Embed Size (px)
Citation preview

Diogo Araujo – Med 92
Imunologia Médica
Segundo Módulo
Antígenos e indução da resposta imune
Prof. Imaculada
Antígeno X Imunógeno: qual a diferença?
- Antígenos = são moléculas capazes de serem reconhecidas por um anticorpo ou receptor de
célula T.
- Imunógeno = é um antígeno capaz de induzir uma resposta imune contra ele.
Epítopo X determinante antigênico: há diferença? Não. Epítopo = determinante antigênico. É a
porção do antígeno que é, de fato, reconhecida pelo receptor do linfócito T ou B. Na natureza,
alguns epítopos são mais frequentes devido à existência de enzimas que clivam os antígenos de
maneira específica ou pela capacidade (ou facilidade) do MHC de apresentar alguns tipos de
epítopos.
[Parênteses]
Quais são as possibilidades de falso negativo em uma imunização?
(Pesquisar! Algo relacionado à formação de defesa a outros antígenos que não ao injetado)
(Talvez seja o seguinte... A aplicação de uma vacina que teve o seu conteúdo alterado - por ser mal
acondicionada, passar do prazo de vacidade, etc - poderá fazer com que o organismo seja
apresentado a outros epítopos que não são os de interesse; assim, haveria resposta imune contra
epítopos que não estariam relacionados com o agente causador da doença)
O que é um hapteno? Trata-se de uma molécula que, sozinha, não consegue desencadear uma
resposta imune. Contudo, ela pode se ligar a uma outra molécula e passar a induzir resposta. São
exemplos de hapteno:
- penicilina: se associa a proteínas sanguíneas e, assim, modifica a estrutura de uma
molécula própria. Logo, essa proteína conjugada ao hapteno leva à formação de

Diogo Araujo – Med 92
resposta imune e memória;
- brincos de cobre: o cobre presente no local onde está o brinco pode funcionar como
hapteno e levar a respostas imunológicas de alergia.
De modo geral, a partir do momento em que há apresentação do imunógeno às células T, haverá
duas possibilidades:
- formação de linfócitos Th1, com produção de anticorpos IgM e, depois, IgG; ou
- formação de linfócitos Th2, com produção de IgE. Caso haja uma segunda exposição ao antígeno,
essa classe de anticorpos produzida em excesso pode levar o indivíduo ao choque anafilático.
São características do antígeno:
- Natureza química: antígenos podem ser proteínas (maioria), lipídeos, carboidratos, DNA
ou RNA.
[Parênteses] Por que é difícil desenvolver memória imunológica contra antígenos que não são
protéicos? E por que as respostas geralmente são lentas?
Isso se deve ao fato dos linfócitos T somente serem capazes de reconhecer antígenos protéicos
apresentados no CPH. Com isso, a única resposta capaz de ser desencadeada contra antígenos não
protéicos é timo-independete (que não leva à memória). Por exemplo: a cápsula do estreptococo é
formada por polissacarídeos. Contudo, o receptor clássico do linfócito T só reconhece sequências
protéicas. Então, as respostas a esse tipo de antígeno não protéico são timo-independentes,

Diogo Araujo – Med 92
incapazes de formar memória. Além disso, a resposta demora porque não há memória prévia. A
chance de fazer infecções graves por esse patógeno é maior. Nas respostas timo-independentes,
somente há produção de IgM.
"Os linfócitos T só conseguem produzir resposta contra antígeno protéico; os outros tipos de
moléculas induzem uma resposta que é muitas vezes ineficiente".
[Parênteses 2] No lúpus, são formados auto-anticorpos contra o DNA celular que se encontra
acoplado a pedacinhos de proteína.
- Requisitos para imunogenicidade: o primeiro requisito para ocorrer a resposta imune é
que a molécula seja estranha ao organismo (não seja própria). Isso porque tudo o que é próprio foi
apresentado aos linfócitos produzidos (selecionados positiva e negativamente lá na medula ou no
timo) ou simplesmente não causa resposta devido aos mecanismos de tolerância periférica ao
antígeno (que serão tratados depois). Além disso, moléculas muito pequenas ou muito grandes
podem não ser reconhecidas pelo sistema imune. Ou seja, o tamanho tem de ser adequado para
ocorrer resposta. E, por fim, a complexidade química faz com que seja maior a chance de uma
molécula de caber na fenda do CPH. Resumindo, são requisitos:
- ser estranho ao organismo;
- ter tamanho ideal para reconhecimento;
- quanto maior a complexidade química, maior a chance de resposta.
- Genética do indivíduo: a genética de cada pessoa influencia na resposta imune. Isso
porque essa resposta depende de fatores do próprio indivíduo, como a capacidade de apresentar
antígenos (envolvendo os tipos de CPHs existentes, os tipos de enzimas presentes no organismo
para clivagem dos antígenos, a capacidade de fagocitose das células). Ou seja, a genética está
relacionada desde as competências individuais da célula, passando pela capacidade do CPH, as
vias intracelulares estimuladas quando se entra em contato com o antígeno, etc. É por isso que
respondemos de maneiras diferentes a um mesmo antígeno.
- Via de administração (ou de entrada) do antígeno: quando uma vacina é aplicada
intravascular, seus antígenos são diluídos na circulação sanguínea. Já na aplicação intradérmica,
ocorre irritação do local, boa absorção dos antígenos, atração de células por substâncias
adjuvantes (presentes na vacina, como o sulfato de alumínio). Quando um antígeno entra por via
oral, ele tende a apresentar baixa imunogenicidade. Isso porque, de maneira geral, a resposta
imune por essa via tende a ser de tolerância. Contudo, há exceções: a vacina da poliomielite é

Diogo Araujo – Med 92
administrada por via oral e consegue ser eficiente; isso acontece porque ela mimetiza a
contaminação natural do microorganismo (por meio de VO, com apresentação por células M à
placa de Peyer, etc). Sempre que se mimetiza a via de contaminação natural se torna a mais
eficiente para a imunização. Resumindo:
- Via mais eficiente: Via intradérmica;
- Via menos eficiente: Via oral (porque tende a ser uma resposta de tolerância);
[Parênteses] Já que ela tende a induzir tolerância aos antígenos, a via oral está sendo utilizada na
pesquisa para tratamento de doenças auto-imunes.
[Parênteses 2] Quando um microorganismo está no intestino delgado, ele pode ser captado pelas
células M presentes na mucosa. Essas células jogam o microorganismo para o interior das placas
de Peyer, que são áreas de linfócitos B, com linfócitos T e macrófagos ao redor. O linfócito da Placa
de Peyer que é estimulado sai por um vaso linfático e se encaminha para as cadeias de linofodos
para-aórticos. Lá, ele se multiplica intensamente e se converte em plasmócito produtor de IgA.
Então, por via sanguínea, volta para uma região de mucosa (como a própria mucosa ileal, por
exemplo) e passa a secretar IgA a todo vapor. Esse IgA atravessa o epitélio e passa a combater os
microorganismos que estejam na superfície, impedindo-os de entrar no organismo. É isso o que
acontece com o vírus da poliomielite, por exemplo.
Quando um antígeno entra em contato com o organismo, o que acontece?
O organismo pode fazer resposta imune, ser indiferente ou induzir tolerância específica para o
antígeno. Mas como isso acontece?
Se o linfócito T for apresentado ao antígeno por meio de CPH com os segundos sinais necessários,
ele será ativado e produzirá resposta imune. Contudo, se o linfócito T reconhecer um antígeno que
não tenha sido apresentado por CPH, esse linfócito entra em estado de deleção clonal ou de
anergia, sendo incapaz de produzir resposta ou levando à tolerância ao antígeno.
É importante lembrar que o linfócito T somente reconhece estruturas lineares. Já o linfócito B é
capaz de reconhecer a estrutura espacial do antígeno (estrutura tridimensional). Geralmente, ele
vê o que está por fora da bactéria ou do vírus.

Diogo Araujo – Med 92
Como ocorre uma resposta timo-dependente? (Isso será abordado com mais detalhes na próxima
aula)
Numa resposta timo-dependente, o linfócito B em contato com um antígeno reconhece a sua
estrutura tridimensional. Em seguida, para ativar o linfócito T, ele faz duas coisas: apresenta um
fragmento desse antígeno ao linfócito T por meio de CPH de classe II; passa um segundo sinal por
meio de B7 e CD28. Assim, o linfócito T passa um estímulo de volta (por CD40) ao linfócito B (por
ligante CD40 também), levando-o à expansão clonal e produção de anticorpos (no início, IgM e,
depois, IgG). Nesse caso, há formação de linfócitos T e B de memória.
As respostas timo-independentes são mais primordiais, mais inespecíficas e incapazes de
produção de memória.
Os experimentos de Zinkernagel e de Nanuni (ou algo parecido com isso) mostraram que os CPHs
estabelecem identidade ao organismo, restrição à espécie. (Ler mais sobre eles se der).
Falando um pouquinho sobre a apresentação de antígenos
A célula dendrítica é a mais eficiente na apresentação de antígenos porque ela produz
constitutivamente (sem precisar de estímulo) a molécula B7, que participa do segundo sinal.
Assim, ela estimula mais facilmente a resposta de linfócitos T virgens.
São dois os sinais necessários para a ativação de um linfócito T:
- CPH + antígeno = primeiro sinal
- B7 (da célula apresentadora de antígeno) + CD28 (no linfócito T) = segundo sinal
Esses dois sinais são evolutivamente importantes porque, assim, não há resposta imune à toa.
O macrófago e o linfócito B são bons apresentadores de antígeno para linfócitos T de memória
(efetores).

Diogo Araujo – Med 92
Quando um macrófago ativa o linfócito T efetor, esse linfócito ativa o macrófago. Isso se dá por
meio de ligação CD40 (do linfócito) com CD40 ligante (do macrófago), o que leva à produção de
um super macrófago (com maior capacidade fagocítica, mais enzimas, etc).
O linfócito que é estimulado pelos dois sinais passa a produzir interleucina 2 e receptores para
essa interleucina 2. Assim, essa IL-2 atuará sobre a própria célula produtora, com expansão clonal,
etc.
Quando a célula dendrítica encontra um antígeno e fica ativada, ela pode passar até 100 horas
apresentando o antígeno, além de aumentar muito o número de moléculas de CPH em sua
superfície.
Quando o macrófago tem de processar um antígeno para depois apresentá-lo, a resposta imune
geralmente é mais eficaz do que se o antígeno tivesse sido previamente processado in vitro.
Como se dá a apresentação de antígeno por meio de CPH de classe I?
O antígeno que vai ser apresentado pelo CPH de classe I está no citoplasma da célula. Essas
proteínas vão ser ubiquitinadas (marcadas molecularmente por ubiquitina) e carreadas até o
proteassoma (que funciona como um tubo para clivagem de proteínas). Então, ao passar no
interior do proteassoma, a proteína é esticada e clivada por sítios enzimáticos em peptídeos de 7 a
10 aminoácidos. Em seguida, a proteína TAP joga esses peptídeos para dentro do retículo
endoplasmático rugoso, onde a molécula de CPH de classe I é formada. Os peptídeos se encaixam
no CPH de classe I e são expostos na membrana.

Diogo Araujo – Med 92
[Parênteses] O que aconteceria em um indivíduo sem TAP?
Indivíduo sem TAP não consegue apresentar antígenos por CPH de classe I. Assim, ele tem
respostas auto-imunes frequentes devido a déficits na apresentação de antígenos no timo por CPH
de classe I, sendo liberados para a circulação periférica linfócitos que reagem contra o próprio.
Mas porque ele não apresenta maior número de infecções virais?
Porque ele conta as respotas imunes de linfócitos NK e de anticorpos (já que os linfócitos B e
linfócitos T CD4 funcionam bem).
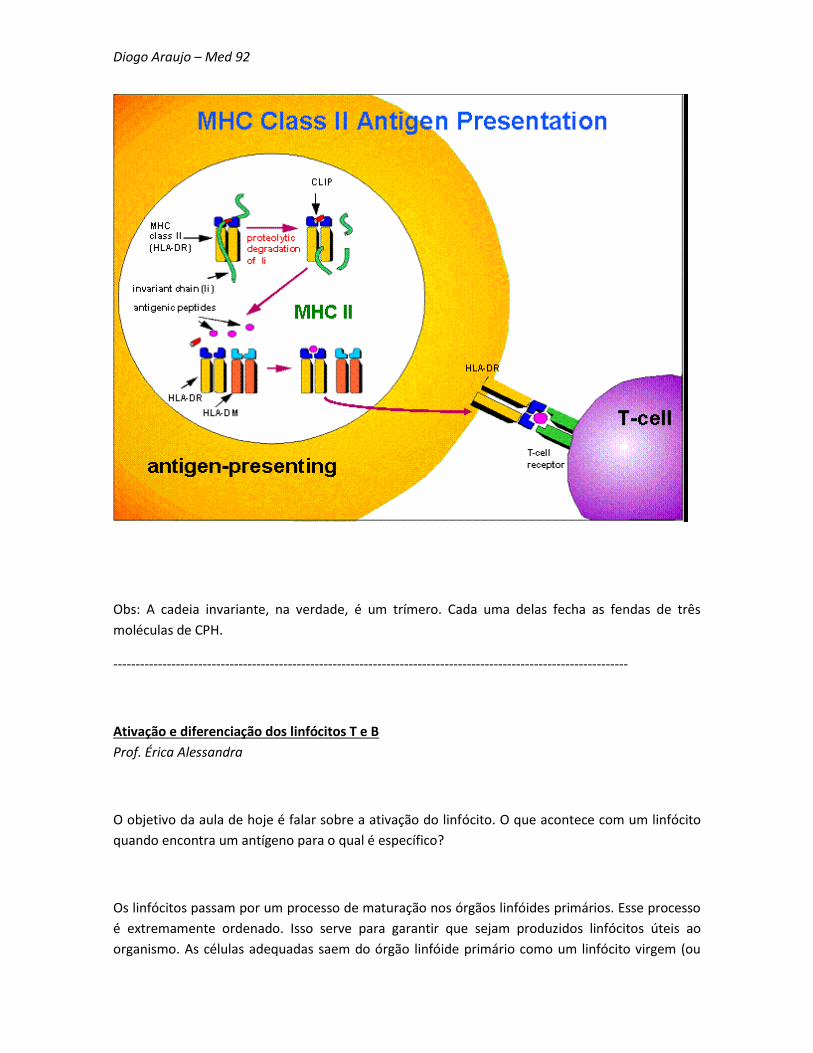
[Parênteses 2] Com relação à apresentação de antígenos pelo CPH de classe II, existe um
mecanismo que é controlado pela chamada "cadeia invariante". Quando o CPH de classe II é
produzido no retículo endoplasmático, a cadeia invariante (molécula protéica) se liga à fenda do
CPH. Assim, eles são colocados em vesículas que vão para o citoplasma. Após a fusão dessa
vesícula com o fagossoma e o lisossoma, as enzimas clivam a porção mais externa da cadeia
invariante, ficando apenas uma parte (chamada de CLIP) no interior da fenda. Nesse momento,
uma outra molécula presente dentro do fagolisossoma, chamada HLA-DM, faz com que o CLIP seja
removido da fenda do CPH. Assim, os peptídeos antigênicos se ligam ao CPH e podem ser
apresentados na membrana.
Imagine o que aconteceria em um indivíduo que não fosse capaz de expressar a cadeia invariante
ou o HLA-DM.
Diogo Araujo – Med 92
Obs: A cadeia invariante, na verdade, é um trímero. Cada uma delas fecha as fendas de três
moléculas de CPH.
-------------------------------------------------------------------------------------------------------------------
Ativação e diferenciação dos linfócitos T e B
Prof. Érica Alessandra
O objetivo da aula de hoje é falar sobre a ativação do linfócito. O que acontece com um linfócito
quando encontra um antígeno para o qual é específico?
Os linfócitos passam por um processo de maturação nos órgãos linfóides primários. Esse processo
é extremamente ordenado. Isso serve para garantir que sejam produzidos linfócitos úteis ao
organismo. As células adequadas saem do órgão linfóide primário como um linfócito virgem (ou

