Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE BIOLOGIA
TAILA DOS SANTOS ALVES
EPIDEMIOLOGIA MOLECULAR DA RESISTÊNCIA A
ANTIMICROBIANOS E FATORES DE VIRULÊNCIA EM
Escherichia coli ISOLADAS DE DÍPTEROS MUSCOIDES EM
PROPRIEDADE LEITEIRA
CAMPINAS
2017

TAILA DOS SANTOS ALVES
EPIDEMIOLOGIA MOLECULAR DA RESISTÊNCIA A ANTIMICROBIANOS E FATORES DE VIRULÊNCIA EM
Escherichia coli ISOLADAS DE DÍPTEROS MUSCOIDES EM PROPRIEDADE LEITEIRA
Dissertação apresentada ao Instituto de Biologia da Universidade Estadual de Campinas como parte dos requisitos exigidos para a obtenção do Título de Mestra em Biologia Animal na Área de Relações Antrópicas, Meio Ambiente e Parasitologia.
Orientador: PROF. DR. DOMINGOS DA SILVA LEITE
CAMPINAS
2017
ESTE ARQUIVO DIGITAL CORRESPONDE À
VERSÃO FINAL DA DISSERTAÇÃO
DEFENDIDA PELA ALUNA TAILA DOS SANTOS
ALVES E ORIENTADA PELO PROF. DR.
DOMINGOS DA SILVA LEITE.


Campinas, 30 de março de 2017.
COMISSÃO EXAMINADORA
Prof. Dr. Domingos da Silva Leite (orientador)
Profa. Dra. Cristina Elisa Alvarez Martinez
Dra. Monique Ribeiro Tiba Casas Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra no processo de vida acadêmica do aluno.

À minha mãe por todo amor, fé e apoio sempre.

Agradecimentos
Ao meu orientador, Prof. Dr. Domingos da Silva Leite, por todos os ensinamentos,
oportunidades, incentivo, confiança, apoio e presença sempre.
Ao Prof. Dr. Márcio Garcia Ribeiro e ao Dr. Gustavo Henrique Batista Lara pela disposição e
auxílio na obtenção das amostras.
À bióloga Ma. Mirtis Maria Giaciani Ferraz pelo auxílio e ensinamentos.
À Profa. Dra. Patrícia Jacqueline Thyssen e a Ma. Natane de Cássia Sibon Purgato pela
identificação das amostras de dípteros.
À Dra. Amanda Keller Siqueira pelo incentivo, disposição, ajuda e ensinamentos.
Ao Prof. Dr. José Luiz Proença Módena, Profa. Dra. Cristina Elisa Alvarez Martinez e Profa.
Dra. Patrícia Jacqueline Thyssen pela participação e sugestões no Exame de Qualificação.
À Dra. Jéssica Wildgrube Bertol pela amizade e apoio.
Profa. Dra. Maria Silvia Viccari Gatti e ao Prof. Dr. Tomomasa Yano pelo apoio e disposição.
Ao Prof. Dr. Wanderley Dias da Silveira e ao Dr. Renato Pariz Maluta pelo auxílio na técnica
de Pulsed-Field Gel Electrophoresis.
À Profa. Dra. Ana Lúcia da Costa Darini, Profa. Dra. Maria Silvia Viccari Gatti e a Dra.
Monique Ribeiro Tiba Casas pelo exame prévio da dissertação.
À Profa. Dra. Cristina Elisa Alvarez Martinez, Dra. Monique Ribeiro Tiba Casas, Profa. Dra.
Fabiana Fantinatti-Garboggini e Prof. Dr. Tomomasa Yano por terem aceitado participar da
banca examinadora.
À Jéssica Gurgel, Taís Baruel Vieira e Guilherme Borelli por toda a ajuda e amizade, e à
Meghi Nogueira de Souza, Luisa de Pontes Ribeiro e Ma. Carolina Lambertini pela amizade e
companheirismo.
A CAPES pela concessão da bolsa de mestrado e a FAPESP pelo apoio financeiro.
À minha família pela compreensão e apoio.

RESUMO
O ambiente rural propicia o desenvolvimento de espécies de dípteros muscoides que podem
atuar como vetores mecânicos de vários patógenos. Dada a possibilidade de veiculação de
bactérias resistentes por meio de moscas, o presente estudo buscou caracterizar a resistência a
antimicrobianos em Escherichia coli isoladas de dípteros muscoides coletados em
propriedades leiteiras, assim como a presença de genes de virulência relacionados à
colibacilose bovina. Musca domestica foi a espécie de díptero muscoide mais abundante nas
duas propriedades estudadas. As cepas de E. coli provenientes das duas propriedades
apresentaram diferenças quanto às frequências de resistência aos antimicrobianos testados por
disco difusão e exibiram frequências baixas ou nulas dos genes de fatores de virulência. A alta
frequência (72%) do grupo filogenético B1 entre as cepas multirresistentes (resistência ≥ a 3
classes de antimicrobianos), aliada a ausência de relação entre presença de multirresistência e
fatores de virulência (p>0,05), indica que a resistência antimicrobiana está associada às
bactérias comensais. A análise de perfil clonal das cepas multirresistentes, realizada por
Pulsed-Field Gel Electrophoresis (PFGE), mostrou ampla diversidade genômica, de modo
que, moscas diferentes podem veicular cepas com perfis genômicos diferentes, porém com os
mesmos genes de resistência antimicrobiana e/ou integron e que uma única mosca pode
veicular tanto cepas quanto clones com genes de resistência distintos. Moscas distintas
apresentaram integron de classe 1 com o mesmo arranjo genético identificado na análise do
sequenciamento. Uma vez que as moscas veiculam cepas de E. coli com perfis
multirresistentes diversos, é possível afirmar que dípteros muscoides contribuem para a
disseminação de genes de resistência a antimicrobianos no ambiente.

ABSTRACT
Rural environment facilitates muscoid diptera development that can act as mechanical vectors
of several pathogens. Given that flies can carry resistant bacteria, this study aimed to
characterize antimicrobial resistance in Escherichia coli isolated from muscoid diptera
collected in dairy farms, as well as the presence of virulence factors genes related to bovine
colibacillosis. Musca domestica was the most collected species in the two studied farms.
Escherichia coli strains from the two farms showed differences in antimicrobial resistance
frequencies tested by disk diffusion and showed low or no frequency to virulence factor
genes. The high frequency (72%) of phylogenetic group B1 among multidrug resistance
strains (resistant ≥ 3 classes), combined with the lack of relationship between the multidrug
resistance and virulence factors (p>0.05), suggest that the antimicrobial resistance is
associated with the commensal bacteria. The clonal profile analysis of the multiresistant
strains, performed by Pulsed-Field Gel Electrophoresis (PFGE), showed wide genomic
diversity, so that different flies may carry unrelated strains, but with the same antimicrobial
resistance genes and/or integron and, a single fly may carry strains and clones with distinct
resistance genes. Different flies presented class 1 integron with the same gene arrangement
identified by sequencing analyze. It is possible to affirm that flies contribute to the spread of
antimicrobial resistance genes in the environment, once they carry strains of E. coli with
diverse multiresistant profiles.

LISTA DE ILUSTRAÇÕES
Figura 1. Região do município de Botucatu-SP, onde estão localizadas as duas propriedades
leiteiras estudadas (denominadas A e B), quanto à resistência antimicrobiana em E. coli
provenientes de dípteros muscoides...................................................................................... 33
Figura 2. Estrutura do integron de classe 1. A região cinza representa o sítio de inserção dos
cassetes gênicos relacionados à resistência antimicrobiana. Reproduzido de Lèvesque et al.
(1995). ................................................................................................................................. 39
Figura 3. Frequência relativa de E. coli resistentes* a cada um dos antimicrobianos testados.
Visualização dos diferentes perfis dos isolados nas duas propriedades leiteiras (A e B). ....... 47
Figura 4. Escherichia coli positiva para o teste de aproximação de disco. O alargamento do
halo de inibição (seta no topo) e o surgimento da “zona fantasma” (seta embaixo) indica a
produção de ESBL (β-lactamase de espectro estendido). ...................................................... 48
Figura 5. Padrão de digestão enzimática de AluI obtido no ensaio de RFLP da região variável
do integron de classe 1 (5’CS-3’CS) de E. coli isoladas de moscas. ...................................... 51
Figura 6. Desenho esquemático da sequência obtida a partir da amplificação dos iniciadores
5’CS e 3’CS. ........................................................................................................................ 52
Figura 7. Desenho esquemático do integron de classe 1 presente em cepas de E. coli isoladas
de dípteros muscoides. ......................................................................................................... 52
Figura 8. Dendrograma obtido pela técnica de PFGE a partir da digestão enzimática com
XbaI do DNA cromossômico de E. coli isoladas de dípteros muscoides coletados em
propriedades leiteiras da região de Botucatu, SP (2015) e os respectivos genes de resistência e
grupos filogenéticos. Padrão gerado por UPGMA, utilizando coeficiente de Dice com
tolerância e otimização de 1%. ............................................................................................. 55
Figura 9. Eletroforese em gel de agarose 0,8% dos plasmídeos extraídos pelo método de lise
alcalina, correspondentes às cepas de E. coli transconjugantes (A) e doadoras (B)................ 57

LISTA DE TABELAS
Tabela 1. Iniciadores utilizados para a detecção dos fatores de virulência, genes de resistência
e integrons em isolados de E. coli provenientes de dípteros muscoides. ................................ 36
Tabela 2. Iniciadores utilizados no esquema proposto por Clermont et al. (2000) e as
combinações de resultados possíveis. ................................................................................... 37
Tabela 3. Iniciadores relacionados a estruturas de integrons de classe 1 utilizados para a
conclusão das sequências de isolados de E. coli provenientes de dípteros muscoides. ........... 38
Tabela 4. Insetos coletados em duas propriedades da região de Botucatu, SP (2015)
organizados por amostra. Espécies e quantidades. ................................................................ 43
Tabela 5. Espécies e abundância de espécies de dípteros muscoides coletados em duas
propriedades leiteiras da região de Botucatu, SP (2015). ...................................................... 44
Tabela 6. Frequência relativa de E. coli resistentes* isoladas da superfície externa de dípteros
muscoides em duas propriedades leiteiras da região de Botucatu, SP (2015). ....................... 47
Tabela 7. Dados referentes ao perfil de resistência*, presença de plasmídeos e grupos
filogenéticos das cepas de E. coli multirresistentes isoladas de dípteros muscoides em
propriedades leiteiras da região de Botucatu, SP (2015). ..................................................... 49
Tabela 8. Cepas de E. coli selecionadas para a avaliação do perfil clonal organizadas de
acordo com o perfil fenotípico de resistência antimicrobiana. ............................................... 53
Tabela 9. Perfil de resistência fenotípica* e genotípica das cepas E. coli doadoras e
transconjugantes(t)
obtidas no ensaio de conjugação. ............................................................ 58
Tabela 10. Resultados para os genes de resistência a partir de DNA plasmidial extraído por
lise alcalina nas cepas de E. coli doadoras e transconjugantes, obtidos por PCR. .................. 59

Sumário
I - Introdução .............................................................................................................. 13
1.1. BOVINOCULTURA ............................................................................................. 14
1.1.1. Uso de antimicrobianos na bovinocultura ............................................................ 15
1.2. DÍPTEROS MUSCOIDES ..................................................................................... 16
1.2.1. Ecologia e biologia das moscas de importância pecuária ..................................... 17
1.2.2. Vetores de bactérias ............................................................................................. 18
1.3. Escherichia coli ...................................................................................................... 21
1.4. RESISTÊNCIA ANTIMICROBIANA ................................................................... 23
1.4.1. Elementos genéticos móveis ................................................................................ 24
1.4.2. Disseminação no ambiente e implicações ............................................................ 26
II – Objetivos ............................................................................................................... 29
2.2. Específico: ............................................................................................................. 30
III - Material e Métodos .............................................................................................. 31
3.1. Áreas de coleta ....................................................................................................... 32
3.2. Obtenção das amostras ........................................................................................... 32
3.3. Isolamento e identificação de E. coli....................................................................... 33
3.4. Caracterização fenotípica da sensibilidade aos antimicrobianos .............................. 34
3.4.1. Sensibilidade microbiana (antibiograma) ............................................................. 34
3.4.2. Produção de ESBLs – Método da aproximação de disco ...................................... 34
3.5. PCR para a pesquisa dos genes de virulência, resistência e integrons ...................... 35
3.5.1. Extração do DNA (método do choque térmico) ................................................... 35
3.5.2. PCR ..................................................................................................................... 35
3.6. Presença de plasmídeos .......................................................................................... 37
3.7. Classificação filogenética de E. coli........................................................................ 37
3.8. Caracterização dos integrons por Restriction Fragment – Length Polymorphism
(RFLP) e sequenciamento ............................................................................................. 38
3.9. Perfil clonal das cepas resistentes ........................................................................... 39
3.10. Ensaio de conjugação ........................................................................................... 39
3.11. Análise estatística ................................................................................................. 40
IV - Resultados ............................................................................................................. 41
4.1. Coleta dos insetos e isolamento de E. coli ............................................................... 42
4.2. Perfil de sensibilidade aos antimicrobianos ............................................................. 45
4.3. Genes relacionados à resistência a antimicrobianos e fatores de virulência ............. 46

4.4. Análises estatísticas ................................................................................................ 48
4.5. Plasmídeos e classificação filogenética de E. coli (Clermont et al., 2000) ............... 49
4.6. Caracterização dos integrons por Restriction Fragment – Length Polymorphism
(RFLP) .......................................................................................................................... 51
4.7. Sequenciamento da região variável do integron de classe 1 ..................................... 51
4.8. Perfil clonal ............................................................................................................ 53
4.9. Conjugação ............................................................................................................ 56
V - Discussão ............................................................................................................... 60
VI – Conclusões ........................................................................................................... 68
VII - Referências Bibliográficas ................................................................................. 70
VIII – Apêndices ......................................................................................................... 86
APÊNDICE A – Resultado do ensaio de disco aproximação para a dectação fenotípica de
ESBL em cepas de E. coli doadoras e transconjugantes obtidas após conjugação. ......... 87
APÊNDICE B – Antibiograma da cepa transconjugante MA17d. Colônias hipermutantes
no interior do halo de inibição de sulfametazol/trimetoprim (SUT). ............................... 88
IX – Anexos ................................................................................................................. 89
ANEXO A – Declaração sobre bioética e biossegurança. .............................................. 90
ANEXO B – Declaração sobre a não infringência da lei nº 9610/98 e do direito autoral de
qualquer editora ............................................................................................................ 91

I - Introdução

Introdução | 14
1.1. BOVINOCULTURA
Os bovinos estão entre as primeiras espécies domesticadas na Europa. Estudos
arqueológicos têm apontado indícios das atividades de criação desde o período Neolítico
(Serjeantson, 2011), com o objetivo de exploração do poder de tração dos animais e produção
de leite (Ebersbach, 2010). A divisão da criação para diferentes finalidades, como para a
obtenção de carne ou leite, data do século XIX (Ebersbach e Schibler, 2003).
Atualmente, cerca de 150 milhões de produtores movimentam o mercado
econômico mundial de leite, sendo a Índia o maior produtor mundial e o Brasil o sexto
(FAO, 2016). No Brasil, segundo o último censo agropecuário realizado em 2006, a criação
de bovinos com aptidão leiteira esteve presente em 25% dos estabelecimentos (IBGE, 2009;
Brasil, 2014), ganhando destaque na economia e na geração de empregos (Zoccal et al.,
2008).
Os sistemas de produção incluem as formas de criação e manejo do gado leiteiro e
são classificados em extensivo, semi-intensivo e intensivo, de acordo com a produtividade do
rebanho e o grau técnico empregado (Embrapa, 2016).
Nos sistemas extensivos os animais são criados soltos no pasto com
suplementação alimentar apenas de sal comum, apresentando menor nível de aplicação
tecnológica, instalações simples e baixa produtividade. Já nos sistemas semi-intensivos, o
gado é manejado em semiconfinamento, recebendo suplementação alimentar a pasto com sal
enriquecido, silagem de milho, sorgo ou capim, geralmente durante a estação seca. Os
sistemas intensivos são destinados à criação de bovinos com elevada produção leiteira que
permanecem confinados constantemente em estábulos. Tais sistemas empregam maiores
níveis de tecnologia, dietas com altos valores nutricionais e mão de obra especializada (Assis
et al., 2005; Sarcinelli et al., 2007; Brasil, 2015).
Em sistemas extensivos e semi-intensivos em que a principal fonte de alimento é
o pasto, práticas de rodízio de pastagens são importantes para manter a qualidade das plantas
forrageiras e o controle de parasitos que apresentam algum estágio do ciclo de vida
dependente do solo (Florião, 2013). A prática consiste na divisão do pasto em piquetes que
são pastejados alternadamente de modo que haja um período de descanso (Martha Júnior et
al., 2003).
Adicionalmente, práticas de manejo que visam o monitoramento da sanidade
animal são fundamentais para a manutenção da produtividade dos rebanhos leiteiros (Florião,
2013). Nesse sentido, inclui-se o uso terapêutico e profilático de antimicrobianos e o controle

Introdução | 15
populacional de artrópodes – insetos e ácaros (Bogard e Stobberingh, 2000; Rutz e Geden,
2010).
1.1.1. Uso de antimicrobianos na bovinocultura
A utilização de antimicrobianos na pecuária é destinada a fins terapêuticos,
profiláticos e como promotores de crescimento (Ventola, 2015).
Nas propriedades leiteiras, a principal razão para a administração de
antimicrobianos consiste no tratamento e prevenção da mastite bovina (Pol e Ruegg, 2007),
uma vez que várias espécies de bactérias, tais como Streptococcus agalactiae, Staphylococcus
aureus e Mycoplasma spp. (contagiosos), Streptococcus uberis, Streptococcus dysgalactiae
subsp. dysgalactiae, Escherichia coli e Klebsiella spp. (ambientais), entre outros, são capazes
de infectar as glândulas mamárias das vacas e produzir enfermidade (Oliver e Murinda, 2012).
O tratamento terapêutico é destinado aos animais com manifestações clínicas de
mastite, enquanto o profilático às vacas em período não lactante (seco), e por esse motivo
popularmente conhecido como “terapia da vaca seca” (Oliver e Murinda, 2012). Ambas as
intervenções são administradas principalmente por via intramamária, mas outras vias também
podem ser utilizadas (Royster e Wagner, 2015).
Além disso, o tratamento com antimicrobianos faz-se necessário em bezerros com
menos de trinta dias de vida que apresentam quadro de diarreia com sinais sistêmicos, uma
vez que independente do agente infeccioso (bactérias, vírus ou protozoários), há aumento do
crescimento bacteriano de coliformes, podendo evoluir para bacteremia e endotoxemia
(Constable, 2009).
Nesse contexto, a administração de antimicrobianos principalmente em doses
subterapêuticas é fonte de constantes debates, pois, apesar de resultar em benefícios como o
aumento da produtividade, redução de patógenos, menor ocorrência de doenças, de morbidade
e mortalidade, gera resíduos de antimicrobianos nos alimentos que podem contribuir para a
seleção de resistência antimicrobiana em patógenos de humanos, devido à ingestão de carne,
leite e derivados. Além disso, a prática colabora para a seleção de resistência em micro-
organismos comensais constituintes da microbiota intestinal de animais saudáveis, que podem
atuar como reservatórios de genes de resistência e na disseminação a outros comensais e
patógenos (Bogard e Stobberingh, 2000; Oliver et al., 2011; Ventola, 2015).
A presença de bactérias comensais resistentes a antimicrobianos parece estar
relacionada à idade dos animais, de forma que animais mais jovens apresentariam maior
quantidade de bactérias resistentes quando comparados aos mais velhos. Nesse sentido,

Introdução | 16
Khachatryan et al. (2004) buscaram relacionar idade e resistência antimicrobiana em bovinos
leiteiros, comprovando, experimentalmente, o pressuposto, e sugerindo que o fenômeno pode
estar associado à dieta já que bezerros que receberam leite apresentaram maior frequência de
cepas resistentes. Este evento pode ser explicado pela associação entre genes de resistência e
genes vantajosos para o metabolismo nutricional das bactérias, no mesmo plasmídeo, como
adesinas e sideróforos. Desse modo, cepas resistentes podem persistir mesmo na ausência de
pressões seletivas (Khachatryan et al., 2004).
1.2. DÍPTEROS MUSCOIDES
A ordem Diptera apresenta mais de 150 mil espécies de moscas e mosquitos que
possuem um par de asas anteriores funcionais e o par de asas posteriores modificados em
halteres, sendo esta a principal sinapomorfia que a distingue das demais ordens da Classe
Insecta (Yeates et al., 2007), de modo que, compreende-se por dípteros muscoides os insetos
popularmente conhecidos como moscas, pertencentes a divisão Schizophora da infraordem
Muscomorpha (McAlpine, 1981 apud Borges, 2006). Nesse contexto, as famílias
Calliphoridae, Sarcophagidae, Muscidae, Faniidae e Anthomyiidae destacam-se devido à
sinantropia e a importância médico-sanitário que apresentam (Linhares, 1979).
Povolný (1971) definiu sinantropia como um fenômeno ecológico baseado na
biocenose, ou seja, a coabitação interativa de espécies diferentes no mesmo ambiente, de
modo que a antropobiocenose acontece quando ambientes modificados pelo homem, através
das habitações humanas e dos animais domésticos, permitem a biocenose, sendo os animais
sinantrópicos componentes espontâneos. Analogamente, dípteros simbovinos são aqueles que
se relacionam com a antropobiocenose através das excretas de ruminantes domésticos
(Povolný, 1971).
A sinantropia em dípteros muscoides é comum (Linhares, 1979). Diferentes
espécies apresentam maior ou menor grau de dependência da associação com o homem de
acordo com as exigências tróficas e ecológicas, sendo fezes e matéria orgânica em
decomposição as principais fontes alimentares de moscas sinantrópicas adultas (Povolný,
1971). Por esse motivo, o ambiente rural, assim como o urbano, demonstra-se propício ao
desenvolvimento dessas espécies de moscas.
Dentre as espécies de importância pecuária destacam-se Musca domestica
(Linnaeus, 1758), Stomoxys calcitrans (Linnaeus, 1758) e Haematobia irritans (Linnaeus,
1758), tanto pela possibilidade de veiculação mecânica e biológica de patógenos, quanto pelo
estresse e consequente perda de peso que ocasionam aos bovinos devido ao hábito

Introdução | 17
hematófago de S. calcitrans e H. irritans (Brito et al., 2008), expondo o gado a micro-
organismos patogênicos e implicando na redução do leite produzido (Rutz e Geden, 2010).
1.2.1. Ecologia e biologia das moscas de importância pecuária
O esterco eliminado pelo gado constitui em criadouros para várias espécies de
moscas e outros artrópodes como besouros, vespas e formigas (Marchiori, 1997; Marchiori et
al., 2001). No entanto, algumas espécies como M. domestica, apresentam comportamento
comunicativo, isto é, a circulação entre ambientes microbiologicamente contaminados (fezes
e matéria orgânica em decomposição) e imediações humanas (fontes de alimentos) (Povolny,
1971).
Musca domestica é cosmopolita e estreitamente associada à antrobiocenose, sendo
troficamente adaptada a alimentos e lixo humano, mas mantém a coprofagia em algumas
situações. Possuem alta capacidade de voo chegando a alcançar cerca de 22 km de distância a
partir do ponto de origem, o que aliado ao comportamento comunicativo marcante da espécie,
permite sua classificação como importante vetor mecânico de uma extensa lista de patógenos
que inclui vírus, bactérias, fungos, protozoários e ovos de helmintos (Greenberg e Povolný,
1971; Greenberg 1971; Linhares, 1979). Indivíduos dessa espécie vivem de 15 a 70 dias e a
fêmea ovipõe em média 120 ovos, sendo que o ciclo de vida apresenta duração de 10 a 14 dias
a depender das condições climáticas (temperatura e umidade) (Brito et al., 2008).
Stomoxys calcitrans são moscas simbovinas de hábito alimentar hematófago,
ativas durante a manhã e no final da tarde, atacam preferencialmente bovinos, mas,
ocasionalmente realizam repasto sanguíneo em outros animais incluindo o homem. A fêmea
ovipõe 40 a 80 ovos a cada postura e o desenvolvimento das larvas ocorre em matéria
orgânica em decomposição fermentada e úmida, e o ciclo de vida consiste em 30 dias quando
em estações de temperaturas elevadas. As fêmeas vivem em média 17 dias e os machos 26
dias (Greenberg, 1973).
Haematobia irritans também são moscas simbovinas hematófagas. As fêmeas
vivem em média 20 dias e ovipõem cerca de 20 ovos entre a massa fecal de bovinos e o solo,
sendo que o ciclo apresenta duração de 10 a 15 dias. Permanecem durante a maior parte do
tempo sobre o hospedeiro bovino, mas o repasto sanguíneo em outros mamíferos como
equinos, ovinos e caninos também pode ocorrer (Brito et al., 2005).

