
CENTRO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE
MESTRADO PROFISSIONAL EM EXERCÍCIO FÍSICO NA PROMOÇÃO DA SAÚDE
RAFAEL MENDES PEREIRA
INFLUÊNCIA DO EXERCÍCIO PRÉ-EXAUSTÃO SOBRE A HIPERTROFIA MUSCULAR DURANTE TREINAMENTO
RESISTIDO EM HOMENS JOVENS
Londrina 2014

ii
RAFAEL MENDES PEREIRA
INFLUÊNCIA DO EXERCÍCIO PRÉ-EXAUSTÃO SOBRE A HIPERTROFIA MUSCULAR DURANTE TREINAMENTO
RESISTIDO EM HOMENS JOVENS
Dissertação apresentada a Universidade Norte do Paraná – UNOPAR, Centro de Ciências Biológicas e da Saúde, como requisito para obtenção do título de Mestre no Programa de Mestrado Profissional em Exercício Físico na Promoção da Saúde.
Orientador: Prof. Dr. Andreo Fernando Aguiar
Londrina 2014

iii
RAFAEL MENDES PEREIRA
INFLUÊNCIA DO EXERCÍCIO PRÉ-EXAUSTÃO SOBRE A HIPERTROFIA MUSCULAR DURANTE TREINAMENTO
RESISTIDO EM HOMENS JOVENS
Dissertação apresentada a Universidade Norte do Paraná – UNOPAR, Centro de Ciências Biológicas e da Saúde, como requisito para obtenção do título de Mestre no Programa de Mestrado Profissional em Exercício Físico na Promoção da Saúde.
BANCA EXAMINADORA
____________________________________
Prof. Dr. Andreo Fernando Aguiar Orientador
____________________________________
Prof. Dr. Cosme Franklim Buzzachera Membro interno
____________________________________
Profa. Dra. Maeli Dal-Pai-Silva Membro externo
Londrina, ____ de ___________ de 2014.

iv
Dedicatória
“Dedico esse trabalho a minha amada esposa Marcele que
me apoiou em todos os momentos, nos difíceis foi minha
sustentação, em todos foi minha eterna parceira”. Obrigado
por todas as alegrias que você me trouxe, em especial pelo
Miguel. Obrigado por você acreditar que tudo isso era
possível antes mesmo que eu pudesse. Toda a dedicatória que
fizer é pouco perto do que você significa pra mim. Te amo
mais que o intervalo de -1 à +1 ( mais que ∞...)”

v
Agradecimentos
Apesar desse trabalho no contexto acadêmico possuir apenas um autor, é de suma
importância ressaltar que outras pessoas colaboraram para a concretização do mesmo,
seja de forma direta ou indireta. Todos os agradecimentos listados abaixo são especiais,
espero não ter me esquecido de ninguém.
Agradeço inicialmente a Deus, pois sem um alicerce na fé acredito ser impossível
seguir em frente. Agradeço em especial a Nossa Senhora Aparecida, a qual tenho imensa
fé e que me guia, me dá forças.
Agradeço ao meu orientador, Prof. Dr. Andreo Fernando Aguiar, pela confiança
depositada desde o início deste projeto, pela sua extrema dedicação e profissionalismo.
Agradeço pelos dois anos de orientação, anos estes muito valiosos e que contribuíram
imensamente para minha formação científico-pessoal. Agradeço também pela amizade
construída. Sou e serei sempre grato a você por essa conquista. Todo sucesso e felicidade
na sua vida.
Agradeço imensamente a minha esposa Marcele Tavares, pois sem a presença dela
com toda certeza nada disso seria possível. Agradeço pelas palavras de incentivo, de
carinho e por sempre acreditar, muitas vezes antes mesmo que eu pudesse acreditar, “Te
amo”.
Agradeço a meu filho Miguel Tavares Mendes Pereira, que mesmo sem saber me
deu muita força para seguir em frente e concretizar esse sonho.
Agradeço a minha mãe Maria Isabel Mendes Pereira, por todo amor e pela vida
dedicada a família, você é e sempre será um exemplo de pessoa para mim.
Agradeço a meu pai Luciano Ribeiro Pereira Filho, por todo orgulho que sentiu de
mim desde que entrei no mestrado. Agradeço pelo exemplo que você passou de força
interior, pois mesmo com o peso da “cruz que carrega” nesses últimos anos nunca
desanimou ou deixou de aproveitar os momentos bons da vida. Você pai é um grande
Homem, amo muito você e a mãe.
Agradeço as minhas irmãs Cristiane e Marcele por todo o carinho e amizade e que
mesmo de forma indireta sempre me apoiaram nessa caminhada.

vi
Agradeço a colega de Mestrado Vanda por toda a ajuda e contribuição nesse
trabalho e por todas as horas que estivemos juntos tentando tornar isso realidade.
Agradeço também aos colegas Jayne, Gisele, Cristiano, pela ajuda durante todo o
processo de coleta de dados.
Agradeço a todos os colegas do Mestrado em Exercício Físico na Promoção da
Saúde pelo apoio e pelas palavras de incentivo.
Agradeço a todos os professores do Mestrado em Exercício Físico na Promoção da
Saúde que participaram, positivamente, na minha formação acadêmico-científica.

vii
“A verdadeira viagem de descobrimento não
consiste em procurar novas paisagens, mas em
ter novos olhos”. (Marcel Proust)

viii
PEREIRA, Rafael Mendes. Influência do exercício pré-exaustão sobre a hipertrofia
muscular durante treinamento resistido em homens jovens. 2014. Dissertação
(Mestrado Profissional em Exercício Físico na Promoção da Saúde) – Centro de Ciências
Biológicas e da Saúde. Universidade Norte do Paraná, Londrina, 2014.
RESUMO
O objetivo do presente estudo foi examinar os efeitos de uma série adicional de exercício
exaustivo imediatamente antes do treinamento hipertrófico tradicional sobre a força
dinâmica máxima, área de secção transversal (AST) e resistência muscular do músculo
quadríceps em homens adultos jovens. Vinte e sete homens saudáveis (idade: 20.6 ± 1.9
anos) realizaram os testes de força dinâmica máxima [uma repetição máxima (1RM)],
resistência muscular localizada e exame de ressonância magnética [magnetic resonance
imaging (MRI)] em dois diferentes momentos: pré (pré-teste) e pós (pós-teste) e um
programa de treinamento resistido de alta intensidade com duração de 8 semanas. O
treinamento foi realizado em uma cadeira extensora bilateral. Após o pré-teste, os
participantes foram divididos em 3 grupos: controle (CO, N = 9), treinamento tradicional
(TR, N = 9) e treinamento pré-exaustão (PE, N = 9). Ambos os grupos TR e PE realizaram
o mesmo protocolo de treinamento (2 vezes na semana, 3 séries de 8-12 repetições à 75%
de 1RM) durante todo período experimental; a única diferença foi que o grupo PE realizou
uma série adicional de exercício exaustivo à 20% de 1RM até a fadiga voluntária, antes
(30seg) de cada sessão de treino. Após o período de treinamento, o grupo PE apresentou
maior aumento (P < 0,05) na força dinâmica máxima, AST e resistência muscular
comparado ao grupo TR. Além disso, nenhuma diferença significante (P > 0,05) foi
observada na massa corporal e consumo de macronutrientes entre os grupos após o período
experimental. Estes resultados sugerem que a inclusão de uma série adicional de exercício
exaustivo à 20% de 1RM, antes do treinamento resistido tradicional, pode ser uma
estratégia adequada para promover efeitos adicionais sobre a força dinâmica máxima,
hipertrofia e resistência muscular do músculo quadríceps em homens jovens.

ix
PEREIRA, Rafael Mendes. 2014. Effects of pre-exhaustion exercise on muscle
hypertrophy during resistance training in young men. Dissertação (Mestrado
Profissional em Exercício Físico na Promoção da Saúde) – Centro de Ciências Biológicas e
da Saúde. Universidade Norte do Paraná, Londrina, 2014.
ABSTRACT
The purpose of this study was to examine the effects of an additional set of exhaustive
exercise before high-intensity resistance training on maximal strength, cross-sectional area
(CSA), and endurance of the quadriceps muscle in young men. Twenty-seven healthy men
(age: 20.6 ± 1.9 years) performed maximal dynamic strength [one repetition maximum
(1RM)], muscular endurance, and magnetic resonance imaging (MRI) tests on 2 separate
occasions (before and after an 8-wk high-intensity resistance training program using a
bilateral knee extensor machine). After baseline testing, the subjects were divided into 3
groups: untrained control (CO, N = 9), traditional training (TR, N = 9), and prior
exhaustive training (PE, N = 9). Both the TR and PE groups trained using the same training
protocol (2 d∙wk-1
; 3 sets of 8-12 repetitions at 75% of 1RM) throughout the 8-wk
experimental period; the only difference was that the PE group performed an additional set
of exhaustive exercise at 20% of 1RM immediately before each training session. After 8
wk, the PE group experienced a greater (P < 0.05) increase in 1RM, CSA, and endurance
than the TR group. Additionally, no changes (P > 0.05) in body mass and daily dietary
intake were observed from pre- to post-test in either group. These results suggest that the
inclusion of a single set of exhaustive exercise at 20% of 1RM immediately before
traditional resistance training can be a suitable strategy for inducing additional beneficial
effects on quadriceps strength, hypertrophy, and endurance in young men.

x
SUMÁRIO
1. INTRODUÇÃO .............................................................................................................. 11
2. OBJETIVOS ................................................................................................................... 14
3. REVISÃO DA LITERATURA ..................................................................................... 15
3.1 Características gerais das fibras musculares .................................................................. 15
3.2 Recrutamento ordenado das fibras musculares .............................................................. 16
3.3 Recrutamento não ordenado das fibras musculares ....................................................... 17
3.4 Relação entre intensidade de esforço e recrutamento das fibras musculares ................. 18
4. METODOLOGIA .......................................................................................................... 21
4.1 Abordagem experimental ............................................................................................... 21
4.2 Sujeitos ........................................................................................................................... 22
4.3 Protocolo de treinamento resistido ................................................................................. 22
4.4 Recordatório alimentar ................................................................................................... 24
4.5 Protocolo de familiarização ............................................................................................ 25
4.6 Força dinâmica máxima (1RM) ..................................................................................... 25
4.7 Ressonância magnética .................................................................................................. 26
4.8 Resistência muscular e sinal eletromiográfico (EMG) .................................................. 27
4.9 Análise estatística ........................................................................................................... 28
5. REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................ 29
6. CAPÍTULO I .................................................................................................................. 36
7. CAPÍTULO II ................................................................................................................. 63

11
1. INTRODUÇÃO
A hipertrofia muscular é caracterizada pelo aumento do tamanho das
fibras/células musculares (Russell et al., 2000), devido ao aumento do número de
miofibrilas adicionadas em paralelo (Paul, 2002). Considerando que existe uma correlação
positiva entre área de secção transversal (AST) do músculo e força produzida, o aumento
da massa muscular é particularmente importante para atletas envolvidos em esportes de
força e potência (ex: atletas de futebol, jogadores de rugby e levantadores de peso), e
bodybuilding, na qual os competidores são avaliados pela quantidade e qualidade de seu
desenvolvimento muscular (Schoenfeld, 2010). O aumento da massa muscular é também
vital para a manutenção ou desenvolvimento da capacidade física e funcional em
indivíduos idosos e praticantes recreacionais de treinamento com pesos. Portanto,
maximizar a hipertrofia muscular tem significativas implicações práticas e clínicas para
diferentes populações associadas com esporte e saúde.
Os mecanismos associados com a hipertrofia envolvem uma complexa interação
de múltiplos fatores anabólicos, incluindo células satélites (CS), hormônios [ex:
testosterona, hormônio do crescimento (GH), e fator de crescimento insulínico (IGF-1)],
citocinas [ex: fator de crescimento de hepatócito (HGF), interleucina-5 (IL-5), interleucina-
6 (IL-6), e fator de crescimento de fibroblasto (FGF)], vias miogênicas intracelulares [ex:
fosfatidilinositol 3-quinase (PI3K), alvo da rapamicina em mamíferos (mTOR) e proteínas
quinases ativadas por mitógeno (MAPK)], e vias de sinalização dependentes de cálcio
(Ca2+
) (para revisão ver Schoenfeld, 2010). A ativação combinada dessas múltiplas vias
metabólicas resulta no aumento da relação entre síntese e degradação protéica,
promovendo maior acúmulo de proteínas miofibrilares (Glass, 2005; Schoenfeld, 2010) e
consequente hipertrofia muscular.
A tensão/sobrecarga mecânica é provavelmente o principal mediador das vias
anabólicas que governam a hipertrofia muscular induzida pelo treinamento resistido
(Vandenburgh, 1987; Goldspink, 2002; Hornberger et al., 2006). A tensão gerada sobre o
tecido muscular é produzida diretamente pela produção de força e alongamento, que são
considerados elementos fundamentais para o desenvolvimento da massa muscular
(Schoenfeld, 2010). A intensidade e duração da carga aplicada sobre os músculos resultam
em pequenas lesões/danos na fibra muscular - um processo natural frequentemente
chamado de ‘miotrauma adaptativo’ (MAD) (Smith, 1999). A fase inicial de reparo do
MAD é caracterizada pela ativação de uma modesta resposta inflamatória (ex: infiltrado de

