Embed Size (px)
Citation preview



Andrea Thompson Da Poian
Debora Foguel
Marílvia Dansa Petretski
Olga Lima Tavares Machado
Volume 1 - Módulos 1 a 3
Bioquímica II
Apoio:

Material Didático
Referências Bibliográfi cas e catalogação na fonte, de acordo com as normas da ABNT.
Copyright © 2005, Fundação Cecierj / Consórcio Cederj
Nenhuma parte deste material poderá ser reproduzida, transmitida e gravada, por qualquer meio eletrônico, mecânico, por fotocópia e outros, sem a prévia autorização, por escrito, da Fundação.
ELABORAÇÃO DE CONTEÚDOAndrea Thompson Da PoianDebora FoguelMarílvia Dansa PetretskiOlga Lima Tavares Machado
COORDENAÇÃO DE DESENHO INSTRUCIONAL Cristine Costa Barreto
DESENHO INSTRUCIONAL E REVISÃOAlexandre Rodrigues AlvesCarmen Irene Correia de OliveiraJosé Meyohas
COORDENAÇÃO DE REVISÃOMaria Angélica Alves
REVISÃO TÉCNICAMarta Abdala
P749b
Poian, Andrea Thompson Da.
Bioquímica 2. v.1/ Andrea Thompson Da Poian. –
Rio de Janeiro : Fundação CECIERJ, 2009.
132p.; 19 x 26,5 cm.
ISBN: 85-89200-75-2
1. Bioquímica. 2. Metabolismo. 3. Bioenergética.
I. Foguel, Debora. II. Petretski, Marílvia Dansa.
III. Machado, Olga Lima Tavares. IV. Título.
CDD: 5722009/1
Fundação Cecierj / Consórcio CederjRua Visconde de Niterói, 1364 – Mangueira – Rio de Janeiro, RJ – CEP 20943-001
Tel.: (21) 2334-1569 Fax: (21) 2568-0725
PresidenteMasako Oya Masuda
Vice-presidenteMirian Crapez
Coordenação do Curso de BiologiaUENF - Milton Kanashiro
UFRJ - Ricardo Iglesias RiosUERJ - Cibele Schwanke
EDITORATereza Queiroz
COORDENAÇÃO EDITORIALJane Castellani
REVISÃO TIPOGRÁFICAEquipe CEDERJ
COORDENAÇÃO DE PRODUÇÃOJorge Moura
PROGRAMAÇÃO VISUALAlexandre d´Oliveira Bruno Gomes Cristiane Matos Guimarães
ILUSTRAÇÃOJefferson CaçadorSalmo Dansa
CAPAEduardo Bordoni
PRODUÇÃO GRÁFICAAndréa Dias FiãesFábio Rapello Alencar
Departamento de Produção

Universidades Consorciadas
Governo do Estado do Rio de Janeiro
Secretário de Estado de Ciência e Tecnologia
Governador
Alexandre Cardoso
Sérgio Cabral Filho
UENF - UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIROReitor: Almy Junior Cordeiro de Carvalho
UERJ - UNIVERSIDADE DO ESTADO DO RIO DE JANEIROReitor: Ricardo Vieiralves
UNIRIO - UNIVERSIDADE FEDERAL DO ESTADO DO RIO DE JANEIROReitora: Malvina Tania Tuttman
UFRRJ - UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIROReitor: Ricardo Motta Miranda
UFRJ - UNIVERSIDADE FEDERAL DO RIO DE JANEIROReitor: Aloísio Teixeira
UFF - UNIVERSIDADE FEDERAL FLUMINENSEReitor: Roberto de Souza Salles


Módulo 1
Aula 1 – Bioenergética - parte I_________________________________ 11 Olga Lima Tavares Machado
Aula 2 – Bioenergética - parte II ________________________________ 15 Olga Lima Tavares Machado
Aula 3 – Conceitos fundamentais do metabolismo __________________ 31 Olga Lima Tavares Machado
Módulo 2
Aula 4 – Fotossíntese I - Princípios gerais _________________________ 41 Marílvia Dansa Petretski
Aula 5 – Fotossíntese II - O processo de fotossíntese _________________ 47 Marílvia Dansa Petretski
Aula 6 – A etapa fotoquímica ou fase clara da fotossíntese ____________ 57 Marílvia Dansa Petretski
Aula 7 – O ciclo de Calvin ou etapa química da fotossíntese ___________ 73 Marílvia Dansa Petretski
Aula 8 – Outros aspectos importantes da fotossíntese________________ 83 Marílvia Dansa Petretski
Módulo 3
Aula 9 – A história da fermentação: do início aos dias de hoje ________ 95 Marílvia Dansa Petretski / Olga Lima Tavares Machado
Aula 10 – Glicólise: oxidação de glicose a piruvato I _______________ 107 Marílvia Dansa Petretski / Olga Lima Tavares Machado
Aula 11 – Glicólise: oxidação de glicose a piruvato II _______________ 119 Marílvia Dansa Petretski / Olga Lima Tavares Machado
Bioquímica II
SUMÁRIO
Volume 1


Introdução ao Metabolismo
Aulas1 - Bioenergética - parte I.
2 - Bioenergética - parte II.
3 - Conceitos fundamentais do metabolismo.
Mód
ulo
1

CEDERJ8
Metabolismo, cujo antepositivo grego metabol – significa
“mudança”, pode ser defi nido como a totalidade dos processos químicos
do organismo. Conseqüentemente podemos admitir que a célula viva seja
uma indústria química em miniatura, onde milhares de reações ocorrem
em nível microscópico.
Você pode pensar no metabolismo celular como um elaborado
mapa rodoviário que representa as reações que ocorrem na célula. As
reações são arranjadas em vias metabólicas intrincadas e bifurcadas, que
transformam moléculas por uma série de etapas. As enzimas aceleram
seletivamente cada etapa no labirinto de reações. Em analogia aos sinais
de trânsito que controlam o fl uxo do tráfego e previnem acidentes,
existem mecanismos que controlam o metabolismo por regulação de
enzimas e, conseqüentemente, mantêm o balanço metabólico, impedindo
defi ciências e excessos de metabólitos.
Poderíamos então começar os nossos estudos apresentando
algumas questões:
1 - Como as enzimas controlam todas estas reações?
2 - Como os seres vivos produzem e armazenam energia?
3 - De onde vem esta energia?
4 - Como um sistema biológico realiza trabalho?
5 - Por que algumas reações químicas liberam e outras
absorvem calor?
6 - Para que servem as vias metabólicas?
Tais questões serão respondidas ao longo de toda a disciplina
Bioquímica II.
Como você pôde observar, energia é o tema central do metabolismo,
por isto é apropriado que comecemos o nosso estudo com uma introdução
à bioenergética, a análise quantitativa de como o organismo ganha e
utiliza energia. A bioenergética é uma parte especial de uma ciência que
estuda as transformações da energia, que é chamada termodinâmica.
INTRODUÇÃO AO METABOLISMO

CEDERJ 9
O módulo Introdução ao metabolismo é composto por três aulas:
Aula 1 – Bioenergética – parte I
Aula 2 – Bioenergética - parte II
Aula 3 – Conceitos fundamentais do metabolismo
Após cada uma das aulas, você verá um resumo da mesma; no
entanto os exercícios; referentes ao módulo serão apresentados após a
Aula 3.


Bioenergética - parte I
Ao fi nal desta aula, você deverá ser capaz de:
• Defi nir conceitos de energia, calor e trabalho.
• Conhecer a primeira lei da termodinâmica.
11AULA
objetivos

Bioquímica II | Bioenergética - parte I
CEDERJ1 2
Uma célula viva é um sistema dinâmico; ela cresce, move, sintetiza
macromoléculas complexas e, seletivamente, direciona substratos para um ou
outro compartimento. Todas estas atividades necessitam de energia; assim, a
célula deve obter energia do meio onde está e deve utilizar esta energia com
a maior efi ciência possível. As plantas a obtêm da luz do sol; animais usam a
energia estocada nas plantas ou em outros animais que consomem.
ENERGIA, CALOR E TRABALHO
Uma palavra que usaremos com freqüência é sistema. Neste
contexto, sistema é uma porção defi nida do espaço que foi selecionada
para o nosso estudo. Pode ser uma simples bactéria, um tubo de ensaio
contendo milhões de células, a Terra ou o universo. Tudo o que estiver
em volta do sistema é o ambiente.
Qualquer sistema pode conter uma certa quantidade de
energia, que chamamos E. Os átomos e moléculas do sistema têm
energia cinética de movimento e energia de vibração e rotação. Além
destas, nós poderíamos incluir toda a energia preservada nas ligações
químicas entre os átomos e a energia de interação não covalente entre
as moléculas.
A menos que um sistema esteja isolado, ele pode, em princípio,
trocar energia com o ambiente e, assim, mudar a sua energia interna.
Esta mudança pode acontecer de duas maneiras. Em uma o calor pode
ser transferido de um sistema para outro. Na outra, o sistema pode
realizar trabalho no seu ambiente ou pode sofrer um trabalho realizado
pelo ambiente. O que denominamos trabalho, neste caso, pode ser, por
exemplo, a contração muscular ou o movimento de um fl agelo de um
protozoário. Certas convenções foram adotadas para descrever estas
trocas de energia:
INTRODUÇÃO

CEDERJ 1 3
AU
LA 1
M
ÓD
ULO
1
Figura 1.1: Esquema geral da fotossíntese.
• Nós denotamos calor pelo símbolo q. Um valor positivo
de q indica que o calor do ambiente foi absorvido pelo
sistema. Um valor negativo de q indica que o sistema
perdeu calor para o ambiente.
• Nós denotamos trabalho pelo símbolo w. Um valor
positivo de w indica que o trabalho é realizado pelo
sistema no seu ambiente. Um valor negativo signifi ca
que o ambiente realizou um trabalho sobre o sistema.
A PRIMEIRA LEI DA TERMODINÂMICA – LEI DA CONSERVAÇÃO DE ENERGIA
Você já deve ter ouvido, em suas aulas de ciências, a seguinte
expressão: “Energia não se cria nem se perde, apenas se transforma”.
A queda d’água de uma cachoeira (energia mecânica) pode acionar
um dínamo (energia elétrica), que por sua vez pode movimentar um
ventilador (energia mecânica), ou acender uma lâmpada (energia
luminosa). Em todas essas transformações a energia não foi criada nem
perdida, apenas transformada.
De onde vem a energia dos reagentes?
A luz do sol é a fonte original de energia. No processo de
fotossíntese, as plantas assimilam a luz do sol através de pigmentos
especiais. Açúcares são sintetizados utilizando essa energia e passam a
ser fonte de energia química que poderá ser utilizada para formação de
outras moléculas. Observe esse processo na Figura 1.1.

Bioquímica II | Bioenergética - parte I
CEDERJ1 4
Em (A) temos a energia cinética transformada somente
em calor;
Em (B) parte da energia cinética é usada para retirar a água (trabalho),
e uma menor quantidade de energia é liberada como calor;
Em (C) a energia cinética estocada no balde d’água suspenso pode
ser usada para dirigir umamáquina hidráulica ou para
qualquer outro trabalho.
Figura 1.2: Transformação de energia.
R E S U M O
Nesta aula, você aprendeu que um sistema pode trocar energia com o
meio ambiente e assim mudar sua energia. Vimos que esta mudança
pode ocorrer na forma de calor ou trabalho. Você deve ter em mente o
seguinte conceito:
• a primeira lei da Termodinâmica é a lei da conservação da energia, ou
seja, energia não se cria, só se transforma.
Veremos, mais adiante, que os seres vivos são transformadores
naturais de energia. Veja exemplos de transformação de energia na fi gura
1.2. Como mencionada anteriormente, a energia de um sistema pode
ser mudada somente na forma de calor ou de trabalho. Tal mudança
da energia interna de um sistema, a qual chamamos ΔE, pode ser
matematicamente representada por:
ΔE = q – w
Essa equação expressa a primeira lei da Termodinâmica. Considere,
por exemplo, alguns processos nos quais uma certa quantidade de calor
é absorvida por um sistema e este realize trabalho sobre o ambiente. No
caso onde q for igual a w, a ΔE será igual a zero.

Bioenergética - parte II
Ao fi nal desta aula, você deverá ser capaz de:
• Analisar as direções das reações químicas a partir do entendimento dos conceitos de entropia, energia livre e da segunda lei da Termodinâmica.
• Conhecer os compostos ricos em energia, ou seja, as fontes de energia dos sistemas biológicos.
• Defi nir e entender a importância das reações acopladas.
2AULA
objetivos

Bioquímica II | Bioenergética - parte II
CEDERJ16
INTRODUÇÃO
AS DIREÇÕES DAS REAÇÕES QUÍMICAS – A SEGUNDA LEI DA TERMODINÂMICA – LEI DA TRANSFERÊNCIA DE ENERGIA
Conceito de entropia
A primeira lei da Termodinâmica não fornece nenhuma
informação sobre o sentido de uma reação química ou sobre o sentido
de transferência de energia. Considere o seguinte exemplo: se colocarmos
um cubo de gelo em um copo d’água à temperatura ambiente, ele derrete.
Por que o inverso não ocorre? Se queimarmos um pedaço de papel, ele
será transformado em CO2 mais água. Mas se misturarmos CO2 com
água não formaremos um pedaço de papel. O tema comum nestes dois
exemplos é este: processos favoráveis ocorrem espontaneamente em
certas direções. Por quê?
A primeira explicação seria a de que os sistemas sempre
caminham para o estado de energia mais baixa. Assim, objetos caem
espontaneamente em virtude do campo gravitacional. Mas tal explicação
não seria adequada para o caso do gelo que se derrete; de fato, energia é
absorvida neste processo. Nós podemos imaginar um outro experimento
muito simples que daria uma indicação clara do que deve estar ocorrendo.
Suponha que nós colocássemos, com muito cuidado, uma camada de
água pura sobre uma solução de sacarose. Depois de algum tempo, nós
verifi caríamos que a solução estaria cada vez mais uniforme, ou seja,
as moléculas de sacarose iriam se distribuir pelo recipiente. Nesse tipo
de transformação nenhuma mudança energética ocorreu. Certamente
o inverso não aconteceria. O que procuramos deixar claro aqui é que
os sistemas formados por moléculas têm uma tendência natural a se
distribuir randomicamente.
Na aula anterior, defi nimos energia, calor e trabalho. Falamos ainda sobre a
primeira lei da Termodinâmica, ou seja, sobre a lei da conservação de energia.
Nesta aula, tentaremos explicar o porquê de algumas reações ocorrerem
espontaneamente e o de outras necessitarem de energia para acontecerem.
Veremos ainda como nosso organismo utiliza reações que liberam energia para
promover reações que necessitam de energia.

CEDERJ 17
AU
LA
MÓ
DU
LO 1
2Figura 2.1: Demonstração das reações espontâneas e não espontâneas.
O grau de distribuição randômica ou grau de desordem é chamado
entropia. Para demonstrarmos a mudança de entropia em um processo
simples, considere a facilidade para desorganizar um quarto e o esforço
para reorganizá-lo. Para isso, veja a Figura 2.1. Diremos então que a
variação de entropia ΔS será a diferença entre a entropia do estado fi nal
(Sf) e a entropia do estado inicial (Si), ou seja,
ΔS = Sf – Si.
Reação espontânea (aumento do grau de desordem, ou seja, aumento da entropia).
Reação não espontânea – requer energia.
Normalmente não medimos a entropia inicial ou fi nal, mas sim
a ΔS. O aumento de entropia é representado por um sinal positivo, e a
redução de entropia por um sinal negativo. Com relação à entropia, dois
aspectos devem ser ressaltados:
a) A entropia total sempre aumenta. A entropia pode aumentar ou
diminuir no sistema ou no ambiente, mas, em todos os processos reais, a
soma entrópica sistema + ambiente é sempre crescente. Pode-se afi rmar
que a entropia do Universo está aumentando continuamente.

Bioquímica II | Bioenergética - parte II
CEDERJ18
b) O signifi cado da quantidade de entropia está associado à
quantidade de energia que não realiza trabalho. Esta representa a energia
inaproveitável em processos reais e é obtida pelo produto da variação da
entropia (ΔS) pela temperatura absoluta do processo. Assim, teremos:
Energia entrópica = TΔS
Esta é também a defi nição formal de entropia.
Para entendermos a segunda lei da Termodinâmica, precisamos
ainda conhecer dois conceitos: entalpia e energia livre.
Conceito de entalpia
A entalpia, que chamamos H, é o conteúdo de energia de um
sistema e pode ser percebido por sua variação, ΔH, que aparece em
qualquer processo. Eis alguns exemplos em Bioquímica:
– Entalpia de reação (ΔHr) – energia liberada ou absorvida quando
ocorre uma reação química.
– Entalpia de solução (ΔHs) – energia liberada ou absorvida
quando uma substância se dissolve.
– Entalpia de formação (ΔHf) – energia liberada ou absorvida
quando se forma um composto.
Conforme o comportamento da entalpia, as reações podem ser
exotérmicas ou endotérmicas. Nas reações exotérmicas, o sistema perde
calor; tal perda é indicada pelo sinal negativo do valor de ΔH. Por
exemplo, quando dissolvemos NaOH em água ocorre aquecimento da
solução em função do calor desprendido, fato que pode ser percebido
pelo tato. Este calor irradia, ou seja, se perde pelo ambiente. Nas reações
endotérmicas, o sistema ganha calor, o que é indicado pelo sinal positivo
de ΔH. Quando dissolvemos o sal NH4Cl em água, ocorre resfriamento
do sistema pelo calor absorvido na dissolução, o que também pode
ser percebido pelo tato. A liberação ou absorção de calor não está
diretamente ligada à realização de trabalho pelo sistema. Este aspecto
será visto ao falarmos de energia livre.
Conceito de energia livre – Energia de Gibbs “G”
As moléculas de uma célula viva possuem energia devido às suas
vibrações, rotações, translações e devido à energia que está estocada
entre as ligações de átomos individuais. Veja uma representação disto
na Figura 2.2.

CEDERJ 19
AU
LA
MÓ
DU
LO 1
2
Na realidade, a energia livre é medida pela sua variação ΔG, e é aquela
capaz de realizar trabalho em processos isotérmicos (processos que
ocorrem à temperatura constante) e isobáricos (processos que ocorrem à
pressão constante). Ela resulta da diferença entre a energia total (entalpia,
ΔH) e a energia inefi caz (quantidade de entropia, TΔS).
ΔG = ΔH - TΔS
Energia que produz trabalho = Energia total - Energia inaproveitável
A equação apresentada é a que rege a segunda lei da Termodinâmica.
Ou seja, um processo ocorrerá espontaneamente quando ΔG for negativo
e não será espontâneo quando ΔG for positivo.
Figura 2.2: Descrição da Energia Livre. A Energia Livre (G) mede a energia da molécula que poderia em princípio ser usada para realizar trabalho à temperatura constante, como ocorre nos seres vivos. G é medido em unidades de quilocalorias/mol ou quilojoules/mol.
Conheça abaixo a defi nição de algumas unidades de medida de energia.
O QUE É UM KJ?
É uma unidade de energia. A unidade básica de energia é o Joule,
nome dado em homenagem a um físico famoso. Um quilojoule
(KJ) corresponde a 1.000 joules. A caloria é uma outra unidade de
energia. Uma caloria corresponde a 4,185 joules. Veja um resumo
destas informações na Figura 2.3.
Unidade básica de energia = joule (J)1quilojoule (KJ) = 1.000 J
1 caloria (cal) = 4,185 J1 caloria alimentar = 1.000 cal = 1kcal = 4.185 J
Figura 2.3: Unidades de energia.

Bioquímica II | Bioenergética - parte II
CEDERJ20
Quando os reagentes possuem uma energia livre menor do que o
produto, como tornar possível a realização da reação?
A solução é usar reagentes que tenham maior energia livre. Veja
alguns exemplos, acompanhando a seqüência de eventos representada
das Figuras 2.4.a a 2.4.d.
Reagentesalternativos
ReagentesProdutos
Ener
gia
livr
ep
adrã
o
DGÉpositivo
0
U
W
YB
D
U+B V+D
V
Reagentesalternativos
Reagentes
Produtos
Ener
gia
livr
ep
adrã
o
DGÉpositivo
0U
W
Y A B C D
A+B C+D
Figura 2.4.a: Os reagentes apresentam energia livre menor do que os produtos. O equilíbrio favorece a formação dos reagentes.
Figura 2.4.b: Os reagentes apresentam energia livre menor do que os produtos. O equilíbrio favorece a formação dos reagentes.

