Embed Size (px)
Citation preview

i
UNIVERSIDADE FEDERAL DE SANTA CATARINA CENTRO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE BOTÂNICA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA VEGETAL
Anatomia do lenho de árvores de espécies de canela, Ocotea Aubl. e Nectandra Rol. Ex. Rottb. (Lauraceae)
ocorrentes no Estado de Santa Catarina
Mestrando: Felipe Bernardino Guimarães
Orientadora: Profa. Dra. Ana Claudia Rodrigues
Co-Orientador: Prof. Dr. Mário Tomazello Filho
Dissertação apresentada ao Programa de Pós- Graduação em Biologia Vegetal da Universidade Federal de Santa Catarina sob a orientação da Profa. Dra. Ana Claudia Rodrigues para obtenção do título de Mestre.
__________________________ __________________________
Mestrando Orientador
Florianópolis, Maio de 2012.

ii
Para minhas preciosas filhas,
Zoê, Olívia e Sofia,
Vida, Árvore e Sabedoria

iii
Agradecimentos
Ao Instituto Brasileiro do Meio Ambiente e Recursos Naturais
Renováveis – IBAMA – pela concessão de licença de capacitação, em especial
a Dalva, ao Liceros e a minha querida Eda, que sempre em atendeu com muita
atenção e carinho; estou certo de que poderei retornar com estes novos
conhecimentos muito aplicáveis em nossas atividades fiscalizatórias.
A Prof. Dra. Ana Claudia Rodrigues, que aceitou o desafio de me
orientar, por sua atenção e curiosidade em aprender junto.
Ao Prof. Dr. Mário Tomazello Filho, pela co-orientação e disponibilização
do Laboratório de Anatomia e Identificação de Madeiras, do Departamento de
Ciências Florestais, da ESALQ/USP.
Aos membros da banca examinadora, Dra. Maria Leonor D’El Rei Souza
(CCB/UFSC), Dr. João de Deus Medeiros (CCB/UFSC) e Dra. Vera Terezinha
Rauber Coradin (LPF/SFB), em especial a Dra. Vera, quem sugeriu o trabalho
com as canelas.
Ao Dr. João Baitello e Dr. Leandro Cézanne de Souza Assis pela
atenção na identificação das exsicatas de Lauraceae.
Ao Dr. Marcos Sobral e Dr. Daniel Barcellos Falkenberg, pela
identificação das plantas extra-lauraceas coletadas nas saídas a campo;
Ao Dr. Hans Georg Richter, que prontamente respondeu minhas dúvidas
com grande atenção e cordialidade.
Ao Prof. Dr. Pablo Riul (UFPB) e em especial ao Prof. Dr. Paulo Horta,
pelo grande apoio dado nas análises estatísticas.
A queridíssima Profa. Dirlei Inês Thiel, que me apresentou uma nova
língua, completamente desconhecida, por suas aulas maravilhosas, que
permitiram descortinar a névoa que pairava sobre os preciosos dados da tese
de H.G.Richter em alemão e pela psicologia com preço bem abaixo da tabela!
Aos casal maravilhoso, Billie e Marcie Miller, por sua disponibilidade em
se dirigir inúmeras vezes a biblioteca em Berkeley, Universidade da Califórnia
em busca de literatura especializada, minha profunda gratidão por seu ato
altruísta, que espero poder recompensar um dia.
A Flávia Santanna, Paulo Licks, Marcelo Reinart, Maycon e Felipe da
ETS Consultoria Ambiental; Ivan Vargas da Desenvix e Sadi Pichetti da Itá

iv
Oeste, pelos contatos e informações preciosas acerca ocorrência de
supressões em andamento e pelo apoio logístico nas coletas a campo.
Aos motosseristas Gerci, Ivanir Apper, Neodir Kulax, Alemão, pelo
manejo das motosserras e atenção na coleta das canelas em campo.
A Baía Sul Madeiras pelos cortes das cunhas e lixação dos discos, em
especial ao Sr. Ademir.
Ao sempre atencioso e dedicado Sr. Emmanuel Bittencourt, Sra.
Verônica e filhas Mirella e Elisângela, pela preparação cuidadosa e paciente
dos corpos de prova.
A Maria A. Bermudez (ESALQ/USP) e Antônio Barbosa (IPT/SP), pelos
auxílios na confecção dos cortes anatômicos e macerados, em especial ao Sr.
Antônio, por toda sua atenção e carinho durante suas explicações.
A Maria das Graças (ESALQ/UPS), por ter sido um anjo na hora certa.
Ao amigo Luiz Henrique Paulli Bianchi, pela confecção dos mapas e das
imagens georreferenciadas, e por todo apoio ao longo destes anos de
convivência no IBAMA em Florianópolis.
Ao CIRAM/EPAGRI pelo fornecimento dos dados meteorológicos, em
especial a Dra. Vera Lúcia, por sua gentileza e atenção.
A Silvia Venturi e ao Dr. Rafael Trevisan, pelo acesso ao herbário e
paciência na inclusão e movimentação das exsicatas.
A Dra. Maria José Miranda, Rafael Pigozzo e ao Sr. Francisco, pelo
recebimento e inclusão das amostras de madeira na xiloteca Dr. Calvino
Mainieri, no Instituto de Pesquisas Tecnológicas de São Paulo.
Ao Laboratório de Fluorescência do departamento de Biologia Celular e
Genática - FLUORBEG/UFSC, especialmente a Chirle Ferreira e Demétrio, dos
quais recebi toda a atenção e orientação durante as análises de fluorescência.
A Elise Galitzki, por sua atenção durante as crises do microscópio
fungado.
Ao Laboratório Central de Microscopia Eletrônica (LCME/UFSC) pela
utilização de instalações e microscópios, em especial a Eliana e Renata Ozório,
pela grande atenção dispensada.
Aos amigos Cláudia Loch e Roque Carmona, por todo apoio e pela
grande amizade, e lógico, pela paciência com minhas amostras de madeira
espalhadas pelo condomínio.

v
Ao Arthur Beck Guimarães e Luiz Pessoa Guimarães, pela acolhida em
Piracicaba.
Aos amigos do mestrado Robson Carlos Avi, Jeovane Warmling, Chirle
Ferreira, Kátia Arenhart Hoss, Halisson Rafael Kedrovski, Rafaela, Graziela,
Taíse, Bruno & Ana, Regina, Francis, Marietu, pela força para continuar, pela
convivência harmoniosa e pelas boas risadas, e claro, por suportarem minha
insegurança e instabilidade nos dias de crise, e aos bolsistas Lucas, Ewerton e
Caiâne, por tanta dedicação da montagem das lâminas das coletas extras.
Aos meus pais, Nelson e Conceição, que de alguma forma, ao longo da
minha vida, me fizeram entender que estudar é preciso, e cá sigo eu, fazendo
isso desde sempre!
A minha vó Zoê Pessoa Guimarães, grande paixão da minha vida, fonte
de inspiração, por sua presença constante, aconselhamento e amor.
A Alessandra Cristina Giuliani, por segurar as pontas nas minhas
ausências e pelo estímulo para continuar.
E por fim, as árvores maravilhosas de canela e imbuia, que ao ceder
suas vidas para a geração de energia, forneceram seus discos para a
realização das análises anatômicas deste trabalho e me abriram janelas para
um mundo novo em anatomia de madeiras. Quiçá possa um dia retribuir à
ciência com conhecimento novo, essas vidas que foram ceifadas.

6
Índice
Página
Resumo 08
Abstract 09
1. Introdução 10
2 - Materiais e Métodos 17
2.1 – Coletas das amostras do lenho das árvores 17
2.2 – Análises macroscópicas do lenho 21
2.3 – Análises microscópicas do lenho 21
2.4 – Análises de caracteres anatômicos quantitativos do lenho 22
2.5 – Análise de agrupamento de caracteres anatômicos
qualitativos do lenho
22
2.6 – Análises estatísticas 25
3 – Resultados 28
3.1 – Descrições macroscópicas do lenho 28
Nectandra lanceolata Nees – Canela-amarela 28
Nectandra megapotamica Mez – Canela-amarela 30
Ocotea porosa (Nees & C. Mart.) Barroso - Imbuia 32
Ocotea puberula (Rich.) Nees – Canela-guaicá 34
Ocotea pulchella (Nees) Mez – Canela-lageana 36
Ocotea sp. - Canela-loura 38
3.2 – Chave dicotômica para diferenciação das espécies pela
estrutura macroscópica do lenho
40
3.3 – Descrições microscópicas do lenho 41
Nectandra lanceolata Nees – Canela-amarela 41
Nectandra megapotamica (Spreng.) Mez - Canela-amarela 43

7
Ocotea porosa (Nees & C. Mart.) Barroso - Imbuia 45
Ocotea puberula (Rich.) Nees – Canela-guaicá 47
Ocotea pulchella (Nees) Mez – Canela-lageana 49
Ocotea sp. – Canela-loura 51
3.4 – Chave dicotômica para diferenciação das espécies pela
estrutura microscópica do lenho
53
3.5 – Análises de caracteres anatômicos quantitativos do lenho 54
3.6 – Análise de agrupamento de caracteres anatômicos
qualitativos do lenho
59
4 – Discussão 77
5 – Considerações finais 100
6 – Referências Bibliográficas 101

8
Resumo
Anatomia do lenho de árvores de espécies de canela, Ocotea Aubl. e
Nectandra Rol. Ex. Rottb. (Lauraceae) ocorrentes no Estado de Santa
Catarina. Lauraceae agrega um grande número de espécies exploradas
comercialmente por sua madeira. Muitas delas, conhecidas popularmente por
“canela” pertencem aos gêneros Ocotea e Nectandra, muito difíceis de serem
separadas, tanto ao nível genérico, quanto ao nível específico, pela anatomia
do lenho. Neste intuito, realizou-se o presente trabalho com o objetivo de
buscar caracteres anatômicos do lenho diagnósticos para a separação das
espécies e dos gêneros. Foram coletadas árvores de canela em quatro
municípios do interior de Santa Catarina: Abdon Batista, Concórdia, Tangará e
Passos Maia. As espécies coletadas foram: Nectandra megapotamica,
Nectandra lanceolata, Ocotea porosa, Ocotea puberula, Ocotea pulchella e
Ocotea sp. As amostras do lenho foram seccionadas e analisadas seguindo os
métodos laboratoriais usuais em anatomia de madeira. Foram realizadas
análises de fluorescência. A partir das descrições macroscópicas e
microscópicas da anatomia do lenho elaboradas, foram confeccionadas chaves
dicotômicas para separação das espécies e realizadas analises estatísticas dos
caracteres anatômicos qualitativos e quantitativos do lenho.
Macroscopicamente, as espécies foram separadas pela cor, cheiro, visibilidade
do parênquima axial sob lente de 10x e arranjo diagonal. Microscopicamente, a
separação das espécies se deu pela presença e abundância de cristais de
oxalato de cálcio nas células dos raios, presença de placas de perfuração
escalariformes, presença, abundância e localização de células
oleíferas/mucilaginosas, largura dos raios e tipo de parênquima axial
paratraqueal. As análises dos caracteres anatômicos quantitativos permitiram a
distinção de quase todas as espécies entre si, mas não foram úteis na distinção
dos gêneros. Foi elaborada uma matriz binária de presença e ausência
considerando-se 35 caracteres anatômicos qualitativos para observação da
similaridade entre 64 espécie considerando-se 114 descrições anatômicas do
lenho de exemplares da família Lauraceae constantes da literatura, sendo 23
espécies de Nectandra, 34 de Ocotea, duas de Pleurothyrium, duas de
Rhodosthemonodaphne, duas de Chlorocardium, e uma de Sextonia. As
análises de similaridade não conseguiram evidenciar relações claras entre os
gêneros e as espécies avaliadas, em virtude de falta de padronização nas
descrições da anatomia do lenho pelos diferentes pesquisadores. Conclue-se
que é possível separar as seis espécies avaliadas no presente estudo pelos
caracteres anatômicos, tanto macroscopicamente quanto microscopicamente,
mas não foi possível esclarecer suas relações de similaridade com outras
espécies de Lauraceae de grupos próximos morfologicamente. Não foram
encontrados caracteres anatômicos diagnósticos que permitissem a distinção
dos gêneros Ocotea e Nectandra entre si. Palavras-chave: Anatomia do lenho,
Ocotea, Nectandra, Santa Catarina.

9
Abstract
Wood anatomy of trees from “canela” species, Ocotea Aubl. e Nectandra
Rol. Ex. Rottb. (Lauraceae) in Santa Catarina, Brazil. Lauraceae includes a
great number of species explored in the timber market due to the great quality
of its wood. Many of these species, known as “canela” belong to the genera
Ocotea and Nectandra, wich are very dificult to distinct considering their quite
homogeneus wood. The present work searched for diagnostic wood anatomical
features that could be helpfull to segregate these plants at specific and generic
levels. Collections of “canela” trees were made in four localities of Santa
Catarina state: Abdon Batista, Concórdia, Tangará and Passos Maia. Six
species were collected: Nectandra megapotamica, Nectandra lanceolata,
Ocotea porosa, Ocotea puberula, Ocotea pulchella and Ocotea sp. Wood
sections and analysis followed usual laboratory methods in wood anatomy.
Fluorescence analyzes were performed. Macroscopic and microscopic
descriptions were used to make dichothomous keys of these species. Statistical
analyzes were carried out using qualitative and quantitative wood characters.
Macroscopically, the species were separated by their color, odor, visibility of
axial parenchyma under 10x magnification and the presence of diagonal
arrangement. Microscopically, the distinction of these species was due to the
presence and abundance of calcium oxalate crystals in ray cells, presence of
scalariform perforations plates, presence, abundance and location of
oil/mucilage cells, rays width and type of paratracheal axial parenchyma. The
quantitative analysis of anatomical characters allowed the distinction of almost
all species, but were not useful in distinguishing the genera. A binary matrix of
presence and absence were prepared considering 35 qualitative anatomical
characters to observe similarity between 64 species considering 114 wood
anatomical descriptions of Lauraceae specimens founded in the literature.
Among these descriptions were 23 species of Nectandra, 34 of Ocotea, 2
species of Pleurothyrium, Rhodosthemonodaphne and Chlorocardium, and one
species of Sextonia. The similarity analyzes failed to show clear relationships
between genera and species evaluated, due to lack of standardization in the
descriptions of these species wood anatomy by different researchers. In
conclusion, it is possible to separate the six species evaluated in this study by
anatomical characters, both macroscopically and microscopically, but it was not
possible to clarify its relations of similarity with other morphologically close
species of Lauraceae. It was not founded clearly diagnostic wood anatomical
features that allowed the distinction between the genera Ocotea and Nectandra.
Key-words: wood anatomy, Ocotea, Nectandra, Santa Catarina.

10
1 - Introdução
A Família Lauraceae Jussieu é composta por árvores ou arbustos,
excepcionalmente trepadeiras, geralmente aromáticos, de distribuição
pantropical, estando bem representada na América, Ásia tropical, Austrália e
Madagascar, sendo rara nas regiões temperadas (Barroso et.al., 2002). Agrega
entre 2.500 a 3.500 espécies, distribuídas em aproximadamente 50 gêneros,
dos quais 20 a 22 ocorrem no Brasil (Barroso et.al., 2002; Baitello et.al., 2003),
com cerca de 390 espécies, habitando diversos ecossistemas, especialmente
as florestas pluviais (Barroso et.al., 2002).
Muitas espécies de Lauraceae são de grande destaque econômico por
seus óleos essenciais na perfumaria, cosmetologia, medicina e culinária, bem
como pela boa qualidade da madeira que produzem, destacando-se sua
utilização na indústria da construção civil – marcenaria, carpintaria, movelaria -
e naval (Kostermans, 1937; Record & Hess, 1942; Bastos, 1943; Brush, 1944;
Stern, 1954; Gemballa, 1955; Ghilardi, E. & Manieri, C. 1964; Rizzini, 1971;
Gottlieb, 1972; Klein, 1974; Loureiro, 1976; Reitz et. al., 1979; Vattimo-Gil,
1979c; Coe-Teixeira,1980; van der Werff & Richter, 1996; Marques, 2001).
Salienta-se, ainda, sua importância na fitofisionomia das formações de florestas
de araucária (Klein, 1974) e seu potencial de utilização em projetos de
reflorestamento (Reitz et. al., 1979).
Barroso et. al. (2002) apresentam uma síntese das características
morfológicas desta família: plantas monóicas, dióicas ou gimnodióicas; folhas
alternas, raramente opostas ou subopostas, glabras ou pilosas, com
indumentos de pêlos simples, unicelulares e células de óleo ou mucilagem no
mesofilo e pecíolos em geral canaliculados; inflorescências axilares ou
terminais, em panículas, tirsos, tirsóides ou botrióides; flores podem
hermafroditas ou unissexuadas, com 6 ou 9 tépalas iguais ou, eventualmente,
com tépalas externas bem menores que as internas; androceu geralmente
constituído de 3-6-9 estames férteis, acompanhados ou não de estaminódios e
dispostos em 3-4 verticilos: 1º e 2º verticilos de estames com anteras introrsas,
extrorso-apicais ou lateralmente extrosas; 3º verticilo de estames com par de
glândulas na base dos filetes e anteras, em geral, extrorsas e o 4º verticilo – o

11
mais interno – formado por estaminódios filiformes, triangulares, cordados,
sagitados ou ausentes; raramente todos os estames apresentam filetes
biglandulosos; anteras bilocelares ou tetralocelares, estas últimas exclusivas da
família; gineceu de ovário livre, mediano, unicarpelar, estilete simples, terminal;
óvulo único, pêndulo, apical ou anátropo; fruto tipo baga, às vezes parcialmente
envolvido pelo hipanto, modificado em cúpula; sementes sem endosperma,
embrião desenvolvido e cotilédones carnosos. A identificação ao nível genérico
e específico das Lauraceas é bastante complexa e difícil (van der Werff,
1991a).
Estes caracteres morfológicos, especialmente dos verticilos de
estames/estaminódios e a abertura das anteras, bem como a morfologia dos
frutos, permitem, quando presentes nos espécimes analisados e com certa
dificuldade, a distinção dos membros desta família ao nível genérico e
específico. Há que ser considerado que, com grande frequência, há
necessidade de identificação de material estéril e no entanto, devido à grande
semelhança observada nos diferentes gêneros, cujas circunscrições estão
baseadas especialmente nas características do androceu (Kostermans, 1957),
são frequentes os problemas na identificação das espécies e na distinção dos
gêneros (Record & Mell, 1924; Record & Hess, 1942; Castiglioni, 1951;
Rohwer, 1993a; van der Werff, 1991a; van der Werff & Richter, 1996).
A última grande proposta de classificação taxonômica para a família
Lauraceae, e mais amplamente aceita, que agrupa os gêneros segundo
caracteres florais, foi realizada por Kostermans (1957) com base no trabalho de
Mez (1889) denominado Lauraceae Americanae. Nela são estabelecidas duas
subfamílias, Lauroideae e Cassythoideae, tendo a primeira cinco tribos:
Perseeae, Cinnamomeae, Litseeae, Cryptocaryeae e Hypodaphneae. Os
gêneros Ocotea Aubl. e Nectandra Rol. Ex. Rottb. foram alocados na subtribo
Cinnamomineae, Tribo Cinnamomeae, juntamente com os gêneros
Pleurothyrium Nees, Cinnamomum Schaeff., Neocinnamomum H. Liu,
Cinnadenia Kosterm., Actinodaphne Ness, Sassafras (Kramer) Boehmer.,
Umbellularia (Nees) Nutt. e Dicypellium Nees & Mart. Posteriormente fora
proposta uma chave dicotômica para os 29 gêneros de Lauraceas do Novo
Mundo por van der Werff (1991a).

12
Van der Werff & Richter (1996) em busca de uma nova classificação da
família levando em conta também caracteres anatômicos do xilema secundário
e da casca apresentados por Richter (1981b), especialmente caracteres de
natureza qualitativa, observam a formação de três grupos, que encontram
correspondência na morfologia das inflorescências, quais sejam: 1 - Tribo
Laureae, que incluiria, entre outros, os gêneros Litsea Lamarck, Lindera
Thunberg, Laurus L. e Sassafras (Kramer) Boehmer; 2 - Tribo Perseeae, que
incluiria os gêneros Ocotea Aubl., Nectandra Rol. Ex. Rottb., Aniba Aubl,
Licaria Aubl, Pleurothyrium Nees, Persea Mil., Cinnamomum Schaeff., Phoebe
Nees e Dehaasia Blume; 3 - Tribo Cryptocaryeae, que incluiria os gêneros
Beilschmiedia Nees, Cryptocarya R. Brown, Endiandra R. Brown e Potameia
Thou. Destes 3 grupos, o primeiro teria como caracteres anatômicos mais
proeminentes a ausência de parênquima marginal e de fibras septadas na
madeira e as fibras floemáticas são sempre presentes na casca; o segundo
sendo caracterizado pela ausência de parênquima marginal e presença de
fibras septadas na madeira e as fibras estão presentes no floema secundário
de quase todos os taxa, com raras exceções; e o terceiro, caracterizado na
madeira pela presença de parênquima marginal, a presença de fibras não-
septadas com pontoações areoladas conspícuas e a presença de placas de
perfuração exclusivamente simples e no floema secundário a ausência de
fibras e a presença de esclereídeos característicos. Os autores salientam que
nem todos os taxa podem ser acomodados satisfatoriamente nestes 3 grupos
com base nos caracteres anatômicos, como p. ex. os gêneros Cinnamomum
Schaeff. e Persea Mil, que aparentam ser transicionais entre o grupo 1 e 2, e o
gênero Mezilaurus Taubert, que apresenta caracteres diagnósticos da madeira
do grupo 2 e caracteres diagnósticos da casca do grupo 3.
Novos gêneros tem sido propostos devido a recentes revisões
taxonômicas, especialmente dos gêneros neotropicais (Assis, 2009) e grandes
gêneros, como Ocotea Aubl. e Nectandra Rol. Ex. Rottb. tem sido
desmembrados. Dentre as contribuições taxonômicas mais significativas,
destacam-se os trabalhos relativos aos gêneros: Aiouea Aubl. (Renner, 1982),
Aniba Aubl (Kubitzki, 1982), Beilschmiedia Nees (Kostermans, 1938),
Dicypellium Nees & Mart. (Rohwer, 1988), Endlicheria Nees (Chanderbali,

13
2004), Systemonodaphne Mez (Rohwer, 1988), Gamanthera van der Werff (van
der Werff, 1991b), Licaria Aubl (Kurz, 2000), Mezilaurus Taubert (van der Werff,
1987), Nectandra Rol. ex. Rottb. (Rohwer, 1993), Ocotea Aubl. (Rohwer, 1986;
Rohwer, 1991; van der Werff, 1999), Sextonia van der Werff (van der Werff,
1997), Persea (Kopp, 1966), Phyllostemonodaphe Kosterm. (Rohwer, 1988),
Pleurothyrium Nees (van der Werff, 1993), Rhodostemonodaphne Rohwer &
Kubitzki (Madriñan, 2004), Urbanodendron Mez (Rohwer, 1988). Algumas
destas contribuições que levaram a propositura de novos gêneros observaram
além dos caracteres morfológicos, também os caracteres anatômicos do xilema
secundário, como nos gêneros Sextonia van der Werff (van der Werff, 1997) e
Paraia Rohwer, Richter & van der Werff (Rohwer et.al, 1991), onde, além da
caracterização microscópica, são mencionados caracteres auxiliares como cor
e densidade, quando da comparação com outros grupos de lauráceas.
Também para as duas espécies do novo gênero Chlorocardium Rohwer,
Richter & van der Werff, desmembradas do gênero Ocotea, são inclusos como
caracteres anatômicos do xilema secundário relevantes para a distinção dos
demais gêneros de lauráceas o tamanho extremamente pequeno das
pontoações inter-vasculares, a ausência de células secretoras
oleíferas/mucilaginosas e a presença de elementos de vasos coronados
(Rohwer et.al, 1991).
Análises filogenéticas com base em dados moleculares atentam para o
monofiletismo da família Lauraceae (Renner, 1999; Renner & Chanderbali,
2000; Chanderbali et. al., 2001), assim como para as tribos Cryptocaryeae e
Cinnamomeae sensu Kostermans (Rohwer, 2000; Chanderbali et. al., 2001).
Entretanto, as mesmas análises apontam para o não-monofiletismo de grandes
gêneros como Endlicheria Nees, Nectandra Rol. ex Rottb, Ocotea Aubl. e
Rhodostemonodaphne Rohwer & Kubitzki (Chanderbali et. al., 2001). Em
virtude da impossibilidade de resolução dos problemas taxonômicos apenas
com dados morfológicos, tem sido sugerido que estudos taxonômicos do grupo
levem em conta não apenas dados morfológicos, mas também dados
anatômicos (Record & Mell, 1924; Richter, 1981a; Richter, 1981b; Rohwer et.al,
1991; van der Werff & Richter, 1996; Quinet, 2001) assim como, a associação
de dados morfológicos e anatômicos com dados moleculares (Assis, 2009).

14
A anatomia do xilema secundário há muito tem sido investigada e
utilizada como uma importante ferramenta para auxiliar a resolução de
problemas taxonômicos (Record, 1934). Descrições anatômicas da madeira de
espécies e gêneros da família Lauraceae e importantes trabalhos sobre
aspectos característicos da estrutura anatômica da madeira da família foram
fornecidos por Knoblauch (1888), Solereder, (1908), Janssonius (1926), Tupper
(1927), Milanez (1930); Dadswell & Record (1936); Record & Hess (1942);
Bastos (1943); Ochioni & Mattos Filho (1947); Botelho (1951); Stern (1954);
Kribs (1959); Castiglioni (1962); van der Slooten et. al., (1970); Loureiro (1976);
Dechamps (1979); Richter (1981a, b; 1985; 1987; 1990); Hernandez (1985);
Carreras & Perez (1988), Feldato et. al., (1989), Richter & Van Wyk (1990);
Rohwer et.al, (1991); Callado (1995), Callado & Costa (1997); León (2000),
Silva & Medeiros (2000), León & Espinoza de Pernía (2000a, b; 2001),
Czarneski et. al. (2001), Oliveira et al. (2001), Silva (2003), Mattos et. al.,
(2003), Oliveira (2005), Loutfy (2009), Marchiori et al., (2009) e Marchiori et al.,
(2010). Merecem grande destaque os trabalhos de Stern (1954) e Richter
(1981b), no qual foram observados padrões anatômicos diagnósticos supra e
infragenéricos e, face à existência de desvios consideráveis destes padrões,
salientam a necessidade de uma reavaliação na hierarquia dos caracteres
utilizados na classificação das Lauraceae.
A estrutura anatômica do xilema da família aparenta ser, em termos
filogenéticos, transicional a avançada, mais do que primitiva (Stern, 1954;
Rohwer, 1993b). A uniformidade dos caracteres anatômicos do xilema
secundário nesta família é bastante evidente (Solereder, 1908; Stern , 1954;
Kostermans, 1957; Richter, 1981b, 1987), o que facilita inclusive a identificação
de fósseis deste grupo (Tupper, 1927). Dentre os caracteres anatômicos mais
significativos do xilema secundário que caracterizam a família Lauraceae
podem ser citados: a porosidade difusa; os vasos nunca exclusivamente
solitários, arredondados ou ovais; as pontoações inter-vasculares sempre
alternas; a presença de parênquima paratraqueal; as fibras com pontoações
localizadas exclusivamente nas paredes radiais; os raios estreitos, geralmente
de 1-4 células de largura; a presença de células oleíferas/mucilaginosas na
maioria das espécies associadas aos parênquimas axial e/ou radial (Richter,

15
1981a, 1981b, 1987, 1990; Metcalfe & Chalk, 1985; Callado & Costa, 1997;
León & Espinoza de Pernía 2000a, 2000b; Oliveira et al. 2001). Padrões e
similaridades anatômicas também tem sido identificados para gêneros (Richter,
1981b, Rohwer et.al, 1991), mas a confusão na taxonômica de alguns grupos,
especialmente Ocotea Aubl. e Nectandra Rol. ex Rottb claramente indicados
como parafiléticos, reflete-se também na dificuldade em observação de
padrões anatômicos nos caracteres da madeira e da casca. Alguns gêneros
possuem características anatômicas bastante específicas, que os distinguem
dos demais gêneros da família, p.ex. os raios homogêneos compostos por
células exclusivamente procumbentes em Hypodaphnis Stapf, raios agregados
em espécies de Cryptocaria R. Brown , anéis porosos somente em espécies de
Sassafras Presl. (Richter, 1987), ausência de células oleíferas/mucilaginosas e
pontoações inter-vasculares muito pequenas em Chlorocardium Rohwer,
Richter & van der Werff e raios estratificados em Aspidostemon Rohwer &
Richter e algumas espécies de Beilschmiedia Nees e Potameia Thou. (Rohwer,
1993b). No entanto, a grande maioria dos gêneros dentro da família
compartilha caracteres anatômicos do xilema secundário, de forma bastante
homogênea, tornando difícil e complexa a identificação tanto ao nível genérico
quanto específico (Castiglioni, 1962; Richter, 1981b, 1987).
No Brasil, contribuições significativas relativas à taxonomia e distribuição
das Lauraceae foram realizadas por Vattimo (1956a,b; 1958; 1959; 1960; 1966;
1976; 1977; 1978a,b), Vattimo-Gil (1979, a, b, c; 1980; 1983), Coe-Teixeira
(1963, 1970, 1980), Pedralli (1985, 1986a, 1986b, 1987a, 1987b 1988, 1996),
Baitello (2001; 2003), Quinet & Andreata (2002), Quinet (2005; 2008), Assis
(2009) Assis & Mello-Silva (2009; 2010a, b). Dados recentes da lista da Flora
do Brasil indicam a presença de 170 espécies de Ocotea Aubl. no país e 46 de
Nectandra Rol. Ex Rottb. (Quinet et. al., 2012). Em Santa Catarina, constam
listadas em Reitz et. al. (1979) nove gêneros da família Lauraceae que
englobam 52 espécies de árvores e arvoretas.
Alheios a toda esta confusão taxonômica, encontram-se os extratores,
comerciantes, consumidores e agentes da lei envolvidos na cadeia produtiva
das espécies de canela exploradas comercialmente, algumas das quais
incluídas em listas de espécies da flora brasileira ameaçadas de extinção (IN
MMA 06/2008), entre elas Ocotea basicordatifolia Vattimo-Gil, Ocotea bragae

16
Coe-Teix., Ocotea catharinensis Mez, Ocotea langsdorffii (Meissn.) Mez,
Ocotea odorifera Rohwer e Ocotea porosa (Nees & Mart.) Barroso. Embora
ferramentas que facilitem a identificação de madeiras estejam sendo
desenvolvidas para auxiliar profissionais que trabalhem nesta área na
identificação de espécies, como a Chave Interativa para Identificação de
Madeiras Comerciais do Brasil (Coradin et. al., 2009), poucos são os dados
disponíveis acerca das espécies de Lauraceas, e constam desta chave apenas
Ocotea porosa (Nees & Mart.) Barroso, Ocotea fragrantissima Ducke,
Mezilaurus itauba (Meissn.) Taubert ex Mez, Mezilaurus lindaviana Schwacke e
Sextonia rubra (Mez) van der Werff.
Pode-se afirmar que o controle da exploração, comercialização e
consumo de madeira de espécies de Lauraceas, que deveria estar sendo
realizado de forma adequada pelos sistemas de controle das autoridades
competentes – Sistema DOF e SISFLORA, está baseado em identificações
equivocadas desde o início da cadeia de custódia, nos Planos de Manejo
Florestal Sustentável (PMFS) e ao longo de toda ela, face a incapacidade dos
entes envolvidos nesta cadeia de custódia em identificarem os táxons desta
família ao nível genérico ou específico.
Espera-se assim, com o presente trabalho, contribuir para o
conhecimento da anatomia do xilema secundário das árvores de algumas
espécies dos gêneros Ocotea Aubl. e Nectandra Rol. Ex Rottb. conhecidas sob
a denominação de canelas ocorrentes no Estado de Santa Catarina.