12
macrófagos e linfócitos), que ativa o processo de crescimento muscular mediado pelas SCs
(Vierck et al., 2000). As SCs são normalmente quiescentes (Cornelison et al., 1997), mas
em resposta ao processo inflamatório induzido pelo MAD, estas células tornam-se ativas,
proliferam, diferenciam, e fundem-se às fibras musculares pré-existentes para: (1) reparar
as miofibrilas danificadas (Charge e Rudnick, 2004; Dhawan et al., 2005) e (2) “doar”
novos mionúcleos para aumentar a capacidade de síntese de proteínas contráteis (Moss et
al., 1970). Estes dois mecanismos celulares tem sido apontados como fatores primários no
processo de hipertrofia muscular.
Considerando que o MAD é essencial para promoção da hipertrofia muscular,
parece lógico pensar que o recrutamento máximo de todos os tipos de fibras (ex: fibras do
tipo I, IIa e IIx) durante as séries de treinamento resistido pode ser favorável para induzir
um maior número de MADs e, consequentemente, expandir a resposta hipertrófica. A
fundamentação teórica que explica a relação entre o ganho hipertrófico e recrutamento das
fibras musculares baseia-se no princípio da ‘falha muscular’. Falha muscular pode ser
definida como o momento na qual o músculo ou grupo muscular não consegue produzir a
força necessária para movimentar/levantar concentricamente uma determinada carga
(Schoenfeld, 2010). É especulado que a continuidade do exercício após falha muscular
pode ser favorável para promover maior recrutamento das unidades motoras (UMs)
(Willardson, 2007) e, consequentemente, maximizar a resposta hipertrófica muscular
(Rooney et al., 1994). Assim, várias técnicas especializadas de treinamento (ex: repetição
forçada, drop set, super série, e cargas negativas) tem sido utilizadas na tentativa de
maximizar o recrutamento de UMs durante as sessões de treinamento resistido (para
revisão ver Schoenfeld, 2011).
Globalmente, o objetivo dessas técnicas é gerar fadiga em um maior número
possível de UMs (ex: tipo I e II) durante as séries de exercícios e, assim, potencializar a
resposta hipertrófica. No entanto, essas técnicas “falham” em promover um recrutamento
preferencial (ou predominante) de um determinado tipo de fibra (ex: tipo I ou II) durante as
séries de exercícios. Prévios estudos têm demonstrado que o aumento global da força e
massa muscular induzido pelo treinamento resistido é resultado de uma maior resposta
hipertrófica das fibras do tipo II (26 a 57%) do que fibras do tipo I (12 a 15%) (Campos et
al., 2002; Kosek et al., 2006; Shepstone et al., 2005; Tesch, 1987). Portanto, a manipulação
do recrutamento das fibras durante as séries de exercício resistido (ex: estimular o
recrutamento das fibras do tipo II) é provavelmente benéfica para aumentar a força e
hipertrofia muscular durante um programa de treinamento resistido de alta intensidade.

13
Assim, a hipótese do presente estudo foi verificar se a fadiga/exaustão das fibras do tipo I
[induzidas por uma série adicional de exercício exaustivo à 20% de uma repetição máxima
(1RM)] antes do treinamento hipertrófico tradicional seria favorável para promover efeitos
adicionais sobre a força e hipertrofia muscular em homens adultos jovens.

14
2. OBJETIVOS
O objetivo deste estudo é apresentar um novo protocolo para maximizar o
recrutamento das fibras musculares (principalmente as fibras do tipo II) durante as séries
de um programa de treinamento hipertrófico. Especificamente, o objetivo deste estudo foi
verificar se a fadiga/exaustão das fibras do tipo I [induzidas por uma série adicional de
exercício exaustivo à 20% de uma repetição máxima (1RM)] antes do treinamento
hipertrófico tradicional seria favorável para promover efeitos adicionais sobre a força e
hipertrofia muscular em homens adultos jovens.
A partir dos dados teóricos e práticos obtidos neste estudo foi elaborado um guia
prático para treinamento hipertrófico em indivíduos adultos jovens. O objetivo primário
deste guia é apresentar um novo protocolo de treinamento resistido, baseado em evidências
científicas, para maximizar o aumento da força e massa muscular em homens adultos
jovens. Esse guia é destinado aos profissionais da área da saúde e esporte que atuam em
todas as áreas relacionadas com a prescrição de exercício físico.

15
3. REVISÃO DA LITERATURA
3.1 Características gerais das fibras musculares
O músculo estriado esquelético é um tecido complexo, versátil e heterogêneo,
constituído por diferentes tipos de fibras musculares. As características genéticas,
morfofisiológicas e bioquímicas distintas dos diferentes tipos de fibras conferem ao tecido
muscular uma ampla diversidade estrutural, metabólica e funcional (Schiaffino e Reggiani,
1994; Campos et al., 2002; Pette e Staron, 2000). Tradicionalmente, as fibras musculares
de mamíferos foram divididas em três discretas categorias, de acordo com o padrão de
reação histoquímica para metabolismo [atividade da adenosina trifosfatase miofibrilar
(mATPase)] e velocidade de contração: (1) fibras oxidativas de contração lenta - tipo I, (2)
fibras oxidativa-glicolíticas de contração rápida - tipo IIA e (3) fibras glicolíticas de
contração rápida - tipo IIB. No entanto, a utilização de métodos mais recentes, como a
imunohistoquímica utilizando anticorpos específicos para isoformas de cadeia pesada da
miosina [do inglês: miosin heavy chain (MHC)] e a análise eletroforética das isoformas de
MHC em fibras isoladas, tem contribuído substancialmente para o entendimento da
diversidade dos tipos de fibras musculares e sua natureza dinâmica.
Coletivamente, estas diversas abordagens metodológicas tem revelado a existência
de fibras musculares que contém uma única isoforma de MHC (nomeadas de fibras puras)
e fibras que expressam duas ou mais isoformas de MHC (nomeadas de fibras híbridas). Em
humanos, os seguintes tipos de fibras puras foram identificados: uma lenta do tipo I,
expressando MHCI, e duas rápidas do tipo II, denominadas tipo IIA e IIX (também
referida como IID), expressando MHCIIa e MHCIIx/IId, respectivamente (Ennion et al.,
1995; Smerdu et al., 1994; Schiaffino e Reggiani, 1994). Além das fibras puras, existem as
fibras “híbridas”, que co-expressam duas ou mais isoformas de miosina. Em humanos,
estas fibras foram classificadas em Tipo ІC (MHCІ>MHCІІa), ІІC (MHCІІa>MHCІ) e
ІІAX (MHCІІa>MHCІІx) (Staron et al., 2012). Vale ressaltar que as fibras musculares de
humanos não expressam a MHCІІb; a classificação prévia de fibras ІІB se equivale às
fibras do tipo ІІX ou ІІD (MHCІІx ou MHCІІd), de acordo com a similaridade da MHC
descrita em ratos (Smerdu et al., 1994). Coletivamente, os diferentes tipos de fibras
conferem ao músculo esquelético uma extensa capacidade para realizar uma variedade de
demandas funcionais (Campos et al., 2002). Enquanto as fibras do tipo I são
predominantemente recrutadas durante esforços prolongados de baixa intensidade, as fibras

16
do tipo IIA e IIX respondem preferencialmente as atividades breves de alta intensidade
(Sargeant, 1999).
3.2 Recrutamento ordenado das fibras musculares
O recrutamento seletivo dos diferentes tipos de fibras de acordo com a natureza
dinâmica da tarefa motora tem despertado interesse de pesquisadores em elucidar os
mecanismos neuromusculares que controlam o recrutamento ordenado das fibras
musculares em diversos padrões de atividades. A base teórica da atual compreensão dos
padrões de recrutamento das fibras musculares foi proposta por Henneman (1957). Em um
estudo com gatos descerebrados este autor observou um recrutamento ordenado dos
motoneurônios (do menor para o maior), de acordo com o aumento do padrão de
estimulação neural; a ordem inversa (do maior para o menor) também foi observada
durante desrecrutamento. Este autor e seus colaboradores encontraram resultados similares
em estudos envolvendo reflexo de estiramento em gatos descerebrados, de modo que
nomearam esta estratégia de recrutamento de "princípio do tamanho" (Henneman et al.,
1965a, b). A base teórica que explica o princípio do tamanho está fundamentada no
tamanho das unidades motoras (UMs) e no diâmetro do axônio dos motoneurônios. As
UMs menores (de menor diâmetro) são formadas por fibras de contração lenta (tipo I),
enquanto as UMs maiores são compostas por fibras de contração rápida (tipo IIA e IIX)
(McPhedran et al., 1965a, b). Além disso, as UMs pequenas apresentam um menor
diâmetro do axônio neural em relação às UMs maiores, resultando em uma menor
velocidade de condução do potencial de ação ao longo dos neurônios (Bawa et al., 1984).
O princípio do tamanho, por conseguinte, prediz que as UMs mais rápidas serão
recrutadas após as UMs mais lentas serem ativadas, e consequentemente, serão as
primeiras a serem desrecrutadas. Este padrão é facilitado pela relação entre os
motoneurônios α (responsável por conduzir a resposta ao músculo esquelético) e seus
respectivos neurônios aferentes Ia (fazem conexão com os motoneurônios α, estimulando-
os). O número de ligações (sinapses) entre os motoneurônios α e os neurônios Ia é
independente do tamanho celular do motoneurônio, o que significa que a densidade
sináptica por unidade de área varia inversamente com o tamanho do motoneurônio (Stein e
Bertoldi, 1981). Portanto, motoneurônios menores recebem maior quantidade de entradas
sinápticas, atingindo seu limiar de despolarização antes dos motoneurônios maiores.

17
Teoricamente, o recrutamento ordenado das UMs apresenta várias vantagens
funcionais, como: (1) o controle simplificado das atividades motoras pelo sistema nervoso
central (Henneman et al., 1974), (2) a preservação das UMs mais rápidas e mais fatigantes
(tipo IIA e IIX) durante tarefas motoras mais intensas e complexas (Henneman e Olson,
1965), e (3) o incremento gradual e suave da força, de modo proporcional ao recrutamento
das UMs para uma determinada atividade motora (Henneman e Olson, 1965; Zajac e
Faden, 1985). Desde que foi proposta pela primeira vez, uma grande quantidade de
evidências experimentais suportam a teoria do “princípio do tamanho” (Milner Brown et
al., 1973; Hoffer et al., 1987; Kier e Curtin, 2002; Hogrel, 2003). No entanto, um número
crescente de evidências indicam que o recrutamento ordenado de UMs pode nem sempre
ocorrer. Prévios estudos em humanos mostraram que os feedbacks auditivo e visual podem
alterar a ordem de recrutamento das UMs (Basmajian, 1963), assim como variações em
entradas proprioceptivas (Wagman et al., 1965) e estímulos cutâneos (Stephens et al.,
1978). Além disso, o rápido encurtamento do músculo extensor curto dos dedos (Grimby e
Hannerz, 1977) e breve alongamento do tríceps sural (Nardone et al., 1989) também
resultam em recrutamento preferencial de UMs mais rápidas.
3.3 Recrutamento não ordenado das fibras musculares
O recrutamento não ordenado das UMs também tem sido relatado a partir dos
resultados de estudos evolvendo a depleção de glicogênio durante atividade supramáxima
de ciclismo (Gollnick et al., 1974) e estimulação elétrica do músculo quadríceps de
humanos (Sinacore et al., 1990). Os resultados destes estudos demonstraram uma marcante
depleção de glicogênio nas fibras de contração rápida (tipo II) quando o músculo foi
submetido a uma intensidade supramáxima de exercício e elevada frequência de
estimulação elétrica, sugerindo um recrutamento preferencial das fibras do tipo II, de
acordo com o estímulo aplicado sobre o músculo esquelético.
Mais recentemente, tem sido sugerida a existência de grupos de UMs dentro de um
músculo individual, que podem ser ativados de forma não ordenada para realizar papéis
funcionais específicos (Loeb, 1985). Estes grupos de UMs foram denominados de "grupos
de tarefa", uma vez que são seletivamente recrutados para diferentes condições cinemáticas
dentro de uma tarefa motora, tal como um passo largo ou um movimento de agarrar (Loeb,
1985; Riek e Bawa 1992; Von Tscharner e Goepfert, 2006; Wakeling e Rozitis, 2004). É,
portanto, provável que outras estratégias de recrutamento, além daquelas previstas pelo

