Embed Size (px)
Citation preview

CLÁUDIA CARNEIRO HECKE KRÜGER
PRODUÇÃO E CARACTERIZAÇÃO QUÍMICA E FISIOLÓGICA DE CASEINOFOSFOPEPTÍDEOS DE
LEITE BOVINO
Tese apresentada ao Programa de Pós-Graduação em Tecnologia de Alimentos, Setor de Tecnologia, Universidade Federal do Paraná, como requisito parcial à obtenção do título de Doutora em Tecnologia de Alimentos. Orientadora: Profª. Drª. Lys Mary Bileski
Cândido Co-orientadora: Profª. Drª. Iara Taborda
Messias
CURITIBA 2006

ii
À memória de meus pais, Paulo e Therezinha;
Ao meu marido, Walter, por todo apoio, amor e compreensão;
Aos meus filhos, Nícolas, Jonathan e Gabriele, pois sua existência é fonte de motivação;
Ao meu Deus, Criador do céu, da terra, do mar e de tudo o que neles existe;
Dedico.

iii
AGRADECIMENTOS
À Profª. Drª Lys Mary Bileski Cândido, por ter acreditado em meu trabalho, pela orientação sempre segura e pela amizade;
À Profª Drª Iara Taborda Messias, pela co-orientação e amizade;
Ao Departamento de Nutrição da Universidade Federal do Paraná;
Ao Programa de Pós-Graduação em Tecnologia de Alimentos;
Ao Conselho Nacional de Pesquisa e Desenvolvimento (CNPq), pelo apoio financeiro;
Ao Hospital e Maternidade Angelina Caron, onde foram desenvolvidos os experimentos com os animais;
Ao Prof. Dr. João Carlos Domingues Repka, pela oportunidade de convívio, ajuda e ensinamentos indispensáveis;
Ao Dr. Sérgio Ioshi, pela contribuição nas análises histopatológicas;
Ao Prof. Dr. Luis Cláudio Fernandes, pelo auxílio nos ensaios com macrófagos;
Ao Prof. Dr. Ricardo Bastos Cunha, pela colaboração na identificação dos peptídeos por MALDI-TOF-MS;
Às bolsistas, Ana Paula e Celine;
À bolsista de iniciação científica, Maria Isabel Vedana, pela grande contribuição à realização deste trabalho;
Ao Claudemir, da PUCPR, pela liofilização dos peptídeos;
Aos técnicos de laboratório, César Aparecido da Silva, Jair José de Lima e Lindamir Tomzack Túlio, pelo auxílio e pronto atendimento todas as vezes que necessitei;
À Else, do Hospital Militar do Paraná, pela realização dos leucogramas e contagens diferenciais;
À Louise e Sandro, pela ajuda no dia da eutanásia dos animais;
Ao Marcus Cândido, pela revisão do abstract;
Ao Renato Nishiara, do Laboratório de Imunopatologia do Hospital de Clínicas, pela colaboração na execução dos experimentos;
Aos membros da banca, pelas correções;
Ao Pastor Ericson, pela amizade, estímulo e orações;
A todos, que de alguma forma contribuíram, direta ou indiretamente, para que este trabalho se efetivasse;
Muito Obrigada!

iv
“The fear of the Lord is the beginning of wisdom and the knowledge of the holy is understanding”
Proverbs of Solomon

v
SUMÁRIO LISTA DE ILUSTRAÇÕES..............................................................................................................vi LISTA DE ABREVIATURAS......................................................................................................... viii RESUMO ......................................................................................................................................... x ABSTRACT .....................................................................................................................................xi 1 ............................................................................................................................. 1INTRODUÇÃO
1.1 JUSTIFICATIVA ..................................................................................................................... ..2 1.2 OBJETIVOS ............................................................................................................................2
2 REVISÃO DE LITERATURA........................................................................................................ 3 2.1 CASEÍNAS ...............................................................................................................................3
2.1.1 Estrutura e Função ............................................................................................................3 2.1.2 Extração e Fracionamento ................................................................................................6 2.1.3 Modificação Enzimática ....................................................................................................9 2.1.4 Obtenção de Petídeos Bioativos.....................................................................................12 2.1.5 Funções dos Peptídeos Bioativos...................................................................................14
2.2 CASEINOFOSFOPEPTÍDEOS ..............................................................................................22 2.2.1 Estrutura ..........................................................................................................................22 2.2.2 Métodos de Obtenção .....................................................................................................25 2.2.3 Funções ...........................................................................................................................29
2.3 RESPOSTA IMUNE NA INTERVENÇÃO NUTRICIONAL....................................................37 2.3.1 Estrutura e Componentes do Sistema Imunitário...........................................................38 2.3.2 Avaliação da Resposta Imunológica...............................................................................45 2.3.3 Função dos Peptídeos do Leite na Resposta Imunológica ............................................46
2.4 O SISTEMA IMUNOLÓGICO E A RESPOSTA INFLAMATÓRIA.........................................48 3 ESTRATÉGIA GERAL ............................................................................................................... 54 4 MATERIAL E MÉTODOS.......................................................................................................... 55
4.1 MATÉRIA PRIMA E REAGENTES ........................................................................................55 4.2 EQUIPAMENTOS ..................................................................................................................55 4.2 METODOLOGIA.....................................................................................................................55
4.2.1 Obtenção dos Caseinofosfopeptídeos............................................................................55 4.2.2 Caracterização dos Caseinofosfopeptídeos ...................................................................57 4.2.3 Atividade Antimicrobiana In Vitro ....................................................................................61 4.2.4 Avaliação da Ação dos Caseinofosfopeptídeos na Resposta Imunológica...................62
5 RESULTADOS E DISCUSSÃO ................................................................................................. 70 5.1 OBTENÇÃO E CARACTERIZAÇÃO DOS CASEINOFOSFOPEPTÍDEOS .........................70
5.1.1 Composição.....................................................................................................................70 5.1.2 Solubilidade .....................................................................................................................80 5.1.3 Seqüenciamento .............................................................................................................82
5.2 ATIVIDADE ANTIMICROBIANA IN VITRO ...........................................................................91 5.3 AVALIAÇÃO DA AÇÃO DOS CASEINOFOSFOPEPTÍDEOS NA RESPOSTA IMUNE......95
5.3.1 Imunidade Inespecífica ...................................................................................................95 5.3.2 Sepse Induzida..............................................................................................................100
6 CONCLUSÕES ........................................................................................................................ 119 CONSIDERAÇÕES FINAIS ........................................................................................................ 121 REFERÊNCIAS ........................................................................................................................... 122

vi
LISTA DE ILUSTRAÇÕES TABELA 1 - CARACTERÍSTICAS FÍSICO-QUÍMICAS DAS CASEÍNAS DO LEITE BOVINO ....... 4 FIGURA 1 - FLUXOGRAMA BÁSICO DE PRODUÇÃO DE CASEÍNA E/OU CASEINATOS...... 7 TABELA 2 - COMPOSIÇÃO TÍPICA DAS CASEÍNAS E CASEINATOS ..................................... 8 FIGURA 2 - PRINCIPAIS FUNÇÕES DOS PEPTÍDEOS BIOATIVOS DERIVADOS DA
CASEÍNA ........................................................................................................................ 15 FIGURA 3 - REPRESENTAÇÃO ESQUEMÁTICA DAS ZONAS ESTRATÉGICAS PRESENTES
NA ESTRUTURA PRIMÁRIA DA Β-CASEÍNA ............................................................... 15 TABELA 3 - ATIVIDADE OPIÓIDE DE Β-CASOMORFINAS HUMANA E BOVINA EM
COMPARAÇÃO À NORMORFINA ................................................................................. 18 FIGURA 4 - ESTRUTURA DA REGIÃO CPP.............................................................................. 23 FIGURA 5 - REGIÕES FOSFORILADAS DE DIFERENTES TIPOS DE CASEÍNAS BOVINAS .. 25 FIGURA 6 - MÉTODOS DE ELABORAÇÃO DE CASEINOFOSFOPEPTÍDEOS....................... 28 FIGURA 7 - APLICAÇÕES POTENCIAIS DOS CASEINOFOSFOPEPTÍDEOS ........................ 30 FIGURA 8 - DIFERENCIAÇÃO E ATIVAÇÃO DOS LINFÓCITOS AUXILIARES (Th)................ 41 FIGURA 9 - DIFERENCIAÇÃO DOS LINFÓCITOS B................................................................. 42 FIGURA 10 - PAPEL DOS MACRÓFAGOS NA INGESTÃO DE PROTEÍNAS ESTRANHAS E
NA APRESENTAÇÃO DE ANTÍGENOS ........................................................................ 43 FIGURA 11 - REPRESENTAÇÃO DIAGRAMÁTICA DOS COMPONENTES PRINCIPAIS DO
SISTEMA IMUNITÁRIO DA MUCOSA INTESTINAL ..................................................... 45 FIGURA 12 - MODELO DAS MEMBRANAS INTERNA E EXTERNA DA E. COLI K-12 ............ 50 FIGURA 13 - LIBERAÇÃO DO LPS POR E. COLI....................................................................... 50 FIGURA 14 - REPRESENTAÇÃO DIAGRAMÁTICA DOS EVENTOS ENVOLVIDOS NA
DETECÇÃO E RESPOSTA AOS ESTÍMULOS FORNECIDOS PELAS ENDOTOXINAS MICROBIANAS NA IMUNIDADE INATA............................................. 51
FIGURA 15 - PATOGÊNESE DO CHOQUE SÉPTICO ............................................................... 52 FIGURA 16 – REPRESENTAÇÃO ESQUEMÁTICA DA ESTRATÉGIA METODOLÓGICA
UTILIZADA PARA REALIZAÇÃO DOS OBJETIVOS DA TESE..................................... 54 TABELA 4 - COMPOSIÇÃO DA DIETA COMERCIAL ................................................................ 63 FIGURA 16 - INJEÇÃO INTRAPERITONEAL COM LPS PARA INDUÇÃO DE CHOQUE
SÉPTICO ........................................................................................................................ 68 TABELA 5 - COMPOSIÇÃO QUÍMICA DOS PEPTÍDEOS ......................................................... 70 TABELA 6 - MINERAIS DOS PEPTÍDEOS................................................................................. 71 FIGURA 17 - ELETROFORESE CAPILAR DE ZONA (ELETROFEROGRAMA) DO CPP-A ...... 72 FIGURA 18 - ELETROFORESE CAPILAR DE ZONA (ELETROFEROGRAMA) DO CPP-B ..... 73 FIGURA 19 - ELETROFORESE CAPILAR DE ZONA (ELETROFEROGRAMA) DO CPP-C..... 73 TABELA 7 - COMPOSIÇÃO AMINOACÍDICA (g AMINOÁCIDO/100g DE PROTEÍNA) DOS
CASEINOFOSFOPEPTÍDOS E DO CASEINATO DE SÓDIO ....................................... 74 TABELA 8 – FORMULAÇÕES ISENTAS DE FENILALANINA .................................................... 77 FIGURA 20 - TEOR DE FENILALANINA DOS CASEINOFOSFOPEPTÍDEOS ........................... 79 TABELA 9 - RELAÇÃO MOLAR ENTRE DADOS DE COMPOSIÇÃO QUÍMICA E CONTEÚDO
AMINOACÍDICO DOS PEPTÍDEOS............................................................................... 79 FIGURA 21 - SOLUBILIDADE DOS PEPTÍDEOS ....................................................................... 81 FIGURA 22 - ESPECTRO DE MASSA POR DESORÇÃO A LASER DO CPP-A......................... 84 FIGURA 23 - ESPECTRO DE MASSA POR DESORÇÃO A LASER DO CPP-B......................... 85 FIGURA 24 - ESPECTRO DE MASSA POR DESORÇÃO A LASER DO CPP-C ........................ 86 QUADRO 1 - MASSAS MOLECULARES DOS PEPTÍDEOS DETECTADOS NO CPP-A E
POSSÍVEIS SEQÜÊNCIAS ............................................................................................ 87

vii
QUADRO 2 - MASSAS MOLECULARES DOS PEPTÍDEOS DETECTADOS NO CPP-B E POSSÍVEIS SEQÜÊNCIAS ............................................................................................ 87
QUADRO 3 - MASSAS MOLECULARES DOS PEPTÍDEOS DETECTADOS NO CPP-C E POSSÍVEIS SEQÜÊNCIAS ............................................................................................ 89
FIGURA 25 – TESTE DE ESTERILIDADE DOS CASEINOFOSFOPEPTÍDEOS......................... 91 TABELA 10 - INIBIÇÃO DO CRESCIMENTO MICROBIANO PELOS
CASEINOFOSFOPEPTÍDEOS....................................................................................... 92 FIGURA 26 - EFEITO DOS CASEINOFOSFOPEPTÍDEOS CONTRA LISTERIA
MONOCYTOGENES ...................................................................................................... 94 FIGURA 27 - FAGOCITOSE DE ZIMOSAN PELOS MACRÓFAGOS PERITONEAIS DOS
RATOS............................................................................................................................ 96 FIGURA 28 - PRODUÇÃO DE ÂNION SUPERÓXIDO (O ) PELOS MACRÓFAGOS
PERITONEAIS DOS RATOS2-
.......................................................................................... 97 FIGURA 29 - PRODUÇÃO DE PERÓXIDO DE HIDROGÊNIO PELOS MACRÓFAGOS
PERITONEAIS DOS RATOS.......................................................................................... 98 FIGURA 30 - VOLUME LISOSSOMAL DOS MACRÓFAGOS PERITONEAIS DOS RATOS ...... 99 FIGURA 31 - CONTAGEM DIFERENCIAL DAS CÉLULAS LEUCOCITÁRIAS ANTES DO
INÍCIO DAS DIETAS (TEMPO = 0) .............................................................................. 100 TABELA 11 - PESO DOS RATOS ALIMENTADOS COM DIETA CONTROLE OU DIETA
ADICIONADA DE CPP, POR UM PERÍODO DE 22 DIAS........................................... 101 FIGURA 32 – EFEITO DE CASEINOFOSFOPEPTÍDEOS SOBRE NÍVEIS PLASMÁTICOS DE
IL-6 APÓS ESTÍMULO COM LPS ................................................................................ 102 FIGURA 33 - CONTAGEM DE LEUCÓCITOS............................................................................ 104 FIGURA 34 - CONTAGEM DE NEUTRÓFILOS.......................................................................... 106 FIGURA 35 - CONTAGEM DE LINFÓCITOS.............................................................................. 107 FIGURA 36 - CONTAGEM DE PLAQUETAS.............................................................................. 108 TABELA 12 - VALORES HISTOLÓGICOS MÉDIOS PARA OS GRUPOS CONTROLE, CPP-A,
CPP-B E CPP-C ........................................................................................................... 109 FIGURA 37 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A
PRESENÇA DE CONGESTÃO .................................................................................... 109 FIGURA 38 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A
PRESENÇA DE EDEMA .............................................................................................. 110 FIGURA 39 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A
PRESENÇA DE HEMORRAGIA ALVEOLAR............................................................... 111 FIGURA 40 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A
PRESENÇA DE INFILTRAÇÃO NEUTROFÍLICA ........................................................ 112 FIGURA 41 - EFEITO DE CASEINOFOSFOPEPTÍDEO (CPP-A) ADMINISTRADO
INTRAPERITONEALMENTE (IP) SOBRE NÍVEIS PLASMÁTICOS DE IL-6 APÓS ESTÍMULO COM LPS................................................................................................................................113
FIGURA 42 - CONTAGEM DE LEUCÓCITOS DOS RATOS...................................................... 115 FIGURA 43 - CONTAGEM DE NEUTRÓFILOS DOS RATOS ................................................... 116 FIGURA 44 - CONTAGEM DE LINFÓCITOS DOS RATOS ....................................................... 117 FIGURA 45 - CONTAGEM DE PLAQUETAS DOS RATOS ....................................................... 118

viii
LISTA DE ABREVIATURAS AAPH - 2,2'-azobis-2-amidinopropano-dihidroclorida AIDS - síndrome da imunodeficiência adquirida Aminoácidos: Ala (A) - alanina Arg (R) - arginina Asn (N) - asparagina Asp (D) - ácido aspártico Cys (C) - cistina Gln (Q) - glutamina Glu (E) - ácido glutâmico Gly (G) - glicina His (H) - histidina Ile (I) - isoleucina Leu (L) - leucina Lys (K) - lisina Met (M) - metionina Phe (F) - fenilalanina Pro (P) - prolina Ser (S) - serina Thr (T) - treonina Trp (W) - triptofano Tyr (Y) - tirosina Val (V) - valina ATCC - American Type Culture Collection C3a - anafilatoxina produzida pela ativação do complemento C5a - anafilatoxina produzida pela ativação do complemento C5b-9 - complexo lítico de membrana CaBP - calcium binding protein CAF - chemically assisted fragmentation CD14 - receptor de superfície que se liga ao complexo LPS-LBP CFP - células formadoras de placas CGP - kappa-caseinoglicopeptídeo CN - caseína ConA - concanavalina A CPP - caseinofosfopeptídeo E/S - relação enzima/substrato ECA, ACE - enzima conversora da angiotensina EDTA - ácido etilenodiamino tetracético ELISA - enzyme linked immuno sorbent assay ESI-CID-MS/MS - electrospray collision induced dissociation FAST - fragmentation analysis and structural time of flight FPLC - fast protein liquid chromatography GH - grau de hidrólise

ix
GMP - glicomacropeptídeo GPI - contrações elétricas induzidas no plexo mioentérico HIV - human imunodeficiency virus HLA - complexo principal de histocompatibilidade HSP - heat shock protein HT -29 - células intestinais tumorais humanas IFN - interferon Ig - imunoglobulina IL - Interleucina IP - intraperitoneal ITAL - Instituto de Tecnologia de Alimentos IUB - União Internacional de Bioquímica LBP - lipopolissacharide binding protein Linfócitos T CD4+ - Linfócitos t auxiliares Linfócitos T CD8+ - Linfócitos t supressores Linfócitos Th - linfócitos t auxiliares Linfócitos Ts, CTL - linfócitos T supressores ou citotóxicos LPS - lipopolissacarídeo MALDI_TOF_MS - espectrometria de massa por desorção a laser MF - microfiltração MIC - concentração inibitória mínima NBT - nitroblue tetrazolium NK - natural killer PBS - tampão fosfato-salina PER - quociente de eficiência protéica PHA - fitohemaglutinina PMA - forbol 12-miristato 13-acetato PSD - post source decay PTN 6.0 - tripsina Novzymes RP - HPLC - cromatografia líquida de alta pressão em fase reversa TECPAR - Instituto de Tecnologia do Paraná TFA - ácido trifluoroacético TGF - fator de transformação de crescimento TLR4 - toll-like receptor four TMB - tetrametilbenzidina TNF - fator de necrose tumoral TOF - time of flight UFC - unidade formadora de colônia

x
RESUMO Introdução: caseinofosfopeptídeos são formados in vivo durante a digestão da caseína e podem ser produzidos in vitro por hidrólise enzimática. Esses peptídeos têm sido apontados por sua bioatividade, podendo ser úteis em alimentos com alegações de propriedades funcionais ou produtos farmacologicamente ativos. Objetivos: avaliar as propriedades funcionais fisiológicas de peptídeos obtidos por hidrólise enzimática de concentrados protéicos de leite bovino. Para atingir esse objetivo, foram elaborados fosfopeptídeos a partir de caseinato de sódio. Métodos: os fosfopeptídeos foram caracterizados do ponto de vista físico-químico e bioquímico. O processamento dos peptídeos foi realizado no Laboratório de Pesquisa e Pós-Graduação do Departamento de Nutrição da Universidade Federal do Paraná. Os peptídeos foram produzidos pela hidrólise tríptica do caseinato de sódio comercial, Lactonat (HV), seguida de precipitação ácida, agregação mineral e precipitação com etanol ou ultrafiltração/diafiltração. Resultados: os produtos obtidos foram liofilizados e denominados CPP-A, CPP-B e CPP-C. Esses produtos se caracterizam como uma mistura de peptídeos derivados da hidrólise das caseínas αS1, αS2, β e κ. A análise da composição centesimal demonstrou os seguintes resultados (em base úmida): CPP-A, 64,28%±0,60 de proteínas e 19,06±0,03% de cinzas; CPP-B, 67,59±0,38% de proteínas, 29,23±0,14% de cinzas; CPP-C, 84,85±0,56% de proteínas, 16,14±0,0003% de cinzas. O rendimento final dos produtos foi de 11,56%, 19% e 45,57% respectivamente para o CPP-A, CPP-B e CPP-C. A análise de aminoácidos revelou redução significativa no teor de fenilalanina para o CPP-A, com redução de 71,89% em relação ao caseinato de sódio. Foram obtidas as seguintes relações molares na caracterização dos produtos: serina/fósforo, 0,973mol/mol no CPP-A, 0,515mol/mol no CPP-B e 0,680mol/mol no CPP-C; nitrogênio/fósforo, 8,082mol/mol no CPP-A, 7,79mol/mol no CPP-B e 16,73mol/mol no CPP-C; nitrogênio/serina, 8,306mol/mol no CPP-A, 15,128mol/mol no CPP-B e 24,611mol/mol no CPP-C. Os peptídeos apresentaram percentuais relativamente elevados de solubilidade tanto em água como em solução salina de tampão-fosfato, pH 7,4. O CPP-C inibiu o crescimento de Staphylococcus aureus, Escherichia coli e Salmonella typhimurium em quase todas as concentrações testadas. A Lysteria monocytogenes foi inibida por todos os peptídeos testados. Os peptídeos CPP-A e CPP-B não alteraram a resposta normal dos macrófagos, indicando que sua administração não prejudica a resposta imune inata. A liberação de IL-6 após administração de dose de 5mg/kg de lipopolissacarídeo via intraperitoneal foi significativamente reduzida para os animais que receberam dieta com 0,14g/rato/dia de CPP-A por um período de 22 dias. Os níveis de IL-6 também se apresentaram reduzidos (p=0,000255) nos animais que receberam o CPP-A via intraperitoneal, concomitante à administração de LPS. Conclusões: a comprovação da ausência de respostas citotóxicas pelos CPP-A e CPP-B, bem como seu potencial efeito protetor da saúde pode ser visto como uma contribuição importante na formulação de novos alimentos e na alegação de propriedade funcional ou de saúde em rótulos de alimentos. Palavras-chave: caseinofosfopeptídeos; espectrometria de massa; antimicrobiano; macrófagos; lipopolissacarídeo.

xi
ABSTRACT Introduction: casein phosphopeptides can be liberated in vivo by normal digestion of casein as well as could be produced in vitro by enzymatic hydrolysis. These peptides were suggested to have biological activity, to be useful in food or pharmaceutical applications. Objective: to study the functional and biological properties of milk protein peptides obtained by enzymatic hydrolysis. For this purpose, phosphopeptides were obtained from sodium caseinate. Methods: peptides were physicochemical and biochemically characterized. Casein phosphopeptides were produced at the Research and Post-Graduation Laboratory of the Department of Nutrition at the Federal University of Parana, Brazil. They were produced by tryptic hydrolysis of commercial sodium caseinate, followed by acid precipitation, mineral aggregation, ethanol precipitation or/and ultrafiltration and diafiltration. The products were lyophilized and named CPP-A, CPP-B and CPP-C. Results: these products were characterized peptide mixtures, derived from hydrolysis of αS1, αS2, β and κ caseins. The centesimal composition analysis demonstrated the following results (wet basis): CPP-A, 64.28%±0.60 of proteins and 19.06±0.03% of ashes; CPP-B, 67.59±0.38% of proteins, 29.23±0.14% of ashes; CPP-C, 84.85±0.56% of proteins, 16.14±0.0003% of ashes. The final yield of the products for CPP-A, CPP-B and CPP-C respectively was 11.56%, 19% and 45.57%. The amino acid analysis disclosed significant reduction in phenylalanin for the CPP-A, with reduction of 71.89% in relation to sodium caseinate. The following molar ratios were obtained in the characterization of the products: Ser/P, 0.973mol/mol in the CPP-A, 0.515mol/mol in the CPP-B and 0.680mol/mol in the CPP-C; N/P, 8.082mol/mol in the CPP-A, 7.79mol/mol in the CPP-B and 16.73mol/mol in the CPP-C; N/Ser, 8.306mol/mol in the CPP-A, 15.128mol/mol in the CPP-B and 24.611mol/mol in the CPP-C. Casein phosphopeptides presented relatively high percentages of solubility in water as in saline solution of PBS, pH 7.4. When tested against Escherichia coli, CPP-A displays antimicrobial activity at 0.2mg/mL and 1mg/mL. CPP-C inhibited Staphylococcus aureus, Escherichia coli and Salmonella typhimurium growth at different concentrations. All tested peptides inhibited Lysteria monocytogenes growth. CPP-A and CPP-B did not modify the response of macrophages, indicating that its administration does not harm the innate immune response. The IL-6 release after administration of 5mg/kg of intraperitoneal LPS was significantly reduced for the animals that had received a diet with 0.14g/rat/day from CPP-A for 22 days. The IL-6 levels were also reduced (p=0,000255) in the animals that had received the intraperitoneal CPP-A, concomitant the LPS administration. Conclusions: the proven absence of cytotoxic response for CPP-A and CPP-B, as well as its potential health-protective effect can be regarded as an important contribution for the food industry, justifying the allegation of functional health property in food labels. Keywords: casein phosphopeptide; mass spectrometry; antimicrobial; macrophages; lipopolysaccharide.

1
INTRODUÇÃO
O aumento na produção de alimentos industrializados tem criado uma demanda
crescente de ingredientes que associem propriedades fisiológicas e funcionais. As
frações protéicas do leite agregam atributos nutricionais, funcionais e fisiológicos, que
podem ser isolados e utilizados pela indústria farmacêutica e de alimentos funcionais.
Alimentos funcionais são entendidos como aqueles que apresentam componentes com
potencial protetor da saúde, especialmente no que se refere às doenças crônicas não
transmissíveis.
O leite é um ingrediente de importância fundamental para a indústria de
alimentos. O fracionamento desta matéria-prima gera uma série de produtos indicados
para inúmeras aplicações, incluindo: soro ácido, caseinatos, co-precipitados protéicos
de caseína e soro, coágulo de caseína, soro doce, concentrados e isolados protéicos
de soro e lactalbumina.
Nos últimos 30 anos, vários peptídeos originados do leite têm sido identificados e
caracterizados. Esses fragmentos protéicos apresentam propriedades bioquímicas
específicas, que incluem atividade opióide, anti-hipertensiva, imunomoduladora, anti-
trombótica e a capacidade de ligar minerais.
Caseinofosfopeptídeos derivam-se da hidrólise da caseína e apresentam
propriedades bioativas. Esses peptídeos são liberados durante a digestão por meio de
proteinases intestinais, melhorando a absorção de cálcio por aumentar sua
biodisponibilidade.
A produção de caseinofosfopeptídeos pode ser realizada pela hidrólise da
caseína, enriquecimento das preparações com íons di e trivalentes, seguida de
processos de purificação e concentração. Os produtos obtidos podem ser considerados
como ingredientes potenciais para receberem alegações de propriedades funcionais.
Portanto, esses produtos devem ser examinados e caracterizados em relação ao seu
efeito benéfico ou possível efeito citotóxico.

2
1.1 JUSTIFICATIVA
A possibilidade de formulação de novos alimentos empregando hidrolisados
protéicos de leite bovino e/ ou seus peptídeos isolados pode ser vista como uma
contribuição importante na prevenção de doenças crônicas e/ou na proteção à saúde.
1.2 OBJETIVOS Objetivo geral: Produção, caracterização e avaliação das propriedades funcionais fisiológicas de
peptídeos obtidos por hidrólise enzimática de concentrados protéicos de leite bovino.
Objetivos específicos: 1. Elaborar fosfopeptídeos a partir de caseinato de sódio
2. Caracterizar os peptídeos do ponto de vista físico-químico
3. Avaliar a atividade antimicrobiana in vitro dos caseinofosfopeptídeos
4. Avaliar o efeito dos caseinofosfopeptídeos na imunidade inespecífica e na indução
de sepse após sua administração em animais de laboratório

3
2 REVISÃO DE LITERATURA As proteínas são componentes importantes dos alimentos, tanto pelo aspecto
nutricional como também pelo valor funcional. Os aminoácidos essenciais ao
funcionamento do organismo são fornecidos pelas proteínas da dieta. Além disso, as
proteínas possuem algumas funções específicas relativas à presença de peptídeos
bioativos em sua seqüência primária, o que as torna agentes promotores de saúde
(LÉONIL et al., 2000).
O leite bovino contém aproximadamente 3,5% de proteínas, que podem ser
classificadas em quatro grupos, de acordo com suas propriedades físico-químicas e
estruturais: a) caseínas; b) proteínas do soro; c) proteínas das membranas dos glóbulos
de gordura; d) enzimas e fatores de crescimento (SGARBIERI, 2005).
2.1 CASEÍNAS
2.1.1 Estrutura e Função As caseínas são as proteínas predominantes no leite da maioria de espécies
mamíferas. Constituem um grupo heterogêneo de fosfoproteínas que estabilizam o
fosfato de cálcio em estruturas micelares. Suas funções biológicas incluem o
fornecimento de fosfato (PO4) e cálcio (Ca2+) para a mineralização de alguns tecidos, o
provimento de aminoácidos essenciais e a formação de peptídeos com atividade
biológica (RASMUSSEN et al., 1999). Compreendem de 2,5 a 3,2% do leite fluido e
aproximadamente 80% das proteínas totais do leite bovino.
Três principais componentes são obtidos a partir da caseína: αS1(50% do total), β
(33%) e κ (15%), existindo ainda quantidades variáveis do componente γ. A análise
elementar da caseína (complexo α, β, κ) revela a seguinte composição: carbono, 53%;
hidrogênio, 7,05%; nitrogênio, 16,65%; enxofre, 0,76%; fósforo, 0,85% (SGARBIERI,
1996).
4
Existem componentes menores no sistema caseína, que são fragmentos C-
terminais da β-caseína produzidos pela ação da enzima plasmina (FOX, 2001).
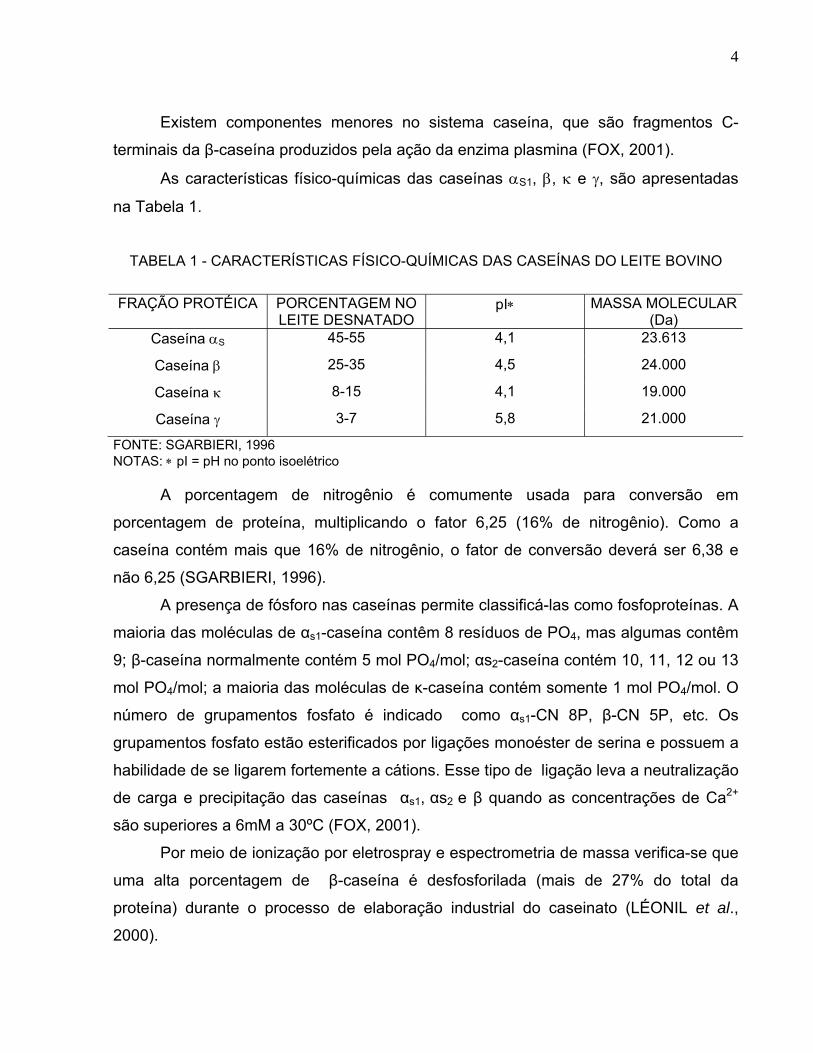
As características físico-químicas das caseínas αS1, β, κ e γ, são apresentadas
na Tabela 1.
TABELA 1 - CARACTERÍSTICAS FÍSICO-QUÍMICAS DAS CASEÍNAS DO LEITE BOVINO
FRAÇÃO PROTÉICA PORCENTAGEM NO
LEITE DESNATADO pI∗ MASSA MOLECULAR
(Da) Caseína αS 45-55 4,1 23.613
Caseína β 25-35 4,5 24.000
Caseína κ 8-15 4,1 19.000
Caseína γ 3-7 5,8 21.000
FONTE: SGARBIERI, 1996 NOTAS: ∗ pI = pH no ponto isoelétrico
A porcentagem de nitrogênio é comumente usada para conversão em
porcentagem de proteína, multiplicando o fator 6,25 (16% de nitrogênio). Como a
caseína contém mais que 16% de nitrogênio, o fator de conversão deverá ser 6,38 e
não 6,25 (SGARBIERI, 1996).
A presença de fósforo nas caseínas permite classificá-las como fosfoproteínas. A
maioria das moléculas de αs1-caseína contêm 8 resíduos de PO4, mas algumas contêm
9; β-caseína normalmente contém 5 mol PO4/mol; αs2-caseína contém 10, 11, 12 ou 13
mol PO4/mol; a maioria das moléculas de κ-caseína contém somente 1 mol PO4/mol. O
número de grupamentos fosfato é indicado como αs1-CN 8P, β-CN 5P, etc. Os
grupamentos fosfato estão esterificados por ligações monoéster de serina e possuem a
habilidade de se ligarem fortemente a cátions. Esse tipo de ligação leva a neutralização
de carga e precipitação das caseínas αs1, αs2 e β quando as concentrações de Ca2+
são superiores a 6mM a 30ºC (FOX, 2001).
Por meio de ionização por eletrospray e espectrometria de massa verifica-se que
uma alta porcentagem de β-caseína é desfosforilada (mais de 27% do total da
proteína) durante o processo de elaboração industrial do caseinato (LÉONIL et al.,
2000).

