Embed Size (px)
Citation preview

0
UNIVERSIDADE ESTADUAL DO CEARA
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
FACULDADE DE VETERINARIA
PROGRAMA DE POS-GRADUACAO EM CIENCIAS VETERINARIAS
ALEXANDRE RODRIGUES DE PAULA JUNIOR
ANÁLISE PROTEÔMICA DO FLUIDO FOLICULAR DE CAPRINOS DA RAÇA
CANINDÉ SUBMETIDOS À ESTIMULAÇÃO HORMONAL OVARIANA
FORTALEZA – CEARÁ
2015

1
ALEXANDRE RODRIGUES DE PAULA JUNIOR
ANÁLISE PROTEÔMICA DO FLUIDO FOLICULAR DE CAPRINOS DA RAÇA
CANINDÉ SUBMETIDOS À ESTIMULAÇÃO HORMONAL OVARIANA
FORTALEZA - CEARÁ
2015
Dissertação apresentada ao Curso de
Mestrado Acadêmico em Ciências
Veterinárias do Programa de Pos-
Graduação em Ciências Veterinárias da
Faculdade de Veterinária da Universidade
Estadual do Ceará, como requisito parcial
para obtenção do título de mestre em
Ciências Veterinárias.
Área de Concentração: Reprodução e
Sanidade Animal.
Orientador: Prof. Dr. Vicente José de
Figueirêdo Freitas

2
Dados Internacionais de Catalogação na Publicação
Universidade Estadual do Ceará
Sistema de Bibliotecas
Paula Junior, Alexandre Rodrigues de Paula Junior. Anàlise proteômica do fluido folicular de caprinos
da raça Canindé submetidos à estimulação hormonal ovariana [recurso eletrônico] / Alexandre Rodrigues de Paula Junior . – 2015.
1 CD-ROM: il.; 4 ¾ pol.
CD-ROM contendo o arquivo no formato PDF do trabalho acadêmico com 59 folhas, acondicionado em caixa de DVD Slim (19 x 14 cm x 7 mm).
Dissertação (mestrado acadêmico) – Universidade Estadual do Ceará, Faculdade de Veterinária, Programa de Pós-Graduação em Ciências Veterinárias, Fortaleza, 2015.
Área de concentração: Reprodução e Sanidade Animal. Orientação: Prof. Dr. Vicente José de Figueirêdo
Freitas.
1. Caprino. 2. Fluido folicular . 3. Proteômica. I. Título.

3

0
A Deus, o qual renova minhas forças todos os dias, me fazendo prosseguir;
A minha família em especial meus pais Alexandre e Vitalina,
minha maior motivação para vencer as adversidades da vida,
A meus avós paternos Antônio de Paula e Maria de Loudes,
em memória.
dedico

1
AGRADECIMENTOS
A Deus, pelo fôlego de vida e pelas bênçãos derramada sobre mim, por renovar minhas
forças todos os dias para que eu vença as adversidades da vida.
Ao PPGCV, pela oportunidade de fazer parte desse grupo seleto de pós-graduandos.
Ao CNPq e CAPES, pelo apoio financeiro permitindo nossa dedicação exclusiva na
realização desse trabalho.
Aos meus familiares, em especial meus pais Alexandre e Vitalina, por serem meus
maiores exemplos de vida, pela educação e ensinamentos prestado a mim, pelo exemplo de
honestidade e caráter e por serem minha maior motivação para que eu chegasse até esse
momento. À minha filha, Ana Leticia, pelo amor e carinho e aos meus irmãos Arlete, Ricardo
e Aline pelo apoio, carinho e incentivo. Ao Ney Rodrigues pela amizade, companheirismo e
todo apoio prestado durante toda minha formação acadêmica e aos meus irmãos de coração
Artur e Edite! Agradeço a todos por fazerem parte da minha vida e contribuírem para realização
desse dia. Amo todos vocês!
À minha namorada Juliana Dantas pelo amor, carinho e compreensão nos momentos
difíceis, me fazendo sorrir e apoio incondicional o que me fez prosseguir diante das
dificuldades.
Ao meu orientador, professor Vicente Freitas, pela oportunidade e confiança na
realização desse trabalho e pela amizade construída durante esses anos, estando sempre disposto
a ajudar.
Ao meu co-orientador, Dr. Mauricio van Tilburg, por todo ensinamento passado, pela
amizade construída, pelas palavras de incentivo sempre acreditando e confiando na minha
capacidade, o que me fez persistir e acreditar que iria dar certo.
Aos professores Arlindo Moura da Universidade Federal do Ceará, Renato Azevedo e
a professora Ana Cristina pertencentes à Universidade de Fortaleza pela grande parceria,
disponibilizando seus laboratórios para realização desse trabalho e pela grande oportunidade de
aprender com pessoas de tamanha competência.
A todos os professores do PPGCV pelos ensinamentos e conselhos que servirão para
formação profissional e pessoal em especial o professor Dárcio Ítalo pelo convívio e amizade
construída e a professora Luciana Melo.
Aos amigos de pós-graduação, em especial Ribrio Ivan, pela amizade, companheirismo,
alegria e simplicidade de sempre. Aos amigos Carlos Henrique e Francisco Carlos pela

2
amizade, apoio e grande contribuição para realização deste trabalho. Aos amigos de sempre
Joanna, Kaká, Deisy, Dowglish e Iana, pessoas que estão sempre dispostas a ajudar.
A toda equipe do LFCR da UECE, pós-graduandos e alunos de iniciação cientifica que
de uma forma ou de outra contribuíram para a realização desse trabalho, pela amizade e
momentos de descontração.
A toda equipe do laboratório de Fisiologia Animal da UFC, em especial Solange,
Révila, Erika, Ana Luiza, Fágner, Taciane e Aderson pela amizade e apoio.
A equipe da professora Ana Cristina na UNIFOR, em especial ao Federico Moreno e
Marina Lobo pelo apoio e grande ajuda durante as atividades realizadas na instituição.
Aos funcionários Selmar, Cesar, Carlos, João e Adriana pelo respeito e prontidão em
ajudar em todos os momentos.
E para finalizar não poderia deixar de agradecer aos animais em especial as cabras da
raça Canindé que contribuíram para a realização desse estudo e que merecem todo nosso
respeito e gratidão.

3
RESUMO
Os caprinos da raça Canindé são animais naturalizados e bem adaptados às condições do
semiárido brasileiro. No entanto, esta raça está em risco de extinção e biotécnicas
reprodutivas, como a produção in vitro de embriões, pode ajudar na conservação da mesma. O
crescimento e a maturação oocitária representam uma série de transformações celulares e
moleculares, de forma sequencial, dos componentes do fluido folicular (FF). A identificação
desses componentes pode ajudar na compreensão dos fenômenos fisiológicos, além de
contribuir com as técnicas de maturação e fecundação in vitro (FIV). O presente estudo teve
como objetivo verificar o perfil de proteínas expressas no fluido de folículos pequenos (< 3
mm), médios (3-4 mm) e grandes (> 4mm) de cabras da raça Canindé. Oito cabras adultas da
raça Canindé receberam um tratamento hormonal de estimulação ovariana e logo em seguida,
através de laparoscopia, as amostras de fluido folicular (FF) foram colhidas por punção com
auxílio de uma bomba de vácuo. As amostras foram submetidas à eletroforese 2D SDS-PAGE
e os spots diferencialmente expressos foram identificados por espectrometria de massa (ESI-
Q-Tof). O estudo verificou que não existiu diferença (P > 0,05) entre os diferentes tamanhos
de folículo quanto à concentração total de proteínas. No entanto, cinco spots diferiram
significativamente, correspondendo a quatro proteínas diferentes. A serotransferrina teve uma
maior expressão nos folículos médios em relação aos pequenos. A zinco‐alfa‐2‐glicoproteina-
like, proteína do complemento C3 e fator B do complemento tiveram uma expressão
aumentada nos folículos grandes em comparação com os médios. Este estudo forneceu a
primeira descrição das proteínas diferencialmente expressas no FF de cabras Canindé
submetidas ao tratamento de estimulação hormonal ovariana. Estudos posteriores sobre a ação
destas proteínas poderão contribuir para o sucesso de programas de FIV na espécie caprina e,
em particular, na preservação da raça Canindé.
Palavras-chave: Caprino, Fluido folicular e Proteômica

4
ABSTRACT
Canindé goats are animals naturalized and well adapted to the conditions of the Brazilian
semiarid region. However, this breed is endangered and reproductive biotechnologies such as
in vitro production of embryos, can help to conserve. The growth and oocyte maturation are a
series of cellular and molecular changes, sequentially, of the components of follicular fluid
(FF). The identification of these components can help in understanding the physiological
phenomena and contribute to the techniques maturation and in vitro fertilization (IVF). This
study aimed to determine the profile of expressed proteins in the fluid of small (<3 mm),
medium (3-4 mm) and large (> 4 mm) follicles of Canindé goats. Eight adult Canindé goats
received an ovarian stimulation treatment and then, they had the follicular fluid (FF)
collected by laparoscopy with the aid of a vacuum pump. The samples were subjected to 2D-
SDS PAGE electrophoresis and the differentially expressed spots were identified by mass
spectrometry ESI-Q-TOF. The study found that there was no difference (P > 0.05) between
the different follicle sizes for total protein concentration. However, five spots were
significantly different, corresponding to four different proteins. The serotransferrin had a
higher expression in the medium follicles in relation to small. Zinc-alpha-2-glycoprotein-like,
C3 complement protein and complement factor B had an increased expression in large
follicles as compared with the medium. This study provided the first description of
differentially expressed proteins in the FF of Canindé goats subjected to treatment of ovarian
hormonal stimulation. Further studies on the action of these proteins may contribute to the
success of IVF programs in goats and in particular the preservation of Canindé breed.
Keywords: goat; folicular fluid, proteomic

5
LISTA DE FIGURAS
REVISÃO DE LITERATURA
Figura 1 Esquema da estacionalidade reprodutiva na espécie caprina …....……….. 15
CAPÍTULO 1
Figura 1 2D-PAGE do fluido folicular de cabras da raça Canindé. Foram aplicadas
500 µg de amostras de proteínas em tira linear com pI 3-10 na primeira
dimensão e separados em 10% gel SDS-PAGE na segunda dimensão. Os
géis foram corados pelo procedimento de azul coloidal...............................
36
Figura 2 Análise computadorizada 2D-PAGE do fluido folicular de cabras da raça
Canindé. (A) Sobreposição dos géis 2D-PAGE de fluido folicular,
destacando as proteínas que apresentaram diferença estatística durante o
crescimento folicular (cores azul, verde e vermelho, correspondem aos
spots dos folículos pequenos (FP), médios (FM) e grandes (FG)
respectivamente). (B) Intensidade dos spots (média ± S.E.M.) em função
do tamanho folicular. (C) Imagens em 3D dos spots diferencialmente
expressos entre os grupos de
folículos.......................................................................................................
36
Figura 3 Análise in silico das interações de proteínas. As proteínas analizadas
foram: a. serotransferrina (TF), b. zinco‐alfa‐2‐glicoproteína‐like
(AZGP1), c. fator B do complemento (CFB), d. C3 do complemento (C3).
Linhas de cores diferentes representam os tipos de evidências para a
associação: (preto) co-expressão, (rosa) experimentos, (verde) textmining
e (azul) banco de dados. Proteína receptora de transferrina (TFRC),
Proteína não caracterizada (CP), Proteína não caracterizada (HEPH),
Soluto carreador da família 40 membro 1 (SLC40A1); Apolipoproteína
A-I (APOA1), Proteína não caracterizada (STEAP3), Próton ATPase do
tipo V 116 kDa subunidade a isoforma 2 (ATP6V0A2), Proteína não
caracterizada (ENSBTAG00000000292), Proteína não caracterizada
(ENSBTAG00000045497), Próton ATPase do tipo V catalítica
subunidade A (ATP6V1A), Proteína homóloga precursora de prolactina-
inducible (ENSBTAG00000006655), Beta-2-microglobulina (B2M),
Fator D do complemento (CFD), Precursor do fator H do complemento
(CFH), Preproteína C3 do complemento (C3), Proteína não caracterizada
(ENSBTAG00000006864), Precursor C4 do complemento (C4A), C2 do
complemento (C2), Proteína não caracterizada (Bt.64039), Proteína não
caracterizada (LOC781004), Fator acelerador de decaimento do
complemento (CD55), Proteína não caracterizada
(ENSBTAG00000005587), Proteína cofator de membrana (LOC616002),
Proteína precursora de cofator de membrana isoforma 5
(ENSBTAG00000005397), Precursor do fator I do complemento (CFI),
Receptor quimiotático da anafilotoxina C5a (C5AR1), Precursor de
plasminogênio (PLG)..…………………………....…….…………...........
37

6
LISTA DE ABREVIATURAS E SIGLAS
2D Bidimensional
3D Tridimensional
ANOVA Análise de Variância
BSA Albumina Sérica Bovina
C3 Proteína C3 do Complemento
C3b Proteína C3b do Complemento
CEUA Comitê de Ética no Uso de Animais
CO2 Dióxido de Carbono
COL Colheita Oocitária por laparoscopia
CR1 Receptor 1 de Complemento
CR2 Receptor 2 de Complemento
DTT Ditiotreitol
eCG Gonadotrofina Coriônica equina
FF Fluido Folicular
FG Folículo Grande
FIV Fecundação in vitro
FM Folículo Médio
FP Folículo Pequeno
FSH Hormônio Folículo Estimulante
g Gravidade
IgA Imunoglobulina A
IgD Imunoglobulina D
IgE Imunoglobulina E
IGF Fator de Crescimento Semelhante à Insulina
IgG Imunoglobulina G
IgM Imunoglobulina M
kDa KiloDalton
LH Hormônio Luteinizante
MAP Acetato de Medroxiprogesterona

7
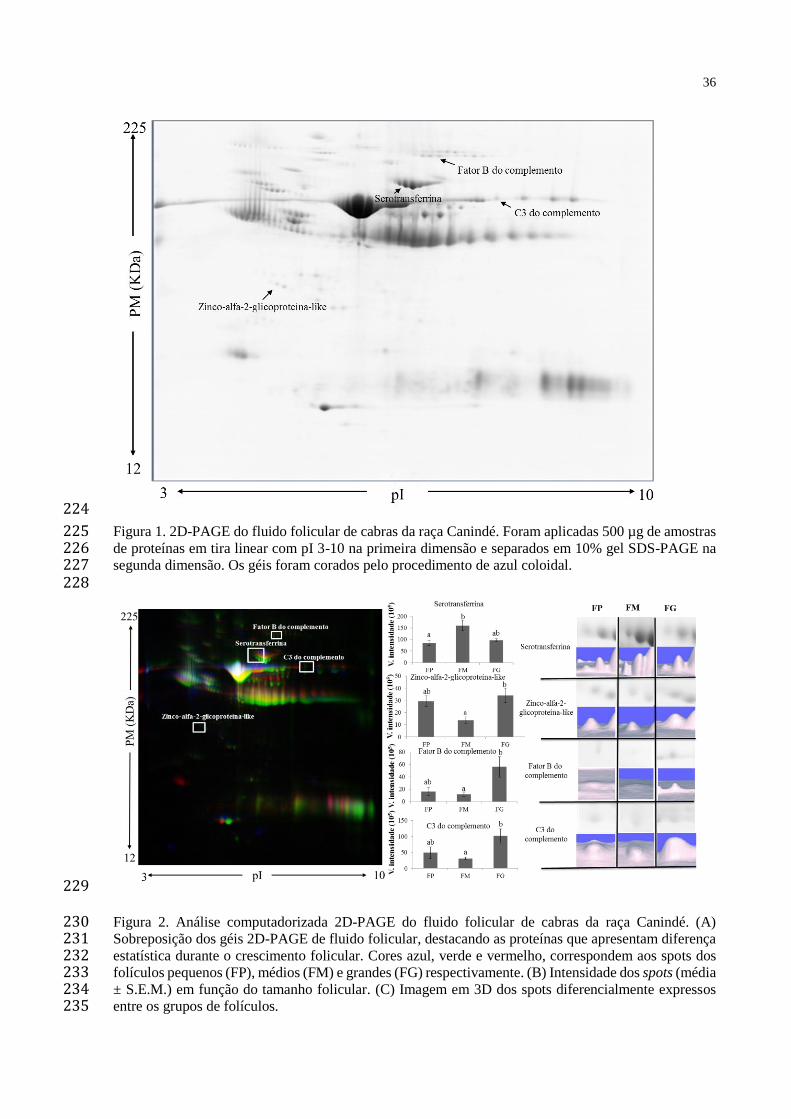
MCP Proteína de Cofator de Membrana
mg Miligrama
min Minuto
MIV Maturação in vitro
mL Mililitro
mm Milímetro
mM Milimolar
mmHg Milímetro de mercúrio
O2 Oxigênio
PAGE Eletroforese em Gel de Poliacrilamida
PBS Tampão Fosfato Salino
pH Potencial hidrogeniônico
pI Ponto isoelétrico
PIV Produção in vitro
RBP Proteína Ligadora de Retinol
ROS Espécies Reativas de Oxigênio
SDS Dodecil-Sulfato de Sódio
SEM Erro Padrão da Média
V Volt
Vh Volts hora
μg Micrograma
μm Micrômetro

