Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO UNIVERSIDADE FEDERAL DO CEARÁ
PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
NÍVEIS DE METIONINA+CISTINA PARA GALINHAS POEDEIRAS LEVES NAS
FASES DE RECRIA E POSTURA
MARCELO HELDER MEDEIROS SANTANA
AREIA – PB
FEVEREIRO - 2014

ii
UNIVERSIDADE FEDERAL DA PARAÍBA
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
UNIVERSIDADE FEDERAL DO CEARÁ
PROGRAMA DE DOUTORADO INTEGRADO EM ZOOTECNIA
NÍVEIS DE METIONINA+CISTINA PARA GALINHAS POEDEIRAS LEVES NAS
FASES DE RECRIA E POSTURA
MARCELO HELDER MEDEIROS SANTANA
Zootecnista
AREIA – PB
FEVEREIRO - 2014

iii
MARCELO HELDER MEDEIROS SANTANA
NÍVEIS DE METIONINA+CISTINA PARA GALINHAS POEDEIRAS
LEVES NAS FASES DE RECRIA E POSTURA
Tese apresentada ao Programa de
Doutorado Integrado em Zootecnia da
Universidade Federal da Paraíba,
Universidade Federal Rural de
Pernambuco e Universidade Federal do
Ceará, como requisito parcial para
obtenção do título de Doutor em
Zootecnia.
Comitê de Orientação:
Prof. Dr. Fernando Guilherme Perazzo Costa – Orientador Principal
Prof. Dra. Patrícia Emília Naves Givisiez
Prof. Dr. Ricardo Romão Guerra
AREIA – PB
FEVEREIRO – 2014

iv
Ficha Catalográfica Elaborada na Seção de Processos Técnicos da Biblioteca Setorial do CCA, UFPB, Campus II, Areia – PB.
S232n Santana, Marcelo Helder Medeiros.
Níveis de metionia+cistina para galinhas poedeiras leves nas fases de recria e postura / Marcelo Helder Medeiros Santana. - Areia: UFPB/CCA, 2014.
127f. : il. Tese (Doutorado em Zootecnia) - Centro de Ciências Agrárias. Universidade Federal da Paraíba, Areia, 2014.
Bibliografia.
Orientadores: Fernando Guilherme Perazzo Costa
1. Avicultura 2. Nutrição animal 3. Galinhas poedeiras -
alimentação I. Costa, Fernando Guilherme Perazzo (Orientador) II.
Título.
UFPB/CCA CDU: 636.5(043.2)

v

vi
DADOS CURRICULARES DO AUTOR
MARCELO HELDER MEDEIROS SANTANA – Nascido em 18 de fevereiro de
1986 na cidade de Campina Grande – PB. Cursou o ensino fundamental e médio nas escolas,
Carl Rogers e Colibri Athenas, na cidade de João Pessoa – PB, concluindo o período escolar
no ano de 2003. No ano seguinte ingressou no Bacharelado em Zootecnia da Universidade
Federal da Paraíba, Campus II, Areia – PB, sendo monitor da disciplina de Genética Animal
durante o ano de 2007 e bolsista PIBIC/CNPq no ano de 2008, graduando-se no início do ano
de 2009. No mesmo ano ingressou no curso de Mestrado do Programa de Pós-Graduação em
Zootecnia da UFPB, Areia – PB, concentrando seus estudos na área de Produção e Nutrição
de Não Ruminantes, obtendo o título de Mestre em Zootecnia em 25 de Fevereiro de 2011.
Em Março de 2011, ingressou no curso de Doutorado em Zootecnia no Programa de
Doutorado Integrado em Zootecnia, UFPB/UFRPE/UFC. Em fevereiro de 2014, submeteu-se
à defesa da Tese para obtenção do título de Doutor em Zootecnia.

vii
Aos meus pais, familiares e amigos
À minha esposa
DEDICO

viii
AGRADECIMENTOS
A Deus, que nos mostra os caminhos a percorrer na vida, nos conforta nos
momentos difíceis e dá sentido a todas as coisas da vida.
Aos meus familiares, em especial os meus pais, irmãos, tios, primos e avó, pela
dedicação na minha educação, ensinamentos e estímulo ao estudo, ao longo dos anos
na minha carreira.
A minha querida esposa Ana Maria Medeiros de A. Santana por todo amor
cedido, que me tranquiliza, me conforta e que me dá forças pra seguir em frente e
enfrentar todas as barreiras da vida.
Aos meus professores e mestres do Ensino Médio, em especial “Tia Leila”,
“Tia Edilma” e “Tia Diva”, por terem iniciado e embasado os meus primeiros passos
na minha carreira profissional, os meus sinceros agradecimentos.
Ao Centro de Ciências Agrárias, da Universidade Federal da Paraíba, por
oferecer toda a estrutura para a minha formação, desde a graduação ao doutorado.
A todos os meus professores da universidade, em especial os meus
orientadores, Professora Patrícia Emília Naves Givisiez e o Professor Fernando
Guilherme Perazzo Costa, pelos ensinamentos, confiança e oportunidades oferecidas
ao longo da minha carreira acadêmica.
Ao Programa de Pós-Graduação em Zootecnia e ao Programa de Doutorado
Integrado em Zootecnia, da Universidade Federal da Paraíba, por toda a estrutura
física e acadêmica, durante a minha formação na pós-graduação.
A CAPES, Adisseo Animal Nutrition e Granja Planalto, pelo apoio a realização
da pesquisa.
Aos membros do Grupo de Estudos em Tecnologias Avícolas (GETA), pelo
companheirismo, ajuda e força para a realização dos trabalhos que envolveram esta
pesquisa.
Aos funcionários do aviário, em especial Josa e Ramalho, pela dedicação e
ajuda para a realização da pesquisa.

ix
Aos meus amigos de universidade, em especial Pessoa, Élcio, Denise (Tuca),
Danielly, Sarah, Rafael e Bruno, pela amizade construída e pela força e dedicação nos
momentos mais difíceis da minha formação.
Aos meus amigos de infância, em especial Luis, Euriko, Renata, Ernani,
Daniel, Paulo Vitor (PV), Larissa, Guilherme, Gabriel, dentre outros, pelo
companheirismo e amizade, que me dá forças e que, apesar da distância, se mantêm
forte e unida.
Aos meus novos colegas de trabalho do IFAC – Sena Madureira, pela força e
companheirismo demonstrados nesse pouco tempo de convivência.
A todas as pessoas que participaram direta ou indiretamente da construção
desse sonho realizado, meus sinceros agradecimentos a todos.

x
SUMÁRIO
LISTA DE TABELAS .................................................................................................. x
LISTA DE FIGURAS................................................................................................. xii
RESUMO GERAL ...................................................................................................... xiv
GENERAL ABSTRACT............................................................................................. xvi
CONSIDERAÇÕES INICIAIS ...................................................................................... 1
CAPÍTULO 1 ................................................................................................................ 3
Referencial teórico .............................................................................................. 3
Introdução ........................................................................................................... 4
Metabolismo de aminoácidos sulfurosos ............................................................ 6
Influência dos níveis de metionina + cistina sobre o desempenho de galinhas
poedeiras na fase de crescimento e postura .................................................................... 9
Influência dos níveis de metionina + cistina sobre a atividade imunológica de
galinhas poedeiras ........................................................................................................ 18
Considerações Finais ........................................................................................ 23
Referências Bibliográficas ................................................................................ 25
CAPÍTULO 2 .............................................................................................................. 33
Níveis de metionina + cistina para galinhas poedeiras leves na fase de recria 33
Resumo ............................................................................................................. 34
Abstract ............................................................................................................. 36
Introdução ......................................................................................................... 38
Material e métodos ........................................................................................... 39
Resultados e discussão ...................................................................................... 44
Conclusões ........................................................................................................ 70
CAPÍTULO 3 .............................................................................................................. 76
Níveis de metionina + cistina para poedeiras leves na fase de postura ............ 76
Resumo ............................................................................................................. 77
Abstract ............................................................................................................. 79
Introdução ......................................................................................................... 80

xi
Material e métodos ........................................................................................... 81
Resultados e discussão ...................................................................................... 85
Conclusões ...................................................................................................... 106
Referências bibliográficas .............................................................................. 107
LISTA DE TABELAS
Capítulo 1 – Referencial Teórico: Níveis de metionina + cistina para poedeiras
leves nas fases de recria e postura
Tabela 1. Composição de aminoácidos essenciais do ovo em pó (mg de
aminoácido/g de proteína) ........................................................................................... 15
Capítulo 2 – Níveis de metionina + cistina para poedeiras leves na fase de recria
Tabela 2. Ingredientes e composição química das dietas experimentais .......... 40
Tabela 3. Efeito dos tratamentos sobre o peso vivo final (PF, g/ave), ganho de
peso (GP, g/ave), consumo de ração (CR, g/ave), conversão alimentar (CA, g/g),
consumo de metionina + cistina (CMC, mg/ave/dia) e consumo de lisina (CLIS,
mg/ave/dia), de poedeiras leves de 13 a 18 semanas de idade ..................................... 44
Tabela 4. Propriedades dos modelos não-lineares utilizados na curva de
crescimento de poedeiras leves de 13 a 18 semanas de idade ...................................... 50
Tabela 5. Efeitos dos tratamentos sobre a atividade da albumina (ALB, g/dL),
alanina aminotransferase (ALT, U/L), aspartato aminotransferase (AST, U/L),
creatinina (CRE, mg/dL), gamma-glutamiltransferase (GGT, U/L) e da proteína sérica
(PTN, g/dL), das aves 13 a 18 semanas de idade ......................................................... 52
Tabela 6. Efeitos dos tratamentos sobre o peso relativo (%) do peso sem
vísceras (PSV), fígado (FIG), baço (BAÇO) e gordura celomática (GCE) das aves com
18 semanas de idade ..................................................................................................... 62
Tabela 7. Efeito residual dos tratamentos sobre a idade ao primeiro ovo (IPO,
dias), peso ao primeiro ovo (PPO, g), número de ovos no período (NOP, total de
ovos), número de dias de produção (NDP, dias), taxa de postura (TXP, %), peso médio
dos ovos (PMO, g) e massa de ovos (MO, g/ave/dia) das de 18 a 29 semanas de idade
...................................................................................................................................... 66

xii
Tabela 8. Efeito residual dos tratamentos sobre a produção de ovos (PR, %),
peso do ovo (PO, g), massa de ovos (MO, g/ave/dia), gravidade específica (GE,
g/mL), espessura de casca (ESPC, mm), percentagem de albúmen (ALB, %), gema
(GEM, %) e casca (CASCA, %), das aves de 34 a 50 semanas de idade .................... 67
Tabela 9. Efeito residual dos tratamentos sobre a produção de ovos (PR, %),
peso do ovo (PO, g), massa de ovos (MO, g/ave/dia), percentagem de albúmen (ALB,
%), gema (GEM, %), casca (CASCA, %), gravidade específica (GE, g/mL) e
espessura de casca (ESPC, mm) das aves de 54 a 70 semanas de idade ...................... 69
Capítulo 3 – Níveis de metionina + cistina para poedeiras leves na fase de postura
Tabela 10. Ingredientes e composição química das dietas experimentais ........ 82
Tabela 11. Efeito dos tratamentos sobre o consumo de ração (CR, g/ave/dia),
produção de ovos (PR, %), peso do ovo (PO, g), massa de ovos (MO, g), conversão
alimentar por massa de ovos (g/g), conversão alimentar por dúzia de ovos (kg/dz),
porcentagem de gema (GEM, %), albúmen (ALB, %) e casca (CASCA, %), espessura
da casca (ESPC, mm), gravidade específica (GE, g/mL), consumo de metionina +
cistina (CMC, mg/ave/dia) e consumo de lisina (CLIS, mg/ave/dia) de poedeiras leves
de 34 a 50 semanas de idade......................................................................................... 86
Tabela 12. Efeito dos tratamentos sobre o peso relativo (%) do peso sem
vísceras (PSV), fígado (FIG), baço (BAÇ) e gordura celomática (GCE) das aves com
50 semanas de idade ................................................................................................... 101
Tabela 13. Efeito residual dos tratamentos sobre a produção de ovos (PR, %),
peso do ovo (PO, g), massa de ovos (MO, g/ave/dia), espessura de casca (ESPC, mm),
percentagem de albúmen (ALB, %), gema (GEM, %) e casca (CASCA, %), das aves
de 54 a 70 semanas de idade....................................................................................... 103
LISTA DE FIGURAS
Capítulo 1 – Referencial Teórico: Níveis de metionina + cistina para poedeiras
leves nas fases de recria e postura
Figura 1. Metabolismo de aminoácidos sulfurosos, colina (CHOL) e betaína
(BET) ............................................................................................................................. 8

xiii
Figura 2. Perfil de crescimento ou ganho de peso semanal de frangas de
reposição ....................................................................................................................... 11
Capítulo 2 – Níveis de metionina + cistina para poedeiras leves na fase de recria
Figura 3. Peso vivo final de poedeiras leves com 18 semanas de idade,
alimentadas com níveis de metionina + cistina ............................................................ 46
Figura 4. Ganho de peso de poedeiras leves com 18 semanas de idade,
alimentadas com níveis de metionina + cistina ............................................................ 47
Figura 5. Conversão alimentar de poedeiras leves com 18 semanas de idade,
alimentadas com níveis de metionina + cistina ............................................................ 47
Figura 6. Consumo de metionina + cistina de poedeiras leves com 18 semanas
de idade, alimentadas com níveis de metionina + cistina ............................................. 48
Figura 7. Curva de crescimento de poedeiras leves de 13 a 18 semanas de
idade, alimentadas com níveis de metionina + cistina e ajustada de acordo com o
modelo de Brody .......................................................................................................... 51
Figura 8. Efeito dos tratamentos sobre o nível de albumina (g/dL) de poedeiras
leves com 18 semanas de idade, alimentadas com níveis de metionina + cistina ........ 54
Figura 9. Efeito dos tratamentos sobre o nível de proteína sérica (g/dL) de
poedeiras leves com 18 semanas de idade, alimentadas com níveis de metionina +
cistina ............................................................................................................................ 54
Figura 10. Fotomicrografias do fígado de poedeiras leves com 18 semanas de
idade suplementadas com níveis de metionina + cistina .............................................. 57
Figura 11. Imagens de microscopia eletrônica do intestino de poedeiras leves
com 18 semanas de idade suplementadas com níveis de metionina + cistina .............. 59
Figura 12. Fotomicrografias do magno de poedeiras leves com 18 semanas de
idade suplementadas com níveis de metionina + cistina e imagem de microscopia
eletrônica do magno ..................................................................................................... 61
Figura 13. Efeito dos tratamentos sobre o peso relativo (%) do baço de
poedeiras leves com 18 semanas de idade, alimentadas com níveis de metionina +
cistina ............................................................................................................................ 63
Figura 14. Efeito residual dos tratamentos sobre a porcentagem de gema de
poedeiras leves de 34 a 50 semanas de idade, alimentadas com níveis de metionina +
cistina ............................................................................................................................ 68

xiv
Capítulo 3 - Níveis de metionina + cistina para poedeiras leves na fase de postura
Figura 15. Efeito dos tratamentos sobre a produção de ovos de poedeiras leves
de 34 a 50 semanas de idade, alimentadas com níveis de metionina + cistina............. 90
Figura 16. Efeito dos tratamentos sobre a massa de ovos de poedeiras leves de
34 a 50 semanas de idade, alimentadas com níveis de metionina + cistina ................. 91
Figura 17. Efeito dos tratamentos sobre a conversão alimentar por massa de
ovos de poedeiras leves de 34 a 50 semanas de idade, alimentadas com níveis de
metionina + cistina ....................................................................................................... 91
Figura 18. Efeito dos tratamentos sobre a conversão alimentar por dúzia de
ovos de poedeiras leves de 34 a 50 semanas de idade, alimentadas com níveis de
metionina + cistina ....................................................................................................... 92
Figura 19. Fotomicrografias do fígado de poedeiras leves com 50 semanas de
idade suplementadas com níveis de metionina + cistina .............................................. 95
Figura 20. Fotomicrografias do intestino delgado de poedeiras leves com 50
semanas de idade suplementadas com níveis de metionina + cistina ........................... 97
Figura 21. Fotomicrografias do magno de poedeiras leves com 50 semanas de
idade suplementadas com níveis de metionina + cistina .............................................. 99
Figura 22. Fotomicrografias do útero de poedeiras leves com 50 semanas de
idade suplementadas com níveis de metionina + cistina ............................................ 100
Figura 23. Efeito residual dos tratamentos sobre a produção de ovos de
poedeiras leves de 54 a 70 semanas de idade, alimentadas com níveis de metionina +
cistina .......................................................................................................................... 105

xv
NÍVEIS DE METIONINA+CISTINA PARA GALINHAS POEDEIRAS LEVES
NAS FASES DE RECRIA E POSTURA
RESUMO GERAL
Objetivou-se com esse trabalho avaliar os níveis de metionina + cistina para galinhas
poedeiras leves na fase de recria (13 a 18 semanas de idade) e na fase de postura (34 a
50 semanas de idade), além do efeito residual nas fases subsequentes. Foram
desenvolvidos seis experimentos, divididos em dois períodos experimentais. No
primeiro período 480 aves foram distribuídas em um delineamento inteiramente
casualizado, com seis tratamentos (níveis de metionina + cistina), 10 repetições, com
oito aves por unidade experimental, para a fase de avaliação das dietas experimentais.
Em seguida foi avaliado o efeito residual da fase anterior no período de produção: pré-
postura (18 a 29 semanas), postura I (34 a 50 semanas) e postura II (54 a 70 semanas),
com oito repetições e oito aves por unidade experimental, e seis repetições e seis aves
por parcela, respectivamente. No segundo período 336 aves foram distribuídas em um
delineamento inteiramente casualizado, com seis tratamentos (níveis de metionina +
cistina), 14 repetições e quatro aves por unidade experimental, para a fase de avaliação
das dietas experimentais. Posteriormente foi avaliado o efeito residual da fase anterior
no período da postura II (54 a 70 semanas), com sete repetições e quatro aves por
unidade experimental. Os tratamentos dietéticos consistiram de um tratamento
controle com base nas exigências de metionina + cistina de acordo com o NRC (1994)
e outras cinco dietas com variações dos níveis de aminoácidos sulfurosos digestíveis,
de acordo com as tabelas brasileiras de exigências nutricionais, equivalentes a 80, 90,
100, 110 e 120% das recomendações nutricionais para a fase de recria e postura.
Foram avaliados dados de desempenho zootécnico e qualidade dos ovos, além de
análise sorológica e histopatológica. Para a fase de recria houve efeito linear crescente
para o peso vivo final, ganho de peso e consumo de metionina + cistina, efeito linear
decrescente para conversão alimentar e efeito quadrático para a atividade da albumina,
níveis de proteínas séricas, porcentagem de gema (postura I) e peso relativo do baço.
As exigências de metionina + cistina estimadas para estas características foram de
91,25% (0,361%), 87,40% (0,346%), 107,79% (0,426%) e 100,44% (0,398%) para

xvi
atividade da albumina, nível de proteína sérica, porcentagem de gema e peso relativo
do baço, respectivamente. Para a fase de postura houve efeito quadrático significativo
para produção e massa de ovos, e conversão por massa e por dúzia de ovos, sendo as
exigências de metionina + cistina estimadas para essas características em 103,25%
(0,691%), 102,91% (0,689%), 110,03% (0,737%) e 109,03% (0,730%),
respectivamente. Também houve efeito quadrático significativo para produção de ovos
(postura II), sendo estimada a exigência de metionina + cistina para essa variável em
105,72% (0,708%). Recomenda-se o nível de metionina + cistina digestível superior a
0,475% (120%) para a fase de recria (13 a 18 semanas de idade), correspondente à
relação metionina + cistina: lisina superior a 98% e o nível de 0,737% de metionina +
cistina digestível, com relação metionina + cistina: lisina de 90% e correspondente ao
consumo de 756,53 mg/ave/dia, para a fase de postura (34 a 50 semanas de idade).
Palavras-chave: aminoácidos sulfurados, análise histopatológica, atividade
enzimática, curva de crescimento, exigências nutricionais

xvii
LEVELS OF METHIONINE AND CYSTINE FOR LIGHT LAYING
HENS AT GROWTH AND POSTURE PHASES
GENERAL ABSTRACT
The objective of this study was to evaluate the levels of methionine and cystine for
light laying hens in the growing phase (13 to 18 weeks old) and the laying phase (34 to
50 weeks old), and the residual effect on subsequent phases. Six experiments were
divided into two experimental periods were developed. In the first period 480 birds
were distributed in a completely randomized design with six treatments (levels of
methionine and cystine), 10 replicates of eight birds per experimental unit for the
assessment phase of the experimental diets. Then the residual effect of the previous
phase has been reported in the period of production: pre - laying (18-29 weeks),
posture one (34-50 weeks) and posture two (54-70 weeks), with eight replicates of
eight birds per experimental unit, and six replicates of six birds per plot, respectively.
In the second period 336 birds were distributed in a completely randomized design
with six treatments (levels of methionine and cystine), 14 replicates of four birds per
experimental unit for the assessment phase of the experimental diets. Subsequently the
residual effect of the previous phase has been reported during the posture two (54-70
weeks), with seven replicates of four birds each. Dietary treatments consisted of a
control treatment based on methionine + cystine according to the NRC (1994) and five
other diets with varying levels of digestible sulfur amino acids, according to the
brazilian tables of nutritional requirements, equivalent to 80, 90, 100, 110 and 120%
of the nutritional recommendations for the rearing and laying. Data production
performance and egg quality were assessed, as well as serological and
histopathological analysis. For the growing phase was increased linearly to the final
live weight, weight gain and consumption of methionine + cystine, linear effect on
feed conversion and quadratic effect on the activity of albumin, serum protein levels,
yolk percentage (posture one) and relative weight of spleen. The methionine + cystine
estimated for these traits were 91,25% (0.361%), 87.40% (0.346%), 107.79%

xviii
(0.389%) and 100.44% (0.398%) to albumin, serum protein level, percentage activity
yolk, and relative weight of spleen, respectively. To the laying phase there was a
significant quadratic effect for production and egg mass, and mass dozen eggs
conversion, with the methionine + cystine estimated for these characteristics in
103.25% (0.691%), 102.91% (0.689%), 110.03% (0.737%) and 109.03% (0.730%),
respectively. There was also significant quadratic effect for egg production (posture
II), estimated the methionine + cystine for this variable at 105.72% (0.708%). The
level of methionine + cystine than 0.475% (120%) for the growing phase (13 to 18
weeks old), corresponding to methionine + cystine is recommended: more than 98%
lysine and 0.737% level of methionine + cystine with methionine and cystine : lysine
ratio of 90% and corresponding to the consumption of 756.53 mg/bird/day for the
laying phase (34-50 weeks old ).
Key words: enzymatic activity, growth curve, histopathology analysis, nutritional
requirements, sulfur amino acids

1
CONSIDERAÇÕES INICIAIS
A avicultura de postura passou por diversas transformações nas últimas
décadas, a partir do surgimento de linhagens mais produtivas, aperfeiçoamento de
equipamentos e instalações, aplicação de técnicas de sanidade e prevenção de
enfermidades e grandes avanços relacionados à nutrição e alimentação das aves. Este
último é responsável por grande parte dos custos de produção na avicultura de postura
e o conhecimento das reais exigências nutricionais dos animais e os valores
nutricionais dos alimentos é extremamente necessário.
A proteína da dieta é caracterizada por ser um dos nutrientes que mais onera no
custo final da ração. Na nutrição de galinhas poedeiras os ingredientes proteicos são
fundamentais para que as aves atinjam índices produtivos satisfatórios e sua eficiência
de utilização depende de diversos fatores, como a porcentagem na ração, composição
em aminoácidos, presença de fatores antinutricionais e a digestibilidade.
Por muitos anos a formulação de rações para aves foi baseada nas exigências
de proteína bruta e, dessa forma, superestimando as reais necessidades nutricionais em
aminoácidos. Como consequência disso, o desbalanceamento dos aminoácidos da
dieta era evidente, provocando queda no desempenho das aves e maior excreção de
poluentes no ambiente de criação.
Atualmente, o conceito de proteína ideal, definido como uma mistura de
aminoácidos com disponibilidade total na digestão e metabolismo (Parsons & Baker,
1994), já está bem difundido na formulação de rações para galinhas poedeiras, graças
ao avanço na indústria de produção de aminoácidos sintéticos. Esse crescimento
possibilitou a redução do preço desses produtos, tornando-o mais acessível e com
maior disponibilidade no mercado. A inclusão dos aminoácidos cristalinos nas rações
de poedeiras proporciona um aumento na eficiência da utilização da proteína dietética,
menor inclusão de fontes proteicas vegetais e, consequentemente, menor excreção de
nitrogênio ao ambiente, principalmente em sistemas de produção avícola em larga
escala.

2
A metionina, primeiro aminoácido limitante nas dietas de galinhas poedeiras,
participa de diversas reações metabólicas e é fundamental para a síntese proteica no
organismo. Em aves poedeiras pode interferir de maneira significativa na produção e
na qualidade dos ovos. Além disso, por ser o principal doador de grupos metil para o
organismo dos animais, sua deficiência ou o excesso na dieta pode provocar sérios
prejuízos metabólicos, como lesões hepáticas e renais, menor desenvolvimento
tecidual e redução da síntese enzimática.
A determinação do nível adequado desse aminoácido para galinhas poedeiras é
essencial, pois interfere diretamente nos custos das dietas e no desempenho produtivo
das aves. As exigências nutricionais de metionina+cistina para poedeiras vêm sendo
determinadas ao longo dos últimos anos, no entanto, ainda divergem. Esse fato pode
ser explicado por mudanças significativas nas linhagens, nutrição, idade e condições
ambientais.
Os dados de pesquisa relativos às exigências nutricionais de aminoácidos
sulfurosos para poedeiras no período de recria e pré-postura ainda são escassos e
bastante conflitantes, principalmente por não existir um consenso nas tabelas quanto à
padronização do período de avaliação experimental e do modelo estatístico utilizado
na pesquisa. Aliado a isso, o conhecimento dos requerimentos das aves em
aminoácidos sulfurosos na fase de recria é fundamental para o ciclo de produção de
ovos, já que nesse período ocorre um intenso desenvolvimento dos órgãos
reprodutivos e o fornecimento de rações com níveis inadequados de metionina +
cistina pode provocar sérios prejuízos fisiológicos.
Diante do exposto, objetivou-se determinar as exigências nutricionais de
metionina + cistina digestível para poedeiras leves durante as fases de recria (13 a 18
semanas de idade) e primeiro ciclo de postura (34 a 50 semanas de idade), além de
avaliar os efeitos dos níveis testados nas fases subsequentes.

3
CAPÍTULO 1
Referencial Teórico
Níveis de Metionina+Cistina para Galinhas Poedeiras Leves nas Fases de
Recria e Postura

4
INTRODUÇÃO
O avanço da avicultura industrial alcançado nas últimas décadas está, em
grande parte, alicerçado no estudo do melhoramento genético dos animais. Atualmente
existem no mercado diversas linhagens de frangos corte e de galinhas poedeiras que
apresentam um alto desempenho produtivo e uma excelente eficiência na conversão de
matérias-primas em produtos nobres para a alimentação humana, como a carne e os
ovos de consumo. Entretanto, o balanço inadequado de nutrientes de uma dieta pode
interferir diretamente no desempenho das aves, já que com a evolução genética da ave,
os níveis nutricionais requeridos pelos animais também foi alterado, sendo necessária
a revisão constante das tabelas de exigências nutricionais.
Os custos com alimentação na avicultura ultrapassam os 70% do custo total de
produção, destacando-se os ingredientes proteicos e energéticos. De acordo com Costa
et al. (2012), os recentes aumentos nos custos de produção, principalmente dos
ingredientes utilizados na alimentação de aves, demandam a adoção de estratégias
nutricionais que proporcionem melhor aproveitamento dos nutrientes, com
consequente melhora da eficiência produtiva.
O desenvolvimento da nutrição animal, acompanhado do melhor conhecimento
do metabolismo proteico e melhor avaliação nutricional dos ingredientes,
possibilitaram a otimização das dietas de galinhas poedeiras, estabelecendo os
requerimentos com base em aminoácidos, reduzindo custos e ocasionando em menor
impacto ao meio ambiente.
Com o avanço da produção de aminoácidos sintéticos pela indústria de rações,
foi possível reduzir a inserção de fontes proteicas tradicionais, principalmente o farelo
de soja. Essa técnica permite a formulação de rações com menor custo, com teores de
proteína bruta inferiores aos preconizados pelas tabelas de exigências nutricionais e
atendendo os requerimentos dos animais em aminoácidos. Segundo Moura (2004), o
conhecimento das reais necessidades em aminoácidos essenciais permite evitar
problemas como a redução no consumo de ração, aumento das perdas energéticas por
incremento calórico e excreção excessiva de ácido úrico, pois diminui o excedente de
aminoácidos circulantes no sangue.

