Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO
BIOLOGIA DE AGENTES INFECCIOSOS E PARASITÁRIOS
DESENVOLVIMENTO DE MÉTODO SOROLÓGICO E MOLECULAR PARA
PESQUISA DE ARENAVÍRUS EM AMOSTRAS BIOLÓGICAS DE ROEDORES
SILVESTRES DA AMAZÔNIA BRASILEIRA
ANA LETÍCIA SCALDELAI BERNARDI
Belém – Pará
2013

2
ANA LETÍCIA SCALDELAI BERNARDI
DESENVOLVIMENTO DE MÉTODO SOROLÓGICO E MOLECULAR
PARA PESQUISA DE ARENAVÍRUS EM AMOSTRAS BIOLÓGICAS
DE ROEDORES SILVESTRES DA AMAZÔNIA BRASILEIRA
Dissertação apresentada ao Curso de Mestrado do
Programa de Pós-Graduação em Biologia de
Agentes Infecciosos e Parasitários do Instituto de
Ciências Biológicas da Universidade Federal do
Pará como requisito para a obtenção do grau de
Mestre em Biologia dos Agentes Infecciosos e
Parasitários.
Orientador: Pedro Fernando da Costa Vasconcelos
Belém – Pará
2013

3
Bernardi, Ana Letícia Scaldelai, 1988
Desenvolvimento de método sorológico e molecular para pesquisa de
arenavírus em amostras biológicas de roedores silvestres da Amazônia
brasileira / Ana Letícia Scaldelai Bernardi. - 2013.
Orientador: Pedro Fernando da Costa Vasconcelos.
Dissertação (Mestrado) - Universidade Federal do Pará, Instituto de
Ciências Biológicas, Programa de Pós-Graduação em Biologia de
Agentes Infecciosos e Parasitários, Belém, 2013.
1. Arenavírus. 2. Arenaviridae. 3. Roedor Amazônia. 4. Teste
imunoenzimático. I. Título.
CDD 22. ed. 579.256

2
FOLHA DE APROVAÇÃO
ANA LETÍCIA SCALDELAI BERNARDI
DESENVOLVIMENTO DE MÉTODO SOROLÓGICO E MOLECULAR PARA
PESQUISA DE ARENAVÍRUS EM AMOSTRAS BIOLÓGICAS DE ROEDORES
SILVESTRES DA AMAZÔNIA BRASILEIRA
Dissertação apresentada ao Programa de Pós – Graduação em Biologia dos Agentes
Infecciosos e Parasitários, do Instituto de Ciências Biológicas da Universidade federal do
Pará, como requisito para a obtenção de grau de Mestre em Biologia dos Agentes Infecciosos
e Parasitários.
Orientador: Prof. Dr. Pedro Fernando da Costa Vasconcelos
Seção de Arbovirologia e Febres Hemorrágicas, IEC.
Banca Examinadora: Dra. Elizabeth Salbé Travassos da Rosa
Seção de Arbovirologia e Febres Hemorrágicas, IEC.
Dra. Daniele de Almeida Medeiros
Seção de Arbovirologia e Febres Hemorrágicas, IEC.
Dra. Lívia Carício Martins
Seção de Arbovirologia e Febres Hemorrágicas, IEC.
Dr. Márcio Teixeira Nunes (Suplente)
Seção de Arbovirologia e Febres Hemorrágicas, IEC
Belém, 26 de junho de 2013.

3
EPÍGRAFE
``Aprender é a única coisa de que a mente
nunca se cansa, nunca tem medo e nunca
se arrepende. ``
Leonardo da Vinci

4
DEDICATÓRIA
Aos meus pais e ao meu irmão,
para que cada minuto de saudade
não tenha sido em vão.

5
AGRADECIMENTOS
Agradeço primeiramente à minha mãe e ao meu pai, por sempre apoiarem as minhas
decisões e por acreditarem em mim. Sem eles minhas conquistas não seriam possíveis.
Ao meu irmão por mesmo distante estar sempre presente.
Ao Ramon Brito, amigo e companheiro incansável, sem o qual nenhuma das vitórias
aqui alcançadas teriam sentido.
À família Brito que me acolheu em Belém, me proporcionando toda a ajuda
necessária.
Ao Dr. Pedro Vasconcelos, por ter me aceitado como orientanda e por ter me
proporcionado o presente trabalho.
À Daniele Medeiros, pela ajuda providencial na parte molecular da dissertação.
À Dra. Elizabeth Salbé, por ceder o laboratório para a realização dos testes de ELISA.
À Darlene Simith, pelo treinamento das técnicas de ELISA e RT-PCR e, pela ajuda
prática durante a parte experimental.
Ao pessoal do IH, por disponibilizarem os antígenos necessários aos ensaios, em
especial o senhor Basílio.
Ao Samir Casseb, por ter ajudado no desenvolvimento dos iniciadores utilizados no
estudo, bem como pelos auxílios diários durante os testes.
A Maria Helena Mendonça, pelo auxílio imprescindível na área da clonagem.
Ao Bruno Tardelli, pelos ensinamentos e ajuda em horas precisas.
Ao pessoal da Biologia Molecular: Alice, Natália, Gregório e Natividade, pelo
companheirismo e ajuda nos momentos necessários.
Aos meus amigos, que mesmo distantes, estão sempre comigo: Juliane, Tamine, Kaoli,
Roger, Fábio, Samantha, Bruno, Daniele Carolina e Kristie.
Ao Instituto Evandro Chagas, por disponibilizar todos os recursos para o
desenvolvimento do projeto.
Enfim, a todos que direta ou indiretamente contribuíram para este trabalho e às
amizades construídas aqui em Belém, muito obrigada.

6
SUMÁRIO
LISTA DE FIGURAS .......................................................................................................... 8
1. INTRODUÇÃO .............................................................................................................. 11
1.1 FAMÍLIA ARENAVIRIDAE................................................................................... 12
1.2 GÊNERO ARENAVÍRUS ..................................................................................... 16
1.2.1 Estrutura Viral ............................................................................................. 16
1.2.2 Ciclo Replicativo ........................................................................................... 17
1.3 ECOLOGIA E INFECÇÃO ................................................................................... 19
1.4 CARACTERÍSTICAS CLÍNICAS DAS FEBRES HEMORRÁGICAS CAUSADAS
POR ARENAVÍRUS ..................................................................................................... 21
1.5 ARENAVÍRUS SUL AMERICANOS CAUSADORES DE FEBRES
HEMORRÁGICAS ........................................................................................................ 22
1.6 DISTRIBUIÇÃO GEOGRÁFICA DOS ARENAVÍRUS NO BRASIL .................. 24
1.7 PROPRIEDADES LABORATORIAIS DOS ARENAVÍRUS ............................... 25
1.8 DIAGNÓSTICO LABORATORIAL ..................................................................... 26
1.9 TRATAMENTO .................................................................................................... 27
1.10 PREVENÇÃO E CONTROLE .............................................................................. 27
1.11 JUSTIFICATIVA .................................................................................................. 28
2. OBJETIVOS ............................................................................................................... 29
2.1 OBJETIVO GERAL .............................................................................................. 29
2.2 OBJETIVOS ESPECÍFICOS ................................................................................. 29
3. MATERIAIS E MÉTODOS ....................................................................................... 30
3.1 AMOSTRAS ......................................................................................................... 30
3.2 TESTE DE ELISA IgG .......................................................................................... 31
3.2.1 Obtenção do antígeno pelo método da sucrose-acetona .............................. 32
3.2.2 Titulação........................................................................................................ 33
3.2.3 Sensibilização ................................................................................................ 33
3.2.4 Adição das amostras ..................................................................................... 34
3.2.5 Adição do conjugado..................................................................................... 35

7
3.2.6 Substrato ....................................................................................................... 35
3.2.7 Interpretação dos resultados ........................................................................ 35
3.3 TRANSCRIÇÃO REVERSA-REAÇÃO EM CADEIA DA POLIMERASE ... 36
3.3.1 Extração do RNA viral ................................................................................. 36
3.3.2 Transcrição Reversa – Reação em Cadeia da Polimerase (RT-PCR) seguida
da reação de Semi-Nested-PCR .................................................................................. 36
3.3.4 Produtos Obtidos .......................................................................................... 38
4. RESULTADOS ........................................................................................................... 39
4.1 ELISA IgG ............................................................................................................ 39
4.2 AMOSTRAS TESTADAS POR RT-PCR .............................................................. 40
4.3 RESULTADOS DO LIMITE DE DETECÇÃO ..................................................... 41
4.3.1 Vírus Amaparí .............................................................................................. 41
4.3.2 Vírus Flexal ................................................................................................... 42
4.3.3 Vírus Sabiá .................................................................................................... 43
4.3.4 Vírus Paraná ................................................................................................. 44
4.3.5 Vírus Junin .................................................................................................... 45
4.3.6 Vírus Guanarito ............................................................................................ 46
4.3.7 Vírus Machupo ............................................................................................. 48
4.3.8 Vírus Latino .................................................................................................. 48
4.3.9 Vírus Inespecíficos ........................................................................................ 49
4.3.10 Limite de detecção mínimo para os arenavírus ........................................... 50
5. DISCUSSÃO ............................................................................................................... 51
6. CONCLUSÃO ............................................................................................................. 57
REFERÊNCIAS ................................................................................................................. 58
ANEXO 1............................................................................................................................ 65

8
LISTA DE FIGURAS
Página
Figura 1 - Estrutura dos arenavírus. ......................................................................... 13
Figura 2 - Classificação genética dos arenavírus. .................................................... 14
Figura 3 - Representação esquemática dos arenavírus. ........................................... 16
Figura 4 - Visão geral do ciclo replicativo dos arenavírus. ..................................... 17
Figura 5 - Mecanismos de replicação, transcrição e tradução do segmento S do
RNA de arenavírus. ................................................................................ 18
Figura 6 - Ciclo de transmissão dos arenavírus. ...................................................... 20
Figura 7 - Distribuição dos arenavírus Sul Americanos. ......................................... 23
Figura 8 - ELISA IgG indireto. ................................................................................. 32
Figura 9 - Placa de ELISA. ........................................................................................ 34
Figura 10 - Adição das amostras. ............................................................................. 34
Figura 11 - Placa de ELISA após a realização da leitura da densidade óptica do
teste. ....................................................................................................... 36
Figura 12 – RT-PCR para o AMV. ........................................................................... 42
Figura 13 – Semi-Nested-PCR do AMAV. .............................................................. 42
Figura 14 – RT-PCR para o FLEV. ........................................................................... 43
Figura 15 – Semi-Nested-PCR do FLEV. ................................................................. 43
Figura 16 – RT-PCR para o SABV. ........................................................................... 44
Figura 17 – Semi-Nested-PCR do SABV. ................................................................. 44
Figura 18 – PCR para o PARV. ................................................................................. 45
Figura 19 – Semi-Nested-PCR do PARV. ................................................................. 45
Figura 20 – PCR para o JUNV. .................................................................................. 46
Figura 21 – Semi-Nested-PCR do JUNV. .................................................................. 46
Figura 22 – PCR para o GTOV. ................................................................................. 47
Figura 23 – Semi-Nested-PCR do GTOV. ................................................................. 47
Figura 24 – PCR e Semi-Nested-PCR para o MACV ............................................... 48
Figura 25 – PCR e Semi-Nested-PCR para o LATV. ............................................... 49
Figura 26 – Semi – Nested-PCR para vírus inespecíficos aos iniciadores................. 49

9
RESUMO
Os arenavírus pertencem a família Arenaviridae, gênero Arenavirus, sendo o grupo do Novo
Mundo compreendido pelos arenavírus nativos das Américas, como os vírus Machupo
(MACV), Junin (JUNV), Guanarito (GTOV) e Sabiá (SABV), sendo estes, os vírus de maior
importância do grupo, pois são a causa de febres hemorrágicas em humanos. Os principais
reservatórios dos arenavírus são os roedores, que ao serem infectados podem desenvolver a
infecção de forma crônica, transmitindo os vírus aos humanos através de aerossóis
provenientes da urina e excretas. A análise de 1.395 amostras de sangue de roedores silvestres
da Amazônia brasileira através da adaptação do teste de ELISA IgG, demonstrou a presença
de arenavírus circulantes na região, indicando a susceptibilidade da população a infecções.
Paralelamente o desenvolvimento de uma técnica de RT-PCR para detecção de arenavírus do
grupo do Novo Mundo, mostrou boa sensibilidade e especificidade, realizadas a partir do
limite de detecção para oito arenavírus: o SABV, Amaparí (AMAV) e Flexal (FLEV), obtidos
a partir da extração de RNA de cérebros de camundongo infectados e, o JUNV, GTOV,
MACV, Paraná (PARV) e Latino (LATV), utilizando cDNA sintético (gBlocks) a partir de
sequências disponíveis no Genbank. Foram feitas diluições seriadas 1:2 e/ou 1:5 do cDNA
sintético e do RNA viral extraído. Orthobunyavírus, Hantavírus e Flavivírus foram utilizados
para verificar a especificidade do teste, demonstrando resultados negativos. Os limites
mínimos de detecção encontrados para os arenavírus foram 0,5pg (PARV), 0,012ng (AMAV),
0,062ng (SABV), 0,062ng (FLEV), 0,062ng (JUNV), 1,56ng (GTOV), 1,56ng (MACV) e
12,5ng (LATV). Considerando que pesquisas focadas em tais vírus no País são muito
precárias, as técnicas desenvolvidas tornam-se ferramentas úteis para futuros estudos sobre
diagnóstico e endemismo viral, os resultados obtidos poderão contribuir para estudos sobre a
circulação dos arenavírus não só no Brasil como em outros países das Américas.

10
ABSTRACT
The arenaviruses belong to the Arenarividae family, genus Arenavirus, being the New
World group comprised by natives arenaviruses of the Americas as the viruses Machupo
(MACV), Junin (JUNV), Guanarito (GTOV) and Sabia (SABV), which are the most
important viruses from the group, they are the cause of hemorrhagic fever in humans. The
main reservoirs of arenaviruses are the rodents, which when infected may develop a chronic
form of the viral disease, transmitting the virus to humans through aerosols from urine and
excretes. Analysis of 1.395 samples from blood of wild rodents from Brazilian Amazon by
adapting an IgG ELISA test, demonstrated de presence of circulating arnaviruses in region,
indicating the susceptibility of local population to infections. At the same time the
development of an RT-PCR for arenaviruses of the New World group showed a good
sensitivity and specificity performed from the detection limit for eight arenaviruses: the
SABV, Amapari virus (AMAV) and Flexal virus (FLEV), followed by RNA genome
extraction from brains of infected mice and, the JUNV, GTOV, MACV, Parana virus (PARV)
and Latino virus (LATV), using synthetic cDNA (gBlocks) from sequences available in
Genbank. Serial dilutions were made in 1:2 and/or 1:5 from synthetic cDNA or viral RNA
extracted. Orthobunyaviruses, hantaviruses and flaviviruses were used to check the specificity
of the test, showing negative results. Minimum detection limits founded for the arenavirures
were 0,5pg (PARV), 0,012ng (AMAV), 0,062ng (SABV), 0,062ng (FLEV), 0,062ng (JUNV),
1,56ng (GTOV), 1,56ng (MACV) and 12,5ng (LATV). Considering that the researches
focuses in these viruses in country are very poor, the techniques employed here are useful
tools for futures studies on viral diagnosis and endemism, the results may contribute to studies
on the circulation of arenaviruses not only in Brazil as in other countries of the Americas.

11
1. INTRODUÇÃO
Os arenavírus foram descritos inicialmente em 1933, quando o vírus da
Coriomeningite Linfocítica (LCMV) foi isolado em amostras de um caso fatal em humano
suspeito de ter ido a óbito devido à encefalite de Saint Louis, vírus causador de grande
epidemia a época (Armstrong & Lillie, 1934). Vários estudos então começaram a ser
realizados com o LCMV. Por volta dos anos de 1960, novos vírus, como os do complexo
Tacaribe, mostraram ter semelhanças morfológicas e sorológicas com tal vírus (Pinheiro et al,
1967; Rowe, Pugh, Webb & Peters, 1970), além da dependência de infecção em roedores, o
que deu origem ao grupo dos arenavírus ( Rowe, Murphy & Bergold, 1970).
Os principais reservatórios para os arenavírus são os membros da ordem Rodentia,
roedores que ao serem infectados, quase sempre são assintomáticos, apresentando a infecção
na forma aguda ou crônica. Presume-se que tais vírus sejam transmitidos aos humanos através
da inalação de aerossóis provenientes das secreções ou excretas de roedores infectados
(Emonet et al., 2009). Enfermidades causadas por arenavírus são comuns em humanos, sendo
graves em alguns casos. Os arenavírus Lassa (LASV), Junín (JUNV), Machupo (MACV),
Guanarito (GTOV) e Sabiá (SABV) são conhecidos por causarem febre hemorrágica severa
na África Ocidental, Argentina, Bolívia, Venezuela e Brasil, respectivamente (Charrel &
Lambalerrie, 2010).
Alguns fatores têm sido associados à transmissão e emergência de novos vírus, tais
como o aumento populacional e a necessidade de expansão da agricultura, por exemplo, são
atividades que favorecem a dispersão dos agentes virais. A extensão do cultivo agrícola para
áreas de floresta pode alterar o ciclo de transmissão zoonótico de determinados vírus,
portanto, a remoção de florestas para o desenvolvimento agrícola torna-se um risco aos
agricultores que são expostos a artrópodes e roedores (Hui, 2006). Na maioria dos casos, a
transmissão de arenavírus aos humanos se dá após incursões de lazer ou atividades agrícolas,
situações em que o homem entra em contato direto com o habitat dos roedores hospedeiros.
Grandes modificações no ambiente ocasionadas por atividades humanas ou mudanças
ecológicas naturais têm implicado no aumento de infecções por arenavírus devido a mudanças
de comportamento dos hospedeiros primários (Charrel & Lambalerrie, 2010). A Febre
Hemorrágica Argentina, causada pelo JUNV, emergiu após a utilização da região dos pampas
para a agricultura, principalmente para a produção de grãos, como o milho, considerado o
fator que atraiu o roedor Calomys musculinus, reservatório natural do vírus (Hui, 2006).

