Embed Size (px)
Citation preview

UNIVERSIDADE DE BRASÍLIA
FACULDADE DE CIÊNCIAS DA SAÚDE
CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE
EFEITOS DA ADMINISTRAÇÃO DO ÓLEO DE COPAÍBA SOBRE O
DESENVOLVIMENTO DA FIBROSE PULMONAR INDUZIDA POR BLEOMICINA
EM CAMUNDONGOS: ASPECTOS MORFOLÓGICOS
HELEMI OLIVEIRA GUIMARÃES DE FREITAS
Dissertação apresentada ao Programa de Pós- graduação em Ciências da Saúde, Faculdade de Ciências da Saúde, Universidade de Brasília, para obtenção do título de Mestre em Ciências da Saúde.
BRASÍLIA
2006

2
EFEITOS DA ADMINISTRAÇÃO DO ÓLEO DE COPAÍBA SOBRE O
DESENVOLVIMENTO DA FIBROSE PULMONAR INDUZIDA POR BLEOMICINA
EM CAMUNDONGOS: ASPECTOS MORFOLÓGICOS
HELEMI OLIVEIRA GUIMARÃES DE FREITAS
Dissertação apresentada ao Programa de Pós- graduação em Ciências da Saúde, Faculdade de Ciências da Saúde, Universidade de Brasília, para obtenção do título de Mestre em Ciências da Saúde.
Orientador: Prof. Dr Ricardo Bentes Azevedo
BRASÍLIA
2006

3
DEDICATÓRIA
Ao Geraldo, meu marido, pelo sempre pontual apoio, vivência, aceitação das
ausências e acima de tudo pelo seu amor.
Ao Giordanne, meu filho, por mostrar que a vida tem motivos para ser bela somente
pelo que ela é, e nada mais.
À Leninne, minha filha, um estímulo para continuar, sempre motivada a lutar pelos
nossos sonhos, minha admiração pela coragem em assumir empreitadas da vida e que por
tantas vezes foi mais mãe do que filha.
Vocês são a minha vida e a minha luz !
“Felicidade é a certeza de que nossa vida não está se passando inutilmente”
Érico Veríssimo.

4
AGRADECIMENTOS
Um agradecimento especial ao meu orientador , Prof. Dr Ricardo Bentes Azevedo,
pelo seu exemplo profissional, pela confiança em mim depositada, pela oportunidade de
cursar esse mestrado e pelo convívio científico.
À Prof. Dra. Maria de Fátima Rodrigues da Silva, pelos vários momentos em que
auxiliou-me na realização deste trabalho, pelo sempre produtivo convívio, as sugestões
sempre úteis, e pelo exemplo de vida sempre demonstrado.
Ao Dr. Silas, e meus colegas do Diagnose, pela colaboração na realização deste
trabalho, pelo respeito e carinho demonstrado ao longo desses muitos anos de convívio.
Aos irmãos Prof. Dr Sérgio e Prof. Ms.Marilucia Zaiden, que sempre me apoiaram e
incentivaram na realização deste e outros trabalhos.
À Prof. Dra.Maria Dolores Lima, pelo apoio e incentivo.
Ao Prof. Dr Gustavo Adolfo Pazzeti, pelas sugestões.
Ao Remo de Castro Russo, pela ajuda nos ensaios bioquímicos.
À Prof. Ms.Umbelina do Rego Leite pelo auxílio nas análises estatísticas.
À Prof. Dra Monica Pereira Garcia, pela ajuda nas capturas das imagens.
Ao pessoal do Laboratório de Morfologia e Genética da UnB, pelos vários momentos
que me auxiliaram na realização deste trabalho.
Aos meus amigos e colegas da Biologia, que me ajudaram a acreditar e realizar este
trabalho.
Aos meus colegas de mestrado, pela cumplicidade nesse ciclo de aprendizagem.
Aos meus pais, irmãos e sobrinhos por participarem de diversos momentos da minha
vida.

5
RESUMO
FREITAS, Helemi Oliveira Guimarães de. Efeito da administração do óleo de copaíba sobre o desenvolvimento da fibrose pulmonar induzida por bleomicina em camundongos: aspectos morfológicos. 2006. 43f. Dissertação de Mestrado (Ciências da Saúde) – UNB – Universidade de Brasília.
A bleomicina é um antibiótico usado para o tratamento de várias neoplasias. Porém esta terapia pode apresentar como efeito colateral a indução de uma intensa resposta inflamatória resultando em fibrose pulmonar. O óleo de copaíba é um importante fitoterápico da medicina popular e vários estudos mostram sua propriedade antiinflamatória. Este trabalho teve como objetivo avaliar o efeito do tratamento com óleo de copaíba sobre o desenvolvimento da fibrose pulmonar induzida pela bleomicina. Camundongos foram tratados com bleomicina (0,5% do peso corporal) diariamente por injeção intraperitoneal concomitante com o tratamento com óleo de copaíba (0,1% do peso corporal) por gavagem diariamente durante 10 dias. Os pulmões foram processados para coloração com HE e ensaios bioquímicos para determinar a concentração de TGFβ e quantificação indireta de neutrófilos e macrófagos por meio da concentração de mieloperoxidase (MPO) e n-acetilglicosaminidase (NAG). Além disso, foi contado o número de células do parênquima e do sangue periférico. O tratamento concomitante com óleo de copaíba e bleomicina atenuou o desenvolvimento da fibrose pulmonar, induzindo apenas um leve processo inflamatório nos pulmões e espessamento dos septos alveolares como observado pela morfometria e histopatologia e dosagem de MPO, NAG e TGF β, quando comparado ao grupo controle.
PALAVRAS-CHAVE
Fibrose pulmonar, bleomicina, óleo copaíba, mieloperoxidase, N acetilglicosaminidase, TGFβ, Histomorfologia.

6
ABSTRACT
FREITAS, Helemi Oliveira Guimarães de. Effects of copaiba oil treatment on bleomycin-induced pulmonary fibrosis in mice: morphological aspects.
Bleomycin is an antibiotic used in the treatment of a variety of neoplasms. A major side-effect of bleomycin therapy is induction of an intense inflammatory response that evolve into pulmonary fibrosis. The copaíba oil is an important phytoterapic used in the popular medicine. Several studies show that this oil copaíba presents anti-inflamatory activity. This study examined the effects of copaiba oil in the development of the fibrosis pulmonary induced by bleomycin. Mice were treated intraperitoneally with bleomycin or with saline solution daily and copaíba oil by gavage for 10 days.The lungs were processed for light histopathological studies and biochemical rehearsals. The copaíba oil concomitant with bleomycin it lessened the development of pulmonary fibrosis as observed by the morphometry, histopathological and content of myeloperoxidase; N-acetylglucosaminidase; and transforming growth factorβ.
KEY WORDS
Copaiba oil; bleomycin; pulmonary fibrosis; myeloperoxidase; N-acetylglucosaminidase; transforming growth factorβ.

7
LISTA DE FIGURAS
FIGURA 1 Estrutura da bleomicina. ........................................................................... 13
FIGURA 2 Estruturas de sesquiterpenos presentes em óleo de copaíba.. .................. 20
FIGURA 3 Estrutura do ácido caurenóico. ................................................................. 20
FIGURA 4 Efeito do tratamento com óleo resina de copaíba e bleomicina sobre o
número de células do parênquima pulmonar. ........................................... 31
FIGURA 5 Fotomicrografia das alterações histológicas dos parênquimas
pulmonares em resposta aos tratamentos. As figuras apresentam cortes
transversais de pulmões corados com Hematoxilina/Eosina, A: SS,
Salina/Salina, B: BO, Bleomicina/Óleo.
32
FIGURA 6 Fotomicrografia das alterações histológicas dos parênquimas
pulmonares em resposta aos tratamentos. As figuras apresentam cortes
transversais de pulmões corados com Hematoxilina/Eosina, A: BS:
Bleomicina/ salina; B: SO, Salina/Óleo. Barra: 50 µm. .
FIGURA 7 Efeito dos tratamentos sobre a quantidade de TGFβ1 produzida no
parênquima pulmonar. .............................................................................. 33
FIGURA 8 Quantificação indireta do acúmulo de neutrófilos no parênquima
pulmonar pela detecção dos níveis da mieloperoxidase. .......................... 34
FIGURA 9 Quantificação indireta do acúmulo de macrófagos no parênquima
pulmonar pela detecção dos níveis de N-acetilglicosaminidase. ............. 35

8
LISTA DE TABELA
TABELA 1 TABELA 1- Comparação entre as massas corpóreas iniciais e finais dos
camundongos após dez dias de tratamento.
29
TABELA 2 Efeito do tratamento com bleomicina e óleo de copaíba sobre o número
de leucócitos totais, absolutos e relativos dos camundongos.
30

9
LISTA DE ABREVIATURAS
CGL – contagem global de leucócitos
Cl- – ânion cloreto
DNA – ácido desoxirribonucléico
EDTA – ácido etilenodiaminotetracético
ERO – Espécie reativa do oxigênio
IL – interleucina
IG – intragástrica
IP – intraperitoneal
MCP – monocyte chemotactic protein
MEC – matriz extracelular
MIP – macrophage inflammatory protein
NO – óxido nítrico
RNA – ácido ribonucléico
RNAm – RNA mensageiro
TGF-b – transforming growth factor-b
TNF – fator de necrose tumoral

10
SUMÁRIO
1 INTRODUÇÃO.....................................................................................................................12
1.1 Fibrose pulmonar idiopática ...............................................................................................12
1.2 Bleomicinas ........................................................................................................................13
1.3 Fibrose pulmonar induzida por bleomicina ........................................................................14
1.4 Células que participam do processo inflamatório na fibrose pulmonar induzida por
bleomicina.........................................................................................................................15
1.5 O TGF -β e a fibrose pulmonar ..........................................................................................17
2 ÓLEO DE COPAIBA............................................................................................................19
3 OBJETIVOS..........................................................................................................................22
3.1 Objetivo geral .....................................................................................................................22
3.2 Objetivos específicos..........................................................................................................22
4 MATERIAIS E MÉTODOS..................................................................................................23
4.1 Animais...............................................................................................................................23
4.2 Tratamentos e grupos experimentais ..................................................................................23
4.3 Grupos experimentais .........................................................................................................23
4.4 Coleta de sangue.................................................................................................................24
4.5 Processamento do material para microscopia óptica..........................................................24
4.6 Morfometria........................................................................................................................25
4.7 Processamento do material para ELISA.............................................................................25
4.8 Ensaio para detecção dos níveis de N-acetilglicosaminidase (NAG) tecidual...................26
4.9 Ensaio para detecção dos níveis de mieloperoxidase (MPO) tecidual ...............................26
4.10 Detecção de TGF-β1 ........................................................................................................27
4.11 Análise estatística .............................................................................................................27
5 RESULTADOS .....................................................................................................................29
5.1 Análise da massa corpórea dos camundongos....................................................................29
5.2 Análise da contagem global e diferencial dos leucócitos do sangue circulante .................30
5.3 Análise da contagem de células do parênquima pulmonar.................................................31
5.4 Aspectos morfológicos do parênquima pulmonar ..............................................................32
5.5 Análise dos mediadores químicos ......................................................................................34

11
5.5.1 Análise do TGFβ1 ...........................................................................................................35
5.5.2 Análise da Mieloperoxidase (MPO) ................................................................................36
5.5.3 Análise da N-acetilglicosaminidase (NAG) ....................................................................37
6 DISCUSSÃO.........................................................................................................................38
7 CONCLUSÕES.....................................................................................................................41
REFERÊNCIAS .......................................................................................................................42

12
1 INTRODUÇÃO
1.1 Fibrose pulmonar idiopática
O termo fibrose pulmonar idiopática, também conhecido como alveolite fibrosante
criptogênica, refere-se a um grupo de doenças pulmonares intersticiais crônicas, fibrosantes,
progressivas, de etiologia desconhecida. Ela é caracterizada histologicamente pela presença
de infiltrado inflamatório, proliferação de fibroblastos, com comprometimento heterogêneo do
interstício, principalmente na matriz septal alveolar, com áreas alternantes de fibrose
intersticial, inflamação, pulmão em favo de mel e pulmão normal (KATZENSTEIN et al.,
1995; FERREIRA, 2000).
Em conseqüência do processo inflamatório há o enrijecimento e redução do tamanho
dos pulmões ocorrendo perda gradativa da capacidade respiratória e dificuldade para realizar
as várias etapas da respiração. Há perda da captação de oxigênio e conseqüente má
oxigenação dos tecidos e órgão vitais do organismo. É referida clinicamente como pneumonia
intersticial ou doença pulmonar intersticial (WITTRAM, 2004).
A fibrose pulmonar apresenta características diferenciadas daquelas de outros órgãos,
a reação que ocorre nos pulmões é resultado de um processo inflamatório prévio após
agressões. A inflamação crônica do parênquima pulmonar continua sendo a idéia mais aceita
para explicar a patogênese da fibrose pulmonar, entretanto evidências de que o pulmão
quando exposto a freqüentes estímulos exógenos (fumaça de cigarros, drogas, fumos,
radiações e poluição) e, estímulos endógenos (infecções e doenças), sofrem danos contínuos
que resultam na fibrose pulmonar, mas os mecanismos são pouco entendidos
(GARANTZIOTIS et al., 2004).
O tratamento para esta doença apresenta maiores chances de sucesso quando
diagnosticada numa fase inicial, na qual as alterações inflamatórias preponderem sobre as
lesões fibróticas irreversíveis. Isso enfatiza a necessidade do reconhecimento e tratamento
precoce dessa condição. Em casos de pacientes com insuficiência respiratória grave, o
transplante pulmonar se apresenta hoje como uma opção terapêutica viável (MARTINEZ,
1998).