5
A proteína ideal, bem definida por Parsons & Baker (1994), é caracterizada
como uma mistura de aminoácidos com disponibilidade total na digestão e
metabolismo e cuja composição seria idêntica às exigências do animal para
manutenção e crescimento. Esses autores ainda relataram que, para ser ideal, uma
proteína ou a combinação destas, devem apresentar todos os aminoácidos em níveis
exatamente requeridos para atender as exigências de manutenção e máxima deposição
de proteína corporal, sem deficiência e sem excesso de aminoácidos.
A proteína corporal é composta por aproximadamente 20 aminoácidos e estes
são requeridos para diversas funções no organismo, como a síntese proteica orgânica,
constituinte de tecidos corporais, imunoglobulinas, hormônios, formação de pele,
penas, músculos, órgãos, além de participarem do metabolismo de outros nutrientes no
organismo. Conceitualmente, os aminoácidos podem ser classificados quanto a sua
capacidade de síntese pelo organismo das aves: os aminoácidos essenciais, que não
podem ser sintetizados pelo organismo ou sua síntese é insuficiente para atender a
demanda metabólica; e os aminoácidos não essenciais, que podem ser sintetizados no
organismo das aves a partir de aminoácidos ou de outros metabólitos.
Dessa forma, a metionina, primeiro aminoácido limitante em rações avícolas, é
o principal doador de grupamentos metil (S-adenosilmetionina) para diversas reações
metabólicas e participa diretamente da síntese proteica (Leeson & Summers, 2001).
Aliado a isto, serve como fonte alternativa de cistina em um processo não-reversível,
e, por esse motivo, segundo Goulart et al. (2011), os requerimentos de metionina e
cistina são usualmente considerados juntos. Esses aminoácidos desempenham um
papel fundamental na estrutura de diversas proteínas e interligando cadeias
polipeptídicas por meio de pontes dissulfeto (Lenningher, 1996).
Como primeiro aminoácido limitante em rações de frangos de corte e
poedeiras, a metionina é caracterizada por interferir diretamente nos índices
produtivos das aves e sua suplementação (aminoácido sintético) resulta em aumento
na eficiência de utilização da proteína. De acordo com Jordão Filho et al. (2006),
rações deficientes em metionina + cistina reduzem a produção e o peso dos ovos, além
de aumentar a deposição de gordura no fígado de poedeiras. A deficiência de
metionina pode causar prejuízos renais e hepáticos (Brumano, 2008), já que as reações

6
de metilação são essenciais no metabolismo da gordura no fígado, evitando assim, a
síndrome do fígado gorduroso.
Assim, a formulação de rações com níveis de aminoácidos sulfurosos em
quantidades requeridas pelas galinhas poedeiras, permite um melhor aproveitamento
do potencial genético das aves e possibilita a produção de alimentos de alta qualidade,
com redução de custos com ingredientes proteicos e melhorando o desempenho
produtivo dos animais.
METABOLISMO DE AMINOÁCIDOS SULFUROSOS
Nas aves a digestão proteica inicia-se no estômago (proventrículo), a partir da
ação de enzimas proteolíticas, ácido clorídrico e pepsinogênio. Este último é
transformado em pepsina (forma ativa) através do conteúdo ácido do estômago (pH
entre 2,0 e 4,0) e pela própria pepsina. As proteínas em meio ácido são desnaturadas e
sofrem a ação direta da pepsina, que hidrolisa as ligações peptídicas entre os
aminoácidos leucina-valina, tirosina-leucina, fenilalanina-tirosina. Em seguida o
alimento vai sendo triturado e digerido, através da motilidade do proventrículo-moela
e as proteínas parcialmente digeridas passam para o duodeno. No intestino, as
proteínas da ingesta sofrem ação de enzimas proteolíticas secretadas pelo pâncreas e
pelo próprio intestino, sendo reduzidos inicialmente para oligo, tri e dipeptídeos e
alguns aminoácidos livres. Finalmente, peptidases de membrana completam a
hidrólise proteica para a absorção de aminoácidos livres pela via porta (Rutz, 2002).
Os principais aminoácidos sulfurosos de relevância para a nutrição animal são
a metionina e a cistina. Esta última é obtida através da oxidação de outro aminoácido
sulfuroso, a cisteína, que faz parte da síntese de enzimas importantes no organismo,
como a ubiquitina ligase, responsável pela síntese de ubiquitina (degradação de
proteínas indesejáveis) e a enzima caspase, fundamental para o processo de apoptose
celular (morte de células que podem desenvolver tumores e doenças). A cisteína é

7
também utilizada para a síntese proteica, formação da pele, penas e pêlos, e sendo
fundamental frente ao estresse e ao status inflamatório (Tesseraud et al., 2008).
Em um processo irreversível, a metionina pode ser uma fonte alternativa de
cistina, apresentando funções importantes nas estruturas de várias proteínas, como
imunoglobulinas e insulina, ligando várias cadeias polipeptídicas via pontes dissulfeto
(Nelson & Cox, 2005). A cistina ainda participa da síntese de glutationa, um
importante antioxidante celular para o organismo dos animais (Tesseraud et al., 2008).
Devido a essa inter-relação, as exigências de metionina e cistina são consideradas
juntas nas tabelas de requerimentos nutricionais para aves.
A metionina é fundamental para o funcionamento normal do organismo de
galinhas poedeiras, pois está envolvida em diversas reações e funções metabólicas
orgânicas, como a participação direta na síntese proteica, em reações de metilação no
organismo, no empenamento, e, além disso, segundo Barbosa et al. (2009), os
aminoácidos sulfurados têm grande importância sobre o tamanho e o peso dos ovos.
Corroborando essa ideia, Pavan et al. (2005) utilizaram três níveis de proteína bruta
(14, 15,5 e 17%) e de aminoácidos sulfurados (0,57, 0,64 e 0,71%) e constataram que
o peso do ovo e a porcentagem de albúmen aumentaram com a elevação dos níveis de
metionina + cistina.
Sob a forma de S-adenosilmetionina, a metionina é exigida para a biossíntese
de várias substâncias envolvidas no crescimento de galinhas poedeiras, como a
cisteína, carnitina, poliaminas, epinefrina, colina e melatonina (Baker et al., 1996). É
também responsável pelo fornecimento de enxofre para a síntese de outros
componentes químicos que apresentam enxofre na sua composição e essencial para a
síntese de taurina, um dos aminoácidos mais abundantes no organismo, que age como
um transmissor neuroinibidor juntamente com a glicina (Wu, 2003).
Outro aspecto que merece destaque no metabolismo da metionina é que esse
aminoácido atua como um agente lipotrópico, através do seu papel como doador de
grupamentos metil ou também pelo envolvimento no metabolismo da colina, betaína,
ácido fólico e da vitamina B12 (Chen et al., 1993). De acordo com Brumano (2008) a
deficiência de metionina pode causar sérios prejuízos renais e hepáticos já que as
reações de metilação são essenciais no metabolismo da gordura no fígado, evitando
assim, a síndrome do fígado gorduroso.

8
Devido à intensa semelhança em algumas funções orgânicas, como a doação de
grupos metil e a ação lipotrófica, há uma forte inter-relação entre a metionina, colina e
betaína. Esta ligação está explicada na Figura 1, na qual a homocisteína, encontrada
num ponto de ramificação do metabolismo de aminoácidos sulfurosos, pode sofrer
transulfuração irreversível à cisteína ou passar pelo processo de remetilação à
metionina via vitamina B12 ou via betaína (dependente de colina) (Pillai et al., 2006).
Figura 1 – Metabolismo de aminoácidos sulfurosos, colina (CHOL) e betaína (BET) (Adaptado
de Pillai et al., 2006).
Os números indicam as enzimas apresentadas a seguir: 1 – Met adenosiltransferase; 2 – Várias enzimas;
3 – S-adenosilhomocisteína hidrolase; 4 – Cistationa β-sintase; 5 – Cistationa γ-liase; 6 – N5, N
10-
metilenotetrahidrofolato redutase; 7 – Met sintase; 8 – Colina desidrogenase; 9 – Betaína aldeído
deidrogenase; 10 – Betaína-Homocisteína metiltransferase.
Devido à intensa relação no metabolismo, a colina e/ou a betaína podem atuar
como fontes poupadoras de metionina, reduzindo os requerimentos nutricionais para
esse aminoácido. Esse efeito poupador está intimamente ligado ao processo de
remetilação da homocisteína à metionina (reciclagem de metionina) no metabolismo

9
de aminoácidos sulfurados (Figura 1), envolvendo enzimas de diversos órgãos, tais
como o fígado, pâncreas e rins.
Entretanto, segundo Pillai et al. (2006) o fluxo de remetilação da homocisteína
varia bastante entre as espécies e está diretamente ligada a atividade da enzima
betaína-homocisteína metil transferase (BHMT). Ainda segundo esses autores,
alterações nos níveis de aminoácidos sulfurados, colina ou betaína resultam em
alterações na atividade dessa enzima, entretanto, o efeito poupador da colina e/ou
betaína sobre o requerimento de metionina ainda não é claro. Pesquisas da década de
80 relatam que cerca de 54% da homocisteína hepática é remetilada à metionina em
mamíferos e que cerca de 38% da homocisteína é remetilada à metionina no fígado de
homens adultos (Finkelstein & Martin, 1984; Storch et al., 1988). Todavia, as
informações sobre o fluxo de remetilação hepática da homocisteína em aves ainda são
escassos.
INFLUÊNCIA DOS NÍVEIS DE METIONINA + CISTINA SOBRE O
DESEMPENHO DE GALINHAS POEDEIRAS NA FASE DE CRESCIMENTO
E POSTURA
Poedeiras em crescimento (frangas de reposição)
A evolução genética alcançada na avicultura moderna proporciona o
surgimento de linhagens cada vez mais produtivas. Esse fato implica na necessidade
em desenvolver dietas balanceadas, a fim de atender com mais precisão os
requerimentos nutricionais das aves. As fases de crescimento e desenvolvimento das
futuras galinhas poedeiras tornam-se essenciais frente a esse desafio, gerado pela
intensa modificação dos materiais genéticos na avicultura de postura.
De acordo com D’Agostini et al. (2012a), a maturidade reprodutiva e o
desempenho produtivo da poedeira moderna estão diretamente relacionados com o
desenvolvimento adequado da ave nas fases anteriores ao período de produção de
ovos. Ainda segundo esses autores, isso ocorre porque fatores que interferem na idade

10
da ave e na maturidade sexual estão diretamente relacionados com o peso corporal e a
uniformidade do lote, alcançados durante as fases iniciais de criação das galinhas
poedeiras.
Ao atingirem a maturidade sexual, as poedeiras devem apresentar peso
corporal satisfatório, de modo a garantir o pleno funcionamento e desenvolvimento
dos órgãos reprodutivos e permitindo a expressão de todo o seu potencial genético.
Segundo Silva et al. (2009), a produção ideal de ovos depende diretamente da forma
como as frangas de reposição foram criadas durante as fases de crescimento, pois o
peso ótimo no início do período produtivo e sua manutenção condicionam o
desempenho da galinha por toda a fase de postura.
A partir do conhecimento da curva de crescimento em aves de reposição, é
possível perceber que cerca de 70% do peso adulto de uma poedeira deve ser atingido
até 12 semanas de idade (Faria & Junqueira, 1998), 82% até 15 semanas e 92% até as
22 semanas de vida, enquanto que os 8% restantes devem ser depositados
posteriormente (Kwakkel, 1992). De acordo com D’Agostini et al. (2012a), aves com
peso abaixo do ideal atrasam o início da postura, reduzindo a produção de ovos; em
contrapartida, aves com peso acima do ideal entram em postura precocemente e
produzem ovos pequenos, prejudicando a rentabilidade do segmento. Da mesma
forma, Leeson & Summers (1997) afirmam que o peso corporal influencia diretamente
a maturidade sexual da ave, ressaltando a importância de otimizar o ganho de peso da
poedeira.
Na Figura 2 encontra-se o perfil de crescimento ou ganho de peso semanal em
frangas de reposição até 24 semanas de idade. De acordo com o esquema proposto por
Bertechini (2012), o crescimento das frangas de reposição segue o modelo fisiológico
multifásico. Na fase de 1 a 5 semanas, ocorre prioridade fisiológica de crescimento
visceral, seguido de crescimento ósseo de 6 a 12 semanas e de 12 a 16 semanas de
idade, o desenvolvimento dos órgãos reprodutivos, com aumento de densidade óssea e
acúmulo de reservas orgânicas para o início de produção.

11
Figura 2 – Perfil de crescimento ou ganho de peso semanal de frangas de reposição (Adaptado
de Bertechini, 2012).
Nesse sentido, a metionina + cistina como primeiro aminoácido limitante para
aves, é fundamental para o desenvolvimento corporal e reprodutivo de galinhas
poedeiras, haja a vista a intensa participação desse aminoácido no processo de síntese
proteica e doação de grupamentos metil no organismo. A intensa modificação do perfil
genético das linhagens de postura fortalece a necessidade de atualizar constantemente
as exigências de metionina + cistina, evitando assim o desequilíbrio aminoacídico da
dieta.
Outro fator relevante é a falta de padronização no período que compreende
cada fase de criação de poedeiras, sendo observado em diversas tabelas diferenças
significativas para estes períodos. Nas tabelas brasileiras de exigências nutricionais
para aves e suínos (Rostagno et al., 2011), as recomendações nutricionais para frangas
de reposição estão divididas em três fases: 1 a 6, 7 a 12 e 13 a 18 semanas de idade. Já
o manual da linhagem Dekalb White (2009) estabelece as exigências para esses
animais para 1 a 5, 6 a 10 e de 11 a 15 semanas de idade. A falta de um consenso entre
as informações contidas nas tabelas de exigências nutricionais para frangas de
reposição pode prejudicar a aplicação de dietas balanceadas para cada fase de criação
dos animais, gerando assim prejuízo econômico aos produtores.

12
De acordo com as tabelas brasileiras de aves e suínos (Rostagno et al., 2011),
as exigências nutricionais de metionina + cistina total para aves de reposição leves é
de 0,710% (0,640% dig.), 0,552% (0,497% dig.) e 0,446% (0,396% dig.), com relação
metionina + cistina/lisina de 73, 80 e 82, para as fases de 1 a 6, 7 a 12 e 13 a 18
semanas de idade, respectivamente. Já para as aves de reposição semipesadas, os
requerimentos de metionina + cistina total para estas fases é de 0,688% (0,619% dig.),
0,543% (0,489% dig.) e 0,431% (0,383% dig.), para as fases de 1 a 6, 7 a 12 e 13 a 18
semanas de idade, respectivamente, com a mesma relação metionina + cistna/lisina
preconizada para aves de reposição leves.
As tabelas oferecidas pelos manuais das linhagens de postura geralmente
apresentam os níveis dos nutrientes recomendados acima do preconizado pelas tabelas
brasileiras de aves e suínos. Por exemplo, o manual da linhagem Dekalb White (2009)
recomenda para frangas de reposição leves os níveis de metionina + cistina digestível
de 0,75%, 0,65% e 0,64%, para as fases de 1 a 5, 6 a 10 e 11 a 15 semanas de idade,
respectivamente.
Algumas pesquisas vêm sendo realizadas ao longo dos últimos anos, com a
finalidade da determinação das exigências nutricionais de metionina + cistina para
poedeiras durante a fase de crescimento, todavia, com resultados ainda contraditórios.
D’Agostini et al. (2012a) determinaram as exigências nutricionais de metionina
+ cistina para frangas (Lohmman LSL e Lohmman Brown) de 13 a 18 semanas de
idade, além dos efeitos residuais durante a fase de postura. Esses autores obsevaram
que os níveis de metionina + cistina oferecidos durante a fase de crescimento da
poedeira não afetaram o desempenho produtivo na fase de postura, além de
estabelecerem as exigências para este aminoácido em 0,679% de metionina + cistina
total (0,611% de met + cis digestível) para aves leves e 0,646% de metionina + cistina
total (0,581% de met + cis digestível) para aves semipesadas.
Este fato implica em afirmar que mesmo recebendo uma dieta deficiente em
metionina + cistina na fase de crescimento, as aves têm a capacidade produtiva
reestabelecida na postura, desde que recebam uma dieta balanceada na fase de
produção, que, segundo Kwakkel et al. (1991), pode ser explicada pela hipótese do
ganho compensatório.

13
Também foi relatado pelos mesmos autores que as frangas de reposição leves
que receberam rações com os menores níveis de metionina + cistina tiveram a postura
retardada e somente atingiram 50% de produção com 161 dias de idade (23 semanas
de idade). Este nível de produção está abaixo do recomendado pelos principais
manuais de criação de linhagens de poedeiras, que preconizam 50% de produção das
aves, em média, entre 21 e 22 semanas de idade.
D’Agostini et al. (2012b) avaliaram a influência dos níveis de metionina +
cistina para frangas de reposição de 7 a 12 semanas de idade, onde observaram
resposta positiva para o aumento no ganho de peso de aves leves e semipesadas, a
partir do aumento da inclusão de metionina + cistina nas rações. Semelhantemente ao
estudo anterior conduzido por D’Agostini et al. (2012a), as frangas semipesadas
apresentaram maior consumo de ração, maior ganho de peso e melhor conversão
alimentar, em comparação com as aves leves, no entanto, a exigência de metionina +
cistina total das aves nesta fase de crescimento foi semelhante para as linhagens
estudadas. O nível recomendado para metionina + cistina total nesse estudo para aves
leves e semipesadas foi de 0,710% (0,639% de met + cis digestível) e de 0,706%
(0,635% de met + cis digestível), respectivamente.
Silva et al. (2009) avaliaram os requerimentos de metionina + cistina para aves
de reposição leves e semipesadas de 1 a 4 semanas de idade, alimentadas com ração
farelada e triturada, e observaram que, independente da linhagem e forma física da
ração, a exigência de metionina + cistina total para as aves nesta fase é de 0,79%, para
um consumo de 146 mg/dia (leves) e 160 mg/dia (semipesadas). Um aspecto relevante
relatado pelos autores é que tanto os menores níveis de aminoácidos sulfurosos,
quanto os mais altos nas dietas das aves, provocaram queda no desempenho produtivo
durante a fase experimental. Outro fator importante observado no estudo foi o
aumento no ganho de penas com a elevação dos níveis de aminoácidos sulfurados nas
dietas experimentais, enfatizando a importância desses aminoácidos com o
empenamento de galinhas poedeiras.
Segundo Baker (1986), as aves mais pesadas exigem maior quantidade de cada
nutriente essencial, em comparação às leves. No entanto, evidências sugerem que as
exigências expressas como porcentagem ou miligrama por quilograma de ração não
variam, ou são inferiores às propostas para poedeiras leves de mesma idade. O que é

14
observado na prática é que as exigências de poedeiras semipesadas são atendidas com
o aumento do consumo de ração.
De acordo com Klasing (1998), a deficiência de um aminoácido essencial se
manifesta na redução do consumo de ração da ave, que torna a deficiência ainda mais
severa. O mesmo acontece a partir da utilização de níveis acima do exigido pela ave
em aminoácidos, tendo influência negativa sobre o consumo de ração, que, segundo
D’Mello (1994), pode provocar sintomas de anorexia em aves que recebem rações
desbalanceadas. D’Agostini et al. (2012b) afirmam que a deficiência de nutrientes nos
estágios iniciais de crescimento de frangas é crítica para o desenvolvimento de tecidos
específicos do corpo da ave, como os músculos, órgãos do trato digestivo e a gordura
no tecido intramuscular.
Poedeiras na fase de produção
O ovo é um alimento completo e rico em proteínas de alto valor biológico,
energia, minerais e vitaminas, além de apresentar baixo custo, quando comparado a
outras fontes alimentares. A comercialização dos ovos de galinhas é realizada em
grande parte sob a forma in natura e em condições de temperatura ambiente, o que
pode comprometer a qualidade do produto. Todavia, nos últimos anos, o
processamento dos ovos vem ganhando mercado, seja na forma líquida ou em pó,
devido a sua praticidade na utilização e no armazenamento.
A indústria vem buscando ao longo dos anos métodos de processamento dos
ovos de galinhas poedeiras, evitando assim perdas significativas durante o processo
produtivo. Devido a esse aumento no processo de industrialização dos ovos de
galinhas poedeiras, os pesquisadores vêm buscando alternativas nutricionais que
permitam melhorar a qualidade do produto, principalmente em relação ao tamanho,
peso e conteúdo de sólidos dos ovos. De acordo com Silva (2006), atualmente as
indústrias exigem ovos com tamanho, peso e frescor bem definidos, além do conteúdo
de sólidos nos componentes dos ovos, que devem ser altos para um maior rendimento
durante o processamento.

15
A metionina, sendo o primeiro aminoácido limitante para aves, em dietas a
base de milho e farelo de soja, tem papel fundamental sobre a produtividade de
galinhas poedeiras leves. Os aminoácidos sulfurosos têm função determinante no
processo de síntese proteica orgânica, além de participarem como os principais
doadores de grupos metil para o organismo e, dessa forma, os níveis desses
aminoácidos nas rações de galinhas poedeiras influenciam tanto de forma quantitativa,
quanto de forma qualitativa a produção de ovos.
Vários fatores podem interferir na qualidade dos ovos de galinhas poedeiras,
como a idade da ave, a genética e, principalmente, a nutrição. Os níveis de proteína e
de aminoácidos presentes na dieta têm influência direta na qualidade e nas
concentrações de proteína ou de sólidos totais dos componentes dos ovos, destacando-
se a metionina lisina e treonina (Shafer et al., 1996; Shafer et al., 1998). Corroborando
essas informações, Sá et al. (2007) e Pavan et al. (2005) afirmam que os níveis de
metionina + cistina podem interferir nos principais parâmetros produtivos em galinhas
poedeiras, como a produção e o peso dos ovos, além das características inerentes à
qualidade interna dos ovos.
Os níveis de aminoácidos essenciais (mg de aminoácido/g de proteína) do ovo
em pó estão descritos na Tabela 1.
Tabela 1 – Composição de aminoácidos essenciais do ovo em pó (mg de aminoácido/g de
proteína)
Aminoácidos essenciais Ovo em pó
Fenilalanina + Tirosina 98,64
Histidina 22,12
Isoleucina 34,64
Leucina 83,9
Lisina 91,44
Metionina + Cistina 40,05
Treonina 53,5
Triptofano nd
Valina 47,52
nd – não determinado
Adaptado de Pires et al. (2006)

16
Como pode ser observado na Tabela 1, a metionina + cistina está presente em
quantidades significativas no conteúdo aminoacídico do ovo em pó, no entanto, outros
aminoácidos estão presentes em níveis superiores aos aminoácidos sulfurosos. Este
fato implica em afirmar que, mesmo não sendo o principal aminoácido presente no
ovo de galinha, a metionina + cistina apresenta papel fundamental para a síntese de
outros aminoácidos, graças a sua intensa participação no metabolismo orgânico para a
síntese proteica.
A importância do perfeito balanço entre os aminoácidos presentes nas dietas de
galinhas poedeiras vem sendo relatado por autores nas pesquisas nos últimos anos.
Pars & Summers (1991) citam que o desbalanço aminoacídico provocado pela
deficiência de aminoácidos sulfurosos causa efeitos negativos sobre o consumo de
alimentos e o desempenho de crescimento em aves de corte. Esse efeito da anorexia
também foi observado por Narváez-Solarte (1996) para poedeiras leves e semipesadas
mantidas com dietas deficientes em metionina + cistina. A elevada deficiência de
metionina na dieta de galinhas poedeiras acarreta na diminuição desse aminoácido na
corrente sanguínea e nesta situação um sinal é enviado ao sistema nervoso central, que
ativa os mecanismos responsáveis pela redução no consumo de alimentos (Austic,
1986).
Da mesma forma, Cao et al. (1995) relataram que a deficiência ou o excesso de
metionina na ração de poedeiras aumenta a degradação de metionina no fígado. No
entanto, de acordo com Koelkebeck et al. (1991), poedeiras de alta produção
apresentam certa tolerância para excessos individuais de aminoácidos em dietas à base
de milho e farelo de soja.
A determinação das exigências de aminoácidos para galinhas poedeiras tem
ocupado espaço significativo na literatura científica, haja vista que atualmente calcular
rações comerciais a partir dos requerimentos dos aminoácidos em vez da proteína
total, se tornou uma prática universal (Sá et al., 2007). Segundo Cupertino et al.
(2009), as exigências nutricionais de metionina + cistina para poedeiras em produção
vêm sendo determinadas ao longo dos anos, embora com pouca analogia entre as
recomendações. Esse fato pode ser explicado pela intensa modificação na genética,
nutrição e no manejo das aves, além dos efeitos da idade, dieta e das condições
ambientais (Waldroup & Hellwig, 1995).

17
De acordo com Jordão Filho et al. (2006), vários fatores podem influenciar os
requerimentos de metionina + cistina, como o teor de proteína da ração, a linhagem, o
ambiente térmico e o teor energético, assim como a presença de fatores
antinutricionais e o processamento da matéria-prima utilizada na ração. Ademais,
segundo Schutte et al. (1994), o nível de metionina + cistina exigido para otimizar a
conversão alimentar é maior que aquele para a maximização da produção de ovos.
Sá et al. (2007) determinaram as exigências nutricionais de aminoácidos
sulfurosos para poedeiras leves e semipesadas durante a fase de postura (34 a 50
semanas de idade). Esses autores observaram efeitos positivos da suplementação de
aminoácidos sulfurados na dieta das aves leves e semipesadas, e recomendaram os
níveis de 0,693 e 0,692% metionina + cistina digestível, respectivamente.
Semelhantemente, Cupertino et al. (2009) avaliaram a exigência nutricional de
metionina + cistina digestíveis para poedeiras leves e semipesadas, de 54 a 70 semanas
de idade e também observaram efeito significativo dos níveis de aminoácidos
sulfurosos sobre as principais variáveis produtivas avaliadas. No entanto, respostas
distintas entre as linhagens foram observadas, sendo que os efeitos da deficiência de
aminoácidos sulfurosos reduziram de forma mais significativa o consumo de ração e
de aminoácidos entre as poedeiras semipesadas, em comparação às aves leves. Os
níveis estimados para aminoácidos sulfurosos digestíveis para galinhas poedeiras leves
e semipesadas foram de 0,645% e 0,655%, correspondente a um consumo de 712 e
723 mg/ave/dia, respectivamente.
Em estudo mais recente, Brumano et al. (2010) avaliaram os níveis de
metionina + cistina digestível para poedeiras leves no período de 42 a 58 semanas de
idade, e observaram aumento no peso dos ovos, a partir da suplementação com
aminoácidos sulfurosos na dieta das galinhas poedeiras. Segundo os autores, essa
elevação no peso médio dos ovos é reflexo do aumento do peso da gema e do
albúmen, que variaram de 15,4 a 17,5 e de 37,8 a 41,4g, respectivamente, além do
aumento da porcentagem desses componentes no ovo.
Esses resultados são semelhantes aos encontrados por Barbosa et al. (2009),
que também observaram aumento linear crescente no peso dos ovos de poedeiras
comerciais leves, alimentadas com níveis de metionina + cistina na ração. Esses

18
autores observaram aumento no peso médio dos ovos de 57,53 a 65,50 g, a partir do
aumento dos níveis de aminoácidos sulfurosos da dieta.
Outros autores (Bertran et al., 1995; Narvaez-Solarte, 1996; Rodrigues et al.,
1996; Novak et al., 2004; Novak et al., 2006) também relataram resposta significativa
na produtividade de galinhas poedeiras ao fornecerem dietas com diferentes níveis de
aminoácidos sulfurosos.
Os níveis de aminoácidos sulfurados para poedeiras no segundo ciclo de
produção também são responsáveis por influenciarem a produtividade dos animais.
Dessa forma, Laurentiz et al. (2005) avaliaram os níveis de aminoácidos sulfurados
totais para poedeiras semipesadas após a muda forçada e foi observado que a
deficiência ocasionada pelo menor fornecimento do aminoácido em estudo provocou
menor consumo de ração e aminoácidos sulfurosos, assim como menor peso corporal
pós-muda. Ainda foi observado nesse estudo que durante a fase de produção, o
fornecimento do menor nível de metionina + cistina total (0,45%) provocou redução
no peso médio dos ovos.
Polese et al. (2012) ao avaliarem a influência dos níveis de aminoácidos
sulfurosos digestíveis para poedeiras em segundo ciclo (75 a 91 semanas de idade)
sobre as principais variáveis produtivas não observaram efeito dos níveis de metionina
+ cistina digestível sobre as características de produção e de qualidade interna e
externa dos ovos de galinhas poedeiras semipesadas pós-muda forçada.
INFLUÊNCIA DOS NÍVEIS DE METIONINA + CISTINA SOBRE A
ATIVIDADE IMUNOLÓGICA DE GALINHAS POEDEIRAS
O avanço do melhoramento genético na avicultura alcançado até os dias de
hoje, possibilitou o aumento da eficiência produtiva das atuais linhagens utilizadas
pelos produtores. No entanto, apesar dessa evolução significativa, a saúde dos animais
preocupa pesquisadores e produtores no mundo inteiro. Os altos índices de ganho de
peso, produção de ovos e precocidade na produção, provavelmente não foram

19
acompanhados pela evolução dos animais, no que se refere às defesas naturais do
organismo. Dessa forma, o aporte de nutrientes presentes nas dietas de aves pode
auxiliar os mecanismos de defesa do organismo, evitando o surgimento de doenças
nos animais.
A imunologia é um campo de estudos referentes às respostas desencadeadas
após a exposição do organismo dos animais a agentes patológicos. Esse sistema no
organismo é capaz de não apenas defender o organismo de agentes patogênicos, mas
também de controlar uma série de eventos importantes para o equilíbrio do organismo,
a sua homeostasia. Com a finalidade de executar essas diversas funções, o organismo
possui células e produz várias moléculas específicas, obtendo assim uma resposta à
entrada de substâncias estranhas, denominada resposta imune. (Morgulis, 2002).
As respostas imunes desencadeadas nos animais são classificadas em dois
tipos, baseadas nos componentes que participam dessas respostas: a imunidade
humoral e a imunidade mediada por células. A primeira é mediada por moléculas,
denominadas anticorpos, responsáveis pelo reconhecimento e eliminação dos
antígenos e que podem ser transferidas através do soro ou do plasma de um indivíduo
imunizado para outro não imunizado. Já a imunidade mediada por células, também
chamada de imunidade celular, envolve as células denominadas linfócitos T, sendo
que a transferência de imunidade de um animal para outro é feita através dessas
células de animais imunizados (Morgulis, 2002).
Os sistemas linfoides e não linfoides constituem as principais estruturas do
sistema imunológico das aves. Entre os componentes linfoides nas aves, a bursa de
Fabrícius e o timo, local de desenvolvimento e diferenciação de linfócitos B e T,
respectivamente, são considerados órgãos linfoides primários, enquanto que o baço é
considerado órgão linfoide secundário. Aliado a isto, estruturas linfoides distribuídas
ao longo do trato intestinal representam uma barreira imunológica. Já as estruturas não
linfoides do sistema imune incluem células que propiciam uma defesa imunológica
não específica para o hospedeiro. As células do sistema fagocítico atuam primeiro
nesta categoria, destacando-se monócitos sanguíneos e macrófagos, que são
importantes devido a sua ampla distribuição nos fluidos corpóreos, órgãos e cavidades
(Rutz et al., 2012).