12
A epidemiologia dos arenavírus em humanos é determinada pela distribuição dos
roedores infectados e seu contato com a população, embora em alguns casos infecções inter-
humanas possam ocorrer (Enria et al, 2005).
Além dos comportamentos peridomiciliares de determinados roedores serem uma
importante fonte de infecção, profissionais em constante contato com roedores infectados, no
campo ou em laboratórios, estão em maior risco de contaminação (Charrel & Lambalerrie,
2010). Casos de infecções adquiridas em laboratório, no Brasil, já foram registrados pelo
vírus Flexal (FLEV) e SABV em 1978 e 1992, respectivamente (Vasconcelos et al., 1993).
1.1 FAMÍLIA ARENAVIRIDAE
Segundo o Comitê Internacional para Taxonomia dos Vírus (International Committe
on Taxonomy of Virus - ICTV), a família Arenaviridae é constituída por um único gênero -
Arenavirus – que recebe este nome devido a aparência de areia (em Latim, arenous) vista nas
partículas virais em microscópio eletrônico. Os ribossomos do hospedeiro, abundantes nestes
vírus, é que conferem esta característica (Fig. 1) (ICTV, 2011; Buchmeier et al., 2007). A
família contém 24 espécies virais (ICTV, 2011) divididas em dois grupos de acordo com suas
propriedades antigênicas: o grupo do Velho Mundo (sorocomplexo Lassa – Coriomeningite
Linfocítica) que inclui vírus nativos da África e em regiões de distribuição do LCMV, e o
grupo do Novo Mundo (sorocomplexo Tacaribe) que inclui vírus nativos das Américas
(Charrel & Lambalerrie, 2010).
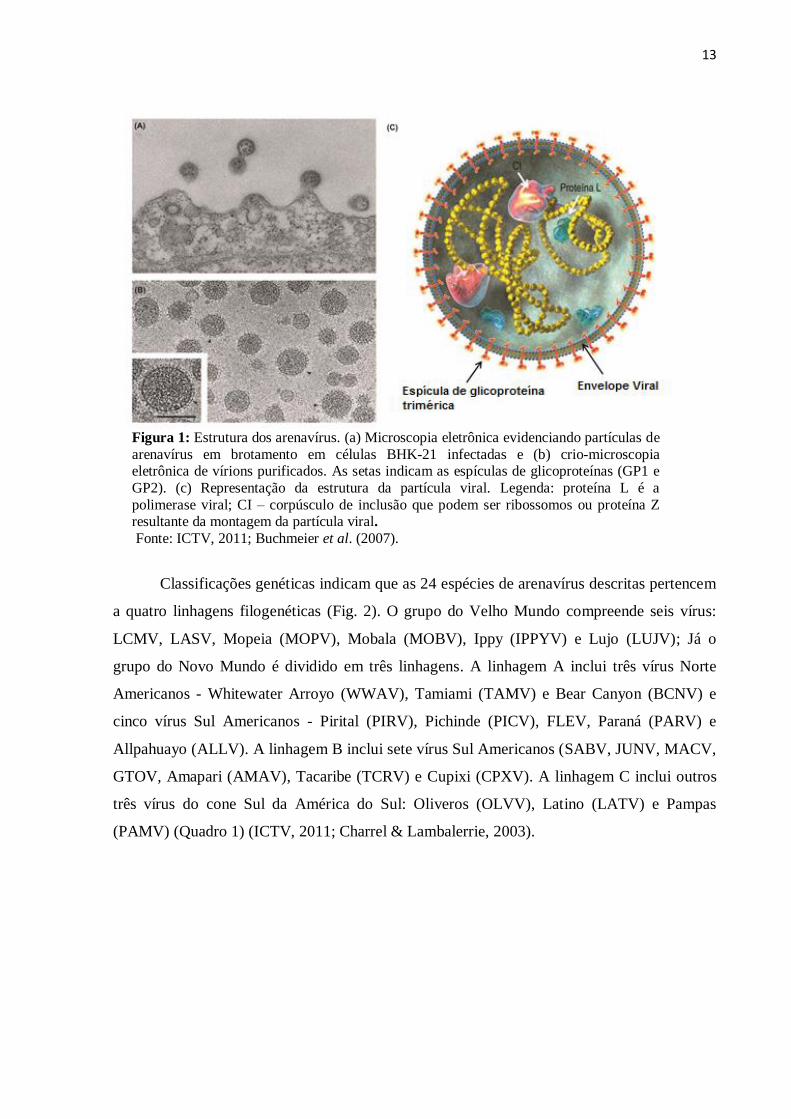
13
Figura 1: Estrutura dos arenavírus. (a) Microscopia eletrônica evidenciando partículas de
arenavírus em brotamento em células BHK-21 infectadas e (b) crio-microscopia eletrônica de vírions purificados. As setas indicam as espículas de glicoproteínas (GP1 e
GP2). (c) Representação da estrutura da partícula viral. Legenda: proteína L é a
polimerase viral; CI – corpúsculo de inclusão que podem ser ribossomos ou proteína Z resultante da montagem da partícula viral.
Fonte: ICTV, 2011; Buchmeier et al. (2007).
Classificações genéticas indicam que as 24 espécies de arenavírus descritas pertencem
a quatro linhagens filogenéticas (Fig. 2). O grupo do Velho Mundo compreende seis vírus:
LCMV, LASV, Mopeia (MOPV), Mobala (MOBV), Ippy (IPPYV) e Lujo (LUJV); Já o
grupo do Novo Mundo é dividido em três linhagens. A linhagem A inclui três vírus Norte
Americanos - Whitewater Arroyo (WWAV), Tamiami (TAMV) e Bear Canyon (BCNV) e
cinco vírus Sul Americanos - Pirital (PIRV), Pichinde (PICV), FLEV, Paraná (PARV) e
Allpahuayo (ALLV). A linhagem B inclui sete vírus Sul Americanos (SABV, JUNV, MACV,
GTOV, Amapari (AMAV), Tacaribe (TCRV) e Cupixi (CPXV). A linhagem C inclui outros
três vírus do cone Sul da América do Sul: Oliveros (OLVV), Latino (LATV) e Pampas
(PAMV) (Quadro 1) (ICTV, 2011; Charrel & Lambalerrie, 2003).

14
Figura 2: Classificação genética baseada na análise de sequências de aminoácidos da
proteína Z, codificada pelo segmento L dos arenavírus. Grupo do Velho Mundo e grupo do Novo Mundo (Linhagens A, B e C).
Fonte: Urata & Yasuda (2012).
15
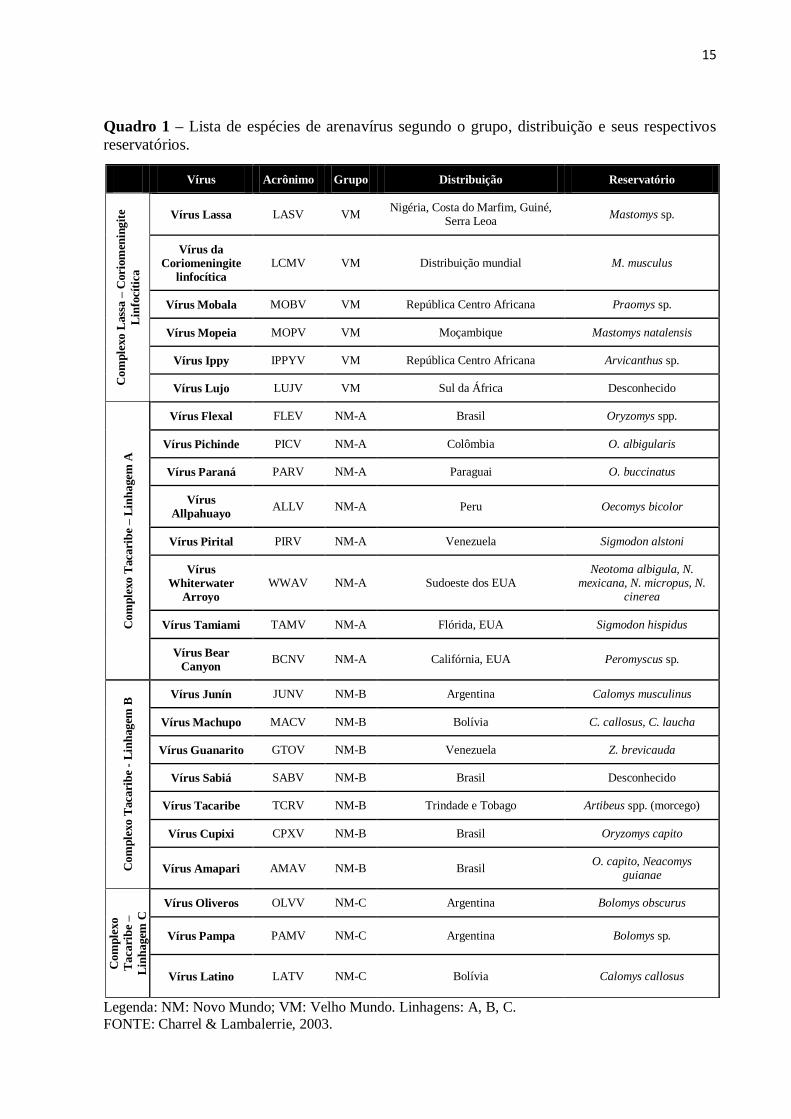
Quadro 1 – Lista de espécies de arenavírus segundo o grupo, distribuição e seus respectivos
reservatórios.
Legenda: NM: Novo Mundo; VM: Velho Mundo. Linhagens: A, B, C.
FONTE: Charrel & Lambalerrie, 2003.
Vírus Acrônimo Grupo Distribuição Reservatório
Com
ple
xo L
ass
a –
Cori
om
enin
git
e
Lin
focí
tica
Vírus Lassa LASV VM Nigéria, Costa do Marfim, Guiné,
Serra Leoa Mastomys sp.
Vírus da
Coriomeningite
linfocítica
LCMV VM Distribuição mundial M. musculus
Vírus Mobala MOBV VM República Centro Africana Praomys sp.
Vírus Mopeia MOPV VM Moçambique Mastomys natalensis
Vírus Ippy IPPYV VM República Centro Africana Arvicanthus sp.
Vírus Lujo LUJV VM Sul da África Desconhecido
Com
ple
xo T
aca
rib
e – L
inh
agem
A
Vírus Flexal FLEV NM-A Brasil Oryzomys spp.
Vírus Pichinde PICV NM-A Colômbia O. albigularis
Vírus Paraná PARV NM-A Paraguai O. buccinatus
Vírus
Allpahuayo ALLV NM-A Peru Oecomys bicolor
Vírus Pirital PIRV NM-A Venezuela Sigmodon alstoni
Vírus
Whiterwater
Arroyo
WWAV NM-A Sudoeste dos EUA Neotoma albigula, N.
mexicana, N. micropus, N. cinerea
Vírus Tamiami TAMV NM-A Flórida, EUA Sigmodon hispidus
Vírus Bear
Canyon BCNV NM-A Califórnia, EUA Peromyscus sp.
Com
ple
xo T
aca
rib
e -
Lin
hagem
B Vírus Junín JUNV NM-B Argentina Calomys musculinus
Vírus Machupo MACV NM-B Bolívia C. callosus, C. laucha
Vírus Guanarito GTOV NM-B Venezuela Z. brevicauda
Vírus Sabiá SABV NM-B Brasil Desconhecido
Vírus Tacaribe TCRV NM-B Trindade e Tobago Artibeus spp. (morcego)
Vírus Cupixi CPXV NM-B Brasil Oryzomys capito
Vírus Amapari AMAV NM-B Brasil O. capito, Neacomys
guianae
Com
ple
xo
Taca
rib
e –
Lin
ha
gem
C Vírus Oliveros OLVV NM-C Argentina Bolomys obscurus
Vírus Pampa PAMV NM-C Argentina Bolomys sp.
Vírus Latino LATV NM-C Bolívia Calomys callosus

16
1.2 GÊNERO ARENAVÍRUS
1.2.1 Estrutura Viral
Os arenavírus são envelopados, possuem estrutura pleomórfica e vírions com tamanho
de 40 a mais de 200 nm de diâmetro (Buchmeier et al., 2007). Seu genoma consiste em dois
segmentos de RNA de fita simples senso negativo, um de tamanho menor (S) e outro maior
(L) (Gómez et al,, 2011). As projeções presentes no envelope dos vírus são complexos
compostos pelas glicoproteínas virais GP1, GP2 e o peptídeo de sinal estável (PSE). As
glicoproteínas de superfície estão alinhadas com proteínas Z subjacentes e
ribonucleoprotéinas (RNP) que formam uma parede bidimensional na membrana interna dos
vírus (Fig. 3) (Buchmeier et al., 2007).
Figura 3: Representação esquemática dos arenavírus. Corte
lateral evidenciando os segmentos genômicos RNA S e RNA L,
glicoproteínas GP1 e GP2, peptídeo de sinal estável (PSE), nucleoproteínas (N), polimerase (L), proteínas Z e ribossomos.
Fonte: Gómez et al. (2011).
A infectividade dos arenavírus é nula quando seu RNA é purificado, sendo a presença
da RNA polimerase dependente de RNA (RdRp), a proteína L, essencial para a replicação de
seu genoma. Diferentes arenavírus apresentam variabilidade no tamanho dos dois segmentos
de RNA, L e S, porém preservam sua estrutura organizacional dentro da família. O ciclo
desses vírus é restrito ao citoplasma da célula hospedeira e, diferente de outros vírus de RNA
de senso negativo, os arenavírus usam uma estratégia de codificação ambisenso que dirige a
síntese de duas proteínas em orientações opostas. As regiões codificantes são separadas por
regiões intergênicas não codificantes (RINC) que formam um grampo estável. O RNA S

17
codifica o precursor do complexo das glicoproteínas virais (GPC) e a nucleoproteína NP,
enquanto o RNA L codifica a RdRp ou L polimerase e a proteína Z (Buchmeier et al., 2007).
Por serem vírus envelopados, os arenavírus tornam-se suscetíveis a solventes
orgânicos e detergentes. Sua infectividade pode ainda ser inativada por temperaturas
superiores a 55oC, luz ultravioleta e radiação gama (Enria et al, 2005).
1.2.2 Ciclo Replicativo
A interação específica com os receptores celulares leva a subsequente entrada do vírus
na célula hospedeira, sendo os receptores celulares determinantes para a infecção e
patogênese. Os arenavírus do Velho Mundo utilizam o receptor α – distroglicana para a
adsorção nas células, já alguns vírus do Novo Mundo do grupo B utilizam como receptor a
transferrina 1 (TfR 1), tais como o JUNV, GTOV, MACV e SABV (Gómez et al., 2011). A
entrada do vírus na célula é feita por endocitose, a posterior fusão entre as membranas viral e
celular é desencadeada pelo meio ácido do endossomo maduro, liberando o material genético
viral no citoplasma celular (Figura 5) (Emonet et al., 2009).
Figura 4: Visão geral do ciclo replicativo dos arenavírus. Entrada por
endocitose através dos receptores α – distroglicana (α –DG) e transferrina
(TrR-1) pelos vírus Lassa (LASV) e Junín (JUNV), respectivamente. EE: Endossomo precoce; LE: Endossomo maduro; mRNA: RNA mensageiro; ER:
retículo endoplasmático; GPC: complexo glicoproteico; vRNA: RNA viral; L:
polimerase; NP: nucleoproteína; Z: proteína estrutural. Fonte: Urata & Yasuda (2012).

18
As sequências terminais 3` e 5`, de 19 nucleotídeos, dos segmentos de RNA S e L são
extremamente conservadas, bem como complementares nos arenavírus, sugerindo que
hibridizações possam ocorrer entre esses segmentos. Acredita-se que essas terminações sejam
essenciais para a replicação e transcrição viral, atuando como sítio de ligação para a
polimerase (Gómez et al., 2011). Assim que a RNP é liberada no citoplasma, a polimerase L
inicia a transcrição do promotor localizado na região terminal 3` do RNA. A transcrição
primária resulta na síntese dos mRNA da NP e L dos segmentos S e L, respectivamente. Em
seguida, a polimerase viral adota a função de replicase e se move através da RINC para gerar
uma nova cópia de toda a extensão do RNA, dando origem a um RNA antigenômico
(agRNA). O novo agRNA servirá de molde para a transcrição dos mRNAs de GP (agS) e Z
(agL). O termo ambisenso se refere a esta situação, onde uma região do segmento S e L é
transcrita em sentido negativo – proteínas NP e L - e outra região transcrita em sentido
positivo a partir do novo RNA – proteínas GP e Z. A fita antigenômica ainda servirá como
molde para a amplificação do RNA genômico correspondente, que dará origem a novas
partículas infecciosas (Fig. 5) (Buchmeier et al., 2007).
Figura 5: Mecanismos de replicação, transcrição e tradução do segmento S do RNA de arenavírus. GP: Glicoproteína; RINC: Região
intergênica não codificante; NP: Nucleoproteína; L: Polimerase;
mRNA: RNA mensageiro.
Fonte: Adapatção de Emonet et al. (2009).
RINC
RINC

19
O complexo GPC origina três proteínas: G1, G2 e o peptídeo de sinal estável (SSP). O
SSP é necessário ao transporte da GPC do retículo endoplasmático (RE) para o complexo de
Golgi, onde a GPC é clivada pela protease celular SKI-1/S1P para formar as subunidades
maduras G1 e G2. Foi demonstrado que a interação entre as proteínas N e Z é necessária para
a montagem dos nucleocapsídeos e glicoproteínas em partículas infecciosas, sendo a proteína
Z de grande importância no brotamento das mesmas (Gómez et al., 2011).
1.3 ECOLOGIA E INFECÇÃO
Os roedores são os reservatórios naturais dos arenavírus, cada espécie viral tem maior
afinidade por determinada espécie de roedor, exceto o vírus Tacaribe, que é associado a uma
espécie de morcego. O LCMV, LASV e vírus relacionados ao grupo do Velho Mundo são
associados aos roedores da família Muridae, subfamília Murinae. Os arenavírus do Novo
Mundo são associados aos roedores da família Cricetidae, subfamília Sigmodontinae (Charrel
& Lambalerrie, 2010; Milazzo et al, 2011).
Atualmente acredita-se que a diversidade dos arenavírus se dá devido ao resultado de
uma longa relação co-evolutiva entre os vírus da família Arenaviridae e os roedores da
família Muridae ou Cricetidae. A infecção crônica do hospedeiro, acompanhada de viremia ou
virúria, aparenta ser crucial para a persistência dos arenavírus em natureza (Charrel &
Lambalerrie, 2003).
A continuidade da infecção entre populações de roedores hospedeiros pode ser por
transmissão horizontal ou vertical. Animais cronicamente infectados geralmente apresentam
crescimento, comportamento e interação intraespecífica normal. Os hospedeiros infectados
podem disseminar grandes quantidades de vírus através da urina, fezes e saliva, o que leva a
contaminação de outros animais, mais comumente por aerossóis (Sabattini et al, 1977). A
transmissão horizontal entre roedores pode ser facilitada por agressões intraespecíficas,
acasalamento e outras atividades que impliquem contato físico. Em algumas associações do
vírus com seu hospedeiro, a transmissão vertical parece ter papel bastante relevante, como
acontece com o PIRV, onde há alta prevalência de espécimes de Sigmodon alstoni infectadas
em idade jovem, sugerindo transmissão uterina ou imediatamente após o parto (Milazzo et al,
2011).
O contato com roedores infectados pode levar a infecção humana através de abrasões
na pele e inalação dos aerossóis de secreções ou excretas desses hospedeiros, uma vez que os
vírus são frequentemente encontrados em amostras de urina e rim (Pinheiro et al, 1977.
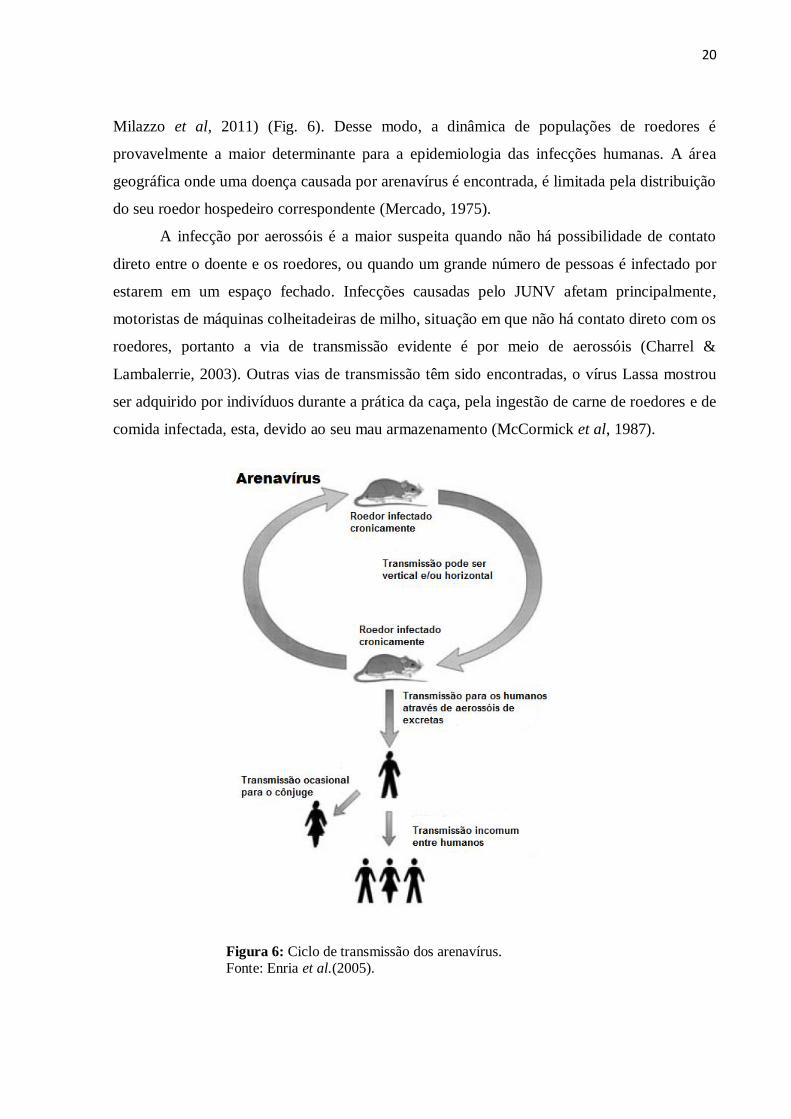
20
Milazzo et al, 2011) (Fig. 6). Desse modo, a dinâmica de populações de roedores é
provavelmente a maior determinante para a epidemiologia das infecções humanas. A área
geográfica onde uma doença causada por arenavírus é encontrada, é limitada pela distribuição
do seu roedor hospedeiro correspondente (Mercado, 1975).
A infecção por aerossóis é a maior suspeita quando não há possibilidade de contato
direto entre o doente e os roedores, ou quando um grande número de pessoas é infectado por
estarem em um espaço fechado. Infecções causadas pelo JUNV afetam principalmente,
motoristas de máquinas colheitadeiras de milho, situação em que não há contato direto com os
roedores, portanto a via de transmissão evidente é por meio de aerossóis (Charrel &
Lambalerrie, 2003). Outras vias de transmissão têm sido encontradas, o vírus Lassa mostrou
ser adquirido por indivíduos durante a prática da caça, pela ingestão de carne de roedores e de
comida infectada, esta, devido ao seu mau armazenamento (McCormick et al, 1987).
Figura 6: Ciclo de transmissão dos arenavírus. Fonte: Enria et al.(2005).

