Embed Size (px)
Citation preview

1
UNIVERSIDADE FEDERAL DE OURO PRETO
NÚCLEO DE PESQUISAS EM CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA
Avaliação dos efeitos antioxidantes e pró-longevidade do
extrato de açaí (Euterpe oleraceae Mart.) no organismo modelo
Caenorhabditis elegans
David Silva Nunes
2011

2
UNIVERSIDADE FEDERAL DE OURO PRETO
NÚCLEO DE PESQUISAS EM CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA
Avaliação dos efeitos antioxidantes e pró-longevidade do
extrato de açaí (Euterpe oleraceae Mart.) no organismo modelo
Caenorhabditis elegans
David Silva Nunes
Orientadora
Riva de Paula Oliveira
Dissertação de Mestrado apresentada ao
Programa de Pós-Graduação em Biotecnologia
da Universidade de Ouro Preto como requisito
parcial para obtenção do Título de Mestre em
Biotecnologia.
Ouro Preto
2011

3
AGRADECIMENTOS RELIGIOSOS
A Deus, por estar sempre ao meu lado guiando meus passos e me socorrendo nos
momentos mais difíceis.

4
AGRADECIMENTOS
Aos meus Pais por apoiar-me e confortar-me e pelo exemplo de vida e às minhas
irmãs Hellem, Sarah e meu irmão Lucas pelo apoio e carinho. Muito obrigada por
estarem comigo mesmo a distancia
A Paula Alves Medeiros por me ajudar nos momentos mais difíceis
A Marilaine Guimaraes Pires por mostrar-me o caminho
Aos amigos do laboratório Franciny, Everton, Igor, Larissa, Patrícia, Ricardo e
Valquíria, Washington, por fazerem do nosso laboratório o mais feliz do planeta e
por momentos inesquecíveis que passamos juntos. Vocês foram importantíssimo,
cada uma com sua peculiaridade só fizeram enriquecer esse trabalho, muito
obrigada por tudo.
Aos amigos da pós-graduação, membros da B.P do mestrado/doutorado Obrigada
pelos momentos inesquecíveis e pela grande amizade que sempre demonstraram.
Os amigos Bruna, Cristiana, Eduardo que estavam sempre prontos a ajudar.
A professora DRa Riva de Paula Oliveira pela orientação e apoio.
A professora DRa Leoneide por ser uma grande amiga.
Aos ilustres desconhecidos aqui eu agradeço a todos os que de certa forma me
ajudaram no período em que esse trabalho foi realizado

5
Resumo:
O alto consumo de comidas e bebidas ricas em flavonóides tem sido associado com o
menor risco de doenças degenerativas crônicas como aterosclerose e câncer devido
tanto às suas atividades antioxidantes diretas de remoção de radicais livres, quanto à sua
capacidade de atuar como antioxidante indireto ativando diferentes vias de sinalização
contra o estresse oxidativo. O Euterpe oleracea Mart. ou açaizeiro é uma palmeira nativa
das regiões amazônicas cuja fruta é rica em flavonóides, especialmente em antocianinas
e proantocianidinas (cianidina-3-glicosídeo e cianidina-3-rutinosídeo). Recentemente,
diversos estudos têm destacado as propriedades antioxidantes, pró-apoptóticas,
antiproliferativas e pró-longevidade do açaí, tanto em ensaios in vitro quanto em modelos
animais. Neste trabalho, o Caenorhabditis elegans foi utilizado como organismo modelo
para testar as atividades antioxidantes in vivo de diferentes extratos de açaí que possam
ser exploradas para aumentar as defesas inatas ao estresse oxidativo e a longevidade.
Através do método do pH diferencial, foi verificado que a quantidade de antocianinas
presentes nos dois extrato de açaí liofilizado (AL-1 e AL-2) apresentaram em torno de
19,2 a 34,7 mg de antocianinas monoméricas/100g de extrato. Para testar se o
tratamento com os extratos de açaí liofilizado (AL-1 e AL-2) aumentam a resistência ao
estresse oxidativo, animais tipo selvagem foram tratados por 48 horas do estágio larval
L1 até L4 com 100 e 200 mg/ml de extrato de açaí e depois submetidos ao estresse
provocado por hidroperóxido de terc-butila (t-BOOH). Animais tratados com 200 mg/ml
não se desenvolvem e permanecem em L1. Já os animais selvagens tratados com 100
mg/ml de AL-1 e AL-2 foram mais resistentes ao estresse oxidativo provocado por 5 mM
de t-BOOH do que os animais não tratados. Por outro lado, o tratamento dos animais tipo
selvagem com 100 mg/ml de AL-1 por 168 horas não aumentou a resistência ao estresse
térmico a 35oC. O tratamento contínuo com 100 mg/ml de AL-1 também não aumentou a
longevidade dos animais tipo selvagem. Apesar de apresentarem uma maior resistência
ao estresse, os animais tratados com 100mg/ml do extrato de açaí por 48 horas não
apresentaram uma diferença significativa quanto aos marcadores bioquímicos (atividade
da catalase) e indicadores de resistência (proteína carbonilada e sulfidrila totais) em
relação aos animais do grupo controle. Para verificar se o aumento da resistência ao
estresse oxidativo promovido pelo tratamento com extrato de açaí depende das vias de
sinalização p38/SKN-1 e DAF-16, os ensaios de resistência ao estresse oxidativo foram
repetidos usando mutantes de deleção sek-1(ku7), a p38 MAPKK, skn-1(RNAi) e daf-
16(mgDf46). Aparentemente, o tratamento com 100 mg/ml AL-1 não aumentou a
resistência ao estresse oxidativo dos mutantes sek-1 e daf-16, sugerindo que este
aumento depende da ativação de SEK-1 e DAF-16. Por outro lado, não foi observada a
indução da localização nuclear nos animais tratados contendo gene repórter DAF-
16::GFP. Estes dados sugerem que o aumento da resistência ao estresse oxidativo
induzido pelo tratamento com extrato de açaí possivelmente depende da p38 MAPK e
DAF-16, porém sem ocorrer ativação da localização nuclear de DAF-16.

6
Abstract
The high consumption of foods and beverages rich in flavonoids has been associated with
lower risk of chronic degenerative diseases such as atherosclerosis and cancer due to both
their direct antioxidant removal of free radicals, for their ability to act as an antioxidant
indirectly by activating different pathways signaling against oxidative stress. Euterpe
oleracea Mart. or açai palm is a native of the Amazon regions whose fruit is rich in
flavonoids, especially anthocyanins and proanthocyanidins (cyanidin-3-glucoside and
cyanidin-3-rutinosídeo). Several recent studies have highlighted the antioxidant properties,
pro-apoptotic, antiproliferative and pro-longevity of acai, both in vitro and in animal
models. In this work, Caenorhabditis elegans was used as a model organism to test the in
vivo antioxidant activities of different extracts of acai that could be exploited to enhance
innate defenses to oxidative stress and longevity. Through the pH differential method, it
was found that the amount of anthocyanins present in açai freeze-dried extract of two (AL-
1 and AL-2) showed around 19.2 to 34.7 mg of anthocyanins monoméricas/100g extract.
To test whether treatment with extracts of freeze-dried açaí (AL-1 and AL-2) increase
resistance to oxidative stress, wild-type animals were treated for 48 hours of larval stage
L1 to L4 with 100 and 200 mg / ml extract acai and then subjected to stress caused by tert-
butyl hydroperoxide (t-Booher). Animals treated with 200 mg / ml do not develop and
remain in L1. Since wild animals treated with 100 mg / ml of AL-1 AL-2 and were more
resistant to oxidative stress caused by 5 mM t-Booher than untreated animals. On the other
hand, treatment of wild-type animals with 100 mg / ml of AL-1 for 168 hours did not
increase the resistance to heat stress at 35 ° C. Continuous treatment with 100 mg / ml of
AL-1 did not increase the longevity of wild-type animals. Despite having a higher
resistance to stress, the animals treated with 100mg/ml of the extract of acai for 48 hours
did not show a significant difference to the biochemical markers (catalase activity) and
indicators of resistance (total sulfhydryl and protein carbonyl) in relation to the control
group. To verify that the increased resistance to oxidative stress promoted by treatment
with acai extract depends on signaling pathways p38/SKN-1 and DAF-16, tests of
resistance to oxidative stress were repeated using deletion mutants of sek-1 (ku7 ), the p38
MAPKK, skn-1 (RNAi) and daf-16 (mgDf46). Apparently, treatment with 100 mg / ml
AL-1 did not increase oxidative stress resistance mutants of sek-1 and daf-16, suggesting
that this increase depends on the activation of SEK-1 and DAF-16. On the other hand, no
significant induction of nuclear localization in treated animals containing reporter gene
DAF-16:: GFP. These data suggest that the increased resistance to oxidative stress induced
by treatment with acai extract possibly depends on p38 MAPK and DAF-16, but occurs
without activation of the nuclear localization of DAF-16.

7
Lista de figuras
FIGURA 1. Ciclo de vida do C. Elegans. Fonte: worm atlas......................................13
FIGURA 2. Regulação de Nrf2/FOXO..........................................................................15
FIGURA 3 Esquema dos genes repórter para DAF-16..........................................30
FIGURA 4 - Localização subcelular de DAF-16::GFP ................................................49

8
Lista de tabelas
TABELA 1 - Conteúdo de antocianinas monoméricas, representado como conteúdo de
cianidina-3-glicosídeomg/100 g).....................................................................................33
TABELA 2 – Análise da longevidade dos animais tipo selvagem tratados com AL-1 a
25oC.................................................................................................................................40
TABELA 3 - Efeito do tratamento com AL-1 sobre a atividade de catalase e os níveis dos
grupos sulfidrilas totais e proteína carbonilada.........................................................43

9
Lista de gráficos
GRÁFICO 1 – Efeito do tratamento com 100 mg/ml de AL na sobrevivência do C.
elegans.............................................................................................................................35
GRÁFICO 2– Efeitos tratamento com açaí na sobrevivência do C elegans em condições
de estresse térmico..........................................................................................37
GRÁFICO 3– Efeitos do tratamento com 100 mg/ml de AL-1 na sobrevivência do C.
elegans.............................................................................................................................39
GRÁFICO 4 - Dosagem dos marcadores bioquímicos e enzimáticos no C. elegans tratado
com açaí...............................................................................................................41
GRÁFICO 5 - Efeito do tratamento com 100 mg/ml de AL-1 na sobrevivência de animais
skn-1(RNAi) em condições de estresse oxidativo...............................................44
GRÁFICO 6 - Efeito do tratamento com 100 mg/ml de AL-1 na sobrevivência de
mutantes sek-1 em condições de estresse oxidativo........................................................46
GRÁFICO 7 - Efeito do tratamento com 100 mg/ml de AL-1 na sobrevivência de
mutantes daf-16 em condições de estresse oxidativo......................................................47

10
ABREVIATURAS
AM antocianinas monomericas
AL-1 Açai liofilizado 100mg/ml
AL-1 Açai liofilizado 100mg/ml
ARE - Elemento de Resposta Antioxidante
CAT - catalase
CYPs - citocromo P450
EpRE - Elemento de Resposta à Eletrófilo
ERK - proteína quinase regulada por sinal extracelular
Eros – Espécies Reativas de Oxigênio
FD- Fator de diluição
FoxO - Forkhead Box
GCS - -glutamilcisteina sintetase
GPx - glutationa peroxidase
GR - glutationa redutase
GSK3β - glicogênio sintase quinase-3β
GST - glutationa S-transferase
H2O2 – Peróxido de Hidrogênio
JNK - c-Jun N-terminal kinase
MAPK - ProteínasAativadas por Mitógenos
Nrf2 - Nuclear factor-erythroid-2-related factor 2
PM- Peso Molecular
PKC - proteína quinase C
SDRs - desidrogenases/redutases de cadeia curta
SOD - superóxido dismutase
t-BOOH - tert-butil peroxide

11
SUMÁRIO
RESUMO....................................................................................................................... .............................v
ABSTRACT..................................................................................................................... .........................vi
LISTA DE FIGURAS................................................................................................................................vii
Lista de
tabelas............................................................................................................................viii
Lista de gráficos.............................................................................................................ix
ABREVIATURAS............................................................................................. ...........................................x
1 INTRODUÇÃO ............................................................................. ..................................................1
1.1 ESTRESSE OXIDATIVO...............................................................................................................2
1.2 MECANISMOS ENZIMÁTICOS DE COMBATE AO ERO.....................................................3
1.2.1 Controle da Expressão Gênica das Enzimas Antioxidantes em Mamífero.................................4
1.3 MECANISMOS NÃO ENZIMÁTICOS DE COMBATE AO ERO............................................5
1.3.1 Antioxidantes Polifenólicos......................................................................................... ..........6
1.4 PROPRIEDADES BIOLÓGICAS DO AÇAÍ................................................................................7
1.4.1 Propriedades Antioxidantes in vitro e ex vivo.....................................................................7
1.4.2 Propriedades Anti-inflamatórias in vitro e in vivo..............................................................8
1.4.3 Propriedades Antiproliferativas in vitro e in vivo...............................................................9
1.4.4 Proteção Contra Estresse Hiperlipídico............................................................................10
1.4.5 Efeitos Benéficos na Saúde Humana............................................................... ..................11
1.5 ORGANIMO MODELO CAENORHABDITIS ELEGANS..................................................................11
1.5.1 Controle da Expressão Gênica das Enzimas Antioxidantes no C. elegans ...................12
1.5.1.1 A via de sinalização de SKN-1/Nrf....................................................................................13
1.5.1.2 A Via de Sinalização de DAF-16........................................................................................16
1.6 C. elegans COMO UM MODELO PARA ESTUDOS DE FITOQUÍMICOS..........................17
2 OBJETIVO ...................................................................................................................................19
2.1 OBJETIVO GERAL................................................................................................................20
2.2 OBJETIVOS ESPECÍFICOS..................................................................................................20
3 METODOLOGIA ..........................................................................................................................21
3.1 CEPAS E LINHAGENS...............................................................................................................22

12
3.2 CRESCIMENTO E MANUTENÇÃO DO C. ELEGANS ..........................................................22
3.3 SINCRONIZAÇÃO CRONOLÓGICA DO C. ELEGANS ........................................................23
3.4 EXTRATOS DE AÇAÍ..................................................................................................................23
3.5 QUANTIFICAÇÃO DE ANTOCIANINAS................................................................................24
3.6 ENSAIOS BIOLÓGICOS .............................................................................................................25
3.6.1 Preparação das Placas NGM Contendo Extrato de Açaí................................................25
3.6.2 Ensaio de Resistência ao Estresse Oxidativo....................................................................25
3.6.3 Ensaio de Resistência ao Estresse Térmico.......................................................................25
3.6.4 Ensaio de Longevidade ................................................................................. .....................26
3.7 DOSAGEM DOS PARÂMETROS BIOQUÍMICOS.................................................................26
3.7.1 Dosagem dos Níveis Totais de Grupos de Sulfidrilas ......................................................27
3.7.2 Dosagem da Atividade da Catalase....................................................................................27
3.7.3 Dosagem dos Níveis de Proteína Carbonilada..................................................................28
3.7.4 Dosagem de Proteínas Totais.............................................................................................29
3.8 LOCALIZAÇÃO SUBCELULAR DE DAF-16..........................................................................30
3.9 ANÁLISE ESTATÍSTICA.............................................................................................................31
4 RESULTADOS..................................................................... .........................................................32
4.1 QUANTIFICAÇÃO DE ANTOCIANINAS.................................................................................33
4.2 ANÁLISE DA RESISTÊNCIA AO ESTRESSE OXIDATIVO DOS ANIMAIS TRATADOS
COM DIFERENTES EXTRATOS DE
AÇAÍ................................................................................................................................................34
4.3 ANÁLISE DA RESISTÊNCIA AO ESTRESSE TÉRMICO DOS ANIMAIS TRATADOS
COM AL-1....................................................................................................................................36
4.4 ANÁLISE DA SOBREVIVÊNCIA EM CONDIÇÕES NORMAIS DOS ANIMAIS .............36
4.5 ANÁLISE DE MARCADORES BIOQUÍMICOS E ENZIMÁTICOS NOS ANIMAIS
TRATADOS COM AL-1...............................................................................................................38
4.6 ANÁLISE DO ENVOLVIMENTO DA VIA DE SINALIZAÇÃO DE SKN-1/P38MAPK NO
AUMENTO DA RESISTÊNCIA AO ESTRESSE OXIDATIVO PROMOVIDO PELO AL-
1........................................................................................................................................................43
4.6.1 Análise da Resistência ao Estresse Oxidativo nos Animais skn-1(RNAi).......................43
4.6.2 Análise da Resistência ao Estresse Oxidativo em mutantes sek-1..................................45

13
4.7 ANÁLISE DO ENVOLVIMENTO DA VIA DE SINALIZAÇÃO DE DAF-16/FOXO NO
AUMENTO DA RESISTÊNCIA AO ESTRESSE OXIDATIVO PROMOVIDO PELO AL-1
..............................................................................................................................................................45
4.7.1.1 Análise da resistência ao estresse oxidativo em mutantes daf-16............................................45
4.7.2 Análise da Localização DAF-16:: GFP......................................................................................48
5 DISCUSSÃO DOS RESULATDOS........................................................................................ .......50
5.1 PROPRIEDADES ANTIOXIDANTES DO EXTRATO DE AÇAÍ..........................................51
5.1.1 Quantificação de Antocianinas nos Extratos de Açaí......................................................52
5.1.2 Ensaios Biológicos...............................................................................................................53
5.1.3 Dosagens Bioquímicas.........................................................................................................58
5.2 POSSÍVEL MECANISMO DE AÇÃO DO EXTRATO AÇAÍ.................................................61
5.2.1 A Via de Sinalização SKN-1/p38 MAPK...........................................................................62
5.2.2 A Via de Sinalização de DAF-16..........................................................................................62
6 CONCLUSÕES..............................................................................................................................65
7 REFERÊNCIAS BIBLIOGRÁFICAS........................................................................................67

14
CAPÍTULO 1
INTRODUÇÃO

15

16
1.1 ESTRESSE OXIDATIVO
O estresse oxidativo é uma conseqüência da produção excessiva de espécies reativas
de oxigênio (ERO), ou uma redução nas defesas antioxidantes do organismo, e ocorre
devido a um desequilíbrio entre os fatores pró-oxidantes e antioxidantes. (FINKEL &
HOLBROOK et al., 2000).
O ERO pode ser gerado de forma endógena durante o metabolismo celular, ou de
forma exógena, como por exposição a drogas e a alimentos (xenobióticos). Esses radicais
danificam importantes macromoléculas celulares tais como DNA, lipídeos, carboidratos e
proteínas (FINKEL & HOLBROOK et al., 2000), portanto ele é considerado como um
fator relevante no desenvolvimento de várias doenças crônicas degenerativas tais como
aterosclerose, diabetes tipo 2 e câncer (COOKE et al., 2003). Além de danificar
macromoléculas, o estresse oxidativo pode causar processos inflamatórios e desempenhar
um papel importante na ativação de oncogenes, bem como destruir genes supressores de
tumor. O estresse oxidativo está envolvido na formação de agregados protéicos que podem
levar ao desenvolvimento de doenças neurodegenerativas como Alzheimer e Parkinson
(FORSBERG et al., 2001). O ERO pode ainda aumentar a inflamação associada a
aterosclerose e artrite reumatóide (GRIFFITHS, 2005; SCHROECKSNADEL et al., 2006).
O sistema de defesa antioxidante, considerado um mecanismo de combate ao ERO,
tem o objetivo primordial de manter o processo oxidativo dentro dos limites fisiológicos e
passíveis de regulação, impedindo que os danos oxidativos se amplifiquem e causem
prejuízos sistêmicos irreparáveis. (KUNDU & SURH, 2010)
Antioxidantes, que são substâncias que atacam e neutralizam o ERO prevenindo os
danos causados por ele, podem ser classificados em dois grupos principais: enzimáticos e
não enzimáticos (RATNAM et al., 2006).