17
2 - Materiais e Métodos
2.1 – Coletas das amostras do lenho das árvores
As coletas foram feitas nos municípios de Passos Maia, Tangará, Abdon
Batista e Concórdia, em Santa Catarina (Fig. 1, 2, 3A e 3B)
Figura 1 – Em destaque, localização dos municípios de coleta das árvores de canelas dos gêneros Ocotea e Nectandra no Estado de Santa Catarina utilizadas no presente trabalho.

18
As áreas de coleta situam-se no oeste, meio-oeste e planalto serrano
catarinense, possuem altitudes, temperaturas médias anuais e pluviosidade
média anual muito similares (Tabela 1). A vegetação dominante é a Floresta
Ombrófila Mista e os bosques de pinhais, nos quais a Araucaria angustifolia
(Bertol.) O. Kze aparece como árvore emergente, são tipicamente
acompanhados das lauráceas que compõem o principal elemento do estrato
das macrofanerófitas (Klein, 1974). As coletas foram realizadas em áreas de
supressão vegetal para instalação da Pequena Central Hidrelétrica (PCH) no
Rio Chapecó, no município de Passos Maia, e PCH de Salto Góes, no rio do
Peixe, no município de Tangará (Fig. 3B), da Usina Hidrelétrica Garibaldi (UHE
Garibaldi), no rio Canoas, em Abdon Batista (Fig. 3A) e do Loteamento Frei
Lency II, em Concórdia (Fig. 2). A floresta suprimida era secundária, em função
de intervenção antrópica no passado, e encontrava-se em estagio médio de
regeneração em Concórdia, Tangará e Abdon Batista e médio a avançado em
Passos Maia . As áreas possuíam autorização de supressão (AUTEX) emitidas
pela FATMA e as coletas de madeira e dos ramos das árvores foram realizadas
em acompanhamento aos trabalhos das equipes de supressão em campo.
Figura 2 – Imagens de satélites georreferenciadas das quatro áreas de coleta das árvores de canela analisadas no presente trabalho. (superior esquerda: Abdon Batista; superior direita: Concórdia; inferior esquerda: Passos Maia; inferior direita: Tangará).

19
Tabela 1 – Localização, altitude e dados meteorológicos dos locais de coleta das amostras do lenho das árvores de canela.
Locais de coleta
Coordenadas geográficas
Altitude (m)
Tº média anual (°C)
Precipitação pluviométrica
anual média (mm)
Abdon Batista¹
27° 38’ 13,57”S 51° 00’ 04,65”W
700
17,5
153,054
Concórdia² 27° 14’ 03,5”S 51° 57’ 48,93”W
750 17,2 168,41
Passos Maia³
26º 42' 06,5"S 51º 55' 12,6"W
1050
16,6
190,170
Tangará⁴
27º 06' 08,0"S 51º 17' 00,8"W
600
17,3
155,5375
1 – Estação meteorológica da EPAGRI em Abdon Batista; 2 – Estação Meteorológica da EPAGRI em Joaçaba; 3 – Estação meteorológica da EPAGRI em Ponte Serrada; 4 - Estação meteorológica da EPAGRI em Videira;
Foram coletados 3 discos do lenho na altura do peito (1,30m) de árvores
de canela (Fig. 3C) das espécies Nectandra lanceolata Nees, Nectandra
megapotamica (Spreng.) Mez, Ocotea puberula (Rich.) Nees, Ocotea pulchella
(Nees) Mez, Ocotea porosa (Nees & C. Mart.) Barroso e Ocotea sp. As alturas
e diâmetros médios das árvores, bem como o número de indivíduos coletados
em cada área estão resumidos na Tabela 2.
As exsicatas foram depositadas no herbário FLOR e encaminhadas para
2 especialistas na família Lauraceae para identificação. As cunhas de madeira
(Fig. 3C e 3D) foram depositadas na Xiloteca do Herbário Flor e na Xiloteca
Calvino Manieri (BCTw) no Instituto de Pesquisas Tecnológicas em São Paulo
(Tabelas 3).
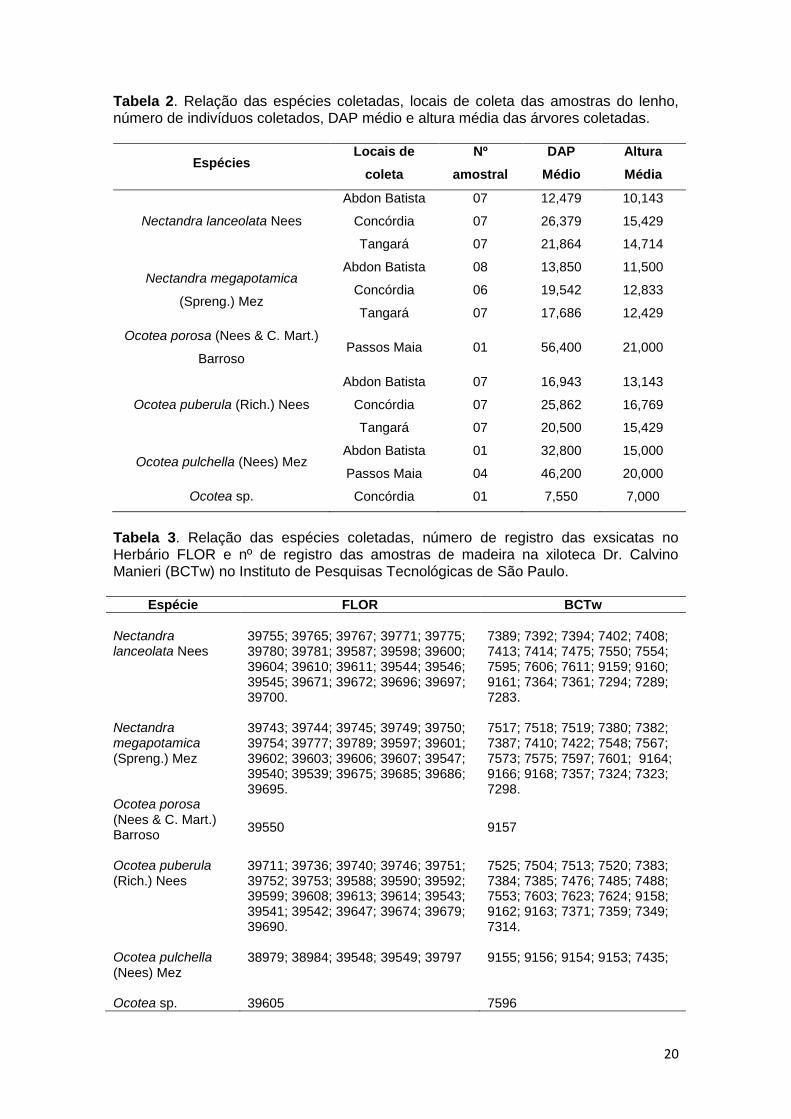
20
Tabela 2. Relação das espécies coletadas, locais de coleta das amostras do lenho, número de indivíduos coletados, DAP médio e altura média das árvores coletadas.
Espécies Locais de
coleta
Nº
amostral
DAP
Médio
Altura
Média
Nectandra lanceolata Nees
Abdon Batista 07 12,479 10,143
Concórdia 07 26,379 15,429
Tangará 07 21,864 14,714
Nectandra megapotamica
(Spreng.) Mez
Abdon Batista 08 13,850 11,500
Concórdia 06 19,542 12,833
Tangará 07 17,686 12,429
Ocotea porosa (Nees & C. Mart.)
Barroso Passos Maia 01 56,400 21,000
Ocotea puberula (Rich.) Nees
Abdon Batista 07 16,943 13,143
Concórdia 07 25,862 16,769
Tangará 07 20,500 15,429
Ocotea pulchella (Nees) Mez Abdon Batista 01 32,800 15,000
Passos Maia 04 46,200 20,000
Ocotea sp. Concórdia 01 7,550 7,000
Tabela 3. Relação das espécies coletadas, número de registro das exsicatas no Herbário FLOR e nº de registro das amostras de madeira na xiloteca Dr. Calvino Manieri (BCTw) no Instituto de Pesquisas Tecnológicas de São Paulo.
Espécie FLOR BCTw
Nectandra lanceolata Nees
39755; 39765; 39767; 39771; 39775; 39780; 39781; 39587; 39598; 39600; 39604; 39610; 39611; 39544; 39546; 39545; 39671; 39672; 39696; 39697; 39700.
7389; 7392; 7394; 7402; 7408; 7413; 7414; 7475; 7550; 7554; 7595; 7606; 7611; 9159; 9160; 9161; 7364; 7361; 7294; 7289; 7283.
Nectandra megapotamica
(Spreng.) Mez
39743; 39744; 39745; 39749; 39750; 39754; 39777; 39789; 39597; 39601; 39602; 39603; 39606; 39607; 39547; 39540; 39539; 39675; 39685; 39686; 39695.
7517; 7518; 7519; 7380; 7382; 7387; 7410; 7422; 7548; 7567; 7573; 7575; 7597; 7601; 9164; 9166; 9168; 7357; 7324; 7323; 7298.
Ocotea porosa (Nees & C. Mart.) Barroso
39550
9157
Ocotea puberula (Rich.) Nees
39711; 39736; 39740; 39746; 39751; 39752; 39753; 39588; 39590; 39592; 39599; 39608; 39613; 39614; 39543; 39541; 39542; 39647; 39674; 39679; 39690.
7525; 7504; 7513; 7520; 7383; 7384; 7385; 7476; 7485; 7488; 7553; 7603; 7623; 7624; 9158; 9162; 9163; 7371; 7359; 7349; 7314.
Ocotea pulchella (Nees) Mez
38979; 38984; 39548; 39549; 39797
9155; 9156; 9154; 9153; 7435;
Ocotea sp. 39605 7596

21
2.2 – Análises macroscópicas do lenho
Os caracteres macroscópicos do lenho foram analisados de acordo
com o proposto por Coradin & Muniz (1991) e Coradin et. al. (2009). As
análises do lenho foram feitas a olho nu ou sob lupa de 10x de aumento, e as
fotografias obtidas com microscópio estereoscópio Leica EZ4D e software LAS
3.0. Realizou-se ainda o teste químico com Cromazurol-S para detecção de
alumínio no lenho.
2.3 – Análises microscópicas do lenho
Foram preparados dois corpos de prova do lenho de cada árvore nas
dimensões de 1,5x1,5x1,5 cm (Fig. 3F), cozidos por 5 a 8 horas (Fig. 3G) em
água com glicerina (5:1) e cortados em micrótomo de deslize Leica (Fig. 3H),
nas faces transversal, longitudinal radial e longitudinal tangencial, na espessura
de 15 a 20 μm. Os cortes foram previamente tratados com hipoclorito de sódio
(20%), corados com Azul de Astra e Safranina (Bukatsch, 1972) e desidratados
em série etílica e acetato de n-butila para montagem de lâminas permanentes
em resina Permount ou Entellan. As análises foram realizadas em microscópio
óptico Leica DM2500 e as medições foram obtidas através do software LAS
3.7. Os testes histológicos realizados foram SUDAN III (Sass, 1951) para
substâncias lipídicas, Cloreto Férrico (Johansen, 1940) para compostos
fenólicos e Fenol para sílica (Johansen, 1940). Os macerados foram
preparados em solução 1:1 de Ácido Acético Glacial e Peróxido de Hidrogênio
segundo Franklin (1945) por 48h em estufa a 60ºC e posteriormente corados
com Safranina (1%). As lâminas semi-permanentes foram montadas em
gelatina glicerinada (Kaiser, 1880). As descrições microscópicas seguiram as
recomendações da IAWA (IAWA Committee, 1989). As análises de
fluorescência foram realizadas através do microscópio Olympus BX41 com
epifluorescência e os filtros para visualização da cor Azul U-MWU2 (excitação:
330 - 385; emissão: 420), da cor Vermelho U-MWG2 (excitação: 510 - 550;
emissão: 590), da cor verde U-MWB2 (excitação: 460 - 490; emissão: 520IF), e
filtro triplo. As imagens foram obtidas por câmara digital colorida com 3.3

22
mpixel QColor 3C da Q-imaging acoplada ao microscópio e analisadas pelo
software Q-capture Pro 5.1 da Q-imaging.
Para as análises em microscopia eletrônia de varredura (MEV), foram
preparados corpos de prova de 0,5 x 05 x 0,5 cm, secos em estufa a 40ºC por
3 dias, aderidos em stubs com fita dupla face de carbono e metalizadas com
ouro no recobridor Leica EM SCD 500. As análises foram realizadas no
microscópio JEOL JSM-6390LV no Laboratório Central de Microscopia
Eletrônica (LCME/UFSC).
2.4 – Análises de caracteres anatômicos quantitativos do lenho
Foram realizadas as mensurações das seguintes estruturas
anatômicas do lenho: diâmetro tangencial e comprimento dos elementos de
vaso, número de vasos por milímetro quadrado, diâmetro tangencial das
pontoações inter-vasculares, altura e largura dos raios número de raios por
milímetro linear, diâmetro mediano, diâmetro do lume, espessura das paredes
e comprimento das fibras, em conformidade com as recomendações da IAWA
(IAWA Committee, 1989) e foram realizadas para os dois corpos de prova de
cada árvore. Nas descrições anatômicas, além dos valores médios, foram
mencionados os menores e maiores valores encontrados, entre parênteses, e
nos resultados, o valor do erro padrão.
2.5 – Análise de agrupamento de caracteres anatômicos qualitativos do
lenho
Para fins de observação da similaridade das espécies de Ocotea e
Nectandra, foi realizada análise de agrupamento com base em uma matriz
binária de presença e ausência (0-1) levando-se em consideração 35
caracteres anatômicos do lenho, dispostos na Tabela 4. As espécies utilizadas
nesta análise foram as 6 espécies avaliadas no presente estudo e outras 114
descrições anatômicas do lenho presentes na literatura, assim identificadas na
Tabela 7: * resultados de análises do presente estudo; 1 - León & Espinoza de
Pernía (2001); 2 - Oliveira et. al. (2001); 3 – León (1999-2000); 4 - Silva &
Medeiros (2000); 5 – Silva (2003); 6 – Oliveira ( 2005); 7 – Castiglioni (1962); 8

23
– Ochioni & Mattos Filho (1947); 9 – Hernandez (1985); 10 – Milanez (1930); 11
- Mattos Filho (1960); 12 – Pinho (1968); 13 – Kostermans et. al. (1969); 14 -
Kribs (1959); 15 – Botelho (1951); 16 – Dechamps (1979), 17 – Stern (1954);
18 – Feldato et. al. (1989); 19 - Carreras & Pérez (1988); 20 – van der Slooten
et. al., 1970; 21 - Marchiori et al., (2009); 22 - Marchiori et al., (2010); 23 -
Mattos et. al., (2003). Ao todo, foram consideradas 114 descrições anatômicas
do lenho. Ressalte-se que algumas destas espécies já foram re-arranjadas em
outros gêneros, como Ocotea rubra Mez e Nectandra rubra (Mez) C. K. Allen,
que foram realocadas para o gênero Sextonia (van der Werff, 1997), Ocotea
rodiei (Schomb. ex. Hook) Mez e Ocotea venenosa Kosterm. & Pinkley
realocadas para o gênero Chlorocardium (Rohwer et. al., 1991), Nectandra
grandis (Mez) Kosterm. e Ocotea cooperi C. K. Allen, realocadas para o gênero
Rhodostemonodaphne (Madriñan, 2004) e Ocotea compactiflora Kosterm. e
Ocotea palmana Mez & J. D. Smith realocadas para o gênero Pleurothyrium
(van der Werff, 1993). Ainda assim, foram mantidas na matriz binária, para que
se pudesse observar seu grau de similaridade junto as demais espécie dos
gêneros Ocotea e Nectandra. Outras espécies incluídas na matriz foram
sinonimizadas, e seus nomes foram atualizadas com base nos dados
disponíveis no TROPICOS e na Lista da Flora do Brasil (Quinet et al., 2012).
Foram elas: Nectandra concinna Nees sinônimo de Nectandra turbacensis
(Kunth) Nees, Nectandra elaiophora Barb. Rodr. e Ocotea barcellensis
(Meissn.) Mez tornaram-se sinônimos de Ocotea cymbarum Kunth., Nectandra
falcifolia (Nees) Castiglioni sinônimo de Nectandra angustifolia (Schrad.) Nees
& Mart., Nectandra nitida Mez sinônimo de Nectandra martinicensis Mez,
Nectandra pichurim (HBK) Mez sinônimo de Nectandra cuspidata Nees & Mart.,
Nectandra pisi Miquel sinônimo de Nectandra globosa (Aubl.) Mez, Nectandra
rigida (H. B. K.) Nees sinônimo de Nectandra reticulata (Ruiz & Pav.) Mez,
Nectandra saligna Nees & Mart. ex. Nees sinônimo de Nectandra
megapotamica (Spreng.) Mez, Ocotea acutangula (Miq.) Mez sinônimo de
Ocotea leucoxylon (Sw.) Laness, Ocotea acutifolia (Nees) Mez sinônimo de
Nectandra acutifolia (Ruiz & Pav.) Mez, Ocotea babosa C. K. Allen sinônimo de
Ocotea puberula (Rich.) Nees, Ocotea calophylla Mez sinônimo de Ocotea
fulvescens Standl. & L.O. Williams, Ocotea caudata (Nees) Mez. sinônimo de
Ocotea cernua (Nees) Mez, Ocotea costulata (Nees) Mez sinônimo de Ocotea

24
aciphylla (Nees), Ocotea cuneata (Griseb.) Gomez sinônimo de Ocotea
cuneifolia (Ruiz & Pav.) Mez, Ocotea lanceolata Nees sinônimo de Ocotea
lancifolia (Schott) Mez, Ocotea suaveolens (Meissn.) Benth ex. Hieron.
sinônimo de Ocotea diospyrifolia (Meissn.) Mez e Ocotea wachenheimii Benoist
tornou-se sinônimo de Ocotea floribunda (Sw.) Mez. Foi excluída da análise a
descrição de Nectandra pardo constante em León & Espinoza de Pernía
(2001), pois esta espécie não existe. Assim, dentre as 114 descrições da
matriz, estão sendo comparadas 64 espécies distintas, sendo duas espécies do
gênero Chlorocardium, 23 espécies do gênero Nectandra, 34 espécies do
gênero Ocotea, duas espécies do gênero Pleurothyrium, duas espécies de
Rhodostemonodaphne e uma espécie de Sextonia.
Tabela 4 – Caracteres qualitativos do lenho utilizados na matriz binária para análise de
agrupamento das espécies de Ocotea e Nectandra.
Nº Caráter
1 Porosidade difusa
2 Camadas de crescimento distintas por zonas fibrosas
3 Poros solitários
4 Poros múltiplos radiais
5 Poros geminados
6 Poros em cachos
7 Arranjo diagonal
8 Pontoações inter-vasculares alternas
9 Pontoações inter-vasculares poligonais
10 Pontoações inter-vasculares ovais ou circulares
11 Pontoações inter-vasculares ornamentadas
12 Pontoações inter-vasculares internamente coalescentes
13 Elementos de vaso coronados
14 Placa de perfuração simples
15 Placa de perfuração escalariforme
16 Placa de perfuração reticulada
17 Tiloses
18 Fibras septadas
19 Parênquima axial paratraqueal escasso
20 Parênquima axial paratraqueal vasicêntrico
21 Parênquima axial paratraqueal unilateral

25
Continuação da Tabela 4...
22 Parênquima axial paratraqueal aliforme
22 Parênquima axial paratraqueal confluente
24 Parênquima em faixas marginais
25 Parênquima apotraqueal difuso
26 Raios unisseriados
27 Raios multisseriados
28 Raios homogêneos
29 Raios heterogêneos
30 Raios fusionados
31 Paredes disjuntivas nas células dos raios
32 Células oleíferas/mucilaginosas associadas aos raios
33 Células oleíferas/mucilaginosas associadas ao Parênquima axial
34 Células oleíferas/mucilaginosas entre as fibras
35 Cristais nas células dos raios
2.6 – Análises estatísticas
As análises dos caracteres anatômicos quantitativos do lenho foram
descritas considerando os valores das médias. A análise de escalonamento
multidimensional foi empregada para se descrever a agregação das espécies
com relação as características anatômicas quantitativas do lenho. As
similaridades e dissimilaridades relativas (SIMPER) e a análise da significância
das similaridades e dissimilaridades observadas (ANOSIM) foram utilizadas
para se avaliar a utilidade das referidas características anatômicas na
separação das espécies entre si, na separação dos gêneros e dos locais de
coleta. A matriz de similaridade utilizada no MDS e ANOSIM foi obtida a partir
do índice de Bray-Curtis, sendo os valores dos respectivos caracteres
anatômicos transformados em Log (X+1).
Para as análises na matriz de dados de presença e ausência (Tabela 7)
dos caracteres anatômicos qualitativos do lenho (Tabela 4) das espécies
analisadas no presente trabalho e dos dados oriundos da literatura utilizou-se o
índice de similaridade de Jaccard (P/A) hierarquizados segundo a análise de
Cluster para construção dos dendrogramas de similaridade. As similaridades e

26
dissimilaridades relativas (SIMPER) e a análise da significância das
similaridades e dissimilaridades observadas (ANOSIM) foram utilizadas para se
avaliar a utilidade das referidas características anatômicas na separação das
espécies entre si, na separação dos gêneros e na avaliação da similaridade
das descrições feitas por um mesmo autor. Foram apresentados gráficos
multidimensionais MDS para as relações inter-genéricas e entre as fontes
bibliográficas dos dados de descrições anatômicas do lenho utilizados. Na
construção da matriz binária. Todas as referidas análises foram realizadas com
o software Primer 6β.

27
Figura 3 – A – área de coleta na UHE Garibaldi, em Abdon Batista (SC); B – área de coleta na PCH Salto Góes, Tangará (SC); C – corte dos discos do tronco das árvores; D – preparação das cunhas de madeira; E – cunhas e discos em processo de secagem natural; F – corpos de prova de madeira; G – cozimento dos corpos de prova de madeira; H – cortes histológicos em micrótomo de deslize.

28
3 – Resultados
3.1 – Descrições macroscópicas do lenho
Nectandra lanceolata Nees – Canela-amarela (Fig. 4A - H)
Características gerais: Cerne/alburno indistintos pela cor; cerne
amarelado, com alteração de cor por foto-oxidação, mudando para
amarronzado-acinzentado (Fig. 4B - C); anéis de crescimento distintos,
individualizados por zonas fibrosas tangencias mais escuras (Fig. 4D); madeira
com brilho nas superfícies transversal e longitudinais (Fig. 4G - H); sem cheiro
perceptível; moderadamente dura ao corte transversal manual; grã revessa;
textura média; figura presente, em faixas causadas pelas camadas de
crescimento (podendo formar "V" ou "U").
Poros: Presentes, visíveis apenas com lente de 10x, pequenos ou
médios, de distribuição difusa, frequência média, predominantemente solitários,
eventualmente múltiplos radiais com menos de 4 vasos, eventualmente
dispostos em arranjo diagonal; formato circular a oval; placas de perfuração
não observadas mesmo com lente de 10x; parte dos vasos obstruído por
tiloses, poucas (Fig. 4E - F).
Parênquima axial: distinto sob lente de 10x., com dificuldade,
paratraqueal escasso ou vasicêntrico (Fig. 4E - F).
Raios: Observados apenas com lente de 10x na superfície transversal
(Fig. 4E) e longitudinal tangencial (Fig. 4G), visíveis a olho nu e bem
contrastados na superfície longitudinal radial (Fig. 4H), finos, baixos, pouco
frequentes.
Estruturas estratificadas: Ausentes.
Estruturas secretoras: Ausentes.
Variantes cambiais: Não observadas.
Testes químicos: Teste de Cromazurol negativo.

29
Figura 4 – Nectandra lanceolata Nees; A – ramo da árvore; B – disco do tronco recém-cortado; C – disco do tronco lixado; D – face transversal: anéis de crescimento; E – vista geral da madeira em corte transversal (10x); F – vista ampliada da madeira em corte transversal (20x); G – vista geral da madeira na face longitudinal tangencial (10x); H – vista geral da madeira na face longitudinal radial da madeira (10x). Escalas: A – 6 cm; B – 10 cm; C – 7 cm ; D – 2 cm ; E a H – 1 mm.

30
Nectandra megapotamica (Spreng.) Mez - Canela-amarela (Fig. 5A - H)
Características gerais: Cerne/alburno pouco distintos pela cor; cerne
amarelado, com alteração de cor por foto-oxidação, mudando para
amarronzado-oliváceo (Fig. 5B - C); anéis de crescimento distintos,
individualizados por zonas fibrosas tangencias mais escuras (Fig. 5D); madeira
com brilho nas superfícies transversal e longitudinais (Fig. 5G - H); com cheiro
perceptível, desagradável, que desaparece com a secagem da madeira,
retornando se a mesma for molhada; moderadamente dura ao corte transversal
manual; grã revessa; textura média; figura presente, em faixas causadas pelas
camadas de crescimento (podendo formar "V" ou "U").
Poros: Presentes, visíveis apenas com lente de 10x., pequenos ou
médios, de distribuição difusa, frequência média; predominantemente solitários,
eventualmente múltiplos radiais com menos de 4 vasos; eventualmente
dispostos em arranjo diagonal; de formato circular a oval; placas de perfuração
não observadas mesmo com lente de 10x.; parte dos vasos obstruídos por
tiloses (Fig. 5E - F)
Parênquima axial: pouco distinto sob lente de 10x., paratraqueal
escasso ou vasicêntrico (Fig. 5E - F).
Raios: Observados apenas com lente de 10x na superfície transversal
(Fig. 5E) e longitudinal tangencial (Fig. 5G), visíveis a olho nu e bem
contrastados na superfície longitudinal radial (Fig. 5H), finos, baixos, pouco
frequentes.
Estruturas estratificadas: Ausentes.
Estruturas secretoras: Ausentes.
Variantes cambiais: Não observadas.
Testes químicos: Teste de Cromazurol negativo.

31
Figura 5 – Nectandra megapotamica (Spreng.) Mez. A – ramo da árvore; B – disco do tronco recém-cortado; C – disco do tronco lixado; D – face transversal: anéis de crescimento; E – vista geral da madeira em corte transversal (10x); F – vista ampliada da madeira em corte transversal (20x); G – vista geral da madeira na face longitudinal tangencial (10x); H – vista geral da madeira na face longitudinal radial (10x). Escalas: A a C– 6 cm; D – 3 cm; E a H – 1 mm.

32
Ocotea porosa (Nees & C. Mart.) Barroso – Imbuia (Fig. 6A - G)
Características gerais: Cerne/alburno distintos pela cor; cerne marrom-
amarelado, com alteração acentuada de cor por foto-oxidação, mudando para
amarronzado-oliváceo (Fig. 6B); anéis de crescimento distintos,
individualizados por zonas fibrosas tangencias mais escuras (Fig. 6C). Madeira
com brilho nas superfícies transversal e longitudinais (Fig. 6F - G), com cheiro
perceptível, agradável, moderadamente dura ao corte transversal manual; grã
direita a revessa; textura média, figura presente em faixas causadas pelas
camadas de crescimento (podendo formar "V" ou "U").
Poros: Presentes, visíveis apenas com lente de 10x, pequenos, de
distribuição difusa, frequência média, predominantemente solitários,
eventualmente múltiplos radiais com menos de 4 elementos, eventualmente em
arranjo diagonal; de formato circular a oval; placas de perfuração não
observadas mesmo com lente de 10x.; parte dos vasos obstruídos por tiloses
(Fig. 6D - E).
Parênquima axial: Não observado, mesmo sob lente de 10x. (Fig. 6D)
Raios: Observados apenas com lente de 10x na superfície transversal
(Fig. 6D) e longitudinal tangencial (Fig. 6F), bem contrastados na superfície
radial (Fig. 6G), finos, baixos, pouco frequentes.
Estruturas estratificadas: Ausentes.
Estruturas secretoras: Ausentes.
Variantes cambiais: Não observadas.
Testes químicos: Teste de Cromazurol negativo.

