Embed Size (px)
Citation preview

Universidade Federal do Rio de Janeiro Instituto de Biologia
Programa de Pós Graduação em Ecologia (PPGE)
Efeito de Peixes Onívoros Sobre a Estrutura e Estabilidade de Cadeias Tróficas Aquáticas
Luciana Silva Carneiro
Rio de Janeiro – RJ
Outubro de 2008
Tese apresentada ao Programa de Pós-Graduação em Ecologia da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do grau de Doutor em Ecologia Orientador: Prof. Dr. Reinaldo Luiz Bozelli

Efeito de Peixes Onívoros Sobre a Estrutura e Estabilidade de Cadeias Tróficas Aquáticas
Luciana Silva Carneiro
__________________________________________________
Profa. Odete Rocha, Dr.
__________________________________________________
Profa. Ana Petry, Dr.
__________________________________________________
Prof. Sidinei Magela Thomaz, Dr.
__________________________________________________
Prof. Érica Caramaschi, Dr.
__________________________________________________
Prof. Reinaldo Luiz Bozelli, Dr.
Rio de Janeiro - RJ
Outubro de 2008
Tese apresentada ao Programa de Pós-Graduação em Ecologia da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do grau de Doutor em Ecologia

Ficha Catalográfica:
Carneiro, Luciana Silva Efeito de peixes onívoros sobre a estrutura e estabilidade de cadeias tróficas aquáticas [Rio de Janeiro 2008] Tese de Doutorado – Universidade Federal do Rio de Janeiro Departamento de Ecologia, UFRJ, 2008. 43p Palavras-chave:
1.Onivoria 2. compartimentos aquáticos 3. cascata trófica 4. comunidades pelágicas 5. meta-análise 6. mesocosmos.

“Tenha em mente que tudo que você aprende
na escola é trabalho de muitas gerações.
Receba essa herança, honre-a, acrescente a ela
e, um dia, fielmente, deposite-a nas mãos de
seus filhos.”
Albert Einstein

iii
Agradecimentos
As 131 páginas que se seguem foram fruto de 11 anos de aprendizado neste grupo
de pesquisa. Então, há muitas pessoas e instituições à agradecer.
Ao meu orientador, Reinaldo Bozelli, por ter sido o incentivo ao estudo da Ecologia.
Compartilhou comigo seu tema de pesquisa, e desde o início, ouviu com interesse todas as
dúvidas e problemas que surgiram durante toda a minha formação. Pela confiança em me
deixar ousar trabalhar com novas idéias e conceitos, correndo riscos inerentes a esta
atitude (Acho que deu certo, né?). Por sua amizade e pela alegria de trabalharmos juntos.
Ao Prof. Dr. Francisco Esteves, graças a seu trabalho árduo desde jovem é que
pude usufruir de uma boa formação dentro da “Família Limnológica”.
Aos membros da pré-banca Prof. Ricardo Iglesias e Prof. Érica Caramaschi pelas
valiosas sugestões.
Aos professores Dr. Sidnei Magela Thomas, Dra. Odete Rocha, Dra. Ana Petry, Dra.
Érica Caramaschi e Dr. Paulo Paiva por terem aceito o convite para participar da banca
avaliadora desta tese de doutorado.
Às professoras Paulina Maria Maia Barbosa e Érica Caramaschi por serem as
grandes incentivadoras da linha de pesquisa que adotei para vida.
Ao Professor José Luiz Attayde (Coca), o irmão científico mais velho e co-orientador,
que é exemplo de entusiasmo pela Ecologia. A semelhança entre nossas linhas de
pesquisa NÃO é mera coincidência, mas sim pura inspiração!
To Professor Mike Vanni who generously let me join his lab group and devoted his
precious time and made “much many” valuable suggestions which indeed helped improve
this study. His body stoichiometry is 1:1 (smartness: friendship). He is the Real McCoy Guy! Meu especial agradecimento àqueles amigos que estiveram envolvidos de corpo e
alma nos cansativos, porém felizes dias de trabalho de campo: Adriana, Adriano, Ellen,
Fred, Jabour, Jayme, Joao Marcelo, Leticia, Leandro (NUPEM), Vinicius, João Leal,
Humberto, Pedro (Lab. Peixe/UFRJ). Obrigada pela ajuda e paciência em aguentar minha
ansiedade!
Obrigada a toda a familia limnológica! Desde 23 de julho de 1997, vocês se
tornaram parte da minha vida! PRIMEIRO A LISTA DOS DINOSSAUROS: Adriana, Ana,

iv
Betina, Chico, Christina, Cláudio, Cléber, Coca, Dinho, Fred, João, Júnior, Kika, Lisiana,
Marquinhos, Mariana, Manuel, Paulina, Reinaldo, Renata, Rose, Sandra, Sérgio e Vinicius.
AGORA A SEGUNDA GERAÇÃO DE DINOSSAUROS: Adriano, Albert, Alexandre,
Andresson, Chiquitito, Coutinho, Fabiana, Fernanda, Humberto, Jabour, Jayme, Juliana,
Letícia, Mário, Murilo, Paloma e Thais. FINALMENTE, AOS FILHOTES DE
DINOSSAUROS, que são tantos, que nem cabem nestas páginas.
Aos imãos planctônicos, Adriana, Ana Cláudia, Jayme, Leticia, Luciana, Monalisa
Paloma, Sandra e Rayanne. Muito obrigada pela troca de experiências no dia a dia!
To my international family for their kidness and affection, and for never letting me feel
that I was away from my parents and country. I will never forget none of you: Lesley Knoll,
Allie Bable, Jen Bobson, Beth Dickman, Leah Jeansen, Albeto Pilati, Mike Vanni, Maria
Gonzalez, Melina Vanni-Gonzalez, Yuta Kawasaky. Thank you so much for making my life
in OHIO awesome!!!!
À amiga Letícia (Let’s Go) que foi meu incentivo para jornada de milhares de horas
em frente ao microscópio! Obrigada pelas 128 amostras de fitoplâncton contadas e medidas
com muito amor e dedicação... Obrigada pela companhia nos fins de semana e noites de
trabalho no Fundão! Obrigada pelo sorriso aberto e pela amizade sincera!
À amiga Adriana (Cãodriana e/ou Mãedriana) porque estamos caminhando juntas e
de mãos dadas há muitos anos. Obrigada pelo companherismo!
Ao amigo Marquinhos (MP) que sempre amenizou meu sofrimento me incentivando
quando experimentos deram errado, chamado para um café de fim de tarde, em conversas
noturnas no MSN ou mesmo tirando minha concentração no trabalho.
Ao Jabouranga, meu querido amigo de todas as horas. Sorrindo ou emburrado, ele
sempre esteve do meu lado para ajudar ou atrapalhar! Obrigada pelo trabalho em
cooperação, pelas horas em frente a televisão e por ser parte da minha família.
Obrigada a toda equipe do Núcleo de Desenvolvimento Sócio Ambiental de Macaé
(NUPEM), especialmente a Lena, Inês, João Marcelo, Sr. Paulo, Prof. Delicius (Vinicius),
Prof. Francisco Esteves. O NUPEM foi minha casa, meu trabalho, meu lazer, meu
descanso! Minha formação como verdadeira Ecóloga se deve aquele pedaço abençoado de
Terra em Macaé.

v
Meu muito obrigada ao Consenho Nacional de Desenvolvimento Tecnológico e
Científico (CNPq) pelas duas modalidades de bolsa que me concedeu para desenvolver
este trabalho (Bolsa de Doutorado e Doutorado Sanduíche). Espero poder retornar ao Brasil
bons frutos de tudo o que foi investido na minha formação.
To the members of Departament of Zoology at Miami University (Oxford, OH), all of
whom I thank for their hospitality and support. I am grateful especially to Dr. Mike Vanni, Dr.
Tom O. Crist, Dr. Maria Gonzalez, Dr. Alberto Pilati, Dr. Maten Hogan, Dr. Jim Stoeckel e
MSc. Stephen Glaholt.
Obrigada a todos os professores do Departamento de Ecologia e do Programa de
Pós-Graduação em Ecologia da UFRJ. Os ensinamentos e influência acadêmica de vocês
me ajudam a exercer esta profissão com sabedoria e carinho.
As amigas que nunca me vêem, mas sempre me ligam para dar apoio: Daniela,
Clarice e Paulinha. Vocês são queridas mesmo à distância!
Toda e qualquer parte deste trabalho, qualquer idéia e toda a inspiração são frutos
do amor incondicional e dedicação que eu tenho recebido do Adriano. Ele é meu amigo,
meu estatistico de plantão, minha fonte de bibliografia, meu conselheiro, meu incentivador,
minha alegria, meu amor! Ao lado dele sei que consigo chegar muito mais longe... Muito
obrigada por você fazer minha vida muito feliz!
Agradeço e dedico este trabalho à minha família. “Seu Jeová” (O PAI), “Mãinha
Salete” (A MÃE), “Ana Cretina” e “Mary Lu” (AS IRMÃS), “Sufiss” (ETERNO NAMORADO),
“Garotinha Insuportável” (A SOBRINHA). Obrigada por tudo, pela educação, pelo apoio,
pela compreensão nas constantes ausências e principalmente pela paciência nas “crises
histéricas”! Obrigada por sempre, sempre, acreditarem em mim!
A todos agradeço, profundamente, e dedico o resultado deste trabalho.
Luciana Silva Carneiro
Rio de Janeiro, Outubro de 2008.

vi
Resumo
As generalizações a respeito do conceito de cascatas tróficas têm sido intensamente
debatidas entre os ecólogos. À onivoria tem sido atribuído o efeito de enfraquecer ou
intensificar a magnitude de cascata trófica; pesquisas recentes sugerem que interações
fracas advindas de interações onívoras promovem estabilidade de redes tróficas. Ao
contrário, o enriquecimento de nutrientes é previsto como um fator desestabilizador em
ecossistemas. O objetivo deste estudo foi testar estas proposições avaliando os resultados
de experimentos publicados a respeito de cascata trófica em comunidades aquáticas de
água doce e os resultados de um experimento conduzido em mesocosmos. Foram
analisados 330 experimentos publicados que manipularam independentemente a presença
de peixes planctívoros e onívoros com o objetivo de avaliar seus efeitos na distribuição de
biomassa a na estabilidade das comunidades planctônicas. Como previsto pela teoria de
cadeias tróficas, a presença de peixes tanto onívoros quanto planctívoros resultou em
significativa diminuição da biomassa do zooplâncton e aumento da biomassa do
fitoplâncton; entretanto o efeito de cascata trófica sobre o fitoplâncton foi significativamente
menor na presença de peixes onívoros. Assim, o efeito das diferentes guildas tróficas de
peixes analisadas não alterou a probabilidade de ocorrência de cascata trófica, no entanto,
fatores como o estado trófico do ambiente e a biomassa de peixes afetaram significativa e
diferentemente o efeito de peixes onívoros e zooplanctívoros sobre a força da cascata.
Além disso, peixes onívoros e zooplanctívoros não apresentaram efeitos significativos sobre
a estabilidade temporal do zooplâncton, mas, ao contrário do previsto pela teoria de cadeias
tróficas, apenas os peixes zooplanctívoros tiveram efeito estabilizador sobre a variabilidade
temporal do fitoplâncton. São apresentados também os resultados de um experimento de
cascata trófica em mesocosmos, que teve duração de 11 semanas, e testou a previsão de
que onívoros multi-cadeia (organismos que se alimentam em diferentes níveis tróficos que
estão imersos em diferentes compartimentos aquáticos) podem atenuar a cascata trófica e
estabilizar a variação temporal da biomassa total e da composição de espécies das
comunidades fito e zooplanctônicas. Foi encontrado suporte parcial aos resultados da meta-
análise. O peixe onívoro atenuou significativamente a cascata trófica e aumentou a
estabilidade temporal das medidas de biomassa e composição das comunidades
planctônicas. Além disso, foi observado que o enriquecimento por nutrientes teve um efeito
contrário, reduzindo significativamente o efeito estabilizador dos peixes onívoros. O
desacordo parcial entre a meta-análise e o experimento (em que um onívoro multi-cadeia foi
utilizado) ressalta a necessidade dos experimentos futuros incorporarem estruturas de
redes tróficas mais complexas dirigidas por predadores multi-cadeia, para promover um
embasamento conceitual mais robusto para análise de redes tróficas em ecossistemas
aquáticos de água doce.

vii
Abstract
The generality of the trophic cascade has been intensely debated among ecologists.
Omnivory has been implicated in both diffusing and intensifying the trophic cascade
strength, as well as, current view holds that weak interactions between species promote
stability of food webs. To the contrary, nutrient enrichment is predicted to destabilize
ecosystems. The aim of this work was to test these propositions by examining the results of
published trophic cascade experiments in freshwater communities and the result of our own
unpublished field mesocosm experiment. In particular, we analyzed the results of 330
published trophic cascade experiments which independently manipulated the presence of
omnivorous and planktivorous fishes and examined their effects on biomass distribution and
stability of planktonic communities. In accordance with the predictions of food web theory,
the presence of both omnivorous and zooplanktivorous fishes consistently reduced
zooplankton biomass and increased phytoplankton biovolume, but the strength of this
trophic cascade was weaker in the presence of omnivorous fishes. Fish trophic guilds did
not affect differently the probability of cascade occurrence, but they have distinct interactive
effects with trophic state and fish biomass on cascade strength. Furthermore, omnivorous
and zooplanktivorous fishes did not affect zooplankton biomass temporal variability, but in
contrast with the predictions of food web theory zooplanktivorous fish stabilized
phytoplankton biomass temporal variability. We therefore presented the results of 11 weeks
study of trophic cascade in experimental mesocosms, to test the predictions that multi-chain
omnivory (i.e., organisms that feed off different trophic levels embedded in multiple aquatic
compartments) can attenuate trophic cascade and stabilize temporal variation on aggregate
community biomass and species composition of phytoplankton and zooplâncton
communities. We found mixed support to the meta-analysis results. Omnivorous fish
attenuate trophic cascade, but significantly increase the temporal stability of aggregate and
compositional metrics of planktonic communities. Additionally, we found that nutrient
enrichment had an opposite effect dampening the effects of omnivorous fish on stability. The
partial disagreement between the meta-analysis and our experiment (i.e., which used a
multi-chain omnivore) claims the need of further research to incorporate more complex food
web structures driven by multi-chain predators to provide a complete conceptual framework
for analysis of reticulate food webs in freshwater ecosystems.

