Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E CONSERVAÇÃO
MESTRADO EM ECOLOGIA E CONSERVAÇÃO
ANÁLISE COMPARATIVA DOS NICHOS ESPACIAL E ALIMENTAR DE ANUROS
EM ÁREAS DE CAATINGA E MATA ATLÂNTICA DE SERGIPE, BRASIL
ADILSON DE OLIVEIRA SILVA
SÃO CRISTOVÃO, SERGIPE
FEVEREIRO, 2014

ii
UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E CONSERVAÇÃO
MESTRADO EM ECOLOGIA E CONSERVAÇÃO
ANÁLISE COMPARATIVA DOS NICHOS ESPACIAL E ALIMENTAR DE ANUROS
EM ÁREAS DE CAATINGA E MATA ATLÂNTICA DE SERGIPE, BRASIL
AUTOR: ADILSON DE OLIVEIRA SILVA
ORIENTADOR: PROF. DR. RENATO GOMES FARIA
SÃO CRISTOVÃO, SERGIPE
FEVEREIRO, 2014

iii
FICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTRAL
UNIVERSIDADE FEDERAL DE SERGIPE
Silva, Adilson de Oliveira S586a Análise comparativa dos nichos espacial e alimentar de anuros
em áreas de Caatinga e Mata Atlântica de Sergipe, Brasil / Adilson de Oliveira Silva ; orientador Renato Gomes Faria. – São
Cristóvão, 2014. 65 f. : il.
Dissertação (mestrado em Ecologia e Conservação) –Universidade Federal de Sergipe, 2014.
O 1. Amphibia. 2. Meio ambiente - Heterogeneidade. 3. Dieta.
4. Mata Atlântica. 5. Caatinga. 6. Sergipe (SE). 7. Nicho
(Ecologia). I. Faria, Renato Gomes, orient. II. Título CDU: 597.8(813.7)

iv
AGRADECIMENTOS

v
A realização deste trabalho foi possível graças à colaboração imprescindível de vários
amigos e familiares aos quais agradeço:
Ao meu orientador professor Renato Gomes Faria, por não medir esforços para
contribuir, querendo sempre o melhor à minha formação, mostrando não só o conhecimento
científico, mas contribuindo muito para a minha formação pessoal.
Ao meu amigo Breno Moura, pelo incentivo e ajuda durante toda a pesquisa,
principalmente nos trabalhos de campo, sem o qual não conseguiria ter realizado o trabalho, e
a sua família que me aceitou de braços abertos em seu lar.
A Anthony Santana e a Lucas França, pela amizade e ajuda durante o curso, assim
também como aos colegas que se mostraram especiais sempre com alegria e disposição
trocando conhecimentos sobre a vida e os temas da Ecologia.
Aos estagiários Dário Junior, Lucas Otávio, Charlles Myller e Mayane Vieira pela
ajuda na triagem do material de campo. Aos integrantes do laboratório de Cordados
(Herpetologia e Ictiologia), pelas horas de descontração e alegria. E a professora Angélica
Bezerra, que muito me motivou a continuar na profissão docente e na pesquisa.
Aos amigos Rodrigo Dantas e Ismael Junior que mesmo não sendo da área me
ajudaram muito durante esse trabalho. Pela disposição em sair de suas casas e ir a campo
viver na pele um dia de ―pesquisador‖.
Ao Programa de Pós-Graduação em Ecologia e Conservação (PPEC/UFS) pelo apoio
logístico para realização das viagens a campo em Poço Redondo, e a Capela. E aos
professores do Programa de Pós-Graduação em Ecologia e Conservação, pelos ensinamentos
transmitidos.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela bolsa
de mestrado, e a Fundação de Apoio à Pesquisa e à Inovação Tecnológica do estado de
Sergipe (FAPITEC/SE) pelos recursos financeiros que facilitaram os trabalhos de campos
realizados nessa dissertação.
Ao professor Geraldo Jorge Barbosa de Moura, que com muito empenho tem me
ajudado nesse longo percurso educacional e da pesquisa na Herpetologia.
Aos meus pais que sempre se preocupavam e me incentivavam da forma que podiam
durante toda a minha formação acadêmica.
A Minha Esposa Thayse, que muito contribuiu com apoio e incentivo durante todo o
meu percurso acadêmico. Pelas noites mal dormidas de preocupação quando eu estava em
campo, e pelo amor e compreensão mesmo quando eu estava ausente.

vi
A minha princesinha Anne Sophie que sempre me confortou nos momentos difíceis e
ao me ver chegar em casa do campo, ou de UFS me recebia com um enorme sorriso e dizia
―Papai... você chegou afinal...‖
A Deus...
Muito Obrigado!

vii
“Nenhum sucesso na vida compensao fracasso no lar...”
David O´Mckey

viii
Sumário
RESUMO ............................................................................................................................... 9
ABSTRACT .......................................................................................................................... 10
INTRODUÇÃO GERAL ...................................................................................................... 11
ESTRUTURA DO HABITAT E DIVERSIDADE DE ESPÉCIES ................................ 11
CAATINGA .................................................................................................................... 12
MATA ATLÂNTICA ..................................................................................................... 12
ANFÍBIOS ....................................................................................................................... 13
OBJETIVO GERAL ............................................................................................................. 15
OBJETIVOS ESPECÍFICOS ............................................................................................... 15
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................... 16
Nicho espacial e alimentar de anuros em áreas de Caatinga e Mata Atlântica de Sergipe,
Brasil ................................................................................................................................... 21
INTRODUÇÃO .................................................................................................................... 23
MATERIAL E MÉTODOS ................................................................................................... 25
RESULTADOS ..................................................................................................................... 31
DISCUSSÕES ...................................................................................................................... 51
CONSIDERAÇÕES FINAIS ................................................................................................ 56
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................... 56

9
RESUMO
Características ambientais distintas podem afetar diretamente a estruturação das
comunidades, pois oferecem recursos e condições variadas, o que permite que em cada área,
grupos diferenciados de espécies se estabeleçam. Nesse estudo duas taxocenoses de anfíbios
anuros foram analisadas entre outubro de 2012 e setembro de 2013, em duas áreas com
diferentes graus de heterogeneidade ambiental, uma na Caatinga (Monumento Natural Grota
do Angico – MNGA) e outra de Mata Atlântica (Refugio da Vida Silvestre Mata do Junco –
RVSMJ), ambas no estado de Sergipe. Foram utilizadas informações sobre os nichos espacial
e alimentar e suas relações com fatores ecológicos e históricos. Foram tomados 1.332
registros (999 - RVSMJ e 333 - MNGA) pertencentes a 28 espécies (22 – RVSMJ e 15 –
MNGA, sendo nove comuns as duas áreas). A sobreposição de nicho espacial e alimentar foi
relativamente alta em ambas as áreas. Nove tipos de microhábitats foram utilizados pelas
espécies sendo os mais comuns, independente da área, solo (45,1%), água (23,7%) e folhiço
(8,6%). As presas mais importantes das dietas foram Formicidae e Coleoptera em ambas as
áreas. Os indivíduos da Mata Atlântica foram relativamente menores que os da Catinga
sugerindo investimentos diferenciados em reprodução e crescimento entre as áreas. Os
resultados obtidos sugerem que as taxocenoses de ambas as áreas são influenciadas por
fatores históricos ou filogenéticos somente na utilização dos recursos espaciais. Suas dietas
são fortemente influenciadas pelas disponibilidades de recursos que o ambiente oferece.
PALAVRAS-CHAVES: amphibia; heterogeneidade ambiental; dieta; uso de microhabitat.

10
ABSTRACT
Distinct environmental characteristics can directly affect community structure, they offer
varying conditions and resources, which allows each area to establish different groups of
species. In this study we analyzed two amphibian assemblages between October 2012 and
September 2013 in two areas with different degrees of environmental heterogeneity, one in
the Caatinga (Monument Natural Grota do Angico - MNGA ) and the other in the Atlantic
Rainforest (Refugio de Vida Silvestre Mata do Junco - RVSMJ), both in the state of Sergipe.
We used information about the spatial and food niches and their relationships with ecological
and historical factors. We analyzed 1,332 records (999 - RVSMJ 333 - MNGA) of 28 species
(22 - RVSMJ and 15 - MNGA, nine common to both areas). The food and space niche
overlap was relatively high in both study sites. Nine types of microhabitats were used by the
species being the most common, regardless of the area, soil (45.1 %), water (23.7 %) and
leaflitter (8.6%). The most important preys in their diets were Formicidae and Coleoptera in
both areas. Individuals in the Atlantic rainforest were relatively smaller than those of the
Caatinga area suggesting differentiated investments between reproduction and growth
between the areas. The results suggest that the assemblages from both areas are influenced by
historical or phylogenetic factors only in the use of space assets. Their diets are strongly
influenced by the availability of resources the environment offers.
KEYWORDS: amphibia, environmental heterogeneity, diet, microhabitat use.

11
INTRODUÇÃO GERAL
ESTRUTURA DO HABITAT E DIVERSIDADE DE ESPÉCIES
As condições estruturais do habitat têm sido consideradas como um importante
determinador da diversidade de espécies em uma comunidade (Mcguinness & Underwood,
1986; Huston, 1994; Fraser, 1998; Moreno-Rueda & Pizarro, 2007; Vasconcelos & Rossa-
Feres, 2008; Vasconcelos et al., 2009; Silva et al., 2011). Logo diferenças nos níveis de
luminosidade, temperatura e umidade apresentadas por ambientes com níveis de
heterogeneidade de habitat variáveis (Li & Reynolds 1995), podem propiciar a ocorrência ou
não de determinadas espécies, pois dão base para a reprodução, nidificação, desenvolvimento
e forrageamento das diferentes espécies de animais (August, 1983; Halffter, 1991; Franklin et
al., 2005).
Relações positivas entre a estrutura do hábitat e a diversidade de espécies já foram
observadas (Van sluys et al., 2007; Silva & Rossa-Feres, 2011), sendo que ambientes mais
heterogênios podem prover as espécies uma maior quantidade de recursos e refúgios, o que
pode alterar padrões no nicho das espécies (Macguiness & Underwood, 1986; Barreto, 2010).
De acordo com a hipotese da variação do nicho populações com nichos mais amplos são mais
variáveis as alterações do ambiente que as populações com um nicho mais estreito, o que
facilitaria a adequação das mesmas a ambientes mais heterogêneos (Van Valen, 1965, Costa
et al., 2008).
Porem, além da heterogeneidade do habitat, outros fatores devem ser considerados ao
tentar explicar os padrões de distribuição das espécies, bem como aspectos de seu nicho
ecológico. Peterson et al., (1999) afirmam que algumas características são mantidas ao longo
da história evolutiva de determinados grupos por certos táxon irmãos, o que pode determinar
suas preferências ecológicas. Assim, uma explicação adequada para os padrões de riqueza e
abundancia de espécies deve incorporar processos evolutivos e biogeográficos em algum
nível, além de considerar as correlações climáticas conhecidas e estrutura do habitat (Wiens,
2004).

12
CAATINGA
A Caatinga é o único bioma exclusivamente brasileiro e está distribuído em toda a
região nordeste do Brasil (exceto para o estado do Maranhão) e norte do estado de Minas
Gerais, abrange uma área de 852.261 km2
e representa 11% do território brasileiro (MMA,
2007), com apenas 1% de seu território atualmente conservado (De Oliveira et al., 2012).
Com solos rasos e pedregosos (Ab’saber, 1974), o bioma Caatinga caracteriza-se pelos climas
árido e semiárido, com longos períodos de seca, e chuvas irregulares concentradas no verão,
apresenta precipitação média anual de 400 a 600 mm (Ab'saber, 1977). Possui temperaturas
anuais elevadas apresentando altos valores de radiação solar, e de evapotranspiração
potencial, contrapondo-se aos baixos índices de umidade relativa do ar, podendo atingir
variações altimetricas superiores a 2000 m acima do nível do mar (Leal et al., 2005;
Camardelli & Napoli, 2012).
A biodiversidade desse bioma é altamente ameaçada devido à exposição da terra para
agricultura e pecuária. Previsões climáticas presumem aumentos de aridez, que possam
representar ameaças adicionais à biodiversidade do bioma (De Oliveira et al., 2012). Embora
o cenário mais comum na Caatinga seja o de savana estépica, que é composto de plantas
espinhosas e decíduas e rios que são em grande parte intermitente e sazonal (MMA, 2007), o
bioma não é homogêneo fisicamente e pode ser dividido em oito ecorregiões menores:
Complexo de Campo Maior, Depressão Sertaneja Meridional, Complexo Ibiapaba – Araripe,
Dunas do São Francisco, Depressão Sertaneja Setentrional, Complexo da Chapada
Diamantina, Planalto da Borborema e Raso da Catarina (Velloso et al., 2002). A altitude varia
de 100 a 500 m, com algumas áreas de 500 a 800 m contendo picos acima de 800 m (Velloso
et al., 2002).
MATA ATLÂNTICA
A Mata Atlântica é composta por florestas ombrófilas (densa, mista, e aberta),
florestas estacionais (semidecidual e decidual), manguezais, restingas, campos e brejos de
altitude e encraves florestais do Nordeste (Tabarelli et al., 2005). É considerada um dos
biomas de maior biodiversidade do mundo, por apresentar alta incidência de endemismo e
grande riqueza de espécies, além de ter sido colocado entre os cinco primeiros “hotspots” da
biodiversidade. (Mittermier et al., 1999).

13
A Mata Atlântica brasileira é uma região heterogênea (1.481.946 km2, que
corresponde 17,4% do território brasileiro), incluindo uma grande variedade de fisionomias
florestais e composições distribuídas através de mais de 3300 km ao longo da costa atlântica
brasileira, do nível do mar até a 2700 m de altitude. Estas florestas são distribuídas em
diferentes condições topográficas e climáticas, abrangendo planícies e regiões de montanha
costeiras com níveis elevados de precipitação que variam entre 1.800 e 3.600 mm/ano
(Metzger, 2009; Felinks et al., 2011). Atualmente sua área total encontra-se reduzida 100 mil
km2. No Nordeste do Brasil a área total de Mata Atlântica é de cerca de 3%, sendo composta
por fragmentos desconectados que se distribuem desde a Bahia até o Ceará (Tabarelli et al.,
2005).
ANFÍBIOS
O Brasil ocupa o primeiro lugar em riqueza de anfíbios do mundo, com 946 espécies
(Segalla et al., 2012), sendo que destas 913 são da ordem Anura, com aproximadamente 370
espécies na Mata Atlântica (286 endemicos) e cerca de 60 na Caatinga (Metzger, 2009;
Albuquerque et al., 2012). Devido ao seu modo de vida que pode ser aquático, misto,
terrestre, arbóreo e fossorial, os habitats desse grupo são bem variados, e nem sempre
dependentes diretamente da água (Dodd, 2010).
Os anfíbios são animais ectotérmicos que apresentam uma característica marcante, a
presença de um tegumento permeável à água. Seus ovos também são altamente suscetíveis à
desidratação e normalmente são depositados na água para garantir o sucesso embrionário e
posterior desenvolvimento larval (Dodd, 2010). Sua reprodução possui uma relação direta
com o tamanho do corpo devido aos vários fatores ecológicos que atuam sobre o
desenvolvimento de machos e fêmeas, com as fêmeas geralmente apresentando tamanhos
maiores já que assim, podem abrigar mais ovos (Howard, 1981; Wollbright, 1989; Wells,
2007).
De acordo com Wells (1977) são observados dois tipos de padrões reprodutivos em
anfíbios: o ―prolongado‖, que não está restrito a uma única estação, e o ―explosivo‖, que
ocorre mediante a disponibilidade de água. Porem apesar dessas classificações a maioria das
espécies ocupa uma posição intermediária entre ambas, se reproduzindo geralmente na
estação quente e úmida do ano, sendo que tanto a riqueza quanto a abundância dessas espécies

