Embed Size (px)
Citation preview

1
UNIVERSIDADE ESTADUAL DO CEARÁ PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
FACULDADE DE VETERINÁRIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
VETERINÁRIAS
JULIÂN DA TRINDADE PONTES
EFEITO DO CORTISOL SOBRE O DESENVOLVIMENTO IN VITRO DE FOLÍCULOS OVARIANOS PRÉ-ANTRAIS CAPRINOS
FORTALEZA - CEARÁ
2015

2
JULIÂN DA TRINDADE PONTES
EFEITO DO CORTISOL SOBRE O DESENVOLVIMENTO IN VITRO DE FOLÍCULOS OVARIANOS PRÉ-ANTRAIS CAPRINOS
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Veterinárias da Faculdade de Veterinária da Universidade Estadual do Ceará, como requisito parcial para obtenção do grau de Mestre em Ciências Veterinárias.
Área de Concentração: Reprodução e Sanidade Animal.
Linha de Pesquisa: Reprodução e Sanidade de Pequenos Ruminantes.
Orientador: Prof. Dr. Claudio Cabral Campello.
FORTALEZA – CEARÁ
2015

3

4
JULIÂN DA TRINDADE PONTES
EFEITO DO CORTISOL SOBRE O DESENVOLVIMENTO IN VITRO DE FOLÍCULOS OVARIANOS PRÉ-ANTRAIS CAPRINOS
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Veterinárias da Faculdade de Veterinária da Universidade Estadual do Ceará, como requisito parcial para obtenção do título de Mestre em Ciências Veterinárias.
Aprovado em :15/12/2015.
BANCA EXAMINADORA

5
AGRADECIMENTOS
Agradeço a Deus, nosso Pai celestial, pelo dom da vida, por suas misericórdias que a
cada manhã se renovam e seu imensurável amor. A cada manhã que nos levantamos é a
oportunidade ideal para louvar e adorar o nome do Senhor, porque o nosso Deus merece
o nosso perfeito louvor e a nossa melhor adoração. Se não fosse por Ele não
existiríamos, por isso é bom nos lembrarmos d´Ele, porque d´Ele e por Ele, para Ele são
todas as coisas (ROMANOS 11.36).
À Universidade Estadual do Ceará (UECE) e ao Programa de Pós-graduação em
Ciências Veterinárias (PPGCV) por minha formação e capacitação profissional.
Agradeço em especial a minha querida esposa Pâmela Katrine da Silva Nogueira que foi
a base mais sólida que eu poderia ter, que me proporcionou todo apoio e ombro amigo
para que eu continuasse forte nesses dois anos de mestrado.
Ao meu pai Juarêz, e a minha mãe Adeliana, professora dedicada que me proporcionou
o melhor ensino que alguém pode dar o viver de uma vida honesta e digna. Agradeço a
minha esposa Pâmela, esposa dedicada que o Senhor preparou para ser a minha eterna
namorada, pelo seu imenso amor, apoio e incentivos e por cuidar de mim todos os dias.
Aos meus irmãos Andréa e Jonatas, pelo amor em família. Agradeço à minha Família,
pelo apoio em todos esses anos de vida, pela educação que com muito amor me deram
ensinamentos na vida acadêmica e na vida pessoal.
Agradeço ao Prof. Dr. Rodrigo Tenório Padilha, pelo incentivo em busca de uma
melhor qualificação profissional e por indicar o LAMOFOPA para realizar este
mestrado. A Prof. Dra. Déborah Padilha e a Dra. Valdevane Araújo pelo apoio durante a
realização deste mestrado.
À Carolina Maside Mielgo, por toda sua contribuição, incentivo, entusiasmo e amizade
durante a finalização deste trabalho!
À professora Maria Helena Tavares Matos e ao Biofov pela contribuição na realização
deste trabalho.
Aos funcionários do PPGCV, em especial a Adriana, ao Sr. João e ao Cesár que desde o
início me acolheram com carinho.

6
Ao meu orientador, professor Cláudio Cabral, por quem tenho tamanha admiração,
obrigada por toda a paciência, confiança e ensinamentos transmitidos.
Ao prof. José Ricardo de Figueiredo e à profa. Ana Paula Ribeiro Rodrigues, por todo
auxílio e incentivo profissional, pela confiança depositada em mim.
Aos membros da banca Dra. Deborah de Melo Magalhães Padilha, Dra. Carolina
Maside Mielgo e Dra. Jamily Bezerra Bruno, pelo auxílio na finalização desse trabalho
e por gentilmente terem aceitado ao convite de participar da minha banca examinadora.
Ao grupo de estudo bíblico formado por irmãos em Cristo pós-graduando do Lamofopa.
A toda equipe do Lamofopa, e aos que aqui não foram citados que contribuíram direta
ou indiretamente para que essa dissertação fosse feita, por toda a amizade e carinho.

7
RESUMO
O objetivo deste estudo foi identificar a expressão do receptor de glicocorticóides em
folículos ovarianos caprino e o efeito do cortisol em diferentes concentrações no cultivo
in vitro de folículos pré-antrais inclusos em tecido ovariano caprino. Fragmentos
ovarianos foram cultivados por 7 dias na ausência ou presenta de cortisol a diferentes
concentrações (0, 1, 5 e 10 ng/ml). Após o cultivo, a identificação da expressão do
receptor de glucocorticoids (NR3C1) em tecido ovariano mediante analises
inmunohistoquímicos assim como a morfologia, viabilidade, ativação e diâmetro
folicular e oocitário foram analisados. Em oócitos, NR3C1 foi fortemente expresso em
folículos primordiais e antrais. Foi observado um aumento progressivo na marcação
para NR3C1 em células da granulosa desde folículos primordiais a antrais,
independentemente do tratamento e tempo do cultivo. Após cultivo, houve uma redução
significativa no percentual de sobrevivência de folículos pré-antrais normais no
tratamento com 10 ng/ml de cortisol em relação aos demais tratamentos. Além disso, o
diâmetro folicular e oocitário diminuíram significativamente em todos os tratamentos
(cortisol 0, 1, 5 e 10 ng/ml) quando comparados com o controle fresco sem diferenças
entre os tratamentos. No entanto, os percentuais de ativação foram significativamente
superiores quando os folículos foram expostos ao cortisol (1, 5 e 10 ng/ml). Em
conclusão, pode-se observar a presença do NR3C1 no oócito e nas células da granulosa
em todas as categorias foliculares, com um papel mais importante nos estádios mais
avançados da foliculogênese. O cultivo in vitro demonstrou que, uma alta concentração
do cortisol (10 ng/ml), exerce um efeito deletério na sobrevivência folicular, reduzindo
o percentual de morfologia normal e o diâmetro folicular e oocitário.
Palavras-chave: Caprino, estresse, cortisol,NR3C1, folículos pré-antrais

8
ABSTRACT
The aim of this study was to identify the glucocorticoid receptor expression in goat
ovarian follicles and the effect of the cortisol in different concentrations on in vitro
culture of preantral follicles included in goat ovarian tissue (in situ). Ovarian fragments
were cultured for 7 days in the absence or presence of different concentrations of
cortisol (1, 5 and 10 ng/ml). After the culture, the expression of the glucocorticoids
receptor (NR3C1) was analysed in ovarian tissue by inmunohistoquimico analysis.
Moreover, the endpoints morphology, viability, activation and follicular and oocyte
diameter were also analyzed. The NR3C1 was strongly expressed in oocytes of antral
and primary follicle. A progressive increase in labeling for NR3C1 in granulosa cells
from primordial follicles to antral was seen regardless of the treatment and the culture
period. After the culture, it was observed a significant (P <0.05) reduction in the
percentage of survival of normal preantral follicles in the treatment with 10 ng/ml
cortisol when compared to the other treatments. Moreover, follicular and oocyte
diameter significantly (P <0.05) decreased in all treatments (cortisol 0, 1, 5 and 10
ng/ml) when compared to the fresh controls. However, there was no differences
between treatments. After 7 days of culture, the activation rate increased (P <0.05)
when the follicles were exposed to cortisol (1, 5 and 10 ng/ml). In conclusion, it was
observed the presence of NR3C1 in the oocyte and granulosa cells in all folicular
categories with a more important role in the late stages of folliculogenesis. The in vitro
culture showed that high concentrations of cortisol (10 ng/ml) exerts a deleterious effect
on follicular survival, reducing the percentage of normal morphology and follicular and
oocyte diameter.
Keywords: Caprine, stress, cortisol, NR3C1, preantral follicles.

9
LISTA DE FIGURAS
Figura 1- Ilustração do ovário mamífero e suas estruturas..............................
Figura 2- Caracterização dos folículos ovarinos................................................
Figura 3- Principais avanços obtidos com a utilização de oócitos oriundos de
folículos pré-antrais de mamíferos cultivados in vitro.....................
16
17
23

10
LISTA DE ABREVIATURAS
ACTH Adrenocorticotropic Hormone (Hormônio Adrenocorticotrófico)
ATP Adenosia Trifosfato
BSA Bovine Serum Albumin (Albumina Sérica Bovina)
CAPES Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
CE Ceará
CG Células da Granulosa
CGP Célula da Granulosa Primordial
CNPq Conselho Nacional de Desenvolvimento Científico e Tecnológico
CO2 Dióxido de Carbono
COCs Cumulus-oocyte complex (Complexo cululus-oocito)
CRH Corticotropin Releasing Hormone (Hormônio Liberador de
Corticotropina)
CT Células da Teca
CYP17 Cytocrome P450 17 - alpha hydroxylase
CYP19A1 (Aromatase)
DAB Diaminobenzina
DNA Desoxynucleic Acid (Ácido Desoxirribonucleico)
Dr. Doutor
Dra. Doutora
EGF Epidermal Growth Factor (Fator de Crescimento Epidermal)
ERK Extracellular Signal-regulate Kinases (Kinases Regulada por
Sinais Extracelulares)

11
E2 Estradiol
FSH Follicle Stimulated Hormone (Hormônio Folículo Estimulante)
FAVET Faculdade de Veterinária
FIG. Figura
GBC Globulin Binding Corticosteroid (Globulina Ligadora de
Corticosteróide)
GnRH Gonadotropin-releasing Hormone (Hormônio Liberador de
Gonadotropina)
GR Glucocorticóide Receptor (Receptor de Glicocorticoids)
H Horas
HC Histologia Clássica
HPA Hipothalamic-pituitary-adrenal (Hipotálamo-Pituitário-Adrenal)
HSP Heta Shock Portein (Porteína de Choque Térmico)
IgG Imunoglobolina G
IHC Immunohistochemistry (Imunohistoquímica)
IVM In vitro Maturation (Maturação in vitro)
LAMOFOPA Laboratório de Manipulação de Oócitos e Folículos Pré-Antrais
LH : Luteinizing Hormone (Hormônio Luteinizante)
MAPK Mitogen-activated Protein Kinases (Kinases Ativados por
Mitógeno)
MEM Minimal Essential Medium (Meio Essencial Mínimo)
MET Microscopia Eletrônica de Transmissão
Mg Miligramas

12
MOIFOPA Manipulação de Oócitos Inclusos em Folículos Ovarianos Pré-
antrais
mRNA Messenger ribonucleic acid (Ácido ribonucléico mensageiro)
MII Metaphase II (Metáfase II)
NA Not Applicable (Não Aplicável)
NR3C1 Nuclear Receptor Subfamily 3, Group C, Member 1 (Receptor
Nuclear Subfamília-3, Grupo –C, Membro -1)
PBS Phospate Buffered Saline (Tampão Fosfato-salino)
pH Potencial de Hidrogênio
RNAm Ribonucleic Acid Messenger (Ácido Ribonucleico mensageiro)
RT-PCR Reverse Transcription Polymerase Chain Reaction (Transcrição
Reversa – Reação em Cadeia da Polimerase)
Vol. Volume
ZP Zona Pelúcida
% Porcentagem
3βHSD 3β – hydroxysteroid dehydrogenase
2D Bidimensional
3D Tridimensional
GDF-9
TCM Tissue Cultered Medium (Meio de Cultivo de Tecido)
p. página
PPGCV Programa de Pós-Graduação em Ciências Veterinárias
PVN Paraventricular Nucleos of Hipothalamus (Núcleo Paraventricular

13
do Hipotálamo)
P34cdc2 Cyclin-dependent Kinase 1 (Ciclina Dependente de Kinase 1)
P<0.05 Probabilidade de Erro Menor do que 5%
SAS Statistical Analysis System (Sistema de Análise Estatística)
SEM Standard Erros of Means (Erro Padrão da Média)
StAR Steroidogenic Acute Regularoty protein (Proteína Reguladora
Aguda da Esteroidogenese)
UECE Universidade Estadual do Ceará
α-MEM Alpha Minimal Essential Medium (Meio Essencial Mínimo
Alpha)
µg Microgramas
µL Micolitros
µm Micrômetro
µM Micromolar
~ Aproximadamente
< Menor
= Igual
> Maior
± Mais ou Menos
≥ Maior ou igual

14
.
Sumário
1 INTRODUÇÃO..........................................................................................................15
2 REVISÃO BIBLIOGRÁFICA..................................................................................16
2.1 OVÁRIO MAMÍFERO.............................................................................................16
2.2 OOGÊNESE E FOLICULOGÊNESE.......................................................................17
2.3 POPULAÇÃO E ATRESIA FOLICULAR..............................................................19
2.4 ESTEROIDOGÊNESE FOLICULAR......................................................................20
2.5 CORTISOL................................................................................................................21
2.6 CULTIVO IN VITRO DE FOLÍCULOS PRÉ-ANTRAIS.......................................22
2.7 SISTEMAS DE CULTIVO.......................................................................................23
2.8 IMPORTÂNCIA DO MEIO DE CULTIVO.............................................................24
2.9 TÉCNICAS PARA AVALIAÇÃO DO CULTIVO IN VITRO...............................25
3 CAPÍTULO I – PAPEL DO CORTISOL NA FOLICULOGÊNESE
OVARIANA................................................................................................................27
4 JUSTIFICATIVA......................................................................................................42
5 HIPÓTESE CIENTÍFICA........................................................................................43
6 OBJETIVOS...............................................................................................................44
6.1 OBJETIVOS GERAL................................................................................................44
6.2 OBJETIVOS ESPECÍFICOS....................................................................................44
7 CAPÍTULO II - IMMUNOLOCALIZATION FOR GLUCOCORTICOID
RECEPTOR AND EFFECT OF CORTISOL ON IN VITRO DEVELOPMENT
OF CAPRINE PREANTRAL FOLLICLES………………………………………...45
8 CONCLUSÃO……………………………………………………………………….65
9 PERSPECTIVA……………………………………………………….…………..66
REFERÊNCIAS…………………………........………………...………………....67

15
1 INTRODUÇÃO
As técnicas de reprodução assistida passaram a ser uma prerrogativa
fundamental para o aumento da eficiência reprodutiva em animais de produção. Nesta
categoria animal, especialmente os ruminantes domésticos como a espécie caprina, a
utilização de biotécnicas reprodutivas estão sendo utilizadas com sucesso para contornar
os baixos índices de reprodução. Esses baixos índices são reflexos de um manejo
inadequado que, por sua vez, expõe cada vez mais os animais a situações de estresse
elevando os níveis de cortisol comprometendo a função reprodutiva (CHAVES et al.,
2013). Uma alternativa para avaliar os possíveis efeitos que o cortisol causa nos ovários
é a MOIFOPA (manipulação de oócitos inclusos em folículos pré-antrais), biotécnica
bastante utilizada para o estudo da foliculogênese, identificando os mecanismos pelos
quais diferentes substâncias atuam no ovário mamífero (FIGUEREDO et al., 2008).
Na reprodução, uma gama de fatores atua no desempenho reprodutivo, como o
estado nutricional, genética do animal e a produção hormonal, entre os hormônios
destaca-se o cortisol. Geralmente caracterizado como o hormônio do estresse, o cortisol
é produzido na zona fasciculada no córtex da glândula adrenal e desempenha um papel
importante em diversos processos fisiológicos como o sistema imunológico, metabólico
e reprodução (MICAHEL et al., 2003). No sistema reprodutivo, a produção excessiva de
cortisol afeta diretamente o eixo hipotalâmico-hipofisário inibindo a secreção do
hormônio liberador de gonadotrofinas - GnRH, como consequência disto, ocorre um
bloqueio na liberação de hormônios hipofisário, hormônio folículo estimulante –FSH e
hormônio luteinizante - LH) (MACFARLANE et al., 2000; BREEN et al., 2005) e,
diretamente nos ovários, o cortisol inibe a secreção de hormônios produzidos
localmente pelos folículos com o estradiol - E2 (BREEN et al., 2005), comprometendo a
qualidade do desenvolvimento folicular.
Dado a importância deste estudo, para uma melhor compreensão da sua
relevância, a seguir serão abordados os aspectos relacionados à estrutura do ovário,
oogênese e foliculogênese, população e atresia folicular, MOIFOPA, cultivo in vitro de
folículos pré- antrais e cortisol.

