Embed Size (px)
Citation preview

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
1
O Modelo de Hodgkin-Huxley
Os mecanismos iônicos responsáveis pela geração de um potencial de ação
foram elucidados pelos trabalhos de Hodgkin e Huxley com o axônio gigante
de lula na primeira metade do Século XX.
Capa do programa da cerimônia de entrega do Prêmio Nobel de Medicina e
Fisiologia de 1963.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
2
Um vídeo do YouTube mostrando como o experimento de HH foi feito está
disponível em: https://www.youtube.com/watch?v=k48jXzFGMc8
O axônio gigante da lula é uma fibra não-mielinizada com um diâmetro em
torno de meio milímetro e vários centímetros de comprimento. Ela é uma das
maiores células de animais conhecidas. Para comparação, as células dos
vertebrados possuem diâmetros de alguns poucos micrômetros.
Por causa disso, o axônio gigante da lula constitui um sistema ideal para a
realização de experimentos eletrofisiológicos.
Para a realização de seus experimentos, Hodgkin e Huxley utilizaram duas
técnicas experimentais, conhecidas como grampeamento espacial e
grampeamento de voltagem (veja a figura abaixo).

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
3
Dois eletrodos finos e longos são inseridos ao longo do axônio e conectados
aos instrumentos mostrados. A resistência dos eletrodos é tão baixa que pode-
se considerar que todos os pontos ao longo do axônio têm o mesmo potencial
elétrico em cada instante de tempo. Portanto, o potencial passa a depender
apenas do tempo e não do espaço, como numa célula pontual (isopotencial).
Dizemos que a célula está sob um grampo espacial.
Um dos eletrodos está emparelhado com um eletrodo externo ao axônio, para
medir a diferença de voltagem V através da membrana. O outro eletrodo é
usado para injetar ou retirar corrente da célula, na quantidade justa para manter
a diferença de potencial através da membrana num valor constante qualquer. A
medida dessa corrente permite o cálculo da condutância (ou da resistência) da
membrana. Esta técnica é chamada de grampo de voltagem, ou fixação de
voltagem, e permite controlar o potencial de membrana, fazendo com que ele
tenha qualquer valor que se queira.
Quando o potencial de membrana é elevado abruptamente do seu valor de
repouso para outro valor, e mantido neste valor, a corrente de membrana Im(t)
apresenta um comportamento como o mostrado a seguir.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
4
A corrente de membrana Im(t) tem três componentes:
1. Um brevíssimo pulso de corrente (de alguns poucos micro-segundos) para
fora da célula. Esta corrente corresponde ao carregamento do capacitor que
constitui a membrana, pois o aumento da voltagem implica num aumento da
carga armazenada na superfície da membrana.
2. Um fluxo de corrente transiente para dentro da célula com duração de 1 a 2
ms. Vários experimentos, como, por exemplo, substituindo-se os íons de
sódio no meio extra-celular por outros íons monovalentes, mostraram que
esta corrente é devida à entrada de íons de sódio na célula.
3. Uma corrente para fora da célula que se manifesta aproximadamente 4 ms
após o início do grampo de voltagem e permanece estável pelo tempo que
durar esse grampo. Estudos com traçadores iônicos revelaram que esta
corrente é devida a íons de potássio. (Em uma escala de tempo de várias
dezenas de milisegundos, esta corrente de potássio também cai para zero
como a de sódio).

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
5
Além dessas três componentes, há também uma pequena corrente constante
para fora (imperceptível na escala do desenho) que corresponde a íons de
cloreto e outros íons que passam pela membrana.
Com base nos seus estudos experimentais, Hodgkin e Huxley postularam o
seguinte modelo fenomenológico para explicar os eventos observados durante
a ocorrência de um potencial de ação no axônio gigante da lula:
1. A corrente de membrana é dada pela soma da corrente capacitiva e de uma
corrente iônica:
dttdVCtItI m
iônicam)()()( += . (1)
2. A corrente iônica é dada pela soma de correntes iônicas para íons
específicos. A corrente de um dado íon é independente das correntes iônicas
dos outros íons. Há três correntes iônicas responsáveis pela geração do
potencial de ação: de sódio, de potássio e de vazamento (ou fuga):
vazKNaiônica IIItI ++=)( (2)
A razão para esta última corrente é que, em seus experimentos, Hodgkin e
Huxley observaram uma corrente adicional além das de sódio e de potássio
cuja condutância não dependia da voltagem; eles atribuíram essa corrente a
um vazamento na membrana, possivelmente devido ao eletrodo.