1
Índice
Resumo ..........................................................................................................................vi
Abstract......................................................................................................................... vii
Introdução Geral ........................................................................................................... 2
Capítulo 1: Diferenças entre os Efeitos de Peixes Onívoros e Zooplanctívoros na
Distribuição de Biomassa e Estabilidade de Teias Tróficas Planctônicas: Uma
Abordagem Meta-Analítica.............................................................................................. 7
Capítulo 2: Efeito da Predação de um Peixe onívoro na Dinâmica e Intensidade de
Cascata Trófica em Comunidades Planctônicas .......................................................... 31
Capítulo 3: Efeitos Diretos e Indiretos da Predação de um Peixe Onívoro na
Estabilidade Temporal da Biomassa e Composição de Espécies de Comunidades
Planctônicas.................................................................................................................. 78
Discussão e Conclusão Geral.................................................................................. 116

2
Introdução Geral
À medida que diversas informações foram reunidas pelos naturalistas durante o
último século, vários conceitos novos emergiram e direcionaram o estudo da Ecologia
moderna. Um deles foi a percepção de que as relações de alimentação ligam as
populações numa entidade funcional reconhecida como comunidade biológica. O primeiro
entre os proponentes desta nova visão foi o ecólogo inglês Charles Elton que desenvolveu
o conceito de cadeias alimentares, uma simples representação de conexões tróficas
verticais na comunidade, que permanece em foco até os dias atuais (Ricklefs 1990).
No último século, duas linhas gerais de investigação emergiram dos estudos de
cadeias alimentares aquáticas. A primeira delas surgiu do interesse na qualidade das
águas, onde os lagos eram considerados sistemas de fluxo unidirecional de energia,
passando dos nutrientes minerais para o fitoplâncton e deste para o zooplâncton até chegar
aos peixes (Straskraba 1965). Apenas a partir da década de 60 surgiram os primeiros
trabalhos sobre a perspectiva reversa, na qual os piscívoros (4o nível trófico) no topo das
cadeias alimentares aquáticas exercem um controle regulatório sobre os peixes
planctívoros (3o nível trófico), reduzindo a pressão de predação sobre o zooplâncton (2o
nível trófico) e conseqüentemente aumentando a pressão de herbivoria sobre o fitoplâncton
(1o nível trófico), modelo que também é conhecido como cascata trófica (Carpenter et al.
1985).
Hrbáček et al. (1961) demonstraram que o fitoplâncton não era apenas reflexo dos
nutrientes do sistema. Em seu trabalho, a presença de peixes em um lago reduziu
significativamente a biomassa de zooplâncton, tendo como resultado aumento da biomassa
de fitoplâncton. Seguiram-se a este, inúmeros experimentos, que independentemente da
escala do sistema (aquários, pequenos lagos, limnocurrais, mesocosmos ou manipulação
de todo o lago) ou da espécie de peixe, obtiveram após a introdução ou a remoção de
carnívoros primários (ex. peixes zooplanctívoros) um expressivo efeito qualitativo sobre
níveis tróficos inferiores (Brooks & Dodson 1965; Lynch & Shapiro 1981).

3
Recentemente, cascata trófica é um fenômeno que tem sido observado em vários
sistemas, incluindo lagos, rios, florestas, pradarias e sistemas marinhos (Power 1990; Brett
& Goldman 1996; Carpenter et al. 2001; Shurin et al. 2002; Borer et al. 2005) e o debate
mais atual tem sido sob quais condições ambientais a cascata trófica é mais freqüente
(Pace et al. 1999; Polis 1999; Polis et al. 2000; Borer et al. 2005).
Inicialmente, em razão dos exemplos de cascatas tróficas mais evidentes terem sido
observados em sistemas aquáticos, vários autores sugeriram que a ocorrência de cascata
trófica era um fenômeno prevalente de ambientes aquáticos (Strong 1992; Polis 1999; Halaj
& Wise 2001). Embora existam muitas evidências empíricas, este modelo de controle
descendente de cadeias tróficas aquáticas tem sido criticado por apresentar pressupostos
espaciais e temporais estáticos (Chase 2003), que ignoram características comuns e
dinamicamente importantes em cadeias tróficas reais, as quais podem potencialmente levar
a padrões diferentes daqueles previstos pela teoria (Polis et al. 1996b). Alguns autores
sugerem que a teoria de cadeias tróficas não acomoda processos dinâmicos cruciais, tais
como competição, ciclagem de nutrientes, heterogeneidade espacial, estrutura de tamanho
das comunidades, mecanismos de defesa contra predação, cadeias de detritivoria e
onivoria (Vanni et al. 1997; Vanni 2002).
Nas últimas décadas, a onivoria tem assumido um papel fundamental no estudo da
ecologia de populações e comunidades (Pimm & Lawton 1978; Polis et al. 1996b; Vanni et
al. 2005). Considerada uma característica comum em muitas teias tróficas, a onivoria reflete
a flexibilidade do predador de adquirir energia de várias fontes (Yodzis 1984). Uma
definição abrangente considera a onivoria como uma estratégia de forrageamento
generalista (Fig. 1A), mas a definição mais comumente aceita é que onívoros alimentam-se
em mais de um nível trófico dentro de uma mesma cadeia (Pimm & Lawton 1978) (Fig. 1B).
No entanto, a estrutura dos ecossistemas raramente pode ser reduzida a apenas uma
cadeia trófica (Polis et al. 1996b) e os predadores, presas e recursos geralmente
ultrapassam o limite de compartimentos, habitats e ecossistemas (Polis 1991), translocando
matéria e energia entre compartimentos (Vanni 2002). Esta visão de acoplamento de

4
habitas foi incorporada à definição de onivoria, considerando que os organismos que se
alimentam em diferentes níveis tróficos estão imersos em compartimentos com múltiplas
vias de transferência de energia. Atualmente, a definição de maior abrangência ecológica
refere-se a onívoros de cadeias múltiplas “multi-chain omnivores” que exploram diferentes
cadeias tróficas baseadas em produtores primários de grupos funcionais e compartimentos
diversos, incluindo detritos (Fig 1C e D) (Polis et al. 1996b). Segundo Vadeboucoeur et al.
(2005), a inclusão deste conceito em trabalhos atuais irá contribuir para o entendimento de
processos ecológicos em escalas temporais e espaciais mais verdadeiras.
Diferentes arquiteturas de cadeias tróficas que incluem interações de predação entre
indivíduos de uma mesma guilda (“intraguild predation”) demonstram reduzir a intensidade
de cascata trófica devido a interações mais fracas e difusas entre os níveis tróficos (Fagan
1997; Leibold et al. 1997; Agrawal 1998; Borer et al. 2005). Além disso, alguns estudos
propõem que predadores onívoros são uma fonte de instabilidade para o sistema predador-
presa, por apresentarem resposta denso-independente às flutuações da presa. Ou seja, o
forrageamento adaptativo, característico de consumidores onívoros capazes de variar a
preferência alimentar em resposta a disponibilidade de recursos, pode manter suas
Generalista/Onívoro Onivoria intra-cadeia
Ecossistema 1 Ecossistema 2
Onivoria inter-cadeia
Figura 1. Modelo esquemático a respeito do desenvolvimento do conceito de onivoria. A) Onívoro definido como um consumidor (C) capaz de forragear sobre múltiplos recursos (R); B) Onivoria intra-cadeia foi originalmente definida como um predador (P) capaz de se alimentar tanto de plantas quanto de animais, mas que agora incorpora definições mais modernas, um organismo que se alimenta em mais de um nível trófico dentro de uma mesma cadeia (Pimm & Lawton 1978). Polis & Strong (1996b) propuseram que a onivoria pode também ocorrer entre cadeias (C) de um mesmo ecossistema ou (D) entre cadeias de ecossistemas distintos. Modificado de Vadeboncoeur et al. (2005).

5
populações em altas densidades independentemente da diminuição populacional de uma
dada presa, podendo portanto levar a supressão da presa de menor densidade (Pimm &
Lawton 1978; Murdoch & Bence 1987; Holt & Polis 1997; Diehl & Feissel 2001; Diehl 2003;
Vanni et al. 2005). Por outro lado, vários autores indicam que a fraca ligação na relação
predador-presa em sistemas sujeitos a onivoria estabiliza drásticas oscilações entre os
consumidores e os recursos. Isto tende a manter a densidade das populações longe da
probabilidade estatística de se tornar extinta, processo que é considerado muito importante
em promover a persistência e a estabilidade em comunidades naturais (McCann & Hastings
1997; McCann et al. 1998a).
Diante da discrepância entre estes dois modelos teóricos se estabeleceu um novo
horizonte de pesquisas na ecologia de cadeias tróficas. A questão mais atual é sobre as
circunstâncias ecológicas em que a onivoria é mantida em comunidades naturais (Diehl
2003). Segundo Rosenheim & Corbett (2003), esta não é uma questão simples de ser
respondida. Vários fatores influenciam as taxas de predação do onívoro, incluindo a
probabilidade de encontro, a probabilidade de ataque, o sucesso de captura e a
probabilidade de consumo. Por sua vez, cada um destes fatores é influenciado pelas
habilidades do predador, da presa e de características do ambiente (Sih 1993).
Muitas pesquisas sobre a interação predador-presa têm demonstrado o importante
papel das defesas ativas da presa (Snyder & Ives 2001), das preferências do predador
(Colfer & Rosenheim 2001), da produtividade do sistema (Holt & Polis 1997; Diehl & Feissel
2001) (HilleRisLambers et al. 2006) e da estrutura de hábitat e de refúgios físicos (MacRae
& Croft 1996; Agrawal & Karban 1997; Finke & Denno 2002; Vanni et al. 2005). Segundo
Rosenzweig (1971) e Gilpin & Rosenzweig (1972), o aumento da produtividade primária em
conseqüência do enriquecimento trófico leva o sistema predador-presa a oscilações tão
drásticas, que aumenta muito a probabilidade das populações se extinguirem. Tais
oscilações na abundância das populações nos níveis tróficos também foram previstas pela
teoria de cascatas tróficas de Carpenter et al. (1985). No entanto, estudos teóricos indicam
que esta desestabilização decorrente do enriquecimento raramente ocorre em cadeias

6
tróficas sujeitas a significante onivoria (Diehl & Feissel 2001). Poucos testes empíricos
desta predição têm sido realizados e seus resultados têm sido contraditórios,
conseqüentemente o efeito de predadores onívoros na estabilidade de comunidades
permanece desconhecido. A instabilidade (variabilidade temporal) de populações e
ecossistemas pode elevar consideravelmente as taxas de extinção (Lande 1993) e reduzem
o valor econômico dos serviços prestados pelos ecossistemas (Armsworth & Roughgarden
2003). Além disso, do ponto de vista da gestão dos recursos hídricos, a suposta fraca
pressão de predação dos peixes onívoros sobre o zooplâncton, torna a implementação de
técnicas de biomanipulação desenvolvidas em cadeias tróficas lineares inoperante para
sistemas onde predominam interações tróficas complexas, tais como as que envolvem um
elevado número de espécies ou a onivoria (Lazzaro 1987; Drenner et al. 1996; Jeppessen
et al. 2005). Sob uma perspectiva de conservação e manejo, torna-se iminente a síntese de
conhecimento sobre como peixes onívoros podem afetar a distribuição de biomassa e
estabilidade de cadeias tróficas planctônicas em ecossistemas aquáticos continentais.
Desta maneira, este trabalho se propõe a testar através de abordagens meta-
analíticas e experimentais a generalidade da hipótese de que peixes onívoros são
responsáveis por enfraquecer a força de cascata trófica e aumentar a estabilidade das
comunidades fitoplanctônicas e zooplanctônicas de ambientes aquáticos continentais.
Além disso, buscou-se entender também se os efeitos da onivoria são dependentes
da densidade de peixes (capítulo I), do estado trófico do sistema (capítulos I e II) e do tipo
de peixe onívoro (capítulos I e II).

