Embed Size (px)
Citation preview

Maiele Bertoldo Lewandowski
LEPTINA: MECANISMOS DE AÇÃO NA OBESIDADE
Santa Maria, RS
2006

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 1
Maiele Bertoldo Lewandowski
LEPTINA: MECANISMOS DE AÇÃO NA OBESIDADE
Trabalho Final de Graduação apresentado ao Curso de Nutrição – Área de Ciências da Saúde,
do Centro Universitário Franciscano, como requisito parcial para obtenção do grau de
Nutricionista – Bacharel em Nutrição.
Orientadora: Cristina Machado Bragança de Moraes
Santa Maria, RS
2006

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 2
Maiele Bertoldo Lewandowski
LEPTINA: MECANISMOS DE AÇÃO NA OBESIDADE
Trabalho final de graduação apresentado ao Curso de Nutrição – Área de Ciência da Saúde, do Centro Universitário Franciscano, como requisito parcial para obtenção do grau de Nutricionista – Bacharel em Nutrição.
____________________________________________________ Cristina Machado Bragança de Moraes – Orientadora (UNIFRA)
_________________________________________________ Viviani Ruffo de Oliveira (UNIFRA)
____________________________________________________ Selvino Luiz Cogo (UNIFRA)
Aprovado em ........ de ...................................... de ...............

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 3
RESUMO
A incidência mundial de obesidade dobrou nos últimos trinta anos e o aumento da prevalência do sobrepeso e da obesidade atribui-se, principalmente, às mudanças nos estilos de vida que incidem sobre a susceptibilidade ou predisposição genética para ser obeso. A leptina é um peptídeo que desempenha importante papel na regulação da ingestão alimentar e no gasto energético, gerando um aumento na queima de energia e diminuindo da ingestão alimentar. As concentrações de leptina são influenciadas pela adiposidade, fatores hormonais e nutricionais. Em pacientes obesos, os níveis de leptina são aumentados, em proporção à gordura corporal e à hiperleptinemia, junto com a escassa resposta do peso corporal pela terapia com leptina recombinante, definindo assim, um estado de resistência à leptina. Considerando o fato de que o hormônio leptina é um marcador da adiposidade corporal, destacando-se como sinalizadora e moduladora do estado nutricional no indivíduo obeso, o objetivo principal deste trabalho foi identificar, através de uma revisão bibliográfica, qual a influência e os mecanismos de ação do hormônio leptina sobre a obesidade e ressaltar a preocupação de diversos autores com a epidemia da obesidade. Palavras-chaves: Obesidade. Leptina. Insulina.
ABSTRACT
The world incidence of obesity bent in the last thirty years and the increase of the prevalence of the overweight and of the obesity it is attributed, mainly, to the changes in the lifestyles that happen on the susceptibility or genetic predisposition to be obese. The leptin is a peptidy that plays important part in the regulation of the alimentary ingestion and in the energy expense, generating an increase in the burning of energy and decreasing of the alimentary ingestion. The leptin concentrations are influenced by the adiposity, hormonal and nutritional factors. In the obese patient, the leptin levels are increased, in proportion to the corporal fat and the hiperleptinemy, with the scarce answer of the corporal weight for the therapy with leptin recombinant, defining like this, a resistance state to the leptin. Considering the fact that the leptin hormone is a marker of the corporal adiposity, standing out as a signalizer and a modulater of the nutricional state in the obese individual, the main objective of this work was to identify, through a bibliographical revision, which the influence and the mechanisms of action of the leptin hormone about the obesity and to point out the several authors' concern with the obesity’s epidemic desease. Keywords: Obesity. Leptin. Insulin.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 4
LISTA DE ILUSTRAÇÕES
Figura 1: Componentes do Balanço Energético ...................................................................... 16
Figura 2: Estrutura da Leptina ................................................................................................. 21

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 5
LISTA DE TABELAS
Tabela 1: Importantes causas e preocupações da obesidade ............................................. 10
Tabela 2: Influência de fatores orgânicos e ambientais nos níveis de leptina ..................... 23

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 6
SUMÁRIO
1 INTRODUÇÃO................................................................................................................ 7
2 REFERENCIAL TEÓRICO .......................................................................................... 9
2.1 OBESIDADE................................................................................................................... 9
2.1.1 Avaliação do Indivíduo Obeso .................................................................................. 14
2.1.2 Balanço Energético .................................................................................................... 15
2.1.3 Substâncias Envolvidas no Controle do Apetite, Ingestão Alimentar e Saciedade .... 17
2.2 LEPTINA....................................................................................................................... 20
2.2.1 Leptina e Obesidade .................................................................................................. 27
2.2.2 Efeito dos Macronutrientes sobre os Níveis de Leptina ............................................ 30
2.2.3 Receptor da Leptina ................................................................................................... 32
2.2.4 Leptina Recombiante ................................................................................................. 33
3 METODOLOGIA .......................................................................................................... 36
4 CONCLUSÃO ................................................................................................................ 37
REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................. 38

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 7
1 INTRODUÇÃO
Devido, em grande parte, à revolução industrial, acompanhada de mudanças nos
estilos de vida (menor atividade física e alimentação com alto teor energético), o peso
corpóreo das populações vem continuamente aumentando. Hoje, a obesidade é considerada
epidêmica e um grave problema de saúde pública mundial, que se sobrepôs aos antigos, como
a desnutrição e doenças infectocontagiosas, inclusive em países emergentes, como é o caso do
Brasil.
No entanto, atribuir este verdadeiro surto epidêmico de obesidade a fatores
nutricionais e comportamentais é ignorar a biologia básica do equilíbrio energético.
Claramente, a vontade de comer e de se exercitar tem uma base bioquímica e hormonal. A
identificação de neurotransmissores, controlados hormonalmente, que interferem no
comportamento alimentar (neuropeptídio Y, por exemplo), hormônios que regulam a
lipogênese (por ex.: insulina) e hormônios que sinalizam a adequação calórica (leptina)
colocam em perspectiva uma nova compreensão dos mecanismos, envolvidos nessa mudança
de padrão corpóreo, e talvez venham a permitir uma abordagem menos empírica e mais
fundamentada em bases fisiopatológicas. Reconhece-se cada vez mais que a obesidade não é
uma condição única, mas um achado comum a várias situações patológicas.
Nos últimos vinte anos, aumentou o conhecimento sobre diversos fatores que
contribuem para o desenvolvimento da obesidade, bem como as conseqüências endócrinas e
metabólicas desta doença. Muito deste conhecimento foi derivado de estudos em modelos de
obesidade animal. Estudos sobre as causas e tratamento da obesidade têm sido desenvolvidos,
em animais que apresentam esta característica, através de lesão neural, alterações endócrinas,
anormalidades genéticas e alterações alimentares.
Atualmente, a obesidade é considerada uma doença de caráter multifatorial que se
integra ao grupo de Doenças Crônicas Não-Transmissíveis (DCNT), que vem atingindo
proporções epidêmicas tanto nos países desenvolvidos quanto naqueles em desenvolvimento.
É considerada uma doença crônica e está relacionada à alta taxa de morbidade e mortalidade.
A obesidade pode ser definida como o acúmulo excessivo de gordura corporal, fator
que traz prejuízos à saúde, destacando-se as dificuldades respiratórias, problemas
dermatológicos e distúrbios do aparelho locomotor, além de favorecer o desenvolvimento de
Diabetes Mellitus tipo II, dislipidemias, doenças cardiovasculares e alguns tipos de câncer.
O acúmulo excessivo do tecido adiposo ocorre devido à ingestão de um aporte
calórico excessivo e crônico de substratos metabolizados, presentes nos alimentos e nas

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 8
bebidas (como: proteínas, carboidratos, lipídios e álcool), em relação ao gasto energético do
organismo (pelo metabolismo basal, efeito termogênico e pela atividade física). Tanto os
hábitos alimentares como o estilo de vida, os fatores sociológicos, as alterações metabólicas e
neuro-endócrinas e os componentes hereditários contribuem para que ocorra o acúmulo de
gordura.
Mesmo levando-se em consideração todos os fatores supracitados, há muitas
evidências de que a genética contribui, expressivamente, na regulação do peso corporal. Dessa
forma, ao longo do tempo, os genes intervêm na manutenção do peso corporal, através de vias
eferentes (leptina, nutrientes, sinais nervosos) e mecanismos centrais (insulina, catecolaminas,
sistema nervoso autônomo).
O interesse pelo tema surgiu devido à leptina ser um hormônio, cujo papel é
importante na regulação da ingestão alimentar, no consumo energético e no peso corporal.
Considerando o fato de que a leptina é um marcador da adiposidade corporal,
destacando-se como sinalizadora e moduladora do estado nutricional no indivíduo obeso,
questionou-se qual a influência e os mecanismos de ação do hormônio leptina sobre a
obesidade, sendo este o principal objetivo deste trabalho. Procurou-se também ressaltar a
preocupação de diversos autores com a epidemia da obesidade.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 9
2 REFERENCIAL TEÓRICO
2.1 OBESIDADE
Obesidade é o resultado de uma complexa interação entre fatores comportamentais,
culturais, genéticos, fisiológicos e psicológicos. Pode, dessa forma, ser classificada em dois
contextos: por determinação genética ou fatores endócrinos e metabólicos, ou, então,
influenciada por fatores externos, sejam eles de origem dietética, comportamental ou
ambiental. Em termos simples, pode ser definida como um estado de desigualdade entre as
calorias ingeridas versus calorias consumidas pelo organismo que levará ao excesso ou
anormal acumulo de gordura (NAMMI et al., 2004; ROMERO; ZANESCO, 2006).
O ganho de peso excessivo ocorre quando a quantidade de energia consumida, através
dos alimentos, é maior do que a energia gasta pelo corpo. Assim, de acordo com Guyton e
Hall (2002, p.754), “para cada 9,3 calorias em excesso que penetram no organismo, ocorre o
armazenamento de 1 grama de gordura”.
A obesidade inexistiu como problema de saúde, pois a escassez de alimentos e a alta
atividade física, praticada pelo homem, fez com que ocorresse seleção dos indivíduos,
portadores de mecanismos orgânicos mais eficientes de estocagem de excesso de energia,
como gordura, para períodos de fome, permitindo, assim, a evolução das sociedades e o seu
surgimento. Acredita-se que este é um dos motivos que tornou a obesidade uma condição
comum na sociedade industrializada e está aumentando rapidamente. É muito provável que a
capacidade para armazenar gordura em tempo de abundância nutricional seja um positivo
traço de seleção há milhares de anos na evolução humana (SPIEGELMAN; FLIER, 2001;
ALMEIDA; FERREIRA, 2005; PARACCHINI; PEDOTTI; TAIOLI, 2005).
A obesidade também é considerada uma doença integrante do grupo de Doenças
Crônicas Não-Transmissíveis (DCNT), as quais são de difícil conceituação, gerando aspectos
polêmicos quanto à sua própria denominação, seja como doenças não-infecciosas, doenças
crônicas não-transmissivéis, sendo esta última a conceituação a mais utilizada (PINHEIRO;
FREITAS; CORSO, 2004).
A incidência mundial de obesidade dobrou nos últimos trinta anos e o aumento da
prevalência do sobrepeso e da obesidade atribui-se, principalmente, às mudanças nos estilos
de vida (como o aumento do consumo de alimentos ricos em gordura e o sedentarismo) que
incidem sobre a susceptibilidade ou predisposição genética para ser obeso. Nesse contexto,
também o fenótipo da obesidade, no qual se distinguem quatro tipos, em função da