3. Os canais iônicos de sódio e de potássio são modelados por condutâncias
ativas, que dependem da voltagem e do tempo, em série com baterias cujas
voltagens são dadas por seus respectivos potenciais de Nernst:

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
6
( )( )íonmmíoníon EtVttVGtI −= )(),()( , (3)
onde íon = Na+ ou K+.
4. O canal iônico de vazamento é modelado por uma condutância passiva Gvaz
em série com uma bateria Evaz (veja a figura a seguir):
Os valores de Gvaz e Evaz são determinados empiricamente para que a
solução numérica do modelo se ajuste bem aos dados experimentais.
O circuito elétrico equivalente ao modelo de Hodgkin-Huxley para a
membrana do axônio gigante da lula é dado pela figura a seguir:

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
7
Notem as posições das baterias: a posição de EK indica que o potencial de
Nernst do potássio é negativo (negativo no interior da célula em relação ao
exterior) e a posição de ENa indica que o potencial de Nernst do sódio é
positivo (positivo no interior da célula em relação ao exterior). A posição da
bateria da corrente de vazamento indica que o seu potencial de reversão é
positivo.
Em seus experimentos de grampeamento de voltagem, Hodgkin e Huxley
aumentavam a voltagem para um certo valor em relação ao repouso e
determinavam o comportamento temporal das correntes dos íons de sódio e
potássio enquanto a voltagem permanecesse constante no valor grampeado.
Para fazer isso, é fundamental conseguir isolar as contribuições de cada
corrente iônica particular para a corrente iônica total.
5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
8
Hodgkin e Huxley conseguiram isso colocando o axônio de lula em um banho
com concentração de sódio muito baixa (ou em que o sódio fosse substituído
por colina, que é um cátion ao qual a membrana é impermeável). Isso fazia
com que a corrente de sódio (INa) fosse praticamente zero, de maneira que toda
a corrente medida era a de potássio (IK). Posteriormente, colocava-se o axônio
em um banho com concentração normal de sódio e media-se a corrente iônica
normal (Iiônica). Subtraindo-se dessa corrente a corrente de potássio determinada
anteriormente podia-se determinar INa.
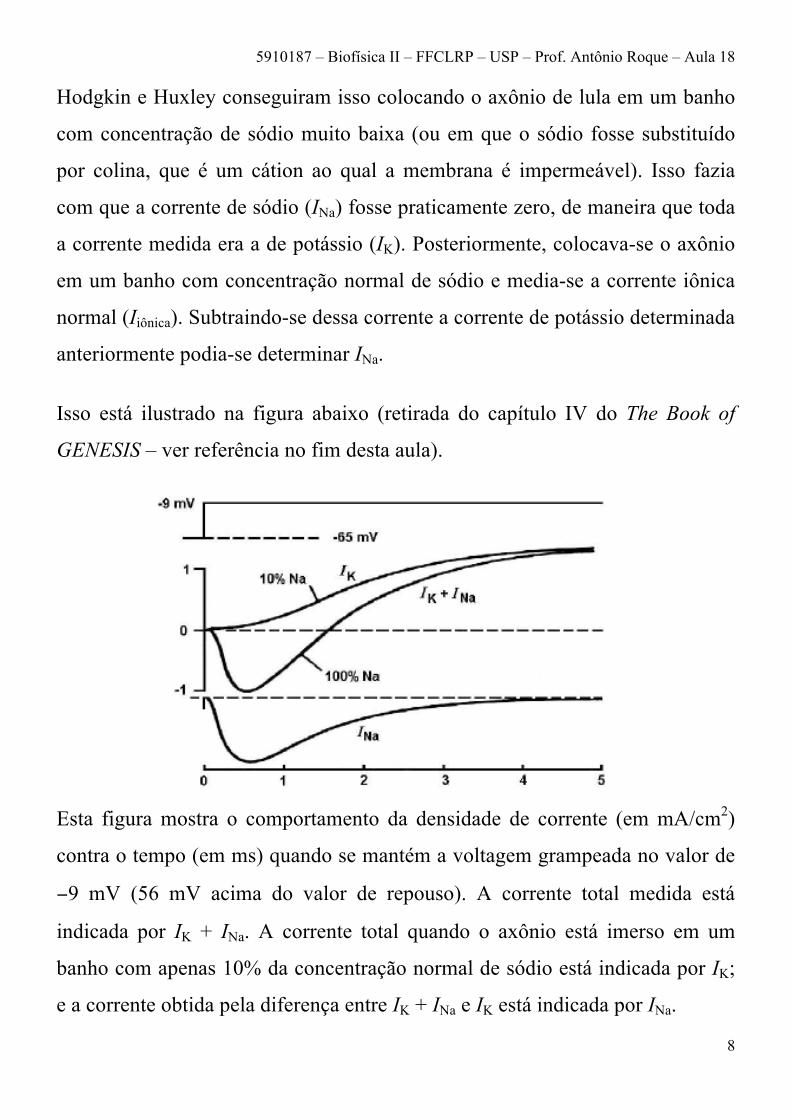
Isso está ilustrado na figura abaixo (retirada do capítulo IV do The Book of
GENESIS – ver referência no fim desta aula).