Introdução | 18
1.2.2. Vetores de bactérias
A relação de dípteros muscoides com excrementos de animais possibilita a
veiculação de bactérias entéricas (Cadavid-Sanchez, 2015). Desde antes da década de 1970 o
tema vem sendo abordado em trabalhos discutidos por Greenberg (1973) que buscaram
relacionar surtos de diarreia aguda, causada por Salmonella, Shigella e E. coli, com a
proliferação de moscas. Além disso, foi elaborado o danger index com o propósito de
representar numericamente a importância sanitária de moscas, levando em consideração a
frequência de visitação a ambientes contaminados e a sinantropia (Linhares, 1979).
Entre os anos 1940 e 2000, as bactérias representaram uma porcentagem
significativa (54,3% incluindo Rickettsia) dos patógenos envolvidos nas Doenças Infecciosas
Emergentes (EID, do inglês Emerging Infectious Diseases), sendo que 22,8% das EID foram
relacionadas a insetos vetores (Jones et al., 2008).
Nas últimas décadas, diversos trabalhos têm sido conduzidos a fim de investigar o
papel das moscas na veiculação de bactérias patogênicas, assim como a disseminação que
desempenham no ambiente. Em razão do comportamento comunicativo de M. domestica ser
bem estabelecido (Povolny, 1971), grande parte das pesquisas buscaram demonstrar o papel
dessa espécie na epidemiologia de doenças infecciosas.
Entre os trabalhos mais notáveis está o de Fontedar et al. (1992) que buscaram
quantificar e caracterizar o perfil microbiano (bactérias, fungos e protozoários) de
M. domestica coletadas em ambiente hospitalar, na Índia, e encontraram que tais moscas são
mais propensas a veicular bactérias potencialmente patogênicas, quando em comparação com
o grupo controle.
Em 1999, Moriya et al., investigaram as origens de um surto de infecções por
E. coli produtora de verotoxina (VTEC) do sorogrupo O157:H7 em uma escola infantil da
zona rural do Japão. Neste trabalho os autores conseguiram encontrar uma forte relação clonal
entre as cepas de E. coli isoladas de M. domestica coletadas na região e as envolvidas no
surto. Ao constatarem que as moscas vinham de uma fazenda próxima, sugeriram, pela
primeira vez, que a contaminação poderia ter acontecido dado o contato das moscas com os
alimentos ingeridos pelas crianças.
Complementando os estudos de Moriya et al. (1999), Kobayashi et al. (1999)
buscaram verificar a via de transmissão de E. coli O157:H7. Para isso, alimentaram as moscas
com bactérias e concluíram que a liberação de E. coli O157:H7 viáveis era possível até três
dias após a ingestão, apontando que a disseminação de bactérias poderia ocorrer não somente
pelo contato com superfícies contaminadas, em que as moscas atuariam como vetores

Introdução | 19
mecânicos, mas também através das excretas e regurgitados, em uma relação mais próxima a
vetores biológicos.
Desse modo, três vias de transmissão de patógenos por moscas têm sido
consideradas: transporte e contato através do exoesqueleto, deposição de fezes e regurgitação
(Rahuma et al. 2005).
Alam e Zurek (2004) buscaram isolar E. coli O157:H7 a partir de moscas
coletadas em vários ambientes de fazenda de gado bovino e apontaram que M. domestica
poderia transportá-las no interior da fazenda e possivelmente a fazendas vizinhas e ambiente
urbano. Posteriormente, Ahmad et al. (2007) elaboraram experimentos a fim de testar a
transmissão de E. coli O157:H7 presente em M. domestica a bezerros, concluindo que a
transmissão era possível.
No Brasil, poucos estudos com essa temática têm sido conduzidos. Destacam-se
os trabalhos de Paraluppi et al. (1996) que pretendiam isolar bactérias patogênicas de moscas
coletadas em feiras livres, associando-as a contaminações de alimentos a venda; Moraes et al.
(2004) que buscaram relacionar isolados de vacas com mastite com os obtidos de
S. calcitrans; Castro et al. (2007, 2008a, 2008b) que apresentaram dados referentes à
microbiota de S. calcitrans com isolados de enterobactérias (patogênicas e não patogênicas),
Pseudomonas, Staphylococcus e Enterococcus; Kappel et al. (2013) que pesquisaram
bactérias presentes na superfície externa de insetos alados (moscas, besouros, borboletas, etc)
coletados em ambiente hospitalar; Almeida et al. (2014) que procuraram isolar e determinar o
perfil de sensibilidade antimicrobiana de E. coli, Salmonella spp. e Sthaphylococcus spp. de
moscas coletadas em fazendas.
Apesar de muito ser conhecido sobre o papel das moscas como vetores de
bactérias patogênicas, poucas pesquisas abordaram a atuação de moscas na disseminação da
resistência antimicrobiana no ambiente, sendo que até o momento não há referências de tais
estudos no Brasil.
Marshall et al. (1990) foram os primeiros a hipotetizar que as moscas em conjunto
com aerossóis e outros animais, poderiam representar uma via de disseminação de genes de
resistência a antimicrobianos entre os animais de uma fazenda, quando buscaram entender as
condições naturais que contribuíam para a disseminação de bactérias e plasmídeos no
ambiente. Adicionalmente, a presença de moscas transportando bactérias resistentes em
outros ambientes, como o hospitalar, pode significar condições sanitárias precárias, além de
contribuir para a disseminação de resistência e de patógenos dentro do hospital, do hospital
para a comunidade e da comunidade para o hospital (Rahuma et al. 2005).

Introdução | 20
O cultivo de bactérias patogênicas resistentes a antimicrobianos, realizado a partir
de moscas coletadas em restaurantes por Macovei e Zurek (2006), colabora para a hipótese
que relaciona a incidência de moscas com eventos de diarreia, o que, aliado à presença de
elementos genéticos móveis ligados a genes de resistência nessas cepas, indica que
M. domestica – alvo do estudo – pode constituir em uma via de disseminação de genes
também nesses ambientes.
Em 2010, Rybaríková et al. (2010), ao pesquisarem resistência antimicrobiana em
isolados de moscas simbovinas e de bezerros, hipotetizaram que as cepas resistentes presentes
nas moscas seriam originárias dos bezerros, uma vez que ambas apresentaram o mesmo perfil
de resistência (fenotípico e genotípico) e clonal, sugerindo que as moscas ao terem contato
com os bezerros adquiririam bactérias resistentes e disseminariam clones pelo ambiente,
comprovando a hipótese de Marshall et al. (1990).
Da mesma forma, Usui et al. (2013, 2015) encontraram clones de E. coli com o
mesmo perfil de resistência antimicrobiana em moscas e em fezes de bezerros e
posteriormente sugeriram que as moscas podem atuar na disseminação de genes de resistência
não só no interior de fazendas mas também entre fazendas. Além disso, cepas resistentes de
E.coli presentes em moscas também poderiam ser transmitidas à comunidade (Blaak et al.,
2014).
Nesse contexto, os animais de produção constituiriam em reservatórios de cepas
bacterianas portadoras de resistência a antimicrobianos, e os insetos sinantrópicos,
principalmente as moscas, atuariam como agentes dispersores de bactérias resistentes e,
consequentemente, dos genes que portam, de modo que, a disseminação do meio rural para o
urbano seria plausível (Zurek e Ghosh, 2014).
Solà-Ginés et al. (2015), ao pesquisarem resistência antimicrobiana em E. coli
isoladas de moscas coletadas em granjas de frangos, discutem que o estudo das bactérias
obtidas por essa via fornecem informações sobre a microbiota dos animais, mas que também
moscas podem atuar introduzindo bactérias portadoras de genes de resistência e virulência
colaborando para que ocorra a colonização dos animais com elementos genéticos diferentes
daqueles já existentes.
Recentemente, Schaumburg et al. (2016) realizaram um levantamento da
resistência em bactérias veiculadas por moscas em áreas rurais e urbanas da cidade de
Münster na Alemanha e compararam com isolados de humanos. Frente à elevada similaridade
dos isolados, sugeriram que haveria uma fonte comum ou transmissão de bactérias das

Introdução | 21
moscas aos humanos. Ressaltaram, porém, que os dados desse estudo não foram suficientes
para indicar a fonte da resistência e o sentido da transmissão.
1.3. Escherichia coli
Escherichia coli é constituinte da microbiota entérica humana e animal. Apesar de
algumas linhagens estarem relacionadas a doenças intestinais e extraintestinais (Clermont et
al., 2000), a maioria não é considerada patogênica (Madigan et al., 2010). As linhagens
patogênicas são divididas em grupos (patotipos), de acordo com as doenças ou toxinas que
produzem: E. coli entero-hemorrágica (EHEC), E. coli enterotoxigênica (ETEC), E. coli
enteropatogênica (EPEC), E. coli enteroinvasiva (EIEC), E. coli enteroagregativa (EAEC),
E. coli que adere difusamente (DAEC) e E. coli extraintestinais (ExPEC) (Kaper et al., 2004).
Escherichia coli entero-hemorrágica (EHEC) é classicamente caracterizada pela
presença de verotoxinas (também denominadas toxinas Shiga-like), codificadas pelos genes
vt1 e/ou vt2, e pela lesão attaching and effacing (A/E), produto da expressão de genes
localizados na ilha de patogenicidade denominada locus of enterocyte effacement (LEE), na
qual se destaca o gene eae, que promove a expressão da intimina, responsável pela adesão e
indução da lesão no tecido intestinal do hospedeiro. As E. coli produtoras de verotoxina
(VTEC) podem ser consideradas um subgrupo de EHEC, pois apresentam apenas vt1 e/ou vt2.
Ruminantes, principalmente bovinos saudáveis, são considerados reservatórios de EHEC, e
infecções em humanos produzem Síndrome Hemolítica Urêmica (HUS) e colite hemorrágica
(HC) (Nataro e Kaper, 1998; Kaper et al., 2004; Piérard et al., 2012).
Já E. coli enterotoxigênica (ETEC) apresenta dois grupos de enterotoxinas, as
enterotoxinas termolábeis (LTs) e as termoestáveis (STs), e podem causar diarreia em
humanos ou animais dependendo dos fatores de colonização que expressam – CFAs, em
humanos e K88 ou K99, em animais (Kaper et al., 2004).
Cepas de E. coli enteropatogênica (EPEC) são caracterizadas pela produção da
lesão A/E devido a expressão dos genes contidos na ilha de patogenicidade LEE e são ligadas
a quadros de diarreia infantil aguda, muitas vezes fatal. São denominadas EPEC típicas as
cepas que possuem plasmídeo EAF (fator de aderência de EPEC) (Kaper et al., 2004).
Escherichia coli enteroinvasiva (EIEC) e Shigella spp. são estreitamente
relacionadas chegando a ser indistinguíveis a nível de espécies, de forma que alguns poucos
testes bioquímicos as diferenciam. Porém, ambas apresentam os mesmos fatores de virulência
essenciais, responsáveis pela penetração das células do epitélio intestinal e apoptose de
macrófagos infectados, gerando colite inflamatória (Kaper et al. 2004).

Introdução | 22
Escherichia coli enteroagregativa (EAEC) não apresenta secreção de
enterotoxinas LT e ST e são caracterizadas por um padrão especial de agregação das células
semelhante a pilhas de tijolos. Podem apresentar citotoxinas e enterotoxinas ShET1, Pic,
EAST1, Pet e fatores de colonização AAFs. Também são relacionadas à diarreia
principalmente em crianças (Nataro e Kaper, 1998; Kaper et al., 2004).
Escherichia coli que adere difusamente (DAEC) possuem padrão de adesão
difuso, envolvendo o fator de colonização F1845 na maioria dos casos. Geralmente estão
envolvidas em episódios diarreicos em crianças de um a cinco anos de idade (Nataro e Kaper,
1998; Kaper et al., 2004).
As E. coli extraintestinais (ExPEC) são aquelas envolvidas em infecções não
intestinais. Compreendem as E. coli uropatogênicas (UPEC), que causam infecção no trato
urinário; E. coli associadas a meningite/sepse (MNEC) que são a principal causa de meningite
em recém nascidos e E. coli patogênica para aves (APEC), responsável por infecções
respiratórias, pericardite e sepse em aves (Kaper et al., 2004).
A microbiota é resultante de milhões de anos de coevolução entre micro-organismos
e o hospedeiro, podendo ser considerada uma relação mutualística, dada a participação em
processos fisiológicos dos hospedeiros que em contrapartida fornecem nichos e nutrientes
necessários à sobrevivência microbiana. No trato gastrintestinal humano a microbiota
contribui para a prevenção da colonização por patógenos, na produção de vitaminas e
auxilia na digestão, além de fazer parte de mecanismos regulatórios do sistema imune do
hospedeiro e do desenvolvimento do tecido linfoide associado ao intestino (Kamada et al.,
2013b).
Bactérias comensais e patogênicas ocupam nichos ecológicos intestinais
semelhantes, de modo que o equilíbrio populacional da microbiota permite a prevenção da
colonização e proliferação de patógenos e patobiontes. Por outro lado, a barreira epitelial do
hospedeiro é fundamental para impedir a disseminação das bactérias comensais pelo
organismo, o que resultaria em choque séptico letal (Kamada et al., 2013a). Porém, Marshall
et al. (2009) sugeriram que a definição de micro-organismos comensais ou patogênicos pode
ser relativa, estando associada à condição física do hospedeiro.
Atualmente, são admitidos sete grupos filogenéticos para E. coli (A, B1, B2, C, D,
E, F) e um pertencente à Escherichia clado I (Clermont et al., 2013), estando os grupos A e
B1 associados às cepas comensais (Clermont et al., 2000), porém não há correspondência
entre os grupos filogenéticos e os patogênicos (Croxen et al., 2013).

Introdução | 23
Cepas de E. coli provenientes de animais de produção frequentemente apresentam
multirresistência a antimicrobianos que pode ser disseminada para a comunidade (Vieira et
al., 2011). Nesse contexto, Verdier et al. (2012) discutiram o papel de E. coli comensal como
reservatório de genes codificadores de resistência a antimicrobianos, assim como a
possibilidade de transferência, e sugeriram que cepas provenientes de animais saudáveis
poderiam constituir em ferramenta de monitoramento.
1.4. RESISTÊNCIA ANTIMICROBIANA
O grupo dos β-lactâmicos está entre os antimicrobianos mais empregados para o
tratamento de diversas infecções em humanos e outros animais. Atua interferindo na síntese
da parede celular bacteriana, sendo a resistência conferida à produção da enzima β-lactamase
que promove a hidrólise do anel β-lactâmico (Aminov e Mackie, 2007). Entre essas enzimas,
as β-lactamases de espectro estendido (ESBLs) têm sido frequentemente estudadas (Pereira et
al., 2003) e são definidas pela capacidade de hidrolisar penicilinas, cefalosporinas de terceira
geração e monobactam sendo inibidas pelo ácido clavulânico, sulbactam e tazobactam (Shah
et al., 2004; Perez et al., 2007).
O isolamento de cepas de E. coli produtoras de β-lactamases em animais e
alimentos de origem animal é frequente, sendo o grupo CTX-M o mais encontrado (Paterson e
Bonomo, 2005; Lee et al., 2010). Adicionalmente, o isolamento de cepas de E. coli e
Klebsiella produtoras de ESBLs tem sido documentado em casos de infecções hospitalares
(Bonnedahl et al., 2009) e em amostras de E. coli isoladas de fezes bovinas, suínas e de aves
(Horton et al., 2011).
Segundo Garcia-Alvarez et al. (2012), a descoberta de cepas de E. coli produtoras
de ESBLs provenientes de fazendas de animais de produção contribuiu para a associação
entre o genótipo presente em E. coli isoladas de infecções animais com amostras
diagnosticadas em humanos. A capacidade de disseminação das ESBLs está relacionada à
localização em elementos genéticos móveis, porém as vias de transmissão parecem ser mais
complexas do que o suposto, e os dados referentes aos reservatórios na comunidade ainda são
escassos, devido à dispersão de genes de β-lactamases no ambiente ser pouco conhecida
(Lachmayr et al. 2009). Assim, a melhor compreensão da complexidade dos processos
envolvidos na disseminação da resistência a antimicrobianos poderia conduzir a um melhor
controle (O’Brien, 2002).
As fluorquinonolonas, derivadas das quinolonas, também são amplamente usadas,
atuam nas topoisomerases DNA girase e topoisomerase IV impedindo a replicação, de modo

Introdução | 24
que a resistência mais comum consiste na mutação dos genes, promovendo a substituição de
aminoácidos nas subunidades Gyr (A e/ou B) da DNA girase e ParC ou ParE na
topoisomerase IV, cuja região geralmente mutada é denominada Região Determinante de
Resistência à Quinolona (QRDR, do inglês Quinolone-Resistance-Determining Region)
(Jacoby, 2005).
Outros mecanismos de resistência a quinolonas como as bombas de efluxo,
proteção do sítio-alvo e alteração dos antimicrobianos envolvem a participação de genes
localizados em plasmídeos (Bolon, 2011). As bombas de efluxo proteicas QepA e OqxAB,
codificadas pelos genes qepA e oqxAB, promovem a expulsão de fluorquinolonas para fora da
célula bacteriana (Hernández et al., 2011). Já as proteínas Qnr protegem as topoisomerases da
atividade das quinolonas, sendo que até o momento, cinco famílias Qnr são conhecidas (genes
qnrA, qnrB, qnrS, qnrC e qnrD) (Hernández et al., 2011). Quanto ao mecanismo de alteração
do antimicrobiano é descrita a inativação enzimática de fluorquinolonas por uma variante da
acetiltransferase AAC(6’)-Ib (Jacoby et al., 2014).
As tetraciclinas correspondem à outra classe de antimicrobianos com ação em
vários micro-organismos (de bactérias gram-positivas e gram-negativas a protozoários).
Atuam inibindo a síntese proteica ao impedir a ligação do RNA transportador ao ribossomo,
de modo que a maioria dos genes que promovem resistência expressam sistemas de efluxo e o
restante uma proteína que protege o ribossomo (Chopra e Roberts, 2001).
1.4.1. Elementos genéticos móveis
De acordo com Frost et al. (2005) “elementos genéticos móveis são segmentos de
DNA que codificam enzimas e outras proteínas que medeiam o movimento do DNA dentro
do genoma (mobilidade intracelular) ou entre células bacterianas (mobilidade intercelular)”.
Os movimentos intracelulares correspondem a eventos de recombinação no
genoma realizados por transposons, que podem se integrar a fagos e plasmídeos, podendo ou
não ser transferidos a outras células através dos mecanismos de transferência horizontal
(HGT, do inglês horizontal gene transfer) (Frost et al., 2005).
Entre os movimentos intercelulares estão os mecanismos de transferência
horizontal de genes: transformação, transdução e conjugação. A transformação necessita de
bactérias naturalmente competentes para ocorrer; a transdução ocorre de maneira
independente da célula bacteriana, uma vez que consiste no empacotamento acidental de
DNA bacteriano no capsídeo do bacteriófago, que ao infectar novas células o transfere; a
conjugação também é um sistema independente, pois necessita da presença de um plasmídeo