18
princípio do tamanho, também sejam utilizadas durante diferentes tarefas motoras. Em
alguns casos, é contraintuitivo para as UMs mais lentas serem recrutadas durante as tarefas
motoras, especificamente aquelas que necessitam produzir elevada força e rápida
velocidade de contração; neste panorama as UMs mais lentas poderiam contribuir muito
pouco para a rápida produção de força (Rome et al., 1988).
Em situações que requerem alto desenvolvimento de força (por ex: ciclistas em
esforços supramáximos), o recrutamento preferencial das UMs mais rápidas seria favorável
devido ao seu rápido tempo de ativação e relaxamento (Burke et al., 1973) e o seu
potencial para a produzir elevados índices de potência mecânica e máxima eficiência em
níveis mais elevados de força (He et al., 2000). Por conseguinte, existem fortes evidências
para sugerir que o recrutamento das UMs com base na demanda mecânica da tarefa motora
possa ocorrer em algumas situações (Lee et al., 2013). Se o recrutamento preferencial das
UMs mais rápidas proporciona uma vantagem mecânica e/ou energética, parece razoável
pensar que exista uma correlação positiva entre o recrutamento de UMs mais rápidas e
maior produção de força durante a contração muscular. Tal fato tem sido demonstrado em
seres humanos durante o ciclismo (Wakeling et al., 2006) e ratos correndo em uma esteira
(Hodson-Tole e Wakeling, 2008). Em ambos os estudos a relação entre taxa de produção
de força e recrutamento das UMs mais rápidas foi mais forte nos músculos com maior
população de fibras rápidas. Em adição, trabalhos anteriores mostraram que o recrutamento
ordenado das UMs é mais difícil de identificar em músculos que apresentam uma
população heterogênea dos tipos de fibras, particularmente quando numerosas fibras de
contração rápida estão presentes (Stephens e Stuart, 1975; Burke e Rymer, 1976; Fleshman
et al., 1981). Os resultados destes estudos sugerem que o padrão de recrutamento das UMs
baseado na demanda mecânica da tarefa motora pode ser mais predominante em
populações de UMs mais rápidas.
3.4 Relação entre intensidade de esforço e recrutamento das fibras musculares
O recrutamento progressivo das UMs durante a tarefa motora parece ser dependente
da intensidade do esforço realizado. Stock et al. (2012) demonstraram um elevado
recrutamento das UMs de alto limiar de estimulação no músculo vasto lateral (composto
predominantemente por fibras do tipo II) após teste isométrico de fadiga. Os autores
relataram que o aumento do recrutamento das UMs mais rápidas é necessário para
compensar o declínio da produção de força durante o teste de fadiga. Adicionalmente, De

19
Luca e Hostage (2010) observaram um recrutamento total das UMs do músculo vasto
lateral de indivíduos jovens, durante exercício isométrico de extensão de pernas em
intensidade correspondente a 95% da contração voluntária máxima (CVM). Neste estudo,
o recrutamento das UMs foi progressivo de acordo com o aumento dos níveis de força
muscular (20, 50, 80 e 100% da CVM), indicando um recrutamento progressivo das UMs
de acordo com o aumento gradual da intensidade de exercício. Este resultado é consistente
com prévios estudos que analisaram os efeitos de uma sessão de treinamento resistido
sobre a depleção de glicogênio do músculo esquelético (Robergs et al., 1991; Tesch et al.,
1998). Robergs et al. (1991) encontraram maior redução dos níveis de glicogênio no
músculo vasto lateral durante exercício de extensão de pernas à 70% de 1RM comparado à
30% de 1RM , resultando em uma diminuição de aprox. 30-40% no conteúdo total de
glicogênio muscular.
Similarmente, Tesch et al. (1998) demonstraram redução de 26% no conteúdo de
glicogênio do músculo vasto lateral de bodybuilders, após uma sessão de treinamento com
pesos (5 séries até a fadiga) envolvendo diferentes modelos de exercícios de extensão de
pernas. Interessantemente, a redução do conteúdo de glicogênio induzida pelo treinamento
resistido de alta intensidade parece ser mais evidente nas fibras de contração rápida do que
nas fibras lentas (Robergs et al., 1991; Tesch et al., 1998), sugerindo um recrutamento
preferencial das UMs rápidas durante tarefas motoras de alta intensidade. Considerando
que o recrutamento preferencial (ou predominante) de um determinado tipo de UM é
diretamente associado à intensidade (sobrecarga) da tarefa motora, parece razoável pensar
que a exaustão total ou parcial (depleção do sistema ATP-CP ou glicogênio) de um
determinado tipo de fibra (por ex: tipo I) poderia promover maior recrutamento de outro
tipo de fibra (por ex: tipo II). Por exemplo, a execução de um exercício prévio de exaustão
com sobrecarga correspondente a 20% de 1RM poderia promover a fadiga das fibras do
tipo I e, consequentemente, favorecer um maior recrutamento das fibras do tipo II durante
as séries de uma sessão de treinamento resistido para hipertrofia. Por conseguinte, o
recrutamento preferencial das fibras do tipo II poderia ser favorável para promover maior
ganho hipertrófico durante as sessões de treino.
Neste sentido, vários modelos empíricos de treinamento resistido (por ex: drop set e
pirâmide) tem sido utilizados na tentativa de maximizar o recrutamento das UMs durante
uma sessão de treino. Estes modelos utilizam diferentes intensidades (sobrecargas) durante
as séries de uma sessão de treino hipertrófico, na tentativa de recrutar o maior número
possível de fibras musculares, e assim, promover maior hipertrofia muscular. No entanto,

20
estes modelos “falham” em promover o recrutamento preferencial de um determinado tipo
de fibra durante as séries de exercício resistido. É provável que a manipulação do
recrutamento de fibras (por ex: estimular o maior recrutamento das fibras do tipo II) seja
benéfica para promover adaptações musculares específicas de acordo com os objetivos do
programa de treinamento.

21
4. METODOLOGIA
4.1 Abordagem experimental
Este estudo é caracterizado como prospectivo comparativo, com delineamento
experimental aleatório, cruzado com grupo controle. O objetivo do presente estudo foi
examinar os efeitos de uma série adicional de exercício exaustivo, imediatamente antes do
treinamento hipertrófico tradicional, sobre a força dinâmica máxima, área de secção
transversal (AST) e resistência muscular do músculo quadríceps em homens adultos
jovens. Para tanto, todos os participantes realizaram os testes de força dinâmica máxima
[uma repetição máxima (1RM)], resistência muscular localizada, e exame de ressonância
magnética [magnetic resonance imaging (MRI)] em dois diferentes momentos [pré (M1) e
pós (M2)] e um programa de treinamento resistido de alta intensidade com duração de 8
semanas], após 3 semanas de familiarização com os equipamentos e exercícios (Figura 1).
Os testes e treinamento foram realizados em uma cadeira extensora bilateral. Após os
testes iniciais (M1), os participantes foram equiparados em relação aos valores de 1RM e
divididos aleatoriamente em 3 grupos: controle (CO, N = 9), treinamento tradicional (TR,
N = 9) e treinamento pré-exaustão (PE, N = 9). Dois dias após o período experimental de 8
semanas, os sujeitos realizaram a mesma bateria de testes (M2), para examinar as possíveis
interações do grupo x tempo (Figura 1).
Figura 1. Desenho experimental

22
4.2 Sujeitos
Participaram deste estudo 27 indivíduos jovens saudáveis (18 – 25 anos) do gênero
masculino, estudantes do curso de graduação em educação física da Universidade Norte do
Paraná (UNOPAR), Londrina-PR. Os critérios para inclusão na amostra foram: (1) não ser
vegetariano, (2) não apresentar histórico de disfunções musculoesquelética, (3) não estar
participando de um programa de treinamento com pesos sistematizado a pelo menos 6
meses, (4) não fazer uso de medicamentos que potencializem ou bloqueiem a ação
muscular, (5) não ter utilizado suplementos ergogênicos e esteroides anabólicos a pelo
menos 6 meses, (6) apresentar a descrição detalhada do recordatório alimentar, e (7) ter
aprovação médica para prática de exercício físico. O recrutamento dos participantes foi
realizado por meio da ampla divulgação do projeto na Instituição (por exemplo: salas de
aula, panfletos e cartazes informativos). Os interessados foram selecionados mediante
entrevista pessoal e anamnese clínica durante uma reunião com os pesquisadores. Todos os
participantes foram previamente informados sobre os objetivos da pesquisa e
procedimentos realizados, e assinaram um termo de consentimento livre e esclarecido
aprovado pelo Conselho de Revisão da Universidade (protocolo n°.
14773113.5.0000.0108). Todos os procedimentos foram realizados de acordo com os
princípios da Declaração de Helsinque de 1964.
4.3 Protocolo de treinamento resistido
Os grupos TR e PE foram submetidos a um similar programa de treinamento
hipertrófico (3 séries de 8-12 repetições à 75% de 1RM, e intervalo de repouso de 1 minuto
entre as séries e exercícios) (American College of Sports Medicine, 2009), por um período
de 8 semanas (2 sessões/semana); a única diferença foi que o grupo PE realizou uma série
adicional de exercício exaustivo à 20% de 1RM, antes de cada sessão de treino (Figura 2).
O treinamento foi realizado em uma cadeira extensora bilateral (Bad Boy Gym, São Paulo,
Brasil), na posição sentada, com amplitude de movimento de 90 a 30° (0° = extensão total
de joelho) (Figura 3). Antes de cada sessão de treino os participantes realizaram um
aquecimento específico para o músculo quadríceps (1 série de 12 repetições, com carga
autosselecionada). O grupo CO permaneceu sem qualquer estímulo de treinamento durante
todo período experimental.
Para a execução da série adicional de exercício exaustivo, os sujeitos foram
instruídos a realizar o maior número possível de repetições à 20% de 1RM, em velocidade

23
moderada (concêntrica: 2s; excêntrica: 2s), até a “falha” voluntária. A série adicional de
exercício exaustivo foi aplicada com o objetivo de induzir a fadiga/exaustão das fibras do
tipo I (American College of Sports Medicine, 2009; Sale, 1987), antes do treinamento
hipertrófico tradicional. As cargas de treinamento foram ajustadas a cada 15 dias, mediante
a aplicação dos testes de 1RM. Ao final de cada sessão de treino, os participantes
realizaram um alongamento específico por um período de 5 minutos. As sessões foram
realizadas sempre no mesmo período do dia, entre 19 e 21 horas.
Figura 2. Sessão de treinamento.

24
Figura 3. Cadeira extensora bilateral
4.4 Recordatório Alimentar
Os participantes foram orientados por nutricionistas para preencher um recordatório
alimentar de três dias da semana (incluindo um dia de final de semana) antes e após as 8
semanas do programa de TR; porções caseiras padronizadas foram utilizadas para a
medição da quantidade de comida e bebida ingeridas. O consumo energético total, a
quantidade e as proporções de macronutrientes foram determinados por meio do programa
de avaliação nutricional (Avanutri, versão 3.1.4., Rio de Janeiro-RJ, Brasil). Os particantes
foram instruídos a manter seus hábitos alimentares durante todo o estudo. A ingestão de
água foi ad libitum.

25
4.5 Protocolo de Familizarização
Todos os participantes (incluindo os controles) foram submetidos a 3 semanas de
familiarização (3x/semana, em dias alternados) com os equipamentos e exercícios, a fim de
minimizar um potencial efeito de aprendizado e estabelecer a confiabilidade dos dados
(Gotshalk et al., 2002). As ssesões incluiram a prática do exercício de extensão de pernas
(3 séries de 8-12 repetições) e testes físicos (1RM e resistência muscular). Cada
participante foi continuamente monitorado e encorajado verbalmente pelos avaliadores
para realizar os testes na máxima intensidade possível. O coeficiente de corrrelação
intraclasse para cada teste foi ≥0.97, indicando a eliminação da curva de aprendizado
(Kraemer et al., 2006). Todas as sessões de familiarização e testes físicos foram realizados
no mesmo local, entre 19 e 21h.
4.6 Força dinâmica máxima (1RM)
Todos os participantes foram submetidos ao teste de 1RM nos momentos pré (M1)
e pós (M2) treinamento. A carga de 1RM foi determinada por meio de um protocolo
padrão, conforme proposto por Baechle e Earle (2008). Antes de iniciar cada teste, os
participantes realizaram um aquecimento muscular específico (1 série de 5-10 repetições)
com carga moderada (aproximadamente 50% da carga a ser utilizada na primeira tentativa
do teste de 1RM). Após 2 minutos de recuperação, os sujeitos foram orientados a realizar
até duas repetições máximas, com carga predeterminada pelos avaliadores. De acordo com
o número de repetições realizadas na primeira tentativa do teste, cargas adicionais foram
acrescentadas até que o participante executasse apenas uma repetição máxima (1RM),
atingindo toda amplitude do movimento. O número máximo de três tentativas foi
estabelecido para determinar a carga de 1RM em um dia de teste; o intervalo entre cada
tentativa foi de 3 a 5 minutos, a fim de garantir a completa recuperação metabólica do
músculo esquelético. Cada participante foi continuamente monitorado e encorajado
verbalmente pelos avaliadores durante todas as tentativas do teste. Além disso, a técnica de
execução do exercício foi padronizada para assegurar a qualidade dos dados coletados.

