Embed Size (px)
Citation preview

57
REVISTA DE BIOLOGIA E CIÊNCIAS DA TERRA ISSN 1519-5228
Volume 12 - Número 1 - 1º Semestre 2012
Análise da relação entre a complexidade vegetacional e as comunidades da macrofauna do solo
Mariana Fregolente1; Janete Brigante2; José Carlos Fogo3; Augusto Hashimoto Mendonça4
RESUMO Este trabalho buscou avaliar a relação entre as comunidades de macrofauna do solo de uma mata e de uma pastagem degradada. Inicialmente foi realizado um levantamento florístico e fitossociológico (simplificado) da vegetação. Para estimar a macrofauna, 45 amostras (25x25x10 cm) foram retiradas de cada área, nas estações seca e chuvosa, (n=180). Os resultados florísticos e fitossociológicos mostraram uma mata em estágio avançado de regeneração e uma pastagem em estágio inicial a médio de regeneração, esta com grande riqueza de herbáceas e arbustivas pioneiras. Quanto à macrofauna associada, os resultados indicaram uma diversidade maior de grupos no solo da mata, independente do período de coleta. Além de uma diversidade menor na pastagem, ocorreu nesta um predomínio de cupins (Isoptera). Pelo método de comparação múltipla de Bonferroni observou-se uma diferença significativa entre os índices de diversidade da mata e do pasto, demonstrando que a estrutura das comunidades de macrofauna edáfica é influenciada pelo estádio sucessional da vegetação. Palavras-chave: Macrofauna, Solo, Cerrado, Estádio de Regeneração.
Analysis of the relationship between vegetation complexity and macrofauna communities in the soil.
ABSTRACT This article examines the relationship between the macrofauna communities in the soil in two areas, one forested and the other a degraded pasture. The study first involved a floristic and phytosociological analysis (simplified) of the vegetation. To assess the macrofauna, 45 soil samples (25x25x10 cm) were taken from each area, in the dry and rainy seasons (n=180). The floristic and phytosociological results revealed a forest in advanced regeneration and a pasture in the initial to middle stage of regeneration, with great richness of herbaceous and pioneer tree species. The results for the associated macrofauna showed a greater diversity of groups in the forest soil, independent of the season. Besides lower diversity in the pasture, there was predominance of termites (Isoptera). According to the Bonferroni multiple comparison method, there was a significant difference between the diversity indices of the forest and pasture, demonstrating that the structure of the macrofauna communities is influenced by the succession stage of the vegetation. Keywords: Macrofauna, Soil, Cerrado, Regeneration State.

58
1 INTRODUÇÃO
A conversão da cobertura florestal nativa para culturas agrícolas provoca inicialmente expressiva mudança na composição vegetacional, sendo que tais mudanças podem perpassar pelo ambiente edáfico e afetar a estrutura e função das comunidades bióticas. Para os componentes originais do solo o efeito será tanto maior quanto mais intensa for a ação destruidora sobre a vegetação nativa, alterando ou eliminando partes e processos ligados à ciclagem de nutrientes (Casagrande & Soares, 2007), e quanto maiores forem as interferências físico-químicas de manejo (subsolagem, gradagem, aração, uso de agrotóxicos e tráfego de máquinas pesadas), de modo que são esperados reflexos negativos sobre as comunidades de organismos do solo (Brigante, 2007).
A regeneração natural, ou mesmo as ações de restauração de ecossistemas degradados tem como objetivo a reconstrução das funções e estruturas destes sistemas, restabelecendo-se os serviços ambientais perdidos. Segundo Carpanezzi (2005) a componente “função” em um ecossistema concentra-se nos serviços ambientais oferecidos por meio de processos, como a regularização do clima ou a proteção do solo. Neste caso, as atenções se voltam para os ciclos de nutrientes, ciclo da água e fluxo de energia. Já a componente “estrutura” se refere aos seres vivos integrantes deste sistema, registrando-se os tipos, suas formas de organização e de inter-relação.
O estudo das flutuações das comunidades da macrofauna do solo em ecossistemas naturais e naqueles alterados pelo homem pode evidenciar possíveis influências sobre esta fauna, da quantidade e qualidade do material vegetal que aporta ao solo, como também do microclima, o que justificaria o seu uso como indicadora da qualidade deste sistema (Stork & Eggleton, 1992; Linden et al., 1994). Além disso, acredita-se que a recuperação da estabilidade do solo e de sua produtividade em longo prazo é dependente da integridade da biota do solo (Whitford, 1996), esta considerada como parte integrante dos sistemas biológicos de regulação mediadores da decomposição, um processo chave para a fertilidade do solo devido
a seus efeitos sobre a humificação e a mineralização da matéria orgânica (Lavelle, 1994).