CEDERJ 21
AU
LA
MÓ
DU
LO 1
2
Figura 2.4.c: Os reagentes apresentam energia livre menor do que os produtos. O equilíbrio favorece a formação dos reagentes.
Figura 2.4.d: Os reagentes apresentam energia livre maior do que os produtos. O equilí-brio favorece a formação dos produtos.
Responda você agora à seguinte questão: uma reação endergônica (ΔH positivo), ou seja,
uma reação que requer energia, pode ocorrer espontaneamente dentro de uma célula?
Se você respondeu que sim, desde que o valor em módulo para a entropia (TΔS) seja maior
do que ΔH, você está acompanhando o nosso raciocínio. Bem, podemos ir um pouco mais adiante,
mas antes precisamos entender como a energia livre de um sistema depende da quantidade dos
vários componentes da mistura de reação.
Reagentesalternativos
Reagentes Produtos
Ener
gia
livr
ep
adrã
o
DGÉpositivo
0
WY
BD
W+B X+D
X
Reagentesalternativos Reagentes
Produtos
Ener
gia
livr
ep
adrã
o
DGÉpositivo
0
Y B
D
Y+B Z+D
Z

Bioquímica II | Bioenergética - parte II
CEDERJ22
Energia livre e concentração das substâncias
Se nós tivermos uma mistura contendo NA moles do componente A e
NB moles do componente B e assim sucessivamente, nós podemos escrever:
G = NAGA + NBGB + NCGC + .....
Ou seja, a energia total do sistema é um somatório das energias
parciais molares ou dos potenciais químicos dos vários componentes.
Para facilitar nossos estudos, químicos e bioquímicos determinaram a
energia livre de alguns componentes de interesse químico/bioquímico
quando a concentração dos mesmos era 1 molar. Denominaram-na
energia do estado fundamental, à qual chamaram G0. Em qualquer
outra situação, a energia desses componentes seria proporcional a essa
energia, conforme podemos expressar na seguinte equação:
GA = GA0 + RT ln[A]
GB = GB0 + RT ln[B],
onde R é a constante absoluta dos gases (8.314 J/mol), T é a
temperatura absoluta expressa em graus Kelvin.
A importância destas equações é que elas nos permitem aplicar
princípios termodinâmicos em problemas práticos. Em particular, elas
nos permitem predizer a direção favorável dos processos reais.
Considere a reação reversível; em equilíbrio:
A + B = C + D
A relação entre energia livre e energia padrão é dada pela equação:
ΔG = ΔG 0 + RT ln [C] x [D]
Em quaisquer concentrações de A, B, C e D quando a reação está
em equilíbrio ΔG = 0, e:
K = [C]eq x [D]eq
[A]eq x [B]eq
K é defi nida como a constante de equilíbrio. Este estado será
atingido quando a velocidade de formação dos produtos C e D for igual
à velocidade de formação (v) dos reagentes A e B.
[A] x [B]

CEDERJ 23
AU
LA
MÓ
DU
LO 1
2
reagentes produtos
No equilíbrio v+ = v- e ΔG = 0 , tem-se
ΔG0 = - RT lnK
V+
V-
4 pares dereagentes
Equilíbrio atingido 8 pares deproduto
Figura 2.5: Representação de uma reação em equilíbrio na qual as concentrações dos produtos e dos reagentes são diferentes.
Agora pense e responda! Será que podemos dizer que no equilíbrio químico a concentração
dos reagentes é igual à concentração dos produtos?
A resposta é não. No equilíbrio químico não é necessário que a concentração dos reagentes seja
igual à concentração do produto. O equilíbrio químico ocorre quando a velocidade da reação
nos dois sentidos é idêntica. No exemplo dado, o equilíbrio ocorre em uma situação em que
existem mais moléculas de produtos do que de reagentes. Veja a Figura 2.5:

Bioquímica II | Bioenergética - parte II
CEDERJ24
E a vida? Será que ela existe quando o equilíbrio químico entre ser vivo e meio ambiente
é atingido?
Não. Somente quando um ser morre é que o equilíbrio químico é atingido. Veja a Figura 2.6.
Figura 2.6: O equilíbrio entre sistema e meio é incompatível com a vida.
COMPOSTOS RICOS EM ENERGIA E REAÇÕES ACOPLADAS
O papel central das mudanças de energia em determinadas
direções favoráveis às reações químicas é de grande importância
para a Bioquímica. Para que uma via metabólica ocorra, ela deve ser
termodinamicamente favorável. Entretanto, reações individuais da via
podem ter um ΔG0 positivo e não serem, portanto, favoráveis. Como
então as reações poderiam ocorrer de maneira efi ciente? Em alguns
casos, in vivo, a chave é encontrada no fato de as concentrações entre
os reagentes e produtos se encontrarem muito distantes do equilíbrio.
Assim, se os reagentes se encontram em concentrações muito elevadas
e se os produtos estiverem sendo continuamente removidos, o valor de
ΔG0 pode se tornar ligeiramente negativo, tornando possível a reação.
Outras vezes, poderemos acoplar reações para tornar possível outras
reações. Veja o tópico a seguir.

CEDERJ 25
AU
LA
MÓ
DU
LO 1
2
Reações acopladas
Embora muitas reações possam ser favorecidas pela remoção
efi ciente dos produtos, existem muitos casos em que isto não é sufi ciente.
Há um outro modo muito importante no qual reações intrinsecamente
desfavoráveis podem acontecer. Suponha por exemplo que nós tenhamos
uma reação endergônica que seja parte essencial de uma via que, na sua
totalidade, poderia gerar energia. Busquemos uma situação do nosso
dia-a-dia para você entender melhor este processo: imagine que você
queira montar uma loja para vender um produto de larga aceitação no
mercado. Com certeza você espera ter lucros quando tudo estiver em
pleno funcionamento. Sem dúvida você deverá gastar alguns recursos
para iniciar seu projeto. Assim funcionam algumas vias metabólicas: você
gasta alguma energia para tornar possível uma das etapas da vias, que é
endergônica, para que outras etapas que forneçam energia cubram esse
“gasto” inicial e ainda liberem energia para outras funções metabólicas.
Vejamos como isto fi ca passo a passo:
A ΔG0 = + 10kJ/mol
C D ΔG0 = - 30kJ/mol
Para que a primeira reação aconteça é preciso energia (reação
endergônica), já a segunda reação libera energia. Dizer que as duas
reações ocorrem de maneira acoplada seria o mesmo que somarmos as
equações. Assim teríamos:
A + C B + D
ΔG0 = (+ 10kJ/mol) + (-30 kJ/mol)
ΔG0 = - 20kJ/mol
Acoplar reações endergônicas com reações exergônicas é um dos
princípios mais importantes em Bioquímica. Tais ligações de processos
favoráveis com processos desfavoráveis são usadas não somente para
inúmeras reações, mas também para o transporte de materiais através de
membranas, transmissão de impulsos nervosos e contração muscular.

Bioquímica II | Bioenergética - parte II
CEDERJ26
Compostos fosfatos de alta energia como estoque de energia
Como vimos, acoplar reações é muito importante nos processos
vitais; por isso existem diversas moléculas que conservam energia em
sua estrutura e, assim, quando clivadas, podem liberar essa energia para
permitir que uma reação endergônica possa acontecer. As moléculas
consideradas “estoque” de energia mais importantes no nosso organismo
são compostos fosforilados, que por hidrólise liberam seu grupo fosfato.
Alguns deles, como o foso-enol-piruvato (PEP), a creatina-fosfato (CP) e
a adenosina tri-fosfato (ATP), quando hidrolisados, liberam uma grande
quantidade de energia (ΔG0 muito negativo). Talvez o mais importante
destes compostos, e que você poderá encontrar com maior freqüência
nos livros, seja o ATP. Por isso a estrutura dessa molécula, bem como
as possíveis reações de hidrólises que ela pode sofrer, são apresentadas
na Figura 2.7.a, 2.7.b e 2.7.c. Como você pode observar, a hidrólise da
molécula de ATP pode proceder de duas maneiras:
1) o fosfato terminal pode ser clivado, formando uma molécula de
adenosina difosfato (ADP) e uma molécula de fosfato inorgânico
(Pi) (Figura 2.7.b);
2) a clivagem pode ocorrer na segunda ligação fosfo-di-éster para
formar uma molécula de adenosina mono-fosfato (AMP) e uma
molécula de pirofosfato inorgânico (PPi) (Figura 2.7.c). A primeira
reação ocorre com maior freqüência in vivo, mas a energia liberada
em ambas as clivagens é a mesma. A molécula de PPi também pode ser
clivada e, neste caso, uma outra parcela de energia pode ser liberada.
A razão pela qual a molécula de ATP é considerada uma das mais
importantes moléculas de transferência de energia é que ela fi ca bem
no meio da escala de potenciais de transferência de energia. Por isso,
quando reações altamente exergônicas acontecem no nosso organismo,
ou mesmo quando a luz incide sobre os organismos fotossintetizantes,
podemos guardar parte dessa energia formando uma molécula de ATP
e, quando necessitamos de energia para a realização de uma reação
exergônica ou para a realização de um trabalho, podemos utilizar a
energia armazenada nessa molécula.

CEDERJ 27
AU
LA
MÓ
DU
LO 1
2
Figura 2.7.a: Estrutura da adenosina trifosfato (ATP).
Figura 2.7.b: Hidrólise do ATP com clivagem do fosfato terminal. • Libera 31 a 55 kJ/mol como calor.• Libera adenosina difosfato e fosfato inorgânico.
Figura 2.7.c: Hidrólise do ATP com clivagem na segunda ligação fosfo-di-éster.• Libera 31 a 55 kJ/mol como calor.• Libera adenosina monofosfato e pirofosfato inorgânico.
NH2
NN
N N
CC
CHCCH
H H H
O
H
OHOH
H C2O O OAdenina
Ribose
Fosfato
O PO- O- O-
O-

Bioquímica II | Bioenergética - parte II
CEDERJ28
Resumindo, podemos dizer: a energia de acoplamento é usada para
permitir que uma reação desfavorável aconteça. O processo substitui a
reação desfavorável por duas reações favoráveis. Na primeira etapa, o
ATP reage para formar um intermediário rico em energia. Na segunda
etapa, o intermediário libera essa energia para formação do produto.
Como as células refazem as moléculas de ATPs gastas?
Elas usam reagentes que possuem mais energia do que o ATP.
Usam, por exemplo, a energia da luz, captada pelos cloroplastos. As
plantas e os animais oxidam açúcares com a ajuda das mitocôndrias,
liberando energia para a formação do ATP. Um detalhamento desses
processos será visto nas aulas de fotossíntese, de glicólise, ciclo de Krebs
e fosforilação oxidativa.
Na próxima aula, após defi nirmos alguns conceitos importantes
para o entendimento do metabolismo, voltaremos a falar da importância
do ATP como molécula transportadora de energia.

CEDERJ 29
AU
LA
MÓ
DU
LO 1
2RESUMO
Nesta aula, você aprendeu os conceitos de entropia, entalpia e energia
livre de Gibbs. Você viu como essas grandezas se relacionam, ou seja, você
aprendeu a segunda lei da Termodinâmica. Você aprendeu a identifi car o
sentido das reações químicas e a importância dos compostos ricos em energia
e das reações acopladas. Vamos resumir esses conceitos.
• O conteúdo de energia de um sistema é denominado entalpia; no entanto,
normalmente medimos sua variação durante uma transformação, ou seja, o
ΔH. Quando ΔH é positivo, a reação é endotérmica, e quando ΔH é negativo,
a reação é exotérmica.
• Entropia é o grau de desordem; normalmente medimos a sua variação
durante um processo, a uma determinada temperatura, ou seja, medimos o
TΔS; a entropia do universo é sempre crescente.
• A energia livre das moléculas é denominada G. Ela é a energia de rotação,
de translação e a energia contida entre as ligações químicas. Neste caso,
também, em Bioquímica, usualmente medimos a sua variação durante uma
reação química, ou seja, medimos o ΔG.
• A segunda lei da Termodinâmica dá informações sobre a espontaneidade de
uma reação química, a qual pode ser visualizada pelo sentido do ΔG. Vimos
que ΔG = ΔH - TΔS; para um ΔG negativo a reação é espontânea e para um
ΔG positivo a reação não é espontânea.
• Se acoplarmos uma reação exotérmica a uma reação endotérmica,
poderemos tornar possível a realização de uma reação termodinamicamente
desfavorável.
• O ATP é o composto rico em energia mais utilizado nos organismos vivos. A
energia contida em suas ligações é utilizada nas reações termodinamicamente
desfavoráveis. Ele é reconstituído às custas da energia solar ou da energia
contida em outras moléculas orgânicas.


Conceitos fundamentais do metabolismo
Ao fi nal desta aula, você deverá ser capaz de:
• Conhecer as funções do metabolismo.
• Dividir o metabolismo em estágios ressaltando a importância de cada um deles.
• Conhecer a organização das vias metabólicas.
• Conceituar catabolismo e anabolismo.
• Identifi car os principais mecanismos de regulação metabólica.
3AULA
objetivos

Bioquímica II | Conceitos fundamentais do metabolismo
CEDERJ32
INTRODUÇÃO
FUNÇÕES DO METABOLISMO
O metabolismo desempenha quatro funções básicas:
1 - Obter energia química do ambiente, por captura de energia solar ou
por degradação de nutrientes ricos em energia.
2 - Converter moléculas de nutrientes (exógenas) em moléculas
características do próprio organismo (endógenas).
3 - Polimerizar precursores monoméricos em produtos poliméricos,
como proteínas, ácidos nucléicos, lipídios, polissacarídeos e outros
componentes celulares.
4 - Sintetizar e degradar biomoléculas requeridas em funções celulares
especializadas.
ESTÁGIOS DO METABOLISMO
O metabolismo pode ser dividido em estágios que refl etem o grau
de complexidade ou tamanho das moléculas geradas
No nível 1, temos as reações químicas de conversão de metabólitos
poliméricos, como polissacarídeos, lipídeos e proteínas, em seus
constituintes monoméricos, como monossacarídeos, ácidos graxos +
glicerol e aminoácidos, respectivamente.
No nível 2 do metabolismo, esses monômeros são quebrados em
intermediários simples, como por exemplo:
O destino dos componentes da dieta, após a digestão e absorção, constitui
o metabolismo intermediário. Assim, ele procura não somente descrever a
via metabólica percorrida individualmente pelas moléculas, mas também
compreender suas inter-relações e os mecanismos que regulam o fl uxo de
metabólitos através das vias metabólicas, envolvendo um amplo domínio.
Glicose Piruvato
Ácidos graxos acetil-CoA

CEDERJ 33
AU
LA
MÓ
DU
LO 1
3
2COH O
2
2
Carregadores de elétronsreduzidos e ATP
GordurasProteínasGlicídeos
Carregadoresde elétronsreduzidos eATP
Aminoácidos Ácidos graxos + glicerolAcúcares simples
(principalmente glicose)
Carregadoresde elétronsreduzidos eATP
Piruvato
Acetil-Co A
Cíclo doácidocítrico
Estágio 3
Estágio 2
Estágio 1
Figura 3.1: Esquema geral dos três estágios do metabolismo.
As reações do metabolismo intermediário não envolvem um
templete de ácido nucléico. A informação necessária para especifi car
cada reação vem da estrutura da enzima que catalisa aquela reação.
Um exemplo de reações metabólicas que dependem de um templete de
ácido nucléico e, portanto, não é considerado metabolismo intermediário,
é a síntese de proteínas.
No nível 3, se tratarmos de organismos aeróbicos (aqueles que
necessitam de oxigênio para sobreviver), a principal via é o ciclo de
Krebs, onde os intermediários do nível 2 sofrem degradação completa
a CO2, H2O.
Por exemplo:
Acetil-CoA CO2 e H2O
Essas relações são esquematizadas na Figura 3.1.
Açúcares simples
(principalmente glicose)

Bioquímica II | Conceitos fundamentais do metabolismo
CEDERJ34
Seus metabólitos são formados seqüencialmente até o produto
fi nal (F). Neste caso, o aumento da concentração do substrato (A), ou o
aumento de qualquer intermediário (B, C, D ou E) resulta no aumento
da concentração do produto (F). Um exemplo de via metabólica linear
é a glicólise, a via de degradação da glicose (veja Aulas 9 e 10). Nesta
via, a glicose é o substrato inicial e o piruvato é o produto fi nal.
Nas vias metabólicas cíclicas, o produto fi nal (F) pode regenerar
o substrato inicial (A). Um exemplo desta via é o ciclo de Krebs (veja
Aulas 12 e 13). Este tipo de via metabólica pode ser representada da
seguinte forma:
ORGANIZAÇÃO DAS VIAS METABÓLICAS
O metabolismo é formado por vias metabólicas que podem ser
lineares ou cíclicas
As vias metabólicas lineares podem ser representadas da
seguinte forma:
A B C D E F
A
D
B
CE
F
As vias cíclicas têm particularidades. Ao contrário das vias lineares, o aumento da
concentração do substrato (A) não resulta necessariamente no aumento da concentração do
produto (F). Na verdade, o aumento da concentração de qualquer um dos metabólitos desencadeia
ativação ou aumento da velocidade da via como um todo.
No metabolismo existem pontos de convergência de vias metabólicas diferentes. A molécula
acetil-CoA é um desses pontos de convergência. Isso signifi ca que várias vias metabólicas têm como
produto fi nal acetil-CoA, ou em alguns casos o produto fi nal da via é convertido nessa molécula.
Chamamos então vias metabólicas convergentes aquelas que têm um produto comum.
A B C D E
A B C D E
H

CEDERJ 35
AU
LA
MÓ
DU
LO 1
3
Figura 3.2: Representação esquemática dos principais aspectos do catabolismo e do metabolismo.
Por outro lado, nas vias metabólicas divergentes, um mesmo
substrato pode sofrer reações distintas que vão determinar a rota
metabólica daquela molécula.
H I J K
D E F G
A B C
Qualquer participante de uma reação metabólica, seja ele substrato,
intermediário ou produto, é chamado metabólito, e as moléculas que
não podem mais ser utilizadas pelo organismo e, portanto, devem ser
eliminadas são denominadas catabólitos. Assim, no nosso organismo as
moléculas de CO2 que eliminamos pela respiração e a molécula de uréia
que eliminamos na urina são exemplos de catabólitos.
CATABOLISMO E ANABOLISMO
O metabolismo pode ainda ser dividido em duas principais
categorias:
Anabolismo – processos que envolvem primariamente a síntese de
moléculas orgânicas complexas. Esses processos necessitam de energia.
Catabolismo – processos relacionados à degradação de substâncias
complexas com concomitante geração de energia.
Veja um resumo destas informações na Figura 3.2.

Bioquímica II | Conceitos fundamentais do metabolismo
CEDERJ36
Esta distinção é arbitrária, especialmente porque muitos substratos
das vias anabólicas são formados como intermediários nos processos
catabólicos e vice-versa. Durante o catabolismo, observamos a oxidação
das moléculas, e durante o anabolismo verifi camos redução das moléculas.
Relembre que redução é um processo no qual ocorre ganho de elétrons
e oxidação envolve perda de elétrons. No nosso organismo existem
moléculas que auxiliam algumas enzimas nos processos de óxido-redução
e, portanto, são denominadas coenzimas. São exemplos de coenzimas a
nicotina adenina di-nucleotídeo (NAD), a fl avino-adenino dinucleotídeo
(FAD). Veja a estrutura de algumas coenzimas, bem como o mecanismo
pelo qual elas transportam moléculas de hidrogênio, na Figura 3.3.
Quando essas coenzimas estão associadas ao hidrogênio, dizemos que
elas estão reduzidas; quando elas perdem esses hidrogênios, dizemos que
a coenzima está oxidada. Na Figura 3.2 você pode observar que durante
o catabolismo ocorre a redução das coenzimas, ou seja, transformação de
NAD e FAD em NADHH e FADH2, respectivamente. Por outro lado, no
anabolismo ocorre a oxidação de NADPHH em NADP. Em resumo, se
quisermos acompanhar os processos de óxido-redução podemos observar
o teor de hidrogênio de uma molécula. Durante a oxidação, a molécula
perde hidrogênios. Observe a Figura 3.3.
C
H
H H
H
Metano
C
H
H OH
H
Metanol
C
H
O
H
Formaldeído
C
C
H
O
OO
HO
Ácido fórmico
OX
IDA
ÇÃ
O
Figura 3.3: Processo de oxidação da molécula de metano à molécula de dióxido de carbono.

CEDERJ 37
AU
LA
MÓ
DU
LO 1
3
Ao contrário da primeira impressão, as vias de síntese e degradação
de uma dada molécula não são as mesmas.
Quase sempre os dois caminhos são bastante distintos um do
outro. Embora apresentem reações enzimáticas e intermediárias comuns,
são vias diferentes por possuírem diferentes enzimas catalisando pelo
menos parte de suas reações.
Essa é uma estratégia para evitar a redundância e o ciclo vicioso.
Enzimas chaves vão ditar a regulação simultânea das vias opostas.
PRINCIPAIS MECANISMOS DE REGULAÇÃO METABÓLICA
Controle dos níveis das enzimas
As concentrações das diversas enzimas intracelulares podem
ser diferentes; assim, enzimas das vias centrais de produção de energia,
como as da via glicolítica (Aula 10), devem ser mais abundantes do que
as que realizam funções limitadas ou especializadas dentro da célula.
Verifi camos ainda que os níveis de uma mesma proteína podem variar
em função das necessidades de um determinado momento. Imagine, por
exemplo, o que ocorre durante a amamentação: durante este período, os
níveis das enzimas envolvidas na síntese da galactose (açúcar do leite)
e da caseína (proteína do leite) estão muito aumentados, retornando a
níveis basais quando a produção do leite não é mais necessária.
Controle da atividade da enzima
Basicamente, a atividade de uma enzima pode ser controlada de
duas maneiras: 1) pela interação com um ligante; 2) por modifi cação
covalente de uma molécula. Ligantes que controlam a atividade de uma
enzima podem ser poliméricos, como por exemplo, interações proteína-
proteína. No entanto, nós nos concentraremos nas interações com
moléculas de baixo peso molecular, principalmente substratos e efetores
alostéricos (veja a aula sobre enzimas na Bioquímica I). Assim, de um
modo geral, quando a concentração de um substrato estiver aumentada
dentro de uma célula, a atividade da enzima que utiliza este substrato
será aumentada e, uma vez formada a quantidade de produto desejado,
mecanismos de inibição desta enzima serão ativados. Muitas vezes, o
produto fi nal de uma seqüência de reações poderá se associar à primeira
enzima da via, inibindo-a.

Bioquímica II | Conceitos fundamentais do metabolismo
CEDERJ38
Modifi cações covalentes de uma enzima representam uma outra
forma efetiva de regulação da atividade de uma determinada enzima.
Você verá que a adição de um grupamento fosfato (fosforilação) a uma
enzima acarretará para algumas delas o aumento de sua atividade;
no entanto, para outras, a fosforilação poderá acarretar inibição da
atividade. Fique atento quando, mais adiante, você for estudar os
mecanismos de ativação das enzimas envolvidas na degradação e
biossíntese do glicogênio (Aulas 27 e 28 desta disciplina)!
Controles através de ligações covalentes normalmente estão
associados com regulações em cascata, ou seja, a modifi cação ativa
uma enzima, a qual ativa uma segunda enzima, que pode ativar uma
terceira enzima, que fi nalmente atua sobre um substrato. Na Figura 3.4,
observamos que cada molécula da primeira enzima ativada pode ativar
dez moléculas da segunda enzima, ou seja, uma amplifi cação do sinal em
dez vezes, e que cada uma dessas pode ativar dez moléculas da terceira
enzima, amplifi cação do sinal em cem vezes, e por fi m cada uma destas
pode ativar dez moléculas da quarta enzima. Teremos no fi nal deste
processo uma amplifi cação de mil vezes do sinal inicial.
Figura 3.4: Reações de ativação enzimática em cascata.
Enzimainativa A
Sinalmodificador
Enzimainativa A
Enzimainativa C
Substrato Produto
Enzimainativa C
Enzimaativa B
Enzimaativa B
amplificação 10 X
amplificação 100 X
amplificação 1000 X

CEDERJ 39
AU
LA
MÓ
DU
LO 1
3
Controle por compartimentalização
Outra estratégia efi ciente é a compartimentalização. Em geral, a
via de síntese de uma dada molécula ocorre em um compartimento celular
distinto daquele onde ocorre a sua via de degradação. Por exemplo, a
via de degradação de ácidos graxos (Aula 24) ocorre nas mitocôndrias,
enquanto a via de síntese de ácidos graxos (Aula 26) ocorre no citoplasma
das mesmas células.
Regulação hormonal
Sobrepondo e permeando os mecanismos que operam dentro
de uma célula estão as mensagens disparadas de um outro tecido ou
órgão. Tais mensagens são transmitidas por hormônios. Um hormônio
é um mensageiro químico, uma substância sintetizada em uma célula
especializada que é transportada pela circulação até uma célula alvo, onde
interage com receptores, resultando em mudanças específi cas na célula
alvo. Estas mudanças são efetivadas por mecanismos regulatórios, tais
como: mudanças na atividade de uma enzima em particular, mudanças na
concentração de uma enzima e mudanças na permeabilidade da membrana
para um substrato em particular. A ação dos hormônios será discutida
em maiores detalhes no último módulo desta disciplina.