7
Capítulo I
Diferenças entre os Efeitos de Peixes Onívoros e Zooplanctívoros na
Distribuição de Biomassa e Estabilidade de Teias Tróficas Planctônicas: Uma
Abordagem Meta-Analítica

8
Introdução
Ecólogos têm há muito reconhecido a importância estrutural e ecológica do papel
que peixes desenvolvem em comunidades e ecossistemas aquáticos (Hrbacek et al. 1961;
Grygierek et al. 1966; Lazzaro 1987; Power 1990; Diehl 1992; Fernando 1994; Flecker
1997; Vanni et al. 1997; Scheffer et al. 2000; Attayde & Hansson 2001a; Vander Zanden &
Vadeboncoeur 2002; Vanni et al. 2005; McIntyre et al. 2007). Através de efeitos
descendentes oriundos da predação (Vanni & Layne 1997; Lazzaro et al. 2003) e
ascendentes mediados pela reciclagem de nutrientes via excreção e bioturbação
(Komarkova 1998; Attayde & Hansson 2001a; McIntyre et al. 2008), peixes podem
significativamente alterar a composição de espécies, a distribuição da biomassa e a
estabilidade de níveis tróficos inferiores (Carpenter et al. 1985; Van De Bund & Van Donk
2004; Vanni et al. 2005). Particularmente o estudo sobre os efeitos de peixes sobre a
dinâmica de cadeias tróficas foi fundamental para o desenvolvimento do conceito de
cascatas tróficas (Carpenter et al. 2001), com um considerável corpo de literatura já
existente a este respeito (sumarizado em Brett & Goldman 1996, 1997; Bell et al. 2003).
Embora atualmente a ocorrência de cascata trófica na natureza seja um fenômeno
já reconhecidamente comprovado (Pace et al. 1999), controvérsia ainda existe em relação à
sua onipresença a todos os tipos de ecossistemas e comunidades (Strong 1992; Polis et al.
2000; Halaj & Wise 2001). Vários estudos têm recentemente tentado sintetizar quais
mecanismos estão relacionados à ocorrência e à força de cascata tróficas (Shurin et al.
2002; Borer et al. 2005; Carpenter et al. 2008). Os resultados têm indicado que uma ampla
gama de fatores ambientais como a produtividade do ambiente (Carpenter et al. 2001;
Chase 2003; Hall et al. 2007; Gruner et al. 2008), diferenças de tamanho entre presa e
predador (Borer et al. 2005; Shurin & Seabloom 2005), heterogeneidade espacial
(Grabowski 2004; Finke & Denno 2006) e diversidade de presas ou predadores (Finke &
Denno 2004; Byrnes et al. 2006; Otto et al. 2008), são aspectos que interferem diretamente
na força e ocorrência de cascata trófica. Um outro tópico que tem recebido considerável
atenção a este respeito é a onivoria (capacidade de se alimentar em mais de um nível

9
trófico). Pelo fato de poder afetar simultaneamente níveis tróficos inferiores, o consumidor
onívoro pode enfraquecer os efeitos descendentes indiretos ao longo da cadeia trófica e
portanto modificar o padrão de força e ocorrência de cascata trófica (Hart 2002; Lancaster
et al. 2005; Vadeboncoeur et al. 2005). Muitos estudos têm destacado a importância de
peixes onívoros em enfraquecer a ocorrência de cascatas tróficas em ambientes pelágicos
(Drenner et al. 1986; Drenner et al. 1987; Lazzaro et al. 1992; Lazzaro et al. 2003; Okun et
al. 2008), mas até o momento nenhum estudo tentou sinteticamente avaliar se diferenças
entre o hábito trófico de peixes onívoros e zooplanctívoros realmente desencadeiam efeitos
distintos sobre a ocorrência e força de cascatas tróficas pelágicas. Além disso, embora
muito abordado em estudos teóricos, pouco se sabe experimentalmente se peixes onívoros
afetam também a estabilidade de cadeias tróficas (Schindler & Scheuerell 2002).
Devido à grande quantidade de estudos experimentais que avaliam os efeitos da
predação por peixes em cadeias tróficas planctônicas (Brett & Goldman 1996, 1997; Bell et
al. 2003) e a necessidade de testar objetivamente a consistência das diferentes hipóteses a
respeito do papel de peixes onívoros sobre a dinâmica de cadeias tróficas aquáticas, foi
utilizada neste trabalho uma abordagem meta-analítica com o objetivo de comparar os
efeitos de peixes zooplanctívoros estritos e onívoros sobre a distribuição e estabilidade
temporal da biomassa de comunidades planctônicas.
Meta-análise é um método estatístico que permite sumarizar, através de revisões
quantitativas, padrões gerais oriundos da análise conjunta de múltiplos experimentos
independentes (Osenberg et al. 1997; Gurevitch & Hedges 1999; Hedges et al. 1999). Ao
contrário dos tradicionais métodos de revisões qualitativas e narrativas, a abordagem meta-
analítica gera conclusões mais confiáveis pois ao invés de se basear em simples contagem
de resultados significativos “vote quoting”, se fundamenta em uma padronizada e rígida
análise de magnitude de efeitos (i.e., diferenças de magnitude entre um tratamento e seu
respectivo controle experimental) calculados de múltiplos experimentos independentes
(Arnqvist & Wooster 1995; Gurevitch & Hedges 1999; Hedges et al. 1999). Desta forma,
revisões meta-analíticas permitem testar de forma mecanicista e generalizada se padrões

10
ecológicos diferem entre si ou se variam consistentemente em relação a outras variáveis
ambientais, uma vez que seus testes levam em consideração a magnitude e direção do
efeito e não apenas a sua ocorrência (Gurevitch & Hedges 1999; Nakagawa & Cuthill 2007).
O objetivo deste trabalho foi testar através de uma abrangente revisão quantitativa
meta-analítica se e como peixes onívoros afetam a biomassa e estabilidade de
comunidades planctônicas diferentemente de peixes zooplanctívoros. Baseado no corpo
teórico e sobre o histórico de informações empíricas a respeito dos efeitos do
comportamento onívoro sobre a dinâmica de cadeias tróficas, nós acreditamos que (1)
peixes onívoros enfraquecerão significativamente a força e ocorrência de cascata trófica
sobre a biomassa fitoplanctônica, (2) peixes onívoros terão um efeito estabilizador sobre a
variabilidade temporal da biomassa fito e zooplanctônica e (3) os efeitos de peixes onívoros
e zooplanctívoros sobre a dinâmica das comunidades planctônicas serão denso-
dependentes das suas biomassas e afetados pelo estado trófico do sistema.

11
Material e Métodos Levantamento bibliográfico, escolha e coleta de dados
O processo de coleta de dados foi iniciado através de um levantamento bibliográfico
de estudos relacionados aos efeitos de peixes onívoros e zooplanctívoros em cadeias
tróficas planctônicas. Foram compilados resultados de 330 experimentos (150
correspondentes a peixes onívoros e 180 a peixes zooplanctívoros) descritos em 109
artigos (Tabela 1). O levantamento foi composto praticamente em sua totalidade de
trabalhos publicados, porém com a inclusão de resultados de um estudo ainda não
publicado (Carneiro, em preparação). Os estudos selecionados foram aqueles que
retratavam experimentos onde peixes onívoros e zooplanctívoros foram adicionados ou
removidos independentemente, e seus efeitos sobre a dinâmica da biomassa do
zooplâncton e fitoplâncton foram quantificados. O levantamento de estudos publicados foi
realizado através de busca na base referencial do Institute of Scientific Information -
Thomson Science Citation Index - (www.isiwebofknowledge.com), utilizando como palavras-
chave: trophic cascade, biomanipulation, pond, enclosure, mesocosm, planktivorous fish,
omnivorous fish e combinações entre elas. O resultado do levantamento realizado na base
referencial do ISI Thomson foi complementado por estudos referenciados por Drenner et al.
(1996) e Bell et al. (2003).
Os critérios para separação das espécies de peixes nas respectivas guildas tróficas
incluiu conhecimentos relacionados à preferência alimentar das espécies; alterações
ontogenéticas dos hábitos alimentares (Schaus et al. 2002); bem como as condições
experimentais capazes de restringir a amplitude trófica das espécies onívoras, tais como
impedimento do acesso ao sedimento de espécies com hábitos bento-pelágicos (Drenner et
al. 1996; Schaus & Vanni 2000), contenção do desenvolvimento de algas perifíticas nas
paredes dos mesocosmos e etc. Desta forma, o critério não foi espécie-específico e uma
mesma espécie de peixe pôde ser classificada como zooplanctívoro ou onívoro
dependendo do seu estágio de vida (estágios larvais foram considerados sempre como
zooplanctívoros) e se a condição experimental permitia a expressão funcional do hábito

12
onívoro (Drenner et al. 1996). Resultados de manipulações experimentais envolvendo um
lago todo, onde a abundância de peixes (onívoros ou zooplanctívoros) foi reduzida através
da adição de piscívoros, só foram incluídos quando a introdução do piscívoro resultou em
uma larga diminuição do estoque destas presas. Além disso, resultados de experimentos
envolvendo todo o lago também não foram considerados quando o lago apresentava
massiva colonização de plantas aquáticas e quando a guilda trófica da espécie ou
comunidade de peixes alvo da redução não se constituía unicamente de onívoros ou
zooplanctívoros.
Os dados foram retirados de gráficos utilizando o software DigitizeIt versão 1.5.8
(http://www.digitizeit.de). A Clorofila-a fitoplanctônica foi usada como medida preferencial da
biomassa do fitoplâncton, entretanto biovolume e fluorescência foram usados quando dados
de clorofila-a não eram fornecidos. A medida de peso seco do zooplâncton foi usada como
estimativa da biomassa zooplanctônica. Quando apenas dados de abundância do
zooplâncton estavam disponíveis, estes foram convertidos para unidade de peso seco
através de relações alométricas de peso-comprimento (Bottrell et al. 1976), mas
freqüentemente foram utilizados os pesos médios de espécies zooplanctônicas publicados
na literatura (Hall et al. 1970; Wetzel & Likens 1991) quando dados do tamanho do
zooplâncton não estavam disponíveis (Brett & Goldman 1996; Bell et al. 2003). Em alguns
casos, quando os dados referentes a toda a comunidade zooplanctônica não estavam
disponíveis, a biomassa de cladóceros e copépodos representou a biomassa total do
zooplâncton (Brett & Goldman 1996; Bell et al. 2003).
Quando os resultados temporais da dinâmica das comunidades fitoplanctônicas e
zooplanctônicas foram disponibilizados, a média temporal e o coeficiente de variação foram
calculados e utilizados nas análises como estimativas de efeito global sobre a quantidade e
variabilidade temporal da biomassa destas comunidades. Para estudos onde a manipulação
dos peixes foi repetida em várias densidades ou conjuntamente com adição de nutrientes,
os tratamentos com a presença do peixe foram contrastados com os respectivos controles
para cada situação, e a biomassa de peixe utilizada, bem como a concentração de fósforo

13
total foram registradas. As informações da biomassa de peixes e da concentração de
fósforo total (i.e. estimativa de estado trófico) foram incluídas na análise pois representam
fatores biológicos e físico-químicos capazes de afetar a intensidade dos efeitos diretos e
indiretos da predação por peixes sobre a dinâmica das comunidades zooplanctônicas e
fitoplanctônicas (Vanni 1987; Drenner et al. 1996; Drenner et al. 1998; Vanni et al. 2005).
Nos trabalhos onde apenas o tamanho e o número de indivíduos de peixes foram
disponibilizados, a biomassa de peixes do tratamento foi estimada utilizando relações
alométricas espécie-específicas de peso-comprimento obtidas através do site
(http://www.fishbase.org). Experimentos onde outros fatores senão a biomassa de peixes e
o estado trófico co-variaram com a presença de peixes, tais como a presença de plantas
aquáticas, não foram considerados neste estudo.
Cálculo da magnitude dos efeitos
A magnitude dos efeitos (“effect size”) da presença de peixes onívoros e
zooplanctívoros na biomassa e estabilidade temporal das comunidades zooplanctônicas e
fitoplanctônicas foi estimada através do logaritmo da razão (“log ratio”) (Osenberg et al.
1997; Gurevitch & Hedges 1999; Hedges et al. 1999). Os efeitos de cada guilda trófica de
peixe sobre a biomassa do zooplâncton e fitoplâncton foram calculados como
{log10(Bp+/Bp-)}, sendo Bp+ a biomassa de zooplâncton ou fitoplâncton na presença de
peixes e Bp- na ausência (controle). Valores positivos indicam que a biomassa da
comunidade em questão aumentou na presença do peixe em relação ao tratamento
controle, valores negativos indicam o oposto. Para o cálculo da magnitude do efeito da
presença dos diferentes tipos de peixes sobre a estabilidade temporal da biomassa
fitoplanctônica e zooplanctônica, foi utilizado como estimativa o coeficiente de variação
(CV). Valores de CV foram incluídos na fórmula {log10(CVp+/CVp-)}, onde CVp+ é a
variabilidade temporal da biomassa do fitoplâncton ou do zooplâncton na presença de
peixes e CVp- na ausência. Valores do logaritmo da razão positivos indicam que a presença
de peixes aumenta a variabilidade temporal da biomassa e valores negativos indicam um
efeito estabilizador dos peixes sobre a variabilidade temporal da biomassa planctônica. O

