Embed Size (px)
Citation preview

1
1. INTRODUÇÃO:
1.1. Escherichia coli enteropatogênica (EPEC):
Escherichia coli são bactérias Gram negativas, anaeróbicas facultativas, que
fazem parte da flora intestinal normal humana, e são importantes na manutenção da
fisiologia intestinal. Essas bactérias foram consideradas inofensivas durante muitos
anos. Contudo, cepas de E. coli causadoras de diarréia, tanto em humanos e
animais domésticos como boi (Fischer et al., 1994), ovelha (Cid et al., 2001), coelho
(Robins-Browne et al., 1994), cachorro (Turk et al., 1998) e porcos (Zhu et al., 1994),
foram descritas entre as décadas de 40 e 50 (Nataro e Kaper, 1998).
Inicialmente, todas as cepas de E. coli que induzem diarréia foram nomeadas
como E. coli enteropatogênicas (EPEC). Estudos posteriores permitiram que as E.
coli enteropatogênicas fossem classificadas em diferentes grupos de acordo com
seus mecanismos de infecção e fatores de virulência produzidos (Kaper, 1994). Os
fatores de virulência são proteínas de adesão, de invasão, e proteínas tóxicas que
caracterizam diversas manifestações clínicas, que vão desde diarréias coleriformes
e colites agudas até disenteria e morte (Nataro e Kaper, 1998; Chen e Frankel,
2004).
Existem hoje seis principais classes de E. coli patogênicas: as
enteropatogênicas (EPEC), as enterohemorrágicas (EHEC), as enterotoxigênicas,
(ETEC), as enteroagregativas (EAEC), as enteroinvasivas (EIEC) e as que aderem
difusamente (DAEC) (Tabela 1).

2
TABELA 1: Comparação entre E. coli enteropatogênicas
E. coli Mecanismos de infecção Fatores de Virulência
EPEC Lesão de Adesão e
desvanecimento do enterócito
(A/E) Attaching/ Effacing.
BfpA (EAF+), EspB,
EspD, EspA, intimina e
Tir, dentre outros.
EHEC Lesão de Adesão e
desvanecimento do enterócito
(A/E) Attaching/ Effacing. Além
de hemorragia e edema da
lâmina própria causada pela
liberação de citotoxina.
Toxina shiga símile/
Verocitotoxina (Stx).
ETEC Colonização da superfície da
mucosa intestinal e liberação de
enterotoxinas.
Toxina Termo estavél
(ST), Toxina Termo
lábel (LT).
EAEC Aderência inicial na mucosa
intestinal, produção de muco e
de citotoxina.
Enterotoxina estável
enteroagregativa
(EAST1)
EIEC Invasão epitelial, rompimento do
vacúolo endocítico,
multiplicação intracelular,
movimento direcional pelo
citoplasma, invasão de células
epiteliais adjacentes.
Plasmídio que codifica
genes de invasão.
DAEC Aderência difusa na membrana
epitelial, elongação das
microvilosidades, sem alteração
no citoplasma.
Hemolisina α, Fator 1
de citotoxicidade
necrosante (CNF1).
Adaptada de Nataro e Kaper,1998.

3
O termo “EPEC” deve ser empregado para espécies de Escherichia coli
diarreicogênicas que não produzem nenhuma enterotoxina, mas que causam a
lesão histopatológica característica determinada pela ligação íntima e
desvanecimento das microvilosidades dos enterócitos (Attaching and Effacing - A/E)
(Nataro e Kaper, 1998) lesão essa que leva a uma pronunciada disfunção na
absorção intestinal.
As EPEC foram responsáveis por surtos de diarréia aguda infantil nas décadas
de 40 e 50 nos países desenvolvidos. Hoje, são a principal causa de diarréia
bacteriana infantil, nos países em desenvolvimento ou em áreas de saneamento
precário, perdendo apenas para o rotavírus em número de pacientes internados
(WHO, 2000). Estudos no Brasil, México, África do Sul e Bangladesh mostraram que
30-40% dos casos de diarréia bacteriana infantil são devido à infecção por EPEC,
sendo a doença responsável por altas taxas de mortalidade. Cerca de 1.000.000 de
crianças-caso/ano (Baldwin, 1998) no mundo e 200.000 somente no Brasil (Gomes
et al., 1991).
Os principais sintomas da doença causada por EPEC são diarréia aquosa, em
vários graus de intensidade, e desidratação. O quadro clínico pode se agravar
rapidamente pela dificuldade na eliminação da bactéria podendo implicar em
infecção do trato urinário e meningite (Chen e Frankel, 2004). Outro fator que pode
contribuir para o agravamento da doença é a falta de um método diagnóstico
diferencial rápido, o que dificulta a detecção, e conseqüentemente, o tratamento
precoce (Vallance e Finlay, 2000).
A diarréia causada por EPEC acomete, freqüentemente, neonatos nas áreas
endêmicas, devido à falta de uma resposta imune adquirida. Os principais fatores de
risco são a ausência do aleitamento ao seio materno, o baixo peso ao nascer,
desnutrição nos primeiros meses de vida, e a residência em áreas de saneamento
precário. Este último sugere que a água utilizada para preparação dos alimentos
seja a principal fonte de contaminação (Fernandes e Medina-Acosta, 2002;
Fernandes e Medina-Acosta, 2004).
Nos países em desenvolvimento a profilaxia para bactéria enteropatogênicas é
dependente de programas de tratamento de água e esgoto, uma vez que a
transmissão é oro-fecal. E os métodos, em geral, utilizados para detecção dessas

4
bactérias, têm duração de aproximadamente 48h, tempo suficiente para o
estabelecimento do processo típico de adesão e destruição das microvilosidades do
epitélio intestinal. Esse processo leva a diarréia e a quadros de forte desidratação,
podendo levar uma criança à morte (Flores, 2001).
Há, então, grande necessidade da produção de imunoprofiláticos e métodos
diagnósticos rápidos contra as bactérias enteropatogênicas, tanto para população
infantil dos países em desenvolvimento, quanto para migrantes ou turistas vindos de
países industrializados. Os últimos são alvo fácil para infecção com enterobactérias
patogênicas, uma vez que têm baixo nível de imunidade adquirida, devido à baixa
incidência da doença em seus países de origem.
1.2. Mecanismo de infecção e Fatores de virulência
Alguns trabalhos propõem que a patogênese de EPEC ocorre em quatro
fases distintas (Knutton et al., 1998; Donnenberg e Kaper, 1992), contudo esse
modelo é artificial e controverso. No primeiro estágio, em condições propícias, EPEC
expressa em sua superfície as adesinas BfpA (Subunidade A dos pili formadores de
feixe de EPEC), a adesina intimina e filamentos curtos de EspA (Proteína A
secretada de EPEC). No segundo estágio, as células bacterianas se aderem ao
epitélio intestinal através dos filamentos de BfpA e EspA, e o sistema de secreção
do tipo III transloca Tir (receptor translocado de intimina) e outras proteínas efetoras
para o citoplasma das células hospedeiras. As proteínas efetoras ativam vias de
sinalização que levam a alteração do citoesqueleto e perda das microvilosidades
locais. No terceiro estágio, os filamentos de EspA são perdidos da superfície
bacteriana, a intimina se liga ao Tir resultando na ligação íntima, com acúmulo de
actina na interface com a bactéria. Na última fase, ocorre a formação do pedestal
típico da lesão de adesão e desvanecimento (A/E) causada pela grande quantidade
de elementos do citoesqueleto no local da adesão por EPEC. Ao final, esse
processo leva a perda eletrolítica e eventual morte celular (Clarke et al., 2003)
(Figura 1).

5
FIGURA 1: Modelo de 4 estágios da patogênese de EPEC. Adaptada de
(Clarke et al., 2003).
Feixe de BfpA Intimina Filamento de EspA
EspB e EspD Actina Tir
EspF
EPECEPEC
EPEC
EPEC
A B C D
EPECEPEC
EPEC
EPEC
A B C D
Microvilos
Enterócitos
1º 2º 3º 4º

6
1.2.1. Adesão inicial
O processo de adesão inicial da bactéria à célula hospedeira é mediado pelo
feixe de pili, formado pela proteína BfpA como demonstrado em experimentos de
infecção in vitro por Hicks et al. (1998) e Cleary et al. (2004). Os genes responsáveis
pela biogênese dos pili de proteína A encontram-se em um megaplasmídio
denominado Fator de Aderência de EPEC (EAF) (Nataro e Kaper, 1998). O feixe de
pili é utilizado pela bactéria na formação de micro-colônias, fenótipo de adesão típico
de EPEC denominada Aderência localizada (LA”Localized Adherence”) (Figura 2)
(Nataro e Kaper, 1998; Rodrigues et al., 2004). Algumas cepas de EPEC,
denominadas atípicas, perdem esse plasmídio, aderem-se difusamente ao epitélio
intestinal e exibem menor virulência e patogenicidade, sendo este fenótipo de
adesão atípico de EPEC denominada aderência difusa (Figura 3) (Nataro e Kaper
1998).
FIGURA 2: Aderência localizada de EPEC.
Fenótipo típico de EPEC em cultura de células epiteliais (Reproduzido com
permissão do Dr. Brett Finlay, University of British Columbia, Canadá, e do Dr.
Michael Donnenberg, Universidade de Maryland, EUA).

7
FIGURA 3: Aderência Difusa de EPEC
Fenótipo atípico de EPEC em cultura de células epiteliais (Reproduzido com
permissão do Dr. Brett Finlay, University of British Columbia, Canadá, e do Dr.
Michael Donnenberg, Universidade de Maryland, EUA).
1.2.2. Ilha de patogenicidade: Lócus de Desvanecimento do Enterócito (LEE)
A capacidade da EPEC de causar a lesão A/E é dependente da ativação de
genes que codificam a síntese dos fatores de virulência de EPEC. Estes estão
localizados no lócus cromossômico LEE (Locus of Enterocyte Effacement) chamado
de Ilha de patogenicidade (PAI - pathogenicity island). Possui 34 Kpb e está dividido
em cinco operons policistrônicos denominados, em ordem, LEE 1, LEE 2, LEE 3,
LEE 5 e LEE 4 (Cleary et al., 2004).
Os genes que codificam um sistema de secreção do tipo III estão no LEE 1, 2,
3; eae, que codifica a adesina intimina, e tir, que codifica o receptor translocado de
intimina, estão no LEE 5; espA, espB e espD, que codificam proteínas secretadas
através do sistema de secreção do tipo III,estão no LEE 4; e o gene que codifica o
regulador ler, que ativa transcrição do LEE 1, 2, 3, 4 e 5, está no LEE 1 (Elliot et al.,
1998).
A região LEE está presente em EPEC e EHEC com alto grau de homologia,
mas não está presente em cepas de E. coli da flora normal, E. coli K-12, ou ETEC.
Seqüências homólogas ao LEE também são encontradas em outras bactérias
enteropatogênicas que causam o fenótipo A/E (Adesão e desvanecimento),
incluindo, REDEC-1 (EPEC de coelho), Hafnia. alvei diarreicogênica e Citrobacter
rodentium (Elliot et al., 1998, Künhe et al., 2004).

8
Mutações em algum dos fatores bacterianos codificados por LEE previnem a
formação da lesão A/E em culturas de células epiteliais, tais como mutantes para
eae, tir ou espA (Cleary et al., 2004). Atualmente, essas mutações tem sido alvo
para o desenvolvimento de vacinas para EPEC, como aqueles mutantes para o
regulador Ler (Zhu et al., 2005) e para eae (Stakenborg et al., 2006). Estes já foram
utilizados como vacinas em coelhos, e mostraram ser imunogênicos e protetores em
testes de desafio com dose letal de EPEC selvagem.
1.2.3. Sistema de secreção do tipo III (TTSS)
O sistema de secreção do tipo III (TTSS) está associado com a virulência de
muitas bactérias Gram negativas que infectam humanos, animais, insetos e plantas.
Sua organização e composição são bastante conservadas (Hueck, 1998), como, por
exemplo, entre a Shigella (Sansonetti et al., 1986), Salmonella (Galan, 1996) e
Yersinia (Rosqvist et al., 1990). Consiste em um corpo basal formado por um
complexo multiprotéico que atravessa ambas membranas bacterianas, e uma
estrutura tipo agulha, projetada para o exterior, que permite a liberação direta de
proteínas efetoras no interior das células hospedeiras (Hueck 1998; Hartland et al.,
2000). Essa organela pode ser visualizada por microscopia eletrônica e é chamada
de complexo agulha ou corpo basal (Sekiya et al., 2001, Ogino et al., 2006). As
principais proteínas formadoras do corpo basal do TTSS em EPEC e EHEC são
EscC, EscV e EscF. As EscC e EscV compõem as estruturas de anéis nas
membranas bacteriana externa e interna, respectivamente (Gauthier et al., 2003) e a
EscF, permanece ancorada na membrana bacteriana através dos anéis interno e
externo (Figura 4) (Sekiya et al.,2001; Ogino et al., 2006).
O TTSS de EPEC e EHEC é o único que possui a extensão de um filamento
para formar o ‘complexo agulha’ (Knutton et al., 1998). A proteína translocada EspA
é a única constituinte desse filamento condutor que liga diretamente a bactéria à
célula hospedeira (Knutton et al., 1998; Hartland et al., 2000; Daniell et al., 2003);
Ogino et al., 2006). Cepas mutantes para EscF não são capazes de expressar EspA
e formar a agulha do TTSS, enquanto que mutantes para EspA não são capazes de
expressar EscF e formar o corpo basal do TTSS, mostrando assim que essas
proteínas podem afetar a forma e a estabilidade do complexo (Sekiya et al.,2001).

9
Os filamentos de EspA são essenciais para translocar para a célula hospedeira as
proteínas EspD (Proteína D Secretada de EPEC), e EspB (Proteína B Secretada de
EPEC), que participam na formação de poros na membrana do enterócito (Knutton
et al., 1998; Hartland et al., 2000; Vallance e Finlay, 2000). Os poros servem de
passagem das proteínas efetoras para a célula hospedeira, como o receptor
translocado de intimina, Tir, envolvido na ligação íntima e transdução de sinal, à
proteína associada a mitocôndra, Map, que interfere no potencial de membrana da
mitocôndria; a proteína F secretada de EPEC, EspF, requerida para ruptura das
junções célula-célula e na morte celular programada; a proteína G secretada de
EPEC, EspG, envolvida na desestabilização dos microtúbulos; e proteína H
secretada de EPEC, EspH envolvida na modulação dos filamentos de actina para a
formação do pedestal (Garmendia et al., 2005).