21
Embora a maioria dos casos de doenças causadas por arenavírus descritos na literatura
sejam por infecção a partir de roedores, a transmissão destes vírus entre humanos já foi
evidenciada. A Febre Hemorrágica Argentina, por exemplo, em geral não é contagiosa, porém
acredita-se que algumas mulheres foram infectadas por seus maridos durante a fase de
convalescência através do contato sexual (Briggiler et al, 1987).
O MACV pode ser transmitido pessoa a pessoa através de fluidos corporais em
pessoas da mesma família ou em infecções hospitalares. Cinco profissionais de saúde que
foram expostos ao caso primário ou a um caso secundário de Febre Hemorrágica Boliviana
(FHB), em um hospital em Cochabamba em 1971, contraíram o MACV (Peters et al, 1974).
Já em 1994, houve seis casos de infecção secundária fatal em seis membros de uma mesma
família, adquiridos a partir de um único caso natural (Kilgore et al, 1995).
O LASV causou infecções hospitalares graves na Nigéria, a disseminação viral foi
atribuída a formação de aerossóis ou a falta de esterilização de equipamentos de injeção
parenteral (Fisher-Hoch et al, 1995).
Os surtos hospitalares, embora raros, possuem a rota parenteral como a mais perigosa
devido à exposição a agulhas contaminadas ou acidentes técnicos e de autópsia. A partir
desses dados, precauções de bio-contenção mostraram-se ser indispensáveis quando se lida
com pacientes e materiais infectados pelos arenavírus (Charrel & Lambalerrie, 2003).
1.4 CARACTERÍSTICAS CLÍNICAS DAS FEBRES HEMORRÁGICAS CAUSADAS
POR ARENAVÍRUS
O período de incubação da doença dura de quatro a 21 dias, com variação média de
seis a 14 dias. Em seguida vem a fase prodrômica, iniciando-se com calafrios, mal-estar,
dores de cabeça, anorexia, mialgias e febre moderada entre 38 e 39° C. Nos dias seguintes, os
pacientes podem apresentar sintomas gastrointestinais, cardiológicos e neurológicos.
Sintomas comuns incluem dor lombar, dor retroorbital, náuseas ou vômitos, dor epigástrica,
fotofobia, tonturas e constipação ou diarréia. Exames físicos demonstraram durante a primeira
semana da doença, a presença de rubores na face, pescoço e parte superior do tórax; congestão
conjuntival com edema periorbital; sangramento espontâneo das gengivas; enantemas no
palato mole com petéquias e pequenas vesículas; petéquias cutâneas nas regiões axilares,
parte superior do tórax e braços; aumento dos linfonodos das regiões cervicais laterais;
irritabilidade, letargia, leves tremores na língua e mãos, ataxia moderada, diminuição nos

22
reflexos musculares e em mulheres, metrorragias leves ou moderadas (Enria & Pinheiro,
2000). Ainda na primeira semana, exames laboratoriais mostram progressiva leucopenia
(1000 a 2000 leucócitos/ mm³) e trombocitopenia (50.000 a 100.000 plaquetas/ mm³), além de
proteinúria e hematúria. Na segunda semana da doença 70 a 80% dos pacientes começam a
melhorar, o restante (20 a 30%) inicia uma fase hemorrágica neurológica. Tais pacientes
podem apresentar sangramento profuso, marcada ataxia, irritabilidade acentuada, intensos
tremores, delírios, convulsões e entrar em coma (Enria & Pinheiro, 2000).
Acredita-se que febre hemorrágica causada por arenavírus ocorre através de um efeito
sobre os macrófagos que induzem a ativação de citocinas. O Interferon – α (IFN-α) e o Fator
de Necrose Tumoral α (TNF-α) são abundantes no soro do paciente e, seus níveis estão
correlacionados com a gravidade da doença. A taxa de letalidade das febres hemorrágicas Sul
Americanas varia entre 15 e 30% dos pacientes dentro de dois a 14 dias após hospitalização.
Anticorpos aparecem de dez a doze dias após a doença instalada, coincidindo com a melhora
clínica, o que sugere que a resposta humoral tem papel importante ao lado da resposta celular
para a recuperação do enfermo. Os sobreviventes passam por uma melhora gradual e lenta,
com perda de cabelo e presença de sulcos transversais nas unhas, características comuns da
recuperação. Não há sequelas a longo prazo, mas a recuperação pode estender-se durante
vários meses (Doyle et al., 1998).
1.5 ARENAVÍRUS SUL AMERICANOS CAUSADORES DE FEBRES
HEMORRÁGICAS
Os arenavírus Sul Americanos (Fig.7) foram identificados na metade do século XX.
Entretanto, acredita-se que os arenavírus circulem silenciosamente em natureza há séculos e
que pelo menos uma das doenças epidêmicas que causaram um colapso na população
mexicana durante o século XVI tenha sido causada por arenavírus (Marrs & Kiracofe, 2000).

23
Figura 7: Distribuição dos arenavírus Sul Americanos. Em destaque, os vírus causadores de febre hemorrágica (GTOV, MOCV, JUNV e
SABV).
Fonte: Instituto Nacional de Enfermidades Virais Humanas - INEVH, Argentina.
Em 1950, foi reconhecida em Buenos Aires, Argentina, uma nova doença hemorrágica
severa, denominada Febre Hemorrágica Argentina (FHA), causada pelo JUNV. A maioria dos
pacientes que apresentaram infecção eram trabalhadores rurais (Greenway et al, 1959). Os
hospedeiros do JUNV são os roedores da espécie Calomys musculinus, amplamente
encontrados na região. A FHA caracteriza-se como uma doença sazonal, apresentando picos
de frequência nos períodos de colheita do milho, com maior incidência no mês de maio (Enria
et al, 2005). A taxa de letalidade dos casos de FHA é de aproximadamente 20% na ausência
de tratamento específico. Estima-se que desde 1950 houve aproximadamente 30.000 casos da
doença sintomática. Durante o período de colheita, 75% dos doentes são agricultores do sexo
masculino, infectados por meio de aerossóis (Charrel & Lambalerrie, 2003). A incidência
anual da doença foi relacionada positivamente com a quantidade populacional de roedores por
região afetada (Mills et al, 1992). Após a introdução da vacina atenuada testada em

24
agricultores em meados de 1980, a epidemiologia da FHA tem sofrido alterações
significativas com drástica redução no número de casos (Enria et al, 2005).
O agente causador da Febre Hemorrágica Boliviana (FHB) foi isolado em 1963, a
partir de cinco amostras de pacientes da cidade de San Joaquim, identificado como MACV
(JOHNSON et al, 1965). Seu reservatório natural é o roedor Calomys callosus, sendo este
arenavírus responsável por grandes surtos de doença hemorrágica grave, com letalidade média
de 20%. Entre os anos de 1962 e 1964, vários surtos de FHB envolveram mais de 1000
pacientes, resultando em 180 mortes. Os picos de incidência ocorreram nas estações de
outono e inverno, correspondente a estação seca (Doyle et al., 1998). Infecções hospitalares
causadas pelo MACV já foram claramente demonstradas, embora a maioria dos casos de FHB
seja pelo contato direto com excretas do C. callosus ou por aerossóis (Charrel & Lambalerrie,
2003).
O GTOV foi originalmente isolado em 1989, quando houve um surto de doença
hemorrágica grave no município de Guanarito, Venezuela. A doença foi designada como
Febre Hemorrágica Venezuelana (FHV) (SALAS et al, 1991). Os roedores Zygodontomys
brevicauda e Sigmodon alstoni são seus hospedeiros principais. Até 2002 cerca de 200
pessoas foram diagnosticadas com a FHV. Neste mesmo ano 18 casos foram registrados,
constatando-se duas mortes (Charrel & Lambalerrie, 2003).
O SABV foi isolado em 1990, mediante de um caso fatal de febre hemorrágica
(Coimbra et al., 1994). Após o isolamento foram relatados dois casos de infecção laboratorial
(Vasconcelos et al., 1993). O reservatório deste vírus ainda não foi identificado, mas
presume-se que seja um roedor encontrado nas proximidades da pequena comunidade de
Sabiá, município de Cotia, estado de São Paulo, onde a única infecção natural conhecida
ocorreu (Buchmeier et al., 2007).
1.6 DISTRIBUIÇÃO GEOGRÁFICA DOS ARENAVÍRUS NO BRASIL
O primeiro arenavírus encontrado no Brasil foi o AMAV isolado em amostras de
roedores capturados na região de Serra do Navio, Amapá, próximo ao Rio Amaparí (Fig.7).
As amostras foram coletadas entre os anos de 1964 e 1966 das espécies de roedores da
espécie Neacomys guinae e Oryzomys capito, reservatórios naturais do AMAV (Pinheiro et
al., 1966; 1967). A presença do vírus foi detectada em amostras de urina dos roedores, o que
evidencia o mecanismo de transmissão a hospedeiros suscetíveis, no entanto, casos de
infecção pelo AMAV em humanos ainda não foram diagnosticados (Pinheiro et al., 1967).

25
O FLEV foi isolado em 1975 a partir de amostras de roedores do gênero Oryzomys
(Pinheiro et al, 1977). Os roedores foram capturados em matas do trecho Itaituba-
Jacareacanga na rodovia Transamazônica, Estado do Pará. Ainda não foi detectada infecção
natural do homem, porém ocorreu um caso de infecção em laboratório, em que o paciente
apresentou sintomas clínicos de febre, calafrios, forte cefaléia, mialgia generalizada, queda de
cabelos, vômitos e diarréia. A doença persistiu por um mês sem alterações nos sintomas
(Pinheiro et al., 1986).
Outro arenavírus também encontrado na região de Serra do Navio no Norte do País na
década de 70 foi o vírus Cupixi, isolado de espécimes de Oryzomys capito. Não há
informações sobre transmissão do vírus para humanos (Charrel et al., 2002).
O arenavírus de maior relevância até o momento para o território nacional é o SABV,
causador de febre hemorrágica severa em humanos. Houve apenas um caso de infecção
natural pelo SABV ocorrido no Estado de São Paulo, em que a paciente apresentou um
quadro grave de febre hemorrágica e veio a óbito no quarto dia após sua internação (Coimbra
et al., 1994). Dois outros casos descritos na literatura derivam de acidentes laboratoriais e,
apesar de terem causado doenças graves não resultaram em morte (Vasconcelos et al., 1993;
Enria & Pinheiro, 2000).
1.7 PROPRIEDADES LABORATORIAIS DOS ARENAVÍRUS
Os arenavírus crescem em várias culturas celulares de vertebrados, como as linhagens
de células VERO E6, BHK-21, de rim de porco, rim de coelho e HeLa. Os arenavírus
apresentam crescimento lento em cultura e, em ensaios de formação de placas levam de
quatro a nove dias em meio semi-sólido para o aparecimento das placas (Enria et al, 2005).
Entre os animais de laboratórios suscetíveis aos arenavírus estão os camundongos,
hamsters e cobaias. A idade do animal hospedeiro, o genótipo do hospedeiro e a via de
inoculação são fatores que interferem no resultado da infecção, os arenavírus do Novo Mundo
em geral não são patogênicos para camundongos adultos, porém a inoculação intracraniana
em camundongos recém-nascidos leva a encefalite (Borden & Nathanson, 1974).
Relações antigênicas entre os arenavírus podem ser evidenciadas através de ensaios
sorológicos. Os testes de fixação do complemento (FC) e de imunofluorescência indireta
(IFI), que reconhecem principalmente epítopos da NP, diferenciaram os arenavírus nos grupos
do Velho e Novo Mundo, mostrando a baixa relação antigênica entre eles (Rowe, Pugh, Webb
& Peters, 1970; Wolff et al, 1978). Por outro lado, ensaios imunoenzimáticos (ELISA), IFI e

26
testes de neutralização (TN), demonstraram fortes relações antigênicas entre os AMAV,
JUNV, MACV e TCRV, além da proximidade entre os vírus PICV, PARV e TAMV
(SANCHEZ et al, 1989; HOWARD et al, 1985).
1.8 DIAGNÓSTICO LABORATORIAL
O isolamento dos arenavírus a partir do soro ou sangue do paciente durante a fase
aguda da doença pode ser feito por inoculação em hamsters ou camundongos recém-nascidos.
Culturas de células Vero também são frequentemente usadas para o isolamento viral. As
técnicas de IFI e FC têm sido pouco utilizadas para diagnóstico, sendo substituídas
principalmente pelo teste de ELISA, devido sua alta sensibilidade (Enria et al, 2005). Vários
ensaios de ELISA foram desenvolvidos, como o ELISA IgG direto para identificação de
arenavírus do Novo mundo como MACV, JUNV, TCRV, AMAV e TAMV, feito em 1981 a
partir da sensibilização de cobaias e o ELISA IgG indireto desenvolvido para a detecção de
anticorpos em roedores infectados (Ivanov et al, 1981; Morales et al, 2002). Mais
recentemente as técnicas de ELISA têm sido aprimoradas, como a técnica de ELISA IgG
indireto a partir de nucleoproteínas recombinantes do JUNV, que mostrou ser eficaz na
detecção de anticorpos de outros arenavírus além do JUNV (Machado et al, 2010). Ensaios
imunoenzimáticos para IgM e IgG são utilizados no diagnóstico das febres hemorrágicas
causadas pelos LASV e MACV (Peters et al, 1973; Branco et al, 2011).
A transcrição reversa seguida da reação em cadeia mediada pela polimerase (Reverse
Transcription Polymerase Chain Reaction- RT-PCR) tem sido aplicada com sucesso e pode
ter papel importante quando há necessidade de diagnóstico etiológico devido à morte do
paciente antes do aparecimento de anticorpos (Enria et al, 2005). A RT-PCR é o método de
escolha para detecção rápida do LASV em amostras de sangue (Demby at al, 1994). Outros
protocolos de RT-PCR convencional já foram desenvolvidos para outros vírus, como o JUNV
e LCMV (Lozano et al, 1995; PARK et al, 1997), além do RT-PCR em tempo real para os
TCRV e LASV (Grajkowska et al, 2009, Branco et al, 2011).
Recentemente, em Serra Leoa, África, foram desenvolvidos testes sorológicos rápidos
para o diagnóstico de infecções pelo LASV. Baseados em dois anticorpos monoclonais para a
proteína NP, os testes podem apresentar o resultado em apenas 20 minutos (Branco et al,
2011).

27
1.9 TRATAMENTO
Em doenças causadas pelo JUNV, na Argentina, o tratamento utilizado até o oitavo dia
após início da infecção é a transfusão de plasma imune. O tratamento diminui a taxa de
mortalidade de 15% a 30% para menos de 1%, porém não possui eficácia quando iniciado
após o oitavo dia da doença instalada (Enria et al, 2005).
O tratamento indicado para outras Febres Hemorrágicas Sul Americanas, tem sido a
ribavirina intravenosa, que tem demonstrado um bom prognóstico em relação aos pacientes
infectados (BARRY et al, 1995, Kilgore et al,1997). A ribavirina é ainda utilizada no
tratamento da Febre Hemorrágica causada pelo LASV (Jahrling et al, 1980) e mostrou ser útil
no tratamento de casos de Febre Hemorrágica Argentina (Enria et al, 2005).
1.10 PREVENÇÃO E CONTROLE
Uma das formas de prevenção da infecção por arenavírus encontrada na Bolívia foi
através de um programa de controle no Departamento de Beni em 1964. A eliminação em
massa de roedores Calomys callosus levou a significante diminuição dos casos de Febre
Hemorrágica Boliviana nas áreas endêmicas (Mercado, 1975). Embora esta forma de controle
tenha sido eficiente, em alguns casos ela torna-se difícil, como em infecções pelo LCMV, em
que seu hospedeiro natural - Mus musculus - tem distribuição mundial, bem como em áreas
onde os reservatórios distribuem-se em campos ou matas próximas a plantações.
A vacina Candid#1, feita a partir de partículas virais do JUNV inativadas, começou a
ser produzida em meados da década de 90, mostrando novos caminhos para a prevenção da
FHA. A eficácia da vacina foi testada em 15.000 trabalhadores rurais que viviam em
constante risco de infecção natural. Posteriormente, mais de 100.000 pessoas foram
imunizadas na Argentina. Um estudo prospectivo mostrou que a vacina teve eficácia igual ou
maior que 84%, apresentando efeitos adversos mínimos. A vacina foi licenciada em 2006 para
uso exclusivo na Argentina (Charrel et al., 2011).
Atualmente não há estudos definitivos comprovando a eficácia da Candid#1 para
outras febres hemorrágicas causadas por arenavírus Sul Americanos. Apesar de esforços para
o desenvolvimento de novas vacinas seguras, ainda não há nenhuma outra disponível e
aprovada pela Organização Mundial de Saúde (Charrel et al., 2011).

28
Pesquisas para o desenvolvimento de outras vacinas para diferentes arenavírus estão
sendo realizadas, porém ainda não existem resultados conclusivos (Geisbert et al, 2005;
Bergthaler et al, 2006).
1.11 JUSTIFICATIVA
Informações sobre o panorama epidemiológico incluindo a circulação dos arenavírus
no Brasil são muito escassas. Poucos estudos têm sido realizados para avaliar áreas
endêmicas desses vírus e quais os riscos que oferecem à população. Devido à gravidade
das infecções causadas por alguns arenavírus incidentes em territórios próximos como o
JUNV, GTOV, MACV e, ainda ao SABV presente no País, pesquisas tornam-se
necessárias para que haja maior segurança e conhecimento que possam subsidiar
providências a serem tomadas em possíveis epidemias.

