Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
INSTITUTO DE MICROBIOLOGIA PROFESSOR PAULO DE GÓES
DEPARTAMENTO DE MICROBIOLOGIA MÉDICA
Tese de Doutorado
Staphylococcus aureus ISOLADOS DE PRÓTESES
ARTICULARES E OUTRAS INFECÇÕES: DIVERSIDADE
GENOTÍPICA E ASPECTOS RELACIONADOS À
RESISTÊNCIA E VIRULÊNCIA
Ricardo Pinto Schuenck
Rio de Janeiro 2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
Ricardo Pinto Schuenck
Staphylococcus aureus ISOLADOS DE PRÓTESES
ARTICULARES E OUTRAS INFECÇÕES: DIVERSIDADE
GENOTÍPICA E ASPECTOS RELACIONADOS À
RESISTÊNCIA E VIRULÊNCIA
Orientadores: Kátia Regina Netto dos Santos
Marcia Giambiagi de Marval
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
INSTITUTO DE MICROBIOLOGIA PROFESSOR PAULO DE GÓES
RIO DE JANEIRO FEVEREIRO DE 2009
Tese de Doutorado apresentada ao Programa de Pós-graduação em Ciências (Microbiologia) do Instituto de Microbiologia Prof. Paulo de Góes, da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências (Microbiologia).

iii
FICHA CATALOGRÁFICA
S385 SCHUENCK, Ricardo Pinto Staphylococcus aureus isolados de próteses articulares e outras infecções: diversidade genotípica e aspectos relacionados à resistência e virulência, 2009. xvi, 131p Tese [Doutorado em Ciências (Microbiologia)] Universidade Federal do Rio de Janeiro, Instituto de Microbiologia Professor Paulo de Góes, 2009 Orientadores: Kátia Regina Netto dos Santos Márcia Giambiagi de Marval 1. Staphylococcus aureus 2. SCCmec 3. resistência antimicrobiana 4. infecções em próteses 5. genes de virulência 6. diversidade genotípica – Teses. I. Santos, Kátia Regina Netto dos (Orientadora). II. Instituto de Microbiologia Professor Paulo de Góes. III. Staphylococcus aureus isolados de próteses articulares e outras infecções: diversidade genotípica e aspectos relacionados à resistência e virulência

iv
Ricardo Pinto Schuenck
Staphylococcus aureus isolados de próteses articulares e outras infecções: diversidade genotípica e aspectos relacionados
à resistência e virulência
Rio de Janeiro, 18 de Fevereiro de 2009
Orientadora (Profa. Kátia Regina Netto dos Santos, Doutora, UFRJ) Membro (Prof. Sergio Eduardo Longo Fracalanzza, Doutor, UFRJ) Membro (Prof. Antônio Carlos Campos Pignatari, Doutor, UNIFESP) Membro (Prof. Raphael Hirata Jr., Doutor, UERJ)
Revisora (Profa. Marinella Silva Laport, Doutora, UFRJ) Suplente (Profa. Simone Aranha Nouér, Doutora, UFRJ) Suplente (Profa. Ana Paula Vieira Colombo, Doutora, UFRJ)

v
Trabalho realizado no Laboratório
de Infecções Hospitalares e no
Laboratório de Microbiologia Molecular
do Departamento de Microbiologia
Médica do Instituto de Microbiologia
Prof. Paulo de Góes, sob orientação das
Profas. Kátia Regina Netto dos Santos e
Marcia Giambiagi de Marval.

vi
AGRADECIMENTOS
Ao final de mais uma importante etapa, gostaria de agradecer as
pessoas que tanto me apoiaram:
À minha orientadora Kátia Regina Netto dos Santos, minha “mãe
científica”, pela orientação, no sentido amplo da palavra, paciência,
incentivo e carinho, que contribuíram, acima de tudo, para o meu
crescimento pessoal. Tive o privilégio, de durante esses quase 10 anos,
estar ao lado de uma pessoa dedicada, exigente e, que acima de tudo, ama
o que faz! Com certeza, não poderia ter escolhido orientadora melhor!
Obrigado por me agüentar todo esse tempo !
À minha co-orientadora Márcia Giambiagi de Marval, por toda sua
imensa ajuda, dedicação e pelas sempre oportunas considerações!
A todos os grandes amigos do Laboratório de Infecções Hospitalares:
Gustavo, Eliezer, Ariane, Natália, Carol, Fred, Milena, André, Aline,
Cristiane, Roberta e Fernanda pela ajuda no trabalho, pelos momentos de
descontração e pela amizade.
Um agradecimento especial a Fernandinha, cuja ajuda, foi
imprescindível para o término desse trabalho. Muito obrigado!
Ao pessoal do Laboratório de Microbiologia Molecular, em especial,
Naira e Elaine, pela ajuda e pelos muitos momentos de descontração.
A todos dos Laboratórios de Micobactérias, Cocos Patogênicos e
Anaeróbios, por toda ajuda durante o desenvolvimento da tese.
Aos amigos da Pós-Graduação: Ivi, Bia, Tati, Carla, Renata, Lili,
Naira, Fabíola, Elaine e Léo. Um agradecimento especial ao Felipe, Marcos

vii
Dornelas, Gustavo e Eliezer, pela importante ajuda e amizade durante
esse tempo.
Aos funcionários do Departamento de Microbiologia Médica,
especialmente, Marlei, Orlando, Joaquim, Marcos, Antônio e Helena, pela
ajuda e por tornarem agradável o dia-a-dia nesse departamento.
À Dra. Simone Nouér, membro da equipe da Comissão de Controle
de Infecção Hospitalar (CCIH) do Hospital Universitário Clementino Fraga
Filho, pela importante ajuda durante o desenvolvimento desse trabalho.
A todos do Laboratório de Microbiologia e da CCIH do Instituto
Nacional de Traumatologia e Ortopedia, em especial, as funcionárias
Francisca, Rita e Maria do Carmo.
À Dra. Thais C. B. S. Souto-Padrón, Coordenadora do Curso de Pós-
Graduação, e aos demais professores do IMPPG.
Aos professores Sérgio Fracalanzza, Walter, Regina e Marinella por
sempre estarem prontos a nos ajudar.
Ao Instituto de Microbiologia, na pessoa da diretora Agnes Marie Sá
Figueiredo.
À FAPERJ pela bolsa a mim concedida durante o desenvolvimento
desse trabalho.
Ao CNPq, FUJB, Pronex e FINEP pelo financiamento dos projetos de
pesquisa desenvolvidos nesse laboratório.
A todos meus verdadeiros amigos Cordeirenses, que mesmo não
entendendo absolutamente nada do que eu fazia, compreendiam a
importância disso para mim.

viii
Ao meu pai (in memorian) que é o responsável direto pelo que sou
hoje e por durante toda a sua vida ter colocado seus filhos acima de tudo
e me ensinado o verdadeiro significado da palavra amar. Amor eterno!
À minha mãe, por muitas vezes se preocupar com meus desejos em
detrimento aos seus e pelo amor incondicional. Se pudesse escolher uma
mãe, seria exatamente como você. Agradeço também por compreender
todos os dias de distância e por tê-la feito vir tantas vezes ao Rio de
Janeiro! Amo você!
Aos meus irmãos, por todo amor e por entenderem minha escolha,
que me mantém distante fisicamente.
Na monografia o agradecimento foi como namorada, no mestrado
como noiva e, agora, como esposa! Gostaria de agradecer a Gisele, que fez
parte de cada passo que dei durante todo esse tempo. Mais do que estar
ao meu lado, você é parte essencial de minha vida. Agradeço por todo seu
amor, ajuda, compreensão pelas muitas horas em que estive ausente e
impaciente, enfim, por ter se mostrado uma verdadeira companheira.

ix
DEDICATÓRIA
Ao meu pai (in memorian) que dedicou cada dia da sua vida a me tornar uma pessoa melhor. À minha mãe, pelo amor incondicional e por ter sido essencial para que eu chegasse até aqui. À minha esposa que me acompanhou em cada passo dessa caminhada e foi acima de tudo uma verdadeira companheira.

x
RESUMO
Staphylococcus aureus isolados de próteses articulares e outras infecções: diversidade genotípica e aspectos relacionados à resistência e virulência.
Ricardo Pinto Schuenck
Orientadoras: Kátia Regina Netto dos Santos
Marcia Giambiagi de Marval Resumo da Tese de Doutorado submetida ao Programa de Pós-Graduação em Ciências (Microbiologia) do Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências (Microbiologia).
Neste trabalho foram realizados dois estudos com 93 amostras de Staphylococcus aureus isoladas de dois hospitais da cidade do Rio de Janeiro. No primeiro, foi realizada a caracterização molecular e determinação das CMIs de oxacilina e vancomicina em 20 amostras de S. aureus resistentes à meticilina não multirresistentes (nMRSA). Destas, 19 apresentaram SCCmecIV e uma SCCmecII. Dezoito amostras apresentaram CMIs <32µg/mL para oxacilina e CMIs para vancomicina entre 1 e 2µg/mL. Foram observados nove genótipos, incluindo o USA300, e 3 STs principais: 1, 5 e 8. Dois novos STs foram descritos: 1203 e 1204. No segundo estudo, foram analisados a diversidade genotípica e os aspectos relacionados à virulência e resistência em 73 amostras, sendo 32 de infecções associadas (IAP) e 41 de não associadas a próteses (INAP). Destas, 45 (61,6%) foram de MSSA e 28 de MRSA. Entre as 28 MRSA, 16 (57%) apresentaram SCCmecIV (8 de IAP e 8 de INAP) e 12 SCCmecIII (5 de IAP e 7 de INAP). A análise por PFGE mostrou 15 genótipos, sendo que cinco (A´-E´), foram responsáveis por 83,5% das infecções. Todas as 12 amostras do genótipo A´ foram relacionadas ao genótipo de MRSA prevalente no Brasil (ST 239-SCCmecIII). Os genótipos B´ e C´ englobaram 20 e 17 amostras, respectivamente, sendo os principais agentes de todas as infecções (IAP e INAP). Nesses dois genótipos, foram encontradas amostras de MRSA SCCmecIV e MSSA. Os genótipos B´ e C´ apresentaram ST 5 e ST 1, respectivamente. O ST 30 foi encontrado nas amostras com o genótipo E´. Mais de 95% das amostras, tanto de IAP quanto de INAP, foram produtoras de biofilme. Os genes de virulência, icaA, fnbA, cna e ebpS estavam amplamente distribuídos, enquanto os genes fnbB, bbp e da PVL foram encontrados associados a genótipos específicos. Não foram observadas diferenças, em relação à virulência e aos genótipos, entre as amostras de IAP e INAP. Neste estudo observamos a emergência de importantes genótipos e demonstramos que as amostras causadoras de infecções em próteses refletem a microbiota existente no hospital. Foram detectados STs, até o momento, não descritos no país (ST 97 e ST 72). A similaridade genética entre amostras de MRSA-SCCmecIV e MSSA de dois genótipos, incluídos no ST 1 e ST 5, nos fez sugerir que essas amostras estariam mais aptas a receberem o cassete mec. Palavras chaves: Staphylococcus aureus – SCCmec – resistência antimicrobiana – infecções em próteses – genes de virulência – diversidade genotípica.
Rio de Janeiro Fevereiro de 2009

xi
ABSTRACT
Staphylococcus aureus isolated from prosthesis and other infections: clonal diversity and resistance and virulence aspects
Ricardo Pinto Schuenck
Orientadoras: Kátia Regina Netto dos Santos
Márcia Giambiagi de Marval Abstract da Tese de Doutorado submetida ao Programa de Pós-Graduação em Ciências (Microbiologia) do Instituto de Microbiologia Professor Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências (Microbiologia). .
In the present work two studies with 93 Staphylococcus aureus isolates obtained from two hospitals in Rio de Janeiro were carried out. In the first study, a molecular characterization and MICs determination to oxacillin and vancomycin in 20 isolates of nonmultirresistant methicillin-resistant Staphylococcus aureus (nMRSA) were performed. Nineteen isolates presented SCCmecIV and one SCCmecII. Eighteen showed MICs to oxacillin of <32µg/mL. MICs to vancomycin ranged from 1 to 2µg/mL. Nine genotypes and three principal STs, 1, 5 and 8, were found. Genotype USA300 was also found among these lineages. Moreover, two new STs were described, STs 1203 e 1204. In the second study we analyzed the clonal diversity, resistance and virulence factors of 73 isolates, 32 recovered from prosthesis related infections (PRI) and 41 from prosthesis not related infections (PNRI). Among 73 isolates, 45 (61,6%) were MSSA and 28 MRSA. Among 28 MRSA, 16 (57%) presented SCCmecIV (8 of PRI and 8 of PNRI) and 12 SCCmecIII (5 of PRI and 7 of PNRI). PFGE clustered all isolates in 15 genotypes. However, five prevalent genotypes (A´ to E´) caused 83.5% of the infections. All 12 isolates of genotype A´ were related to the MRSA prevalent in Brazil (ST 239-SCCmecIII). Genotypes B´ and C´ included 20 and 17 isolates, respectively. These two genotypes were found among MRSA SCCmecIV and MSSA isolates, presenting ST 5 (genotype B´) and ST 1 (genotype C´). ST 30 was found among isolates from genotype E´. At least 95% of isolates produced biofilm. Virulence genes, icaA, fnbA, cna and ebpS were extensively distributed, and fnbB, bbp and PVL genes were associated with specific genotypes. There was no difference between PRI and PNRI regarding the prevalence of virulence genes and genotypes found. Our results show the presence of important international lineages of MRSA-SCCmecIV in our institutions and STs not described in Brazil (ST 97 e ST 72). The genetic similarity observed between MRSA SCCmecIV and MSSA isolates that were included in genotypes B´ and C´ (STs 5 and 1, respectively) indicates that some lineages are more adapted to receive cassette mec.
Key words: Staphylococcus aureus – SCCmec – microbial resistance - prosthesis infection – virulence genes - genotypic diversity
Rio de Janeiro Fevereiro de 2009

xii
ÍNDICE
1 Introdução
1.1 O Gênero Staphylococcus................................................................ 2
1.2 Staphylococcus aureus.................................................................... 3
1.3 Fatores de virulência ..................................................................... 4
1.4 Aspectos gerais das infecções associadas a próteses articulares causadas por Staphylococus aureus.......................................................... 11
1.5 Resistência à meticilina em Staphylococcus aureus......................... 14
1.6 Epidemiologia das infecções por Staphylococcus aureus resistentes à meticilina............................................................................................... 21
1.7 Justificativa do presente estudo...................................................... 29
2 Objetivos............................................................................................... 32
3 Material e Métodos
3.1 Locais de estudo............................................................................. 34
3.2 Amostras bacterianas..................................................................... 34
3.3 Amostras-controle.......................................................................... 35
3.4 Definições utilizadas neste estudo................................................... 36
3.5 Identificação bacteriana.................................................................. 36
3.5.1 Confirmação da identificação das amostras de Staphylococus aureus...................................................................................................... 37
3.6 Determinação da susceptibilidade aos antimicrobianos................... 38
3.7 Tipagem do SCCmec por PCR-multiplex nas amostras de Staphylococcus aureus resistentes à meticilina.......................................... 40
3.7.1 Liberação do DNA................................................................... 40
3.7.2 PCR-multiplex para detecção do tipo de SCCmec.................... 42
3.8 Análise do perfil de fragmentação do DNA cromossômico após tratamento com enzima de restrição e separação por eletroforese em gel de campo pulsado (PFGE).........................................................................
45
3.9 Caracterização das amostras de Staphylococcus aureus através da análise dos fragmentos de restrição de genes que codificam isenzimas (MLFRT)....................................................................................................
47
3.10 Caracterização das amostras de Staphylococcus aureus através de tipagem por seqüênciamento de multilócus enzimático (MLST).................. 49
3.11 Detecção de genes que codificam fatores de virulência bacteriana. 50
3.12 Método quantitativo de análise da produção de biofilme................ 52

xiii
3.13 Análise estatística......................................................................... 54
4 Resultados............................................................................................ 55
5 Discussão.............................................................................................. 86
6 Conclusões............................................................................................ 110
7 Referências bibliográficas...................................................................... 112
8 Anexo.................................................................................................... 130

xiv
LISTA DE ABREVIATURAS
A – Adenina
ATCC – “American type culture collection”
C – Citosina
CA-MRSA – Staphylococcus aureus resistente à meticilina de origem comunitária
CC – Complexo clonal
CCIH – Comissão de controle de infecção hospitalar
CDC – Centro de Prevenção Controle de Doenças, Estados Unidos
CLSI – “Clinical and Laboratory Standards Institute”
CMI(s) – Concentração (ões) mínima (s) inibitória (s)
oC – Graus centígrados
DNA – Ácido desoxirribonucléico
dNTPS – Desoxirribonucleotídeos trifosfatados
D.O. – Densidade ótica
EDTA – Ácido etilenodiaminotetracético
G – Guanina
h – Horas
HA-MRSA - Staphylococcus aureus resistente à meticilina de origem hospitalar
HIV – Vírus da imunodeficiência humana
HUCFF – Hospital Universitário Clementino Fraga Filho
IAP – Infecções associadas a prótese
INAP - Infecções não associadas a prótese
INTO – Instituto Nacional de Traumatologia e Ortopedia
IS – Seqüência de inserção
kb – Quilobase (s)
M - Molar
µg – Micrograma (s)
mg – Miligrama (s)
min - Minutos
mL – Mililitro (s)
µL – Microlitro (s)
MLFRT – Polimorfismo dos fragmentos de restrição de genes que codificam
isoenzimas (“Multilocus fragment restriction type”)
MLST – Tipagem por seqüenciamento de multilocus enzimático (“Multilocus
sequence typing”)

xv
MRSA – Staphylococcus aureus resistente à meticilina
MSCRAMMs – Componentes da superfície microbiana que reconhecem moléculas
da matriz extracelular
MSSA - Staphylococcus aureus sensível à meticilina
mM - Milimolar
NNISS – “National nosocomial infection surveillance system”
nMRSA - Staphylococcus aureus resistente à meticilina não multirresistente
OSPC – Clone Oceania Pacífico Sul
pb – Par (es) de base (s)
PBP – Proteína ligadora de penicilina
PCR – Reação em cadeia da polimerase
PFGE – Eletroforese de campo elétrico pulsado
pH – Potencial de hidrogênio
PVL – Leucocidina de Panton-Valentine
p/v – Peso por volume
RFT – “Restriction fragment type”
SCCmec – Cassete cromossômico mec de Staphylococcus
SCN – Staphylococcus coagulase-negativo
s – Segundo (s)
ST – “Sequence type”
T – Timina
TBE – Tris – Borato – EDTA
TE – Tris – EDTA
Tn – Transpósons
TSA – Ágar tripcaseína de soja
TSB – Caldo tripcaseína de soja
UFC – Unidade formadora de colônia
UI – Unidade internacional
UTI – Unidade de terapia intensiva
UFRJ – Universidade Federal do Rio de Janeiro
V – Volts
v/v – Volume por volume

xvi
VISA – Staphylococcus aureus com resistência intermediária à vancomicina
VRSA – Staphylococcus aureus resistentes à vancomicina
X g – Força gravitacional
1
1 - Introdução

2
1.1 O Gênero Staphylococcus
Membros do gênero Staphylococcus apresentam-se como cocos
Gram-positivos, com 0,5 a 1,5µm de diâmetro, isoladamente, aos pares,
em tétrades, em pequenas cadeias (3 ou 4 células) ou irregulares, na
forma de cachos. Esses microrganismos são imóveis, resistentes à
bacitracina, não formam esporos, são anaeróbios facultativos,
normalmente, produzem a enzima catalase, são capazes de crescer em até
10% de NaCl e sua temperatura ótima de crescimento se encontra entre
30oC e 37oC. Pertencem à família Staphylococcaceae e em seu genoma
apresentam baixo conteúdo G-C (HOLT et al., 1994; BANNERMAN &
PEACOCK, 2007).
O gênero Staphylococcus é composto por 41 espécies e 24
subespécies (EUZEBY, 2008). Essas espécies estão amplamente
distribuídas na natureza e colonizam, principalmente, mucosas, pele e
glândulas da pele de mamíferos e aves (JARLOV,1999).
Baseado na capacidade de coagular o plasma, através da ação da
enzima coagulase, as espécies do gênero são classificadas em coagulase-
positivas e coagulase-negativas. A espécie S. aureus subespécie aureus
(referida apenas como S. aureus) é a principal representante do grupo das
espécies coagulase-positivas. Essa espécie é reconhecida desde 1883 como
agente de infecções, tendo sido responsável, nessa era pré-antibiótico, por
taxas de mortalidade em bacteriemias de até 82% (SMITH & JARVIS,
1999). Hoje, essa espécie continua sendo considerada como patogênica e
de maior importância entre os estafilococos em infecções humanas, tanto
de origem comunitária quanto de origem hospitalar.

3
Os Staphylococcus coagulase-negativos (SCN) constituem um grupo
onde é encontrada a maior parte das espécies do gênero Staphylococcus
(JARLOV et al., 1999). Até 1975, os SCN eram agrupados juntos e
distinguidos do S. aureus por sua inabilidade em coagular plasma
sanguíneo. Eles foram considerados, durante muito tempo,
microrganismos saprófitas, porém, sua importância como patógenos tem
sido reconhecida nas últimas décadas, principalmente, devido ao aumento
no uso de próteses, cateteres e dispositivos invasivos, associado ao
crescente número de pacientes imunocomprometidos (BANNERMAN &
PEACOCK, 2007). Esses fatores trouxeram os SCN para a vanguarda dos
patógenos hospitalares. Contudo, como essas bactérias são habitantes
normais da pele e mucosas, um dos principais desafios é distinguir as
amostras de SCN com significância clínica das amostras contaminantes
(VON EIFF, PETERS & HEILMANN, 2002). Entre as espécies de SCN,
aquelas que têm sido mais isoladas de infecções humanas, especialmente
de bacteriemias, infecções relacionadas aos dispositivos médicos e
infecções urinárias são: S. epidermidis, S. haemolyticus, S. hominis e S.
saprohyticus (ING, BADDOUR & BAYER, 1997).
1.2 Staphylococcus aureus
As infecções por S. aureus muitas vezes são precedidas por
colonização, principalmente nasal, que ocorre em cerca de 25% dos
indivíduos sadios (VANDENBERGH et al., 1999; SCANVIC et al., 2001). A
narina anterior funciona como reservatório primário de S. aureus em

4
humanos (CASEWEL & HILL, 1986). Esse microrganismo pode ser
encontrado colonizando outros sítios, como axilas e períneo, porém,
havendo eliminação do microrganismo das narinas, normalmente, há
eliminação também em outros sítios, indicando ser a narina o principal
sítio de colonização da espécie (KLUYTMANS et al., 1997). A partir desse
local, o microrganismo pode colonizar pele, mucosas e outras regiões
(REAGAN et al., 1991). Além disso, indivíduos que são portadores nasais
podem contaminar as mãos de profissionais de saúde e/ou outros
pacientes e contribuir para a dispersão do microrganismo no ambiente ao
seu redor (BOYCE, 1996).
1.3 Fatores de virulência
Devido à variedade de mecanismos de virulência conhecidos, S. aureus é
considerado um dos patógenos humanos mais versáteis (ARCHER, 1998). S.
aureus pode causar desde infecções cutâneas superficiais, como impetigo,
celulite e abscessos, até infecções invasivas decorrentes da invasão direta
dos tecidos, como bacteriemia, endocardite, pneumonia e meningite (SMITH
& JARVIS, 1999). Além disso, esse patógeno causa síndromes clínicas
relacionadas à produção de toxinas, incluindo intoxicação alimentar
associada à ingestão de enterotoxina estafilocócica pré-formada, síndrome da
pele escaldada e síndrome do choque tóxico estafilocócico (BANNERMAN &
PEACOCK, 2007).
Inúmeros fatores de virulência atuam na patogenia bacteriana.
Durante a etapa de colonização está envolvido um conjunto de adesinas

5
que são capazes de se ligar aos componentes da matriz extracelular do
hospedeiro (tabela 1), que são designadas pela sigla MSCRAMMs
(“Microbial Surface Components Recognizing Adhesive Matrix Molecules”)
e reconhecidas como importantes receptores para aderência estafilocócica
(PROJAN & NOVICK, 1997). A maioria dessas MSCRAMMs possui em
comum a característica de estarem ancoradas, covalentemente, à
peptideoglicana da parede celular bacteriana através de ligações de
transpeptidação. Essas proteínas, freqüentemente, apresentam um sinal
peptídico, C-terminal, do tipo LPXTG, o qual é reconhecido por uma
enzima denominada sortase. Essa enzima é responsável pelo ancoramento
de proteínas de superfície a peptídeoglicana. A sortase é uma proteína
localizada na membrana do S. aureus que cliva os peptídeos entre a
treonina e a glicina do domínio LPXTG e catalisa a formação de uma ponte
amida entre o radical carboxil da treonina e o grupamento amina da ponte
cruzada do peptideoglicana (TON-THAT et al., 1999). Entre as MSCRAMMs
de S. aureus estão proteínas que se ligam ao fibrinogênio, como o fator
clumping A e B (ClfA e ClfB), proteínas que se ligam à elastina (Ebp), ao
colágeno (Cna), ao ácido siálico ósseo (Bbp) e proteínas que se ligam à
fibronectina A e B (FnBPA e FnBPB), que são reconhecidas como
importantes invasinas capazes de promover a entrada do S. aureus em
vários tecidos (FOSTER & HOOK, 1998; FOWLER et al., 2000; TUNG et
al., 2000). Essas MSCRAMMs têm papel essencial no início das infecções
endovasculares, e daquelas relacionadas a dispositivos médicos, entre
outras. Diferentes amostras de S. aureus podem apresentar conjuntos
diversos de MSCRAMMs, o que pode explicar a capacidade apresentada

6
por algumas cepas em causar determinadas infecções (PATTI et al. 1994;
FOSTER & HOOK, 1998).
Tabela 1. Principais adesinas de ligação à matriz extracelular do hospedeiro encontradas em Staphylococcus aureus.
Gene Função da proteína codificada Referência
Cna ligação ao colágeno PATTI et al., 1994
clfA/clfB fator clumping – ligação ao fibrinogênio PATTI et al., 1994
fnbA/fnbB ligação à fibronectina MENZIES, 2003
Ebp ligação à elastina FOSTER et al., 1998
Bbp ligação ao ácido siálico ósseo TUNG et al., 2000
Spa proteína A (adesina) e ligação a porção Fc do anticorpo
HARTLEIB et al., 2000
Uma vez aderido ao tecido, S. aureus é capaz de se estabelecer e
persistir, utilizando vários mecanismos. Esse microrganismo é capaz de
formar uma estrutura complexa denominada biofilme, tanto em
componentes biológicos quanto em superfícies inanimadas (CONSTERTON
et al, 1999). Costerton e colaboradores (1999) definiram biofilme como
uma comunidade estruturada de células bacterianas embebidas em uma
matriz polimérica, produzidas por estas bactérias, e aderidas em uma
superfície biótica ou abiótica.
O biofilme atua como barreira física à ação do sistema imunológico
do paciente e tem se tornado um grave problema na área de saúde, pois,
está intrinsecamente relacionado ao aumento da resistência dos
microrganismos aos antimicrobianos. Além disso, está associado a

7
infecções relacionadas a dispositivos médicos e a dificuldade de
tratamento dessas infecções, uma vez que as células embebidas no
biofilme (sésseis) encontram-se menos expostas aos antimicrobianos, e
adicionalmente, neste microambiente há maior concentração de
nutrientes, como carbono e nitrogênio (DONLAN, 2001). El-Azizi e
colaboradores (2005) ao compararem a susceptibilidade à vancomicina e
quinupristina/dalfopristina em amostras de S. aureus e S. epidermidis, no
biofilme e em condições planctônicas, observaram que todas as células
crescidas em condições planctônicas foram sensíveis aos antimicrobianos
testados, enquanto no biofilme, grande parte das células era resistente a
concentrações superiores a 1g/mL, de ambas as drogas.
A formação de biofilme em Staphylococcus é um processo
multifatorial e complexo que pode ser subdividido em duas fases distintas:
(1) uma fase de rápida aderência da bactéria à superfície biótica ou
abiótica e (2) uma fase mais prolongada de acúmulo de células, que
envolve proliferação, adesão entre células bacterianas e a produção de
uma matriz levando a formação do biofilme maduro (HEILMANN et al.,
1996). A aderência inicial a superfícies depende não apenas da natureza
do material do polímero, mas também das características da superfície
celular bacteriana (GÖTZ, 2002). Essa interação inicial envolve forças
físico-químicas não específicas, como força de van der Waal’s, interações
hidrofóbicas e polaridade (VON EIFF, PETERS & HEILMANN, 2002). S.
aureus também pode se ligar às superfícies via proteínas da matriz
extracelular do hospedeiro uma vez que após a implantação do dispositivo
médico, o polímero rapidamente torna-se recoberto por plasma e proteínas

8
da matriz extracelular, como fibronectina, fibrinogênio, vitronectina,
colágeno e outras (DICKINSON & BISNO, 1989).
Na fase de acúmulo, adesinas polissacarídicas têm papel importante
na interação entre as células bacterianas (GÖTZ, 2002). Vários
polissacarídeos têm sido estudados, porém, a maioria dos estudos tem
mostrado que o principal composto presente nessa fase de formação de
biofilme é a adesina intercelular polissacarídica, referida como PIA, sendo
sua biossíntese dependente de genes do operon icaADBC (ARCIOLA,
BALDASSARRI & MONTANARO, 2001). A estrutura desse principal
polissacarídeo é formada por uma glicosaminoglicana, com ligações β-1,6
com pelo menos 130 resíduos de 2-deoxi-2-amino-D-glicopiranosil, onde
80 a 85% são N-acetilados, sendo 15-20% desses resíduos deacetilados e
assim, carregados positivamente (MACK et al., 1996).
O operon ica é composto pelo gene icaR (regulatório) e pelos genes
ica ADBC (biossíntese de PIA). Esse operon foi, inicialmente, descrito em S.
epidermidis e mais tarde foi observada sua presença também em S. aureus
(CRAMTON et al., 1999). Apesar das funções das proteínas IcaA, B, C e D
não estarem completamente elucidadas, acredita-se que IcaA e D têm
atividade de glicosil transferase, atuando na síntese dos oligômeros e
utilizando como substrato UDP-N-acetilglicosamina, enquanto IcaC
estaria envolvida na externalização do polissacarídeo crescente e IcaB
seria responsável pela reação de deacetilação (GERKE et al., 1998). O gene
icaR que está localizado à montante do operon ica codifica o repressor
desse operon (HEILMANN et al., 1996).

9
Em S. aureus, outros fatores, além da PIA, parecem ter papel
importante na formação do biofilme. Alguns estudos demonstram que
amostras de S. aureus que sofreram deleção no lócus ica foram capazes de
continuar produzindo biofilme, revelando que há outros mecanismos
envolvidos nessa produção, independente do operon ica (CUCARELA et al.,
2001; FITZPATRICK et al., 2005; O´NEILL et al., 2008).
A manutenção da bactéria no biofilme requer mecanismos
adaptativos ao modo séssil de vida. Dentro do biofilme a bactéria
desenvolve uma organizada e complexa comunidade, onde os canais
servem como um rudimentar sistema circulatório (BOLES, THOENDEL &
SINGH, 2004). A liberação de moléculas sinalizadoras, que é finamente
regulada por um sistema “quorum sensing”, induz a bactéria a responder
às mudanças, alterando seu perfil de expressão gênica. Além disso, esse
microambiente fornece condições ótimas para transferência de genes
extra-cromossomiais (DUNNE et al., 2002). Beenken e colaboradores
(2004) analisando a expressão global dos genes de S. aureus em biofilme e
nas fases exponencial e pós-exponencial de culturas planctônicas, através
de “microarray”, identificaram 48 genes em que a taxa de expressão foi
aumentada pelo menos duas vezes mais no biofilme. Concomitantemente,
foi observado que 84 genes tiveram sua expressão diminuída pela metade
nas culturas planctônicas.
Os S. aureus também possuem outras características que o permitem
evadir do sistema imune do hospedeiro (FOSTER et al., 2005). Um dos
principais fatores é uma microcápsula, presente em mais de 90% das
amostras clínicas de S. aureus que atua como fator antifagocitário, sendo

10
os tipos CP5 e CP8, os mais encontrados (JOHN & BARG, 1996; LEE &
LEE, 2000).
Durante o processo infeccioso S. aureus é capaz de produzir várias
enzimas, como proteases e lipases que permitem que o patógeno invada e
destrua os tecidos, migrando para outros sítios (LOWY, 1998). Além disso,
essa espécie produz várias toxinas, como a da síndrome do choque tóxico
(TSST), esfoliatinas e enterotoxinas que causam síndromes severas
associadas a essas toxinas (DINGES et al., 2000).
A leucocidina de Panton Valentine (PVL) é uma toxina cuja importância
vem sendo destacada devido ao isolamento cada vez maior de amostras de S.
aureus que a produzem. A PVL tem se destacado como um importante fator
de virulência, devido a constante presença dessa proteína em amostras de S.
aureus resistentes à meticilina causadoras de infecções comunitárias e
também hospitalares (DAVIS et al., 2006; BARTELS et al., 2007). Essa
toxina, codificada pelos genes lukS-PV e lukF-PV, consiste em duas proteínas
secretadas separadamente, conhecidas como componentes S e F, que atuam
sinergicamente para lisar células polimorfonucleares, monócitos e
macrófagos (PREVÓST et al., 1995). Os genes que a codificam encontram-se
inseridos em bacteriófagos que possuem a capacidade de integrar-se no
cromossomo de S. aureus (KANECO & KAMIO, 2004). Amostras MRSA
produtoras de PVL estão fortemente associadas com infecções cutâneas e
pneumonias necrotisantes (GILLET et al., 2002; YAMASAKI et al., 2005).