8
SUMÁRIO
1 INTRODUÇÃO....................................................................................................... 13
2 REVISÃO DE LITERATURA.............................................................................. 15
2.1 Ciclo Reprodutivo da Cabra................................................................................ 15
2.2 Foliculogenese na Cabra...................................................................................... 17
2.3 Fluido Folicular.................................................................................................... 18
2.4 Proteínas do Fluido Folicular.............................................................................. 20
2.4.1 Albumina............................................................................................................ 21
2.4.2 Imunoglobulina G............................................................................................... 22
2.4.3 Transferrina......................................................................................................... 23
2.4.4 Proteína C3 do Complemento.............................................................................. 23
3 JUSTIFICATIVA................................................................................................... 25
4 HIPÓTESE CIENTÍFICA..................................................................................... 26
5 OBJETIVOS............................................................................................................ 27
5.1 Objetivo Geral...................................................................................................... 27
5.2 Objetivos Específicos........................................................................................... 27
6 CAPÍTULO 1........................................................................................................... 28
7 CONCLUSÕES....................................................................................................... 47
8 PERSPECTIVAS.................................................................................................... 48
REFERÊNCIAS BIBLIOGRÁFICAS..................................................................... 49

13
1 INTRODUÇÃO
A caprinocultura vem há algum tempo ganhando destaque no cenário mundial e isso se
deve, entre alguns pontos, também aos avanços obtidos no uso de biotécnicas reprodutivas na
espécie. No Brasil, o rebanho caprino possui um efetivo de aproximadamente 8,8 milhões de
animais, sendo que 91,5% são criados na região Nordeste do país (IBGE, 2013). Nesta região,
também encontra-se um grupo de raças naturalizadas, as quais constituem um importante
recurso genético por suas características de rusticidade e prolificidade. Dentre estas raças
destaca-se a Canindé, por sua maior habilidade leiteira, quando em comparação com as outras
raças naturalizadas brasileiras (EMBRAPA-CNPC, 1994; EGITO; MARIANTE;
ALBUQUERQUE, 2002). No entanto, cerca de 75% do rebanho nordestino são de animais
mestiços resultante de cruzamentos indiscriminados entre raças naturalizadas e raças exóticas
especializadas (MADRUGA et al., 2008), fato que tem promovido a descaracterização e o risco
de extinção das raças naturalizadas (EGITO; MARIANTE; ALBUQUERQUE, 2002).
O uso de biotécnicas reprodutivas, tais como a produção in vitro (PIV) de embriões,
pode ser uma ferramenta de grande importância para multiplicação e preservação de material
genético das populações ameaçadas de extinção (HOLT; BENNETT; VOLOBOUEV, 1996).
A PIV de embriões possibilita a produção de um maior número de descendentes de animais
com elevado potencial genético em um menor intervalo de tempo (COGNIÉ et al., 2003), além
da possibilidade de utilização de fêmeas pré-púberes, senis, gestantes e, até mesmo, post-
mortem (BALDASSARRE, 2007).
A PIV de embriões em caprinos tem evoluído em qualidade, porém as taxas de
blastocisto ainda são consideradas baixas, com variação de 10 a 20% (ROMAGUERA et al.,
2011), enquanto que esses índices em bovinos são de 35% a 50% (MOROTTI et al., 2014). As
taxas de sucesso da produção in vivo de embriões, independentemente da espécie, ainda são
bastante superiores do que em embriões produzidos in vitro (MERTON et al., 2003), devido
principalmente à complexidade metabólica envolvida no microambiente folicular
(LONERGAN & FAIR, 2014). Por isso a importância de estudos moleculares que visem uma
melhor compreensão dos eventos biológicos que ocorrem dentro dos folículos ovarianos.
O fluido de folículos ovarianos é composto de várias proteínas biologicamente ativas as
quais podem afetar eventos como a maturação folicular e a ovulação (RICHARDS, 1994). Da
mesma maneira, os meios utilizados na PIV de embriões em qualquer espécie devem levar em
conta a composição do fluido folicular (FF), uma vez que as substâncias existentes no mesmo

14
possuem ação importante nos aspectos fisiológicos, bioquímicos e metabólicos da maturação
nuclear e citoplasmática do oócito (WATSON, 2007; GOTTARDI & MINGOTI, 2009). Desta
forma, estudos proteômicos do FF poderão fornecer informações valiosas para o entendimento
dos processos fisiológicos envolvidos na maturação oocitária e desenvolvimento folicular.
Estudos do proteoma do FF já foram descritas em bovinos (MORTARINO et al., 1999;
MANIWA et al., 2005), suínos (BIJTTEBIER et al. 2009), caninos (FAHIMINIYA et al. 2010)
e em equinos (FAHIMINIYA et al. 2011). Entretanto, até o presente momento não existem
relatos sobre a expressão de proteínas do FF de caprinos. Dessa forma, a identificação de
proteínas do fluido de folículos ovarianos caprinos poderá contribuir para uma melhor
compreensão dos mecanismos envolvidos no crescimento e maturação oocitária, podendo gerar
informações que irão contribuir para melhoria dos índices de PIV de embriões nessa espécie.

15
2 REVISÃO DE LITERATURA
2.1 O Ciclo Reprodutivo da Cabra
As cabras são animais poliéstricos estacionais, com estação reprodutiva determinada
principalmente por uma combinação de fatores genéticos e ambientais. Diversos elementos
climáticos, mas sobretudo o fotoperíodo, regulam o ciclo reprodutivo destes animais
(FONSECA, 2005). Em latitudes superiores a 25°, a estação reprodutiva em caprinos é
estacional, ocorrendo durante o período de menor luminosidade do ano, particularmente no
outono e inverno, enquanto que nos dias longos (primavera e verão) a atividade sexual diminui,
com animais em anestro (Figura 1). À medida que se aproxima da linha do equador, esta
estacionalidade é reduzida ou praticamente deixa de existir (MORAES et al., 2008). Desta
forma, em zonas tropicais e subequatoriais, desde que haja aporte nutricional adequado, cabras
irão ciclar durante todo o ano. Esta ciclicidade também é fortemente influenciada pelo fator
raça, uma vez que caprinos de raças naturalizadas como Canindé e Moxotó apresentam
atividade reprodutiva durante todo o ano, o que não acontece com caprinos de raças leiteiras
especializadas (Saanen, Alpina e Toggenburg), mesmo em áreas próximas aos trópicos
(FONSECA, 2005).
FIGURA 1 - Esquema da estacionalidade reprodutiva na espécie caprina.
Fonte: GRANADOS (2006).

16
De maneira geral, as cabras estarão aptas à reprodução quando os ovários assumirem
simultaneamente a esteroidogênese e a gametogênese, sendo capaz de liberar um oócito e de
manifestar comportamento sexual completo. Isso tem início durante a puberdade, evento que
em caprinos ocorre por volta de 6 a 7 meses de idade (FREITAS et al., 2004), porém esta idade
pode ser influenciada por diversos fatores, dos quais o peso corpóreo é fundamental para que
cabras tenham seu primeiro ciclo estral que é descrito como o intervalo entre dois estros
sucessivos, tendo na fêmea caprina uma duração média de 21 dias e apresentando uma fase
luteínica de 17 dias e uma fase folicular de quatro dias (FONSECA, 2005). Durante o ciclo
estral ocorrem mudanças morfológicas (crescimento e recrutamento folicular), bioquímicas
(maturação folicular) e fisiológicas (regulação endócrina) que levam à ovulação (FATET;
PELLICER-RUBIO; LEBOEUF, 2011).
A fase folicular corresponde à onda de desenvolvimento caracterizada pela sequência
de eventos dependentes de gonadotrofina, ou seja: crescimento, recrutamento, seleção e
dominância do folículo (DRIANCOURT, 2001). O Hormônio Folículo Estimulante (FSH)
estimula o crescimento folicular, uma vez que logo após a formação do antro, os folículos
tornam-se dependentes de gonadotrofinas para, posteriormente, serem recrutados a se
desenvolverem, o que caracteriza a emergência da onda folicular (ADAMS et al., 2008). Em
cabras, o número de ondas foliculares pode variar de três a quatro por ciclo (GINTHER & KOT,
1994; CRUZ; RONDINA; FREITAS, 2005). Após o recrutamento folicular, somente dois ou
três folículos irão atingir 4 mm de diâmetro e serão selecionados para entrar na fase de
dominância (FATET; PELLICER-RUBIO; LEBOEUF, 2011).
Os folículos dominantes produzem estradiol e inibina, sendo esta última responsável
por modular a liberação de FSH pela adenohipófise (DRIANCOURT, 2001).
Concomitantemente, tais folículos adquirem mais receptores para o hormônio luteinizante
(LH), quando comparado aos menores. Sob a influência do FSH, os folículos atingem a fase
pré-ovulatória, chegando de 6-7 mm de diâmetro na espécie caprina (VAN DEN HURK;
ZHAO, 2005). Após o pico de LH, ocorre a ovulação e inicia-se a fase luteínica com formação
do corpo lúteo e aumento dos níveis de progesterona (DRIANCOURT, 2001).

17
2.2 A Foliculogenese na cabra
A foliculogênese é um processo complexo caracterizado pela formação, crescimento e
maturação dos folículos ovarianos. É um evento iniciado ainda na vida pré-natal na maioria das
espécies, iniciando com a gênese do folículo primordial e terminando com a formação do
folículo de DeGraaf ou pré-ovulatório (VAN DEN HURK; ZHAO, 2005). O folículo é
considerado a unidade morfofuncional do ovário, constituído por um oócito circundado por
células somáticas (células da granulosa e tecais), sendo responsável por oferecer um
microambiente favorável para o crescimento e maturação do oócito (CORTVRINDT; SMITZ,
2001), além de produzir hormônios e peptídeos (ADASHI, 1994). Durante o desenvolvimento,
o folículo sofre alterações morfológicas devido ao crescimento oocitário, diferenciação e
multiplicação das células da granulosa, bem como o aparecimento da cavidade antral. Por esse
motivo são classificados em folículos pré-antrais ou não-cavitários (primordiais,
intermediários, primários e secundários) e antrais ou cavitários (terciários e de DeGraaf ou pré-
ovulatórios) (FIGUEIREDO et al., 2008).
Em fêmeas caprinas, a foliculogênese é iniciada por volta do 62º dia de vida fetal com
a formação dos folículos primordiais. Estes são os primeiros folículos encontrados no ovário e
consistem de um oócito imaturo circundado por uma camada de células somáticas planas,
conhecidas como células da pré-granulosa (BEZERRA et al., 1998). Na transição de folículos
primordiais para primários, o oócito imaturo é circundado por células da granulosa planas e
cúbicas, sendo classificado como folículo intermediário. Os primários por sua vez contêm uma
única camada de células da granulosa de formato cúbico circundando o oócito. Enquanto que
os folículos secundários, possuem duas ou mais camadas de células da granulosa cúbicas ao
redor do oócito (FIGUEIREDO et al., 2008).
Com o crescimento dos folículos secundários e a intensa proliferação das células da
granulosa, ocorre a formação de uma cavidade repleta de fluido, caracterizando os folículos
antrais. O início da formação do antro em caprinos, ocorre quando os folículos atingem cerca
de 130 μm de diâmetro, ainda em fase fetal, por volta de 110 dias de gestação (BEZERRA et
al., 1998). Os folículos terciários ainda possuem um oócito imaturo que sofreram uma série de
transformações biomoleculares até chegar a folículo pré-ovulatório (FIGUEIREDO et al.,
2008).
O mecanismo de formação do antro folicular provém do metabolismo das células da
granulosa, produzindo substâncias de alto peso molecular como os glicosaminoglicanos e os

18
proteoglicanos que geram um gradiente osmótico, resultando no acúmulo de fluido entre as
células da granulosa (CLARKE et al., 2006). A produção de fluido no antro folicular é
intensificada pelo aumento da vascularização e permeabilidade dos vasos sanguíneos e síntese
de células somáticas durante o desenvolvimento folicular.
2.3 O Fluido Folicular
Os folículos ovarianos passam por uma série de transformações morfológicas durante
seu desenvolvimento e ao atingirem cerca de 0,2 mm de diâmetro, na maioria dos mamíferos,
as células da granulosa se organizam em camadas formando uma cavidade, denominada de
antro folicular que será repleto de fluido (MONNIAUX et al., 2009). Após a formação do antro,
esteróides, fatores de crescimento e o FSH, irão regular a formação e o crescimento de folículos,
enquanto que fatores locais induzirão uma intensa atividade proliferativa nas células da
granulosa estimulando o influxo de fluido no antro (ANGELUCCI et al., 2006).
O FF é resultante da secreção do metabolismo das células somáticas (granulosa e teca)
e do transudado sanguíneo que se acumula no antro por difusão dos capilares (GOSDEN et al.,
1988; FORTUNE, 1994). Assemelhando-se muito com o plasma sanguíneo, contendo
constituintes específicos como esteróides, metabólitos, polissacarídeos, proteínas, espécies
reativas de oxigênio (ROS) e enzimas que auxiliam principalmente no crescimento e na
maturação das células foliculares e do oócito (EDWARDS, 1974).
Este fluido é o meio essencial para o crescimento e maturação das células somáticas e
germinativas ovarianas, contendo substâncias envolvidas na diferenciação e proliferação de
células da granulosa, qualidade oocitária e ruptura da parede folicular, pré-requisito para que
ocorra a ovulação (FAHIMINIYA et al., 2011). Este líquido ovariano fornece o microambiente
ideal para o crescimento e maturação folicular, possuindo ação importante nos aspectos
fisiológicos, bioquímicos e metabólicos da maturação nuclear e citoplasmática do oócito
(WATSON, 2007; GOTTARDI & MINGOTI, 2009), tornando-o competente para se tornar,
posteriormente, um embrião.
A qualidade do oócito é finamente programada por sinalizações parácrinas locais e
autócrinas durante a foliculogênese (HILLIER, 2009), onde o FF é a forma pela qual os
mediadores de sinalização são transportados para fora do folículo, bem como dentro do folículo,
entre vários tipos de células. Essa comunicação intrafolicular é fundamental para o crescimento
e a perfeita maturação oócitaria (ZAMAH et al 2015). Por isso, estudos tem sido direcionados