29
2. OBJETIVOS
2.1 OBJETIVO GERAL
Desenvolver testes de ELISA e RT-PCR, visando a detecção da circulação de arenavírus
em amostras de roedores silvestres capturados na Amazônia brasileira.
2.2 OBJETIVOS ESPECÍFICOS
- Desenvolver um ensaio imuno-enzimático (ELISA) para detecção de anticorpos da
classe IgG em amostras de roedores silvestres, utilizando antígenos de sucrose-acetona do
SABV (arenavírus brasileiro);
- Detectar anticorpos da classe IgG para arenavírus em amostras de sangue de roedores
capturados na área de abrangência da Amazônia brasileira, pelo ensaio de ELISA;
- Desenvolver a técnica de RT-PCR e Semi-Nested-PCR para a detecção do genoma
viral de arenavírus do complexo Tacaribe;
- Tentativa de detecção do genoma viral pela técnica de RT-PCR em vísceras de
roedores sorologicamente positivas pelo teste de ELISA IgG.
30
3. MATERIAIS E MÉTODOS
3.1 AMOSTRAS
As amostras de sangue de roedores silvestres utilizadas neste estudo pertencem a
Seção de Arbovirologia e Febres Hemorrágicas do Instituto Evandro Chagas (IEC), e foram
coletadas durante estudos ecoepidemiológicos para a investigação de hantavirose realizados
pelo Serviço de Vigilância em Saúde (SVS) do Ministério da Saúde (MS), com a participação
do IEC e das secretarias de saúde dos estados e/ou municípios em questão. Foram analisadas
1.395 amostras de sangue de roedores capturados entre os anos de 2001 e 2010 através do
método de ELISA (Quadro 2). Posteriormente, as vísceras dos roedores soropositivos para
arenavírus foram submetidas à técnica de RT-PCR. O estudo foi autorizado pela Direção do
IEC (Anexo 1).
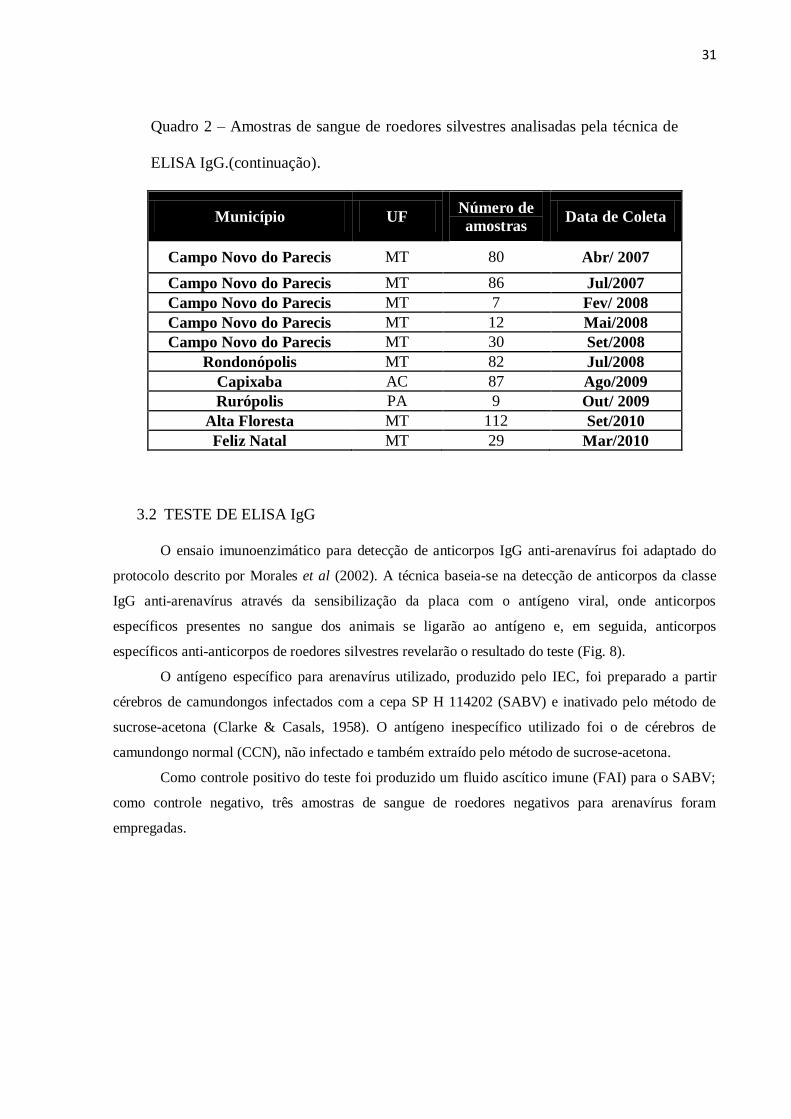
Quadro 2 – Amostras de sangue de roedores silvestres analisadas pela técnica de
ELISA IgG.
Município UF Número de amostras Data de Coleta
Tangará da Serra MT 106 Nov/ 2001
Anajatuba MA 92 Mai/ 2003
Itacoatiara AM 7 Ago/ 2004
Anajatuba MA 119 Mai/ 2005
Alto Paraíso RO 120 Jul/ 2005
Altamira PA 28 Nov/2005
Campo Novo do Parecis MT 105 Dez/2005
Campo Novo do Parecis MT 48 Set/2006
Campo Novo do Parecis MT 166 Out/ 2006
Campo Novo do Parecis MT 29 Nov/2006
Campo Novo do Parecis MT 41 Fev/2007
31
Quadro 2 – Amostras de sangue de roedores silvestres analisadas pela técnica de
ELISA IgG.(continuação).
Município UF Número de
amostras Data de Coleta
Campo Novo do Parecis MT 80 Abr/ 2007
Campo Novo do Parecis MT 86 Jul/2007
Campo Novo do Parecis MT 7 Fev/ 2008
Campo Novo do Parecis MT 12 Mai/2008
Campo Novo do Parecis MT 30 Set/2008
Rondonópolis MT 82 Jul/2008
Capixaba AC 87 Ago/2009
Rurópolis PA 9 Out/ 2009
Alta Floresta MT 112 Set/2010
Feliz Natal MT 29 Mar/2010
3.2 TESTE DE ELISA IgG
O ensaio imunoenzimático para detecção de anticorpos IgG anti-arenavírus foi adaptado do
protocolo descrito por Morales et al (2002). A técnica baseia-se na detecção de anticorpos da classe
IgG anti-arenavírus através da sensibilização da placa com o antígeno viral, onde anticorpos
específicos presentes no sangue dos animais se ligarão ao antígeno e, em seguida, anticorpos
específicos anti-anticorpos de roedores silvestres revelarão o resultado do teste (Fig. 8).
O antígeno específico para arenavírus utilizado, produzido pelo IEC, foi preparado a partir
cérebros de camundongos infectados com a cepa SP H 114202 (SABV) e inativado pelo método de
sucrose-acetona (Clarke & Casals, 1958). O antígeno inespecífico utilizado foi o de cérebros de
camundongo normal (CCN), não infectado e também extraído pelo método de sucrose-acetona.
Como controle positivo do teste foi produzido um fluido ascítico imune (FAI) para o SABV;
como controle negativo, três amostras de sangue de roedores negativos para arenavírus foram
empregadas.

32
Figura 8: ELISA IgG indireto. Na placa de ELISA (1) é feita a sensibilização com o antígeno (2), em seguida, a
amostra de sangue é colocada e se esta contiver anticorpos
específicos IgG, os mesmos se ligarão ao antígeno formando um complexo antígeno-anticorpo (3). Para a
detecção da reação, um anti- anticorpo de roedor
conjugado com a enzima peroxidase é adicionado ligando-
se ao complexo (4), a revelação é feita pela adição do substrato da enzima (peróxido de hidrogênio e
cromógeno), que ao se ligar a peroxidase forma um
produto colorido (5). Fonte: Adaptação de http:.. www.leinco.com.indirect_elisa.
3.2.1 Obtenção do antígeno pelo método da sucrose-acetona
Camundongos albinos suíços recém-nascidos foram inoculados intra-cranialmente com
0,02mL de uma suspensão de cérebro de camundongo infectado com o SABV. Os
camundongos inoculados foram examinados duas vezes ao dia para observar sinais de doença
e, quando apresentaram estes sinais foram levados para o laboratório, onde os cérebros foram
removidos assepticamente, colocados em uma placa de Petri, pesados e guardados à
temperatura de –70 °C, até o preparo do antígeno.

33
Para a extração do antígeno, os cérebros já pesados foram macerados e, em seguida, foram
adicionados quatro volumes de solução aquosa de sucrose a 8,5 %, a qual foi colocada aos
poucos, misturando bem até que a suspensão ficasse homogênea. Foram colocados lentamente
junto ao homogeneizado vinte volumes de acetona pura gelada, sendo agitado e deixado em
repouso durante cinco minutos a 4 °C. Em seguida a decantação, o sobrenadante foi
desprezado e foi reposto o mesmo volume de acetona. Após agitação vigorosa, a mistura foi
deixada uma hora em repouso a 4°C. A seguir foi feita a decantação e descarte do
sobrenadante. O precipitado foi seco durante uma hora em bomba para vácuo. Em seguida foi
hidratado com solução de cloreto de sódio a 0,85 %, em volume igual a duas vezes o peso
inicial dos cérebros. Foi deixado overnight a 4°C para completa hidratação do sedimento.
Após isso, a solução foi centrifugada a 8.000 rpm durante quinze minutos a 4 °C, sendo o
sobrenadante decantado, obtendo dessa forma, o antígeno que foi mantido à temperatura de
-70 °C até o seu uso.
3.2.2 Titulação
Para definir as concentrações utilizadas no teste, foi previamente realizada uma titulação com
diferentes concentrações dos antígenos e do conjugado, sendo os soros mantidos na
concentração de 1:100. As concentrações testadas para o antígeno foram 1:50, 1:100, 1:200,
1:400, 1:800 e 1:1600, já as concentrações para o conjugado foram 1:400, 1:800, 1:1000,
1:2000 e 1:4000. A partir desta diluição definiu-se como título padrão para os antígenos
(específico e inespecífico) a concentração de 1:200 e, como título para o conjugado definiu-se
a concentração de 1:1000. O controle positivo para o teste apresentou absorbância de 0,904 e
os controles negativos em torno de 0,006, após a correção óptica.
3.2.3 Sensibilização
Em uma placa de ELISA foram adicionados 100µL do antígeno específico em cada
poço, em fileiras alternadas. Do mesmo modo foram colocados 100µL do antígeno
inespecífico em cada poço das fileiras alternadas vazias. Para o teste, os antígenos foram
diluídos 1:200 em solução salina tamponada (PBS) pH 7,4 e a placa incubada overnight a 4°C
em câmara úmida (Fig. 9).

34
Figura 9: Placa de ELISA. Sensibilização com o antígeno do
arenavírus Sabiá (A) e com o antígeno inespecífico CCN (I).
3.2.4 Adição das amostras
A placa de ELISA sensibilizada foi lavada seis vezes com PBS contendo Tween 20
(0,1%) - tampão de lavagem. Posteriormente, foram colocados os controles positivo e
negativos e as amostras de sangue de roedores, todos diluídos a 1:100 em solução de PBS
contendo Tween 20 (0,1%) com leite desnatado a 5%. Foram adicionados 100 µL dos
controles positivo e negativo e, igual volume das amostras de roedores em dois poços, um
sensibilizado com o antígeno do SABV (específico) e outro com o antígeno inespecífico
(CCN). Após a adição dos soros a placa foi incubada em câmara úmida, em estufa a 37°C por
60 minutos.
Figura 10: Adição das amostras. Na placa, primeiramente coloca-se o
controle positivo (C+) e os três controles negativos (C-), em seguida inicia-se a adição das amostras de sangue a serem analisadas (S1-S44).

35
3.2.5 Adição do conjugado
Após a incubação a placa foi novamente lavada seis vezes com o tampão de lavagem
para retirada dos anticorpos não ligados e, em seguida 100µL do conjugado Anti-IgG de
Peromyscus leucopus (Kirkegaard and Perry Laboratories -KPL) foram adicionados a cada
poço da placa na diluição de 1:1000. O conjugado foi diluído em PBS contendo Tween 20
(0,1%) com leite desnatado a 5%. A placa foi incubada em câmara úmida a 37°C durante 60
minutos.
3.2.6 Substrato
Após uma hora a placa foi lavada seis vezes com o tampão de lavagem e, em seguida,
100µL de substrato ABTS (2,2´ azino- di[ 3- ethyl-benzothiazoline sulfonate (6)] – KPL) e
peróxido de hidrogênio foram adicionados em cada poço da placa e esta, foi incubada por 30
minutos a 37°C. Em seguida, foi realizada a leitura da densidade óptica em comprimento de
onda usando filtro de 405 nm e a placa colocada novamente na estufa a 37°C. Passados 15
minutos nova leitura foi realizada.
3.2.7 Interpretação dos resultados
A análise dos resultados foi realizada através da diferença entre as densidades ópticas
(DO) obtidas nos poços sensibilizados com o antígeno viral e com o antígeno inespecífico
(Fig. 11). As amostras que apresentaram valores iguais ou acima de 0,200 foram considerados
positivos, entre 0,100 e 0,200, limítrofes e menores que 0,100, negativos (Rodrigues et al,
2002).

36
Figura 11: Placa de ELISA após a realização da leitura da densidade óptica do teste.
3.3 TRANSCRIÇÃO REVERSA-REAÇÃO EM CADEIA DA POLIMERASE
3.3.1 Extração do RNA viral
As amostras das vísceras dos roedores silvestres que apresentaram positividade no
teste de ELISA IgG foram maceradas e as suspensões obtidas , submetidas à extração de RNA
pelo kit Pure Link RNA Mini Kit, InvitrogenTM
, seguindo as recomendações do fabricante. Os
procedimentos foram realizados dentro do Laboratório de Nível de Biossegurança 3 (NB3).
3.3.2 Transcrição Reversa – Reação em Cadeia da Polimerase (RT-PCR) seguida
da reação de Semi-Nested-PCR
As amostras biológicas passaram por dois processos para a detecção do genoma viral.
A primeira etapa foi a produção de um DNA complementar (cDNA) ao RNA viral através da
Transcrição Reversa (RT) seguida da amplificação do cDNA gerado pela PCR. Objetivando
aumentar a sensibilidade do teste, foi realizado um Semi-Nested-PCR. Os iniciadores
necessários aos procedimentos foram desenhados com base em sequências genômicas
completas do segmento S, gene N, de arenavírus do grupo Tacaribe disponíveis na base de
dados do GenBank (Quadro 3).
Como controle positivo do teste utilizou-se o genoma do SABV e para o controle
negativo o RNA total extraído de cérebros de camundongos normais (CCN).
37
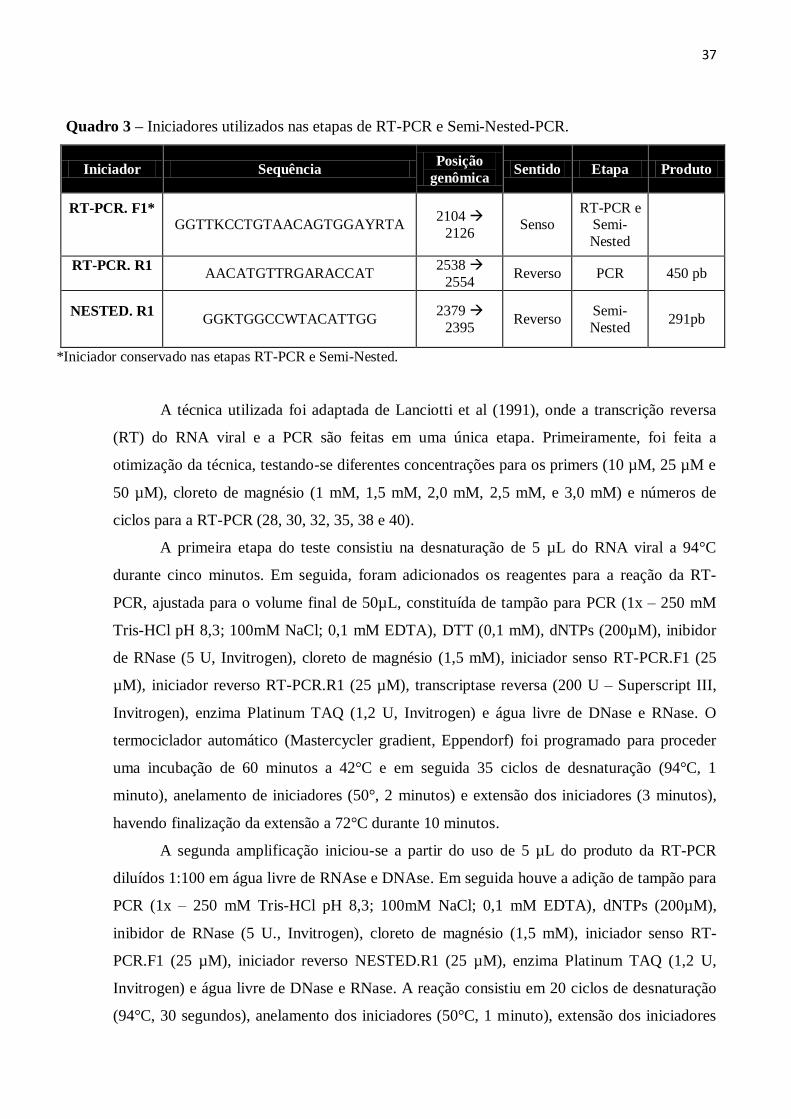
Quadro 3 – Iniciadores utilizados nas etapas de RT-PCR e Semi-Nested-PCR.
Iniciador Sequência Posição
genômica Sentido Etapa Produto
RT-PCR. F1*
GGTTKCCTGTAACAGTGGAYRTA 2104
2126 Senso
RT-PCR e Semi-
Nested
RT-PCR. R1
AACATGTTRGARACCAT
2538
2554 Reverso PCR 450 pb
NESTED. R1
GGKTGGCCWTACATTGG
2379
2395 Reverso
Semi-
Nested 291pb
*Iniciador conservado nas etapas RT-PCR e Semi-Nested.
A técnica utilizada foi adaptada de Lanciotti et al (1991), onde a transcrição reversa
(RT) do RNA viral e a PCR são feitas em uma única etapa. Primeiramente, foi feita a
otimização da técnica, testando-se diferentes concentrações para os primers (10 µM, 25 µM e
50 µM), cloreto de magnésio (1 mM, 1,5 mM, 2,0 mM, 2,5 mM, e 3,0 mM) e números de
ciclos para a RT-PCR (28, 30, 32, 35, 38 e 40).
A primeira etapa do teste consistiu na desnaturação de 5 µL do RNA viral a 94°C
durante cinco minutos. Em seguida, foram adicionados os reagentes para a reação da RT-
PCR, ajustada para o volume final de 50µL, constituída de tampão para PCR (1x – 250 mM
Tris-HCl pH 8,3; 100mM NaCl; 0,1 mM EDTA), DTT (0,1 mM), dNTPs (200µM), inibidor
de RNase (5 U, Invitrogen), cloreto de magnésio (1,5 mM), iniciador senso RT-PCR.F1 (25
µM), iniciador reverso RT-PCR.R1 (25 µM), transcriptase reversa (200 U – Superscript III,
Invitrogen), enzima Platinum TAQ (1,2 U, Invitrogen) e água livre de DNase e RNase. O
termociclador automático (Mastercycler gradient, Eppendorf) foi programado para proceder
uma incubação de 60 minutos a 42°C e em seguida 35 ciclos de desnaturação (94°C, 1
minuto), anelamento de iniciadores (50°, 2 minutos) e extensão dos iniciadores (3 minutos),
havendo finalização da extensão a 72°C durante 10 minutos.
A segunda amplificação iniciou-se a partir do uso de 5 µL do produto da RT-PCR
diluídos 1:100 em água livre de RNAse e DNAse. Em seguida houve a adição de tampão para
PCR (1x – 250 mM Tris-HCl pH 8,3; 100mM NaCl; 0,1 mM EDTA), dNTPs (200µM),
inibidor de RNase (5 U., Invitrogen), cloreto de magnésio (1,5 mM), iniciador senso RT-
PCR.F1 (25 µM), iniciador reverso NESTED.R1 (25 µM), enzima Platinum TAQ (1,2 U,
Invitrogen) e água livre de DNase e RNase. A reação consistiu em 20 ciclos de desnaturação
(94°C, 30 segundos), anelamento dos iniciadores (50°C, 1 minuto), extensão dos iniciadores