11
1.4 Aspectos gerais das infecções associadas a próteses articulares
causadas por Staphylococcus aureus
Implantes de próteses articulares estão entre os principais avanços
da cirurgia e medicina nas últimas décadas (STECKELBERG & OSMON,
2000). Contudo, esses avanços vieram acompanhados de maior
susceptibilidade às infecções, apesar das taxas ainda serem relativamente
baixas. Segundo Steckelberg e Osmon (2000) a incidência de infecções em
artroplastias de joelho e quadril é de 5,9/1.000 pacientes-ano, nos dois
primeiros anos pós-operatório, e de 2,3/1000 pacientes-ano, entre 2 e 10
anos após a cirurgia. Na Inglaterra, o Serviço Nacional de Vigilância de
Infecções Hospitalares divulgou as taxas de infecções em próteses de
joelho e quadril, entre 1997 e 2005, tendo sido observados percentuais de
1% e 1,8%, respectivamente (http://www.hpa.org.uk/infections/topics_az/
surgical_site_infection/all_97_05_ssi.pdf). Essas infecções ocasionam alta
morbidade devido a dor, perda da prótese, necessidade de nova cirurgia, e
em muitos casos, pode levar a óbito. Além disso, o custo com o reimplante
é 3 a 4 vezes maior do que o implante original e há prolongado uso de
antimicrobianos (STECKELBERG & OSMON, 2000). Sculco e
colaboradores (1995) demonstraram que o custo com cada episódio de
infecção em próteses, nos Estados Unidos, excede os $50.000,00.
Em relação aos principais agentes causadores de infecções em
próteses articulares os Staphylococcus, sobretudo, as espécies S. aureus e
S. epidermidis, possuem um papel essencial, uma vez que, normalmente,
são encontrados em mais de 70% dessas infecções (EHLRICH et al., 2004;
MORAN et al., 2007). Moran e colaboradores (2007) analisando infecções

12
de próteses articulares, em 112 pacientes, demonstraram que S. aureus
estava presente em 52% e os SCN em 47% dessas infecções, sendo que em
algumas delas havia a presença de ambos os patógenos.
Microrganismos podem colonizar a prótese, diretamente no
momento da colocação do implante através da manipulação pela equipe
médica, através da via hematogência durante uma bacteriemia, ou por
contigüidade de sítios infectados (LIDWELL et al., 1983). Muitas vezes,
devido ao longo período existente entre a realização do implante e o
surgimento dos sintomas e/ou diagnóstico, torna-se difícil determinar a
causa da infecção (TSUKAYAMA, ESTRADA & GUSTILO, 1996). As
infecções que ocorrem em próteses são classificadas de acordo com o
período em que apareceram os sintomas, após a colocação do implante,
em: i) infecção rápida, aquela que ocorre até três meses após implante da
prótese, sendo, em geral, adquirida durante a colocação da prótese; ii)
infecção retardada, que ocorre entre três meses e dois anos após a
colocação do implante, podendo ser adquirida durante a colocação da
prótese, sendo causada por microrganismos menos virulentos, como S.
epidermidis; iii) infecção tardia, que ocorre após 24 meses de implantação
da prótese, sendo predominantemente adquirida por via hematogênica
(TRAMPUZ & ZIMMERLI, 2005). Vários trabalhos sugerem que a maioria
das infecções em próteses articulares é adquirida na sala de cirurgia, já
que diversos tipos de profilaxia pré-operatória, incluindo antimicrobianos
sistêmico e local, diminuem significativamente a incidência dessas
infecções (HILL et al., 1981; LIDWELL et al., 1982; STECKELBERG &
OSMON, 2000). Moram e colaboradores (2007) analisando infecções em

13
próteses articulares, entre 1998 e 2003, mostraram que 69% das
infecções ocorreram nos três primeiros meses após o implante.
Normalmente, são utilizados os seguintes critérios para caracterizar
os casos como infecção de próteses (pelo menos um devendo estar
presente): i) duas ou mais culturas do fragmento do tecido ou do aspirado
da prótese devem ser positivas para o mesmo microrganismo, ii) formação
de secreção observada no sítio cirúrgico, iii) inflamação aguda com
infecção observada em exame histopatológico (STECKELBERG & OSMON,
2000).
No Brasil, não há conhecimento do número preciso de implantes de
próteses que é realizado anualmente. Na Alemanha, 2,5 milhões de
dispositivos médicos, como, cateteres, próteses articulares e marcapassos
são utilizados por ano. Ainda assim, é esperado um aumento nesse
número devido ao crescimento da expectativa de vida da população, pela
realização de exercícios de alto impacto, obesidade, entre outros (MACK et
al., 2004). Nos Estados Unidos são feitos, aproximadamente, 250.000
implantes de próteses de joelho e 450.000 de quadril, anualmente e,
levando em conta que a taxa média de infecções nesses implantes seja de
1%, cerca de 7.000 artroplastias são infectadas anualmente (EHLRICH et
al., 2004; KURTZ et al., 2005).
O principal fator da patogênese bacteriana relacionado a próteses
articulares é a produção de biofilme. Como foi dito anteriormente, o
biofilme age protegendo a bactéria da ação antimicrobiana. Dois
mecanismos têm sido propostos para explicar como as bactérias tornam-
se resistentes na presença do biofilme: (i) os antibióticos não conseguiriam

14
alcançar bactérias presentes nas camadas mais profundas do biofilme,
devido a dificuldade destes em penetrarem na camada de polissacarídeo;
(ii) as bactérias das camadas mais profundas estariam com o metabolismo
sensivelmente alterado, o que as tornariam resistentes aos
antimicrobianos (CONSTERTON et al., 1999)
1.5 Resistência à meticilina em Staphylococcus aureus
Com a introdução das penicilinas semi-sintéticas, como meticilina e
oxacilina, em 1959, ocorreu um grande avanço no tratamento das
infecções estafilocócicas, causadas por amostras produtoras de
penicilinases, diminuindo, consideravelmente, os índices de mortalidade
por essas infecções (JOHN & BARG, 1996). Porém, já no início da década
de 60, surgiram as primeiras amostras de S. aureus resistentes a essas
drogas na Inglaterra (JEVONS, 1961 apud ITO et al., 2004). Na década
seguinte, amostras de S. aureus resistentes à meticilina emergiram como
principais causadoras de infecções hospitalares em várias partes do
mundo (BENNER & KAYSER, 1968; ZECHOVSKY, 1974).
O surgimento da resistência à meticilina em S. aureus deveu-se a
aquisição do gene mecA, cujo tamanho é de 2,1kb e é responsável pela
produção de uma proteína ligadora de penicilina (PBP) alterada, de 78
kDa, chamada PBP 2a (ITO et al., 2003). Essas PBPs são enzimas que
atuam como transpeptidases durante a biossíntese da parede celular
bacteriana. Em amostras de S. aureus sensíveis à meticilina (MSSA -
“methicillin-suceptible Staphylococcus aureus”) os antimicrobianos β-
lactâmicos ligam-se às PBPs nativas e impedem a síntese da camada de

15
peptideoglicana. Como esses antimicrobianos possuem baixa afinidade
pela PBP2a, a síntese da parede celular ocorre normalmente nas amostras
resistentes (MRSA - “methicillin-resistant Staphylococcus aureus”)
(CHAMBERS, 1993).
O gene mecA está inserido em um cassete cromossômico mec de
estafilococos (“staphylococcal cassete chromosome mec”, SCCmec), um
elemento que pode variar de 20,9 a 66,9kb de DNA. Até o momento foram
descritos sete tipos de elementos SCCmec, classificados de I a VII (tabela
2; figura 1) (ITO et al. 2001; DAUM et al., 2002; ITO et al, 2004; OLIVEIRA
et al., 2006; HIGUCHI et al. 2008). Normalmente, os tipos I, IV, V, VI e VII
determinam resistência apenas aos β-lactâmicos, enquanto os tipos II e III
determinam resistência a várias classes de antimicrobianos, devido aos
elementos genéticos adicionais integrados a esses cassetes, tais como:
plasmídeos (pUB110, pI258 e pT181) e transpósons (Tn554 e ΨTn554). O
plasmídeo pUB110 apresenta o gene ant(4´) que confere resistência a
vários aminoglicosídeos, enquanto o pI258 confere resistência às
penicilinas e aos metais pesados, e o pT181 resistência à tetraciclina. Os
elementos genéticos móveis atuam como potenciais regiões “hot spot” (alta
freqüência de recombinação), contribuindo para a variação na composição
do cassete. Além disso, esses elementos possuem seqüências de inserção
(IS) como, IS431 e IS1272 e os genes responsáveis pela regulação do gene
mecA: mecI e mecR1 (ITO et al., 2001; LECLERCQ, 2002; ITO et al., 2003;
OLIVEIRA et al., 2006). A proteína MecI atua na repressão dos genes mecA
e mecR1 enquanto a MecR1 é uma proteína transmembranar com
capacidade de reconhecer os β-lactâmicos. Na presença do

16
antimicrobiano, MecR1 sofre clivagem e o domínio metaloprotease
intracelular torna-se ativo. Esse domínio cliva a proteína MecI que está
ligada à região promotora do mecA, permitindo assim, a expressão do gene
mecA e, conseqüentemente, a síntese da PBP2a (BERGER-BACHI &
ROHER, 2002). As seqüências de inserção IS431 e IS1272 podem se
inserir na região do mecI e mecR1 e, como resultado, ocorre a indução do
gene mecA (KATAYAMA et al., 2001). Já foram descritas, até o momento,
cinco classes distintas de complexo do gene mec (A, B, C, D e E) em
Staphyloccoccus, das quais, três, foram descritas para S. aureus: classe A
(mecI-mecR1-mecA-IS431), classe B (IS1272-∆mecR1-mecA-IS431) e classe
C (IS431-mecA-∆mecR1-IS431) (ITO et. al., 2001, OLIVEIRA, MILHEIRIÇO
& DE LENCASTRE, 2006; DEURENBERG & STOBBERINGH, 2008).
O SCCmec também é caracterizado pela presença de genes que
codificam as recombinases, denominados ccr, que estão presentes em
todos os elementos SCCmec (tabela 2). Sua função é integrar e excisar o
SCCmec no genoma bacteriano em um sítio específico denominado sítio de
ligação do SCCmec (attBscc), que está localizado na região terminal 3´ de
uma seqüência de leitura aberta ou orf (“open reading frame”), cuja função
não é conhecida (ITO et al., 1999). São descritos quatro alotipos (1, 2, 3 e
4) para os genes ccrA e ccrB e três para o gene ccrC (1, 2 e 8). Os tipos de
SCCmec são determinados pela composição do complexo mec e dos genes
ccr (tabela 2). As regiões que não fazem parte desses sítios são
denominadas regiões J (do inglês junkiard). Cada SCCmec é dividido em
três regiões J que, baseado em sua posição, apresentam-se,
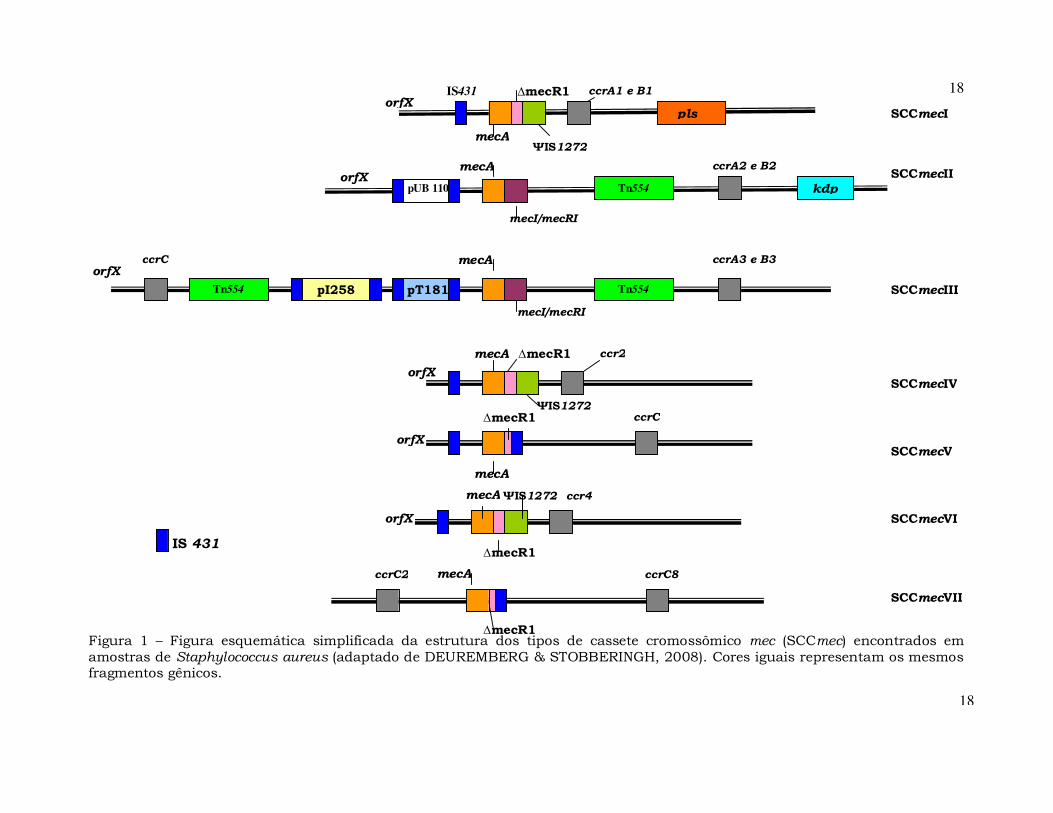
17
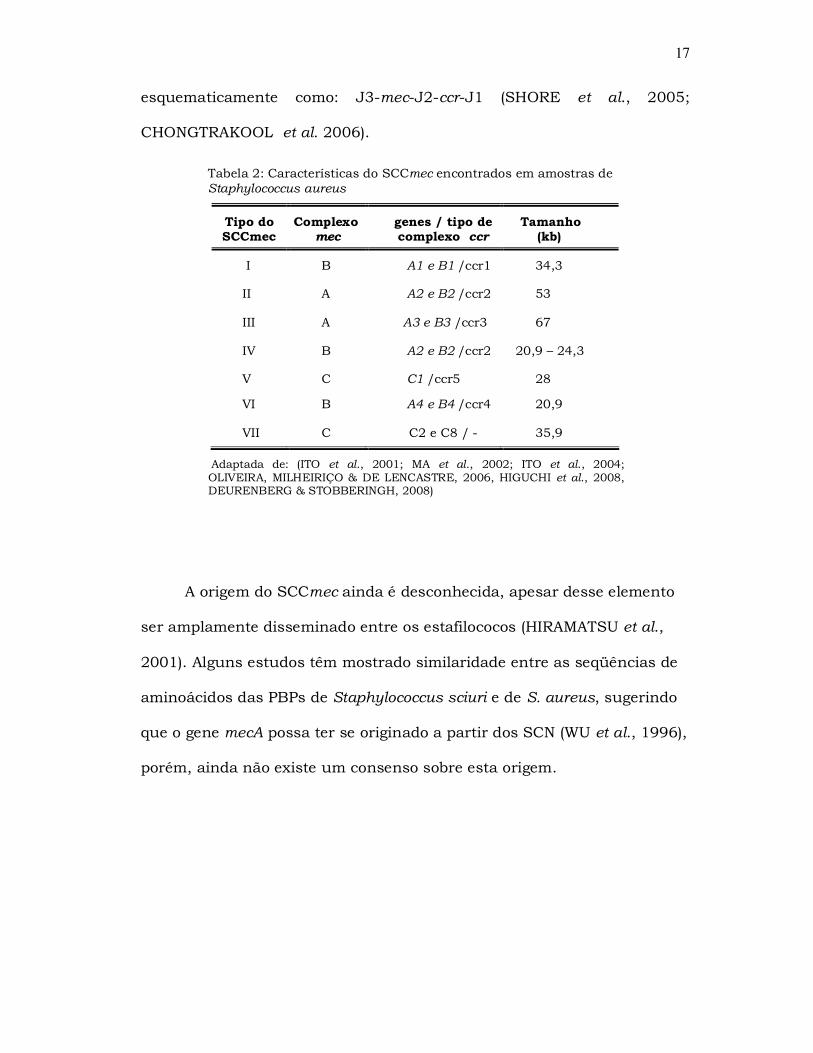
esquematicamente como: J3-mec-J2-ccr-J1 (SHORE et al., 2005;
CHONGTRAKOOL et al. 2006).
Tabela 2: Características do SCCmec encontrados em amostras de Staphylococcus aureus
Tipo do SCCmec
Complexo mec
genes / tipo de complexo ccr
Tamanho (kb)
I B A1 e B1 /ccr1 34,3
II A A2 e B2 /ccr2 53
III A A3 e B3 /ccr3 67
IV B A2 e B2 /ccr2 20,9 – 24,3
V C C1 /ccr5 28
VI B A4 e B4 /ccr4 20,9
VII C C2 e C8 / - 35,9
Adaptada de: (ITO et al., 2001; MA et al., 2002; ITO et al., 2004; OLIVEIRA, MILHEIRIÇO & DE LENCASTRE, 2006, HIGUCHI et al., 2008, DEURENBERG & STOBBERINGH, 2008)
A origem do SCCmec ainda é desconhecida, apesar desse elemento
ser amplamente disseminado entre os estafilococos (HIRAMATSU et al.,
2001). Alguns estudos têm mostrado similaridade entre as seqüências de
aminoácidos das PBPs de Staphylococcus sciuri e de S. aureus, sugerindo
que o gene mecA possa ter se originado a partir dos SCN (WU et al., 1996),
porém, ainda não existe um consenso sobre esta origem.
18
IS 431 Figura 1 – Figura esquemática simplificada da estrutura dos tipos de cassete cromossômico mec (SCCmec) encontrados em amostras de Staphylococcus aureus (adaptado de DEUREMBERG & STOBBERINGH, 2008). Cores iguais representam os mesmos fragmentos gênicos.
IS431
pUB 110 Tn554
mecA
∆mecR1
ΨIS1272
ccrA1 e B1
pls orfX
orfX
orfX
orfX
orfX
orfX
ccrA2 e B2
mecI/mecRI
kdp
pI258 pT181
ccrA3 e B3 ccrC
Tn554 Tn554
mecA
mecA
mecA
mecA
mecA
mecA
SCCmecI
SCCmecII
SCCmecIII
SCCmecIV
SCCmecV
SCCmecVI
SCCmecVII
∆mecR1
ΨIS1272
ccr2
ccrC
ccr4
ccrC2 ccrC8
mecI/mecRI
∆mecR1
∆mecR1
ΨIS1272
∆mecR1
18

19
As taxas de resistência à meticilina em S. aureus têm sido bastante
variadas nos últimos anos. Em um estudo realizado por Tiermesma e
colaboradores (2004), onde foi analisada a resistência à meticilina em S.
aureus em diversos países da Europa, foram encontradas taxas menores
que 1%, em países como Holanda e Suécia, porém, em outras localidades,
como França (33,1%), Portugal (34,7%), Itália (40,9%), Inglaterra (41,5%) e
Grécia (44%) as taxas de resistência foram superiores a 30%. No Brasil,
um trabalho realizado por Sader e colaboradores (2004), em hospitais do
Rio de Janeiro, São Paulo, Porto Alegre, Brasília e Florianópolis,
envolvendo 1.516 amostras, coletadas de janeiro de 1997 a dezembro de
2001, mostrou uma taxa média de 38% de resistência à meticilina entre
as amostras de S. aureus. Vivoni e colaboradores (2006) analisando
amostras de S. aureus isoladas de infecções em pacientes do Hospital
Universitário Clementino Fraga Filho, no Rio de Janeiro, encontraram
uma taxa de 37% de resistência à meticilina nas amostras de S. aureus
analisadas.
A terapêutica utilizada em infecções causadas por amostras MRSA
ainda é bastante limitada. Algumas drogas começaram a ser utilizadas
nos últimos anos, tais como, linezolida, quinupristina/dalfopristina e
tigeciclina (AKSOY & UNAL, 2007). Contudo, a vancomicina continua
sendo o principal antimicrobiano de escolha para o tratamento dessas
infecções. Essa droga é administrada endovenosamente e pode induzir
efeitos colaterais devido a sua toxicidade (HIRAMATSU et al., 2001).

20
Apesar da existência de relatos esporádicos de amostras clínicas de
SCN resistentes à vancomicina, desde a década de 80, o interesse sobre a
possibilidade de aquisição de resistência à vancomicina em estafilococos
aumentou somente após o primeiro relato de amostras de S. aureus com
resistência intermediária a essa droga, em 1996, no Japão (HIRAMATSU
et al., 1997) e, em 1997, nos Estados Unidos (CDC, 1997). Desde então, o
isolamento de amostras clínicas conhecidas como VISA (“vancomycin
intermediate Staphylococus aureus”) tem sido relatado em vários países
(KANTZANOU et al., 1999; ROTUN et al., 1999; MARCHESE et al., 2000).
Em 2002, nos Estados Unidos, foi relatado o isolamento de uma amostra
de S. aureus com resistência plena à vancomicina (chamada VRSA -
“vancomycin resistant Staphylococcus aureus) com concentração mínima
inibitória de 32µg/mL. Além disso, foi detectado nessa amostra o gene
vanA, indicando que essa resistência tenha sido adquirida por
transferência a partir da amostra de Enterococcus faecalis, que foi isolada
do mesmo sítio do VRSA (CHANG et al., 2003). No Brasil, Oliveira e
colaboradores (2001) publicaram o primeiro relato de isolamento de
amostras VISA em São Paulo. Nesse estudo, foram isoladas cinco
amostras VISA entre 140 amostras MRSA, isoladas no período de um ano,
sendo quatro amostras provenientes de pacientes internados na unidade
de queimados, que tinham utilizado vancomicina por 30 dias, e a quinta
amostra isolada de um paciente da unidade de ortopedia com sete dias de
tratamento com a droga. Esse fato vem reforçar a preocupação com o uso
continuado de vancomicina e, conseqüentemente, com a questão da

21
emergência de amostras de Staphylococcus resistentes à vancomicina no
ambiente hospitalar.
1.6 Epidemologia das infecções por Staphylococcus aureus
resistentes à meticilina
A primeira amostra de MRSA (NTCT 10442) foi isolada na Inglaterra
em 1960 e apresentava o SCCmec tipo I (ITO et al., 2001). Esse clone,
agora denominado clone Arcaico, disseminou-se pelo mundo na década de
60. Em 1982, outra amostra de MRSA (N315) foi isolada no Japão e
apresentou o SCCmecII. Esse clone foi chamado de Nova Iorque/Japão e
também se disseminou para vários países, sendo inclusive, o genótipo
predominante nas infecções por MRSA nos Estados Unidos. Três anos
mais tarde, em 1985, foi isolada uma amostra de MRSA na Nova Zelândia
(85/2082) que apresentou o SCCmecIII (ITO et al., 2001). Já na década de
1990 e 2000 vários clones de MRSA, com SCCmec IV, também começaram
a se disseminar por vários países (OKUMA et al., 2002; HUANG et al.,
2007; AMORIN et al., 2008) e, em 2004, na Austrália, foi descrita uma
amostra de MRSA (WIS) com SCCmecV (ITO et al., 2004). O SCCmecVI foi
observado apenas em amostras de Portugal e o protótipo dessas amostras
é chamado HDE288 (OLIVEIRA et al., 2006) e o SCCmecVII foi,
recentemente, descrito na amostra TSG-17 em Taiwan (HIGUCHI et al.,
2008).
No início da década de 1990, vários estudos utilizando técnicas de
tipagem molecular, em grandes coleções de S. aureus, começaram a
elucidar aspectos da epidemiologia desse patógeno. Duas teorias sugerem

22
a evolução clonal das amostras MRSA. A primeira, chamada de teoria
clonal simples, sugere que todos os MRSA teriam um ancestral em comum
e que o SCCmec teria sido inserido somente uma vez nesse ancestral e ele
teria se disseminado (KREISWIRTH et al., 1993). A outra teoria, chamada
multiclonal, sugere que o SCCmec foi introduzido várias vezes, em
diferentes linhagens de S. aureus (ENRIGHT et al., 2002). Vários estudos
indicam que esta última é a teoria mais provável (MUSSER & KAPUR,
1992; FITZGERALD et al, 2001). Musser e Kapur (1992) publicaram
resultados da análise de 254 amostras de MRSA, isoladas entre 1961 e
1992, em nove países de quatro continentes. Foi observado que o gene
mecA estava inserido em amostras de diversas linhagens de S. aureus de
diferentes origens, tendo esses autores sugerido que a aquisição desse
gene seria um evento de transmissão horizontal entre membros da
espécie, que teria ocorrido repetidas vezes em amostras de MSSA,
originando diversas linhagens de MRSA (MUSSER & KAPUR, 1992). O
estudo foi baseado na avaliação do perfil eletroforético de isoenzimas
(“Multilocus Enzyme Electrophoresis”, MLEE), responsáveis pela
manutenção celular e que apresentam pouca variação entre os membros
de uma mesma espécie (BOERLIN, 1997).
O estudo da epidemiologia de infecções por S. aureus teve grande
avanço com a aplicação da técnica de análise dos perfis de fragmentação
do DNA cromossômico, após o tratamento com enzimas de restrição e de
separação por PFGE (“Pulsed Field Gel Electrophoesis”). Em 1995, a
publicação de uma proposta de padronização da interpretação dos
resultados, obtidos pelo emprego da técnica, veio facilitar a utilização da

23
mesma em estudos epidemiológicos envolvendo amostras isoladas de uma
mesma localidade, em períodos curtos de tempo (TENOVER et al., 1995). A
técnica de PFGE passou a ser muito utilizada em grupos de amostras de
S. aureus responsáveis por surtos hospitalares (CLANCY et al., 2005;
LARSSEN et al., 2005).
Clones epidêmicos de MRSA são conhecidos por sua facilidade de
transmissão, longa persistência, rápida disseminação intra e inter-
hospitalar e facilidade de cruzar barreiras geográficas (PAPAKYRIACOU et
al., 2000). A tabela 3 mostra os principais clones de MRSA disseminados
pelo mundo. O clone Ibérico foi primeiramente descrito na Espanha, em
1989 (DOMINGUEZ et al., 1994) e, desde então, tem sido isolado em
Portugal (SANCHES et al., 1995), na Itália e Reino Unido (MATO et al.,
1998), Alemanha (WITTE et al., 1994), Bélgica, Suíça e França (DEPLANO
et al., 2000) e Estados Unidos (EUA) (ROBERTS et al., 1998b) entre
outros. O clone Húngaro foi descrito como extensivamente disseminado
nos hospitais da Hungria (SANCHES et al., 1998), tendo sido também
encontrado na Tailândia (CRISOSTOMO et al., 2001). O clone Nova
Iorque/Japão foi identificado como sendo predominante em hospitais de
Nova Iorque (ROBERTS et al. 1998a), Nova Jérsei, Pensilvânia e
Connecticut (ROBERTS et al., 2000), além de ter sido descrito em um
hospital localizado em Tóquio (AIRES DE SOUZA et al., 2000). O clone
pediátrico foi, inicialmente, descrito em um hospital pediátrico de Portugal
em 1992 (SÁ-LEÃO et al., 1999). Sua ocorrência também foi relatada na
Polônia (LESKI et al., 1998), EUA (ROBERTS et al., 1998b), Argentina
(CORSO et al., 1998) e Colômbia (GOMES et al., 2001), entre outros

24
países. O clone brasileiro (CB) foi inicialmente descrito por Teixeira e
colaboradores (1995) e encontrado em vários hospitais do Brasil
(TEIXEIRA, LOURENÇO & FIGUEIREDO, 1996; SANTOS et al., 1999),
Argentina, Uruguai e Chile (AIRES et al., 2001) e Portugal (DE SOUZA et
al., 1998). Esse clone encontra-se disseminado em vários países do
mundo.
Apesar da técnica de PFGE ser utilizada em inúmeros estudos, a
mesma pode se mostrar inadequada para avaliação de amostras coletadas
em longos períodos, bem como, em estudos de epidemiologia em nível
nacional ou global. Além da limitação dos critérios de interpretação, com
relação ao período de tempo e localidade onde foram coletadas as
amostras, outro problema é a dificuldade de comparação dos resultados
obtidos pelos diferentes laboratórios (TENOVER et al., 1995).
No ano de 2000, Enright e colaboradores descreveram a aplicação da
técnica de tipagem por seqüenciamento de multilócus enzimáticos
(“multilocus sequence typing”, MLST) na caracterização de amostras
MSSA e MRSA. Através dessa técnica, sete genes (arcC, aroE, glpF, gmk,
tpi, yqiL e pta) que codificam enzimas de manutenção celular são
seqüenciados, após amplificação através da técnica de reação em cadeia
da polimerase (PCR). Cada nova seqüência caracteriza-se por um novo
alelo. Da mesma forma que na eletroforese de isoenzimas, a combinação
desses alelos forma o perfil alélico (“sequence type”, ST) de cada amostra
(ENRIGHT et al., 2000). Os resultados obtidos com o uso do MLST vêm
sendo utilizados para a construção de um banco de dados que permite a
comparação de amostras de S. aureus isoladas de várias partes do mundo.

25
Em 2003, Diep e colaboradores (2003) descreveram uma nova
metodologia para tipagem de amostras de S. aureus, baseada na técnica
de MLST, denominada polimorfismo dos fragmentos de restrição de genes
que codificam isoenzimas (“multilocus restriction fragment type”, MLRFT).
Nessa metodologia são analisados os mesmos sete genes de manutenção
celular avaliados no MLST, porém, como alternativa ao seqüenciamento, é
realizada a digestão enzimática dos produtos obtidos por PCR. Esses
produtos digeridos são, então, analisados em eletroforese e os diferentes
perfis, obtidos pela avaliação dos sete genes analisados, são considerados
como diferentes tipos de fragmentação (RFTs - “restriction fragment
types”).
A associação do MLST com tipagem do SCCmec determinou uma
nova proposta de nomenclatura para os clones epidêmicos de MRSA. As
amostras são classificadas de acordo com o ST, fenótipo de resistência à
oxacilina e tipo de SCCmec. Por exemplo, o clone brasileiro é representado
como ST 239-MRSAIIIA e o clone Nova Iorque/Japão é caracterizado como
ST 5-MRSAII (OLIVEIRA, TOMASZ & DE LENCASTRE, 2001).
Em 2002, Enright e colaboradores analisaram, por MLST, 359
amostras comunitárias e hospitalares de MRSA, isoladas de 20 países.
Essa análise revelou a prevalência de cinco complexos clonais (CC),
compostos pelo mesmo ST ou STs relacionados (5 ou mais loci
semelhantes entre as amostras), que foram: CC 8, CC 5, CC 30, CC 45 e
CC22.
Os STs e complexos clonais dos principais clones distribuídos pelo
mundo estão descritos na tabela 3.

26
Tabela 3. Características dos principais clones de Staphylococcus aureus resistentes à meticilina disseminados pelo mundo.
Genótipo Sequence type (ST)
Complexo clonal (CC)
Tipo de SCCmec
Distribuição geográfica
Arcaico 250 8 I Austrália., Dinamarca, Alemanha., Suíça, Uganda, Reino Unido, Estados Unidos
Ibérico 247 8 I Bélgica, Croácia, República Tcheca, Dinamarca, Finlândia, França, Alemanha, Itália, Polônia,
Holanda, Portugal, Eslovênia, Espanha, Suécia, Suíça, Reino Unido, Estados Unidos
Irlandês 1 8 8 II Austrália, Irlanda, Estados Unidos, Reino Unido
Nova Iorque /Japão
5 5 II Austrália., Bélgica, Canadá, Dinamarca, Finlândia, França, Alemanha., Irlanda, Japão, Coréia, México, Singapura, Suíça, Uruguai,
Reino Unido, Estados Unidos
EMRSA-16 36 36 II Austrália., Bélgica, Canadá, Dinamarca, Grécia, Irlanda, México, Noruega, Espanha, Suíça,
Suécia, Reino Unido, Estados Unidos
Brasileiro/ Húngaro
239 8 III Argélia, Austrália, Brasil, China, Chile, República Tcheca, Finlândia, Alemanha., Grécia, Índia, Coréia, Mongólia, Holanda, Polônia, Portugal,Cingapura, Eslovênia, Espanha, Sri-Lanka, Suécia, Tailândia,
Uruguai, Reino Unido, Vietnã
Berlim 45 45 IV Armênia, Austrália., Bélgica, Finlândia, Alemanha, Hungria, Holanda, Noruega, Espanha, Suécia, Suíça, Estados Unidos
Pediátrico 5 5 IV Argélia, Austrália, Brasil, Colômbia, Dinamarca, França, Coréia, Noruega, Polônia, Portugal, Espanha, Uruguai, Reino Unido, Estados
Unidos
E-MRSA - 2/6 8 8 IV Austrália, Bélgica, Finlândia, França, Alemanha, Irlanda, Holanda, Noruega, Taiwan,
Reino Unido, Estados Unidos
E-MRSA- 15 22 22 IV Austrália, Bélgica, República Tcheca, Dinamarca, Alemanha, Irlanda, Kwait, Nova Zelândia, Noruega, Portugal, Singapura,
Espanha, Suíça, Reino Unido
Adaptado de DEURENBERG et al., 2007.
Em relação às amostras MSSA, apesar de sua importância como
agentes de infecções hospitalares e comunitárias, poucos são os estudos
envolvendo tipagem molecular. Contudo, o emprego da técnica de PFGE
na tipagem de MSSA tem mostrado uma grande variedade de perfis
eletroforéticos (ENRIGHT et al., 2000; NETTO DOS SANTOS et al., 2001).