Considerando a diversidade de invertebrados do solo e suas múltiplas funções nos ecossistemas terrestres, pode-se inferir que o conhecimento do papel que a biota edáfica pode prestar nos processos de regeneração natural de sistemas vegetacionais degradados torna-se estratégico nos dias atuais, como resposta à crescente demanda por ações de recuperação. Da mesma forma, os estudos sobre a estrutura de sistemas vegetacionais naturais são de grande importância, pois oferecem a possibilidade de conhecimento das relações bióticas, além de permitir testar teorias ecológicas e fornecer subsídios indispensáveis para o manejo da restauração ecológica. Assim, o objetivo deste trabalho foi avaliar a abundância e a diversidade da macrofauna do solo em dois sistemas vegetacionais em estádios diferenciados de regeneração natural, buscando detectar possíveis relações entre a complexidade vegetacional e os grupos da macrofauna invertebrada. 2 MATERIAL E MÉTODOS
O estudo ocorreu na região Centro-Norte do Estado de São Paulo, na bacia hidrográfica do Alto Jacaré-Guaçu, margem direita do Ribeirão do Lobo, município de Itirapina. A região está assentada sobre um mosaico de formações geológicas constituídas de arenitos amarelados e rosados e intrusões e derrames basálticos da Formação Botucatu, além de material de origem continental flúvio-lacustre originada da Formação Bauru, sendo que a bacia hidrográfica do ribeirão do Lobo encontra-se na Província das Cuestas Basálticas (Villela & Mattos 1975; Souza, 1977; Almeida, 1974). Quanto aos tipos de solo destacam-se o solo Hidromórfico, o Latossolo Vermelho Amarelo, o Regosol e as manchas de Latossolo Vermelho, férrico (Souza, 1977; Embrapa, 1999). A área de estudo está assentada sobre mancha de Latossolo Vermelho, férrico. A classificação Koëppen registra um clima do tipo Cwb, temperado úmido, com inverno seco e temperatura média do mês mais quente inferior a 22°C (Gonçalves, 1994). A vegetação nativa

59
característica da região tem forte associação com as formações areníticas já citadas, com vegetação predominantemente de Cerrado em grande parte da área. Apenas nas depressões, onde ocorrem variações litológicas expressivas e solos mais férteis, os cerrados são substituídos por matas mais densas (Villela & Mattos, 1975). No presente estudo as amostras de vegetação e solo foram extraídas de uma área de 2,2 ha de uma área em processo de regeneração natural, e de uma área de 3,0 ha de mata secundária do tipo Floresta Estacional Semidecidual. Histórico de uso das áreas
Até 1960 a área que hoje se encontra em processo de regeneração natural era ocupada por culturas mistas, em ciclos rotacionados de milho, arroz e feijão. Após 1960 a área passou por vários ciclos de abandono, de uso para pastagem e de usos para cultivos de milho, arroz, feijão. O último uso, de 1990 a 2000, foi para estes cultivos, e desde então a área foi abandonada para regeneração natural sendo, no entanto, eventualmente utilizada como pastagem. Durante todo esse período foram aplicados apenas adubos químicos e em nenhum momento foram utilizados pesticidas agrícolas. Com relação aos usos da área de mata secundária, a mesma não sofreu intervenções humanas significativas nestes últimos 50 anos. Caracterização da vegetação
O delineamento experimental da vegetação foi feito no período de novembro/2007 a março/2008 e utilizou o método de Avaliação Ecológica Rápida, caracterizado por ser um levantamento flexível, acelerado e direcionado das espécies e fisionomias vegetacionais (Sayre et. al., 2000). O método consistiu de caminhadas aleatórias pelas áreas, coletando-se espécies botânicas inéditas (florística), com paradas constantes para medidas de alguns parâmetros fitossociológicos. Foram recolhidas amostras para a montagem de exsicatas e posterior identificação por meio da morfologia comparada (Lorenzi, 2000; Lorenzi, 2002a,b; Lorenzi & Matos, 2002; Durigan et al., 2004; Souza & Lorenzi, 2005; Sobral & Jarenkow,
2006; Ramos et al, 2008). Para a taxonomia das Angiospermas seguiu-se o sistema Angiosperm Phylogeny Group II (APG II, 2003). Os grupos sucessionais foram identificados com base em Kageyama & Viana (1992) e Gandolfi et al. (1995).