Esta figura mostra o comportamento da densidade de corrente (em mA/cm2)
contra o tempo (em ms) quando se mantém a voltagem grampeada no valor de
−9 mV (56 mV acima do valor de repouso). A corrente total medida está
indicada por IK + INa. A corrente total quando o axônio está imerso em um
banho com apenas 10% da concentração normal de sódio está indicada por IK;
e a corrente obtida pela diferença entre IK + INa e IK está indicada por INa.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
9
Outra maneira de repetir esse experimento é usando um bloqueador de canais
iônicos. Existem toxinas e agentes farmacológicos capazes de bloquear
seletivamente apenas alguns tipos de canais iônicos. Um exemplo é a
tetrodotoxina (TTX), capaz de bloquear apenas os canais de sódio.
Quando se faz um experimento como o descrito acima e se consegue isolar a
corrente devida apenas a um íon, a condutância do íon pode ser determinada da
seguinte maneira. Supondo que apenas um tipo de íon pode passar através da
membrana, a equação para o potencial de membrana torna-se:
),())(,()(injrev tIEVtVG
dttdVC m
m =−+
ou
),())(,()(injrev tIEVtVG
dttdVC m
mm +−−=
onde Iinj(t) é a corrente que tem que ser injetada pelo eletrodo dentro da célula
para manter o potencial constante no valor grampeado e Erev é o potencial de
reversão do único íon presente. Como a corrente injetada faz com que o
potencial de membrana permaneça constante, dVm/dt = 0 e o lado esquerdo da
equação anterior deve ser nulo. Pode-se então escrever:
).)(()( revinj EVtGtI m −= (4)
Esta equação indica que o comportamento temporal da corrente injetada é o
mesmo da condutância do íon que está fluindo através da membrana. Como os
valores de Vm e de Erev são conhecidos, ela permite que se determine G(t).