Diogo Araujo – Med 92
seja, que ainda não entrou em contato com os antígenos nos órgãos linfóides periféricos). Lá, eles
não estabelecem uma morada definitiva: sofrem um processo de recirculação. Assim, as células
interagem com diversas células do organismo nos mais diversos locais para aumentar a chance de
encontrar o antígeno para o qual é específico. Se o antígeno não for encontrado, o linfócito morre.
O linfócito, quando entra em contato com o antígeno, sofre ativação e se multiplica. Esse
fenômeno de multiplicação é chamado de expansão clonal. Essa multiplicação é necessária
porque é preciso que se combata demais antígenos presentes no organismo e para acompanhar a
taxa de multiplicação do microorganismo.
Depois da expansão, o linfócito ainda não tem o fenótipo que dá capacidade para combate ao
antígeno. Pra isso, ele tem de passar pela etapa de diferenciação (adquirir a capacidade de
combater o organismo). Assim, passam a ser linfócitos T e B EFETORES (atuam combatendo o
microorganismo).
Ex: quando linfócitos T de memória são ativados, eles geram linfócitos T efetores que migram para
o local a fim de eliminar os antígenos por meio da ativação de macrófagos teciduais (graças à
liberação de interferon gama), tornando-os super macrófagos.
A atuação das células efetoras faz com que os antígenos sejam destruídos e a sua quantidade seja
diminuída. No final, o antígeno pode ser eliminado ou sobrar em pequenas quantidades. Então, as
células que resultaram da expansão clonal sofrem apoptose. Porém, algumas sobrevivem e
tornam-se células de memória.
Por que é importante eliminar a grande massa de linfócitos resultante da expansão clonal? Porque
esses linfócitos, por estarem ativos, ficariam o tempo inteiro produzindo citocinas e estimulando
os macrófagos. Assim, poderiam causar danos ao "próprio".
Além disso, alguns linfócitos ativados podem produzir TNF, que é deletério para o organismo
quando não é necessário. Logo, esses linfócitos precisam ser eliminados após a resposta ao
antígeno (fase de declínio de sua existência). O TNF induz inflamação e causa inapetência (perda
de função). Leva à perda de peso, à febre e quadros de choque (bradicardia e hipotensão).
Linfócito T pode ser ativado nos linfonodos, no baço e no TLAM (tecido linfóide associado à
mucosa). Nesses locais, isso acontece porque existe a célula apresentadora de antígeno.

Diogo Araujo – Med 92
Uma vez que o antígeno esteja presente no organismo, ele pode:
- ser capturado por uma célula apresentadora de antígeno, que se destaca do tecido e migra até o
linfonodo;
- o próprio antígeno pode migrar diretamente para o linfonodo e ser apresentado por células que
estejam nesse local;
- podem ser carreados por via hematogênica para o baço.
A ativação dos linfócitos T depende de dois sinais (primeiro e segundo sinais). A célula
apresentadora de antígeno tem de expressá-lo associado ao CPH (primeiro sinal); além disso, ela
precisa apresentar moléculas B7, que atuam como coestimuladoras ao se ligarem ao CD28 do
linfócito T (segundo sinal).
Por que é necessário ter dois sinais (via CPH e B7)?
Porque, se isso não existisse, poderia haver respostas excessivas de linfócitos auto-reativos contra
o "próprio". Isso é um mecanismo de segurança.
As células apresentadoras de antígeno expressam grande quantidade de moléculas
coestimuladoras porque há contato constante com o antígeno. As células dendríticas, por
exemplo, são ótimas apresentadoras de antígenos justamente porque têm muitos CPHs e
moléculas B7 em sua superfície.
Após o reconhecimento do primeiro e do segundo sinal, o linfócito entra em um processo de
expansão clonal, expressando receptor de il-2 e expressando a própria il-2. Esse é um processo
autócrino. Só depois que há expressão do receptor do il-2, há início da divisão celular (expansão
clonal). Além disso, essa célula é induzida a sobreviver porque ela mesma começa a expressar
moléculas do tipo Bcl (xL e 2).
Após a expansão clonal, inicia-se a etapa de diferenciação, com formação de linfócitos T efetores e
de memória.
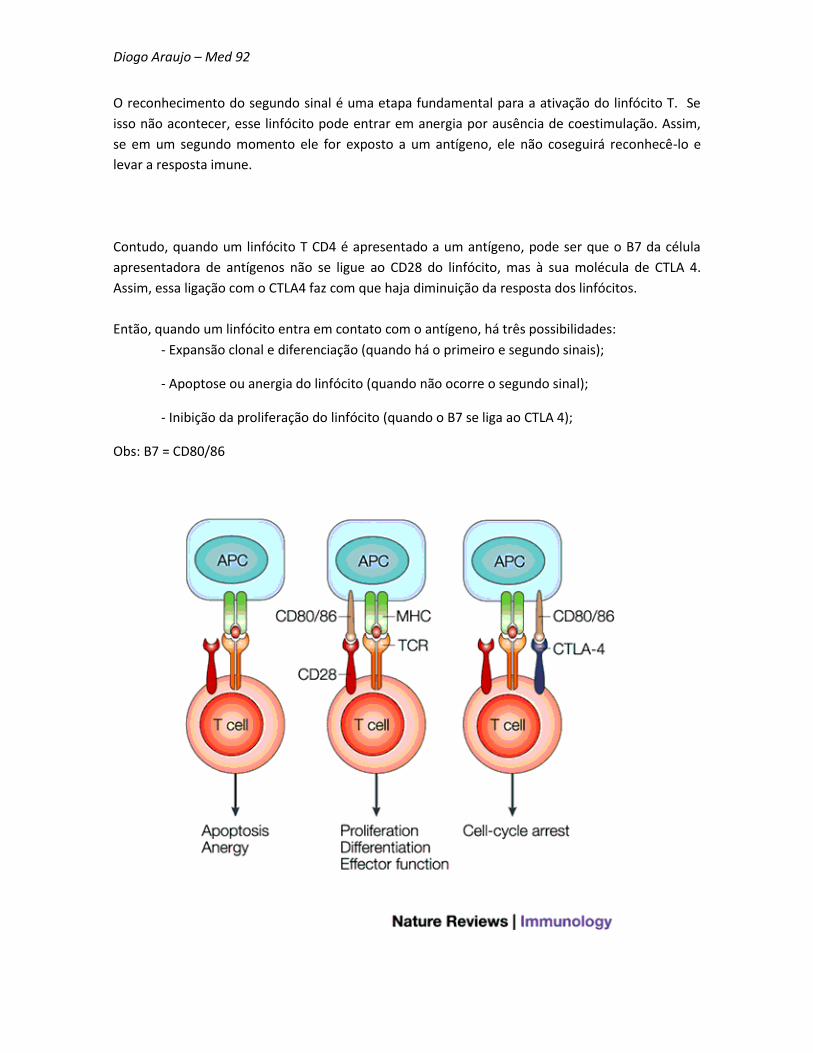
Diogo Araujo – Med 92
O reconhecimento do segundo sinal é uma etapa fundamental para a ativação do linfócito T. Se
isso não acontecer, esse linfócito pode entrar em anergia por ausência de coestimulação. Assim,
se em um segundo momento ele for exposto a um antígeno, ele não coseguirá reconhecê-lo e
levar a resposta imune.
Contudo, quando um linfócito T CD4 é apresentado a um antígeno, pode ser que o B7 da célula
apresentadora de antígenos não se ligue ao CD28 do linfócito, mas à sua molécula de CTLA 4.
Assim, essa ligação com o CTLA4 faz com que haja diminuição da resposta dos linfócitos.
Então, quando um linfócito entra em contato com o antígeno, há três possibilidades:
- Expansão clonal e diferenciação (quando há o primeiro e segundo sinais);
- Apoptose ou anergia do linfócito (quando não ocorre o segundo sinal);
- Inibição da proliferação do linfócito (quando o B7 se liga ao CTLA 4);
Obs: B7 = CD80/86

Diogo Araujo – Med 92
O macrófago, quando entra em contato com o antígeno, reconhece padrões moleculares de
patógenos (ou seja, moléculas que normalmente estão expressas em patógenos de um modo
geral). Esse reconhecimento leva à expressão de mais moléculas coestimulatórias na superfície.
Contudo, a fagocitose de células próprias não leva a essa expressão de moléculas coestimulatórias
(já que não possuem padrões moleculares de patógenos). Assim, quando um macrófago fagocita
uma hemácia ou uma célula velha, por exemplo, não há inflamação no local. : )
[Parênteses] Adjuvantes da formulação de vacinas estimulam a inflamação e, assim, fazem com
que as células apresentadoras de antígenos expressem mais moléculas coestimuladoras. Se fosse
administrado só o antígeno, a vacina poderia até mesmo induzir que os linfócitos específicos se
tornassem anérgicos (devido à falta de expressão de coestimuladores). Cada adjuvante tem um
papel que leva à expressão de coestimuladores.
Depois que o linfócito reconhece os dois sinais, há expressão do CD40 ligante no linfócito T. A
ligação do CD40 ligante ao CD40 da CAA. Isso amplifica a capacidade da célula apresentadora de
antígenos de ativar linfócitos T CD4 e CD8.
Há, também, expressão do CD69 (provoca a retenção do linfócito no local onde está acontecendo
a ativação da célula).
À medida que o linfócito vai respondendo ao antígeno, ele passa a expressar o CTLA-4, que tem
função supressora.
Podemos dizer que quando o CD28 se liga na molécula B7, o linfócito é ativado. Quando o CTLA-4
se liga na molécula B7, o linfócito tem a sua função suprimida. Esse é um balanço de sinais que
rege a permanência da função do linfócito.

Diogo Araujo – Med 92
O que acontece dentro do linfócito T para que ele passe a expressar il-2, seu receptor, CD69 e
CTLA-4? Quais são as vias internas de sinalização?
[Não é preciso decorar essas cascatas]
A ligação do antígeno ao RLT (receptor do linfócito T) faz com que a Lck (que está na cauda
citoplasmática do CD4 ou CD8) fosforile as cadeias zeta e as moléculas de CD3. Essa fosforilação
forma pontos de ancoragem de proteínas intracitoplasmáticas (da família ZAP-70). Essas enzimas
fosforilam a LAT (trasmembrana). A GRB2 e a Sos se ancoram na LAT e levam à ativação da via das
MAP cinases e da via do fosfatidilinositol.
Na via das MAP cinases, há por fim, a expressão do fator de transcrição AP-1, que vai ao núcleo do
linfócito e estimula a transcrição dos genes importantes para a ativação do linfócito.
A via do fosfatidilinositol se dá por ativação da fosfolipase C. O fosfatidilinositol é clivado em IP3 e
DAG. A presença de IP3 leva à ativação de reservas de cálcio intracelular. A presença da DAG
estimula a PKC, que ativa o NF kappa B, que vai para o núcleo estimular a transcrição de genes. A
ação dos íons cálcio sobre a calmodulina/calcinerina torna-a ativa, que cliva a NFAT e a torna
ativa. Ela vai para o núcleo e estimula a transcrição de genes.
A ciclosporina atua inibindo a calcineurina e, consequentemente, a formação de NFAT ativo, com
baixa produção de proteínas essenciais para a ativação de linfócitos.
Resumindo:
A presença do primeiro e do segundo sinal leva à ativação das vias do fosfatidilinositol (que forma
os fatores de transcrição NF kappa beta e NFAT) e das MAP cinases (que forma o fator de
transcrição AP-1).
Se somente há o primeiro sinal, não há expressão de cofatores e eles não levam à formação de
proteínas que ativem o linfócito.
Resumindo todas as etapas de ativação do linfócito T CD4:
1. Primeiro e segundo sinal: apresentação de antígeno por CPH de classe II ao TCR do
linfócito T CD4; segundo sinal com B7 se ligando a CD28 do linfócito;
2. Expressão de fatores de transcrição no linfócito: NF kappa beta, NFAT e AP-1;

Diogo Araujo – Med 92
3. Ativação e expansão clonal frente à expressão de IL-2, de seu receptor e da ação
autócrina;
4. Expressão de Bcl (xL e 2), que leva à sobrevivência dos clones;
5. Diferenciação dos linfócitos: formação de linfócitos T efetores e de memória;
6. Linfócito T passa a expressar CD40 em sua superfície. Ele se liga ao CD 40 ligante da célula
apresentadora de antígeno, que se torna mais capaz de ativar linfócitos T CD4 e T CD8;
7. Expressão de CD 69, que leva à retenção dos linfócitos ao local onde foram ativados;
8. À medida que a resposta ocorre, há maior expressão de CTLA-4, fazendo com que haja
declínio da resposta.
A ativação do linfócito T CD4 pode fazer com que ele se diferencie em subpopulações: Th1, Th2 ou
Th17. Isso vai depender de fatores locais como:
- Tipo de antígeno;
- Quantidade de antígeno;
- Tipo de célula apresentadora do antígeno envolvida;
- Genética do indivíduo;
- Citocinas presentes no meio.
A célula Th1 surge mais quando há vírus no meio, além da presença de Il-12 e INF gama. Assim,
quando ela surge, passa a produzir mais INF gama, que ativa macrófagos e leva à produção de
anticorpos IgM e IgG.
A célula Th2 surge frente à presença de helmintos e de Il-4 no meio. Então, ela passa a produzir Il-
4, Il-5 e Il-13, que induzem linfócitos B a secretarem IgE, além de ativarem os eosinófilos e
estimularem as secreções mucosas.
A célula Th17 surge quando há Il-1, Il-6, TGF beta, bactérias e fungos. Ela libera Il-17 e Il-22, que
induzem inflamação.