33
Figura 6 – Ocotea porosa (Nees & C. Mart.) Barroso. A – vista geral da árvore; B – disco do tronco recém-cortado; C – face transversal: anéis de crescimento; D – vista geral da madeira em corte transversal (10x); E – vista ampliada da madeira em corte transversal (20x); F – vista geral da madeira na face longitudinal tangencial (10x); G – vista geral da madeira na face
longitudinal radial (10x). Escalas: A – aprox. 2 m; B – 15 cm; C – 4 cm ; D a G – 1 mm.

34
Ocotea puberula (Rich.) Nees – Canela-guaicá (Fig. 7A - H)
Características gerais: Cerne/alburno indistintos pela cor; cerne
branco-amarelado-acinzentado, com alteração acentuada de cor por foto-
oxidação, mudando para amarronzado-acinzentado (Fig. 7B - C); anéis de
crescimento distintos, individualizados por zonas fibrosas tangencias mais
escuras (Fig. 7D); madeira com brilho nas superfícies longitudinais (Fig. 7G -
H); sem cheiro perceptível; macia ao corte transversal manual; grã direita a
revessa; textura média; figura presente, em faixas causadas pelas camadas de
crescimento (podendo formar "V" ou "U").
Poros: Presentes, visíveis apenas com lente de 10x.; pequenos ou
médios, de distribuição difusa; frequência média; solitários , frequentemente
geminados e múltiplos radiais com menos de 4 vasos, ou mais raramente com
até 7 elementos, dispostos em padrão não definido, de formato circular a oval;
placas de perfuração não observadas mesmo com lente de 10x.; vasos
eventualmente obstruídos por tiloses (Fig. 7E - F).
Parênquima axial: Não observado, mesmo sob lente de 10x. (Fig. 7E).
Raios: Observados apenas com lente de 10x na superfície transversal e
longitudinal tangencial (Fig. 7G), visíveis a olho nu e bem contrastados na
superfície longitudinal radial (Fig. 7H), finos, baixos, pouco frequentes.
Estruturas estratificadas: Ausentes.
Estruturas secretoras: Ausentes.
Variantes cambiais: Não observadas.
Testes químicos: Teste de Cromazurol negativo.

35
Figura 7 – Ocotea puberula (Rich.) Nees. A – ramo da árvore; B – tronco recém-cortado; C – disco do tronco lixado; D – face transversal: anéis de crescimento; E – vista geral da madeira em corte transversal (10x); F – vista ampliada da madeira em corte transversal (20x); G – vista geral da madeira na face longitudinal tangencial (10x); H – vista geral da madeira na face
longitudinal radial (10x). Escalas: A – 7 cm; B – 6 cm; C– 5 cm; D – 1 cm; E a H – 1 mm.

36
Ocotea pulchella (Nees) Mez – Canela-lageana (Fig. 8A - G)
Características gerais: Cerne/alburno pouco distintos pela cor; cerne
marrom-amarelado, com alteração acentuada de cor por foto-oxidação,
mudando para amarronzado (Fig. 8B); anéis de crescimento distintos,
individualizados por zonas fibrosas tangencias mais escuras (Fig. 8C). Madeira
com brilho nas superfícies transversal e longitudinais (Fig. 8F - G); com cheiro
perceptível, agradável; Moderadamente dura ao corte transversal manual; grã
direita a revessa; textura média; figura presente em faixas causadas pelas
camadas de crescimento (podendo formar "V" ou "U").
Poros: Presentes, visíveis apenas com lente de 10x. pequenos ou
médios, de distribuição difusa; frequência média; vasos predominantemente
solitários e eventualmente múltiplos radiais com menos de 4 vasos, sem
padrão definido de disposição; de formato circular a oval; placas de perfuração
não observadas mesmo com lente de 10x.; parte dos vasos obstruídos por
tiloses (Fig. 8D - E);
Parênquima axial: Não observado, mesmo sob lente de 10x. (Fig. 8D)
Raios: Observados apenas com lente de 10x na superfície transversal e
longitudinal tangencial (Fig. 8F), bem contrastados na superfície radial (Fig.
8G), finos, baixos, pouco frequentes.
Estruturas estratificadas: Ausentes.
Estruturas secretoras: Ausentes.
Variantes cambiais: Não observadas.
Testes químicos: Teste de Cromazurol negativo.

37
Figura 8 – Ocotea pulchella (Nees) Mez. A – vista geral da árvore; B – disco do tronco recém-cortado; C – face transversal: anéis de crescimento; D – vista geral da madeira em corte transversal (10x); E – vista ampliada da madeira em corte transversal (20x); F – vista geral da madeira na face longitudinal tangencial (10x); G – vista geral da madeira na face longitudinal
radial (10x). Escalas: A – aprox. 2 m; B – 9 cm; C – 2 cm ; D a G – 1 mm;

38
Ocotea sp. - Canela-loura (Fig. 9A - G)
Características gerais: Cerne/alburno indistintos pela cor; cerne
marrom acinzentado (Fig. 9B); anéis de crescimento distintos individualizadas
por zonas fibrosas tangencias mais escuras (Fig. 9C); madeira com brilho nas
superfícies transversal e longitudinais (Fig. 9F - G); sem cheiro perceptível;
moderadamente dura ao corte transversal manual; grã direita; textura fina;
figura presente em faixas causadas pelas camadas de crescimento (podendo
formar "V" ou "U").
Poros: Presentes; visíveis apenas com lente de 10x; pequenos, de
distribuição difusa; frequência média; vasos predominantemente solitários e
eventualmente múltiplos radiais, com menos de 4 vasos, eventualmente em
arranjo diagonal; de formato circular a oval; placas de perfuração não
observadas mesmo com lente de 10x.; parte dos vasos obstruídos por tiloses
(Fig. 9D - E).
Parênquima axial: Não observado, mesmo sob lente de 10x (Fig. 9D).
Raios: Observados apenas com lente de 10x na superfície transversal
(Fig. 9D) e longitudinal tangencial (Fig. 9F), bem contrastados na superfície
radial (Fig. 9G), finos, baixos, pouco frequentes.
Estruturas estratificadas: Ausentes.
Estruturas secretoras: Ausentes.
Variantes cambiais: Não observadas.
Testes químicos: Teste de Cromazurol negativo.

39
Figura 9 – Ocotea sp. A – ramo herborizado; B – disco do tronco; C – face transversal: anéis de crescimento; D – vista geral da madeira em corte transversal (10x); E – vista ampliada da madeira em corte transversal (20x); F – vista geral da madeira na face longitudinal tangencial (10x); G – vista geral da madeira na face longitudinal radial (10x). Escalas: A – 4 cm; B – 1 cm; C – 0,8 cm ; D a G – 1 mm.

40
3.2 – Chave dicotômica para diferenciação das espécies pela estrutura
macroscópica do lenho
1 – Madeira com odor característico....................................................................2
2 – Odor característico agradável, cerne marrom-amarelado, parênquima
axial indistinto sob lente de 10x...................................................................3
3 – Cerne/alburno distintos pela cor, poros eventualmente em
arranjo diagonal ..........................................................Ocotea porosa
3’ – Cerne/alburno pouco distintos pela cor, poros sem arranjo
definido ....................................................................Ocotea pulchella
2’ – Odor característico desagradável, cerne amarelado, parênquima axial
distinto sob lente de 10x, escasso ou vasicêntrico........................................
...........................................................................Nectandra megapotamica
1’ – Madeira sem odor característico..................................................................4
4 – Cerne amarelado, parênquima axial observado sob lente de 10x,
paratraqueal escasso ou vasicêntrico.......................Nectandra lanceolata
4’ – Cerne branco-amarelado-acinzentado, parênquima axial não
observado sob lente de 10x........................................................................5
5 – Poros pequenos, predominantemente solitários, eventualmente
dispostos em arranjo diagonal, textura fina........................Ocotea sp.
5’ – Poros pequenos ou médios, solitários ou frequentemente
geminados ou múltiplos radiais, sem arranjo definido, textura média
..................................................................................Ocotea puberula

41
3.3 – Descrições microscópicas do lenho
Nectandra lanceolata Nees – Canela-amarela (Fig. 10 A - J)
Camadas de crescimento distintas, demarcadas pelo espessamento
das paredes e achatamento radial das fibras (Fig 10A).
Porosidade difusa, (05) 12 (30) vasos/mm²; elementos de vaso
predominantemente solitários, ou eventualmente geminados, em pequenas
cadeias radiais de 3 a 4 elementos ou em cachos, eventualmente em arranjo
diagonal, secção circular a oval, comprimento (230,69) 526,76 (949,54) μm,
diâmetro tangencial (40,85) 109,57 (203,16) μm (Fig. 10A); tiloses presentes;
placas de perfuração simples ou escalariformes (Fig. 10I); pontoações inter-
vasculares grandes, (6,51) 11,10 (17,33) μm de diâmetro, alternas, de formato
circular ou oval, areoladas, abertura elíptica e inclusa (Fig. 10D), internamente,
não-coalescentes (Fig. 10J); eventualmente, presença de depósitos de
extrativos ornamentando algumas aréolas (Fig. 10E); pontoações raio-
vasculares arredondadas ou alongadas no sentido radial ou axial, com bordas
distintas ou muito reduzidas a aparentemente simples; pontoações
parênquimo-vasculares similares as pontoações raios-vasculares.
Fibras septadas (Fig. 10B), comprimento (381,88) 987,72 (2199,13) μm,
diâmetro (13,28) 26,77 (46,45) μm, lume (9,83) 15,56 (23,21) μm, paredes com
(3,35) 5,60 (13,65) μm de espessura, mais espessadas no lenho outonal.
Parênquima axial paratraqueal escasso ou vasicêntrico (Fig. 10A), de 2
a 3 células.
Raios (4) 8 (13)/mm, unisseriados ou multisseriados (Fig. 10B), (2) 3 (4)
células de largura, altura de (74,75) 326,35 (1149,93) μm, largura de (12,24)
38,01 (72,82) μm, formados por células procumbentes na região central e 1 a 4
(7) fileiras de células quadradas ou eretas nas margens; presença eventual de
raios fusionados; cristais aciculares e rombóides quando presentes nas células
procumbentes, pouco abundantes (Fig. 10F - G).
Células oleíferas frequentes, associadas principalmente ao parênquima
radial, eventualmente associadas ao parênquima axial ou entre as fibras (Fig.
10C e H); conteúdo dos idioblastos fluorescente sob filtros verde, vermelho e
triplo (Fig. 10H).

42
Figura 10 – Nectandra lanceolata Nees; A – face transversal (4x); B – face longitudinal tangencial (10x); C – face longitudinal radial (10x); D – pontoações (40x); E – pontoação com extrativos (7.000x); F – cristais (setas) nas células procumbentes dos raios (40x); G – cristal em célula do raio (3.000x); H – idioblastos com conteúdo fluorescente sob filtro triplo (40x); I – placa de perfuração escalariforme (1.000x); J – vista interna do elemento de vaso: pontoações inter-vasculares não-coalescentes (2.000x). Escalas: A – 100 μm; B – 125 μm; C – 100 μm ; D – 50 μm; E – 2 μm; F – 50 μm; G – 5 μm; H – 50 μm ; I e J – 10 μm.

43
Nectandra megapotamica (Spreng.) Mez Canela-amarela (Fig.11A - K)
Camadas de crescimento distintas, demarcadas pelo espessamento
das paredes e achatamento radial das fibras (Fig. 11A).
Porosidade difusa, (05) 16 (40) vasos/mm²; elementos de vaso
predominantemente solitários, geminados ou em pequenas cadeias radiais de
3 a 4 elementos ou mais raramente em cachos, eventualmente em arranjo
diagonal, secção circular a oval, comprimento (245,98) 507,61 (853,03) μm,
diâmetro tangencial (34,40) 94,42 (170,08) μm (Fig. 11A); placas de perfuração
simples ou escalariformes (Fig. 11D), 1-6 barras; tiloses presentes; pontoações
inter-vasculares médias, (6,06) 9,59 (17,29) μm de diâmetro, alternas, de
formato circular ou oval (Fig. 11B), areoladas, abertura elíptica e inclusa (Fig.
11F), internamente não-coalescentes (Fig. 11G); pontoações raio-vasculares
arredondadas ou alongadas no sentido radial ou axial, com bordas distintas ou
muito reduzidas a aparentemente simples (Fig. 11E); pontoações parênquimo-
vasculares similares as pontoações raios-vasculares (Fig. 11H).
Fibras septadas (Fig. 11B), comprimento (333,44) 941,00 (1664,56) μm,
diâmetro (10,31) 24,65 (46,58) μm, lume (7,84) 14,02 (20,25) μm, paredes com
(3,14) 5,31 (9,77) μm de espessura, mais espessadas no lenho outonal (Fig.
11A).
Parênquima axial paratraqueal escasso ou vasicêntrico de 4 a 6 células
(Fig. 11A), eventualmente confluente.
Raios (2) 8 (12)/mm, unisseriados ou comumente multisseriados (Fig.
11B), 2-3 células de largura, altura de (87,93) 289,50 (913,90) μm, largura de
(8,52) 29,59 (70,74) μm, formados por células procumbentes na região central
e 1 a 4 fileiras de células quadradas ou eretas nas margens (Fig. 11C);
presença eventual de raios fusionados; cristais aciculares, alongados,
rombóides e tabletóides (Fig. 11J e K) frequentemente presentes nas células
procumbentes, eretas e quadradas dos raios.
Células oleíferas frequentes, associadas ao parênquima radial, ao
parênquima axial ou entre as fibras (Fig. 11A - C e I); conteúdo dos idioblastos
fluorescente sob filtros verde (Fig. 11I), vermelho e triplo.
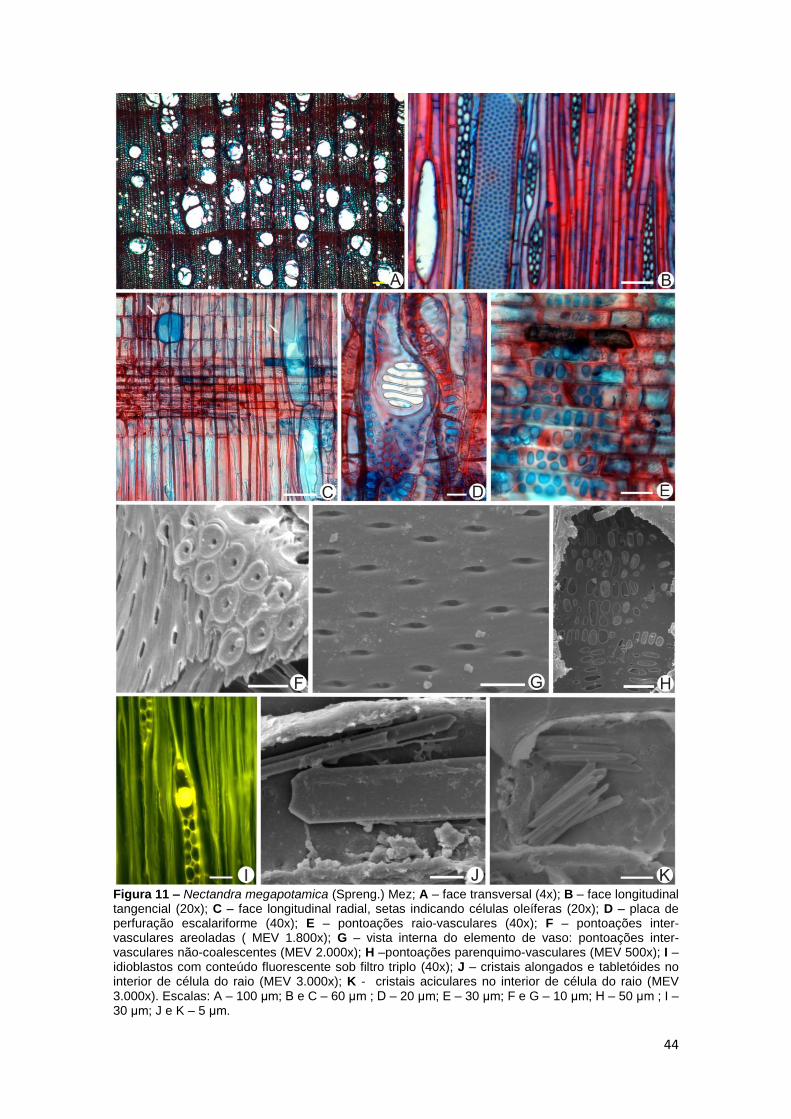
44
Figura 11 – Nectandra megapotamica (Spreng.) Mez; A – face transversal (4x); B – face longitudinal tangencial (20x); C – face longitudinal radial, setas indicando células oleíferas (20x); D – placa de perfuração escalariforme (40x); E – pontoações raio-vasculares (40x); F – pontoações inter-vasculares areoladas ( MEV 1.800x); G – vista interna do elemento de vaso: pontoações inter-vasculares não-coalescentes (MEV 2.000x); H –pontoações parenquimo-vasculares (MEV 500x); I – idioblastos com conteúdo fluorescente sob filtro triplo (40x); J – cristais alongados e tabletóides no interior de célula do raio (MEV 3.000x); K - cristais aciculares no interior de célula do raio (MEV 3.000x). Escalas: A – 100 μm; B e C – 60 μm ; D – 20 μm; E – 30 μm; F e G – 10 μm; H – 50 μm ; I – 30 μm; J e K – 5 μm.

45
Ocotea porosa (Nees & C. Mart.) Barroso - Imbuia (Fig. 12A - I)
Camadas de crescimento distintas, demarcadas pelo espessamento
das paredes e achatamento radial das fibras (Fig. 12A).
Porosidade difusa, (11) 15 (23) vasos/mm²; elementos de vaso
predominantemente solitários, geminados ou em pequenas cadeias radiais de
3 elementos ou eventualmente em cachos, secção circular a oval (Fig. 12A),
comprimento (243,49) 473,38 (735,14) μm, diâmetro tangencial (54,54) 90,375
(125,17) μm,; placas de perfuração simples; espessamentos internos
escalariformes presentes; tiloses presentes; pontoações inter-vasculares
médias, (7,82) 10,04 (12,18) μm de diâmetro, alternas (Fig. 12E), de formato
circular ou oval, areoladas, abertura elíptica e inclusa (Fig. 12F); internamente
coalescentes (Fig. 12G); pontoações raio-vasculares arredondadas ou
alongadas no sentido radial ou axial, com bordas distintas ou muito reduzidas a
aparentemente simples; pontoações parênquimo-vasculares similares as
pontoações raios-vasculares (Fig. 12H).
Fibras septadas (Fig. 12B-C), comprimento (632,85) 950,14 (1241,93)
μm, diâmetro (16,98) 23,49 (47,10) μm, lume (10,70) 11,44 (12,18) μm,
paredes com (4,22) 6,02 (8,70) μm de espessura, mais espessadas no lenho
outonal (Fig. 12A).
Parênquima axial paratraqueal escasso ou mais raramente vasicêntrico
de 1 a 2 células (Fig. 12A).
Raios (8) 10 (13)/mm, unisseriados ou multisseriados, 2-3 células de
largura (Fig. 12B), altura de (140,93) 329,04 (579,07) μm, largura de (18,05)
33,45 (56,39) μm, formados por células procumbentes na região central e 1 a 2
fileiras de células quadradas ou eretas nas margens (Fig. 12C); presença
eventual de raios fusionados; cristais não observados.
Células oleíferas associadas ao parênquima radial (Fig. 12B), axial (Fig.
12C) e entre as fibras; conteúdo dos idioblastos fluorescente sob filtros verde,
vermelho e triplo (Fig. 12I).

46
Figura 12 – Ocotea porosa (Nees & C. Mart.) Barroso ; A – face transversal (4x); B – face longitudinal tangencial (10x); C – face longitudinal radial (10x); D – placa de perfuração simples (MEV 500x); E – pontoações inter-vasculares (40x); F – detalhe de pontoações inter-vasculares areoladas (100x); G – vista interna da parede do elemento de vaso com pontoações inter-
vasculares coalescentes (MEV 1.500x); H – pontoações parenquimo-vasculares (MEV 2.000x); I – face longitudinal tangencial: idioblasto com conteúdo fluorescente sob filtro triplo(40x); Escalas: A – 250 μm; B e C – 100 μm; D e E – 50 μm; F, G e H – 10 μm; I – 30 μm.

47
Ocotea puberula (Rich.) Nees – Canela-guaicá (Fig. 13A - K)
Camadas de crescimento distintas, demarcadas pelo espessamento
das paredes e achatamento radial das fibras.
Porosidade difusa, (05) 15 (29) vasos/mm²; elementos de vaso
predominantemente solitários e geminados ou frequentemente em cadeias
radiais de 3 a 7 elementos ou eventualmente em cachos, secção circular a oval
(Fig. 13A), comprimento (178,39) 515,78 (947,29) μm, diâmetro tangencial
(33,64) 106,74 (205,46) μm; placas de perfuração simples ou escalariformes
(Fig. 13F), 1-6 barras; tiloses presentes (Fig. 13H); pontoações inter-vasculares
grandes (Fig. 13D), (6,67) 11,29 (20,56) μm de diâmetro, alternas, de formato
circular ou oval, areoladas, abertura elíptica e inclusa (Fig. 13G); internamente,
não-coalescentes; pontoações raio-vasculares arredondadas ou alongadas no
sentido radial ou axial, com bordas distintas ou muito reduzidas a
aparentemente simples (Fig. 13E); pontoações parênquimo-vasculares
similares as pontoações raios-vasculares.
Fibras septadas (Fig. 13B), comprimento (331,03) 916,93 (1651,72) μm,
diâmetro (13,11) 27,94 (46,18) μm, lume (12,52) 16,90 (21,66) μm, paredes
com (3,40) 5,52 (10,67) μm de espessura, mais espessadas no lenho outonal.
Parênquima axial paratraqueal escasso, ou eventualmente vasicêntrico
delgado (Fig. 13A), de 2-3 células.
Raios (4) 7 (13)/mm, unisseriados ou comumente multisseriados (Fig.
13B), 2-4 células de largura, altura de (93,96) 326,70 (1233,35) μm, largura de
(12,82) 32,55 (68,69) μm, formados por células procumbentes na região central
e 1 a 3 fileiras de células quadradas ou eretas nas margens (Fig. 13C);
presença eventual de raios fusionados; cristais aciculares, rombóides ou
piramidais raramente presentes nas células procumbentes dos raios (Fig. 13I).
Células oleíferas pouco frequentes, associadas ao parênquima radial
(Fig. 13C, J e K); conteúdo dos idioblastos fluorescente sob filtros verde (Fig.
13J), vermelho e triplo (Fig. 13K).

48
Figura 13 – Ocotea puberula (Rich.) Nees; A – face transversal (4x); B – face longitudinal tangencial (40x); C – face longitudinal radial (20x); D – pontoações inter-vasculares (40x); E – pontoações raio-vasculares (40x); F – placa de perfuração escalariforme (20x); G – detalhe de pontoações inter-vasculares areoladas (MEV 3.000x); H – tiloses (MEV 300x); I – cristal em célula procumbente do raio (MEV 5.000x); J - face transversal: idioblasto com conteúdo fluorescente sob filtro verde (40x); K - face longitudinal radial: idioblasto com conteúdo fluorescente sob filtro triplo (40x). Escalas: A – 100 μm; B – 50 μm; C – 50 μm; D – 25 μm; E – 50 μm; F – 100 μm; G – 5 μm; H – 50 μm; I – 5 μm; J e K – 50 μm.

49
Ocotea pulchella (Nees) Mez – Canela-lageana (Fig. 14A - K)
Camadas de crescimento distintas, demarcadas pelo espessamento
das paredes e achatamento radial das fibras (Fig. 14A).
Porosidade difusa, (05) 13 (28) vasos/mm²; elementos de vaso
predominantemente solitários, geminados ou em pequenas cadeias radiais de
3 a 4 elementos ou eventualmente em cachos, secção circular a oval (Fig.
14A), comprimento (237,46) 523,27 (934,48) μm, diâmetro tangencial (56,57)
114,74 (208,22) μm; placas de perfuração simples (Fig. 14E); espessamentos
internos escalariformes presentes; tiloses presentes (Fig. 14F); pontoações
inter-vasculares médias, (6,20) 8,78 (12,52) μm de diâmetro, alternas (Fig.
14D), de formato circular ou oval, areoladas, abertura elíptica e inclusa (Fig.
14G); internamente coalescentes (Fig. 14I - J); pontoações raio-vasculares
arredondadas ou alongadas no sentido radial ou axial, com bordas distintas ou
muito reduzidas a aparentemente simples; pontoações parênquimo-vasculares
similares as pontoações raios-vasculares.
Fibras septadas (Fig. 14B), comprimento (481,06) 933,62 (1417,70) μm,
diâmetro (13,94) 26,12 (39,61) μm, lume (5,79) 14,31 (17,94) μm, paredes com
(3,78) 5,90 (13,51) μm de espessura, mais espessadas no lenho outonal (Fig
14A).
Parênquima axial paratraqueal escasso ou vasicêntrico de 3 a 4 células
(Fig. 14A).
Raios (5) 8 (12)/mm, raramente unisseriados ou multisseriados (Fig
14B), 3-6 células de largura, altura de (133,37) 374,52 (845,83) μm, largura de
(20,20) 48,79 (92,95) μm, formados por células procumbentes na região central
e 1 a 3 fileiras de células quadradas ou eretas nas margens (Fig. 14C);
presença eventual de raios fusionados; areias de cristais eventualmente
presentes nas células dos raios (Fig. 14K).
Células oleíferas muito frequentes, associadas ao parênquima radial,
axial ou entre as fibras (Fig. 14B - C); conteúdo dos idioblastos fluorescente
sob filtros verde, vermelho e triplo (Fig. 14H).

50
Figura 14 – Ocotea pulchella (Nees) Mez; A – face transversal (4x); B – face longitudinal tangencial (10x); C – face longitudinal radial (10x); D – elementos de vasos e pontoações inter-vasculares (40x); E – placas de perfuração simples (MEV 500x); F – tiloses (MEV 250x); G – detalhe de pontoações inter-vasculares areoladas (MEV 3.000x); H – face longitudinal tangencial: idioblasto vinculado ao raio com conteúdo fluorescente sob filtro triplo (40x); I – vista interna da parede do elemento de vaso com pontoações inter-vasculares coalescentes (MEV 500x); J – detalhe das câmaras das pontoações inter-vasculares coalescentes (MEV 4.000x); K – areia de cristais em célula procumbente do raio (MEV 5.000x). Escalas: A – 100 μm; B – 250 μm; C – 100 μm; D e E – 50 μm; F – 100 μm; G – 5 μm; H e I – 50 μm; J e K – 5 μm.

51
Ocotea sp. – Canela-loura (Fig. 15A - I)
Camadas de crescimento distintas, demarcadas pelo espessamento
das paredes e achatamento radial das fibras.
Porosidade difusa, (15) 19 (23) vasos/mm²; elementos de vaso
predominantemente solitários, e mais raramente geminados ou em pequenas
cadeias radiais de 3 a 4 elementos ou mais raramente em cachos,
eventualmente em arranjo diagonal, secção circular a oval (Fig. 15A),
comprimento (275,52) 500,05 (805,48) μm, diâmetro tangencial (34,83) 66,96
(96,38) μm; placas de perfuração simples (Fig. 15D); tiloses presentes;
pontoações inter-vasculares grandes, (7,0) 10,32 (12,85) μm de diâmetro,
alternas (Fig. 15E), de formato circular ou oval, areoladas, abertura elíptica e
inclusa, internamente eventualmente coalescentes (Fig. 15F); pontoações raio-
vasculares arredondadas ou alongadas no sentido radial ou axial, com bordas
distintas ou muito reduzidas a aparentemente simples; pontoações
parênquimo-vasculares similares as pontoações raios-vasculares (Fig. 15G).
Fibras septadas (Fig. 15B), comprimento (688,32) 1019,68 (1351,07)
μm, diâmetro (19,80) 26,19 (34,25) μm, lume (14,11) 15,52 (16,94) μm,
paredes com (3,42) 5,33 (7,91) μm de espessura, mais espessadas no lenho
outonal.
Parênquima axial paratraqueal escasso (Fig. 15A).
Raios (5) 8 (10)/mm, unisseriados ou comumente multisseriados (Fig.
15B), 2-3 células de largura, altura de (147,35) 356,47 (760,97) μm, largura de
(23,75) 41,85 (65,32) μm , formados por células procumbentes na região
central e 1 a 3 fileiras de células quadradas ou eretas nas margens (Fig. 15C);
presença eventual de raios fusionados; cristais aciculares, alongados e
rombóides frequentemente presentes nas células procumbentes dos raios (Fig.
15H).
Células mucilaginosas muito frequentes, longas (>500 μm), associadas
ao parênquima radial (Fig. 15B), axial ou frequentemente entre as fibras (Fig.
15C); conteúdo dos idioblastos não-fluorescente sob filtros verde, vermelho e
triplo (Fig. 15I).