5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
10
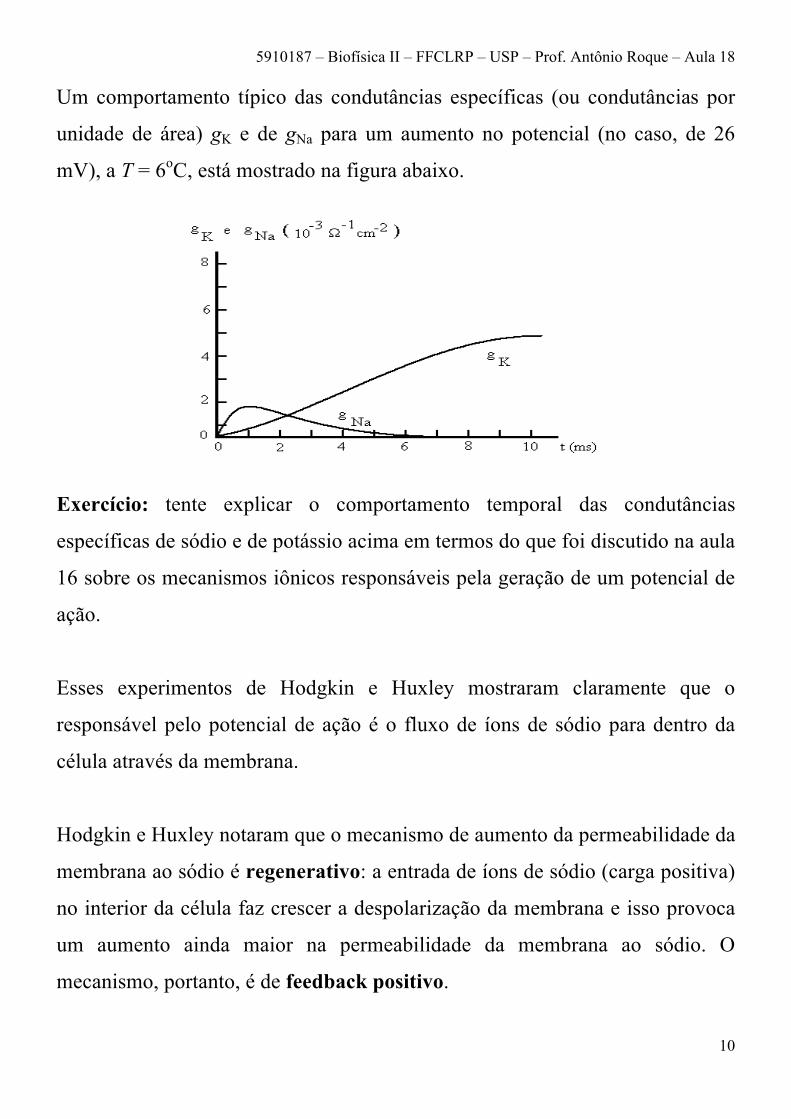
Um comportamento típico das condutâncias específicas (ou condutâncias por
unidade de área) gK e de gNa para um aumento no potencial (no caso, de 26
mV), a T = 6oC, está mostrado na figura abaixo.
Exercício: tente explicar o comportamento temporal das condutâncias
específicas de sódio e de potássio acima em termos do que foi discutido na aula
16 sobre os mecanismos iônicos responsáveis pela geração de um potencial de
ação.
Esses experimentos de Hodgkin e Huxley mostraram claramente que o
responsável pelo potencial de ação é o fluxo de íons de sódio para dentro da
célula através da membrana.
Hodgkin e Huxley notaram que o mecanismo de aumento da permeabilidade da
membrana ao sódio é regenerativo: a entrada de íons de sódio (carga positiva)
no interior da célula faz crescer a despolarização da membrana e isso provoca
um aumento ainda maior na permeabilidade da membrana ao sódio. O
mecanismo, portanto, é de feedback positivo.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
11
Após a identificação experimental do papel das condutâncias de potássio e
sódio, Hodgkin e Huxley tiveram que encontrar equações capazes de fitar o
comportamento temporal dessas condutâncias para cada valor de grampo de
voltagem usado.
A figura abaixo (também retirada do The Book of GENESIS) mostra o resultado
de vários experimentos dando a variação temporal da condutância específica do
potássio para diferentes valores da voltagem grampeada. Os círculos abertos
indicam os dados experimentais e as linhas sólidas indicam as curvas obtidas
por Hodgkin e Huxley para fitar os dados.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
12
Observe que o valor final (chamado de valor estacionário) da condutância
específica aumenta com o valor da voltagem grampeada (os valores de
voltagem dados no gráfico são medidos em relação ao potencial de repouso da
célula). Observe também que a taxa de crescimento da condutância específica
em direção ao valor estacionário (a inclinação da curva) aumenta com a
voltagem grampeada.
Observando seus dados, Hodgkin e Huxley pensaram em fitar as curvas de
condutância específica por uma função envolvendo uma exponencial, tal como
⎟⎠⎞
⎜⎝⎛ −=
−)(e1)(),( V
t
KK VgtVg τ,
com gK e τ dependentes do valor da voltagem. gK(V) daria o valor estacionário
para o qual a condutância específica tenderia a uma dada voltagem grampeada
e τ(V) determinaria quão rapidamente a condutância específica cresceria para
esse valor estacionário.