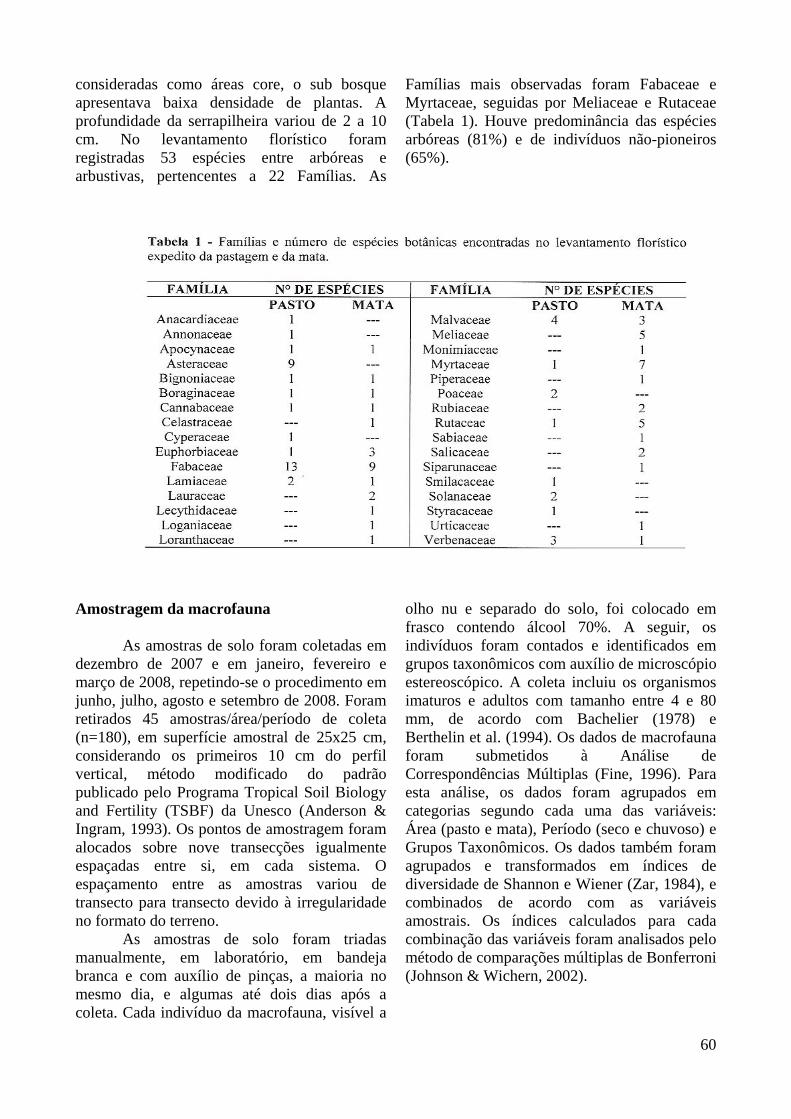
De acordo com as orientações da Resolução Conjunta SMA IBAMA/SP Nº 1, de 17 de fevereiro de 1994, a vegetação da pastagem apresentou estágio inicial a médio de regeneração, mesclando fisionomias campestres e savânicas. Nos trechos campestres predominaram estratos herbáceos e arbustos, abertos, e alturas entre 0,20 m a 0,90 m, com frequentes espécies heliófilas, incluindo forrageiras exóticas como o Capim-braquiária (Brachiaria decumbens Staf.). Nesta formação não foram observadas epífitas e as trepadeiras foram predominantemente herbáceas. Nos trechos savânicos ocorreu estrato lenhoso aberto, apresentando plantas com alturas variáveis entre 1,5 m e 4 m. O Diâmetro à Altura do Peito (DAP a 1,30 m do solo) foi de até 8,1 cm, apresentando pequeno produto lenhoso. As epífitas foram pouco abundantes e ocorreram trepadeiras herbáceas. Foram registradas 47 espécies entre arbóreas e arbustivas, distribuídas em 20 famílias botânicas (Tabela 1). Houve predominância de espécies herbáceas, trepadeiras e ervas que, juntas, corresponderam a 38% dos indivíduos amostrados. As arbustivas representaram 28% e as arbóreas, 34%. As Famílias predominantes foram Fabaceae e Asteraceae, e o grupo sucessional mais representativo foi o das pioneiras, representando 87% das espécies encontradas
Pela mesma referência legal, a vegetação da mata foi caracterizada como uma Mata Estacional em estádio médio a avançado de regeneração, com fisionomia florestal fechada, ocorrendo distribuição contínua de copas, altura do dossel entre 8 e 18 m e indivíduos arbóreos emergentes a 22 m. O DAP médio dos troncos foi superior a 20 cm e boa quantidade de material lenhoso. Próximo das bordas as trepadeiras estavam presentes em grande quantidade, com predomínio das lenhosas, e registrou-se expressiva diversidade de formas de vida, com árvores, arbustos (incluindo espécies umbrófilas), ervas terrícolas, trepadeiras, epífitas e palmáceas. Nas áreas mais declivosas
60
consideradas como áreas core, o sub bosque apresentava baixa densidade de plantas. A profundidade da serrapilheira variou de 2 a 10 cm. No levantamento florístico foram registradas 53 espécies entre arbóreas e arbustivas, pertencentes a 22 Famílias. As
Famílias mais observadas foram Fabaceae e Myrtaceae, seguidas por Meliaceae e Rutaceae (Tabela 1). Houve predominância das espécies arbóreas (81%) e de indivíduos não-pioneiros (65%).
Amostragem da macrofauna
As amostras de solo foram coletadas em dezembro de 2007 e em janeiro, fevereiro e março de 2008, repetindo-se o procedimento em junho, julho, agosto e setembro de 2008. Foram retirados 45 amostras/área/período de coleta (n=180), em superfície amostral de 25x25 cm, considerando os primeiros 10 cm do perfil vertical, método modificado do padrão publicado pelo Programa Tropical Soil Biology and Fertility (TSBF) da Unesco (Anderson & Ingram, 1993). Os pontos de amostragem foram alocados sobre nove transecções igualmente espaçadas entre si, em cada sistema. O espaçamento entre as amostras variou de transecto para transecto devido à irregularidade no formato do terreno.
As amostras de solo foram triadas manualmente, em laboratório, em bandeja branca e com auxílio de pinças, a maioria no mesmo dia, e algumas até dois dias após a coleta. Cada indivíduo da macrofauna, visível a
olho nu e separado do solo, foi colocado em frasco contendo álcool 70%. A seguir, os indivíduos foram contados e identificados em grupos taxonômicos com auxílio de microscópio estereoscópico. A coleta incluiu os organismos imaturos e adultos com tamanho entre 4 e 80 mm, de acordo com Bachelier (1978) e Berthelin et al. (1994). Os dados de macrofauna foram submetidos à Análise de Correspondências Múltiplas (Fine, 1996). Para esta análise, os dados foram agrupados em categorias segundo cada uma das variáveis: Área (pasto e mata), Período (seco e chuvoso) e Grupos Taxonômicos. Os dados também foram agrupados e transformados em índices de diversidade de Shannon e Wiener (Zar, 1984), e combinados de acordo com as variáveis amostrais. Os índices calculados para cada combinação das variáveis foram analisados pelo método de comparações múltiplas de Bonferroni (Johnson & Wichern, 2002).