17
1.2 MECANISMOS ENZIMÁTICOS DE COMBATE AO ERO
Os antioxidantes enzimáticos são produzidos endogenamente e fazem parte de um
complexo sistema de detoxificação de três fases, altamente conservado entre eucariotos. As
proteínas de detoxificação envolvidas neste processo são classificadas em Fase I, II e III.
(SARKADI et al., 2006).
A citocromo P450 monooxigenase (CYP) é a principal enzima envolvida na Fase I
do metabolismo xenobiótico. As CYPs oxidam os xenobióticos por hidroxilação. Uma rede
de enzimas detoxificadoras, tais como superóxido dismutase (SOD), catalase (CAT),
glutationa peroxidase (GPx) e glutationa transferase (GT) fazem parte da Fase II. A SOD
age transformando dois ânions radicais superóxidos em um peróxido de hidrogênio,
podendo ocorrer de três formas dependendo do metal associado a ela, sendo cobre (Cu) e
zinco (Zn) no citoplasma e manganês (Mn) na matriz mitocondrial de eucariotos e ferro
(Fe) em bactérias Perez et al., 2009. A CAT tem a função de proteger as células
catalisando a decomposição do peróxido de hidrogênio em oxigênio molecular e água sem
produção de radicais livres (McCORD, 1986; SIES, 1993; RATNAM et al., 2006). A GPx
é uma enzima localizada no citosol e na matriz mitocondrial, que reduz o peróxido de
hidrogênio e hidropeptídeos orgânicos utilizando a glutationa (GSH). A GSH atua como
co-substrato da GPx, com propriedade doadora de elétrons, podendo ser regenerada através
da glutationa redutase (GR) com transferência de hidrogênio do NADPH. Neste processo
são transferidos dois hidrogênios dos grupamentos sulfidrila para os peróxidos,
transformando-os em álcool e/ou água, resultando em glutationa dissulfeto (GSSG). A GPx
geralmente ocorre associada ao selênio (Se). Os principais locais de ação da GPx são o
fígado e os eritrócitos, podendo ocorrer no coração, pulmões e músculos. Finalmente na
Fase III de detoxificação, os conjugados tóxicos são bombeados para fora da célula por
transportadores cassetes de ligação de ATP (Transpotador ABC) ou outros transportadores
(SARKADI et al., 2006).

18
1.2.1 Controle da Expressão Gênica das Enzimas Antioxidantes em Mamíferos
A condição de estresse oxidativo na célula induz várias respostas fisiológicas e
patológicas. Estas respostas, em última análise, causam mudanças no perfil transcricional
de forma a influenciar o destino da célula e a progressão de uma doença. Nas últimas
décadas, vários fatores de transcrição e vias de sinalização foram identificados pelo seu
papel crítico na resposta ao estresse oxidativo (MA et al., 2010). Dentre estes fatores,
podemos citar as proteínas Nrf e FoxO.
A ativação transcricional das enzimas da Fase II é principalmente mediada pelo
elemento cis atuante denominado ARE (Antioxidant Response Element) que é reconhecido
pelo fator de transcrição do tipo zíper de leucina básico (bZIP) Nrf (Nrf1 e Nrf2) (ITOH et
al., 1997). Em condições normais, Nrf localiza-se no citoplasma ligado a uma proteína
associada à actina chamada Keap1 (ITOH et al., 1999). Durante o estresse oxidativo, Nrf
se desliga de Keap1 e transloca-se para o núcleo da célula ativando a expressão dos genes
detoxificadores da Fase II (ITOH et al., 1999; ZIPPER et al., 2002). Esta dissociação de
Nrf é alcançada através da mudança conformacional em Keap1 causada pela oxidação, da
modificação de cisteínas essenciais (ITOH et al., 1999) ou pela remoção de Zn de
cisteínas coordenadoras na sua estrutura (DINKOVA-KOSTOVA et al., 2005). Portanto os
ativadores de Nrf2 são eletrofílicos e têm a capacidade de oxidar ou alquilar grupos tióis
ou de quelar íons metais (KLUTH et al., 2007).
A importância da ativação da resposta contra o estresse oxidativo e contra compostos
xenobióticos pela proteína Nrf2 foi demonstrada em camundongos “knock-out” Nrf2-/-.
Estes animais não foram capazes de ativar os genes da Fase II no trato digestório e foram
mais sensíveis ao estresse oxidativo e drogas tóxicas (ITOH et al., 1997; CHAN et al.,
2001; HAYES, 2001). Os camundongos mutantes para ambas as proteínas Nrf1 e Nrf2
(Nrf1-/-;Nrf2 -/-) morreram antes de nascer, e as células isoladas destes camundongos
foram hipersensíveis ao estresse oxidativo (LEUNG et al., 2003). Evidências in vivo
mostraram que camundongos deficientes para Nrf2 não respondem à proteção dos
antioxidantes e são mais susceptíveis à carcinogênese (RAMOS-GOMES et al., 2001).
As proteínas FoxO são fatores de transcrição conhecidos por seu papel nas respostas
celulares ao estresse ambiental e fisiológico, cuja atividade é regulada por modificação
pós-transcricional, incluindo a fosforilação. Um único gene FoxO está presente em

19
drosofila (Dfoxo) e C. elegans (daf-16) e 4 estão presentes em camundongos e humanos
(FoxO1, FoxO3, FoxO4 e FoxO6) (LANDIS & MURPHY, 2010; YEN; NARASIMHAN;
TISSENBAUM, 2011). Em resposta à via da insulina/IGF-1 e a outros fatores de
crescimento, as proteínas quinases Akt e SGK fosforilam FoxO (com exceção dos FoxO6)
em três locais conservados. Esta fosforilação promove a ligação de FoxO às proteínas 14-
3-3 no citoplasma impedindo sua localização nuclear. Em resposta ao estresse oxidativo, a
via JNK (c-Jun N-terminal kinase) fosforila diretamente FoxO, ativando a sua localização
nuclear (MA, 2010; YEN; NARASIMHAN; TISSENBAUM, 2011). O aumento da
ativadade transcricional de FoxO ativa a expressão de catalase e SOD2, enzimas que são
reguladas por FoxO aumentando a detoxificação de ERO (MA, 2010).
1.3 MECANISMOS NÃO ENZIMÁTICOS DE COMBATE AO ERO
Os antioxidantes não enzimáticos, exceto os de baixos peso molecular, são obtidos
de fontes dietéticas. Estes são classificados em várias classes, das quais os polifenóis são a
maior parte. As outras classes incluem vitaminas, carotenóides, compostos
organosulfurados, minerais e cofatores que desempenham um papel importante na
manutenção da saúde humana (RATNAM et al., 2006).
A glutationa, o tocoferol, o ácido ascórbico e os carotenóides, são substâncias que
podem participar diretamente em processos fisiológicos, bioquímicos e celulares
neutralizando os radicais livres ou prevenindo as reações químicas deflagradas por eles
(SHARONI et al. 2004; MASELLA et al, 2005). Além destas características, antioxidantes
diretos são consumidos ou quimicamente modificados no processo de ação e devem ser
repostos ou regenerados após sua ação.
Entre os minerais destacam-se o zinco, cobre, selênio e magnésio. Já entre as
vitaminas, destaca-se a vitamina C que tem ação removedora de ERO e também
regeneradora da vitamina E (tocoferol). Como a vitamina C é hidrossolúvel, possui maior
ação no plasma sanguíneo, enquanto a vitamina E, lipossolúvel, tem maior ação em
membranas celulares (MANELA-AZULAY et al., 2003)

20
1.3.1 Antioxidantes Polifenólicos
Os polifenóis são a maior classe de antioxidantes da dieta que compreendem uma
variedade de compostos, divididos em várias classes (ácido hidrobenzóico, antocianinas,
proantocianinas, flavanóis, flavanonas, isoflavonas, flavonóides etc), que ocorrem em
frutas e vegetais, vinhos e chás, chocolate e outros produtos do cacau (MANACH et al.,
2004).
O alto consumo de comidas e bebidas ricas em polifenóis, especialmente os
flavonóides, tem sido associado com o menor risco de doenças degenerativas crônicas
como aterosclerose e câncer (KLUTH et al., 2007). Muitos dos efeitos fisiológicos dos
flavonóides têm sido atribuídos à sua capacidade antioxidante direta de remoção de
radicais livres. Os flavonóides são substâncias fenólicas de baixo peso molecular e
possuem em sua estrutura química anéis aromáticos, tendo como base o 2-fenil-
benzopirano (C6-C3-C6) (AHERNE & O’BRIEN, 2002). O efeito antioxidante direto dos
flavonóides se deve à capacidade destes compostos em neutralizar os radicais livres (ânion
superóxido, radicais hidroxilas e peroxilas) ou quelar íons metálicos (cobre e ferro). Os
flavonóides apresentam grupos hidroxilas em sua estrutura, portanto eles podem inibir as
reações de oxidação das espécies reativas de oxigênio, por doarem átomos de hidrogênio às
espécies, estabilizando-as e transformando-se em quinonas (substâncias pardas). Ao quelar
íons metálicos ocorrem à inibição da Reação de Fenton, uma forma endógena de produzir
espécies reativas (ENIO, 2003).
Além da atividade antioxidante, os flavonóides também podem apresentar
propriedades anti-inflamatória, anticancerígena, antiproliferativa e pró-apoptótica
(MASELLA et al., 2005). Os flavonóides regulam a atividade de enzimas antioxidantes,
modulam receptores nucleares, atuam sobre a expressão gênica e vias de sinalização
celular (MOSKAUG, 2004). Apesar de intensa investigação, os mecanismos específicos,
pelo qual estes compostos alteram a saúde humana permanece incerto. Recentemente foi
descrito que os flavonóides também podem atuar como antioxidantes indiretos (HU et al.,
2006; JUGE; MITHEN; TRAKA, 2007). O modo de atuação destes compostos é a
ativação transcricional dos genes das enzimas detoxificantes da Fase II. Dentre os
antioxidantes indiretos, já foi demonstrado que vários compostos fitoquímicos são potentes
indutores de Nrf2 e das enzimas de detoxificação da Fase II, aumentando a capacidade

21
antioxidante da célula (DINKOVA-KOSTOVA et al., 2005; SPORN & LIBY et al., 2005
OSBURN & KENSLER, 2008).
1.4 PROPRIEDADES BIOLÓGICAS DO AÇAÍ
O Euterpe oleracea Mart é uma espécie de palmeira nativa das regiões amazônicas
conhecida como açaizeiro, que tem ganhado grande destaque nas últimas décadas, em
consequência das suas propriedades antioxidantes e nutricionais. O açaí, fruto oriundo
dessa palmeira, tem uma importância econômica devido ao seu valor nutricional entre as
populações indígenas da Amazônia. A polpa é rica em lipídeos essenciais e em certos
minerais como o cálcio e o potássio, que podem contribuir para garantir o crescimento e
bom funcionamento do corpo humano em geral, uma vez que esses nutrientes participam
de várias reações metabólicas importantes no organismo (MENEZES et al., 2008). O açaí
liofilizado pode ser considerado como uma excelente alternativa de conservação da polpa
do fruto, devido à presença de importantes componentes nutricionais encontrados no
mesmo. Por ser altamente calórico é um excelente alimento a ser incorporado nas refeições
de indivíduos com baixo peso, em especial a faixa etária infantil. Além disso, polpa de açaí
liofilizada é uma alternativa para suprir deficiência nutricional de adultos e adolescentes
afetados pelos hábitos alimentares do século XXI, que normalmente consomem refeições
pobres em macronutrientes e em minerais (SANTOS et al.,2008)
1.4.1 Propriedades Antioxidantes in vitro e ex vivo
Em relação as suas propriedades antioxidantes, já foi demonstrado que o açaí é rico
em polifenóis, especialmente em flavonóides do tipo antocianinas e proantocianas (DEL
POZO-INSFRAN et al., 2004; SCHAUSS et al., 2006). Dentre os principais componentes
fitoquímicos do açaí, as antocianinas cianidina 3-glicosídio e cianidina 3-rutinosídio, são
os compostos encontrados em maior abundância (DEL POZO-INSFRAN et al., 2004;
SCHAUSS et al., 2006). Foi demonstrada a capacidade antioxidante in vitro da polpa de
açaí para radicais peroxil (ROO-) e peroxinitrito (ONOO
-) e hidroxila (
-OH)
(LICHTENTHALER et al., 2005). A correlação desta capacidade antioxidante do açaí é

22
positiva para as frações de antocianinas e os compostos fenólicos totais (SANTOS et al.,
2008). Spada et al. (2008) também demostraram in vitro que a polpa do açaí possui
atividade antioxidante para o radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) e atividades
semelhantes a da SOD e CAT. Jensen et al. (2008) analisaram os efeitos antioxidantes da
bebida MonaVie cujo principal componente é o açaí. Esta bebida contém altos níveis das
antocianididas cianidina 3-rutenosídeo, e cianidina 3-glicosídeo e diminui a produção de
ERO em células polinucleares incubadas com H2O2. A capacidade antioxidante também foi
avaliada ex vivo em tecidos isolados do córtex cerebral, hipocampo e cerebelo de ratos. O
pré-tratamento com açaí reduziu o dano oxidativo induzido por H2O2 em lipídios e
proteínas e a atividade das enzimas SOD e CAT, restabelecendo níveis basais semelhantes
ao grupo controle em todos os tecidos avaliados. É interessante ressaltar porém que nos
tecidos tratados somente com açaí, nenhum dos parâmetros avaliados se alteraram
(SPADA et al., 2009). Guerra et al. (2011) demonstraram através de experimento com
ratos, que o açaí foi capaz de modular a produção de EROs em neutrófilos e enzimas
antioxidantes como a glutationa e a proteína carbonilada no figado de animais diabéticos.
1.4.2 Propriedades Anti-inflamatórias in vitro e in vivo
A polpa de açaí também apresenta propriedade anti-inflamatória, como demonstrado
pela sua habilidade de inibir a atividade das ciclooxigenases COX-1 e COX-2 em cultura
de células (SCHAUSS et al., 2006). De acordo com Noratto et al. (2011), o extrato de açaí
também é capaz de proteger células endoteliais do estresse induzido por glicose, reduzindo
a expressão de interleucina -6 e -8. Ainda segundo o autor, a expressão de moléculas de
adesão e ativação de NF-kB foram inibidas nas células tratadas com açaí e induzidas com
lipopolisacarídeo (NORATO et al., 2011).
A presença do extrato de açaí em cigarros teve um efeito protetor contra o enfisema
em camundongos (MOURA et al., 2011). Os camundongos submetidos à fumaça de
cigarro e extrato de açaí apresentaram menor nível de estresse oxidativo devido ao
aumento na atividade de GPx e de GSH, em relação ao grupo exposto somente com
fumaça de cigarro. Houve também uma redução da atividade da mieloperoxidase (MPO),
uma enzima marcadora do influxo de neutrófilo e dano pulmonar (MOURA et al., 2011).

23
1.4.3 Propriedades Antiproliferativas in vitro e in vivo
Além dos efeitos antioxidantes e anti-inflamatórios, várias pesquisas apontaram que
frações de polifenóis, extraídas da polpa de açaí, demonstraram atividade antiproliferativa
e pró-apoptótica em células leucêmicas HL-60, através de mecanismo mediado pela
caspase-3 (DEL POZO-INSFRAN et al., 2006; SCHAUSS et al., 2006). Ding et al. (2006)
demonstraram que a cianidina 3-glicosídio (C3G) possui propriedades quimiopreventivas e
quimioterapêuticas. Em cultura de células, a C3G foi capaz de eliminar espécies reativas
de oxigênio induzidas por radiação ultravioleta, enquanto o estudo in vivo demonstrou que
o tratamento com C3G diminuiu o número de tumores de pele malignos e benignos
induzidos por tetradecanoilforbol-13-acetato (TPA) (MATIAS et al., 2008). Os efeitos
antiproliferativos de extratos obtidos a partir da polpa de açaí também foram avaliados em
cultura de células de adenocarcinoma humano HT-29 onde apresentaram efeito inibitório
de até 90,7% sobre a proliferação celular (PACHECO-PALENCIA et al., 2008). Hogan et
al. (2010) mostraram que o extrato enriquecido em antocianinas obtido a partir da polpa de
açaí apresentou grande capacidade antiproliferativa em células do glioma cerabral C6. A
análise de fragmentação do DNA indicou que este efeito pode ser devido à indução da
apoptose nas células C6, após 48 horas de tratamento com 100 e 200 ug/mL do extrato. Os
mesmos resultados não foram observados em células de câncer de mama MDA-468,
sugerindo que a atividade antiproliferativa do extrato pode ser específica para
determinados tipos celulares como as células C6. Ribeiro et al. (2010) observaram através
do ensaio de cometa que o tratamento de ratos com açaí não teve nenhum efeito
genotóxico nos micronúcleos de células da medula e nos eritrócitos e o tratamento foi
capaz de reduzir o dano no DNA provocado por doxorubicina (DXR) nas células.
No entanto, Spada et al. (2008) mostraram que apesar da polpa do açaí possuir
atividade antioxidante, também apresentou efeitos mutagênicos em altas concentrações em
linhagens de Saccharomyces cerevisiae.

24
1.4.4 Proteção Contra Estresse Hiperlipídico
Além dos efeitos protetores contra o estresse oxidativo, o tratamento com extrato de
açaí também exerce efeitos benéficos contra o estresse induzido por dietas hiperlipídicas
(SOUZA et al., 2010; SUN et al., 2010; OLIVEIRA et al., 2010).
A suplementação com 2% da polpa de açaí na dieta de ratos alimentados com dietas
padrão e hipercolesterolêmica melhorou o perfil lipídico e de biomarcadores do estresse
oxidativo no soro. Animais hipercolesterolêmicos apresentaram redução dos níveis séricos
de colesterol total e colesterol não HDL após seis semanas de suplementação com a polpa
de açaí. Além disso, a suplementação com açaí reduziu os níveis de proteínas carboniladas,
aumentou a concentração de grupos sulfidrila livres e ligados a proteína e a atividade da
enzima paraoxonase no soro de ratos alimentados com ambas as dietas. A suplementação
com açaí também reduziu significativamente a atividade da superóxido dismutase apenas
nos ratos hipercolesterolêmicos. Em camundongos submetidos à dieta hiperlipídica, o
tratamento com extrato de açaí reduziu o peso corporal, os níveis de triglicérides e
colesterol total no plasma (OLIVEIRA et al., 2010). O tratamento também diminuiu os
níveis de glicose, a resistência a insulina e intolerância a glicose dos camundongos
submetidos à dieta hiperlipídica. Estes resultados sugerem que o açaí é uma fonte
nutricional alternativa para prevenção da síndrome metabólica (OLIVEIRA et al., 2010).
Usando a Drosophila melanogaster como modelo, Sun et al. (2010) observaram que
moscas tratadas com dieta hipercalórica suplementada com açaí tiveram seu tempo de vida
médio e máximo aumentados significativamente, quando comparadas àquelas apenas
submetidas à dieta hipercalórica. Esse efeito pró-longevidade parece estar associado a um
aumento na resposta ao estresse e a diminuição da expressão da enzima fosfoenolpiruvato
carboxiquinase (PEPCK), enzima chave do controle da gliconeogênese e gliceroneogênese
e modulada pelo fator de transcrição FoxO. Além disso, observou-se que a polpa de açaí
aumentou a sobrevivência de moscas tratadas com RNAi para sod1 (superóxido
dismutase), principal enzima citosólica envolvida na eliminação de radicais superóxido da
célula, sugerindo que o açaí pode aliviar o dano oxidativo in vivo. Análises moleculares
demonstraram que a suplementação com açaí em moscas alimentadas com dieta
hipercalórica aumentou a transcrição de enzimas envolvidas em funções antioxidantes e de
resposta ao estresse, entre elas a glutationa-S-transferase.