20
Os efeitos dos níveis de nutrientes sobre as respostas imunes em frangos de
corte e galinhas poedeiras vêm sendo avaliadas ao longo dos últimos anos,
destacando-se principalmente a utilização de fontes orgânicas de minerais e
suplementação vitamínica. No entanto, os estudos ainda são escassos no que se refere
ao efeito dos aminoácidos na imunocompetência de aves de produção. Segundo Rutz
et al. (2012), a deficiência crônica severa de micronutrientes (vitaminas e minerais)
são mais debilitantes ao sistema imunológico das aves do que a energia e o nível
proteico da dieta.
Características de desempenho e função imune de aves que sofreram estresse
por calor são melhoradas com a suplementação de vitamina C (Pardue et al., 1985),
vitamina E (El-Boushy, 1988) e piridoxina (Blalock et al., 1984). De acordo com Rutz
et al. (2012), as vitaminas participam no metabolismo como imunomoduladores para
melhorar as funções imunológicas e a resistência a infecções em aves e outros animais
domésticos.
Do mesmo modo, minerais complexados a moléculas orgânicas também vêm
sendo avaliados como alternativas nutricionais frente ao status imunológico de aves de
produção. De acordo Figueiredo Júnior et al. (2012), os minerais na forma orgânica
são absorvidos pelos carreadores intestinais de aminoácidos e peptídeos e não por
transportadores intestinais clássicos de minerais, evitando assim a competição entre
minerais pelos mesmos mecanismos de absorção.
Dentre os minerais orgânicos utilizados em ensaios experimentais para avaliar
o papel desses micronutrientes no sistema imunológico, destacam-se o zinco e o
selênio. Este último é componente da glutationa peroxidase, que tem como principal
função evitar a oxidação dos tecidos e manter a integridade das membranas das
células. Já o zinco é um micromineral intimamente ligado ao metabolismo de
aminoácidos, além de estar envolvido em diversas reações enzimáticas, já que,
segundo Underwood (1999), mais de 200 proteínas contendo zinco em sua estrutura
apresentam papeis biológicos no organismo dos animais.
No entanto, as respostas desencadeadas pelo sistema imunológico da ave
também podem estar diretamente ligadas ao suprimento adequado de proteína e
aminoácidos na dieta. Wu et al. (1999) afirmaram que diversos estudos indicaram que
a deficiência de proteína dietética, com consequente redução na concentração de

21
aminoácidos plasmáticos, pode comprometer o funcionamento do sistema imune.
Dessa forma, há um grande interesse dos pesquisadores em avaliar o verdadeiro papel
dos aminoácidos na função imune de mamíferos, aves, peixes e outras espécies
animais (Roch, 1999; Calder, 2006; Grimble, 2006; Kim et al., 2007).
Nas últimas décadas, pesquisadores do mundo inteiro vêm buscando entender
os mecanismos celulares e moleculares, que envolvem o papel dos aminoácidos nas
respostas imunes. O entendimento dessas funções metabólicas pode ajudar no
desenvolvimento de estratégias eficazes para melhorar a saúde e prevenir doenças
infecciosas nos animais (Li et al., 2007).
Vários aminoácidos essenciais e não essenciais estão envolvidos nas diversas
rotas metabólicas que envolvem as respostas imunes nas aves, destacando-se a
arginina, asparagina, aminoácidos de cadeia ramificada, aspartato, glutamato,
glutamina, glicina, histidina, lisina, fenilalanina, tirosina, serina, prolina e aminoácidos
sulfurosos. Estes últimos participam diretamente na doação de grupamentos metil e
como fonte de enxofre para diversas reações metabólicas.
Tsiagbe et al. (1987) observaram que a suplementação de metionina em dietas
a base de milho e farelo de soja aumentam a produção de anticorpos e elevam a
estimulação mitogênica provocada pela fitohematoglutinina. Outras pesquisas
revelaram que a cisteína pode exercer efeitos específicos na modulação da resposta
imune em frangos de corte (Takahashi et al., 1997; Konashi et al., 2000). De acordo
com Grimble (2006), o consumo adequado de metionina e cistina são importantes para
a síntese de proteínas do sistema imune.
Diversos trabalhos nas últimas décadas determinaram as exigências
nutricionais de aminoácidos sulfurosos para galinhas poedeiras, porém, baseados em
resultados de desempenho produtivo, como ganho de peso, conversão alimentar e
produção de ovos. Segundo Swain & Johri (2000), as necessidades de metionina para
uma ótima produção de anticorpos em aves de produção são maiores do que para um
ótimo crescimento corporal.
Segundo Grimble (2006), o sistema imune dos animais, agindo contra
organismos invasores, abrange uma série complexa de células e moléculas. Entretanto,
o mecanismo da variação da disponibilidade tecidual de aminoácidos sulfurados e os

22
produtos de seu metabolismo, interagindo com as respostas imunes, ainda são
desconhecidos.
De acordo com Rubin (2007), a suplementação dietética com aminoácidos
sulfurados e a quebra das proteínas teciduais são necessárias para a síntese de
proteínas e dos peptídeos envolvidos em um funcionamento normal do sistema
imunológico. Todavia, os estudos avaliando o consumo elevado de aminoácidos
sulfurosos sobre a função imune em animais de produção são escassos.
Através da S-adenosilmetionina, a metionina doa grupamentos metil que
participam da metilação do DNA e proteínas e regulação da expressão gênica (Wu et
al., 2006). Devido as poliaminas serem importantes para a proliferação e diferenciação
dos linfócitos (Flynn et al., 2002), a metionina pode desenvolver um papel importante
como constituinte direto das proteínas. Ademais, a metionina age como substrato para
a síntese da colina, assim como da fosfatidilcolina e acetilcolina, que são essenciais
para as funções nervosas do organismo e para o metabolismo dos leucócitos (Kim et
al., 2007).
Outra forte relação dos aminoácidos sulfurosos com a atividade imunológica de
galinhas poedeiras se dá através da síntese de glutationa. De acordo com Malmezat et
al. (2000), a cisteína é precursora da glutationa em células animais e seu metabolismo
é significativamente alterado em resposta a processos infecciosos. Outro fator
importante citado por Wu et al. (2004) é que a síntese desse importante antioxidante
celular é influenciado pelo consumo dietético de aminoácidos sulfurosos e, dessa
forma, observa-se uma correlação positiva entre a via de transulfuração e as
concentrações de glutationa no fígado, baço e músculos (Malmezat et al., 2000).
A glutationa destrói radicais livres e outras substâncias oxidativas (radicais
hidroxila, lipoperóxidos, peróxido de hidrogênio), além de participar do processo de
detoxificação de substâncias tóxicas ao organismo dos animais (Fang et al., 2002).
Existem evidências que as concentrações intracelulares de GSH desempenham um
papel importante na regulação de vias de sinalização celular, em resposta aos desafios
imunológicos (Fratelli et al., 2005).
De acordo com Rubin (2007), a síntese de glutationa, que é realizada
principalmente no fígado, utiliza com enzima precursora a gama-glutamil cisteína

23
sintetase. Assim, a conversão de cisteína à glutationa é influenciada fortemente pela
taxa de utilização e do transporte de glutationa dentro e entre as células do corpo, ou
seja, a síntese desse antioxidante celular é um processo conduzido pela demanda
orgânica, desde que a cisteína esteja disponível. Em condições de baixa
disponibilidade de cisteína, a síntese proteica será mantida e a síntese de sulfato,
taurina e glutationa diminuída. As mudanças na disponibilidade de taurina e glutationa
podem influenciar significativamente a função imune dos animais.
A deficiência dietética de aminoácidos sulfurosos em galinhas poedeiras pode
reduzir a atividade do sistema imunológico, principalmente através da diminuição da
produção e proliferação de linfócitos (Obled et al., 2004) e redução da atividade
antioxidante da glutationa. A suplementação com aminoácidos sulfurosos em dietas de
galinhas poedeiras pode melhorar o efeito das respostas imunológicas, no entanto, é
necessário evitar o excesso desses aminoácidos na dieta, devido aos possíveis efeitos
negativos do imbalanço aminoacídico, com consequente redução na ingestão e
utilização de outros aminoácidos.
CONSIDERAÇÕES FINAIS
As alterações metabólicas da deficiência e do excesso de aminoácidos
sulfurosos nas dietas de poedeiras em crescimento e durante a fase de produção são
evidentes. Esse imbalanço provoca queda no desempenho produtivo e redução da
qualidade dos ovos destinados ao consumo humano.
O intenso trabalho dos geneticistas na avicultura de postura nas últimas
décadas e a grande variação dos materiais genéticos utilizados nos atuais sistemas de
criação reforça a necessidade da constante avaliação dos requerimentos em
aminoácidos sulfurosos em todas as fases de criação de galinhas poedeiras. No
entanto, os estudos referentes à determinação das exigências de aminoácidos
sulfurosos para galinhas poedeiras durante a fase de crescimento e pré-postura ainda
são escassos na literatura.

24
O correto estabelecimento do nível de aminoácidos sulfurosos nas dietas de
aves de postura permite um melhor desempenho dos animais, desde a produção e
qualidade dos ovos ao melhor desenvolvimento de órgãos reprodutivos e ativação do
sistema imune e a atividade antioxidante do organismo.
Em consequência disso, os melhores resultados obtidos a campo permitem uma
maior rentabilidade e lucratividade da atividade para os produtores, sempre na busca
por dietas com menores custos e sem prejudicar a eficiência produtiva dos animais e a
qualidade dos produtos gerados.

25
REFERÊNCIAS BIBLIOGRÁFICAS
ALBINO, L.F.T.; ROSTAGNO, H.S.; SANT’ANNA, R. et al., Determinação dos
valores de aminoácidos metabolizáveis e proteína digestiva de alimentos para aves.
Revista Brasileira de Zootecnia, v.21, p.1059-1068, 1992.
AUSTIC, R.E. Biochemical description of nutritional effects. In: FISHER, C.
Nutrient requirements of poultry and nutritional research, BOORMAN, K.N.
(Eds.). London: Butterworths, 1986. p.59-77.
BAKER, D.H. Problems and pitfalls in animal experiments designed to establish
dietary requirements for essential nutrients. Journal of Nutrition, v.116, p.2339-
2349, 1986.
BAKER, D.H.; FERNANDEZ, S.R.; WEBEL, D.M. et al. Sulfur amino acid
requirement and cystine replacement value of broiler chicks during the period three
to six weeks post-hatching. Poultry Science, v.75, p.37-42, 1996.
BARBOSA, M.J.B.; CARDOZO, R.M.; SOUZA, V.L.F. et al. Níveis de metionina +
cistina no desempenho de poedeiras comerciais leves com 45 semanas de idade.
Revista Brasileira de Saúde e Produção Animal, v.10, p.1032-1039,2009.
BLALOCK, J.L.; THAXTON, J.P.; GARLICH, J.D. Humoral immunity in chicks
experiencing marginal vitamin B-6 deficiency. Journal of Nutrition, v.114, p.312-
322, 1984.
BRUMANO, G. Níveis de metionina + cistina digestíveis em rações para poedeiras
leves nos períodos de 24 a 40 e de 42 a 58 semanas de idade. 2008, 103f. Tese
(Doutorado), Universidade Federal de Viçosa, UFV, Viçosa – MG.
CALDER, P.C. Branched-chain amino acid and immunity. Journal of Nutrition,
v.136, p.288–293, 2006.
CAO, Z.; CAI, F.L.; COON, C. The methionine and cystine metabolism and
requirement of laying. Poultry Science, v.74, p.105, 1995.

26
CHEN, P.; JOHNSON, P.; SOMMER, T. et al. Multiple ubiquitin-conjugating
enzymes participate in the in vivo degradation of the yeast MATα2 repressor. Cell,
v.74, p.357-369, 1993.
COSTA, F.G.P.; SILVA, J.H.V.; FIGUEIREDO-LIMA, D.F. et al [2012]. Novos
avanços da nutrição de aves. Engormix. Disponível em: <www.engormix.com>.
Acesso em: Out. 30, 2013.
CUPERTINO, E.S.; GOMES, P.C.; ROSTAGNO, H.S. et al. Exigência nutricional de
metionina + cistina digestíveis para galinhas poedeiras de 54 a 70 semanas de
idade. Revista Brasileira de Zootecnia, v.38, p.1238-1246, 2009.
D’AGOSTINI, P.; GOMES, P.C.; MELLO, H.H.C. et al. Exigência de metionina +
cistina para frangas de reposição na fase de recria de 13 a 18 semanas de idade.
Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.64, p.1691-1698,
2012a.
D’AGOSTINI, P.; GOMES, P.C.; CALDERANO, A.A. et al. Exigência de metionina
+ cistina para frangas de reposição na fase de recria de 7 a 12 semanas de idade.
Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.64, p.1699-1706,
2012b.
D’MELLO, J.P.F. Amino acid imbalances, antagonisms and toxicities. In: D’MELLO,
J.P.L. (Ed.) Amino acids in farm animal nutrition. Edinburgh: The Scottish
Agrigultural College, CAB Internacional, 1994. p.63-97.
EL-BOUSHY, A.R. Vitamin E affects viability, immune response of poultry.
Feedstuffs, v.60, p.20-26, 1988.
FANG, Y.Z.; YANG, S; WU, G. Free radicals, antioxidants, and nutrition. Nutrition,
v.8, p.872–879, 2002.
FARIA, D.E.; JUNQUEIRA, O.M. Nutrição e alimentação de poedeiras comerciais.
In: SIMPÓSIO SOBRE NUTRIÇÃO ANIMAL E TECNOLOGIA DA
PRODUÇÃO DE RAÇÕES, 1998, Campinas-SP. Anais...Campinas-SP, 1998,
p.77-95.

27
FIGUEIREDO JÚNIOR, J.P.; GIVISIEZ, P.E.N.; COSTA, F.G.P. et al.
Microminerais orgânicos na nutrição de poedeiras. Revista Eletrônica Nutritime.
v.9, p.1833-1843, 2012.
FLYNN, N.E.; MEININGER, C.J.; HAYNES, T.E. et al. The metabolic basis of
arginine nutrition and pharmacotherapy. Biomedicine Pharmacotherapy. v.56,
p.427–438, 2002.
FRATELLI, M.; GOODWIN, L.O.; OROM, U.A. et al. Gene expression profiling
reveals a signaling role of glutathione in redox regulation. Proceedings of National
Academy of Sciences USA, v.102, p.13998–14003, 2005.
GRIMBLE, R.F. The effects of sulfur amino acid intake on immune function in
humans. Journal of Nutrition, v.136, p.1660-1665, 2006.
JORDÃO FILHO, J.; SILVA, J.H.V.; SILVA, E.S. et al. Exigências nutricionais de
metionina + cistina para poedeiras semipesadas do início de produção até o pico de
postura. Revista Brasileira de Zootecnia, v.35, p.1063-1069, 2006 (supl.).
KIM, S.W.; MATEO, R.D.; YIN, Y.L. et al. Functional amino acids and fatty acids for
enhancing production performance of sows and piglets. Asian-Australasian
Journal Animal Science, v.20, p.295–306, 2007.
KOELKEBECK, K.W.; BAKER, D.H.; HAN, Y. et al. Research note: effect of excess
lysine, methionine, threonine or tryptophan on production performance of laying
hens. Poultry Science, v.70, p.1651-1653, 1991.
KONASHI, S.; TAKAHASHI, K.; AKIBA, Y. Effects of dietary essential amino acid
deficiencies on immunological variables in broiler chickens. British Journal of
Nutrition, v.83, p.449-456, 2000.
KLASING, K.C. Comparative avian nutrition. New York: CAB Internacional,
1998. 350p.
KWAKKEL, R.P. Nutritional studies on body development and performance in
laying-type pullets and hens: a multiphasic approach. In: WOURLD’S POULTRY
CONGRESS, 19, 1992, Amsterdam, Proceedings... Amsterdam: 1992, p.480-484.

28
LAURENTIZ, A.C.; FILARDI, R.S.; RODRIGUES, E.A. et al. Níveis de aminoácidos
sulfurados totais para poedeiras semi-pesadas após a muda forçada. Ciência Rural,
v.35, p.164-168, 2005.
LEESON, S.; SUMMERS J.D. Nutrition of the chicken. 4.ed. Guelph: University
Books, 2001. 591p.
LENNINGHER, A.L. Princípios de bioquímica. 2.ed. São Paulo: Sarvier, 1996.
839p.
LI, P.; YIN, Y.; LI, D. et al. Amino acids and immune function. British Journal of
Nutrition, v.98, p.237–252, 2007.
MALMEZAT, T.; BREUILLÉ, D.; POUYET, C. et al. Methionine transulfuration is
increased during sepsis in rats. American Journal of Physiology, v.279, p.1391–
1397, 2000.
MORGULIS, M.S. Imunologia aplicada. In: MACARI, M., FURLAN, R.L.,
GONZALES, E. (Ed.) Fisiologia aviária aplicada a frangos de corte. 2.ed.
Jaboticabal: FUNEP/UNESP, 2002. p.231-245.
MOURA, A.M.A., Conceito da proteína ideal aplicada na nutrição de aves e suínos.
Revista Eletrônica Nutritime, v.1, p.31-34, 2004.
NARVÁEZ-SOLARTE, W.V. Exigências em metionina + cistina para poedeiras
leves e semipesadas. 1996. 57f. Dissertação (Mestrado em Zootecnia) –
Universidade Federal de Viçosa, Viçosa, MG.
NELSON, D.L.; COX, M.M. Principles of Biochemistry, 4 ed. Freeman Publishers,
New York, 2005.
NOVAK, C.; YAKOUT, H.; SCHEIDELER, S. The combined effects of dietary
lysine and total sulfur amino acid level on egg production parameters and egg
components in Dekalb delta laying hens. Poultry Science, v.83, p.977-984, 2004.
NOVAK, C.; YAKOUT, H.; SCHEIDELER, S. The effect of dietary protein level and
total sulfur amino acid:lysine ratio on egg production parameters and egg yield in
Hy-Line W-98 hens. Poultry Science, v.85 (suppl.12), p.2195-2206, 2006.

29
OBLED, C.; PAPET, I.; BREUILLE, D. Sulfur-containing amino acids and
glutathione in diseases. In: Metabolic & Therapeutic Aspects of Amino Acids in
Clinical Nutrition, LA Cynober Editor. Boca Raton, FL: CRC Press, 2.ed. 2004, p.
667–687.
PARDUE, S.L.; THAXTON, J.P.; BRAKE, J. Role of ascorbic acid in chicks exposed
to high environmental temperature. Journal Applied Physiology, v.58, p.1511-
1516, 1985.
PARS, J.F.; SUMMERS, J.D. The effects of minimizing amino acid excess in broiler
diets. Poultry Science, v.70, p.1540-549, 1991.
PARSONS, C. M., BAKER, D. H., The concept and use of ideal proteins in the
feeding of nonruminants. In: 31ª REUNIÃO ANUAL DA SOCIEDADE
BRASILEIRA DE ZOOTECNIA, 1994, Maringá-PR, Anais...Maringá-PR, 1994,
p.120-128.
PAVAN, A.C.; MÓRI, C.; GARCIA, E.A. et al. Níveis de proteína bruta e de
aminoácidos sulfurados totais sobre o desempenho, a qualidade dos ovos e a
excreção de nitrogênio de poedeiras de ovos marrons. Revista Brasileira de
Zootecnia, v.34, p. 568-574, 2005.
PILLAI, P.B.; FANATICO, A.C.; BEERS, K.W. et al. Homocysteine remethylation in
young broilers fed varying levels of methionine, choline and betaine. Poultry
Science, v.85, p. 90–95, 2006.
PIRES, C.V.; OLIVEIRA, M.G.A.; ROSA, J.C. et al. Qualidade nutricional e escore
químico de aminoácidos de diferentes fontes proteicas. Ciência e Tecnologia de
Alimentos, v.26, p.179-187, 2006.
POLESE, C., NUNES, R.V., VILELA, C.G. et al. Quantidade de metionina + cistina
digestível para poedeiras semipesadas de 75 a 91 semanas de idade. Arquivo
Brasileiro de Medicina Veterinária e Zootecnia, v.64, p.1682-1690, 2012.
ROCH, P. Defense mechanisms and disease prevention in farmed marine
invertebrates. Aquaculture, v.172, p.125–145, 1999.

30
RODRIGUES, P.B.; BERTECHINI, A.G.; OLIVEIRA, B.L. et al. Fatores nutricionais
que influenciam a qualidade do ovo no segundo ciclo de produção. I. Níveis de
aminoácidos sulfurosos totais. Revista da Sociedade Brasileira de Zootecnia,
v.25, p.248-260, 1996.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. Tabelas brasileiras para
aves e suínos. 2011. 3ed. UFV-MG. 252 p.
RUTZ, F.; BERMUDEZ, V.L.; PAN, E.A. et al. [2012]. Impacto da nutrição
vitamínica sobre a resposta imunológica das aves. Engormix. Disponível em:
<www.engormix.com> Acesso em: Out. 23, 2013.
SÁ, L.M.; GOMES, P.C.; ALBINO, L.F.T., et al. Exigência nutricional de metionina
+ cistina digestível para galinhas poedeiras no período de 34 a 50 semanas de idade.
Revista Brasileira de Zootecnia, v.36, p.1837-1845, 2007.
SCHUTTE, J.B.; JONG, J.; BERTRAM, H.L. Requirements of the laying hen for
sulfur amino acids. Poultry Science, v.73, p.274-280, 1994.
SHAFER, D.J.; CAREY, J.B.; PROCHASKA, J.F. Effect of dietary methionine intake
on egg component yield and composition. Poultry Science, v.75, p.1080-1085,
1996.
SHAFER, D.J.; CAREY, J.B.; PROCHASKA, J.F. Dietary methionine effects on egg
component yield, composition, functionally and texture profile analysis. Poultry
Science, v.7, p. 1056-1062, 1998.
SILVA, M.F.R. Desempenho, qualidade dos ovos e balanço de nitrogênio de
poedeiras comerciais alimentadas com diferentes níveis de proteína bruta,
metionina e lisina. 2006, 107f, Tese (Doutorado). Faculdade de Zootecnia e
Engenharia de Alimentos da Universidade de São Paulo. Pirassununga-SP.
SILVA, E.S.; SILVA, J.H.V.; BERTECHINI, A.G. et al. Exigência de metionina +
cistina para aves de reposição leves e semipesadas de 1 a 4 semanas de idade
alimentadas com rações farelada e triturada. Revista Brasileira de Zootecnia,
v.38, p.500-507, 2009.

31
SWAIN, B.K.; JOHRI, T.S. Effect of supplemental methionine, choline and their
combinations on the performance and immune response of broilers. British
Poultry Science, v.41, p.83-88, 2000.
TAKAHASHI, K.; OHTA, N.; AKIBA, Y. Influences of dietary methionine and
cystine on metabolic responses to immunological stress by escherichia coli
lipopolyssacharide injection, and mitogenic response in broiler chickens. British
Journal Nutrition, v.78, p.815-821, 1997.
TESSERAUD, S.; COUSTARD, S.M.; COLLIN, A. et al. Role of sulfur amino acids
in controlling nutrients metabolism and cell finctions: implications of nutrition.
British Journal of Nutrition, p. 1-8, 2008.
TSIAGBE, V.K.; COOK, M.E.; HARPER, A.E. et al. Enhanced immune responses in
broiler chick fed methionine supplemented diets. Poultry Science. v.66, p.1147-
1154, 1987.
UNDERWOOD, E.J. The mineral nutrition of livestock. CABI. Wallingford. 1999,
614 pp.
WALDROUP, P.W.; HELLWIG, H.M. Methionine and total sulfur amino acid
requirements influenced by stage of production. Journal of Applied Poultry
Science, v.4, p.283-292, 1995.
WU, G. Interrelationship among methionine, choline and betaine in channel
catfish – Ictalurus punctatus. 2003, 45f. Dissertação (Mestrado) – Auburn
University, Alabama, USA.
WU, G.; FLYNN, N.E.; FLYNN, S.P. et al. Dietary protein or arginine deficiency
impairs constitutive and inducible nitric oxide synthesis by young rats. Journal of
Nutrition, v.129, p.1347–1354, 1999.
WU, G.; FANG, Y.Z.; YANG, S. et al. Glutathione metabolism and its implications
for health. Journal of Nutrition, v.134, p.489–492, 2004.
WU G.; BAZER, F.W.; WALLACE, J.M. et al. Intrauterine growth retardation:
implications for the animal sciences. Journal Animal Science. v.84, p.2316–2337,
2006.

32

33
CAPÍTULO 2
Níveis de Metionina+Cistina para Galinhas Poedeiras Leves na Fase de
Recria

34
Níveis De Metionina+Cistina Para Galinhas Poedeiras Leves Na Fase De
Recria
RESUMO
Objetivou-se com esse trabalho avaliar os níveis de aminoácidos sulfurosos nas dietas
de galinhas poedeiras leves durante a fase de recria (13 a 18 semanas de idade). Foram
distribuídas 480 aves da linhagem Dekalb White com 13 semanas de idade em um
delineamento inteiramente casualizado, com seis tratamentos, 10 repetições e oito aves
por unidade experimental. As dietas consistiram em um tratamento controle,
formulada para atender as exigências nutricionais de aminoácidos sulfurosos de acordo
com o NRC (1994) e outras cinco dietas que foram estabelecidas de acordo com as
recomendações das tabelas brasileiras de aves e suínos (Rostagno et al., 2005), exceto
para metionina + cistina, que consistiram em cinco variações do nível desse
aminoácido (80, 90, 100, 110 e 120%), resultando nos níveis de 0,317 0,356; 0,396;
0,436 e 0,475% de metionina + cistina digestível. A pesquisa teve a duração de 329
dias e foi dividida em quatro fases experimentais: recria (13 a 18 semanas de idade),
pré-postura (19 a 29 semanas de idade), postura I (34 a 50 semanas de idade) e postura
II (54 a 70 semanas de idade). No primeiro período da pesquisa foram avaliados os
níveis de metionina + cistina entre os tratamentos e nas demais fases foi avaliado o
efeito residual dos níveis testados na fase de recria. Foram avaliados dados de
desempenho zootécnico e de qualidade dos ovos, sorológicos e histopatológicos das
aves. Houve efeito linear crescente para o peso vivo final, ganho de peso e consumo
de metionina + cistina, efeito linear decrescente para conversão alimentar e efeito
quadrático para a atividade da albumina, níveis de proteínas séricas, porcentagem de
gema (postura I) e peso relativo do baço. As exigências de metionina + cistina
digestível estimadas para estas características foram de 91,25% (0,361%), 87,40%

35
(0,346%), 107,79% (0,426%) e 100,44% (0,398%) para atividade da albumina, nível
de proteína sérica, porcentagem de gema e peso relativo do baço, respectivamente.
Baseado nos resultados obtidos e nas respostas biológicas das aves recomenda-se a
utilização de níveis superiores a 0,475% de metionina + cistina digestível,
correspondente à relação metionina + cistina: lisina digestível superior a 98% para
galinhas poedeiras leves de 13 a 18 semanas de idade.
Palavras-chave: aminoácidos sulfurosos, curva de crescimento, desempenho
produtivo, exigência nutricional

36
Levels Of Methionine And Cystine For Laying Hens At Growth Phase
ABSTRACT
The objective of this study was to evaluate the levels of sulfur amino acids in the diets
of laying hens hens during the rearing period (13-18 weeks old). 480 birds of Dekalb
White with 13 weeks of age were distributed in a completely randomized design with
six treatments, 10 replicates and eight birds each. The diets consisted in a control
treatment, formulated to meet the nutritional requirements of sulfur amino acids
according to the NRC (1994) and five other diets that have been established in
accordance with the recommendations of the brazilian poultry and swine tables
(Rostagno et al. 2005), except for methionine and cystine, which consisted of five
variations in the level of this amino acid (80, 90, 100, 110 and 120%) , resulting in
levels of 0.317, 0.356, 0.396, 0.436 and 0.475% of methionine + cystine. The study
lasted 329 days and was divided into four experimental stages: growth phase (13 to 18
weeks of age), pre-posture (19 to 29 weeks of age) posture one (34 to 50 weeks of age)
and posture two (54-70 weeks old). In the first survey period were evaluated levels of
methionine and cystine between treatments and the remaining stages the residual
effect of the levels tested in the growing phase was evaluated. Data production
performance and egg quality, serological and histological were evaluated. Increased
linearly for the final body weight, weight gain and intake of methionine and cystine,
linear effect on feed conversion and quadratic effect on the activity of albumin, serum
protein levels, yolk percentage (posture one) and relative weight of spleen. The
methionine + cystine estimated for these traits were 91,25% (0.361%), 87.40%
(0.346%), 107.79% (0.426%) and 100.44% (0.398%) to albumin activity, levels of
serum protein, yolk percentage and relative weight of spleen, respectively. Based on

37
these results obtained and the biological responses of birds is recommended using
levels above 0.475% of methionine + cystine, corresponding to methionine + cystine:
lysine digestible relationship more than 98%, for light laying hens with 13-18 weeks
old.
Key words: growth curve, nutritional requirements, productive performance, sulfur
amino acids

38
INTRODUÇÃO
A proteína dietética, ao lado do conteúdo energético da dieta, é caracterizada
por ser um dos nutrientes que mais onera o custo final das rações para galinhas
poedeiras. Nesse contexto, como primeiro aminoácido limitante para poedeiras, em
dietas a base de milho e farelo de soja, a metionina desenvolve papel importante na
produtividade dos animais e o fornecimento de níveis exatamente requeridos por estes
nas dietas tem impacto significativo nos resultados econômicos.
Dentre as principais funções dos aminoácidos sulfurosos, destacam-se: a
participação na doação de radicais metil para diversas reações metabólicas no
organismo, participa diretamente da síntese proteica orgânica, destacando-se a síntese
de proteínas do sistema imune (Grimble, 2006), regulação da expressão gênica (Wu et
al., 2006), é substrato para a síntese de colina e, assim, fosfatidilcolina e acetilcolina,
que são essenciais para a função nervosa e no metabolismo de leucócitos (Kim et al.,
2007).
Durante a fase de crescimento em galinhas poedeiras, há um intenso
desenvolvimento de órgãos reprodutivos e outros tecidos. O fornecimento de dietas
com deficiência em metionina nessa fase podem afetar o ganho de peso dos animais e
a maturidade sexual. Segundo D’Agostini et al. (2012), os fatores que interferem na
idade da ave e na maturidade sexual estão diretamente relacionados com o peso
corporal e a uniformidade do lote na fase de recria. Esses autores ainda afirmam que
aves com peso abaixo do ideal retardam o início da postura, reduzindo a produção de
ovos. Em contrapartida, aves com excesso de peso iniciam a postura precocemente e
produzem ovos pequenos, o que também não é viável.