Bioquímica II | Conceitos fundamentais do metabolismo
CEDERJ40
RESUMO
Nesta aula, você aprendeu que o metabolismo é o conjunto de reações que
ocorre dentro da célula, tendo como funções obter energia química do ambiente;
converter moléculas de nutrientes (exógenas) em moléculas características do
próprio organismo (endógenas); sintetizar e degradar biomoléculas requeridas
em funções celulares especializadas.
O metabolismo pode ser dividido em três estágios; no nível 1 ocorrem as reações
de conversão de metabólitos poliméricos em seus constituintes monoméricos;
no nível 2 estes monômeros são quebrados em intermediários simples tais
como moléculas de acetil coA; no nível 3 essas moléculas são transformadas em
catabólitos para excreção.
As vias metabólicas podem ser lineares ou cíclicas, convergentes ou divergentes.
As reações de degradação, normalmente convergentes e associadas com liberação
de energia, e a formação de coenzimas reduzidas fazem parte do catabolismo,
enquanto as reações dependentes de energia e com utilização de coenzimas
reduzidas fazem parte do anabolismo.
O controle do metabolismo é feito por regulação dos níveis e das atividades das
enzimas. Este controle é norteado por regulações alostéricas, por modulações
covalentes, por compartimentalização e por regulação hormonal.

Fotossíntese I - Princípios gerais
4AULA

Bioquímica II | Fotossíntese I - Princípios gerais
CEDERJ4 2
INTRODUÇÃO Um ecossistema consiste do conjunto de organismos de uma determinada área,
mais os fatores abióticos com os quais eles interagem; em outras palavras, uma
comunidade e o seu ambiente físico. Podemos considerar como um ecossistema
um terrário ou um lago, uma fl oresta ou a biosfera inteira.
Um ecossistema mantém-se através de dois processos que serão discutidos
tomando-se a biosfera como ecossistema referência. São eles:
A energia necessária para a manutenção da vida entra no
ecossistema na forma de luz solar ou energia luminosa. Esta energia é
convertida em energia química através da fotossíntese. Os organismos
capazes de realizar a fotossíntese são chamados autotrófi cos.
• Fluxo de energia e
• Ciclos biogeoquímicos.
Fotossíntese:
Energia luminosa Energia química
Os autotrófi cos convertem parte da energia solar que chega à
superfície da Terra em energia química, e esta energia química pode ser
estocada nas células ou convertida em trabalho celular. Como vimos
anteriormente, este processo é baseado em transdução de energia.
Energia luminosa Energia química Trabalho celular
Normalmente, na natureza, os elementos químicos, como carbono,
nitrogênio etc. são reciclados entre os componentes abióticos e bióticos
do ecossistema.
Organismos fotossintéticos ou fotossintetizantes adquirem estes
elementos na forma inorgânica, do ar e solo, e os assimilam para a
construção de moléculas orgânicas, algumas das quais serão consumidas
por animais.
Os elementos retornam na forma inorgânica ao solo e ar através
do metabolismo de plantas e animais e por outros organismos (como
bactérias e fungos) que degradam lixo orgânico e organismos mortos
(os decompositores).
Figura 4.1: Um gafanhoto se alimentando de uma folha. Esta imagem repre-senta a relação de inter-dependência metabólica entre os organismos.

CEDERJ 4 3
AU
LA 4
M
ÓD
ULO
2
Figura 4.2: O ciclo do carbono. Vegetais e outros organismos convertem CO2 e água em glicose e O2 através do processo de fotossíntese. Animais e outros organismos heterotrófi cos utilizam glicose e O2 no processo de respiração celular, devolvendo CO2 e H2O à atmosfera.
Fluxo de energia e reciclagem química estão relacionados porque
ambos ocorrem por transferência de substâncias através dos diferentes
níveis alimentares (trófi cos) do ecossistema. Assim, o carbono que hoje
é parte do corpo de um animal amanhã poderá estar na forma de CO2.
Este gás será utilizado por vegetais e o carbono voltará à sua forma
orgânica como amido, por exemplo. Esta molécula (que você conhece
da Bioquímica I) é fonte de alimento para muitos animais e, assim, o
carbono voltará a fazer parte da constituição de algum consumidor de
amido. Não há desperdício. A matéria é reciclada.
Entretanto, a energia (diferente da matéria) não pode ser
reciclada, um ecossistema deve sobreviver graças a um contínuo infl uxo
de energia nova de uma fonte externa (o sol).
Nossas próximas aulas falam de como se dá esse fl uxo de energia
que mantém a biosfera. Temos por objetivo mostrar a interdependência
entre os diferentes processos usados pelos organismos para obtenção de
energia e como essa interdependência é crucial para o equilíbrio biológico
no nosso planeta.
+CO +H O2 2
Glicose +O2
respiração
fotossíntese

Bioquímica II | Fotossíntese I - Princípios gerais
CEDERJ4 4
O CICLO DO CARBONO
Na natureza, CO2 e H2O são combinados para formar carboidratos
na fotossíntese.
CO2 + H2O Carboidratos (fotossíntese)
Tanto nos próprios vegetais como nos herbívoros, estes
carboidratos são reoxidados para gerar CO2 e H2O, no processo
conhecido como respiração celular. Parte da energia é aprisionada na
forma de ATP.
Carboidratos CO2 + H2O (respiração celular)
ATP
LUZ
Os vegetais adquirem CO2 da atmosfera, por difusão através dos
estômatos de suas folhas, e o incorporam à matéria orgânica que será
parte de sua biomassa. Parte desse material orgânico torna-se fonte de
carbono para os organismos heterotrófi cos (consumidores). A respiração
de autotrófi cos e heterotrófi cos retorna o CO2 para a atmosfera (veja
a Figura 4.2).
O ciclo do carbono é relativamente rápido porque as plantas
têm alta demanda de CO2, mas o gás está presente na atmosfera a
uma concentração relativamente baixa (~0,03%). A cada ano, plantas
removem 1/7 do CO2 da atmosfera, e isto é equilibrado pela respiração,
que devolve esse CO2 à atmosfera. Parte do carbono pode ser desviado
desse ciclo por um longo período de tempo. Isto acontece quando ele é
acumulado em madeira e outros materiais orgânicos duráveis. O processo
de decomposição desses materiais eventualmente recicla esse carbono
para a atmosfera na forma de CO2, mas o fogo pode oxidar o material
orgânico acumulado em madeira a CO2 mais rapidamente.
Pense na situação atual: o CO2 fi xado na forma de combustíveis fósseis retorna à
atmosfera rapidamente, certo? Ao mesmo tempo, diminui a cobertura vegetal. O que
acontece com o nível de CO2 atmosférico? Você pode imaginar as conseqüências disso?
!

CEDERJ 4 5
AU
LA 4
M
ÓD
ULO
2
Alguns processos, entretanto, podem remover carbono por
milhões de anos. Em alguns ambientes, o material orgânico é acumulado
mais rapidamente do que os decompositores podem quebrar. Sob estas
condições, no passado, estes depósitos formaram carvão e petróleo, os
combustíveis fósseis.
FLUXO DE ENERGIA
A cada dia a Terra é bombardeada por 1022 joules de energia solar,
o equivalente à energia de 100 milhões de bombas atômicas do tamanho
da que foi lançada sobre Hiroshima na 2ª Guerra Mundial. A maior parte
dessa energia é absorvida, desviada ou refl etida pela atmosfera ou pela
superfície da Terra. Da radiação (luz visível, que atinge folhas e algas),
somente 1% é convertido em energia química pela fotossíntese – a efi ciência
varia com o tipo de planta, o nível de luz e outros fatores. Mesmo assim,
isto é sufi ciente para a produção de, aproximadamente, 170 bilhões de
toneladas de matéria orgânica por ano, globalmente. Vamos discutir aqui
como acontece essa conversão.
A fotossíntese, portanto, não somente fornece carboidratos para obtenção de energia
por plantas e animais, mas é também o principal caminho pelo qual o carbono retorna
à biosfera e é a principal fonte de O2 na atmosfera terrestre.
!
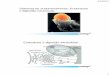
EVOLUÇÃO, OXIGÊNIO E FOTOSSÍNTESE
Acredita-se que, antes do surgimento dos organismos
fotossintéticos, a atmosfera terrestre era pobre em O2 e rica em CO2.
Organismos pré-fotossintéticos devem ter utilizado moléculas ricas
em energia, presentes no meio e sintetizadas abioticamente. Sem a
fotossíntese, estas fontes de energia teriam se esgotado e teria sido
impossível a sobrevivência de organismos vivos. Os registros fósseis
sugerem que fotossintetizantes apareceram há 3 bilhões de anos.
A conversão gradual, de atmosfera primitiva não oxidante a
uma atmosfera terrestre oxidante, permitiu o boom de organismos com
metabolismo aeróbico e a evolução de animais (veja a Figura 4.3).

Bioquímica II | Fotossíntese I - Princípios gerais
Níveis de oxigênio na atmosfera
(%)
Bilhões de anos
Formação da terra
Formação de oceanos
e continentes
Primeiras células vivas
Primeiras células
fotossintetizante
Liberação de O2 a partir da
fotólise da água
Respiração celular
Origem das células
eucarióticas fotossintetizantes
Primeirasplantas
e animais multicelula
Primeirasvertebrados
Hoje
20
10 Rápido acúmulo de O2 pela saturação do Fe12
nos oceanos
0 1 2 3 4
CEDERJ4 6
Figura 4.3: Seqüência de eventos evolutivos e sua relação com os níveis de O2 atmosférico.
Hoje a fotossíntese representa a última e única fonte de energia
direta ou indireta para a maior parte dos organismos vivos (exceto
bactérias terosófi las, que usam H2S e H2 como fonte de energia).
Você viu nesta aula a importância da Fotossíntese para a manutenção da vida. Nas próximas
aulas você verá os detalhes deste processo.

Fotossíntese II –O processo de fotossíntese 5AULA

Bioquímica II | Fotossíntese II - O processo de fotossíntese
CEDERJ48
INTRODUÇÃO
LOCALIZAÇÃO
Cloroplastos
Em todas as plantas e algas, a fotossíntese está localizada em
estruturas especializadas chamadas cloroplastos. Em plantas, a maior
parte dos cloroplastos é encontrada em células mesófi las, logo abaixo da
superfície (cutícula) da folha (veja a Figura 5.1); cada célula contém entre
20 a 50 dessas organelas. As algas também têm cloroplastos, mas em
geral apresentam um único cloroplasto muito grande em cada célula.
Como as mitocôndrias, os
cloroplastos são organelas semi-
autônomas: carregam seu próprio DNA,
que codifi ca algumas de suas proteínas, e
os ribossomos necessários para a tradução
de seus RNAs mensageiros. Existem
evidências de que cloroplastos evoluíram
de organismos unicelulares similares a
cianobactérias. Como os procariontes
fotossintetizantes não possuem cloroplastos,
mas possuem membranas que desempenham
o mesmo papel das membranas de
cloroplastos, as cianobactérias lembram
cloroplastos de vida livre.
Os cloroplastos são formados por uma membrana externa
livremente permeável e uma membrana interna seletivamente permeável
(veja as Figuras 5.2 e 5.3). A membrana interna envolve um material
análogo à matriz mitocondrial – que é chamado estroma.
Na aula passada, falamos da importância da fotossíntese para a vida no planeta
e já sabemos um pouco sobre os eventos evolutivos que culminaram na vida
como a conhecemos hoje. Agora falaremos dessa via metabólica, sua localização
celular, o que ocorre em cada uma das etapas e, é claro, um pouco da história.
Vamos destacar as enzimas chaves quando chegar a hora. Não se preocupe com
os nomes pontualmente, mas prefi ra sempre entender o processo e a lógica
metabólica e fi siológica. Os nomes virão como conseqüência.
Figura 5.1: Microscopia óptica de campo claro de células vegetais. Os cloroplastos aparecem como pequenos círculos em tom mais escuro.
Cloroplastos
Você conhece mito-côndrias e cloroplastos do ensino médio e também das aulas de Biologia Celular.
!

CEDERJ 49
AU
LA
MÓ
DU
LO 2
5
Mergulhados no estroma estão os tilacóides, estruturas formadas
por membranas que lembram sacos. Estes estão organizados empilhados
como moedas, formando unidades chamadas grana.
Grana são irregularmente conectados por extensões dos tilacóides
chamadas lamela. A membrana dos tilacóides envolve um espaço interior
chamado lúmen do tilacóide.
A divisão de trabalho no cloroplasto é única. Absorção de luz
e todas as reações que dependem de luz ocorrem na membrana dos
tilacóides. As outras reações ocorrem no estroma.
Singular = granumPlural = grana
Figura 5.2: Estrutura de um cloroplasto.
Granum
Membrana Interna
EstromaTilacóide
Membrana Externa
Lamela
Figura 5.3: Microscopia eletrônica de transmissão de uma célula vegetal (A) e um cloroplasto em destaque (B).
Parede Celular
Espaçocom ar
Núcleo
Cloroplasto
Vacúolo
Citoplasma
Membrana plasmática
Grana
Membrana do tilacóide
Estroma
Membranas interna eexterna do cloroplasto
Grânulode amido
a b

Bioquímica II | Fotossíntese II - O processo de fotossíntese
CEDERJ50
AS ETAPAS
A fotossíntese é descrita, de uma maneira bem geral, como:
onde (CH2O)n representa um carboidrato qualquer.
CO2 + H2O (CH2O)n + O2,
luz
Entretanto, o processo pode ser dividido em duas etapas:
1) Reações do claro ou fase clara ou etapa fotoquímica.
2) Reações do escuro ou fase escura ou etapa química.
Figura 5.4: Equação geral do processo de fotossíntese.
CO2 + H2O + energia solar açúcar + CO2

CEDERJ 51
AU
LA
MÓ
DU
LO 2
5
A etapa fotoquímica
Nesta etapa, a molécula de água é quebrada. O oxigênio de duas
moléculas de água se combinam e formam O2. Os hidrogênios restantes
se associam com moléculas chamadas aceptores de elétrons. Neste caso,
o aceptor é o NADP (Nicotinamida adenina dinucleotídeo fosfato), cuja
forma reduzida é representada por NADPH. (veja a Figura 5.5).
Figura 5.5: Nicotinamida Adenina dinucleotideo fosfato (NADP).
O
O
O
O
H
H
H
H
H
H
H
H
H
HOHO
HO
C
N+N+
N
CH2
CH2
NH2
NH2
O
O
P
P
O-
O-
O
O
OPO H3 22-
N
N+
Assim, já na primeira etapa, a célula libera oxigênio (pela quebra
da água) e forma um composto de alta energia (o NADPH). Na etapa
fotoquímica há também a síntese de outra molécula de alta energia,
conhecida como ATP (adenosina trifosfato). Essa etapa pode, então, ser
representada da seguinte maneira:
Dizemos então que, nesta fase, há conversão de energia solar em
energia química (NADPH e ATP).
H2O + NADP + ADP + Pi ½ O2 + H+ + NADPH + ATP
luz
RESUMINDO
Na etapa fotoquímica, a energia luminosa é utilizada para quebrar
a molécula de água e sintetizar NADPH e ATP.

Bioquímica II | Fotossíntese II - O processo de fotossíntese
CEDERJ52
Esquema 1 ATP + NADPH + CO2 (CH2O)n + NADP + ADP + Pi
Esquema 2 ATP + NADPH + 6CO2 C6H12O6 + NADP + ADP+ Pi
Etapa química ou Ciclo de Calvin
A etapa química não depende diretamente da energia solar.
Ela utiliza os produtos da etapa anterior para formar carboidratos.
Lembre-se sempre de que sintetizar moléculas custa caro. A etapa
química é uma fase de síntese de glicose e, portanto, precisa de energia.
Esta energia é transferida pelas moléculas de alta energia produzidas
na etapa fotoquímica (NADPH e ATP). Além de energia, é preciso
matéria-prima para a construção. A matéria-prima é o CO2, que fornece
os elementos necessários à construção do esqueleto carbonado. A seguir,
representamos um esquema da etapa química, no qual o carboidrato
construído é (CH2O)n (esquema 1) e um esquema em que este carboidrato
é a glicose (esquema 2).
Dizemos que esta fase é a fase de fi xação do CO2 em uma molécula
orgânica.
RESUMINDO
Na etapa química, O CO2 é assimilado e fi xado, para sua posterior
conversão em uma molécula orgânica. Este processo de síntese
requer NADPH e ATP, sintetizados na etapa fotoquímica.
Assim, somando-se as duas etapas:
Etapa fotoquímica H2O + NADP+ + ADP + Pi O2 + ATP + NADPH
Etapa química ATP + NADPH + CO2 NADP+ + ADP + Pi + glicose
Temos:
6 H2O + 6 CO
2 C6H12O6 + 6 O2
luz
luz
Lembre-se de que para somar as duas etapas podemos cortar os componentes que aparecem nos dois lados das reações. Portanto, na soma, NADP+, ADP, Pi, ATP e NADPH podem ser eliminados.
!

CEDERJ 53
AU
LA
MÓ
DU
LO 2
5
HISTÓRICO
Até 1930, o modelo da fotossíntese especulava que o dióxido
de carbono era quebrado e então a água era adicionada ao carbono
resultante.
1 - CO2 C + O2
2 - C + H2O CH2O
Neste modelo, era proposto que o O2 liberado durante a
fotossíntese vinha como resultado da quebra do CO2. Este modelo caiu
por terra graças a Cornelis Bernardus van Niel, um estudante (na época)
que investigava a fotossíntese em bactérias que produziam carboidratos
a partir de CO2 mas não liberavam oxigênio.
Van Niel concluiu que, pelo menos em bactérias, o CO2 não é
quebrado em carbono + O2.
Esse grupo de bactérias requer H2S (sulfeto de H) e não H2O para
a fotossíntese, formando glóbulos amarelos de enxofre como produto
de excreção.
Assim:
CO2 + 2H2S CH2O + H2O + 2S
Van Niel ponderou que a bactéria deveria quebrar H2S e usar o
hidrogênio para produzir açúcar. Ele generalizou que todos os organismos
fotossintetizantes requerem uma fonte de hidrogênio, mas que essa fonte
pode variar:
Em sulfobactérias:
CO2 + 2H2 CH2O + H2O + 2S
Em plantas:
CO2 + 2H2O CH2O + H2O + O2
Esquema geral:
CO2 + H2X CH2O + H2O + 2X
Cornelis Bernardus van Niel (1897-
1985) era um engenheiro químico
que trabalhava como microbiologista
estudando taxonomia de bactérias.
Trabalhou na estação marinha Hopkins,
da Universidade de Stanford, na
Califórnia, até 1962. Retirado de http:
//www.asmusa.org/mbrsrc/archive/pdfs/
530287p75.pdf

Bioquímica II | Fotossíntese II - O processo de fotossíntese
CEDERJ54
Desta forma, Van Niel levantou a hipótese de que as plantas
quebram água como fonte de hidrogênio, liberando O2 como um produto
desta quebra.
Nos vinte anos seguintes, a hipótese de Van Niel foi confi rmada
graças ao uso de 18O – um isótopo pesado do oxigênio, utilizado como um
traçador para mostrar o destino dos átomos de oxigênio na fotossíntese.
Esses experimentos mostraram que o O2 gerado na fotossíntese é 18O
somente se a H2O é a fonte do traçador. Se o 18O foi introduzido na planta
na forma de CO2, a marcação não aparece no oxigênio produzido.
Experimento 1: CO2 + 2H2O CH2O + H2O + O2
Experimento 2: CO2 + 2H2O CH2O + H2O + O2
É importante lembrar que os elétrons associados ao hidrogênio
têm uma energia potencial muito maior em moléculas orgânicas do que
na água. A energia é estocada em açúcares e outras moléculas orgânicas
na forma de elétrons de alta energia.

CEDERJ 55
AU
LA
MÓ
DU
LO 2
5R E S U M O
Você viu nesta aula que a fotossíntese ocorre no cloroplasto das células
vegetais e que ela pode ser dividida em etapa fotoquímica e etapa
química. Na primeira etapa, a molécula de água é quebrada e os
hidrogênios associam-se a aceptores de elétrons. O oxigênio molecular
(O2), liberado na atmosfera, também é resultado desta etapa. Nesta fase
são sintetizados NADPH e ATP. Na etapa seguinte, estas moléculas serão
a fonte de energia para a síntese de glicose.
Nas próximas duas aulas, você verá cada uma dessas etapas
separadamente. Vamos discutir como cada uma acontece e qual a bateria
enzimática responsável pela catálise das reações. Na Aula 6, veremos a
etapa fotoquímica – aquela que depende de luz. Você conhecerá o papel
da clorofi la e de outros pigmentos neste processo. Na aula seguinte (Aula
7), veremos como a célula vegetal utiliza o ATP e o NADPH, gerados na
primeira fase, para sintetizar glicose e outros compostos. Lembre que os
exercícios só virão no fi nal do módulo. Então... estamos prontos?