19
para melhor compreensão dos mecanismos de sinalização intrafolicular, envolvidos durante o
desenvolvimento folicular. Estes eventos podem ser mediados por pequenas moléculas solúveis
através de junções gap ou sinais lipídicos (BECKER et al., 2011; VACCARI et al., 2009), mas
a maioria dos componentes envolvidos na sinalização intrafolicular identificados, até o
momento, são moléculas peptídicas (HILLIER, 2009; OTSUKA, 2010). Este fluido biológico
pode fornecer informações valiosas sobre os processos de desenvolvimento oocitário, tendo em
vista a íntima relação entre FF e oócito,
Diante da complexidade dos eventos biológicos envolvidos no desenvolvimento
folicular e da tentativa de otimizar as biotécnicas reprodutivas como a PIV de embriões,
melhorando as etapas de maturação, fertilização e cultivo in vitro, estudos moleculares, tais
como a proteômica do FF podem fornecer informações importante sobre os componentes
envolvidos durante a foliculogênese. As primeiras análises em FF visando comparar os padrões
de proteínas de folículos maduros e imaturos de pacientes submetidos a fecundação in vitro
(FIV) foram realizados em humanos (SPITZER et al., 1996). Posteriormente, Anahory et al.
(2002) identificaram três proteínas: tioredoxinaperoxidase 1, transtirretina e RBP (retinol-
binding protein), em fluido de folículos maduros de mulheres estimuladas hormonalmente.
Angelucci et al., (2006) em abordagem proteômica, identificaram 210 proteínas em FF
de mulheres normo-ovulatórias e estimuladas hormonalmente, das quais um grande número
eram proteínas de fase aguda, levantando a hipótese de que a ovulação é comparável a um
evento inflamatório. Estudos também demostram que a maioria das proteínas do sistema
complemento estão presentes no FF de mulheres submetidas à estimulação ovariana,
apresentando um envolvimento na função imunológica inata no folículo, possuindo papel
importante no desenvolvimento e maturação do oócito (JARKOVSKA et al., 2010; YOO et al.,
2013).
Análises proteômicas também foram realizadas no FF de fêmeas domésticas. Em
bovinos realizaram-se estudos do perfil de proteínas do fluido de folículos ovarianos normais e
císticos (MORTARINO et al., 1999). Maniwa et al., (2005) identificaram oito proteínas a mais
em cistos foliculares de vacas, em relação ao perfil proteico de folículos ovarianos normais. A
proteína 4 de ligação ao retinol (RBP-4) foi mais expressa em fluido de folículos císticos,
quando comparada aos folículos normais, apresentando-se como um possível biomarcador para
cisto folicular em suíno (SUN et al., 2011).
Em equinos, analisou-se a expressão de proteínas em três estágios de desenvolvimento
folicular: início da dominância, dominante e pré-ovulatório, os quais não apresentaram

20
diferenças significativas entre si. No entanto, foram encontradas 30 proteínas diferencialmente
expressas entre FF e soro sanguíneo (FAHIMINIYA et al., 2011). Já em caninos, verificou-se
que expressão de algumas proteínas podem estar relacionadas com o pico de LH
(FAHIMINIYA et al., 2010).
É evidente que o FF está intimamente relacionado ao desenvolvimento e maturação do
oócito e que este fluido pode fornecer informações para uma melhor compreensão do processo
que ocorre durante a maturação folicular (ZAMAH et al., 2015). No entanto, até o momento
não existem estudos sobre a proteômica do FF que contemple a espécie caprina.
2.4 Proteínas do Fluido Folicular
Durante a foliculogenese os folículos tornam-se mais permeáveis às proteínas do
plasma, resultando em um maior número de proteínas do sangue atravessando a barreira
hemato-folicular. Assim, são bastante notáveis as semelhanças entre a composição proteica do
FF e do sangue, apesar dos processos seletivos de transporte (MOOS et al., 2009). No entanto,
estudos demostram que o conteúdo de proteínas totais do FF é superior ao do plasma sanguíneo
em éguas, mas foi significativamente menor nas vacas (DESJARDINS; KIRTON; HAFS,
1966; GERARD et al, 1998).
A capacidade das proteínas de atravessar esta barreira hemato-folicular é baseada tanto
no peso molecular bem como nas características de carga (HESS; CHEN; LARSEN, 1998).
Estudos mostraram que esta barreira é permeável para proteínas de peso inferior a 500 kDa
(GOSDEN et al., 1988), no entanto as proteínas plasmáticas de tamanhos acima de 100 kDa,
são encontradas em concentrações progressivamente mais baixas no FF do que no plasma
(MANARANG-PANGAN & MENGE,1971; CLARKE et al., 2006).
As proteínas plasmáticas contribuem com boa parte da composição de todos fluidos
corporais. Porém os constituintes e as concentrações de proteínas derivadas do plasma em
fluidos corporais irão depender do peso molecular, carga, solubilidade, permeabilidade
microvascular e da estrutura molecular do compartimento (ZAMAH et al., 2015).
As proteínas de fluído detectadas em folículos humanos estão envolvidas com os
processos de metabolismo (19%), processos celulares (14%), comunicação celular (11%) e
respostas imunitárias (11%) (ZAMAH et al., 2015). Em éguas, as proteínas do FF estão
envolvidas tanto na imunidade e defesa (14%) quanto no transporte (11%), sendo as mais
abundantes. O sistema complemento e proteínas de coagulação do sangue em conjunto

21
representam 17% das proteínas (FAHIMINIYA et al., 2011). Quanto a classificação sobre a
localização subcelular, estudos demostraram que 56% e 85% das proteínas do FF de humanos
e equinos, respectivamente estão localizados na região extracelular (FAHIMINIYA et al., 2011;
ZAMAH et al., 2015).
A maioria das proteínas plasmáticas identificadas no FF são de fase aguda da inflamação
(ANGELUCCI et al., 2006) quando presentes em superfícies celulares, respondem a um
processo fisiológico inflamatório por meio de mecanismos de reconhecimento e de ativação a
proteínas estranhas, lesão tecidual, apoptose e necrose. Algumas proteínas apresentam grande
abundância no FF de humanos tais como: albumina, IgG, antitripsina, IgA, transferrina,
haptoglobina, fibrinogênio, alfa-2 macroglobulina, alfa-1 glicoproteína ácida, IgM,
apolipoproteína AI e AII e complemento C3 (JARKOVSKA et al., 2010; AMBEKAR et al.,
2013). Podemos destacar algumas, como: albumina, imunoglobulina G, transferrina e a proteína
C3 do complemento, proteínas de alta abundância e relacionadas com os processos
reprodutivos.
2.4.1 Albumina
A albumina é a principal proteína do plasma, sendo de fundamental importância para a
regulação da pressão osmótica coloidal do sangue (SHAMAY et al., 2005), agindo ainda no
transporte de hormônios tireoidianos e lipossolúveis, ácidos graxos livres, bilirrubina não
conjugada, fármacos e drogas, além de controlar o pH do sangue. No entanto, uma das
propriedades mais interessante é a capacidade de ligar-se reversivelmente a uma grande
variedade de substâncias, transportando-as para diversos órgãos através do sistema circulatório
(GÓES FILHO, 2005).
Esta proteína é altamente expressa no FF da maioria das espécies, sendo considerada de
grande abundância, o que prejudica a expressão de outras proteínas menos abundantes em géis
2D-PAGE (FAHIMINIYA et al., 2011; AMBEKAR et al., 2013). Estudos demostraram que
após a depleção de albumina no FF de éguas, foi possível identificar uma nova proteína em um
ponto ocupado inicialmente pela albumina (FAHIMINIYA et al., 2011).
As concentrações de albumina no FF não diferiram durante o desenvolvimento folicular
de búfalas, porém os níveis desta proteína no FF foram significativamente maiores, quando
comparados aos do plasma sanguíneo. Isto sugere ser uma proteína de grande importância nas
funções fisiológicas do folículo, incluindo crescimento e maturação (ARSHAD et al., 2005),

22
atuando também, como um sequestrador de ROS, protegendo o oócito contra o estresse
oxidativo (OTSUKI et al., 2012).
A albumina também tem sido utilizada em cultivos in vitro, principalmente a albumina
sérica bovina (BSA), por atuar como um quelante de metais pesados (MEHTA & KIESSLING,
1999) e por sua propriedade surfactante, prevenindo a aderência das células às superfícies
plásticas ou de vidro. Estudos têm demonstrado que a manutenção da estrutura dos folículos
durante o cultivo in vitro favorece a sobrevivência folicular (JIN et al, 2010).
2.4.2 Imunoglobulina G
As imunoglobulinas são glicoproteínas produzidas pelos plasmócitos em resposta a um
imunógeno construídas por uma estrutura básicas compostas de duas cadeias leves idênticas
(23kDa) e duas cadeias pesadas idênticas (50-70kDa). Baseado nas diferenças estruturais da
composição de suas cadeias pesadas, as imunoglobulinas são divididas em cinco classes (IgG,
IgM, IgA, IgD e IgE), onde IgG é a principal imunoglobulina da circulação sanguínea e de
espaços extravasculares (MESQUITA JÚNIOR et al., 2010).
As IgG´s são divididas em 4 subclasses: IgG1, IgG2, IgG3 e IgG4 com diferentes
atividades funcionais, sendo ativação da cascata do complemento uma das mais importantes,
realizada principalmente pela IgG1 e IgG3 (SCHROEDER & CAVACINI, 2010).
A IgG juntamente com albumina são consideradas proteínas de alta abundância e
identificadas no FF de humanos (ANGELUCCI et al., 2006), suínos (BIJTTEBIER et al., 2009),
caninos (FAHIMINIYA et al., 2010) e equinos (FAHIMINIYA et al., 2011). Estudos
demostram que a expressão dessas proteínas no FF de equinos não diferiu significativamente
durante o desenvolvimento folicular (FAHIMINIYA et al., 2011).
A presença de imunoglobulinas nos fluidos, pode estar associado a processos
infecciosos, inflamatórios e a tratamentos hormonais de indução a ovulação que podem
produzir uma resposta autoimune (CURTIS et al., 1991; RHIM et al., 1992). Estudos
demostram que o FF de humanos contém anticorpos capazes de induzir a reação acrossomal no
espermatozoide e inibir a ligação entre espermatozoides-zona pelúcida no oócito, tendo as IgGs
participação nesses processos (MARÍN-BRIGGILER et al., 2003).

23
2.4.3 Transferrina
As transferrinas são proteínas pertencente à família das glicoproteínas de ligação a
metais, responsáveis pelo transporte de ferro a partir de locais de absorção e degradação da
partícula heme para os de armazenamento e utilização. Podendo ter papel adicional na
estimulação da proliferação celular. Sua síntese, ocorre principalmente no fígado
(SCHAEFFER et al., 1987), podendo também ser sintetizada pelo ovário, através das células
da granulosa e transportada ao oócito por endocitose (BRIGGS et al., 1999).
A transferrina é constituída por uma única cadeia polipeptídica. Ela já foi identificada
no FF da maioria dos mamíferos (AMBEKAR et al., 2013), apresentando importante papel no
transporte de ferro no folículo, além de estimular o desenvolvimento folicular, agindo como um
fator de crescimento, reduzindo as gerações de ROS que provocam atresia folicular (DE JONG;
VAN DIJK; VAN EIJK, 1990; TILLY, 1998). A transferrina aumenta gradativamente no
interior das células da granulosa e oócito em estádios mais avançados do desenvolvimento
folicular e no cumulus com a ampliação do antro folicular (BRIGGS et al., 1999).
Estudos tem identificado a expressão da transferrina no FF de espécies domésticas
(FAHIMINIYA et al., 2010; FAHIMINIYA et al., 2011) e em mulheres, verificou-se um
aumento progressivo desta proteína nas células da granulosa e oócito, a medida que o folículo
se desenvolvia (BRIGGS et al., 1999). Possuindo alta afinidade com fator de crescimento
semelhante à insulina (IGF) que desempenha papel fundamental na foliculogênese, atuando
principalmente em folículos antrais, na seleção do folículo dominante (DRIANCOURT, 2001).
2.4.4 Proteína C3 do Complemento
A proteína C3 do complemento é sintetizada principalmente no fígado por hepatócitos
(ALPER et al., 1969), sendo o componente plasmático mais abundante do sistema
complemento. Este é composto por mais de 30 proteínas, tanto plasmática, quando presentes
em superfícies celulares, que respondem por meio de mecanismos de reconhecimento e de
ativação a proteínas estranhas, lesão tecidual, apoptose e necrose (JARKOVSKA et al., 2010).
Proteínas do complemento também participam em diversos processos, tais como angiogênese,
regeneração de tecidos e metabolismo de lipídeos.
As proteínas Fator B e C3 do complemento foram encontradas no FF de humanos,
caninos e equinos, através de análises proteômica (FAHIMINIYA et al., 2010; FAHIMINIYA

24
et al., 2011; YOO et al., 2013). A proteína C3 do complemento possui genes com ação
regulatória sobre estrógenos, glicocorticóides e hormônio da tireóide (VIK et al., 1991),
desempenhando papel importante no processo reprodutivo, tendo a C3 ação durante o processo
de fecundação do espermatozoide ao oócito. Estudos demostraram a expressão de receptores
do complemento CR1 e CR3 em oócitos humanos. O espermatozoide possui uma protease que
realiza a clivagem de C3 em C3b, que por sua vez liga-se à proteína de cofator de membrana
(MCP) do espermatozoide, facilitando a fecundação mediante a ligação da C3b ao seu epítopo
presente na zona pelúcida do oócito (ANDERSON; ABBOTT; JACK, 1993; YOO et al., 2013).
Estudos também demostraram que oócitos que clivaram após a FIV eram oriundos de
folículos em que o FF apresentava concentrações significativamente mais elevadas de proteína
C3 do complemento, quando comparados aos não clivados ou não fecundados (GONZALES et
al., 1992). Outros estudos também descrevem que os níveis de proteína C3 do complemento no
FF de mulheres jovens estão presentes em maior quantidade quando comparado ao de adultas
(HASHEMITABAR et al., 2014), podendo ser regulada por hormônios esteroides, em
particular o estrógeno, dando suporte aos processos fisiológicos de pré-implantação
embrionária (LEE et al., 2009).

25
3 JUSTIFICATIVA
Os caprinos da raça Canindé apresentam padrão racial definido, sendo uma das raças
naturalizadas que mais se destacam no Brasil, por sua prolificidade, rusticidade e maior
habilidade leiteira quando comparadas a outras raças naturalizadas no Nordeste. No entanto, o
cruzamento indiscriminado entre raças exóticas e naturalizadas tem provocado impactos
consideráveis, descaracterizando este padrão racial adquirido ao longo dos séculos, além de
provocar sério risco de extinção às raças naturalizadas. Segundo Gutiérrez et al. (2003), o
conhecimento da diversidade genética é a base para o estabelecimento de programas de seleção
e conservação. Diante disso, inúmeros grupos vêm desenvolvendo estudos que objetivam a
preservação destas raças naturalizadas, para que o material genético não seja perdido de forma
definitiva. Neste âmbito, podemos destacar a importância do uso, sobretudo das modernas
biotécnicas reprodutivas na tentativa de propagação e conservação dessas raças.
Dentre as biotécnicas reprodutivas, a PIV de embriões exerce papel importante na
conservação de diferentes espécies, dando suporte à sobrevivência das populações existentes
através de manipulações de gametas, pois permite a otimização e multiplicação de fêmeas, além
da utilização de animais pré-púberes, senis e gestantes, quando utilizada conjuntamente com a
colheita oocitária por laparoscopia ou COL (BALDASSARRE et al., 2002). Entretanto, muitos
problemas ainda necessitam ser resolvidos para que esta técnica possa alcançar resultados
compatíveis com a produção esperada. Uma vez que em caprinos as técnicas de reprodução
assistida, como a MIV de oócitos, a FIV dos oócitos maturados e o desenvolvimento
embrionário inicial in vitro têm uma menor taxa de sucesso quando comparados a bovinos. Isso
pode ser devido à falta de conhecimento dos processos biológicos complexos que envolvem o
desenvolvimento folicular, tendo o FF intima relação com o crescimento e maturação do oócito.
Diante do exposto, estudos moleculares como a proteômica do FF de caprinos são necessários
para uma melhor compreensão dos processos fisiológicos que ocorrem durante o
desenvolvimento folicular. Estes estudos devem fornecer informações importantes para a
compreensão dos mecanismos fisiológicos que ocorrem em diferentes espécies, inclusive em
caprinos.

26
4 HIPÓTESE CIENTÍFICA
O proteoma do fluido folicular apresenta alterações em função do desenvolvimento dos
folículos em cabras da raça Canindé, previamente submetidas a tratamento hormonal de
estimulação ovariana.

27
5 OBJETIVOS
5.1 Objetivo Geral
o Verificar a expressão de proteínas do fluido de folículos pequenos, médios e grandes,
oriundos de cabras da raça Canindé, submetidas à estimulação hormonal ovariana.
5.2 Objetivos Específicos
o Realizar uma abordagem proteômica do fluido de folículos de diferentes tamanhos em
cabras da raça Canindé, submetidas à estimulação hormonal ovariana.
o Identificar as proteínas diferencialmente expressas do fluido de folículos de diferentes
tamanhos em cabras da raça Canindé, submetidas à estimulação hormonal ovariana.