10
Membrana da célula hospedeira
ME
MI
EspA
Filamento de EspA
AE
AI
EscF
Tir, EspB, EspD
EscN
PP
EspB, EspD
Membrana da célula hospedeira
ME
MI
EspA
Filamento de EspA
AE
AI
EscF
Tir, EspB, EspD
EscN
PP
Membrana da célula hospedeira
ME
MI
EspA
Filamento de EspA
AE
AI
EscF
Tir, EspB, EspD
EscN
PP
EspB, EspD
FIGURA 4: Modelo Esquemático do sistema de secreção do tipo III de EPEC
Primeiro os anéis interno e externo são formados, e em seguida EscF é polimerizada
através dos anéis. EspA é secretada e ligada no topo do complexo agulha de onde
inicia sua polimerização e formação da agulha desse complexo (filamento). Através
dessa estrutura em forma de tubo as proteínas EspB, EspD e Tir são translocadas
para a célula hospedeira, onde EspB e EspD formam poros para que Tir e outras
proteínas efetoras cheguem ao citoplasma. ME: Membrana Externa, MI: Membrana
Interna, PP: Periplasma, AE: Anel Externo da porção ancorada do sistema de
secreção do tipo III de EPEC, AI: Anel Interno da porção ancorada do sistema de
secreção do tipo III de EPEC. Adaptada de Sekiya et al. (2001).
1.2.4. Aderência íntima
Dentre as proteínas efetoras que a bactéria transloca através do TTSS para o
citoplasma do enterócito, está a proteína Tir de 90 kDa. Esta insere-se na membrana
servindo de receptor para a proteína intimina de 94 kDa, que está presente na
membrana externa bacteriana. A ligação Tir-intimina estabelece uma adesão íntima
entre a bactéria e o enterócito, com conseqüente perda localizada das
microvilosidade das células do epitélio intestinal. Com a translocação de proteínas
efetoras para a célula hospedeira e a adesão íntima entre a bactéria e o enterócito, a
bactéria torna-se capaz de modular algumas vias de sinalização na célula

11
hospedeira (Kenny et al., 1997). Dessa forma, EPEC pode iniciar a formação do
pedestal e mediar seu efeito patogênico na célula hospedeira enquanto permanece
na superfície extracelular.
1.2.5. Transdução de sinal
Ao se fixar na membrana da célula hospedeira a proteína Tir é fosforilada,
ativando a via de sinalização da fosfolipase C (PLC), o que leva a formação de
fosfatidil inositol (IP), aumento no fluxo de cálcio intracelular e ativação de proteína
quinase C (PKC). Como conseqüência à sinalização intracelular, ocorre
reorganização do citoesqueleto e formação de um pedestal na interface com a
bactéria, caracterizando a típica lesão A/E (Moon et al., 1983; Dytoc et al., 1994;
DeVinney et al., 1999; Campellone e Leong, 2003) (Figura 5).
A lesão A/E causada pela infecção por EPEC provoca danos celulares que
levam a diarréia. Estes são caracterizados pela perda da absortividade através das
microvilosidades, pela redução das junções do tipo oclusão entre as células, pelo
aumento da permeabilidade paracelular, com conseqüente perda de importantes
eletrólitos como Cl- e HCO3-, e pelo processo inflamatório mediado pela infiltração
de neutrófilos (Vallance e Finlay, 2000) (Figura 5).
Dano tecidual
Recrutamento de Neutrófilos
Alteração de Cl- e HCO3
-
Secreção iônica
Diarréia
DiminuiçãoIntegridadedas junções
Tight
Perda da superfície de absorção
DiminuiçãoDa
Resistência
H2O H2OCl- Cl- EPEC
Dano tecidual
Recrutamento de Neutrófilos
Alteração de Cl- e HCO3
-
Secreção iônica
Diarréia
DiminuiçãoIntegridadedas junções
Tight
Perda da superfície de absorção
DiminuiçãoDa
Resistência
H2O H2OCl- Cl- EPEC
FIGURA 5: Esquema dos danos celulares que levam à diarréia na infecção por
EPEC. Adaptado de Vallance e Finlay, 2000.

12
1.3. A Proteína EspA
Como mencionado anteriormente, os genes que codificam as proteínas
secretadas encontram-se no lócus cromossômico LEE, que também inclui todos os
outros componentes formadores do TTSS, além das proteínas efetoras. Dentre as
proteínas secretadas codificadas por LEE está EspA, uma proteína de
aproximadamente 25 kDa, que é a formadora da agulha do TTSS (Chen e Frankel,
2004).
Daniell e colaboradores (2003) mostraram, em experimentos de microscopia
eletrônica dos filamentos de EspA, que a forma com que esses filamentos se
organizam é muito semelhante ao arranjo da flagelina na formação do flagelo.
Porém, eles diferem na região central, sugerindo que o filamento de EspA forme um
canal central por onde as proteínas da bactéria são liberadas no citoplasma do
enterócito.
A formação dos filamentos de EspA ocorre por adição de muitas subunidades
de EspA no topo de crescimento do filamento, e as proteínas efetoras são
translocadas através de um canal central do filamento de EspA como confirmado no
trabalho de Crepin e colaboradores 2005. Além disso, mostraram que o aumento da
concentração intracelular das subunidades de EspA resulta em filamentos mais
longos, sugerindo que a quantidade de EspA produzida pela bactéria constitui um
fator limitante para definir o comprimento do filamento.
A secreção de muitas proteínas do TTSS é regulada pela presença de
chaperoninas citoplasmáticas. Creasey e colaboradores (2003) identificaram uma
interação entre EspA e EspB com uma proteína citoplasmática que, além de impedir
a formação de agregados de EspA intracelulares, mostrou regular a expressão, o
nível de transcrição e a secreção da EspA, sendo chamada de chaperoninas CesAB.
Em termos funcionais o TTSS é chamado de injectiossomo, uma organela
especializada na translocação, para o interior do enterócito, de proteínas efetoras
que são necessárias para que ocorra a adesão íntima, subversão, transdução de
sinal e formação da lesão A/E (Kenny et al., 1997; Knutton et al., 1998) (Figura 6).

13
Membrana da célula do hospedeiro
Membrana bacteriana externa
Membrana bacterianainterna
Espaço Periplasmático
Membrana da célula do hospedeiro
Membrana bacteriana externa
Membrana bacterianainterna
Espaço Periplasmático
FIGURA 6: Esquema representando o aparato do sistema de secreção do tipo III de
EPEC/EHEC. Estão representadas as proteínas que formam o sistema de secreção
do tipo III e as proteínas que são translocadas. Em destaque a proteína EspA, a
principal proteína que compõe o sistema de secreção do tipo III, formadora do
filamento translocador que liga a bactéria à célula hospedeira. Adaptada de
(Garmendia et al., 2005).
Além de mediar a translocação de proteínas efetoras, os filamentos de EspA
podem funcionar como adesinas, como demonstrado em estudos in vitro com
células epiteliais, em que EPEC deficiente para os genes que codificam BfpA e
intimina foram capazes de estabelecer adesão inicial através dos filamentos de
EspA, porém, não tão eficientemente quanto a cepa selvagem. Os filamentos de
EspA funcionaram como um tipo de adesina estabelecendo uma ligação transiente
entre a bactéria e a célula hospedeira (Hartland et al., 2000; Cleary et al., 2004).
Além disso, Moreira e colaboradores (2006) mostraram, em substratos abióticos,

14
que EspA e BfpA de EPEC estão envolvidas na formação de biofilme, o que pode
justificar a alta resistência dessa bactéria a antibióticos.
A grande importância e contribuição dos filamentos de EspA para a lesão A/E
foram comprovados em estudos in vitro com cultura de CaCo-2 (modelo de células
epiteliais) e cepa de EPEC deficiente para o gene espA, em que essa cepa foi
incapaz de induzir a típica lesão A/E (Cleary et al., 2004). Resultados semelhantes
foram obtidos por Abe e colaboradores em 1998, utilizando cepas de REPEC (EPEC
de coelho) deficiente para espA na infecção experimental de coelhos.
No entanto, utilizando microscopia de fluorescência e anticorpos policlonais
anti-EspA marcados com diferentes fluorocromos, Knutton e colaboradores (1998)
observaram que a expressão dos filamentos de EspA in vitro é transiente,
demonstrando que, após o início da formação da lesão A/E, os filamentos de EspA
vão diminuindo, gradualmente, até não serem mais detectados na lesão A/E
madura. (Figura 7). Esses dados sugerem uma aplicação profilática importante para
anticorpos anti –EspA. Embora, estes não previnam a formação dos filamentos de
EspA, como descrito anteriormente (La Ragione et al., 2005), podem, de alguma
forma, impedir a formação da lesão A/E, seja por inibição da função de
‘ancoramento’ dos filamentos de EspA com a membrana da célula hospedeira,
através da prevenção da interação de EspA com outras proteínas efetoras, ou por
estabilizar os filamentos, prevenindo seu desarranjo, que é o pré-requisito para que
ocorra a adesão íntima.

15
FIGURA 7: Foto micrografia de fluorescência. Anticorpos marcados em vermelho
reconhecem actina, nas células epiteliais, e anticorpos marcados em verde
reconhecem EspA, em EPEC, mostrando que a presença dos filamentos de EspA é
transiente, barra = 5µm (Knutton et al., 1998).
Os filamentos de EspA também são essenciais para desencadear a hemólise
de células vermelhas do sangue em experimentos in vitro, uma vez que EPEC
mutantes para EspA não foram capazes de induzir a lise, assim como mutantes para
EspD. Entretanto, mutantes para EspB ainda induziram fraca hemólise. Esse é um
modelo de estudo simples que sugere a necessidade dos filamentos de EspA para
desencadear a lesão A/E (Shaw et al., 2001), e mostra que, a formação dos
filamentos de EspA é dependente de EspD, como confirmado por Crepin et al.
(2005a).
Trabalhos de caracterização de EspA em diferentes sorotipos de EPEC
mostraram a importância dessa proteína na patogênese de EPEC e o seu possível
uso em diagnóstico e profilaxia, devido a alta similaridade entre as seqüências de
aminoácidos das diferentes cepas de EPEC, e entre EPEC e EHEC (Neves et al.,
1998 e 2003). Análises comparativas (Tabela 2) das seqüências de aminoácidos da
proteína EspA de EPEC de coelho (REPEC sorotipo O103:H2) com a de outras
bactérias enteropatogênicas revelam alta similaridade da seqüência de aminoácidos
entre diferentes cepas de E. coli eteropatogênicas e diferentes sorotipos de variados
hospedeiros específicos.

16
TABELA 2: Porcentagem de similaridade entre a seqüência de aminoácidos da
proteína EspA de REPEC (0103:H2) com as diferentes E.coli enteropatogênicas
E.coli Sorotipo Hospedeiro Similaridade
Protéica %
Nº de acesso
REPEC “0103:H2” Coelho Af200363
STEC “026:H” Bovino 100 AJ277443
EPEC “0111:H2” Ovelha 99 AJ225018
EPEC “0128:H2” Ovelha 99 AJ225021
EPEC -- Cachorro 98 U65681
EPEC -- Suíno 98 AF064683
EHEC “0103:H2” Bovino 94 AJ303141
EPEC “0127:H6 Humano 91 Z54352
EHEC “0127:H7” Humano 91 AF022236
EPEC “055:H7” Ovelha 90 AJ225220
EPEC “055:H6” Ovelha 90 AJ225019
BLAST (National Center for Biotechnology Information, NCBI, NIH, Bethesda, MD).
(Análise feita para este trabalho).
Estudos recentes mostraram que a proteína EspA de EPEC e EHEC de
diferentes cepas possui a extremidade N e C terminal com alta similaridade,
formando a região central do filamento, e uma região entre as extremidades que é
hipervariável, exposta e imunodominante. Além disso, esse estudo mostrou que
pequenos peptídeos, inseridos na região hipervariável, são apresentados na
superfície do filamento de EspA e são acessíveis para anticorpos. Essa observação
pode ter um considerável impacto para o futuro uso do filamento de EspA como
‘peptide display’ (Crepin et al., 2005b).
O polimorfismo encontrado na região central da EspA indica que há interações
entre os filamentos de EspA e o sistema imune do hospedeiro, o que faz da EspA
um possível alvo para o desenvolvimento de vacina (Crepin et al., 2005b). O estudo
de La Ragione e colaboradores (2005) reforça essa idéia, uma vez que anticorpos
policlonais anti-EspA demonstraram reconhecer todos fragmentos de EspA com
exceção dos fragmentos correspondentes aos aminoácidos 129-171 e 150-192,

17
onde o último compõe a região conservada do filamento, ou seja, a região não
exposta.
A proteína EspA demonstrou estimular a produção de IgG e IgA humanos, uma
vez que anticorpos anti-EspA são detectados em soros de pacientes acometidos por
EPEC (Martinez et al., 1999), e em colostros de mães que residem em regiões
endêmicas para EPEC (Loureiro et al., 1998). Sendo assim, a detecção de
anticorpos anti- EspA em pacientes abre a possibilidade de diagnóstico específico e
profilaxia de pessoas acometidas por E. coli enteropatogênicas. Em nosso grupo de
pesquisa, trabalhos buscando a caracterização e diagnóstico para EPEC já foram
realizados, tanto utilizando BfpA recombinante com soro de pacientes e colostro das
mães (Fernandes et al., 2002 a e b) quanto BfpA e EspB recombinante com
coproanticorpos (Fernandes et al., 2003), em testes de Western blot.
1.4. Resposta imune, vacinas e imunobiológicos contra E. coli
enteropatogênicas
Para avaliar a resposta imune contra REPEC Ramirez e colaboradores (2005)
infectaram coelhos com REPEC selvagem e com mutantes para intimina e EspA.
Em geral, observaram um aumento da migração de leucócitos polimorfonuleares e a
expressão de citocinas proinflamatórias como IL-6, IL-8, TNF-α sendo secretadas
por linfócitos da Placa de Peyer e baço. Essas características indicam que a
colonização por REPEC dirige a resposta imunológica para Th1. Durante a infecção
com REPEC mutante para EspA, houve um aumento dos níveis de expressão das
citocinas IL-8 e TNF-α, e fraco aumento de IL-10.
Diversos trabalhos visam determinar a presença e a especificidade de
anticorpos protetores contra proteínas essenciais para o estabelecimento da
infecção por E. coli enteropatogênicas. A resposta imune contra EPEC e EHEC é
detectada no leite de lactantes com a presença de altos níveis de anticorpos IgA
contra as proteínas EspA, EspB, intimina (Loureiro et al., 1998; Noguera-Obenza et
al., 2003) e BfpA (Loureiro et al., 1998; Fernandes et al., 2002b). Também foram
detectados, no soro de crianças, altos níveis de anticorpos IgG anti-EspA, EspB,
intimina (Martinez et al., 1999), BfpA (Fernandes et al., 2002a) e Tir (Li et al., 2000).
Anticorpos anti-BfpA e anti-EspB também foram encontrados em fezes de crianças

18
alimentadas naturalmente e artificialmente (Fernandes et al., 2003). Carbonare e
colaboradores (2003) também analisaram anticorpos do soro e saliva de crianças
residentes em área endêmica para EPEC e encontraram anticorpos reativos para as
adesinas (BfpA e intimina) e para as proteínas secretadas de EPEC (EspA e EspB).
Comprovada a imunogenicidade de diversas proteínas de EPEC e EHEC, o
seu uso para produção de imunobiológicos tornou-se alvo de estudos para muitos
pesquisadores. Em busca de um método diagnóstico rápido para E. coli
enteropatogênicas, anticorpos anti-EspB foram produzidos e usados em testes de
aglutinação reversa passiva em látex. Estes anticorpos foram capazes de detectar
fatores de virulência tanto EPEC quanto EHEC (Lu et al., 2002). Com o mesmo
objetivo, anticorpos policlonais e monoclonais recombinantes anti-EspA e intimina
foram capazes de reconhecer essas proteínas de EHEC, em testes de Western blot
( Kühne et al., 2004). Recentemente, La Ragione e colaboradores (2005), por meio
de experimentos de fluorescência, demonstraram que anticorpos policlonais e
monoclonais anti-EspA e intimina inibem a reorganização do citoesqueleto de
células epiteliais HEp-2 induzida pela infecção por EHEC, e que anticorpos anti-
EspA da cepa EHEC O157:H7 interferem na reorganização de actina de 13 dos 21
sorotipos de EPEC testados, mostrando que houve proteção cruzada.
O uso de anticorpos IgY, produzidos na gema do ovo de galinhas
hiperimunizadas, anti-intimina, Tir, EspA, EspB e EspD foi proposto por Girard e
colaboradores (2006) como uma alternativa promissora na prevenção da lesão A/E
causada por EPEC. Nesse estudo, IgY anti-EspA inibiu a aderência de EPEC
humana em cultura de órgãos in vitro (IVOC).
Uma outra abordagem que vem sedo explorada por alguns pesquisadores é o
uso de proteínas recombinantes, ou mesmo E. coli enteropatogênicas mutantes,
como vacinas. Potter e colaboradores (2004) utilizaram proteínas secretadas (Esps
e Tir) para imunizarem bovinos. A resposta imune induzida foi suficiente para manter
baixos níveis de bactérias nas fezes após o desafio com EHEC. A vacinação de
coelhos com REPEC mutantes para o regulador (ler) do lócus LEE conferiu proteção
aos coelhos após desafio com cepa selvagem de REPEC (Zhu et al., 2005).
Resultados semelhantes foram descritos com EPEC mutante para EspB e Tir
(Boullier et al., 2003).