14
CV foi utilizado no presente estudo devido à sua utilização freqüente em estudos que
abordam estabilidade de ecossistemas (Tilman 1996; Cottingham et al. 2001) e por fornecer
uma medida adimensional passível de comparações entre dados coletados de múltiplos
estudos (Ives et al. 2000).
A utilização do logaritmo da razão como uma estimativa comparável da magnitude
do efeito das diferentes guildas tróficas sobre as comunidades planctônicas deveu-se ao
fato de que a mesma confere significados biológicos claros através de mudanças
proporcionais na variável resposta, além de satisfazer propriedades estatísticas
paramétricas como distribuição normal dos valores (Hedges et al. 1999). Entretanto, a
variância ao redor do logaritmo da razão é afetada pela variância temporal da variável
resposta e o seu número de replicação amostral, o que representa uma potencial
desvantagem para contrastar médias temporais de experimentos realizados em diferentes
escalas de tempo e/ou que contem com desigual número de amostragens (Osenberg et al.
1997). Uma alternativa para corrigir tal problema é calcular a magnitude do efeito utilizando
índices que produzem magnitude de efeitos padronizada pela variância conjunta das
médias envolvidas no cálculo (Gurevitch & Hedges 1999). Entretanto, tais índices exigem
replicação nos tratamentos dos experimentos. Considerando que vários experimentos
utilizados neste estudo não contaram com réplicas e muitos outros apresentaram os
resultados apenas em valores médios sem fornecer medidas de variância, a não utilização
do logaritmo da razão reduziria consideravelmente o número de experimentos utilizados
neste estudo, repercutindo em perda do poder de generalização e possivelmente uma
amostragem tendenciosa (Englund et al. 1999). Embora o uso do logaritmo da razão possa
por um lado diminuir o poder de detectar diferenças estatísticas (aumento do erro tipo II),
ele garante uma estimativa não tendenciosa da magnitude média dos efeitos (Gurevitch &
Hedges 1999; Hedges et al. 1999). Além disso, um recente estudo de revisão meta-
analítico, que utilizou resultados de mais de 90 estudos empíricos, demonstrou que a
duração experimental não afeta a força de cascata trófica originada pela predação por
peixes em cadeias tróficas planctônicas (Bell et al. 2003), o que refuta a existência de

15
potenciais inadequações relacionadas ao uso do logaritmo da razão no cálculo da
magnitude dos efeitos no presente trabalho.
Análises estatísticas Diferenças globais (independentes do estado trófico ou da biomassa de peixes)
entre o efeito de peixes onívoros e zooplanctívoros sobre a distribuição e estabilidade da
biomassa das comunidades fitoplanctônicas e zooplanctônicas foram testadas contrastando
os valores médios e seus respectivos ±95% intervalos de confiança. Este método de
comparação é amplamente indicado para testes de significância em estudos de revisão
meta-analíticos, devido a sua insensibilidade à diferenças do número de réplicas entre os
tratamentos e facilidade de interpretação (Nakagawa & Cuthill 2007). Valores sobrepostos
de ±95% intervalos de confiança indicam que os tratamentos não diferem significativamente
entre si e o caso específico de estudos que testam hipóteses através de medidas de
magnitude do efeito, a sobreposição do ±95% intervalo de confiança com o eixo x, denota
que a magnitude do efeito do tratamento sobre a variável em questão não difere
significativamente de zero (Gurevitch & Hedges 1999; Nakagawa & Cuthill 2007).
Para testar se a biomassa de peixes e o estado trófico afetaram diferencialmente os
efeitos da presença de peixes onívoros e zooplanctívoros sobre as comunidades
planctônicas, foi utilizada uma análise de covariância (ANCOVA), onde as magnitudes dos
efeitos foram consideradas como a variável dependente (ANCOVAS separadas para cada
magnitude de efeito), o estado trófico e a biomassa de peixes como variáveis contínuas ou
co-variáveis e as guildas tróficas como variável categórica. Foi utilizado um nível de
significância de α = 0.05 para todos os testes.

16
Tabela1: Lista de referências dos estudos experimentais utilizados nesta meta-análise.
Referência Comunidade analisada
Duração experimental
Unidade experimental
Guilda trófica
1 Albright, et al., (2004) Fito 2 anos Lago inteiro Zoo 2 Acunã et al., (2008) Fito/Zoo 3 semanas Enclosure Zoo 3 Arcifa, et al., (1986) Fito/Zoo 2 meses Enclosure Zoo 4 Attayde & Hansson (2001a) Fito/Zoo 5 semanas Enclosure Zoo 5 Attayde & Hansson (2001b) Fito/Zoo 5 semanas Enclosure Zoo 6 Baca & Drenner (1995) Fito/Zoo semanas/anos Mesocosmo/Lago inteiro Oni 7 Beklioglu & Moss (1995) Fito/Zoo 4 semanas Enclosure Zoo 8 Beklioglu & Moss (1996) Fito/Zoo 4 semanas Enclosure Oni 9 Beklioglu & Moss (1998) Fito/Zoo 5 semanas Enclosure Oni/ Zoo
10 Bell et al., (2003) Fito/Zoo 4 anos Poça Zoo 11 Bertolo et al., (1999) Fito/Zoo 3 meses Enclosure Zoo 12 Bertolo et al., (1999) Fito/Zoo 4 semanas Enclosure Zoo 13 Bertolo et al., (2000) Fito/Zoo 2 meses Enclosure Zoo 14 Burke &Bayne (1986) Fito/Zoo 7 meses Poça Oni 15 Byers & Vinyard (1990) Fito/Zoo 2 semanas Mesocosmo Oni 16 Carneiro et al., (Em Preparação) Fito/Zoo 11 semanas Enclosure Oni 17 Carpenter et al., (2001) Fito/Zoo 7 anos Lago inteiro Zoo 18 Christoffersen et al., (1993) Fito/Zoo 5 meses Enclosure Zoo 19 Crisman & Beaver (1990) Fito/Zoo 4 semanas Enclosure Oni 20 Dettmers & Stein (1996) Fito/Zoo 3 meses Enclosure/Lago inteiro Oni 21 Devries & Stein (1992) Fito/Zoo 2 semanas Enclosure/Lago inteiro Zoo 22 Diana et al., (1991) Fito/Zoo 5 meses Lago inteiro Oni 23 Drenner et al., (1990) Fito/Zoo 3 semanas Mesocosmo Zoo 24 Drenner (1982) Zoo 3 meses Poça Oni 25 Drenner et al., (1986) Fito/Zoo 2 meses Mesocosmo Oni 26 Drenner et al., (1987) Fito 1 semana Mesocosmo Oni 27 Drenner et al., (1989) Fito/Zoo 4 semanas Mesocosmo Zoo 28 Drenner et al., (1996) Fito/Zoo 1 semana Mesocosmo Oni 29 Drenner et al.,(1998) Fito 1 mês/18 meses Mesocosmo/Lago inteiro Oni 30 Elhigzi et al., (1995) Fito/Zoo 2 meses Poça Oni 31 Elser & Carpenter (1988) Fito/Zoo 2 anos Lago inteiro Zoo 32 Faafeng et al., (1990) Fito/Zoo 4 semanas Enclosure Zoo 33 Figueredo &Giani (2005) Fito 4 semanas Enclosure Oni 34 Findlay et al., (1994) Fito/Zoo 5 anos Lago inteiro Oni 35 Giussani et al., (1990) Fito/Zoo 3 anos Lago inteiro Oni 36 Goad (1984) Fito/Zoo 2 semanas Enclosure Zoo 37 Hall et al., (1970) Fito/Zoo 5 meses Poça Zoo 38 Hambright (1994) Fito/Zoo 3 anos Lago inteiro Zoo 39 Hambright et al., (1986) Fito/Zoo 4 meses Poça Zoo 40 Hanazato et al., (1990) Zoo 4 semanas Enclosure Zoo 41 Hansson & Carpenter (1993) Fito/Zoo 3 meses Enclosure Zoo/Oni 42 Hansson et al., (1998) Fito/Zoo 4 meses Mesocosmo Oni 43 Horppila & Kairesalo (1992) Fito/Zoo 4 semanas Enclosure Zoo/Oni 44 Horppila &Kairesalo (1990) Fito 6 semanas Enclosure Oni 45 Horppila et al., (1998) Fito 5 anos Lago inteiro Oni 46 Hurlbert &Mulla (1981) Fito/Zoo 10 meses Poça Oni 47 Hurlbert et al., (1972) Fito/Zoo 3 meses Mesocosmo Oni 48 Khan et al., (2003) Fito/Zoo 4 semanas Poça Oni 49 Lancaster & Drenner (1990) Fito/Zoo 4 semanas Mesocosmo Oni 50 Laws & Weisburd (1990) Fito 10 meses Poça Oni 51 Lazzaro et al., (1992) Fito/Zoo 4 semanas Mesocosmo Zoo/Oni 52 Leibold (1989) Fito/Zoo 4 semanas Enclosure Zoo 53 Lu et al., (2002) Fito/Zoo 8 semanas Enclosure Oni 54 Lynch (1979) Fito/Zoo 6 semanas Enclosure Zoo 55 Lynch & Shapiro (1981) Fito/Zoo 8 semanas Enclosure Zoo 56 Markosova & Jezek (1993) Fito/Zoo 4 semanas Enclosure Zoo 57 Mattson (1998) Fito 5 meses Poça Oni 58 Matveev et al., (2000) Fito/Zoo 5 semanas Mesocosmo Oni/ Zoo 59 Mazumder et al., (1990) Fito/Zoo 4 meses Enclosure Zoo 60 Mcqueen et al., (1989) Fito/Zoo 5 anos Lago inteiro Zoo 61 Mcqueen et al., (1992) Fito/Zoo 4 meses Enclosure Zoo 62 Meijer et al., (1990) Fito/Zoo 4 meses Poça Oni 63 Meijer et al., (1990) Fito/Zoo 1 ano Lago inteiro Oni

17
Tabela1: Lista de referências dos estudos experimentais utilizados nesta meta-análise.
Referência Comunidade analisada
Duração experimental
Unidade experimental
Guilda trófica
64 Nowlin & Drenner (2000) Fito/Zoo 4 semanas Mesocosm Zoo/Oni 65 Okun & Mehner (2005) Zoo 5 semanas Enclosure Oni/Zoo 66 Okun et al., (2007) Fito/Zoo 5 semanas Enclosure Oni 67 Ortega-Mayagoita et al., (2002) Fito/Zoo 3 semanas Microcosmo Zoo 68 Otsuki et al., (2000) Fito/Zoo 6 semanas Mesocosmo Oni 69 Parker et al., (2001) Fito/Zoo 2 anos Lago inteiro Zoo 70 Perez-Fuentetaja et al., (1996a) Zoo 4 semanas Enclosure Zoo 71 Perez-Fuentetaja et al., (1996b) Fito/Zoo 4 semanas Enclosure Oni 72 Persson (1997) Fito/Zoo 4 semanas Mesocosmo Zoo 73 Persson et al., (1993) Fito/Zoo 3 anos Lago inteiro Oni 74 Post & Mcqueen (1987) Fito/Zoo 4 meses Enclosure Zoo 75 Proulx et al., (1996) Fito 8 semanas Enclosure Zoo 76 Qin & Culver (1995) Fito/Zoo 4 semanas Enclosure Zoo 77 Qin & Culver (1996) Fito/Zoo 7 semanas Poça Zoo 78 Qin & Threlkeld (1990) Fito/Zoo 8 semanas Mesocosmo Zoo/Oni 79 Radke et al., (2002) Fito/Zoo 4 semanas Enclosure Oni 80 Ramcharan et al., (1996) Fito/Zoo 9 semanas Enclosure Zoo 81 Ranta et al., (1987) Fito/Zoo 6 semanas Poça Zoo 82 Reinertsen et al., (1990) Fito/Zoo 3 anos Lago inteiro Zoo 83 Rejas et al., (2005) Fito/Zoo 2 semanas Enclosure Zoo 84 Richardson (1990) Fito/Zoo 4 semanas Mesocosmo Zoo/Oni 85 Riemann et al., (1990) Fito/Zoo 2 anos Lago inteiro Oni 86 Rondel et al., (2007) Fito/Zoo 4 semanas Enclosure Oni/Zoo 87 Schaus & Vanni (2000) Fito/Zoo 2 semanas Enclosure Oni/Zoo 88 Schindler (1992) Fito/Zoo 4 semanas Enclosure Zoo 89 Sondergaard et al., (1990) Fito/Zoo 3 anos Lago inteiro Oni 90 Spencer & King (1984) Fito/Zoo 5 meses Poça Zoo 91 Starling (1993) Fito/Zoo 4 semanas Enclosure Oni 92 Starling &Rocha (1990) Fito/Zoo 5 semanas Enclosure Zoo 93 Stephen et al., (1998) Fito/Zoo 5 semanas Enclosure Zoo 94 Tang et al., (2002) Fito/Zoo 4 semanas Enclosure Oni 95 Tatrai et al, (1990) Zoo 3 meses Enclosure Oni 96 Tatrai et al., (1985) Fito/Zoo 3 semanas Enclosure Oni 97 Threlkeld (1988) Fito/Zoo 6 semanas Mesocosmo Zoo 98 Turner & Mittelbach (1990) Fito/Zoo 10 semanas Poça Zoo 99 Turner & Mittelbach (1992) Fito/Zoo 5 semanas Enclosure Zoo
100 Vakkilainen et al., ( 2004) Zoo 6 semanas Enclosure Zoo/Oni 101 Van De Bund et al., (2004) Fito 6 semanas Enclosure Zoo/Oni 102 Vanni (1987) Fito/Zoo 6 semanas Enclosure Zoo 103 Vanni & Findlay (1990b) Fito/Zoo 3 meses Enclosure Zoo 104 Vanni et al., (1990) Fito/Zoo 2 anos Lago inteiro Zoo 105 Vanni et al., (1997) Fito/Zoo 6 semanas Enclosure Zoo 106 Vinebrooke et al., (2001) Zoo 6 semanas Enclosure Oni 107 Vinyard et al., (1988) Fito/Zoo 3 semanas Mesocosmo Oni 108 Watson et al., (2003) Fito/Zoo 2 semanas/3 anos Mesocosmo/Lago inteiro Oni 109 Williams & Moss (2003) Fito/Zoo 5 semanas Enclosure Oni
Oni = Peixes onívoros; Zoo = Peixes zooplanctívoros estritos; Oni/Zoo = Experimento que manipulou independentemente peixes onívoros e zooplanctívoros estritos.