Você já sabe o básico sobre fotossíntese. Agora vamos descrever a etapa fotoquímica – processo pelo qual a energia solar é capturada pelo cloroplasto – as moléculas envolvidas e a seqüência de reações. Na aula seguinte (Aula 7), explicaremos como ocorre a etapa química.
A etapa fotoquímica oufase clara da fotossíntese 6AULA
obje
tivo
s

Bioquímica II | A etapa fotoquímica ou fase clara da fotossíntese
CEDERJ58
COMO AS REAÇÕES DA FASE CLARACAPTURAM ENERGIA SOLAR?
Como vimos anteriormente, a primeira etapa da fotossíntese
transforma energia luminosa em energia química, como ATP e NADPH.
Isso acontece graças a um conjunto organizado de moléculas presentes
na membrana do tilacóide, presente nos cloroplastos. O evento mais
importante, que é a origem de todo o processo, é a fotólise da água.
A energia do sol é uma forma de energia conhecida como ENERGIA
ELETROMAGNÉTICA OU RADIAÇÃO. A energia eletromagnética viaja em ondas.
O comprimento de onda varia de menos de um nanômetro (para raios
gama) até mais de 1km (para ondas de rádio). Essa variação é conhecida
como espectro eletromagnético. A radiação conhecida como luz visível
(400 a 700nm de comprimento de onda) é o segmento mais importante
para a vida.
EN E R G I A
E L E T RO M A G N É T I C A O U R A D I A Ç Ã O
A luz do sol é uma forma de energia eletromagnética ou radiação.
Figura 6.1: Espectro eletromagnético. O olho humano é capaz de perceber apenas os comprimentos de onda correspondentes ao espectro visível. A maior parte do espectro eletromagnético é invisível.

CEDERJ 59
AU
LA
MÓ
DU
LO 2
6
Não esqueça!
A quantidade de energia é inversamente relacionada ao comprimento de onda da luz.
!
A radiação também pode ser vista como partículas chamadas
quanta ou fótons. A quantidade de energia é inversamente relacionada
ao comprimento de onda da luz.
Assim, um fóton de luz violeta tem duas vezes mais energia que
o fóton de luz vermelha.
A mesma parte do espectro que pode ser vista – a luz visível – é
também a radiação que dirige a fotossíntese. Azul e vermelho são os dois
comprimentos de onda mais efi cientemente absorvidos pela clorofi la
(Figura 6.2) e, portanto, as cores mais usadas como energia para as
reações da fotossíntese.
CLOROFILA a
CLOROFILA b
R
R
= CH3
= CH
O
CH2
CH2
CH2
CH2
CH2
CH3 CH3
CH3
CH3
CH3
CHCH
CH
CH
CH
CH2
C
C
C
H C3
H C3
H C2
HC
H
H
H
O
O
O O
O
N N
N N
Mg
R
CH3 CH3
CH2 CH2
CH2
CH2
CH2
CH2
CH CH
CH3
Figura 6.2: A molécula de clorofi la. Note o anel porfi rínico semelhante ao heme da hemoglobina (que você viu em Bioquímica I). No caso da clorofi la, o átomo central do anel é o magnésio. O grupo na posição representada por R diferencia as várias clorofi las. Observe a longa cauda de hidrocarboneto. Como você classifi caria essa molécula? Fonte: fai.unne.edu.ar/biologia/planta/fotosint.htm
Substâncias que absorvem luz visível são chamadas pigmentos.
Diferentes pigmentos absorvem diferentes comprimentos de onda, e o
comprimento de onda absorvido desaparece, enquanto os outros são
refl etidos. Se o pigmento absorve todos os comprimentos de ondas
ele aparece preto. A habilidade de um pigmento para absorver vários
comprimentos de onda de luz pode ser medida colocando-se uma solução
do pigmento em um instrumento chamado espectrofotômetro.
Então pense... por que a clorofi la é verde?

Bioquímica II | A etapa fotoquímica ou fase clara da fotossíntese
CEDERJ60
O pigmento chave quando se fala de fotossíntese é a clorofi la
(Figura 6.2). Isso porque apenas a clorofi la pode participar diretamente
das reações da fase clara que convertem energia solar em química. Mas
outros pigmentos podem absorver luz e transferir a energia para a
clorofi la, que então iniciará as reações.
Alguns pigmentos encontrados nos cloroplastos:
• Clorofi la a
• Clorofi la b
• β- Caroteno
• Ficoeritrina
• Ficocianina
Figura 6.3: O espectro de absorção de luz de diferentes pigmentos.
Ab
sorç
ão
Clorofila a
Clorofila b Ficoeritrina Ficocianina
�- Caroteno
violeta azul verde amarelo vermelho
Comprimento de onda (nm)
250 300 350 400 450 500 550 600 650 700

CEDERJ 61
AU
LA
MÓ
DU
LO 2
6
COMO OS PIGMENTOS ESTÃO ORGANIZADOS?
Os pigmentos estão organizados na membrana tilacóide em
conjuntos de poucas centenas (~300) de moléculas de pigmentos. Essas
membranas são ricas em glicolipídios e contêm grande quantidade
de proteína. Os pigmentos estão ligados a algumas dessas proteínas.
Clorofi las também interagem com lipídios de membrana por suas caudas
hidrofóbicas.
Estes conjuntos de pigmentos e proteínas estão organizados em
fotossistemas. Existem dois fotossistemas na membrana tilacóide:
• Fotossistema I
• Fotossistema II
COMO OS FOTOSSISTEMAS TRABALHAM?
Em cada um dos fotossistemas há uma única clorofi la capaz de
iniciar a reação. A localização dessa clorofi la no fotossistema é chamada
centro de reação (alguns autores sugerem a existência de um par de
moléculas de clorofi las a especializadas no centro de reação).
Robert Emerson e Willian Arnold, em 1930, mostraram que,
quando o sistema fotossintético da alga Chlorella operava na efi ciência
máxima, somente uma molécula de O2 era produzida para cada 2.500
moléculas de clorofi la.
A maior parte dessas moléculas não está diretamente engajada no
processo fotoquímico, mas atua como moléculas antenas ou clorofi las
antenas ou, ainda, complexo antena, absorvendo fótons e transferindo
energia para os centros de reação. Em outras palavras:
A energia de um fóton absorvido por qualquer molécula do
complexo antena viaja pelo sistema de uma forma randômica, aleatória.
Eventualmente (~10-10seg) a energia encontra o seu caminho para a
molécula de clorofi la a no centro de reação.

Bioquímica II | A etapa fotoquímica ou fase clara da fotossíntese
CEDERJ62
A clorofi la do centro de reação difere das outras moléculas de
clorofi la porque se encontra em um ambiente diferente, de forma que
o seu nível de energia (no estado excitado) é um pouco menor. Desta
forma, ela atua como uma armadilha para a energia absorvida pelos
outros pigmentos do fotossistema.
Complexoantena Centro de
reação
Energiaredutora
Energia luminosa
Figura 6.4: A organização das moléculas de clorofi la nos fotossistemas. A maioria das moléculas de clorofi la absorve luz e transfere a energia para o complexo antena, também chamado molécula antena ou proteína antena.

CEDERJ 63
AU
LA
MÓ
DU
LO 2
6
Cada complexo antena possui aproximadamente 300 moléculas
de clorofi las.
A energia é absorvida pelo centro de reação e os elétrons da clorofi la
passam de um estado fundamental para um estado excitado. Estes elétrons
de alta energia são então transferidos dos centros de reação para uma
cadeia de tranportadores até o NADP para formar o NADPH.
Figura 6.5: Seqüência de excitação do elétron da molécula aceptora pela luz solar.
ener
gia
Moléculaaceptora deelétrons e-
ener
gia
ener
gia
Estadofundamental
e-e-
e-
Estadoexcitado
Fóton
e-

Bioquímica II | A etapa fotoquímica ou fase clara da fotossíntese
CEDERJ64
AS REAÇÕES NOS FOTOSSISTEMAS
Os dois fotossistemas realizam diferentes reações e dependem
de luz de diferentes comprimentos de onda.
O fotossistema I pode ser excitado por luz de comprimentos
de onda mais curtos que 700nm e o fotossistema II requer luz de
comprimentos de onda mais curtos que 680nm para sua excitação.
Fotossistema I - 700nm
Fotossistema II - 680nm
Os dois fotossistemas operam no cloroplasto para produzir
NADPH (poder redutor), ATP e O2, porque estão interligados por uma
cadeia de transporte de elétrons. Os dois fotossistemas são estruturalmente
distintos no cloroplasto. No centro de reação do fotossistema I, uma
clorofi la especializada é chamada P700; e no fotossistema II a clorofi la
é chamada P680.
Ambas absorvem luz vermelha e são moléculas idênticas. Suas
poucas diferenças estão associadas com diferentes proteínas de membrana
que afetam sua distribuição de elétrons e, portanto, apresentam pequenas
diferenças no espectro de absorção.
A absorção de luz nos centros de reação dos fotossistemas I e II
produz um estado excitado de alta energia em suas clorofi las. Esse evento
dispara uma série de REAÇÕES REDOX.
RE A Ç O E S RE D O X
Reações redox ou reações de
oxidação e redução são reações de
transferência de elétrons. Lembre-
se: redução é ganho de elétrons
e oxidação é perda de elétrons.

CEDERJ 65
AU
LA
MÓ
DU
LO 2
6
ATP
fótonfóton
Estroma
Membranatilacóide
Quinone
NADPRedutaseFerredoxina
Citocromobf
plastocianina
e
e
Tilacoide
Gradiente deprótons usadospara gerar
2 H O
O 4H 2H
e-
ePSIIP680
NADPH
PSIP700
2
2+ +
–
––
+
Figura 6.6: As reações luminosas da fotossíntese produzem ATP e NADPH. A energia requerida nesta etapa é derivada da oxidação da água (fotólise). Em plantas esse processo envolve dois centros de rea-ção fotossintéticos ligados em série por uma cadeia transportadora de elétrons. O fotossistema II (FSII) usa energia luminosa para oxidar a água a O2 e então passa os elétrons para o fotossistema I (FSI). No fotos-sistema I o elétron é excitado novamente pela luz e é usado para reduzir a ferredoxina. A ferredoxina reduzida então reduz NADP+ a NADPH.
Em ambos os fotossistemas, a primeira etapa é a transferência do
elétron excitado pela luz do centro de reação para a cadeia de transporte
de elétrons.
A fonte desse elétron é a água, e o destino fi nal é a molécula de
NADP+, que será reduzida a NADPH.
Durante as reações luminosas, existem duas possíveis rotas para
o fl uxo de elétrons.
Fluxo cíclico – produz apenas ATP
Fluxo não cíclico – produz ATP, NADPH, O2
Antes de falar dos tipos de fl uxos de elétrons, vamos apresentar
os componentes do transporte de elétrons da fotossíntese na ordem em
que estão presentes na membrana interna do cloroplasto, que é a ordem
de funcionamento da cadeia transportadora:
1. Fotossistema II (P680) - conjunto de pigmentos ligados
a proteínas.
2. Plastoquinona (PQ) - Chamada plastoquinona pela
primeira vez em 1959 por Crane, por ser uma quinona
encontrada em grandes quantidades no cloroplasto de
vegetais superiores.
No cloroplasto
Elétron de baixa energia + luz = elétron de alta

Bioquímica II | A etapa fotoquímica ou fase clara da fotossíntese
CEDERJ66
Reação:
PQ PQH2
O
O
H C3
H C3
CH3 CH3CH3
CH3
OH
H C3
H C3
CH3 CH3 CH3
CH3
OH
Figura 6.7: Plastoquinona oxidada (A) e reduzida (B).
3. Citocromo bf - recebe os elétrons da plastoquinona.
O complexo protéico é formado por quatro subunidades:
um citocromo f de 33kDa, um citocromo b563 de 23kDa
(dois hemes), uma proteína ferro-enxofre (Fe-S) de 20kDa
e uma proteína de 17kDa.
4. Plastocianina (Pc) - também é uma proteína integral de
membrana que apresenta uma única cadeia polipeptídica
de 10 500 Da. Esta proteína tem um centro constituído
de um átomo de cobre envolvido por duas pontes de
nitrogênio e duas pontes de enxofre, que lhe confere uma
geometria tetraédrica.
5. Fotossistema I (P700) - conjunto de pigmentos ligados
a proteínas.
6. Ferredoxina - é uma proteína ferro-enxofre solúvel,
presente no estroma do cloroplasto.
Figura 6.8: Plastocianina. Fonte: http://www.biologie. uni-hamburg.de/b-online/e17/plasto.htm
Cys
Cys S
S
Fe
S
S
Fe
S
S
Cys
Cys
Cys
Cys S
S
Fe S
S
Fe
S
S
Cys
CysFe
SS
Fe
A B C
Figura 6.9: Ferredoxina. (A) complexo ferro-enxofre de uma proteína Fe2S2 - presente em ferredoxinas vegetais;(B) complexo ferro-enxofre de proteínas Fe4S4 - presentes em ferredoxinas de bactérias; (C) estrutura tridimensional de uma ferredoxina.

CEDERJ 67
AU
LA
MÓ
DU
LO 2
6
7. NADP redutase - último componente da cadeia
transportadora de elétrons, a NADP redutase é a enzima
que catalisa a reação:
NADP+ NADPH + H+, usando 2 e- e 2 H+ da FdH2
(ferredoxina reduzida).
O fl uxo cíclico de elétrons
O fl uxo cíclico de elétrons é o mais simples. Envolve somente
o fotossistema I e gera ATP, mas não produz NADPH nem O2 . Ele é
chamado cíclico porque os elétrons excitados que saem da clorofi la do
centro de reações retornam.
Figura 6.10: Ferredoxina Fe4S4. Fonte:http://www.life.uiuc.edu/crofts/bioph354/4fe4s.html
Figura 6.11: Transporte cíclico. Quando os níveis de NADPH são altos, os níveis de NADP+ são baixos, e os elétrons na ferredoxina reduzida não têm para onde ir. Nessas condições, a ferredoxina transfere seus elétrons para o citocromo bf, onde eles serão transferidos novamente para a clorofi la do fotossistema I. Este processo é chamado transporte de elétrons cíclico e permite o bombeamento de prótons e a síntese de ATP, mas não fornece NADPH.
Para o lúmen do
e-
2H+
e-
tilacóide
fóton
ferredoxina
Pod
er r
edu
tor
Citocromoo bf
pc
(P700)(P700)
(P700)(P700)

Bioquímica II | A etapa fotoquímica ou fase clara da fotossíntese
CEDERJ68
Quando pigmentos do complexo antena do fotossistema I absorvem
luz, a energia atinge, no centro de reação, a clorofi la P700. Isto excita dois
elétrons da P700. Os elétrons de alta energia são aprisionados por um aceptor
primário de elétrons para uma ferroproteína – ferredoxina (Fd).
A ferredoxina passa os elétrons para a plastoquinona (Pq). Daí os
elétrons vão para um complexo de dois citocromos. Estes citocromos doam
seus elétrons para outra proteína, plastocianina (Pc), que contém cobre.
Figura 6.12: Seqüência de transferência de elétrons na etapa fotoquímica da fotossíntese. Fd = ferredoxina; PQ = plastoquinona; Pc = plastocianina.
A cada etapa da cadeia de transporte, os elétrons perdem energia
potencial. Esse caminho, energeticamente “morro abaixo”, é usado pela
cadeia de transporte para bombear prótons H+ através da membrana
tilacóide. O gradiente de prótons gerado ativa a ATP-sintase, a enzima
que catalisa a síntese de ATP, que veremos mais adiante.
Certos transportadores de elétrons somente transportam elétrons
na companhia de íons H+, que são levados de um lado da membrana
tilacóide para o lado oposto.
O processo é similar à fosforilação oxidativa da mitocôndria
(que veremos na Aula 16).
Nos cloroplastos, a síntese de ATP é um processo de fosforilação.
Ela é chamada fotofosforilação por ser dirigida por energia luminosa.
No caso da fotofosforilação cíclica há somente síntese de ATP.
Nenhum NADPH é gerado.

CEDERJ 69
AU
LA
MÓ
DU
LO 2
6
O fl uxo não cíclico de elétrons
No caso do fl uxo não cíclico de elétrons, os dois fotossistemas
cooperam. Elétrons passam da H2O para o NADP+ para formar
NADPH.
Como no fl uxo cíclico, a luz excita um par de elétrons da clorofi la
P700. Entretanto, os elétrons não retornam, mas são armazenados como
elétrons de alta energia na forma de NADPH. Este será utilizado na fase
escura como doador de elétrons para a síntese de carboidrato.
NADP+ NADPH
ADP + Pi ATP
H2O 1/2O2 + 1H+
Elétrons ejetados da clorofi la P680 são repostos por elétrons
removidos da água. Os elétrons excitados fl uem pela cadeia de transporte
de elétrons até a clorofi la P700 do fotossistema I, fornecendo energia
para a síntese de ATP. A iluminação do fotossistema I leva os elétrons
novamente para um estado de alta energia. Os elétrons excitados passam
por uma aceptor primário – a ferrodoxina, e fi nalmente ao NADP+
redutase, que catalisa a formação de NADPH.
Figura 6.13: Fluxo não cíclico de elétrons na etapa fotoquímica da fotossíntese. Neste caso, ATP e NADPH são formados.
Pod
er r
edu
tor
Ferredoxina
Redutase
NADP+
NA
DPH
Citocromoo bf
fóton
(P660)(P660)
(P660)(P660)
fóton
(P700)(P700)
(P700)(P700)
2H O2
e-e-O +4H2 +

Bioquímica II | A etapa fotoquímica ou fase clara da fotossíntese
CEDERJ70
A SÍNTESE DE ATP
Tanto nas mitocôndrias quanto nos cloroplastos, o ATP é gerado
por um mecanismo básico chamado quimiosmótico.
A membrana tilacóide bombeia H+ do estroma (onde ocorreu
a hidrólise da água) para dentro do tilacóide, que funciona como
reservatório de H+. Os prótons concentrados no lúmen do tilacóide geram
um gradiente de concentração de prótons (gradiente eletroquímico).
Esses prótons encontram uma única saída para voltar ao estroma: passar
através de uma ATP-sintase. A ATP-sintase usa a energia de retorno dos
prótons para sintetizar ATP. A ATP-sintase produz ATP no estroma e é
utilizada no ciclo de Calvin que ali acontece.
F (80 A )1 °
F (50 A )0 °
Talo 50 A°
Canal transportador de H+
Superfície voltadapara a matriz
Outras superfícies
a ab
ab b
F (80 A )1 °
F (50 A )0 °
VISTA PERSPECTIVADA VISTA LATERALSIMPLIFICADA

CEDERJ 71
AU
LA
MÓ
DU
LO 2
6RESUMO
Você viu nesta aula que a primeira etapa da fotossíntese transforma energia
luminosa em energia química, como ATP e NADPH. O evento mais importante, que
é a origem de todo o processo, é a fotólise da água. O pigmento chave, quando
se fala de fotossíntese, é a clorofi la. Mas outros pigmentos podem absorver luz e
transferir a energia para a clorofi la, que então iniciará as reações.
Esses conjuntos de pigmentos e proteínas estão organizados em fotossistemas:
o fotossistema I e o fotossistema II, em plantas e cianobactérias. A fotólise
da água, promovida pela fotooxidação do fotossistema II, libera elétrons
que fl uem através de um complexo citocromo bf, passam pelo fotossistema
I e fi nalmente para o NADP+. O fl uxo dos elétrons pode ser não cíclico,
resultando na redução do NADP+, ou cíclico, no qual os prótons adicionais
são transferidos para o lúmen do tilacóide. Os prótons liberados pela
oxidação da água e aqueles transferidos para o lúmen do tilacóide geram
um gradiente transmembrana de prótons, que é usado pela ATP-sintase do
cloroplasto para realizar a fosforilação do ADP (síntese de ATP).
Na próxima aula você verá como ocorre a etapa química, ou ciclo de Calvin. É neste
ciclo que o CO2 será utilizado como matéria-prima para a síntese de moléculas
orgânicas, especialmente glicose. Lembre que os exercícios serão apresentados
no fi nal do módulo. Portanto, não perca a visão geral da fotossíntese.


O ciclo de Calvin ouetapa química da fotossíntese 7AULA

Bioquímica II | O ciclo de Calvin ou etapa química da fotossíntese
CEDERJ7 4
Na aula passada, nós falamos da etapa fotoquímica ou fase clara da fotossíntese.
Você viu como o oxigênio é produzido e ATP e NADPH são sintetizados e o
papel da luz do sol neste processo. Agora, vamos falar da etapa seguinte,
chamada etapa química ou ciclo de Calvin ou ainda fase escura da fotossíntese.
Antes, lembre-se de que você já viu, na aula 05, que na etapa química o CO2
atmosférico é fi xado em carboidratos.
A estequiometria global do ciclo de Calvin é:
INTRODUÇÃO
18 ATP 18 ADP + 18 Pi
6 CO2 + 12 NADPH C6H12O6 + 12 NADP+
UM POUCO DE HISTÓRIA
A etapa química da fotossíntese acontece através de uma série de
reações cíclicas que deram a Melvin Calvin, um bioquímico americano, o
Prêmio Nobel de Química em 1961. Tal etapa foi elucidada por Calvin,
James Bassham e Andrew Benson entre 1946 e 1953. Estes pesquisadores
descreveram o destino metabólico do CO2 marcado com 14C (14CO2)
em uma cultura de células de algas. Os primeiros experimentos em que
o 14CO2 era incubado com as células de algas por um minuto ou mais
indicaram que estas sintetizavam uma mistura complexa de metabólitos,
dentre eles açúcares e aminoácidos.
14CO2 + células de algas em cultura 5 minutos
ou mais
Açúcares, aminoácidos e
outros compostos orgânicos
Entretanto, se o tempo de incubação das algas com o 14CO2
era reduzido para cinco segundos, a análise do material sintetizado
mostrava que o primeiro composto radioativo estável formado era o
3-fosfoglicerato (3PG).