27
A epidemiologia das infecções por MRSA vem sofrendo alterações
nos últimos anos, uma vez que está cada vez mais comum o isolamento de
amostras MRSA não multirresistentes (chamadas nMRSA – “non-
multirresistant oxacillin-resistant Staphylococcus aureus”) nos hospitais
(OKUMA et al., 2002; TRINDADE et al., 2005). Esse fato era observado
apenas em amostras MRSA isoladas de infecções de origem comunitária
(CA-MRSA – “community-acquired meticillin resistant Staphylococcus
aureus”). As primeiras amostras de CA-MRSA foram descritas na
população indígena na Austrália (UDO et al., 1993). Desde então, há
registro de infecções por CA-MRSA em diversas partes do mundo (OKUMA
et al., 2002; MA et al., 2005; RIBEIRO et al., 2005). Essas infecções
acometem, principalmente, população indígena, crianças em creches,
grupos militares e atletas (SHAHIN et al., 1999; GROOM et al., 2001; CDC,
2003). No Brasil, o primeiro relato de infecções causadas por CA-MRSA foi
realizado por Ribeiro e colaboradores (2005). Essas amostras
apresentavam o SCCmecIV, ST 30 e os genes da PVL. Além disso, três das
quatro amostras apresentavam o mesmo perfil pela técnica de PFGE.
Essas amostras comunitárias são diferentes das amostras
tradicionalmente encontradas nos hospitais, ou seja, freqüentemente,
apresentam resistência apenas aos β-lactâmicos, o SCCmecIV ou V e os
genes da PVL (GILLET et al., 2002; OKUMA et al., 2002; DIETRICH et al.,
2004).
Nos últimos anos, essas características apontadas anteriormente
para as amostras de CA-MRSA vêm sendo encontradas em amostras
isoladas de infecções hospitalares. Muitos estudos têm demonstrado a

28
emergência de amostras de nMRSA com SCCmec IV em hospitais de todo
mundo (TRINDADE et al., 2005; VAN DER MEE-MARQUET et al., 2006;
BARTELS et al., 2007; AMORIN et al., 2008). A grande preocupação em
relação a essas linhagens de amostras que emergem como MRSA não
multirresistentes e com SCCmec curtos é que elas parecem apresentar um
maior equilíbrio entre a resistência e virulência, demonstrando vantagens
adaptativas, tais como, tempo de geração menor, tendência de maior
facilidade de transmissão dos elementos de resistência e CMIs de oxacilina
próximas ao “ponto de corte”, o que pode acarretar em dificuldade na
detecção de sua resistência, facilitando a sua disseminação (GILLET et al.,
2002; OKUMA et al., 2002; ITO et al., 2004). Dados que ratificam a
preocupação com essas novas amostras podem ser verificados pela
substituição, em hospitais de diversos países, dos clones de MRSA
endêmicos por essas linhagens. Além disso, essas amostras estão sendo
responsáveis pelo aumento das infecções por MRSA em países cujas taxas
eram inferiores a 1%. Bartels e colaboradores (2008) relataram que na
Dinamarca, entre 1980 e 2002, as taxas de isolamento eram inferiores a
100 amostras de MRSA por ano. A partir de 2003 os números de infecções
por MRSA foram de 243 casos em 2003, 549 em 2004 e 864 em 2005.
Esse grupo caracterizou 143 amostras de MRSA isoladas de infecções
ocorridas em Copenhagen, entre 2003 e 2004, e observou que 71% dos
casos de infecções por MRSA foram caracterizadas como comunitárias.
Nesse estudo foram identificados 16 STs, sendo os prevalentes ST 8, ST
30 e ST 80. Além disso, 86% das amostras apresentavam SCCmecIV e
44% tinham os genes da PVL. Amorim e colaboradores (2007) analisando

29
280 amostras MRSA isoladas, entre 2003 e 2005, de um hospital de
Portugal, onde o clone ST 239-SCCmecIII era endêmico, verificaram que a
maioria das amostras era resistente apenas aos β-lactâmicos e que 80,3%
das amostras apresentaram ST 22 e SCCmecIV. Apenas 11,8% das
amostras apresentaram ST 239 e SCCmecIII, características do clone
prevalente até então, demonstrando claramente a substituição de um
clone pelo outro.
1.7 Justificativas do presente estudo
1 – Entre 2005 e 2006 houve uma alteração no perfil de resistência
aos antimicrobianos nas amostras de S. aureus resistentes à meticilina
isoladas no Hospital Universitário Clementino Fraga Filho, da UFRJ. Ao
longo desses dois anos, foram isoladas amostras de MRSA com grande
sensibilidade aos demais antimicrobianos. Nesse hospital, antes desse
período, era observada a prevalência do clone brasileiro (ST 239 e
SCCmecIII), cuja característica principal é a resistência a múltiplos
antimicrobianos. Outro aspecto relevante é o fato do Laboratório de
Microbiologia do hospital apresentar dificuldade na correta detecção da
resistência à oxacilina nessas amostras. Assim, para compreender essa
alteração na resistência e responder outras questões pertinentes,
realizamos um estudo epidemiológico e da resistência à oxacilina e à
vancomicina nas amostras em questão.

30
2 – A alta prevalência de S. aureus nas infecções associadas a
dispositivos médicos relatada em diversos estudos, torna cada vez mais
importante a caracterização molecular dessas amostras, bem como a
análise de aspectos genotípicos e fenotípicos relacionados a sua
virulência. Além disso, na literatura mundial e, sobretudo no Brasil, são
extremamente escassos trabalhos que apresentem dados moleculares de
amostras de MRSA causadores de infecções em próteses. Estudos de
epidemiologia molecular podem fornecer dados de crucial importância
para a identificação e caracterização de genótipos prevalentes, bem como
de aspectos relevantes relacionados a virulência nessas amostras. Esse
tipo de análise pode ser uma importante ferramenta para auxiliar na
compreensão desse tipo de infecção e direcionar futuras terapias com o
objetivo de diminuir a incidência de infecções associadas a próteses por
essa espécie bacteriana.

31
2 - OBJETIVOS

32
O objetivo geral desse estudo foi avaliar características relacionadas
a epidemiologia molecular, a resistência e a virulência em amostras de
Staphylococcus aureus isoladas de infecções associadas e não associadas
a próteses articulares em dois hospitais da cidade do Rio de Janeiro.
Os objetivos específicos foram:
a – Determinar a susceptibilidade aos antimicrobianos utilizados na
prática clínica e as concentrações mínimas inibitórias de oxacilina e
vancomicina.
b – Determinar o tipo de cassete cromossômico mec (SCCmec).
c – Avaliar a diversidade genotípica das amostras através da técnica de
eletroforese em campo pulsado (PFGE), “multilocus fragment restriction
type” (MLFRT) e “multilocus sequence typing” (MLST).
d – Detectar a presença de diferentes genes relacionados à virulênca
bacteriana nas amostras isoladas de próteses articulares e de outras
infecções e correlacionar com a clonalidade dessas amostras.
e – Verificar a formação de biofilme nas amostras isoladas de próteses
articulares e de outras infecções.

33
3 – MATERIAL E MÉTODOS

34
3.1 – Locais de obtenção das amostras
a) Hospital Universitário Clementino Fraga Filho (HUCFF). O HUCFF
é um hospital de ensino que está ligado a Universidade Federal do Rio de
Janeiro e possui 490 leitos, sendo 10 de terapia intensiva. Em geral, 1.200
pacientes são admitidos mensalmente nas diversas especialidades
médicas existentes.
b) Instituo Nacional de Traumatologia e Ortopedia (INTO). O INTO é
uma instituição especializada em atendimento cirúrgico, não presta
serviços de emergência e está localizado na cidade do Rio de Janeiro. Os
pacientes são encaminhados ao INTO por outras unidades de saúde. O
INTO possui 144 leitos, sendo 15 de terapia intensiva e oferece serviços
médicos em 13 especialidades ortopédicas.
3.2 Amostras bacterianas
No total, foram analisadas 93 amostras de S. aureus isoladas dos
dois hospitais citados anteriormente, sendo 20 amostras de MRSA não
multirresistentes, obtidas de infecções ocorridas em pacientes do HUCFF,
no período de Fevereiro de 2005 a Março de 2006. As outras 73 amostras
foram de S. aureus resistentes ou sensíveis à oxacilina, isoladas,
continuamente, de infecções em pacientes internados no INTO, sendo 32
de infecções associadas a próteses articulares (IAP) e 41 de infecções não
associadas a próteses (INAP), no período de Agosto de 2005 a Setembro de
2007.

35
3.3 Amostras-controle
Para os testes utilizados nesse estudo foram empregadas as
amostras-controle listadas na tabela 4.
Tabela 4. Amostras-controle utilizadas no presente estudo.
Espécie Amostra Descrição Referência
Staphylococcus aureus ATCC 25923 Padrão para antibiograma
Controle dos testes fenotípicos de identificação
CLSI
S. aureus ATCC 29213 Padrão para teste de diluição em ágar CLSI
S. aureus ATCC 33591 Padrão para teste de triagem em ágar com oxacilina
CLSI
S. aureus 52a Amostra clínica resistente à oxacilina (gene mecA+)
SCHUENCK et al. (2006)
S. aureus 63a Amostra clínica com SCCmecIII / Clone Brasileiro
VIVONI et al (2006)
S. aureus Mu50 Amostra SCCmecII / Clone Nova Iorque-Japão HIRAMATSU et al (1997)
S. aureus 522a Amostra clínica com SCCmecIV / Clone Pediátrico
VIVONI et al (2006)
S. aureus 523a Amostra clínica com os genes da PVL VIVONI et al (2006)
S. aureus ATCC 12600 Amostra tipo - Controle de testes fenotípicos de identificação
-
Staphylococcus epidermidis
ATCC 14990 Amostra tipo - Controle de testes fenotípicos de identificação
-
S. epidermidis ATCC 35984 Amostra produtora de biofilme (icaA +) ARCIOLA et al., 2001
S.epidermidis ATCC 12228 Amostra não produtora de biofilme (icaA-) -
Enterococcus faecalis ATCC 29212 Controle de testes fenotípicos de identificação CLSI
Micrococcus luteus ATCC 10240 Amostra tipo - Controle de testes fenotípicos de identificação
-
ATCC - American Type Culture Collection; CLSI – “Clinical and Laboratory Standards Institute”

36
3.4 Definições utilizadas neste estudo
No presente estudo, infecção de prótese foi caracterizada quando
observados os seguintes critérios: (i) inflamação aguda no local de
implantação da prótese e (ii) quando duas ou três culturas do fragmento
do tecido adjacente a prótese eram positivas para o mesmo
microrganismo.
Para diferenciar infecções comunitárias (IC) de infecções
hospitalares (IH) por MRSA foi utilizada a definição recomendada pelo
CDC (BUCK et al., 2005). A classificação das IC foi baseada nos seguintes
critérios:
- diagnóstico em até 48h após a admissão hospitalar;
- ausência de histórico de infecção ou colonização por MRSA;
- não ter, no período de um ano, histórico de hospitalização,
admissão em “home-care”, diálise, cirurgia, implantes de cateteres ou
outros dispositivos médicos.
3.5 Identificação bacteriana
As amostras foram previamente identificadas pelos Laboratórios de
Bacteriologia do HUCFF e do INTO, através do sistema automatizado
Vitek® (Biomerieux, Paris, França) e Microscan – AutoScan®4 (Siemens,
Mϋnchen, Alemanha), respectivamente. A confirmação da identificação
das amostras de S. aureus foi realizada segundo metodologia descrita por
MacFaddin (1976) e Smibert & Krieg (1994), com modificações, conforme
descrito a seguir.

37
3.5.1 Confirmação da identificação das amostras de Staphylococcus
aureus
As amostras foram cultivadas em ágar-sangue (Plast Labor LTDA,
Rio de Janeiro) e incubadas a 35oC, por até 72 horas, para análise da
pureza, morfologia colonial e presença de hemólise. Posteriormente, foram
realizados os seguintes testes:
a) Produção da enzima catalase
A partir de cultura bacteriana em ágar TSA (“Trypticase Soy Agar”,
Difco Laboratories, Le Pont de Claix, França) foram transferidas 3 colônias
para uma lâmina de vidro que continha uma solução de peróxido de
hidrogênio a 3% (v/v). A produção de bolhas foi indicativa de reação
positiva. As amostras de S. aureus ATCC 12600 e Enterococcus faecalis
ATCC 29212 foram utilizadas como controles positivo e negativo,
respectivamente.
b) Susceptibilidade à bacitracina
Uma suspensão bacteriana com uma turbidez equivalente à 0,5 da
escala McFarland (~108 UFC/mL) foi preparada em salina 0,9% (p/v) e
semeada, confluentemente, com auxílio de um “swab” em agar Müeller-
Hinton (Difco Laboratories). Posteriormente, foi depositado sobre o meio,
um disco impregnado de bacitracina 0,04U (CECON, São Paulo, Brasil) e a
placa foi incubada a 35oC por 18h. Um halo menor ou igual a 10mm foi
indicativo de resistência à bacitracina. As amostras utilizadas como

38
controles do teste foram S. aureus ATCC 12600 (resistente) e Micrococcus
luteus ATCC 10240 (sensível).
c) Produção de fator “clumping”
A produção de “fator clumping” foi verificada utilizando-se o sistema
comercial “Slidex Staph Plus” (Biomerieux), conforme instruções do
fabricante. A amostra utilizada como controle-positivo foi S. aureus ATCC
12600 e a amostra de S. epidermidis ATCC 14990 como controle-negativo.
d) Produção da enzima coagulase
A partir do crescimento bacteriano em meio ágar-sangue 3 colônias
foram transferidas para um tubo contendo plasma de coelho (Biocampo,
Rio de Janeiro, Brasil), diluído a 1:2 (v/v) em solução salina 0,9% (p/v).
Foi realizada incubação a 37oC, por 4h, em banho de imersão. Em
seguida, foi verificada a produção ou não de coágulo. Nos testes que ainda
se apresentavam negativos, uma nova leitura foi realizada após 24h de
incubação. Amostras de S. aureus ATCC 12600 e S. epidermidis ATCC
14990 foram utilizadas como controles positivo e negativo,
respectivamente.
3.6 Determinação da susceptibilidade aos antimicrobianos
a) Teste de difusão a partir do disco (Clinical and Laboratory
Standards Institute [CLSI] 2006a)
O perfil de susceptibilidade das amostras foi determinado frente a
15 antimicrobianos. As amostras foram inicialmente semeadas em ágar

39
sangue, por 18-24h, a 35oC. O crescimento microbiano foi então diluído
em solução salina 0,9% (p/v) para obtenção do padrão de turvação 0,5 da
escala de McFarland (~108 UFC/mL), e inoculado em placa de ágar
Müeller-Hinton (Difco Laboratories), com auxílio de um “swab”, de forma a
obter um crescimento confluente. Os discos foram depositados sobre o
meio e a leitura dos halos de inibição foi feita após 18h de incubação a
35oC, exceto para oxacilina, cefoxitina e vancomicina, cujo período de
incubação foi de 24h.
As amostras foram testadas frente aos seguintes antimicrobianos:
ciprofloxacina (5µg), clindamicina (2µg), cloranfenicol (30µg), eritromicina
(15µg), gentamicina (10µg), penicilina G (10UI), rifampicina (5µg),
sulfametoxazol/trimetoprima (25µg), teicoplanina (30µg), tetraciclina
(30µg), vancomicina (30µg) (Cecon, Rio de Janeiro, Brasil), cefoxitina
(30µg), linezolida (30µg), mupirocina (5µg) e oxacilina (1µg) (Oxoid).
Concomitantemente ao teste de difusão a partir do disco foi
realizado o teste de indução de resistência à clindamicina por macrolídeos
(teste D), conforme determinação do CLSI (2006a). Os discos de
clindamicina e eritromicina foram colocados a 20mm de distância,
margem a margem, e a análise da indução da resistência à clindamicina
por macrolídeos foi verificada pelo surgimento de um “achatamento” no
halo de inibição da clindamicina, próximo ao disco de eritromicina.
A interpretação do teste foi realizada conforme o CLSI (2007) para os
antimicrobianos em geral e de acordo com Fuchs, Jones e Barry (1990)

40
para mupirocina. A amostra padrão S. aureus ATCC 25923 foi utilizada
como controle do teste.
b) Determinação da concentração mínima inibitória (CLSI, 2006b)
A determinação das concentrações mínimas inibitórias (CMIs) de
oxacilina (Sigma-Aldrich Company, St. Louis, EUA) e vancomicina (Sigma-
Aldrich Company) foi realizada pelo método de diluição em ágar, segundo
o CLSI (2006b). As amostras bacterianas foram inicialmente semeadas em
agar-sangue, por 18-24h, a 35oC. O crescimento microbiano foi então
diluído em salina 0,9% (p/v) para obtenção do padrão de turvação 0,5 da
escala de McFarland (~108 UFC/mL). Em seguida, foi realizada uma
diluição 1:10 (v/v) em salina 0,9% e as amostras foram transferidas para
o replicador de “Steers”. Posteriormente, as amostras (~104 UFCs/mL)
foram inoculadas nas placas contendo agar Mueller-Hinton (Difco
Laboratories), 2% de NaCl (Reagen, Rio de Janeiro, Brasil) e concentrações
crescentes do antimicrobiano, variando de 0,25 a 256µg/mL.
Posteriormente, as placas foram incubadas por 24h à 35oC. A leitura do
teste foi realizada de acordo com os valores do CLSI (2007). Foi utilizada a
amostra S. aureus ATCC 29213 como controle do teste.
c) Teste de triagem em ágar contendo 6µg/mL de oxacilina ou
4µg/mL de vancomicina (NCCLS – 2006b)
O crescimento microbiano em ágar sangue foi diluído em solução
salina 0,9% para obtenção do padrão de turvação 0,5 da escala de
McFarland (~108 UFC/mL) e inoculado em um quadrante da placa de ágar

41
Müeller-Hinton (Difco) com 6µg/mL de oxacilina e 4% de NaCl (Difco), com
auxílio de um “swab” de forma a obter um crescimento confluente. As
amostras foram incubadas por 24h a 35oC. A leitura do teste foi realizada
de acordo com o NCCLS (2007). Foram utilizadas as amostras S. aureus
ATCC 29213 (sensível) e ATCC 33591 (resistente) como controles do teste.
Para o teste de triagem em 4µg/mL de vancomicina, foram
realizados os mesmos procedimentos descritos anteriormente, porém, as
amostras foram inoculadas no meio BHI (“Brain Heart Infusion”, Difco)
com 4µg/mL de vancomicina. As amostras foram incubadas por 24h a
35oC. O crescimento de mais de uma colônia na superfície do meio foi
indicativo de resultado positivo, isto é, susceptibilidade reduzida à
vancomicina. A amostra S. aureus ATCC 25923 (amostra sensível à
vancomicina) e a amostra S. aureus Mu50 (resistência intermediária à
vancomicina) foram utilizadas como controles do teste.
3.7 Tipagem do SCCmec por PCR-multiplex nas amostras de S.
aureus resistentes à meticilina
3.7.1 Liberação do DNA
A liberação do DNA foi realizada de acordo com Pitcher, Sauders e
Owen (1989). Cinco colônias bacterianas, obtidas após crescimento em
ágar-sangue, foram inoculadas em 3mL de caldo TSB (Oxoid) e incubadas
por 18h, a 35oC. A seguir, 1mL dessa suspensão foi centrifugada
(centrífuga modelo 5417R, Eppendorf, Hamburgo, Alemanha) (5min, 8000

42
X g) e o sedimento suspenso em 1,5mL de tampão TE (Tris 10 mM, EDTA
1 mM, pH 7,8). A suspensão foi novamente centrifugada (5min, 8000 X g)
e ao precipitado foram adicionados 100µL de solução de lise (50mg/mL de
lisozima [Sigma-Aldrich Company] em tampão TE, pH 7,8 e 5% de BRIJ
0,5% [v/v] (Sigma-Aldrich Company). A essa solução foram adicionados
5µL de lisostafina (1mg/mL) comercial (Sigma-Aldrich Company) e após
rápida agitação esta solução foi incubada por 1h, a 37oC em banho de
imersão. Após esse período, foram adicionados 500µL de isotiocianato de
guanidina 5M (p/v) (Life Technologies, California, EUA). As suspensões
celulares foram agitadas manualmente e então incubadas a temperatura
ambiente, por 10 min. Posteriormente, os lisados foram resfriados no gelo,
por 10 min, e a seguir adicionados 250µL de acetato de amônio (Vetec, Rio
de Janeiro, Brasil) a 7,5M (p/v). Após agitação manual os tubos foram
mantidos por 10min no gelo. Após esse período uma solução de 500µL
composta de clorofórmio/álcool isoamílico (24:1 v/v – Vetec) foi adicionada
e a mistura agitada. Posteriormente, essa mistura foi centrifugada por
10min, a 13000 X g. A fase aquosa, foi então, transferida para um novo
tubo, onde foram adicionados 430µL de isopropranol (Vetec). Os tubos
foram invertidos repetidas vezes, por 1 min, para misturar as soluções.
Após centrifugação a 7000 X g, por 20 seg o DNA precipitado foi então
lavado 5 vezes com etanol gelado (Merck) a 70% (v/v). Por fim, o DNA foi
suspenso em 50 µL de tampão TE, incubado por 16h, a 37oC e então
estocado a -20ºC.

43
3.7.2 PCR-multiplex para detecção do tipo do SCCmec.
A determinação do tipo de SCCmec presente nas amostras
resistentes à oxacilina foi realizada de acordo com Oliveira e de Lencastre
(2002).
A técnica baseia-se na presença de diferentes fragmentos de DNA
específicos dos diferentes tipos de SCCmec. O conjunto de
oligonucleotídeos utilizados (IDT, Coralville, EUA), a especificidade dos
mesmos e o tamanho dos produtos amplificados estão descritos tabela 5.
Tabela 5 – Seqüência, tamanho do amplicon e especificidade dos oligonucleotídeos utilizados na reação de PCR-multiplex para tipagem do SCCmec em amostras de Staphylococcus resistentes à meticilina.
Oligonucleotídeo Seqüência (5’-3’) Amplicon (pb)
Especificidade (tipo de SCCmec)
CIF2 F2
CIF2 R2
TTGGAGTTGCTGATGAAGAAGG
ATTTACCACAAGGACTACCAGC
495 I
KDP F1
KDP R1
AATCATCTGCCATTGGTGATGC
CGAATGAAGTGAAAGAAAGTGG
284 II
MECI P2
MECI P3
ATCAGACTTGCATTCAGGC
GCGGTTTCAATTCACTTGTC
209 II, III
DCS F2
DCS R1
CATCCTATGATAGCTTGGTC
CTAAATCATAGCCATGACCG
342 I, II, IV
RIF4 F3
RIF4 R9
GTGATTGTTCGAGATATGTGG
CGCTTTATCTGTATCTATCGC
243 III
RIF5 F10
RIF5 R13
TTCTTAAGTACACGCTGAATCG
GTCACAGTAATTCCATCAATGC
414 III
IS431 P4
pUB110 R1
CAGGTCTCTTCAAGATCTACG
GAGCCATAAACACCAATAGCC
381 IA
IS431 P4
pT181 R1
CAGGTCTCTTCAAGATCTACG
GAAGAATGGGGAAAGCTTCAC
303 IIIA

44
Como controles internos da reação de PCR-multiplex foram
utilizados os oligonucleotídeos MRS1 e MRS2 (IDT) (DEL VECCHIO et al.,
1995) para amplificação de um fragmento do gene mecA de 154pb.
O volume de 50µL para realização da PCR-multiplex, foi composto
por: 1µL (~30ng) de DNA, tampão da enzima 1X (10mM de Tris-HCl,
25mM de KCl), 2 mM de MgCl2, 200µM de cada deoxinucleotídio
trifosfatado (dNTPS - dATP, dGTP, dCTP e dTTP) (Biotools, Madrid,
Espanha), 1,5U de Taq DNA polimerase (Biotools), 0,8µM dos
oligonucleotídeos CIF2 F2, CIF2 R2, MECI P2, MECI P3, RIF5 F10, RIF5
R13, pUB110 R1 e pT181 R1, 1,6µM dos oligonucleotídeos DCS F2, DCS
R1 e IS431 P4 e 0,4µM dos oligonucleotídeos KDP F1, KDP R1, RIF4 F3 e
RIF4 R9.
As condições da PCR-multiplex foram as seguintes: desnaturação
inicial a 94°C, por 4min, seguida de 30 ciclos de 94°C, por 30s, 53°C por
30s e 72°C, por 1min, e, em seguida, extensão final de 4min, a 72°C. Após
a realização do ciclo, os produtos amplificados foram analisados por
eletroforese em gel de agarose (Invitrogen, Carlsbad, Estados Unidos) a 2%
em TBE 1x (Tris 0,89M, ácido bórico 0,89M, EDTA 2,5mM, pH 8,2), a 100
V, por 1h e 20min. Posteriormente, o gel foi corado com solução de
brometo de etídio (0,5µg/mL) e a imagem capturada sob luz ultravioleta
em um fotodocumetador (Vilber Lourmat, Marne-la-Valleé, França). Como
padrão de DNA para a corrida eletroforética foi utilizado o marcador 100pb
DNA ladder (Biotools).

45
3.8 Análise do perfil de fragmentação do DNA cromossômico após
tratamento com enzima de restrição e separação por eletroforese em
gel de campo pulsado (PFGE).
A análise do perfil de fragmentação do DNA cromossômico das
amostras isoladas nesse estudo foi realizada após separação, por PFGE,
dos fragmentos gerados após tratamento com a enzima de restrição SmaI .
Foi utilizada a técnica descrita por Vivoni e colaboradores (2005). As
amostras, inicialmente, foram cultivadas em ágar-sangue a 35oC, por 48h.
Posteriormente, cinco colônias isoladas foram inoculadas em 5mL de caldo
TSB (Oxoid) e incubadas durante 4h, a 37oC em banho de imersão. A
seguir, 1mL desta suspensão foi centrifugado (7000 X g / 5min) e o
sedimento suspenso em 250µL de tampão PIV (NaCl 1M, Tris-HCl 10mM,
pH 7,6). A esta suspensão foi adicionado o mesmo volume de agarose de
baixo ponto de fusão (“Low Melting Point Agarose”, IBI Technical, New
Heaven, EUA) a 2%, dissolvida em tampão PIV e mantida a 58oC. Após
homogeneização, a agarose foi distribuída em moldes que foram mantidos
a 4oC por cerca de 10 min. Posteriormente, os blocos de agarose foram
colocados em 2mL de solução de lise EC (Tris-HCl 6mM, NaCl 1M, EDTA
100mM, 0,5% de Brij 58 e 0,5% lauril sarcosinato de sódio; pH final 7,5)
contendo 50.000U de lisozima (Sigma-Aldrich Company) e 50U de
lisostafina (Sigma-Aldrich Company) e incubados a 37oC, sob agitação
lenta, durante 18h. Após este período, os tubos foram resfriados a 4°C e a
solução foi substituída por 2mL de solução ESP (EDTA 0,4M pH 9,5, 1%
(p/v) de lauril sarcosinato de sódio) contendo 0,1mg/mL de proteinase K
(Sigma-Aldrich Company), sendo essa solução incubada a 50oC, em

46
banho de imersão, sob leve agitação, durante 18h. Ao final desta fase, os
blocos de agarose foram mantidos em nova solução ESP (sem proteinase)
a 4oC. A digestão do DNA cromossômico foi realizada a partir de um bloco
de agarose, lavado cinco vezes em tampão TE a 37oC, sendo as quatro
primeiras lavagens de 1h cada e a última de 18h. Após este processo, o
bloco de agarose foi transferido para uma solução contendo 250µL do
tampão específico da enzima de restrição SmaI (New England Biolabs,
Ipswich, Inglaterra) e incubado a 25oC, por 4h. Em seguida, o bloco de
agarose foi novamente transferido para um novo tampão da enzima de
restrição contendo 20U da enzima SmaI e incubado a 25oC, durante 18h.
Posteriormente, foi removida a solução contendo a enzima e o bloco de
agarose foi fundido a 70oC e aplicado no gel de agarose (Invitrogen) a 1%
em tampão TBE 0,5x. O gel foi submetido a eletroforese em campo
pulsado (CHEF DR III, Bio-Rad), utilizando um tempo de pulso crescente
de 1 a 35s, durante 23h, a 6V/cm, 13oC, com ângulo de 120o. O gel foi
corado com brometo de etídio (0,5µg/mL), por 40min e descorado por 1h
com água destilada. Posteriormente, a imagem foi capturada sob luz
ultravioleta em um fotodocumetador (Vilber Lourmat). Como padrão de
tamanho molecular de DNA foi utilizado o marcador 50-1.000Kb Lambda
Ladder PFGE Marker (New England BioLabs).
Os padrões de bandas foram analisados através da comparação
visual entre as amostras e foram classificados de acordo com os critérios
de Tenover e colaboradors (1995) e através da análise automatizada pelo
programa GelCompar II, versão 3,5 (Applied Maths, Bélgica), usando o
coeficiente Dice de similaridade e o método de “Unweitgthed Pair Group

47
Method using Arithmetic Averages” (UPGMA) para análise dos
agrupamentos.
3.9 Caracterização das amostras de S. aureus através da análise do
polimorfismo dos fragmentos de restrição de genes que codificam
isoenzimas (MLRFT) (DIEP et al., 2003).
Após o crescimento das amostras de S. aureus em ágar-sangue, a
liberação do DNA bacteriano foi realizada como descrito no item 3.7.1.
Inicialmente foram realizadas as reações de PCR para amplificação dos
sete genes “housekeeping”, que codificam enzimas metabólicas essenciais
(tabela 6). As reações foram realizadas em um volume final de 25µL
contendo: 1µL de DNA (~30ng), tampão da enzima 1X (10mM de Tris-HCl,
25mM de KCl), 5mM de MgCl2, 0,2µM de cada oligonucleotídeo, 200µM de
cada dNTP e 1U de Taq DNA polimerase (Biotools). As condições de
amplificação foram: um ciclo inicial de 94°C/2min, seguido de 30 ciclos de
94°C/1min, 55°C/1min, 72°/1min e um ciclo final de extensão de
72°/5min. Após a realização dos ciclos, os produtos amplificados foram
analisados por eletroforese em gel de agarose (Invitrogen) a 1% em TBE 1X
a 100V por 1h e 20min. Posteriormente, o gel foi corado com solução de
brometo de etídio (0,5µg/mL) e a imagem capturada sob luz ultravioleta
em um fotodocumetador (Vilber Lourmat). Como padrão de DNA para a
corrida eletroforética foi utilizado o marcador 100pb DNA ladder (Biotools).
Após a amplificação, os produtos foram submetidos à digestão
enzimática, nas condições apropriadas a cada enzima, segundo
determinações dos fabricantes (Promega, Madison, EUA e Biotools). Para

48
cada reação de digestão foram utilizadas 3,5U de cada enzima de restrição
e 3µL do produto de PCR. Após a etapa de restrição os fragmentos obtidos
foram separados através de eletroforese em gel de agarose (Invitrogen) a
4%, exceto para os produtos da fragmentação do gene glpF pela enzima
Tsp5091 que foram separados em gel de poliacrilamida. Os possíveis
alelos descritos para cada gene estão apresentados na tabela 6.
A eletroforese em gel de agarose (Invitrogen), feito em tampão TBE
1X, foi realizada a 50V durante 3h. O padrão de tamanho molecular
utilizado foi de 50pb (Biotools).
O gel de poliacrilamida foi preparado utilizando acrilamida
(Pharmacia Biotech, New Jersey, Estados Unidos) e bis-acrilamida (Biorad)
na proporção 29:1 (p/v) na concentração de 20%, TBE 1X, 0,7% (p/v) de
persulfato de amônio (Sigma-Aldrich Company) e 0,7% (v/v) de solução
TEMED (GE Healthcare). As amostras foram submetidas a eletroforeses
em cuba vertical a 150V, por 3h. O padrão de tamanho molecular
utilizado foi de 10pb (Biotools).
A visualização dos géis foi realizada através do emprego de
transiluminador de luz UV e a imagem, capturada por um fotocumentador
(Vilber Lourmat)

49
Tabela 6. Oligonucleotídeos, enzimas de restrição e alelos descritos utilizados nas técnicas de MLST e MLFRT.
Lócus (gene) oligonucleotídeo (5´-3´)a Enzima de restriçãob
Alelos descritosb
Carbamato quinase (arcC)
TTGATTCACCAGCGCGTATTGTC
AGGTATCTGCTTCAATCAGCG
HinfI A, B e C
Shiquimato desidrogenase (aroE)
ATCGGAAATCCTATTTCACATTC
GGTGTTGTATTAATAACGATATC
AluI e CfoI A, B, C e D
Glicerol quinase (glpF)
CTAGGAACTGCAATCTTAATCC
TGGTAAAATCGCATGTCCAATTC
Tsp5091 A, B, C, D, E, F e G
Guanilato quinase (gmk)
ATCGTTTTATCGGGACCATC
TCATTAACTACAACGTAATCGTA
CfoI A, B e C
Fosfato acetil transferase (pta)
GTTAAAATCGTATTACCTGAAGG
GACCCTTTTGTTGAAAAGCTTAA
RsaI A, B e C
Triose fosfato isomerase (tpi)
TCGTTCATTCTGAACGTCGTGAA
TTTGCACCTTCTAACAATTGTAC
BbuI e MboI A, B, C e D
Acetil coenzima A acetil transferase
(yqiL)
CAGCATACAGGACACCTATTGGC
CGTTGAGGAATCGATACTGGAAC
VspI e DdeI A, B e C
a – ENRIGHT et al., 2000; b – DIEP et al., 2003.
3.10 Caracterização das amostras de S. aureus através da tipagem
por seqüenciamento de multilócus enzimáticos (MLST) (ENRIGHT et
al., 2000)
As amostras de S. aureus foram selecionadas para realização do
MLST, segundo os resultados obtidos após as análises por PFGE e/ou
MLFRT. Nessa técnica foi realizada PCR para amplificação dos mesmos
genes e nas mesmas condições descritas no item anterior (3.8). Após a
amplificação dos fragmentos dos genes, esses produtos foram purificados
utilizando o sistema comercial “GTX PCR and band purification” (GE

50
Healthcare, Buckinghamshire, Inglaterra), de acordo com as
especificações do fabricante.
Os produtos purificados foram seqüenciados pelo Centro de Estudo
do Genoma Humano da Universidade de São Paulo utilizando sistema
MegaBACE 1000 com 96 capilares (GE Healthcare). As reações de
seqüenciamento foram realizadas utilizando o sistema comercial
“DYEnamic ET Dye Terminator (com Thermo Sequenase™ II DNA
Polimerase)”.
As seqüências obtidas foram analisadas no sítio
http://saureus.mlst.net/ pela comparação com o banco de dados para
determinação dos alelos e do ST (combinação dos sete alelos) de cada
amostra.
3.11 – Detecção de genes que codificam fatores de virulência
bacteriana
Foram pesquisados os seguintes genes relacionados a virulência
bacteriana em S. aureus: icaA (codifica o polissacarídeo de adesão
intercelular), cna (proteína ligadora de colágeno), bbp (proteína ligadora de
sialoproteína óssea), ebpS (proteína ligadora de elastina), fnbB (proteína B
ligadora de fibronectina), fnbA (proteína A ligadora de fibronectina) e os
genes lukS e lukF (leucocidina de Panton Valentine).
A liberação do DNA bacteriano foi realizada conforme descrito no
item 3.7.1. A amplificação foi realizada para cada gene separadamente.
Cada reação de PCR foi realizada em um total de 25µL compostos por:
1µL de DNA (~30ng), tampão da enzima 1X (10mM de Tris-HCl, 25mM de

51
KCl), 2 mM de MgCl2, 200µM de cada dNTP (dATP, dGTP, dCTP e dTTP)
(Biotools), 1U de Taq DNA polimerase (Biotools).
Os oligonucleotídeos e as concentrações utilizadas, os tamanhos dos
fragmentos amplificados e os ciclos de amplificação estão descritos na
tabela 7.
Após a realização dos ciclos, os produtos amplificados foram
analisados por eletroforese em gel de agarose (Invitrogen) a 1% em TBE 1X
a 100V por 1h e 20min. Posteriormente, o gel foi corado com solução de
brometo de etídio (0,5µg/mL) e a imagem capturada sob luz ultravioleta
em um fotodocumetador (Vilber Lourmat). Como padrão de DNA para a
corrida eletroforética foi utilizado o marcador 100pb DNA ladder (Biotools).