28
6 CAPÍTULO 1
Análise diferencial de proteínas do fluido folicular de cabras da raça Canindé
submetidas ao tratamento de estimulação hormonal ovariana
Differential analysis of proteins in follicular fluid of Canindé goats previously submitted to an
ovarian hormonal stimulation treatment
Submetido em 07 de junho de 2015 ao periódico Arquivo Brasileiro de Medicina
Veterinária e Zootecnia (Qualis A2)

29
Análise diferencial de proteínas do fluido folicular de cabras da raça Canindé 1
submetidas ao tratamento de estimulação hormonal ovariana 2
Differential analysis of proteins in follicular fluid of Canindé goats previously submitted to an 3
ovarian hormonal stimulation treatment 4
A.R. Paula Junior1, M.F. van Tilburg2, M.D.P. Lobo3, A.C.O. Monteiro-Moreira3, R.A. 5
Moreira3, J.M.G. Souza-Fabjan4, A.A. Araujo1, L.M. Melo1, D.I.A. Teixeira1, A.A. Moura5, 6
V.J.F. Freitas1 7
1 Programa de Pós-graduação em Ciências Veterinárias, Universidade Estadual do Ceará, Fortaleza-CE, Brasil 8 2 Laboratório de Bioquímica Humana, Universidade Estadual do Ceará, Fortaleza-CE, Brasil 9
3 Departamento de Farmácia, Universidade de Fortaleza, Fortaleza-CE, Brasil 10 4 Universidade Federal Fluminense, Rio de Janeiro-RJ, Brasil 11
5 Departamento de Zootecnia, Universidade Federal do Ceará, Fortaleza-CE, Brasil 12
13 RESUMO 14
O presente estudo teve como objetivo verificar o perfil de proteínas expressas no fluido de 15
folículos pequenos (< 3 mm), médios (3-4 mm) e grandes (> 4mm) de cabras da raça Canindé. 16
Oito cabras adultas da raça Canindé receberam um tratamento hormonal de estimulação 17
ovariana e logo em seguida, através de laparoscopia, as amostras de fluido folicular (FF) foram 18
colhidas por punção com auxílio de uma bomba de vácuo. As amostras foram submetidas à 19
eletroforese 2D SDS-PAGE e os spots diferencialmente expressos foram identificados por 20
espectrometria de massa (ESI-Q-Tof). O estudo verificou que não existiu diferença (P > 0,05) 21
entre os diferentes tamanhos de folículos quanto à concentração de proteínas totais. No entanto, 22
cinco spots diferiram significativamente, correspondendo a quatro proteínas diferentes. A 23
serotransferrina teve uma maior expressão nos folículos médios em relação aos pequenos. A 24
zinco‐alfa‐2‐glicoproteina-like, proteína do complemento C3 e fator B do complemento 25
tiveram uma expressão aumentada nos folículos grandes em comparação com os médios. Este 26
estudo forneceu a primeira descrição das proteínas diferencialmente expressas no FF de cabras 27
Canindé submetidas ao tratamento de estimulação hormonal ovariana. Estudos posteriores 28
sobre a ação destas proteínas poderão contribuir para o sucesso de programas de FIV na espécie 29
caprina e, em particular, na preservação da raça Canindé. 30
Palavras-chave: caprino; fluido folicular; proteômica 31
32
33

30
34
ABSTRACT 35
This study aimed to determine the profile of expressed proteins in the fluid of small (<3 mm), 36
medium (3-4 mm) and large (> 4 mm) follicles of Canindé goats. Eight adult Canindé goats 37
received an ovarian stimulation treatment and then, they had the follicular fluid (FF) collected 38
by laparoscopy with the aid of a vacuum pump. The samples were subjected to 2D-SDS PAGE 39
electrophoresis and the differentially expressed spots were identified by mass spectrometry 40
ESI-Q-TOF. The study found that there was no difference (P > 0.05) between the different 41
follicle sizes for total protein concentration. However, five spots were significantly different, 42
corresponding to four different proteins. The serotransferrina had a higher expression in the 43
medium follicles in relation to small. Zinc-alpha-2-glycoprotein-like, C3 complement protein 44
and complement factor B had an increased expression in large follicles as compared with the 45
medium. This study provided the first description of differentially expressed proteins in the FF 46
of Canindé goats subjected to treatment of ovarian hormonal stimulation. Further studies on 47
the action of these proteins may contribute to the success of IVF programs in goats and in 48
particular the preservation of Canindé breed. 49
Keywords: goat; folicular fluid, proteomic 50
51
INTRODUÇÃO 52
Quando os europeus descobriram o Brasil, há mais de 500 anos atrás, os colonizadores 53
portugueses trouxeram os primeiros ruminantes para o país recém-descoberto e, desde então, 54
estes animais formaram raças denominadas de “naturalizadas”. Ao longo dos anos, a seleção 55
natural foi responsável pelo aparecimento de características morfológicas e funcionais 56
necessárias para a adaptação às condições ambientais no Brasil. No entanto, ao início do século 57
XX, a importação de raças comerciais começaram a substituir gradualmente as raças 58
naturalizadas, de tal forma que estas últimas estão agora em risco de extinção (Mariante et al., 59
2009). A raça caprina Canindé, está entre as raças em risco de extinção e é encontrada 60
principalmente no Nordeste do Brasil. Estes caprinos são criados de forma extensiva e 61
atualmente estima-se que existam pouco mais de 3000 animais. Alguns esforços estão sendo 62
realizados para evitar o desaparecimento desta raça, inclusive pelo uso de biotécnicas 63
reprodutivas, como por exemplo a fecundação in vitro (FIV) para produção de embriões e 64
posterior transferência. No entanto, na espécie caprina, a FIV ainda possui alguns entraves (p. 65

31
ex., falhas na maturação oocitária) os quais são responsáveis pela baixa produção de embriões 66
(revisado por Paramio e Izquierdo, 2014). 67
O fluido folicular (FF) é um dos constituintes do folículo ovariano que fornece um 68
microambiente biológico ideal para o desenvolvimento do oócito. O acúmulo de FF ocorre 69
durante a formação da cavidade antral no folículo e se deve principalmente à secreção 70
produzida pela atividade metabólica das células da teca e granulosa e por difusão de proteínas 71
dos capilares sanguíneos para o antro (Gosden et al., 1988). Em humanos, nos programas de 72
FIV, os ovários são hormonalmente estimulados para colheita de um maior número de oócitos 73
a serem fecundados. Na realidade, pouco se conhece sobre a qualidade dos oócitos nestes 74
programas, em que a morfologia é quase o único critério utilizado na prática médica diária 75
(Virant-Klun e Krijgsveld, 2014). Entre os poucos estudos sobre o tema, Powell et al. (2010) 76
objetivaram descobrir possíveis marcadores de qualidade em oócitos suínos. Os autores 77
verificaram 16 proteínas abundantes que foram identificados no proteoma, as quais foram 78
diferencialmente expressas nos oócitos de elevada ou baixa qualidade. 79
Estudos com diferentes abordagens em proteômica já foram realizados para identificação das 80
proteínas no FF de várias espécies domésticas, tais como em bovinos (Maniwa et al., 2005), 81
suínos (Bijttebier et al., 2009), caninos (Fahiminiya et al., 2010) e equinos (Fahiminiya et al., 82
2011). Entretanto, ao nosso conhecimento, não existe nenhum relato sobre a proteômica do FF 83
na espécie caprina. Portanto, o presente estudo teve por objetivo verificar possíveis variações 84
da expressão proteica no FF, oriundo de folículos de diferentes tamanhos, em cabras da raça 85
Canindé, as quais receberam previamente um tratamento hormonal de estimulação ovariana. 86
87
MATERIAL E MÉTODOS 88
Foram utilizadas oito cabras da raça Canindé, com idade entre um e três anos e pesando de 20 89
a 30 kg. Estes animais foram mantidos nas instalações do Laboratório de Fisiologia e Controle 90
de Reprodução (Universidade Estadual do Ceará), localizado na cidade de Fortaleza (3°47'38"S 91
e 38º33'29"W). Os animais foram mantidos em regime de manejo intensivo, recebendo feno de 92
Tifton (Cynodon dactylon) nos cochos e concentrado comercial uma vez ao dia, além de água 93
e sal mineral ad libitum. Todos os procedimentos foram aprovados pelo Comitê de Ética no 94
Uso de Animais (CEUA) (n° 3246402/2014) e obedecendo o guia para uso ético de animais em 95
pesquisa (ASAB, 2006). 96
Os animais receberam um tratamento hormonal de estimulação ovariana, o qual consistiu de 97
esponjas intravaginais de 60 mg de acetato de medroxiprogesterona (Progespon, Coopers Saúde 98

32
Animal, Cotia, Brasil) por 10 dias. No sétimo dia de tratamento, foi aplicada por via 99
intramuscular, 75 µg de d-cloprostenol (Prolise, ARSA, Buenos Aires, Argentina). Neste 100
momento, foi iniciada a estimulação ovariana com administração de 120 mg de pFSH 101
(Folltropin-V; Bioniche, Belleville, Canadá), os quais foram divididos em cinco doses 102
decrescentes (30/30, 20/20 e 20), administradas por via intramuscular em intervalos de 12 h 103
(Sousa et al., 2011). 104
Trinta e seis horas antes da colheita do FF, as cabras foram submetidas a um jejum hídrico e 105
alimentar de 36 h. As esponjas foram removidas e as fêmeas anestesiadas com 0,048 mg/kg de 106
sulfato de atropina (Atropina 1%, Fagra, Mairiporã, Brasil) e 20 mg/kg de tiopental (Tiopentax 107
2,5%, Cristália, Itapira, Brasil) por via endovenosa. Posteriormente, os animais foram mantidos 108
em plano anestésico profundo com 3% de isoflurano (Isoforine, Cristália) em equipamento de 109
anestesia inalatória. Os animais foram submetidos à laparoscopia (Karl Storz, Tuttlingen, 110
Alemanha). Os folículos foram puncionados e o FF colhido através de sistema de aspiração, 111
constituído por uma agulha de 22G e uma bomba de vácuo (WTA, Cravinhos, Brasil), a qual 112
foi regulada a uma pressão constante de 50 mmHg. Os folículos foram mensurados com o 113
auxílio da agulha de aspiração folicular que possui tamanho de cinco milímetros e classificados 114
de acordo com Khan et al. (2005) em pequenos (< 3 mm), médios (3-4 mm) e grandes (> 4 115
mm). Cada grupo de folículos era puncionado separadamente para não ocorrer mistura de FF. 116
Após a colheita, os tubos contento o FF foram centrifugados a 3000 g por 20 min e a 4o C, o 117
sobrenadante foi colhido e colocado em tubos de 1,5 mL, os quais foram devidamente 118
identificados e armazenados a -80°C para posterior análise. 119
A quantificação proteica foi realizada de acordo com o método de Bradford (Bradford, 1976). 120
Um volume do FF contendo 500 μg de proteínas foi adicionado a uma solução de rehidratação 121
contendo ureia (7 M), tioureia (2 M), anfólitos livres 0,8% (IPG buffer, 3-10), CHAPS (2% 122
w/v), ditiotreitol (DTT) 16 mM e traços de azul de bromofenol. A solução foi adicionada às 123
canaletas da bandeja de hidratação e incubada com tiras de 18 cm e faixa de pH 3-10 linear 124
(IPGs; GE Healthcare Lifesciences, Piscataway, EUA) por 17 h. Para focalização isoelétrica 125
(IEF), utilizou-se o equipamento ETTANTMIPGphorIITM (GE Healthcare Lifesciences), com as 126
seguintes condições: 250 V durante 2 h, 500 V durante 1 h, 800 V por 1 h, 1000 V por 1 h, 127
10000 V por 22000 Vh e 10000 por 40000 Vh, totalizando 67200 Vh. Após a focalização, as 128
tiras foram incubadas no tampão de equilíbrio, por duas vezes, contendo (75 mM Tris-HCl, pH 129
8,8; glicerol 29,3%, ureia 6 M, dodecil sulfato de sódio (SDS) 2%, azul de bromofenol (traços) 130
e água ultrapura (q.s.p. 200 mL). Numa primeira etapa, as tiras foram mantidas, sob leve 131

33
agitação, em solução de equilíbrio (10 mL/tira) adicionada de 2% de DTT, por 20 min, sendo, 132
em seguida, incubadas por mais 20 min com a solução de equilíbrio contendo 2,5% de 133
iodoacetamida. Logo após, as proteínas foram separadas em géis de poliacrilamida acrescido 134
de dodecil sulfato de sódio (SDS-PAGE 10%), em sistema vertical (ETTANTMDALTsix, GE 135
Healthcare Lifesciences). Os géis foram colocados em tampão de corrida contendo (0,25 M) de 136
Tris Base, (2 M) glicina e (1 %) SDS, que foi conduzido como se segue: 15 mA/gel durante 1 137
h e 40 mA/gel por mais 5-6 h, adicionados de marcadores moleculares Amersham™ Full-Range 138
Rainbow™ variando de 12 a 225 kDa, com base em sua massa molecular (GE Healthcare 139
Lifesciences) (O’Farrel, 1975; van Tilburg et al., 2013). 140
Após a separação, as proteínas foram visualizadas utilizando-se o método Coomassie coloidal 141
(Candiano et al., 2004) com modificações (van Tilburg et al., 2013). Para tanto, os géis foram 142
colocados em solução de fixação (ácido fosfórico 2% e etanol 30%, em água) por 143
aproximadamente 12 h. Sequencialmente os géis foram lavados três vezes por 20 min, em uma 144
solução de ácido fosfórico 2%, em água. Após a última lavagem os géis permaneceram em 145
contato com uma solução contendo ácido fosfórico 2%, etanol 18% e sulfato de amônio 15% 146
em água, por 30 min. Em seguida, adicionou-se a esta solução 2% do volume total de 147
Coomassie Blue G-250. O corante permaneceu em contato com os géis por cerca de 5 dias. 148
Após este período, a solução contendo o corante foi desprezada, e os géis lavados com água 149
destilada. As imagens dos géis foram digitalizadas utilizando o Image Scanner II (GE 150
Healthcare Lifesciences, USA) e analisadas utilizando o aplicativo PDQuest versão 8.0.1 (Bio-151
Rad Laboratories). A intensidade dos spots de cada perfil foi avaliada quanto à normalidade de 152
sua distribuição, por meio do teste Kolmogorov-Smirnov, bem como de sua assimetria e 153
curtose. Transformações logarítmicas foram realizadas para ajuste da normalidade da 154
intensidade dos spots. Os spots foram avaliados por meio de ANOVA, e as médias foram 155
comparadas entre os tratamentos por meio de teste de Tukey (P < 0,05), utilizando o software 156
GraphPad Prism 5. 157
As proteínas separadas por eletroforese 2D que diferiram significativamente entre os folículos 158
pequenos, médios e grandes foram submetidas à digestão com tripsina conforme descrito 159
previamente (Moura et al., 2006; van Tilburg et al., 2013). Os spots selecionados foram 160
cortados individualmente dos géis em pedaços de aproximadamente 1 mm3 e completamente 161
descorados com 400 μL de solução contendo acetonitrila 50% e bicarbonato de amônio 50% 162
(25 mM, pH 8,0). A solução foi retirada e acrescentado 400 μL da mesma solução, repetindo o 163
procedimento por três vezes. Os spots foram desidratados com 200 μL de acetonitrila 100% 164