5
Somente as caseínas αs2 e κ contêm cisteína. Esse resíduo forma pontes
dissulfeto intermoleculares. A ausência de cisteína ou cistina nas caseínas αs1 e β
permite uma maior flexibilidade nessas moléculas. As caseínas, especialmente a β-
caseína, são proteínas de estruturas abertas e com predominância de estruturas
primárias (randomizadas) o que se deve, em parte, aos altos teores de prolina,
distribuída em toda a cadeia polipeptídica, que previne a formação de estruturas
secundárias (DICKINSON, 1999).
A falta da estruturação secundária e terciária torna as caseínas mais
susceptíveis à desnaturação e contribui para sua elevada atividade de superfície, que
lhes confere boa capacidade de formação de espumas e emulsões e facilita a
proteólise. As caseínas são relativamente hidrofóbicas. Os resíduos hidrofóbicos,
polares e carregados não se apresentam uniformemente distribuídos, mas ocorrem
como áreas hidrofóbicas ou hidrofílicas, tornando-as anfipáticas. Sua alta
hidrofobicidade explica a propensão dos hidrolisados de caseína ao aparecimento de
amargor. Todas as caseínas exibem polimorfismo genético, o qual envolve a
substituição de um ou mais aminoácidos (FOX, 2001).
As moléculas de caseína apresentam elevada afinidade por cátions bi- ou
trivalentes em função da presença de resíduos serina fosforilados. Os cátions também
podem se ligar a grupamentos carboxílicos (ácidos glutâmico e aspártico), grupamentos
fenólicos (resíduo tirosil), grupamentos sulfidrílicos (resíduo cisteinil) e grupamentos
imidazólicos (resíduos histidil). A associação de minerais à caseína leva à formação de
ingredientes com propriedades funcionais diferenciadas, de acordo com o íon
associado (GAUCHERON et al., 1997).
As micelas de caseína se apresentam suspensas na fase líquida do leite, com um
diâmetro médio de aproximadamente 120 nm, contendo 93% de proteínas e 7% de sais
inorgânicos, principalmente cálcio e fosfato. A estrutura das micelas não está bem
estabelecida. A estrutura proposta por WALSTRA (1999), segundo SGARBIERI (2005),
apresenta as seguintes características: a) a micela apresenta-se essencialmente
esférica, contudo sua superfície não se apresenta lisa; b) é formada de unidades
menores denominadas submicelas, contendo principalmente caseína; c) as submicelas

6
variam em composição, existindo particularmente dois tipos principais, isto é, um tipo
formado pelas caseínas αs, β e κ e outro formado pelas caseínas αs e κ; d) as
submicelas parecem permanecer ligadas por aglomerados (clusters) de fosfato de
cálcio; e) dessa forma, as submicelas se agregam até a formação completa da micela,
em que a caseína κ se posiciona superficialmente; f) a porção C-terminal da caseína κ
(glicopeptídio) projeta-se para fora da superfície da micela, formando uma camada
esponjosa que previne, por repulsões estéricas e eletrostáticas, qualquer agregação
posterior de submicelas. Os fatores que podem alterar a estrutura do sistema micelar
são a acidificação química ou biológica, a adição de cálcio ou etanol ou a combinação
desses agentes (CREAMER et al., 1998; KRUIF, 1999).
Do ponto de vista nutricional, as proteínas do leite apresentam um Quociente de
Eficiência Protéica (PER) elevado. A caseína é considerada proteína referência,
apresentando um PER acima de 2,5 e o caseinato de sódio apresenta um PER de 2,6
(MING, 2000).
Um estudo de avaliação nutricional de caseinato de sódio e de coágulo de
caseína produzidos no Instituto de Tecnologia de Alimentos (ITAL, Campinas), revelou
valores de PER de 3,15 para o caseinato de sódio e de 3,65 para a caseína coagulada
enzimaticamente (BORGES et al., 1999).
Além do fornecimento de aminoácidos necessários ao crescimento do neonato, o
sistema micelar das caseínas possui importância fisiológica, pois está relacionado à
prevenção de uma calcificação patológica das glândulas mamárias. A inclusão de cálcio
e fósforo nas micelas de caseína apresenta-se na forma de "nanoclusters", permitindo a
retenção e transporte desses importantes e pouco solúveis minerais ao recém-nascido
sem o perigo da precipitação do cálcio nas glândulas mamárias (MAZA, 1998).
2.1.2 Extração e Fracionamento
Há 2 processos básicos de produção de caseínas em escala industrial: a
precipitação isoelétrica e a coagulação enzimática. O esquema geral de fabricação da
caseína e dos caseinatos está representado na Figura 1, sendo que modificações nos

7
agentes utilizados nas diversas etapas levam à obtenção de produtos diferenciados em
composição (Tabela 2) e aplicação funcional.
O coágulo de caseína, obtido por enzimas, é usado principalmente na
elaboração de queijos. A caseína isoelétrica, obtida pela precipitação no ponto
isoelétrico, geralmente é convertida para a forma de sal (caseinato). Neste caso, um
álcali (sódio, cálcio ou potássio) é adicionado ao produto, que então é desidratado.
FIGURA 1 - FLUXOGRAMA BÁSICO DE PRODUÇÃO DE CASEÍNA E/OU CASEINATOS
Leite Desnatado Pasteurizado
Coagulação/Precipitação
Dessoragem
Lavagem
Ajuste de pH
Secagem
Caseína / Caseinatos

8
TABELA 2 - COMPOSIÇÃO TÍPICA DAS CASEÍNAS E CASEINATOS
CASEÍNA
ISOELÉTRICA
COÁGULO DE
CASEÍNA
CASEINATO
DE SÓDIO
CASEINATO
DE CÁLCIO
PROTEÍNA (%) 87,3 80,6 90,4 90,5
UMIDADE (%) 9,6 11,0 4,6 4,6
CINZAS (%) 1,8 7,8 3,8 3,7
GORDURA (%) 1,2 0,5 1,1 1,1
LACTOSE (%) 0,1 0,1 0,1 0,1
SÓDIO (%) <0,01 <0,01 1,2 0,01
CÁLCIO(%) 0,02 2,8 0,03 1,3
FÓSFORO (%) 0,7 1,6 0,8 0,8
pH 4,6 7,1 6,8 6,8
FONTE: Adaptado de HUFFMAN & HARPER, 1999.
Os caseinatos de sódio e potássio são solúveis em pH neutro, possuem
elevadas propriedades de superfície e estabilidade ao congelamento. Formam soluções
de alta viscosidade quando em concentrações acima de 10%. O caseinato de cálcio
apresenta alta dispersibilidade e baixa viscosidade. O coágulo de caseína e os
caseinatos de sódio e cálcio podem ser usados em produtos de panificação, alimentos
infantis, alimentos para atletas, na elaboração de emulsões, coberturas, pós para
bebidas, sopas e sobremesas. A utilização de coágulo de caseína e de caseinatos na
elaboração de queijos leva a uma melhor hidratação e rápida dispersão dos
componentes, tendo como resultado uma melhor textura do produto final. Além das
aplicações alimentícias, o caseinato de sódio pode ser usado na fabricação de papéis e
adesivos (MANN, 1991; USDEC, 1997; HUFFMAN; HARPER, 1999; KRÜGER, 2002).
Dois novos processos têm sido propostos para a desestabilização das micelas
de caseína. O primeiro denomina-se crioprecipitação e tem como princípio o
aproveitamento do efeito de temperaturas negativas na estabilidade micelar. O
congelamento a -10ºC do leite com posterior armazenamento leva ao abaixamento do
pH para valores próximos a 5,8, devido à precipitação do fosfato de cálcio. O segundo

9
processo se baseia na baixa estabilidade da caseína quando o leite é acidificado a pH
6,0 e misturado a 10 a 15% de etanol (DAMODARAN; PARAF, 1997).
As micelas de caseína também podem ser separadas do soro pelo uso de
membranas de microfiltração (MF). A microfiltração de leite integral ou desnatado em
membranas com porosidade variando de 0,1 a 0,2µm, permite a coleta de um
permeado com composição semelhante ao soro doce, porém mais cristalino e de
qualidade bacteriológica satisfatória. Este soro doce pode sofrer purificação posterior
por ultrafiltração gerando um isolado protéico de soro com 96% de proteínas. Como
retentado se obtém o fosfocaseinato de cálcio micelar ou caseína micelar
(DAMODARAN; PARAF, 1997; MAUBOIS; LEONIL; SABOYA, 2002). Essa é uma
tecnologia emergente e promissora para a indústria alimentícia, pois a caseína micelar
não apresenta a eletronegatividade causada pela clivagem do glicomacropeptídeo.
Além disso, a caseína micelar pode ser considerada um ingrediente potencial para
fracionamento posterior.
As micelas de caseína também podem ser sedimentadas por centrifugação a
100.000xg por 1 hora, processo que pode ser realizado em nível laboratorial. Micelas
de caseína obtidas como retentado de ultrafiltração podem ser sedimentadas por
centrifugação a 75.000xg por 1 hora a 50ºC. O sobrenadante resultante tem a
composição de um concentrado protéico de soro com 35% de proteínas (FOX, 2001).
IMAFIDON, FARKYE & SPANIER (1997) fazem uma revisão detalhada das
técnicas de isolamento e purificação das frações caseínicas. A separação da β-caseína
tem sido proposta tanto por troca iônica como por processos de membranas. A fração
apresenta funcionalidade superior ao caseinato de sódio.
Do ponto de vista industrial, maior interesse tem sido dado ao fracionamento da
β-caseína, glicomacropeptídeo e caseinofosfopeptídeo, pois esses produtos
apresentam aplicações potenciais para a indústria de alimentos para fins especiais.
2.1.3 Modificação Enzimática
As proteínas podem ser modificadas intencionalmente por reações químicas,
reações catalisadas por enzimas, transformações físicas ou alterações genéticas.

10
Essas modificações podem ocorrer in vitro ou in vivo (GONZÁLEZ-TELLO et al., 1994;
SGARBIERI, 1996; IMAFIDON et al., 1997).
O uso de enzimas na manipulação de proteínas garante maior especificidade,
controle e segurança nos hidrolisados obtidos do que a utilização de meios químicos. A
hidrólise enzimática de proteínas produz peptídeos com pesos moleculares variáveis e
alterações na estrutura nativa da proteína. Previne reações indesejáveis devido à sua
especificidade de ação, utiliza condições moderadas de tratamento, menor energia no
processo, sendo possível sua inativação após o uso. Os hidrolisados obtidos por via
enzimática geralmente apresentam menor conteúdo de sais do que os obtidos por
hidrólise ácida ou alcalina (LÖFFLER, 1986; DEESLIE; CHERYAN, 1988; GONZÁLEZ-
TELLO et al., 1994; IMAFIDON et al., 1997).
Como resultado da hidrólise, há aumento na solubilidade das proteínas, melhoria
das propriedades interfaciais e redução na viscosidade (TURGEON; GAUTHIER;
PAQUIN, 1992; CARNEIRO, 1997).
O grau de hidrólise (GH) é definido como o número de ligações peptídicas
hidrolisadas em relação ao número total de ligações peptídicas de uma determinada
proteína. Proteínas intactas têm um valor GH = 0% e uma proteína completamente
hidrolisada tem valor GH = 100%. O grau de hidrólise é determinado pelas condições
do processo (relação enzima:substrato, tempo, temperatura, pH) e influenciado pela
natureza da atividade enzimática (PANYAM; KILARA, 1996).
As proteases têm sido classificadas de diferentes formas, mas a classificação
mais satisfatória é a adotada pela Comissão de Enzimas (CE) da IUB: proteases
contendo serina, contendo cisteína, metaloproteases e proteases ácidas (proteases
aspárticas). Em relação ao local de atuação na molécula de proteína são classificadas
como exo e endopeptidases (CÂNDIDO, 1998).
Os hidrolisados podem ser classificados de acordo com o grau de hidrólise e
com a aplicação dos mesmos. Assim, se agrupam em três grandes blocos: a)
hidrolisados com baixo grau de hidrólise, entre 1 e 10%, para melhoria de propriedades
funcionais; b) hidrolisados com graus de hidrólise variável, geralmente alto, para
utilização como aromatizantes; c) hidrolisados extensivos, com grau de hidrólise

11
superior a 10%, para uso em alimentos para fins especiais. Estes podem se subdividir
em hidrolisados para utilização em suplementos protéicos e hidrolisados com
composição definida para o tratamento de enfermidades e síndromes específicas
(VIOQUE et al., 2001)
Os hidrolisados têm sido utilizados como suplementos protéicos na elaboração
de dietas para alimentação enteral de bebês e adultos enfermos, fórmulas
hipoalergênicas para lactentes e lactantes, suplementos para praticantes de atividade
física, dietas para idosos e para controle de peso. Hidrolisados com composição
definida têm sido usados em erros metabólicos congênitos, como na fenilcetonúria, em
que se propõem as remoções dos aminoácidos aromáticos e em enfermidades
hepáticas, onde há necessidade de uma alta razão de Fischer (ramificados/aromáticos).
Finalmente, devido à sua alta solubilidade, digestibilidade e boa absorção intestinal, os
hidrolisados extensivos podem ser usados em pacientes com atividade gastrointestinal
deficiente, como no caso da doença de Crohn. Portanto, esses produtos devem possuir
um adequado balanço osmótico, hipoalergenicidade, sabor agradável, baixa
concentração salina, valor nutritivo igual ou similar ao concentrado original
(GONZÁLEZ-TELLO et al., 1994; SINGH; DALGLEISH, 1998; NEKLYUDOV; IVANKIN;
BERDUTINA, 2000; VIOQUE et al., 2001; PACHECO et al., 2002; CÂNDIDO;
SGARBIERI, 2003).
Tradicionalmente, proteases têm sido aplicadas no amaciamento de carnes
utilizando-se papaína. A modificação enzimática mais clássica do leite se refere à
clivagem da κ-caseína, nos resíduos Phe105-Met106 pela ação da renina e a proteólise
subseqüente que ocorre na maturação dos queijos. Enzimas também agem na
prevenção da turbidez da cerveja, na produção de detergentes e em fermentações de
alimentos orientais. Vários estudos têm demonstrado a aplicação de hidrolisados
protéicos solúveis elaborados a partir de proteínas vegetais ou de pescados (LÖFFLER,
1986; FENNEMA, 1993; CÂNDIDO; SGARBIERI, 2003).
A produção industrial de hidrolisados de caseína teve seu início a partir da
década de 70. A matéria-prima mais freqüentemente empregada tem sido o caseinato
de sódio e, dependendo da aplicação é selecionado o tipo de enzima proteolítica a ser

12
usado. Muitas vezes, o hidrolisado é ultrafiltrado para eliminação de enzimas e de
peptídeos de alto peso molecular (BRULE et al., 1980).
A hidrólise seletiva das proteínas do leite, com a finalidade de produzir
hidrolisados protéicos ou peptídeos com propriedades fisiológicas ativas, tem sido
pesquisada por vários autores (ROKKA et al., 1997; CHU; MACLEOD; OZIMEK, 1996;
YAMAMOTO; TAKANO, 1999; FITZGERALD; MEISEL, 1999; LEDOUX, et al., 1999).
Após o processo de hidrólise por uma enzima específica, há necessidade da
caracterização dos peptídeos obtidos. Vários métodos podem ser utilizados e incluem
eletroforese em gel de poliacrilamida uni ou bidimensional, focalização isoelétrica,
imunoeletroforese, cromatografia líquida de alta pressão em fase reversa (RP-HPLC),
eletroforese capilar e espectrometria de massa do tipo MALDI-TOF. Este último é
composto por um ionizador (MALDI, matrix assisted laser desorption/ionization), um
analisador (TOF, time of flight) e um detector que registra o tempo de deslocamento
(time of flight) dos vários peptídeos ionizados. Como os íons menores “voam” mais
rápido que os maiores, a razão m/z pode ser calculada a partir dos tempos de vôo
registrados, após calibração do instrumento, permitindo assim determinar as massas de
cada peptídeo. Este método pode ser associado ao seqüenciamento aminoacídico por
degradação de Edman (KANEKANIAN, GALLAGHER, EVANS, 2000; PACHECO et al.,
2002).
2.1.4 Obtenção de Petídeos Bioativos
Substâncias bioativas de origem alimentar são definidas como componentes dos
alimentos consumidos que podem vir a exercer atividades regulatórias no organismo
humano, independentemente de suas funções nutricionais (MEISEL, 1999).
Baseados no conceito acima, vários pesquisadores têm considerado diferentes
substâncias bioativas como ingredientes promotores da saúde, a fim de utilizá-los em
alimentos com alegações de propriedades funcionais, que podem ser consumidos como
parte da dieta diária e em produtos farmacologicamente ativos.
Vários estudos estão sendo conduzidos na determinação de substâncias
bioativas, para aplicação em alimentos formulados:

13
- O uso de microorganismos probióticos na prevenção de infecções do trato
gastrointestinal e no restabelecimento da microflora (VAUGHAN; MOLLET,
1999);
- A utilização de oligossacarídeos e catequinas do chá no controle da
hiperglicemia de pacientes diabéticos (SHIMIZU, 1999);
- Peptídeos bioativos derivados das prolaminas de cereais, capazes de
prevenir, in vitro, as alterações causadas pelos mesmos cereais em pacientes
portadores de doença celíaca (SILANO; VICENZI, 1999);
- Os efeitos antioxidantes dos flavonóides (HEMPEL et al., 1999; PFORTE et
al., 1999);
- Lipídios bioativos do leite bovino (MOLKENTIN, 1999);
- A capacidade das isoflavonas em prevenir e/ou tratar doenças crônico-
degenerativas como a osteoporose, câncer e doenças coronarianas
(BEDANI; ROSSI, 2005).
- O potencial farmacológico dos extratos etanólicos de própolis: anticâncer,
anti-inflamatório, antibiótico, antioxidante, antifúngico e antiviral (CHOI et al.,
2006).
As proteínas, especialmente as caseínas, que sofreram digestão in vivo, in vitro
ou durante o processamento dos alimentos podem ser consideradas substâncias
bioativas. Provavelmente, esses peptídeos não possuem um potencial similar ao de
drogas comumente utilizadas no tratamento de doenças, porém podem regular alguns
processos orgânicos quando ingeridos diariamente. Em alguns casos, a expressão do
potencial biológico ocorre num sinergismo entre os peptídeos e substâncias não
protéicas (SHANBACHER et al., 1998).
Há uma série de peptídeos bioativos que compõem a estrutura primária das
caseínas, porém que não apresentam atividade biológica enquanto a proteína se
mantém intacta. Sua ativação depende da ocorrência de proteólise que pode acontecer
durante o processamento do alimento protéico ou no decorrer de sua digestão.
No caso do leite, a proteólise pode acontecer de forma natural, pela presença de
enzimas ou bactérias ácido-lácticas. Ocorre nos processos de fermentação para

14
elaboração de produtos de valor agregado ou na maturação dos queijos. Os
caseinofosfopeptídeos são constituintes naturais dos queijos e o processo de
maturação leva ao aparecimento de peptídeos com atividade anti-hipertensiva. Leites
fermentados com a utilização de Lactobacillus helveticus e Saccharomyces cerevisae
produzem β-casoquininas e β-caseína (169 – 175), um peptídeo com moderada
atividade inibidora da enzima conversora da angiotensina (ECA), com atividade no
sistema cardiovascular (MEISEL; BOCKELMANN, 1999; GOBETTI et al., 2002).
No processo digestivo, após a ingestão de leite se detecta uma considerável
quantidade de proteínas do soro no jejuno e somente traços de caseína intacta. A
diferença no tempo de esvaziamento gástrico entre proteínas do soro e frações
caseínicas se dá pela alta solubilidade das primeiras. O coágulo de caseína formado no
meio ácido estomacal permanece sofrendo hidrólise e é liberado na forma de
peptídeos, com potencial atividade biológica como o glicomacropeptídeo (GMP),
peptídeos com atividade opióide (casomorfinas) e fragmentos fosfopeptídicos (TOMÉ;
DEBABBI, 1998).
A maioria dos peptídeos derivados de caseína que apresentam atividade
biológica é produzida in vitro pelo uso de proteinases pancreáticas, especialmente
tripsina. Combinações de endo-proteinases também podem ser utillizadas, incluindo
quimotripsina, pepsina, termolisina, pancreatina, carboxipetidase entre outras. Essas
enzimas podem ser de origem microbiana, vegetal ou animal (GOBETTI et al., 2002).
2.1.5 Funções dos Peptídeos Bioativos
Os peptídeos com atividade fisiológica derivada da caseína podem exercer
funções em diferentes sistemas do organismo, conforme esquema apresentado na
Figura 2. Algumas seqüências peptídicas são consideradas “zonas estratégicas” por
apresentarem mais de uma atividade biológica. Estas zonas são parcialmente
protegidas contra a proteólise, de maneira que a atividade fisiológica é preservada
durante a digestão ou processamento. A Figura 3 representa esta “multi-
funcionalidade”.
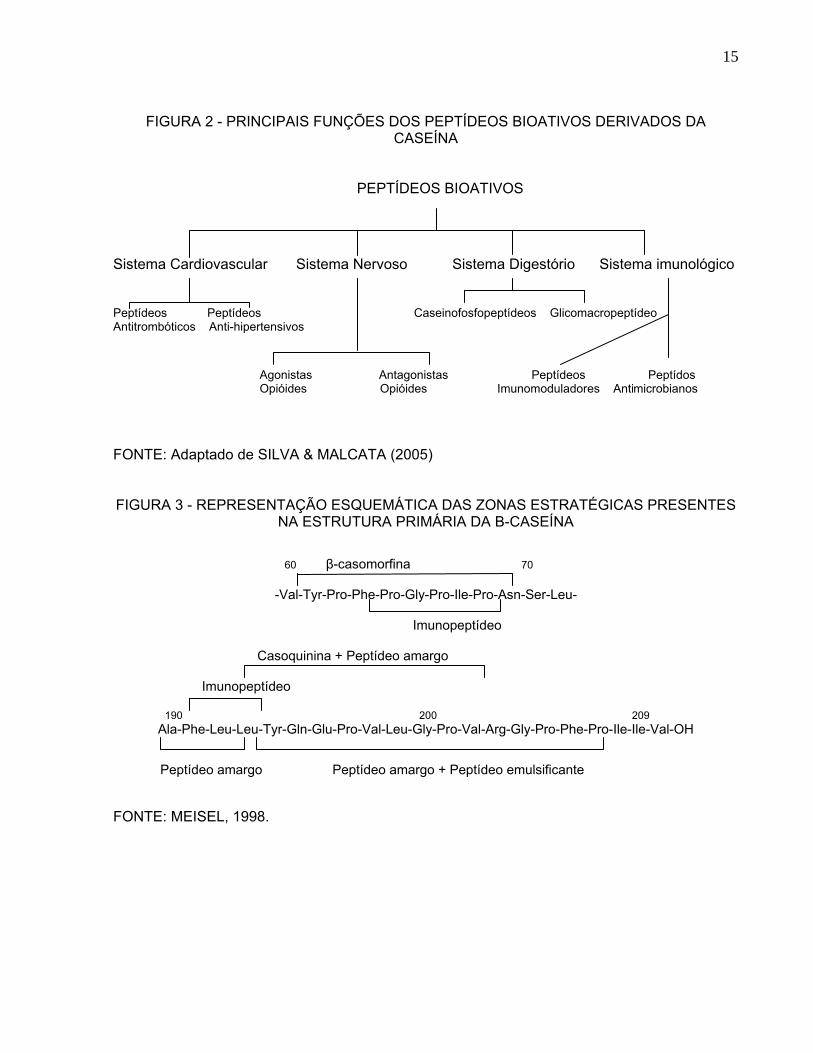
15
FIGURA 2 - PRINCIPAIS FUNÇÕES DOS PEPTÍDEOS BIOATIVOS DERIVADOS DA CASEÍNA
PEPTÍDEOS BIOATIVOS
Sistema Cardiovascular Sistema Nervoso Sistema Digestório Sistema imunológico
Peptídeos Peptídeos Caseinofosfopeptídeos Glicomacropeptídeo Antitrombóticos Anti-hipertensivos
Agonistas Antagonistas Peptídeos Peptídos Opióides Opióides Imunomoduladores Antimicrobianos
FONTE: Adaptado de SILVA & MALCATA (2005)
FIGURA 3 - REPRESENTAÇÃO ESQUEMÁTICA DAS ZONAS ESTRATÉGICAS PRESENTES NA ESTRUTURA PRIMÁRIA DA Β-CASEÍNA
60 β-casomorfina 70
-Val-Tyr-Pro-Phe-Pro-Gly-Pro-Ile-Pro-Asn-Ser-Leu-
Imunopeptídeo
Casoquinina + Peptídeo amargo
Imunopeptídeo
190 200 209 Ala-Phe-Leu-Leu-Tyr-Gln-Glu-Pro-Val-Leu-Gly-Pro-Val-Arg-Gly-Pro-Phe-Pro-Ile-Ile-Val-OH Peptídeo amargo Peptídeo amargo + Peptídeo emulsificante
FONTE: MEISEL, 1998.

16
2.1.5.1 Sistema Cardiovascular
Alguns autores apontam semelhanças entre os mecanismos envolvidos na
formação do coágulo de leite e no processo de coagulação do sangue. Existem
algumas homologias entre os resíduos 106 - 116 da κ-caseína e os resíduos 400 - 411
do fibrinogênio humano. Um peptídeo denominado casopiastrina foi obtido pela
hidrólise tríptica da κ-caseína e demonstrou atividade antitrombótica por impedir a
ligação do fibrinogênio às plaquetas (SMACHI; GOBETTI, 2000).
As casoplatelinas são peptídeos derivados da clivagem da κ-caseína (f106 –
116, f106 – 112, f 113 – 116) que possuem atividade inibidora da agregação plaquetária
(JOLLÉS et al., 1986).
A regulação da pressão sanguínea é parcialmente dependente do sistema
renina-angiotensina. A renina age sobre a angiotensina liberando a angiotensina I. Esta
se converte em angiotensina II, um hormônio vasoconstritor, pela ação do sistema da
enzima conversora da angiotensina (ECA ou ACE). A angiotensina II inativa a
bradiquinina (um vasodilatador) aumentando a produção de aldosterona que diminui a
excreção renal e aumenta a retenção de líquidos pelo organismo (SILVA; MALCATA,
2005).
A enzima conversora da angiotensina (ECA ou ACE) se localiza em vários
tecidos. A inibição da ECA pode influenciar diferentes sistemas de regulação do
organismo envolvidos na modulação da pressão arterial, defesa imune e atividade do
sistema nervoso (MEISEL, 1998).
Uma característica estrutural da maioria dos peptídeos inibidores de ECA e de
alguns peptídeos imunomoduladores é a presença de um resíduo Arg carboxi-terminal
(MEISEL; GÜNTHER, 1998)
São encontrados peptídeos inibidores de ECA a partir de algumas fontes
vegetais, como o glúten, caseínas humanas e bovinas e a partir de veneno de cobra
(SMACHI; GOBETTI, 2000).
A inibição da ECA é registrada em peptídeos obtidos a partir de αs1-caseína (f23
– 34, 23 – 27, 294 – 299), β-caseína bovina (f177 – 183), β-caseína humana (f43 – 52)

17
e κ-caseína (f63 – 65). Esses peptídeos são conhecidos como casoquininas
(SCHANBACHER et al., 1998).
2.1.5.2 Sistema Nervoso
Peptídeos derivados da caseína podem desempenhar funções regulatórias no
sistema nervoso. São conhecidos como peptídos opióides, podendo exercer atividades
agonistas ou antagonistas.
Estudos farmacológicos, bioquímicos e de comportamento têm estabelecido a
existência de pelo menos três tipos de receptores opióides no sistema nervoso central e
periférico dos mamíferos. Esses receptores opióides interagem com moléculas
protéicas que se ligam a eles e que podem ser de origem endócrina ou exócrina (BITRI,
2004).
Os peptídeos opióides “típicos” são a encefalina, a endorfina e a dinorfina que
são peptídeos derivados da propiomelanocortina, da proencefalina e da prodinorfina.
Essas substâncias apresentam uma seqüência N-terminal semelhante: Tyr-Gly-Gly-
Phe. Os peptídeos “atípicos” com atividade opióide são derivados das proteínas lácteas
e apresentam seqüências N-terminais diferentes dos “típicos”. Entretanto, ambos
grupos possuem algumas semelhanças estruturais, como a presença de um resíduo
tirosina amino terminal e de resíduos aromáticos (tirosina ou fenilalanina) na 3ª ou 4ª
posição. Peptídos com ausência de tirosina não apresentam atividade opióide (MEISEL,
1998).
Os peptídeos agonistas opióides derivados da caseína possuem de 5 a 10
resíduos e são conhecidos como casomorfinas ou exorfinas. Ligam-se aos receptores
opióides nas células intestinais e outros tecidos. São obtidos a partir da β-caseína (f60-
70 – β-casomorfina) e αs1-caseína (f90 – 96 – α-casomorfina). A morficetina é o
peptídeo com maior potencial opióide. Pertence ao grupo da β-casomorfina e apresenta
a seguinte seqüência N-terminal: Tyr-Pro-Phe-Pro. Estudos têm demonstrado que as
casomorfinas são responsáveis pela produção de analgesia, aumento do tempo de
trânsito intestinal, efeitos antidiarréicos, aumento na absorção de aminoácidos e

18
eletrólitos e estímulo da secreção de insulina e somastostatina (MEISEL; SCHLIMME,
1990; KORHONEN et al., 1998; SCHANBACHER et al., 1998; SHAH, 2000).
BRANTL (1985) seqüenciou β-casomorfinas humanas e avaliou sua atividade
opióide, tendo verificado que as β-casomorfinas bovinas apresentam um potencial
opióide mais intenso (Tabela 3).
A penetração de peptídeos pela barreira hemato-encefálica em adultos é restrita,
porém há evidências de que sob determinadas condições alguns oligopeptídeos podem
ultrapassar essa barreira. UMBACH et al. (1985) verificaram a presença de β-
casomorfina imunoreativa no plasma de bezerros e apontam a possibilidade de
receptores opióides do sistema nervoso central apresentarem maior afinidade pela β-
casomorfina do que pela morfina.
TABELA 3 - ATIVIDADE OPIÓIDE DE Β-CASOMORFINAS HUMANA E BOVINA EM
COMPARAÇÃO À NORMORFINA
SUBSTÂNCIA GPI*
β-casomorfina -4 : (Tyr-Pro-Phe-Pro) 14,3
β-casomorfina -4 humana: (Tyr-Pro-Phe-Val) 56,2
β-casomorfina -5: (Tyr-Pro-Phe-Pro-Gly) 2,0
β-casomorfina -5 humana: (Tyr-Pro-Phe-Val-Gly) 33,1
Normorfina 0,1
* Os valores indicam concentrações µM, causando 50% de inibição das contrações elétricas induzidas no plexo mioentérico do íleo de roedores por preparação longitudinal do músculo (GPI). BRANTL (1985)
A hidrólise das proteínas lácteas também leva à formação de peptídeos opióides
com atividade antagonista. Estes se denominam casoxinas, quando obtidos a partir das
caseínas. Surgem em concentrações mais elevadas em leite proveniente de animais
com mastite e promovem aceleração no trânsito intestinal. As casoxinas A e B são
derivadas da κ-caseína, sendo que a casoxina A corresponde ao fragmento 35 – 41 e a
casoxina B ao fragmento 58 – 61. A casoxina C (f 25 – 34) é obtida da hidrólise tríptica
da κ-caseína e apresenta o maior potencial biológico dentre as demais. A casoxina D é
formada a partir da αs1-caseína. Algumas casoxinas são metoxiladas durante os

19
processos de isolamento e purificação e se tornam mais ativas (MEISEL;
BOCKELMAN, 1999; SILVA; MALCATA, 2005).
2.1.5.3 Sistema Imunológico
Os peptídeos do leite apresentam propriedades reguladoras no desenvolvimento
do sistema imunológico. Essas propriedades podem ser classificadas em dois grandes
grupos: a imunomodulação e a atividade antimicrobiana.
Sabe-se que o leite contém inúmeras substâncias que protegem a criança contra
infecções (COSTE et al., 1992; SMACHI; GOBETTI, 2000).
Estudos experimentais com animais demostraram que o concentrado protéico de
soro de leite apresenta atividade anticarcinogênica. O concentrado protéico de soro
induziu o aumento de glutationa nos tecidos, diminuindo o volume tumoral,
provavelmente devido ao efeito estimulador da glutationa sobre a resposta imunológica
(BOUNOUS, 2000).
MONTAGNIER; OLIVIER e PASQUIER (1998), estudaram o efeito do
ImmunocalTM, produto do soro de leite patenteado no Canadá, em crianças com
síndrome da imunodeficiência adquirida (AIDS), e constataram que o tratamento é bem
tolerado. A utilização do concentrado de soro melhorou o estado nutricional dos
pacientes, com reflexo no peso e nas medidas antropométricas.
MORENO (2002) investigou o papel imunomodulador do concentrado protéico de
soro doce (obtido no Instituto de Tecnologia de Alimentos ITAL-Campinas) em crianças
com AIDS, e constatou que a suplementação nutricional com um concentrado protéico
rico em cisteína aumenta os níveis de glutationa nos eritrócitos e induz a melhora da
relação entre os linfócitos T CD4+/CD8+. O hemograma e a liberação de intermediários
reativos do oxigênio mantiveram-se sem modificações. Não foram observadas
alterações nos marcadores de estresse oxidativo das crianças envolvidas no ensaio
clínico, situação semelhante aos resultados obtidos com adultos infectados pelo HIV
suplementados com as proteínas do soro de leite bovino. As crianças suplementadas
apresentaram diminuição da ocorrência de episódios infecciosos.

20
DIAS et al. (1999) investigaram o efeito das proteínas do lactosoro no sistema
imunológico de camundongos. Os autores utilizaram animais recém-desmamados, que
receberam dietas durante 21 dias contendo 20% de proteína e água ad libitum. Após 15
dias, os animais foram injetados com 5x106 hemácias de carneiro, lavadas com solução
salina. No quinto dia após a injeção, os animais foram sacrificados e os baços e fígados
retirados. Nos testes, que verificam o numero de células formadoras de placas (CFP),
características da existência de resposta imune humoral, os camundongos tratados com
um produto de soro de leite patenteado no Canadá (Immunocal), apresentaram valores
significativamente mais elevados de CFP (95,85x103), seguidos dos camundongos
tratados com concentrados protéicos de soro ácido (obtido no ITAL). Por outro lado, os
camundongos tratados com concentrado protéico de soro doce (obtido no ITAL)
apresentaram os valores mais elevados de glutationa (8,4µmol/g de tecido).
Imunopeptídeos, obtidos da αS1, β-caseína e α-lactalbumina estimulam a
atividade fagocitária e também apresentam papel protetor contra infecções por
Klebsiella pneumoniae em camundongos. Esses peptídeos podem estimular a
proliferação e maturação dos linfócitos T e de células natural killer na proteção de
neonatos contra bactérias entéricas (SHAH, 2000; SMACHI; GOBETTI, 2000).
COSTE et al. (1992) produziram hidrolisados de β-caseína bovina utilizando
quimosina. Os peptídeos apresentavam a seqüência 193 - 209. Os autores avaliaram a
proliferação celular de células de baço de ratos não sensibilizados e células de
linfonodos inguinais de ratos sensibilizados com ovalbumina. A atividade biológica dos
peptídeos mostrou-se mais efetiva na ativação celular dos linfonodos.
Em 1996, WONG et al. comunicaram que a β-caseína bovina era capaz de
aumentar significativamente a resposta proliferativa de linfócitos ovinos, induzidos por
mitógenos.
Os mecanismos pelos quais os peptídeos lácteos exercem efeitos
imunomoduladores não estão completamente definidos. A presença do resíduo Arg nas
regiões C- ou N- terminais dos peptídeos é apontada como um elo de ligação entre o
peptídeo e receptores de membrana da superfície de células de defesa imunológica
(MEISEL, 1998).

21
Há evidências de que neonatos exibem peptídeos antibacterianos formados após
a ingestão do leite. Ainda não existem estudos in situ que comprovem esta hipótese
(MALKOSKI et al., 2001).
Peptídeos antimicrobianos têm sido identificados em frações de leite bovino,
apresentando um provável papel regulador da flora intestinal (ZUCHT et al., 1995).
Um peptídeo denominado caseicidina, obtido pela digestão da caseína com
quimosina em pH neutro possui atividade inibidora do crescimento de Staphylococcus
spp., Sarcina spp., Bacillus subtilis, Diplococcus pneumoniae e Streptococcus
pyogenes. O fragmento derivado da αs1-caseína (f1 - 23) denominado isracidina
demonstrou atividade bacteriostática contra S. aureus e Candida albicans e foi capaz
de proteger ovinos e bovinos de infecções nas glândulas mamárias (LAHOV;
REGELSON, 1996).
RECIO & VISSER (1999) identificaram dois peptídeos, obtidos da α-s2 caseína
com atividade antibacteriana contra uma série de microorganismos.
A casoplatelina é um decapeptídeo derivado da κ-caseína (fragmento 127 – 137)
com atividade na função plaquetária e provável atividade antimicrobiana (RIZZELLO et
al., 2005).
2.1.5.4 Sistema Digestório
O trato gastrintestinal pode ser considerado como uma porta de entrada para
nutrientes e como um sítio de ação para substâncias de origem alimentar que possuem
atividade biológica. Alguns produtos apresentam efeito local, outros apresentam efeito
sistêmico ao longo do sistema digestório. Podem agir como antimicrobianos, ligarem-se
a patógenos ou toxinas, seqüestrarem nutrientes microbianos, se associarem a
receptores de mucosa e renovarem o crescimento das células (HOERR; BOSTWICK,
2000).
As caseínas possuem atividade biológica relacionada ao transporte de cálcio,
zinco, cobre, ferro e fosfato. Os caseinofosfopeptídeos atuam como carreadores de
minerais.

22
O glicomacropeptídeo (GMP), que se forma na clivagem da κ-caseína, possui
uma composição aminoacídica singular. O produto não apresenta resíduos aromáticos
e é rico em resíduos ramificados, sendo uma opção interessante para a elaboração de
fórmulas alimentícias destinadas a pacientes com doenças hepáticas e desordens do
metabolismo (SILVA; MALCATA, 2005).
NAKAJIMA et al. (2005) avaliaram o efeito preventivo do GMP contra infecções
intestinais, tendo verificado que o peptídeo se liga a diversas bactérias impedindo sua
aderência em células intestinais.
2.2 CASEINOFOSFOPEPTÍDEOS
O termo caseinofosfopeptídeo (CPP) foi introduzido na década de 50 para
descrever peptídeos fosforilados derivados da caseína que apresentam a propriedade
de melhorar a calcificação de crianças portadoras de raquitismo. Essa influência de
peptídeos no metabolismo mineral partiu das observações de Mellander que incubava
caseína com pepsina e suco pancreático, obtendo uma fração peptídica resistente à
degradação posterior por outras enzimas. Os peptídeos obtidos apresentavam elevado
conteúdo de resíduos fosfoserina e aumentavam o balanço de cálcio de 39 a 78% em
neonatos raquíticos. Desde então, verificou-se cientificamente que os
caseinofosfopeptídeos possuíam a habilidade de ligar microelementos como Ca, Mg e
Fe e também, elementos-traço, como Zn, Ba, Cr, Ni, Co e Se (FITZGERALD, 1998;
SCHOLZ-AHRENS; SCHREZENMEIR, 2000).
2.2.1 Estrutura
Os primeiros estudos sobre a hidrólise da caseína, em 1895, demonstraram que
a utilização de extratos pancreáticos permitia a quebra da molécula e a obtenção de
compostos inorgânicos de fósforo. Mais tarde, verificou-se que os compostos
fosforilados derivados da caseína formavam sais insolúveis com vários metais sendo
denominados “fosfopeptonas” (RIMINGTON, 1941; KITTS; YUAN, 1992).

23
LOWNDES et al. (1941) obtiveram um polipeptídeo a partir da caseína e com uso
de tripsina, com uma relação N:P de 4:1. A caracterização desta fosfopeptona foi
realizada pelos métodos então disponíveis, revelando a presença de serina e um ácido
dicarboxílico, que aparentemente seria o ácido hidroxiglutâmico.
Estudos subseqüentes verificaram que os caseinofosfopeptídeos consistem
numa mistura de peptídeos de diferentes pesos moleculares formados in vivo quando a
caseína é degradada pelas enzimas proteolíticas no trato digestivo. Esses peptídeos,
em sua estrutura primária, contêm uma seqüência de aminoácidos carregada
negativamente em pH fisiológico, constituída pelos resíduos de ácido glutâmico e serina
fosforilada (Ser(P)-Ser(P)-Ser(P)-Glu-Glu), cuja conformação está representada na
Figura 4. (ERBA; CIAPPELANO; TESTOLIN, 2001).
FIGURA 4 - ESTRUTURA DA REGIÃO CPP
A partir de então, vários estudos têm demonstrado que a digestão da caseína
eleva a biodisponibilidade do cálcio. Os caseinofosfopeptídeos formam complexos
solúveis com fosfato de cálcio em pH alcalino. Esses complexos previnem a
precipitação do fosfato de cálcio e aumentam a concentração do cálcio solúvel in vitro e
no lúmen do intestino delgado (KITTS et al., 1992).