52
Tabela 7 – Oligonucleotídeos, tamanho dos fragmentos e ciclo de amplificação utilizados para detecção de genes de virulência nas amostras de Staphylococcus aureus analisadas.
oligonucleotídeo (5´-3´) / concentração na reação gene amplicon (pb)
ciclo referência
FNBA1 – GATTATTAACGCAGCAGTAG/ 0,4µM FNBA2 – GATAATTTAATGCCAGAGC / 0,4µM
fnbA
1362
1
PEACOCK et al., 2002
ICAF - TCTCTTGCAGGAGCAATCAA/ 0,4µM ICAR - TCAGGCACTAACATCCAGCA/ 0,4µM
icaA
188
2
ARCIOLA et al., 2001
FNBB1 – GTAACAGCTAATGGTCGAATTGATACT/ 0,4µM FNBB2 – CAAGTTCGATAGGAGTACTATGTTC/ 0,4µM
fnbB
524
3
TRISTAN et al, 2003
BBP1 – AACTACATCTAGTACTCAACAACAG/ 0,8µM BBP2 – ATGTGCTTGAATAACACCATCATCT/ 0,8µM
bbp
575
3
TRISTAN et al, 2003
EBP1 – CATCCAGAACCAATCGAAGAC/ 0,4µM EBP2 – CTTAACAGTTACATCATCATGTTTATCTTT/ 0,4µM
ebpS
186
3
TRISTAN et al, 2003
CNA1 – GTCAAGCAGTTATTACACCAGAC/ 0,8µM CNA2 – AATCAGTAATTGCACTTTGTCCACT/ 0,8µM
cna
423
3
TRISTAN et al, 2003
PVL1-ATCATTAGGTAAAATGTCTGGACATGATCCA/ 0,8µM PVL2-GCATCAATGTATTGGATAGCAAAAGC/ 0,8µM
luk-S e F
433
4
LINA et al.,
1999
ciclo 1 – 30 ciclos de 1min a 94°C / 1min a 50°C / 2min a 72°C ciclo 2 – 5min a 94°C e 50 ciclos de 30s a 94°C / 30s a 55,5°C / 30s a 72°C e 1min 72°C ciclo 3 – 5min a 94°C e 30 ciclos de 1min a 94°C / 1min a 55°C / 1min a 72°C e 10min 72°C ciclo 4 – 30 ciclos de 30s a 94°C / 30s a 55°C / 30s a 72°C.
3.12 Método quantitativo de análise da produção de biofilme
Para a análise da formação de biofilme foi utilizado o método e
interpretação descritos por Stepanovic e colaboradores (2000).
As culturas bacterianas foram crescidas em TSB (Difco) a 35°C, por
18-24h. Em seguida, foi realizada uma diluição (1:100v/v) desse
crescimento em novo caldo TSB suplementado com 1% de glicose (Merck,
New Jersey, EUA). Posteriormente, foram distribuídos, em quadriplicata,
200µL da suspensão bacteriana em poços de uma placa de microtitulação
de 96 poços (modelo 92096 TPP – “Techno Plastic Products”, Trasadingen,

53
Suíça), e essa foi incubada a 35°C, por 24h, e em seguida, o meio foi
desprezado para retirada das células planctônicas. Os poços foram
lavados quatro vezes com 200µL de tampão PBS (“Phosphate Buffer
Saline” – pH 7,2). Em seguida, a placa permaneceu por 1h, a 60ºC, para
fixação das células aderidas. Em seguida, foram adicionados 200µL de
cristal violeta, a 0,1% (p/v), em cada poço e a placa foi mantida à
temperatura ambiente, por 45min. Após esse período foi realizada uma
lavagem em água corrente e a placa mantida, por 30 min, a 35°C, para
secagem. Em seguida, foram colocados 200µL etanol (Merck) a 70% em
cada poço da placa.
Para análise da produção de biofilme foi avaliada a absorbância das
células aderidas, utilizando-se um leitor de ELISA, modelo 680 (Bio-Rad)
em comprimento de onda de 570nm.
As amostras de S. epidermidis ATCC 35984 (produtora de biofilme) e
S. epidermidis ATCC 12228 (não produtora de biofilme) foram utilizadas
como controles do teste.
As amostras foram classificadas em quatro categorias: não
produtoras ou fracas, moderadas ou fortes produtoras de biofilme,
fundamentadas no valor da densidade ótica (DO), obtida a partir do
biofilme bacteriano formado. Para cada amostra analisada foi
determinado um valor (chamado de N), que correspondeu a média
aritmética dos valores encontrados nos quatro poços. O valor da
densidade ótica final (DOf) considerado para determinar se as
amostras produziam ou não biofilme foi obtido calculando-se a

54
média aritmética dos quatro poços da amostra controle-negativo (S.
epidermidis ATCC 12228) acrescida de três vezes o valor do desvio-
padrão encontrado para essa amostra, ou seja, DOf = P/4 + (3 x DP),
onde P é a soma algébrica dos valores das DOs nos 4 poços do
controle-negativo e DP é o desvio-padrão observado nesses 4 poços.
A tabela 8 mostra as quatro classificações possíveis.
Tabela 8 – Critérios e classificações utilizados na análise da produção de biofilme nas amostras de Staphylococcus aureus.
Critérios Classificações
N < DOf Amostras não produtoras de biofilme
DOf < N ≤ 2DOf Amostras fracas produtoras de biofilme
2DOf < N < 4DOf Amostras moderadamente produtoras de biofilme
N > 4DOf Amostras fortes produtoras de biofilme
N= média aritmética dos quatro poços da amostra testada; DOf – média aritmética dos quatro poços da amostra controle-negativo (S. epidermidis ATCC 12228) acrescida de três vezes o valor do desvio-padrão encontrado para essa amostra.
3.13 – Análise estatística Neste estudo foi utilizado o teste do Qui-quadrado para analisar se
houve diferença quanto a produção de biofilme, resistência aos
antimicrobianos e presença dos genes de virulência entre as amostras
isoladas de próteses e de outras infecções. Foi estabelecida significância
em valores de 5% (p<0,05).

55
4 - RESULTADOS

56
Os resultados obtidos no presente estudo foram divididos em duas
partes, sendo uma envolvendo a justificativa 1, referente a estudos
realizados em amostras MRSA não multirresistentes (nMRSA) isoladas de
infecções no HUCFF, e a outra relacionada com a justificativa 2, referente
a estudos realizados em amostras de S. aureus isoladas de infecções
associadas e não associadas a próteses, ocorridas no INTO.
Estudo I – Resultados do estudo envolvendo 20 amostras de nMRSA
isoladas no HUCFF.
4.1 – Amostras de nMRSA e aspectos relacionados aos pacientes do
HUCFF.
Nesse estudo foram avaliadas 20 amostras de MRSA, que foram
detectadas como sensíveis a pelo menos seis classes diferentes de
antimicrobianos pelo laboratório do HUCFF, tendo sido denominadas
nMRSA. Esse tipo de amostra começou a emergir no HUCFF, no início de
2005, e surpreendeu a equipe de CCIH e os profissionais do laboratório de
bacteriologia devido a grande sensibilidade frente aos antimicrobianos,
mesmo sendo resistentes a oxacilina. Esse tipo de perfil fenotípico era
diferente daquele encontrado nas amostras MRSA isoladas até então na
instituição. No período de 01/2005 a 03/2006, 30 amostras de nMRSA
foram isoladas de 27 pacientes no HUCFF. Desse total, 20 amostras de 17
pacientes foram recuperadas e analisadas.

57
As características relacionadas aos 17 pacientes de onde foram
isoladas as 20 amostras de nMRSA estão apresentadas na tabela 9.
A idade média dos pacientes, sendo 10 do sexo masculino e 7 do
sexo feminino, foi de 46 anos (variando de 15 a 79 anos). Em relação aos
tipos de infecção (hospitalar ou comunitária), quatro foram caracterizadas
como comunitárias e 16 como hospitalares. Dos quatro pacientes que
apresentaram infecções comunitárias (pacientes 4, 11, 16 e 17), três não
tinham quaisquer fatores de risco para aquisição de MRSA e um fazia uso
de antimicrobianos (paciente 17). Um desses pacientes era de origem
americana, que se encontrava no Brasil a turismo. Esse paciente (número
4) apresentou infecção cutânea e foi encaminhado ao HUCFF. As
principais infecções apresentadas pelos pacientes foram: bacteriemia (6
pacientes) e infecções cutâneas e de tecidos moles (4 pacientes, sendo em
três casos infecções causadas por CA-MRSA). Os principais fatores de
risco para aquisição de MRSA foram: uso de antimicrobianos, prévia
hospitalização e realização de procedimentos invasivos. Hipertensão
arterial e diabetes foram as principais comorbidades encontradas nos
pacientes analisados.
Dos 17 pacientes que apresentaram infecções por MRSA, oito (47%)
evoluíram para óbito, sendo que seis apresentaram bacteriemia e um
paciente apresentou pneumonia.
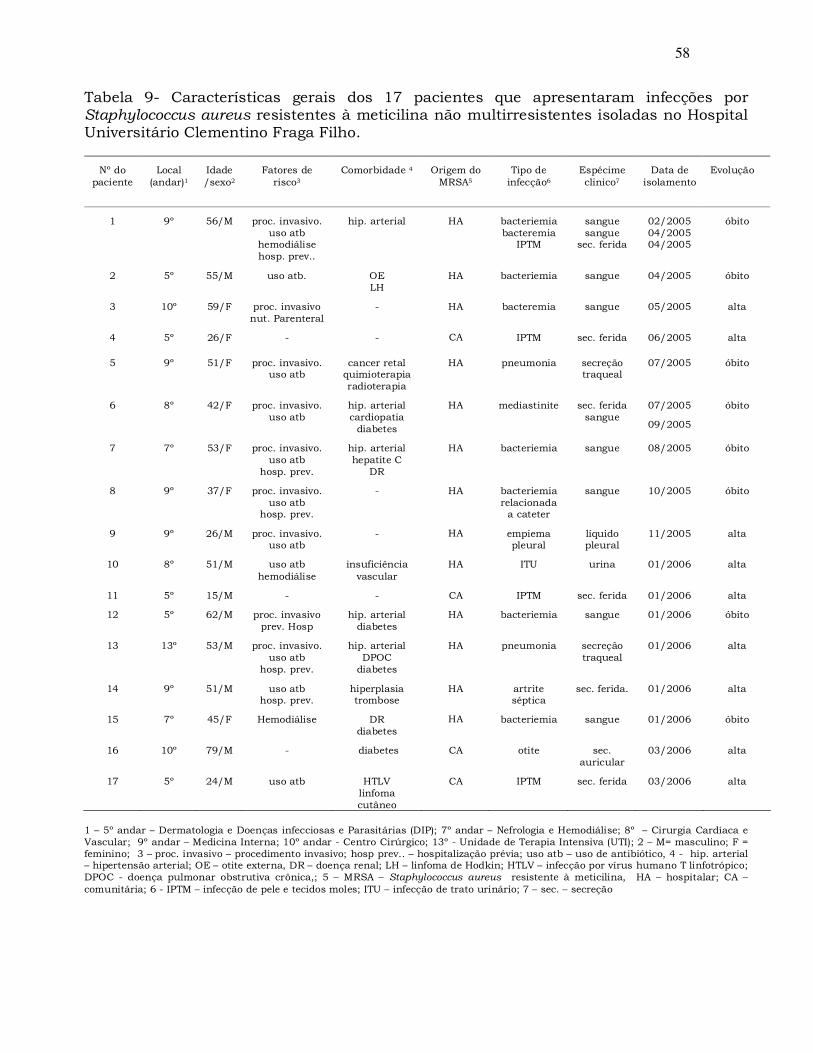
58
Tabela 9- Características gerais dos 17 pacientes que apresentaram infecções por Staphylococcus aureus resistentes à meticilina não multirresistentes isoladas no Hospital Universitário Clementino Fraga Filho.
Nº do paciente
Local (andar)1
Idade/sexo2
Fatores de risco3
Comorbidade 4 Origem do MRSA5
Tipo de infecção6
Espécime clinico7
Data de isolamento
Evolução
1 9º 56/M proc. invasivo. uso atb
hemodiálise hosp. prev..
hip. arterial HA bacteriemia bacteremia
IPTM
sangue sangue
sec. ferida
02/2005 04/2005 04/2005
óbito
2 5º 55/M uso atb. OE LH
HA bacteriemia sangue 04/2005 óbito
3 10º 59/F proc. invasivo nut. Parenteral
- HA bacteremia sangue 05/2005 alta
4 5º 26/F - - CA IPTM sec. ferida 06/2005 alta
5 9º 51/F proc. invasivo. uso atb
cancer retal quimioterapia radioterapia
HA pneumonia secreção traqueal
07/2005 óbito
6 8º 42/F proc. invasivo. uso atb
hip. arterial cardiopatia diabetes
HA mediastinite sec. ferida sangue
07/2005
09/2005
óbito
7 7º 53/F proc. invasivo. uso atb
hosp. prev.
hip. arterial hepatite C
DR
HA bacteriemia sangue 08/2005 óbito
8 9º 37/F proc. invasivo. uso atb
hosp. prev.
- HA bacteriemia relacionada a cateter
sangue 10/2005 óbito
9 9º 26/M proc. invasivo. uso atb
- HA empiema pleural
líquido pleural
11/2005 alta
10 8º 51/M uso atb hemodiálise
insuficiência vascular
HA ITU urina 01/2006 alta
11 5º 15/M - - CA IPTM sec. ferida 01/2006 alta
12 5º 62/M proc. invasivo prev. Hosp
hip. arterial diabetes
HA bacteriemia sangue 01/2006 óbito
13 13º 53/M proc. invasivo. uso atb
hosp. prev.
hip. arterial DPOC
diabetes
HA pneumonia secreção traqueal
01/2006 alta
14 9º 51/M uso atb hosp. prev.
hiperplasia trombose
HA artrite séptica
sec. ferida. 01/2006 alta
15 7º 45/F Hemodiálise DR diabetes
HA bacteriemia sangue 01/2006 óbito
16 10º 79/M - diabetes CA otite sec. auricular
03/2006 alta
17 5º 24/M uso atb HTLV linfoma cutâneo
CA IPTM sec. ferida 03/2006 alta
1 – 5º andar – Dermatologia e Doenças infecciosas e Parasitárias (DIP); 7º andar – Nefrologia e Hemodiálise; 8º – Cirurgia Cardíaca e Vascular; 9º andar – Medicina Interna; 10º andar - Centro Cirúrgico; 13º - Unidade de Terapia Intensiva (UTI); 2 – M= masculino; F = feminino; 3 – proc. invasivo – procedimento invasivo; hosp prev.. – hospitalização prévia; uso atb – uso de antibiótico, 4 - hip. arterial – hipertensão arterial; OE – otite externa, DR – doença renal; LH – linfoma de Hodkin; HTLV – infecção por vírus humano T linfotrópico; DPOC - doença pulmonar obstrutiva crônica,; 5 – MRSA – Staphylococcus aureus resistente à meticilina, HA – hospitalar; CA – comunitária; 6 - IPTM – infecção de pele e tecidos moles; ITU – infecção de trato urinário; 7 – sec. – secreção

59
4.2 – Testes de susceptibilidade, determinação dos tipos de SCCmec e
detecção do gene da PVL nas amostras nMRSA isoladas do HUCFF.
O perfil de resistência aos 15 antimicrobianos testados, os
resultados das concentrações mínimas inibitórias (CMIs) para oxacilina e
vancomicina, bem como a determinação dos tipos de SCCmec e da
presença dos genes da PVL estão apresentados na tabela 10.
Das 20 amostras analisadas, 12 (60%) foram resistentes à
eritromicina, 8 (40%) à ciprofloxacina, 6 (30%) ao cloranfenicol, 6 (30%) à
clindamicina, duas (10%) à gentamicina, uma (5%) a rifampicina e uma
outra a mupirocina. Todas as amostras foram sensíveis a vancomicina,
teicoplanina, linezolida, sulfametoxazol/trimetoprima e tetraciclina.
Em relação às CMIs de oxacilina, 18 (90%) amostras apresentaram
valores <32µg/mL, sendo que duas apresentaram CMI de 4µg/mL. Apenas
duas amostras apresentaram CMIs elevadas para oxacilina (128µg/mL).
Em relação a vancomicina, as CMIs variaram entre 1 e 2µg/mL, sendo que
16 (80%) amostras apresentaram CMI de 2µg/mL, como apresentado na
tabela 10.
No teste de triagem com oxacilina, somente as duas amostras com
CMIs de 4µg/mL apresentaram crescimento não confluente no meio.
Todas as amostras de nMRSA avaliadas não foram capazes de crescer no
ágar com 4µg/mL de vancomicina.
Em relação ao tipo de SCCmec, das 20 amostras analisadas, 19
apresentaram o tipo IV e apenas uma apresentou o tipo II (tabela 10).

60
A presença dos genes da PVL foi observada em três amostras, sendo
duas relacionadas a infecções comunitárias (amostras 526 e 601) e uma
relacionada a infecção hospitalar (551) (tabela 10).
4.3 – Caracterização molecular das amostras de nMRSA isoladas no
HUCFF.
A caracterização genotípica por análise da fragmentação do DNA
cromossômico (PFGE) revelou a presença de 14 pulsotipos em 9 genótipos
distintos (A até I), demonstrando uma grande variedade clonal entre as 20
amostras. Os pulsotipos prevalentes foram o A (5 amostras) e o D (3
amostras), como demonstrado na tabela 10 e figuras 2A e 2B. Em relação
ao MLFRT, todas as amostras foram agrupadas em 6 RFTs distintos. A
associação entre as técnicas de MLFRT e PFGE demonstrou que o RFT
AAAAAAA englobou todas as amostras do genótipo A, o RFT AAACCAC
agrupou as amostras do genótipo B, o RFT BBBBBAB as amostras do
genótipo C e o RFT AAACCAA incluiu quatro diferentes genótipos, D, E, F
e G. Os RFTs únicos CAAAABC e CAAACAC englobaram os genótipos H e
I, respectivamente.
Para a realização da técnica de MLST foram selecionadas 7 amostras
representativas, uma de cada RFT, sendo que para o RFT AAACCAA foram
analisadas duas amostras: uma que apresentou o SCCmecII e outra o
SCCmecIV. A análise por MLST demonstrou os seguintes resultados: a
amostra 633 (genótipo A e RFT AAAAAAA) apresentou o ST 1 (Complexo
Clonal 1 – CC 1); a amostra 517 (genótipo B e RFT AAACCAC) apresentou
o ST 72 (CC 8); a amostra 601 (genótipo C e RFT BBBBBAB) apresentou

61
perfil alélico 2-2-161-2-3-6-2, demonstrando a alteração de um
nucleotídeo na seqüência do gene gmk e foi incluída em um novo ST 1203,
que é um variante de um lócus (SLV) do ST 30, porém, a amostra foi
classificada em um complexo clonal único. As amostras 664 (SCCmecIV) e
603 (SCCmecII) apresentaram o mesmo RFT AAACCAA, porém, na
primeira foi observado o ST 5, enquanto a amostra 603 apresentou o perfil
alélico 1-4-1-4-144-1-10 (com alteração na seqüência nucleotídica no gene
pta), um SLV do ST 5, e foi classificada em um novo ST, o ST 1204 (CC 5).
A amostra 563 (genótipo H e RFT CAAAABC) foi classificada como ST 97
(CC 97), enquanto a amostra 526 (genótipo I e RFT CAAACAC) apresentou
o ST 8 (CC 8).

62
Tabela 10 – Características gerais das 20 amostras de MRSA não multirresistentes isoladas no Hospital Universitário Clementino Fraga Filho
Nº da amostra
Nº do paciente/ origem do MRSA 1
Perfil de resistência além dos ß-lactâmicos 2
CMI (µg/mL) 3 OXA VAN
Tipo de SCCmec
genes da PVL
Pulsotipo
Clonalidade RFT 4 ST 5 CC6
527 5 / HA CIP – CLI – CLO – ERI – GEN 16 2 IV - A1 USA400 AAAAAAA
530 3 / HA CIP – CLI – CLO – ERI – GEN 32 2 IV - A1 AAAAAAA
548 7 / HA CIP – CLI – CLO – ERI 32 2 IV - A1 AAAAAAA
608 10 / HA CIP – CLI – ERI 8 2 IV - A1 AAAAAAA
633 16 / CA CIP – CLI – CLO – ERI 128 1 IV - A2 AAAAAAA 1 1
517, 518, 519
1 / HA -
8 2
IV
-
B
CA-MRSA AAACCAC 72 8
551 8 / HA - 4 2 IV + C1 CA-MRSA BBBBBAB
601 11 / CA - 4 1 IV + C2 BBBBBAB 1203 Único
646 12 / HA CLO 8 1 IV - D1 AAACCAA
664 17 / CA CIP – ERI –MUP 8 2 IV - D2 AAACCAA 5 5
598 15 / HA ERI 8 2 IV - D3 AAACCAA
603 13 / HA CIP – CLI – CLO – ERI 32 2 II - E1 AAACCAA 1204 5
604 14 / HA - 8 1 IV - E2 USA800 AAACCAA
528, 529 6 / HA ERI 32 2 IV - F AAACCAA
520 2 / HA CIP – ERI 8 2 IV - G AAACCAA
563 9 / HA RIF 128 2 IV - H relacionados a MSSA
CAAAABC 97 97
526 4 / CA ERI 16 2 IV + I USA300 CAAACAC 8 8
1 – MRSA – Staphylococcus aureus resistentes à meticilina, HA – hospitalar, CA – comunitária; 2 - CIP – ciprofloxacina; CLI – clindamicina; CLO – cloranfenicol; ERI – eritromicina; GEN – gentamicina; MUP – mupirocina; RIF - rifampicina; 3 – concentração minima inibitória de oxacilina (OXA) e vancomicina (VAN); 4 - RFT restriction fragment type obtido por MLFRT, ordem dos genes – arcC-aroE- glpF-gmk-pta-tpi-yqiL; 5–sequence type obtidos por MLST; 6 – CC – complexo clonal; - indica ausência; + indica presença.

63
48,5
97
145,5
194
242,5
kb
339,5
.
Figura 2. A – Genótipos obtidos pela fragmentação do DNA cromossômico com a enzima SmaI das 20 amostras de MRSA não multirresistentes analisadas nesse estudo. B – Dendograma obtido por análise computadorizada dos genótipos encontrados.
Nº da
amostra
Pulsotipo
A1
A1
A1
A1
A2
H
D1
D2
D3
I
C1
C2
B B
B
E1
E2
F
F
G
a)
b)
Pulsotipo
Nº da amostra

64
Estudo II – Resultados do estudo envolvendo 73 amostras de S. aureus
isoladas continuamente, no período de 08/2005 a 09/2007, de infecções
associadas e não associadas a próteses articulares.
4.4 – Amostras de S. aureus isoladas no INTO
Foram analisadas 73 amostras de S. aureus, sensíveis e resistentes à
oxacilina, sendo 32 isoladas de infecções associadas a próteses articulares
(IAP) de joelho e quadril e 41 de infecções não associadas a próteses (INAP).
As amostras isoladas de INAP foram obtidas a partir de infecção de sítio
cirúrgico (21 amostras), bacteriemia (10), osteomielite (7), infecção urinária
(2) e pneumonia (1). Apenas uma amostra de cada paciente foi analisada no
presente estudo.
4.5 – Resultados da susceptibilidade antimicrobiana nas amostras de S.
aureus isoladas de IAP e INAP no INTO
O percentual de resistência aos antimicrobianos nas 73 amostras de S.
aureus isoladas de IAP (32) e INAP (41) está apresentado na figura 3. Em
relação a oxacilina, 13 (40,6%) amostras de IAP e 15 (36,6%) de INAP foram
resistentes a esse antimicrobiano. Em relação aos outros antimicrobianos o
número de amostras resistentes isoladas de IAP e INAP foram os seguintes,
respectivamente: sulfametoxazol/trimetoprima, 6 (18,7%) e 8 (19,5%);
rifampicina, 3 (9,4%) e 8 (19,5%); cloranfenicol, 7 (21,9%) e 9 (21,9%);
gentamicina, 8 (25%) e 11 (26,8%); ciprofloxacina, 12 (37,5) e 11 (26,8%);
clindamicina, 13 (40,6%) e 12 (29,3%); tetraciclina, 9 (28,1%) e 12 (29,3%) e

65
0
10
20
30
40
50
60
70
80
90
100
% d
e r
esis
tên
cia
VAN TEIC LIN MUP SUT RIF CLO GEN CIP CLI TET OXA CEF ERI PEN
Antimicrobianos
Amostras de IAP
Amostras de INAP
eritromicina, 21 (65,6%) e 19 (46,3%). Todas as amostras de INAP foram
resistentes à penicilina, enquanto 28 (87,5%) amostras de IAP também
foram resistentes a essa droga. Todas as 73 amostras foram sensíveis à
vancomicina, teicoplanina e linezolida. Apenas duas (4,9%) amostras foram
resistentes à mupirocina, e ambas eram sensíveis à oxacilina e isoladas de
INAP. Para todos os antimicrobianos não houve diferença estatisticamente
significativa no perfil de resistência antimicrobiana observado entre as
amostras isoladas de IAP e INAP (p>0,05).
Não foi observada a indução de resistência à clindamicina por
macrolídeos entre as 73 amostras analisadas.
Figura 3 – Perfil de resistência das amostras de S. aureus isoladas de infecções associadas (IAP) e não associadas a próteses (INAP). VAN – vancomicina; TEIC – teicoplanina; LIN – linezolida; MUP – mupirocina; SUT – sulfametoxazol/trimetoprima; RIF – rifampicina; CLO – cloranfenicol; GEN – gentamicina; CIP – ciprofloxacina; CLI – clindamicina; TET – tetraciclina; OXA – oxacilina; CEF – cefoxitina; ERI – eritromicina; PEN – penicilina. Não houve diferença estatisticamente significativa, pelo teste qui-quadrado (p>0,05), na resistência aos antimicrobianos testados, entre as amostras isoladas de IAP e INAP.