CEDERJ 7 5
AU
LA 7
M
ÓD
ULO
2
14CO2 + células de algas em cultura 5 segundos 2 moléculas de
3-fosfoglicerato
A análise do composto marcado com 14CO2 mostrava que ele era
inicialmente marcado no seu grupo carboxila. Calvin sugeriu que o 3PG
era formado pela carboxilação de um composto de dois carbonos. Este
composto foi, por muito tempo, procurado por vários pesquisadores, mas
nunca encontrado. A reação de carboxilação que realmente ocorre envolve
uma pentose chamada ribulose 1,5-bisfosfato (RuBP).
O CICLO DE CALVIN
A carboxilação de uma pentose tem como produto dois compostos
de três carbonos cada: as moléculas de 3-fosfogliceratos. A rota completa
envolve a carboxilação da pentose, a formação de carboidratos como
produto principal da via e a regeneração de RuBP. Para que isso ocorra,
existem várias reações catalisadas por diferentes enzimas.
Mas não esqueça, no ciclo...
1- O CO2 é aprisionado na forma de um carboxilato e
reduzido, o que resulta na síntese de carboidratos.
2- A molécula aceptora de CO2 (ribulose 1,5-fosfato) é
regenerada.
O CO2 difunde-se no estroma do cloroplasto, onde é adicionado ao
grupamento carbonil da ribulose 1,5-bisfosfato (RuBP), reação catalisada
pela enzima ribulose 1,5-Bisfosfato Carboxilase/Oxigenase – RuBisCO
(veja a Figura 7.2).
CH O2
CHOH
CHOH
CH O2
C O
P
P
Figura 7.1: Ribulose 1,5 - bisfosfato (RuBP).
A molécula aceptora
de CO2 é a ribulose
1,5-bisfosfato.

Bioquímica II | O ciclo de Calvin ou etapa química da fotossíntese
CEDERJ7 6
Figura 7.2: Estrutura tridimensional da RuBisCO. A. Esta enzima é composta por oito grandes subunidades e nove subunidades menores. Esta imagem mostra quatro subunidades de cada tipo. Ela fi xa CO2 catalisando a reação da ribulose 1,5-bisfosfato com o CO2, produzindo duas moléculas de 3-fosfoglicerato. RuBisCO é a proteína mais abundante do mundo. Esta abundância, assim como a fi xação de CO2, é fundamental para todas as formas de vida; B. Detalhe de A mostrando as subunidades maiores da RuBisCO que ligam o substrato da enzima e íons magnésio.
B
A
Esta enzima é uma das mais importantes enzimas conhecidas e
certamente a mais abundante – 40 milhões de toneladas de RuBisCO
no mundo. Como o seu nome diz, ela alterna a atividade carboxilase e
oxigenase. Mas isso nós discutiremos mais tarde, ainda nesta aula.
A incorporação de CO2 em 3-fosfoglicerato representa o processo
de fixação de CO2 em si; as reações subseqüentes convertem o 3-
fosfoglicerato em gliceraldeído 3-fosfato. Isto acontece de uma forma
similar à gliconeogênese (que você verá na aula 30), exceto por uma
única característica: as reações requerem NADPH e não NADH, como
na gliconeogênese.
CH O2
C
C
C
CH O2
O
OHOH
OHH
P
P
COOH
C
CH O2
OHH
P
COOH
C
CH O2
OHH
P
CO2
Rubisco
Ribulose 1,5-bisfosfato
2 moléculas de
3-fosfogliceratoReação 1
CO2

CEDERJ 7 7
AU
LA 7
M
ÓD
ULO
2
Após serem formadas, as duas moléculas de 3-fosfoglicerato são
convertidas a 1,3-bisfosfoglicerato (BPG), uma molécula duplamente
fosforilada. Esta reação é catalisada pela enzima fosfoglicerato quinase.
A reação de fosforilação requer a hidrólise de uma molécula de ATP para
cada BPG formado. Lembre que uma ribulose 1,5-bisfosfato, ao sofrer
carboxilação, foi capaz de gerar duas moléculas de 3-fosfoglicerato. Logo,
serão gastos 2 ATPs por ciclo.
CH2P O CH
OH
CO
O—CH2P O CH
OH
CO
O
ATP ADP + Pi
P
1,3 - bisfosfoglicerato3 - fosfoglicerato
ATP ADP
Cada molécula de 1,3-bisfosfoglicerato formada pode ser convertida
a gliceraldeído 3-fosfato (GAP). A enzima que catalisa esta reação é a
gliceraldeído 3-fosfato desidrogenase.
Você lembra, das aulas de Bioquímica I, o que fazem as enzimas denominadas
desidrogenases?
!
Reação 2
Neste caso, a enzima reoxida o NADPH gerado durante a fase
clara da fotossíntese.
1,3 - bisfosfoglicerato
NADPH NADP+Reação 3
CH2P O CH
OH
CO
O PCH2P O CH
OH
CO
H
gliceraldeído 3-fosfato

Bioquímica II | O ciclo de Calvin ou etapa química da fotossíntese
CEDERJ7 8
A conversão de 3-fosfoglicerato em gliceraldeído-3fosfato e, a
seguir, em glicose é a etapa da fotossíntese conhecida como ciclo de
Calvin, fase escura ou ainda etapa química. Neste ponto, uma molécula
de CO2 foi incorporada e duas moléculas de G3P foram produzidas às
custas de 2 ATPs e 2 NADPHs. Entretanto, seis moléculas de CO2 são
necessárias para produzir uma molécula de glicose (porque a glicose é
uma molécula de seis carbonos). Portanto, as seis moléculas de CO2 irão
formar doze moléculas de 3-fosfoglicerato que, então, serão convertidas
em doze moléculas de gliceraldeído 3P.
O gliceraldeído 3P pode ser drenado do ciclo para uso em vias
biossintéticas.
Quando gliceraldeído 3-P é formado, ele pode ter dois destinos:
1) Produção de açúcares de 6 carbonos (glicose).
2) Regeneração de ribulose 1,5-bisfosfato.
O resumo das reações pode ser visto na fi gura 7.3.
Ribulose 1,5-bisP
Ribulose-5P
6
6ATP
6
12
12NADPH
12
Regeneraçãoda ribulose 1,5-bisP
6
ATP
CO2
Síntese de glicose
3-fosfoglicerato 12 x 1,3-bisPglicerato
Gliceraldeído 3P ou(ou DHAP)
de
Gliceraldeído 3P ou(ou DHP)
6
Frutose-6P
Gicose 1P
Frutose-6P
2
Figura 7.3: Resumo das reações do ciclo de Calvin. O ciclo ocorre para cumprir duas funções: uma é a síntese de glicose e outras moléculas orgânicas; a outra é a regeneração da ribulose 1,5-bisfosfato, molécula essencial para reiniciar o ciclo.

CEDERJ 7 9
AU
LA 7
M
ÓD
ULO
2
PRODUÇÃO DE GLICOSE
Seis moléculas de CO2 podem gerar doze moléculas de gliceraldeído
3-fosfato (G3P). As doze moléculas de gliceraldeído-3-fosfato podem seguir
vários destinos diferentes e, portanto, ser substrato para a síntese de muitas
outras moléculas orgânicas ou regenerar o aceptor inicial de CO2 (ribulose
1,5-bisfosfato).
Duas dessas moléculas de G3P são utilizadas para produzir uma
glicose-fosfato (via gliconeogênese), e as outras dez são usadas para
regenerar as seis moléculas iniciais de ribulose 1,5-bisfosfato. Estas são
necessárias para ligar outras seis moléculas de CO2 e reiniciar o ciclo.
Das dez G3P envolvidas na regeneração da RBP, quatro formarão duas
moléculas de frutose 6-fosfato.
Muitas das reações do ciclo de Calvin são comuns à via das pentoses-
fosfato (Aula 20). As duas moléculas de frutose 6-fosfato reagem com 6
G3P. Gliceraldeído 3P pode ainda ser isomerizado a dihidroxiacetona-
fosfato (DHAP). As enzimas transcetolase, aldolase, fosfatase e outra
transcetolase geram intermediários combinando G3P e DHAP. Estes
intermediários incluem xilulose 5-fosfato e ribulose 5-fosfato. A ribulose
5P pode ser convertida a ribulose 1,5-bisfosfato, cuja reação requer um
ATP por molécula convertida. A conversão de seis moléculas de ribulose
1,5-bisfosfato, necessárias para fi xar 6 CO2, requer seis moléculas de
ATP. A síntese de uma molécula de glicose requer dezoito ATPs e doze
NADPHs.
Assim, o gliceraldeído 3-P, um dos produtos mais importantes
da fotossíntese, é usado em uma grande variedade de rotas metabólicas,
tanto dentro como fora do cloroplasto. Como você viu, ele pode, por
exemplo, ser convertido a frutose 6-fosfato. A frutose 6-fosfato pode ser
convertida a glicose 1-fosfato pela ação da fosfoglicose isomerase e pela
fosfoglicomutase. A glicose 1-fosfato é uma molécula importante, pois é
precursora de carboidratos complexos característicos de vegetais. Entre eles
destaca-se a sacarose, o principal açúcar de transporte para fornecimento
de carboidratos para células não fotossintetizantes; o amido, o principal
polissacarídeo de reserva; e a celulose, o principal componente estrutural
de paredes celulares. Na síntese de todas essas substâncias, a glicose 1-P é
ativada pela formação de ADP-, CDP-, GDP- ou UDP-glicose, dependendo
da espécie e da via metabólica. A unidade de glicose é, então, transferida
para o terminal não redutor de uma cadeia crescente de polissacarídeo,
como ocorre na síntese do glicogênio.
Figura 7.4: Algas fotossin-tetizantes. Fonte: http: //old.jccc.net/~pdecell/photosyn/photoframe.html
CH2P O CH
OH
CO
H
Gliceraldeído-3P
Figura 7.5
A síntese dog l i cogên io você verá nas Aulas 26 e 27.
!

Bioquímica II | O ciclo de Calvin ou etapa química da fotossíntese
CEDERJ8 0
REGENERAÇÃO DA RIBULOSE 1,5-BISFOSFATO
A seguir, temos as possíveis reações do ciclo de Calvin até a regeneração
de ribulose 1,5-bisfosfato. Note que esta seqüência possibilita a síntese de
várias moléculas orgânicas, com diferentes números de carbonos.
1) 2X Gliceraldeído 3P 2Dihidroxicetona P
2) Gliceraldeído 3P + Dihidroxicetona-P Frutose 1,6-bisfosfato
3)Frutose 1,6-bisfosfato Frutose 6P + Pi
4) Frutose 6P + Gliceraldeído 3P Eritrose 4P + Xilulose 5P
5) Eritrose 4P + Dihidroxicetona Sedoheptulose 1,7-bisfosfato
6)Sedoheptulose 1,7-bisfosfato Sedoheptulose 7P + Pi
7) Sedoheptulose 7P + Gliceraldeído 3P Ribulose 5P + Vilulose 5P
8) Ribulose 5P Ribulose 5P
9) 2 Xilulose 5P 2 Ribulose 5P
10) 3 Ribulose 5P + 3ATP 3 Ribulose 1,5-bisP + ADP
= 5 Gliceraldeído 3P + 3ATP 3Ribulose 1,5-bisP + 3ADP
2X 3C 2X 3C
3C 3C 6C +
6C 6C
6C 3C 4C 5C
3C 4C 7C
7C 7C
7C + 3C 5C
5C 5C
2X 5C 2X 5C
3X 5C 3X 5C
3X 5C
5C
5X 3C

CEDERJ 8 1
AU
LA 7
M
ÓD
ULO
2
TRANSPORTE DE TRIOSES FOSFATO
As trioses fosfato (3-fosfoglicerato, 1,3-bisfosfoglicerato, gliceraldeído
3-fosfato, dihidroxiacetona-fosfato) são críticas para o metabolismo de
açúcares em plantas. No estroma do cloroplasto, as trioses fosfato são
usadas como substrato para a gliconeogênese (síntese de glicose).
Sacarose
Citoplasma
Estroma docloroplasto
ATP ADP NADH NAD+
1,3BPG
Pi
G3PPi
PiATP
ADP
DHAP
NADP+
1,3BPG G3P
3-PG
NADPH
Figura 7.6: Transporte de trioses fosfato.
Trioses fosfato são também transportadas do estroma do cloroplasto
para o citoplasma da célula, por transportadores específicos. Estes
transportam Pi do citoplasma para o estroma, onde ele será necessário
para a síntese de ATP, enquanto transfere dihidroxiacetona fosfato (DHAP)
para o citoplasma, onde ele pode ser convertido a 3-PG (produzindo ATP
e NADPH). DHAP pode também ser usado no citoplasma para a síntese
de sacarose (veja a Figura 7.7).
Síntese de sacarose – a reação de UDP-glicose com frutose 6P é catalisada pela
enzima sacarose 6P sintase.
!

Bioquímica II | O ciclo de Calvin ou etapa química da fotossíntese
CEDERJ8 2
Figura 7.7: ATP e NADPH podem ser transferidos do cloroplasto para o citoplasma da célula vegetal por trans-portadores específi cos. Estas moléculas são usadas no citoplasma como fonte de energia e equivalentes redutores para converter as trioses fosfato em sacarose, que é transportada através do sistema circulatório do vegetal para tecidos não fotossintéticos (como as raízes) onde será usada no metabolismo energético.
R E S U M O
Nesta aula, você viu que as reações da fase escura usam ATP e NAPH produzidos
na fase clara para realizar a síntese de carboidratos a partir de CO2. O primeiro
evento do ciclo de Calvin é a fi xação do CO2 através de sua reação com a ribulose
1,5-bisfosfato para formar, em última análise, duas moléculas de gliceraldeído
3P (GAP). As reações remanescentes do ciclo regeneram o aceptor inicial de
CO2, ribulose 1,5-bisfosfato. A próxima aula será a última do módulo, e nela
falaremos de fotorrespiração e regulação da fotossíntese. Finalmente, teremos
também os exercícios e auto-avaliação do módulo. Vamos então à Aula 8.
UDP-Glicose
Frutose 6-P
HOCH2
H H
HO
OH
HOH
OHHO
OO P O
O
O—O—
CH2
O
O N
HN
O
OH OH
H HHH
HOCH2
CH OPO2 32 —
H
HOHHO
HOH
O

Outros aspectosimportantes da fotossíntese 8AULA

Bioquímica II | Outros aspectos importantes da fotossíntese
CEDERJ84
OUTROS ASPECTOS IMPORTANTES DA FOTOSSÍNTESE
Você viu na aula passada o ciclo de Calvin. Até aqui, as aulas
deste módulo concentraram-se em mostrar as etapas da fotossíntese, os
objetivos de cada etapa e as moléculas envolvidas como matéria-prima,
intermediários, enzimas, ou produtos. A Aula 8 é a última do módulo e
aqui discutiremos o papel da RuBisCO como oxigenase, participando do
processo conhecido como FOTORRESPIRAÇÃO. Conheceremos ainda algumas
plantas que apresentam desvios metabólicos para aumentar a efi ciência da
fotossíntese. Finalmente, discutiremos um pouco de como este processo,
tão importante para a vida, é regulado.
Fotorrespiração
A RuBisCO é uma enzima peculiar. Ela pode comportar-se como
uma oxigenase quando a concentração de O2 na atmosfera é alta e a
concentração de CO2 baixa.
Neste caso, a ribulose 1,5-bisfosfato reage com o oxigênio e é
convertida em fosfoglicolato e 3-fosfoglicerato (fi gura 8.1).
Figura 8.1: A dupla atividade da RuBisCO.
FO T O R RE S P I R A Ç Ã O
Uma série de reações que ocorrem nas plantas quando a RuBisCO funciona como oxigenase.
Fosfoglicolato pode ser convertido a 3-fosfoglicerato por uma
série de reações que requerem ATP e O2. Como O2 é consumido e CO2
é produzido, este processo é chamado fotorrespiração. Esta reação
desperdiça 3-fosfoglicerato e ATP, porque o processo não resulta em
fi xação de carbono.
CH2
CH2 OCCC
HH
OHOH
O
O
HHO CC
Ch2 O
O–
O
O– OCCCH2 O
H OH
CH2 OC
O– O
O– OCCH OHCH2 O
+
+
CarboxilaseCo2
Oxigenase
Grupo fosfato (Po )2–3=
2-fosfoglicolato 3-fosfoglicerat
RuBisCO(ribulosebisfosfato carboxilase/oxigenase)
2-moléculas de fosfoglicerato
Ribulose 1,5bisfosfato
P
P
P
P
P
P
P
O2

CEDERJ 85
AU
LA
MÓ
DU
LO 2
8
Desta forma, a enzima inicia a via
conhecida como fotorrespiração, onde a
ribulose 1,5-fosfoglicerato é convertida a
fosfoglicolato (Figura 8.2).
Figura 8.2: A localização de cada etapa da fotorrespiração.
-
Cloroplastos
Peroxissomas(microcorpos)
Mitocôndrias
Ciclo deCalvin
Ribulose1,5 -bisP
RuBisCO
fosfoglicolato
Glioxilato
Glicina
Glicina
O2O2 CO2
3 PG ADPP1
O2O2
H2O2O2
CO +NH2 3
Serine
Serine
ATP
Glicolato Glicerato
O fosfoglicolato é então desfosforilado e passa para os pero-
xissomos, onde ele é oxidado, formando peróxido de hidrogênio e
glioxilato.
Fosfoglicolato H2O2 + Glioxilato
O H2O2 tóxico é quebrado pela catalase e o glioxilato é transa-
minado, produzindo glicina.
A glicina entra na mitocôndria e duas moléculas são convertidas
em uma serina. Neste processo há descarboxilação e desaminação: CO2
e amônia são liberados. A serina novamente é convertida a glicerato. No
cloroplasto, sofre fosforilação pelo ATP, formando 3-fosfoglicerato.
Assim, neste processo:
1. A ribulose 1,5-bisfosfato não pode ser usada no ciclo de Calvin.
2. A fi xação de CO2 é revertida, pois o O2 é consumido e CO2
é liberado.
3. ATP é gasto desnecessariamente.

Bioquímica II | Outros aspectos importantes da fotossíntese
CEDERJ86
Plantas C4
Portanto, até hoje se acredita que a fotorrespiração trabalha
contra a fotossíntese.
Isto acontece com a maior parte dos vegetais e tem-se trabalhado
no sentido de criar em laboratório, por engenharia genética, plantas que
não tenham esse comportamento.
Algumas plantas, entretanto, ao longo da evolução, conseguiram
uma solução para esse problema. Esta solução não é uma enzima modi-
fi cada. Na verdade, as plantas chamadas plantas C4 desenvolveram uma
rota metabólica que ajuda a conservar o CO2. Esta rota é chamada Ciclo
C4 ou Hatch - Slack. Este ciclo, encontrado em plantas como o milho e
a cana-de-açúcar, é importante em plantas tropicais que são expostas a
intensa luz solar e altas temperaturas.
Embora fotorrespiração ocorra sempre, em maior ou menor grau,
em todas as plantas, é mais ativa sob condições de muita luminosidade,
altas temperaturas e baixa concentração de CO2.
Plantas C4 concentram sua fotossíntese em uma camada de células
que se localiza logo abaixo das células mesófi las, as células da bainha da
lâmina foliar. As células mesófi las, neste caso, mais diretamente expostas
ao CO2, contêm as enzimas do ciclo C4.
A denominação C4 se deve à síntese de um composto de quatro
carbonos, o oxaloacetato.
A enzima chave, fosfoenol piruvato carboxilase, fi xa CO2, con-
densando-o com o fosfoenol piruvato para formar oxalacetato.
Para plantas C4, ver também: http://www.icb.ufmg.br/prodabi/grupo5/Photosynthesis/c4/c4.htmlExplore a página: tem muita coisa legal sobre fotossíntese.
!
As células mesófi las e as células da bainha (vascular) formam a camada mediana ou mais interna da folha.
!

CEDERJ 87
AU
LA
MÓ
DU
LO 2
8
Figura 8.3: Metabolismo de fi xação de CO2 em plantas C4.
Plantas CAM
Outra modifi cação de fi xação de carbono é encontrada em plan-
tas suculentas (armazenadoras de água), incluindo plantas de gelo,
cactos, que são adaptados para climas muito secos. Essas plantas abrem
seus estômatos durante a noite e fecham durante o dia, exatamente
o inverso de como outras plantas se comportam. Estômatos fechados
ajudam a preservar água durante o dia, mas também impedem a entrada
de CO2. Durante a noite, quando seus estômatos abrem, essas plantas
tomam CO2 e o incorporam em uma variedade de ácidos orgânicos.
Este modo de fi xação de CO2 é chamado metabolismo do ácido cras-
suláceo (em inglês, CAM), por causa da família Crassulacea, onde foi
primeiro descrito.
Regulação da fotossíntese
Plantas possuem dois sistemas dependentes de luz para regular o
ciclo de Calvin. O primeiro sistema tem como alvo a RuBisCO, ativando
a enzima em resposta à luz.
CO2
Co2
CO2
Pi COO
COO-
COO
COO-Pi
COO
PEPcarboxilase
CH2
CH2
CH2
COO
Ch3
C O
Oxaloacetato
NADPH+H+
CH OH
C O PO32
NADP+
Malate
C O
fosfoenolpiruvato
AMP+PPi ATP+Pi
piruvato
Malate NADP+
NADPH
Células da bainha
Ciclode
Calvin+

Bioquímica II | Outros aspectos importantes da fotossíntese
CEDERJ88
Fotofosforilação bombeia prótons do estroma para o lúmen do tilacóide
e Mg+2 do lúmen do tilacóide para o estroma. Este bombeamento causa
um aumento no pH e na concentração de Mg do estroma. Os dois efeitos
ativam a RuBisCO.
O segundo sistema responde ao estado redox do estroma
dependente de luz. Durante a fotofosforilação, os elétrons da água são
usados para reduzir a ferredoxina. Parte da ferredoxina reduzida é usada
para reduzir tiorredoxina. Tiorredoxina reduzida pode reduzir diversas
enzimas chaves do ciclo de Calvin, levando à ativação destas enzimas.
Figura 8.4: Regulação da fotossíntese. Os níveis de prótons e Magnésio no estroma do cloroplasto modulam a atividade da RuBisCO. O estado redox das enzimas do ciclo de Calvin é outro importante mecanismo de regulação das enzimas.
Estroma do
cloroplasto
Ativação da
RuBisCO
Elétrons gerados por
fotofosforilação
Tioredoxina
reduzida
Ferredoxina
reduzida
Ferredoxina
oxidada
Tiorredoxina
oxidadaH+
Mg2+
[ ]Mg2+
[ ]H+
2e–
2H+
2e–
2H+
SHSH
SH
SH S
S
S
S
citoplasma
Lúmen
do
tilacóide
Enzimas do ciclode calvin oxidadas(inativas)
Enzimas do ciclode calvin reduzidas(ativas)
Destino dos produtos fotossintéticos
A fotossíntese é um processo onde a energia fl ui dos fótons às
moléculas orgânicas. As reações luminosas capturam a energia solar e
a usam para produzir ATP e transferir elétrons da água até o NADP+.
No ciclo de Calvin, ATP e NADPH são usados para sintetizar açúcar a
partir de CO2. A energia que entra no cloroplasto como luz solar torna-
se energia química estocada nas ligações das moléculas de açúcar.
O açúcar produzido nos cloroplastos sustenta a planta inteira
com energia química e esqueletos de carbono para sintetizar todas as
principais moléculas orgânicas das células. Aproximadamente 50%
do material orgânico produzido na fotossíntese, entretanto, são con-
sumidos como combustível para a respiração celular na mitocôndria
das células vegetais.