24
HAN; SHIN E BYUN (2000) identificaram uma nova variante genética da β-
caseína (β-caseína-H ou β-CN-H) com substituição do 25º resíduo (região CPP). Em
ratos, o efeito da solubilização do cálcio é aumentado em 23% comparado com outras
variantes genéticas.
A elevada afinidade das moléculas de caseína por cátions bi e trivalentes é
atribuída à sua fosforilação. A extensão da fosforilação das caseínas é dependente do
tipo de caseína em questão. Por exemplo, a caseína bovina αs2 possui 13 grupos
fosfato, enquanto a κ-caseína apresenta somente um grupamento. O fósforo aparece
ligado às caseínas através de ligações monoéster nos resíduos serina, criando um
domínio ácido favorável à ligação com metais. A seqüência de aminoácidos na cadeia
peptídica também é outro fator a definir o grau de fosforilação da molécula, isto é, uma
seqüência tripla de resíduos de aminoácidos aniônicos: SerP-SerP-SerP-Glu-Glu
(FITZGERALD, 1998; KITSS, 2005).
A Figura 5 apresenta as regiões fosforiladas encontradas nas caseínas de leite
bovino.
A presença de sítios fosforilados conduz a um aumento na hidrofilicidade e na
mobilidade daquele determinado sítio peptídico. Aproximadamente 80% das regiões
fosforiladas da caseína se apresentam com a estrutura β-pregueada (ZHANG; OTANI,
2003)
HUQ, CROSS e REYNOLDS (2004) investigaram a estrutura da αs1-caseína f59
– 79 utilizando ressonância nuclear magnética. Observaram a existência de alto grau de
flexibilidade e mobilidade. Esse achado se correlaciona bem com a habilidade do
peptídeo em se ligar a uma variedade de íons.
MIQUEL et al. (2006) hidrolisaram fórmulas infantis com pepsina e pancreatina,
simulando o processo fisiológico e obtiveram caseinofosfopeptídeos com massas
moleculares na faixa de 1400 a 9600Da. Em aproximadamente 50% dos CPPs
identificados obtiveram a seqüência SerP-SerP-SerP-Glu-Glu, responsável pela ligação
com minerais.

25
FIGURA 5 - REGIÕES FOSFORILADAS DE DIFERENTES TIPOS DE CASEÍNAS BOVINAS
αs1-caseína gln-met-glu-ala-glu-ser(P)-ile-ser(P)-ser(P)-ser(P)-glu-glu-ile-val-pro-asn-ser(P)val-glu-gln-lys f(59 – 79) val–pro-asn-ser(P)-ala-glu-glu-arg f(112 – 119) αs2-caseína glu-his-val-ser(P)-ser(P)-ser(P)-glu-glu-ser-ile-ile-ser(P)-gln-glu f(5 - 18) asn-pro-ser(P)-lys-glu-asn f(29 - 34) gly-ser(P)-ser(P)-ser(P)-glu-glu-ser(P)-ala-glu-val f(55 - 64) gln-leu-ser(P)-thr-ser(P)-glu-glu-asn-ser-lys-lys-thr-val-asp-met-glu-ser(P)-thr-glu-val-phe f(127 - 147) β-caseína ile-val-glu-ser(P)-ser(P)-ser(P)-glu-glu-ser-ile-lys variante A f(12 - 23) ile-val-glu-ser(P)-lys-ser(P)-glu-glu-ser-ile-lys variante B f(12 - 23) κ-caseína glu-ala-ser(P)-pro-glu-val-ile f(147 - 153)
2.2.2 Métodos de Obtenção
Caseinofosfopeptídeos têm sido produzidos in vitro por meio da utilização de
enzimas pancreáticas na digestão da caseína. Estudos in vivo também têm
demonstrado a formação de caseinofosfopeptídeos a partir de caseína em animais
experimentais (McDONAGH; FITZGERALD, 1998; BOUHALLAB et al., 1999; PÈRES et
al., 1999; MEISEL; BOCKELMANN, 1999; OTANI et al., 2000; PARK; ALLEN, 2000).
Os caseinofosfopeptídeos podem ser produzidos a partir de diferentes enzimas
de origem bacteriana, fúngica, vegetal ou animal, mas a habilidade de solubilizar o
cálcio diferencia-se com o método de obtenção. Endoproteinases de origem microbiana
resultaram na produção de CPPs de melhor qualidade e menores custos (McDONAGH;
FITZGERALD, 1998).

26
A partir de 1950, vários métodos foram estabelecidos para a produção de
fosfopeptídeos derivados da caseína, em escala laboratorial (PETERSON; NAUMAN;
McMEEKIN, 1958; MANSON & ANNAN, 1971; NAITO & SUZUKI, 1974). Entretanto,
esses métodos eram executados para produzir caseinofosfopeptídeos para fins de
pesquisa e para confirmar sua presença e pureza. Alguns reagentes utilizados, como
BaCl2, eram impróprios para uso em ingredientes alimentícios e as operações difíceis
de serem convertidas para a escala industrial.
BRULE et al. (1980) propuseram um método de obtenção de fosfocaseinatos a
partir da hidrólise enzimática, seguida da agregação de cátions divalentes ao
hidrolisado e de ultrafiltração. Apontam os produtos obtidos como ingredientes
terapêuticos a serem incluídos em dietas para alimentação oral ou enteral.
Em 1987, REYNOLDS propôs a extração de fosfopeptídeos a partir de solução
de caseinato de sódio digerida com tripsina por 1 hora, a 37ºC e pH 5. Neste método,
ajusta-se o pH para 4,7 e removendo-se o precipitado resultante. Adiciona-se BaCl2
(0,25% p/v) seguido de igual volume de etanol absoluto e o precipitado resultante é
removido e seco. O precipitado é dissolvido 10 vezes e a solução acidificada. Adiciona-
se igual volume de acetona e o precipitado é novamente removido e seco. O
precipitado é então dissolvido em água e acidificado para pH 2 com HCl. O precipitado
resultante é removido e descartado e o sobrenadante ajustado para pH 3,5 com NaOH
e adiciona-se volume igual de acetona. Coleta-se o precipitado resultante, redissolve-se
em água e adiciona-se H2SO4. O sobrenadante é dialisado e liofilizado. Obtém-se uma
mistura de cinco fosfopeptídeos.
A utilização de uma matriz de quitosana para adsorver os fosfopeptídeos é
preconizada em uma patente (KOIDE et al., 1994). A adsorção dos fosfopeptídeos
começa no momento em que a solução de caseína digerida e com pH na faixa de 2,5 a
4,5 entra em contato com a quitosana. Pode-se eluir os 10 a 20% de fosfopeptídeos
que ficam adsorvidos após a lavagem da matriz com água pela adição de 0,2 a 1mol/L
de NaCl. Após, os peptídeos podem ser facilmente separados por métodos
convencionais de separação sólido-líquido. Pode-se usar centrifugação, decantação,
filtração ou filtração por membranas. Para a adsorção e desorção dos fosfopeptídeos

27
com quitosana, pode-se usar uma coluna empacotada com a mesma, tanque ou
fermentador usado rotineiramente na indústria láctea.
REYNOLDS, RILEY e ADAMSON (1994) descreveram um método de purificação
de fosfopeptídeos a partir da precipitação com cálcio e etanol de digestos trípticos de
caseína. Os peptídeos fosforilados passam por processos de purificação pelo uso de
FPLC.
McDONAGH e FITZGERALD (1998) obtiveram fosfopetídeos a partir de
hidrolisados de caseinato de sódio utilizando agregação de cloreto de cálcio em pH
neutro seguido de precipitação destes agregados com etanol, conforme segue: o
sobrenadante resultante da precipitação a pH 4,6 é elevado para o pH 7,0 com o
acréscimo de solução 2N de NaOH. Adiciona-se CaCl2 até a concentração final de 1%
(p/v) e as soluções são deixadas por 1 hora à temperatura ambiente. Acrescenta-se
etanol a 50% (v/v) e o precipitado resultante é coletado por centrifugação a 6000xg por
10 minutos.
Os métodos de obtenção dos caseinofosfopeptídeos são, em sua maioria,
patenteados. FITZGERALD (1998), em sua revisão propõe a elaboração dos
caseinofosfopeptídeos, conforme apresentado na Figura 6.
PÉRÈS et al. (1999) isolaram β-caseína de caseinato de sódio comercial por
solubilização a frio (pH 4,5 e 4 ºC), seguida de cromatografia de troca iônica. Obtiveram
o caseinofosfopeptídeo (β-caseína, fragmento 1 – 25) por hidrólise tríptica e
precipitação com cálcio e etanol. Os autores ligaram Fe ao caseinofosfopeptídeo
adicionando solução de FeCl2 (4.10-2M; pH 5,3; 30 minutos e 25ºC). O excesso de ferro
(não ligado) foi descartado por ultracentrifugação e diafiltração numa membrana de
celulose regenerada com um “cutt off” de 3.000Da. O complexo obtido foi liofilizado.
Verificou-se que 1 mol do peptídeo obtido foi capaz de ligar 4 moles de Fe.
PARK e ALLEN (2000) separaram a caseína de leite integral utilizando a
precipitação no ponto isoelétrico, seguida de separação das frações por coluna de troca
aniônica. Os autores prepararam hidrolisados utilizando tripsina (Sigma) e GSE SP 446
(Novo Nordisk). Separaram, então, os caseinofosfopeptídeos, pelo emprego de coluna
QAE-Sephadex A 25, que foi equilibrada com 50mM de tampão Tris-HCl, pH 8,0. As

28
frações αs e β-caseína hidrolisadas foram colocadas na coluna com uma taxa de fluxo
de 3mL/min., monitorando-se o eluído a 280nm.
FIGURA 6 - MÉTODOS DE ELABORAÇÃO DE CASEINOFOSFOPEPTÍDEOS
agregação de CaCl2ultrafiltração/diafiltração
desidratação ou liofilização
CPP
agregação de CaCl2precipitação c/ etanol
separação cromatográfica
remoção do material não peptídico(centrifugação/ultrafiltração)
digestão por endoproteinase(pH 7,0 - 8,0)
Caseinato de sódio
ELLEGARD et al. (1999) propuseram um método de obtenção de
caseinofosfopeptídeos em escala industrial utilizando hidrólise tríptica, precipitação
ácida, diafiltração e cromatografia de troca aniônica. O produto resultante se caracteriza
como de alta pureza e boa funcionalidade.
OTANI et al. (2000) prepararam caseinofosfopeptídeos a partir de solução de
caseína a 15% (p/v) de pH 7,0 tratada com tripsina bovina (Novo Industry,
caseína:tripsina = 100:0,01, p/p) por 2 horas a 60ºC. O pH da solução de caseína
tratada é ajustado para 4,5 e o precipitado formado removido por ultrafiltração.

29
Adiciona-se cloreto de cálcio e etanol ao filtrado até uma concentração final de 1,1%
(p/v) e 50% (v/v), respectivamente. O precipitado formado é então coletado e
desidratado. O produto obtido consiste de 12% (p/p) de caseinofosfopeptídeos dos
resíduos 1 – 32 da caseína bovina αs2 e dos resíduos 1 – 28 da caseína bovina β.
Os caseinofosfopeptídeos têm sido produzidos em escala industrial no Japão,
por meio de hidrólise tríptica da caseína. São considerados alimentos funcionais e
utilizados para elevar a biodisponibilidade de cálcio no trato intestinal (OTANI;
WATANABE; TASHIRO, 2001).
2.2.3 Funções
Os caseinofosfopeptídeos são formados in vivo durante a digestão da caseína.
Com o objetivo de avaliar a distribuição do CPP no trato digestivo de ratos (Sprague-
Dawley), TAKANORI et al. (1995) coletaram o conteúdo do trato digestivo após 4 e 10
horas da ingestão de caseína. Os autores observaram as maiores taxas no íleo após 4
horas (5,2%) sendo que houve redução significativa no conteúdo após 10 horas (1,4%).
Também foi observada a presença de CPP no ceco e cólon intestinal.
A presença de caseinofosfopeptídeos também foi observada no íleo de adultos
humanos após algumas horas da ingestão de leite. Como o trato epitelial intestinal
representa a porta de entrada para os componentes dos alimentos, várias preparações
de caseinofosfopeptídeos foram testadas em relação à possível citotoxicidade. Não
foram observadas respostas citotóxicas dos caseinofosfopeptídeos com o uso de
modelos citoquímicos recomendados pela literatura (HARTMANN et al., 2000;
HARTMANN; MEISEL, 2002).
PARK e ALLEN (2000) utilizaram um modelo animal para verificar o potencial
alergênico dos caseinofosfopeptídeos. Verificaram que os fosfopeptídos derivados da
hidrólise tríptica da αs-caseína possuem imunogenicidade significativamente menor do
que a αs-caseína intacta ou seus hidrolisados.
Ratos que receberam injeção intraperitoneal de caseinofosfopeptídeos não
apresentaram diferença significativa nos níveis de IgE em relação a animais não
imunizados, indicando baixa alergenicidade do produto (OTANI et al, 2000).

30
A antigenicidade de uma proteína tem sido associada à hidrofilicidade,
mobilidade e propensão à formação de estruturas β-pregueadas. Esses requisitos são
atendidos pelos sítios fosforilados das β-caseínas, indicando uma alta possibilidade
dessas regiões se localizarem nos epítopos ou muito próximas a eles. ZHANG e OTANI
(2003) num estudo com camundongos confirmaram que os anticorpos reconhecem as
regiões fosforiladas da β-caseína como epítopos intactos ou parte deles. Os anticorpos
reconheciam a seqüência Ser(P)-Ser(P)-Ser(P), mas não mostraram reatividade com a
β-caseína desfosforilada ou com peptídeos contendo dois resíduos serina fosforilados,
intercalados por qualquer resíduo não-fosforilado (Ser(P)-X-Ser(P)).
Desta forma, segmentos fosforilados contendo dois resíduos serina provenientes
da β-caseína ou fosfopeptídos derivados da hidrólise tríptica da αs-caseína podem ser
utilizados na elaboração de fórmulas hipoalergênicas.
Várias formulações alimentícias têm sido citadas como carreadores em potencial
para os fosfopeptídeos. Estas incluem: produtos de confeitaria, alimentos matinais,
produtos de panificação e confeitaria, sorvetes e sobremesas congeladas, leite e
produtos lácteos, sucos, chocolates, chás e maionese (FITZGERALD, 1998).
As principais aplicações dos caseinofosfopeptídeos estão resumidas na Figura 7.
FIGURA 7 - APLICAÇÕES POTENCIAIS DOS CASEINOFOSFOPEPTÍDEOS
CASEINOFOSFOPETÍDEOS
Suplementação mineral Efeito anticariogênico Humanização do leite bovino Imunomodulador Antioxidante

31
2.2.3.1 Suplementação Mineral
Os fosfopeptídeos possuem um alto conteúdo de cargas negativas e sítios de
ligação fosfoseril ou carboxílicos que os tornam eficientes na formação de complexos
solúveis com cátions divalentes. São relatados complexos formados entre
fosfopeptídeos e diversos minerais (VEGARUD; LANGSRUD E SVENNING, 2000).
Quando são comparados os níveis intestinais de isótopos de Ca45 entre animais
que recebem diferentes fontes protéicas em suas dietas (caseína, glúten, gelatina,
caseína desfosforilada, albumina de ovo, soro de leite ou isolado protéico de soja) se
torna claro que a fosforilação da caseína é um pré-requisito indispensável para o efeito
significativamente maior dessa proteína na solubilidade do mineral (SCHOLZ-AHRENS;
SCHREZENMEIR, 2000).
O leite e os produtos lácteos representam uma excelente fonte de cálcio. As
formas básicas de absorção de cálcio pelo homem são: um sistema dependente de
vitamina D localizado no duodeno e jejuno. Neste, o fator chave é uma proteína
("calcium-binding protein" - CaBP) que facilita a difusão do mineral pelas membranas,
participando do transporte ativo. Esta proteína é estimulada pela vitamina D ou por seu
metabólito ativo (1,25 - (OH)2 - D3). Outra forma de absorção se faz por meio de um
sistema passivo, independente de vitamina D localizado no íleo. O transporte passivo
constitui o maior canal de absorção do mineral derivado da dieta. Existe também, um
sistema pinocítico de absorção, importante em crianças. Vários fatores dietéticos
podem afetar a absorção do mineral: presença de fitatos, oxalatos, compostos
fenólicos, lactose, fósforo e vitamina D. Por exemplo, têm sido bem documentado que a
presença de fitatos em isolados protéicos de soja reduz a absorção do mineral por
complexar com o cálcio, formando quelatos, altamente insolúveis (KITTS et al., 1992).
Quando diferentes doses de CPP são incubadas in vitro, elevadas quantidades
de cálcio são transportadas do íleo numa relação dose-dependente, sendo que a
absorção ocorre pelo sistema passivo. Além disso, a formação de complexos solúveis
entre o peptídeo e o mineral previne a precipitação do fosfato de cálcio e, como

32
conseqüência, aumenta a quantidade de cálcio solúvel (SCHOLZ-AHRENS;
SCHREZENMEIR, 2000).
A avaliação da habilidade de CPP (contendo 80% de peptídeos fosforilados e
obtida pela remoção seletiva dos não-fosforilados) em solubilizar o cálcio foi comparada
com hidrolisado protéico de soro de leite. Uma solução de cloreto de cálcio foi
preparada e teve seu pH elevado para 7,4 com a adição de fosfato de potássio, tendo
sido adicionada ao CPP ou ao hidrolisado de soro. O aumento progressivo da solução
de CPP levou à prevenção da precipitação de fosfato de cálcio em pH neutro, sendo
que 150mg de CPP ligaram 100mg de cálcio. O hidrolisado de soro de leite não
solubilizou o cálcio (STEINJS, 2001).
Embora a aplicação potencial dos CPP se relacione à sua habilidade em
solubilizar minerais, são observadas diferenças de solubilidade em diferentes estudos
realizados. Essas disparidades podem ser atribuídas a diferenças na metodologia
utilizada para medir extrínsica e intrínsicamente a absorção de cálcio (KITTS et al.,
1992; MEISEL; OLEIMAN, 1998)
A utilização do cálcio absorvido na mineralização óssea é um método alternativo
para avaliar a biodisponibilidade do mineral (KITTS; YUAN, 1992)
As investigações do papel do CPP em melhorar a absorção de cálcio também
apresentam resultados contraditórios. Estudos em suínos em crescimento e em ratas
lactentes apontam que a suplementação de CPP não foi capaz de influenciar a
mineralização óssea. Por outro lado, a adição de CPP às dietas de ratas
ovariectomizadas preveniu o declínio na densidade mineral óssea. Galinhas normais e
raquíticas tiveram aumento da absorção do mineral. Ratos alimentados com dieta
contendo 200g/kg de caseína tiveram a formação de 5 mg de CPP (aproximadamente
1,6µmol de CPP de β-caseína), que evitou a precipitação de 20µmol de cálcio no íleo
distal. O caseinofosfopeptídeo também foi capaz de aumentar a incorporação de Ca2+
aos espermatozóides facilitando sua penetração nos oócitos de suínas (SATO et al.,
1991; TSUCHITA et al., 1996; SCHOLZ-AHRENS; SCHREZENMEIR, 2000;
FERRARETTO et al., 2001)

33
Provavelmente as discrepâncias entre resultados se devem a diferenças no
delineamento experimental, estado dos animais ou composição química dos
caseinofosfopeptídeos utilizados.
BOUHALLAB et al. (2002) estudaram a influência de diferentes métodos de
preparação de caseinofosfopeptídeos na absorção de ferro. Concluíram que a eficiência
do CPP em aumentar a absorção de ferro depende de sua origem e propriedades
estruturais. Caseinofosfopeptídeos produzidos pela digestão de β-caseína resultaram
em índices maiores de absorção do mineral do que CPP produzido a partir de caseína
integral e de frações obtidas de αs-caseína.
A biodisponibilidade do ferro é melhorada pela ligação com os
caseinofosfopeptídeos por diversos mecanismos: a hidrólise protéica melhora a
absorção do mineral, provavelmente por melhorar sua solubilidade, que se correlaciona
com sua melhor absorção. A ligação do ferro ao CPP forma complexos solúveis e
resistentes a alterações no pH e força iônica, que ocorrem no trato digestório. Frações
de CPP não digeridos são encontradas nas fezes permitindo um maior contato entre o
ferro solúvel e a mucosa. O ferro também pode ser absorvido por endocitose, ligado ao
CPP, sendo os peptídeos fragmentados dentro do enterócito (AÏT-OUKHATAR et al.,
1997; PÉRÈS et al., 1999; YEUNG; GLAHN; MILLER, 2002).
FERRARETTO et al. (2001) investigaram as interações entre o CPP e as células
intestinais, utilizando o cultivo de células tumorais humanas (HT-29). Sugerem que a
ação do CPP no fluxo de cálcio ocorra pela formação de canais próprios ou pelo
carreamento do mineral via endocitose.
Algumas evidências epidemiológicas, clínicas e de experimentação animal
apontam uma possível relação entre a suplementação de cálcio e a redução nos níveis
de pressão arterial. O íon Ca2+ possui atividade na regulação do tônus da musculatura
vascular. KITTS et al. (1992) avaliaram o papel da ingestão de cálcio ou de
fosfopeptídeos que melhoram a biodisponibilidade do mineral no desenvolvimento da
hipertensão. Os autores concluíram que o uso dessa suplementação não leva a uma
redução significativa nos níveis arteriais de ratos espontaneamente hipertensos.

34
A maioria dos dados apresentados na literatura sobre os efeitos do CPP na
absorção mineral se baseia em estudos in vitro, in situ e de cultivo celular. Muitos foram
realizados com roedores. Ao se realizar um balanço entre os resultados apresentados,
se conclui que há elevada evidência de que estes peptídeos melhoram a absorção
mineral. Entretanto, há necessidade de estudos longitudinais em humanos, com
monitoramento de dados relativos a mineralização óssea.
Um estudo realizado avaliou pacientes acima de 50 anos que receberam
suplementação de 1200mg/dia de cálcio bioativo (cálcio ligado ao CPP), num período
de 34 meses. Houve redução significativa nas fraturas ósseas e diminuição dos custos
médicos diretos (HANSEN, 1995).
2.2.3.2 Efeito Anticariogênico
O consumo de produtos lácteos e leite está associado à redução no
aparecimento de cáries dentárias. Essas estão relacionadas à desmineralização
dentária gerada por ácidos orgânicos produzidos por bactérias odontopatogênicas
durante a fermentação dos açúcares da dieta. Fosfopeptídeos possuem também a
habilidade de prevenir a formação de cálculos dentários, que constituem um depósito
de fosfato de cálcio na forma de hidroxiapatita. Além disso, seqüestram minerais
antibacterianos como Zn, Cu, Sn, Ag e Al. Agem como agentes tampões contra a placa
dentária ácida. Devido a essa variedade de aplicações odontológicas, os CPPs podem
ser utilizados em produtos para a higiene dental (FITZGERALD, 1998).
REYNOLDS et al. (1995) utilizaram um modelo animal para comparar a atividade
do CPP com o flúor na redução de cáries. Constataram que o uso de CPP (0,5 – 1%
p/v), duas vezes ao dia nos dentes é equivalente ao uso de 500ppm de flúor.
A aderência de S. sobrinus e S. mutans à película salivária foi reduzida pela
incorporação de CPP. Provavelmente se dá a formação de um biofilme que aumenta a
remineralização dos dentes e impede a adesão bacteriana (SCHÜPBACH et al., 1996)
O CPP pode ser incorporado a uma série de produtos para higiene oral. Após 3
horas do uso de goma de mascar de xilitol contendo CPP ainda é possível se detectar a

35
presença do peptídeo na superfície dos dentes. O uso de enxagüatório bucal contendo
CPP aumenta o nível de cálcio e fosfato na região oral. Pasta dentária com CPP
aumenta a remineralização impedindo a formação de placas dentárias. O uso de pasta
dental contendo CPP também reduz a sensibilidade em indivíduos que sofrem de
elevada sensibilidade dos dentes em sua higiene oral (YAMAGUCHI et al., 2006).
Os derivados de leite constituem o grupo de alimentos com maior atividade
anticariogênica. WALKER et al. (2006) verificaram que a adição de CPP aumenta
significativamente a habilidade do leite bovino em remineralizar lesões dentais in situ.
2.2.3.3 Humanização do Leite Bovino
O leite bovino possui uma relação entre fósforo orgânico/fósforo total de 0,34,
enquanto no leite humano esta relação é de 0,83. BRULE et al. (1980) sugerem a
incorporação de CPP às formulações destinadas a lactentes como uma forma de
aproximá-las do leite humano.
A fosfatase alcalina é uma fosfatase específica que se liga às microvilosidases
apicais da borda em escova e que tem como função melhorar a absorção de fosfato. A
fosfatase alcalina intestinal de bezerros é capaz de hidrolisar resíduos de fosfoserina,
mas possui baixa atividade em hidrolisar a caseína intacta. A enzima hidrolisa somente
10% dos resíduos de fosfoserina presentes numa dispersão de caseína intacta
incubada a 37ºC por 60 minutos. Já, quando a mesma quantidade de enzima age sobre
αs1- ou β-caseínas, hidrolisa 50% dos resíduos fosfoserina em 30 minutos (YEUNG;
GLAHN; MILLER, 2002).
2.2.3.4 Modulação do Sistema Imunológico
Em 1998, MEISEL e GÜNTHER em comunicação prévia, relatam que até aquele
momento não haviam evidências na literatura de que fosfopeptídeos pudessem exercer
atividades imunomodulatórias. Entretanto, estudo realizado pelos mesmos autores

36
demonstrou que tanto os caseinofosfopeptídeos como seus derivados desfosforilados
possuem efeitos imunomoduladores.
HATA; UEDA; OTANI (1999) avaliaram a atividade imunoestimuladora de
fosfopeptídeos derivados da caseína em culturas de células de baço, timo e intestino de
ratos. Os autores apontam que o resíduo 59 - 79 da αs1-caseína, derivada de hidrólise
tríptica, é responsável pelo aumento da produção de imunoglobulinas nas células de
baço desses animais.
KIHARA e OTANI (2000) produziram caseinofosfopeptídeos com diversas
seqüências e observaram que aqueles cuja matéria-prima recebeu tratamento térmico
antes do processo apresentaram atividade mitogênica sobre células de baço de ratos.
Esses peptídeos também aumentaram as respostas proliferativas nas células, induzidas
por lipopolissacarídeos e concanavalina-A.
OTANI et al. (2000) estudaram o efeito da administração oral de
caseinofosfopeptídeo em camundongos. Os resultados obtidos pelo grupo indicam um
aumento na produção de imunoglobulina A (IgA) pela mucosa intestinal nos animais
estudados.
Em 2001, OTANI; WATANABE; TASHIRO, investigaram o centro ativo dos
caseinofosfopeptídeos responsável pela proliferação de linfócitos e produção de IgA,
utilizando o fragmento 1-28 da β-caseína. Os autores avaliaram a proliferação de
linfócitos B estimulados com LPS derivada de Salmonella typhimurium e de linfócitos T
pelo uso de PHA e concanavalina A, bem como a produção de IgA secretória. Os
ensaios foram realizados em células de baço de camundongos. Os autores concluíram
que os fosfopeptídeos que contêm a seqüência SerP- X – SerP são responsáveis pela
atividade imunoregulatória. Propuseram então a utilização do tripeptídeo SerP- X –
SerP como ingrediente alimentício adjuvante no estímulo da produção de IgA.
Indivíduos que ingeriram tortas contendo CPP por um período de 15 dias
apresentaram valores aumentados de IgA secretória, sendo que a resposta foi maior
naqueles indivíduos que raramente consumiam leite e seus derivados (KITAMURA;
OTANI, 2002).

37
Alguns peptídeos antibacterianos são liberados após a ingestão do leite,
contribuindo na função do sistema imunológico dos neonatos. O glicomacropeptídeo
(GMP) tem sido apontado com capacidade de inibir a aderência de Streptococcus
mutans à película salivária e de inibir o crescimento de patógenos orais como o
Streptococcus mutans e Porphyromonas gingivalis, assim como de Escherichia coli.
Esta forma ativa do GMP foi designada kapacina e possui homologia estrutural com
peptídeos catiônicos anfipáticos presentes em vertebrados (MALKOSKI et al., 2001).
2.2.3.5 Antioxidante
Os caseinofosfopeptídeos exibiram atividade antioxidante primária e secundária
em modelo que avaliou a degradação da desoxiribose. Os caseinofosfopeptídeos
também protegeram os lipossomas de soja da oxidação gerada pela adição de íons
ferro e de radicais livres como a AAPH (2,2’-azobis-2-amidinopropano-dihidroclorida).
Esses estudos iniciais sobre o papel antioxidante dos caseinofosfopeptídeos apontam
efeitos benéficos da sua presença no intestino após o consumo de leite, contribuindo
provavelmente para a redução do stress oxidativo e manutenção da saúde intestinal
(KITTS, 2005).
2.3 RESPOSTA IMUNE NA INTERVENÇÃO NUTRICIONAL
A interdependência entre as disciplinas de Nutrição e Imunologia foi reconhecida
formalmente a partir de 1970 quando medidas imunológicas foram introduzidas como
parte da avaliação nutricional. Atualmente, a desnutrição protéico-calórica é
reconhecida como a maior causa mundial de imunodeficiências. Também ocorrem
alterações do sistema imunológico na maioria das doenças crônicas nas quais a dieta
possui um papel de prevenção e/ou tratamento, tais como artrite reumatóide, diabetes
tipo 1, doença celíaca, câncer e doenças cardiovasculares (FIELD, 2000).
Proteínas, carboidratos, lipídios, vitaminas e minerais são essenciais para a
manutenção e o crescimento dos seres humanos. Alterações qualitativas e quantitativas

38
nesses nutrientes podem afetar significativamente as funções do sistema imunológico.
Células desse sistema, quando ativadas, sintetizam e secretam moléculas. Assim, uma
deficiência ou um excesso de nutrientes específicos pode influenciar a síntese dessas
moléculas, alterando dessa forma a regulação da resposta imune (STITES; TERR,
1992).
Com relação a defesa inata, a influência da desnutrição sobre o muco epitelial é
consistente com o risco de infecção e translocação de bactérias. A imunidade mediada
por células é afetada em lactentes de baixo peso ao nascimento. Na resposta imune
humoral, a expansão clonal e a produção de anticorpos requerem rápida síntese
protéica, sendo que a restrição de aminoácidos inevitavelmente interfere nessa função
(BOUNOUS; BATIST; GOLD, 1989; NESTLÉ, 2001).
2.3.1 Estrutura e Componentes do Sistema Imunitário
A resposta imune é desencadeada pela introdução de um antígeno e geralmente
culmina com a sua eliminação. Existem dois tipos de resposta imune. A resposta imune
inata, inespecífica ou natural que está presente desde o nascimento, não possui
especificidade nem memória e se utiliza de barreiras físicas (por exemplo, pele íntegra
e membranas mucosas íntegras), barreiras químicas (por exemplo, ácido gástrico e
enzimas digestivas), células fagocíticas e do sistema complemento. O sistema
complemento consiste de pelo menos 30 proteínas plasmáticas e associadas às
membranas celulares. O complemento desempenha importante papel na iniciação da
resposta inflamatória, solubilizando e transportando os complexos imunes, modulando a
produção de imunoglobulinas, opsonizando os microorganismos patogênicos e
induzindo a sua lise direta pela formação do complexo lítico de membrana (C5b-9). Ao
contrário dos mecanismos de defesa inespecíficos, a imunidade adquirida ou adaptativa
é específica para o antígeno invasor, apresenta memória e se baseia nas respostas
específicas de linfócitos T e B. A imunidade específica pode ser adquirida naturalmente
por infecção ou artificialmente por imunização. A imunidade específica é dividida em
imunidade humoral e imunidade mediada por células (STITES; TERR, 1992,
CUNNINGHAM-RUNDLES, 1998).

39
Os linfócitos são células capazes de reconhecer e responder a antígenos.
Apresentam-se como células circulantes no sangue e linfa, surgindo como coleções de
células dos órgãos linfóides (timo, baço e linfonodos) ou organizadas em estruturas
linfóides associadas às mucosas. Os principais tipos são os linfócitos T, B e as células
NK (ou natural killer). Assim como os monócitos, os linfócitos são células
mononucleares (CALDER, 1998).
As células T são originárias da medula óssea e diferenciam-se no timo em
distintas populações, os linfócitos T auxiliares (Th - T helper), CD4+ e os linfócitos T
citotóxicos ou supressores (CTL ou Ts), CD8+. Os linfócitos T auxiliares (Th) são
constituídos por duas subpopulações, Th1 e Th2 que se distinguem funcionalmente
pelo tipo de citocinas que produzem. (CALDER, 1998; ELIA; SOUZA, 2001).
As células CD8+ possuem atividade citolítica e representam o principal
mecanismo de defesa contra patógenos intracelulares. Estão relacionadas com o
fenômeno de apoptose (morte celular programada). A maioria das células CD8+
reconhece peptídeos endógenos antigênicos apresentados pelas moléculas de classe I
do HLA e apresentam atividade citotóxica. Observa-se também a existência de
respostas de células CD8+ em conseqüência da apresentação de antígenos exógenos,
sendo que este tipo de resposta é efetivo no tratamento de algumas doenças do
sistema imune. O estudo de peptídeos análogos aos peptídeos exógenos que ativam as
células CD8+ é interessante pois esse tipo de apresentação leva essas células à
produção de citocinas como o IFN-γ, que é um inibidor potente dos eventos
dependentes de Th2 (Figura 8), inclusive da produção de imunoglobulina E (IgE)
(TOTSUKA et al., 1998).
O evento central, tanto na resposta humoral como na mediada por células,
depende da ativação e da expansão clonal de linfócitos T auxiliares (Th). A interação do
linfócito Th com o antígeno dá início a uma seqüência de reações bioquímicas que
induzem a expressão do receptor de IL-2 e a secreção de IL-2. Em resposta à IL-2, a
célula Th ativada prolifera e se diferencia em célula de memória ou célula efetora. A
resposta do tipo Th1 compreende uma produção aumentada de IFN-γ e IL-2, que
ativam predominantemente linfócitos T citotóxicos e macrófagos, capazes de atuar

40
diretamente sobre o antígeno ou de causar lesão na célula infectada ou tumoral. Os
macrófagos ativados, por sua vez, produzem citocinas pró-inflamatórias tais como a IL-
1 e TNF-α, que favorecem a reação do tipo de hipersensibilidade tardia e também
produzem IL-12 que estimula linfócitos Th1 em um mecanismo de retroalimentação
positiva. Na resposta do tipo Th2 ocorre um aumento na produção de IL-4, IL-5 e IL-6,
que favorecem a imunidade humoral, estimulando a proliferação e diferenciação de
linfócitos B e a conseqüente produção de anticorpos. Além disso, a resposta do tipo
Th2 é capaz de inibir paralelamente a resposta Th1, através do aumento na produção
de IL-10 e de IL-13, responsáveis pela retroalimentação negativa sobre linfócitos Th1 e
macrófagos (Figura 8).
O papel dos anticorpos na defesa imunológica é de neutralização e eliminação
do antígeno. Os anticorpos pertencem a diferentes classes de imunoglobulinas,
dependendo do tipo de estímulo apresentado (Figura 9).
As células NK ou natural killer caracterizam-se por não expressarem marcadores
de superfície, como os linfócitos T e B. Possuem a habilidade de destruir células
neoplásicas ou células infectadas por vírus, sem prévia estimulação antigênica. São
ativadas pela IL-2, IL-12, IFN-γ ou TNF-α (CALDER, 1998).
O sistema fagocítico mononuclear é composto por células de uma mesma
linhagem, cuja função primária é a de fagocitose. Todas as células desse sistema
originam-se da medula óssea, adquirindo diferentes formas, após a maturação. A
primeira célula a deixar a medula óssea e entrar na corrente sanguínea, ainda não
completamente diferenciada, é o monócito. Nos tecidos, após maturar, passa a se
chamar macrófago ou histiócito. Dentre as funções dos macrófagos destacam-se a
fagocitose de partículas estranhas e a função de células apresentadoras de antígenos
(Figura 10). Os macrófagos também produzem intermediários reativos de oxigênio
(peróxido de hidrogênio, superóxido e óxido nítrico) mediadores derivados dos lipídios
(prostaglandinas e leucotrienos), citocinas e fatores de crescimento. Os compostos
produzidos pelos macrófagos agem na eliminação de células neoplásicas e
microbianas, na regulação das funções de linfócitos T e B e da resposta inflamatória.
Entretanto, esses mesmos produtos podem estar envolvidos em doenças auto-imunes,

41
inflamatórias e no desenvolvimento da aterosclerose. Por exemplo, espécies reativas de
oxigênio e citocinas derivadas dos macrófagos foram encontradas em tecidos de uma
variedade de doenças auto-imunes, inflamatórias e em placas ateroscleróticas. Estudos
epidemiológicos com esquimós sugerem que a utilização de óleo de peixe pode resultar
de alguma forma na supressão da atividade dos macrófagos, pois essas populações
apresentam uma incidência muito baixa de doenças inflamatórias, auto-imunes e
coronarianas. Esses dados indicam que há possibilidade de intervenções nutricionais
ou farmacológicas que atuem no processo de ativação dos macrófagos (YAQOOB,
CALDER, 1995; ELIA; SOUZA, 2001).
FIGURA 8 - DIFERENCIAÇÃO E ATIVAÇÃO DOS LINFÓCITOS AUXILIARES (Th)
FONTE: CALDER, 1998. NOTAS: CTL, linfócitos citotóxicos; IFN-gama, interferon-gama; IL, interleucina; NK, natural killer; TGF-beta, fator de transformação do crescimento-beta, + , promove, - suprime; - - - →, modula, →, produz.

42
FIGURA 9 - DIFERENCIAÇÃO DOS LINFÓCITOS B
FONTE: CALDER, 1998. NOTAS: IFN-gama, interferon-gama; Ig, imunoglobulina; IL, interleucina; TGF-beta, fator de transformação do crescimento-beta, +, promove; - - - →, modula, ──, produz.
Os eosinófilos são granulócitos derivados da medula óssea que possuem um
núcleo bilobulado e grânulos citoplasmáticos com enzimas lisossomais, proteína
catiônica e neurotoxinas. Essas proteínas apresentam propriedades tóxicas para
helmintos, células tumorais e para as próprias células do hospedeiro. O citoplasma dos
eosinófilos também contém grânulos com corpos lipídicos que apresentam atividade
pró-inflamatória. São fontes de citocinas, que estão estocadas nos grânulos
citoplasmáticos, juntamente com as proteínas catiônicas. O mecanismo de liberação
dessas citocinas parece estar relacionado a degranulação seletiva da célula. Algumas
das principais citocinas liberadas pelos eosinófilos incluem: IL-3, IL-5, fatores de
crescimento (TGF-α, TGF-β – transforming growth factors), citocinas imunoregulatórias
(IL-2, IL-4 e IFN-γ) e pró-inflamatórias (IL-1, IL-6, IL-8, TNF-α). A ativação eosinofílica
representa, assim, importante função efetora no processo inflamatório (ELIA; SOUZA,
2001).