Diogo Araujo – Med 92
E quanto aos linfócitos T CD8? Como eles são ativados?
Eles são apresentados a antígenos por quaisquer células do organismo (exceto espermatozóides e
hemácias) através do CPH de classe I. Então, ele apresenta ação citotóxica sobre a célula,
liberando perforinas e granzimas.
Contudo, o linfócito T CD4 do tipo Th1 pode estimular essa citotoxicidade do linfócito T CD8. Isso
se dá através de:
- secreção de citocinas, que atuam na membrana do linfócito T CD8 como um segundo sinal; ou
- estimulação de células apresentadoras de antígenos, que irão ativar mais linfócitos T CD8. (Achei
isso estranho... Como uma célula apresentadora de antígenos, que tem CPH de classe II, poderia
estimular linfócitos T CD8?) :P
Falando um pouquinho sobre os linfócitos T de memória...
Os linfócitos T de memória sobrevivem por anos após a eliminação do antígeno, já que possuem
altos níveis das moléculas antiapoptóticas Bcl-2 e Bcl-xL. Respresetam cerca de 50% dos linfócitos
de pessoas com mais de 50 anos. A il-7 e a il-15 são as responsáveis pela autorrenovação dessas
células.
Essas células são menos dependentes de coestimulação para responder ao antígeno, combatendo-
o com maior eficiência.
Existem os linfócitos T de memória:
- centrais (presentes nos linfonodos), que têm alta proliferação e baixa capacidade efetora;
-efetores, que estão localizados nos tecidos periféricos, com baixa capacidade de proliferação mas
grande capacidade efetora.
A exposição ao antígeno estimula a sobrevivência das células de memória.
Existem dois mecanismos descritos para a formação do linfócito T de memória: por conversão de
um efetor ou independentemente. (Esses mecanismos serão vistos com detalhe posteriormente)

Diogo Araujo – Med 92
Falando um pouco de linfócitos B...
Os linfócitos B são ativados nos mesmos locais que os linfócitos T (linfonodos, TLAM, baço). Os
folículos linfóides que são as regiões de linfócitos B.
Ele é capaz de reconhecer antígenos protéicos, polissacarídicos e lipídicos.
Se ele reconhece antígenos proteicos, ele precisará do auxilio de linfócito T CD4. Ou seja, será uma
resposta timo-dependente. Se não houver esse auxílio, não há resposta. Se houver, o linfócito B é
ativado e passa a produzir IgM, IgG, IgA e IgE (dependendo do tipo de linfócito T CD4 que é
formado - se Th1 ou Th2). Nesse processo, ocorrem a troca de classe e a maturação da afinidade
(clones com maior afinidade pelo antígeno vão sobreviver, devido à produção de proteínas
antiapoptóticas). Isso gera plasmócitos de longa vida (memória), que ficam na medula óssea
produzindo anticorpos por anos e anos.
Se a resposta é realizada contra um antígeno polissacarídico ou lipídico, haverá uma resposta
timo-independente. Nesse caso, o linfócito só produzirá IgM e pouquinho IgG, convertendo-se em
plasmócito de vida curta (que não é capaz de formar memória).
Olhando a curva de anticorpos, vemos que há uma resposta primária ao antígeno, por meio de
produção de IgM e pouco IgG. Se a resposta foi contra um antígeno protéico, quando a exposição
for repetida, os linfócitos B de memória serão ativados produzindo a resposta secundária, com
grande produção de IgG, IgA ou IgE. O isotipo do anticorpo vai depender do tipo de antígeno
apresentado (helminto, microorg de mucosa, etc).
Vamos supor que um antígeno protéico consiga chegar ao folículo linfático de um linfonodo. O que
aconteceria?
As células apresentadoras de antígeno ou o antígeno em si podem entrar em contato com as
células T CD4 e ativá-las (pelos mecanismos vistos anteriormente). Contudo, esses linfócitos T CD4
se encontram ao redor dos folículos linfáticos, distantes dos linfócitos B.
Para acontecer uma resposta timo-dependente contra esse antígeno proteico, seria necessário
que esse linfócito T CD4 entrasse em contanto com os linfócitos B.
Então, o linfócito T CD4 ativado migra para a periferia do folículo linfático, onde ele se encontra
com o linfócito B (que também migrou pra lá). Lá, eles se encontram, se reconhecem, se ativam e

Diogo Araujo – Med 92
vão juntos para ao interior do folículo linfóide.
Os linfócitos B podem se converter em plasmócitos de longa vida (que vão para a medula) e
células B de memória (que podem permanecer no linfonodo ou ir para outros locais). Já os
linfócitos T permanecem no folículo linfático, de modo a ativar mais linfócitos B, promover troca
de classe, etc.
Mas como exatamente que os linfócitos B e T interagem entre si?
São dois mecanismos:
- O linfócito B apresenta antígeno para o linfócito T CD4 a partir de CPH de classe II. O segundo
sinal é a ligação do ligante do CD40 (que é expresso pelo linfócito T) com o CD40 do linfócito B.
- Passagem de citocinas (INF gama) do linfócito T CD4 para o linfócito B. Assim, ele entra em
proliferação e maturação de afinidade.
Os três subgrupos de linf T CD4 são capazes de auxiliar o linfócito B nesse processo.
Na resposta timo-independente, o linfócito B entra em contato com antígenos multivalentes (com
subunidades regularmente repetidas; um lipídeo, por exemplo). Isso faz com que várias moléculas
de Ig que funcionam como receptores possam se ligar a cada uma dessas subunidades e ativar o
linfócito B. Assim, esse linfócito pode formar plasmócito produtor de IgM.
Se são necessários dois sinais para que uma célula T se ative, será que o mesmo acontece com o
linfócito B na resposta timo-independente?
Sim. O primeiro sinal se dá através da ligação do antígeno ao receptor do linfócito B (BCR). O
segundo sinal ocorre através da ligação do antígeno a uma molécula Toll ou CR2 presente na
membrana.

Diogo Araujo – Med 92
Como é essa ligação com o CR2?
Ocorre por meio de um fragmento resultante da cascata do complemento: o C3d.
E quanto ao receptor Toll?
O antígeno também pode se ligar ao receptor do tipo Toll diretamente, sem o auxílio de
fragmento do complemento.
Tanto a sinalização via CR2 quanto via receptor Toll levam à proliferação e à diferenciação.
A existência dos dois sinais para ativação serve para garantir que o linfócito B seja ativado
somente em momentos adequados.
A descoberta de cooperação entre linfócitos T e B foi importante para o desenvolvimento de
vacinas.
A associação de carbohidratos a proteínas para a produção de vacinas (vacinas conjugadas) faz
com que a parte protéica ative o linfócito T e a não protéica, o linfócito B. A ativação mútua faz

Diogo Araujo – Med 92
com que haja memória imunológica contra um microorganismo que só desencadearia resposta
timo-independente.
Isso acontece nas repostas contra pneumococo, meningococo e Haemophilus.
Alguns microorganismos apresentam moléculas que são chamados de superantígenos. Ou seja, ele
é capaz de ativar INESPECIFICAMENTE vários clones de linfócitos simultaneamente, tanto linfócitos
B quanto T.
Isso porque esse superantígeno se liga a regiões que não são as de reconhecimento de antígeno
tanto no receptor do linfócito T quanto no receptor do linfócito B.
Logo, como esse superantígeno age de maneira inespecífica e simultânea para vários linfócitos, ele
causa ativação policlonal de linfócitos T e B.
E isso é ruim porque:
- nesse processo, linfócitos auto-reativos podem sofrer expansão, levando a doenças auto-imunes;
- pode haver imunossupressão temporária, devido à indução da morte de muitos linfócitos ao
mesmo tempo;
-------------------------------------------------------------------------------------------------------------------
MONITOR MONOGRAFIA
Levi Bezerra Sena
81794520
-----------------------------------------------------------------------------------------------------------------
Respostas mediadas pelos linfócitos T
Prof. Imaculada

Diogo Araujo – Med 92
Existem as fases de indução e efetora. A de indução consiste no processamento e na apresentação
do antígeno. A fase efetora compreende a resposta que o linfócito terá após ter sido ativado.
Tipos de respostas mediadas pelos linfócitos T:
- Linfócitos T CD4 exercem função auxiliar (na maior parte das vezes) ou citotóxica. Antígeno
processado é apresentado a esse tipo de linfócito. Assim, ele fica ativado e entra na fase efetora.
Passa a secretar citocinas para ativar macrófago, estimular inflamação ou funcionar como auxiliar
para o linfócito B produzir anticorpos ou para o linfócito TCD8 ser citotóxico.
- Linfócitos T CD8 exercem função citotóxica (na maior parte das vezes) ou auxiliar. Ele é efetor por
contato célula-célula, matando a célula alvo (qualquer célula que esteja infectada e que esteja
apresentando antígenos não-próprios na sua superfície).
Conclusões de um certo experimento:
- Proteção contra bactérias intracelulares não pode ser dada por anticorpos;
- Mas pode ser dada por linfócitos T CD8;
- Linfócitos T (CD4) sozinhos, in vitro, não conseguem levar o microorganismo extracelular à morte.
Ou seja, eles dependem de outros elementos para conseguirem matar microorganismos. Isso
porque o braço efetor da resposta do linfócito T CD4 é o macrófago. Ou seja, é o macrófago que
mata o microorganismo que não está presente dentro da célula.
A resposta dos linfócitos se dá nos órgãos linfóides secundários. Os antígenos têm de ser
capturados e levados até lá. Após ter sido ativado, o linfócito tem de ser jogado no sangue ou na
linfa para ir exercer função no local onde está acontecendo a infecção.
Esse linfócito chega até o local de infecção por quimiotaxia. Citocinas aumentam a permeabilidade
do vaso, sendo que leucócitos passam a migrar para esse local. E as quimiocinas levam ao
recrutamento dos linfócitos, que expressam receptores para reconhecimento de ligantes
presentes no endotélio da área de infecção.
Existem os linfócitos T CD8 tipo 1 e tipo 2, assim como linfócitos Th1 e Th2.

Diogo Araujo – Med 92
FALANDO DE LINFÓCITOS T CD4
O linfócito Th2 :
- produz il-4, il-5 e il-13 que fazem: ativação de linfócitos B (para produção de IgE); ativação de
eosinófilos e aumento de secreção mucosa;
- produz il-10, que inibe o Th1.
- inibe o macrófago graças à ação pela via da arginase.
O linfócito Th1:
- produz INF gama, que ativa macrófagos e estimula linfócitos B a secretarem IgM e IgG;
- inibe a resposta Th2 também através da produção de INF gama.
Bactérias extracelulares não levam à uma polarização evidente do tipo de linfócito T auxiliar.
Contudo, quando se trata de bactérias intracelulares, essa polarização pode acontecer. Na
hanseníase, por exemplo, que tem patógeno intracelular, há formação de uma população de
linfócitos T auxiliares que são ou Th1 OU Th2.
A resposta polarizada totalmente para um lado (Th1 ou Th2) nunca é benéfica para o organismo.
Se polariza para a forma de Th1, no caso da hanseníase, surge uma forma tuberculosa da
hanseníase. A resposta é tão forte que leva à morte do bacilo de Hansen, mas mata e destrói
células do organismo também. São formadas lesões de pele bem evidentes. É forma lepromatosa.

Diogo Araujo – Med 92
Por outro lado, se polarizar para o lado Th2, há inibição total das funções do macrófago (devido à
via da arginase). Como o Mycobacterium leprae fica dentro do macrófago, devido à supressão, o
patógeno passa a se multiplicar intensamente. O indivíduo pode passar a ter até 2kg de bactérias
no corpo, formando placas na pele. Essa é a forma tuberculóide (ou virchowiana).
O ideal é fazer uma resposta tendendo para o Th1 a fim de matar o microorganismo, mas com
presença de Th2 também. O ideal é não polarizar!
Mas o que determina que o linfócito T CD4 virgem forme linfócitos Th1 ou Th2?
- Antígeno: dependendo das característica do antígeno, há uma resposta. A maioria dos
organismos intracelulares (como vírus) induzem resposta Th1. Larvas de Schistosoma, por
exemplo, induzem reposta Th2.
- Estado de ativação da célula apresentadora de antígeno: se for macrófago forte produtor de INF
gama, tenderá para Th1. Se for uma célula dendrítica, a resposta tenderá para Th2.
- Células presentes no local: presença de mastócitos secretores de Il-4 podem levar o linfócito T
CD4 virgem a virar Th2. A presença de células NK ou de macrófagos produtores de INF gama leva à
formação de Th1.
- Citocinas: após um macrófago ter fagocitado um antígeno, ele passa a produzir il-12. Essa il-12
estimula células NK que produzem INF gama. Essa citocina, por sua vez, atua sobre o macrófago
inicial estimulando a expressão de B7 e produção de Th1.
- Genética do indivíduo: são as vias metabólicas de ativação que a célula irá utilizar
preferencialmente dentro da célula. Se for T bet e STAT 1, vira Th1. Se for GATA-3 ou STAT6, vira

Diogo Araujo – Med 92
Th2.
Para uma criança ser alérgica, ela tem de tender para a via Th2 uma vez que tenha entrado em
contato com o antígeno. Essa formação de Th2 será favorecida se houver um microambiente rico
em Il-4. Th2 estimula os linfócitos B a produzirem IgE e IgA.
Esse IgE se liga a receptores presentes na membrana de eosinófilos e mastócitos. Quando há uma
segunda exposição ao antígeno, esse IgE leva essas células à degranulação, promovendo alergia,
choque anafilático, etc.
O primeiro fator de melhora da criança contra a alergia é anatômico: crescimento dos brônquios.
Além disso, com o passar do tempo, essa criança tende a desviar para respostas imunes do tipo
Th1.
Papel da IL-12 e da IFN gama na diferenciação da Th1
O próprio microorganismo em contato com a CAA, a célula NK (que produz INF gama) e a ligação
de linfócito T CD4 por meio de CD40/ligante do CD40 ativam a CAA a produzir il-12. Assim, essa
CAA expressa IL-12 que atua sobre células T CD4 virgens a se tornarem linfócitos Th1. Esse tipo de
linfócito secreta INF gama que estimula o macrófago a matar o microorganismo (efetor).
Como a Th1 tem ação efetora?
- Ao se ligar ao macrófago (que fagocitou o microorganismo), há ligação CD40/ligante do CD40.
Assim, os macrófagos se sentem mais estimulados a matarem os microorganismos do meio em
que estão. O macrófago ativado é aquele que tem sua expressão de receptores aumentada para
facilitar fagocitose; expressa mais CHP de classe 2 para facilitar apresentação de antígenos;
aumenta toda a produção de suas moléculas microbicidas (radicais de nitrogênio e oxigênio; lisa
membrana, destrói proteína, DNA, RNA).
Como Th1 tem função auxiliar?
- Ele auxilia o linfócito B a produzir anticorpos, uma vez que ele secreta INF gama e estimula o
linfócito B a produzir IgM e IgG (resposta timo-dependente). Essas Ig atuam como opsoninas.
- Ele auxilia o linfócito T CD8 a se transformar em linfócito citolítico ativado para matar a célula
infectada.