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 10
distribuição anatômica da gordura corporal (global, andróide, ginóide, visceral), é
influenciado pela base genética e por fatores ambientais (LOPES et al., 2004; CARNEIRO,
2005).
Identificar a etiologia da obesidade não parece ser simples e objetivo. A epidemia da
obesidade não está restrita à determinada raça, etnia ou grupo socioeconômico. Apesar de os
indivíduos com excesso de peso serem, rotineiramente, considerados pessoas com distúrbio
de comportamento ou de personalidade, há fatores que fazem um papel crítico no
desenvolvimento da obesidade pela genética ou pela suscetibilidade metabólica. Existem
vários estudos que mostram que os fatores genéticos são predominantes no surgimento da
obesidade e que a predisposição genética para a obesidade está acima dos fatores ambientais.
Alguns dados sugerem que a hereditariedade pode estar presente em 50 a 90% dos casos. A
ação da influência ambiental leva a um aumento na energia ingerida ou a uma diminuição da
energia gasta com baixa atividade física e, portanto, há um aumento da probabilidade de
favorecer a obesidade. Comportamentos sedentários, particularmente assistir à televisão ou
andar de carro também contribuem para o risco de obesidade, além de condições médicas, tais
como doenças hereditárias raras e uma desigualdade hormonal (por ex.: hipotireodismo),
idade, fatores genéticos, etnia e gênero (Tabela 1) (NAMMI et al., 2004; PINHEIRO;
FREITAS; CORSO; 2004; CARNEIRO, 2005; PARACCHINI; PEDOTTI; TAIOLI, 2005).
Tabela 1 Importantes causas e precauções da obesidade
Obesidade
Causas Precaução
Estilo de Vida Sedentário Atividade Física
Alimentos Disponíveis Controle da Dieta
Dieta com alto teor de gordura Terapia Comportamental
Hereditariedade Medicamentos
Drogas que causam ganho de peso Consulta
Fonte: NAMMI et al., 2004.
Segundo Nammi et al. (2004), a obesidade pode ser descrita como “Nova Síndrome do
Mundo”. Os autores apresentam dados estatísticos e revelam que o problema da obesidade
tem aumentado de 12 para 20%, em homens, e de 16 para 25%, em mulheres, nos últimos dez
anos. Pacientes obesos têm sido associados a aumento do risco de morbidade e mortalidade,

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 11
se comparados a indivíduos com peso corporal ideal. Uma modesta redução de peso, 5-10%
do peso inicial, está associada a um significativo progresso das condições de co-morbidade.
A obesidade está associada a condições médicas que podem causar prejuízos à saúde e
morte prematura, as quais são: artrites, defeitos no nascimento, várias formas de câncer,
doenças cardiovasculares, diabetes, hipertensão, infertilidade, trombose venosa grave e função
respiratória debilitada, doença hepática, pancreatites, dispnéia do sono e derrame
(PARACCHINI; PEDOTTI; TAIOLI, 2005).
Para Fisberg, Cintra e Oliveira (2005, p.11):
O relatório da IOTF – Internacional Obesity Task Force – (2003), para a Organização Mundial da Saúde, estima que aproximadamente 10% dos indivíduos entre 5 e 17 anos de idade, apresentam excesso de gordura corporal, sendo que 2% a 3% são obesos; o que correspondia no ano 2000 a 155 milhões de crianças com excesso de peso, sendo 30 a 45 milhões de crianças obesas ao redor do mundo.
Romero e Zanesco (2006, p.86) colocam que, de acordo com a Organização Mundial
da Saúde (OMS), “o número de obesos entre 1995 e 2000 passou de 200 para 300 milhões,
perfazendo quase 15% da população mundial. Estimativas mostram que, em 2025, o Brasil
será o quinto país no mundo a ter problemas de obesidade em sua população”.
No Brasil, a obesidade é um evento recente, principalmente como problema de Saúde
Pública. A partir da análise de relatos da Era Paleolítica sobre ‘homens corpulentos’, Pinheiro,
Freitas e Corso (2004) afirmam que o grau epidêmico da obesidade nunca esteve tão elevado
como na atualidade. E ressaltam que esse detalhe torna-se ainda mais relevante, quando se
verifica que o aumento é, proporcionalmente, mais elevado na classe mais baixa, apesar de
estar distribuído em todas as regiões do país e nos diferentes estratos socioeconômicos da
população. O autor acrescenta que, em todas as regiões do Brasil, parcelas significativas da
população adulta apresentam sobrepeso e obesidade. A situação mais crítica é verificada na
Região Sul, onde 34% dos homens e 43% das mulheres apresentam algum grau de excesso de
peso (aproximadamente 5 milhões de adultos de ambos os sexos). Já na Região Sudeste, mais
de 10 milhões de adultos têm sobrepeso e cerca de 3 milhões e meio apresentam obesidade.
O comportamento da obesidade tem variado no país ao longo do tempo. De acordo
com Monteiro, Conde e Castro (2003), entre 1975 e 1989, o risco de obesidade se elevava
juntamente com os níveis de escolaridade, tendendo a ser máximo para homens e mulheres
com maior escolaridade. Porém, em um segundo período analisado, de 1989 a 1997, o maior
aumento na obesidade foi verificado entre indivíduos sem escolaridade, tendo sido constatada

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 12
estabilidade ou mesmo diminuição da doença nos estratos femininos de média ou de alta
escolaridades (ALMEIDA; FERREIRA, 2005).
É claro que as tendências da obesidade não são limitadas a um determinado grupo
étnico ou região. Tem se verificado um aumento constante desta doença na América do Norte,
América Latina, África do Sul, Malásia e nações do Pacífico. Em 1980, a prevalência da
obesidade, em adultos americanos, era de 14,5%. No final da década passada, 1 de cada 4
americanos eram obesos. Atualmente, o número de pessoas com IMC>30 kg/m² é estimado
em 250 milhões, ou 7% da população adulta do mundo. Porém, há uma grande variação entre
países e populações, com prevalência de menos de 5% na China, no Japão e em certos locais
da África, enquanto a população urbana de Samoa apresenta mais de 75% de ocorrência da
obesidade (ALMEIDA; FERREIRA, 2005; OLIVEIRA et al., 2004; VIEIRA et al., 2005).
Recentemente, a obesidade está sendo considerada a mais importante desordem
nutricional. As mudanças epidemiológicas, demográficas e socioeconômicas ocorridas
permitiram mudanças nos padrões alimentares. Por isso, nas últimas décadas, verifica-se um
processo de transição nutricional no Brasil, caracterizada pela diminuição progressiva da
desnutrição e aumento da obesidade (PEREIRA; FRANCISCHI; LANCHA, 2003).
A Transição Nutricional integra os processos de Transição Demográfica e
Epidemiológica. De acordo com Pinheiro, Freitas e Corso (2004), a Transição Nutricional “é
um processo de modificações seqüenciais no padrão de nutrição e consumo, que acompanham
alterações econômicas, sociais e demográficas, e do perfil de saúde populacional”.
Há três hipóteses que tentam explicar o aumento da obesidade, as quais são objetos de
estudos: (1) possibilidade de populações apresentarem-se, geneticamente, mais suscetíveis à
obesidade, associando este fator com as influências ambientais; (2) aumento da obesidade em
países desenvolvidos e em desenvolvimento, a rápido e intenso declínio de gasto energético
dos indivíduos, pelo sedentarismo, pela diminuição do consumo de fibras e aumento do
consumo de gorduras e açúcares; (3) obesidade seria resultado de uma desnutrição energético-
protéica precoce, seqüela da desnutrição (PINHEIRO; FREITAS; CORSO; 2004).
Almeida e Ferreira (2005, p.190-191) definem as características do predomínio da
obesidade em quatro itens:
1.Correlação negativa com o estrato socioeconômico. Dieta pouco saudável, estresse psicossocial, baixo nível de educação e de orientação em saúde contam para essa relação. 2.Populações urbanas tendem a uma maior prevalência de obesidade, o que seria justificado por hábitos mais sedentários (como dependência de transporte motorizado e trabalhos sedentários), dieta com mais carboidratos complexos, maior

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 13
teor de gordura, mais alimentos processados e de fast food, e pela exposição a uma grande quantidade de propagandas de comida (transição nutricional). 3. Há um aumento da prevalência de obesidade com a idade até cerca de 60 anos. Após isso, a tendência é haver um declínio em razão de uma diminuição na massa magra, influenciando o IMC, e da provável maior mortalidade em indivíduos obesos. 4. Minorias étnicas são predispostas a desenvolver obesidade e doenças associadas quando expostas a um balanço energético positivo e hábito da vida urbana moderna.
Na Política Nacional de Alimentação e Nutrição (PNAN), a obesidade é apontada
como evento de controle prioritário; como conseqüência, foi publicado, pelo Ministério da
Saúde, o Plano Nacional para Promoção da Alimentação Adequada e Peso Saudável, com
mensagens de incentivo a hábitos de vida e alimentação saudável, principalmente para
promoção da saúde. Os serviços de saúde começam a se organizar para implementar
propostas e estratégias de atenção primária para a obesidade (PINHEIRO; FREITAS;
CORSO, 2004).
Em 1995, o número de mortes em adultos, atribuídas a excesso de peso, foi de 1
milhão, o dobro das atribuídas à desnutrição. É natural concluir que esse problema de saúde
pública que traz consigo uma gama de comorbidades e morte, causa enorme impacto na
economia mundial e que é urgente implantar medidas de prevenção no âmbito populacional
(ALMEIDA; FERREIRA, 2005).
A co-existência de obesidade, em vários membros da mesma família, aponta para a
participação da herança genética no desenvolvimento da obesidade. Segundo Lopes et al.
(2004), a probabilidade de que os filhos sejam obesos, quando os pais o são, é de 50% a 80%.
O autor confirma essa hipótese, por existirem indivíduos com alteração na termogênese, no
metabolismo basal ou na ativação simpática, e ainda por constatar que os fatores genéticos
podem modificar os efeitos da atividade física sobre o peso e a composição corporal.
Baseado na análise do estudo, realizado por Boejerson, em 1976, com 40 gêmeos
monozigóticos e 60 dizigóticos, Schneider e Meyer (2005) afirmam que os fatores genéticos
são de grande importância na determinação da obesidade. Apresentam também, através do
estudo feito por Stunkard, em 1986, com 540 adultos adotados, que existe uma forte relação
do índice de massa corporal (IMC) com os pais biológicos, o que não ocorre com os pais
adotivos. Dessa forma, evidencia-se que a obesidade humana e a gordura corpórea são
altamente hereditárias.
Uma criança obesa, nos primeiros seis meses de vida, tem 2,3 vezes mais
probabilidade de se tornar um adulto obeso, enquanto que adolescentes obesos dos 10 aos 13
anos têm 6,5 vezes mais probabilidade de se tornarem adultos obesos (DAMIANI, 2000).

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 14
Levando-se em consideração todas as afirmações ora referidas, é evidente que a
genética contribui, substancialmente, na regulação do peso corporal, influenciada pela dieta
ocidental e, principalmente, pelo estilo de vida sedentário. Através de estudos, realizados em
animais, especialmente em camundongos, geneticamente obesos, foram descobertos vários
genes que estão relacionados com a obesidade. Portanto, “o peso corpóreo é regulado por uma
interação complexa entre hormônios e neuropeptídios, sob o controle principal de núcleos
hipotalâmicos” (RODRIGUES; SUPLICY; RADOMINSKI, 2003, p.398).
2.1.1 Avaliação do Indivíduo Obeso
Em vários estudos, a obesidade é definida, baseada em medidas antropométricas,
principalmente altura, peso e circunferência da cintura. Cada medida fornece informações do
grau e distribuição da obesidade. Sujeitos adultos são definidos como pré-obesos, se o Índice
de Massa Corporal (IMC) estiver entre 25-39,9 Kg/m² e, se o IMC estiver acima de 30 Kg/m²,
o indivíduo é considerado obeso. Alguns estudos utilizam-se da relação cintura-quadril (RCQ)
com uma áspera aproximação da distribuição da gordura corporal. Nos idosos, a obesidade
tem sido descrita como um excesso de gordura com a perda de massa magra (CUPPARI,
2005; PARACCHINI; PEDOTTI; TAIOLI, 2005).
A composição corporal também pode ser estimada usando uma variedade de métodos
tecnológicos, incluindo a densitometria com emissão de raios-X de dupla energia (DEXA),
pletismografia, pesagem hidrostática e bioimpedância (NAMMI et al., 2004).
O IMC não descreve a ampla variação que ocorre na composição corporal de
indivíduos, desconsiderando idade, relação entre IMC e indicadores de composição corporal,
como, por exemplo, a gordura corporal. Assim, estes critérios podem significar pouca
especificidade, em termos de associação de risco de saúde, entre diferentes indivíduos ou
populações (PINHEIRO; FREITAS; CORSO; 2004).
Existe uma boa correlação entre o IMC e a porcentagem de gordura corporal. A
porcentagem de gordura é maior em mulheres do que homens. De acordo com Nammi et al.
(2004), a razão para isto pode ser que, em mulheres, o excesso de gordura corporal é
usualmente distribuída como gordura subcutânea e é principalmente periférica (coxas,
nádegas, peitos), uma vez que, em homens, há um excesso de armazenamento de gordura
corporal, em cavidade abdominal, como gordura abdominal subcutânea que, conforme os
autores (p.425), a porcentagem de gordura pode ser calculada da seguinte maneira:

