Embed Size (px)
Citation preview

UM
inho |
2015
Universidade do Minho
Escola de Ciências
Ana Gabriela Pereira Barros
Caracterização das fontes de matéria
orgânica que suportam a produção de
ictioplâncton no estuário do Rio Minho.
Dezembro de 2015
Ana G
abriela
Pere
ira B
arr
os
Ca
racte
riza
çã
o d
as f
on
tes d
e m
até
ria
org
ân
ica
qu
e s
up
ort
am
a p
rod
uçã
o d
e icti
op
lân
cto
n n
o e
stu
ári
o d
o R
io M
inh
o.


Ana Gabriela Pereira Barros
Caracterização das fontes de matéria
orgânica que suportam a produção de
ictioplâncton no estuário do Rio Minho.
Dezembro de 2015
Dissertação de MestradoMestrado em Ecologia
Trabalho efetuado sob a orientação da
Doutora Sofia Ester e Sousa de Aguilar Dias
e da
Professora Doutora Fernanda Maria Fraga
Mimoso Gouveia Cássio
Universidade do Minho
Escola de Ciências

iii
Não sou nada. Nunca serei nada.
Não posso querer ser nada. À parte isso, tenho em mim todos os sonhos do mundo.
Fernando Pessoa

iv
AGRADECIMENTOS
A Ester Dias, o meu sincero agradecimento, por já fora de horas, ter aceite orientar-me.
Agradeço toda a paciência, até nas questões mais elementares. Agradeço por nunca ter desistido de
mim e por fazer-me acreditar. Agradeço a exigência, por obrigar-me a superar-me. Por fim, agradeço a
amizade.
Ao CIIMAR – Centro de Investigação Interdisciplinar Marinha e Ambiental (Universidade do
Porto), o meu agradecimento por permitirem a realização desta tese, disponibilizando todo o apoio
logístico necessário durante o período de investigação.
Ao Aquamuseu do Rio Minho e colaboradores, em especial ao Professor Doutor José Carlos
Antunes, pelo apoio logístico fundamental para a realização das amostragens e por toda a
disponibilidade e paciência que sempre demonstrou.
Aos investigadores do Grupo Estuarine Ecology and Biological Invasions do CIIMAR, Ronaldo
Sousa, Martina Llarri, Vanessa Modesto, Allan Souza e em especial ao Jacinto Cunha, por toda a
prontidão em ajudar e tirar dúvidas sempre que precisei.
Aos meus amigos, em especial ao Stefano Araújo, Helena Machado e Filipa Martins, por todo o
apoio e por toda a paciência que tiveram ao ouvir-me lamentar cada vez que aparecia uma pedra no
caminho.
Aos meus colegas, em especial à Ana Lages e à Maria Luís Miranda, pelo apoio e
companheirismo.
Aos meus pais, que sem a sua insistência não teria embarcado nesta viagem. Agradeço pelo
apoio e confiança em todas as fases desta etapa, sem eles nada disto seria possível.
Ao meu irmão, por existir, que só isso chega.

v
RESUMO
Barros, A. G. P. Caracterização das fontes de matéria orgânica que suportam a produção de
ictioplâncton no estuário do Rio Minho. Characterization of organic matter sources that support the
production of ichthyoplankton in the Minho river estuary. 2015. Dissertação (Mestrado em Ecologia) –
Universidade do Minho, Campus Gualtar, Braga – Portugal, 2015.
Uma das funções das áreas berçário é fornecer as condições ideais para a sobrevivência e
crescimento dos estágios iniciais de desenvolvimento de diversas espécies de peixe. O objectivo
principal deste estudo consistiu em avaliar a importância dos habitats estuarinos, e dos ecossistemas
adjacentes, para a produção da biomassa do ictioplâncton. Era esperado que com o aumento do
caudal, existisse um aumento no contributo de fontes de matéria orgânica (MO) de origem alóctone e
durante períodos de caudal baixo, aumentasse a contribuição das fontes de MO de origem autóctone.
Para tal, foi identificado e quantificado, o tipo e origem da MO assimilada pelo ictioplâncton no estuário
do rio Minho, através da utilização dos isótopos estáveis de carbono (C) e azoto (N). Foi também
avaliada a resposta funcional das larvas de peixe, face às variações naturais na disponibilidade de
alimento ao longo do gradiente estuarino de salinidade e face às variações do caudal. Os valores dos
rácios de C(δ13C: 13C/12C) e N (δ15N: 15N/14N) das larvas de peixe analisadas revelam que a sua energia
provém essencialmente de fontes autóctones, isto é, produzidas localmente. As variações do caudal,
provocaram alterações na qualidade da MO particulada (MOP) disponível, nomeadamente, com o
aumento do caudal, aumentou o contributo de MO com origem terrestre no estuário (C/NMOP> 10), e
com a diminuição do caudal, terá aumentado o contributo de fitoplâncton para o MOP (C/NMOP≈7). No
entanto, esta alteração não foi registada nos tecidos das larvas dos peixes, sugerindo que o caudal não
terá um papel determinante na utilização dos diferentes tipos de fontes de MO. Em todo o caso, o
contributo do ecossistema terrestre foi notório (até 49%), pois registou-se a sua contribuição para a
produção da biomassa das larvas, durante todo o período de estudo, e ao longo do gradiente de
salinidade. O contributo do ecossistema marinho foi mais reduzido (até 57%) e estará mais confinado
às regiões mais próximas da foz. Verificou-se ainda que, as larvas de peixe no estuário do rio Minho,
utilizam quer energia proveniente das cadeias tróficas pelágicas (e.g. MOP), quer da cadeia trófica
bentónica (e.g. epilíton e MOS). A distribuição e abundância de ictioplâncton, foi também um tema
deste estudo. As famílias mais abundantes foram, Gobiidae n.i. (18.8%) e Ammodytes tobianus
(48.7%). Este estudo mostra assim que, apesar dos ecossistemas estuarinos funcionarem como áreas
de berçário para o desenvolvimento de várias espécies de peixes, os ecossistemas adjacentes
subsidiam as cadeias tróficas nas quais estas larvas de peixe se alimentam.
Palavras-chave: ictioplâncton, estuários, δ15N, δ13C, alóctone, autóctone

vi

vii
ABSTRACT
Barros, A. G. P. Caracterização das fontes de matéria orgânica que suportam a produção de
ictioplâncton no estuário do Rio Minho. Characterization of organic matter sources that support the
production of ichthyoplankton in the Minho river estuary. 2015. Dissertação (Mestrado em Ecologia) –
Universidade do Minho, Campus Gualtar, Braga – Portugal, 2015.
Nursery areas are critical for the survival and growth of early fish larval stages. The main goal of
this study was to evaluate the importance of estuarine habitats, and adjacent ecosystems, for the
ichthyoplankton biomass. It was expected that with the increase of the river flow, there would be an
increase in the contribution of alochthonous sources, and that during low river inflow the importance of
autochthonous sources would increase. In order to accomplish this goal, the organic matter (OM)
assimilated by the fish larvae in the Minho estuary, was identified and quantified using carbon (C) and
nitrogen (N) stable isotopes. It was also evaluated the functional response of fish larvae to the natural
variability in food availability throughout the estuarine salinity gradient and due to variations in the river
flow. The C(δ13C: 13C/12C) and N (δ15N: 15N/14N) values of the fish larvae revealed that they relied
essentially on autochthonous sources, i.e., local photosynthetic production. The quality of particulate
OM (POM) changed with the river inflow; with increasing river inflow, the contribution of terrestrial-
derived OM also increased (C/NPOM> 10), whereas the decrease in the river inflow lead to an increased
contribution of phytoplankton to the POM pool (C/NPOM≈7). However, these changes were not reflected
on the fish larvae tissues, suggesting that river inflow has a minor role on the type of OM assimilated by
fish larvae. Nonetheless, there was a relevant contribution of the terrestrial ecosystem (to 49%) to the
fish larvae food web in time and along the estuarine salinity gradient. The contribution of the marine
ecosystem (to 57%) was more limited to the stations closer to the river mouth. It was also possible to
verify that the fish larvae from the Minho river estuary rely on both pelagic (e.g. POM), and benthic food
webs (e.g. epilithon, SOM). Distribution and abundance of ichthyoplankton were also a theme of the
present study. Gobiidae n.i. (18.8%) and Ammodytes tobianus (48.7%), were the most abundant
species. Therefore, this study shows that, although estuaries may act as important nursery areas for
fish species, their food webs are subsidized by the adjacent ecosystems.
Keywords: ichthyoplankton, estuaries, stable isotopes, δ15N, δ13C, allochthonous, autochthonous

viii

ix
ÍNDICE
Pág ina
AGRADECIMENTOS iv
RESUMO v
ABSTRACT vii
LISTA DE FIGURAS xi
LISTA DE TABELAS xv
LISTA DE ABREVIATURAS xvii
1 In trodução 1
2 Object ivos 5
3 Mater ia l e Métodos 6
3.1 Área em estudo 6
3.2 Amostragem 6
3.3 No laboratório 8
3.4 Análise de dados 9
4 Resul tados 11
4.1 condições ambientais 11
4.2 Abundância de ictioplâncton 13
4.3 Valores isotópicos das fontes de matéria orgânica 16
4.4 Valores isotópicos do ictioplâncton 20
5 Discussão 29
5.1 Abundância larvar 29
5.2 Variabilidade espacial e temporal das fontes de energia que suportam a biomassa das larvas 31
6 Conclusão 34
7 Referências b ib l iográf icas 36
8 Anexos 45

x

xi
LISTA DE FIGURAS
Figura 1 – localização das estações de amostragem ao longo do estuário do Rio Minho.
Figura 2 – Valores de concentração de clorofila (µg.L-1) e salinidade recolhidos no fundo e na
superfície, nas estações de 1 a 6 (E1 a E6), entre Janeiro e Julho de 2015, no estuário do rio Minho.
Figura 3 - Caudal médio no rio Minho medido entre 1991 e 2005 (linha sólida; estação Foz do Mouro -
SNIRH, 2015) e durante o ano de 2015 (linha picotada; estação N015 - Confederación Hidrográfica del
Miño-Sil, 2015).
Figura4–Análise de cluster realizada para os valores de caudal medidos no estuário do rio Minho entre
Janeiro e Dezembro de 2015 (Confederación Hidrográfica del Miño-Sil, 2015). Linhas a cinzento
indicam os grupos não separados (a a< 0.05) pelo SIMPROF.
Figura 5 – Média (± DP) dos valores de δ13C e δ15N (‰), do ictioplâncton, ajustado para dois níveis
tróficos de fraccionamento (+0.8 ‰ δ13C; +5.9 ‰ δ15N) e das potenciais fontes de material orgânica
recolhidas entre Janeiro e Abril (caudal elevado) de 2015 em E1, E2, E3, E4 e EAD (correspondente às
estações de água doce – 5 e 6): Parablenius gattorugine (Pg), Ammodytes tobianus (A), Lipophrys
pholis (Lf), Lepadogaster purpurea (Lp), Centrolabrus exoletus (Ce), Platichthys flesus (Pf),
Pomatoschistus microps (Pm), Gobiidae n.i. (G), Solea solea (Ss), Blennidae n.i. (B), Dicentrarchus
labrax (Dl), Atherina presbyter (Ap), Pomatoschistus sp. (P), Cyprinidae n.i. (C). As fontes de matéria
orgânica (MO) incluem a MO no sedimento (MOS), MO particulada do fundo (MOPf) e da superfície
(MOPs), Epilíton (Epi), vegetação aquática emerge (VAE), vegetação aquática submersa (VAS), plantas
terrestres (T), detritos (D) Fucus sp. (F), plantas de via fotossintética C3 (C3) e C4 (C4) e macroalgas
(MA).
Figura 6 –Média (± DP) dos valores de δ13C e δ15N (‰), do ictioplâncton, ajustado para dois níveis
tróficos de fraccionamento (+0.8 ‰ δ13C +5.9 ‰ δ15N) e das potenciais fontes de material orgânica
recolhidas entre Maio e Julho de 2015 (caudal baixo) em E1, E2 , E3, E4 e EAD (correspondente às
estações de água doce – 5 e 6): Lipophrys pholis (Lf), Platichthys flesus (Pf), Pomatoschistus microps
(Pm), Gobiidae n.i. (G1 e G2), Dicentrarchus labrax (Dl), Atherina presbyter (Ap), Cyprinidae n.i. (C),
Lepadogaster purpurea (Lp), Symphodus melops (Sm), Labridae n.i. (L), Gobius niger (Gn),
Lepadogaster sp. (Lsp), Syngnathidae n.i. (Sn), Syngnathus acus (Sa) e Labrus mixtus (Lm). As fontes
de matéria orgânica (MO) incluem a MO no sedimento (MOS), a MO particulada do fundo (MOPf) e da

xii
superfície (MOPs), epilíton (Epi), vegetação aquática emerge (VAE), vegetação aquática submersa
(VAS), plantas terrestres (T), detritos (D), Fucus sp. (F), plantas de via fotossintética C3 (C3) e C4 (C4)
e macroalgas (MA).
Figura 7 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o
caudal elevado, correspondente aos meses de Janeiro, Fevereiro e Março, na estação 1, baseado no
modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As fontes de
matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica particulada (MOP) e matéria
orgânica no sedimento (MOS).
Figura 8 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o
caudal baixo, correspondente aos meses de Abril, Maio, Junho e Julho, na estação 1, baseado no
modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As fontes de
matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica particulada (MOP), matéria orgânica
no sedimento (MOS) e plantas de origem terrestres.
Figura 9 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o
caudal elevado, correspondente aos meses de Janeiro, Março e Abril, na estação 2, baseado no
modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As fontes de
matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica particulada (MOP), matéria orgânica
no sedimento (MOS), vegetação aquática emerge (VAE) e plantas de via fotossintética C4.
Figura 10 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o
caudal elevado, correspondente aos meses de Janeiro, Fevereiro e Março e Abril, na estação 3,
baseado no modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As
fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica particulada (MOP) e
matéria orgânica no sedimento (MOS).
Figura 11 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o
baixo, correspondente aos meses de Maio, Junho e Julho na estação 3, baseado no modelo de mistura
de isótopos estáveis com intervalos de credibilidade Bayesianos. As fontes de matéria orgânica são:
epilíton (Epi), macroalgas, matéria orgânica particulada (MOP), matéria orgânica no sedimento (MOS) e
detritos.

xiii
Figura 12 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o
caudal elevado, correspondente aos meses de Janeiro, Fevereiro e Março e Abril, nas estações 5 e 6,
baseado no modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As
fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica particulada (MOP), matéria
orgânica no sedimento (MOS), vegetação aquática emerge (VAE) e vegetação aquática submerge (VAS).
Figura 13 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o
caudal baixo, correspondente aos meses de Maio, Junho e Julho nas estações 5 e 6, baseado no
modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As fontes de
matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica partículada (MOP), matéria orgânica
no sedimento (MOS) e plantas de origem terrestre.