16
2 REVISÃO DE LITERATURA
2.1 OVÁRIO MAMÍFERO
O ovário é o principal órgão do sistema reprodutor feminino, o qual é
constituído de duas regiões distintas: a medular e a cortical. Na maioria das espécies a
região medular localiza-se mais internamente no ovário, sendo composta de tecido
conjuntivo, nervos, artérias e veias, responsáveis pela sustentação e nutrição deste órgão
(SILVA, 2005). O córtex ovariano por sua vez, é a porção mais externa do ovário,
considerado a porção funcional, sendo composta por células germinativas (oócitos) e
somáticas (células da granulosa - CG, da teca – CT e do estroma), as quais interagem e
organizam a formação dos folículos ovarianos (MCGEE; HSUEH, 2000). Em
decorrência da ovulação, o folículo ovariano sofre transformação na sua estrutura dando
origem ao corpo lúteo, estrutura também presente no córtex. O ovário desempenha duas
importantes funções fisiológicas, a saber: gametogênica e endócrina. A função
gametogênica está relacionada à produção de oócitos maturos (ovulação) aptos a serem
fecundados (BARNETT et al., 2006) enquanto que a função endócrina esta relacionada
a produção de hormônios, fatores de crescimento e peptídeos (HIRSHFIELD, 1991). A
interação destas duas funções é essencial para a manutenção da fertilidade.
Figura 1: Ilustração do ovário mamífero e suas estruturas. Adaptado de
http://www.angelfire.com/rings/juls/page2.html.

17
2.2 OOGÊNESE E FOLICULOGÊNESE
Segundo VAN DEN HURK; ZHAO, (2005) a oogênese consiste na formação e
diferenciação das células germinativas primordiais (CGP) culminando com a formação
do oócito haploide fecundado. A foliculogênese por sua vez, pode ser compreendida
pelo processo de formação e desenvolvimento (crescimento e maturação) folicular, o
qual se inicia na vida intrauterina/pré-natal em ruminantes e primatas (BEZERRA et al.,
1998). Este processo é decorrente da diferenciação das células germinativas primordiais
(CGP) e formação dos folículos primordiais culminando com o estágio de folículos pré-
ovulatórios (VAN DEN HURK; ZHAO, 2005).
O folículo ovariano é a unidade morfológica e funcional do ovário de
mamíferos, é responsável por proporcionar um ambiente ideal capaz de assegurar a
viabilidade, nutrição e desenvolvimento (crescimento e maturação) no oócito nele
incluso (CORTVRINDT; SMITZ, 2001) e produzir hormônios e peptídeos (ADASH,
1994). No interior dos folículos ovarianos encontra-se um oócito imaturo circundado
por células somáticas (granulosas e tecais), estes folículos podem ser classificados em
pré-antrais ou não cavitários (primordiais, transição, primários e secundários) e antrais
ou cavitários (terciários ou pré-ovulatórios) de acordo com o grau de evolução.
(FIGUEIREDO et al., 2008).
Figura 2: Caracterização dos folículos ovarinos: Folículos pré-antrais (1-3). Folículo
primordial (1). Folículo primário (2). Folículo secundário (3). Folículos terciário (4).
Folículos pré-ovulatório (5).

18
Os folículos primordiais representam o pool de reserva dos folículos ovarianos,
sendo que estes folículos representam 95% da população folicular do ovário. No interior
dos folículos primordiais encontra-se um oócito imaturo em estado de quiescência
circundado por uma camada de células da pré-granulosa deforma pavimentosa
(JUENGEL et al., 2002). As características morfológicas que marcam o início do
crescimento de folículos primordiais são: aumento do diâmetro oocitário, proliferação
das células da granulosa e transformação do formato destas células de pavimentoso para
cúbico. Durante esta fase, os folículos que apresentam células da granulosa de ambos os
formatos pavimentoso e cúbico são denominados intermediários ou de transição
(BARNETT et al., 2006). A fase de transição de transição de folículo primordial para
folículo primário caracteriza a ativação folicular e, alguns estudos tem demostrados que
é uma fase pouco dependente de gonadotrofinas, sendo sua regulação mediada por
fatores intraovarianos (VAN DEN HURK & ZHAO, 2005).
Os folículos primários são o primeiro estádio de crescimento após a saída do
pool de reserva, são morfologicamente caracterizados pela presença de uma única
camada completa de células da granulosa de forma cúbica em torno do oócito (SILVA,
2005). Neste estádio de desenvolvimento há o aparecimento da Zona Pelúcida (ZP), a
qual é formada por três glicoproteínas denominadas ZP1, ZP2 e ZP3 produzidas pelo
próprio oócito, com função de protegê-lo. Este evento é uma característica notável neste
estádio de desenvolvimento (RANHIN et al., 2001).
Os folículos secundários são caracterizados pelo aumento do oócito e
proliferação das células da granulosa, as quais formam duas ou mais camadas completas
de células com formato cúbico. Neste estádio também surgem as primeiras células da
teca (SILVA et al., 2004). As células da granulosa apresentam uma extensiva rede de
comunicações denominadas junções do tipo Gap. Essas junções do tipo Gap são canais
membranários que promovem a interação entre o oócito e as células da granulosa. As
junções do tipo Gap são compostas por proteínas denominadas Conexinas, ocorrem em
locais de próxima aposição celular, são estruturas de comunicação intercelular que
permitem a passagem de nutrientes, íons inorgânicos, segundos mensageiros e pequenos
metabólitos entre as células até chegar ao oócito que esta em constante crescimento
(KIDDER; MHAWI, 2002). Sabe-se que a interação entre o oócito e as células da
granulosa via junções do tipo Gap, é essencial para o crescimento do oócito, maturação
citoplasmática, comunicação intercelular, e inclusive para a liberação de fatores como o

19
GDF-9 e BMP originários do oócito, que tem ação anti-apoptótica sobre as células da
granulosa (HUSSEIN et al., 2005).
Os folículos antrais são formados a partir de uma intensa proliferação e
organização em várias camadas das células da granulosa resulta no crescimento dos
folículos, como consequência um aumento da atividade metabólica por estes tipos
celulares culmina com a produção em grandes quantidades de substâncias, que se
acumulam entre as células da granulosa caracterizando a formação de uma cavidade
repleta de líquido denominada antro folicular (DRIANCOURT, 2001). O fluido
folicular que preenche esta cavidade contém água, eletrólitos, proteínas séricas e alta
concentração de hormônios esteróides secretados pelas células da granulosa
(BARNETT et al., 2006).
2.3 POPULAÇÃO FOLICULAR E ATRESIA
A população de folículos ovarianos vária de entre espécies e entre indivíduos em
função de inúmeros fatores como: raça (CAHILL; MARIANA; MAULÉON, 1979),
idade (ERICKSON, 1966; RÜSSE, 1983), produção hormonal (PETERS, 1976),
genética (ERICKSON, 1966) e status reprodutivo e nutricional (ERICKSON;
REYNOLDS; MURPHREE, 1976; SCARAMUZZI et al., 1993). É sabido que o total
de folículos presentes no ovário é definido na vida intrauterina em ruminantes
(KNIGHT; GLISTER, 2006) e poucos dias após o nascimento em roedores (OJEDA et
al., 2000), sendo de 285.000 em bovinos (SILVA-SANTOS et al., 2011), 33.000 em
ovinos (AMORIM et al., 2000), 45.000 em caprinos (LUCCI et al., 1999) e
aproximadamente 2.000.000 na mulher (ERICKSON, 1986).
Apesar do grande população folicular presente no ovário mamífero, é sabido
que aproximadamente 99,9% dos folículos são eliminados pelo processo fisiológico
conhecido como atresia folicular, diminuindo assim o potencial reprodutivo das fêmeas.
A atresia ocorre por um processo de morte celular programada conhecido por apoptose
(TSAFIRI; BRAW, 1984). O destino final dos folículos ovarianos, sendo ovulação ou
atresia, é dependente de um balanço entre diferentes fatores endócrinos, parácrinos e
autócrinos que promovem a sobrevivência e aqueles que induzem a apoptose (HSU;
HSUEH, 2000).
Diante disso, visando evitar a enorme perda folicular que ocorre naturalmente in
vivo pela atresia, nas últimas décadas têm sido desenvolvidos vários sistemas de cultivo

20
in vitro de folículos pré-antrais e antrais que possibilitam o estudo dos fatores que
controlam a atresia e o crescimento folicular.
2.4 ESTEROIDOGÊNESE FOLICULAR
A produção de hormônios esteróides no ovário é importante para assegurar o
desenvolvimento normal do folículo. Sendo assim, o controle da produção hormonal
pelo ovário é orquestrado pela interação entre folículos e oócitos com a hipófise, nos
quais os hormônios de origem ovariana exercem uma retro-alimentação negativa ou
positiva sobre a produção e secreção de gonadotrofinas pela hipófise. A ação dos
esteroides nas suas células-alvo ocorre através receptores nucleares específicos, as
quais já foram localizados nos ovários (estroma, CG e CT) ovinos (JUENGEL et al.,
2006)
A biossíntese de esteróides é iniciada a partir do colesterol, sendo o precursor
desses hormônios. Deste modo, as células foliculares especializaram-se em sintetizar
o colesterol, uma vez que estas células expressam as enzimas do complexo
enzimático P450. A proteína StAR é responsável pelo transporte do colesterol da
membrana externa da mitocôndria para a interna, onde se localiza a enzima
desmolase, participante da conversão do colesterol em pregnenolona (GIOMETTI et
al., 2009). A pregnonelona sob ação da enzima 3β hidroxidesidrogenase é convertida
em progesterona que pode servir de substrato para a produção de androstenediona
via enzima 17,20-liase ou apresentar ação biológica (VALDEZ et al., 2005;
MIZRACHI E AUCHUS, 2009). A androstenediona quando não é destinada a
exercer sua função, pode ser convertida em testosterona pela enzima 17β- redutase,
que por sua vez, converter-se em dihidrotestosterona pela enzima 5α-redutase ou
penetrar nas células da granulosa ou estradiol pela enzima aromatase (DRUMMOND
E FINDLAY, 1999; YARAK et al., 2005; STOCCO, 2008). O estradiol também
pode ser produzido por uma segunda via, na qual a androstenediona sofre
aromatização nas células da granulosa e é convertida em estrona, e esta em estradiol,
pela enzima 17β-redutase (YARAK et al., 2005).
O estradiol – E2 juntamente com o estriol e estrona compõem os hormônios
estrogênios sendo o mais potente deles. Os folículos começam a sintetizar estradiol nos
estádios de pré-antrais avançados (Drummond e Findlay, 1999). E a ação desses

21
hormônios são mediadas são mediadas nos tecidos específicos por, pelo menos, dois
tipos de receptores (ER) denominados ERα e ERβ (Tomic et al., 2007). Altos níveis de
estradiol normalmente acarretam uma redução na secreção de FSH nas células da
granulosa em momentos próximos a ovulação, sendo então relacionados à redução da
fertilidade. Devido o inicio do processo de ovulação, inicia-se a luteinização das células
induzindo a produção de progesterona. A progesterona, por sua vez, é secretada pelas
células da granulosa e teca e células estroma ovariano, além de células luteais. Duas
forma de receptores de progesterona são descritos, a PRA e PRB (GAVA et al., 2004).
Existe ainda, a hipótese de a progesterona exerce uma ação luteotrópica local,
estimulando a sua própria produção e liberação (ROTHCHILD et al., 1991).
Desta forma, estradiol e progesterona são importantes hormônios reguladores da
foliculogênese, que atuam de forma isolada ou conjunta com outras hormônios atuando
diretamente nas células via receptores.
2.5 CORTISOL
O cortisol é produzido na zona fasciculada no córtex da glândula adrenal.
Pertencente à classe dos hormônios esteróides, os glicocorticoides, com o principal
representante o cortisol, diferem dos outros hormônios esteróides por apresentarem
receptores específicos. A biodisponibilidade do cortisol na corrente sanguínea se dá
através da associação com proteínas plasmáticas (ROSNER, 1991). Sendo que 96%
deste hormônio encontra-se ligado a glubulinas ligadoras de corticosteróides (CGB) ou
Transcortina e a albumina e dessa forma o cortisol encontra-se inativo. Apenas 4% do
cortisol encontra-se na forma livre (desassociado de proteínas) caracterizando-o como
biologicamente ativo (MICHAEL, 2003).
As ações do cortisol são mediadas através do receptor de glicocorticoides
(glucocorticoids receptor - GR) presente no interior das células. O GR pertencente à
superfamília de receptores nucleares – NR3C1 e já foram detectados na maioria dos
tecidos (ADCOCK, 2000), inclusive em tecido ovariano de camundongas (ILLERA et
al., 2005). É sabido que a biodisponibilidade do cortisol depende da ação da enzima
11β-hidroxiesteróide desidrogenase (11β-HSD) tipo 1 e 2, que tornam o cortisol ativo
ou inativo no interior das células. Quando ligado ao seu receptor, o cortisol é
translocado para o núcleo onde atua como fator de transcrição.