Diogo Araujo – Med 92
O teste intradérmico (como no caso da tuberculina) mede a capacidade de resposta Th1. É uma
reação de hipersensibilidade retardada porque demorou 48 horas. Significa que o indivíduo tem
linfócitos de memória para o antígeno que está sendo apresentado. E são linfócitos de memória
porque, se fossem somente linfócitos virgens, a resposta somente viria após uma semana.
Teste de Fernandes: é como o PPD da tuberculose, mas aplicado à hanseníase. Tem de ser feita
com duas leituras: uma de 48 horas e outra de 40 dias depois (que mede a resposta Th2).
Na região de reação do PPD, há uma expressão histopatológica de granuloma. Ou seja, há uma
coleção de células do sistema imune organizadas e estruturadas, que se forma em resposta à
estimulação antigênica crônica, no contexto de fatores mediados pelo macrófago. É uma resposta
demorada.
No granuloma é encontrada uma bainha de linfócitos circundando uma região (centro) de
macrófagos que fagocitaram e morreram. Ao redor, há macrófagos muito ativados que podem
formar células gigantes de Langerhans. Depois, uma bainha de linfócitos que, na maior parte das
vezes, é do tipo Th1.
Como o Th2 atua?
- Inibe Th1 (através da produção de il-10);
- Estimula linfócito B a produzir anticorpos da classe IgE e IgA. O IgE se liga à superfície de
mastócitos. Quando entra em contato com um alérgeno (mastócito) ou helminto (eosinófilo), a
célula se degranula. Faz também defesa contra parasitas;
- Estimula produção de IgG (pouco) por linfócitos B por meio de Il-4;
- Inibe o macrófago através da via da arginase.

Diogo Araujo – Med 92
O linfócito Th17 e Treg são subpopulações recentemente descobertas.
Th17 é produzido pela presença de il-6 e TGF beta, usando a via de ativação ROR gama t. A célula,
então, passa a produzir il-17 e il-22, combatendo bactérias extracelulares.
O linfócito Treg é produzido pela presença de Il-2 e TGF beta, usando a via de ativação FOXp3. Faz
regulação negativa do sistema imunitário.
FALANDO DE LINFÓCITOS T CD8
A fase de indução consiste na apresentação do antígeno por meio de CPH de classe I.
A fase efetora do linfócito T CD8 se dá por contato célula-célula. Mas como ele mata a célula
irregular? São três os mecanismos envolvidos na morte:
- FAS
- Granzima
- Perforina
Uma vez ativado, ele vira linfócito T CD8 citolítico. Ele mata especificamente a célula infectada. Na
maior parte das vezes, ele induz a morte por apoptose. Como não há necrose/estouro da célula,
não há liberação de tantos fatores que estimulam a inflamação e a sintomatologia clínica.
A programação de morte da célula (apoptose) se dá por meio da via do FAS/ligante do FAS (que
ativa caspases dentro da célula) e da liberação de perforinas e granzimas.
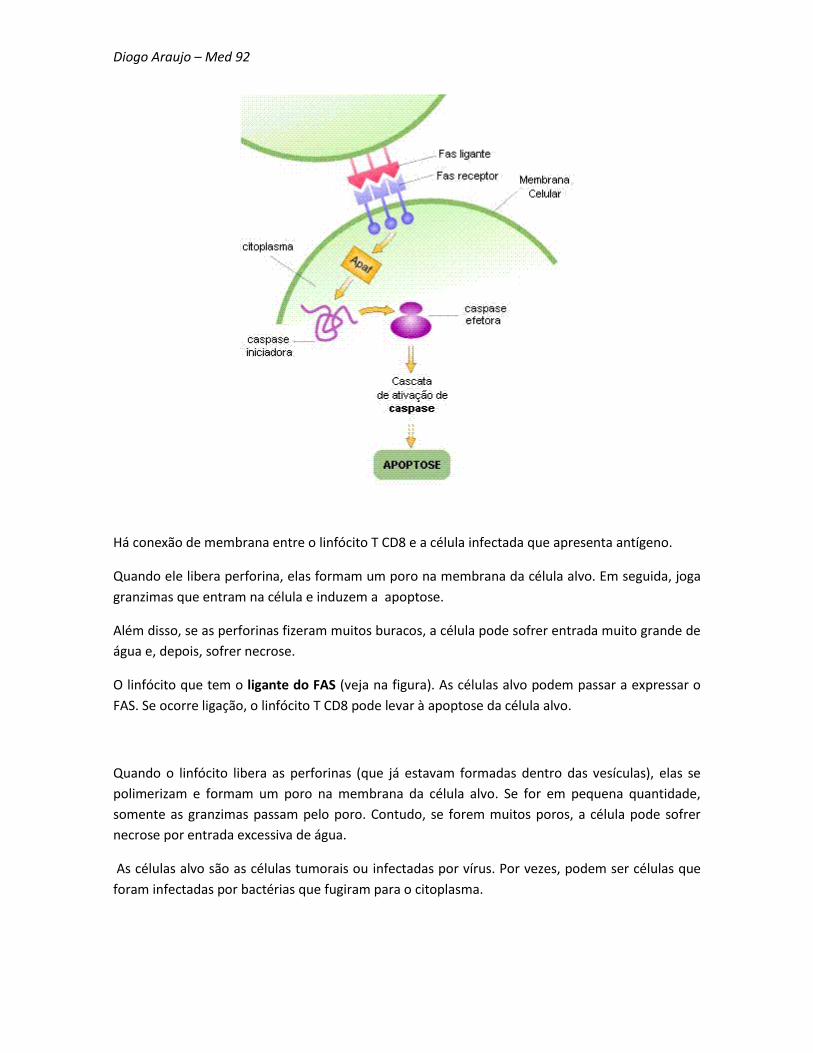
Diogo Araujo – Med 92
Há conexão de membrana entre o linfócito T CD8 e a célula infectada que apresenta antígeno.
Quando ele libera perforina, elas formam um poro na membrana da célula alvo. Em seguida, joga
granzimas que entram na célula e induzem a apoptose.
Além disso, se as perforinas fizeram muitos buracos, a célula pode sofrer entrada muito grande de
água e, depois, sofrer necrose.
O linfócito que tem o ligante do FAS (veja na figura). As células alvo podem passar a expressar o
FAS. Se ocorre ligação, o linfócito T CD8 pode levar à apoptose da célula alvo.
Quando o linfócito libera as perforinas (que já estavam formadas dentro das vesículas), elas se
polimerizam e formam um poro na membrana da célula alvo. Se for em pequena quantidade,
somente as granzimas passam pelo poro. Contudo, se forem muitos poros, a célula pode sofrer
necrose por entrada excessiva de água.
As células alvo são as células tumorais ou infectadas por vírus. Por vezes, podem ser células que
foram infectadas por bactérias que fugiram para o citoplasma.

Diogo Araujo – Med 92
FALANDO UM POUQUINHO SOBRE CÉLULA NK (QUE NÃO É LINFÓCITO T)
A célula NK não tem o receptor do linfócito T. Então, ela não é tão específica. Mas possui
mecanismos de destruição muito semelhantes aos do linfócito T CD8. Ela aparece no início de uma
resposta a uma infecção viral.
A célula NK age diferente do linfócito T CD8 no sentido de que, se não houver CPH de classe I com
antígeno próprio, a célula NK mata. Já o linfócito T CD8 não mata se não houver CPH de classe I.
----------------------------------------------------------------------------------------------------------------
Regulação da resposta imuntária
Profª Érica Alessandra
Como vimos na aula anterior, existe uma dinâmica na resposta imunológica, com as fases:
- Expansão clonal;
- Diferenciação;
- Ação contra o antígeno;
- Eliminação do antígeno (por respostas celulares ou humorais);
- Apoptose de clones ;
- Permanência de linfócitos de memória.
Em cada uma dessas etapas, há mecanismos de regulação. Isso é importante porque, regulando
cada uma das etapas, a ação do sistema imune é adequada e sem excessos (matando o antígeno
sem causar danos ao próprio).
Mas quais são os mecanismos envolvidos nessa modulação da resposta imune?

Diogo Araujo – Med 92
REGULAÇÃO POR ANTÍGENO
Os antígenos ajudam a regular a resposta imunológica. De que forma?
"Se ele está presente, há resposta imunológica."
Como?
O linfócito T, ao reconhecer uma célula que esteja apresentando um antígeno, produz il-2 e, assim,
sofre proliferação. Ao final da resposta, se o sistema imune elimina os antígenos, o linfócito fica
privado da estimulação de que ele precisa para sobreviver. Então, ele entra em apoptose a partir
da ativação interna de caspases. Essa é a chamada morte passiva ou morte por negligência:
quando antígeno é eliminado, faltam estímulos para fazer com que os linfócitos permaneçam
vivos.
Em contrapartida, se o antígeno fica presente no meio por um tempo muito prolongado e, mesmo
após o ataque inicial do sistema imune, ele permanece vivo, há um estímulo excessivo dos
linfócitos que o combatem. Esses linfócitos acabam expressando o Fas e o ligante do Fas. Assim,
caspases-B são ativadas dentro dos linfócitos e eles entram em apoptose. Essa é a morte celular
induzida por ativação.
Se não houvesse esses sistemas de apoptose, a cada ativação, aumentaria muito a quantidade de
linfócitos no organismo, podento criar mesmo linfócitos que o atacassem. A forma de controlá-los
é justamente pela indução da apoptose.
Pessoas e animais que têm defeitos na via do FAS e do ligante do FAS apresentam doenças. A
síndrome linfoproliferativa autoimune (ALPS) é um quadro clínico em que não se consegue
expressar o Fas. Assim, são mantidos muitos linfócitos vivos na circulação do indivíduo. Ele
desenvolve uma linfoadenomegalina não maligna, além de esplenomegalia e uma série de
citopenias autoimunes (pela manutenção de células auto-reativas na circulação do indivíduo).
REGULAÇÃO POR MOLÉCULAS
CTLA-4: via de regulação inibitória
CTLA-4 significa Antígeno-4 do Linfócito T Citotóxico.
O linfócito virgem possui baixíssima concentração de CTLA-4 na sua membrana.

Diogo Araujo – Med 92
Se, em uma apresentação de antígeno a um linfócito T virgem, existe o primeiro sinal (CPH e
antígeno) mas não existe o segundo (B7 se ligando em CD28), a célula T virgem entra em anergia
ou apoptose.
No entanto, se essa célula T virgem é apresentada a um antígeno com coestimulação (CD28 do
linfócito ligando a B7 da célula apresentadora de antígeno), essa célula T entra em proliferação,
diferenciação e função efetora.
Depois que está ativa, essa célula passa a matar os antígenos.
Contudo, ela também aumenta a sua expressão de CTLA-4. Essa molécula se liga ao B7 da célula
apresentadora de antígeno. Uma vez que isso anconteça, o linfócito T ativo deixa de produzir il-2 e
entra em modo de repouso.
Essa via permite que, enquanto alguns linfócitos são ativados, outros sejam "desligados", havendo,
então, uma população adequada de linfócitos.
O CTLA-4 faz com que a célula entre em repouso tanto por modificação de vias intracelulares do
linfócito quanto por inibição da ligação do B7 ao CD28 (que promove ativação).
A via do CTLA-4 também é importante para a contenção de células auto-reativas. Isso porque o
tecido próprio que está sendo agredido não apresenta coestimulação, fazendo com que esses
linfócitos auto-reativos parem de se proliferar.
Essa via está sendo utilizada para tratamento de doenças em que há ativação excessiva de
linfócitos. Para isso, utiliza-se uma molécula semelhante ao CTLA-4 (é o CTLA-4Ig). Essa molécula,
ao se ligar ao B7 da célula apresentadora de antígenos, faz com que não haja ativação dos
linfócitos responsáveis pela patogenia.
A artrite reumatóide é um exemplo de doença que poderia ser tratada com CTLA-4Ig.
Um outro exemplo de terapia que explora essa via é o uso do anticorpo anti-CTLA-4, com função
de bloquear o CTLA-4. Assim, essa via seria inibida, o que é interessante no contexto dos tumores
(quadro em que se deseja manter ativos os linfócitos que combatem tumores).
Anticorpos

Diogo Araujo – Med 92
Os anticorpos se ligam aos antígenos e fazem com que eles sejam removidos através de
fagocitose. Assim, há menor taxa de ativação de linfócitos B e T.
Eles também participam da regulação negativa por anticorpos. Como é isso?
Quando um linfócito B produz IgG, esse anticorpo se liga a receptores presentes na superfície do
próprio linfócito que o produziu.
Assim, um antígeno que esteja presente no meio e que vá se ligar ao linfócito, acaba grudando
tanto no receptor do linfócito B quanto na IgG que estava na membrana. A ligação à IgG faz com
que as enzimas necessárias para a ativação do linfócito B não atuem. Assim, a ativação do linfócito
B é suprimida.
Essa via é importante porque ela inibe que muitos anticorpos sejam produzidos pelo linfócito B.
Anticorpos em excesso são danosos para os tecidos (é o que acontece na glomérulo nefrite, por
exemplo).
CD22
O CD22 uma molécula presente na membrana do linfócito B. Quando é ativada, ela faz inibição do
linfócito B. Contudo, não se conhecem os ligantes do CD22.
A proteína SHP-1 está ligada na cauda do CD22. Os camundongos knockout para SPH-1
desenvolvem autoimunidade grave com ativação descontrolada de linfócitos B e produção de
autoanticorpos.
Revisando:
Até agora, vimos como mecanismos de regulação:
- O próprio antígeno;
- via do CTLA-4;
- atuação de anticorpos; e
- via do CD22.