25
1.4.5 Efeitos Benéficos na Saúde Humana
Apesar de grande parte dos estudos sobre os efeitos do extrato de açaí in vivo serem
em organismos modelos, alguns estudos já foram realizados em humanos também
(JENSEN et al., 2008; MERTENS-TALCOO et al., 2008; UDANI et al., 2011; JENSEN
et al., 2011). Jensen et al. (2008) fizeram um ensaio randomizado, duplo-cego com doze
indivíduos saudáveis, no qual o consumo de 120 mL da bebida a base de açaí causou um
aumento da capacidade antioxidante do plasma, e inibição da peroxidação lipídica no soro,
após intervalo de duas horas do consumo. Outro estudo clínico realizado em 12 humanos
saudáveis demonstrou que o consumo de uma única dose de suco e polpa de açaí (7mL/Kg
de peso) foi capaz de aumentar a capacidade antioxidante do plasma em até 2,3 e 3 vezes.
As antocianinas foram quantificadas no plasma após um período de 0 a 12 horas do
consumo das bebidas, atingindo o pico de concentração plasmática após 2,2 horas para
polpa e 2.0 horas para o suco clarificado (MERTENS-TALCOTT et al. 2008).
Udani et al. (2011) mostraram em um estudo piloto realizado com 10 adultos com
sobrepeso que a ingestão de 100 mg de polpa de açaí duas vezes ao dia durante um mês
promoveu a redução da glicemia de jejum e níveis os insulina. Houve também uma
redução no colesterol total bem como no LDL-colesterol (UDANI et al., 2011). Tendo em
vista os efeitos anti-inflamatórios do açaí, um estudo piloto foi feito com 14 pacientes com
dores e dificuldade de locomoção para verificar se o consumo de suco contento na sua
porção maior açaí (MonaVie) estaria associado com alívio da dor (JENSEN et al., 2011).
Além de melhorar significantemente a dor e a execução de atividades diárias, o consumo
deste suco, também melhorou o status antioxidante do soro, reduziu a peroxidação lipídica
e a proteína C reativa, um marcador inflamatório (JENSEN et al., 2011).
1.5 ORGANIMO MODELO CAENORHABDITIS ELEGANS
Caenorhabditis elegans é um nematóide terrestre de vida livre da família
Rhabditidae. Nas últimas décadas, o C. elegans tem sido utilizado como organismo

26
modelo por mais de 200 laboratórios mundiais em estudos de biologia do
desenvolvimento, genética, envelhecimento e de ecotoxicologia, levando a muitas
descobertas importantes para os mamíferos, incluindo o homem. Uma das principais
razões para isto é devido à grande semelhança que existe entre C. elegans e mamíferos,
tanto a nível celular quanto molecular. Cerca de 60-80% dos genes homólogos aos
humanos foram identificados no C. elegans (The C. elegans Sequencing Consortium,
1998; KALETTA E HENGARTER, 2006).
Além da alta homologia com o genoma dos mamíferos, este organismo possui uma
série de características que o torna muito eficiente como modelo genético: é fácil de
cultivar e possui um ciclo de vida curto. O C. elegans é facilmente cultivado em
laboratório em placas de petri contendo ágar semeadas com a bactéria Escherichia coli
como alimento. Os vermes atingem a maturidade com 2,5 dias a 25º C e tem um tempo de
vida curto de 20 dias a 25oC. O ciclo de vida do animal consiste de um período de
desenvolvimento embrionário dentro do ovo, quatro estágios larvais e finalmente o adulto
(Figura 1). O ciclo de vida deste organismo inicia-se como uma única célula, que dá
origem, através de divisões celulares repetidas, há 558 células que formam o pequeno
verme dentro da casca do ovo. Após eclodir, divisões posteriores resultam no crescimento
e na maturação sexual do verme. Após a muda final, o verme adulto hermafrodita começa
a produzir os seus próprios ovos. Em condições adversas, tais como escassez de alimento
ou estresse oxidativo, a larva L1 de C. elegans pode entrar em um estágio alternativo de
diapausa. Neste estado, os animais são hipometabólicos e altamente resistentes ao estresse,
com capacidade de sobreviver por longos períodos de várias semanas até meses (Figura 1).
Outra característica importante é que o C. elegans é pequeno e transparente, permitindo
que suas células possam ser acompanhadas individualmente e que os genes repórteres
ligados ao GFP (Green Fluorescent Protein) possam ser observados no organismo ao vivo.
1.5.1 Controle da Expressão Gênica das Enzimas Antioxidantes no C. elegans
Assim como os mamíferos, o C. elegans possui o sistema de detoxificação de Fase I
(citocromo p450), Fase II (AN et al., 2003; MENZEL et al., 2005) e Fase III (OLIVEIRA
et al., 2009). No C. elegans, a resistência ao estresse oxidativo é controlado, em parte, por
dois principais fatores de transcrição: p38/SKN-1/Nrf e DAF-2/Ins/IGF-1 que ativam a

27
expressão dos genes de detoxificação.
1.5.1.1 A via de sinalização de SKN-1/Nrf
O SKN-1 é uma proteína bZIP que atua de forma análoga ao fator Nrf de mamíferos
e promove resistência ao estresse e longevidade (WALKER et al., 2001; AN &
BLACKWELL, 2003). Em condições normais, SKN-1 localiza-se constitutivamente no
núcleo dos neurônios ASI, correspondente ao hipotálamo, que detectam ou regulam a
ingestão de comida (YOU et al., 2008). Nestes neurônios, SKN-1 é necessário para o
aumento da longevidade induzida pela restrição calórica (BISHOP & GUARENTE, 2007).
FIGURA 1. Ciclo de vida do C. Elegans. Fonte: worm atlas
Na ausência de estresse, a atividade de SKN-1 parece ser controlada tanto no
citoplasma quanto no núcleo. No citoplasma, SKN-1 é regulado por várias vias de
sinalização. SKN-1 é negativamente regulado através da fosforilação pela enzima
glicogênio sintetase quinase (GSK-3) prevenindo sua acumulação no núcleo (AN et al.,
2005). Foi também demonstrado que a via da IIS (via de sinalização da insulina) regula

28
diretamente o fator de transcrição SKN-1. DAF-2 é homólogo ao receptor do fator de
crescimento semelhante à insulina (IGF-1) em mamíferos. DAF-2 ativa as enzimas
fosfatidilinosiol 3-quinase AGE-1 (PI3K), PDK-1, e AKT-1/2 quinases que antagonizam o
fator de transcrição SKN-1 (TULLET et al., 2009), assim como DAF-16, o fator ortólogo
de FoxO (HENDERSON & JOHNSON, 2001; LEE et al., 2003). Assim como nos
mutantes daf-2, mutações em SKN-1 dirigidas para os sítios de fosforilação da AKT
resultam na localização nuclear constitutiva de SKN-1. A deleção de skn-1 em mutantes
daf-2 suprime os fenótipos da resistência ao estresse oxidativo e a longevidade, associados
a este último. Além disso, o envelhecimento é retardado nos animais quando SKN-1 é
superexpresso transgenicamente, e quando possuem o sítio de fosforilação para AKT-1/2
mutados na ausência de daf-16. Estes dados mostram que a via de IIS inibe diretamente
SKN-1 em paralelo a DAF-16 e que SKN-1 contribui para a resistência ao estresse e
longevidade, que são observados quando ocorre a redução da atividade de IIS.
Recentemente, foi demonstrado que a via de sinalização ERK MAPK fosforila diretamente
SKN-1 e ativa a localização nuclear deste (OKUYAMA et al., 2010). A inibição da
expressão dos genes da via ERK diminui a longevidade de animais tipo selvagem, mas não
de mutatnes daf-16 e skn-1 sugerindo que a longevidade controlada pela via ERK MAPK
depende de SKN-1 e DAF-16 (OKUYAMA et al., 2010).
Em condições normais SKN-1 associa-se à proteína WDR-23 e ao complexo
protéico da ubiquitina ligase CUL-4/DDB-, sugerindo que SKN-1 seja degradado pelo
sistema proteassômico (CHOE et al., 2009). Também foi demonstrado que WDR-23 regula
negativamente a expressão de genes de detoxificação de Fase II (HOSEGAWA & MIWA,
2010).
Já em condições de estresse, SKN-1 acumula-se no núcleo das células do intestino, o
maior órgão de detoxificação no C. elegans, e ativa a transcrição genes alvos em resposta a
vários estresses (AN & BLACKWELL et al., 2003; OLIVEIRA et al., 2009). A
localização nuclear de SKN-1 em condições de estresse no núcleo requer a sua fosforilação
pela PMK-1, uma quinase conservada, pertencente à via de sinalização da p38 MAPK
(MAPK -Mitogen-Activated Protein Kinase). SEK-1, a MAPK quinase que fosforila PMK-
1 é necessária para ativar a localização de SKN-1 no núcleo e a expressão de gcs-1
(INOUE et al., 2005). Os mutantes com perda de função skn-1 e sek-1 são mais sensíveis
ao estresse oxidativo, reforçando a importância de SEK-1-SKN-1 na defesa contra o
29
estresse oxidativo (AN & BLACKEWELL, 2003; INOUE et al., 2005). Além disso,
mutantes skn-1 possuem um tempo de vida reduzido em cerca de 25-30%, o que é
consistente com associações feitas entre estresse oxidativo e longevidade (AN &
BLACKWELL, 2003) (Figura 2).
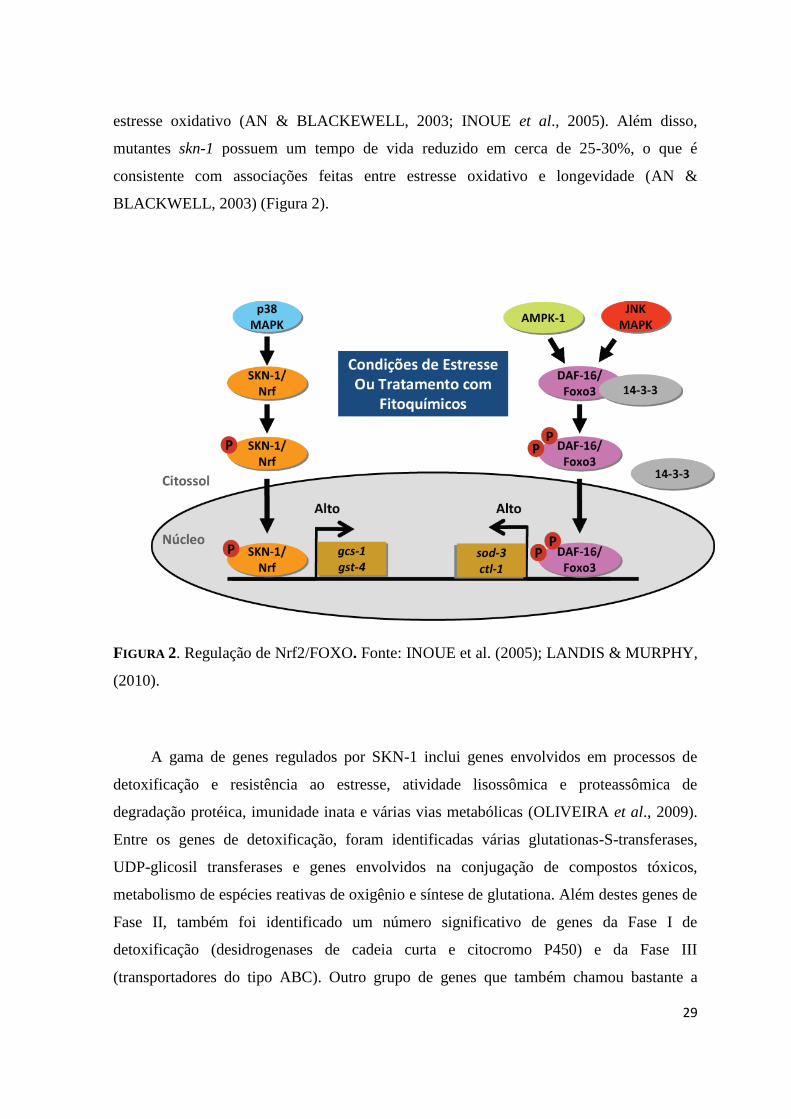
FIGURA 2. Regulação de Nrf2/FOXO. Fonte: INOUE et al. (2005); LANDIS & MURPHY,
(2010).
A gama de genes regulados por SKN-1 inclui genes envolvidos em processos de
detoxificação e resistência ao estresse, atividade lisossômica e proteassômica de
degradação protéica, imunidade inata e várias vias metabólicas (OLIVEIRA et al., 2009).
Entre os genes de detoxificação, foram identificadas várias glutationas-S-transferases,
UDP-glicosil transferases e genes envolvidos na conjugação de compostos tóxicos,
metabolismo de espécies reativas de oxigênio e síntese de glutationa. Além destes genes de
Fase II, também foi identificado um número significativo de genes da Fase I de
detoxificação (desidrogenases de cadeia curta e citocromo P450) e da Fase III
(transportadores do tipo ABC). Outro grupo de genes que também chamou bastante a

30
atenção foi os genes envolvidos nas vias lisossômicas e proteassômicas de degradação de
proteínas sugerindo que SKN-1 também teria um possível papel na reciclagem de proteínas
danificadas (OLIVEIRA et al., 2009). Em resposta ao estresse provocado por arsenato de
sódio (As), foi observada a expressão dependente de SKN-1 de vários genes de
detoxificação sendo que a maioria deles é também expressos em condições normais. Por
outro lado o tratamento com t-BOOH ativa a expressão de genes de detoxificação e
metabolismo que são na sua maioria SKN-1 independentes. Estas observações sugerem
que SKN-1 tem um papel importante tanto em condições normais como durante o estresse
(OLIVEIRA et al., 2009).
1.5.1.2 A Via de Sinalização de DAF-16
Em C. elegans, a via de sinalização da insulina/IGF-1 controla vários processos
biológicos tais como longevidade, reserva lipídica, reprodução e resposta ao estresse,
termotolerância, resistência a patógenos, metabolismo e autofagia. (LEE et al., 2003). Esta
via é iniciada pelo receptor DAF-2, o homólogo do receptor do fator de crescimento
semelhante à insulina (IGF-1) em mamíferos. Quando DAF-2 é ativado fosforila a
fosfoinositil 3-kinase, AGE-1, gerando PIP3, que por sua vez recruta as kinases AKT-1,
AKT-2, SGK-1 e PDK-1 para a membrana plasmática onde PDK-1 fosforila AKT e SGK-
1. O complexo AKT-1/AKT-2/SGK-1 fosforila o fator de transcrição DAF-16,
sequestrando-o no citoplasma e então prevenindo a ativação ou repressão de genes alvos
no núcleo. Inibidores desta cascata incluem DAF-18, o fosfoinositil 3-fosfatase PTEN, que
antagoniza AGE-1 pela defosforilação de PIP3, e PPTR-1, uma subunidade reguladora da
PP2A que defosforila AKT-1 (LANDIS & MURPHY, 2010). O papel desta via de
sinalização na longevidade e metabolismo é conservado em C. elegans, Drosophila e
mamíferos. A via da DAF-2 insulina/IGF1 regula a expressão de várias enzimas de
detoxificação, tais como superóxido dismutase (sod-3), catalases (ctl-1 e ctl-2) (MURPHY
et al., 2003). Mutantes com alelo hipomórfico de daf-2 são mais resistentes ao estresse
oxidativo e possuem um tempo de vida prolongado em 40% comparado com nematóides
do tipo selvagem, reforçando a teoria do dano oxidativo do radical livre e o envolvimento
da mitocôndria e do ERO no processo de envelhecimento (VANFLETEREN & DE
VREESE 1995; HONDA & HONDA, 1999).

31
Além da via DAF-2/Ins/IGF1, DAF-16 também pode ser regulado pela interação
com a s proteínas 14-3-3, AMPK e JNK. Similar a sinalização de FoxO de mamíferos, as
proteínas 14-3-3 interagem com DAF-16 e regulam sua distribuição
nuclear/citoplasmática. A proteína kinase ativada por AMP (AMPK) ativa DAF-16 pela
fosforilação direta. A família JNK é um subgrupo da superfamília MAPK e está associada
com a regulação de processos biológicos críticos como o desenvolvimento, apoptose e
sobrevivência celular. Em C. elegans, a via de sinalização JNK é ativada por diferentes
estresses tais como estresse térmico e oxidativo. JNK interage fisicamente e fosforila DAF-
16 em um sítio diferente dos sítios de fosforilação de AKT, e esta fosforilação resulta em
um aumento da translocação nuclear de DAF-16 (MA, 2010).
1.6 C. elegans COMO UM MODELO PARA ESTUDOS DE FITOQUÍMICOS
Como apresentado acima, o C. elegans é um excelente modelo para identificação
rápida de compostos com propriedades biológicas e apresenta grandes vantagens em
relação à cultura de células, pois é um organismo multicelular que permite que a ação dos
compostos seja avaliada de uma forma sistêmica, em um organismo. Além disso, o cultivo
e os experimentos realizados com este organismo são muito mais baratos comparados com
camundongos ou outro modelo mamífero. Isto se deve principalmente porque no caso do
C. elegans, o requerimento de animais não é limitante, pois eles crescem rapidamente em
um sistema simples e barato, onde podem ser gerados milhares de animais em alguns
poucos dias. Além disso, o fato do seu genoma já ser conhecido e da disponibilidade de
mutantes para a maioria dos seus genes, o C. elegans, permite que os mecanismos da
resposta ao estresse oxidativo sejam facilmente investigados in vivo.
Devido a todas estas características, vários compostos que aumentam a longevidade e
a resistência ao estresse oxidativo têm sido estudados neste organismo. Em especial
atenção, dois estudos relataram o efeito de extratos ricos em polifenóis e antocianinas no
C. elegans (WILSON et al., 2006; PUN et al., 2010). Extratos de mirtilo “blueberry” ricos
em polifenóis (WILSON et al., 2006) aumentaram o tempo médio de vida em 28% e o
máximo em 14%. Além disso, o extrato de mirtilo aumentou a termotolerância do C.
elegans, mas não a resistência ao estresse oxidativo. A análise indicou que os polifenóis do

32
extrato de mirtilo parecem atuar através de uma via de sinalização da proteína quinase
dependente da Ca2+
/calmodulina (CaMKII). Já foi demonstrado que Casca de Pinheiro,
rico em antocianinas (CP) diminui os níveis de EROs em doses muito baixas e muito altas,
mas aumenta a concentração de EROs em doses intermediárias. O maior decréscimo nos
níveis de EROs foi observada a 5 e 10 CP lg / ml, que foram as mesmas doses, que
provocaram significativo aumento do tempo de vida do organismo C. elgans.(PUN et al.,
2010)
O extrato de Ginkgo biloba, complexo rico glicosídeos e terpenóides
(KAMPKÖTTER et al., 2007), aumentou a resistência ao estresse oxidativo em 33% (WU
et al., 2002). O resveratrol, polifenol abundante nas cascas de uva (BASS et al., 2007),
prolongou o tempo de vida de uma maneira dose-dependente (VISWANATHAN, 2005). A
quercetina, flavonóide abundante em plantas comestíveis, aumentou a resistência ao
estresse oxidativo e o tempo de vida médio em 18% no C. elegans (PIETSCH et al., 2009).
Tomando em consideração todas estas informações, o presente projeto teve como
proposta estudar os efeitos do açaí no organismo multicelular C. elegans. Além disso,
foram investigados os possíveis mecanismos envolvidos na resposta aos estresses oxidativo
e térmico e na longevidade dos animais tratados com açaí.

33
CAPÍTULO 2
OBJETIVO

34
2.1 OBJETIVO GERAL
Avaliar os efeitos antioxidantes e pró-longevidade do extrato de açaí (Euterpe oleracea
Mart.) no organismo modelo Caenorhabits elegans
2.2 OBJETIVOS ESPECÍFICOS
- Quantificar antocianinas de extratos obtidos da polpa de açaí liofilizado (AL);
- Avaliar os efeitos do tratamento com extrato de açaí liofilizado na sobrevivência do
C. elegans em condições de estresse e em condições normais;
- Dosar marcadores bioquímicos de estresse oxidativo (catalase, grupos sulfidrilas
totais, proteína carbonílica);
- Verificar o envolvimento de SKN-1 e DAF-16 na resposta antioxidante do C.
elegans tratado com extrato de açaí.