26
4.7 Ressonância magnética
A área de secção transversal (AST) do músculo quadríceps da perna dominante foi
mensurada nos momentos pré (M1) e pós (M2) treinamento, por meio do exame de
Ressonância Magnética (Signa Horizon LX-GE scanner, GE Medical System, Milwaulkee,
WI, USA) (Figura 4). O exame de ressonância magnética é considerado ‘padrão ouro’ para
mensuração da AST do músculo quadríceps (Mitsiopoulos et al., 1998), além de ser livre
de radiação. Durante o exame, os indivíduos foram posicionados no interior do campo
estacionário na posição supina, considerando a área medial da coxa (ponto médio entre o
trocânter maior e epicôndilo lateral do fêmur) como ponto de referência (Esmarck et al.,
2001). Para mensurar a AST do quadríceps, 6 imagens transaxiais foram obtidas do ponto
médio da ‘coxa’, com ângulo flip de 90°, de acordo com os seguintes parâmetros: tempo de
repetição = 600 ms e tempo de eco = 13 ms. A coleta dos dados foi obtida utilizando um
campo de visão = 18 mm, o que consiste em uma matriz de 384/224 pixels. A espessura do
corte foi de 60 mm e espaçamento foi de 10 mm. As imagens transaxiais foram transferidas
para um software de análise de imagens (Axiovision v3.0; Carl Zeiss, Jena, Alemanha), na
qual foi determinada as AST do m. quadríceps. A AST de cada porção (vasto medial, vasto
intermédio, vasto lateral e reto femoral) foi mensurada três vezes consecutivas e a média
aritmética das três medidas considerada como a AST do quadríceps (em cm2) (Esmarck et
al., 2001). As análises foram realizadas pelo mesmo avaliador, de modo randomizado,
duplo-cego, mediante a realização de sorteio. Análises anteriores revelaram um coeficiente
de correlação intraclasse (CCI) ≥0.96 para as medidas da AST do m. quadríceps.
Figura 4. Exame de ressonância magnética

27
4.8 Resistência muscular e sinal eletromiográfico (EMG)
O teste de resistência muscular localizada (RML) foi realizado nos momentos pré
(M1) e pós (M2) treinamento. Para tanto, os sujeitos foram orientados a realizar o maior
número possível de repetições à 60% de 1RM até a “falha” voluntária (Campos et al.,
2002). Durante o teste, sinais eletromiográficos de superfície foram registrados do músculo
vasto lateral (VL) a partir de um eletrodo ativo de superfície pré-amplificado (ganho:
1000) (modelo DE-2.3; Delsys, Inc., Wellesley, MA) a uma taxa de amostragem de 2000
Hz e um software de análise eletromiográfica (EMGworks 4.0.5, Delsys System, Boston,
MA) (Figura 5). Previamente à colocação do eletrodo, a pele foi tricotomizada e limpa com
álcool a 70%. O eletrodo foi posicionado perpendicularmente às fibras do músculo vasto
lateral, próximo ao centro do ventre muscular, de acordo com as recomendações da
SENIAM (do inglês: Surface EMG for Non-Invasive Assessment of Muscles ou
eletromiografia de superfície para avaliação não invasiva de músculos). O eletrodo de
referência foi fixado no processo estilóide direito (região do cotovelo). Os sinais EMG
foram filtrados com filtro digital de 20 a 500 Hz e tratados com rotinas do programa
MATLAB 11.0 (Mathworks®, South Natick, MA, USA). A partir dos sinais EMG
correspondente a segunda e penúltima contração no teste de RML, os dados de RMS (do
inglês: Root Mean Square) foram coletados em sucessivas janelas de tempo de 250 ms
(512 pontos) (50% de sobreposição) durante a execução dos movimentos de extensão e
flexão de pernas. Os valores de RMS foram normalizados pelo número máximo de
repetições (NmáxRep) executadas no teste de RML, utilizando a seguinte equação: %RMS
= [(RMS/NmáxRep) x 100%]. O valor de %RMS foi utilizado como parâmetro para
determinar o índice de trabalho muscular nos momentos pré e pós-treinamento.
Especificamente, um menor valor de %RMS durante o teste de RML indicaria maior
eficiência muscular (um menor recrutamento de fibras seria requerido para suportar a
demanda funcional durante o teste de RML).

28
Figura 5. Posicionamento do eletrodo (A) e captação do sinal eletromiográfico durante o teste de
RML (B).
4.9 Análise Estatística
Os dados foram expressos em média ± DP. Para garantir a confiabilidade dos
dados, os procedimentos estatísticos foram realizados após um estudo preliminar das
variáveis para determinar a normalidade e igualdade de variância entre os grupos. Testes
de ANOVA [grupo (3) x tempo (2)] para medidas repetidas foram utilizados para avaliar
os dados sobre tempo e entre os grupos. As diferenças específicas entre os grupos foram
detectadas pelo teste post-hoc de Bonferroni. O nível de significância α foi de 0.05. Todas
as análises foram realizadas através do software Statistica 7.0 (StatSoft, Tulsa, OK).

29
5. REFERÊNCIAS BIBLIOGRÁFICAS
1. American College of Sports Medicine position stand. Progression models in resistance
training for healthy adults. Med Sci Sports Exerc 2009; 41: 687-708
2. Baechle TR, Earle RW. Resistance training and spotting techniques. In: Earle R,
Baechle T (eds) Essentials of strength and conditioning: national strength and
conditioning association, 3rd edn. Human Kinetics, Champaign, 2008; 326–376
3. Basmajian JV. Control and training of individual motor units. Science 1963; 141:440–
441
4. Bawa P, Binder MD, Ruenzel P, Henneman E. Recruitment order of motoneurons in
stretch reXexes is highly correlated with their axonal conduction velocity. J
Neurophysiol 1984; 52:410–420
5. Burke RE, Levine DN, Tsairis P, Zajac FEIII. Physiological types and histochemical
proWles in motor units of the cat gastrocnemius. J Physiol 1973; 234:723–748
6. Burke RE, Rymer WZ. Relative strength of synaptic input from short-latency pathways
to motor units of deWned type in cat medial gastrocnemius. J Neurophysiol 1976;
39:447–458
7. Campos GE, Luecke TJ, Wendeln HK, Toma K, Hagerman FC, Murray TF, Ragg KE,
Ratamess NA, Kraemer WJ, Staron RS. Muscular adaptations in response to three
different resistance-training regimens: specificity of repetition maximum training
zones. Eur J Appl Physiol 2002; 88: 50–60
8. Charge´ SB, Rudnicki MA. Cellular and molecular regulation of muscle regeneration.
Physiol Rev 2004; 84: 209–238
9. Cornelison DD, Wold BJ. Single-cell analysis of regulatory gene expression in
quiescent and activated mouse skeletal muscle satellite cells. Dev Biol 1997; 191:
270–283
10. De Luca CJ, Hostage EC. Relationship between firing rate and recruitment threshold
of motoneurons in voluntary isometric contractions. J Neurophysiol 2010; 104(2):
1034-46

30
11. Dhawan J, Rando TA. Stem cells in postnatal myogenesis: molecular mechanisms of
satellite cell quiescence, activation and replenishment. Trends Cell Biol 2005; 15:
666–673.
12. Ennion S, Sant’ana Pereira J, Sargeant AJ, Young A, Goldspink G. Characterization
of human skeletal muscle fibres according to the myosin heavy chains they express. J
Muscle Res Cell Mot 1995; 16(1): 35-43
13. Esmarck B, Andersen JL, Olsen S, Richter EA, Mizuno M, Kjaer M. Timing of
postexercise protein intake is important for muscle hypertrophy
with resistance training in elderly humans. J Physiol 2001; 535: 301–311
14. Fleshman JW, Munson JB, Sypert GW, Friedman WA. Rheobase, input resistance, and
motor-unit type in medial gastrocnemius motoneurons in the cat. J Neurophysiol 1981;
46:1326–1338
15. Glass DJ. Skeletal muscle hypertrophy and atrophy signaling pathways. Int J Biochem
Cell Biol 2005; 37: 1974–1984
16. Goldspink G. Gene expression in skeletal muscle. Biochem Soc Trans 2002; 30: 285–
290
17. Gollnick PD, Piehl K, Saltin B. Selective glycogen depletion pattern in human muscle
Wbres after exercise of varying intensity and at varying pedalling rates. J Physiol
1974; 241:45–57
18. Gotshalk LA, Volek JS, Staron RS, Denegar CR, Hagerman FC, KraemerWJ. Creatine
supplementation improves muscular performance in older men. Med Sci Sports Exerc
2002; 34: 537–543
19. Grimby L, Hannerz J. Firing rate and recruitment order of toe extensor motor units in
diVerent modes of voluntary contraction. J Physiol 1977; 264:865–879
20. He Z-H, Bottinelli R, Pellegrino MA, Ferenczi MA, Reggiani C. ATP consumption and
eYciency of human single muscle fibers with different myosin isoform composition.
Biophys J 2000; 79:945– 961
21. Henneman E. Relation between size of neurons and their susceptibility to discharge.
Science 1957; 126:1345–1347

31
22. Henneman E, Clamann HP, Gillies JD, Skinner RD. Rank order of motoneurons
within a pool: law of combination. J Neurophysiol 1974; 37:1338–1349
23. Henneman E, Olson CB. Relations between structure and function in the design of
skeletal muscles. J Neurophysiol 1965; 28:581–598
24. Henneman E, Somjen G, Carpenter DO. Excitability and inhibitability of motoneurons
of diVerent sizes. J Neurophysiol 1965a; 28:599–620
25. Henneman E, Somjen G, Carpenter DO. Functional significance of cell size in spinal
motoneurons. J Neurophysiol 1965b; 28:560–580
26. Hodson-Tole EF, Wakeling JM. Motor unit recruitment patterns 2: the inXuence on
myoelectric intensity and muscle fascicle strain rate. J Exp Biol 2008; 211: 1893–1902
27. Hogrel JY. Use of surface EMG for studying motor unit recruitment during isometric
linear force ramp. J Electromyogr Kinesiol 2003; 13: 417–423
28. Hoffer JA, Loeb GE, Marks WB, O’Donovan MJ, Pratt CA, Sugano N. Cat hindlimb
motoneurons during locomotion. I. Destination, axonal conduction velocity, and
recruitment threshold. J Neurophysiol 1987; 57: 510–529
29. Hornberger TA, Chien S. Mechanical stimuli and nutrients regulate
rapamycinsensitive signaling through distinct mechanisms in skeletal muscle. J Cell
Biochem 2006; 97: 1207–1216
30. Kier WM, Curtin NA. Fast muscle in squid (Loligo pealei): contractile properties of a
specialized muscle fibre type. J Exp Biol 2002; 205: 1907–1916
31. Kosek DJ, Kim JS, Petrella JK, Cross JM, Bamman MM. Efficacy of 3 days/wk
resistance training on myofiber hypertrophy and myogenic mechanisms in young vs.
older adults. J Appl Physiol 2006; 101: 531–544
32. Kraemer WJ, Ratamess NA, Fry AC, French DN. Strength training: development and
evaluation of methodology. In: Physiological assessment of human fitness. Eds: Maud
P.J. and Foster, C.Champain, IL: Human Kinetics 2006
33. Lee SS, de Boef Miara M, Arnold AS, Biewener AA, Wakeling JM. Recruitment of
faster motor units is associated with greater rates of fascicle strain and rapid changes
in muscle force during locomotion. J Exp Biol 2013; 216(Pt 2): 198-207