18
Resultados
Os peixes zooplanctívoros e onívoros induziram cascata trófica no fitoplâncton (Fig.
2A), ou seja, levaram a um aumento da biomassa total nos tratamentos. Os valores médios
do logaritmo da razão entre os tratamentos e os controles foram sempre significativamente
maiores que zero, no entanto, a intensidade de cascata trófica desencadeada por peixes
onívoros foi significativamente menor. Além disso, o efeito positivo dos peixes onívoros na
biomassa do fitoplâncton foi significativamente maior à medida que a concentração de
fósforo total aumentou (Fig. 2B). Este resultado indica que peixes onívoros são menos
efetivos em reduzir cascata trófica em condições de alta trofia. Por outro lado, peixes
zooplanctívoros tiveram efeito de induzir cascata independentemente do estado trófico do
sistema (Fig. 2B). Considerando os efeitos interativos entre a biomassa de peixes e as
estratégias de forrageamento (Fig. 2C), apenas os peixes zooplanctívoros levaram a um
aumento da biomassa do fitoplâncton em função do seu aumento de biomassa. A biomassa
de peixes onívoros não apresentou nenhum efeito interativo na resposta do fitoplâncton.
Peixes onívoros e zooplanctívoros reduziram significativamente a biomassa do
zooplâncton (Fig. 2D). Este efeito foi ligeiramente mais negativo na presença de peixes
onívoros, no entanto a diferença entre ambos não foi significativa. Os peixes
zooplanctívoros exerceram uma pressão de predação menor no zooplâncton à medida que
a concentração de fósforo total aumentou, mas este resultado não ocorreu para peixes
onívoros (Fig. 2E). O aumento de biomassa dos peixes onívoros teve um efeito significativo
na redução da biomassa do zooplâncton, e o mesmo resultado não foi observado para
peixes zooplanctívoros (Fig. 2F).
Peixes zooplanctívoros estabilizaram a dinâmica temporal da biomassa do
fitoplâncton, e os peixes onívoros apresentaram a mesma tendência, no entanto, seu efeito
não foi significativo (Fig. 3A). O efeito de peixes zooplanctívoros e onívoros apresentou uma
tendência de desestabilização da biomassa fitoplanctônica ao longo de um gradiente de
concentração de fósforo total; no entanto esta tendência não foi significativa para ambos

19
(Fig. 3B). A variação da biomassa de peixes zooplanctívoros e onívoros não influenciou
significativamente a estabilidade temporal da biomassa fitoplanctônica (Fig. 3C).
O efeito de peixes zooplanctívoros e onívoros teve tendência de desestabilizar a
dinâmica temporal da biomassa do zooplâncton, mas esta tendência não foi significativa
para zooplanctívoros e foi marginalmente não significativa para onívoros (Fig. 3D). A
estratégia de forrageamento dos peixes não alterou a estabilidade temporal do zooplâncton
ao longo de um gradiente de concentração de fósforo (Fig. 3E). Considerando a variação da
biomassa dos peixes, o aumento da biomassa de onívoros teve um efeito significativo na
estabilização temporal da biomassa do zooplâncton, já a variação da biomassa de
zooplanctívoros em nada afetou a dinâmica do zooplâncton (Fig. 3F).
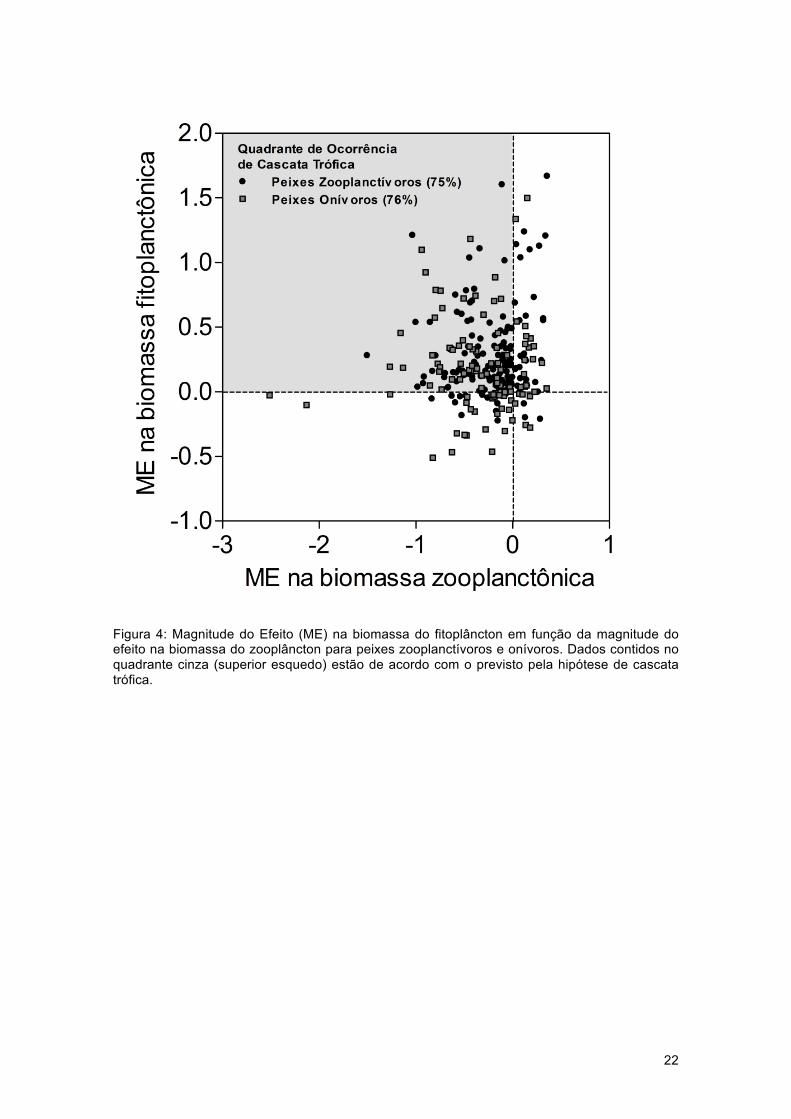
Nós também avaliamos qual porcentagem do aumento na biomassa do fitoplâncton
(magnitude de efeitos >1) estaria associada a uma redução da biomassa do zooplâncton
(magnitude de efeitos <1), considerando as diferentes estratégias de forrageamento dos
peixes. O resultado demonstra que a porcentagem de ocorrência de cascata trófica foi
bastante similar entre estudos com peixes onívoros (76%) e peixes zooplanctívoros (75%)
(Fig. 4), o que indica que a ocorrência de cascata não é afetada pela estratégia de
forrageamento dos peixes.

20
Figura 2: Magnitude do efeito (ME) de peixes zooplanctívoros e onívoros sobre a biomassa fito e zooplanctônica analisadas (A e D) de forma global, (B e F) em relação ao gradiente de trofia e (C e F) em relação à biomassa de peixes. Valores negativos indicam que a biomassa das comunidades foi menor na presença do que na ausência de peixes. Figuras A e D apresentam valores médios da magnitude do efeito calculado pelo log da razão do coeficiente de variação da biomassa do fitoplâncton e zooplâncton na presença e na ausência de peixes zooplanctívoros ou onívoros. Barras de erro são intervalos de confiança de ±95%. Efeitos da presença de peixes são estatisticamente significativos sobre a biomassa das comunidades quando as barras de erro não sobrepõem o eixo x. Tratamentos são diferentes entre si quando não houver sobreposição entre suas barras de erros. Figuras B, C, E e F cada ponto representa a magnitude do efeito de experimentos individuais.

21
Figura 3: Magnitude do efeito (ME) de peixes zooplanctívoros e onívoros sobre a variabilidade temporal da biomassa fito e zooplanctônica analisadas (A e D) de forma global, (B e F) em relação ao gradiente de trofia e (C e F) em relação à biomassa de peixes. Valores negativos indicam que a variabilidade temporal das comunidades foi menor na presença do que na ausência de peixes (i.e. efeito estabilizador). Figuras A e D apresentam valores médios da magnitude do efeito calculado pelo log da razão do coeficiente de variação da biomassa do fitoplâncton e zooplâncton na presença e na ausência de peixes zooplanctívoros ou onívoros. Barras de erro são intervalos de confiança de ±95%. Efeitos da presença de peixes são estatisticamente significativos sobre a biomassa das comunidades quando as barras de erro não sobrepõem o eixo x. Tratamentos são diferentes entre si quando não houver sobreposição entre suas barras de erros. Figuras B, C, E e F cada ponto representa a magnitude do efeito de experimentos individuais.
22
Figura 4: Magnitude do Efeito (ME) na biomassa do fitoplâncton em função da magnitude do efeito na biomassa do zooplâncton para peixes zooplanctívoros e onívoros. Dados contidos no quadrante cinza (superior esquedo) estão de acordo com o previsto pela hipótese de cascata trófica.

23
Discussão
Neste trabalho nós sintetizamos os resultados de quase três décadas de
experimentos que manipularam diferentes estratégias de forrageamento de peixes na
distribuição de biomassa e estabilidade de comunidades zooplanctônicas e
fitoplanctônicas. Mais especificamente, nosso objetivo foi responder qual o papel de
interações onívoras em teias alimentares aquáticas, uma questão que até hoje foi
debatida apenas à luz de pesquisas teóricas (Pimm & Lawton 1978; Polis et al. 1989;
Diehl 1993; Fagan 1997; McCann & Hastings 1997; McCann et al. 1998a). Embora
existam outros trabalhos meta-analíticos que avaliaram o efeito de cascata trófica em
diferentes ecossistemas (Brett & Goldman 1996; Shurin et al. 2002; Bell et al. 2003;
Borer et al. 2005), este é o primeiro que mede o efeito da predação de peixes na
estabilidade de cadeias aquáticas planctônicas.
Os resultados da análise dos 330 experimentos demonstraram que a
ocorrência de cascata trófica não variou de acordo com a estratégia alimentar dos
peixes (Fig. 4), no entanto a intensidade foi significativamente enfraquecida pela
onivoria (Fig. 2A). A zooplanctivoría e a onivoria reduziram a biomassa do zooplâncton
de modo similar, mas a resposta da biomassa fitoplanctônica foi diferente. A resposta
mais intensa do fitoplâncton à zooplanctivoría deveu-se provavelmente à estratégia
destes peixes que na sua maioria são predadores visuais de águas abertas, cuja
eficiência na remoção do macrozooplâncton, traduz-se numa forte resposta do
fitoplâncton. Ao reduzirem o macrozooplâncton, os peixes zooplanctívoros liberam o
microzooplâncton da competição, permitindo que seu crescimento compense parte da
perda de biomassa total do zooplâncton (Vanni 1986a). Além disso, a dominância de
espécies menores como rotíferos tem efeito muito pouco eficiente no controle do
fitoplâncton, ao mesmo tempo em que suas mais elevadas taxas metabólicas reciclam
e liberam nutrientes em maior quantidade e mais rapidamente, subsídio que pode
garantir um crescimento ainda maior para o fitoplâncton (Vanni 1986a, 2002).