35
CAPÍTULO 3
METODOLOGIA

36
3.1 CEPAS E LINHAGENS
As cepas utilizadas neste projeto foram obtidas através do Caenorhabditis Genetics
Center (CGC), University of Minesota, EUA (http://www.cbs.umn.edu/cgc) ou do Dr. T.
Keith Blackwell do Joslin Diabetes Center, Boston, EUA. Foram utilizadas a cepa tipo
selvagem N2 Bristol, as cepas mutantes de deleção “knockout” sek-1(km4), e daf-
16(mgDf47) e a cepa transgênica com gene repórter N2 zIs356[DAF-16::GFP]. Também
foram usados animais “knockedown” para skn-1 obtidos por RNA interferente (RNAi)
(TULLET et al., 2008).
A incorporação do RNA fita dupla (dsRNA) nos vermes é feita através de um sistema
bacteriano. Para tal, um fragmento exônico do gene alvo é clonado no plasmídeo pL4440
(distribuído gratuitamente por Andrew Fire) que é específico para síntese de dsRNA por
indução com IPTG (KAMATH et al., 2001). Para o silenciamento de skn-1, o plasmídeo foi
construído através da subclonagem de um fragmento obtido por PCR do cDNA de skn-1
(WALKER et al., 2001) do plasmídeo pPD129.36 (cedido gentilmente por Andrew. Fire). O
plasmídeo pL4440 sem nenhum inserto foi usado como controle negativo do RNAi. O
plasmídeo recombinante foi transformado em E. coli HT115 e a cultura foi então semeada
em placas de petri com NGM contendo IPTG. Os vermes tipo selvagem no estágio larval 1
(L1) foram então cultivados nestas placas e utilizados nos experimentos após 48h a 20oC
quando atingiram estágio L4. A confirmação da redução da expressão de skn-1 foi feita
deixando os mesmos animais descritos acima atingirem a fase adulta. Na ausência de skn-1
os embriões colocados por eles não eclodem.
3.2 CRESCIMENTO E MANUTENÇÃO DO C. ELEGANS
As cepas foram mantidas congeladas até o momento de uso. O congelamento foi
transferindo animais L1 para uma solução 1:1 de M9 (22 mM KH2PO4; 42 mM Na2HPO4;
85,5 mM NaCl; 1 mM MgSO4) e solução de congelamento (50 mM Tampão Fosfato; 100
mM NaCL; 30% de glicerina). Alíquotas desta mistura foram transferidas para tubos de 1,5
mL e colocadas no freezer -80oC. Quando necessário, as cepas utilizadas foram

37
descongeladas e transferidas para o meio NGM (Nematode Growth Médium) (BRENNER,
1974) semeadas com E. coli OP50 como alimento e incubada a 20°C, o que permitiu o
desenvolvimento normal do ciclo de vida. A manutenção das cepas foi feita transferindo
animais em diferentes estágios para novas placas à medida que a camada de bactéria ficava
escassa.
3.3 SINCRONIZAÇÃO CRONOLÓGICA DO C. ELEGANS
Para os experimentos de resistência ao estresse oxidativo, térmico e longevidade, os
animais tiveram sua idade sincronizada. A sincronização para a todas as cepas foi feita
através da lise alcalina. Este método baseia-se no tratamento de hermafroditas adultos
grávidos com solução de lise (50% de hipoclorito de sódio; 2,5 mM NaOH). Os embriões
resistentes a este tratamento foram coletados e colocados em meio líquido M9 de um dia
para o outro na ausência de alimento. Isto permite que os embriões eclodam, mas
permaneçam estacionários em L1. Os animais L1 foram então transferidos para placas
NGM semeadas com E. coli OP50 por 48h a 20°C, tempo necessário para o C. elegans
chegar a L4.
3.4 EXTRATOS DE AÇAÍ
Neste projeto foram utilizados extratos de açaí. Esses extratos consistem em um pó
liofilizado que foi gentilmente cedido pela empresa Liotécnica Alimentos LTDA (Embu,
SP), sendo o açaí liofilizado (AL-1) código 120.044.032 e o açaí liofilizado (AL-2) código
120.044.022. Este pó liofilizado foi usado para preparar o extrato de açaí AL através da sua
diluição em solução basal para 100 mg/ml como descrito posteriormente no item
3.6.1.Preparação das Placas NGM Contendo Extrato de Açaí

38
3.5 QUANTIFICAÇÃO DE ANTOCIANINAS
O conteúdo de antocianinas dos extratos utilizados foi quantificado através do
método do pH diferencial (COHEN et al., 2006, adaptação de GIUSTI & WROSLTAD,
2001), sendo a concentração de antocianinas monoméricas calculada e representada como
concentração de cianidina-3-glicosídeo, antocianina que, juntamente com a cianidina-3-
rutinosídeo, é encontrada em maior abundância no açaí (DEL POZO-INSFRAN et al.,
2004; SCHAUSS et al., 2006).
Foram preparadas duas soluções tampão: cloreto de potássio – ácido clorídrico pH
1,0 (0,025 M) e acetato de sódio pH 4,5 (0,4 M). O extrato liofilizado passou por uma
prévia etapa de extração em água destilada. As amostras foram diluídas nas soluções
tampão até fator de diluição igual a 20 de forma que a densidade óptica não ultrapassasse o
máximo de 1.200 nm. As medidas de absorbâncias foram efetivadas em máximo de
absorção na região visível 510 nm e em 700 nm, sendo utilizada água destilada como
branco.
A diferença de absorbância foi calculada de acordo com a fórmula 1:
A = (Aλmáz_vis – A700nm)pH 1,0 - (Aλmáz_vis – A700nm)pH 4,5
A concentração de antocianinas monoméricas foi calculada de acordo com a fórmula 2:
AM = A x PM x FD x 102
ε x L
onde AM representa a concentração de antocianinas monoméricas (mg.100g-1
); A
representa a diferença de absorbância calculada pela fórmula 1; PM, o peso molecular da
cianidina-3-glicosídeo (449,2); FD, o fator de diluição da amostra nas soluções tampão; ε a
absortividade molar da cianidina-3-glicosídeo (26900); e L representa o caminho óptico
(1,0cm).

39
3.6 ENSAIOS BIOLÓGICOS
3.6.1 Preparação das Placas NGM Contendo Extrato de Açaí
Para o tratamento com o extrato AL-1 e AL-2, 2 gramas de açaí foram ressuspensos
com 20ml de solução basal, esterilizado por filtração, obtendo-se 5ml de extrato líquido, e
então acrescido de 50µl de solução estéril de 10mM MgSO4. Essas misturas foram
utilizadas para ressuspender o pellet de 1ml de E. coli OP50 com OD 1 que foi então
semeado em placas contendo meio NGM. Para as placas de controle do tratamento, foi
utilizada apenas 1ml de E. coli OP50 com OD 1, ressuspensa em quantidade proporcional
de solução basal e MgSO4. Em caso das placas não serem usadas imediatamente, elas
foram mantidas a 4oC por no máximo 1 semana.
3.6.2 Ensaio de Resistência ao Estresse Oxidativo
Para o tratamento, larvas L1 sincronizadas foram colocadas em placas contendo ou
não açaí e deixadas crescer por 48h a 20°C até o estágio L4. Para testar se o tratamento
com açaí promove um aumento na resistência ao estresse oxidativo, os animais tratados ou
não com açaí foram transferidos para solução de M9 contendo 5 e 7,5mM de tert-butil
hidroperoxido (t-BOOH), um análogo do peróxido de hidrogênio mais estável em meio
líquido. Cada experimento foi realizado em placas de 24 poços, onde 50 animais por grupo
foram colocados em 5 poços com 10 animais por poço. A viabilidade dos animais foi
analisada após 6, 9, 12 e 15 horas usando uma lupa estereoscópica para contar o número de
animais mortos, que são determinados pela ausência de batimento faríngeo e de
movimento quando provocados com uma alça de fio de platina (LITHGOW et al., 1995).
3.6.3 Ensaio de Resistência ao Estresse Térmico
Para testar a resistência ao estresse térmico, vermes com idade sincronizada foram
mantidos em placas de NGM com ou sem extrato de açaí até o quinto dia de vida adulta.
Em seguida, cerca de 60 animais de cada grupo foram transferidos para três novas placas
contendo 20 animais em cada e mantidos a 35°C. A sobrevivência foi verificada em 6, 9 e

40
12 horas, usando lupa estereoscópica. Animais que morreram por dissecação nas laterais
das placas foram desconsiderados nas contagens.
3.6.4 Ensaio de Longevidade
Para os ensaios de longevidade as placas de NGM foram preparadas com 10 mM
FUdR (5-Fluoro-2'-deoxyuridine). FUdR é normalmente fosforilado nas células em
fluorodeoxiuridilato, um inibidor competitivo da enzima timidilato sintetase que converte
deoxiuridilato em timidilato. Desta forma a replicação do DNA é rapidamente
interrompida uma vez que esta é a única via de síntese de timidilato nas células. Após
atingirem a fase adulta, as únicas células a se duplicarem no C. elegans são as
germinativas. Assim, animais adultos mantidos em placas contendo FUdR não produzem
nenhuma progênie e quaisquer embriões produzidos nesta condição não se desenvolvem.
Após a solidificação das placas NGM acrescidas de FUdR, estas foram semeadas com E.
coli OP50 com ou sem extrato de açaí como descrito anteriormente.
Os ensaios de longevidade foram iniciados com 60 animais por grupo com idade
sincronizada no estádio larval L4. Estes são cultivados por todo seu tempo de vida em
placas NGM com FUdR com ou sem extrato de açaí a 25°C. Os 60 animais são divididos
em 3 placas contendo 20 animais em cada. A sobrevivência dos vermes foi verificada
diariamente. Os dados foram submetidos no programa estatístico GraphPad Prisma versão
5 para construção dos gráficos de sobrevivência e cálculo de valores p (Log-Rank)
(TULLET et al., 2008).
3.7 DOSAGEM DOS PARÂMETROS BIOQUÍMICOS
Além da análise da resistência ao estresse também foi verificada a capacidade
antioxidante dos C. elegans tratados e não tratados com extrato de açaí através da medida
dos níveis de proteína carbonilada, grupo sulfidrilas totais e da atividade da enzima
catalase. As medidas de catalase e sulfidrilas totais foram feitas a partir de uma cultura de
5.000 animais e de proteína carbonilada de uma cultura de 10.000 vermes, todos

41
sincronizados em estágio L4. Após a lise e trituração dos animais, o extrato foi submetido
às análises citadas acima. Estes experimentos foram repetidos pelo menos 2 vezes.
3.7.1 Dosagem dos Níveis Totais de Grupos de Sulfidrilas
Foi utilizado o método de Ellman para a execução destas técnicas (SEDLAK &
LINDSAY, 1968). Este método baseia-se na reação de grupos tióis com o ácido 5,5’-ditio-
bis-2-nitro-benzóico (DTNB) formando um ânion amarelo do ácido 5-tio-2-nitrobenzóico
que é medido espectrofotometricamente a 412nm.
Para cada 120µL de amostra, foi adicionado 720µL de metanol, 150µL de Tris-HCl,
pH8,2, 50µL de DTNB (ácido 6,5 ditiobis 2-nitrobezóico). Esta mistura foi centrifugada a
10.000g durante 15 minutos à temperatura ambiente. As absorbâncias das amostras foram
lidas em 412nm, a 25°C. A concentração de sulfidrila foi feita através da regressão linear
usando cisteína como padrão. Todas as concentrações foram obtidas em mol/proteína.
3.7.2 Dosagem da Atividade da Catalase
Determinação da atividade da enzima catalase, foi realizada baseada na sua
capacidade de converter o peróxido de hidrogênio (H2O2) em água e oxigênio molecular,
conforme descrito por Aebi, (1984). 5000 animais L4 foram homogeneizados com 0,5mL
de 100mM tampão fosfato pH 7,2 e em seguida centrifugado por 10 minutos à 4ºC. O
sobrenadante foi retirado e usado como amostra biológica. Em um tubo de polipropileno
colocaram-se 50L de 100mM tampão fosfato pH 7,2 e 40L de água destilada. Em
seguida foi adicionado 100L da amostra e 810L de 10mM H2O2. As absorbâncias das
amostras foram lidas a 240nm a cada minuto, durante cinco minutos. 1U de catalase é
equivalente a hidrólise de 1 mol de H2O2 (ε = 39,4 L.mol-1
.cm-1
) por minuto (AEBI,
1984). Usualmente a atividade dessa enzima é representada em Unidade por mL de
amostra. Calculamos a atividade da catalase segundo a lei de Lambert Beer. A absorbância
utilizada nessa expressão é o delta obtido das cinco absorbâncias lidas (absorbância final –
absorbância inicial / 4).

42
3.7.3 Dosagem dos Níveis de Proteína Carbonilada
A oxidação de proteína por ERO leva à formação de derivados carbonilados. Estes
podem ser mensurados por métodos sensíveis, particularmente aqueles que utilizam o 2,4-
dinitrofenilhidrazina (DNP). O DNP reage com grupos carbonílicos gerando a hidrazona
correspondente, a qual pode ser analisada espectrofotomicamente. A determinação da
concentração sérica de proteína carbonilada foi realizada conforme descrito por Levine et
al. (1994). 10000 animais sincronizados em L4 foram homogeneizados com 0,5mL de 50
mM tampão fosfato pH 6,7 contendo 1mM EDTA. Em seguida centrifugado a 10000g, por
10 minutos à 4ºC. O sobrenadante foi retirado e usado como amostra biológica. Para cada
amostra utilizou-se dois tubos de polipropileno, um foi denominado de Amostra (A) e
outro de Controle (C). Transferiu-se 400L de homogeneizado para todos os tudo (amostra
(A) / controle (C)). Em seguida foi adicionado aos tubos 400L de TCA 10% e misturado
no vórtex, logo após, foram centrifugados (tubo A e C) à 5000g por 10minutos a 4°C. O
próximo passo foi adicionar ao tubo A 500L de DNPH (2,4-dinitrofenilhidrazina) e no
tubo C 500L de 2,5M HCL. Ambos foram mantidos no escuro à temperatura ambiente
por um período de 30 minutos, e a cada 15 minutos eram misturados no vórtex. Em
seguida foram adicionados 500L de 10% ácido tricloroácetico (TCA) em cada tubo,
misturado no vórtex e centrifugados à 5000g por 10 minutos a 4°C. Depois de
centrifugados, o sobrenadante dos tubos foi descartado e 1mL de mistura de etanol com
acetato de etila (1:1) foi adicionado aos tubos e misturados no vórtex. Uma nova
centrifugação foi realizada. Em seguida o sobrenadante dos tubos A e C foram descartados
e à estes foram adicionados mais 1mL da mistura etanol e acetato de etila, foram
misturados no vórtex e novamente centrifugados. No final das centrifugações, o
sobrenadante dos tubos A e C foram novamente descartados e adicionado em ambos 1mL
de 6% SDS, misturados no vórtex e centrifugados à 10000g por 10 minutos a 4°C.
Finalmente o sobrenadante dos tubos foram retirados e transferidos para cubeta, onde
foram lidos no espectrofotômetro a 370nm. A concentração de proteína carbonilada foi
determinada utilizando a equação: ..bCA , onde A é a subtração da absorbância do tubo
A (amostra) pela absorbância do tubo C (controle), C é a concentração, b é o caminho
óptico e ε é o coeficiente de extinção molar. O conteúdo de proteína carbonilada foi
calculado usando o coeficiente de extinção molar de 22 000 M-1 cm-1 e expresso por nmol

43
de proteína carbonilada formada por mg de proteína. Para se obter a concentração de
proteína carbonilada em relação à concentração de proteínas totais este parâmetro foi
determinado pelo método de Lowry (descrito a seguir).
3.7.4 Dosagem de Proteínas Totais
Este método foi descrito a primeira vez por Lowry et al. (1951). É baseado nas
ligações das proteínas, que em meio alcalino, com os íons cobre (Cu2+
) formam uma cor
azul que é dependente em partes, do índice de tirosina e triptofano da amostra, já que os
íons cobre catalisam a oxidação de aminoácidos aromáticos.
Em um tudo de polipropileno, foram pipetados 100µL de amostra ou o padrão de
albumina. O branco, usado para zerar o espectrofotômetro, foi feito apenas com 100µL de
água destilada. Posteriormente foi adicionado 1mL do reagente C em todos os tubos. A
mistura foi levada ao vórtex e mantida a temperatura ambiente por 15 minutos. Em
seguida, foi adicionado em cada tubo, 100µL do reagente D. O volume foi misturado e
incubado a temperatura ambiente, no escuro, por 30 minutos. A leitura foi feita em
espectrofotômetro a 660nm.
Para o cálculo de proteínas totais foi feito um gráfico expressando a concentração do
padrão de albumina (Eixo Y) X absorbância do padrão de albumina (Eixo X). Após
regressão linear, foi determinada a equação da reta com a seguinte característica:
Concentração = a X Absorbância + b. Esta equação foi utilizada para determinar a
concentração de proteínas totais nos homogenatos de C. elegans. Todas as concentrações
foram obtidas em mg/mL.
Os reagentes utilizados e forma de preparo estão descritos a seguir. Reagente A: Foi
dissolvido 0,25g de sulfato de cobre e 0,5 de citrato de sódio em 100mL de água destilada.
Reagente B: Foi dissolvido 5g dr carbonato de sódio e 1g de hidróxido de sódio em 250mL
de água destilada. Reagente C: Foi adicionado 1mL do reagente A em 50mL do reagente
B. Reagente D: Foi dissolvido 1mL de Folin-Ciocateau em 1mL de água destilada.

44
3.8 LOCALIZAÇÃO SUBCELULAR DE DAF-16
Para investigar se o extrato de açaí ativa a localização nuclear dos fatores de
transcrição DAF-16, foram utilizadas as cepas transgênicas N2 zIs356[DAF-16::GFP] e
TJ356, contendo os genes repórteres daf-16::gfp (AN et al., 2003; HENDERSON &
JOHNSON, 2001). A seleção dos animais transgênicos daf-16::gfp foi feita através da
presença marcador rol-6, um gene defectivo do colágeno que provoca nos animais
portadores o fenótipo de se arrastem em círculos ao invés de se arrastarem de maneira
sinuosa. A cepa N2 zIs356[DAF-16::GFP] é uma linhagem também integrada de um
plasmídeo contendo uma fusão traducional da sequência genômica do promotor e do gene
DAF-16 com o gene GFP (HENDERSON & JOHNSON, 2001). Esta linhagem
transgênica codifica a isoforma DAF-16a2 que é expressa na maioria das células do corpo
do C. elegans, com exceção das células da faringe (HENDERSON & JOHNSON, 2001).
Após o tratamento com extrato açaí, os animais transgênicos nos estágio de L2 a L4
foram colocados em lâminas contendo uma fina camada de 2% de agarose, imobilizados
com 2% de azida sódica e cobertos com uma lamínula. A localização subcelular dos genes
repórteres foi analisada através de um microscópio de fluorescência Leika TM5000,
disponível no Núcleo de Pesquisa Básica (NUPEB) da UFOP, usando aumento ocular de
10 vezes e filtro para excitação de 365nm. A localização foi classificada como citossólica,
intermediária e nuclear de acordo com a localização predominante do GFP. Baixa indução
corresponde à ausência de localização nuclear. Indução média corresponde à concentração
de localização nuclear na região anterior e/ou posterior do intestino. Indução alta
corresponde à localização nuclear através de todo o intestino.
FIGURA 3 Esquema dos genes repórter para DAF-16
Linha sólida representa DNA genômico, caixa de cor azul representa região não
traduzida, caixa vermelha representa região codificantes e caixa verde representa região
codificante para proteína fluoresente verde (GFP). Fonte Esquema: Adaptado de
(HENDERSON & JOHNSON, 2001).

45
3.9 ANÁLISE ESTATÍSTICA
Para os ensaios de resistência ao estresse oxidativo e térmico e para as quantificações
de marcadores bioquímicos, as análises estatísticas foram realizadas através do teste t de
Student’s sendo que p ≤ 0,05 foi considerado estatisticamente significante. As curvas de
sobrevivência Kaplan-Meier foram criadas usando o software GraphPad Prisma versão 5 e
o valor de p foi calculado através do Log-Rank test.

46
CAPÍTULO 4
RESULTADOS

47
4.1 QUANTIFICAÇÃO DE ANTOCIANINAS
O açaí é um fruto muito utilizado na indústria alimentícia de diferentes maneiras,
seja na forma de polpa congelada ou até mesmo o produto liofilizado. A primeira pergunta
feita neste trabalho foi de verificar o conteúdo de antocianinas monoméricas presentes
nestes tipos das amostras de açaí (Tabela 1). A quantificação das antocianinas foi realizada
em dois extratos liofilizados da Liotécnica Alimentos, AL-1 código120.044.032 e AL-2
código 120.044.022
A dosagem das antocianinas monoméricas na amostra AL-1 foi feita duas vezes com
um ano de diferença entre elas (05/03/2010 e 10/04/2011). Os resultados obtidos foram
muito próximos (15,8 e 14,5 mg/100g) indicando que a quantidade de antocianinas não
foram perdidas com tempo.
TABELA 1 - Conteúdo de antocianinas monoméricas, representado como conteúdo
de cianidina-3-glicosídeomg/100 g)
AMOSTRAS LIOFILIZADO
AL-1 (05/03/2010) 15,8
AL-1 (10/04/2011) 14,6
AL-2 (06/05/2011) 34,7

48
4.2 ANÁLISE DA RESISTÊNCIA AO ESTRESSE OXIDATIVO DOS ANIMAIS
TRATADOS COM DIFERENTES EXTRATOS DE AÇAÍ
Tendo em vista que os extratos AL-1 e AL-2 apresentaram antocianinas
monoméricas, foi investigado em seguida se animais tipo selvagem tratados com os
extratos apresentariam um aumento da resistência ao estresse oxidativo.
Para isso estes foram mantidos por 48 horas (de L1 a L4) em 100 ou 200mg/ml de
AL-1 e AL-2 e posteriormente foi verificada a sobrevivência deles quando incubados com
5 e 7,5 mM de t-BOOH. Os animais tratados com 200mg/ml não se desenvolveram até L4
e ficaram estacionados no estágio L1, indicando que esta concentração de extrato de açaí é
tóxica. O mesmo não foi observado com os animais tratados com 100 mg/ml sendo que
eles chegaram ao estágio L4 em 48h como esperado para animais tipo selvagem mantidos a
20oC. Portanto, todos os experimentos feitos a partir deste ponto foram feitos usando
apenas a concentração de 100mg/ml de extrato.
O Gráfico 1 mostra a fração de sobreviventes em diferentes tempos dos animais
tratados com 100 mg/ml de AL-1 ou AL-2. Foi observado que a sobrevivência dos animais
tratados com AL-1 e AL-2 foram significativamente maior (p ≤ 0,05) após 9 e 12 horas em
5 mM de t-BOOH (Gráfico 1A). Já a fração de animais tratados com AL-1 e AL-2 em 7
mM não apresentaram aumento da resistência ao estresse (Gráfico 1B)

49
A
B.
GRÁFICO 1 – Efeito do tratamento com 100 mg/ml de AL na sobrevivência do C.
elegans em condições de estresse oxidativo. A sobrevivência dos animais em (A) 5mM e
(B) 7,5mM de t-BOOH foram verificadas em tempos determinados. O gráfico acima
representa a média de três experimentos. A barra de erro representa o SEM. O símbolo *
indica resultado significante p ≤ 0,05 através do teste de Student sobreviveram em 7,5 mM
não foi significativamente maior em relação aos animais não tratados (Gráfico 1B).