52
Figura 15 – Ocotea sp.; A – face transversal (4x); B – face longitudinal tangencial (10x); C – face longitudinal radial: idioblastos entre as fibras (10x); D – placas de perfuração simples (MEV 600x); E – elemento de vaso e pontoações inter-vasculares alternas (40x); F – vista interna do elemento de vaso: pontoações inter-vasculares eventualmente coalescentes (MEV 2.500x); G – pontoações parênquimo-vasculares (40x); H – cristais nas células procumbentes dos raios (MEV 2.000x); I – face longitudinal radial: idioblastos entre as fibras sem conteúdo fluorescente sob filtro triplo(10x); Escalas: A a C – 100 μm; D – 20 μm e E – 50 μm; F – 10 μm; G – 50 μm; H – 10 μm e I – 100 μm.

53
3.4 – Chave dicotômica para separação das espécies pela estrutura
microscópica do lenho
1 – Presença abundante de cristais aciculares, alongados ou rombóides nas
células dos raios..................................................................................................2
2 – Parênquima axial paratraqueal escasso ou vasicêntrico; placas de
perfuração simples ou frequentemente escalariformes; células oleíferas
pouco maiores que as células do parênquima axial ou radial ao qual se
encontram associadas.......................................Nectandra megapotamica
2’– Parênquima axial paratraqueal escasso; placas de perfuração simples;
células mucilaginosas muito frequentes e muito maiores que as células do
parênquima radial ou axial aos quais estão associadas, ou muito longas
(>500μm), e mais frequentemente dispostas entre as fibras;....Ocotea sp.
1’ – Ausência de cristais aciculares, alongados ou rombóides nas células dos
raios ou, se presentes, pouco freqüentes/abundantes........................................3
3 – Placas de perfuração simples ou escalariformes; pontoações inter-
vasculares grandes.....................................................................................4
4 – Células oleíferas pouco frequentes, associadas ao parênquima
radial;.......................................................................Ocotea puberula
4’ – Células oleíferas frequentes, associadas ao parênquima radial
ou ao parênquima axial e entre as fibras;.........Nectandra lanceolata
3’ – Placas de perfuração exclusivamente simples; pontoações inter-
vasculares médias.......................................................................................5
5 – Parênquima axial paratraqueal escasso ou mais raramente
vasicêntrico de 1 a 2 células; raios multisseriados
predominantemente de 2 a 3 células de largura;....... Ocotea porosa
5’ – Parênquima axial paratraqueal escasso ou frequentemente
vasicêntrico de 3 a 4 células; raios multisseriados
predominantemente de 3 a 6 células de largura;.....Ocotea pulchella

54
3.5 – Análises de caracteres anatômicos quantitativos do lenho
As médias dos valores e o erro padrão obtidos da mensuração dos
caracteres anatômicos do lenho nas lâminas permanentes e nos macerados
estão expostos na Tabela 5.
Os valores de similaridades e dissimilaridades e os níveis de significância
das análises entre as espécies estão dispostos na Tabela 6.
Os valores de similaridades e dissimilaridades e os níveis de significância
das análises entre os locais de coleta estão dispostos na Tabela 7.
O escalonamento multidimencional das análises de similaridade de Bray-
Curtis dos caracteres anatômicos quantitativos do lenho estão dispostos por
comparações interespecíficas (Fig. 16), intergenéricas (Fig. 17) e por locais de
coleta (Fig. 18).

55
Tabela 5 – Caracteres anatômicos quantitativos do lenho mensurados nas amostras das árvores das espécies coletadas.
Características anatômicas
Ocotea porosa
(Nees & C. Mart.)
Barroso
Ocotea pulchella
(Nees) Mez Ocotea puberula
(Rich.) Nees Ocotea sp.
Nectandra lanceolata
Nees
Nectandra megapotamica
(Spreng.) Mez
Média Erro padrão Média Erro padrão Média Erro padrão Média Erro padrão Média Erro padrão Média Erro padrão
Diâmetro tangencial
do vaso (μm) 90,37 ± 2,14 114,74 ± 3,42 106,74 ± 2,58 66,96 ± 1,50 109,58 ± 2,67 94,42 ± 2,24
Vasos/mm² 15 ± 0,79 13 ± 0,89 15 ± 1,09 19 ± 0,69 12 ± 0,88 16 ± 1,05
Comprimento do vaso (μm) 473,38 ± 22,07 523,27 ± 23,68 515,78 ± 21,54 500,05 ± 20,89 526,77 ± 21,50 507,61 ± 20,20
Diâmetro tangencial da
pontoação intervascular (μm) 10,04 ± 0,11 8,78 ± 0,10 11,29 ± 0,14 10,32 ± 0,15 11,11 ± 0,13 9,59 ± 0,11
Altura do raio (μm) 329,04 ± 12,32 374,52 ± 14,96 326,70 ± 16,26 356,47 ± 17,70 326,35 ± 15,88 289,50 ± 13,10
Largura do raio (μm) 33,45 ± 0,81 48,79 ± 1,38 32,55 ± 0,82 41,85 ± 1,11 38,01 ± 0,97 29,59 ± 0,75
Raios/mm linear 10 ± 0,47 8 ± 0,37 7 ± 0,40 8 ± 0,37 8 ± 0,40 8 ± 0,41
Comprimento da fibra (μm) 950,14 ± 27,08 933,62 ± 28,37 916,93 ± 30,73 1019,68 ± 32,43 987,72 ± 33,72 941,00 ± 31,99
Diâmetro da fibra (μm) 23,48 ± 1,17 26,12 ± 0,80 27,94 ± 0,91 26,19 ± 0,76 26,77 ± 0,92 24,65 ± 0,85
Espessura das parede da
fibra (μm) 6,02 ± 0,21 5,90 ± 0,19 5,52 ± 0,16 5,33 ± 0,13 5,60 ± 0,16 5,31 ± 0,15
Lume da fibra (μm) 11,44 ± 0,91 14,31 ± 0,89 16,90 ± 0,87 15,52 ± 0,91 15,56 ± 0,77 14,02 ± 0,84

56
Tabela 6 – Análise de ANOSIM e SIMPER das características anatômicas quantitativas do lenho das espécies estudadas nas relações entre espécies. As similaridades intra-específicas estão na diagonal, em negrito. As dissimilaridades inter-específicas estão acima da diagonal e os valores p do ANOSIN estão abaixo da diagonal. As características anatômicas quantitativas do lenho que mais contribuíram para as semelhanças e diferenças observadas e os valores relativos (em porcentagem, entre parênteses) estão destacados abaixo dos respectivos valores de similaridade e dissimilaridade.
Espécies Nectandra
lanceolata
Nees
Nectandra
megapotamica
(Spreng.) Mez
Ocotea porosa
(Nees & C. Mart.)
Barroso
Ocotea pulchella
(Nees) Mez
Ocotea puberula
(Rich.) Nees
Ocotea sp.
Nectandra
lanceolata
Nees
93,28
e(48,06);b(25,76);
c(15,25);a(5,35)
6,99
e(40,36);c(23,71);
b(20,07);a(6,81)
5,92
e(38,1);b(24,22);
c(19,55);a(8,21)
6,99
e(42,71);c(24,39);
b(19,9);d(4,32)
6,88
e(43,64);c(22,45);
b(20,92);a(5,62)
6,45
e(36,81);c(21,01);
b(18,25);a(15,91)
Nectandra
megapotamica
(Spreng.) Mez 0,1
93,79
e(49,02);b(26,51);
c(14,38);a(4,74)
5,30
e(34,71);c(27,31);
b(21,60);a(6,24);
d(3,81)
7,70
e(34,32);c(30,37);
b(17,32);a(7,23);
d(6,36)
6,67
e(38,01);c(25,03);
b(20,75);a(7,52);
6,65
e(37,01);c(28,21);
b(15,02);a(10,32)
Ocotea porosa
(Nees & C.
Mart.)
Barroso
2,5 53,8
97,14
e(48,74);b(24,57);
c(16,93)
6,15
e(34,74);b(22,61);
c (20,78);a (9,94);
d (6,30)
5,56
e(34,98);b(24,86);
c(20,21);a(8,81);
d(3,16)
4,29
e(40,31);b(16,52);
c(15,89);a(13,56);
d(4,89)
Ocotea
pulchella
(Nees) Mez 0,1 0,1 6,1
93,67
e (44,84); b (25,46);
c(18,12); a(5,66)
7,05
e(38,32);c(25,52);
b(19,78);d(5,85);
a(5,56)
6,30
e(42,71);a(18,39);
b(17,57);c(13,04)
Ocotea
puberula
(Rich.) Nees 0,1 0,1 1,2 0,1
93,46
e(46,72);b(26,30);
c(15,91);a(5,27)
6,82
e(40,09);c(20,7);
b(16,78);a(14,28)
Ocotea sp.
0,3 6,6 33,3 6,1 0,6
97,32
e(49,39);b(23,67);
c(17,65)
a – diâmetro dos elementos de vaso; b – comprimento dos elementos de vaso; c – altura dos raios; d – largura dos raios; e – comprimento das fibras;

57
Tabela 7 – Análise de ANOSIM e SIMPER das características anatômicas quantitativas do lenho das espécies estudadas nas relações entre locais de coleta.
As similaridades intra-locais estão na diagonal, em negrito. As dissimilaridades entre os locais estão acima da diagonal e os valores p do ANOSIN estão
abaixo da diagonal. As características anatômicas quantitativas do lenho que mais contribuíram para as semelhanças e diferenças observadas e os valores
relativos (em porcentagem, entre parênteses) estão destacados abaixo dos respectivos valores de similaridade e dissimilaridade.
Locais de coleta Abdon Batista Concórdia Passos Maia Tangará
Abdon Batista
97,95
e(16,93);b(14,98);
c(13,56);a(10,80)
2,04
f(15,08);d(13,68);
a(11,36);c(10,48);g(10,36)
2,51
d(20,44);f(12,66);
c(11,63);g(10,73); a(10,53)
2,21
f(16,56);d(13,92);
c(12,41);a(10,80); g(9,41)
Concórdia 1,1
98,10
e(16,42);b(14,91);
c(13,74);a(11,05)
2,25
d(18,43);g(13,41);
f(12,33);h(9,27)
2,05
f(16,25);d(12,65);
c(11,42);g(11,17)
Passos Maia 0,2 0,1
97,88
e(16,25);b(14,84);
c(14,00);a (11,21)
2,40
d(17,25);f(13,75);
g (12,44);c (11,11);
Tangará 1 25,3 1
97,81
e (16,41) b (14,99);
c(13,63); a(11,13)
a – diâmetro dos elementos de vaso; b – comprimento dos elementos de vaso; c – altura dos raios; d – largura dos raios; e – comprimento das fibras; f – vasos/mm²; g - lume das fibras; h – raios/mm linear
.

58
Figura 16 – Escalonamento multidimencional da distribuição dos valores de similaridade nas
relações interespecíficas, das características anatômicas quantitativas do lenho das espécies
analisadas.
Figura 17 – Escalonamento multidimencional da distribuição dos valores de similaridade nas
relações inter-genéricas, das características anatômicas quantitativas do lenho das espécies
analisadas.

59
Figura 18 – Escalonamento multidimencional da distribuição dos valores de similaridade nas
relações entre os locais de coleta, das características anatômicas quantitativas do lenho das
espécies analisadas.
3.6 – Análise de agrupamento de caracteres anatômicos qualitativos do
lenho
A partir dos dados dispostos na matriz binária (Tabela 8), foram obtidos
as semelhanças inter-específicas das espécies ora estudadas (Tabela 9) e os
dendrogramas de similaridade (CLUSTER) baseados no índice de similaridade
de Jaccard (Fig. 19 – 23). Os valores SIMPER para as relações de
similaridade e dissimilaridade estabelecidas entre os gêneros, bem como as
características anatômicas que mais contribuíram nestas relações estão
expostos na Tabela 10. As relações se mostraram significativas, com valores
de significância inferiores a 0,05, entre os gêneros: Nectandra e
Rhodostemonodaphne; Nectandra e Sextonia; Nectandra e Chlorocardium;
Ocotea e Rhodostemonodaphne; Ocotea e Sextonia; Ocotea e Chlorocardium.
Não houve significância nas demais relações inter-genéricas. Os
escalonamentos multidimencionais (MDS) das espécies da matriz binária são
apresentados por gênero (Fig. 24) e por fonte da descrição anatômica do lenho
das espécies na literatura (Fig. 25).

60
Tabela 8 – Matriz binária de presença (1) e ausência (0) de caracteres qualitativos do lenho utilizada na construção dos dendrogramas de similaridade pelo
índice de Jaccard.
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Chlorocardium rodiei (R.H. Schomb.)
Rohwer, H.G. Richt. & van der Werff
(14)
1 0 1 1 1 0 0 1 0 1 0 0 1 1 0 0 1 0 1 1 0 1 0 0 0 1 1 1 1 0 0 0 0 0 0
Chlorocardium rodiei (R.H. Schomb.)
Rohwer, H.G. Richt. & van der Werff
(16)
1 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 0 1 0 1 0 1 0 0 0 1 1 1 1 0 0 0 1 1 0
Chlorocardium venenosum (Kosterm. &
Pinkley) Rohwer, H.G. Richt. & van der
Werff (13)
1 1 1 1 1 1 0 1 1 1 0 0 1 1 0 0 0 0 1 1 0 1 1 0 0 1 1 0 1 0 0 0 0 0 0
Nectandra acutifólia (Ruiz & Pav.) Mez
(1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 1 0 0 0 1 1 1 1 0 1 1 1 0 0
Nectandra acutifolia (Ruiz & Pav.) Mez
(9) 1 1 1 1 1 1 0 1 1 0 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 1 0 1 0 0 0
Nectandra amazonum Nees (1) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 1 0 1 1 0 1 0 1 1 0 0 1 1 1 1 0 1 1 1 1 0
Nectandra angustifolia (Schrad.) Nees &
Mart. (7) 1 1 1 1 1 0 0 1 1 1 0 0 0 1 1 0 0 1 1 1 0 0 0 0 0 1 1 0 1 1 0 1 1 0 0
Nectandra angustifolia (Schrad.) Nees &
Mart. (9) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 0 0 0
Nectandra cissiflora Nees (1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 0 1 0 1 1 0 0 1 1 1 1 0 1 1 0 0 0
Nectandra coriacea (Sw.) Griseb. (1) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 0 0 0 1
Nectandra coriacea (Sw.) Griseb. (17) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0
Nectandra cuspidata Nees & Mart. (1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 1 1 0 0 1 1 1 1 0 1 1 1 1 1

61
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Nectandra cuspidata Nees & Mart. (16) 1 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 0 1 0 1 0 0 0 1 1 1 1 0 0 1 1 1 0
Nectandra cuspidata Nees & Mart. (7) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 1 0 1 0 0 0
Nectandra cuspidata Nees & Mart. (9) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 0 0 0
Nectandra globosa (Aubl.) Mez (1) 1 1 1 1 1 1 0 1 0 1 1 0 0 1 1 0 1 1 1 1 0 1 1 0 0 1 1 1 1 0 1 1 1 1 0
Nectandra globosa (Aubl.) Mez (14) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0
Nectandra globosa (Aubl.) Mez (16) 1 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 1 0 0 0 1 1 1 1 0 0 1 1 1 0
Nectandra globosa (Aubl.) Mez (17) 1 0 1 1 1 1 0 1 1 0 0 0 0 1 1 0 1 1 0 1 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0
Nectandra grandiflora Nees (1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 1 1 0 1
Nectandra hihua (Ruiz & Pav.) Rohwer
(1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 0 1 0 1 1 0 0 1 1 1 1 0 1 1 1 1 1
Nectandra lanceolata Nees (*) 1 1 1 1 1 1 1 1 0 1 1 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 1 0 1 1 1 1
Nectandra lanceolata Nees (21) 1 1 1 1 1 1 0 1 1 1 1 0 0 1 1 0 1 1 0 1 0 0 0 0 0 1 1 0 1 1 0 1 1 0 0
Nectandra lanceolata Nees (7) 1 1 1 1 1 0 0 1 1 0 0 0 0 1 0 0 0 1 1 1 0 0 0 0 0 1 1 0 1 1 0 0 1 1 0
Nectandra lanceolata Nees (9) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 0 0 0
Nectandra laurel Klotzsch ex Nees (1) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 1 1 1 1 0 1 0 0 0 1 1 1 1 0 1 1 1 1 0
Nectandra leucantha Nees & Mart. (2) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 1 0 0 0 0 1 1 0 1 0 0 0 0 0 0
Nectandra martinicensis Mez (1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 1 1 0 0 1 1 0 1 0 1 1 1 0 1
Nectandra megapotamica (Spreng.) Mez
(*) 1 1 1 1 1 1 1 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 1 0 0 1 1 0 1 1 0 1 1 1 1

62
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Nectandra megapotamica (Spreng.) Mez
(21) 1 1 1 1 1 1 0 1 0 1 1 1 0 1 1 0 1 1 0 1 0 1 1 0 1 1 1 0 1 1 0 1 1 0 1
Nectandra megapotamica (Spreng.) Mez
(7) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 1 1 1 0 0 1 1 0 0 1 1 0 1 1 0 1 1 0 1
Nectandra megapotamica (Spreng.) Mez
(9) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 1 0 1
Nectandra membranacea (Sw.)
Griseb. (1) 1 0 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 0 1 0 1 1 0 0 0 1 1 1 0 1 1 1 1 1
Nectandra membranacea (Sw.)
Griseb. (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 0 0 1 0 0 1 1 0 1 1 0 0 0 0 0
Nectandra nitidula Nees & Mart. (12) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0
Nectandra oppositifolia Nees & Mart.
(22) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 1 1 1 1 0 0 0 0 0 1 1 1 0 1 1 0 1 1 0 0
Nectandra oppositifolia Nees & Mart. (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 0 0 1 0 0 1 1 0 1 0 0 0 0 0 0
Nectandra puberula (Schott) Nees (1) 1 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 1 1 0 0
Nectandra puberula (Schott) Nees (2) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 1 1 1 0 0 1 1 0 1 0 0 1 1 0 0
Nectandra reticulata (Ruiz & Pav.) Mez
(1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 1 1 0 0 1 1 0 1 0 1 1 1 0 1
Nectandra reticulata (Ruiz & Pav.) Mez
(2) 1 1 1 1 1 1 1 1 1 1 0 0 0 1 1 0 0 1 0 1 0 1 1 0 0 1 1 0 1 1 0 0 0 0 0
Nectandra sanguinea Rol. Ex Rottb. (14) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0
Nectandra sp.(20) 1 1 1 1 1 0 0 1 0 1 0 1 0 1 0 0 0 1 1 0 0 0 0 0 0 1 1 1 1 1 0 1 0 0 0

63
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Nectandra surinamensis Rottb. Ex
Bercht. & J. Presl (1) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 0 1 0 1 1 0 0 1 1 1 1 0 1 1 1 0 0
Nectandra turbacensis (Kunth) Nees (1) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 1 1 1 1 0 1 1 0 0 1 1 1 1 0 1 0 0 0 1
Nectandra turbacensis (Kunth) Nees (1) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 1 1 0 0 1 1 1 1 0 1 1 1 1 0
Nectandra turbacensis (Kunth) Nees
(14) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0
Ocotea aciphylla (Nees & Mart.) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 1 1 0 0 1 1 1 1 0 1 1 1 1 0
Ocotea aciphylla (Nees & Mart.) Mez (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 0 0 1 1 0 1 1 0 0 0 0 1 1 0 1 1 0 1 1 0 0
Ocotea aciphylla (Nees) Mez (15) 1 1 1 1 1 0 0 1 1 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 1 0 0 1 1 1 0
Ocotea aciphylla (Nees) Mez (16) 1 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 0 1 0 1 0 0 0 1 1 0 1 0 0 1 1 1 0
Ocotea adenotrachelium (Nees) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 1 0 0 0 1 1 1 1 0 1 0 0 0 0
Ocotea amplissima Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 1 1 0 0
Ocotea austinii C.K. Allen (20) 1 1 1 1 1 0 0 1 1 1 0 1 0 1 0 0 1 1 1 0 0 0 0 0 0 1 1 1 1 1 0 1 0 0 0
Ocotea bofo Kunth (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 1 1 0 1
Ocotea bullata (Burch.) E. Meyer in
Drege (14) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 0 0 0 0 0 0 1 1 1 1 0 0 1 0 0 0
Ocotea canaliculata (Rich.) Mez (15) 1 1 1 1 1 0 0 1 1 0 0 1 0 1 0 0 1 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 1 1 0
Ocotea canaliculata (Rich.) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 1 1 1 0 0 1 1 1 1 0 1 1 1 1 1
Ocotea caracasana (Nees) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 1 1 1 0 1 1 1 0 1

64
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Ocotea catharinensis Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 1 1 1 1 1
Ocotea catharinensis Mez (4) 1 1 1 1 1 0 1 1 0 1 0 1 0 1 0 0 1 1 1 0 0 0 0 0 0 1 1 1 1 1 0 1 0 1 0
Ocotea cernua (Nees) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 1 0 0 0 1 1 1 1 0 1 1 1 1 1
Ocotea cernua (Nees) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 1 1 0 0 1 1 0 1 0 1 1 1 1 1
Ocotea corymbosa (Meissn.) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 1 1 0 1 1 0 0 1
Ocotea cuneifolia (Ruiz & Pav.) Mez (19) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 1 1 1 1 1 0 0 1 0 0 0
Ocotea cymbarum Kunth (14) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0
Ocotea cymbarum Kunth (3) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 0 0 1 1 0 1 0 1 1 0 0 1 1 1 1 0 1 1 1 1 1
Ocotea diospyrifolia (Meissn.) Mez (23) 1 1 1 1 1 0 0 1 0 1 0 1 0 1 0 0 1 1 1 1 0 0 1 0 0 1 1 0 1 1 0 0 1 1 0
Ocotea diospyrifolia (Meissn.) Mez (9) 1 1 1 1 1 1 0 1 1 0 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0
Ocotea diospyrifolia (Meissn.) Mez (9) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 1 0 1 0 0 0
Ocotea divaricata (Nees) Mez (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 1 0 0 0 1 1 1 0 1 1 0 1 1 1 0
Ocotea elegans Mez (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 0 0 0 1 0 1 1 0 0 0 0 1 1 0 1 1 0 1 1 1 0
Ocotea floribunda (Sw.) Mez (14) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 1 0 1 0 0 1 1 0 0
Ocotea floribunda (Sw.) Mez (16) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 0 1 1 1 0
Ocotea fragrantissima Ducke (8) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 1 0 1 0 1 0 0 0 0 0 0 1 1 1 1 1 0 1 1 0 0
Ocotea fulvescens Standl. & L.O.
Williams (3) 1 0 1 1 1 1 0 1 1 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 0 0 0 0
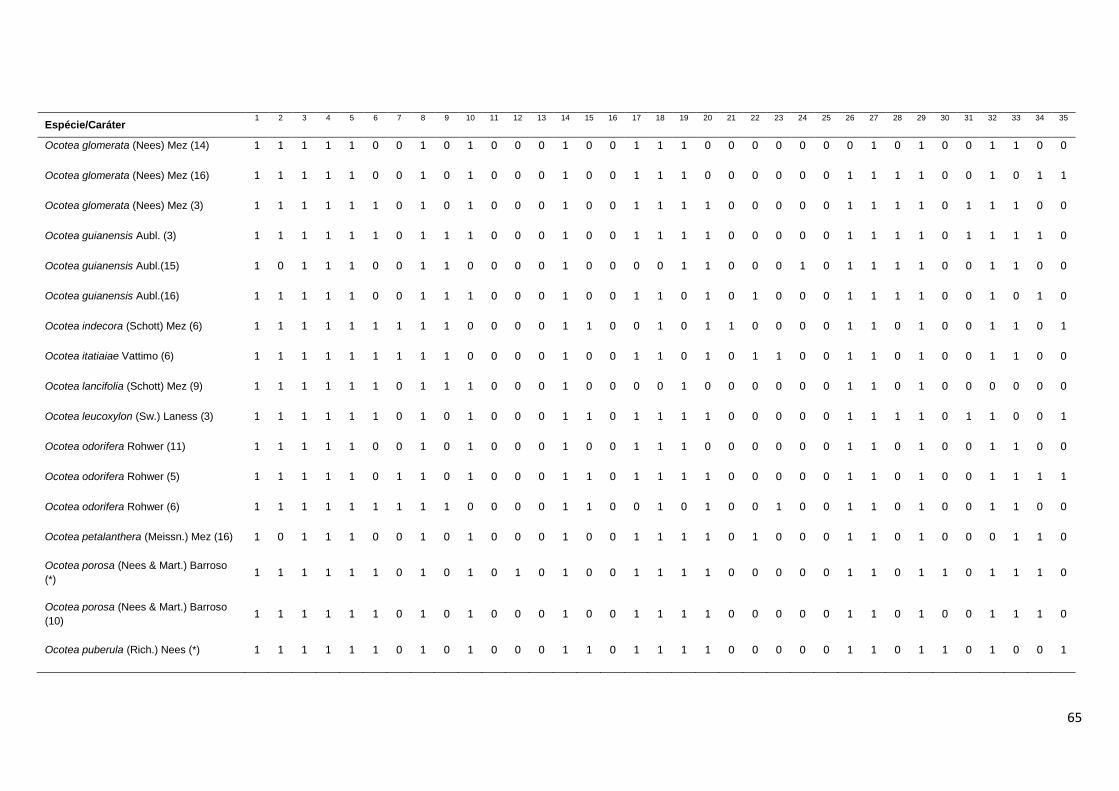
65
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Ocotea glomerata (Nees) Mez (14) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 0 0 0 0 0 0 0 1 0 1 0 0 1 1 0 0
Ocotea glomerata (Nees) Mez (16) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 0 0 0 0 0 0 1 1 1 1 0 0 1 0 1 1
Ocotea glomerata (Nees) Mez (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 1 1 0 0
Ocotea guianensis Aubl. (3) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 1 1 1 0
Ocotea guianensis Aubl.(15) 1 0 1 1 1 0 0 1 1 0 0 0 0 1 0 0 0 0 1 1 0 0 0 1 0 1 1 1 1 0 0 1 1 0 0
Ocotea guianensis Aubl.(16) 1 1 1 1 1 0 0 1 1 1 0 0 0 1 0 0 1 1 0 1 0 1 0 0 0 1 1 1 1 0 0 1 0 1 0
Ocotea indecora (Schott) Mez (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 1 0 0 0 0 1 1 0 1 0 0 1 1 0 1
Ocotea itatiaiae Vattimo (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 0 0 1 1 0 1 0 1 1 0 0 1 1 0 1 0 0 1 1 0 0
Ocotea lancifolia (Schott) Mez (9) 1 1 1 1 1 1 0 1 1 1 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0
Ocotea leucoxylon (Sw.) Laness (3) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 1 1 0 1 1 0 0 1
Ocotea odorifera Rohwer (11) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0
Ocotea odorifera Rohwer (5) 1 1 1 1 1 0 1 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 0 1 1 1 1
Ocotea odorifera Rohwer (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 0 0 1 0 0 1 1 0 1 0 0 1 1 0 0
Ocotea petalanthera (Meissn.) Mez (16) 1 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 1 0 0 0 1 1 0 1 0 0 0 1 1 0
Ocotea porosa (Nees & Mart.) Barroso
(*) 1 1 1 1 1 1 0 1 0 1 0 1 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 0 1 1 0 1 1 1 0
Ocotea porosa (Nees & Mart.) Barroso
(10) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 0 1 1 1 0
Ocotea puberula (Rich.) Nees (*) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 1 0 1 0 0 1