Diogo Araujo – Med 92
Existe interferência mútua entre as subpopulações de linfócitos T?
Durante a resposta imunológica dos linfócitos T, a população de linfócitos T auxiliares se diferencia
em Th17 (bactérias, fungos), Th2 (helmintos e microorganismos extracelulares), Th1
(microorganismos intracelulares), Treg (que tem papel de regulação da resposta).
Em uma lesão cutânea por leishmaniose, por exemplo, o linfócito Th1 é o responsável por ativar o
macrófago, para que ele produza radicais de oxigênio e nitrogênio para matar a Leishmania.
Contudo, nesse mesmo local, as outras subpopulações de linfócitos T auxiliares estão presentes, a
fim de regular a atividade do linfócito Th1.
Obs: Na forma cutâneo-mucosa, a resposta Th1 é muito forte.
O linfócito Treg diminui a atividade tanto de Th1 quanto de Th2.
Relembrando:
- A população Th2 produz citocinas como a il-10 e TGF beta para diminuir a atividade da população
Th1.
- A Th1 produz INF gama para inibir a Th2.
- Além disso, Th1 e Th2 juntas inibem a ação da Th17 (por meio de il-4 e INF gama).
Na leishmaniose, o indivíduo que faz a forma muco-cutânea possui uma ativação de população
Th1 muito importante, com altos níveis de TNF alfa e IFN gama nos tecidos, com grande ativação
de macrófagos.
Na forma visceral da leishmaniose, há indícios de que o indivíduo polariza para Th2, não
conseguindo combater o parasito intracelular. Há baixa produção de INF gama nesse caso.
Linfócitos T reguladores
Existem várias populações de linfócitos Treg. A mais bem caracterizada é aquela que é CD4 +,
FOXP3 +, CD25+ (uma das cadeias do receptor para il-2) e CTLA-4 +.

Diogo Araujo – Med 92
Essa população é dependente de Il-2 para se desenvolver.
Esses linfócitos Treg podem ser gerados no timo ou na periferia. Mas como?
Durante o processo de formação de linfócitos T no timo, aqueles que se ligam a auto-antígenos
podem sofrer apoptose ou virar linfócito Treg (não se sabe ainda quais são os mecanismos
responsáveis por fazê-lo sofrer apoptose ou virar Treg).
Para ser Treg, ele ganha o receptor FOXP3 ao entrar em contato com o antígeno na periferia ou no
timo. Isso dá funcionalidade ao linfócito Treg.
Mas como o linfócito Treg atua depois de ter sido formado? São dois os mecanismos:
- Ele pode fazer inibição da ativação de células T virgens, por meio da inibição da célula
apresentadora de antígeno.
- Ele pode atuar diretamente sobre as células T efetoras, diminuindo a sua função.
[Cada um desses mecanismos será descrito em detalhes um pouco mais adiante]
Indivíduos com deficiência de FOXP3 desenvolvem uma doença chamada IPEX (Endocrinopatia e
enteropatia autoimunes) devido à ausência de linfócito Treg. Nesse caso, são duas as explicações:
- Por não conseguirem expressar a FOXP3, os linfócitos T auto-reativos no timo não são
convertidos em Treg e, assim, podem continuar vivos e cair na circulação;
- A ausência de linfócitos Treg conduz para ação excessiva dos outros linfócitos.
Se há pouco linfócito Treg:
- o patógeno é eliminado, mas a ação excessiva dos linfócitos causa dano tecidual.
Se há muito linfócito Treg:
- o patógeno permanece, há maior transmissão da doença, que pode destruir o
hospedeiro e, por vezes, levar à reativação da doença.

Diogo Araujo – Med 92
Se há número normal de linfócitos Treg:
- existe proteção imunológica e, ao mesmo tempo, controle de repostas excessivas
contra o próprio. Algumas vezes, o microorganismo permanece no indivíduo, que se
torna um reservatório da doença.
Como o linfócito Treg tem função regulatória?
Por mecanismos dependentes de contato célula-célula
A presença de CTLA-4 no linfócito Treg pode fazer com que esse linfócito se ligue à B7 da célula
apresentadora de antígeno, ocupando o espaço para ativação de outras células.
Por produção de fatores supressivos solúveis
Além disso, ele pode passar à celula apresentadora de antígenos uma série de substâncias
supressoras, como a il-10, o TGF beta e a il-35.
Competição
Vimos que a il-2 atua como molécula importante para a formação de linfócitos Treg. Mas essa
citocina também atua na proliferação das outras subpopulações de linfócitos T CD4. Assim, o fato
da il-2 atuar sobre a célula Treg e não sobre as demais subpopulações pode ser encarado como um
"roubo" de il-2. Esse roubo implicaria em menor formação de linfócitos T efetores, o que seria
uma maneira de regular a resposta.
Então, até agora, vimos que são mecanismos de regulação:
- presença do antígeno;
- via do CTLA-4;
- ação do anticorpo;
- via do CD 22;
- ação dos linfócitos Treg.

Diogo Araujo – Med 92
Existe um outro mecanismo de modulação da resposta imune que é a TOLERÂNCIA. Ela será
estudada à parte na próxima aula.
HIPÓTESE DA REDE IDIOTÍPICA
Cobaia exposta a antígenos, sete dias depois, produz anticorpos contra o antígeno. Esses são os
chamados anticorpos idiotípicos.
Mas, 14 dias depois, há produção de anticorpos contra os anticorpos idiotípicos. Esses são os
anticorpos antidiotípicos. Representam a imagem do antígeno, porque ele encaixa na região de
reconhecimento do antígeno.
"Uma conseqüência imediata das interações idiotípicas é noção da imagem interna de um
antígeno: se o antígeno é reconhecido pelo anticorpo idiotípico e este pelo anticorpo
antidiotípicos, esse anticorpo antidiotípico pode apresentar uma estrutura similar ao
antígeno e é conseqüentemente uma imagem interna do antígeno em questão." Ou seja:
Anticorpo antidiotípico = antígeno.
Parte dessa teoria não é verdade. Mas parte existe sim.
Quais são as possíveis funções desses anticorpos antidiotípicos?
A sua presença leva à modulação das respostas imunitárias (celular e humoral). Isso porque os
anticorpos anti-idiotípicos, por lembrarem os antígenos, serviriam de estímulo para a manutenção
da resposta imune de células de memória.
--------------------------------------------------------------------------------------------------------------------
Tolerância imunológica
Profª. Érica Alessandra

Diogo Araujo – Med 92
É mais uma forma de regulação do sistema imune.
Compreende uma série de mecanismos que levam à tolerância à microbiota, autoantígenos, etc.
Por que o nosso sistema imune não reage contra a microbiota presente no trato gastrointestinal?
Isso se deve à tolerância. : )
A tolerância é a não reatividade imunológica específica resultante de um contato prévio com um
antígeno. Ela é importante porque garante a não reatividade a antígenos próprios (endógenos) e
inócuos (microbiota). Caso contrário, se não houvesse essa tolerância, a cada vez que nos
alimentássemos, haveria reação contra os antígenos, com inflamação do sistema digestório.
Seríamos um ser o tempo todo doente e inflamado.
Essa questão de tolerância iniciou com os estudos de Dr. Ray Owen (1945). Ele observou que em
gêmeos dizigóticos bovinos que tinham placentas compartilhadas apresentavam trocas de células
entre si. Então, após o nascimento, eles apresentavam dois tipos de eritrócitos distintos. Eram
quiméricas hematopoiéticas. E o interessante é que não havia rejeição das hemácias que não
faziam parte do organismo do animal. E, depois de crescido, era possível pegar a pele de um
animal e enxertar no outro sem haver rejeição.
Dr. Petter Medawar (1950) também fez uma demonstração semelhante. Ele pegou camundongos
neonatos da linhagem CBA e dividiu em dois grupos. Em um dos grupos, ele não fez nada. No
outro, assim que o camundongo nasceu, transferiu células esplênicas da linhagem B. Depois de
adultos, os camundongos dos dois grupos foram submetidos a enxertos da linhagem B. Somente
aqueles do grupo que tinha sido exposto a células da linhagem B foram capazes de aceitar o
enxerto. Quanto eram transplantados com pele da linhagem C, eles também rejeitavam o enxerto.
São as bases da tolerância imunológica:
- É um processo específico induzido por um antígeno. Esse antígeno que induz tolerância contra
ele é chamado de antígeno tolerogênico.
- O processo de tolerância se inicia durante o período de vida intra-uterino e é mantido por toda a
vida.
- A tolerância pode ser alcançada tanto para antígenos endógenos (próprios) e não próprios (como
a microbiota e os alimentos, por exemplo).

Diogo Araujo – Med 92
- Conta com a participação de linfócitos T e B.
- Conta com múltiplos mecanismos para acontecer. Mas... Quais são eles?
São dois mecanismos básicos:
- a tolerância central (que acontece na medula ou no timo);
- a tolerância periférica (que acontece nos sítios de residência dos linfócitos; ou seja, linfonodos,
intestino, TLAM, etc.)
Os mecanismos de tolerância central, de maneira isolada, não são 100% efetivos. Durante a
tolerância central, algumas células conseguem escapar e serem jogadas na periferia. Assim, elas
podem entrar em atividade e causar lesão tecidual.
Vamos discutir um pouco sobre os mecanismos de tolerância a antígenos próprios.
TOLERÂNCIA DO LINFÓCITO T
A mecanismos de tolerância central do linfócito T
Durante o processo de formação de linfócitos no timo, há seleção negativa de linfócitos T auto-
reativos. Linfócitos auto-reativos que se ligam muito fortemente a auto-antígenos podem entrar
em apoptose e serem eliminados. Além disso, como vimos anteriormente, ao invés de sofrer
apoptose, esse linfócito pode se tornar um linfócito Treg, que cai na periferia e vai ajudar nos
mecanismos de regulação da resposta imune.
Os genes AIRE e FOXP3 são importantíssimos para a tolerância central.
O gene AIRE codifica a proteína AIRE (regulador autoimune) que é um fator de transcrição
expresso por células dendríticas e epiteliais tímicas responsáveis pela expressão de autoantígenos
no timo (isso está envolvido na remoção de linfócitos T auto-reativos).
Caso contrário não exista o gene AIRE e, consequentemente, a expressão de auto-antígenos no

Diogo Araujo – Med 92
timo, há a síndrome poligradular autoimune (APS-1), em que o indivíduo apresenta
hipoparatireioidismo, doença de Addison, diabetes insulino-dependente, vitiligo, alopécia,
ceratopatia e candidíase, por exemplo.
O AIRE também tem papel na manutenção da estrutura da queratina. Assim, sua falta leva a
alterações ungueais, por exemplo.
Nos macrófagos, o AIRE também é importante para a via de sinalização responsável pelo
reconhecimento da cândida. É por isso que sua falta implica em maior ocorrência de candidíase.
O FOXP3 (Forkhead Box Protein 3) é importantíssimo para que o linfócito T auto-reativo passe a
ser linfócito Treg (como descrito na aula anterior). Relembrando os mecanismos de ação dos
linfócitos Treg:
- supressão da função linfocitária, com a secreção de citocinas supressoras (il-10,
TGF beta, il-35). Elas fazem com que a divisão celular do linfócito seja mais difícil.
Isso quebra a fase de proliferação de linfócitos T.
- consumo da il-2, que é indispensável para a proliferação e manutenção de linfócitos
que estão respondendo a antígenos. Assim, a célula fica privada de um estímulo
necessário para sobrevivência, podendo entrar em apoptose.
- pode promover citólise, por meio de secreção de granzima e apoptose pela ativação
da cascata do FAS. (WTF?! Esse mecanismo nem havia sido citado na aula anterior!).
- expressão de moléculas de superfície que irão agir em receptores de outros linfócitos
por contato célula-célula, fazendo-o parar no ciclo celular. Um exemplo é a galectina-1.
Os linfócitos T reg também atuam sobre as células apresentadoras de antígenos, já que são elas
que vão participar da ativação de linfócitos T:
- Via ligação CTLA-4 com B7, com diminuição da capacidade de coestimulação pela
célula apresentadora de antígeno (CAA).
- por meio da molécula LAG-3, que se liga ao MHC de classe II, fazendo com que a
CAA fique imatura, com baixa apresentação de antígenos também por ocupação
física do local de apresentação de antígenos.
- por meio de degradação de moléculas extracelulares (como o ATP) que seriam capazes
de ativar a CAA. Assim, essas células ficam menos ativadas e são menos capazes de fazer
coestimulação.
- uma série de moléculas que fazem com que o linfócito Treg se ligue à superfície

Diogo Araujo – Med 92
da CAA, ocupando o sítio de ligação de outras moléculas que teriam potencial
estimulatório.
Lembrar da doença em que há falta da FOXP3: Endocrinopatia e enteropatia autoimunes (IPEX).
Mecanismo de tolerância periférica do linfócito T
Como nem todos os autoantígenos são expressos no timo e há eventual saída de linfócitos auto-
reativos do timo, essas células acabam indo para a periferia, podendo causar lesões teciduais.
Se há contato adequado entre um linfócito auto-reativo e um antígeno próprio, ele poderia ser
ativado, sofrer proliferação e agredir o tecido.
Contudo, esse linfócito pode encontrar mecanismos que o levam à tolerância. A falta de
coestimuladores durante a sua apresentação a auto-antígenos, por exemplo, poderia ser um fator
que o levasse a não agir contra estruturas do próprio.
Explicando direitinho:
Na resposta imunitária normal, o antígeno imunogênico aumenta o nível de moléculas de CPH de
classe II e coestimulatórias na superfície da CAA.
Contudo, quando se trata de auto-antígenos, os antígenos endógenos são apresentados na
ausência de coestimulação (porque essa CAA estava em repouso, quase sem expressar
coestimuladores). O linfócito T auto-reativo que é apresentado a esse antígeno sem coestimulação
acaba se tornando anérgico, ou seja, não responsivo. Esse mecanismo acontece muito durante a
formação inicial de linfócitos nas crianças.
A falta de coestimulação pode também fazer com que esse linfócito T passe a expressar CTLA-4 em
sua superfície, o que o faria se tornar anérgico. Ratinhos sem CTLA-4, por exemplo, morrem
precocemente por ter proliferação maciça de linfócitos auto-reativos.
Além da via do CTLA-4, existe a morte celular induzida por ativação. Nela, linfócitos que entram em
contato com auto-antígenos expressos por CAA mas sem o segundo sinal, passam a expressar Fas
e ligante do Fas. Assim, são levados à apoptose.
TOLERÂNCIA DO LINFÓCITO B