50
4.3 ANÁLISE DA RESISTÊNCIA AO ESTRESSE TÉRMICO DOS ANIMAIS
TRATADOS COM AL-1
As propriedades antioxidantes do AL também foram avaliadas através de um ensaio
de resistência ao estresse térmico. Para avaliar a sobrevivência em condições de estresse
térmico, os animais foram tratados com 100 mg/mL de AL-1 desde o estágio L1 até o
quinto dia de vida adulta, ou seja por 168 horas. No 5º dia de vida adulta, os animais já
terminaram a maior parte da ovoposição e com isto eles eram menos ágeis para escapar da
placa nas condições de estresse. Após este período os animais foram submetidos ao
estresse térmico por elevação de sua temperatura de cultivo de 20°C para 35°C e
asobrevivências dos animais foi verificada 6, 9 e 12 horas após o inicio do estresse. O
Gráfico 2 mostra que o tratamento com 100 mg/mL de AL-1 não foi capaz de aumentar a
resistência ao estresse térmico dos animais tipo selvagem em nenhum dos tempos
analisados.
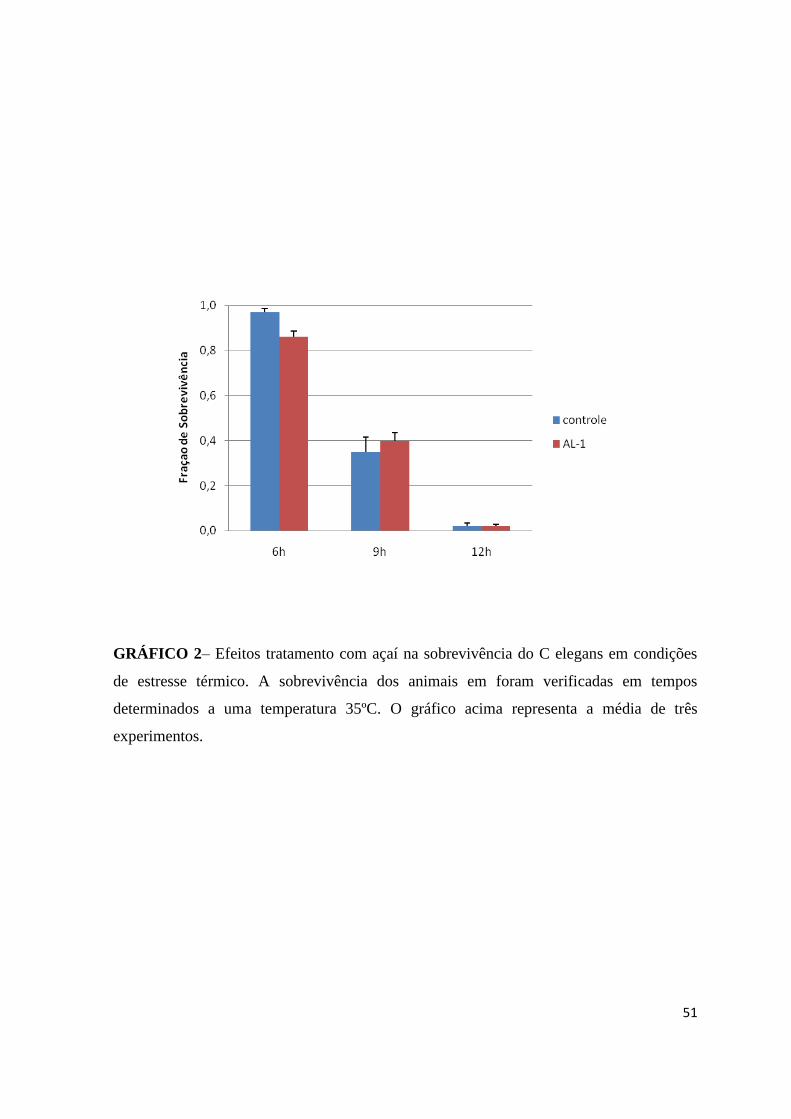
51
GRÁFICO 2– Efeitos tratamento com açaí na sobrevivência do C elegans em condições
de estresse térmico. A sobrevivência dos animais em foram verificadas em tempos
determinados a uma temperatura 35ºC. O gráfico acima representa a média de três
experimentos.

52
4.4 ANÁLISE DA SOBREVIVÊNCIA EM CONDIÇÕES NORMAIS DOS
ANIMAIS TRATADOS COM AL-1
O terceiro ensaio biológico usado para avaliar a capacidade antioxidante do AL in
vivo foi a análise da longevidade dos animais em condições normais. Neste experimento,
os animais tipo selvagem foram tratados desde L1 por todo o seu tempo de vida com 100
mg/mL de AL-1 a 25oC e o número de animais vivos foram contados diariamente.
O Gráfico 3 e Tabela 2 mostram os resultados obtidos de um experimento de
longevidade realizado a 25oC. Pode-se observar tanto no gráfico como na tabela, que o
tratamento com AL-1 não aumentou nem o tempo de vida médio e o nem máximo dos
animais tipo selvagem. A 25oC, o tempo de vida médio foi de 14 dias para os animais do
grupo controle e 15 dias para o grupo tratado sendo esta diferença não significante. O
tempo de vida máximo foi de 20 dias a 25oC tanto para o grupo controle quanto para o
grupo tratado (Tabela 2).
4.5 ANÁLISE DE MARCADORES BIOQUÍMICOS E ENZIMÁTICOS NOS
ANIMAIS TRATADOS COM AL-1
Uma vez que o tratamento com AL foi capaz de aumentar a resistência ao estresse
oxidativo no C.elegans, o próximo passo foi verificar se tratamento também estava
associado a uma melhora do status antioxidante in vivo no C. elegans. Para isso foi dosada
a grupos sulfidrilas totais atividade da catalase como marcadores antioxidante e os níveis
de proteína carbonilada como marcador de estresse.
Os resultados apresentados no Gráfico 4 mostram que não existe uma diferença
significativa entre animais controle e tratados com AL-1 em nenhum dos parâmetros
dosados. Devido à variação entre experimentos, os resultados foram apresentados na forma
de níveis relativos onde em cada experimento, o valor obtido para o grupo controle foi
considerado como 1 e o valor obtido para o grupo tratamento foi convertido por razão.
Após esta conversão foi obtida a média dos níveis relativos das triplicatas experimentais
(Tabela 3).

53
0 5 10 15 20 250.0
0.2
0.4
0.6
0.8
1.0controle
AL-1
Dias
Fra
çao
de S
ob
reviv
ên
cia
GRÁFICO 3– Efeitos do tratamento com 100 mg/ml de AL-1 na sobrevivência do C.
elegans. A sobrevivência dos animais foi verificada diariamente. As curvas de
sobrevivência Kaplan-Meier foram criadas usando o GraphPad Prism versão 5. O gráfico
acima representa a média de dois experimentos.
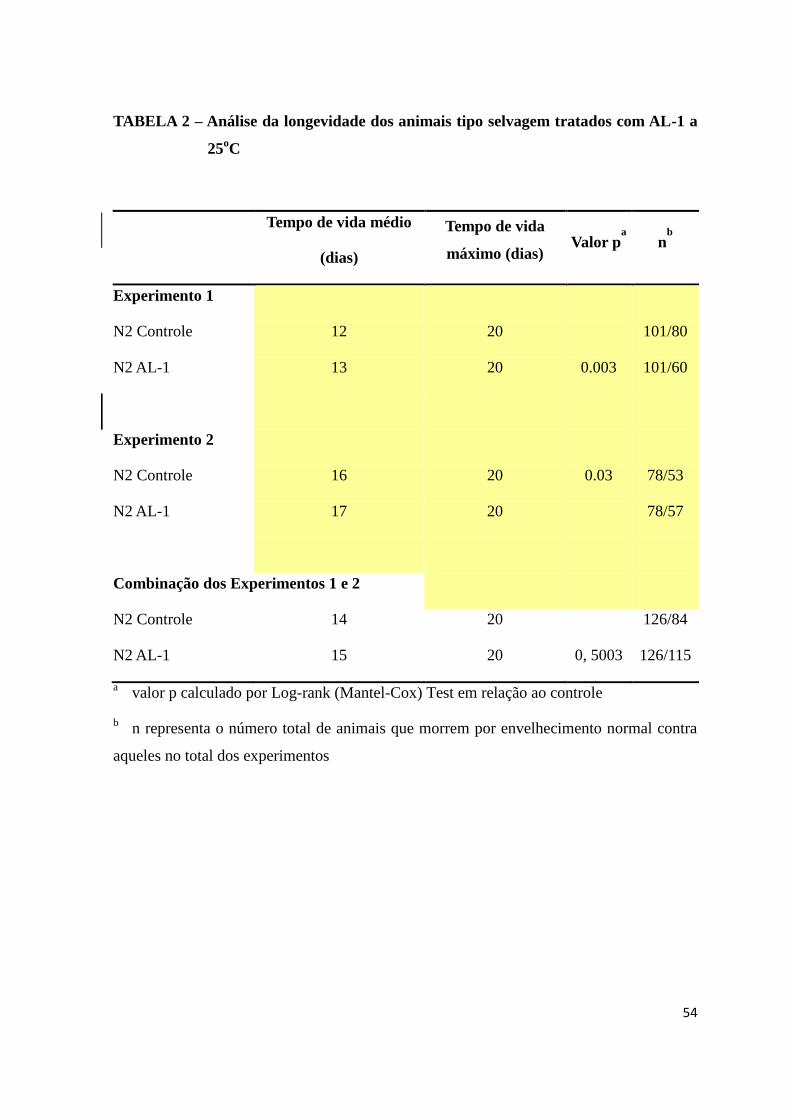
54
TABELA 2 – Análise da longevidade dos animais tipo selvagem tratados com AL-1 a
25oC
Tempo de vida médio
(dias)
Tempo de vida
máximo (dias) Valor p
a
nb
Experimento 1
N2 Controle 12 20 101/80
N2 AL-1 13 20 0.003 101/60
Experimento 2
N2 Controle 16 20 0.03 78/53
N2 AL-1 17 20 78/57
Combinação dos Experimentos 1 e 2
N2 Controle 14 20 126/84
N2 AL-1 15 20 0, 5003 126/115
a valor p calculado por Log-rank (Mantel-Cox) Test em relação ao controle
b n representa o número total de animais que morrem por envelhecimento normal contra
aqueles no total dos experimentos
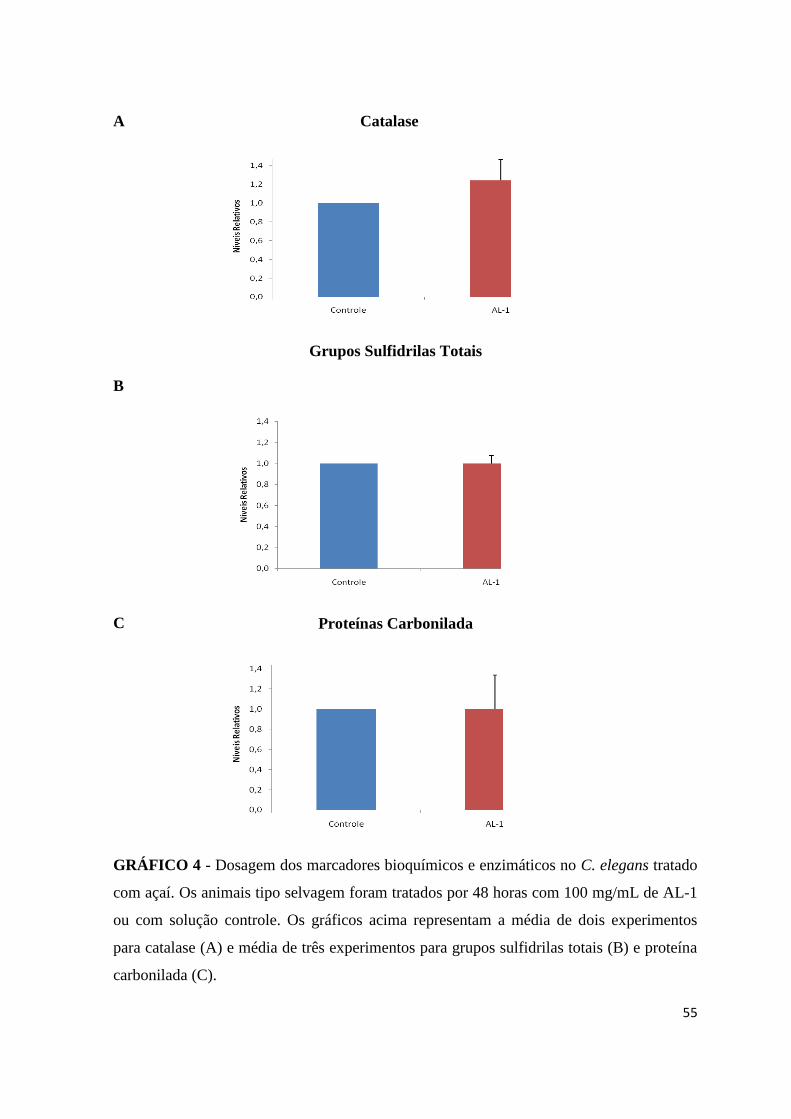
55
A Catalase
B
C
GRÁFICO 4 - Dosagem dos marcadores bioquímicos e enzimáticos no C. elegans tratado
com açaí. Os animais tipo selvagem foram tratados por 48 horas com 100 mg/mL de AL-1
ou com solução controle. Os gráficos acima representam a média de dois experimentos
para catalase (A) e média de três experimentos para grupos sulfidrilas totais (B) e proteína
carbonilada (C).
Grupos Sulfidrilas Totais
Proteínas Carbonilada
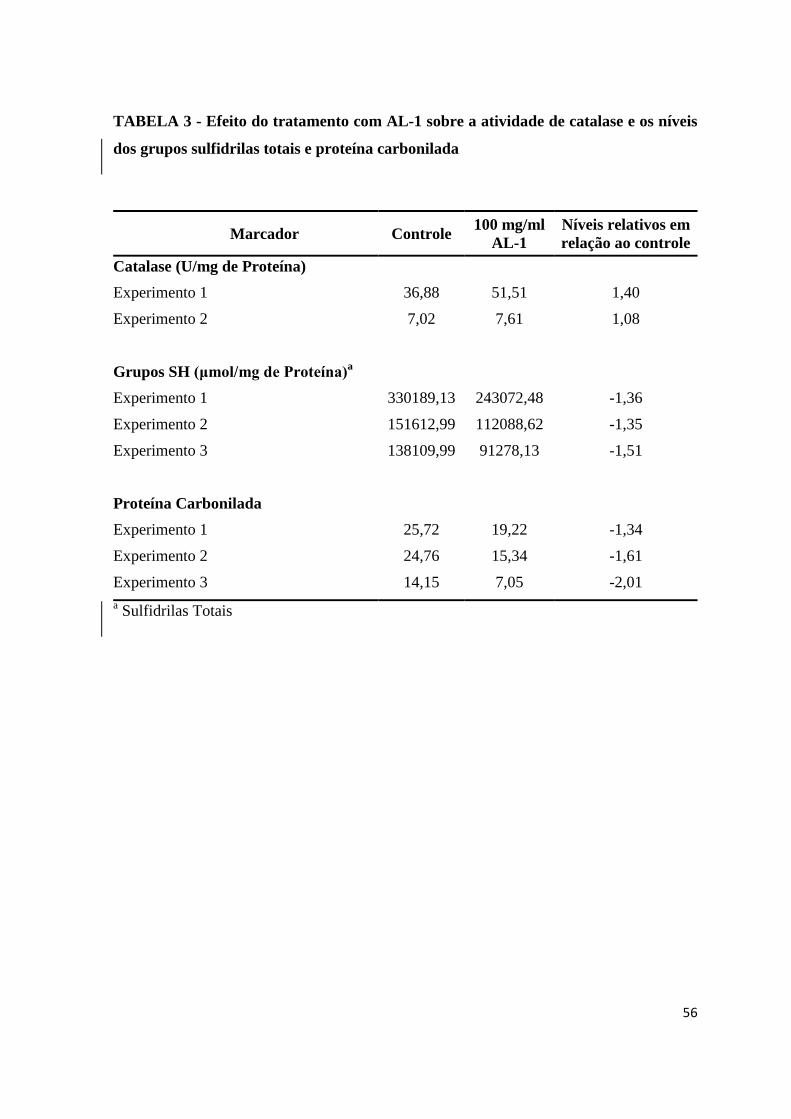
56
TABELA 3 - Efeito do tratamento com AL-1 sobre a atividade de catalase e os níveis
dos grupos sulfidrilas totais e proteína carbonilada
Marcador Controle 100 mg/ml
AL-1
Níveis relativos em
relação ao controle
Catalase (U/mg de Proteína)
Experimento 1 36,88 51,51 1,40
Experimento 2 7,02 7,61 1,08
Grupos SH (μmol/mg de Proteína)a
Experimento 1 330189,13 243072,48 -1,36
Experimento 2 151612,99 112088,62 -1,35
Experimento 3 138109,99 91278,13 -1,51
Proteína Carbonilada
Experimento 1 25,72 19,22 -1,34
Experimento 2 24,76 15,34 -1,61
Experimento 3 14,15 7,05 -2,01
a Sulfidrilas Totais

57
4.6 ANÁLISE DO ENVOLVIMENTO DA VIA DE SINALIZAÇÃO DE SKN-
1/P38MAPK NO AUMENTO DA RESISTÊNCIA AO ESTRESSE
OXIDATIVO PROMOVIDO PELO AL-1
As vias de sinalização celulares para genes que afetam a resistência ao estresse
oxidativo são conservadas entre C. elegans e humanos. O aumento da resistência ao
estresse oxidativo no C. elegans promovido pelo tratamento com 100 mg/mL de AL-1
oferece uma oportunidade de usar o poderoso sistema genético do C. elegans para testar o
mecanismo de ação do açaí.
4.6.1 Análise da Resistência ao Estresse Oxidativo nos Animais skn-1(RNAi)
Assim, o próximo passo foi verificar se o aumento da resistência ao estresse
oxidativo promovido pelo tratamento com AL-1 depende da ativação do fator de
transcrição SKN-1. Para tal, os ensaios de resistência ao estresse oxidativo foram repetidos
usando mutantes com expressão reduzida para SKN-1 (knockdown) obtidos através do
silenciamento gênico (RNAi). Após 48 horas de tratamento com AL-1 os animais foram
transferidos para solução com 5 e 7,5 mM de t-BOOH e a sobrevivência foi analisada em
horários pré estabelecidos. Os animais controle(RNAi) tratados com AL-1 não
apresentaram um aumento da resistência ao estresse oxidativo estatisticamente
significativo em relação aos animais controle(RNAi) não tratados (Gráfico 6). Portanto não
foi possível concluir neste caso se SKN-1 está envolvido no aumento da resistência ao
estresse oxidativo promovido pelo tratamento com AL-1.

58
A
B
GRÁFICO 5 - Efeito do tratamento com 100 mg/ml de AL-1 na sobrevivência de animais
skn-1(RNAi) em condições de estresse oxidativo. A sobrevivência dos animais em (A)
5mM e (B) 7,5mM de t-BOOH foram verificadas em tempos determinados. O gráfico
acima representa a média de três experimentos. A barra de erro representa o SEM.

59
4.6.2 Análise da Resistência ao Estresse Oxidativo em mutantes sek-1
Em condições de estresse oxidativo, a ativação e localização nuclear de SKN-1
dependem da via p38 MAPK (INOUE et al., 2005). SEK-1, a MAPK quinase que fosforila
PMK-1, o ortólogo da p38 de mamíferos, é necessário para que SKN-1 acumule-se no
núcleo das células intestinais. Para verificar se o aumento da resistência ao estresse
oxidativo promovido pelo tratamento com AL-1 também depende da via p38 MAPK, os
ensaios foram repetidos usando mutantes de deleção sek-1(km4).
Observa-se no Gráfico 6 que mutantes sek-1 tratados com AL-1 não apresentam
uma maior resistência ao t-BOOH como os animais tipo selvagem. Ou seja, na ausência de
SEK-1 os animais perderam o aumento da resistência ao estresse oxidativo promovida pelo
tratamento com açaí. Estes resultados apresentados acima sugerem que a via de sinalização
p38 MAPK está envolvida no aumento da resistência ao estresse oxidativo promovido por
AL-1.
4.8 ANÁLISE DO ENVOLVIMENTO DA VIA DE SINALIZAÇÃO DE DAF-
16/FOXO NO AUMENTO DA RESISTÊNCIA AO ESTRESSE OXIDATIVO
PROMOVIDO PELO AL-1
4.7.1 Análise da resistência ao estresse oxidativo em mutantes daf-16
Para verificar se o aumento da resistência ao estresse oxidativo promovido pelo
tratamento com AL-1 também depende da ativação do fator de transcrição DAF-16 os
ensaios de resistência ao estresse oxidativo foram repetidos usando mutantes de deleção
daf-16(mgDf47). Após 48 horas de tratamento com 100 mg/mL de AL-1 os animais foram
transferidos para solução com 5 e 7,5mM de t-BOOH e a sobrevivência foi analisada após
6, 9, 12 e 15 horas.
O resultado apresentado no Gráfico 7 mostra que os mutantes daf-16 tratados com
AL-1 não apresentaram um aumento da resistência ao estresse oxidativo como os animais
tratados tipo selvagens tiveram. Este resultado sugere que o aumento da resistência ao
estresse oxidativo promovido por AL-1 também depende de DAF-16.