19
Recentemente, a proteção de coelhos contra REPEC 8+/0103 foi realizada
com vacinação oral com EPEC mutante para intimina, porém, não foi eficiente para
proteger totalmente os coelhos desafiados com REPEC de outros bio-sorogrupos
(Stakenborg et al., 2006).
2. Justificativa
A proteína EspA, secretada por E. coli enteropatogênicas via sistema de
secreção do tipo III, é a subunidade formadora do filamento que liga a bactéria à
célula hospedeira. Ela permite a translocação de proteínas efetoras para o interior
da célula hospedeira, desencadeando a adesão íntima, subversão, transdução de
sinal e formação da lesão patognomônica de adesão e desvanecimento. Desta
forma, a proteína EspA é de grande importância no estabelecimento da infecção.
A EspA é considerada uma proteína imunogênica alvo de estudos de
profilaxia experimental, podendo ser utilizada na produção de uma vacina protetora
contra EPEC e EHEC de vários sorotipos, pois, a exposição natural a EPEC resulta
em produção de IgG e IgA específicas prevalente em indivíduos nas áreas
endêmicas.
A produção de proteína EspA de EPEC, de forma recombinante, e de
anticorpos anti-EspA oferecem algumas vantagens, pois a similaridade estrutural da
EspA das diferentes E. coli enteropatogênicas e as altas freqüências de anticorpos
anti–EspA, sugerem que uma proteção cruzada seria possível contra múltiplos
sorotipos de maneira eficaz no bloqueio da formação da lesão A/E. Portanto, a
produção de EspA de forma recombinante e de anticorpos anti-EspA recombinante
abrem a possibilidade para o desenvolvimento de métodos diagnóstico mais rápidos,
baratos e eficientes, além de sua aplicação em profilaxia e terapia.

20
3. OBJETIVOS
Clonar e expressar o gene espA de EPEC em sistema de expressão bacteriano
pQE-30.
Produzir e purificar proteína EspA recombinante.
Produzir anticorpos policlonais anti- EspA recombinante em coelhos hiperimunes.
Avaliar a antigenicidade da proteína EspA recombinante com IgG policlonais anti-
EspA produzidos em coelhos, e com IgG e IgA séricas de crianças assintomáticos
de 0 a 2 anos.

21
4. MATERIAIS E MÉTODOS
4.1. Estratégia metodológica
A seqüência codificadora espA foi amplificada por meio da técnica de PCR
utilizando iniciadores específicos desenhados para conter sítios das enzimas Bam HI
e Hind III. A seqüência amplificada foi clonada no vetor de clonagem comercial
pGEM-T-Easy®. A construção recombinante obtida, pGEM/espA, foi digerida com as
enzimas de restrição Bam HI e Hind III, para obtenção do inserto relativo à
seqüência espA com as extremidades compatíveis, para ligação com o vetor de
expressão bacteriano pQE-30, também digerido com Bam HI e Hind III. Tanto o
vetor como a seqüência espA foram purificados e ligados em condições e
concentrações apropriadas. As construções recombinantes pQE-30/espA foram
utilizadas para transformar E. coli M15 (pREP4). Após a seleção de clones contendo
pQE-30/espA e pREP4, foi feita a indução da expressão da proteína EspA com
IPTG. A proteína EspA foi purificada por coluna de afinidade Ni-NTA-Sepharose,
utilizada na imunização de coelhos, e testada quanto sua antigenicidade. Anticorpos
policlonais de coelhos imunizados foram analisados in vitro por ELISA e Western
blot. A proteína EspA recombinante também foi utilizada em testes de captura
(ELISA e Western blot) para IgG e IgA anti-EspA de crianças assintomáticas
menores de 2 anos. Anticorpos anti-EspA foram utilizados em ensaios de inibição da
hemólise por EPEC e REPEC.

22
FIGURA 8: Esquema representativo da Estratégia metodológica
PCR
espA
+
VVeettoorr ppGGEEMM--TT
eeaassyy
pGEM/espA
Bam HI Hind III
pGEM/espA VVeettoorr ppGGEEMM--
TT eeaassyy
+
espA
Hind III
Bam HI
Vetor pQE-30 VVeettoorr ppQQEE--3300 ddiiggeerriiddoo
+
Transformação
pQE-30/espA
E.coli M15
Expressão
Purificação
NNii--NNTTAA--SSeepphhaarroossee HHiissEEssppAA
IImmuunniizzaaççããoo ddee ccooeellhhooss
AAnnttiiccoorrppooss
-- EElliissaa -- BBlloott -- IInniibbiiççããoo ddaa hheemmóólliissee

23
4.2. Material Biológico
4.2.1. Células
TABELA 3: Células utilizadas
Célula Cepa Fonte
Escherichia coli DH5-α Invintrogen
Escherichia coli M15 Qiagen
Escherichia coli Enteropatogênica
humana (hEPEC)
B171 Cedida por Wilmar Dias da
Silva – LBR/UENF
Escherichia coli Enteropatogênica
de coelho (rEPEC)
REPEC-1 Cedida por João Ramos
Costa Barros - UERJ
4.2.2 Coelhos
Para o experimento de imunização com EspA recombinante foram utilizados
dois coelhos albinos pesando 2Kg, obtidos de criador local. Todo o procedimento
experimental foi realizado dentro dos padrões éticos e de acordo com as leis de
proteção de animais de laboratório.
4.2.3 Soro e anticorpos
Amostras de sangue foram coletadas de 100 crianças, assintomáticas para
diarréia, de 0 a 2 anos de idade, atendidas pela Drª Regina Célia de Souza Campos
Fernandes no Centro da Criança e do Adolescente da cidade de Campos dos
Goytacazes. O sangue foi coagulado a temperatura ambiente e separado por
centrifugação a 12.000x g e o soro mantido a -20 ºC até o uso. Todo o procedimento
foi realizado dentro dos padrões éticos e com o consentimento dos pais.
Para obtenção do sangue dos coelhos foi feita a sangria total por um médico
veterinário, o sangue foi coagulado a temperatura ambiente e separado por
centrifugação a 12.000x g e o soro mantido a -20 ºC até o uso.
Os anticorpos anti-His, anti-IgG e anti-IgA peroxidase foram adquiridos da
Sigma-Aldrich, EUA e mantidos a -20 ºC até o uso.

24
4.3. Amplificação da região codificadora espA
O DNA de EPEC EAF + cepa B171 sorotipo O111:NM (Riley et al., 1990),
extraído pelo método TENS (Solução Tris, EDTA, NaOH, SDS) (Sambrook, 1989) foi
usado para amplificação da região codificadora da EspA. Foram utilizados
iniciadores específicos desenhados de acordo com pesquisa de seqüências
disponíveis no banco de seqüências do Genebank (número de acesso: AF200363)
Forward 5 ’CGC CGC GGA TCC GCG ATG GAT ACA TCA ACT GCA ACA - 3’ e
Reverse 5’ CGC CCC AAG CTT GGG TTA TTT ACC AAG GGA TAT TGC – 3’. As
regiões sublinhadas correspondem aos sítios alvos para as enzimas de restrição
Bam HI e Hind III, engenheiradas para facilitar a clonagem direcionada. O produto
da amplificação esperado seria de 579 pb de acordo com pesquisa no banco de
dados (nº acesso: AF200363). Para 50 µL da reação de PCR foram utilizados de 10-
20 ng de DNA molde, 25 pmol de cada iniciador, mistura de deoxinucleotídeos na
concentração final de 0,2 mM (Biotools), e 1U da polimerase (Biotools) diluídos em
tampão da própria enzima 1X concentrado (Biotools com 2mM MgCl2), outras
concentrações de iniciadores, dNTPs e MgCl2 foram testadas até se chegar a
25pmoles, 25mM, 2,5mM respectivamente que foram as concentrações ótimas. As
condições de amplificação foram 95 ºC, 5 min (1 ciclo); 94 ºC, 1 min, 49 ºC, 2.5 min,
72 ºC, 2.5 min (30 ciclos); 72 ºC, 15 min (1 ciclo). Ao final da PCR as amostras foram
analisadas em gel de agarose contendo 0.8% de brometo de etídio.
4.4. Purificação de fragmentos de DNA a partir de géis de agarose
A região amplificada (espA) foi retirada do gel de Agarose 0,8%, tratado com
brometo de etídio, com a utilização do Kit de extração de DNA Concert Rapid Gel
Extraction System, (GIBCO BRL, Life Technologies).
4.5. Preparação de células DH5-α e M15 (PREP4) competentes
As células foram cultivadas em Erlenmeyer contendo 50 mL de meio LB (Luria
Broth) até DO600nm= 0,5 - 0,7 e centrifugadas a 4 ºC a 2.060 x g durante 5 min; o
sobrenadante foi descartado e o pellet dissolvido em 25 mL de 50 mM CaCl2 gelado,

25
misturado gentilmente, e colocado em gelo durante 20 min. Essa mistura foi
centrifugada a 4 ºC a 2.060xg durante 5 min, o pellet dissolvido em 3,5 mL de 50
mM/ 15% glicerol/ CaCl2. Foram feitas alíquotas de 100 µL, congeladas em
nitrogênio líquido e estocados a -70 ºC.
4.6. Clonagem da região codificadora espA e transformação de E.coli DH5-α
O DNA referente ao amplicon purificado foi diretamente ligado, utilizando 0,1U
T4 ligase® (Promega), ao vetor de clonagem pGEM-T Easy® (Promega). O produto
da ligação foi utilizado para a transformação de E. coli da cepa DH5-α® (Invitrogen)
pelo método de choque térmico (3 minutos 42 ºC, 15 minutos 0 ºC). Os
transformantes foram selecionados em meio LB sólido contendo ampicilina (100
µg/mL), 0,5 mM IPTG e X- Gal (80 µg/mL). Neste sistema as colônias transformadas
com as construções positivas crescem com coloração branca, enquanto as
negativas crescem com coloração azul. A coloração branca se dá quando o gene
para a β-galactosidase é interrompido pelo inserto que possui o gene de interesse. A
β-galactosidase expressa pela colônia cliva o substrato cromogênico X-Gal presente
no meio de cultura. As colônias brancas foram selecionadas e crescidas em meio LB
líquido por 16h contendo ampicilina (100 µg / mL). O DNA das bactérias foi extraído
pelo método TENS e digerido com 0,1U da enzima Bam HI e Hind III® (Promega). O
fragmento restrito de 579 pb liberado do vetor de 3015pb foi purificado a partir do gel
utilizando Kit de extração de DNA (GIBCO BRL®).
4.7. Análise dos clones por PCR
A validação dos possíveis clones (pGEM-Teasy/espA) foi feita pela reação da
PCR , nas condições descritas no item 4.5, utilizando o DNA extraídos das colônias
brancas. As amostras positivas foram selecionadas e utilizadas para obtenção do
fragmento para clonagem no vetor de expressão. Os clones positivos foram
chamados de pLUA-04(1, 2, 4, 8 e 10).

26
4.8. Subclonagem da região codificadora espA no vetor de expressão pQE-
30
O amplicon espA purificado, de 579 pb, foi ligado ao vetor de expressão pQE-
30 ® (Qiagen) linearizado com Bam HI ® (Promega) e Hind III® (Promega).A
quantificação desses DNA foi feita em visualização eletroforética comparativa com
marcador de DNA ג hind. O vetor pQE-30 possui o gene bla que confere resistência
ao β-lactâmico ampicilina. A ligação foi feita nas proporções 1:1 e 1:3 vetor: inserto,
utilizando 0,1U de enzima ligase (®Promega, EUA) em tampão próprio da ligase, 1X
concentrado, contendo ATP. O volume da reação foi de 20 µL e a incubação a 16 ºC
por 16h. As E. coli competentes, cepa M15 (pREP4), foram transformadas, por
choque térmico, com a construção originada da ligação da região codificante
amplificada com o vetor de expressão (Sambrook, 1989). O plasmídio pREP4
contém o gene nptll que confere resistência ao antibiótico canamicina, e contém o
gene lac Iq, que codifica um repressor da expressão do operador lac, presente no
vetor de expressão pQE-30® (Qiagen). Bactérias co-tranformadas com o vetor de
expressão recombinante e pREP4 foram selecionadas em meio LB contendo
ampicilina (100 µg/mL) e canamicina (25 µg/mL). O DNA plasmidial das colônias
obtidas na transformação foi extraído pelo método TENS (Sambrook, 1989) e para a
confirmação da clonagem foi feito digestão com as enzimas Bam HI® (Promega) e
Hind III® (Promega), seguida de reação da PCR nas condições descritas no item 4.5.
Os clones positivos foram chamados de pLUA-052.6, 8.3, 8.5, 10.3 e 10.6.
4.9. Validação da clonagem por seqüenciamento
A validação do processo de clonagem foi realizada por seqüenciamento total
em ambas as direções do inserto com DNA do clone pLUA-0510.3. O
seqüenciamento foi feito por encomenda (Genemed Synthesis, Califórnia, EUA).
4.10. Teste da expressão da proteína His 6EspA
Os clones pQE-30/espA em M15 (pREP4), confirmados PCR e
sequenciamento foram testados para expressão da proteína recombinante. Colônias
isoladas foram utilizadas como pré-inóculo, cultivadas em tubo Falcon de 30 mL
contendo 5 mL de meio LB líquido com ampicilina (100 µg/mL) e canamicina (25

27
µg/mL), a 37 ºC durante 16 h, sob agitação constante, em 250 rpm. Novos inóculos
foram feitos a partir desta cultura na diluição 1:20 em erlenmeyer de 4 mL contendo
2 L de meio LB e foram crescidos até DO600nm = 0,6. Alíquotas foram retiradas a
cada hora para análise. A expressão do transgene foi induzida, durante 3h, com a
adição de 1,0 mM de IPTG, o indutor sintético que reprime lac Iq. Assim, a expressão
de Iac foi liberada permitindo a transcrição do transgene inserido no pQE-30. A
indução também foi testada utilizando 0,7 mM, 0,5 mM, 0,3 mM, 0,1 mM e 0, 05 mM
de IPTG.
A figura 9 mostra um esquema representativo do vetor de expressão pQE-30®
(Qiagen), mostrando o sítio múltiplo de clonagem com os respectivos sítios das
enzimas de restrição, onde foi inserido o gene que codifica para EspA, e uma
seqüência que codifica para um hexâmero de histidina. Os sítios das enzimas de
restrição Bam HI e Hind III, inseridos na seqüência gênica espA, flanqueiam o sítio
múltiplo de clonagem no vetor pQE-30. Amostra de material induzido e não induzido
foram analisadas,quanto a indução, em SDS-PAGE segundo Laemmli (1972).
A proteína EspA recombinante foi expressa como uma proteína de fusão a um
hexâmero de histidina e foi denominada His6-EspA. A proteína de fusão tem 206
aminoácidos, sendo que 193 são correspondentes a seqüência para EspA e 13
aminoácidos extras para facilitar a purificação em coluna de afinidade, como
ilustrado na figura 10 abaixo.