38
(72°C, 2 minutos), além da finalização da extensão (72°C, 10 minutos). Esta etapa também foi
realizada com volume final da reação ajustado para 50 µL.
3.3.3 Sensibilidade e especificidade
A sensibilidade RT-PCR seguida da Semi-Nested-PCR foi determinada através das
diluições seriadas em base 2 e 5, sendo o procedimento realizado em duplicata.
Para a avaliação da especificidade dos iniciadores da técnica, foram utilizados os
genomas de oito arenavírus pertencentes ao grupo Tacaribe. O limite de detecção foi
verificado através do teste com o SABV, AMAV e FLEV extraídos de amostras de cérebro de
camundongo infectado, e com os vírus Junín, Paraná, Latino, Guanarito, e Machupo, estes
com genomas sintéticos na forma de cDNA (gBlocksTM
, IDT). Tais genomas foram
desenhados com base no GenBank e suas sequências construídas a partir das regiões que os
iniciadores desenvolvidos se anelam, tal estratégia foi utilizada devido a indisponibilidade de
tais materiais genéticos no laboratório. Como os vírus sintetizados são fitas de DNA, estes
não passaram pelo processo da RT, apenas pela PCR. Para a determinação da especificidade
ainda foram paralelamente testados os Dengue-1, Febre Amarela, Bussuquara, Hantavírus –
Castelo dos Sonhos, Bushbush, Benfica, Apeu, Icoaraci e Belterra.
As concentrações finais utilizadas para o limite de detecção foram de 200ng, 100ng,
50ng, 25ng, 12,5ng, 6,25ng, 3,125ng, 1,56ng, 0,312 ng, 0,062ng, 0,012ng, 0,002ng, 0,5pg e
0,09pg. As concentrações finais do SABV, AMAV, e FLEV foram medidas a partir do RNA
total extraído, já as dos arenavírus sintetizados foram medidas a partir do cDNA puro. A
quantificação do material genético foi feita pelo aparelho Qubit® 2.0 Fluorometer,
InvitrogenTM.
3.3.4 Produtos Obtidos
Os produtos obtidos na Semi-Nested-PCR foram submetidos a análise por eletroforese
em gel de agarose 1,5%, imerso em tampão TAE (1x) e corado com SYBR safe DNA gel
stain, Invitrogen. Os amplicons foram comparados ao padrão do peso molecular 100 bp DNA
Mass Leader (Invitrogen), visualizados em transluminador com emissão de raios UV e
fotografados para documentação.

39
4. RESULTADOS
4.1 ELISA IgG
Dentre as 1.395 amostras de soro de roedores testadas pela técnica de ELISA IgG, 31
amostras foram positivas, representando 2,2% do total analisado. As amostras positivas
pertencentes ao Estado do Mato Grosso representaram 96,77%, sendo apenas uma dessas
amostras proveniente do Estado de Rondônia (Quadro 4).
Qu
ad
ro 4
– A
most
ras
posi
tivas
par
a ar
enav
írus
atra
vés
do t
este
de
EL
ISA
IgG
.
40
4.2 AMOSTRAS TESTADAS POR RT-PCR
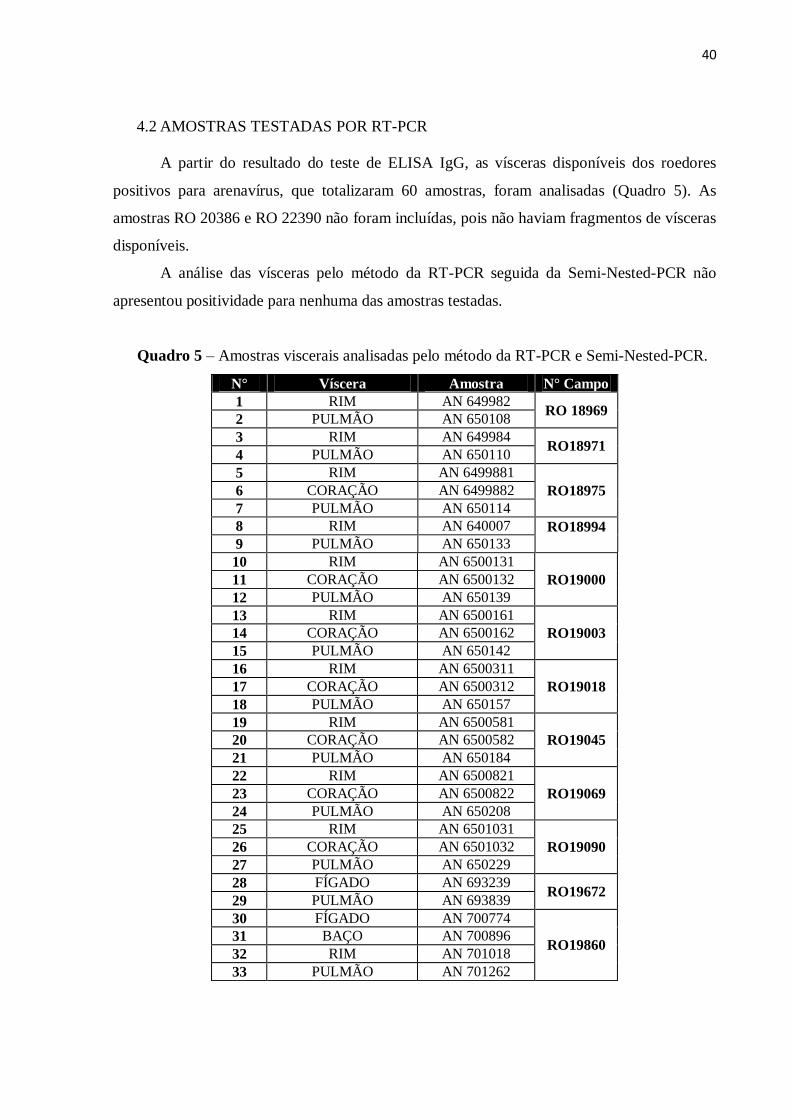
A partir do resultado do teste de ELISA IgG, as vísceras disponíveis dos roedores
positivos para arenavírus, que totalizaram 60 amostras, foram analisadas (Quadro 5). As
amostras RO 20386 e RO 22390 não foram incluídas, pois não haviam fragmentos de vísceras
disponíveis.
A análise das vísceras pelo método da RT-PCR seguida da Semi-Nested-PCR não
apresentou positividade para nenhuma das amostras testadas.
Quadro 5 – Amostras viscerais analisadas pelo método da RT-PCR e Semi-Nested-PCR.
10 RIM AN 6500131
RO19000
11 CORAÇÃO AN 6500132
12 PULMÃO AN 650139
13 RIM AN 6500161
RO19003
14 CORAÇÃO AN 6500162
15 PULMÃO AN 650142
16 RIM AN 6500311
RO19018
17 CORAÇÃO AN 6500312
18 PULMÃO AN 650157
19 RIM AN 6500581
RO19045
20 CORAÇÃO AN 6500582
21 PULMÃO AN 650184
22 RIM AN 6500821
RO19069
23 CORAÇÃO AN 6500822
24 PULMÃO AN 650208
25 RIM AN 6501031
RO19090
26 CORAÇÃO AN 6501032
27 PULMÃO AN 650229
28 FÍGADO AN 693239 RO19672
29 PULMÃO AN 693839
30 FÍGADO AN 700774
RO19860 31 BAÇO AN 700896
32 RIM AN 701018
33 PULMÃO AN 701262
N° Víscera Amostra N° Campo
1 RIM AN 649982 RO 18969
2 PULMÃO AN 650108
3 RIM AN 649984 RO18971
4 PULMÃO AN 650110
5 RIM AN 6499881
RO18975
6 CORAÇÃO AN 6499882
7 PULMÃO AN 650114
8 RIM AN 640007 RO18994
9 PULMÃO AN 650133
41
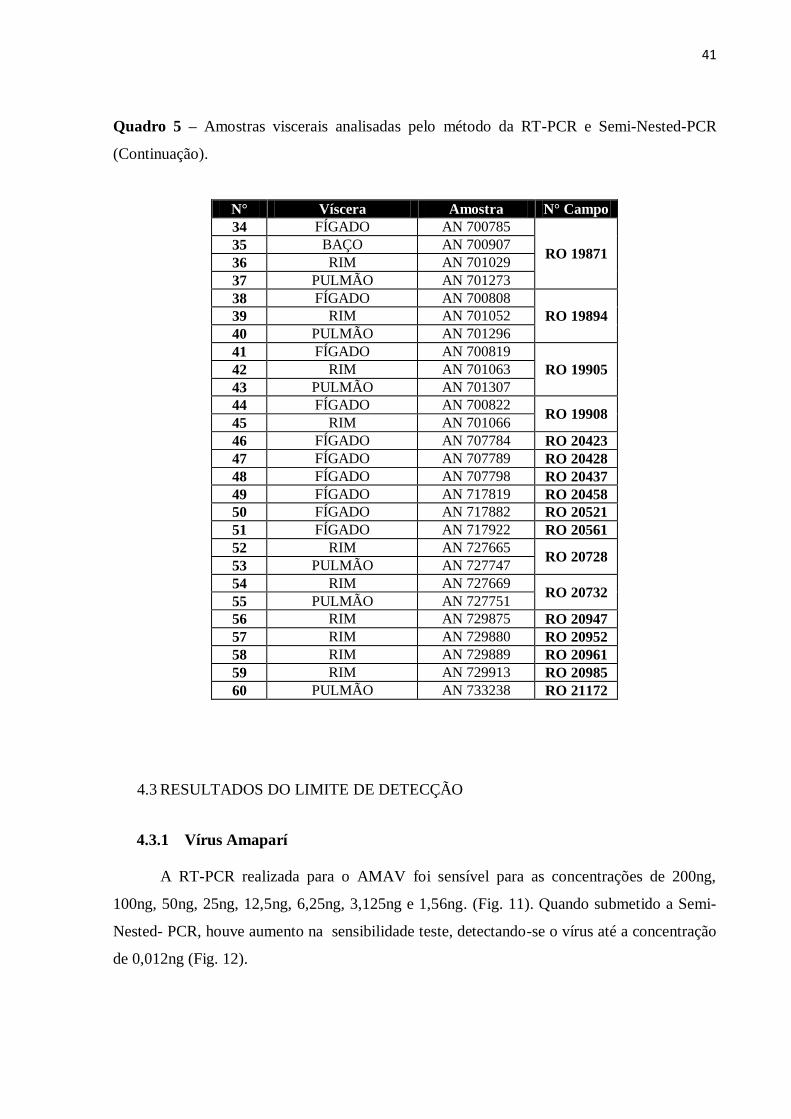
Quadro 5 – Amostras viscerais analisadas pelo método da RT-PCR e Semi-Nested-PCR
(Continuação).
N° Víscera Amostra N° Campo
34 FÍGADO AN 700785
RO 19871 35 BAÇO AN 700907
36 RIM AN 701029
37 PULMÃO AN 701273
38 FÍGADO AN 700808
RO 19894 39 RIM AN 701052
40 PULMÃO AN 701296
41 FÍGADO AN 700819
RO 19905 42 RIM AN 701063
43 PULMÃO AN 701307
44 FÍGADO AN 700822 RO 19908
45 RIM AN 701066
46 FÍGADO AN 707784 RO 20423
47 FÍGADO AN 707789 RO 20428
48 FÍGADO AN 707798 RO 20437
49 FÍGADO AN 717819 RO 20458
50 FÍGADO AN 717882 RO 20521
51 FÍGADO AN 717922 RO 20561
52 RIM AN 727665 RO 20728
53 PULMÃO AN 727747
54 RIM AN 727669 RO 20732
55 PULMÃO AN 727751
56 RIM AN 729875 RO 20947
57 RIM AN 729880 RO 20952
58 RIM AN 729889 RO 20961
59 RIM AN 729913 RO 20985
60 PULMÃO AN 733238 RO 21172
4.3 RESULTADOS DO LIMITE DE DETECÇÃO
4.3.1 Vírus Amaparí
A RT-PCR realizada para o AMAV foi sensível para as concentrações de 200ng,
100ng, 50ng, 25ng, 12,5ng, 6,25ng, 3,125ng e 1,56ng. (Fig. 11). Quando submetido a Semi-
Nested- PCR, houve aumento na sensibilidade teste, detectando-se o vírus até a concentração
de 0,012ng (Fig. 12).
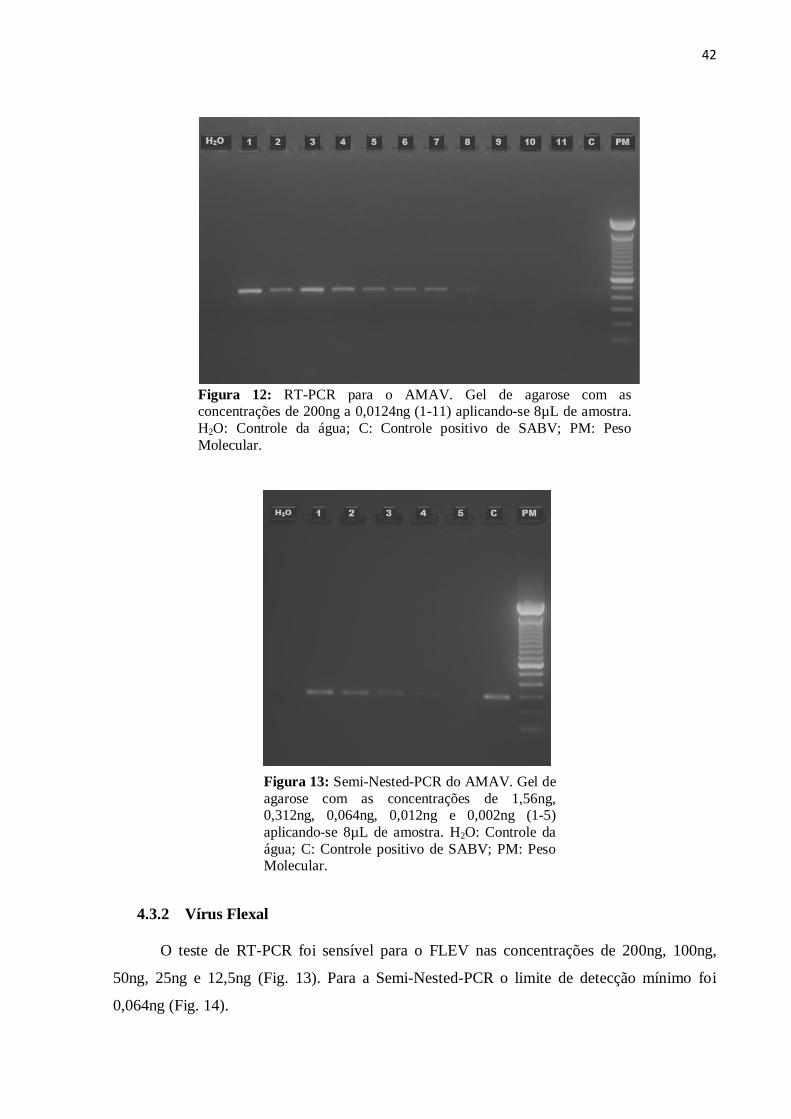
42
Figura 12: RT-PCR para o AMAV. Gel de agarose com as concentrações de 200ng a 0,0124ng (1-11) aplicando-se 8µL de amostra.
H2O: Controle da água; C: Controle positivo de SABV; PM: Peso
Molecular.
Figura 13: Semi-Nested-PCR do AMAV. Gel de
agarose com as concentrações de 1,56ng, 0,312ng, 0,064ng, 0,012ng e 0,002ng (1-5)
aplicando-se 8µL de amostra. H2O: Controle da
água; C: Controle positivo de SABV; PM: Peso Molecular.
4.3.2 Vírus Flexal
O teste de RT-PCR foi sensível para o FLEV nas concentrações de 200ng, 100ng,
50ng, 25ng e 12,5ng (Fig. 13). Para a Semi-Nested-PCR o limite de detecção mínimo foi
0,064ng (Fig. 14).
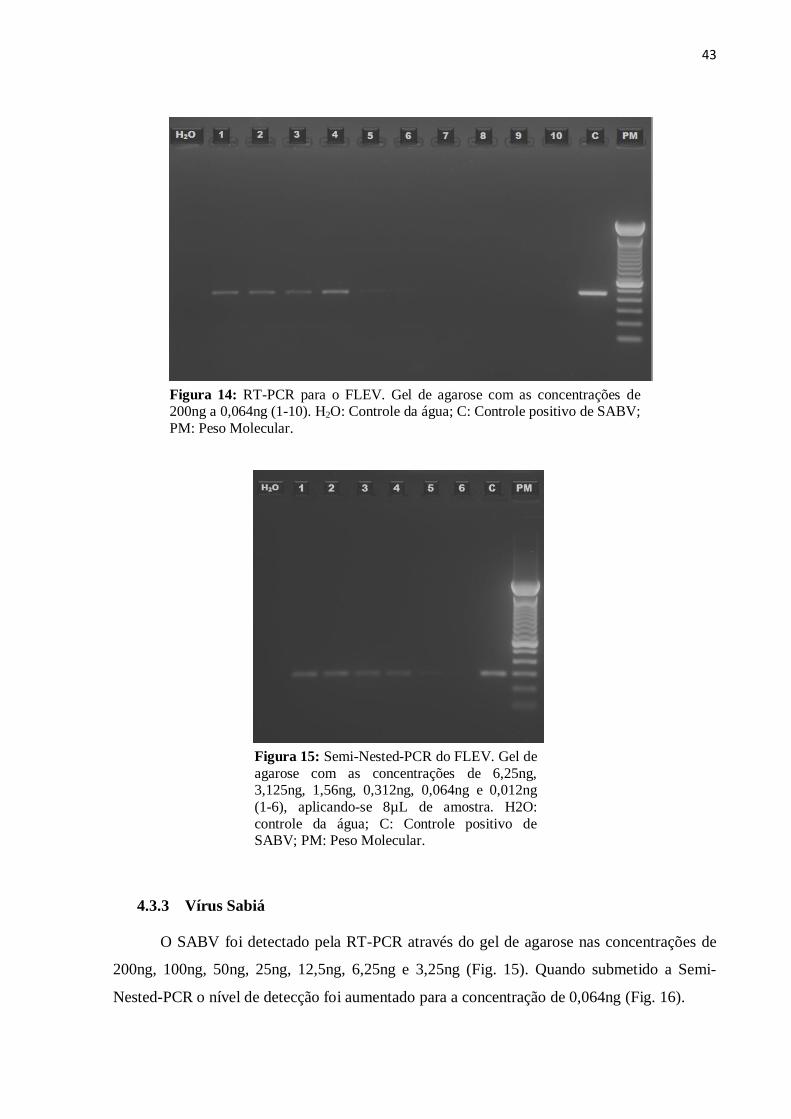
43
Figura 14: RT-PCR para o FLEV. Gel de agarose com as concentrações de 200ng a 0,064ng (1-10). H2O: Controle da água; C: Controle positivo de SABV;
PM: Peso Molecular.
Figura 15: Semi-Nested-PCR do FLEV. Gel de
agarose com as concentrações de 6,25ng, 3,125ng, 1,56ng, 0,312ng, 0,064ng e 0,012ng
(1-6), aplicando-se 8µL de amostra. H2O:
controle da água; C: Controle positivo de SABV; PM: Peso Molecular.
4.3.3 Vírus Sabiá
O SABV foi detectado pela RT-PCR através do gel de agarose nas concentrações de
200ng, 100ng, 50ng, 25ng, 12,5ng, 6,25ng e 3,25ng (Fig. 15). Quando submetido a Semi-
Nested-PCR o nível de detecção foi aumentado para a concentração de 0,064ng (Fig. 16).
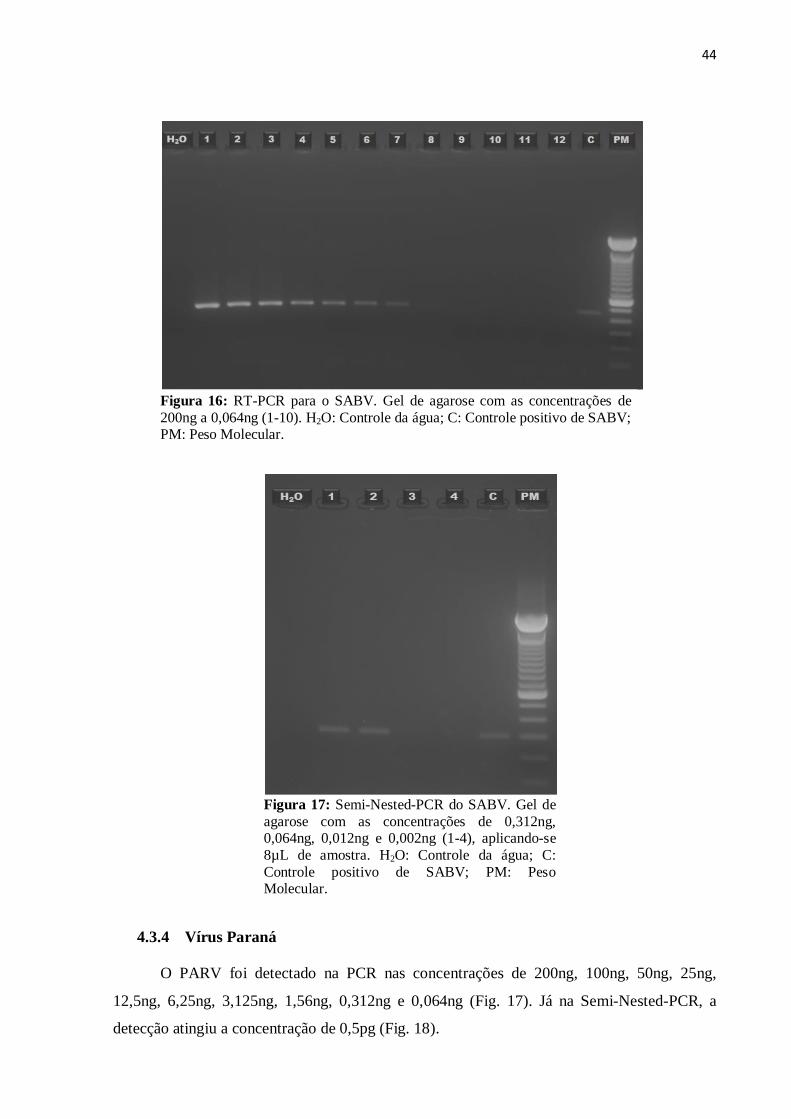
44
Figura 16: RT-PCR para o SABV. Gel de agarose com as concentrações de
200ng a 0,064ng (1-10). H2O: Controle da água; C: Controle positivo de SABV; PM: Peso Molecular.
Figura 17: Semi-Nested-PCR do SABV. Gel de
agarose com as concentrações de 0,312ng, 0,064ng, 0,012ng e 0,002ng (1-4), aplicando-se
8µL de amostra. H2O: Controle da água; C:
Controle positivo de SABV; PM: Peso Molecular.
4.3.4 Vírus Paraná
O PARV foi detectado na PCR nas concentrações de 200ng, 100ng, 50ng, 25ng,
12,5ng, 6,25ng, 3,125ng, 1,56ng, 0,312ng e 0,064ng (Fig. 17). Já na Semi-Nested-PCR, a
detecção atingiu a concentração de 0,5pg (Fig. 18).
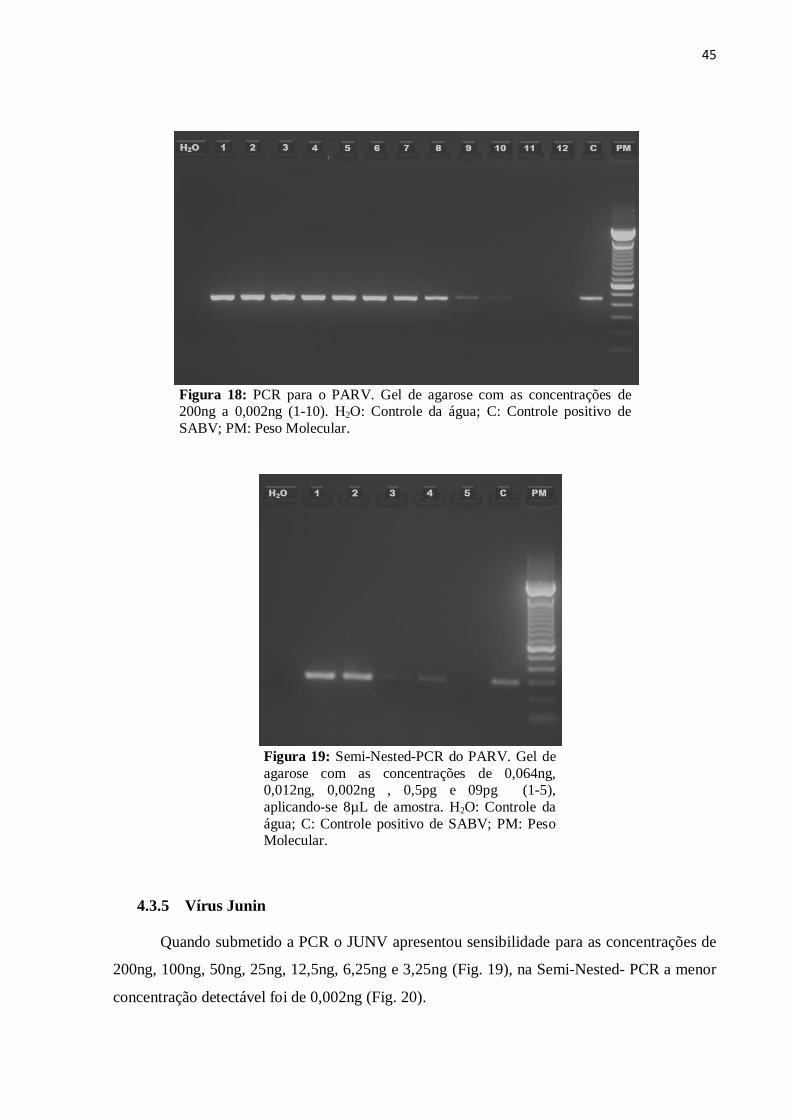
45
Figura 18: PCR para o PARV. Gel de agarose com as concentrações de 200ng a 0,002ng (1-10). H2O: Controle da água; C: Controle positivo de
SABV; PM: Peso Molecular.
Figura 19: Semi-Nested-PCR do PARV. Gel de
agarose com as concentrações de 0,064ng, 0,012ng, 0,002ng , 0,5pg e 09pg (1-5),
aplicando-se 8µL de amostra. H2O: Controle da
água; C: Controle positivo de SABV; PM: Peso Molecular.
4.3.5 Vírus Junin
Quando submetido a PCR o JUNV apresentou sensibilidade para as concentrações de
200ng, 100ng, 50ng, 25ng, 12,5ng, 6,25ng e 3,25ng (Fig. 19), na Semi-Nested- PCR a menor
concentração detectável foi de 0,002ng (Fig. 20).
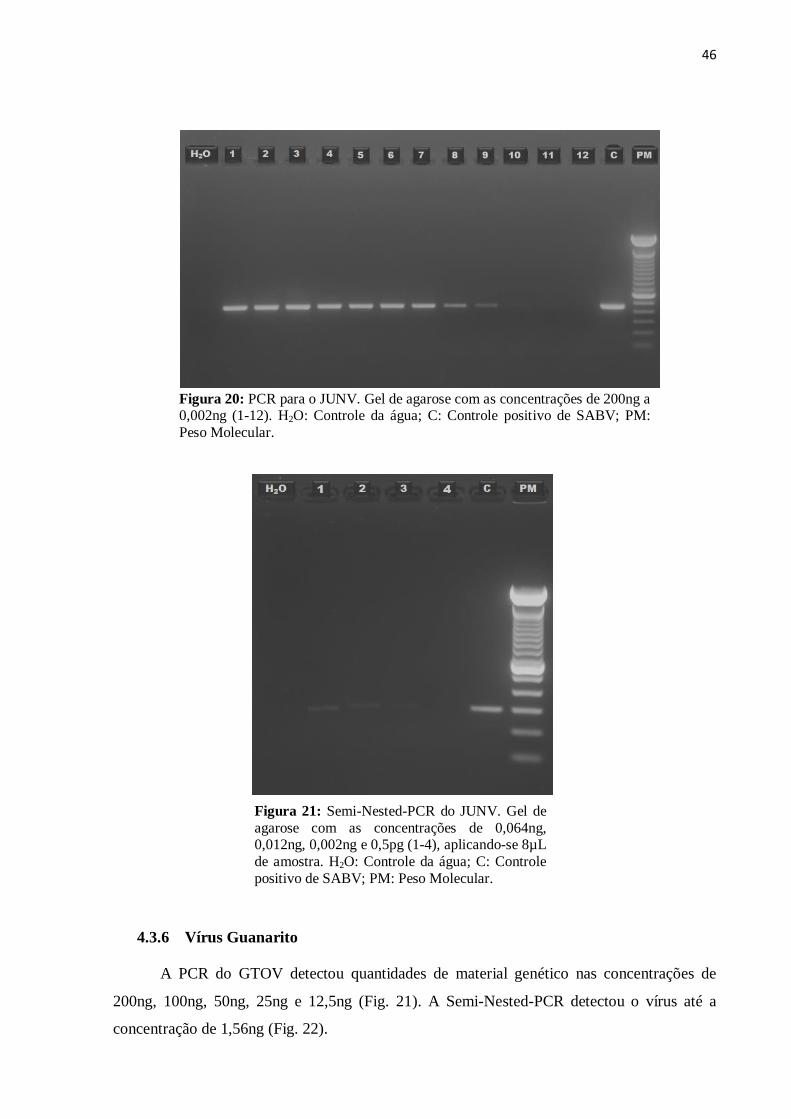
46
Figura 20: PCR para o JUNV. Gel de agarose com as concentrações de 200ng a 0,002ng (1-12). H2O: Controle da água; C: Controle positivo de SABV; PM:
Peso Molecular.
Figura 21: Semi-Nested-PCR do JUNV. Gel de
agarose com as concentrações de 0,064ng, 0,012ng, 0,002ng e 0,5pg (1-4), aplicando-se 8µL
de amostra. H2O: Controle da água; C: Controle
positivo de SABV; PM: Peso Molecular.
4.3.6 Vírus Guanarito
A PCR do GTOV detectou quantidades de material genético nas concentrações de
200ng, 100ng, 50ng, 25ng e 12,5ng (Fig. 21). A Semi-Nested-PCR detectou o vírus até a
concentração de 1,56ng (Fig. 22).
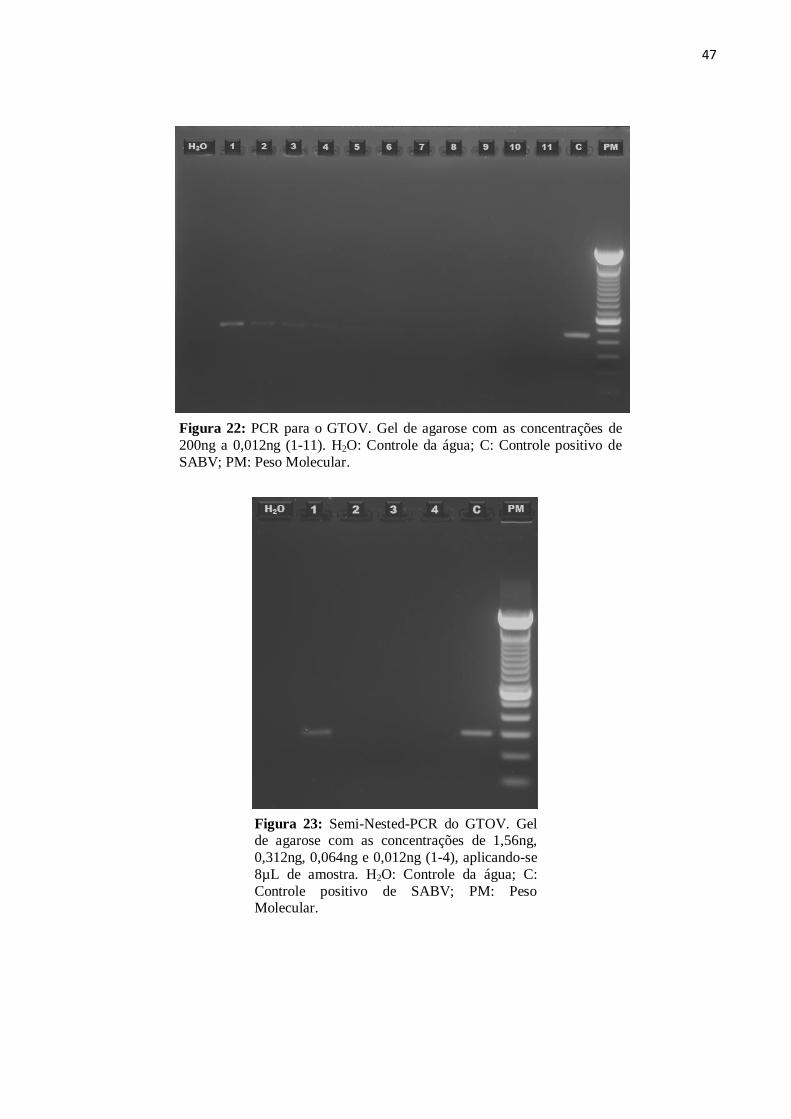
47
Figura 22: PCR para o GTOV. Gel de agarose com as concentrações de
200ng a 0,012ng (1-11). H2O: Controle da água; C: Controle positivo de
SABV; PM: Peso Molecular.
Figura 23: Semi-Nested-PCR do GTOV. Gel de agarose com as concentrações de 1,56ng,
0,312ng, 0,064ng e 0,012ng (1-4), aplicando-se
8µL de amostra. H2O: Controle da água; C:
Controle positivo de SABV; PM: Peso Molecular.