xiv

xv
LISTA DE TABELAS
Tabela 1– Valores dos rácios isotópicos de carbono (C) e azoto (N), de fontes de matéria
orgânica tipicamente encontradas em estuários.
Tabela 2- Número total de indivíduos capturados no estuário do rio Minho entre Janeiro e
Julho de 2015.Abundância, em número de indivíduos por 100 m3 (±Desvio padrão) e
abundância relativa (%), por espécie. Designação do seu habitat e ciclo de vida (Antunes e
Rodrigues, 2004; fishbase.org).
Tabela 3 – Número de espécies capturadas por estação de amostragem e por período de
amostragem, abundância de indivíduos por 100 m3 (±Desvio padrão) por estação de
amostragem e por período de amostragem.
Tabela 4 - Média (± Desvio padrão) dos valores (‰) de 13C e 15N, das potenciais fontes de
material orgânica, por estação e período de amostragem: material orgânica particulada (MOP);
material orgânica no sedimento (MOS); Plantas terrestres (Terrestres); plantas de via
fotossintética C4 (C4): vegetação aquática submersa (VAS) e emergente (VAE).

xvi

xvii
LISTA DE ABREVIATURAS
C – Carbono
N - Azoto
13C – Isótopos de carbono
15N – Isótopos de azoto
δ13C – Rácios de carbono
δ15N – Rácios de azoto
CO2 – Dióxido de carbono
h – Horas
ºC – Graus centigrados
Km -Quilómetros
m - Metros
Km2 – Quilómetros quadrados
HCL – Ácido clorídrico
µm – Micrómetro
L – Litros
ind/100m3 – Indivíduos por metro cúbico
µg.L-1 – microgramas por litro

xviii

1
1 INTRODUÇÃO
Os estuários são ecossistemas costeiros que desempenham diversas funções
económicas importantes, incluindo o transporte, o turismo e a drenagem de resíduos de
atividades domésticas, industriais e agrícolas (Heip e Herman, 1995; Raz-Guzman e Huidobro,
2002). Para além disto, são ecossistemas altamente produtivos (Costanza et al., 1997), com
elevada relevância ecológica, pois funcionam como locais de desova para várias espécies de
peixes migradores como Alosa alosa (sável), Petromyzon marinus (lampreia marinha) (Antunes
e Rodrigues, 2004; Mota e Antunes, 2011), e como locais de alimentação e refúgio para
várias espécies de aves, peixes e crustáceos marinhos (Reis, 2009). Os estuários são
ecossistemas heterogéneos, constituídos por diversos tipos habitats que incluem, sapais,
canaviais, canais subtidais e intertidais, fanerogâmicas marinhas, habitats definidos pela
salinidade ou pela profundidade da coluna de água, que se interconectam entre si através do
movimento da água (e nutrientes) ou de animais (Pihl et al., 2002; Vanni et al., 2004;
Sheaves, 2009; Hofman et al., 2015). Apesar da elevada produtividade dos habitats
estuarinos, o ictioplâncton é geralmente reduzido quer em diversidade, quer em biomassa
(Elliot et al.. 1990; Whitfield, 1994). Tal pode dever-se ao elevado stress fisiológico a que os
organismos estão sujeitos devido às oscilações ambientais estuarinas (e.g. salinidade, caudal,
turbidez, luz, etc; McLusky e Elliot, 2004), ou como consequência do reduzido tempo de vida
dos estuários, do ponto de vista geológico (Whitfield, 1994). No entanto, as comunidades de
ictioplâncton estuarino são variáveis quer na composição específica quer nos seus padrões de
distribuição (Harris et al., 1999), que estão dependentes das épocas de reprodução de cada
espécie e das flutuações ambientais (Whitfield, 1994; Harris e Cyrus, 1995), havendo uma
tendência para que as larvas atinjam o seu pico de abundância durante a primavera e o verão
dentro dos estuários (Whitfield, 1989; Harris et al., 1999; Young e Potter, 2003). Contudo, os
estuários desempenham um papel vital na sobrevivência e desenvolvimento de várias espécies
de peixes, constituindo importantes áreas berçário para as fases larvares de inúmeras
espécies costeiras, tais como, Solea solea (Linguado), Solea senegalensis (Linguado do
Senegal), Platichthys flesus (Solha-das-pedras) e Dicentrarchus labrax (Robalo) (Vasconcelos et
al., 2008; Freitas et al., 2009). Um berçário pode ser descrito como uma zona restrita onde
juvenis de uma dada espécie passam um determinado período do seu ciclo de vida, e durante
o qual estão espacialmente e temporalmente separados dos adultos da mesma espécie (Beck

2
et al., 2001). Segundo Beck et al., (2001) um habitat funciona como berçário, para uma
espécie, se a sua contribuição por unidade de área para a população dos indivíduos que
recrutam para a população adulta é maior, em média, do que a produção noutros habitats
onde os juvenis dessa espécie também ocorrem.
A função de um habitat como berçário, implica assim que exista uma ligação entre
diferentes habitats e/ou ecossistemas através do movimento de matéria orgânica (MO) e
nutrientes, ou através do movimento dos peixes (Sheaves, 2009). No entanto, enquanto que
existem vários estudos sobre a dinâmica das fases larvares de peixes em estuários (e.g.
Rakocinski et al., 1996; Harris et al., 1999; Faria et al., 2006), poucos estudos se debruçaram
sobre a importância que o contributo de diferentes habitats têm para o crescimento e
sobrevivência das fases larvares nas áreas de berçário (e.g. Hoffman et al., 2015), ao
contrário do que acontece com os juvenis (e.g. Vinagre et al., 2008; Vinagre et al., 2011; Le
Pape et al., 2012).
Visto que os estuários são amplamente considerados como locais importantes para a
alimentação dos estágios iniciais de desenvolvimento de diversas espécies de peixe (Elliot e
McLusky, 2002), é determinante identificar as principais origens das fontes de matéria
orgânica que suportam a biomassa das fases larvares, a fim de compreender qual o seu grau
de dependência dos habitats estuarinos. No entanto, identificar as fontes de energia que
suportam os consumidores em estuários pode ser difícil, dada a multiplicidade de fontes e de
origens que estas podem ter. Podem ter origem autóctone (i.e. produção fotossintética local)
ou alóctone (i.e. vegetação de origem terrestre ou ripícola, fitoplâncton com origem noutros
habitats). Uma ferramenta útil para fazer a identificação das fontes de energia que suportam
as cadeias tróficas estuarinas, e compreender a sua dinâmica, são os isótopos estáveis de
carbono (C) e azoto (N). A sua utilização em estudos ecológicos, tem aumentado ao longo dos
anos, devido ao facto de apresentarem algumas vantagens em relação a outras técnicas como
a análise conteúdos estomacais ou observações diretas. Enquanto que os isótopos estáveis
permitem identificar os alimentos assimilados pelos tecidos dos consumidores, os conteúdos
estomacais ou as observações diretas, apenas permitem identificar o que foi ingerido
(Pasquaud et al., 2007). Assim, os isótopos estáveis de C e N fornecem uma informação
integrada no espaço e no tempo da MO disponível no estuário e que foi incorporada nos
componentes estruturais e nas reservas energéticas dos consumidores (Peterson e Fry,
1987). Os rácios de C(δ13C=13C/12C) são usados para determinar as fontes de matéria

3
orgânica que suportam a biomassa dos consumidores, uma vez que os valores dos
consumidores se assemelham aos da sua dieta (Currin et al., 1995). Os rácios isotópicos de
N(δ15N=15N/14N) complementam a informação dada pelo C e permitem determinar o nível
trófico dos consumidores (Minagawa e Wada, 1984; Peterson e Fry, 1987). Assim, os rácios
isotópicos de um consumidor refletem a sua dieta apresentando um fracionamento trófico
médio (i.e. a diferença entre os rácios isotópicos do consumidor e da sua dieta) de +0.4‰
δ13C e +3.2‰ δ15N por nível trófico (Vander Zander e Rasmussen, 2001).
Uma outra vantagem da utilização dos isótopos estáveis é dada pelo facto de,
geralmente, as fontes primárias de energia apresentarem rácios isotópicos distintos. Por
exemplo, as plantas ripícolas que utilizam a via fotossintética C3 têm um rácio de carbono de
cerca de -28 ‰ (Tabela 1), uma vez que ocorre um fracionamento de cerca de -21 ‰ sobre o
CO2 atmosférico (δ13C; -7 ‰; Smith e Epstein, 1971). Por outro lado, as plantas que utilizam a
via fotossintética C4 (e.g. Spartina sp.) são mais enriquecidas em 13C (δ13C: -10 a -14 ‰;
Tabela 1) em virtude da redução do fraccionamento (Smith e Epstein, 1970; Fry e Sherr,
1984). Em sistemas de água doce, onde o carbono inorgânico dissolvido é empobrecido em
13C (δ13C <-10 ‰), o fitoplâncton pode ser distinguido da vegetação ripícola (Hoffman e Bronk,
2006). Em geral, os microfitobentos são mais enriquecidos em 13C do que o fitoplâncton na
coluna de água, devido à existência de uma camada limite difusora entre o sedimento e a
coluna de água que reduz o fraccionamento isotópico (France, 1995).
No entanto, vários factores podem fazer variar a qualidade e disponibilidade das
diferentes fontes de MO nos ecossistemas estuarinos. O balanço hidrológico entre a água que
vem dos rios e a que entra nos estuários, proveniente do mar, influencia a circulação de
nutrientes e de MO dentro do estuário. Com o aumento do caudal, aumenta a quantidade de
MO com origem alóctone no estuário (Hoffman et al., 2008). Por outro lado, como diminui o
tempo de residência, a disponibilidade de fitoplâncton no estuário diminui (Sin et al., 1999).
Pelo contrário, em períodos de caudal baixo, o tempo de residência aumenta o que permite
que a biomassa de fitoplâncton produzida localmente se acumule, estando por isso, mais
disponível para ser consumida (Hoffman e Bronk, 2006). Os ciclos diários de maré, bem como
o efeito dos ciclos lunares, podem distribuir a MO e os nutrientes de formas distintas ao longo
do gradiente estuarino de salinidade (Riley et al., 2004). Os eventos climáticos extremos
podem ainda alterar a magnitude e a direção do material que é transferido entre habitats e
ecossistemas (Valiela e Bartholomew, 2015).

4
Fontes de matér ia orgânica δ13C δ15N Referências
P lantas r ip íco las -26 a -30 -4 a 4 Peterson e Fry , 1987; C loern et a l . ,2002
Plantas vasculares emergentes -28 a -30 6 a 10 Cloern et a l . , 2002; Hof fman e Bronk,
2006
Vegetação aquát ica submersa -19 a -22 2 a 8 Cloern et a l . , 2002; Hof fman et a l . ,
2010
Spart ina spp. (C4) -10 a -14 2 a 6 Fry e Sherr , 1984
Diatomáceas bentónicas (Sa l in idade <5)
-18 a -22 6 a 7 Cloern et a l . , 2002; Deegan e Garr i t t ,
1997 Diatomáceas bentónicas
(Sa l in idade >5) -14 a -24 7 a 9 Dias et a l . , 2014
F i top lâncton de água doce Var iáve l 0 a 8 Hof fman e Bronk, 2006
F i top lâncton estuar ino -18 a -26 5 a 10 Cloern et a l . 2002
Microf i tobentos (MPB) -21 a -24 8 a 14 França et a l . , 2011; D ias et a l . , 2014
Sedimento (MOS) -19 a -28 0,2 a 7 França et a l . , 2011; Novais , 2013; Dias
et a l . , 2014
Tabela 1– Valores dos rácios isotópicos de carbono (C) e azoto (N), de fontes de matéria orgânica tipicamente encontradas em
estuários.

5
2 OBJECTIVOS
O principal objectivo deste estudo foi o de identificar a importância que os habitats
estuarinos e os ecossistemas adjacentes (terrestre e marinho) têm para a produção de
biomassa de ictioplâncton no estuário do rio Minho. Neste estudo, apenas se consideraram
para análise, as larvas de peixe. Para tal, foram identificadas e caracterizadas, através da
análise de isótopos estáveis de C e N, as principais fontes basais de energia neste estuário
que poderiam suportar as cadeias tróficas onde se alimentam as larvas de peixe. Procurou-se
também avaliar a resposta funcional das larvas de peixe, quando sujeitas à variabilidade
natural da matéria orgânica (MO) existente num ecossistema estuarino ao longo do gradiente
estuarino de salinidade e face às oscilações provocadas pelos caudais. Os principais
resultados esperados com este estudo seriam que o ictioplâncton dependeria
predominantemente da cadeia trófica pelágica, e que o caudal teria um papel fundamental no
tipo e origem das fontes assimiladas. Assim, esperava-se que durantes os meses de caudal
mais elevado, houvesse um aumento da contribuição de MO com origem terrestre na cadeia
trófica estuarina, , com particular relevância nas zonas de salinidade mais baixa. Por outro
lado, nos meses de caudal mais baixo, como o tempo de residência aumenta, esperar-se-ia
um maior contributo de fontes autóctones, nomeadamente de fitoplâncton, para a produção
do ictioplâncton, bem como um aumento do contributo de MO com origem marinha nas
cadeias tróficas estuarinas, sobretudo nas zonas de maior influência salina.

6
3 MATERIAL E MÉTODOS
3.1 ÁREA DE ESTUDO
O rio Minho localiza-se no noroeste da Península ibérica (NO-Europa; Fig. 1) e é um
dos rios, em Portugal, menos intervencionados e com enorme relevância para a conservação
de espécies piscícolas migratórias (ICNB, 2015). Possui uma extensão de 300 km, uma área
de 17 080 km2, sendo que apenas 5% desta se localizada em Portugal (Ferreira et al., 2003).
O estuário é mesotidal, com marés que variam entre 0.7 e 3.7 m (Alves 1996), com um
comprimento de aproximadamente 40 Km e uma área total de 23 km2 (Ferreira et al., 2003).
A largura máxima do estuário é de cerca de 2 km perto da foz, diminuindo até cerca de 10 m
para montante (Ferreira et al., 2003). A sua profundidade média é de 2.6 m e a profundidade
máxima é de cerca de 26 metros (Antunes et al., 2011).O estuário estende-se até Valença e
reúne um conjunto de habitats de elevada importância ecológica tais como, bancos de vasa e
de areia, sapais, matas ripícolas, caniçais e juncais (ICNB, 2015). O estuário é um local
importante de passagem migratória para diversas espécies de aves migratórias, sobretudo nas
áreas de caniçal e de mancha de floresta aluvial (ICNB, 2015). Devido à sua importância
como área de abrigo e alimentação para aves, o estuário foi classificado como uma área
Natura 2000 e como uma área importante para aves (BirdLife International, 2015). O estuário
do rio Minho é também uma zona de elevada relevância económica, onde se praticam
diversas atividades, como a pesca, a caça, a agricultura e a pecuária (ICNB, 2015).
3.2 AMOSTRAGEM
Para avaliar a importância do ecossistema estuarino e dos ecossistemas adjacentes
para a produção de ictioplâncton no estuário do rio Minho, foram realizadas amostragens,
quinzenalmente, entre Janeiro e Julho de 2015, em seis estações fixas situadas ao longo do
gradiente de salinidade do estuário (Fig. 1). As amostragens foram realizadas quinzenalmente,
dado que as larvas de peixe apresentam geralmente taxas de crescimento rápido, chegando
rapidamente ao equilíbrio isotópico, após uma mudança de dieta isotopicamente diferente
(turnover). De uma foram geral, as larvas demoram, entre 10 a 20 dias para que 90% do seu

7
tecido assimile uma nova dieta (Herzka, 2005; Hoffman et al., 2011). O período de
amostragem foi selecionado por forma a que as amostragens ocorressem em períodos de
caudal contrastantes, uma vez que a sua variação influencia a disponibilidade e variabilidade
de alimento (Sin et al., 1999, Hoffman e Bronk 2006).
As recolhas foram realizadas em seis estações fixas ao longo do gradiente de
salinidade do estuário: as estações 1 e 2 (sapal do rio Coura) estão sujeitas a uma maior
influência marinha, as estações 3 e 4 encontram-se numa zona meso a polihalina e as
estações 5 e 6 são oligohalinas.
Em cada estação, e sempre que possível, foram recolhidas as potenciais fontes de
matéria orgânica para a biomassa do ictioplâncton: Estas incluíram: matéria orgânica
particulada (MOP), matéria orgânica no sedimento (MOS), plantas aquáticas (emergentes e
submersas),plantas terrestres, detritos, epilíton e macroalgas. Em cada estação foram
recolhidas amostras de água da superfície e do fundo (cerca de 0.5 m a cima do fundo), para
determinação da concentração de clorofila (Chla: µg. L-1) e composição isotópica da MOP
Figura 1 – Localização das estações de amostragem no estuário do Rio Minho.