No entanto, uma exponencial simples como a da função acima não daria um
bom ajuste. A figura abaixo mostra por quê. Uma curva de condutância
específica tem inclinação (derivada) nula em t = 0, enquanto que uma
exponencial tem inclinação não nula.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
13
Existem muitas maneiras de se gerar uma função matemática que se pareça
melhor com a curva experimental de uma condutância específica. Uma delas é
usando o fato de que para x pequeno (1 – e-x) é aproximadamente igual a x
(mostre isso a partir da expansão da função exponencial em série de Taylor).
A função x elevada a uma potência se aproxima da origem com inclinação nula
(veja o gráfico esboçado a seguir). Por outro lado, para x → ∞, (1 – e-x) → 1
assim como (1 – e-x)α, onde α é um expoente qualquer.
Isto implica que uma função que envolva um termo do tipo (1 – e-t/τ)α pode ser
capaz de fitar a curva experimental da condutância específica: para t → 0 ela se
aproxima da origem com inclinação nula e para t → ∞ ela tende a 1.
Hodgkin e Huxley propuseram uma expressão do tipo a seguir para descrever o
comportamento da condutância específica do potássio para cada valor
grampeado de V: α
τ⎥⎦⎤
⎢⎣⎡ ⎟
⎠⎞⎜
⎝⎛ −=
−
∞
t
KK eVngtVg 1)(),( .

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
14
Nesta expressão, Kg é o maior valor possível que a condutância específica do
potássio pode atingir durante a aplicação do grampo de voltagem por um longo
tempo. Vamos chamar este valor de condutância específica máxima do
potássio.
Já o valor n∞α (V ) é um número entre 0 e 1 e determina a fração da condutância
específica máxima Kg que será atingida em t → ∞ para o valor de voltagem
grampeada V. Portanto, o valor estacionário da condutância específica para um
dado valor de voltagem grampeada V é dado por ).(VngKα∞
Após muitas tentativas, Hodgkin e Huxley obtiveram um bom ajuste para os
seus dados com um expoente α = 4 na expressão proposta (este valor foi o
menor expoente capaz de fornecer uma concordância aceitável da equação com
os dados experimentais).
Desta maneira, para cada valor de V, o comportamento experimental da
condutância específica do potássio pode ser bem descrito empiricamente pela
expressão:
gK (V, t) = gKn4 (V, t),
(5)
onde
n(V, t) = n∞(V ) 1− e−tτ n (V )#
$%
&'(.
(6)

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
15
Note que a constante τ está agora indicada com um sub-índice n, τ = τn. Isto foi
feito para indicar que ela é relativa à variável n sendo usada para o potássio,
pois aparecerão outras variáveis do tipo de n e outras constantes de tempo
quando a condutância específica do sódio for modelada mais para a frente.
Note que, para um dado valor de V, a função n(V, t) em (6) é solução da
equação diferencial,
τ n (V )dndt= n∞(V )− n, (7)
para constantes τn(V) e n∞(V).
A variável n(V, t) é chamada de variável de ativação do potássio. A equação
diferencial acima descreve como a ativação do potássio aumenta até atingir o
valor estacionário para um dado valor de voltagem.
Para completar a modelagem fenomenológica da condutância específica do
potássio, Hodgkin e Huxley tiveram que determinar como os parâmetros τn(V)
e n∞(V) dependem de V. Isto foi feito a partir de fittings de dados experimentais
como os mostrados anteriormente. As equações obtidas por eles para τn(V) e
n∞(V) serão mostradas mais adiante.
A modelagem da variação temporal da condutância do sódio para cada valor de
grampo de voltagem foi feita de maneira similar por Hodgkin e Huxley. Porém,
olhando para as curvas experimentais a seguir, dando o comportamento de gNa
contra t, vemos que o comportamento de gNa é mais complicado que o de gK.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
16
A condutância específica do sódio inicialmente sobe em resposta ao aumento
do potencial, mas depois decai enquanto o potencial fica mantido num valor
alto.
O gráfico acima mostra as respostas da condutância específica do sódio, gNa, a
aumentos em V do tipo degrau de diferentes valores, obtidos pelo grampo de
voltagem e indicados pelos números à esquerda (em mV). As curvas suaves
são os ajustes fornecidos pelas equações do modelo de Hodgkin-Huxley. As
escalas verticais à direita estão em unidades de mS/cm2.