39
Mesmo com a devida importância dessa fase, poucos trabalhos são encontrados
na literatura sobre as exigências de aminoácidos sulfurosos para poedeiras na fase de
recria. Esse fato pode ser explicado pelo alto custo e ao tempo necessário para a
realização de pesquisas com esses animais (Rostagno et al., 1996)
Diante do exposto, objetivou-se com esta pesquisa determinar as exigências
nutricionais de aminoácidos sulfurosos para frangas de reposição na fase de recria (13
a 18 semanas de idade), assim como avaliar os efeitos desses aminoácidos no
desempenho produtivo nas fases subsequentes (pré-postura, postura I e postura II).
MATERIAL E MÉTODOS
O estudo foi realizado no Setor de Avicultura, do Departamento de Zootecnia,
da Universidade Federal da Paraíba, Câmpus II, Areia-PB. Esse município está
localizado a 112 km de João Pessoa e situado na microrregião do Brejo Paraibano
(6°58’12’’ de latitude sul e 35°42’15’’ de longitude oeste). Apresenta como
característica climática tropical semi-úmido, estação chuvosa no período outono-
inverno, precipitação pluviométrica de aproximadamente 1500 mm, temperatura anual
entre 22 e 30ºC e umidade relativa do ar entre 75 e 87%.
A pesquisa teve a duração de 329 dias e foi subdividida em quatro fases
experimentais: recria (13 a 18 semanas de idade), pré-postura (19 a 29 semanas de
idade), postura I (34 a 50 semanas de idade) e postura II (54 a 70 semanas de idade).
Na primeira fase da pesquisa foram avaliados os níveis de metionina + cistina entre os
tratamentos e nas demais foi avaliado o efeito residual dos níveis testados na fase de
recria.
Período de Avaliação das Dietas Experimentais

40
Nesse período foram utilizadas 480 aves da linhagem Dekalb White, com idade
inicial de 13 semanas. As aves foram alojadas em gaiolas de produção, com
comedouro do tipo calha e bebedouro do tipo sparkcup, para fornecimento de ração e
água à vontade, respectivamente.
Os animais foram distribuídos em um delineamento inteiramente casualizado,
composto por seis tratamentos e dez repetições, com oito animais por unidade
experimental. As dietas experimentais (Tabela 2) foram elaboradas da seguinte forma:
um tratamento controle, formulada para atender as exigências nutricionais seguindo as
recomendações do NRC (1994); e os demais tratamentos foram estabelecidos de
acordo com as recomendações das tabelas brasileiras de aves e suínos (Rostagno et al.,
2005), exceto para metionina + cistina, que consistiram em cinco variações do nível
desse aminoácido (80, 90, 100, 110 e 120%), resultando nos níveis de 0,317 0,356;
0,396; 0,436 e 0,475% de metionina + cistina digestível. As dietas foram
suplementadas com DL-Metionina-99% (0; 0,039; 0,078; 0,117; 0,156%), em
substituição ao farelo de glúten de milho, para atingir os níveis dietéticos.
Tabela 2 – Ingredientes e composição química das dietas experimentais.
Ingredientes NRC Rostagno et al, 2005
(1994)* 80% 90% 100% 110% 120%
Milho
77,653 79,847 79,847 79,847 79,847 79,847
Farelo de Soja, 45% 18,518 15,212 15,212 15,212 15,212 15,212
Fosfato Bicálcico 2,994 1,180 1,180 1,180 1,180 1,180
Sal
0,332 0,329 0,329 0,329 0,329 0,329
Cloreto de Colina
0,070 0,070 0,070 0,070 0,070 0,070
Premix Vitamínico2
0,050 0,050 0,050 0,050 0,050 0,050
Premix Mineral1
0,050 0,050 0,050 0,050 0,050 0,050
Antioxidante3
0,010 0,010 0,010 0,010 0,010 0,010
Carbonato de Potássio 0,00 0,005 0,005 0,005 0,005 0,005
Calcário
0,323 1,123 1,123 1,123 1,123 1,123
Farelo de Glúten de Milho, 60% 0,00 2,124 2,085 2,046 2,007 1,968
DL-Metionina 0,00 0,00 0,039 0,078 0,117 0,156
Total 100 100 100 100 100 100
Composição Química
(D-Digestível) Unidade
Proteína Bruta % 15 14 14 14 14 14

41
Cálcio % 0,800 0,780 0,780 0,780 0,780 0,780
Fósforo Disponível % 0,646 0,310 0,310 0,310 0,310 0,310
Energia Metabolizável kcal/kg 2900 2900 2900 2900 2900 2900
Arginina-D % 0,670 0,531 0,531 0,531 0,531 0,531
Isoleucina-D % 0,400 0,372 0,372 0,372 0,372 0,372
Lisina-D % 0,450 0,483 0,483 0,483 0,483 0,483
Met + Cis-D % 0,420 0,317 0,356 0,396 0,436 0,475
Treonina-D % 0,370 0,333 0,333 0,333 0,333 0,333
Triptofano-D % 0,110 0,106 0,106 0,106 0,106 0,106
Sódio % 0,150 0,150 0,150 0,150 0,150 0,150
Cloro % 0,243 0,244 0,244 0,244 0,244 0,244
Potássio % 0,544 0,500 0,500 0,500 0,500 0,500
Balanço Eletrolítico mEq/kg 135,87 124,36 124,36 124,36 124,36 124,36
* Ração formulada com base em aminoácidos totais. 1Premix mineral inorgânico por kg de produto: Mn, 20 g; Fe,
10 g; Zn, 13,7 g; Cu, 2,5 g; Se, 0,063 g; I, 0,19 g; e veículo q.s.p., 500 g .2 Premix vitamínico por kg de ração: Vit.
A - 15.000.000 Ul, Vit. D3 - 1.500.000 Ul, Vit. E - 15.000 Ul, Vit.B1 - 2,0 g, Vit.B2-4,0 g, Vit B6 - 3,0 g, Vit.B12 -
0,015 g, Ácido nicotínico - 25 g, Ácido pantotênico- 10 g, Vit.K3 - 3,0 g, Ácido fólico- 1,0 g, Bacitracina de zinco
- 10 g, Selênio - 250 mg.3Antioxidante BHT - 10 g, e veículo. q.s.p. - 1.000 g.
As variáveis avaliadas foram: peso vivo final (g/ave), ganho de peso (g/ave),
consumo de ração (g/ave), conversão alimentar (g/g), consumo de metionina + cistina
(mg/ave), consumo de lisina (mg/ave), peso sem vísceras (g/ave), peso do fígado
(g/ave), peso da gordura celomática (g/ave), peso do baço (g/ave), análise sorológica
da alanina aminotransferase ((U/L), aspartato aminotransferase (U/L), gamma-
glutamiltransferase (U/L), creatinina (mg/dL), albumina (g/dL) e proteína sérica
(g/dL). Ademais, foi realizada análise histopatológica do fígado, intestino delgado e
magno, além da mensuração da curva de crescimento no período de 13 a 18 semanas
de idade.
Os animais e as rações foram pesados no início e no final do período
experimental para a mensuração do peso vivo final, ganho de peso, consumo de ração,
conversão alimentar, consumo de metionina + cistina e consumo de lisina.
Para as análises sorológicas, histopatológicas e as mensurações dos órgãos,
cinco aves por unidade experimental foram abatidas no final do período experimental.
As análises sorológicas foram realizadas por meio do equipamento VetTest Blood
Chemistry Analyzer. Já para as análises histopatológicas, fragmentos do intestino
delgado, fígado e magno foram imersos em substância fixadora metacarn (60%

42
metanol, 30% clorofórmio e 10% ácido acético), por 12 horas e posteriormente
transferidos para álcool 70%.
Para a microscopia óptica foi feita a inclusão dos fragmentos dos órgãos em
paraplast, sendo realizados cortes seriados dos mesmos de 5µm de espessura. Para a
análise histológica, foram utilizadas as colorações, hematoxilina e eosina, periodic
acid Schiff (PAS) e tricômio de Masson. As fotomicrografias foram obtidas através de
microcâmera acoplada a um microscópio Olympus BX-51 e as imagens digitalizadas
no software KS 400.3 (Zeiss).
Para a mensuração da curva de crescimento, todas as aves das unidades
experimentais foram pesadas semanalmente durante o período de 13 a 18 semanas de
idade. Os dados coletados foram submetidos aos modelos equacionais propostos Von
Bertalanffy, Brody, Gompertz, Logístico e Richards, utilizando-se o programa
estatístico computacional GOSA, obtendo-se o melhor ajuste as equações
matemáticas. Foi utilizado o critério de informação de AKAIKE (AIC) para a escolha
do modelo matemático com melhor ajuste aos dados de peso e idade das aves na curva
de crescimento.
Avaliação do Efeito Residual
Após o período de avaliação das dietas experimentais, as aves foram
alimentadas com água e ração à vontade, de acordo com as exigências nutricionais
para cada fase, seguindo as recomendações de Rostagno et al. (2005). Desse modo, foi
permitido avaliar o efeito residual dos níveis de metionina + cistina do período de 13 a
18 semanas de idade sobre as fases subsequentes.
Para a fase de pré-postura (19 a 29 semanas de idade), 384 aves selecionadas
aleatoriamente nos tratamentos foram distribuídas em oito repetições, com oito
animais por unidade experimental. Nesse período foram avaliadas as seguintes
variáveis: idade ao primeiro ovo (dias), peso do primeiro ovo (g), número de ovos por
período (total de ovos), número de dias de produção (dias), taxa de postura (%), peso
dos ovos e massa de ovo (g/ave/dia).

43
Ao iniciar a produção de ovos, os mesmos eram anotados para a determinação
da idade ao primeiro ovo e posteriormente pesados em balança analítica, para a
determinação do peso do primeiro ovo, assim como a determinação do número de
ovos por período e o número de dias de produção.
A taxa de postura foi determinada através da relação entre o total de ovos do
período e o número de aves corrigido pela mortalidade, sobre o número de dias de
produção, multiplicado por 100. O peso médio dos ovos foi obtido por meio da relação
entre o peso total dos ovos sobre o número de ovos do período. A massa de ovos foi
determinada pela multiplicação da produção pelo peso dos ovos.
Nas fases subsequentes (postura I e postura II) as variáveis analisadas foram:
produção de ovos (%), peso dos ovos (g), massa de ovo (g/ave/dia), percentagem de
gema (%), albúmen (%) e casca (%), gravidade específica (g/mL) e espessura de casca
(mm).
A produção de ovos foi determinada pela relação do número de ovos
produzidos e o número de aves corrigido pela mortalidade, por cada período e
multiplicado por 100. A massa de ovos foi calculada multiplicando-se a produção e o
peso dos ovos. Para o cálculo do peso médio e dos componentes internos dos ovos, os
mesmos foram pesados individualmente em balança analítica. As percentagens de
gema e albúmen foram obtidas através da relação entre o peso médio da gema e do
albúmen sobre o peso médio do ovo, multiplicando-se por 100.
As cascas dos ovos foram secas em estufa a 55-60ºC, por um período de 24
horas e pesadas em balança digital de precisão para a obtenção do peso médio. A
porcentagem da casca foi determinada pela relação entre o peso médio da casca sobre
o peso médio do ovo e multiplicado por 100. Já a espessura da casca foi obtida por
meio da utilização de micrômetro digital Mitutoyo de 0-25 mm, com precisão de 0,001
mm. A gravidade específica foi determinada pelo método de flutuação salina, proposto
por Hamilton (1982). Os ovos foram imersos em quinze soluções salinas (NaCl),
variando entre 1,070 a 1,090 g/mL, com gradiente de 0,0025 entre elas.
Análises Estatísticas

44
As análises estatísticas das variáveis avaliadas foram realizadas por meio do
programa estatístico computacional SAS (SAS Institute, 2011). O tratamento controle
(NRC, 1994) foi comparado com os demais tratamentos (variações dos níveis de
metionina + cistina, de acordo com Rostagno et al., 2005), através do teste Dunnett a
5% de probabilidade. O requerimento de metionina + cistina digestível foi estimado
através de análise de regressão, de modo a considerar o valor do R2 e a resposta
biológica das aves.
RESULTADOS E DISCUSSÃO
Variáveis de Desempenho Zootécnico
De acordo com a Tabela 3, foi verificado efeito significativo (P<0,05) entre os
tratamentos para todas as variáveis em estudo, exceto para consumo de ração, quando
submetidos ao teste Dunnet.
Tabela 3 – Efeito dos tratamentos sobre o peso vivo final (PF, g/ave), ganho de peso (GP, g/ave), consumo de
ração (CR, g/ave), conversão alimentar (CA, g/g), consumo de metionina + cistina (CMC, mg/ave/dia) e
consumo de lisina (CLIS, mg/ave/dia), de poedeiras leves de 13 a 18 semanas de idade.
Tratamento MET:LIS*
PF GP CR CA CMC CLIS
NRC, 1994 93 1207,36 350,30 2207,80 6,32 220.78 236.55
Rostagno et
al. (2005)
80% 66 1209,17 351,16 2131,47 6,08 160.24¥ 244.15
90% 74 1215,09 361,28 2121,64 5,88 179.83¥ 243.99
100% 82 1235,62¥ 378,18¥ 2174,54 5,75 205.03¥ 250.07¥
110% 90 1240,53¥ 382,73¥ 2132,83 5,58 221.41 245.28
120% 98 1253,60¥ 395,87¥ 2129,85 5,38¥ 240.88¥ 244.93
Regressão
Rostagno et al. (2005)
L** L** ns L** L** ns
C.V.(%)
1,53 4,33 3,17 4,89 3,13 3,14

45
*Relação metionina+cistina:lisina (%). C.V.(%): Coeficiente de variação; ¥: médias diferentes pelo teste Dunnett
a 5% de probabilidade; L**: Efeito significativo ao modelo de regressão linear a 1% de probabilidade
As aves que receberam as dietas com os níveis recomendados pelo NRC (1994)
apresentaram menor peso final e ganho de peso, quando comparado com os animais
alimentados com as dietas com 100, 110 e 120% dos níveis de metionina + cistina
digestível preconizados por Rostagno et al. (2005). As aves alimentadas com o nível
de 120% das exigências deste aminoácido, recomendado pelas tabelas brasileiras de
exigências nutricionais, apresentou melhor conversão alimentar, quando comparado ao
tratamento controle.
Estes resultados podem ser explicados pelo melhor balanço aminoacídico das
dietas formuladas com base em aminoácidos digestíveis, quando comparado com
dietas que estabelecem suas exigências em aminoácidos totais. A tabela de exigência
nutricional do NRC (1994), além dos aminoácidos sulfurosos, recomenda níveis de
aminoácidos essenciais abaixo dos requerimentos das aves, quando considerados os
valores de aminoácidos digestíveis. Dessa forma, possivelmente as aves alimentadas
com o tratamento controle, ou seja, com a dieta do NRC (1994), tiveram redução no
desempenho devido a menor disponibilidade de aminoácidos na dieta, já que os
mesmos são essenciais às funções normais do organismo das aves.
A formulação de ração com base na composição em aminoácidos digestíveis
contribui para maior precisão no fornecimento de aminoácidos, pois considera a
capacidade da ave em aproveitar os aminoácidos dos alimentos e superar eventuais
diferenças na digestibilidade dos ingredientes (Rombola et al., 2008). No entanto, os
resultados de estudos referentes aos critérios de formulação (total ou digestível) para
poedeiras ainda são controversos. Casartelli et al. (2004), utilizando rações formuladas
pelos conceitos de aminoácidos totais e digestíveis, observaram que as formulações
com aminoácidos totais promoveram melhores resultados de consumo de ração e
conversão alimentar.
Também foi observado efeito significativo (P<0,05) entre os tratamentos para
as variáveis, consumo de metionina + cistina e consumo de lisina. As aves que
receberam a dieta controle apresentaram um maior consumo de aminoácidos
sulfurosos, quando comparado com as aves alimentadas com as dietas com 80, 90 e

46
100% das exigências desses aminoácidos das tabelas brasileiras de exigências
nutricionais, enquanto que os animais alimentados com a dieta com 120% de
metionina + cistina digestível apresentaram maior consumo de aminoácidos
sulfurosos, quando comparado com a dieta controle.
Mesmo não sendo observado efeito significativo (P<0,05) entre os tratamentos
para o consumo de ração, o maior consumo de aminoácidos sulfurosos das aves
alimentadas com a dieta controle pode ser explicado pelo maior consumo de ração dos
animais, quando comparado aos tratamentos com 80, 90 e 100% das exigências
nutricionais de metionina + cistina digestível recomendadas por Rostagno et al.
(2005).
De acordo com a análise de regressão, foi observado comportamento linear
significativo (P<0,01) para as variáveis, peso vivo final, ganho de peso, conversão
alimentar e consumo de metionina + cistina. Seguem abaixo, as figuras e as equações
correspondentes para cada variável analisada.
Figura 3 – Peso vivo final de poedeiras leves com 18 semanas de idade, alimentadas com
níveis de metionina + cistina.
Ŷ = 1,143x + 1116,5
R² = 0,966
1200
1210
1220
1230
1240
1250
1260
80 90 100 110 120
Pes
o v
ivo
fin
al
(g)
Níveis de metionina + cistina digestível (Rostagno et al., 2005)

47
Figura 4 – Ganho de peso de poedeiras leves com 18 semanas de idade, alimentadas com níveis
de metionina + cistina.
Figura 5 – Conversão alimentar de poedeiras leves com 18 semanas de idade, alimentadas com
níveis de metionina + cistina.
Ŷ = 1,108x + 262,97
R² = 0,9792
340
350
360
370
380
390
400
80 90 100 110 120
Ga
nh
o d
e P
eso
(g
)
Níveis de metionina + cistina digestível (Rostagno et al., 2005)
Ŷ= -0,017x + 7,434
R² = 0,9955
5,20
5,40
5,60
5,80
6,00
6,20
80 90 100 110 120
Co
nv
ersã
o a
lim
enta
r (g
/g)
Níveis de metionina + cistina digestível (Rostagno et al., 2005)

48
Figura 6 – Consumo de metionina + cistina de poedeiras leves com 18 semanas de idade,
alimentadas com níveis de metionina + cistina.
Houve efeito linear crescente (P<0,01) para as variáveis, peso vivo final, ganho
de peso e consumo de metionina + cistina, em poedeiras leves alimentadas com níveis
de metionina + cistina digestível, de 13 a 18 semanas de idade. De acordo com as
Figuras 3, 4, 5 e 6, é possível observar que a suplementação com aminoácidos
sulfurosos melhorou o desempenho do crescimento das aves durante a fase de recria,
no entanto, não foi possível estimar a exigência nutricional para este período. Desta
forma, a exigência de metionina + cistina digestível para poedeiras leves de 13 a 18
semanas de idade é superior ao nível de 0,475% na dieta.
D’Agostini et al. (2012) avaliaram as exigências de metionina + cistina para
frangas de reposição na fase de recria (13 a 18 semanas de idade) e observaram que a
exigência desse aminoácido, estimada para a variável de ganho de peso foi de 0,502%,
com base em aminoácidos digestíveis, resultado superior ao sugerido no presente
estudo (0,475%).
O manual da linhagem Dekalb White (2009) sugere como recomendação
nutricional para metionina + cistina digestível o nível de 0,640%, para a fase de 11 a
15 semanas, valor também superior ao sugerido no estudo (0,475%). A nova tabela
brasileira de exigências nutricionais para aves e suínos (Rostagno et al., 2011)
recomenda o mesmo nível de metionina + cistina digestível da edição anterior (ano de
Ŷ= 2,028x - 1,382
R² = 0,996
160
175
190
205
220
235
250
80 90 100 110 120
Co
nsu
mo
de
met
ion
ina
+ c
isti
na
(m
g)
Níveis de metionina + cistina digestível (Rostagno et al., 2005)

49
2005), sugerindo o nível de 0,396%, correspondente ao nível de 100% desse
aminoácido no estudo.
Da mesma forma, houve efeito positivo da suplementação de aminoácidos
sulfurosos sobre a conversão alimentar das aves durante o período experimental, sendo
que os maiores níveis desses aminoácidos na dieta proporcionaram os melhores
resultados. Esses resultados estão de acordo com os dados encontrados por D’Agostini
et al. (2012), que observaram efeito linear decrescente dos níveis de metionina +
cistina, sobre a conversão alimentar de poedeiras Lohmann LSL, de 13 a 18 semanas
de idade.
Segundo Silva et al. (2009), o atendimento das exigências dos aminoácidos
para aves de reposição durante a fase de crescimento é uma etapa crítica na otimização
do desempenho na fase de produção. Os níveis mais baixos de metionina + cistina
digestível nas dietas das poedeiras em crescimento proporcionaram os piores
resultados de desempenho nesse estudo, haja vista a intensa participação dos
aminoácidos sulfurosos como doadores de grupamentos metil para diversas reações
metabólicas, assim como na participação direta na síntese proteica.
De acordo com Baker et al. (1996), a metionina sob a forma de S-
adenosilmetionina é exigida para a biossíntese de substâncias envolvidas no período
crescimento das aves, como a cisteína, carnitina, poliaminas, epinefrina, colina e
melatonina e, dessa forma, a deficiência desse aminoácido observada nos tratamentos
com os menores níveis de suplementação, pode explicar o baixo desempenho dos
animais.
Não houve efeito dos tratamentos sobre o consumo de ração das aves durante o
período experimental. Mesmo sendo considerado um dos aminoácidos mais tóxicos
para as aves, quando consumido em excesso, os níveis mais altos de metionina +
cistina nas dietas não foram capazes de depreciar o consumo de ração no período.
Jordão Filho et al. (2006) afirmam que as aves possuem a habilidade de reduzir
a ingestão de alimentos para compensar o excesso de aminoácidos da ração. Esse
mecanismo é utilizado como estratégia em situações de alta ingestão de misturas
desbalanceadas, que podem causar lesões patológicas e, geralmente é percebida de 2 a
3 dias do início da alimentação das aves com uma dieta desbalanceada (Park & Austic,

50
1998). Dessa forma, o maior nível de metionina + cistina (0,475%) não foi suficiente
para ativarem esses mecanismos fisiológicos de redução no consumo de ração durante
o período experimental.
Curva de Crescimento
Os dados da curva de crescimento das aves no período de 13 a 18 semanas de
idade obtiveram melhor ajuste ao modelo proposto por Brody, que apresentou o menor
AIC (1610,565), de acordo com a Tabela 4. Esses dados estão representados na Figura
7, que demonstra a curva média de crescimento das aves no período de 13 a 18
semanas de idade.
Os piores ajustes foram observados nos modelos de Richards e Logistic,
apresentando o AIC de 1612,620 e 1611,064, respectivamente, valores superiores aos
do modelo matemático selecionado.
Tabela 4 – Propriedades dos modelos não-lineares utilizados na curva de crescimento de poedeiras leves de 13 a
18 semanas de idade
MODELO EQUAÇÃO AIC
Von Bertalanffy Y = A(1− Be−Kt
) 3 + ε 1610,650
Brody Y = A(1- Be -Kt
) + ε 1610,565
Gompertz Y = Ae -Be-Kt
+ ε 1610,747
Logistic Y = A(1+ Be-Kt
) -1 + ε 1611,064
Richards Y = A(1- Be-Kt
)M
+ ε 1612,620
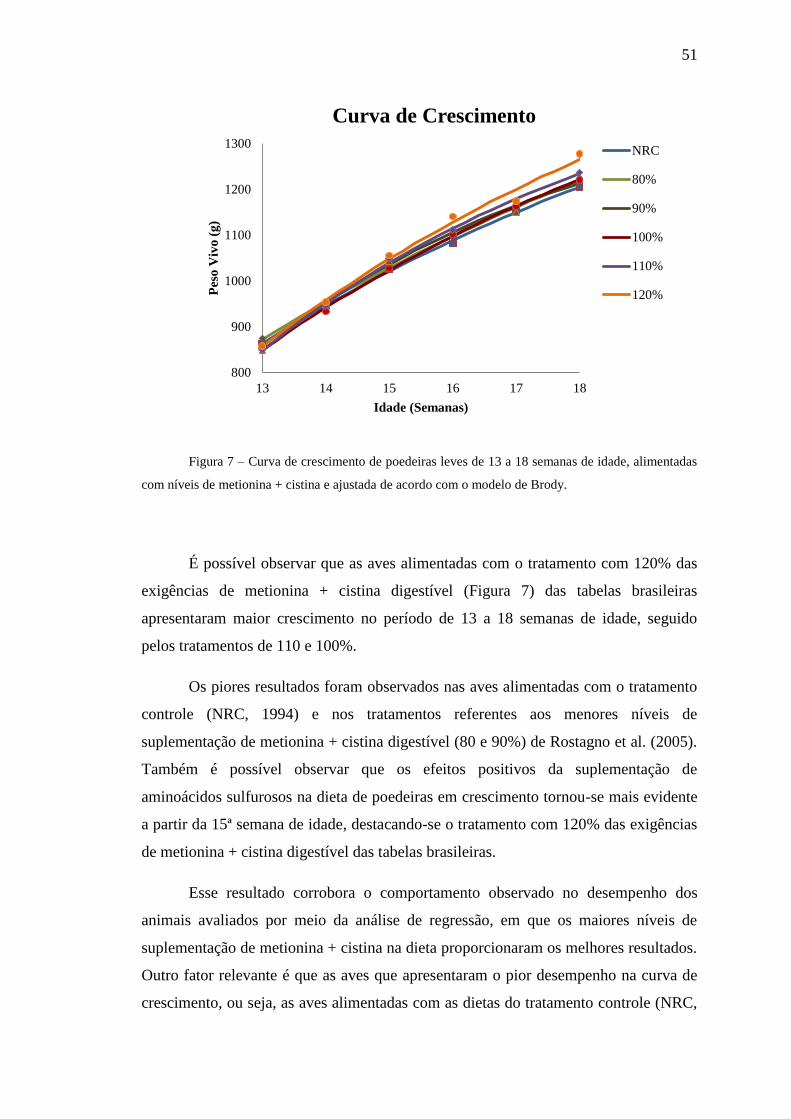
51
Figura 7 – Curva de crescimento de poedeiras leves de 13 a 18 semanas de idade, alimentadas
com níveis de metionina + cistina e ajustada de acordo com o modelo de Brody.
É possível observar que as aves alimentadas com o tratamento com 120% das
exigências de metionina + cistina digestível (Figura 7) das tabelas brasileiras
apresentaram maior crescimento no período de 13 a 18 semanas de idade, seguido
pelos tratamentos de 110 e 100%.
Os piores resultados foram observados nas aves alimentadas com o tratamento
controle (NRC, 1994) e nos tratamentos referentes aos menores níveis de
suplementação de metionina + cistina digestível (80 e 90%) de Rostagno et al. (2005).
Também é possível observar que os efeitos positivos da suplementação de
aminoácidos sulfurosos na dieta de poedeiras em crescimento tornou-se mais evidente
a partir da 15ª semana de idade, destacando-se o tratamento com 120% das exigências
de metionina + cistina digestível das tabelas brasileiras.
Esse resultado corrobora o comportamento observado no desempenho dos
animais avaliados por meio da análise de regressão, em que os maiores níveis de
suplementação de metionina + cistina na dieta proporcionaram os melhores resultados.
Outro fator relevante é que as aves que apresentaram o pior desempenho na curva de
crescimento, ou seja, as aves alimentadas com as dietas do tratamento controle (NRC,
800
900
1000
1100
1200
1300
13 14 15 16 17 18
Pes
o V
ivo
(g
)
Idade (Semanas)
Curva de Crescimento
NRC
80%
90%
100%
110%
120%

52
1994) e dos tratamentos com 80 e 90% de metionina + cistina digestível, irão levar
mais tempo para atingir o peso ideal para o período de produção.
De acordo com Kwakkel (1999), as empresas fornecedoras de material
genético enfatizam a importância das aves atingirem o chamado “peso alvo”, que as
tornariam aptas à produção. No entanto, as reações fisiológicas entre o peso alvo e a
produção não são tão claras.
Dessa forma, diversos modelos de crescimento das aves de reposição vêm
sendo estudados nos últimos anos e a configuração da curva de crescimento corporal
passou a fornecer informações adicionais sobre o subsequente desempenho das aves.
Sendo assim, o estudo das curvas de crescimento é importante para entender como as
aves crescem e podem depositar os seus tecidos, possibilitando a elaboração de
programas nutricionais mais adequados (Neme et al., 2006).
Análise Sorológica
Houve efeito significativo (P<0,05) entre o tratamento controle (NRC, 1994) e
os demais tratamentos (Rostagno et al., 2005) para todas as variáveis analisadas, de
acordo com o teste Dunnett (Tabela 5).
Tabela 5 – Efeitos dos tratamentos sobre a atividade da albumina (ALB, g/dL), proteína sérica (PTN, g/dL),
aspartato aminotransferase (AST, U/L), alanina aminotransferase (ALT, U/L), gamma-
glutamiltransferase (GGT, U/L) e creatinina (CRE, mg/dL) das aves de 13 a 18 semanas de idade.
Tratamento MET:LIS* ALB PROT AST ALT GGT CRE
NRC, 1994 93 1,562 3,48 123,4 3,4 28,4 0,508
Rostagno et al. (2005)
80% 66 2,334¥ 4,74¥ 175,2¥ 3,8 37,6¥ 0,654¥
90% 74 2,384¥ 4,92¥ 157,8 3,2 37,2 0,626¥
100% 82 2,414¥ 4,56¥ 176,4¥ 5,4¥ 32,2 0,676¥
110% 90 2,258¥ 4,44¥ 156 5,2 33,6 0,744¥
120% 98 1,852 3,74 124,2 4,4 28,6 0,604¥
Regressão Rostagno et al. (2005) Q* Q* ns ns ns ns
C.V. (%) 16,273 13,319 15,48 21,84 12,17 12,412 *Relação metionina+cistina:lisina (%). C.V.(%): Coeficiente de variação; ¥: médias diferentes pelo teste Dunnett
a 5% de probabilidade; Q*: Efeito significativo ao modelo de regressão quadrática a 5% de probabilidade.