8
(δ13CCOP, δ15NNP, C/N molar). A água foi pré-crivada (150 µm) e filtrada através de filtros
Whatman GF/F, previamente calcinados. Os filtros foram mantidos congelados até análise
Todas as plantas, detritos, macroalgas e sedimento, foram recolhidos à mão. As
amostras de epilíton foram raspadas de seixos submersos nas margens dos locais de
amostragem.
O ictioplâncton foi recolhido durante as marés altas de marés vivas. Para tal,
realizaram-se arrastos com uma rede de malha de 200 µm (Hydro-Bios), a uma velocidade
constante, durante três minutos. Foram recolhidas três réplicas por estação de amostragem.
As amostras foram preservadas em etanol a 70%, ainda no campo, para posterior identificação
e análise.
Em cada estação de amostragem foram também medidos os parâmetros abióticos
temperatura, potencial oxidação/redução, pH, percentagem de oxigénio, concentração de
oxigénio, condutividade, salinidade, turbidez e sólidos dissolvidos (TDS), utilizando uma sonda
multiparamétrica YSI EXO 2.
3.3 NO LABORATÓRIO
Para análise de Chla, os filtros foram colocados, durante 24h, numa solução de
acetona a 90% e posteriormente analisados num espectrofotómetro Spectronic® 20 Genesys.
A concentração de Chla (µg.L-1) foi determinada segundo o método Lorenzen (1967).
Para a análise de isótopos estáveis da MOP e do epilíton, os filtros foram secos numa
estufa a 60 ℃, durante 24h. Posteriormente, foi realizada uma subamostragem do filtrado,
para acidificação, numa câmara de vácuo, com ácido clorídrico concentrado. A acidificação foi
necessária para a remoção do carbono inorgânico, uma vez que este iria influenciar os valores
finais de δ13C (Lorrain et al., 2003).
Para a determinação dos rácios isotópicos da matéria orgânica no sedimento (MOS:
COS, NS), as amostras foram secas numa estufa a 60 ℃, durante pelo menos 48h.
Posteriormente, estas amostras foram reduzidas a pó num moinho de bolas (Retsch MM200).
Uma subamostra de sedimento foi depois acidificada através da aplicação direta, gota a gota,
de HCl a 10%, para remoção do carbono inorgânico.

9
As plantas e as macroalgas foram lavadas com água ultra pura, para remoção de
epífetos, secas numa estufa a 60 ℃, e posteriormente reduzidas a pó com um moinho de
bolas (Retsch MM200).
O ictioplâncton foi triado e identificado até ao nível taxonómico mais baixo possível,
utilizando para tal bibliografia adequada (e.g. Nichols, 1971, Nichols, 1976, Ré et al., 2008).
Todas as larvas foram medidas, com recurso ao software Leica® Application Suite V4.6
acoplado a uma lupa (Leica® S8AP0). Para posterior análise isotópica, as amostras foram
secas numa estufa a 60 ℃, durante 24 horas.
Os rácios dos isótopos estáveis de C e N foram determinados com recurso a um
espectrómetro de massa de razões isotópicas (Thermo scientific Delta V Advantage IRMS via
conflo IV interface (Marinnova, Universidade do Porto)). As razões isotópicas são expressas em
notação δ:
δX : δX = (RAmostra / RPadrão – 1) x 103
onde, X representa os isótopos estáveis de C ou N,R é o rácio 13C/12C ou15N/14N e Pee Dee
Belamite e o ar são os padrões utilizados para a análise de δ13C e δ15N, respectivamente. O
erro analítico, foi de ±0.1‰ para o δ13C e δ15N.
3.4 ANÁLISE DE DADOS
Foram efectuadas análises multivariadas para ver se existiam agrupamentos nos dados quer
entre estações quer ao longo do tempo. Foi efectuada uma análise de cluster, usando a
distância de similaridade Euclideana, para verificar de que forma os meses de amostragem se
agrupavam de acordo com os valores dos caudais. Foi também efectuada uma análise two-
way PERMANOVA, para verificar se existiriam diferenças temporais ou espaciais nos valores de
δ13C e δ15N das larvas de peixes recolhidas. . A significância estatística da variância (α=0.05)
foi testada utilizando 9999 permutações dos resíduos num modelo reduzido. As análises
foram efectuadas no PRIMER (v.6.1.6, PRIMER-E) com a análise da variância multivariada
baseada em permutações (PERMANOVA) + 1.0.1 (Anderson et al., 2008).
Para quantificar a contribuição de cada uma das fontes de matéria orgânica para a
biomassa do ictioplâncton foi usado um modelo de mistura de isótopos estáveis. O modelo de

10
mistura estima a contribuição proporcional de cada fonte de alimento para o tecido do
consumidor, segundo as seguintes equações:
δ13CM=fA.δ
13CA+fB.δ13CB+fC. δ
13CC
δ15NM=fA.δ
15NA+fB.δ15NB+fC. δ
13N
fA + fB + fC = 1
onde δM corresponde à composição isotópica do tecido do consumidor M e ƒA , ƒB e
ƒC são as contribuições das fontes de matéria orgânica A, B e C. O modelo de mistura
utilizado foi um modelo que usa a inferência Bayesiana (Stable Isotope Analysis in R- SIAR)e
que produz uma distribuição de probabilidades que representa a probabilidade com que uma
determinada fonte contribui para a biomassa do consumidor (Parnell et al., 2010). OSIAR
permite incluir não só o valor médio dos isótopos de cada fonte e do fraccionamento trófico,
mas permite também a inclusão da sua variabilidade (desvio padrão). O SIAR vai produzir uma
gama de soluções possíveis para o problema do modelo de mistura, ao qual são atribuídos
intervalos de credibilidade (CIs) (Parnell et al., 2010). Para a execução dos modelos, foram
selecionadas as fontes mais prováveis através de uma análise gráfica dos valores de δ13C e
δ15N dos consumidores e fontes. As fontes com uma contribuição inferior a 10% para a
biomassa dos consumidores, foram excluídas dos modelos.
Os valores de δ13C das larvas de peixe, foram corrigidos para os lípidos, segundo o
modelo proposto por Hoffman e Sutton (2010; Eq.6). Os valores de δ13C e δ15N foram ainda
corrigidos para o etanol (+0.4‰δ13C, +0.6‰ δ15N; Feuchtmayer & Grey, 2003). Para os
modelos, os valores de δ13C e δ15N foram ajustados para dois níveis tróficos, uma vez que as
larvas de peixe, são consumidores secundários: +0.8± 1.3‰ δ13C, +5.9 ± 1.0‰ δ15N (Vander
Zander and Rasmussen, 2001).
11
4 RESULTADOS
4.1 CONDIÇÕES AMBIENTAIS
As estações de amostragem apresentaram variações ambientais ao longo do tempo.
As estações de 1 a 3 são de maior influência marinha, com salinidades que variaram entre
2.7 (Janeiro) e 37.6 (Julho). A estação 4 é uma zona de transição de salinidade, com valores
a variarem entre 0.2 (Janeiro) e 7.8 (Março). As estações 5 e 6, são consideradas estações de
água doce, uma vez que apresentam sempre valores de salinidade inferiores ou iguais 0.05 e
portanto, são consideradas estações de água doce, não existindo diferenças entre os valores
medidos à superfície e no fundo (Fig. 2). Os valores de salinidade entre E1 e E4 foram, de
uma forma geral, superiores no fundo em relação à superfície (Fig. 3).
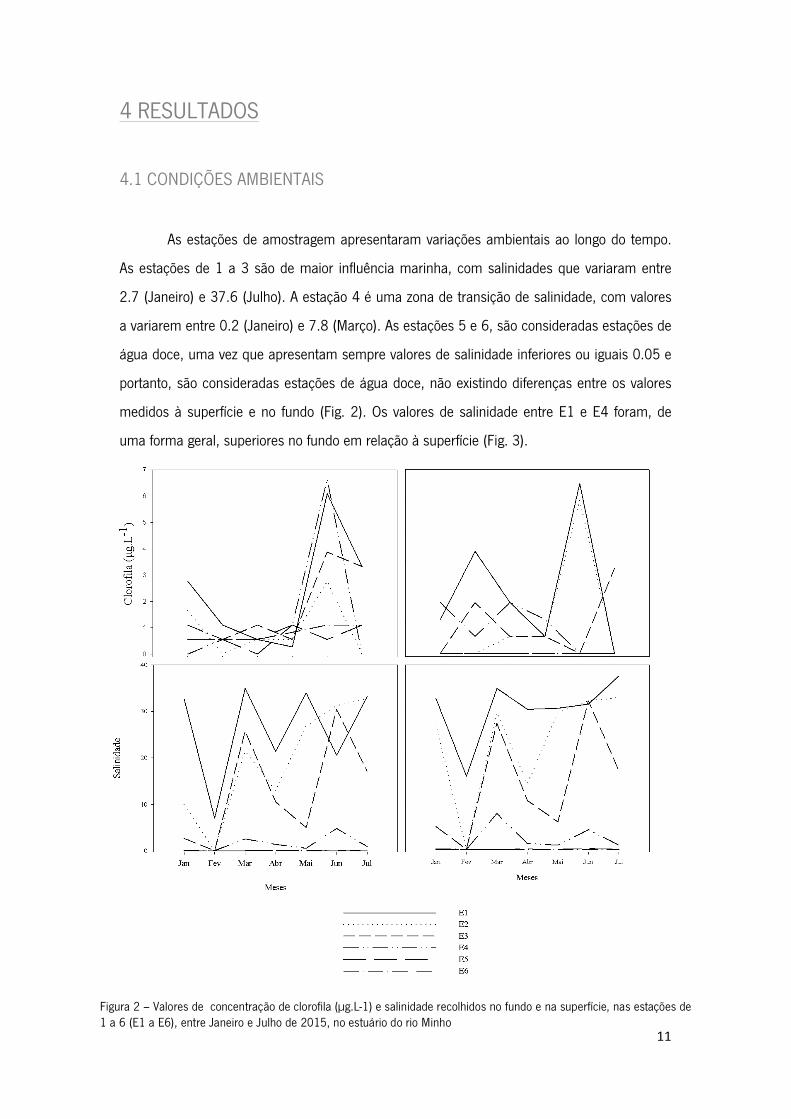
Figura 2 – Valores de concentração de clorofila (µg.L-1) e salinidade recolhidos no fundo e na superfície, nas estações de 1 a 6 (E1 a E6), entre Janeiro e Julho de 2015, no estuário do rio Minho

12
A clorofila apresenta valores superiores nos meses de Junho e Julho, tendo o valor
mais elevado (6.65µg.L-1) sido registado em E1, no mês de Junho e à superfície e o valor mais
baixo (0µg.L-1), foi registado em E4 nos meses de Fevereiro e Abril.
A salinidade foi sempre mais alta nas estações mais próximas da foz (E1 e E2), e
mais baixa nas estações E5 e E6. Junto à foz, os valores de salinidade variaram entre 7
(Fevereiro) e 34 (Julho) em E1, e entre 0.1 (Fevereiro) e 33 (Julho) em E2 (Fig. 2). As
estações E5 e E6 são consideradas estações de água doce, uma vez que a salinidade foi
sempre inferior a 0.5, não existindo diferenças entre os valores medidos à superfície e no
fundo (Fig. 2). As estações E3 e E4 apresentaram valores de salinidade mais variáveis. Na E3,
os valores de salinidade variaram entre 0.05 (Fevereiro) e 32 (Julho) (Fig. 2) e na E4, a
salinidade variou entre 0.04 (Fevereiro) e 1.4 (Abril) (Fig. 2). Os valores de salinidade entre E1
e E4 foram, de uma forma geral, superiores no fundo em relação à superfície (Fig. 3).
Os valores médios do caudal observados durante o período de amostragem, foram, de
uma forma geral, mais baixos do que a média dos valores registados entre 1991 e 2005 ,
com exceção do mês de Fevereiro de 2015 (Fig. 3). No entanto, a tendência observada é
mesma. A partir de Setembro começa a haver um aumento do caudal, sendo os valores
máximos atingidos entre Janeiro e Fevereiro (Fig. 3). Entre Março e Abril ocorre um
decréscimo acentuado nos valores do caudal, que se mantêm baixos até ao final do verão (Fig.
3).
Figura 3 - Caudal médio no rio Minho medido entre 1991 e 2005 (linha sólida; estação Foz do Mouro- SNIRH, 2015) e durante o ano de 2015 (linha picotada; estação N015- Confederación Hidrográfica del Miño-Sil, 2015).

13
A análise de cluster separou os meses, com base nos valores do caudal, em dois
grandes grupos (Fig. 4). Um grupo é constituído pelos meses em que os valores de caudal são
tendencialmente mais baixos, entre o final da primavera e o final do outono (Maio a
Novembro; Fig. 3) e outro grupo é constituído pelos meses de inverno e primavera, quando os
caudais são mais elevados e variáveis (Dezembro a Abril; Fig. 3) (Fig. 4). Uma vez que os
sistemas tendem a ser biogeoquimicamente estáveis em períodos de caudais semelhantes
(Hoffman e Bronk, 2006), com algumas reservas para os meses de inverno, para facilitar a
análise e interpretação dos dados, optou-se por dividir o período de amostragem em período
de caudal elevado (Janeiro a Abril) e período de caudal baixo (Maio a Julho).
4.2 ABUNDÂNCIA DE ICTIOPLÂNCTON
Durante o período de amostragem foram capturadas 611 larvas de peixe no
estuário do rio Minho, pertencentes a 13 famílias (Tabela 2). Os grupos mais abundantes
foram, Gobiidae n.i. (60.2 ± 67.8 ind/100m3), seguido de Ammodytes tobianus (48.7
ind/100m3), Syngnathus spp. (21.1 ± 25.3 ind/100m3) e Cyprinidae n.i. (20.3 ± 17.4
ind/100m3) (Tabela 2).
Figura4–Análise de cluster realizada para os valores de caudal medidos no estuário do rio Minho entre Janeiro e Dezembro de 2015 (Confederación Hidrográfica del Miño-Sil, 2015). Linhas a cinzento indicam os grupos não separados (a α< 0.05) pelo SIMPROF.