60
A
B
GRÁFICO 6 - Efeito do tratamento com 100 mg/ml de AL-1 na sobrevivência de
mutantes sek-1 em condições de estresse oxidativo. A sobrevivência dos animais em (A)
5mM e (B) 7,5mM de t-BOOH foram verificadas em tempos determinados. O gráfico
acima representa a média de três experimentos. A barra de erro representa o SEM. O
símbolo * indica resultado significante p ≤ 0,05 através do teste t de Student´s.

61
A
B
GRÁFICO 7 - Efeito do tratamento com 100 mg/ml de AL-1 na sobrevivência de
mutantes daf-16 em condições de estresse oxidativo. A sobrevivência dos animais em (A)
5mM e (B) 7,5mM de t-BOOH foram verificadas em tempos determinados. O gráfico
acima representa a média de três experimentos. A barra de erro representa o SEM. O
símbolo * indica resultado significante p ≤ 0,05 através do teste t de Student´s.

62
4.7.2 Análise da Localização DAF-16:: GFP
O envolvimento de DAF-16 no aumento da resistência ao estresse oxidativo
promovido por AL-1 também foi avaliado através da ativação da sua localização nuclear.
A localização subcelular de DAF-16 foi realizada utilizando linhagem transgênica
contendo uma fusão traducional do gene DAF-16::GFP (HENDERSON; JOHNSON,
2001). Em condições normais, DAF-16::GFP se encontra no citoplasma de quase todas as
células do corpo do C. elegans. Em resposta aos vários agente estressores, DAF-16::GFP
acumula-se no núcleo destas células (LANDIS;MURPHY,2010.
O tratamento dos animais transgênicos com AL-1 por 48 horas não foi capaz de
induzir a localização nuclear de DAF-16::GFP (Figura 4).
Os resultados relacionados à sobrevivência de mutantes em condições de estresse
acima sugerem que AL-1 aumenta a resistência ao estresse oxidativo de uma maneira
SKN/p38 MAPK e DAF-16 dependente. No entanto as análises de genes repórteres
realizadas não evidenciaram essa localização.

63
FIGURA 4 - Localização subcelular de DAF-16::GFP em animais tipo selvagem
tratados com AL-1. GFP, proteína fluorescente verde; DIC, contraste de interferência
diferencial.

64
CAPÍTULO 5
DISCUSSÃO DOS
RESULTADOS

65
5.1 PROPRIEDADES ANTIOXIDANTES DO EXTRATO DE AÇAÍ
A flora brasileira é rica em espécies com princípios ativos de importância
terapêutica, com potencial não apenas para a utilização na medicina natural como
também na agricultura natural no controle integrado de pragas e doenças de plantas
cultivadas. Muitas espécies de plantas medicinais contêm fenóis, quinonas, saponinas,
flavanóides e terpenóides em quantidades apreciáveis de repelir insetos, também exercer
atividades antioxidantes, antiinflamatórias, antifungicidas e etc (SILVA & VIZZOTO,
1996). Desta forma, a flora brasileira é alvo de grande interesse pelas indústrias
farmacêuticas, na tentativa de descoberta de novos medicamentos para o arsenal
terapêutico ou agropecuário. Mas para identificar um composto natural com
propriedades biológicas de interesse é necessário investigar as suas propriedades tanto
in vitro assim como avaliar as suas funções em sistemas biológicos in vivo.
O estresse oxidativo contribui para a etiologia de várias doenças degenerativas
como isquemia e processos de envelhecimento, então foi sugerido que compostos que
eliminem os EROS, seja por ação direta ou indireta, podem prevenir ou ao menos
retardar estes processos. A identificação de uma classe de compostos químicos que
retardam o envelhecimento em muitas espécies é altamente desejável tanto como
instrumento para futuras pesquisas, mas também como potencial terapêutico para
retardar as doenças relacionadas à idade.
O açaí (Euterpe olerace Mart.) é uma fruta original da Amazônia rica em
antioxidantes polifenólicos, flavanóides e antocianinas principalmente cianidina 3-
glicosídio e cianidina 3-rutinosídio (DEL POZO-INSFRAN et al., 2004; SCHAUSS et
al., 2006). Vários trabalhos têm demonstrado que açaí tem efeitos antioxidantes tanto in
vitro (SPADA et al., 2008,) quanto in vivo (JENSEN et al., 2008; SPADA et al., 2009,
de MOURA et al., 2011) e que esta capacidade antioxidante está associada as suas
frações de antocianinas e dos compostos fenólicos totais (SANTOS et al., 2008). Apesar
dos vários estudos in vivo, apenas um deles demonstrou suas propriedades antioxidantes
e pró-longevidade de maneira sistêmica usando a Drosophila melanogaster. Nesse
contexto, o principal objetivo deste trabalho foi estabelecer o C. elegans como modelo
para avaliar os efeitos antioxidantes globais do extrato de açaí no C. elegans.

66
5.1.1 Quantificação de Antocianinas nos Extratos de Açaí
Devido à crescente comercialização e ao consumo do extrato de açaí, e à sua
constante indicação como fonte de antioxidantes, o primeiro objetivo deste trabalho foi
o de quantificar as antocianinas presentes no extrato de açaí de origem industrial por
processo de liofilização.
Neste trabalho, o método de pH diferencial foi utilizado para quantificar as
antocianinas em dois extratos liofilizados da Liotécnica Alimentos, AL-1 codigo
120.044.32 e AL-2 codigo 120.044.022. Como demonstrado na Tabela 1, a
concentração de antocianinas monoméricas foram próximos para AL-1 medida com um
ano de diferença de 15,8 mg/100 g em 05/03/2010 e 14,6 mg/100 g em 10/04/2011,
indicando que as antocianinas neste processamento são estáveis. Apenas a quantidade
de antocianinas monoméricas da amostra AL-2 foi superior (34,7 mg/100 mg) em
relação a AL-1. Os resultados obtidos neste presente trabalho estão de acordo com
dados já publicados na literatura. Teixeira; Estringheta; Oliveira (2008) e Kuskoski et
al., (2006) encontraram respectivamente 19,62mg/100g e 22,8mg/100g de antocianinas
de várias polpas congeladas. Por outro lado, valores bem mais elevados de 92,8 mg/100
g foi observado por Albarici et al.(2007).
Já foi documentada que existe uma grande variação nas concentrações de
antocianinas em diferentes extratos de açaí sendo encontrado valores desde 13,75 a
228,77 mg/100 g (COHEN et al., 2006). A justificativa em parte para esta grande
variação se deve ao fato das antocianinas serem pigmentos muito instáveis que podem
ser degradados durante o processamento e a estocagem. Outros fatores também podem
influenciar na composição dos frutos e de sua polpa, como época da safra, tipo de solo,
influência das marés, qualidade da água de inundação dos solos, localização espacial.
Em síntese, acredita-se que os fatores acima mencionados podem exercer influências
sobre a composição dos frutos e da polpa e, portanto sobre a qualidade do produto final
comercializado. Tais fatores devem ser responsáveis pela distinção de populações de
diferentes procedências, mas não se sabe a real contribuição de cada um (FARIAS
NETO et al., 2005). Apesar disto, nossos resultados indicam que a antocianina presente
no extrato de açaí liofilizado AL-1 é relativamente estável. Dessa forma seria

67
interessante não só dosar antocianinas encontradas com a capacidade antioxidante in
vitro utilizando ensaio de DPPH e TROLOX.
5.1.2 Ensaios Biológicos
O C. elegans é um organismo modelo já bem estabelecido para estudos de
longevidade assim como para a avaliação dos efeitos biológicos tanto de antioxidantes
fitoquímicos (PIETSCH et al, 2009; MENSACK et al., 2010; MATORELL et al., 2011)
quanto sintéticos (HOSEGAWA et al., 2010).
Como até o momento, nenhum estudo utilizando açaí foi realizado no C. elegans,
a primeira etapa do trabalho foi estabelecer as condições ideais para o tratamento do C.
elegans com este extrato. Inicialmente foram testadas a adição de 100 mg/ml e 200
mg/ml de AL-1 ao meio de cultura E. coli que serve como alimento para o nematodo.
Foi observado que o desenvolvimento dos animais tratados com 100 mg/ml de AL-1 de
L1 até L4 ocorreu normalmente. Por outro lado, o tratamento com 200 mg/ml foi tóxico
pois os animais não se desenvolveram e ficaram estacionados no estágio L1. Portanto,
todos os experimentos de resistência ao estresse e longevidade realizados para avaliar
capacidade antioxidante do AL no C. elegans foram feitos usando apenas a
concentração de 100mg/ml de extrato.
Spada et al. (2008) observaram que o tratamento com 5, 10 e 15% de polpa de
açaí em Saccharomyces cerevisiae tem um efeito mutagênico devido ao aumento de
linhagens mutantes revertentes tratadas em relação ao controle. Apesar disto, os autores
sugerem que o risco de efeitos mutagênicos nos seres humanos é baixo, uma vez que as
concentrações testadas são muito mais altas do que as usadas no consumo humano
(SPADA et al., 2008). Apoiando este argumento, Ribeiro et al. (2010), utilizando a
contagem de células micronucleadas e ensaio do DNA cometa, não observaram
respectivamente nenhum efeito citotóxico ou genotóxico em camundongos tratados com
3,33, 10 ou 16,67 g/Kg peso corporal, doses estas que se aproximam ao consumo de
açaí pelo homem.
Para criar as condições de estressse oxidativo, foi utilizado o tert-butil
hidroperóxido, um análogo do peróxido de hidrogênio mais estável em meio liquido que
ataca proteínas e lipídeos e remove GSH (MATHEWS et al., 1994). Normalmente o

68
tratamento do C. elegans com 5 e 7,5 mM de t-BOOH causa uma redução da
sobrevivência dos animais tipo selvagem para cerca de 42% e 21% após 6 a 9 horas
respectivamente (Gráfico 1 e 2). Foi observado que o tratamento por 48h com 100
mg/ml dos extratos de açaí liofilizado (AL) aumentou a sobrevivência do C. elegans em
t-BOOH. O tratamento do C. elegan com 100 mg/ml do AL aumentou para quase 75%
(p<0,05) a fração de sobreviventes após 9 horas em 5 mM t-BOOH (Gráfico 1A) mas
não em 7,5 mM (Gráfico 1B).
Uma variedade de ensaios in vitro já mostrou que a polpa de açaí tem uma alta
capacidade antioxidante, especialmente contra radicais superóxido e peroxil
(SCHAUSS et al., 2006). Dentre os ensaios in vitro utilizando culturas de células,
Jensen et al (2008) observaram que células polimornucleares incubadas com H2O2 e
tratadas com bebida MonaVie, cujo principal componente é o açaí, tem uma menor
produção de ERO. Dentre os estudos in vivo, já foram demonstrados que a
suplementação da polpa de açaí na dieta melhora o perfil lipídico e os marcadores de
estresse oxidativo de ratos hipercolesterolêmicos (SOUZA et al., 2009), aumenta a
longevidade de Drosophila tratado com dieta hiperlipídica (SUN et al, 2010), modula a
produção de ERO em neutrófilos e aumenta a produção de GSH no fígado de ratos
(GUERRA et al., 2011). Além disso, ensaios clínicos em indivíduos saudáveis
mostraram que o consumo de açaí resulta em um aumento significativo da capacidade
antioxidante do plasma (JENSEN et al., 2008). Apesar de todos estes dados, o resultado
apresentado nesta dissertação é o primeiro que mostra de uma maneira sistêmica que o
tratamento com açaí aumenta a sobrevivência de um animal em condições de estresse
oxidativo.
Este efeito antioxidante do tratamento de compostos ricos em polifenóis no C.
elegans também já foi descrito por Pun et al. (2009). Neste trabalho, estes
pesquisadores observaram que o tratamento com extratos da casca do pinheiro, que é
rico em polifenóis e procianidinas, aumenta a resistência do C. elegans ao paraquat,
herbicida que gera radicais superóxido. Os autores mostraram que estes extratos
possuem atividade antioxidante através do ensaio TEAC (Trolox Equivalent.
Antioxidant Capacity) e sugerem que o aumento da resistência se deve a capacidade do
extrato de pinheiro de remover o OH-.

69
Por outro lado, Wilson et al. (2006) não observaram nenhuma melhora na
sobrevivência do C. elegans tratado com extrato de mirtilo quando subemetidos a H2O2
e paraquat, apesar deste extrato conter uma mistura de vários polifenóis tais como
antocianinas, proantocianidinas e ácidos clorogênicos. Neste mesmo trabalho, eles
observaram que o extrato de mirtilo aumenta a resistência ao estresse térmico e
longevidade do C. elegans e que a fração das procianidinas é a responsável por estes
efeitos benéficos (WILSON et al., 2006).
Os estudos listados acima sobre açaí nos levam a pensar que o efeito antioxidante
do açaí se deve aos seus compostos polifenólicos, especialmente as antocianidinas 3-
glicosídeo e 3-rudinosídeo. Entretanto, isto não parece ser exclusivo para os polifenóis
uma vez que Wilson et al. (2006) não observou o mesmo efeito com extrato de mirtilo.
Neste cenário é possível supor que deva existir uma ação sinérgica para proporcionar os
efeitos benéficos e/ou para minimizar os efeitos tóxicos ou adversos de constituintes
individuais dos extratos de frutas e vegetais (YU et al., 2010). Uma possível resposta a
essa questão seria a utilizaçao do corantes H2DCF-DA (utilizado para quantificar EROs
na célula).
O efeito benéfico do tratamento com 100 mg/ml e tóxico com 200 mg/ml de
AL-1 é uma questão bastante interessante que sugere que este possui um efeito do tipo
hormético. Hormese ou efeito hormético é o termo utilizado quando uma determinada
condição tem efeitos opostos em doses altas e em doses baixas. Já tem sido bastante
documentado o efeito hormético de plantas medicinais e de fitoquímicos específicos
(RATTNAM, 2006; SON; CAMANDOLA; MATTSON, 2008).
Tem sido sugerido que o mecanismo geral de ação dos fitoquímicos seja através da
ativação de vias de resposta celular adaptativa ao estresse. De uma perspectiva
evolutiva, as propriedades nocivas de fitoquímicos desempenham um papel importante
na dissuasão de insetos e outras pragas fitófagas. No entanto, quando ingerido em doses
sub-tóxicas, os fitoquímicos induzem uma resposta celular moderada ao estresse. As
vias “horméticas” ativadas por fitoquímicos incluem diferentes quinases e fatores de
transcrição que induzem a expressão de genes que codificam enzimas antioxidantes,
proteínas chaperonas, fatores neurotróficos e outras proteínas citoprotetoras. Exemplos
específicos de tais vias incluem as vias de sinalização de NF-B, Nrf-2/ARE, FOXO,
sirtuinas e outras (RATTNAM, 2006; SON; CAMANDOLA; MATTSON, 2008).

70
Quando ingeridos polifenóis também podem modular uma variedade de enzimas de
respostas a xenobióticas tais com a citocromo P450 (McPHERSON; TINGLE;
FERGUSON, 2001). É muito comum a geração de compostos mais tóxicos pela P450
(VERMEULEN, 1996). A produção de tais compostos secundários mais tóxicos
poderia explicar estes efeitos deletérios do tratamento com maiores concentrações do
extrato de açaí.
Tem sido postulado que compostos que possuem propriedades antioxidantes em
sistemas biológicos podem conferir proteção a diferentes condições de estresse. Assim,
além da resistência ao estresse oxidativo também foi avaliado se o AL-1 aumenta a
resistência ao estresse térmico. O C. elegans se desenvolve normalmente em
temperaturas de 16oC a 25
oC sendo que a sobrevivência da população começa a
declinar rapidamente acima de 25oC. Ensaios de termotolerância em C. elegans são
realizados com bastante frequência para avaliar o efeito de compostos e extratos de
plantas (WILSON et al., 2006; BENEDETTI et al., 2008 KAMPKÖTTER et al., 2007;
KAMPKÖTTER et al., 2008; RATTNAM, 2006; SON; CAMANDOLA; MATTSON,
2008; PIETCSH et al., 2009). No presente estudo, o tratamento com 100 mg/ml de AL-
1 não aumentou a sobrevivência do C. elegans a 35oC. Nenhum trabalho publicado até o
momento avaliou o efeito do açaí na resistência ao choque térmico a fim de
comparação.
Estes resultados sugerem que o potencial antioxidante do açaí é mais eficiente
para proteção contra danos gerados por estresse oxidativo do que térmico. Enquanto que
o ERO gerado no estresse oxidativo é capaz de destruir macromoléculas, a resposta
celular ao estresse térmico causa primariamente a desnaturação e agregação protéica.
Muitos efeitos fenotípicos e morfológicos deste estresse podem ser explicados pela
agregação de proteínas e desequilíbrio da homeostase protéica (HASLBECK et al.,
2004). Assim, é possível propor que os efeitos do açaí sejam específicos para aliviar a
injúria do estresse oxidativo e não do térmico uma vez que os polifenóis são capazes de
neutralizar cátions diretamente. Mas ao contrário do nosso resultado, Wilson et al.
(2006) observaram que a fração de proantocianidina do extrato de mirtilo aumenta a
resistência ao estresse térmico, mas não oxidativo, do C. elegans.
Os efeitos benéficos de compostos polifenólicos de frutas e vegetais são
principalmente extrapolados a partir de estudos in vitro ou in vivo de suplementação

71
dietética de curta duração. Devido ao custo e duração, relativamente pouco se conhece
se os polifenóis dietéticos são benéficos de uma forma sistêmica para os seres vivos,
particularmente em respeito à longevidade.
Neste sentido, o C. elegans é um excelente modelo para ensaios de longevidade,
pois além da alta homologia com genoma de mamíferos, experimentos de longevidade
podem ser realizados em algumas semanas a baixo custo. Neste trabalho, o C. elegans
foi tratado com 100 mg/ml de AL-1 por todo seu tempo de vida a partir do estágio L1.
Mas não foi observado um aumento da longevidade, nem no tempo de vida médio e
máximo do C. elegans tratado com AL-1 em relação ao controle (Gráfico 3).
Até o presente, apenas um artigo na literatura investigou o efeito do açaí na
longevidade que foi realizado na Drosophila melanogaster. Assim como observado
nesta dissertação, a suplementação de 2% do extrato de açaí na dieta padrão da
Drosophila não promoveu um aumento da longevidade delas (SUN et al., 2010).
Entretanto, moscas fêmeas alimentadas com uma dieta hiperlipídicas e com 2% do
extrato de açaí tiverem um aumento do seu tempo de vida médio (SUN et al., 2010).
Esta ausência de efeito pró-longevidade com a suplementação do açaí na dieta
padrão se parece com a do resveratrol, um composto polifenólico abundante na uva. Já
foi documentado que a suplementação com o resveratrol não aumentou a longevidade
da Drosophila, do C. elegans (BASS et al., 2007) e nem do camundongo (BAUR et al.,
2006) submetidos a dietas padrão. Entretanto, o resveratrol prolonga a longevidade de
camundongos alimentados com dieta hiperlipídica (BAUR et al., 2006). Apesar de não
aumentar a longevidade dos camundongos com dieta padrão, o resveratrol parece
melhorar a saúde de maneira geral, pois foi observado um efeito benéfico nas funções
cognitivas destes camundongos (PERSON et al., 2008).
Por outro lado, vários trabalhos têm descrito que extratos de plantas ricos em
polifenóis, tais como mirtilo (WILSON et al., 2006), casca de pinheiro (PUN et al.,
2009) e maçã (SUNAGAWA et al., 2010) são capazes de aumentar a longevidade do C.
elegans. Extrato de mirtilo aumentou o tempo de vida médio do C. elegans em 28%.
Este tratamento também retardou o declínio de várias funções relacionadas ao
envelhecimento tais como batimento faríngeo e acúmulo de lipofuscina (WILSON et
al., 2006). Os efeitos pró-longevidade assim como o aumento da termotolerância foram
atribuídos a fração de procianidinas do extrato de mirtilo (WILSON et al., 2006). A

72
casca de pinheiro, além de aumentar a resistência ao estresse oxidativo, aumentou o
tempo de vida médio e máximo do C. elegans (PUN et al., 2009). Sunagawa et al.
(2010) observaram que os polifenóis e as procianidinas da maçã são capazes de
aumentar o tempo de vida médio em 12,0 e 12,1% respectivamente. Entretanto o
tratamento do C. elegans com a (-)-epicatequina, uma das principais procianidina da
maçã, não teve nenhum efeito na longevidade dos animais (SUNAGAWA et al., 2010).
Esta variabilidade dos efeitos dos extratos de plantas ricos em polifenóis na
modulação da longevidade do C. elegans chama a atenção por dois pontos. O primeiro é
de que a atribuição de atividade antioxidante in vitro a estes extratos não
necessariamente irá ter efeitos biológicos in vivo. Na verdade muitos antioxidantes in
vitro mostram pouca ou nenhuma capacidade antioxidante in vivo (GRUBER et al.,
2009; PUN et al., 2009). O segundo ponto é de que a presença de outros componentes
nos extratos de plantas deve exercer uma influência nos efeitos benéficos sobre as
diferentes funções biológicas (YU et al., 2010).
5.1.3 Dosagens Bioquímicas
Baixos e constantes níveis de EROS são indispensáveis em muitos processos
bioquímicos, incluindo apoptose celular (GHOSH & MYERS, 1998). Em contraste,
altas concentrações ou inadequada remoção de EROS resultam em um quadro de
estresse oxidativo, o qual pode gerar danos a macromoléculas biológicas e severos
distúrbios metabólicos. Para que se predomine o equilíbrio homeostático e as funções
biológicas sejam preservadas, é indispensável que organismo controle a presença de
ambos, oxidantes e antioxidantes, continuamente. Os sistemas biológicos
desenvolveram a capacidade de promover resposta adaptativa que podem compensar o
estresse oxidativo, como por exemplo, um sistema antioxidante altamente complexo,
que envolve uma variedade de componentes endógenos e exógenos com a função de
neutralizar espécies reativas de oxigênio (SIES,1993).
Pelo fato do AL ter propiciado um significante efeito antioxidante in vivo no C.
elegans, foi avaliado também seu efeito sobre a atividade de sulfidrilas totais, catalase e
de proteína carbonilada .