Para fitar o comportamento transiente de gNa observado na figura acima,
Hodgkin e Huxley propuseram que a condutância específica do sódio deve
obedecer a uma equação do tipo:
),,(),(),( 3 tVhtVmgtVg NaNa = (8)

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
17
onde Nag é o máximo valor que a condutância específica do sódio pode atingir
e m e h são funções de V e t com valores entre 0 e 1, de maneira que o produto
m3h determina a fração da condutância específica máxima Nag que é atingida
em t → ∞ para um dado valor do grampo de voltagem V.
Observe que a maneira de modelar a condutância específica do sódio é análoga
à usada para modelar a condutância específica do potássio, só que agora são
usadas duas variáveis (m e h) para descrever o comportamento temporal de Nag
ao invés de apenas uma (n) como no caso de Kg .
A função m é chamada de variável de ativação do sódio e é uma função
exponencialmente crescente de V e t. A função h é chamada de variável de
inativação do sódio e é uma função exponencialmente decrescente de V e t.
As variáveis m e h obedecem equações diferenciais similares àquela para a
variável n do potássio:
mVmdtdmVm −= ∞ )()(τ e ,)()( hVh
dtdhVh −= ∞τ (9)
onde τm(V), m∞(V), τh(V) e h∞(V) têm a mesma interpretação que as variáveis
τn(V) e n∞(V) para o potássio.
Combinando tudo o que foi visto acima, o modelo de Hodgkin-Huxley é
caracterizado pela seguinte equação,
,)(
),(AtI
tVJdtdVC inj
mm
m +−= (10)

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
18
onde Cm é a capacitância específica da membrana, Iinj(t) é a corrente injetada na
célula e A é a área da superfície da membrana da célula. A divisão por A foi
feita porque a equação acima está escrita em termos de grandezas específicas
(por unidade de área).
A densidade de corrente de membrana Jm é composta por três componentes
(corrente de sódio, de potássio e de vazamento):
( ) ( ) ( ) ( ).),(, 433
1VmVKmkNamNa
kkm EVgEVngEVhmgtVJtVJ −+−+−==∑
= (11)
As variáveis de ativação m e n e de inativação h, por sua vez, obedecem às
seguintes equações diferenciais:
τ n (V )dndt= n∞(V )− n, (12)
τm (V )dmdt
=m∞(V )−m, (13)
τ h (V )dhdt= h∞(V )− h, (14)
Por razões que serão vistas na próxima aula, é costume reescrever cada uma
destas três equações em termos de duas novas variáveis, αi e βi, onde i = n, m,
h.
Exercício: Mostre que as equações acima para n, m e h podem ser reescritas
como:

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
19
( ) ,)(1)( nVnVdtdn
nn βα −−= (15)
dmdt
=αm (V ) 1−m( )−βm (V )m, (16)
( ) ,)(1)( hVhVdtdh
hh βα −−= (17)
com as variáveis αi e βi, i = n, m, h, definidas por
;)()(
)(VVnV
nn τ
α ∞= ;)()(
)(VVmV
mm τ
α ∞= ;)()(
)(VVhV
hh τ
α ∞= (18)
;)()(1
)(VVnV
nn τ
β ∞−= ;
)()(1
)(VVmV
mm τ
β ∞−= .
)()(1
)(VVhV
hh τ
β ∞−= (19)
As dependências das variáveis, αi e βi com o potencial de membrana V foram
determinadas empiricamente por Hodgkin e Huxley, resultando nas expressões:
11010exp
1001,0)(−⎟⎠
⎞⎜⎝
⎛ −−
=VVVnα e ⎟
⎠
⎞⎜⎝
⎛ −=80
exp125,0)( VVnβ (potássio) (20)
11025exp
251,0)(−⎟⎠
⎞⎜⎝
⎛ −−
=VVVmα e ⎟
⎠
⎞⎜⎝
⎛ −=18
exp4)( VVmβ (sódio) (21)
⎟⎠
⎞⎜⎝
⎛ −=20
exp07,0)( VVhα e 1
1030exp
1)(+⎟⎠
⎞⎜⎝
⎛ −=
VVhβ (sódio). (22)
A voltagem V que aparece nestas equações é o potencial de membrana medido
em relação ao potencial de repouso do axônio gigante da lula, em milivolts
(isto é, no repouso V = 0 nas equações acima).