14
A abundância média foi superior nos meses de caudal baixo (16.3 ± 19.7
ind/100m3), sendo que a estação onde os valores foram mais elevados, foi E4 (34.4 ± 35.1
ind/100m3) (Tabela 2). A diversidade específica também foi superior nos meses de caudal
baixo (16 ± 8.2). A diversidade diminui entre E1 e E6, tanto nos meses de caudal mais baixo
(entre 28 e 5), como nos meses de caudal mais alto (entre 9 e 1) (Tabela 3).
Ordem Familia Espécies Habitat Ciclo de vida Abundância
(ind/100m3) Abundância relativa (%)
Perciformes
Ammodytidae Ammodytes tobianus Marinho oportunista Demersal 48.65 16.11
Moronidae Dicentrarchus labrax Marinho oportunista Demersal 3,12 (2.3) 0.97
Sparidae Sparidae Estuarino/Marinho - 6,40 (0.2) 1.99
Gobiidae
Pomatoschistus spp. Estuarino/Marinho Demersal 10,54 (8.9) 3.28
Gobiidae n.i. - Demersal 60,24 (67.8) 18.77
Pomatoschistus microps
Estuarino Demersal 15,72 (13.4) 4.90
Gobius niger Marinho Demersal 13.71 4.27
Gobiesocidae
Lepadogaster purpurea
Marinho Demersal 6,19 (3.5) 1.93
Gobiesocidae spp. Marinho Demersal 1.91 0.59
Lepadogaster n.i. Marinho Demersal 6,58 (0.4) 2.05
Blenniidae
Blenniidae n.i. Marinho Demersal 5,36 (3.3) 1.67
Lipophrys pholis Marinho ocasional Demersal 4,91 (1.7) 1.53
Parablennius gattorugine
Marinho Demersal 1.91 0.59
Labridae
Centrolabrus exoletus Marinho Demersal 8,23 (8.9) 2.56
Labridae spp. Marinho Demersal 10,31 (3.9) 3.21
Symphodus melops Marinho Demersal 9,69 (3.3) 3.02
Labrus mixtus Marinho Demersal 14.55 4.53
Pleuronectiformes Pleuronectidae Platichthys flesus marinho oportunista Pelágico 7,23 (4.8) 2.25
Pleuronectiformes Soleidae Solea solea marinho oportunista Pelágico 8,01 (8.8) 2.52
Atheriniformes Atherinidae Atherina presbyter marinho oportunista Pelágico/Neritica 6,08 (3.9) 1.89
Cypriniformes Cyprinidae Cyprinidae n.i. Dulciaqui ́cola Demersal 20,30 (17.4) 6.32
Clupeiformes Clupeidae Clupeidae Marinho adventicio Pelágico 5,79 (3.6) 1.80
Syngnathiformes Syngnathidae Syngnathus spp. Marinho oportunista /Marinha
adventicia Demersal 21,13 (25.3) 6.58
Não identificado Não
identificado n.i. - - 17,49 (21.3) 5.57
Numero total de individuos 611
Tabela 2- Número total de indivíduos capturados no estuário do rio Minho entre Janeiro e Julho de 2015.Abundância, em número de
indivíduos por 100 m3 (±Desvio padrão) e abundância relativa (%), por espécie. Designação do seu habitat e ciclo de vida (Antunes e
Rodrigues, 2004; fishbase.org).

15
Relativamente ao habitat, das 23 espécies identificadas, 11 são marinhas, 5 marinhas
oportunistas, 3 marinhas e estuarinas, 1 marinha adventícia, 1 marinha oportunista e marinha
adventícia 1 estuarina e 1 dulciaquícola (Antunes e Rodrigues, 2004). As espécies com habitat
marinho oportunista foram as que apresentaram uma maior distribuição no estuário,
encontrando-se representadas em todas as estações amostradas nos meses de caudal mais
baixo e em quase todas (E1, E2, E3 e E6) nos meses de caudal mais alto. As espécies com
ciclo de vida marinho adventício foram aquelas representadas num menor número de
estações no estuário, tendo sido apenas capturadas em E1 e nos meses de caudal mais baixo.
As espécies marinhas encontram-se apenas em E1, E2 e E3, as espécies estuarinas em E2,
E3 e E4 e as espécies dulciquícolas em E4, E5 e E6. A única espécie que apresentou uma
distribuição ao longo de todo o gradiente de salinidade estudado, foi a Platichthys flesus
(Anexo 1), encontrando-se representada em E1, E2, E3 e E6 nos meses de caudal mais alto e
em E1, E3, E4 e E5 nos meses de caudal mais baixo. Isto deve-se ao facto de P.flesus ter uma
elevada tolerância às oscilações ambientais existentes nos ambientes estuarinos (Daverat et
al., 2012). As espécies que apresentaram uma distribuição mais restrita, isto é, que foram
encontradas apenas numa estação foram Ammodytes tobianus (E1), Gobiesocidae sp. (E1),
Parablenius gattorugine (E1), Gobius niger (E1), Clupeidea n.i. (E1) e Labrus mixtus (E3).
Caudal Estação Número de
espécies por estação
Média do número de espécies por
per íodo de
amostragem (±DP)
Abundância ( ind/100m 3)
(±DP) por estação
Abundância
( ind/100m 3) (±DP)
por per íodo de amostragem
Al to
1 9
3.5 (3.5)
10.28 (14.9)
11.59 (10.0)
2 4 17.81 (26.7)
3 6 5.94 (7.85)
4 1 2.98 (1)
5 0 -
6 1 2.24 (1)
Ba ixo
1 28
16 (8.2)
10.79 (9.6)
16.26 (19.7)
2 18 26.15 (49.1)
3 21 27.71 (51.2)
4 14 34.44 (35.1)
5 10 15.82 (16.4)
6 5 10.27 (8.8)
Tabela 3 – Número de espécies capturadas por estação de amostragem e por período de amostragem, abundância de indivíduos por
100 m3 (±Desvio padrão) por estação de amostragem e por período de amostragem.

16
A espécie P. flesus apresentou valores mais elevados de abundância nos meses de
caudal mais baixo (8.7± 3.6 ind/100m3), do que nos meses de caudal mais elevado (4.1 ±
2.6 ind/100m3) (Anexo 1). Nos meses de caudal mais elevado, os valores de abundância vão
diminuindo entre E1 (7.6 ± 1.0 ind/100m3) e E3 (2.03 ± 1.0 ind/100m3), apresentando um
ligeiro aumento em E6 (2.2 ± 1.0 ind/100m3) (Tabela 3). Nos meses de caudal mais baixo, os
valores vão diminuindo entre E1 (11.9 ± 3.3 ind/100m3) e E5 (3.5 ± 2.4 ind/100m3) (Tabela
3).
4.3 VALORES DOS ISÓTOPOS DAS FONTES DE MATÉRIA ORGÂNICA
Os valores médios (±DP)de δ13C da matéria orgânica particulada (MOP) foram mais
elevados em E1, quer nos meses de caudal alto (δ13C: -24.6 ± 1.0‰), quer nos meses de
caudal baixo (δ13C: -22.6 ± 0.5‰).Os valores médios (±DP) de δ15N foram mais elevados em
E4 (δ15N: 5.5 ± 0.9‰ ), nos meses de caudal alto e em E3 (δ15N: 6.1 ± 0.6‰), nos meses de
caudal baixo. De uma maneira geral, nos meses de caudal mais baixo, a MOP estava mais
enriquecida em 13C(δ13C: -25.5 ± 2.3‰ ), do que nos meses de caudal mais elevado (δ13C: -
26.5 ± 1.4‰). Os valores médios (±DP) de δ15N de MOP, foram idênticos nos meses de caudal
mais elevado e nos meses de caudal mais baixo, onde a média foi de 5.3± 0.3‰. A média
(±DP) dos valores de δ13CMOP e δ15NNP foram superiores no fundo (δ13C: -24.9 ± 2.1‰ ; δ15N:
5.5 ± 1,5‰ ) do que à superfície (δ13C: 26.6 ± 2.3‰; δ15N: 5.3± 1.6‰ ). Nos meses de caudal
mais baixo, a média (±SD) de MOP C:NMOP foi de 4.6 ± 1.6‰ à superfície e de 4.9 ± 1.5‰ no
fundo; nos meses de caudal mais alto, a média (±SD) de C:NMOP foi de 13.3 ± 5.4‰ à
superfície e de 12.6 ± 5.1‰ no fundo (Tabela 4).
Os valores médios (±DP) de δ13C do epilíton foram mais elevados em E1 (δ13C: -18.6
± 0.1‰), nos meses de caudal elevado e em E2 (δ13C:-15.6 ± 1.1‰), nos meses de caudal
baixo. Os valores médios (±DP) de δ15N foram mais elevados em E1 (δ15N: 4.2 ± 0.5‰), nos
meses de caudal baixo, e entre E3 e E4(δ15N: 6.5 ± 2.4‰), nos meses de caudal alto. De uma
maneira geral, nos meses de caudal mais baixo, o epilíton encontrava-se mais enriquecido em
13C(δ13C: -18.3 ± 2.2‰ ), do que nos meses de caudal mais alto (δ13C: -23.5 ± 3.1‰ ). Os
valores médios (±DP) de δ15N do epilíton, foram mais elevados nos meses de caudal alto
(δ15N: 7.7 ± 2.7‰) em relação aos meses de caudal baixo, onde a média foi de 6.9 ± 1.4‰
(Tabela 4).

17
Os valores médios (±DP) de δ13C de matéria orgânica no sedimento (MOS) foram
mais elevados em E1 (δ13C: -20.9 ± 2.8‰), nos meses de caudal baixo e entre E3 e E4 (δ13C: -
25.5 ± 0.4‰), nos meses de caudal alto. Os valores médios (±DP) de δ15N foram mais
elevados nas estações de água doce -EAD- (δ15N: 6.0 ± 1.6‰ ), nos meses de caudal alto e em
E2 (δ15N: 5.0 ± 0.8‰), nos meses de caudal baixo. De uma maneira geral, nos meses de
caudal mais baixo, a MOS encontrava-se mais enriquecida em 13C(δ13C: -24.7± 2.2‰ ), do que
nos meses de caudal mais alto (δ13C: -25.9 ± 0.4‰). Os valores médios (±DP) de δ15N de
MOS foram mais elevados nos meses de caudal mais baixo (5.0± 0.8‰) em relação aos
meses de caudal mais alto, onde a média foi de 3.6± 2.02‰ (Tabela 4).
Os valores médios (±DP) de δ13C da vegetação aquática emergente (VAE) foram mais
elevados em EAD, quer nos meses de caudal baixo (δ13C: -21.7 ± 0.02‰), quer nos meses de
caudal alto (δ13C: -30 ± 1.2‰). Os valores médios (±DP) de δ15N foram mais elevados em EAD
(δ15N: 8.2 ± 1.2‰ ), nos meses de caudal alto e em E4 (δ15N: 7.6 ± 1.4‰), nos meses de
caudal baixo. De uma maneira geral, nos meses de caudal mais baixo, a VAE encontrava-se
mais enriquecida em 13C(δ13C: -25.7 ± 3.8‰ ), do que nos meses de caudal mais alto (δ13C: -
29.3 ± 1.6‰ ). Os valores médios (±DP) de δ15N da VAE, foram mais elevados nos meses de
caudal mais alto (δ15N: 7.3 ± 1.1‰) em relação aos meses de caudal mais baixo, onde a
média foi de 7.2 ± 0.5‰ (Tabela 4).
Os valores médios (±DP) de δ13C da vegetação aquática submersa (VAS) foram mais
elevados em E2 (δ13C: -13.9 ± 0.04‰), nos meses de caudal baixo e em EAD (δ13C: -27.5 ±
4.5‰), nos meses de caudal alto. Os valores médios (±DP) de δ15N foram mais elevados em
E3 e E4 (δ13C: 10.02 ± 1.0‰ ), nos meses de caudal alto e em E3 (δ13C: 10.02 ± 1.0‰ ), nos
meses de caudal baixo. De uma maneira geral, nos meses de caudal mais baixo, a VAS
encontrava-se mais enriquecida em 13C(δ13C: -23.4 ± 1.0‰ ), do que nos meses de caudal
mais alto (δ13C: -33.4 ± 5.2‰ ). Os valores médios (±DP) de δ15N da VAS, foram mais
elevados nos meses de caudal mais alto (δ15N: 9.1 ± 1.6‰) em relação aos meses de caudal
mais baixo, onde a média foi de 8.3 ± 1.3‰ (Tabela 4).
Os valores médios (±DP) de δ13C das plantas de origem terrestre foram mais
elevados em EAD, tanto nos meses de caudal mais baixo (δ13C: -27.5 ± 0.04‰), como nos
meses de caudal mais alto (δ13C: -29.8 ± 1.4‰). Os valores médios (±DP) de δ15N foram mais
elevados em EAD, nos meses de caudal alto (δ15N: 5.8 ± 1.3‰ ), e em E3 (δ13C: 7.3 ± 1.3‰),
nos meses de caudal baixo.

18
De uma maneira geral, nos meses de caudal mais baixo, as plantas de origem
terrestre encontravam-se mais enriquecidas em 13C (δ13C: -28.8 ± 1.3‰ ), do que nos meses
de caudal mais alto (δ13C: -30.04 ± 0.2‰ ). Os valores médios (±DP) de δ15N das plantas de
origem terrestre, foram mais elevados nos meses de caudal mais alto (δ15N: 5.7 ± 1.0‰) em
relação aos meses de caudal mais baixo, onde a média foi de 2.8 ± 3.2‰ (Tabela 4).
Os valores médios (±SD) de δ13C das macroalgas, nos meses de caudal mais alto,
foram mais enriquecidos em 13C (δ13C: -16.4 ± 1.0‰ ) do que nos meses de caudal mais baixo
(δ13C: -17.2 ± 0,8‰). Os valores médios (±DP) de δ15N das macroalgas, foram mais elevados
nos meses de caudal alto (δ15N: 9.1 ± 0.6‰) em relação aos meses de caudal baixo, onde a
média foi de 6.9 ± 0.5‰ (Tabela 4).