43
FIGURA 10 - PAPEL DOS MACRÓFAGOS NA INGESTÃO DE PROTEÍNAS ESTRANHAS E NA APRESENTAÇÃO DE ANTÍGENOS
FONTE: PATHMAKANTHAN; HAWKEY, 2000. NOTAS: HLA, complexo principal de histocompatibilidade.
As citocinas são proteínas solúveis, com peso molecular entre 5 e 50 KDa,
produzidas pelos leucócitos e outros tipos celulares, que desempenham importante
funções na comunicação intercelular, incluindo a ativação de receptores específicos de
membrana, a proliferação e diferenciação celular, a quimiotaxia e a produção de
imunoglobulinas. Após serem secretadas, as citocinas se ligam a receptores específicos
na superfície das células do sistema imunológico, exercendo suas diferentes funções.
Em estudo experimental, observou-se uma correlação positiva entre a
biodisponibilidade de glutamina e a produção de citocinas. Camundongos alimentados
com dieta rica em glutamina tiveram um aumento expressivo na produção de citocinas
pelos macrófagos. Esse efeito sugere que a administração oral de glutamina possa
melhorar a resposta imunológica contra bactérias, vírus, parasitas e tumores (YAQOOB;
CALDER, 1998; WELLS et al, 1999).

44
Algumas citocinas desempenham um papel importante nos processos
inflamatórios. A interleucina-6 (IL-6), IL-1β e o fator de necrose tumoral (TNF-α) são
citocinas pró-inflamatórias produzidas em resposta à inflamação sistêmica. São
produzidos basicamente por macrófagos ativados, mas também por linfócitos e células
endoteliais. São responsáveis por vários sinais fisiológicos que acompanham a
endotoxemia. O TNF-α regula a resposta de fase aguda, induz a produção de outras
citocinas imunoregulatórias; a expressão de moléculas do complexo maior de
histocompatibilidade (HLA) de classes I e II e de moléculas de adesão; ativa células T,
B, NK e neutrófilos; promove quimiotaxia de monócitos, neutrófilos e granulócitos para
áreas de reatividade imune e aumenta a síntese de proteínas de fase aguda. A IL-6
regula a síntese de proteínas de fase aguda pelo fígado e ativa células T e B. A IL-6
pode modular diferentes eventos biológicos, tais como a diferenciação celular, a
proliferação e a apoptose. As células intestinais produzem IL-6 em resposta à
exposição de bactérias e infecções virais, demonstrando que esta citocina também
apresenta um papel de regulação do sistema imune intestinal (YAQOOB; CALDER,
1995; KITTS; NAKAMURA, 2006)
O sistema imunológico das mucosas também constitui uma parte importante do
sistema de defesa, sendo a imunoglobulina A (IgA) o componente principal, o qual se
encontra em secreções e superfícies mucosas. O epitélio intestinal é constantemente
invadido por antígenos derivados de patógenos e da própria dieta e inicia a interação
desses com o sistema imunológico. A maioria dos linfócitos T encontrada nas células
intraepiteliais consiste do fenótipo citotóxico, enquanto na lâmina própria se encontram
as células Th, B e monócitos/macrófagos. As células M são células de transporte,
isentas de microvilosidases e aptas a fagocitar antígenos solúveis e microorganismos.
As placas de Peyer são encontradas na lâmina própria (Figura 11). As células epiteliais
do intestino estão envolvidas no transporte da IgA da lâmina própria até o lúmen
intestinal e produzem várias citocinas imunoregulatórias importantes, que influenciam a
secreção local de Igs pelas células B. Esses linfócitos B expressam principalmente IgA
(60% na mucosa brônquica e até 85% na mucosa intestinal), além de IgG, IgM e IgE
(KITTS; NAKAMURA, 2006).

45
FIGURA 11 - REPRESENTAÇÃO DIAGRAMÁTICA DOS COMPONENTES PRINCIPAIS DO SISTEMA IMUNITÁRIO DA MUCOSA INTESTINAL
FONTE: PATHMAKANTHAN; HAWKEY, 2000.
2.3.2 Avaliação da Resposta Imunológica
Um dos testes mais utilizados para avaliar a função dos linfócitos é a proliferação
das células em resposta a sinais mitogênicos. Utiliza-se a concanavalina A (Con A) e a
fitohemaglutinina (PHA), para se estimular linfócitos T e o lipopolissacarídeo (LPS), para
linfócitos B. A resposta proliferativa dos linfócitos é medida pelo aumento no número de
células através da incorporação de timidina radioativamente marcada no DNA dessas
células ou por meio do uso de ensaios colorimétricos (LOOSDRECHT et al., 1994;
CALDER, 1998; FIELD, 2000).
A medição de citocinas circulantes ou secretadas, quando bem conduzida,
permite uma excelente correlação com os estudos de resposta proliferativa
(CUNNINGHAM-RUNDLES, 1998).
A função dos linfócitos e a produção de citocinas derivadas das células T
também podem ser avaliadas após indução de sepse por endotoxinas bacterianas
(LANZA-JACOBY; FLYNN; MILLER, 2001).

46
2.3.3 Função dos Peptídeos do Leite na Resposta Imunológica
Devido ao fato das proteínas do leite representarem a única fonte protéica na
alimentação dos neonatos, algumas evidências apontam que seus peptídeos possuem
uma ação local na regulação da função imune gastrintestinal (COSTE et al., 1992;
SMACHI; GOBETTI, 2000). Os primeiros meses de vida se caracterizam como um
período crítico no desenvolvimento de tolerância oral a algumas moléculas presentes
nos alimentos e na obstrução à entrada de antígenos derivados de patógenos. O
sistema imunitário da mucosa intestinal é desafiado constantemente a discriminar
patógenos de antígenos não patogênicos derivados da dieta e de microorganismos da
flora normal. Portanto, peptídeos com propriedades bioativas podem gerar a supressão
da resposta imunológica a alguns produtos, dos quais se requer a tolerância oral e a
indução do sistema imunológico, como um sinal de alerta para a presença de antígenos
(BALDI et al., 2005).
O estudo do fenômeno fisiológico de tolerância oral aponta para implicações
terapêuticas futuras em doenças relacionadas com anormalidades no mecanismo de
tolerância, além de permitir novas abordagens para o desenvolvimento de vacinas orais
para diversas doenças inflamatórias sistêmicas (ELIA; SOUZA, 2001).
A supressão da proliferação celular também possui aplicações clínicas
importantes na melhoria do quadro clínico de pacientes que sofrem de doenças auto-
imunes, como artrite reumatóide, psoríase, esclerose múltipla, entre outras (PABLO;
CIENFUEGOS, 2000).
Vários trabalhos realizados com proteínas do leite apontam a presença de
peptídeos imunomoduladores, tanto com atividade supressora como indutora do
sistema imunológico.
OTANI & MONNAI (1995) verificaram que o κ-caseinoglicopeptídeo (resíduos
106 – 169, CGP) foi capaz de inibir a resposta proliferativa dos linfócitos de baço de
ratos quando estimulados por mitógenos. Em 1996, OTANI; HORIMOTO E MONNAI
propuseram que a atividade supressora do CGP ocorra por dois mecanismos: produção

47
de um componente inibitório que reage com o anticorpo anti-IL-1ra e supressão do
receptor de IL-2 nos linfócitos Th.
KAYSER & MEISEL (1996) apontam que β-casomorfina-7 e a β-casomorfina-10,
podem inibir a proliferação de linfócitos de sangue periférico humano, quando em
baixas concentrações.
HATA; HIGASHIYAMA; OTANI (1998) digeriram a αS1-caseína com tripsina
bovina, obtendo peptídeos com efeito inibitório na proliferação de células de baço de
camundongos, quando estimuladas por concanavalina A.
Existem indicações de que o papel dos hidrolisados de caseína na imunidade
seja dependente do nível de amadurecimento do sistema imunológico. BALDI et al.
(2005) examinaram o efeito de dois peptídeos sintéticos, correspondentes aos resíduos
191 – 193 e 63 – 68 da β-caseína, na expressão do complexo principal de
histocompatibilidade de classe II (HLA) por neutrófilos de sangue de suínos. Para esse
modelo experimental, os autores concluíram que os peptídeos se apresentam efetivos
no período de tempo que corresponde à imaturidade do sistema imunológico e não
quando o sistema imunológico adquire sua plenitude funcional.
BERTHOU et al. (1987) purificaram um digesto de caseína humana até a
obtenção de um tripeptídeo: Gly-Leu-Phe. Tanto este peptídeo natural como o similar
sintético elevou a atividade fagocítica de macrófagos peritoneais de ratos.
Dois peptídeos sintéticos (Tyr-Gly e Tyr-Gly-Gly) tiveram seu efeito proliferativo
avaliado com o emprego de ConA e utilizando linfócitos periféricos humanos. Ambos
demonstraram proliferação, sendo que o dipeptídeo teve um resultado
significativamente mais elevado. Os peptídeos Tyr-Gly e Tyr-Gly-Gly são seqüências
parciais da estrutura primária da κ-caseína e da α-lactalbumina, respectivamente. Esses
peptídeos foram usados para imunoterapia de pacientes infectados pelo HIV: um
extrato de leucócitos dialisado, obtido de doadores normais teve a inclusão dos
peptídeos. Após um período de duas semanas os pacientes apresentaram redução nas
manifestações clínicas (KAYSER; MEISEL, 1996)
Camundongos alimentados com caseinofosfopeptídeos por via oral exibem
aumento na produção de IgA em resposta a antígenos via peritoneal (OTANI; KIHARA;

48
PARK, 2000). O efeito estimulatório do CPP na produção de IgA ocorre também após a
ingestão oral de LPS (OTANI; NAKANO; KAWAHARA, 2003).
OTANI; WATANABE; TASHIRO (2001) avaliaram a proliferação celular dos
linfócitos T e B usando mitógenos. Ao final do período de cultivo, coletaram os
sobrenadantes para medição dos níveis de imunoglobulinas. Os resultados obtidos
indicam que fosfopeptídeos contendo a seqüência SerP-X-SerP aumentam a
proliferação dos linfócitos e a produção de imunoglobulina A, indicando o tripeptídeo
como adjuvante na produção de IgA.
2.4 O SISTEMA IMUNOLÓGICO E A RESPOSTA INFLAMATÓRIA
A inflamação é uma resposta normal do hospedeiro contra agentes infecciosos.
A sepse é caracterizada pela produção excessiva de mediadores inflamatórios e pela
excessiva ativação de células inflamatórias. Os mediadores inflamatórios promovem a
injúria dos tecidos, as endotoxinas produzidas pelas bactérias destroem os tecidos pela
ativação da cascata de coagulação, da cascata do complemento, da injúria de vasos ou
pela liberação do ácido araquidônico ou óxido nítrico. A principal conseqüência desta
resposta inflamatória é o comprometimento de muitos órgãos e o quadro de choque
com evolução para a síndrome da imunodeficiência de múltiplos órgãos, que é
acompanhada de alta mortalidade (GLAUSER, 2000).
Os componentes da parede bacteriana são os principais ativadores da resposta
do hospedeiro: as endotoxinas dos microorganismos gram-negativos (principalmente o
lipídio A), o ácido teicóico, o ácido lipoteicóico e as peptidoglicanas dos
microorganismos gram-positivos (GLAUSER, 2000).
Os lipopolissacarídeos bacterianos consistem basicamente de 3 partes: o lipídio
A (ou endotoxina), um oligossacarídeo central e um polissacarídeo distal (antígeno-O)
(Figura 12). O lipídio A ou endotoxina é uma “âncora” hidrofóbica do lipopolissacarídeo,
sendo um componente das membranas externas da maioria das bactérias gram-
negativas. Existem aproximadamente 106 resíduos de endotoxina em uma única célula
de Escherichia coli. As moléculas do lipídio A são detectadas em níveis picomolares

49
pelo receptor do sistema imunológico inato presente nos macrófagos e nas células
endoteliais. Este receptor recentemente foi identificado e denominado TLR4 (toll-like
receptor 4) (RAETZ; WHITFIELD, 2002).
O lipopolissacarídeo (LPS) é liberado da membrana externa das bactérias
durante a divisão celular, apoptose ou como resultado do tratamento com antibióticos
contra a infecção bacteriana (Figura 13). Após sua liberação, o lipopolissacarídeo é
reconhecido pelas células fagocíticas (monócitos e macrófagos), ativando-as. Como
resultado, há um aumento na atividade fagocítica e indução da secreção de citocinas
pró-inflamatórias. Embora essas citocinas possuam um papel fundamental na resposta
inflamatória local, sua produção excessiva gera o choque séptico (ROSENFELD;
PAPO; SHA, 2006).
O mecanismo de ativação dos macrófagos pelo LPS se inicia pela ligação do
LPS (através de sua porção tóxica, o lipídio A) a uma proteína específica, denominada
LPS-LBP (LPS-binding protein) no plasma, acelerando a ligação do LPS ao CD14, um
receptor primário de LPS, que é expresso na maioria dos macrófagos. O complexo
LPS-CD14 inicia a sinalização intracelular para interagir com o receptor de membrana,
TLR-4, resultando na ativação dos macrófagos, conforme representado na Figura 14.
A ativação dos macrófagos conduz à biossíntese de diversos mediadores
inflamatórios, como o TNF-α, a IL-1 β e a IL-6 e coestimula a produção de algumas
moléculas necessárias para a resposta imune adaptativa (RAETZ; WHITFIELD, 2002).

50
FIGURA 12 - MODELO DAS MEMBRANAS INTERNA E EXTERNA DA E. COLI K-12
FONTE: RAETZ; WHITFIELD, 2002 NOTAS: LPS, lipopolissacarídeo da superfície de E.coli.
FIGURA 13 - LIBERAÇÃO DO LPS POR E. COLI
NOTAS: LPS, lipopolissacarídeo da superfície de E.coli; LBP, LPS binding protein . FONTE: RAETZ; WHITFIELD, 2002

51
FIGURA 14 - REPRESENTAÇÃO DIAGRAMÁTICA DOS EVENTOS ENVOLVIDOS NA DETECÇÃO E RESPOSTA AOS ESTÍMULOS FORNECIDOS PELAS ENDOTOXINAS
MICROBIANAS NA IMUNIDADE INATA
FONTE: GLAUSER, 2000. NOTAS: LPS, lipopolissacarídeo; LBP, LPS binding protein; CD14, receptor de superfície dos monócitos que se liga ao complexo LPS-LBP; TLR-4, receptor que se liga ao complexo CD14-LPS, resultando na ativação celular.
A presença de citocinas pró-inflamatórias leva a anorexia, perda de peso,
alterações no metabolismo de lipídios por suprimirem a atividade da enzima lipase
lipoprotéica favorecendo a lipólise, aumentam o “turnover” de proteínas, os níveis de
cortisol, glucagon e adrenalina (hormônios catabólicos), reduzem os níveis de insulina
(hormônio anabólico) e favorecem o quadro de resistência periférica à insulina
(PIZATO, 2004)
A Figura 15 representa a complexidade dos eventos envolvidos na endotoxemia.

52
FIGURA 15 - PATOGÊNESE DO CHOQUE SÉPTICO
Quimiotaxia Radicais superóxido
Complemento Enzimas lisossomais
Neutrófilos
Moléculas de adesão
A ativação do complemento conduz à liberação de anafilatoxinas, especialmente
C3a e C5a, que são responsáveis por vasodilatação, aumento da permeabilidade
vascular, agregação plaquetária e ativação de granulócitos, sendo que este é um fator
desencadeante da síndrome respiratória. Os neutrófilos ativados formam aglomerados
e aderem ao endotélio vascular. Também ocorre liberação de derivados do ácido
araquidônico, subprodutos citotóxicos do oxigênio e enzimas lisossomais, produzindo
efeitos vasculares e endoteliais. Os neutrófilos ativados contribuem às injúrias
vasculares e teciduais. O Fator XII (Fator de Hageman) desempenha um papel
importante na patogênese do choque séptico, sendo ativado pelo lipídio A. Após sua
ativação, conduz ao consumo dos fatores de coagulação, disseminando a coagulação
intravascular. Sua ativação também leva à conversão da pré-quallikreína em
LPS
LBP
Células monocíticas
Citocinas
Mediadores lipídicos
Fator tissular
Sistema de coagulação Fibrinólise
Extravasamento capilar Acúmulo de neutrófilos Febre Alterações metabólicas Alterações hormonais Vasodilatação Coagulação intravascular
Sídrome séptica Hipotensão Falhas de múltiplos órgãos Morte
Fator XII
Bradiquinina
Células endoteliais

53
quallikreína. Esta, cliva o quininogênio em bradiquinina, que se caracteriza como um
potente agente hipotensor (GLAUSER et al., 1991).
A apoptose celular em pacientes com sepsis varia de acordo com o tipo celular.
Encontra-se aumentada para linfócitos do sangue e baço, não se altera para monócitos
circulantes e apresenta-se diminuída para neutrófilos do sangue e macrófagos
alveolares (ANNANE; BELISSANT; CAVAILLON, 2005).
A inalação de LPS, em humanos, conduz à ativação, migração de neutrófilos e
superprodução de mediadores inflamatórios. A instilação intratraqueal de LPS é
responsável por inflamação pulmonar e lesão pulmonar aguda, sendo que esta se
caracteriza pela presença de infiltrados neutrofílicos e aumento da permeabilidade
alvéolo-capilar (FERNÁNDEZ-MARTÍNEZ, et al., 2004; JANSSON; ERIKSSON; WANG,
2005)
A administração intravenosa de Escherichia coli ou de endotoxina em ratos
rapidamente (< 20 minutos) desencadeia hipertensão pulmonar e lesão pulmonar
aguda. A avaliação histológica de pulmões de ratos que receberam LPS via intravenosa
ou intraperitoneal revela freqüentemente os seguintes achados: edema pulmonar,
edema perivascular, ativação e infiltração de neutrófilos, infiltração de macrófagos,
extravasamento do fluido do espaço vascular, resultando em hemoconcentração,
hemorragia severa no interstício e alvéolos, hipertensão pulmonar, destruição da
arquitetura pulmonar e fibrose (SILLIMAN et al., 1998; CLAVIJO et al., 2000; LEE et al.,
2001; AKCA et al., 2005; KOKSEL et al., 2005; KONO et al., 2005; ZHANG; SUN,
2005).
A administração de endotoxina de bactérias gram-negativas a ratos resulta em
hipotensão sistêmica, aumento do hematócrito, redução do número de leucócitos
circulantes (polimorfonucleares), monócitos e plaquetas. Essas alterações podem ser
parcialmente atribuídas à ativação do complemento e geração de anafilatoxinas pela
endotoxina (SMEDEGARD; CUI; HUGLI, 1989).

54
3 ESTRATÉGIA GERAL A Figura 16 apresenta a estratégia geral utilizada neste trabalho.
FIGURA 16 – REPRESENTAÇÃO ESQUEMÁTICA DA ESTRATÉGIA METODOLÓGICA UTILIZADA PARA REALIZAÇÃO DOS OBJETIVOS DA TESE
Avaliações
Caseinato de sódio
Hidrólise Tríptica
CPP-A
CPP-B
CPP-C
Caracterização Físico-Química
Atividade antimicrobiana in vitro
Inespecífica
Sepse induzida
Imunidade

55
4 MATERIAL E MÉTODOS
4.1 MATÉRIA PRIMA E REAGENTES Utilizou-se caseinato de sódio (LACTONAT HV, Lactoprot Deutschland GmbH),
cedido pela empresa Colorcon do Brasil Ltda., com a seguinte composição química:
proteína, 87,5%; umidade, 5,8%; cinzas, 5,3%; gordura, 1,7%, lactose, 0,3% e pH 6,6.
Os reagentes utilizados, de grau analítico, foram os disponíveis comercialmente
de empresas como: Merck, Sigma, BD Pharmigen, Pharmacia, etc. A enzima PTN 6.0
foi gentilmente cedida pela empresa Novozymes.
4.2 EQUIPAMENTOS Agitadores; homogeneizadores; balança analítica; banho termostático;
liofilizador; equipamento para eletroforese capilar; sistema de ultrafiltração;
espectrofotômetros; digestor e destilador de Kjeldahl; estufas; capela de fluxo laminar;
centrífugas; demais equipamentos e material de uso corrente em laboratório.
4.2 METODOLOGIA
4.2.1 Obtenção dos Caseinofosfopeptídeos
Após levantamento bibliográfico dos métodos existentes na literatura para
obtenção de caseinofosfopeptídeos, optou-se pelo agrupamento destes em três tipos
de processos. Estes processos foram reproduzidos no Laboratório de Pesquisa e Pós-
Graduação do Departamento de Nutrição da Universidade Federal do Paraná. Os
parâmetros foram cuidadosamente ajustados para a otimização dos processos citados
no trabalho. Os produtos foram liofilizados na Planta Piloto de Secagem da Pontifícia
Universidade Católica do Paraná.

56
Os peptídeos foram produzidos pela hidrólise tríptica do caseinato de sódio,
seguida de precipitação ácida, agregação mineral e precipitação com etanol (OTANI et
al., 2000), agregação de cloreto de cálcio (REYNOLDS, 2002) ou agregação de sulfato
de zinco (FITZGERALD, 1998), conforme processos descritos abaixo. Os produtos
foram liofilizados e denominados respectivamente, CPP-A, CPP-B e CPP-C.
4.2.1.1 Processo CPP-A:
Dissolveram-se 228,57g de caseinato de sódio em 2000mL de água nanopura para
produzir solução a 10% de proteínas. O pH inicial da solução foi de 6,32. Ajustou-se o
pH para 8 com NaOH 0,5N. Adicionou-se tripsina (PTN 6.0) dissolvida em água na
relação E/S 1:100. Procedeu-se à hidrólise, sob leve agitação, à temperatura de 50ºC e
pH 8. O pH foi mantido em 8 durante a hidrólise pelo acréscimo de NaOH 0,5N,
segundo o método de pH-stat (ADLER-NIESSEN, 1979). A reação de hidrólise ocorreu
até não haver mais o consumo de base. Após, ajustou-se o pH para 4,64 com HCl 2M
O precipitado insolúvel que se formou foi removido por centrifugação a 3000xg por 10
minutos. Ao sobrenadante foram adicionados CaCl2 a 1% e etanol a 50%, com o
objetivo de promover a agregação do cálcio ao peptídeo e conseqüente precipitação. O
precipitado resultante foi coletado por centrifugação a 4000xg por 15 minutos.
Redissolveu-se o precipitado em água destilada. O produto foi congelado, liofilizado e
denominado CPP-A.
4.2.1.2 Processo CPP-B:
Dissolveram-se 228,57g de caseinato de sódio em 2000mL de água para obtenção
de solução a 10% de proteínas. O pH da solução foi ajustado para 8 com NaOH 2M.
Adicionou-se tripsina (PTN 6.0) dissolvida em água numa relação E/S 1:100. O produto
foi submetido à hidrólise a 50ºC por 4 horas, sem adição de base. Após a hidrólise o pH
foi reduzido para 4,7 com HCl 2M. O precipitado insolúvel que se formou foi removido
por centrifugação a 3000xg por 10 minutos. Ajustou-se o pH para 7 com NaOH 0,5M.

57
Adicionou-se CaCl2 a 1% e o produto foi submetido à agregação por 1 hora. A solução
foi diafiltrada em membrana de 10kDa pelo acréscimo de 3 volumes de solução de
CaCl2 a 1%. O pH do retentado foi reduzido para 3,55. O produto foi diafiltrado com
água e concentrado em membrana de 1kDa. Após, elevou-se o pH para 7. O produto foi
congelado, liofilizado e denominado CPP-B.
4.2.1.3 Processo CPP-C:
Dissolveram-se 228,57g de caseinato de sódio em 2000mL de água para obtenção
de solução a 10% de proteínas. O pH da solução foi ajustado para 8 com NaOH 2M.
Adicionou-se tripsina (PTN 6.0) dissolvida em água numa relação E/S 1:100. O produto
foi submetido à hidrólise a 50ºC por 4 horas, sem adição de base. Após a hidrólise o pH
foi reduzido para 4,7com HCl 2M. O precipitado insolúvel que se formou foi removido
por centrifugação a 3000g por 10 minutos. Ajustou-se o pH para 7 com NaOH 0,5M.
Acrescentou-se 1,1% de ZnSO4. A solução foi mantida por 1 hora sob agitação suave
para promover a agregação de zinco ao peptídeo. Após, foi ultrafiltrada em membrana
de 10 kDa, diafiltrada com 1 volume de solução 38mM de ZnSO4 e concentrada. O
produto foi então liofilizado e denominado CPP-C.
4.2.2 Caracterização dos Caseinofosfopeptídeos 4.2.2.1 Composição Centesimal
A determinação de nitrogênio protéico foi realizada pelo método semi-micro
Kjeldahl, utilizando-se o fator 6,38 para conversão em proteína (AOAC, 1990). Cinzas
foram dosadas pelo método de incineração em mufla a 450ºC até peso constante
(AOAC, 1990). Umidade por meio de secagem em estufa com circulação de ar forçada
a 105ºC até peso constante (AOAC, 1990).

58
4.2.2.2 Minerais
Para determinação dos minerais, realizou-se abertura seca das amostras, que
foram pré-calcinadas em fogareiro, calcinadas em forno mufla a 550ºC até a obtenção
de cinzas brancas ou acinzentadas. Após adicionou-se 25mL de HNO3 25%, 1mL de
KCl 20% e os cadinhos foram colocados em banho–maria e deixados em aquecimento
por cerca de 1 hora. As soluções foram filtradas utilizando papel filtro qualitativo faixa
preta, para balão volumétrico de 100mL. Os teores de Ca, Na e Zn foram determinados
por espectrofotometria de absorção atômica em equipamento Varian modelo SpectrAA-
200 (AOAC, 1990). A dosagem de P foi realizada pelo método colorimétrico vanado-
molíbdico, que se baseia na formação de um complexo de adição do fósforo ao
reagente vanado-molíbdico, de coloração amarela (procedimento 22.042-2045 – AOAC,
1990).
4.2.2.3 Aminoácidos
O método de eletroforese capilar de zona baseia-se na migração diferencial de
compostos iônicos ou ionizáveis, em um campo elétrico (TAVARES, 1996). Na
eletroforese capilar de zona essa separação ocorre por diferença na razão carga-
massa. As moléculas eluem-se no capilar de sílica fundida por ordem de carga positiva
decrescente (TOMÁS-BARBERÁN, 1994). Para dosagem de aminoácidos, as amostras
foram hidrolisadas a 110°C em presença de HCl 6M por 22 horas. Ao término deste
tempo, evapora-se o ácido (evaporador à vácuo) elevando-se o pH a 8 e o volume a
20ml. Previamente à análise, procede-se à filtração em membranas de 0,22µm.
A análise foi realizada sob as seguintes condições: equipamento modelo PNA 402,
tampão: Borato, pH=9 e T=15oC em colunas: 67cm / 65cm / 375um / 50um. A Injeção
da amostra foi de 15 seg. e 5KV. Empregou-se voltagem de 15KV com ~11,2 uA. A
detecção foi efetuada a: 400nm/500nm com laser a 10mW e 500KHz .

59
4.2.2.4 Solubilidade
Determinada pelo método de MORR et al. (1985). Foram preparadas dispersões
protéicas a 1% com agitação mecânica em 50 mL de água destilada e em PBS pH 7,4.
A dispersão foi agitada por 1 hora. A dispersão foi centrifugada por 30’ a 20000xg e
temperatura de 5ºC. A fração sobrenadante resultante foi filtrada em papel de filtro
Whatman nº 1. O teor de proteína do filtrado foi determinado pelo método semi-micro-
Kjedahl. A porcentagem de proteína solúvel foi dada pela equação:
100(g%)/100 amostra da proteína de teor x (g) amostra da peso
(g) tesobrenadan de mL 50 em proteína de ãoconcentraç(%) xS =
4.2.2.5 Espectrometria de Massa por Desorção a Laser (MALDI-TOF-MS)
As amostras foram dissolvidas em água nanopura na concentração de 5pmol/µL.
Retirou-se um volume de 10µL de cada uma das mostras dissolvidas e se realizou a
dessalinização com colunas ZipTips C18 (Millipore, Billerica, MA). As amostras foram
ressuspensas em 10µL de ácido trifluoroacético (TFA 0,1%), dos quais 1µL foi aplicado
na placa de MALDI de aço inoxidável da Bruker Daltonics, junto com 1µL de solução de
ácido α-ciano-4-hidroxicinâmico 10mg/mL, acetonitrila 50% e TFA 0,1% (matriz de
MALDI).
Os espectros de MALDI-TOF-MS foram obtidos em um espectrômetro de massa
Reflex IV (Bruker Daltonics, Bremen, Germany), com laser UV de 337nm e intensidade
média de 12%, no modo refletor positivo (voltagem do refletor igual a 19,8kV) e
extração retardada (delayed extraction), usando faixa de massa de 0,3 a 3,5kDa. Cada
espectro foi calibrado externamente com uma mistura-padrão de peptídeos PeptideMix
(Bruker Daltonics, Bremen, Germany).
A identificação dos precursores de caseína, dos peptídeos encontrados nos
espectros de MALDI-TOF-MS, foi realizada inicialmente com o auxílio do programa
Paws (MacOS). Com este programa, as massas obtidas são comparadas com as

60
seqüências das caseínas (α-s1-casein, α-s2-casein, β-casein and κ-casein),
considerando um erro de 0,5 Da para cada peptídeo. As possíveis seqüências
correspondentes a cada massa foram então comparadas com as seqüências das
caseínas com o auxílio das ferramentas do Expasy (www.expasy.org).
4.2.2.6 Seqüenciamento por MS/MS
Devido ao fato das amostras consistirem numa mistura de peptídeos, realizou-se
inicialmente cromatografia líquida de alta eficiência em fase reversa (RP-HPLC). Foram
obtidas novas frações, contendo um ou mais peptídeos.
Foram dissolvidos exatamente 1,2mg da amostra em 200µL de TFA 0,1% (v/v),
agitado em vortex por 1 minuto e centrifugado a 14000rpm por 10 minutos. A amostra
foi analisada por RP-HPLC utilizando uma coluna C18 (Vydac 218TP54, 4.6mm ×
250mm, Separations Group, USA) sob um gradiente de 0 a 100% de acetonitrila em 85
minutos com uma taxa de fluxo de 0,8mL/minuto. A absorção em UV foi monitorada a
216nm.
Para confirmação dos peptídeos identificados por MALDI-TOF-MS foram
realizados experimentos de seqüenciamento utilizando técnicas de ESI-CID-MS/MS
(electrospray collision induced dissociation) e MALDI PSD (post source decay).
As frações liofilizadas isoladas por HPLC foram dissolvidas em acetonitrila 50% e
ácido acético 50% na concentração de 5pmol/µL e analisadas por espectrometria de
massa em um instrumento API300 equipado com fonte de nanospray - Applied
Biosystems (Foster City, CA).
Os dados obtidos foram interpretados com o auxílio do Programa BioMultiView
v1.2. Os experimentos de PSD foram realizados em um espectrômetro de massa Reflex
IV (Bruker Daltonics, Bremen, Germany), operando no modo refletor positivo e
utilizando o acessório FAST (Fragmentation Analysis and Structural TOF). Os dados
foram analisados com o auxílio do programa BioTools v. 2.0. Para alguns peptídeos,
cuja fragmentação não foi observada pelo método PSD convencional, foi realizado o
seqüenciamento por CAF (chemically assisted fragmentation) usando o kit de
seqüenciamento Ettan CAF MALDI, GE Healthcare (Chalfont St. Giles, UK).

61
4.2.3 Atividade Antimicrobiana In Vitro
Antes da execução dos experimentos, realizou-se teste de esterilidade das
amostras. Este teste foi realizado sete dias antes da inoculação final, com o objetivo de
verificar possível contaminação prévia da matéria-prima.
Soluções dos peptídeos CPP-A, CPP-B e CPP-C foram preparadas em
duplicata, em solução salina de tampão fosfato (PBS) estéril, pH 7,4, na concentração
de 125mg/mL. Foram inoculados 0,9mL destas soluções em kit HEMOPROV I com
seringa de 1mL e agulha previamente esterilizadas. Os frascos foram mantidos em
estufa a 37ºC por uma semana, quando se deu a avaliação dos resultados.
A atividade antimicrobiana foi avaliada pelo teste de susceptibilidade através do
método de microdiluição em caldo. O método de microdiluição em caldo permite avaliar
de forma quantitativa a atividade in vitro de determinado antimicrobiano frente a um
isolado bacteriano. Esta técnica é realizada através de placas de microdiluição estéreis
contendo quantidades pré-determinadas do meio de cultura adicionado de diluições
crescentes dos antimicrobianos. Nestas diluições aplicam-se inóculos de supensões
bacterianas previamente padronizadas (ISENBERG, 1998).
A escolha das cepas se deu de acordo com pesquisa de microorganismos mais
freqüentemente envolvidos em surtos de infecção e/ou intoxicação alimentar, de acordo
com dados da Secretaria da Saúde do Estado do Paraná (disponível em
<http\\:www.saúde.gov.pr.br>).
As bactérias Gram-positivas Staphylococcus aureus (ATCC 6538) e Listeria
monocytogenes (ATCC 7644) e as Gram-negativas Escherichia coli (ATCC 8739) e
Salmonella typhimurium (ATCC 14028) foram pré-cultivadas aerobicamente a 37ºC em
meio Mueller-Hinton. Após, foram suspensas neste mesmo meio e a atividade
antimicrobiana foi testada pela inibição do crescimento. Soluções dos peptídeos foram
preparadas em triplicata, em solução salina de tampão fosfato (PBS) estéril, pH 7,4, nas
seguintes concentrações: 125mg/mL, 25mg/mL, 5mg/mL, 1mg/mL e 0,2mg/mL. O
ensaio foi realizado em placas de cultura de 24 poços. O inóculo foi preparado pela

62
diluição exponencial das células em meio de crescimento, até que o inóculo contivesse
3 x 106UFC/mL para a Listeria monocytogenes e 1,5 x 107UFC/mL para as demais
bactérias, de acordo com a escala de McFarland.
Em cada poço foram adicionados 1800µL das soluções de peptídeos, contendo
os peptídeos CPP-A, CPP-B, CPP-C e 200µL do inóculo bacteriano. Os controles
consistiam de todos os componentes, com exceção dos peptídeos. O crescimento
microbiano foi avaliado pelo aumento na absorbância após 24 horas de incubação a
37ºC. O valor da A620 nm das culturas controle (sem o peptídeo) foi tomado como 100%.
Após avaliação, as amostras que obtiveram redução no crescimento microbiano,
comparativamente ao controle, foram novamente semeadas em placas com ágar
Mueller-Hinton, por 48 horas a 37ºC.
4.2.4 Avaliação da Ação dos Caseinofosfopeptídeos na Resposta Imunológica
4.2.4.1 Animais
Foram utilizados 54 ratos albinos machos (Rattus norvegicus), da linhagem
Wistar, com 30 dias pós-desmame. Os animais foram adquiridos do Instituto de
Tecnologia do Paraná (TECPAR). O biotério possuía temperatura e umidade
controladas com ciclos de luzes acesas e apagadas de 12 horas. O projeto seguiu os
Princípios Éticos adotado pelo Colégio Brasileiro de Experimentação Animal (COBEA).
4.2.4.2 Dietas
Os animais foram distribuídos em nove grupos de seis e receberam dieta
comercial (Nuvilab CR1, Nuvital, Curitiba) suplementada com 0,14g/rato/dia de CPP
(dieta com CPP) ou somente dieta comercial (dieta controle) por um período de 22 dias.
A composição detalhada da dieta comercial está demonstrada na Tabela 4.

63
TABELA 4 -COMPOSIÇÃO DA DIETA COMERCIAL
COMPOSIÇÃO DIETA CONTROLE*(%)
UMIDADE (máx.) 12,5
PROTEÍNA BRUTA (min.) 22
EXTRATO ETÉREO (min.) 4
MATERIAL MINERAL (máx.) 10
MATÉRIA FIBROSA (máx.) 8
CÁLCIO (max.) 1,4
FÓSFORO (min.) 0,8
* Por kg apresenta não menos que: vit. A, 12000 UI; D3, 1800 UI; E, 30 mg; K3, 3 mg; B1, 5 mg; B2, 6 mg; B6, 7 mg; B12, 20 µg, niacina, 60 mg; ácido pantotênico, 20 mg; ácido fólico, 1 mg; biotina, 0,05 mg; colina, 600 mg; ferro, 50 mg; zinco, 60 mg; cobre, 10 mg; iodo, 2 mg; manganês, 60 mg; selênio, 0,05 mg; cobalto, 1,5 mg; DL metionina, 300 mg; lisina, 100 mg.
4.2.4.3 Imunidade Inespecífica
a) Obtenção de Macrófagos
Para este ensaio, foram selecionados os animais que receberam a
suplementação com os peptídeos (CPP-A, CPP-B e CPP-C) por um período de 22 dias.
Estes grupos não foram submetidos à indução de endotoxemia. Após a eutanásia, os
animais tiveram a pele da região abdominal cuidadosamente removida e foram
injetados 10 mL de tampão fosfato, pH 7,4 (PBS) estéril na cavidade peritoneal. Após, a
cavidade foi aberta e o fluido aspirado com auxílio de pipeta de Pasteur estéril. Estas
células foram centrifugadas a 2500xg, 4ºC por 5 minutos. O sobrenadante foi
descartado e os macrófagos foram ressuspendidos em PBS. As células foram então
incubadas em placas de cultura, por 2 horas e após, lavadas três vezes com PBS para
remoção das células não-aderentes.