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 15
porcentagem de gordura corporal = 1.2 (IMC) + 0.23 (idade) – 10.8 (gênero) – 5.4; colocar no
lugar de gênero = 1 para homem e 0 para mulher
Quando se estuda a relação da adiposidade com a obesidade, é preciso considerar o
número e o tamanho das células adiposas. Estudos transversais e longitudinais de
desenvolvimento de células de tecido adiposo mostraram que, de 2 a 10 anos de idade,
crianças obesas exibem um número significativamente maior de células adiposas do que seus
pares não-obesos da mesma idade. O número de células adiposas foi significativamente maior
em crianças obesas desde os dois anos e essa elevação persistiu até os 16 anos (SCHNEIDER;
MEYER, 2005).
2.1.2 Balanço Energético
O peso corporal é uma função do balanço energético e de nutrientes ao longo de um
período de tempo. O peso, a composição corpórea e o estoque de energia, como triglicerídio
no tecido adiposo, são determinados pela interação entre fatores genéticos, ambientais e
psicossociais. Esta influência, ultimamente, age na mudança da equação do balanço
energético, determinado pela ingestão de macronutrientes, pelo gasto energético e pela
termogênese dos alimentos (BOUCHARD, 2000; SPIEGELMAN; FLIER, 2001).
O balanço energético positivo que ocorre quando o valor calórico ingerido é superior
ao gasto, é importante por contribuir para o desenvolvimento da obesidade, promovendo
aumento nos estoques de energia e peso corporal. O início da manutenção de um balanço
calórico positivo, relativo às necessidades do organismo, pode ser conseqüência tanto de
aumento na ingestão calórica, como redução no total calórico gasto, ou dos dois fatores
combinados (PEREIRA, FRANCISCHI; LANCHA, 2003).
O Sistema Nervoso Central influencia o balanço energético e o peso corporal, através
de três mecanismos: (1) efeitos no comportamento, incluindo alimentação e atividade física;
(2) efeitos na ativação do Sistema Nervoso Autônomo, que regula a energia gasta e outros
aspectos do metabolismo; (3) efeitos no sistema neuroendócrino, incluindo a secreção de
hormônios, tais como o hormônio do crescimento, tireóide, cortisol, insulina e hormônios
esteróides. Portanto, existem vários fatores que atuam e interagem na regulação da ingestão de
alimentos e de armazenamento de energia, contribuindo para o surgimento e a manutenção da
obesidade. Entre eles, fatores neuronais, endócrinos e adipocitários, bem como fatores
intestinais (SPIEGELMAN; FLIER, 2001; HALPERN; RODRIGUES; COSTA, 2004).
Supondo que um indivíduo não tenha problema de absorção de nutrientes, a energia

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 16
armazenada somente aumentará se a energia ingerida exceder o total de energia, gasta pelo
corpo. A energia consumida é exigida para atividade física, metabolismo basal e termogênese.
A atividade física refere-se a todo movimento voluntário do indivíduo, enquanto que o
metabolismo basal engloba os processos bioquímicos, necessários para sustentar a vida. A
termogênese está relacionada à energia dissipada, na forma de calor, em resposta às mudanças
ambientais, tais como a exposição ao frio e alterações na dieta. Não há evidências de que os
indivíduos obesos apresentam a taxa metabólica basal do repouso diminuída. Além do mais,
alguns estudos mostraram que o aumento da massa corpórea total eleva a taxa do
metabolismo basal para retardar um ganho de peso excessivo. Logo, um indivíduo obeso
possui TMB mais elevada do que um indivíduo magro da mesma altura. Essa observação
sugere que existe um mecanismo de manutenção do peso corpóreo e suporta a teoria de que o
comportamento não é o único determinante da obesidade (SPIEGELMAN; FLIER, 2001;
CARNEIRO, 2005).
A regulação do comportamento alimentar pode ser divido em sistemas de controle a
curto e longo prazos (Figura 1).
Figura 1: Componente do Balanço Energético Fonte: Flier (2004, p.338)
Controle a curto prazo envolve o início e o término das refeições. O principal
determinante do tamanho da refeição é a saciedade, uma resposta de fatores neural e

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 17
endócrino, tais como distensão e a liberação da colecistocinina no intestino, gerada durante a
refeição. A longo prazo, há sinais que refletem o estado das reservas energéticas, tais como a
gordura, derivada do hormônio leptina, que proporcionam informações ao Sistema Nervoso
Central e levam à regulação do comportamento alimentar, para promover energia
homeostática. Estes dois sistemas se interrelacionam (SPIEGELMAN; FLIER, 2001).
2.1.3 Substâncias Envolvidas no Controle do Apetite, Ingestão Alimentar e Saciedade
NEUROPEPTÍDIO Y (NPY) E PROTEÍNA RELACIONADA À AGOUTI (AGRP)
O NPY é um peptídeo de 36 aminoácidos, concentrado principalmente no hipotálamo,
uma região crucial para a regulação do apetite que tem emergido como uma possível chave
neurotransmissora, candidata à regulação do balanço energético (SPIEGELMAN; FLIER,
2001; NAMMI et al., 2004; HERMSDORFF; VIEIRA; MONTEIRO, 2006).
O NPY tem um papel importante no sistema orexígeno, promovendo aumento da
ingestão alimentar, diminuição do gasto energético e aumento da lipogênese em animais. A
redução dos níveis de leptina e insulina ativa os neurônios produtores de NPY no núcleo
arqueado do hipotálamo. Entretanto, não foram demonstradas diferenças nos níveis de NPY
no plasma nem no líquor entre humanos de peso normal e obesos (RODRIGUES; SUPLICY;
RADOMINSKI, 2003).
O NPY não age isoladamente no controle do peso. O AGRP é sintetizado no núcleo
arqueado apenas nos neurônios que expressam NPY e antagoniza os efeitos anorexígenos do
hormônio estimulador de melanócitos (α-MSH) sobre os receptores de melanocortina. A
expressão gênica do NPY, bem como a do AGRP, são ativadas durante o jejum, levando
certos autores a postular que os neurônios que expressam ambos os peptídeos representam
uma unidade funcional no núcleo arqueado. A queda dos níveis de leptina, ao final de um
período de jejum, levaria a um aumento da ingestão alimentar, gerada pela estimulação do
NPY e o bloqueio do efeito anorexígeno do α-MSH pelo AGRP (NEGRÃO; LICINIO, 2000;
RODRIGUES; SUPLICY; RADOMINSKI, 2003).
O gene da proteína relacionada à agouti (AGRP - agouti-related protein) é expresso
no núcleo arqueado do hipotálamo, nas mesmas células que expressam o neuropeptídio Y,
sendo sua expressão inibida pela leptina (SPIEGELMAN; FLIER, 2001).
GRELINA
Este hormônio é responsável pelo aumento da ingestão alimentar, seu principal local

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 18
de síntese é o intestino. Em obesos, os níveis plasmáticos de grelina estão diminuídos
(WEIGLE et al. 2005).
COLECISTOCININA (CCK)
No início dos anos 1970, descobriu-se que a CCK, um peptídeo intestinal, atuava na
promoção da saciedade. Evidências demonstram que a saciedade prandial é atribuída
predominantemente à ação da CCK, liberada pelas células do trato gastrintestinal, em resposta
à presença de gordura e proteína. A CCK, além de inibir a ingestão alimentar, também induz a
secreção pancreática, a secreção biliar e a contração vesicular (HALPERN; RODRIGUES;
COSTA, 2004).
FATOR DE TRANSCRIÇÃO COCAÍNA-ANFETAMINA DEPENDENTE (CART)
O CART foi descoberto em 1998 e atua como um anorexígeno, sendo que sua
expressão gênica, no núcleo arqueado, diminui durante o jejum e aumenta após a
administração de leptina (RODRIGUES; SUPLICY; RADOMINISKI. 2003).
MELANOCORINA 4 (MCR4)
A ativação do receptor da MCR4 pelo MSH (melanocyte-stimulating hormone) reduz
a quantidade ingerida, enquanto que a supressão de MCR4 sinaliza, através deste receptor,
pelo antagonista endógeno AGRP ou antagonistas farmacológicos, o aumento do volume da
alimentação e diminuição da resposta da leptina (SPIEGELMAN; FLIER, 2001).
PRÓ-ÓPIO-MELANOCORTINA (POMC)
É um peptídeo anorexígeno. Um dos principais papéis da leptina no SNC é ativar os
neurônios do núcleo arqueado do hipotálamo que expressam a POMC. Este pró-hormônio dá
origem à corticotrofina (ACTH), às melanocortinas-MSH (α, β e ϒ ) e a β-endorfina (FLIER,
2004; CARNEIRO, 2005).
FATOR DE NECROSE TUMORAL
O TNF-alfa modula, positivamente, a secreção de leptina pelos adipócitos, tendo-se
demonstrado uma associação independente entre a leptinemia e o nível circulante do receptor
solúvel do TNF-alfa, um indicador sensível de ativação desta interleucina (SPIEGELMAN;
FLIER, 2001).

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 19
PEPTÍDIO YY
Este é outro peptídeo que atua como inibidor da ingestão alimentar. É expresso pelas
células da mucosa intestinal e sugere-se que a regulação seja neural, já que seus níveis
plasmáticos aumentam quase que imediatamente após a ingestão alimentar. Indivíduos obesos
apresentam menor elevação dos níveis de PYY pós-prandial, especialmente em refeições
noturnas, levando a uma ingestão calórica maior (HALPERN; RODRIGUES; COSTA, 2004).
INSULINA
Insulina é um produto das células β pancreáticas e é a mestre metabólica interruptora
entre os estados de alimentação e jejum. O sistema de transporte da insulina, através da
barreira hemato-encefálica, foi descrita, e a administração de doses farmacêuticas de insulina,
dentro do SNC, suprime a quantidade ingerida em roedores e primatas subumanos e regula a
expressão dos neuropeptídios hipotalâmicos que influenciam o apetite. Ratos com mutações
no receptor da leptina apresentam uma redução na resposta na administração da insulina
centralmente. A ação da leptina pode influenciar o caminho de ação da insulina no cérebro
(FLIER, 2004).
A insulina elevada pode estimular o tecido adiposo a produzir leptina, inibindo a
produção e a secreção do neuropeptídio Y, levando a uma redução da ingestão alimentar e
aumento do gasto energético. Independente da adiposidade, a insulina pode ser um
determinante da concentração de leptina. O efeito da insulina sobre a produção de leptina
diminui com a resistência a insulina e com o aumento da adiposidade (UEHARA; ROSA;
CARMO, 2006).
Segundo Mueler et al. (1998 apud UEHARA; ROSA; CARMO, 2006, p.10), “a
secreção da leptina é diretamente proporcional à quantidade de glicose transportada e
metabolizada pelo tecido adiposo”.
A leptina, produzida no tecido adiposo, exerce efeito inibitório na insulina, estimula a
capitação da glicose e aumenta a lipólise. No músculo esquelético, aumenta a capacitação da
glicose, a síntese do glicogênio e da oxidação dos ácidos graxos na presença da leptina. A
secreção da insulina parece ser modulada pela leptina. Por outro lado, insulina estimula a
produção de leptina nos adipócitos, indicando a existência de um “eixo adipoinsulinar”. Tem
sido bem demonstrado que a secreção da leptina é diretamente proporcional ao tamanho e
número de adipócitos e que a expressão do gene ob RNAm é estimulada pela insulina. A fome
suprime a expressão do gene ob RNAm e a realimentação reverte isto (CEDDIA et al., 1998;
UEHARA; ROSA; CARMO, 2006).