32
34. Loeb GE. Motoneurone task groups: coping with kinematic heterogeneity. J Exp Biol
1985; 115: 137–146
35. McPhedran AM, Wuerker RB, Henneman E. Properties of motor units in a
heterogeneous pale muscle (M. Gastrocnemius) of the cat. J Neurophysiol 1965a; 28:
85–99
36. McPhedran AM, Wuerker RB, Henneman E. Properties of motor units in a
homogeneous red muscle (Soleus) of the cat. J Neurophysiol 1965b; 28: 71–84
37. Milner-Brown HS, Stein RB, Yemm R. The orderly recruitment of human motor units
during voluntary isometric contractions. J Physiol 1973; 230: 359–370
38. Mitsiopoulos N, Baumgartner RN, Heymsfield SB, Lyons W, Gallagher D, Ross R.
Cadaver validation of skeletal muscle measurement by magnetic resonance imaging
and computerized tomography. J Appl Physiol 1998; 85: 115–122
39. Moss FP, Leblond CP. Satellite cells are the source of nuclei in muscles of growing
rats. Anat Rec 1970; 170: 421–435
40. Nardone A, Romano C, Schieppati M. Selective recruitment of high-threshold human
motor units during voluntary isotonic lengthening of active muscles. J Physiol 1989;
409: 451–471
41. Paul AC, Rosenthal N. Different modes of hypertrophy in skeletal muscle fibers. J Cell
Biol 2002; 18: 156: 751–760
42. Pette D, Staron RS. Myosin isoforms, muscle fiber types, and transitions. Microsc Res
Tech 2000; 50(6): 500–509
43. Riek S, Bawa P. Recruitment of motor units in human forearm extensors. J
Neurophysiol 1992; 68: 100–108
44. Robergs RA, Pearson DR, Costill DL, Fink WJ, Pascoe DD, Benedict MA, Lambert
CP, Zachweija JJ. Muscle glycogenolysis during differing intensities of weight-
resistance exercise. J Appl Physiol 1991; 70(4):1700–1706
45. Rome LC, Funke RP, Alexander RM, Lutz GJ, Aldridge H, Scott F, Freadman M. Why
animals have different muscle fibre types. Nature 1988; 335:824–827
46. Rooney KJ, Herbert RD, Balnave RJF. Fatigue contributes to the strength training
stimulus. Med Sci Sport Exerc 1994; 26: 1160–1164

33
47. Russell BD, Motlagh D, Ashley WW. Form follows functions: how muscle shape is
regulated by work. Journal of Applied Physiology 2000; 88: 1127–1132
48. Sale DG. Influence of exercise and training on motor unit activation. Exerc Sport Sci
Rev 1987; 15: 95–151
49. Sargeant AJ. Neuromuscular determinants of human performance. In Physiological
Determinants of Human Exercise Tolerance (ed. B. J. Whipp and A. J. Sargeant), pp.
13–28. London: The Physiological Society/Portland Press 1999
50. Schiaffino S, Reggiani C. Myosin isoforms in mammalian skeletal muscle. J Appl
Physiol 1994; 77(2): 493–501
51. Schoenfeld BJ. The mechanisms of muscle hypertrophy and their application to
resistance training. J Strength Cond Res 2010; 24: 2857–2872
52. Schoenfeld BJ. The use of specialized training techniques to maximize muscle
hypertrophy. J Strength Cond Res 2011; 33: 60–65
53. Shepstone TN, Tang JE, Dallaire S, Schuenke MD, Staron RS, Phillips SM. Short-term
high- vs. low-velocity isokinetic lengthening training results in greater hypertrophy of
the elbow flexors in young men. J Appl Physiol 2005; 98: 1768–1776
54. Sinacore DR, Delitto A, King DS, Rose SJ. Type II fiber activation with electrical
stimulation: a preliminary report. Phys Ther 1990; 70(7):416–22
55. Smerdu V, Karsch-Mizrachi I, Campione M, Leinwand L, Schiaffino S. Type IIx
myosin heavy chain transcripts are expressed in type IIb fibers of human skeletal
muscle. Am J Physiol 1994; 267(6): 1723–1728
56. Smith LL. Cytokine hypothesis of overtraining: a physiological adaptation to excessive
stress? Med Sci Sports Exerc 1999; 32: 317– 333
57. Staron RS, Herman JR, Schuenke MD. Misclassification of hybrid fast fibers in
resistance-trained human skeletal muscle using histochemical and
immunohistochemical methods. J Strength Cond Res 2012; 26(10): 2616–22
58. Stein RB, Bertoldi R. Unique contributions of slow and fast extensor muscles to the
control of limb movements. In: Desmedt JE (ed) Motor unit types, recruitment and
plasticity in health and disease. Karger, Basel, 1981; pp 85–96

34
59. Stephens JA, Garnett R, Buller NP. Reversal of recruitment order of single motor units
produced by cutaneous stimulation during voluntary muscle contraction in man.
Nature 1978; 272:362–364
60. Stephens JA, Stuart DG. The motor units of cat medial gastrocnemius: speed–size
relations and their significance for the recruitment order of mortor units. Brain Res
1975; 91:177–195
61. Stock MS, Beck TW, Defreitas JM. Effects of fatigue on motor unit firing rate versus
recruitment threshold relationships. Muscle Nerve 2012; 45(1): 100–119
62. Tesch, P.A., LL. Ploutz-Snyder, L. Ystrom, M.J. Castro, and G.A. Dudley. Skeletal
muscle glycogen loss evoked by resistance exercise. J Strength and Cond Res 1998;
12(2):67–73
63. Tesch PA Acute and long-term metabolic changes consequent to heavy-resistance
exercise. Med Sci Sports Exerc 1987; 26: 67–89
64. Vandenburgh HH. Motion into mass: How does tension stimulate muscle growth?
Med Sci Sports Exerc 1987; 19: S142–S149
65. Vierck J, O’Reilly B, Hossner K, Antonio J, Byrne K, Bucci L, Dodson M. Satellite cell
regulation following myotrauma caused by resistance exercise. Cell Biol Int 2000; 24:
263–272
66. Von Tscharner V, Goepfert B. Estimation of the interplay between groups of fast and
slow muscle fibers of the tibialis anterior and gastrocnemius muscle while running. J
Electromyogr Kinesiol 2006; 16: 188–197
67. Wagman IH, Pierce DS, Burger RE. Proprioceptive influence, involitional control of
individual motor units. Nature 1965; 207: 957–958
68. Wakeling JM, Rozitis AI. Spectral properties of myoelectric signals from different
motor units in the leg extensor muscles. J Exp Biol 2004; 207: 2519–2528
69. Wakeling JM, Uehli K, Rozitis AI. Muscle fibre recruitment can respond to the
mechanics of the muscle contraction. J R Soc Interface 2006; 3: 533–544
70. Willardson JM. The application of training to failure in periodized multiple-set
resistance exercise programs. J Strength Cond Res 2007; 21: 628–631

35
71. Zajac FE, Faden JS. Relationship among recruitment order, axonal conduction
velocity and muscle-unit properties of type-identiWed motor units in cat plantaris
muscle. J Neurophysiol 1985; 53: 1303–1322

36
6. CAPÍTULO I
A single set of exhaustive exercise before high-intensity
resistance training increases quadriceps strength, hypertrophy
and endurance in young men
Este artigo foi submetido para publicação no International Journal of Sports Medicine
Abstract
The purpose of this study was to examine the effects of an additional set of exhaustive
exercise before high-intensity resistance training on maximal strength, cross-sectional area
(CSA), and endurance of the quadriceps muscle in young men. Twenty-seven healthy men
(age: 20.6 ± 1.9 years) performed maximal dynamic strength [one repetition maximum
(1RM)], muscular endurance, and magnetic resonance imaging (MRI) tests on 2 separate
occasions (before and after an 8-wk high-intensity resistance training program using a
bilateral knee extensor machine). After baseline testing, the subjects were divided into 3
groups: untrained control (CO, N = 9), traditional training (TR, N = 9), and prior
exhaustive training (PE, N = 9). Both the TR and PE groups trained using the same training
protocol (2 d∙wk-1
; 3 sets of 8-12 repetitions at 75% of 1RM) throughout the 8-wk
experimental period; the only difference was that the PE group performed an additional set
of exhaustive exercise at 20% of 1RM immediately before each training session. After 8
wk, the PE group experienced a greater (P < 0.05) increase in 1RM, CSA, and endurance
than the TR group. Additionally, no changes (P > 0.05) in body mass and daily dietary
intake were observed from pre- to post-test in either group. These results suggest that the
inclusion of a single set of exhaustive exercise at 20% of 1RM immediately before
traditional resistance training can be a suitable strategy for inducing additional beneficial
effects on quadriceps strength, hypertrophy, and endurance in young men.

37
Introduction
Skeletal muscle hypertrophy is characterized by increased muscle fiber/cell size [32] due to
an increase in the number of myofibrils, which are added in parallel [28]. Given that a
positive correlation exists between muscle cross-sectional area (CSA) and muscular
strength [24], hypertrophy is particularly important to athletes who are involved in strength
and power sports (e.g., football linemen, rugby players, and powerlifters) and
bodybuilding, where competitors are judged on both the quantity and quality of their
muscle development [35]. Increased muscle mass is also vital for maintaining or
developing physical fitness and functional capacity in recreational lifters and older adults.
Therefore, maximizing muscle hypertrophy has significant practical and clinical
implications for a variety of populations associated to sports and health.
The steps associated with hypertrophy involve a complex interaction of multiple
anabolic factors, including satellite cells (SCs), hormones [e.g., testosterone, growth
hormone (GH), and insulin-like growth factor (IGF-I)], cytokines [e.g., hepatocyte growth
factor (HGF), interleukin-5 (IL-5), interleukin-6 (IL-6), and fibroblast growth factor
(FGF)], intracellular myogenic pathways [e.g., phosphatidylinositol 3-kinase (PI3K)-Akt-
mammalian target of rapamycin (mTOR) and mitogen-activated protein kinase (MAPK)],
and calcium (Ca2+
)-dependent signaling pathways [for a review, see Schoenfeld (35)]. The
combined activation of these multiple anabolic pathways results in a higher protein
synthesis/degradation ratio, leading to a greater accumulation of myofibrillar proteins
[11,35] and, consequently, hypertrophy.
Mechanical tension/overload is perhaps the primary mediator of the anabolic
pathways that govern exercise-induced muscle hypertrophy [40,12,17]. Mechanical tension
is directly produced by force generation and stretch, which are considered key elements in
developing muscle hypertrophy [35]. Mechanical tension (e.g., the intensity and duration

38
of the load applied) results in a mild degree of damage/injury to muscle fibers, which is a
natural process often called ‘adaptive microtrauma’ (AMT) [38]. The initial phase of AMT
repair is characterized by the activation of a mild inflammatory response (e.g., infiltration
of macrophages and lymphocytes), which activates the SC-mediated muscle growth
process [41]. Adult SCs are normally quiescent [7], but in response to AMT-induced
inflammation, they become activated, proliferate, differentiate, and fuse to pre-existing
fibers to: (1) repair damaged myofibers [6,9] and (2) donate extra myonuclei to increase
the capacity of contractile protein synthesis [26]. These 2 cellular mechanisms have been
advocated as key factors in the process of generating muscle hypertrophy.
Because AMT is essential to SC-induced muscle hypertrophy, maximal recruitment
of all muscle fiber types (e.g., type I, IIa, and IIx fibers) during resistance training bouts
may generate a higher number of AMTs in a wide spectrum of muscles fibers and,
consequently, expand the hypertrophic response. The theoretical foundation that explains
the relationship between hypertrophic gain and fiber recruitment is based on the principle
of muscle failure. Muscular failure can be defined as the point during a set when muscles
can no longer produce the force necessary to concentrically lift a given load [35]. Training
to muscle failure is hypothesized to activate a larger number of motor units (MUs) with
continued exercise [42] and further stimulate hypertrophy [31]. In this context, several
specialized training techniques (e.g., forced repetitions, drop sets, supersets, and heavy
negatives) have been used in an attempt to optimize the recruitment of MUs during
resistance training bouts [for a review, see Schoenfeld (36)].
Globally, the aim of these techniques is to fatigue more MUs (e.g., type I and II)
during exercise sets and thus further stimulate hypertrophy. However, these techniques fail
to promote a preferential (or predominant) recruitment of a specific fiber type (e.g., type I
or type II) during the exercise sets. Previous studies have shown that the global increase in

39
muscle strength and mass in response to progressive resistance training is the result of a
greater hypertrophic effect on type II fibers (26 to 57%) than on type I fibers (12.5 to 15%)
[5,19,37,39]. Therefore, manipulating fiber recruitment during exercise sets (e.g.,
stimulating the recruitment of type II fibers) is likely beneficial for increasing strength and
hypertrophy during a high-intensity resistance training (RT) program. Hence, we
hypothesized that fatigue/exhaustion of type I fibers [induced by a single set of prior
exhaustive exercise at 20% of one repetition maximum (1RM)] before traditional RT
would promote a greater global recruitment of muscle fibers (principally, type II fibers)
and thus further stimulate muscle strength and hypertrophy in young men.