66
4.6 – Determinação dos tipos de SCCmec nas amostras de MRSA
isoladas de IAP e INAP no INTO
A tipagem do SCCmec foi realizada nas 28 amostras que foram
resistentes à oxacilina, através do teste de difusão a partir do disco, sendo 13
amostras isoladas de IAP e 15 de INAP.
Nas 45 amostras que se apresentaram sensíveis à oxacilina no teste de
difusão a partir do disco foi realizada a confirmação da sensibilidade, através
da PCR, utilizando-se os oligonucleotídeos MRS1 e MRS2, utilizados para a
detecção do gene mecA. Esse gene não foi encontrado em nenhuma das 45
amostras sensíveis analisadas.
Entre as 28 amostras MRSA, 16 (57,1%) apresentaram o SCCmecIV
enquanto 12 (42,9%) apresentaram o SCCmecIII. Quando avaliadas em
relação ao tipo de infecção, verificamos que entre as 13 amostras isoladas de
IAP, 8 (61,5%) apresentaram o SCCmec tipo IV, enquanto 5 (38,5%)
apresentaram o tipo III. Nas 15 amostras isoladas de INAP, 8 (53,3%)
apresentaram o SCCmec tipo IV e 7 (46,7%) o tipo III (tabela 11; figura 4).
Tabela 11 – Tipos de SCCmec detectados nas amostras MRSA isoladas de infecções associadas e não associadas à próteses no INTO.
IAP – infecções associadas a próteses; INAP – infecções não associadas a próteses.
Nº ( % de amostras) Tipos de SCCmec
(Nº/% de amostras) Amostras isoladas de IAP
Amostras isoladas de INAP
III (12/42,9) 5 (38,5) 7 (46,6)
IV (16/57,1) 8 (61,5) 8 (53,4)
Total (28/100) 13 (100) 15 (100)
kb Tipos de SCCmec III II IV IV III III IV

67
4.7 – Associação entre a resistência antimicrobiana e os tipos de
SCCmec em 28 amostras MRSA isoladas no INTO.
Os perfis de susceptibilidade antimicrobiana, entre os grupos de
amostras MRSA com os tipos III e IV de SCCmec, foram comparados com o
objetivo de verificar se havia diferença no padrão de resistência entre eles, já
que a prevalência de amostras com esses dois tipos de cassetes foi
semelhante nas amostras de próteses e de outras infecções. Esta associação
está apresentada na figura 5.
Todas as amostras de ambos os grupos foram sensíveis à vancomicina,
teicoplanina, mupirocina e linezolida. As amostras com SCCmecIII também
foram resistentes à tetraciclina, sulfametoxazol/trimetoprima, gentamicina,
ciprofloxacina, clindamicina e eritromicina. Além disso, 9 (75%) amostras
foram resistentes à rifampicina e 7 (58,3%) ao cloranfenicol. Em relação às
500pb
100pb
Figura 4 - Eletroforese em gel de agarose dos tipos de SCCmec detectados por PCR-multiplex em amostras MRSA. Linha 1 - padrão de tamanho de DNA (100pb ladder); Linhas 2 a 4 – amostras de S. aureus 63a, Mu50 e 522, controles dos tipos III, II e IV de SCCmec, respectivamente; Linhas 5 e 8 – amostras clínicas com SCCmec IV; linhas 6 e 7 - amostras clínicas com SCCmecIII; linha 9 - S. aureus ATCC 29213 - controle sensível à oxacilina.
1 2 3 4 5 6 7 8 9
300pb

68
0
10
20
30
40
50
60
70
80
90
100
% d
e r
es
istê
nc
ia
VAN TEIC LIN MUP TET SUT RIF GEN CIP CLI ERI CLO
Antimicrobianos
Amostras SCCmecIII Amostras SCCmecIV
amostras com SCCmecIV, todas foram sensíveis à tetraciclina,
sulfametoxazol/trimetoprima e rifampicina. Em relação aos outros
antimicrobianos foi observado que 6 (37,5%) amostras foram resistentes à
gentamicina, 6 (37,5%) ao cloranfenicol, 9 (56,2%) à ciprofloxacina, 9 (56,2%)
à clindamicina e 11 (68,7%) à eritromicina. Exceto para o cloranfenicol,
houve diferença significativa na taxa de resistência encontrada para todos os
antimicrobianos avaliados, quando os dois grupos de MRSA, com SCCmecIII
e IV, foram comparados.
Figura 5 – Associação entre resistência antimicrobiana e os tipos de SCCmec detectados nas amostras de MRSA isoladas no INTO; * - diferença significativa pelo teste qui-quadrado (p<0,05) na resistência a esses antimicrobianos entre os dois grupos.
* * *
*
* * *
Amostras com SCCmecIII Amostras com SCCmecIV g g

69
4.8 - Concentrações mínimas inibitórias (CMIs) de oxacilina e
vancomicina e associação com os tipos de SCCmec nas 28 amostras de
MRSA isoladas de IAP e INAP no INTO.
Como um dos principais problemas para os laboratórios clínicos na
detecção de amostras de MRSA é a existência de CMIs de oxacilina muito
próximas ao “ponto de corte”, realizamos a determinação das mesmas nas 28
amostras MRSA, com interesse principal nas amostras com SCCmecIV, que,
geralmente, são aquelas que apresentam essa característica. Outro
importante fator que também foi analisado refere-se à CMI de vancomicina,
uma vez que, vem sendo relatado o aumento de seus valores em muitos
hospitais, comprometendo a resposta clínica do paciente nas infecções por
MRSA.
As CMIs de oxacilina e vancomicina foram determinadas para as 12
amostras de MRSA, com SCCmecIII (5 de IAP e 7 de INAP) e para as 16
amostras com SCCmecIV (8 de IAP e 8 de INAP). Em relação à oxacilina todas
as amostras com SCCmecIII apresentaram CMIs >256µg/mL enquanto que
as amostras com SCCmecIV mostraram CMIs variando de 8 a 256µg/mL,
como apresentado na tabela 12. É importante destacar que, apesar do
número reduzido de amostras, entre aquelas com SCCmecIV, 5 (62,5%)
amostras isoladas de INAP apresentaram CMIs <32µg/mL, enquanto entre
aquelas amostras isoladas de IAP, 7 (87,5%) delas tiveram CMIs >64µg/mL.
Em relação à vancomicina, 20 amostras apresentaram CMI de 1µg/mL
(9 amostras com SCCmecIII e 11 com SCCmecIV), 4 amostras apresentaram
CMI de 2µg/mL (2 amostras SCCmecIII e 2 com SCCmecIV) e 4 apresentaram

70
CMI <0,5µg/mL (1 amostra com SCCmecIII e 3 com SCCmecIV). Não houve
diferença significativa em relação a resistência à vancomicina entre as
amostras com SCCmec tipo III e IV (p>0,05).
Tabela 12 – Associação entre os tipos de SCCmec e concentrações mínimas inibitórias de oxacilina e vancomicina nas 28 amostras de MRSA associadas e não associadas a próteses.
CMI de oxacilina (µg/mL)
CMI de vancomicina (µg/mL)
SCCmec / Nº de
amostras
Tipo de infecção (Nº de amostras)
4 8 16 32 64 128 256 >256 >0,5 1 2 4 8 16
Tipo III (12) IAP (5) - - - - - - - 5 1 4 - - - -
INAP (7) - - - - - - - 7 - 5 2 - - -
Tipo IV (16) IAP (8) - - 1 - 3 2 2 - 3 4 1 - - -
INAP (8) - 1 - 4 1 2 - - - 7 1 - - -
IAP – infecções associadas a próteses; INAP - infecções não associadas a próteses; CMI – concentração mínima inibitória.
4.9 – Presença de genes de virulência e análise da produção de biofilme
nas 73 amostras de S. aureus isoladas de IAP e INAP no INTO
A presença de genes de virulência e a formação de biofilme foram
comparadas entre as amostras isoladas de IAP e INAP. Todas as 73 amostras,
independente da origem, apresentaram o gene icaA. Em relação às amostras
isoladas de IAP, 31 (96,9%) apresentaram o gene fnbA, 24 (75%) o gene ebpS,
21 (65,6%) o gene cna, 5 (15,6%) o gene fnbB, 1 (3,1%) o gene bbp e 1 (3,1%)
o gene da PVL. Nas amostras isoladas de INAP, 33 (80,5%) apresentaram o
gene fnbA, 29 (70,7%) o gene ebpS, 28(68,3%) o gene cna, 7 (17,1%) o gene
fnbB, 6 (14,6%) o gene bbp e 6 (14,6%) o gene da PVL. Não houve diferença

71
100
100100
96,9
80,5
87,7
75 70,7
72,6
65,6 68
,367,
1
15,6
17,1
16,4
3,1
14,6
9,6
3,1
14,6
9,6
0
10
20
30
40
50
60
70
80
90
100
% d
e a
mo
str
as
icaA fnbA ebpS cna fnbB bbp pvl
genes de virulência
Amostras de IAP (n=32) Amostras de INAP (n=41) total (n=73)
estatisticamente significativa, em relação à presença dos genes de virulência,
entre os dois tipos de infecções (IAP e INAP). Os resultados da detecção dos
genes de virulência estão apresentados nas figuras 6 e 7
Figura 6 – Percentual de positividade para diferentes genes de virulência nas 73 amostras de S. aureus isoladas de infecções associadas (IAP) e não associadas a próteses (INAP) no INTO. Não houve diferença estatisticamente significativa, pelo teste qui-qudrado (p>0,05), em relação à presença dos genes de virulência entre as amostras isoladas de IAP e INAP.

72
Figura 7 – Eletroforese em gel de agarose dos genes de virulência pesquisados neste estudo. Linha 1 - padrão de tamanho de DNA (100pb ladder); Linhas 2 a 8 – Diferentes amostras clínicas de S. aureus que apresentaram os genes fnbA (1362 pb), fnbB (524 pb), bbp (575 pb) ebpS (186), da PVL (433 pb), cna (423 pb) e icaA (188pb).
Em relação à produção de biofilme, utilizando a metodologia descrita
por Stepanovic e colaboradores (2000), os seguintes valores aproximados de
DO foram considerados para classificação das amostras: valor de DO menor
ou igual a 0,6, para amostras não produtoras de biofilme; DO entre 0,6 e
<1,2, para amostras fracas produtoras; DO entre 1,2 e <2,4, para moderadas
produtoras e acima de 2,4 para amostras fortes produtoras de biofilme.
Entre as 32 amostras isoladas de IAP, 31 (96,9%) produziram biofilme,
sendo 5 fortes, 23 moderadas e 3 fracas produtoras. Nas 41 amostras
isoladas de INAP, 39 (95,1%) produziram biofilme, sendo 3 fortes, 30
moderadas e 6 fracas produtoras. O resultado geral da produção de biofilme
1 2 3 4 5 6 7 8
600pb
100pb
300pb
kb
1kb
Genes de virulência fnbA fnbB bbp ebpS PVL cna icaA
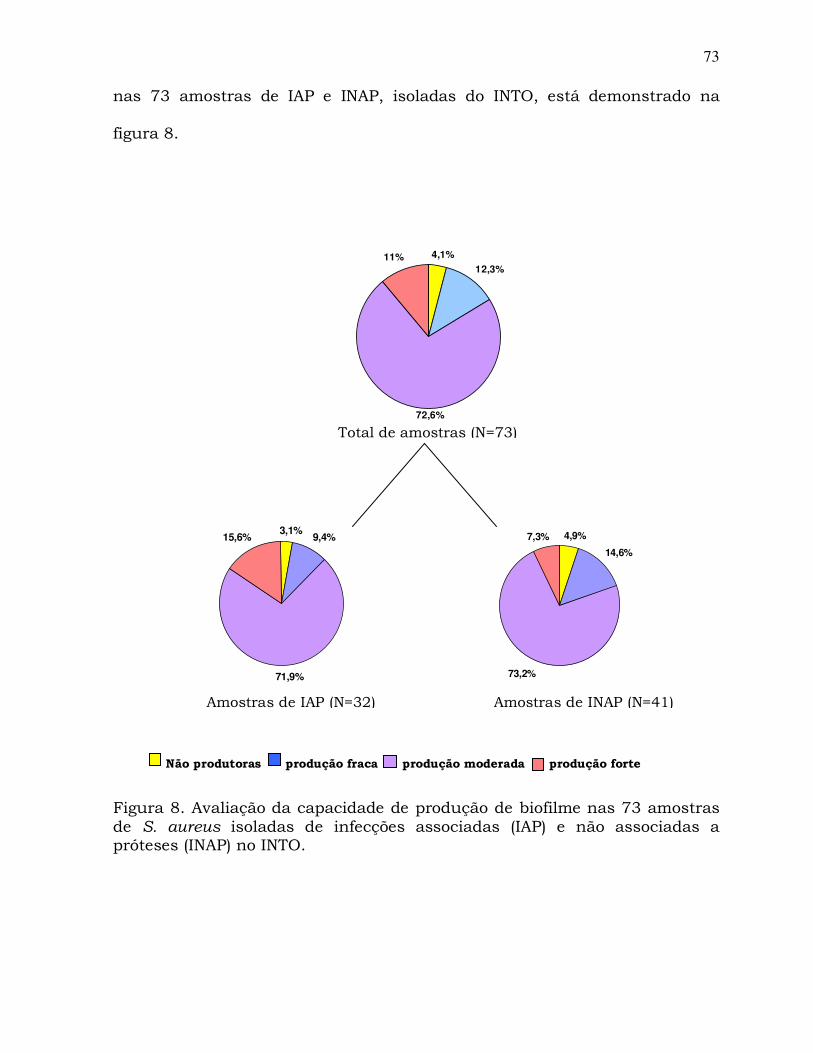
73
4,9%
14,6%
73,2%
7,3%3,1%
9,4%15,6%
71,9%
4,1%
12,3%
11%
72,6%
nas 73 amostras de IAP e INAP, isoladas do INTO, está demonstrado na
figura 8.
Figura 8. Avaliação da capacidade de produção de biofilme nas 73 amostras de S. aureus isoladas de infecções associadas (IAP) e não associadas a próteses (INAP) no INTO.
Amostras de IAP (N=32) Amostras de INAP (N=41)
Total de amostras (N=73)
Não produtoras produção fraca produção moderada produção forte

74
4.10 – Caracterização molecular das 73 amostras de S. aureus isoladas
de IAP e INAP no INTO.
Com o objetivo de determinar a diversidade genotípica das amostras de
S. aureus causadoras de IAP e de INAP, no INTO, e verificar se havia
prevalência e/ou diferença de genótipos nesses dois grupos, foram realizados
os testes de PFGE e MLST.
A caracterização genotípica através da análise da fragmentação do DNA
cromossômico (PFGE) revelou 15 genótipos distintos (A´ a P´) entre as
amostras de MRSA e MSSA avaliadas. Nas tabelas 13 e 14 estão descritas as
características gerais das amostras, incluindo os genótipos encontrados.
Apenas 5 genótipos (A´ a E´) englobaram 61 (83,5%) das 73 amostras de S.
aureus isoladas.
A tabela 13 mostra as características gerais das 28 amostras MRSA
isoladas de IAP e INAP. Todas as 12 amostras MRSA, com SCCmecIII,
isoladas nesse estudo estão representadas pelo genótipo A´ (figura 9) e foram
geneticamente relacionadas ao clone multirresistente, prevalente no Brasil.
As 12 amostras desse genótipo foram incluídas em 11 pulsotipos distintos,
demonstrando que apesar de serem relacionados, não existiu a disseminação
de um pulsotipo específico no hospital. A amostra desse grupo (número 530)
selecionada para análise por MLST apresentou o ST 239, similar ao descrito
para o clone brasileiro multirresistente.
O genótipo B´ foi prevalente nas amostras de infecções no INTO e
apresentou 14 pulsotipos distintos (B´1 a B´14). Esse genótipo (figura 10) foi
encontrado em 20 (27,4%) amostras, sendo seis MRSA que apresentaram o

75
SCCmecIV (tabela 13), e 14 amostras de MSSA (tabela 14). Entre as amostras
resistentes foram observados cinco pulsotipos, sendo dois em amostras
isoladas de IAP (pulsotipos B´3 e B´4) e quatro de INAP (pulsotipos B´3, B´10,
B´13 e B´14). Um mesmo pulsotipo (B´3) foi observado nos dois tipos de
amostras, isoladas de IAP e INAP. As amostras selecionadas para realização
do MLST foram: 4948 (MRSA SCCmecIV; pulsotipo B´3) e 2807 (MSSA;
pulsotipo B´6), ambas isoladas de IAP, e as amostras 2638 (MRSA SCCmecIV;
pulsotipo B´10), 2520 (MRSA SCCmecIV; pulsotipo B´13), e 3210 (MSSA;
pulsotipo B´1), que foram isoladas de INAP. Essas amostras foram escolhidas
como representativas do genótipo B´ e pela diferença na susceptibilidade à
oxacilina. Todas apresentaram perfil alélico 1-4-1-4-12-1-10 e foram
alocadas no ST 5 (CC 5).
Dentro do genótipo C´ (figura 11) foram incluídas 17 amostras (23,3%),
distribuídas em nove pulsotipos distintos (C´1 a C´9). Entre elas, nove eram
de MRSA com SCCmecIV, sendo seis isoladas de IAP e três de INAP, e foram
incluídas em cinco pulsotipos. As amostras do pulsotipo C´1 foram isoladas
de IAP (três amostras) e INAP (uma amostra). Entre as oito amostras de
MSSA, quatro foram isoladas de IAP e outras quatro de INAP, e foram
incluídas em cinco pulsotipos. Assim como, entre as amostras de MRSA,
também foi observado, entre as de MSSA pulsotipos iguais em amostras de
IAP (pulsotipo C´4 - duas amostras) e de INAP (pulsotipo C´3 – duas
amostras), além do pulsotipo C´5, que foi encontrado nos dois grupos de
infecção. As amostras do genótipo C´ selecionadas para realização do MLST
foram: 3216 (MRSA SCCmecIV; pulsotipo C´1), 5398 (MSSA; pulsotipo C´4 e)

76
e 2879 (MRSA SCC mecIV; pulsotipo C´7), que foram isoladas de IAP e a
amostra 5917 (MRSA SCCmecIV; pulsotipo C´2), que foi isolada de INAP.
Todas as amostras foram incluídas no ST 1 (perfil alélico 1-1-1-1-1-1-1),
integrando o CC 1.
Em conjunto, os genótipos B´ e C´ foram responsáveis por 59,4% das
amostras de IAP e 44% das INAP .
Oito amostras foram incluídas no genótipo D´, sendo três isoladas de
IAP e cinco de INAP, e foram incluídas em seis pulsotipos. Todas as amostras
desse genótipo foram sensíveis à oxacilina e quatro amostras (3210-C, 3474,
4511 e 4855) de dois pulsotipos (D´1 e D´4), apresentaram os genes da PVL.
Três amostras isoladas de INAP apresentaram o mesmo pulsotipo D´1. Foi
realizado o MLST em duas amostras, 4511 (pulsotipo D´1) e 3210-C
(pulsotipo D´4), que representaram os dois pulsotipos de amostras positivas
para os genes da PVL. Nas duas amostras foi observado o perfil alélico 3-1-1-
4-1-1-1, o qual não apresentou correspondência com nenhum outro descrito
no banco de dados. A similaridade encontrada foi com o ST 188, cujo perfil
alélico é 3-1-1-8-1-1-1-1, sendo, então, um variante em um lócus (gene gmk)
do ST 188 (SLV 188).
O genótipo E´ englobou quatro amostras e todas elas eram de amostras
isoladas de INAP. Dessas quatro amostras, três eram MSSA e uma MRSA
com SCCmecIV. Das quatro amostras, três apresentaram os genes da PVL,
sendo duas amostras de MSSA (amostra 3651, pulsotipo E´1 e amostra 324,
pulsotipo E´2) e uma MRSA (amostra 1970, pulsotipo E´3). Foi realizado o

77
MLST dessas três amostras e todas apresentaram perfil alélico 2-2-2-2-6-3-2,
sendo representantes do ST 30 (CC 30).
Na figura 12 estão apresentados os pulsotipos de todas as sete
amostras que apresentaram os genes da PVL. Dessas sete amostras, quatro
apresentaram o genótipo D´ e três o genótipo E´.
Todos as outras 12 amostras apresentaram 10 genótipos distintos (F´ a
P´) e todas elas eram MSSA, como mostrado nas tabelas 13 e14.

78
Tabela 13 – Características gerais das 28 amostras MRSA isoladas de infecções associadas e não associadas a próteses no INTO.
1 – IAP – infecções associadas a próteses, INAP – infecções não associadas a próteses; 2 – CMI- concentração mínima inibitória, OXA – oxacilina, VAN – vancomicina; 3 – ST – sequence type; 4 – CC – complexo clonal
Tipo de infecção / Nº de amostras1
Nº da amostra
Data de isolamento
CMI (µg/mL)2 OXA VAN
Tipo de SCCmec
Pulsotipo (PFGE)
ST3 CC4
IAP (13) 178 01/2006 >256 1 III A´1
530 02/2006 >256 0,5 III A´2 239 8
590 02/2006 >256 1 III A´3
499 02/2006 >256 1 III A´4
6452 12/2006 >256 1 III A´5
4948 12/2005 128 2 IV B´3 5 5
6769 08/2007 16 1 IV B´4
3671 09/2005 256 0,5 IV C´6
3216 08/2005 128 0,5 IV C´1 1 1
445 01/2006 256 0,5 IV C´1
468 02/2006 64 1 IV C´8
2879 06/2006 64 1 IV C´7 1 1
510 01/2007 64 0,5 IV C´1
INAP (15) 3679-3 09/2005 >256 1 III A´7
3478 09/2005 >256 1 III A´11
4144 10/2005 >256 2 III A´6
4122 10/2005 >256 2 III A´8
5238 12/2005 >256 1 III A´10
1140 03/2006 >256 1 III A´7
3200 07/2006 >256 1 III A´9
2520 05/2006 32 1 IV B´13 5 5
2638 06/2006 32 1 IV B´10 5 5
1509 03/2007 32 1 IV B´3
3211 04/2007 32 1 IV B´14
4435 11/2005 64 2 IV C´1
5917 11/2006 128 1 IV C´2 1 1
795 02/2007 128 1 IV C´2
1970 05/2006 8 1 IV E´3 30 30

79
Tabela 14 – Características gerais das 45 amostras de S. aureus sensíveis à oxacilina isoladas de infecções associadas e não associadas a próteses no INTO.
Tipo de infecção / Nº de amostras1
Nº da amostra
Data de isolamento
Pulsotipo (PFGE)
ST2 CC3
IAP (19) 4341 10/2005 B´1
4840 12/2005 B´1
791 02/2006 B´5
2195 05/2006 B´2
2807 06/2006 B´6 5 5
710 02/2007 B´2
7856 09/2007 B´7
1183 03/2006 C´9
2733 06/2006 C´4
5398 10/2006 C´4 1 1
6562 08/2007 C´5
4233 10/2005 D´5
2405 05/2006 D´6
3210-C 04/2007 D´4 SLV 188 1
6436 08/2007 F´1
4608 11/2006 H´
3676-2 09/2005 I´
3473 07/2006 J´
4700 11/2005 L´
INAP (26) 727 02/2006 B´12
490 02/2006 B´5
1572 04/2006 B´8
3460 07/2006 B´1
3210 07/2006 B´1 5 5
4412 09/2006 B´9
1496 03/2007 B´11
4715 11/2005 C´10
3346-1 08/2005 C´3
024 01/2006 C´5
1215 03/2006 C´3

80
continuação da tabela 14.
3630 09/2005 D´2
3884 10/2005 D´3
3474 07/2006 D´1
4511 09/2006 D´1 SLV 188 1
4855 09/2006 D´1
3651 08/2006 E´1 30 30
5218 10/2006 E´4
324 01/2007 E´2 30 30
5210 10/2006 F´2
2389 05/2006 G´1
4976 10/2006 G´2
4318 10/2005 M´
4104 08/2006 N´
2032 05/2006 O´
3670-2 09/2005 P´
1 – IAP – infecções associadas a próteses, INAP – infecções não associadas a próteses; 2 – ST – sequence type; 3 – CC – complexo clonal.

81
Figura 9 – Pulsotipos do genótipo A´ obtidos por PFGE após fragmentação do DNA genômico com a enzima SmaI. Amostras de MRSA com SCCmecIII. Amostra 63a - representante do genótipo do clone brasileiro.M – marcador molecular.
48,5
97
145,5
339,5
kb
M M M

82
Figura 10 – Pulsotipos representativos do genótipo B´, obtidos por PFGE após fragmentação do DNA genômico com a enzima SmaI. Amostras de MRSA com SCCmecIV: 6769, 4948, 3211, 2638 e 2520; amostras de MSSA: 791, 7856, 2195, 2807, 3210, 3460, 490, 1572 e 4412. M – marcador molecular.
Figura 11 - Pulsotipos representativos do genótipo C´ obtidos por PFGE após fragmentação do DNA genômico com enzima a SmaI. Amostras de MRSA com SCCmecIV: 445, 3216, 510, 2879, 468, 3671, 4435, 795 e 5917; amostras de MSSA: 5398, 2733, 6562, 1215, 3346-1, 024 e 4715.M – marcador molecular.
48,5
97
145,5
kb
242,2
339,5
48,5
97
145,5
kb
242,2
339,5
M M M
M M M
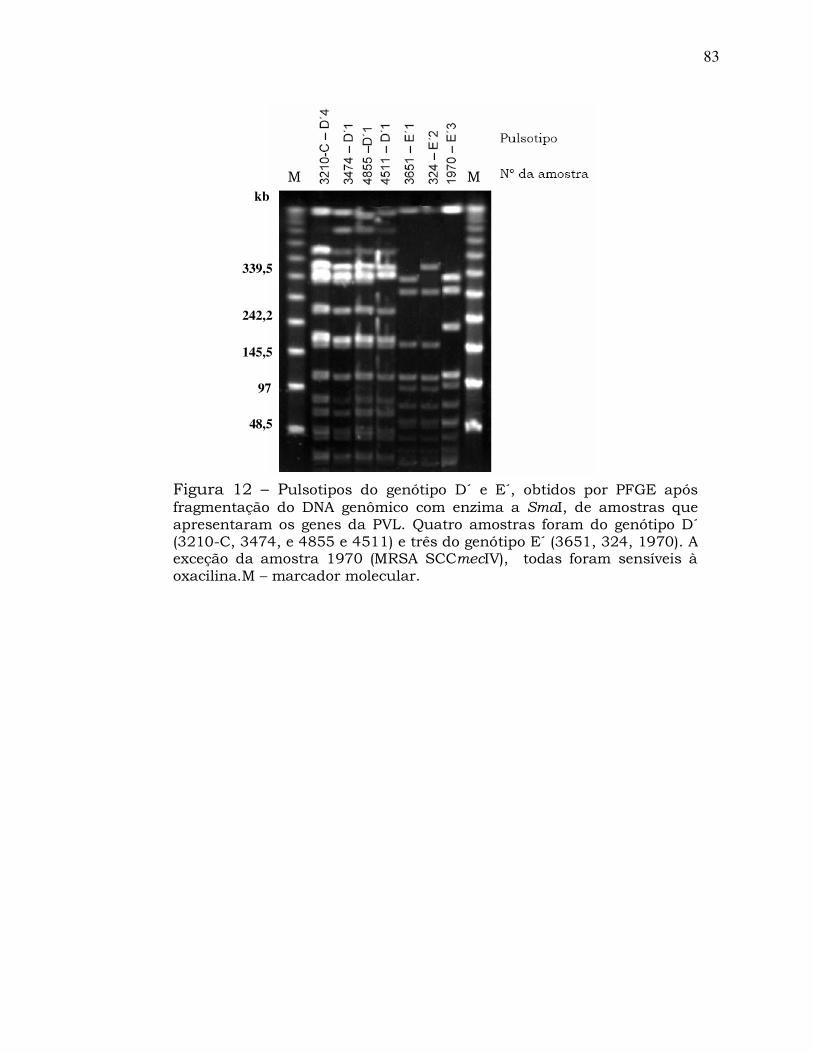
83
Figura 12 – Pulsotipos do genótipo D´ e E´, obtidos por PFGE após fragmentação do DNA genômico com enzima a SmaI, de amostras que apresentaram os genes da PVL. Quatro amostras foram do genótipo D´ (3210-C, 3474, e 4855 e 4511) e três do genótipo E´ (3651, 324, 1970). A exceção da amostra 1970 (MRSA SCCmecIV), todas foram sensíveis à oxacilina.M – marcador molecular.
48,5
97
145,5
kb
242,2
339,5
M M

84
4.11 – Relação entre os principais perfis genotípicos encontrados e
a presença de genes de virulência
A tabela 15 mostra a associação entre os genótipos prevalentes e a
presença de genes de virulência nas 73 amostras de S. aureus isoladas de
IAP e INAP, no INTO. À exceção do genótipo D´, todos os outros genótipos e
seus pulsotipos apresentaram, no máximo, duas combinações (perfis) de
genes de virulência. As 12 amostras do genótipo A´ (SCCmecIII) apresentaram
os genes cna, fnbA, fnbB e ica e não apresentaram os genes bbp, ebpS e da
PVL. Apenas no genótipo A´ foi encontrado o gene fnbB. Em relação ao
genótipo B´, em 19 amostras que apresentaram SCCmecIV, das 20
analisadas, só foram detectados os genes fnbA, ebpS e ica, exceto a amostra
de MSSA que, além destes genes, também apresentou o gene cna. No
genótipo C´, onde se encontrou a maior parte das amostras de MRSA com
SCCmecIV, das 17 amostras, 16 apresentaram os genes cna, fnbA, ebpS e ica
e apenas uma de MSSA não apresentou o gene cna. No genótipo D´ foi
observada a maior variação de combinação dos genes. Entre as oito
amostras, seis combinações distintas de genes de virulência foram
detectadas. Quatro amostras tinham os genes da PVL. Em relação ao
genótipo E´, todas as quatro amostras apresentaram os genes cna, fnbA, bbp,
ebpS e ica e três apresentaram o gene da PVL (1970, 3651 e 324). Somente
amostras desse genótipo apresentaram o gene bbp. O perfil 3 de virulência foi
observado entre amostras dos genótipos B’, C’ e D’. Em geral, os genes ica
(100%), fnbA (87,7%), cna (72,6%) e ebpS (75,3%) estavam presentes em
grande parte das amostras.
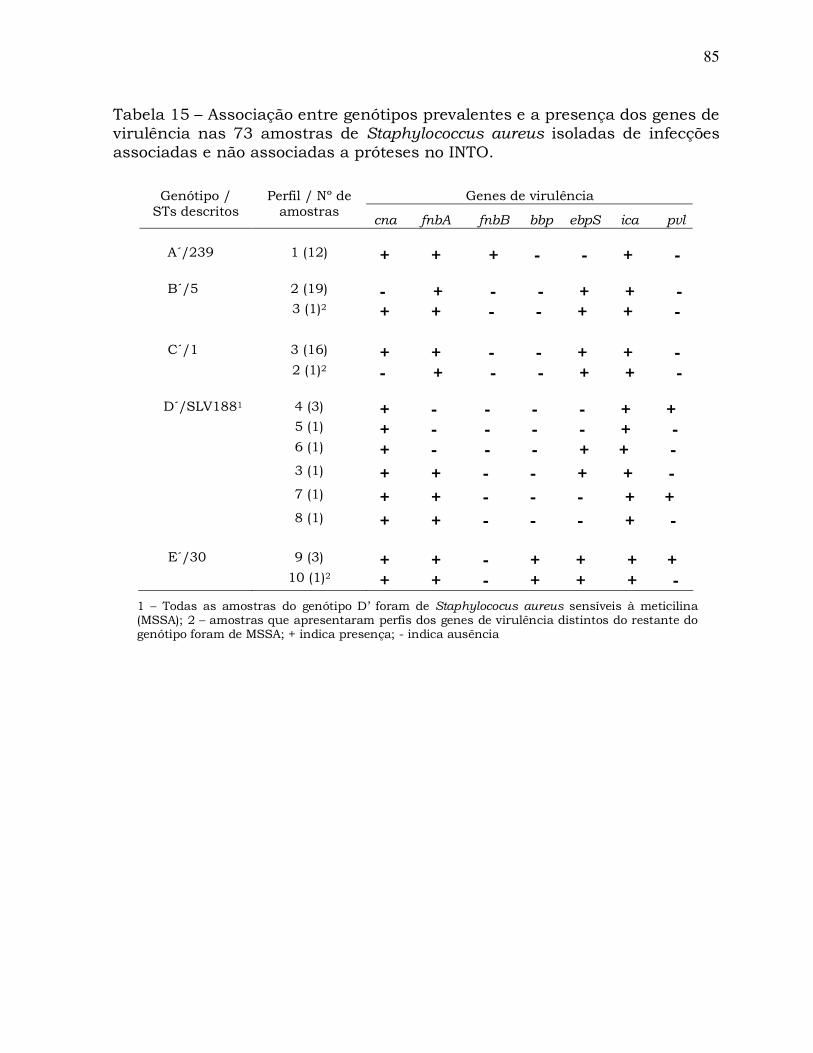
85
Tabela 15 – Associação entre genótipos prevalentes e a presença dos genes de virulência nas 73 amostras de Staphylococcus aureus isoladas de infecções associadas e não associadas a próteses no INTO.
Genes de virulência Genótipo / STs descritos
Perfil / Nº de amostras
cna fnbA fnbB bbp ebpS ica pvl
A´/239 1 (12) + + + - - + -
2 (19) - + - - + + - B´/5
3 (1)2 + + - - + + -
C´/1 3 (16) + + - - + + - 2 (1)2 - + - - + + -
D´/SLV1881 4 (3) + - - - - + + 5 (1) + - - - - + -
6 (1) + - - - + + -
3 (1) + + - - + + -
7 (1) + + - - - + +
8 (1) + + - - - + -
E´/30 9 (3) + + - + + + + 10 (1)2 + + - + + + -
1 – Todas as amostras do genótipo D’ foram de Staphylococus aureus sensíveis à meticilina (MSSA); 2 – amostras que apresentaram perfis dos genes de virulência distintos do restante do genótipo foram de MSSA; + indica presença; - indica ausência

86
5 - DISCUSSÃO

87
A espécie S. aureus está entre aquelas de maior prevalência em
infecções, tanto de origem hospitalar quanto comunitária. A presença de uma
grande variedade de fatores de virulência, bem como sua capacidade de
desenvolver ou adquirir mecanismos de resistência a diversas classes de
antimicrobianos, torna desafiadora e extremamente relevante a realização de
estudos que permitam a compreensão desses mecanismos.
Grande parte das amostras de S. aureus, isoladas de diversas partes do
mundo, é resistente à oxacilina (DIEKEMA et al., 2001). Em hospitais, a
multirresistência é comum entre as amostras, assim como, um limitado
número de genótipos é responsável pela maioria das infecções (SANTOS et al,
1999; OLIVEIRA et al., 2002; TIEMERSMA et al., 2004). No Brasil, um
genótipo de MRSA, com SCCmecIII e incluído no ST 239, tem sido o principal
agente de infecções hospitalares, sendo a multirresistência sua principal
característica (OLIVEIRA et al., 2001, VIVONI et al., 2006). A prevalência de
amostras de MRSA, apresentando resistência múltipla aos antimicrobianos,
tem sido demonstrada em estudos anteriores de nosso grupo, inclusive entre
amostras isoladas no HUCFF (SANTOS et al., 1996; SCHUENCK et al., 2004,
VIVONI et al., 2006). Contudo, entre fevereiro de 2005 e março de 2006, foi
observada a emergência de amostras MRSA não multirresistentes (nMRSA)
nesse hospital. Dados da CCIH revelaram que esse tipo de amostra foi
responsável por 14% das infecções por MRSA no ano de 2004, mas que esse
percentual tinha aumentado quatro vezes mais (56%) em 2005. Para
compreender melhor essa alteração na epidemiologia das infecções por
amostras MRSA, isoladas no HUCFF, avaliamos 20 amostras de nMRSA,

88
obtidas entre fevereiro de 2005 e março de 2006, através de PFGE, MLRFT e
MLST. Além disso, determinamos o tipo de SCCmec, as concentrações
mínimas inibitórias de oxacilina e vancomicina e verificamos a presença dos
genes da PVL nessas amostras de nMRSA.
Amostras MRSA de origem comunitária (CA-MRSA), geralmente,
possuem SCCmecIV, e podem apresentar heterorresistência à oxacilina.
Outra característica importante é que essas amostras possuem um tempo de
geração menor do que outras que apresentam cassetes mec maiores e, além
disso, possuem CMIs para oxacilina próximas ao “ponto de corte” (OKUMA
et al. 2002). Entre as 20 amostras analisadas em nosso estudo, 19
apresentaram SCCmecIV e, em 18 destas, as CMIs obtidas variaram de 4 a
32µg/mL, características similares às encontradas em CA-MRSA.
Quanto ao tipo de SCCmec, amostras hospitalares, geralmente,
apresentam os tipos I, II ou III, diferentes daquelas de origem comunitária,
que apresentam, normalmente, os tipos IV ou V (DIETRICH et al., 2004,
KILLIC et al., 2006, UDO et al., 2008). Esse quadro vem sendo alterado na
última década, sendo cada vez mais comum o isolamento de amostras com
SCCmecIV, no ambiente hospitalar (WANNET et al., 2005; FOSSUM &
BUKHOLM, 2006; UDO et al., 2008). Em São Paulo, Trindade e
colaboradores (2005) avaliando amostras isoladas de infecções de corrente
sanguínea, demonstraram a presença de amostras com o SCCmecIV entre
elas. Uma característica importante dessas amostras emergentes é a
presença de um tipo SCCmec de tamanho reduzido e, uma presença mais
freqüente de genes de virulência, como o da PVL. Essas características