66
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Ocotea puberula (Rich.) Nees (16) 1 0 1 1 1 1 0 1 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 1 1 1 1 0 0 0 1 1 0
Ocotea puberula (Rich.) Nees (21) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 0 1 0 0 1
Ocotea puberula (Rich.) Nees (3) 1 0 1 1 1 1 0 1 1 1 0 0 0 1 1 0 0 1 1 1 0 0 0 0 0 1 1 1 1 0 1 0 0 0 0
Ocotea puberula (Rich.) Nees (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 1 1 0 1 0 0 1 0 0 1 1 0 1 0 0 0 0 0 0
Ocotea puberula (Rich.) Nees (9) 1 1 1 1 1 1 0 1 1 0 0 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 0 0 0
Ocotea pulchella (Nees & Mart.) Mez (*) 1 1 1 1 1 1 0 1 0 1 0 1 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 0 1 1 0 1 1 1 1
Ocotea pulchella (Nees & Mart.) Mez
(21) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 1 0 1 0 1 1 0 0 1 1 0 1 0 0 1 1 0 0
Ocotea pulchella (Nees & Mart.) Mez (9) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 0 0 0
Ocotea schomburgkiana (Nees) Mez
(14) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 1 0 1 0 0 1 1 0 0
Ocotea schomburgkiana (Nees) Mez
(16) 1 1 1 1 1 0 0 0 0 1 0 0 0 1 0 0 1 1 0 1 0 1 0 0 0 1 1 1 1 0 0 1 1 1 0
Ocotea sp. (*) 1 1 1 1 1 1 1 1 0 1 0 0 1 1 0 0 1 1 1 0 0 0 0 0 0 1 1 0 1 1 0 1 0 1 1
Ocotea tenuiflora (Nees) Mez (6) 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1 0 1 0 1 1 1 0 1 1 0 1 1 0 1 1 0 0
Ocotea usambarensis Engl.(14) 1 1 1 1 1 1 0 1 0 1 0 0 0 1 1 0 1 0 1 1 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0
Pleurothyrium palmanum (Mez & Donn.
Sm.) Rohwer (17) 1 1 1 1 1 1 0 1 1 0 0 0 0 1 1 0 1 1 0 1 0 0 0 0 0 1 1 0 1 0 0 0 1 0 0
Pleurothyrium parviflorum Ducke (3) 1 1 1 1 1 0 0 1 1 1 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 1 1 0 1 0 1 1 1 0 1
Rhodostemonodaphne grandis (Mez)
Rohwer (1) 1 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 0 0 1 0 1 1 0 0 1 1 0 1 0 0 0 0 0 1
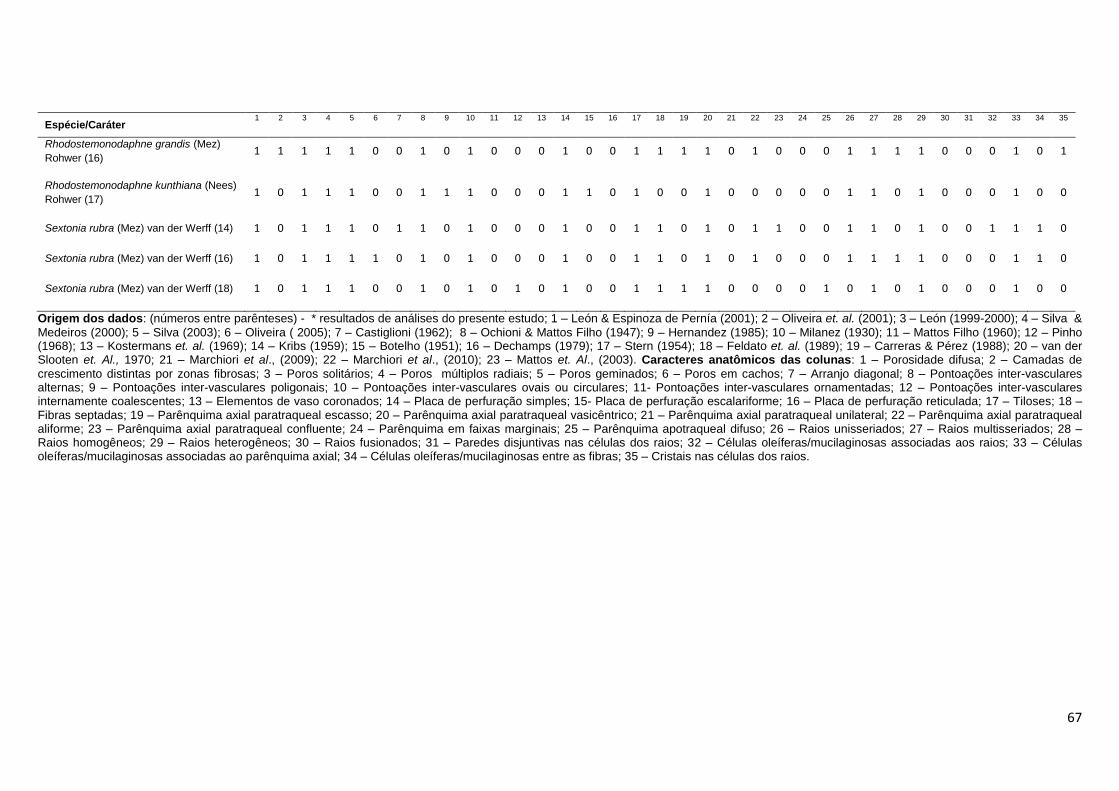
67
Espécie/Caráter 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
Rhodostemonodaphne grandis (Mez)
Rohwer (16) 1 1 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1 1 0 1 0 0 0 1 1 1 1 0 0 0 1 0 1
Rhodostemonodaphne kunthiana (Nees)
Rohwer (17) 1 0 1 1 1 0 0 1 1 1 0 0 0 1 1 0 1 0 0 1 0 0 0 0 0 1 1 0 1 0 0 0 1 0 0
Sextonia rubra (Mez) van der Werff (14) 1 0 1 1 1 0 1 1 0 1 0 0 0 1 0 0 1 1 0 1 0 1 1 0 0 1 1 0 1 0 0 1 1 1 0
Sextonia rubra (Mez) van der Werff (16) 1 0 1 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 0 1 0 1 0 0 0 1 1 1 1 0 0 0 1 1 0
Sextonia rubra (Mez) van der Werff (18) 1 0 1 1 1 0 0 1 0 1 0 1 0 1 0 0 1 1 1 1 0 0 0 0 1 0 1 0 1 0 0 0 1 0 0
Origem dos dados: (números entre parênteses) - * resultados de análises do presente estudo; 1 – León & Espinoza de Pernía (2001); 2 – Oliveira et. al. (2001); 3 – León (1999-2000); 4 – Silva &
Medeiros (2000); 5 – Silva (2003); 6 – Oliveira ( 2005); 7 – Castiglioni (1962); 8 – Ochioni & Mattos Filho (1947); 9 – Hernandez (1985); 10 – Milanez (1930); 11 – Mattos Filho (1960); 12 – Pinho (1968); 13 – Kostermans et. al. (1969); 14 – Kribs (1959); 15 – Botelho (1951); 16 – Dechamps (1979); 17 – Stern (1954); 18 – Feldato et. al. (1989); 19 – Carreras & Pérez (1988); 20 – van der Slooten et. Al., 1970; 21 – Marchiori et al., (2009); 22 – Marchiori et al., (2010); 23 – Mattos et. Al., (2003). Caracteres anatômicos das colunas: 1 – Porosidade difusa; 2 – Camadas de
crescimento distintas por zonas fibrosas; 3 – Poros solitários; 4 – Poros múltiplos radiais; 5 – Poros geminados; 6 – Poros em cachos; 7 – Arranjo diagonal; 8 – Pontoações inter-vasculares alternas; 9 – Pontoações inter-vasculares poligonais; 10 – Pontoações inter-vasculares ovais ou circulares; 11- Pontoações inter-vasculares ornamentadas; 12 – Pontoações inter-vasculares internamente coalescentes; 13 – Elementos de vaso coronados; 14 – Placa de perfuração simples; 15- Placa de perfuração escalariforme; 16 – Placa de perfuração reticulada; 17 – Tiloses; 18 – Fibras septadas; 19 – Parênquima axial paratraqueal escasso; 20 – Parênquima axial paratraqueal vasicêntrico; 21 – Parênquima axial paratraqueal unilateral; 22 – Parênquima axial paratraqueal aliforme; 23 – Parênquima axial paratraqueal confluente; 24 – Parênquima em faixas marginais; 25 – Parênquima apotraqueal difuso; 26 – Raios unisseriados; 27 – Raios multisseriados; 28 – Raios homogêneos; 29 – Raios heterogêneos; 30 – Raios fusionados; 31 – Paredes disjuntivas nas células dos raios; 32 – Células oleíferas/mucilaginosas associadas aos raios; 33 – Células oleíferas/mucilaginosas associadas ao parênquima axial; 34 – Células oleíferas/mucilaginosas entre as fibras; 35 – Cristais nas células dos raios.

68
Tabela 09 – Matriz de similaridade obtida pelo índice de Jaccard (valores abaixo da diagonal), a partir dos dados anatômicos qualitativos do lenho oriundos da matriz binária de ausência e presença, para as espécies analisadas no presente estudo
Espécies
Nectandra lanceolata
Nees
Nectandra megapotamica
(Spreng.) Mez
Ocotea porosa
(Nees & C. Mart.)
Barroso
Ocotea puberula
(Rich.) Nees
Ocotea pulchella
(Nees) Mez Ocotea sp.
Nectandra lanceolata
Nees
Nectandra megapotamica
(Spreng.) Mez 92,00
Ocotea porosa
(Nees & C. Mart.)
Barroso
80,00 80,00
Ocotea puberula
(Rich.) Nees 83,33 83,33 78,26
Ocotea pulchella
(Nees) Mez 84,00 84,00 95,45 82,61
Ocotea sp. 83,33 83,33 78,26 81,82 82,61

69
Tabela 10 – Análise de ANOSIM e SIMPER nas relações entre gêneros a partir da similaridade dada pelo índice de Jaccard quando da análise das
características anatômicas qualitativas do lenho das espécies constantes da matriz binária de presença e ausência. As similaridades intra-genéricas estão na
diagonal, em negrito. As dissimilaridades intra-genéricas estão acima da diagonal e os valores p do ANOSIN estão abaixo da diagonal. As características
anatômicas qualitativas do lenho que mais contribuíram para as semelhanças e diferenças observadas e os valores relativos (em porcentagem, entre
parênteses) estão destacados abaixo dos respectivos valores de similaridade e dissimilaridade.
Gêneros Nectandra Ocotea Rhodostemonodaphne Sextonia Pleurothyrium Chlorocardium
Nectandra
80,09
a(6,32); c(6,32);
d(6,32); e(6,32);
h(6,32); n(6,32); r(6,32);
cc(6,32); ee(6,32);
bb(6,04)
20,57
o(7,01); dd(6,08);
i(6,03); ii(5,75);
jj(5,75); v(5,60);
f(5,38); q(5,31);
gg(5,31); x(5,31)
24,89
hh(8,75); f(8,00); r(7,71);
b(7,20); kk(6,36);
o(6,25); s(6,25); v(5,90);
i(5,30); x(5,19)
25,03
b(9,42); o(7,42);
hh(6,43); f(6,13); jj(6,12);
s(5,85); v(5,64); x(4,90);
dd(4,87); i(4,48)
19,02
i(7,99); j(7,05); hh(7,03);
s(6,97); gg(6,78); o(6,78);
f(6,77); kk(6,75); v(5,60);
x(5,32)
26,90
hh(8,01); o(7,09); m(6,75);
r(6,75); b(6,50); v(6,07);
f(5,90); q(5,65); dd(5,58);
ii(5,57)
Ocotea 18,5
79,52
a(6,65); c(6,65);
d(6,65); e(6,65);
cc(6,65); ee(6,65);
h(6,42); n(6,40);
r(5,73); bb(5,49)
25,75
hh(9,29); f(7,34); r(7,33);
b(7,19); s(6,58); v(6,52);
kk(6,36); dd(5,25);
i(5,10); o(5,02)
24,42
b(10,00); hh(6,96);
v(6,58); s(6,44); f(6,22);
jj(5,89); dd(5,35); g(4,32);
l(4,24); x(4,23)
19,34
i(8,65); hh(7,11); j(7,08);
s(7,07); o(7,01); f(6,84);
kk(6,78); gg(6,78);
dd(6,03); jj(5,70)
27,18
hh(8,70); v(8,19); m(6,84);
b(6,63); r(6,60); q(5,95);
f(5,78); ii(5,68); dd(5,42);
jj(4,88)
Rhodostemonodaphne 1 2,7
79,61
a(7,72); c(7,72); d(7,72);
e(7,72); h(7,72); j(7,72);
n(7,72); q(7,72); t(7,72);
bb(7,72)
21,09
r(9,69); kk(9,16); jj(9,10);
v(6,35); x(6,23); s(6,21);
dd(6,09); l(4,91);
aa(4,91); bb(4,91)
20,75
b(9,61); r(9,61); i(9,10);
v(9,10); f(7,25); j(7,25);
o(7,04); kk(7,04); s(6,82);
gg(6,65)
20,58
m(9,76); q(9,76); kk(9,66);
dd(8,42); s(8,30); ii(8,12);
x(6,58); i(6,58); r(6,48);
b(6,36)
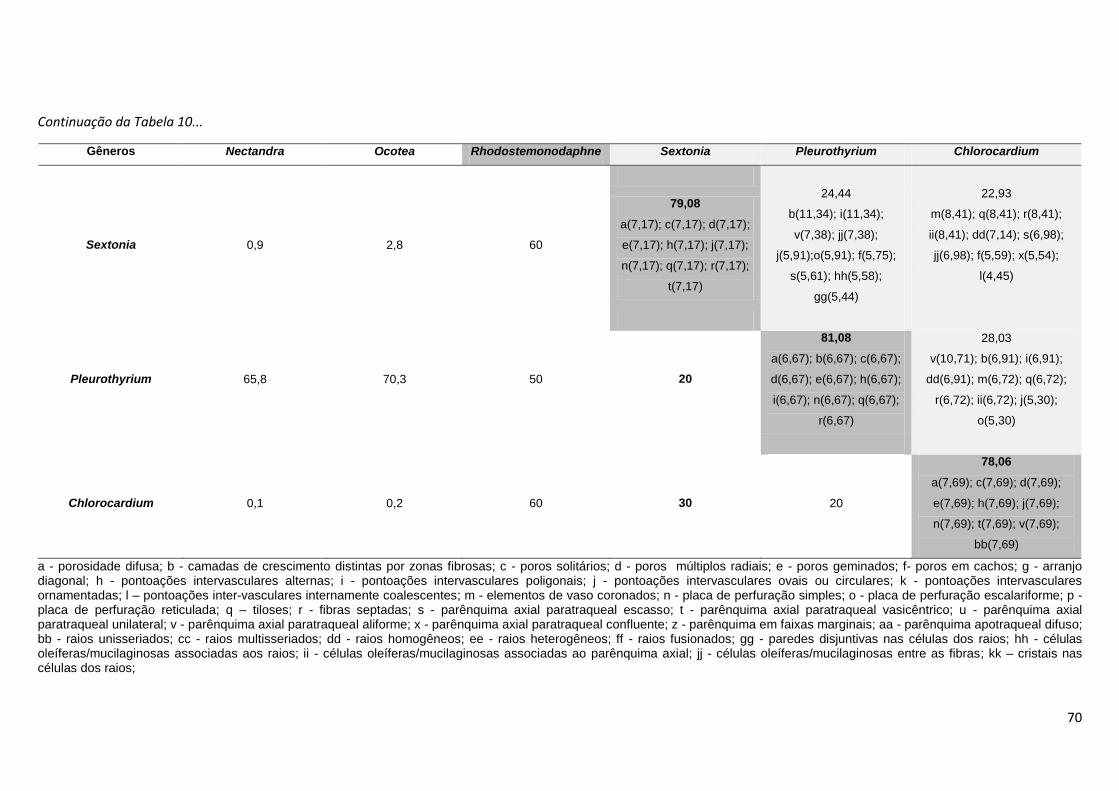
70
Continuação da Tabela 10...
Gêneros Nectandra Ocotea Rhodostemonodaphne Sextonia Pleurothyrium Chlorocardium
Sextonia 0,9 2,8 60
79,08
a(7,17); c(7,17); d(7,17);
e(7,17); h(7,17); j(7,17);
n(7,17); q(7,17); r(7,17);
t(7,17)
24,44
b(11,34); i(11,34);
v(7,38); jj(7,38);
j(5,91);o(5,91); f(5,75);
s(5,61); hh(5,58);
gg(5,44)
22,93
m(8,41); q(8,41); r(8,41);
ii(8,41); dd(7,14); s(6,98);
jj(6,98); f(5,59); x(5,54);
l(4,45)
Pleurothyrium 65,8 70,3 50 20
81,08
a(6,67); b(6,67); c(6,67);
d(6,67); e(6,67); h(6,67);
i(6,67); n(6,67); q(6,67);
r(6,67)
28,03
v(10,71); b(6,91); i(6,91);
dd(6,91); m(6,72); q(6,72);
r(6,72); ii(6,72); j(5,30);
o(5,30)
Chlorocardium 0,1 0,2 60 30 20
78,06
a(7,69); c(7,69); d(7,69);
e(7,69); h(7,69); j(7,69);
n(7,69); t(7,69); v(7,69);
bb(7,69)
a - porosidade difusa; b - camadas de crescimento distintas por zonas fibrosas; c - poros solitários; d - poros múltiplos radiais; e - poros geminados; f- poros em cachos; g - arranjo diagonal; h - pontoações intervasculares alternas; i - pontoações intervasculares poligonais; j - pontoações intervasculares ovais ou circulares; k - pontoações intervasculares ornamentadas; l – pontoações inter-vasculares internamente coalescentes; m - elementos de vaso coronados; n - placa de perfuração simples; o - placa de perfuração escalariforme; p - placa de perfuração reticulada; q – tiloses; r - fibras septadas; s - parênquima axial paratraqueal escasso; t - parênquima axial paratraqueal vasicêntrico; u - parênquima axial paratraqueal unilateral; v - parênquima axial paratraqueal aliforme; x - parênquima axial paratraqueal confluente; z - parênquima em faixas marginais; aa - parênquima apotraqueal difuso; bb - raios unisseriados; cc - raios multisseriados; dd - raios homogêneos; ee - raios heterogêneos; ff - raios fusionados; gg - paredes disjuntivas nas células dos raios; hh - células oleíferas/mucilaginosas associadas aos raios; ii - células oleíferas/mucilaginosas associadas ao parênquima axial; jj - células oleíferas/mucilaginosas entre as fibras; kk – cristais nas células dos raios;

71
Figura 19 – Dendrograma de similaridade dos caracteres anatômicos qualitativos, com base no índice de similaridade de Jaccard, com identificação do fator gênero.
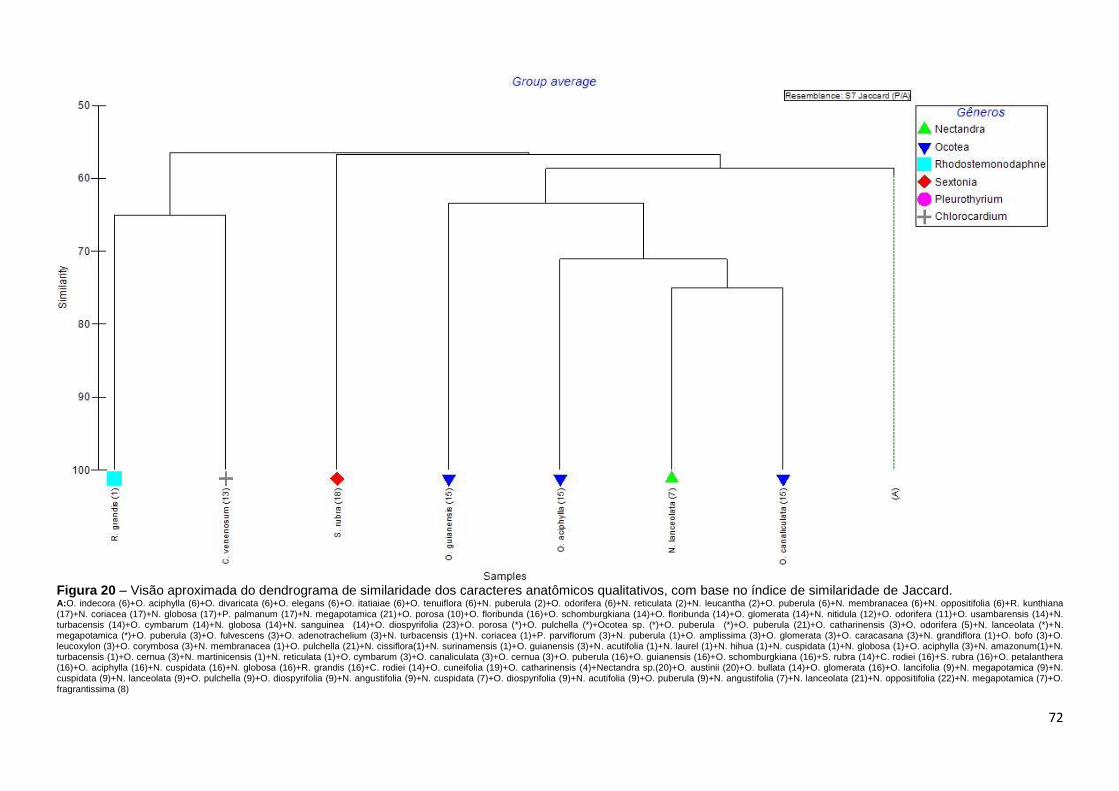
72
Figura 20 – Visão aproximada do dendrograma de similaridade dos caracteres anatômicos qualitativos, com base no índice de similaridade de Jaccard. A:O. indecora (6)+O. aciphylla (6)+O. divaricata (6)+O. elegans (6)+O. itatiaiae (6)+O. tenuiflora (6)+N. puberula (2)+O. odorifera (6)+N. reticulata (2)+N. leucantha (2)+O. puberula (6)+N. membranacea (6)+N. oppositifolia (6)+R. kunthiana (17)+N. coriacea (17)+N. globosa (17)+P. palmanum (17)+N. megapotamica (21)+O. porosa (10)+O. floribunda (16)+O. schomburgkiana (14)+O. floribunda (14)+O. glomerata (14)+N. nitidula (12)+O. odorifera (11)+O. usambarensis (14)+N. turbacensis (14)+O. cymbarum (14)+N. globosa (14)+N. sanguinea (14)+O. diospyrifolia (23)+O. porosa (*)+O. pulchella (*)+Ocotea sp. (*)+O. puberula (*)+O. puberula (21)+O. catharinensis (3)+O. odorifera (5)+N. lanceolata (*)+N. megapotamica (*)+O. puberula (3)+O. fulvescens (3)+O. adenotrachelium (3)+N. turbacensis (1)+N. coriacea (1)+P. parviflorum (3)+N. puberula (1)+O. amplissima (3)+O. glomerata (3)+O. caracasana (3)+N. grandiflora (1)+O. bofo (3)+O. leucoxylon (3)+O. corymbosa (3)+N. membranacea (1)+O. pulchella (21)+N. cissiflora(1)+N. surinamensis (1)+O. guianensis (3)+N. acutifolia (1)+N. laurel (1)+N. hihua (1)+N. cuspidata (1)+N. globosa (1)+O. aciphylla (3)+N. amazonum(1)+N. turbacensis (1)+O. cernua (3)+N. martinicensis (1)+N. reticulata (1)+O. cymbarum (3)+O. canaliculata (3)+O. cernua (3)+O. puberula (16)+O. guianensis (16)+O. schomburgkiana (16)+S. rubra (14)+C. rodiei (16)+S. rubra (16)+O. petalanthera (16)+O. aciphylla (16)+N. cuspidata (16)+N. globosa (16)+R. grandis (16)+C. rodiei (14)+O. cuneifolia (19)+O. catharinensis (4)+Nectandra sp.(20)+O. austinii (20)+O. bullata (14)+O. glomerata (16)+O. lancifolia (9)+N. megapotamica (9)+N. cuspidata (9)+N. lanceolata (9)+O. pulchella (9)+O. diospyrifolia (9)+N. angustifolia (9)+N. cuspidata (7)+O. diospyrifolia (9)+N. acutifolia (9)+O. puberula (9)+N. angustifolia (7)+N. lanceolata (21)+N. oppositifolia (22)+N. megapotamica (7)+O. fragrantissima (8)

73
Figura 21 – Visão aproximada do dendrograma de similaridade dos caracteres anatômicos qualitativos, com base no índice de similaridade de Jaccard. A:R. grandis (1)+C. venenosum (13); B:O. guianensis (15)+O. aciphylla (15)+N. lanceolata (7)+O. canaliculata (15); C:N. megapotamica (21)+O. porosa (10)+O. floribunda (16)+O. schomburgkiana (14)+O. floribunda (14)+O. glomerata (14)+N.
nitidula (12)+O. odorifera (11)+O. usambarensis (14)+N. turbacensis (14)+O. cymbarum (14)+N. globosa (14)+N. sanguinea (14)+O. diospyrifolia (23)+O. porosa (*)+O. pulchella (*)+Ocotea sp. (*)+O. puberula (*)+O. puberula (21)+O. catharinensis (3)+O. odorifera (5)+N. lanceolata (*)+N. megapotamica (*)+O. puberula (3)+O. fulvescens (3)+O. adenotrachelium (3)+N. turbacensis (1)+N. coriacea (1)+P. parviflorum (3)+N. puberula (1)+O. amplissima (3)+O. glomerata (3)+O. caracasana (3)+N. grandiflora (1)+O. bofo (3)+O. leucoxylon (3)+O. corymbosa (3)+N. membranacea (1)+O. pulchella (21)+N. cissiflora(1)+N. surinamensis (1)+O. guianensis (3)+N. acutifolia (1)+N. laurel (1)+N. hihua (1)+N. cuspidata (1)+N. globosa (1)+O. aciphylla (3)+N. amazonum(1)+N. turbacensis (1)+O. cernua (3)+N. martinicensis (1)+N. reticulata (1)+O. cymbarum (3)+O. canaliculata (3)+O. cernua (3)+O. puberula (16)+O. guianensis (16)+O. schomburgkiana (16)+S. rubra (14)+C. rodiei (16)+S. rubra (16)+O. petalanthera (16)+O. aciphylla (16)+N. cuspidata (16)+N. globosa (16)+R. grandis (16)+C. rodiei (14)+O. cuneifolia (19)+O. catharinensis (4)+Nectandra sp.(20)+O. austinii (20)+O. bullata (14)+O. glomerata (16)+O. lancifolia (9)+N. megapotamica (9)+N. cuspidata (9)+N. lanceolata (9)+O. pulchella (9)+O. diospyrifolia (9)+N. angustifolia (9)+N. cuspidata (7)+O. diospyrifolia (9)+N. acutifolia (9)+O. puberula (9)+N. angustifolia (7)+N. lanceolata (21)+N. oppositifolia (22)+N. megapotamica (7)+O. fragrantissima (8)

74
Figura 22 – Visão aproximada do dendrograma de similaridade dos caracteres anatômicos qualitativos, com base no índice de similaridade de Jaccard. A:R. grandis (1)+C. venenosum (13); B:O. guianensis (15)+O. aciphylla (15)+N. lanceolata (7)+O. canaliculata (15); C:O. indecora (6)+O. aciphylla (6)+O. divaricata (6)+O. elegans (6)+O. itatiaiae (6)+O. tenuiflora (6)+N. puberula (2)+O. odorifera (6)+N. reticulata (2)+N. leucantha (2)+O. puberula (6)+N. membranacea (6)+N. oppositifolia (6)+R. kunthiana (17)+N. coriacea (17)+N. globosa (17)+P. palmanum (17); D:O. cuneifolia (19)+O. catharinensis (4)+Nectandra sp.(20)+O. austinii (20)+O. bullata (14)+O. glomerata (16)+O. lancifolia (9)+N. megapotamica (9)+N. cuspidata (9)+N. lanceolata (9)+O. pulchella (9)+O. diospyrifolia (9)+N. angustifolia (9)+N. cuspidata (7)+O. diospyrifolia (9)+N. acutifolia (9)+O. puberula (9)+N. angustifolia (7)+N. lanceolata (21)+N. oppositifolia (22)+N. megapotamica (7)+O. fragrantissima (8)
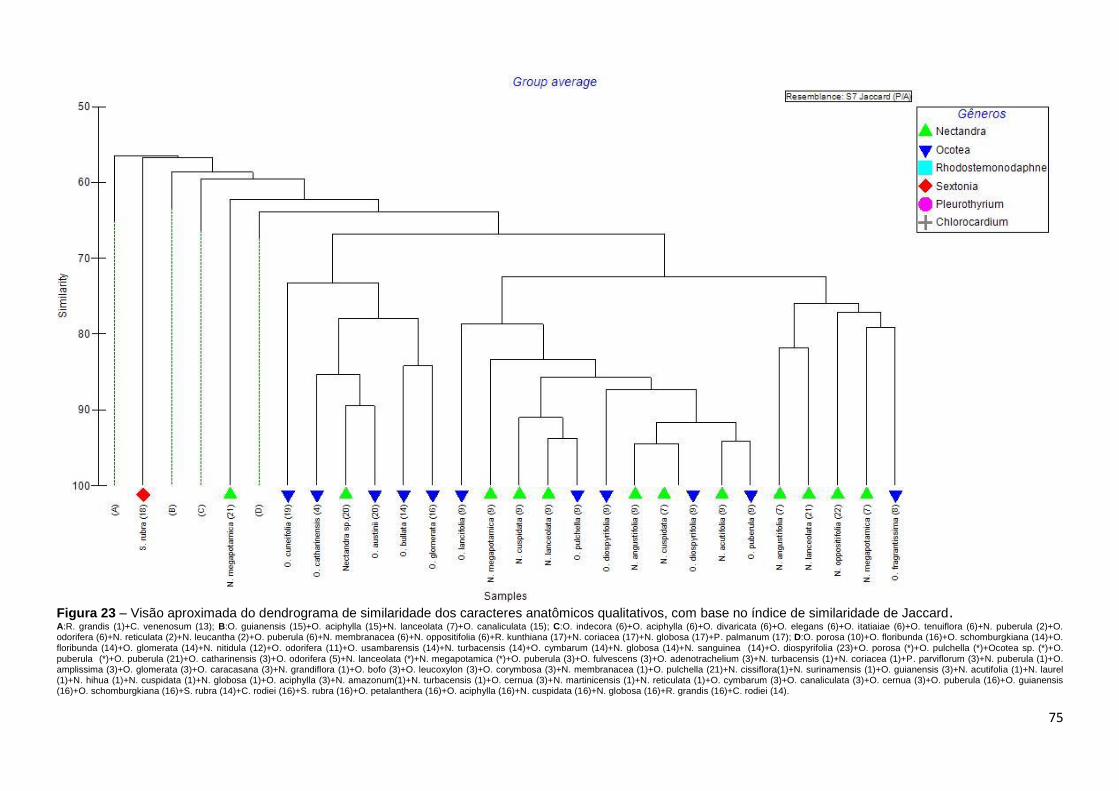
75
Figura 23 – Visão aproximada do dendrograma de similaridade dos caracteres anatômicos qualitativos, com base no índice de similaridade de Jaccard. A:R. grandis (1)+C. venenosum (13); B:O. guianensis (15)+O. aciphylla (15)+N. lanceolata (7)+O. canaliculata (15); C:O. indecora (6)+O. aciphylla (6)+O. divaricata (6)+O. elegans (6)+O. itatiaiae (6)+O. tenuiflora (6)+N. puberula (2)+O. odorifera (6)+N. reticulata (2)+N. leucantha (2)+O. puberula (6)+N. membranacea (6)+N. oppositifolia (6)+R. kunthiana (17)+N. coriacea (17)+N. globosa (17)+P. palmanum (17); D:O. porosa (10)+O. floribunda (16)+O. schomburgkiana (14)+O. floribunda (14)+O. glomerata (14)+N. nitidula (12)+O. odorifera (11)+O. usambarensis (14)+N. turbacensis (14)+O. cymbarum (14)+N. globosa (14)+N. sanguinea (14)+O. diospyrifolia (23)+O. porosa (*)+O. pulchella (*)+Ocotea sp. (*)+O. puberula (*)+O. puberula (21)+O. catharinensis (3)+O. odorifera (5)+N. lanceolata (*)+N. megapotamica (*)+O. puberula (3)+O. fulvescens (3)+O. adenotrachelium (3)+N. turbacensis (1)+N. coriacea (1)+P. parviflorum (3)+N. puberula (1)+O. amplissima (3)+O. glomerata (3)+O. caracasana (3)+N. grandiflora (1)+O. bofo (3)+O. leucoxylon (3)+O. corymbosa (3)+N. membranacea (1)+O. pulchella (21)+N. cissiflora(1)+N. surinamensis (1)+O. guianensis (3)+N. acutifolia (1)+N. laurel (1)+N. hihua (1)+N. cuspidata (1)+N. globosa (1)+O. aciphylla (3)+N. amazonum(1)+N. turbacensis (1)+O. cernua (3)+N. martinicensis (1)+N. reticulata (1)+O. cymbarum (3)+O. canaliculata (3)+O. cernua (3)+O. puberula (16)+O. guianensis (16)+O. schomburgkiana (16)+S. rubra (14)+C. rodiei (16)+S. rubra (16)+O. petalanthera (16)+O. aciphylla (16)+N. cuspidata (16)+N. globosa (16)+R. grandis (16)+C. rodiei (14).