28
espA
FIGURA 9: Esquema representativo do vetor de expressão pQE-30 (Qiagen)
NH2 ATg AgA ggA TCg 6XHis ggA TCC NNN 579 pb(EspA)
FIGURA 10: Esquema representativo da sequência da proteína His6EspA
4.11. Purificação de His6EspA expressa em células M15 por cromatocrafia de
afinidade em Ni-NTA-Agarose
Após o período de indução, a cultura foi centrifugada a 2800x g por 30 min a
4º C. O meio foi descartado, o pellet ressuspendido em 20-40 mL de tampão de lise
desnaturante (10 mM Tris pH 8,0, 100 mM NaH2PO4, 7 M Uréia, 2 M Tiouréia) e
levado para ultrasom (10 Watts (RMS)). O lisado foi centrifugado a 11.600x g
durante 5 min e o sobrenadante transferido para outro tubo. Amostras de 5 mL de
Ni-Sepharose (50% resina e 50% de etanol a 30%) foram adicionados em coluna
Bam HI
13 aminoácidos de fusão
*
*

29
de 15 mL. Esta foi lavada três vezes com 1 mL de tampão de equilíbrio (10 mM Tris
pH 8,0, 100 mM NaH2PO4, 7 M Uréia, 2 M Tiouréia) deixando o lisado em contato
com a resina durante 40 min, sob agitação leve, a temperatura ambiente. A
quantidade de His6EspA induzida foi determinada previamente em SDS-PAGE
(Laemmli, 1970), e 1 mL do lisado foi passado na coluna por vez, uma vez que a
capacidade da coluna é de 5-10 mg de proteína por mL de resina. Após a coleta do
material não retido a coluna foi lavado com intuito de eliminar as proteínas
contaminantes. A lavagem foi feita 10 vezes com 5 mL de tampão de lavagem (10
mM Tris pH 8,0, 100 mM NaH2PO4, 7 M Uréia, 2 M Tiouréia) e 5 vezes com tampão
de lavage (10 mM Tris pH 8,5, 100 mM NaH2PO4, 7 M Uréia, 2 M Tiouréia) . Após as
lavagens o material ligado à coluna foi eluído com tampão de eluição (10 mM Tris
pH 4,5, 100 mM NaH2PO4, 7 M Uréia, 2 M Tiouréia). Todos os lavados e eluídos
foram coletados de 5 em 5 mL. Imediatamente à eluição, a resina foi lavada três
vezes com 5 mL da solução de lavagem após uso em condições desnaturantes e
guardada em etanol 30% a 4ºC. A proteína purificada passou pelo processo de
diálise para dessalinização (4 vezes em 4L de solução de diálise) e por
concentração pelo processo de sublimação (liofilização). Todas as amostras
coletadas foram aplicadas em gel desnaturante de acrilamida (SDS-PAGE) a 15%
para visualização da proteína His6EspA. A proteína purificada foi quantificada por
ácido biscínconínico (Redinbaugh e Turley, 1986) e por densitometria ( Bozzo e
Retamal , 1991).
4.12. Produção de anticorpos policlonais anti-His6EspA em coelho
A imunização de dois coelhos albinos, pesando cerca de 2 Kg cada, para a
produção de anticorpos policlonais contra a proteína His6EspA seguiu o esquema de
imunização descrito por Harlon e Lane (1998). A primeira imunização foi realizada
com 150 µg de proteína, emulsificada com 500 µL de adjuvante completo de
Freünd® (Sigma-Aldrich, EUA). As aplicações foram subcutâneas na região cervical
do animal. Após 15 e 30 dias foram feitos reforços aplicando a quantidade de 100 µg
antígeno emulsificado com 500 µL de adjuvante incompleto de Freünd® (Sigma-
Aldrich, EUA) . Após 15 dias do último reforço foi feita a sangria total dos animais via
punção cardíaca, por um Veterinário. O sangue foi coagulado, centrifugado, e o soro

30
coletado foi aliquotado e estocado a -20º C para uso posterior em testes de
interação antígeno-anticorpo.
4.13. Purificação de IgG anti-His 6EspA de coelho
A IgG do soro de coelho pré-imunizado e imunizado foi purificada em coluna
de afinidade de proteína A (segundo as especificações do fabricante -® Sigma-
Aldrich, EUA) utilizando o Kit Protein A Antibody Purification KitR . A IgG total
purificada foi quantificada por ácido biscinconínico (Redinbaugh e Turley, 1986).
4.14. Testes de antigenicidade da His6EspA
A antigenicidade da His6EspA foi avaliada por Western blot/imunoblot
segundo metodologia descrita (Towbin et al., 1979). A proteína EspA (4 µg)
recombinante purificada por afinidade foi fracionada por SDS-PAGE (Laemmli, 1970)
e eletrotransferidas para membrana de nitrocelulose de 0,22 µm (Bio-Rad
Laboratories, Richmond, EUA). As membranas foram bloqueadas com 1% de leite
desnatado em tampão TST-20 (Tris, Sal (NaCl), Tween-20)(ANEXO) por 16 h a 4 ºC.
Os anticorpos utilizados para detectar His6EspA foram: monoclonal anti-His,
anticorpos policlonais anti-EspA produzidos em coelho (nesse trabalho) e soros (n=
100) de pacientes pediátricos assintomáticos diluídos em TST-20 com 0,3% de leite
desnatado, a 1:6.000, 1:10.000, 1:500, respectivamente. As membranas foram
incubadas com os anticorpos diluídos durante 4 h. Os reagentes para detecção dos
anticorpos imobilizados foram proteína A conjugada a peroxidase (Amersham
Pharmacia Biotech, EUA), na diluição 1:6.000, anticorpo monoclonal de coelho anti-
IgG humana conjugada a peroxidase (® Sigma-Aldrich, EUA ) e anticorpo
monoclonal de coelho anti-IgA humana conjugada a peroxidase (® Sigma-Aldrich,
EUA), na diluição de 1:40.000, em TST-20 com 0,3% de leite desnatado. O tempo
de incubação foi de 2 h. Entre cada etapa as membranas foram lavadas 3 vezes
com TST-20. As membranas foram reveladas pela adição de uma solução do
cromógeno diaminobenzidina (DAB) (® Sigma-Aldrich, EUA). A reação foi bloqueada
com água destilada.
A antigenicidade da His-EspA foi avaliada por testes de ELISA segundo
metodologia descrita (Coligan, 1991). No ensaio de ELISA, utilizando placa de 96

31
poços, foram incubados 50 µL/poço de uma solução de 2 µg/mL da proteína
His6EspA recombinante em tampão carbonato/bicarbonato (ANEXO), durante 16 h a
4º C. Após a sensibilização, a placa foi bloqueada com 1% de leite em pó desnatado
(diluído em PBS 1X, Tween-20 0,05%) por uma hora a 37º C. Logo após foram
adicionados os soros (anti- HisEspA recombinante) dos diferentes coelhos diluídos
em log 2 a partir de 1:100 até 1:3,276,800 e de pacientes de 0 a 2 anos de idade
diluídos a 1:500, ambos em 0,2% de leite/Tween-20 0,05% em PBS e incubados por
duas horas. Foi utilizada proteína A conjugada a peroxidase, na diluição de
1:30.000, ou IgG de cabra anti-IgG de coelho conjugada a peroxidase, na diluição
1:40.000. Para os soros humanos foram utilizadas IgG de coelho anti-IgG e IgA
humana conjugada a peroxidase. A revelação ocorreu após 25 min, na presença de
2 mM de orto-fenil-diamina (® Sigma-Aldrich, EUA) e 0,006% de peróxido de
hidrogênio. A reação foi parada com a adição de 20 µL de 0,1 N de ácido sulfúrico. A
leitura das reações colorimétricas foi realizada em leitor de placas Multiskan,
Labsystems Uniscience, a 492 nm. Para esse ensaio consideramos o End Point
sendo diluição que apresenta uma D.O. duas vezes maior a do controle negativo.
4.15. Ensaio de hemólise e inibição de hemólise
A atividade hemolítica foi determinada como descrito em (Shaw et al., 2001;
Ribeiro e Medina-Acosta, 2003) com adaptações. O sangue de um indivíduo
saudável foi coletado em vacuum tube contendo heparina como anticoagulante. As
hemácias foram sedimentadas por centrifugação por 10 min 2.000 x g 20 ºC e
lavadas três vezes em PBS (Tampão fosfato salino) (ANEXO) de alta força iônica ou
IGP (Tampão Isotônico Glicose Fosfato) (ANEXO) de baixa força iônica. As células
foram reconstituídas em PBS ou em IGP para 20% (v/v) volume de célula
empacotada. Essa suspensão foi diluída 1:20 em PBS ou em IGP, e desta
suspensão 100 µL/ poço a uma placa contendo 96 poços .
As EPEC (EAF+) B171 (O111:NM) (cepa patogênica para humano), REPEC-1
(cepa patogênica para coelho) e DH5-α (Invitrogem) (cepa não patogênica) foram
crescidas em DMEM contendo 10% de soro fetal bovino inativado, a 37 ºC durante
16 h sob agitação de 170 rpm. Um mililitro de cada cultura foi transferido para tubo
de microcentrífuga e foram sedimentadas por centrifugação por 10 min a 2.000 x g a

32
20ºC. As células foram lavadas com PBS ou IGP e recontituídas em 500 µL de PBS
ou IGP. Dessa solução 100µL/poço foram adicionados em placa de 96 poços
juntamente com as hemácias para indução de hemólise. A cepa de E. coli DH5-α, foi
utilizada como um dos controles negativo. Um segundo controle negativo foi feito
com hemácia e PBS ou hemácia e IGP. O controle de hemólise total foi feito
incubando hemácias com 1% de Tween-20 em água destilada.
Para os testes de inibição da hemólise 50 µg ou 100 µg de IgG de coelho
imunizados com EspA, purificada utilizando o Kit Protein A Antibody Purification KitR
(Sigma Co., St. Louis, EUA), foram incubadas com as bactérias durante 25 min
antes de acrescentar as hemácias diluídas em PBS ou IGP.
Todas as amostras foram incubadas durante 1 h a 37 ºC e centrifugadas
durante 5 min por 1.500 x g a 20 ºC. O sobrenadante foi transferido para outros
poços e a absorbância medida a 405nm. Todas as amostras foram feitas em
duplicatas.
A atividade hemolítica foi calculada como porcentagem, usando a fórmula:
% de hemólise=[ valor da média D.O. 405nm da amostra teste] - [valor da média D.O.
405nm com tampão] / [valor da média D.O. 405nm amostra tratada com Tween-20] -
[valor da média D.O. 405nm com tampão] X 100%.

33
5. RESULTADOS
5.1. Amplificação da região codificadora para EspA
Utilizamos DNA extraído de EPEC (EAF+) B171 (O111:NM) para amplificação
da região codificante da EspA. Foram utilizados iniciadores específicos, dNTPs e
MgCl2 em diferentes concentrações até se encontrar a melhor, que foi 25pmoles,
25mM, 2,5mM respectivamente. Foi usado o pQE-30 como controle negativo e o
megaplasmídio, que contém a região codificadora bfpA, como controle positivo.
Para visualizar o resultado os 25 µL da reação da PCR foram aplicados em gel
de agarose 0,8% com brometo de etídio (Figura 11 A). Após estabelecimento das
condições ótimas, a PCR foi repetida para ter o fragmento em grande quantidade
para a clonagem (Figura 11 B).
Figura 11: Visualização eletroforética da amplificação da espA
Gel de agarose 0,8% contendo 0,03% de brometo de etídio. A: Amplificação da
espA com iniciadores específicos. M: marcador λ Hind de 564 pb a 23.000 pb; 1:
espA amplificada; 2: controle negativo pQE-30; 3: controle positivo com pLUA-013 4:
controle positivo com DNA de EPEC. B: 1-8 amostras da amplificação da espA em
grande quantidade para a ligação ao pGEM- Teasy. M: marcador 1kb. As setas
indicam a posição relativa do amplicom de 579pb.
579pb 579pb
1 2 3 4 5 6 7 8 M A B M 1 2 3 4

34
5.2. Ligação do Amplicon no vetor de clonagem
Foi realizada a ligação do amplicom ao vetor pGEM-Teasy, (®Kit Promega).
Células DH5-α competentes foram transformadas com o produto da ligação e
plaqueadas em meio LB sólido contendo ampicilina 100 µg/mL, IPTG 80µg/Ml e X-
Gal 16 mM para selecionar as colônias brancas. As colônias selecionadas foram
crescidas em 5mL de meio LB líquido contendo ampicilina 100 µg/mL por 16 h. O
DNA de 10 colônias brancas foi extraído pelo método de TENS.
5.3. Validação dos possíveis clones (pGEM-Teasy/espA)
Foi feita uma PCR para validação dos possíveis clones (pGEM-Teasy/espA) e
seis dos dez possíveis clones (1, 2, 3, 4, 8 e 10) foram positivos para espA (Figura
12).
Foi realizada uma digestão com as enzimas Bam HI ®(Promega) e Hind
III®(Promega) dos DNA das colônias (1, 2, 3, 4, 8 e 10) positivas por PCR para
linealizar o vetor de clonagem e deixar o inserto espA com extremidades coesivas,
prontas para serem ligadas no vetor de expressão pQE-30. Um gel de agarose 0.8%
foi feito e nele aplicado toda a amostra das digestões. Foi possível visualizar a
liberação do fragmento correspondente a espA somente na digestão dos DNA das
colônias 1, 2, 4, 8, e 10, de onde a banda correspondente ao inserto espA foi
extraída do gel utilizando o Kit da Gibco (Figura 13). As colônias positivas receberam
o nome de pLUA-041, 2, 4, 8 ou 10.
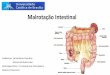
35
Figura 12: Visualização eletroforética do produto da PCR para validar clones
pGEM/espA
Gel de agarose 0,8% contendo 0,03% de brometo de etídio. M: marcador λ Hind de
564 pb a 23.000 pb; 1-10: DNA de colônias 1-10 de pGEM/espA; C+: controle
positivo com DNA de EPEC; C- : controle negativo com pQE-30.
Figura 13: Visualização eletroforética da digestão para confirmação de clones
pGEM/espA e purificação de espA.
Gel de agarose 0,8% contendo 0,03% de brometo de etídio. M: marcador λ Hind de
564 pb a 23.000 pb; 1-7: DNA de clones pGEM/espA positivos por PCR e digeridos
com Bam HI e Hind III .
579pb
500pb
M 1 2 3 4 5 6 7
M 1 2 3 4 5 6 7 8 9 10 C+ C-
579pb