Mês Estação
MOP Epi l í ton Macroalgas MOS Detr i tos Terrestres C4 VAS VAE Fucus sp.
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
δ13C
(‰)
δ15N
(‰)
Ca
ud
al
alt
o
1 -24.6
(1)
5.2
(1)
-18.6
(0.1)
4.2
(0.5)
-15,88
(2,7)
9.5
(1,3)
-26.0
(1.5)
1.7
(0.3) - - - - - - - - - - - -
2 -26.0
(1.5)
5.1
(0.1)
-27.2
(1)
5.7
(1)
-17,56
(0,50)
8.5
(1)
-26.1
(1.5)
1.2
(0.3) - - - -
-14.1
(1.2)
5.0
(0.7) - -
-27.2
(1)
5.7
(1) - -
3 -26.1
(0.2)
5.5
(1)
-23.3
(1.6)
12.3
(6)
-15,88
(2,7)
9.5
(1.3)
-25.5
(0.4)
4.44
(0.2) -32.8
0.13
(1)
-30.1
(0.1)
5.6
(3.6) - -
-36.4
(1.5)
10.02
(0.9)
-30.4
(0.8)
7.6
(1.8) - -
4 -27.7
(0.8)
5.5
(1)
-23.8
(2)
7.9
(1) - -
-25.5
(0.4)
4.44
(0.2) -32.8
0.13
(1)
-30.1
(0.1)
5.6
(3.6) - -
-36.4
(1.5)
10.02
(0.9)
-30.4
(0.8)
7.6
(1.8) - -
5 e 6 -28.0
(1)
5.4
(1)
-24.8
(3)
8.4
(1.7) - -
-26.4
(0.6)
5.9
(1.6)
-29.9
(0.5)
4.9
(0.9)
-29.8
(1.4)
5,76
(1,4) - -
-27.5
(4.5)
7,27
(1.7)
-29.2
(1.2)
8.2
(1.2) - -
Média/Caudal
e levado
-26.5
(1.4)
5.3
(0.2)
-23.5
(3.1)
7.7
(2.7)
-16.4
(1)
9.1
(0.6)
-25.9
(0.4)
2.9
(2.6)
-31.8
(1.7)
1.7
(2.7)
-30.0
(0.2)
5.65
(0.1)
-33.4
(5.2)
9.10
(1.6)
-29.3
(1.6)
7.3
(1.1)
Ca
ud
al
ba
ixo
1 -22.6
(0.5)
4.7
(0.02)
-16.1
(1.5)
5.04
(0.8)
-16.7
(0.6)
6.5
(1.2)
-20.9
(2.8)
5.0
(1.2)
-28.9
(0.7)
3.9
(2.8)
-27.9
(2.6)
2,97
(2,4) - - - - - - - -
2 -24.0
(1.3)
5.01
(1.6)
-15.6
(1.1)
7.2
(0.4)
-17.8
(0.04)
7.2
(0.1)
-24.6
(0.3)
6.2
(0.4) - - - - - -
-13.9
(0.04)
8.17
(0.1)
-25.9
(1.3)
7.2
(1.9)
-13.9
(0.1)
9.0
(0.1)
3 -25.3
(1.6)
6.1
(0.6)
-19.9
(0.4)
6.5
(2.4) - -
-26.1
(0.4)
4.8
(0.5)
-32.8
(1)
0.1
(1)
-30.2
(0.7)
7,25
(1,3) - -
-36.4
(1.5)
10.02
(0.9)
-30.4
(0.9)
7.5
(1.8) - -
4 -27.7
(1.1)
5.2
(0.3)
-19.9
(0.4)
6.5
(2.4) - -
-26.1
(0.4)
4.8
(0.5)
-28.7
(0.1)
4.6
(0.02)
-29.6
(0.1)
0,15
(0,02) - - -19.5
6.9
(0.02)
-28.6
(0.5)
7.6
(1.4) - -
5 e 6 -27.7
(0.5)
5.6
(1.7)
-19.8
(1.4)
9.0
(2.3) - -
-25.7
(0.8)
4.1
(0.6)
-30.2
(0.04)
1.5
(0.03)
-27.5
(0.04)
0,84
(0,07) - -
-23.8
(0,7)
8.2
(0.3)
-21.6
(0.02)
6.4
(0.1) - -
Média/Caudal
baixo
-25.5
(2.3)
5.3
(0.4)
-18.3
(2.2)
6.9
(1.4)
-17.2
(0.8)
6.9
(0.5)
-24.7
(2.2)
4.9
(0.8)
-30.6
(2.1)
2.5
(2.1)
-28.8
(1.3)
2.80
(3.2)
-14.1
(1.2)
5.0
(0.7)
-23.4
(9.6)
8.3
(1.3)
-26.7
(3.8)
7.2
(0.5)
-13.9
(0.1)
9.0
(0.1)
Tabela 4- Média (±Desvio padrão) dos valores (‰) de δ13C e δ15N, das potenciais fontes de material orgânica, por estação e período de amostragem: material orgânica particulada (MOP); material
orgânica no sedimento (MOS); Plantas terrestres (Terrestres); plantas de via fotossintética C4 (C4): vegetação aquática submersa (VAS) e emergente (VAE).

20
4.4 VALORES ISOTÓPICOS DO ICTIOPLÂNCTON
Figura 5 – Média (± Desvio padrão) dos valores de δ13C e δ15N (‰), do ictioplâncton, ajustado para dois níveis tróficos de fraccionamento (+0.8 ‰ δ13C; +5.9 ‰ δ15N) e das potenciais fontes de material orgânica recolhidas entre Janeiro e Abril (caudal elevado) de 2015 em E1,
E2, E3, E4 e EAD (correspondente às estações de água doce – 5 e 6): Parablenius gattorugine (Pg), Ammodytes tobianus (A), Lipophrys
pholis (Lf), Lepadogaster purpurea (Lp), Centrolabrus exoletus (Ce), Platichthys flesus (Pf), Pomatoschistus microps (Pm), Gobiidae n.i. (G), Solea solea (Ss), Blenniidae n.i. (B), Dicentrarchus labrax (Dl), Atherina presbyter (Ap), Pomatoschistus sp. (P), Cyprinidae n.i. (C). As fontes de matéria orgânica (MO )incluem a MO no sedimento (MOS), MO particulada do fundo (MOPf) e da superfície (MOPs), Epilíton (Epi), vegetação aquática emergentes (VAE), vegetação aquática submersa (VAS), plantas terrestres (T), detritos (D) Fucus sp. (F), plantas de via fotossintética C3 (C3) e C4 (C4) e macroalgas (MA).

21
Figura 6 –Média (± Desvio padrão) dos valores de δ13C e δ15N (‰), do ictioplâncton, ajustado para dois níveis tróficos de
fraccionamento (+0.8 ‰ δ13C +5.9 ‰ δ15N) e das potenciais fontes de material orgânica recolhidas entre Maio e Julho de 2015
(caudal baixo) em E1, E2 , E3, E4 e EAD (correspondente às estações de água doce – 5 e 6): Lipophrys pholis (Lf), Platichthys flesus
(Pf), Pomatoschistus microps (Pm), Gobiidae n.i. (G1 e G2), Dicentrarchus labrax (Dl), Atherina presbyter (Ap), Cyprinidae n.i. (C),
Lepadogaster purpurea (Lp), Symphodus melops (Sm), Labridae n.i. (L), Gobius niger (Gn), Lepadogaster sp. (Lsp), Syngnathidae n.i.
(Sn), Syngnathus acus (Sa) e Labrus mixtus (Lm). As fontes de matéria orgânica (MO) incluem a MO no sedimento (MOS), a MO
partículada do fundo (MOPf) e da superfície (MOPs), epilíton (Epi), vegetação aquática emergente (VAE), vegetação aquática submersa
(VAS), plantas terrestres (T), detritos (D), Fucus sp.(F), plantas de via fotossintética C3 (C3) e C4 (C4) e macroalgas (MA).

22
0
10
20
30
40
50
60
70
80
90
100
Proporção(%)
Fontes
Caudalalto
Não foram observadas diferenças significativas nos valores globais de δ13C e δ15N
das larvas capturadas nem entre períodos diferentes de caudal (Pseudo-F= 2.57; P > 0.05),
nem entre estações de amostragem (Pseudo-F= 4.20; P > 0.05). No entanto, apesar de não
haver, na generalidade uma alteração no tipo de fontes basais de energia que contribuíram
para a biomassa das larvas de peixe, entre períodos de amostragem, o mesmo já não se
verificou entre estações de amostragem (Figs. 5 e 6). Enquanto que, nas estações junto à foz,
as macroalgas estão disponíveis e são aparentemente utilizadas, nas estações mais a
montante, parece haver uma predominância na disponibilidade e assimilação material com
origem em plantas vasculares (Figs. 5 e 6). De uma maneira geral, os valores médios de δ13C
são mais elevados nos meses de caudal mais alto (-19.1 ± 2.2‰), do que nos meses de
caudal mais baixo (-20.1 ± 2.4‰) e os valores médios (±DP) de δ15N, foram superiores nos
meses de caudal baixo (6.4 ± 3.1‰), do que nos meses de caudal alto (4.7 ± 2.2‰ ).
Figura 7 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o caudal elevado, correspondente aos meses de
Janeiro, Fevereiro e Março, na estação 1, baseado no modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As
fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica partículada (MOP) e matéria orgânica no sedimento (MOS).
Pla t i ch thys
f lesus
Cent ro lab rus
exoletus
Lepadogaster
purpurea
L ipophrys
pho l is
Parab lennius
ga tto rugine
Ammodytes
tobianus

23
0102030405060708090100
Epilíton
Macroalgas
MOP
SOM
Terrestre
Epilíton
Macroalgas
MOP
SOM
Terrestre
Epilíton
Macroalgas
MOP
SOM
Terrestre
Epilíton
Macroalgas
MOP
SOM
Terrestre
Epilíton
Macroalgas
MOP
SOM
Terrestre
Epilíton
Macroalgas
MOP
SOM
Proporção(%)
Fontes
Caudalbaixo
Em E1, nos meses de caudal alto, as larvas de peixe analisadas parecem estar a
alimentar-se de organismos que se alimentam de uma mistura de fontes na coluna de água e
no sedimento e que incluem as macroalgas (MA), matéria orgânica particulada (MOP), matéria
orgânica no sedimento (MOS) e epilíton (Epi) (Fig. 5-E1). Nos meses de caudal mais baixo, a
mistura de fontes basais utilizada, parece ser semelhante. No entanto, os valores médios de
δ15N de P. flesus (2.9 ± 0.7‰) e de S. melops (2.6 ± 0‰), sugerem que estes poderão estar a
alimentar-se de organismos que utilizarão também material com origem terrestre ou detritos
(Fig. 6-E1). Os resultados do modelo de mistura apoiam esta interpretação. Assim, nos meses
de caudal alto, o epilíton foi a fonte com maior contributo para a biomassa deP. flesus (11-
95%) e de L. purpurea (11-68%; Fig. 7). As macroalgas tiveram também um importante
contributo para a biomassa de L. purpurea (17-55%) e A. tobianus (6-52%;Fig. 6). No período
de caudal mais baixo, e de acordo com os resultados do SIAR, o epilíton foi novamente uma
fonte relevante, com contributo até 50% para a biomassa das larvas analisadas (Fig.8). A
matéria orgânica com origem terrestre revelou-se um contributo importante para a biomassa
de P. flesus (0-31%) e S. melops (0-39%;Fig. 8).
Pla t i ch thys f le sus Symphodus
me lops
Lepadogaster
purpurea
L ipophrys pho l is Gob ius n iger Labr idae sp.
Figura 8 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o caudal baixo, correspondente aos meses de
Abril, Maio, Junho e Julho, na estação 1, baseado no modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As
fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica partículada (MOP), matéria orgânica no sedimento (MOS) e plantas
de origem terrestres.
24
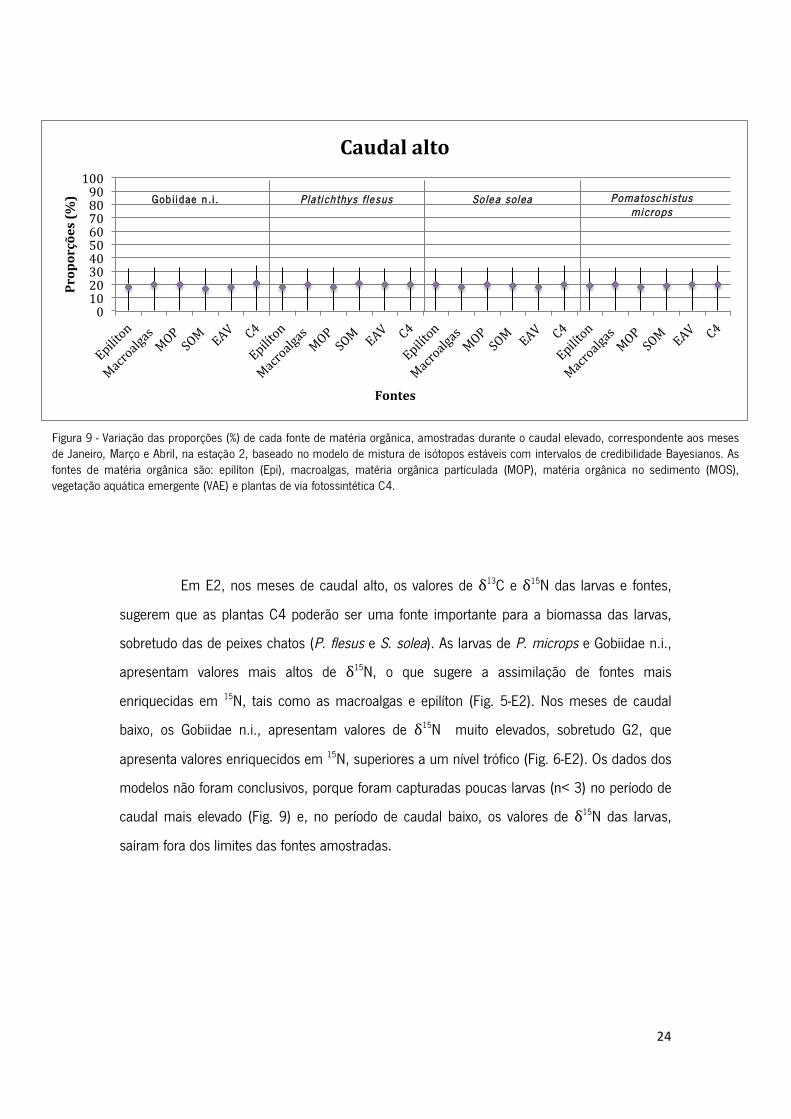
Em E2, nos meses de caudal alto, os valores de δ13C e δ15N das larvas e fontes,
sugerem que as plantas C4 poderão ser uma fonte importante para a biomassa das larvas,
sobretudo das de peixes chatos (P. flesus e S. solea). As larvas de P. microps e Gobiidae n.i.,
apresentam valores mais altos de δ15N, o que sugere a assimilação de fontes mais
enriquecidas em 15N, tais como as macroalgas e epilíton (Fig. 5-E2). Nos meses de caudal
baixo, os Gobiidae n.i., apresentam valores de δ15N muito elevados, sobretudo G2, que
apresenta valores enriquecidos em 15N, superiores a um nível trófico (Fig. 6-E2). Os dados dos
modelos não foram conclusivos, porque foram capturadas poucas larvas (n< 3) no período de
caudal mais elevado (Fig. 9) e, no período de caudal baixo, os valores de δ15N das larvas,
saíram fora dos limites das fontes amostradas.
0
10
20
30
40
50
60
70
80
90
100
Proporções(%)
Fontes
Caudalalto
Gob iidae n . i . Pla t i ch thys f le sus So lea solea Pomatoschis tus
microps
Figura 9 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o caudal elevado, correspondente aos meses
de Janeiro, Março e Abril, na estação 2, baseado no modelo de mistura de isótopos estáveis com intervalos de credibilidade Bayesianos. As
fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica partículada (MOP), matéria orgânica no sedimento (MOS),
vegetação aquática emergente (VAE) e plantas de via fotossintética C4.