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 20
Além da leptina, a insulina também informa a adequação das reservas energéticas ao
hipotálamo. Estes dois hormônios são importantes para o controle do peso corporal a longo
prazo (RODRIGUES; SUPLICY; RADOMINSKI, 2003).
2.2 LEPTINA
A descoberta da leptina ocorreu, em 1994, pelo grupo do Dr. Friedman da
Universidade Columbia de Nova York (RODRIGUES; SUPLICY; RADOMINSKI, 2003).
Segundo Negrão e Licinio (2000), a descoberta concretizou-se através de investigações com
modelos experimentais, especialmente com camundongos que tiveram a obesidade induzida
por dieta, proporcionando estudos do controle homeostático de energia.
A leptina, termo de origem grega (leptos = magro), é um sinal endócrino que informa
o tamanho das reservas energéticas para o sistema regulador da alimentação, substratos
utilizados e o balanço energético, ligando a energia, utilizada como combustível, com a
quantidade. Isto pode, portanto, ter um papel fisiológico no mecanismo que adapta o
comportamento alimentar às necessidades energéticas, de acordo com os estoques de energia
(CHAPELOT et al., 2000; FRIED et al., 2000; LOPES et al., 2004).
A leptina é uma proteína composta por 167 aminoácidos e possui uma estrutura,
semelhante às citocinas, que consiste em um complexo de quatro hélices (Figura 2). Seu gene
está localizado no cromossomo 7alfa31.3. Após a sua produção, a leptina é secretada na
corrente sanguínea, ligando-se a proteínas, sendo que a porcentagem de ligação é maior em
indivíduos magros (aproximadamente 45%) do que em obesos (aproximadamente 20%),
sendo transportada até o cérebro, onde estimula ou inibe a liberação de vários
neurotransmissores. A leptina plasmática cruza a barreira hemato-encefálica, através de um
transporte ativo, e age sobre os receptores nas regiões lateral e medial do hipotálamo, para
diminuir o apetite e aumentar o gasto energético por meio da estimulação simpática. A
entrada da leptina no cérebro acontece por um mecanismo de transporte saturável, talvez por
transcitose, mediado por receptor, através da membrana hemato-encefálica ou via líquido
cefálico-raquidiano. Esta idéia é sustentada pela presença de grande número de receptores de
leptina Ob-R nos microvasos cerebrais, os quais, embora não sejam capazes de expressar o
gene da leptina, são capazes de ligar e internalizar este hormônio (BRANDS et al., 2000;
CISTERNAS, 2002; LOPES et al., 2004; PARACCHINI; PEDOTTI; TAIOLI, 2005;
ROMERO; ZANESCO, 2006).
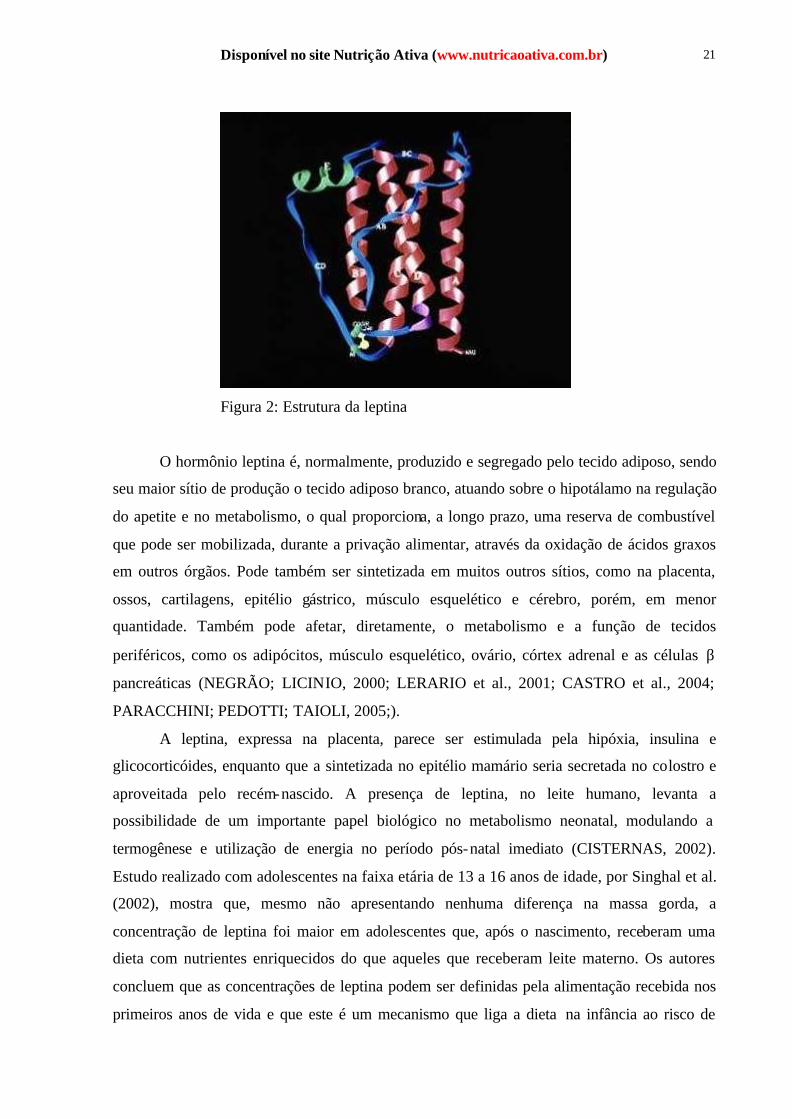
Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 21
Figura 2: Estrutura da leptina
O hormônio leptina é, normalmente, produzido e segregado pelo tecido adiposo, sendo
seu maior sítio de produção o tecido adiposo branco, atuando sobre o hipotálamo na regulação
do apetite e no metabolismo, o qual proporciona, a longo prazo, uma reserva de combustível
que pode ser mobilizada, durante a privação alimentar, através da oxidação de ácidos graxos
em outros órgãos. Pode também ser sintetizada em muitos outros sítios, como na placenta,
ossos, cartilagens, epitélio gástrico, músculo esquelético e cérebro, porém, em menor
quantidade. Também pode afetar, diretamente, o metabolismo e a função de tecidos
periféricos, como os adipócitos, músculo esquelético, ovário, córtex adrenal e as células β
pancreáticas (NEGRÃO; LICINIO, 2000; LERARIO et al., 2001; CASTRO et al., 2004;
PARACCHINI; PEDOTTI; TAIOLI, 2005;).
A leptina, expressa na placenta, parece ser estimulada pela hipóxia, insulina e
glicocorticóides, enquanto que a sintetizada no epitélio mamário seria secretada no colostro e
aproveitada pelo recém-nascido. A presença de leptina, no leite humano, levanta a
possibilidade de um importante papel biológico no metabolismo neonatal, modulando a
termogênese e utilização de energia no período pós-natal imediato (CISTERNAS, 2002).
Estudo realizado com adolescentes na faixa etária de 13 a 16 anos de idade, por Singhal et al.
(2002), mostra que, mesmo não apresentando nenhuma diferença na massa gorda, a
concentração de leptina foi maior em adolescentes que, após o nascimento, receberam uma
dieta com nutrientes enriquecidos do que aqueles que receberam leite materno. Os autores
concluem que as concentrações de leptina podem ser definidas pela alimentação recebida nos
primeiros anos de vida e que este é um mecanismo que liga a dieta na infância ao risco de

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 22
desenvolver a obesidade na vida adulta.
A função da leptina vai além da atividade de regulação do peso corpóreo, pois também
participa da fisiologia da puberdade e da reprodução, informando ao cérebro se as reservas
energéticas são suficientes para sustentar o início da puberdade e da reprodução; é
responsável por estimular a oxidação de ácidos graxos e a captação de glicose, impedindo
assim o acúmulo de gordura nos tecidos não-adiposos, causando prejuízo nas suas funções,
devido à “lipotoxidade”, causada pelo excesso de triglicerídios nos tecidos, como músculos,
fígado, coração e pâncreas. É provável que a leptina interfira, indiretamente, sobre a massa
óssea, através da sua relação com a massa gordurosa, fonte importante de esteróides sexuais
no climatério. Além destes fatores, a leptina tem papel fundamental sobre os sistemas
imunológico, hematopoiético e cardiovascular (NEGRÃO; LICINIO, 2000; MINOKOSHI et
al., 2002; COSTA et al., 2003; RODRIGUES; SUPLICY; RADOMINSKI, 2003; AZEVEDO
et al., 2004; ROMERO; ZANESCO, 2006; HERMSDOFF; VIEIRA; MONTEIRO, 2006).
A leptina participa, ativamente, na regulação do metabolismo da glicose e da gordura.
Assim, diminui agudamente a insulinemia e glicemia no camundongo ob/ob, antes de se
observar uma efetiva perda de peso., no camundongo de cepa nativa, também estimula a
gliconeogênese e metabolismo da glicose. Este hormônio estimula a lipólise, altera o
conteúdo de gordura no músculo esquelético e tem a capacidade de estimular a síntese
hepática de ácidos graxos (CISTERNAS, 2002).
Para Negrão e Licínio (2000), existem fatores que resultam na diminuição da
expressão do gene da leptina, como o jejum, o exercício físico moderado, o frio, a
testosterona, catecolaminas e a ativação do Sistema Nervoso Simpático, influenciando a
queda da concentração de leptina durante o jejum, vista como sinal que informaria ao cérebro
da necessidade de ajustes neuro-hormonais de modo a garantir um metabolismo mais
eficiente. Entretanto, alimentação pós-jejum, glicocorticóides e a insulina estimulam a
transcrição do gene, levando à produção do hormônio leptina. Situações de estresse, impostas
ao corpo, como jejum prolongado e exercícios físicos intensos, provocam a diminuição dos
níveis circulantes de leptina, comprovando, dessa maneira, a atuação do Sistema Nervoso
Central na inibição da liberação de leptina pelos adipócitos (Tabela 2) (NEGRÃO; LICINIO,
2000; ROMERO; ZANESCO, 2006).

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 23
Tabela 2 Influência de fatores orgânicos e ambientais nos níveis de leptina
Situações Níveis de leptina
Ganho de peso Aumentados
Insulina Aumentados
Glicocorticóides Aumentados
Infecções agudas Aumentados
Citoquinas inflamatórias Aumentados
Perda de peso Diminuídos
Jejum Diminuídos
Estimulação adrenérgica Diminuídos
Hormônio do crescimento (GH) Diminuídos
Hormônio tireoidianos Diminuídos
Melatonina Diminuídos
Fumo Diminuídos
Fonte: Romero e Zanesco (2006)
Na infância e adolescência, há diferença nos níveis plasmáticos de leptina entre os
sexos: nas meninas, os níveis de leptina aumentam progressivamente de acordo com a idade,
com o ganho de peso e com gordura corporal, enquanto que, nos meninos, ocorre uma
diminuição progressiva. Isso devido às diferenças hormonais, em que a testosterona tem uma
correlação negativa com níveis de leptina. Tanto em indivíduos eutróficos como em obesos, a
síntese da leptina plasmática é 3 vezes superior em mulheres do que em homens, sendo as
primeiras relativamente mais resistentes aos efeitos da leptina. Isso pode explicar o aumento
da suscetibilidade aos distúrbios, tanto alimentares, como do peso corporal. A expressão dessa
proteína é 2 a 3 vezes maior no tecido subcutâneo do que no visceral, particularmente em
mulheres do que nos homens, devido aos adipócitos subcutâneos serem maiores do que os
viscerais (NEGRÃO; LICINIO, 2000; PÉREZ, 2000; WAJCHENBERG, 2000; OLIVEIRA et
al., 2004; HERMSDOFF; VIEIRA; MONTEIRO, 2006).
Entre os adultos dos Estados Unidos, as concentrações de leptina sérica são maiores
em mulheres do que em homens e são maiores em negros do que em brancos,
independentemente das medidas antropométricas da gordura corporal. No estudo realizado
por Ruhl et al. (2004), em que se analisou a relação ente adiposidade, raça e sexo, identificou-