34
durante 5 min e secos no concentrador (Concentrator plus, Eppendorf). As proteínas foram 165
incubadas por, aproximadamente, 20 horas, a 37ºC, com solução contendo tripsina (Promegacat 166
# V5111, Madison, EUA), ácido acético 50 mM (solução estoque 200 ng/μL) e diluída com 167
bicarbonato de amônio 50 mM (concentração final 20 ng/μL). Os peptídeos foram extraídos 168
por meio de duas lavagens com solução de ácido fórmico 5%/acetonitrila 50%, por 30 min. Os 169
peptídeos foram separados, inicialmente, por cromatografia líquida em sistema NanoUPLC, 170
acoplado a um espectrômetro de massa Synapt G1 HDMS (Waters Corporation, Milford, EUA). 171
Para os procedimentos cromatográficos iniciais, foi utilizada uma coluna Waters Symmetry 172
300TM (C-18, filme de 5 μm; 0,3 mm × 5 mm) para dessalinização e pré-concentração. Após 173
lavagens por 3 min com solvente, os peptídeos foram então eluídos em um gradiente de 174
concentração para a coluna analítica de fase reversa nanoACQUITY HSS T3 (C-18, 5 mm, 175
0,075 mm x 150 mm). A cromatografia na coluna de fase reversa foi realizada utilizando-se um 176
gradiente linear durante 40 min. A medida que os peptídeos iam sendo eluídos, os mesmos eram 177
ionizados para posterior aquisição de dados no espectrômetro de massas. 178
O espectrômetro de massas foi operado a fim de obter espectros MS/MS dos peptídeos trípticos 179
em modo dependente de dados (data-dependent acquisition; DDA) para o íon precursor usando 180
o reconhecimento de estado de carga e limite de intensidade como critérios de seleção, através 181
da utilização do aplicativo MassLynx 4.1. De forma a se obter os dados MS/MS, uma varredura 182
a cada 1,5 seg foi executada para dados de relação massa/carga (m/z) entre 400-1500. A partir 183
de cada varredura, até três íons mais intensos, baseando-se nos critérios de seleção já descritos, 184
foram escolhidos para obtenção dos espectros resultantes da dissociação induzida por colisão 185
(CID) na presença de argônio. O espectrômetro de massa foi calibrado utilizando fragmentos 186
de íons do peptídeo Glu-1-fibrinopeptídeo B (Glu-Fib), conforme necessário para manter a 187
precisão em 10 partes por milhão. 188
Os espectros iônicos resultantes (6-8 seg) foram processados utilizando o aplicativo Protein 189
Lynx Global Server 2.1 e convertidos em arquivos de lista de picos (peak list files; PKL). Esses 190
últimos foram utilizados para a pesquisa em banco de dados realizada através da ferramenta 191
MASCOT (www.matrixscience.com) no banco de dados NCBInr, utilizando o modo MS/MS 192
ion search. As análises das interações proteína-proteína, foram realizadas através do banco de 193
dados STRING 9.0 (Snel et al., 2000). A rede de análise só foi avaliada para proteínas 194
diferencialmente expressas em diferentes tamanhos de folículos. 195
196
197

35
RESULTADOS 198
Entre os diferentes tamanhos de folículos não foram encontradas variações significativas (P > 199
0,05) quanto à concentração de proteínas totais (mg/mL) presentes no FF dos folículos 200
pequenos (45,3 ± 3,1), médios (44,2 ± 3,3) e grandes (45,1 ± 2,3). Foram analisados 24 géis, 201
sendo oito géis de cada grupo, que apresentaram uma média de 155,4 ± 4,5 spots por gel (Fig. 202
1). 203
Dos 155 spots analisados, cinco foram diferencialmente expressos (P < 0,05) de acordo com o 204
tamanho do folículo, correspondendo a quatro proteínas diferentes. Estas foram identificadas 205
como serotransferrina, zinco‐alfa‐2‐glicoproteina‐like, fator B do complemento e proteína C3 206
do complemento (Tab. 1). Não foram observadas proteínas consistentemente expressas em 207
apenas um tamanho de folículo. 208
A Fig. 2A mostra a sobreposição dos géis referentes ao fluido de folículos pequenos, médios e 209
grandes. A serotransferrina apresentou um aumento significativo entre folículos pequenos e 210
médios (Fig. 2B). A zinco‐alfa‐2‐glicoproteina‐like se comportou de forma contrária à 211
serotransferrina, apresentado uma redução na sua expressão quando comparada o folículo 212
pequeno com o folículo médio (Fig. 2B). As proteínas fator B do complemento e proteína C3 213
do complemento se comportaram de forma semelhante durante o desenvolvimento folicular, 214
aumentando os níveis de expressão com o crescimento folicular, sendo significativamente mais 215
expressos em folículos grandes, quando comparados a folículos médios (Fig. 2B). É possível 216
observar (Fig. 2C) a representação tridimensional dos spots diferencialmente expressos entre 217
os grupos foliculares. De acordo com a análise da rede de proteínas, a serotransferrin (TF) 218
apresentou interações com um conjunto de 11 proteínas (Fig. 3a). A zinco‐alfa‐2‐glicoproteína‐219
like (AZGP1) está associada à beta-2-microglobulina e uma proteína homóloga a prolactina-220
inducible (Fig. 3b). O fator B de complemento (CFB) está associada a 10 diferentes proteínas, 221
incluindo a proteína do complemento C3(C3) (Fig. 3c). E finalmente a C3, que está associada 222
a 10 proteínas (Fig. 3d). 223
36
224
Figura 1. 2D-PAGE do fluido folicular de cabras da raça Canindé. Foram aplicadas 500 µg de amostras 225 de proteínas em tira linear com pI 3-10 na primeira dimensão e separados em 10% gel SDS-PAGE na 226 segunda dimensão. Os géis foram corados pelo procedimento de azul coloidal. 227 228
229
Figura 2. Análise computadorizada 2D-PAGE do fluido folicular de cabras da raça Canindé. (A) 230 Sobreposição dos géis 2D-PAGE de fluido folicular, destacando as proteínas que apresentam diferença 231 estatística durante o crescimento folicular. Cores azul, verde e vermelho, correspondem aos spots dos 232 folículos pequenos (FP), médios (FM) e grandes (FG) respectivamente. (B) Intensidade dos spots (média 233 ± S.E.M.) em função do tamanho folicular. (C) Imagem em 3D dos spots diferencialmente expressos 234 entre os grupos de folículos. 235

37
236 237
238 Figura 3. Análise in silico das interações de proteínas. As proteínas analizadas foram: a. serotransferrina 239 (TF), b. zinco‐alfa‐2‐glicoproteína‐like (AZGP1), c. fator B do complemento (CFB), d. C3 do 240 complemento (C3). Linhas de cores diferentes representam os tipos de evidências para a associação: 241 (preto) co-expressão, (rosa) experimentos, (verde) textmining e (azul) banco de dados. Proteína 242 receptora de transferrina (TFRC), Proteína não caracterizada (CP), Proteína não caracterizada (HEPH), 243 Soluto carreador da família 40 membro 1 (SLC40A1); Apolipoproteína A-I (APOA1), Proteína não 244 caracterizada (STEAP3), Próton ATPase do tipo V 116 kDa subunidade a isoforma 2 (ATP6V0A2), 245 Proteína não caracterizada (ENSBTAG00000000292), Proteína não caracterizada 246 (ENSBTAG00000045497), Próton ATPase do tipo V catalítica subunidade A (ATP6V1A), Proteína 247 homóloga precursora de prolactina-inducible (ENSBTAG00000006655), Beta-2-microglobulina 248 (B2M), Fator D do complemento (CFD), Precursor do fator H do complemento (CFH), Preproteína C3 249 do complemento (C3), Proteína não caracterizada (ENSBTAG00000006864), Precursor C4 do 250 complemento (C4A), C2 do complemento (C2), Proteína não caracterizada (Bt.64039), Proteína não 251 caracterizada (LOC781004), Fator acelerador de decaimento do complemento (CD55), Proteína não 252 caracterizada (ENSBTAG00000005587), Proteína cofator de membrana (LOC616002), Proteína 253 precursora de cofator de membrana isoforma 5 (ENSBTAG00000005397), Precursor do fator I do 254 complemento (CFI), Receptor quimiotático da anafilotoxina C5a (C5AR1), Precursor de plasminogênio 255 (PLG). 256 257
DISCUSSÃO 258
Este estudo consiste na primeira caracterização de proteínas do FF expressas diferencialmente 259
em folículos de diferentes tamanhos, colhidos in vivo na especie caprina. A obtenção de 260
informações sobre a expressão das proteínas no FF poderá contribuir para a compreensão dos 261

38
fenômenos fisiológicos e, posteriormente, incrementar as taxas de sucesso em biotécnicas 262
reprodutivas, tais como a FIV. 263
A expressão de algumas proteínas são modificadas durante o desenvolvimento folicular e 264
estudos tem evidenciado que a barreira hemato-folicular é permeável a proteínas com massa 265
até 500 kDa (Gosden et al., 1988). Dessa forma, a presença de proteínas no FF, pode estar 266
associada ao aumento da vascularização folicular e permeabilidade dos vasos sanguíneos e/ou 267
aumento da síntese em células somáticas do folículo, intensificando a produção de fluido no 268
antro durante o desenvolvimento folicular (Fortune, 1994; Hanrieder et al., 2008). No presente 269
estudo, não foram observadas variações na concentração total de proteínas entre os diferentes 270
tamanhos foliculares e também nenhuma proteína foi consistetemente expressa em um único 271
grupo de folículos. Isso se deve provavelmente à presença de proteínas de grande abundância 272
no FF e soro sanguíneo, como albumina (Fahiminiya et al., 2011), que é de fundamental 273
importância para a regulação osmótica coloidal dos fluidos corporais, realizando assim o 274
equilibrio das proteínas entre FF e o plasma sanguineo. 275
As transferrinas são proteínas responsáveis pelo transporte de ferro e exercem um papel 276
adicional na estimulação da proliferação celular. Sua síntese ocorre principalmente no fígado 277
(Schaeffer et al., 1987), mas também é sintetizada pelo ovário através das células da granulosa 278
e transportada ao oócito por endocitose (Briggs et al., 1999). No folículo, a transferrina realiza 279
a função de transporte de ferro, além de estimular o desenvolvimento folicular, agindo como 280
um fator de crescimento e reduzindo a geração de espécie reativa de oxigenio (ROS) que 281
provocam atresia folicular (De Jong et al., 1990; Tilly, 1998). No ovário de humanos e 282
camundongos, a transferrina aumenta gradativamente sua expressão no interior das células da 283
granulosa e oócito em estágios mais avançados do desenvovimento folicular e no cumulus, com 284
a ampliação do antro folicular (Briggs et al., 1999). 285
Em nosso estudo observamos o aumento na expressão da serotransferrina em folículos médios 286
quando comparados a folículos pequenos, provavelmente devido ao aumento da proliferação 287
das células da granulosa durante o desenvolvimento folicular. Esses resultados corrobaram com 288
aqueles encontrados no FF de éguas (Rocha, 2014). 289
No interactoma da serotransferrina podemos destacar a interação com apolipoproteina-AI 290
(APOA1) encontrada em FF de mulheres (Angelucci et al., 2006). A APOA1 é responsável 291
pelo transporte de colesterol e triglicerídeos, tendo importante relação com o desenvolvimento 292
folicular devido ao estímulo mitogênico e angiogênico (Von Otte et al., 2006). A proteína 293
APOA1 também pode atuar como transportador para os precursores de esteróides, além de 294

39
possuir efeitos antioxidantes que protegem o oócito de lesão tóxica, aumentando assim o 295
potencial de fecundação (Von Wald et al., 2010). 296
A zinco‐alfa‐2‐glicoproteina‐like (AZPG1 ou ZAG) apresenta efeito lipolítico que resulta em 297
um aumento no AMPc intracelular (Russell et al., 2004). Embora as funções reprodutivas da 298
AZPG1 não sejam bem elucidadas, o aumento significativo na sua expressão de folículos 299
médios para grandes pode significar um envolvimento da mesma na maturação oocitária. 300
Outras proteínas diferencialmente expressas encontradas no FF de cabras foram o fator B do 301
complemento e a proteína C3 do complemento, que pertecem ao sistema do complemento. Esse 302
sistema é composto por mais de 30 proteínas, tanto plasmáticas quanto presentes em superfícies 303
celulares (Jarkovska et al., 2010). Em nosso estudo, duas proteínas do sistema do complemento 304
aumentaram significativamente a sua expressão quando da comparação entre folículos médios 305
e grandes. Em mulheres, o insucesso da FIV pode estar relacionada aos níveis de algumas 306
proteínas, incluindo a proteína C3 do complemento (Estes et al., 2009). 307
Também foi possível observar um comportamento semelhante do Fator B com a proteína C3 308
do complemento. Esse comportamento pode ser justificado pelo fato da proteína fator B do 309
complemento ser parte da via alternativa do sistema complemento que participa do processo de 310
iniciação da ativação desta via para gerar C3 ou C5 convertase. 311
A proteína C3 também possui uma ação importante durante o processo de fecundação. 312
Proteases do acrossoma de espermatozóides clivam a C3 do meio em C3b, que por sua vez liga-313
se a proteína cofator de membrana (LOC616002) presente no espermatozoide. A proteína C3 314
ligada à membrana espermática vai mediar a interação do espermatozóide como oócito através 315
dos receptores do complemento CR1 e CR3 expressos no oócito. Dessa maneira, as proteínas 316
do sistema complemento facilitam a interação espermatozóide-oócito (Anderson, Abbott e 317
Jack, 1993). Em caprinos, já foi demostrado que oócitos submetidos à maturação e posterior 318
fecundação in vitro apresentam capacidade de desenvolvimento inferior em comparação aos 319
maturados in vivo (Cognié et al., 2003). Portanto, parece evidente que o processo de 320
competência oocitária é modulado por diferentes substâncias presentes no FF, tendo as 321
proteínas uma importante participação. 322
323
CONCLUSÃO 324
Este estudo forneceu a primeira descrição das proteínas diferencialmente expressas no fluido 325
folicular colhido in vivo de cabras da raça Canindé submetidas à tratamento de estimulação 326

40
hormonal ovariana, que poderá contribuir para o sucesso de programas de FIV na espécie 327
caprina e, em particular, na preservação da raça Canindé. 328
329
AGRADECIMENTOS 330
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e à Coordenação 331
de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pelo suporte financeiro para bolsa 332
de estudos e apoio ao projeto. Os autores gostariam também de agradecer ao MSc. Carlos 333
Henrique Sousa de Melo, docente do Instituto Superior de Teologia Aplicada (INTA) pela 334
importante participação na realização e no treinamento em laparoscopia para colheita do FF. 335
336
REFERÊNCIAS 337
ANDERSON, D.J.; ABBOTT, A.F.; JACK, R.M. The role of complement component C3b and 338
its receptors in sperm-oocyte interaction. Proc. Natl. Acad. Sci., v.90, p.10051-10055, 1993. 339
ANGELUCCI, S.; CIAVARDELLI, D.; DI GIUSEPPE, F. et al. Proteome analysis of human 340
follicular fluid. Biochim. Biophys. Acta, v.1764, p.1775-1785, 2006. 341
ASSOCIATION FOR THE STUDY OF ANIMAL BEHAVIOUR (ASAB). Guidelines for the 342
treatment of animals in behavioral research and teaching. Anim. Behav., v.71, p.245-253, 2006. 343
BIJTTEBIER, J.; TILLEMAN, K.; DHAENENS, M. et al. Comparative proteome analysis of 344
porcine follicular fluid and serum reveals that excessive alpha (2) macroglobulin in serum 345
hampers successful expansion of cumulus-oocyte complexes. Proteomics, v.9, p.4554-4565, 346
2009. 347
BRADFORD, M.M. A rapid and sensitive method for the quantization of microgram quantities 348
of protein utilizing the principle of dye binding. Anal. Biochem., v.72, p.248-254, 1976. 349
BRIGGS, D.A.; SHARP, D.J.; MILLER, D. et al. Transferrin in the developing ovarian follicle: 350
evidence for de-novo expression by granulosa cells. Mol. Hum. Reprod., v.5, p.1107-1114, 351
1999. 352
CANDIANO, G.; BRUSCHI, M.; MUSANTE, L. et al. Blue silver: A very sensitive colloidal 353
Coomassie G-250 staining for proteome analysis. Electrophoresis, v.25, p.1327-1333, 2004. 354
COGNIÉ, Y.; BARIL, G.; POULIN, N. et al. Current status of embryo technologies in sheep 355
and goat. Theriogenology, v.59, p.171-188, 2003. 356