13
1.2 Bleomicinas
As bleomicinas pertencem a uma família de glicopeptídeos antibióticos com
atividade tumoricida que foram isoladas inicialmente do fungo Streptomyces verticillus. Elas
apresentam efeitos antineoplásicos e são utilizadas de forma isolada ou em associação com
outros quimioterápicos para o tratamento de diversos tipos de cânceres como linfomas, câncer
de útero, carcinomas de cabeça e pescoço, testicular e peniano (SUNTRES; SHEK, 1997).
Entretanto, seu uso clínico é limitado devido a seus efeitos colaterais, sendo um dos mais
graves o desenvolvimento da fibrose pulmonar, visto que de 30 a 40% dos pacientes que são
tratados com bleomicina são passíveis de desenvolver a doença (SUNTRES; SHEK, 1997;
MISHRA et al., 2000; SELMAN et al., 2001).
A estrutura da bleomicina é composta por uma molécula central ligada ao metal que
contém um cromóforo primidínico ligado a uma proprionamida, uma cadeia lateral amida β-
aminoalanina e os açucares L-glicose e 3- 0 carbamoil-d-manose. A bleomicina forma
complexos equimolares com diversos metais como o ferro e cobre e ao se ligar ao DNA
provoca sua ruptura.
FIGURA 1 – Estrutura da Bleomicina. Fonte: Almeida et al. (2005).

14
1.3 Fibrose pulmonar induzida por bleomicina
A injúria pulmonar induzida por bleomicina, semelhante à fibrose pulmonar
idiopática, é caracterizada pela inflamação do trato respiratório inferior, dano em células
epiteliais do tipo I, proliferação de fibroblastos e deposição de proteínas da matriz
extracelular. Os eventos inflamatórios que causam a fibrose pulmonar são iniciados quando
células epiteliais alveolares e outras células residentes liberam mediadores químicos que
promovem o recrutamento de células inflamatórias. Assim ocorre o acúmulo de neutrófilos,
eosinófilos, linfócitos, mastócitos, monócitos e macrófagos alveolares (CROUCH, 1990;
McANULTY; LAURENT, 1995; COSTABEL; KING, 2001).
Os mecanismos de destruição de células tumorais pela bleomicina são diferentes dos
mecanismos de toxicidade pulmonar. O pulmão é seletivamente afetado pelo bleomicina por
ser um órgão que não contém enzimas hidrolases as quais hidrolizam o grupo aminoalanina
da bleomicina evitando que aconteça a ligação de seus metabólitos a metais como o ferro, na
presença do oxigênio, formando a bleomicina ativada a qual oxida lipídios, hidrolisa ligações
aminas de proteínas e causa danos a DNA e RNA. A bleomicina ativada também pode se
decompor para gerar radicais hidroxilas que reagem de maneira rápida e inespecífica com
qualquer molécula que encontrar (FILDERMAN et al., 1998; LAZO; HUMPHREYS, 1993;
LAZO; HUMPHREYS, 1993; CHEN; STUBBE, 2005).
Animais como os camundongos, semelhante ao homem, apresentam esta enzima em
órgãos como fígado, baço e rins e não as apresentam nos pulmões sendo, portanto, modelo
experimental para pesquisas sobre os mecanismos de desenvolvimento desta doença
(ADAMSON; BOWDEN, 1974).
Vários estudos têm sido realizados com o intuito de esclarecer o mecanismo de ação
da bleomicina no desenvolvimento da fibrose pulmonar. Fichtner et al., (2004) demonstraram
que a bleomicina induz o aumento da expressão de moléculas que estão envolvidos na
patogênese da fibrose pulmonar como interleucinas e moléculas de adesão intercelular das
células do endotélio pulmonar.
O início da inflamação parece se iniciar quando a bleomicina interage com o DNA
levando à alterações fibroproliferativas via a ação em conjunto de várias citocinas tais como
TNF, TGFβ, interleucinas, havendo o acúmulo do colágeno nos espaços alveolares e capilares
dos pulmões resultando na redução da troca gasosa nos pulmões (CHEN, STUBBE, 2005).
A bleomicina também promove a depleção de antioxidantes endógenos e assim
exacerbando a injúria tissular mediada por oxidantes (ATZORI et al., 2004).

15
1.4 Células que participam do processo inflamatório na fibrose pulmonar induzida por
bleomicina
As reações inflamatórias são desencadeadas por vários estímulos e pela migração de
vários tipos celulares, os vasos sanguíneos sofrem alterações que visam facilitar o movimento
dessas células da circulação para o local da lesão. É um processo que envolve a ligação dos
leucócitos circulantes às células endoteliais e sua migração através do endotélio. O
recrutamento dessas células é feito por quimiotaxia.
Os mediadores químicos originam se do plasma ou de células, ligam-se a receptores
específicos na célula alvo e podem estimular a liberação de outros mediadores. Eles possuem
vida curta, tem efeitos em um ou diversos alvos ou ainda, efeitos diferentes em células,
também, diferentes e finalmente, tem potencial de causar efeitos danosos (RUFINO, SILVA
JR., 2006).
Dentre as células envolvidas na inflamação algumas estão normalmente presentes
nos tecidos, enquanto outras têm acesso a partir do sangue.
Os macrófagos podem ser ativados por uma variedade de estímulos, incluindo as
citocinas secretadas pelos linfócitos T. Macrófagos ativados secretam uma variedade de
produtos biologicamente ativos que, se não forem controlados, resultam na lesão tecidual e
fibrose por proliferação dos fibroblastos e deposição de colágeno. Os mastócitos também
estão presentes nas reações inflamatórias e podem produzir citocinas que contribuem para a
fibrose (ROBBINS, 2000).
Os neutrófilos encontram-se envolvidos em todas as fases da inflamação e são os
iniciadores do processo inflamatório nos tecidos. Existem mecanismos para sua identificação
e neutralização, muitos deles podem ser ativados por restos celulares após algum trauma,
mesmo sem infecção, e apresentarem uma ação nociva para o organismo (RUFINO, SILVA
JR., 2006).
Para isso os grânulos dos neutrófilos contêm lisozima, colagenase, gelatinase,
lactoferrina, histaminase e fosfatase alcalina. Os radicais livres derivados do oxigênio podem
ser liberados pelos leucócitos no meio extracelular após a exposição a quimiocinas. A
liberação desses mediadores em maiores quantidades pode ser danosa para os tecidos. A
influência dos radicais livres derivados do oxigênio em qualquer reação inflamatória depende
do equilíbrio entre a produção e a inativação desses metabólitos pelas células. Após a lesão
ocorre a expressão de proteínas responsáveis pela interação entre células endoteliais dos

16
capilares e os neutrofilos, as moléculas de adesão, ocorrendo a diapedese e a sua deposição no
espaço intersticial levará à produção de polipeptideos responsáveis pela quimiotaxia de novas
células inflamatórias (SGARBI, 2006; DUNSMORE et al, 2006).
Quando o neutrófilo é ativado, ele expressa suas moléculas de adesão da família das
L-selectinas e passa a consumir ativamente oxigênio em suas mitocôndrias, esse processo é
chamado de explosão oxidante (burst oxidativo das mitocôndrias) e esse consumo não é
direcionado para o metabolismo da célula e, sim, para a produção de espécies reativas de
oxigênio. Uma vez nos tecidos os neutrófilos sofrem desgranulação com a liberação para o
citoplasma da enzima mieloperoxidase. A enzima mieloperoxidase atua na presença de ânion
superóxido (ERO identificada como O-2) e do ânion cloreto (Cl-) para formar o ácido
hipoclorídico. O ácido hipoclorídrico apresenta intensa ação tóxica sobre as bactérias, sendo,
também, lesiva para as células do tecido. Outra ação dos neutrófilos nos tecidos é mediada
pela “elastase”, uma enzima do grupo das metaloproteinases (proteínas envolvidas nos
processos de lise tecidual), que é armazenada em grânulos não-azurófilos do citoplasma dos
neutrófilos. A mieloperoxidase pode ser usada como um índice de infiltração de neutrófilos
nos tecidos (SGARBI, 2006; DUNSMORE et al, 2006).
A infiltração dos neutrofilos é seguida pela migração dos macrófagos que produzem
substâncias relacionadas à modulação das respostas inflamatórias, secretam a maioria das
citocinas importantes da inflamação.
Os neutrófilos e macrófagos contêm grânulos de lisossomos que, quando liberados
podem contribuir com a reação inflamatória (RUFINO; SILVA JR, 2006).
Foi constato em estudos realizados com seres humanos mostram que a dosagem de
elastase do plasma sanguíneo e ensaios imunohistoquímicos de indivíduos com fibrose
pulmonar indicam um aumento da infiltração destas células no pulmão, mas o seu papel no
processo inflamatório que desencadeia a fibrose pulmonar não é esclarecido( OBAYASHI;
YAMADORI, 2006)
Intercrinas ou quimiocinas são citocinas que ativam leucócitos induzidas por
estímulos pró-inflamatórios como lipopolissacarídeos, TNF ou IL-1. Elas apresentam em sua
estrutura quatro resíduos de cisteína que formam duas pontes dissulfeto e são classificadas em
duas subfamílias. A subfamília CC a qual inclui quimiocinas beta como a MIP (macrophage
inflammatory protein); e a subfamília CXC que inclui quimiocina alfa que apresenta os
resíduos de cisteína separados por um aminoácido intervenientes como, por exemplo, a MCP
(monocyte chemotactic protein).
Estas moléculas modulam a quimiotaxia, proliferação e expressão de citocinas em

17
subgrupos de leucócitos. Elas são potentes atrativos para leucócitos circulantes durante a
inflamação. Por exemplo, as MIP-1α, MIP-1β e MIP-2 apresentam propriedades
quimioatrativas para neutrófilos.
Macrófagos, células epiteliais, células musculares lisas e fibroblastos liberam MIP-
1α a qual é aumentada na fibrose pulmonar idiopática devido ao aumento de sua produção por
células alveolares epiteliais tipo II e macrófagos (SMITH et al., 1995).
Pensava-se inicialmente que apenas fibroblastos residentes nos pulmões mediavam a
fibrose pulmonar, mas em trabalhos recentes Phillips et al. (2004) demonstraram, em modelo
murino de fibrose pulmonar induzida por bleomicina, que um grupo de fibrócitos
(CD45+colI+CXC4+) circulantes também participam do desenvolvimento da fibrose. Estes
fibrócitos migram para o pulmão em resposta à quimiocina CXCL12 e contribuem para o
aumento da deposição de colágeno. Esta migração foi inibida por anticorpos anti-CXCL12
atenuando a fibrose pulmonar (PHILLIPS et al., 2004).
Estudos realizados por Hashimoto et al. (2004) demonstraram que o processo
proliferativo dos fibroblastos não se origina unicamente no pulmão, mas tem a participação de
células oriundas da medula óssea. Alterações estruturais e quantitativas no parênquima
pulmonar podem ser observadas durante o esse processo, provocando um remodelamento das
vias aéreas e, conseqüentemente comprometendo o funcionamento pulmonar.
Quando ocorre ativação e proliferação de fibroblastos no parênquima pulmonar,
aparece nos septos alveolares feixes de miofilamentos de actina, essas células passam a ser
classificadas como células intersticiais contráteis ou miofibroblastos(CROUCH, 1990;
MCKAY; SHARMA, 2002).
1.5 O TGF -β e a fibrose pulmonar
O TGFβ tem sido considerado como a principal citocina fibrogênica. Foi constato
que o TGFβ aumenta a síntese e inibe a degradação de vários componentes da MEC por
diminuir as proteases e aumentar os inibidores de proteases, estimula a quimiotaxia de
fibroblastos e a produção de colágeno e fibronectina , aumenta a síntese de integrinas e pode
transformar fibroblastos em miofibroblastos. Em conjunto esses efeitos favorecem a
fibrinogênese, e esse fator parece estar envolvido no desenvolvimento de fibrose em uma
variedade de condições inflamatórias como na fibrose pulmonar induzida por bleomicina
(MATTOS, 2002).