14
são fortemente correlacionadas com as variáveis climáticas encontradas em cada bioma
(Rossa-Feres & Jim 2001; Santos et al., 2005).
Ao se trabalhar com anfíbios anuros, deve-se levar em consideração fatores como
temperatura e pluviosidade por serem de vital relevância na influência do comportamento
desses organismos (Prado & Pombal Jr, 2005; Araujo & Almeida-Santos, 2013), sendo que os
locais com um nível de umidade mais elevada irão suportar mais modos reprodutivos do que
locais mais secos e, consequentemente, apresentam uma maior diversidade filogenética (Silva
et al., 2012).
Outro fator relevante então seria o tamanho corpóreo que é um traço morfológico
fundamental, importante em um contexto fisiológico, ecológico e social (Avise, 2000).
Variações na morfologia dos indivíduos, particularmente em relação ao tamanho corpóreo,
são comuns a várias espécies (Dodd, 2010; Castellano & Giacoma,1998; Castellano et al.,
1998). Alterções morfológicas em escala geográfica são comuns em algumas espécies animais
e normalmente estão associadas a particularidades de cada localidade como distintas
condições climáticas afetando as taxas de crescimento (Riha & Berven, 1991; Castellano &
Giacoma,1998; Castellano et al., 1998; Schäuble, 2004). Schäuble (2004) realizou estudos
com as variações morformetricas entre indivíduos das populações de Limnodynastes
tasmaniensis and L. peronii, espécies distribuídas amplamente no território australiano e
constatou que o clima e as variações latitudinais influenciaram muito na morfologia das
espécies em questão.
Características ambientais tão contrastantes como as apresentadas por biomas
diferentes como os citados anteriormente afetam diretamente as comunidades de anfíbios,
animais ectotérmicos que apresentam uma característica marcante, a presença de um
tegumento permeável à água. Assim os locais com nível de umidade mais elevada irão
suportar mais modos reprodutivos do que locais mais secos e, consequentemente, apresentam
uma maior diversidade filogenética (Silva et al., 2012). Já em áreas áridas e semiáridas as
altas temperaturas e a perda de água por evaporação rápida, muitas vezes acompanhada por
uma fonte de água limitada a alguns períodos do ano, fazem com o que a reprodução, o uso de
habitat e a dieta desses organismos sejam afetados (Bentley, 1966; Wells, 2007). Por isso, as
variáveis ambientais e climáticas são então de suma importância ao estudo de comunidades de

15
anfibios, uma vez que nenhum sistema ecológico pode ser estudado isoladamente do ambiente
no qual está inserido (Wells, 2007).
OBJETIVO GERAL
Avaliar como a heterogeneidade ambiental e fatores climáticos influenciam
parâmetros ecológicos como o nicho trófico e espacial de anfíbios anuros em remanescentes
de Caatinga e Mata Atlântica do estado de Sergipe.
OBJETIVOS ESPECÍFICOS
i. Comparar a forma de utilização dos recursos espaciais e alimentares pelos anuros entre
as áreas estudadas;
ii. Avaliar a influência dos parâmetros climáticos sobre a riqueza e abundância das
populações em cada bioma
iii. Verificar a influência de fatores ecológicos e históricos na forma de utilização dos
recursos espaciais e alimentares nas taxocenose estudadas;
iv. Investigar possíveis variações na morfologia das espécies comuns aos dois biomas;

16
REFERÊNCIAS BIBLIOGRÁFICAS
Ab’sáber, A. N. (1974). O Domínio Morfoclimático das Caatingas Brasileiras.
Geomorfologia, 43.
Ab’saber, A. N. (1977). Os domínios morfoclimáticos na América do Sul. Primeira
aproximação. Geomorfologia 52, 1–21.
Albuquerque, U. P. ; Araújo, E; El-Deir, A. C. A. ; Lima, A. L. A. ; Souto, A. ; Bezerra, B. M.
; Ferraz, E. M. N.; Freire, E. ; Sampaio, E.; Las-Casas, F.; Moura, G. J. B.; Pereira, G.;
Melo, J.; Ramos, M.; Rodal, M.; Schiel, N.; Lyra-Neves, R.; Alves, R.; Azevedo-
Júnior, S.; Telino Júnior, W.; Severi, W. (2012). Caatinga Revisited: Ecology and
Conservation of an Important Seasonal Dry Forest. The Scientific World Journal, 1-18.
Araujo, C. O., Almeida-Santos, S. M. (2013). Composição, riqueza e abundância de anuros
em um remanescente de Cerrado e Mata Atlântica no estado de São Paulo. Biota
Neotropica 13.
August, P. V. (1983). The role of habitat complexity and heterogeneity in structuring tropical
mammal communities. Ecology 64, 1495-1507.
Barreto, C. C. (2010). Heterogeneidade espacial do habitat e diversidade específica:
implicações ecológicas e métodos de mensuração. Oecologia Australis 7, 121-154.
Bentley, P. J. (1966). Adaptations of Amphibians to arid environments. Science 152, 619-623.
Camardelli, M., Napoli, M. F. (2012). Amphibian Conservation in the Caatinga Biome and
Semiarid Region of Brazil. Herpetologica 68, 31-47.
Castellano, S. & Giacoma, C. (1998). Stabilizing and directional female choice for male
callsin the European green toad. Animal Behavior 56, 275-287.
Castellano, S., Giacoma, C., Dujsebayeva, T., Odierna, G. e Balletto, E. (1998).
Morphometrical and acoustical comparison between diploid and tetraploid green
toads. Biological Journal of the Linnean Society 63, 257- 281

17
Costa, G., Mesquita, D. O., Colli, G., Vitt, L. (2008). Niche Expansion and the Niche
Variation Hypothesis: Does the Degree of Individual Variation Increase in
Depauperate Assemblages?. American Naturalist 172, 868-877.
De Oliveira, G. et al. (2012). Conserving the Brazilian semiarid (Caatinga) biome under
climate change. Biodiversity and Conservation 21, 2913–2926
Dodd, C. K. (2010). Amphibian ecology and conservation: a handbook of techniques. [S.l.]
Oxford University Press New York.
Felinks, B., Pardini, R., Dixo, M., Follner, K., Metzger, J. P., Henle, K. (2011). Effects of
species turnover on reserve site selection in a fragmented landscape. Biodiversity and
Conservation 20, 1057-1072.
Felinks, B.; Pardini, R. ; Dixo, M. ; Follner, K. ; Metzger, J. P. ; Henle, K. 2011. Effects of
species turnover on reserve site selection in a fragmented landscape. Biodiversity and
Conservation 20, 1057-1072.
Franklin E., Magnusson, W. E., Luizão, F. J. (2005). Relative effects of biotic and abiotic
factors on the composition. Applied Soil Ecology 29, 259-273.
Halffter, G. (1991) Historical and ecological factors determining the geographical distribution
of beetles (Coleoptera: Scarabaeidade: Scarabaeinae). Folia Entomologica Mexicana
82, 195-238.
Howard, R. D. 1981. Sexual dimorphism in bullfrogs. Ecology 62, 303-310.
Huston, M. A. (1994). Biological Diversity, The coexistence of species on changing
landscapes. Cambridge University Press, 681p.
Leal, I. R., Silva, J. M. C., Tabarelli, M., Lacher, J. R. T. (2005). Changing the course of
biodiversity conservation in the Caatinga of Northeastern Brazil. Conservation
Biology 19, 701-706.
Li H. & Reynolds J. F. (1995). On definition and quantification of heterogeneity. Oikos 73,
280–284.

18
Mcguinness, K. A. & Underwood, A. J. (1986). Habitat structure and the nature of
communities on intertidal boulders. Journal of Experimental Marine Biology and
Ecology 104. 97-123
Metzger, J. P. (2009). Conservation issues in the Brazilian Atlantic forest. Biological
Conservation 142, 1138-1140.
Metzger, J. P. (2009). Conservation issues in the Brazilian Atlantic forest. Biological
Conservation 142, 1138-1140.
Mittermeier, R. A., Myers, N., Gil, P. R.; Mittermeier, C. G. (1999). Hotspots: earth’s
biologically richest and most endangered terrestrial ecoregions. Mexico City:
CEMEX, 431p.
MMA (Ministério do Meio Ambiente). (2007). Áreas prioritárias para conservação, uso
sustentável e repartição de benefícios da biodiversidade brasileira. Atualização —
Portaria MMA No. 9, de 23 de janeiro de 2007. Ministério do Meio Ambiente,
Secretaria de Biodiversidade e Florestas, série biodiversidade, Distrito Federal, Brazil
Moreno-Rueda, G. & Pizarro, M. (2007). The relative influence of climate, environmental
heterogeneity, and human population on the distribution of vertebrate species richness
in south-eastern Spain. Acta Oecologica 32, 50-58.
Peterson, A.T., Soberón, J., Sánchez-Cordero, V. (1999). Conservatism of ecological niches
in evolutionary time. Science 285, 1265–1267.
Prado, G. M., Pombal, J. P. (2005). Distribuição espacial e temporal dos anuros em um brejo
dareserva biológica de duas bocas, sudeste do brasil. Arquivos do Museu Nacional 63,
685-705.
Riha V. F. & Berven K. A. (1991). An analysis of latitudinal variation in the larval
development of the wood frog (Rana sylvatica). Copeia, 209–221.
Rossa-Feres, D. C., Jim, J. (2001). Similaridade do sítio de vocalização em uma comunidade
de anfíbios anuros na região noroeste do Estado de São Paulo, Brasil. Revista
Brasileira de Zoologia 18, 439-454.

19
Santos, T. G.; Rossa-Feres, D. C.; Casatti, L. (2007). Diversidade e distribuicão espaço-
temporal de anuros em região com pronunciada estação seca no sudeste do Brasil.
Iheringia: Série Zoologia 97, 37–49.
Schäuble, C. S. (2004). Variation in body size and sexual dimorphism across geographical
and environmental space in the frogs Limnodynastes tasmaniensis and L. peronei.
Biological Journal of the Linnean Society 82, 39–56.
Segalla, M. V.; Caramaschi, U.; Cruz, C. A. G.; Garcia, P. C. A.; Grant, T.; Haddad, C. F. B e
Langone, J. (2012). Brazilian amphibians – List of species. Accessible at
http://www.sbherpetologia.org.br. Sociedade Brasileira de Herpetologia. Captured
on 13/05/2013.
Silva, F. R., Almeida Neto, M., Prado, V. H. M., Haddad, C. F. B., Rossa-Feres, D. C. (2012).
Humidity levels drive reproductive modes and phylogenetic diversity of amphibians in
the Brazilian Atlantic Forest. Journal of Biogeography 39, 1720-1732.
Silva, F. R., Rossa-Feres, D. C. (2011). Influence of terrestrial habitat isolation on the
diversity and temporal distribution of anurans in an agricultural landscape. Journal of
Tropical Ecology 27, 327-331.
Silva, R. A., Martins, I. A., Rossa-Feres, D. C. (2011). Environmental heterogeneity: Anura
diversity in homogeneous environments. Revista Brasileira de Zoologia, 28, 610-618.
Tabarelli, M., Pinto, L.P., Silva, J.M.C., Hirota, M.M., Bede, L.C. (2005). Desafios e
oportunidades para a conservação da biodiversidade na Mata Atlântica brasileira.
Megadiversidade: Desafios e oportunidades para a conservação da biodiversidade
brasileira, 132-138.
Van Sluys, M., Vrcibradic, D., Alves, M. A. S., Bergallo, H. G., Rocha, C. F. D.
(2007).Ecological parameters of the leaf-litter frog community of an Atlantic
Rainforest area at Ilha Grande, Rio de Janeiro state, Brazil. Austral Ecology 32, 254-
260.
Van Valen, L. (1965). Morphological variation and width of ecological niche. American
Naturalist 99, 377–389.

20
Vasconcelos, T. S. & Rossa-Feres, D. C. (2008). Habitat heterogeneity and use of physical
and acoustic space in anuran communities in southeastern Brazil. Phyllomedusa 7,
127-142.
Vasconcelos, T. S., Santos, T. G., Rossa-Feres, D. C. & Haddad, C.F.B. (2009). Influence of
the environmental heterogeneity of breeding ponds on anuran assemblages from
southeastern Brazil. Canadian Journal Zoology 87, 699-707.
Velloso, A. L.; Sampaio, E. V. S. B.; Giulietti, E. M.; Barbosa, M. R. V.; Castro, A. A. J. F.;
Queiroz, l. P.; Fernandes, A.; Oren, D. C.; Cestaro, L. A.; Carvalho, A. J. E.;Pareyn, F.
G. C.; Silva, F. B. R.; Miranda, E. E.;Keel, S. E Gondim, r. S. (2002). Ecorregiões do
Bioma Caatinga. Recife: APNE/TNC. 75p.
Wells, K. D. (2007). The Ecology and Behavior of Amphibians.The University of Chicago
Press.Chicago and London.p. 25.
Wiens, J. J. (2004). Speciation and ecology revisited:phylogenetic niche conservatism and the
origin of species. Evolution 58, 193–197.
Wiens, J. J. (2007). Global patterns of species richness and diversification in amphibians. The
American Naturalist 170, 86–106.
Woolbright, L. L. 1989. Sexual dimorphism in Eleutherodactylus coqui: selection pressures
and growth rates. Herpetologica 45, 68-74.

21
ARTIGO A SER SUBMETIDO À HERPETOLOGICAL JOURNAL
Nicho espacial e alimentar de anuros em áreas de Caatinga e Mata Atlântica de Sergipe,
Brasil
ADILSON DE OLIVEIRA SILVA1,2
E RENATO GOMES FARIA1,3
1Programa de Pós graduação em Ecologia e Conservação - PPEC, Universidade Federal de
Sergipe, São Cristóvão, Sergipe, Brazil, 40100-000
2Corresponding author: [email protected]
3Departamento de Biologia, Universidade Federal de Sergipe, São Cristóvão, Sergipe, Brazil

22
Resumo — Características ambientais distintas podem afetar diretamente a estruturação das
comunidades, pois contribuem com recursos e condições variadas, o que permite que em cada
área, grupos diferenciados de espécies se estabeleçam. Nesse estudo duas taxocenoses de
anfíbios anuros foram analisadas entre outubro de 2012 e setembro de 2013, em duas áreas
com diferentes graus de heterogeneidade ambiental, uma na Caatinga (Monumento Natural
Grota do Angico – MNGA) e outra de Mata Atlântica (Refugio da Vida Silvestre Mata do
Junco – RVSMJ), ambas no estado de Sergipe. Foram utilizadas informações sobre os nichos
espacial e alimentar e suas relações com fatores ecológicos e históricos. Foram tomados 1.332
registros (999 - RVSMJ e 333 - MNGA) pertencentes a 28 espécies (21 – RVSMJ e 15 –
MNGA, sendo dez comuns as duas áreas). A sobreposição de nicho espacial e alimentar foi
relativamente alta em ambas as áreas. Nove tipos de microhábitats foram utilizados pelas
espécies sendo os mais comuns, independente da área, solo (45,1%), água (23,7%) e folhiço
(8,6%). As presas mais importantes das dietas foram Formicidae e Coleoptera em ambas as
áreas. Os indivíduos da Mata Atlântica foram relativamente menores que os da Catinga
sugerindo investimentos diferenciados em reprodução e crescimento entre as áreas. Os
resultados obtidos sugerem que as taxocenoses de ambas as áreas são influenciadas por
fatores históricos ou filogenéticos somente na utilização dos recursos espaciais. Suas dietas
são fortemente influenciadas pelas disponibilidades de recursos que o ambiente oferece.
Palavras-Chave: Amphibia; Heterogeneidade Ambiental; Dieta; Uso de microhabitat.
Abstract — Distinct environmental characteristics can directly affect community structure,
they offer varying conditions and resources, which allows each area to establish different
groups of species. In this study we analyzed two amphibian assemblages between October
2012 and September 2013 in two areas with different degrees of environmental heterogeneity
, one in the Caatinga (Monument Natural Grota do Angico - MNGA ) and the other in the
Atlantic Rainforest (Refugio de Vida Silvestre Mata do Junco - RVSMJ ) , both in the state of
Sergipe . We used information about the spatial and food niches and their relationships with
ecological and historical factors. We analyzed 1,332 records (999 - RVSMJ 333 - MNGA) of
28 species (21 - RVSMJ and 15 - MNGA, ten common to both areas). The food and space
niche overlap was relatively high in both study sites. Nine types of microhabitats were used
by the species being the most common , regardless of the area , soil (45.1 %) , water (23.7 %)
and leaflitter (8.6%). The most important prey diets were Formicidae and Coleoptera in both