36
5.4. Preparação do vetor de expressão pQE-30
Células DH5- α competentes foram transformadas com pQE-30, plaqueadas
em meio LB contendo ampicilina (100 µg/mL). Algumas colônias foram selecionadas
e crescidas em 5mL de meio LB com ampicilina (100 µg/mL) por 16 h.
Os DNAs dessas colônias foram extraídos pelo método de TENS. Foi feita a
digestão do pQE-30 extraído, com as enzimas Bam HI ®(Promega) e Hind
III®(Promega). Depois da digestão foi feita uma precipitação utilizando 0.3M de NaCl
e 2,5 V de etanol e um gel de agarose 0.8% foi feito para visualização do vetor
extraído, digerido e precipitado (Figura 14).
Figura 14: Visualização eletroforética da digestão e precipitação do vetor de
expressão pQE-30.
Gel de agarose 0,8% contendo 0,03% de brometo de etídio. M: marcador de 100pb
a 1517pb. 1 e 2 vetor pQE-30 linearizado pelas enzimas Bam HI e Hind III, e
precipitado.
Vetor pQE-30 linear
M 1 2
3500pb
500pb

37
O inserto espA obtido da digestão de clones positivos pGEM/espA foi
purificado do gel pelo Kit da Qiagen. Após a purificação, um gel de agarose
0.8% foi feito para quantificação do vetor e inserto purificado (Figura 15).
Figura 15: Visualização eletroforética do vetor de expressão pQE-30 e o inserto
espA com extremidades Bam HI e Hind III purificados.
Gel de agarose 0,8% contendo 0,03% de brometo de etídio. 1: pQE-30 linearizado;
2-6: espA purificada de 5 diferentes clones pLUA-04(1, 2, 4, 8 e 10); M: marcador de
100pb a 1517pb.
5.5. Ligação do inserto espA no vetor de expressão Uma vez digeridos com as enzimas Bam HI e Hind III, o inserto extraído do
gel via Kit, e o vetor pQE-30 precipitado foram quantificados em torno de 5 ng/µL e
40 ng/µL, respectivamente. Diferentes reações de ligação vetor – inserto foram
feitas: 1:1 e 1:3 com o fragmento espA purificado dos clones pLUA-04(1, 2, 4, 8 e 10),
sendo a concentração do vetor foi sempre de 50ng/µL. A reação de autoligação
(sem inserto) também foi realizada como controle.
As E. coli da cepa M15 foram transformadas por choque térmico, com o
produto de cada ligação, plaqueadas em meio LB contendo ampicilina (100 µg/mL) e
canamicina (25 µg /mL) e crescidas por 16 h a 37 ºC.
3400pb 1 2 3 4 5 6
579pb
1 2 3 4 5 6 M

38
5.6. Validação dos clones por PCR
Uma PCR foi também realizada com o DNA dos clones pLUA-052.6, 8.3, 8.5, 10.3 e
10.6. O DNA de EPEC foi utilizado como controle positivo e pQE-30 foi usado como
controle negativo (Figura 16).
FIGURA 16: Visualização eletroforética da reação da PCR para validação da
clonagem de espA no vetor pQE-30.
Gel de Agarose 0.8% contendo 0,03% de brometo de etídio. M: marcador λ Hind de
564 pb a 23.000 pb. Nos poços 1-5 DNA de cinco clones; C-: controle negativo com
pQE-30; C+: controle positivo com DNA de EPEC.
5.7. Teste da expressão da proteína His6espA
Trinta colônias foram selecionas, crescidas em meio LB contendo ampicilina
(100 µg/mL) e canamicina (25 µg/mL) e incubadas por 16 h a 37 ºC.
Como as células M15 contém o plasmídio pREP4 (que contem um gene para
resistência a canamicina), juntamente com pQE-30/espA (que contem gene para
resistência a ampicilina), colônias que crescerem em meio contendo os dois
antibióticos possuem ambos os plasmídios. Cultivos positivos para ambos os
plasmídios, foram incubadas com 1 mM de IPTG para a expressão da proteína
recombinante EspA. Cinco cultivos expressaram EspA recombinante e foram
M 1 2 3 4 5 C- C+
579pb

39
chamados de pLUA-052.6, 8.3, 8.5, 10.3 e 10.6 (Figura 17). Esse mesmo procedimento de
indução da expressão foi repetido, porém utilizando diferentes concentrações do
indutor IPTG, a fim de encontrar a sua concentração mínima ainda capaz de induzir,
e dessa forma evitar desperdício (Figura 18). O DNA desses cultivos foi extraído
pelo método TENS e armazenados para fim de manutenção. Foi realizado um
ensaio de Western blot com o lisado total das colônias positivas, utilizando anticorpo
monoclonal anti-hexâmero de histidina como anticorpo primário (Figura 19).
FIGURA 17: Visualização eletroforética da expressão da proteína EspA
recombinante
SDS-PAGE com gel de acrilamida/bisacrilamida a 15% corado com azul de
Comassie R-250. Nas raias 1, 3, 5, 7 e 9 são amostras de lisado de cultura não
induzida; nas raias 2, 4, 6, 8 e 10 são amostras de lisado de cultura induzida durante
3h com IPTG. A seta representa a banda de proteína EspA super expressa.
25 kDa
1 2 3 4 5 6 7 8 9 10

40
NI 1 2 3 4 5 6
Induzidos
25 kDa
NI 1 2 3 4 5 6
Induzidos
25 kDa
FIGURA 18: Visualização eletroforética da indução da expressão da proteína
EspA recombinante com diferentes concentrações de IPTG
SDS-PAGE com gel de acrilamida/bisacrilamida a 15% corado com azul de
Comassie R-250. NI: referente ao lisado não induzido, 1-6: referente aos lisados
induzidos, respectivamente, com 1 mM, 0,7 mM, 0,5 mM, 0,3 mM, 0,1 mM e 0,05
mM de IPTG.

41
FIGURA 19: Detecção de EspA recombinante por Western blotting
Lisado total de proteína EspA recombinante foi aplicada num gel de acrilamida 15%
e transferido para membrana de nitrocelulose onde IgG anti-His reconheceu o
hexâmero de histidina fusionado à proteína EspA. M: marcador low range (BioRad);
1, 3, 5, 7 e 9: são amostras de lisado de cultura induzidas durante 3 h com IPTG; 2,
4, 6 e 8: são amostras de lisado de cultura não induzida. A seta indica a proteína
EspA super expressa.
20,7
M (kDa) 1 2 3 4 5 6 7 8 9 107
81
48,7
33,8
27
20,7

42
5.8. Validação do clone pLUA-0510.3 por sequenciamento
A partir do resultado da PCR foram feitas extrações de DNA para se obter
quantidades suficientes, e de boa qualidade, a serem precipitados e enviados para
sequenciamento. O sequenciamento foi feito por encomenda (Genemed Synthesis,
Califórnia, EUA). O clone pLUA-0510.3 foi seqüenciado totalmente, em ambas as
direções do inserto (Figura 20).
CCCTTCCGGCGGAACATTTCACACAGAATTCATTAAAGAGGAGAAATTAACTATGATGATGATGAGAGGATCGCCCCATCACCATCACCATCACATCACCATCACCATCACATCACCATCACCATCACATCACCATCACCATCACGGATCCGGATCCGGATCCGGATCCGCGGCGGCGGCGATGGATACATCAACTGCAACATCAGTTGCTAGTGCGAACGCGAGTACTTCGACATCGACAGTCTATGACTTAGGCAGTATGTCGAAAGACGAAGTAGTTCAGCTATTTAATAAAGTCGGTGTTTTTCAGGCTGCGCTTCTCATGTTTGCCTATATGTATCAGGCACAAAGCGATCTGTCGATTGCAAAGTTTGCTGATATGAATGAGGCATCTAAGGAGTCAACCACAGCCCAAAAAATGGCTAATCTTGTGGATGCTAAAATTGCTGATGTTCAGAGTAGTTCTGACAAGAATAAGAAAGCCAAACTTCCTCAAGAAGTGATTGACTATATAAATGATCCTCGCAATGACATTACAGTAAGTGGTATTAGCGATCTAAATGCTGAATTAGGCGCTGGTGATTTGCAAACGGTGAAGGCCGCTATTTCGGCCAAATCGAATAACTTGACCACGGTAGTGAATAATAGCCAGCTTGAAATACAGCAAATGTCAAATACGTTAAACCTATTAACGAGTGCACGTTCTGATATTCAGTCACTGCAATACAGAACTATTTCAGCAATATCCCTTGGTAAATAACCCAAGCTTAAGCTTAAGCTTAAGCTTAATTAGCTGAGCTTGGACTCCTGTTGATAGATCCAGTAATGAC
CTCAGAACTCCATCTGGATTTGTTCAGAACGCTCGGTTGCCGCCGGGCGTTTTTTATTGGTGAGAATCCAAGCTAGCTGGGCGAGATTTTCAGGAGCTAAAGGAAGCTAAAATGGAGAAAAAAATCACTGGATATACCACCGTTGATATATCCCAATGGCATCGTAAGAACATTTTGAGGCATTTCAGTCAGTTGCTCATGTACCTATACCAGACCGTTCAGCTGGATATACGCTTTTTAAGACCGTAAAGAAAAATAGCACAGTTTATCGGCTTTATCACATCTGCCCGCTGATGATGCTCATCGATTTCGTATGGCATGAGGACGGTGAGCTGTGTAATTGGATAGTGTTCACCTTGTTCACCGTTTCATGGAGCACTGGAACGTTTCATCGCCTTGATGTAATCCCACGACGATTCGGCGCGAACTTATGTC
FIGURA 20: Seqüência total do clone pLUA-05 10.3
A região em vermelho esta indicando a seqüência parcial do vetor pQE-30; em preto
o códon inicial; em lilás o hexâmero de histidina; em verde o sítio para restrição Bam
Hl; em alaranjado o sítio para restrição Hind III; em roxo um aminoácido adicional
marcador de identificação; e em azul a seqüência referente a EspA.

43
Ao desenhar os iniciadores, tomamos como referência a seqüência de espA
da REPEC sorotipo “O103:H2”, porém, a reação da PCR foi realizada utilizando o
DNA da EPEC humana (B171) sorotipo “O111:NM”. Comparando o resultado desse
sequenciamento com a seqüência de espA da REPEC (“O103:H2”) vimos que
possui 100% de homologia com a espA de EPEC humana (“O111:NM”).
5.9. Purificação da EspA recombinante em coluna de Ni- Sepharose
Foi montada uma coluna com 5 mL de resina Ni-Sepharose que foi
equilibrada com 10 mL de tampão de lise pH = 8,0. Foi aplicado 1 mL do lisado total
acrescido de 1mL do tampão de lise pH 8,0 e incubado durante 40 min. Uma única
porção de material não retido foi coletada. A primeira lavagem foi feita com 50 mL
(10x de 5 mL) de tampão de lise pH 8,0. A segunda lavagem foi feita com 25 mL (5x
de 5 mL) de tampão pH 8,5, e a eluição foi feita com 50 mL de tampão pH 4,5 (10x
de 5 mL) (Figuras 21 e 22).
FIGURA 21: Visualização da purificação da proteína EspA recombinante em
coluna de afinidade Ni- Sephaose.
SDS-PAGE com gel de acrilamida/bisacrilamida a 15% corado com azul de
Comassie R-250. Raia 1: lisado total; 2: não retido; 3: lavado em pH 8,0; 4: lavado
em pH 8,5; 5-10: Eluído 1, 2, 3, 4, 5 e 6 em pH 4,5.
1 2 3 4 5 6 7 8 9 10
25 kDa
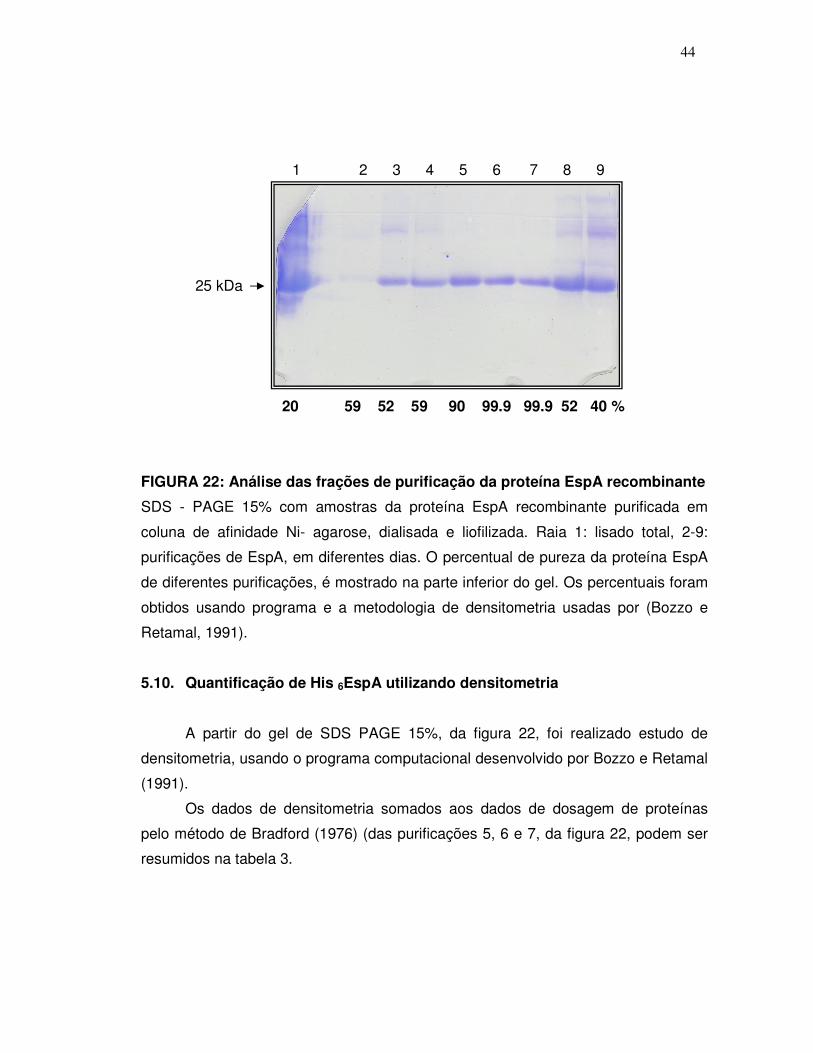
44
20 59 52 59 90 99.9 99.9 52 40 %
FIGURA 22: Análise das frações de purificação da proteína EspA recombinante
SDS - PAGE 15% com amostras da proteína EspA recombinante purificada em
coluna de afinidade Ni- agarose, dialisada e liofilizada. Raia 1: lisado total, 2-9:
purificações de EspA, em diferentes dias. O percentual de pureza da proteína EspA
de diferentes purificações, é mostrado na parte inferior do gel. Os percentuais foram
obtidos usando programa e a metodologia de densitometria usadas por (Bozzo e
Retamal, 1991).
5.10. Quantificação de His 6EspA utilizando densitometria
A partir do gel de SDS PAGE 15%, da figura 22, foi realizado estudo de
densitometria, usando o programa computacional desenvolvido por Bozzo e Retamal
(1991).
Os dados de densitometria somados aos dados de dosagem de proteínas
pelo método de Bradford (1976) (das purificações 5, 6 e 7, da figura 22, podem ser
resumidos na tabela 3.
1 2 3 4 5 6 7 8 9
25 kDa