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 24
se que as concentrações de leptina são maiores em mulheres negras do que em mulheres
brancas, entretanto, essa diferença significativa não há entre homens brancos e homens
negros. A elevada concentração de leptina em mulheres é explicada pela alta porcentagem de
gordura corporal. Não há uma explicação pela alta taxa de leptina em negros através da
porcentagem de gordura corporal.
Em média, a liberação de leptina, por grama de tecido adiposo, é duas vezes maior em
obesos do que em sujeitos magros, porque o tamanho das células adipócitas é usualmente
aumentado 2-4 vezes em obesos; quando expressados por células gorduras, a secreção da
leptina está elevada para 7 vezes maior em obesos do que em sujeitos magros. Em adição, um
aumento no número de células adiposas, particularmente na obesidade extrema,
indubitavelmente contribui para o aumento de leptina sérica (FRIED et al., 2000).
Níveis de leptina plasmática aumentam com o ganho de peso e diminuem com a perda
de peso, correspondente ao papel da leptina como um sinal do tamanho dos estoques dos
tecidos adiposos. Porém, sobrepondo-se à regulação da secreção da leptina pelos níveis de
obesidade, sinais agudos nutricionais e hormonais modulam a velocidade da secreção da
leptina com um tempo e direção, abrangendo efeitos sobre aproximadamente 7h. O ritmo
diurno da leptina é vinculado ao momento escolhido da alimentação, pois um atraso de 6h na
refeição produz uma fase similar que muda o perfil da leptina plasmática. Além disso,
freqüente amostra de sangue revela que níveis da leptina plasmática apresentam pulsos, de
aproximadamente 30 pulsos por um período de 24h. A leptina apresenta um ritmo circadiano
e o pico de sua secreção ocorre durante a noite e nas primeiras horas da manhã, e sua meia-
vida plasmática é de 30 minutos. A velocidade de secreção da leptina é lenta. Assim se, após
uma ingestão alta de alimentos, for mantida prolongadamente, poderá dar lugar a maior
síntese de leptina (FRIED et al., 2000; WAJCHENBERG, 2000; CISTERNAS, 2002;
ROMERO; ZANESCO, 2006).
A maior amplitude do pulso, verificada nas mulheres, em comparação aos homens, é o
único fator que contribui para os valores mais altos de leptina das mulheres. Esses achados
sugerem, para Negrão et al. (1998), “que picos individuais de secreção de leptina, em
mulheres, contêm mais moléculas de leptina que em homens”. Se considerar que a meia-vida
plasmática da leptina é idêntica em homens e mulheres, pode-se afirmar que exista um
dimorfismo sexual na produção da leptina. Para poder manter um peso normal, as mulheres
parecem requerer maior liberação de leptina por pulso, o que pode indicar que as mulheres
são mais resistentes às ações da leptina que os homens.
A superalimentação aguda (120 Kcal/Kg) aumenta a leptina plasmática para 40% após

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 25
cinco horas. Comparada com uma dieta hiperlipídica (60% de calorias), uma hipolipídica
(20%), dieta hiperglicídica produz elevados picos de leptina durante a noite, sem afetar níveis
de leptina pela manhã (FRIED et al., 2000).
Com a elevação da leptina, ocorre uma redução, na ingesta alimentar, e um aumento
do gasto energético, indicando que há, como afirma Wajchenberg (2000, p.16), “uma forte
correlação positiva entre os níveis circulantes de leptina e a quantidade de gordura corpórea
nos homens, sugerindo que a secreção da leptina é um reflexo da hipertrofia gordurosa”.
Portanto, a quantidade de gordura corporal é o principal determinante dos níveis de circulação
deste hormônio. Entretanto, além da adiposidade, os níveis de leptina são influenciados por
fatores nutricionais e hormonais. A adiposidade tem sido relatada como principal fator
determinante de leptinemia, correlacionada em estudos com a massa total de gordura e o
índice de massa corporal (IMC) (LÖNNERDAL; HAVEL, 2000; PARACCHINI;
PEDOTTI; TAIOLI, 2005; HERMSDORFF; VIEIRA; MONTEIRO, 2006).
As taxas sangüíneas de leptina, em roedores, elevam-se poucas horas após ingestão
alimentar, diferentemente do homem, no qual só acontece após uma ingestão alimentar
exagerada, mantida por vários dias. Contudo, diminui em ambas espécies, após poucas horas
de inanição ou jejum, indicando que a leptina não só seria regulada pela massa de tecido
adiposo, mas também por outros fatores; neste caso, por alterações agudas do balanço
energético (CISTERNAS, 2002).
Praticamente nada é conhecido sobre o(s) caminho(s) celular(es), envolvidos na
secreção da leptina, por ex.: se a secreção da leptina por si só é regulada (aumenta/diminui em
reposta para um secretagogo) ou é apenas constitutivelmente secretada por razão proporcional
à sua síntese. O rápido efeito da estimulação β-adrenérgica, na liberação da leptina do tecido
adiposo in vitro e in vivo, sugere a possibilidade que a secreção de leptina por si só é regulada
pela AMPc. Um recente relato sugere que as catecolaminas podem regular diretamente a
secreção da leptina. Adicionalmente, o fator de necrose tumoral, é um importante regulador
parácrino da leptina, o qual aumenta a sua liberação (FRIED et al., 2000).
Segundo Flier (2004), na década passada, houve o consenso de que existe um sistema
fisiológico para controlar a homeostase dos estoques de energia, em resposta às variações do
acesso à nutrição e à demanda da energia consumida. A curto prazo, há sinais que controlam o
início e o término da refeição, e os a longo prazo, que controlam as estoques energéticos.
Ambos os sinais convergem para o cérebro, mais especificamente ao hipotálamo, onde são
integrados, e a direção e magnitude da resposta são determinadas.
O cortisol elevado em restrições alimentares inibe a secreção diurna de leptina, via

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 26
redução da sensibilidade insulínica, e o efeito da insulina no metabolismo de glicose no tecido
adiposo. Tais efeitos são coerentes com o ciclo circadiano da leptina, com maiores
concentrações por volta de uma hora e menores em torno de nove horas da manhã.
Particularmente, a leptina age de modo conspícuo em quatro peptídeos, produzidos em
neurônios do núcleo arqueado: o neuropeptídio Y (NPY), a pró-opiomelanocorticotropina
(POMC) e o fator de transcrição cocaína-anfetamina dependente (CART). Embora seja um
modelo incompleto, postula-se que a leptina supra a atividade dos neurônios que produzem
NPY/AGRP (efeito orexígeno) e que ela estimule a atividade de neurônios, produtores de
POMC ou CART (efeito anorexígeno). Outros hormônios podem alterar os níveis séricos e
cerebroespinhais da leptina, como os estrógenos, estimuladores e catecolaminas, e andrógenos
(testosterona), inibidores da secreção da leptina. Citocinas, tais como interleucina 1, 6 e o
fator de necrose tumoral-α, também interferem na expressão do RNA mensageiro. Inúmeras
linhas de evidência, derivadas principalmente de estudos com modelos de roedores, sugerem
que a insulina influencia, diretamente, a secreção da leptina (FRIED et al., 2000; NEGRÃO;
LICINIO, 2000; HERMSDORFF; VIEIRA; MONTEIRO, 2006).
O agente hipoglicemiante, toglitazone, diminui a síntese de leptina, como também a
exposição ao frio e catecolaminas, deprimem sua expressão gênica, talvez através da ativação
dos receptores β-adrenérgicos. O efeito do toglitazone e das catecolaminas nos níveis de
leptina, provavelmente, seja devido a uma ação direta no gene ob. Além disso, o toglitazone,
o qual melhora a sensibilidade à insulina, produz efeitos opostos na leptina e na fome.
Entretanto, isto pode depender de dois fatores: (1) saciedade, a qual consiste na ausência da
motivação de ingestão, que termina quando a próxima refeição é iniciada, e, (2) satisfação, o
sinal que interrompe a refeição. A saciedade produzida por uma refeição é, deste modo,
associada pela medida do intervalo até o espontâneo pedido para a próxima refeição; a
satisfação está envolvida na medida da energia ingerida da refeição (por ex.: o tamanho da
refeição). A ausência de alguma mudança na concentração sanguínea de leptina, durante e
após refeições isoenergéticas que diferem na sua composição ou na patabilidade, tem sido
proposta como um argumento contra um efeito da leptina na satisfação. Entretanto, em ratos,
a administração de leptina foi demonstrada para reduzir o tamanho da refeição, mas não um
espontâneo intervalo entre as refeições, sugerindo que a leptina afeta a satis fação, mas não a
saciedade. Isto também demonstra que o pré-tratamento com colecistocinina aumenta o
aferente gástrico vagal sensitivamente para a leptina, favorecendo seu papel na quantidade de
alimentos ingeridos durante uma refeição. A relação inversa entre leptina plasmática e o
momento pré-prandial, induzindo salivação, direciona para o efeito da leptina na quantidade

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 27
de alimento ingerido, pois aumento da salivação foi positivamente associado à fome e a
reflexo da depleção energética. A recente descoberta que a leptina inibe os neurônios LH
também suporta, a curto prazo, o efeito da leptina no molde da refeição (CHAPELOT et al.,
2000; SPIEGELMAN; FLIER, 2001; CISTERNAS, 2002).
Brandão et al. (2003) revelam que o melhor parâmetro, a ser usado para estimar os
níveis de leptina, em ambos os sexos, é a % da massa gorda e IMC. Os valores de leptina, em
indivíduos magros, são <17 ng/mL e em obesos são <59 ng/mL. Isso demonstra que as
concentrações séricas de leptina são maiores em obesos. Concentrações elevadas de leptina,
associadas à obesidade, implicam quadro de resistência à ação da leptina (MARTINS;
MOREIRA; PIEROSAN, 2003; UEHARA; ROSA; CARMO, 2006).
Acreditava-se que o aumento dos níveis de leptina seria capaz de prevenir a obesidade
pela inibição da fome e estímulo da termogênese, através de uma ação no cérebro. Entretanto,
as mutações do receptor de leptina (Ob-Rb) provocam o aparecimento precoce de obesidade
em roedores (SPIEGELMAN; FLIER, 2001; CISTERNAS, 2002).
2.2.1 Leptina e Obesidade
A mutação do gene da leptina, em ratos, foi primeiramente descoberta em 1950: ratos
ob/ob obesos demonstraram um disparate na mutação do códon 105 do rato original,
resultando na ausência na produção de leptina. Esta mutação causa obesidade, hiperfagia,
extrema resistência à insulina, infertilidade, hipogonadismo, hipercortisolismo e uma massa
óssea duas a três vezes maior que o animal selvagem. A administração de leptina restabelece a
condição normal. A estrutura do gene da leptina é preservada em todos os mamíferos: a
leptina humana e a leptina de ratos compartilham 84% da seqüência idêntica. O gene ob é
expresso somente no tecido adiposo, principalmente o branco, mas também no tecido adiposo
marrom, em proporção ao tamanho e número dos adipócitos. A produção do recentemente
clonado gene ob provoca uma acentuada redução do peso corporal, da porcentagem de
gordura corporal e a quantidade ingerida, seguida do aumento na energia consumida e
reduzindo a concentração sérica de glicose e insulina em ratos obesos (CEDDIA et al., 1998;
BORBA; KULAK; CASTRO, 2003; PARACCHINI; PEDOTTI; TAIOLI, 2005).
A deficiência de leptina e defeitos no receptor da leptina, em humanos, são muito
raros. Em média, a leptina sérica é usualmente elevada, na obesidade, e é positivamente
relacionada ao índice de massa corporal, porcentagem de gordura corporal e à massa gorda.
Porém, vários laboratórios têm notado um largo aumento dos valores da leptina sérica em