18
O TGFβ, amplamente encontrado em mamíferos, é uma proteína produzida por
plaquetas, células endoteliais, linfócitos e macrófagos e é sintetizada na forma de proteínas
precursoras, as quis são proteoliticamente clivadas para liberar o fator de crescimento e um
segundo componente latente, esses fatores possuem funções inibitórias e estimulatórias.
A participação do TGF-β sobre a patogênese da fibrose tem sido demonstrada em
ensaios onde o TGF-β ativado foi introduzido em células de pulmão de rato e provocou uma
severa e prolongada fibrose caracterizada pela extensiva deposição de colágeno, fibronectina
e elastina na matriz extracelular além do surgimento de fibroblastos com fenótipo de
miofibroblastos (SIME et al., 1997). Após o dano das células endoteliais e infiltração de
células inflamatórias no interstício, os fibroblastos são ativados e ocorre a deposição de
colágeno. A bleomicina é capaz de ativar fibroblasto de maneira direta e indireta por meio de
citocinas tais como o TNF- e IL-1 (MOSELEY; HEMKEN; HUNNINGHAKE, 1986;
SUGERMAN et al., 1985; SCHMIDT et al., 1982).
Em pulmão normal de rato as cópias de RNAm para TGF-β são abundantes no
epitélio bronqueoalveolar.Quando ratos são tratados com bleomicina a expressão de RNAm
para TGF-β é aumentada predominantemente em macrófagos alveolares, mas também em
células do endotélio pulmonar, células mesenquimais, fibroblastos e células mesoteliais,
enquanto a expressão de RNAm para TGF-β2 e TGF-β3 não é afetada. Assim a produção
local e liberação de TGF-β neste caso é feita principalmente por macrófagos alveolares,
contribuindo a patogênese da fibrose pulmonar. Em biópsia pulmonar de pacientes com
fibrose pulmonar observam-se focos de fibroblastos ativados que expressam colágeno e
fibronectina e macrófagos alveolares que aumentam a expressão de mRNA e da proteína
TGF-β (BROEKELMANN et al., 1991).
Li et al. (2006) verificaram que a utilização de EM703, um derivado da eritromicina
caracterizada por não apresentar propriedades antibióticas e com propriedades
antiinflamatórias, atenua a fibrose e a infiltração de macrófagos e neutrófilos no espaço aéreo
e inibiu a produção de colágeno produzido por fibroblastos induzida por TGFβ.
O TGF-ß é um mediador que desempenha um papel central no processo de produção
do colágeno pelos fibroblastos e a utilização de anticorpos anti TGF-ß resulta na prevenção da
fibrose pulmonar induzida por bleomicina (GIRI; HYDE; HOLLINGER, 1993).
A administração de bleomicina em animais estimula vários tipos celulares a produzir
TGF-ß no pulmão tais como macrófagos, fibroblastos, células endoteliais e eosinófilos
(PHAN et al., 1991; ZHANG; FLANDERS; PHAN, 1995).

19
2 ÓLEO DE COPAIBA
As copaíbas são árvores do gênero Copaifera L. (Leguminosae-Caesalpinoideae),
conhecidas também como pau d´óleo, apresentam mais de 60 espécies catalogadas. Este
gênero ocorre no norte da América do Sul, principalmente nos estados brasileiros do Pará e
Amazonas e na região Centro Oeste.
Dentre as espécies mais abundantes, destacam-se: C. officinalis L. (norte do
Amazonas, Roraima, Colômbia, Venezuela e São Salvador), C. guianensis Desf. (Guianas),
C. reticulata Ducke, C. multijuga Hayne (Amazônia), C. confertiflora Bth (Piauí), C.
langsdorffii Desf. (Brasil, Argentina e Paraguai), C. coriacea Mart. (Bahia), C. cearensis
Huber ex Ducke (Ceará), (VEIGA Jr.; PINTO, 2001).
O óleo de copaíba é apresenta uma coloração que varia do amarelo ao marrom, sendo
que para estudos farmacológicos os óleos escuros e viscosos são os mais utilizados. As
espécies botânicas mais freqüentemente utilizadas na produção de óleo são: C. reticulata
(70%), C. guianensis (10%), C. multijuga (5%) e C. officinalis (5%) (VEIGA Jr.; PINTO,
2002).
O óleo de copaíba é um importante fitoterápico da medicina popular, indicado para
uma grande variedade de propriedades farmacológicas. Sua utilização como antiinflamatório e
cicatrizante está documentada desde o século XVI, quando colonizadores das Américas
relataram que as índias aplicavam esse óleo nos ferimentos dos guerreiros e umbigos de
recém-nascidos e, atualmente, é comercializado em farmácias e lojas de produtos naturais de
todo o país (MACIEL et al., 2002; VEIGA JR; PINTO, 2002).
A este óleo são também atribuídas propriedades como: estimulante, diurético,
laxante, expectorante, cicatrizante, antifúngico, antitetânico, anti-hemorrágico, anti-reumático,
tratamento de bronquites, doenças de origem sifilítica, moléstias de infecções dos sistemas
pulmonar e urinário, e ainda combate diferentes tipos de câncer ( VEIGA Jr.; PINTO, 2002;
MACIEL et al., 2002).
A composição do óleo resina varia tanto entre as espécies ou quanto dentro de uma
mesma espécie. Por exemplo, análises de duas amostras do óleo resina de copaífera multijuga
mostram que predomina na composição de ambas os sesquiterpenos α copaeno e β cariofileno
como principal componente e o diterpeno ácido copálico porém variando suas concentrações
(CASCON; GILBERT, 2000).
20
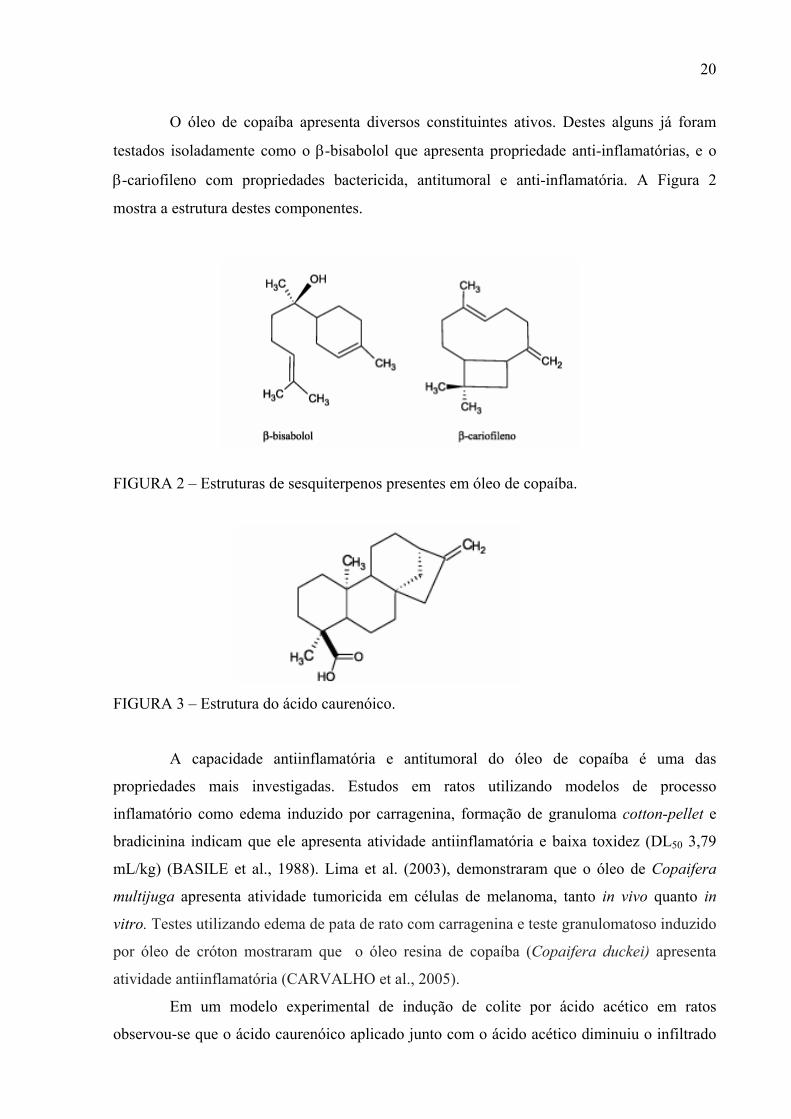
O óleo de copaíba apresenta diversos constituintes ativos. Destes alguns já foram
testados isoladamente como o β-bisabolol que apresenta propriedade anti-inflamatórias, e o
β-cariofileno com propriedades bactericida, antitumoral e anti-inflamatória. A Figura 2
mostra a estrutura destes componentes.
FIGURA 2 – Estruturas de sesquiterpenos presentes em óleo de copaíba.
FIGURA 3 – Estrutura do ácido caurenóico.
A capacidade antiinflamatória e antitumoral do óleo de copaíba é uma das
propriedades mais investigadas. Estudos em ratos utilizando modelos de processo
inflamatório como edema induzido por carragenina, formação de granuloma cotton-pellet e
bradicinina indicam que ele apresenta atividade antiinflamatória e baixa toxidez (DL50 3,79
mL/kg) (BASILE et al., 1988). Lima et al. (2003), demonstraram que o óleo de Copaifera
multijuga apresenta atividade tumoricida em células de melanoma, tanto in vivo quanto in
vitro. Testes utilizando edema de pata de rato com carragenina e teste granulomatoso induzido
por óleo de cróton mostraram que o óleo resina de copaíba (Copaifera duckei) apresenta
atividade antiinflamatória (CARVALHO et al., 2005).
Em um modelo experimental de indução de colite por ácido acético em ratos
observou-se que o ácido caurenóico aplicado junto com o ácido acético diminuiu o infiltrado

21
de células inflamatórias e edema da mucosa intestinal sugerindo seu efeito antiinflamatório
(PAIVA et al., 2003).
Veiga e colaboradores (2001) investigaram a composição de oito amostras de óleo de
copaíba comercializados. Foi observado que os óleos que continham vários sesquiterpenos e
diterpenos apresentaram maior eficiência para diminuir a formação de edema induzido por
carragenina em pata de rato sugerindo que estes componentes são responsáveis pela ação
antiiflamatória.
Estudos in vitro utilizando fibroblastos (V79) de pulmão de hamster chinês sugerem
que um dos componentes obtido do óleo de Copaifera langsdorfii, o ácido caurenóico, um
diterpeno (figura 2), apresenta atividade genotóxica quanto utilizado em altas concentrações
(30 e 60 µg/mL) (CAVALCANTE et al., 2006).
A atividade antitumoral da Copaífera multijuga foi demonstrada em bioensaios com
carcinma CMI em camundongo onde observou-se a redução dos nódulos de metástase no
tecido pulmonar pela administração in vivo e in vitro do óleo (LIMA, et al, 1999)
Recentemente, os pesquisadores Oswaldo de Freitas e Mônica Freiman de Souza
Ramos da USP de São Paulo mostraram que a capacidade antiinflamatória do óleo reside em
seus componentes voláteis e que é mais potente que o fármaco diclofenaco. Neste trabalho os
pesquisadores conseguiram isolar e encapsular componentes voláteis do óleo de copaíba e
mostraram sua ação antiinflamatória em camundongos com pleurisia induzida e em edemas
de patas (Agencia USP de notícias, 2006) (dados não publicados).