41
DE JONG, G.; VAN DIJK, J.P.; VAN EIJK, H.G. The biology of transferrin. Clin. Chim. Acta, 357
v.190, p.1-46, 1990. 358
ESTES, S.J.; YE, B.; QIU, W. et al. A proteomic analysis of IVF follicular fluid in women ≤ 359
32 years old. Fertil. Steril., v.92, p.1569-1578, 2009. 360
FAHIMINIYA, S.; REYNAUD, K.; LABAS, V, et al. Steroid hormones content and proteomic 361
analysis of canine follicular fluid during the preovulatory period. Reprod. Biol. Endocrinol., 362
v.8, p.1-14, 2010. 363
FAHIMINIYA, S.; LABAS, V.; DACHEUX, J.L., et al. Improvement of 2D-PAGE resolution 364
of human, porcine and equine follicular fluid by means of hexapeptide ligand library. Reprod. 365
Domest. Anim., v.46, p.561-563, 2011. 366
FORTUNE, J.E. Ovarian follicular growth and development in mammals. Biol. Reprod., v.50, 367
p.225-232, 1994. 368
GOSDEN, R.G.; HUNTER, R.H.; TELFER, E. et al. Physiological factors underlying the 369
formation of ovarian follicular fluid. J. Reprod. Fert., v.82, p.813-825, 1988. 370
HANRIEDER, J.; NYAKAS, A.; NAESSEN, T. et al. Proteomic analysis of human follicular 371
fluid using an alternative bottom-up approach. J. Proteome Res., v.7, p.443-449, 2008. 372
JARKOVSKA, K.; MARTINKOVA, J.; LISKOVA L. et al. Proteome mining of human 373
follicular fluid reveals a crucial role of complement cascade and key biological pathways in 374
women undergoing in vitro fertilization. J. Proteome Res., v.9, p.1289-1301, 2010. 375
KHAN, M.H., AGARWAL, S.K., MAJUMDAR, A.C., HOQUE, M., SHANKAR, U. Effect 376
of exogenous administration of insulin on follicular dynamics in goat. Indian J. Anim. Sci. v.75, 377
p.1140–1143, 2005. 378
MANIWA, J.; IZUMI, S.; ISOBE, N. et al. Studies on substantially increased proteins in 379
follicular fluid of bovine ovarian follicular cysts using 2-D PAGE and MALDI-TOF MS. 380
Reprod. Biol. Endocrinol., v.3, p.1-9, 2005. 381
MARIANTE, A.S.; ALBUQUERQUE, M.S.M.; EGITO, A.A. et al. Present status of the 382
conservation of livestock genetic resources in Brazil. Livest. Sci., v.120, p.204-212, 2009. 383
MOURA, A.A.; KOC, H.; CHAPMAN, D.A. et al. Identification of accessory sex gland fluid 384
proteins as related to fertility indexes of dairy bulls: a proteomic approach. J. Androl., v.27, 385
p.201-211, 2006. 386

42
O'FARREL, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem., 387
v.250, p.4007-4021, 1975. 388
PARAMIO, M.T.; IZQUIERDO, D. Current status of in vitro embryo production in sheep and 389
goats. Reprod. Domest. Anim., v.49, p.37-48, 2014. 390
POWELL, M.D.; MANANDHAR, G.; SPATE, L. et al. Discovery of putative oocyte quality 391
markers by comparative ExacTag proteomics. Proteomics, v.4, p.337-351, 2010. 392
ROCHA B.P.L.O. Perfil protéico do fluido folicular durante a foliculogênese da égua. 2014, 78 393
p. Tese (Doutorado em reprodução animal) - Universidade Federal do Rio Grande do Sul, Porto 394
Alegre. 2014. 395
RUSSELL, S.T.; ZIMMERMAN, T.P.; DOMIN, B.A. et al. Induction of lipolysis in vitro and 396
loss of body fat in vivo by zinc-alpha2-glycoprotein. Biochim. Biophys. Acta, v.36, p.59-68, 397
2004. 398
SCHAEFFER, E.; LUCERO, M.A.; JELTSCH, J.M. et al. Complete structure of the human 399
transferrin gene. Comparison with analogous chicken gene and human pseudogene. Gene, v.56, 400
p.109-116, 1987. 401
SNEL, B.; LEHMANN, G.; BORK, P. String: a web-server to retrieve and display the 402
repeatedly occurring neighborhood of a gene. Nucleic Acids Res., v.28, p.3442-3444, 2000. 403
SOUSA, F.C.; MELO, C.H.S.; TELES FILHO, A.C.A.; AVELAR, S.R.G.; MOURA, A.A.A.; 404
MARTINS, J.A.M.; FREITAS, V.J.F.; TEIXEIRA, D.I.A. Ovarian follicular response to 405
different hormonal stimulation treatments in Canindé goats. Anim. Reprod. Sci., v.125, p.88-406
93, 2011. 407
TILLY, J.L. Molecular and genetic basis of normal and toxicant-induced apoptosis in female 408
germ cells. Toxicol. Lett., v.102-103, p.497-501, 1998. 409
VAN TILBURG, M.F.; RODRIGUES, M.A.; MOREIRA, R.A. et al. Membrane-associated 410
proteins of ejaculated sperm from Morada Nova rams. Theriogenology., v.79, p.1247-1261, 411
2013. 412
VIRANT-KLUN, I.; KRIJGSVELD, J. Proteomes of animal oocytes: what can we learn for 413
human oocytes in the in vitro fertilization programme? BioMed Res. Inter., article ID 856907, 414
11 pages, doi:10.1155/2014/856907. 415

43
VON OTTE, S.; PALETTA, J.R.; BECKER, S. et al. Follicular fluid high density lipoprotein-416
associated sphingosine 1-phosphate is a novel mediator of ovarian angiogenesis. J. Biol. Chem., 417
v.281, p.5398-5405, 2006. 418
VON WALD, T.; MONISOVA, Y.; HACKER, M.R. et al. Age-related variations in follicular 419
apolipoproteins may influence human oocyte maturation and fertility potential. Fertil. Steril., 420
v.93, p.2354-2361, 2010. 421
422
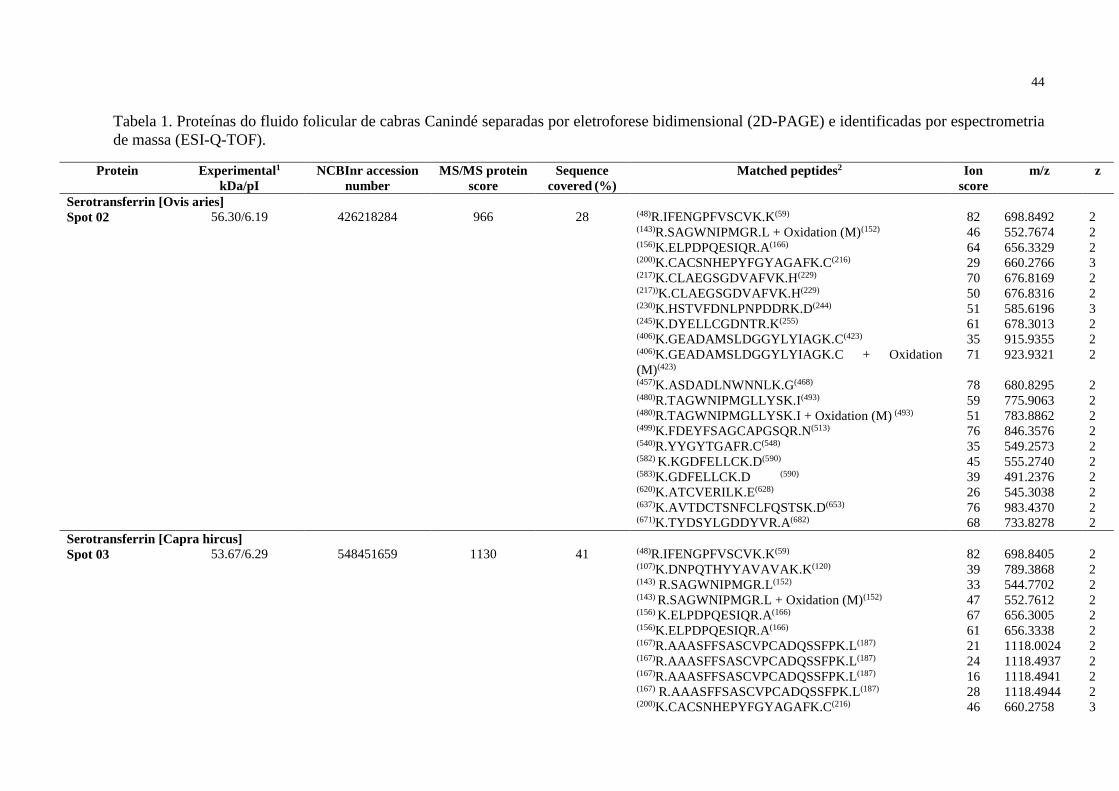
44
Tabela 1. Proteínas do fluido folicular de cabras Canindé separadas por eletroforese bidimensional (2D-PAGE) e identificadas por espectrometria
de massa (ESI-Q-TOF).
Protein Experimental1
kDa/pI
NCBInr accession
number
MS/MS protein
score
Sequence
covered (%)
Matched peptides2 Ion
score
m/z z
Serotransferrin [Ovis aries]
Spot 02 56.30/6.19 426218284 966 28 (48)R.IFENGPFVSCVK.K(59) 82 698.8492 2
(143)R.SAGWNIPMGR.L + Oxidation (M)(152) 46 552.7674 2
(156)K.ELPDPQESIQR.A(166) 64 656.3329 2
(200)K.CACSNHEPYFGYAGAFK.C(216) 29 660.2766 3
(217)K.CLAEGSGDVAFVK.H(229) 70 676.8169 2
(217))K.CLAEGSGDVAFVK.H(229) 50 676.8316 2
(230)K.HSTVFDNLPNPDDRK.D(244) 51 585.6196 3
(245)K.DYELLCGDNTR.K(255) 61 678.3013 2
(406)K.GEADAMSLDGGYLYIAGK.C(423) 35 915.9355 2
(406)K.GEADAMSLDGGYLYIAGK.C + Oxidation
(M)(423)
71 923.9321
2
(457)K.ASDADLNWNNLK.G(468) 78 680.8295 2
(480)R.TAGWNIPMGLLYSK.I(493) 59 775.9063 2
(480)R.TAGWNIPMGLLYSK.I + Oxidation (M) (493) 51 783.8862 2
(499)K.FDEYFSAGCAPGSQR.N(513) 76 846.3576 2
(540)R.YYGYTGAFR.C(548) 35 549.2573 2
(582) K.KGDFELLCK.D(590) 45 555.2740 2
(583)K.GDFELLCK.D (590) 39 491.2376 2
(620)K.ATCVERILK.E(628) 26 545.3038 2
(637)K.AVTDCTSNFCLFQSTSK.D(653) 76 983.4370 2
(671)K.TYDSYLGDDYVR.A(682) 68 733.8278 2
Serotransferrin [Capra hircus]
Spot 03 53.67/6.29 548451659 1130 41 (48)R.IFENGPFVSCVK.K(59) 82 698.8405 2
(107)K.DNPQTHYYAVAVAK.K(120) 39 789.3868 2
(143) R.SAGWNIPMGR.L(152) 33 544.7702 2
(143) R.SAGWNIPMGR.L + Oxidation (M)(152) 47 552.7612 2
(156) K.ELPDPQESIQR.A(166) 67 656.3005 2
(156)K.ELPDPQESIQR.A(166) 61 656.3338 2
(167)R.AAASFFSASCVPCADQSSFPK.L(187) 21 1118.0024 2
(167)R.AAASFFSASCVPCADQSSFPK.L(187) 24 1118.4937 2
(167)R.AAASFFSASCVPCADQSSFPK.L(187) 16 1118.4941 2
(167) R.AAASFFSASCVPCADQSSFPK.L(187) 28 1118.4944 2
(200)K.CACSNHEPYFGYAGAFK.C(216) 46 660.2758 3
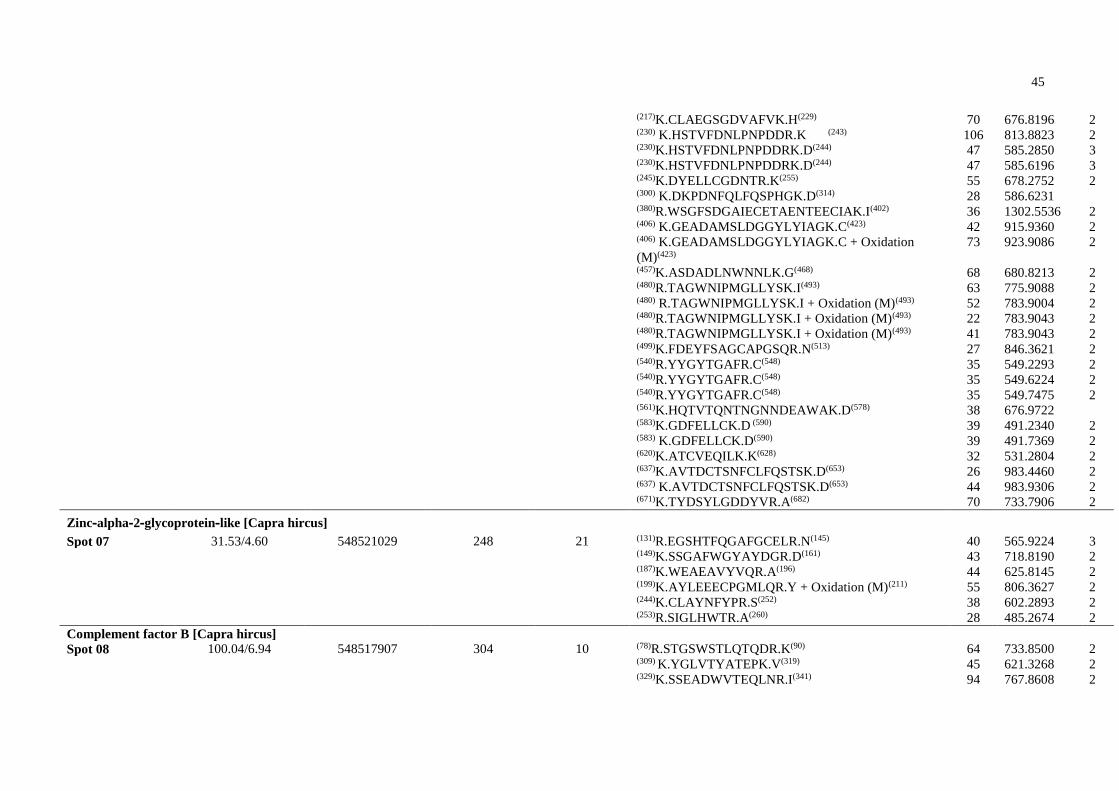
45
(217)K.CLAEGSGDVAFVK.H(229) 70 676.8196 2
(230) K.HSTVFDNLPNPDDR.K (243) 106 813.8823 2
(230)K.HSTVFDNLPNPDDRK.D(244) 47 585.2850 3
(230)K.HSTVFDNLPNPDDRK.D(244) 47 585.6196 3
(245)K.DYELLCGDNTR.K(255) 55 678.2752 2
(300) K.DKPDNFQLFQSPHGK.D(314) 28 586.6231
(380)R.WSGFSDGAIECETAENTEECIAK.I(402) 36 1302.5536 2
(406) K.GEADAMSLDGGYLYIAGK.C(423) 42 915.9360 2
(406) K.GEADAMSLDGGYLYIAGK.C + Oxidation
(M)(423)
73 923.9086 2
(457)K.ASDADLNWNNLK.G(468) 68 680.8213 2
(480)R.TAGWNIPMGLLYSK.I(493) 63 775.9088 2
(480) R.TAGWNIPMGLLYSK.I + Oxidation (M)(493) 52 783.9004 2
(480)R.TAGWNIPMGLLYSK.I + Oxidation (M)(493) 22 783.9043 2
(480)R.TAGWNIPMGLLYSK.I + Oxidation (M)(493) 41 783.9043 2
(499)K.FDEYFSAGCAPGSQR.N(513) 27 846.3621 2
(540)R.YYGYTGAFR.C(548) 35 549.2293 2
(540)R.YYGYTGAFR.C(548) 35 549.6224 2
(540)R.YYGYTGAFR.C(548) 35 549.7475 2
(561)K.HQTVTQNTNGNNDEAWAK.D(578) 38 676.9722
(583)K.GDFELLCK.D (590) 39 491.2340 2
(583) K.GDFELLCK.D(590) 39 491.7369 2
(620)K.ATCVEQILK.K(628) 32 531.2804 2
(637)K.AVTDCTSNFCLFQSTSK.D(653) 26 983.4460 2
(637) K.AVTDCTSNFCLFQSTSK.D(653) 44 983.9306 2
(671)K.TYDSYLGDDYVR.A(682) 70 733.7906 2
Zinc‐alpha‐2‐glycoprotein‐like [Capra hircus]
Spot 07 31.53/4.60 548521029 248 21 (131)R.EGSHTFQGAFGCELR.N(145) 40 565.9224 3
(149)K.SSGAFWGYAYDGR.D(161) 43 718.8190 2
(187)K.WEAEAVYVQR.A(196) 44 625.8145 2
(199)K.AYLEEECPGMLQR.Y + Oxidation (M)(211) 55 806.3627 2
(244)K.CLAYNFYPR.S(252) 38 602.2893 2
(253)R.SIGLHWTR.A(260) 28 485.2674 2
Complement factor B [Capra hircus]
Spot 08 100.04/6.94 548517907 304 10 (78)R.STGSWSTLQTQDR.K(90) 64 733.8500 2
(309) K.YGLVTYATEPK.V(319) 45 621.3268 2
(329)K.SSEADWVTEQLNR.I(341) 94 767.8608 2
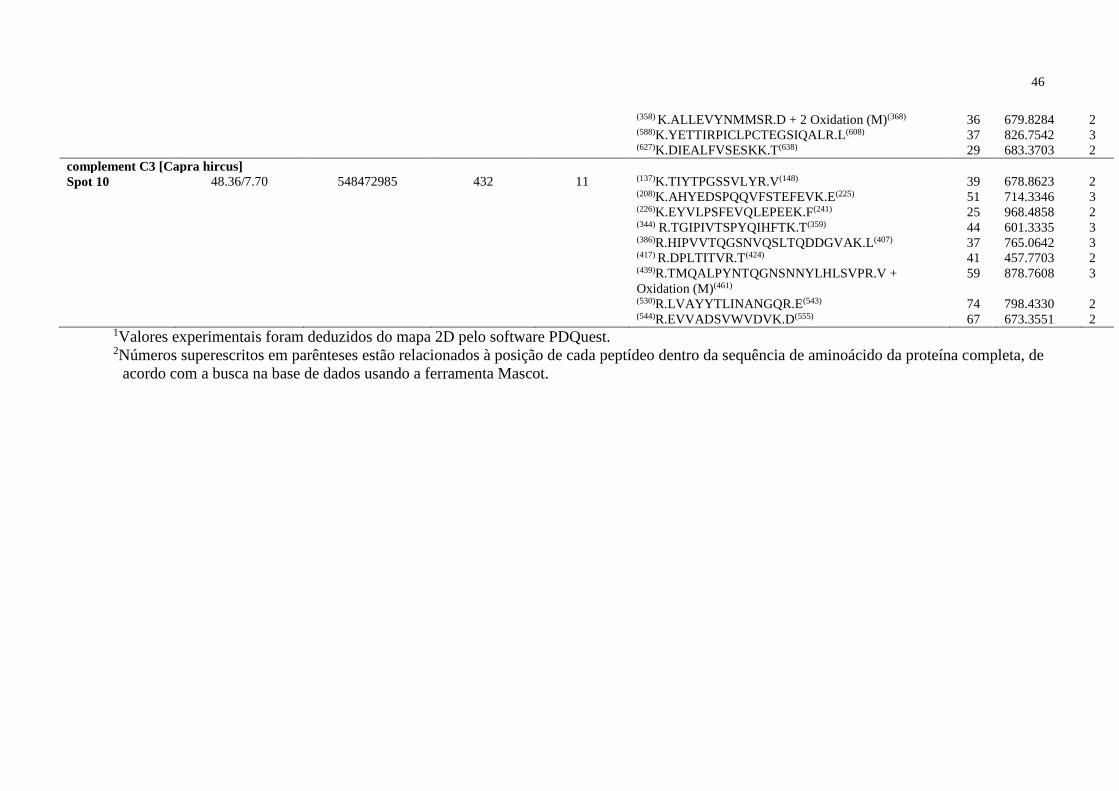
46
(358) K.ALLEVYNMMSR.D + 2 Oxidation (M)(368) 36 679.8284 2
(588)K.YETTIRPICLPCTEGSIQALR.L(608) 37 826.7542 3
(627)K.DIEALFVSESKK.T(638) 29 683.3703 2
complement C3 [Capra hircus]
Spot 10 48.36/7.70 548472985 432 11 (137)K.TIYTPGSSVLYR.V(148) 39 678.8623 2
(208)K.AHYEDSPQQVFSTEFEVK.E(225) 51 714.3346 3
(226)K.EYVLPSFEVQLEPEEK.F(241) 25 968.4858 2
(344) R.TGIPIVTSPYQIHFTK.T(359) 44 601.3335 3
(386)R.HIPVVTQGSNVQSLTQDDGVAK.L(407) 37 765.0642 3
(417) R.DPLTITVR.T(424) 41 457.7703 2
(439)R.TMQALPYNTQGNSNNYLHLSVPR.V +
Oxidation (M)(461)
59 878.7608 3
(530)R.LVAYYTLINANGQR.E(543) 74 798.4330 2
(544)R.EVVADSVWVDVK.D(555) 67 673.3551 2 1Valores experimentais foram deduzidos do mapa 2D pelo software PDQuest. 2Números superescritos em parênteses estão relacionados à posição de cada peptídeo dentro da sequência de aminoácido da proteína completa, de
acordo com a busca na base de dados usando a ferramenta Mascot.