48
4.3.7 Vírus Machupo
Quando feita a PCR para o MACV, não foi possível a sua detecção, porém quando
submetido a Semi-Nested-PCR, o vírus pode ser detectado até a concentração de 1,56ng (Fig.
23).
Figura 24: PCR e Semi-Nested-PCR para o MACV. Em a: PCR das concentrações de 200ng a 1,56ng (1-8); b: Semi-Nested-PCR com as concentrações de 6,25ng, 3,12ng, 1,56ng e
0,064ng (1-5). H2O: Controle da água; C: Controle positivo de SABV; PM: Peso Molecular.
4.3.8 Vírus Latino
Bandas fracas apareceram na PCR do LATV, aparecendo apenas nas concentrações de
200ng e 100ng. A Semi-Nested- PCR detectou o material genético até a concentração de
12,5ng (Fig. 24).
49
Figura 25: PCR e Semi-Nested-PCR para o LATV. Em a: PCR das concentrações de
200ng a 1,56ng (1-8); b: Semi-Nested-PCR com as concentrações de 25ng, 12,5ng, 6,25ng 3,25ng e 1,56ng (1-5). H2O: Controle da água; C: Controle positivo de SABV; PM: Peso
Molecular.
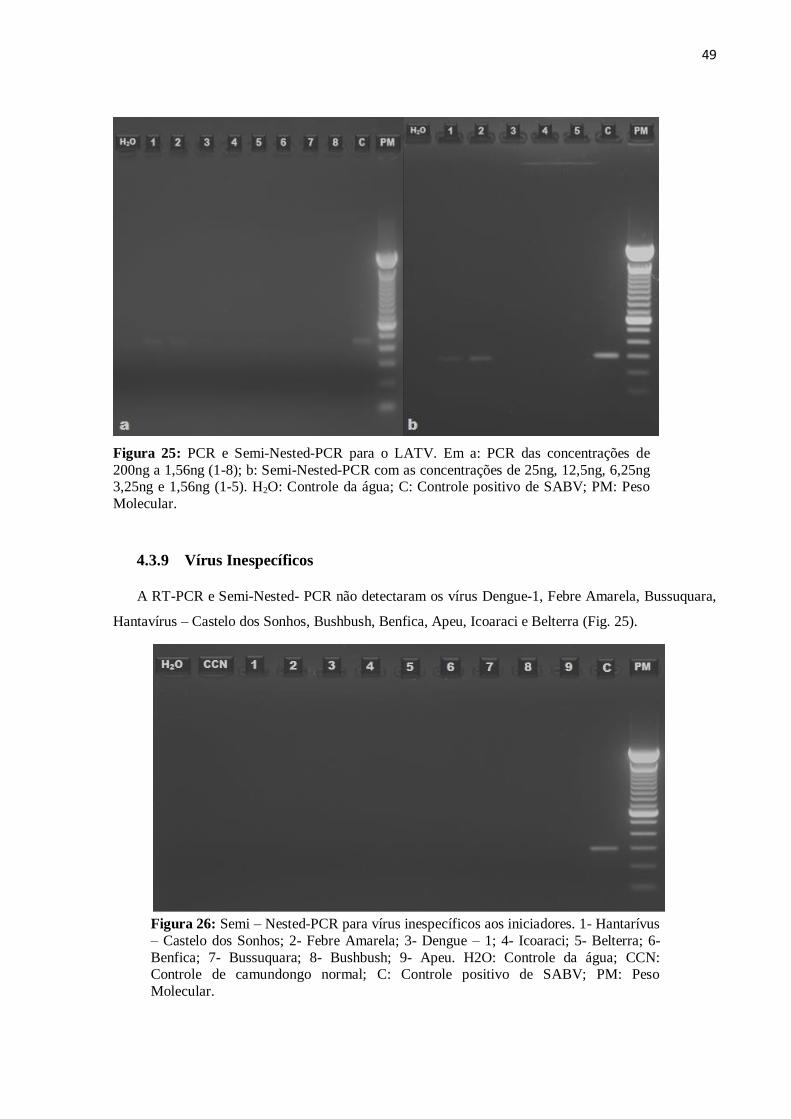
4.3.9 Vírus Inespecíficos
A RT-PCR e Semi-Nested- PCR não detectaram os vírus Dengue-1, Febre Amarela, Bussuquara,
Hantavírus – Castelo dos Sonhos, Bushbush, Benfica, Apeu, Icoaraci e Belterra (Fig. 25).
Figura 26: Semi – Nested-PCR para vírus inespecíficos aos iniciadores. 1- Hantarívus
– Castelo dos Sonhos; 2- Febre Amarela; 3- Dengue – 1; 4- Icoaraci; 5- Belterra; 6-
Benfica; 7- Bussuquara; 8- Bushbush; 9- Apeu. H2O: Controle da água; CCN: Controle de camundongo normal; C: Controle positivo de SABV; PM: Peso
Molecular.
50
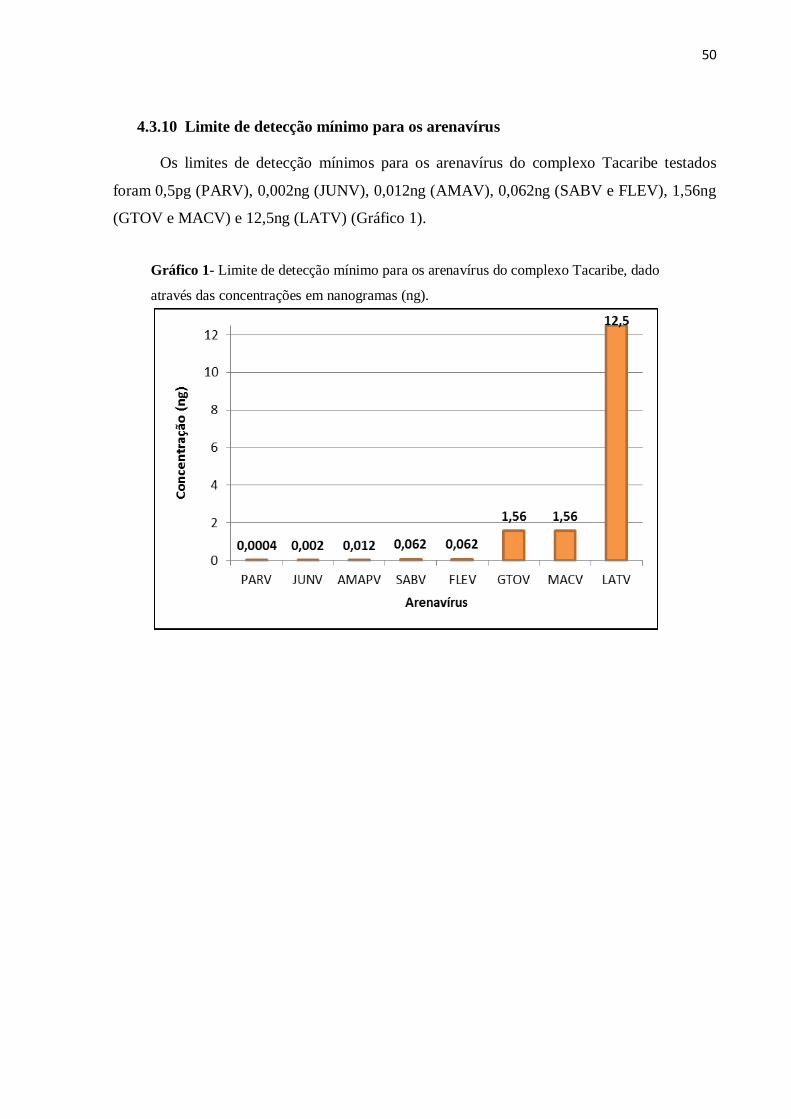
4.3.10 Limite de detecção mínimo para os arenavírus
Os limites de detecção mínimos para os arenavírus do complexo Tacaribe testados
foram 0,5pg (PARV), 0,002ng (JUNV), 0,012ng (AMAV), 0,062ng (SABV e FLEV), 1,56ng
(GTOV e MACV) e 12,5ng (LATV) (Gráfico 1).
Gráfico 1- Limite de detecção mínimo para os arenavírus do complexo Tacaribe, dado
através das concentrações em nanogramas (ng).