73
A enzima catalase participa do sistema endógeno de defesa antioxidante, sendo
responsável, em conjunto com a enzima superóxido dismutase, pela neutralização de
radicais superóxido, nocivos ao organismo, transformando-os em água e oxigênio
molecular (SCHAUSS, 2006). A análise da atividade da catalase in vivo no C. elegans
mostrou que o tratamento com AL-1 não causou nenhuma alteração em relação aos
animais não tratados. Estes resultados foram semelhantes aos resultados observados in
vivo por Guerra et al. (2011) e de Souza et al (2010). Nestes trabalhos, a suplementação
de 2% de açaí na dieta normal de ratos não alterou significativamente os níveis de
atividade da catalase. Spada et al. (2009) também não observaram nenhuma diferença
estatística na atividade da catalase nos tecidos do cortex cerebral, hipocamo e cerebelo
de ratos tratados ex vivo com açaí.
Além da análise das atividades da catalase como um marcador antioxidante, foram
também analisados os níveis de sulfidrilas totais e proteína carbonilada como
marcadores de estresse oxidativo no C. elegans. Os radicais sulfidrilas representam
todos os grupos tióis encontrados na glutationa, em proteínas e em compostos de baixo
peso molecular. Os tióis podem ser oxidados quando o estresse oxidativo está elevado,
tendo assim seus níveis diminuídos (WISDOM et al .,1991). Já os grupos carbonílicos
representam o dano oxidativo às proteínas, podendo constituir análise do estado
oxidativo do organismo. O tratamento com 100 mg/ml de AL-1 não proporcionou
alterações significantes para ambos os parâmetros. Spada et al. (2009) também relatou
que o tratamento ex vivo com 40% de açaí nos homogenatos do tecido nervoso não
alterou os níveis de proteína carbonilada. Por outro lado, Guerra et al. (2011) e de Souza
et al. (2010) observaram uma redução significativa de proteína carbonilada e aumento
de grupos SH totais ou GSH no fígado de ratos suplementados com 2% de açaí na dieta
normal.
A capacidade de extratos de plantas em alterar marcadores de estresse oxidativo é
bastante variável na literatura. Por exemplo, Pun et al. (2009) observaram que diferentes
extratos de plantas com comprovada capacidade antioxidante in vitro foram capazes
tanto de aumentar quanto diminuir os níveis de proteína carbanilada no C. elegans.
Em nossa opinião, a justificativa para não ter sido observada quaisquer variações
nestas dosagens é porque o açaí não altera a homestose do animal crescendo em
condições normais, porém este mesmo tratamento é capaz de aumentar a defesa

74
oxidante do animal perante a um desafio tal como com t-BOOH. Uma proposta
interessante para confirmar esta hipótese seria tratar previamente o C. elegans com açaí
e então submetê-los a uma condição de estresse para então realizar as dosagens
bioquímicas. Se o tratamento com açaí tiver algum efeito nos marcadores bioquímicos,
poderia ser esperado um aumento dos níveis de sulfidrilas totais e reduzisse os níveis de
proteína carbonilada em comparação aos animais somente submetidos ao estresse
oxidativo.
Dentro desta linha de raciocínio, vários trabalhos já demostraram que a
suplementação de açaí associada a uma condição de estresse é capaz de melhorar vários
marcadores bioquímicos. Spada et al. (2009) observaram que no homogenato do córtex
cerebral, hipocampo e cerebelo de ratos tratados com H2O2 a atividade de SOD, CAT e
os níveis de proteína carbonilada estão aumentados e quando são tratados previamente
com a polpa do açaí os valores encontrados retornam para níveis semelhantes ao do
grupo controle. De Moura et al. (2011) mostraram que a quantidade de GSH no pulmão
de camundongos com efisema pulmonar é reduzida em relação ao controle e que a
ingestão de fumaça de cigarro associada com açaí retorna os níveis de GSH semelhante
ao grupo controle. De Souza et al. (2010) relata que a suplementação de 2% de açaí na
dieta hipercolesterolêmica reduz os níveis de proteína carbonilada e aumenta os níveis
de sulfidrilas totais no fígado de ratos quando comparados com aqueles submetidos a
dieta hipercolesterolêmica. Guerra et al. (2011) também observaram uma redução dos
níveis de proteina carbonilada no fígado de ratos diabéticos tratados com 2% de açaí em
relação ao grupo de ratos diabéticos. Finalmente, Sun et al. (2010) mostraram que
moscas crescidas em dietas hiperlipídicas e suplementadas com açaí tem um tempo de
vida médio maior que as moscas alimentadas somente com a dieta hiperlipídica.

75
5.2 POSSÍVEL MECANISMO DE AÇÃO DO EXTRATO DE AÇAÍ
As propriedades biológicas de extratos de plantas e flavonóides têm sido
intensamente investigadas in vitro e em sistemas ex vivo e in vivo de mamíferos. Estes
compostos e extratos podem agir como anti- ou pró-oxidantes influenciando o sistema
redox celular e/ou modulando vias de sinalização (WILLIAMS et al. 2004). Por esta
razão, os mecanismos de sua ação biológica ainda são objetos de intensa investigação.
Neste sentido, o C. elegans é um excelente modelo não só para determinar o potencial
protetor dos flavonóides e extratos vegetais, bem como para investigar os mecanismos
moleculares da sua ação (WU et al., 2002;. SMITH; LUO, 2003, 2004, GUTIERREZ-
ZEPEDA et al., 2005, BROWN; EVANS; LUO, 2006, WILSON et al., 2006;
KAMPKÖTTER et al., 2007).
No presente estudo, foi mostrado que o tratamento de C. elegans com AL
aumentou a sua sobrevivência em estresse oxidativo, porém sem aumentar a resistência
em condições de estresse térmico e em condições normais. Considerando que os
antioxidantes podem atuar diretamente devido a sua capacidade de captar radicais das
substâncias ou indiretamente através da modulação de diferentes vias de sinalização
levando a um aumento da resistência ao estresse oxidativo, este trabalho se propôs
também a tentar elucidar o possível mecanismo de ação da atividade protetora do AL.
Em C. elegans dois fatores de transcrição, DAF-16 e SKN-1, promovem a expressão de
enzimas antioxidantes ou de detoxificação.
5.2.1 A Via de Sinalização SKN-1/p38 MAPK
No C. elegans, a conservada via de sinalização SKN-1/p38 MAPK é responsável
pela ativação de genes de Fase II de detoxificação durante estresse oxidativo e na
regulação da longevidade em condições normais, de restrição calórica e de sinalização
reduzida da insulina (TULLET et al., 2009).
Para investigar se o aumento da resistência ao estresse observado no C. elegans
tratado com AL foi dependente da via de SKN-1/p38 MAPK, os ensaios de resistência
ao estresse oxidativo foram repetidos usando os mutantes skn-1(RNAi) e sek-1(km4)
(Gráficos 5 e 6). Os resultados obtidos com os mutantes skn-1(RNAi) foram

76
inconclusivos. Apesar deste experimento ter sido realizados várias vezes, em nenhuma
delas o tratamento com açaí aumentou a sobrevivência em condições de estresse do
grupo controle (Gráfico 5). Uma possível razão de não se observar o efeito protetor do
açaí neste experimento pode ser devido ao procedimento de RNAi. Nos experimentos
de RNAi, foi utilizada a linhagem de E. coli HT115 enquanto que nos outros
experimentos os C. elegans foram crescidos na linhagem OP50. Já foi descrito que a
linhagem HT115 é menos patogênica para o C. elegans (PARTRIDGE & GEMS, 2007;
SHUHEISO et al., 2011). O crescimento do C. elegnas em uma bactéria menos
patogênica pode estabelecer um novo estado de homeostase no animal e assim, o
aumento da resistência ao estresse observado nos animais mantidos em OP50 não é
mais observado nos animais mantidos em HT115. Para elucidar esta questão é
importante repetir estes experimentos com o mutante de delação skn-1(zu67) que são
mantidos em OP50 para comparar os resultados.
Os resultados de resistência ao estresse oxidativo obtidos com os mutantes sek-
1(km4) mostraram que os mutantes tratados não tiveram a sobrevivência aumentada
(Gráfico 6). Este resultado sugere que o aumento da resistência ao estresse oxidativo
promovido por AL depende da ativação da via p38 MAPK. Estes dados são ainda muito
preliminares, pois ainda falta realizar outros experimentos para corroborar este dado.
Entre os experimentos que pretendemos realizar está a análise da resistência ao estresse
oxidativo com o mutante skn-1, análise da localização nuclear da linhagem transgênica
SKN-1B/C::GFP, e a análise da ativação da expressão de genes regulados por SKN-1
tais como gcs-1 e gst-4.
5.2.2 A Via de Sinalização de DAF-16
O fator de transcrição DAF-16/FoxO ativa a expressão de genes que aumentam a
longevidade e a resistência ao estresse (MURPHY et al., 2003).
Para investigar se o aumento da resistência ao estresse observado no C. elegans
tratado com AL ativa a via de DAF-16/FoxO, os ensaios de resistência ao estresse
oxidativo foram repetidos usando o mutante de deleção daf-16(mgDf47). Os resultados
obtidos indicam que o aumento da resistência ao estresse oxidativo promovido pelo com
AL depende de DAF-16 (Gráfico 7).

77
Outra estratégia utilizada para verificar o envolvimento de DAF-16 foi através da
análise da localização nuclear da linhagem transgênica DAF-16::GFP tratada com AL.
Apesar da resistência ao estresse oxidativo parecer depender de DAF-16, não foi
observada a indução de DAF-16::GFP com o tratamento de AL-1. Uma explicação para
os resultados aparentemente contraditórios pode ser devido ao tempo usado para
analisar este transgênico não ter sido o mais adequado, 48 horas desde o estágio L1.
Vários autores analisaram a localização subcelular de DAF-16::GFP apenas após 1 a 24
horas após a indução (PRZYBYSZ et al., 2009; POWOLNY et al., 2011). Já foi
demonstrado que a quercetina é capaz de aumentar a localização nuclear de DAF-
16::GFP após 72 horas com o tratamento de quercetina (KAMPKÖTTER et al., 2007;
KAMPKÖTTER et al., 2008). Além disso, alguns autores têm sugerido que fatores de
transcrição podem ter sua atividade modulada, mas sem necessariamente aumentar a sua
concentração no núcleo (WANG et al., 2010). Outra possível explicação pode ser
devido à dependência de co-fatores necessários a ação de fatores de transcrição. Já foi
demonstrado que a translocação de DAF-16 pode não ser efetiva se cofatores nucleares
estiverem faltando (HENDERSON; BONAFÈ; JOHNSON, 2006).
Da mesma maneira que foi sugerido para investigar o envolvimento de SKN-1 no
tratamento com AL, é importante também avaliar a indução de genes efetores de DAF-
16 para corroborar os resultados. Sun et al. (2010) observaram que o tratamento com
açaí aumentou a expressão do gene l(2)elf, que codifica uma proteína de heat shock e
dois genes de detoxificação gstd1 e mtnA e diminuiu a expressão do gene para piruvato
carboxiquinase pepck da via da gliconeogênese. Wilson et al. (2006) mostraram que o
extrato de mirtilo além de retardar o declínio de várias funções relacionadas ao
envelhecimento também impediu o aumento da expressão de genes para proteínas heat
shock e aumentou a longevidade do C. elegans. O aumento da longevidade induzida
pelo extrato de mirtilo parece depender via de sinalização da proteína quinase
dependente de Ca(+2)-Calmodulina (CaMKII), que é importante para a resistência ao
estresse osmótico (WILSON et al., 2006).
Alimentos funcionais são um dos segmentos que mais crescem dentro da indústria
de alimentos. Este mercado é movido pelo aumento do tempo de vida da população,
aumento do custo de assistência á saúde, avanços tecnológicos na produção de
alimentos e consciência dos benefícios de uma dieta saudável. Apesar do

78
desenvolvimento de drogas e fármacos ser necessário para tratar problemas de saúde
específicos, a intervenção nutricional através de alimentos funcionais pode ser uma
maneira efetiva de melhorar a qualidade de vida. Conseqüentemente, estudos sobre
ingredientes alimentares com propriedades antioxidantes é de grande interesse. Assim, é
importante avaliar estas propriedades em sistemas biológicos in vivo. O C. elegans se
tornou um organismo modelo central na pesquisa de longevidade e estresse oxidativo.
Os motivos que levaram a isto se devem ao fato de ser um organismo multicelular com
o genoma já completamente seqüenciado e possuir vias de metabólicas conservadas
entre mamíferos, o que permite explorar os mecanismos genéticos que controlam a
longevidade e resistência ao estresse. Neste contexto, este trabalho destaca-se por ser o
primeiro a avaliar as propriedades antioxidantes do açaí através da abordagem global
dos seus efeitos em organismo como um todo mostrando que este aumenta a resistência
ao estresse oxidativo e que é possivelmente através de SKN-1 e DAF-16.

79
CAPÍTULO 6
CONCLUSÕES

iii
- O extrato de açaí industrializado liofilizado (AL) apresenta quantidades de
anoticianinas comparável com já publicados
- O crescimento do C. elegans em 200 mg/ml de AL foi tóxico enquanto que o
tratemento com 100 mg/ml permitiu o desenvolvimento normal.
- O tratamento do C. elegans com 100 mg/ml de AL aumenta a resistência ao
estresse oxidativo,
- O tratamento do com 100 mg/ml de AL não aumenta a sobrevivência do C.
elegans nem condições de estresse térmico e nem em condições normais.
- O aumento da resistência ao estresse oxidativo promovido pelo AL foi dependente
das vias de sinalização p38 e DAF-16, uma vez que o tratamento com AL não
aumentou a sobrevivência dos mutantes sek-1 e daf-16 em condições de estresse
oxidativo.
- As análises dos genes repórteres DAF-16::GFP realizadas até o momento não
corroboraram estas observações.

iv
CAPÍTULO 7
REFERÊNCIAS
BIBLIOGRÁFICAS

v
REFERÊNCIAS BIBLIOGRÁFICAS
AEBI, H. Catalase in vitro. Methods Enzymol, v. 105, p. 121-126, 1984.
AHERNE, S.A.; O’BRIEN, N.M. Dietary flavonols: chemistry, food content and
metabolism. Nutrition, v. 18, p. 75-81, 2002.
ALBARICI, T.R.; VALETA, A.C. Efeitos da temperatura nas antocianinas do açaí.
Comunicado técnico, p. 1517-4786, 2007.
AN, J.H. et al. Regulation of the Caenorhabditis elegans oxidative stress defense
protein SKN-1 by glycogen synthase kinase-3. Proceedings of the National Academic
of Science, v. 102, p. 16275-16280, 2005.
AN, J.H.; BLACKWELL, T.K. SKN-1 links C. elegans mesendodermal specification to
a conserved oxidative stress response. Genes & Development, v. 17, p. 1882-1893,
2003.
BASS, T. M. et al. Effects of resveratrol on lifespan in Drosophila melanogaster and
Caenorhabditis elegans. Mechanisms of Ageing and Development, v. 128, n. 10, p.
546-552, 2007.
Baur, J.A. et al. Resveratrol improves health and survival of mice on a high calorie
diet. Nature, v. 444, n. 7117, p. 337-42, 2006.
BENEDETTI, M.G. et al. Compounds that confer thermal stress resistance and
extended lifespan. Experimental Gerontology. v. 43, p. 882–891, 2008.
BISHOP, N.A, GUARENTE,L. Two neurons mediate diet-restriction-induced longevity
in C. elegans. Nature, v. 447, p. 545-549, 2007.
BRENNER, S. The genetics of Caenorhabditis elegans. Genetics, v. 77, p. 71-94, 1974.
BROWN, M. K.; EVANS, J. L.; LUO, Y. Beneficial effects of natural antioxidants
EGCG and [alpha]-lipoic acid on life span and age-dependent behavioral declines in
Caenorhabditis elegans. Pharmacology Biochemistry and Behavior, v. 85, n. 3, p.
620-628, 2006.
Chan, K., X. D. Han, et al. (2001). An important function of Nrf2 in combating oxidative
stress: detoxification of acetaminophen. Proc Natl Acad Sci U S A, v. 98, n. 8, p. 4611-
6, 2001.
CHOE, K.P.; PRZYBYSZ, A.J.; STRANGE, K. The WD40 repeat protein WDR-23
functions whit the CUL4/DDB1 ubiquitin ligase to regulate nuclear abundance and
activity of SKN-1 Caenorhabditis elegans. Mol Cell Biol, v. 29, n. 10, p. 2704-15, 2009.

vi
CHOI, S.K. et al. Promotion of Met-tRNAiMet binding to ribosomes by yIF2, a
bacterial IF2 homolog in yeast. Science, v. 280, p. 1757–1760, 1998.
COHEN, K.O.; MONTE, D.C “Quantificação do teor de antocianinas totais da polpa de
açaí de diferentes populações de açaizeiro”. Embrapa Amazônia Oriental. Boletim de
Pesquisa e Desenvolvimento, v. 60, p. 1517-2228, 2006.
COOKE, M.S.et al. "Oxidative DNA damage: mechanisms, mutation, and disease."
Faseb Journal, v. 17, n. 10, p. 1195-214, 2003.
DEL POZO-INSFRAN, D., C. H. Brenes, et al. "Phytochemical composition and
pigment stability of Acai (Euterpe oleracea Mart.)." J Agric Food Chem. v. 52, n.6, p.
1539-45, 2004.
DEL POZO-INSFRAN, D., S. S. Percival, et al. Acai (Euterpe oleracea Mart.)
polyphenolics in their glycoside and aglycone forms induce apoptosis of HL-60 leukemia
cells. J Agric Food Chem. v.54, n. 4, p. 1222-9. 2006.
Ding, M., Shi, X, et al “Cyanidin-3-glucoside, a Natural Product Derived from
Blackberry, Exhibits Chemopreventive and Chemotherapeutic Activity”. The Journal
of Biological Chemistry, v. 281, n. 25, p. 17359-68, 2006.
DINKOVA-KOSTOVA, A.T. et al. Direct evidence that sulfhydryl groups of Keap1 are
the sensors regulating induction of phase 2 enzymes that protect against carcinogens
and oxidants. Proceedings of the National Academic of Science, v. 99, p. 11908–
11913, 2002.
DINKOVA-KOSTOVA, A.T. et al. Extremely potent triterpenoid inducers of the phase
2 response: correlations of protection against oxidant and inflammatory stress.
Proceedings of the National Academy of Sciences, v. 102, p. 4584-9, 2005.
ENIO, C.V. Alimentos Funcionais. Revista de Medicina de Minas Gerais, v. 13, n. 4,
p. 260-262, 2003.
GUERRA, J.F.C. et al. Dietery açaí modulates ROS production by neutrophilis and
gene expression of liver antioxidant exzinmes in rats. J Clin Biochem Nutr, v.49, n X
p. 1-7, 2011.
FARIAS NETO, J.T.de. et al. Variabilidade genética em progênies jovens de açaizeiro.
Cerne. v. 11, n.4, p. 336-41, 2005.
FINKEL, T.; HOLBROOK, N.J. Oxidants, oxidative stress and the biology of ageing.
Nature, v. 408, n. 6809, p. 239-247, 2000.
FORSBERG, H. et al. The role of the yeast plasma membrane SPS nutrient sensor in
the metabolic response to extracellular amino acids. Molecular Microbiology, v. 42, n.
1, p. 215-228, 2001.