40
Methods
Experimental approach
A prospective, 3 group, matched, double-blind, randomized controlled study with repeated
measures was performed to examine the effects of an additional set of exhaustive exercise
(for more details see description in the resistance training protocol section) immediately
before high-intensity RT [1] on the maximal dynamic strength (1RM), CSA, and
endurance of the quadriceps muscle in young men (Fig. 1). For this purpose, all subjects
performed 1RM, muscular endurance, and magnetic resonance imaging (MRI) tests on 2
separate occasions [before (M1) and after (M2) an 8-wk resistance training program] after
a 3-wk familiarization period (3 non-consecutive d per wk, for a total of 9 sessions) with
the physical tests to minimize potential learning effects. The 1RM and muscle endurance
tests were performed using a bilateral knee extensor machine. After baseline testing (M1),
the subjects were matched according to 1RM values and then randomly assigned in a
double-blind fashion to 1 of 3 groups: untrained control (CO, N=9), traditional training
(TR, N=9), and prior exhaustive training (PE, N=9). Two days after the 8-wk experimental
period, the subjects completed the same battery of tests (M2) to examine possible group-
by-time interactions (Fig. 1).
Figure 1. Experimental design.

41
Subjects
Apparently healthy 18- to 25-yr-old men were invited to attend a meeting aimed at
explaining the purpose and details of the study protocol. To qualify as participants, the
subjects were required to 1) not be vegetarian, 2) have not ingested any ergogenic
supplement or anabolic steroids for the 6 months prior to the start of study, 3) have not
ingested any medication that could affect muscle growth or the ability to train intensely
during the study, 4) not be involved in the practice of systematized physical activity (i.e.,
2-3 d∙wk-1
) for the 6 months prior to the start of study, 5) have a detailed description of
their lifestyle and daily food intake, and 6) have medical approval for the practice of
physical exercise. Twenty-seven men [mean (SD): age 20.6 (1.9) yr, height 174.8 (6.0) cm,
and body mass 72.7 (9.7) kg] who met these criteria volunteered to participate in the study.
The physical characteristics of the CO, TR, and PE groups at baseline are presented in
Table 1. All subjects were informed of the procedures, risks, and benefits of the
investigation and signed an informed consent document approved by the Institutional
Review Board of the University (protocol no: 14773113.5.0000.0108). All procedures
were performed according to the principles outlined in the 1964 Declaration of Helsinki.

42
Table 1. Baseline characteristics
CO (N = 9) TR (N = 9) PE (N = 9)
Age (yr) 20.0 ± 1.8 20.9 ± 2.0 21.0 ± 1.9
Body mass (kg) 75.0 ± 8.8 73.7 ± 9.4 69.8 ± 11.2
Height (cm) 176.4 ± 8.1 173.8 ± 6.9 174.4 ± 2.2
BMI (kg∙m2) 24.1 ± 2.0 24.4 ± 2.5 22.9 ± 3.6
1RM (kg) 106.4 ± 2.6 107.4 ± 3.9 106.6 ± 4.5
Values are mean ± SD.
There were no differences between the groups
BMI: Body mass index
1RM: one repetition maximum
Resistance training protocol
Both the TR and PE groups trained under the same training regime (2 d∙wk-1
; 3 sets of 8-12
repetitions at 75% of 1RM, with 1 min rest between sets) during an 8-wk RT program
designed to promote muscle hypertrophy [1]. The only difference in the training protocol
was that the PE group performed an additional set of exhaustive exercise immediately
before each training session (Fig. 2). The exercise used for training was a bilateral knee
extension in a seated position using a commercial knee extensor machine (Bad Boy Gym
equipment, São Paulo, Brazil), with a range of motion of 90 to 30° (0° = full knee
extension). Each training session began with a specific warm-up exercise (1 set of 12
repetitions with a self-selected load) for the quadriceps muscle. For the additional set of
exhaustive exercise, the subjects were instructed to perform as many repetitions as possible
at 20% of 1RM until failure. We used a light load of 20% of 1RM during the prior
exhaustive exercise because previous studies reported that training to failure at a relatively
low intensity (up to 30% of 1RM) primarily recruits slow-twitch motor units [1,33]. The
prior exhaustive exercise was designed to induce the fatigue/exhaustion of the type I fibers

43
before the high-intensity RT training. Qualified personnel supervised each participant
individually during every workout. Each subject received a training logbook, in which the
researchers recorded the weekly training load (weight). The training load was adjusted
every 15 d according to a 1RM test. At the end of each session, the muscles exercised were
stretched for approximately 5 min. The sessions were performed between 7 and 9 pm.
Figure 2. Training session design. The numbers in the boxes indicate the exercise intensity (% of
1RM)
Nutrient intake
Under the supervision of nutritionists, the subjects completed 3-day dietary intake records
(including 1 weekend day) before and after the 8-wk RT program; standard portions were
used to assess the amount of food and drink consumed. The total energy intake and
macronutrient amounts were calculated using software for nutritional assessment
(Avanutri, version 3.1.4, Rio de Janeiro-RJ, Brazil). Participants were instructed to
maintain their habitual daily diet Water intake was ad libitum.

44
Familiarization protocol
All subjects (including the controls) completed a 3-wk orientation program (3 non-
consecutive d per wk, for a total of 9 sessions) before M1 for familiarization with the
equipment and exercise to minimize any potential learning effects and establish the
reliability of the testing protocols [15]. The sessions consisted of repeated performance of
knee extension exercise (3 sets of 8-12 repetitions) and physical tests (1RM and muscular
endurance). Maximal effort in each test was requested during the last 5 sessions to reduce
any learning effects and to make the data consistent. The intraclass correlation coefficients
were ≥0.97 for each test, indicating the elimination of the learning curve for the subjects
[22]. All familiarization sessions and physical tests were performed at the same location,
between 7 and 9 pm.
Maximal dynamic strength
Maximal dynamic strength was assessed using a 1RM standard testing protocol as
previously documented by Baechle and Earle [3]. The 1RM test was preceded by a set of
warm-up exercise (5-10 repetitions) at approximately 50% of the load to be used in the
first attempt of the 1RM test. After 2 min of rest, the 1RM attempts were performed with a
progressively increasing load for each attempt and were separated by 3- to 5-min rest
intervals. The subjects were permitted 2 to 3 attempts to determine the 1RM value. 1RM
was defined as the greatest load lifted through a full range of motion before 2 failed
attempts at a given load. Verbal encouragement was provided during all 1RM attempts.
The exercise execution technique was standardized and continuously monitored in an
attempt to assure the quality of the data.

45
MRI
The CSA of the quadriceps muscle of the dominant side was measured before (M1) and
after (M2) the 8-wk training period via MRI using a 1.5 T Signa Horizon LX-GE scanner
(GE Medical System, Milwaukee, WI, USA). MRI was used because it provides the most
accurate in vivo measurement of a muscle’s cross section [25] and is radiation free. The
subjects had not been engaged in any testing or training 2 d prior to the MRI scanning. The
subjects were placed within the stationary, external field in the supine position with the
mid-point of the thigh (half-way between trochanter major and epicondylus lateralis of the
femur) as the point of reference [10]. To determine the CSA, 6 transaxial images were
obtained at midthigh with a flip angle of 90° with the following parameters: repetition time
= 600 ms and echo time = 13 ms. Each data set was obtained with a field of view = 18 mm
consisting of a 384/224 pixel matrix. The slice thickness was 60 mm, and the interslice gap
was 10 mm. The transaxial images were analyzed using the software Axiovision v3.0
software (Carl Zeiss, Jena, Germany). All 4 heads of the quadriceps femoris muscle were
outlined manually 3 times for analysis (Fig. 4), and the CSA was determined as the
average of the 3 analyses [10]. Previous analysis revealed an intra-rater reliability level
greater than 0.96.
Muscular endurance tests and electromyographic (EMG) signal recordings
A local muscular endurance test was performed before (M1) and after (M2) the 8-wk
experimental period. Briefly, the subjects were instructed to perform as many repetitions as
possible at 60% of 1RM until failure [5]. During the test, surface EMG signals were
recorded from the vastus lateralis (VL) muscle using a preamplified (gain: 1000) active
surface electrode (Model DE-2.3; Delsys, Inc., Wellesley, MA) at a sampling rate of 2000
Hz and EMGworks 4.0.5 analyses software (Delsys System, Boston, MA). The subject’s
skin was prepared by removing the superficial dead skin and was sterilized with an alcohol

46
swab. The electrode was placed on a location near the center of the belly of the muscle
according to the recommendations of SENIAM (Surface EMG for Non-Invasive
Assessment of Muscles), and the reference electrode was fixed at the right styloid process.
The EMG signals were filtered with a band-pass digital filter between 20 and 500 Hz using
Matlab 11.0 routines (Mathworks®, South Natick, MA, USA). From the EMG signals
corresponding to the second- and before last-contractions of the endurance test, a moving
RMS method was executed on successive 250-ms (512 points) time windows (50%
overlapped) to obtain the RMS average values during the leg flexion-extension exercise.
The RMS values were normalized by the maximal number of repetitions (NmáxRep)
performed during the endurance test, according to the following equation: %RMS=
[(RMS/NmáxRep) × 100%]. The %RMS values were used as the pre- and post-training
muscular work index. Specifically, a lower %RMS value during the endurance test at 60%
of 1RM indicated greater muscle efficiency (lower fiber recruitment was required to
support the functional demand of the endurance exercise until failure).

47
Statistical analysis
Data are expressed as the mean ± SD. To ensure data reliability, the statistical procedures
were performed after a preliminary study of the variables to determine the normality and
equality of the variance between groups; the statistical power was 80% for the assessed
comparisons. On the basis of a statistical power of 0.80, a moderately large effect size
(0.35), and an overall level of significance of 0.05, 9 subjects were required in each group.
A one-way ANOVA was used to determine differences in the baseline measurements
between the groups for the body mass, age, height, BMI, and 1RM variables. A 3 (group:
CO vs. TR vs. PE) x 2 (time: pre- and post-test) ANOVA with repeated measures was used
to evaluate the data across time and between groups for the 1RM, CSA, muscle endurance,
and nutrient intake (dependent variables). When significant differences were confirmed
with ANOVA, multiple comparisons testing were performed using Bonferroni post-hoc
analysis to identify these differences. The level of significance was set at P ≤ 0.05.
Statistical analyses were performed using Statistica 7.0 software (StatSoft, Tulsa, OK).