51
5. DISCUSSÃO
Estudos anteriores demonstraram que as características antigênicas dos arenavírus
permitem seu agrupamento em vírus do Novo Mundo e do Velho Mundo (Rowe, Pugh, Webb
& Peters, 1970). Testes de IFI, FC e ELISA já demonstraram serem eficazes para a
identificação cruzada entre arenavírus do Grupo Tacaribe (Sanchez et al, 1989; Casals et al,
1975; Rowe, Pugh, Webb & Peters, 1970). Antígenos tratados pelo método de sucrose-
acetona (Kuno et al, 1991; Clarke e Casals, 1958), foram utilizados por Vasconcelos et al
(1993) para o diagnóstico específico do soro de paciente infectado com o SABV através dos
métodos de FC e pela detecção de anticorpos IgM pelo Ensaio Imunoenzimático de Captura
(MAC – ELISA). Outros testes de ELISA baseados no uso de antígenos tratados pelo método
de sucrose- acetona são muito utilizados no diagnóstico de enfermidades virais, como as
causadas pelos vírus Dengue (Nunes et al, 2011), Oropouche (Vasconcelos et al, 1989 e
Mayaro (Casseb, 2010).
Para o teste de ELISA, desenvolvido neste trabalho, não foi possível a formação de um
painel composto de amostras de roedores silvestres positivos para arenavírus. Para a melhor
avaliação do teste seria necessária uma quantidade considerável dessas amostras que
permitiriam a análise da sensibilidade e especificidade do ensaio. Além das dificuldades para
dispor de amostras de roedores silvestres positivas, a captura de tais roedores e manejo de
colônias também tornaram-se inviáveis para este projeto devido a necessidade de autorização
do Instituto Brasileiro do Meio Ambiente e Recursos Naturais Renováveis (IBAMA) e o curto
tempo após o desenvolvimento do teste para a defesa desta dissertação. Além disso, a maioria
das amostras testadas pertencem ao gênero Calomys sp., roedores não característicos da
região Norte do Pará.
Morales et al (2002) realizou a captura de Calomys musculinus e Calomys laucha para
avaliar um teste de ELISA IgG indireto para roedores infectados com o vírus Junin, , que após
submetidos a testes para arenavírus e mantidos em quarentena foram utilizados como modelo
para a inoculação do vírus e empregados como um painel com soro e sangue positivos. O teste
de ELISA desenvolvido apresentou sensibilidade e especificidade de 100% e
reprodutibilidade de 87,9% em comparação a IFI, que apresentou sensibilidade de apenas
53%. O teste realizado a partir de antígenos do JUNV também mostrou sensibilidade em
reações cruzadas quando testado com o soro do roedor Neacromys benefactus, reservatório
natural dos vírus Oliveros e Pampas. Os autores ainda comprovaram que não há diferenças

52
estatísticas significantes entre os resultados obtidos pelo teste de ELISA IgG realizado com
soro ou com o sangue total dos roedores, o que facilita o trabalho de campo, já que a obtenção
do sangue total dos animais é mais fácil. Esses resultados reforçam os obtidos no teste de
ELISA desenvolvido no presente estudo, com o antígeno do SABV, pois, apenas o sangue
total dos roedores estava disponível para a avaliação do teste.
A realização de testes sorológicos adicionais FC e TN para comparação com os
resultados obtidos no ELISA desenvolvido neste estudo, ficaram impossibilitados devido à
não disponibilidade de soros nas amostras testadas. Testes de neutralização utilizados na
avaliação do vírus Amaparí realizados por Pinheiro et al. (1966), apresentaram resultados
insatisfatórios quando feitos a partir de vírus coletados de cérebros de camundongos devido a
mortes irregulares dos animais. Vários camundongos sobreviveram às inoculações
intracranianas iniciais do vírus e apenas na sétima passagem, a dose de letalidade (LD50)
chegou a 2,5 log, indicando que, a presença de baixos títulos de anticorpos para arenavírus
podem influenciar no resultado do teste. A maior sensibilidade do ELISA já foi evidenciada
em relação a outros ensaios, como a FC (Ivanov et al, 1981) e, tem sido utilizada como
padrão para a detecção de infecções causadas por arenavírus (Morales et al, 2002).
O desenvolvimento do ensaio de ELISA IgG deste estudo mostrou a presença de
arenavírus circulantes na Amazônia Brasileira. Dentre as 1.395 amostras analisadas, 31
(2,2%) apresentaram positividade. Dados comparativos sobre a pesquisa epidemiológica de
roedores infectados com arenavírus no Brasil são inexistentes, porém os resultados obtidos
entram em consenso com o estudo realizado na Argentina por Mills et al.(1991), que, através
dos testes de ELISA e isolamento viral, analisou a prevalência do JUNV em roedores de áreas
endêmicas e não endêmicas do país. A prevalência do estudo variou de 0,4% a 3,7%, sendo
que, as áreas endêmicas para o JUNV apresentaram de duas a três vezes maior positividade do
que as áreas não endêmicas. Esses resultados corroboram com os obtidos no presente trabalho
e mesmo não havendo no Brasil áreas consideradas endêmicas para arenavírus acredita-se
que, há a circulação de tais vírus, sendo de primordial atenção áreas próximas a plantações.
Para uma melhor avaliação do teste de ELISA IgG desenvolvido, a adaptação de
outros antígenos de arenavírus se faz necessária. Antígenos produzidos em cultura de células
(Mattar et al, 2011) e antígenos de proteínas recombinantes (Machado et al, 2010) poderiam
ser utilizados na sensibilização da placa para posterior comparação com os resultados obtidos
pelas diferentes metodologias. Outra alternativa seria levar a técnica desenvolvida para ser

53
testada em centros de referência para a avaliação de amostras disponíveis de roedores
silvestres sabidamente positivas.
A confirmação do teste de ELISA IgG seria realizada pela técnica de RT-PCR
desenvolvida para os arenavírus sul americanos, porém, todos os testes realizados foram
negativos. Padrões de infecção persistente nos hospedeiros ainda não são bem definidos. Em
Calomys musculinus, a resposta pelo JUNV é dividida, sendo que a metade dos hospedeiros
silvestres torna-se crônica, com viremia persistente e, a outra metade elimina a infecção (Mills
et al, 1992). Porém, mesmo quando existe a cronicidade da infecção, há períodos em que a
excreção do vírus não é detectada (Pinheiro et al, 1977). A ausência de material genético viral
pode ter ocorrido devido a não cronicidade nos roedores estudados, ou mesmo pela baixa
carga viral presente nos tecidos desses roedores. Outro ponto a ser levado em consideração é
o armazenamento das amostras, que podem resultar em variações de temperatura durante os
anos de estocagem e, como os arenavírus são vírus de RNA, estes se degradam facilmente
com o acondicionamento inadequado, o que pode justificar os resultados negativos pela RT-
PCR. Por outro lado, a ausência de detecção do genoma pela RT- Semi- Nested – PCR
desenvolvida pode ser devido ao cruzamento sorológico por ELISA entre diferentes
arenavírus, o que não se observa por RT-PCR. Assim, é possível que os soros positivos por
ELISA IgG sejam devidos a um arenavírus ainda não isolado, e portanto, por RT-PCR o
resultado negativo para esses arenavírus desconhecidos é esperado.
Os arenavírus apresentam uma grande variabilidade genética entre si devido a vários
fatores, dentre eles, a alta taxa de mutação da polimerase RdRp, o rearranjo genético e
possivelmente as recombinações ocorridas durante a evolução desses vírus, contribuindo para
sua grande diversidade. Acredita-se que, a alta frequência de erros de transcrição seja a
principal razão para a diversidade evolutiva dos arenavírus (Zapata & Salvato, 2013). A
análise filogenética a partir de sequências parciais da NP dividiram as linhagens A, B e C dos
arenavírus do Novo Mundo, que apresentaram divergências de 61% entre as linhagens A e B,
39% entre B e C, e 55% entre A e C (Archer & Rico-Hesser, 2002).
Apesar de todos os arenavírus do complexo do Novo Mundo apresentarem
divergências, eles continuam no mesmo sorocomplexo quando classificados através de soros
imunes, isso significa que, embora haja grande divergência genética entre as linhagens virais,
existem sítios antigênicos comuns que podem ser detectados por ensaios sorológicos (Archer
& Rico-Hesser, 2002). As variabilidades genéticas são obstáculos quando se busca
desenvolver iniciadores para vários tipos de arenavírus. Na literatura é rara a descrição de

54
técnicas de RT-PCR sensíveis a mais de um tipo viral e geralmente os iniciadores são
desenvolvidos para diagnósticos de arenavírus endêmicos, como ocorre com os vírus Lassa e
Junin (Demby, 1994; Lozano, 1995). Uma técnica de RT- PCR para detecção do gênero
Arenavírus foi desenvolvida com base em regiões conservadas das extremidades do segmento
S do RNA genômico, porém o teste codifica todo o segmento S (3,4 Kb), o que acaba
dificultando o diagnóstico viral (Gunther et al, 2000).
Os iniciadores desenvolvidos neste trabalho apresentaram uma boa sensibilidade para
os vírus avaliados. Os menores limites de detecção foram obtidos com os vírus Amaparí,
Junin e Paraná, apresentando valores de 0,012ng, 0,002ng e 0,5pg, respectivamente. Para o
FLEV e SABV a RT-PCR seguida da Semi-Nested detectou até 0,062ng a partir do RNA
total. Menor sensibilidade foi encontrada para os vírus MACV e GTOV, com detecção de
1,56ng e para o LATV, onde a Semi-Nested detectou até a 12,5ng de material genético. Como
os vírus Sabiá, Flexal e Amaparí foram obtidos de amostras de RNA total, a quantidade real
de detecção para o genoma de tais vírus é possivelmente menor do que o estipulado no estudo,
o que indica que a técnica tem boa sensibilidade para os arenavírus brasileiros.
Os outros cinco arenavírus analisados a partir dos cDNAs sintetizados apresentaram
quantidades da sequência alvo puras, sendo possível uma análise mais fidedigna da
quantidade de material genético detectável no teste, porém, para a melhor validação da técnica
a reprodutibilidade do teste deve ser avaliada, sendo necessária, para isto, a análise das
amostras a partir do RNA. Como a aquisição de amostras de arenavírus provenientes de
outros países é difícil, sendo necessários rigorosos critérios de biossegurança, uma alternativa
para a obtenção dos RNAs de tais vírus seria a clonagem das sequências de cDNA alvo e sua
posterior transformação em RNA pela enzima transcriptase reversa.
Estudos com iniciadores degenerados demonstraram sensibilidade de até 1ng de
material genético viral para os Orthopoxvirus a Parapoxvírus, desenvolvidos para Nested-
Multiplex PCR (Abrahão et al, 2009), demonstrando que, os valores obtidos para os
arenavírus deste trabalho estão dentro dos valores sensíveis admissíveis. O resultado da
detecção molecular do LATV foi consideravelmente inferior aos outros arenavírus, contudo,
quando se trabalha com degeneração de iniciadores, principalmente no caso desses vírus, é
esperado que a sensibilidade seja discrepante entre eles. Como a técnica desenvolvida neste
trabalho tem o objetivo de identificar arenavírus sul americanos em geral, o limite de detecção
mínimo para o LATV ainda é aceitável.

55
A especificidade da técnica molecular empregada foi avaliada a partir do teste com os
vírus inespecíficos Dengue-1, Febre Amarela, Bussuquara, Hantavírus (Castelo dos Sonhos),
Bushbush, Benfica, Apeu, Icoaraci e Belterra. Nenhum dos vírus foi detectado, o que
evidencia a qualidade dos iniciadores utilizados para evitar resultados falso-positivos.
A maioria das amostras positivas no teste de ELISA IgG indireto foram provenientes
do Estado do Mato Grosso. Acredita-se, desta forma, na hipótese de que os vírus circulantes
na região sejam o Machupo e Latino ou outro a eles relacionado, já que o estado faz fronteira
com a Bolívia, país endêmico para estes arenavírus. Além disso, quase todos os hospedeiros
da amostragem deste trabalho são do gênero Calomys sp., reservatório do MACV e LATV.
Considerando-se que, todas as amostras analisadas pela RT- PCR foram negativas,
pode-se correlacionar a menor eficiência dos iniciadores para estes dois vírus, que poderiam
não ter sido detectados devido a quantidade de material genético presente nas amostras ser
inferior aos limites detectáveis, ou a um vírus relacionado e ainda não isolado.
Das amostras positivas obtidas, 30 são oriundas de Campo Novo do Parecis, Mato
Grosso, um dos estados que apresentam alto índice de produção agropecuária no Brasil. Nos
últimos quarenta anos, o Brasil desenvolveu diversos núcleos de ocupação e modernização de
fronteiras agrícolas, sendo o município de Campo Novo do Parecis/MT um núcleo de alta
produção agrícola, especializado na produção de grãos, como arroz, milho e soja. O rápido
desenvolvimento agrícola desta região transformou drasticamente a paisagem, substituindo o
cerrado por pastos e plantações. (Portela, 2009). A rápida modificação do cenário é um fator
determinante para a adaptação de roedores que antes habitavam apenas as matas, o que
culmina na transmissão para o homem de doenças antes deconhecidas, como é o caso dos
arenavírus (Hui, 2006). Com todas as condições favoráveis para emergirem na população de
regiões com características como essas, os arenavírus tornam-se grandes candidatos a
possíveis epidemias, por isso, a vigilância em regiões onde há abundância de alimento para
roedores silvestres são de grande relevância.
A RT-PCR realizada com iniciadores degenerados para os arenavírus sul americanos é
uma ferramenta bastante útil para estudos sobre a circulação dos mesmos em determinadas
regiões e até mesmo para a descoberta de novos arenavírus circulantes. Como a técnica nos
permite a detecção de vários arenavírus, uma análise sobre a dispersão de tais vírus entre os
diferentes países sul americanos pode ser feita através dela. No Brasil, não existem
publicações concisas sobre o panorama da prevalência de roedores infectados com arenavírus,
os iniciadores degenerados desenvolvidos neste trabalho serão de grande auxílio para futuros

56
estudos sobre o endemismo dos arenavírus em diferentes regiões do País, sendo um
instrumento efetivo para a coleta de dados que poderão dar suporte a criação de políticas de
prevenção e controle de doenças causadas por esses vírus.

57
6. CONCLUSÃO
- O teste de ELISA IgG indireto desenvolvido para a análise das amostras dos roedores
silvestres da Amazônia Brasileira mostrou capacidade de detectar anticorpos para arenavírus
brasileiros;
- Foi desenvolvido um protocolo para detecção de genomas de arenavírus do Novo
Mundo usando a técnica de RT-PCR seguida da Semi-Nested – PCR.;
- A técnica de RT-Semi-Nested-PCR desenvolvida apresentou alta sensibilidade, tendo
detectado genoma de arenavírus do Novo Mundo até valores de 0,5pg de material genético
dependendo da espécie viral;
- Todos os arenavírus testados foram detectados, sendo o vírus Latino o que apresentou
níveis menores de sensibilidade;
- A especificidade da RT-PCR foi comprovada para diferentes vírus inespecíficos (não
arenavírus), demonstrando que, os iniciadores específicos desenvolvidos não produzem
resultados falso-positivos.
- Os resultados preliminares obtidos neste trabalho servirão como referência para outros
estudos no Brasil, visando o diagnóstico e o esclarecimento sobre a circulação dos arenavírus
no País.