23
areas. Individuals in the Atlantic rainforest were relatively smaller than those of the Caatinga
area suggesting differentiated investments between reproduction and growth between the
areas. The results suggest that the assemblages from both areas are influenced by historical or
phylogenetic factors only in the use of space assets. Their diets are strongly influenced by the
availability of resources the environment offers.
Keywords: Amphibia, Environmental heterogeneity, Diet, Microhabitat use.
INTRODUÇÃO
Ambientes podem diferir em grau de complexidade oferecendo recursos e condições
variados, o que permite que em cada área, grupos diferenciados de espécies se estabeleçam.
Ambientes mais heterogêneos podem prover as espécies uma maior quantidade de recursos e
refúgios, reduzindo assim perdas catastróficas para os competidores, predadores, e estresse
ambiental, além de possibilitar uma percepção e utilização diferenciada do microhabitat por
organismos de diferentes classes (Tews et al., 2004; Barreto, 2010). Assim, a estrutura do
habitat tem sido considerada por vários autores como um importante determinador da
abundância e riqueza de espécies em uma comunidade (Mcguinness & Underwood, 1986;
Huston, 1994; Fraser, 1998; Moreno-Rueda & Pizarro, 2007; Vasconcelos & Rossa-Feres,
2008; Vasconcelos et al., 2009, Silva et al., 2011). Pois, ambientes mais heterogêneos
apresentam diferentes níveis de pluviosidade, temperatura, e luminosidade, influenciando
diretamente o nicho (temporal, espacial e alimentar) de determinadas espécies (Prado &
Pombal Jr, 2005; Araujo & Almeida-Santos, 2013).
Desta maneira, entender como esses fatores podem influenciar na estrutura das
comunidades, além de caracterizar as possíveis causas que fazem com que determinadas
espécies possam coexistir em certas condições ambientais e não em outras, são aspectos
críticos nos estudos ecológicos (Dodd, 2010; Silva & Rossa-Feres, 2011). No entanto, apesar
da heterogeneidade do habitat explicar muito dos padrões de distribuição e da ecologia das
espécies, há outros pontos que também devem ser levados em consideração, como o fator
histórico ou filogenético (Brooks & Mclennan, 1991), a partilha e disponibilidade de recursos
(Torrez-Orosco, et al., 2002), e a morfologia dos indivíduos (Avise, 2000).
Peterson et al., (1999) afirmam que algumas características são mantidas por certos
táxon irmãos, determinando suas preferências ecológicas. Essas características são então

24
retidas ao longo da história evolutiva de determinados grupos (Wiens, 2007). Assim, uma
explicação adequada para os padrões de riqueza e abundancia de espécies deve incorporar
processos evolutivos e biogeográficos em algum nível, além de considerar as correlações
climáticas conhecidas (Wiens, 2004).
A relação entre a disponibilidade e a partilha de recursos nas comunidades é outro
fator determinante a ser considerado. Alimento, espaço e tempo são as três categorias mais
comumente partilhadas pelas populações (Torrez-Orosco, et al., 2002), sendo de suma
importância compreender como as dinâmicas ambientais interferem nesses três aspectos do
nicho (Popielarz & Neal, 2007).
Tão importante quanto à partilha de recursos o tamanho corpóreo também é um
elemento significativo, pois é um traço morfológico fundamental, fazendo-se relevante em um
contexto fisiológico, ecológico e social (Avise, 2000). Assim, variações na morfologia dos
indivíduos, particularmente em relação ao tamanho corpóreo, são comuns dentro de muitas
espécies (Castellano & Giacoma,1998; Castellano et al., 1998; Dodd, 2010) e normalmente
estão associadas a particularidades de cada localidade, como condições climáticas distintas e
estrutura da vegetação (Riha & Berven, 1991; Schäuble, 2004; Juncá et al., 2008).
Logo, analisar os fatores que influenciam o nicho é uma ferramenta importante no
entendimento dos fenômenos ecológicos e evolutivos que determinam a coexistência das
espécies e os mecanismos que governam a especiação e a evolução dos ecossistemas (Alley,
1982; Popielarz & Neal, 2007; Costa et al., 2008). No entanto, em se tratando de anfíbios
existe muita controvérsia acerca da influência desses parâmetros e em especial da
heterogeneidade ambiental sobre o nicho das espécies, sendo que alguns autores apontam uma
influência negativa (Vasconcelos & Rossa-Feres, 2005; Santos et al., 2007), e outros afirmam
que populações de anfíbios que ocupam ambientes com características distintas podem ter
aspectos de sua biologia afetados por essas variações ambientais, sendo que uma mesma
espécie pode ter seu ciclo de vida influenciado de diferentes maneiras, dependendo das
condições que lhe forem impostas (Dunham & Overall, 1994; Schäuble, 2004; Prado &
Pombal, 2005; Araujo & Almeida-Santos, 2013).
Anfíbios possuem várias características fisiológicas e anatômicas que os tornam
dependentes de água (Dodd, 2010). A presença de um tegumento permeável, por exemplo, é
uma característica marcante do grupo que faz com que ambientes que ofereçam uma maior

25
disponibilidade de agua e umidade tenham maior riqueza e diversidade de espécies, como é o
caso das florestas tropicais úmidas (Azevedo-Ramos & Galatti, 2002; Araujo & Almeida-
Santos, 2013). Os ovos dos anfíbios são também altamente suscetíveis à desidratação, logo
ambientes mais secos suportam menos modos reprodutivos e consequentemente limitam a
riqueza e a diversidade de espécies nesses locais (Bentley, 1966; Wells, 2007; Dodd, 2010;
Young et al., 2011).
O estudo de organismos em ambientes contrastantes podem trazer informações
importantes a respeito das respostas desses às condições locais. Prates & Navas (2009)
sugerem que em ambientes com condições climáticas e estruturais diferenciadas como é o
caso dos biomas Caatinga (menor umidade e sítios reprodutivos mais temporários) e Mata
Atlântica (maior umidade e sítios reprodutivos mais permanentes), a forma de uso do habitat e
a morfologia dos indivíduos são fatores essenciais para sobrevivência das espécies nos locais
menos favoráveis. Em áreas mais áridas como as verificadas para a Caatinga, por exemplo, os
anfíbios anuros possuem adaptações ecofisiológicas, que minimizam a perda de água, têm
desenvolvimento mais rápido das larvas, capacidade de estivação e maior amplitude de nicho
(mais generalistas) do que organismos que habitam áreas mais úmidas (Mata Atlântica)
(Navas, et al., 2004; Tejedo et al., 2010).
Com base no disposto foram escolhidas duas áreas com características climáticas e
estruturais bem distintas para verificar a ação de fatores locais no uso dos recursos espaciais e
alimentares por anfíbios anuros. A influência de fatores históricos no padrão de uso de
recursos pelas espécies avaliadas foi também investigada. E por último procurou-se verificar
se espécies ocorrentes nas duas áreas diferiam em características morfológicas.
MATERIAL E MÉTODOS
Áreas de estudo
O estudo foi realizado de outubro de 2012 a setembro de 2013, em duas Unidades de
Conservação (UC) do estado de Sergipe, Brasil: Monumento Natural Grota do Angico
(37°40’W; 09°39’S - MNGA) e Refúgio da Vida Silvestre Mata do Junco (37°03’W; 10°32’S
- RVSMJ) sendo que em cada área foram amostrados seis corpos d´água (Figura 1).
O MNGA está inserido no domínio morfoclimático da Caatinga, apresentando uma
área de 2.182 ha e altitude média de 100 m. O clima é árido, limitado por espaços semiáridos

26
(classe BShw de Köppen), com altas temperaturas (amplitude anual: 20 – 40°C) e baixa
precipitação (em geral menos de 500 mm por ano). A região apresenta uma floresta de baixa
estatura com árvores e arbustos espinhosos (vegetação xerófila e esparsa). O regime
pluviométrico é irregular, normalmente concentrado em quatro meses (abril a agosto), seguido
por um período seco (Santos & Andrade, 1992). As condições climáticas locais caracterizam
uma elevada evapotranspiração e a presença quase exclusiva de rios temporários (excetuando
o Rio São Francisco) e acúmulos de água de tamanho e duração variáveis chamados de
tanques ou cacimbas.
O Refúgio de Vida Silvestre Mata do Junco está localizado na porção sul do município
de Capela - Sergipe, possuindo uma área total de 1520 hectares (Santos, 2007). Os fragmentos
florestais da Mata do Junco abrigam remanescentes de Mata Atlântica com estratificação bem
definida e importantes espécies vegetais (Santos, 2007) com um dossel fechado, temperatura
média anual em torno de 24,9ºC e a média anual pluviométrica é de 1372 mm. Sua
hidrografia está associada ao abastecimento aquífero da bacia do Rio Japaratuba, através do
Rio Lagartixo que tem sua nascente, grande parte do seu curso e de seus afluentes
alimentados pelas nascentes que surgem dentro da mata.
Coleta de dados
Para as amostragens foram adotadas campanhas mensais de três noites consecutivas,
em cada área com intervalos de dois dias entre elas. Em cada localidade foram amostrados
seis corpos d`água. O método utilizado foi o de busca ativa limitada por tempo (18 a 24 h).
Para cada espécime observado foram tomadas as seguintes informações: data, área, hábitat
(e.g. lagoa, brejo, riacho) e substrato utilizado (e.g. caule de árvore, solo, água).
Caracterização dos ambientes
Cada corpo d´água foi caracterizado mensalmente quanto ao: i) hidroperíodo: retenção
de água; ii) tamanho do corpo: em m2; iii) profundidade máxima: em cm; iv) altura da
vegetação na margem dos corpos d´água: (1) somente vegetação com até 30 cm de altura, (2)
categoria anterior mais vegetação com 31 a 60 cm, (3) as duas categorias anteriores mais
vegetação com 61 a 90 cm, (4) as três anteriores mais categoria de 91 a 120 cm, e (5) todas as
categorias anteriores; v) porcentagem de vegetação aquática na superfície do corpo d´água:
(1) sem vegetação; (2) até 30%; (3) entre 31% e 60%; (4) acima de 60%; vi) porcentagem de

27
cobertura vegetal emergente ereta na superfície do corpo d´água: (1) sem vegetação; (2) até
30%; (3) entre 31% e 60%; (4) acima de 60%; vii) perfil das margens: plana, inclinada ou em
barranco; viii) tipo de margens: seca, úmida ou alagada.
A
B
Figura 1 Mapa das unidades de conversão estaduais no estado de Sergipe – Brasil. A - Refugio da Vida Silvestre
Mata do Junco (RVSMJ) e B Monumento Natural Grota do Angico (MNGA).

28
Riqueza e diversidade
A eficiência da amostragem para cada área foi verificada através das curvas de
rarefação de espécies. Dois estimadores não-paramétricos (Chao 1 e Chao 2) foram adotados,
considerando espécies versus indivíduos de cada espécie. Para obtenção das curvas foi
utilizado o programa EstimateSWin 7.5.2 (Colwell, 2006), empregando 10.000
aleatorizações. Foi considerado o estimador com menor desvio padrão.
A diversidade alfa em cada área foi calculada através do índice de Shannon Wiener
(H’) (Pielou 1966, Magurran 1988) e a diversidade beta (diferença da composição de espécies
entre as áreas) pelo inverso do índice de Jaccard (1-Cj) (Krebs, 1999), ambos utilizando o
programa PAST 1.80 (Hammeret al., 2001). A influência dos montantes de precipitação sobre
a abundância total das espécies foi avaliada através do Coeficiente de Correlação de Spearman
(Legendre & Legendre, 1998).
Morfometria
Para as análises morfométrica e da dieta foram coletados, sempre que possível cinco
indivíduos, de cada espécie por mês em cada área (Licença SEMARH-SE 032.000-
01144/2012-1). Os animais foram sacrificados com doses elevadas de anestésico (lidocaína
2%), fixados em formalina 10% e conservados em álcool 70%. Todos os animais foram
depositados na Coleção Herpetológica da Universidade Federal de Sergipe (CHUFS).
Vinte medidas foram tomadas com um paquímetro digital Mitutoyo® (precisão de
0,01 mm) seguindo Freitas et al., (2008) sendo elas: comprimento rostro-cloacal (CRC);
largura e altura do corpo (LC e ACO); comprimento, largura e altura da cabeça (CC, LCA e
ACA); diâmetro do olho (DO); distâncias inter-ocular, olho-narina e inter-nasal (DIO, DON e
DI); comprimentos e larguras do braço (CB e LB), antebraço (CAB e LAB), coxa (CCX e
LCX) e perna (CPE e LPE) e comprimentos da mão e do pé (CM e CP).
Todas as variáveis morfométricas foram log10- transformadas visando aproximar a
amostra de uma distribuição normal e reduzir possíveis efeitos de escala. Variações na
morfologia das espécies foram inicialmente investigadas por duas Análises de Componentes
Principais (ACP), uma para cada área. Espécies comuns as duas áreas tiveram o primeiro
fator (isométrico –―tamanho‖), gerado de cada análise de componentes principais,
comparados por análise de variância (ANOVA). Esse fator foi adotado por acumular a maior

29
parte das variações morfológicas presentes no grupo de variáveis trabalhadas. Todas as
análises morfométricas foram realizadas no programa Systat 12.0 (Wilkinson,1990)
Dieta
Em laboratório os estômagos foram retirados e os seus conteúdos identificados com o
auxílio de lupa e bibliografia especializada (Buzzi, 2002) até o menor nível taxonômico
possível, normalmente ordem. As presas encontradas inteiras foram medidas quanto aos seus
maiores comprimento e largura (mm) com o auxílio de um paquímetro digital (0,01 mm) e
seus volumes estimados usando a fórmula do volume de um elipsóide (Magnusson et al.,
2003): Volume= (π.comprimento.largura2).
Um índice relativo de importância (IRI) foi calculado para verificar com quanto cada
item alimentar participa na dieta dos anfíbios, através da seguinte equação (Gadsden &
Palacios-Orona, 1997): IRI=F%(N%+V%) onde F é a frequência, N o número e V o volume
de presas, todos em porcentagem.
Larguras e sobreposições de nicho
As larguras de nicho espacial e trófico foram calculadas através do inverso do índice
de diversidade Simpson (Simpson, 1949):
onde i é a categoria de microhábitat ou de presa, n é o número de categorias e p é a proporção
da categoria i.
Para caracterizar as espécies (em cada área) como especialistas e generalistas quanto
aos nichos espacial e alimentar, foi utilizado o índice de Levins (1968) padronizado.
Conforme a equação:
onde: Ba é o índice de Levins padronizado, B refere-se ao inverso do índice de Simpson e N
ao número de possíveis categorias de recurso. Este índice varia de 0 a 1, sendo que os valores
próximos a 0 indicam nicho espacial ou trófico mais restrito e próximos a 1 mais amplo. Os

30
resultados de Ba ≥0,5 foram atribuídos às espécies generalistas e Ba<0,1 às especialistas
(Krebs 1999).
As sobreposições de nicho espacial e alimentar das taxocenoses foram calculadas
através do índice de sobreposição de Pianka (1973)
onde p representa a proporção da categoria de microhabitat ou de recurso alimentar, e j e k
representam as espécies comparadas. As sobreposições (Øjk) podem variar de 0 (sem
similaridade ou sobreposição) a 1 (similaridade ou sobreposição completa). Os dados foram
processados no módulo de sobreposição de nicho do programa ECOSIM (Gotelli &
Entsminger, 2010). Para as simulações foi adotado o algoritmo de randomização RA3, ideal
para detectar padrões de sobreposição de nichos não aleatórios (Gotelli & Entsminger, 2010).
Influência filogenética na dieta e uso do hábitat
O método de ordenação filogenética canônica (CPO) (Giannini, 2003) é uma forma de
comparar a conservação filogenética sobre os aspectos ecológicos das espécies no caso do
presente trabalho, dos anfíbios anuros. Ele é baseado na análise de correspondência canônica
(Ter Braak, 1986). E consiste em obter uma estimativa dos efeitos históricos sobre um
conjunto de variáveis biológicas através de métodos de ordenação. Para isso, foram
construídas duas matrizes para cada tipo de avaliação, no caso, dieta e uso do microhábitat,
uma com dados filogenéticos (X) e outra com dados ecológicos (Y). A matriz filogenética X
foi construída com base em uma árvore filogenética preexistente das espécies envolvidas e é
composta por uma combinação de indicadores binários (0 e 1) para cada grupo monofilético.
O número 1 é assinalado para a espécie que pertence a um dado grupo, e o 0 para as que não
pertencem àquele grupo. Isso foi feito com todas as espécies até que se obtivesse uma matriz
que representasse a relação entre as espécies e os clados monofiléticos que as agrupam. A
árvore filogenética utilizada para a matriz X foi construída segundo Pyron & Wiens (2011). A
segunda Matriz, Y, consiste nas proporções adotadas de cada categoria de recurso (presas ou
substrato) por cada espécie. As duas áreas foram avaliadas separadamente.