22
3 OBJETIVOS
3.1 Objetivo geral
Verificar se óleo de copaíba (C. multijuga) pode se administrado juntamente com a
bleomicina, inibir ou mesmo atenuar a resposta inflamatória pulmonar, evitando o
aparecimento da fibrose pulmonar e avaliar se o óleo, por si só, pode induzir um processo
inflamatório pulmonar.
3.2 Objetivos específicos
• Analisar morfologicamente e quantificar morfometricamente as alterações que
ocorrem no parênquima pulmonar em função da administração do óleo de copaíba e/ou
bleomicina.
• Verificar os efeitos do óleo de copaíba e/ou bleomicina sobre a modulação de
TGF-β1.
• Verificar os efeitos do óleo de copaíba e/ou bleomicina sobre a quantidade de
neutrófilos e macrófagos no parênquima pulmonar por meio da quantificação indireta de
NAG e MPO.

23
4 MATERIAIS E MÉTODOS
4.1 Animais
Camundongos Swiss, machos, com peso entre 30 e 35 gramas, foram obtidos do
Biotério do Instituto de Química do Estado de Goiás (IQUECO) em Goiânia e mantidos,
durante o experimento, no biotério do Departamento de Genética e Morfologia da UnB. Eles
receberam água e ração ad libitum e ciclo de 12 horas dia e noite.
4.2 Tratamentos e grupos experimentais
Tratamento com bleomicina: 100 µL de uma solução de 7,5 mg/ kg de peso corporal
de sulfato de bleomicina (Blenoxane®, Brystol Meyers-Squibb), ou solução salina 0,9%,
foram injetados em camundongos por via intraperitoneal (IP), por dez dias.
Suplementação com óleo resina de copaíba (Copaifera multijuga): animais
receberam óleo de copaíba, ou solução salina 0,9%, por via intragástrica (IG) nas doses de
0,1% relativo ao peso corporal, por dez dias (dose previamente definidas de acordo com
experimentos pilotos realizados).
Todos os animais foram tratados entre sete e nove horas da manhã, durante dez dias e
mortos por deslocamento cervical entre sete e nove horas do primeiro dia após o término do
tratamento. Os tratamentos intragástricos e intraperitoneais foram realizados no mesmo
período (dez dias).
4.3 Grupos experimentais
Os animais foram divididos nos seguintes grupos:
1- Grupo controle – Salina/salina (SS) - animais receberam solução salina pelas vias
IG e IP.
2- Grupo bleomicina/óleo de copaíba (BO) - animais tratados com sulfato de
bleomicina IP e suplementados com óleo de copaíba IG em uma dose de 0,1% relativa à
massa corporal.
3- Grupo bleomicina/salina (BS) - animais tratados com bleomicina IP e salina por
via IG.

24
4- Grupo óleo de copaíba/salina (OS) - animais tratados com solução salina por via
IP e suplementados com óleo de copaíba em uma dose de 0,1% relativa à massa corporal por
via IG.
4.4 Coleta de sangue
Após anestesia, o sangue foi coletado por punção cardíaca para contagem total e
diferencial de leucócitos periféricos, feito esfregaço e posteriormente diluído em EDTA.
Para a contagem total dos leucócitos foi utilizado 20 µL de sangue diluído em 400
µL da solução de Turk e feito a contagem em câmara de Neubauer. Foram contados todas as
células dos quatro retículos da câmara de Neubauer; e aplicado a fórmula (CGL/4.50).
A contagem diferencial dos leucócitos foi feita em esfregaço sanguíneo, corado pelo
corante Wright-Giemsa. Foram contados 500 células por animal para determinação das
populações de monócitos, linfócitos, neutrófilos e eosinófilos; e aplicado a fórmula (X = CGL
x número de células/500). A análise foi realizada em microscópio de luz em.
Em seguida o animal recebeu uma dose excessiva de anestésico e foi morto por
deslocamento cervical.
4.5 Processamento do material para microscopia óptica
Após a morte dos animais, os pulmões foram coletados e os pulmões direitos foram
congelados em nitrogênio líquido e os pulmões esquerdos foram fixados por imersão durante
duas a três horas em paraformoldeído. Depois de fixado, o material foi colocado para
desidratação em banhos de concentrações crescentes de álcool etílico (70%, 80%, 90% e três
vezes 100%) durante uma hora para cada banho. Em seguida foi submetido a uma mistura de
álcool-xilol (1:1) por uma hora, diafanizado em três banhos de xilol de 40 minutos cada e
posteriormente incluído em três banhos de parafina de uma hora cada. As amostras foram
então emblocadas individualmente em parafina usando-se barras de Leuckhart. Os blocos
foram cortados com espessura de cinco micrômetros em micrótomo Spencer ‘820’ e os cortes
colocados em estufa, por 24 horas, para que aderissem às lâminas. Foram feito três
colorações, Hematoxilina/Eosina, Tricrômico de Masson e Taenzer-Unna (orceína). Para HE
as lâminas foram colocadas em uma seqüência de banhos, de um minuto cada, de: xilol puro
(três vezes), álcool/xilol (1:1), álcool etílico 100%, 90%, 80% e 70%, Hematoxilina, água
corrente, Eosina, álcool 70%, 80%, 90%, álcool/xilol (1:1), xilol puro (três vezes) Para o

25
Tricômica de Masson as lâminas foram colocadas em uma seqüência de banhos, de um
minuto cada, de xilol puro (três vezes), álcool/xilol (1:1), álcool etílico 100%, 90%, 80% e
70%, Hematoxilina Férrica, água corrente, Ponceau, água corrente, ácido fosfotúngstico-
ácido fosfomolíbdico, azul de anilina, álcool 70%, 80%, 90%, álcool/xilol (1:1), xilol puro
(três vezes). Para o Taenzer-Unna (orceína) as lâminas foram colocadas em uma seqüência
de banhos, de um minuto cada, de xilol puro (três vezes), álcool/xilol (1:1), álcool etílico
100%, 90%, 80% e 70%, orceína, álcool 70%, solução saturada de ácido pícrico, água
corrente, álcool 70%, 80%, 90%, álcool/xilol (1:1), xilol puro (três vezes), e montadas em
meio Bálsamo sintético do Canadá. Posteriormente foram analisadas e fotomicrografadas em
microscópio Axioskop da Zeiss.
4.6 Morfometria
Foram cortados três lâminas (corte semi-seriado) de cada bloco de parafina contendo
os três fragmentos de pulmão dos diferentes grupos, experimentais e controle, de modo a
obter de cada grupo experimental uma coleção de 18 lâminas. Foram escolhidos as três
laminas de três animais de cada coleção, uma do início, outra intermediaria e a terceira da
parte final da coleção. Os cortes foram quadriculados com a ajuda de uma fotocópia em
transparência com um dado não viciado de 20 faces foram sorteados três quadrantes de cada
corte, que foram então fotografados em microscópio de luz Zeiss Axioskop acoplado a uma
câmera CCD-IRIS, utilizando uma objetiva de 40x. Desse modo, a área do tecido pulmonar
captura em cada fotografia era a mesma para todos os quadrantes, ou seja, 77.289 mm². As
imagens fotografadas tiveram o número de núcleos totais de cada imagem contado com o
auxilio do programa ImagePro 4.0, e assim determinada a densidade celular de cada grupo
experimental. Para a determinação da densidade nuclear foram contados os núcleos de células
epiteliais alveolares, como pneumócitos do tipo I e II, células inflamatórias, macrófagos
alveolares e células endoteliais de capilares.
4.7 Processamento do material para ELISA
Foram pesados 100 mg do pulmão direito e homogeneizados em 0.9 mL de uma
solução para extração de citocinas (NaCl 0.4 M, NaPO4 10 mM, PMSF 0,1 mM, cloreto de
benzetonio 0.1 mM, EDTA 10 mM, tween 20 0.05%, 0.5% de BSA, 20 mM KI, aprotinina),
numa relação de 10% peso/volume, e posteriormente centrifugados a 3000 RPM durante 15

26
minutos a 4º C. O sobrenadante foi utilizado para detecção de citocinas e quimiocinas
teciduais pelo método de ELISA. A detecção das atividades de NAG e MPO foram feitas no
pellet, após processamento das amostras para estes ensaios. Desta maneira, o pellet foi
ressuspendido em 2 mL de tampão fosfato (0.1 M NaCl, 0.02 M NaPO4, 0.015 M NaEDTA;
pH 4.7), homogeneizado por 1 minuto; 1,5 mL de NaCl 0.2% foi adicionado e
homogeneizado novamente; 1,5 mL de NaCl 1.6% com 5% de glicose foi adicionado,
homogeneizado e vortexado. Após esta etapa, o volume total (1.6 mL) foi dividido em dois
tubos, 0.8 mL para cada tubo, para NAG e MPO respectivamente. Estas amostras foram
centrifugadas a 4º C, por 15 minutos, a uma velocidade de 10.000 RPM e o sobrenadante foi
posteriormente descartado. Para o ensaio de NAG, as amostras foram ressuspendidas e
homogeneizadas em 0.8 mL de salina 0.9% com 0.1% v/v de Triton X-100, centrifugadas a 4º
C, por 10 minutos a 1.500 RPM e o ensaio foi feito com o sobrenadante destas amostras. Para
o ensaio de MPO, as amostras foram ressuspendidas e homogeneizadas em 0.8 mL de tampão
fosfato (0.05 M Na3PO4, 0.5% HETAB; pH 5.4), congeladas em nitrogênio líquido e então
descongeladas, sendo este processo repetido por 3 vezes. Após esta etapa, as amostras para
MPO foram centrifugadas a 4º C, por 15 minutos a 10.000 RPM e o sobrenadante utilizado
para o ensaio.
4.8 Ensaio para detecção dos níveis de N-acetilglicosaminidase (NAG) tecidual
A reação foi iniciada pela adição de 100 µL de p-nitrofenil-N-acetil-β-D-
glicosaminidina (Sigma), diluídos em tampão citrato/fosfato (acido cítrico 0,1 M, Na2HPO4
0,1 M, pH 4.5) na concentração final de 2.24 mM, a 100 µL de sobrenadante das amostras
previamente processadas e diluídas em tampão citrato/fosfato, em placas de 96 poços, por 10
minutos a 37º C. Ao final desta etapa adicionou-se 100 µL de tampão glicina 0.2 M (pH 10.6)
para término da reação. As placas de 96 poços foram lidas em leitor de ELISA (Emax,
Molecular Devices) à 405 nm. O conteúdo de macrófagos foi calculado com base na curva
padrão de NAG, feita pela coleta de macrófagos peritoneais recolhidos de animais
estimulados com 3 mL de tioglicolato 3% (dados não mostrados).
4.9 Ensaio para detecção dos níveis de mieloperoxidase (MPO) tecidual
O ensaio utiliza 25 µL de 3,3’-5,5’-tetrametilbenzidina (TMB, Sigma) dissolvidos
em dimetilsulfoxido (DMSO, Merck) com concentração final de 1.6mM, 100 µL de tampão
fosfato (0.05 M Na3PO4, 0.5% H-TAB; pH 5.4) com H2O2 diluído numa concentração final de