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 28
alguns níveis de gordura corporal. Alguns sujeitos obesos estão expostos, particularmente, a
níveis superiores ou inferiores de leptina que estão de acordo com sua massa de gordura,
sugerindo que outro mecanismo, e não a massa gorda, por si, regula a leptina sérica. Este
ponto parte para a possibilidade que uma diferenciação na regulação da secreção da leptina,
contribuindo para manter o peso corporal elevado em alguns sujeitos obesos (FRIED et al.,
2000).
Estima-se que a presença de altos níveis de leptina, no plasma, seriam indicativos de
uma resistência a este hormônio, seja pela alteração da sua síntese e/ou secreção, por
alterações no transporte cerebral, ou por anomalias nos receptores e ou posterior sinalização; é
também sugerido que as taxas deste hormônio no líquor estariam deprimidas. Os mecanismos
pelos quais o aumento de tecido adiposo é traduzido em aumento da concentração sérica de
leptina, envolvem tanto o número de células adiposas quanto a indução do RNAm ob.
Indivíduos obesos apresentam um aumento do número de células adiposas, o que significa
uma maior quantidade de RNAm ob, encontrada em seus adipócitos, do que em sujeitos
eutróficos. Todavia, a concentração sérica de leptina não é dependente somente do tamanho
do tecido adiposo, uma vez que a redução de 10% do peso corporal provoca diminuição de
cerca de 53% de leptina plasmática, sugerindo que outros fatores, além da adiposidade
tecidual, estão envolvidos na regulação de sua produção (CISTERNAS, 2002; ROMERO;
ZANESCO, 2006).
O estudo dos níveis de leptina sérica, em relação às várias medidas de adipócitos,
demonstrou que a obesidade não é caracterizada pela deficiência de leptina, mas pela
hiperleptinemia, encontrada em pacientes obesos. A incapacidade de tal elevação dos níveis
de leptina para alterar o estado de obesidade destas pessoas, podem ser relacionadas com a
“resistência à leptina”, uma incapacidade da leptina em entrar no fluido cérebro-espinhal, para
alcançar as regiões hipotalâmicas que regulam o apetite, ou pode simplesmente refletir a
quantidade de tecido adiposo nessas pessoas. Indivíduos obesos apresentam elevados níveis
plasmáticos de leptina, cerca de cinco vezes maiores do que aqueles encontrados em sujeitos
magros (PARACCHINI; PEDOTTI; TAIOLI, 2005; ROMERO; ZANESCO, 2006).
A hiperleptinemia, encontrada em pessoas obesas, é atribuída a alterações no receptor
de leptina ou a uma deficiência em seu sistema de transporte na barreira hemato-encefálica,
fenômeno denominado resistência à leptina. Existem dois mecanismos identificados. O
primeiro pode envolver um defeito no transporte da leptina, através da barreira hemato-
encefálica, para sítios de ação dentro do SNC, visando regular o balanço energético. A
existência de um transporte defeituoso é ressaltada pela potencial administração da leptina no

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 29
cérebro (opondo-se à periférica) em ratos, com dieta induzida para obesidade (FLIER, 2004;
ROMERO; ZANESCO, 2006).
A perda da sensibilidade à insulina (resistência à insulina) pode permitir um balanço
dos estoques energéticos negativos, culminando na obesidade. Outra hipótese é compatível
com estudos em humanos demonstrando que, diferentemente de ratos ob/ob, a obesidade não
é um problema pela falta de leptina, já que elevados níveis de leptina são encontrados em
sujeitos obesos. Portanto, a obesidade pode ocorrer devido à interrupção no sinal da leptina
nos seus sítios alvos, localizados central ou perifericamente (CEDDIA et al., 1998; MARS et
al., 2005).
Estudos da relação liquórica/leptina sérica, na obesidade humana, sugerem que a
resistência poderia resultar de um defeito de transporte da leptina ao sistema nervoso central.
A resistência à leptina também poderia ocorrer por um defeito pós-receptor, levando à falha
na ativação dos mediadores neuroendócrinos, reguladores do peso corporal. Postula-se que
um acúmulo excessivo de leptina, a curto prazo, poderia levar a uma “down-regulation” dos
receptores centrais e a um reajuste do seu efeito inibitório sobre o apetite. Desta maneira, uma
concentração supranormal de leptina seria necessária para o mesmo efeito inibitório sobre o
apetite. Outra possibilidade é a de que haja uma insuficiência do sistema de transporte da
leptina para dentro do cérebro, pois pacientes obesos têm uma diminuição das concentrações
liquóricas de leptina, quando comparadas às concentrações plasmáticas do hormônio
(NEGRÃO; LICINIO, 2000; RODRIGUES; SUPLICY; RADOMINSKI, 2003).
Steinberg et al. (2002) foram os primeiros a fornecer evidências de que a leptina
aumenta a oxidação de ácidos graxos no músculo esquelético de indivíduos magros, o que já
não ocorre em obesos, apontando para o desenvolvimento de resistência à leptina no músculo
esquelético de obesos. Na obesidade, o aumento dos níveis de leptina pode ocorrer devido à
hiperinsulinemia crônica.
Mais importante do que o papel antiobesidade, antes atribuído à leptina, é que este
hormônio é responsável pelo fornecimento da informação ao cérebro de que o organismo está
passando fome. Com a ausência da leptina, o cérebro tem a sensação de fome, apesar da
obesidade. Deficiência da leptina leva à fome, supressão da energia consumida e inibe os
componentes da reprodução, sendo que todos estes eventos ocorrem para adaptar-se à falta de
alimento (FLIER, 2004).
Em crianças obesas, têm sido verificados níveis aumentados de leptina. Alguns
estudos avaliaram a relação da leptina com a sensibilidade à insulina e constataram que,
independente da obesidade, da idade, do sexo, da relação circunferência cintura-quadril,

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 30
houve associação inversa, isto é, quanto menor a sensibilidade à insulina, maior o nível de
leptina (OLIVEIRA et al., 2004). Sudi et al. (2000) observaram que o principal determinante
para a variação dos níveis de leptina, em crianças e adolescentes obesos, é o IMC e não a
insulina basal e o índice de resistência à insulina.
A importância genética, na etiologia da obesidade, também tem sido foco de pesquisa
em todo o mundo. A identificação e seqüenciamento do gene ob, que codifica o peptídeo
leptina, e a descoberta que o defeito, neste gene, parece ser a simples causa da obesidade em
ratos ob/ob, tem gerado considerável interesse no estudo da genética da obesidade
(PEREIRA, FRANCISCHI; LANCHA, 2003).
2.2.2 Efeito dos Macronutrientes sobre os Níveis de Leptina
Ainslie et al. (2000) realizaram estudo com ratas, divididas em dois grupos: um, com
uma dieta hiperlipídica e outro, com dieta controle. O peso corporal, a quantidade de energia,
a composição corporal e as concentrações de leptina, no jejum, foram comparados, após a 4ª e
14ª semana do tratamento dietético. Constataram que, após 4 semanas, a gordura abdominal
foi 38% maior no grupo com a dieta hiperlipídica do que as do grupo controle. Porém, a
concentração de leptina plasmática foi 24% menor em animais alimentados com a dieta
hiperlipídica, resultando em uma significativa diminuição das concentrações plasmáticas por
unidade de massa gorda abdominal que dos animais controles. Da 4ª para a 14ª semana, o
grupo da dieta hiperlipídica ganhou 12Kg ou mais de peso. Após 14 semanas, as
concentrações de leptina plasmática, por unidade de massa gorda abdominal, foi 27% menor
em ratos com dieta hiperlipídica. Portanto, a curto prazo, uma moderada dieta hiperlipídica é
associada com menor circulação de leptina do que o esperado, a qual correlaciona com maior
peso corporal. Logo, uma dieta hiperlipídica pode contribuir para ganho de peso pela redução
da secreção no tecido adiposo. Uma dieta, rica em gordura, pode contribuir para um ganho de
peso, não somente devido ao fornecimento de substrato para acumulação de triacilglicerol,
mas também pela quantidade ingerida como um resultado da redução na secreção da leptina
pelo tecido adiposo.
O consumo de pré-carga de carboidrato ou lipídio tem efeitos diferentes sobre a
leptinemia, sendo que a ingestão de lipídio resulta em menor secreção de leptina, o que
aumenta a ingestão energética e o risco de obesidade. Concentrações de leptina na circulação
(24h) são inferiores, quando sujeitos consomem uma refeição hiperlipídica do que quando
eles consomem uma refeição hiperglicídica. Mudanças na leptina, em resposta ao jejum,

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 31
alimentação e aos macronutrientes da dieta são provavelmente relacionadas com a insulina
mediadora do metabolismo da glicose no tecido adiposo (LÖNNERDAL; HAVEL, 2000;
HERMSDORFF; VIEIRA; MONTEIRO, 2006).
As dietas hiperlipídicas, para Uehara, Rosa e Carmo (2006), podem afetar a entrada de
glicose nos adipócitos, estimulada pela insulina. Assim, o aumento da lipólise pode reduzir a
síntese de leptina. Tem-se demonstrado que as dietas hiperlipídicas diminuem a saciedade,
levando ao aumento da ingestão de alimentos, favorecendo o ganho de peso. Isso pode ocorrer
devido a uma redução na concentração de leptina. Fisiologicamente, dentre todos os outros
macronutrientes, os lipídios são os que apresentam a maior densidade energética e a maior
capacidade de estoque no organismo. No entanto, alguns autores acreditam que o tipo de
lipídio, ingerido na dieta, também pode influenciar o acúmulo de adiposidade e a forma de
agir sobre a lipólise e a entrada de glicose nas células adiposas, visto que alguns trabalhos
mostram significativa correlação entre percentual de gordura saturada e monoinsaturada,
ingerida na alimentação. Além disso, a duração do tratamento dietético também influencia o
efeito da gordura: por exemplo, o consumo de dietas hiperlipídicas durante 12 dias não afeta
os níveis de leptina. Contudo, durante 5 meses, os níveis de leptina elevam-se, o que pode ser
explicado pelo desenvolvimento de resistência à leptina (AINSLIE et al. 2000; PEREIRA,
FRANCISCHI; LANCHA, 2003; UEHARA; ROSA; CARMO, 2006).
Alterações agudas no balanço energético regulam a expressão da leptina e os
correspondentes níveis circulantes. Desta forma, um aumento na ingestão de carboidrato
induz a uma elevação da leptina (secundária a hiperinsulinemia) em aproximadamente 40%,
nas primeiras 12 horas, na ausência de alterações do peso corporal. Entretanto, a ingestão
isocalórica de gordura provoca uma redução dos níveis de leptina. O jejum prolongado leva a
uma queda na leptinemia, em desproporção com as variações da massa de tecido adiposo. Por
conseguinte, em condições de equilíbrio energético, a leptina é um índice estático da
quantidade de triglicerídios no tecido adiposo e, em situações de balanço energético, em
equilíbrio não-estável, a leptina pode ser regulada agudamente de uma maneira independente
dos depósitos de triglicerídios, servindo como um sensor do balanço energético
(WAJCHENBERG, 2000).
A longo prazo, dietas hiperlipídicas podem induzir à hiperalimentação e ganho de
peso. Entretanto, o mecanismo pelo qual isto ocorre é desconhecido. Ratos com dietas bem
estabelecidas que induzem à obesidade apresentam hiperleptinemia, são ainda hiperfágicos e
gastam menos energia, sugerindo que dieta hiperlipídica pode causar ganho de peso pela
limitada ação da leptina (AINSLIE et al., 2000).