22
Em níveis basais, o cortisol é essencial para preservar a homeostasia e o
equilíbrio interno de funções como o sistema imunológico, metabólico e reprodutivo
(ANDERSEN, 2002). No entanto, o aumento nos níveis plasmáticos desse hormônio é
oriundo de condições estressantes. Em animais estressados, a função reprodutiva é
comprometida, uma vez que o aumento dos níveis de cortisol inibe a secreção de GnRH
no hipotálamo, bem como a liberação de FSH, além de reduzir a frequência do pulso
LH, bem como uma diminuição no pico pré-ovulatório de LH em ovelhas
(MACFARLANE et al., 2000; BREEN et al., 2005). A nível folicular, o cortisol pode
inibir a secreção de estradiol, isso contribuirá para interrupção do desenvolvimento
folicular e ovulação (BREEN et al., 2005). No tocante a morfologia folicular, em
animais estressados ocorre uma redução no diâmetro de folículos dominantes
acarretando danos a sua estrutura, porém alguns folículos podem completar seu
desenvolvimento chegando a ovular oócitos subférteis (LUCY et al., 2001).
Portanto, o cortisol é um importante hormônio ligado ao estresse e sua influência
na reprodução tem sido pesquisada, no entanto os efeitos que esse hormônio faz na
reprodução ainda não são totalmente conhecidos, podendo atuar em vários níveis do
sistema reprodutivo.
2.6 CULTIVO IN VITRO DE FOLÍCULOS PRÉ-ANTRAIS
O cultivo in vitro de folículos pré-antrais, também conhecido como “Ovário
Artificial”, é uma importante etapa da MOIFOPA que vem sendo largamente
empregada com o intuito de avaliar o efeito de diferentes substâncias, em diferentes
concentrações e em diferentes fases do desenvolvimento folicular, desde a ativação até a
produção de oócitos maturados (FIGUEIREDO et al., 2011). Sendo assim, o cultivo in
vitro destaca-se como uma importante técnica para elucidar mecanismos que controlam
a foliculogênese e atresia folicular.
A MOIFOPA consiste no isolamento ou regate de folículos pré-antrais do
ambiente ovariano, seguido da conservação por curto (resfriamento) ou longo período
(congelamento) e/ou cultivo folicular, visando-se a estocagem, bem como o
crescimento, maturação e fecundação in vitro dos oócitos inclusos em folículos
ovarianos pré-antrais (FIGUEIREDO et al., 2008). Atualmente, esta biotécnica tem
contribuído para elucidar a foliculogênese inicial, no entanto, para alcançar o objetivo
final com a produção de crias vivas é necessário garantir crescimento folicular,

23
maturação oocitária adequada e posterior fecundação dos oócitos pré-antrais através de
um sistema de cultivo eficiente. Com uma perspectiva futura desta técnica, a MOIFOPA
poderá contribuir para a multiplicação de animais de alto valor zootécnico ou em via de
extinção através do fornecimento de um grande número de oócitos provenientes de um
mesmo animal, os quais poderão ser destinados aos programas de produção in vitro de
embriões ou clonagem e posterior transferência de embriões (FIGUEIREDO et al.,
2008).
Figura 3: Principais avanços obtidos com a utilização de oócitos oriundos de folículos
pré-antrais de mamíferos cultivados in vitro. (Chaves et al., 2010)
2.7 SISTEMAS DE CULTIVO
Para alcançar sucesso no cultivo in vitro de folículos é necessário o uso de
métodos que permitam o isolamento folicular adequado sem que haja danos na estrutura
folicular. Além disso, o sistema de cultivo adotado tem como objetivo promover um
crescimento in vitro adequado que possibilite a manutenção da viabilidade de folículos
pré-antrais e a obtenção de oócitos aptos à fertilização, diversos sistemas de cultivo têm
sido desenvolvidos para atender a esse objetivo (VAN DEN HURK et al., 2000).
Atualmente dois sistemas de cultivos são comumente empregados em estudos no cultivo
in vitro de folículos pré-antrais, no entanto a escolha do sistema de cultivo é
determinada de acordo com os objetivos a serem estudados.

24
O sistema de cultivo in situ é caracterizado pelo cultivo de folículos pré-antrais
inclusos no próprio tecido ovariano, utilizando pequenos fragmentos do córtex ovariano
ou ainda utilizando o ovário inteiro. Este sistema torna-se bastante prático uma vez que
permite a manutenção da integridade tridimensional natural dos folículos, bem como a
interação destes com as células do estroma ovariano. No modelo isolado os folículos
podem ser cultivados de maneira bidimensional, aderidos diretamente à placa de cultivo
ou a uma monocamada de substrato e ainda na forma tridimensional (DEMEESTERE et
al., 2005). Este sistema de cultivo possibilita acompanhar de forma individual os
folículos durante o cultivo, favorecendo a perfusão do meio e o fornecimento de
nutrientes.
Existe ainda o sistema de cultivo em dois passos, neste modelo é efetuado
inicialmente o cultivo dos folículos no córtex ovariano (in situ), maximizando a
ativação folicular e o crescimento dos folículos primordiais até o estádio de secundários
e em sequência o isolamento e o cultivo dos folículos secundários isolados (TELFER et
al.,2008). Utilizando o modelo de cultivo em dois passos, possibilitou avanços obtidos
no cultivo in vitro de folículos pré-antrais, com resultados satisfatórios obtidos na
espécie murina, com a obtenção de crias viáveis a partir do cultivo de oócitos oriundos
de folículos pré-antrais (O’BRIEN et al., 2003). No entanto, nas espécies domésticas, as
taxas de maturação e produção de embriões oriundos do cultivo in vitro de folículos
pré-antrais ainda são variáveis. Em caprinos, por exemplo, foram obtidos apenas três
embriões após a fertilização in vitro de oócitos oriundos de folículos pré-antrais
cultivados in vitro (SARAIVA et al., 2010; MAGALHÃES et al., 2011).
2.8 IMPORTÂNCIA DO MEIO DE CULTIVO
O sucesso no desenvolvimento, crescimento e maturação folicular in vitro
depende de vários fatores, com destaque para a composição do meio no qual serão
cultivados os folículos. Diferentes meios comerciais podem influenciar o cultivo in vitro
de folículos ovarianos, estimulando a sobrevivência e o crescimento folicular (LIMA et
al., 2012). Dentre os meios podemos destacar o meio essencial mínimo (MEM) e suas
modificações (α-MEM, MEM Glutamax, etc.), o meio de cultivo de tecido (TCM 199),
além de outros meios como o McCOy’s, Waymouth, Leibowitz e Menezo B2 (ROCHA,
2013). A maioria dos meios de cultivo de base é suplementada com tampões,
antibióticos, antioxidantes e fontes protéicas (FIGUEIREDO et al., 2008). Outras

25
substâncias também são adicionadas ao meio de cultivo como: insulina, transferrina,
selênio, hormônio folículo estimulante (MAGALHÃES et al., 2009), proteína
morfogenética óssea-7 (ARAÚJO et al., 2011) e albumina sérica bovina (RODRIGUES
et al., 2010). Sendo assim, avaliar o efeito de diversas substâncias, por exemplo, o
cortisol o desenvolvimento folicular poderá contribuir para elucidação da
foliculogênese.
2.9 TÉCNICAS PARA AVALIAÇÃO DO CULTIVO IN VITRO
Atualmente existem várias técnicas disponíveis que permitem a avaliação da
qualidade folicular. As principais técnicas utilizadas permitem observar a ativação
folicular, mudanças na morfologia do folículo e oócito e produção de hormônios e
esteroides pelo folículo durante o crescimento in vitro. Para cada parâmetro a ser
avaliado existe uma técnica específica, porém a associação de mais de uma técnica
permite observar de forma mais precisa a qualidade folicular (MATOS et al., 2007).
As principais técnicas utilizadas para detectar alteração ou degeneração folicular
são: Histologia Clássica (HC) e Microscopia Eletrônica de Transmissão (MET).
A Histologia Clássica (HC) permite verificar mudanças na morfologia do
citoplasma e do núcleo, mais especificamente permite verificar o número e a mudança
no formato das células da granulosa através da ativação folicular e avaliar a integridade
do oócito e das células da granulosa. A Histologia Clássica é uma técnica quantitativa,
pois possibilita avaliar um grande número de folículos (MATOS et al., 2004), porém,
não é possível avaliar a integridade das organelas citoplasmáticas ou atividade
enzimática dos folículos ovarianos (GOSDEN, 2000).
No tocante a integridade das organelas citoplasmáticas ou avaliar a atividade
enzimática dos folículos ovarianos utiliza-se a Microscopia Eletrônica de Transmissão
(MET) uma técnica qualitativa que permite a avaliação de mudanças ultra-estruturais
corridas durante a atresia folicular, sendo destacada sua importância após conservação
in vitro de folículos pré-antrais caprinos e ovinos, mostrando que folículos considerados
normais, após a avaliação histológica, podem apresentar alterações degenerativas na sua
ultraestrutura (MATOS et al., 2004; SANTOS et al., 2006).
Existem outras técnicas que permitem a avaliação da viabilidade folicular e
apoptose com o uso de microscopia de fluorescência e corantes vitais. Esta técnica

26
utiliza marcadores fluorescentes que, ao serem excitados com radiação de baixo
comprimento de onda, absorvem energia e emitem luz de comprimento de onda maior
(JUNQUEIRA; CARNEIRO, 2004). Por ser uma técnica rápida, confiável e prática para
avaliação da viabilidade folicular antes ou após estudos de cultivo in vitro
(CORTVRINDT; SMITZ, 2001; LOPES et al., 2009), a microscopia de fluorescência
vem sendo comumente adotada para análise de folículos pré-antrais e oócitos após a
maturação in vitro (MIV) (ROSSETTO et al., 2009; BRUNO et al., 2009; CELESTINO
et al., 2011; MAGALHÃES-PADILHA et al., 2012). A calceína acetoximetil (calceína-
AM), o etídio homodímero-1 (EthD-1) e o Hoescht 33342 são os corantes mais
utilizados na microscopia de fluorescência.
Outras técnicas que vêm sendo bastante utilizadas são as técnicas de biologia
molecular, que podem ser realizadas antes, durante e após o cultivo in vitro de folículos
pré-antrais, com o intuito de identificar e quantificar o local de atuação e produção de
cada substância (ligante e receptor) envolvidas nas diferentes etapas do
desenvolvimento folicular. Uma técnica bastante utilizada é a Reação de Transcriptase
Reversa em Cadeia de Polimerase em tempo real (qRT-PCR) (KREUZER; MASSEY,
2002) que permite uma análise da quantificação relativa da expressão gênica em
determinado tecido ou amostra biológica, utilizando um sistema fluorescente em
plataforma capaz de detectar a luz oriunda da reação de amplificação de um
determinado gene no momento real da amplificação (BUSTIN, 2002). Através desta
técnica e da RT-PCR convencional, tornou-se possível identificar diferentes
substâncias, como hormônios e fatores de crescimento, presentes em folículos ovarianos
caprinos (CELESTINO et al., 2011; FROTA et al., 2011; ALMEIDA et al., 2012;
MAGALHÃES et al., 2012).
Além das técnicas citadas, a dosagem hormonal também é uma ferramenta que
auxilia na avaliação do cultivo in vitro. Hormônios esteroides, tais como progesterona, e
estradiol podem ser mensurados em meio de descarte, ou seja, meios retirados durante
as trocas, podendo ou não, serem avaliados utilizando radioimunoensaio ou ensaio
imunoenzimático (enzime-linked immunosorbent assay- ELISA).

27
Capitulo I
PAPEL DO CORTISOL NA FOLICULOGÊNESE NOS
MAMÍFEROS

28
Resumo Os glicocorticóides são hormônios essenciais para a manutenção do equilíbrio de
funções vitais para o organismo como a função imunológica e especialmente a
reprodutiva. O cortisol é o principal glicocorticoide produzido na glândula adrenal, sua
produção e liberação segue o ritmo circadiano, resultando em níveis basais. Porém
frente a situações de estresse que provoca um aumento acentuado nas concentrações de
cortisol, desta forma o excesso desse hormônio é prejudicial para todas as funções do
organismo. Em tecidos alvos, o cortisol é ativado pela enzima 11βHSD e desta forma,
se liga ao receptor de glicocorticóide presente no citoplasma das células. Após a ligação
com o receptor, o cortisol é translocado para o núcleo onde atua como fator de
transcrição. Alguns indícios sugerem que o receptor de glicocorticóide esta presente no
ovário, uma vez que esse que o cortisol foi identificado no fluido folicular, bem como a
sua associação com a maturação oocitária, resultado da presença de maiores
concentrações de cortisol com a maturação de oócitos em humanos. Porém, grandes
concentrações de cortisol podem interromper o desenvolvimento folicular,
principalmente a esteroidogênese folicular, suprimindo a produção de hormônios
esteróides pelos folículos. Além disso, A longa exposição ao estresse pode interromper
a função reprodutiva. Desta forma, visto a importância deste hormônio para o
organismo, principalmente o sistema reprodutivo, são necessários mais estudos visando
elucidar o papel do cortisol na função reprodutiva.

29
INTRODUÇÃO
Na reprodução, uma gama de fatores atua no desempenho reprodutivo, como o
estado nutricional, genética do animal e a produção hormonal, entre os hormônios
destaca-se o cortisol. Geralmente caracterizado como o hormônio do estresse o cortisol
é produzido na zona fasciculada no córtex da glândula adrenal e desempenha um papel
importante em diversos processos fisiológicos como o sistema imunológico, metabólico
e reprodução (MICAHEL et al., 2003). No sistema reprodutivo, a produção excessiva de
cortisol afeta diretamente o eixo hipotalâmico-hipofisário inibindo a secreção d
hormônio liberador de gonadotrofinas - GnRH, como consequência disto, ocorre um
bloqueio na liberação de hormônios hipofisário, hormônio folículo estimulante –FSH e
hormônio luteinizante - LH (MACFARLANE et al., 2000; BREEN et al., 2005) e
diretamente nos ovários, o cortisol inibe a secreção de hormônios produzidos
localmente pelos folículos com o estradiol - E2 (BREEN et al., 2005) comprometendo a
qualidade do desenvolvimento folicular.
A detecção de níveis de cortisol no fluído folicular (FATEH, 1989; ACOSTA,
2005) e a presença de receptores de glicocorticoides em células ovarianas de
camundongas (ILLERA et al., 2005) são indicativo da participação deste hormônio no
processo da foliculogênese ovariana. Com base nas informações apresentadas, a
presente revisão objetivou abordar de maneira geral alguns aspectos relacionando a
influencia dos glicocorticóides, em especial o cortisol, caracterizando sua síntese,
receptores, mecanismo de ação e enfatizando sua ação no sistema reprodutivo feminino.
CORTISOL
O cortisol (C21H30O5) é o principal hormônio representante da classe dos
hormônios glicocorticóides, este hormônio é sintetizado na zona fasciculada no córtex
da glândula suprarrenal. Após a produção do cortisol, sua liberação na corrente
sanguínea é regida sob controle neuroendócrino através do eixo hipotalâmico-
hipofisário-adrenal (HPA) e sua disponibilidade se dá através da ligação com proteínas
plasmáticas, dentre elas as globulinas ligadoras de corticosteróides (CGB) ou
Transcortina, a albumina e por último, pode ser encontrado na forma livre, a qual é
considerada a fração biologicamente ativa (ESCHER et al., 1997; ANDERSEN, 2003).
Em células alvos a ação dos glicocorticóides é mediada pelo receptor de

30
glicocorticóides (GR) um membro da superfamília dos receptores nucleares (NR3C1)
(GRIEKSPOOR et al., 2007)
Os glicocorticóides são hormônios esteróides essenciais para manter a
homeostasia, comumente envolvidos no controle de numerosas funções fisiológicas
vitais, tanto através da regulação do metabolismo celular genômico e do ciclo celular,
bem como através de efeitos não genômicos sobre o metabolismo das células e
sinalização (SCHONEVELD et al., 2004; OGIAS et al., 2009; BORSKI, 2000). Entre
essas funções, a saber, o metabolismo, ações anti-inflamatórias e imunossupressoras e o
sistema reprodutivo. Níveis endógenos de cortisol são controlados pela ativação do eixo
HPA, enquanto que níveis intracelulares deste hormônio além de estarem sob controle
deste eixo também são controlados pelas enzimas 11-β hidroxiesteróide desidrogenase
(11βHSD) que ativa (11βHSD1) ou inativa (11βHSD2).
LOCAL DE PRODUÇÂO
Sendo um dos principais hormônios que regem funções fisiológicas vitais, a
síntese de cortisol pela glândula adrenal esta sob controle do eixo hipotálamo-hipófise-
adrenal que depois de ativado tem como produto final a produção de cortisol. Devido ao
intenso metabolismo, o organismo torna-se carente deste hormônio. A síntese de
cortisol ocorre naturalmente de acordo com a necessidade do organismo ou em
decorrência a distúrbios envolvendo os sistemas que controlam sua produção.
Em um estado de homeostase, o organismo produz inúmeras substâncias em
níveis adequados para manter a capacidade de se adaptar ao meio. Em níveis adequados
as catecolaminas ativam neurônios parvocelulares situados no núcleo paraventricular do
hipotálamo (PVN). Estes neurônios estão relacionados no controle da atividade
simpática e cardiovascular, bem como, na liberação de hormônios que atuam na hipófise
anterior. Posteriormente a ativação dos neurônios do núcleo paraventricular no
hipotálamo, ocorre à produção do hormônio liberador de corticotropina (CRH). Este por
sua vez, controla a atividade do eixo hipotálamo-hipófise-adrenal pela ação na hipófise
anterior controlando a liberação do hormônio adrenocorticotrófico (adrenocorticotropic
hormone-ACTH) produzido pelas células corticotróficas da hipófise anterior. Ao ser
liberado o ACTH chega via corrente sanguínea na glândula adrenal e atua sobre a região
cortical iniciando a síntese e liberação de glicocorticóides em especial o cortisol,
estimulando também o aumento dessa região mantendo a integridade da mesma.