Introdução | 25
conjugativo ou de elementos conjugativos integrados ao cromossomo como os transposons
conjugativos (Frost et al., 2005).
Desse modo, é possível distinguir três elementos genéticos de importância para a
disseminação de resistência a antimicrobianos – plasmídeos, integrons e transposons (Stokes e
Gillings, 2011).
Por definição, plasmídeos são elementos genéticos extracromossomais, capazes de
autorreplicação e que não carregam genes essenciais para o metabolismo bacteriano
(Carattoli, 2009). Podem ser classificados de acordo com os grupos de incompatibilidade
(Inc), com base no fato que plasmídeos com o mesmo sistema de replicação são incompatíves
e, portanto, não se propagam na mesma célula (Carattoli, 2009).
Nos plasmídeos dois conjuntos de genes são necessários para a conjugação, os
genes de mobilidade (MOB) e do complexo de formação do par de acoplamento (MPF)
associado a membrana, que produz um sistema de secreção tipo IV (T4SS), de modo que
plasmídeos conjugativos são aqueles que apresentam complexo MPF próprio. Os plasmídeos
mobilizáveis possuem MOB, mas não apresentam MPF e podem utilizá-lo de outros
elementos genéticos presentes na célula bacteriana a fim de realizar conjugação, enquanto os
não mobilizáveis não possuem nenhum dos dois conjuntos de genes (MOB e MPF) (Smillie et
al., 2010).
Os plasmídeos geralmente apresentam genes de resistência a antimicrobianos o
que, aliado à conjugação e a possibilidade de agregação de outros elementos genéticos
(integrons e transposons), demonstra o papel significativo que desempenham na disseminação
dessa resistência (Carattoli, 2013).
Os integrons foram descritos pela primeira vez por Stokes e Hall (1989). São
elementos genéticos que promovem a integração de cassetes gênicos através de recombinação
sítio-específica e são constituídos pelo gene da integrase (enzima que atua na integração do
cassete gênico), um sítio de recombinação e o promotor (Koczura et al., 2013; Ravi et al.,
2014). Podem ser divididos em dois grupos, os integrons de resistência e os superintegrons
(Fluit e Schmitz, 2004).
Integrons de resistência não são naturalmente móveis, pois a integrase atua apenas
na movimentação dos cassetes gênicos, porém, frequentemente estão associados a sequências
de inserção, transposons e plasmídeos conjugativos que garantem a mobilidade e
disseminação dos cassetes de resistência a antimicrobianos. São classificados em classes de
acordo com a similaridade genética da integrase (gene intI), sendo que até o momento cinco

Introdução | 26
classes são conhecidas (classe 1, 2, 3, 4 e 5). Os integrons de classe 1 são os mais comuns e os
de classe 4 e 5 presentes em Vibrio spp. (Mazel, 2006; Ravi et al., 2014).
Os superintegrons localizam-se no cromossomo bacteriano e são grandes, uma
vez que possuem mais de 20 cassetes gênicos com funções variadas (Fluit e Schmitz, 2004;
Mazel, 2006).
A estrutura clássica dos integrons de classe 1, no sentido 5’ a 3’ da sequência,
compreende a integrase (intI1), o sítio de recombinação (attI), os cassetes gênicos com seus
respectivos sítos de recombinação (attC) e uma região 3’ conservada apresentando os genes
qacΔE e sul1, que conferem resistência a quaternário de amônio e sulfonamidas,
respectivamente (Mazel, 2006). No entanto, integrons de classe 1 não usuais apresentam o
gene sul3 na região 3’ conservada (Sáenz et al., 2010).
Acredita-se que os integrons de classe 1 tenham surgido a partir da captura do
integron de classe 1 cromossomal de betaproteobactérias ambientais, por um transposon da
família Tn402, evento que tem favorecido a disseminação de genes de resistência no ambiente
e possibilitado o uso dessa classe de integron como indicativo de poluição gerada pela
atividade humana (Gillings et al., 2015).
Já os transposons são caracterizados por um fragmento de DNA com duas
sequências de inserção nas extremidades (Mazel, 2006), sendo que um grupo particular de
transposons apresenta capacidade de conjugação (transposons conjugativos). Tais transposons
apresentam características de transposons clássicos e de bacteriófagos. Semelhante a
transposons clássicos, transposons conjugativos excisam e integram ao DNA de plasmídeos
conjugativos quando apresentam forma circular fechada e são transferidos por conjugação,
apesar de não se replicarem. Por outro lado, apresentam características de excisão e integração
semelhante a fagos temperados (Salyers et al., 1995).
1.4.2. Disseminação no ambiente e implicações
Resultados obtidos em análise retrospectiva, quanto à susceptibilidade
antimicrobiana em E. coli indicaram que a seleção de mecanismos de resistência estaria
temporalmente relacionada ao uso de antimicrobianos na medicina humana e veterinária
(Tadesse et al., 2012).
Nesse sentido, Marshall et al. (2009) ressaltaram que o uso de antimicrobianos na
pecuária, inclusive como aditivo na alimentação, tem contribuído para a seleção de genes de
resistência em bactérias comensais, localizados em elementos móveis como transposons e
plasmídeos. Isso merece atenção especial, pois indica a possibilidade de transferência gênica

Introdução | 27
entre bactérias provenientes de diferentes animais, as associadas aos humanos ou entre
comensais e patógenos. Dessa forma, o estudo da resistência antimicrobiana em bactérias não
patogênicas é fundamental para a melhor compreensão da distribuição ambiental dos genes,
dada a possibilidade de transferência a bactérias patogênicas (Lachmayr et al., 2009).
Devido ao contato que algumas espécies de dípteros muscoides estabelecem com
as fezes de bovinos, torna-se notável a possibilidade de veiculação de bactérias resistentes a
antimicrobianos, fato que contribui para a alocação das moscas nas rotas de disseminação de
resistência antimicrobiana (Rybaríková et al., 2010). Isto, aliado à distribuição de E. coli em
hospedeiros animais ou humanos e no ambiente, e à versatilidade patógeno/comensal da
espécie, permite que estudos direcionados à abordagem de cepas comensais como
reservatórios genéticos, assim como a contribuição com a emergência de fatores de virulência
e resistência sejam conduzidos (Tenaillon et al., 2010). Além disso, trabalhos desenvolvidos
com E. coli podem contribuir para a orientação de estudos em outras bactérias do mesmo
nicho (O’Brien, 2002).
No ambiente, a participação das moscas na disseminação geralmente não é
considerada, sendo associada a alimentos de origem animal, pelo contato direto com animais
ou através de rotas ambientais (Costa et al., 2013), que geralmente incluem os animais
domésticos, seus excrementos e os vegetais cultivados, de modo que um fator importante para
a dispersão de bactérias resistentes pode ser a aplicação de esterco como fertilizante
(Khachatourians, 1998; Marshall et al., 2009; Costa et al., 2013). Isto, aliado ao emprego de
antimicrobianos na pecuária, colocaria as fazendas como fontes de genes e de bactérias
resistentes a antimicrobianos, de modo que entender a contribuição da prática agropecuária
para a disponibilização de genes de resistência no ambiente é essencial (Wichmann et al.,
2014). No entanto, apesar de vários estudos terem como tema a resistência antimicrobiana no
ambiente, análises mais abrangentes sobre os elementos genéticos e o perfil clonal das cepas
presentes na disseminação são secundárias (Novais et al., 2013).
Considerando que a ecologia de genes de resistência antimicrobiana e de
virulência no ambiente não é bem compreendida (Macovei e Zurek, 2006), a abordagem de
dípteros muscoides como vetores mecânicos de bactérias resistentes a antimicrobianos
contribuiria para o desenvolvimento de estudos mais completos sobre a disseminação da
resistência antimicrobiana, não só em termos epidemiológicos clássicos como também
moleculares, uma vez que a relação de dípteros muscoides com fezes de bovinos possibilita a
veiculação de bactérias da microbiota intestinal desses animais.

Introdução | 28
A possibilidade de classificação de E. coli em comensais ou em potenciais
patógenos permite que estudos sobre resistência antimicrobiana e fatores de virulência
relacionados a esses grupos sejam conduzidos, uma vez que E. coli não patogênica pode
significar um importante reservatório de genes de resistência antimicrobiana, sendo a
proximidade entre o ambiente animal e humano um provável fator para a troca de genes entre
bactérias de origens distintas (Liebana et al., 2006).
Assim, estudos de resistência antimicrobiana em isolados animais são
fundamentais para a melhor compreensão do papel dos hospedeiros, e das espécies
ecologicamente associadas a eles, nos processos de disseminação e de eventos de
transferência horizontal dos genes de resistência a antimicrobianos entre cepas comensais e
patogênicas. Nesse sentido, este projeto propõe o isolamento, identificação e a caracterização
fenotípica e genotípica da resistência de E. coli isoladas de dípteros muscoides em
propriedade rural leiteira; a detecção de fatores de virulência relacionados à colibacilose
bovina e a avaliação da capacidade de disseminação dos genes de resistência através da
presença de plasmídeos, ensaios de conjugação e perfil clonal das cepas isoladas.

II – Objetivos

Objetivos | 30
2.1. Objetivo Geral: Caracterizar a resistência fenotípica e genotípica a antimicrobianos e
fatores de virulência em E. coli isoladas de dípteros muscoides presentes em propriedade
leiteira.
2.2. Específicos:
1. Isolar e identificar cepas de E. coli em amostras de dípteros muscoides e determinar o
perfil de sensibilidade fenotípica dos isolados frente aos principais antimicrobianos
administrados em terapia veterinária e humana, assim como a produção fenotípica de
ESBLs;
2. Identificar por PCR os principais genes de resistência, integrons e fatores de
virulência associados à colibacilose em bovinos;
3. Verificar a presença de plasmídeos nas cepas resistentes;
4. Caracterizar os integrons por Restriction Fragment- Length Polymorphism (RFLP) e
sequenciamento
5. Verificar a possibilidade de transferência dos integrons e genes de resistência através
de ensaios de conjugação;
6. Determinar por PCR os grupos filogenéticos das cepas resistentes;
7. Avaliar o perfil clonal das cepas resistentes através de Pulsed Field Gel
Electrophoresis (PFGE);

III - Material e Métodos

Material e Métodos| 32
3.1. Áreas de coleta
A coleta das moscas foi realizada com a colaboração do Prof. Dr. Márcio Garcia
Ribeiro - Departamento de Higiene Veterinária e Saúde Pública (DHVSP) da Faculdade de
Medicina Veterinária e Zootecnia (FMVZ) - UNESP, Botucatu-SP, em dois
estabelecimentos rurais, aqui designados por A e B, localizados na região de Botucatu – SP,
com sistema semi-intensivo de produção leiteira (Figura 1).
A propriedade A caracteriza-se pela produção de leite destinada à
comercialização, apresentando maior densidade de animais (número de animais por área),
com sistema de rotação de pastagem e ordenha mecânica realizada duas vezes ao dia. Os
animais são mantidos em sistema de semiconfinamento e os bezerros são separados de
acordo com a idade. A suplementação alimentar é realizada no estábulo com ração e feno.
A propriedade B apresenta produção de leite para autoconsumo e criação de gado
de corte, sendo que para a realização do presente trabalho apenas o gado leiteiro foi
contabilizado. A propriedade apresenta menor densidade de animais devido a maior área
destinada à pastagem, com ordenha manual realizada uma vez ao dia. A suplementação
alimentar é realizada no estábulo com ração, feno e silagem de milho.
As propriedades distam aproximadamente 35 km.
3.2. Obtenção das amostras
As moscas foram coletadas utilizando puçá entomológico previamente
desinfetado com solução de hipoclorito de sódio a 10% por 30 minutos, a fim de evitar a
interferência de contaminantes presentes anteriormente no equipamento. Foi dada
preferência às moscas que estavam mais próximas aos animais em lactação (vacas e
bezerros). As coletas foram realizadas em junho de 2015, durante o período da manhã por
duas horas (7h30 às 9h30 na propriedade A e das 10h às 12h na propriedade B) com
tentativas a cada 3 minutos, em média.
As moscas obtidas em cada captura foram transferidas para tubos tipo falcon
estéreis e inativadas por congelamento a -10 ºC por 30 minutos.

Material e Métodos| 33
Figura 1. Região do município de Botucatu-SP, onde estão localizadas as duas propriedades
leiteiras estudadas (denominadas A e B), quanto à resistência antimicrobiana em E. coli
provenientes de dípteros muscoides. Fonte: Elaborado com Google Maps (https://maps.google.com.br/)
3.3. Isolamento e identificação de E. coli
Aos tubos contendo as moscas foram adicionados 2 mL de caldo EC
(Escherichia coli). Os tubos foram incubados a 37 ºC por 24 horas e posteriormente, as
culturas foram semeadas em meio ágar MacConkey, com e sem a adição de ampicilina
(50 μg/mL), e incubadas a 37 ºC por 24 horas. As moscas foram removidas do meio líquido,
lavadas com álcool 70 °GL e mantidas sob refrigeração, também em álcool 70 °GL, para
posterior identificação das espécies.
Cinco colônias típicas de E. coli cultivadas em ágar MacConkey com adição de
ampicilina foram selecionadas por meio das características morfológicas das colônias e da
diferenciação bioquímica a partir das seguintes provas: fermentação de glicose, produção de
gás, urease, L-triptofanase, produção de ácido sulfídrico, lisina descarboxilase, motilidade,
indol e citrato. Para a realização das provas foi utilizado o conjunto de meios
EPM/MILi/Citrato (Toledo et al., 1982 a, b).
As cepas foram estocadas em meio semissólido (ágar nutriente 1,15%; caldo
nutritivo 0,4%) e congeladas (Caldo BHI - Brain Heart Infusion; 20% glicerol).

Material e Métodos| 34
A identificação das espécies de dípteros foi realizada com a colaboração da Profa.
Dra. Patrícia Jacqueline Thyssen - Departamento de Biologia Animal, Instituto de Biologia –
UNICAMP.
3.4. Caracterização fenotípica da sensibilidade aos antimicrobianos
3.4.1. Sensibilidade microbiana (antibiograma)
O perfil de sensibilidade microbiana foi realizado mediante a técnica de difusão
com discos (Bauer et al., 1966; CLSI, 2012), frente aos antimicrobianos: ampicilina (10 g),
amoxicilina/ácido clavulânico (30 g), cefalexina (30 g), cefoperazona sódica (75 g),
ceftiofur (30 g), ceftriaxona (30 g), cloranfenicol (30 g), enrofloxacina (5 g),
ciprofloxacina (5 g) gentamicina (10 g), sulfametoxazol/trimetoprim (25 g) e tetraciclina
(30 g) (Ribeiro, 2008; Winn Jr. et al., 2008). Como controle de sensibilidade foi utilizada a
linhagem de E. coli ATCC 25922 e a interpretação dos resultados de acordo com o disposto
pelo Clinical and Laboratory Standards Institute (CLSI, 2012).
A técnica consiste no preparo de uma suspensão bacteriana na escala 0,5 de Mc
Farland em solução de NaCl 0,15 M estéril, a partir de colônias com características idênticas,
isoladas em meio ágar triptona de soja (TSA). Posteriormente, a suspensão de cada amostra
foi espalhada na superfície de placas contendo ágar Mueller Hinton com auxílio de swabs
estéreis. Em seguida, os discos correspondentes aos antimicrobianos a serem testados foram
aplicados sobre o meio, obedecendo a uma distância mínima de 20 mm entre os discos e
incubados a 37 °C por 24 horas. As cepas que apresentaram perfil intermediário foram
consideradas resistentes.
3.4.2. Produção de ESBLs – Método da aproximação de disco
A detecção de ESBL foi realizada conforme o método de aproximação de discos
(Jarlier et al.,1988), utilizando discos de aztreonam (30 g), ceftriaxona (30 g), cefotaxima
(30 g), ceftazidima (30 g) e amoxicilina/ácido clavulânico (30 μg). Para a execução da
técnica foram escolhidos os isolados de E. coli que apresentaram perfil resistente, no
antibiograma, aos β-lactâmicos (cefalosporinas) de terceira geração empregando os mesmos
procedimentos do item anterior. Os discos de aztreonam, ceftriaxona, cefotaxima e
ceftazidima foram dispostos a 20 mm do disco de amoxicilina/ácido clavulânico ao centro.
Considera-se o resultado positivo quando há alargamento do halo de inibição ou surgimento

Material e Métodos| 35
de uma faixa de inibição entre os halos de amoxicilina/ácido clavulânico e os antimicrobianos
testados, conhecida como “zona fantasma”.
3.5. PCR para a pesquisa dos genes de virulência, resistência e integrons
3.5.1. Extração do DNA (método do choque térmico)
O método baseado em Chapman et al. (2001) consistiu na fervura por 10 minutos
da suspensão bacteriana, preparada a partir do crescimento confluente obtido em placas de
TSA em 200 L de água ultrapura estéril, seguido de congelamento a -10 °C por 30 minutos.
Após o descongelamento (temperatura ambiente) as amostras foram centrifugadas a
12.000 rpm (9.676,8 g) (centrífuga Sorvall® MC12) por 5 minutos e o sobrenadante
transferido para um novo tubo.
3.5.2. PCR
A detecção dos genes de interesse (Tabela 1) foi realizada pela Reação em Cadeia
da Polimerase (PCR), primeiramente utilizando pool de DNA de cinco amostras1. Em cada
reação foram adicionados 1,5 U Taq polimerase, 1,5 L de Tampão 10X para PCR, 200 M
de dNTP, 2 mM de MgCl2, 0,5 µL dos iniciadores em concentrações ótimas, água ultrapura
estéril qsp., em um volume final de 15 μL e 2 L de DNA bacteriano obtido por choque
térmico. Cerca de 5 µL dos produtos da PCR adicionados de 1-2 µL de tampão de amostra
(azul de bromofenol 0,25%; sacarose em água 40% w/v) foram submetidos à eletroforese em
gel de agarose em tampão TAE 1X (Tris 2 M; Ácido acético 0,04 M; EDTA 0,01 M em pH
8,0) a 100 V e 200 mA por 50 minutos. Posteriormente, o gel foi incubado em solução de
Brometo de Etídeo a 1 g/mL por 10 minutos e visualizado em transiluminador de luz UV e
as imagens foram capturadas e fotografadas com o sistema Image Master® VDS (Pharmacia
Biotech).
A programação das PCRs (termociclador Techne TC-312) foi realizada da
seguinte forma: desnaturação inicial a 94ºC por cinco minutos, 30 ciclos (94ºC por 45
segundos, temperatura de anelamento por 45 segundos, 72ºC por 45 segundos) e extensão
final a 72ºC por sete minutos.
1 Os resultados positivos foram repetidos para a amostra de DNA de cada bactéria contida no pool.

Material e Métodos| 36
Tabela 1. Iniciadores utilizados para a detecção dos fatores de virulência, genes de resistência
e integrons em isolados de E. coli provenientes de dípteros muscoides.
Iniciador Produto Sequência (5’-3’) T* pb
** Ref.
Fatores de virulência
eae intimina GACCCGGCACAAGCATAAGG
CCACCTGCAGCAACAAGAGC 63 ºC 384
Yu e Kaper,
1992
vt1 verotoxina AAGTTGCAGCTCTCTTTGAATA
TGCAAACAAATTATCCCCTGAG 50 ºC 364
Ojeniyi et al.,
1994 vt2 verotoxina
GGGCAGTTATTTTGCTGTGGA GTATCTGCCTGAAGCGTAA
50 ºC 386
hlyA hemolisina AACAAGGATAAGCACTGTTCTGGCT ACCATATAAGCGGTCATCCCGTCA
63 ºC 1177 Yamamoto et
al., 1995
K99 fator de
colonização TATTATCTTAGGTGGTATGG
GGTATCCTTTAGCAGCAGTATTTC 50 ºC 314
Franck et al.,
1998
LTII toxina
termolábil AGATATAATGATGGATATGTATC
TAACCCTCGAAATAAATCTC 48 °C 300
Schultsz et al.,
1994
STa toxina
termoestável TCCGTGAAACAACATGACGG ATAACATCCAGCACAGGCAG
48 °C 244 So e McCarty,
1980
β-lactâmicos
ampC AmpC β-
lactamases*** CCCCGCTTATAGAGCAACAA TCAATGGTCGACTTCACACC
61 °C 634 Féria et al.,
2002
blaTEM ESBL TCGGGGAAATGTCGCG
TGCTTAATCAGTGAGGCACC 61 °C 972
Cao et al.,
2002 blaSHV ESBL TTATCTCCCTGTTTAGCCACC GATTTGCTGATTTCGCTCGG
61 °C 795
blaCTX-M ESBL CGATGTGCAGTACCAGTAA
TTAGTGACCAGAATCAGCGG 60 °C 585
Batchelor et al., 2005
blaCMY-4 AmpC β-
lactamases**** GATTCCTTGGACTCTTCAG
TAAAACCAGGTTCCCAGATAGC 56 ºC 1800
Stapleton et al., 1999
Quinolonas
gyrA mutação DNA
girase TACACCGGTCAACATTGAGG
TTAATGATTGCCGCCGTCGG 64 ºC 648
Oram e Fisher,
1992
parC Mutação
topoisomerase
IV
AAACCTGTTCAGCGCCGCATT
GTGGTGCCGTTAAGCAAA 55 ºC 395
Vila et al.,
1996
Tetraciclina
tetA sistema efluxo GTAATTCTGAGCACTGTCGC
CTGTCCTGGACAACATTGCTT 62 °C 937
Guardabassi et
al., 2000
Integrons
intI1 integrase
classe 1 GGGTCAAGGATCTGGATTTCG
ACATGGGTGTAAATCATCGTC 62 ºC 483
Mazel et al.,
2000
5’CS 3’CS
região dos
cassetes GGCATCCAAGCAGCAAG AAGCAGACTTGACCTGA
60 ºC V§ Lévesque et
al., 1995
intI2 integrase
classe 2 CACGGATATGCGACAAAAAGGT GTAGCAAACGAGTGACGAAATG
62 ºC 788 Mazel et al.,
2000
attI2-F
orfX-R região dos
cassetes GACGGCATGCACGATTTGTA
GATGCCATCGCAAGTACGAG 58 ºC V§
Machado et al.,
2005
T*: Temperatura de anelamento; pb
**: tamanho do produto em pares de bases; Ref.: Referência;
***: gene
cromossomal; ****: gene plasmidial; V§: variável; ESBL: β-lactamase de espectro estendido