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 32
Estudo, realizado por Weigle et al. (2005), conc lui que um aumento de proteína na
dieta de 15% para 30% da energia, com quantidade de carboidrato constante, produz uma
contínua diminuição na quantidade de calorias ingeridas que podem ser mediadas pelo
aumento da sensibilidade da leptina no Sistema Nervoso Central e resulta em uma
significativa perda de peso. Este efeito anoréxico da proteína pode contribuir para a perda de
peso, produzida por dietas com pouco carboidrato.
2.2.3 Receptor da Leptina
O gene do receptor da leptina, em humanos, está localizado no cromossomo 1p31. Os
receptores da leptina são incluídos na família das citocinas e possuem várias formas
identificadas, mas uma, em especial, encontra-se em grande quantidade no hipotálamo,
especificamente no núcleo hipotalâmico arqueado, o que o indica como local de importante
ação da leptina. Estes receptores têm três partes funcionais: (1) Extracelular que interage com
a proteína leptina e que é idêntica a todas as isoformas dos receptores ob-r; (2) Intracelular
que, quando ativada, estimula os eventos celulares, sendo que seu tamanho e seu domínio
determinam a ação que a leptina exercerá sobre a célula alvo; (3) Transmembrana que serve
para “ancoramento” do receptor à membrana celular (PARACCHINI; PEDOTTI; TAIOLI, 20
05; HERMSDORFF; VIEIRA; MONTEIRO, 2006).
Os domínios extracelular e transmembrana são idênticos entre a pequena e a longa
isoformas; as diferenças ocorrem devido às mudanças no comprimento do domínio
citoplasmático. As seis isoformas do receptor caracterizam-se por compartilhar o mesmo
domínio de ligação extracelular, porém somente o receptor Ob-Rb (forma longa) contém um
domínio intracelular que é capaz de transmitir o sinal de ligação, junto à leptina, para dentro
da célula. Este receptor Ob-Rb ativa o sistema JAK (Janus quinase/STAT) – transdutores de
sinais e ativadores da transcrição – de transdução após a ligação do hormônio ao receptor
(CISTERNAS, 2002; PARACCHINI; PEDOTTI; TAIOLI, 2005).
Há pelo menos seis variantes das isoformas do receptor da leptina (a- f) do Ob-R. O
Ob-Ra foi a forma originalmente clonada e esta tem sido postulada pelo transporte da leptina
através da barreira hemato-encefálica. Duas outras variantes, Ob-Rc e Ob-Rd, têm sido
implicadas na remoção da leptina da circulação, e a Ob-Re, é considerado um receptor solúvel
que transporta proteína periférica. O Ob-Rf é, principalmente, encontrado em tecidos
envolvidos com o sistema imunológico, mas sua função ainda não está bem esclarecida. Estes

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 33
variantes contrastam com a forma Ob-Rb, o qual possui um longo domínio intracelular,
essencial para a transdução do sinal intracelular (CEDDIA et al., 1998).
Existem dois tipos de receptores para a leptina: o ObRb, de cadeia longa, o qual possui
um número maior de aminoácidos, com maior expressão no hipotálamo, e os receptores de
cadeia curta, ObRa, encontrados no pâncreas e, mais especificamente, nas células α e δ das
ilhotas de Langerhans. A leptina age através dos receptores ObRb em duas populações
distintas de neurônios no núcleo arqueado. Uma população coexpressa os orexígenos,
neuropeptídio Y e AgRP, e a leptina age reduzindo suas expressões. A outra população
coexpressa peptídeos anorexígenos, cocaína e transcrição da anfetamina (CART) e α-MSH
(derivada da proopiomelanocortina – POMC), e a leptina aumenta a sua expressão. A leptina
suprime dois peptídeos orexígenos e aumenta dois peptídeos anorexígenos (SPIEGELMAN;
FLIER, 2001; ROMEO; ZANESCO, 2006).
Estudos em ratos demonstraram que a forma longa é a mais importante para transmitir
o sinal da leptina para as células, localizadas predominantemente no hipotálamo e não maior
em outros tecidos. Análise dos moldes, expressos pelas duas formas (longa e a curta) dos
receptores da leptina, demonstrou que os receptores são expressos na maior parte dos tecidos
(cérebro, ovário, rim, baço, intestinos, tecido adiposo marrom e branco, coração, pulmão,
fígado, músculo esquelético, adrenais, glândula pituitária, órgãos reprodutores masculinos e
femininos, glândula mamária e sistema imunológico), com a forma longa sendo prevalente no
hipotálamo. Também pode ser encontrada na medula da adrenal, na zona interna da medula
do rim, ilhas pancreáticas e tecido adiposo, ao passo que as formas pequenas são expressas em
todo o corpo, especialmente nos rins e pulmões (CEDDIA et al., 1998; PARACCHINI;
PEDOTTI; TAIOLI, 2005).
O receptor de leptina humano também estimula alguns hormônios hipotalâmicos,
mesmo na ausência da leptina, portanto, o defeito é mais severo quando a função do receptor
é perdida do que na do ligante (RODRIGUES; SUPLICY; RADOMINSKI, 2003).
2.2.4 Leptina Recombinante
Recentemente, ensaio clínicos com leptina recombinante fecharam um ciclo de
investigações que começaram com modelos animais de obesidade e chegaram até o
tratamento da obesidade em humanos. A leptina recombinante foi administrada, com sucesso,
numa paciente obesa com deficiência do hormônio, devido a uma mutação do gene ob.
Espera-se que, com o tratamento da leptina recombinante, o paciente apresente perda de peso,

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 34
embora ainda não se tenha nenhum relato disso. Em pacientes obesos, os níveis de leptina são
aumentados, em proporção à gordura corporal e à hiperleptinemia, junto com a escassa
resposta do peso corporal pela terapia com leptina recombinante, definindo assim, um estado
de resistência à leptina (NEGRÃO; LICINIO, 2000; FLIER, 2004).
Existe uma homologia de 84% entre o gene da leptina humana e o gene encontrado em
camundongos. Logo a seguir, passou-se a produzir leptina recombinante e demonstrou-se que
a leptina, administrada em camundongos da cepa ob/ob, levava a uma perda de peso e reduzia
a ingestão calórica. Uma pequena queda, na produção de leptina, seria percebida pelo sistema
de regulação do peso corporal, com aumento da massa de gordura, a fim de restaurar os níveis
de leptina em um novo set point. Pessoas obesas com níveis relativamente baixos de leptina,
com ou sem mutações, poderiam se beneficiar do uso da leptina recombinante (NEGRÃO;
LICINIO, 2000; RODRIGUES; SUPLICY; RADOMINSKI, 2003).
Segundo Hukshorn, Plantenga e Saris (2003), o aumento das evidências sugerem que
a queda das concentrações de leptina, observadas durante o jejum, agem como um sinal
periférico de fome, que serve para conservar energia em face as reservas limitadas. Em
extensão desta hipótese é que a leptina exógena deve afetar a regulação energética durante
restrição severa. Em um estudo, realizado pelo autor, com um grupo de obesos em tratamento
com leptina recombinante (PEG-OB – pegylated polyethylene glycol protein) e um grupo
controle, mostrou que o tratamento com a leptina recombinante, após 46 dias, ocorreu perda
de peso (14.6 ± 0.8 comparada com 11.8 ± 0.9 Kg) e redução do apetite, em relação ao grupo
controle. Entretanto, não houve significativa mudança na composição corporal e na energia
consumida. Isso mostra que, com a redução da concentração de leptina, durante estado de
fome, ocorre um aumento no apetite em humanos. Este fato também foi observado em um
estudo, realizado por Platenga et al. (2001), com a leptina recombinante administrada
subcutaneamente em homens obesos, ao mostrar que este medicamento modificou o apetite,
mas também não houve mudança na composição corporal, energia consumida ou perda
relativa de massa corporal, comparados com um grupo placebo. Isto sugere que PEG-OB tem
atividade biológica mais central do que periférica.
Em ratos e camundongos magros e obesos, a administração intraperitoneal ou
subcutânea de leptina foi associada com dose-dependente a qual reduz a quantidade inferida
de alimento e peso corporal. O aumento da energia consumida durante o tratamento com
leptina foi observado na restrição alimentar em ratos magros. Os autores concluem que a
leptina controla a termoregulação da energia gasta quando alimentos são escassos, porém
modifica a quantidade ingerida, maior do que a energia gasta, quando se tem alimentos em

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 35
abundância. Estas observações sugerem que a leptina recombinante humana foi potencial no
tratamento da obesidade humana. Além disso, foi sugerido que a administração exógena da
leptina cruza a barreira hemato-encefálica em humanos, embora se espere que a alta
concentração de leptina sérica deva saturar o sistema de transporte. A existência de um
transporte que não se sature foi proposto, e mais do que um caminho pode estar envolvido no
transporte de leptina no cérebro. Desta maneira, a administração exógena da leptina pode ser
estudada para determinar se o aumento da concentração de leptina no fluido cerebroespinhal
resulta em perda de peso em pessoas obesas (PLANTENGA et al., 2001).
No final de 1999, foi publicado o primeiro estudo clínico da leptina recombinante, em
pacientes com obesidade e controles de peso normal, por um grupo patrocinado pela Amgen
Inc. Tanto pacientes com obesidade quanto os controles tiveram uma perda de peso
proporcional a doses crescentes de leptina num período de quatro semanas. Os pacientes
obesos receberam a droga por um total de 24 semanas e mantiveram, em média, uma perda de
peso ao longo deste período. Oito pacientes obesos perderam, em média, 7,1Kg ao longo de
24 semanas, recebendo a dose máxima da droga (0,30mg/kg), porém os autores observaram
uma considerável variabilidade da resposta entre indivíduos. A perda de peso foi constatada,
principalmente, na forma de massa de gordura, conforme observado no paciente com a
mutação do gene da leptina. Além da grande variabilidade na resposta à leptina, outra
limitação do estudo foi o volume administrado do peptídeo (8mL/dia), levando a reações
inflamatórias importantes no local da aplicação da droga (HEYMSFIELD et al., 1999).
A deficiência de leptina, em ratos, tem reduzido claramente a energia consumida e a
eficiência do metabolismo, causando obesidade mesmo sem haver uma hiperalimentação.
Além de serem espécies diferentes, ratos e humanos apresentam deficiência de leptina. Eles
são unidos pelo acentuado aumento do apetite e conseqüente obesidade, assim como pela
falha para iniciar a puberdade; ambas as anormalidades são dramaticamente suprimidas pela
terapia com leptina recombinante (FLIER, 2004).