27
0.003% v/v, e 25 µL de sobrenadante das amostras de tecidos previamente processadas. A
reação se inicia pela adição de tetrametilbenzidina as amostras em placas de 96 poços, a 37º C
por 5 minutos. Após esta etapa, adicionou-se H2O2 diluída em tampão fosfato e encubou-se a
37º C por 5 minutos. Ao final desta etapa adicionou-se 100 µL de H2SO4 4M para término da
reação. As reações foram lidas em leitor de ELISA (Emax, Molecular Devices) à 450 nm. O
conteúdo de neutrófilos foi calculado com base em curva padrão de MPO, feita pela coleta de
neutrófilos peritoneais recolhidos de animais estimulados com 3 mL de caseína 5% (dados
não mostrados).
4.10 Detecção de TGF-β1
Para a dosagem TGF-beta1 foi utilizado kit específico (BD OptEIA) onde foram
seguidas as instruções do fabricante. Basicamente, 100 µL/poço do anticorpo de captura (5,5
µL/mL) diluído em PBS estéril foram adicionados à cada placa (NUNC FLAT, 96-well,
FALCON). Estas foram vedadas e incubadas a 4ºC overnight. O conteúdo de cada placa foi
retirado e esta foi lavada 3 vezes (300 µL/poço) com um tampão de lavagem (PBS/TWEEN
20 0,05%). Após este procedimento, adicionou-se 300 µL/poço do tampão de bloqueio (BSA
1% em PBS). As placas foram incubadas à temperatura ambiente por no mínimo uma hora.
Repetiu-se o procedimento de lavagem e adicionou-se 100 µL/poço das amostras, lavado
bronco-alveolar ou tecido, e padrões, diluídos em tampão de diluição (BSA 0,1% em PBS
1x). As placas foram novamente incubadas overnight. Repetiu-se o procedimento de lavagem
e foram adicionados 100µL/poço do anticorpo de detecção biotinilado (5,55 µL/mL) diluído
em tampão de diluição (o mesmo das amostras e padrões). As placas foram incubadas à
temperatura ambiente por 2 horas. Após mais um procedimento de lavagem, adicionou-se 100
µL/poço de estreptavidina – HRP 1:200 em tampão de diluição e incubou-se as placas à
temperatura ambiente por 20 minutos. Novamente as placas foram lavadas e adicionou-se 100
µL/poço do substrato OPD (em tampão citrato pH 5 com adição de H2O2). As placas foram
incubadas por 20-30 minutos, à temperatura ambiente, protegidas da luz. Após este período
observou-se a formação de cor e foram adicionados 50 µL/poço de H2SO4 1M para
interromper a reação. As placas foram lidas em leitor de ELISA (Emax, Molecular Devices) à
492 nm.
4.11 Análise estatística

28
Os dados foram analisados estatisticamente por análise de variância (One-way
ANOVA), seguida do teste Newman-Keuls, e as diferenças foram consideradas
estatisticamente significativas se p < 0.05. Os dados foram expressos em média ± EPM. As
análises estatíticas e os gráficos foram elaborados utilizando-se o software GraphPad Prism
3.0.
A determinação das diferenças estatísticas nas freqüências dos diferentes tipos
celulares, na contagem total e diferencial sangüínea e na densidade celular foi feita por análise
de variância com teste estatístico de PLSD de Fischer (para N regular em todos os grupos) ou
Scheffe (para N irregular em alguns grupos), com o programa estatístico Statview. O nível de
significância adotado em todos os testes foi de 5% (p < 0,05).

29
5 RESULTADOS
5.1 Análise da massa corpórea dos camundongos
Realizada a análise de variância dos pesos iniciais dos camundongos, constatou-se
que não existe diferença estatística significativa entre os grupos, o que demonstra
homogeneidade entre as unidades experimentais. Os valores médios do peso inicial e desvio
padrão podem ser visualizados na tabela 1.
TABELA 1- Comparação entre as massas corpóreas iniciais e finais dos camundongos
após dez dias de tratamento.
Media do peso inicial
Media do peso final
Diferença P
Grupo Salina/Salina – SS 31.83±2.40 31.00±1.78 0.83 0,010 Grupo -Salina/Óleo – SO 29.33±1.86 27.00±0.89 2.33 0,005 Grupo -Bleomicina/Salina – BS 32.17±2.48 29.67±1.36 2.50 0,010 Grupo -Bleomicina/Óleo – BO 28.50±2.48 24.67±1.36 3.83 0,006 Salina/Salina – SS; Salina/Óleo – SO; Bleomicina/Salina – BS; Bleomicina/Óleo – BO.
A comparação dos valores médios do peso dos camundongos, no início e ao final do
tratamento, mostra que ocorreu perda de massa corpórea em todos os grupos testados.
Entretanto vale ressaltar que os camundongos que receberam tratamento com Salina/Salina
(SS) apresentaram a menor perda de massa corpórea, que inclusive difere significativamente
da maior perda de massa corpórea registrada nos camundongos tratados com Bleomicina/Óleo
(BS) (TABELA 1).
Entre os camundongos tratados com BS e aqueles tratados com SO não se registrou
diferença estatística significativa na perda de massa corpórea; como também não se constatou
diferença estatística entre esses tratamentos e o tratamento considerado controle.
Os resultados mostram que os animais dos grupos que receberam o óleo de copaíba
emagreceram mais que os animais dos outros grupos.
30
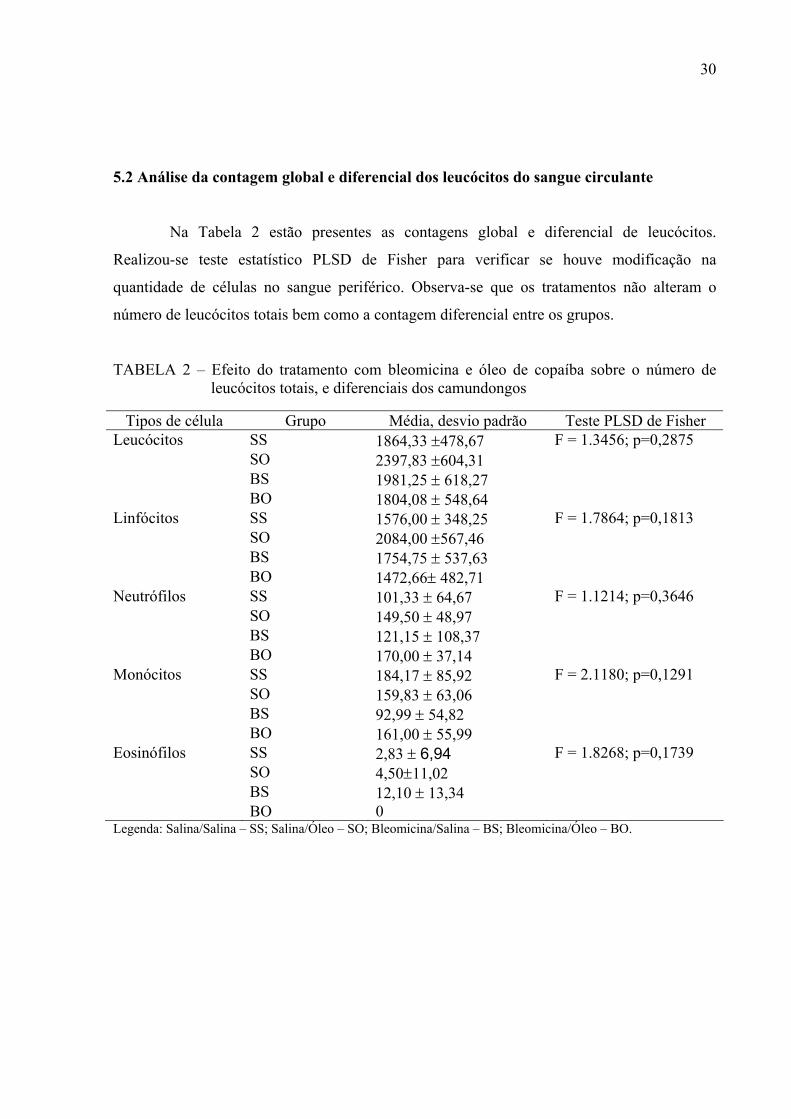
5.2 Análise da contagem global e diferencial dos leucócitos do sangue circulante
Na Tabela 2 estão presentes as contagens global e diferencial de leucócitos.
Realizou-se teste estatístico PLSD de Fisher para verificar se houve modificação na
quantidade de células no sangue periférico. Observa-se que os tratamentos não alteram o
número de leucócitos totais bem como a contagem diferencial entre os grupos.
TABELA 2 – Efeito do tratamento com bleomicina e óleo de copaíba sobre o número de leucócitos totais, e diferenciais dos camundongos
Tipos de célula Grupo Média, desvio padrão Teste PLSD de Fisher
Leucócitos SS 1864,33 ±478,67 F = 1.3456; p=0,2875 SO 2397,83 ±604,31 BS 1981,25 ± 618,27 BO 1804,08 ± 548,64 Linfócitos SS 1576,00 ± 348,25 F = 1.7864; p=0,1813 SO 2084,00 ±567,46 BS 1754,75 ± 537,63 BO 1472,66± 482,71 Neutrófilos SS 101,33 ± 64,67 F = 1.1214; p=0,3646 SO 149,50 ± 48,97 BS 121,15 ± 108,37 BO 170,00 ± 37,14 Monócitos SS 184,17 ± 85,92 F = 2.1180; p=0,1291 SO 159,83 ± 63,06 BS 92,99 ± 54,82 BO 161,00 ± 55,99 Eosinófilos SS 2,83 ± 6,94 F = 1.8268; p=0,1739 SO 4,50±11,02 BS 12,10 ± 13,34 BO 0 Legenda: Salina/Salina – SS; Salina/Óleo – SO; Bleomicina/Salina – BS; Bleomicina/Óleo – BO.
31
S/S B/S S/O B/O0.00.51.01.52.02.53.03.54.04.5
Núm
ero
de c
élul
as/m
m2
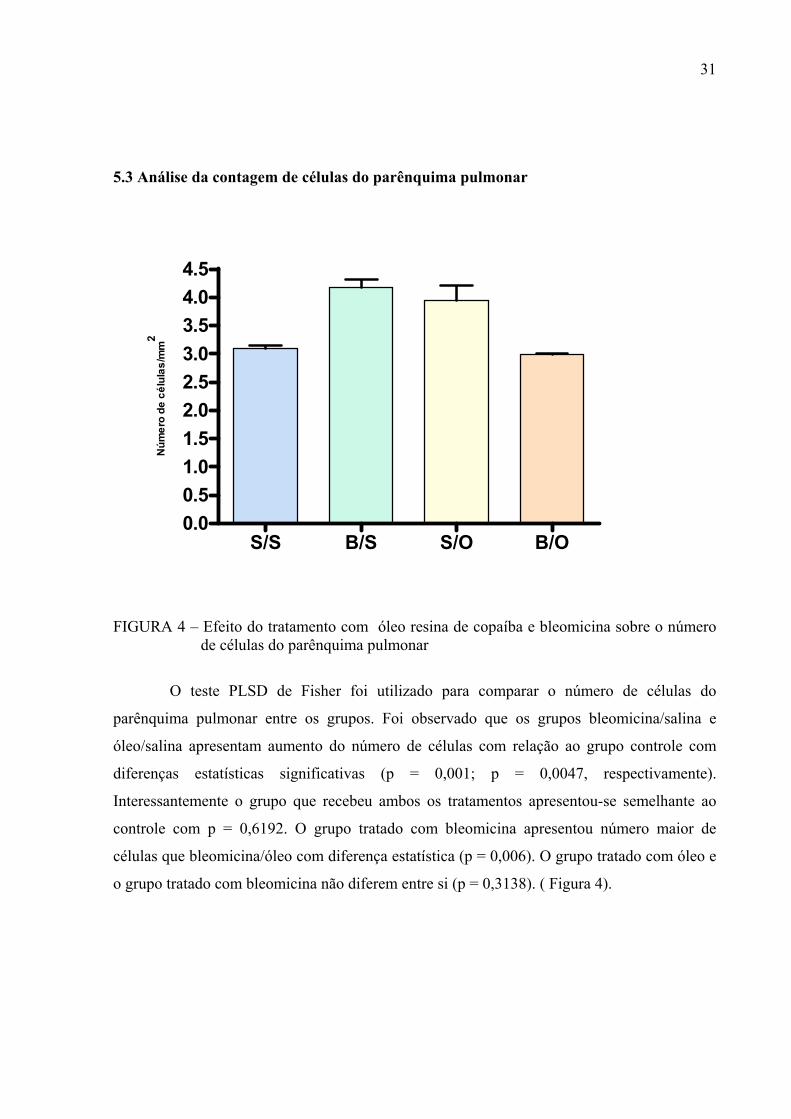
5.3 Análise da contagem de células do parênquima pulmonar
FIGURA 4 – Efeito do tratamento com óleo resina de copaíba e bleomicina sobre o número
de células do parênquima pulmonar
O teste PLSD de Fisher foi utilizado para comparar o número de células do
parênquima pulmonar entre os grupos. Foi observado que os grupos bleomicina/salina e
óleo/salina apresentam aumento do número de células com relação ao grupo controle com
diferenças estatísticas significativas (p = 0,001; p = 0,0047, respectivamente).
Interessantemente o grupo que recebeu ambos os tratamentos apresentou-se semelhante ao
controle com p = 0,6192. O grupo tratado com bleomicina apresentou número maior de
células que bleomicina/óleo com diferença estatística (p = 0,006). O grupo tratado com óleo e
o grupo tratado com bleomicina não diferem entre si (p = 0,3138). ( Figura 4).
32
5.4 Aspectos morfológicos do parênquima pulmonar
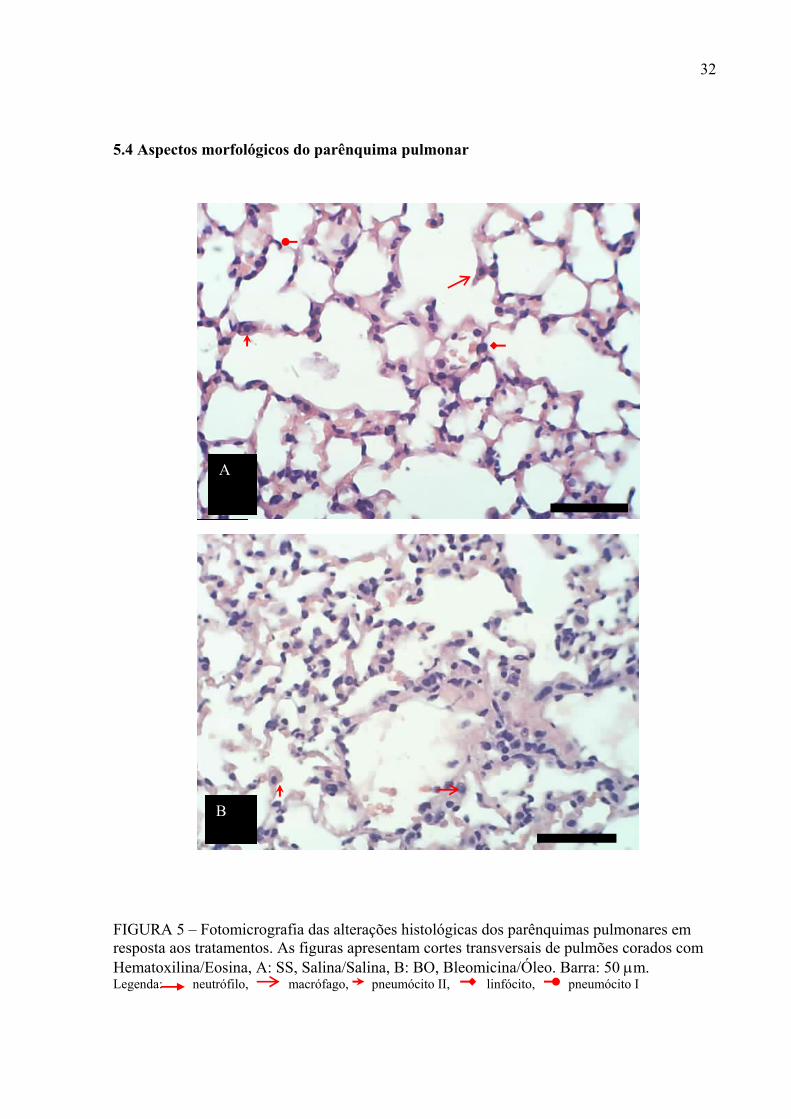
FIGURA 5 – Fotomicrografia das alterações histológicas dos parênquimas pulmonares em resposta aos tratamentos. As figuras apresentam cortes transversais de pulmões corados com Hematoxilina/Eosina, A: SS, Salina/Salina, B: BO, Bleomicina/Óleo. Barra: 50 µm. Legenda: neutrófilo, macrófago, pneumócito II, linfócito, pneumócito I
A
B
A
33
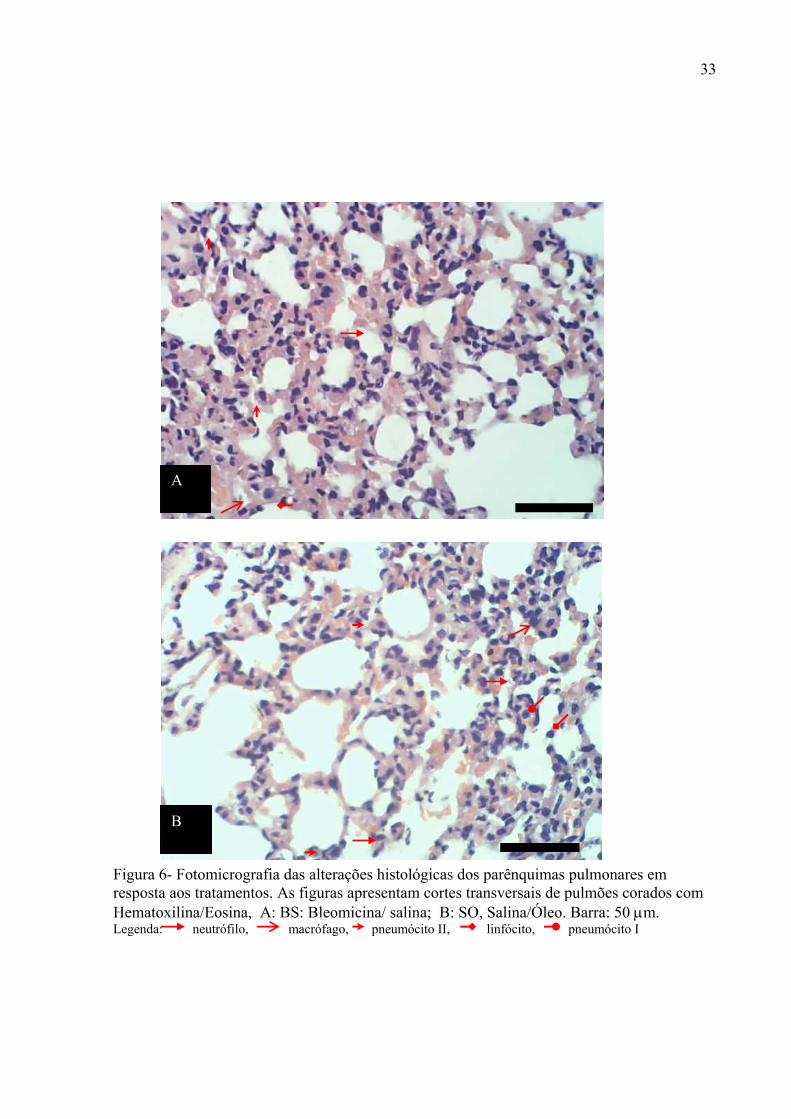
Figura 6- Fotomicrografia das alterações histológicas dos parênquimas pulmonares em resposta aos tratamentos. As figuras apresentam cortes transversais de pulmões corados com Hematoxilina/Eosina, A: BS: Bleomicina/ salina; B: SO, Salina/Óleo. Barra: 50 µm. Legenda: neutrófilo, macrófago, pneumócito II, linfócito, pneumócito I
A
B