53
O nível sérico de albumina das aves alimentadas com o nível de metionina +
cistina do tratamento controle foi significativamente menor (P<0,05), quando
comparado com os animais que receberam os níveis de aminoácidos sulfurosos das
tabelas brasileiras, com exceção para o tratamento com 120% das exigências de
metionina + cistina digestível. De acordo com Schmidt et al. (2007), a albumina, que é
sintetizada no fígado, representa de 40 a 50% da proteína plasmática total das aves,
sendo que seus teores normais podem variar de 0,8 a 2,0 g/dL, valores próximos aos
encontrados nesse estudo.
A redução dos níveis de albumina sérica pode ser ocasionada por diversos
fatores. Aves inoculadas com cepa virulenta do vírus da doença de Newcastle
provocaram diminuição nos níveis de albumina e, consequentemente, diminuição da
concentração das proteínas totais. Outra possível causa para a redução do nível de
albumina pode ser ocasionada por lesões em vísceras, principalmente do sistema
digestório, com ulcerações intestinais e hemorragias (Schmidt et al., 2007). Ademais, a
anorexia, causada pela diminuição na ingestão de alimentos, também pode reduzir as
concentrações séricas de albumina em aves (Kaneko et al., 1997).
O comportamento dos resultados dos níveis séricos de albumina nas aves
alimentadas com níveis de metionina + cistina é semelhante aos dados obtidos dos
níveis de proteínas totais séricas, sendo observado comportamento quadrático
significativo (P<0,05), entre os tratamentos correspondentes aos níveis de metionina +
cistina digestível de Rostagno et al. (2005), conforme as figuras 8 e 9. Os níveis de
metionina + cistina digestível recomendados, a partir da derivada das equações das
funções obtidas para albumina e proteínas séricas, é de 91,25 e 87,40%,
correspondente ao nível de 0,324 e 0,311% de metionina + cistina digestível,
respectivamente.

54
Figura 8 – Efeito dos tratamentos sobre o nível de albumina (g/dL) de poedeiras leves com 18
semanas de idade, alimentadas com níveis de metionina + cistina.
Figura 9 – Efeito dos tratamentos sobre o nível de proteína sérica (g/dL) de poedeiras leves
com 18 semanas de idade, alimentadas com níveis de metionina + cistina.
A albumina é um dos principais componentes das proteínas séricas, fato este
que pode explicar o comportamento semelhante entre essas duas variáveis.
Os níveis mais elevados de albumina e proteínas séricas nas aves alimentadas
com os tratamentos correspondentes aos níveis de metionina + cistina das tabelas
brasileiras podem ser explicados pelo início da produção de ovos. Segundo Schmidt et
Ŷ = -0,0008x2 + 0,146x - 4,3476
R² = 0,9741
0
0,5
1
1,5
2
2,5
3
80 90 100 110 120
Alb
um
ina
(g
/dL
)
Níveis de metionina + cistina digestível (Rostagno et al., 2005)
y = -0,001x2 + 0,1923x - 3,68
R² = 0,955
3,5
4
4,5
5
5,5
80 90 100 110 120
Pro
teín
a S
éric
a (
g/d
L)
Níveis de metionina + cistina digestível (Rostagno et al., 2005)
91,25
%
87,40
%

55
al. (2007), a produção de ovos pode afetar as concentrações de proteínas totais,
incluindo a albumina. Galinhas poedeiras antes da postura podem apresentar
hiperproteinemia induzida por estrógenos (Campbell, 2004), fato que pode explicar os
níveis elevados de albumina e proteínas séricas das aves. Souza et al. (2013) não
observaram efeito da suplementação com níveis de aminoácidos sulfurosos para
codornas de corte sobre o nível sérico de albumina.
As aves alimentadas com os tratamentos com 80 e 100% das exigências de
metionina + cistina digestível apresentaram maior nível da enzima aspartato
aminotransferase, quando comparado com o tratamento controle. Para a atividade da
alanina aminotransferase, apenas o tratamento com 100% das exigências nutricionais
de metionina + cistina digestível diferiu do tratamento controle.
De acordo com Kaneko et al. (1997), o aumento significativo da aspartato
aminotransferase sugere lesão hepática grave e difusa, além de distúrbios musculares,
pois a aspartato aminotransferase não é hepato-específica. No entanto, para que seja
considerada lesão hepática considerável, os valores dessa enzima devem ser superiores
a 275 U/L (Schmidt et al., 2007) e, dessa forma, não foi observada lesão no fígado das
aves que receberam os níveis de metionina + cistina.
Souza et al. (2013) avaliaram as atividades enzimáticas séricas (aspartato
aminotransferase e alanina aminotransferase) de codornas de corte alimentadas com
níveis de metionina + cistina: lisina digestível durante a fase inicial e não observaram
efeito dos tratamentos sobre os níveis das enzimas estudadas. Essas enzimas não
possuem atividade específica para o fígado, sendo encontrada atividade em outros
órgãos, como o coração e músculo, e por esse fato, não podem ser utilizadas
exclusivamente como indicador de lesões hepáticas.
De acordo com Borsa et al. (2006), os valores de referência de perfil
bioquímico em aves comerciais são escassos na literatura mundial, não havendo
padronização quanto a atividade produtiva, linhagem, período, região, nutrição, entre
outros fatores, dificultando a interpretação dos resultados de pesquisas nessa área.
As aves alimentadas com o tratamento com 80% das exigências de metionina +
cistina digestível apresentaram maior atividade da enzima gamma-glutamiltransferase,
quando comparado com o tratamento controle. A atividade sérica elevada dessa

56
enzima se dá pelo aumento da produção e liberação pelo tecido hepatobiliar (Meyer et
al., 1995). Provavelmente, a deficiência de aminoácidos sulfurosos pode ter provocado
a elevação da atividade da gamma-glutamiltransferase, nas aves alimentadas com a
dieta com 80% das exigências de metionina + cistina digestível.
Segundo Schmidt et al. (2007), os valores de GGT entre 0-10 U/L são
considerados normais, no entanto, as diferenças nas metodologias para determinação
da GGT podem contribuir nas variações nos valores de referência dessa enzima
(Lumeij, 1997). Além disso, os níveis de GGT elevados encontrados nesse estudo
podem ser explicados pela idade das aves, já que animais jovens apresentam valores
elevados dessa enzima.
De acordo com os resultados de GGT, AST e ALT obtidos no estudo, não foi
possível observar lesões hepáticas significativas, com o aumento dos níveis de
metionina + cistina na dieta.
Os animais que receberam as dietas com a variação dos níveis de metionina +
cistina (80 a 120% de metionina + cistina digestível) apresentaram maior nível sérico
de creatinina, quando comparados com os animais do tratamento controle. Não foi
observado efeito significativo (P<0,05) dos níveis de aminoácidos sulfurosos sobre a
atividade da creatinina nas aves, após a análise de regressão. No entanto, os níveis de
metionina + cistina nas dietas não foram capazes de ocasionar uma deficiência na
funcionalidade renal das aves.
Batista et al. (2013) avaliaram os efeitos da suplementação de lisina e arginina
sobre os níveis séricos de creatinina e ácido úrico de codornas de corte e também não
observaram efeito significativo dos tratamentos sobre as variáveis analisadas. Esses
autores ainda afirmam que altos níveis de creatinina indicam deficiência na
funcionalidade renal e valores baixos podem ser explicados pela baixa ingestão de
proteínas. Da mesma forma, Rojas (2012) também não observou efeito significativo
dos níveis de glicerina + serina e do excesso de treonina sobre os níveis séricos de
creatinina em frangos de corte na fase inicial.

57
Análises Histológicas
De acordo com a Figura 10 (C e D), é possível observar que as aves
alimentadas com 100 e 110% das exigências de metionina + cistina digestível das
tabelas brasileiras, apresentaram menor acúmulo de glicogênio hepático, com menor
positividade para coloração periodic acid Schiff. Todavia, os animais alimentados com
as dietas com os menores níveis de metionina + cistina digestível (Figura, 10 B), não
apresentaram redução no acúmulo de glicogênio hepático.
Figura 10 – Fotomicrografias do fígado de poedeiras leves com 18 semanas de idade
suplementadas com níveis de metionina + cistina. A) Imagem representando o tratamento controle
(NRC, 1994); B) Imagem representando os tratamentos 80 e 90% de metionina + cistina digestível
(Rostagno et al., 2005); C) Imagem representando o tratamento com 100% de metionina + cistina
digestível (Rostagno et al., 2005); D) Imagem representando o tratamento com 110% de metionina +
cistina digestível (Rostagno et al., 2005); E) Imagem representando o tratamento com 120% de
metionina + cistina digestível (Rostagno et al., 2005). Coloração periodic acid Schiff. Aumento de
100x.
Segundo Lehnninger et al. (1995), o glicogênio hepático é uma fonte de
carboidrato fundamental para o organismo, servindo como fonte de energia e
A
)
B
)
C
)
D
)
E
)

58
fornecedor de glicose para a manutenção da homeostase glicêmica. Possivelmente, a
deficiência de aminoácidos sulfurosos reduziu a deposição de glicogênio hepático, que
é resultado da sua conversão a α-cetobutirato e consequente transformação em energia,
sendo acumulada a nível hepático. O menor acúmulo de glicogênio nos animais dos
tratamentos com 100 e 110% das exigências de metionina + cistina digestível das
tabelas brasileiras também pode estar atribuído ao amadurecimento mais precoce do
sistema reprodutivo, com deposição de energia excedente em outros tecidos.
Aliado a isso, observa-se a presença de vacúolos citoplasmáticos de lipídios
nos hepatócitos das aves alimentadas com o tratamento com 100% de metionina +
cistina digestível. No entanto, não foi possível observar esse efeito nos tratamentos
com os menores níveis de metionina + cistina digestível (Figura 10, B), assim como
não foi evidenciado o processo de esteatose hepática em nenhum dos tratamentos.
Uma maior presença de lipídios nos hepatócitos pode indicar o início da
síndrome do fígado gorduroso, ocasionado pela deficiência de aminoácidos sulfurosos.
A metionina pode atuar no metabolismo orgânico como um agente lipotrópico, tanto
pela doação de grupamentos metil, como também pelo envolvimento no metabolismo
da colina, betaína, ácido fólico e da vitamina B12 (Chen et al., 1993). Pompeu et al.
(2011) não observaram efeito significativo sobre a histopatologia de frangos de corte
machos alimentados com níveis crescentes de colina, resultados semelhantes aos
relatados por Wolford & Murphy (1972), que não detectaram nenhuma alteração no
conteúdo de gordura no fígado de aves que receberam dietas suplementadas com
vitaminas lipotrópicas.
Provavelmente, a deficiência de metionina + cistina observada no tratamento
com 100% das exigências desse aminoácido das tabelas brasileiras, pode ter
ocasionado o início do acúmulo de gordura no fígado, já que, segundo Brumano
(2008), as reações de metilação são essenciais no metabolismo da gordura no fígado,
evitando a síndrome do fígado gorduroso nas aves.
De acordo com Jordão Filho et al. (2006), rações deficientes em metionina
aumentam a deposição de gordura no fígado. De acordo com os dados de desempenho
zootécnico, poedeiras na fase de crescimento (13 a 18 semanas de idade) possuem uma
exigência para metionina + cistina digestível acima do valor de 20% superior ao nível
recomendado por Rostagno et al. (2005).

59
De acordo com as imagens de microscopia eletrônica de varredura do intestino
delgado das poedeiras leves com 18 semanas de idade (Figura 11), é possível observar
aumento na altura das vilosidades intestinais, a partir da suplementação com metionina
+ cistina nas dietas. A imagem representando as aves alimentadas com o tratamento
com 120% de metionina + cistina digestível (Rostagno et al., 2005) demonstra maior
altura das vilosidades intestinais, ilustrado pelo linha vermelha na imagem (Figura 8,
C), evidenciando o efeito benéfico desse aminoácido no aumento da área de superfície
de absorção do intestino delgado.
Figura 11 – Imagens de microscopia eletrônica de varredura do intestino delgado de poedeiras
leves com 18 semanas de idade suplementadas com níveis de metionina + cistina. A) Imagem
representando os tratamentos, controle (NRC, 1994), 80 e 90% de metionina + cistina digestível
(Rostagno et al., 2005); B) Imagem representando os tratamentos com 100 e 110% de metionina +
cistina digestível (Rostagno et al., 2005); C) Imagem representando o tratamento com 120% de
metionina + cistina digestível (Rostagno et al., 2005).
Resultados semelhantes foram encontrados por Soares et al. (2008), que
observaram aumento na altura das vilosidades intestinais e maior profundidade de
cripta em frangos de corte alimentados com dietas com níveis de metionina + cistina
digestível, na fase pré-inicial. Semelhantemente, Lima et al. (2012) também
observaram maior largura das vilosidades intestinais em galinhas poedeiras leves
alimentadas com níveis de triptofano digestível.
Segundo Gomide Junior et al. (2004), o desenvolvimento intestinal está
relacionado com o consumo de nutrientes e consequente aumento do diâmetro do
intestino. Aliado a isso, o desenvolvimento da mucosa intestinal consiste no aumento
da altura e densidade dos vilos, correspondendo a um aumento em número de suas
A B C

60
células epiteliais (enterócitos, células caliciformes e enteroendócrinas) (Maiorka et al.,
2002). Com o aumento das vilosidades e, consequentemente, maior área de contato do
alimento com as células absortivas (enterócitos), ocorre aumento da absorção
intestinal (Aptekmann et al., 2001; Gomide Júnior et al., 2004).
A presença de agentes tróficos, neste caso os aminoácidos sulfurosos,
provavelmente estimularam o desenvolvimento da mucosa intestinal, a partir do
processo de síntese de novas células epiteliais e sendo fator determinante para o
equilíbrio do turnover proteico, pois, de acordo com Rezende e Beletti (2008), o
desenvolvimento das vilosidades intestinais em galinhas poedeiras ocorre
intensamente até a nona semana de idade.
Segundo Tarachi & Yamauchi (2000), na presença de nutrientes, o lúmen
intestinal apresenta maior desenvolvimento, traduzido pela altura dos vilos, número de
células em mitose e extrusão, sugerindo que o estímulo primário para o
desenvolvimento da mucosa são as características químicas dos nutrientes.
O magno das poedeiras leves com 18 semanas de idade, alimentadas com os
tratamentos com 110 e 120% das exigências de metionina + cistina digestível
apresentaram um aumento no número de glândulas funcionais secretoras de albúmen
(setas), além de maior presença de dobras secundárias do órgão (Figura 12, C), que
aumenta a área epitelial do magno.

61
Figura 12 – Fotomicrografias do magno de poedeiras leves com 18 semanas de idade
suplementadas com níveis de metionina + cistina e imagem de microscopia eletrônica do magno. A)
Imagem de microscopia eletrônica do magno de poedeiras leves com 18 semanas de idade, com
destaque para as dobras do órgão; B) Imagem representando os tratamentos, controle (NRC, 1994), 80,
90 e 100% de metionina + cistina digestível (Rostagno et al., 2005); C) Imagem representando os
tratamentos com 110 e 120% de metionina + cistina digestível (Rostagno et al., 2005). Coloração
hematoxilina-eosina (100x). Setas: estoques de albúmen; Pontas de setas: dobras secundárias do magno.
Estes resultados indicam um desenvolvimento mais precoce do trato
reprodutivo das aves alimentadas com os maiores níveis de suplementação com
aminoácidos sulfurosos na dieta. Aliado a isso, o aumento da área epitelial do magno
facilita a passagem do ovo em formação no oviduto, já que o epitélio desse órgão é
rico em células caliciformes que produzem muco.
Efeito semelhante foi encontrado por Lima et al. (2012), que observaram
aumento na produção de albúmen pelo magno de poedeiras leves suplementadas com
níveis de triptofano digestível. Segundo Novak et al. (2004), a síntese de proteína no
tecido do magno, local de produção do albúmen, pode ser afetada pela concentração
de aminoácidos no sangue. Dessa forma, possivelmente a maior concentração de
B C
A

62
aminoácidos sulfurosos no sangue permitiu um melhor desenvolvimento das pregas do
magno, com consequente aumento no número de células secretoras de albúmen nas
aves alimentadas com os maiores níveis de metionina + cistina na dieta.
Peso dos Órgãos
Houve efeito quadrático significativo (P<0,05) entre os níveis de metionina +
cistina digestível para o peso do baço de poedeiras leves com 18 semanas de idade.
Para as demais variáveis, não houve efeito significativo (P>0,05) entre os tratamentos,
tanto pelo teste Dunnet, quanto pela análise de regressão (Tabela 6).
Tabela 6 - Efeitos dos tratamentos sobre o peso relativo (%) do peso sem vísceras (PSV), fígado (FIG), baço
(BAÇO) e gordura celomática (GCE) das aves com 18 semanas de idade.
*Relação metionina + cistina: lisina (%); ns: não significativo; Q*: Efeito significativo ao modelo de regressão
quadrática a 5% de probabilidade; C.V. (%): Coeficiente de variação.
O nível de metionina + cistina digestível estimado pela derivada da equação do
gráfico (Figura 13), para o peso do baço das aves com 18 semanas de idade é de
100,44%, correspondente ao nível de 0,398% na dieta. O aumento do peso do baço das
aves suplementadas com níveis de metionina + cistina pode estar atribuído à maior
modulação da resposta imune das aves, já que esse órgão é caracterizado como uma
das principais glândulas responsáveis pela atividade imunológica das aves, juntamente
com a Bursa de Fabricius.
Tratamento
MET:LIS* PSV BAÇO FIG GCE
NRC, 1994 93 89.26 0.22 2.36 3.24
Rostagno et al.
(2011)
80% 66 90.51 0.22 2.40 3.60
90% 74 89.21 0.28 2.28 4.16
100% 82 91.49 0.28 2.20 4.38
110% 90 90.58 0.26 2.12 3.74
120% 98 90.67 0.24 2.30 4.12
Regressão Rostagno et al. (2011) ns Q* ns ns
C.V. (%) 0.98 11.02 4.52 10.91

63
Segundo Rubin (2007), a suplementação dietética com aminoácidos sulfurosos
e a quebra das proteínas teciduais são necessárias para a síntese de proteínas e dos
peptídeos envolvidos no funcionamento normal do sistema imunológico. No entanto, o
mecanismo de modulação da resposta imune frente à suplementação com metionina +
cisitina para galinhas poedeiras ainda é controverso.
Figura 13 – Efeito dos tratamentos sobre o peso relativo (%) do baço de poedeiras leves com
18 semanas de idade, alimentadas com níveis de metionina + cistina.
De acordo com Grimble (2006), o mecanismo da variação da disponibilidade
tecidual de aminoácidos sulfurosos e os produtos de seu metabolismo, interagindo com
as respostas imunes, ainda são desconhecidos. Esse fato pode estar atribuído às
diversas interrelações dos aminoácidos sulfurosos com a modulação da resposta imune
nas aves.
Segundo Tsiagbe et al. (1987) a suplementação de metionina em dietas a base
de milho e farelo de soja aumentaram a produção de anticorpos e elevaram a
estimulação mitogênica, provocada pela fitohematoglutinina.
Outra forte relação dos aminoácidos sulfurosos com a atividade imunológica de
galinhas poedeiras se dá através da síntese de glutationa., já que a cisteína, obtida
através da metionina, é precursora da glutationa em células animais e seu metabolismo
y = -0,000129x2 + 0,025914x - 1,024000
R² = 0,865546
0,2000
0,2100
0,2200
0,2300
0,2400
0,2500
0,2600
0,2700
0,2800
0,2900
80 90 100 110 120
Pes
o d
o b
aço
Níveis de metionina + cistina digestível (Rostagno et al., 2005)
100,44
%

64
é significativamente alterado em resposta a processos infecciosos (Malmezat et al.
2000). Outro fator importante é que a síntese desse importante antioxidante celular é
influenciado pelo consumo dietético de aminoácidos sulfurosos (Wu et al. 2004) e,
dessa forma, observa-se uma correlação positiva entre a via de transulfuração e as
concentrações de glutationa no fígado, baço e músculos (Malmezat et al., 2000).
Ademais, segundo Fang et al. (2002), a glutationa destrói radicais livres e outras
substâncias oxidativas (radicais hidroxila, lipoperóxidos, peróxido de hidrogênio),
além de participar do processo de detoxificação de substâncias tóxicas ao organismo
dos animais.
Segundo Rubin (2007), a síntese de glutationa no fígado utiliza como enzima
precursora a gama-glutamil cisteína sintetase. Dessa forma, a conversão de cisteína à
glutationa é altamente influenciada pela taxa de utilização e do transporte de glutationa
dentro e entre as células do corpo, ou seja, a síntese desse antioxidante celular é um
processo conduzido pela demanda orgânica, desde que a cisteína esteja disponível.
Outra forma de estímulo da resposta imune a partir da suplementação com
aminoácidos sulfurosos se dá pela participação ativa da metionina como doadora de
grupamentos metil. Segundo Wu et al. (2006), sob a forma da S-adenosilmetionina, a
metionina doa grupamentos metil que participam da metilação do DNA e proteínas,
além da regulação da expressão gênica. Aliado a isso, a metionina age como substrato
para a síntese da colina, assim como da fosfatidilcolina e acetilcolina, que são
essenciais para as funções nervosas do organismo e para o metabolismo dos leucócitos
(Kim et al., 2007).
Assim, a deficiência de aminoácidos sulfurosos nas dietas de galinhas
poedeiras pode reduzir a atividade do sistema imunológico, haja vista a grande
participação desses aminoácidos na resposta imune. A suplementação com
aminoácidos sulfurosos em dietas de galinhas poedeiras pode melhorar o efeito das
respostas imunológicas e consequentemente aumentar o peso do baço. No entanto, é
necessário evitar o excesso desses aminoácidos na dieta, devido aos possíveis efeitos
negativos do imbalanço aminoacídico, com consequente redução na ingestão e
utilização de outros aminoácidos.
Para as demais variáveis, as diferentes bases de formulação das rações
(aminoácidos totais vs aminoácidos digestíveis), assim como os níveis de metionina +

65
cistina digestível, não foram capazes de alterar o peso relativo do fígado e da gordura
celomática das aves. No entanto, mesmo não apresentando efeito significativo
(P>0,05), o peso do fígado das aves alimentadas com os tratamentos com 100 e 110%
de metionina + cistina digestível foi menor, quando comparado com os demais
tratamentos. Esse resultado pode ser explicado pelo menor acúmulo de glicogênio
hepático das aves desses tratamentos, representados pela menor positividade à
coloração periodic acid Schiff.
De acordo com Bertechini (2012), o desenvolvimento dos órgãos vitais e o
crescimento visceral das frangas de reposição ocorrem principalmente de 1 a 5
semanas de idade. Esse fato pode explicar a ausência de diferença significativa entre
os tratamentos para o peso relativo do fígado das aves com 18 semanas de idade.
O fígado é o principal órgão responsável pelo metabolismo e distribuição dos
nutrientes no organismo. Dessa forma, o fluxo de nutrientes nesse órgão pode
aumentar a sua taxa metabólica e consequentemente alterar o seu peso. No entanto, os
níveis de aminoácidos sulfurosos avaliados nesse estudo não foram capazes de
modificar de forma significativa o metabolismo e o peso relativo do fígado das aves no
período experimental.
Avaliação do Efeito Residual
Não foi observado efeito estatístico significativo (P>0,05) dos tratamentos
oferecidos no período experimental (13 a 18 semanas de idade) sobre as variáveis
inerentes ao início de produção de ovos (19 a 29 semanas de idade), tanto pelo teste
Dunnet, quanto pela análise de regressão (Tabela 7).

66
Tabela 7 - Efeito residual dos tratamentos sobre a idade ao primeiro ovo (IPO, dias), peso ao primeiro ovo (PPO,
g), número de ovos no período (NOP, total de ovos), números de dias de produção (NDP, dias), taxa
de postura (TXP, %), peso médio dos ovos (PMO, g) e massa de ovos (MO, g/ave/dia) das aves de 19
a 29 semanas de idade.
Tratamento IPO PPO NOP NDP TXP PMO MO
NRC 1994 148.38 41.88 230.25 59.63 54.66 58.47 31.99
Rostagno et al. (2005)
80% 148.13 44.00 256.88 59.88 56.86 59.60 33.88
90% 143.13 42.50 246.88 64.88 51.96 58.51 30.35
100% 140.25 41.88 244.63 67.75 48.19 58.33 28.11
110% 140.75 41.75 260.38 67.25 50.87 58.05 29.55
120% 139.75 41.75 250.50 68.25 48.95 58.68 28.76
Regressão
Rostagno et al. (2005) ns ns ns ns ns ns ns
C.V.(%) 6.27 8.47 25.25 13.91 19.85 2.42 19.86
C.V.(%): Coeficiente de variação; ns: não significativo.
Os níveis de aminoácidos sulfurosos, presentes nas dietas das aves no período
experimental, não foram capazes de afetar a precocidade sexual das aves, assim
como a taxa de produção no início da postura. D’Agostini et al. (2012), em estudo
semelhante, avaliaram a suplementação de metionina + cistina na fase de recria (13 a
18 semanas de idade) para poedeiras leves e os efeitos residuais nas fases
subsequentes. Esses autores observaram que o fornecimento do menor nível de
aminoácidos sulfurosos (0,399% de metionina + cistina total) retardou o início da
postura nas aves, atingindo 50% de produção com 161 dias. Também foi relatado que
as aves que foram alimentadas com níveis superiores a 0,469% de metionina +
cistina total, atingiram 50% de produção aos 150 dias de idade, ou seja, com onze
dias de antecedência.
Ao final do período de avaliação das dietas experimentais, todas as aves
receberam ração basal de acordo com as exigências nutricionais estabelecidas por
Rostagno et al. (2005) e possivelmente, apresentaram recuperação metabólica na fase
subsequente avaliada. Dessa forma, é possível afirmar que a restrição nutricional
(aminoácidos sulfurosos) nesse estudo, provocada no período experimental, não afeta
o desempenho das aves no início de produção.