76
Figura 24 – Escalonamento multidimencional da distribuição dos valores de similaridade nas relações inter-genéricas, das características anatômicas qualitativas do lenho das espécies analisadas na matriz binária de presença e ausência.
Figura 25 – Escalonamento multidimencional da distribuição dos valores de similaridade nas relações das características anatômicas qualitativas do lenho das espécies analisadas na matriz binária de presença e ausência com a literatura da qual a informação fora retirada (autores da descrição).

77
4 – Discussão
A importância da anatomia do lenho é evidente para a comercialização de
madeira, uma vez que a identificação anatômica é indispensável à lisura da
transação (Araújo & Mattos Filho, 1980). No entanto, o grupo de árvores
conhecido popularmente por “canela” representa hoje um grande desafio para
os anatomistas quanto se trata de tentar separar as espécies pelas
características da madeira. As dificuldades existentes entre os especialistas do
meio acadêmico na distinção dos gêneros e das espécies desta família são
enormes e, ao longo da cadeia de custódia das madeiras deste grupo, desde
os profissionais que elaboram os planos de manejo, passando pelos extratores,
transportadores, comerciantes, consumidores e agentes de fiscalização, a
identificação, quando realizada, é uníssona: canela. Para os dois maiores
laboratórios de identificação do país, quais sejam, Laboratório de Produtos
Florestais – LPF, ligado ao Serviço Florestal Brasileiro em Brasília, e
Laboratório de Anatomia de Madeiras, vinculado ao Instituto de Pesquisas
Tecnológicas de São Paulo, que recebem um grande fluxo de amostras de
madeira para identificação anatômica, oriundos especialmente de apreensões
resultantes de ações fiscalizatórias em empresas ligadas ao ramo madeireiro,
realizadas por agentes públicos de fiscalização, os laudos de identificação para
materiais deste grupo de Lauraceas comumente informam: Canela – Nome
científico: Ocotea/Nectandra. Ainda assim, correm o risco de não estar
informando adequadamente sequer o gênero, tendo em vista os recentes
rearranjos de espécies que tem sido propostos e os novos gêneros criados em
virtude das revisões taxonômicas que tem sido levadas a cabo para os grandes
gêneros da família Lauraceae.
Macroscopicamente, as espécies de canela dos gêneros Ocotea e
Nectandra apresentam lenho muito similares, sendo muito difícil a sua
distinção, tanto ao nível genérico, quanto ao nível específico. Entre os
caracteres macroscópicos do lenho que puderam ser considerados na tentativa
de diferenciação das espécies analisadas na presente pesquisa estão: cor,
odor, disposição dos elementos de vaso e parênquima axial.
A coloração amarelada ou amarronzada, com nítida foto-oxidação durante
a secagem, e o brilho perceptível nas faces longitudinais do lenho parecem ser

78
comuns neste grupo. Observou-se que a madeira das espécies de Nectandra
ora analisadas tendem mais para a cor amarela, especialmente quando recém
cortadas, e escurecem pouco, tornando-se amarronzadas após secas, e
difíceis de serem distintas entre si, considerando-se apenas este caráter. Já as
espécies de Ocotea porosa e O. pulchella apresentaram cerne marrom desde
o momento do corte e, quando secas a coloração acentuou-se e escureceu,
adquirindo, a primeira, um tom mais oliváceo. O. puberula se apresentou mais
clara, esbranquiçada, no momento do corte, escurecendo para marrom
acinzentado após a secagem, muito similar a cor de Ocotea sp., o que torna
inviável a distinção destas últimas pela cor. Na literatura, as espécies de
Ocotea e Nectandra não possuem grande variação na cor, sendo descritas
como madeiras amareladas, castanho-amareladas, marrom-amareladas,
amarelo-esverdeadas, amarelo-oliváceas, marrom-douradas ou amarelo-
grisáceo (Kribs, 1959; Loureiro & Silva, 1968; var der Slooten et. al.;1970,
Dechamps, 1979; Mainieri, 1983). Apesar da subjetividade que a observação
da cor do lenho possa fornecer ao nível macroscópico, este caráter tem sido
utilizado em chaves dicotômicas para identificação anatômica de madeiras,
como em Mainieri (1983), Coradin et. al. (1991), Zenid & Ceccantini (2007),
Coradin et. al. (2009b), inclusive como elemento principal de entrada na chave,
como em Camargos (1996).
A presença de odores característicos, agradáveis ou desagradáveis no
lenho, como o observado para O. porosa, O. pulchella e N. megapotamica, é
comum ao grupo das canelas e foi, possivelmente, um dos caracteres mais
importantes para separação das espécies. Esses odores são amplamente
relatados inclusive nos nomes populares, por vezes chulos, para espécies
deste grupo, como: canela-fedorenta, canela-fedida, canela-bosta, canela-
merda, canela-cheirosa, canela-de-cheiro, canela-de-mau-cheiro, canela-
cânfora, casca-cheirosa, catinga-de-negro, louro-cânfora, louro-cheiroso, pau-
cravo, entre muitos outros (Camargos et. al., 2001). Ochioni & Mattos Filho
(1947) menciona o cheiro de “funcho” para O. fragrantissima. Vattimo (1956)
indica a presença de aroma de anis para O. puberula e cheiro “nauseabundo”
para O. spectabilis (Meissn) Mez. Mattos Filho (1960) indica a presença de
cheiro muito característico para O. odorifera, variáveis porém entre os
espécimes analisados. Mainieri (1971) indica cheiro ativo e agradável para O.

79
cymbarum e a ausência de cheiro para Sextonia rubra. Loureiro & Silva (1968)
indicam cheiro “aromático ativo” para Ocotea cymbarum. Kribs (1959) descreve
cheiro agradável para O. schomburgkiana, O. floribunda, O. glomerata,
resinoso-agradável para O. cymbarum, Nectandra globosa, N. sanguinea e N.
turbacensis, odor desagradável para O. bullata e odor de cânfora para O.
usambarensis. Rizzini (1971) destaca o cheiro característico, resinoso e
agradável de O. porosa, resinoso de O. catharinensis, odor de sassafrás e odor
de canela para as variedades fisiológicas de O. odorífera. Usa o termo
“perfumada” para S. rubra, “aromática” para O. aciphylla, “fétida” para N.
cissiflora Nees (ali descrita como N. myriantha Meissn.). Indica odor
desagradável para N. megapotamica e não indica a presença de odores para
N. puberula, N reticulata e O. pulchella. Castiglioni (1962) refere-se ao odor de
N. megapotamica como desagradável e penetrante, a N. angustifolia como
pronunciado e agradável, N. lanceolata e N. cuspidata com odores pouco
pronunciados. Rohwer (1986) diante a dificuldade de diferenciar espécimes de
herbário de O. porosa e O. catharinensis, recebeu de Richer comunicação
pessoal indicando que o cheiro de O. porosa é “perfumado” e de O.
catharinensis quase insuportavelmente fétido. Neste contexto, necessário se
faz destacar as madeiras do gênero Aniba, como Aniba canelilla (HBK) Mez,
Aniba rosaeodora Ducke e Aniba duckei Kostermans, todas com cheiro muito
agradável, pronunciado, e cuja exploração econômica, especialmente das duas
últimas, para extração de compostos químicos para a indústria de perfumaria
quase levou-as à extinção (Bastos, 1943; Loureiro & Silva, 1968; Loureiro,
1976).
A disposição dos vasos arranjados diagonalmente, como observado no
lenho das árvores de N. lanceolata, N. megapotamica, O. porosa e Ocotea sp.
não deve ser considerada um caráter diagnóstico forte, como se observa em
Calophyllum brasiliense Cambess. e Caraipa densifolia Mart. (Clusiaceae) e
em Myrtaceae do gênero Eucalyptus (Coradin et al. 2009a), pois existiu grande
variação entre as amostras analisadas, e eventualmente dentro das mesmas
amostras. Dentre as madeiras de Lauraceas comerciais mais comuns no país,
Sextonia rubra parece ter este caráter mais bem definido (Kribs, 1959; Coradin
et al. 2009a,b), apesar de não ter sido relatado por Dechamps (1979) e Feldato
et. al. (1989).

80
A distinção macroscópica do parênquima axial no lenho das espécies de
Ocotea e Nectandra estudadas também se mostrou uma tarefa árdua e
questionável. Macroscopicamente, com auxílio de lupa de 10x, é muito difícil
distinguir a presença de parênquima axial e se este é paratraqueal escasso ou
vasicêntrico, sobretudo se for vasicêntrico pouco abundante ou delgado,
especialmente em razão da presença de células oleíferas/mucilaginosas
vinculadas ao parênquima axial, o que leva a uma indefinição sobre a real
abrangência deste em torno dos poros. No presente estudo, se pôde observar
sob lente de 10x, parênquima axial paratraqueal escasso ou vasicêntrico em
Nectandra lanceolata e N. megapotamica. Nas demais espécies, sob esta
ótica, o parênquima axial não se mostrou distinto. Kribs (1959) também indica
como indistinto sob lente de 10x o parênquima axial de Ocotea cymbarum, O.
bullata, O. glomerata, O. schomburgkiana, O. floribunda e O. usambarensis N.
globosa, N. sanguinea, N. turbacensis, e dificilmente visto sob lente para
Chlorocardium rodiei. Loureiro & Silva (1968) descrevem como praticamente
indistinto, mesmo com auxílio de lentes, o parênquima axial de O. cymbarum.
Feldato et. al. (1989) e Coradin et al. (2009a,b) afirmam ser indistinto sob lente
o parênquima axial de Sextonia rubra e Kribs (1959) indica para esta mesma
espécie como indistinto sem lentes. A chave de identificação de madeiras do
IPT, elaborada por Zenid & Ceccantini (2007) considera todas as espécies de
Ocotea e Nectandra com parênquima axial indistinto, mesmo sob lente.
No que tange aos caracteres anatômicos microscópicos do lenho, o gênero
Nectandra pôde ser assim caracterizado por León & Espinoza de Pernía
(2001), quando da análise de 18 espécies distintas: porosidade difusa, poros
sem padrão definido de disposição, pontoações raio-vasculares distintas das
pontoações inter-vasculares, com bordos reduzidos a aparentemente simples,
arredondadas ou alongadas horizontalmente e fibras septadas. As células
oleíferas/mucilaginosas estavam presentes em 83% das espécies e cristais nas
células dos raios em 55% das espécies, variando entre cristais aciculares,
alargados, tabletóides, isodiamétricos, fusiformes e areia de cristais. Similar
percentual fora encontrado para as espécies que apresentaram placas de
perfuração simples e escalariforme concomitantemente. Caracterização muito
semelhante foi feita para o gênero Ocotea por León (1999-2000) após
avaliação do lenho de 17 espécies, entre as quais 82% apresentaram células

81
oleíferas/mucilaginosas, 59% apresentaram cristais nas células dos raios e
35% placas de perfuração simples e escalariformes.
As características comuns encontradas no lenho das 6 espécies analisadas
no presente estudo foram: camadas de crescimento distintas pela presença de
fibras de paredes mais espessas e achatamento radial das fibras, porosidade
difusa, poros predominantemente solitários, geminados, em pequenas cadeias
radiais ou eventualmente em cachos, pontoações inter-vasculares alternas,
circulares ou ovaladas, areoladas, de abertura elíptica e inclusa, pontoações
raio-vasculares e parênquimo-vasculares distintas das pontoações inter-
vasculares, com bordos reduzidos a aparentemente simples, arredondadas ou
alongadas horizontalmente, fibras septadas, parênquima axial paratraqueal
escasso, raios frequentemente multisseriados, raramente unisseriados,
eventualmente fusionados, heterogêneos, com células procumbentes na região
central e eretas ou quadradas em fileiras marginais e a presença de células
oleíferas/mucilaginosas, vinculadas ao parênquima radial, axial ou entre as
fibras. Dentre as características do lenho que apresentaram variação entre
estas espécies, estão: vasos em arranjo diagonal, presença de placas de
perfuração simples e escalariformes, tamanho das pontoações inter-vasculares
- médias ou grandes, espessamentos na parede dos elementos de vaso,
tornando coalescentes a abertura das pontoações inter-vasculares
internamente, presença de parênquima vasicêntrico ou eventualmente
confluente e presença de cristais de oxalato de cálcio nas células do
parênquima radial.
Segundo Richter (1987), a presença de placas de perfuração
escalariformes, em “grau limitado”, seria o comumente esperado para as
espécies de Ocotea e Nectandra. No presente estudo, foram encontradas
placas de perfuração escalariformes nas espécies Nectandra lanceolata, N.
megapotamica e Ocotea puberula – 50% das espécies analisadas. De todas as
espécies analisadas na matriz binária (Tabela 8), 18 espécies de Nectandra
(78%), 15 espécies de Ocotea (44%), uma de Pleurothyrium e 1 de
Rhodostemonodaphne apresentaram placas escalariformes. Desta forma,
pode-se concluir que, embora a presença de placas de perfuração apenas
simples seja predominante entre as espécies deste grupo, a frequência de
espécies com placas simples e escalariformes, é maior do que sugeriu Richter

82
(1987), sendo mais comum em espécies do gênero Nectandra do que no
gênero Ocotea. A presença de placas de perfuração reticulada foi referida
apenas para N. oppositifolia (Marchiori et. al., 2010)
Os depósitos de extrativos em pontoações inter-vasculares simulando
ornamentações, como observadas em N. lanceolata devem ser analisados com
critério e cuidado para que não sejam confundidas com ornamentações
verdadeiras. Os depósitos de extrativos são eliminadas quando os cortes são
tratados com hipoclorito (Richter, comunicação pessoal). De acordo com León
& Espinoza de Pernía (2001), que observaram a presença de pontoações
ornamentadas apenas em Nectandra globosa, as ornamentações não são
comuns neste grupo de plantas, mas podem ocorrer ocasionalmente (Richter,
1987).
Espessamentos internos nos elementos de vaso foram observados no
presente estudo em Ocotea pulchella, O. porosa e Ocotea sp. e, em função
desses espessamentos, 2 ou 3 pontoações inter-vasculares encontram-se
coalescentes internamente numa câmara alongada. Situação similar foi
descrita para O. canaliculata (Botelho, 1951), Nectandra sp. e Ocotea austinii
(van der Slooten et. al., 1970), N. megapotamica (Machiori et. al., 2009), O.
diospyrifolia (Mattos et. al., 2003 ) e Sextonia rubra (Feldato et. al. 1989). Estes
espessamentos não foram mencionados por Milanez (1930) quando da
descrição do lenho de O. porosa nem por Hernández (1985), quando da
descrição do lenho de O. pulchella. Cabe salientar que não se tratam de
espessamentos espiralados, ou espiral-reticulados, como mencionado para O.
catharinensis por Silva & Medeiros (2000). Estes, segundo León & Espinoza de
Pernía (2001) são ausentes nos gêneros Ocotea e Nectandra respectivamente.
Richter (comunicação pessoal) entende que estes espessamentos não são
espiralados, que são comuns em várias espécies do grupo e ainda, que não
possuem nenhum valor diagnóstico, mas que devem ser mencionados nas
descrições das espécies.
Dentre as espécies analisadas, nenhuma apresentou gomas. A presença
de gomas foi observada apenas por León (1999-2000) em O. aciphylla e O.
catharinensis.
No que tange às pontoações raio-vasculares, todas as espécies analisadas
no presente estudos enquadram-se no tipo “II” proposto por Richter (1981b),

83
apresentando-se arredondadas ou alongadas no sentido radial ou axial, com
bordas distintas ou muito reduzidas a aparentemente simples, semelhante às
observadas nas espécies de Ocotea, Nectandra e outros gêneros da família
descritos na literatura (Castiglioni, 1962; Hernandez, 1985; Callado & Costa,
1997; León, 1999-2000; León & Espinoza de Pernía, 2000a; León & Espinoza
de Pernía, 2000b; León & Espinoza de Pernía, 2001; Oliveira et. al., 2001).
Nas espécies do presente estudo, o comprimento médio das fibras variou
entre 916 e 1019 μm e o diâmetro médio das fibras entre 23 e 27 μm. A
espessura das paredes de delgadas a grossas, variou conforme o
posicionamento das fibras, no lenho primaveril (mais delgadas) ou no lenho
outonal, região limítrofe a camada de crescimento (mais espessas). Não foram
observadas fibras de paredes muito grossas. As seis espécies analisadas
apresentaram fibras septadas. Segundo Richter (1987), a septação das fibras é
caráter comum aos gêneros Ocotea e Nectandra, o que também foi constatado
para a maioria das espécies analisadas por Hernandez (1985), León (1999-
2000) e León & Espinoza de Pernía (2001). Fibras não-septadas são descritas
apenas para Ocotea lancifolia (Hernandez, 1985), O. guianensis (Botelho,
1951), O. fragrantissima (Ochioni & Mattos Filho, 1947), O. usambarensis
(Kribs, 1959), e Rhodostemonodaphne grandis (León & Espinoza de Pernía,
2001), R. kunthiana (Stern, 1954), Chlorocardium rodiei (Kribs, 1959) e C.
venenosum (Kostermans et. al., 1969).
Analisado sob microscopia de luz, o parênquima axial paratraqueal é
comum às espécies de Ocotea e Nectandra, e segundo Richter (1987) pode
ser escasso, vasicêntrico, aliforme curto ou longo, unilateral e eventualmente
confluente. Observando as variações deste caráter constatadas no presente
estudo nas diferentes amostras de uma mesma espécie, não pôde ser
considerado, por si só, um caráter determinante para diagnóstico ou
diferenciação interespecífica. Da literatura analisada para a construção da
matriz binária, observou-se, no que tange ao parênquima axial, o seguinte
quadro: no gênero Nectandra, 73,9% apresentaram paratraqueal escasso,
91,3% paratraqueal vasicêntrico, 8,7% paratraqueal unilateral, 60,9%
paratraqueal aliforme, 56,5% paratraqueal confluente, nenhuma espécie
apresentou parênquima axial em faixas marginais e 8,7% apotraqueal difuso;
para as espécies do gênero Ocotea, estas proporções foram, respectivamente,

84
79,4%, 82,3%, 11,8%, 32,3%, 29,4%, 5,9% e 5,9%. Cabe destacar que a
presença de idioblastos entre as fibras gera confusão entre os autores na
designação destas como parênquima axial apotraqueal difuso.
Todas as espécies analisadas apresentaram raios heterogêneos, com
células procumbentes na região central e 1 a 4 (7) fileiras de células quadradas
ou eretas nas margens, similares as descrições para espécies dos gêneros
Ocotea e Nectandra encontradas na literatura (Kribs, 1959; Castiglioni, 1962;
Dechamps, 1979; Hernandez, 1985; León, 1999-2000; León & Espinoza de
Pernía, 2001; Oliveira et. al., 2001; Silva & Medeiros, 2003; Marchiori et. al.,
2009). Em média, raios mais frequentes em O. porosa - 10/mm linear - e
menos frequentes em O. puberula - 7/mm linear, e mais baixos e estreitos em
N. megapotamica - 289,5 μm de altura e 29,5 μm de largura (2-3 células) e
mais altos e largos em O. pulchella – 374,5 μm de altura e 48,8 μm de largura
(3-6 células).
A presença de cristais aciculares, alongados, rombóides e tabletóides nas
células procumbentes dos raios foi observada no presente estudo em
abundância em N. megapotamica, e aciculares, rombóides e ráfides em Ocotea
sp. Em N. lanceolata e O. puberula foram observados em algumas amostras,
com menor frequência e abundância, cristais aciculares, rombóides e
piramidais e em O. pulchella, areia de cristais, só observáveis sob MEV. Em O.
porosa, não foram observados cristais. Milanez (1930) também não registra a
presença de cristais em O. porosa. Cristais em N. megapotamica também são
registrados por Castiglioni (1962), Hernandez (1985) e Marchiori et. al. (2009).
As descrições de N. lanceolata de Castiglioni (1962), Hernandez (1985) e
Marchiori et. al. (2009) não mencionam a presença de cristais nas células do
raio desta espécie. Marchiori et. al. (2009) também observou cristais nas
células de O. puberula, mas Oliveira (2005), Hernandez (1985) e Dechamps
(1979) analisando a mesma espécie, não os observaram, em discrepância com
os resultados do presente trabalho. Para O. pulchella, também não havia sido
mencionada anteriormente a presença de cristais por Hernandez (1985) e
Marchiori et. al. (2009).
Segundo Richter (1980, 1987), a presença e a distribuição de compostos
inorgânicos, como sílica e cristais, é de grande importância taxonômica neste
grupo, indicando que pouco mais de 50% das espécies de lauráceas possuem

85
cristais de oxalato de cálcio. Na literatura, são relatados cristais nas células dos
raios nas outras espécies do grupo: Nectandra coriacea, N. cuspidata, N.
grandiflora, N. hihua, N. martinicensis, N. membranacea, N. reticulata, N.
turbacensis e Rhodostemonodaphne grandis (León & Espinoza de Pernía,
2001); Ocotea bofo, O. canaliculata, O. caracasana, O. catharinensis, O.
cernua, O. corymbosa, O. cymbarum, O. leucoxylon e Pleurothyrium
parviflorum (León, 1999-2000); O. indecora (Oliveira, 2005); O. odorifera (Silva,
2003) e O. glomerata (Dechamps, 1979).
A presença de raios fusionados foi eventual em todas as espécies
analisadas. Raios fusionados foram observados por Oliveira et. al. (2001) para
N. leucantha e N. rigida, por Castiglioni (1962) para N. falcifolia, N. lanceolata,
N. pichurim e N. saligna. Não foram observados raios agregados nas espécies
estudadas. Segundo Richter (1987) esta característica está restrita na família a
um pequeno grupo de espécies do gênero Cryptocarya mas no entanto,
Oliveira et. al. (2001) descrevem a presença deste caráter para N. rigida. Não
foi constatado, nas espécies ora analisadas, a presença de parênquima radial
disjuntivo. León (1999-2000) constatou a presença de parênquima radial
disjuntivo em todas as espécies de Ocotea analisadas, caráter este não
mencionado anteriormente para este grupo de lauráceas. León & Espinoza de
Pernía (2000a, b, 2001) registraram, a posteriori, esta característica para
espécies de outros gêneros desta família, como Aniba, Beilschmiedia e
Nectandra, respectivamente. Não há menção a presença deste caráter nos
estudos realizados por Stern (1954), Kribs (1959), Dechamps (1979), Richter
(1981b, 1987), Hernandez (1985), Silva & Medeiros (2000), Oliveira et. al.
(2001), Silva (2003), Oliveira (2005) Marchiori et. al. (2009) e Marchiori et. al.
(2010).
Constatou-se a presença de células oleíferas/mucilaginosas no lenho de
todas as espécies analisadas no presente estudo, o que é uma característica
comum aos membros desta família (Richter, 1977, 1980, 1981b; 1987). Dentre
estas, todas apresentaram conteúdo oleoso no interior dos idioblastos, a
exceção de Ocotea sp., cujos idioblastos apresentaram conteúdo mucilaginoso.
Merece destaque a grande quantidade dos idioblastos em O. pulchella, e o
tamanho relativamente grande destes em Ocotea sp., comumente maiores que
500 μm, muito similares aos encontrados por Richter (1981b) em O.

86
paucinervia. Oliveira (2005) não havia detectado a presença destas células em
O. puberula, em contradição ao presente estudo. Hernandez (1985) as
encontrou nesta mesma espécie, vinculadas apenas ao parênquima radial. A
ocorrência de idioblastos é citado para várias espécies de Ocotea e Nectranda
entretanto, Richter (1987) afirma que, dentro da família, pouco significado
taxonômico pode ser atribuído à localização, forma, tamanho e conteúdo
destas células.
As análises de fluorescência indicaram a presença de compostos auto-
fluorescentes sob filtros azul, vermelho e triplo nas células oleíferas das
espécies estudadas, a exceção de Ocotea sp., cujas células mucilaginosas não
se mostraram auto-fluorescentes. Não foi possível assim diferenciar estas
espécies através desta ferramenta. Não há dados disponíveis na literatura
acerca da utilização de fluorescência do xilema secundário para diferenciação
de espécies.
As análises estatísticas demonstraram que os caracteres quantitativos
do lenho apresentaram grande similaridade intra-específica e no entanto, se
mostraram úteis na diferenciação das espécies analisadas entre si, com um
nível de significância global (p) de 0,1% e R global de 0,251. As diferenças
observadas foram significativas nas relações entre as espécies (>0,05), exceto
nas relações estabelecidas entre O. porosa e Ocotea sp. entre si e entre estas
duas espécies e N. megapotamica e O. pulchella, possivelmente em função do
número amostral baixo (n=1). As dissimilaridades foram maiores entre O.
pulchella e N. megapotamica e O. pulchella e O. puberula. Os caracteres
anatômicos quantitativos do lenho que contribuíram na dissimilaridade entre as
espécies foram, em ordem de importância: comprimento das fibras,
comprimento dos elementos de vaso, altura dos raios e diâmetro tangencial
dos vasos (Tabela 6 e Fig. 16)
. Na comparação entre os gêneros, as análises estatísticas
demonstraram que os caracteres quantitativos do lenho apresentaram grande
similaridade intra-genérica: 97,95% para Ocotea e 97,87% para Nectandra (Fig.
17). Os fatores que mais contribuíram para esta similaridade foram
comprimento das fibras, comprimento dos elementos de vaso, altura dos raios,
diâmetro tangencial dos vasos e largura dos raios. A dissimilaridade entre os
gêneros foi de 2,15%, com nível de significância global de 2,1%. Também

87
evidenciou-se grande similaridade, acima de 97%, quando da comparação de
semelhança entre os locais de coleta, com nível de significância global (p) de
0,1% e R global de 0,082, exceto para a relação entre Concórdia e Tangará (p
= 25,3). Estas semelhanças levam em consideração que, a maior parte das
amostras é de apenas 3 espécies (O. puberula, N. megapotamica e N.
lanceolata) Ressalte-se que as condições climáticas das quatro localidades de
coleta - temperatura, precipitação anual e altitude - também eram muito
similares. Vários autores na literatura já utilizaram-se de caracteres anatômicos
qualitativos do lenho na relação entre táxons, tanto ao nível genérico quanto ao
nível específico (Baas, 1973; Baas & van der Graff, 1974; Noshiro & Baas,
2000; Liu & Noshiro, 2003; Lens et. al. 2007; Lens et. al., 2008; Boura et. al.
2011)
As análises de agrupamento por similaridade baseadas na matriz binária
de presença e ausência de caracteres anatômicos qualitativos do lenho,
permitiram a elaboração de algumas considerações acerca das relações de
proximidade observadas entre as espécies em função dos caracteres
anatômicos do lenho. São comparadas estas proximidades/agregações com as
relações estabelecidas entre estas espécies, baseadas em caracteres
morfológicos utilizados na definição/delimitação das mesmas para com outras
entidades taxonômicas próximas, explicitados por Rohwer após a revisão
taxonômica dos gêneros Ocotea (Rohwer, 1986) e Nectandra (Rohwer, 1993a).
As quatro descrições de N. megapotamica (presente trabalho;
Castiglioni,1962; Hernandez, 1985 e Machiori et al. 2009) apresentaram-se
relativamente distantes no dendrograma de similaridade, e o melhor índice de
similaridade nas relações entre as descrições foi de 82% na relação entre a
descrição de Castiglioni (1962) e Hernandez (1985), ambas analisando lenho
oriundo da Argentina. A similaridade na relação obtida entre a descrição do
presente trabalho, com coleta na região oeste de planalto serrado de Santa
Catarina e a de Machiori et al. 2009, com material oriundo do norte do Rio
Grande do Sul foi de apenas 75%. A espécie mais próxima de N.
megapotamica é N. angustifolia (Rowher, 1993a). As duas descrições de N.
angustifolia constantes da matriz binária (Castiglioni,1962 e Hernandez, 1985)
obtiveram índice de similaridade entre si de 80%. O maior percentual de