CEDERJ 89
AU
LA
MÓ
DU
LO 2
8Figura 8.5: Síntese de sacarose a partir de UDP-glicose.
Tecnicamente, células verdes são a única parte autotrófi ca da
planta. O resto da planta depende de moléculas orgânicas exportadas
pelas folhas nos sistemas vasculares. Na maioria das plantas, carboidratos
são transportados das folhas na forma de sacarose, um dissacarídeo
(Figura 8.5). Quando chega às células não fotossintetizantes, a sacarose
é usada na respiração celular e em uma grande variedade de vias
anabólicas. Uma considerável parte da glicose é usada para produzir
celulose. Celulose é a molécula orgânica mais abundante na planta e
provavelmente na superfície do planeta.
O excesso de glicose é usado na síntese de amido que é estocado em folhas (no próprio
cloroplasto) e em células de armazenagem como raízes, tubérculos e frutos. Isto tudo é substrato
oxidativo para heterotrófi cos, incluindo humanos.
Em uma escala global, a produtividade do cloroplasto é prodigiosa. Estima-se que a fotossín-
tese produza 160 bilhões de toneladas métricas de carboidratos por ano (ton métrica = 1000 Hg).
Nenhum outro processo químico na Terra tem essa produtividade e nenhum é mais impor-
tante para a manutenção da vida no planeta.
CH OH2
H H
H
H
H
H H
H H
HH
H
H
HH
H H
H
H
CH OH2
CH OH2
CH OH2
HOHO
H
HOH
HOHO
HOHO
H H
O
O
O
O
OO
O
O
O
O
O
OH OH
OH
OHOH
OH
OH
OH OH
H
CH2
CH2
HOCH2
HOCH2
HOCH2
UDP P
P
UDP
Pi
UDP-glicose Frutose-6-fosfato
Sacarose 6-fosfato sintase
Sacarose 6-fosfato
Sacarose 6-fosfato fosfatase
Sacarose
1 2
1 2
1 2+

Bioquímica II | Outros aspectos importantes da fotossíntese
CEDERJ90
R E S U M O
Nessa aula falamos de fotorrespiração, um desvio metabólico da fotossíntese que
ocorre quando a RuBisCO passa a trabalhar como oxigenase. Geralmente isso
acontece em altas temperaturas e quando a concentração de O2 na atmosfera é
alta e a concentração de CO2 baixa. Mostramos que algumas plantas, como as
C4 e as CAM, apresentam adaptações metabólicas que otimizam a fotossíntese,
aumentando a concentração de CO2 nas células. Além disso, nesta aula discutimos
a regulação da fotossíntese. Lembre que a velocidade do ciclo de Calvin é regulada
por dois sistemas: um é a concentração de prótons e magnésio no estroma do
cloroplasto que ativa a RuBisCO; o outro são as reações de oxi-redução promovidas
pelos elétrons e prótons da cadeia transportadora. Falamos, fi nalmente, do destino
da glicose gerada na fotossíntese, mais especifi camente da síntese de sacarose
e formação de amido.
Com isso terminamos o módulo Fotossíntese. Espero que você
tenha gostado, entendido e, acima de tudo, percebido a importância
deste processo bioquímico. Nas próximas aulas falaremos de
fermentação e de respiração celular. Esses dois processos formam, com
a fotossíntese, o pacote básico do metabolismo energético. Como você
já percebeu desde as primeiras aulas de fotossíntese, eles caminham
em paralelo e são interdependentes. Esta interdependência é a base do
fenômeno biológico.
Antes de continuar, faça os exercícios de fi xação que estão a seguir.

CEDERJ 91
AU
LA
MÓ
DU
LO 2
8
EXERCÍCIOS DE FIXAÇÃO
1. O que é fotossíntese?
2. Que afi rmativa(s) sobre a fotossíntese não é correta?
a) As reações luminosas são dirigidas pela energia derivada da oxidação da água.
( )
b) Fotossistema II usa energia luminosa para oxidar água a O2 e então passa os
eletrons para o fotossistema I. ( )
c) No fotossistema I, o elétron é outra vez excitado pela luz e é usado para reduzir
a ferredoxina. ( )
d) A produção de ATP e NADPH é dirigida pela energia que é derivada da absorção
de luz. ( )
e) Ferredoxina reduzida reduz NADP+ a NADPH. ( )
3. Ao contrário da fosforilação oxidativa, a fotofosforilação não envolve complexos
protéicos ligados à membrana. Falso ou verdadeiro? Por quê?
4. Quando a cadeia transportadora de elétrons está em pleno funcionamento, o
pH do lúmen do tilacóide será (maior que, menor que
ou igual a) o pH do estroma.
5. Analisando as equações da fotossíntese e da quimiossíntese abaixo, o que se
pode concluir da origem do oxigênio molecular formado?
Fotossíntese: H2O + CO2 glicose + O2 + Energia
Quimiossíntese: H2S + CO2 glicose + 2S + Energia
6. O que não acontece na fosforilação cíclica?
a) produção de ATP;
b) atividade do centro de reação do FSI;
c) o transporte de elétrons na membrana fotossintética;
d) utilização de energia luminosa;
e) formação de NADPH.

Bioquímica II | Outros aspectos importantes da fotossíntese
CEDERJ92
7. O reagente DCMU, 3-(3,4-diclorofenil)-1,1-dimetil urea, inibe a transferência de
elétrons para a plastoquinona no fotossistema II. Que efeito fi nal você esperaria
encontrar, se fosse administrado DCMU a uma suspensão de cloroplastos iluminada?
8. Por que nossos olhos detectam como verde estruturas contendo clorofi la?
9. Suponha um breve pulso de 14CO2 em uma planta verde.
a) Trace o caminho percorrido pelo 14C até a síntese de frutose 1,6-bisfosfato,
mostrando que os átomos de carbono em cada intermediário devem carregar a
marcação em cada ciclo.
b) Todas as moléculas de frutose 1,6-bisfosfato apresentarão dois átomos de
carbono marcados?
10. As plantas C3 e C4 são mantidas juntas numa caixa selada e iluminada, a planta
C4 viceja, enquanto a planta C3 adoece e, eventualmente, morre. Explique.
RESPOSTAS DOS EXERCÍCIOS DE FIXAÇÃO
1. O que é fotossíntese?
É a via metabólica de síntese de glicose a partir de CO2 e H2O que ocorre em células
especializadas com energia derivada do sol (energia luminosa).
2. Que afi rmativa (s) sobre a fotossíntese não é correta?
a) As reações luminosas são dirigidas pela energia derivada da oxidação da água
( x ).
b) Fotossistema II usa energia luminosa para oxidar água a O2 e, então passa os
elétrons para o fotossistema I ( ).
c) No fotossistema I, o elétron é outra vez excitado pela luz e é usado para reduzir
a ferredoxina ( ).
d) A produção de ATP e NADPH é dirigida pela energia que é derivada da absorção
de luz ( ).
e) Ferredoxina reduzida reduz NADP+ a NADPH ( ).
3. Ao contrário da fosforilação oxidativa, a fotofosforilação não envolve complexos
protéicos ligados à membrana. Falso ou verdadeiro? Por quê?
Errado. Os fotossistemas, assim como os transportadores, são proteínas.

CEDERJ 93
AU
LA
MÓ
DU
LO 2
8
4. Quando a cadeia transportadora de elétrons está em pleno funcionamento, o
pH do lúmen do tilacóide será menor que (maior que, menor que ou igual a) o
pH do estroma.
5. Analisando as equações da fotossíntese e da quimiossíntese abaixo, o que se
pode concluir da origem do oxigênio molecular formado?
Fotossíntese: H2O + CO2 glicose + O2 + Energia
Quimiossíntese: H2S + CO2 glicose + 2S + Energia
O oxigênio liberado na fotossíntese se origina da molécula de água.
6. O que não acontece na fosforilação cíclica?
a) produção de ATP.
b) atividade do centro de reação do FSI.
c) o transporte de elétrons na membrana fotossintética.
d) utilização de energia luminosa.
e) formação de NADPH.
7. O reagente DCMU, 3-(3,4-diclorofenil)-1,1-dimetil urea, inibe a transferência
de elétrons para a plastoquinona no fotossistema II. Que efeito fi nal você
esperaria encontrar, se fosse administrado DCMU a uma suspensão de
cloroplastos iluminada?
A inibição do transporte de elétrons da plastoquinona para o fotossistema II
interrompe o início do transporte de elétrons, não permitindo que ocorra a síntese
de ATP nem a redução de NADPH.
8. Por que nossos olhos detectam como verde estruturas contendo clorofi la?
Porque a clorofi la absorve os comprimentos de onda azul e vermelho. Nossos olhos
detectam os comprimentos de onda restantes. A percepção dos comprimentos de
onda transmitidos é verde.
9. Suponha um breve pulso de 14CO2 em uma planta verde.
a) Trace o caminho percorrido pelo 14C até a síntese de frutose 1,6-bisfosfato,
mostrando que átomos de carbono em cada intermediário devem carregar a
marcação em cada ciclo.
Para responder, veja a ordem dos intermediários do ciclo de Calvin e observe suas
estruturas. Nem todas são mostradas na aula. Pesquise nos livros de bioquímica
do pólo.

Bioquímica II | Outros aspectos importantes da fotossíntese
CEDERJ94
b) Todas as moléculas de frutose 1,6-bisfosfato apresentarão dois átomos de
carbono marcados?
Discuta com seus colegas e seu tutor, procure nos livros, siga a pista dada para a
pergunta anterior.
10. As plantas C3 e C4 são mantidas juntas numa caixa selada e iluminada; a planta
C4 viceja, enquanto a planta C3 adoece e eventualmente morre. Explique.
Plantas C4 são mais efi cientes na utilização do CO2.

A história da fermentação:do início aos dias de hoje 9AULA

Bioquímica II | A história da fermentação: do início aos dias de hoje
CEDERJ96
Desde a Antigüidade, o processo de fermentação era conhecido
e muito utilizado na fabricação de vinhos, queijos e pães. Aristóteles
descreveu quatro etapas na formação do vinho: a infância, quando a uva
estava na parreira; a adolescência, com o suco de uva; a maturidade com
o vinho; e fi nalmente a velhice e morte, quando o vinho se transformava
em vinagre.
Uma das primeiras definições do processo fermentativo foi
elaborada em 1684, por Willis:
A fermentação é um movimento intestino de partículas ou
princípios de cada corpo, tendendo para a perfeição ou para a
transformação em outro. As partículas elementares postas em
movimento, devido à sua própria natureza ou ocasionalmente
vibrando maravilhosamente, são aprisionadas e transformam-se
em outras: as sutis e mais ativas esforçam-se na tentativa de escapar
suavemente, mas estando entrelaçadas com outras mais espessas
são impedidas de fazê-lo. Também as mais espessas são mantidas
unidas pelo intento e expansão das mais sutis e são enfraquecidas até
que cada uma alcance sua própria grandeza e exaltação. Elas fi xam
em si a devida perfeição ou completam as alterações e mutações
designadas pela natureza.
Até o início do século XIX, a conservação dos alimentos era
feita utilizando-se basicamente a técnica da salmoura. Nessa época, o
Império Napoleônico estava em expansão. Entretanto, um dos maiores
empecilhos para a conquista de novos territórios era a alimentação de
seus soldados. Napoleão, ofereceu, então, um prêmio de 12000 francos
para quem inventasse uma maneira efi ciente de conservar os alimentos
por um longo período de tempo.
Nesse contexto, destacou-se o francês Mr. Nicolas Appert, dono
de uma loja de secos e molhados, que exportava alimentos para diversas
localidades e pessoas, inclusive para o Czar da Rússia. Sua técnica de
conservação de alimentos, herdada de seu pai, consistia em adicionar
grandes quantidades de açúcar às frutas e ferver os alimentos, que
deveriam imediatamente ser tampados e selados com parafi na.
Figura 9.1: Capa do livro de Mr. Appert, A arte de preservar alimentos de origem animal e veg-etal, 1810.

CEDERJ 97
AU
LA
MÓ
DU
LO 3
9
Suas receitas foram publicadas em um livro (Figura 9.1), considerado
por muitos como o primeiro livro de Bioquímica.
Hoje o processo fermentativo é amplamente
conhecido e mundialmente utilizado em escala
industrial. A cerveja é um exemplo de produto obtido
a partir deste processo, onde o organismo fermentador
é uma levedura.
A IMPORTÂNCIA DA HISTÓRIA DA FERMENTAÇÃO NA HISTÓRIA DA BIOQUÍMICA
Com base no livro de Mr. Appert, um físico-químico da época
chamado Gay-Lussac fez a seguinte experiência (Figura 9.2):
Aqueceu duas vasilhas com caldo de carne e cobriu apenas uma
delas com uma camada de mercúrio.
Em qual das vasilhas você esperaria que ocorresse fermentação?
Por quê?
Na metade do século XIX a qualidade dos vinhos na França era
muito baixa. Já se tinha conhecimento, na época, de que a qualidade
do vinho dependia do tipo de uva utilizada, bem como do repique (uma
porção de um vinho de boa qualidade já fermentado). A adição do
repique acelerava o processo de fermentação e uniformizava o paladar
do vinho.
Na tentativa de solucionar o problema da qualidade do vinho
de paladar ruim, uma vinícola francesa contratou Louis Pasteur, um
químico simples e muito religioso.
Ao analisar o vinho ao microscópio, Pasteur observou a
existência de grande quantidade de “glóbulos castanhos”, alguns
inclusive se dividindo (Figura 9.3).
Figura 9.2: O experimento de Gay-Lussac. Em A a vasilha com caldo de carne; em B a vasilha com caldo de carne coberta por uma camada de mercúrio.
A B
Figura 9.3: Glóbulos castanhos observados por Pasteur ao analisar o vinho ao microscópio.
Pense sobre isso!
Acesse http://perso.magic. f r / o r m e r r y /newindex.html para saber mais sobre Nicolas Appert e suas importantes contribuições para a indústria de alimentos.e http://members.nbci.com/_XMCM/cervas/fabricacao.htm) se você quer saber mais sobre como se fabrica cerveja.
!

Bioquímica II | A história da fermentação: do início aos dias de hoje
CEDERJ98
Pasteur lançou mão de suas experiências anteriores como cientista
para tentar desvendar os segredos da fermentação. Antes de ser contratado
pela fábrica de vinhos, Pasteur havia se dedicado ao estudo da polarização
da luz por ácidos orgânicos e observou, usando um polarímetro, que ácidos
orgânicos extraídos de tecidos de seres vivos eram capazes de desviar o
plano da luz polarizada. Ao contrário, os ácidos orgânicos sintéticos não
eram capazes de desviar o plano da luz polarizada.
Pasteur concluiu que os ácidos orgânicos eram constituídos de uma
MISTURA RACÊMICA e que somente os seres vivos eram capazes de transformá-los
em um grupo homogêneo de substâncias capazes de desviar o plano da luz.
Pense sobre isso!
Isolando ácidos orgânicos de um repique e colocando-os em um
polarímetro, Pasteur observou que esses compostos eram capazes de
desviar a luz.
O que você concluiria no lugar de Pasteur?
Assim, Pasteur propõe que os glóbulos castanhos são organismos
vivos e que somente na presença destes pode haver fermentação. Pasteur
associou não só a fermentação, mas também outros processos à presença
de organismos vivos e formulou, assim, a Teoria Vitalista.
Surgem imediatamente críticas ferozes à sua teoria. A mais
contundente e irônica foi escrita por um renomado químico da época
chamado Liebig:
MI S T U R A R A C Ê M I C A
Mistura racêmica é uma mistura de substâncias capazes de desviar o plano de luz polarizado para a esquerda (levrógiras) com substâncias capazes de desviar o plano de luz polarizado para a direita (dextrógiras).
SOLUÇÃO DO SEGREDO DA
FERMENTAÇÃO ALCOÓLICA
O levedo de cerveja, quando disperso em água, quebra-se em um número
infi nito de pequenas esferas. Se estas esferas são transferidas para uma
solução aquosa de açúcar, elas desenvolvem-se em pequenos animais.
Estes animais são dotados de uma espécie de trompa de sucção com a
qual eles devoram o açúcar da solução. A digestão é reconhecível clara
e imediatamente por causa da descarga de excrementos. Estes animais
evacuam álcool etílico dos seus intestinos e dióxido de carbono dos seus
órgãos urinários. Deste modo pode-se observar que um fl uido especifi camente mais claro é
exudado de seu ânus e ascende verticalmente, ao passo que um jato de dióxido de carbono
é ejetado em intervalos muito curtos dos seus genitais enormemente grandes.
(WOEHLER E LIEBIG,1893).
Pense sobre isso!

CEDERJ 99
AU
LA
MÓ
DU
LO 3
9
Essa passagem mostra que havia uma forte resistência à idéia
de que a fermentação era um processo biológico que dependia do
metabolismo dos “glóbulos” observados por Pasteur. Ao contrário,
Liebig e boa parte da comunidade científi ca da época acreditavam
que a fermentação era, na verdade, um processo meramente químico,
decorrente da reação entre substâncias albuminóides (nitrogenadas),
presentes na superfície dos “glóbulos”, com o açúcar do meio, na
presença de ar.
Para responder às críticas, Pasteur mostrou que os “glóbulos”
presentes no meio contendo açúcar eram capazes de promover a
fermentação, mesmo na ausência de ar (Figura 9.4).
Com os resultados obtidos a partir das experiências realizadas na
ausência de oxigênio, Pasteur afi rma que a fermentação pode ocorrer
mesmo em condições de anaerobiose, descrevendo pela primeira vez
La vie sans l’air (“A vida sem ar”).
Foram necessários mais de cinqüenta anos para que fossem
acrescentados novos conhecimentos à área da fermentação.
Dois irmãos, Hans e Edward Büchner, após terem sido expulsos de
laboratórios de pesquisa por serem muito desastrados, resolveram abrir
em sociedade uma pequena fábrica para produzir um xarope contra a
tuberculose. Hans era microbiologista e Edward, botânico e químico; este
último tendo inclusive trabalhado com o renomado químico Adolf von
Bayer. Ambos decidiram investir em um negócio que lhes fosse rentável.
No início do século XIX, as principais doenças eram a sífi lis e a
tuberculose. Por não haver tratamento efetivo para ambas as doenças,
possuí-las signifi cava sentença de morte para o paciente. Era costume, na
época, internar os portadores de tuberculose em sanatórios localizados
nas montanhas, onde respiravam ar puro, repousavam e eram obrigados
a beber uma cerveja escura e amarga. Na opinião de muitos especialistas,
o levedo depositado no fundo da cerveja produzia antitoxinas capazes de
combater os microorganismos causadores da tuberculose. Entretanto, para
o total efeito terapêutico eram necessários vários litros dessa cerveja.
Valendo-se disto, os irmãos Büchner resolveram preparar um
xarope dessa levedura e poupar o sofrimento dos doentes, que poderiam
adquirir um xarope com extrato concentrado de antitoxinas, dispensando
assim a ingestão dos litros de cerveja amarga.
Figura 9.4: Resultado do experimento de Pasteur. Foi medido CO2 em função do tempo de fermentação na presença (c/O2) ou na ausên-cia (s/O2) de oxigênio.
s/O2
c/O2c/O
pro
du
zid
o2
Tempo (min)

Bioquímica II | A história da fermentação: do início aos dias de hoje
CEDERJ100
Para preparar o xarope, eles montaram uma estratégia que
consistia em:
1 – fazer crescer grandes quantidades de levedura em frascos contendo
caldo de açúcar (leveduras em cultura);
2 – isolar as leveduras do meio de cultura através de centrifugação
(leveduras íntegras isoladas);
3 – romper as leveduras (a ruptura das células era realizada através de
um processo trabalhoso que utilizava uma espécie de prensa, chamada
prensa francesa, com a capacidade de gerar até 300 atmosferas) (extrato
de levedura);
4 – e, por fi m, fi ltrar o extrato de leveduras para obter um concentrado
livre de células (extrato livre de células).
Para conservar o xarope, recorreram ao clássico livro de Mr. Appert
sobre conservar os alimentos e adicionaram grandes quantidades de açúcar
(xarope de levedura).
Seria esse procedimento efi ciente para conservar o xarope?
Para surpresa dos irmãos Büchner, o xarope continuou a fermentar.
Baseando-se nas experiências de Pasteur, eles pensaram prontamente
que se tratava de uma contaminação com células intactas. Para resolver
o problema, passaram o xarope pela prensa francesa diversas vezes e
certifi caram-se, utilizando um microscópio, de que não havia mais células
íntegras. No entanto, o xarope continuava a fermentar.
Por quê ?
Hans, o microbiologista, cansado de trabalhar sem obter
resultado, desfez a sociedade com o irmão. Edward, consciente de que
a fermentação obtida com o xarope não era por erro experimental,
continuou seu trabalho. Após repetir inúmeras vezes o processo de quebra
das leveduras, Edward teve certeza de que a presença de célula íntegra de
levedura não era necessária para que a fermentação ocorresse. E, assim,
submeteu sua descoberta à comunidade científi ca.
Edward Büchner (Figura 9.5) recebeu, em 1907, o 1º Prêmio
Nobel de Química por seu trabalho Der Einfl uss des Sauerstoffs auf
Gärungen (A infl uência do oxigênio na fermentação), publicado em
1885, e o seguinte comentário de Bayer, seu antigo chefe:
“Esse rapaz, apesar de não ter nenhum talento para química, vai
fi car famoso.”
Que conseqüências os resultados acima proporcionaram à teoria
Vitalista de Pasteur e à pesquisa bioquímica?
Pense sobre isso!
Figura 9.5: Edward Buchner. 1º Prêmio Nobel de Química. Der Einfluss des Sauerst-offs auf Gärungen (A infl uência do oxigênio na fermentação), pub-licado em 1885.
Pense sobre isso!
Pense sobre isso!