Material e Métodos| 37
3.6. Presença de plasmídeos
A extração de DNA plasmidial foi realizada pelo método de lise alcalina
(Birnboim e Doly, 1979) nas cepas que apresentaram multirresistência (perfil fenotípico
resistente a três ou mais classes de antimicrobianos) (Magiorakos et al., 2012). Os plasmídeos
foram submetidos à eletroforese em gel de agarose a 0,8% em tampão TBE (Tris 0,89 M;
ácido bórico 0,89 M; EDTA 0,25 M em pH 8,0) a 50 V e 100 mA durante 2 horas.
Posteriormente o gel foi visualizado e fotografado conforme o item 3.5.2. Como controles da
extração foram utilizadas as cepas de E. coli DH10B com um plasmídeo inserido (positivo),
J53 (negativo) e V517.
3.7. Classificação filogenética de E. coli
As cepas que apresentaram multirresistência foram atribuídas aos grupos
filogenéticos A, B1, B2 e D de acordo com o esquema de PCR multiplex elaborado por
Clermont et al. (2000). As reações foram realizadas de acordo com o exposto no item 3.5.2.,
utilizando 1 L de mix contendo os três pares de iniciadores nas mesmas concentrações e
programação dos ciclos de acordo com o proposto no trabalho citado. As cepas alocadas nos
grupos A e B1 são consideradas comensais e nos grupos B2 e D patogênicas.
Dados referentes aos iniciadores e a interpretação dos resultados estão expostos na
Tabela 2.
Tabela 2. Iniciadores utilizados no esquema proposto por Clermont et al. (2000) e as
combinações de resultados possíveis.
Iniciador Sequência (5’-3’) pb* Cepas
controles** Combinações/Grupos
ChuA GACGAACCAACGGTCAGGAT
TGCCGCCAGTACCAAAGACA 279 O157:H7 - - - + + +
YjaA TGAAGTGTCAGGAGACGCTG
ATGGAGAATGCGTTCCTCAAC 211 K12C600 - + - + + -
TspE4C2 GAGTAATGTCGGGGCATTCA CGCGCCAACAAAGTATTACG
152 2348/69 - - + - + +
A A B1 B2 B2 D
pb*: tamanho do produto em pares de bases; **controles positivos dos iniciadores

Material e Métodos| 38
3.8. Caracterização dos integrons por Restriction Fragment – Length Polymorphism
(RFLP) e sequenciamento
Os produtos obtidos na PCR com os primers 5’CS e 3’CS, correspondente à região
variável do integron de classe 1, foram submetidos ao ensaio de RFLP utilizando 10 U da
enzima AluI, 10 µL do produto de PCR, 2 µL do tampão da enzima e água ultrapura estéril
qsp., para o volume final de 30 µL. As reações foram incubadas a 37 °C por 1 hora e
inativadas a 65 °C por 20 minutos. Os perfis das bandas obtidas por eletroforese a 2,5% de
agarose a 100 V e 200 mA por 1 hora foram analisados com o programa de código aberto
GelJ (Heras et al., 2015).
O sequenciamento foi realizado em laboratório prestador de serviços utilizando
BigDye Terminator v.3.1 Cycle Sequencing Kit e sequenciador ABI-PRISM 3700 DNA
Analyzer (Applied Biosystems, USA) a partir dos produtos de PCR, primeiramente da região
5’CS-3’CS. Os produtos foram purificados com Kit AxyPrep PCR Clean Up®
(Axygen
Biotechnology) conforme normas do fabricante. As amostras foram preparadas e enviadas
para sequenciamento de acordo com as especificações do serviço.
Para a conclusão da sequência correspondente aos genes cassetes e da estrutura do
integron de classe 1 foram realizados novos PCRs (Tabela 3) ligando os fragmentos
correspondentes aos genes amplificados pelos iniciadores (Figura 2), de forma a combinar
diferentes pares. Os produtos das reações foram encaminhados para sequenciamento.
A montagem dos contigs foi realizada com o software Sequencher 4.7 (Gene
Codes Corporation) e as sequências foram analisadas utilizando as ferramentas BLAST e
ORF Finder (National Center for Biotecnology Information - NCBI) visando à identificação
dos cassetes gênicos relacionados à resistência antimicrobiana, a identidade dos isolados com
os depositados no banco de dados e a estrutura dos integrons presentes nos isolados.
Tabela 3. Iniciadores relacionados a estruturas de integrons de classe 1 utilizados para a
conclusão das sequências de isolados de E. coli provenientes de dípteros muscoides.
Iniciador Produto Sequência (5’-3’) T* pb** Ref.
intI1 integrase classe 1
GGGTCAAGGATCTGGATTTCG
ACATGGGTGTAAATCATCGTC 62 °C 483 Mazel et al., 2000
5’CS
3’CS
região dos cassetes
GGCATCCAAGCAGCAAG
AAGCAGACTTGACCTGA 60 °C V§
Lévesque et al.,
1995
dfrA7/dfrA17 resistência a trimetoprim
CAGAAAATGGCGTAATCG TCACCTTCAACCTCAACG
55 °C 345 Frech et al., 2003
qac resistência a quaternário de amônio
GGCTGGCTTTTTCTTGTTATCG TGAGCCCCATACCTACAAAGC
62 °C 287 Mazel et al., 2000
sul1 resistência a sulfonamidas
TGGTGACGGTGTTCGGCATTC GCGAGGGTTTCCGAGAAGGTG
63 °C 789 Sáenz et al., 2004
T*: Temperatura de anelamento; pb
**: tamanho do produto em pares de bases; Ref.: Referência;V
§: variável

Material e Métodos| 39
Figura 2. Estrutura do integron de classe 1. A região cinza representa o sítio de inserção dos
cassetes gênicos relacionados à resistência antimicrobiana. Reproduzido de Lèvesque et al.
(1995).
3.9. Perfil clonal das cepas resistentes
As cepas multirresistentes foram selecionadas para a avaliação do perfil clonal de
acordo com a sensibilidade fenotípica que apresentaram, dando preferência para os perfis
semelhantes ao das cepas portadoras de integrons. A técnica de eletroforese em campo
pulsado (PFGE) foi realizada conforme o protocolo padronizado para a rede PulseNet (Ribot
et al., 2006). Os blocos (plugs) contendo o DNA das amostras a serem testadas foram
digeridos com a enzima XbaI (NEB). Após a digestão foram transferidos para gel de agarose
para PFGE a 1% (Bio-Rad) e submetidos à eletroforese em TBE 0,5X realizada em
equipamento CHEF-DR III (Bio-Rad) a 6 V e ângulo de 120° com tempo de pulso inicial de
2,2 segundos e final de 54,2 segundos, por 19 horas. Posteriormente, o gel foi incubado em
solução de Brometo de Etídeo a 1g/mL por 30 minutos e descorados em água ultrapura por
30 minutos para a remoção do “fundo” gerado pelo Brometo de Etídeo. O gel foi fotografado
e a imagem capturada pelo sistema Image Master® VDS (Pharmacia Biotech).
As fotos dos géis foram analisadas com o programa GelComparII 5.1(Applied
Mayhs) utilizando o método Unweighted Pair Group Method Using Arithmetic Average
(UPGMA), utilizando coeficiente de Dice com tolerância e otimização de 1%.
3.10. Ensaio de conjugação
A possibilidade de transferência dos genes de resistência e dos integrons foi
avaliada através de ensaio de conjugação baseado no método apresentado por Soufi et al.
(2009). As cepas que apresentaram resultado positivo na PCR para os integrons de classe 1
foram utilizadas como doadoras no ensaio de conjugação. Para o ensaio, a cepa de E. coli J53
resistente a azida sódica foi utilizada como receptora e se mostrou sensível a tetraciclina, o
antimicrobiano escolhido para a seleção dos transconjugantes.

Material e Métodos| 40
As bactérias foram cultivadas em 2 mL de caldo LB (triptona 10%; extrato de
levedura 5%; NaCl 10% w/v) a 37 °C por 18 horas. Posteriormente, alíquotas das culturas das
cepas doadoras e receptora, foram transferidas para um mesmo tubo na razão 1:10 (v/v) e
incubadas a 37 °C por 18 horas. Após, 100 µL das culturas contendo doadora + receptora
foram semeadas em placas de Mueller Hinton contendo azida sódica (100 µg/mL), tetraciclina
(100 µg/mL) e azida sódica combinada com tetraciclina (100 µg/mL). As placas foram
incubadas a 37 °C por até 72 horas para a visualização das colônias transconjugantes.
Em seguida, uma colônia transconjugante representante de cada amostra foi
escolhida aleatoriamente e estocada por congelamento para a confirmação das marcas
transferidas, de modo que novos ensaios de sensibilidade microbiana, PCR e extração de
plasmídeos foram realizados utilizando as cepas transconjugantes. As cepas doadoras foram
utilizadas como controle na PCR e extração de plasmídeos.
3.11. Análise estatística
A associação entre a presença/ausência de cepas multirresistentes aos
antimicrobianos e a presença/ausência de integron de classe 1 e fatores de virulência foram
realizadas com teste Exato de Fisher com intervalo de confiança 95%. A ocorrência de cepas
fenotipicamente resistentes e a presença dos genes de resistência testados foram analisadas
por regressão logística múltipla. Todos os testes foram realizados utilizando o programa
BioEstat 5.3 (Instituto Mamirauá).

IV - Resultados

Resultados | 42
4.1. Coleta dos insetos e isolamento de E. coli
Na Propriedade A foram coletadas 57 moscas, das quais foram isoladas da
superfície externa, 135 cepas de E. coli (MA). Já na Propriedade B foram coletados 39
insetos, sendo que 63 cepas de E. coli foram isoladas da superfície externa (MB). No entanto
é importante destacar que as moscas coletadas em cada tentativa (varredura de puçá) foram
acondicionadas no mesmo tubo, de modo que o número de indivíduos no tubo foi variável.
Além disso, não foram todas as amostras que apresentaram crescimento de E. coli e para as
que apresentaram, um número de cinco colônias foi fixado para a continuidade do trabalho.
Por esse motivo, cálculos de frequência de isolamento por espécies de dípteros muscoides não
foram possíveis de serem realizados. Porém, efetuando-se uma média simples e considerando
os dados não agrupados, foram isoladas em média 2,4 cepas de E. coli por mosca, na
Propriedade A e 1,6 cepas de E. coli por inseto na Propriedade B. Os dados referentes a cada
amostra (espécies e/ou famílias dos insetos e isolados de E. coli) estão expostos na Tabela 4.
Os resultados referentes à identificação das espécies de dípteros muscoides estão
expostos na Tabela 5, sendo que um indivíduo da família Cicadellidae e um da família
Formicidae não foram contabilizados por não pertencerem ao grupo de dípteros muscoides.
As espécies mais abundantes na Propriedade A foram Musca domestica (Linnaeus, 1758),
Fannia spp. e Stomoxys calcitrans (Linnaeus, 1758), respectivamente. Na Propriedade B, os
dípteros mais abundantes foram M. domestica, Hippelates spp. e Acalyptratae, e Fannia spp.
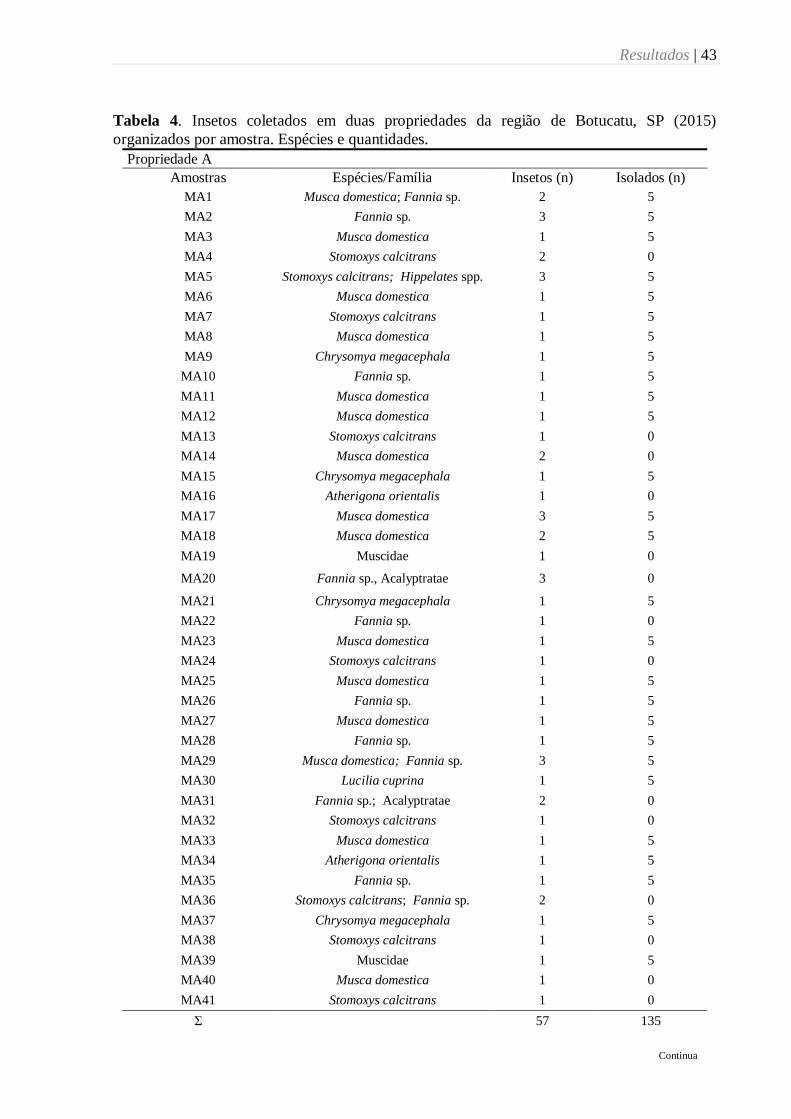
Resultados | 43
Tabela 4. Insetos coletados em duas propriedades da região de Botucatu, SP (2015)
organizados por amostra. Espécies e quantidades.
Propriedade A
Amostras Espécies/Família Insetos (n) Isolados (n)
MA1 Musca domestica; Fannia sp. 2 5
MA2 Fannia sp. 3 5
MA3 Musca domestica 1 5
MA4 Stomoxys calcitrans 2 0
MA5 Stomoxys calcitrans; Hippelates spp. 3 5
MA6 Musca domestica 1 5
MA7 Stomoxys calcitrans 1 5
MA8 Musca domestica 1 5
MA9 Chrysomya megacephala 1 5
MA10 Fannia sp. 1 5
MA11 Musca domestica 1 5
MA12 Musca domestica 1 5
MA13 Stomoxys calcitrans 1 0
MA14 Musca domestica 2 0
MA15 Chrysomya megacephala 1 5
MA16 Atherigona orientalis 1 0
MA17 Musca domestica 3 5
MA18 Musca domestica 2 5
MA19 Muscidae 1 0
MA20 Fannia sp., Acalyptratae 3 0
MA21 Chrysomya megacephala 1 5
MA22 Fannia sp. 1 0
MA23 Musca domestica 1 5
MA24 Stomoxys calcitrans 1 0
MA25 Musca domestica 1 5
MA26 Fannia sp. 1 5
MA27 Musca domestica 1 5
MA28 Fannia sp. 1 5
MA29 Musca domestica; Fannia sp. 3 5
MA30 Lucilia cuprina 1 5
MA31 Fannia sp.; Acalyptratae 2 0
MA32 Stomoxys calcitrans 1 0
MA33 Musca domestica 1 5
MA34 Atherigona orientalis 1 5
MA35 Fannia sp. 1 5
MA36 Stomoxys calcitrans; Fannia sp. 2 0
MA37 Chrysomya megacephala 1 5
MA38 Stomoxys calcitrans 1 0
MA39 Muscidae 1 5
MA40 Musca domestica 1 0
MA41 Stomoxys calcitrans 1 0
Σ
57 135
Continua

Resultados | 44
Propriedade B
Amostras Espécies/Família Insetos (n) Isolados (n)
MB1 Fannia sp. 1 5
MB2
Hippelates sp., Fannia sp.,
Drosophilidae, Acalyptratae,
Cicadellidae
12 5
MB3 Musca domestica 1 5
MB4 Faniidae 1 5
MB5 Hippelates sp. 1 0
MB6 Musca domestica 2 3
MB7 Stomoxys calcitrans 1 5
MB8 Musca domestica 1 5
MB9 Musca domestica 1 5
MB10 Musca domestica 1 5
MB11 Musca domestica 1 5
MB12 Stomoxys calcitrans 1 0
MB13 Musca domestica; Stomoxys
calcitrans; Acalyptratae 5 5
MB14 Hippelates sp.; Formicidae 4 5
MB15 Musca domestica 1 0
MB16 Toxomerus sp. 1 0
MB17 Sepsidae; Acalyptratae 2 5
MB18 Musca domestica; Euxesta sp. 2 0
Σ 39 63
n: quantidade; Σ: somatório; MA: moscas coletadas na Propriedade A; MB: moscas coletadas na Propriedade B.
Tabela 5. Espécies e abundância de espécies de dípteros muscoides coletados em duas
propriedades leiteiras da região de Botucatu, SP (2015).
Propriedade A
Família Espécie Abundância
Muscidae Musca domestica (Linnaeus, 1758) 20
Stomoxys calcitrans (Linnaeus, 1758) 10
Atherigona orientalis (Schiner) 2
Não identificado 2
Fanniidae Fannia spp. 14
Calliphoridae Chrysomya megacephala (Fabricius, 1794) 4
Lucilia cuprina (Wiedemann, 1830) 1
Chloropidae Hippelates spp. 2
Acalyptratae ----- 2
Continua
Continuação Tabela 4
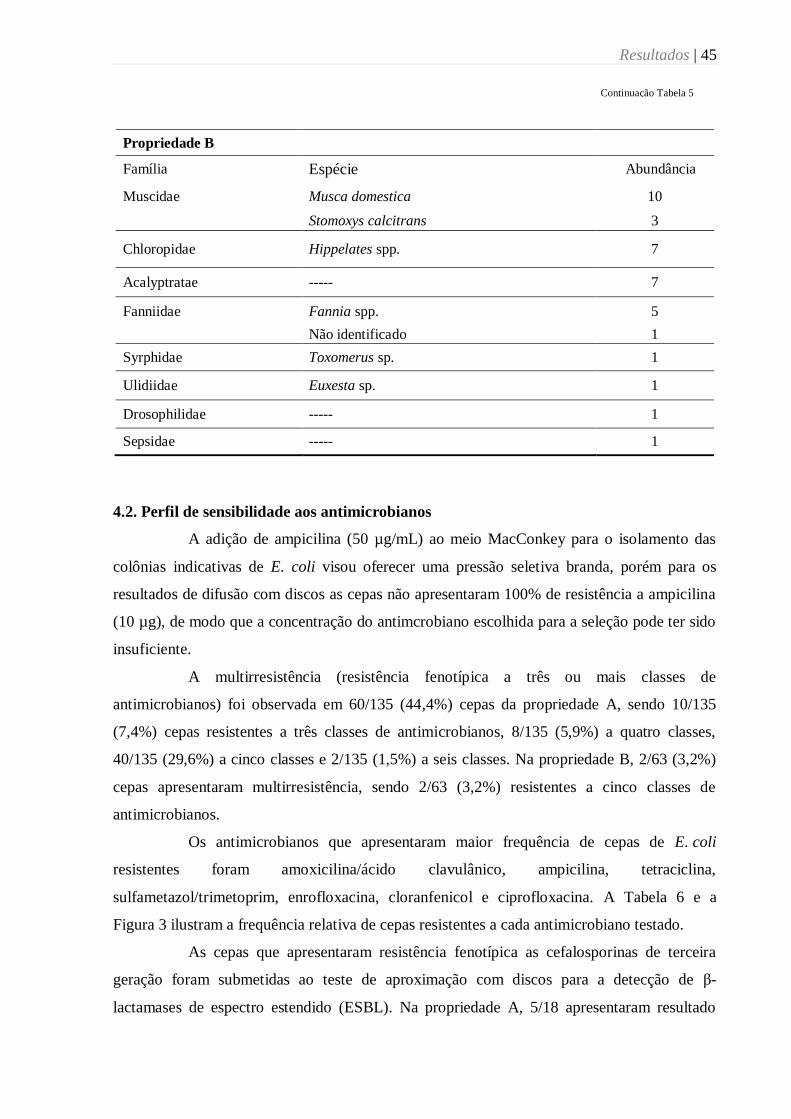
Resultados | 45
Propriedade B
Família Espécie Abundância
Muscidae Musca domestica 10
Stomoxys calcitrans 3
Chloropidae Hippelates spp. 7
Acalyptratae ----- 7
Fanniidae Fannia spp. 5
Não identificado 1
Syrphidae Toxomerus sp. 1
Ulidiidae Euxesta sp. 1
Drosophilidae ----- 1
Sepsidae ----- 1
4.2. Perfil de sensibilidade aos antimicrobianos
A adição de ampicilina (50 µg/mL) ao meio MacConkey para o isolamento das
colônias indicativas de E. coli visou oferecer uma pressão seletiva branda, porém para os
resultados de difusão com discos as cepas não apresentaram 100% de resistência a ampicilina
(10 µg), de modo que a concentração do antimcrobiano escolhida para a seleção pode ter sido
insuficiente.
A multirresistência (resistência fenotípica a três ou mais classes de
antimicrobianos) foi observada em 60/135 (44,4%) cepas da propriedade A, sendo 10/135
(7,4%) cepas resistentes a três classes de antimicrobianos, 8/135 (5,9%) a quatro classes,
40/135 (29,6%) a cinco classes e 2/135 (1,5%) a seis classes. Na propriedade B, 2/63 (3,2%)
cepas apresentaram multirresistência, sendo 2/63 (3,2%) resistentes a cinco classes de
antimicrobianos.
Os antimicrobianos que apresentaram maior frequência de cepas de E. coli
resistentes foram amoxicilina/ácido clavulânico, ampicilina, tetraciclina,
sulfametazol/trimetoprim, enrofloxacina, cloranfenicol e ciprofloxacina. A Tabela 6 e a
Figura 3 ilustram a frequência relativa de cepas resistentes a cada antimicrobiano testado.
As cepas que apresentaram resistência fenotípica as cefalosporinas de terceira
geração foram submetidas ao teste de aproximação com discos para a detecção de β-
lactamases de espectro estendido (ESBL). Na propriedade A, 5/18 apresentaram resultado
Continuação Tabela 5