61
3 RESULTADOS E DISCUSSÃO
Abundância e riqueza da macrofauna
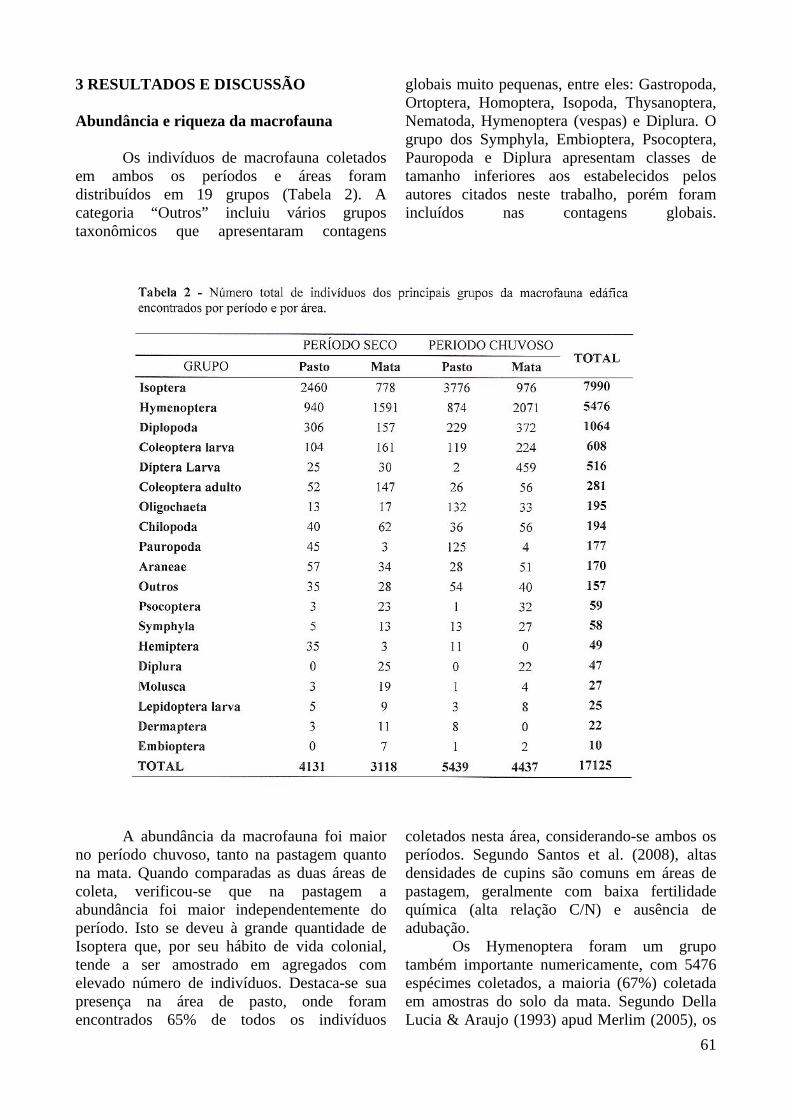
Os indivíduos de macrofauna coletados em ambos os períodos e áreas foram distribuídos em 19 grupos (Tabela 2). A categoria “Outros” incluiu vários grupos taxonômicos que apresentaram contagens
globais muito pequenas, entre eles: Gastropoda, Ortoptera, Homoptera, Isopoda, Thysanoptera, Nematoda, Hymenoptera (vespas) e Diplura. O grupo dos Symphyla, Embioptera, Psocoptera, Pauropoda e Diplura apresentam classes de tamanho inferiores aos estabelecidos pelos autores citados neste trabalho, porém foram incluídos nas contagens globais.
A abundância da macrofauna foi maior
no período chuvoso, tanto na pastagem quanto na mata. Quando comparadas as duas áreas de coleta, verificou-se que na pastagem a abundância foi maior independentemente do período. Isto se deveu à grande quantidade de Isoptera que, por seu hábito de vida colonial, tende a ser amostrado em agregados com elevado número de indivíduos. Destaca-se sua presença na área de pasto, onde foram encontrados 65% de todos os indivíduos
coletados nesta área, considerando-se ambos os períodos. Segundo Santos et al. (2008), altas densidades de cupins são comuns em áreas de pastagem, geralmente com baixa fertilidade química (alta relação C/N) e ausência de adubação.
Os Hymenoptera foram um grupo também importante numericamente, com 5476 espécimes coletados, a maioria (67%) coletada em amostras do solo da mata. Segundo Della Lucia & Araujo (1993) apud Merlim (2005), os

62
Hymenoptera preferem explorar ambientes que possuem alimentos de mais alta qualidade, embora estejam presentes em ambientes diversificados (Damasceno, 2005).
A predominância de insetos sociais (Isoptera e Hymenoptera) também foi observada por Merlim (2005) e Baretta (2007), o que pode estar relacionado com o método de amostragem utilizado.
Os Oligochaeta destacaram-se no pasto, no período chuvoso, seguido pela mata, também no período chuvoso. Fragoso & Lavelle (1992) encontraram, em florestas do México, uma relação entre as populações de Oligochaeta e as variações sazonais, com valores máximos ocorrendo na estação úmida. Além disso, eles migram para camadas mais profundas do solo durante a estação seca. Sendo assim, a profundidade da coleta, de apenas 10 cm, pode ter desfavorecido a coleta deste grupo durante o período seco. Ainda, segundo Baretta (2007), o método do monólito de dimensões 25x25cm não amostra os Oligochaeta de forma adequada. O uso de monólitos maiores (40x40cm) e formaldeído diluído garantiriam uma melhor caracterização de sua população (Baretta et al., 2007).
A maior parte dos Coleoptera adultos e de suas larvas foi coletada em amostras da mata. Os adultos apareceram com maior freqüência no período seco, enquanto a maior parte das larvas foi encontrada no período chuvoso. Isto pode ser explicado pelo fato de os adultos de Coleoptera preferirem solos não excessivamente úmidos, embora seu desenvolvimento possa ocorrer sob estas circunstâncias (Ekschimitt et al., 1997, apud Damasceno, 2005).
Os Diplopoda foram o terceiro grupo mais expressivo numericamente, sendo que no período seco foram encontrados mais indivíduos deste grupo no pasto do que na mata. De acordo com Correia et al. (2005) eles são classificados como saprófagos. Pimentel et al. (2006) obtiveram resultados relacionando um maior teor de carbono orgânico à ocorrência de espécies saprófagas e predadoras. Assim, a presença de Diplopoda pode significar alta disponibilidade de matéria orgânica.
Os Chilopoda e os Araneae não apresentaram grandes variações de populações nem entre os períodos nem entre as áreas. Segundo Damasceno (2005), a presença de altas
densidades de predadores, como os dos grupos Araneae e Chilopoda, pode significar uma maior estruturação da teia alimentar do solo. Além disso, esses grupos são mais vulneráveis que outros insetos à dissecação porque sua cutícula geralmente é mais permeável e o sistema de trocas gasosas não é fechado (Wolters & Ekschmitt, 1997 apud Ducatti, 2002). Sendo assim, sua presença pode não estar relacionada apenas à disponibilidade de alimentos, mas também às condições climáticas, como umidade e sombreamento.