25
0
20
40
60
80
100
Proporção
Fontes
Caudalbaixo
Figura 10 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o caudal elevado, correspondente aos
meses de Janeiro, Fevereiro e Março e Abril, na estação 3, baseado no modelo de mistura de isótopos estáveis com intervalos de
credibilidade Bayesianos. As fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica partículada (MOP) e matéria
orgânica no sedimento (MOS).
Athe r ina presbyte r Lepadogaster sp. Labrus mix tus
Figura 11 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o caudal baixo,
correspondente aos meses de Maio, Junho e Julho na estação 3, baseado no modelo de mistura de isótopos estáveis com
intervalos de credibilidade Bayesianos. As fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica
particulada (MOP), matéria orgânica no sedimento (MOS) e detritos.
0
10
20
30
40
50
60
70
80
90
100
Epilíton
Macroalgas
MOP
SOM
Epilíton
Macroalgas
MOP
SOM
Epilíton
Macroalgas
MOP
SOM
Epilíton
Macroalgas
MOP
SOM
Epilíton
Macroalgas
MOP
SOM
Epilíton
Macroalgas
MOP
SOM
Epilíton
Macroalgas
MOP
SOM
Proporções(%)
Fontes
Caudalalto
Lepadogaster
pupurea
Blenn i idae n . i . Dicen tra rchus
labrax
Athe r ina
presbyter
Pomatoschis tus
sp.
Cent ro lab rus
exoletus
P la t i ch thys
f lesus

26
Em E3, nos meses de caudal mais alto, os valores de δ13C das larvas e fontes,
sugerem que as macroalgas poderão ser uma fonte importante para a biomassa das larvas
(Fig. 5-E3). Nos meses de caudal baixo, existem quatro grupos de taxa, quanto aos rácios
isotópicos. Um grupo será constituído pelas larvas de P. microps e S. acus, que estarão a
utilizar uma mistura de epilíton ou plantas vasculares e MOP; um grupo mais enriquecido em
15N, constituído pelos Gobiidae n.i e Syngnathidae n.i., que se estará a alimentar de outras
larvas ou fontes mais enriquecidas não amostradas; um terceiro grupo que se estará a
alimentar de uma mistura de epilíton, MOP, MOS, detritos, e que é constituído pelos taxa
Lepadogaster sp., S. melops e A. presbyter (Fig. 11); e um quarto grupo de indivíduos que
apresenta valores mais elevados de δ13C em relação às fontes capturadas, indicando a
assimilação de MO mais enriquecida em 13C do que aquela amostrada (Fig. 6-E3). Os
resultados do modelo de mistura apoiam esta interpretação. Assim, nos meses de caudal alto,
as macroalgas foram a fonte com maior contributo para a biomassa de L. purpuria (6-57%) e o
epilíton para a biomassa de P. flesus (0-56%) (Fig. 9). Nos meses de caudal baixo, os dados
do modelo não foram conclusivos, porque foram capturadas poucas larvas (n< 3) (Fig. 10).
Em E4, nos meses de caudal mais alto, não foi possível identificar as fontes de
matéria orgânica assimiladas pelas larvas de peixe, pois todas apresentaram valores de δ13C e
de δ15N mais elevados do que as fontes amostradas (Fig. 5-E4). Nos meses de caudal baixo,
existem dois grupos de taxa, quanto aos rácios isotópicos. Um grupo será constituído pelas
larvas de Cyprinidae n.i. e Gobiidae n.i., que estarão a utilizar uma mistura de plantas
vasculares, detritos, MOP e MOS; um segundo grupo constituído por D. labrax e P. flesus, que
se estão a alimentar de epilíton ou plantas vasculares de origem aquática. Os dados dos
modelos não foram conclusivos, porque foram capturadas poucas larvas (n< 3) no período de
caudal mais elevado (Fig. 11).

27
0
20
40
60
80
100
Proporções(%)
Fontes
Caudalbaixo
Em EAD, nos meses de caudal alto, as larvas analisadas parecem estar a alimentar-
se de organismos que se alimentam de uma mistura de fontes na coluna de água e no
sedimento e que incluem MOP, Epi, VAS, VAE, detritos e MOS (Fig. 5-EAD). A espécie P. flesus
não apresenta correspondência isotópica com nenhuma das fontes de matéria orgânica, pois
apresentaram valores de δ13C e de δ15N mais elevados do que as fontes amostradas (Fig. 5-
EAD). Nos meses de caudal baixo as larvas de A. presbyter e P. flesus que estarão a utilizar
0
20
40
60
80
100
Epilíton MOP SOM VAE VAS
Prporção(%)
Fontes
Caudalalto
Cyprinidaen.i.
Figura 12 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o caudal elevado, correspondente
aos meses de Janeiro, Fevereiro e Março e Abril, nas estações 5 e 6, baseado no modelo de mistura de isótopos estáveis com
intervalos de credibilidade Bayesianos. As fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica particulada
(MOP), matéria orgânica no sedimento (MOS), vegetação aquática emergente (VAE) e vegetação aquática submerge (VAS).
Platichthysflesus Atherinapresbyter
Figura 13 - Variação das proporções (%) de cada fonte de matéria orgânica, amostradas durante o caudal baixo, correspondente
aos meses de Maio, Junho e Julho nas estações 5 e 6, baseado no modelo de mistura de isótopos estáveis com intervalos de
credibilidade Bayesianos. As fontes de matéria orgânica são: epilíton (Epi), macroalgas, matéria orgânica partículada (MOP),
matéria orgânica no sedimento (MOS) e plantas de origem terrestre.

28
uma mistura de epilíton, plantas vasculares, MOP ou MOS. Os dados dos modelos não foram
conclusivos, porque foram capturadas poucas larvas (n< 3) no período de caudal mais baixo
(Fig. 13).

29
5 DISCUSSÃO
5.1 ABUNDÂNCIA LARVAR
Verificou-se que um pequeno número de taxa, como Gobiidae n.i e Ammodytes
tobianus, apresentou um elevado número de indivíduos, enquanto que a maioria dos taxa
identificados apresentou, de uma maneira geral, valores de abundância baixos ao longo do
período de estudo. Esta é uma característica comum na generalidade dos estuários (Harris e
Cyrus, 1995; Whitfield, 1999; Ramos et al., 2006), visto que poucas espécies têm a
capacidade de sobreviver e de se desenvolver em ambientes com a enorme variabilidade
ambiental existente nos ecossistemas estuarinos, ou pelo facto dos estuários serem
ecossistemas recentes do ponto de vista geológico (McLusky, 1989).
Algumas espécies foram capturadas durante todo o ano como Platichthys flesus,
Lepadogaster purpurea, Solea solea, Lipophrys pholis, Atherina presbyter, Centrolabrus
exoletus e espécies pertencentes ao género Pomatoschistus, enquanto que outras só
apareceram em determinadas períodos. Os taxa capturados exclusivamente durante os meses
de caudal alto foram a Ammodytes tobianus e Gobiesocidae sp. e os taxa capturados apenas
durante os meses de caudal baixo foram os Cyprinidae sp., Dicentrarchus labrax, Parablennius
gattorugine, Labridae sp., Pomatoschistus microps, Lepadogaster sp., Gobius niger,
Syngnathus sp., Sparidea n.i., Clupeidea n.i., Symphodus melops e Labrus mixtus. Gobiidae
n.i. foi o taxon mais frequente em todas as estações de amostragem, à exceção de E6, que
foi dominada por indivíduos do género Cyprinidae. A dominância de Gobiidae n.i. pode ser
explicada pelo facto de algumas espécies deste género, como Pomatoschistus microps, terem
a capacidade de desovar várias vezes ao logo do ciclo reprodutivo e por apresentarem uma
grande capacidade de adaptação a diferentes condições abióticas (Bouchereau e Guelorget,
1998; Souza et al., 2014a). Outra explicação, prende-se com o facto de possuírem ovos
demersais, (reduzindo o risco de mortalidade associado com a dispersão descontrolada de
ovos e larvas para fora do estuário) (Whitfield, 1990), ovos e larvas bentónicas (organismos
mais desenvolvidos e menos dependentes do alimento disponível, quando eclodem) (Faria et
al., 2006) e logo depois da eclosão assentam-se no substrato, o que dificulta a sua captura
(Nonaka, 2000).
A abundância larvar e a riqueza específica foram superiores nos meses em que o
caudal foi mais baixo (abundância: 16.3 ± 19.7 ind/100m3; riqueza específica: 16 ± 8.2),

30
comparativamente aos registados nos meses de caudal mais alto (abundância: 11.6 ± 10.0
ind/100m3; riqueza específica: 3.5 ± 3.5). Vários factores poderão explicar os resultados
encontrados. Em primeiro lugar, quando o caudal é mais elevado, ocorre uma diminuição da
temperatura e da salinidade da água, o que poderá dificultar o recrutamento das espécies
marinhas para o estuário (Ramos et al., 2007). Por outro lado, a disponibilidade de alimento
poderá também influenciar a distribuição do ictioplâncton. É no final da Primavera e durante o
Verão, que aumenta a produtividade de zooplâncton (o alimento preferencial das larvas; Ré,
1999), na costa portuguesa e nos seus estuários (Cunha, 1993; Cabeçadas et al., 1999).
Embora o zooplâncton não tenha sido quantificado, estudos realizados neste ecossistema
indicam que, de uma forma geral, a diversidade e abundância de zooplâncton é mais elevada
entre o final da Primavera e durante o Verão (Vieira et al., 2015).
A estação que apresentou maiores valores de abundância larvar foi E4 (32.3 ± 34.8
ind/100m3). Esta estação situa-se na zona de transição de salinidade no estuário, podendo
coincidir com a zona de turbidez máxima do estuário. No entanto, são necessários estudos
que o comprovem. As zonas de turbidez máxima têm sido identificadas como importantes
áreas de berçário (Boynton et al., 1997; North e Houde, 2003; Winkleretal et al., 2003), uma
vez que a turbidez desempenha um papel relevante na sobrevivência de peixes estuarinos
(Kattenfeld, 1997; Whitfield, 1999), pois apresentam uma elevada produção/retenção de
zooplâncton (Kimmerer et al., 1998; Roman et al., 2001).
A estação que apresentou maior riqueza específica foi E1 (18.5 ± 13.4) devido à
captura de um grande número de espécies com origem marinha. Como o ictioplâncton tende
a entrar no estuário por transporte passivo, isto é, por ação da corrente (Ré, 1999), e as
amostragens decorreram durante a enchente da maré, é esperada uma maior riqueza
específica nesta zona. A estação que apresentou menor riqueza específica foi E6 (3.0 ± 2.8), o
que pode dever-se ao facto de existir pouca diversidade de espécies dulciaquícolas, no
estuário do rio Minho (Sousa et al., 2008).O número de taxa (espécies e géneros) identificados
durante este estudo foi inferior ao encontrado em estudos realizados noutros ecossistemas
nomeadamente no estuário do rio Lima (50 taxa; Ramos et al., 2006), Mondego (28 taxa;
Ribeiro, 1991) ou Guadiana (34 taxa; Faria et al., 2006). Apesar de existirem diferenças nas
condições de amostragem, uma explicação possível para estas diferenças poderá dever-se às
diferenças nos valores dos caudais (Whitfield e Harrison, 2003; Faria et al., 2006), que no
estuário do rio Minho é superior (300 m3s-1; Ferreira et al., 2003), em relação ao rio Guadiana

31
(180 m3s-1; Faria, 2006) e ao rio Lima (70 m3s-1; Ramos, 2007). O elevado caudal poderá
dificultar a retenção das larvas no interior do estuário, que são exportadas para a zona
costeira(Faria et al., 2006). No entanto, espera-se que a longo praxo, com o aumento do
caudal, possa haver um aumento da abundância de peixes na zona costeira adjacente ao
estuário. Tal foi observado na espécie Engraulis encrasicolus (anchova), no estuário do rio
Guadiana (Chícharo et al., 2001).
5.2 VARIABILIDADE ESPACIAL E TEMPORAL DAS FONTES BASAIS DE ENERGIA
QUE SUPORTAM A BIOMASSA DAS LARVAS
Os valores de δ13C e δ15N das larvas analisadas revelam que a sua energia provém de
diferentes tipos de fontes e com diferentes origens. A contribuição das fontes autóctones foi
superior nas estações mais junto à foz e nos meses de caudal baixo (28.7%) em relação aos
meses de caudal alto (25.8%). A contribuição da matéria orgânica com origem terrestre foi
superior nas estações mais afastadas da foz e a contribuição da matéria orgânica com origem
marinha foi superior nas estações mais próximas da foz. Para além disto, algumas larvas (eg.
P. flesus) apresentaram valores de δ13C que não eram consistentes mais enriquecidos em 13C
do que com os valores das fontes amostradas, sugerindo que a sua chegada a essas estações
terá sido recente. Assim, verificou-se que as larvas no estuário do rio Minho, dependem
bastante de fontes com uma origem diferente daquela onde foram capturadas.
O contributo de matéria orgânica com origem marinha e terrestre terá sido mais
elevado nos meses de caudal mais alto (25% e 22%, respetivamente) do que nos meses de
caudal mais baixo (12% e 28%, respetivamente). Apesar de não ter sido determinada a
quantidade de carbono com origem terrestre que terá entrado no estuário durante o período
do estudo, os valores de C/N da matéria orgânica particulada (MOP) fornecem informação
relativamente à sua qualidade (Hedges et al., 1986; Hedges et al., 1997). Assim, os valores
de C/N do MOP durante o período de caudal alto variaram entre 11 (E3) e 14 (E1) nas
estações junto à foz e de ca. 14 nas estações de água doce (E5 e E6), indicando uma forte
contribuição da matéria orgânica de origem terrestre para o MOP (C/N > 10; Hedges et al.,
1986; Hedges et al., 1997). Pelo contrário, no período de caudal mais baixo, os valores de
C/N do MOP variaram entre 5 e 6 ao longo do estuário. Como estes valores se aproximam do
rácio de Redfield (≈7), o MOP poderá ter tido uma forte contribuição de fitoplâncton, durante

32
este período (Hedges et al., 1986; Hedges et al., 1997). É esperado que, quando o caudal
baixa, o tempo de residência aumente, aumentado assim a acumulação de fitoplâncton
produzido localmente (Sin et al., 1999). Para além disto, os valores de Chla, indicam que terá
havido um aumento na disponibilidade de fitoplâncton a partir de Abril 2015, tendo-se atingido
os valores mais elevados de Chla, na maioria das estações, em Junho 2015 (entre 13.7µg. L-
1 e 21.1µg. L-1 à superfície e entre 4.4µg. L
-1 e 5.5µg. L-1 no fundo). No entanto, este
aumento na disponibilidade de fitoplâncton, não se refletiu nos tecidos das larvas capturadas,
pois não houve um aumento generalizado da proporção de MOP assimilada neste período. Era
também esperado que o contributo de matéria orgânica com origem marinha fosse superior
nos meses de caudal baixo, mas tal não se refletiu nos tecidos das larvas amostradas (caudal
alto: 25.3%; caudal baixo: 12.0%).
Contrariamente ao esperado, durante o período de caudal mais baixo, houve uma
importante contribuição de plantas, e seus detritos, para a biomassa das larvas,
nomeadamente para a P. flesus (0-30%) e S. melops (0-40%) em E1, para L. mixtus e
Lepadogaster sp. (0-50%) em E3, e para P. flesus (0-50%) nas estações de água doce. Apesar
do zooplâncton ser geralmente considerado a presa preferencial das larvas de peixes, algumas
espécies poderão alimentar-se de invertebrados bentónicos (e.g. larvas de insecto), o que
permitirá a transferência da matéria orgânica com origem terrestre para os níveis tróficos
superiores (Hoffman et al., 2015). Embora este tipo de presas não esteja descrito para as
larvas de P. flesus, esta espécie sofre metamorfose na fase larvar onde, entre outras coisas,
adquire um hábito demersal (Summers, 1979). Como o estuário do rio Minho é um estuário
de baixa profundidade (Antunes et al. 2011), estas presas poderão estar mais disponíveis para
serem capturadas.. Contudo, à semelhança do que foi observado noutros ecossistemas (Cole
et al., 2011) estudos recentes no estuário do rio Minho indicam que, também o zooplâncton
poderá ter a capacidade para assimilar matéria orgânica com origem terrestre (Dias, comm.
pess).
O epilíton e a matéria orgânica no sedimento (MOS) foram as fontes de MO que mais
contribuíram, e de uma forma constante, para a biomassa das larvas de peixe capturadas.
Assim, estes resultados indicam que há uma forte dependência destas espécies, pelo menos
na fase larvar, da cadeia trófica bentónica do estuário. A razão para este facto poderá estar
relacionada com a aparente baixa disponibilidade de fitoplâncton na coluna de água neste
ecossistema. Estudos anteriores revelaram que a concentração média anual de Chla é de 1.3