45
TABELA 4: Demonstrativo da purificação da proteína EspA recombinante
5 6 7 Lisado
Proteína solúvel a 2,65 mg 2,35 mg 2,25 mg 3,72 mg
Produção 71,2% 63,2% 60,5% ----
Rendimento b 17,7 mg/g
peso úmido
15,7 mg/g
peso úmido
15 mg/g
peso úmido
25 mg/g
peso úmido
Eficiência 70,8% 62,8% 60% ----
Homogeneidade 99,9% 99,9% 90% ----
a Proteína total por mL. b A partir de 4 L de cultura foi obtido 2,4 g de peso úmido de
células que foi diluído em 16 mL de tampão de lise. A quantidade relativa de proteína
total solúvel presente em 1 mL é 18,6 mg. Os cálculos foram realizados considerando
um nível de expressão de 20% que representa 3,72 mg do lisado total.
5.11. Imunização de coelhos com proteína EspA recombinante
A proteína EspA recombinante, 99,9% homogênea, foi utilizada para o
processo de imunização.
5.12. Avaliação da especificidade do anticorpo policlonal de coelho contra
EspA recombinante
Para avaliar a especificidade do anticorpo policlonal anti-EspA produzido em
coelhos foi sensibilizada uma placa de ELISA, com 2 µg/mL de EspA recombinante e
incubada com o anticorpo policlonal anti-EspA de coelho nas diluições em log 2 a
partir de 1:100 até 1:3,276,800. Consideramos o anticorpo ainda reativo (End Point)
numa diluição com Densidade Óptica (D.O.) duas vezes a do controle negativo (soro
pré-imune), e o título foi de 1:400,000.
Com o objetivo de visualizar a reatividade do anticorpo de coelho contra
EspA, 4 µg de EspA recombinante foi aplicada por poço de um gel SDS – PAGE
15% e transferida para uma membrana de nitrocelulose. Essa membrana foi cortada
em tiras, uma foi incubada com IgG de coelho anti-His, (controle positivo), e a outra

46
foi incubada com IgG de coelho anti-EspA, ambas durante 3 h. Por último, a
membrana foi incubada com proteína A peroxidase por mais 2 h e a reação foi
revelada. Anticorpo anti-EspA produzido em coelho mostrou ser reativo para EspA
recombinante, e na mesma intensidade do controle positivo (Figura 23).
1 2 3 4 5 6
25 kDa
1 2 3 4 5 6
25 kDa
FIGURA 23: Avaliação da reatividade do anticorpo anti-EspA produzido em
coelho.
Uma montagem para comparação onde 1 e 2 são SDS-PAGE 15% corado com
Comassie blue. 1: lisado bacteriano, 2: Proteína EspA recombinante pura, 3, 4, 5:
proteína EspA pura transferida para membrana de nitrocelulose, 6: lisado bacteriano
transferido para membrana de nitorcelulose. 3: proteína EspA recombinante pura,
anticorpo monoclonal anti-Hexâmero de Histidina na diluição 1:6.000, 4: proteína
EspA recombinante pura, anticorpo anti-EspA produzido em coelho na diluição
1:10000, 5: Proteína EspA recombinante pura, soro de paciente pediátrico na
diluição de 1:500, 6: lisado bacteriano, anticorpo anti-EspA produzido em coelho na
diluição 1:10.000.

47
5.13. Estudo da soro prevalência de IgG e IgA séricas anti-EspA de crianças
de 0 a 2 anos.
Para avaliar a soro prevalência de IgG e IgA séricas, placas de ELISA foram
sensibilizadas com 2 µg/mL de EspA recombinante, e incubada com soro de 100
crianças de 0 a 2 anos numa diluição de 1:500. Consideramos soro positivo para
EspA o soro que na diluição 1:500 teve D.O. duas vezes a do controle negativo (sem
anticorpo primário). Dos 100 soro analisados 75 foram positivos para IgG e 23 para
IgA (Figura 24).
0 20 40 60 80 100
0-6 meses
6-12 meses
12-18 meses
18-24 meses
0-24 meses
Fai
xa E
tári
a (m
eses
)
Porcentagem (%)
IgA
IgG
FIGURA 24: Soro prevalência de IgG e IgA séricas anti-EspA recombinante em
lactentes.
O gráfico mostra a porcentagem de crianças soro positivas para IgG e IgA anti-EspA
em diferentes faixas etárias.
23/100 75/100
40/49 10/49
5/12 7/12
27/36 8/36
1/3
0/3

48
Depois de realizada a seleção dos soros positivos para EspA recombinante,
foi avaliada a especificidade desses soros para EspA através da sua titulação. Mais
placas de ELISA foram sensibilizadas com 2 µg/mL de EspA recombinante e 75
soros positivos para IgG e 23 soros positivos para IgA foram diluídos 1:100 a
1:3.200. Foi considerado ainda positivo o soro diluído que apresentou uma D.O duas
vezes maior a do controle negativo chamado de “End Point”. A reatividade dos soros
nas outras diluições foram consideradas End Point de 1 a 6 de acordo com o grau
da reatividade com EspA recombinante (Figuras 25 e 26).
ELISASoroprevalência IgG anti-EspA
0
1
2
3
4
5
6
7
0 6 12 18 24 30
Faixa Etária (meses)
En
d P
oin
ts
FIGURA 25: Especificidade da IgG anti-EspA
O gráfico mostra a relação da especificidade da IgG anti-EspA com a faixa etária dos
pacientes soro positivos.

49
ELISA Soroprevalência de IgA anti-EspA
0
1
2
3
4
5
6
0 5 10 15 20 25 30
Faixa Etária (meses)
En
d P
oin
ts
FIGURA 26: Especificidade da IgA anti-EspA
O gráfico mostra a relação da especificidade da IgG anti-EspA com a faixa etária dos
pacientes soro positivos.
Para podermos visualizar a especificidade de soros de crianças de 0 a 2 anos anti-
EspA recombinante foram feitos dois géis preparativos de acrilamida contendo
aproximadamente 4 µg de EspA recombinante e transferidos para membranas de
nitrocelulose. Depois de bloqueadas com 1% de leite bovino, foram secas e cortadas
em tiras. Os soros que no ELISA foram mais positivos para IgG ou IgA foram
incubados com diferentes tiras. A IgG anti-Human foi utilizado como anticorpo
secundário (Figuras 27 e 28).

50
FIGURA 27: Imunodetecção de IgG anti-EspA de crianças assintomáticas.
SDS – PAGE 15% preparativo, contendo HisEspA purificada, foi transferida a uma
membrana de nitrocelulose, depois cortada em tiras. 1-12: soros das crianças (1, 2,
9, 13, 17, 41, 73, 74, 81, 85, 89, 99) de até 2 anos de idade (1:500) e revelado com
IgG de cabra anti-IgG humano conjugada a peroxidase (1:40.000).
1 2 3 4 5 6 7 8 9 10 11 12
+ + + + + + + + + + + +
25 kDa

51
FIGURA 28: Imunodetecção de IgA anti-EspA de crianças assintomáticas.
SDS – PAGE 15% preparativo contendo HisEspA purificada foi transferida para
membrana de nitrocelulose, depois cortada em tiras. 1-12: soros das crianças (4, 5,
6, 9, 15, 17, 41, 45, 51, 54, 74, 81) de até 2 anos de idade (1:500) e revelado com
IgG de cabra anti-IgA humano conjugada a peroxidase (1:40.000).
1 2 3 4 5 6 7 8 9 10 11 12
+ - + + + + + + + + + -
25 kDa

52
5.14. Ensaio de hemólise e inibição da hemólise
TABELA 5: Porcentagem de hemólise de hemácias com diferentes tratamentos
EPEC REPEC DH5-α (c-)
Hemácia+IGP 47% 43,2% 2,5%
Hemácia+PBS 0,85% 1,2% 4,7%
Hemácia+IGP+50µg IgG anti-EspA
38,1% 38,5% ----
Hemácia+IGP+100µg IgG anti-EspA
4,6% 5,5% ----
Hemácia+PBS+50µg IgG anti-EspA
1% 0,3% ----
Hemácia+PBS+100µg IgG anti-EspA
0,06% 0,26% ----
Hemácia+IGP+100 µg
IgG pré-imune
---- -0,09 ----
Hemácia+PBS+100 µg
IgG pré-imune
---- 0,9% ----
As cepas de EPEC e REPEC utilizadas nesses ensaios mostraram induzir a
lise de hemácias quando em solução com IGP (baixa força iônica), e lise
extremamente baixa quando em solução com PBS, pois, em baixa força iônica a
aproximação da bactéria (dos filamentos de EspA) com as hemácias fica facilitada,
sendo o ensaio mais sensível. A cepa de E. coli DH5-α foi utilizada como controle
negativo por não ser patogênica, portando, não possui filamentos de EspA, que é o
que medeia a lise (Shaw et al., 2001).
O teste de inibição da hemólise se baseia em inibir o contato dos filamentos
de EspA utilizando anticorpos anti-EspA produzidos em coelho. Ao utilizarmos 50 µg
de IgG anti-EspA, a inibição da hemólise foi apenas de 8,9% em comparação ao
ensaio somente com EPEC, e de 4,7% em comparação ao ensaio somente com
REPEC. Com utilização de 100 µg de IgG anti-EspA, houve inibição de 42,4% em
comparação ao ensaio somente com EPEC, e de 37,7% em comparação ao ensaio
somente com REPEC.
Contudo, ao utilizarmos IgG pré-imune como controle negativo da inibição, a
hemólise não foi observada.

53
6. Discussão
Dada a alta morbidade e mortalidade causada por EPEC, principalmente em
crianças de até 2 anos de idade, existe interesse em desenvolver estratégias para
prevenir, diagnosticar e tratar pessoas acometidas por EPEC.
Uma estratégia que vem sendo estudada é o desenvolvimento de uma vacina
baseada em fatores de virulência de EPEC e EHEC. Para isso, a identificação e
caracterização desses antígenos devem ser estudadas no sentido de determinar sua
antigenicidade e imunogenicidade.
Com o objetivo de tornar a profilaxia, o diagnóstico e o tratamento da doença
diarréica causada por EPEC mais rápidos e eficazes, focalizamos nossos estudos
na proteína EspA que está presente tanto em EPEC quanto em EHEC formando a
agulha do sistema de secreção do tipo III. Ela é essencial para translocar proteínas
efetoras para a membrana e citoplasma das células hospedeiras e para
desencadear a transdução de sinal, adesão íntima, redução local das
microvilosidades e formação de um pedestal, que juntos caracterizam a lesão
patognomônica A/E.
Acredita-se, que ao bloquear a proteína EspA, a lesão A/E será inibida e,
conseqüentemente, a diarréia, como demonstrado in vitro para EspA e in vivo para
intimina por Girard e colaboradores (2006).
Alguns trabalhos avaliaram a resposta imune para EspA, utilizando para isso
extratos totais de EPEC (Loureiro et al., 1998; Martinez et al., 1999 ), extratos
bacterianos expressando proteínas recombinantes de EHEC (Li et al., 2000) e EPEC
(Loureiro et al., 1998) e proteínas recombinantes de EHEC semi purificadas ou
purificadas (Li et al., 2000; Noguera-Obenza et al., 2003).
No nosso trabalho foi utilizado o sistema de expressão pQE-30 para produzir
EspA recombinante acrescida da seqüência NH2-fMet-Arg-Gly-Ser-(His6)-Gly-Ser-
NNN para facilitar a purificação em coluna de afinidade (Figuras 9 e 10 ). O nível de
expressão obtido foi de 20% da proteína EspA em relação ao total de proteínas do
lisado bacteriano, similar aos resultados do nosso grupo para BfpA e EspB (Flores et
al., 2002). A eficiência do processo de purificação chegou a 70,8% , com pureza de
90 a 99,9% em diferentes purificações (Tabela 3).

54
A proteína recombinante obtida com alta homogeneidade (99,9%) foi utilizada
para imunização de coelhos. Anticorpos anti-EspA foram obtidos em altos títulos
1:400.000 como determinado por ELISA.
Os anticorpos anti-EspA produzidos em coelhos, em teste de western blot,
reagiram fortemente com a proteína de 25 kDa equivalente a EspA recombinante,
porém, também reagiram com uma proteína de massa molecular maior. Essa
proteína provavelmente é um dímero de EspA, pois, no ensaio para imunodetecção
de IgG anti-EspA de algumas das 100 crianças assintomáticas de 0 a 2 anos de
idade, também houve o mesmo padrão de bandas.
Um dos objetivos do presente estudo foi determinar se crianças,
assintomáticas para a doença causada por EPEC, possuíam anticorpos séricos anti-
EspA. Para avaliar a reatividade de IgG e IgA séricas anti-EspA, testes de ELISA e
Western blot foram realizados utilizando o soro de 100 crianças entre 0 e 2 anos de
idade. Dos soros em estudo, 75% foram positivos para IgG e 23% para IgA séricas,
o que indica que EspA é uma proteína bastante imunogênica. A alta prevalência de
principalmente IgA sérica na faixa de 12-18 meses de idade como representado na
figura 24, sugere que esses anticorpos não sejam maternais e que essas crianças
sofreram exposição antigênica natural, uma vez que residem em área endêmica
para EPEC (Campos dos Goytacazes), e estão em uma faixa etária onde o sistema
imune é capaz de responder a estímulos antigênicos. Além da soro prevalência,
analisamos também por ELISA os níveis desses anticorpos das crianças que foram
consideradas positivas para IgG e IgA séricas anti-EspA. Esses resultados são
mostrados nas figuras 25 e 26 onde mostram que os níveis desses anticorpos foram
bastante variados, o que pode ser devido ao fator nutricional, a maior ou menor
exposição antigênica e as diferentes idades, pois até os 12 meses os títulos são
baixos, sugerindo que a resposta seja ainda incipiente, e que essas crianças podem
ser mais susceptíveis, uma vez que ainda dependem de anticorpos maternais para
proteção. Não podemos afirmar se esses anticorpos são efetivamente protetores,
particularmente IgA, que está relacionada à proteção de mucosas. Porém, com
esses dados, somados aos resultados de um trabalho anterior do nosso grupo
(Fernandes et al., 2002b), é possível fazer algumas inferências. Embora o número
de crianças na faixa etária de 0 a 6 meses seja restrito, essas crianças não
apresentaram IgA anti-EspA, mas a partir dos 12 meses a maioria já apresenta,