Com relação aos Molusca, eles apresentaram-se em números mais expressivos apenas na mata (85% dos indivíduos desse grupo coletados). Quase não foram coletados espécimes no pasto. Resultados semelhantes foram obtidos por Brigante (2000), que observou 81% desses organismos ocorrendo no solo da mata, em comparação com pastagens.
Análise de Correspondências Múltiplas da macrofauna
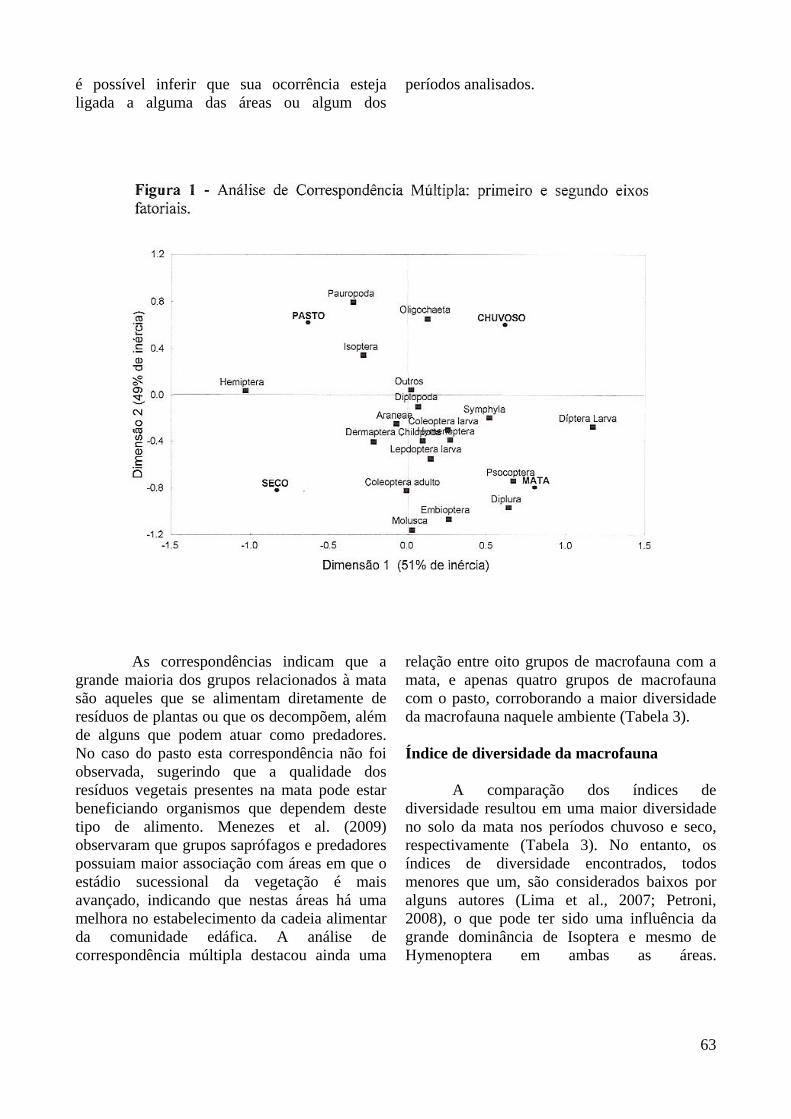
A Análise de Correspondências Múltiplas mostrou que existe uma fraca associação entre a área amostrada e o período de coleta. Dentre os grupos de macrofauna houve destaque para as seguintes associações: i) os grupos Diptera larva (saprófago/predador/macrofitófago) e Symphyla (alimenta-se de resíduos de plantas) apresentaram forte associação com a mata/chuvoso; ii) o grupo Hemiptera (macrofitófago) correlacionou-se destacadamente na área de pasto/seco; iii) os grupos Pauropoda, Oligochaeta (saprófago) e Isoptera (saprófago/predador/macrofitófago) com menos intensidade, associaram-se com a área de pasto/chuvoso; iv) os grupos Molusca, Embioptera, Coleoptera adulto (saprófago/predador) e Lepdoptera larva (saprófago/predador/macrofitófago) com menos intensidade associaram-se com mata/seco; v) os grupos Diplura e Psocoptera (ambos alimentam-se de resíduos de plantas), associaram-se de modo semelhante na mata/chuvoso/seco (Figura 1). Os grupos que não apresentaram correspondência com nenhuma área ou período de coleta (como Araneae, Diplopoda, Larva de Coleoptera, Outros) não se podem ter sua presença relacionada a estes fatores, ou seja, não
63
é possível inferir que sua ocorrência esteja ligada a alguma das áreas ou algum dos
períodos analisados.
As correspondências indicam que a grande maioria dos grupos relacionados à mata são aqueles que se alimentam diretamente de resíduos de plantas ou que os decompõem, além de alguns que podem atuar como predadores. No caso do pasto esta correspondência não foi observada, sugerindo que a qualidade dos resíduos vegetais presentes na mata pode estar beneficiando organismos que dependem deste tipo de alimento. Menezes et al. (2009) observaram que grupos saprófagos e predadores possuiam maior associação com áreas em que o estádio sucessional da vegetação é mais avançado, indicando que nestas áreas há uma melhora no estabelecimento da cadeia alimentar da comunidade edáfica. A análise de correspondência múltipla destacou ainda uma
relação entre oito grupos de macrofauna com a mata, e apenas quatro grupos de macrofauna com o pasto, corroborando a maior diversidade da macrofauna naquele ambiente (Tabela 3). Índice de diversidade da macrofauna A comparação dos índices de diversidade resultou em uma maior diversidade no solo da mata nos períodos chuvoso e seco, respectivamente (Tabela 3). No entanto, os índices de diversidade encontrados, todos menores que um, são considerados baixos por alguns autores (Lima et al., 2007; Petroni, 2008), o que pode ter sido uma influência da grande dominância de Isoptera e mesmo de Hymenoptera em ambas as áreas.