31
No interior do córtex adrenal na zona fasciculada a síntese de cortisol assim
como todos os hormônios esteróides inicia com uma molécula precursora de colesterol é
convertido em diferentes esteróides. O primeiro passo da esteroidogênese ocorre dentro
da mitocôndria quando o colesterol presente no interior das células da zona fasciculada
é transportado para interior da membrana mitocondrial pela proteína StAR
(steroidogenic acute regulatory proteis-StAR), o colesterol é convertido a pregnonelona
pela enzima P450 de clivagem de cadeia lateral (side-chain cleavage P450 - P450scc)
uma enzima mitocondrial membro da superfamília de enzimas citocromo P450 de gene
CYP11A1. A seguir a enzima 17α-hidroxilase (CYP17A1) atua sobre a pregnonelona
desta vez fora da mitocôndria, no retículo endoplasmático, convertendo-a em 17α-
hidroxipregnonelona que por sua vez é degradada em 17-hidroxiprogesterona em
resposta a ação da 3β-hidroxiesteróide desidrogenase (3β-HSD). Seguindo a via
esteroidogênica da síntese de cortisol, a formação de 11-Deoxicortisol é resultado da
ação da enzima 21-hidroxilase (CYP21) e por último, como resultado final a enzima
11β-hidroxilase (CYP11B1) presente nas células da zona fasciculada atua promovendo
a hidroxilação do carbono na posição 11 (C11) do composto 11-Deoxicortisol formando
cortisol.
Figura 1: Síntese de cortisol.

32
RECEPTOR DE GLICOCORTICÓIDES
Os glicocorticóides diferenciam-se dos demais hormônios esteróides por
apresentarem receptores específicos que são encontrados em quase todos tecidos. Nas
células alvos existem numerosos sítios de ligação dos glicocorticóides que variam de
2.000 a 30.000 por célula (ADCOCK, 2000), possibilitando ampla ação em diversas
funções fisiológicas. Estes hormônios unem-se a seus receptores presentes no citosol.
Após a entrada na célula sem a necessidade de um receptor transmembranário,
características dos hormônios lipossolúveis. Os glicocorticóides posteriormente são
translocados para o núcleo, onde atuam como fatores de transcrição.
Os receptores de glicocorticoides são proteínas intracelulares pertencente à
superfamília de receptores nucleares, mais especificamente denominados receptor
nuclear da superfamília 3, grupo C, membro 1 (nuclear receptor 3, group C, member 1 –
NR3C1). Esta classe compreende três domínios funcionais: um domínio N-terminal, um
domínio de ligação ao DNA e um domínio de ligação com o ligante C-terminal
(OAKLEY AND CIDLOWSKI, 2011). No citoplasma, na ausência do ligante, o
receptor de glicocorticóides está associado a proteínas de choque-térmico (heat-chock
protein – HSP), característica atribuída aos receptores de hormônios esteróides
(AGARWAL AND MIRSHASHI, 1999). As HSP estão comumente presente no
citoplasma não apenas associadas ao receptor de glicocorticóides, como também a
outras proteínas. Um complexo proteico é formado quando o receptor de
glicocorticóides é associado às proteínas HSP-90 e HSP70 (PRATT et al., 1993;
ADCOCK, 2000) e imunofilinas que participam da sinalização dos glicocorticóides
(DAVIES et al., 2002). As proteínas de choque térmico atuam retendo o receptor de
glicocorticóides no citoplasma, além de manter a capacidade ligação com o hormônio.
Após a ligação do cortisol no seu receptor, um complexo receptor-ligante é
formado e transloca-se para o núcleo onde se liga a elementos de resposta ao hormônio
na região promotor de gene alvo para atuar como um fator de transcrição.
11β-HIDROXISTEROIDE DESIDROGENASE
O acesso do cortisol biologicamente ativo no receptor de glicocorticóides nas
células alvo é regulado por duas enzimas desidrogenases, 11β-Hidroxisteróide
Desidrogenase tipo 1 e tipo 2. A ativação do cortisol é regulada pela enzima tipo 1 (11β-

33
HSD1), uma enzima de baixa afinidade para o cortisol que predominantemente atuar
como uma redutase dependente de NADPH catalisando a conversão de cortisona (uma
forma inativa do cortisol) em cortisol. Em contraste, a enzima tipo 2 (11βHSD2) atua
inativando cortisol convertendo-o em cortisona por ter uma atividade NAD+
dependente de alta afinidade para o cortisol (AGARWAL et al., 1994; ALBISTON et
al., 1994). A enzima 11β-HSD1 é altamente expressa no fígado, pulmão, tecido adiposo,
SNC e principalmente nos ovários, também nesses locais há uma expressão significante
dos receptores de glicocorticóides (SECKL AND WALKER, 2001). Em quanto que a
enzima 11β-HSD2 é expressa em tecidos alvos de aldosterona como cólon, nefrons
distal e glândulas sudoríparas. No entanto ambos os mRNAs das duas enzimas 11β-
HSD1 e 11β-HSD2 foram identificados no ovário bovino (TETSUKA et al., 2003), e a
sua expressão e atividade têm sido associados com o desenvolvimento e maturação
folicular (TETSUKA et al., 2010; THURSTON et al., 2007).
Após a ligação com o hormônio, ocorre uma alteração conformacional no
receptor de glicocorticóides que induz à dissociação do receptor de glicocorticóides com
as proteínas de choque-térmico, formando um complexo hormônio-receptor. Após a
formação do complexo hormônio-receptor, este é translocado para o núcleo. Uma vez
no núcleo, ocorre a dimerização do GR antes da sua ligação com o DNA (MARCEL et
al., 2003) seguido da ligação a elementos responsivos a glicocorticoides (GRES)
estimulando a expressão de genes alvos.
AÇÃO DO CORTISOL NA FOLICULOGÊNESE
Altas concentrações de glicocorticóides, especialmente o cortisol configuram a
perda da homeostasia pelo organismo, deste modo, um quadro de estresse é
caracterizado. Geralmente associado como hormônio do estresse, no tocante a
reprodução o cortisol tem a capacidade de atuar de três formas: 1 hipotálamo, 2 hipófise
ou diretamente nas gônadas (ANDERSEN, 2003), porém, independente da via de
atuação, o cortisol exerce efeitos diretos na foliculogênese o que pode levar a supressão
da atividade gonadal, bem como a esterilidade e perda do estimulo sexual.
Na ausência de condições ideais para a reprodução, a influência do estresse
reflete no baixo desempenho reprodutivo em animais de reprodução. Pequenos
ruminantes como caprinos e ovinos representam uma fonte de lucro para o Nordeste
brasileiro, devido a grande concentração desses animais nessa região (IBGE, 2010),

34
porém são mais facilmente afetados pelos efeitos deletérios que o estresse pode causar
oriundo de inúmeras fontes como alimentação e manejo inadequado e altas temperaturas
(CHAVES et al., 2013; OZAWA, et al., 2005).
Levando em consideração que o processo de reprodução dos animais domésticos
está sobre controle do eixo hipotalâmico-hipofisário-gonadal, para obter sucesso é
necessário que os mecanismos envolvidos nesse eixo estejam em completa sincronia
nos eventos relacionados. Nesse sentido, é necessário desenvolver mais estudos sobre
como o cortisol oriundo do estresse pode interferir a função reprodutiva.
Diversos estudos tem demonstrado que o cortisol implica em várias funções
ovarianas como, maturação, esteroidogênese e diferenciação das células da granulosa
(GONZÁLES et al., 2010; BREEN et al., 2005 – ovinos, ANDERSEN et al., 2002 -
humanos). O aumento das concentrações desse hormônio em situações de estresse
durante fases críticas do ciclo estral pode interferir nos estímulos hormonais indutores
do comportamento sexual e da ovulação (PAPARGARIS et al., 2011).
Efeito dos glicocorticoides na esteroidogênese
Durante o estresse, ocorre a ativação do eixo hipotalâmico-hipofisário-adrenal,
esta ativação desencadeia mudanças em quase todo o sistema endócrino, como
resultados podemos observar que em ovinos altos níveis de cortisol inibem a secreção
pulsátil do hormônio liberador de gonadotrofinas (GnRH) pelo hipotálamo
(MACFARLANE et L., 2000), consequentemente, quantidades reduzidas de GnRH
exercem pouco efeito na hipófise, reduzindo a secreção do hormônio folículo
estimulante (FSH) que aliado ao efeito inibidor cortisol na hipófise torna insuficiente a
quantidade de FSH para promover o recrutamento dos folículos durante a fase folicular.
Uma vez que se instala o estresse há pouca oferta de FSH, assim, existe uma
menor mobilização de folículos em crescimento durante a nova onda folicular. Há estes
folículos dar-se-á a característica de pouco produtores de estradiol, uma vez que o
próprio cortisol inibe a produção de estradiol pelos folículos em crescimento
contribuindo para a interrupção do desenvolvimento folicular (BREEN et al., 2005). A
produção reduzida de estradiol torna-se insuficiente para o surgimento do LH, foi
demostrado que o cortisol inibe o feedback positivo do estradiol sobre a indução do pico
de LH (WAGENMAKER et al., 2009). Em adição, o comportamento sexual é

35
prejudicado através da inibição das ações do estradiol para estimular o estro não
havendo receptividade sexual na ovelha (PAPARGARIS et al., 2011).
Efeito dos glicocorticoides na foliculogênese e maturação
A maturação oocitária tem por objetivo a produção de um oócito meioticamente
competente, apto a ser fertilizado. Nas espécies mamíferas a maturação é decorrente de
um crescimento em condições adequadas do oócito no folículo ovariano, porém este
processo é extremamente sensível podendo resultar em falhas na maturação por ocasião
de alterações fisiológicas (MAO et al., 2014). No âmbito da maturação in vitro, alguns
estudos têm se concentrado na utilização de substâncias que possam aumentar as taxas
de maturação in vitro, por exemplo, fatores de crescimento e hormônios (ARAÚJO et
al., 2010; MAGALHÃES et al., 2009).
Desde a década de 1980, alguns estudos se propuseram a investigar os efeitos
que os glicocorticóides podem causar na maturação oocitária, bem como a relação
destes hormônios com este evento final da foliculogênese. Um estudo pioneiro em
mulheres foi observado que após a aspiração folicular para a obtenção de oócitos para
fertilização in vitro, após a obtenção de oócitos maturados a concentração de cortisol no
fluido folicular foi duas vezes maior em relação ao fluido folicular de folículos
contendo oócitos imaturos, sugerindo uma correlação positiva entre o cortisol e a
maturação oocitária (FATEH, 1989). Semelhantemente, HARLOW, (1997) observou
que durante o surgimento da onda pré-ovulatória de LH há um aumento na concentração
de cortisol, deste modo o cortisol pode exercer um papel fisiológico na maturação e
ovulação.
Os mecanismos pelo qual o cortisol pode atuar na maturação ainda não são
claros, porém algumas hipóteses são levantadas como uma possível forma de ação deste
hormônio. Durante a retomada da meiose é necessário um fornecimento de energia
através da beta-oxidação de ácidos graxos livres presente nas células do cúmulos, assim
sendo, os lipídios são uma grande fonte de energia para a maturação (VALSANGKAR
et al., 2013). Mulheres em tratamento para fertilização in vitro, o cortisol promoveu a
mobilização de moléculas de lipídios presente em células do cúmulos como fonte de
energia, uma vez que este hormônio tem propriedades lipofílicas em outros tecidos
(CARMAN, 2012). Nesse estudo foi observada uma correlação positiva entre os níveis

36
de cortisol no fluido folicular com a maturação oocitária, portanto o cortisol no fluido
folicular pode promover o fornecimento de energia necessária para a retomada da
meiose através da beta-oxidação de ácidos graxos livres (SIMERMAN et al., 2015).
Estudos anteriores sugeriram que altos níveis de cortisol podem atuar em outros
mecanismos no controle da maturação Na maturação de oócitos suínos, cortisol e a
dexametasona inibiram a retomada da meiose (YANG et al, 1999). Durante a retomada
da meiose o fator promotor da maturação (MPF) é ativado através da desfosforilação da
tirosina p34cdc2 e síntese da ciclina B (NURSE, 1990). Deste modo, foi observado a
influência desses glicocorticóides na expressão da p34cdc2 e ciclina B1 durante a
maturação. Verificou-se que a exposição a 1 µg/ml de dexametasona reduziu i nível da
ciclina B1, mas não teve efeito na p34 cdc2. A exposição a dexametasona também
reduziu o complexo p34cdc2-ciclina B1, sugerindo que a ação dos glicocorticóides na
maturação pode ser atribuída a diminuição deste complexo.
Sobre um possível efeito dos glicocorticoides em outras vias de sinalização, na
maturação de oócitos ovinos, cortisol e dexametasona diminuíram o percentual de
oócitos maturados através da inibição da ativação das cinases reguladas por sinais
extracelulares (ERKS), ERK-1 e ERK-2 na presença de altas concentrações
(GONZÁLES et al., 2010). ERK-1 e 2 são membros da família das proteínas quinases
ativadas por mitógenos (MAPKs), as MAPKs são importantes reguladores da maturação
do oócito associada à organização dos microtúbulos e condensação da cromatina (FAN
and SUN, 2004). Para testar a hipótese de que os glicocorticoides afetam a maturação
oocitária, GONZÁLES (2010), utilizou oócitos de ovelhas para avaliar o efeito in vitro
do cortisol e dexametasona na via MAP kinase durante a maturação oocitária. Ambos os
glicocorticoides inibiram a ativação das ERK-1 e 2 durante o período de maturação,
resultando na diminuição do percentual de oócitos maturados in vitro na presença de
altas concentrações, sugerindo que níveis fisiológicos de glicocorticoides não acometem
danos à maturação do oócito.
Em conclusão, nota-se a importância do cortisol para diversos sistemas do
organismo incluindo a reprodução. Em níveis fisiológicos, este hormônio contribui para
equilíbrio natural do organismo preservando a homeostasia, especialmente na função
reprodutiva, o cortisol pode participar do processo de maturação oocitária. No entanto, o
aumento das concentrações de cortisol ocasionado por um estímulo estressante, acarreta