34
As figuras 5 e 6 mostram os efeitos dos tratamentos sobre o parênquima pulmonar
dos camundongos. Os animais controle SS apresentou o parênquima pulmonar normal, com
alvéolos revestidos por um epitélio constituído por dois tipos de células: pneumócitos tipo I e
pneumócitos tipo II. Os septos interalveolares estão revestidos por epitélio alveolar e se
mostram ora extremamente delgado, organizado apenas pelas células epiteliais alveolares e a
lâmina basal e ora um pouco mais espesso incluindo elementos do tecido conjuntivo como
fibras elásticas, finas fibrilas colágenas, fibroblastos e raros macrófagos (Figura 5A).
Os animais tratados com bleomicina o parênquima pulmonar apresenta aspecto típico
do estágio inicial de fibrose com áreas focais de desorganização do tecido, com grande
quantidade de células inflamatórias como neutrófilos e macrófagos, dispersos pelo
parênquima e próximos aos vasos sanguíneos, congestão septal, edema e hemorragia intra
alveolar, ruptura dos septos, macrófagos livres no espaço alveolar, e vasos congestos.
Apresentam também algumas áreas com afilamento dos septos e dilatação alveolar (Figura
5B).
Os animais tratados com óleo de copaíba apresentaram o parênquima pulmonar
semelhante aos dos animais BS, é observado aumento da quantidade de células inflamatórias
mononucleares e polimorfonucleares com espessamento dos septos interalveolares, edema
agudo, hemorragia, vasos congestos, estreitamento alveolar na região central e dilatação
alveolar na periferia lobular (Figura 6A).
Os animais tratados com bleomicina e óleo apresentou áreas com parênquima normal
alternados com áreas com espessamento septal com presença de fibras elásticas e fibras
colágenas, fibroblastos e raros macrófagos e polimorfonucleares (Figura 6B).
5.5 Análise dos mediadores químicos
Efeito dos tratamentos sobre a quantidade de TGFβ1, MPO e NAG sobre o
parênquima pulmonar.
35
S/S B/S S/O B/O0
1000
2000*
**
* P < 0.05
TGF-β 1
(pic
ogra
ms)
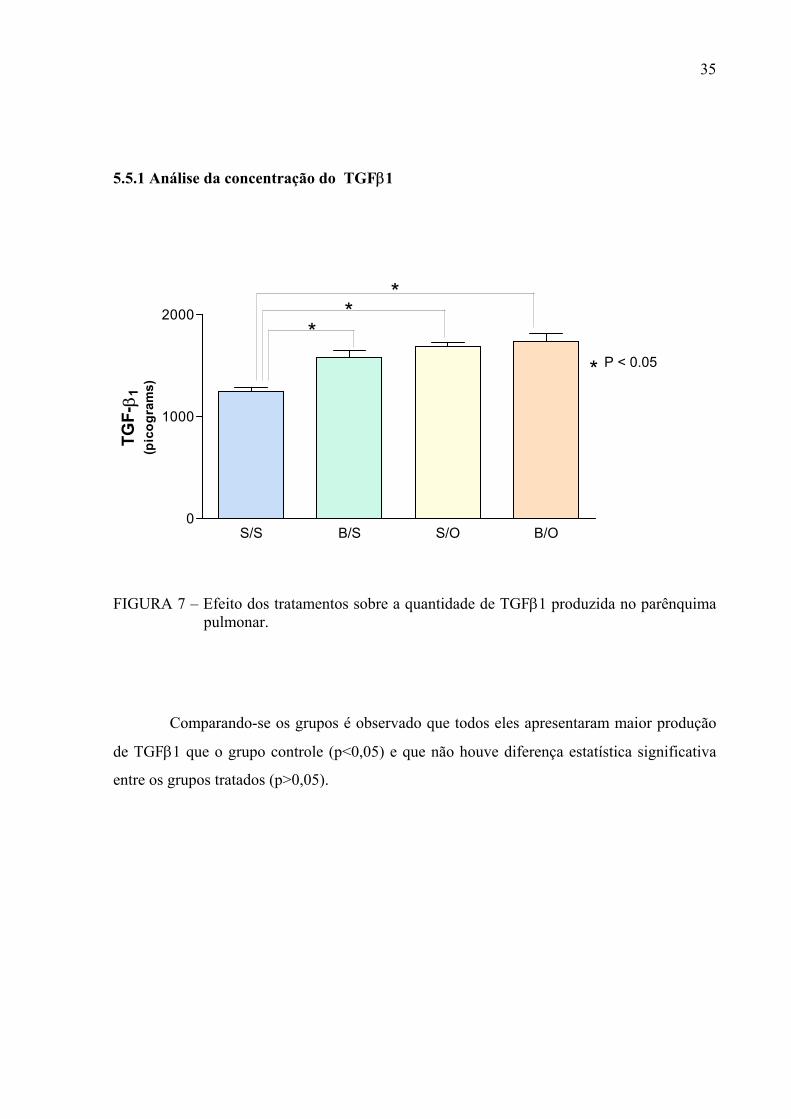
5.5.1 Análise da concentração do TGFβ1
FIGURA 7 – Efeito dos tratamentos sobre a quantidade de TGFβ1 produzida no parênquima
pulmonar.
Comparando-se os grupos é observado que todos eles apresentaram maior produção
de TGFβ1 que o grupo controle (p<0,05) e que não houve diferença estatística significativa
entre os grupos tratados (p>0,05).
36
S/S B/S S/O B/O0
250
500
750
1000
1250
***
P < 0.01P < 0.05
**** * *
Neu
trof
ilos
(núm
ero
de c
el x
105 ) /
100m
g te
cido
pul
mon
ar
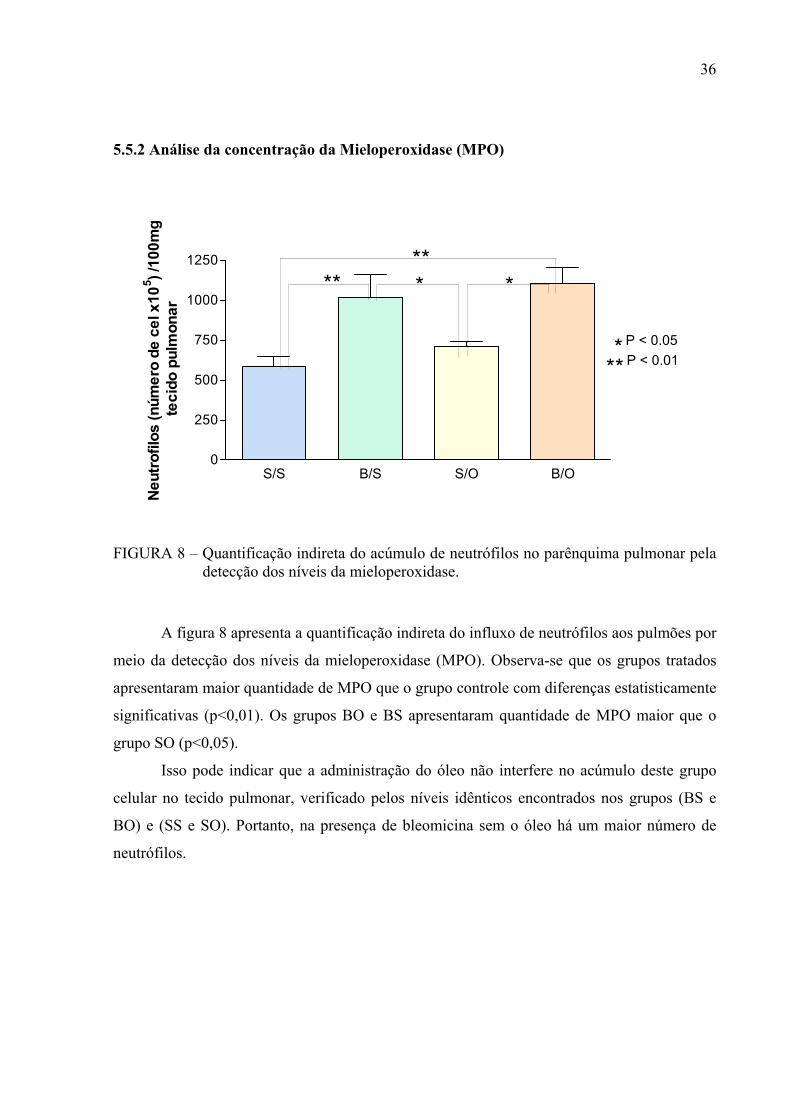
5.5.2 Análise da concentração da Mieloperoxidase (MPO)
FIGURA 8 – Quantificação indireta do acúmulo de neutrófilos no parênquima pulmonar pela
detecção dos níveis da mieloperoxidase.
A figura 8 apresenta a quantificação indireta do influxo de neutrófilos aos pulmões por
meio da detecção dos níveis da mieloperoxidase (MPO). Observa-se que os grupos tratados
apresentaram maior quantidade de MPO que o grupo controle com diferenças estatisticamente
significativas (p<0,01). Os grupos BO e BS apresentaram quantidade de MPO maior que o
grupo SO (p<0,05).
Isso pode indicar que a administração do óleo não interfere no acúmulo deste grupo
celular no tecido pulmonar, verificado pelos níveis idênticos encontrados nos grupos (BS e
BO) e (SS e SO). Portanto, na presença de bleomicina sem o óleo há um maior número de
neutrófilos.
37
S/S B/S S/O B/O0
10
20
30
40
50
60
70*****
***** P < 0.001
P < 0.01
Mac
rofá
gos
(núm
ero
de c
els
x104 ) /
100m
gte
cido
pul
mon
ar
5.5.3 Análise da N-acetilglicosaminidase (NAG)
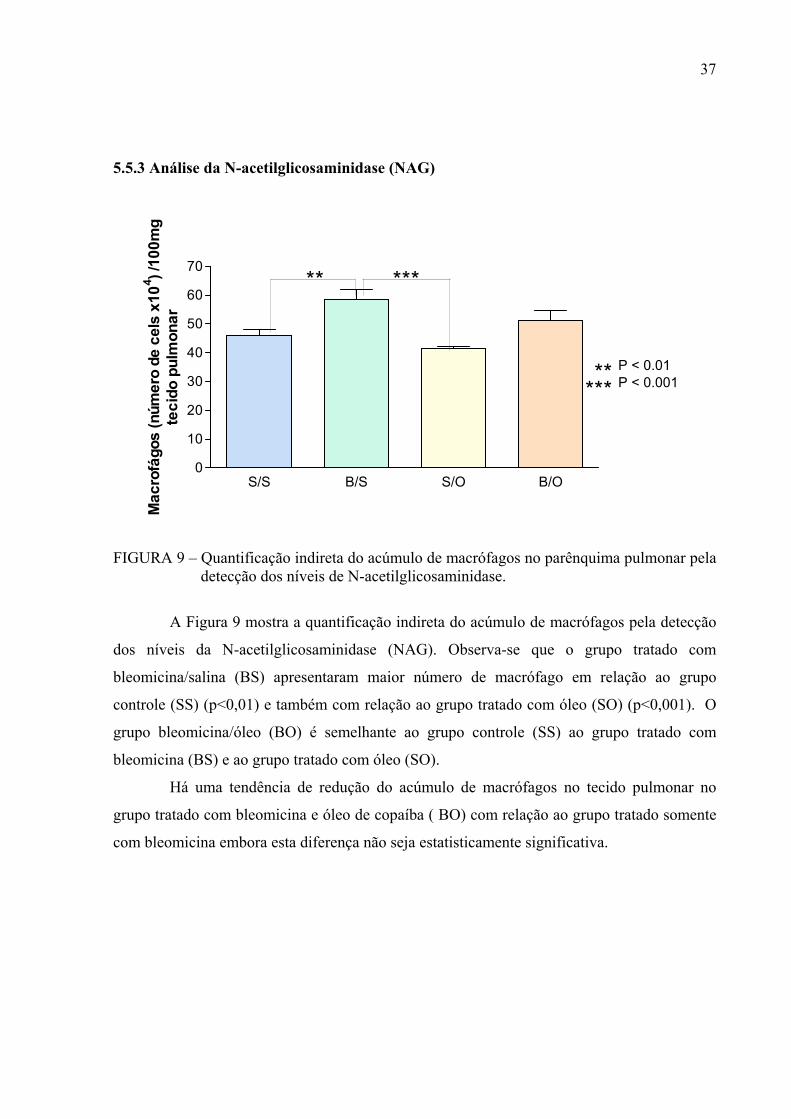
FIGURA 9 – Quantificação indireta do acúmulo de macrófagos no parênquima pulmonar pela detecção dos níveis de N-acetilglicosaminidase.
A Figura 9 mostra a quantificação indireta do acúmulo de macrófagos pela detecção
dos níveis da N-acetilglicosaminidase (NAG). Observa-se que o grupo tratado com
bleomicina/salina (BS) apresentaram maior número de macrófago em relação ao grupo
controle (SS) (p<0,01) e também com relação ao grupo tratado com óleo (SO) (p<0,001). O
grupo bleomicina/óleo (BO) é semelhante ao grupo controle (SS) ao grupo tratado com
bleomicina (BS) e ao grupo tratado com óleo (SO).
Há uma tendência de redução do acúmulo de macrófagos no tecido pulmonar no
grupo tratado com bleomicina e óleo de copaíba ( BO) com relação ao grupo tratado somente
com bleomicina embora esta diferença não seja estatisticamente significativa.