89
poderiam conferir uma vantagem adaptativa a esses clones e ajudar a
disseminá-los (OKUMA et al., 2002).
Outro aspecto relevante, é a dificuldade de detecção da resistência à
oxacilina em algumas dessas amostras de nMRSA. Vandenesch e
colaboradores (2003) analisando 81 amostras de CA-MRSA com SCCmecIV,
observaram que as CMIs variavam de 4 a 64µg/mL, semelhante ao observado
no presente estudo. Esse fato representa um grande problema para os
laboratórios clínicos, uma vez que esses valores de CMIs se aproximam aos
de sensibilidade. Adicionalmente, o ágar triagem com 6µg/mL também pode
fornecer resultados de falsa sensibilidade. Os resultados obtidos com a
utilização de ágar com oxacilina confirmam essa dificuldade, uma vez que
duas amostras com CMI de 4µg/mL apresentaram crescimento de apenas 2 a
5 UFCs no meio, característico de heterorresistência à oxacilina, um dos
principais motivos para a detecção errônea de MRSA (VELASCO et al., 2005).
No presente estudo, a resistência à eritromicina foi freqüente entre as
amostras (12/20), seguida por ciprofloxacina (8/20), clindamicina (6/20) e
cloranfenicol (6/20). Esses fatos vem sendo observados em amostras MRSA
carreando o SCCmecIV (BARTELS et al., 2007). Em relação a vancomicina
80% das amostras apresentaram CMIs de 2µg/mL. Já foi observada a
ocorrência de falha terapêutica no tratamento de infecções de corrente
sanguínea por MRSA, em amostras com CMI aumentada (1 e 2µg/mL), em
comparação com amostras com CMIs reduzidas (<0,5) (SAKOULAS et al,
2004). Contudo, mesmo com o desenvolvimento de novas drogas como
tigeciclina e daptomicina, a vancomicina continua a ser a droga mais

90
utilizada no tratamento de infecções por amostras MRSA. Logo, a diminuição
de sua atividade pode tornar o tratamento dessas infecções ainda mais difícil.
A técnica de PFGE agrupou as 20 amostras de nMRSA isoladas no
HUCFF, em 9 pulsotipos diferentes (A até I), demonstrando uma ampla
variedade clonal entre elas. Essa diversidade de perfis genotípicos entre
amostras com SCCmecIV foi também descrita por vários outros autores
(DIETRICH et al., 2004; NIMMO et al., 2006; BARTELS et al., 2007). Dados
similares aos nossos foram encontrados por Bartels e colaboradores (2007),
que também observaram uma emergência de amostras nMRSA, com
SCCmecIV e geneticamente distintas, entre 2003 e 2004, em um hospital na
Dinamarca. De acordo com Robinson e Enright (2003) esta elevada
diversidade pode ser devido não apenas a disseminação de determinados
genótipos, mas também a transmissão do SCCmecIV para amostras de MSSA
apresentando diferentes genótipos. Trindade e colaboradores (2005)
observaram a emergência de amostras nMRSA SCCmecIV, em infecções de
corrente sanguínea, em um hospital em São Paulo, porém, os autores
verificaram a prevalência de um genótipo causando estas infecções.
No presente estudo, cinco das 20 amostras nMRSA avaliadas
apresentaram o genótipo A, considerado prevalente no estudo. Essas
amostras foram isoladas de cinco pacientes e apresentaram resistência a um
maior número de drogas, entre todas as outras amostras avaliadas. Todas
foram resistentes à eritromicina, clindamicina e ciprofloxacina. A prevalência
desse genótipo no hospital pode ter favorecido a aquisição de outros genes de
resistência. Todas as amostras do genótipo A apresentaram o RFT AAAAAAA,

91
e uma dessas amostras (633) foi selecionada e analisada por MLST, sendo
incluída no ST 1 (CC 1). Esse ST é similar ao do genótipo denominado WA-1,
que foi inicialmente descrito em amostras de CA-MRSA na Austrália (UDO et
al., 1993). A similaridade também foi observada em relação ao genótipo
USA400, associado a infecções comunitárias nos Estados Unidos
(MCDOUGAL et al., 2003). Entre 1999 e 2000, amostras apresentando
características semelhantes também foram encontradas causando infecção
no HUCFF (VIVONI et al, 2006). Porém, essas amostras carreavam os genes
da PVL e eram sensíveis à oxacilina. Ribeiro e colaboradores, em 2007,
analisando amostras isoladas de dois hospitais nacionais também
encontraram amostras que apresentaram ST 1 e eram semelhantes aos
genótipos WA-1/USA400. Em nosso estudo, as amostras de nMRSA
relacionadas ao WA-1/USA400 não apresentaram os genes da PVL e foram
resistentes à meticilna. Como esse genótipo já havia sido detectado em
amostras de MSSA isoladas no HUCFF (VIVONI et al., 2006), anteriormente,
uma hipótese seria que estas amostras tenham incorporado o SCCmecIV e
perdido os genes da PVL.
O RFT AAACCAA foi aquele mais freqüente (7/20) entre as amostras
nMRSA avaliadas, tendo sido associado a 4 genótipos: D, E, F e G. Uma
amostra representativa desse RFT (664 - genótipo D) foi analisada por MLST
e apresentou perfil alélico 1-4-1-4-12-1-10, sendo incluído no ST 5 (CC 5).
Amostras de S. aureus apresentando SCCmecIV, com genótipo similar por
PFGE, e mesmo RFT, já tinham sido descritas anteriormente no HUCFF
(VIVONI et al, 2006). Contudo, a maioria dessas amostras (9/11) não

92
apresentava o cassete de resistência. Esse fato pode indicar uma tendência a
aquisição de SCCmec por essas linhagens. Uma outra amostra (604), com o
mesmo RFT e do genótipo E, apresentou um perfil por PFGE idêntico ao do
USA800 (COCKFIELD et al, 2007), um MRSA de origem hospitalar descrito
como clone pediátrico (SÁ-LEÃO et al., 1999). Miranda e colaboradores
(2007) também encontraram amostras de MRSA com esse genótipo em um
estudo envolvendo dois hospitais brasileiros, indicando a emergência desse
perfil genômico também entre amostras MRSA isoladas no Brasil.
O complexo clonal 5 (CC 5) tem agrupado amostras de diferentes
SCCmec, como os tipos I, II e IV (ENRIGHT et al., 2002). As amostras de
nMRSA 664 e 603, isoladas no presente estudo, apresentaram os tipos IV e II
de SCCmec, respectivamente, porém o mesmo RFT AAACCAA foi detectado,
o que indicou que havia uma relação entre elas. O MLST classificou a
amostra 664 no ST 5 e a amostra 603 em um ST que variou em um lócus
(gene pta) em relação ao ST 5, recebendo uma nova denominação de ST
1204, sendo portanto, um SLV (do inglês, “single lócus variant”) do ST 5.
As amostras nMRSA de números 517, 518 e 519, que foram isoladas
de infecções hospitalares em diferentes sítios do mesmo paciente,
apresentaram o RFT-AAACCAC e o ST 72 (CC 8). Não encontramos descrição
desse ST no Brasil, porém, amostras com ST 72 e SCCmecIV têm sido
associadas a infecções comunitárias na Coréia (KIM et al., 2007) e Suécia
(FANG et al., 2008). Outro ST descrito entre as amostras analisadas foi o ST
97 (CC 97), relacionado à amostra 563. No banco de dados do MLST existe
pouca informação a respeito da distribuição desse ST no mundo e os que

93
estão descritos são referentes a amostras de MSSA. A causa da emergência
desses novos genótipos em nossa instituição não está clara, porém, pode
estar relacionada ao fato das amostras de nMRSA possuírem vantagens
adaptativas em comparação com as amostras multirresistentes (LAURENT et
al., 2001).
Geralmente associadas apenas a infecções hospitalares, no final da
década de 90, amostras de MRSA foram também isoladas de infecções
comunitárias (CA-MRSA), na Austrália (HEROLD et al., 1998) e Estados
Unidos (GROOM et al., 2001). A partir de então, houve relatos em diversos
locais pelo mundo (MA et al., 2005; DIETRICH et al, 2007). No Brasil, o
primeiro relato de CA-MRSA foi realizado por Ribeiro e colaboradores (2005).
Essas amostras foram isoladas de abscessos cutâneos e artrite séptica, em
um hospital de Porto Alegre, Rio Grande do Sul, apresentavam o SCCmecIV
e estavam relacionadas ao genótipo do sudoeste do pacífico, denominado,
OSPC (do inglês, “Oceania Southwest Pacific clone”) (NIMMO et al, 2000). Em
nosso trabalho, foram observadas quatro amostras MRSA isoladas de
infecções de origem comunitária, sendo uma delas (amostra 526) isolada de
uma turista americana de 26 anos, que se encontrava no Rio de Janeiro.
Essa amostra foi geneticamente idêntica ao genótipo USA300, relacionado,
principalmente com casos, de infecções comunitárias nos Estados Unidos e
mais recentemente, isoladas também de infecções hospitalares (HUANG et
al., 2006; PATEL et al., 2007). Essa amostra apresentou o RFT CAAACAC e
foi incluída no ST 8, e também apresentou os genes da PVL, características

94
comuns no genótipo USA300. Esse é o primeiro relato da presença de
amostra USA300, carreando o gene da PVL em hospital do Rio de Janeiro.
Em nosso estudo, as quatro amostras de CA-MRSA foram isoladas de
quatro pacientes (4, 11, 16 e 17). Em três deles não havia a presença de
fatores de risco para aquisição de MRSA e nem alguma comorbidade
associada, o que está de acordo com os critérios observados para pacientes
com CA-MRSA (MILLAR et al., 2007). O paciente 17, contudo, estava fazendo
uso de antimicrobiano e apresentava infecção viral. De fato, tanto as
amostras MRSA de origem comunitária quanto de origem hospitalar
apresentaram características em comuns, tais como: SCCmecIV e o fenótipo
de não multirresistência. Além disso, os mesmos STs, como o ST 1 e 5 foram
identificados para os dois tipos de amostras. Em adição, a apresentação
clínica foi a mesma para ambos os pacientes, com infecção por HA e CA-
MRSA. Portanto, amostras com características de CA-MRSA devem ser
consideradas como potencial causa de infecções hospitalares.
Dos quatro pacientes com infecção por CA-MRSA três apresentaram
infecções cutâneas, sendo em dois deles, por amostras com genes da PVL. A
outra amostra do estudo que também apresentava esses genes foi isolada de
sangue, em paciente com bacteriemia relacionada a cateter. Esses dados
confirmam estudos anteriores que demonstram serem mais freqüentes
amostras PVL positivas em infecções comunitárias do que hospitalares (LINA
et al, 1999; RIBEIRO et al, 2005; FANG et al, 2008).
Além da amostra 526, isolada de uma turista americana, uma outra
amostra de CA-MRSA (601) também carreava os genes da PVL. Essa amostra

95
apresentou o genótipo C, RFT BBBBBAB, e foi incluída no novo ST 1203
(SLV 30, CC não definido). Vivoni e colaboradores (2006) observaram em
estudo anterior, no HUCFF, a presença de amostras MSSA com genes da PVL
causando infecções. Essas amostras também eram do mesmo RFT
mencionado anteriormente e do ST 30. As outras duas amostras de CA-
MRSA (633 e 664) não apresentaram os genes da PVL e foram incluídas nos
genótipos A (ST 1) e D (ST 5), similares aos genótipos USA400 e USA800,
importantes agentes de infecções comunitárias e hospitalares,
respectivamente (SÁ-LEÃO et al., 1999; MCDOUGAL et al., 2002; RIBEIRO et
al, 2007).
Na segunda parte deste estudo, caracterizamos aspectos relacionados à
resistência e virulência em amostras de S. aureus, isoladas de infecções
associadas a próteses e outras infecções no INTO. S. aureus e S. epidermidis
são responsáveis por mais de 70% das infecções relacionadas a próteses
(EHLRICH et al., 2004; MORAN et al., 2007). Contudo, no Brasil são muito
escassos dados ou informações sobre estudos realizados com amostras
isoladas de próteses articulares (ERCOLLE & CHIANCA, 2002).
No período de 08/2005 a 10/2007 foram isoladas 73 amostras de S.
aureus no INTO, sendo 32 amostras isoladas de próteses articulares e 41 de
outras infecções. Diferente do estudo realizado no HUCFF, as amostras
isoladas no INTO foram obtidas consecutivamente de infecções por S. aureus
durante o período relatado.
Das 73 amostras avaliadas, 28 foram identificadas como MRSA (sendo
13 de próteses e 15 de outras infecções). Dezesseis (57,1%) carreavam o

96
SCCmecIV e 12 (42,9%) o SCCmecIII. Foi observado que houve prevalência de
amostras com SCCmecIV, tanto nas amostras de MRSA isoladas de IAP
quanto nas isoladaas de INAP. Essas amostras foram não multirresistentes
(nMRSA), ao contrário do observado para amostras com SCCmecIII.
Informações obtidas junto a CCIH do INTO demostraram que esse tipo de
amostra, nMRSA, não havia sido isolada no hospital até então.
Assim como entre as amostras isoladas no HUCFF, aquelas de MRSA
com SCCmecIV isoladas no INTO, apresentaram altas taxas de resistência à
eritromicina, ciprofloxacina e clindamicina. Outros trabalhos têm mostrado,
nos últimos anos, um aumento nas taxas de resistência aos antimicrobianos
não β-lactâmicos, sobretudo à eritromicina e à clindamicina, entre amostras
de MRSA com SCCmecIV (TRINDADE et al., 2005; KILIC et al., 2006; PARK et
al., 2007). Em nosso estudo, particularmente em relação as quinolonas,
mais de 50% das amostras com SCCmecIV foram resistentes. Kilic e
colaboradores (2006) analisando a resistência antimicrobiana em 423
amostras de MRSA com SCCmecIV, isoladas de variadas infecções em
adultos, observaram taxas de resistência à eritromicina, clindamicina e
levofloxacina de 90,1%, 13,2% e 16,8%, respectivamente. Esses autores
fazem uma associação desses resultados com o rápido aumento na
resistência a quinolonas ocorrido, em amostras com SCCmecII no passado, e
sugerem que o mesmo pode acontecer com as amostras MRSA que carreiam
o SCCmecIV.
Em nosso estudo, as taxas de resistência à gentamicina observadas
entre as amostras com SCCmecIV foi de 37,5%. Esses dados são conflitantes

97
com os de outros trabalhos que demonstraram serem essas amostras, em
geral, sensíveis a essa droga (LAURENT et al., 2001; KILIC et al., 2006;
MIRANDA et al., 2007). Inclusive, Laurent e colaboradores (2001) sugerem
que a gentamicina serviria como marcador para amostras com SCCmecIV. É
importante destacar, contudo, que, em nosso estudo, as amostras SCCmecIV
se agruparam em apenas dois genótipos, o que pode ter contribuído para
influenciar as elevadas taxas de resistência detectadas. Ainda assim, a
presença de resistência, sobretudo a antimicrobianos que não estão
associados com a resistência em amostras MRSA com SCCmecIV, como a
ciprofloxacina e gentamicina, pode retratar uma tendência nessas linhagens,
caracterizadas como ST 1 e ST 5. Provavelmente, esse fato está associado a
maior exposição aos antimicrobianos, e a presença, cada vez maior, dessas
amostras no ambiente hospitalar, o que facilitaria a aquisição de novos
elementos de resistência.
Em relação às CMIs de oxacilina em amostras SCCmecIV, mais de 60%
apresentaram valores entre 8 e 64µg/mL, confirmando que essas amostras
apresentam CMIs próximas aos “ponto de corte”. Como foi dito
anteriormente, esses valores dificultam a detecção, in vitro, dessas amostras
de MRSA, o que pode levar a resultados falso-negativos. Analisando as CMIs
das 16 amostras de MRSA com SCCmecIV, isoladas de IAP e INAP,
observamos que sete (87,5%), das oito amostras isoladas de IAP
apresentaram CMIs >64µg/mL, enquanto entre as oito amostras de INAP,
cinco (62,5%) apresentaram CMIs <32µg/mL. Apesar do pequeno número de
amostras, essas CMIs mais elevadas entre amostras isoladas de IAP, podem

98
refletir a exposição a concentrações subinibitórias da droga, provavelmente,
devido ao biofilme formado sobre o material implantado.
Quanto a vancomicina, 85,7% das amostras MRSA apresentaram CMIs
de 1 ou 2µg/mL. Esses resultados também foram observados para as
amostras do HUCFF, demonstrando que está havendo uma tendência de
aumento nas CMIs para vancomicina, entre as amostras MRSA (SAKOULAS
et al, 2004), o que pode reduzir as possibilidades terapêuticas no tratamento
de infecções por esse patógeno.
Todas as amostras de MRSA com SCCmecIII, analisadas no presente
estudo, foram relacionadas ao genótipo prevalente no Brasil e, como era
esperado, todas as amostras apresentaram multirresistência antimicrobiana.
Todas as CMIs nessas amostras foram >256µg/mL para oxacilina,
confirmando dados anteriores (SCHUENCK et al., 2004; VIVONI et al., 2006)
Nesse estudo, 100% (73) das amostras analisadas apresentaram o gene
icaA e mais de 95% das amostras, tanto de IAP quanto de INAP, produziram
biofilme em placas de poliestireno. Muitos estudos demonstraram que a
maioria das amostras clínicas de S. aureus apresenta o operon ica (PEACOCK
et al., 2002; FRANK et al., 2004; ROHDE et al., 2007; O´DONNEL et al.,
2008). Rohde e colaboradores (2007), analisando 18 amostras de S. aureus,
isoladas de próteses de joelho e quadril, mostraram que todas as amostras
carreavam o gene icaA e também produziam biofilme. O´Donnel e
colaboradores (2008) também detectaram o operon ica em 100% das
amostras isoladas de portadores nasais (n=20) e naquelas isoladas de
infecções (n=26). Diferente dos estudos citados, alguns autores têm

99
comentado que, apesar das altas taxas de detecção do operon ica, muitas
vezes as amostras não formam biofilme, in vitro (FOWLER et al., 2001;
KNOBLOCH et al., 2002). Esse fato pode estar relacionado com as distintas
metodologias empregadas, incluindo diferentes concentrações de
suplementos, como, glicose e/ou sacarose. Contudo, é possível que a
expressão, em agumas amostras, ocorra, preferencialmente, in vivo, sobre
condições mais apropriadas. Enfim, os autores, em geral, concordam que a
presença do operon ica e a produção de biofilme parecem ser fatores
essenciais para o estabelecimento das infecções em próteses articulares.
Logo, a presença do operon ica, na maioria das amostras clínicas de S.
aureus avaliadas, é de relevância e, medidas que evitem ou dificultem a
chegada do patógeno ao local do implante e sua ligação ao dispositivo médico
devem ser consideradas.
S. aureus é um microrganismo altamente versátil com grande
capacidade de colonizar e estabelecer infecções em várias partes do
organismo. O primeiro passo para o estabelecimento dessas infecções é a
adesão ao tecido do hospedeiro. No presente estudo, investigamos a presença
de genes relacionados a adesão, e verificamos que os genes fnbA, ebpS e cna
foram encontrados em mais de 65% das amostras, tanto nas isoladas de IAP
quanto nas de INAP, enquanto os genes fnbB e bbp foram associados a
genótipos específicos, sendo detectados em menos de 20% das amostras
avaliadas.
Já foi demonstrado que a fibronectina é um importante mediador da
adesão de S. aureus a diversos materiais utilizados em próteses ortopédicas,

100
como, titânio e alumínio (DELMI et al., 1994). As proteínas FnBPA e FnBPB,
codificadas pelos genes fnbA e fnbB, respectivamente, não apenas
apresentam capacidade de se ligar à fibronectina, como também tem
capacidade de se ligar ao fibrinogênio (WANN et al., 2000), o que demonstra a
importância dessas proteínas na adesão bacteriana. Peacock e colaboradores
(2000) analisando a presença dos genes fnbA e fnbB em 163 amostras de S.
aureus, isoladas de implantes ortopédicos, endocardite, artrite séptica,
osteomielite e narina anterior, demonstraram que 162 (99,4%) apresentavam
o gene fnbA e 127 (77,9%) o gene fnbB. Analisando, in vitro, a ligação à
fibronectina por essas amostras, os autores observaram que um número
maior de amostras isoladas de implantes ortopédicos se ligava a essa
proteína quando comparado a amostras isoladas de outros sítios. Arciola e
colaboradores (2005) analisando a prevalência dos genes fnbA e fnbB em 191
amostras de S. aureus isoladas de infecções ortopédicas, demonstraram a
presença desses genes em mais de 98% das amostras. Em nosso estudo,
96% das amostras de IAP e 80,5% das amostras de INAP apresentaram o
gene fnbA Ao contrário do observado nos estudos anteriormente citados, o
gene fnbB, foi detectado em apenas 12 (16,4%) amostras, todas de MRSA
com SCCmecIII e todas similares ao genótipo prevalente nos hospitais
brasileiros. Amaral e colaboradores (2005), em estudos desenvolvidos, in
vitro, verificaram que amostras desse genótipo apresentavam maior
habilidade para se ligar, persistir e invadir células epiteliais. Segundo Gordon
e Lowy (2008) a presença de domínios de ligação a fibronectina mais
acessíveis parece ser necessária para um alto nível de invasão dessas

101
amostras. Booth e colaboradores (2001) encontraram resultados similares
aos nossos quando analisaram a presença de diversos genes de virulência em
405 amostras de S. aureus, isoladas de diversas infecções em vários hospitais
dos Estados Unidos e, em um da Alemanha, observaram a prevalência (65%)
de cinco genótipos entre essas amostras. O gene fnbA estava amplamente
disseminado, sendo encontrado em mais de 91% das amostras. Porém, o
gene fnbB foi encontrado em 20,1% das amostras e todas pertenciam a um
genótipo específico. Contudo, diferente dos nossos resultados, todas as
amostras eram de MSSA, isoladas de infecções de corrente sanguínea.
Devemos ressaltar que na literatura não foram encontrados estudos
relacionando o gene fnbB com tipos de SCCmec ou complexos clonais.
Em relação ao gene cna, Booth e colaboradores (2001) também
observaram que sua prevalência estava relacionada a genótipos específicos,
pois, em três dos cinco genótipos, esse gene foi encontrado em mais de 90%
das amostras, enquanto que em um outro genótipo esse gene não foi
detectado. Outros estudos têm demonstrado taxas variadas em relação à
presença desse gene (TRISTAN et al. 2003; ROHDE et al., 2007; BABA-
MOUSSA et al., 2008). Rohde e colaboradores (2007) relataram a presença do
gene cna em 22% das amostras de S. aureus isoladas de próteses de joelho e
quadril, enquanto Arciola e colaboradores (2005) ao analisarem 191
amostras de S. aureus, isoladas de infecções ortopédicas, observaram taxas
de 98% para esse gene. Em nosso estudo esses genes estavam presentes em
67,1% das 73 amostras. Entre os cinco principais genótipos observados,
apenas entre amostras do genótipo B´ esse gene não foi encontrado.

102
Excetuando o genótipo B´, em geral, todas as outras amostras dos genótipos
prevalentes (A´, C´, D´ e E´) apresentaram esse gene, não sendo possível
associá-lo a genótipos específicos.
Outra importante proteína de matriz que atua na ligação do S. aureus
aos tecidos é a elastina. Essa proteína apresenta a função de manter a
integridade estrutural de tecidos que requerem elasticidade, como pulmões e
vasos sanguíneos (DOWNER et al., 2002). Nesse estudo, o gene ebpS foi
encontrado em 75% das amostras isoladas de IAP e 70,7% das amostras de
INAP. Taxas semelhantes em relação à presença do gene ebpS foram
encontradas por outros autores. Rohde e colaboradores (2007), relataram a
presença desse gene em 71% das amostras de S. aureus isoladas de próteses
de joelho e quadril. Tristan e colaboradores (2003) comparando amostras
isoladas de endocardite, osteomielite/artrite séptica e colonização nasal
encontraram taxas de 61%, 62% e 55%, respectivamente. Da mesma forma
que o gene fnbA, o gene ebpS parece estar amplamente distribuído entre
amostras clínicas de S. aureus. Contudo, sua ausência foi observada
especificamente entre amostras do genótipo A´, onde foram incluídas as
amostras relacionadas ao genótipo de MRSA prevalente no Brasil, que
apresenta SCCmecIII e pertence ao CC 8.
Em nosso estudo, o gene bbp, que codifica uma proteína ligadora de
sialoproteína óssea, foi encontrado em apenas sete amostras (9,6%). Esse
gene estava presente em todas as quatro amostras do genótipo E´, o qual foi
associado apenas a amostras de INAP. As outras três amostras que
apresentaram esse gene foram dos genótipos G´ e H´, associados a amostras

103
de IAP e INAP, respectivamente. Tristan e colaboradores (2003) observaram
que o gene bbp estava mais associado com amostras isoladas de osteomielite
e artrite séptica, quando comparado com amostras de colonização nasal e
endocardite. Essa relação encontrada se deve a capacidade da proteína Bbp
ligar-se à sialoproteína óssea (TUNG et al., 2000). Baba-Moussa e
colaboradores (2008) ao analisar a presença de genes de virulência, em 100
amostras de S. aureus, isoladas de infecções do trato urinário, não
encontraram o gene bbp. Por outro lado, Rohde e colaboradores (2007)
detectaram 27% de amostras de S. aureus carreando esse gene, entre 18
amostras isoladas de próteses de quadril e joelho. Em nosso estudo, das sete
amostras isoladas de osteomielite, apenas uma apresentou esse gene. Como
infecções associadas a próteses têm relação direta com a estrutura óssea, era
esperada uma presença significativa entre as amostras de IAP, porém, não foi
o observado no presente estudo.
A análise da diversidade clonal das 73 amostras de S. aureus isoladas
no INTO mostrou que apenas cinco genótipos eram prevalentes e foram
responsáveis por 83,5% das infecções.
As amostras do genótipo A´ foram relacionadas ao genótipo de MRSA
prevalente no Brasil. A amostra representativa desse grupo, selecionada para
o MLST, apresentou ST 239, confirmando essa relação. Esse genótipo é
amplamente disseminado em hospitais nacionais, como demonstrado em
vários estudos (TEIXEIRA et al., 1995; SANTOS et al., 1999; VIVONI et al.,
2005). As principais características associadas a esse genótipo são a
multirresistência antimicrobiana e a diversidade de fatores de virulência,

104
incluindo a produção de biofilme, o que pode ter permitido que essas
amostras se disseminassem por diversos países (AMARAL et al., 2005;
DEURENBERG et al., 2006). Em nosso estudo, das oito amostras fortes
produtoras de biofilme, quatro eram desse genótipo, confirmando os relatos
anteriores.
Os dois principais genótipos detectados (B´ e C´) e que foram
prevalentes, tanto em infecções de prótese quanto em outras infecções,
englobaram amostras MRSA com SCCmecIV e amostras MSSA. O genótipo B´
apresentou uma grande quantidade de pulsotipos (14) demonstrando uma
variedade de eventos genéticos envolvendo essa estirpe. Ainda assim, o perfil
encontrado para a presença dos genes de virulência foi idêntico em 19 das 20
amostras avaliadas (6 MRSA e 14 MSSA). Em todas as cinco amostras em
que foi realizado o MLST, foi encontrado o ST 5 (CC 5). Algumas amostras,
isoladas no HUCFF, e avaliadas na primeira parte do estudo, também
pertenciam ao mesmo CC 5. A comparação entre as amostras mostrou
grande similaridade entre elas, sendo o pulsotipo B´7, isolado no INTO,
idêntico ao pulsotipo D3, isolado do HUCFF. Essas amostras são
relacionadas ao genótipo USA800 (clone pediátrico), amplamente
disseminado em vários hospitais dos Estados Unidos e Europa
(DEURENBERG et al., 2006, MCDOUGAL et al., 2003). Sola e colaboradores
(2006), analisando amostras de MRSA isoladas em Córdoba, Argentina,
demonstraram que as amostras relacionadas ao USA800 (SCCmecIV; ST 5)
estavam substituindo o genótipo predominante naquela região (SCCmecIII;

105
ST 239). Nosso estudo mostrou a emergência de amostras relacionadas ao
genótipo USA800 nas duas instituições de saúde avaliadas.
No genótipo C´ foi incluída a maior parte das amostras SCCmecIV,
isoladas tanto de IAP quanto de INAP. Nesse genótipo foram observadas nove
amostras de MRSA e 8 de MSSA. Em relação à presença de genes de
virulência, da mesma forma que no genótipo B´, todas, exceto uma,
apresentaram o mesmo perfil. As quatro amostras desse genótipo que foram
submetidas ao MLST apresentaram o ST 1. Esse mesmo ST foi associado ao
genótipo A, prevalente nas amostras de nMRSA isoladas no HUCFF. Os
pulsotipos C´1 e C´7 das amostras do INTO foram idênticas aos pulsotipos A1
e A2, respectivamente, encontrados nas amostras do HUCFF. Esses
genótipos são similares ao WA-1/USA400 que, como foi discutido
anteriormente, é um importante agente de infecções comunitárias na
Austrália (UDO et al., 1993) e Estados Unidos (McDOUGAL et al., 2003) e
foram descritos no Brasil.
A presença, em dois hospitais distintos, desses genótipos de MRSA
SCCmecIV, com ST 1 e 5, demonstra uma nova realidade nas infecções por
esse patógeno, uma vez que genótipo de MRSA prevalente no Brasil, vêm
sendo substituído ou co-existindo com essas novas estirpes. Quando
introduzidas nos hospitais, as amostras com tipos de SCCmec de tamanhos
mais reduzidos, podem apresentar algumas vantagens em relação àqueles
genótipos de MRSA tipicamente hospitalares, uma vez que já foi observado
que tais amostras apresentam menor tempo de geração e presença de fatores

106
de virulência adicionais, em especial os genes da Leucocidina de Panton
Valentine (BABA et al., 2002; MIRANDA et al., 2007).
Nos genótipos D´ e E´ foram incluídas todas as sete amostras que
apresentaram os genes da PVL. Apenas uma amostra (número 1970) era
MRSA com SCCmecIV, enquanto outras duas do mesmo genótipo E´ eram
MSSA. O MLST demonstrou que elas pertenciam ao ST 30, enquanto as
amostras do genótipo D´ foram classificadas em um SLV do ST 188. Vivoni e
colaboradores (2006) analisando amostras de S. aureus, isoladas do HUCFF,
entre 1999 e 2000, também encontraram, em amostras de MSSA carreando
os genes da PVL, o ST 188 e o ST 432, um SLV do 188. Os mesmos autores
encontraram, entre 11 amostras de um mesmo genótipo, oito que carreavam
os genes da PVL. Dentre essas amostras o MLST foi realizado em duas delas,
e os STs encontrados foram, ST 30 e ST 642, um SLV do ST 30. E assim,
como em nosso estudo, dentre as amostras que apresentaram o ST 30 havia
uma de MRSA, porém, o SCCmec não foi tipável. Ribeiro e colaboradores
(2007) relataram a presença de amostras de CA-MRSA, com ST 30 e genes da
PVL, em amostras isoladas no Rio de Janeiro e Porto Alegre. O ST 30 está
amplamente relacionado com amostras que possuem os genes da PVL,
sobretudo em amostras de CA-MRSA. Vandenesh e colaboradores (2003), em
um estudo envolvendo 117 amostras de CA-MRSA, isoladas da Europa,
Estados Unidos e Oceania demonstraram que das 17 amostras isoladas da
Oceania, 13 apresentavam o ST 30 e eram representantes do chamado
genótipo do Sudoeste do Pacífico (OSPC). Além disso, todas as 13 amostras
apresentaram os genes da PVL. Nossos dados se enquadram no modelo

107
evolutivo proposto para as linhagens do CC 30, descritas por Robinson e
colaboradores (2005) e Diep e colaboradores (2006). Esses autores propõem
um modelo baseado no SCCmec, genes da PVL e genes de virulência. De
acordo com esse modelo, haveria um ancestral comum, de MSSA - ST 30 que
teria recebido os genes da PVL, e mais recentemente, o SCCmecIV. Assim, as
amostras de S. aureus com ST 30, genes da PVL e o SCCmecIV, serviriam
como reservatórios para a transferência, tanto do SCCmec, quanto dos genes
da PVL. Este fato torna ainda mais importante a caracterização das
amostras envolvidas em infecção nas instituições de saúde, para controlar,
não apenas a disseminação dessas amostras, mas a transmissão desses
fatores a outras linhagens.
A associação entre os genes de virulência e os STs encontrados
demonstra que as amostras com o ST 30 apresentaram o perfil com maior
número de genes de virulência (perfil 10- presença dos genes icaA, cna, fnbA,
fnbB, bbp, ebpS e da PVL). Além disso, somente amostras do ST 30
apresentaram o gene bbp.
Assim como Arciola e colaboradores (2005) demonstraram que as
condições geradas por implantes não alteram a população de S. aureus no
ambiente, em nosso estudo, não foram observadas características específicas
entre os dois grupos de amostras, de infecções associadas (IAP) e não
associadas a próteses (INAP).
Apesar do principal genótipo de MRSA isolado de hospitais, estar
relacionado com amostras multirresistentes com SCCmecIII e ST 239, nesse
estudo, ficou claramente demonstrada a entrada de novos genótipos de

108
MRSA não multirresistentes, nos dois hospitais analisados. O aumento das
infecções por amostras MRSA observado em todo o mundo pode ter relação
com dois fatores: a disseminação de linhagens já existentes e/ou a bem
sucedida conversão de determinadas linhagens de MSSA em amostras de
MRSA, pela aquisição do SCCmec (NOTO et al., 2008). Noto e colaboradores
(2008) em um estudo que analisou a região de inserção do SCCmec (chamada
attB) no cromossomo de diferentes linhagens de MSSA demonstraram que
alguns grupos de amostras possuem maior capacidade de adquirir o
SCCmec, pela presença de seqüências de DNA similares às encontradas
nesse cassete, o que facilitaria uma possível recombinação e inserção no
cromossomo. Em nosso estudo, as amostras de MRSA com SCCmecIV e de
MSSA incluídas nos mesmos genótipos e STs foram encontradas em
infecções ocorridas no INTO. Analisando os pulsotipos desses dois grupos,
verificamos que, em algumas amostras, a diferença era inferior a três
bandas. Essa similaridade pode ser indicativa de uma pré-disposição desses
genótipos em receberem o cassete mec. Futuros estudos devem ser realizados
para melhor avaliação dessas amostras, em especial a região attB.
Os resultados desse estudo mostram que é cada vez mais importante a
realização de uma vigilância ativa nos hospitais nacionais, visando à
detecção e caracterização de amostras MRSA, não somente quanto aos tipos
de SCCmec, como também em relação aos genótipos, já que vem ocorrendo
uma mudança mundial na epidemiologia das infecções por S. aureus. É
importante ressaltar ainda a necessidade de informar aos profissionais de
saúde sobre a emergência dessas novas linhagens, para que medidas de

109
controle apropriadas para prevenção de sua transmissão, dentro das
instituições de saúde, sejam adotadas.