33
µg.L-1 (Brito et al., 2012). Os picos de Chla observados em Junho, poderão refletir a
ocorrência de um bloom de fitoplâncton.
Foi possível também observar que, algumas espécies, apresentavam valores de δ13C e
δ15N mais elevados do que as fontes amostradas, sugerindo que, ou as fontes possíveis não
foram todas amostradas, ou o local de captura não foi o mesmo onde se alimentaram do
material assimilado. Este facto é particularmente evidente sobretudo quando se analisam os
valores de δ13C para as espécies L.purpurea, D.labrax, C.exoletus, P.flesus e Blenniidae n.i.,
que apresentaram valores entre 1 a 8‰ mais enriquecidos em 13C do que a fonte mais
enriquecida (MA; 16.8 ± 0.9‰). Os tecidos das larvas de peixe demoram entre 10 a 20 dias
para que ocorra o turnover de 90% do seu tecido (Hoffman et al., 2011). Assim, existem duas
possibilidades para os resultados observados: ou houve deslocação de MO proveniente de
uma cadeia trófica mais enriquecida em 13C para as zonas amostradas, ou houve uma
deslocação recente das larvas para esses locais. O facto de as larvas apresentarem
constantemente valores mais enriquecidos em 13C, suporta a hipótese de que estas se
deslocaram para esses locais e ainda não assimilaram a MO aí existente. No caso da P.
flesus, que apresentou valores mais enriquecidos em 13C do que todas fontes amostradas,
sobretudo nas estações de água doce, sugere que estas terão migrado de uma zona de maior
salinidade, como a zona costeira para o interior do estuário, e posteriormente, terão migrado
para as zonas de água doce (Bos, 1999). No entanto, apesar de apenas terem sido analisadas
larvas com a vesícula vitelina aparentemente reabsorvida, não se pode descartar a hipótese de
que ainda exista algum efeito materno nos valores dos rácios de isótopos estáveis dos seus
tecidos, ou que a progenitora tenha migrado recentemente da costa, para se reproduzir na
zona de água doce do estuário (sea-run mother hypothesis), pois estudos recentes sugerem
que a desova, nesta espécie, poderá ocorrer em zonas de água salobra ou mesmo doce
(Daverat et al., 2012).

34
6 CONCLUSÃO
Com este estudo foi possível pela primeira vez, inferir qual a importância dos habitats
estuarinos, e dos ecossistemas adjacentes, para o crescimento e desenvolvimento das larvas
de peixes numa importante área berçário, o estuário do rio Minho. Foi também possível, pela
primeira vez, identificar e avaliar a abundância e distribuição da comunidade de larvas de
peixes neste ecossistema. Verificou-se que o caudal teve um papel determinante na
abundância e diversidade específica, uma vez que estas foram superiores nos meses de
caudal mais baixo. Este facto que pode ser explicado pela existência de um aumento de
alimento, temperatura e salinidade, durante este período. Abundância no estuário do rio Minho
ficou marcada pela dominância de indivíduos da família Gobiidae.
As larvas de peixe neste ecossistema dependem predominantemente de fontes de
matéria orgânica (MO) autóctones (e.g. epilíton, MOS). Ao contrário do esperado, o caudal não
teve um papel determinante no tipo de fontes basais de energia que suportaram a biomassa
larvar. Foi possível constatar que, o ecossistema terrestre tem um contributo importante para
as cadeias tróficas nas quais as larvas de peixe se alimentam, visto que foi assimilada MO
com origem terrestre, independentemente da sua maior ou menor disponibilidade. Durante os
meses de caudal mais baixo, quando a disponibilidade de fitoplâncton foi maior verificou-se
que a MO com origem terrestre, contribui para a biomassa das espécies capturadas ao longo
do gradiente de salinidade estuarino, e não só nas estações de água doce, como seria
expectável. Da mesma forma, não foi observado um aumento da contribuição de fontes com
origem marinha (por exemplo, as macroalgas, ou de MOP enriquecida em 13C), com a
diminuição do caudal. No entanto, verificou-se que o ecossistema marinho, subsidia as
cadeias tróficas da região mais próxima da foz.
Foi possível também observar que as larvas de peixe, no estuário do rio Minho,
dependem não só da cadeia trófica pelágica, através do consumo de matéria orgânica
particulada (MOP), mas também são dependentes da cadeia trófica bentónica, uma vez que o
epilíton e o MO no sedimento (MOS) foram as fontes que, de uma maneira geral, tiveram um
maior contributo para a sua biomassa.
Desta forma, conclui-se que, apesar dos habitats do estuário do rio Minho
funcionarem como um importante local para o crescimento e desenvolvimento de várias

35
espécies de peixes, as suas cadeias tróficas são subsidiadas pelos ecossistemas adjacentes.
Assim, qualquer plano de gestão ou conservação neste estuário, terá que incluir também, os
ecossistemas marinho, mas sobretudo o terrestre, no planeamento.

36
7 REFERÊNCIAS BIBLIOGRÁFICAS
Abrahams, M., Kattenfeld, M. (1997). The role of turbidity as a constraint on predator-prey
interactions in aquatic environments. Behavioral Ecology Sociobiology 40:169-174.
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance.
Austral Ecology 26: 32-46.
Anderson, M. J., Gorley, R. N., Clarke, K. R. (2008). Plymouth: primer-E; 2008. PERMANOVA+
for PRIMER: guide to software and statistical methods. 214.
Antunes, C., Rodrigues, H. (2004). Guia Natural do Rio Minho: Os Peixes. Aquamuseu do Rio
Minho. Vila Nova de Cerveira – Portugal. 86 pgs.
Antunes, C., Araújo, M. J., Braga, C., Roleira, A., Carvalho, R., Mota, M. (2011). Valorização
dos recursos naturais da bacia hidrográfica do rio Minho. Final report from the project Natura
Miño-Minho, Centro interdisciplinar de Investigação Marinha e Ambiental, Universidade do
Porto.
Altominho.pt – disponível em http://www.altominho.pt/gca/?id=858; consultado em
03/10/2015.
Alves, A. M. (1996). Causas e processos da dinâmica sedimentar na evolução atual do litoral
do Alto Minho. Tese de doutoramento, Universidade do Minho: Portugal, 442 pgs.
Beck, M. W., Heck Jr. K. L., Able, K. W., Childers, D.L., Eggleston, D. B., Gillanders, B. M.,
Halpern, B., Hays, C. G., Hoshino, K., Minello, T. J., Orth, R. J., Sheridan, P. F., and Weinstein,
M. P. (2001). The identification, conservation, and management of estuarine and marine
nurseries for fish and invertebrates. BioScience 51: 633–641
Boehlert, G. W., Mundy, B. C. (1988). Roles of behavioral and physical factors in larval and
juvenile fish recruitment to estuarine nursery areas. American Fisheries Society Symposium 3,
51–67.
Bos, A. R. (1999). Tidal transport of flounder larvae (Pleuronectes flesus) in the Elbe River,
Germany. Arch. Fish. Mar. Res. 47, 47–60.
Bouchereau, J. L., Guelorget, O. (1998). Comparison of three Gobiidæ (Teleostei) life history
strategies over their geographical range. Oceanologica Acta 21(3):503–517. doi:10.1016/
S0399-1784(98)80034-0.
Boynton, W. R., Boicourt, W., Brandt, S., Harding, L., Holliday, Y., Jech, M., Kemp, M.,
Lascara, C., Leach, S., Madden, A., Roman, M., Sanford, S., Smith, E. (1997). Interactions

37
between physics and biology in the estuarine turbidity maximum (ETM) of Chesapeake Bay,
USA. ICES CM 1997/S:11.
Brito, A. C., Brotas, V., Caetano, M., Coutinho, T. P., Bordalo, A. A., Icely, J., Neto, J. M.,
Serôdio, J., Moita, T. (2012). Defining phytoplankton class boundaries in Portuguese
transitional waters: an evaluation of the ecological quality status according to the water
framework directive. Ecological Indicators 19:5–14. doi:10.1016/j.ecolind.2011.07.025.
Cabeçadas, G., Nogueira, M., Brogueira, M.J. (1999). Nutrient dynamics and productivity in
three European estuaries. Marine Pollution Bulletin 38 (12), 1092-1096.
Chícharo, M. A., Esteves, E., Andrade, J. P., Morais, P. (2001). Effects of alterations in fresh
water supply on the abundance and distribution of Engalius encrasicolus in the Guadiana
estuary and adjacent coastal areas of South Portugal. Ecohydrology and Hydrology 3, 341-345.
Cloern, J. E., Canuel, E. A., Harris. D. (2002). Stable carbon and nitrogen isotope composition
of aquatic and terrestrial plants of the San Francisco Bay estuarine system. Limnology and
Oceanography 47:713–729.
Costa-Dias, S., Freitas, V., Sousa, R., Antunes, C. (2010). Factors influencing epibenthic
assemblages in the Minho Estuary (NW Iberian Peninsula). Marine Pollution Bullerin 61:240-
246.
Costanza, R., d'Arge, R., de Groot, R., Farber, S., Grasso, M., Hannon, B., Limburg, K.,
Naeem, S., O'Neill, R., Paruelo, J., Raskin, R., Sutton, P., van den Belt, M. (1997). The value of
the world's ecosystem services and natural capital. Nature 387:253-260. In: Ramos, S.
(2007). Icthyoplankton of the Lima estuary (NW-Portugal): Ecology of the early life stages of
Pleuronectiformes. Tese de Mestrado, Universidade do Porto: Porto, 228 pgs.
Cunha, M. E. (1993). Variabilidade estacional do zooplâncton na plataforma continental
portuguesa. Boletim UCA Universidade do Algarve 1, 229 e 241. In: Faria, A., Morais, P.,
Chícharo, M. A. (2006). Ichthyoplankton dynamics in the Guadiana estuary adjacent coastal
area, South-East Portugal. Estuarine, Coastal and Shelf Science 70, 85-97.
Currin, C. A., Newell, S. Y., Paerl, H. W. (1995). The role of standing dead Spartina alterniflora
and benthic macroalgae in salt marsh food webs: considerations based on multiple stable
isotope analysis. Marine Ecology Progress Series 121, 99-116.
Deegan, L. A., Garritt, R. H. (1997). Evidence for spatial variability in estuarine food webs.
Marine Ecology Progress Series 147: 31–47.
Deverat, F., Morais, P., Dias, E., Babaluk, J., Martin, J., Eon, M., Fablet, R., Peycheran, C.,

38
Antunes, C. (2012). Plasticity of European flounder life history patterns discloses alternatives to
catadromy. 465:267-280.
Dias, E., Morais, P., Antunes, C., Hoffman, J. C. (2014). Linking terrestrial and benthic
estuarine ecosystems: organic matter sources supporting the high secondary production of a
non-indigenous bivalve. Biological invasions, DOI 10.1007/s10530-014-0655-8.
Elliott, M., O’Reilly, M. G., Taylor, C. J. L. (1990). The Forth estuary: a nursery and
overwintering area for North Sea fishes. Hydrobiologia 195: 89-103.
Faria, A., Morais, P., Chícharo, M. A. (2006). Ichthyoplankton dynamics in the Guadiana
estuary adjacent coastal area, South-East Portugal. Estuarine, Coastal and Shelf Science 70,
85-97.
Ferreira, J. G., Simas, T., Nobre, A., Silva, M. C., Schifferegger, K., Lencart-Silva, J. (2003).
Identification of sensitive areas and vulnerable zones in transitional and coastal Portuguese
systems. Application of the United States National Estuarine Eutrophication Assessment to the
Minho, Lima, Douro, Ria de Aveiro, Mondego, Tagus, Sado, Mira, Ria Formosa and Guadiana
systems. INAG/IMAR Technical Report.
Fishbase – disponível em http://www.fishbase.org/TrophicEco/FishEcoList.php?ve_code=13;
consultado em 10/12/2015.
Feuchtmayer, H., Grey, J. (2003). Effect of preparation and preservation procedures on carbon
and nitrogen stable isotope determinations from zooplankton. Rapid Communications in Mass
Spectrometry 17: 2605–2610.
Fisher, L., Pereira, L. E., Vieira, J. (2011). Peixes estuarinos e costeiros. “Ed.2”. Rio grande,
Portugal.
França, S., Vasconcelos, R. P., Tanner, S., Máguas, C., Costa, M. J., Cabral, H. N. (2011).
Assessing food web dynamics and relative importance of organic matter for fish species in two
Portuguese estuaries: A stable isotope approach. Marine Environmental Research 72 204-215.
France, R. L. (1995). Carbon-13 enrichment in benthic compared to planktonic algae: food
web implications. Marine Ecology Progress Series 124:307-312.
Freitas, V., Costa-Dias, S., Campos, J., Bio, A., Santos, P., Antunes, C. (2009). Patterns in
abundance and distribution of juvenile flounder, Platichthys flesus, in Minho estuary (NW
Iberian Peninsula). Aquatic Ecology 43, 1143–1153.
Fry, B., Sherr, E. B. (1984). 13C measurements as indicators of carbon flow in marine and
freshwater ecosystems. Contributions in Marine Science 27: 13-47

39
Harris, S. A., Cyrus, D. P. (1995). Occurrence of larval fishes in the St. Lucia estuary, KwaZulu-
Natal, South Africa. South African Journal of Marine Science 16:333-350.
Harris, S. A., Cyrus, D. P., Beckley, L. E. (1999). The larval fish assemblage in nearshore
coastal waters off the St Lucia estuary, South Africa. KwaZulu-Natal, South Africa. South
African Journal of Marine Science 49:789-811.
Hedges, J. I., Clark, W. A., Quay P. D., Richey, J. E., Devol A. H., Santos U. M. (1986).
Compositions and fluxes of particulate organic material in the Amazon River. Limnology and
Oceanography 31:717-738.
Heip, C. H. R., Herman, P. M. J. (1995). Major biological processes in European tidal
estuaries: a synthesis of the JEEP-92 project. Hydrobiologia 311:1-7.
Herzka, S. Z. (2005). Assessing connectivity of estuarine fishes based on stable isotope ratio
analysis. Estuarine, Coastal and Shelf Science 64:58-69.
Hoffman, J. C., Bronk, D. A. (2006). Interannual variation in stable carbon and nitrogen
isotope biogeochemistry of the Mattaponi River, Virginia. Limnology and Oceanography
51:2319–2332.
Hoffman, J. C., Bronk, D. A., Olney, J. E. (2008). Organic matter supporting lower food web
production In the tidal freshwater portion of the York river estuary, Virginia. Coastal and
estuarine research federation.
Hoffman, J. C., Sutton, T. T. (2010). Lipid correction for carbon stable isotope analysis of
deep-sea fishes. Deep sea Research Part I: Oceanography Research Papers 57:956–964.
Hoffman, J. C., Cotter, A. M., Peterson, G. S. & Kelly, J. R. (2011). Rapid stable isotope
turnover of larval fish in a Lake Superior coastal wetland: implications for diet and life history
studies. Aquatic Ecosystem Health and Management 14:403-413.
Hoffman, J. C., Kelly, J. R., Peterson, G. S., Cotter, A. M. (2015). Landscape-Scale food webs
of fish nursery habitat along a river-coast mixing zone. Estuarine and Coasts 38: 1335-1349.
Kimmerer, W. J., Burau, J. R., Bennett, W. A. (1998). Tidally oriented vertical migration and
position maintenance of zooplankton in a temperate estuary. Limnology and Oceanography
43:1697–1709
Leggett, W. C., Deblois, E. (1994). Recruitment in marine fishes: is it regulated by starvation
and predation in the egg and larval stages? Netherlands Journal of Sea Research 32 (2), 119-
134. In: Faria, A., Morais, P., Chícharo, M. A. (2006). Ichthyoplankton dynamics in the
Guadiana estuary adjacent coastal area, South-East Portugal. Estuarine, Coastal and Shelf