37
danos na função reprodutiva, principalmente na esteroidogênese folicular e o
desenvolvimento do folículo, além disso, ocorrendo estresse por um longo período o
cortisol pode até suprimir a função reprodutiva.
REFERÊNCIAS
ACOSTA, T.J.; TETSUKA, M.; MATSUI, M.; SHIMIZU, T.; BERISHA, B.;
SCHAMS, D.;
MIYAMOTO, A. In vivo evidence that local cortisol production increases in
the preovulatory follicle of the cow. J Reprod Dev v. 51, p.483–489, 2005
ADCOCK, I.M. Molecular Mechanisms of Glucocorticosteroid Actions. Pulmonary
Pharmacology & Therapeutics. vol. 13, p. 115–126. 2000.
AGARWAL, A.K.; MONDER, C.; ECKSTEIN, B.; WHITE, P.C. Cloning and
�expression of rat cDNA encoding corticosteroid 11 -dehydrogenase. Journal of
Biological Chemistry 264 18939–18943. 1998
AGARWAL, A.K.; MUNE, T.; MONDER, C.; WHITE, P.C. NAD+- dependent
�isoform of 11 -hydroxysteroid dehydrogenase. Journal of Biological Chemistry 269
25959–25962. 1994
ALBISTON, AL.; OBEYESEKERE, V.R.; SMITH, R.E.; KROZOWSKI, Z.S. Cloning
�and tissue distribution of the human 11 -hydroxysteroid dehydrogenase type 2 enzyme.
Molecular and Cellular Endocrinology. v. 105, p.11:17, 1994
ANDERSEN, C.Y. Possible new mechanism of cortisol action in female
reproductive organs: physiological implications of the free hormone
hypothesis. J. Endocrinol. v. 173. P. 211–217. 2002
VALDEVANE R. ARAÚJO, V.R.; SILVA, C.M.G.; MAGALHÃES, D.M.; SILVA,
G.M.; BÁO, S;N.;
SILVA, J.R.V.; FIGUEIREDO, J.R.; RODRIGUES, A.P.R. Effect of Bone
Morphogenetic Protein-7 (BMP-7) on in
vitro survival of caprine preantral follicles. Pesq. Vet. Bras. v. 30. p. 305:310, 2010

38
BORSKI, R.J. Nongenomic membrane actions of glucocorticoids in vertebrates. Trends
Endocrinol Metab. v. 11, p.427:36, 2000
BREEN, K.M.; BILLINGS, H.J.; WAGENMAKER, E.R.; WESSINGER, E.W.;
KARSCH, F.J. Endocrine basis for disruptive
effect of cortisol on preovulatory events. Endocrinology, v. 146,
n. 4, p. 2107-2115, 2005.
CHEN, W.Y.; YANG, J.G.; LI, P.S. Effect of dexamethasone on the expression of
p34cdc2 and cyclin B1 in pig oocytes in vitro. Molecular Reproduction and
Development. v. 56, p.74:79, 2000
DAVIES, T.H.; NING, Y.M.; SÁNCHEZ, E.R. A new fi rst step in activation of steroid
receptors: hormone-induced switching of fkbp51 and fkbp52 immunophilinso. The
Journal of Biological Chemistry. v. 277, p. 4597:4600, 2002
ESCHER G.; GALLI, E. VISHWANATH, B.S.; FREY, B.; FELIX, F.J. Tumor
necrosis factor alpha and interleukin i beta enhance the cortisol/cortisone suttle. Journal
of Experimental Medicine. v. 186, p. 189:198, 1997
FAN, H.Y.; SUN, Q.Y. Involvement of mitogen-activated protein kinase cascade during
oocyte maturation and fertilization in mammals. Biology of Reproduction v.70,
p.535;:47, 2004
FATEH, M; BEN-RAFAEL, Z; BENADIVA, C.A; MASTROIANNI, L. JR;
FLICKINGER, G.L. Cortisol levels in human follicular fluid. Fertil Steril. Vol. 51. P.
538-41. 1989
GONZÁLEZ, R; RUIZ-LEÓN, Y; GOMENDIO, M; ROLDAN, E.R.S. The effect of
glucocorticoids on ERK-1/2 phosphorylation during maturation of lamb oocytes and
their subsequent fertilization and cleavage ability in vitro. Reproductive Toxicology. V
29, p. 198–205, 2010.
GRIEKSPOOR, A.; ZWART, W.; NEEFJES, J.; MICHALIDES, R. Visualizing the
action of steroid hormone receptors in living cells. Nucl. Recept. Signal. v. 5, p. 003.
2002

39
HARLOW, C.R.; JENKINS, J.M.; WINSTON, R.M. Increased follicular fluid total and
free cortisol levels during the luteinizing hormone surge. Fertil Steril. v. 68(1), p.48:53.
1997
HUANG, T.J.; LI, P.S. Dexamethasone inhibits luteinizing hormone-induced synthesis
of steroidogenic acute regulatory protein in cultured rat preovulatory follicles. Biology
of Reproduction v. 64, p.163:170, 2001.
ILLERA,J.C.; G. SILVÁN, C.; MARTÍNEZ, M.; BLASS, A.; PEÑA, L. The effect of
dexamethasone on disruption of ovarian steroid levels and receptors in female rats. J.
Physiol. Biochem., 61 (3), 429-438, 2005.
MACFARLANE, M. S.; BREEN, K. M.; SAKURAI, H.; ADAMS,
B. M.; ADAMS, T. E. Effect of duration of infusion of stress-like
concentrations of cortisol on follicular development and the preovulatory surge of LH in
sheep. Animal Reproduction Science,
v. 63, p. 167-175, 2000.
MAGALHÃES, D.M.; ARAÚJO, V.R.; LIMA-VERDE, I.B.; MATOS, M.H.T.;
SILVA, R.C.; LUCCI, C.M.; BÁO, S.N.; CAMPELLO, C.C.; FIGUEIREDO, J.R.
Different FollicleStimulating Hormone (FSH) sources influence caprine preantral
follicle viability and development in vitro. Braz. J. Vet. Res. Anim. Sci. 5, 378-386.
2009
MAO, L.; LOU, H.; LOU, Y.; WANG, N.; JIN, F. Behaviour of cytoplasmic organelles
and cytoskeleton during oocyte maturation. Reprod Biomed Online. 28: 284-299. 2014
MICHAEL, A.E.; THURSTON, L.M.; RAE, M.T. Glucocorticoid metabolism and
reproduction: a tale of two enzymes. Reproduction. v. 126, p.425:441, 2003
NURSE, P. (1990). Universal control mechanism regulating onset of M-phase. Nature,
344:503-508
OAKLEY, R.H.; CIDLOWSKI, J.A. Cellular Processing of the Glucocorticoid
Receptor Gene and Protein: New Mechanisms for Generating Tissue-specific Actions of
Glucocorticoids. J Biol Chem. v. 286, p. 3177:3184, 2011

40
ONARD, J.L.M.; SCHONEVELD, INGRID, C. GAEMERS, WOUTER H. LAMERS.
Mechanisms of glucocorticoid signalling. Biochimica et Biophysica Acta. v. 1680, p.
114:128, 2004
OZAWA, M.; TABAYASHI, D.; LATIEF, T.A.; SHIMIZU, T.; OSHIMA, I; KANAI,
Y. Alterations in follicular dynamics and steroidogenic abilities induced by heat stress
during follicular recruitment in goats. Reproduction. v. 129, p. 621:630, 2005
PAPARGARIS, M.M; RIVALLAND, E.T .A; HEMSWORTH, P. H; MORRISEY, A.
D; TILBROOK, A. J. Acute and chronic stress-like levels of cortisol inhibit the
oestradiol stimulus to induce sexual receptivity but have no effect on sexual attractivity
or proceptivity in female sheep. Hormones and Behavior. V. 60. N.4. p.336-345. 2011
PRATT, W.B. The role of heat shock proteins in regulating the function, folding, and
trafficking of the glucocorticoid receptor. J. Biol. Chem. 268:21455- 21458, 1993.
ROSNER W. Plasma steroid-binding proteins. Endocrinol Metab Clin North Am.
20(4):697-720, 1991
SCHONEVELD, O.J.; GAEMERS, I.C.; LAMERS, W.H. Mechanisms of
glucocorticoid signalling. Biochim Biophys Acta. v. 1680, p.114:128, 2004
SECKL, J.R.; WALKER, B.R Minireview: 11b-Hydroxysteroid Dehydrogenase Type
1— A Tissue-Specific Amplifier of Glucocorticoid Action. Endocrinology. v. 142, p.
1371;1376, 2001
SIMERMAN, A.A.; HILL, D.L.; GROGAN, T.R.; ELASHOFF, D.; CLARCK, N.J.;
GOLDSTEIN, E.H.; MANRRIQUEZ, A.N.; CAZENBALK, G.D.; DUMESIC, D.A.
Intrafolliclar cortisol levels inversely correlate with cumulus cell lipid content as a
possible energy source during oocyte meiotic resumption in women undergoing ovarian
stimulation for in vitro fertilization. Fertility and Sterility®. v. 103, p.249-257, 2015
TETSUKA, M.; YAMAMOTO, S.; HAYASHIDA, N.; HAYASHI, K.G.; HAYASHI,
M.; ACOSTA, T.J.; MIYAMOTO, A. Expression of 11 beta-hydroxysteroid
dehydrogenases in bovine follicle and corpus luteum. J Endocrinol. v. 177, p.445–452,
2010

41
THURSTON, L.M.; ABAYASEKARA, D.R.; MICHAEL, AE. 11 Betahydroxysteroid
dehydrogenase expression and activities in bovine granulosa cells and corpora lutea
implicate corticosteroids in bovine ovarian physiology. J Endocrinol. v. 193, p. 299:310,
2007
TOMLINSON, J.W.; STEWART, P.M. �Cortisol metabolism and the role of 11 -
hydroxysteroid dehydrogenase. Best Practice and Research Clinical Endocrinology and
Metabolism. v. 15, p.61;78, 2001
VALSANGKAR, D.; STEPHEN, M.D. A Requirement for Fatty Acid Oxidation in the
Hormone-Induced Meiotic Maturation of Mouse Oocytes. BIOLOGY OF
REPRODUCTION. v. 89, 43 .p. 1:9, 2013
WAGENMAKER, E.R; BREEN, K.M.; OAKLEY, A.E.; TILBROOK, A.J.; KARSCH,
F.J. Psychosocial stress inhibits amplitude of gonadotropin-releasing hormone pulses
independent of cortisol action on the type II glucocorticoid receptor. Endocrinology.
v. 150, p. 762–769. 2009.

42
4 JUSTIFICATIVA
a. Escolha da espécie:
A caprinocultura tem grande importância no contexto da pecuária brasileira. A
região Nordeste ganha destaque com a expansão da caprinocultura concentrando mais
de 90% do plantel nacional, consolidando-se como a maior região produtora de carnes,
subprodutos e animais de alto valor zootécnico oriundos dessa espécie.
A espécie caprina apresenta melhor tolerância a nas regiões semi-áridas como o
Nordeste, porém o manejo reprodutivo inadequado aliado a temperaturas elevadas
refletem em um baixo desempenho reprodutivo dessa espécie.
b. Relevância do protocolo:
Estudos in vitro avaliando o efeito dos glicocorticóides, naturais e sintéticos
possibilitaram identificar os mecanismos pelos quais cortisol e dexametasona atuam
durante a maturação oocitária de folículos crescidos in vivo, elucidando o efeito dos
glicocorticoides durante a maturação no final da foliculogênese. Entretanto, na
foliculogênese pré-antral, folículos primários e secundários, não existe relato de como
os glicocorticoides atuam durante o desenvolvimento. Deste modo, o cultivo in vitro do
córtex ovariano em diferentes concentrações de cortisol poderá fornecer informações
importante de como esse hormônio atua nessa fase da foliculogênese.
c. Originalidade do trabalho:
Até o presente momento não existe relato da ação dos glicocorticóides na
foliculogênese pré-antral de caprinos. Portanto, a originalidade deste trabalho foi à
investigação dos efeitos do cortisol durante o desenvolvimento de folículos pré-antrais
caprinos cultivados in vitro visando elucidar como esse hormônio atua na
foliculogênese.

43
5 HIPÓTESES
O cortisol atua na função ovariana mediante o receptor de glicocorticóides
presente nos folículos ovariano em todas as categorias foliculares, além disso, esse
hormônio afeta o desenvolvimento e a viabilidade de folículos pré-antrais de uma forma
concentração-dependente.

44
6 OBJETIVOS
6.1 OBJETIVO GERAL
Identificar a imunolocalização do receptor dos glicocorticoides NR3C1 assim
como avaliar o efeito do hormônio cortisol sobre o desenvolvimento in vitro de
folículos ovarianos pré-antrais inclusos em tecido ovariano caprino.
6.2 OBJETIVOS ESPECÍFICOS
- Avaliar a expressão da proteína do receptor de glicocorticoides, NR3C1 em
folículos ovarianos incluso no tecido ovariano caprino;
- Verificar efeito do hormônio cortisol em diferentes concentrações (0, 1, 5 e 10
ng/ml) na ativação, morfologia e a viabilidade folicular bem como no diâmetro folicular
e oocitário de folículos ovariano caprino cultivados por 7 dias.

45
Capitulo II
IMMUNOLOCALIZATION FOR GLUCOCORTICOID RECEPTOR AND
EFFECT OF CORTISOL ON IN VITRO DEVELOPMENT OF CAPRINE
PREANTRAL FOLLICLES
PONTES, J.T.1,*; MASIDE, C.1; LIMA, L.F.1; MAGALHÃES-PADILHA, D.M.2;
MATOS, M.H.T.3; FIGUEIREDO, J.R.1; CAMPELLO, C.C.1
1Laboratory of Manipulation of Oocytes Enclosed in Preantral Follicles (LAMOFOPA), Faculty, State University of Ceará, Av. Paranjana, 1700, Campus do Itaperi, Fortaleza, 60740-000, CE, Brazil.
2Postgraduate biotechnology, Potiguar University / Laureate International Universities, Av. Senador Salgado Filho, 1610, Lagoa Nova, Natal, 59056-000, CE, Brazil
3Nucleus of Biotechnology Applied to Ovarian Follicle Development, Federal University of San Francisco Valley, Petrolina, PE, Brazil
*Correspondence should be addressed to: Faculty of Veterinary Medicine. Programa de Pós-Graduação em Ciências Veterinárias (PPGCV). Laboratório de Manipulação de Oócitos e Folículos Pré-Antrais (LAMOFOPA). Universidade Estadual do Ceará (UECE) Av. Paranjana, 1700, Campus do Itaperi. Fortaleza – CE – Brasil. CEP: 60740 903 Tel.: +55.85. 3101.9852; Fax: +55.85.3101.9840 E-mail address: [email protected]

46
Abstract
The aim of this study was to evaluate the immunolocalization for glucocorticoid
receptor (NR3C1) in goat ovarian follicles and the effect of cortisol on in vitro
development of caprine preantral follicles. Ovarian fragments were cultured for 7 days
under different cortisol concentrations (0, 1, 5 and 10 ng/ml). Before and after culture,
the protein expression of NR3C1 was analyzed in ovarian tissue by
immunohistochemical analysis. Moreover, the end points follicular morphology,
viability, activation as well as follicular and oocyte diameter were also analyzed. The
NR3C1 was strongly expressed in oocytes of primary and antral follicles. A progressive
increase of immunostaining for NR3C1 in granulosa cells from primordial to antral
follicles was observed regardless of the treatment. After culture, it was observed a
reduction in the rate of normal preantral follicles rate in the 10 ng/ml cortisol treatment
when compared to the other treatments (P <0.05). Moreover, follicular and oocyte
diameter significantly decreased in all treatments (cortisol 0, 1, 5 and 10 ng/ml)
compared to the fresh control. After culture, the activation rate increased when the
follicles were exposed to 1, 5 and 10 ng/ml cortisol compared to the fresh control (P
<0.05). In conclusion, it was observed the presence of NR3C1 in the oocyte and
granulosa cells in all follicular categories. The in vitro culture showed that high cortisol
concentrations (10 ng/ml) exerts a deleterious effect on follicular survival, reducing the
percentage of normal follicles as well as follicular and oocyte diameter.
Keywords: Caprine, stress, cortisol, NR3C1, preantral follicles.