24
A maioria dos estudos com peixes onívoros incluiu espécies filtradoras
generalistas representadas principalmente por ciprinídeos (e.g. Cyprinus carpio,
Hypophthalmichthys molitrix, Rutilus rutilus, Abramis brama, Oreochromis niloticus) e
clupeídeos (Dorosoma cepedianum) que se alimentam de macrozooplâncton,
microzooplâncton, fitoplâncton e detritos (Fernando 1994). Ao reduzirem a biomassa
não apenas do macrozooplâncton (i.e. cladóceros e copépodos), mas também do
microzooplâncton (i.e. rotíferos e náuplios), os resultados mostraram um efeito global
ligeiramente mais negativo dos peixes onívoros sobre a biomassa total do zooplâncton
(Fig. 2D). Este resultado não se traduziu num aumento maior da biomassa do
fitoplâncton (Fig. 2A), porque peixes onívoros também exercem forte pressão de
predação direta sobre essa comunidade, com taxas de consumo maiores sobre algas
de grande tamanho (Vinyard et al. 1988; Smith 1989; Drenner et al. 1996; Okun et al.
2008).
A análise do efeito dos peixes onívoros sobre a comunidade fitoplanctônica fica
ainda mais interessante ao analisarmos os resultados sob uma perspectiva de
gradiente de trofia dos experimentos. Quanto maior o grau de trofia do sistema, mais
pronunciado foi o efeito positivo dos peixes onívoros na biomassa do fitoplâncton (Fig.
2B). A hipótese sobre estado trófico de lagos e interação com peixes proposta por
McQueen et al. (1986) prevê que o aumento da biomassa do fitoplâncton pela
presença de peixes será mais intensa em sistemas oligotróficos. Os resultados deste
estudo meta-analítico não foram consistentes com esta hipótese.
Os peixes onívoros são reconhecidos por atuarem como eficientes
regeneradores e recicladores de nutrientes através de translocação de sedimentos do
fundo para coluna d’agua (i.e. bioturbação) e através de excreção, respectivamente
(Vanni 2002). Ao consumirem matéria orgânica dos sedimentos, parte dos nutrientes
ingeridos são excretados em uma razão estequiométrica nitrogênio:fósforo mais baixa
e portanto mais apropriada a assimilação de outros organismos. Desta forma, os
peixes onívoros podem aumentar a biomassa do fitoplâncton agindo como verdadeiras

25
“bombas de nutrientes” que estavam imobilizados no sedimento (Schaus et al. 1997;
Vanni & Layne 1997).
Alguns trabalhos demonstram que a porção de nutriente incorporada aos
sedimentos é cada vez maior, à medida que o grau de trofia dos sistemas aumenta
(Sondergaard et al. 2003; Vanni et al. 2006), provendo aos onívoros filtradores e
detritívoros um potencial ainda maior de agirem como bombas de nutrientes. De fato, a
ocorrência desse mecanismo tem sido demonstrada empiricamente em estudos
experimentais (Drenner et al. 1996) e de observação realizados ao longo de
gradientes tróficos em ecossistemas lacustres naturais colonizados por peixes
onívoros. Desta forma, fica compreensível o mecanismo pelo qual a magnitude do
efeito de peixes onívoros na biomassa do fitoplâncton pode aumentar com os estado
trófico do ambiente (Fig 2B). Outra evidência que contribui para acreditarmos que o
aumento da biomassa do fitoplâncton dá-se primariamente pelo efeito ascendente da
bioturbação e excreção de peixes onívoros, é o efeito invariável que a predação de
peixes onívoros teve na biomassa zooplanctônica ao longo de um gradiente trófico
(Fig. 2E).
O efeito de peixes zooplanctívoros sobre a biomassa do zooplâncton
decresceu ao longo de um gradiente de trofia (Fig. 2E). Nesse caso, a predação do
macrozooplâncton pelos peixes, concomitante ao aumento de trofia do sistema, pôde
favorecer o crescimento do microzooplâncton e compensar minimamente a redução na
biomassa total do zooplâncton. Com isso, sistemas de alta trofia, dominados por
microzooplâncton e onde o impacto de predação foi presumivelmente pequeno, não
tiveram correspondente aumento significativo da biomassa do fitoplâncton (Fig. 2B).
Além disso, a presença dos peixes zooplanctívoros pode não ter surtido o efeito de
cascata trófica, uma vez que em condições meso-eutróficas a proliferação de algas
menos palatáveis (e.g. blooms de cianobactérias) pode minimizar a susceptibilidade
do fitoplâncton à predação pelo zooplâncton (McQueen et al. 1986; Brett & Goldman
1996; Cottingham et al. 2004; Moss et al. 2004; Rondel et al. 2008).

26
Verificar efeitos sinérgicos da biomassa de peixes onívoros e zooplanctívoros
na distribuição de biomassa fito e zooplanctônicas tem implicações práticas muito
importantes do ponto de vista da biomanipulação de ecossistemas. Conhecer os
efeitos da estocagem de peixes amplia o entendimento sobre a ocorrência e a
magnitude de cascata trófica, tanto via remoção do zooplâncton quanto via
translocação de nutrientes entre compartimentos, e pode prevenir os efeitos negativos
sobre a qualidade da água. A reunião de todos os trabalhos que possuíam
informações sobre a biomassa dos peixes possibilitou-nos concluir que o aumento da
biomassa de peixes onívoros teve um efeito significativo na redução da biomassa do
zooplâncton, porém o mesmo resultado não foi observado para peixes zooplanctívoros
(Fig. 2F).
A estratégia de predação ativa dos zooplanctívoros utilizando a visão é
extremamente eficiente em reduzir a biomassa do macrozooplâncton prontamente
(Confer & Blades 1975). Sendo assim, mesmo baixas densidades de zooplanctívoros
podem causar reduções extremas na biomassa do zooplâncton, de modo que um
aumento na biomassa de peixe não se reflete necessariamente em um aumento na
taxa de predação per capita. Apesar da resposta invariável do zooplâncton ao
aumento da biomassa dos peixes zooplanctívoros (Fig. 2F), observamos um
correspondente aumento na biomassa do fitoplâncton (Fig. 2C). Este aumento do
fitoplâncton não pode ser atribuído a nenhum mecanismo de controle ascendente
relacionado ao aumento de biomassa dos peixes (e.g. regeneração e excreção de
nutriente), uma vez que a biomassa de peixes e a concentração de fósforo não foram
significativamente correlacionados ao longo dos múltiplos experimentos analisados (p
> 0.05). Alternativamente, podemos explicar tal aumento do fitoplâncton por um efeito
descendente indireto, onde o aumento da biomassa de peixes zooplanctívoros
representou maior risco de predação para o zooplâncton, diminuindo suas taxas de
herbivoria por mecanismos comportamentais e não pela redução de sua densidade
[i.e. cascata trófica comportamental, (Werner & Peacor 2003)]. Tais efeitos não letais

27
na eficiência de herbivoria ocasionada pela presença de predadores é amplamente
documentada em vários tipos de ecossistemas e comunidades, incluindo cadeias
tróficas pelágicas (Peacor & Werner 1997; Wojdak & Luttbeg 2005; Stief & Holker
2006).
A biomassa dos peixes onívoros reduziu de forma significativa a do biomassa
do zooplâncton (Fig 2F). A diferença no efeito da biomassa de onívoros e
zooplanctívoros ocorreu provavelmente porque predadores generalistas filtradores são
menos eficientes na captura de presas do que os predadores zooplanctívoros visuais
(Drenner & McComas 1980). O modo de captura por sucção e não visual da maioria
dos predadores onívoros faz com que o aumento da sua eficiência de predação seja
função exclusiva da probabilidade de encontro com sua presa. Por esta razão, o
aumento de biomassa do predador reflete-se proporcionalmente em um aumento na
probabilidade de captura da presa. No entanto, a redução do zooplâncton não se
reverteu em um aumento significativo do fitoplâncton, porque os peixe onívoros
também exerceram pressão de predação sobre esta comunidade (Fig. 2C).
Os estudos reunidos nesta meta-análise mostram que a adição de um
predador à comunidade gerou uma tendência de desestabilização na biomassa do
zooplâncton, independentemente da estratégia de forrageamento deste predador (Fig.
3D). Mesmo assim, a biomassa de herbívoros mais variável traduziu-se em
significativa estabilidade da biomassa fitoplanctônica em comunidades sujeitas à
zooplanctivoria (Fig. 3A). Este resultado é contrário a alguns modelos teóricos que
prevêem que a onivoria estabiliza teias tróficas porque aumenta o número de
interações tróficas fracas (McCann & Hastings 1997; McCann et al. 1998a; Kuijper et
al. 2003; Emmerson & Yearsley 2004; Vadeboncoeur et al. 2005). Interessantemente,
a predação por onívoros nesta síntese mostrou ser um mecanismo ligeiramente mais
forte na redução da biomassa zooplanctônica (Fig. 2D). Segundo alguns estudos, a
resposta denso-independente de predadores onívoros à flutuação de suas presas

28
pode gerar supressão mais intensa sobre elas (Pimm & Lawton 1978; Holt & Polis
1997; Diehl 2003).
Diante deste resultado, a questão mais interessante é saber qual foi o
mecanismo de estabilização desencadeado pelos peixes zooplanctívoros. Esta
abordagem não foi elaborada para testar mecanismos específicos e sim hipóteses
específicas, e portanto, estudos adicionais serão necessários para esclarecer de forma
consistente o padrão observado. Entretanto, algumas hipóteses podem ajudar a dirigir
futuras investigações. A predação visual dos zooplanctívoros, muito seletiva para o
macrozooplâncton, porém pouco eficaz na redução do microzooplâncton e de
espécies muito evasivas (Confer & Blades 1975), teve um efeito global não diferente
dos peixes onívoros, porém mais ameno sobre a biomassa total do zooplâncton (Fig.
2D). Esta sutil diferença, foi suficiente para produzir um efeito de cascata trófica
significativamente maior na biomassa do fitoplâncton em relação aos peixes onívoros
(Fig. 2A) e também pode ter sido fundamental para diminuir significativamente a
estabilidade temporal do fitoplâncton (Fig. 3A). Esta hipótese considera o fato de que
em razão dos peixes zooplanctívoros terem provocado um efeito ligeiramente mais
fraco sobre o controle da biomassa zooplanctônica em relação aos onívoros, este
efeito foi suficiente para manter o fitoplâncton sob certo nível de controle descendente
capaz de evitar grandes oscilações temporais na biomassa desta comunidade. Alguns
trabalhos têm demonstrado que uma moderada pressão de consumo pode funcionar
prevenindo dominância e extinção de espécies, bem como impedindo picos de
crescimento descontrolado e assim produzir uma estabilidade temporal mais
consistente (Worm et al. 2002).
Outro fato intrigante que foi contrariado pelos resultados deste estudo e que
tem sido proposto por vários estudos teóricos é o de que a onivoria, ao reduzir a
cascata trófica, produziria efeitos responsáveis por estabilizar drásticas oscilações
entre os consumidores e os recursos (Polis et al. 1996b; McCann & Hastings 1997;
McCann et al. 1998a; Thompson & Gese 2007). Entretanto, os resultados desta

29
síntese de trabalhos empíricos não corroboram esta predição. De fato, um recente
trabalho meta-analítico que considerou vários ecossistemas em sua abordagem, não
encontrou nenhuma relação óbvia de susceptibilidade entre a força de cascata trófica
e o efeito de predadores na estabilidade temporal de produtores primários e de
herbívoros (Halpern et al. 2005). A falta de mecanismos aparentes para elucidar este
padrão levou os autores a proporem que fatores determinantes da distribuição de
biomassa entre níveis tróficos podem ser diferentes daqueles responsáveis por
determinar a variância da biomassa. Este fato coloca em dúvida a relação de
acoplamento causal entre a ocorrência de cascata trófica e a instabilidade temporal,
sugerida como um mecanismo estabilizador oriundo da onivoria. Um fato importante
que deve ser considerado é o nível de generalização dos modelos teóricos que podem
ser eficientes para preverem os efeitos de peixes onívoros em condições mais
controladas e restritas, mas não ao longo de uma ampla e heterogênea escala de
condições ecológicas tais como as abordadas neste estudo. Os dados utilizados neste
estudo para o cálculo de estabilidade refletem processos ecológicos em escalas
temporais reais (i.e. os estudos analisados levaram em consideração várias gerações
de herbívoros e produtores primários), tendo sido improvável a ocorrência de uma
análise experimental tendenciosa.
Ao mostrar que peixes onívoros enfraquecem a cascata trófica mas
aparentemente não afetam a estabilidade temporal de cadeias tróficas aquáticas em
um conjunto significativo de experimentos publicados na literatura, estes resultados,
dentro de seus limites de abrangência (e.g., cadeias tróficas planctônicas de
ecossistemas lacustres), podem refletir um importante grau de generalização e
chamarem a atenção para revisões de conceitos que tangem a dinâmica do
funcionamento de teias tróficas além, também de, importantes ramificações práticas e
de conservação. Uma importante constatação é que peixes onívoros, ao promoverem
uma teia trófica reticulada e portanto com maior conectividade entre níveis tróficos,
não necessariamente promovem a maior estabilidade destes. Outra é que, de fato,

30
estratégias de biomanipulação que, baseadas no conceito de cascata trófica, utilizam
a introdução de piscívoros para indiretamente promover a redução da biomassa
fitoplanctônica, podem não prosperar se a comunidade de peixes planctívoros for
dominada por onívoros. Além disso, várias das mais bem sucedidas espécies
invasoras de peixes de ecossistemas aquáticos dulcícolas ao redor do mundo são
onívoras e portanto sua introdução pode modificar significativamente a dinâmica
natural de ecossistemas aquáticos lacustres.