88
similaridade na relação entre esta espécie e N. megapotamica foi de 89,4%,
ambas descritas por Hernandez (1985). Pelas análises dos dados da matriz, N.
megapotamica apresentou-se mais próxima de N. lanceolata, na relação entre
as descrições (ambas) do presente trabalho, com índice de similaridade de
92% (Fig. 22).
Nectandra reticulata é a entidade central de um complexo de espécies
que inclui, entre outras, N. laurel e N. oppositifolia (Rowher, 1993a). No
entanto, com base nos dados oriundos da matriz binária, as 2 descrições desta
espécie apresentaram um índice de similaridade entre si de 65%, mantendo-se
relativamente afastadas no dendrograma (Fig. 19). De igual maneira,
mantiveram-se distantes as duas descrições (Oliveira, 2005 e Machiori et al.
2010) de N. oppositifolia mantendo entre si apenas 52% de similaridade. A
melhor relação de semelhança dentro deste grupo de espécies tidas como
próximas foi de 85% de similaridade (Fig. 21) - se deu entre N. reticulata
(Oliveira et. al., 2001) e N. oppositifolia (Oliveira, 2005), seguida da relação
entre N. reticulata e N. laurel (León & Espinoza de Pernía, 2001), com 80,7%
de similaridade. A descrição de N. reticulata de León & Espinoza de Pernía,
(2001) apresentou grande similaridade (95,6%) com a descrição de N.
martinicensis, realizada pelo mesmo autor (Fig. 22).
Nectandra membranacea possui maior proximidade com N. cuspidata
(Rowher, 1993a). As duas descrições de N. membranacea (León & Espinoza
de Pernía, 2001 e Oliveira et. al., 2005) mantiveram entre si similaridade de
apenas 44,4%. Entre as quatro descrições de N. cuspidata (León & Espinoza
de Pernía, 2001; Castiglioni ,1962; Hernandez ,1985 e Dechamps, 1979), a
maior similaridade foi de 88,8% entre as descrições de Castiglioni (1962) e
Hernandez (1985), ambas analisando lenho oriundo da Argentina, ainda sob o
nome já sinonimizado de N. pichurim. A maior similaridade evidenciada na
relação entre N. membranacea e N. cuspidata foi de 84%, nas amostras
descritas por León & Espinoza de Pernía (2001). Na análise da matriz binária,
N. membranacea apresentou maior relação de similaridade (94,4%) com N.
oppositifolia (Fig. 21) nas descrições de Oliveira (2005). Já as descrições
anatômicas de N. cuspidata apresentaram maior proximidade (96%) com O.
aciphylla, N. globosa e N. hihua nas descrições de León & Espinoza de Pernía

89
(2001) e León (1999-2000). A relação de similaridade também foi alta (ca. 95%
- Fig.22) entre N. cuspidata, O. aciphylla e N. globosa descritas por Dechamps
(1979).
As duas descrições de N. coriacea (Stern, 1954 e León & Espinoza de
Pernía, 2001) obtiveram índice de similaridade de 75%, mantendo-se
relativamente distantes entre si. A espécie mais próxima desta, segundo
Rohwer (1993a) é N. salicifolia (Humboldt, Bonpland & Kunth) Nees, que não
foi analisada na presente matriz binária. Pelos dados da matriz, a relação de
maior similaridade de N. coriacea (86,9%) foi mantida com N. turbacensis
(Fig.22) ambas descrições de León & Espinoza de Pernía (2001).
As três descrições de N. turbacensis, duas das quais obtidas de León &
Espinoza de Pernía, (2001), e outra de Kribs (1959) obtiveram índice de
similaridade entre si de apenas 73,9%. As maiores similaridades desta espécie
se deram com N. amazonum (91,3%), também descrita por León & Espinoza
de Pernía, (2001) e O. aciphylla (91,6%) na descrição de León (1999-2000). As
descrições de Kribs (1959) para N. turbacensis, O. cymbarum, N. globosa e N.
sanguinea.foram idênticas (Fig. 22). Segundo Rohwer (1993a), a delimitação e
N. turbacensis se dá por caracteres morfológicos que em sua maioria não são
constantes e as espécies próximas a este táxon não possuem dados
anatômicos do lenho disponíveis e, portanto, não foram incluídas na matriz
binária de presença e ausência.
As espécies mais próximas de N. cissiflora, quais sejam: N. viburnoides
Meissn., N. pearcei Mez e N. subbullata Rohwer não tiveram suas descrições
analisadas na presente matriz binária. A melhor relação de similaridade de N.
cissiflora foi estabelecida com N. surinamensis (95,4% - Fig. 22), ambas
descrições de León & Espinoza de Pernía, (2001). Esta última foi sugerida por
Rohwer (1993a) como sinônimo de Ocotea guianensis.
Nectandra nitidula é uma espécie bem definida, e tem maior
aproximação com N. grandiflora (Rohwer, 1993a), A similaridade obtida entre
estas dias espécies, com base nas descrições anatômicas do lenho obtidas
respectivamente em Pinho (1968) e León & Espinoza de Pernía, (2001) foi de
76%. A descrição anatômica do lenho de N. nitidula foi idêntica (100% de

90
similaridade – Fig. 22) a de Ocotea odorifera realizada por Mattos Fillho (1960).
Já N. grandiflora também obteve similaridade de 100% (Fig. 22) com O. bofo,
descrita por León (1999-2000), e 95,2% de similaridade com O. amplissima, O.
glomerata e O. caracasana, descritas pelo mesmo autor.
Nectandra leucantha é muito próxima a N. hihua (Rohwer , 1993a). A
similaridade entre estas duas espécies, nas descrições anatômicas do lenho
feitas por Oliveira et al. (2001) e León & Espinoza de Pernía, (2001) foi de
apenas 51,8%. A maior relação de similaridade de N. leucantha foi com N.
oppositifolia (88,8%), N. membranacea e O. puberula (84,2% - Fig. 21), nas
descrições de Oliveira (2005). Já N. hihua obteve maior similaridade com N.
cuspidata (96% - Fig. 22) também descrita por León & Espinoza de Pernía,
(2001).
Nectandra puberula é muito próxima a N. lanceolata, N. venulosa
Meissn., N. barbellata Coe-Teix. e N. warmingii Meissn. (Rohwer, 1993a). Estas
três últimas não foram analisadas na matriz binária de presença e ausência,
impossibilitando comparações entre os lenhos. As duas descrições de N.
puberula analisadas (León & Espinoza de Pernía, 2001 e Oliveira et al. 2001)
obtiveram apenas 50% de similaridade, restando distantes no dendrograma
(Fig. 21). Já as quatro descrições de N. lanceolata analisadas (presente
estudo; Castiglioni,1962; Hernandez, 1985 e Machiori et al., 2009)
apresentaram melhor índice de similaridade (80%) entre as descrições do
presente estudo e de Machiori et al. (2009), cujas análises se deram de
material oriundo de região muito próxima – oeste de Santa Catarina e Norte do
Rio Grande do Sul. Entre N. puberula e N. lanceolata, a maior similaridade foi
de apenas 68%. As maiores similaridades obtidas para as descrições de N.
puberula foram a análise oriunda de León & Espinoza de Pernía (2001) com O.
amplissima e O. glometara (90%), nas descrições de León (1999-2000). As
maiores similaridades de N. lanceolata foram detectadas entre a descrição de
Hernandez (1985) e a descrição de O. pulchella do mesmo autor (93,7% - Fig.
23). Segundo Rohwer (1993a), N. lanceolata também é muito próxima a N.
weddelli Meissn., cuja descrição anatômica do lenho não esteve disponível na
literatura, impossibilitando comparações.

91
Nectandra globosa, espécie cuja delimitação morfológica é difícil
(Rohwer, 1993a), tem proximidades com N. baccans (Meissn.) Mez, N.
matogrossensis Coe-Teix., N. aurea Rohwer, N. riparia Rohwer e N. ruforamula
Rohwer, cujas descrições anatômicas do lenho não se encontraram disponíveis
na literatura para comparações. As quatro descrições disponíveis – Stern
(1954), Kribs (1959), Dechamps (1979) e León & Espinoza de Pernía, (2001)
apresentaram maior similaridade entre si de ca de 76%.. Kribs (1959) forneceu
a N. globosa a mesma descrição que a fornecida para N. sanguinea, N.
turbacensis e O. cymbarum. Excluindo-se esta similaridade de 100% já
explicitada anteriormente, as maiores agregações foram na relação com O.
aciphylla 96% nas descrições de León & Espinoza de Pernía (2001) e León
(1999-2000) e de 88 a 92% de similaridade nas descrições de León & Espinoza
de Pernía, (2001) com N. acutifolia, N. amazonum, N. cuspidata e N. laurel,
todas descrições do mesmo autor (Fig. 22).
Nectandra acutifolia possui proximidade com N. acuminata (Nees)
Maacbride, N. canescens Nees, N. crassiloba Rohwer, N. fulva Rohwer e N.
kruggi Mez (Rohwer, 1993a), espécies cujas descrições anatômicas do lenho
não estavam disponíveis para comparação. Entre as duas descrições
analisadas (León & Espinoza de Pernía, 2001 e Hernandez, 1985) a
similaridade foi de apenas 62,5%. As maiores similaridades foram encontradas
na relação da descrição de León & Espinoza de Pernía (2001) com O. aciphylla
(91,6%) descrita por León (1999-2000) e na descrição de Hernandez (1985)
com O. puberula, O. diopspyrifolia e N. cuspidata (Ca. 94%) em descrições do
mesmo autor (Fig. 23).
As duas descrições de O. cymbarum (León, 1999-2000 e Kribs, 1959)
apresentaram entre si similaridade de 61,5%, dispondo-se distantes no
dendrograma de similaridade. A descrição de Kribs para O. cymbarum foi
idêntica a descrição fornecida para N. globosa. N. sanguinea e N. turbacensis
(Fig. 22). A descrição de León (1999-2000) agregou-se com O. canaliculata e
O. cernua também descritas pelo mesmo autor (Fig. 22). Na relação de O.
cymbarum com O. fragrantissima, a similaridade foi de apenas 72,7%, nas
descrições de Ochioni & Mattos Filho (1947) e Kribs (1959).

92
As espécies O. elegans, O. odorifera e O. indecora pertencem a um
complexo de espécies em torno de O. indecora (Rohwer, 1986; Assis, 2009).
Na matriz binária de caracteres qualitativos foram analisadas as descrições
anatômicas do lenho de O. indecora e O. elegans, ambas oriundas de Oliveira,
(2005) e três descrições de O. odorifera (Mattos Filho, 1960; Silva, 2003 e
Oliveira, 2005). A similaridade entre O. indecora e O. elegans foi de 81%, e
estas mantiveram-se relativamente próximas, juntamente com outras
descrições de Oliveira (2005) para O. aciphylla e O. divaricata (Fig. 21). A
descrição de Oliveira (2005) obteve maior similaridade (90%) com N. puberula,
(Fig. 21) na descrição de Oliveira et al. (2001). Entre O. indecora e O. odorifera
a maior similaridade foi de 85,7%, e entre O. elegans e O. odorifera foi de 77%.
As três descrições O. odorifera mantiveram-se relativamente distantes entre si
no dendrograma e a maior similaridade entre as três descrições - 76% - foi
obtida entre as descrições de Silva (2003) e Mattos Filho (1960). A descrição
de O. odorifera de Mattos Filho (1960) agregou-se com N. nitidula (100% de
similaridade – Fig. 22) descrita por Pinho (1968) e a descrição de Silva (2003)
apresentou maior similaridade (87,5%) com as descrições do presente estudo
de N. lanceolata e N. megapotamica.
Segundo Rohwer (1986) O.porosa é muito similar O. catharinensis,
quando herborizadas. Mas recebeu de Richter rica diferenciação pela madeira:
enquanto a primeira apresenta um perfume muito agradável, a segunda
apresenta um odor fétido quase insuportável. Na análise de similaridade da
anatomia do lenho pela matriz binária, as duas descrições de O. porosa (dados
do presente estudo e de Milanez (1930) apresentaram-se relativamente
próximas no dendrograma, com similaridade de 90% (Fig. 22). A descrição de
O. porosa, considerando-se os dados do presente estudo, se apresentou muito
similar (95%) a outra espécie descrita no presente estudo, O. pulchella (Fig.
22). As duas descrições de O. catharinensis (León, 1999-2000 e Silva &
Medeiros, 2000) apresentaram-se distantes no dendrograma, com similaridade
de apenas 61% (Fig. 22 e 23). A descrição de Silva & Medeiros (2000)
apresentou-se muito próxima da descrição de Nectandra sp. e O. austinii (van
der Slooten et. al. 1970) e a descrição de León (1999-2000) muito próxima de
O. odorifera descrita por Silva (2005).

93
Ocotea aciphylla encabeça um outro complexo de espécies (Rohwer,
1986), das quais apenas O. amplissima encontra-se representada com a
descrição anatômica de caracteres qualitativos do lenho na matriz binária. As
quatro descrições de O. aciphylla (León, 1999-2000; Oliveira, 2005; Botelho,
1951 e Dechamps, 1979) apresentaram-se distantes no dendrograma, e a
maior similaridade (70%) foi obtida entre as descrições de León (1999-2000) e
Dechamps (1979). A forma descrita por Oliveira (2005) manteve-se muito
próxima de outras três espécies distintas descritas pelo mesmo autor (Fig. 21).
A descrição de León (1999-2000) foi a que mais se aproximou de O.
amplissima, descrita pelo mesmo autor, apresentando 83% de similaridade. O.
amplissima apresentou 100% de similaridade com O. glomerata, também
descrita por León (Fig. 22).
Ocotea guianensis encabeça outro complexo de espécies, que inclui,
entre outras, O. glomerata, e O. canaliculata (Rowher, 1986). As três
descrições de O. guianensis (Botelho, 1951; Dechamps, 1979 e León, 1999-
2000) apresentaram-se distantes no dendrograma, e a maior similaridade
(78%) foi obtida entre as descrições de Dechamps (1979) e León (1999-2000).
As duas descrições de O. canaliculata (León, 1999-2000 e Botelho, 1951)
apresentaram apenas 59% de similaridade. As três descrições de O. glomerata
(Kribs, 1959; Dechamps, 1979 e León, 1999-2000) também apresentaram-se
distantes no dendrograma, e a maior similaridade (75%) foi obtida entre as
descrições de Kribs (1959) e León (1999-2000). Entre estas espécies, a maior
proximidade por similaridade (ca. 91%) se deu entre O. glomerata e O.
guianensis, ambas descrições de León (1999-2000).
Ocotea austinii pertence ao grupo de O. insularis (Meiss.) Mez, que não
teve outros representantes analisados na matriz binária. A descrição de van der
Slooten et. al., (1970) apresentou-se próxima (90% de similaridade) a
Nectandra sp., também descrita pelo mesmo autor (Fig. 23).
As duas descrições de O. schomburgkiana (Kribs, 1959 e Dechamps,
1979) apresentaram-se distantes no dendrograma, com apenas 70% de
similaridade. A descrição de Kribs (1959) fora idêntica a de O. floribunda, e a
descrição de Dechamps (1979) apresentou cerca de 85% de similaridade com

94
O. guianensis, descrita pelo mesmo autor. As duas descrições de O. floribunda
(Kribs, 1959 e Dechamps, 1979) apresentaram 84% de similaridade. Esta
espécie, segundo Rohwer (1986), encabeça um complexo de espécies que
inclui O. lancifolia. A descrição de Hernandez (1985) para O. lancifolia
apresentou apenas 57% de similaridade com as descrições de O. floribunda.
Ocotea pulchella pertence a um complexo de espécies, entre elas O.
tristis (Nees) Mez, O. semicompleta (Nees) Mez, O. montana (Meissn.) Mez e
O. pomaderroides (Meissn.) Mez, nenhuma das quais fora analisada na matriz
binária. As três descrições de O. pulchella (dados do presente estudo;
Hernandez, 1985 e Machiori et al. 2009) apresentaram-se distantes no
dendrograma (Fig. 22 e 23), e a maior similaridade obtida entre estas
descrições foi de 68% comparando-se os dados do presente estudo com os de
Hernandez (1985) e de Machiori et al. (2009). A descrição de Hernandez
(1985) para O. pulchella apresentou-se muito similar (93,7%) a N. lanceolata
descrita pelo mesmo autor.
Ocotea puberula tem como espécie mais próxima O. schwackeana Mez
(Rowher, 1986), ausente da análise na matriz binária. Duas, dentre as seis
descrições de O. puberula (dados do presente estudo; Dechamps, 1979;
Hernandez, 1985; León, 1999-2000; Oliveira, 2005 e Machiori et al. 2009)
apresentaram-se muito próximas entre si (95% de similaridade): dados do
presente estudo e Machiori et al. 2009, ambas coletadas em regiões muito
próximas geograficamente (Fig. 22) A única diferença foi a constatação da
presença de raios fusionados no presente estudo, não mencionados por
Machiori et al. 2009 em sua descrição. As demais descrições de O. puberula
mostraram-se distantes no dendrograma. A descrição de Dechamps (1979)
apresentou-se próxima a outras duas descrições do mesmo autor – O.
guianensis e O. schomburgkiana (Fig. 22). A descrição de Hernandez (1985)
agregou-se próxima a N. acutifolia, também descrita pelo mesmo autor (Fig.
23) e a descrição de Oliveira (2005) muito próxima a N. membranacea e N.
oppositifolia, ambas descritas pelo mesmo autor (Fig. 21). A descrição de León
(1999-2000) apresentou grande similaridade (ca. 95%) com O. fulvescens,
descrita pelo mesmo autor.

95
Ocotea adenotrachelium (León, 1999-2000) é espécie próxima de O.
albopunctulata Mez (Rohwer, 1986), ausente da matriz binária, para fins de
comparação da anatomia do lenho. O. adenotrachelium posicionou-se próxima
a duas espécies de Nectandra (Fig. 22) descritas por León & Espinoza de
Pernía (2001). Ocotea corymbosa pertence ao complexo de espécies que inclui
O. urbaniana Mez, O. polyantha (Nees) Mez e O. confertiflora (Meissn.) Mez
(Rohwer, 1986), também ausentes da matriz binária. A amostra analisada por
León (1999-2000) aproximou-se de O. leucoxylon, analisada pelo mesmo autor,
com cerca de 90% de similaridade.
Ocotea cernua encabeça um dos maiores complexos “informais” de
espécies definidos por Rohwer (1986). Entre as espécies deste grupo está O.
divaricata, cuja descrição anatômica do lenho realizada por oliveira (2005)
encontra-se na matriz binária. As duas descrições de O. cernua, ambas
oriundas de León (1999-2000) apresentaram-se próximas no dendrograma,
com ca. de 92% de similaridade (Fig. 22) e muito distantes de O. divaricata,
com apenas 55% de similaridade.
Ocotea petalanthera e O. bofo não tiveram suas relações de
proximidade estabelecidas por Rohwer (1986), que analisou amostras com
flores incompletas. Ocotea bullata e O. usambarensis, espécies do velho
mundo, também não tiveram suas relações de proximidade mencionadas por
Rohwer (1986). Nos dendrogramas gerados a partir dos dados da matriz
binária, todas elas aparecem próximas a outras espécies descritas pelos
mesmos autores
As três descrições de O. diospyrifolia (Hernandez, 1985 e Mattos et. al.,
2003) apresentaram-se distantes no dendrograma, e a maior similaridade
(83%) foi obtida entre as duas descrições de Hernandez (1985) para a mesma
espécie.
Segundo Rowher (1986), Ocotea rubra apresentava-se bastante isolada
entre as espécies deste gênero, o que inclusive favoreceu sua migração para o
novo gênero Sextonia (van der werff, 1997). A madeira desta espécie é
facilmente diferenciável de outras Lauraceas, especialmente pela cor, róseo-
avermelhada, incomum no grupo (Fedalto et. al.,1989, Coradin et. al., 2009).

96
Tal isolamento entre as Ocotea também fora observado na análise da matriz
binária, ficando bastante evidente nos dendrogramas (Fig. 20 e 22) As três
descrições de Sextonia rubra (Kribs, 1959, Dechamps, 1979 e Fedalto et.
al.,1989) apresentaram menor similaridade com relação as outras espécies
analisadas, e duas delas (Kribs, 1959 e Dechamps, 1979) mantiveram-se
próximas entre si (76%), e de Chlorocardium rodiei – 88%., ambas descritas
por Dechamps (1979). A descrição de Fedalto et. al.,1989 manteve-se
relativamente próxima a Chlorocardium venenosum.e Rhodostemonodaphne
grandis (Fig. 20)
As duas espécies de Pleurothyrium presentes na análise da matriz
binária, P. palmanum e P. parviflorum não pertencem a um grupo de espécies
próximas (Rohwer, 1986). Na análise de similaridade, com base nas
características anatômicas do lenho, mantiveram entre si apenas 68% de
similaridade. A melhor relação de agregação de P. palmanum foi de 88,8%
(Fig. 21) com N. globosa, em descrição do mesmo autor (Stern, 1954). Já P.
parviflorum obteve 86% de similaridade com N. grandiflora, em descrição de
León & Espinoza de Pernía (2001) e O. bofo em descrição de León (1999-
2000).
As espécies de Rhodostemonodaphne, R. grandis e R. kunthiana,
também não mantém entre si relacionamento próximo (Rohwer, 1986). Na
matriz de similaridade, obtiveram entre si 66% de similaridade. As duas
descrições de R. grandis (León & Espinoza de Pernía, 2001 e Dechamps,
1979) obtiveram entre si 70% de similaridade, apresentando-se distantes no
dendrograma (Fig. 20 e 22) e relativamente aproximadas das espécies de
Chlorocardium.
As espécies do gênero Chlorocardium. C. rodiei e C. venenosum, até
recentemente pertenciam ao gênero Ocotea, ao grupo de O. dendrodaphne
Mez (Rohwer, 1986) e foram rearranjadas para este novo gênero (Rowher et al,
1991). Dentro deste grupo de Ocotea, mantinham relações de proximidade com
O. fragrantissima, O cymbarum, O staminea (Griseb.) Mez, O. dendrodaphne
Mez e O. veraguensis (Meissn.) Mez, (Rohwer, 1986) estas três últimas
ausentes da matriz de similaridade, impossibilitando comparações. As relações

97
de similaridade entre as duas descrições de C. rodiei (Kribs, 1959 e Dechamps,
1979) foi de 68%, mantendo-as relativamente próximas no dendrograma (Fig.
22). As espécies mais próximas de Chlorocardium foram Sextonia rubra –
88,8% de similaridade, especialmente a descrita por Dechamps (1979) Na
relação de C. rodiei com as espécies próximas no grupo Ocotea, a maior
similaridade (66,6%) foi obtida com O. cymbarum, em descrição de (León
1999-2000). C. venenosum obteve 65% de similaridade (Fig. 20) com R.
grandis descrita por León & Espinoza de Pernía (2001).
A análise SIMPER dos dados da matriz binária ao nível de gênero
(Tabela 10), demonstrou que os caracteres qualitativos do lenho das espécies
do presente trabalho e daquelas oriundas da literatura apresentaram a
similaridade intra-genérica de 80,09% para Nectandra e 79,52% para Ocotea.
Para Nectandra, os fatores que mais contribuíram para esta similaridade foram,
em ordem de importância: porosidade difusa, poros solitários, poros múltiplos
radiais, poros geminados, pontoações intervasculares alternas, placa de
perfuração simples, fibras septadas raios multisseriados e raios unisseriados.
Em Ocotea, os fatores que contribuíram para a similaridade foram, em ordem
de importância: porosidade difusa, poros solitários, poros múltiplos radiais,
poros geminados, raios multisseriados, raios heterogêneos, pontoações
intervasculares alternas, placa de perfuração simples, fibras septadas e raios
unisseriados. Observe-se que os fatores que contribuíram para a similaridade
das espécies dentro dos gêneros Ocotea e Nectandra foram os mesmos, com
alteração na ordem de importância dada aos caracteres dos raios, que se
apresentaram de forma mais uniforme nas espécies no que tange a
multisseriação e a heterogeneidade. A dissimilaridade entre os gêneros Ocotea
e Nectandra foi de 20,57 % e os fatores que mais contribuíram para estas
dissimilaridades foram: placas de perfuração escalariformes, raios
homogêneos, pontoações inter-vasculares poligonais, células
oleíferas/mucilaginosas ao parênquima axial e às fibras, parênquima axial
paratraqueal aliforme, poros em cachos, presença de tiloses, paredes
disjuntivas nas células dos raios e parênquima axial paratraqueal confluente.
Ressalte-se que o teste ANOSIN demonstrou que as diferenças observadas
entre os gêneros não foram significativas (p = 18,5).

98
As relações inter-genéricas que se mostraram estatisticamente
significativas a partir dos dados qualitativos foram entre os gêneros: Nectandra
e Rhodostemonodaphne, Nectandra e Sextonia, Nectandra e Chlorocardium e
ainda entre os gêneros Ocotea e Rhodostemonodaphne, Ocotea e Sextonia e
Ocotea e Chlorocardium. As relações de similaridade e dissimilaridade de
Pleurothyrium com os outros gêneros analisados não se mostraram
significativas. O escalonamento multidimensional (MDS) dessa relação entre os
gêneros Ocotea, Nectandra, Chlorocardium, Sextonia, Rhodostemonodaphne e
Pleurothyrium pode ser observado na Fig. 24.
Destaque deve ser dado aos caracteres que mais contribuíram na
dissililaridade de 26,9% (Tabela 10) do gênero Chlorocardium para com o
gênero Ocotea: células oleíferas/mucilaginosas associadas aos raios, placa de
perfuração escalariforme, elementos de vaso coronados e fibras não-septadas.
Estes caracteres, aliados a outras características anatômicas do lenho que não
foram inclusas na matriz binária, como as pontoações em tamanho muito
pequeno e as pontoações parenquimo-vasculares unilateralmente compostas,
foram justamente as características anatômicas que auxiliaram Rohwer et. al.
(1991) quando da propositura do gênero Chlorocardium, retirando as duas
espécies (C. rodiei e C. venenosum) do gênero Ocotea.
As análises estatísticas dos caracteres anatômicos qualitativos avaliados
permitem concluir que as : (i) todas as espécies analisadas apresentaram ao
menos 50% de similaridade; (ii) não existiram grupos bem definidos de
espécies similares exclusivamente dos gêneros Ocotea ou do gênero
Nectandra, com base nestas características anatômicas; (iii) distanciaram-se
espécies iguais, cujas descrições anatômicas são oriundas de diferentes fontes
da literatura; (iv) agregaram-se espécies distintas cujas descrições anatômicas
são oriundas da mesma fonte na literatura;
Para testar a hipótese de que as relações estabelecidas entre as
espécies estavam sendo influenciadas pela excssiva uniformidade na descrição
das espécies pelo mesmo autor e falta de padronização das descrições entre
os autores destas, realizou-se a análise SIMPER de similaridade dos dados da
tabela binária de presença e ausência entre as descrições anatômicas dos
lenhos incluindo-se como fator de agregação a fonte dos dados, i.e. autor (es)
da informação anatômica ora prestada. Os resultados das similaridades “intra-

99
literárias” foram: análises do presente estudo – 90,94%; León & Espinoza de
Pernía (2001) – 88,64%; Oliveira et. al. (2001) – 87,92%; León (1999-2000) –
87,14%; Oliveira (2005) – 87,79%; Castiglioni (1962) – 84,88%; Hernandez
(1985) – 91,97%; Kribs (1959) – 87,27%; Botelho (1951) – 79,58%; Dechamps
(1979) – 87,80%; Stern (1954) – 87,01%; van der Slooten et. al., 1970 –
94,44%; Marchiori et al. (2009) – 86,27%. Os demais autores apresentaram a
descrição de apenas 1 espécie. O escalonamento multidimensional MDS da
relação estre as descrições e os autores fontes dessas descrições pode ser
observado na Fig. 25. A análise visual desta figura deixou claro que houve
grande agregação entre as espécies, mesmo de diferentes gêneros, que foram
analisadas pelo mesmo(s) autor(es). Alguns dos caracteres analisados na
matriz evidenciam esta questão, como p.ex., a presença de paredes disjuntivas
nas células dos raios, que só foram descritas para as espécies analisadas por
León (2000) e León & Espinoza de Pernía (2001), caráter este cuja observação
é muito difícil, e a declinação pela presença ou ausência do caráter pode ser
uma questão de interpretação (Richter, comunicação pessoal). Outras
características podem ter sido negligenciadas nas descrições da anatomia do
lenho ao longo do tempo, já que algumas análises são anteriores a década de
70. Entre estas características, a presença de cristais nas células dos raios,
cuja importância na taxonomia das espécies é mais recente (Richter, 1981a).
Cabe observar que as amostras do lenho analisadas no presente trabalho
aparecem agrupadas, com grande similaridade, o que reforça a tese de
agregação em função da homogeneidade na análise do pesquisador.
Também deve ser levado em consideração que, para a maioria das
espécies analisadas - 57,1%, as descrições anatômicas do lenho foram
baseadas em apenas uma amostra de madeira, 32,4% baseadas em duas a
quatro amostras e apenas 10,5% baseadas cinco ou mais amostras do lenho.
Ainda, não pode ser desprezada a possibilidade de existência de erros
nas identificações taxonômicas das espécies cujos dados são oriundos de
literatura que, na grande maioria, não mencionam os registros dos vouchers ou
números de registro das amostras nas xilotecas, impedindo a checagem das
identificações e possíveis re-análises por taxonomistas.