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
20
As condutâncias específicas máximas para cada um dos três canais e os
respectivos potenciais de reversão também foram determinados
experimentalmente por Hodgkin e Huxley e seus valores são:
120=Nag mS/cm2; 36=Kg mS/cm2; 3,0=Vg mS/cm2; (23)
ENa = 115 mV; EK = −12 mV; Ev = 10,163 mV; Cm = 1 µF/cm2. (24)
A solução numérica das equações do modelo de Hodgkin e Huxley mostra o
aparecimento de potenciais de ação para correntes acima de um valor limiar,
conforme observado experimentalmente.
O exemplo a seguir é de uma simulação das equações de Hodgkin e Huxley
com um potencial de repouso Vrep = −65mV e uma densidade de corrente
injetada Jinj = 10 µA/cm2 com duração de 1 ms aplicada em t = 0.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
21
Para entender o modelo de Hodgkin-Huxley, é conveniente fazer uma análise
dos comportamentos de n(t), m(t) e h(t) quando a voltagem é fixada em um
dado valor (simulando um experimento de grampeamento de voltagem). Esses
comportamentos são mostrados na figura a seguir:
Observe que:
- As variáveis de ativação do potássio (n, em azul) e do sódio (m, em preto)
são inicialmente pequenas e aumentam com o tempo de maneira
exponencial, porém a variável m aumenta muito mais rapidamente que a
variável n.
- Já a variável de inativação do sódio (h, em vermelho) tem um valor inicial
mais alto e depois decai para zero. Quando h = 0, a corrente de sódio está
completamente inativa.
O mecanismo de geração de um potencial de ação pode ser entendido a partir
de uma análise da figura acima.

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
22
Inicialmente, existe um balanço entre a corrente de Na+ para dentro e a corrente
de K+ para fora. No repouso, m é pequena e h é grande, de maneira que o termo
m³h é pequeno, mas não nulo. A variável de ativação do K+, n, tem um valor
intermediário entre m e h, mas n4 é também pequeno.
Após o início do pulso de voltagem, a despolarização aumenta m rapidamente e
os canais de Na+ se abrem. Após alguns milissegundos, h vai a zero e a
condutância do Na+ se inativa. Enquanto isso, n aumenta e a condutância do K+
cresce, atingindo um valor estacionário alto. Após o fim do pulso, n, m e h
retornam aos seus valores de repouso.
A chave para se entender o comportamento do potencial de membrana gerado
pelas variáveis m, h e n está nas suas respectivas constantes de tempo, τm, τh e
τn.
Observando a figura acima, vemos que as variáveis de ativação do potássio (n)
e de inativação do sódio (h) levam muito mais tempo para atingir seus valores
estacionários (indicados pelo subíndice ∞) do que a variável de ativação do
sódio (m). Como o comportamento temporal das três variáveis é exponencial,
do tipo e−t/τ, as constantes de tempo associadas às variáveis n e h são maiores
do que a constante de tempo associada à variável m.
Isto pode ser mais bem entendido observando as figuras abaixo, que mostram
como as constantes de tempo e as variáveis de ativação e inativação dependem
da voltagem no modelo de Hodgkin-Huxley.