31
RESULTADOS
Riqueza, abundância e diversidade de espécies
Foram tomados 1.332 registros (999 - RVSMJ e 333 - MNGA) de anfíbios anuros
pertencentes a 28 espécies (21 – RVSMJ e 15 – MNGA, sendo dez comuns as duas áreas;
Tabela 1). As espécies mais abundantes no RVSMJ foram Hypsiboas albomarginatus
(14,4%), Rhinela jimi (11,5%), Leptodactylus latrans (9,9%) e Hypsiboas faber (9,2%) e para
o MNGA Physalaemus albifrons (29,7%), Pleurodema diplolister (16,82%), Scinax
pachicrus (14,4%), Scinax x-signatus (8,1%) e Rhinela granulosa (7,2%) (Tabela 1).
As riquezas de espécies (Chao1) encontradas em cada área de estudo foram 21,00 ±
0,25 espécies para RVSMJ e de 15,50 ± 1,30 espécies para o MNGA (Figuras 2 e 3). As
diversidades totais das áreas de estudo foram de H’=2,68 para o RVSMJ e H’=2,13 para o
MNGA. A diversidade beta (1-C) foi de 0,69.
Figura 2 Curva de rarefação de espécies observadas e obtidas através do estimador não paramétrico Chao 1 a partir
do número de indivíduos registrados na Refugio da Vida Silvestre Mata do Junco, Sergipe, Brasil, no período de
Outubro de 2012 a Setembro de 2013.
Figura 3 Curva de rarefação de espécies observadas e obtidas através do estimador não paramétrico Chao 1 a partir do
número de indivíduos registrados no Monumento Natural grota do Angico, Sergipe, Brasil, no período de Outubro de
2012 a Setembro de 2013.

32
Tabela 1 Microhábitat e amplitude de nicho espacial para as espécies de anfíbios do Refúgio de Vida Silvestre Mata do Junco e Monumento Natural Grota do Angico,
Sergipe, Brasil. So = solo, Ag = agua, Ar = arvores, PAq = planta aquática, GAr = galho de arvore, Fo = folhiço, Ro = rocha, CHer = caule de herbácea, Gr = gramínea.
Taxón Bioma Microhabitat
N Largura do nicho
So Ag Ar PAq GAr Fo Ro CHer Gr B Levins Padronizado
Brachycephalidae I. ramagii Ma 7 0 0 0 0 24 0 1 6 38 3,03 0,25
I. vinhai Ma 0 0 0 0 0 22 0 0 1 23 1,09 0,01
Bufonidae
R. granulosa Ma 21 1 0 0 0 2 0 0 0 24 1,29 0,04
R. granulosa Ca 15 2 0 0 0 5 0 0 0 22 2,62 0,2
R. jimi Ma 61 47 0 0 0 2 0 0 0 110 2,06 0,13
R. jimi Ca 3 0 0 0 0 0 0 0 0 3 2 0,13
Cycloramphidae
P. renalis Ma 2 0 0 0 0 14 0 0 0 16 1,28 0,03
P. cristiceps Ca 13 0 0 0 0 0 0 1 0 14 2,14 0,14
Hylidae
C. greeningi Ma 0 2 0 1 1 0 0 0 0 4 2,67 0,21
C. greningii Ca 0 0 0 0 6 2 0 0 0 8 2,46 0,18
D. branneri Ma 0 1 0 2 3 0 0 11 7 24 3,13 0,27
D. minutus Ma 0 2 0 13 1 0 0 13 10 39 3,43 0,3
H. albomarginatus Ma 5 21 0 12 42 0 0 55 1 136 3,51 0,31
H. crepitans Ma 4 6 0 0 4 0 0 3 0 17 3,75 0,34
H. crepitans Ca 0 0 0 0 1 0 0 0 0 1 1 0
H. faber Ma 14 55 0 0 0 0 0 23 0 92 2,26 0,16
H. raniceps Ma 8 25 0 1 1 0 0 32 6 73 3,04 0,26
P. nordestina Ma 2 0 0 0 3 0 0 4 0 9 2,79 0,22
P. nordestina Ca 1 3 1 0 8 0 2 0 0 15 2,96 0,25
S. x-signatus Ma 3 0 0 0 0 0 0 0 0 3 2 0,13
S. x-signatus Ca 14 0 1 0 2 10 0 0 0 27 2,83 0,23
S. pachicrus Ca 1 3 0 2 2 13 27 0 0 48 2,86 0,23

33
Tabela 2 (Continuação) Microhábitat e amplitude de nicho espacial para as espécies de anfíbios do Refúgio de Vida Silvestre Mata do Junco e Monumento Natural Grota do
Angico, Sergipe, Brasil. So = solo, Ag = agua, Ar = arvores, PAq = planta aquática, GAr = galho de arvore, Fo = folhiço, Ro = rocha, CHer = caule de herbácea, Gr =
gramínea.
Taxón Bioma Microhabitat
N Largura do nicho
So Ag Ar PAq GAr Fo Ro Cher Gr B Levins Padronizado
Leiuperidae
P. cuvieri Ma 11 1 0 0 0 16 0 0 1 29 2,22 0,15
P. albifrons Ca 78 17 0 0 0 0 0 0 0 95 2,34 0,17
P. diplolister Ca 50 6 0 0 0 0 0 0 0 56 2,21 0,15
Leptodactylidae
L. natalensis Ma 19 43 0 0 0 9 0 0 7 78 2,57 0,2
L. trogodytes Ma 23 2 0 0 0 0 0 0 0 25 1,17 0,02
L. trogodytes Ca 1 1 0 0 0 0 0 0 0 2 2,67 0,21
L. latrans Ma 43 50 0 0 0 1 0 0 4 98 2,2 0,15
L. latrans Ca 7 2 0 0 0 0 0 0 0 9 2,41 0,18
L. fuscus Ma 37 21 0 0 0 1 0 1 10 70 2,56 0,2
L. fuscus Ca 15 6 0 0 0 21 0 0 0 42 2,51 0,19
L. vastus Ma 28 47 0 0 0 0 0 0 0 75 1,91 0,11
Microhylidae
D. mulleri Ma 1 0 0 0 0 0 0 0 0 1 1 0
D. mulleri Ca 4 0 0 0 0 1 0 0 0 5 2,38 0,17
Pipidae
P. carvalhoi Ca 1 0 0 0 0 0 0 0 1 1 1 0

34
Não houve correlação significativa entre os montantes de precipitação mensais e a
abundância de indivíduos das duas áreas rma= 0,190; p= 0,157 e rca = 0,006; p = 0,804 e os
modelos propostos respondem muito pouco do que foi observado (Figura 4).
Figura 4. Regressão Linear simples. A - Refugio da Vida Silvestre Mata do Junco (RVSMJ) e B -Monumento
Natural Grota do Angico (MNGA).
A
B

35
Uso do espaço
A Tabela 2 apresenta um resumo das características estruturais dos corpos d’água
avaliados em cada área. Nove tipos de microhábitats foram utilizados pelas espécies sendo os
mais comuns, independente da área, solo (45,1%), água (23,7%) e folhiço (8,6%). Os
substratos mais adotados pelas espécies do RVSMJ foram água (33,4%), solo (29,3%) e
herbáceas (14,2%) e para o MNGA solo (60,1%), água (14,1%) e folhiço (9,3%) (Tabela 1).
As larguras de nicho (B) espacial dos anuros do RVSMJ variaram de 1,09 (I. vinhai) a
3,75 (H. crepitans). Maiores especializações no uso dos microhábitats (BA) foram verificadas
para I. vinhai (0,01), P. renalis (0,03) e R. granulosa (0,04) e o uso mais generalizado dos
substratos para H. crepitans (0,34), H. albomarginatus (0,31), D. minutus (0,30) e D. branneri
(0,27) (Tabela 1). Em relação ao MNGA, as larguras de nicho (B) espacial variaram de 2,00
(R. jimi) a 2,96 (P. nordestina), com as maiores especializações (BA) para P. nordestina
(0,25), S. x-signatus (0,23) e S. pachicrus (0,23) e o uso mais generalizado para R. jimi
(0,13), P. cristiceps (0,14) e P. diplolister (0,15) (Tabela 1).
A sobreposição de nicho espacial média observada para o RVSMJ foi de 0,41
(variando de 0 a 0,99), sendo essa maior do que a média esperada ao acaso (P<0,000001)
(Tabela 3). As maiores sobreposições foram entre L. troglodytes e R. granulosa (0,99) e L.
latrans e R.jimi (0,99). Já para o MNGA a sobreposição média observada foi de 0,53 (0 a
0,99), também maior que a verificada ao acaso (P<0,000001). As maiores sobreposições
foram entre P. albifrons e L. latrans e entre R. jimi e P. cristceps (ambas 0,99) (Tabela 4).
Dieta
Foram analisados 313 estômagos (227 – RVSMJ e 86 – MNGA), desses 63 (46 –
RVSMJ e 17 MNGA) estavam vazios ou em estado avançado de digestão. Foram encontradas
21 categorias de presas (16 RVSMJ e 17 MNGA). As presas mais importantes (IRI)
encontradas no RVSMJ foram Formicidae (20733,23), Coleoptera (13892,22) e Orthoptera
(5424,32) (Tabela 5). Já no MNGA os itens mais importantes foram Formicidae (20910,21),
Coleoptera (15329,2) e larva de insetos (11212,9) (Tabela 6).

36
Tabela 3 Caracteristicas estruturais presentes nos corpos d´água estudados em ambas as áreas. Estratos de vegetação: N = nenhum; HR = herbáceas rasteiras; HE = herbáceas
eretas; Ab = arbustos; Av = árvores. Perfis de Margem: P = plana; I = inclinada; B = barranco. Tipos de margem: S = seca; U = úmida; A = alagada.
Corpo d’água Tamanho
(m²)
Estratos de
vegetação
Perfis de
margem
Tipos de
margem
Vegetação
aquática
(%)
Plantas
emergente
(%)
Altura da
vegetação
(cm)
Profundidad
e (cm)
Duraçã
o
(Meses)
RVSMJ
lagoa seca 500 HR P S 0 0 0 90 12
borda 1 16 HR,HE, AB, AV P, B S 0 0 700 150 6
borda 2 12 HR,HE, AB P, B S 0 0 700 60 3
borda 3 8 HR, HE, AB P, B S 0 0 400 40 2
brejo saae 78 HR P U, A 0 90 30 10 12
Riacho 400 HR,HE, AB, AV P, I U 10 5 1200 200 12
MNGA
lagoa da vacaria 12,3
HR, HE, AB,
AV I, B S 0 0 200 100 12
lagoa da casa 8,9 HR,HE, AB I, B S 0 10 60 0 3
lagoa da sede 6 HR, HE, AB I S 0 0 60 0 1
lagoa da trilha 10 HR, HE, AB I S 0 0 60 0 1
lagoa da estrada 30
HR, HE, AB,
AV P S 35 15 200 0 5
Riacho 400 AB, AV P S 5 0 400 0 4

37
Tabela 4 Sobreposição de Nicho espacial entre os pares de espécies coletadas no Refugio da Vida Silvestre Mata do Junco, Sergipe, Brasil no período de Outubro de 2012 a
Setembro de 2013. Corythomantis greeningi (Cg), Dendropsophus branneri (Db), Dendropsophus minutus (Dmi), Dermatonotus muelleri (Dmu), Hypsiboas albomarginatus
(Ha), Hypsiboas creptans (Hc), Hypsiboas faber (Hf), Hypsiboas raniceps (Hr), Ischnocnema ramagii (Ir), Ischnocnema vinhai (Iv), Leptodactylus natalensis (Ln),
Leptodactylus troglodytes (Lt), Leptodactylus latrans (Ll), Leptodactylus fuscus (Lf), Leptodactylus vastus (Lv), Physalaemus cuvieri (Pc), Phyllomedusa nordestina (Pn),
Proceratophrys renalis (Pr), Rhinella granulosa (Rg), Rhinella jimi (Rj), Scinax x-signatus (Sx). Em negrito os valores de sobreposição maiores que 0,9.
Espécies Ir Iv Rg Rj Pr Cg Db Dmi Há Hc Hf Hr Pn Pc Ln Lt Ll Lf Lv
Ir
0,62 0,46 0,33 0,64 0 0,39 0,35 0,06 0,21 0,11 0,21 0,18 0,75 0,37 0,41 0,32 0,53 0,22
Iv
0,09 0,02 0,98 0 0,02 0,02 0 0 0 0 0 0,82 0,19 0 0,01 0,03 0
Rg
0,79 0,23 0,03 0 0 0,07 0,48 0,26 0,21 0,36 0,64 0,45 0,99 0,72 0,86 0,54
Rj
0,13 0,52 0,04 0,06 0,25 0,78 0,75 0,52 0,28 0,48 0,87 0,81 0,99 0,97 0,95
Pr
0 0 0 0 0,06 0,03 0,02 0,05 0,88 0,23 0,14 0,11 0,14 0,07
Cg
0,21 0,34 0,57 0,74 0,73 0,51 0,22 0,04 0,72 0,07 0,59 0,39 0,7
Db
0,85 0,76 0,42 0,37 0,74 0,72 0,03 0,14 0 0,08 0,27 0,71
Dm
0,61 0,29 0,31 0,61 0,48 0,02 0,15 0 0,09 0,16 0,06
Há
0,76 0,57 0,77 0,88 0,05 0,31 0,09 0,28 0,23 0,08
Hc
0,84 0,76 0,67 0,29 0,78 0,51 0,81 0,72 0,31
Hf
0,86 0,36 0,17 0,88 0,31 0,81 0,62 0,82
Hr
0,65 0,14 0,62 0,24 0,57 0,49 0,88
Pn
0,21 0,14 0,36 0,25 0,33 0,61
Pc
0,23 0,56 0,43 0,53 0,19
Ln
0,46 0,92 0,79 0,33
Lt
0,73 0,88 0,96
Ll
0,93 0,97
Lf
0,84
Lv

38
Tabela 5 Sobreposição de Nicho espacial entre os pares de espécies coletadas no Monumento Natural Grota do Angico, Sergipe, Brasil no período de Outubro de 2012 a
Setembro de 2013. Rhinella granulosa (Rg), Rhinella jimi (Rj) Proceratophrys cristiceps (Pcr), Corythomantis greeningi (Cg), Hypsiboas creptans (Hc), Phyllomedusa
nordestina (Pn), Scinax x-signatus (Sx), Scinax pachicrus (Sp), Physalaemus albifrons (Pa), Pleurodema diplolister (Pd) Dermatonotus muelleri (Dmu), Leptodactylus
latrans (Ll), Leptodactylus fuscus (Lf). Em negrito os valores de sobreposição maiores que 0,9.
Espécies Rg Rj Pcr Cg Hc Pn Sx Sp Pa Pd Ll Lf Dm
Rg
0,94 0,93 0,09 0 0,21 0,94 0,17 0,94 0,94 0,93 0,92 0,98
Rj
0,99 0 0 0,11 0,8 0,03 0,97 0,99 0,96 0,92 0,97
Pcr
0 0 0,11 0,8 0,03 0,97 0,98 0,95 0,92 0,96
Cg
0,94 0,32 0,29 0,19 0 0 0 0 0,07
Hc
0,33 0,11 0,06 0 0 0 0 0
Pn
0,13 0,31 0,3 0,21 0,35 0,43 0,1
Sx
0,28 0,78 0,8 0,77 0,79 0,92
Sp
0,05 0,04 0,05 0,06 0,13
Pa
0,99 0,99 0,98 0,94
Pd
0,98 0,96 0,96
Ll
0,99 0,93
Lf
0,91
Dm