55
sugerindo a presença de uma janela imunológica nos primeiros meses de vida, o
que pode determinar uma maior susceptibilidade. Essa janela imunológica pode ser
explorada para o estudo de proteção através do acompanhamento de crianças
nessa faixa etária, soro negativas para IgA anti-EspA, e a evolução para casos de
diarréia causada por EPEC. Dessa forma, poderíamos analisar se a soro conversão
está relacionada com a diminuição dos casos de diarréia causada por EPEC. Com a
mesma soroteca utilizada neste trabalho, Fernandes e colaboradores (2002b)
demonstraram que 2 das 3 crianças que estavam na faixa etária de 0 a 6 meses,
tinham anticorpos IgA anti-BfpA (outro fator de virulência importante de EPEC),
diferente do encontrado para EspA nesse trabalho. De acordo com essas
informações, essas crianças possivelmente foram expostas naturalmente a EPEC,
sendo que inicialmente o sistema imune respondeu para a proteína BfpA, que é mais
imunogênica, e somente a partir dos 12 meses quando o sistema imune já está mais
maduro há uma resposta para EspA. Contudo, para saber se esses anticorpos são
realmente protetores, seria importante saber se essas crianças que soro convertem
são ou não acometidas por EPEC.
Além disso, a exposição natural a EspA parece estimular boa resposta
imune, que é refletida pelas altas porcentagens de crianças assintomáticas que
possuem tanto IgG quanto IgA séricas anti-EspA. O trabalho de Noguera-Obenza e
colaboradores (2003) demonstrou alta prevalência de IgA anti-EspA de EHEC no
colostro, evidenciando sua capacidade protetora, uma vez que IgA secretada é o
fator primário para prevenir a colonização do epitélio intestinal por enteropatógenos
em neonatos amamentados em seio materno. Em nosso grupo de pesquisa, o
estudo da presença de anticorpos IgA anti-BfpA de colostro e fezes dos neonatos
mostrou que os anticorpos encontrados nas fezes não perderam sua atividade,
sugerindo sua capacidade protetora e o possível uso na imunoterapia oral para
prevenir e/ou tratar a infecção por EPEC típica na infância (Fernandes et al., 2002b;
Fernandes et al., 2003). Esses dados mostram a importância de usar a proteína
EspA recombinante como reagente de captura em estudos epidemiológicos da
doença diarréica por EPEC, e em estudos de desafio, para avaliação da sua
capacidade protetora .
Com o resultado do sequenciamento do clone pLUA-05 demonstrou-se que a
seqüência de aminoácidos da EspA de REPEC sorotipo “O103:H2” é idêntica a

56
EspA de EPEC (B171) sorotipo “O111:NM” que utilizamos para amplificação do DNA
no início dos experimentos (Figura 20). Esse dado é de grande importância para uso
futuro de coelho como modelo animal em experimentos in vivo utilizando a proteína
EspA ou anticorpos anti-EspA na inibição da lesão causada por EPEC.
Diante dos dados da soro prevalência e do seqüenciamento podemos afirmar
que a proteína EspA recombinante é antigenicamente idêntica a proteína EspA de
EPEC e também de REPEC.
O presente trabalho demonstrou que EPEC e REPEC são capazes de causar
hemólise como descrito previamente para EPEC e EHEC (Shaw et al., 2001).
Embora condições isotônicas sejam necessárias para prevenir a hemólise
espontânea, foi mostrado que quando a sensibilidade do ensaio é aumentada (feita
com IGP, solução isotônica, de baixa força iônica) EPEC e REPEC induziam forte
atividade hemolítica, enquanto em PBS essa atividade foi indetectável. Isso pode
ser explicado pelo fato de baixas forças iônicas permitirem maior interação da
bactéria com a hemácia (Ribeiro et al., 2003).
Testes utilizando anticorpos anti-EspA in vitro já demonstraram que não há
inibição da formação dos filamentos de EspA (La Ragione et al., 2005), mas que de
alguma forma a lesão A/E é inibida, seja por inibição da função de ‘ancoramento’
dos filamentos de EspA com a membrana da célula hospedeira, ou por prevenir a
interação de EspA com outras proteínas efetoras. Além disso, esses anticorpos
podem estabilizar os filamentos de EspA prevenindo seu desarranjo, que é pré-
requisito para a adesão íntima. Para demonstrar uma propriedade imunobiológica
dos anticorpos anti-EspA, também foram realizados testes de inibição da hemólise,
que foram eficientes com 100 µg de IgG de coelho anti-EspA (Tabela 4). Contudo,
houve inibição não esperada quando utilizamos 100 µg de IgG pré-imune (controle
negativo para inibição). Esse resultado pode ser explicado pelo fato da utilização de
IgG total e não IgG purificada por afinidade com EspA, uma vez que a produção
desses anticorpos foi feita em coelhos e estes são coprofágicos, eles podem ter
desenvolvido uma resposta imune contra E. coli não patogênica. Dessa forma,
anticorpos pré-imunes podem estar reconhecendo outras proteínas de E. coli e
formando agregados, impedindo que a bactéria lise a hemácia, dando um resultado
falso positivo para inibição. Acreditamos que para termos um controle negativo da
inibição mais confiável, um ensaio de absorção deva ser realizado anteriormente ao

57
ensaio da inibição, onde o pool de IgG pré-imune e o pool de IgG imune devam ser
incubados com bactéria não patogênica DH5-α. Anticorpos IgG que reconheçam
proteínas de E. coli DH5-α formarão agregados que poderão ser precipitados por
centrifugação. Dessa forma, teremos menos anticorpos inespecíficos no
sobrenadante da amostra de anticorpos pré-imune e IgG anti-EspA no
sobrenadante da amostra de anticorpos imune.
Alguns outros testes imunobiológicos podem ser vislumbrados com os
anticorpos anti-EspA de coelho produzidos nessa tese. Como foi demonstrado por
Moreira e colaboradores (2006), as proteínas BfpA e EspA medeiam a formação de
biofilme, uma vez que nosso grupo já produziu anticorpos policlonais tanto para Bfpa
(Flores et al., 2002) quanto para EspA, é possível utilizá-los em um teste simples de
inibição da formação de biofilme em diferentes substratos.
Anticorpos anti-EspA também podem ser utilizados para co-imunoprecipitar
EspA e EspB, pois, como demonstrado previamente (Hantland et al., 2000) há forte
interação entre essas proteínas.
É importante para nosso grupo de pesquisa a produção de EspA
recombinante e de anticorpos anti-EspA recombinante para que, em breve, possam
ser utilizados em ensaios de proteção in vivo. A proteína EspA poderá ser utilizada
como vacina e posterior desafio com REPEC ou os anticorpos produzidos contra
EspA poderão ser utilizados na imunoprofilaxia, como demonstrado com anticorpos
IgY anti-intimina na inibição in vivo da diarréia em porcos desafiados com PEPEC
(E.coli Enteropatogênica de Porcos) (Girard et al., 2006). Além disso, como o grupo
já produziu outras proteínas recombinantes de EPEC como EspB e BfpA (Flores et
al., 2002) seria importante avaliar o potencial vacinal de um coquetel contendo todas
essas proteínas recombinantes.
Anticorpos produzidos anti-EspA recombinante também poderão ser utilizados
como ferramenta para o diagnóstico da infecção por EPEC e EHEC no
desenvolvimento de Kits baseados na aglutinação como descrito para EspB (Lu et
al., 2002), são testes que podem ser utilizados no campo, rápidos, precisos e de
baixo custo.
De acordo com os resultados discutidos, acredita-se que a proteína EspA
recombinante e anticorpos anti-EspA recombinante possam ser excelentes
ferramentas para o desenvolvimento de formulações vacinais, para o diagnóstico

58
rápido de aplicação no campo e para estudos de epidemiologia da doença diarréica
aguda em crianças menores de 2 anos.
7. Conclusões:
A quantidade, na ordem de miligramas, da proteína EspA recombinante foi
produzida com alto nível de expressão e purificação;
A proteína EspA recombinate foi imunogênica para coelhos. Os anticorpos de coelho
anti-EspA mostraram uma titulação de 1:400000 por ELISA;
O fator de virulência EspA mostrou ser imunogênico estimulando a produção de IgG
e IgA em crianças assintomáticas de até dois anos de idade;
A proteína EspA recombinante pode ser usada como candidata a vacina, para a
imunodiagnose e estudos de epidemiologia na infecção por EPEC.

59
8. REFERÊNCIAS BIBLIOGRÁFICAS
Abe, A., Heczko, U., Hegele, R.G. and Finlay, B.B. (1998) Two enteropathogenic
Escherichia coli type III secreted proteins, EspA and EspB, are virulence factors.” J.
Exp. Med 188(10): 1907–1916.
Baldwin, T.J. (1998) Pathogenicity of enteropathogenic Escherichia coli. J. Med.
Microbiol 47: 283-93.
Boullier, S., Nougayrède, J. P., Marchès, O, Tasca, C., Boury, M., Oswald, E., De
Rycke, J. and Milon, A. (2003) Genetically engineered enteropathogenic Escherichia
coli strain elicits a specific immune response and protects against a virulent
challenge. Microbes and Infection 5: 857-867.
Bozzo, S., and Retamal, C. (1991) Gels unidimensionales. Un Nuevo método
densitometrico para computadores personales. Arch.Biol. Med. Exp. 24: 181.
Bradford, M. M. (1976) “A rapid and sensitive method for the qualitation of microgram
quantities of protein the principle of protein-dye binding”. Analyt. Biochem 72, 248-
254.
Campellone, K.G. and Leong, J.M. (2003) Tails of two Tirs: actin pedestal formation
by enteropathogenic E. coli and enterohemorrhagic E. coli O157: H7. Curr. Opin.
Microbiol. 6: 82–90.
Carbonare C.B., Carbonare S.B., Carneiro-Sampaio M.M.S. (2003) Early acquisition
of serum and saliva antibodies reactive to enteropathogenic Echerichia. Coli
virulence-associated proteins by infants living in an endemic area. Pediatr Allergy
Immunol 14: 222-228.
Chen, D.H. and Frankel, G. (2004) Enteropathogenic Escherichia coli: unravelling
pathogenesis. FEMS Microbiology Reviews 29: 83-98.

60
Cid, D., Ruiz-Santa-Quiteria, J.A., Marin, I., Sanz, R., Orden, J.A., Amils, R. and
de la Fuente, R. (2001) Association between intimin (eae) and EspB gene subtypes
in attaching and effacing Escherichia coli strains isolated from diarrhoeic lambsand
goat kids. Microbiology 147:2341–2353.
Clarke, S. C., (2001) Diarrhoeagenic Echerichia coli-an emerging problem?
Diagnostic Microbiology and Infectious Disease 41: 93-98.
Clarke, S.C., Haigh, R.D., Freestone, P.P.E. and Williams, P.H. (2003). Virulence of
Enteropathogenic Escherichia coli, a Global Pathogen. Clinical Microbiology Reviews
16 (3): 365–378.
Cleary, J., Lai, L.C., Shaw, R.K., Straatman-Iwanowska, A., Donnenberg, M.S.,
Frankel, G. and Knutton, S. (2004) Enteropathogenic Escherichia coli (EPEC)
adhesion to intestinal epithelial cells: role of bundle-forming pili (BFP), EspA
filaments and intimin. Microbiology 150: 527–538.
Coligan, J.E., Kruisbuk, A..M., Margulies, D.H., Sherach, E.M. and Strober, W.
(1991). Current Protocols in Immunology. Ed. John Wiley and Sons. Cap. 2; unid.
2.1; pág. 2.1.3-2.1.6.
Creasey, E.A., Friedberg, D., Shaw, R.K., Umanski, T., Knutton, S., Rosenshine, I.
and Frankel, G. (2003) CesAB is an enteropathogenic Escherichia coli chaperone for
the type-III translocator proteins EspA and EspB. Microbiology 149: 3639–3647.
Crepin, V. F., R. Shaw, C. M. Abe, S. Knutton, and G. Frankel (2005) Polarity of
enteropathogenic Escherichia coli EspA filament assembly and protein secretion. J.
Bacteriol 187 (8): 2881-2889.
Crepin, V. F., Shaw, R., Abe, C. M., Knutton, S. and Frankel G. (2005b) Molecular
Basis of Antigenic Polymorfism of EspA Filaments: Development of a Peptide Display
Technology. J. Mol .biol 350: 42-52.

61
Daniell, S. J., Kocsis, E., Morris, E., Knutton, S., Booy, F.P. and Frankel, G. (2003)
3D structure of EspA filaments from enteropathogenic Escherichia coli. Molecular
Microbiology 49 (2),301–308.
De Vinney, R., Knoechel, D.G. and Finlay, B.B. (1999) Enteropathogenic Escherichia
coli: cellular harassment. Current Opinion in Microbiology 2: 83-88.
Donnenberg, M.S. and Kaper, J.B. (1992) Enteropathogenic Escherichia coli. Infect.
Immun. 60: 3953-3961.
Dytoc, M, Fedorko, L., Sherman, P.M. (1994) Signal transduction in human epithelial
cells infected with attaching and effacing Escherichia coli in vitro. Gastroenterology
106:1150-1161.
Elliott, S.J., Wainwright, L.A., McDaniel, T.K., Jarvis, K.G., Deng, Y.K., Lai, L.C.,
McNamara, B.P., Donnenberg, M.S. and Kaper, J.B. (1998) The complete sequence
of the locus of enterocyte effacement (LEE) from enteropathogenic Escherichia coli
E2348/69. Mol. Microbiol. 28, 1–4.
Fernandes, R.C.S.C. and Medina-Acosta, E. (2002a) Diagnóstico da infecção por
Escherichia coli enteropatogênica. Pediatria Atual 15(3): 20-23.
Fernandes, R.C.S.C., Flores, V.M.Q., Macedo, Z.S. and Medina-Acosta, E. (2002b)
Antibody responsiveness to the entheropathogenic Escherichis coli bundle-forming
pilus subunit A in lactating mothers and asyntomactic children. Eur J Pediatr 161:
471-474.
Fernandes, R.C.S.C., Flores, V.M.Q., Macedo, Z.S. and Medina-Acosta, E. (2003)
Coproantibodies to the enteropathogenic Escherichia coli vaccine candidates BfpA
and EspB in breastfed and artificially fed children. Vaccine 21: 1725–1731.
Fernandes, R.C.S.C. and Medina-Acosta, E. (2004) Uso de antimicrobianos no
tratamento da diarréia aguda infantil. Pediatria Atual 17 (2): 8-12.