CEDERJ 101
AU
LA
MÓ
DU
LO 3
9
A FERMENTAÇÃO E A VIA GLICOLÍTICA
Paralelamente à descoberta dos irmãos Büchner, de
que a fermentação ocorria mesmo na ausência de vida,
Arthur Harden e William John Young, dois pesquisadores
ingleses, foram contratados por uma fábrica de cerveja para
estudar a fermentação.
Em 1903, eles estabeleceram a relação de 1:1 entre gás
carbônico e etanol produzidos pela fermentação do açúcar.
Utilizando o extrato de levedura na presença de glicose e
medindo a produção de CO2, eles observaram que a fermentação ocorria
durante algum tempo, sendo porém interrompida após certo período.
Eles observaram também que a fermentação alcoólica da glicose,
por suco de levedura (o extrato de levedura dos irmãos Büchner), era
muito aumentada pela adição de mais suco de levedura, o qual tinha sido
previamente fervido e fi ltrado. Este líquido fervido, por si só, era incapaz
de fermentar quando em presença de glicose. Observe o Gráfi co 9.1. Ele
representa o andamento da fermentação ao longo do tempo.
Gráfi co 9.1: Medida de dióxido de carbono em função do tempo de fermentação.Curva A: na presença de 25 mL de suco de levedura + 25 mL de água + 5g de glicose; Curva B: na presença de 25 mL do mesmo suco de levedura que na curva A + 25 mL de suco de levedura fervido + 5g de glicose.
O que sugere esse resultado?
Harden e Young submeteram o suco de levedura fervido à DIÁLISE
e assim separaram duas frações distintas (Tabela 9.1). Uma fração era
composta de moléculas pequenas que passaram pela membrana de diálise.
Outra fração era composta de moléculas grandes, que não passaram na
membrana de diálise e, portanto, foram chamadas resíduo.
DIÁLISE
Diálise é um procedimento através
do qual se separam moléculas grandes de moléculas pequenas.
A diálise baseia-se na capacidade de estas
moléculas atravessarem os poros de uma
membrana. O limitante, então, será
o tamanho, ou seja, moléculas pequenas passam e moléculas grandes não passam
pelos poros. Para que esta seletividade seja efi ciente, você pode
imaginar o tamanho que estes poros devem ter?
Figura 9.6: Uma placa de estrada na Inglaterra anunciando Young e Harden. Para onde esses dois levaram a Bioquímica?
Pense sobre isso!
B
Dió
xid
o d
e ca
rbo
no
Tempo em horas
0 10 20 30 40 60 70 9080 100
120011001000900800700600500400300200
0100
A
50

Bioquímica II | A história da fermentação: do início aos dias de hoje
CEDERJ102
No Água(mL)
Suco fervido
(mL)
Resíduo(mL)
Dialisado(mL)
Tempo(horas)
Dióxido de car-bono
(grama)
1
2
2500
250
0250
00
0025
00
000
025
484848
4848
0,2530,5610,264
0,1540,251
Levando-se em consideração os resultados, que característica
teria, então, o ativador da fermentação?
Como resultado de um grande número de tentativas de isolar o
constituinte do suco de levedura responsável pelo aumento da
fermentação, observou-se que quando ácido fosfórico na forma
de um fosfato solúvel estava presente era produzido um aumento
na taxa de fermentação. O efeito da adição de fosfatos solúveis ao
suco de levedura foi então examinado e encontrou-se uma evolução
inicial rápida bem marcada de dióxido de carbono produzido.
Além disso, os sucos de levedura usados invariavelmente contêm
fosfatos, precipitáveis por mistura magnésia; nesse ponto não
pode haver nenhuma dúvida de que o fenômeno inicial é devido à
presença destes (HARDEN e YOUNG, 1906).
Com estas descobertas, Harden e Young afi rmaram que o processo
fermentativo necessitava tanto da fração residual da diálise do suco não
fervido, que chamaram zimazes (proteínas), quanto da fração dialisável
do suco de levedura, que eles chamaram cozimazes. A observação
de fosfatos durante o processo fermentativo levou–os a supor que as
cozimazes fossem compostas por fosfato.
Adicionando sucessivamente Pi (fosfato inorgânico) a um extrato
de leveduras, observaram que a retomada da fermentação ocorria até
certo ponto, e a partir daí tanto a adição de Pi como a adição de glicose
não mais ativavam o processo. Por outro lado, a adição de mais suco de
levedura fervido ainda era capaz de ativar a fermentação (Gráfi co 9.2).
Tabela 9.1: Diálise do suco de levedura fervido: 25 mL de suco de levedura+ 5 gramas glicose
Pense sobre isso!

CEDERJ 103
AU
LA
MÓ
DU
LO 3
9
Gráfi co 9.2: Efeito da adição de fosfato na fermentação.Mistura da reação: 25 mL de suco de levedura + 20 mL de água + 5 g de glicose.Curva A: representa a fermentação na ausência de fosfato adicionado;Curva B e C: representam o efeito da adição sucessiva de duas quantidades de 5 mL de fosfato de sódio (0,3 M).
Qual seria então o destino do fosfato inorgânico?
Utilizando um sal de prata, Harden e Young foram capazes de
isolar, por precipitação, um composto fosforilado com seis carbonos
(6C) e dois fosfatos, e identifi caram como sendo frutose-1,6-bisfosfato,
denominado Éster de Harden e Young, o primeiro intermediário da via
glicolítica identifi cado. Na época, os autores fi caram em dúvida se o
composto era uma hexose difosfatada ou duas trioses monofosfatadas.
Como você faria para provar que este composto realmente
participa do processo fermentativo?
A seguir, Otto Meyerhoff, um pesquisador alemão, trabalhando
com músculo de rã, peito de pombo e de coelho, mostrou que o músculo
se contraía na ausência de oxigênio e isso resultava na produção de
ácido lático.
Esse processo pode ser ativado por duas frações do extrato de
músculo, uma termossensível (fração sensível ao calor, também chamada
zimazes) e outra termorresistente (fração insensível ao calor, chamada
cozimazes). Mas este não foi o único paralelo entre a fermentação da
levedura e os músculos do pombo: Meyerhoff testou Pi nos músculos
e obteve os mesmos resultados de Harden e Young. Assim, sabendo
que a glicose é consumida pelo extrato de levedura na fermentação,
resultando em etanol e dióxido de carbono, e que a adição de glicose em
macerados de músculo de coelho resultava no aparecimento de lactato,
Otto Meyerhoff levantou a possibilidade de as proteínas envolvidas nos
dois processos serem também semelhantes.
Que experiência você faria para testar esta hipótese?
Pense sobre isso!
Pense sobre isso!
Pense sobre isso!
Dió
xid
o d
e ca
rbo
no
Tempo em minutos
120
130
110
100
90
80
70
60
50
40
30
20
0
10
10 20 30 40 60 70 9080 100 110 120 130 140 15050
A
B
C

Bioquímica II | A história da fermentação: do início aos dias de hoje
CEDERJ104
É uma fração termossensível, porque o aquecimento a 50º C por
somente um minuto resulta num considerável grau de inativação.
(...) O ativador tem as propriedades de uma enzima. Eu proponho,
portanto, designá-la “hexoquinase”, pois ela atua especifi camente
em hexoses fermentáveis. (...) Com certas variações, a velocidade
inicial de formação de ácido lático é proporcional à quantidade de
hexoquinase. Levedura é o único material do qual foi possível isolar
o ativador. (...) A procura da “hexoquinase” em leveduras lisadas
foi baseada na idéia de que extratos de levedura apresentam uma
esterifi cação rápida das hexoses fermentáveis, com subseqüente
fermentação. Extrato de músculos, ao contrário, não apresenta
qualquer acumulação de ésteres. Esta diferença deve ser devida
ao fato de o fator essencial para a esterifi cação da glicose ser mais
concentrado em leveduras. A validade dessa hipótese é ilustrada por
observações simultâneas da formação de ácido lático e esterifi cação
de hexoses em extratos de músculo ativados (MEYERHOFF,
1930).
Já nessa época, se conhecia o efeito letal de alguns compostos,
seja em bactérias e leveduras, como também em mamíferos. Entretanto,
o mecanismo de ação de tais compostos ainda não era conhecido.
Nesse contexto, alguns pesquisadores decidiram testar o efeito
desses venenos durante o processo fermentativo e, após análise dos
produtos, obtiveram o seguinte resultado:
DROGA EFEITO
1. Fluoreto de sódio Acúmulo de 2-fosfoglicerato e a seguir 3-fosfoglicerato.
2. Cloro-acetal-fosfato (CAP) Acúmulo de dihidroxicetona fosfato e frutose 1,6-bisfosfato.
3. Iodoacetato Um súbito acúmulo de gliceraldeído 1,3-bisfosfato seguido do seu rápido
desaparecimento e de um grande acúmulo de frutose 1,6- bisfosfato.
A partir destes dados, deduza as possíveis etapas envolvidas na
degradação da glicose.
Pista 1: Uma via metabólica pode ser descrita como
A B C D
Onde A é o substrato ou matéria-prima da via, B e C são os
Células lisadas são células cujas membranas foram rompidas. Isso pode ocorrer por uma grande variedade de técnicas, como congelamento/descongelamento, pressão, choque osmótico etc.
Pense sobre isso!
Tabela 9.2: Efeito de diferentes inibidores em reações da via glicolítica.

CEDERJ 105
AU
LA
MÓ
DU
LO 3
9
intermediários e D é o produto fi nal.
Se um determinado inibidor interrompe a reação B D, qual
dos intermediários será acumulado? O que não será mais produzido?
Utilize esta lógica para cada uma das linhas da tabela e encontre
as reações isoladamente.
Pista 2: A glicólise é uma via de degradação ou quebra da glicose
e, portanto, os produtos são moléculas menores. Observe as reações
isoladamente. Consulte as fórmulas das moléculas, veja quais delas são
maiores e quais são menores. Observe também o grau de semelhança
entre elas.
Se você chegou até aqui, entendeu como uma via metabólica
pode ser desvendada. Na próxima aula mostraremos a glicólise na sua
totalidade. Agora, a maior parte da via você já conhece. Então vai ser
fácil compreender o que falta. Vamos lá?
Para saber mais, visite estes endereços:Harden e Young: http://ir2lcb.cnrs-mrs.fr/~athel/harden.htmEdward Büchner: http://ir2lcb.cnrs-mrs.fr/~athel/buchner.htm#buch;http://ir2lcb.cnrs-mrs.fr/~athel/buchner.htmLouis Pasteur: http://www.woodrow.org/teachers/chemistry/institutes/1992/Pasteur.htmlhttp://www.pasteur.net/louis/index.htmBaseado no material organizado pelo departamento de Bioquímica Médica, CCS, UFRJ.
H d Y htt //i 2l b f / th l/h d ht


Glicólise: oxidaçãode glicose a piruvato I 10A
UL
A

Bioquímica II | Glicólise: oxidação de glicose a piruvato I
CEDERJ108
Na aula anterior, você conheceu a história da Glicólise e soube como se chegou
ao conhecimento disponível sobre esta via metabólica. Agora podemos falar um
pouco mais sobre ela, suas funções, seus intermediários, enzimas, cofatores etc.
Essa é uma via importante, pois a glicose (que você já conhece da Bioquímica I)
é, quantitativamente, o principal substrato oxidável na maioria dos seres vivos:
dos microorganismos ao homem. A glicólise é a principal via para a utilização da
glicose e ocorre no citosol da maioria das células, sendo considerada, portanto,
uma via universal. Ela pode ocorrer se o oxigênio estiver disponível (aerobiose)
ou na total ausência dele (anaerobiose).
A GLICOSE COMO SUBSTRATO OXIDÁVEL
Quase todas as células são capazes de atender às suas demandas
energéticas apenas a partir de glicose. Esse açúcar é imprescindível para
algumas células e tecidos, como hemácias e tecido nervoso, por constituir
o único substrato que eles são capazes de utilizar para obter energia.
Você viu nas nossas aulas iniciais que a energia contida nas moléculas
oxidáveis, como a glicose, são derivadas em última análise da radiação
solar. Esse tipo de energia é transformado em energia química durante
a síntese de moléculas orgânicas (principalmente glicose) que ocorre na
fotossíntese. Os organismos chamados heterotrófi cos são capazes de
utilizar tal energia presente nas ligações químicas das moléculas orgânicas
para realizar suas atividades celulares. Mas essa utilização não ocorre de
forma direta: primeiro a energia precisa ser convertida em ATP.
INTRODUÇÃO
CO2 + H2O (CH2O)n piruvato
Luz solar ADP + Pi ATP
fotossíntese glicólise
Uma das vias metabólicas de extração de energia, acoplada à
síntese de ATP, é a glicólise, também chamada via glicolítica. Nela,
a glicose é oxidada (degradada, quebrada) e, durante esse processo
oxidativo, a célula é capaz de utilizar a energia liberada para sintetizar
duas moléculas de ATP.
Esquema 10.1

CEDERJ 109
AU
LA 1
0 M
ÓD
ULO
3
A GLICOSE COMO INDICADOR METABÓLICO
O nível sangüíneo de glicose é chamado glicemia. Variações
na glicemia são indicadores do estado fisiológico do organismo.
Assim, quando a glicemia está baixa (hipoglicemia), as células têm
um comportamento metabólico distinto daquele que ocorre quando a
glicemia está alta (hiperglicemia).
Hiperglicemia indica um estado fi siológico de fartura e, portanto,
de armazenamento ou biossíntese (anabolismo – como você viu nas aulas
de introdução ao metabolismo).
Hipoglicemia indica um estado fi siológico de privação, mesmo
que momentânea, e as células respondem derivando seu metabolismo
para a degradação (catabolismo).
Os níveis sangüíneos de glicose são mantidos dentro de uma
faixa estreita, graças a diferentes vias metabólicas de síntese de glicose
(gliconeogênese) ou armazenamento de glicose na forma de glicogênio
(glicogenogênese), em contraposição a vias de oxidação de glicose
(glicólise). Esse processo, que procura manter os níveis de glicose no sangue
constantes, chama-se homeostase e é fi namente regulado por hormônios.
Pequenas variações nesse nível indicam, para as células, o que fazer.
Glossário:
Glicogênio – um polímero de glicose.
Gliconeogênese ou neo-glicogênese – síntese de glicose a partir de precur-sores não glicídicos.
Glicogenogênese – sín-tese de glicogênio.
Glicólise – quebra da glicose.
Glicogenólise – quebra do glicogênio.
Figura 10.1: A homeostase de glicose é mantida pelas vias de síntese e de degradação.
Glicogênio, amido
Fotossíntese
GlicoseGlicose
Gliconeogênese
Co eluz solar
2 Piruvatoe
Alanina
ATP NADPH eribose-5-P
Glicólise e ciclode Krebs
Via daspentoses-fosfato

Bioquímica II | Glicólise: oxidação de glicose a piruvato I
CEDERJ110
GLICÓLISE – ASPECTOS GERAIS
Na via glicolítica, uma molécula de glicose (seis carbonos
– Figura 10.2) é quebrada, produzindo duas moléculas de três carbonos
denominadas piruvato. É um processo oxidativo no qual duas moléculas
de NAD+ (Figura 10.3) são reduzidas a duas moléculas de NADH + H+.
Figura 10.3: Nicotinamida adenina dinucleotídeo – NAD. O NAD pode estar na sua forma oxidada NAD+ ou na sua forma reduzida NADH.
Para que esta via ocorra, existem etapas que consomem energia
(através da hidrólise de ATP - Figura 10.4): etapas endergônicas, onde a
molécula de glicose é preparada para ser clivada; e etapas que sintetizam
ATP: etapas exergônicas, onde moléculas de ATP são formadas a partir
da energia liberada durante o processo oxidativo.
Figura 10.4: Adenosina trifosfato – ATP. Em destaque, a ligação fosfoéster entre a adenosina e o fosfato e as ligações fosfoanidras, entre fosfatos.
Figura 10.2: Glicose ou α-D- glicopiranose (projeção de Haworth).
H H HO O
R
+HC CNH2 NH2
2H+
2e-
+N
O-O
O
O
P
O
O-
O
O H
OH OH
H HHP
CH2
O
H
OH OH -
H H
CH2
..N
NADHReduzido
NAD+
(oxidado)
Adenina
NH2
N
H
N
N
N
ATP
AMP
Adenosina
ADP
OO
O O O
O
OO
OP P
Ligaçõesfosfoanidras
fosfoéster
O
O
P
H
OH OH -
H H
CH2
H
NH2
N
N
N
N
�� �
HHO
HO
H OH
OH H OH
H
CH2OH
23
4
5
6
1

CEDERJ 111
AU
LA 1
0 M
ÓD
ULO
3
No processo completo, associando-se consumo e formação de
ATP, temos um saldo líquido de duas dessas moléculas. Vale ressaltar
que, na realidade, conservamos parte da energia liberada nesse processo
oxidativo na formação de ligações éster–fosfato (ligações ricas em energia)
entre a molécula de ADP e fosfato inorgânico (Pi). Uma visão geral deste
processo é apresentada na Figura 10.5.
Como produtos desta via temos, além de ATP e NADH, duas
moléculas de piruvato. O destino deste piruvato depende da célula e da
presença ou ausência de oxigênio, como veremos mais adiante.
Figura 10.5: Resumo da via glicolítica. Uma molécula de glicose (seis carbonos) é quebrada em duas moléculas de piruvato (três carbonos). Neste processo, a energia liberada é utilizada para a síntese de duas moléculas de ATP. O produto fi nal (piruvato) pode ter des-tinos diferentes (lactato, CO2 e H2O ou etanol e CO2), como veremos mais adiante.
Glicólise
Glicose
2 piruvato
Frutose - 1,6 -bisfosfato
2NAD+2ADP+ 2Pii
2NADH2ATP
Lactato CO +H O
2222 Etanol
+ CO22

Bioquímica II | Glicólise: oxidação de glicose a piruvato I
CEDERJ112
A VIA GLICOLÍTICA PASSO A PASSO
A glicólise pode ser dividida em dois estágios. O estágio 1 é chamado
estágio de investimento; e o estágio 2, o de pagamento ou de síntese de
ATP. Nesta aula, nós iremos mostrar o primeiro estágio apenas. Na aula
seguinte você verá a última fase, o estágio 2.
Estágio 1: Neste primeiro estágio da via glicolítica, são gastas duas
moléculas de ATP, sendo a molécula de glicose (seis carbonos) convertida
em duas moléculas de três carbonos (gliceraldeído 3-fosfato).
Glicose 2 x gliceraldeído 3P
A glicose entra na via glicolítica por fosforilação, na hidroxila do
carbono 6, formando uma molécula de glicose 6-fosfato (G6P).
Glicose + ATP Glicose 6-fosfato + ADPReação 1:
Resumo do estágio 1:

CEDERJ 113
AU
LA 1
0 M
ÓD
ULO
3
Esta reação é dependente de energia, sendo, portanto, acoplada à
clivagem de uma molécula de ATP e é catalisada pela enzima hexoquinase.
(Figura 10.6).
Figura 10.6: Hexoquinase e seu mecanismo de ação. A seta indica o substrato (glicose) em (a) se aproximando do sítio ativo. Em (b) a mudança conformacional induzida pela ligação do substrato (fi t induzido).
Entretanto, nas células do parênquima hepático e nas ilhotas
pancreáticas, tal reação é catalisada pela enzima glicoquinase, cuja
atividade é induzida e afetada por mudanças no estado nutricional. A
hexoquinase é inibida de maneira alostérica pelo produto de sua reação,
a G6P. A hexoquinase está presente em baixas concentrações em todos
os tecidos extra-hepáticos e possui alta afi nidade (baixo Km) por seu
substrato, a glicose. Sua função é permitir que a glicose seja utilizada pelos
tecidos, mesmo quando os níveis de glicose no sangue estejam baixos.
No momento em que uma molécula de glicose entra na célula e é
fosforilada, ela permanece dentro da célula mesmo contra um gradiente
de concentração. A hexoquinase pode catalisar a fosforilação dos
isômeros α (alfa) e β (beta) da glicose, bem como de outros açúcares de
seis carbonos. A função da glicoquinase é remover glicose do sangue após
a alimentação. Ao contrário da hexoquinase, a glicoquinase tem um alto
Km por glicose e opera em condições ótimas quando os níveis de glicose
do sangue são elevados, sendo específi ca para este tipo de açúcar.
A glicose 6-fosfato, além de ser a molécula utilizada nas próximas
etapas da via glicolítica, é um componente importante na junção de diversas
outras vias metabólicas, tais como gliconeogênese, via das pentoses,
glicogenogênese e glicogenólise, como estudaremos mais adiante.
Para relembrar os conceitos de enzima alostérica e Km – ver as aulas de enzimas da Bioquímica I.
!
a b

Bioquímica II | Glicólise: oxidação de glicose a piruvato I
CEDERJ114
EN Z I M A I N D U Z Í V E L
Uma enzima cuja síntese pode ser aumentada (induzida) em uma determinada situação metabólica. Em contraposição, utilizamos o termo enzima constitutiva, cujo nível é constante, independente da situação metabólica.
Além disso, essa primeira reação da glicólise tem conseqüências
importantes para a célula: ao ser fosforilada, a glicose não pode mais
sair da célula, pois os mecanismos de transporte dessa molécula não
servem para aquela que já foi fosforilada. Isto mantém o nível de glicose,
na célula, sempre baixo em relação à concentração extracelular. Como
o transporte de glicose é dependente da concentração (ver difusão
facilitada e glicose, em Biologia Celular I), a tendência da glicose é
sempre entrar na célula.
Outra conseqüência desta primeira reação é indicar o caminho
metabólico que a glicose vai seguir. A fosforilação no carbono 6 indica
que a glicose será degradada na via glicolítica. Como veremos mais
tarde, a glicose pode receber diferentes “etiquetas” que indicam outros
destinos metabólicos.
Na segunda reação da glicólise, a glicose 6P é convertida em
frutose 6P (Fru 6P). Essa é uma reação do tipo isomerização aldose-cetose
e é catalisada por uma fosfo-hexoisomerase (reação 2).
Glicose 6P Frutose 6PReação 2:
Na etapa seguinte, também dependente da energia de clivagem de
ATP, a molécula de frutose 6P é fosforilada na hidroxila do carbono 1,
formando uma molécula de Frutose 1-6-bisfosfato (Fru 1,6-bisP) (reação
3). Tal reação é catalisada pela enzima fosfofrutoquinase (PFK). (Veja
reação 3 e Figura 10.7). A PFK é tanto uma enzima alostérica quanto
uma ENZIMA INDUZÍVEL, cuja atividade é considerada o principal ponto de
regulação da velocidade da via glicolítica.