110
6 – CONCLUSÕES

111
Entre as principais conclusões observadas nesse estudo, podemos citar:
• Os altos valores de CMI para vancomicina (1 e 2µg/mL), observados
nas amostras de MRSA, tanto no HUCFF (100%), quanto no INTO
(85,7%), podem refletir a sua ampla utilização em infecções por MRSA,
aumentando a possibilidade de ocorrência de falha terapêutica com o
uso desse antimirobiano.
• A análise das amostras de MRSA, isoladas nos dois hospitais
estudados, mostrou que o genótipo de MRSA multirresistente
(SCCmecIII; ST 239), predominante nos hospitais nacionais, vem sendo
substituído por linhagens de MRSA não multirresistentes. Pudemos
observar a emergência de três linhagens de MRSA com SCCmecIV, ST
1, ST 5 e ST 30, em ambos os hospitais do Rio de Janeiro.
• A similaridade genética detectada entre amostras MRSA-SCCmecIV
(comunitárias e hospitalares) e MSSA, no INTO, com a inclusão da
maioria delas em duas grandes linhagens, ST 1 e ST 5, indica que as
mesmas podem ter migrado dos hospitais para comunidade ou vice-
versa.

112
• Neste estudo detectamos STs, não descritos no país até o momento,
tais como: ST 97 e ST 72, entre amostras de MRSA com SCCmecIV,
isoladas do HUCFF, assim como, descrevemos dois novos STs (ST 1203
e ST 1204).
• Detectamos uma amostra do genótipo USA300 (PVL + e resistente à
eritromicina) em um hospital no Rio de Janeiro. Esse fato é relevante,
pois, esse genótipo mostrou rápida capacidade de disseminação na
comunidade e hospitais americanos.
• A presença do gene icaA em todas as amostras de S. aureus isoladas
no INTO demonstra seu grande potencial para a produção de biofilme,
como demonstrado, in vitro, em mais de 90% das amostras avaliadas.
Os genes de virulência, icaA (100%), fnbA (87,7%), ebpS (70,7%) e cna
(67,1%) estavam amplamente distribuídos entre as amostras, enquanto
os genes fnbB (16,4%), bbp (9,6%) e da PVL (9,6%) foram encontrados
associados a genótipos específicos.

113
7 – REFERÊNCIAS BIBLIOGRÁFICAS

114
AIRES DE SOUZA, M., MIRAGAIA, M., SANCHES, I. S., AVILA, S., ADAMNSON, I., CASAGRANDE, S. T., BRANDILEONE, M. C., PALACIO, R., DELL´ACQUA, L., HORTAL, M., CAMOU, M., ROSSI, A., VELAZQUEZ-MEZA, M. E., ECHANIZ-AVILES, G., SOLORZANO-SANTOS, F., HEITMANN, I. & de LENCASTRE, H. Three-year assessment of methicillin-resistant Staphylococus aureus clones in Latin America from 1996 to 1998. J. Clin. Microbiol., 39, 2197-2205, 2001. AIRES DE SOUZA, M., de LENCASTRE, H., SANTOS, S., KIKUCHI, I., TOTSUKA, K. & TOMASZ, A. Similarity o antibiotic resistance patterns and molecular typing properties of methicillin-resistant Staphylococcus aureus isolates widely spread in hospitals in New York City and in a hospital in Tokio, Japan. Microbiol. Drug Resist. 6, 253-258, 2000. AKSOY, D.Y. & UNAL, S. New antimicrobial agents for the treatment of Gram-positive bacterial infections. Clin. Microbiol. Infect. 14, 411-420, 2008. AMARAL, M.M., COELHO, L.R., FLORES, R.P., SOUZA, R.R., SILVA-CARVALHO, M.C., TEIXEIRA, L.A., FERREIRA-CARVALHO, B.T., FIGUEIREDO, A.M. The predominant variant of the Brazilian epidemic clonal complexo of methicillin-resistant Staphylococcus aureus has an enhanced ability to produce biofilm and to adhere to and invade airway epithelial cells. J. Infect. Dis. 192, 801-810, 2005. AMORIM, M.L., FARIA, N.A., OLIVEIRA, D.C., VASCONCELOS, C., CABEDA, J.C., MENDES, A.C., CALADO, E., CASTRO, A.P., RAMOS, M.H., AMORIM, J.M. & DE LENCASTRE, H. Changes in the clonal nature and antibiotic resistance profiles of methicillin-resistant Staphylococcus aureus isolated associated with spread of the EMRSA-15 clone in a tertiary care portuguese hospital. J. Clin. Microbiol. 45, 2881-2888, 2007. ARCHER, G.L. Staphylococcus aureus: a well-armed pathogen. Clin. Infect. Dis. 26, 1179-1181, 1998. ARCIOLA, C.R.; BALDASSARI, L. & MONTANARO, L. Presence of icaA and icaD genes and slime production in a collection of staphylococcal strains from catheter-associated infections. J. Clin. Microbiol. 39: 2151-2156, 2001 ARCIOLA, C.R., CAMPOCCIA, D., GAMBERINI, S., BALDASSARRI, L., MONTANARO, L. Prevalence of cna, fnbA and fnbB adhesin genes among Staphylococcus aureus isolates from orthopedic infections associated to different types of implant. FEMS Microbiol. 246, 81-86, 2005. ARCIOLA, C. R., COLLAMATI, S., DONATI, E. & MONTANARO, L. A rapid PCR method for the detection of slime-producing strains of Staphylococcus epidermidis and S. aureus in periprosthesis infections. Diagn. Mol. Pathol. 10, 130-137, 2001. BABA-MOUSSA, L., ANANI, L., SCHEFTEL, J.M., COUTURIER, M., RIEGEL, P., HAIKOU, N., HOUNSOU, F., MONTEIL, H., SANNI, A. & PRÉVOST, G. Virulence factors produced by strains of Staphylococcus aureus isolated from urinary tract infections. J. Hosp. Infect. 68, 32-38, 2008. BANNERMAN, T. L.A & PEACOCK, S. J.Staphylococcus, Micrococcus and other catalase-positive cocci. In. Manual of Clinical Microbiology, Murray, P. R.; Baron, E. J.; Jorgensen, J. H., Landry, M. L. & Pfaller, M. A. (eds), 9th ed, ASM Press. Washington, D.C. p. 390-411. 2007. BARTELS, M.D., BOYE, K., LARSEN, A.R., SKOV, R. & WESTH, H. Rapid increase of genetically diverse methicillin-resistant Staphylococcus aureus, Copenhagen, Denmark. Emerg. Infect. Dis. 13, 1533-1540, 2007.

115
BEENKEN, K. E., DUNMAN, P. M., McALEESE, F., MACAPAGAL, D., MURPHY, E., PROJAN, S. J., BLEVINS, J. S. & SMELTZER, M. S. Global gene expression in Staphylococcus aureus biofilm. J. Bacteriol. 186, 4665-4684, 2004. BENNER, E. J. & KAYSER, F. H. Growing clinical significance of methcillin-resistant Staphylococcus aureus. Lancet 2:741-744 BERGER-BACHI, B. & ROHRER, S. Factors influencing methicillin resistance in staphylococci. Arch. Microbiol. 178:165-171, 2002 BOERLIN, P. Applications of multilocus enzyme electrophoresis in medical microbiology. J. Med. Methods. 28, 221-231, 1997. BOLES, B. R., THOENDEL, M. & SINGH, P. K. Self-generated diversity produces "insurance effects" in biofilm communities. Proc. Natl. Acad. Sci. 101, 6630-16635, 2004. BOOTH, M.C., PENCE L.M., MAHASRESHTI, P., CALLEGAN, M.C. & GILMORE, M.S. Clonal Associations among Staphylococcus aureus Isolates from Various Sites of Infection. Infect. Immun. 69, 345-352, 2001. BOYCE, J.M. 1996. Coagulase-negative staphylococci. In: Mayhall, C.G. (ed.). Hospital Epidemiology and Infection Control. Willians & Wilkins Company, Baltimore, USA. p. 306-334. BUCK, J. M., K. COMO-SABETTI, K. H. HARRIMAN, R. N. DANILA, D. J. BOXRUD, A. GLENNEN, & D R. LYNFIELD. Community-associated methicillin-resistant Staphylococcus aureus, Minnesota, 2000-2003. Emerg. Infect. Dis. 11,1532-1538, 2005. CASEWELL, M. W. & HILL, R. L. R. Elimination of nasal carriage of Staphylococcus aureus with mupirocin (“pseudomonic acid”) - a controlled trial. J. Antimicrob. Chemother. 17, 365-372, 1986. CENTER FOR DISEASE CONTROL AND HOSPITAL EPIDEMIOLOGY (CDC). Staphylococcus aureus with reduced susceptibility to vancomycin – United States. Morb. Mortal. Wkly. Rep. 46, 765-766, 1997. CENTERS FOR DISEASE CONTROL AND PREVENTION (CDC). Public Health Dispatch: Outbreaks of community-associated methicillin-resistant Staphylococcus aureus skin infections - Los Angeles County, California, 2002-2003. Morb. Mortal. Wkly. Rep. 52, 88, 2003. CHAMBERS, H. F. Detection of methicillin-resistant staphylococci. Infect Dis. Clin. North. Am. 7, 425-433, 1993. CHANG, S., SIEVERT, D. M., HAGEMAN, J. C., BOULTON, M. L., TENOVER F.C., DOWNES, F. P., SHAH, S., RUDRIK, J. T., PUPP, G. R.., BROWN W. J., CARDO, D., FRIDKIN, S. K. & THE VANCOMYCIN-RESISTANT Staphylococcus aureus INVESTIGATIVE TEAM. Infection with vancomycin-resistant Staphylococcus aureus containing the vanA resistance gene. N. Engl. J. Med. 348, 1342-1347, 2003. CHONGTRAKOOL, P., ITO, T., MA, X.X., KONDO, Y., TRAKULSOMBOON, S., TIENSASITORN, C., JAMKLANG, M., CHAVALIT, T., SONG, J.H. & HIRAMATSU, K. Staphylococcal cassette chromosome mec (SCCmec) typing of methicillin-resistant Staphylococcus aureus strains isolated in 11 Asian countries: a proposal for a new

116
nomenclature for SCCmec elements. Antimicrob. Agents Chemother. 50,1001–1012, 2006. CLINICAL AND LABORATORY STANDARDS INSTITUTE - CLSI 2006a Performance standards for antimicrobial disk susceptibility test – eighth ed. Approved standards-nine edition: M2-A9 . Wayne, Pensylvania, USA. CLINICAL AND LABORATORY STANDARDS INSTITUTE - CLSI 2006b. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically – sixth ed. Approved standards – seventh edition: M7-A7. Wayne, Pensylvania, USA CLINICAL AND LABORATORY STANDARDS INSTITUTE - CLSI 2007. Performance standards for antimicrobial susceptibility testing – eightheenth information supplement. M100-S18. Wayne, Pensylvania, USA. COCKFIELD, J. D., S. PATHAK, J. EDGEWORTH, & J. A. LINDSAY. Rapid determination of hospital-acquired methicillin-resistant Staphylococcus aureus lineages. J. Med. Microbiol. 56, 614-619,2007. COSTERTON, J. W., STWART, P. S. & GREENBERG, E.P. Bacterial biofilms: a common cause of persistent infection. Science. 284,1318-1322, 1999. CRAMTON, S.E., GERKE, C., SCHNELL, N.F., NICHOLS, W.W. & GÖTZ, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect Immun. 67, 5427-5433, 1999. CRISOSTOMO, M. I., WESTH, H., TOMASZ A., CHUNG, M., OLIVEIRA, D. C. & de LENCASTRE H. The evolution of methicillin resistance in Staphylococcus aureus: similarity of genetics backgrounds in historically early methicillin-susceptible and –resistant isolates and contemporary epidemic clones. Proc. Natl. Acad. Sci. 98, 9865-9870, 2001. CUCARELLA, C., TORMO, M. A., ÚBEDA, C., TROTONDA, M. P.MÓNZON, M., PERIS, C., AMOREMA, B. LASA, I. & PENADÉS, J. R. Role of biofilm-associated protein bap in the pathogenesis of bovine Staphylococcus aureus. Infect. Immun. 72, 2177-2185, 2001. DAUM, R.S., ITO, T., HIRAMATSU, K., HUSSAIN, F., MONGKOLRATTANOTHAI, K., JAMKLANG, M. & BOYLEVAVRA, S. A novel methicillinresistance cassette in communityacquired methicillinresistant Staphylococcus aureus isolates of diverse genetic backgrounds. J. Infec. Dis., 186, 1344-1347, 2002. DAVIS, S.L., RYBAK, M.J., AMJAD, M., KAATZ, G.W. & MCKINNON, P.S. Characteristics of patients with healthcare-associated infection due to SCCmec type IV methicillin-resistant Staphylococcus aureus. Infect. Control. Hosp. Epidemiol. 27, 1025-1031, 2006. DE SOUZA, M.A.; SANCHES, I.S.; FERRO, M.L.; VAZ, M.J.; SARAIVA, Z.; TENDEIRO, T.; SERRA, J. & DE LENCASTRE, H. Intercontinental spread of a multidrug-resistant methicillin resistant Staphylococcus aureus clone. J. Clin. Microbiol., 36: 2590-2596, 1998. DELMI, M., VAUDAUX, P., LEW, D.P., & VASEY, H. Role of fibronectin on staphylococcal adhesion to metallic surfaces used as models of orthopaedic devices. J. Orthop. Res. 12, 432-438, 1994. DEPLANO, A., WITTE, W., VAN LEEUWEN, W. J., BRUN, Y. & STRUELENS, M. J. Clonal dissemination of epidemic methicillin-resistant Staphylococus aureus in Belgium and neighboring countries. Clin. Microbiol. Infect. 6, 239-245, 2000.

117
DEURENBERG, R.H. & STOBBERINGH, E.E. The evolution of Staphylococcus aureus. Infection, Genetics and Evolution.8, 747-763, 2008. DEURENBERG, R.H., VINK, C., KALENIC, S., FRIEDRICH, A.W., BRUGGEMAN, C.A. & STOBBERINGH, E.E. The molecular evolution of methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 13, 222-235, 2007. DICKINSON, G. M. & BISNO, A. L. Infections associated with indwelling devices: concepts of pathogenesis; infections associated with intravascular devices. Antimicrob. Agents Chemother. 33, 597-601, 1989. DIEKEMA, D.J., PFALLER, M.A., SCMITZ, F.J., SMAYEVSKY, J., BELL, J., JONES, R.N., BEACH, M. & SENTRY GROUP. Survey of infections due to Staphylococcus species: frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, Latin America, Europe, and the Western Pacific Region for the SENTRY antimicrobial surveillance program, 1997-1999. Clin. Infect. Dis. 32, S114-132, 2001. DIEP, B.A., CARLETON, H.A., CHANG, R.F., SENSABAUGH G.F. & PERDREAU-REMINGTON F. Roles of 34 Virulence Genes in the Evolution of Hospital – and Community-Associated Strains of Methicilin-Resistant Staphylococcus aureus. J. Infect. Dis. 193, 1495-1503, 2006. DIEP, B.A., PERDREAU-REMINGTON, F. & SENSABAUGH, G.F. Clonal characterization of Staphylococcus aureus by multilocus restriction fragment typing, a rapid screening approach for molecular epidemiology. J. Clin. Microbiol. 41, 4559-4564, 2003. DIETRICH, D.W, AULD, D. B. & MERMEL, L. A.. Community-acquired methicillin-resistant Staphylococcus aureus in southern New England children. Pediatrics. 113, 347-352, 2004. DINGES, M.M., ORWIN, P.M. & SCHILIEVERT, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Review. 13, 16-34, 2OOO. DOMINGUEZ, M. A., de LENCASTRE, H., LINARES, J. & TOMAZS, A. Spread and maintenence of a dominant methicillin-resistant Staphylococcus aureus (MRSA) clone during an outbreak of MRSA disease in a Spanish hospital. J. Clin. Microbiol. 32, 2081-2087, 1994. DONLAN, R.M. Biofilm formation: a clinically relevant microbiological process. Clin. Infect. Dis. 33, 1387-1392, 2001. DOWNER, R., ROCHE, F., PARK, P.W., MECHAM, R.P. & FOSTER, T.J. The Elastin-binding Protein of Staphylococcus aureus (EbpS) Is Expressed at the Cell Surface as an Integral Membrane Protein and Not as a Cell Wall-associated Protein. J. Biol. Chem. 277, 243-250, 2002. DUNNE, W.M. Bacterial adhesion: seen any good biofilms lately? Clin. Microbiol. Rev. 15, 155-166, 2002. EHRLICH, G. D., HU, F. Z., LIN, QIAO, L., CONSTERTON, J. W. & POST, J. C. Intelligent implants to battle biofilms. ASM News. 70, 127-133, 2004. EL-AZIZI, M., RAO, S., KANCHANAPOOM, T. & KHARDORI, N. In vitro activity of vancomycin, quinupristin/dalfopritin, and linezolid against intact and disrupted biofilms of staphylococci. Ann. Clin. Microbiol. Antimicrob. 4, 2, 2005.

118
ENRIGHT, M.C., DAY, N.P.J., DAVIES, C.E., PEACOCK, S.J & SPRATT, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin Microbiol. 38, 1008-1015, 2000. ENRIGHT, M.C., ROBINSON, D.A., RANDLE, G., FEIL, E.J., GRUNDMANN, H. & SPRATT, B.G. The evolutionary history of methicilin-resistant Staphylococcus aureus (MRSA). Proc. Natl. Acad. Sci. 99, 7687-7692, 2002. ERCOLE, F.F. & CHIANCA, T.C.M. Infecção de sítio cirúrgico em pacientes submetidos a artroplastias de quadril. Rev. Latino-am Enfermagem. 10, 157-165, 2002. EUZÉBY, J.P. List of Prokaryotic names with Standing in Nomenclature - Genus Staphylococcus. http://www.bacterio.cict.fr/s/staphylococcus.html 20/12/2008. FANG, H., G. HEDIN, G. LI, AND C. E. NORD. Genetic diversity of community-associated methicillin-resistant Staphylococcus aureus in southern Stockholm, 2000-2005. Clin. Microbiol. Infect. 14, 370-376, 2008. FITZGERALD, J.R., STURDEVANT, D.E., MACKIE, S.M., GILL, S.R. & MUSSER, J.M. Evolutionary genomics of Staphylococcus aureus: insights into the origin of methicillin-resistant strains and the toxic shock syndrome epidemic. Proc. Natl. Acad.Sci. U.S.A. 98, 8821–8826. 2001. FITZPATRICK, F., HUMPHREYS, H. & O'GARA, J.P. Evidence for icaADBC-independent biofilm development mechanism in methicillin-resistant Staphylococcus aureus clinical isolates. J. Clin. Microbiol., 43, 1973-1976, 2005. FOSSUM, A. E., & G. BUKHOLM. Increased incidence of methicillin-resistant Staphylococcus aureus ST80, novel ST125 and SCCmecIV in the south-eastern part of Norway during a 12-year period. Clin. Microbiol. Infect. 12, 627-633, 2006. FOSTER, T.J. Immune evasion by Staphylococci. Nature. 3, 948-958, 2005. FOSTER, T.J. & HÖÖK, M. Surface protein adhesins of Staphylococcus aureus. Trends Microbiol., 6, 484-488, 1998. FOWLER, V.G.Jr, FEY, P.D., RELLER, L.B., CHAMIS, A.L., COREY, G.R. & RUPP, M.E. The intercellular adhesion locus ica is present in clinical isolates of Staphylococcus aureus from bacteremic patients with infected and uninfected prosthetic joints. Med. Microbiol. Immunol. 189, 127-131, 2001. FRANK, K.L., HANSSEN, A.D. & PATEL, R. icaA is not a useful diagnostic marker for prosthetic joint infection. J. Clin. Microbiol. 42, 4846-4849, 2004. FUCHS, P. C., JONES, R. N. & BARRY, A. Interpretative criteria for disk diffusion susceptibility testing of mupirocin, a topical antibiotic. J. Clin. Microbiol. 28, 608-609, 1990. GERKE C., KRAFT, A., SUSSMUTH R., SCHWEITZER O. & GÖTZ, F. Characterization of the N-acetylglucosaminyltransferase activity involved in the biosynthesis of the Staphylococcus epidermidis polysaccharide intercellular adhesin. J. Biol. Chem. 17, 18586-18593, 1998. GILLASPY, A.F., PATTI, J.M., PRATT, F.L.Jr., IANDOLO, J.J. & SMELTZER, M.S. The Staphylococcus aureus collagen adhesin-encoding gene (cna) is within a discrete genetic element. Gene. 196, 239-248, 1997.

119
GILLET, Y., ISSARTEL, B., VANHEMS, P., FOURNET, J.C., LINA, G., BES, M., VANDENESCH, F., PIÉMONT, Y., BROUSSE, N., FLORET, D. & ETIENNE, J. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet. 359, 753-779, 2002. GOMES, A. R., SANCHES, I. S.,AIRES, D. S., CASTANEDA, E. & DE LECANSTRE, H. Molecular epidemiology of methicillin-resistant Staphylococcus aureus in Colombian hospitals: dominance of a single unique multidrug resistant clone. Microb. Drug Resist. 7, 23-32, 2001. GORDON, R.J. & LOWY, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect. Dis. 46, S350-S359, 2008. GÖTZ, F. Staphylococcus and biofilms. Mol. Microbiol. 43, 1367-1378, 2002. GROOM, A.V., WOLSEY, D.H., NAIMI, T.S., SMITH, K., JOHNSON, S., BOXRUD, D., MOORE, K.A. & CHEEK, J.E. Community-acquired methicillin-resistant Staphylococcus aureus in a rural American Indian community. JAMA. 286, 1201–1205, 2001. GRUNDMANN, H., AIRES-DE-SOUZA, M., BOYCE, J., TIEMERSMA, E. Emergence and resurgence of meticillin-resistant Staphylococcus aureus as a public-health threat. Lancet. 368, 874-885, 2006. HARTLEIB, J., KOHLER, N., DICKINSON, R.B., CHHATWAL, G.S., SIXMA, J.J., FOSTER, T.J., PETERS, G., KEHREL, B.E. & HERMANN, M. Protein A is the von Willebrand factor binding protein on Staphylococcus aureus. Blood. 96, 2149-2156, 2000. HEILMANN, C., SCHWEITZER, O., GERKE, C., VANITTANAKOM, N., MACK, D., GÖTZ, E. Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. Mol. Microbiol. 20, 1083-1091,1996. HEROLD, B.C., IMMERGLUCK, L.C., MARANAN, M.C., LAUDERDALE, D.S., GASKIN, R.E., BOYLE-VAVRA, S., LEITCH, C.D. & DAUM, R.S. Community-acquired methicillin-resistant Staphylococcus aureus in children with no identified predisposing risk. JAMA. 279, 593-598, 1998. HIGUCHI, W., TAKANO, T., TENG, L., YAMAMOTO, T. Structure and especific detection of staphylococcal cassette chromosome mec type VII. Biochemical and Biophysical Research Communications. 377, 752-756, 2008. HILL, C.R., FLAMANT, R., MAZAS, F. & EVRARD, J. Prophylatic cefazolin versus placebo in total hip replacement. Report of multicentre double-mind randomised trial. Lancet. I, 795-796, 1981. HIRAMATSU, K., CHUI, L., KURODA, M. & ITO, T. The emergence and evolution of methicillin-resistant Staphylococcus aureus. Trends Microbiol. 9, 486-493, 2001. HIRAMATSU, K., HANAKI, H., INO, T., YABUTA, K., OGURI, T. & TENOVER, F.C. Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility. J. Antimicrob. Chemother. 40, 135-136, 1997. HOLT, G.H., KRIEG, N.R., SNEATH, P.H.A., STALEY, J.T. & WILLIANS, S.T. 1994. Gram-positive cocci. In: Bergey’s Manual of Determinative Bacteriology - 9th ed. Willians & Wilkins, Baltimore, USA p.532-558.

120
HUANG, H., FLYNN, N. M., KING, J. H., MONCHAUD, C., MORITA, M. & COHEN, S. H. Comparisons of community-associated methicillin-resistant Staphylococcus aureus (MRSA) and hospital-associated MSRA infections in Sacramento, California. J. Clin. Microbiol. 44, 2423-2427, 2006. HUANG, Y. H., TSENG, S.P., HU, J.M., TSAI, J.C., HSUEH P.R. &. TENG, L.J. Clonal spread of SCCmec type IV methicillin-resistant Staphylococcus aureus between community and hospital. Clin. Microbiol. Infect. 13, 717-724, 2007. HUDSON, M. C., RAMP, W. K. & FRANKENBURG, K. P. Staphylococcus aureus adhesion to bone matrix and bone-associated biomaterials. FEMS Microbiol. 173, 279-284, 1999. HUEBNER, J. & GOLDMANN, D.A. Coagulase-negative staphylococci: role as pathogens. Annu. Rev. Med. 50, 223-236, 1999. ING, H.B., BADDOUR, L.M. & BAYER, A.S. 1997. Bacteremia and infective endocarditis: pathogenesis, diagnosis, and complications. In: The Staphylococci in Human Disease, Crossley, K.B. & Archer, G.L. (eds), Churchill Livingstone, New York, USA. p.331-355. ITO, T., KATAYAMA, Y., ASADA, K., MORI, N., TSUTSUMIMOTO, K. TIENSATORN, C. & HIRAMATSU, K. Structural comparison of three types of staphylococcal cassette chromosome mec integrated in the chromosome in methicillin- resistant Staphylocccus aureus. Antimicrob. Agents Chemother. 45, 1323-1336, 2001. ITO, T., KATAYAMA, Y. & HIRAMATSU, K. Cloning and nucleotide sequence determination of the entire mec DNA of pre-methicillin resistant Staphylococcus aureus N315. Antimicrob. Agents Chemother. 43, 1449-1458, 1999. ITO, T., MA, X. X., TAKEUCHI, F., OKUMA, K., YUZAWA, H. & HIRAMATSU, K. Novel type V staphylococcal cassete chromosome mec driven by a novel cassete chromosome recombinase, ccrC. Antimicrob. Agents Chemother. 48, 2637-2651, 2004. ITO, T., OKUMA, K., MA, X.X., YUZAMA, H. & HIRAMATSU, K. Insights on antibiotic resistance of Staphylococcus aureus from its whole genome: genomic island SCC. Drug Resist. Updat. 6, 41-52, 2003. JARLOV, J.O. Phenotypic characteristics of coagulase-negative staphylococci: typing and antibiotic susceptibility. APMIS. 107, 1S-42S, 1999. JEVONS, M.P. Celbenin-resistant Staphylococci. BMJ. 1, 124–125, 1961. JOHN, J. F. & BARG N. L. 1996.Staphylococcus aureus. In: Hospital Epidemiology and Infection Control. Mayhall, C. G. (ed.) 1st ed. Williams & Wilkins Company, Baltimore, USA p. 271-289. KANECO, J. & KAMIO, Y. Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: structures, pore-forming mechanism and organization of the genes. Biosci. Biotechnol. Biochem. 68, 981-1003, 2004. KANTZANOU, M., TASSIOS, P.T., TSELINI-KOTSOVILI, A., LEGAKIS, N.J. & VATAPOULOS, A. Reduced susceptibility to vancomycin of nosocomial isolates of methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 43, 729-731, 1999. KATAYAMA, Y., ITO, T. & HIRAMATSU K. Genetic organization of the chromosome region surrounding mecA in clinical staphylococcal strains: role of IS431-mediated mecI

121
deletion in expression of resistance in mecA-carrying, low-level methicillin-resistant Staphylococcus haemolyticus. Antimicrob. Agents. Chemother. 45:1955-1963 KATAYAMA, Y., ITO, T. & HIRAMATSU, K. A new class of genetic element, staphylococcus cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob. Agents. Chemother. 44, 1549-1555, 2000. KILIC, A., LI, H., STRATTON, C.W. & TANG, Y-W. Antimicrobial susceptibility patterns and staphylococcal cassette chromosome mec types of, as well as panton-valentine leukocidin occurrence among, methicillin-resistant Staphylococcus aureus isolates from children and adults in middle Tennessee. J. Clin. Microbiol. 44, 4436-4440, 2006. KIM, E.S., SONG, J.S., LEE, H.J. , CHOE, P.G., PARK, K.H., CHO, J.H., PARK, W.B., KIM, S.H., BANG, J.H., KIM, D.M., PARK, K.U., SHIN, S., LEE, M.S., CHOI, H.J., KIM, E.C., OH, M.D., KIM, H.B. & CHOE, K.W. A survey of community-associated methicillin-resistant Staphylococcus aureus in Korea. J. Antimicrob. Chemother. 60, 1108-1114, 2007. KLUYTMANS, J., VAN BELKUM, A. & VERBRUGH, H. Nasal carriage of Staphylococcus aureus: epidemiology, underlying mechanisms, and associated risks. Clin. Microbiol. Rev. 10, 505-520, 1997. KNOBLOCH, J.K., HORSTKOTTE, M.A., ROHDE, H. & MACK, D. Evaluation of different detection methods of biofilm formation in Staphylococcus aureus. Med. Microbiol. Immunol., 191, 101-106, 2002. KONDO, Y., ITO, T., MA, X.X., WATANABE, S., KREISWIRTH, B.N., ETIENNE, J. & HIRAMATSU, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: rapid identification system for mec, ccr, and major differences in junkyards regions. Antimicrob. Agents Chemother. 51, 264-274, 2007. KREISWIRTH, B., KORNBLUM, J., ARBEIT, R.D., EISNER, W., MASLOW, J.N., MCGEER, A., LOW, D.E. & NOVICK, R.P., Evidence for a clonal origin of methicillin resistance in Staphylococcus aureus. Science. 259, 227–230, 1993. KURTZ, S., MOWAT, F., ONG., K., CHAN, N., LAU, E. & HALPEM, M. Prevalence of primary and revision total hip and knee arthroplasty in the United States from 1990 through 2002. J. Bone Joint Surg (Am). 87, 1487-1497, 2005. LARSSEN, K.W., JACOBSEN, T., BERGH, K., TVETE, P., KVELLO, E. & SCHEEL, O. Outbreak of methicillin-resistant Staphylococcus aureus in two nursing homes in Central Norway. J. Hosp. Infect. 60, 312-316, 2005. LAURENT, F., LELIÈVRE, H., CORNU, M., VANDENESCH, F., CARRET, G. ETIENNE, J. & FLANDROIS, J.P. Fitness and competitive growth advantage of new gentamicin-susceptible MRSA clones spreading in French hospitals. J. Antimicrob. Chemother. 47, 277-283, 2001. LECLERCQ, R. Mechanisms of resistance to macrolides and lincosamides: nature of the resistance elements and their clinical implications. Clin. Infect. Dis. 34, 482–492, 2002. LEE, C. Y. & LEE, J. C. 2000. Staphylococcal capsule. In: Gram-Positive Pathogens. Fischetti, V.A.; Novick, R.P.; Ferretti, J.J.; Portnoy, D.A. & Rood, J.I. (eds), 1st edition. ASM Press, Washington, D.C., USA. p 361-366. LIDWELL, O. M., LOWBURY, E. J. L., WHYTE, W., BLOWERS, R., STANLEY, S. J & LOWE. Airbone contamination of wounds in joint replacement operations: the relationship to sepsis rates. J. Hosp. Infect. 4, 111-113, 1983.