vii
GHOSH, J.; MYERS CE. Inhibition of arachidonate 5-lipoxygenase triggers massive
apoptosis in human prostate cancer cells. Proc Natl Acad Sci U S A. v. 95, n. 22, p.
13182-7, 1998
GRIFFITHS, M.M.; WILLIAMS JR., R.C. In vitro peripheral blood and synovial fluid
lymphocyte interactions. Arthritis and Rheumatism, 2005.
GRUBER, J. et al. Deceptively simple but simply deceptive - Caenorhabditis elegans
lifespan studies: Considerations for aging and antioxidant effects. FEBS Letters, v.
583, n. 21, p. 3377-3387, 2009.
GUTIERREZ-ZEPEDA, A. et al. Soy isoflavone glycitein protects against beta
amyloid-induced toxicity and oxidative stress in transgenic Caenorhabditis elegans.
BMC Neuroscience, v. 25, n. 6, p. 54, 2005.
HASLBECK, M. et al. Hsp42 is the general small heat shock protein in the cytosol of
Saccharomyces cerevisiae. EMBO Journal, v. 23, n. 3, p. 638-649, 2004.
HASEGAWA K, MIWA J. Genetic and cellular characterization of Caenorhabditis
elegans mutants abnormal in the regulation of many phase II enzymes. Plos one, v. 5, n
.6, p.1-10, 2010.
HAYES, M. et al. The Cap'n'Collar basic leucine zipper transcription factor Nrf2 (NF-
E2 p45-related factor 2) controls both constitutive and inducible expression of intestinal
detoxification and glutathione biosynthetic enzymes. Cancer Research, v. 61, n. 8, p.
3299-307, 2001.
HENDERSON, S. T.; JOHNSON, T. E. daf-16 integrates developmental and
environmental inputs to mediate aging in the nematode Caenorhabditis elegans.
Current Biology, v. 11, n. 24, p. 1975-1980, 2001.
HENDERSON, S.T.; BONAFÈ, M.; JOHNSON, T.E. daf-16 protects the nematode
Caenorhabditis elegans during food deprivation. Journal of Gerontology, v. 61, n. 5,
p. 444-60, 2006.
HOGAN, S. et al. Antiproliferative and antioxidant properties of anthocyanin-rich
extract from Açai. Food Chemistry, v. 118, p. 208-214. cccccccccc2010
1 HONDA, Y.; HONDA, S. The daf-2 gene network for longevity regulates oxidative
stress resistance and Mn-superoxide dismutase gene expression in Caenorhabditis
elegans. The FASEB Journal, v. 13, p. 1385-1393, 1999.
HU, R. et al. Identification of Nrf2-regulated genes induced by chemopreventive
isothiocyanate PEITC by oligonucleotide microarray. Life Science, v. 79, p. 1944–
1955, 2006.
Itoh, K., T. Chiba, et al. . "An Nrf2/small Maf heterodimer mediates the induction of
phase II detoxifying enzyme genes through antioxidant response elements." Biochem
Biophys Res Commun, v.236, n. 2, p. 313-22, 1997.

viii
Itoh, K., N. Wakabayashi, et al. Keap1 represses nuclear activation of antioxidant
responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes
Dev, v. 13, n. 1, 76-86, 1999.
INOUE, H. et al. The C. elegans p38 MAPK pathway regulates nuclear localization of
the transcription factor SKN-1 in oxidative stress response. Genes and Development,
v. 19, p. 2278-2283, 2005.
JENSEN, G.S. et al. Pain Reducion and Improvement in Ranger of Motion After Daily
Consuption of an Açai (Euterpe oleracea Mart.) Pulp-Fortified Polyphenolic-Rich fruit
and Berry Juice Blend. J Med Food, v. XX, n. X, p. 1-10, 2011
JENSEN, G.S. et al. In Vitro and in Vivo Antioxidant and Anti-inflammatory Capacities
of an Antioxidant-Rich Fruit and Berry Juice Blend. Results of a Pilot and Randomized,
Double-Blinded, Placebo-Controlled, Crossover Study, J. Agric. Food Chem, v. 56, n.
18, p. 8326–8333, 2008.
JUGE, N.; MITHEN, R.F.; TRAKA, M. Molecular basis for chemoprevention by
sulforaphane: a comprehensive review. Cellular and Molecular Life Science, v. 9, p.
1105–1127, 2007.
KALETTA, T.; HENGARTNER, M. O. Finding function in novel targets: C. elegans as
a model organism. Nature Reviews Drug Discovery, v. 5, n. 5, p. 387-399, 2006.
KAMATH, R.S. et al. Effectiveness of specific RNA-mediated interference through
ingested doublestranded RNA in Caenorhabditis elegans. Genome Biology, v. 2, n. 1, p.
1-10, 2001.
KAMPKÖTTER, A. et al. Increase of stress resistance and lifespan of Caenorhabditis
elegans by quercetin. Comparative Biochemistry and Physiology Part B: Biochemistry
and Molecular Biology, v. 149, n. 2, p. 314-323, 2008.
KAMPKÖTTER, A. et al. The Ginkgo biloba extract EGb761 reduces stress sensitivity,
ROS accumulation and expression of catalase and glutathione S-transferase 4 in
Caenorhabditis elegans. Pharmacological ResearchI, v. 55, p. 139-147, 2007.
KUSKOSKI, E.M.; ASUERO, A.G.; MORALES, M.T.; FETT, R. Frutos tropicais
silvestres e polpas de frutas congeladas: atividade antioxidante, polifenóis e
antocianinas. Ciência Rural, v.36, n.4, p 1283-1287, 2006.
KUNDU, J.; SURH, Y.-J. Nrf2-Keap1 Signaling as a Potential Target for
Chemoprevention of Inflammation-Associated Carcinogenesis. Pharmaceutical
Research, v. 27, n. 6, p. 999-1013, 2010.

ix
KLUTH, D., A. et al. "Modulation of pregnane X receptor- and electrophile responsive
element-mediated gene expression by dietary polyphenolic compounds. Free Radic Biol
Med, v. 42, n3, p.315-25.2007.
LANDIS, J.N.; MURPHY, C.T. Integraiqwtion of diverse inputs in the regulation of
Caenorhabditis elegans DAF-16/FOXO. Developmental Dynamics, v. 239, n. 5, p.
1405-12, 2010.
LEE, J.M. et al. Identification of the NFE2-related factor-2-dependent genes conferring
protection against oxidative stress in primary cortical astrocytes using oligonucleotide
microarray analysis. The Journal of Biological Chemistry, v. 278, p. 12029-12038,
2003.
LEVINE, R.L. et al. Carbonyl assays for determination of oxidatively modified
proteins. Methods Enzymol, v. 233, p. 346-356, 1994.
Leung, L., M. Kwong, et al. Deficiency of the Nrf1 and Nrf2 transcription factors results
in early embryonic lethality and severe oxidative stress." J Biol Chem, v. 278, n. 48, p.
48021-9.2003.
LICHTENTHALER, R. et al. Total oxidant scanveng capacities of Euterper olereacea
Mart. (Açaí) fruits. Int J Food Sci Nutri, v. 56, n. 1, p. 53-56, 2005.
2 LITHGOW, G.J. et al. Thermotolerance and extended life-span conferred by
single-gene mutations and induced by thermal stress. Proceedings of the National
Academic of Science, v. 92, n. 16, p. 7540-4, 1995.
LOWRY, et al. Protein measurement with the Folin phenol reagent. The Journal of
Biological Chemistry, v. 193, p. 265-275, 1951.
MA, Q. Transcriptional responses to oxidative stress: Pathological and toxicological
implications. Pharmacology & Therapeutics, v. 125, p. 376–393, 2010.
Manela-Azulay, M. et al. Vitamina C. An bras Dermatol, v.78, n.3, p.265-274, 2003.
MANACH, C. et al. Polyphenols: food sources and bioavailability, American Journal
of Clinical Nutrition, v. 79, p. 727–747, 2004.
MARTORELL P. et al. Use of Saccharomyces cerevisiae and Caenorhabditis elegans as
model organisms to study the effect of cocoa polyphenols in the resistance to oxidative
stress. J Agric Food Chem. v. 59, n. 5, p. 2077-85, 2011
MATIAS, G.S. et al. Correlação entre atividade antioxidante e compostos bioativos de
polpas comerciais de açaí (Euterpe oleracea Mart). ALAN, vol.58, no.2, p.187-192.
2008
MATHEWS W.R. et al. Lipid peroxidation as molecular mechanism of liver cell injury
during reperfusion after ischemia. Free Radic Biol Med. V. 16, n. 6, p.763-70, 1994.

x
MASELLA, R. Novel mechanisms of natural antioxidant compounds in biological
systems: involvement of glutathione and glutathione-related enzymes. Journal of
Nutritional Biochemistry, v. 16, p. 577-586, 2005.
MENEZES, E.M.S.; TORRES. A.T.; SRUR. A.U.S. Valor nutricional da polpa de
açaí (Euterpe oleracea Mart) liofilizada. Acta Amazonica, v. 38, n.2, p. 311-316, 2008.
MENZEL, R., M. Rodel, "CYP35: xenobiotically induced gene expression in the
nematode Caenorhabditis elegans." Arch Biochem Biophys. v. 438, n.1, p. 93-102,
2005.
MENSACK.M.M. et al. Characterization of low molecular weight chemical fractions of
dry bean (Phaseolus vulgaris) for bioactivity using Caenorhabditis elegans longevity
and metabolite fingerprinting. J Agric Food Chem, v. 58, n.11, p. 6697-705, 2010.
MERTENS-TALCOTT, U.S. et al. Pharmacokinetics of Anthocyanins and Antioxidant
Effects after the Consumption of Anthocyanin-Rich Açai Juice and Pulp (Euterpe
oleracea Mart.) in Human Healthy Volunteers. J. Agric. Food Chem, v. 56, n. 17, p.
7796–7802, 2008.
McCORD, J.M. Superoxide dismutase: rationale for use in reperfusion injury and
inflammation. Free Radical Biology and Medicine, v. 2, p. 307–310, 1986.
McCORD, J.M. The evolution of free radicals and oxidative stress. The American
Journal of Medicine, v. 108, p. 652– 659, 2000.
McPHERSON, R.A.; TINGLE, M.D.; FERGUSON, L.R. Contrasting effects of acute
and chronic dietary exposure to 2-amino-3-methyl-imidazo[4,5-f]quinoline (IQ) on
xenobiotic metabolising enzymes in the male Fischer 344 rat: implications for
chemoprevention studies. European Journal of Nutrition, v. 40, n. 1, p. 39-47, 2001.
MOSKAUG et al. Molecular imaging of the biological effects of quercetin and
quercetin-rich foods. Mechanisms of Ageing and Development, v. 125, p. 315-324,
2004.
MOURA, R.S. et al. Addition of açaí (Euterpe oleracea) to cigarettes has a protective
effect against emphysema in mice. Food and Chemical Toxicology, p. 2-9, 2011.
MURPHY, C. T. et al. Genes that act downstream of DAF-16 to influence the lifespan
of Caenorhabditis elegans. Nature, v. 424, n. 6946, p. 277-283, 2003.
NORATTO, G.D. et al. Polyphenolics from Açai (Euterpe oleracea. Mart.) and RED
Muscadine Grape (Vitis rotundifolia) Protect Human Umbilical Vascular Endothelial
Cells (HUVEC) from Glucose and Lipopolysaccharide (LPS)-Induced Inflammation
and Target MicroRNA-126. J Agric Food Chem 2011

xi
OKUYAMA, T. et al. The ERK-MAPK pathway regulates longevity through SKN-1
and insulin-linke signaling in Caenorhabditis elegans. J Biol Chem, v. 285, n. 39, p.
30274-81, 2010.
OLIVEIRA, et al. Effects of an extract obtained from of Euteroe oleracea Mart. In the
compenntes of metabolic syndrome induced in C57BL/6J mice fed a hihh-fat diet. J
Cardiovasc Pharmacol, v. 56, n. 6, p. 619-26, 2010.
OLIVEIRA, R. P. et al. Condition-adapted stress and longevity gene regulation by
Caenorhabditis elegans SKN-1/Nrf. Aging Cell, v. 8, p: 524–541, 2009.
OSBURN, W.O.; KENSLER, T.W. Nrf2 signaling: an adaptive response pathway for
protection against environmental toxic insults. Mutation Research, v. 659, p. 31-9.
2008.
PACHECO-PALENCIA, L. A.; MERTES-TALCOTT.S.; TALCOTE, S. T. Chemical
composition, properties, and thermal stability of a phytochemical enriched oil Acai
(Euterpe oleracea Mart). J Agric Food Chem, v. 56. N. 12, p. 4631-6, 2008.
PARTRIDGE.; L, GEMS D. Benchmarks for ageing studies. Nature, v. 450, p. 165-
167, 2007.
PÉREZ. V et al. Is the Oxidative Stress Theory of Aging Dead?. Biochim Biophys
Acta, n. 1790, v. 10, p. 1005–1014, 2009.
PIETSCH, K. Quercetin mediated lifespan extension in Caenorhabditis elegans is
modulated by age-1, daf-2, sek-1 and unc-43. Biogerontology, v. 10, p. 565-578, 2009.
POWOLNY, A.A. et al. The garlic constituent diallyl trisulfide increases the lifespan of
C. elegans via skn-1 activation. Experimental Gerontology, v. 46, n. 6, p. 441-52,
2011.
PRZYBYSZ, A.J. et al. Increased age reduces DAF-16 and SKN-1 signaling and the
hormetic response of Caenorhabditis elegans to the xenobiotic juglone. Mechanismis
of Ageing and Development, v. 130, n. 6, p. 357-69, 2009.
PUN, P.B. et al. Ageing in nematodes: do antioxidants extend lifespan in Caenorhabitis
elegan?. Biogerontology. v. 11, n. 1, p. 17-30, 2009.
RAMOS-GOMEZ, M. et al. Sensitivity to carcinogenesis is increased and
chemoprotective efficacy of enzyme inducers is lost in nrf2 transcription factor-
deficient mice, Proceedings of the National Academic of Science, v. 98, p. 3410–
3415, 2001.
RATNAM, D.V. et al. Role of antioxidants in prophylaxis and therapy: A
pharmaceutical perspective. Journal of Controlled Release, v. 113, p. 189–207, 2006.
RIBEIRO, J.C. et al. Evaluation of the genotoxic and antigenotoxic effectos after acute
and subacute treatements with açaí pulp (Euterpe oleracea Mart.) on mice using the

xii
erythrocytes micronucleus test and tehe comet assay. Mutation Research, v. 695, p22-
28, 2010
SARKADI, B. et al. Human multidrug resistance ABCB and ABCG transporters:
participation in a chemoimmunity defense system. Physiologycal Reviews, v. 86, p.
1179-1236, 2006.
SANTOS,G.M. et al. Correlação entre atividade antioxidante e compostos bioativos de
polpas comerciais de açaí (Euterpe oleracea Mart). Archivos Latinoamericanos de
Nutricion, v. 58, n. 2, p187-192, 2008.
SCHAUSS, A. et al. Phytochemical and nutrient composition of the freeze-dried
Amazonian palm berry, Euterpe oleraceae Mart.(Acai). Journal of Agricultural and
Food Chemistry, v. 54, n. 22, p. 8604-10, 2006.
SCHROECKSNADEL, K. et al. Tryptophan degradation increases with stage in
patients with rheumatoid arthritis. Clinical Rheumatology, v. 25, p. 334–337, 2006.
SEDLAAK, J.; LINDSAY, R.H. et al. Estimation of total, protein-bound, and
nonprotein sulfhydryly groups in tissue with Ellmam’s reagent. Anal Biochem, v. 25,
n.1, p. 192-205, 1968.
SHARONI, Y. et al. Carotenoids and transcription. Archives of Biochemistry and
Biophysics, v. 430, p. 89-96, 2004.
SIES, H. Strategies of antioxidant defense. Europen Journal of Biochemistry, v. 215,
p. 213-219, 1993.
SILVA JR., A.A.; VIZZOTTO, V.J. Plantas medicinais, aromáticas e fitoprotetoras.
Agropecuária Catarinense, v.9, n.1, p.5-8, 1996.
SMITH, J.V.; LUO, Y. Elevation of oxidative free radicals in Alzheimer's disease
models can be attenuated by Ginkgo biloba extract EGb 761. Journal of Alzheimer’s
Disease, v.5, n. 4, p. 287-300, 2003.
SON, T.G.; CAMANDOLA, S.; MATTSON, M.P. Hormetic Dietary Phytochemicals.
NeuroMolecular Medicine, v. 10, p. 236–246, 2008.
SOUZA. M.O. et al. Diet supplementation with acai(Euterpe oleracea Mart.) pulp
improves biomarkes of oxidativo stress and the serum lipid profile in rats.Nutrion. v.
26, n 7-8, p. 804-10, 2009.
SUN, X. et al. “Açaí palm fruit (Euterpe oleracea Mart.) pulp improves survival of flies
on a high fat diet”. Exp. Gerontol, v. 45, n. 3, p. 243-51, 2010.
SPADA, P.D. et al. Frozen fruit pulp of Euterpe oleraceae Mart. (Acai) prevents
hydrogen peroxide-induced damage in the cerebral cortex, cerebellum, and
hippocampus of rats. Journal of Medicinal Food, v. 12, n. 5, p. 1084-1088, 2009.

xiii
SPADA, P.D. et al. Antioxidant, Mutagenic, and Antimutagenic Activity of Frozen
Fruits. Journal of Medicinal Food. v. 11, n. 1, p. 144-151, 2008.
SPORN, M. B. and K. T. Liby. Cancer chemoprevention: scientific promise, clinical
uncertainty. Nat Clin Pract Oncol. v.2, n.10, p. 518-25. 2005.
SUNAGAWA, T. et al. Procynidinis from Apples (Malus pumila Mill.) Extende the
Lifespan of Caenorhabits elegans. Biol and Pharmacological, v.77, n. 2, p. 122-
127,2011
TEIXEIRA , L.N; STRIGHETA, P.C; OLIVEIRA, F.A. Comparaçao de métodos para
quantificação de antocianinas. Revista Ceres, v. 55, n. 4, p. 297-304, 2008.
TULLET, J.M. et al. Direct inhibition of the longevity-promoting factor skn-1 by
insulin-like signaling in C. elegans. Cell biology, v. 132, p. 1025-1038, 2008.
VANFLETEREN, J.R.; DE VREESE, A.The gerontogenes age-1 and daf-2 determine
metabolic rate potential in aging Caenorhabditis elegans. FASEB Journal, v. 9, p.
1355-1361, 1995.
VERMEULEN, P.B. et al. Quantification of angiogenesis in solid human tumours: an
international consensus on the methodology and criteria of evaluation. European
Journal of Cancer, v. 32A, n. 14, p. 2474-84, 1996.
VISWANATHAN, M. et al. A role for SIR-2.1 regulation of ER stress response genes
in determining C. elegans life span. Devlopmental Cell, v. 9, p. 605-615, 2005.
WALKER, A.K. et al. A conserved transcription motif suggesting functional parallels
between C. elegans SKN-1 and Cap 'n' Collar-related bZIP proteins. The Journal of.
Biological.Chemistry, v. 275, p. 22166-22171, 2000.
WANG. J. et al. RNAi screening implicates a SKN-1-dependent transcriptional
response in stress resistance and longevity deriving from translation inhibition. PLoS
Genet. v. 6, n. 8, p.1-17, 2010
WILLIAMS, R. J.; SPENCER, J. P. E.; RICE-EVANS, C. Flavonoids: antioxidants or
signalling molecules? Free Radical Biology and Medicine, v. 36, n. 7, p. 838-849,
2004.
WISDOM. S.J,; WILSON. R,; MCKILLOP, J.H,; WALKER J.J. Antioxidant systems
in normal pregnancy and in pregnancy-induced hypertension. , v. 165, n. 6, p. 1701-4,
1991
WU, J. et al. Effects of the narrow band gap on the properties of InN. Physical Review,
v. 66, n. 2002.
UDANI, J.K.; SINGH, B.B.;SINGH, V.J.; BARRETT. M.L. Effects of Açai (Euterpe
oleracea Mart.) berry preparation on metabolic parameters in a healthy overweight
population: A pilot study. Nutrition Journal, v. 10, n. 45, p. 1-7, 2011.

xiv
YEN, K.; NARASIMHAN, S.D.; TISSENBAUM, H.A. DAF-16/Forkhead box O
transcription factor: many paths to a single Fork(head) in the road. Antioxididants
e Redox Signaling, v. 14, n. 4, p. 623-34, 2011.
YOU. Y. et al. Insulin, cGMP, and TGF-β Signal Regulate Food Intake and Quiescence
in C. elegans: A Model for Satiety. Cell metabolism, v. 7, n. 3, 249-257, 2008.
YU, Y. et al. Cinnamomum cassia Bark in Two Herbal Formulas Increases Life Span in
Caenorhabditis elegans via Insulin Signaling and Stress Response Pathways. Plos one,
v. 5, n. 2, p. 1-10, 2010.
ZIPPER, L. M. AND R. T. MULCAHY. The Keap1 BTB/POZ dimerization function is
required to sequester Nrf2 in cytoplasm." J Biol Chem, v.277, n.39, p 36544-52. 2002.





