47
7 CONCLUSÕES
Este estudo fornece a primeira descrição das proteínas diferencialmente expressas no
fluido de folículos pequenos, médios e grandes, colhido in vivo de cabras da raça Canindé
submetidas a estimulação hormonal.
Quatro proteínas (serotransferrina, zinco‐alfa‐2‐glicoproteina-like, proteína do
complemento C3 e fator B do complemento) presentes no FF foram expressas diferencialmente,
aumentando sua quantidade de acordo com o tamanho do folículo.

48
8 PERSPECTIVAS
Finalmente, os resultados obtidos neste estudo apresentam-se como um primeiro passo
na tentativa de melhorar os índices da produção in vitro de embriões em cabras da raça Canindé.

49
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, G. P.; JAISWAL, R.; SINGH, J.; MALHI, P. Progress in understanding ovarian
follicular dynamics in cattle. Theriogenology, v. 69, n. 1, p. 72-80, 2008.
ADASHI, E. Y. Endocrinology of the ovary. Human Reproduction, v. 9, p. 815-827, 1994.
ALPER, C. A.; JOHNSON, A. M.; BIRTCH, A. G.; MOORE, F. D. Human C3: evidence for
the liver as the primary site of synthesis, Science, v. 163, p. 286-288, 1969.
AMBEKAR, A. S.; NIRUJOGI, R. S.; SRIKANTH, S. M.; CHAVAN, S.; KELKAR, D. S.;
HINDUJA, I.; ZAVERI, K.; PRASAD, T. S.; HARSHA, H. C.; PANDEY, A.; MUKHERJEE,
S. Proteomic analysis of human follicular fluid: A new perspective towards understanding
folliculogenesis. Journal of Proteomics, v. 87, p. 68-77, 2013.
ANAHORY, T.; DECHAUD, H.; BENNES, R.; MARIN, P.; LAMB, N. J.; LAOUDJ, D.
Identification of new proteins in follicular fluid of mature human follicles. Electrophoresis, v.
23, p. 1197-1202, 2002.
ANDERSON, D. J.; ABBOTT, A. F.; JACK, R. M. The role of complement component C3b
and its receptors in sperm-oocyte interaction. Proceedings of the National Academy of Sciences,
v. 90, p. 10051-10055, 1993.
ANGELUCCI, S.; CIAVARDELLI, D.; DI GIUSEPPE, F.; ELEUTERIO, E.; SULPIZIO, M.;
TIBONI, G. M.; GIAMPIETRO, F.; PALUMBO, P.; DI ILIO, C. Proteome analysis of human
follicular fluid. Biochimica et Biophysica Acta, v. 1764, p. 1775-1785, 2006.
ARSHAD, H. M.; AHMAD, N.; ZIA-UR-RAHMAN, SAMAD, H. A.; AKHTAR, N.; ALI, S.
Studies on some biochemical constituents of ovarian follicular fluid and peripheral blood in
buffaloes. Pakistan Veterinary Journal, v. 25, p. 189-193, 2005.
ASSOCIATION FOR THE STUDY OF ANIMAL BEHAVIOUR (ASAB). Guidelines for the
treatment of animals in behavioral research and teaching. Animal Behavior, v. 71, p. 245-253,
2006.
BALDASSARRE, H. Aplicaciones prácticas de la produccion de embriones in vitro en la
especie caprina. Revista Brasileira de Reprodução Animal, v. 31, p. 261-267, 2007.
BALDASSARRE, H.; WANG, B.; KAFIDI, N.; KEEFER, C.; LAZARIS, A.; KARATZAS,
C.N. Advances in the production and propagation of transgenic goats using laparoscopic ovum
pick-up and in vitro embryo production technologies. Theriogenology, v. 57, p. 275-284, 2002.

50
BECKER, S.; VON OTTE, S.; ROBENEK, H.; DIEDRICH, K.; NOFER, J. R. Follicular fluid
high-density lipoprotein-associated sphingosine 1-phosphate (S1P) promotes human granulosa
lutein cell migration via S1P receptor type 3and small G-protein RAC1. Biology of
Reproduction, v. 84, p. 604-612, 2011.
BEZERRA, M. B.; RONDINA, D.; LIMA, A. K. F.; OLIVEIRA, L. C.; CECCHI, R.; LUCCI,
C. M.; GIORGETTI, A.; FIGUEIREDO, J. R. Aspectos quantitativos e qualitativos da
foliculogênese na fase pré-natal na espécie caprina. Ciência Animal, v. 8, p. 47-56, 1998.
BIJTTEBIER, J.; TILLEMAN, K.; DHAENENS, M.; DEFORCE, D.; VAN SOOM, A.;
MAES, D. Comparative proteome analysis of porcine follicular fluid and serum reveals that
excessive alpha (2) macroglobulin in serum hampers successful expansion of cumulus-oocyte
complexes. Proteomics, v. 9, p. 4554-4565, 2009.
BRADFORD, M. M. A rapid and sensitive method for the quantization of microgram quantities
of protein utilizing the principle of dye binding. Analytical Biochemistry, v. 72, p. 248-254,
1976.
BRIGGS, D. A.; SHARP, D. J.; MILLER, D.; GOSDEN, R. G. Transferrin in the developing
ovarian follicle: evidence for de-novo expression by granulosa cells. Molecular Human
Reproduction, v. 5, n. 12. p. 1107-14, 1999.
CANDIANO, G.; BRUSCHI, M.; MUSANTE, L.; SANTUCCI, L.; GHIGGERI, G. M.;
CARNEMOLLA, B.; ORECCHIA, P.; ZARDI, L.; RIGHETTI, P. G. Blue silver: A very
sensitive colloidal Coomassie G-250 staining for proteome analysis. Electrophoresis, v. 25, p.
1327-1333, 2004.
CLARKE, H. G.; HOPE, S. A.; BYERS, S.; RODGERS, R. J. Formation of ovarian follicular
fluid may be due to the osmotic potential of large glycosaminoglycans and proteoglycans.
Reproduction, v. 132, p. 119-131, 2006.
COGNIÉ, Y.; BARIL, G.; POULIN, N.; MERMILLOD, P. Current status of embryo
technologies in sheep and goat. Theriogenology, v. 59, p. 171-188, 2003.
CORTVRINDT, R.; SMITZ, J. In vitro follicle growth: Achievements in mammalian species.
Reproduction in Domestic Animals, v. 36, p. 3-9, 2001.

51
CRUZ, J. F.; RONDINA, D.; FREITAS, V. J. F. Ovarian follicular dynamics during anoestrus
in Anglo-nubian and Saanen goats raised in tropical climate. Tropical Animal Health and
Production, v. 37, p. 395-402, 2005.
CURTIS, P.; KEITH, E.; BURFORD, G.; SHAW, R. W.; AMSO, N. Assessment of the
relevance of zona pellucida antibodies in serum and cervical mucus in patients who have
fertilization failure during in vitro fertilization. Fertility and Sterility, v. 56, p. 1124–1127,
1991.
DE JONG, G.; VAN DIJK, J. P.; VAN EIJK, H. G. The biology of transferrin. Clinica Chimica
Acta, v. 190, p. 1-46, 1990.
DESJARDINS, C.; KIRTON, K. T.; HAFS, H. D. Some chemical, immunochemical and
electrophoretic properties of bovine follicular fluid. Reproduction, v. 11, p. 237-244, 1966.
DRIANCOURT, M. A. Regulation of ovarian follicular dynamics in farm animals: Implications
for manipulation of reproduction. Theriogenology, v. 55, p. 1211-1239, 2001.
EDWARDS, R. G. Follicular fluid. Journal of Reproduction & Infertility, v. 37, p. 189-219,
1974.
EGITO, A. A., MARIANTE, A. S. E.; ALBUQUERQUE, M. S. M. Programa brasileiro de
conservação de recursos genéticos animais. Archivos de Zootecnia, v. 51, p. 39-52, 2002.
EMBRAPA-CNPC. Documentos, 23. In: SEMANA DA CAPRINOCULTURA E DA
OVINOCULTURA TROPICAL BRASILEIRA, 1., Sobral, 1994. Anais da 1 Semana da
Caprinocultura e da Ovinocultura Tropical Brasileira. Brasilia, DF, EMERAPA-SPI, 1994. 154
p.
ESTES, S. J.; YE, B.; QIU, W.; CRAMER, D.; HORNSTEIN, M. D.; MISSMER, S. A. A
proteomic analysis of IVF follicular fluid in women ≤32 years old. Fertility and Sterility, v.
92, n. 5, p. 1569-1578, 2009.
FAHIMINIYA, S.; LABAS, V.; DACHEUX, J. L.; GÉRARD, N. Improvement of 2D-PAGE
resolution of human, porcine and equine follicular fluid by means of hexapeptide ligand library.
Reproduction in Domestic Animals, v. 46, p. 561-563, 2011.
FAHIMINIYA, S.; REYNAUD, K.; LABAS, V.; BATARD, S.; CHASTANT-MAILLARD,
S.; GERARD, N. Steroid hormones content and proteomic analysis of canine follicular fluid
during the preovulatory period. Reproductive Biology and Endocrinology, v. 8, p. 1-14, 2010.

52
FATET, A.; PELLICER-RUBIO, M. T.; LEBOEUF, B. Reproductive cycle of goats. Animal
Reproduction Science, v. 124, p. 211-219, 2011.
FIGUEIREDO, J. R.; RODRIGUES, A. P. R.; AMORIM, C. A.; SILVA, J. R. V. Manipulação
de oócitos inclusos em folículos ovarianos pré-antrais - MOIFOPA. In: Gonçalves, P. B. D.;
Figueiredo, J. R.; Freitas, V. J. F. Biotécnicas aplicadas à reprodução animal. São Paulo: Roca,
2ª ed., 2008. p. 303-327.
FONSECA, J. F. Estratégias para o controle do ciclo estral e superovulação em ovinos e
caprinos. In: CONGRESSO BRASILEIRO DE REPRODUÇÃO ANIMAL, 16., 2005,
Goiânia. Anais: Palestras...Goiânia, 2005.
FORTUNE, J. E. Ovarian follicular growth and development in mammals. Biology of
Reproduction, v. 50, p. 225-232, 1994.
FREITAS, V. J. F.; LOPES-JUNIOR, E. S.; RONDINA, D.; SALMITO-VANDERLEY, C. S.
B.; SALLES, H. O.; SIMPLICIO, A. A.; BARIL, G.; SAUMANDE, J. Puberty in Anglo-
Nubian and Saanen female kids raised in the semi-arid of North-eastern Brazil. Small Ruminant
Research, v. 53, p. 167-172, 2004.
GERARD, N.; DUCHAMP, G.; GOUDET, G.; BEZARD, J.; MAGISTRINI, M.; PALMER,
E. A high molecular-weight preovulatory stage-related protein in equine follicular fluid and
granulosa cells. Biology of Reproduction, v. 58, p. 551-557, 1998.
GINTHER, O. J.; KOT, K. Follicular dynamics during the ovulatory season in goats.
Theriogenology, v. 42, p. 987-1001, 1994.
GÓES FILHO, L. S. Caracterização e estudos cinéticos de albumina tratada com espécies
reativas derivadas de óxidos de nitrogênio: espectroscopia de absorção e fluorescência. 2005.
84 f. Dissertação (Mestrado em Física) – Pontifícia Universidade Católica do Rio de Janeiro,
Rio de Janeiro. 2005.
GONZALES, J.; LESOURD, S.; VAN DREDEN, P.; RICHARD, P.; LEFEBVRE, G.;
BROUZES, D. V. Protein composition of follicular fluid and oocyte cleavage occurrence in
vitro fertilization (IVF). Journal of Assisted Reproduction and Genetics, v. 9, p. 211-216, 1992.
GOSDEN, R. G.; HUNTER, R. H.; TELFER, E.; TORRANCE, C.; BROWN, N. Physiological
factors underlying the formation of ovarian follicular fluid. Journal of Reproduction and
Fertility, v. 82, p. 813-825, 1988.