40
Science 70, 85-97.
Le Pape, O., Modéran, J., Beaunée, G., Riera, P., Nicolas, D., Savoye, N., Harmelin-Vivien, M.,
Darnaude, A. M., Brind´Amour, A., Le Bris, H., Cabral, H., Vinagre, C., Pasquaud, S., França,
S., Kostecki, C. (2012). Sources of organic matter for flatfish juveniles in coastal and estuarine
nursery grounds: A meta-analysis for the common sole (Solea solea) in contrasted systems of
Western Europe. Journal of Sea Research.
Lorenzen, C. J. (1967). Determination of chlorophyll and pheo-pigments: spectrophotometric
equations. Limnology and Oceanography 12:343-346.
Lorrain, A., Savoye, N., Chauvaud, L., Paulet, Y., Naulet, N. (2003). Decarbonation and
preservation method for the analysis of organic C and N contents and stable isotope ratios of
low-carbonated suspended particulate material. Analytica Chimica Acta 491:125-133.
McLusky. (1989). The Estuarine Ecosystem.“ed.2”. Blackie and Son, 215 pp.
McLusky, D.S., Elliott, M., 2004. The Estuarine Ecosystem. Oxford University Press, Oxford,
214 pp.
Minagawa, M., Wada, E. (1984). Stepwise enrichment of 15N along food chains: further
evidence and the relation between 15N and animal age. Geochimica er Cosmochimica Acta Vol.
58:1135-1140.
Miranda, A., Cal, R. M., Iglesias, J. (1990). Effect of temperature on the development of eggs
and larvae of sardine Sardina pilchardus Walbaum in captivity. Journal of Experimental Marine
Biology and Ecology 140, 69-77.
Morais, P., Dias, E., Babaluk, J., Antunes, C. (2011). The migration patterns of the European
flounder Platichthys fleus (Linnaeus, 1758) (Pleuronectidae, Pisces) at the southern limit of its
distribution range: Ecological implications and fishery management. Journal of Sea Research
65:235-246.
Mota, M., Antunes, C. (2011). First report on the status of Allis shad (Alosa alosa) in the Minho
River (Northwestern Iberian Peninsula). Journal of applied ichthyology 27;56-59.
Nichols, J. H. (1971). Pleuronectidae. Identification Sheets on Fish Eggs and Larvae, number
4-6. Conseil International pour l’Exploration de la Mer, Danemark, 18 pp.
Nichols, J. H. (1976). Soleidae. Identification Sheets on Fish Eggs and Larvae, number 150-
151. Conseil International pour l’Exploration de la Mer, Danemark, 10 pp.
Nonaka, R. H., Matsuura, Y., Suzuki, K. (2000). Seasonal variation in larval fish assemblages
in the Abrolhos. Bank region off eastern Brazil. Fishery Bulletin. v. 98, p. 767-784.In: Mota, E.

41
M. (2013). Distribuição e abundância do ictioplâncton na plataforma continental (42-37 W, 48-
21 S) no nordeste do Brasil. Tese de Mestrado, Universidade Federal do Ceará: Fortaleza, 71
pgs.
Norcross, B. L., Shaw, R. F. (1984). Oceanic and estuarine transport of fish eggs and larvae: a
review. Transactions of the American Fisheries Society 113, 153–165.
North, E. W., Houde, E. D. (2003). Linking ETM physics, zooplankton prey, and fish early-life
histories to striped bass Morone staxatilis and white perch M. americana recruitment. Marine
Ecological Progress Series 260:219-236.
Novais, A. (2013). Análise comparative das diferentes fontes de recursos utilizadas por uma
espécie invasora (Corbicula fluminea) e por espécies nativas simpátricas. Tese de mestrado,
Universidade do Minho: Braga, 109 pgs.
Parnell, A. C., Inger, R., Bearhop, S., Jackson, A. L. (2010). Source partitioning using stable
isotopes: coping with too much variation. Plos One 5:1–5.
Pasquaud, S., Lobrym J., Ellie, P. (2007). Facing the necessity of describing estuarine
ecosystems: a review of food web ecology study techniques. Hydrobiologia 588:159-172.
PBHL. (2002). http://www.dre.pt/pdfgratis/2002/03/057B00.PDF
Peterson, B. J., Fry, B. (1987) Stable isotopes in ecosystem studies. Annual Review Ecology,
Evolution and Systematics 18:293–320.
Pihl, L., Cattrijsse, A., Codling, I., Mathieson, S., McLusky, D. S., Roberts, C. (2002). Habitat
use by fishes in estuaries and other brackish areas. In: Elliott, M., Hemingway, K.L. (Eds.),
Fishes in Estuaries. Blackwell Science.
Post, D. M. (2002). Using stable isotopes to estimate trophic position: Models, Methods and
Assumptions. Ecology, 83:703-718, Ecological Society of America.
Rakocinski, C. F., Lyczkowski-Shultz, J., Richardson, S. L. (1996). Ichthyoplankton assemblage
structure in Mississippi sound as revealed by canonical correspondence analysis. Estuarine,
coastal and shelf science 43:237-257.
Ramos, S., Cowen, R. K., Ré, P., Bordalo, A. A. (2006). Temporal and spatial distributions of
larval fish assemblages in the Lima estuary, Portugal. Estuarine, coastal and shelf science.
Ramos, S. (2007). Icthyoplankton of the Lima estuary (NW-Portugal): Ecology of the early life
stages of Pleuronectiformes. Tese de Mestrado, Universidade do Porto: Porto, 228 pgs.
Raz-Guzman, A., Huidobro, L. (2002). Fish communities in two environmentally different
estuarine systems of Mexico. Journal of Fish Biology 61:182-195.

42
Ré, P. (1999). Ictioplâncton estuarino da Península Ibérica. Guia de identificação dos ovos e
estados larvares planctónicos. Câmara Municipal de Cascais, Cascais.
Ré, P. (2008). Early stages of marine fishes occurring in the Iberian Peninsula. IPIMAR/IMAR:
282pp.
Reis, P. A. P. (2009). Estudo geoquiímico de metais em sedimentos do sepal dos rios Minho e
Coura. Tese de Mestrado, Universidade do Porto: Porto, 89 pgs.
Ribeiro, R. (1991). Ictioplâncton do estuário do Mondego e Resultados. Revista de Biologia:
Actas do 1º Encontro de Planctologistas Portugueses 4, 233e244. In: Ramos, S., Cowen, R.
K., Ré, P., Bordalo, A. A. (2006). Temporal and spatial distributions of larval fish assemblages
in the Lima estuary, Portugal. Estuarine, coastal and shelf science.
Riley, R. H., Townsend, C. R., Raffaelli, D. A., Flecker, A. S. (2004). Sources and effects of
subsidies along the stream-estuary continuum. In: Polis, G. A., Power, M. E., Huxel, G. R. (eds)
Food webs at the landscape level. Pp.241-245. University of Chicago Press. Chicago.
Roman, M. R., Holliday, D. V., Sandford, L. P. (2001). Temporal and spatial patterns of
zooplankton in the Chesapeake Bay turbidity maximum. Marine Ecology Progress Series
213:215-227.
Sheaves, M. (2009). Consequences of ecological connectivity: the coastal ecosystem mosaic.
Marine Ecology Progress Series 391:107–115.
Sin, Y., Wetzel, R. L., Anderson, I.C. (1999). Spatial and temporal characteristics of nutrient
and phytoplankton dynamics in the York River estuary, Virginia: analyses of long-term data.
Estuaries 22:260–275.
Smith, B. N., Epstein, S. (1970). Biogeochemistry of the stable isotopes of hydrogen and
carbon in salt marsh biota. Plant Physiology 46:738–742.
Smith, B. N., Epstein, S. (1971). Two categories of 13C/12C ratios for higher plants. Plant
Physiology 47:380–384.
Souza, A. T., Dias, E., Campos, J., Marques, J. C., Martins, I. (2014a). Structure, growth and
production of a remarkably abundant population of the common goby, Pomatoschistus
microps (Actinopterygii: Gobiidae). Environmental Biology Fishes 97:701-715.
Souza, A. T., Dias, E., Marques, J. C., Antunes, C. (2014b). Population structure, production
and feeding habitat of the goby Pomatoschistus minutus (Actinopterygii: Gobiidae) in the Minho
estuary (NW Iberian Peninsula). Environmental Biology Fishes DOI 10.1007/s10641-014-
0259-2.

43
Sousa, R., Costa Dias, S., Guilhermino, L., Antunes, C. (2008). Minho River tidal freshwater
wetlands: threats to faunal biodiversity. Aquatic Biology 3:237-250.
Summers, R.W. (1979). Life cycle and population ecology of the flounder Platichthys flesus (L.)
in the Ythan estuary, Scotland. J. Nat. Hist. 13, 703–723.
Valiela, I., Bartholomew, M. (2015). Land-Sea Coupling and Global Forcing: Following Some os
Scott Nixon´s Challenges. Estuaries and Coasts 38:1189-1201.
Vander Zander, M. J., Rasmunssen, J. B. (2001). Variations in δ15N and δ13C trophic
fractionation: Implications for aquatic food web studies. Limnology and Oceanography,
46:2061-2066.
Vasconcelos, R. P., Reis-Santos, P., Tanner, S., Maia, A., Latkoczy, C., Gunther, D., Costa, M.
J., Cabral, H. (2008). Evidence of estuarine nursery origin of five coastal fish species along the
Portuguese coast through otolith elemental fingerprints. Estuarine, Coastal and Shelf Science
317-327.
Vieira, L. R., Guilhermino, L., Morgado, F. (2015). Zooplankton structure and dynamics in two
estuaries from the Atlantic coast in relation to multi-stressors exposure. 167:347-367.
Vinagre, C., Salgado, J., Costa, M. J., Cabral, H. N. (2008). Nursery fidelity, food web
interactions and primary sources of nutrition of the juveniles of Solea solea and S.
senegalensis in the Tagus estuary (Portugal): A stable isotope approach. Estuarine, Coastal
and Shelf science 76:255-264.
Vinagre, C., Salgado, J., Cabral, H. N., Costa, M. J. (2011). Food web structure and habitat
connectivity in fish estuarine nurseries – impact of river flow. Estuaries and Coasts 34:663-
674.
Whitfield, A.K. (1990). Life-history styles of fishes in South African estuaries. Environmental
Biology of Fishes 28, 295-308.
Whitfield, A.K. (1989). Ichthyoplankton in a southern African surf zone: nursery area for the
postlarvae of estuarine associated fish species? Estuarine, Coastal and Shelf Science 29:533-
547.
Whitfield, A. K. (1994). An estuary-association classification for the fishes of southern Africa.
South African Journal of Science 90:411-417.
Whitfield, A. K. (1999). Ichthyofaunal assemblages in estuaries: a South African study. Reviews
in Fish Biology and Fisheries 9:151-186.
Whitfield, A. K., Harrison, T. D. (2003). River flow and fish abundance in a South African

44
estuary. Journal of Fish Biology 62:1467-1472.
Winkler, G., Dodson, J. J., Bertrand, N., Thivierge, D., Vincent, W. F. (2003). Trophic coupling
across the St. Lawrence River estuarine transition zone. Marine Ecology Progress Series
251:59-73.
Young, G. C., Potter, I. C. (2003). Do the characteristics of the ichthyoplankton in an artificial
and a natural entrance channel of a large estuary differ? Estuarine, Coastal and Shelf Science
56:765-779.

45
Anexo I – Abundância de espécies
Espécies
Caudal alto Total/Caudal
alto
Caudal baixo Total/Caudal
baixo E1 E2 E3 E4 E5 E6 E1 E2 E3 E4 E5 6
Ammodytes
tobianus 48.65 - - - - - 8.11 (18.9) - - - - - - -
Platichthys flesus 7.64 4.45 2.03 - - 2.24 2.73 (2.9) 11.96
(3.3) - 9.71 (6.8) 9.46 (5.5) 3.53 (1.7) - 5.78 (4.9)
Lepadogaster
purpurea 11.46 - 4.06 - - - 2.59 (4.6) 4.56 (3.2) - 6.30 - - - 1.81(±3.0)
Gobiesocidae sp. 1.91 - - - - - 0.32 (0.8) - - - - - - -
Lepadogaster sp.
- - - - - - 6.86 - 6.30 - - - 2.19(±3.6)
Solea solea 1.91 - - - - - 0.32 (0.8) - 14.29 - - - - 2.38(±6.4)
Blenniidae n.i. 1.91 - 4.06 - - - 0.99 (1.7) 7.32(3.7) 6.52 7.27 - - - 3.52(±3.8)
Lipophrys pholis 3.82 - 4.06 - - - 1.31 (2) 6.86 - - - - - 1.14 (3.1)
Parablennius
gattorugine - - - - - - - 1.91 - - - - - 0.32 (0.9)
Atherina
presbyter - - 2.03 - - - 0.34 (0.8) - - 6.30(±0) - - 9,91(±1) 2.70 (2.8)
Centrolabrus
exoletus - - - - - - - 1.91 - 14.55(±0) - - - 2.74 (6.3)
Labridae sp. - - - - - - - 10.11 10.40 (5.5) - - - - 3.42 (5.6)
Symphodus
melops - - - - - - - 8.33 13.48 7.27 - - - 4.85 (5.8)
Labrus mixtus - - - - - - - - - 14.55 - - - 2.42 (6.5)
Pomatoschistus
sp. - 4.45 - - - - 0.74 (1.8) 8.33 10.82 (9.2) 5.59 (1.01) 27.91 - - 8.77 (10.5)
Gobiidae n.i. - - - 2.98 - - 0.50 (1.2) 20.45
(17.04) 82.97 (89.5)
118.85
(102.3)
66.21
(38.02) 31.68 (1) - 35.14 (31.2)
Pomatoschistus
microps - - - - - - - - 9.42 11.42 (4.5)
23.16
(22.5) - - 7.33 (9.6)
Gobius niger - - - - - - - 13.71 - - - - - 2.29 (6.1)
Cyprinidae n.i. - - - - - - - - - - 25.38 18.60
(20.9) - 7.33 (12.3)
Dicentrarchus
labrax - - - - - - - - 5.80 2.01 1.55 - - 1.56 (2.4)
Sparidea n.i. - - - - - - - 6.0 - 6.30 - - - 2.13 (3.5)
Clupeidea n.i. - - - - - - - 5.79 (3.6) - - - - - 0.97 (2.6)
Syngnathus sp. - - - - - -
- - - 38.99 - - 3.26 7.04 (17.4)
Tabela 1- Abundância das espécies, por estação e por período de amostragem em número de indivíduos por 100 m3 (± Desvio padrão).





