62
Flores, V.M.Q. (2001) Expressão de Antígenos de Escherichia coli Enteropatogênica
(EPEC) em Sistemas Heterólogos. Tese (Biociências e Biotecnologia) – Campos dos
Goytacazes – Rj, Universidade Estadual do Norte Fluminense – UENF, 113p.
Flores, V.M.Q., Fernandes, R.C.S., Macedo, Z.S. and Medina-Acosta, E. (2002)
Expression and purification of the recombinant enteropathogenic Escherichia coli
vaccine candidates BfpA and EspB. Protein Expression And Purification 25(1): 16-
22.
Fischer, J., Maddox, C., Moxley, R., Kinden, D. and Miller, M. (1994) Pathogenicity of
a bovine attaching effacing Escherichia coli isolate lacking Shiga-like toxins. Am. J.
Vet. Res. 55, 991–999.
Galan, J.E. (1996) Molecular genetic bases of Salmonella entry into host cells. Mol
Microbiol 20: 263–271.
Garmendia, J., Frankel, G. and Crepin, V.F. (2005) Enteropathogenic and
Enterohemorrhagic Escherichia coli Infections:Translocation, Translocation,
Translocation. Infection And Immunity 73 (5): 2573–2585.
Gauthier, A., Puente, J. and Finlay, B. (2003) Secretin of the enteropathogenic
Escherichia coli type III secretion system requires components of the type III
apparatus for assembly and localization. Infect. Immun 71: 3310-3319.
Girard, F., Batisson I., Martinez, G., Breton, C., Harel, J. and Fairbrother, J.M. (2006)
Use of virulence factor-specific egg yolk-derived immunoglobulins as a promising
alternative to antibiotics for prevention of attaching and effacing Escherichia coli
infections. FEMS Immunol Med Microbiol 46: 340–350.
Gomes, T.A., Rassi, V, MacDonald, K.L., Ramos, S.R., Trabulsi, L.R., Vieira M.A.,
Guth, B.E., Candeias, J.A., Ivey, C. and Toledo, M.R. (1991). Enteropathogens

63
associeted with acute diarrheal disease in urban infants in São Paulo, Brazil. J Infect
Dis 164: 331-7.
Harlon, E. and Lane, D. (1998) Antibodies – A laboratory manual, Editora Cold
Spring Harbor Laboratory, capítulo 13.
Hartland, L.E., Daniell, S.J., Delahay, R.M., Neves, B.C, Wallis, T., Shaw, R.K., Hale,
C., Knutton, S. and Frankel, G. (2000) The type III protein translocation system of
enteropathogenic Escherichia coli involves EspA-EspB protein interactions.
Molecular Microbiology 35(6): 1483±1492.
Hicks, S., Frankel, G., Kaper, J.B, Dougan, G. and Phillips, A.D. (1998) Role of
intimin and bundle-forming pili in enteropathogenic Escherichia coli adhesion to
pediatric intestinal tissue in vitro. Infect Immun 66: 570–1578.
Hueck, J. C. (1998). “Type III Protein Secretion Systems in Bacterial Pathogens of
Animals and Plants.” Microbiology and Molecular Biology Reviews 62(2): 379–433
Kaper, J. B. (1994) Molecular pathogenesis of enteropathogenic Escherichia coli,. In
Miller, V. L., Kaper, J. B., Portnoy, D. A., and Isberg, R. R. (Eds.), Molecular Genetics
of Bacterial Pathogenesis. American Society of Microbiology 12:173-195.
Kenny, B., DeVinney, R., Stein, M., Reinscheid, D.J., Frey, E.A. and Finlay, B.B.
(1997) Enteropathogenic E. coli (EPEC) transfers its receptor for intimate adherence
into mammalian cells. Cell 91: 511-520.
Knutton, S., Rosenshine, I., Pallen, M.J., Nisan, I., Neves, B.C., Bain, C., Wolff, C.,
Dougan, G. and Frankel G. (1998) A novel EspA-associated surface organelle of
enteropathogenic Escherichia coli involved in protein translocation into epithelial
cells. EMBO J 17: 2166±2176.

64
Kühne, S.A., Hawes, W.S., La Ragione, R.M., Woodward, M.J., Whitelam, G.c. and
Gough, K.C. (2004) Isolation of recombinant antibodies against EspA and intimin of
Escherichia coli O157:H7. J. Clin.Microbiol 42: 2966–2976.
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 227(259): 680-685.
La Ragione, R.M., Patel, S., Maddison, B., Woodward, M.J., Best, A., Whitelam, G.C.
and Gough, K.C. (2005) Recombinant anti-EspA antibodies block Escherichia coli
O157:H7-induced attaching and effacing lesions in vitro. Microbes and Infection 8 (2):
426-33.
Li, Y., Frey, E., Mackenzie, A.M, Finlay, B.B. (2000) Human response to E. coli
O157:H7 infection: antibodies to secreted virulence factors. Infect Immun
68(9):5090–5.
Loureiro, I., Frankel, G., Adu-Bobie, j., Dougan, g., Trabulsi, L.R., Carneiro-Sampaio
and Magda, M.S. (1998) Human colostrums contains IgA antibodies reactive to
enteropathogenic Escherichia coli virulence-associated proteins: intimin, BfpA, EspA
and EspB. J. Pediatr. Gastroenterol. Nutr 27, 166-171.
Lu, Y., Toma, C., Honma, Y. and Iwanaga, M. (2002). Dtection of EspB using
reversed passive látex agglutination: application to determination of
enteropathogenic Escherichi coli. Diagnostic Microbiology and Infectious Disease 43:
7-12.
Manjarrez-Hernandez, H.A., Gavilanes-Parra, S., Chavez-Berrocal, E., Navarro-
Ocan, A., Cravioto, A. (2000) Antigen Detection in Enteropathogenic Escherichia coli
Using Secretory Immunoglobulin A Antibodies Isolated from Human Breast Milk.
Infection and Immunity 68(9): 5030–5036.
Martinez, M.B., Taddei, C.R., Ruiz-Tagle, A., Trabulsi, L.R. and Girón, J.A. (1999)
Antibody Response of Children with Enteropathogenic Escherichia coli Infection to

65
the Bundle-Forming Pilus and Locus of Enterocyte Effacement–Encoded Virulence
Determinants. Infectious Diseases 179: 269-274.
Moon, H. W., Whipp, S.C., Argenzio, R.A., Levine, M.M. and Giannella, R.A. (1983)
Attaching and effacing activities of rabbit and human enteropathogenic Escherichia
coli in pig and rabbit intestines. Infect. Immun. 41:1340–1351.
Moreira, C.G., Palmer, K., Whiteley, M., Sircili, M.P., Trabulsi, L.R., Castro, A.F.P.
and Sperandio, V. (2006) Bundle-Forming Pili and EspA Are Involved in Biofilm
Formation by Enteropathogenic Escherichia coli. Journal of Bacteriology 188 (11):
3952–3961.
Nataro, J. P., and Kaper, J. B. (1998) Diarrheagenic Escherichia coli. Clin.
Microbiol. Rev 11:142–201.
Neves, B.C., Knutton, S., Trabulsi, L.R., Sperandio, V., Kaper, J.B., Dougan, G. and
Frankel, G. (1998) Molecular and ultrastructural characterisation of EspA from
different enteropathogenic Escherichia coli serotypes. FEMS Microbiology Letters
169: 73-80.
Neves, B.C., Shaw, R.K., Frankel, G. and Kunutton, S. (2003). Polymorphisms
within EspA Filaments of Enteropathogenic and Enterohemorrhagic Escherichia coli.
Infection and Immunity 71(4): 2262–2265.
Noguera-Obenza, M., Ochoa, T.J., Gomez, H.F., Guerrero, M.L., Herrera-Isua, I.,
Morrow, A.L., Ruiz-Palacios, G., Pickering, L.K., Guzman, C.A. and Cleary, T.G.
(2003) Human Milk Secretory Antibodies against Attaching and Effacing Escherichia
coli Antigens. Emerging Infectious Diseases 9 (5): 545-551.
Ogino, T., Ohno, R., Sekiya, K., Kuwae, A., Matsuzawa, T., Nonaka, T., Fukuda, H.,
Imajoh-Ohmi, S. and Abe, A. 2006 Assembly of the Type III Secretion Apparatus of
Enteropathogenic Esccherichia coli. Journal of Bcter. 188(8): 2801-2811.

66
Potter, A.A., Klashinsky, S., Li, Y., Frey, E., Townsend, H., Rogan, D., Erickson, G,
Hinkley, S., Klopfenstein, T., Moxley, R., Smith, D.R. and Finlay, B.B. (2004)
Decreased shedding of Escherichia coli O157:H7 by cattle following vaccination with
type III secreted proteins. Vaccine 22:362–369.
Ramirez, K., Hurta, R., Oswald, E., Garcia-Tovar, C., Hernandez, J.M. and Nvarro-
Garcia, F. (2005) Role of EspA and Intimin in Expression of Proinflammatory
Cytokines from Enterocytes and Lynphocytes by Rabbit Enteropathogenic
Escherichia coli-Infected Rabbits. Infection and Immunity 73 (1): 103-113.
Rendinbaugh, M.G. and Turley, R.B. (1986) Adaptation of the bicinchoninic acid
protein assay for use with microtiter plates and sucrose gradient fractions. Anal
Biochem 153(2):267-71.
Ribeiro, P.D. and Medina-Acosta, E. (2003) Prevention of lethal murine candidiasis
using HP (2-20), an antimicrobial peptide derived from the N-terminus of Helicobacter
pylori ribosomal protein L1. Peptides: 24: 1807-1814.
Riley, L.W., Junio, L.N. and Schoolnik, G.K. (1990) HeLa cell invasion by a strain of
enteropathogenic Escherichia coli that lacks the O-antigenic polysaccharide. Mol.
Microbiol. 4: 1661-1666.
Robins-Browne, R.M., Tokhi, A.M., Adams, L.M. and Bennett-Wood, V. (1994) Host
specificity of enteropathogenic Escherichia coli from rabbits: lack of correlation
betweenadherence in vitro and pathogenicity for laboratory animals. Infect. Immun.
62, 3329–3336.
Rodrigues, J., Thomazini, C.M., Morelli, A. and De Batista, G.C.M. (2004) Reduced
Ethiological Role for Enteropathogenic Escherichia coli in Cases of Diarrhea in
Brazilian Infants. Journal of Clinical Microbiology 42 (1) 398–400.

67
Rosqvist, R., Forsberg, A., Rimpilainen, M., Bergman, t. and Wolf-Watz, H. (1990)
The cytotoxic protein YopE of Yersinia obstructs the primary host defence. Mol
Microbiol 4: 657–667.
Sambrook, J. F., Fritsch, E. and Maniatis, T. (1989) Molecular cloning: a laboratory
manual. New York, Col Spring Habor Laboratory.
Sansonetti, P.J., Ryter, A., Clerc, P., Maurelli, A.T. and Mounier, J. (1986)
Multiplication of Shigella flexneri within HeLa cells: lysis of the phagocytic vacuole
and plasmid-mediated contact hemolysis”. Infect Immun 51: 461–469.
Sekiya, k., Ohishi, M., Ogino, T., Tamano, K., Sasakawa, C. and Abe A. (2001)
Supermolecular structure of the enteropathogenic Escherichia coli type III secretion
system and its direct interaction with the EspA-sheath-like structure. PNAS 98(20):
11638-11643.
Shaw, R.K., Daniell, S., Ebel, F., Frankel, G. and Knutton, S. (2001) EspA filament-
mediated protein translocation into red blood cells. Cellular Microbiology 3(4): 213-
222.
Stakenborg, T., Vandekerchove, D., Mariën, J., Laevens, H., Imberechts, H. and
Peeters, J. (2006) Protection of rabbits against enteropathogenic Escherichia coli
(EPEC) using an intimin null mutant. BMC Veterinary Research 2:22.
Towbin, H., Staehelin, T. and Gordon, J. (1979) Electrophoretic transfer of proteins
from polyacrylamide gels to nitrocellulose sheets: procedure and some applications.
Proc Natl Acad Sci USA 76(9): 4350-4354.
Turk, J., Maddox, C., Fales, W., Ostlund, E., Miller, M., Johnson, G., Pace, L.,
Turnquist, S. and Kreeger, J. (1998) Examination for heat-labile, heat-stable, and
Shiga-like toxinsand for the eaeA gene in Escherichia coli isolates obtained from

68
dogs dying with diarrhea: 122 cases (1992–1996). J. Am. Vet. Med. Assoc. 212,
1735–1736.
Vallance, B.A. and Finlay, B.B. (2000) Exploitation of host cells by enteropathogenic
Escherichia coli. PNAS 97(16): 8799–8806.
Zhu, C., Harel, J., Jacques, M., Desautels, C., Donnenberg, M.S., Beaudry, M.,
Fairbrother, J.M. (1994) Virulence propertiesand attaching-effacing activity of
Escherichia coli O45 from swine postweaning diarrhea. Infect. Immun. 62: 4153–
4159.
Zhu, C., Feng, S., Thate, T.E., Kaper, J.B. and Boedeker, E.C. (2005) Towards a
vaccine for attaching/effacing Escherichia coli: a LEE encoded regulator (ler) mutant
of rabbit enteropathogenic Escherichia coli is attenuated, immunogenic, and protects
rabbits from lethal challenge with the wild-type virulent strain. Vaccine 24(18): 3845-
55.
Fonte de Financiamento: FAPERJ/Cientista do Nosso Estado E-26/151.991/2004. CAPES - programa de bolsas de Mestrado

69
ANEXO

70
Meio de cultura LB:
Triptona………………………………………………………10,0 g
Extrato de levedo…………………………………………… 5,0 g
NaCl…………………………………………………………..10,0 g
H2O……………………………………………………………800 ml
Ajustar o pH para 7,0 com NaOH
Completar água para 1000 mL.
Meio sólido – Agar ...........................................................15,0 g
Esterilizar por autoclavagem por 20 minutos
Soluções:
TENS
SDS...................................................................................10%
NaOH................................................................................0,2 M
Tris HCl pH 8,0.................................................................100 mM
EDTA pH 8,8………………………………………………....50 mM
TE
Tris HCl pH 7,5..................................................................10 mM
EDTA pH 8,0……………………………………………….....1 mM
TE RNAse
Tris HCl pH 7,5..................................................................10 mM
EDTA pH 8,0……………………………………………….....1 mM
RNAse………………………………………………………….20 µg/ml
Tampão de lise desnaturante
Tris HCl pH 8,0....................................................................10 mM
NaH2PO4..............................................................................100 mM
Uréia.....................................................................................7 M
Tiouréia.................................................................................2 M

71
Tampão de lavagem
Tris HCl pH 8,0 ou 8,5 .............................................................. 10 mM
NaH2PO4...................................................................................100 mM
Uréia...........................................................................................7 M
Tiouréia.......................................................................................2 M
Tampão de eluição
Tris HCl pH 4,5 .........................................................................10 mM
NaH2PO4....................................................................................100 mM
Uréia...........................................................................................7 M
Tiouréia.......................................................................................2 M
Tampão de lavagem da coluna após uso em condições desnaturantes
Ácido acético...............................................................................0,2 M
Glicerol........................................................................................30%
Solução para diálise
Tris pH 8,0..................................................................................150 mM
NaCl…………………………………………………………………..50 mM
Solução reveladora de ELISA
Ácido cítrico (0,1 M)…………………………………………..26 mM
Na2HPO4 (0,2 M)................................................................28 mM ml
H2O2.................................................................................... 0,03% (vol/vol)
OPD ...................................................................................0,4 mg/ml
Quantidades referentes para 25 ml de solução reveladora.
Tampão TST-20 (10X)
Tris-HCl pH 7.4(12,11 g/l).................................................100 mM
NaCl (87,1 g/l) ……………………………………………….1,5 M
Tween-20 (5mL/l) ……………………………………………0,5%

72
Solução reveladora de Westernblot
Tris HCl pH 7,4 .................................................................40 mM
Imidazol..............................................................................10mM
H2O2....................................................................................0,03% (vol/vol)
DAB.....................................................................................1mg/ml
PBS
Fosfato de sódio pH 7,0.............................................................9 mM
NaCl...........................................................................................150 mM
IGP
Tampão fosfato de potássio pH 7,0.........................................1 mM
Glicose......................................................................................287 mM









![Higiene Intestinal.[1]](https://img.document.onl/doc/110x75/55cf9da3550346d033ae8004/higiene-intestinal1.jpg)