100
5 – Considerações finais
Os lenhos das árvores das espécies de Ocotea e Nectandra ora
analisados, bem como os dados disponíveis na literatura, apresentam alto grau
de similaridade.
Não foram encontrados caracteres anatômicos qualitativos e
quantitativos diagnósticos do xilema secundário que permitissem a distinção
entre os gêneros Ocotea e Nectandra.
A análise macroscópica de caracteres anatômicos do lenho de espécies
de Ocotea e Nectandra não permite a clara distinção das espécies. Caracteres
organolépticos, especialmente cor e odores, devem ser usados em conjunto
com os caracteres anatômicos, para auxiliar nesta distinção. Ainda assim, há
um certo grau de subjetividade nestes caracteres, que podem facilmente levar
ao cometimento de erros.
O conjunto de caracteres anatômicos microscópicos do lenho podem
auxiliar na distinção das espécies; entre os mais relevantes, estão: a presença
de placas de perfuração escalariformes, o tipo de parênquima axial
paratraqueal, a presença, abundância e os tipos de cristais nas células dos
raios e a presença e abundância de células oleíferas ou mucilaginosas e sua
vinculação aos parênquimas axial, radial ou disposição entre as fibras.
Caracteres anatômicos quantitativos do lenho, como diâmetro tangencial
e comprimento de elementos de vasos, altura dos raios e comprimento das
fibras podem ser úteis na distinção/segregação das espécies mas não
permitiram a segregação dos gêneros Ocotea e Nectandra.
O conteúdo das células oleíferas das espécies analisadas se mostrou
auto-fluorescente sob filtros de luz verde, vermelho e triplo e não fluorecente
nas células mucilaginosas.
A falta de padronização nas 114 descrições de caracteres anatômicos
qualitativos do lenho oriundas de diferentes fontes literárias das 23 espécies de
Nectandra, 34 espécies de Ocotea, duas de Chlorocardium, duas de
Pleurothyrium, duas de Rhodostemonodaphne e uma de Sextonia pode ter
levado a distorções nas análises de similaridades e dissimilaridades destas
espécies entre si.

101
6 – Referências Bibliográficas
Araújo, P. A. M. & Mattos Filho, A. de. 1980. A importância da anatomia do
lenho para a comercialização da madeira. Rodriguésia 32 (53): 315-318.
Assis, L. C. S. 2009. Sistemática e filosofia: filogenia do complexo Ocotea
e revisão do grupo Ocotea indecora (Lauraceae). Tese de Doutorado.
Universidade de São Paulo, São Paulo. 268p.
Assis, L. C. S. & Mello-Silva, R. de. 2009. Three new species of Ocotea
(Lauraceae) from the Brazilian atlantic forest. Rodriguésia 60 (3): 641-649.
Assis, L. C. S. & Mello-Silva, R. de. 2010a. Two new species of Ocotea
(Lauraceae) from the Brazilian restinga. Novon 20 (2): 123-128.
Assis, L. C. S. & Mello-Silva, R. de. 2010b. Three new species of Ocotea
(Lauraceae) from the campus rupestres of Brazil. Brittonia 62: 86-94.
Baas, P. 1973. The wood anatomical range Ilex (Aquifoliaceae), and its
ecological and phylogenetic significance. Blumea 21: 193 – 258.
Baas, P. & van der Graff, N. A. 1974. Wood structure in relation to latitudinal
and altitudinal distribuition. Iawa Bulletin 4: 3-5.
Baitello, J. B. 2001. Novas espécies de Lauraceae para a flora brasileira. . Acta
Bot. Bras. 15 (3): 445-450.
Baitello, J. B., Hernández, F. G. L., Moraes, P. L. R.de, Esteves, R. &
Marcovino, J. R. 2003. Lauraceae. In: Wanderley, M. G. L., Shepherd, G.
J., Giulietti, A. M. & Melhen, T. S. Flora Fanerogâmica do Estado de São
Paulo. São Paulo: FAPESP. v. 3. p. 149-223.
Barroso, G. M., Peixoto, A. L., Ichaso, C. L. F., Guimarães, E. F. & Costa, C. G.
2002. Sistemática de Angiospermas do Brasil. v. 1. 2ª ed. Editora UFV,
Viçosa. 309p.
Bastos, A. M. 1943. Os paus rosa da indústria da essência. Rodriguésia 6
(16): 45-63.

102
Berlyn, G. P. & Miksche, J. P. 1976. Botanical microtechnique and
cytochemistry. Ames (Iowa):The Iowa State University Press.
Botelho, E. A. 1951. Fichas anatômicas de espécies vegetais brasileiras.
Bol. INT, Rio de Janeiro. v. 2: 65-71.
Boura, A.; Le Péchon, T. & Thomas, R. 2011. Wood anatomy of the Mascarene
Dombeyoideae: systematic and ecological implications. IAWA Journal 32
(4): 493-519.
Brush, W. D. 1944. Foreign Woods. Forest Service - US Dept. of Agriculture:
Washington. 10p.
Bukatsch, F. 1972. Bemerkungen zur Doppeffärbung Astrablau-Safranin.
Mikrokosmos 61 (8): 255.
Callado, C. H. & Costa, C. G. 1997. Wood anatomy of some Anaueria and
Beilschmiedia species (Lauraceae). IAWA Journal 18 (3): 247-259.
Callado, C. H. 1995. Anatomia do lenho de quatro espécies do gênero
Beilschmiedia Nees (Lauraceae). Dissertação de Mestrado. Universidade
Federal do Rio de Janeiro, Rio de Janeiro. 96p.
Camargos, J. A. A. 1996. Madeiras comerciais de Mato Grosso: chave de
identificação. Brasília: Ibama. 82 p.
Camargos, J. A. A.; Coradin, V. T. R.; Czarneski, C. M..; Oliveira, D. &
Meguerditchian, I. 2001. Catálogo de árvores do Brasil. Brasília: IBAMA.
896p.
Carreras, R & Pérez, E. 1988. Descripción anatomica de la madera de ocho
espécies forestales. Revista Forestal Baracoa 18: 17-37.
Castiglioni, J. A. 1951. Lauraceas Argentinas (1). I. Genero Nectandra. Boletin
de La Sociedad Argentina de Botanica 4 (1; 2): 66-94.
Castiglioni, J. A. 1962. El leño secondario de las especies argentinas de
Nectandra. Revista de Investigaciones Forestales 3 (1): 1-15.

103
Chanderbali, A. S. Endlicheria (Lauraceae). 2004. Flora Neotropica,
Monograph 91. New York Botanical Garden, New York. 141p., ilust.
Chanderbali, A. S.; van der Werf, H. & Renner, S. 2001. Phylogeny and
historical biogeography of Lauraceae: evidence of the chloroplast and
nuclear genomes. Ann. Missouri Bot. Gard. 88: 104-134.
Coe-Teixeira, B. 1963. Lauraceas do estado de São Paulo I: Beilschmiedia,
Endlicheria e Aniba. Sec. Agri. Do Estado de São Paulo. bl. 1. 1-29.
Coe-Teixeira, B. 1970. Algumas espécies novas de Lauraceas da Amazônia.
Bol. Mus. Paraense Emílio Goeldi- Botânica 34: 1-21.
Coe-Teixeira, B. 1980. Lauráceas do gênero Ocotea do estado de São Paulo.
Rodriguésia 32 (52): 55-190.
Coradin, V. T. R. & Muñiz, G. M. B. 1991. Normas e Procedimentos em
Estudos de Anatomia de Madeira: I. Angiospermae. II. Gimnospermae.
LPF. 15: 1-19.
Coradin, V. T. R.; Marques, M. H. B.; Camargos, J. A. A. & Muñiz, G. B.; 1991.
Chave de identificação das principais madeiras da Floresta Nacional
do Tapajós. Brasília: IBAMA. 51p.
Coradin, V. T. R.; Camargos, J. A. A.; Marques, L. F. & Silva Júnior, E. R.
2009a. Madeiras similares ao mogno (Swietenia macrophylla King.):
chave ilustrada para identificação anatômica em campo. Brasília: SFB.
28p.
Coradin, V. T. R.; Camargos, J. A.A; Pastore, T. C. M. & Christo, A. G. 2009b.
Madeiras comerciais do Brasil: chave interativa de identificação baseada
em caracteres macroscópicos. Versão 1.0.
Czarneski, C. M., Freitas, G. F. de., Coradin, V. T. R. & Camargos, J. A. A.
2001. Estudo anatômico comparativo do lenho de quatro espécies da
família Lauraceae ocorrentes na Amazônia. Brasil Florestal 70: 48-60.
Dadswell, H. E. & Record, S. J. 1936. Identification of Woods with conspicuous
rays. Tropical Woods 48: 1-30

104
Dechamps, R. 1979. Etude anatomique de bois d’Amérique du Sud. I.
Acanthaceae à Lecythidaceae. Ann. Mus. Roy. Afr. Cent., Tervuren. Sér.
IN-8, Sci. econ, nº 10: 332p.
Fedalto, L. C.; Mendes, I. C. A. & Coradin, V. T. R. 1989. Madeiras da
Amazônia: descrição do lenho de 40 espécies ocorrentes na Floresta
Nacional do Tapajós. Brasília: IBAMA. 156p.
Franklin, G. L. 1945. Preparation of thin sections of synthetic resins and Wood-
resin composites, and a new macerating method for Wood. Nature 155 . n.
3924. p. 51.
Gemballa, G. 1955. Contribuição para a caracterização da essência de
Ocotea pretiosa Mez: essência do sassafrás brasileiro. Tese de
Doutorado, Faculdade Nacional de Farmácia da Universidade do Brasil, Rio
de Janeiro. 181p.
Ghilardi, E. & Manieri, C. 1964. Caracterização e aplicações das madeiras
nacionais no estado de São Paulo. Silvicultura em São Paulo 3 (3). 311-
332.
Gottlieb, O. R. 1972. Review article chemosystematics of the Lauraceae
Phytochemistry 11: 1537-1570.
Hernandez, H. 1985. Estudio del xilema secundário de las Lauraceae
argentinas. Lilloa 2 (38): 41-71.
IAWA Committee. 1989. In: Wheeler, E. A.; BASS, P. & GASSON, P. E. (eds.)
List of microscopic feature of hardwood identification. IAWA Bulletin 10 (3):
219-332.
Instrução Normativa do Ministério do Meio Ambiente nº 06/2008. Lista Oficial
das Espécies da Flora Brasileira Ameaçadas de Extinção. Anexo I
Janssonius, H. H. 1926. Mucilage cells and oil cells in the woods of the
Lauraceae. Tropical Woods 6: 3-4.
Johansen, D. A. 1940. Plant Microtechnique. New York, McGraw - Hill Book
Company, XI. 523 p.

105
Kaiser, E. 1880. Verfahren zur Herstellung einer tadellosen Glycerin-Gelatine.
Bot. Zentralb. p. 25-26.
Klein, R. M. 1974. Importância e fidelidade das Lauraceae na “Formação de
Araucária” do Estado de Santa Catarina. Ínsula 7: 1-19.
Knoblauch, R. 1888. Anatomia des halzes der Laurineen. In: Solereder, H.
Systematic Anatomy of the Dicotyledons. Vol II. 705-706.
Kopp, e. l. 1966. A taxonomic revision of the genus Persea in the western
hemisphere (Perseae-Lauraceae). Mem. New York. Bot. Gard. 14: 1-120.
Kostermans, A. J. G. H. 1957. Lauraceae. Reinwardtia 4 (2): 193-256.
Kostermans, A. J. G. H. 1937. Revision of the Lauraceae II. The genera
Endlicheria, Cryptocarya (American species) and Licaria. Recueil des
Travaux Botaniques Néerlandais 34: 500-609.
Kostermans, A. J. G. H. 1938. Revision of the Lauraceae V. Recueil des
Travaux Botaniques Néerlandais 35: 834-931.
Kostermans, A. J.G. H., Homer V. Pinkley & William L. 1969. A new Amazonian
arrow poison: Ocotea venenosa. Botanical Museum Leaflets, Harvard
University 22(7): 241 - 252.
Kribs, D. A. 1959. Commercial foreign woods on the American market.
Pennsylvania State Univ. 203p.
Kubitzki, K. 1982. Aniba. In: Flora Neotropica 31. New York Botanical Garden,
New York. 1-84.
Kurz, H. 2000. Revision der gattung Licaria (Lauraceae). Mitt. Inst. Allg. Bot.
28/29: 89-221.
Lens, F.; Endress, M. E.; Baas, P.; Jansen, S. & Smets, E. 2008. Wood
anatomy of Rauvolfioideae (Apocynaceae): a search for meaningful non-
DNA characters at the tribal level. Amer. Journal of Botany 95: 1199-
1215.

106
Lens, F.; Schönenberger, J.; Baas, P.; Jansen, S. & Smets, E. 2007. The role
of wood anatomy in phylogeny reconstruction of Ericales. Cladistics 23: 1-
26.
León, W. J. 1999-2000. Anatomía del leño de 17 especies del género Ocotea
Aublet (Lauraceae). Pittieria 1 (29-30): 53-65.
León, W. J. & Espinoza de Pernía, N. 2000a. Estudio anatômico del leño de
ocho especies del género Aniba Aublet (Lauraceae). Revista Forestal
Venezolana 44 (1): 37-46.
León, W. J. & Espinoza de Pernía, N. 2000b. Estudio anatômico del leño de
siete especies del género Beilschmiedia Nees (Lauraceae). Revista
Forestal Venezolana 44 (1): 47-56.
León, W. J. & Espinoza de Pernía, N. 2001. Estudio anatômico del leño de 18
especies del género Nectandra Rol. ex Rottb. (Lauraceae). Revista
Forestal Venezolana 45 (1): 33-44.
Liu, J. & Noshiro, S. 2003. Lack of latitudinal trends in wood anatomy of
Dodonea viscosa (Sapindaceae), a species with a worldwide distribution.
Amer. Journal of Botany 90(4): 532-539.
Loureiro, A. A. & Silva, M. F. da. 1968. Catálogo das madeiras da Amazônia.
SUDAM: Belém. v.1. 433p.
Loureiro, A. A. 1976. Estudo anatômico macro e microscópico de 10 espécies
do Gênero Aniba (Lauraceae) da Amazônia. Acta Amazonica 6 (2): 5 - 85.
Loutfy, M. H. A. 2009. Wood anatomy and its implications on the taxonomy of
Apollonias Nees (Lauraceae). Feddes Repertorium 120 (1-2.): 75-90
Madriñan, S. 2004. Lauraceae: Rhodostemodaphne. Flora Neotropica,
Monograph 92. New York Botanical Garden, New York. 102p., ilust.

107
Mainieri, C. 1971. 25 madeiras da Amazônia de valor comercial,
caracterização macroscópica, usos comuns e índices qualificativos.
IPT: São Paulo. publ. nº 798. 33p.
Mainieri, C. 1983. Manual de Identificação das principais madeiras
comerciais brasileiras – São Paulo: IPT. 241p.
Marchiori, J. N. C.; Muñiz, G. I. B. de & Santos, S. R. dos. 2009. Madeiras do
Rio Grande do Sul 1: descrição microscópica de 33 espécies nativas.
Santa Maria: s.n. 80p.
Marchiori, J. N. C.; Muñiz, G. I. B. de & Santos, S. R. dos. 2010. Madeiras do
Rio Grande do Sul 2: descrição microscópica de 35 espécies nativas.
Santa Maria: Anaterra. s.n. 80p.
Marques, A. C. 2001. Importância econômica da família Lauraceae Lindl.
Floresta e Ambiente 1: 195 – 206.
Mattos Filho, A. de. 1960. A estrutura do lenho de sassafrás. Anu. Bras. Econ.
Flor. 12: 289-296.
Mattos, P. P. de; Teixeira, L. L.; Seitz, R. A.; Salis, S. M. de & Botosso, P. C.
2003. Anatomia de madeiras do Pantanal mato-grossense:
(características microscópicas). EMPRABA: Colombo. 182p.
Metcalfe, C. & Chalk, L. 1985. Anatomy of the Dicotyledons. Vol II 2 ed.
Claredon Press. Oxford. 297p.
Mez, C. 1889. Lauraceae Americanae. Jahrbuch des Königlichen
Botanischen Gartens und des Botanischen Museums zu. Berlin 5: 1-
556.
Milanez, F. R. 1930. A estructura do lenho da embuia. Min. Agri. Ind. Com.
Rio de Janeiro. 17p.

108
Noshiro, S. & Baas, P. 2000. Latitudinal trends in Wood anatomy within species
and genera: case study in Cornus s.l. (Cornaceae). Amer. Journal of
Botany 87 (10): 1495-1506.
Ochioni, P. & Mattos Filho, A. 1947. Estudo anatômico do lenho secundário do
puchury-rana Ocotea fragrantissima Ducke. Rodriguésia 10 (21): 1-12.
Oliveira, C. W. de, Callado, C. H. & Marquete, O. 2001. Anatomia do lenho de
espécies do gênero Nectandra Rol. ex Rottb. (Lauraceae). Rodriguésia 52
(81): 125-134.
Oliveira. C. W. de. 2005. Anatomia da casca e do lenho de espécies de
Nectandra Rol. ex Rottb. e Ocotea Aubl. (Lauraceae) – Dissertação de
mestrado. Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Rio de
Janeiro. 92p.
Pedralli, G. 1985. A família Lauraceae Lindley no Rio Grande do Sul, Brasil:
gêneros Persea Clus. Ex Mill. e Phoebe Nees. Roessléria 7 (1): 23-42.
Pedralli, G. 1986a. A família Lauraceae Lindley no Rio Grande do Sul, Brasil:
gênero Nectandra Rol. Ex Rottb. Iheringia 35: 133-149.
Pedralli, G. 1986b. O gênero Ocotea Aublet (Lauraceae) no Estado do Rio
Grande do Sul, Brasil. XXXVII Congresso Nacional de Botânica.
Universidade Federal de Ouro Preto/Minas Gerais. Brasil, p. 426-466;
Pedralli, G. 1987a. A família Lauraceae Lindley no RS, Brasil: gêneros
Endlicheria Nees, Laurus L. e Cryptocarya R. BR. . Acta Bot. Bras. 1(1):
27-41
Pedralli, G. 1987b. Lauraceas: Nectandra In: Reitz, R. (ed.). Flora Ilustrada
Catarinense. Herbário Barbosa Rodrigues: Itajaí, 93p.
Pedralli, G. 1988. Chave para os gêneros da família Lauraceae Lindley
ocorrentes no estado do Rio Grande do Sul, Brasil. Albertoa 1 (12): 137-
140.

109
Pedralli, G. 1996. Uma análise das observações de Rohwer sobre Lauráceas. 6
Nectandra da flora ilustrada Catarinense. Sellowia 45-48: 109-114.
Pinho, R. A. de. 1968. Estudo anatômico do lenho secundário de três espécies
florestais do estado de São Paulo. Arq. de Bot. do Estado de São Paulo.
v. IV. 137-140.
Quinet, A. & Andreata, R. H. P. 2002. Lauraceae Jussieu na Reserva Ecológica
de Macaé de Cima, município de Nova Friburgo, Rio de Janeiro, Brasil.
Rodriguésia 53 (82): 59-121.
Quinet, A. 2001. Lauraceae Jussieu na Reserva Ecológica de Macaé de
Cima, Município de Nova Friburgo, Rio de Janeiro, Brasil. Dissertação
de Mestrado. Universidade Federal do Rio de Janeiro, Rio de Janeiro.
146p.
Quinet, A. 2005. Sinopse taxonômica da família Lauraceae no estado do Rio de
Janeiro, Brasil. Acta Bot. Bras. 19 (3): 563-572.
Quinet, A. 2008. Uma nova espécie de Ocotea (Lauraceae) para o estado do
Espírito Santo, Brasil. Rodriguesia 59 (2): 339-342.
Quinet, A.; Baitello, J.B. & Moraes, P.L.R. de. 2012. Lauraceae. In: Lista de
Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro.
(http://floradobrasil.jbrj.gov.br/2012/FB008440)
Record, S. J. & Hess, R. W. 1942. American timbers of the family Lauraceae.
Tropical Woods 69: 7-33.
Record, S. J. & Mell, C. D. 1924. Timbers of Tropical America New Haven,
Yale Univers. Press. 610p.
Record, S. J. 1934. Rôle of wood anatomy in taxonomy. Tropical Woods 37: 1-
9.
Reitz, R.; Klein, M. R. & Reis, A. 1979. Madeiras do Brasil: Santa Catarina.
Lunardelli: Florianópolis. 320 p.

110
Renner, S. S. 1999. Circunscription and phylogeny of the Laurales: evidence
from molecular and morphological data. Amer. Journal of Botany 86:
1301-1315.
Renner, S. 1982. Aiouea. In: Flora Neotropica 31. New York Botanical Garden,
New York. p. 85-116.
Renner, S. S. & Chanderbali, A. S. 2000. What is the relationship among
Hernandiaceae, Lauraceae and Monnimiaceae and why is this so difficult to
answer? Int. J. Plant. Sci. 161: 109-119.
Richter, H. G. & Van Wyk, A. E. 1990. Wood and bark anatomy of Lauraceae.
IV: Dahlgrenodendron J. J. M. Van der Merwe & Van Wyk. IAWA Bull. 11:
173-182.
Richter, H. G. 1977. Differencial staining of oil and mucilage in idioblasts of
Lauraceae. IAWA Bull. 4: 76.
Richter, H. G. 1980. Occurrence, morphology and taxonomic implications of
crystalline and siliceous inclusions in the secondary xylem of the Lauraceae
and related families. Wood Science and Technology 14: 35-44.
Richter, H. G. 1981a. Wood and bark anatomy of Lauraceae I: Aniba Aublet.
IAWA Bull. 2: 79-87.
Richter, H. G. 1981b. Anatomie des sekundären Xylems und der Rinde der
Lauraceae. Sonderbände des Naturwiss. Vereins Hamburg 5: 1-148.
Richter, H. G. 1985. Wood and bark anatomy of Lauraceae. II: Licaria Aublet.
IAWA Bull. n. s. Vol. 6: 187-199.
Richter, H. G. 1987. Mature Secondary Xylem. In: Metcalfe, C. R. 1987.
Anatomy of the Dicotyledons 2ª. ed. Oxford, Claredon Press. III. 162-
171p.
Richter, H. G. 1990. Wood and bark anatomy of Lauraceae. III: Aspidostemon
Rohwer & Richter. IAWA Bull. 11: 47-56.

111
Rizzini, C. T. 1971. Plantas do Brasil. Árvores e Madeiras Úteis do Brasil:
Manual de Dendrologia Brasileira. São Paulo. Edgar Blücher/EDUSP.
294 p.
Rohwer, J. G. 1986. Prodomus einer Monographie der Gattung Ocotea Aubl.
(Lauraceae), sensu lato. Mitteilungen aus dem Institut fur Allegemeine
Botanik Hamburg 20. 278p.
Rohwer, J. G. 1988. The genera Dicypellium, Phyllostemonodaphne,
Systemonodaphne and Urbanodendron (Lauraceae). Bot. Jahrb. Syst.
110: 157-171.
Rohwer, J. G. 1991. Borderline cases between Ocotea, Nectandra and Phoebe
(Lauraceae): the “marginal” species of the Ocotea helicterifolia-group
including the Ocotea heydeana-group. Bot. Jahrb. Syst. 112: 365-397.
Rohwer, J. G. 1993a. Lauraceae (Nectandra). Flora Neotropica, Monograph
60. New York Botanical Garden, New York. 332p., ilust.
Rohwer, J. G. 1993b. Lauraceae. In: Kubitzki, K., Rohwer, J. G. & Bittrich, V.
(eds.). The Familias and Genera of Vascular Plants II. Springer-Verlag,
Berlin. p. 366-391.
Rohwer, J. G. 2000. Toward a phylogenetic classification of the Lauraceae:
evidence from matK sequences. Systematic Botany, 25 (1): 60-71.
Rohwer, J. G.; Richter, H. G. & van der Werf, H. 1991. Two new genera of
neotropical Lauraceae and critical remarks on the generic delimitations.
Ann. Missouri Bot. Gard. 78: 388-400.
Saas, J. E. 1951. Botanical microtechnique. The Iowa State College Press:
Ames. 391p.
Silva, M. da. 2003. Anatomia da madeira de Ocotea odorifera (Vell.) Rohwer
(Lauraceae). Dissertação de Mestrado. Universidade Federal de Santa
Catarina, Florianópolis. 86p.

112
Silva M. da. & Medeiros, J. D. 2000. Anatomia da madeira de Ocotea
catharinensis Mez, Lauraceae (Canela-Preta). Ínsula 29: 67 – 87.
Solereder, H. 1908. Laurinae. In: Systematic anatomy of the dicotyledons.
Oxford, Clarendo Press. Vol. II. 702-706
Stern, W. L. 1954. Comparative anatomy of xylem and phylogeny of Lauraceae.
Tropical Woods 100: 1-75.
Tupper, W. W. 1927. A comparative study of Lauraceous woods. Amer.
Journal of Botany 14: 520-525.
Van der Slooten, H. J.; Acosta-contreras I. & Aas, P. S. 1970. Madeiras
latinoamericanas. IV. Nectandra sp., Ocotea austinii, Persea sp. aff.
vesticula, Persea schiedeana. Turrialba 20 (2): 223 – 232.
Van der Werff, H. & Richter, H. G. 1996. Toward an improved classification of
Lauraceae. Ann. Missouri Bot. Gard. 83: 409-418.
Van der Werff, H. 1987. A revision of Mezilaurus (Lauraceae). Ann. Missouri
Bot. Gard. 74: 153-182.
Van der Werff, H. 1991a. A key to the genera of Lauraceae in the New World.
Ann. Missouri Bot. Gard. 78: 377-387.
Van der Werff, H. 1991b. Gamanthera (Lauraceae), a new genus from Costa
Rica. Ann. Missouri Bot. Gard. 78: 401-408.
Van der Werff, H. 1993. A revision of the genus Pleurothyrium (Lauraceae).
Ann. Missouri Bot. Gard. 80: 39-118.
Van der Werff, H. 1997. Sextonia, a new genus of Lauraceae from South
America. Novon 7: 436-439.
Van der Werff, H. 1999. New taxa and combinations in the Ocotea helicterifolia
(Lauraceae) species group. Novon 9: 571-583.
Vattimo, I. de. 1956a. Lauraceae do Itatiaia. Rodriguésia 18-19 (30-31): 41-86.

113
Vattimo, I. de. 1956b. O gênero Ocotea Aubl. no sul do Brasil I. Espécies de
Santa Catarina e do Paraná. Rodriguésia 18-19 (30-31): 265-350.
Vattimo, I. de. 1958. Seis novas espécies brasileiras do gênero Ocotea.
Arquivos do Serviço Florestal 10: 109-123.
Vattimo, I. de. 1959a. Flora da cidade do Rio de Janeiro (Lauraceae).
Rodriguésia 21/22 (33- 34): 157-175.
Vattimo, I. de. 1959b. O gênero Ocotea Aubl. (Lauraceae) no sul do Brasil II.
Espécies dos estados de São Paulo e Rio Grande do Sul. Arquivos do
Jardim Botânico do Rio de Janeiro 17: 199-228.
Vattimo, I. de. 1960a Duas novas Lauraceae brasileiras. Rodriguésia 23-24
(35-36): 253-255.
Vattimo, I. de. 1960b. O gênero Ocotea Aubl. no nordeste do Brasil
(Lauraceae). Rodriguésia 23-24 (35-36): 241-252.
Vattimo, I. de. 1966. Lauraceae do estado da Guanabara. Rodriguésia 25 (37):
75-122.
Vattimo, I. de. 1976. Estudos sobre Ocotea Aubl., Phyllostemonodaphne
Kosterm. e Licaria Aubl. (Lauraceae). Rodriguésia 28 (41): 121-127.
Vattimo, I. de. 1977. Três novas espécies de Lauraceae brasileiras.
Rodriguésia 29 (42): 127-131.
Vattimo, I. de. 1978. Contribuição ao conhecimento da distribuição geográfica
das Lauraceae I. Rodriguésia 29 (44): 269-305.
Vattimo, I. de. 1978. Contribuição ao conhecimento da distribuição geográfica
das Lauraceae II. Rodriguésia 30 (47): 83-103.
Vattimo-Gil, I. de. 1979a. Contribuição ao conhecimento da distribuição
geográfica das Lauraceae III. Rodriguésia 31 (48): 7-57.
Vattimo-Gil, I. de. 1979b. Contribuição ao conhecimento da distribuição
geográfica das Lauraceae IV. Rodriguésia 31 (49): 5-15.

114
Vattimo-Gil, I. de. 1979c. Contribuição ao conhecimento da distribuição
geográfica das Lauraceae V – Novas localidades de ocorrência nos
estados do Paraná e Rio Grande do Sul. Rodriguésia 31 (50): 37-65.
Vattimo-Gil, I. de. 1980. Contribuição ao conhecimento da distribuição
geográfica das Lauraceae VI. Rodriguésia 32 (53): 9-32.
Vattimo-Gil, I. de. 1983. Contribuição ao conhecimento da distribuição
geográfica das Lauraceae VIII. Rodriguésia 35 (57): 7-28.
Vattimo-Gil, I. de. 1996. In: Marques, M. do C. M. Espécies Coletadas no
Estado do Rio de Janeiro Depositadas no Herbário RB. Rio de Janeiro.
107p.
Zenid, G.J. & Ceccantini, G.C.T. 2007. Identificação macroscópica de
madeiras. São Paulo: IPT. 51p.










![Billie Warren Chai - [Noiva Por Encomenda 03] - A Noiva de Josh (Julia Historicos 1621.3)](https://img.document.onl/doc/110x75/577cd14d1a28ab9e78941a49/billie-warren-chai-noiva-por-encomenda-03-a-noiva-de-josh-julia-historicos.jpg)


