64
A partir da análise de Comparações
Múltiplas de Bonferroni, seis combinações diferentes foram testadas quanto à hipótese de igualdade dos índices de diversidade, assumindo nível de confiança global de 95%: Pasto/Seco-Mata/Seco; Pasto/Seco-Pasto/Chuvoso; Pasto/Seco-Mata/Chuvoso; Mata/Seca-Pasto/Chuvoso; Mata/Seca-Mata/Chuvoso e Pasto/Chuvoso-Mata/Chuvoso. Os resultados comparativos indicaram que para a mata nos dois períodos amostrados não está rejeitada a hipótese de igualdade entre os índices de diversidade, indicado por uma linha que liga as duas combinações (Figura 2). Para o pasto nos
dois períodos, houve uma rejeição da hipótese de igualdade. Os resultados indicam que não é possível estabelecer uma relação direta entre abundância e diversidade. Segundo Moço et al. (2005), altas densidades podem reduzir a diversidade em função do predomínio de poucos grupos. Quando se avaliou diretamente a diversidade de grupos de macrofauna da mata e do pasto, observou-se que o valor p resultou menor que 0,05 o que significa que os índices de diversidade da mata e do pasto diferem significativamente (Tabela 4), sendo o da mata maior que o do pasto.
Os resultados da análise de correspondências múltiplas e da comparação dos índices de diversidade mostraram que as comunidades da macrofauna do solo diferiram nas áreas estudadas, indicando que o estádio
sucessional da vegetação influencia a composição dessas comunidades. Resultados semelhantes foram encontrados por Menezes et al. (2009).

65
De acordo com diversos autores
(Townsendo et al., 2006; Correia & Andrade, 1999; Podgaisk et al., 2007), a heterogeneinade espacial de um ambiente está diretamente relacionado à riqueza de espécies devido ao fato de que ambientes mais diversificados proporcionam um maior número de microambientes e de microclimas, além de maior variedade de recursos, possibilitando a colonização de várias espécies da macrofauna com diferentes estratégias de sobrevivência. Assim, segundo Leitão-Lima & Teixiera (2002), quanto mais diversificada for a cobertura vegetal, maior o número de nichos a serem colonizados, resultando em maior diversidade das comunidades da fauna do solo. Além disso, uma maior complexidade estrutural aérea implica em igual complexidade radicular, representando incrementos nutricionais e físicos que favorecem o aumento da diversidade da biota (Wardle & Giller, 1996; Hole, 1980).
No entanto, o sistema com vegetação menos complexa apresentou grande parte dos grupos da mata, apesar da menor abundância. Isto pode ser devido ao fato de que gramíneas como as braquiárias (Brachiaria sp), embora promovam interferências negativas ao infestar as áreas em processo de regeneração natural, apresentam alta produtividade de matéria orgânica, o que é favorável para a atividade
microbiana edáfica (Tauk-Tornisielo, 2007; Casagrande & Soares, 2007) e para o incremento de comunidades da macrofauna.
Mesmo considerando certa representatividade dos grupos, o solo da área em regeneração natural ainda apresenta sinais de desequilíbrio das comunidades de macrofauna, evidenciado pelo elevado número de Isoptera, indicando que este grupo não está sofrendo predação ou mesmo que as condições do solo nesta área o favorecem competitivamente e desfavorecem seus predadores.
A análise temporal mostrou que, de maneira geral, as maiores quantidades de indivíduos foram observadas no período chuvoso, independente da área ser pasto ou mata. Isso pode significar que a macrofauna edáfica é afetada pela estação seca, quer seja por limitação de recursos, água ou temperatura, que os leva a sincronizar estádios de resistência, dormência, ou outra estratégia biológica para sobreviver nos períodos mais restritivos à sua natureza. CONCLUSÕES
O solo da área de pasto em regeneração
mostrou ser ainda um ambiente seletivo, favorecendo alguns grupos, que têm seu número

66
elevado na comunidade, e desfavorecendo outros, que têm seu número reduzido ou estão ausentes, demonstrando que a estrutura das comunidades de macrofauna edáfica é influenciada pelo estádio sucessional da vegetação. Apesar da diferença ecológica entre as áreas comparadas, a composição da macrofauna edáfica do pasto – com a presença expressiva de grupos importantes para os ciclos de nutrientes e para a estrutura física do solo, como Oligochaeta, Hymenoptera e Isoptera – pode influenciar a qualidade do solo desta área, em um nível que não foi estudado neste trabalho. Embora o período chuvoso tenha se mostrado o mais favorável para a macrofauna em termos de número de indivíduos, em termos de diversidade foi constatada a hipótese de igualdade nos índices de diversidade entre os períodos. Portanto, a estação seca afetou a população da macrofauna, mas não interferiu na composição das espécies de sua comunidade.
AGRADECIMENTOS
Os autores agradecem o apoio à pesquisa oferecido pelo Centro de Recursos Hídricos e Ecologia Aplicada (CRHEA/SHS/EESC/USP); ao Sr. Edgar Nori por viabilizar o estudo da vegetação em sua propriedade; à Profa. Dra. Suzana Trivinho Strixino pela grande ajuda na identificação de alguns dos indivíduos da macrofauna edáfica. REFERÊNCIAS BIBLIOGRÁFICAS ALMEIDA, F.F.M. Fundamentos geológicos do relevo paulista. 14.ed. São Paulo, Instituto de Geografia-USP - Série Teses e Monografias, 1974. 99p. ANDERSON, J.M. & INGRAM, J.S.I. (eds). Tropical soil biology and fertility – a handbook of methods. 2.ed. C. A. B. International, 1993. 171p. APG II. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnaean Society, 141:399-436, 2003.