53
GOTTARDI, F. P.; MINGOTI, G. Z. Maturação de oócitos bovinos e influência na aquisição
da competência para o desenvolvimento do embrião. Revista Brasileira de Reprodução Animal,
v. 33, n. 2, p. 82-94, 2009.
GRANADOS, L. B. C. Aspectos gerais na reprodução de caprinos e ovinos. 1. ed. Campos dos
Goytacazes: Projeto PROEX/UENF, 2006. 54p.
GUTIÉRREZ, J. P.; ALTARRIBA, J.; DÍAZ, C.; QUINTANILLA, R.; CAÑÓN, J.;
PIEDRAFITA. J. Pedigree analysis of eight Spanish beef cattle breeds. Genetics, Selection and
Evolution, v. 35, p. 43-63, 2003.
HANRIEDER, J.; NYAKAS, A.; NAESSEN, T.; BERGQUIST, J. Proteomic analysis of
human follicular fluid using an alternative bottom-up approach. Proteome Research, v. 7, p.
443-449, 2008.
HASHEMITABAR, M.; BAHMANZADEH, M.; MOSTAFAIE, A.; ORAZIZADEH, M.;
FARIMANI, M.; NIKBAKHT, R. A Proteomic analysis of human follicular fluid: comparison
between younger and older women with normal FSH levels. International Journal of Molecular
Sciences, v. 15, p. 17518-17540, 2014.
HESS, K. A.; CHEN, L.; LARSEN, W. J. The ovarian blood follicle barrier is both charge- and
size-selective in mice. Biology of Reproduction, v. 58, p. 705-11, 1998.
HILLIER, S. G. Paracrine support of ovarian stimulation. Molecular Human Reproduction, v.
15, p. 843-850, 2009.
HOLT, W. V.; BENNETT, P. M.; VOLOBOUEV, V. Genetic resource banks in wildlife
conservation. Journal of Zoology, v. 238, p. 531-544, 1996.
IBGE, 2013. Instituto Brasileiro de Geografia e Estatística. Produção da pecuária municipal.
http://www.ibge.gov.br.
JARKOVSKA, K.; MARTINKOVA, J.; LISKOVA, L.; HALADA, P.; MOOS, J.; REZABEK,
K.; GADHER, S.; KOVAROVA, H. Proteome mining of human follicular fluid reveals a
crucial role of complement cascade and key biological pathways in women undergoing in vitro
fertilization. Journal of Proteome Research, v. 9, p. 1289-1301, 2010.

54
JIN, S. Y.; LEI, L.; SHIKANOV, A.; SHEA, L. D.; WOODRUFF, T. K. A novel two-step
strategy for in vitro culture of early-stageovarian follicles in the mouse. Fertility and Sterility,
v. 93, p. 2633-2639, 2010.
KHAN, M.H., AGARWAL, S.K., MAJUMDAR, A.C., HOQUE, M., SHANKAR, U. Effect
of exogenous administration of insulin on follicular dynamics in goat. The Indian Journal of
Animal Sciences, v.75, p.1140-1143, 2005.
LEE, Y. L.; CHEONG, A. W. Y.; LEE, W. N. K. F.; YEUNG, W. S. B. Regulation of
complement-3 protein expression in human and mouse oviducts. Molecular Reproduction and
Development, v. 76, p. 301-308, 2009.
LONERGAN, P.; FAIR, T. The ART of studying early embryo development: progress and
challenges in ruminant embryo culture. Theriogenology, v. 81, p. 49-55, 2014.
MADRUGA, M. S.; TORRES, T. S.; CARVALHO, F. F.; QUEIROGA, R. C.; NARAIN, N.;
GARRUTTI E, D.; SOUZA NETO, M. A.; MATTOS, C. W.; COSTA, R. G. Meat quality of
Moxotó and Canindé goats as affected by two levels of feeding. Meat Science, v. 80, p. 1019-
1023, 2008.
MANARANG-PANGAN, S.; MENGE, A. C. Immunologic studies on human follicular fluid.
Fertility and Sterility, v. 22, p. 367-372, 1971.
MANIWA, J.; IZUMI, S.; ISOBE, N.; TERADA, T. Studies on substantially increased proteins
in follicular fluid of bovine ovarian follicular cysts using 2-D PAGE and MALDI-TOF MS.
Reproductive Biology and Endocrinology, v. 3, p. 1-9, 2005.
MARIANTE, A. S.; ALBUQUERQUE, M. S. M.; EGITO, A. A.; MCMANUS, C.; LOPES,
M. A.; PAIVA, S. R. Present status of the conservation of livestock genetic resources in Brazil.
Livestock Science, v. 120, p. 204-212, 2009.
MARÍN-BRIGGILER, C. I.; VAZQUEZ-LEVIN, M. H.; GONZALEZ-ECHEVERRÍA, F.;
BLAQUIER, J. A.; MIRANDA, P. V.; TEZÓN, J. G. Effect of Antisperm Antibodies Present
in Human Follicular Fluid upon the Acrosome Reaction and Sperm–Zona pellucida Interaction.
American Journal of Reproductive Immunology, v. 50, p. 209-219, 2003.
MEHTA, T. S.; KIESSLING, A. A. Development potential of mouse embryos conceived in
vitro and cultured inethylenediaminetetraacetic acid with or without amino acids or serum.
Biology of Reproduction, v. 43, p. 600-606, 1990.

55
MERTON, J. S.; ROOS, A. P. W.; MULLAART, E.; RUIGH, L.; KAAL, L.; VOS, P. L. A.
M.; DIELEMAN, S. J. Factors affecting oocyte quality and quantity in commercial application
of embryo technologies in the cattle breeding industry. Theriogenology, v. 59, p. 651-674, 2003.
MESQUITA JÚNIOR, D.; ARAÚJO, J. A. P.; CATELAN, T. T. T.; SOUZA, A. W. S.;
CRUVINEL, W. M.; ANDRADE, L. E. C.; SILVA, N. P. Sistema Imunitário – Parte II
Fundamentos da resposta imunológica mediada por linfócitos T e B. Revista Brasileira de
Reumatologia, v. 50, n. 5, p. 552-580, 2010.
MONNIAUX, D.; CARATY, A.; CLEMENT, F.; DALBIES-TRAN, R.; DUPONT, J.;
FABRE, S.; GÉRARD, N.; MERMILLOD, P.; MONGET, P.; UZBEKOVA, S.
Développement folliculaire ovarien et ovulation chez les mammifères. Inra Productions
Animales, v. 22, p. 59-76, 2009.
MOOS, J.; FILOVA, V.; PAVELKOVA, J.; MOOSOVA, M.; PEKNICOVA, J.; REZABEK,
K. Follicular fluid and serum levels of inhibin A and pregnancy-associated plasma protein A in
patients undergoing IVF. Fertility and Sterility, v. 91, p. 1739-1744, 2009.
MORAES, J. C. F.; SOUZA, C. J. H.; GONÇALVES, P. B. D.; FREITAS, V. J. F.; LOPES
JÚNIOR, E. S. Controle do estro e da ovulação em ruminantes. In: Gonçalves, P. B. D;
Figueiredo, J. R.; Freitas, V. J. F. Biotécnicas aplicadas à reprodução animal. São Paulo: Roca,
2ª ed., 2008. p. 303-327.
MOROTTI, F.; SANCHES, B. V.; PONTES, J. H.; BASSO, A. C.; SIQUEIRA, E. R.;
LISBOA, L. A.; SENEDA, M. M. Pregnancy rate and birth rate of calves from a large-scale
IVF program using reverse-sorted semen in Bos indicus, Bos indicus-taurus, and Bos taurus
cattle. Theriogenology, v. 81, p. 696-701, 2014.
MORTARINO, M.; VIGO, D., MAFFEO, G.; RONCHI, S. Two-dimensional polyacrylamide
gel electrophoresis map of bovine ovarian fluid proteins. Electrophoresis, v. 20, p. 866-869,
1999.
MOURA, A. A.; KOC, H.; CHAPMAN, D. A.; KILLIAN G. J. Identification of accessory sex
gland fluid proteins as related to fertility indexes of dairy bulls: a proteomic approach. Journal
of Andrology, v. 27, p. 201-211, 2006.
O'FARREL, P. H. High resolution two-dimensional electrophoresis of proteins. Journal of
Biological Chemistry, v. 250, p. 4007-4021, 1975.
OTSUKA, F. Multiple endocrine regulation by bone morphogenetic protein system. Endocrine
Journal, v. 57, p. 3-14, 2010.

56
OTSUKI, J.; NAGAI, Y.; MATSUYAMA, Y.; TERADA, T.; ERA, S. The influence of the
redox state of follicular fuid albumin on the viability of aspirated human oocytes. Systems
Biology Reproductive Medicine, v. 58, p. 149-153, 2012.
PARAMIO, M. T.; IZQUIERDO, D. Current status of in vitro embryo production in sheep and
goats. Reproduction in Domestic Animals, v. 49, p. 37-48, 2014.
POWELL, M. D.; MANANDHAR, G.; SPATE, L.; SUTOVSKY, M.; ZIMMERMAN, S.;
SACHDEV, S. C.; HANNINK, M.; PRATHER, R. S.; SUTOVSKY, P. Discovery of putative
oocyte quality markers by comparative ExacTag proteomics. Proteomics, v. 4, p. 337-351,
2010.
RHIM, S. H.; MILLAR, S. E.; ROBEY, F.; LUO, A-M.; LOU, Y-H.; YULE, T.; ALLEN, P.;
DEAN, J.; TUNG, K. S. K. Autoimmune disease of the ovary induced by a ZP3 peptide from
the mouse zona pellucida. The Journal of Clinical Investigation, v. 89, p. 28-35, 1992.
RICHARDS, J. S. Hormonal control of gene expression in the ovary. Endocrinology Review,
v. 15, p. 725-751, 1994.
ROCHA, Bianca do Prado Lima Petrucci. Perfil protéico do fluido folicular durante a
foliculogênese da égua. 2014, 78 p. Tese (Doutorado em reprodução animal) - Universidade
Federal do Rio Grande do Sul, Porto Alegre, 2014.
ROMAGUERA, R.; MOLL, X.; MORATÓ, R.; ROURA, M.; PALOMO, M. J.; CATALÁ, M.
G.; JIMÉNEZ-MACEDO, A. R.; HAMMAMI, S.; IZQUIERDO, D.; MOGAS, T.; PARAMIO,
M. T. Prepubertal goat oocytes from large follicles result in similar blastocyst production and
embryoploidy than those from adult goats. Theriogenology, v. 76, p. 1-11, 2011.
RUSSELL, S. T.; ZIMMERMAN, T. P.; DOMIN, B. A.; TISDALE, M. J. Induction of lipolysis
in vitro and loss of body fat in vivo by zinc-alpha2-glycoprotein. Biochimica et Biophysica
Acta, v. 1636, p. 59-68. 2004.
SCHAEFFER, E.; LUCERO, M. A.; JELTSCH, J. M.; PY, M. C.; LEVIN, M. J.; CHAMBON,
P.; COHEN, G. N.; ZAKIN, M. M. Complete structure of the human transferrin gene.
Comparison with analogous chicken gene and human pseudogene. Gene, v. 56, p. 109-116,
1987.
SCHROEDER, H. W.; CAVACINI, L. Structure and function of immunoglobulins. Journal of
Allergy and Clinical Immunology, v. 125, n. 2, p. S41-S52, 2010.

57
SHAMAY, A.; HOMANS, R.; FUERMAN, Y.; LEVIN, I.; BARASH, H.; SILANIKOVE, N.;
MABJEESH, S. J. Expression of albumin in nonhepatic tissues and its synthesis by the bovine
mammary gland. Journal of Dairy Science, v. 88, n. 2, p. 569-576, 2005.
SNEL, B.; LEHMANN, G.; BORK, P. String: a web-server to retrieve and display the
repeatedly occurring neighborhood of a gene. Nucleic Acids Research, v. 28, p. 3442-3444,
2000.
SOUSA, F.C.; MELO, C.H.S.; TELES FILHO, A.C.A.; AVELAR, S.R.G.; MOURA, A.A.A.;
MARTINS, J.A.M.; FREITAS, V.J.F.; TEIXEIRA, D.I.A. Ovarian follicular response to
different hormonal stimulation treatments in Canindé goats. Animal Reproduction Science,
v.125, p.88-93, 2011.
SPITZER, D.; MURACH, K. F.; LOTTSPEICH, F.; STAUDACH, A.; ILLMENSEE, K.
Different protein patterns derived from follicular fluid of mature and immature human follicles.
Human Reproduction, v. 11, p. 798-807, 1996.
SUN, Y. L.; PING, Z. G.; LI, C. J.; SUN, Y. F.; YI, K. L.; CHEN, L.; LI, X. Y.; WANG, X.
L.; ZHOU, X. Comparative proteomic analysis of follicular fluids from normal and cystic
follicles in sows. Reproduction in Domestic Animals, v. 46, p. 889-895, 2011.
TILLY, J. L. Molecular and genetic basis of normal and toxicant-induced apoptosis in female
germ cells. Toxicology Letters, v. 102-103, p. 497-501, 1998.
VACCARI, S.; WEEKS, J. L.; HSIEH, M.; MENNITI, F. S.; CONTI, M. Cyclic GMP
signaling is involved in the luteinizing hormone-dependent meiotic maturation of mouse
oocytes. Biology of Reproduction, v. 81, p. 595-604, 2009.
VAN DEN HURK, R.; ZHAO, J. Formation of mammalian oocytes and their growth,
differentiation and maturation within ovarian follicles. Theriogenology, v. 63, p. 1717-1751,
2005.
VAN TILBURG, M. F.; RODRIGUES, M. A.; MOREIRA, R. A.; MORENO, F. B.;
MONTEIRO-MOREIRA, A. C.; CÂNDIDO, M. J.; MOURA, A. A. Membrane-associated
proteins of ejaculated sperm from Morada Nova rams. Theriogenology, v. 79, p. 1247-1261,
2013.

58
VIK, D. P.; AMIGUET, P.; MOFFAT, G. J.; FEY, M.; AMIGUET-BARRAS, F.; WETSEL,
R. A. Structural features of the human C3 gene: íntron/exon organization, transcriptional start
site, and promoter region sequence. Biochemistry, v. 30, p. 1080-1085, 1991.
VIRANT-KLUN, I.; KRIJGSVELD, J. Proteomes of animal oocytes: what can we learn for
human oocytes in the in vitro fertilization programme? BioMed Research International, article
ID 856907, 11 pages, doi:10.1155/2014/856907, 2014.
VON OTTE, S.; PALETTA, J. R.; BECKER, S.; KÖNIG, S.; FOBKER, M.; GREB, R. R.;
KIESEL, L.; ASSMANN, G.; DIEDRICH, K.; NOFER, J. R. Follicular fluid high density
lipoprotein-associated sphingosine 1-phosphate is a novel mediator of ovarian angiogenesis.
Journal of Biological Chemistry, v. 281, p. 5398-5405, 2006.
VON WALD, T.; MONISOVA, Y.; HACKER, M. R.; YOO, S. W.; PENZIAS, A. S.;
REINDOLLAR, R. R.; USHEVA, A. Age-related variations in follicular apolipoproteins may
influence human oocyte maturation and fertility potential. Fertility and Sterility, v. 93, p. 2354-
2361, 2010.
WATSON, A. J. Oocyte cytoplasmic maturation: A key mediator of oocyte and embryo
developmental competence. Journal of animal science, v. 85, p. E1-3, 2007.
YOO, S. W.; BOLBOT, T.; KOULOVA, A.; SNEERINGER, R.; HUMM, K.; DAGON, Y.;
USHEVA, A. Complement factors are secreted in human follicular fluid by granulosa cells and
are possible oocyte maturation factors. Journal of Obstetrics and Gynaecology Research, v. 39,
p. 522-527, 2013.
ZAMAH, A. M.; HASSIS, M. E.; ALBERTOLLE, M. E.; WILLIAMS, K. E. Proteomic
analysis of human follicular fluid from fertile women. Clinical Proteomics, v. 5, n. 1, p. 1-12,
2015.





