64
b) Índice de Fagocitose
Foram retiradas alíquotas de 0,1mL das suspensões de macrófagos e
adicionadas a placas com 96 poços para cultivo celular. O material ficou em repouso por
40 minutos, para a aderência das células. Foram adicionados 10µL de “zimosan”
(Sigma, 1x108 partículas/mL) a cada perfuração. O “zimosan” é uma preparação de
células de levedura (Saccharomyces cerevisiae) que conduz ao processo de fagocitose.
A incorporação do “zimosan” pelos macrófagos é classificada como uma fagocitose não
específica. Essa característica de não especificidade a distingue da incorporação de
antígenos mediada por receptores de superfície (CARVALHO et al., 2000).
O material foi incubado por 30 minutos com solução formol-cálcio de Baker (4%
formaldeído, 2% cloreto de sódio, 1% acetato de cálcio) por 30 minutos. As células
foram lavadas por centrifugação (435xg por 5 minutos). O “zimosan” foi solubilizado
pela adição de 0,1mL de álcool acidificado (10% ácido acético, 40% etanol em água
destilada) aos poços. Após 30 minutos, realizou-se a leitura de absorbância a 550nm.
c) Produção de Superóxido
A produção de superóxido foi estimada pelo ensaio de redução do NBT (nitroblue
tetrazolium), um composto amarelo que se torna insolúvel e de cor azul no estado
reduzido (MADHAVI; DAS, 1994). A utilização de PMA (forbol 12-miristato 13-acetato)
torna o método seletivo para superóxido, pois este composto estimula a atividade do
sistema NADPH-oxidase (que é dormente em células não desafiadas
imunologicamente). Uma vez organizado e funcional na membrana, este complexo
enzimático catalisa a oxidação do composto NADPH (produzido pela via pentose
fosfato a partir de glicose) a NADP+, sendo que os elétrons liberados nesta oxidação
são utilizados para a redução do oxigênio molecular ao ânion radical superóxido, O2-
(HALLIWELL; GUTTERIDGE, 1989; CADENAS, 1995).

65
Alíquotas de 0,45mL de solução de macrófagos foram suspensas em PBS e
incubadas por 1 hora a 37ºC na presença de 0,03mL de PMA e 0,1% de NBT. A reação
foi interrompida pela adição de 0,45mL de ácido acético glacial. A mistura foi
centrifugada a 2500xg por 30 segundos e o NBT reduzido presente no sedimento foi
solubilizado em 900µL de ácido acético a 50%. Os restos celulares foram sedimentados
por nova centrifugação (30 segundos) e a absorbância do sobrenadante determinada a
560nm.
d) Produção de Peróxido de Hidrogênio
Para processar os microorganismos fagocitados, os macrófagos precisam
produzir substâncias com alta capacidade lítica, como é o caso dos produtos envolvidos
no burst respiratório (aumento da captação de O2). Dentre estes produtos, encontra-se
o peróxido de hidrogênio, que é o resultado do aumento da captação de oxigênio,
seguida da redução do oxigênio em ânion superóxido (O2-) pela enzima NADPH
oxidase, e a conseqüente dismutação (espontânea ou enzimática) do ânion superóxido
que forma então, o peróxido de hidrogênio (PICK; MIZEL, 1981).
A produção de peróxido de hidrogênio foi realizada conforme método de PICK e
MIZEL (1981). Esse ensaio se baseia na reação que ocorre entre o vermelho de fenol
e o peróxido de hidrogênio, na presença de peroxidase (horseradish peroxidase).
Alíquotas de 100µL de solução de macrófagos e 10µL de éster de forbol 12-miristato
13-acetato (PMA – 20µM) foram colocados em placas de microdiluição, no escuro (para
impedir a foto-oxidação), por um período de 1 hora a 20ºC. Após esse período, o
sobrenadante foi descartado por inversão e foram adicionados 100µL da solução de
vermelho de fenol contendo peroxidase e zimosan. A solução foi incubada por 30
minutos e realizada leitura com absorbância de 620nm.

66
e) Avaliação do Volume do Lisossoma
A incorporação do corante catiônico vermelho, que se concentra nas células
lisossomais, corando-as, foi utilizada para avaliar o volume do sistema lisossomal dos
macrófagos (BONATTO et al., 2004).
Adicionou-se 20µL de solução de vermelho neutro a 3% dissolvida em PBS a
0,1mL de solução de macrófagos. A suspensão foi incubada por 30 minutos em poços
de microdiluição. As células foram lavadas por duas vezes com PBS por centrifugação
(435xg por 5 minutos). O vermelho neutro foi solubilizado pela adição de 0,1mL de
ácido acético a 10% e solução de etanol. Incubou-se por 30 minutos. A absorbância foi
lida a 550nm.
4.2.4.4 Sepse Induzida
Duas condições experimentais foram estudadas: (1) administração
intraperitoneal de LPS após dieta via oral de CPP e (2) injeção intraperitoneal de LPS
após administração intraperitoneal de CPP.
O Experimento 1 seguiu o delineamento abaixo:
Grupo 1: Grupo controle
Grupo 2: Grupo controle + LPS
Grupo 3: CPP A
Grupo 4: CPP A + LPS
Grupo 5: CPP B
Grupo 6: CPP B + LPS
Grupo 7: CPP C
Grupo 8: CPP C + LPS

67
Após o período de 22 dias foi realizada a indução de endotoxemia por meio de
injeção intraperitoneal de lipopolissacarídeo (LPS) de Escherichia coli (sorotipo
0111:B4; Sigma Chemical, St. Louis, MO) (Figura 16), em dose de 5mg/kg. Após
tratamento com LPS, os animais retornaram às suas gaiolas e foram monitorados nas 6
horas subseqüentes. A toxicidade foi avaliada pela observação de sinais adversos,
como: prostração, pilo-ereção e blefarite sanguinolenta. Foram avaliados também a
taxa e tempo de sobrevivência. Os animais foram submetidos a eutanásia por
exanguinação por punção intracardíaca seis horas após a injeção de LPS. Foram
realizados leucogramas e determinados os níveis de interleucina 6 (IL-6) no plasma. Os
pulmões dos animais submetidos ao LPS foram removidos, fixados em tampão
formalina-fosfato e embebidos em parafina. Cortes histológicos foram corados com
hematoxilina e eosina para avaliação. Na avaliação histológica, as variáveis analisadas
foram: presença de leucócitos, congestão, necrose perivascular, fibrose septal, edema
alveolar, hemorragia alveolar e colapso alveolar. As variáveis foram quantificadas por
uma escala de 4 pontos: 0, ausente; 1, discreto; 2, moderado; 3, intenso.
No Experimento 2, preparou-se solução a 5% de CPP-A, dissolvido em tampão
fosfato-salina (PBS, estéril, pH 7,4). Animais adultos, com peso variando entre 280 a
305g, receberam o peptídeo (CPP-A) via intraperitoneal (IP) (0,75g/kg). Em seguida,
induziu-se a sepse por injeção intraperitoneal de lipopolissacarídeo (LPS) de
Escherichia coli (sorotipo 0111:B4; Sigma Chemical, St. Louis, MO), em dose de
5mg/kg. O grupo controle recebeu somente a injeção de LPS. Os animais foram
submetidos a eutanásia após 6 horas da administração de LPS. Foram realizados
leucogramas e determinados os níveis de interleucina 6 (IL-6) no plasma.

68
FIGURA 16 - INJEÇÃO INTRAPERITONEAL COM LPS PARA INDUÇÃO DE CHOQUE
SÉPTICO
NOTAS: LPS, Lipopolissacarídeo.
4.2.4.5 Determinação dos Níveis de IL-6 Para determinação da produção de citocina, o sangue dos ratos foi coletado em
tubos contendo EDTA, para separação do plasma. Na determinação da interleucina 6,
utilizou-se kit específico para ELISA (BD OptEIATM, Pharmigen).
As amostras foram pipetadas sobre as escavações de placas de 96 poços
(Maxisorp F-96, Nunc) cobertas previamente com anticorpo de captura específico para
a interleucina (anticorpo monoclonal anti-rato IL-6), seguido de diversas lavagens e da

69
adição de diluente (tampão fosfato com 10% de soro fetal bovino, pH 7,0). Após a
colocação das amostras nas escavações, as placas foram incubadas por duas horas à
temperatura ambiente. Realizaram-se de cinco lavagens totais. Realizou-se então a
adição do anticorpo de detecção (anticorpo monoclonal anti-rato IL-6 biotinilado) e
incubação por uma hora a temperatura ambiente. Após remoção do excesso do
segundo anticorpo, adicionou-se a enzima peroxidase (avidin-horseradish peroxidase
conjugate). Esta se ligou ao anticorpo de detecção para completar a reação. Após a
lavagem para remoção de toda enzima não ligada, colocou-se solução de substrato
(TMB, tetrametilbenzidina e peróxido de hidrogênio) e as amostras foram incubadas por
30 minutos a temperatura ambiente, no escuro. Após esta etapa, foi adicionada solução
2N de ácido sulfúrico (stop solution), para interromper a reação. Realizou-se leitura a
450 nm e os valores obtidos para as amostras foram comparados com curva padrão,
que utilizou IL-6 liofilizada de rato.

70
5 RESULTADOS E DISCUSSÃO
5.1 OBTENÇÃO E CARACTERIZAÇÃO DOS CASEINOFOSFOPEPTÍDEOS
5.1.1 Composição
A análise química dos produtos está apresentada na Tabela 5,
comparativamente ao caseinato de sódio. O conteúdo protéico do CPP-C é semelhante
ao teor do caseinato, indicando que foram removidos poucos peptídeos da proteína
original. Em conseqüência do alto conteúdo mineral dos peptídeos, os valores de cinzas
são superiores aos do caseinato. O teor de umidade pode ser considerado elevado
para produtos cuja secagem foi realizada por liofilização, provavelmente devido à alta
higroscopicidade e elevada capacidade de absorção de água pelos hidrolisados.
TABELA 5 -COMPOSIÇÃO QUÍMICA DOS PEPTÍDEOS
PRODUTO PROTEÍNAS
(%) UMIDADE
(%) CINZAS (%)
Caseinato* 87,5 5,8 5,3 CPP-A 64,28±0,60 15,72±0,03 19,06±0,03 CPP-B 67,59±0,38 6,66±0,29 29,23±0,14 CPP-C 84,85±0,56 4,89±0,02 16,14±0,0003
NOTAS: *Lactoprot, HV (dados fornecidos pelo fabricante) Os dados estão apresentados como média ± desvio padrão da média para amostras analisadas em triplicata.
O conteúdo mineral dos peptídeos está listado na Tabela 6. Os teores
encontrados para cálcio, sódio e fósforo estão de acordo com valores apresentados na
literatura (ELLEGARD et al., 1999). O CPP-A apresentou um conteúdo de fósforo mais
elevado, indicando maior rendimento na obtenção dos fosfopeptídeos.
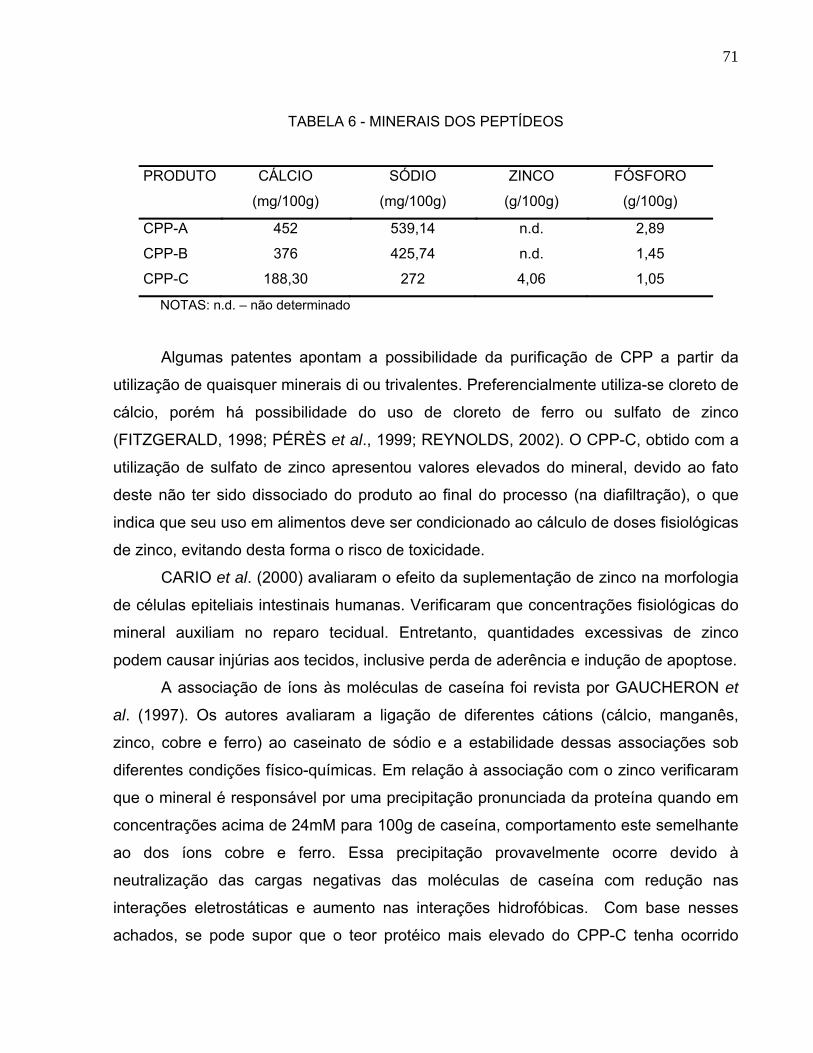
71
TABELA 6 - MINERAIS DOS PEPTÍDEOS
PRODUTO CÁLCIO
(mg/100g)
SÓDIO
(mg/100g)
ZINCO
(g/100g)
FÓSFORO
(g/100g)
CPP-A 452 539,14 n.d. 2,89
CPP-B 376 425,74 n.d. 1,45
CPP-C 188,30 272 4,06 1,05
NOTAS: n.d. – não determinado
Algumas patentes apontam a possibilidade da purificação de CPP a partir da
utilização de quaisquer minerais di ou trivalentes. Preferencialmente utiliza-se cloreto de
cálcio, porém há possibilidade do uso de cloreto de ferro ou sulfato de zinco
(FITZGERALD, 1998; PÉRÈS et al., 1999; REYNOLDS, 2002). O CPP-C, obtido com a
utilização de sulfato de zinco apresentou valores elevados do mineral, devido ao fato
deste não ter sido dissociado do produto ao final do processo (na diafiltração), o que
indica que seu uso em alimentos deve ser condicionado ao cálculo de doses fisiológicas
de zinco, evitando desta forma o risco de toxicidade.
CARIO et al. (2000) avaliaram o efeito da suplementação de zinco na morfologia
de células epiteliais intestinais humanas. Verificaram que concentrações fisiológicas do
mineral auxiliam no reparo tecidual. Entretanto, quantidades excessivas de zinco
podem causar injúrias aos tecidos, inclusive perda de aderência e indução de apoptose.
A associação de íons às moléculas de caseína foi revista por GAUCHERON et
al. (1997). Os autores avaliaram a ligação de diferentes cátions (cálcio, manganês,
zinco, cobre e ferro) ao caseinato de sódio e a estabilidade dessas associações sob
diferentes condições físico-químicas. Em relação à associação com o zinco verificaram
que o mineral é responsável por uma precipitação pronunciada da proteína quando em
concentrações acima de 24mM para 100g de caseína, comportamento este semelhante
ao dos íons cobre e ferro. Essa precipitação provavelmente ocorre devido à
neutralização das cargas negativas das moléculas de caseína com redução nas
interações eletrostáticas e aumento nas interações hidrofóbicas. Com base nesses
achados, se pode supor que o teor protéico mais elevado do CPP-C tenha ocorrido
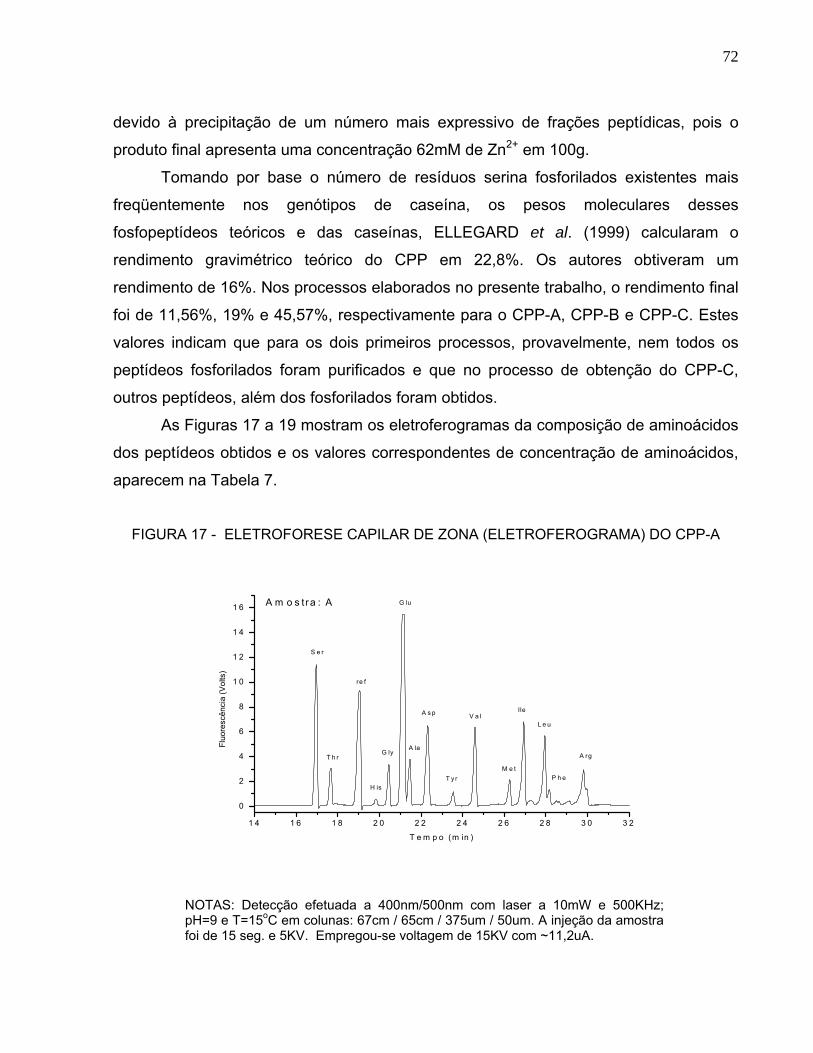
72
devido à precipitação de um número mais expressivo de frações peptídicas, pois o
produto final apresenta uma concentração 62mM de Zn2+ em 100g.
Tomando por base o número de resíduos serina fosforilados existentes mais
freqüentemente nos genótipos de caseína, os pesos moleculares desses
fosfopeptídeos teóricos e das caseínas, ELLEGARD et al. (1999) calcularam o
rendimento gravimétrico teórico do CPP em 22,8%. Os autores obtiveram um
rendimento de 16%. Nos processos elaborados no presente trabalho, o rendimento final
foi de 11,56%, 19% e 45,57%, respectivamente para o CPP-A, CPP-B e CPP-C. Estes
valores indicam que para os dois primeiros processos, provavelmente, nem todos os
peptídeos fosforilados foram purificados e que no processo de obtenção do CPP-C,
outros peptídeos, além dos fosforilados foram obtidos.
As Figuras 17 a 19 mostram os eletroferogramas da composição de aminoácidos
dos peptídeos obtidos e os valores correspondentes de concentração de aminoácidos,
aparecem na Tabela 7.
FIGURA 17 - ELETROFORESE CAPILAR DE ZONA (ELETROFEROGRAMA) DO CPP-A
1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0 3 2
0
2
4
6
8
1 0
1 2
1 4
1 6 A m o s tra : A
A rg
P h e
L e u
Ile
M e t
V a l
T y r
A s p
A la
G lu
G ly
H is
re f
T h r
S e r
Fluo
resc
ênci
a (V
olts
)
T e m p o (m in )
NOTAS: Detecção efetuada a 400nm/500nm com laser a 10mW e 500KHz; pH=9 e T=15oC em colunas: 67cm / 65cm / 375um / 50um. A injeção da amostra foi de 15 seg. e 5KV. Empregou-se voltagem de 15KV com ~11,2uA.

73
FIGURA 18 - ELETROFORESE CAPILAR DE ZONA (ELETROFEROGRAMA) DO CPP-B
1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0 3 2
0
2
4
6
8
1 0
1 2
1 4
1 6
Ile
A m o s tr a : D
A rg
P h e
L e u
M e t
V a l
T y r
A s p
A la
G lu
G ly
H is
re f
T h r
S e r
Fluo
resc
ênci
a (V
olts
)
T e m p o ( m in )
NOTAS: Detecção efetuada a 400nm/500nm com laser a 10mW e 500KHz; pH=9 e T=15oC em colunas: 67cm / 65cm / 375um / 50um. A injeção da amostra foi de 15 seg. e 5KV. Empregou-se voltagem de 15KV com ~11,2uA.
FIGURA 19 - ELETROFORESE CAPILAR DE ZONA (ELETROFEROGRAMA) DO CPP-C
1 4 1 6 1 8 2 0 2 2 2 4 2 6 2 8 3 0 3 2
0
2
4
6
8
1 0
1 2
1 4
Ile
A m o s t r a : F
A r g
P h e
L e u
M e t
V a l
T y r
A s p
A la
G lu
G ly
H is
r e f
T h r
S e r
Fluo
resc
ênci
a (V
olts
)
T e m p o ( m in )
NOTAS: Detecção efetuada a 400nm/500nm com laser a 10mW e 500KHz; pH=9 e T=15oC em colunas: 67cm / 65cm / 375um / 50um. A injeção da amostra foi de 15 seg. e 5KV. Empregou-se voltagem de 15KV com ~11,2uA.

74
TABELA 7 - COMPOSIÇÃO AMINOACÍDICA (g AMINOÁCIDO/100g DE PROTEÍNA) DOS CASEINOFOSFOPEPTÍDOS E DO CASEINATO DE SÓDIO
AMINOÁCIDOS
(g/100g de ptn.) CASEINATO DE SÓDIO
PEPTÍDEOS CPP-A CPP-B CPP-C
Serina
4,91 8,49 5,35 4,12
Treonina
3,61 3,08 3,68 3,35
Histidina
2,83 0 1,59 2,61
Glicina
1,6 1,51 1,45 1,18
Àcido glutâmico
21,3 29,49 24,77 17,95
Alanina
2,78 2,23 2,2 1,91
Ácido aspártico
6,23 8,07 7,21 5,86
Tirosina
4,98 1,63 1,96 2,67
Valina
5,74 5,8 5,99 4,43
Metionina
2,3 2,58 2,14 2,08
Isoleucina
4,65 8,04 6,69 4,43
Leucina
8,35 6,28 8,56 6,49
Fenilalanina
4,59 1,29 2,04 3,28
Arginina 3,24 0,76 2,92 2,93

75
A composição aminoacídica do caseinato de sódio revela altos teores de tirosina
e fenilalanina. A comparação do perfil de aminoácidos dos peptídeos obtidos e do
caseinato de sódio revela redução expressiva nos teores de fenilalanina e tirosina. Os
peptídeos CPP-A e CPP-B apresentam altos teores de serina, ácido glutâmico,
asparagina e isoleucina. Não foram observadas perdas elevadas de aminoácidos nos
peptídeos. O triptofano não foi determinado, pois o aminoácido é destruído durante a
hidrólise ácida utilizada na preparação das amostras.
BERNARDI et al. (2003) verificaram que o uso do evaporador a vácuo para
eliminar o HCl promove perdas de treonina e serina, relacionadas principalmente à
destruição por calor. A neutralização com solução padrão de citrato/NaOH diminui
essas perdas, contudo a inclusão de sais interfere com a determinação por eletroforese
capilar, sendo que por esta razão não foi adotada.
A análise quantitativa de aminoácidos por eletroforese capilar apresenta
desafios, incluindo a separação entre os compostos. Observa-se uma certa dificuldade
na separação de ácido glutâmico e alanina (algumas vezes os resultados aparecem
como a soma dos dois aminoácidos) e leucina com fenilalanina.
A detecção da cisteína (Cys), lisina (Lys) e arginina (Arg) por fluorescência
induzida a laser também apresenta alguns problemas. Nestes hidrolisados a Cys ocorre
como Cys-Cys (cistina) e não como cisteína livre. Por consequência, na reação de
derivatização, formam-se dois fluoróforos por molécula Cys-Cys, uma em cada amina
primária. Estes sistemas não fluorescem porque cada fluoróforo se encontra muito
próximo ao outro. Ocorre um fenômeno chamado "quenching", onde um rouba a
energia do outro, o que provoca diminuição na fluorescência (como se pode ver pelo
tamanho dos picos nos eletroferogramas). O mesmo acontece com a Lys, pois ela tem
uma segunda amina primária na cadeia lateral (pK=10,7). Com a Arg o problema não
ocorre porque o pK da amina primária da cadeia lateral é 12,1, este valor elevado
impede que a reação de derivatização ocorra com esta amina (da cadeia lateral).
No processo de elaboração dos peptídeos, após a realização da digestão tríptica,
reduziu-se o pH para 4,64 no CPP-A e 4,7 nos demais produtos e os precipitados

76
obtidos foram removidos por centrifugação. Esse procedimento tem como objetivo
principal remover os peptídeos hidrofóbicos do digesto protéico. Sabe-se que as
proteínas podem ser precipitadas pela indução de modificações no pH, força iônica,
constante dielétrica e temperatura. LÉONIL et al. (1994) verificaram que a precipitação
de peptídeos hidrofóbicos ocorre pela redução do pH a valores inferiores ao pH 5. Os
autores sugerem que a diminuição brusca da solubilidade dos peptídeos, nessa faixa de
pH, independente de seus pontos isoelétricos, seja responsável pela destruição das
associações entre peptídeos que os mantêm em solução. Como conseqüência, ocorre
um aumento nas interações hidrofóbicas, promovendo o fenômeno de precipitação
desses peptídeos.
A análise de aminoácidos revelou redução expressiva no teor de fenilalanina
(Phe) para o CPP-A (Figura 20). O valor obtido para este produto representou redução
de 71,89% em relação ao caseinato de sódio, 51,89% e 26,61% em relação a estudos
de desenvolvimento de hidrolisados com baixo teor de fenilalanina (LOPEZ-BAJONERO
et al., 1991; MONTEIRO, 2003). Estes trabalhos utilizaram carvão ativo na adsorção do
aminoácido, o que pode ocasionar perdas de aminoácidos essenciais, devido à baixa
especificidade do carvão.
Os resultados obtidos apontam os caseinofosfopeptídeos como uma alternativa
importante na elaboração de formulações a serem utilizadas em pacientes
fenilcetonúricos. A fenilcetonúria é uma das doenças causadas por erros inatos do
metabolismo. É de herança autossômica recessiva e resulta da deficiência da
fenilalanina hidroxilase hepática, que converte o aminoácido fenilalanina em tirosina. A
fenilcetonúria clássica tem uma incidência internacional média de 1:11000 recém
nascidos vivos. A manifestação clínica mais severa é o atraso no desenvolvimento
neuropsicomotor. A detecção precoce e o controle dietético rigoroso são essenciais
para esses pacientes. A dieta deve ser suplementada com fórmulas que podem ser
elaboradas a partir de misturas de aminoácidos ou produtos derivados de proteínas,
dos quais se tenha retirado a fenilalanina (LOPEZ-BAJONERO et al., 1991).
Diversas formulações isentas de fenilalanina estão disponíveis no mercado,
conforme se verifica na Tabela 8. São isentas por se constituírem em misturas de

77
aminoácidos, acrescidas de carboidratos, vitaminas, minerais, e outros ingredientes,
dependendo de cada formulação. Normalmente apresentam-se em forma de pó e
devem ser reconstituídas em água de acordo com a quantidade recomendada pelo
profissional de saúde que acompanha o paciente. O Lofenalac é um hidrolisado
protéico de caseína que contém aproximadamente 75 mg Phe/100g de produto.
TABELA 8 – FORMULAÇÕES ISENTAS DE FENILALANINA
FABRICANTE PRODUTO PROTEÍNA
(g/100g)
IDADE*
Mead Johnson Lofenalac 14,0
Phenyl-free 1 16,2
Phenyl-free 2 22,0
Phenyl-free 2 40,0
Vitaffix Rilla 1 60,0 Até 12 meses
Rilla 2 60,8 1 a 8 anos
Rilla 3 60,8 Mais de 8 anos
Support PKU 1 50,3 Até 12 meses
PKU 2 66,8 1 a 8 anos
PKU 3 68,0 Mais de 8 anos
XP Analog 13,0 Até 12 meses
XP Maxamaid 25,0 1 a 8 anos
XP Maxamum 39,0 Mais de 8 anos
NOTAS: *Idade: recomendação do fabricante Fonte: http://www.supportnet.com.br/produtos/erros_finilcetonuria/index.htm, http://www.vittafix.com.br/produtos_bod.htm, http://www.meadjohnson.com/professional/indivdescr.html#metabolics De acordo com a Resolução RDC nº 269, de 22 de setembro de 2005 (ANVISA,
2005), que trata da ingestão diária recomendada (IDR) de proteína, vitaminas e
minerais, crianças de 0 a 6 meses devem ingerir 9,1g de proteína por dia (ptn/d); até 11
meses, 11g ptn/d; de 1 a 3 anos, 13g ptn/d; de 4 a 6 anos, 19g ptn/d; e de 7 a 10 anos,
34g ptn/d.

78
A dieta de pacientes fenilcetonúricos deve conter entre 250 a 500mg Phe/dia,
enquanto que para não portadores a recomendação é de 2500mg Phe/dia (BRASIL,
2002).
A vantagem do uso dos caseinofosfopeptídeos em relação aos hidrolisados
tratados para eliminação de fenilalanina, é que o CPP não apresenta perda de
aminoácidos que possam comprometer o valor nutricional.
Além disso, a produção de caseinofosfopeptídeos a partir da digestão da caseína
eleva a biodisponibilidade de cálcio (KITTS et al., 1992). A ingestão de caseína ou β-
caseína produz o aparecimento in vivo de CPP no trato intestinal de ratos e associa-se
a uma maior concentração de cálcio solúvel no lúmen do íleo distal (SATO et al., 1991).
Estudos do estado mineral ósseo de crianças e adultos com fenilcetonúria revelaram
retardo na maturação do esqueleto e osteoporose (McMURRY et al., 1992; AL-
QADREH et al., 1998), sugerindo que os caseinofosfopeptídeos podem ser utilizados
para melhorar o perfil de mineralização óssea desses pacientes.
O teor de Phe do CPP-A é de 1,29g de Phe/100g proteína. Apesar de
aparentemente elevado, considerando que a recomendação de proteína para uma
criança de menos de três anos é de 13g ptn/dia, uma formulação à base deste peptídeo
respeitaria a ingestão recomendada de Phe ainda que o CPP fosse a única fonte de
proteína na dieta.
Além disto, o uso de hidrolisados protéicos contendo peptídeos de cadeia curta é
altamente desejado em formulações. A absorção de peptídeos de cadeia curta é
considerada mais eficiente quando comparada a uma quantidade equivalente de
aminoácidos livres. Isto se deve à disponibilidade de sistemas específicos de transporte
de peptídeos e a subseqüente quebra em aminoácidos por ação das peptidases
citoplasmáticas nos enterócitos, antes do transporte à circulação. Por outro lado,
peptídeos são menos hipertônicos que misturas de aminoácidos, facilitando a absorção
de outros componentes da dieta e eliminando problemas osmóticos. Além disto, devido
à instabilidade química e à insolubilidade em água, alguns aminoácidos como
glutamina, tirosina e cisteína não podem ser facilmente administrados em forma livre.
(CLEMENT, 2000)

79
FIGURA 20 - TEOR DE FENILALANINA DOS CASEINOFOSFOPEPTÍDEOS
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
CPP-A CPP-B CPP-C CASEÍNA * **
Phe
(g/1
00g
prot
eína
)
NOTAS: * , ** Hidrolisados de caseína tratados com carvão ativo para remoção da Phe (*MONTEIRO, 2003; **LOPEZ-BAJONERO et al., 1991).
A relação entre os dados de composição química e o conteúdo de serina permite
descrever alguns parâmetros de qualidade dos caseinofosfopeptídeos (Tabela 9).
TABELA 9 - RELAÇÃO MOLAR ENTRE DADOS DE
COMPOSIÇÃO QUÍMICA E CONTEÚDO AMINOACÍDICO DOS PEPTÍDEOS
AMOSTRA Ser/P* N/P N/Ser
CPP-A 0,973 8,082 8,306
CPP-B 0,515 7,790 15,128
CPP-C 0,680 16,73 24,611
NOTAS: *mol/mol

80
A relação molar entre serina/fosfato próxima de 1 indica que todos os resíduos
serina estão fosforilados.
TAKANORI et al. (1995) encontraram uma relação molar entre serina e fósforo
de 0,96, em caseinofosfopeptídeos obtidos das fezes de ratos que foram alimentados
com caseína, indicando que praticamente todos os resíduos de serina estavam
fosforilados. Valores baixos na relação molar entre nitrogênio/fósforo e nitrogênio/serina
indicam que os fosfopeptídeos apresentam, respectivamente, elevada fosforilação e
alto conteúdo de serina. Os valores obtidos nos processos realizados apontam o CPP-A
como um produto que apresenta teor mais elevado de serina fosforilada. LIHME et al.
(1998) obtiveram as seguintes relações molares na caracterização de fosfopeptídeos
obtidos por troca iônica: Ser/P, 0,84 a 0,93mol/mol; N/P, 6,71 a 6,92mol/mol; N/Ser,
7,00 a 7, 56mol/mol. Os autores comparam os valores obtidos com um processo de
obtenção de fosfopeptídeos com uso de etanol, (CPP III), cujos valores são os
seguintes: Ser/P, 1,06mol/mol; N/P, 10,05mol/mol; N/Ser, 9,58 mol/mol.
5.1.2 Solubilidade
A solubilidade de um material protéico descreve a capacidade do mesmo formar
soluções coloidais. O conceito de solubilidade para proteínas define a proporção de
nitrogênio (ou proteína) de uma amostra protéica que se solubiliza a partir de um
determinado procedimento de solubilização. A adição de sais pode modificar a
solubilidade protéica pela estabilização ou desdobramento da estrutura da proteína e
pela modificação na carga líquida da mesma (PILOSOF; BARTHOLOMAI, 2000).
A Figura 21 apresenta a solubilidade dos peptídeos. Ao se realizar a
comparação do perfil de solubilidade de cada produto em relação ao tratamento
realizado (dissolução em água ou PBS), se verificou que para o CPP-A não houve
diferença significativa (p= 0,400035). Esse achado pode se relacionar ao fato desse
produto ter sido obtido pela adição de etanol. Para os demais produtos houve diferença
significativa entre os tratamentos (CPP-B, p= 0,042463; CPP-C, p= 0,013688).
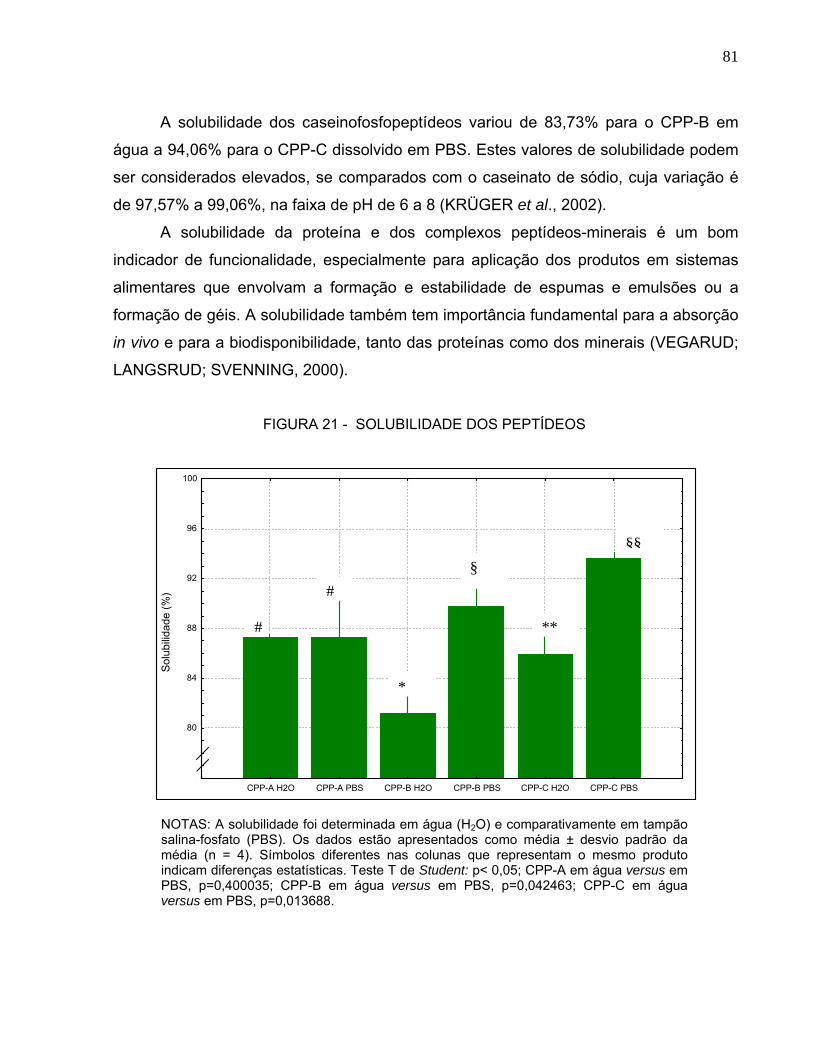
81
A solubilidade dos caseinofosfopeptídeos variou de 83,73% para o CPP-B em
água a 94,06% para o CPP-C dissolvido em PBS. Estes valores de solubilidade podem
ser considerados elevados, se comparados com o caseinato de sódio, cuja variação é
de 97,57% a 99,06%, na faixa de pH de 6 a 8 (KRÜGER et al., 2002).
A solubilidade da proteína e dos complexos peptídeos-minerais é um bom
indicador de funcionalidade, especialmente para aplicação dos produtos em sistemas
alimentares que envolvam a formação e estabilidade de espumas e emulsões ou a
formação de géis. A solubilidade também tem importância fundamental para a absorção
in vivo e para a biodisponibilidade, tanto das proteínas como dos minerais (VEGARUD;
LANGSRUD; SVENNING, 2000).
FIGURA 21 - SOLUBILIDADE DOS PEPTÍDEOS
Sol
ubilid
ade
(%)
76
80
84
88
92
96
100
§ #
# *
NOTAS: A solsalina-fosfato média (n = 4)indicam diferenPBS, p=0,400versus em PBS
CPP-A H2O C
ubilidade foi(PBS). Os d. Símbolos ças estatíst
035; CPP-B, p=0,01368
PP-A PBS C
*
determinadaados estãodiferentes nicas. Teste T em água 8.
PP-B H2O CPP-B PBS CP
em água (H2O) e comp apresentados como mas colunas que repres de Student: p< 0,05; C
versus em PBS, p=0,0
*
P-C H2O CPP-C
arativamenteédia ± desvioentam o mesPP-A em água42463; CPP-C
§§
PBS
em tampão padrão da mo produto versus em em água

82
5.1.3 Seqüenciamento
A digestão tríptica do caseinato de sódio, seguida de processos de separação
permitiu a obtenção de 3 produtos que se caracterizam como uma mistura de peptídeos
derivados da hidrólise das caseínas αS1, αS2, β e κ (KRÜGER et al., 2005).
Os picos encontrados nos espectros de massa estão demonstrados nas Figuras
22 a 24 e foram comparados com seqüências conhecidas das caseínas bovinas,
tomando como base seu peso molecular (FERRANTI et al., 2004). A confirmação das
seqüências foi feita por espectrometria de massa e, quando necessário, as amostras
foram analisadas por seqüenciamento automatizado, utilizando um kit para
seqüenciamento de peptídeos, conforme descrito nos métodos. Essas análises
permitiram a determinação da seqüência de aminoácidos da maioria dos fragmentos
obtidos. A caracterização completa dos peptídeos das três amostras está descrita nos
Quadros 1 a 3.
A técnica de MALDI-TOF-MS pode ser utilizada na identificação de resíduos
fosforilados. A análise por PSD (post source decay) identifica o resíduo fosforilado na
seqüência peptídica. Esse processo é relativamente simples no caso da fosforilação da
tirosina, pois a fosfotirosina é detectada como um resíduo correspondente à soma da
massa da tirosina com o grupamento fosforil. Entretanto, o resíduo serina fosforilado é
mais lábil, sendo perdido facilmente durante a fragmentação, resultando na formação
de um resíduo deidroalanina no lugar da fosfoserina, o que conduz a formação de um
pico de intensidade mais elevada (TALBO et al., 2001). Optou-se então, pela utilização
das técnicas de seqüenciamento já mencionadas, sendo que a presença de peptídeos
fosforilados foi deduzida a partir da verificação da existência de resíduos fosfoserina
nos fragmentos obtidos.
A análise dos espectros resultou na identificação dos seguintes peptídeos
fosforilados: αs1-caseína (f62-73) no CPP-B, αs1-caseína (f106-115) no CPP-A e CPP-C,
αs1-caseína (f100-115), no CPP-B, αs1-caseína (f109-139) no CPP-C e αs2-caseína
(f151 -165) no CPP-B. Entretanto, não foram detectados peptídeos fosforilados da β-
caseína.