38
6 DISCUSSÃO
A fibrose pulmonar induzida por bleomicina em camundongos é um modelo
experimental utilizado em pesquisas por serem capazes de desenvolver a doença de maneira
semelhante ao observado em seres humanos (ADAMSON; BOWDEN, 1974). No presente
trabalho, a fibrose pulmonar induzida pela bleomicina ficou evidenciada histologicamente
pelo aumento do número de células do parênquima pulmonar bem como pelo espessamento
da parede alveolar pela deposição de colágeno e bioquimicamente pelo aumento da migração
celular por meio da concentração de mieloperoxidase (MPO), n-acetilglicosaminidase (NAG),
TGFβ.
O desenvolvimento da fibrose pulmonar induzida por bleomicina inicia–se por
edema pulmonar, exsudato intra-alveolar e infiltração dos septos alveolares por células
mononucleares e discreta hiperplasia de pneumócitos tipo II seguida do aumento do tecido
fibroso pela proliferação de fibroblastos, perda do equilíbrio no remodelamento da matriz
extracelular levando à deposição de colágeno com espessamento dos septos alveolares e grau
variável de inflamação (AZAMBUJA et al., 2005).
A migração de células como os neutrófilos e macrófagos são importantes para o
desenvolvimento da fibrose pulmonar. Os macrófagos são fontes de TGFβ, uma potente
citocina pró-fibrótica, que desempenha um papel central no estabelecimento da doença.
Dentre seus efeitos está o estímulo para a migração de linfócitos e fibroblastos para o local da
injúria (BRANTON; KOOP, 1999). Além disso, Broekelmann, et al. (1991) demonstraram
que o TGFβ é responsável pelo estimulo para a produção de colágeno, fibronectina, e actina
de músculo liso semelhante ao que é encontrado em uma cicatrização de feridas, ocasionando
a fibrose pulmonar.
A comparação entre as massas corpóreas dos grupos tratados mostra que aqueles que
receberam bleomicina e os que receberam óleo emagreceram igualmente. Porém, o grupo que
recebeu os dois tratamentos foi o que apresentou maior perde de massa corpórea. Estudos
relatam que a bleomicina causa emagrecimento em animais tratados pela perda de apetite

39
levando à deficiência nutricional (TAOOKA et al., 1997).
Neste trabalho foi utilizado o óleo resina em sua forma bruta talvez por isso tenha
havido perturbações gastrointestinais nos camundongos levando-os ao emagrecimento.
A capacidade antiinflamatória do óleo resina de copaíba sobre a fibrose pulmonar foi
testada frente a administração do óleo resina concomitante com a bleomicina. O grupo tratado
apresentou uma redução do processo inflamatório caracterizado pela diminuição do
recrutamento celular, porém, houve uma maior produção da enzima mieloperoxidase
indicando um aumento do número de neutrófilos.
O aumento da atividade mieloperoxidase observada neste estudo sugerem também o
aumento do stress oxidativo elicitado pela bleomicina e pelo óleo de copaíba.
Quanto ao número de macrófagos nossos resultados indicaram que o óleo
administrado juntamente com a bleomicina reduziu o acúmulo de macrófagos no tecido
pulmonar sugerido que o óleo pode modular a quimiotaxia deste tipo celular para o
parênquima pulmonar e estimular a produção de TGFβ.
Dentre as propriedades medicinais atribuídas ao óleo resina de copaíba a ação
antiiflamatória é uma das mais investigadas. Estudos utilizando modelos de inflamação como
edema de pata de rato induzido por carragenina e bradicinina bem como teste granulomatoso
induzido por óleo de cróton mostram seu efeito antiinflamatório (VEIGA et al., 2000). A
utilização do ácido caurenóico presente no óleo de copaíba quando utilizado concomitante
com ácido acético para indução de colite mostrou a diminuição de infiltrado de células
inflamatórias e de edema da mucosa intestinal (PAIVA et al., 2003). Além disso, a medicina
popular utiliza óleo de copaíba como antiinflamatório de vias aéreas superiores (TAPPIN et
al., 2003).
Recentemente, pesquisadores da USP conseguiram isolar e encapsular componentes
voláteis deste óleo resina. Testes com estas cápsulas em pleurisia induzida em camundongo e
também em edema de pata mostraram que são estes os componentes que apresentam ação
antiinflamatória. Além disso, mostrou também que a ação antiinflamatória da fração volátil é
mais potente que a do diclofenaco, um fármaco muito utilizado (Agencia USP de notícias,
2006).
Curiosamente, em nosso trabalho, quando camundongos saudáveis foram tratados
com o óleo resina observou-se maior recrutamento celular e regiões focais de fibrose e
enfizema, que puderam ser visualizados na microscopia óptica e análise morfométrica,
sugerindo, portanto, que o óleo por si pode desencadear uma resposta inflamatória nos
pulmões contrariando o que está dito na literatura até o presente momento.