Diogo Araujo – Med 92
A tolerância de linfócito B deve ser dada contra proteínas, polissaracídeos, linpídeos e ácidos
nucleicos, já que o receptor do linfócito B pode reconhecer todos esses tipos de antígenos.
Mecanismos de tolerância central do linfócito B
Na medula, durante a selação negativa, se esse linfócito reconhece autoantígeno, ele pode entrar
em apoptose ou fazer reedição do receptor do linfócito B (ou seja, sofrer pequenas mutações no
gene do BCR para tentar ser selecionado). Se ele for útil, ele pode ir para a circulação. Caso
contrário, pode passar por outros processos de reedição ou sofrer apoptose (isso foi estudado
com detalhes no primeiro módulo).
Esses dois eventos são a deleção clonal (apoptose) e a reedição do receptor do linfócito B.
Contudo, esses mecanismos não são muito fortes, sendo que linfócitos B auto-reativos são
produzidos constantemente, devendo ser regulados negativamente na periferia.
Mecanismos de tolerância periférica do linfócito B
O processo de tolerância do linfócito B acontece em órgãos linfóides periféricos. São três os
fatores envolvidos: a anergia do linfócito (que não é ativado), a sua exclusão do folículo e
consequente apoptose.
Como os linfócitos T auto-reativos sofrem os mecanismos de tolerância central e periférica, eles
não ficam presentes ao redor dos folículos de órgãos linfáticos para ativar linfócitos B auto-
reativos. Assim, esses linfócitos B ficam anérgicos (não são ativados), sofrem exclusão folicular
(ficam à margem do folículo) e podem sofrer apoptose.
Tudo isso que foi dito é para antígeno próprios. Contudo, há necessidade de tolerância para
antígenos exógenos.
INDUÇÃO DE TOLERÂNCIA A ANTÍGENOS EXÓGENOS

Diogo Araujo – Med 92
Existem mecanismos de tolerância central e periférico para antígenos exógenos.
Contudo, essa tolerância central só acontece em nível experimental. Não é comum nos indivíduos.
Os mecanismos periféricos são os mais comuns para a tolerância a antígenos exógenos.
Um exemplo é a indução de tolerância oral. Ou seja, toda vez em que ingerimos algo (alimentos,
bactérias, antígenos derivados de microorganismos patogênicos) preferencialmente induzimos
tolerância a esse antígeno. Isso pode acontecer em qualquer mucosa.
Células epiteliais, macrófagos e células mesenquimais estão respondendo aos antígenos ingeridos
e comensais presentes na luz. Essas células produzem TGF-B e PGE2 (prostaglandina E2), que
atuam sobre as células apresentadoras de antígeno, inibindo-as. Assim, quando a CAA encontra o
linfócito T virgem para um antígeno, pela ausência de coestimulação, pode haver formação de
linfócito Th3 ou linfócito Treg. Esses linfócitos fazem supressão local e sistêmicas de resposta
contra esse microorganismo.
Esses linfócitos Treg atuam secretando citocinas que causam tolerância sistêmica e local. Já os
linfócitos Th3 produzem il-4, que estimula a produção de IgA pelos linfócitos B. (Achei isso
estranho, porque, se há produção de IgA, essas células Th3 não seriam inibitórias da função
imune, né). :P
No caso de microorganismos patogênicos presentes na luz, há produção de uma série de citocinas
(il-1, il-5, il-8, CCL5) que levam à ativação forte das células apresentadoras de antígeno. Desse
modo, as células T CD4 podem se diferenciar em Th1 ou Th2, levando a maior resposta local e
sistêmica.
Fatores que favorecem a tolerância:
- quantidade do antígeno: altas quantidades favorecem tolerância;
- persistência: aquele antígeno que permanece no organismo por tempo prolongado tende a
causar tolerância (isso estaria relacionado com aquela morte induzida por ativação?);
- porta de entrada: entrada via oral tende a fazer tolerância, por exemplo;
- presença de adjuvantes: antígenos sem adjuvantes são tolerogênicos;
- propriedades das células apresentadoras de antígenos: baixos níveis de coestimuladores e
citocinas.
-------------------------------------------------------------------------------------------------------------------

Diogo Araujo – Med 92
Citocinas
Prof. Érica Alessandra
Citocinas são polipeptídeos (proteínas!) produzidos em resposta a microorganismos e outros
antígenos, que medeiam e regulam a resposta imunitária.
Todas as citocinas têm propriedades gerais. São elas:
- Toda secreção de citocina é um evento breve e autolimitado. A célula produz a citocina, ela age
e a célula pára de produzir. Ela é produzida no RER, vai para o Golgi, é colocada em vesículas que
são eliminadas por exocitose, indo atuar no meio extracelular. Ela pode agir sobre a própria célula
que a produziu (autócrina), atuar em células ao lado (parácrina) ou cair na corrente sanguínea
(endócrina, atuando como um hormônio).
- A ação da citocina não é aleatória. Ou seja, ela atua sobre receptores específicos. Existem 5
grupos de receptores: receptores de citocina tipo 1, tipo 2, família do receptor do TNF, receptor da
superfamília das imunoglobulinas (il-1 se liga nesse receptor) e receptor de sete alfa-hélices
(quimiocinas).
- As citocinas alteram a expressão gênica das células-alvo, levando a novas funções e, algumas
vezes, à proliferação (dependendo da citocina). A ligação do TNFalfa, por exemplo, ao seu receptor
promove a ativação do NF kappa Beta (fator de transcrição), que leva à produção de citocinas,
moléculas de adesão, radicais de oxigênio. Ou seja, esse fator induz inflamação. Ele é o
responsável por iniciar a inflamação.
- Podem influenciar a síntese e as ações de outras citocinas. Quando o tecido é infectado por
microorganismos, a primeira célula a ser encontrada é o macrófago. Ele produz uma citocina
importante que é a il-12. Essa citocina atua sobre a célula NK, que produz INF gama como
resposta. Essa molécula ativa o macrófago, o que resulta na produção de TNF, il-1 e quimiocinas
que levam à migração de células inflamatórias para o local. Isso faz com que a resposta imunitária
seja uma resposta coordenada.
- Podem ser pleiotrópicas, redundantes, sinérgicas ou antagônicas. Peliotropismo indica que uma
mesma citocina é capaz de agir sobre diversos pontos do sistema imunitário, determinando em
cada local uma função diferente. A redundância é a convergência de atuação de citocinas sobre
uma mesma célula, influenciando uma mesma função. A sinergia é a atuação conjunta de citocinas
de modo que o efeito seja multiplicado. O antagonismo é a ação de citocinas sobre a mesma célula

Diogo Araujo – Med 92
determinando funções opostas.
Citocinas que medeiam e regulam a imunidade inata
TNF ALFA
É produzido principalmente por fagócitos mononucleares ativados, além de células T ativadas,
células NK e mastócitos.
O ativador mais potente de TNF alfa é o lipopolissacarídeo bacteriano (presente em bactérias
gram-negativas).
Ela apresenta sinergismo com a linfotoxina (produzida por linfócitos T).
Atua em dois tipos de receptores.
Como havia sido citado anteriormente, o TNF atua em seu receptor e aumenta a expressão do
NFkappaB, que é um fator de transcrição. Isso leva à expressão de citocinas, moléculas de adesão
e radicais de oxigênio e nitrogênio que promovem a inflamação tecidual.
Além disso, durante o processo de ativação do receptor, ao invés de ativar a expressão de NK
kappa beta, o TNF pode resultar em aumento da molécula FADD, que ativa caspases que
conduzem à apoptose. Assim, a ação do TNF alfa pode levar à inflamação do tecido ou a morte das
células.
O TNF faz ações no corpo dependendo da sua quantidade produzida. Se as quantidades são baixas,
a ação do TNF é local, com inflamação local, produzindo alteração endotelial, atração de fagócitos
e aumento de sua capacidade de fagocitose.
Se as quantidades são moderadas, há efeito sistêmico, agindo no hipotálamo e causando febre.
Além disso, atua no fígado, aumentando as proteínas de fase aguda, como a proteína C reativa
(essas proteínas atuam como opsoninas, auxiliando a fagocitose do microorganismo). Também
age na medula óssea, estimulando a produção de leucócitos.
Se as quantidades são muito altas, o indivíduo pode entrar em choque. Infecções por gram-
negativos podem conduzir ao choque. Isso porque o TNF em excesso atua causando baixo débito
cardíaco, baixa resistência vascular, formação de trombos e hipoglicemia por efeito hepático.

Diogo Araujo – Med 92
Assim, quantidades normais de TNF alfa são importantes. Mas quantidades exageradas podem
levar a prejuízos e ao óbito.
INTERLEUCINA-1
Produzida por fagócitos mononucleares ativados. Ou seja, são fagócitos que foram ativados pelo
microorganismo e pela própria produção de TNF. Pode ser produzida também por neutrófilos,
queratinócitos e células endoteliais.
LPS (lipopolissacarídeo bacteriano) e TNF alfa induzem sua produção.
Existem as interleucinas 1 alfa e beta. São produzidas por mecanismos diferentes. A alfa é
produzida e secretada sem sofrer mudanças em sua estrutura. A beta é produzida e sofre ação do
inflamassoma, tornando-se ativada antes de ser secretada. Contudo, possuem as mesmas funções.
A il-1 tem sinergismo com il-6 e TNF alfa (lembre-se que são exatamente essas as citocinas que,
em conjunto, estimulam a formação de linfócitos Th17).
A il-1 possui praticamente os mesmos efeitos locais e sistêmicos que o TNF alfa.

Diogo Araujo – Med 92
- Em baixas quantidades, promove inflamação local, com migração de leucócitos, mudanças no
endotélio, etc.
- Em quantidades moderadas, produz febre, aumenta proteínas de fase aguda e estimula a
produção de leucócitos.
Contudo, quando em grandes quantidades, o choque séptico só acontece quando há ação
sinérgica da il-1 com outras citocinas. A il-1 isoladamente não é capaz de gerar choque séptico.
INTERLEUCINA-12
Produzida por fagócitos mononucleares ativados e células dendríticas.
LPS e bactérias intracelulares, vírus, ligação CD40/CD40 ligante e presença de INF gama estimulam
sua produção.
Tem fuções parecidas com outras interleucinas, como a il-18, 23 e 27.
A il-12 faz ativação de linfócitos T virgens, estimulando-as a se diferenciarem em linfócitos Th1.
Além disso, ativa células NK e linfócitos T CD8. Assim, essas células secretam o INF gama, que atua
sobre o macrófago, aumentando o seu poder (mais enzimas, mais radicais, mais receptores, mais
CPH). Isso é essencial em algumas doenças como na leishmaniose.
Ela também atua sobre a NK e sobre a TCD8 aumentando a sua ação citolítica, com mais granzimas
e perforinas e, consequentemente, maior capacidade de induzir a morte do microorganismo. A
falta de il-12 prejudica a defesa contra microorganismos intracelulares.
A il-12 se liga em receptor do tipo JAK/STAT. Ele se dimeriza e, como está associado à JAK, é
fosforilado. Isso traz para a membrana celular as proteínas STAT. Elas são fosforiladas e se
dimerizam também. Então, vão para o núcleo da célula, onde atuam como fator de transcrição
para a produção de uma série de proteínas.
São vários os tipos de receptores do tipo JAK/STAT. Contudo, os receptores que são ativados são
os do tipo 1 e 2. A STAT 4 que é utilizada quando há a ação da il-12.

Diogo Araujo – Med 92
As citocinas da imunidade inata possuem relações entre si.
A ação do LPS sobre o macrófago faz com que ele produza a TNF (é a citocina mais precocemente
produzida). Assim, esse TNF ativa o endotélio dos vasos, com migração de leucócitos para o local.
Quando essas células chegam, elas já encontram o TNF, sendo ativadas.
Esse TNF atua sobre o próprio macrófago, estimulando a produção de il-1. Então, essa citocina
surge mais tardiamente que o TNF. Assim, há atuação sobre o endotélio, estimulando ainda mais a
migração de células inflamatórias, que são ativadas.
Essas células ativadas (por microorganismo, TNF e il-1) produzem il-12, que ativa linfócito T,
formando células Th1. Além disso, ativa células NK e T CD8. Essas células produzem INF gama. Essa
citocina ativa ainda mais o macrófago inicial, a fim de que ele elimine o microorganismo inicial.
Então, há uma sequência durante as fases iniciais de ataque ao microorganismo invasor:
1. Presença de LPS do microorganismo aumenta produção de TNF alfa;
2. Vasodilatação, migração de leucócitos e ativação;
3. TNF atua sobre macrófago, que libera il-1;
4. Mais vasodilatação, migração e ativação de células;
5. Il-1 atua sobre macrófago, que libera il-12;
6. Há formação de células Th1 e ativação de células NK e T CD8;
7. Essas três células produzem INF gama, que ativa ainda mais o macrófago inicial.
Nem sempre o microorganismo consegue ser eliminado. Isso porque esse microorganismo possui
mecanismos que o fazem ser capaz de escapar das nossas defesas. A leishmania, por exemplo,
produz uma série de proteínas que bloqueiam a produção de óxido nítrico dentro do macrófago.
Assim, ela consegue se multiplicar.O vírus Epstein Barr produz uma proteína semelhante à il-10,
que suprime a capacidade fagocítica do macrófago e as funções do linfócito T.
INTERFERON TIPO 1

Diogo Araujo – Med 92
Existem dois interferons do tipo 1: INF alfa (produzido por fagócitos mononucleares) e INF beta
(produzido por células infectadas).
O interferon atua sobre receptor tirosina quinase.
O maior estímulo é a infecção viral.
Essas citocinas atuam sobre a própria célula ou sobre células vizinhas. Isso tem a função de
bloquear a replicação viral nas células vizinhas.
Eles induzem na célula um estado chamado de "estado antiviral". Ou seja, se o vírus penetrar
nessa célula, ele não conseguirá fazer transcrição gênica. Isso porque, quando o interferon tipo 1
atua sobre a célula, ele bloqueia mecanismos que permitam a transcrição gênica.
Além disso, o interferon do tipo I aumenta a expressão de moléculas CPH de classe I nas células
infectadas, estimulando a ação do linfócito T CD8.
Contudo, alguns tipos de vírus fazem justamente o oposto: inibem a produção de CPH de classe I
na célula, diminuindo o reconhecimento por linfócito T CD8 e "anulando" e efeito do interferon do
tipo 1.
Contudo, o interferon do tipo I também ativa linfócitos NK. Assim, devido à presença de proteínas
virais na superfície da célula, há maior chance de lise da célula infectada por esse linfócito.
Além disso, o interferon do tipo I ativa células Th1, que combatem microorganismos
intracelulares.
INTERLEUCINA-10
Ela é uma citocina com funções supressoras.
Produzida por fagócitos mononucleares (retroalimentação negativa) e por linfócitos Th2.
Ela tem duas funções principais:
- inibir macrófago: reduz a produção de il-12, a expressão de coestimuladores e de CPH de classe II
nas células apresentadoras de antígeno;
- inibir formação de células Th1.