83
Os resultados parecem controversos se comparados com a Tabela 9 que
relaciona a composição química com a análise de aminoácidos, indicando um alto grau
de fosforilação para o CPP-A e, para os demais produtos, valores médios.
Sabe-se que existem dificuldades na localização de resíduos fosforilados. Uma
das estratégias que podem ser empregadas consiste na separação da fosfoproteína ou
peptídeo em pequenos fragmentos. Utiliza-se preferencialmente a composição em
aminoácidos do que a seqüência para a identificação do resíduo (MEYER et al., 1986).
Na análise dos dados se verificou que os segmentos N-teminais não foram
identificados nas caseínas αs2, β e κ. No caso da αs1-caseína, que apresenta uma
porção N-terminal hidrofóbica e rica em prolina, os fragmentos 23-37 foram identificados
no CPP-A e CPP-C. Na seqüência da β-caseína esse fato se torna mais evidente, pois
o segmento N-terminal, com elevada densidade de carga e bastante hidrofílico,
apresenta quatro radicais fosfato entre os resíduos 15 – 19 e um resíduo na posição 35
(SGARBIERI, 1998), que não foram identificados.
MINKIEWICZ et al. (2000) verificaram que durante a ionização para os ensaios
de espectrometria pode ocorrer fragmentação de alguns peptídeos com liberação de
íons e elevação de até 15Da nas massas observadas.
Em relação ao tamanho dos peptídeos obtidos, os espectros indicam pesos
moleculares coincidindo com outros trabalhos de obtenção de caseinofosfopeptídeos
(ELLEGARD et al., 1999), sendo que para o CPP-A e o CPP-B os fragmentos variaram
de 5 a 20 resíduos de aminoácidos e para o CPP-C de 6 a 30 resíduos.
Um dos peptídeos cuja seqüência foi deduzida a partir dos dados de MS é o
fragmento 128 – 137 da κ-caseína, que compõem quase toda a estrutura da
casoplatelina (f 127 – 137), análoga ao fibrinogênio e com provável ação antimicrobiana
(RIZZELLO et al., 2005).
Praticamente todos os peptídeos identificados podem ser classificados como
peptídeos trípticos, clivados pela tripsina nos resíduos Lys e Arg. A ausência de
peptídeos quimotrípticos demonstra a baixa quantidade de quimiotripsina na enzima
utilizada (PTN 6.0, Novozymes, Denmark).

84
FIGURA 22 - ESPECTRO DE MASSA POR DESORÇÃO A LASER DO CPP-A
NOTAS: Espectro obtido em espectrômetro de massa Reflex IV, com laser UV de 337nm e intensidade média de 12%, no modo refletor positivo (voltagem do refletor igual a 19,8kV) e extração retardada (delayed extraction), usando faixa de massa de 0,3 a 3,5kDa.

85
FIGURA 23 - ESPECTRO DE MASSA POR DESORÇÃO A LASER DO CPP-B
NOTAS: Espectro obtido em espectrômetro de massa Reflex IV, com laser UV de 337nm e intensidade média de 12%, no modo refletor positivo (voltagem do refletor igual a 19,8kV) e extração retardada (delayed extraction), usando faixa de massa de 0,3 a 3,5kDa.

86
FIGURA 24 - ESPECTRO DE MASSA POR DESORÇÃO A LASER DO CPP-C
NOTAS: Espectro obtido em espectrômetro de massa Reflex IV, com laser UV de 337nm e intensidade média de 12%, no modo refletor positivo (voltagem do refletor igual a 19,8kV) e extração retardada (delayed extraction), usando faixa de massa de 0,3 a 3,5kDa.

87
QUADRO 1 - MASSAS MOLECULARES DOS PEPTÍDEOS DETECTADOS NO CPP-A E
POSSÍVEIS SEQÜÊNCIAS
Amostra Peptídio
(m/z)
Seqüência deduzida a partir
dos dados de MS
Seqüência
Confirmada por MS/MS
Caseína
Identificada
CPP-A
1104,47*
KTKVIPYVR − α - s2 caseína
(212 – 220)
CPP-A
1104,47*
QTEDELQDK − β − caseína
(55 – 63)
CPP-A 1267,52
#
YLGYLEQLLR YLGYLEQLLR α - s1 caseína
(106 – 115)
CPP-A
1759,73
HQGLPQEVLNENLLR HQGLPQEVLNENLLR α - s1 caseína
(23 – 37)
CPP-A
1867,72
VKEAMAPKHKEMPFPK − β - caseína
(113 – 128)
CPP-A
2281,02*
QPLPPTVMFPPQSVLSLSQSK − β - caseína
(164 – 184)
CPP-A
2281,02*
QTPVVVPPFLQPEVMGVSKVK − β - caseína
(94 – 114)
NOTAS:1) As massas marcadas com um asterisco (*) correspondem aos peptídeos isobáricos, que apresentam duas possibilidades de seqüência com base nos dados de MS. As massas marcadas com sinal jogo da velha (#) correspondem aos peptídeos fosforilados; 2) Os experimentos para dedução das sequências foram realizados em um espectrômetro de massa Reflex IV, operando no modo refletor positivo e utilizando o acessório FAST (Fragmentation Analysis and Structural TOF). Os dados foram analisados com o auxílio do programa BioTools v. 2.0. Para alguns peptídeos, cuja fragmentação não foi observada pelo método PSD convencional, foi realizado o seqüenciamento por CAF (chemically assisted fragmentation) usando kit de seqüenciamento. QUADRO 2 - MASSAS MOLECULARES DOS PEPTÍDEOS DETECTADOS NO CPP-B E
POSSÍVEIS SEQÜÊNCIAS
Amostra Peptídio
(m/z)
Seqüência deduzida a partir
dos dados de MS
Seqüência
Confirmada por MS/MS
Caseína
Identificada
CPP-B
751,44
YPVEPF YPVEPF β - caseína
(129 – 133)
continua

88
Amostra Peptídio
(m/z)
Seqüência deduzida a partir
dos dados de MS
Seqüência
Confirmada por MS/MS
CPP-B
977,54
TKLTEEEK −
CPP-B
1139,40
AIPPKKNQDK −
CPP-B
1383,80
LLYQEPVLGPVR LLYQEPVLGPVR
CPP-B 1476,55
#
ESTEDQAMEDIK −
CPP-B
1494,59
QGPIVLNPWDQVK −
CPP-B
1759,61
HQGLPQEVLNENLLR HQGLPQEVLNENLLR
CPP-B 1771,61
#
KKTVDMESTEVFTKK −
CPP-B
1867,52
VKEAMAPKHKEMPFPK −
CPP-B 1951,56
#
DVPSERYLGYLEQLLR −
CPP-B
2280,66*
QPLPPTVMFPPQSVLSLSQSK −
CPP-B
2280,66*
QTPVVVPPFLQPEVMGVSKVK −
CPP-B
2794,68
KLTEEEKNRLNFLKKISQRYQK −
NOTAS: 1) As massas marcadas com um asterisco (*) correspondem aos pepapresentam duas possibilidades de seqüência com base nos dados de MS. As msinal jogo da velha (#) correspondem aos peptídeos fosforilados; 2) Os experimentos para dedução das sequências foram realizados em um esmassa Reflex IV, operando no modo refletor positivo e utilizando o acessório FASTAnalysis and Structural TOF). Os dados foram analisados com o auxílio do progra2.0. Para alguns peptídeos, cuja fragmentação não foi observada pelo método PSfoi realizado o seqüenciamento por CAF (chemically assisted fragmentation) seqüenciamento.
continuação
Caseína
Identificada
α - s2 caseína
(166 – 173)
κ - caseína
(128 – 137)
β - caseína
(206 – 217)
α - s1 caseína
(62 – 73)
α - s2 caseína
(116 – 128)
α - s1 caseína
(23 – 37)
α - s2 caseína
(151 – 165)
β - caseína
(113 – 128)
α - s1 caseína
(100 – 115)
β - caseína
(164 – 184)
β - caseína
(94 – 114)
α - s2 caseína
(167 – 188)
tídeos isobáricos, que assas marcadas com
pectrômetro de (Fragmentation ma BioTools v.
D convencional, usando kit de

89
QUADRO 3 - MASSAS MOLECULARES DOS PEPTÍDEOS DETECTADOS NO CPP-C E POSSÍVEIS SEQÜÊNCIAS
Amostra Peptídio
(m/z) Seqüência deduzida a partir
dos dados de MS Seqüência
Confirmada por MS/MS Caseína
Identificada
CPP-C 634,39+ LNFLK LNFLK α - s2 caseína
(176 – 180)
CPP-C 751,37+ YPVEPF YPVEPF β - caseína
(129 – 134)
CPP-C
827,47
NQFYQK − α - s2 caseína
(101 – 106)
CPP-C 830,45+ AVPYPQR AVPYPQR β - caseína
(192 – 198 )
CPP-C 910,47+ EGIHAQQK − α - s1 caseína
(140 – 147)
CPP-C 960,00+ PIGSENSEK PIGSENSEK α - s1 caseína
(200 – 208)
CPP-C
977,51
TKLTEEEK − α - s2 caseína
(166 – 173)
CPP-C 994,57+ QEPVLGPVR QEPVLGPVR β - caseína
(209 – 217)
CPP-C
1057,49
QVLSNTVPAK QVLSNTVPAK κ- caseína
(98 – 107)
CPP-C 1267,56
#
YLGYLEQLLR YLGYLEQLLR α - s1 caseína
(106 – 115)
CPP-C
1759,58
HQGLPQEVLNENLLR HQGLPQEVLNENLLR α - s1 caseína
(23 – 27)
CPP-C 2104,97+ TDAPSFSDIPNPIGSENSEK TDAPSFSDIPNPIGSENSEK
α - s1 caseína
(189 – 208)
CPP-C
2280,61*
QPLPPTVMFPPQSVLSLSQSK − β - caseína
(164 – 184)
CPP-C
2280,61*
QTPVVVPPFLQPEVMGVSKVK − β - caseína
(94 – 114)
continua

90
CPP-C
2794,60
KLTEEEKNRLNFLKKISQRYQK − α - s2 caseína
(167 – 188)
CPP-C 3833,69
#
YLEQLLRLKKYKVPQLEIVPNSAEERLHSMK − α - s1 caseína
(109 – 139)
continuação
NOTAS: 1) As massas marcadas com um asterisco (*) correspondem aos peptídeos isobáricos, que apresentam duas possibilidades de seqüência com base nos dados de MS. As massas marcadas com sinal jogo da velha (#) correspondem aos peptídeos fosforilados; 2) As massas marcadas com sinal de adição (+) correspondem a peptídeos que não foram detectados nos primeiros dados de MS obtidos por MALDI-TOF, mas foram isolados posteriormente por cromatografia líquida de alta performance (HPLC) e seqüenciados por ESI-MS/MS. 3) Os experimentos para dedução das sequências foram realizados em um espectrômetro de massa Reflex IV, operando no modo refletor positivo e utilizando o acessório FAST (Fragmentation Analysis and Structural TOF). Os dados foram analisados com o auxílio do programa BioTools v. 2.0. Para alguns peptídeos, cuja fragmentação não foi observada pelo método PSD convencional, foi realizado o seqüenciamento por CAF (chemically assisted fragmentation) usando kit de seqüenciamento.

91
5.2 ATIVIDADE ANTIMICROBIANA IN VITRO
O resultado do teste de esterilidade das amostras, que detecta possíveis
contaminações da matéria-prima utilizada para os experimentos, foi analisado após 72
horas e após sete dias da inoculação dos peptídeos em frascos Hemoprov I. Observou-
se imediata turvação do CPP-A no momento da inoculação das amostras, sendo que
esta não se modificou no período de avaliação. Não foram observadas quaisquer
alterações nos frascos, conforme se pode visualizar na Figura 25.
FIGURA 25 – TESTE DE ESTERILIDADE DOS CASEINOFOSFOPEPTÍDEOS
NOTAS: Caseinofosfopeptídeos em solução salina de PBS estéril, pH 7,4, na concentração de 125mg/mL. Foram inoculados 0,9mL destas soluções em kit HEMOPROV I.
Neste trabalho se verificou que os três produtos elaborados apresentaram
atividade antimicrobiana.
A Tabela 10 apresenta a porcentagem da inibição do crescimento pelos
peptídeos nos ensaios realizados com Escherichia coli, Staphylococcus aureus e
Salmonella typhimurium.

92
TABELA 10 - INIBIÇÃO DO CRESCIMENTO MICROBIANO PELOS CASEINOFOSFOPEPTÍDEOS
% DA INIBIÇÃO DO CRESCIMENTO2AMOSTRA QUANTIDADE1
(mg/mL) E. coli S.
aureus
S.
typhimurium
CPP-A 1,0 53±2 NI3 NI
CPP-A 0,2 10±6 NI NI
CPP-B 125 NI 53±14 27±2
CPP-B 25 NI NI 5±3
CPP-C 125 15±8 87±6 94±7
CPP-C 25 73±0,8 71±6 89±1
CPP-C 5 84±1 41±11 48±2
CPP-C 1 30±3 NI 16±1
NOTAS: 1- Quantidade do peptídeo no ensaio antimicrobiano 2- % média da inibição do crescimento ± desvio padrão (n = 3), avaliado pelo aumento na
absorbância após 24 horas de incubação a 37ºC. O valor da A620 nm das culturas controle (sem o peptídeo) foi tomado como 100%
3- NI = nenhuma inibição A inibição do crescimento de Escherichia coli pelo CPP-A ocorreu nas
concentrações de 0,2 e 1mg/mL. Nas demais concentrações, não houve inibição do
crescimento. O CPP-B reduziu o crescimento de Staphylococcus aureus e Salmonella
typhimurium, respectivamente em 53% e 27%, quando numa concentração de
125mg/mL. O CPP-C inibiu o crescimento de Escherichia coli e Salmonella typhimurium
a partir de 1mg/mL. A inibição do crescimento de Staphylococcus aureus ocorreu a
partir de uma concentração de 5mg/mL.
A Figura 26 apresenta os resultados obtidos na inibição do crescimento da
Listeria monocytogenes. Em todos os peptídeos estudados houve diferença significativa
entre o controle (amostra sem adição do peptídeo) e as amostras com adição dos três
caseinofosfopeptídeos.
A concentração inibitória mínima (MIC) da atividade dos microorganismos é
definida como a concentração mínima na qual há um declínio significativo na
absorbância (DEVIENNE; RADDI, 2002). No presente trabalho está apresentado o

93
comportamento dos inóculos frente aos peptídeos, além dos valores inibitórios mínimos.
Foram realizados repiques das negativas, sendo que as placas de cultivo apresentaram
crescimento escasso. Desta forma se pode notar que o comportamento da Listeria
monocytogenes nem sempre se apresenta linear, sendo que no caso do CPP-B há um
efeito bacteriostático inicial que não se perpetua quando as doses de peptídeo são
aumentadas. Estes dados foram confirmados por repetições, sendo o MIC o método de
escolha para determinação da atividade antimicrobiana.
O CPP-C foi obtido com a utilização de sulfato de zinco. A presença do zinco no
produto pode ser responsável pela potencialização do efeito antimicrobiano.
O peptídeo denominado kappacina, derivado da κ-caseína, também denominado
glicomacropeptídeo (GMP) fosforilado demonstrou atividade inibidora do crescimento
contra bactérias oportunistas da via oral (Streptococcus mutans e Porphyromonas
gingivalis) e contra Escherichia coli (MALKOSKI et al., 2001). DASHPER et al. (2005)
ligaram cátions divalentes à kappacina e verificaram que o metal Zn2+ permitiu uma
melhoria na propriedade antibacteriana. A ligação de cátions divalentes ao peptídeo
provavelmente altera sua conformação, podendo facilitar a ligação do peptídeo à
membrana celular da bactéria.
O íon zinco também é capaz de reduzir o crescimento e o metabolismo de
bactérias por interagir com grupos sulfidrílicos das enzimas bacterianas, inibindo sua
atividade. O zinco é um mineral bacteriostático, contudo quando em altas
concentrações, apresenta atividade bactericida (PHAN et al., 2004).

94
FIGURA 26 - EFEITO DOS CASEINOFOSFOPEPTÍDEOS CONTRA LISTERIA MONOCYTOGENES
A
B
0,0
0,1
0,1
0,2
0,2
0,3
0,3
Abso
rbân
cia
(650
nm
)
0 0,2 1 5 25 125Concentração de CPP-A (mg/mL)
a a
bab
a
c
0,0
0,1
0,2
0,3
0,4
0,5
0,6
Abso
rbân
cia
( 650
nm
)
0 0,2 1 5 25 125
Concentração de CPP-B (mg/mL)
c
d d
c
b
a
C
0,0
0,1
0,1
0,2
0,2
0,3
0,3
Abso
rbân
cia
(650
nm
)
0 0,2 1 5 25 125Concentração de CPP-C (mg/mL)
b b
cc
b
a
NOTAS: Os dados estão apresentados como valores médios obtidos para análises realizadas em triplicata. A concentração inicial (0 mg/mL do CPP) se refere ao controle, sendo que colunas com letras diferentes desta, apresentam resultados estatísticos diferentes. ANOVA com teste de Tukey para comparação das médias: p<0,05; A = CPP-A, p=0,0001; B = CPP-B, p=0,0001; C = CPP-C, p=0,0001

95
O mecanismo de inibição do crescimento das bactérias pelos peptídeos testados
não é conhecido, porém algumas possibilidades da interação peptídeos/bactérias
podem ser apontadas.
Alguns peptídeos antibacterianos têm sido isolados e caracterizados. Geralmente
possuem uma alta porcentagem de resíduos básicos em sua estrutura primária,
facilitando a interação entre as cargas positivas do peptídeo e as cargas negativas da
membrana bacteriana (MALKOSKI et al., 2001).
Peptídeos antimicrobianos geralmente possuem resíduos prolina e fosforilação
e/ou glicosilação em peptídeos aniônicos. Cita-se também a presença de resíduos
cisteína ou de pontes dissulfeto (MALKOSKI et al., 2001).
Os caseinofosfopeptídeos testados se apresentam como uma mistura de
peptídeos com pontos isoelétricos variáveis, havendo alguns peptídeos catiônicos e
também peptídeos aniônicos fosforilados, tais como o fragmento 62 - 73 da αs1-caseína,
presente no CPP-B, com pI 3,83. O resíduo prolina também aparece em alguns
segmentos, como é o caso do fragmento 113 – 128 da β-caseína com pI 9,53.
(KRUGER et al., 2005). Embora estas seqüências não possuam atividade
antimicrobiana descrita, atendem parcialmente às características dos peptídeos
antimicrobianos citados por MALKOSKI et al., 2001. Dentre os peptídeos obtidos
também está o fragmento 128 – 137 da κ-caseína no CPP-B, semelhante à
casoplatelina (f 127 – 137) e com provável ação antimicrobiana (RIZZELLO et al.,
2005).
5.3 AVALIAÇÃO DA AÇÃO DOS CASEINOFOSFOPEPTÍDEOS NA RESPOSTA IMUNE
5.3.1 Imunidade Inespecífica
5.3.1.1 Índice de Fagocitose
Os macrófagos desempenham um papel fundamental no reconhecimento,
internalização e degradação de substâncias estranhas. Possuem receptores de

96
superfície, que reconhecem os componentes moleculares comuns a muitos patógenos
bacterianos. O processo fagocítico é benéfico ao organismo na defesa contra
constituintes microbianos, mas também pode contribuir para a patogênese de algumas
doenças inflamatórias (CARVALHO et al., 2000).
A Figura 26 apresenta os resultados obtidos na ativação dos macrófagos
peritoneais pelo “zimosan”. Os animais que receberam a suplementação de CPP-A e
CPP-B não diferiram do controle. O índice de fagocitose do grupo CPP-C se apresentou
significativamente menor do que o controle (p = 0,01178), sendo que os demais
peptídeos não diferiram entre si.
A utilização de partículas de “zimosan” demonstrou que os peptídeos CPP-A e
CPP-B não alteram a atividade dos macrófagos na capacidade de promover a
fagocitose. O CPP-C induziu a uma diminuição da capacidade fagocítica, sugerindo uma
possível interferência na eliminação de patógenos bacterianos ou virais.
FIGURA 27 - FAGOCITOSE DE ZIMOSAN PELOS MACRÓFAGOS
PERITONEAIS DOS RATOS
Índi
ce d
e Fa
goci
tose
,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
controle CPP-A CPP-B C
*
NOTAS: Grupos controle ou com suplementação de peptídeos (CPP-A, um período de 22 dias. Índice de fagocitose expresso em absorbância/2dados apresentados se referem às médias ± desvio padrões da média (n de teste de Tukey para comparação das médias: * p< 0,05 versus Controle
PP-C
1CPP-B e CPP-C) por x105 células/poço. Os = 5). ANOVA, seguida .

97
5.3.1.2 Produção de superóxido
A produção de superóxido não apresentou diferença entre os grupos
(p=0,059855). Ao se observar a Figura 28, verifica-se que os animais alimentados com o
CPP-C apresentaram valores mais elevados na produção deste radical tóxico e que
aqueles animais alimentados com o CPP-B apresentaram os menores valores.
Embora a produção de superóxido e peróxido de hidrogênio pelos macrófagos
dos animais não tenha sido superior ao controle, os dados obtidos indicam que os
peptídeos avaliados não alteraram a imunidade inata.
FIGURA 28 - PRODUÇÃO DE ÂNION SUPERÓXIDO (O2-) PELOS
MACRÓFAGOS PERITONEAIS DOS RATOS
Prod
ução
de
Supe
róxi
do
0,74
0,80
0,86
0,92
0,98
1,04
1,10
controle CPP-A CPP-B CPP-C
NOTAS: Grupos controle ou com suplementação de peptídeos (CPP-A, CPP-B e CPP-C) por um período de 22 dias. Produção de ânion superóxido expressa em absorbância/2x105 células/poço. Os dados apresentados se referem às médias ± desvio padrões da média (n = 5). ANOVA, p< 0,05.

98
5.3.1.3 Produção de peróxido de hidrogênio
Quase imediatamente após um patógeno ser englobado por um macrófago
ativado, uma bateria de enzimas degradativas e outras substâncias tóxicas são
produzidas. Oxidases geram radicais tóxicos do oxigênio e de peróxido de hidrogênio.
A produção de peróxido de hidrogênio pelos macrófagos peritoneais não
apresentou diferença entre o controle e os peptídeos testados (p = 0,388661), conforme
Figura 29.
FIGURA 29 - PRODUÇÃO DE PERÓXIDO DE HIDROGÊNIO PELOS
MACRÓFAGOS PERITONEAIS DOS RATOS
Pro
duçã
o de
Per
óxid
o de
Hid
rogê
nio
0,00
0,06
0,12
0,18
0,24
0,30
0,36
controle CPP-A CPP-B CPP-C
NOTAS: Grupos controle ou com suplementação de peptídeos (CPP-A, CPP-B e CPP-C) por um período de 22 dias. Produção de peróxido de hidrogênio expressa em absorbância/2x105 células/poço. Os dados apresentados se referem às médias ± desvio padrões da média (n = 5). ANOVA, p< 0,05.
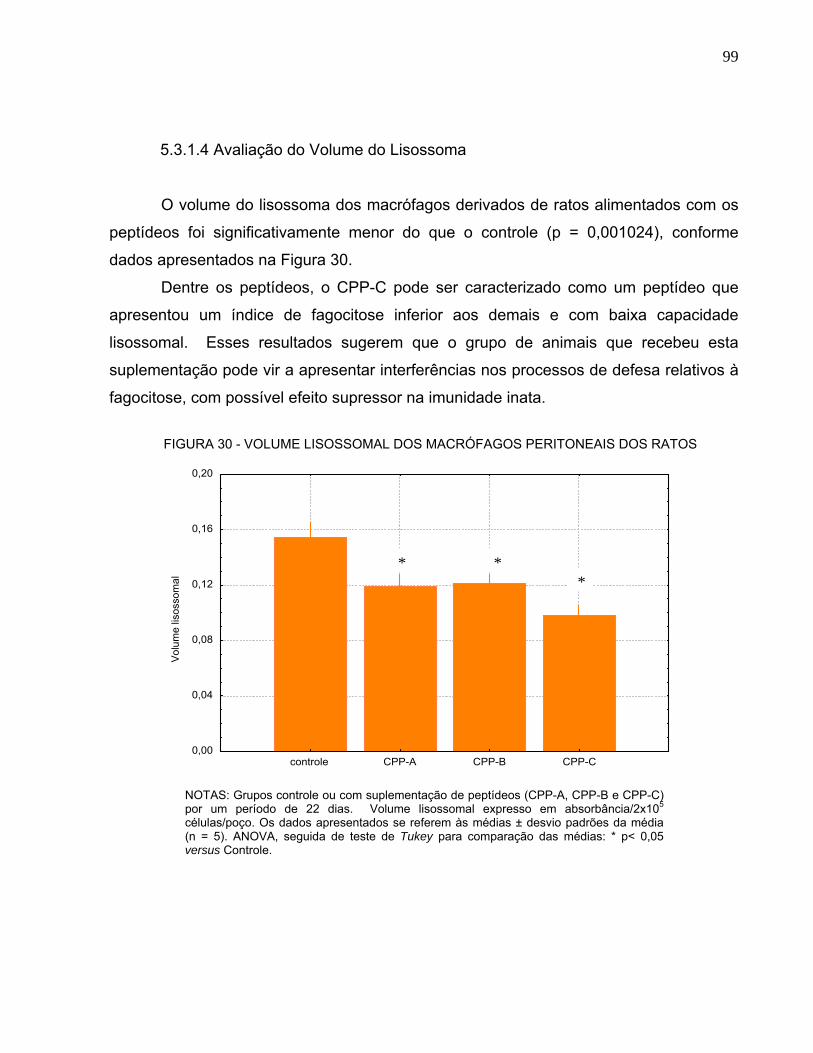
99
5.3.1.4 Avaliação do Volume do Lisossoma
O volume do lisossoma dos macrófagos derivados de ratos alimentados com os
peptídeos foi significativamente menor do que o controle (p = 0,001024), conforme
dados apresentados na Figura 30.
Dentre os peptídeos, o CPP-C pode ser caracterizado como um peptídeo que
apresentou um índice de fagocitose inferior aos demais e com baixa capacidade
lisossomal. Esses resultados sugerem que o grupo de animais que recebeu esta
suplementação pode vir a apresentar interferências nos processos de defesa relativos à
fagocitose, com possível efeito supressor na imunidade inata.
FIGURA 30 - VOLUME LISOSSOMAL DOS MACRÓFAGOS PERITONEAIS DOS RATOS
Vol
ume
lisos
som
al
0,00
0,04
0,08
0,12
0,16
0,20
controle C
* *
NOTAS: Grupos controle ou com suplepor um período de 22 dias. Volucélulas/poço. Os dados apresentados(n = 5). ANOVA, seguida de teste deversus Controle.
PP-A CP
mentação de pepme lisossomal e
se referem às mé Tukey para com
P-B C
*
tídeos (CPP-A,xpresso em adias ± desvio pparação das m
PP-C
CPP-B e CPP-C) bsorbância/2x105 adrões da média édias: * p< 0,05

100
5.3.2 Sepse Induzida
Antes do início dos Experimentos 1 e 2, os animais foram submetidos a punção
intracardíaca para realização de leucograma. Os dados apresentados na Figura 31 se
referem à contagem diferencial das células leucocitárias. Os valores encontrados estão
dentro dos padrões de normalidade, de acordo com HARKNESS e WAGNER (1993).
Os autores apontam os seguintes valores hematológicos para ratos: leucócitos totais, 6
a 17 x103/mm3; neutrófilos, 9 a 34%; linfócitos 65 a 85%; eosinófilos, 0 a 6%;
monócitos, 0 a 5%; basófilos, 0 a 1,5%; plaquetas, 500 a 1300 x103/mm3.
FIGURA 31 - CONTAGEM DIFERENCIAL DAS CÉLULAS LEUCOCITÁRIAS
ANTES DA INTRODUÇÃO DAS DIETAS (TEMPO = 0)
%
0
20
40
60
80
100
Eosinófilos Bastonetes Segmentados Linfócitos Monócitos Basófilos
NOTAS: Os dados apresentados se referem às médias ± desvio padrões da média.

101
5.3.2.1 Experimento 1: Administração Intraperitoneal de LPS Após Dieta Via Oral
de CPP
A Tabela 11 apresenta a comparação entre o ganho de peso dos ratos que
receberam a dieta controle e as dietas com adição de CPP. Os pesos finais dos grupos
com adição de CPP-B foram mais elevados, porém sem diferença estatística. Houve
homogeneidade entre as variâncias para ganho de peso, comprovada pelo teste de
Levene (p = 0,44928). OTANI et al. (2000) forneceram caseinofosfopeptídeo (CPP-III) a
camundongos, não encontrando diferenças significativas entre o ganho de peso de
dietas acrescidas de CPP-III (0,1 a 1g CPP por 100g de dieta) e dietas controle.
TABELA 11 - PESO DOS RATOS ALIMENTADOS COM DIETA CONTROLE OU DIETA
ADICIONADA DE CPP, POR UM PERÍODO DE 22 DIAS
DIETA
PESO CORPÓREO (g)
INÍCIO Dieta controle 198.58±9.79 Adição de CPP-A 203.5±20.10 Adição de CPP-B 201.25±12.05 Adição de CPP-C 202.92±13.17 FINAL Dieta controle 291.32±13.14 Adição de CPP-A 295.21±17.33 Adição de CPP-B 302.05±17.33 Adição de CPP-C 289.51±14.72 NOTAS: Os dados apresentados se referem às médias ± desvio padrões da média (n = 12 animais por grupo). ANOVA: p< 0,05
Os animais receberam uma dose de 5 mg/kg de LPS, que induz a uma alta taxa
de mortalidade. A sobrevivência foi de 100% para os grupos controle, CPP-A e CPP-C
e de 83% para o grupo CPP-B, num período de 6 horas.
Após 90 minutos de injeção intraperitoneal de endotoxina, a IL-6 já pode ser
detectada no plasma, mesmo em baixas doses de LPS (5µg/kg) (LENCZOWSKI et al.,
1997). Baixas doses de LPS (10 ou 100µg/kg) induzem à elevação de citocinas pró-
inflamatórias, porém não geram disfunções hepáticas. É necessário uma dose de
1mg/kg de LPS para induzir alterações na função normal do fígado (BENO et al., 2001).
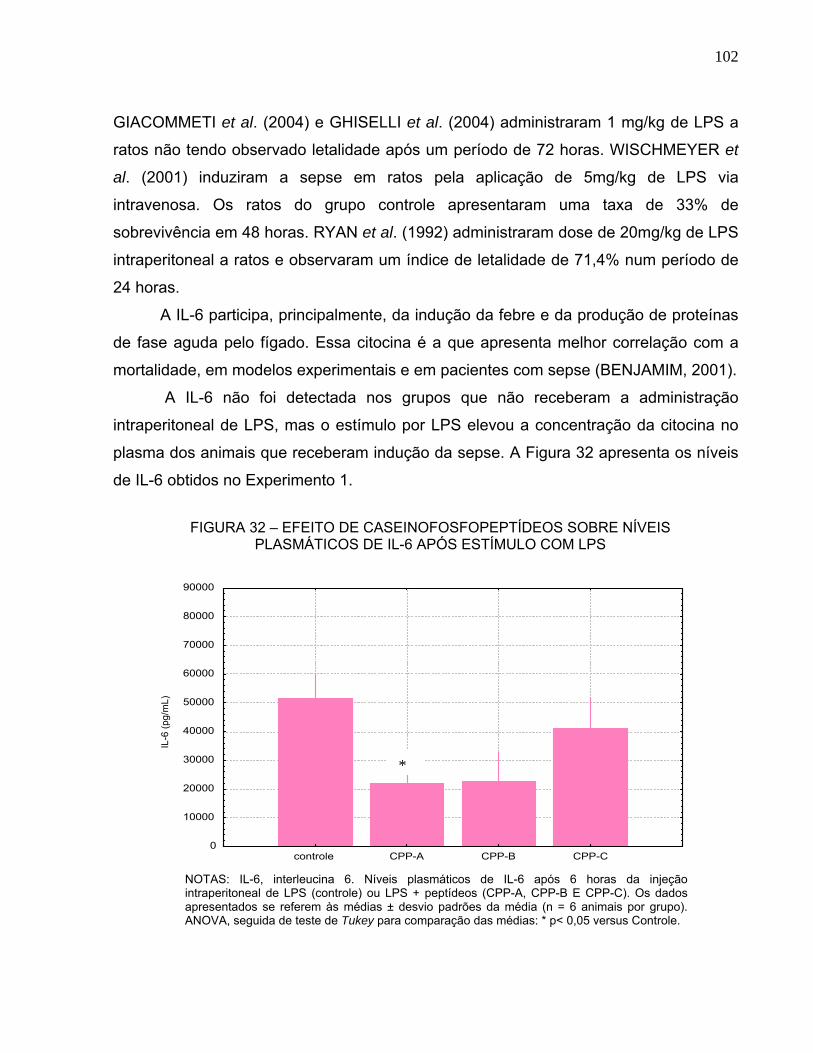
102
GIACOMMETI et al. (2004) e GHISELLI et al. (2004) administraram 1 mg/kg de LPS a
ratos não tendo observado letalidade após um período de 72 horas. WISCHMEYER et
al. (2001) induziram a sepse em ratos pela aplicação de 5mg/kg de LPS via
intravenosa. Os ratos do grupo controle apresentaram uma taxa de 33% de
sobrevivência em 48 horas. RYAN et al. (1992) administraram dose de 20mg/kg de LPS
intraperitoneal a ratos e observaram um índice de letalidade de 71,4% num período de
24 horas.
A IL-6 participa, principalmente, da indução da febre e da produção de proteínas
de fase aguda pelo fígado. Essa citocina é a que apresenta melhor correlação com a
mortalidade, em modelos experimentais e em pacientes com sepse (BENJAMIM, 2001).
A IL-6 não foi detectada nos grupos que não receberam a administração
intraperitoneal de LPS, mas o estímulo por LPS elevou a concentração da citocina no
plasma dos animais que receberam indução da sepse. A Figura 32 apresenta os níveis
de IL-6 obtidos no Experimento 1.
FIGURA 32 – EFEITO DE CASEINOFOSFOPEPTÍDEOS SOBRE NÍVEIS PLASMÁTICOS DE IL-6 APÓS ESTÍMULO COM LPS
IL-6
(pg/
mL)
0
10000
20000
30000
40000
50000
60000
70000
80000
90000
controle C
NOTAS: IL-6, interleucina 6. Níveiintraperitoneal de LPS (controle) ou Lapresentados se referem às médias ±ANOVA, seguida de teste de Tukey pa
*
PP-A CPP-B CPP-C
s plasmáticos de IL-6 após 6 horas da injeção PS + peptídeos (CPP-A, CPP-B E CPP-C). Os dados desvio padrões da média (n = 6 animais por grupo). ra comparação das médias: * p< 0,05 versus Controle.