48
RESULTS
Participant characteristics. All participants (CO, N = 9; TR, N = 9; PE, N = 9) who began
the 8-wk RT program completed the study. The baseline characteristics of the subjects are
presented in Table 1. All groups had similar (P > 0.05) baseline physical characteristics. In
addition, no significant (P > 0.05) differences in the daily dietary intakes were observed
among the groups before and after the 8-wk intervention period (Table 2).
Table 2. Dietary analyses
CO (N = 9) TR (N = 9) PE (N = 9)
Carbohydrate (%)
Pre 65.5 ± 2.2 64.8 ± 4.6 64.6 ± 3.1
Post 65.9 ± 0.8 64.2 ± 5.3 63.0 ± 5.3
Protein (%)
Pre 17.4 ± 1.4 17.9 ± 1.8 19.1 ± 2.3
Post 17.6 ± 1.8 19.8 ± 5.6 21.5 ± 5.3
Fat (%)
Pre 17.1 ± 2.7 16.6 ± 3.8 16.3 ± 2.1
Post 16.5 ± 2.1 16.0 ± 2.3 15.5 ± 1.9
There were no differences between the groups
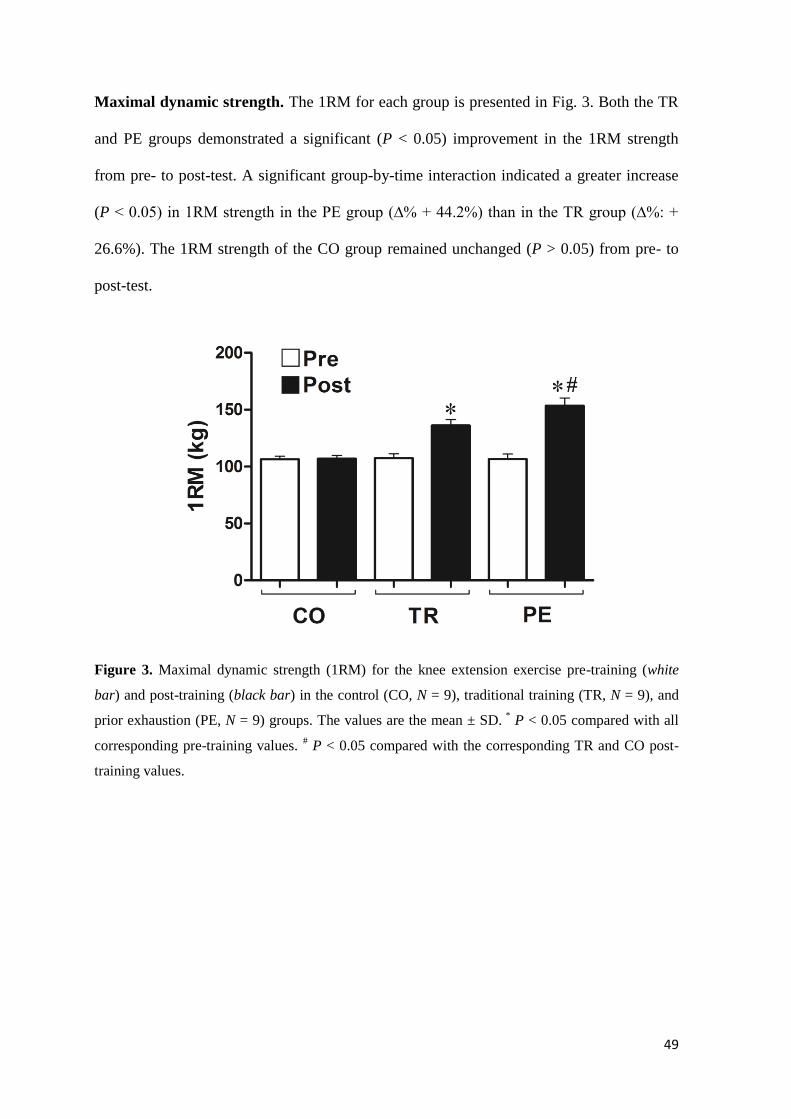
49
Maximal dynamic strength. The 1RM for each group is presented in Fig. 3. Both the TR
and PE groups demonstrated a significant (P < 0.05) improvement in the 1RM strength
from pre- to post-test. A significant group-by-time interaction indicated a greater increase
(P < 0.05) in 1RM strength in the PE group (∆% + 44.2%) than in the TR group (∆%: +
26.6%). The 1RM strength of the CO group remained unchanged (P > 0.05) from pre- to
post-test.
Figure 3. Maximal dynamic strength (1RM) for the knee extension exercise pre-training (white
bar) and post-training (black bar) in the control (CO, N = 9), traditional training (TR, N = 9), and
prior exhaustion (PE, N = 9) groups. The values are the mean ± SD. *
P < 0.05 compared with all
corresponding pre-training values. # P < 0.05 compared with the corresponding TR and CO post-
training values.

50
Quadriceps CSA. A representative MRI axial image from each group is shown in Fig. 4,
and the corresponding data are presented in Fig. 5A and B. Both the TR and PE groups
demonstrated a significant (P < 0.05) increase in the total quadriceps CSA from pre- to
post-test. A significant group-by-time interaction indicated a greater increase (P < 0.05) in
total CSA (4 heads of the quadriceps) in the PE group (∆%, VL: +16.7%; VM: +16.9%;
VI: +13.4%; RF: +28.4%) than in the TR group (∆%, VL: +10.2%; VM: +9.3%; VI:
+8.0%; RF; +18.4%). The total quadriceps CSA of the CO group remained unchanged (P >
0.05) from pre- to post-test.
Figure 4. MRI axial images of the right thigh from a subject from the control (CO, N = 9),
traditional training (TR, N = 9), and prior exhaustion (PE, N = 9) groups pre-training (A, C, and E)
and post-training (B, D, and F). RF, rectus femoris; VM, vastus medialis; VL, vastus lateralis; VI,
vastus intermedius. Note the greater increase in the quadriceps cross section (as indicated by the
VL thickness - dashed line) from pre- to post-test in the PE group compared with the TR group.
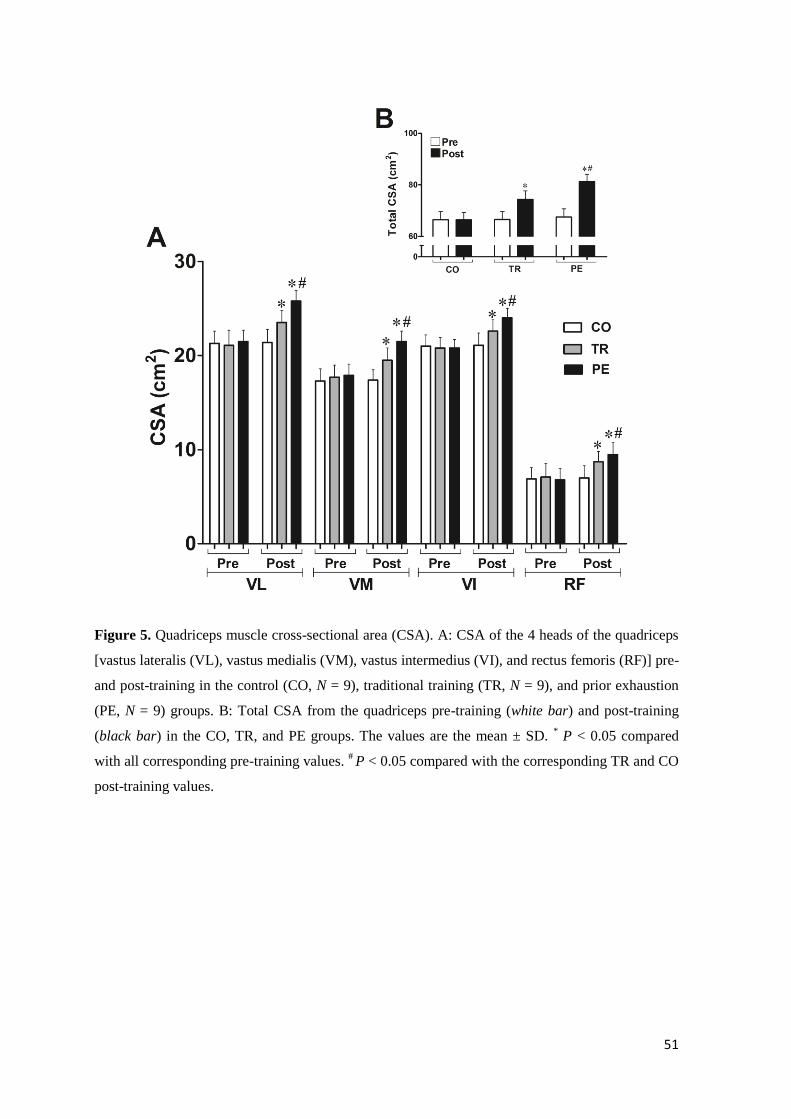
51
Figure 5. Quadriceps muscle cross-sectional area (CSA). A: CSA of the 4 heads of the quadriceps
[vastus lateralis (VL), vastus medialis (VM), vastus intermedius (VI), and rectus femoris (RF)] pre-
and post-training in the control (CO, N = 9), traditional training (TR, N = 9), and prior exhaustion
(PE, N = 9) groups. B: Total CSA from the quadriceps pre-training (white bar) and post-training
(black bar) in the CO, TR, and PE groups. The values are the mean ± SD. * P < 0.05 compared
with all corresponding pre-training values. # P < 0.05 compared with the corresponding TR and CO
post-training values.

52
Muscle endurance and efficiency. A significant group-by-time interaction (P < 0.05) was
observed for muscle endurance (number of total repetitions) and muscle work index
(%RMS) (Fig. 6A and B). The PE group had a significantly greater number of total
repetitions (∆%, TR: +20.2% vs. PE: +36.9%, P < 0.05) and lower %RMS values (∆%,
TR: –30.8% vs. PE: –50.5%, P < 0.05) at post-test than the TR group. The number of
repetitions and %RMS in the CO group remained unchanged (P > 0.05) from pre- to post-
test.
Figure 6. Maximal number of repetitions (A) and muscular work index (%RMS) (B) for the knee
extension exercise at 60% of 1RM pre-training (white bar) and post-training (black bar) in the
control (CO, N = 9), traditional training (TR, N = 9), and prior exhaustion (PE, N = 9) groups.
%RMS= [(RMS/NmáxRep) × 100%]. The values are the mean ± SD. *
P < 0.05 compared with all
corresponding pre-training values. # P < 0.05 compared with the corresponding TR and CO post-
training values.

53
DISCUSSION
This study was mainly designed to determine whether the inclusion of a single set of
exhaustive exercise at 20% of 1RM immediately before traditional resistance training
would provide additional beneficial effects on the maximal dynamic strength (1RM knee
extension), CSA, endurance, and muscle efficiency of the quadriceps muscle in young
men. We hypothesized that the fatigue/exhaustion of type I fibers induced by prior
exhaustive exercise would promote a greater global recruitment of muscle fibers
(principally type II fibers) during traditional training sets and, thus, further stimulate
muscle performance and adaptations. Our results confirm this hypothesis by showing for
the first time that 1RM knee extension, CSA, endurance, and muscle efficiency were
significantly enhanced by the inclusion of a single set of exhaustive exercise immediately
before traditional resistance training.
The traditional hypertrophic training protocol used in this investigation was
originally designed in accordance with the ACSM Position Stand guidelines on
progression models in resistance training for healthy adults [1]. The design seems to be
incompatible with including a set of exhaustive exercise at 20% of 1RM before traditional
resistance training because the muscle adaptations are more likely to be attributed to the
greater training volume than to the prior exhaustive exercise. However, the greater training
volume in the PE group compared with that in the TR group was intentional because we
are particularly interested in exhausting type I fibers, regardless of the training volume.
Our study is limited because we did not perform muscle biopsies to confirm the
fatigue/exhaustion of type I fibers (e.g., depletion of the ATP-PC system); however,
evidence from previous studies confirms that training to failure at a relatively low intensity
(up to 30% of 1RM) primarily recruits slow-twitch motor units [1,33]. Therefore, despite a
higher training volume in the PE group, the additional set of exhaustive exercise indeed

54
induced fatigue in the type I fibers before the traditional training. To our knowledge, this
study is the first to use a training strategy to manipulate the recruitment of a specific fiber
type (e.g., type IIa) during sets of traditional resistance training.
The greater increase in 1RM and hypertrophy observed in the PE group compared
with that in the TR group is similar to previous findings from studies involving different
specialized training techniques (e.g., drop set and forced repetitions), which necessarily
include additional sets during the training routines [for a review, see Schoenfeld (36)].
Although these techniques are often based on empirical evidence, the techniques and
results are widely used by coaches, athletes, and sports scientists in an attempt to maximize
muscle hypertrophy. The drop sets technique (also known as descending sets), for
example, involves performing an exercise set to muscular failure with a given load and
then immediately reducing the load to perform another set until subsequent failure. This
technique is thought to stimulate greater muscular growth by inducing greater MU fatigue
[42] and GH secretion [13]. Goto et al. [13] analyzed the effects of including a low-
intensity set (50% of 1RM) immediately after the performance of a high-intensity set on
GH levels. The authors found a significant increase in the GH levels that was associated
with the additional low-intensity drop set In a later study [14], the same authors also
showed that the addition of a drop set to a standard strength training protocol resulted in a
greater increase in the 1RM leg press and muscle CSA compared with a strength training
protocol alone. The results of our study together with those from Goto et al. [13,14]
indicate that the greater work volume from an additional set can be beneficial to promote
additional effects on muscle strength and hypertrophy. In fact, Goto et al. [13] found a
significant positive correlation between GH secretion and the work volume (r = 0.80; P <
0.05) of the additional set, suggesting that work volume is an important factor in enhancing
GH secretion and consequently increasing muscle strength and hypertrophy. This effect

55
appears to be consistent with the classical principle of resistance training programs for
strength and hypertrophy, in which higher-volume, multiple-set protocols result in greater
responses for GH [8,27], testosterone [20,21], hypertrophy [23,43], and strength [29,30,43]
compared with the responses to single set protocols.
Although there is no question that the greater work volume from an additional set
can induce beneficial effects on muscle strength and hypertrophy, it is not clear why the
additional set of exhaustive exercise prior to traditional training elicited such benefits. A
possible explanation for this result is that the prior fatigue of the type I fiber induced by the
prior exhaustive exercise at 20% of 1RM could maximize the recruitment of type II fibers
during the subsequent training sets, resulting in a greater total hypertrophic response. This
hypothesis is supported by the principle of muscle fiber fatigue; when a specific group of
MUs becomes fatigued, a progressively greater number of MUs are recruited (principally
the highest threshold MUs - type II) to continue activity [42], further stimulating strength
and hypertrophy [31]. Previous studies have shown that the global increase in muscle
strength and mass in response to progressive resistance training results from a greater
hypertrophic effect on type II fibers (26 to 57%) than on type I fibers (12.5 to 15%)
[5,19,37,39]. Furthermore, type II muscle fibers appear to be more sensitive to GH
stimulation [2], indicating that the increased recruitment of type II fibers could favor the
hypertrophy response. Thus, it seems logical to think that the greater recruitment of type II
fibers in the PE group should result in greater increases in 1RM (+17.6%) and total CSA
(+8.5%) compared with those in the traditional training group (TR group). From a practical
perspective, our results reveal that the manipulation of fiber-type recruitment using
additional sets with light loads (e.g., 20% of 1RM) before traditional training sets can be a
suitable strategy for promoting muscular strength and hypertrophy in young subjects.