Resultados | 46
positivo para os antimicrobianos ceftazidima e aztreonam (Figura 4). Apesar de algumas
cepas de E. coli terem apresentado resultado sugestivo de produção de ESBL para
cefotaxima, tais resultados foram considerados negativos, uma vez que o método utilizado já
emprega distâncias reduzidas de 20 mm entre os discos de amoxicilina/ácido clavulânico e as
cefalosporinas de terceira geração e aztreonam (Bradford, 2001; Rawat e Nair, 2010)
(Figura 4; Apêndice A). Uma cepa da propriedade B foi resistente a cefoperazona, porém não
apresentou resultados positivos para a produção de ESBL.
4.3. Genes relacionados à resistência a antimicrobianos e fatores de virulência
Os genes relacionados à resistência a antimicrobianos foram selecionados para
estudo considerando a frequência com que são abordados na literatura ou a uma marca
específica almejada, no caso do gene blaCMY-4, relacionado à carbapenemase em E. coli
(Stapleton et al., 1999).
Os isolados da Propriedade A apresentaram genotipos positivos para os seguintes
genes e região gênica: intI1 (10/135), 5’CS-3’CS (25/135), sendo integron de classe 1 (intI +
5’CS-3’CS) presente em 8/135, ampC (6/135), blaTEM (49/135), blaCTX-M (15/135), parC
(38/135), gyrA (67/135), tetA (20/135). No total, 89/135 (66%) cepas foram positivas a pelo
menos um gene relacionado à resistência. Entre os fatores de virulência pesquisados houve
genótipo positivo para eae (4/135), vt1 (7/135), hlyA (11/135), K99 (5/135), LTII (13/135),
STa (5/135). Todas as sete E. coli positivas para vt1 foram isoladas de M. domestica.
Quanto à Propriedade B, não foram encontradas cepas positivas para o gene da
integrase (intI), no entanto, 5/63 cepas amplificaram a região 5’CS-3’CS. Houve genótipo
positivo para ampC (5/63), parC (17/63) e gyrA (7/63), sendo que 19/63 (30,2%) foram
positivas a pelo menos um gene relacionado à resistência. Não houve genótipo positivo a
nenhum dos fatores de virulência pesquisados.
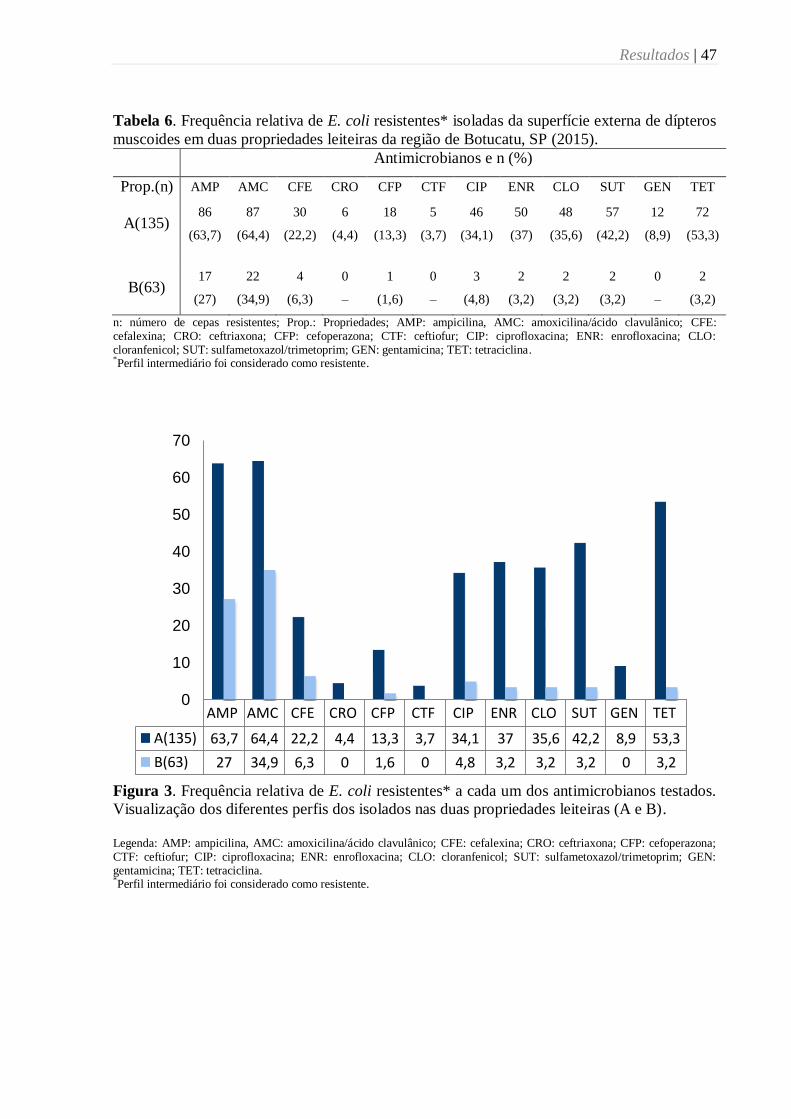
Resultados | 47
Tabela 6. Frequência relativa de E. coli resistentes* isoladas da superfície externa de dípteros
muscoides em duas propriedades leiteiras da região de Botucatu, SP (2015).
Antimicrobianos e n (%)
Prop.(n) AMP AMC CFE CRO CFP CTF CIP ENR CLO SUT GEN TET
A(135) 86
(63,7)
87
(64,4)
30
(22,2)
6
(4,4)
18
(13,3)
5
(3,7)
46
(34,1)
50
(37)
48
(35,6)
57
(42,2)
12
(8,9)
72
(53,3)
B(63) 17
(27)
22
(34,9)
4
(6,3)
0
–
1
(1,6)
0
–
3
(4,8)
2
(3,2)
2
(3,2)
2
(3,2)
0
–
2
(3,2)
n: número de cepas resistentes; Prop.: Propriedades; AMP: ampicilina, AMC: amoxicilina/ácido clavulânico; CFE: cefalexina; CRO: ceftriaxona; CFP: cefoperazona; CTF: ceftiofur; CIP: ciprofloxacina; ENR: enrofloxacina; CLO: cloranfenicol; SUT: sulfametoxazol/trimetoprim; GEN: gentamicina; TET: tetraciclina. *Perfil intermediário foi considerado como resistente.
Figura 3. Frequência relativa de E. coli resistentes* a cada um dos antimicrobianos testados.
Visualização dos diferentes perfis dos isolados nas duas propriedades leiteiras (A e B).
Legenda: AMP: ampicilina, AMC: amoxicilina/ácido clavulânico; CFE: cefalexina; CRO: ceftriaxona; CFP: cefoperazona; CTF: ceftiofur; CIP: ciprofloxacina; ENR: enrofloxacina; CLO: cloranfenicol; SUT: sulfametoxazol/trimetoprim; GEN: gentamicina; TET: tetraciclina. *Perfil intermediário foi considerado como resistente.
AMP AMC CFE CRO CFP CTF CIP ENR CLO SUT GEN TET
A(135) 63,7 64,4 22,2 4,4 13,3 3,7 34,1 37 35,6 42,2 8,9 53,3
B(63) 27 34,9 6,3 0 1,6 0 4,8 3,2 3,2 3,2 0 3,2
0
10
20
30
40
50
60
70

Resultados | 48
Figura 4. Escherichia coli positiva para o teste de aproximação de disco. O alargamento do
halo de inibição (seta no topo) e o surgimento da “zona fantasma” (seta embaixo) indica a
produção de ESBL (β-lactamase de espectro estendido).
4.4. Análises estatísticas
Os testes exato de Fisher e regressão logística múltipla foram realizados para a
análise dos dados obtidos com as cepas de E. coli isoladas de dípteros muscoides capturados
na propriedade A. Considerando as 135 cepas e o valor de p<0,05 como estatisticamente
significativo, os resultados demonstraram associação entre multirresistência e a presença de
integron (p=0,0012) e multirresistência e a presença do gene para o fator de virulência K99
(p=0,0158). Porém a presença do gene vt1 foi associada à ausência de multirresistência
(p=0,0171). Os fatores de virulência eae, hlyA, LTII e STa não apresentaram nenhuma
associação (p>0,05).
A regressão logística múltipla foi efetuada a fim de verificar a presença dos genes
de resistência antimicrobiana em relação à resistência fenotípica a cada um dos
antimicrobianos testados, de modo que a resistência genotípica consistiu na variável
dependente (Y) e a resistência fenotípica às variáveis independentes (X). Os genes testados
foram blaTEM, blaCTX-M, ampC e tetA, sendo que houve relação significativa entre blaTEM e
resistência a ampicilina (p=0,0242), cloranfenicol (p=0,0179) e tetraciclina (p= 0,0056). O
gene blaCTX-M apresentou relação significativa com resistência a cefoperazona sódica
(p=0,0458). Os genes ampC e tetA não apresentaram relação com a resistência a nenhum dos
antimicrobianos testados.

Resultados | 49
4.5. Plasmídeos e classificação filogenética de E. coli (Clermont et al., 2000)
Os isolados que apresentaram multirresistência (60 da Propriedade A e dois da
Propriedade B descritos no item 4.2) foram submetidos à extração de plasmídeos e a
classificação filogenética. No total, 57/62 (91,9%) cepas de E. coli apresentaram plasmídeo
visível nas condições estabelecidas pela técnica de lise alcalina, 45/62 (72,6%) das cepas
foram classificadas no grupo filogenético B1, do restante, 9/62 (14,5%) foram alocadas no
grupo A, 6/62 (9,7%) no grupo D, 1/62 (1,6%) em B2 e 1/62 (1,6%) não foi possível de ser
tipada pelo método. Detalhes sobre a resistência de cada isolado, presença de plasmídeo e
grupo filogenético estão apresentados na Tabela 7.
Tabela 7. Dados referentes ao perfil de resistência*, presença de plasmídeos e grupos
filogenéticos das cepas de E. coli multirresistentes isoladas de dípteros muscoides em
propriedades leiteiras da região de Botucatu, SP (2015).
Isolado Perfil de resistência Presença de
plasmídeo**
Grupo
filogenético
MA1c AMP/AMC/CFE/ENR/SUT/TET Sim B1
MA1d AMP/CIP/ENR/SUT/TET Sim A
MA2a AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET Sim A
MA2b AMP/CFE/CIP/ENR/CLO/SUT/TET Sim B1
MA2c AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET Sim B1
MA2d AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET Sim B1
MA2e AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA3a AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET Sim B1
MA3b AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA3c AMP/AMC/CFE/CIP/ENR/CLO/SUT/GEN/TET Sim NT
MA3e AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/GEN/TET Sim B1
MA5a AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA5b AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA5c AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA5d AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA5e AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET Sim B1
MA6a AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET Sim B1
MA6b AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA6d AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA7c AMP/AMC/CFE/SUT/TET Sim B1
MA7e AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET Sim D
MA8b AMP/AMC/CIP/ENR/CLO/SUT/TET Sim A
MA9a AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA9b AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA9c AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA9d AMP/AMC/CIP/ENR/CLO/SUT/TET Sim A
Continua.
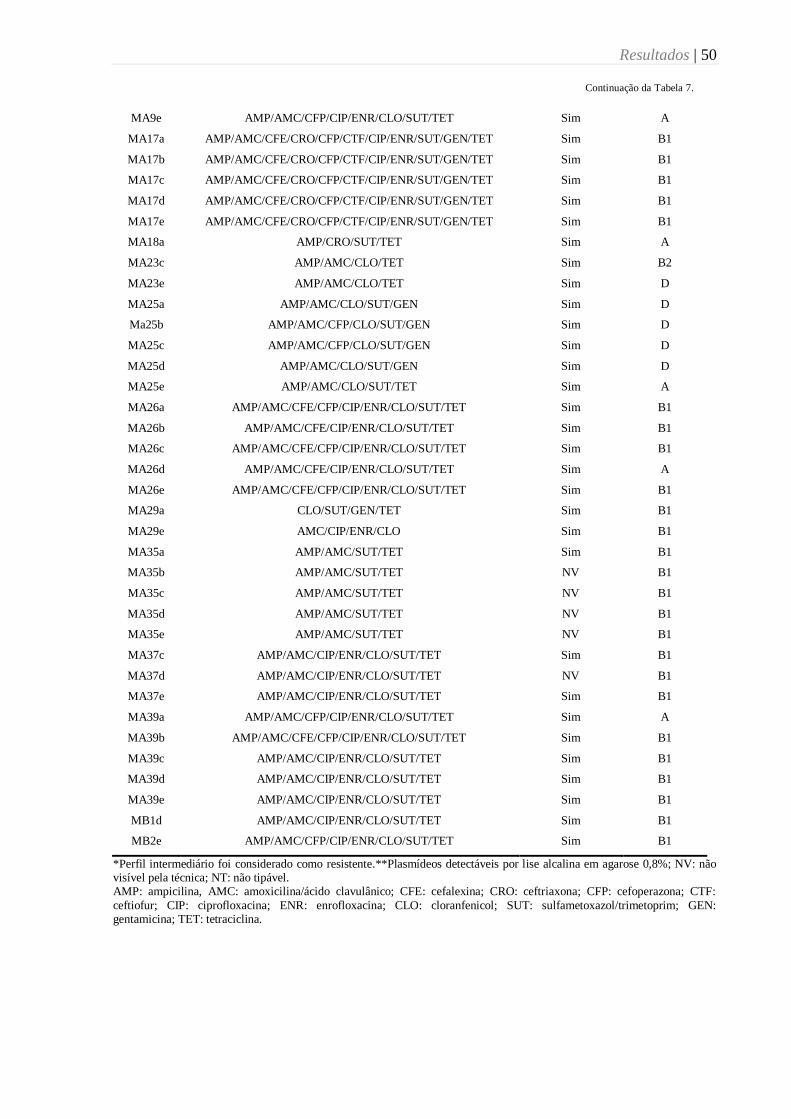
Resultados | 50
MA9e AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET Sim A
MA17a AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET Sim B1
MA17b AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET Sim B1
MA17c AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET Sim B1
MA17d AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET Sim B1
MA17e AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET Sim B1
MA18a AMP/CRO/SUT/TET Sim A
MA23c AMP/AMC/CLO/TET Sim B2
MA23e AMP/AMC/CLO/TET Sim D
MA25a AMP/AMC/CLO/SUT/GEN Sim D
Ma25b AMP/AMC/CFP/CLO/SUT/GEN Sim D
MA25c AMP/AMC/CFP/CLO/SUT/GEN Sim D
MA25d AMP/AMC/CLO/SUT/GEN Sim D
MA25e AMP/AMC/CLO/SUT/TET Sim A
MA26a AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET Sim B1
MA26b AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET Sim B1
MA26c AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET Sim B1
MA26d AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET Sim A
MA26e AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET Sim B1
MA29a CLO/SUT/GEN/TET Sim B1
MA29e AMC/CIP/ENR/CLO Sim B1
MA35a AMP/AMC/SUT/TET Sim B1
MA35b AMP/AMC/SUT/TET NV B1
MA35c AMP/AMC/SUT/TET NV B1
MA35d AMP/AMC/SUT/TET NV B1
MA35e AMP/AMC/SUT/TET NV B1
MA37c AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA37d AMP/AMC/CIP/ENR/CLO/SUT/TET NV B1
MA37e AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA39a AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET Sim A
MA39b AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET Sim B1
MA39c AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA39d AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MA39e AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MB1d AMP/AMC/CIP/ENR/CLO/SUT/TET Sim B1
MB2e AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET Sim B1
*Perfil intermediário foi considerado como resistente.**Plasmídeos detectáveis por lise alcalina em agarose 0,8%; NV: não visível pela técnica; NT: não tipável. AMP: ampicilina, AMC: amoxicilina/ácido clavulânico; CFE: cefalexina; CRO: ceftriaxona; CFP: cefoperazona; CTF:
ceftiofur; CIP: ciprofloxacina; ENR: enrofloxacina; CLO: cloranfenicol; SUT: sulfametoxazol/trimetoprim; GEN: gentamicina; TET: tetraciclina.
Continuação da Tabela 7.

Resultados | 51
4.6. Caracterização dos integrons por Restriction Fragment – Length Polymorphism
(RFLP)
Oito cepas de E. coli da Propriedade A foram positivas para integron de classe 1
(positivas na PCR para o gene da integrase de classe 1 intI1 e para a região variável 5’CS-
3’CS). O produto da PCR para a região 5’CS-3’CS apresentou um tamanho aproximado de
750 pb e foi submetido à digestão enzimática por AluI com o objetivo de caracterizar a região
variável onde são inseridos os cassetes gênicos no integron e, assim, auxiliar nas análises do
sequenciamento dessas mesmas amostras. As bandas geradas pela digestão de cada um dos
fragmentos apresentaram tamanhos aproximados a 750 pb, 450 pb e 300 pb (Figura 5).
O fragmento de 750 pb corresponde ao produto de PCR que não foi completamente digerido
pela enzima.
Figura 5. Padrão de digestão enzimática de AluI obtido no ensaio de RFLP da região variável
do integron de classe 1 (5’CS-3’CS) de E. coli isoladas de moscas. M: Marcador de peso molecular 1 kb (Fermentas); 1: MA17a; 2: MA17b; 3: MA17c; 4: MA17d; 5: MA17e; 6:
MA26a; 7: MA26b; 8: MA26d.
4.7. Sequenciamento da região variável do integron de classe 1
A amplificação da região 5’CS-3’CS, para as oito amostras de E. coli positivas
oriundas das moscas coletadas na propriedade A, gerou produtos de tamanhos aproximados a
750 pb. Visando primeiramente obter as sequências iniciais da região 5’CS e 3’CS, conforme
o ilustrado no esquema apresentado por Lèvesque et al. (1995) (Figura 2), os produtos das
PCRs foram enviados para sequenciamento. As análises das sequências obtidas demonstraram
a presença de um único gene cassete com identidade de 100% com o gene dfrA7 –
diidrofolato redutase tipo VII que confere resistência ao antimicrobiano trimetoprim em todas
as oito amostras. A Figura 6 ilustra o esquema elaborado inicialmente.

Resultados | 52
Figura 6. Desenho esquemático da sequência obtida a partir da amplificação dos iniciadores
5’CS e 3’CS. 59-be: 59- base element (sítio de recombinação do gene cassete)
Posteriormente, novos PCRs foram realizados utilizando os iniciadores dos genes
estruturais do integron de classe 1 (intI1, 5’CS-3’CS, qac, sul1) e do gene cassete
correspondente (dfrA7) de maneira combinada, buscando propor a estrutura completa dos
integrons presentes nas amostras.
Os produtos sequenciados corresponderam às combinações intI1R/dfrA7R,
5’CS/dfrA7R, 5’CS/qacR, dfrA7F/qacR, 3’CS/dfrA7F e intIF/intIR.
A análise dos contigs resultou em uma sequência consenso única para as oito
amostras, contendo a integrase de classe 1 com aproximadamente 1200 pb. O gene dfrA7
apresentou tamanho de 564 pb com start códon alternativo “GTG” e stop códon “TAA”
compartilhado com o sítio de recombinação 59-base element (59-be) também conhecido como
attC, apresentando 143 pb (Figura 7).
Figura 7. Desenho esquemático do integron de classe 1 presente em cepas de E. coli isoladas
de dípteros muscoides.

Resultados | 53
4.8. Perfil clonal
Para a avaliação do perfil clonal, os isolados de E. coli foram organizados em seis
grupos de acordo com o perfil de resistência fenotípico que apresentaram (Tabela 8). No total
foram analisadas 43 E. coli multirresistentes com perfil igual ou semelhante às cepas positivas
para integron ou àqueles obtidos para as duas cepas da Propriedade B. Foram gerados 36
pulsotipos, a partir dos perfis de macrorrestrição do DNA genômico, analisados segundo os
critérios estabelecidos por Tenover et al. (1995), representados na Figura 8 pelos números 1
a 36.
As cepas que apresentaram 100% de similaridade genômica foram: MA9a e
MA37c (pulsotipo 3), MA3a e MA2e (pulsotipo 12), MA2c e MA3b (pulsotipo 13), MB2e,
MA39b e MA39d (pulsotipo 24) e MA17a e MA17c (pulsotipo 34). A cepa MB1d apresentou
94% de similaridade com as cepas MA6a e MA2d.
Quanto as cepas com integrons MA17a, MA17c, MA17d e MA17e, formaram um
cluster com similaridade maior que 94%. O pulsotipo de MA26a com MA2a também
apresentaram 94% de similaridade e MA26b com MA5a, 98%. A cepa MA26d apresentou
80% de similaridade com as outras cepas de MA26, e a cepa MA17b, não foi relacionada a
nenhum dos outros pulsotipos obtidos.
Tabela 8. Cepas de E. coli selecionadas para a avaliação do perfil clonal organizadas de
acordo com o perfil fenotípico de resistência antimicrobiana.
Cepas Resistência* Genes de resistência
Grupo I
MA7e AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET blaTEM / ampC
MA2d AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET blaTEM / tetA
MA26aint AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET blaTEM / blaCTX-M / ampC / tetA
MA26c AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET blaTEM / blaCTX-M / ampC /tetA
MA26e AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET ampC/ tetA
MA39b AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET blaCTX-M
Grupo II
MA17aint AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET blaTEM / blaCTX-M
MA17bint AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET blaTEM / blaCTX-M
MA17cint AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET blaCTX-M
MA17dint AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET blaTEM / blaCTX-M
MA17eint AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET blaCTX-M
Continua.