103
Houve homogeneidade entre as variâncias para níveis de IL-6, comprovada pelo
teste de Levene (p = 0,275211). O grupo que recebeu CPP-B aparentemente
apresentou um efeito imunomodulador, porém a IL-6 não foi significativamente
reduzida. Os valores são estatisticamente inferiores para o CPP-A.
O CPP-C não apresentou efeito redutor significativo nas concentrações de IL-6.
Este produto tinha como um de seus diferenciais o acréscimo de zinco à formulação.
Sabe-se que o zinco é um mineral importante em vários processos metabólicos, sendo
essencial para a atividade de mais de 300 enzimas. Possui também um papel
fundamental na manutenção da estabilidade e permeabilidade das membranas
celulares. A deficiência de zinco pode se expressar na involução do timo, depleção de
timócitos, redução na atividade proliferativa dos linfócitos T periféricos, resposta
humoral a antígenos e atividade de células NK (natural killer). As funções imunológicas
que se apresentam diminuídas, tanto em humanos como em animais experimentais
submetidos a dietas com teores insuficientes do mineral, podem ser corrigidas por sua
suplementação (OBMINSKA-DOMORADZKA; SZCZYPKA; DEBOWYJ, 2002).
Entretanto, CHANG et al. (2006) verificaram que concentrações fisiológicas de zinco em
células mononucleares de sangue periférico humano obtidas de indivíduos sadios não
apresentam funções imunológicas ou biológicas, mas concentrações farmacológicas
consideradas máximas (100µM) ou valores superiores alteraram ambas as funções.
Concentrações superiores a 100µM reduziram a viabilidade celular e a partir de 100µM
a expressão de citocinas foi aumentada. A concentração de IL-6 foi aumentada de
50,9±8,9pg/mL sem adição do mineral ou em dose fisiológica de 30µM para
356±23,6pg/mL a 100µM e 701±66,5 a 300µM. Os autores também avaliaram a indução
da apoptose por eletroforese de fragmentação do DNA e citometria de fluxo, tendo
verificado que o mineral induz a morte celular programada. Após 24 horas de
tratamento, o percentual de células apoptóticas se elevou de 11,6% a 30µM de Zn2+
para 23,19% a 100µM e 95,2% a 1000µM.
Em dietas destinadas a ratos, existem algumas variações nos valores apontados
pela literatura para a administração de zinco. São consideradas dietas deficientes,
aquelas contendo 0,8 a 5mg/kg de peso corpóreo; dietas adequadas ou suficientes

104
contêm de 25 a 40mg/kg. Em suplementação, os valores variam de 54 a 155mg/kg. As
concentrações fisiológicas variam de 6,25 a 50µM, sendo que doses acima de 100µM
induzem a apoptose (KAGANDA; TATSUHIRO; HIROO, 2003; PASKI et al., 2003;
ROTH, 2003). O CPP-C foi adicionado à dieta dos animais em valores de
0,14g/rato/dia, o que corresponde a uma concentração de 86,8µM de zinco. Esta
concentração está acima dos valores fisiológicos, podendo ser considerada como uma
dose farmacológica, que pode ter estimulado a elevação dos níveis de IL-6 (CHANG et
al., 2006), comparativamente aos resultados obtidos pelos demais
caseinofosfopeptídeos.
O leucograma dos animais submetidos à administração de LPS indica alterações
hematológicas, com redução na contagem de leucócitos (25% dos valores normais),
linfócitos (45% dos valores normais) e plaquetas (31% da normalidade). A contagem de
neutrófilos foi aumentada em 282% em relação aos valores normais.
Os valores obtidos na contagem de leucócitos estão demonstrados na Figura 33.
FIGURA 33 - CONTAGEM DE LEUCÓCITOS
Leuc
ócito
s (K
/uL)
0
1
2
3
4
5
6
CONTROLE CPP-A CPP-B CPP-C
NOTAS: Contagem de leucócitos em K/µL após 6 horas da injeção intraperitoneal de LPS (controle) ou LPS + peptídeos (CPP-A, CPP-B e CPP-C). Os dados apresentados se referem às médias ± desvio padrões da média (n = 6 animais por grupo). Kruskal-Wallis. p< 0,05.

105
Na avaliação deste parâmetro foi realizada estatística não paramétrica, pois os
grupos não apresentaram homogeneidade nas variâncias. Não houve diferença
significativa entre os grupos na contagem de leucócitos (p = 0,5398) com teste de
Kruskal-Wallis.
KITAJIMA et al. (1995) avaliaram a relação entre o tratamento com LPS (1 mg/kg
de peso corpóreo) e as alterações hematológicas de ratos. Verificaram um decréscimo
tempo-dependente dos leucócitos (80% do controle), linfócitos (40% do controle) e
plaquetas (35% do controle) e um aumento nos neutrófilos (330% do controle) e
monócitos (650% do controle).
A administração de endotoxina causa um decréscimo na contagem de
leucócitos, seguida de significativa neutrofilia (FIJEN et al., 2000). Dentre as
manifestações hematológicas também pode ocorrer leucocitose, sendo que o
prognóstico da leucopenia é pior do que da leucocitose (PEREIRA et al. 1998).
A contagem de neutrófilos (Figura 34), realizada pela soma das contagens
diferenciais de bastonetes e segmentados, apresentou homogeneidade nas
variâncias, segundo teste de Levene (p = 0,421301) e a ANOVA não registrou
diferença significativa entre os grupos (p = 0,804072). Embora sem diferença
estatística, o grupo CPP-B apresentou os menores resultados. Um achado constante
nas contagens diferenciais foi o desvio nuclear à esquerda, indicando um aumento no
número de neutrófilos bastonetes e caracterizando um quadro de infecção aguda.
Esse achado se apresentou em todos os grupos estudados.
Um achado comum na endotoxemia é o aumento no número de bastonetes (>
10%) com linfopenia (PEREIRA et al. 1998).
Em todos os grupos estudados se verificou um aumento importante na contagem
de neutrófilos, o que indica que a dose de 5 mg/kg de LPS administrada aos animais
conduz a um quadro infeccioso agudo. Os peptídeos não influíram nesse achado
hematológico.

106
FIGURA 34 - CONTAGEM DE NEUTRÓFILOS
N
eutró
filos
(%)
0
10
20
30
40
50
60
70
80
90
100
CONTROLE CPP-A CPP-B CPP-C
NOTAS: Contagem de neutrófilos em %, correspondente a soma das contagens diferenciais de segmentados e bastonetes após 6 horas da injeção intraperitoneal de LPS (controle) ou LPS + peptídeos (CPP-A, CPP-B e CPP-C). Os dados apresentados se referem às médias ± desvio padrões da média (n = 6 animais por grupo). ANOVA, p< 0,05.
A Figura 35 apresenta os resultados obtidos na análise de linfócitos. O teste de
Levene para linfócitos demonstrou homogeneidade nas variâncias (p = 0,636841) e,
segundo a análise de variâncias, não houve diferença significativa entre os grupos.
Todos os grupos demonstraram um comportamento semelhante com redução de
aproximadamente 45% dos valores obtidos no início do experimento, sendo que os
peptídeos não influíram nestes resultados. O grupos CPP-A e CPP-B apresentaram
redução de 38% e 36% dos linfócitos encontrados nos animais no início do
experimento, indicando uma redução não significativa, porém expressiva.

107
FIGURA 35 - CONTAGEM DE LINFÓCITOS
Linf
ócito
s (%
)
0
10
20
30
40
50
60
70
CONTROLE CPP-A CPP-B CPP-C
NOTAS: Contagem de linfócitos em %, após 6 horas da injeção intraperitoneal de LPS (controle) ou LPS + peptídeos (CPP-A, CPP-B e CPP-C). Os dados apresentados se referem às médias ± desvio padrões da média (n = 6 animais por grupo). ANOVA, p< 0,05.
Os valores obtidos na contagem de plaquetas foram homogêneos (Levene, p =
0,194727) e sem diferença significativa na ANOVA (p = 0,080607), conforme se verifica
na Figura 36.
Mesmo com o quadro clínico de um paciente com disfunção de múltiplos órgãos
resolvido, a plaquetopenia pode persistir, retornando aos valores normais apenas três
ou quatro semanas após o início do quadro (PEREIRA et al. 1998).
Alguns autores descrevem a avaliação do escore de gravidade do paciente com
disfunção de múltiplos órgãos pela pontuação de sinais orgânicos e achados
bioquímicos e hematológicos, sendo que a contagem de plaquetas é um dos
indicadores da gravidade do quadro e sua redução é utilizada no prognóstico da doença
(LE GALL et al., 1996).
Embora os valores não tenham diferido entre os grupos, se observou que no
grupo CPP-B a contagem de plaquetas apresentou valores mais elevados, o que pode
indicar uma tendência de proteção do processo inflamatório pela administração do
peptídeo.

108
FIGURA 36 - CONTAGEM DE PLAQUETAS
P
laqu
etas
(K/u
L)
0
75
150
225
300
375
450
525
CONTROLE CPP-A CPP-B CPP-C
NOTAS: Contagem de leucócitos em K/µL após 6 horas da injeção intraperitoneal de LPS (controle) ou LPS + peptídeos (CPP-A, CPP-B e CPP-C).Os dados apresentados se referem às médias ± desvio padrões da média (n = 6 animais por grupo). ANOVA, p< 0,05.
Devido às injúrias que afetam os pulmões na endotoxemia, este órgão foi
selecionado para avaliação histopatológica. A endotoxina age nos pulmões causando
alterações estruturais na microcirculação, induzindo a destruição da barreira sangüínea,
edema alveolar e intersticial. Essas alterações iniciam o processo de infiltração de
neutrófilos e macrófagos e a hemorragia alveolar. A liberação de citocinas pró-
inflamatórias, mediadores reativos do oxigênio e metabólitos do ácido araquidônico
também contribuem na patofisiologia da injúria pulmonar (KOKSEL et al., 2005).
Os valores histológicos médios para os grupos estão apresentados na Tabela 12.
Como não houve homogeneidade nas variâncias, foi realizada a análise não-
paramétrica dos dados. No teste de Kruskal-Wallis não foram encontradas diferenças
entre os grupos (p=0,6003). Contudo, houve prevalência dos seguintes sinais:
infiltração leucocitária, congestão, edema alveolar, hemorragia e colapso alveolar. As
microfotografias, apresentadas nas Figuras 37 a 40, demonstram alguns destes

109
achados. Na Figura 37 evidencia-se a congestão pela presença de aglomerados
celulares. O edema está demonstrado na vacuolização do parênquima pulmonar
(Figura 38). Na Figura 39 é possível a observação de hemácias no parênquima,
caracterizando a presença de hemorragia alveolar. A figura 40 registra a presença de
neutrófilos, com infiltração neutrofílica.
TABELA 12 - VALORES HISTOLÓGICOS MÉDIOS PARA OS GRUPOS CONTROLE,
CPP-A, CPP-B E CPP-C
Grupo Leucócitos Congestão Necrose
PerivascularFibrose Septal
Edema alveolar
Hemorragia alveolar
Colapso alveolar
Controle 1,5 1,2 0 0,0 0,3 1,2 1,0CPP-A 1,5 2 0 0,0 1,0 1,5 1,2CPP-B 1,2 1,5 0 0,2 0,7 1,0 1,2CPP-C 1,4 1,2 0 0 0,6 1,2 0,8Quantificação: 0, ausente; 1, discreto; 2, moderado; 3, intenso. Os dados apresentados se referem às médias (n = 6 animais por grupo). Kruskal-Wallis: p< 0,05.
FIGURA 37 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A PRESENÇA DE CONGESTÃO
NOTAS: Pulmões dos animais removidos após 6 horas da administração intraperitoneal de LPS; fixados em tampão formalina-fosfato e embebidos em parafina. Cortes histológicos corados com hematoxilina e eosina. Microscopia com aumento de 400x.

110
FIGURA 38 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A PRESENÇA DE EDEMA
NOTAS: Pulmões dos animais removidos após 6 horas da administração intraperitoneal de LPS; fixados em tampão formalina-fosfato e embebidos em parafina. Cortes histológicos corados com hematoxilina e eosina. Microscopia com aumento de 200x.

111
FIGURA 39 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A
PRESENÇA DE HEMORRAGIA ALVEOLAR
NOTAS: Pulmões dos animais removidos após 6 horas da administração intraperitoneal de LPS; fixados em tampão formalina-fosfato e embebidos em parafina. Cortes histológicos corados com hematoxilina e eosina. Microscopia com aumento de 200x.

112
FIGURA 40 - ANÁLISE HISTOPATOLÓGICA DE PULMÃO DE RATOS, DEMONSTRANDO A PRESENÇA DE INFILTRAÇÃO NEUTROFÍLICA
NOTAS: Pulmões dos animais removidos após 6 horas da administração intraperitoneal de LPS; fixados em tampão formalina-fosfato e embebidos em parafina. Cortes histológicos corados com hematoxilina e eosina. Microscopia com aumento de 400x.

113
5.3.2.2 Experimento 2: Injeção Intraperitoneal de LPS Após Administração
Intraperitoneal de CPP.
No Experimento 2, verificou-se o efeito protetor dos peptídeos num modelo de
choque tóxico por LPS. Uma dose de 5mg/kg de LPS foi administrada concomitante à
administração de CPP-A (0,75g/kg). A sobrevivência dos ratos foi avaliada num período
de 6 horas, sendo de 100% tanto nos ratos controle como nos animais que receberam o
CPP-A. Os ratos submetidos ao choque apresentaram blefarite sanguinolenta, assim
como prostração. Não houve diferença desses sinais entre os que receberam o CPP e
os que não receberam. Os animais que receberam o peptídeo via intraperitoneal
tiveram uma redução significativa na produção de IL-6 (Figura 41).
FIGURA 41 - EFEITO DE CASEINOFOSFOPEPTÍDEO (CPP-A) ADMINISTRADO
INTRAPERITONEALMENTE (IP) SOBRE NÍVEIS PLASMÁTICOS DE IL-6 APÓS ESTÍMULO COM LPS
IL-6
(pg/
mL)
0
10000
20000
30000
40000
50000
60000
70000
80000
90000
CONTROLE C
*
NOTAS: Os dados apresentados se referem às médias(n=6 animais por grupo). Teste T de Student. * p< 0,05 ve
PP-A IP
± desvio padrões da média rsus Controle.

114
WISCHMEYER et al. (2001) administraram doses de glutamina de 0,15 a 0,75
g/kg a ratos submetidos à indução de choque séptico pela injeção intravenosa de LPS
(5mg/kg). Os autores confirmaram o papel protetor da glutamina contra as injúrias
geradas pela endotoxemia, sendo que o mecanismo de proteção envolve a expressão
de proteínas de choque térmico (HSP, heat shock proteins). Elevações na expressão
das HSP se associam a reduções na liberação de citocinas pró-inflamatórias. Além
disso, verificaram que a glutamina é capaz de conferir maior proteção quando
administrada no início da sepsis do que como um pré-tratamento.
O CPP-A foi o peptídeo selecionado para o Experimento 2, pois foi aquele que
gerou uma redução significativa na expressão de IL-6, após sua inclusão na dieta por
um período de 22 dias. Entretanto, os resultados obtidos na liberação de IL-6, por esse
peptídeo, quando administrado no início da sepsis foram inferiores aos resultados para
o mesmo peptídeo no Experimento 1, assemelhando-se ao papel terapêutico da
glutamina.
A avaliação do leucograma dos animais submetidos ao Experimento 2 indica
alterações hematológicas, com redução na contagem de leucócitos (70% dos valores
normais), linfócitos (28% dos valores normais) e plaquetas (55% da normalidade). A
contagem de neutrófilos foi aumentada em 347% em relação ao tempo 0 do
experimento. Os valores obtidos estão demonstrados nas Figuras 42 a 45. A análise
estatística foi realizada pelo teste T de Student, sendo que não houve diferença
significativa entre os grupos na contagem de leucócitos (p = 0,56047). Embora não se
apresente diferença significativa, os valores de leucócitos para o grupo CPP-A IP se
apresentam bem superiores e próximos dos valores normais, com redução de 30%
contra uma redução de 70% dos animais do grupo controle, indicando um possível
efeito protetor.

115
FIGURA 42 - CONTAGEM DE LEUCÓCITOS DOS RATOS
Le
ucóc
itos
(K/u
L)
0
2
4
6
8
10
12
CONTROLE CPP-A IP
NOTAS: Contagem de leucócitos em K/µL, após 6 horas da injeção intraperitoneal de LPS (controle) ou LPS + CPP-A intraperitoneal (0,75g/kg). Os dados apresentados se referem às médias ± desvio padrões da média (n = 6 animais por grupo). Teste T de Student, p< 0,05.

116
A contagem de neutrófilos, realizada pela soma das contagens diferenciais de
bastonetes e segmentados, não registrou diferença significativa entre os grupos (p =
0,174026), conforme dados apresentados na Figura 43.
FIGURA 43 - CONTAGEM DE NEUTRÓFILOS DOS RATOS
Neu
trófil
os (%
)
0
10
20
30
40
50
60
70
80
90
CONTROLE CPP-A IP
NOTAS: Contagem de neutrófilos em %, correspondente a soma das contagens diferenciais de bastonetes e segmentados, após 6 horas da injeção intraperitoneal de LPS (controle) ou LPS + CPP-A intraperitoneal (0,75g/kg). Os dados apresentados se referem às médias ± desvio padrões da média (n = 6 animais por grupo). Teste T de Student, p< 0,05.

117
Na contagem de linfócitos não houve diferença significativa (p = 0,139370), pelo
teste T de Student. Os valores estão representados na Figura 44. Embora não se
caracterize a diferença estatística, o grupo controle apresentou uma redução na
contagem de linfócitos de 49% dos valores obtidos no tempo zero do experimento e o
grupo que recebeu o peptídeo teve uma redução de 28%.
FIGURA 44 - CONTAGEM DE LINFÓCITOS DOS RATOS
Linf
ócito
s (%
)
0
10
20
30
40
50
60
70
CONTROLE CPP-A IP
NOTAS: Contagem de linfócitos em %, após 6 horas da injeção intraperitoneal de LPS (controle) ou LPS + CPP-A intraperitoneal (0,75g/kg). Os dados apresentados se referem às médias ± desvio padrões da média (n = 6 animais por grupo). Teste T de Student, p< 0,05.

118
A contagem de plaquetas apresentou diferença significativa entre o grupo
controle e o grupo CPP-A (p = 0,015088), conforme Figura 45, indicando um provável
papel protetor do peptídeo.
FIGURA 45 - CONTAGEM DE PLAQUETAS DOS RATOS
Plaq
ueta
s (K
/uL)
0
150
300
450
600
CONTROLE CPP-A IP
*
NOTAS: Contagem de plaquetas em K/µL, após 6 horas da injeç(controle) ou LPS + CPP-A intraperitoneal (0,75g/kg). Os dados apmédias ± desvio padrões da média (n = 6 animais por grupo). Testversus Controle (p = 0,015088).
ão intraperitoneal de LPS resentados se referem às e T de Student, * p< 0,05

119
6 CONCLUSÕES
Os resultados obtidos e apresentados neste trabalho permitem concluir que:
1. É possível a obtenção de misturas de peptídeos fosforilados pela utilização dos
métodos preconizados na literatura.
2. Os peptídeos apresentaram percentuais relativamente elevados de solubilidade,
indicando baixa desnaturação nos processos de obtenção, com conseqüente
manutenção das propriedades fisiológicas. O perfil aminoacídico apresentado indica
que não foram observadas perdas elevadas de aminoácidos nos peptídeos. A análise
dos espectros e o seqüenciamento posterior indicam a obtenção de três produtos que
se caracterizam como uma mistura de peptídeos derivados da hidrólise das caseínas
αS1, αS2, β e κ. A correlação entre os dados de composição química, análise de
aminoácidos e técnicas de seqüenciamento empregadas denota a necessidade da
otimização da metodologia de MALDI-TOF-MS para peptídeos fosforilados.
3. Os três produtos apresentaram diferentes graus de atividade antimicrobiana, sendo
que a presença do zinco no produto CPP-C sugere que o íon desempenhou um papel
fundamental na redução do crescimento microbiano.
4. Os peptídeos CPP-A e CPP-B não alteraram a resposta normal dos macrófagos,
indicando que sua administração não prejudica a resposta imune inata. Num modelo
experimental de endotoxemia, os níveis da interleucina 6 se apresentaram diminuídos,
pela administração do CPP-A via oral ou intraperitoneal, sugerindo que este peptídeo
possui um papel imunomodulador pela redução no processo inflamatório dos animais.
Os resultados obtidos neste trabalho são inovadores, sugerem que estes
peptídeos apresentam bioatividade e apontam novas perspectivas para o emprego de
caseinofosfopeptídeos em alimentos.
A comprovação da ausência de respostas citotóxicas pelos CPP-A e CPP-B,
bem como seu potencial efeito protetor da saúde pode ser visto como uma contribuição

120
importante na formulação de novos alimentos e na alegação de propriedade funcional
ou de saúde em rótulos de alimentos.

121
CONSIDERAÇÕES FINAIS Este trabalho teve como objetivo a obtenção, caracterização e avaliação da
atividade biológica de caseinofosfopeptídeos de leite bovino. Pode-se verificar que a
distribuição dos peptídeos e conseqüente atividade biológica foram afetadas pela forma
de obtenção do CPP. Conhecimento adicional da forma de atuação das seqüências
verificadas em cada CPP poderia ser obtido com o isolamento destas frações e
posterior avaliação da atividade. Para exemplificar: uma seqüencia similar a do
peptídeo casoplatelina foi detectada neste trabalho, e segundo a literatura apresenta
atividade antimicrobiana. Com o isolamento destas frações seria possível estudar o
mecanismo da propriedade fisiológica. Esta seria uma proposta de caráter acadêmico e
poderia colaborar no conhecimento da relação estrutura-função de peptídeos bioativos.
Do ponto de vista do agronegócio, a otimização da metodologia de obtenção de CPP,
fisiologicamente ativo, e isento de toxicidade, poderia contribuir para agregação de
valor aos produtos lácteos e para a segurança alimentar.

122
REFERÊNCIAS AÏT-OUKHATAR, N.; BOUHALLAB, S.; BUREAU, F.; ARHAN, P.; MAUBOIS, J.L.; DROSDOWSKY, M.A.; BOUGLÉ, D.L. Bioavailability of caseinophophopeptide bound iron in the young rat. The Journal of Nutritional Biochemistry, Stonehan, v.8, n. 4, p. 190 – 194, , 1997. AKCA, T.; CANBAZ, H.; TATAROGLU, C.; CAGLIKULEKCI, M.; TAMER, L.; COLAK, T.; KANIK, H.; BILGIN, O.; AYDIN, S. The effect of N-acetylcysteine on pulmonary lipid peroxidation and tissue damage. Journal of Surgical Research, New York, v. 129, n. 1, p. 38 – 45, 2005. AL-QADREH, A.; SCHULPIS, K.H.; ATHANASOPOULOU, H. MENGRELI, C.; SKARPALEZOU, A.; ZOSKAKI, I. Bone mineral status in children with phenylketonuria under treatment. Acta Pediatric, Madrid, v. 87, p. 1162-1166, 1998. ANNANE, D.; BELISSANT, E.; CAVAILLON, J.M. Septic shock. The Lancet, London, n. 9453, v. 365, p. 63 - 78, 2005. ANVISA. Resolução RDC nº 269, de 22 de setembro de 2005. Regulamento técnico sobre a ingestão diária recomendada (IDR) de proteína, vitaminas e minerais. Disponível em: http://e-legis.anvisa.gov.br/leisref/public/showAct.php?id=18828&word=. Consultado em: 12/08/2006. ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS. HORWITZ W. (Ed.) Official methods of analysis of the Association of Analytical Chemists. 15 ed. Arlington : AOAC Inc., 1990. v. 2, p. 1167 – 1174. BALDI, A.; IOANNIS, P.; CHIARA, P.; ELEONORA, F.; ROUBINI, C.; VITTORIO, D.O. Biological effects of milk proteins and their peptides with emphasis on those related to the gastrointestinal ecosystem. Jouranl of Dairy Research. Cambridge, v. 72, Special Issue, p. 66 – 72, 2005. BEDANI, R.; ROSSI, E.A. Isoflavonas: bioquímica, fisiologia e implicaçõs para a saúde. Boletim do CEPPA. Curitiba, v. 23, n. 2, p. 231 – 264. 2005.
BENJAMIM, C.F. Atualização sobre mediadores e modelos experimentais de sepse. Medicina, ribeirão Preto, v. 34, p. 18 – 26, 2001. BENO, D.W.A.; VHING, M.R.; GOTO, M.; CHEN, Y.; JIYA-MAPA-SERNA, V.A.; KIMURA, R.E. Staphylococcal enterotoxin B potentials LPS induced hepatic dysfunction in chronically catheterized rats. American Journal of Gastrointestinal and liver Physiology, . Bethesda, v. 280, n. 5, p 866 – 872, 2001. BERNARDI, C.R.; LUIZ, M.T.B.; ZANOTTO, D.L.; GUIDONI, A.L. Preparo de hidrolisados protéicos para a análise de aminoácidos. Ciência e Tecnologia de Alimentos, Campinas, v. 23, n. 3, p. 317 – 322, 2003.

123
BERTHOU, J.; MIGLIORE-SAMOUR, D.; LIFCHITZ, A.; DELETTRÉ, J.; FLOC’H, F.; JOLLÉS, P. Immunostimulating properties and three-dimensional structure of two tripeptides from human and cow caseins. FEBS Letters, Amsterdam, v. 218, n. 1, p. 55 – 58, 1987. BITRI, L. Optimization study for the production of na opiod-like preparation from bovine casein by mild acidic hydrolysis. International Dairy Journal. Barking, v. 14, n. 6, p. 535 – 539, 2004. BONATTO, S.J.R.; FOLADOR, A.; AIKAWA, J.; YAMAZAKI, R.; PIZATTO, N.; OLIVEIRA, H.H.P; ECCHI, R.; CURI, R.; CLDER, P.C.; FERNANDES, L.C. Lifelong exposure to dietary fish oil alters macrophage responses in Walker 256 tumor beating-rats. Cellular Immunology, v. 231, p. 56 – 62, 2004. BORGES, P.Z.; SGARBIERI, V.C.; JACOBUCCI, H.B.; DIAS, N.F.G. Avaliação nutricional de concentrado protéico de soro de leite, caseinato de sódio e coágulo de caseína, comparados à caseína comercial. Anais do III Simpósio Latino-Americano de Ciência e Tecnologia de Alimentos, Campinas: 16 a 19/11. Resumo. p. 113,114. 1999.
BOUHALLAB, S.; OUKHATAR, N.A.; MOLLÉ, D.; HENRY, G.; MAUBOIS, J.L.; ARHAN, P.; BOUGLÉ, D.L. Sensitivity of beta-casein phosphopeptide-iron complex to digestive enzymes in ligated segment of rat duodenum. Journal of Nutritional Biochemistry. New York, v. 10, p. 723 - 727, 1999. BOUHALLAB, S.; CINGA, V.; OUKHATAR, N.; BUREAU, F.; NEUVILLE, D.; ARHAN, P.; MAUBOIS, J.L.; BOUGLÉ, D. Influence of various phosphopeptides of caseins os iron absorption. Journal of Agricultural and Food Chemistry. Easton, v. 50, n. 24, p. 7127 - 7130, 2002. BOUNOUS, G. Whey protein concentrate (WPC) and glutathione modulation in cancer treatment. Anticancer Research, Philadelphia, v. 20, n. 6C, p. 4785 – 4792, 2000. BOUNOUS, G.; BATIST, G.; GOLD, P. Immunoenhancing property of dietary whey protein in mice: role of glutathione. Clinical and Investigative Medicine. Toronto, v. 12, n. 3, p. 154 – 161, 1989. BRANTL, V. Novel opiod peptides derived from human β-casein: human β-casomorphin. European Journal of Pharmacology. Amsterdam, v. 106, n. 1, p. 213 – 214, 1985. BRASIL. Ministério da Saúde. Portaria nº 847 de 06 de novembro de 2002. Protocolo clínico e diretrizes terapêuticas – Fenilcetonúria. Disponível em: http://dtr2001.saude.gov.br/sas/PORTARIAS/PORT2002/PT-847.htm. Consultado em: 12/08/2006. BRULE, G.; ROGER, L.; FAUQUANT, J.; PIOT, M. Procedé de traitement d'une matière à base de caséine contenant des phosphocaséinates de cations monovalents ou leurs dérivés, produits obtenus et apllications. Word Patent nº 8002281, 1980.

124
CADENAS, E. Mechanisms of oxygen activation and reactive oxygen species detoxification. In: AHMAD, S. (ed.). Oxidative stress and antioxidant defenses in biology. Chapman & Hall : New York, 1995. p. 18 – 25. CALDER, P.C. Dietary fatty acids and lymphocyte functions. Proceedings of the Nutrition Society, Cambridge, v. 57, p. 487 – 502, 1998. CÂNDIDO, L. M. B. Obtenção de concentrados e hidrolisados protéicos de tilápia do Nilo (Oreochromus niloticus): composição, propriedades nutritivas e funcionais.Tese de Doutorado. Universidade Estadual de Campinas, 1998. 207 p. CÂNDIDO, L.M.B; SGARBIEIRI, V.C. Enzymatic hydrolysis of Nile Tilapia (Oreochromus niloticus) myofibrilar proteins: effects on nutritional and hydrophilic properties. Journal of the Science of Food and Agriculture, United Kingdom, v. 83, p. 937-944, 2003. CARIO, E.; JUNG, S.; HARDER d’HEUREUSE, J.; SCHULTE, C.; STURM, C.; WIEDENMANN, B.; GOEBELL, H.; DIGNASS, A.U. Effects of exogenous zinc suoolementation on intestinal epithelial repair in vitro. European Journal of Clinical Investigation, Oxford, v. 30, n. 5, p. 419 – 428, 2000. CARNEIRO, J.G.M. Características funcionais de concentrados protéicos de soro de leite de cabras. Tese de Doutorado. Universidade Estadual de Campinas, 1997. 127 p. CARVALHO, M.D.T.; TOBIAS, V.E.; VENDRAME, C.M.V.; SHIMAKURRO, A.F.M.; GIDLUND, M.; QUINTÃO, E.C.R. Zymosan phagocytosis by mouse peritoneal macrophage is incresead by apoHDl- and not by intact HDL- covered particles. Braszilian Journal of Medical and Biological Research, . Ribeirão Preto, v. 33, n. 3, p. 313 – 316, 2000. CHANG, K.L.; HUNG, T.C.; HSIEH, B.S.; CHEN, Y.H.; CHEN, T.F.; CHENG, H.L. Zinc at pharmacologic concentrations affects cytokine expression and induces apoptosis of human peripheral blood mononuclear cells. Nutrition, New York, v. 22, n. 5, p. 465 – 474, 2006. CHOI, Y.M.; NOH, D.O.; CHO, S.Y.; SUH, H.J.; KIM, K.M.; KIM, J.M. Antioxidant and antimicrobial activities of propolis from several regions of Korea. LWT. v. 39, p. 756 – 761, 2006. CHU, L. MACLEOD, A., OZIMEK, L. Isolation of glycomacropeptide from sodium caseinate hydrolysate solution by ultrafiltration. Milchwissenschaft, Nurnberg, v. 51, n. 6, p. 303 – 306, 1996. CLAVIJO, L.C.; CARTER, M.B.; MATHESON, P.J.; WILLS-FRANK, L.A.; WILSON, M.A.; WEAD, W.B.; GARRISON, R.N. Platelet-activating factor and bacteremia-induced pulmonary hypertension. Journal of Surgical Research, New York, v. 88, n. 2, p. 173 – 180, 2000. CLEMENT, A. Enzymatic protein hydrolysates in human nutrition. Trends in Food Science and Nutrition, Cambridge, v. 11, n. 7, p. 254-262, 2000.

125
COSTE, M.; ROCHET, V.; LÉONIL, J.; MOLLÉ, D.; BOUHALLAB, S.; TOMÉ, D. Identification of C-terminal peptides of bovine beta-casein that enhance proliferation of rat lymphocytes. Immunology Letters. Amsterdam, V. 33, n. 1, p. 41 - 46, 1992. CREAMER, L.K.; PLOWMAN, J.E.; LIDDELL, M.J.; SMITH, M.H.; HILL, J.P. Micelle stability: κ-casein structure and function. Journal of Dairy Science, Champaign, v . 81, n. 11, p. 3004 - 3012, 1998. CUNNINGHAM-RUNDLES, S. Analytical methods for evaluation of immune response in nutrient intervention. Nutrition Reviews. New York, V. 56, n. 1, p. S27 - S37, 1998. DAMODARAN, S.; PARAF, A. Food proteins and their applications. New York : Marcel Dekker, Inc., 1997. DASHPER, S.G.; O’BRIEN-SIMPSON, N.M.; CROSS, K.J.; PAOLINI, R.A.; HOFFMANN, B.; CATMULL, D.V.; MALKOSKI, M.; REYNOLDS, E. Divalent metal cátions increase the activity of the antimicrobial peptide kappacin. Antimicrob. Agents Chemother., Bethesda, v. 49, p. 2322 – 2328, 2005. DEESLIE, W. D.; CHERYAN, M., Functional properties of soy protein hydrolysates from a continuous ultrafiltration reactor. Journal of Agriculture and Food Chemistry, Washington, v. 36, n. 1, p. 26 - 31, 1988. DEVIENNE, K.F.; RADDI, M.S.G. Screening for antimicrobial activity of natural products using a microplate photometer. Braz. J. Microbiol., São Paulo, v.33, p.166-168, 2002. DIAS, N.F.G.P.; SGARBIERI, V.C.; RANGEL, H.A.; JACOBUCCI, H.B; BORGES, P.Z.; PACHECO, M.T.B.; TANIKAWA, C. Efeito das proteínas do lactosoro no sistema imunológico de camundongos. In: III Simpósio Latino Americano de Ciências de Alimentos, 1999, Campinas. Livro de Programa e Resumos do III SLACA. Campinas : Unicamp, 1999. p. 116. DICKINSON, E. Caseins in emulsions: interfacial properties and interactions. International Dairy Journal, Cambridge, v. 9, n. 3 - 6, p. 305-312. 1999. ELIA, C.C.S.; SOUZA, H.S.P. Imunologia da Mucosa Intestinal: da Bancada ao Leito. Editora Atheneu : São Paulo, 2001. ELLEGARD, K.H.; GAMMELRLGARD-LARSEN, C.; SORENSEN, E.S.; FEDOSOV, S. Process scale chromatographic isolation, characterizatrion and identification of tryptic bioactive casein phosphopeptides. International Dairy Journal. Cambridge, v. 9, p. 639 – 652, 1999. ERBA; CIAPPELANO; TESTOLIN. Effect of caseinphosphopeptides on inhibition of calcium intestinal absorption to phosphate. Nutrition Research, New York, v. 21, n. 4, p. 649 – 656, 2001. ERROS inatos do Metabolismo. Disponível em: http://www.supportnet.com.br/produtos/erros_finilcetonuria/index.htm. Consultado em: 12/08/2006.

126
FENILCETONÚRIA. O que é? Disponível em: http://www.vittafix.com.br/produtos_bod.htm. Consultado em: 12/08/2006. FENNEMA,O.R. Química de los alimentos. Zaragoza : Editorial Acribia , 1993. p. 415 – 536. FERNÁNDEZ-MARTINEZ, E.; MORALES-RÍOS, M.S.; PÉREZ-ÁLVAREZ, V.; MURIEL, P. Immunomodulatory effects of thalidomide analogs on LPS-induced plasma and hepatic cytokines in the rat. Biochemical Pharmacology, New York, v 68, p. 1321 – 1329, 2004. FERRANTI, P.; TRAISCI, M.V.; PICARIELLO, G.; NASI, A.; BOSCHI, V.; SIERVO, M.; FALCONI, C.; CHIANESE, L.; ADDEO, F. Casein proteolysis in human milk: tracing the pattern of casein breakdown and the formation of potential bioactive peptides. Journal of Dairy Research, Cambridge, v. 71, p. 74 – 87, 2004. FERRARETO, A.; SIGNORILE, A.; GRAVAGHI, C.; FIORILLI, A.; TETTAMANTI, G. Casein phosphopeptides influence calcium uptake by cultured human intestinal HT-29 tumor cells. Journal of Nutrition. Bethesda, v. 131, n. 6, p. 1655 – 1661, 2001. FIELD, C.J. Use of T cell function to determine the effect of physiologically active food components. American Journal of Clinical Nutrition, New York, v. 71, Suppl., p. 1720S – 1725S, 2000. FIJEN, J.W.; KOBOLD, A.C.M.; BOER, P.; JONES, C.R.; WERF, T.S.; TERVAERT, J.W.C.; ZIJLSTRA, J.G.; TULLEKEN, J.E. Leukocyte activation and cytokine production during experimental human endotoxemia. European Journal of Internal Medicine, Oxford, v. 11, n. 2, p. 89 – 95, 2000. FITZGERALD, R.J. Potencial uses of caseinophosphopeptides. International Dairy Journal. Great Britain, v. 8, n. 5/6, p. 451 – 457, 1998. FITZGERALD, R.J.; MEISEL, H. Lactokiinins: Whey protein-derived ACE inhibitory peptides. Nahrung, Weinheim, v. 43, n. 3, p. 165 - 167, Sept. 1999. FOX, P.F. Milk proteins as food ingredients. International Journal of Dairy Technology. Huntingdon, v. 54, n. 2, p. 41 – 55, 2001. GAUCHERON, F.; GRAET, Y.L.; BOYAVAL, E.; PIOT, M. Binding of cations to casein molecules: importance of physicochemical conditions. Milchwissenschaft. Nurnberg, v. 52, n. 6, p. 322 – 327, 1997. GIACOMETTI, A.; CIRIONI, O.; GHISELLI, R.; MOCCHEGIANI, F.; DÁMATO, G.; CIRCO, R.; ORLANDO, F.; SKERLAVAJ, B.; SILVESTRI, C.; SABA, V.; ZANETTI, M.; SCALISE, G. título. American Journal of Respiratory and Critical Care Medicine, . New York, v. 169, n. 2, p 187 – 194, 2004. GHISELLI, R.; GIACOMETTI, A.; CIRIONI, O.; MOCHEGIANI, F.; ORLANDO, F.; D’AMATO, G.; SISTI, V.; SCALISE, G.; SABA, V. Cecropin B enhances betalactams activities in experimental rat models of gram-negative septic shock. Annals of Surgery, Philadelphia v. 239, n. 2, p. 251 – 256, 2004.