58
REFERÊNCIAS
ARCHER, M.A., RICO-HESSER, R. High genetic divergence and recombination in
arenaviruses from the Americas. Virology 304: 274-281, 2002.
ABRAHÃO, J.S., LIMA, L.S., ASSIS, F.L., ALVES, P.A., FERNANDES, A.T.S., COTA,
M.M.G., FERREIRA, V.M., CAMPOS, R.K., MAZUR, C., LOBATO, Z.I.P., TRINDADE,
G.S., KROON, E.G. Nested-multiplex PCR detection for Orthopoxvirus and Parapoxvirus
directly from exanthematic clinical samples. Virology Journal 6: 140-155, 2009.
ARMSTRONG, C., LILLIE R.D. Experimental lymphocytic choriomeningitis of monkeys
and mice produced by a virus encountered instudies of the 1933 St. Louis encephalitis
epidemic. Public Health Reports 49: 1019-1027, 1934.
BARRY, M., RUSSI, M., ARMSTRONG, L., GELLER, D., TESH, R., DEMBRY, L.,
GONZALEZ, J.P., KHAN, A.S., PETERS, C.J. Brief report: treatment of a laboratory-
acquired Sabiá virus infection. The New England Journal of Medicine 333: 294-296, 1995.
BERGTHALER, A., GERBER, N.U., MERKLER, D., HORVATH, E., TORRE, J.C.,
PINSCHEWER, D.D. Envelope Exchange for the Generation of Live-Attenuated Arenavirus
Vaccines. PLoS Pathogens 2: e51, 2006.
BORDEN, E.C., NATHANSON, N. Tacaribe virus infection of the mouse: an
immunopathologic disease model. Laboratory Investigation 30: 465-473, 1974.
BRANCO, L.M., MATT, BOISEN, M.L., ANDERSEN, K.G., GROVE, J.N., MOSES, L.M.,
MUNCY, I.J., HENDERSON, L.A., SCHIEFFELLIN, J.S., ROBINSON, J.E., BANGURA,
J.J., GRANT, D.S., RAABE, V.N., FONNIE, M., SABETI, P.C., GARRY, R.F. Lassa
hemorrhagic fever in a late term pregnancy from northern sierra leone with a positive
maternal outcome: case report. Virology Journal 8: 404, 2011.
BRIGGILER, A. ENRIA, D., FEUILLADE, M. R., MAIZTEGUI, J. Contagio interhumano e
infección clínica con virus Junin en matrimonios residentes en el área endemica de fiebre
hemorrágica argentina. Medicina 47: 565, 1987.
BUCHMEIER, M. J. TORRE, J., PETERS, C.J. Arenaviridae: The viruses and their
replication. In: Fields Virology. Knipe, D.M. & Howley, P.M. 5.ed. Philadelphia, Lippincott
Williams & Wilkins, 2007. P. 1791–1827.
CASALS, J.; BUCKLEY, S.M.; CEDENO, R. Antigenic properties of arenaviruses. Bulletin
of the World Health Organization 72: 421-427, 1975.

59
CASSEB, R.C. Soroprevalência de anticorpos e padronização do teste de ELISA
sanduíche indireto para 19 tipos de arbovírus em herbivírus domésticos. Tese (Doutorado
em Biologia de Agente Infecciosos e Parasitários) – Belém, Universidade federal do Pará,
2010. 188p.
CLARKE, D. H., CASALS, J. Techinique for hemagglutination and hemagglutination
inhibition with arthropod-borne viruses. The American Journal of Tropical Medicine and
Hygiene 7: 561-573, 1958.
CHARREL, R.N., COUTARD, B., BARONTI, C., CANARD, B., NOUGAIREDE, A.,
FRANGEUL, A., MORIN, B., JAMAL, S., SCHMIDT, C.L., HILGENFELD, R., KLEMPA,
B., LAMBALLERIE, X. Arenaviruses and hantaviruses: From epidemiology and genomics to
antivirals. Antiviral Research 90: 102-114, 2011.
CHARREL, R.N., FELDMANN, H. FULHORST, C.F., KHELIFA, R., CHESSE, R.,
LAMBALLERIE, X. Phylogeny of New World arenaviruses based on the complete coding
sequences of the small genomic segment identified an evolutionary lineage produced by
intrasegmental recombination. Biochemical and Biophysical Research Communications
296: 1118- 1124, 2002.
CHARREL, R.N., LAMBALLEIRE, X. Arenavirus other than Lassa virus. Antiviral
Research 57: 89-100, 2003.
CHARREL, R.N., LAMBALLERIE, X. Zoonotic aspects of arenavirus infections.
Veterinary Microbiology 140: 213-220, 2010.
COIMBRA, T.L.M., NASSAR, E.S., BURATTINI, M.N., SOUZA, L.T.M., FERREIRA,
I.B., TRAVASSOS DA ROSA, A.P.A., VASCONCELOs, P.F.C., PINHEIRO, F.P., LEDUC,
J.W., RICO-HESSE, R., GONZALEZ, J., JAHRLING, P.B., TESH, R.B. New arenavirus
isolated in Brazil. Lancet 343: 391-392, 1994.
DEMBY, A.H., CHAMBERLAIN, J., BROWN, D.W., CLEGG, C.S. Early diagnosis of
Lassa fever by reverse transcription-PCR. Journal of Clinical Microbiology 32: 2898-
28903, 1994.
DOYLE, T.J., BRYAN, R.T., PETERS, C.J. Viral hemorrhagic fevers and hantavirus
infections in the Americas. Emerging Infectious Diseases 12: 95-110, 1998.
EMONET, S.F., TORRE, J.C., DOMINGO, E., SEVILLA, N. Arenavirus genetic diversity
and its biological implications. Infection, Genetics and Evolution 9: 417-429, 2009.
ENRIA, D.A., PINHEIRO, F. Rodent-borne emerging viral zoonosis. Hemorragic fevers and
hantavirus infections in South America. Infectious Disease Clinics of North America 14:
167-184, 2000.

60
ENRIA, D.A., MILLS, J.N., FLICK, R., BOWEN, M.D., BAUSCH, D., SHIEH, W.J.,
PETERS, C.J. Arenavirus Infections. In: Tropical Infectious Diseases: Principles,
Pathogens, and Practice, 2nd
ed. GUERRANT, R.L., WALKER, D.H., WELLER, P.F. New
York: Elsevier, 2005. P. 734-755.
FISHER- HOCK, S.P., TOMORI, O., NASIDI, A., PEREZ-ORONOZ, G.I., FEKILE, Y.,
HUTWAGNER, L., MCCORMICK, J.B. Review of cases of nosocomial Lassa Fever in
Nigeria: the high price of poor medical practice. BMJ 311: 857-859, 1995.
GEISBERT, T.W., JONES, S., FRITZ, E.A., SHURTLEFF, A.C., GEISBERT, J.B.,
LIEBSCHER, R., GROLLA, A., STROHER, U., FERNANDO, L., DADDARIO, K.M.,
GUTTIERI, M.C., MOTHE´ , B.R., LARSEN, T., HENSLEY, L.E., JAHRLING, P.B.,
FELDMANN, H. Development of a New Vaccine for the Prevention of Lassa Fever. PLoS
Medicine 2: e183, 2005.
GÓMEZ, R.M., GIUSTI, C.J., VALLDUVI, M.M.S., FRIK, J., FERRER, M.F.,
SCHATTNER, M. Junín vírus. A XXI century update. Microbes Infect 13: 303-311, 2011.
GRAJKOWSKA, L.T., PEDRAS-VASCONCELOS, J.A., SAUDER, C., VERTHELYI, D.,
PUIG, M. High-throughput real-time PCR for early detection and quantitation of arenavirus
Tacaribe. Journal of Virological Methods 159: 239-243, 2009.
GREENWAY, D.J., RUGIERO, H.S., PARODI, M., RIVERO, E., BARRERA, J.M.,
GARZON, F., BOXACA, M. METTLER, N., GUERRERO, L.B., NOTA, N. Epidemic
Hemorrhagic Fever in Argentina. Public Health Reports 74: 1011-1014, 1959.
GUNTHER, S., EMMERICH, P., LAUE, T., KUHLE, O., ASPER, M., JUNG, A.,
GREWING, T., MEULEN, J., SCHMITZ, H. Imported Lassa Fever in Germany: Molecular
Characterization of a New Lassa virus strain. Emerging Infectious Disease 5: 466-476,
2000.
HOWARD, C.R., LEWICKI, H., ALLISON, L., SALTER, M., BUCHMEIER, M.J.
Properties and Characterization of Monoclonal Antibories to Tacaribe Virus. The Journal of
General Virology 66: 1383-1395, 1985.
HUI, E.K. Reasons for the increase in emerging and re-emerging viral infectious diseases.
Microbes and Infection 8: 905-916, 2006.
International Committee on Taxonomy of Viruses (ICTV) - Virus Taxonomy: 2011
Release. Disponível em: <http://www.ictvonline.org/virusTaxonomy.asp?bhcp=1 >. Acesso
em: 21/11/2012.
IVANOV, A.P., BASHKIRTSEV, TKACHENKO, E.A. Enzyme-Linked Immunosorbent
Assay for Detection of Arenaviruses. Archives of Virology 67: 71-74, 1981
.

61
JAHRLING, P.B., HESSE, R.A., EDDY, G.A., JOHNSON, K.M., CALLIS, R.T.,
STEPHEN, E.L. Lassa virus infection of rhesus monkeys: pathogenesis and treatment with
ribavirin. Journal of Infectious Diseases 141: 580-509, 1980.
JOHNSON, K.M., WIEBENGA, N.H., MACKENZIE, R.B., KUNS, M.L., TAURASO,
N.M., SHELOKOV, A., WEBB, P.A., JUSTINES, G., BEYE, H.K. Virus Isolation from
Human Cases of Hemorrhagic Fever in Bolivia. Experimental Biology and Medicine 118:
113-118, 1965.
KILGORE, P.E., KSIAZEK, T.G., ROLLIN, P.E., MILLS, J.N., VILLAGRA, M.R.,
MONTENEGRO, M.J., COSTALES, M.A., PAREDES, L.C., PETERS, C.J. Treatment of
Bolivian hemorrhagic fever with intravenous ribavirin. Clinical Infectious Diseases 24:
718-722, 1997.
KILGORE, P.E., PETERS, J.C., MILLS, J.N., ROLLIN, P.E., ARMSTRONG, L., KHAN,
A.S., KSIAZEK, T.G. Prospects for the control of Bolivian hemorrhagic fever. Emerging
Infectious Diseases 1: 97-100, 1995.
KUNO, G.; GÒMEZ, I.; GUBLER, D.J. An ELISA procedure for the diagnosis of dengue
infections. Journal of Virologic Methods 33: 101-113, 1991.
LANCIOTTI, R.S., CALISHER, A. H., GUBLER, D.J., CHANG, G, VORNDAM, V. Rapid
Detection and Typing of Dengue Viruses from Clinical Samples by Using Reverse
Transcriptase- Polymerase Chain Reaction. Journal of Clinical Microbiology 30: 545-551,
1992.
LOZANO, M.E., ENRÍA, D., MAIZTEGUI, J.I., GRAU, O., ROMANOWISK, V. Rapid
Diagnosis of Argentine Hemorrhagic Fever by Reverse Transcriptase PCR-Based Assay.
Journal of Clinical Microbiology 33: 1327-1332, 1995.
MACHADO, A.M., FIGUEIREDO, G.G., CAMPOS, G.M., LOZANO, M.E., MACHADo,
A.R.S.R., FIGUEIREDO, L.T.M. Standardization of an ELISA test using a recombinant
nucleoprotein from the Junin virus as the antigen and serological screening for arenavirus
among the population of Nova Xavantina, State of Mato Grosso. Revista da Sociedade
Brasileira de MedicinaTropical 43: 229-233, 2010.
MARRS, J. S., KIRACOFE, J.B. Was the Huey Cocoliztli a Hemorrhagic fever? Medical
History 44: 341-362, 2000.
MATTAR, S, GUZMAN, C., ARRAZOLA, J., SOTO, E., BARRIOS, J., PINI, N., LEVIS,
S., SALAZAR-BRAVO, J., MILLS, J.N. Antibody to Arenavirus in rodents, Caribbean,
Colombia. Emerging Infectious Disease 17: 1315-1317, 2011.

62
MCCORMIK, J.B., WEBB,P.A., KREBS, J.W., JOHNSON, K.M., SMITH, E.S. A
prospective study of the epidemiology and ecology of Lassa fever. The Journal of Infectious
Diseases 155: 437-444, 1987.
MERCADO, R. Rodent control programmes in areas affected by Bolivian haemorrhagic
fever. Bulletin of the World Health Organization 52: 691-696, 1975.
MILAZZO, L.M., CAJIMAT, M.N.B., DUNO, G., DUNO, F., UTRERA, A., FULHORST,
C.F. Transmission of Guanarito and Pirital Viruses among Wild Rodents, Venezuela.
Emerging Infectious Diseases 17: 2209-2215, 2011.
MILLS, J.N., ELLIS, B.A., MCKEE, K.T.JR., KSIAZEK, T.G., ORO, J.B., MAIZTEGUI,
J.I., KSIAZEK, T.G., PETERS, C.J., CHILDS, J.E. Junin virus activity in rodents from
endemic and nonendemic loci in central Argentina. The American Society of Tropical
Medicine and Hygiene 44: 589-597, 1991.
MILLS, J.N., ELLIS, B.A., MCKEE, K.T.JR., CALDERON, G.E., MAIZTEGUI, J.I.,
NELSON, G.O., KSIAZEK, T.G., PETERS, C.J., CHILDS, J.E. A longitudinal study of Junin
virus activity in the rodent reservoir of Argentine hemorrhagic fever. The American Journal
of Tropical Medicine and Hygiene 47: 749-763, 1992.
MORALES, M.A., CALDERÓN, G.E., RIERA, L.M., AMBROSIO, A.M., ENRÍA, D.A.,
SABATTINI, M.S. Evaluation of an enzyme-linked immunosorbent assay for detection of
antibodies to Junin virus in rodents. Journal of Virological Methods 103: 57-66, 2002.
NUNES, M.R.T., NETO, J.P.N., CASSEB. S.M.M., NUNES, K.N.B., MARTINS, L.C.,
RODRIGUES, S.G., MATHEUS, S., DUSSART, P., CASSEB, L.M.N., VASCONCELOS,
P.F.C. Journal of Virological Methods 171: 13-20, 2011.
PARK, J.Y., PETERS, C.J., ROLLIN, P.E., KSIAZEC, T.G., GRAY, B., WAITES, K.B.,
STHEPENSEN, C.B. Development of a reverse transcription-polymerase chain reaction assay
for diagnosis of lymphocytic choriomeningitis virus infection and its use in a prospective
surveillance study. Journal of Medical Virology 51: 107-114, 1997.
PETERS, C.J. KUHENE, R.W., MERCADO, R.R., LE BOW, R.H., SPERTZEL, R.O.,
WEBB, P.A. Hemorrhagic fever in Cochabamba, Bolivia, 1971. American Journal of
Epidemiology 99: 425-433, 1974.
PETERS, C.J., WEBB, P.A., JOHNSON, K.M. Measuremet of antibodies to Machupo virus
by the indirect fluorescent technique. Proceedings of the Society for Experimental Biology
and Medicine 142: 526-531, 1973.
PINHEIRO, F.P., BENSABATH, G., ANDRADE, A.H.P., WOODALL, J.P. Isolamento no
Território Federal do Amapá de um vírus novo relacionado sorologicamente os agentes

63
responsáveis pelas febres hemorrágicas da Argentina e da Bolívia. Revista do Serviço
Especial de Saúde Pública 13: 87-94, 1967.
PINHEIRO, F.P., SHOPE, R.E., ANDRADE, A.H.P., BENSABATH,G., CACIOS, G.V.,
CASALS, J. Amaparí, a new vírus of the Tacaribe group from rodents and mites of Amapá
Territory, Brazil. Procedeedings of the Society for Experimental Biology and Medicine
122: 531-535, 1966.
PINHEIRO, F.P., TRAVASSOS DA ROSA, A.P.A., FREITAS, R.B., TRAVASSOS DA
ROSA, J.F.S., VASCONCELOS, P.F.C. Aspectos clínico-epidemiológicos. In: Instituto
Evandro Chagas; 50 anos de contribuição às ciências biológicas e à medicina tropical.
Belém, Fundação Serviços de Saúde Pública, 1986. P. 375-408.
PINHEIRO, F.P., WOODALL, J.P., TRAVASSOS DA ROSA, A.P.A., TRAVASSOS DA
ROSA, J.F. Studies on arenviruses in Brazil. Medicina (Buenos Aires) 37: 175-181, 1977.
PORTELA, A.L.S., PARPINELLI, N.L. Caracterização de uma cidade pequena inserida no
contexto do agronegócio mato-grossense. In: Encontro Nacional de Geografia Agrária., XIX,
2009, São Paulo, Anais: 2009, p. 1-18.
REED, L. J., MUENCH H. A simple method of estimating fifty percent endpoints. American
Journal of Hygiene 27: 493-497, 1938.
RODAS, J.D., SALVATO, M.S. Tales of mice and men: Natural history of arenaviruses.
Revista Colombiana de Ciencias Pecuarias 19: 382-399, 2006.
RODRIGUES, E.M.S., DAL-FABBRO, A.L., SALOMÃO, R., FERREIRA, I.B., ROCCO,
I.M., FONSECA, B.A.L. Epidemiologia da infecção pela dengue em Riberão Preto, SP,
Brasil. Revista de Saúde Pública 2: 160-165, 2002.
ROWE, W.P., MURPHY, F.A., BERGOLD G.H. Arenoviruses: proposed name for a newly
defined virus group. Journal of Virology 5: 651-652, 1970.
ROWE, W.P., PUGH, W.E., WEBB, P.A., PETERS, C.J. Serological relationship of the
Tacaribe complex of viruses to lymphocytic choriomeningitis virus. Journal of Virology 5:
289-292, 1970.
SABATTINI, M.S., RIOS, L.E.G., DIAZ, G., VEJA, R. Infecction Natural y Experimental de
Roedores com Virus Junin. Medicina 37: 149-161, 1977.
SALAS, R., PACHECO, M.E., RAMOS, B., TAIBO, M.E., JAIMES, E., Vasquez, C.,
QUERALES, J., de MAZIONE, N., GODOY, O., BETANCOURT, A., ARAOZ, F.,
BRUZUAL, R., GARCIA, J., TESH, R.B., RICO-HESSE, R., SHOPS, R.E. Venezuelan
Hemorrhagic Fever. Lancet 338: 1033-1036, 1991,

64
SANCHEZ, A., PYFAT, D.Y., KENYON, R.H., PETERS, C.J., MCCORMICK, J.B.,
KILEY, M.P. Junin Vírus Monoclonal Antibodies: characterization and cross-reactivity with
other arenaviruses. The Jounal of General Virology 70: 1125-1132, 1989.
SANGER, F., NICKLEN, S., COULSON, A.R. DNA sequencing with chain-terminating
inhibitors. Proceedings of the National Academy of Sciences 74: 5463-5467, 1977.
URATA, S., YASUDA, J. Molecular Mechanism of Arenavirus Assembly and Budding.
Viruses 4: 2049-2079, 2012.
VASCONCELOS, P.F.C., TRAVASSOS DA ROSA, J.F.S., GUERREIRO, S.C.,
DÉGALLIER, N., TRAVASSOS DA ROSA, E.S., TRAVASSOS DA ROSA, A.P.A.
Primeiro registro de epidemias causadas pelo vírus Oropouche nos estados do Maranhão e
Goiás, Brasil. Revista do Instituto de Medicina Tropical de São Paulo 31: 271-278, 1989.
VASCONCELOS, P.F.C., TRAVASSOS DA ROSA, A.P.A., RODRIGUES, S.G., TESII, R.,
TRAVASSOS DA ROSA, J.F.S., TRAVASSOS DA ROSA, E.S. Infecção humana adquirida
em laboratório causada pelo vírus SP H 114202 (Arenavirus: Família Arenaviridae): Aspectos
clínicos e laboratoriais. Revista do Instituto de Medicina Tropical de São Paulo 35: 521-
525, 1993.
WOLFF, H., LANGE, J.V.; WEBB, P.A. Interrelationships among arenaviruses measured by
indirect immunofluorescence. Intervirology 9: 344-350, 1978.
ZAPATA, J.C., SALVATO, M.S. Arenavirus variations due to the host-specific adaptation.
Viruses 5: 241-278, 2013.

65
ANEXO 1





