47
INTRODUCTION
Goat production is economically and socially important activity with an
increasing interest in milk, meat and skin production around the world. Although, goats
are well adapted to many different environments, improper handling and feeding
practices may produce stress reducing their productivity and profitability (18).
Conditions of stress lead to an increase of cortisol inducing suppression of reproductive
system function (15, 28) with subsequent economic impact on the livestock.
Cortisol, commonly known as hormone stress, is produced in the adrenal gland
and is responsible for the regulation of several physiological processes, including
metabolism, immunological response, and female reproductive function. High level of
cortisol in response to stress, acts directly or indirectly on the reproductive functionality
resulting in low reproductive rates (11, 28). The rise of cortisol acts on the
hypothalamic -pituitary suppressing GnRH secretion which in turn inhibits of FSH
release and reduce the LH surge frequency in sheep (7,20). Furthermore, excessive
cortisol levels can inhibit the estradiol secretion, inducing the disruption of the follicular
development and ovulation (7).
In physiological conditions, several studies have demonstrated the role of
cortisol on the ovary during the oocyte maturation as well as granulosa cell
differentiation and steroidogenesis (7, 13 sheep 3 - human). Moreover, this hormone is
associated with oocyte maturation in human because human follicular fluid from
matured oocytes had greater cortisol level than immature oocytes (12). In addition,
cortisol increases lipid metabolism in pre-ovulatory follicle and promoting meiotic
resumption in human (26).

48
Despite numerous studies reporting the role of cortisol in reproduction, it is
unclear the effect of this hormone on the early folliculogenesis (primordial and primary
follicles). In addition, at least for goats, there is no information on the localization of
cortisol receptor in the ovarian follicles. Therefore, the aims of this study were to
investigate the immunolocatization for cortisol receptor, NR3C1, and the effect of
cortisol on the survival and activation of caprine preantral follicles cultured in vitro.
MATERIAL AND METHODS
Chemicals
Unless mentioned otherwise, all chemicals used in the present study were
purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Source of ovaries
Ovaries (n=10) from 5 adults, mixed-breed goats between 1 to 3 years old were
collected at a local slaughterhouse. Immediately after slaughter, the ovaries were
washed in 70% ethanol, followed by two washes in washing medium consisting of
Minimum Essential Medium (MEM) buffered with 25mM HEPES (MEM-HEPES)
supplemented with 100 µg/ml penicillin and 100 µg/ml streptomycin. Ovaries were
transported to the laboratory within 1 hour in 50 ml tubes containing wash medium at 4
ºC (10).
Immunohistochemical localization of glucocorticoid receptor (NR3C1)
In the laboratory, before culture (fresh control) and after 7 days of culture, all of
the ovarian fragments cultured with different concentration of cortisol (0, 1, 5, 10
ng/ml) were fixed in 4% paraformaldehyde for 12 h, dehydrated, and embedded in
paraffin. For detections of NR3C1, immunohistochemistry was performed according to

49
the protocol described previously by Sales et al. (24). Briefly, sections of 5 µm from
each block were obtained and mounted in poly- L-lysine slides and dried overnight at
37 ºC, deparaffinised in xylene and rehydrated in graded ethanol series, followed by
antigen retrieval with citric acid buffered solution (pH 6.0) and Tween 20. After
blocking solution endogenous peroxidase activity with 3% hydrogen peroxide and
methanol for 10 min, the sections were incubated for 1 h with blocking solution (25 ml
PBS containing 1.25% bovine serum albumin and 3% Triton X) at room temperature
and overnight with rabbit polyclonal primary antibody (NR3C1) (1:800 Abcam Inc.,
USA) at 4 °C. After repeated washing in PBS for 5 min, slides were incubated with
secondary antibody Rabbit anti-rabbit IgG (1:500 Abcam Inc., USA) for 30 minutes at
room temperature followed by washing in PBS for 5 minutes. Then the sections were
incubated with horseradish peroxidase-conjugated avidin (ABC kit, Vector
Laboratories, Burlingame, CA, USA). The positive reactions were visualized with 3,3’ -
diaminobenzidine tetrahydrochloride (DAB) solution. Finally, the sections were
counterstained with hematoxylin, dehydrated, and mounted in Depex. Replacing the
primary antibody for IgG of the same species in which the primary antibody was
produced performed negative controls. The preantral follicles were classified as
primordial (oocyte surrounded by one layer of squamous granulosa cells), primary
(oocyte surrounded by one layer of cuboidal granulosa cells), or secondary (oocyte
surrounded by two or more layers of cuboidal granulosa cells). The presence of small
antral follicles (presence of cavity filled with follicular fluid and well-developed layers
of granulosa cells) was also noted. In the various follicular compartments (oocyte,
granulosa or theca cells), the immunostaining was classified as absent (−), weak (+),
moderate (++) or strong (+++).

50
In vitro culture of goat preantral follicle
In vitro culture of caprine follicles was performed according to the protocol
described previously by Chaves et al. (10). Briefly, in the laboratory, the surrounding
fatty tissues and ligaments were stripped from the ovaries. Ovarian cortical slices
(3x3x1 mm) were cut from the ovarian surface using a surgical blade under sterile
conditions and subsequently placed in holding medium consisting of MEM-HEPES
with antibiotics (100 mg/mL penicillin and 100 mg/mL streptomycin). Subsequently,
half the ovarian fragments were randomly fixed for histological analysis (fresh control
or noncultured fragments). The remaining fragments were cultured individually in vitro
in 24-well culture dishes (Corning, Corning, NY, USA) containing 1mL culture
medium at 39◦C in presence of 5% CO2 in air for 7 days. The basic culture medium
consisted of α-MEM (pH 7.2–7.4) supplemented with 10 ng/ml insulin, 5.5 ng/ml
transferrin, 5 ng/ml selenium, 2 mM glutamine, 2 mM hypoxanthine and 1.25 mg/ml
bovine serum albumin This medium was called MEM+ and was supplemented with
different cortisol (Hydrocortisone) concentrations (0, 1, 5 and 10 ng/ml). Before (fresh
control) and after culture, the samples were fixed and destined for histological analysis.
Fresh medium was prepared immediately before use. Every 2 days, the culture medium
was replaced by fresh medium. Before (fresh control) and after in vitro culture for 7
days, ovarian fragments were processed as described below to evaluate the morphology
of goat preantral follicles.
Morphological analysis of goat preantral follicles
Ovarian fragments from each treatment, including the fresh control or non-
cultured fragments, were fixed individually in 4% paraformaldehide for 12 h.
Subsequently, fragments were dehydrated in a graded series of ethanol, clarified with

51
xylene and embedded in paraffin wax (8). Tissues were sectioned serially at a thickness
of 7μm and sections were stained using standard protocols with periodic acid-Schiff and
haematoxylin (PAS staining system; Sigma). Sections were examined by light
microscopy (Zeiss, Jena, Germany) at 400× magnification.
Preantral follicles were counted and evaluated in the section where the oocyte
nucleus was visible. According to the developmental stage, preantral follicles were
classified as primordial (oocyte surrounded by one layer of flattened pre-granulosa
cells) or developing, with the latter category further subdivided into intermediate
(oocyte surrounded by one layer of flattened pregranulosa and cuboidal granulosa cells),
primary (oocyte surrounded by one layer of cuboidal granulosa cells) or secondary
(oocyte surrounded by two or more layers of cuboidal granulosa cells). Each follicle
was evaluated according to the following morphological parameters: (1) integrity of
oocyte and granulosa cells; (2) the presence or absence of pyknotic bodies; (3)
ooplasmic retraction; and (4) the organisation of granulosa cells. Based on this
evaluation, preantral follicles were classified as normal, when a morphologically normal
oocyte with a non-pyknotic nucleus was surrounded by granulosa cells organised in
discrete layers. Degenerated follicles were defined as those with a retracted oocyte, with
a pyknotic nucleus and/or surrounded by disorganised granulosa cells, which were
detached from the basement membrane. The tissue was performed by a single player
using the same optical microscope (Nikon, Tokyo, Japan). Slide analysis was made
blind reading (reviewing the slides without knowledge of treatment group).
Primordial follicle activation and growth
To evaluate primordial follicle activation and growth, only intact follicles with a
visible oocyte nucleus were recorded and the proportion of primordial and developing

52
follicles was calculated for fresh control and for all treatments where the ovarian
fragments were cultured for 7 days with different cortisol concentrations (0, 1, 5 and 10
ng/ml). In addition, for each follicle category, oocyte and follicle diameters were
measured using an ocular micrometer and the average of the minor and major axes was
reported as oocyte and follicle diameter, respectively.
Statistical analisys
Data were initially submitted to Shapiro-Wilk and Bartlett tests to verify
normality of residues distribution and homoscedasticity, respectively. Confirmed both
requirements underlying analysis of variance (ANOVA), it was performed according to
a completely randomized design using General Linear Model Procedure (GLM) of SAS
(2002) and Student-Newman-Keuls (SNK) test was applied to compare means when a
significant effect of treatment (cortisol concentration) was present. Results were
expressed as mean ± standard deviation and differences were considered significant
when p <0.05.
RESULTS
Immunohistochemical localization of glucocorticoid receptor (NR3C1) in the caprine
ovarian tissue
The results from immunohistochemistry demonstrated the presence of the
NR3C1 in all ovarian follicular categories in caprine, varying only the intensity and
location. There was no difference in the intensity of immunoreaction NR3C1 among the
treatments. However, when it was evaluated the follicular structures in preantral follicle,
the oocyte of primordial follicles showed a moderate immunostaining in the cytoplasm
and strong immunoreaction in the nucleus; a weak immunostaining in the oocyte
(cytoplasm and nucleus) of primary follicles and a moderate immunostaining in the

53
oocytes of secondary follicles (cytoplasm and nucleus) was detected. Regarding to the
granulosa cells, there was a progressive increase of immunostaning for NR3C1 in
primordial, primary and secondary follicles, which were classified as absent, weak and
moderate immunoreaction, respectively (Tab. 1). In antral follicles, there was a strong
immunoreaction in the oocyte (cytoplasm and nucleus) as well in the granulosa cells. In
theca cells, it was not detected the presence of NR3C1 in the secondary follicles,
however, the antral follicles had a weak immunoreaction.
Table. 1: Relative intensity of immunohistochemical staining for NR3C1 in caprine
ovarian follicles.
Follicular category
Follicular
structure
Primordial Primary Secondary Antral
Oocyte +++ + ++ +++
Granulosa - + ++ +++
Theca NA NA - +
NA, not applicable. (-) absent; (+) weak; (++) moderate; (+++) strong immunoreaction.
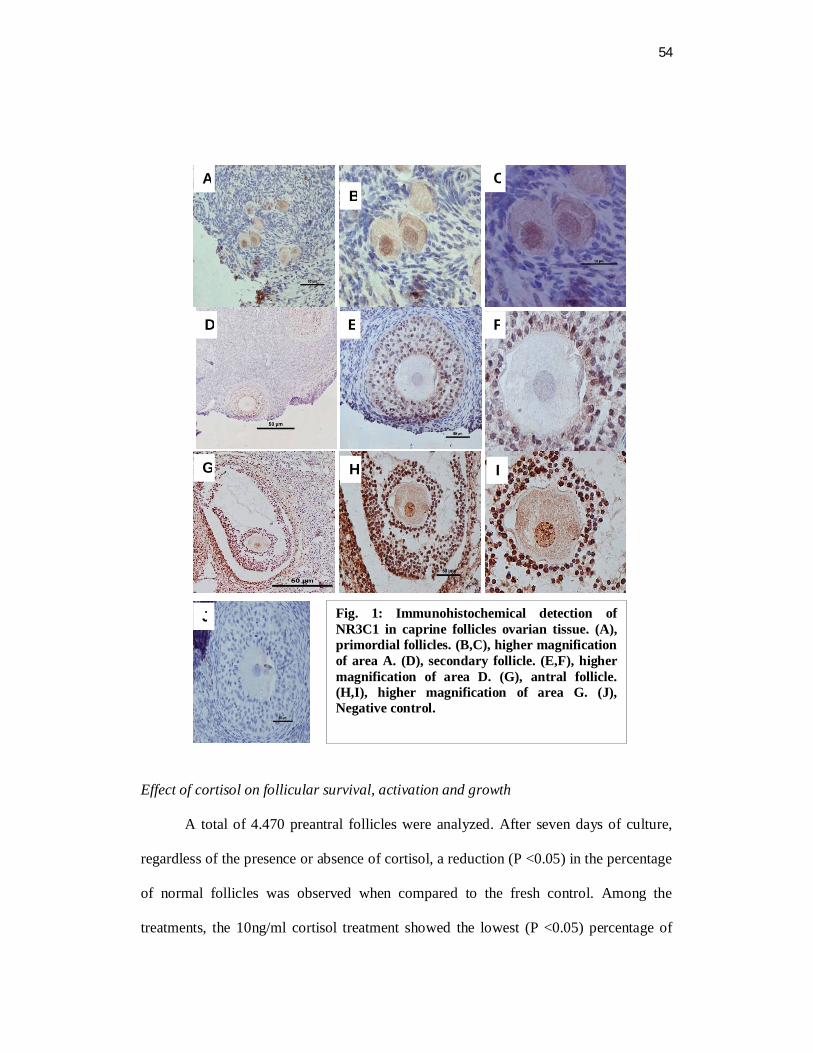
54
A B
C
F E D
G H I
Effect of cortisol on follicular survival, activation and growth
A total of 4.470 preantral follicles were analyzed. After seven days of culture,
regardless of the presence or absence of cortisol, a reduction (P <0.05) in the percentage
of normal follicles was observed when compared to the fresh control. Among the
treatments, the 10ng/ml cortisol treatment showed the lowest (P <0.05) percentage of
J Fig. 1: Immunohistochemical detection of NR3C1 in caprine follicles ovarian tissue. (A), primordial follicles. (B,C), higher magnification of area A. (D), secondary follicle. (E,F), higher magnification of area D. (G), antral follicle. (H,I), higher magnification of area G. (J), Negative control.
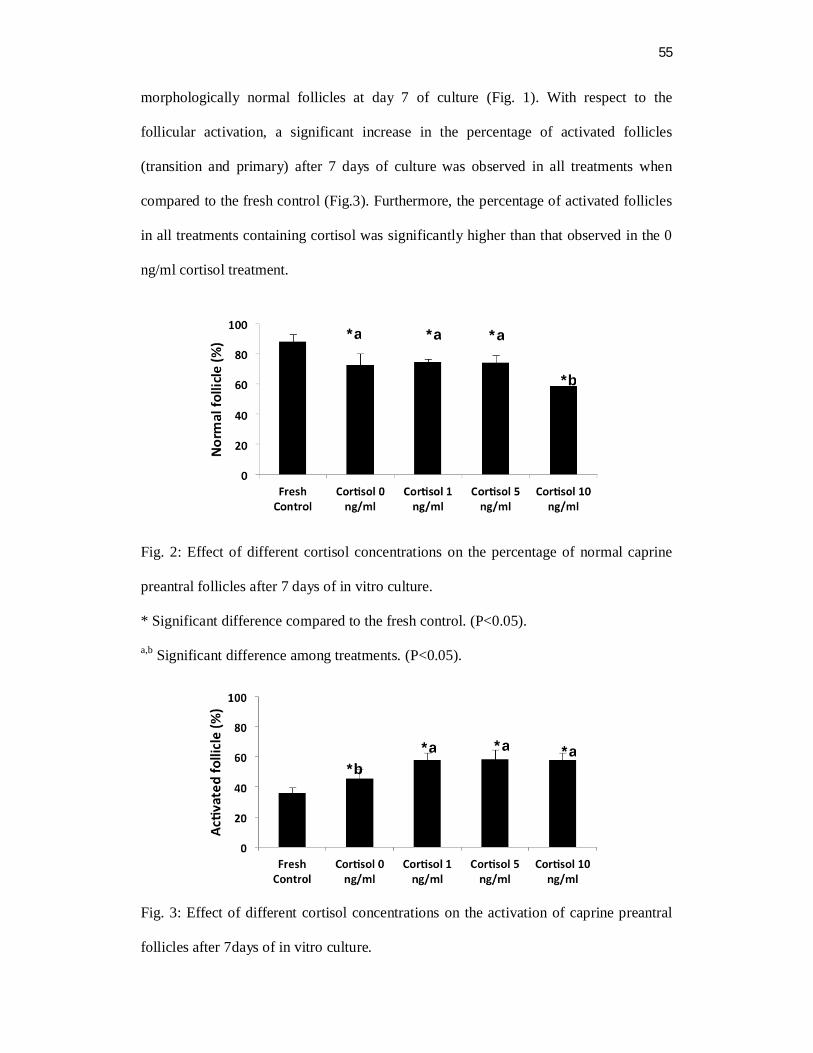
55
morphologically normal follicles at day 7 of culture (Fig. 1). With respect to the
follicular activation, a significant increase in the percentage of activated follicles
(transition and primary) after 7 days of culture was observed in all treatments when
compared to the fresh control (Fig.3). Furthermore, the percentage of activated follicles
in all treatments containing cortisol was significantly higher than that observed in the 0
ng/ml cortisol treatment.
Fig. 2: Effect of different cortisol concentrations on the percentage of normal caprine
preantral follicles after 7 days of in vitro culture.
* Significant difference compared to the fresh control. (P<0.05).
a,b Significant difference among treatments. (P<0.05).
Fig. 3: Effect of different cortisol concentrations on the activation of caprine preantral
follicles after 7days of in vitro culture.
*a *a *a
*b
*b
*a *a *a