Resultados | 54
Grupo III
MA26bint AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET blaCTX-M / ampC / tetA
MA26dint AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET ampC / tetA
MA2a AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET blaTEM / tetA
MA2c AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET blaTEM / tetA
MA3a AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET blaTEM
MA5e AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET blaTEM
Grupo IV
MB1d AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA2e AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM / tetA
MA3b AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA5a AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM
MA5b AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM
MA5c AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM
MA5d AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM
MA6b AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM
MA6d AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM
MA8b AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA9a AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM / tetA
MA9b AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM / tetA
MA9c AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM / blaCTX-M / tetA
MA9d AMP/AMC/CIP/ENR/CLO/SUT/TET blaTEM / tetA
MA37c AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA37d AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA37e AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA39c AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA39d AMP/AMC/CIP/ENR/CLO/SUT/TET N
MA39e AMP/AMC/CIP/ENR/CLO/SUT/TET N
Grupo V
MB2e AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET N
MA39a AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET blaCTX-M
MA9e AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET blaTEM / blaCTX-M / tetA
MA6a AMP/AMC/CFP/CIP/ENR/CLO/SUT/TET blaTEM
Grupo VI
MA3e AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/GEN/TET blaTEM
MA3c AMP/AMC/CFE/CIP/ENR/CLO/SUT/GEN/TET N
*Perfil intermediário foi considerado como resistente. AMP: ampicilina, AMC: amoxicilina/ácido clavulânico; CFE: cefalexina; CRO: ceftriaxona; CFP: cefoperazona; CTF: ceftiofur; CIP: ciprofloxacina; ENR: enrofloxacina; CLO: cloranfenicol; SUT: sulfametoxazol/trimetoprim; GEN: gentamicina; TET: tetraciclina. N: negativo. int: cepas com integron.
Continuação Tabela 8
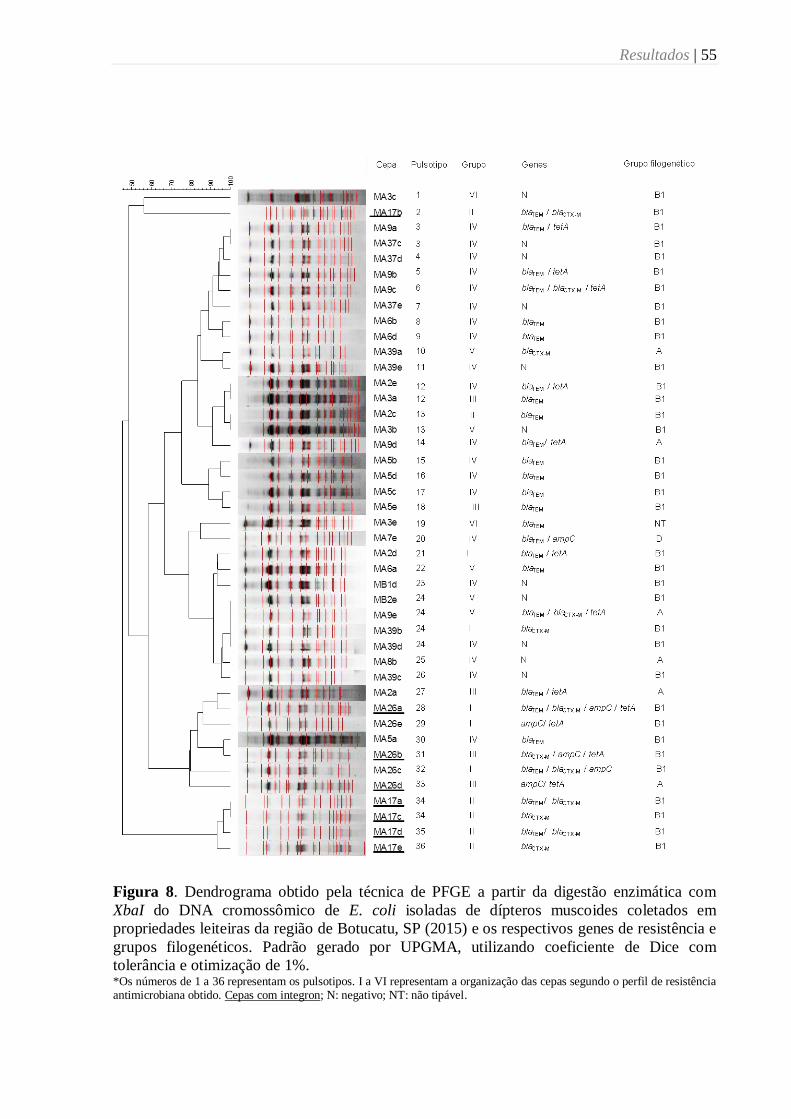
Resultados | 55
Figura 8. Dendrograma obtido pela técnica de PFGE a partir da digestão enzimática com
XbaI do DNA cromossômico de E. coli isoladas de dípteros muscoides coletados em
propriedades leiteiras da região de Botucatu, SP (2015) e os respectivos genes de resistência e
grupos filogenéticos. Padrão gerado por UPGMA, utilizando coeficiente de Dice com
tolerância e otimização de 1%. *Os números de 1 a 36 representam os pulsotipos. I a VI representam a organização das cepas segundo o perfil de resistência antimicrobiana obtido. Cepas com integron; N: negativo; NT: não tipável.

Resultados | 56
4.9. Conjugação
As oito cepas de E. coli que apresentaram integrons foram selecionadas como
doadoras para o ensaio de conjugação in vitro, uma vez que um dos objetivos propostos foi
verificar a transferência desses elementos genéticos, assim como dos genes de resistência a
antimicrobianos presentes nessas cepas.
Dada a possibilidade da localização cromossomal dos integrons, os
antimicrobianos estruturalmente relacionados a esses elementos, como as sulfonamidas, não
puderam ser utilizados na seleção das cepas transconjugantes obtidas após a conjugação.
Portanto, o antimicrobiano selecionado para esse fim foi a tetraciclina, uma vez que é
amplamente administrada em animais de produção e a resistência em cepas bacterianas é bem
documentada (Marshall e Levy, 2011). Além disso, alguns trabalhos já descreveram relação
entre genes de resistência a tetraciclinas e integrons de classe 1, uma vez que ambos podem
estar localizados no mesmo elemento móvel (Agersø e Sandvang, 2005).
A fim de confirmar a transferência dos genes relacionados à resistência
antimicrobiana, os testes fenotípicos foram repetidos para as cepas doadoras e
transconjugantes, assim como as PCRs utilizando DNA total de ambas. Os resultados
referentes às cepas doadoras e transconjugantes estão apresentados na Tabela 9.
Adicionalmente, os plasmídeos das cepas transconjugantes e doadoras foram purificados pela
técnica da lise alcalina (Figura 9) e as PCRs, anteriormente realizadas com DNA total, foram
repetidas utilizando o DNA plasmidial extraído pela técnica (Tabela 10). Os plasmídeos
extraídos das cepas transconjugantes não puderam ser nitidamente visualizados nas condições
estabelecidas pela técnica (Figura 9).
É possível verificar que a resistência fenotípica aos antimicrobianos testados
apresenta genes ligados a elementos conjugativos, uma vez que as colônias transconjugantes
adquiriram perfis resistentes. Apesar de o perfil de resistência ter se mantido nas
transconjugantes de MA17a, MA17b, MA17c, MA17d e MA17e, estas apresentaram as
medidas dos halos de inibição diferentes da doadora (exceto MA17e), significando perfis
intermediários de resistência, mas que foram considerados no trabalho todo, para
contabilização, como perfis resistentes.
Já as cepas MA26a, MA26b e MA26d apresentaram resultados divergentes entre
as cepas doadoras e suas respectivas transconjugantes quanto a produção de ESBL (Tabela 9).
Esses resultados são complementados com as imagens dos testes de disco aproximação
presentes no Apêndice A.

Resultados | 57
A cepa doadora MA26b não foi contabilizada no item 4.2 pois não foi resistente a
cefalosporinas de terceira geração, porém apresentou resultado positvo para a produção de
ESBL em aztreonam (Tabela 9; Apêndice A) .
Os resultados expostos nas Tabelas 9 não incluem o gene ampC, pois a cepa
receptora E. coli J53, utilizada no ensaio de conjugação, apresentou amplificação desse gene
na PCR, o que pode ser devido a origem constitutiva de ampC em E. coli (Peter-Getzlaff et
al., 2011), apesar de as cepas de E. coli analisadas no presente estudo terem apresentado
baixas frequências de detecção (Propriedade A, 6/135 e Propriedade B, 5/63).
Interessantemente, o gene tetA foi amplificado apenas nas cepas transconjugantes
de MA17b e MA17d, e a região variável 5’CS-3’CS não foi dectada na cepa MA26a doadora,
apesar desta ser comprovadamente uma das cepas positivas para integron de classe 1.
Figura 9. Eletroforese em gel de agarose 0,8% dos plasmídeos extraídos pelo método de lise
alcalina, correspondentes às cepas de E. coli transconjugantes (A) e doadoras (B). 1: E. coli V517; 2: MA17a; 3: MA17b; 4: MA17c; 5: MA17d; 6: MA17e; 7: MA26a; 8: MA26b; 9: MA26d; 10:
E. coli J53; 11: MA17a; 12: MA17b; 13: MA17c; 14: MA17d; 15: MA17e; 16: MA26a; 17: MA26b; 18:
MA26d.
A B

Resultados | 58
Tabela 9. Perfil de resistência fenotípica* e genotípica das cepas E. coli doadoras e transconjugantes(t)
obtidas no ensaio de conjugação.
Cepa Resistência fenotípica Genes de resistência
DD** DA§
(ESBL)
intI 5CS-3CS dfrA7 qac sul1 blaTEM blaCTX-M tetA
DNA total
MA17a AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET CAZ/ATM + + + + + - + +
MA17at AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET ATM + + + + + - + +
MA17b AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET CAZ/ATM + + + + + + + -
MA17bt AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET ATM + + + + + - + +
MA17c AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET CAZ/ATM + + + + + + + +
MA17ct AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET ATM + + + + + - + +
MA17d AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET CAZ/ATM + + + + + + + -
MA17dt AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET ATM + + + + + - + +
MA17e AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET CAZ/ATM + + + + + - + +
MA17et AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/SUT/GEN/TET ATM + + + + + - + +
MA26a
AMP/AMC/CFE/CFP/CIP/ENR/CLO/SUT/TET N + - + + + + + +
MA26at AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/CLO/SUT/TET ATM/CTX + + + + + + + +
MA26b AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET ATM + + + + + + + +
MA26bt AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/CLO/SUT/TET CAZ + + + + + + + +
MA26d AMP/AMC/CFE/CIP/ENR/CLO/SUT/TET N + + + + + + + +
MA26dt AMP/AMC/CFE/CRO/CFP/CTF/CIP/ENR/CLO/SUT/TET N + + + + + + + +
J53 Sensível a todos os antimicrobianos testados N - - - - - - - -
*Perfil intermediário foi considerado como resistente.**DD: disco difusão/antibiograma; § DA:disco aproximação; ESBL: β-lactamase de espectro estendido; N: negativo; AMP: ampicilina,
AMC: amoxicilina/ácido clavulânico; CFE: cefalexina; CRO: ceftriaxona; CFP: cefoperazona; CTF: ceftiofur; CIP: ciprofloxacina; ENR: enrofloxacina; CLO: cloranfenicol; SUT: sulfametoxazol/trimetoprim; GEN: gentamicina; TET: tetraciclina; CAZ: ceftazidima; ATM: aztreonam; CTX: Cefotaxima; SUT: presença de colônias hipermutantes no interior do halo de 21mm de diâmetro.
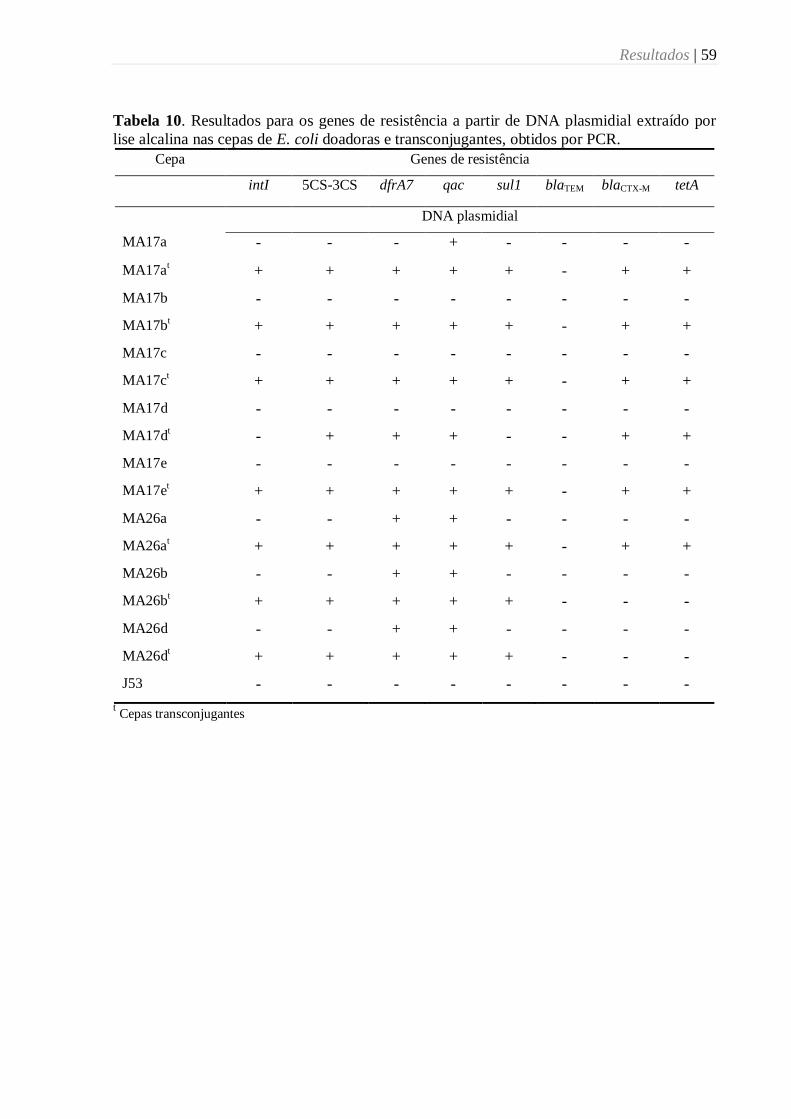
Resultados | 59
Tabela 10. Resultados para os genes de resistência a partir de DNA plasmidial extraído por
lise alcalina nas cepas de E. coli doadoras e transconjugantes, obtidos por PCR.
Cepa Genes de resistência
intI 5CS-3CS dfrA7 qac sul1 blaTEM blaCTX-M tetA
DNA plasmidial
MA17a - - - + - - - -
MA17at + + + + + - + +
MA17b - - - - - - - -
MA17bt + + + + + - + +
MA17c - - - - - - - -
MA17ct + + + + + - + +
MA17d - - - - - - - -
MA17dt - + + + - - + +
MA17e - - - - - - - -
MA17et + + + + + - + +
MA26a
- - + + - - - -
MA26at + + + + + - + +
MA26b - - + + - - - -
MA26bt + + + + + - - -
MA26d - - + + - - - -
MA26dt + + + + + - - -
J53 - - - - - - - -
t Cepas transconjugantes

V - Discussão

Discussão | 61
O ambiente é considerado fonte de resistência antimicrobiana, uma vez que os
micro-organismos presentes nas comunidades bióticas do solo e água naturalmente
apresentam genes de resistência devido à produção de antibióticos por outras bactérias e
fungos, além da constante pressão seletiva a qual estão submetidos devido a resíduos de
antimicrobianos depositados no ambiente, advindos da utilização na medicina humana,
veterinária e na agricultura (Allen et al., 2010; Costa et al., 2013).
Nesse sentido, vários autores têm proposto esquemas que buscam conectar as
fontes de resistência antimicrobianas não naturais às vias de disseminação no ambiente
(Khachatourians, 1998; Marshall, 2009; Cantas et al., 2013; Costa et al., 2013; Berglund,
2015).
Considerando que nenhum dos trabalhos consultados inclui a veiculação de
bactérias multirresistentes por moscas nas rotas de disseminação ambiental de resistência
antimicrobiana, este estudo buscou caracterizar a resistência de E. coli isoladas de moscas em
ambiente de ordenha, a fim de propor que bactérias constituintes da microbiota de animais
eliminadas em excrementos podem ser transportadas por moscas, fato ainda pouco discutido
na literatura (Marshall et al., 1990; Rahuma et al., 2005; Macovei e Zurek, 2006; Rybaríková
et al., 2010; Usui et al., 2013; Wang, 2013; Blaak et al., 2014; Zurek e Ghosh, 2014; Solà-
Ginés et al., 2015; Usui et al., 2015).
Entre as espécies de dípteros muscoides coletadas, Musca domestica foi a espécie
mais abundante nas duas propriedades estudadas, o que já era esperado devido a elevada
sinantropia que apresenta (Povolný, 1971).
A maioria das cepas de E. coli multirresistentes (68%) isoladas da superfície
externa de cada grupo de moscas, apresentaram resistência a cinco classes de antimicrobianos
– β-lactâmicos, quinolonas, fenicois, inibidores de folato e tetraciclinas ou β-lactâmicos,
quinolonas, inibidores de folato, aminoglicosídeo e tetraciclina. Isso aliado à produção de
enzimas β-lactamases de espectro estendido (ESBLs), detectada em um grupo de cepas a
partir da sinergia entre amoxicilina/ácido clavulânico e ceftazima e aztreonam, indica que tais
cepas estão expostas a pressões seletivas geradas pelos principais antimicrobianos utilizados
na medicina veterinária e humana.
Nesse contexto, destaca-se a resistência a quinolonas, uma vez que a resistência
selecionada para um antimicrobiano do grupo implica em resistência a todos, pois apresentam
o mesmo mecanismo de ação, de modo que a administração não terapêutica em animais é
considerada ilegal em muitos países (Webber e Piddock, 2001), além de serem amplamente
usadas no tratamento de infecções bacterianas na medicina humana (Lastours et al., 2014).

Discussão | 62
Porém, estudos conduzidos na Suécia indicam que a existência de E. coli resistente a
quinolonas parece ser comum em amostras fecais de bovinos, principalmente bezerros (Duse
et al., 2016).
Nesse contexto, Rybaríková et al. (2010), discutiram sobre a possibilidade de
disseminação de cepas oriundas dos bezerros no ambiente, e Costa et al. (2013) alertaram
quanto ao desenvolvimento de resistência cruzada para os antimicrobianos usados na
veterinária, agricultura e tratamento humano, possívelmente devido a similaridade química
que possuem. Segundo Costa et al. (2013), a resistência antimicrobiana encontrada em
bactérias comensais, prediz a resistência em patógenos.
Desse modo, os resultados deste trabalho podem contribuir para confirmação da
hipótese apresentada por Solà-Ginés et al. (2015) que bactérias provenientes de moscas
podem refletir as condições da microbiota dos animais.
Os isolados das Propriedades A e B diferiram quanto à frequência relativa de
resistência fenotípica e genotípica, uma vez que os resultados obtidos para a Propriedade A
foram maiores em relação à Propriedade B.
Em contrapartida, quanto aos genes cromossomais gyrA, parC e ampC era
esperado que todas as cepas de E. coli testadas apresentassem amplificação na PCR, já que
correspondem a sequências de genes constitutivos bacterianos (gyrA e parC) (Jacoby, 2005)
e de E. coli (ampC) (Peter-Getzlaff et al., 2011). No entanto, das 198 cepas isoladas da
Propriedade A e B, 124 foram negativas para gyrA, 143 para parC e 187 para ampC. Tais
resultados podem significar diferenças nas enzimas em relação às de referência e devem ser
estudadas em maiores detalhes.
Quanto aos fatores de virulência investigados, todas as sete amostras de E. coli
positivas para verotoxina (vt1) detectadas foram isoladas de M. domestica, analogamente aos
estudos de Moriya et al. (1999), e de maneira geral, as amostras apresentaram frequência de
genes de virulência relativamente baixos para as cepas de E. coli isoladas da Propriedade A e
ausentes para a Propriedade B.
A ausência de associação entre multirresistência e virulência, aliada à baixa
frequência ou ausência de genes de virulência nas E. coli estudadas, sugere que a
multirresistência está direcionada às cepas comensais. Este dado é sustentado pela
classificação de mais de 72% das cepas multirresistentes no grupo filogenético B1,
conhecidos como comensais, sendo o mais frequente em E. coli de animais (Tenaillon et al.,
2010).

Discussão | 63
Nesse contexto, Beceiro et al. (2013) apresentaram um extenso trabalho de
revisão, discutindo os mecanismos e as vantagens evolutivas da seleção de genes de
resistência antimicrobiana e de virulência em bactérias, no qual argumentaram que a
vantagem evolutiva bacteriana constituiria na cosseleção de resistência e virulência, de modo
que os casos de resistência elevada e virulência diminuída, levaria a um processo de seleção
mais longo até que alguns genes de virulência apresentassem vantagens suficientes para serem
fixados na população. Considerando esses mecanismos os autores concluem alertando para a
necessidade de práticas de monitoramento aliando resistência antimicrobiana e virulência em
cepas e clones.
As diferenças de frequência de resistência e fatores de virulência entre as duas
propriedades pode ser explicada, de acordo com Lara (2016)2, pelo manejo dos bovinos em
cada propriedade. A maior aglomeração de animais na Propriedade A pode facilitar a
circulação e a manutenção de cepas multirresistentes e/ou patogênicas entre os animais,
implicando em surtos mais frequentes e na necessidade de intervenção antibiótica em doses
mais levadas, de modo que as moscas poderiam desempenhar um papel importante na
disseminação dessas cepas nas propriedades leiteiras.
A presença de integron de classe 1 nos isolados de E. coli foi considerada quando
houve genotipo positivo para intI1 e 5’CS-3’CS. Porém, alguns isolados apresentaram apenas
intI1 ou apenas a região variável 5’CS-3’CS. É possível que esses isolados apresentem
organização não frequente do integron de classe 1, mas a ausência do gene sul3 (confere
resistência a sulfonamidas) associado a esse evento, indica que outras organizações, diferentes
das estudadas por Sáenz et al. (2010) podem estar presentes.
As oito cepas de E. coli selecionadas para sequenciamento apresentaram o mesmo
arranjo do integron de classe 1, com um único cassete gênico, dfrA7, inserido. A presença de
start códon alternativo “GTG” e sítio de recombinação (59-base element) com 143 pb, parece
ser variável entre dfrA7 de diferentes origens, já que trabalhos caracterizando esses genes e o
presente estudo, apresentaram diferenças no tamanho do sítio de recombinação, o que pode
estar relacionado a mobilização desses genes (Ojo et al., 2002; Kumar et al., 2015).
Os genes dfr estão relacionados à resistência ao antimicrobiano trimetoprim, uma
vez que expressam variantes da enzima diidrofolato redutase que não são sensíveis ao
fármaco (Sköld, 2001). O mecanismo de ação de trimetoprim consiste na analogia química
entre o antimicrobiano e o ácido fólico, que interage com a enzima diidrofolato redutase,
2 Lara, G.H.B. (Médico Veterinário, presta atendimento às propriedades estudadas – Faculdade de Medicina
Veterinária e Zootecnia, Unesp – Botucatu, SP. Comunicação pessoal, 2016).