BACHELIER, G. La faune des sols, son écologie et son action. (Documentations Techniques, 38). Orstom, 1978. 391p. BARETTA, D. et al., Earthworm populations sampled using collection methods in atlantic forests with Araucaria angustifolia. Sci. agric. (Piracicaba, Braz.), 64: 384-392, 2007. BARETTA, D. Fauna do solo e outros atributos edáficos como indicadores da qualidade ambiental em áreas com Araucaria angustifolia no Estado de São Paulo. Tese (Doutorado em Agronomia) - Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 2007. 158p. BERTHELIN, J.; LEVAL, C.; TOUTAIN, F. Biologie des sols. Rôle des organisms dans l’altérations et humification. In: Bonneau, M.; Souchier, B. (coord.). Pédologie. 2 – Constituants et proprietés du sol. Paris, Masson, 1994. p. 143-237. BRIGANTE, J. 2007. Adequação ambiental de áreas de pastagem. Requisitos de Qualidade na Bovinocultura de Corte. Santos, F. A. P.; Moura, J. C.; Pedroso de Faria, V. Anais... 6º Simpósio sobre Bovinocultura de Corte. Piracicaba – FEALQ. ISBN: 978-85-7133-051-1. p. 39-64. BRIGANTE, J. Comparação de algumas comunidades de macrofauna e microrganismos de solo encontradas em áreas de mata e pastagens, em um Latossolo. (Tese de Doutorado). Universidade Federal de São Carlos, 2000. 122p. CARPANEZZI, A.A. Fundamentos para a reabilitação de ecossistemas florestais. In: GALVÃO, A.P.M & PORFÍRIO-DA-SILVA, V. (Eds. Téc.). Restauração florestal – fundamentos e estudos de caso. Colombo, Embrapa Florestas, 2005. p. 27-45. CASAGRANDE, J.C. & SOARES, M.R. Recuperação de solos degradados: interação solo – planta. In: BARBOSA, L.M. & JÚNIOR, N.A.S. (org.), A botânica no Brasil: Pesquisa, ensino e políticas públicas ambientais. São Paulo, Percepção Design, 2007. p 53-57.

67
CORREIA, M.E.F; ANDRADE, A.G. Formação de serrapilheira e ciclagem de nutrientes. In: SANTOS, G.A; CAMARGO, F.A. de. Fundamentos da matéria orgânica do solo: ecossistemas tropicais e subtropicais. Porto Alegre, Genesis, 1999. p.209-214. CORREIA, M.E.F.; FARIA, S.M.; CAMPELLO, E.F. & FRANCO, A.A. Organização da comunidade de macroartrópodos edáficos em plantios de eucalipto e leguminosas arbóreas. In: CONGRESSO BRASILEIRO DE CIÊNCIA DO SOLO, 25., Viçosa, 1995. DAMASCENO, A.C.F. Macrofauna edáfica, regeneração natural de espécies arbóreas, lianas e epífitas em florestas em processo de restauração com diferentes idades no Pontal do Paranapanema. Piracicaba. Dissertação (Mestrado em Recursos Florestais) – Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo. 2005, 107p. DUCATTI, F. Fauna edáfica em fragmentos florestais e em áreas reflorestadas com espécies da Mata Atlântica. Dissertação (Mestrado em Recursos Florestais) – Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, 2002. 70p. DURIGAN, G.; BAITELLO, J.B; FRANCO, G.A.D.C.; SIQUEIRA, M.F. Plantas do Cerrado Paulista – imagens de uma paisagem ameaçada. São Paulo, Páginas e Letras Editora e Gráfica, 2004. 475p. EMBRAPA. Centro Nacional de Pesquisa de Solos (Rio de Janeiro, RJ). Sistema brasileiro de classificação de solos (SIBCS). Brasília, Embrapa Produção de informação; Rio de Janeiro, Embrapa Solos, 1999. 412p. FINE, J. Introdução aos Métodos estatísticos multidimensionais a partir de exemplos. São Carlos, DEs-UFSCar & Rede PRESTA, 1996. FRAGOSO, C. & LAVELLE, P. Earthworm communities of tropical rain forests. Soil Biology and Biochemistry, 24:1397-1408, 1992.
