level Eepression of the B. microplus Bm 86 antigen in the yeast Pichia
Pastoris forming highly immunogenic particles for cattle. J. Biotechnol., 33: 135-146.
RODRÍGUEZ, M.; PENICHET, M.; MOURIS, A. E.; LABARTA, V.; LORENZO, L.; RUBIERA, R.;
CORDOVÉS, C.; SÁNCHEZ, P. A.; RAMOS, E.; SOTO, A.; CANALES, M.; PALENZUELA, D.;
TRIGUERO, A.; LLEONART, R.; HERRERA, L. & DE LA FUENTE, J. 1995a. Control of Boophilus
microplus Population in Grazing Vaccinated with a Recombinant Bm 86 Antigen Preparation. Vet.
Parasitol., 57: 339-349.
RODRÍGUEZ, M., MASSARD, C. L., FONSECA, A. H., RAMOS, N. F., MACHADO, H., LABARTA, V. &
DE LA FUENTE, J. 1995b. Effect of vaccination with a recombinant Bm 86 antigen preparation on
natural infestation of Boophilus microplus in grazing dairy and beef pure and cross breed cattle in Brazil.
Vaccine, 13 (18): 1804-1808.
SAUER, J. R.; MCSWAIN, J. L.; BOWMAN, A S. & ESSENBERG, R. C. 1995. Tick salivary gland
physiology. Annu. Rev. Entomol., 40: 245-267.
SHARP, P. J.; MCINERNEY, B. V.; SMITH, D. R; TURNBULL, I. F.; KEMP, D. H.; RAND, K. N. &
COBON, G. S. 1990. Chromatography and generation of specific antisera to synthetic peptides from a
protective Boophilus microplus antigen. J. Chromatogr., 512: 189-202.
SUTHERLAND, G. B. & EWEN, A. B. 1974. Fecundity decrease in mosquitoes ingesting blood from
specifically sensitized mammals. J. Insect Physiol., 20(4): 655-660.
TATCHELL, R. J. 1964. Digestion in the tick Argas persicus. Parasitology, 54: 423-440.
TIZARD, I. 1995. Imunologia Veterinária. 4a edição. Interamericana, México. 558p.

22TORO-ORTIZ, R. D.; DA SILVA VAZ, I.; GONZALES, J. C. & MASUDA, A. 1997. Monoclonal antibodies
against Boophilus microplus and their effects on tick reproductive effeciency. Vet. Parasitol., 69: 297-306.
TRACEY-PATTE, P. D.; KEMP, D. H. & JHONSTON, L. A. Y. 1987. Boophilus microplus, passage of bovine
immunoglobulins and albumin across the gut of cattle ticks feeding on normal or vaccinated cattle.
Research Vet. Sci., 43: 287-290.
TRAGER, W. 1939. Further observations on acquired immunity to the tick Dermacentor variabilis Say. J.
Parasitol., 25: 137-139.
TURNBULL, I. F.; SMITH, D. R. J.; SHARP, P. J.; COBON, G. S. & HYNES, M. J. 1990. Expression and
secretion in Aspregillus nidulans and A. niger. of a cell surface glycoprotein from the cattle tick,
Boophilus microplus, by using the fungal and S promoter system. Appl. And Env. Microbiol., 56 (9) :
2847 - 2852.
URIOSTE, S.; HALL, L. R.; TELFORD, S. R. & TITUS, R. G. 1994. Saliva of the lyme disease vector, Ixodes
dammini, blocks cell activation by a nonprostaglandin E2-dependent mechanism. J. Exp. Med., 180: 1077-
1085.
WEBSTER, K. A.; RANKIN, M.; GODDARD, N.; TARRY, D. W. & COLES, G. C. 1992. Immunological and
feeding studies on antigens derived from the biting fly Stomoxys calcitrans. Vet. Parasitol., 44(1-2): 143-
150.
WIKEL, S. K. & OSBURN, R. L. 1982. Immune responsiveness of the bovine to repeated low-level infestations
with Dermacentor andersoni. Ann. Trop. Med. Parasitol., 76: 405-414.
WIKEL, S. K. 1985. Resistence to ixodid ticks infestation induced by administration of tick tissue culture cells.
Ann. Trop. Ned. Parasitol., 79: 513-518.
WIKEL, S. K. 1988. Immunological control of hematophagous arthropod vectors: Utilization of novel antigen.
Vet. Parasitol., 29: 235-264.
WIKEL, S. K. 1996. Host immunity to ticks. Anna. Rev. Entomol., 41: 1-22.
WILLADSEN, P. & WILLIAMS, P. G. 1976. Isolation and partial characterization of an antigen from the cattle
tick, Boophilus microplus. Immunochemistry, 13: 591-597.
WILLADSEN, P.; WILLIAMS, P. G.; ROBERTS, J. A. & KERR, J. D. 1978. Responses of Cattle to Allergens
from Boophilus microplus. Intern. J. Parasitol., 8: 89-95.
WILLADSEN, P. 1980. Immunity to Ticks. Adv. Parasitol., 18: 293- 313.
WILLADSEN, P. 1987. Immunological approaches to the control of ticks. Intern. J. Parasitol., 17(2): 671-677..
WILLADSEN, P.; MCKENNA, R. V. & RIDING, G. A. 1988. Isolation from the cattle tick, Boophilus
microplus, of antigenic material capable of eliciting a protective immunological response in the bovine
host. Intern. J. Parasitol., 18(2): 183-189.
WILLADSEN, P. & KEMP, D. H. 1988. Vaccination with “Concealed” antigens for tick control. Parasitology
Today, 4 (7) 196-198.

23WILLADSEN, P.; RIDING, G. A.; MCKENNA, R. V.; KEMP, D. H.; TELLAM, R. L.; NIELSEN, J. N.;
LAHNSTEIN, J.; COBON, G. S. &. GOUGH, J. M. 1989. Immunologic control of a parasitic arthropod,
identification of a protective antigen from Boophilus microplus. J. Immunol., 143(4):1346-1351 .
WILLADSEN, P. & MC KENNA, R. V. 1991. Vaccination with “concealed” antigens: Myth or reality ?
Parasite Immunol., 13: 605-616.
WILLADSEN, P.; BIRD, P. E.; COBON, G. S. & HUNGERFORD, J. 1995. Commercialization of a
recombinant vaccines against Boophilus microplus. Parasitology, 110: S 43-50.
WHEELER, C. M.; COLEMAN, J. L. & BENACH, J. L. 1991. Salivary gland antigens of Ixodes dammini are
Glycoproteins that have interspecies cross-reactivity. J. Parasitol., 77(6): 965-973.