67
No entanto, esse comportamento pode variar de acordo com o tipo de restrição
nutricional. Araújo et al. (2008) observaram que o aumento da fibra, provocado pela
inclusão do farelo de trigo na dieta de poedeiras semipesadas na fase de recria, reduz
a taxa de crescimento das frangas e retarda o início da postura. Dessa forma, é
necessário que se busque constantemente informações acerca dos efeitos do
fornecimento de dietas na fase de crescimento das frangas, sobre a precocidade
sexual e produtividade dos animais no início da postura.
Houve efeito estatístico significativo (P<0,05) para a espessura da casca entre o
tratamento controle e os tratamentos com 90, 100 e 110% dos níveis de metionina +
cistina digestível das tabelas brasileiras, para o período de avaliação do efeito
residual de 34 a 50 semanas de idade (Tabela 8). Ademais, houve efeito quadrático
significativo (P<0,05) para a percentagem de gema entre os tratamentos com os
níveis de metionina + cistina digestível das tabelas brasileiras, para o mesmo
período.
Tabela 8 - Efeito residual dos tratamentos sobre a produção de ovos (PR, %), peso de ovo (PO, g), massa de
ovos (MO, g/ave/dia), gravidade específica (GE, g/mL), espessura de casca (ESPC, mm),
percentagem de albúmen (ALB, %), gema (GEM, %) e casca (CASCA, %), das aves de 34 a 50
semanas de idade.
Tratamento PR PO MO GE ESPC ALB GEM CASCA
NRC 1994 96.11 66.19 63.61 1.0844 509.16 60.97 26.36 9.70
Rostagno et al. (2005)
80% 93.77 66.58 62.41 1.0835 496.17 61.93 25.56 9.52
90% 95.48 65.34 62.34 1.0846 490.51¥ 61.65 25.76 9.48
100% 92.99 65.78 61.16 1.0842 491.94¥ 61.03 26.52 9.61
110% 95.81 65.83 63.07 1.0833 492.13¥ 60.84 26.77 9.57
120% 96.71 66.30 64.12 1.0834 500.99 61.46 26.01 9.55
Regressão
Rostagno et al. (2005) ns ns ns ns ns ns Q* ns
C.V.(%) 2.91 2.13 3.03 0.17 2.11 1.35 2.93 2.68
C.V.(%): Coeficiente de variação; ¥: médias diferentes pelo teste Dunnett a 5% de probabilidade; Q* Efeito
significativo ao modelo de regressão quadrática a 5% de probabilidade.
As aves alimentadas com os tratamentos com 90, 100 e 110% dos níveis de
metionina + cistina digestível apresentaram menor espessura de casca, quando
comparadas com as aves que receberam o tratamento controle. No entanto, as aves que
receberam o tratamento com 120% das exigências de metionina + cistina digestível

68
não diferiram estatisticamente das aves que foram alimentadas com o tratamento
controle, pelo teste Dunnet.
A menor espessura da casca das aves alimentadas com os tratamentos com 90,
100 e 110% dos níveis de metionina + cistina pode ser explicada pelo aumento no
tamanho dos ovos. Esses resultados são semelhantes aos encontrados por Cupertino et
al. (2009), que também observaram redução no percentual de casca, à medida que
aumentaram os níveis de aminoácidos sulfurosos nas dietas de galinhas poedeiras
leves, e, da mesma forma, sendo atribuída a justificativa da redução da qualidade da
casca, devido ao aumento do tamanho dos ovos.
O nível estimado de metionina + cistina digestível, a partir da derivada da
equação referente à porcentagem de gema, no período de 34 a 50 semanas, foi de
107,79%, correspondente ao nível de 0,426% de metionina + cistina digestível na
ração (Figura 14).
Figura 14 – Efeito residual dos tratamentos sobre a porcentagem de gema de poedeiras leves de
34 a 50 semanas de idade, alimentadas com níveis de metionina + cistina.
Esses resultados demonstram que as aves alimentadas com os maiores níveis de
aminoácidos sulfurosos durante o período de avaliação das dietas experimentais
apresentaram maior percentual de gema durante a postura, devido à maior
disponibilidade de aminoácidos para a formação da gema dos ovos. Esses resultados
y = -0,0017x2 + 0,3665x + 7,1909
R² = 0,7635
25,2
25,4
25,6
25,8
26
26,2
26,4
26,6
26,8
27
80 90 100 110 120
Po
rce
nta
gem
de
ge
ma
Níveis de metionina + cistina digestível (Rostagno et al., 2005)
107,79
%

69
são semelhantes aos encontrados por diversos pesquisadores (Shafer et al., 1998;
Harms & Russel, 2003; Novak & Scheideler, 2004), que comprovaram aumento nos
componentes internos dos ovos (gema e albúmen), ocasionado pelo aumento do nível
de metionina da dieta.
Não houve efeito significativo (P>0,05) dos níveis de metionina + cistina da
fase de recria para nenhuma das variáveis avaliadas no período de 54 a 70 semanas de
idade das aves (postura II) (Tabela 9).
Tabela 9 - Efeito residual dos tratamentos sobre a produção de ovos (PR, %), peso de ovo (PO, g), massa de ovos
(MO, g/ave/dia), percentagem de albúmen (ALB, %), gema (GEM, %) e casca (CASCA, %),
gravidade específica (GE, g/mL) e espessura de casca (ESPC, mm) das aves de 54 a 70 semanas de
idade.
Tratamento PR PO MO GE ESPC ALB GEM CASCA
NRC 1994 88.38 67.01 59.23 1.0779 491.19 60.03 27.41 9.57
Rostagno et al. (2005)
80% 86.78 66.49 57.70 1.0764 488.02 60.71 26.87 9.20
90% 90.29 64.95 58.60 1.0774 478.44 59.94 27.88 9.30
100% 88.53 65.45 57.95 1.0789 476.61 60.68 26.94 9.21
110% 88.35 66.05 58.36 1.0769 477.07 59.94 27.10 9.24
120% 89.46 66.43 59.43 1.0768 479.61 60.94 27.87 9.23
Regressão
Rostagno et al. (2005) ns ns ns ns ns ns ns ns
C.V.(%) 3.51 2.46 4.16 0.23 2.91 2.10 5.66 4.27
C.V.(%): Coeficiente de variação; ns: não significativo.
Esses resultados demonstram a capacidade de recuperação metabólica das aves
que receberam dietas deficientes em aminoácidos sulfurosos durante o período de
avaliação experimental, a partir do momento em que é fornecida uma dieta basal,
atendendo as exigências nutricionais das aves nas fases subsequentes. Segundo
D’Agostini et al. (2012), a inclusão de níveis de metionina + cistina na fase de recria
afeta as características produtivas das aves nesta idade, no entanto, não mantém seus
efeitos nas aves durante a fase de postura. Esses autores ainda afirmam que as aves
alimentadas com deficiência de aminoácidos sulfurosos na fase de recria podem
recuperar seu desempenho produtivo, caso recebam dietas adequadas durante a fase de
produção.

70
CONCLUSÕES
Recomendam-se níveis superiores a 0,475% de metionina + cistina digestível e
relação metionina + cistina:lisina superior a 98, para aves de reposição leves de 13 a
18 semanas de idade.
REFERÊNCIAS BIBLIOGRÁFICAS
APTEKMANN, K. P. et al. Morphometric analysis of the intestine of domestic quails
(Coturnix coturnix japonica) treated with different levels of dietary calcium. Anat.
Hist. Embryol. 30:277–280. 2001.
ARAÚJO, D.M.; SILVA, J.H.V.; ARAÚJO, J.A. et al. Farelo de trigo na alimentação de
poedeiras semipesadas na fase de recria. Revista Brasileira de Zootecnia, v.37, p.67-
72, 2008.
BAKER, D.H.; FERNANDEZ, S.R.; WEBEL, D.M. et al. Sulfur amino acid requirement
and cystine replacement value of broiler chicks during the period three to six weeks
post-hatching. Poultry Science, v.75, p.737-42, 1996.
BATISTA, E.; ZANCANELA, V.; FURLAN, A.C. et al. Níveis séricos de creatinina e
ácido úrico em codornas de corte dos 15 a 35 dias de idade suplementadas com
diferentes níveis de lisina e arginina digestível. In: SIMPÓSIO INTERNACIONAL
DE COTURNICULTURA, 2013, Lavras-MG. Anais...Lavras-MG, 2013.

71
BORSA, A.; KOHAYAGAWA, A.; BORETTI, L.P. et al. Níveis séricos de enzimas de
função hepática em frangos de corte de criação industrial clinicamente saudáveis.
Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.58, p.675-677, 2006.
BRUMANO, G. Níveis de metionina + cistina digestíveis em rações para poedeiras
leves nos períodos de 24 a 40 e de 42 a 58 semanas de idade. 2008, 103f. Tese
(Doutorado), Universidade Federal de Viçosa, UFV, Viçosa – MG.
CAMPBELL, T.W. Clinical Chemistry of Birds. In: THRALL, M.A. Veterinary
Hematology and Clinical Chemistry. Philadelphia, Lippincott, Williams & Wilkins,
2004, p.479-492.
CASARTELLI, E.M.; JUNQUEIRA, O.M.; FILARDI, R.S. et al. Formulações de rações
para poedeiras comerciais utilizando-se diferentes recomendações de aminoácidos
totais e digestíveis. Revista Brasileira de Ciência Avícola, v.6, p.25, 2004 (supl.).
CHEN, P.; JOHNSON, P.; SOMMER, T. et al. Multiple ubiquitin-conjugating enzymes
participate in the in vivo degradation of the yeast MATα2 repressor. Cell, v.74, p. 357-
369, 1993.
CUPERTINO, E.S.; GOMES, P.C.; ROSTAGNO, H.S. et al. Exigência nutricional de
metionina + cistina digestíveis para galinhas poedeiras de 54 a 70 semanas de idade.
Revista Brasileira de Zootecnia, v.38, p.1238-1246, 2009.
D’AGOSTINI, P.C.; MELLO, H.H.C.; CALDERANO, A.A. et al. Exigência de
metionina + cistina para frangas de reposição na fase de recria de 13 a 18 semanas de
idade. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.64, p.1691-
1698, 2012.
JORDÃO FILHO, J.; SILVA, J.H.V.; SILVA, E.S. et al. Exigências nutricionais de
metionina + cistina para poedeiras semipesadas do início de produção até o pico de
postura. Revista Brasileira de Zootecnia, v.35, p.1063-1069, 2006 (supl.).
GOMIDE JUNIOR, M.H.; STERZO, E.V.; MACARI, M. et al. Use of scanning electron
microscopy for the evaluation of intestinal epithelium integrity. Revista Brasileira de
Zootecnia, v.33, p.1500-1505, 2004.

72
GRIMBLE, R.F. The effects of sulfur amino acid intake on immune function in humans.
Journal of Nutrition, v.136, p.1660–1665, 2006.
HAMILTON, R.M.G. Methods and factors that affect the measurement of egg shell
quality. Poultry Science, v.61, p.2002-2039, 1982.
HARMS, R.H.; RUSSEL, G.B. Performance of commercial laying hens fed diets with
various levels of methionine. Journal of Applied Poultry Research, v.12, p.449-455,
2003.
KANEKO, J.J.; HARVEY, J.W.; BRUSS, M.L. Clinical Biochemistry of Domestic
Animals, 5th ed., San Diego, Academic Press, 1997, 932p.
KIM, S.W.; MATEO, R.D.; YIN, Y.L. et al. Functional amino acids and fatty acids for
enhancing production performance of sows and piglets. Asian-Australasian Journal
of Animal Science, v.20, p.295–306, 2007.
KWAKKEL, R.P. Rearing the layer pullet – A multiphasic approach. In: WISEMAN, J.;
GARNSWORTHY, P.C. (Eds.) Recent development in poultry nutrition. 2.ed.
Nottingam: Nottingam University Press, 1999. p.227-249.
LEHNINGER, A. L.; NELSON, D. L; COX, M. M. Princípios de bioquímica. São
Paulo: Sarvier, 1995. 839 p.
LIMA, M.R., COSTA, F.G.P., GUERRA, R.R. et al. Digestible tryptophan:lysine ratio
for laying hens. Revista Brasileira de Zootecnia. v.41, p.2203-2210, 2012.
LUMEIJ, J.T. Avian Clinical Biochemistry. In: KANEKO, J.J.; HARVEY, J.W.;
BRUSS, M.L. Clinical Biochemistry of Domestic Animals. 5th edition. San Diego,
Academic Press, 1997. 932p.
MANUAL DE MANEJO DAS POEDEIRAS DEKALB WHITE. Granja Planalto. 5 ed.
Minas Gerais: Uberlândia, 2009. 42p.
MEYER, D.J.; COLES, E.H.; RICH,L, J. Medicina de laboratório veterinário:
interpretação e diagnóstico. São Paulo: Roca, 1995. 308 p.
NATIONAL RESEARCH COUNCIL. Nutrient requirements of poultry. 9 ed.
Washington, D.C.: National Academy Press, 1994. 157p.

73
NEME, R.; SAKOMURA, N.K.; FUKAYAMA, E.H. et al. Curvas de crescimento e de
deposição dos componentes corporais em aves de postura de diferentes linhagens.
Revista Brasileira de Zootecnia, v.35, p.1091-1100, 2006 (supl.).
NOVAK C.; YAKOUT, H.; SCHEIDELER, S. The combined effects of dietary lysine
and total sulfur amino acid level on egg production parameters and egg components in
dekalb delta laying hens. Poultry Science, v.83, p.977-984, 2004.
PARK, B,C.; AUSTIC, R.E. Changes in hepatic branched-chain áketo acid
dehydrogenase activity in response to isoleucine imbalance in growing chickens.
Journal of nutrition of Biochemistry, v.9, p.687-696, 1998.
PIRES, C.V.; OLIVEIRA, M.G.A.; ROSA, J.C. et al. Qualidade nutricional e escore
químico de aminoácidos de diferentes fontes proteicas. Ciência e Tecnologia de
Alimentos, v.26, p.179-187, 2006.
POMPEU, M.A.; LARA, L.J.C.; BAIÃO, N.C. et al. Suplementação de colina em dietas
para frangos de corte machos na fase inicial de criação. Arquivo Brasileiro de
Medicina Veterinária e Zootecnia, v.63, p.1446-1452, 2011.
ROJAS, I.C.O. Estratégias nutricionais para redução dos níveis proteicos de dietas
para frangos de corte na fase inicial. 2012. 89f. Dissertação (Mestrado em
Zootecnia) – Programa de Pós-Graduação em Zootecnia/Universidade Estadual de
Maringá, Maringá-PR.
ROMBOLA, L.G.; FARIA, D.E.; DEPONTI, B.J. et al. Fontes de metionina em rações
formuladas com base em aminoácidos totais ou digestíveis para frangas de reposição
leves e semipesadas. Revista Brasileira de Zootecnia, v.37, p.1990-1995, 2008.
ROSTAGNO, H.S.; BARBARINO JR., P.; BARBOZA, W. Exigências nutricionais das
aves determinadas no Brasil. In: SIMPÓSIO INTERNACIONAL SOBRE
EXIGÊNCIAS NUTRICIONAIS DE AVES E SUÍNOS, 1996, Viçosa.
Anais...Viçosa: Universidade Federal de Viçosa, 1996, p.361-388.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. et al. Tabelas brasileiras para
aves e suínos: composição de alimentos e exigências nutricionais. 2 ed. Viçosa:
Universidade Federal de Viçosa, 2005. 186p.

74
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. et al. Tabelas brasileiras para
aves e suínos: composição de alimentos e exigências nutricionais. 3 ed. Viçosa:
Universidade Federal de Viçosa, 2011. 252p.
RUTZ, F. Proteínas: Digestão e Absorção. In: MACARI, M.; FURLAN, R.L.;
GONZALES, E.; Fisiologia Aviária Aplicada a Frangos de Corte. 2 ed. Jaboticabal:
FUNEP/UNESP, 2002, p.135-141.
SAS. SAS/STAT 9.3 User’s Guide. Cary, NC: SAS Institute Inc. 2011, 8621p.
SCHMIDT, E.M.S.; LOCATELLI-DITTRICH, R.; SANTIN, E. et al. Patologia clínica
em aves de produção – uma ferramenta para monitorar a sanidade avícola – Revisão.
Archives of Veterinary Science, v 12, p.9-20, 2007.
SHAFER, D.J.; CAREY, J.B.; PROCHASKA, J.F. Dietary methionine intake on egg
component yield, composition, functionality and texture profile analysis. Poultry
Science, v.77, p.1056-1062, 1998.
SILVA, E.L.; SILVA, J.H.V., BERTECHINI, A.G. et al. Exigência de metionina +
cistina para aves de reposição leves e semipesadas de 1 a 4 semanas de idade
alimentadas com rações farelada e triturada. Revista Brasileira de Zootecnia, v.38,
p.500-507, 2009.
SOARES, A.D.M., GIVISIEZ, P.E.N., SANTOS, E.G. et al. Efeito dos níveis de
metionina + cistina digestíveis da dieta pré-inicial sobre a morfometria intestinal de
frangos de corte. In: CONGRESSO BRASILEIRO DE ZOOTECNIA, 2008. João
Pessoa-PB. Anais... João Pessoa-PB, 2008.
SOUZA, C.S., FERNANDES, A., MORAES, G.H.K. et al. Atividades enzimáticas
séricas de codornas de corte (Coturnix coturnix coturnix) alimentadas com diferentes
níveis de metionina + cistina:lisina digestível nas rações na fase inicial. In: SIMPÓSIO
INTERNACIONAL DE COTURNICULTURA, 2013. Lavras-MG. Anais... Lavras-
MG, 2013.
TARACHAI, P. E.; YAMAUCHI, K. Effects of luminal nutrient absorption, intraluminal
physical stimulation and intravenous parenteral alimentation on the recovery reponses
of duodenal villus morphology following feed withdrawal in chickens. Poultry
Science, v.79, p.1578-1585, 2000.

75
WYSS, M.; KADDURAH-DAOUK, R. Creatine and creatinine metabolism.
Physiological Reviews, v.80, p.1107–1213, 2000.
WOLFORD, J.H.; MURPHY, D. Effect of diet on fatty liver-hemorrhagic syndrome
incidence in laying chickens. Poultry Science, v. 51, p. 2087-2094, 1972.
WU G.; BAZER, F.W.; WALLACE, J.M. et al. Intrauterine growth retardation:
implications for the animal sciences. Journal of Animal Science, v.84, p.2316–2337,
2006.

76
CAPÍTULO 3
Níveis de Metionina+Cistina para Galinhas Poedeiras Leves na Fase de
Postura

77
Níveis de Metionina + Cistina para Poedeiras Leves na Fase de Postura
RESUMO
Objetivou-se com esse trabalho avaliar os níveis de aminoácidos sulfurosos nas dietas
de galinhas poedeiras leves de 34 a 50 semanas de idade. 336 aves da linhagem
Dekalb White foram distribuídas em um delineamento inteiramente casualizado,
composto por seis tratamentos, 14 repetições e quatro aves por unidade experimental.
As dietas consistiram em um tratamento controle, formulada para atender as
exigências nutricionais de aminoácidos sulfurosos de acordo com o NRC (1994) e
outras cinco dietas que foram estabelecidas de acordo com as recomendações das
tabelas brasileiras de aves e suínos (Rostagno et al., 2011), exceto para metionina +
cistina, que consistiram em cinco variações do nível desse aminoácido (80, 90, 100,
110 e 120%), resultando nos níveis de 0,536; 0,603; 0,670; 0,737 e 0,804% de
metionina + cistina digestível. A pesquisa teve a duração de 224 dias e foi dividida em
duas fases experimentais: postura I (34 a 50 semanas de idade) e postura II (54 a 70
semanas de idade). No primeiro período da pesquisa foram avaliados os níveis de
metionina + cistina entre os tratamentos e na fase posterior foi avaliado o efeito
residual dos níveis testados na fase de postura I. Foram avaliados dados de
desempenho zootécnico e de qualidade dos ovos e histopatológicos. Houve efeito
quadrático significativo para produção e massa de ovos, e conversão por massa e por
dúzia de ovos, sendo as exigências de metionina + cistina estimadas para essas
características em 103,25% (0,691%), 102,91% (0,689%), 110,03% (0,737%) e
109,03% (0,730%), respectivamente. Também houve efeito quadrático significativo
para produção de ovos (postura II), sendo estimada a exigência de metionina + cistina
para essa variável em 105,72% (0,708%). Os níveis de metionina + cistina
influenciaram significativamente os tecidos analisados no estudo. Recomenda-se o
nível de metionina + cistina digestível de 0,737%, com relação metionina + cistina:
lisina de 90% e correspondente ao consumo de 756,53 mg/ave/dia, para galinhas
poedeiras leves durante a fase de postura (34 a 50 semanas de idade).

78
Palavras-chave: aminoácidos sulfurosos, exigências nutricionais, produção de ovos,
qualidade de ovos.

79
Levels of Methionine + Cistine for Light Laying Hens at Production Phase
ABSTRACT
The objective of this study was to evaluate the levels of sulfur amino acids in the diets
of laying hens of 34-50 weeks of age. 336 Dekalb White birds were distributed in a
completely randomized design with six treatments, 14 replicates of four birds each.
The diets consisted in a control treatment, formulated to meet the nutritional
requirements of sulfur amino acids according to the NRC (1994) and five other diets
that have been established in accordance with the recommendations of the brazilian
poultry and swine tables (Rostagno et al., 2011), except for methionine and cystine,
which consisted of five variations in the level of this amino acid (80, 90, 100, 110 and
120%), resulting in levels of 0.536, 0.603, 0.670, 0.737 and 0.804% of methionine +
cystine. The study lasted 224 days and were divided into two experimental phases:
posture one (34 to 50 weeks of age) and posture two (54 to 70 weeks old). In the first
survey period the levels of methionine and cystine between treatments and later stage
were evaluated the residual effect of the tested levels has been reported in the laying
phase one. Data production performance and egg quality, serological and histological
were evaluated. There was a significant quadratic effect for production and egg mass,
and mass and dozen conversion per eggs, with the methionine + cystine estimated for
these characteristics in 103.25% (0.691%), 102.91% (0.689%), 110.03% (0.737%) and
109.03% (0.730%), respectively. There was also significant quadratic effect for egg
production (posture two), estimated the methionine + cystine for this variable in
105.72% (0.708%). The methionine + cystine levels significantly influenced the
tissues analyzed in the study. The level of 0.737% of digestible methionine + cystine,
with 90% of methionine + cystine: lysine relationship, and 756.53 mg/bird/day of
consumption, is recommended to light laying hens in the posture phase (34 to 50
weeks old).
Key words: egg quality, egg production, nutritional requirements, sulfur amino acids.

80
INTRODUÇÃO
A produção de ovos de galinha no Brasil, de acordo com dados do IBGE
(2013), foi de 668,3 milhões de dúzias no primeiro trimestre de 2013 e apresentando-
se constante nos últimos três anos. Desse total da produção de ovos, aproximadamente
99% é destinado ao cosumo interno e apenas 1% dos ovos de galinha são exportados
(Ubabef, 2012). Esse fato pode estar atribuído aos baixos índices de industrialização
desses produtos, que viabilizaria a sua comercialização ao exterior.
Nesse contexto, a melhor qualidade desse produto é fundamental para a
manutenção desses índices de produção, assim como na busca por novos mercados
consumidores. Grande parte dessa melhoria na qualidade dos ovos de consumo está
atribuída ao fornecimento de dietas balanceadas, respeitando o atendimento das
exigências de todos os nutrientes, nas diversas fases de criação.
A metionina, primeiro aminoácido limitante nas dietas de galinhas poedeiras, a
base de milho e farelo de soja, são essenciais para a manutenção da produção de ovos
em um ciclo de postura e diretamente ligados à qualidade do produto, já que a
metionina é fundamental no processo de síntese proteica no organismo.
Os aminoácidos são importantes no desempenho produtivo de poedeiras, pois
estão diretamente ligados à produção de ovos, com a conversão e eficiência alimentar
e à melhoria na utilização do nitrogênio, evitando assim maiores níveis de excreção ao
meio ambiente. Além disso, são componentes essenciais dos ovos e constituem as
moléculas proteicas presentes no albúmen e na gema (Leeson & Summers, 2001).
Ademais, o imbalanço aminoacídico da dieta pode provocar inúmeros prejuízos
metabólicos, como maior excreção de nitrogênio, lesões hepáticas e renais, além de
um menor desenvolvimento de vilosidades intestinais e das células do trato
reprodutivo de galinhas poedeiras.

81
Dessa forma, objetivou-se com este trabalho avaliar os níveis de metionina +
cistina nas dietas de galinhas poedeiras leves em postura, assim como os efeitos dos
tratamentos testados na fase subsequente.
MATERIAL E MÉTODOS
O estudo foi realizado no Setor de Avicultura, do Departamento de Zootecnia,
da Universidade Federal da Paraíba, Câmpus II, Areia-PB. Esse município está
localizado a 112 km de João Pessoa e situado na microrregião do Brejo Paraibano
(6°58’12’’ de latitude sul e 35°42’15’’ de longitude oeste). Apresenta como
característica climática tropical semi-úmido, estação chuvosa no período outono-
inverno, precipitação pluviométrica de aproximadamente 1500 mm, temperatura anual
entre 22 e 30ºC e umidade relativa do ar entre 75 e 87%.
A pesquisa teve a duração de 224 dias e foi dividida em duas fases: período de
avaliação das dietas experimentais (34 a 50 semanas de idade, postura 1); e avaliação
dos efeitos dos tratamentos oferecidos na postura 1, sobre o desempenho dos animais
na fase subsequente (54 a 70 semanas de idade, postura 2).
Período de Avaliação das Dietas Experimentais
Nesse período foram utilizadas 336 aves da linhagem Dekalb White, com 34
semanas de idade. As aves foram alojadas em gaiolas de produção, com comedouro do
tipo calha e bebedouro do tipo sparkcup, para fornecimento de ração e água à vontade,
respectivamente. O programa de luz adotado para o período experimental foi de 17
horas de luz (natural + artificial).
Os animais foram distribuídos em um delineamento inteiramente casualizado,
composto por seis tratamentos e quatorze repetições, com quatro animais por unidade
experimental. As dietas experimentais (Tabela 10) foram elaboradas da seguinte
forma: um tratamento controle, formulada para atender as exigências nutricionais

82
seguindo as recomendações do NRC (1994); e os demais tratamentos foram
estabelecidos de acordo com as recomendações das tabelas brasileiras de aves e suínos
(Rostagno et al., 2011), exceto para metionina + cistina, que consistiram em cinco
variações do nível desse aminoácido (80, 90, 100, 110 e 120%), resultando nos níveis
de 0,536; 0,603; 0,670; 0,737 e 0,804% de metionina + cistina digestível. As dietas
foram suplementadas com DL-Metionina-99% (0,093; 0,160; 0,227; 0,294; 0,361%),
em substituição ao amido, para atingir os níveis dietéticos.
Tabela 10 – Ingredientes e composição química das dietas experimentais.
Ingredientes NRC Rostagno et al, 2011
(1994)* 80% 90% 100% 110% 120%
Milho
66,797 71,675 71,675 71,675 71,675 71,675
Farelo de Soja 16,077 13,360 13,360 13,360 13,360 13,360
Calcário 9,654 9,587 9,587 9,587 9,587 9,587
Fosfato Bicálcico 1,195 1,158 1,158 1,158 1,158 1,158
Sal
0,435 0,523 0,523 0,523 0,523 0,523
Cloreto de Colina
0,070 0,070 0,070 0,070 0,070 0,070
Premix Vitamínico2
0,100 0,100 0,100 0,100 0,100 0,100
Premix Mineral1
0,050 0,050 0,050 0,050 0,050 0,050
Farelo de Glúten de Milho 60% 4,281 2,642 2,642 2,642 2,642 2,642
L-Lisina 0,211 0,222 0,222 0,222 0,222 0,222
L-Treonina
0,000 0,103 0,103 0,103 0,103 0,103
L-Triptofano
0,000 0,040 0,040 0,040 0,040 0,040
DL-Metionina 0,179 0,093 0,160 0,227 0,294 0,361
Amido de Milho
0,936 0,361 0,294 0,227 0,160 0,093
Total 100 100 100 100 100 100
Composição Química
(D- Digestível) Unidade
Proteína Bruta % 18,800 16,500 16,500 16,500 16,500 16,500
Cálcio % 4,060 4,020 4,020 4,020 4,020 4,020
Fósforo Disponível % 0,310 0,300 0,300 0,300 0,300 0,300
Energia Metabolizável kcal/kg 2800,0 2800,0 2800,0 2800,0 2800,0 2800,0
Arginina D % 0,880 0,736 0,736 0,736 0,736 0,736
Isoleucina D % 0,810 0,559 0,559 0,559 0,559 0,559
Lisina D % 0,860 0,736 0,736 0,736 0,736 0,736
Met + Cis D % 0,730 0,536 0,603 0,670 0,737 0,804
Treonina D % 0,590 0,559 0,559 0,559 0,559 0,559

83
Triptofano D % 0,159 0,169 0,169 0,169 0,169 0,169
Sódio % 0,190 0,225 0,225 0,225 0,225 0,225
Cloro % 0,303 0,355 0,355 0,355 0,355 0,355
Potássio % 0,532 0,580 0,580 0,580 0,580 0,580
Balanço Eletrolítico mEq/kg 0,419 0,450 0,450 0,450 0,450 0,450
* Ração formulada com base em aminoácidos totais. 1
Premix mineral inorgânico por kg de produto: Mn, 20g;
Fe, 10g; Zn, 13,7g; Cu, 2,5g; Se, 0,063g; I, 0,19g; e veículo q.s.p., 500g. 2
Premix vitamínico por kg de ração:
vit. A – 15.000.000Ul, vit. D3 – 1.500.000Ul, vit. E – 15.000Ul, vit.B1 – 2,0g, vit.B2 – 4,0g, vit. B6 – 3,0g,
vit.B12 – 0,015g, Ácido nicotínico – 25g, Ácido pantotênico – 10g, vit. K3 – 3,0g, Ácido fólico – 1,0 g,
Bacitracina de zinco – 10g, Selênio – 250mg, Antioxidante BHT – 10g, e veículo. q.s.p. – 1.000g.
As variáveis estudadas foram: consumo de ração (g/ave), produção de ovos
(%), peso dos ovos (g), massa de ovos (g), conversão alimentar por massa de ovos
(g/g), conversão alimentar por dúzia de ovos (g/g), porcentagem de gema (%), casca
(%) e albúmen (%), espessura da casca (mm), consumo de metionina + cistina
(mg/ave), consumo de lisina (mg/ave), peso sem vísceras (g/ave), peso do fígado
(g/ave), peso da gordura celomática (g/ave) e peso do baço (g/ave).
Ademais, foi realizada análise histopatológica do fígado, rins, intestino
delgado, magno e útero. Para tanto, fragmentos desses órgãos foram imersos em
substância fixadora metacarn (60% metanol, 30% clorofórmio e 10% ácido acético),
por 12 horas e posteriormente transferidos para álcool 70%.
Para a microscopia óptica foi feita a inclusão dos fragmentos dos órgãos em
paraplast, sendo realizados cortes seriados dos mesmos de 5µm de espessura. Para a
análise histológica, foram utilizadas as colorações, hematoxilina e eosina, periodic
acid Schiff (PAS) e tricômio de Masson. As fotomicrografias foram obtidas através de
microcâmera acoplada a um microscópio Olympus BX-51 e as imagens digitalizadas
no software KS 400.3 (Zeiss).
As rações foram pesadas no início e no final de cada ciclo de avaliação
experimental (28 dias) para a mensuração do consumo de ração, conversão alimentar
por massa e por dúzia de ovos, consumo de metionina + cistina e consumo de lisina.
Diariamente, os ovos eram coletados em cada unidade experimental para a
mensuração da produção de ovos. A conversão alimentar por dúzia de ovos foi
calculada dividindo-se o consumo de ração pelo número de dúzias de ovos produzidos.
Já a conversão por massa de ovos foi obtida pela divisão do consumo de ração pela