39
Tabela 5 Índice Relativo de Importância de categorias de presas na dieta da taxocenose de anfíbios no Refugio da Vida Silvestre Mata do Junco, Sergipe, Brasil. Hypsiboas albomarginatus
(Ha), Hypsiboas creptans (Hc), Hypsiboas faber (Hf), Hypsiboas raniceps (Hr), Ischnocnema ramagii (Ir), Leptodactylus natalensis (Ln), Leptodactylus latrans (Ll), Leptodactylus fuscus
(Lf), Leptodactylus vastus (Lv), Physalaemus cuvieri (Pc), Proceratophrys renalis (Pr), Rhinella granulosa (Rg), Rhinella jimi (Rj).
Categoria das Presas
Espécies
D. m. H. a. H. c. H. f. H. r. I. r. L. l. L. n. L. f. L.v. P. c. P. r. R. g. R. j.
Acarina 0,0 15,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Araneae 123,2 92,4 0,0 10,2 0,0 0,0 0,0 438,9 22,1 31,3 0,0 0,0 0,0 0,3
Blattaria 0,0 0,0 0,0 0,0 0,0 0,0 0,0 15,9 183,3 0,0 0,0 277,2 0,0 0,0
Coleoptera 1345,4 88,0 150,2 460,1 2848,1 71,6 5341,4 894,0 278,3 144,6 770,2 45,8 123,4 1331,3
Dermaptera 0,0 0,0 0,0 0,0 0,0 0,0 0,0 33,6 0,0 0,0 0,0 0,0 0,0 0,0
Diplopoda 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 486,2 0,0 0,0 0,0 12,0
Diptera 0,0 151,5 0,0 0,0 13,9 0,0 25,7 28,9 53,6 0,0 0,0 0,0 0,0 0,0
Hymenoptera 0,0 0,0 0,0 0,0 0,0 0,0 0,0 6,9 26,6 20,0 0,0 0,0 0,0 0,5
Formicidae 58,9 604,1 310,4 213,3 328,7 87,9 140,3 83,1 406,6 543,8 1231,4 38,3 3751,0 12935,5
Hemiptera 59,2 158,8 0,0 0,0 55,2 0,0 46,0 228,2 52,7 30,9 0,0 0,0 0,0 119,8
Isoptera 0,0 0,0 0,0 0,0 0,0 1051,2 104,7 0,0 0,0 0,0 0,0 0,0 87,9 0,0
Larva de Inseto 0,0 445,3 38,1 20,5 40,3 0,0 348,7 233,4 0,0 98,6 0,0 0,0 0,0 3,7
Lepidoptera 0,0 0,0 0,0 27,6 82,4 0,0 65,6 0,0 0,0 0,0 0,0 0,0 0,0 2,4
Ortoptera 0,0 50,5 382,0 794,1 100,6 48,5 438,8 847,3 138,1 238,6 500,4 1867,7 0,0 17,7
Mollusca 0,0 0,0 0,0 0,0 0,0 0,0 0,0 141,8 0,0 38,0 0,0 0,0 0,0 0,8
Vertebrado 0,0 0,0 0,0 0,0 0,0 0,0 20,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Material vegetal 0,0 0,0 0,0 9,9 22,8 0,0 364,1 524,2 94,0 0,0 0,0 214,3 1,4 4,5
Material mineral 0,0 0,0 0,0 0,0 0,0 0,0 0,0 8,5 10,4 0,0 0,0 0,0 0,0 1,8
Largura do nicho numérico 3,00 4,52 1,67 2,79 3,67 1,53 5,34 10,12 5,90 4,47 2,36 3,60 1,40 1,25
Largura do nicho
volumétrico 1,48 3,78 2,87 2,23 2,05 1,79 2,91
5,00 3,70 2,18 2,46 1,50 1,06 1,49

40
Tabela 6 Índice Relativo de Importância das categorias de presas na dieta da taxocenose de anfíbios do Monumento Natural Grota do Angico, Sergipe, Brasil. Leptodactylus
latrans (Ll), Leptodactylus fuscus (Lf), Physalaemus albifrons (Pa), Proceratophrys cristiceps (Pcr), Pleurodema diplolister (Pd), Rhinella jimi (Rj), Rhinella granulosa
(Rg), Scinax pachicrus (Sp), Scinax x-signatus (Sx).
Categoria das Presas Espécies
Ll Lf Pa Pcr Pd Rj Rg Sp Sx
Acarina 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 126,2
Aranae 0,0 23,8 2143,5 0,0 0,0 0,0 0,0 1675,5 138,3
Blattaria 213,8 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Coleoptera 4737,1 1702,7 0,0 2600,0 2610,6 49,7 628,4 2638,2 362,5
Diptera 0,0 35,0 0,0 0,0 141,3 0,0 0,0 710,4 0,0
Hymenoptera 173,9 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Formicidae 0,0 387,1 3876,6 904,2 0,0 2193,1 11788,2 0,0 1761,6
Hemiptera 0,0 190,4 0,0 0,0 396,6 59,2 3,4 0,0 0,0
Homoptera 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Isoptera 0,0 857,0 0,0 0,0 0,0 613,8 229,9 0,0 0,0
Larva de Inseto 2685,7 158,2 624,8 326,2 3809,6 0,0 295,7 293,8 3024,0
Lepidoptera 460,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Mantodea 0,0 0,0 0,0 0,0 277,6 88,5 0,0 0,0 0,0
Ortoptera 0,0 0,0 452,4 2369,8 0,0 278,1 0,0 0,0 0,0
Quilopoda 0,0 0,0 0,0 52,4 0,0 0,0 0,0 0,0 0,0
Scorpionida 460,7 0,0 0,0 0,0 0,0 36,3 0,0 0,0 0,0
Mollusca 0,0 196,5 0,0 73,2 0,0 347,8 0,0 0,0 0,0
Mat. Vegetal 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Mat. Mineral 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Largura do nicho numérico 3,92 1,82 1,94 3,80 3,06 3,12 1,46 3,00 3,86
Largura do nicho volumétrico 2,84 3,69 2,90 3,57 2,98 3,10 1,91 2,83 2,39

41
As larguras de nicho alimentar para os anuros amostrados no RVSMJ variaram de 1,25
(R.jimi) a 10,12 (L. natalensis) para número e de 1,06 (R. granulosa) a 5,00 (L. natalensis)
para volume de presas ingeridas (Tabela 5). No MNGA a largura de nicho variou 1,46 (P.
diplolister) a 3,92 (L. latrans) para número e 1,91 (R. granulosa) a 3,69 (L.fuscus) para
volume de presas ingeridas (Tabela 6).
A média de sobreposição de nicho alimentar observada no RVSMJ foi 0,44 sendo
significativamente maior que a esperada ao acaso (P < 0,00001). No MNGA a média foi 0,42
e também significativamente maior que a simulada (P<0,000001). Com relação à
sobreposição dos itens alimentares utilizados, os valores variaram de 0 a 0,99 para a RVSMJ
sendo as maiores sobreposições verificadas entre R.jimi e R. granulosa (0,99) e entre L.
latrans e H. raniceps (0,99) (Tabela 7). Já no MNGA, a sobreposição variou de 0 até 0,94,
sendo as mais altas entre R. jimi e R. granulosa (0,95) e entre P. diplolister e L. latrans (0,88)
(Tabela 8).
Morfometria
Informações sobre a morfometria das espécies das duas áreas estão presentes nas
Tabelas 9 e 10. Foram analisados 345 registros (96 para o MNGA – e 249 para o RVSMJ). Os
resultados da Análise de Componentes Principais (ACP) indicam variações na morfologia das
espécies que compõem cada taxocenose. Os dois primeiros componentes acumularam 88,73%
da variação explicada pelo conjunto de componentes da ACP para a área do RVSMJ e 83%
para o MNGA. Na área do RVSMJ valores positivos estão associados a maiores CRC,
comprimentos do antebraço e da mão e largura do antebraço (Tabela 11). Na área do MNGA
valores positivos estão associados ao CRC e comprimentos da mão e do antebraço (Tabela
11).
Espécies comuns as duas áreas apresentaram diferenças quanto ao primeiro
componente principal (eixo isométrico - tamanho) (L. latrans - F1,30 = 61.796; P < 0,0001; L.
fuscus- F1,26 = 62.615; P < 0,0001; R. granulosa- F1,20 = 7.799; P = 0,011; R. jimi- F1,27 =
16.561; P < 0,0001). Em todos os casos, espécimes maiores foram verificados para a área do
MNGA (Figura 5).

42
Figura 5 Analise de variância entre o primeiro componente da ACP do indivíduos comuns as duas áreas. A = L. latrans,
B = L. fuscus, C = R. granulosa e D = R. jimi.
Least Squares Means
angico junco
AREA$
-1
0
1
2
3
CP
1
Least Squares Means
angico junco
AREA$
-2
-1
0
1
CP
1
Least Squares Means
angico junco
AREA$
-2
-1
0
1
CP
1
Least Squares Means
angico junco
AREA$
-1
0
1
2
3
4
CP
1
A B
C D

43
Tabela 7 Sobreposição de Nicho alimentar entre os pares de espécies coletadas no Refugio da Vida Silvestre Mata Junco, Sergipe, Brasil no período de Outubro de 2012 a
Setembro de 2013. Dendropsophus minutus (Dm), Hypsiboas albomarginatus (Ha), Hypsiboas creptans (Hc), Hypsiboas faber (Hf), Hypsiboas raniceps (Hr), Ischnocnema
ramagii (Ir), Leptodactylus latrans (Ll), Leptodactylus natalensis (Ln), Leptodactylus fuscus (Lf), Leptodactylus vastus (Lv), Physalaemus cuvieri (Pc), Proceratophrys
renalis (Pr), Rhinella granulosa (Rg), Rhinella jimi (Rj), Em negrito os valores de sobreposição maiores que 0,9.
Espécies Dm Há Hc Hf Hr Ir Ll Ln Lf Lv Pc Pr Rg Rj
Dm
0,16 0,00 0,96 0,99 0,07 0,99 0,65 0,54 0,22 0,53 0,02 0,76 0,15
Ha
0,58 0,29 0,21 0,07 0,17 0,31 0,67 0,65 0,69 0,80 0,76 0,77
Hc
0,90 0,38 0,10 0,37 0,66 0,77 0,70 0,09 0,75 0,61 0,63
Hf
0,54 0,09 0,06 0,08 0,62 0,50 0,70 0,84 0,24 0,28
Hr
0,08 0,99 0,64 0,59 0,27 0,60 0,06 0,15 0,22
Ir
0,07 0,07 0,11 0,70 0,12 0,05 0,09 0,09
Ll
0,69 0,55 0,23 0,54 0,11 0,06 0,13
Ln
0,59 0,37 0,54 0,63 0,08 0,12
Lf
0,67 0,92 0,34 0,74 0,78
Lv
0,74 0,32 0,69 0,71
Pc
0,35 0,82 0,85
Pr
0,02 0,02
Rg
1,00
Rj

44
Tabela 8 Sobreposição de nicho alimentar entre os pares de espécies coletadas no Monumento Natural Grota do Angico, Sergipe, Brasil, no período de Outubro de 2012 a
Setembro de 2013. Leptodactylus latrans (Ll), Leptodactylus fuscus (Lf), Physalaemus albifrons (Pa), Proceratophrys cristiceps (Pcr), Pleurodema diplolister (Pd), Rhinella
jimi (Rj), Rhinella granulosa (Rg), Scinax pachicrus (Sp), Scinax x-signatus (Sx). Em negrito os valores de sobreposição maiores que 0,9.
Espécies Ll Lf Pa Pc Pd Rg Sp Sxs Rj
Ll
0,78 0,07 0,66 0,89 0,06 0,75 0,51 0,02
Lf
0,19 0,67 0,57 0,25 0,73 0,26 0,34
Pa
0,29 0,11 0,86 0,26 0,57 0,83
Pcr
0,47 0,29 0,59 0,27 0,33
Pd
0,05 0,54 0,76 0,02
Rg
0,05 0,53 0,95
Sp
0,18 0,02
Sx
0,47
Rj

45
Tabela 9 Estatística descritiva dos caracteres morfológicos dos anfíbios anuros do Monumento Natural Grota do Angico, Sergipe - Brasil. Medidas lineares são dadas em
milímetros. CRC = Comprimento rostro-cloacal, LC = Largura do corpo, ACO = Altura do corpo, CC = Comprimento da cabeça, LCA = Largura da cabeça, ACA = Altura da
cabeça, DO = Diâmetro do olho, DIO = Distância inter-ocular, DON = Distância olho-narina, DIN = Distância inter-nasal, CB = Comprimento do braço, LB = Largura do
braço, CAB = Comprimento do antebraço, LAB = Largura do antebraço, CCX = Comprimento da coxa, LCX = Largura da coxa, CPE = Comprimento da perna, LPE =
Largura da perna, CM = Comprimento da mão, CP = Comprimento do pé. Dermatonotus muelleri (Dmu), Leptodactylus troglodytes (Lt), Leptodactylus fuscus
(Lf),Leptodactylus latrans (Ll), Physalaemus albifrons (Pa), Proceratophrys cristiceps (Pcr), Pleurodema diplolister (Pd), Phyllomedusa nordestina (Pn), Rhinella granulosa
(Rg), Rhinella jimi (Rj), Scinax pachicrus (Sp), Scinax x-signatus (Sx), Pipa carvalhoi (Pc) Hypsiboas creptans (Hc), Corythomantis greeningi (Cg).
Dmu Lt Lf Ll Pa Pcr Pd Pn Rg Rj Sp Sx Pc Hc Cg
n=4 n=2 n=9 n=5 n=17 n=14 n=7 n=2 n=11 n=2 n=11 n=9 n=1 n=1 n=1
CRC 54,8±3,4 36,81±0 44,5±3,3 78,3±1,6 28,06±1,7 47,6±4,3 31,4±4,8 39,8±0,9 45,2±4,5 100,1±6,9 31,3±2,9 37,4±2,3 46,7 56,5 49,0
LC 35,8±3,7 13,7±0,9 20,6±3,2 32,8±0,5 14,06±2,7 29,3±4,4 14,8±1,5 12,5±0,9 23,8±2,5 64,6±1,9 14,8±1,1 17,3±1,3 23,3 21,5 18,9
ACO 15,06±1,1 5,09±0,3 8,8±1,3 15,5±0,7 5,6±0,9 12,6±1,9 7,2±1,2 6,5±0,8 8,2±1,2 24,3±0,1 6,2±1,1 7,2±0,4 7,5 9,4 7,8
CC 12,7±2,8 14,01±1,1 15,1±1,8 23,3±2,2 7,3±0,7 16,4±1,5 10,3±1 11,5±0,7 12,9±1, 28,3±0,1 10,±1,21 12,4±0,8 11,0 26,4 20,1
LCA 16,5±1,7 11,5±0,6 14,4±2,2 23,3±1 9,4±1 20,4±1,6 10,8±2,7 11,5±0,2 135,5±395 34,2±2,9 9,8±0,9 11,47±1,2 13,7 17,5 15,0
ACA 7,7±1,8 4,23±0,2 5,9±0,4 11,05±0,2 3,8±0,5 9,2±1,5 4,7±1,2 5,3±0,9 6,09±0,7 12,7±1,1 3,8±0,7 4,7±0,3 4,4 7,7 3,9
DO 3,6±0,4 3,6±0,6 4,3±0,3 5,8±1,5 2,4±0,4 4,9±0,5 3,5±0,6 3,8±0,3 5,04±0,5 9,2±0,3 3,28±0,3 3,7±0,3 2,8 4,8 4,9
DIO 6,9±0,9 4,7±0,8 5,05±1 9,2±1,2 4,5±0,5 7,8±0,8 4,4±0,8 5,84±0 7,4±0,7 13,2±1,1 4,5±0,4 4,5±0,7 6,5 8,0 8,3
DON 3,3±0,7 3,35±0,8 3,9±0,5 6,3±0,3 2,7±0,5 4,04±0,4 2,6±0,6 2,76±0,1 3,5±0,6 6,06±0 3,5±0,4 4,1±0,2 4,0 5,5 7,1
DI 2,70,2 2,8±0,5 2,9±0,6 5,1±0,9 2,1±0,4 3,6±0,5 2,6±0,3 4,6±3 3,1±0,5 4,11±0,9 2,8±0,4 3,19±o,3 3,1 3,3 3,4
CB 14,2±1,5 8,4±1,5 9,1±1,9 20,1±0,6 7,1±0,9 12,7±1,3 8,6±1,9 10,5±0,5 11,2±1,7 20,3±0,6 7,5±0,9 8,7±0,4 9,7 11,2 10,0
LB 5,9±1,8 1,7±0,6 3,3±0,7 6,9±1 2,1±0,4 3,8±1,2 2,2±0,5 1,75±0 3,1±1,1 7,6±0 2,17±0,4 2,29±0,4 2,7 3,3 2,3
CAB 11,4±1 6,3±1,1 8,4±0,8 13,6±0,3 5,1±0,3 9,7±1,1 5,6±1,2 9,37±0,6 10,4±1,2 21,5±0,9 6,64±0,9 7,23±0,5 11,3 11,1 9,1
LAB 5,8±1,1 2,2±1 3,6±0,6 6,4±0,8 2,03±0,4 3,4±0,5 2,3±0,5 2,4±0,3 3,0±0,4 8,005±1 2,28±0,2 2,47±0,1 2,8 4,3 3,5
CCX 19,8±1,3 12,2±2 17,6±2,5 29,8±0,7 11,5±1,6 17,2±1,2 12,07±1,7 15,1±1,6 13,3±1,5 34,59±5,2 14,3±1,8 15,8±1 20,7 27,2 20,3
LCX 8,6±0,7 6,4±0,6 8,6±1,3 14,7±2 3,9±0,7 6,9±1,4 4,9±1,5 3,1±0,3 5,2±0,7 15,5±2,1
4,2±0,7 5,04±0,3 8,4 10,0 6,4
CPE 15,8±1,3 17,1±5,3 17,8±1,1 31,0±0,9 10,1±0,9 15,4±1,7 11,5±2,2 14,9±0,2 11,6±2,8 29,1±0,5 14,6±2 16,3±1,3 21,7 25,3 23,7
LPE 6,5±0,8 4±0,8 5,9±0,5 10,4±1,5 2,7±0,5 5,01±0,8 3,6±0,7 2,69±0 4,7±0,6 11,7±0,4 4,1±0,4 4,4±0,2 6,7 5,7 3,3
CM 14,04±0,3 7,2±1,1 8,9±0,7 16,08±0,7 5,4±0,4 12,5±0,6 7,8±1,1 10,2±1,5 9,7±1,4 22,3±2,7 7,62±0,8 8,9±0,4 9,3 15,4 13,4
CP 26,4±1,3 22,2±1 30,1±2,5 49,7±2,1 16,9±1,4 23,4±2,3 18,3±3,2 21,9±1,6 24,01±2,8 49,6±2,2 20,6±2 23,4±1,1 28,5 38,2 33,6