127
GLAUSER, M.P. Pathophysiologic basis of sepsis: considerations for future strategies of intervention. Critical Care Medicine, Baltimore, v. 28, n. 9, Suppl., p. 54 – 58, 2000. GLAUSER, M.P.; ZANETTI, G.; BAUMGARTNER, J.D.; COHEN, J. Septic shock: pathogenesis. The Lancet. London, V. 338, p. 732 – 739, 1991. GOBBETTI, M.; STEPANIAK, L.; DE ANGELIS, M.; CORSETTI, A.; DI CAGNO, R. Latent bioactive peptides im milk proteins: proteolytic activation and significance in dairy processing. Critical Reviews in Food Science and Nutrition. Boca Raton,. v. 42, n. 3, p. 223 – 239, 2002. GONZÁLEZ-TELLO, P.; CAMACHO, F.; JURADO, E.; PÁEZ, M. P.; GUADIX, E. M. Enzymatyc hydrolysis of whey proteins: II. Molecular weight range. Biotechnology and Bioengineering, New York, v. 44, n. 4, p. 529 - 532, 1994. HALLIWELL, B. GUTTERIDGE, J.M.C. Free radicals in biology and medicine. Clarendon Pres : Oxford, 2. ed., 1989. HAN, S.K.; SHIN, Y.C.; BYUN, H.D. Biochemical, molecular and physiological characterization of a new β-casein variant detected in Korean cattle. Animal Genetics. Oxford, v. 31, n, p. 49 – 51, 2000. HANSEN, M. Effect of casein phosphopeptides on zinc and calcium absorption in experimental models and humans. Ph.D. thesis, The Royal Veterinary and Agricultural University, Denmark, 1995. HARKNESS, J.E.; WAGNER, J.E. Biologia e Clínica de Coelhos e Roedores. São Paulo : Editora Roca, 3ª edição, 1993. HARTMANN, R.; GÜNTHER, S.; MARTIN, D.; MEISEL, H.; PENTZIEN, A.K.; SCLIMME, E.; SCHOLZ, N. Cytochemical model systems for the detection and characterization of potentially bioactive milk components. Kieler Milchwirstschaftliche Forschungsberichte. Hildesheim v. 52, n. 1, p. 61 – 85, 2000. HARTMANN, R.; MEISEL, H. Cytochemical assessment of phosphopeptides derived from casein as potential ingredients for functional food. Nahrung/Food. Weinheim, v. 46, n. 6, p. 427 – 431, 2002. HATA, I.; HIGASHIYAMA, S.; OTANI, H. Identification of a phosphopeptide in bovine αs1-casein digest as a factor influencing proliferation and immunoglobulin production in lymphocyte cultures. Journal of Dairy Research. United Kingdom, v. 65, p. 569 – 578, 1998. HATA, I.; UEDA, J.; OTANI, H. Immunostimulatory action of a commercially available casein phosphopeptide preparation, CPP-III, in cell cultures. Milchwissenschaft, Nurnberg, v. 54, n. 1, p. 429 - 432, 1999. HEALTH care professionals product informations. Disponível em: http://www.meadjohnson.com/professional/indivdescr.html#metabolics. Consultado em: 12/08/2006.

128
HEMPFEL, J.; PFORTE, H.; RAAB, B.; ENGST, W.; BÖHM, H.; JACOBASCH, G. Flavonols and flavones of parsley cell suspension culture change the antioxidative capacity of plasma in rats. Nahrung, Weinheim, v. 43, n. 3, p. 201 - 204, 1999. HOERR, R.A.; BOSTWICK, E.F. Bioactive proteins and probiotic bacteria: modulators of nutritional health. Nutrition. New York, v. 16, n. 7/8, p. 711 - 713, 2000. HUFFMAN, L.M.; HARPER, W.J. Maximizing the value of milk through separation technologies. Journal of Dairy Science, Champaign, v . 82, n. 10, p. 2238 - 2244, 1999 HUQ, N.L.; CROSS, K.J.; REYNOLDS, E.C. Molecular modeling of the multiphophorylated casein phosphopetide αs1-casein (59 – 79) based on NMR constraints. Journal of Dairy Technology. Barking,, v. 71, n. , p. 28 – 32, 2004. IMAFIDON, G. I.; FARKYE, N. Y.; SPANIER, A .M. Isolation, purification, and alteration of some functiona groups of major milk proteins: a review. Critical Reviews in Food Science and Nutrition, Boca Raton, v. 37, n.7, p.663-689, 1997. ISENBERG, H.D. Essential Procedures for Clinical Microbiology. ASM Press : Washington, 1998. p. 216 – 223. JANSSON, A.H.; ERIKSSON, C.; WANG, X. Effects of budesonide and N-acetylcysteine on acute lung hyperinflation, inflammation and injury in rats. Vascular Pharmacology, New York, v. 43, p. 101 – 111, 2005. JOLLÈS, P.; LÉVY-TOLEDANO, S.; FIAT, A.M.; SORIA, C.; GILLESSEN, D.; THOMAIDIS, A.; DUNN, F.W.; CAEN, J.P. Analogy between fibrinogen and casein: Effect of an undecapeptide isolated from kappa-casein on platelet function. European Journal of Biochemistry, Berlin, v. 158, n. 2, 1986. KAGANDA, J.; TATSUHIRO, M.; HIROO, S. Development of iron deficiency decreases zinc requirement of rat. Journal of Nutritional Science and Vitaminology, . Tokyo, v. 49, n. 4, p 234 – 240, 2003. KANEKANIAN, A.; GALLAGHER, J.; EVANS, E.P. Casein hydrolysis and peptide mapping. International Journal of Dairy Technology. Barking,, v. 53, n. 1, p. 1 – 5, 2000. KAYSER, H.; MEISEL, H. Stimulation of human peripheral blood lymphocytes by bioactive peptides derived from bovine milk proteins. FEBS Letters. Amsterdam, V. 383, n. 1 - 2, p. 18 - 20, 1996. KIHARA, Y.; OTANI, H. Preparation of casein phosphopeptides from different kinds of pasteurized milks and their action on proliferative responses of mouse pleen cells in cell cultures . Milk Science, Munchen, v. 49, n. 2, p. 73 - 79, 2000. KITAJIMA, S.; TSUDA, M.; ESHITA, N.; MATSUSHIMA, Y.; SAITOH, M.; MOMMA, J.; KUROKAWA, Y. Lipopolysaccharide-associated elevation of serum and urinary

129
nitrite/nitrate levels and hematological changes in rats. Toxicology Letters, Amsterdam, v. 78, n. 2.; p. 135 – 140, 1995. KITAMURA, H.; OTANI, H. Fecal IgA levels in healthy persons who ingested cakes with or without bovine casein phsphopeptides. Milchwissenschaft, Nurnberg, v. 57, n. 11/12, 2002. KITTS, D.D. Antioxidant properties of casein-phosphopetides. Trends in Food Science and Technology. Cambridge, v. 16, n. 12, p. 549 – 554, 2005. KITTS, D.D.; NAKAMURA, S. Calcium –enriched casein phosphopeptide stimulates release of IL-6 cytokine in human epithelial intestinal cell line. Journal of Dairy Research, Cambridge, v. 73, p. 44 – 48, 2006. KITTS, D.D.; YUAN, Y.V. Caseinophosphopeptides and calcium biovailability. Trends in Food Science and Technology, Cambridge, v. 3, n .2, p. 31 - 35, 1992. KITTS, D.D.; YUAN, Y.V.; NAGASAWA, T.; MORYAMA, Y. Effects of casein, casein phosphopeptide and calcium intake on ileal Ca disappearance and temporal systolic blood pressure in spontaneously hypertensive rats. British Journal of Nutrition. Cambridge, v. 68, p. 765 – 781, 1992. KOIDE, K.; FUKUSHIMA, T.; KUWATA, T.; ITOYAMA, K.; MIYAZAWA, F. Method for separation and concentration of phosphopeptides. United States Patent, nº US005290685, 1994. KOKSEL, O.; OZDULGER, A.; TAMER, L.; CINEL, L.; ERCIL, M.; DEGIMENCI, U.; UNLU, S.; KANIK, A. Effects of caffeic acid phenethyl ester on lipopolysaccharide-induced luna injury in rats. Pulmonary, Pharmacology and Therapeutics, , 2005 in press p. 1 -6. KONO,H.; FUJII, H.; AMEMIYA, H.; ASAKAWA, M.; HIRAI, Y.; MAKI, A.; TSUCHIYA, M.; MATSUDA, M.; YAMAMOTO, M. Role of Kupffer cells in luna injury in rats administered endotoxin. Journal of Surgical Research. New York, v. 129, p. 176 – 189, 2005. KORHONEN, H.; PIHLANTO-LEPPÄLÄ, A.; RANTAMÄKI ,P.; TUPASELA, T. Impact of processing on bioactive proteins and peptides. Trends in Food Science and Technology., Cambridge, v. 9, p. 307 – 319, 1998. KRÜGER, C.C.H. Propriedades funcionais de concentrados protéicos de leite bovino. Dissertação de Mestrado. Universidade Federal do Paraná, Curitiba, 2002, 77p. KRÜGER, C.C.H.; CENI, G.C.; SGARBIERI, V.C.; CÂNDIDO, L.M.B. Propriedades hidrofílicas de concentrados protéicos de leite bovino. Revista da Sociedade Brasileira de Ciência e Tecnologia de Alimentos. Campinas, p.122 - 127, 2002. KRÜGER, C. C. H. ; ANDRADE, A. ; CUNHA, R. B. ; SGARBIERI, V.C. ; BOUHALAB, S. ; CÂNDIDO, L.M.B. . Analysis of casein phosphopeptides by matrix-assisted laser desorption ionization mass spectrometry. In: INTRAFOOD 2005 Innovations in Traditional Foods, 2005, Valencia. INTRAFOOD 2005, 2005. p. P1.17.

130
KRUIF, A.G. Casein micelle interactions. International Dairy Journal. Great Britain, v. 9, n. 3, p. 183 – 188, 1999. LAHOV, E.; REGELSON, W. Antibacterial and immunostimulating casein-derived substances from milk: caseicidin, isracidin peptides. Food and Chemical Toxicology. Oxford, v. 34, n. 1, p. 131 – 145, 1996. LANZA-JACOBY, S.; FLYNN, J.T.; MILLER, S. Parenteral supplementation with a fish oil emulsion prolongs survival and improves lymphocyte function during sepsis. Nutrition, New York, v. 17, n. 2, p. 112 – 116, 2001. LEDOUX, N.; MAHÉ, S.; DUBARRY, M.; BOURRAS, M.; BENAMOUZIG, R.; TOMÉ, D. Intraluminal immunoreactive caseinomacropeptide after milk protein ingestion in humans. Nahrung, Weinheim, v. 43, n. 3, p. 196 - 200, 1999. LEE, R.P.; WANG, D.; KAO, S.J.; CHEN, H.I. The lung is the majopr site that produces nitric oxide to induce acute pulmonary oedema in endotoxin shock. Clinical and Experimental Pharmacology, Carlton, v. 28, p. 315 – 320, 2001. LE GALL, J.R.; KLAR, J.; LEMESHOW, S.; SAULNIER, F.; ALBERTI, C.; ARTIGAS, A.; TERES, D. The logistic organ dysfunction system. A new way to assess organ dysfunction in the intensive care unit. JAMA, Chicago, v. 276, p. 802 – 810, 1996. LENCZOWSKI, M.J.P.; VAM-DAM, A.M.; POOLE, S.; LARRICK, J. W.; TILDERS, F.J.H. Role of circulating endotoxin and interleukin-6 in the ACTH and corticosterone response to intraperitoneal LPS. American Journal of Regulatory, Integrative and Comparative Physiology, . Bethesda, v. 276, n. 6, p. 1870 – 1877, 1997. LÉONIL, J.; GAGNAIRE, V.; MOLLÉ, D.; PEZENNEC, S.; BOUHALLAB, S. Application of chromatography and mass spectrometry to the characterization of food proteins and derived peptides. Journal of Chromatography A, Arlington, v. 881, n. 1, p. 1-21, 2000. LÉONIL, J.; MOLLÉ, D.; BOUHALLAB, S.; HENRY, G. Precipitation of hydrophobic peptides from tryptic casein hydrolysate by salt and pH. Enzyme and Microbial Technology. New York, V. 16, p. 591 – 594, 1994. LIHME, A.O.F.; AAGESEN, M.I.; GAMMELGAARD-LARSEN, C.; ELLEGAARD, K.H. Method of isolating biomolecules by ion exchange. Patent Publication, United States nº US5780593, 1998. LÖFFLER, A. Proteolytic enzymes: sources and applications. Food Technology, Chicago, v. 42, n. 1, p. 63 - 70, 1986. LOOSDRECHT, A.A.; BEELEN, R.H.J.; OSSENKOPPELE, G.J. BROEKHOVEN, M.G.; LANGENHUIJSEN, M.M.A.C. A tetrazolium based colorimetric MTT assay to quantitate human monocyte mediated cytotoxicity against leukemic cells from cell lines and patientes with acute myeloid leukemia. Journal of Immunological Methods.Amsterdam, v. 174, p. 311 - 320, 1994.

131
LOPEZ-BAJONERO, L.J.; LARA–CALDERON, P.; GALVEZ-MARISCAL, A.; VELÁSQUEZ-ARELLANO, A.; LOPEZ-MUNGUIA, A. Enzymatic production of a low-phenylalanine product from skim powder and caseinate. Journal of Food Science. Chicago, v. 56, n. 4, p. 938 – 942, 1991. LOWNDES, J.; MACAA, T.J.R.; PLIMMER, H.A. Analysis of proteins: caseo-phosphopeptone. The Biochemical Journal. London, v. 35, n. 3, p. 315 – 320, 1941. MADHAVI, N.; DAS, V.N. Effects of N3 and N6 fatty acids on the survival of vineristine sensitive and resistant human cervical carcinoma cells, in vitro. Cancer Letters, v. 84, p. 31 – 41, 1994. MALKOSKI, M.; DASPER, S.G.; O’BRIEN-SIMPSON, N.M.; TALBO, G.H.; MACRIS, M.; CROSS, K.J.; REYNOLDS, E.C. Kappacin, a novel antibacterial peptide from bovine milk. Antimicrobial Agents and Chemotherapy, Bethesda, v. 45, p. 2309-2315, 2001. MANN, E.J. Casein and caseinates. Dairy Industries International London, v.56, n. 2, p. 13 - 14, 1991. MANSON, W.; ANNAN, W.D. The structure of a phosphopeptide derived from β-casein. Archives of Biochemistry and Biophysics, New York, v. 145, n. 1, p. 15 – 26, 1971. MAUBOIS, J.L.; LEONIL, J.; SABOYA, L.V. Milk-derived bioactive peptides: technological challenges. Boletim SBCTA. Campinas, v. 36, n. 2, p. 116 – 121, 2002. MAZA, G. Functional Foods: Biochemical and Processing Aspects. Lancaster : Technomic Publishing CO, Inc., 1998, 460 p. McDONAGH, D.; FITZGERALD, R.J. Production of caseinophosphopeptides from sodium casseinate using a range of commercial protease preparations. International Dairy Journal. Great Britain, v. 8, n. 1, p. 39 – 45, 1998. McMURRY, M.P.; CHAN, G.M.; LEONARD, C.O.; ERNST, S.L. Bone mineral status in children with phenylketonuria relationship to nutricional intake and phenylalanine control. American Journal Clinical Nutrition, Bethesda, v. 55, p. 997-1004, 1992. MEISEL, H.; BOCKELMANN, W. Bioactive peptides encrypted in milk proteins: proteolytic activation and thropho-functional properties. Antonie van Leeuwenhoek. Netherlands, v. 76, n. 1 -4, p. 207 – 215, 1999. MEISEL, H.; GÜNTHER, S. Food proteins as precursors of peptides modulating human cell activity (Short communication). Nahrung. Weinheim, v 42, n. 3/4, p. 175 – 176, 1998. MEISEL, H.; OLIEMAN, C. Estimation of calcium-binding constants of casein phophopetides by capillary electrophoresis. Analytica Chimica Acta. Amsterdam, V. 372, n. ½, p. 291 – 297, 1998. MEISEL, H.; SCHLIMME, E. Milk proteins: precursors of bioactive peptides. Trends in Food Science and Technology, Cambridge, v. 1, n. 2, p. 41 - 48, 1990.

132
MEISEL, H. Overview on milk protein-derived peptides. International Dairy Journal. Great Britain, v. 8, n.5/6, p. 363 – 373, 1998. MEISEL, H. Bioactive substances of food origin: a challenge for food and nutrition science. Nahrung, Weinheim, v. 43, n. 3, p. 147, Sept. 1999. MEYER, H.E.; HOFFMANN-POSORSKE, E.; KORTE, H.; HEILMEYER, M.G. sequence analysis of phosphoserine-containing peptides. FEBS Letters. Amsterdam, V. 204, n. 1, p. 61 – 66, 1986. MING, P. Descrição da indústria de produtos lácteos dos Estados Unidos. Leite e Derivados, São Paulo, n. 12, ano IX, p. 64 – 68, 2000. MINKIEWICZ, P.; SLANG, C.J.; DZIUBA, J.; VISSER, S.; MIODUSZEWSKA, H. Identification of peptides obtained via hydrolysis of bovine casein by chimosin using HPLC and mass spectrometry. Milchwissenschaft, Nurnberg, v. 55, n. 1, p. 14 – 17, 2000. MIQUEL, E.; ALEGRÍA, A.; BARBERÁ. R.; FARRÉ, R. Casein phosphopeptides released by simulated gastrointestinal digestion of infant formulas and their potential in mineral binding. International Dairy Journal. Great Britain, v. 16, n. 9, p. 992 – 1000, 2006. MOLKETIN, J. Bioactive lipids ocurring in bovine milk. Nahrung, Weinheim, v. 43, n. 3, p. 185 - 189, 1999. MONTAGNIER, L.; OLIVIER, R.; PASQUIER, C. Oxidative stress in cancer, AIDS, and neurodegenerative diseases. New York : Marcel Dekker, 1998 MONTEIRO, L.T.B. Desenvolvimento de uma formulação protéica com teor reduzido de fenilalanina. Curitiba. Dissertação de mestrado. Universidade Federal do Paraná, 2003. MORENO, Y.M.F. Influência das proteínas de soro de leite bovino no estado nutricional, composição corporal e sistema imunológico em um coorte de crianças com síndrome de imunodeficiência adquirida (AIDS). Dissertação de mestrado. Universidade Estadual de Campinas, 2002. 109 p. MORR, C.V.; GERMAN, B.; KINSELLA, J.E.; REGENSTEIN, J.M.; VAN BUREN, J.M.; KILARA, A.; LEWIS, B.A.; MANGINO, M.E. Acollaborative study to develop a standardized food protein solubility procedure. Journal of Food Science, Chicago, v. 50, n. 6, p. 1715 - 1718, 1985. NAITO, H.; SUZUKI, H. Further evidene for the formation in vivo of phosphopeptide in the intestinal lúmen from dietary β-casein. Agricultural and biological chemistry, Tokyo, v. 38, n. 8, p. 1543 – 1545, 1974. NAKAJIMA, K.; TAMURA, N.; KOBAYASHI-HATTORI, K.; YOSHIDA, T.; HARA-KUDO, Y.; IKEDO, M.; SUGITA-KONISHI, Y.; HATTORI, M. Prevention of intestinal infection by glycomacropeptide. Bioscience, Biotechnology and Biochemistry. Tokyo, v. 69, n. 12, p. 2294 – 2301, 2005.

133
NEKLYUDOV, A.D; IVANKIN, A.N.; BERDUTINA, A.V. Properties and uses of protein hydrolysates. Applied Biochemistry and Microbiology. New York, v. 36, n. 5, p. 452 – 459, 2000. NESTLÉ NUTRITION WORKSHOP. Nutrição, imunidade e infecção na infância. Séries Programa Pediátrico n.º 45, 2001. OBMINSKA-DOMORADZKA, B.; SZCZYPKA, M.; DEBOWY, J. Thymomimetic drugs and zinc supplementation on the celular immune response in hydrocortisone-suppressed mice. Journal of Veterinary Medicine, Berlin, v. 49, p. 469 – 475, 2002. OTANI, H.; HORIMOTO, Y.; MONNAI, M. Suppression of interleukin-2 receptor expresión on mouse CD4+ T cells by bovine kappa-caseinoglycopeptide. Bioscience, Biotechnology and Biochemistry. Tokyo, v. 60, n. 6, p. 1017 – 1019, 1996. OTANI, H.; KIHARA, Y.; PARK, M. The immunoenhancing property of a dietary casein phosphopeptide preparation in mice. Food and Agricultural Immunology. Abingdon, V. 12, p. 165 – 173, 2000. OTANI, H.; KITAMURA, H.; PARK, M.; OSHIDA, T.; KUSUHARA, S.; SAWADA, K. Enhancement of intestinal IgA levels in piglets by oral administration of a commercially available casein phosphopeptide preparation. Milchwissenschaft, Nurnberg, v. 55, n. 8, p. 429 - 432, 2000. OTANI, H.; MONNAI, M. Induction o fan Interleukin-1 receptor antagonst-like component produced from mouse soleen cells by bovine kappa-caseinoglycopeptide. Bioscience, Biotechnology and Biochemistry. Tokyo, V. 59, n. 6, p. 1166 – 1168, 1995. OTANI, H.; NAKANO, K.; KAWAHARA, T. Stimulatory effect of a dietary casein phosphopeptide prepartion on the mucosal IgA response of mice too rally ingested lipopolysaccharide from Salmonella typhimurium. Bioscience, Biotechnology and Biochemistry. Tokyo, V. 67, n. 4, p. 729 - 735, 2003. OTANI, H.; WATANABE, T.; TASHIRO, Y. Effects of bovine beta-casein (1 – 28) and its chemically synthesized partial fragments on proliferative responses and immunoglobulin production in mouse spleen cell cultures. Bioscience, Biotechnology and Biochemistry. Tokyo, V. 65, n. 11, p. 2489 - 2495, 2001. PABLO, M.A.; CIENFUEGOS, G.A. Modulatory effects of dietary lpids on immune system functions. Immunology and Cell Biology, Adelaide, v. 78, p. 31 – 39, 2000. PACHECO, M.T.B.; AMAYA-FARFAN,J.; SGARBIERI, V.C. Partial characterization of a whey protein concentrate and its enzyme hydrolysates. Journal of Food Biochemistry, Trumbull, v. 26, p. 327 – 338, 2002. PANYAM, D; KILARA, A. Enhancing the functionality of food proteins by enzymatic modification. Trends in Food Science and Technology, Cambridge, v. 7, n.4, p. 120 - 125, 1996.

134
PARK, O.J.; ALLEN, J.C. Antigenicity of casein phosphopeptides prepared with immobilized glutamic acid-specific endopeptidase or trypsin. Nutrition Research, Tarrytown, v. 20, n. 3, p. 359 - 370, 2000. PASKI, S.C.; COVERY, L.; KUMMER, A.; XU, Z. Role of melallothionein in regulating the abundance of histochemically reactive zinc in rat tissues. Canadian Journal of Physiology and Pharmacology. Ottawa, v. 81, n. 8, p. 815 – 824, 2003. PATHMAKANTHAN, S.; HAWKEY, C.J. A lay doctor’s guide to the inflammatory process in the gastrointestinal tract. Post-Graduation Medicine Journal, v. 76, p. 611 – 617, 2000. PEREIRA, G.A.J.; MARSON, F.; ABERD, M.; OSLINI, F.; SOUZA, S.H.; BASILE-FILHO, A.; Fisiopatologia da sepse e suas implicações terapêuticas. Medicina, Ribeirão Preto, v. 31, p. 349 – 362, 1998. PÉRÈS, J.M.; BOUHALLAB, S.; BUREAU, F.; NEUVILLE, D.; MAUBOIS, J.L.; DEVROEDE, G.; ARHAN, P.; BOUGLÉ, D. Mechanisms of absorption of caseinophosphopeptide bound iron. Journal of Nutritional Biochemistry. Stoneham, V. 10, p. 215 - 222, 1999. PETERSON, R.F.; NAUMAN, L.W.; MCMEEKIN, T.L. The separation and amino acid composition of a pure phosphopetone prepared from beta-casein by the action of trypsin. Journal of American Chemical Society. Easton, V. 80, p. 95 – 99, 1958. PFORTE, H.; HEMPEL, J.; JACOBASCH, G. Distribution pattern of a flavonoid extract in the gastrointestinal lumen and wall of rats. Nahrung, Weinheim, v. 43, n. 3, p. 205 - 208, 1999. PHAN, T.N.; BUCKNER, T.; SHENG, J.; BALDECK, J.D.; MARQUIS, R.E. Physiologic actions of zinc related to inhibition of acid and alkali production by oral streptococci in suspensions and biofilms. Oral Microbiology and Imunnology. Copenhagen, v. 19, p. 31 – 38, 2004. PICK , E.; MIZEL, M. Rapid microassays for the measurement of superoxide and hydrogen peroxide production by macrophages in culture using an automatic enzyme immunoassay reader. Journal of Immunological Methods. Amsterdam, V. 46, p. 211 – 226, 1981. PILOSOF, A.M.R.; BARTHOLOMAI, G.B. Caracterizacion Functional y Structural de Proteinas. Buenos Aires : Eudeba, 2000. 181 p. PIZATO, N.M.P. Efeito da dieta com diferentes proporções de ácidos graxos N-6:N-3 sobre o crescimento tumoral, caquexia e sistema imunitário em ratos portadores do tumor de Walker 256. Tese de Doutorado. Universidade Federal do Paraná, Curitiba, 2005. 93p. RAETZ, C.R.H.; WHITFIELD, C. Lipopolysaccharide endotoxinas. Annual Review of Biochemistry, Palo Alto, v. 71, p. 635 – 700, 2002. RASMUSSEN, L.K.; JOHNSEN, L.B.; TSIORA, A.; SORENSEN, E.S.; THOMSEN, J.K.; NIELSEN, N.C.; JAKOBSEN, H.J.; PETERSEN, T.E. Disulphide-linked caseins and micelles. International Dairy Jounal., Great Britain, v.6. n. 3, p. 215 - 218, 1999.

135
RECIO, I.; VISSER, S. Identification of two distinct antibacterial domains within the sequence of bovine αs2-casein. Biochimica et Biophysica Acta. Amsterdam, V. 1428, n. 2 - 3, p. 314 - 326, 1999. REYNOLDS, E.C. The prevention of subsurface demineralization of bovine enamel and change in plaque composition by casein in na intra-oral model. Journal of Dental Research. Alexandria, v. 66, p. 1120 – 1127, 1987. REYNOLDS, E.C. Production of phosphopeptide from casein. United States Patent nº US 6448374, 2002. REYNOLDS, E.C.; CAIN, C.J.; WEBBER, F.L..; BLACK, C.L..; RILEY, P.F..; JOHNSON, I.H.; PERICH, J.W. Anticariogenicity of calcium phosphate complexes of tryptic casein phosphopeptides in the rat. Journal of Dental Research. Alexandria, v. 74, n. 6, p. 1272 – 1279, 1995. REYNOLDS, E.C.; RILEY, P.F.; ADAMSON, N.J.; A selective-precipitation purification procedure for multiple-phosphoseryl containing peptides and methods for their identification. Analytical Biochemistry. V. 217, n. 2, p. 277 – 284, 1994. RIMINGTON, C. Note on the amino-acids present in phosphopeptone. The Biochemical Journal. v. 35, n. 3, p. 321 - 327, 1941. RIZZELLO, C.G.; LOSITO, I.; GOBETTI, M.; CARBONARA, T.; BARI, M.D.; ZAMBONIN, P.G. Antibacterial activities of peptides from the water-soluble extracts of Italian cheese varietes. Journal of Dairy Science. Champaign, V. 88, n. 7, p. 2348 – 2360, 2005. ROKKA, T.; SYVÄOJA, E.; TUOMINEN, J.; KORHONEN, H. Release of bioactive peptides by enzymatic proteolysis of Lactobacillus GG fermented UHT milk. Milchwissenschaft, Nurnberg, v. 52, n. 12, p. 675 - 678, 1997. ROSENFELD, Y.; PAPO, N.; SHA, Y. Endotoxin (lipopolysaccharide) neutralization by innate immunity host-defense peptides. The Journal of Biological Chemistry, Bethesda, v. 281, n. 3, p. 1636 – 1643, 2006. ROTH, H.P. Development of alimentary zind deficiency in growing rats is retarde at low dietary protein levels. Journal of Nutrition, . New York, v. 133, n. 7, p. 2294 – 2301, 2003. RYAN, A.J. FLANAGAN, S.W.; MOSELEY, P.L.; GISOLFI, C.V. Acute heat stress protects rats against endotoxin shock, Journal of Applied Physiology, v. 73, n. 4, p. 1517 – 1522, 1992. SATO, R.; SHINDO, M.; GUNSHIN, H.; NOGUCHI, T.; NAITO, H. Characterization of phosphopeptide derived from β-casein: an inhibitor to intra-intestinal precipitation of calcium phosphate. Biochimica et Biophysica Acta, v. 1077, p. 413 – 415, 1991.

136
SCHANBACHER, F.L.; TALHOUK, R.S.; MURRAY, F.A.; GHERMAN, L.I.; WILLET, L.B. Milk-borne bioactive peptides. International Dairy Journal. Greait Britain, v. 8, n.5/6, p. 393 – 403, 1998. SCHOLZ-AHRENS, K.E.; SCHREZENMEIR, J. Effects of bioactive substances im milk on mineral and trace element metabolism with special reference to casein phopsphopeptides. British Journal of Nutrition, Cambridge, V. 84, suppl. 1, p. S147 – S153, 2000. SCHÜPBACH, P.; NEESER, J.R.; GOLLIARD, M.; ROUVET, M.; GUGGENHEIM, B. Incorporation of caseinoglycomacropeptide and caseinophosphopetide into the salivary pellicle inhibits adherence of mutans streptococci. Journal of Dental Research. Alexandria, v. 75, n. 10, p. 1779 – 1788, 1996. SGARBIERI, V. C. Proteínas em alimentos protéicos: Propriedades, Degradações, Modificações. São Paulo : Livraria Varela, 1996. 547 p. SGARBIERI, V.C. Revisão: Propriedades estruturais e físico-químicas das proteínas do leite. Brazilian Journal of Food Technology. Campinas, v. 8, n. 1, p. 43 – 56, 2005. SHAH, N.P. Effects of milk-derived bioactives: an overview. British Journal of Nutrition. Cambridge, v. 84, suppl. 1, p. S3 – S10, 2000. SHIMIZU, M. Modulation of intestinal functions by food substances. Nahrung, Weinheim, v. 43, n. 3, p. 154 - 158, 1999. SILANO, M.; VINCENZI, M.D. Bioactive antinutritional peptides derived from cereal prolamins: a review. Nahrung, Weinheim, v. 43, n. 3, p. 175 - 184, 1999 SILLIMAN, C.C.; VOELKEL, N.F.; ALLARD, J.D.; ELZI, D.J.; TUDER, R.M.; JOHNSON, J.L. Plasma and lipids from stored packed red blood cells cause acute lung injury in na animal model. Journal of Clinical Investigation, New York, v. 101, n. 7, p. 1458 – 1467, 1998. SILVA, A.V.; MALCATA, F.X. Caseins as source of bioactive peptides. International Dairy Journal. Great Britain, v. 15, n. , p. 1 – 15, 2005 SINGH, A.M.; DALGLEISH, D. G. The emulsifying properties of hydrolyzates of whey proteins. Journal of Dairy Science, Champaign, v. 81, n.4, p. 918 – 924, 1998. SMACHI, E.; GOBBETTI, M. Bioactive peptides in dairy products: synthesis and interaction with proteolytic enzymes. Food Microbiology. London, V. 17, p. 129 - 141, 2000. SMEDEGARD, G.; CUI, L.X.; HUGLI, T.E. Endotoxin-induced schock in the rat. A role for C5a. American Journal of Pathology, Bethesda, v. 135, n. 3, p. 489 – 497, 1989. STEIJNS, J.M. Milk ingredients as nutraceuticals. International Journal of Dairy Technology. Barking, v. 54, n. 8, p. 81 – 88, 2001 STITES, D.P.; TERR, A.I. Imunologia Básica. Rio de Janeiro : Prentice-Hall do Brasil Ltda., 1992.

137
TALBO, G.H.; SUCKAU, D.; MALKOSKI, M.; REYNOLDS, E.C. MALDI-PSD-MS analysis of the phophorilation sites of caseinomacropeptide. Peptides. V. 22, n. 7, p. 1093 – 1098, 2001. TAKANORI, K.; IWASAKI, R.; TANAKA, M.; KIRIYAMA, S. Caseinophosphopeptides (CPP) in feces and contents in digestive tract of rats fed casein and CPP preparations. Bioscience, Biotechnology, Biochemistry. Tokyo, v. 59, n. 1, p. 26 – 30, 1995. TAVARES, M. F. M. Eletroforese capilar básicos. Química Nova, v. 19; n.2, 1996, p. 173-181. TOMÁS-BARBERÁN, F. A. Capillary electrophoresis: a new technique in the analysis of plant secondary metabolites, 1994 TOMÉ, D.; DEBABBI, H. Physiological effects of milk protein components. International Dairy Journal. , Great Britain, v. 8, n.5/6, p. 383 – 392, 1998. TOTSUKA, M.; KAKEHI, M.; KOHYAMA, M.; HACHIMURA, S.; HISATSUNE, T.; KAMINOGAWA, S. Enhancement of antigen-specific IFN-γ production from CD8+ T cells by a single amino-acid-substituted peptide derived from bovine αs1-casein. Clinical Immunology and Immunopathology, New York, v. 88, n. 3, p. 277 – 286, 1998. TURGEON, S.L.; GAUTHIER, S.F.; PAQUIN, P. Emulsifying property of whey peptide fraction as a funtion of pH and ionic strength. Journal of Food Science, Chicago, v. 57, n. 3, p. 601 - 604, 634, 1992. TSUCHITA, H.; GOTO, T; SHIMIZU, T.; YONEHARA, Y.; KUWATA, T. Dietary casein phosphopeptides prevent bone loss in aged ovariectomized rats. Journal of Nutrition. Bethesda, v. 126, n. 1, p. 86 – 93, 1996. UMBACH, M.; TESCHEMACHER, H.; PRAETORIUS, K.; HIRSCHHÃUSER, R.; BOSTEDT, H. Demosntration of a β-casomorphin immunoreactive material in the plasma of newborn calves after milk intake. Regulatory Peptides. Amsterdam, v. 12, , p. 223 – 230, 1985. USDEC - U.S. DAIRY EXPORT COUNCIL. Manual de referência para produtos de soro dos EUA. Arlington, 1997. VAUGHAN, E.E; MOLLET, B. Probiotics in the new millennium. Nahrung, Weinheim, v. 43, n. 3, p.148 - 153, 1999. VEGARUD, G.E.; LANGSRUD, T.; SVENNING, C. Mineral-binding milk proteins and peptides; occurrence, biochemical and technological characteristics. British Journal of Nutrition. Cambridge, v. 84, suppl. 1, p. S91 – S 98, 2000. VIOQUE, J; CLEMENTE, A.; PEDROCHE, J.; YUST, M.M.; MILLÁN, F. Obténcion y aplicaciones de hidrolizados protéicos. Grasas y Aceites. Sevilla, v. 52, n. 2, p. 132 – 136, 2001. WALKER, G.; CAI, F.; SHEN, P.; REYNOLDS, C.; WARB, B.; FONE, C.; HONDA, S.; KOGANEI, M.; ODA, M.; REYNOLDS, E. Increased remineralization of tooth enamel by milk

138
containing added casein phophopeptide-amorphous calcium phosphate. Journal of Dairy Research, United Kingdom, v. 73, p. 74 – 78, 2006. WALSTRA, P. Casein sub-micelles: do they exist? International Dairy Journal. Barking,, v. 9, n. , p. 189 – 192, 1999. WELLS, S.M.; KEW, S.; YAQOOB, P.; WALLACE, F.A.; CALDER, P.C. Dietary glutamine enhances cytokine production by murine macrophages. Nutrition, New York, v. 15, n. 11/12, p. 881 – 884, 1999. WISCHMEYER, P.E.; KAHANA, M.; WOLFSON, R.; REN, H.; MUSCH, M.M.; CHANG, E.B. Glutamine induces heat shock protein and protects against endotoxic shock in the rat. Journal of Applied Physiology. V. 90; n. 6, p. 2403 – 2410, 2001. WONG, C.W.; SEOW, H.F.; LIU, A.H.; HUSBAND, A.J.; SMITHERS, G. W.; WATSON, D.L. Modulation of immune responses by bovine beta-casein. Immunology and Cell Biology v. 74, n. 4, p. 323 – 329, 1996. YAQOOB, P.; CALDER, P. Effects of dietary lipid manipulation upon inflammatory mediator production by murine macrophages. Celular Immunology, v. 163, p. 120 – 128, 1995. YAQOOB, P.; CALDER, P. Cytokine production by human peripheral blood mononuclear cells: differential sensitivity to glutamine availability. Cytokine, New York, v. 10, n. 10, p. 790 – 794, 1998. YAMAGUCHI, K.; MIYAZAKI, M.; TAKAMIZAWA, T.; INAGE, H.; MOORE, B.K. Effect of CPP-ACP paste on mechanical properties of bovine enamel as determined by an ultrasonic device. Journal of Dentistry, Bristol, v. 34, n. 3, p. 230 – 236, 2006. YEUNG, A.C.; GLAHN, R.P.; MILLER, D.D. Effects of iron source on iron availability from casein and casein phosphopetides. Journal of Food Science. Chicago, v. 67, n. 3, p. 1271 – 1275, 2002. YAMAMOTO, N.; TAKANO, T. Antihypertensive peptides derived from milk proteins. Nahrung, Weinheim, v. 43, n. 3, p. 159 – 164, 1999. ZHANG, F.M.; OTANI, H. Immunogenicity and antigenicity of casein phosphopeptides. Milchwissenschaft. Nurnberg, v. 58, n. ½, p. 9 – 13, 2003. ZHANG, H.; SUN, G.Y. LPS induces permeability in lung microvascular endothelium via AT1 receptor. Archives of Biochemistry and Biophysics, v. 441, p. 75 – 83, 2005. ZUCHT, H.D.; RAIDA, M.; ADERMANN, K.; MÄGERT, H.J.; FORSSMANN, W.G. Casocidin-I: a casein-αs2 derived peptide exhibits antibacterial activity. FEBS Letters, Amsterdam, v. 372, n. 2 - 3, p. 185 - 188, 1995.





