84
massa de ovos produzidos. Esta última foi obtida multiplicando-se a produção pelo
peso dos ovos.
Os ovos de cada parcela foram pesados em balança digital de três dígitos
(0,001g), para o cálculo do peso médio dos ovos, assim como os componentes internos
dos ovos (gema e albúmen), que também foram pesados individualmente e
separadamente em balança digital de três dígitos (0,001g), para mensuração das
porcentagens de gema e albúmen.
As cascas dos ovos, previamente identificadas, foram secas em estufa a 55-
60ºC por um período de 24 horas e posteriormente pesadas em balança digital com
precisão de 0,01g, para a obtenção da porcentagem das cascas. A espessura da casca
foi obtida através da utilização de um micrômetro digital Mitutoyo de 0-25 mm, com
precisão de 0,001 mm. Para as análises histopatológicas e as mensurações dos órgãos,
cinco aves por unidade experimental foram abatidas no final do período experimental.
Avaliação do efeito residual
Após o período de avaliação das dietas experimentais na postura 1, as aves
foram alimentadas com água e ração à vontade, de acordo com as exigências
nutricionais recomendadas por Rostagno et al. (2011). Desse modo, foi permitido
avaliar o efeito residual dos níveis de metionina + cistina do período de 34 a 50
semanas de idade sobre a fase subsequente (54 a 70 semanas de idade).
Para a avaliação do efeito residual, 168 aves foram selecionadas aleatoriamente
nos tratamentos em sete repetições, com quatro animais por unidade experimental. As
variáveis avaliadas nesse período foram: produção de ovos (%), peso dos ovos (g),
massa de ovos (g), porcentagem de gema (%), casca (%) e albúmen e espessura de
casca (mm). Para a determinação dessas variáveis foi utilizada a mesma metodologia
empregada na fase de avaliação das dietas experimentais.
Análises Estatísticas

85
As análises estatísticas das variáveis analisadas foram realizadas por meio do
programa computacional SAS (SAS Institute, 2011). O tratamento controle (NRC,
1994) foi comparado com os demais tratamentos (variações dos níveis de metionina +
cistina digestível, de acordo com Rostagno et al., 2011), através do teste de médias
Dunnett a 5% de probabilidade. O requerimento de metionina + cistina digestível foi
estimado através de análise de regressão, de modo a considerar o valor do R2 e a
resposta biológica das aves.
RESULTADOS E DISCUSSÃO
Variáveis de Desempenho Zootécnico
Houve efeito significativo (P<0,05) para as seguintes variáveis de desempenho
zootécnico: produção, peso, massa e conversão alimentar por massa de ovos, consumo
de metionina + cistina e consumo de lisina, quando submetidos ao teste Dunnet
(Tabela 11). Já para as demais variáveis, não houve diferença estatística significativa
entre o tratamento controle (NRC, 1994) e os demais tratamentos.
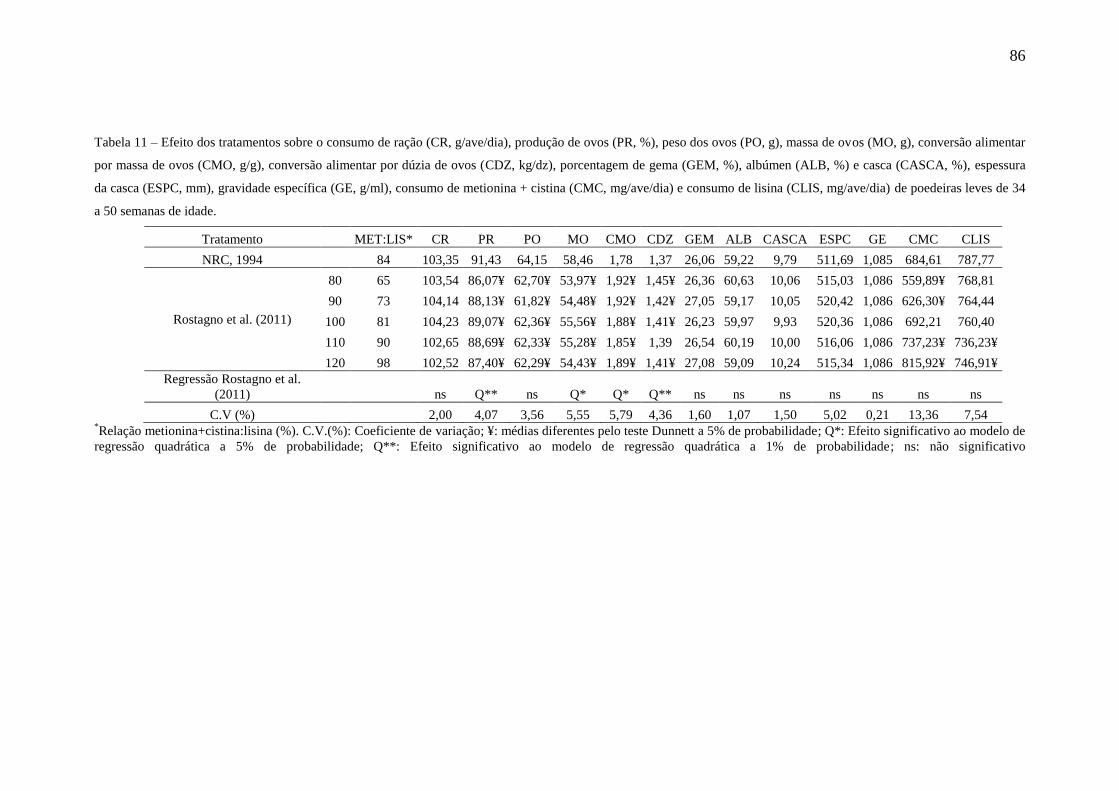
86
Tabela 11 – Efeito dos tratamentos sobre o consumo de ração (CR, g/ave/dia), produção de ovos (PR, %), peso dos ovos (PO, g), massa de ovos (MO, g), conversão alimentar
por massa de ovos (CMO, g/g), conversão alimentar por dúzia de ovos (CDZ, kg/dz), porcentagem de gema (GEM, %), albúmen (ALB, %) e casca (CASCA, %), espessura
da casca (ESPC, mm), gravidade específica (GE, g/ml), consumo de metionina + cistina (CMC, mg/ave/dia) e consumo de lisina (CLIS, mg/ave/dia) de poedeiras leves de 34
a 50 semanas de idade.
Tratamento MET:LIS* CR PR PO MO CMO CDZ GEM ALB CASCA ESPC GE CMC CLIS
NRC, 1994 84 103,35 91,43 64,15 58,46 1,78 1,37 26,06 59,22 9,79 511,69 1,085 684,61 787,77
Rostagno et al. (2011)
80 65 103,54 86,07¥ 62,70¥ 53,97¥ 1,92¥ 1,45¥ 26,36 60,63 10,06 515,03 1,086 559,89¥ 768,81
90 73 104,14 88,13¥ 61,82¥ 54,48¥ 1,92¥ 1,42¥ 27,05 59,17 10,05 520,42 1,086 626,30¥ 764,44
100 81 104,23 89,07¥ 62,36¥ 55,56¥ 1,88¥ 1,41¥ 26,23 59,97 9,93 520,36 1,086 692,21 760,40
110 90 102,65 88,69¥ 62,33¥ 55,28¥ 1,85¥ 1,39 26,54 60,19 10,00 516,06 1,086 737,23¥ 736,23¥
120 98 102,52 87,40¥ 62,29¥ 54,43¥ 1,89¥ 1,41¥ 27,08 59,09 10,24 515,34 1,086 815,92¥ 746,91¥
Regressão Rostagno et al.
(2011) ns Q** ns Q* Q* Q** ns ns ns ns ns ns ns
C.V (%) 2,00 4,07 3,56 5,55 5,79 4,36 1,60 1,07 1,50 5,02 0,21 13,36 7,54 *Relação metionina+cistina:lisina (%). C.V.(%): Coeficiente de variação; ¥: médias diferentes pelo teste Dunnett a 5% de probabilidade; Q*: Efeito significativo ao modelo de
regressão quadrática a 5% de probabilidade; Q**: Efeito significativo ao modelo de regressão quadrática a 1% de probabilidade; ns: não significativo

87
As aves alimentadas com a dieta controle (NRC, 1994) apresentaram melhor
desempenho produtivo, quando comparado com os animais que receberam os níveis
crescentes de metionina + cistina digestível estabelecido pelas tabelas brasileiras de
Rostagno et al. (2011), expressados pela maior produção, peso e massa de ovos, além
de menor conversão alimentar por massa de ovos.
Os níveis de arginina, isoleucina e lisina digestível da dieta estabelecida pelo
NRC (1994) estão acima dos valores preconizados pelas tabelas brasileiras de aves e
suínos. Provavelmente, as dietas formuladas com base nas exigências nutricionais das
tabelas brasileiras (níveis de metionina + cistina digestível) não atenderam os
requerimentos desses aminoácidos para as aves, acarretando em menor desempenho
produtivo, quando comparado com os animais que receberam a dieta com base em
aminoácidos totais.
As linhagens de poedeiras comerciais atuais apresentam melhor capacidade de
conversão de nutrientes da ração na produção, peso e massa de ovos. Em detrimento a
isso, os requerimentos nutricionais das aves elevam-se, com o intuito de dar suporte
aos altos índices produtivos. Possivelmente, as aves alimentadas com a dieta
formulada com base nas exigências nutricionais do NRC (1994) apresentaram melhor
aproveitamento dos nutrientes da dieta, expressadas pela melhor produção, peso,
massa e conversão alimentar por massa de ovos.
Também foi possível observar que os níveis elevados dos aminoácidos
essenciais, das dietas do NRC (1994), principalmente a arginina, lisina e isoleucina,
não foram capazes de provocar redução no desempenho ocasionada pelo possível
excesso de nitrogênio da dieta. De acordo com Koelkebeck et al. (1991), poedeiras de
alta produção apresentam certa tolerância para excessos de aminoácidos em dietas à
base de milho e farelo de soja.
Segundo Rombola et al. (2008), a formulação de rações com base na
composição em aminoácidos digestíveis contribui para maior precisão no
fornecimento de aminoácidos, já que permite considerar a capacidade da ave em
aproveitar os aminoácidos dos alimentos e superar eventuais diferenças na
digestibilidade dos ingredientes.

88
A efetividade das dietas com base em aminoácidos digestíveis já está
comprovada para frangos de corte, a partir do desenvolvimento de diversas pesquisas
ao longo dos últimos anos (Rostagno et al., 1995; Maiorka et al., 2004; Araújo et al.,
2006). No entanto, essa efetividade não está comprovada para poedeiras comerciais
durante a fase de postura e os resultados ainda são controversos (Silva et al., 2000;
Casartelli et al., 2004).
O menor rendimento das aves alimentadas com as dietas formuladas com base
nas tabelas de Rostagno et al. (2011) pode ser explicado pela possível deficiência
aminoácidos essenciais nas dietas, como a lisina, arginina e isoleucina. Segundo
Jordão Filho et al. (2006), os aminoácidos essenciais são exigidos pelo organismo das
galinhas poedeiras para atender três necessidades básicas: mantença, formação de
tecidos corporais e deposição de proteína para o ovo. Ainda segundo esses autores,
erros na concentração dos aminoácidos essenciais nas rações podem comprometer o
desempenho produtivo das aves, destacando-se a metionina, em que a sua deficiência
pode provocar a redução na produção e no peso dos ovos.
O consumo de aminoácidos sulfurosos pelas aves alimentadas com o
tratamento controle (NRC, 1994) foi significativamente superior (P<0,05) ao consumo
desse aminoácido para os animais alimentados com os tratamentos 80 e 90% de
metionina + cistina digestível, enquanto que não diferiu do tratamento com 100% e foi
significativamente menor com relação aos tratamentos com 110 e 120% de metionina
+ cistina digestível das tabelas brasileiras.
Esses resultados podem ser explicados pela ausência de diferença significativa
(P>0,05) para o consumo de ração entre os tratamentos, já que o nível de metionina +
cistina digestível do tratamento controle (0,657%) é superior aos níveis dos
tratamentos com 80 e 90%, semelhante ao tratamento com 100% e inferior aos níveis
dos tratamentos com 110 e 120% de metionina + cistina digestível.
O consumo de lisina digestível dos animais alimentados com a dieta controle
(NRC, 1994) foi significativamente superior (P<0,05) às aves alimentadas com as
dietas com os níveis 110 e 120% de aminoácidos sulfurosos estabelecidos por
Rostagno et al. (2011).

89
O nível de lisina digestível do tratamento controle (0,756%) é semelhante ao
nível desse aminoácido das tabelas brasileiras (0,736%). Mesmo não havendo
diferença significativa entre os tratamentos para o consumo de ração, os animais
alimentados com os tratamentos com 110 e 120% de metionina + cistina digestível
apresentaram menor consumo de ração, quando comparado com o tratamento controle,
explicando assim o menor consumo de lisina digestível para esses tratamentos.
As diferentes bases de formulação de ração (aminoácidos toais vs aminoácidos
digestíveis), assim como os níveis de metionina + cistina digestível (Rostagno et al.,
2011), não foram capazes de afetar significativamente o consumo de ração dos
animais. A ausência de diferença significativa (P>0,05) para o consumo de ração nesse
estudo corrobora os resultados encontrados em outros estudos que avaliaram os níveis
de metionina + cistina para galinhas poedeiras (Sá et al., 2007; Cupertino et al., 2009;
Varela, 2009; Polese et al., 2012).
No entanto, o comportamento do consumo de ração é bastante controverso em
pesquisas que avaliaram os níveis crescentes de aminoácidos sulfurosos nas dietas de
galinhas poedeiras, podendo ser demonstrado tanto pelo decréscimo no consumo de
ração (Jordão Filho et al., 2006), quanto pelo aumento no consumo de alimentos
(Narvaez Solarte, 1996; Harms et al., 1998; Barbosa et al., 2009; Brumano et al.,
2010). O consumo de ração pode ser afetado por diversos fatores, como a temperatura
ambiente, umidade relativa do ar, níveis dietéticos dos nutrientes, estresse, dentre
outros, fato que pode explicar a grande variação de respostas dos animais relacionadas
ao consumo de alimentos.
Os níveis de aminoácidos sulfurosos não foram capazes de alterar a qualidade
dos ovos das aves nesse estudo, sendo expressados pela ausência de diferença
estatística significativa (P>0,05) entre os tratamentos dietéticos, tanto para o teste
Dunnet, quanto para os dados submetidos à análise de regressão linear e quadrática.
Esses resultados estão de acordo aos encontrados por Jordão Filho et al. (2006), Sá et
al. (2007), Valera (2009) e Polese et al. (2012), que também não observaram efeito
significativo dos níveis de metionina + cistina sobre a qualidade dos ovos de galinhas
poedeiras.
Por outro lado, Mendonça Jr & Lima (1999), Barbosa et al. (2009) e Brumano
et al. (2010) encontraram influência significativa dos níveis aminoácidos sulfurosos

90
sobre a qualidade dos ovos de galinhas poedeiras. Mesmo não estando presente em
quantidades significativas nos componentes internos dos ovos de galinha (Pires et al.,
2006), os aminoácidos sulfurosos participam diretamente da síntese proteica orgânica
e podem influenciar a qualidade dos ovos de poedeiras, no entanto, os níveis dietéticos
trabalhados nesse estudo não foram suficientes para ocasionar mudanças significativas
na qualidade dos componentes internos e externos dos ovos.
Com base nos dados da análise de regressão, houve efeito quadrático
significativo (P<0,01) para produção (Figura 15) e conversão alimentar por dúzia de
ovos (Figura 18), além de efeito quadrático (P<0,05) para as variáveis, massa (Figura
16) e conversão alimentar por massa de ovos (Figura 17).
Figura 15 – Efeito dos tratamentos sobre a produção de ovos de poedeiras leves de 34 a 50
semanas de idade, alimentadas com níveis de metionina + cistina.
y = -0,0057x2 + 1,1779x + 28,512
R² = 0,9983
85,5
86
86,5
87
87,5
88
88,5
89
89,5
80 90 100 110 120
Pro
du
ção
de
ov
os
(%)
Níveis de metionina + cistina digestível (Rostagno et al., 2011)
103,25%

91
Figura 16 – Efeito dos tratamentos sobre a massa de ovos de poedeiras leves de 34 a 50
semanas de idade, alimentadas com níveis de metionina + cistina.
Figura 17 – Efeito dos tratamentos sobre a conversão alimentar por massa de ovos de poedeiras
leves de 34 a 50 semanas de idade, alimentadas com níveis de metionina + cistina.
y = -0,0029x2 + 0,6001x + 24,464
R² = 0,863
53,6
53,8
54
54,2
54,4
54,6
54,8
55
55,2
55,4
55,6
55,8
80 90 100 110 120
Ma
ssa
de
ov
os
(g)
Níveis de metionina + cistina digestível (Rostagno et al., 2011)
y = 0,0000643x2 - 0,0141571x + 2,6520000
R² = 0,6518883
1,84
1,85
1,86
1,87
1,88
1,89
1,9
1,91
1,92
1,93
1,94
80 90 100 110 120
Co
nv
ersã
o a
lim
enta
r p
or
ma
ssa
de
ov
os
(g/g
)
Níveis de metionina + cistina digestível (Rostagno et al., 2011)
110,03%
102,91%

92
Figura 18 – Efeito dos tratamentos sobre a conversão alimentar por dúzia de ovos de poedeiras
leves de 34 a 50 semanas de idade, alimentadas com níveis de metionina + cistina.
A partir da derivada das equações obtidas nas análises acima, o nível de
metionina + cistina digestível estimado para produção de ovos, massa de ovos,
conversão por massa de ovos e conversão por dúzia de ovos é de 103,25% (0,691% de
metionina + cistina digestível), 102,91% (0,689% de metionina + cistina digestível),
110,03% (0,737% de metionina + cistina digestível) e 109,03% (0,730% de metionina
+ cistina digestível), respectivamente.
Como pôde ser observado, as exigências de metionina + cistina digestível para
as variáveis relacionadas à conversão alimentar são superiores às variáveis de
produção e massa de ovos. Esses resultados estão de acordo a afirmação de Schutte et
al. (1994), que verificaram que o nível de metionina + cistina exigido para otimizar a
conversão alimentar das aves é maior que aquele para a maximização da produção de
ovos.
Com base nos resultados de desempenho zootécnico obtidos nesse estudo e nas
respostas biológicas dos animais frente aos níveis de aminoácidos sulfurosos
avaliados, estima-se o nível de metionina + cistina digestível correspondente à
conversão alimentar por dúzia de ovos, ou seja, 0,730% de metionina + cistina
digestível, para galinhas poedeiras leves de 34 a 50 semanas de idade.
y = 0,000064x2 - 0,013957x + 2,156000
R² = 0,931548
1,38
1,39
1,4
1,41
1,42
1,43
1,44
1,45
1,46
80 90 100 110 120
Co
nv
ersã
o a
lim
enta
r p
or
dú
zia
de
ov
os
(g/g
)
Níveis de metionina + cistina digestível (Rostagno et al., 2011)
109,03%

93
Os valores estimados para aminoácidos sulfurosos nesse estudo estão acima do
nível preconizado no manual da linhagem Dekalb White (2009), que recomenda
0,660% de metionina + cistina digestível durante a fase de postura. Da mesma forma,
o nível de aminoácidos sulfurosos estimado nesse estudo são superiores aos
encontrados em pesquisas semelhantes ao longo dos últimos anos.
Nesse sentido, tem-se observado em pesquisas recentes um aumento
significativo nos requerimentos nutricionais em aminoácidos sulfurosos para galinhas
poedeiras em produção. Esse fato está principalmente atribuído ao melhoramento
genético, que tornaram as aves mais eficientes e com maior capacidade produtiva,
devendo sempre serem revistas e atualizadas as tabelas de exigências nutricionais de
galinhas poedeiras em produção. Corroborando esse fato, Sá et al. (2007) afirmaram
que as diferenças nos valores da exigência em metionina para poedeiras tornam-se
evidentes, principalmente com as mudanças na genética, nutrição e no manejo que
aves estão submetidas, além dos efeitos da idade, do tipo da dieta e das condições
ambientais.
Togashi et al. (2002) determinaram o nível de 0,565% de metionina + cistina
para poedeiras semipesadas em postura alimentadas com rações contendo Levedura
Seca (Saccharomyces cerevisae). Sá et al. (2007) e Cupertino et al. (2009), estimaram
as exigências de aminoácidos sulfurosos para galinhas poedeiras leves em produção e
recomendaram o nível de 0,693 e 0,655% de metionina + cistina digestível,
respectivamente.
Mais recentemente, Brumano et al. (2010) avaliaram os níveis de metionina +
cistina digestível para poedeiras leves no período de 42 a 58 semanas de idade e
estimaram o nível de aminoácidos sulfurosos para a fase em 0,775%, valor próximo ao
estimado nesse estudo (0,730%).
De acordo com Waldroup & Hellwig (1995), os requerimentos em
aminoácidos sulfurosos para galinhas poedeiras apresentam um pico para produção de
ovos no período que compreende entre a 51ª e a 58ª semana de idade, provavelmente
pelo aumento no tamanho dos ovos. Na presente pesquisa, o período de avaliação das
dietas experimentais situou-se próximo à fase do pico de exigência de metionina para
poedeiras e, dessa forma, pode explicar o nível estimado nesse estudo que foi

94
significativamente superior aos valores preconizados nas principais tabelas de
exigências nutricionais.
As aves alimentadas com os menores níveis de aminoácidos sulfurosos (80 e
90%), assim como os animais que receberam o tratamento com 120% de metionina +
cistina digestível, tiveram menor produção, peso e massa de ovos, além de pior
conversão alimentar. Dessa forma, pode-se inferir que o imbalanço aminoacídico
provocado tanto pela deficiência, quanto pelo excesso de aminoácidos sulfurosos na
dieta, provoca prejuízos na produção de ovos de galinha.
Os resultados encontrados nesse estudo são semelhantes aos relatados por
Jordão Filho et al. (2006), que observaram redução no peso dos ovos de poedeiras
semipesadas nos níveis extremos de metionina + cistina, que foram explicados pelos
autores devido a deficiência e o excesso de aminoácidos sulfurosos da ração sobre esta
variável.
De acordo com D’Mello (2003), a metionina tem um papel chave no
metabolismo como doador de grupamentos metil ativos. Aliado a isto, tem função
importante como precursora da homocisteína e cisteína, que é fundamental para o
organismo na síntese de diversas enzimas envolvidas no metabolismo proteico. De
acordo com Jordão Filho et al. (2006), erros nas concentrações de aminoácidos das
rações de poedeiras, principalmente metionina e lisina, podem comprometer o
desempenho produtivo, expressados pela redução na produção e no peso dos ovos,
além de aumentar a deposição de gordura no fígado.
O desbalanço aminoacídico provocado pela deficiência de metionina causa
efeitos negativos sobre o consumo de alimentos, a taxa de crescimento e sobre a
produtividade de frangos de corte e galinhas poedeiras leves e semipesadas (Pars &
Summers, 1991; Narváez Solarte, 1996).
Dentre os aminoácidos essenciais, os aminoácidos sulfurosos são considerados
um dos mais tóxicos para não ruminantes, principalmente por conter enxofre em sua
composição. Dessa forma, possivelmente, o excesso de metionina + cistina da dieta
com 120% das exigências nutricionais para esse aminoácido, pode ter provocado
toxidez aos animais, reduzindo principalmente a produção e a eficiência alimentar no
período.

95
Análises Histológicas
A partir das imagens das fotomicrografias do fígado das aves (Figura 19), é
possível perceber que as aves alimentadas com o tratamento controle (NRC, 1994) e
com a dieta com 120% das exigências de metionina + cistina digestível, apresentaram
desorganização parenquimal hepática, em função de um maior acúmulo de vacúolos
citoplasmáticos nas células do fígado.
Figura 19 – Fotomicrografias do fígado de poedeiras leves com 50 semanas de idade suplementadas
com níveis de metionina + cistina. A) Imagem representando o tratamento controle (NRC, 1994); B) Imagem
representando os tratamentos 80 e 90% de metionina + cistina digestível (Rostagno et al., 2011); C) Imagem
representando o tratamento com 100 e 110% de metionina + cistina digestível (Rostagno et al., 2011); D)
Imagem representando o tratamento com 120% de metionina + cistina digestível (Rostagno et al., 2011).
Coloração de periodic acid Schiff. Barras: 400µm.

96
Aliado a isto, os animais alimentados com o tratamento controle (NRC, 1994) e
as aves que receberam o nível de 120% de metionina + cistina digestível, apresentaram
maior acúmulo de glicogênio hepático, representado pela maior positividade a
coloração de periodic acid Schiff.
Em contrapartida, o fígado das aves dos tratamentos com 80, 90, 100 e 110%
das exigências de metionina + cistina digestível não apresentaram indícios de esteatose
ou desorganização parenquimal hepática, além de ser observado menor acúmulo de
glicogênio hepático, com menor positividade a coloração periodic acid Schiff, quando
comparado com os demais tratamentos.
Esses dados estão de acordo com os resultados encontrados em trabalho
realizado por Bunchasak & Silapasorn (2005), que relataram aumento na gordura
hepática de aves de postura que receberam altos níveis de metionina na dieta. Esses
autores ainda afirmaram que o aumento da gordura do fígado das aves pode ter sido
causado pela maior síntese de estrogênio no ovário, em detrimento às altas taxas de
produção de ovos, corroborando os dados desse estudo, onde foram observados
maiores índices de produção de ovos no tratamento controle (NRC, 1994).
As células hepáticas estão envolvidas em processos de síntese proteica,
secreção biliar, armazenamento de glicogênio, biotransformação e detoxificação e no
metabolismo de aminoácidos, lipídios e glicose, dentre outras funções (Takashima &
Hibiya, 1995). Provavelmente, os níveis de metionina + cistina alteraram o
metabolismo hepático, evidenciado pelo maior acúmulo de glicogênio hepático dos
animais do tratamento controle (NRC, 1994) e as aves que receberam o nível de 120%
de metionina + cistina digestível.
Apesar de instalada esteatose hepática moderada nesses tratamentos,
provavelmente os efeitos desse distúrbio metabólico não interferiram diretamente
sobre os resultados de desempenho zootécnico, já que os melhores índices produtivos
foram encontrados nos animais que receberam a dieta controle (NRC, 1994). Aliado a
isso, nessa idade os animais já possuem todos os órgãos e sistemas desenvolvidos e, o
excedente de energia, pode ter sido estocado no fígado e utilizado para outras funções,
como a produção de ovos (Bunchasak & Silapasorn, 2005).

97
O fígado é um órgão capaz de armazenar diversas substâncias no organismo,
dentre elas lipídios e glicogênio. A síntese de glicogênio no organismo é o processo
pelo qual a glicose é polimerizada a glicogênio, que é armazenada nos hepatócitos e
funcionam como depósito de energia acessível à célula.
Segundo Graciano (2009), as reservas de glicogênio nos animais podem ser
afetadas por desvios metabólicos, principalmente em função da dieta. Dessa forma, um
possível excesso de aminoácidos nos animais alimentados com a dieta controle (NRC,
1994), ocasionado pelo maior nível de proteína bruta da dieta e dos níveis elevados
dos principais aminoácidos essenciais dietéticos, pode explicar o maior nível de
glicogênio hepático dos animais desse tratamento.
Com relação às imagens das fotomicrografias do intestino delgado das aves
(Figura 20) alimentadas com níveis de aminoácidos sulfurosos, é possível observar
que os animais que receberam os níveis mais baixos de metionina + cistina digestível
(Figura 20A) apresentaram menor largura das vilosidades intestinais.
Figura 20 – Fotomicrografias do intestino delgado de poedeiras leves com 50 semanas de idade
suplementadas com níveis de metionina + cistina. A) Imagem representando os tratamentos com 80 e 90% de
metionina + cistina digestível (Rostagno et al., 2011); B) Imagem representando os tratamentos com 100, 110 e
120% de metionina + cistina digestível (Rostagno et al., 2011) e o tratamento controle (NRC, 1994), os quais
apresentaram vilosidades com maior largura (barra vertical) e mais ramificações (pontas de seta). Coloração de
hematoxilina-eosina. Barras: 400µm.