56
* Significant difference compared to the fresh control. (P<0.05).
a,b Significant difference among treatments. (P<0.05).
The oocyte and follicular diameters in the fresh control and in the ovarian tissue
cultured are shown (table 2). There was a decrease (P <0.05) in the follicular and oocyte
diameters in all treatments when compared to the fresh control. However, no significant
difference (P >0.05) in follicular and oocyte diameters was observed among the cultured
treatments.
Table 2: Effect of different cortisol concentrations on follicular and oocyte diameters of
caprine preantral follicles during in vitro culture.
Treatment Follicle Oocyte
FRESH CONTROL 36,95 ± 6,93 24,09 ± 3,19
CORTISOL 0 ng/ml 30,62 ± 5,20* 19,44 ± 3,28*
CORTISOL 1 ng/ml 31,72 ± 6,37* 19,37 ± 3,83*
CORTISOL 5 ng/ml 29,71 ± 6,49* 17,94 ± 2,99*
CORTISOL 10 ng/ml 26,77 ± 6,06* 17,72 ± 3,72*
* Significant difference compared to the fresh control. (P<0.05).
DISCUSSION
The caprine species is adapted to different environments, but due to high
temperatures, improper handling and feeding suffer due to stress, causing a rise in
cortisol concentration (18) disturbing ovarian function, such as oocyte maturation as
well as granulosa cell differentiation and steroidogenesis (7, 13- sheep, 3 - humans).
Despite numerous studies reporting the role of cortisol in reproduction, it is unclear the
effect of this hormone on the early folliculogenesis (primordial and primary follicles).

57
The current study showed for the first time the immunolocalization for NR3C1
in goat ovarian follicles. After the receptor is bound to cortisol, this protein is activated
and translocated to the nucleus acting as transcription factor (15). The mRNA for
glucocorticoid receptor has been detected in other species (27), using the RT-PCR.
However, in goats, there was no studied about glucocorticoid receptor in the ovary. The
current study was the first one that demonstrated the presence of glucocorticoid receptor
NR3C1 in the caprine ovary. NR3C1 was expressed in all follicular categories and
compartments, especially in the oocyte (nucleus and cytoplasm) and in the granulosa
cells of secondary and antral follicles. The NR3C1 expression increased in advanced
stages of folliculogenesis. In bovine, mRNA was detected for the glucocorticoid
receptor in all follicular categories, both in granulosa and theca cells (27). Furthermore,
in primordial follicles, our results showed that NR3C1 was expressed with a strong
immunoreaction in the oocyte, suggesting an important role for the maintenance of
follicular quiescence or activation. In the secondary follicles, granulosa cells express the
protein receptor for glucocorticoids, however it was not possible to classify the intensity
of the immunostaining in the theca cells, probably due to the reduction in cytoplasmic
volume during the histology procedure (2).
The present study was the first to demonstrate the effect of different
concentrations of cortisol on in vitro culture of goat preantral follicles. After the culture,
it was observed that cortisol affected in concentration-dependent manner the follicular
survival and growth. Regarding the survival, the exposure to 10 ng/ml cortisol resulted
in a lower percentage of normal preantral follicles when compared to the other
treatments and the fresh control. One explanation for this fact is that the glucocorticoids
may deregulate ovarian steroidogenesis, inhibiting the biosynthesis of steroids in
granulosa cells, probably by a decreased expression of StAR protein (16, 6). In cattle,

58
during induction of stress by ACTH administration, there was a decrease in the
expression of P450 aromatase and CYP17 enzyme, reducing the production of estradiol
and androstenedione (5), and therefore contribute to the reduction of follicle viability.
The data from the current study were consistent with previous in vivo studies about the
effect of cortisol caused by the stress. In sheep, the cortisol was correlated with the
inhibition of follicular recruitment and selection phase in the first half of the follicular
phase (29).
In the current study, a significant increase in the percentage of activated follicles
after in vitro culture was observed in all treatments in the presence of cortisol. This
suggests that this hormone can act affecting primordial follicle category and promoting
their activation. Moreover, a strong immunoreaction in oocytes of primordial follicles
was observed in the present study.
One of the alternatives to assess the growth of follicles after in vitro culture is
the measurement of follicle and oocyte diameter. In the present study, it was observed a
significant reduction in the follicle and oocyte diameter of preantral follicles cultured in
the presence of cortisol when compared to the fresh control. It is known that in the
preovulatory follicle, the stress induces a reduction in the diameter of the dominant
follicle (19), however, even with damage caused by stress, the follicles continue to grow
and may ovulate subfertile oocytes (19).
In conclusion, it was observed the presence of NR3C1 in the oocyte and
granulosa cells in all follicular categories. The in vitro culture showed that high cortisol
concentrations (10 ng/ml) exerts a deleterious effect on follicular survival, reducing the
percentage of normal follicles as well as follicular and oocyte diameter. percentage of
normal.

59
References
1. Adcock IM (2000) Molecular Mechanisms of Glucocorticosteroid Actions.
Pulmonary Pharmacology & Therapeutics 13:115–126
2. Almeida AP, Saraiva MVA, Alves Filho JG, Silva GM, Gonçalves RFB, Brito IR,
Silva AWB, Lima AKF, Cunha RMS, Silva JRV, Figueiredo JR (2011) Gene
expression and Immunolocalization of Fibroblast Growth Factor 2 in the Ovary and Its
Effects on the In vitro Culture Of Caprine Preantral Ovarian Follicle. Reprod. Dom.
Anim. 10:1439-0531
3. Andersen CY (2002) Possible new mechanism of cortisol action in female
reproductive organs: physiological implications of the free hormone
hypothesis. J. Endocrinol. 173:211–217
4. Badinga L, Thacher WW, Diaz T, Drost M, Wolfenson D (1993) Effect of
environmental heat stress on follicular development and steroidogenesis in lactating
Holstein cows. Theriogenology, 39:797-810
5. Biran D, Braw-Tal R, Gendelman M, Lavon Y, Roth Z (2015) ACTH administration
during formation of preovulatory
follicles impairs steroidogenesis and angiogenesis in
association with ovulation failure in lactating cows. Domestic Animal Endocrinology.
53:52-59
6. Breen K.M, Billings HJ, Debus N, Karsch FJ (2004) Endotoxin inhibits the surge
secretion of gonadotropin-releasing hormone via a prostaglandin-independent pathway.
Endocrinology. 145:221–7

60
7. Breen KM, Billings HJ, Wagenmaker ER, Wessinger EW, Karsch FJ (2005)
Endocrine basis for disruptive
effect of cortisol on preovulatory events. Endocrinology. 146:2107-2115
8. Bruno JB, Lima-Verde IB, Martins FS, Matos MHT, Lopes CAP, Maia-JR JE, Báo
SN, Nobre Junior HV, Maia FD, Pessoa C, Moraes MO,
Silva JRV, Figueiredo JR, Rodrigues APR (2008) Característica histológica, ultra-
estrutural e produção de nitrito de folículos pré-antrais
caprinos cultivados in vitro na ausência ou presença de soro. Arq. Bras. Med. Vet.
Zootec. 60:1329-1337
9. Charmandari E, Tsigos C, Chrousos GP (2005) Endocrinology of the stress response.
Annu Rev Physiol. 67:259–284
10. Chaves RN, Martins FS, Saraiva MVA, Celestino JJH, Lopes CAP, Correia JC,
Lima-Verde IB, Matos MHT, Báo SN, Name KPO, Campello CC, Silva JRV,
Figueiredo JR (2008) Chilling ovarian fragments during transportation improves
viability and growth of goat preantral follicles cultured in vitro. Reprod. Fert. Dev.
20:640–647
11. Chaves, Ricardo De Macêdo et al . Influência das estações seca e chuvosa na
capacidade de desenvolvimento de oócitos e produção in vitro de embriões da espécie
caprina. Ciênc. anim. bras. Goiânia, v. 14, n. 1, p. 135-142, Mar. 2013. Available from
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1809-
68912013000100017&lng=en&nrm=iso, access on 19 Aug. 2015.
http://dx.doi.org/10.5216/cab.v12il.9375.
12. Fateh M, Ben-Rafael Z, Benadiva CA, Mastroianni LJR, Flickinger GL (1989)
Cortisol levels in human follicular fluid. Fertil Steril. 51:538-41

61
13. González R, Ruiz-León Y, Gomendio M, Roldan ERS (2010) The effect of
glucocorticoids on ERK-1/2 phosphorylation during maturation of lamb oocytes and
their subsequent fertilization and cleavage ability in vitro. Reproductive Toxicology.
29:198–205
14. Gougeon A (1996) Regulation of ovarian follicular development in primates: facts
and hypotheses. Endocr Rev. 17:121–54
15. Griekspoor A, Zwart W, Neefjes J, Michalides R (2002) Visualizing the action of
steroid hormone receptors in living cells. Nucl. Recept. Signal. 5:003
16. Huang TJ, Li PS 2001 Dexamethasone Inhibits Luteinizing Hormone-Induced
Synthesis of Steroidogenic Acute Regulatory Protein in Cultured Rat Preovulatory
Follicles. Biology of Reproduction. 64:163-170
17. Illera JC, Silván GM, MartínezM , Blass A, Peña L (2005) The effect of
dexamethasone on disruption of ovarian steroid levels and receptors in female rats. J.
Physiol. Biochem. 61:429-438
18. Kaushish SK, et al (1987) Effect of heat and water restriction on physiological
responses of Beetal and Black Bengal goats. Indian Journal of Animal Science. 57:461-
65
19. Lucy MC, (2001) Estratégias de manejo de vacas leiteiras para melhoria dos índices
reprodutivos durante o verão. Anais do V curso "Novos enfoques na produção e
reprodução de bovinos, p.12-18
20. Macfarlane MS, Breen KM, Sakurai H, Adams BM, Adams TE (2000) Effect of
duration of infusion of stress-like

62
concentrations of cortisol on follicular development and the preovulatory surge of LH in
sheep. Animal Reproduction Science. 63:167-175
21. Papargaris MM, Rivalland ET, Hemsworth PH, Morrisey AD, Tilbrook AJ (2011)
Acute and chronic stress-like levels of cortisol inhibit the oestradiol stimulus to induce
sexual receptivity but have no effect on sexual attractivity or proceptivity in female
sheep. Hormones and Behavior. 60:336-345
22. Reddy TE, Pauli F, Sprouse RO, Neff NF, Newberry KM, Garabedian MJ, et al
(2009) Genomic determination of the glucocorticoid response reveals unexpected
mechanisms of gene regulation. Genome Research. 19:2163-71
23. Ristić N, Nestorović N, Manojlović-Stojanoski M, Filipović B, Šošić-Jurjević B,
Milosević V, Sekulić M (2008) Maternal Dexamethasone Treatment Reduces Ovarian
Follicle Number in Neonatal Rat Offspring. J. Microsc. 232:549-557
24. Sales AD, Brito IR, De Lima LF, Lobo CH, Duarte AB, et al (2014). Expression and
localization of Aquaporin 3 (AQP3) in folliculogenesis of ewes. Acta Histochem.
116:831–837
25. Schaaf MJ, Cidlowski JA (2002) Molecular mechanisms of glucocorticoid action
and resistance. J Steroid Biochem Mol Biol. 83:37–48
26. Simerman AA, Hill DL, Grogan TR, Elashoff D, Clarck NJ, Goldstein EH,
Manrriquez AN, Cazenbalk GD, Dumesic DA (2015) Intrafolliclar cortisol levels
inversely correlate with cumulus cell lipid content as a possible energy source during
oocyte meiotic resumption in women undergoing ovarian stimulation for in vitro
fertilization. Fertility and Sterility® 103:249-257.

63
27. Tetsuka M, Nishimoto H, Myamoto A, Okuda K, Hamano S (2010) Gene
Expression of 11β-HSD and Glucocorticoids Receptor in the Bovine (Bos Taurus)
Follicle During Follicular Maturation and Atresia:The Role of Follicular Stimulating
Hormone. Journal of Reproduction and Development. 56:616-622
28. Tilbrook1 AJ,Turner AI, Clarke IJ (2000) Effects of stress on reproduction in non-
rodent mammals: the role of glucocorticoids and sex differences. Review of
Reproduction. 5:105-113
29. Wagenmaker ER, Breen KM, Oakley AE, Tilbrook AJ, Karsch FJ (2009)
Psychosocial stress inhibits amplitude of gonadotropin-releasing hormone pulses
independent of cortisol action on the type II glucocorticoid receptor. Endocrinology.
150:762–769

64
8 CONCLUSÃO
Em conclusão, através da imunohistoquímica, pode-se observar a presença da
proteína para o receptor de glicocorticóides NR3C1 no ovário caprino em todas as
categorias foliculares mediando à ação do cortisol na foliculogênese caprina, com um
papel mais importante nos estádios mais avançados da foliculogênese. No entanto, o
cortisol, em altas concentrações (10 ng/ml), exerce efeito deletério na sobrevivência
folicular durante o desenvolvimento in vitro, reduzindo o percentual de morfologia
normal, bem como o diâmetro folicular e oocitário. Além disso, nos estádios iniciais, o
cortisol, independente da concentração, foi capaz de promover a ativação de folículos
pré-antrais cultivados in vitro.

65
9 PERSPECTIVAS
Devido à importância dos glicocorticóides na reprodução, mais estudos são
necessários para avaliar o efeito desses hormônios na foliculogênese in vitro, em
especial na esteroidogênese, uma vez que o cortisol exerce efeitos diretos nas enzimas
esteroidogênicas.