31
Capítulo II
Efeito da Predação de um Peixe Onívoro na Dinâmica e Intensidade de
Cascata Trófica em Comunidades Planctônicas

32
Introdução
A natureza dos fatores responsáveis por regular a estrutura e funcionamento
de teias tróficas tem sido uma área de considerável debate dentro da ecologia (Pimm
1980; Pimm et al. 1991; Borer et al. 2006; Hillebrand et al. 2007). Um importante tópico
emergente deste cenário foi a teoria de cascatas tróficas, a qual prevê que a biomassa
de cada nível trófico, em uma determinada cadeia, é inversamente e diretamente
relacionada à biomassa dos níveis tróficos sobrejacentes e subjacentes a ele
(Oksanen 1991; Brett & Goldman 1996; Carpenter et al. 2001). Grande parte dos
estudos que originaram a proposição desta teoria se remete a ecossistemas aquáticos
(Strong 1992; Carpenter et al. 2001; Shurin et al. 2006) envolvendo cadeias tróficas
pelágicas, tendo o fitoplâncton como produtor primário, zooplâncton como herbívoro e
peixes como predadores (McQueen et al. 1986).
Embora a ocorrência de cascatas tróficas devido a manipulações da presença
ou do estoque de peixes seja atualmente um fato comprovado tanto
experimentalmente (Brett & Goldman 1996, 1997; Bell et al. 2003) quanto em estudos
de observação (Vanni et al. 1990) realizados em ecossistemas aquáticos continentais,
o conhecimento sobre fatores responsáveis por determinar a dinâmica temporal bem
como a força da cascata trófica ainda é permeado de considerável incerteza (Shurin et
al. 2002; Borer et al. 2005; Shurin & Seabloom 2005). Um fator que tem merecido
considerável atenção tanto teórica quanto experimental, é como o hábito trófico (i.e.
peixes zooplanctívoros ou onívoros) pode afetar a força da cascata trófica (Pace et al.
1999). Um considerável número de evidências sustenta o fato de que peixes
zooplanctívoros visuais, por exibirem predação direcionada a organismos
zooplanctônicos de maior tamanho exercem forte efeito indireto no aumento da
biomassa fitoplanctônica pela redução da herbivoria imposta pelo zooplâncton,
ocasionando cascatas tróficas de grande magnitude (McQueen et al. 1986; Vanni
1986b; McQueen et al. 1989; Vanni & Findlay 1990; Vanni et al. 1997). Por outro lado,
considerando que peixes onívoros filtradores se alimentariam tanto do zooplâncton

33
quanto também do produtor primário (fitoplâncton), vários estudos propuseram-se a
testar a hipótese de que a predação por peixes onívoros poderia ser um fator
amenizador da intensidade de cascatas tróficas uma vez que a diminuição da força de
interação entre o predador onívoro e o zooplâncton, aliada ao fato do peixe também se
alimentar do produtor primário, diminuiria seus efeitos positivos indiretos sobre a
biomassa algal, reduzindo a intensidade da cascata trófica (Drenner et al. 1982;
Drenner et al. 1984b; McCann et al. 1998a; McCann et al. 1998b; Scheffer 1998;
Lazzaro et al. 2003).
Embora uma considerável parte destes trabalhos de certa forma suporte a
hipótese de que a onivoria reduz a força de cascatas tróficas (Drenner et al. 1996),
parte da variação observada nestes experimentos não pôde ser explicada apenas por
diferenças entre as duas guildas tróficas de peixes, revelando que a força da cascata
trófica pode ser um fenômeno regulado de forma interativa por múltiplos fatores
ambientais (Brett & Goldman 1996, 1997; Agrawal 1998; Borer et al. 2005). Dentre
estes, o estado trófico do ecossistema (McQueen et al. 1986; Drenner et al. 1996;
Drenner et al. 1998) bem como a heterogeneidade da composição intratrófica (Attayde
& Hansson 2001b; Hansson et al. 2004) têm sido apontados como importantes fatores
capazes de modificar tanto a ocorrência quanto a intensidade de cascatas tróficas em
ecossistemas pelágicos (Vakkilainen et al. 2004; Van de Bund et al. 2004; Rondel et
al. 2008). Adicionalmente, os experimentos que buscam avaliar o papel de peixes
onívoros sobre a força de cascata trófica têm sido realizados basicamente com peixes
onívoros-planctívoros filtradores (e.g. tilápia, carpa etc) que consomem tanto a
biomassa zooplanctônica como também a biomassa fitoplanctônica (Drenner et al.
1996). Entretanto, o número de espécies de peixes nos quais a onivoria não
necessariamente encontra-se relacionada ao consumo de algas planctônicas, mas sim
ao consumo de perifíton, detritos e etc, é imensamente maior, principalmente na
região tropical (Fernando 1994). Tal fato impede que generalizações consistentes em

34
relação aos efeitos da onivoria sobre a dinâmica e força de cascata trófica pelágica
sejam feitas.
Uma inovadora proposição funcional do hábito onívoro, conhecida como
onivoria de cadeias múltiplas (“multi-chain omnivory”) (Polis et al. 1996b;
Vadeboncoeur et al. 2005), propõem que onívoros que utilizam subsídios oriundos de
níveis tróficos pertencentes a cadeias alimentares distintas e portanto espacialmente
e/ou funcionalmente desacoplados (i.e. produtor primário bentônico, herbívoro
planctônico, detritos alóctones) podem, através de complexos mecanismos indiretos,
modificar a dinâmica de interações tróficas e o fluxo de energia responsáveis por
determinar padrões de regulação trófica entre as diferentes comunidades e
compartimentos aquáticos (Huxel et al. 2002; Vadeboncoeur et al. 2002; Vander
Zanden & Vadeboncoeur 2002; McCann et al. 2005; Vander Zanden et al. 2005).
Desta forma, peixes onívoros não filtradores poderiam afetar a força de cascata trófica
pelágica sobre o fitoplâncton se por exemplo, o forrageamento sobre o perifíton e/ou
invertebrados bentônicos contribuir para a diminuição da taxa de predação sobre o
zooplâncton (alterando a pressão de herbivoria sobre o fitoplâncton) ou interferir nas
relações competitivas por nutrientes entre as algas pelágicas e o perifíton.
Neste estudo, através de experimentos de campo realizados em mesocosmos,
foi avaliado se a presença de um peixe onívoro-zooplanctívoro não filtrador afeta a
dinâmica temporal e a força de cascata trófica em comunidades pelágicas (fitoplâncton
e zooplâncton), e se estes efeitos são dependentes do estado trófico (disponibilidade
de nutrientes).
A seguinte hipótese geral foi testada:
Os efeitos da predação pelo peixe onívoro-zooplanctívoro não filtrador sobre a
dinâmica temporal da cascata trófica bem como sobre sua intensidade são
dependentes do estado trófico e da identidade (heterogeneidade) funcional das
assembléias fitoplanctônicas e zooplanctônicas.

35
Para testar esta hipótese a dinâmica individual (e.g. biomassa de diferentes
grupos funcionais e taxonômicos) e agregada das comunidades fitoplanctônicas e
zooplanctônicas foram comparadas em relação aos efeitos individuais e interativos da
presença de peixe e adição de nutrientes ao longo de 11 semanas.

36
Materiais e Métodos Área de Estudo
Este trabalho foi realizado na lagoa Cabiúnas, localizada no Parque Nacional
da Restinga de Jurubatiba, Rio de Janeiro – Brasil (22º 15’ S, 41º 40’ O) (Fig. 1a).
Seus limites físicos estão contidos no domínio geomorfológico da planície de
sedimentação aluvial estando sua gênese referente ao período terciário (Soffiati 2000).
A lagoa Cabiúnas é uma típica lagoa costeira permanecendo separada do oceano por
uma barra de areia, possui uma área de aproximadamente 0,35 km2 e profundidade
média de 2,0 m (Panosso et al. 1998). Suas águas se caracterizam por serem húmicas
e ligeiramente ácidas (pH 6,3) com temperatura média anual de 23,6º e transparência
medida a partir do disco de Secchi de aproximadamente 0,5 m (Petrucio 1998). Seu
formato dendrítico confere uma alta relação perímetro:volume, permitindo o
desenvolvimento de uma extensa região litorânea densamente colonizada pela
Figura 1: (a) Fotografia aérea da Lagoa Cabiúnas localizada no Parque Nacional da Restinga de Jurubatiba. A seta indica o local (b) onde os mesocosmos foram instalados próximo da região litorânea. (c) Detalhe dos mesocosmos.
A
B
C

37
macrófita aquática Typha domingensis (Pers). Pode ser classificada como oligo-
mesotrófica, com concentrações médias de fósforo total (PT) e nitrogênio total (NT) de
0,35 e 55 µM, respectivamente e biomassa média fitoplanctônica, estimada a partir da
concentração de clorofila-a, de 4 µg/L (Enrich-Prast et al. 2004). A lagoa apresenta
elevada biodiversidade no que se refere à fauna e flora pelágica possuindo o maior
número de táxons zooplanctônicos (Castelo-Branco 1998), fitoplanctônicos (de Mello &
Suzuki 1998) e ictiológicos (Caramaschi et al. 2004) documentados para as lagoas
costeiras da região. A maior parte da ictiofauna é composta por peixes de hábitos
onívoros associados à região litorânea da lagoa (Caramaschi et al. 2004).
Estrutura dos mesocosmos e desenho experimental
Dezesseis mesocosmos cilíndricos (2,0 m de diâmetro × 2,4 m de altura)
construídos de polietileno translúcido (300 mm de espessura) foram instalados
próximos à região litorânea da lagoa em local sem a presença de macrófitas aquáticas
(Fig. 1b e 2). Cada mesocosmo foi equipado com um anel de ferro em cada
extremidade para permitir estabilidade estrutural e o acoplamento de bóias e âncoras.
Os mesocosmos foram abertos para a atmosfera e para o sedimento, isolando
completamente a coluna d’água interna da água da lagoa, mas permitindo que trocas
gasosas e a incidência luminosa fossem próximas às condições naturais além do
acesso irrestrito dos peixes ao sedimento. Para evitar a fuga de peixes, a extremidade
inferior de cada mesocosmo foi firmemente enterrada (≈ 0,1 m) dentro do sedimento e
ancorada com placas de concreto. Todos os mesocosmos foram colocados em
profundidades de aproximadamente 2 m, de maneira que a parte superior
permanecesse 0,3 m acima do nível d’água evitando a entrada de água da lagoa pela
ação de ondas (Fig. 2). Desta forma a profundidade dos mesocosmos correspondeu à
profundidade média da lagoa Cabiúnas, sendo o volume médio dos mesocosmos
nesta profundidade de aproximadamente 6300 L. Após a instalação, cada mesocosmo

38
foi inspecionado visualmente, não sendo detectada a presença de peixes em nenhum
deles.
O desenho experimental consistiu de um arranjo fatorial do tipo 2x2 com dois
níveis do fator nutriente (sem ou com adição) e dois níveis do fator peixe (ausência ou
presença) resultando nos seguintes tratamentos: CTRL (sem peixe e sem adição de
nutrientes), PEIXE (presença de peixe sem a adição de nutrientes), NUT (ausência de
peixe com a adição de nutrientes) e PEIXE+NUT (presença de peixe e adição de
nutrientes) (Fig. 2). Desta forma foi possível avaliar os efeitos independentes e
interativos dos fatores principais (i.e. peixe e nutrientes) sobre as variáveis analisadas.
Cada tratamento foi replicado 4 vezes e cada réplica igualmente distribuída em 4
diferentes blocos para diluir potenciais efeitos da heterogeneidade ambiental de forma
homogênea entre os tratamentos (Fig. 2). Entretanto, para detectar eventuais efeitos
da heterogeneidade sobre as variáveis mensuradas, os tratamentos só foram
implementados uma semana após a instalação dos mesocosmos (semana 1), período
no qual foram realizadas coletas para avaliar a homogeneidade das condições iniciais
entre os diferentes tratamentos.
Procedimento Experimental
O experimento ocorreu ao longo de 11 semanas, de maio a agosto de 2005.
Semanalmente após a semana 1, nitrogênio (NH4NO3) e fósforo (KH2PO4 e K2HPO4)
foram adicionados conjuntamente de forma a estabelecer uma concentração final
desejada de 200 µM de nitrogênio e 40 µM de fósforo aos tratamentos que recebiam
adição de nutrientes, resultando em uma razão N:P = 5:1. Os nutrientes foram
adicionados diluindo a quantidade específica desejada em um balde contendo água do
próprio mesocosmo. Em seguida a água era retornada para o mesocosmo e
cuidadosamente homogeneizada com o auxílio de um remo. Esta metodologia foi
repetida em períodos intercalados à coleta das variáveis, o que além de permitir a