GANDOLFI, S.; LEITÃO FILHO, H.F. & BEZERRA, C.L.F. Levantamento florístico e caráter sucessional das espécies arbustivo-arbóreas de uma floresta mesófila semidecídua no Município de Guarulhos, SP. Revista Brasileira de Biologia, 55: 753-767, 1995. GONÇALVES, J.M.M. Caracterização das coberturas de alteração e suas aplicações na região entre os municípios de Analândia, Itirapina e São Carlos (SP). Rio Claro, 1994. (Dissertação de Mestrado) – UNESP. HOLE, F. D., Effects of animals on soil. Geoderma, 25:75-112, 1980. JOHNSON, R.A. & WICHERN, D.W. Applied multivariate statistical analysis, 5st. Englewood Cliffs, Prentice Hall, 2002. KAGEYAMA, P.Y. & VIANA, V.M. Tecnologia de sementes e grupos ecológicos de espécies arbóreas tropicais. In: SIMPÓSIO BRASILEIRO SOBRE TECNOLOGIA DE SEMENTES FLORESTAISP, 2., Atibaia, 1989. Anais. São Paulo, Instituto Florestal (Série Documentos), 1992. p.197-215. LAVELLE, P. Correspondência do autor. Soil fauna and soil processes: Adaptative strategies that determine ecosystem function. Laboratoire d’Ecologie des Sols Tropicaux, Centre ORSTOM, 93143. Bondy Cedex, France, 1994. LEITÃO-LIMA, P. da S.; TEIXEIRA, L. B. Macrofauna do solo em capoeiras natural e enriquecidas com leguminosas arbóreas. Belém, Embrapa-CPATU, 2002. 3 p. (Comunicado técnico, 62). LIMA, S.S.; AQUINO, A.M.; LEITE, L.F.; SILVA, P.H.; CASTRO, A.A. & OLIVEIRA, F. Diversidade da macrofauna edáfica em agroflorestas de diferentes estádios sucessionais. Revista Brasileira de Agroecologia, 2:1029-1033, 2007. LINDEN, D.R., HENDRIX, P.F., COLEMAN, D.C. Faunal indicators of soil quality. In: DORAN, J.W. et al. (Ed). Defining soil quality for a sustainable environment. Madison, Soil

68
Science Society of American, 1994. p. 91-106. (Special Publication, 35). LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 4.ed. Volume 1. Nova Odessa, Instituto Plantarum, 2002a. 352p. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 2.ed. Volume 2. Nova Odessa, Instituto Plantarum, 2002b. 368p. LORENZI, H. Plantas daninhas do Brasil: terrestres, aquáticas, parasitas e tóxicas. 3.ed. Nova Odessa, Instituto Plantarum, 2000. 608p. LORENZI, H.; MATOS, F. J. A. Plantas medicinais do Brasil: nativas e exóticas. Nova Odessa, Instituto Plantarum, 2002. 512p. MENEZES C. E. G. et al. Macrofauna edáfica em estádios sucessionais de Floresta Estacional Semidecidual e pastagem mista em Pinheiral (RJ). Revista Brasileira de Ciência do Solo, 33:1647-1656, 2009. MERLIM, A. O. Macrofauna edáfica em ecossistemas preservados e degradados de araucária no parque estadual de Campos do Jordão. Piracicaba, 2005. Dissertação (Mestrado em Ecologia de Agrossistemas) – Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo. 89p. MOÇO, M.K.S.; GAMA-RODRIGUES, E.F.; GAMA-RODRIGUES, A.C.; & CORREIA, M.E.F. Caracterização da fauna edáfica em diferentes coberturas vegetais na região norte Fluminense. Revista Brasileira de Ciência do Solo, 29:555-564, 2005. PETRONI, D.M. Diversidade de famílias de Coleoptera em diferentes fragmentos florestais no município de Londrina, PR-Brasil. Londrina, Dissertação (Mestrado em Ciências Biológicas) – Universidade Estadual de Londrina. 2008. 61p. PIMENTEL, M. S., et al. Atributos Biológicos do solo sob manejo orgânico de cafeeiro, pastagem e floresta em região do médio
Paraíba Fluminense-RJ. Coffee Science, 1:85-93, 2006. PODGAISK, L. R.; OTT, R.; GANADE, G. Ocupação de microhábitats artificiais por invertebrados de solo em um fragmento florestal no sul do Brasil. Neotropical Biology and Conservation, 2:71-79, 2007. RAMOS, V.S.; DURIGAN, G.; FRANCO, G.F.; SIQUEIRA, M.F. de. Árvores da floresta estacional semidecidual: guia de identificação de espécies. São Paulo, EDUSP, 2008. 277p. RESOLUÇÃO CONJUNTA SMA/IBAMA/SP 01. Define vegetação primária e secundária nos estágios pioneiro, inicial, médio e avançado de regeneração de Mata Atlântica em cumprimento ao disposto no art. 6º do Decreto nº 750, de 10 de Fevereiro de 1993, na Resolução CONAMA 10 de 10 de Outubro de 1993. DOU de 03 de fevereiro de 1994, Seção I, v. 132, fasc. 24, 1994. SANTOS, G. G. et al., Macrofauna edáfica associada a plantas de cobertura em plantio direto em um latossolo vermelho do cerrado. Pesquisa Agropecuária Brasileira, 43:115-122, 2008. SAYRE, R.; ROCA, E.; SEDAGHATKISH, G.; YOUNG, B.; KEEL, S.; ROCA, R. & SHEPPARD, S. Nature in focus: rapid ecological assessment. Washington, D. C., EUA, The Nature Conservancy, 2000. 182 p. SOBRAL, M. & JARENKOW, J.A. (Org.). Flora arbórea e arborescente do Rio Grande do Sul, Brasil. São Carlos, Editoras Rima e Novo Ambiente, 2006. 350p. SOUZA, M.H.A.O. Alguns aspectos ecológicos da vegetação na região perimetral da Represa do Lobo (Brotas-Itirapina, SP). Tese (Doutorado). Instituto de Biociências – USP, 1977. 369p. SOUZA, V.C. & LORENZI, H. Botânica sistemática: Guia ilustrado para identificação das famílias de Angiospermas da floresta brasileira, baseado em APG II. Nova Odessa, Instituto Plantarum, 2005. 640p.

69
STORK, N.E. & EGGLETON, P. Invertebrates as determinants and indicators of soil quality. American Journal of Alternative Agriculture, 7:2-6, 1992. TAUK-TORNISIELO, S. M. Os fungos nos processos de decomposição da matéria orgânica nos ambientes terrestres. In: BARBOSA, L. M. & JÚNIOR, N. A. S. A botânica no Brasil: Pesquisa, ensino e políticas públicas ambientais. São Paulo, Percepção Design, 2007. p. 177-181. TOWSEND, C.R.; BEGON, M. e HARPER, J.L. Fundamentos de Ecologia. Porto Alegre, Artmed, 2006. 592 p. VILLELA, S. M. & MATTOS, A. Hidrologia Aplicada. São Paulo, Mc Graw-Hill do Brasil, 1975. 245p. WARDLE, D. A. & GILLER, K. E., The quest for a contemporary ecological dimension to soil biology. Soil Biology and Biochemistry, 28:1549-1554, 1996. WHITFORD, W. G., The importance of the biodiversity of soil biota in arid ecosystems. Biodiversity and Conservation, 5:185-195, 1996. ZAR, J.M. Bioestatistical analysis. Englewood Cliffs, Prentice-Hall, 1984.





