40
Os grupos tratados com bleomicina apresentaram maior quantidade de neutrófilos,
mostrado, pelos níveis de MPO. Essas células podem migrar para os pulmões pela
quimiotaxia provocada pelos efeitos da bleomicina (SATO et al., 1999), no entanto, o grupo
tratado com óleo resina de copaíba apresentou os níveis de MPO semelhante ao grupo
controle (SS), sugerindo que o óleo resina não apresenta por si só um menor efeito
inflamatório.
Neste trabalho, tanto a bleomicina quanto o óleo elevaram a quantidade de TGFβ
quando comparados ao controle, sugerindo que esse mediador seja um dos responsáveis pelo
processo inflamatório nessa fase do desenvolvimento da fibrose pulmonar. Em pequena
quantidade o TGFβ1 é benéfico numa resposta inflamatória, mas em níveis elevados pode
contribuir para a patogênese da fibrose (BRANTON; KOOP, 1999). Estes dados indicam que
a administração do óleo resina concomitante com o bleomicina modula a resposta imune
colaborando para a diminuição da resposta inflamatória.
Este é o primeiro estudo relatando o efeito do óleo resina de copaíba sobre o pulmão
de camundongos saudáveis e com fibrose pulmonar. Os estudos que existem sobre o efeito
antiinflamatório do óleo são realizados em modelos de edema, diferente de fibrose pulmonar,
o que não é comparável, e também, estudo utilizando outros óleos essenciais com os mesmos
componentes também não relatam seus efeitos sobre o parênquima pulmonar.

41
7 CONCLUSÕES
- A utilização do óleo resina de copaíba a 0,1% da massa corpórea concomitante
com a indução da fibrose pulmonar pela bleomicina, durante 10 dias, não impede
desenvolvimento da fibrose pulmonar, mas atenua o seu desenvolvimento;
- O óleo resina de copaíba administrado sozinho na dose de 0,1% da massa corpórea,
durante 10 dias, estimula o surgimento de processo inflamatório nos pulmões;
- Sugere-se que o próximo passo a ser pesquisado seja utilizar frações isoladas do
óleo de copaíba com propriedades antiinflamatórias para investigar sua ação sobre a fibrose
pulmonar induzida por bleomicina.

42
REFERÊNCIAS
BASILE, A. C.; SERTIE, J. A.; FREITAS, P. C.; ZANINI, A. C. Anti-inflammatory activity of oleoresin from Brazilian Copaifera. Ethonopharmacol. Journal of ethnopharmacology. Ireland, v. 22, n. 1, p. 101- 109, 1988.
BROEKELMANN., T. J; LIMPER.;A. H.; COLBY.; T. V.; MCDONALD, J.A. Transforming growth factor beta 1 is present at sites of extracellular matrix gene expression in human pulmonary fibrosis. Proc Natl Acad Sci. USA, Aug, ano 1, v. 88, n°15, p.6642-6, 1991. CAVALCANTI, B. C.; COSTA-LOTUFO, L.V. ; MORAES, M. O.; BURBANO, R. R.; SILVEIRA, E. R.; CUNHA, K.M.A.A.; . RAO V.S.N.; MOURA D.J.; ROSA R.M. D.; HENRIQUES, J.A.P. PESSOA, C. Genotoxicity evaluation of kaurenoic acid, a bioactive diterpenoid present in Copaiba oil. Food and Chemical Toxicology v.44, p. 388–392. Disponível em: www.elsevier.com/locate/foodchemtox. 2006. CARVALHO, J. C. T.; CASCON, V. ; POSSEBON, L. S.; MORIMOTO, M. S.; CARDOSO L.G.V. Topical Antiinflammatory and Analgesic Activities of Copaifera duckei Dwyer.Phytother. Res. v19, p.946–950). Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/ptr.1762. 2005.
COSTABEL, U.; KING T, E. International consensus statement on idiopathic pulmonary fibrosis. Eur Respir J. Stanford University, USA, v.17, p.163–167, 2001.
COTRAN, R S.; KUMAR, V.; COTRAN, R.S. Robbins/Patologia estrutural e funcional. 6ªed., São Paulo: Guanabara-Koogan, 2000.
CROUCH, E. C. Pathobiology of pulmonary fibrosis. Am J Physiol- Lung, Stanford University Libraries, USA, vol. 259, p. 159–184, 1990.
DUNSMORE,S.E.; ROES,J.; CHUA,J.F.; SEGAL,A.W.; MUTSAERS,E.S.; LAURENT,G.J. Evidence that neutrophil elastase-deficient mice are resistant to bleomycin induced fibrosis. Chest. Salt Lake City, Utah, vol. 120, p. 35-36, 2001.

43
FILDERMAN, A. E., GENOVESE, L. A., LAZO, J. S. Alteration in pulmonary protective enzymes following systemic bleomycin treatment in mice. Biochem. Pharmacol. Elsevier, v.37, p.1111 – 1116, 1988.
GIRI, S. N.; HYDE, D. M.; HOLLINGER, M. A. Effect of antibody to transforming growth factor ß on bleomycin induced accumulation of lung collagen in mice. Thorax. Tavistock Square, London, v.48, p.959-966.1993. HIRANO, E. S.; MANTOVANI, M, MORANDIN, R. C. Isquemia e reperfusão hepática total em condições de normalidade e sob estado de choque hemorrágico controlado: efeitos no sequestro de neutrófilos no rim do rato. Acta Cir Bras. São Paulo, jul-ago.2004. Disponível em: http://www.scielo.br/acb. Acesso em 20 jun.2005.
KATZENSTEIN, A-LA, MYERS, J. Idiopathic pulmonary fibrosis: clinical relevance of pathologic classification. Am J Respir Crit Care Med. Stanford University Libraries, v.157, p.1301-1315. 1998.
LAZO, J. S.; HUMPHREYS, C. J. Lack of metabolism as the biochemical basis of bleomycin-induced pulmonary toxicity (bleomycin hydrolase/high-pressure liquid chromatography/lung fibrosis/fluorescence/aminopeptidase). Proc Natl Acad Sci. USA, Vol. 80, p. 3064-3068, 1983.
LEONARD, S. S.; MOWREY, K.; PACK, D.; SHI, X.; CASTRANOVA, V.; KUPPUSAMY, P.; VALLYATHAN, V. In vivo bioassays of acute asbestosis and its correlation with ESR spectroscopy and imaging in redox status. Mol Cell Biochem. London, v. 234-235, (1-2), p.369-77. 2002.
LI, Y. J. et al. EM703 improves bleomycin-inducede pulmonary fibrosis in mice by hte inhibition OF TGFβ signaling in lung fibroblasts. Respiratory Research. London, v.7, p. 2, 2006.
LIMA, S. R. M.; FERNANDES, P. D.; VEIGA JR., V. F.; PINTO, A. C. III Reunião da Sociedade Latino-Americana de Fitoquímica, Gramado, Brasil, 1999.
MARTINEZ, J. A. B. Doenças intersticiais pulmonares. Medicina. Ribeirão Preto, v.31, p.247-256, abr./jun.1998.
MATTOS, M.D.; KIMURA,E.T.; SILVA,M.R.R.; EGAMI,M.I.; SEGRETO,R.A.; SEGRETO,H.R.C.Ativação da proteína TGFβI latente em pulmão irradiado in vivo. Rev. Assoc. Med. Bras. São Paulo, v. 48, p.329-340, 2002.

44
MCANULTY, R. J.; LAURENT, G.J. Collagen and its regulation in pulmonary fibrosis. In Lung Biology in Health and Disease:Pulmonary Fibrosis, Phan SH, Thrall RS (org.). Marcel Dekker: New York, 135–171. 1995.
MOSELEY, P. L.; HEMKEN, C.; HUNNINGHAKE, G. W. Augmentation of fibroblast proliferation by bleomycin. J Clin Invest. Columbia University, v. 78, p.1150-1154, 1986. OBAYASHI, Y.; YAMADORI, I.; FUJITA, J.; YOSHINOUCHI, T.; UEDA, N.; TAKAHARA, J. The role of neutrophils in the pathogenesis of idiopathic pulmonary fibrosis. Chest; v.112, p.1338-1343, 1997.
PAIVA, L. A. F.;GURGEL, L. A.; SILVA, R. M.; TOMÉ, A. R.; GRAMOSA, N. V.; SILVEIRA, E. R.; SANTOS, F. A.; RAO, V. S. N. Anti-inflammatory effect of kaurenoic acid, a diterpene from Copaifera langsdorffii on acetic acid-induced colitis in rats. Vascular Pharmacology. USA, v. 39, p. 303-307, 2003.
PHAN, S. H.; GHARAEE-KERMANI, M.; WOLBER, F.; Stimulation of rat endothelial cell transforming growth factor-ß production by bleomycin. J Clin Invest. Columbia University, v.87, p.148-154. 1991.
PHILLIPS,R.J.;BURDICK,M.D.;HONG,K.;LUTZ,M.A.;MURRAY, L.A.; XUE,Y.Y.; BELPERIO,J.A.; KEANE, M.P.; STRIETER1,R.M.Circulating fibrocytes traffic to the lungs in response to CXCL12 and mediate fibrosis. J. Clin. Invest. Columbia University, vol. 114, p. 438–446, 2004.
RUFINO,R.; SILVA,J.R.L.; Cellular and biochemical bases of chronic obstructive pulmonary disease. J Bras Pneumol. Ribeirão Preto, São Paulo, v. 32, p. 241-248, 2006.
SCHMIDT, J. A.; MIZEL, S. B.; COHEN, D. Et al. Interleukin-1: a potential regulator of fibroblast proliferation. J Immunol. Maryland, v.128, p.2177-2182, 1982.
SGARBI, M. W.; SILVA Jr, B.A.; NETO, H. J. S. Importância da resposta inflamatória sistêmica (SIRS) no prognóstico dos pacientes politramatizados. Ver. Bras. Ortop. v.41, n. 1/2, p. 1-6, 2006.
SMITH, R. E.; STRIETER, R. M.; ZHANG, K.; PHAN, S. H.; STANDFORD, T. J.; LUKACS, N. W. ; KUNKEL, S.L. A role for C-C chemokines in fibrotic lung disease. J. Leukoc Biol. England, v. 57, n 5, p. 782-787, 1995.

45
STARK, J. M, STARK, M. A.; COLASURDO, G. N.; LEVINE, A. M. Decreased bacterial clearance from the lungs of mice following primary respiratory syncytial virus infection. J Med Virol. USA,v.78(6), p.829-38. 2006.
SUGERMAN, B. J.; AGGARWAL, B. B.; FIGARI, I. S. et al. Recombinant human tumor necrosis factor : effects on proliferation of normal and transformed cells in vitro. Science. USA, v.230, p.943-945, 1985.
VEIGA, F.; ZUNINO, L.; CALIXTO, J. B.; PATITUCCI, M. L.; PINTO, C. A. Phytochemical and antioedematogenic studies of commercial copaíba oils avaliable in Brazil. Phytotherapy Research. England, v. 15, p. 476-480, 2001. VEIGA JUNIOR, Valdir F.; PINTO, Angelo C.. The Copaifera L. genus. Quím. Nova., São Paulo, v. 25, n. 2, 2002. Available from: <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-40422002000200016&lng=en&nrm=iso>. Access on: 07 Dec 2006. doi: 10.1590/S0100-40422002000200016.
ZHANG, K.; FLANDERS, K. C.; PHAN, S. H. Cellular localization of transforming growth factor-ß expression in bleomycin-induced pulmonary fibrosis. Am J Pathol. Stanford University Libraries', v. 147, p.352-361,1995.