Discussão | 64
impedindo a redução do ácido diidrofólico e consequentemente a síntese de timina em
procariotos (Sköld, 2001).
Considerando que as cepas de E. coli portadoras de dfrA7 foram isoladas de
moscas diferentes (MA17, M. domestica e MA26 Fannia sp.), e que possuem pulsotipos
distintos, é possível sugerir que dfrA7 pode apresentar disseminação regional, do mesmo
modo que o discutido por Labar et al. (2012) que encontraram altas frequências de integron de
classe 1 com uma única cópia desse cassete gênico em E. coli isoladas de humanos saudáveis
na Nigéria.
Ainda segundo Labar et al. (2012), o arranjo contendo apenas dfrA7 no integron
não é comum na literatura, e não há registros desse cassete gênico em isolados no Brasil.
A avaliação do perfil clonal revelou elevada diversidade cromossômica entre as
cepas de E. coli multirresistentes, dentre os quais destaca-se a presença de três perfis
genéticos apresentando 100% de similaridade (MB2e, MA39b e MA39d) provenientes de
insetos coletados em propriedades diferentes e que exibiram diferentes perfis de resistência
(fenotípico e genotípico), representadas pelo pulsotipo 24. Este pulsotipo apresentou
similaridade maior que 90% com os pulsotipos 21 (MA2d), 22 (MA6a), 23 (MB1d), 25
(MA8d) e 26 (MA39c), exibindo perfis de resistência diferentes entre si.
A presença de isolados clones em propriedades distintas e relativamente distantes
(35 km entre Propriedade A e B), com perfis de resistência antimicrobiana distintos, indica
que cepas podem ser disseminadas no ambiente assim como os genes de resistência.
As cepas MA39b e MA39d, apesar de serem isoladas de uma única mosca
(família Muscidae) e apresentarem perfil genético 100% similar (pulsotipo 24), são diferentes
quanto aos genes de resistência que possuem e à sensibilidade aos antimicrobianos testados.
Diferentemente, as cepas MA26d e MA26e, isoladas de uma única mosca (Fannia sp.),
apresentaram similaridade menor que 80%, mas os mesmos genes de resistência
Diante disso e das discussões realizadas anteriormente, é possível notar que uma
única mosca pode veicular cepas clones com genes de resistência distintos, ou que moscas
diferentes podem veicular cepas não relacionadas, porém com os mesmos genes – como
ocorrido com as cepas portadoras de integron de classe 1.
Nesse sentido, a investigação de integrons teve por objetivo estabelecer um
referencial para os estudos de disseminação molecular da resistência a antimicrobianos, de
modo que os resultados obtidos com PFGE, aliados as diferenças e similaridades dos perfis de
sensibilidade antimicrobiana, suportam a interpretação que genes de resistência e integrons

Discussão | 65
podem ser disseminados não apenas por meio de cepas clones, mas também ao serem
transferidos por elementos genéticos conjugativos.
Ensaios de conjugação são importantes para avaliar a capacidade de disseminação
de genes por essa via, devido à associação com elementos conjugativos que podem estar
presentes no genoma bacteriano.
Os resultados obtidos e apresentados na Tabela 9 indicam que o integron de classe
1 e os outros genes de resistência presentes nas cepas testadas, com exceção de blaTEM, podem
ser transferidos por conjugação, assim como aqueles envolvidos nos perfis de resistência
fenotípicos e na produção de ESBLs. No entanto, alguns resultados atípicos foram observados
nas cepas transconjugantes. Interessantemente, as cepas transconjugantes de MA26a, MA26b
e MA26d, expressaram no teste de difusão com discos, além da resistência anteriormente
verificada para as doadoras, resistência a ceftriaxona e ceftiofur (MA26a), e a ceftriaxona,
cefoperazona sódica e ceftiofur (MA26b e 26d). Adicionalmente, no ensaio de aproximação
de discos, a cepa transconjugante de MA26a produziu ESBL diante dos antimicrobianos
aztreonam e cefotaxima, sendo que na cepa doadora a produção de ESBL não havia sido
identificada, e a cepa transconjugante MA26b para ceftazidima, porém, a produção de ESBL
havia sido identificada apenas para aztreonam na cepa doadora.
A partir disso, é possível propor que os resultados obtidos podem estar
relacionados à regulação da expressão de genes de resistência aos β-lactâmicos de terceira
geração nas cepas doadoras, uma vez que tais genes poderiam estar sobre o efeito de
repressores e/ou que tiveram a expressão induzida com a ocorrência da conjugação.
Segundo Depardieu et al. (2007), a expressão de resistência antimicrobiana é
constantemente regulada, pois a manutenção desses genes em ambientes favoráveis
significaria gasto energético desnecessário, e diante desse tema elaboraram uma revisão dos
mecanismos envolvidos na regulação gênica da resistência. Uma das formas de regulação
apresentadas pelos autores, e que são relacionadas aos resultados deste trabalho, diz respeito
às funções de regulação que podem ser desempenhadas por elementos de inserção e integrons,
uma vez que os elementos de inserção podem conter nas extremidades sequências parciais ou
completas de promotores que conduzem a expressão de genes vizinhos.
São conhecidos vários elementos de inserção portadores de promotores que atuam
na transcrição de genes de ESBLs, dentre os quais se destaca o elemento ISEcp1, responsável
pela regulação da expressão de fenótipos resistentes a cefotaxima, aztreonam e ceftazidima,
em enterobactérias e Pseudomonas aeruginosa, afetando principalmente os genes blaCTX-M
(Depardieu et al., 2007). Adicionalmente, é conhecida a associação de ISEcp1 com a

Discussão | 66
mobilização de blaCTX-M por transposição e como os genes blaCTX-M podem estar localizados
downstream de integrons de classe 1, é possível relacioná-los também a esses elementos
(Poirel et al., 2008), sendo que, no Brasil, o elemento de inserção ISEcp1 já foi associado a
blaCTX-M em enterobactérias de origem humana (Minarini et al., 2009).
Nesse sentido, é plausível que as cepas transconjugantes que apresentaram
produção de ESBLs apresentem genes regulados por ISEcp1. Porém, estudos mais
minunciosos precisam ser realizados a fim de comprovar tal hipótese.
Por outro lado, os plasmídeos das cepas transconjugantes, extraídos por lise
alcalina, não puderam ser visualizados nitidamente nas condições estabelecidas pela técnica,
enquanto que aqueles extraídos das cepas doadoras sim (Figura 9), de modo que PCRs a partir
do DNA plasmidial extraído foram realizados. No entanto, alguns genes não foram
amplificados a partir do DNA plasmidial de cepas doadoras, mas somente das
transconjugantes (Tabela 10).
Diante disso, duas situações são possíveis: a) os genes estudados estão localizados
no DNA cromossômico na forma de elementos conjugativos integrados ao cromossomo
(como transposons conjugativos ou epissomos), de modo que foram transferidos através da
conjugação e permaneceram na conformação circular covalentemente fechada após o
processo, sendo os genes detectados por PCR, mas os plasmídeos não; b) os genes estão
localizados em megaplasmídeos e os plasmídeos de tamanhos menores purificados por lise
alcalina e visualizados em eletroforese comum não correspondem aos elementos onde os
genes estudados estão localizados.
Os elementos conjugativos integrados ao cromossomo (ICE, do inglês integrative
and conjugative element) são grandes, apresentando tamanho de 20 kb até 500 kb, pois
carregam todos os genes necessários a conjugação, podendo portar elementos de inserção,
transposons e integrons. Vários estímulos podem estar envolvidos na indução da conjugação
desses elementos, tais como resposta SOS, contato entre células e indução dependente do
fenótipo, de modo que é conhecido que tetraciclina pode induzir conjugação em ICE (Johnson
e Grossman, 2015).
A extração de plasmídeos pela técnica de lise alcalina é amplamente conhecida e
utilizada para a detecção de plasmídeos em várias situações, porém, o tamanho dos
plasmídeos é um fator limitante da técnica, uma vez que megaplasmídeos podem ser
renaturados juntamente com o DNA cromossomal (Pedraza e Díaz, 2002), além de as
condições oferecidas por equipamentos de eletroforese convencional não permitirem a

Discussão | 67
visualização de plasmídeos grandes, sendo necessária a digestão do DNA com enzima S1 e
separação dos fragmentos por PFGE (Barton et al., 1995).
Devido às características gerais dos ICEs e as limitações da técnica utilizada, é
possível propor que ambas as hipóteses (a e b) anteriormente apresentadas são aceitáveis e
podem ter acontecido em conjunto, pois, ICEs são grandes e, portanto não detectáveis em
eletroforese convencional quando em estado não integrado ao cromossomo.
Outra situação incomum ocorrida durante a execução do trabalho foi a
inconstância na amplificação de algumas regiões como 5’CS-3’CS, do gene tetA e blaTEM,
quando réplicas de PCR da mesma amostra apresentaram resultados distintos, ou quando a
cepa doadora não amplificou o gene transferido à transconjugante. Tais eventos podem estar
relacionados à inespecificidade dos iniciadores usados frente às amostras testadas ou resultado
da atuação de elementos móveis associados a esses casos.
Apesar de resistência a antimicrobianos em cepas bacterianas ser um tema
mundialmente difundido e amplamente estudado, ainda são poucos os trabalhos direcionados
aos mecanismos envolvidos na resistência de cepas de origem animal, as práticas veterinárias
e às pressões ambientais envolvidas na seleção dos micro-organismos resistentes.
No presente estudo, são apresentados resultados inéditos no Brasil referente à
possibilidade de disseminação de E. coli comensais multirresistentes por meio de moscas,
dado que fornece subsídios para a inclusão desses insetos nas rotas de disseminação ambiental
de resistência antimicrobiana, o que é favorecido pelo comportamento sinantrópico e
comunicativo das moscas.

VI – Conclusões

Conclusões | 69
Diante dos resultados obtidos é possível concluir que espécies de dípteros
muscoides podem veicular, em ambiente de ordenha, cepas de E. coli não patogênicas
multirresistentes a antimicrobianos com e sem integrons e cepas de E. coli potencialmente
patogênicas não multirresistentes, de modo que dípteros muscoides podem contribuir para a
disseminação de resistência antimicrobiana, associada a E. coli, entre os animais e o ambiente
ao veicularem cepas e/ou clones, que apresentam diversidade de perfis de resistência
fenotípico e genotípico.

VII - Referências Bibliográficas

Referências Bibliográficas| 71
Agersø Y, Sandvang D. Class 1 integrons and tetracycline resistance genes in Alcaligenes,
Arthrobacter, and Pseudomonas spp. isolated from pigsties and manured soil. Appl. Environ.
Microbiol. 2005; 71(12): 7941-7947.
Ahmad A, Nagaraja TG, Zurek L. Transmission of Escherichia coli O157:H7 to cattle by
house flies. Prev. Vet. Med. 2007; 80:74–81.
Alam MJ, Zurek L. Association of Escherichia coli O157:H7 with houseflies on a cattle farm.
Appl. Environ. Microbiol. 2004; 70(12): 7578–7580.
Allen HK, Donato J, Wang HH, Cloud-Hansen KA, Davies J, Handelsman J. Call of the wild:
antibiotic resistance genes in natural environments. Nature Rev. Microbiol. 2010; 1-9.
Almeida JL, Giuffrida R; Andrade RAP; Chaves MP. Muscoid Diptera as potential vectors of
bacterial agents on dairy farms in the northern region of Paraná, Brazil. Semin: Ciênc. Agrár.
2014; 35(6): 3127-3138.
Aminov RI, Mackie RI. Evolution and ecology of antibiotic resistance genes. FEMS
Microbiol. Lett. 2007; 271: 147-161.
Assis AG, Stock LA, Campos OF, Gomes AT, Zoccal R, Silva MR. Sistemas de produção de
leite no Brasil. Circular Técnica 85. Embrapa Gado de Leite. 2005; 1-5.
Barton BM, Harding GP, Zuccarelli AJ. A general method for detecting and sizing large
plasmids. Anal. Biochem. 1995; 226(2): 235-240.
Batchelor M, Hopkins K, Threlfall EJ, Clifton-Hadley FA, Stallwood AD, Davies RH, et al.
blaCTX-M genes in clinical Salmonella isolates recovered from humans in England and Wales
from 1992 to 2003. Antimicrob. Agents Chemother. 2005; 49: 1319-1322.
Bauer AW, Kirby WMM, Sherres JC, Turck M. Antibiotic susceptibility testing by a
standardized single disk method. Am. J. Clin. Pathol. 1966; 45 (4): 493-496.
Beceiro A, Tomás M, Bou G. Antimicrobial resistance and virulence: a successful or
deleterious association in the bacterial world? Clin. Microbiol. Rev. 2013; 26(2): 185-230.
Berglund B. Environmental dissemination of antibiotic resistance genes and correlation to
anthropogenic contamination with antibiotics. Infect. Ecol. Epidemiol. 2015; 5: 1-10.

Referências Bibliográficas| 72
Birnboim HC, Doly J. A rapid alkaline extraction procedure for screening recombinant
plasmid DNA. Nucleic Acids Res. 1979; 7(3): 1513-1523.
Blaak H, Hamidjaja RA, Hoek AHAM, Heer L, Husman AMR, Schets FM. Detection of
extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli on flies at poultry
farms. Appl. Environ. Microbiol. 2014; 80(1): 239-246.
Bogard AE, Stobberingh EE. Epidemiology of resistance to antibiotics. Links between
animals and humans. Int. J. Antimicrob. Agents. 2000; 14: 327-335.
Bolon MK. The newer fluoroquinolones. Med. Clin. North Am. 2011; 95(4):793-817.
Bonnedahl J, Drobni M, Gauthier-Clerc M, Hernandez J, Granholm S, Kayser Y, et al.
Dissemination of Escherichia coli with CTX-M Type ESBL between humans and yellow-
legged gulls in the south of France. Plos One. 2009; 4: 1-6.
Borges MAZ. Flutuação populacional de dípteros muscoides (Diptera: Muscomorpha),
parasitoides e foréticos predadores Igarapé, MG. Belo Horizonte. Tese [Doutorado em
Ciência Animal] - Escola de Veterinária da UFMG; 2006.
Bradford PA. Extended-spectrum β-lactamases in the 21st century: characterization,
epidemiology, and detection of this important resistance threat. Clin. Microbiol. Rev. 2001;
14(4): 933-951.
Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Bem estar animal e sistema de
produção de gado leiteiro. In: Código sanitário de animais terrestres – OIE 2014. 2015; Cap
7.11.
Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Plano mais pecuária. Brasília,
2014. [acesso em 29 maio 2016]. Disponível em: www.agricultura.gov.br/mais-pecuaria
Brito LG, Borja GEM, Oliveira MCS, Netto FGS. Mosca-dos-chifres: aspectos bio-
ecológicos, importância econômica, interações parasito-hospedeiro e controle. Comunicado
Técnico, 302. Porto Velho : Embrapa Rondônia ; 2005.
Brito LG, Oliveira MCS, Giglioti R, Barbieri FS, Netto FGS, Chagas ACS, et al. Manual de
identificação, importância e manutenção de colônias estoque de dípteras de interesse
veterinário em laboratório. Porto Velho: Embrapa Rondônia; 2008.

Referências Bibliográficas| 73
Cadavid-Sanchez IC, Amat E, Gomez-Piñerez LM. Enterobacteria isolated from synanthropic
flies (Diptera, Calyptratae) in Medellín, Colombia. Caldasia. 2015; 37(2): 319–332.
Cantas L, Shah SQA, Cavaco LM, Manaia CM, Walsh F, Popowska M, et al. A brief multi-
disciplinary review on antimicrobial resistance in medicine and its linkage to the global
environmental microbiota. Front. Microbiol. 2013; 4(96): 1-17.
Cao V, Lambert T, Nhu DQ, Loan HK, Nguyen KH, Arlet G, et al. Distribution of extended-
spectrum β-lactamases in clinical isolates of Enterobacteriaceae in Vietnam. Antimicrob.
Agents Chemother. 2002; 46: 3739-3743.
Carattoli A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013; 303: 298–
304
Carattoli A. Resistance plasmids families in Enterobacteriaceae. Antimicrob. Agents
Chemother. 2009; 53(6): 2227–2238.
Castro BG, Souza MMS, Bittencourt AJ. Aerobic bacterial microbiota in Stomoxys calcitrans:
preliminary studies in Brazil Rev. Bras. Parasitol. Vet. 2007; 16(4): 193-197.
Castro BG, Souza MMS, Bittencourt AJ. Isolamento de espécies enterobacterianas em
Stomoxys calcitrans. Ciência Rural. 2008a; 38(9): 2654-2657.
Castro BG, Souza MMS, Bittencourt AJ. Microbiota bacteriana em segmentos de mosca do
estábulo Stomoxys calcitrans no Brasil: primeiro relato de espécies. Arq. Bras. Med. Vet.
Zootec. 2008b; 60(3): 1029-1031.
Chapman PA, Ellin M, Ashton R, Shafique W. Comparison of culture, PCR end
immunoassays for detecting Escherichia coli O157H7 following enrichment culture and
immunomagnetic separation performed on naturally contaminated raw meat products. Int. J.
Food Microbiol. 2001; 68: 11-20.
Chopra I, Roberts M. Tetracycline antibiotics: mode of action, applications, molecular
biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001; 65(2):
232–260.
Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli
phylogenetic group. Appl. Environ. Microbiol. 2000; 66: 4555-4558.

Referências Bibliográficas| 74
Clermont O, Christenson JK, Denamur E, Gordon DM. The Clermont Escherichia coli phylo-
typing method revisited: improvement of specificity and detection of new phylo-groups.
Environ. Microbiol. Rep. 2013; 5: 58-65.
CLSI. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial
disk susceptibility tests; approved standard - 11th ed. CLSI document M02-A11. Wayne, PA,
USA: 2012.
CLSI. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial
susceptibility testing - 2nd Informational supplement. CLSI document M100-S22. Wayne,
PA, USA: 2012.
Constable PD. Treatment of calf diarrhea: Antimicrobial and ancillary treatments. Vet. Clin.
Food Anim. 2009; 25: 101–120.
Costa PM, Loureiro L, Matos AJF. Transfer of multidrug-resistant bacteria between
intermingled ecological niches: the interface between humans, animals and the environment.
Int. J. Environ. Res. Public Health. 2013; 10: 278-294.
Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M, Finlay BB. Recent advances
understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013; 26(4):822-
880.
Depardieu F, Podglajen I, Leclercq R, Collatz E, Courvalin P. Modes and modulations of
antibiotic resistance gene expression. Clin. Microbiol. Rev. 2007; 20(1): 79–114.
Duse A, Waller KP, Emanuelson U, Unnerstad HE, Persson Y, Bengtsson B. Occurrence and
spread of quinolone-resistant Escherichia coli on dairy farms. Appl. Environ. Microbiol.
2016; 82(13): 3765-3773.
Ebersbach R. My farmland – our livestock forms of subsistence farming and forms of sharing
in peasant communities. In: M. Benz. The principle of sharing. segregation and construction
of social identities at the transition from foraging to farming. Studies in early near eastern
production, subsistence, and environment. Berlin. 2010. p. 159-182
Ebersbach R, Schibler J. A criação de animais domésticos. Os genes e a alimentação. Nestlé,
Foundation Alimentarium e Ministério da Cultura: Coimbra, 2003. 200p. [acesso em 25 set
2014]. Disponível em: http://triplov.com/hist_fil_ciencia/genes/index.htm

Referências Bibliográficas| 75
Embrapa. Sistemas de Produção Embrapa Gado de Leite. [homepage da internet].
[acesso em 27 dez 2016]. Disponível em:
https://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Leite/LeiteCerrado/infra/15.html
FAO. Food and Agriculture Organization of the United Nations [homepage da internet]. Dairy
production and products: Milk production. 2016. [acesso em 27 dez 2016]. Disponível em:
www.fao.org/agriculture/dairy-gateway/milk-production/.
Féria C, Ferreira E, Correia JD, Gonçalves J, Caniça M. Patterns and mechanisms of
resistance to β-lactams and β-lactamase inhibitors in uropathogenic Escherichia coli isolated
from dogs in Portugal. J. Antimicrob. Chemother. 2002; 49: 77-85.
Florião MM. Boas práticas em bovinocultura leiteira com ênfase em sanidade preventiva.
Niteroi: Programa Rio Rural; 2013.
Fluit AC, Schmitz FJ. Resistance integrons and super-integrons. Clin. Microbiol. Infect. 2004;
10(4): 272-88.
Fontedar R, Banerjee U, Singh S, Shriniwas, Verma AK. The housefly (Musca domestica) as
a Carrier of pathogenic microorganisms in the hospital environment. J. Hosp. Infect. 1992;
20: 209-215.
Franck SM, Bosworth BT, Moon HW. Multiplex PCR for enterotoxigenic, attaching and
effacing, and Shiga toxin-producing Escherichia coli strains from calves. J. Clin. Microbiol.
1998; 36: 1795-1797.
Frech G, Kehrenberg C, Schwarz S. Resistance phenotypes and genotypes of multiresistant
Salmonella. J. Antimicrob.l Chemother. 2003; 51: 180-182.
Frost LS, Leplae R, Summers AO, Toussaint A. Mobile genetic elements: the agents of open
source evolution. Nat. Rev. Microbiol. 2005; 3(9): 722-732.
Garcia-Alvarez L, Dawson S, Cookson B, Hawkey P. Working across the veterinary and
human health sectors. J. Antimicrob Chemother. 2012; 67 (Suppl 1): i37-i49.
Gillings MR, Gaze WH, Pruden A, Smalla K, Tiedje JM, Zhu YG. Using the class 1 integron-
integrase gene as a proxy for anthropogenic pollution. ISME J. 2015; 9: 1269-1279.

Referências Bibliográficas| 76
Greenberg B. Flies and disease: Biology and disease transmission. Volume 2. New Jersey:
Princeton University Press; 1973. p. 20-94.
Greenberg B. Fly organism associations with systematic list of fly species. In: Greenberg B.
Flies and disease: Ecology, classification and biotic associations. v.1. New Jersey: Princeton
University Press; 1971. p. 83-856.
Greenberg B, Povolný D. Bionomics of flies. In: Greenberg B. Flies and disease: Ecology,
classification and biotic associations. v.1. New Jersey: Princeton University Press; 1971. p.
57-83.
Guardabassi L, Dijkshoorn L, Collard JM, Olsen JE, Dalsgaard A. Distribution and in-vitro
transfer of tetracycline resistance determinants in clinical and aquatic Acinetobacter strains. J.
Med. Microbiol. 2000; 49: 929-936.
Hernández A, Sánchez MB, Martínez JL. Quinolone resistance: much more than predicted.
Front. Microbiol. 2011; 2: 22.
Horton RA, Randall LP, Snary EL, Cockrem H, Lotz S, Wearing H, et al. Fecal carriage and
shedding density of CTX-M Extended-Spectrum-Lactamase-Producing Escherichia coli in
cattle, chickens, and pigs: implications for environmental contamination and food production.
Appl. Environ. Microbiol. 2011; 77: 3715–3719.
IBGE – Instituto Brasileiro de Geografia e Estatística. Censo agropecuário 2006. Brasil,
Grandes Regiões e Unidades da Federação. Rio de Janeiro; 2009. [acesso em 29 maio 2016].
Disponível em: www.biblioteca.ibge.gov.br
Jacoby GA. Mechanisms of resistance to quinolones. Clin. Infect Dis. 2005; 41: S120-S126.
Jacoby GA, Strahilevitz J, Hooper DC. Plasmid-mediated quinolone resistance. Microbiol.
Spectr. 2014; 2(2): 1-42.
Jarlier V, Nicolas M, Fournier G, Phillipon A. ESBLs conferring transferable resistance to
newer β-lactam agents in Enterobacteriaceae; hospital prevalence and susceptibility patterns.
Rev. Infect Dis. 1988; 10: 867–78.
Johnson CM, Grossman AD. Integrative and conjugative elements (ICEs): What they do and
how they work. Annu. Rev. Genet. 2015; 49: 577–601.