46
Tabela 10 Estatística descritiva dos caracteres morfológicos dos anfíbios anuros do Refúgio de Vida Silvestre Mata do Junco, Sergipe - Brasil. Medidas lineares são dadas em
milímetros. CRC = Comprimento rostro-cloacal, LC = Largura do corpo, ACO = Altura do corpo, CC = Comprimento da cabeça, LCA = Largura da cabeça, ACA = Altura da
cabeça, DO = Diâmetro do olho, DIO = Distância inter-ocular, DON = Distância olho-narina, DIN = Distância inter-nasal, CB = Comprimento do braço, LB = Largura do
braço, CAB = Comprimento do antebraço, LAB = Largura do antebraço, CCX = Comprimento da coxa, LCX = Largura da coxa, CPE = Comprimento da perna, LPE =
Largura da perna, CM = Comprimento da mão, CP = Comprimento do pé. Dendropsophus branneri (Db), Dendropsophus minutus (Dmi), Hypsiboas albomarginatus (Ha),
Hypsiboas creptans (Hc), Hypsiboas faber (Hf), Hypsiboas raniceps (Hr), Ischnocnema ramagii (Ir), Leptodactylus latrans (Ll), Leptodactylus natalensis (Ln).
Db Dmi Ha Hc Hf Hr Ir Lf Ll Ln
n=8 n=12 n=20 n=6 n=12 n=22 n=15 n=19 n=27 n=28
± dp
CRC 17,51 ±1 23,7±1,9 45,6±8,3 59,3±4,8 94,2±11,7 67,8±,17,7 23,6±13,2 39,68±3,8 76,5±13,7 37,8±10,7
LC 5,1±0,1 9,37±0,7 15,23±3,6 22,7±3,3 37,46±7,5 25,86±8,2 8,29±4,9 15,90±3,3 33,3±6,9 16,07±5,7
ACO 2,75±0,2 4,63±0,8 7,04±1,9 9,66±2,6 17,25±5,8 12,01±4,8 4,46±2,9 8,04±1,2 17,84±5,1 8,58±2,3
CC 5,04±0,3 6,71±1,9 14,44±3,1 22,2±4,5 33,84±2,8 20,24±6,8 8,15±3,7 14,40±2 27,36±4,9 13,25±4,4
LCA 5,93±0,2 6,13±1,8 14,37±2,4 15,18±5 33,46±5,7 19,44±7,1 8,05±3,2 14,76±5,4 22,52±7,3 12,28±4
ACA 2,93±0,2 3,89±0,6 6,59±1,7 11,95±6,6 14,42±6 9,03±3,6 3,88±1,6 6,48±2,4 15,84±6,1 5,63±1,6
DO 2,59±0,1 2,45±0,3 4,33±0,8 5,37±0,3 9,37±2,5 6,1±1,8 2,62±0,9 3,54±0,7 9,92±15,3 3,88±1,4
DIO 4,68±1 3,63±1,4 7,66±1,4 8,79±1 16,72±4,4 10,87±3,1 5,06±1,8 5,25±0,9 49,07±21,1 5,53±1,2
DON 2,15±0,3 2,38±0,3 4,49±0,8 5,65±0,2 9,52±2,4 5,48±1,8 2,91±1 3,48±1 8,37±11,2 3,55±0,9
DI 1,75±01 2,05±0,2 3,30±1,8 3,62±0,3 6,41±2,3 4,80±1,4 2,04±1,1 2,59±0,7 4,703±1,2 2,84±0,6
CB 3,87±0,3 4,86±0,4 9,61±2,2 12,31±1,4 21,78±4,5 12,75±3,7 4,17±2,7 8,18±1,6 21±19,1 8,50±3,5
LB 0,84±0,1 1,34±0,4 2,70±1,4 2,85±0,5 6,54±3 3,51±1,1 1,46±0,8 17,97±66,8 7,90±4,5 2,81±1,7
CAB 3,21±0,4 4,63±0,8 8,62±1,8 12,74±1,9 17,46±3 11,89±3,4 5,09±2,3 7,32±1 13,84±2,8 7,21±1,9
LAB 1,46±0,2 1,33±0,4 2,81±0,6 5,64±2,4 7,97±2,3 4,95±1,7 1,55±0,8 2,73±0,6 7,42±2,7 3,10±1,5
CCX 8,59±0,5 10,48±1,9 23,54±4,8 30,67±4,9 45,05±5,2 33,24±8,5 11,04±6,5 16,87±1,8 32,30±5,2 15,08±4,6
LCX 2,17±0,1 2,79±0,6 5,08±1 10,18±3 14,48±2,7 10,69±3,6 3,26±1,9 7,05±1,2 14,7±3,2 7,03±2,3
CPE 9,35±0,4 11,92±0,7 24,02±4,2 30,72±4,6 44,62±6,2 35,43±7,7 11,91±6,8 18,64±2,2 31,19±4 15,43±4,2
LPE 2,37±0,3 2,72±0,4 4,38±1,2 7,62±2,6 10,12±2,5 6,89±3 2,67±1,6 5,20±1 10,22±2,3 5,31±1,6
CM 3,86±0,2 6,36±0,8 11,69±2,7 17,75±3 28,73±4,4 18,47±6,1 6,48±3,8 8,46±0,9 16,02±2,9 8,78±1,8
CP 12,85±1,1 16,98±1,7 30,89±6,3 41,8±3,8 57,97±16,1 44,63±11,9 16,75±9,3 28,89±3,4 50,77±6,2 26,67±8,3

47
Tabela 10 (Continuação) Estatística descritiva dos caracteres morfológicos dos anfíbios anuros do Refúgio de Vida Silvestre Mata do Junco, Sergipe - Brasil. Medidas
lineares são dadas em milímetros. CRC = Comprimento rostro-cloacal, LC = Largura do corpo, ACO = Altura do corpo, CC = Comprimento da cabeça, LCA = Largura da
cabeça, ACA = Altura da cabeça, DO = Diâmetro do olho, DIO = Distância inter-ocular, DON = Distância olho-narina, DIN = Distância inter-nasal, CB = Comprimento do
braço, LB = Largura do braço, CAB = Comprimento do antebraço, LAB = Largura do antebraço, CCX = Comprimento da coxa, LCX = Largura da coxa, CPE =
Comprimento da perna, LPE = Largura da perna, CM = Comprimento da mão, CP = Comprimento do pé. Leptodactylus troglodytes (Lt), Leptodactylus vastus (Lv),
Physalaemus cuvieri (Pc), Proceratophrys renalis (Pr), Rhinella granulosa (Rg), Rhinella jimi (Rj), Scinax x-signatus (Sx). Ischnocnema vinhai (Iv), Dermatonotus muelleri
(Dm), Corythomantis greeningi (Cg).
Lt Lv Pc Pr Rg Rj Sx Iv Cg Dm n=8 n=13 n=9 n=7 n=11 n=27 n=1 n=1 n=1 n=1
CRC 49,34±5,2 133,4±32,4 36,79±40,1 49,5±11,1 50,9±19,5 117±30 36,2 22,3 36,8 52,3
LC 19,98±2,2 57,40±18 16,1±19,8 26,4±7,3 26,41±10,4 65,8±21,8 12,6 8,2 14,2 29,9
ACO 9,23±1,3 27,41±9,1 8,4±8 12,7±2,9 10,34±3,4 28,4±10,7 5,7 3,7 5,8 9,8
CC 18,93±2,4 45,1±10,5 13,52±15,7 21,2±5,6 14,9±4,6 33,3±9,2 12,2 8,2 15,1 15,8
LCA 13,10±4,6 45,5±15,6 13±15,2 23,5±7,5 13,79±6,5 42,1±15,3 11,7 8,2 11,2 7,7
ACA 11,73±5,6 24,5±7,1 6,4±6,2 11,8±3,7 8,52±5,3 19,6±6,5 1,6 4,0 2,9 17,7
DO 4,31±0,7 12,2±3,3 3,8±3,7 5,5±1,8 4,5±1,1 11,2±4,3 3,7 2,6 3,7 3,6
DIO 6,15±0,7 18,18±5,2 5,8±4,2 8,8±2,4 6,9±2 16,4±5 3,6 4,9 6,3 5,8
DON 4,34±0,7 10,16±3,1 3,3±1,7 4,8±1,1 3,5±1,2 7,4±2,9 3,7 2,8 5,3 2,7
DI 3,65±0,5 8,44±2,7 3,4±2,7 3,8±1 2,5±1 6,9±3,2 3,3 1,8 2,6 2,4
CB 11,60±1 30±9,6 9,1±8,6 13,7±3,3 12,2±3,1 29,5±11,3 9,8 3,8 7,5 13,2
LB 3,25±0,4 11,82±4,5 2,3±2,8 4,5±1,7 3,4±1,4 8,5±4 2,3 1,3 1,7 4,4
CAB 9,24±1,1 24±6,1 7,4±6,6 11,5±3,1 10,5±3,4 24,9±8 7,5 5,2 6,8 11,3
LAB 3,95±1 12,3±3,7 3,2±4,5 3,9±1,1 3,7±1,5 10,08±3,9 2,5 1,5 2,6 4,1
CCX 17,8±3,6 48,9±11,9 14,05±14,1 17,6±2,4 14,7±6 41,6±12,9 16,0 10,6 15,2 20,0
LCX 9,25±1,1 23,9±7,1 7,2±7,8 8,38±1,4 6,1±3,1 19,6±6,6 5,3 3,2 4,8 8,8
CPE 17,77±1,8 46±12,9 14,6±10,8 16±2,3 14,4±4,3 38,8±13,2 37,7 10,6 17,8 13,4
LPE 6,02±1,3 16,4±4,4 4,4±4,1 5,2±1,2 4,2±1,7 13,1±6,8 5,1 2,3 2,5 3,5
CM 10,63±0,9 29,2±7,2 9,3±8 12,1±2,7 10,9±3,3 27,1±8,8 9,8 5,8 10,1 10,6
CP 28,90±1,3 74,6±17,5 23,4±22,6 25,4±6,2 24,8±7,3 61,5±17,7 22,7 15,8 25,2 24,3

48
Tabela 11 Resultados da Análise de Componentes Principais (ACP) em ambas as áreas. Refugio da Vida
Silvestre Mata do Junco – RVSMJ, Monumento Natural Grota d Angico – MNGA, Sergipe - Brasil.
Variáveis
Componentes
RVSMJ MNGA
1 2 1 2
Comprimento rostro
cloacal
0.977 0.074 0.977 0.036
Largura do corpo 0.948 0.205 0.892 0.284
Altura do corpo 0.944 0.191 0.902 0.242
Comprimento da cabeça 0.952 0.080 0.917 -0.100
Largura da cabeça 0.900 -0.037 0.593 0.458
Altura da cabeça 0.891 0.213 0.881 0.300
Diâmetro do olho 0.930 -0.128 0.869 0.143
Distância inter-ocular 0.828 -0.316 0.828 0.304
Distância olho-narina 0.893 -0.346 0.822 -0.449
Distância inter-nasal 0.893 -0.346 0.822 -0.449
Comprimento do braço 0.932 0.093 0.929 0.183
Largura do braço 0.867 0.142 0.750 0.058
Comprimento do
antebraço
0.975 0.047 0.937 0.113
Largura do antebraço 0.961 0.056 0.925 0.064
Comprimento da coxa 0.918 -0.081 0.917 -0.243
Largura da coxa 0.943 0.141 0.886 -0.164
Comprimento da perna 0.921 -0.137 0.791 -0.436
Largura da perna 0.947 0.091 0.885 -0.106
Comprimento da mão 0.962 -0.020 0.939 0.101
Comprimento do pé 0.933 0.008 0.917 -0.277
Variância Explicada pelos Componentes
17.167 0.580 15.239 1.396
Porcentagem Total de Variação Explicada
85.834 2.900 76.197 6.978

49
Efeito histórico
No RVSMJ a influência de fatores históricos para o uso de microhabitat foi
significativa nos clados Hylidae (C), Cruciobatrachia (J), Hylinae (G), Brachycephalidae e
Athesphatanura (A/B), Dendropsophi (I), e Leptodactylidae (K) (Tabela 12). Já no MNGA o
uso de microhabitat foi influenciado pelos fatores históricos nas junções que representam os
táxons Hylidae (C), Cruciobatrachia (F) e Hylinae (D) (Tabela 12; Figura 6). Não houve
influencia significativa dos efeitos históricos sobre dieta das espécies.
D C
B
A
Figura 6 Grupos utilizados para ordenação filogenética canônica para os dados de dieta (A – Refúgio de Vida Silvestre
Mata do Junco - RVSMJ e C – Monumento Natural Grota do Angico - MNGA) e microhabitat (B - RVSMJ e D - MNGA).
Filogenia baseada em Pyron & Wiens 2011.

50
Tabela 12. Efeitos históricos na ecologia dos anuros do Refúgio de Vida Silvestre Mata do Junco (RVSMJ) e
Monumento Natural Grota do Angico (MNGA), Sergipe - Brasil. Resultados das permutações do teste de Monte
Carlo para os grupos (definidos na Figura 4) para as matrizes de dieta e microhabitat. Porcentagem de variação
explicada (em relação à variação total, sem restrições), e F e P são dados para cada variável (9999 permutações
realizadas) para cada matriz principal.
Grupo (s) Variação Variação % F P
Dieta (RVSMJ)
A/B 0,270 20,9 2.545 0,0766
L 0,209 20,6 1,872 0,0601
I 0,152 17,4 1,306 0,1847
K 0,150 12,8 1,283 0,2745
F 0,126 - 1,054 0,3884
J 0,125 - 1,045 0,3969
G 0,115 - 0,954 0,5125
H 0,107 - 0,884 0,6048
C 0,101 - 0,828 0,6741
D 0,086 - 0,703 0,7755
E 0,076 - 0,614 0,7558
Microhabitat (RVSMJ)
C 0,469 92,80 8,892 0,0001
G 0,373 7,20 6,445 0,0003
J 0,327 - 5,400 0,0005
H 0,266 - 4,133 0,0058
P 0,221 - 3,301 0,0151
A/B 0,218 - 3,246 0,0134
I 0,207 - 3,050 0,0290
K 0,196 - 2,869 0,0286
M 0,166 - 2,375 0,0558
D 0,137 - 1,901 0,1018
N 0,110 - 1,492 0,1803
E 0,109 - 1,487 0,1744
Q 0,082 - 1,100 0,3385
O 0,075 - 0,993 0,3970
F 0,065 - 0,849 0,5072
L 0,054 - 0,706 0,6151
Dieta (MNGA)
G 0,251 28,70 1,346 0,2197
F 0,227 20,70 1,196 0,2821
A/B 0,196 16,40 1,009 0,4804
C 0,164 13,60 0,825 0,6789
D 0,122 - 0,596 0,8830
E 0,098 - 0,470 0,9458
Microhabitat (MNGA)
C 0,468 47,70 6,555 0,0014
F 0,377 27,10 4,695 0,0016
D 0,354 15,20 4,276 0,0100
G 0,308 6,50 3,532 0,0034
E 0,207 - 2,120 0,0737
H 0,207 - 2,128 0,0535
J 0,090 - 0,830 0,4313
I 0,086 - 0,783 0,4689
K 0,062 - 0,550 0,6402
A/B 0,050 - 0,444 0,4221