98
Já os animais que receberam o tratamento controle (NRC, 1994) e os níveis
com 100, 110 e 120% de metionina + cistina digestível das tabelas brasileiras,
apresentaram maior largura das vilosidades intestinais.
O aumento da largura e da altura das vilosidades está diretamente ligado a uma
maior área de absorção de nutrientes no intestino delgado, já que este órgão é o
principal sítio de digestão e captação de nutrientes no organismo. Vários ingredientes
vêm sendo utilizados na nutrição com função trófica sobre a mucosa intestinal,
destacando-se os aminoácidos sintéticos. Nesse sentido, a metionina, primeiro
aminoácido limitante para aves de produção em dietas a base de milho e farelo de soja,
e elemento chave no processe de síntese proteica no organismo, tem participação
direta no desenvolvimento intestinal.
Soares et al. (2008) observaram melhor desenvolvimento das estruturas do
intestino delgado de pintos de corte com sete dias de idade e suplementados com
metionina na dieta. Em contrapartida, Zavarize et al. (2011) não observaram
modificações morfológicas representativas na mucosa intestinal de galinhas poedeiras
leves submetidas ao estresse por calor e termoneutralidade e suplementadas com
glutamina na dieta.
No entanto, de acordo com Rezende e Beletti (2008), independentemente do
segmento intestinal considerado, as vilosidades tendem a ter a sua altura e largura
intensamente aumentadas até a nona semana de vida e, após essa etapa, existe uma
tendência de se estabilizarem.
Dessa forma, possivelmente a deficiência de aminoácidos sulfurosos provocou
uma redução na capacidade do processo de renovação das células epiteliais do
intestino delgado das aves alimentadas com os menores níveis de metionina + cistina,
havendo redução da largura do vilo, quando comparado com os demais tratamentos.
De acordo com a Figura 21, todas as imagens demonstram a presença de
glândulas ativas secretoras de albúmen no magno das aves de todos os tratamentos.
Todavia, os animais que receberam as dietas com 80 e 90% das exigências de
metionina + cistina digestível (Figura 21B) apresentaram quantidade reduzida de
células secretoras de albúmen no magno, quando comparado com as aves que
receberam o tratamento controle (Figura 21A) e os animais dos tratamentos com 100,

99
110 e 120% das exigências de metionina + cistina digestível, os quais apresentaram
maior quantidade de glândulas ativas em comparação aos demais tratamentos.
Figura 21 – Fotomicrografias do magno de poedeiras leves com 50 semanas de idade suplementadas
com níveis de metionina + cistina. A) Imagem representando o tratamento controle (NRC, 1994) B) Imagem
representando os tratamentos com 80 e 90% de metionina + cistina digestível (Rostagno et al., 2011); C)
Imagem representando os tratamentos com 100, 110 e 120% de metionina + cistina digestível (Rostagno et al.,
2011). Asterisco representa glândulas ativas do magno. Coloração de hematoxilina-eosina. Barras: 200µm.
Efeito semelhante foi relatado por Lima et al. (2012), que observaram maior
atividade das glândulas secretoras de albúmen no magno e consequente maior
produção de albúmen em galinhas poedeiras leves suplementadas com triptofano
digestível na dieta.
O aumento das pregas do magno, aliado a uma maior atividade desse órgão e
maior estoque de albúmen, permite a produção de ovos em um período mais curto,
aumento a produtividade e o peso e massa de ovos (King & McLelland, 1979). Dessa
forma, o melhor desenvolvimento do órgão e o maior estoque de albúmen,
principalmente nos animais alimentados com a dieta controle e com os maiores níveis
de metionina + cistina digestível das tabelas brasileiras, permitiram uma maior
produção, peso e massa de ovos, além de uma melhor conversão alimentar, conforme
demonstrado nos dados de desempenho zootécnico.
As fotomicrografias do útero das aves não demonstraram mudanças
histológicas significativas entre os tratamentos, no entanto, pode-se observar que as

100
aves dos tratamentos com 110 e 120% de metionina + cistina digestível apresentaram
uma maior largura das pregas uterínicas (Figura 22).
Figura 22 – Fotomicrografias do útero de poedeiras leves com 50 semanas de idade suplementadas com
níveis de metionina + cistina. A) Imagem representando os tratamentos com 80 e 90% de metionina + cistina
digestível (Rostagno et al., 2011); C) Imagem representando os tratamentos com 100, 110 e 120% de
metionina + cistina digestível (Rostagno et al., 2011) e o tratamento controle (NRC, 1994). Coloração de
hematoxilina-eosina (barra de 100µm).
Corroborando esses dados, Lima et al. (2012) observaram que a suplementação
de triptofano na dieta de galinhas poedeiras leves em postura proporciona um maior
desenvolvimento e elevada quantidade de pregas secundárias no útero das aves, além
de hiperplasia do órgão.
De acordo com King & McLlland (1979), o maior desenvolvimento das pregas
do útero, também proporciona a produção de ovos mais rapidamente, já que o
processo de formação e deposição dos cristais de carbonato de cálcio na casca dos
ovos se dá com mais eficiência. No entanto, essas mudanças no desenvolvimento do
útero das aves não foram suficientes para alterar a qualidade da casca dos ovos, não
sendo encontradas diferenças significativas para porcentagem e espessura de casca, e
gravidade específica.
Peso dos Órgãos

101
Não foi verificado efeito estatístico significativo (P>0,05) dos tratamentos
dietéticos sobre o peso dos órgãos das aves com 50 semanas de idade, não sendo
observadas diferenças entre as diferentes formulações, aminoácidos total vs digestível,
pelo teste Dunnett, assim como entre os níveis de metionina + cistina digestível, pela
análise de regressão (Tabela 12).
Tabela 12 - Efeitos dos tratamentos sobre o peso relativo (%) do peso sem vísceras (PSV), fígado (FIG),
baço (BAÇ) e gordura celomática (GCE) das aves com 50 semanas de idade.
Tratamento MET:LIS* PSV BAÇO FIG GCE
NRC, 1994 93 70,82 0,100 2,58 3,03
Rostagno et al.
(2011)
80% 66 71,16 0,119 2,66 2,22
90% 74 70,16 0,105 2,79 2,23
100% 82 72,27 0,094 2,73 1,98
110% 90 71,08 0,098 2,65 2,41
120% 98 72,33 0,091 2,73 2,16
Regressão Rostagno et al. (2011) ns ns ns ns
C.V. (%) 1,18 9,87 2,77 15,64
*Relação metionina + cistina: lisina (%); ns: não significativo; C.V. (%): Coeficiente de variação.
A ausência de diferença estatística significativa para essas variáveis,
principalmente o fígado e o baço, pode ser explicada pelo fato do crescimento e
desenvolvimento desses órgãos ocorrerem principalmente nas primeiras semanas de
vida da ave, onde é possível observar diferenças nos pesos dos órgãos a partir da
manipulação dos níveis dos nutrientes dietéticos.
Segundo Bertechini (2012), o crescimento das frangas de reposição segue o
modelo fisiológico multifásico, sendo que na fase de 1 a 5 semanas ocorre prioridade
fisiológica de crescimento visceral (órgãos vitais), seguido de crescimento ósseo de 6
a 12 semanas e de 12 a 16 semanas de idade o desenvolvimento dos órgãos
reprodutivos, com aumento de densidade óssea e acúmulo de reservas orgânicas para o
início de produção.
Dessa forma, o aumento no peso dos órgãos viscerais em galinhas poedeiras é
mais evidente durante a fase de crescimento e nas semanas que antecedem a postura.
Segundo Saki et al. (2011), a metionina pode influenciar as características de carcaça e

102
órgãos viscerais, principalmente o fígado, sendo que altos níveis de metionina
aumentaram a atividade metabólica desse órgão em poedeiras durante a fase de
crescimento. Do mesmo modo, Hazelwood et al. (1986) relataram que o fígado de
poedeiras aumenta de peso nos períodos que antecedem a fase de postura.
Já o baço é um órgão diretamente ligado à atividade imunológica das aves e os
níveis de aminoácidos dietéticos também podem influenciar a atividade metabólica e
consequentemente, o peso desse órgão. De acordo com Carvalho (2012), os
aminoácidos sulfurosos podem exercer diversas funções no organismo das aves, dentre
elas a melhoria no sistema imune das aves. No entanto, segundo Kidd (2004), as
exigências nutricionais para as respostas imunes nos animais diferem daquelas para o
crescimento e deposição tecidual, e este fato, pode explicar a ausência de diferença
significativa entre os tratamentos para o peso do baço das aves.
As diferentes formas de formulação da ração (aminoácidos totais vs
aminoácidos digestíveis), assim como os níveis de metionina + cistina digestível das
tabelas brasileiras não foram capazes de alterar o acúmulo de gordura corporal das
aves durante o período experimental.
Silva Júnior et al. (2005) observaram que os níveis de metionina + cistina
afetaram de forma significativa a deposição de gordura celomática em frangos de corte
aos 42 dias de idade, sendo atribuído ao maior acúmulo de gordura no corpo ao
desequilíbrio aminoacídico das dietas com excesso de aminoácidos sulfurosos. Esses
autores ainda afirmaram que esse desequilíbrio limita o acúmulo de tecido magro e
direciona as calorias para os adipócitos, aumentando o acúmulo de gordura na
cavidade abdominal das aves.
Outro fato que pode explicar a ausência de diferença significativa entre os
tratamentos para a porcentagem de gordura celomática das aves é a grande variação
entre as médias, destacada pelo valor elevado do coeficiente de variação. Como pôde
ser observado na Tabela 11, os animais alimentados com a dieta controle apresentaram
maior porcentagem de gordura celomática (efeito numérico), quando comparado com
os demais tratamentos. Aliado a isto, esses animais também apresentaram maior
acúmulo de glicogênio hepático (Figura 18), provavelmente ocasionado pelo excesso
de aminoácidos na dieta das aves.

103
Avaliação do Efeito Residual
Houve efeito residual significativo (P<0,05) dos níveis de metionina + cistina
entre o tratamento controle e os demais tratamentos oferecidos no período de 34 a 50
semanas de idade, sobre a produção e massa de ovos, casca e espessura de casca, das
aves no período de produção de 54 a 70 semanas de idade (Tabela 13).
Tabela 13 - Efeito residual dos tratamentos sobre a produção de ovos (PR, %), peso de ovo (PO, g), massa de
ovos (MO, g/ave/dia), espessura de casca (ESPC, mm), percentagem de albúmen (ALB, %), gema
(GEM, %) e casca (CASCA, %), das aves de 54 a 70 semanas de idade.
Tratamento PR PO MO ESPC ALB GEMA CASCA
NRC, 1994 82,51 67,55 55,66 512,39 58,39 28,66 8,65
Rostagno et al. (2011)
80% 85,90 67,58 58,01 541,71 59,98 27,96 9,05
90% 89,40 65,41 58,57 530,64 59,63 27,33 9,13
100% 92,76¥ 67,37 62,41 530,42 58,80 28,58 8,98
110% 94,01¥ 68,65 64,37¥ 541,71 60,48 27,68 6,57
120% 90,31 67,74 61,20 559,82¥ 56,40 27,04 6,13¥
Regressão Rostagno et al.
(2011) Q* ns ns ns ns ns ns
C.V (%) 3,54 1,58 5,32 2,94 2,47 2,35 16,82
C.V.(%): Coeficiente de variação; ¥: médias diferentes pelo teste Dunnett a 5% de probabilidade; Q*: Efeito
significativo ao modelo de regressão quadrática a 5% de probabilidade; ns: não significativo
As aves alimentadas com as dietas dos tratamentos com 100 e 110% de
metionina + cistina digestível das tabelas brasileiras apresentaram maior produção de
ovos, quando comparadas com as aves que receberam a dieta controle (NRC, 1994).
Da mesma forma, a massa de ovos das aves alimentadas com a dieta do tratamento
com 110% de metionina + cistina digestível foi superior à massa das aves alimentadas
com o tratamento controle.
Esses resultados demonstram uma perda da capacidade produtiva das aves
alimentadas com a dieta controle (NRC, 1994) durante o período de avaliação do
efeito residual das dietas oferecidas de 34 a 50 semanas de idade. Essas aves, durante

104
o período de avaliação das dietas experimentais, apresentaram melhor desempenho
produtivo, quando comparado com as aves que receberam os níveis de aminoácidos
sulfurosos das tabelas brasileiras, no entanto, não mantiveram esse desempenho
quando receberam uma ração basal (Rostagno et al., 2011) de 54 a 70 semanas de
idade.
Essa redução no desempenho pode ser explicada pela mudança na dieta das
aves do tratamento controle (NRC, 1994), para a dieta formulada com base nas
exigências nutricionais das tabelas brasileiras (Rostagno et al., 2011). Esta última
possui o nível de proteína bruta, assim como os níveis dos principais aminoácidos
essenciais dietéticos, abaixo dos valores recomendados pela tabela do NRC (1994).
Outra hipótese para a redução no desempenho das aves alimentadas com a
dieta controle (NRC, 1994) é a possível toxicidade ocasionada pelo excesso de
aminoácidos dietéticos. De acordo com Rutz (2002), a toxicidade se aplica a condições
nas quais um efeito adverso é causado devido à suplementação excessiva de um
aminoácido, nesse caso, vários dos aminoácidos essenciais da dieta. Ainda segundo
esses autores, sob o ponto de vista prático, dentre os aminoácidos potencialmente
tóxicos, a metionina é o que apresenta maior preocupação.
Dessa forma, a possível toxicidade do excesso do conteúdo proteico da dieta
citada acima só apresentou efeitos negativos sobre o desempenho produtivo das aves
após o período de avaliação das dietas experimentais.
O excesso de aminoácidos da dieta é catabolizado no organismo e podem
seguir diversas rotas metabólicas. Segundo Rutz (2002), os aminoácidos liberados
após o catabolismo podem ser utilizados na síntese proteica ou desviados para a
produção de ATP. Após a remoção do nitrogênio da cadeia, o restante da cadeia
aminoacídica (ceto-ácido) pode entrar na cadeia glicolítica ou ciclo de Krebs. Essa
cadeia pode ser metabolizada para produzir dióxido de carbono e formar ATP, ou pode
ser usada como intermediária para a formação de glicose. Esse fato está evidenciado
pelo maior acúmulo de glicogênio hepático nas aves alimentadas com a dieta controle
(NRC, 1994) no período de avaliação das dietas experimentais (34 a 50 semanas de
idade).
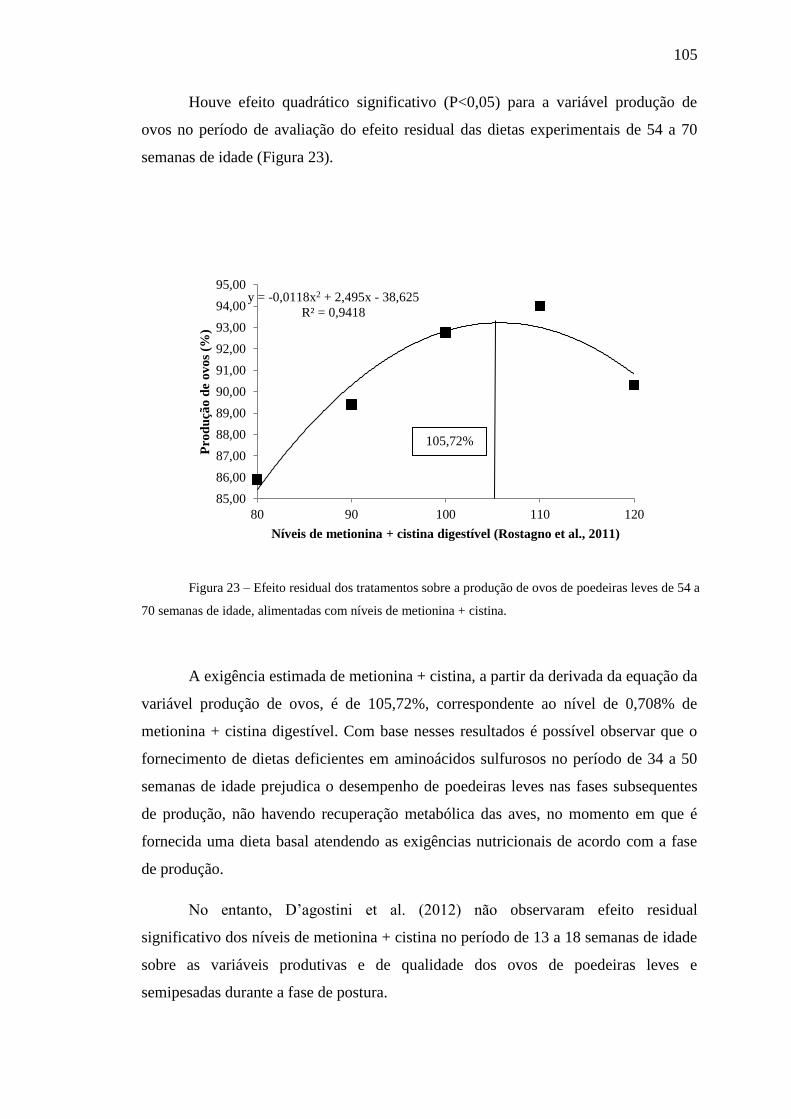
105
Houve efeito quadrático significativo (P<0,05) para a variável produção de
ovos no período de avaliação do efeito residual das dietas experimentais de 54 a 70
semanas de idade (Figura 23).
Figura 23 – Efeito residual dos tratamentos sobre a produção de ovos de poedeiras leves de 54 a
70 semanas de idade, alimentadas com níveis de metionina + cistina.
A exigência estimada de metionina + cistina, a partir da derivada da equação da
variável produção de ovos, é de 105,72%, correspondente ao nível de 0,708% de
metionina + cistina digestível. Com base nesses resultados é possível observar que o
fornecimento de dietas deficientes em aminoácidos sulfurosos no período de 34 a 50
semanas de idade prejudica o desempenho de poedeiras leves nas fases subsequentes
de produção, não havendo recuperação metabólica das aves, no momento em que é
fornecida uma dieta basal atendendo as exigências nutricionais de acordo com a fase
de produção.
No entanto, D’agostini et al. (2012) não observaram efeito residual
significativo dos níveis de metionina + cistina no período de 13 a 18 semanas de idade
sobre as variáveis produtivas e de qualidade dos ovos de poedeiras leves e
semipesadas durante a fase de postura.
y = -0,0118x2 + 2,495x - 38,625
R² = 0,9418
85,00
86,00
87,00
88,00
89,00
90,00
91,00
92,00
93,00
94,00
95,00
80 90 100 110 120
Pro
du
ção
de
ov
os
(%)
Níveis de metionina + cistina digestível (Rostagno et al., 2011)
105,72%

106
Dessa forma, pode-se concluir que os efeitos do fornecimento de dietas
deficientes em aminoácidos sulfurosos sobre as variáveis produtivas no período de
postura, dependem do estágio em que são fornecidas as dietas, ou seja, a deficiência de
metionina + cistina digestível no período de 34 a 50 semanas de idade tem efeito
negativo sobre a produtividade no período subsequente (54 a 70 semanas de idade).
Também foi verificado efeito estatístico significativo (P<0,05) entre o
tratamento controle (NRC, 1994) e tratamento com 120% das exigências nutricionais
de metionina + cistina digestível das tabelas brasileiras para as variáveis porcentagem
e espessura de casca. A porcentagem da casca dos ovos das aves alimentadas com a
dieta com 120% de metionina + cistina digestível foi significativamente menor,
quando comparado com as aves que receberam a dieta controle (NRC, 1994). Aliado a
isto, a espessura da casca das aves que receberam o tratamento com 120% de
metionina + cistina digestível foi significativamente maior, quando comparado com as
aves do tratamento controle.
Esses resultados podem ser explicados pela maior largura das pregas uterínicas
(Figura 22) dos animais do tratamento com 120% das exigências de metionina +
cistina digestível, devido ao melhor desenvolvimento do órgão e expressado pela
maior espessura da casca.
Em contrapartida, D’Agostini et al. (2012) não observaram efeito residual
significativo sobre a qualidade da casca de poedeiras leves em postura, alimentadas
com níveis de metionina + cistina digestível na fase de recria (13 a 18 semanas de
idade).
CONCLUSÕES
Recomenda-se o nível de 0,730% de metionina + cistina digestível, com
relação metionina + cistina:lisina de 99% e correspondente ao consumo aproximado
de metionina + cistina de 751,24 mg/ave/dia, para poedeiras leves de 34 a 50 semanas
de idade.

107
REFERÊNCIAS BIBLIOGRÁFICAS
ARAÚJO, L.F.; JUNQUEIRA, O.M.; ARAÚJO, C.S.S. et al. Different criteria for feed
formulation base non digestible amino acids for broilers. Brazilian Journal of
Poultry Science, v.8, p.201-205, 2006.
BARBOSA, M.J.B.; CARDOZO, R.M.; SOUZA, V.L.F. et al. Níveis de metionina +
cistina no desempenho de poedeiras comerciais leves com 45 semanas de idade.
Revista Brasileira de Saúde e Produção Animal, v.10, p.1032-1039, 2009.
BERTECHINI, A. G., Nutrição de Monogástricos, 2ed. Lavras-MG. UFLA, 2012. 373p.
BRUMANO, G.; GOMES, P.C.; DONZELE, J.L. Níveis de metionina + cistina
digestível para poedeiras leves no período de 24 a 40 semanas de idade. Revista
Brasileira de Zootecnia, v.39, p.1228-1236, 2010.
BUNCHASAK, C.; SILAPASORN, T. Effects of adding methionine in low-protein diet
on production performance, reproductive organs and chemical liver composition of
laying hens under tropical conditions. International Journal of Poultry Science, v.4,
p.301-308, 2005.
CARVALHO, C.B. Níveis de metionina+cistina e suas relações com a lisina em
rações para poedeiras leves no período de 79 a 95 semanas de idade. 2012. 55f.
Tese (Doutorado em Zootecnia) – Universidade Federal de Viçosa, Viçosa.
CASARTELLI, E.M.; JUNQUEIRA, O.M.; FILARDI, R.S. et al. Formulações de rações
para poedeiras comerciais utilizando-se diferentes recomendações de aminoácidos
totais e digestíveis. Revista Brasileira de Ciência Avícola, v.6, p.25, 2004 (supl.).
CUPERTINO, E.S.; GOMES, P.C.; ROSTAGNO, H.S. et al. Exigência nutricional de
metionina + cistina digestíveis para galinhas poedeiras de 54 a 70 semanas de idade.
Revista Brasileira de Zootecnia, v.38, p.1238-1246, 2009.

108
D’AGOSTINI, P.C.; MELLO, H.H.C.; CALDERANO, A.A. et al. Exigência de
metionina + cistina para frangas de reposição na fase de recria de 13 a 18 semanas de
idade. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.64, p.1691-
1698, 2012.
D’MELLO, J.P.F. Amino Acids in Animal Nutrition. Wallingford: CABI International,
2003. 526p.
HARMS, R.H.; RUSSEL, G.B.; HERLOW, H. et al. The influence of methionine on
commercial laying hens. Journal of Applied Poultry Reseach, v.7, p.45-52, 1998.
HAZELWOOD, R.L. Carbohydrate metabolism. In: STURKIE, P.D. Avian Physiology.
New York: Spring-Verlag, 1986, p.303-325.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA – IBGE. Indicadores
IBGE: Estatística da Produção Pecuária. 2013, 43p.
JORDÃO FILHO, J.; SILVA, J.H.V.; SILVA, E.S. et al. Exigências nutricionais de
metionina + cistina para poedeiras semipesadas do início de produção até o pico de
postura. Revista Brasileira de Zootecnia, v.35, p.1063-1069, 2006 (supl.).
KIDD, M.T. Nutritional modulation of immune function in broilers. Poultry Science,
v.83, p.650-657, 2004.
KING A.S.; MCLELLAND, J. Form and function in birds. London: Academic Press,
1979. 493p.
KOELKEBECK, K.W., BAKER, D.H., HAN, Y., et al. Research note: effect of excess
lysine, methionine, threonine, or tryptophan on production performance of laying
hens. Poultry Science, v.70, p.1651-1653, 1991.
LEESON, S., SUMMERS, J.D., CASTON, L.J. Response of layers to low nutrient
density diets. Journal applied Poultry Research, v.10, p.46-52. 2001.
LIMA, M.R.; COSTA, F.G.P.; GUERRA, R.R. et al. Digestible tryptophan:lysine ratios
for laying hens. Revista Brasileira de Zootecnia, v.41, p. 2012.

109
MAIORKA, A.; DAHLKE, F.; SANTIN, E. et al. Effect of energy levels of diets
formulated on total or digestible amino acids basis on broiler performance. Brazilian
Journal of Poultry Science, v.6, n.2, p.87-91, 2004.
MANUAL DE MANEJO DAS POEDEIRAS DEKALB WHITE. Granja Planalto. 5 ed.
Minas Gerais: Uberlândia, 2009. 42p.
MENDONÇA JR. C.X.; LIMA, F.R. Efeito dos níveis de proteína e de metionina da dieta
sobre o desempenho de galinhas poedeiras após a muda forçada. Journal Veterinary
Research and Animal Science, v.36, p.332-338, 1999.
NARVÁEZ-SOLARTE, W.V. Exigências em metionina+cistina para poedeiras leves
e semipesadas. 1996. 57f. Dissertação (Mestrado em Zootecnia) - Universidade
Federal de Viçosa, Viçosa, MG, 1996.
NATIONAL RESEARCH COUNCIL. Nutrient requirements of poultry. 9 ed.
Washington, D.C.: National Academy Press, 1994. 157p.
PARS, J.F.; SUMMERS, J.D. The effects of minimizing amino acid excess in broiler
diets. Poultry Science, v.70, p.1540-549, 1991.
PIRES, C.V.; OLIVEIRA, M.G.A.; ROSA, J.C. et al. Qualidade nutricional e escore
químico de aminoácidos de diferentes fontes proteicas. Ciência e Tecnologia de
Alimentos, v.26, p.179-187, 2006.
POLESE, C.; NUNES, R.V.; VILELA, C.G. et al. Quantidade de metionina + cistina para
poedeiras semipesadas de 75 a 91 semanas de idade. Arquivo Brasileiro de
Medicina Veterinária e Zootecnia, v.64, p.1682-1690, 2012.
REZENDE, B.J.; BELETTI, M.E. Análise histomorfométrica do desenvolvimento do
intestino delgado de galinhas poedeiras. In: XII Seminário de Iniciação Científica,
2008, Uberlândia-MG. Anais... Uberlândia: Universidade Federal de Uberlândia,
2008.
ROMBOLA, L.G.; FARIA, D.E.; DEPONTI, B.J. et al. Fontes de metionina em rações
formuladas com base em aminoácidos totais ou digestíveis para frangas de reposição
leves e semipesadas. Revista Brasileira de Zootecnia, v.37, p.1990-1995, 2008.

110
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. et al. Tabelas brasileiras para
aves e suínos: composição de alimentos e exigências nutricionais. 3 ed. Viçosa:
Universidade Federal de Viçosa, 2011. 252p.
RUTZ, F. Proteínas: Digestão e Absorção. In: MACARI, M.; FURLAN, R.L.;
GONZALES, E.; Fisiologia Aviária Aplicada a Frangos de Corte. 2 ed. Jaboticabal:
FUNEP/UNESP, 2002, p.135-141.
SÁ, L.M.; GOMES, P.C.; ALBINO, L.F.T. et al. Exigência nutricional de metionina +
cistina digestível para galinhas poedeiras no período de 34 a 50 semanas de idade.
Revista Brasileira de Zootecnia, v.36, p.1837-1845, 2007.
SAKI, A.A.; MIRZAAGHATABAR, F.; ZAMANI, P. et al. Energy utilization by
chickens fed various levels of balanced methionine. Global Veterinaria, v.7, p.276-
282, 2011.
SAS. SAS/STAT 9.3 User’s Guide. Cary, NC: SAS Institute Inc. 2011, 8621p.
SCHUTTE, J.B.; DE JONG, J. Requirement of the laying hen for sulfur amino acids.
Poultry Science, v.73, p.274-280, 1994.
SILVA, J.H.V.; MUKAMI, F.; ALBINO, L.F.T. Uso de rações à base de aminoácidos
digestíveis para poedeiras. Revista Brasileira de Zootecnia, v.29, p.1446-1451, 2000.
SILVA JUNIOR, R.G.C.; LANA, G.R.Q.; RABELLO, C.B.V. et al. Exigências de
metionina + cistina para frangos de corte machos de 1 a 21 e de 22 a 42 dias de idade,
em clima tropical. Revista Brasileira de Zootecnia, v.34, p.2399-2407, 2005 (supl.).
SOARES, A.D.M., GIVISIEZ, P.E.N., SANTOS, E.G. et al. Efeito dos níveis de
metionina + cistina digestíveis da dieta pré-inicial sobre a morfometria intestinal de
frangos de corte. In: CONGRESSO BRASILEIRO DE ZOOTECNIA, 2008. João
Pessoa-PB. Anais... João Pessoa-PB, 2008.
TAKASHIMA, F.; HIBIYA, T. An atlas of fish histology. Normal and Pathological
features. 2ed. New York:Fisher, 1995, 195p.
TOGASHI, C.K.; FONSECA, J.B.; SOARES, R.T.B.N. Determinação de níveis de
metionina + cistina para poedeiras semipesadas alimentadas com rações contendo

111
Levedura Seca (Saccharomyces cerevisae). Revista Brasileira de Zootecnia, v.31,
p.1426-1433, 2002 (supl.).
UNIÃO BRASILEIRA DE AVICULTURA – UBABEF [2012]. Destino da produção
brasileira de ovos em 2012. Ubabef, 2012. Disponível em:
<http://www.ubabef.com.br/estatisticas/ovoses/Destino_da_Producao_Brasileira_de_
Ovos_em_2012> Acesso em: Dez. 03, 2013.
VARELA, E.V. Níveis nutricionais de metionina + cistina digestíveis em poedeiras
Hy Lyne W36 com base no conceito de proteína ideal. 2009, 34f. Dissertação
(Mestrado), Universidade Estadual de Maringá, UEM, Maringá – PR.
WALDROUP, P.W.; HELLWIG, H.M. Methionine and Total Sulfur Amino Acid
requirements influenced by stage of production. Journal of Applied Poultry
Research, v.4, p.283-292, 1995.
ZAVARIZE, K.C.; SARTORI, J.R.; PEZZATO, A.C. et al. Glutamina na dieta de
poedeiras leves submetidas ao estresse por calor e à termoneutralidade. Ciência
Animal Brasileira, v.12, p. 400-406, 2011.





