Referências Bibliográficas| 77
Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in
emerging infectious diseases. Nature. 2008; 451: 990-993.
Kamada N, Chen GY, Inohara N, Núñez G. Control of pathogens and pathobionts by the gut
microbiota. Nat. Rev. Immunol. 2013a; 14: 85-690.
Kamada N, Seo SU, Chen GY, Núñez. Role of the gut microbiota in immunity and
inflammatory disease. Nat Rev. Immunol. 2013b; 13: 321-335.
Kaper JB, James P, Nataro JP, Mobley HLT. Pathogenic Escherichia coli. Nat. Rev.
Microbiol. 2004; 2: 123-140.
Kappel HB, Oliveira AG, Silva PR, Pelli A. Non-biting flying insects as carriers of
pathogenic bacteria in a Brazilian hospital. Rev. Soc. Bras. Med. Trop. 2013; 46(2):234-236.
Khachatourians GG. Agricultural use of antibiotics and the evolution and transfer of
antibiotic-resistant bacteria. J. Can. Med. Assoc. 1998; 159: 1129-1136.
Khachatryan AR, Hancock DD, Besser TE, Call DR. Role of calf-adapted Escherichia coli in
maintenance of antimicrobial drug resistance in dairy calves. Appl. Environ. Microbiol. 2004;
70(2): 752-757.
Kobayashi M, Sasaki T, Saito N, Tamura K, Suzuki K, Watanabe H, et al. Houseflies: not
simple mechanical vectors of enterohemorrhagic Escherichia coli O157:H7. Am. J. Trop.
Med. Hyg. 1999; 61(4): 625-629.
Koczura R, Mokracka J, Barczak A, Krysiak N, Kaznowski A. Association between the
presence of class 1 integrons, virulence genes, and phylogenetic groups of Escherichia coli
isolates from river water. Microb. Ecol. 2013; 65: 84-90.
Kumar A, Tiwary BK, Kachhap S, Nanda AK, Chakraborty R. An Escherichia coli strain,
PGB01, isolated from feral pigeon faeces, thermally fit to survive in pigeon, shows high level
resistance to trimethoprim. Plos One. 2015; 10(3): 1-21.
Lachmayr KL, Kerkhof LJ, Di Rienzo G, Cavanaugh CM, Ford TE. Quantifying nonspecific
TEM β-Lactamase (blaTEM) genes in a wastewater stream. Appl. Environ. Microb. 2009;
75(1): 203-211.

Referências Bibliográficas| 78
Lastours V, Bleibtreu A, Chau F, Burdet C, Duval X, Denamur E, et al. Quinolone-resistant
Escherichia coli from the faecal microbiota of healthy volunteers after ciprofloxacin exposure
are highly adapted to a commensal lifestyle. J. Antimicrob. Chemother. 2014; 69(3): 761-768.
Lee S, Yu JJ, Park K, Eun-Jee O, Kim SY, Park YJ. Phylogenetic groups and virulence
factors in pathogenic and commensal strains of Escherichia coli and their association with
blaCTX-M. Ann. Clin. Lab. Sci. 2010; 40: 361-367.
Lévesque C, Piche L, Larose C, Roy PH. PCR mapping of integrons reveals several novel
combinations of resistance genes. Antimicrob. Agents Chemother. 1995; 39:185-195.
Liebana E, Batchelor M, Hopkins KL, Clifton-Hadley FA, Teale CJ, Foster A, et al.
Longitudinal farm study of Extended-Spectrum β-Lactamase-mediated resistance. J. Clin.
Microbiol. 2006; 44(5): 1630-1634.
Linhares AX. Sinantropia de dípteros muscoides de Campinas. Campinas. Dissertação
[Mestrado em Ciências Biológicas] - Universidade Estadual de Campinas; 1979.
Machado E, Cantón R, Baquero F, Galán JC, Rollán A, Peixe L, et al. Integron content of
extended-spectrum-β-lactamase-producing Escherichia coli strains over 12 years in a single
hospital in Madrid, Spain. Antimicrob. Agents Chemother. 2005; 49: 1823-1829.
Macovei L, Zurek L. Ecology of antibiotic resistance genes: Characterization of Enterococci
from houseflies collected in food settings. Appl. Environ. Microbiol. 2006; 72(6): 4028-4035.
Madigan MT, Martinko JM, Dunlap PV, Clark DP. Microbiologia de Brock. 12ed. Porto
Alegre: Artmed; 2010. p. 1055.
Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, et al.
Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international
expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol.
Infect. 2012; 18(3): 268-281.
Marchiori CH. Dípteros muscoideos associados a fezes frescas de gado bovino e seus
parasitoides, nos municípios de Uberlândia-MG e Itumbiara-GO. Campinas. Tese [Doutorado
em Ciências Biológicas] - Universidade Estadual de Campinas; 1997.
Marchiori CH, Oliveira AT, Linhares AX. Artrópodes associados a massas fecais bovinas no
sul do estado de Goiás. Neotrop. Entomol. 2001; 30(1): 19-24.

Referências Bibliográficas| 79
Marshall BM, Levy SB. Food animals and antimicrobials: impacts on human health. Clin.
Microbiol. Rev. 2011; 24(4): 718-733.
Marshall BM, Ochieng DJ, Levy SB. Commensals: Underappreciated Reservoir of Antibiotic
Resistance. Microbe. 2009; 4: 231-238.
Marshall BM, Petrowsk D, Levy SB. Inter and intraspecies spread of E. coli in a farm
environment in the absence of antibiotic usage. Proc. Natl. Acad. Sci. USA. 1990; 87: 6609-
6613.
Martha Júnior GB, Barioni LG, Vilela L, Barcellos AO. Área do piquete e taxa de lotação no
pastejo rotacionado. Comunicado Técnico 101. Planaltina: Embrapa; 2003
Mazel D, Dychinco B, Webb VA, Davies J. Antibiotic resistance in the ECOR collection:
integrons and identification of a novel aad gene. Antimicrob. Agents Chemother. 2000; 44:
1568-1574.
Mazel D. Integrons: agents of bacterial evolution. Nat. Rev. Microbiol. 2006; 4: 608-620.
Minarini LAR, Poirel L, Trevisani NAC, Darini ALC, Nordmann P. Predominance of CTX-
M–type extended-spectrum β-lactamase genes among enterobacterial isolates from outpatients
in Brazil. Diagn. Microbiol. Infect. Dis. 2009; 65: 202–206.
Moraes APR, Badini PV, Souza MMS; Bittencourt AJ. Avaliação da capacidade de Stomoxys
calcitrans (Linnaeus, 1758) em carrear bactérias envolvidas nas etiologias das mastites de
municípios do Rio de Janeiro. Rev. Bras. Parasitol. Vet. 2004; 13: 143-149.
Moriya K, Fujibayashi T, Yoshihara T, Matsuda A, Sumi N, Umezaki N, et al. Verotoxin-
producing Escherichia coli O157:H7 carried by the housefly in Japan. Med. Vet. Entomol.
1999; 13: 214-216.
Nataro JP, Kaper JB. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998; 11(1): 142–
201.
Novais C, Freitas AR, Silveira E, Antunes P, Silva R, Coque TM, et al. Spread of multidrug-
resistant Enterococcus to animals and humans: an underestimated role for the pig farm
environment. J. Antimicrob. Chemother. 2013; 68: 2746-2754.

Referências Bibliográficas| 80
O’Brien TF. Emergence, spread, and environmental effect of antimicrobial resistance: how
use of an antimicrobial anywhere can increase resistance to any antimicrobial anywhere else.
Clin. Infect. Dis. 2002; 34(Suppl 3): S78-84.
Ojeniyi B, Ahrens P, Meyling A. Detection of fimbrial and toxin genes in Escherichia coli
and their prevalence with piglets with diarrhea: the application of colony hybridization assay
polymerase chain reaction and phenotype assays. J. Vet. Med. 1994; 41: 49-59.
Ojo KK, Waturangi DE, Odelola HA, Suwanto A, Schwarz S. Identification of a cassette-
borne dfrA7-like gene that shows a 97 bp extension at the 3'-end of the reading frame. J.
Antimicrob. Chemother. 2002; 49(3):573-574.
Oliver SP, Murinda SE. Antimicrobial resistance of mastitis pathogens. Vet. Clin. North. Am.
Food Anim. Pract. 2012; 28: 165-185.
Oliver SP, Murinda SE, Jayarao BM. Impact of antibiotic use in adult dairy cows on
antimicrobial resistance of veterinary and human pathogens: a comprehensive review.
Foodborne Pathog. Dis. 2011; 8(3): 337-355.
Oram M, Fisher LM. 4-Quinolone resistance mutations in the DNA gyrase of Escherichia coli
clinical isolates identified by using the polymerase chain reaction. Antimicrob. Agents
Chemother. 1992; 35: 387-389.
Paraluppi ND, Vasconcelos JC, Aquino JS, Castellón EG, Silva MSB. Calliphoridae (Diptera)
in Manaus: IV. Bacteria isolated from blowflies collected in street markets. Acta Amaz. 1996;
26: 93-96.
Paterson DL, Bonomo RA. Extended-spectrum β-lactamases: a clinical update. Clin.
Microbiol. Rev. 2005; 18(4): 657-686.
Pedraza RO, Díaz JCR. In-well cell lysis technique reveals two new megaplasmids of 103.0
and 212.6 MDa in the multiple plasmid-containing strain V517 of Escherichia coli. Lett.
Appl. Microbiol. 2002; 34(2):130-133.
Pereira AS, Carmo Filho JR, Tognim MCB, Sader HS. Avaliação da acurácia de testes
laboratoriais para detecção de amostras de Klebsiella pneumoniae produtora de β-lactamase
de espectro estendido. J. Bras. Patol. Med. Lab. 2003; 39: 301-308.

Referências Bibliográficas| 81
Perez F, Endimiani A, Hujer KM, Bonomo RA. The continuing challenge of ESBL. Curr.
Opin. Pharmacol. 2007; 7: 459-469.
Peter-Getzlaff S, Polsfuss S, Poledica M, Hombach M, Giger J, Böttger EC, et al. Detection
of AmpC beta-lactamase in Escherichia coli: comparison of three phenotypic confirmation
assays and genetic analysis. J. Clin. Microbiol. 2011; 49(8): 2924-2932.
Piérard D, De Greve H, Haesebrouck F, Mainil J. O157:H7 and O104:H4 Vero/Shiga toxin-
producing Escherichia coli outbreaks: respective role of cattle and humans. Vet. Res. 2012;
43(13): 1-12.
Poirel L, Naas T, Nordmann P. Genetic support of extended-spectrum β-lactamases. Clin.
Microbiol. Infect. 2008; 14 (Suppl. 1): 75–81.
Pol M, Ruegg PL. Treatment practices and quantification of antimicrobial drug usage in
conventional and organic dairy farms in Wisconsin. J. Dairy Sci. 2007; 90: 249-261.
Povolný D. Synanthropy. In: Greenberg B. Flies and disease: Ecology, classification and
biotic associations. Volume 1. New Jersey: Princeton University Press; 1971. p. 17-54.
Rahuma N, Ghenghesh KS, Ben Aissa R, Elamaari A. Carriage by the housefly (Musca
domestica) of multiple-antibiotic-resistant bacteria that are potentially pathogenic to humans,
in hospital and other urban environments in Misurata, Libya. Ann. Trop. Med. Parasitol.
2005; 99(8): 795-802.
Ravi A, Avershina E, Ludvigsen J, L’Abée-Lund TM, Rudi K. Integrons in the intestinal
microbiota as reservoirs for transmission of antibiotic resistance genes. Pathogens. 2014; 3:
238-248.
Rawat D, Nair D. Extended-spectrum β-lactamases in gram negative bacteria. J. Glob. Intect
Dis. 2010; 2(3): 263-274.
Ribeiro MG. Princípios terapêuticos na mastite em animais de produção e de companhia. In:
Andrade, S.F. (Ed). Manual de Terapêutica Veterinária. 3ed. São Paulo: Rocca; 2008. p. 759-
771.
Ribot EM, Fair MA, Gautom R, Cameron DN, Hunter SB, Swaminathan B, et al.
Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia

Referências Bibliográficas| 82
coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006; 3(1): 59-
67.
Royster E, Wagner S. Treatment of mastitis in cattle. Vet. Clin. Food. Anim. 2015; 31: 17–46.
Rutz DA, Geden CJ. Pest management recommendations for dairy cattle. [Monografia na
internet]. A Cornell and Penn State Cooperative Extension Publication; 2010 [acesso em 15
set 2016]. Disponível em: http://extension.psu.edu/publications/xf0275/view.
Rybaríková J, Dolejská M, Materna D, Literák I, Cízek A. Phenotypic and genotypic
characteristics of antimicrobial resistant Escherichia coli isolated from symbovine flies, cattle
and sympatric insectivorous house martins from a farm in the Czech Republic (2006-2007).
Res. Vet. Sci. 2010; 89: 179-183.
Sáenz Y, Briñas L, Domínguez E, Ruiz J, Zarazaga M, Vila J, et al. Mechanisms of resistance
in multiple-antibiotic-resistant Escherichia coli strains of human, animal, and food origins.
Antimicrob. Agents Chemother. 2004; 48: 3996-4001.
Sáenz Y, Vinué L, Ruiz E, Somalo S, Martínez S, Rojo-Bezares B, et al. Class 1 integrons
lacking qacED1 and sul1 genes in Escherichia coli isolates of food, animal and human origin.
Vet. Microbiol. 2010; 144: 493-497.
Salyers AA, Shoemaker NB, Stevens AM, Li LY. Conjugative transposons: an unusual and
diverse set of integrated gene transfer elements. Microbiol. Rev. 1995; 59(4): 579–590.
Sarcinelli MF, Venturini KS, Silva LC. Produção de bovinos – Tipo leite. [Boletim Técnico].
Universidade Federal do Espírito Santo. 2007. [acesso em 23 dez 2016]. Disponível em:
www.agais.com/telomc/b00407_leite_bovinodeleite.pdf.
Schaumburg F, Onwugamba FC, Akulenko R, Peters G, Mellmann A, Köck R, et al. A
geospatial analysis of flies and the spread of antimicrobial resistant bacteria. J. Med.
Microbiol. 2016; 306(7): 566-571.
Schultsz C, Pool GJ, Van Ketel R, De Wever B, Speelman P, Dankert J. Detection of
enterotoxigenic Escherichia coli in stool samples by using nonradioactively labeled
oligonucleotide DNA probes and PCR. Clin. Microbol. 1994; 32: 2393-2397.

Referências Bibliográficas| 83
Serjeantson D. Review of aminal remains from the Neolithic and Early Bronze Age of
Southern Britain (4000BC–1500BC). Research Department Report Series. 2011. Portsmouth,
UK: English Heritage
Shah AA, Hansan F, Ahmed S, Hameed A. Characteristics, epidemiology and clinical
importance of emerging strains of Gram-negative bacilli producing extended-spectrum β-
lactamases. Res. Microbiol. 2004; 155: 409-421.
Sköld O. Resistance to trimethoprim and sulfonamides. Vet. Res. 2001; 32: 261-273.
Smillie C, Garcillán-Barcia MP, Francia MV, Rocha EP, de La Cruz F. Mobility of plasmids.
Microbiol. Mol. Biol. Rev. 2010; 74(3): 434-452.
Solà-Ginés M, González-López JJ, Cameron-Veas K, Piedra-Carrasco N, Cerdà-Cuéllar M,
Migura-Garcia L. Houseflies (Musca domestica) as vectors for extended-spectrum β-
lactamase-producing Escherichia coli on Spanish broiler farms. Appl. Environ. Microbiol.
2015; 81(11): 3604-3611.
So M, McCarty B J. Nucleotide sequence of the bacterial transposon Tn1681 encoding a heat-
stable (ST) toxin and its identification in enterotoxigenic Escherichia coli strains. Proc. Natl.
Acad. Sci. USA. 1980; 77: 4011-4015.
Soufi L, Abbassi MS, Sáenz Y, Vinué L, Somalo S, Zarazaga M, et al. Prevalence and
diversity of integrons and associated resistance genes in Escherichia coli isolates from poultry
meat in Tunisia. Foodborne Pathog. Dis. 2009; 6(9): 1067-1073.
Stapleton PD, Shannon KP. French GL. Carbapenem resistance in Escherichia coli associated
with plasmid-determined CMY-4-lactamase production and loss of an outer membrane
protein. Antimicrob. Agents Chemother. 1999; 43: 1206-1210.
Stokes HW, Gillings MR. Gene flow, mobile genetic elements and the recruitment of
antibiotic resistance genes into gram-negative pathogens. FEMS Microbiol. Rev. 2011; 35(5):
790-819.
Stokes HW, Hall RM. A novel family of potentially mobile DNA elements encoding site-
specific gene-integration functions: integrons. Mol Microbiol. 1989; 3(12): 1669-1683.

Referências Bibliográficas| 84
Tadesse DA, Zhao S, Tong E, Ayers S, Singh A, Bartholomew MJ, et al. Antimicrobial drug
resistance in Escherichia coli from humans and food animals, United States,1950–2002.
Emerg. Infect. Dis. 2012; 18: 741-749.
Tenaillon O, Skurnik D, Picard B, Denamur E. The population genetics of commensal
Escherichia coli. Nat. Rev. Microbiol. 2010; 8: 207-217.
Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, et al.
Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel
electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 1995; 33(9): 2233-2239.
Toledo MRF, Fontes CF, Trabulsi LR. EPM: modificação do meio de Rugai e Araújo para a
realização simultânea dos testes de produção de gás a partir de glicose, H2S, uréase e
triptofano desaminase. Rev. Microbiol. 1982a; 13: 309-315.
Toledo MRF, Fontes CF, Trabulsi LR. MILi: um meio para a realização dos testes de
motilidade, indol e lisina descarboxilase. Rev. Microbiol. 1982b; 13: 230-235.
Usui M, Iwasa T, Fukuda A, T. Sato T, Okubo T, Tamura Y. The role of flies in spreading the
extended- spectrum β-lactamase gene from cattle. Microb. Drug Resist. 2013; 19:415-420.
Usui M, Shirakawa T, Fukuda A, Tamura Y. The role of flies in disseminating plasmids with
antimicrobial-resistance genes between farms. Microb. Drug Resist. 2015; 21(5): 562-569.
Ventola CL. The antibiotic resistance crisis. Part 1: Causes and threats. P&T. 2015; 40(4):
277-283.
Verdier K, Nyman A, Greko C, Bengtsson B. Antimicrobial resistance and virulence factors
in Escherichia coli from Swedish dairy calves. Acta Vet. Scand. 2012; 54: 1-10.
Vieira AR, Collignon P, Aarestrup FM, McEwen SC, Hendriksen RS, Hald T, et al.
Association between antimicrobial resistance in Escherichia coli isolates from food animals
and blood stream isolates from humans in Europe: An ecological study. Foodborne Pathog.
Dis. 2011; 8: 1295-1301.
Vila J, Ruiz J, Goni P, De Anta MT. Detection of mutations in parC in quinolone-resistant
clinical isolates of Escherichia coli. Antimicrob. Agents Chemother. 1996; 40: 491-493.

Referências Bibliográficas| 85
Wang W. Houseflies as potential vectors for antibiotic resistant bacteria. Columbus. Thesis
[Master of Science in the Graduate School of The Ohio State University] - Ohio State
University ; 2013.
Webber M, Piddock LJV. Quinolone resistance in Escherichia coli. Vet. Res. 2001; 32: 275-
284.
Wichmann F, Udikovic-Kolic N, Andrew S, Handelsman J. Diverse antibiotic resistance
genes in dairy cow manure. MBio. 2014; 5: 1-9.
Winn Jr WC, Allen SD, Janda WM, Koneman EW, Procop G, Schreckenberger PC, Woods,
G. Diagnóstico Microbiológico. Texto e Atlas Colorido. 6ed. Guanabara Koogan: Rio de
Janeiro, 2008. p. 1565.
Yamamoto S, Tsukamoto T, Kurazono H, Takeda Y, Yoshida O. Distribution of virulence
factors in Escherichia coli isolated from urine of cystitis patient. Microbiol. Immunol. 1995;
39: 401-404.
Yeates DK, Wiegmann BM, Courtney GW, Meier R, Lambkin C, Pape T. Phylogeny and
systematics of Diptera: Two decades of progress and prospects. Zootaxa. 2007; 1668: 565–
590.
Yu J, Kaper, J.B. Cloning and characterization of the eae gene of enterohaemorragic
Escherichia coli O157:H7. Mol. Microbiol. 1992; 6: 411-417.
Zoccal R, Carneiro AV, Junqueira R, Zamagno MA. A nova pecuária leiteira brasileira. In:
Barbosa SBP, Batista AMV, Monardes H. III Congresso Brasileiro de Qualidade do Leite.
CCS Gráfica e Editora: Recife. 2008; 1: 85-9.
Zurek L, Ghosh A. Insects represent a link between food animal farms and the urban
environment for antibiotic resistance traits. Appl. Environ. Microbiol. 2014; 80(12): 3562-
3567.

VIII – Apêndices

Apêndices| 87
APÊNDICE A – Resultado do ensaio de disco aproximação para a dectação fenotípica
de ESBL em cepas de E. coli doadoras e transconjugantes obtidas após conjugação.
Cepas doadoras: A: MA17a; B: MA17b; C: MA17c; D: MA17d; I: MA17e; J: MA26a; K: MA26b; L: MA26d.
Cepas transconjugantes: E: MA17a; F: MA17b; G: MA17c; H: MA17d; M: MA17e; N: MA26a; O: MA26b; P:
MA26d.
A B C D
E F G H
I J K L
M N O P

Apêndices| 88
APÊNDICE B – Antibiograma da cepa transconjugante MA17d. Colônias
hipermutantes no interior do halo de inibição de sulfametazol/trimetoprim (SUT).

IX – Anexos

Anexos| 90
ANEXO A – Declaração sobre bioética e biossegurança.

Anexos| 91
ANEXO B – Declaração sobre a não infringência da lei nº 9610/98 e do direito autoral de
qualquer editora





