CEDERJ 115
AU
LA 1
0 M
ÓD
ULO
3
Frutose 6P Frutose 1,6-bisfosfatoReação 3:
Figura 10.7: Fosfofrutoquinase (PFK). A PFK é uma enzima chave da glicólise. Preste atenção nas características da enzima e da reação que ela catalisa. Esta reação é um importante ponto de regulação da via, como veremos na próxima aula.
A seguir, a molécula de frutose 1-6-bisfosfato sofre uma clivagem
pela ação da enzima aldolase, gerando duas moléculas isômeras, que
possuem três carbonos: gliceraldeído 3P (G3P) e dihidroxiacetona
fosfato (DHAP). Pela ação de uma triose-isomerase específi ca, a DHAP
é convertida em G3P. Assim, iniciaremos os passos seguintes com duas
moléculas de G3P.
Fique atento. Em alguns livros você vai encontrar notações diferentes para as moléculas. A frutose 1,6-bisfosfato pode ser encontrada como frutose 1,6-bifosfato ou frutose 1,6-difosfato; a fosfofrutoquinase pode ser encontrada como fosfo-frutose-quinase ou ainda fosfofrutocinase, junto ou separado.
!
Frutose 1,6-bisfosfato dihidroxiacetona-P + gliceraldeído 3PReação 4:
ΔG10 = 23.8 kJ/mol
O P O CH2
O
O
- 6
-
HH HO
HOH
OH
O CH2 OH5
4 3
2
1 ATP ADPMg2+
26
HH HO
HOH
OH
O CH
5
4 3
2
2CH1
O P O
O
O-
-O P O
O
O-
-
Frutose 6-fosfato frutose 1,6-bisfosfato
Gº = -14,2 kJ/mol
O P O CH 2
O
OO
CH O P O
O
O
-
-
OH
HO
H
HH OH
2
2
34
5
6
CH O P O
C O
C H OH
2
2
-
-O
OCH O P O2
HCOH O
O
-
-
C
O H
+
Frutose 1,6-bisfosfatoGliceraldeído
3-fosfatoDihidroxiacetona
3-fosfato
aldolase

Bioquímica II | Glicólise: oxidação de glicose a piruvato I
CEDERJ116
dihidroxiacetona-P gliceraldeído 3PReação 5:
Até aqui uma molécula de seis carbonos (a glicose) foi parcialmente
quebrada em duas moléculas de três carbonos (gliceraldeído 3-P). Note
que as duas moléculas geradas são fosforiladas e apresentam a metade
do número de carbonos que a molécula original de glicose.
Se você prestou atenção, provavelmente reparou que até aqui não
ocorreu síntese de ATP. Pelo contrário, a célula gastou duas moléculas
de ATP (reações 1 e 3). Por este motivo, essa primeira fase é chamada
etapa de investimento ou etapa preparatória. Tal etapa é fundamental,
pois nela a célula coloca à disposição do processo a energia já
existente, e esse investimento possibilita, em etapas posteriores, não
só a recuperação do investimento inicial, como também a síntese de
mais ATP. Isto acontece através de um conjunto de reações conhecido
por etapa de pagamento ou etapa de conversão de energia ou simplesmente
etapa de síntese de ATP.
ΔG10 = 7,5 kJ/mol
CH OH2
C O
CH O P O
O
O -
-2
HCOH
CH O P O2
O
O-
-
CO H
Triose fosfatoisomerase
Dihidroxiacetonafosfato
Gliceraldeído3-fosfato

CEDERJ 117
AU
LA 1
0 M
ÓD
ULO
3
Figura 10.8: Resumo das reações da etapa de investimento ou etapa preparatória da glicólise. Nessa fase, os eventos principais são a fosforilação da glicose e sua conversão a gliceraldeído-3P.
Clivagem de um açúcar-fosfato de seis
carbonos em dois açúcares- fosfato de
três carbonos.
Lembre-se de que esse assunto não acabou. Na próxima aula, você
verá o estágio 2 da glicólise, e como a célula usa a energia da quebra
da glicose, para sintetizar ATP. Não esqueça que, embora por motivos
didáticos os dois estágios estejam separados, o processo é um só.


Glicólise: oxidaçãode glicose a piruvato II 11A
UL
A

Bioquímica II | Glicólise: oxidação de glicose a piruvato II
CEDERJ120
INTRODUÇÃO Nesta aula continuaremos a falar da glicólise. Dessa vez, exploraremos a etapa
de pagamento ou de síntese de ATP. Lembre-se de que:
• Na primeira fase, dois ATPs foram hidrolisados para fosforilar a molécula
de glicose.
• Os produtos da primeira fase são duas moléculas de gliceraldeído 3P, um
açúcar fosforilado de três carbonos.
É daí que partiremos agora. Recorde-se de que, com uma molécula de glicose,
duas moléculas de gliceraldeído 3P são geradas e, portanto, a partir delas, duas
moléculas de todos os intermediários são produzidas também.
Estágio 2: Nesse último estágio da glicólise, as duas moléculas de
três carbonos (G3P) são convertidas em duas moléculas de piruvato.
2 Gliceraldeído 3-fosfato 2 Piruvato
A molécula de G3P é oxidada e ao mesmo tempo fosforilada,
formando uma molécula de 1,3-bisfosfoglicerato. Observe que, nessa
etapa, uma molécula de fosfato inorgânico (Pi) é incorporada à molécula
de G3P no entanto não há consumo de qualquer molécula de ATP
(reação 6).
De onde vem então a energia para formar uma ligação covalente
entre o G3P e o Pi?
Resumo do estágio 2:
Reação 6: O HC
HCOH
CH OPO2 32–
P+
O–
O
HO O–
OC
HCOH
CH OPO2 32–
P
O–
O
O O–
NAD+
NADH + H+ +
Gliceraldeído3-fosfato
desidrogenaseFosfato
inorgânicoGliceraldeído
3-fosfato1,3
bifosfoglicerato
�G´ = 6,3 kJ/mol0
Bifosfoglicerato

CEDERJ 121
AU
LA 1
1 M
ÓD
ULO
3
Parte da energia que foi liberada no processo oxidativo
(transformação de gliceraldeído em glicerato) foi conservada na formação
de uma molécula de 1,3-BISFOSFOGLICERATO, que possui, então, um potencial
energético maior do que o G3P. Nesta reação, uma molécula de NAD+
é reduzida a NADH + H+. Esta é uma reação típica, catalisada por uma
classe de enzimas denominadas desidrogenases ligadas ao NAD+.
Na etapa seguinte, a energia conservada na molécula de 1,3-
bisfosfoglicerato (BPG) será utilizada na formação de uma molécula de
ATP (ligação éster-fosfórica entre ADP + Pi), sendo o BPG convertido,
então, em 3-fosfoglicerato (3PG).
1,3-B I S F O S F O G L I -C E R A T O
É a primeira molécula de alta energia formada
na glicólise.
Reação 7:
Essa reação é catalisada pela enzima fosfoglicerato quinase. Uma
vez que duas moléculas de triose fosfato são formadas por molécula de
glicose, duas moléculas de ATP são geradas nesse estágio. Este processo de
formação de ATP é um exemplo de fosforilação em nível de substrato.
Pela ação de uma fosfoglicerato mutase, o 3PG será convertido
em 2-P-Glicerato (2PG).
OC
HCOH
CH OPO2 32–
P
O–
O
O O–
1,3bifosfoglicerato �G´ = –18,5 kJ/mol0
O
P
P
Rib Adenina
+
ADP
OC
HCOH
CH OPO2 32–
O–
O
P
P
Rib Adenina
P
O–
–O O
ATP
3–fosfoglicerato
+fosfoglicerato
quinase
Mg2+

Bioquímica II | Glicólise: oxidação de glicose a piruvato II
CEDERJ122
Reação 8:
A etapa subseqüente é catalisada pela enolase e envolve a
desidratação e redistribuição da energia dentro da molécula. A
proximidade do grupamento funcional hidroxila com o íon fosfato
favorece a formação de um enol-fosfato, o fosfoenolpiruvato (PEP),
que é também considerado um composto de alta energia.
Reação 9:
O passo seguinte é também uma fosforilação em nível do substrato.
O fosfoenolpiruvato transfere seu grupo fosfato para o ADP, formando
ATP. O produto é o piruvato, e a reação é catalisada pela piruvato
quinase.
FO S F O E N O L P I R U-V A T O
É a segunda molécula de alta energia formada na glicólise.
3-fosfoglicerato
O—
O
HC
CH
C
2 P O—
O—
O
O
OHMg2+
fosfogliceratomutase
O—
O
HC
CH
C
2
P O—
O—
O
OH
O
2-fosfoglicerato
G´°= 4,4 kJ/mol
H C O P O
HO CH 2 O
OCOO
-
-
-
C O P O
CH2 O
OCOO
-
-
-H O2
2-fosfoglicerato fosfoenolpiruvato
enolase
G´º= 7,5 kJ/mol
OC
C
Ch2
O–
Fosfoenolpiruvato
�G´ = –31,4 kJ/mol0
O
P
P
Rib Adenina
+
OC
C
Ch3
O
O
P
P
Rib Aden
P
O–
–O O
ATP
Piruvato
+Piruvatoquinase
Mg , K2+ +
P
O–
O
O O–
O
Fosfoenolpiruvato

CEDERJ 123
AU
LA 1
1 M
ÓD
ULO
3
Devemos lembrar que, nesse segundo estágio, partimos de duas
moléculas de gliceraldeído 3P e, portanto, duas moléculas de piruvato,
duas de NADH + H+ e quatro moléculas de ATP foram formadas. Ao
calcularmos o rendimento energético em termos de formação de ATPs,
veremos que foram gastas duas moléculas no primeiro estágio da via e
que foram ressintetizadas quatro moléculas, ou seja ocorreu formação
líquida de duas moléculas de ATP (Figura 11.1).
Figura 11.1: Fase de pagamento da glicólise. Também chamada etapa de conservação de energia ou, simplesmente, etapa de síntese de ATP.
Fase de pagamento
Conversão oxidativa de gliceraldeído 3-fosfato em piruvato e a formação acoplada de ATP e NADH.
!

Bioquímica II | Glicólise: oxidação de glicose a piruvato II
CEDERJ124
UTILIZAÇÃO DO PIRUVATO E REOXIDAÇÃO DO NADH
Já vimos no início da aula que o piruvato pode ter vários destinos.
O estado redox do tecido e a presença ou ausência de O2 determinam
qual via deverá ser seguida.
1o destino – Quando em condições anaeróbicas, o piruvato é
reduzido pelo NADH, formando lactato (na fermentação lática) ou
etanol e CO2 (na fermentação alcoólica).
Fermentação lática
Na fermentação lática a reação é catalisada pelo complexo
enzimático piruvato desidrogenase. Nessas condições, a molécula de
NAD+ é regenerada, estando pronta para ser reutilizada em um outro
ciclo da via glicolítica.
Piruvato + NADH + H+ L(+)-Lactato + NAD+
A fermentação lática ocorre em alguns microorganismos e também
no músculo de vertebrados e invertebrados.
Fermentação alcoólica
Na fermentação alcoólica a conversão de piruvato em etanol e CO2
envolve duas reações sucessivas. A primeira reação é a conversão de
piruvato em acetaldeído, uma reação de descarboxilação, na qual o CO2
é liberado. Esta reação é catalisada pela piruvato descarboxilase.
piruvato acetaldeído
CO2
piruvatodescarboxilase
A segunda reação é a reoxidação do NADH a NAD+. Esta última
reação é catalisada pela enzima álcool desidrogenase. Os produtos fi nais
da fermentação alcoólica são, portanto, o etanol, uma molécula de dois
carbonos, CO2 e NAD+.
acetaldeído etanol
NADH NAD+
álcool desidrogenase

CEDERJ 125
AU
LA 1
1 M
ÓD
ULO
3
2o destino – Quando em condições aeróbicas, o piruvato é
transferido para as mitocôndrias e, após conversão em acetil-Coenzima A
(Acetil-CoA), é oxidado em CO2 no ciclo do ácido cítrico. Os equivalentes
do NADH + H+ produzidos na via glicolítica são também levados para
as mitocôndrias, onde serão transferidos para aceptores específi cos,
em um processo que veremos mais tarde e que é denominado cadeia
transportadora de elétrons.
Piruvato + NAD+ + CoA Acetil-CoA + NADH + H+ + CO2
REGULAÇÃO DA VIA GLICOLÍTICA
Embora a maioria das reações da via glicolítica
seja reversível, três delas são marcadamente
exergônicas e são consideradas irreversíveis do ponto
de vista fi siológico. Essas reações são catalisadas
pela HEXOQUINASE E GLICOQUINASE, FOSFOFRUTOQUINASE
e PIRUVATO QUINASE e são consideradas os principais
sítios de regulação da glicólise.
A regulação da via ocorre segundo
o princípio da máxima economia. A via será
inibida se o organismo estiver em um estado fi siológico energeticamente
favorável, ou seja, quando o balanço ATP/ADP for positivo. Em outras
palavras, se a célula tem muito ATP, as vias metabólicas de síntese de
ATP serão inibidas. Em baixas concentrações de ATP (altas concentrações
de ADP), a via será ativada. Entretanto, não se engane... Não existe
nenhuma situação fi siológica em que a via esteja parada. Ela pode
funcionar a uma velocidade maior ou menor, mas qualquer inibidor
ou situação metabólica que paralise a via glicolítica certamente levará
o organismo à morte.
GLICOQUINASE E HEXOQUINASE
glicose + ATP glicose –6P + ADP + Pi
FOSFOFRUTOQUINASE
frutose 6-P + ATP frutose 1,6-bisfosfato + ADP + Pi
PIRUVATUVATO QUINASE
fosfoenolpiruvato + ADP + Pi piruvato + ATP
Enzimas chaves da via glicolítica:HEXOQUINASE FOSFOFRUTO-QUINASE PIRUVATO QUINASE
!

Bioquímica II | Glicólise: oxidação de glicose a piruvato II
CEDERJ126
Tabela 11.1: Reguladores das enzimas chaves da via glicolítica.
Enzima Inibidor (-) Ativador (+)
Hexoquinase
Fosfofrutoquinase
Piruvato quinase
Glicose 6-fosfato; ATP
ATP; citrato AMP; ADP
O detalhamento dessa regulação será visto após estudarmos outras
vias metabólicas, uma vez que os processos metabólicos ocorrem de
maneira integrada.
R E S U M O
A glicólise é uma seqüência de dez reações catalisadas por enzimas e por
meio das quais uma molécula de glicose é convertida em duas moléculas de
piruvato, com a produção de duas moléculas de ATP e duas de NADH. A via
glicolítica pode ser dividida em uma etapa de investimento e uma etapa de
pagamento. Na primeira etapa, a célula investe dois ATPs e quebra a glicose em
duas moléculas de gliceraldeído 3P. Na segunda etapa, duas moléculas de alta
energia são produzidas: 1,3-bisfosfoglicerato e fosfenolpiruvato. A formação
dessas moléculas é fundamental para a síntese de quatro moléculas de ATP.
Além de ATP, dois NAD+ são reduzidos a NADH, para cada molécula de glicose
quebrada. A via glicolítica é regulada principalmente pela carga energética da
célula. O indicador da carga energética é a relação entre a concentração de
ATP e a concentração de ADP e/ou AMP. Os principais pontos de regulação são
as enzimas hexoquinase, fosfofrutoquinase e piruvato quinase, todas as três
metabolicamente irreversíveis. A via trabalha tanto em condições aeróbicas
quanto em condições anaeróbicas.

CEDERJ 127
AU
LA 1
1 M
ÓD
ULO
3
Nas três últimas aulas, você conheceu uma das vias metabólicas
mais importantes do metabolismo energético. Não perca de vista a
totalidade dessas aulas, pois, mais importante que saber detalhes da via
metabólica, é a compreensão do seu sentido fi siológico, ou seja, para
que ela serve. Isso vai orientar você quanto ao que precisar ser fi xado
realmente. Não deixe de entender os conceitos de máxima economia, de
moléculas de alta energia, de acoplamento entre reações. Tais conceitos
são universais quando se trata de Bioquímica e entendê-los facilita a
compreensão dessa e de cada uma das outras vias metabólicas que você
verá na Bioquímica II.
O próximo módulo chama-se respiração celular. Ele fala das vias
complementares à glicólise que ocorrem na presença de oxigênio (ciclo
de Krebs ou ciclo do ácido cítrico e cadeia transportadora de elétrons).
Portanto, não esqueça que a glicólise também é parte desse processo.
EXERCÍCIOS
De 1 a 6, assinale a(s) afi rmativa(s) correta(s):
1. Sobre os aspectos gerais da via glicolítica:
a) na via glicolítica são produzidas seis moléculas de ATP;
b) duas moléculas de piruvato são formadas por cada molécula de glicose que entra
na via;
c) ocorrem duas reações de condensação na via;
d) a regulação alostérica é importante para esta via;
e) a via glicolítica só ocorre em organismos anaeróbicos.
2. A glicólise apresenta duas etapas:
a) ambas requerem ATP para ativação;
b) somente uma é catalisada pela aldolase;
c) somente uma produz piruvato;
d) ambas são catalisadas por quinases.

Bioquímica II | Glicólise: oxidação de glicose a piruvato II
CEDERJ128
3. Qual enzima não é importante para a via glicolítica?
a) aldolase;
b) enolase;
c) piruvato carboxilase;
d) piruvato quinase.
4. A conversão de gliceraldeído 3P em 1,3-bisfosfoglicerato.
a) requer ATP;
b) é inibida por frutose 1,6 bi-P;
c) é catalisada por uma desidrogenase NAD+ dependente;
d) é uma reação de condensação.
5. A produção de frutose 1,6-bisP.
a) libera energia;
b) é uma etapa comprometida na via glicolítica;
c) é catalisada pela hexoquinase;
d) é inibida por ATP.
6. A produção de ATPs na glicólise.
a) ocorre em duas reações separadas;
b) ocorre na ausência de 2P-glicerato;
c) requer a produção de lactato;
d) requer a produção de piruvato.

CEDERJ 129
AU
LA 1
1 M
ÓD
ULO
3
7 . Com relação ao processo de fermentação alcoólica (etanólica), podemos dizer que:
a) as reações da glicólise são diferentes daquelas que ocorrem nos organismos que
produzem lactato;
b) o acetaldeído é um composto intermediário nesta transformação;
c) NAD+ é convertido em NADH;
d) grandes quantidades de oxigênio são necessárias para que tal processo ocorra.
8 . Estabeleça os aspectos comparativos entre as enzimas hexoquinase e glicoquinase.
9. Como você explicaria a existência de uma mesma via metabólica presente desde
os microorganismos até os seres mais complexos que conhecemos?
10. Você reparou que todos os compostos da via glicolítica, exceto a molécula
de piruvato, encontram-se fosforilados? Pense/especule sobre uma razão
para este fato.

Bioquímica II | Glicólise: oxidação de glicose a piruvato II
CEDERJ130
RESPOSTAS DOS EXERCÍCIOS AVALIATIVOS
Assinale a(s) afi rmativa(s) correta(s)
1. Sobre os aspectos gerais da via glicolítica.
a) na via glicolítica são produzidas seis moléculas de ATP:
b) duas moléculas de piruvato são formadas por cada molécula de glicose que
entra na via; CORRETA
c) ocorrem duas reações de condensação na via;
d) a regulação alostérica é importante para esta via; CORRETA
e) a via glicolítica só ocorre em organismos anaeróbicos.
2. A glicólise apresenta duas etapas e:
a) ambas requerem ATP para ativação;
b) somente uma é catalisada pela aldolase;
c) somente uma produz piruvato; CORRETA
d) ambas apresentam reações catalisadas por quinases. CORRETA
3. Qual dessas enzimas não é importante para a via glicolítica?
a) enolase;
b) aldolase;
c) piruvato carboxilase; CORRETA
d) piruvato quinase.

CEDERJ 131
AU
LA 1
1 M
ÓD
ULO
3
4. A conversão de gliceraldeído 3P a 1,3-bisfosfoglicerato:
a) requer ATP; CORRETA
b) é inibida por frutose-1,6 bisP;
c) é catalisada por uma desidrogenase NAD+ dependente;
d) é uma reação de condensação.
5. A produção de frutose 1,6-bisP:
a) libera energia;
b) é a reação fi nal na via glicolítica;
c) é catalisada pela hexoquinase;
d) é inibida por ATP. CORRETA
6. A produção de ATPs na glicólise:
a) ocorre em duas reações separadas; CORRETA
b) ocorre na ausência de 2P-glicerato;
c) requer a produção de lactato;
d) requer a produção de piruvato.
7 . Com relação ao processo de fermentação alcoólica (etanólica), podemos dizer que:
a) as reações da glicólise são diferentes daquelas que ocorrem nos organismos que
produzem lactato;
b) o acetaldeído é um composto intermediário nesta transformação; CORRETA
c) NAD+ é convertido em NADH;
d) grandes quantidades de oxigênio são necessárias para que este processo ocorra.

Bioquímica II | Glicólise: oxidação de glicose a piruvato II
CEDERJ132
8 . Estabeleça os aspectos comparativos entre as enzimas hexoquinase e glicoquinase.
Ambas são enzimas que catalisam a reação de fosforilação de glicose em glicose
6-P. Ambas são inibidas pelo produto. Hexoquinase é a isoforma presente na
maior parte dos tecidos extra-hepáticos, enquanto a glicoquinase é a isoforma
encontrada no fígado. As isoformas têm diferentes afi nidades pelo substrato
(glicose). Enquanto a hexoquinase é uma enzima generalista, que catalisa a
fosforilação de hexoses, como glicose, manose e frutose, a glicoquinase é bastante
específi ca para glicose. A hexoquinase possui alta afi nidade (baixo Km) por seu
substrato glicose, enquanto a glicoquinase possui baixa afi nidade.
9. Como você explicaria a existência de uma mesma via metabólica presente desde
os microorganismos até os seres mais complexos que conhecemos?
Falar de relações evolutivas entre os organismos.
10. Você reparou que todos os compostos da via glicolítica, exceto a molécula
de piruvato, encontram-se fosforilados? Pense/especule sobre uma razão
para este fato.
Falar de organização celular, fosfato como etiqueta de endereçamento.