Diogo Araujo – Med 92
A produção exacerbada de TNF alfa, il-12 e IFN gama podem levar a quadros de doença, como a
coagulação intravascular disseminada.Na leishmaniose mucocutânea, por exemplo, há aumento
exagerado de TNF alfa e ativação muito acentuada de macrófagos, que danificam o tecido.
Assim, é preciso ter mecanismos de contrarregulação da resposta. A IL-10 atua nesse papel.
O anticorpo monoclonal anti-TNF alfa bloqueia essa citocina, diminuindo a sua ligação no receptor
e induzindo melhora do quadro de dor e inflamação. Esse anticorpo, então, poderia ser utilizado
no tratamento de artrite reumatóide, por exemplo. Contudo, esse fármaco pode induzir
imunossupressão, com susceptibilidade a infecções secundárias.
Citocinas que medeiam e regulam a imunidade adaptativa
INTERLEUCINA 2
Produzida por linfócitos T CD4 e TCD8 que entram em contato com o antígeno.
Elas se ligam a receptores do tipo I (com as cadeias alfa, beta e gama) e sinaliza com a JAK1 e
JAK3/STAT5.
Nos linfócitos inativos ou em repouso não conseguem proliferar na presença de il-2 porque só
apresentam cadeia beta e gama no receptor.
Uma vez que eles são apresentados a antígenos, passam a produzir a cadeia alfa, formando um
trímero. Assim, o receptor fica com alta afinidade pela il-2. Dessa forma, pequenas quantidades de
il-2 já dão ao linfócito a capacidade de se ligar ao receptor, estimulando a sua proliferação .
A cadeia gama é muito importante porque é por ela que faz sinalização.
Quando o indivíduo tem um defeito na cadeia gama, há defeito na reposta à il-2.

Diogo Araujo – Med 92
A il-2 faz:
- proliferação de linfócitos T;
- desenvolvimento de células Treg;
- proliferação e ativação de células NK, havendo sinergismo com a il-15 e a il-21.
INTERLEUCINA 4
Produzida por linfócitos da subpopulação Th2, mastócitos e basófilos.
Atua em receptores do tipo I (alfa gama) e JAK3 ou 4/STAT 6.
Possui sinergismo com a il-13.
Ela faz:
- diferenciação e proliferação de células Th2 (defesa contra helmintos; alergias);
- promove a troca de isótipos (de modo a formar IgE e IgG);
- inibe a diferenciação de linfócitos virgens para a população Th1.
A il-13 também promove a troca de isótipo para IgE e determinados tipos de IgG.
Além disso, ela estimula a cicatrização (estimulando fibroblastos e macrófagos) e aumento da
produção de muco.
INTERLEUCINA-5
Produzida por linfócitos da subpopulação Th2 e mastócitos.
Atua em receptores tipo I e JAK2/STAT3.

Diogo Araujo – Med 92
Ela ativa o eosinófilo, fazendo com que ele expresse mais receptores do IgE e enzimas.
Aumenta a formação de eosinófilos na medula óssea.
INTERFERON GAMA
Produzido por linfócitos da subpopulação Th1, CD4, CD8 e NK.
Atua sobre receptores do tipo II.
Importantíssimo na defesa contra microorganismos intracelulares.
Ele faz:
- ativação de macrófago, fazendo mais fagocitose, matando microorganismos, apresentando mais
antígenos;
- ativação de linfócitos T CD4 virgens e transformação em células Th1;
- inibição do desenvolvimento da subpopulação Th2;
- estimula linfócitos B, fazendo com que eles produzam anticorpos fixadores do complemento
(IgM ou IgG) e anticorpos que atuem como opsoninas.
TGF BETA
Produzida por linfócitos T estimulados por antígeno, linfócitos Treg e fagócitos mononucleares
ativados.
Inibe linfócitos efetores (contribui para diminuir a ativação linfócitaria).
Estimula a diferenciação do linfócitos Th17.
Estimula troca de isótipo para IgA (importante pra mucosas).
Faz reparo tecidual, estimulando a produção de colágeno e a angiogênese. Risco de formação de

Diogo Araujo – Med 92
quelóides.
Indivíduo asmático tem desenvolvimento Th2, com produção de IgE. Essa IgE se liga a mastócitos e
basófilos do sistema respiratório. Quando há antígeno que se liga a ao menos duas moléculas de
IgE, há degranulação celular, o que provoca broncoespasmo, aumento da secreção de muco e
inflamação crônica. Il-4, il-5, il-8, il-10, il-13 e TNF alfa estão entre as citocinas envolvidas.
QUIMIOCINAS
São citocinas que induzem a migração de leucócitos.
Atuam sobre o receptor de quimiocina (sete alfa hélices), que são acoplados à proteína G.
A ativação da proteína G leva a várias cascatas: MAP quinases, via do ácido aracdônico, via do PKC,
etc. Com isso, há migração dos leucócitos para o local onde existem essas quimiocinas. Como?
Devido à expressão de receptores e ligantes. O leucócito, que está no interior do vaso, se liga às
selectinas endoteliais, ligantes de integrinas e quimiocinas. Em seguida, há margeamento e
rolamento dos leucócitos. A ligação do receptor à quimiocina promove maior afinidade das
integrinas pelo seu receptor.
Há mudança do citoesqueleto da célula, com achatamento de sua estrutura. Com ajuda das
ICAMs, há diapedese e passagem dos leucócitos para o subendotélio.
As células migram obedecendo o gradiente de quimiocinas. É uma movimentação orientada. Há
projeção da região anterior da célula, devido à reorganização do citoesqueleto. Então, a região
anterior e ventral fica fixa. A parte posterior se solta e é puxada para frente. Assim, a célula se
desloca até que ela encontre o antígeno.
Organização dos órgãos linfóides
O linfócito virgem consegue residir no linfonodo porque ele expressa receptores para quimiocinas
específicas do local.
Isso acontece também para os linfócitos de memória, efetores, etc.

Diogo Araujo – Med 92
Orienta o padrão de migração das células do sistema imune
A célula dendrítica ativada deixa de expressar o receptor de quimiocina que a fazia ficar no seu
local e passa a expressar o receptor CCR7. Assim, ela é capaz de migrar para o linfonodo. Lá, ela
ativa linfócitos T.
Ou seja, para migrar, a célula precisa mudar o padrão de receptores de ligação a quimiocinas.
-------------------------------------------------------------------------------------------------------------------
Sistema Complemento
Prof. Imaculada
Sistema do complemento não resiste ao aquecimento (aquecimento a 36°C por 30 minutos para
inativá-lo). Já o anticorpo é termoestável.
O sistema complemento é um manismo de defesa da imunidade inata.
É formado por moléculas presentes de modo inativo no soro.
A ativação é dada por três maneiras:
- direta (via alternativa, dada pelo próprio microorganismo);
- por molécula de anticorpo (via clássica);
- por proteína de ligação à manose (via das lecitinas).

Diogo Araujo – Med 92
VIA CLÁSSICA
Foi a primeira a ser descrita. Depende dos anticorpos. Essa via é um dos braços efetores da
imunidade humoral. Por utilizar anticorpo, é mais específica e possui um efeito de amplificação da
resposta e da sua eficiência.
[Não precisa saber todos os detalhes da via]
A proteína C1q é a maior. A C1r e C1s se encontram dentro da C1q. Quando ocorre a ligação do
anticorpo (porção Fc) na coroa radiada da C1q, há uma modificação da molécula, sendo que a C1r
passa a funcionar como enzima da C1s.
A proteína C1 não é ativada naturalmente pelo anticorpo porque, para que ocorra ativação de C1,
a molécula de Cq1 tem de se ligar a pelo menos dois anticorpos. E essa ligação é facilitada quando
os anticorpos estão aderidos a uma superfície (como a de uma bactéria). Assim, a proteína C1
passa a ser ativada principalmente quando esses anticorpos estão sobre uma superfície. Isso evita
ativação do complemento sem a presença de microorganismo.

Diogo Araujo – Med 92
A IgG e a IgM são capazes de ativar C1. A ativação de C1 pela IgM circulante não se dá
naturalmente porque, quando a IgM está circulante, a sua porção Fc não está aberta para a ligação
à coroa radiada do C1. Esse sítio somente se abre quando esse IgM se liga ao antígeno, com
mudança conformacional.
Os IgGs ligados ao antígeno se ligam ao C1. Esse C1, pela parte C1q, se liga ao C4 , clivando-o em
C4a e C4b. A parte do C4b fica ligado à membrana. O C2 é clivado pela C4, sendo formado um
complexo C42b (também chamado de C3 convertase). Assim, esse complexo cliva o C3 (que é o
ponto de convergência das vias).
No final, será formado um poro na membrana da célula.
VIA ALTERNATIVA
No soro, a C3 circulante sofre clivagem espontânea. Se a porção C3b encontra a superfície de uma
bactéria, ela se liga a essa superfície (caso não ocorra, ela é eliminada).
A porção do C3b fica ligada à membrana e, com o fator B (cofator), forma a C3 convertase. A
properdina estabiliza essa convertase, que cliva mais moléculas de C3.
O que garante que esse processo se continue na superfície da bactéria e não caia na circulação
sanguínea é o fato de que a porção maior (C3b, por exemplo) precisa que o seu radical tiol esteja
ligado à membrana. Se ele não estiver ligado à membrana, essa proteína é perdida.
VIA DA LECITINA
O microorganismo presente no meio possui manose em sua superfície. A proteína de ligação à
manose, circulante em condições fisiológicas, se liga à superfície da bactéria, servido de ponto de
início para a ativação da via. A partir daí, a proteína C1 se liga à proteína de ligação à manose,
dando início à mesma cascata que a via alternativa (mas sem a presença de anticorpo).
As vias alternativa e da lecitina são as duas primeiramente ativadas quando há contato com o
organismo. A via clássica, que depende de anticorpo, vem posteriormente, conferindo maior
especificidade ao processo, além de maior ampliação do sinal.

Diogo Araujo – Med 92
Vimos que as três vias convergem para a formação de C3.
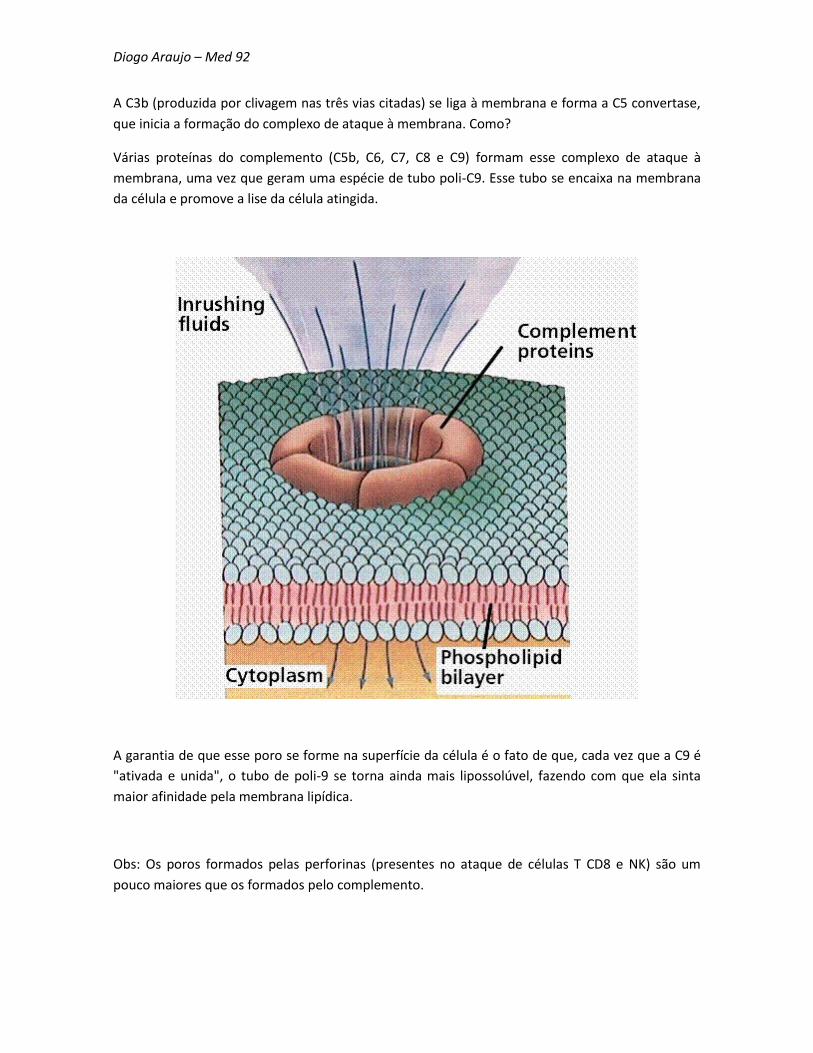
Diogo Araujo – Med 92
A C3b (produzida por clivagem nas três vias citadas) se liga à membrana e forma a C5 convertase,
que inicia a formação do complexo de ataque à membrana. Como?
Várias proteínas do complemento (C5b, C6, C7, C8 e C9) formam esse complexo de ataque à
membrana, uma vez que geram uma espécie de tubo poli-C9. Esse tubo se encaixa na membrana
da célula e promove a lise da célula atingida.
A garantia de que esse poro se forme na superfície da célula é o fato de que, cada vez que a C9 é
"ativada e unida", o tubo de poli-9 se torna ainda mais lipossolúvel, fazendo com que ela sinta
maior afinidade pela membrana lipídica.
Obs: Os poros formados pelas perforinas (presentes no ataque de células T CD8 e NK) são um
pouco maiores que os formados pelo complemento.

Diogo Araujo – Med 92
A formação de poros faz com que o conteúdo do meio externo (água) entre na bactéria. Isso
acarreta em lise celular.
Resumindo: o início da reação se dá na superfície do patógeno. As enzimas formadas ficam
aderidas à superfície do patógeno, garantindo que o processo não se espalhe pela corrente
sanguínea. O início da cascata pode ser desendadeado por Ig, manose ou pelo C3b que se auto-
clivou. Por fim, a ativação das enzimas em série (cascata) culmina com uma via de ativação
comum (C3), que resulta na formação de poro na superfície da célula (tubo de poli-C9).
O que garante que não haja destruição de células próprias?
Existem mecanismos de defesa contra o sistema complemento. As nossas células possuem
mecanismos de inativação de cada ponto da cascata de ativação do complemento.
Por exemplo, existe a molécula inibidora do C1, que se liga às moléculas de C1r e C1s impedindo a
ligação e ativação do C1. A falta desse fator leva ao edema angioneurótico hereditário. Em
indivíduos com esse distúrbio, a compressão de uma região do corpo leva à formação de edema,
graças à fácil ativação do C1 e inflamação resultante.
Outro exemplo é o DAF/CR1, que impede a ligação do C2b ao C4b durante as vias clássica e
alternativa.
A hemoglobinúria paroxística noturna é uma doença causada pela deficiência genética do DAF ou
do CD59.
Existe também a proteína S, que inibe que o complexo de formação do poli-C9 (tubo) se forme.