5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
23
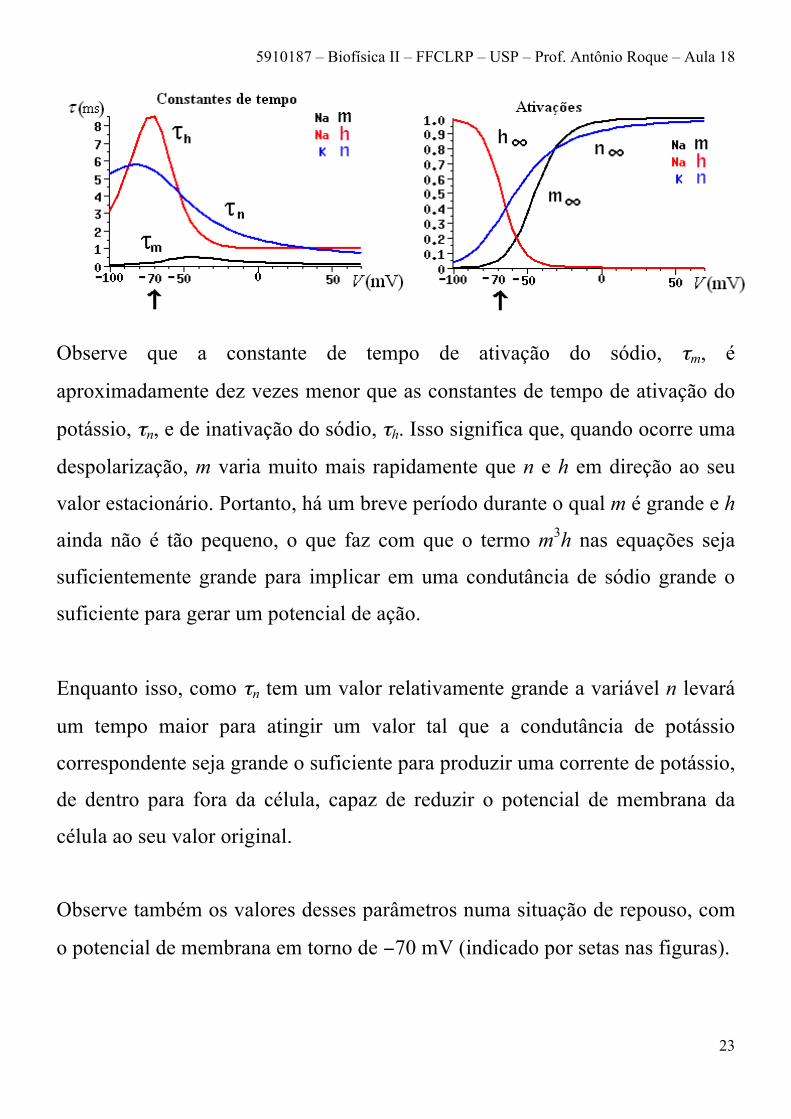
Observe que a constante de tempo de ativação do sódio, τm, é
aproximadamente dez vezes menor que as constantes de tempo de ativação do
potássio, τn, e de inativação do sódio, τh. Isso significa que, quando ocorre uma
despolarização, m varia muito mais rapidamente que n e h em direção ao seu
valor estacionário. Portanto, há um breve período durante o qual m é grande e h
ainda não é tão pequeno, o que faz com que o termo m3h nas equações seja
suficientemente grande para implicar em uma condutância de sódio grande o
suficiente para gerar um potencial de ação.
Enquanto isso, como τn tem um valor relativamente grande a variável n levará
um tempo maior para atingir um valor tal que a condutância de potássio
correspondente seja grande o suficiente para produzir uma corrente de potássio,
de dentro para fora da célula, capaz de reduzir o potencial de membrana da
célula ao seu valor original.
Observe também os valores desses parâmetros numa situação de repouso, com
o potencial de membrana em torno de −70 mV (indicado por setas nas figuras).

5910187 – Biofísica II – FFCLRP – USP – Prof. Antônio Roque – Aula 18
24
Note que o valor de h aumenta com a hiperpolarização de célula e que existe
uma faixa de valores de hiperpolarização tal que m se mantém diferente de
zero.
Isto implica que uma possível estratégia para se despolarizar a célula é,
primeiro, hiperpolarizá-la um pouco e depois despolarizá-la.
Quando a célula é hiperpolarizada, o valor da variável de inativação do sódio,
h, cresce – um processo chamado de desinativação. Posteriormente, quando
ocorre a despolarização, a variável de ativação do sódio, m, cresce rapidamente
e h se mantém em um valor relativamente alto por um certo tempo (por causa
da sua constante temporal grande). Isto implica que o produto m3h pode chegar
a atingir valores maiores neste caso do que quando ocorre apenas uma
despolarização a partir do repouso.
Referências:
- Bower, J.M. and Beeman, D. (1998) The book of GENESIS (2nd ed.),
Springer-Verlag, New York. Disponível gratuitamente na internet em:
http://genesis-sim.org/GENESIS/bog/bog.html.
- Hodgkin, A. L. and Huxley, A. F., A quantitative description of membrane
current and its application to conduction and excitation in nerve. Journal of
Physiology, London, 117: 500-544, 1952.
- Nicholls, J.G., Martin, A.R., Wallace, B.G. and Fuchs, P.A. (2001) From
Neuron to Brain (4th ed.), Sinauer, Sunderland, MA.