51
DISCUSSÕES
Riqueza de espécies e abundância
A maior riqueza de espécies foi verificada para o RVSMJ, o que se justifica pelas
condições locais que oferecem ambientes mais diversificados e com menores restrições de
água durante o ano. Em geral áreas de Mata Atlântica mantêm maiores riquezas de espécies
(em média 27 ± 5,2 espécies - Juncá, 2006; Santos & Santos, 2010; Santos, 2010; Silva &
Moura, 2010) quando comparadas a áreas de Caatinga (em média 18 ± 3,5 espécies - Borges-
Nojosa & Santos, 2005; Vieira et al., 2007, Silva e Santos, 2011). Os valores verificados
nesse estudo diferem um pouco dos conhecidos para cada área, onde foram registradas 34
espécies para a RVSMJ (Morato et al. 2011; Ferreira & Faria, 2011) e 19 espécies para o
MNGA (dados não publicados CHUFS). Isso indica que algumas espécies provavelmente não
utilizam os açudes amostrados em ambas as áreas. Os resultados podem estar relacionados ao
fato que as informações já existentes para cada área agrupam dados que extrapolam os corpos
d’água trabalhados aqui. Contudo os valores estimado e observado das espécies de anuros
para as duas áreas indicam que o esforço amostral foi adequado.
Nas duas áreas houve a predominância de representantes das famílias Lepdoctylidae e
Hylidae, o que é esperado para as áreas neotropicais (Duellman, 1978; Vieira et al. 2007).
Entre as espécies registradas em cada área, nove são comuns aos dois biomas, sendo elas D.
muelleri, R. jimi, R. granulosa, L. latrans, L. troglodytes, L. fuscus, C. greeningi e H. creptans
S. x-signatus. Essas espécies são amplamente distribuídas ao longo dos biomas brasileiros,
com exceção de C. greeningi que é uma espécie com distribuição restrita ao nordeste do
Brasil (Borges-Nojosa & Skuk, 2004; Reynolds, et al., 2004; Arzabe & Heyer, 2004; Heyer et
al., 2010; La Marca et al., 2013).
Uso do espaço
Dentre os ambientes trabalhados o uso mais diversificado dos recursos espaciais foi
em geral verificado para as espécies ocorrentes no RVSMJ. Características locais como a
grande disponibilidade de água, a diversidade de microhábitats e os estratos florestais
abundantes, a maior parte do ano, podem gerar maiores possibilidades às espécies que ali
vivem e contribuir para larguras de nicho mais amplas (Titon et al., 2010; Kubisch et al.,
2012). O período de desenvolvimento do estudo foi de grande escassez de água para a área do
MNGA, o que restringiu muito os recursos locais às espécies, refletindo no uso mais restrito

52
de microhábitats na área. Possivelmente em anos de maiores precipitações e retenção de água
nas lagoas, novos recursos podem tornar-se disponíveis as espécies o que poderia gerar outras
respostas.
Os representantes da família Hylidae foram os que utilizaram de forma mais ampla os
substratos disponíveis em cada área, pois ocuparam praticamente todos os disponíveis no
ambiente em especial a vegetação marginal e as plantas emergentes ao corpo d´água, o que é
corroborado por Donnelly & Guyer (1994). Grandes sobreposições no uso de recursos, como
as observadas para essas espécies nas duas áreas, podem ser indicativas de um
compartilhamento de recursos e uma falta de concorrência, ou de modo alternativo, que há
forte concorrência, mas que ainda não levou a divergência no uso dos recursos (Gotelli &
Graves, 1996).
Maiores sobreposições, independente da área, foram observadas principalmente para
as espécies do gênero Leptodactylus, Proceratophrys e Rhinela que ocuparam o solo e a água.
Espécies desses gêneros estão historicamente associadas ao uso do solo, que por si só poderia
gerar os altos valores de sobreposição (Vitt & Caldwell, 2013). A própria forma de
categorização pode levar a falsas interpretações, visto que não integra uma visão temporal
e/ou mesmo espacial mais refinada. Na área do RVSMJ, por exemplo, L. vastus e L.
natalensis adotaram as mesmas categorias de recursos espaciais (solo úmido ou água), porém
a primeira espécie foi observada principalmente nas lagoas marginais a reserva, e já segunda
foi exclusiva da área brejosa. Outro exemplo, P. renalis e I. vinhai, também adotaram o solo,
porém estiveram associados ao folhiço no interior da mata. A maioria dos registros de R. jimi,
L. latrans, L. fuscus e L. vastus, para o RVSMJ, foi tomada para lagoa seca, que apresentou
grande proporção e retenção de água em todo o período do trabalho. Apesar dessas espécies
utilizarem categorias semelhantes de recursos espaciais elas ocuparam regiões diferenciadas
da lagoa. Rhinela jimi foi mais frequente na região mais antropizada e exposta da lagoa, já as
demais espécies predominaram na margem oposta, onde o acesso era mais restrito.
Outro fato que chamou atenção foi a alternância das espécies mais abundantes, que
ocuparam o solo (L. vastus, L. latrans, R. granulosa e R. jimi), incluindo também alguns
hilídeos (H. faber e H. raniceps), nos períodos de menor precipitação. Assim a forma de
distribuição das espécies, bem como a alternância observada pode contribuir para a

53
coexistência das mesmas e reduzir possíveis pressões competitivas entre elas. Mecanismos
semelhantes já foram sugeridos por Kopp et al. (2010).
Devido à baixa precipitação no MNGA durante o período de estudo, apenas duas
lagoas retiveram água por mais de dois meses, limitando muito os recursos espaciais
disponíveis às espécies, e contribuindo assim para os altos valores de sobreposição e menores
larguras de nicho. Apesar do uso semelhante do espaço por P. albifrons e P. diplolister e
pelas espécies dos gêneros Leptodactylus e Rhinella, as duas primeiras só foram visualizadas
após períodos de maior precipitação e retenção de água. A expressiva maioria das
visualizações dessas espécies foram de juvenis recém metamorfoseados presentes em fendas
no solo úmido na lagoa da estrada, assim que água evaporava.
Apesar das diferenças estruturais entre as áreas, algumas espécies mantiveram certa
semelhança no uso do microhabitat. Contudo apesar da influência dos parâmetros ambientais
sobre o nicho das espécies é preciso considerar também a pressão seletiva exercida no
passado. Zimmerman & Simberloff (1996) mostraram que a estrutura da comunidade de
anuros, quanto ao tipo de habitat utilizado para a reprodução e desenvolvimento, é
determinada principalmente por restrições filogenéticas de espécies colonizadoras em cada
área, em vez de pressões ambientais ou competitivas. Esses autores sugerem que
diferenciações nos padrões de nicho dos organismos podem ter ocorrido em algum momento
no passado e parecem ter uma grande interferência no uso do microhabitat por anuros adultos,
pelo menos em fluxos permanentes. Logo nesse estudo muito dos padrões de uso do habitat
foi explicado pelo efeito histórico ou filogenético das populações, como é o caso dos táxons
Phyllomedusinae, Hylidae, Brachycephalidae e Athesphatanura, Dendropsophi e Leiuperidae.
Os indivíduos dessas famílias tem seu nicho espacial influenciado mais pelos efeitos
históricos que pelos fatores ambientais que lhes são impostos.
Dieta
Apesar das diferenças climáticas e fitofisionômicas entre as áreas de estudo, houve
sobreposição alimentar entre os indivíduos de ambas as áreas, mais comum para as do
RVSMJ, principalmente entre as espécies de solo. Os itens mais consumidos foram
Formicidae, Coleoptera e Orthoptera, o que sugere que apesar das diferenças nas
características ambientais e na disponibilidade de itens alimentares, as espécies de ambas as
áreas tem preferencias alimentares similares.

54
Algumas famílias apresentaram uma dieta bastante especializada, conforme observado
em Bufonidae que apesar de ter se alimentado de várias categorias de presas, Formicidae foi o
item alimentar mais consumido em ambas as áreas. Essa preferência por formigas em
bufonídeos já foi observada em outros trabalhos, principalmente devido aos seus hábitos de
forrageio (Ferreira & Teixeira, 2009; Batista et al., 2011), contudo as presas secundarias
diferiram em cada área analisada, o que sugere uma disponibilidade diferenciada de
artrópodes. Os representantes dessa família (R. jimi e R. granulosa) não só tiveram as maiores
amplitudes de nicho em cada área, mas também foram as de maiores sobreposições nas duas
localidades.
Já os representantes da família Leptodactylidae apresentaram dietas bem variadas e
consequentemente maiores amplitudes de nicho, o que é comum ao grupo (Solé, et al., 2009).
Leptodactylus natalensis foi a espécie com a maior variedade de categorias ingeridas sendo
considerado um animal de dieta bem generalista, o que também foi comprovado por estudos
similares em áreas de Mata Atlântica (Ferreira et al., (2007).
No RVSMJ, Leptodactylus latrans e H. raniceps também apresentaram alta
sobreposição sendo coleóptera o item mais importante de suas dietas, porem os demais itens
variaram de importância, o que é comum para áreas de Mata Atlântica (Solé et al., 2009;
Ferreira et al., 2007). Esse resultado é pouco esperado, uma vez que historicamente as
famílias em que estão incluídos possuem hábitos bem diferenciados, sendo Leptodactylidae
um grupo de hábitos terrestres (Solé, et al., 2009; Heyer et al., 2013; Vitt & Caldwell, 2013) e
Hylidae, em geral, mais arborícola (La Marca et al., 2004; Vitt & Caldwell, 2013). As
diferenças de ocupação do espaço por representantes destas famílias poderiam reduzir as
sobreposições em suas dietas, por proverem recursos alimentares diferenciados em cada
estrato. Entretanto, devido a ausência de estratificação vegetal na lagoa seca (área com maior
riqueza e abundancia de espécies no RVSMJ), espécies arborícolas adotaram ambientes
alternativos, coincidentes com as que ocuparam o solo, o que poderia justificar as altas
sobreposições entre as mesmas. O uso de categorias de presas semelhantes foi também
verificado entre H. faber, H raniceps e D. minutus, essas ocuparam locais distintos,
minimizando os efeitos que uma espécie teria sobre as outras. As duas primeiras foram
encontradas na lagoa seca e D. minutus exclusivamente nas lagoas da borda.

55
Os resultados sugerem que a dieta das espécies em ambas áreas é uma característica
variável, superando o sinal filogenético, sendo que pouca variação na utilização do itens
alimentares pode ser explicado pelo contexto histórico. Isso indica que os hábitos alimentares
das espécies nessas áreas são provavelmente influenciados diretamente pelo que está
disponível no ambiente.
Morfometria
Pequenas diferenças nos tamanhos e formas foram observadas entre as populações de
L. latrans, L. fuscus, R. granulosa e R. jimi. Alguns autores afirmam que essas variações
morfológicas são comuns em algumas espécies de vertebrados tais como anfíbios, repteis e
aves e normalmente estão associadas a particularidades de cada localidade (Castellano &
Giacoma, 1998; Santiago-Alarcon et al., 2006; Juncá et al., 2008; Kaliontzopoulou et al.,
2010), sendo que possivelmente na maioria dos casos pressões diferenciadas (particularidades
ambientais e/ou climáticas, diferenças nos tipos de presas consumidas e predação) são
impostas sobre as populações podendo ser essa a fonte das alterações (Riha & Berven, 1991;
Schneider et al., 1999; Castellano et al., 2000).
Leptodactylus latrans, L. fuscus, R. granulosa e R. jimi da área de Mata Atlântica
apresentaram-se ligeiramente menores que os espécimes da Caatinga. Resultados semelhantes
foram observados para P. nordestina em trabalho desenvolvido nas mesmas áreas do presente
estudo (Caldas, 2011). Provavelmente diferenças climáticas e fitofisionômicas das duas áreas,
nos níveis de alteração do ambiente, taxas de predação e ou mesmo no número de
competidores possam contribuir para as variações observadas. Possivelmente a escassez de
água na Caatinga pode levar a um maior investimento em crescimento, uma vez que nem
sempre os corpos d’água estão disponíveis para a reprodução. O maior tamanho poderia
compensar, em termos da produção de um número maior de ovos, nos períodos de maior
precipitação e retenção de água nas lagoas. Como a água é mais constante nos ambientes de
Mata Atlântica, menores restrições à reprodução seriam impostas a esses animais,
possibilitando assim um número maior de desovas por animal.
Alternativamente, níveis diferenciados de degradação nos ambientes podem interferir
nas taxas de crescimento dos indivíduos, sendo essas possivelmente menores nos locais de
maior distúrbio. Alguns autores afirmam que mudanças na qualidade do habitat refletem
negativamente na vida dos indivíduos, sobretudo em seu crescimento (Neckel-Oliveira, 2003;

56
Neckel-Oliveira & Gascon, 2006). A maioria dos espécimes do RVSMJ, utilizados nas
comparações morfométricas, é proveniente da lagoa seca, corpo d´água com maior grau de
alteração entre aqueles estudados, o que talvez justifique em parte o menor tamanho dos
indivíduos para a área. O mesmo padrão foi verificado por Caldas (2011) para P. nordestina.
Alterações na vegetação nativa podem interferir na quantidade e qualidade de abrigos,
disponíveis para adultos e girinos, e na abundância de microrganismos importantes na
alimentação das larvas. Estes fatores poderiam favorecer a sobrevivência de espécimes
menores em localidades mais alteradas como sugerido por Juncá et al., (2008), que relataram
variações na morfometria de C. greeningi em populações de diferentes localidades da
Caatinga baiana.
CONSIDERAÇÕES FINAIS
A diversidade de anfíbios foi maior na área do RVSMJ do que no MNGA, o que já era
esperado devido ás condições ambientais presentes em cada bioma. As duas taxocenoses
apresentaram alta sobreposição quanto ao uso dos recursos espaciais e alimentares.
Aparentemente o que mais influenciou na sobreposição espacial em ambas as áreas foram os
fatores históricos, pois apesar das diferenças impostas pelo ambiente a maioria das espécies
tendeu a conservação de aspectos fundamentais de seus nichos. O mesmo não ocorreu com os
itens alimentares, pois as espécies se mostraram muito suscetíveis à utilização do que estava
disponível no ambiente, do que seguir um padrão filogenético.
A influência dos parâmetros ambientais também foi notada na morfologia das espécies
que são comuns aos dois biomas, pois os indivíduos do RVSMJ são ligeiramente menores que
os do MNGA, o que indica uma diferenciação nos investimentos em crescimento,
provavelmente devido a escassez de água na Caatinga e níveis diferenciados de degradação
entre as áreas. Quando há restrição de água, os indivíduos investem seus recursos mais em
crescimento que em reprodução, pois ter um corpo maior pode abrigar mais ovos e dessa
forma compensar restrições de água. Níveis maiores de degradação podem contribuir também
para indivíduos menores.
REFERÊNCIAS BIBLIOGRÁFICAS
Alley, T. R. (1982). Competition theory, evolution, and the concept of ecological niche. Acta
Biotheoretica 31, 165-179.

57
Araujo, C. O. & Almeida-Santos, S. M. (2013).Composição, riqueza e abundância de anuros em um
remanescente de Cerrado e Mata Atlântica no estado de São Paulo. Biota Neotropica 13, 265-
275.
Arzabe, C. & Heyer, R. 2004. Leptodactylus troglodytes. 2006 IUCN Red List of Threatened
Species. Downloaded on 24 January 2014.
Arzabe, C. (1999). Reproductive activity pattern of anurans in two different altitudinal sites within
the Brazilian Caatinga. Revista Brasileira de Zoologia16, 851-864.
Avise J. C. (2000). Phylogeography: the history and formation of species. Cambridge, MA: Harvard
University Press.
Azevedo-Ramos, C., & Galatti, U. (2002). Patterns of amphibian diversity in Brazilian Amazonia:
conservation implications. Biological Conservation 103, 103-111.
Barreto, C. C. (2010). Heterogeneidade espacial do habitat e diversidade específica: implicações
ecológicas e métodos de mensuração. Oecologia Australis 7, 121-154.
Batista, R. C., De-Carvalho, C. B., Freitas, E. B., Franco, S. C., Batista, C. C., Coelho, W. A.& Faria,
R. G. (2011). Diet of Rhinella schneideri (Werner, 1894) (Anura: Bufonidae) in the Cerrado,
Central Brazil. Herpetology Notes 4, 17-21.
Bentley,P. J. (1966).Adaptations of Amphibians to arid environments. Science 152, 619-623.
Borges-Najosa, D. & Skuk, G. (2004). Corythomantis greeningi. In: IUCN 2013. IUCN Red List of
Threatened Species. Version 2013.2. Downloaded on 24 January 2014.
Brooks, D. R. & Mclennan, D. A. (1991). Phylogeny, Ecology, and Behavior: a Research Program
in Comparative Biology. Chicago: University of Chicago Press.
Buzzi, Z.J. (2002). Entomologia Didática. 4 ed. Editora UFPR, Curitiba.
Castellano S., Giacoma C. & Dujsebayeva T. (2000). Morphometric and advertisement call
geographic variation in polyploidy green toads. Biological Journal of the Linnean Society 70,
341–360.
Castellano, S. & Giacoma, C. (1998). Morphological variation of the green toad, Bufo viridis, in
Italy: a test of causation. Journal of Herpetology32, 540-550.