56
It is important to note that an increase in training volume from the additional set
seems to be effective only when the training intensity (workload) is sufficient to promote
an adaptive stimulus of strength and hypertrophy. Campos et al. [5] have reported that an
8-wk traditional training program involving 2 sets of 20–28 repetitions (high-volume, low-
intensity) did not result in muscle fiber hypertrophy. In contrast, a low-volume, moderate-
intensity protocol (3 sets of 9–11 repetitions) promoted a significant increase in the CSA of
the 3 major muscle fiber types (I, IIa, and IIx) [5]. Additionally, previous studies showed
that gains in muscle strength are inhibited when muscular endurance training is performed
concurrently [4,16,34], suggesting that an increase in training volume can decrease the
training intensity and, thus, suppress the training effects on muscle strength. In our study,
the subjects who performed prior exhaustive exercise at 20% of 1RM to failure (PE group)
were able to complete the total number of repetitions (range of 8-12 repetitions) required
for each of the 3 sets of traditional hypertrophic training; that is, the prior exhaustive
exercise did not affect the intensity of the subsequent training sets. The maintenance of
exercise intensity even with the volume increase from an additional set could explain the
benefits of the protocol presented in this study. Thus, the combination of prior exhaustive
exercise (emphasizing increased total work) and traditional resistance training
(emphasizing mechanical loading) used in our study appears to be the most effective
combination for promoting an additional increase in muscle strength and hypertrophy. The
training protocol presented here highlights that the variation in training volume and its
interaction with intensity are key factors for maximizing strength and hypertrophy.
Additionally, the PE group experienced greater increases in muscle endurance and
work efficiency compared with the TR group from pre-to post-training. These results are
perhaps not surprising considering the previously published data on resistance training in
humans [5,14,18]. The results of these studies show that high-volume, low-intensity

57
protocols result in a greater increase in muscular endurance than low-volume, high-
intensity protocols. Thus, the greater increase in training volume in the PE group compared
with the TR group could, at least partially, explain the additional increase in muscle
endurance after the 8-wk resistance training program. It is important to note that a
considerable increase in training volume (e.g., multiple sets with 15-20 repetitions or
more) generally suppresses the training effects on strength and hypertrophy [5] due to the
reduction in training intensity. However, in our study, the greater training volume in the PE
group further promoted muscle endurance without affecting the additional gains in 1RM
and hypertrophy. As a result, the combination of these effects promoted greater muscular
work efficiency (as indicated by the lowest muscular work index) during the muscle
endurance test in the PE group compared with the group TR. This result indicates that this
novel training protocol can also improve muscle fiber recruitment during repeated,
maximal contraction activities. Together, our results provide new insight into the muscular
adaptations that occur in response to increased resistance training volume by showing for
the first time that the inclusion of a single set of exhaustive exercise at a light load (20% of
1RM) before high-intensity resistance training can be a suitable strategy for promoting
muscle endurance, strength, and hypertrophy in young men.
In conclusion, our results show that the inclusion of a single set of exhaustive
exercise at 20% of 1RM immediately before traditional resistance training promote
additional beneficial effects on quadriceps maximal strength, hypertrophy, and endurance
in young subjects. To define the usefulness of this protocol in other populations (e.g.,
women and older subjects) and muscle groups (e.g., muscles of the upper body
extremities), future studies should be performed to clarify the physiological (e.g., potential
for inducing overtraining) and metabolic (e.g., GH, testosterone, lactate, and cortisol

58
levels) responses to prolonged training (> 8 wk) and to determine how including this set
with other types of exercise affects a periodized training program.
Acknowledgments
We address a special thanks to Jayne Borim, Giseli Lombardi and Christiano Papi for their
great help in the physical tests and training.
Conflict of interest
No conflicts of interest, financial or otherwise, are declared by the author(s).

59
REFERENCES
1. American College of Sports Medicine. American College of Sports Medicine position
stand. Progression models in resistance training for healthy adults. Med Sci Sports
Exerc 2009; 41: 687–708
2. Aroniadou-Anderjaska V, Lemon PW, Gilloteaux J. Effects of exogenous growth
hormone on skeletal muscle of young female rats. Tissue Cell 1996; 28: 719–24
3. Baechle TR, Earle RW. Resistance training and spotting techniques. In: Earle R,
Baechle T (eds) Essentials of strength and conditioning: national strength and
conditioning association, 3rd edn. Human Kinetics, Champaign, 2008; 326–376
4. Bell GJ, Petersen SR, Wessel J, Bagnall K, Quinney HA. Physiological adaptations to
concurrent endurance training and low velocity resistance training. Int J Sports
Med 1991; 12: 384–390.
5. Campos GE, Luecke TJ, Wendeln HK, Toma K, Hagerman FC, Murray TF, Ragg KE,
Ratamess NA, Kraemer WJ, Staron RS. Muscular adaptations in response to three
different resistance-training regimens: specificity of repetition maximum training
zones. Eur J Appl Physiol 2002; 88: 50–60
6. Charge´ SB, Rudnicki MA. Cellular and molecular regulation of muscle regeneration.
Physiol Rev 2004; 84: 209–238
7. Cornelison DD, Wold BJ. Single-cell analysis of regulatory gene expression in
quiescent and activated mouse skeletal muscle satellite cells. Dev Biol 1997; 191:
270–283
8. Craig B, Kang H. Growth hormone release following single versus multiple sets of
back squats: Total work versus power. J Strength Cond Res 1994; 8: 270–275
9. Dhawan J, Rando TA. Stem cells in postnatal myogenesis: molecular mechanisms of
satellite cell quiescence, activation and replenishment. Trends Cell Biol 2005; 15:
666–673.
10. Esmarck B, Andersen JL, Olsen S, Richter EA, Mizuno M, Kjaer M. Timing of
postexercise protein intake is important for muscle hypertrophy
with resistance training in elderly humans. J Physiol 2001; 535: 301–311

60
11. Glass DJ. Skeletal muscle hypertrophy and atrophy signaling pathways. Int J Biochem
Cell Biol 2005; 37: 1974–1984
12. Goldspink G. Gene expression in skeletal muscle. Biochem Soc Trans 2002; 30: 285–
290
13. Goto K, Sato K, Takamatsu K. A single set of low intensity resistance exercise
immediately following high intensity resistance exercise stimulates growth hormone
secretion in men. J Sports Med Phys Fitness 2003; 43: 243–249
14. Goto K, Nagasawa M, Yanagisawa O, Kizuka T, Ishii N, Takamatsu K. Muscular
adaptations to combinations of high- and low-intensity resistance exercises. J Strength
Cond Res 2004; 18: 730–737
15. Gotshalk LA, Volek JS, Staron RS, Denegar CR, Hagerman FC, KraemerWJ. Creatine
supplementation improves muscular performance in older men. Med Sci Sports Exerc
2002; 34: 537–543
16. Hickson RC. Interference of strength development by simultaneously training for
strength and endurance. Eur J Appl Physiol Occup Physiol 1980; 45: 255–263
17. Hornberger TA, Chien S. Mechanical stimuli and nutrients regulate
rapamycinsensitive signaling through distinct mechanisms in skeletal muscle. J Cell
Biochem 2006; 97: 1207–1216
18. Huczel HA, Clarke DH. A comparison of strength and muscle endurance in strength-
trained and untrained women. Eur J Appl Physiol. 1992; 64:467–470
19. Kosek DJ, Kim JS, Petrella JK, Cross JM, Bamman MM. Efficacy of 3 days/wk
resistance training on myofiber hypertrophy and myogenic mechanisms in young vs.
older adults. J Appl Physiol 2006; 101: 531–544
20. Kraemer WJ, Marchitelli L, Gordon SE, Harman E, Dziados JE, Mello R, Frykman P,
McCurry D, Fleck SJ. Hormonal and growth factor responses to heavy resistance
exercise protocols. J Appl Physiol 1990; 69: 1442–1450
21. Kraemer WJ, Gordon SE, Fleck SJ, Marchitelli LJ, Mello R, Dziados JE, Friedl K,
Harman E, Maresh C, Fry AC. Endogenous anabolic hormonal and growth factor
responses to heavy resistance exercise in males and females. Int J Sport Med 1991; 12:
228–235

61
22. Kraemer WJ, Ratamess NA, Fry AC, French DN. Strength training: development and
evaluation of methodology. In: Physiological assessment of human fitness. Eds: Maud
P.J. and Foster, C.Champain, IL: Human Kinetics 2006
23. Krieger JW. Single vs. multiple sets of resistance exercise for muscle hypertrophy: A
meta-analysis. J Strength Cond Res 2010; 24: 1150–1159
24. Maughan RJ, Watson JS, Weir J. Strength and cross-sectional area of human skeletal
muscle. J Physiol 1983; 338: 37–49
25. Mitsiopoulos N, Baumgartner RN, Heymsfield SB, Lyons W, Gallagher D, Ross R.
Cadaver validation of skeletal muscle measurement by magnetic resonance imaging
and computerized tomography. J Appl Physiol 1998; 85: 115–122
26. Moss FP, Leblond CP. Satellite cells are the source of nuclei in muscles of growing
rats. Anat Rec 1970; 170: 421–435
27. Mulligan SE, Fleck SJ, Gordon SE, Koziris LP. Influence of resistance exercise
volume on serum growth hormone and cortisol concentrations in women. J Strength
Cond Res 1996; 10: 256–262
28. Paul AC, Rosenthal N. Different modes of hypertrophy in skeletal muscle fibers. J Cell
Biol 2002; 18: 156: 751–760
29. Rhea MR, Alvar BA, Burkett LN. Single versus multiple sets for strength: a meta-
analysis to address the controversy. Res Q Sport Exerc. 2002; 73: 485–488
30. Rhea MR, Alvar BA, Burkett LN, Ball SD. A meta-analysis to determine the dose
response for strength development. Med Sci Sports Exerc. 2003; 35: 456–464
31. Rooney KJ, Herbert RD, Balnave RJF. Fatigue contributes to the strength training
stimulus. Med Sci Sport Exerc 1994; 26: 1160–1164
32. Russell BD, Motlagh D, Ashley WW. Form follows functions: how muscle shape is
regulated by work. Journal of Applied Physiology 2000; 88: 1127–1132
33. Sale DG. Influence of exercise and training on motor unit activation. Exerc Sport Sci
Rev 1987; 15: 95–151
34. Sale DG, Jacobs I, MacDougall JD, Garner S. Comparison of two regimens of
concurrent strength and endurance training. Med Sci Sports Exerc 1990; 22: 348–356

62
35. Schoenfeld BJ. The mechanisms of muscle hypertrophy and their application to
resistance training. J Strength Cond Res 2010; 24: 2857–2872
36. Schoenfeld BJ. The use of specialized training techniques to maximize muscle
hypertrophy. J Strength Cond Res 2011; 33: 60–65
37. Shepstone TN, Tang JE, Dallaire S, Schuenke MD, Staron RS, Phillips SM. Short-term
high- vs. low-velocity isokinetic lengthening training results in greater hypertrophy of
the elbow flexors in young men. J Appl Physiol 2005; 98: 1768–1776
38. Smith LL. Cytokine hypothesis of overtraining: a physiological adaptation to excessive
stress? Med Sci Sports Exerc 1999; 32: 317– 333
39. Tesch PA Acute and long-term metabolic changes consequent to heavy-resistance
exercise. Med Sci Sports Exerc 1987; 26: 67–89
40. Vandenburgh HH. Motion into mass: How does tension stimulate muscle growth?
Med Sci Sports Exerc 1987; 19: S142–S149
41. Vierck J, O’Reilly B, Hossner K, Antonio J, Byrne K, Bucci L, Dodson M. Satellite cell
regulation following myotrauma caused by resistance exercise. Cell Biol Int 2000; 24:
263–272
42. Willardson JM. The application of training to failure in periodized multiple-set
resistance exercise programs. J Strength Cond Res 2007; 21: 628–631
43. Wolfe BL, LeMura LM, Cole PJ. Quantitative analysis of single- vs. multiple-set
programs in resistance training. J Strength Cond Res 2004; 18: 35–47

63
7. CAPÍTULO II
GUIA PRÁTICO
Recommended

