Diogo Araujo – Med 92
Contudo, alguns patógenos desenvolveram estratégias que impedem a ativação do complemento.
Isso é resultado da utilização de genes e mecanismos de defesas humanos ao longo da evolução. O
Schistosoma mansoni é um exemplo.
Quais as funções do complemento?
- O C3b se liga ao microorganismo, sendo que existem receptores para a porção C3b do
complemento nas células fagocitárias. Assim, esse C3b possui papel de facilitar a fagocitose, de
modo que ele atua como opsonina. É um potente amplificador de uma resposta inata celular.
- C3a e C3b são potentes quimiotáticos. Haverá, então, recrutamento de leucócitos (como os
neutrófilos) para o local onde se encontra o microorganismo, estimulando a sua destruição. Em
situações em que há formação de imunocomplexos que se depositam, haverá maior ativação do
complemento, formação dessas moléculas quimiotáticas e infiltração de neutrófilos nas paredes
de um órgão, havendo lesão tecidual.
- formação de poros: citólise.
- os imunocomplexos formados se ligam ao complemento. Como a hemácia possui receptores
para o complemento e para a porção Fc de anticorpos, se liga a esse
imunocomplexo/complemento, transportando-o. No fígado, a passagem das hemácias pelos
sinusóides (em fila indiana) porporciona o contato das células de Kupffer (macrófago) com a sua
superfície. Essa superfície é limpa pelos macrófagos. Assim, pacientes que têm deficiência da
atividade do sistema complemento possuem dificuldade de remover imunocomplexos. O lúpus
eritematoso (quadro em que há vasculite por deposição de imunocomplexos) é um exemplo.
- remoção de colesterol: o excesso de colesterol deve ser removido do sangue circulante. Uma das
formas de removê-lo é através da produção de anticorpos anti-colesterol, formando
imunocomplexos que também são removidos pela hemácia quando há ligação do complemento. É
importante lembrar que os hepatócitos permitem entrada de colesterol. Contudo, há limite. Já os
macrófagos colocam colesterol para o seu interior sem limites. Assim, formam-se macrófagos
muito cheios de colesterol. Essa é a base para a formação de placas ateroscleróticas.
- papel de vasodilatação pelo C5. Além disso, há maior permeabilidade dos vasos e degranulação
de basófilos.
- atuam como moléculas de aderência, que realizam a ponte quando há apresentação de antígeno
para o linfócito T.
- função de ponte entre célula alvo e a célula citolítica (TCD8 ou NK). Essa é a função ADCC.
- aumento da ativação de linfócitos B e manutenção da memória imunológica. Os patógenos

Diogo Araujo – Med 92
podem ser mantidos em células imunes presentes em gânglios linfáticos (células foliculares
dendríticas). Essas células se mantêm expressando antígenos do patógeno, sendo que os linfócitos
B de memória são apresentados a eles. O complemento intermedeia esse processo.
- na ativação timo-independente do linfócito B, o complemento pode servir como fator necessário
para a formação do segundo sinal. É a ligação micróbio/C3d/CR2.
A ocorrência de meningite meningocóccica por duas vezes ou mais é fator indicativo de teste para
análise da função do C9.
A deficiência do sistema complemento é rara e seu tratamento, difícil.
-------------------------------------------------------------------------------------------------------------------
Imunidade Anti-Infecciosa
Prof. Imaculada
IMUNIDADE ANTI-VIRAL
São exemplos de doenças causadas por vírus: sarampo, hepatite, mononucleose, varicela.
A infecção pode acontecer por via respiratória, por via sanguínea, etc.
Pode evoluir para câncer, insuficiência hepática, etc.
Sarampo e varicela geralmente ocorrem nas crianças.
A primeira defesa contra a infecção viral é a célula NK, que lisa as células infectadas pelo vírus.
O interferon alfa e o beta são produzidos por quaisquer células do organismo que são infectadas
por vírus. Qualquer molécula que tenha função antiviral é chamada de interferon. O interferon age
na célula vizinha, impedindo a síntese protéica. Assim, quando o vírus entra nessa célula vizinha,
ele não consegue se replicar devido à baixa síntese protéica.
O macrófago tem um papel importante porque ele representa uma ponte entre o sistema inato e
adquirido. Assim, devido à sua ação, ocorre ativação de linfócitos T e B, de modo que há produção
de anticorpos, as principais moléculas de defesa contra vírus. Isso porque os anticorpos se ligam

Diogo Araujo – Med 92
ao vírus, fazem neutralização, facilitam fagocitose, etc.
O anticorpo neutraliza e impede que o vírus infecte outra célula. Um exemplo é a IgA, que inibe a
entrada do vírus por mucosas. A IgG facilita opsonização do vírus, sua neutralização, etc. IgM
facilita aglutinação viral (por ter estrutura pentamérica, um só IgM pentamérico se liga a vários
vírus), o que facilita a fagocitose por macrófagos. Além disso, IgG e IgM ativam a via clássica do
sistema complemento.
Linfócito T CD8 é importante também. Ele é a célula do sistema imune adquirido que atua contra
as células infectadas (juntamente com o linfócito NK). Ele se liga ao CPH de classe I (que apresenta
proteínas virais), joga perforinas e granzimas sobre a célula e faz lise.
Ficar ou não doente por uma infecção viral depende dos mecanismos inatos de defesa (potente
capacidade de produzir interferon, potente capacidade de lise das células NK e TCD8, existência de
anticorpos prévios), da virulência (capacidade de multiplicação, de replicação) e do número de
partículas (carga viral).
A carga e a resposta das células NK e T CD8 e de interferon também determinam a evolução do
quadro. A forte resposta T CD8 pode conduzir a hepatite fulminante, por exemplo. O aumento de
interferon diminui muito a infecção viral de outras células. Podemos dizer, então, que é o
equilíbrio entre o microorganismo e a defesa que determina a evolução do quadro (a
sintomatologia, a gravidade, etc).
E quais são os mecanismos de defesa dos vírus?
Vírus podem fazer latência. O vírus da herpes é um exemplo. Ele pode ser transmitido por via
respoiratória, podendo ficar latente na medula óssea. Quando há uma situação de estresse, ele
pode ser reativado.
O vírus H1N1 possui as proteínas Hemaglutinina e Neuraminidase do tipo 1. Pequenas mutações
nessas moléculas podem fazer com que os anticorpos já produzidos não sejam mais eficientes
contra o vírus. Isso faz com que, de tempos em tempos, haja epidemias de certas doenças
causadas por esses vírus. Essa mutação faz parte da chamada variação antigênica. A maioria dos
microorganismos utiliza esse mecanismo. A cada multiplicação, aumenta o risco do
microorganismo fazer mutação.
[Parênteses] o Mycobacterium tuberculosis também possui esse mecanismo de variação
antigênica. É por isso que o tratamento requer uma série de medicamentos (esquema RIP).

Diogo Araujo – Med 92
Os vírus (como o Epistein Barr e o citomegalovírus) impedem o proteassoma de funcionar. Assim,
ele diminui a sua apresentação a linfócitos.
O vírus da herpes simples inibe a proteína TAP, que tem a função de passar as proteínas
processadas pelo proteassoma para o interior do retículo endoplasmático. Há outros víruso que
inibem a síntese de MHC dentro do retículo endoplasmático.
Resumindo imunidade celular e humoral contra vírus:
- Linfócitos NK, linfócitos T CD8, macrófagos;
- Anticorpos e interferons;
Resumindo mecanismos de defesa do vírus:
- variação antigênica (influenza, rinovirus, VIH);
- inibição do processamento antigênico (bloqueio do TAP, remoção de moléculas de CPH do RER).
(Herpes simplex, citomegalovirus).
- Produção de homólogos de receptores de citocinas; (poxvirus, citomegalovirus)
- Produção de citocinas imunodepressoras; (Epstein Barr)
- Infecção de células imunocompetentes (nesse caso, as crianças são mais gravemente acometidas,
devido ao fato de que ela ainda não teve tempo de produzir defesas contra algumas doenças). (VIH
e vírus do sarampo).
IMUNIDADE PARA BACTÉRIAS EXTRA-CELULARES
São exemplos: estreptococos, diplococos, gram negativos, estafilococos.
Diogo Araujo – Med 92
São exemplos de doenças: impetigo bolhoso, abscesso amigdaliano, meningite bacteriana,
penumonia bacteriana.
São patógenos que induzem a formação de pus (piogênicos). Isso porque é o neutrófilo um dos
maiores responsáveis pelo combate a bactérias extracelulares. Restos de neutrófilos mortos e
bactérias lisadas formam o pus.
As bactérias extracelulares as grandes responsáveis pelas infecções bacterianas encontradas em
ambiente hospitalar.
Lesões de continuidade na pele facilitam a entrada desses patógenos.
O sistema complemento também tem grande importância. Isso porque ele faz quimiotaxia para os
neutrófilos, além de opsonizar e facilitar a fagocitose.
O grande amplificador da ação dos neutrófilos é o anticorpo. Eles são muito importantes, visto que
atuam até mesmo no processo de ADCC.
A deficiência de neutrófilos, de complemento e de anticorpos levam a um mesmo quadro de
maior susceptibilidade à infecção por bactérias extra-celulares.

Diogo Araujo – Med 92
Então, podemos dizer que são os principais fatores na defesa contra bactérias extracelulares:
- Celular: neutrófilos.
- Humoral: anticorpos e sistema complemento.
São mecanismos de evasão do sistema por bactérias extra-celulares:
- variação antigênica; (Neisseria gonorrhoeae, Escherichia coli, Salmonella typhimurium)
- Inibição da atuação do complemento através da produção da molécula DAF; (S. aureus;
streptococcus do grupo A);
- Resistência à fagocitose. Isso reduz muito a opsonização por bactérias e pelo complemento.
Pneumococcus é um exemplo de agente que faz esse mecanismo, sendo o agente causador mais
frequente de pneumonias em adultos;
- Inibição de radicais de oxigênio. Um exemplo é o Staphylococcus catalase positivo. A catalase
cliva os radicais de oxigênio.
IMUNIDADE PARA BACTÉRIAS/MICROORGANISMOS INTRACELULARES
São exemplos: micobactérias (causadoras da tuberculose e da hanseníase, por exemplo),
espiroquetas, protozoários.
A célula da imunidade inata mais importante é o macrófago (que precisa estar ativado). Em
contrapartida, com relação à imunidade adquirida, pode-se destarcar o linfócito CD4 (que ativa o
macrófago por meio de interferon; esse macrófago produz então muitos radicais de oxigênio, mais
moléculas de CPH, mais fatores de coestimulação, etc).
O linfócito CD8 também é importante porque ele é o responsável por detectar e lisar células
infectadas por bactérias intracelulares. Pelo mesmo motivo, a célula NK é muito importante
(principalmente no começo da infecção). Após os estágios iniciais, o linfócito TCD8 passa a ser
mais importante.

Diogo Araujo – Med 92
[Divagando] Num animal normal, há infecção, mas depois de alguns dias, ele consegue controlá-la,
diminuindo o número de bactérias intracelulares no organismo. Quando o organismo é knockout
para interferon gama, a bactéria se multiplica descontroladamente e o indivíduo morre em poucos
dias, já que essa citocina é a responsável por ativar os macrófagos. Num indivíduo knockout para
il-12, há intensa proliferação bacteriana, apesar desta ser de menor intensidade que na ausência
de interferon gama. Isso acontece porque, apesar de não haver il-12 (e a sua produção por células
NK estar prejudicada), o INF gama consegue ser produzido por outras células (como linfócitos Th1,
por exemplo).
O CD4 é a célula da imunidade adquirida mais importante para a defesa, estimulando a a
fagocitose e a lise das bactérias pelo macrófago (apesar do linfócito CD8 também ser importante,
como mencionado anteriormente).
A forma cutânea ou mucocutânea da leishmaniose depende da cepa da Leishmania e da resposta
do indivíduo ao microorganismo.
Resumindo os principais mecanismos de defesa contra microorganismos intracelulares:
- Celular: macrófago, linfócito T CD4, linfócito T CD8 e linfócito NK;
- Humoral: INF gama (por estimular os macrófagos) e il-12.
São mecanismos de escape dos microorganismos intracelulares:
- Inibição da formação do fagolisossoma (fusão entre fagossoma e lisossoma);
- Ruptura da membrana do fagolisossoma e escape apara o citoplasma;
- Inibição de radicais de oxigênio.
Mecanismos de escape gerais dos protozoários:
- Variação antigênica;
- Resistência adquirida ao complemento e ao linfócito T CD8;
- Inibição da resposta imune do hospedeiro;

Diogo Araujo – Med 92
- Eliminação de antígenos.
IMUNIDADE CONTRA HELMINTOS
É impossível o sistema imunitário fazer defesa contra um verme adulto. A defesa é feita na fase de
larva. Se não houver eficiência nessa resposta, ela vira um verme adulto. A partir daí, o máximo
que o organismo consegue fazer é irritar a parede do intestino em que a larva está aderida, numa
tentativa de fazê-la se desligar da mucosa. Mas o que ocorre é a mudança de lugar pela larva.
[Parênteses] O esquistossoma vive nas veias do plexo mesentérico inferior, fazendo com que
metade dos ovos (cerca de 300 por dia) sejam levados para o intestino (metade) ou para demais
órgãos do corpo. Quando o ovo chega no fígado, por exemplo, por ser muito grande, não consegue
entrar nos sinusóides. Então, fica preso, dando origem a um granuloma no local, que
eventualmente evolui para fibrose da região e, consequentemente, dificuldade na circulação
sanguínea da região. O aumento da pressão do sangue dentro das veias causa esplenomegalia,
assim como a má circulação gera aumento do calibre das veias da circualção colateral, formando
até varizes esofagianas (que podem se romper durante a alimentação e causar hemorragias
graves).
Obs: nomeia-se "imunidade concomitante" como a incapacidade de se defender contra o verme
adulto e realização de ataque apenas contra novos parasitas.
A produção de IgE, feita quando existe infecção por helmintos, promove o mecanismo de ADCC,
facilitando a destruição das larvas dos parasitas.
São fatores imunológicos importantes na defesa contra helmintos:
- Celular: Eosinófilos, neutrófilos, monócitos, linfócitos Th2.
- Humoral: IgE.

Diogo Araujo – Med 92
São mecanismos de escape do Schistosoma mansoni (diz-se que ele é o "mestre" em escapar do
sistema imune):
- Clivagem da porção Fc de anticorpos ligados ao parasito;
- Capacidade de restaurar a superfície lesionada por uma célula de defesa;
- Produção de susbtância inibitórias para o sistema imune;
- Produção de DAF, que inibe o complemento.
---------------------------------------------------------------------------------------------------------------