122
LIDWELL, O. M., LOWBURY, E. J. L., WHYTE, W., BLOWERS, R., STANLEY, S. J & LOWE. Effect of ultraclean air in operating roons on deep sepsis in the joint after total hip or knee replacement: a randomized study. Br. Med J. 250, 99-102, 1982. LINA, G., PIÉMONT, Y., GODALL-GAMOT, F., BES, M. PETER, M.O., GAUDUCHON, V., VANDENESH, F. & ETIENNE J. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumoniae. Clin. Infect. Dis. 29, 1128-1132, 1999. LODISE, T.P., MILLER, C.D. , GRAVES, J., EVANS, A., GRAFFUNDER, E., HELMECKE M. & STELLRECHT K. Predictors of high vancomycin MIC values among patients with methicillin-resistant Staphylococcus aureus bacteraemia. J. Antimicrob. Chemother. 62, 1138-1141, 2008. LOWY, F.D. Medical Progress: Staphylococcus aureus infections. N. Engl. J. Med. 339, 520-532, 1998. MA, X.X., GALIANA, A., PEDREIRA, W., MOWSZOWICZ, M., CHRISTOPHERSEN, I., bhnMACHIAVELLO, S., LOPE, L., BENADERET, S., BUELA, F., VICENTINO, W., ALBINI, M., BERTAUX, O., CONSTENLA, I., BAGNULO, H., LIOSA, L., ITO, T. & HIRAMATSU, K. Community-acquired methicillin-resistant Staphylococcus aureus, Uruguay. Emerg. Infect. Dis., 11, 973-976, 2005. MAC FADDIN, J.F. Biochemical tests for identification of medical bacteria. Baltimore: Williams & Wilkins, 312p, 1976. MACK, D., BECKER, P., CHATTERJJE, I., DOBINSKY, S., KNOBLOCH, J.K.M., PETERS, G., HOLDE, H. & HERRMANN, M. Mechanism of biofilm formation in Staphylococcus epidermidis and Staphylococus aureus: functional molecules, regulatory circuits, and adaptative responses. Intern. J. Med. Microbiol. 294, 203-212, 2004. MACK, D., HAEDER, M., SIEMSSEN, N. & LAUFS, R. Association of biofilm production of coagulase negative staphylococci with expression of a specific polysaccharide intercellular adhesin. J. Infect. Dis. 174, 881-884, 1996. MARCHESE, A., BALISTRERI, G., TONOLI, E., DEBBIA, E.A. & SCHITO, G.C. Heterogeneous vancomycin resistance in methicillin-resistant Staphylococcus aureus strains in a large Italian hospital. J. Clin. Microbiol. 38, 866-869, 2000. MATO, R., SANTOS, S., VENDITII, M., PLATT, D. J., CHUNG, M. & DE LENCASTRE, H. Spread of the multiresistant Iberian clone of methicillin-resistant Staphylococcus aureus (MRSA) to Italy and Scotland. Microbial. Drug Resist. 4, 107-112, 1998. MCDOUGAL, L.K., STEWARD, C.D., KILLGORE, G.E., CHAITRAM, J.M., MCALLISTER, S.K. & TENOVER, F.C. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: establishing a national database. J. Clin. Microbiol. 41, 5113-5120, 2003. MENZIES, B.E. The role of fibronectin binding proteins in the pathogenesis of Staphylococcus aureus infections. Curr. Opin. Infect. Dis. 16, 225-229, 2003. MILLAR, B.C., LOUGHREY, A., ELBORN, J.S. & MOORE, J.E. Proposed definitions of community-associated meticillin-resistant Staphylococcus aureus (CA-MRSA). J. Hosp. Infect. 67, 109-113, 2007.

123
MIRANDA, O.P., SILVA-CARVALHO, M.C. , RIBEIRO, A., PORTELA, F., CORDEIRO, R.P. , CAETANO, N., VIDAL, C.F. & FIGUEIREDO A.M.S. Emergence in Brazil of methicillin-resistant Staphylococcus aureus isolates carrying SCCmecIV that are related genetically to the USA800 clone. Clin. Microbiol. Infect. 13, 1165-1172, 2007. MONTANARO, L., CAMPOCCIA, D. & ARCIOLA, C.R. Advancements in molecular epidemiology of implant infections and future perspectives. Biomaterials. 28, 5155-5168, 2007. MORAN, E., MASTERS, S., BERENDT, A.R., MCLARDY-SMITH, P., BYREN, I. & ATKINS, B.L. Guiding empirical antibiotic therapy in orthopaedics: the microbiology of prosthetic joint infection managed by debridement, irrigation and prosthesis retention. J. Infect. 55, 1-7, 2007. MUSSER, J.M. & KAPUR, V. Clonal analysis of methicillin-resistant Staphylococcus aureus from intercontinental sources: association of the mec gene with divergent phylogenetic lineages implies dessemination by horizontal transfer and recombination. J. Clin. Microbiol. 30, 2058-2063, 1992. NATIONAL NOSOCOMIAL INFECTIONS SURVEILLANCE SYSTEM (NNISS) REPORT. 2001. Data summary from January 1992-June 2001, issued August 2001. Centers for Disease Control and Prevention. Atlanta, Georgia, USA. NATIONAL NOSOCOMIAL INFECTIONS SURVEILLANCE SYSTEM (NNIS) REPORT. 1996. Data summary from October 1986-April 1996, issued May 1996. Centers for Disease Control and Prevention. Atlanta, Georgia, USA. NETTO-DOS-SANTOS, K.R., DE SOUZA, F.L., TEIXEIRA, L.M. & GONTIJO FILHO, P.P. Typing of Staphylococcus aureus from surgical site infections: comparison of pulsed field electrophoresis (PFGE) and PCR technique using extragenic palindromic (rep) and Tn916-shine-Dalgarno (tnSD) target sequences. Int. J. Med. Microbiol. 291, 231-236, 2001. NIMMO, G.R., COOMBS, G.W., PEARSON, J.C., O'BRIEN, F.G., CHRISTIANSEN, K.J., TURNIDGE, J.D, GOSBELL, I.B., COLLIGNON, P. & MCLAWS, M.L. Methicillin-resistant Staphylococcus aureus in the Australian community: an evolving epidemic. Med. J. Aust. 184, 384-388, 2006. NOSOCOMIAL INFECTION NATIONAL SURVEILLANCE SERVICE. Surveillance of surgical site infection in England, October 1997-September 2005. http://www.hpa.org.uk/infections/topics_az/surgical_site_infection/all_97_05_SSI.pdf. Acesso em Novembro 2008. NOTO, M. J., KREISWIRTH, B.N., MONK, A.B. & ARCHER, G.L. Gene Acquisition at the Insertion Site for SCCmec, the Genomic Island Conferring Methicillin Resistance in Staphylococcus aureus. J. Bacteriol. 190, 1276-1283, 2008. O’DONNELL, S., HUMPHREYS, H. & HUGHES D. Distribution of virulence genes among colonizing and invasive isolates of methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 14, 625-626, 2008. OKUMA, K., IWAKAWA, K., TURNIDGE, J.D. , GRUBB, W.B. , BELL, J.M., O'BRIEN, F.G., COOMBS, G.W., PEARMAN, J.W. , TENOVER, F.C. , KAPI, M. , TIENSASITORN, C., ITO, T. & HIRAMATSU, K. Dissemination of new methicillin-resistant Staphylococcus aureus clones in the community. J. Clin. Microbiol. 40, 4289-4294, 2002.

124
OLIVEIRA, D.C. & DE LENCASTRE, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother.46, 2155-2161, 2002. OLIVEIRA, G.A., DELL`AQUILA, A.M., MASIERO, R.L., LEVY, C.E., GOMES, M.S., CU, L., HIRAMATSU, K. & MAMIZUKA, E.M. Isolation in Brazil of nosocomial Staphylococcus aureus with reduced susceptibility to vancomycin. Infect. Control. Hosp. Epidemiol. 22, 443-448, 2001. OLIVEIRA, D.C., MILHEIRIÇO, C. & DE LENCASTRE, H. Redefining a structural variant of staphylococcal cassette chromosome mec, SCCmec type VI. Antimicrob. Agents Chemother. 50, 3457-3459, 2006. OLIVEIRA, D. C., TOMASZ, A. & DE LENCASTRE, H. Secrets of success of a human pathogen: molecular evolution of pandemic clones of methicillin-resistant Staphylococcus aureus. Lancet Infect. Dis. 2, 180:189, 2002. OLIVEIRA, D.C., TOMASZ, A. & DE LENCASTRE, H. The evolution of pandemic clones of methicillin-resistant Staphylococcus aureus: identification of two ancestral genetic backgrounds and the associated mec elements. Microb. Drug. Resist. 7, 349-361, 2001. O’NEILL, E., POZZI, C., HOUSTON, P., SMYTH, D., HUMPHREYS, H., ROBINSON, D.A. & O’GARA, J.P. Association between methicillin susceptibility and biofilm regulation in Staphylococcus aureus isolated from device-related infections. J. Clin. Microbiol. 45, 1379-1388, 2007. PALAZZO, I. C. V., ARAÚJO, M. L. C. & DARINE, A. L. C. First report of vancomycin-resistant staphylococci isolated from healthy carriers in Brazil. J. Clin. Microbiol. 43, 179-185, 2005. PAPAKYRIACOU, H., VAZ, D., SIMOR, A., LOUIE, M. & MCGAVIN, M.J. Molecular analysis of the acessory gene regulator (agr) locus and balance of virulence factor expression in epidemic methicillin-resistant Staphylococcus aureus. J. Infect. Dis. 181, 990-1000, 2000. PARK, J.Y., JIN, J.S., KANG, H.Y., JEONG, E.H., LEE, J.C., LEE, Y.C., SEOL, S.Y., CHO, D.T. & KIM, J. A comparison of adult and pediatric methicillin-resistant Staphylococcus aureus isolates collected from patients at a University Hospital in Korea. J. Microbiol. 45, 447-452, 2007. PARK, P.W., ROSENBLOOM, J., ABRAMS, W.R., ROSENBLOOM, J. & MECHAM, R.P. Molecular cloning and expression of the gene for elastin-binding protein (ebpS) in Staphylococcus aureus. J. Biol. Chem., 271, 15803-15809, 1996. PATEL, M., KUMAR, R.A., STAMM, A.M., HOESLEY, C.J., MOSER, S.A. & WAITES, K.B. USA300 genotype community-associated methicillin-resistant Staphylococcus aureus as a cause of surgical site infections. J. Clin. Microbiol. 45, 3431-3433, 2007. PEACOCK S.J., DAY N.P.J., THOMAS M.G., BERENDT, A.R., & FOSTER, T.J. Clinical Isolates of Staphylococcus aureus Exihibit Diversity in fnb Genes and Adhesion to Human Fibronectin. Jornual of Infection. 41, 23-31, 2000. PEACOCK, S.J., MOORE, C.E., JUSTICE, A., KANTZANOU, M., STORY, L., MACKIE, K., O’NEILL, G. & DAY, N.P.J. Virulent Combinations of Adhesin and Toxin Genes in Natural Populations of Staphylococcus aureus. Infect. Immun. 70, 4987-4996, 2002.

125
PETER, G., LOCCI, R. & PULVERER. Microbial colonization of prosthetic devices II. Scaning electron microscopy of naturally infect intravenous catheters. Zentralbl. Baketeriol. Mikrobiol. Hyg. B. 173, 293-299, 1981. PITCHER, D.G., SAUDERS, N.A. & OWEN, R.J. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol. 8, 151-156, 1989. PREVOST, G., COUPPIE, P., PREVOST, P., GAYET, S., PETIAU, P., CRIBIER, B., MONTEIL, H. & PIEMONT, Y. Epidemiological data on Staphylococcus aureus strains producing synergohymenotropic toxins. J. Med. Microbiol. 42, 237–245, 1995. PROJAN, S.J. & NOVICK, R.P. The molecular basis of pathogenicity. In: The Staphylococci in human disease. Crossley, K.B. & Archer, G.L. (ed). Churchill Livingstone, New York, USA, p. 55-81. 1997. REAGAN, D.R., DOEBBELING, B.N., PFALLER, M.A., SHEETZ, C.T., HOUSTON, A.J., HOLLIS R.J. & WENZEL, R.P. Elimination of coincident Staphylococcus aureus nasal and carriage with intranasal application of mupirocin calcium ointment, Ann. Inten. Med. 114, 101-106, 1991. RIBEIRO, A., CORONADO, A.Z. , SILVA-CARVALHO, M.C., FERREIRA-CARVALHO, B.T., DIAS, C., ROZENBAUM, R., DEL PELOSO, P.F., LEITE, C.C.F., TEIXEIRA, L.A. & FIGUEIREDO A.M.S. Detection and characterization of international community-acquired infections by methicillin-resistant Staphylococcus aureus clones in Rio de Janeiro and Porto Alegre cities causing both community- and hospital-associated diseases. Diagn. Microbiol. Infect. Dis. 59, 339-345, 2007. RIBEIRO, A., DIAS, C., SILVA-CARVALHO, M.C., BERQUÓ, L. FERREIRA, F.A., SANTOS, K.R.N., FERREIRA-CARVALHO, B.T. & FIGUEIREDO A.M. First report of infection with community-acquired methicillin-resistant Staphylococcus aureus in South America. J. Clin. Microbiol. 43, 1985-1988, 2005. ROBERTS, R.B., CHUNG, M., DE LENCASTRE, A., HARGRAVE, J., TOMASZ, A., NICOLAU, D.P., JOHN, J.F.Jr. & KORZENIOWSKI, O. Distribuition of methicillin-resistant Staphylococcus aureus clones among health care facilities in Connecticut, New Jersey, and Pennsylvania. Microbial. Drug Resist. 6, 245-251, 2000. ROBERTS, R.B., DE LENCASTRE, A., EISNER, W., SEVERINA, E.P., SHOPSIN, B., KREISWIRTH, B.N. & TOMASZ, A. Molecular epidemiology of methicillin-resistant Staphylococcus aureus in 12 New York hospitals. J. Hosp. Infect. 178, 164-171, 1998a. ROBERTS, R.B., TENNENBERG, A.M., EISNER, W., HARGRAVE, J., DRUSIN, L.M., YURT, R. & KREISWIRTH, B.N. Outbreak in a New York teaching hospital burn center caused by Iberian epidemic clone of MRSA. Microbial. Drug Resist. 4, 175-183, 1998b. ROBINSON, D. A. & ENRIGHT, M. C. Evolutionary models of the emergence of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 47, 3926-3934, 2003.
ROBINSON, D.A, KEARNS, A.M, HOLMES, A., MORRISON, D., GRUNDMANN, H., EDWARDS, G., O'BRIEN, F.G., TENOVER, F.C., MCDOUGAL, L.K., MONK, A.B., ENRIGHT, M.C. Re-emergence of early pandemic Staphylococcus aureus as a community-acquired meticillin-resistant clone. Lancet. 365, 1256-1258, 2005. ROCHE, F.M., MASSEY, R., PEACOCK, S.J., DAY, N.P.J., VISAI, L., SPEZIALE, P., LAM, A., PALLEN, M. & FOSTER T.J. Characterization of novel LPXTG-containing proteins of

126
Staphylococcus aureus identified from genome sequences. Microbiology. 149, 643-654, 2003. ROHDE, H., BURANDT, E.C., SIEMSSEN, N., FROMMELT, L., BURDELSKI, C., WURSTER, S., SCHERPE, S., DAVIES, A.P., HARRIS, L.G., HORSTKOTTE, M.A., KNOBLOCH, J.K.-M., RAGUNATH, C., KAPLAN, J.B., MACK, D. Polysaccharide intercellular adhesin or protein factors in biofilm accumulation of Staphylococcus epidermidis and Staphylococcus aureus isolated from prothetic hip and knee joint infections. Biomaterials., 28, 1711-1720, 2007. ROTUN, S.S., MCMATH, V., SCHOONMAKER, D.J., MAUPIN, P.S., TENOVER, F.C., HILL, B.C. & ACKMAN, D. M. Staphylococcus aureus with reduced susceptibility to vancomycin isolated from a patient with fatal bacteremia. Emerg. Infect. Dis. 5: 147-149, 1999. SADER, H.S., GALES, A.C., PFALLER, M.A., MENDES, R.E., ZOCCOLI, C., BARTH, A., JONES, R.N. & THE SENTRY PARTICIPANTS GROUP. Pathogen frequency and resistance patterns in brazilian hospitals: summary of results from three years of th SENTRY antimicrobial surveillance program. Braz. J. Infect. Dis. 5, 200-214, 2001. SADER, H.S., JONES, R.N., GALES A.C., SILVA J.B., PIGNATARI A.C. & THE SENTRY PARTICIPANTS GROUP (LATIN AMERICA). SENTRY antimicrobial surveillance program report: Latin American and Brazilian results for 1997 through 2001. Braz. J. Infect. Dis. 8, 25-79, 2004. SAKOULAS, G., MOISE-BRODER, P.A., SCHENTAG, J., FORREST, A., MOELLERING Jr, R.C. & ELIOPOULOS, G.M. Relationship of MIC and bactericidal activity to efficacy of vancomycin for treatment of methicillin-resistant Staphylococcus aureus bacteremia. J. Clin. Microbiol. 42, 2398-2402, 2004. SA-LEAO, R., SANTOS, S.I., DIAS, D., PERES, I., BARROS, R.M. & DE LENCASTRE, H. Detection of na archaic clone of Staphylococcus aureus with low-level resistance to methicillin in a pediatric hospital in Portugal and in international samples: relics of a formely widely disseminated strain? J. Clin. Microbiol.37, 1913-1920, 1999. SANCHES, I.S., AIRES DE SOUZA, M., SOBRAL, L., CALHEIROS, I., FELICIO, L., PEDRA, I. & DE LENCASTRE, H. Multidrug-resistant iberian epidemic clone of methicillin-resistant Staphylococcus aureus endemic in a hospital in northern Portugal. Microb. Drug. Resist. 1, 299-306, 1995 SANCHES, I.S., SARAIVA, Z.C., TENDEIRO, T.C., SERRA, J.M., DIAS, D.C. & DE LENCASTRE, H. Extensive intra-hospital spread of a methicillin-resistant staphylococcal clone. Int. J. Infect. Dis. 3, 26-31, 1998. SANTOS, K.R.N., TEIXEIRA, L.M., LEAL, G.S., FONSECA, L.S. & GONTIJO FILHO, P.P. DNA typing of methicillin-resistant Staphylococcus aureus: isolates and factors associated with nosocomial acquisition in two Brazilian university hospitals. J. Med. Microbiol. 48, 17-23, 1999. SCANVIC, A., DENIC, F., GAILLON, S., GIRY, P., ANDREMONT, A. & LUCET, J. Duration of colonization by methicillin-resistant Staphylococcus aureus after hospital discharge and risk factors for prolonged carriage. Clin. Infect. Dis. 32, 1393-1398, 2001. SCHUENCK, R.P., DADALTI, P., SILVA, M.G., FONSECA, L.S. & SANTOS, K.R.N. Oxacilin- and mupirocin-resistant Staphylococcus aureus : in vitro activity of silver sulphadiazine and cerium nitrate in hospital strains. J. Chemother. 16, 453-458, 2004.

127
SCHUENCK, R.P., LOURENÇO, M.C., IÓRIO, N.L., FERREIRA, A.L., NOUÉR, S.A. & SANTOS, K.R. Improved and rapid detection of methicillin-resistant Staphylococcus aureus nasal carriage using selective broth and multiplex PCR. Res. Microbiol. 157, 971-975, 2006. SCULCO, T.P. The economic impacto f infected joint arthroplasty. Orthopedics. 18, 871-873, 1995. SHAHIN, R., JOHNSON, I. L., JAMIESON, F., MCGEER, A., TOLKIN, J, FORD-JONES, E. L. Methicillin-resistant Staphylococcus aureus carriage in a child care center following a case of disease. Toronto Child Care Center Study Group. Arch. Pediatr. Adolesc. Méd. 153:864-868. SHORE, A., ROSSNEY, A.S., KEANE, C.T., ENRIGHT, M.C. & COLEMAN, D.C. Seven novel variants of the staphylococcal chromosomal cassette mec in methicillin-resistant Staphylococcus aureus isolates from Ireland. Antimicrob. Agents Chemother.49, 2070–2083, 2005. SMELTZER, M.S., GILLASPY, A.F., PRATT, F.L. & THAMES, M.D. Comparative evaluation of use of can, fnbA, fnbB, and hlb for genomic fingerprinting in the epidemiological typing of Staphylococcus aureus. J. Clin. Microbiol. 35, 2444-2449, 1997. SMIBERT, R. & KRIEG, N.R. Methods for general and molecular bacteriology. In: Phenotypic characterization. Gerhardt, P.; Murray, R.G. E.; Wood, W. A. & Krieg, N. R. (eds.) ASM Press. Washington, D.C., USA, p. 611-651. 1994 SMITH, T. & JARVIS, W. Antimicrobial resistance in Staphylococcus aureus. Microb. Infect. 1, 795-805, 1999. SOLA, C., CORTES, P., SAKA, H.A., CORDOBA MRSA COLLABORATIVE STUDY GROUP, VINDEL, A. & BOCCO, J.L. Evolution and molecular characterization of methicillin-resistant Staphylococcus aureus epidemic and sporadic clones in Cordoba, Argentina. J. Clin. Microbiol. 44, 192-200, 2006. STECKELBERG, J. M. & OSMON, R. D. Prosthetic joint infection. In: Waldvogel, F. A. and Bisno, A. L.(eds). Infections associated with indwelling medical devices. 3th edition. ASM Press. Washington D. C. p. 173-210. 2000 STEPANOVIC, S., VUKOVIC, D., DAKIC, I., SAVIC, B. & SVABIC-VLAHOVIC, M. A modified microtiter-plate for quantification of staphylococcal biofilm formation. J. Microbiol. Methods., 40, 175-179, 2000. TEIXEIRA, L.A., LOURENÇO, M. & FIGUEIREDO, A. Emergence of a methicillin-resistant Staphylococcus aureus clone related to the Brazilian epidemic clone III:B:A causing invasive disease among AIDS patients in a Brazilian hospital. Microb. Drug Resist. 2,393-399, 1996. TEIXEIRA, L. A., RESENDE, C. A., ORMONDE, L. R., ROSENBAUM, R., FIGUEIREDO, A.M., DE LENCASTRE, H. & TOMAZS, A. Geographic spread od epidemic multiresistant Staphylococcus aureus clone in Brazil. J. Clin. Microbiol. 33, 2400-2404, 1995. TENOVER, F.C., ARBEIT, R.D., GOERING, R.V., MICKELSEN, P.A., MURRAY, B.E., PERSING, D.H. & SWAMINATHAN, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 33, 2233-2239, 1995.

128
TENOVER, F.C. & GAYNES, R.P. The epidemiology of Staphylococcus infections. In: Gram-Positive Pathogens. Fischetti, V.A.; Novick, R.P.; Ferretti, J.J.; Portnoy, D.A. & Rood, J.I. (eds), 1st edition. ASM Press, Washington, D.C., USA. p 414-421. 2000. TIERMESMA, E.W., BRONZWARR, S.L.A.M., LYYTIKAINEN, O., DEGENER, J.E., SCHRIJNEMAKERS, P., BRUNISMA, N., MONEN, J., WITTE, W., GRUNDMAMN, H. & EUROPEAN ANTIMICROBIAL RESISTANCE SURVEILLANCE SYSTEM PARTICIPANTS. Methicilli-resistant Staphylococcus aureus in Europe, 1999-2002. Emerg. Infect. Dis. 10, 1627-1634, 2004. TON-THAT, H., LIU, G., MAZMANIAN, S.K., FAULL, K.F. & SCHNEEWIND, O. Purification and characterization of sortase; the transpeptidase that cleaves surface proteins of Staphylococcus aureus at the lpxtg motif. Proc. Natl. Acad. Sci. 96,12424-12429, 1999. TRAMPUZ, A. & ZIMMERLI, W. Prosthetic joint infections: update in diagnosis and treatment. Swiss. Med. Wkly. 135, 243-251, 2005. TRINDADE, P.A., PACHECO, P.L., COSTA, S.F., ROSSI, F., BARONE, A.A., MAMIZUKA, E.M. & LEVIN, A.S. Prevalence of SCCmec type IV in nosocomial bloodstream isolates of methicillin-resistant Staphylococus aureus. J. Clin. Microbiol. 43, 3435-3437, 2005. TRISTAN, A., YING, L., BES, M., ETIENNE, J., VANDENESCH, F. & LINA, G. Use of multiplex PCR to identify S. aureus adhesions involved in human haematogenous infections. J. Clin. Microbiol. 41, 4465-4467, 2003. TSUKAYAMA, D. T., ESTRADA, R. & GUSTILO, R. B. Infection after total hip arthroplasty. A study of the treatment of one hundred and six infections. J. Bone Joint Surg. Am. 78, 512-23, 1996. TUNG, H., GUSS, B., HELLMAN, U., PERSSON, L., RUBIN, K. & RYDÉN, C. A bone sialoprotein-binding protein from Staphylococcus aureus: a member of the staphylococcal Sdr family. Biochem. J. 345, 611-619, 2000. UDO, E.E., PANIGRAHI, D. & JAMSHEER, A.E. Molecular typing of methicillin-resistant Staphylococcus aureus isolated in a Bahrain hospital. Med. Princ. Pract. 17, 308-314, 2008. UDO, E.E., PEARMAN, J.W. & GRUBB, W.B. Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia. J. Hosp. Infect. 25, 97-108, 1993. VANDENBERGH, M.F.Q., YZERMAN, E.P.F., BELKUM, A., BOELENS, H.A.M., SIJMONS, M. & VERBRUGH, H.A. Follow-up of Staphylococcus aureus nasal carriage after 8 years: redefining the persistent carrier state. J. Clin. Microbiol. 37, 3133-3140, 1999. VANDENESCH, F., NAIMI, T., ENRIGHT, M.C., LINA, G. NIMMO, G.R., HEFFERNAN, H., LIASSINE, N., BES, M., GREENLAND, T., REVERDY, M.E. & ETIENNE, J. Community-acquired methicillin-resistant Staphylococcus aureus carrying panton-valentine leukocidin genes: worldwide emergence. Emerg. Infect. Dis. 9, 978-984, 2003. VAN DER MEE-MARQUET, N., EPINETTE, C., LOYAU, J., ARNAULT, L., DOMELIER, A., LOSFELT, B., GIRARD, N., QUENTIN, R. & THE BLOODSTREAM INFECTION STUDY GROUP OF THE RELAIS D’HYGIÉNE DU CENTRE. Staphylococcus aureus strains isolated from bloodstream infections changed significantly in 2006. J. Clin. Microbiol. 45, 851-857, 2007.

129
VELASCO, D., DEL MAR TOMAS, M., CARTELLE, M., BECEIRO, A,, PEREZ, A., MOLINA, F., MOURE, R. & VILLANUEVA R, BOU G. Evaluation of different methods for detecting methicillin (oxacillin) resistance in Staphylococcus aureus. J Antimicrob Chemother. 55:379-382. 2005 VIVONI, A.M., DIEP, B.A., MAGALHÃES, A.C.G., SANTOS, K.R.N., RILEY, L.W., SENSABAUGH, G.F. & MOREIRA B.M. Clonal composition of Staphylococcus aureus isolates at a Brazilian university hospital: identification of international circulating lineages. J. Clin. Microbiol. 44, 1686-1691, 2006. VIVONI, A.M., NETTO-DOS-SANTOS, K.R., DE-OLIVEIRA, M.P., GIAMBIAGI-DEMARVAL, M., FERREIRA, A.L.P., RILEY, L.W. & MOREIRA, B.M. Mupirocin for controlling methicillin-resistant Staphylococcus aureus: lessons from a decade of use at a university hospital. Infect. Control. Hosp. Epidemiol. 26, 662-667, 2005. VON EIFF, C. PETERS, G. & HEILMANN, C. Pathogenesis of infection due to coagulase-negative staphylococci. Lancet Infect. Dis. 2, 677-685, 2002. WANN, E.R., GURUSIDDAPPA S. & HOOK M. The Fibronectin-binding MSCRAMM FnbpA of Staphylococcus aureus Is a Bifunctional Protein That Also Binds to Fibrinogen. J. Biol. Chem. 275, 13863-13871, 2000. WANNET, W.J.B., SPALBURG, E., HECK, M.E.O.C., PLUISTER, G.N., TIEMERSMA, E., WILLEMS, R.J.L., HUIJSDENS, X.W., NEELING, A.J. DE & ETIENNE, J. Emergence of virulent methicillin-resistant Staphylococcus aureus strains carrying panton-valentine leucocidin genes in the Netherlands. J. Clin. Microbiol. 43, 3341-3345, 2005. WITTE, W., CUNY, C., BRAULKE, C. & HEUCK, D. Clonal dissemination of two MRSA strains in Germany. Epidemiol. Infect. 113, 67-73, 1994. WU, S., PISCITELLI, C., DE LENCASTRE, H. & TOMASZ, A. Tracking the evolutionary origin of the methicillin resistance gene: cloning and sequencing of a homologue of mecA from a methicillin susceptible strain of Staphylococcus sciuri. Microb. Drug Resist. 2, 435-441,. 1996. YAMASAKI, O., KANEKO, J., MORIZANE, S., AKIYAMA, H., ARATA, J., NARITA, S., CHIBA, J., KAMIO, Y. & IWATSUKI, K. The association between Staphylococcus aureus strains carrying panton-valentine leukocidin genes and the development of deep-seated follicular infection. Clin. Infect. Dis. 40, 381-385, 2005. ZAMAM, R. & DIBB, W.L. Methicillin resistant Staphylococcus aureus isolated in Saudi Arabia: epidemiology and antimicrobial resistance patterns. J. Hosp. Infect. 26, 297-300, 1994. ZHANG, Y.L., WANG, N.P., LAI, F.C., LI, Q. & LI, Z.Q. Survey on drug resistance of Staphylococcus aureus to commonly used antibiotics. Chemother. 4, 251-253, 2003. ZIMMERLI, W. & OCHSNER, P.E. Management of infection associated with prosthetic joints. Infection. 31, 99-107, 2003. ZIMMERLI, W., TRAMPUZ, A. & OCHSNER, P.E. Prosthetic-joint infections. N. Engl. J. Med. 351, 1645-1654, 2004.

130
ANEXO Artigos submetidos e em preparação relacionados a presente tese:
Artigo 1 - Schuenck, R. P.; Nouér, S. A.; Winter, C. O.; Cavalcante, F. S.;
Scotti , T. D.; Ferreira, A. L. P.; Giambiagi deMarval, M. & Santos, K. R. N.
Polyclonal Presence of Nonmultiresistant Methicillin–Resistant
Staphylococcus aureus Isolates Carrying SCCmec IV in Healthcare-
Associated Infections in a Hospital in Rio de Janeiro, Brazil. (Artigo
submetido ao Diagnostic Microbiology and Infectious Disease)
Artigo 2 - Schuenck, R. P.; Cavalcante, F. S., Emery, E.; Giambiagi deMarval,
M. & Santos, K. R.N. Prevalence of fnbA, fnbB, cna, bbp, ebpS and PVL
genes among Staphylococcus aureus isolates from orthopedic and other
infections in a Brazilina hospital. (Manuscrito em preparação).
Artigo 3 - Schuenck, R. P.; Cavalcante, F. S., Emery, E.; Giambiagi deMarval,
M. & Santos, K. R.N. Emergence of lineages ST 1, 5 and 30 among
Staphylococcus aureus isolates from orthopedical Hospital in Rio de
Janeiro. (Manuscrito em preparação).

131
Polyclonal Presence of Nonmultiresistant Methicillin–Resistant
Staphylococcus aureus Isolates Carrying SCCmec IV in
Healthcare-Associated Infections in a Hospital in Rio de Janeiro, Brazil
Ricardo Pinto Schuenck 1; Simone Aranha Nouér 2, 3; Carolina de Oliveira Winter 1;
Fernanda Sampaio Cavalcante 1; Tatiana Dantas Scotti 3; Adriana Lúcia Pires
Ferreira 3; Márcia Giambiagi-de Marval 1; Kátia Regina Netto dos Santos 1*
Departamento de Microbiologia Médica, Instituto de Microbiologia Prof. Paulo de
Góes 1 and Faculdade de Medicina 2 and Hospital Universitário Clementino Fraga
Filho 3, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
Running title: nMRSA SCCmec IV in healthcare-associated infections
*Corresponding author. Mailing address: Laboratório de Infecções Hospitalares,
Departamento de Microbiologia Médica, Instituto de Microbiologia Prof. Paulo de
Góes, CCS, Bloco I, UFRJ, Cidade Universitária, Rio de Janeiro, RJ, Brazil, CEP:
21941-590. Phone: 55-21-2560-8344, Fax: 55-21-2560-8028. E-mail:

132
Abstract
Changing in epidemiology of methicillin-resistant S. aureus (MRSA) was
observed due to the emergence of infections by nonmultiresistant MRSA (nMRSA) in
our hospital in Rio de Janeiro, Brazil. Clinical characterization and molecular
analysis of 20 nMRSA isolates recovered from 17 patients, between February 2005
and March 2006 was performed. The analysis included SCCmec (staphylococcal
cassette chromosome mec), pulsed field gel electrophoresis, multilocus restriction
fragment, and multilocus sequence typing. Minimum inhibitory concentrations
(MICs) for oxacillin and vancomycin, and presence of PVL genes were also
investigated. All but one of the 20 isolates presented SCCmec type IV. PFGE
clustered all isolates into nine genotypes. MIC <16µg/ml to oxacillin was found for
65% of the isolates, while 80% exhibited MIC of 2µg/ml for vancomycin. PVL
encoding genes were observed in three isolates. Polyclonal presence of nMRSA
SCCmec IV was observed in our institution, including community and healthcare-
associated isolates, belonged to the sequence types (STs) 1 (Clonal Complex-CC1),
ST5 (CC5), ST8 and ST72 (CC8), ST97 (CC97) and 2 STs singletons (SLV5 and
SLV30).
Key words: methicillin-resistant Staphylococcus aureus, SCCmec IV, PFGE, MLRFT,
MLST
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