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 36
3 METODOLOGIA
No presente trabalho, foi realizada uma revisão bibliográfica, utilizando, como
instrumento de pesquisa, obras, artigos e periódicos científicos nacionais e internacionais,
sobre a relação entre o hormônio leptina e a obesidade, produzidos nos últimos cinco anos,
disponíveis em bancos de dados, tais como: Medline, Periódicos Capes, Scielo, Pubmed, entre
outros. Lima (2004) afirma que a pesquisa bibliográfica é caracterizada pela consulta de
fontes diversificadas sobre um tema específico, com o objetivo de concretizar não apenas um
estudo de caráter descritivo, mas atingir o nível analítico da questão.
Optou-se pela Revisão Bibliográfica, porque uma pesquisa, realizada em seres
humanos, em relação à taxa de leptina produzida e à obesidade, não seria viável, devido ao
tempo disponível para a realização do Trabalho Final de Graduação. Para se obter resultados
concretos e científicos, seriam necessários anos de pesquisa e material apropriado para
investigação, através da análise bioquímica em exames laboratoriais.
O estudo da obesidade em humanos, provavelmente, responderia a muitas dúvidas. No
entanto, pesquisas com humanos têm óbvias limitações éticas, financeiras, além do estudo em
animais permitir grande quantidade de pesquisas e resultados. Além disso, animais de
laboratório podem ser mantidos em condições rigidamente controladas, consumindo dieta
controlada e mantidos livre de patógenos e germes. O fato de animais de laboratório também
se tornarem obesos espontaneamente, alimentando-se de ração comercial, ou através de outras
manipulações, abriu novas áreas para pesquisa na área da obesidade.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 37
4 CONCLUSÃO
Hábitos da cultura humana moderna, tais como alimentação inadequada e
sedentarismo, atingem também países em desenvolvimento como o Brasil, onde a
denominada transição nos padrões nutricionais, com decorrente redução da desnutrição e
aumento da obesidade, considerada uma epidemia mundial, já podem ser identificadas. Isso se
torna um problema de saúde pública, uma vez que a obesidade é um dos principais fatores de
risco para inúmeras doenças prevalentes na sociedade moderna.
Os recentes achados, envolvendo a descoberta da leptina, produzida pelo adipócito,
abrem novos campos de estudo para o controle da obesidade, principalmente nas áreas de
nutrição e metabolismo. Portanto, o aprofundamento dos conhecimentos sobre esse peptídeo
torna-se de grande relevância na manutenção e preservação da qualidade de vida da
população, podendo proporcionar novas abordagens terapêuticas no tratamento da obesidade.
Constata-se que as perspectivas do uso da leptina recombinante em humanos devem
ser vistas com parcimônia. Por um lado, a visível perda de peso, observada no paciente com
deficiência congênita da leptina, reforça a importância desse hormônio na regulação do peso e
do apetite. Por outro lado, os resultados do estudo clínico em pacientes obesos sugerem que
existam pessoas com diferentes graus de suscetibilidade aos efeitos da leptina. Seguramente,
ainda há muito que se investigar sobre a complexidade da ação da leptina. Por exemplo, é
preciso esclarecer qual a importância de todos os peptídeos que atuam no circuito
hipotalâmico que regula o peso sobre a leptina. Finalmente, a identificação dos fatores de
transcrição, envolvidos na regulação do gene da leptina, poderão ser alvos de drogas que
facilitem a sua ação na obesidade.
Conclui-se que o hormônio leptina é um dos fatores envolvidos no desenvolvimento
da obesidade, em grande parte, pela resistência à leptina. Entretanto, maior número de
trabalhos científicos são necessários para o esclarecimento das alterações séricas da leptina no
estado de jejum e pós-prandiais.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 38
REFERÊNCIAS BIBLIOGRÁFICAS AINSLIE, Deborah A et al. Short-term, high-fat diets lower circulating leptin concentrations in rats. The American Journal of Clinical Nutrition, USA, v.71, p.438-442, 2000. ALMEIDA, Bianca de; FERREIRA, Sandra R.G. Epidemiologia. In: CLAUDINO, Angélica de Medeiros; ZANELLA, Maria Teresa. Transtornos Alimentares e Obesidade . Baueri, SP: Manole, 2005, p.185. ASSOCIAÇÃO BRASILEIRA DE NORMAS TÉCNICAS. NBR 6023. Informação e documentação: referências: elaboração. Rio de Janeiro, 2002. ______. NBR 14724. Informação e documentação: trabalhos acadêmicos: apresentação. 2.ed. Rio de Janeiro, 2005. ______. NBR 10520: Informação e documentação: citações em documentos: apresentação. Rio de Janeiro, 2002. AZEVEDO, George Dantas de et al. Níveis séricos de leptina e densidade mineral óssea em mulheres na pós-menopausa. Revista Brasileira de Ginecologia e Obstetrícia, Rio de Janeiro, v.26, n.6, p.429-433, 2004. BORBA, Victória Z.C; KULAK, Carolina A. Moreira; CASTRO, Marise Lazaretti. Controle neuroendócrino da massa óssea: mito ou verdade? Arquivos Brasileiro de Endocrinologia e Metabologia, São Paulo, v.47, n.4, p.453-457, ago. 2003. BOUCHARD, C. Physical activity and obesity. Human Kinetics, Champaign, 2000. BRANDÃO, C.M.A et al. Serum leptin concentration during puberty in healthy non-obese adolescents. Brazilian Journal of Medical and Biological Research, São Paulo, v.36, n.10, p.1293-1296, 2003. BRANDS, J.E. et al. Role of sympathetic nervous system and neuropeptides in obesity hypertension. Brazilian Journal of Medical and Biological Research, São Paulo, v.33, n.6, p.605-618, 2000. CARNEIRO, Gláucia. Patogênese. In: CLAUDINO, Angélica de Medeiros; ZANELLA, Maria Teresa. Transtornos Alimentares e Obesidade . Baueri, SP: Manole, 2005, p.203.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 39
CASTRO, Flávia Cipriano et al. Associação entre a antropometria e a leptina circulante nos compartimentos materno, fetal e placentário, na gravidez normal. Revista Brasileira de Ginecologia e Obstetrícia, Rio de Janeiro, v.26, n.9, p.691-970, 2004. CEDDIA, R. B et al. Pivotal role of leptin in insulin effects. Brazilian Journal of Medical and Biological Research, São Paulo, v.31, n.6, p.715-722, 1998. CENTRO UNIVERSITÁRIO FRANCISCANO. Orientações para Elaboração e Apresentação de Projetos de Pesquisa, Monografias e Dissertações. Santa Maria, RS: 2006. Disponível em: <http//www.unifra.br/utilitarios/arquivos/Normas_UNIFRA_vfinal6a. pdf>. Acesso em: nov. 2006. CHAPELOT, Didier et al. An endocrine and metabolic definition of the intermeal in humans: evidence for a role of leptin on the prandial pattern through fatty acid disposal. The American Journal of Clinical Nutrition, USA, v.72, p.421-431, 2000. CISTERNAS, J.R. Fisiologia do tecido adiposo e leptina. In: DOUGLAS, Carlos Roberto. Tratado de Fisiologia aplicado à Nutrição. São Paulo: Robe Editorial, 2002, p.805. COSTA, Elaine M. Frade et al. Genética molecular do eixo hipotálamo-hipófise-gonodal. Arquivos Brasileiros de Endocrinologia e Metabologia, São Paulo, v.47, n.4, ago. 2003. DAMIANI, Durval. Obesidade na infância e adolescência – um extraordinário desafio. Arquivos Brasileiros de Endocrinologia e Metabologia, São Paulo, v.44, n.5, p.363-365, out., 2000. DI, Marzo V et al. Leptin- regulated endocannabinoids are involved in maintaining food intake. Nature , v.12, n.410, p.822-825, abr., 2001. FILIER, Jeffrey. Obesity Wars: molecular progress confronts na expanding epidemic. Cell Press, v.116, p.337-350, jan. 2004. FISBERG, Mauro; CINTRA, Isa de Pádua; OLIVEIRA, Cecília Lacroix. Epidemiologia e diagnósticos da obesidade: abordagem inicial. In: FISBERG, Mauro. Atualização em obesidade na infância e adolescência. São Paulo: Atheneu, 2005. FRIED, Susan K et al. Regulation of leptin production in humans. The Journal of Nutrition, p.3127S-3131S, 2000.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 40
GUYTON, Arthur C.; HALL, John E. Tratado de fisiologia médica. Rio de Janeiro: Guanabara Koogan, 2002. HALPERN, Zuleika; RODRIGUES, Mariana Del Bosco; COSTA, Roberto Fernandes da. Determinantes fisiológicos do controle do peso e apetite. Revista de Psiquiatria Clínica, v.31, n.4, p.150-153, 2004. HERMSDORFF, Helen Hermana Miranda; VIEIRA, Maria Aparecida de Queiroga Milagres; MONTEIRO, Josefina Bressan Resende. Leptina e sua influência na patofisiologia de distúrbios alimentares. Revista de Nutrição, São Paulo, v.19, n.3, p.369-379, maio/jun. 2006. HEYMSFIELD, S.B et al. Recombinant leptin for weight loss in obese and lean adults: a randomized, controlled, dose-escalation trial. Jama, v.282, p.1568-1575, 1999. HUKSHORN, Chris J; PLANTENGA, Magriet S. Westerterp; SARIS, Wim HM. Pegylated human recombinant leptin (PEG-OB) causes additional weight loss in severely energy-restricted, overweight men. The American Journal of Clinical Nutrition, USA, v.77, p.771-776, 2003. LERARIO, D.D.G. et al. Influence of dexamethasone and weight loss on the regulation of serum leptin levels in obese individuals. Brazilian Journal of Medical and Biological Research, São Paulo, v.34, n.4, p.479-487, 2001. LIMA, Manolita Correia. Monografia: a engenharia da produção acadêmica. São Paulo: Saraiva, 2004. LÖNNERDAL, Bo; HAVEL, Peter J. Serum leptin concentrations in infants: effects of diet, sex, and adiposity. The American Journal of Clinical Nutrition, USA, v.72, p.484-489, 2000. LOPES, Ivã Marques et al. Aspectos genéticos da obesidade. Revista de Nutrição, São Paulo, v.17, n.3, p.327-338, jul./set. 2004. MARS, Monica et al. Decreases in fasting leptin and insulin concentrations after acute energy restriction and subsequent compensation in food intake. The American Journal of Clinical Nutrition(CHAPELOT et al., 2000), USA, v.81, p.570-577, 2005. MARTINS, Cristina; MOREIRA, Silvania de Moura; PIEROSAN, Simone Regina. Interações Droga & Nutriente. 2. ed. Curitiba, PR: Nutroclínica, 2003.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 41
MINOKOSHI, Y. et al. Leptin stimulates fatty-acid oxidation by activating AMP-activated protein kinase. Nature , USA, v.17, n.415, p.339-343, jan. 2002. MONTEIRO, C.A; CONDE, W.L; CASTRO, I.R.R. A tendência cambiante da relação entre escolaridade e risco de obesidade no Brasil (1975-1997). Caderno de Saúde Pública, v.19, n.1; p.S67-S75, 2003. NAMMI, Srinivas et al. Obesity: an overview on its current perpectives and treatment options. Nutrition Journal, v.3, n.3, p.424-451, abr. 2004. NEGRÃO, André B.; LICINIO, Julio. Leptina: o diálogo entre adipócitos e neurônios. Arquivos Brasileiros de Endocrinologia e Metabologia, São Paulo, v.44, n.3, p.205-214, jun. 2000. ______ et al. Diferenças na secreção diária de leptina em homens e mulheres: possíveis implicações na neurologia dos transtornos alimentares. Revista de Psiquiatria Clínica, v.25, n.4, p.184-190, 1998. OLIVEIRA, Cecília Lacroix et al. Obesidade e síndrome metabólica na infância e adolescência. Revista de Nutrição, Campinas, SP, v.17, n.2, p.237-245, abr./jun., 2004. PARACCHINI, Valentina; PEDOTTI, Paola; TAIOLI, Emanuela. Genetics of leptin and obesity: a huge review. American Journal of Epidemiology, USA, v.162, n.2, p.101-114, jul. 2005. PEREIRA, Luciana O.; FRANCISCHI, Rachel P. de; LANCHA JÚNIOR, Antonio H. Obesidade: hábitos nutricionais, sedentarismo e resistência à insulina. Arquivos Brasileiros de Endocrinologia e Metabologia, São Paulo, v.47, n.2, p.111-127, abr. 2003. PERÉZ B., Francisco et al. Asociación obesidad y leptina em três poblaciones aborígenes de Chile. Revista Médica do Chile, Santiago, v.128, n.1, jan. 2000. PINHEIRO, Anelise Rizzolo de Oliveira; FREITAS, Sérgio Fernando Torres de; CORSO, Arlete Catarina Tittoni. Uma abordagem epidemiológica da obesidade. Revista de Nutrição, São Paulo, v.17, n.4, p.523-533, out./dez. 2004. PLATENGA, Margriet S. Westerterp et al. Effects of weekly administration of pegylated recombinant human OB protein on appetite profile and energy metabolism un obese men. The American Journal of Clinical Nutrition, USA, v. 74, p.426-434, 2001.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 42
RODRIGUES, Adriane M.; SUPLICY, Henrique L.; RADOMINSKI, Rosana B. Controle neuroendócrino do peso corporal: implicações na gênese da obesidade. Arquivos Brasileiros de Endocrinologia e Metabologia, São Paulo, v.47, n.4, p.398-409, ago. 2003. ROMERO, Carla Eduarda Machado; ZANESCO, Angelina. O papel dos hormônios leptina e grelina na gênese da obesidade. Revista de Nutrição, São Paulo, v.19, n.1, p.85-91, jan./fev. 2006. RUHL, Constance E et al. Serum leptin concentrations and body adipose measures in older black and white adults. The American Journal of Clinical Nutrition, USA, v.80, p.576-583, 2004. SCHNEIDER, Patrícia; MEYER, Flávia. Relação entre crescimento do tecido adiposo e obesidade em idades precoces. Nutrição em Pauta, São Paulo, v.13, n.74, p.21-26, set./out. 2005. SINGHAL, Atul et al. Early nutrition and leptin concentrations in later life. The American Journal of Clinical Nutrition, USA, v.75, p. 993-999, 2002. SPIEGELMAN, Bruce M; FLIER, Jeffrey S. Obesity and the regulation of energy balance. Cell Press, v.104, p.531-543, fev. 2001. STEINBERG, G.R et al. Leptin increases FA oxidation in lean but not obese human skeletal muscle: evidence of peripheral leptin resistance. Am. J. Physiol. Endocrinol. Metab., v.283, n.1, p.187-192, 2002. SUDI, K et al. Insulin and insulin resistance index are not independent determinants for the variation in leptin in obese children and adolescents. J Pediatr Endocrinol Metab, v.13, n.7, p.923-932, 2000. UEHARA, Sofia Kimi; ROSA, Glorimar; CARMO, Maria das Graças Tavares do. Atualização sobre a Síndrome Metabólica. Revista de Metabolismo e Nutrição, Porto Alegre, RS, v.8, n.1, p.6-14, 2006. VIEIRA, Victoria J et al. Elevated atopy in healthy obese women. The American Journal of Clinical Nutrition, USA, v.82, p.504-509, 2005. WAJCHENBERG, Bernardo Léo. Tecido adiposo como glândula endócrina. Arquivos Brasileiros de Endocrinologia e Metabologismo, São Paulo, v.44, n.1, p.13-20, fev. 2000.

Disponível no site Nutrição Ativa (www.nutricaoativa.com.br) 43
WEIGLE, David S et al. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. The American Journal of Clinical Nutrition, USA, v.82, p.41-48, 2005.





