66
REFERÊNCIAS
ACOSTA, T.J.; TETSUKA, M.; MATSUI, M.; SHIMIZU, T.; BERISHA, B.;
SCHAMS, D.;
MIYAMOTO, A. In vivo evidence that local cortisol production increases in
the preovulatory follicle of the cow. J Reprod Dev v. 51, p.483–489, 2005
ADCOCK, I.M. Molecular Mechanisms of Glucocorticosteroid Actions. Pulmonary
Pharmacology & Therapeutics. vol. 13, p. 115–126. 2000.
AGARWAL, A.K.; MONDER, C.; ECKSTEIN, B.; WHITE, P.C. Cloning and
�expression of rat cDNA encoding corticosteroid 11 -dehydrogenase. Journal of
Biological Chemistry 264 18939–18943. 1998
AGARWAL, A.K.; MUNE, T.; MONDER, C.; WHITE, P.C. NAD+- dependent
�isoform of 11 -hydroxysteroid dehydrogenase. Journal of Biological Chemistry 269
25959–25962. 1994
ALBISTON, AL.; OBEYESEKERE, V.R.; SMITH, R.E.; KROZOWSKI, Z.S. Cloning
and tissue distribution of the human 11�-hydroxysteroid dehydrogenase type 2 enzyme.
Molecular and Cellular Endocrinology. v. 105, p.11:17, 1994
ALMEIDA, A.P.; SARAIVA, M.V.A.; ALVES FILHO, J.G.; SILVA, G.M.;
GONÇALVES, R.F.B.; BRITO, I.R.; SILVA, A.W.B.; LIMA, A.K.F.; CUNHA,
R.M.S.; SILVA, J.R.V.; FIGUEIREDO, J.R. Gene expression and Immunolocalization
of Fibroblast Growth Factor 2 in the Ovary and Its Effects on the In vitro Culture Of
Caprine Preantral Ovarian Follicle. Reprod. Dom. Anim. v. 10, p. 1439:0531, 2011
ANDERSEN, C.Y. Possible new mechanism of cortisol action in female
reproductive organs: physiological implications of the free hormone
hypothesis. J. Endocrinol. v. 173. P. 211–217. 2002
ARAÚJO, V.R.; SILVA, C.M.G.; MAGALHÃES, D.M.; SILVA, G.M.; BÁO, S;N.;
SILVA, J.R.V.; FIGUEIREDO, J.R.; RODRIGUES, A.P.R. Effect of Bone
Morphogenetic Protein-7 (BMP-7) on in
vitro survival of caprine preantral follicles. Pesq. Vet. Bras. v. 30. p. 305:310, 2010

67
BADINGA, L.; THACHER, W.W.; DIAZ, T.; DROST, M.; WOLFENSON, D. Effect
of environmental heat stress on follicular development and steroidogenesis in lactating
Holstein cows. Theriogenology. v. 39, p.797:810, 1993
BIRAN, D.; BRAW-TAL, R.; GENDELMAN, M.; LAVON, Y.; ROTH, Z. ACTH
administration during formation of preovulatory
follicles impairs steroidogenesis and angiogenesis in
association with ovulation failure in lactating cows. Domestic Animal Endocrinology. v.
53, p.52:59, 2015
BORSKI, R.J. Nongenomic membrane actions of glucocorticoids in vertebrates. Trends
Endocrinol Metab. v. 11, p.427:36, 2000
BREEN, K.M.; BILLINGS, H.J.; DEBUS, N.; KARSCH, F.J. Endotoxin inhibits the
surge secretion of gonadotropin-releasing hormone via a prostaglandin-independent
pathway. Endocrinology. v. 145, p. 221:7, 2004
BREEN, K.M.; BILLINGS, H.J.; WAGENMAKER, E.R.; WESSINGER, E.W.;
KARSCH, F.J. Endocrine basis for disruptive
effect of cortisol on preovulatory events. Endocrinology, v. 146,
n. 4, p. 2107-2115, 2005.
BRUNO, J.B.; LIMA-VERDE, I.B.; MARTINS, F.S.; MATOS, M.H.T.; LOPES,
C.A.P.; MAIA-JR, J.E.; BÁO, S.N.; NOBRE JUNIOR, H.V.; MAIA, F.D.; PESSOA,
C.; MORAES, M.O.;
SILVA, J.R.V.; FIGUEIREDO, J.R.; RODRIGUES, A.P.R. Característica histológica,
ultra-estrutural e produção de nitrito de folículos pré-antrais
caprinos cultivados in vitro na ausência ou presença de soro. Arq. Bras. Med. Vet.
Zootec. v. 60, p. 1329:1337, 2008
CHARMANDARI, E.; TSIGOS, C.; CHROUSOS, G.P. Endocrinology of the stress
response. Annu Rev Physiol. v. 67, p. 259:284, 2005
CHAVES, R.N.; MARTINS, F.S.; SARAIVA, M.V.A.; CELESTINO, J.J.H.; LOPES,
C.A.P.; CORREIA, J.C.; LIMA-VERDE, I.B.; MATOS, M.H.T.; BÁO, S.N.; NAME,
K.P.O.; CAMPELLO, C.C.; SILVA, J.R.V.; FIGUEIREDO, J.R. Chilling ovarian

68
fragments during transportation improves viability and growth of goat preantral follicles
cultured in vitro. Reprod. Fert. Dev. v. 20, p. 640:647, 2008
CHAVES, R.M. et al . Influência das estações seca e chuvosa na capacidade de
desenvolvimento de oócitos e produção in vitro de embriões da espécie caprina. Ciênc.
anim. bras. Goiânia, v. 14, n. 1, p. 135-142, Mar. 2013. Available from
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1809-
68912013000100017&lng=en&nrm=iso, access on 19 Aug. 2015.
http://dx.doi.org/10.5216/cab.v12il.9375.
CHEN, W.Y.; YANG, J.G.; LI, P.S. Effect of dexamethasone on the expression of
p34cdc2 and cyclin B1 in pig oocytes in vitro. Molecular Reproduction and
Development. v. 56, p.74:79, 2000
DAVIES, T.H.; NING, Y.M.; SÁNCHEZ, E.R. A new fi rst step in activation of steroid
receptors: hormone-induced switching of fkbp51 and fkbp52 immunophilinso. The
Journal of Biological Chemistry. v. 277, p. 4597:4600, 2002
ESCHER G.; GALLI, E. VISHWANATH, B.S.; FREY, B.; FELIX, F.J. Tumor
necrosis factor alpha and interleukin i beta enhance the cortisol/cortisone suttle. Journal
of Experimental Medicine. v. 186, p. 189:198, 1997
FAN, H.Y.; SUN, Q.Y. Involvement of mitogen-activated protein kinase cascade during
oocyte maturation and fertilization in mammals. Biology of Reproduction v.70,
p.535;:47, 2004
FATEH, M; BEN-RAFAEL, Z; BENADIVA, C.A; MASTROIANNI, L. JR;
FLICKINGER, G.L. Cortisol levels in human follicular fluid. Fertil Steril. Vol. 51. P.
538-41. 1989
GONZÁLEZ, R; RUIZ-LEÓN, Y; GOMENDIO, M; ROLDAN, E.R.S. The effect of
glucocorticoids on ERK-1/2 phosphorylation during maturation of lamb oocytes and
their subsequent fertilization and cleavage ability in vitro. Reproductive Toxicology. V
29, p. 198–205, 2010
GOUGEON, A. Regulation of ovarian follicular development in primates: facts and
hypotheses. Endocr Rev. v. 17, p. 121:54, 1996

69
GRIEKSPOOR, A.; ZWART, W.; NEEFJES, J.; MICHALIDES, R. Visualizing the
action of steroid hormone receptors in living cells. Nucl. Recept. Signal. v. 5, p. 003.
2002
HARLOW, C.R.; JENKINS, J.M.; WINSTON, R.M. Increased follicular fluid total and
free cortisol levels during the luteinizing hormone surge. Fertil Steril. v. 68(1), p.48:53.
1997
HUANG, T.J.; LI, P.S. Dexamethasone inhibits luteinizing hormone-induced synthesis
of steroidogenic acute regulatory protein in cultured rat preovulatory follicles. Biology
of Reproduction v. 64, p.163:170, 2001.
ILLERA,J.C.; G. SILVÁN, C.; MARTÍNEZ, M.; BLASS, A.; PEÑA, L. The effect of
dexamethasone on disruption of ovarian steroid levels and receptors in female rats. J.
Physiol. Biochem., 61 (3), 429-438, 2005
KAUSHISH, S.K., et al. Effect of heat and water restriction on physiological responses
of Beetal and Black Bengal goats. Indian Journal of Animal Science. v. 57, p, 461:65,
1987
LUCY, M.C. Estratégias de manejo de vacas leiteiras para melhoria dos índices
reprodutivos durante o verão. Anais do V curso "Novos enfoques na produção e
reprodução de bovinos, p.12-18, 2001
MACFARLANE, M. S.; BREEN, K. M.; SAKURAI, H.; ADAMS,
B. M.; ADAMS, T. E. Effect of duration of infusion of stress-like
concentrations of cortisol on follicular development and the preovulatory surge of LH in
sheep. Animal Reproduction Science,
v. 63, p. 167-175, 2000.
MAGALHÃES, D.M.; ARAÚJO, V.R.; LIMA-VERDE, I.B.; MATOS, M.H.T.;
SILVA, R.C.; LUCCI, C.M.; BÁO, S.N.; CAMPELLO, C.C.; FIGUEIREDO, J.R.
Different FollicleStimulating Hormone (FSH) sources influence caprine preantral
follicle viability and development in vitro. Braz. J. Vet. Res. Anim. Sci. 5, 378-386.
2009

70
MAO, L.; LOU, H.; LOU, Y.; WANG, N.; JIN, F. Behaviour of cytoplasmic organelles
and cytoskeleton during oocyte maturation. Reprod Biomed Online. 28: 284-299. 2014
MICHAEL, A.E.; THURSTON, L.M.; RAE, M.T. Glucocorticoid metabolism and
reproduction: a tale of two enzymes. Reproduction. v. 126, p.425:441, 2003
NURSE, P. (1990). Universal control mechanism regulating onset of M-phase. Nature,
344:503-508
OAKLEY, R.H.; CIDLOWSKI, J.A. Cellular Processing of the Glucocorticoid
Receptor Gene and Protein: New Mechanisms for Generating Tissue-specific Actions of
Glucocorticoids. J Biol Chem. v. 286, p. 3177:3184, 2011
ONARD, J.L.M.; SCHONEVELD, INGRID, C. GAEMERS, WOUTER H. LAMERS.
Mechanisms of glucocorticoid signalling. Biochimica et Biophysica Acta. v. 1680, p.
114:128, 2004
OZAWA, M.; TABAYASHI, D.; LATIEF, T.A.; SHIMIZU, T.; OSHIMA, I; KANAI,
Y. Alterations in follicular dynamics and steroidogenic abilities induced by heat stress
during follicular recruitment in goats. Reproduction. v. 129, p. 621:630, 2005
PAPARGARIS, M.M; RIVALLAND, E.T .A; HEMSWORTH, P. H; MORRISEY, A.
D; TILBROOK, A. J. Acute and chronic stress-like levels of cortisol inhibit the
oestradiol stimulus to induce sexual receptivity but have no effect on sexual attractivity
or proceptivity in female sheep. Hormones and Behavior. V. 60. N.4. p.336-345. 2011
PRATT, W.B. The role of heat shock proteins in regulating the function, folding, and
trafficking of the glucocorticoid receptor. J. Biol. Chem. 268:21455- 21458, 1993.
REDDY, T.E.; PAULI, F.; SPROUSE, R.O.; NEFF, N.F.; NEWBERRY, K.M.;
GARABEDIAN, M.J.; et al. Genomic determination of the glucocorticoid response
reveals unexpected mechanisms of gene regulation. Genome Research. v. 19, p.
2163:71, 2009
RISTIĆ, N.; NESTOROVIĆ, N.; MANOJLOVIĆ-STOJANOSKI, M.; FILIPOVIĆ, B.;
ŠOŠIĆ-JURJEVIĆ, B.; MILOSEVIĆ, V.; SEKULIĆ, M. Maternal Dexamethasone
Treatment Reduces Ovarian Follicle Number in Neonatal Rat Offspring. J. Microsc. v.
232, p. 549:557, 2008

71
SALES, A.D.; BRITO, I.R.; DE LIMA, L.F.; LOBO, C.H.; DUARTE, A.B.; et al
Expression and localization of Aquaporin 3 (AQP3) in folliculogenesis of ewes. Acta
Histochem. 116:831–837, 2014
SCHAAF, M.J.; CIDLOWSKI, J.A. Molecular mechanisms of glucocorticoid action
and resistance. J Steroid Biochem Mol Biol. 83:37–48, 2002
ROSNER W. Plasma steroid-binding proteins. Endocrinol Metab Clin North Am.
20(4):697-720, 1991
SCHONEVELD, O.J.; GAEMERS, I.C.; LAMERS, W.H. Mechanisms of
glucocorticoid signalling. Biochim Biophys Acta. v. 1680, p.114:128, 2004
SECKL, J.R.; WALKER, B.R Minireview: 11b-Hydroxysteroid Dehydrogenase Type
1— A Tissue-Specific Amplifier of Glucocorticoid Action. Endocrinology. v. 142, p.
1371;1376, 2001
SIMERMAN, A.A.; HILL, D.L.; GROGAN, T.R.; ELASHOFF, D.; CLARCK, N.J.;
GOLDSTEIN, E.H.; MANRRIQUEZ, A.N.; CAZENBALK, G.D.; DUMESIC, D.A.
Intrafolliclar cortisol levels inversely correlate with cumulus cell lipid content as a
possible energy source during oocyte meiotic resumption in women undergoing ovarian
stimulation for in vitro fertilization. Fertility and Sterility®. v. 103, p.249-257, 2015
TETSUKA, M.; YAMAMOTO, S.; HAYASHIDA, N.; HAYASHI, K.G.; HAYASHI,
M.; ACOSTA, T.J.; MIYAMOTO, A. Expression of 11 beta-hydroxysteroid
dehydrogenases in bovine follicle and corpus luteum. J Endocrinol. v. 177, p.445–452,
2010
THURSTON, L.M.; ABAYASEKARA, D.R.; MICHAEL, AE. 11 Betahydroxysteroid
dehydrogenase expression and activities in bovine granulosa cells and corpora lutea
implicate corticosteroids in bovine ovarian physiology. J Endocrinol. v. 193, p. 299:310,
2007
TILBROOK, A.J.; TURNER, A.I.; CLARKE, I.J. Effects of stress on reproduction in
non-rodent mammals: the role of glucocorticoids and sex differences. Review of
Reproduction. v. 5, p.105:113, 2000

72
TOMLINSO �N, J.W.; STEWART, P.M. Cortisol metabolism and the role of 11 -
hydroxysteroid dehydrogenase. Best Practice and Research Clinical Endocrinology and
Metabolism. v. 15, p.61;78, 2001
VALSANGKAR, D.; STEPHEN, M.D. A Requirement for Fatty Acid Oxidation in the
Hormone-Induced Meiotic Maturation of Mouse Oocytes. BIOLOGY OF
REPRODUCTION. v. 89, 43 .p. 1:9, 2013
WAGENMAKER, E.R; BREEN, K.M.; OAKLEY, A.E.; TILBROOK, A.J.; KARSCH,
F.J. Psychosocial stress inhibits amplitude of gonadotropin-releasing hormone pulses
independent of cortisol action on the type II glucocorticoid receptor. Endocrinology.
v. 150, p. 762–769. 2009.





