58
Castellano, S., Giacoma, C., Dujsebayeva, T., Odierna, G. & Balletto, E. (1998). Morphometrical and
acoustical comparison between diploid and tetraploid green toads. Biol. Journal of the
Linnean Society 63, 257- 281.
Colwell, R. K. (2006). EstimateS 7.5.2. Statistical estimation of species richness and shared
species from samples.Version 7. Acessado em 18/09/2013.
Costa, G. C., Mesquita, D. O., Colli, G. R. & Vitt, L. J. (2008). Niche Expansion and the Niche
Variation Hypothesis: Does the degree of individual variation increase in depauperate
assemblages? The American Naturalist 172, 868-877.
Dodd, C. K. (2010). Amphibian ecology and conservation: a handbook of techniques. [S.l.] Oxford
University Press New York.
Donnelly, M. A. & Guyer, C. (1994). Patterns of reproduction and habitat use in an assemblage of
Neotropical hylid frogs. Oecologia 98, 291- 302.
Duellman, W.E. (1978). The biology of an equatorial herpetofauna in Amazonian Ecuador.
Miscellaneous Publications Museum of Natural History University of Kansas 65, 1–352
Dunham, A. E. & Overall, K. L. (1994). Population Responses to Environmental Change: Life
History Variation, Individual-based Models, and the Population Dynamics of Short-lived
Organisms. Integrative and Comparative Biology 34, 382-396.
Ferreira, A. S. & Faria, R. G. (2011) First record of Lithobates palmipes (Spix, 1824) (Anura:
Ranidae) for the State of Sergipe, Brazil. Herpetology Notes 4, 139-140.
Ferreira, R. B. & Teixeira, R. L. (2009). Feeding patterns and use of reproductive habitat of the
Striped toad (Rhinella crucifer) from southeastern Brazil. Acta Herpetologica,. Acta
Herpetologica 4, 125-134.
Ferreira, R. B., Dantas, R. B.& Teixeira, R. L. (2007). Reproduction and ontogenetic diet shifts in
Leptodactylus natalensis (Anura, Leptodactylidae) from southeastern Brazil. Boletim do
Museu de Biologia Mello Leitão 22, 45-55.
Fraser, R. H. (1998). Vertebrate species richness at the mesoscale: relative roles of energy and
heterogeneity.Global Ecology and Biogeography Letters 7, 215-220.

59
Freitas, E. B., De Carvalho, C. B., Faria, R. G., Batista, R. C., Batista, C. C., Coelho, W, A.,
Bocchiglieri, A. (2008). Nicho ecológico e aspectos da história natural de Phyllomedusa
azurea (Anura: Hylidae, Phyllomedusinae) no Cerrado do Brasil Central. Biota Neotropica 8,
101-110.
Gadsden, H.E. & Palacios-Orona, L.E. (1997). Seasonal dietary patterns of the Mexican Fringe-Toed
Lizard (Uma paraphygas). Journal of Herpetology 31:1–9.
Giannini, N. P. (2003). Canonical phylogenetic ordination. Systematic Biology, 52, 684–695.
Gotelli, N. J & Graves, G. R. (1996). Null models in ecology. Smithsonian Intitution press,
Washington.
Gotelli, N.J & Entsminger, G. L. (2010). EcoSim: Null models software for ecology. Version 7.
Acquired Intelligence Inc. & Kesey-Bear. Jericho, VT 05465. Acessado em 18/06/2013
Kopp, K., Signorelli, L. & Bastos, R. P. (2010). Distribuição temporal e diversidade de modos
reprodutivos de anfíbios anuros no Parque Nacional das Emas e entorno, Estado de Goiás,
Brasil. Iheringia. Série Zoologia 100, 192-200.
Hammer O., Harper, D. A. T.& Ryan, P. D. (2001). PAST: Paleontological statistics software
package for education and data analysis. Palaeontologia Electronica 4, 1-9.
Heyer, R., Langone, J., La Marca, E., Azevedo-Ramos, C., Tada, I., Baldo, D., Lavilla, E., Scott, N.,
Aquino, L., Hardy, J. (2010). Leptodactylus latrans. In: IUCN 2013. IUCN Red List of
Threatened Species. Version 2013.2. Downloaded on 02 February 2014.
Huston, M. A. (1994). Biological Diversity, The coexistence of species on changing
landscapes. Cambridge University Press, 681p.
Juncá, F. A. (2006): Diversidade e uso de hábitat por anfíbios anuros em duas localidades de Mata
Atlântica, no norte do estado da Bahia. Biota Neotropica 6, 1–17.
Juncá, F. A., Carneiro, M.C.L. & Rodrigues, N. N. (2008). Is a dwarf population of Corythomantis
greeningi Boulenger, 1896 (Anura, Hylidae) a new species?. Zootaxa 1686, 48–56.

60
Kaliontzopoulou, A.; Carretero, M. A. & Sillero, N. (2010). Geographic patterns of morphological
variation in the lizard Podarcis carbonelli, a species with fragmented distribution.
Herpetological Journal 20, 41–50.
Krebs, C. J. (1999). Ecological Methodology. 2ed. Menlo Park: Addison Wesley Longman.
Kubisch, E., Piantoni, C., Williams, J., Scolaro, A., Navas, C. A., Ibargüengoytía, N. R. (2012).Do
Higher Temperatures Increase Growth in the Nocturnal Gecko (Gekkota: Phyllodactylidae)?
A Skeletochronological Assessment Analyzed at Temporal and Geographic Scales. Journal of
Herpetology 46, 587-595.
La Marca, E., Azevedo-Ramos, C., Silvano, D., Scott N., Aquino, L., Faivovich J., 2004. Hypsiboas
raniceps. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2013.2..
Downloaded on 28 January 2014.
La Marca, E., Azevedo-Ramos, C., Silvano, D., Solís, F., Ibáñez, R., Jaramillo, C., Fuenmayor, Q.,
Hardy, J. (2010). Hypsiboas crepitans. In: IUCN 2013. IUCN Red List of Threatened
Species. Version 2013.2. Downloaded on 24 January 2014.
Legendre, P. & Legendre, L. (1998). Numerical Ecology. Developments in Environmental Modelling
20. 2º ed.Elsevier Science B.V.
Levins, R. (1968). Evolution in changing environments. New Jersey, Princeton University Press,
IX+120p.
Magnusson, W.E., Lima, P.A., Silva, W.A.& Araujo, M.C. (2003). Use of geometric forms to
estimate volume of invertebrates in ecological studies of dietary overlap. Copeia 1, 13-19.
Magurran, A.E. (1988). Ecological diversity and its measurement. New Jersey: Princenton
University Press.
Mcguinness, K. A., Underwood, A. J. (1986). Habitat structure and the nature of communities on
intertidal boulders. Journal of.Experimental Marine Biology and Ecology 104, 97-123.
Morato,S. A. A., Lima, A. M. X., Staut, D. C. P., Faria, R. G., Alves, J. P. A., Gouveia, S. F.,
Scupino, M. R. C., Gomes, R., Silva, M. J. (2011). Amphibians and Reptiles of the Refúgio
de Vida Silvestre Mata do Junco, municipality of Capela, state of Sergipe, northeasten Brazil.
Check List (São Paulo. Online) 7, 756-762.

61
Moreno-Rueda, G. & Pizarro, M. (2007). The relative influence of climate, environmental
heterogeneity, and human population on the distribution of vertebrate species richness in
south-eastern Spain. Acta Oecologica 32, 50-58.
Morrison, C., & Hero, J. M. (2003). Geographic variation in life‐history characteristics of
amphibians: a review. Journal of Animal Ecology 72, 270-279.
Navas, C., Antoniazzi, M., JARED, C. (2004). A preliminary assessment of anuran physiological and
morphological adaptation to the Caatinga, a Brazilian semi-arid environment. International
Congress Series 1275, 298-305.
Neckel-Oliveira, S. & Gascon, C. (2006). Abundance, body size and movement patterns of a tropical
treefrog in continuous and fragmented forests in the Brazilian Amazon. Biological
Conservation 28, 308–315.
Neckel-Oliveira, S. (2003). Effects of landscape change on clutches of Phyllomedusa tarsius, a
neotropical treefrog. Biological Conservation 118, 109–116.
Pazinato, D. M. M. ; Trindade, A. D. O. ; Oliveira, S. V. D; Cappellari, L. H. (2011). Dieta de
Leptodactylus latrans (Steffen, 1815) na Serra do Sudeste, Rio Grande Do Sul, Brasil.
Biotemas (UFSC) 24, 147-151.
Peterson, A.T. Soberón, J. Sánchez-Cordero, V. (1999). Conservatism of ecological niches in
evolutionary time. Science 285, 1265–1267.
Pianka, E.R. (1973). The structure of lizard communities.Annual Review of Ecology and Systematics
4, 53-74.
Pielou, E. C. (1966). Shannon's Formula as a Measure of Specific Diversity: Its Use and Misuse. The
American naturalist 100, 463-465.
Pombal Jr., J. P. & Haddad, C. F. B. (2005). Estratégias e modos reprodutivos de anuros (Amphibia)
em uma poça permanente na Serra de Paranapiacaba, Sudeste do Brasil. Papéis Avulsos de
Zoologia (São Paulo) 45, 215-229.
Popielarz, P. A., & Neal, Z. P. (2007). The niche as a theoretical tool. Annual Review of
Sociology33, 65-84.

62
Prado, G. M.& Pombal Jr., J.P. (2005). Distribuição espacial e temporal dos anuros em um brejo da
reserva biológica de duas bocas, sudeste do brasil. Arquivos do Museu Nacional, Rio de
Janeiro 63, 685-705.
Prates, I. & Navas, C. A. (2009). Cutaneous Resistance to Evaporative Water Loss in Brazilian
Rhinella (Anura: Bufonidae) from Contrasting Environments. Copeia 2009, 618-622.
Pyron, R. A. & Wiens, J. J. (2011). A large-scale phylogeny of Amphibia including over 2800
species, and a revised classification of extant frogs, salamanders, and caecilians. Molecular
Phylogenetics and Evolution61, 543-583.
Reynolds, R., Caramaschi, U., Mijares, A., Acosta-Galvis, A., Heyer, R., Lavilla, E., Hardy, E.
2004. Leptodactylus fuscus. In: IUCN 2013. IUCN Red List of Threatened Species. Version
2013.2. Downloaded on 24 January 2014.
Riha V. F. & Berven K. A. (1991). An analysis of latitudinal variation in the larval development of
the wood frog (Rana sylvatica). Copeia 1991, 209–221.
Rossa-Feres, D. C.& JIM, J. (2001). Similaridade do sítio de vocalização em uma comunidade de
anfíbios anuros na região noroeste do Estado de São Paulo, Brasil. Revista Brasileira de
Zoologia18, 439-454,
Santiago-Alarcon, D., Tanksley, S. M. & Parker, P. G. (2006). Morphological variation and genetic
structure of Galapagos dove (Zenaida galapagoensis) populations: issues in conservation for
the Galapagos bird fauna. The Wilson Journal of Ornithology 118, 194–207.
Santos, A. F. & Andrade, J. A. (1992). Caracterização e delimitação do semi-árido sergipano.
Aracaju; UFS.
Santos, M. J. S. (2007). Mata do Junco (Capela – SE): Identidade Territorial e Gestão de Conflitos
Ambientais. NESA/UFS, São Cristóvão.
Santos, T. G.; Rossa-Feres, D. C.; Casatti, L. (2007). Diversidade e distribuicão espaço-temporal de
anuros em região com pronunciada estação seca no sudeste do Brasil. Iheringia: Série
Zoologia 97, 37–49.

63
Schäuble, C. S. (2004). Variation in body size and sexual dimorphism across geographical and
environmental space in the frogs Limnodynastes tasmaniensis and L. peronii. Biological
Journal of the Linnean Society 82, 39–56.
Schneider C. J., Smith T. B., Larison B. & Moritz C. (1999). A test of alternative models of
diversification in tropical rainforests: Ecological gradients vs. rainforest refugia. Proceedings
of the National Academy of Sciences 96, 13869–13873.
Silva, F. R. & Rossa-Feres, D. C. (2011). Influence of terrestrial habitat isolation on the diversity and
temporal distribution of anurans in an agricultural landscape. Journal of Tropical Ecology 27,
327-331.
Silva, R. A., Martins, I. A. & Rossa-Feres, D. C. (2011). Environmental heterogeneity: Anura
diversity in homogeneous environments. Revista Brasileira de Zoologia 28, 610-618.
Simpson, E.H. (1949). Measurement of diversity. Nature 163:688.
Solé, M., Dias, I. R., Rodrigues, E. A. S., Silva Junior, E. M. S., Branco, S. M. J., Cavalcante, K. P.,
Rödder, D. (2009). Diet of Leptodactylus ocellatus (Anura: Leptodactylidae) from a cacao
plantation in southern Bahia, Brazil. Herpetology Notes 2, 9-15.
Tejedo, M., Marangoni, F., Pertoldi, C., Richter-Boix, A., Laurila, A., Orizaola, G., Gomez-Mestre, I.
(2010). Contrasting effects of environmental factors during larval stage on morphological
plasticity in post-metamorphic frogs.Climate research 43.
TerBraak, C. J. F. (1986). Canonical correspondence analysis: a new eigenvector method for
multivariate direct gradient analysis. Ecology 67, 1167–1179.
Tews, J., Brose, U., Grimm, V., Tielborger, K., Wichmann, M.C., Schwager, M. & Jeltsch,
F. (2004) Animal species diversity driven by habitat heterogeneity/ diversity: the importance
of keystone structures. Journal of Biogeography 31, 79–92.
Titon, B., Navas, C. A., Gomes, F. R. (2010). Water balance and locomotor performance in three
species of neotropical toads that differ in geographical distribution. Comparative
Biochemistry and Physiology. Part A, Molecular & Integrative Physiology 156, 129-135.

64
Toft, C. A. (1980). Feeding ecology of thriteen syntopic species of Anurans in a seasonal tropical
environment. Oecologia 15, 131-141.
Toft, C. A. (1981). Feeding ecology of Panamanian litter anurans: patterns in diet and foraging mode.
Journal of Herpetology 15, 139-144.
Torres-Orozco, R. E., Jimenez-Sierra, C. L., Vogt, R. C., & Benitez, J. L. V. (2002). Neotropical
tadpoles: spatial and temporal distribution and habitat use in a seasonal lake in Veracruz,
Mexico. Phyllomedusa 1, 81-91.
Vasconcelos, T. S. & Rossa-Feres, D. C. (2005). Diversidade, distribuição espacial e temporal de
anfíbios anuros (Amphibia, Anura) na região noroeste do Estado de São Paulo. Biota
Neotropica 5, 1-14.
Vasconcelos, T.S. & D.C. Rossa-Feres. (2008). Habitat heterogeneity and use of physical and
acoustic space in anuran communities in southeastern Brazil. Phyllomedusa 7, 127-142.
Vasconcelos, T.S.; Santos, T.G.; Rossa-Feres, D.C., Haddad, C.F.B. (2009). Influence of the
environmental heterogeneity of breeding ponds on anuran assemblages from southeastern
Brazil. Canadian Journal Zoology 87, 699-707.
Vieira, W.L.S., Arzabe, C., Santana, G.G. (2007). Composição e distribuição espaço-temporal de
anuros no Cariri paraibano, Nordeste do Brasil. Oecologia Brasiliensis 11, 383-396.
Vitt, L. J. & Caldwell, J. P. (2013). Herpetology: an introductory biology of amphibians and reptiles.
Access Online via Elsevier.
Wells, K. D. (2007). The Ecology and Behavior of Amphibians.The University of Chicago
Press.Chicago.
Wiens, J. J. (2004). Speciation and ecology revisited: Phylogenetic niche conservatism and the origin
of species. Evolutionv 58, 193–197.
Wiens, J. J. (2007). Global patterns of species richness and diversification in amphibians. The
American Naturalist 170, 86–106.
Wilkinson, L. (1990). SYSTAT: The System for Statistics. Systat Inc., Evanston, Illinois.

65
Young, K. M., Cramp, R. L., White, C. R., & Franklin, C. E. (2011). Influence of elevated
temperature on metabolism during aestivation: implications for muscle disuse atrophy. The
Journal of Experimental Biology 214, 3782-3789.
Zimmermann, B. L. & Simberloff , D. (1996). An historical interpretation of habitat use by frogs in a
Central Amazonian Forest. Journal of Biogeography 23:27-46






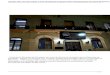















![[2014] - Plegados concatenados diferenciados](https://img.document.onl/doc/110x75/57906d061a28ab68748e2180/2014-plegados-concatenados-diferenciados.jpg)