39
homogeneização dos nutrientes minimiza o processo de decantação do séston, que é
mais acelerado em experimentos de mesocosmos pela redução do hidrodinamismo da
coluna d’água (Stephen et al. 2004). Considerando que a dinâmica de nutrientes pode
ser diferente em réplicas de um mesmo tratamento em experimentos de campo, a
determinação semanal das concentrações das formas inorgânicas de nitrogênio (N-
NH4 e N-NO3) e fósforo (P-PO4), permitiram calcular a quantidade necessária de N e P
a ser adicionada em cada mesocosmo semanalmente, de forma a manter as
concentrações finais homogeneamente próximas aos limites desejados. Embora estas
concentrações finais reflitam a de um ambiente hipereutrófico, elas foram escolhidas
com base em um experimento piloto realizado previamente na lagoa Cabiúnas, onde
foi demonstrado que após 48 horas, havia a redução de aproximadamente 70% e 85%
do N e P adicionados, respectivamente. De acordo com vários estudos esta redução
deve-se principalmente à absorção/complexação de nutrientes pelo sedimento
(Schindler et al. 1997; Carpenter et al. 2001). Portanto, a adição de quantidades
menores de nutrientes a sistemas experimentais onde o contato do sedimento com a
Ctrl Peixe Nut Nut + Peixe
N + P
N + P
Ctrl Peixe
NutNut+
Peixe
Arranjo espacial dos tratamentos
Nut
Peixe
Ctrl Peixe
NutNut+
PeixeCtrlPeixe
Sedimento
Nível d ’água
Flutuadores
Anel de ferro
Nut+
PeixeNut
Nut+
Peixe
Ctrl
Fig. 2 - Representação esquemática do desenho experimental adotado e detalhes estruturais dos mesocosmos.

40
coluna d`água existe pode prejudicar o objetivo da manipulação, pela redução dos
nutrientes, devido a fatores geoquímicos. Entretanto, as concentrações utilizadas
neste experimento não se apresentaram tóxicas às comunidades aquáticas e estão
dentro da amplitude de variação daquelas utilizadas em experimentos que buscaram
avaliar os efeitos do enriquecimento e predação por peixes sobre comunidades
planctônicas (Stephen et al. 2004).
A presença de peixes foi manipulada através da adição de 40 indivíduos
adultos (comprimento padrão 3 - 3,5 cm, 0,5 g/ind) da espécie Hyphessobrycon
bifasciatus (Ellis 1911) (Characidae - Fig. 3). A densidade final de peixes nos
mesocosmos foi de 13 ind/m2. Os indivíduos de H. bifasciatus foram cuidadosamente
capturados com o auxílio de redes de arrasto na região litorânea próxima ao local da
realização do experimento, e imediatamente acondicionados em tanques rede
instalados na própria lagoa Cabiúnas.
Figura 3: Indivíduo adulto de Hyphessobrycon bifasciatus. A espécie é comumente encontrada na região litorânea da lagoa Cabiúnas (Macaé, RJ) e possui hábitos onívoros, alimentando-se de perifíton, detritos orgânicos, zooplâncton, invertebrados aquáticos e insetos terrestres. (Crédito da foto - www.fishbase.org).
Os peixes foram mantidos nestas condições por 24 horas para recuperação do
estresse da captura. Apenas indivíduos ativos e aparentemente saudáveis foram
utilizados no experimento. A espécie H. bifasciatus foi escolhida pelo fato de possuir
hábitos onívoros (Sánchez-Botero 2005) (alimenta-se de zooplâncton, invertebrados
bentônicos, perifíton, detritos e etc.) além de apresentar ampla distribuição nas lagoas
_______ 1 cm

41
costeiras da região Norte Fluminense (Caramaschi et al. 2004) e em ecossistemas de
água doce na América do Sul (www.fishbase.org). Embora a densidade de peixes não
tenha sido registrada ao final do experimento, inspeções diárias permitiram detectar
indivíduos mortos que foram substituídos imediatamente por peixes saudáveis
mantidos em tanques-rede na lagoa. Entretanto a mortalidade de peixes foi baixa
durante o experimento (< 10%) e não relacionada a algum dos tratamentos onde a
presença de peixe foi manipulada, garantindo, portanto, a densidade de peixes em
níveis desejáveis ao longo do experimento.
Amostragem e Análise das Variáveis
Amostragens semanais, conduzidas consecutivamente entre a 1ª e 7ª semana
e uma última na 11ª semana, foram realizadas totalizando 8 amostragens ao longo do
experimento. Para evitar efeitos relacionados a variações diárias das variáveis
analisadas, as amostragens foram sempre conduzidas entre 10:00 h e 12:00 h. As
variáveis analisadas foram íon amônio e nitrato (nitrogênio inorgânico dissolvido –
NID), ortofosfato, oxigênio dissolvido, temperatura da água, biovolume fitoplanctônico
e biomassa zooplanctônica. Com exceção das medidas de oxigênio dissolvido e
temperatura que foram feitas na superfície, meio e fundo através de um oxímetro (YSI-
95), as demais amostragens foram realizadas de forma integrada ao longo da coluna
d’água. Para cada mesocosmo, amostras de água da superfície e fundo foram
coletadas com auxílio de garrafa tipo Van Dorn e integradas em um balde de onde
foram retiradas amostras de 100 ml para determinação do biovolume fitoplanctônico
(contagem de células) e 50 ml para determinação das concentrações de nutrientes. O
zooplâncton foi coletado através de 1 arrasto vertical (1,6 m) realizado com uma rede
de plâncton de 65 µm. O fitoplâncton foi fixado com lugol e o zooplâncton em solução
açucarada de formalina a 4%. As concentrações do íon amônio foram determinadas
pelo método colorimétrico do indofenol azul, conforme proposto por Koroleff (1978) e
as concentrações de nitrato medidas através de um analisador de nutrientes (FIA),

42
pelo método de redução do nitrato a nitrito em coluna de cádmio (Golterman et al.
1978). Em laboratório células fitoplanctônicas foram contadas através de microscopia
invertida e identificadas até o nível de espécie ou gênero e medidas de acordo com
sua conformação geométrica (Hillebrand et al. 1999). O volume celular do fitoplâncton
foi calculado através de aproximações a formas geométricas sólidas similares. As
algas foram em seguida agrupadas e classificadas em suas respectivas classes.
Organismos zooplanctônicos foram contados, medidos (apenas náuplios) e
identificados a nível específico através de microscópio estereoscópico. Os organismos
foram agrupados em cladóceros, copépodes (Calanoida e Cyclopoida), náuplios e
rotíferos. Para determinação da biomassa de microcrustáceos (copépodes e
cladóceros), os primeiros 20 exemplares (somente adultos para copépodes) de cada
espécie contados foram separados em cadinhos espécie-específicos, secos em estufa
a 60 oC por 24 horas e em seguida pesados em microbalança Metler modelo UMT2
para a determinação da biomassa por indivíduo. A biomassa de adultos de
microcrustáceos foi calculada multiplicando o peso médio por indivíduo de cada
espécie pela sua respectiva densidade na amostra. Para a determinação da biomassa
de náuplios foram utilizadas equações alométricas (peso x comprimento) segundo
Culver et al. (1985). Para a determinação da biomassa de rotíferos, a densidade de
cada espécie foi multiplicada por seu respectivo peso médio obtido da literatura e a
biomassa de todas as espécies somadas (Wetzel & Likens 1991).
Análises dos Dados
Para detectar efeitos independentes e interativos da adição dos fatores
nutrientes e peixes sobre a dinâmica da biomassa das comunidades fitoplanctônica e
zooplanctônica, foi adotada uma rotina hierárquica de análises de variância.
Inicialmente foram conduzidas análises de variância multivariadas com medidas
repetidas (RM-MANOVA) para analisar os efeitos globais dos fatores manipulados
sobre as comunidades zooplanctônicas e fitoplanctônicas separadamente. Peixes e

43
nutrientes foram tratados como fatores categóricos principais, o tempo (semanas) foi
considerado como fator repetido e a biomassa total e dos grupos (zooplâncton) e o
biovolume total e das classes (fitoplâncton) como variáveis dependentes. A utilização
da MANOVA é indicada para casos onde múltiplas variáveis dependentes que mantém
interdependência são analisadas (Scheiner 2001). Seu uso é especialmente
importante em análises que incluam o tempo como fator repetido, pois é mais robusta
a problemas de circularidade associados à RM-ANOVA, além de prevenir erro do Tipo
I associado à ocorrência de múltiplos testes (Maxwell & Delaney 2004). O índice de
Pillai-Trace foi utilizado como critério estatístico em todas as MANOVAs, pois é o teste
mais indicado para avaliar significância em relação a múltiplas variáveis
interdependentes (Scheiner 2001). Em seguida, como a MANOVA testa os efeitos
independentes e interativos dos fatores categóricos sobre todas as variáveis
dependentes simultaneamente, fatores que apresentaram resultados significativos da
MANOVA foram analisados por RM-ANOVAs fatoriais (com correções de Huyn-Feldt
para o termo repetido) para cada variável dependente, para avaliar os efeitos
principais e interativos da presença de peixes e nutrientes sobre a dinâmica temporal
de cada variável resposta separadamente. Por fim para detectar diferenças
específicas entre cada tratamento (CTRL, NUT, PEIXE e PEIXE+NUT), análises de
contraste pareadas foram realizadas para cada variável dependente. Considerando
que 6 comparações pareadas podem ser feitas entre os quatro tratamentos, o valor de
a inicialmente selecionado como P = 0,05, foi corrigido pelo procedimento seqüencial
de Bonferroni para o ajuste de múltiplas comparações (Rice 1989). Neste
procedimento, para cada variável os valores de P obtidos das 6 comparações
pareadas entre os tratamentos são classificados hierarquicamente do menor para o
maior valor e em seguida o menor valor de P obtido é comparado ao menor valor de P
ajustado calculado como a/k, onde k é o número de comparações pareadas existentes
(neste caso 0,05/6 = 0,008). O resultado é considerado significativo se o menor valor
de P obtido da análise de contraste for menor que o menor valor de P ajustado. Em

44
seguida sucessivamente os crescentes valores de P obtidos são comparados com os
crescentes valores de a calculados como a/k-1, a/k-2,..., até que todas as
comparações sejam efetuadas. RM-ANOVAs fatoriais foram feitas para detectar
diferenças entre os tratamentos em relação à temperatura, oxigênio dissolvido, NID e
ortofosfato. Para todas as análises tanto em relação às comunidades quanto às
variáveis fisico-químicas, a primeira semana (semana pré-tratamento) não foi incluída
no modelo. Análises de variância simples (ANOVA) foram feitas para todas as
variáveis (bióticas e abióticas) com apenas os dados da 1 semana (antes da
implementação dos tratamentos) para avaliar se haviam diferenças prévias entre os
mesocosmos que compreenderiam os tratamentos.
Para analisar como a presença de peixes afetou globalmente (ao longo de todo
o experimento) a relação entre a biomassa zooplanctônica e o biovolume
fitoplanctônico, regressões lineares foram feitas considerando todos os tratamentos
conjuntamente bem como separadamente enfocando o efeito da presença de peixes
sob condições de disponibilidade de recursos variada. Todos os dados foram
transformados pelo Log10 para redução da heterogeneidade das variâncias. Todas as
análises de variância foram feitas através do programa STATISTICA 7.0 (StatSoft
2001).
Para estimar uma medida integrativa referente à intensidade de cada fator (i.e.
peixe e nutriente) sobre a distribuição da biomassa dos níveis tróficos planctônicos, a
intensidade dos efeitos da presença de peixes e da adição de nutrientes (effect sizes)
sobre grupos individuais e sobre a biomassa zooplanctônica e fitoplanctônica total
foram calculadas como a diferença padronizada entre as médias (D) conforme
proposto por Gurevitch & Hedges (1993), de acordo com a seguinte equação:
JS
D CT χχ ˆˆ −=
Onde XT e Xc são respectivamente as médias do tratamento manipulado e seu
respectivo controle. S é o desvio padrão agrupado e J corrige os desvios gerados pela

45
diferença de tamanho amostral. Uma vez que a variância geralmente aumenta com o
aumento da média, é vantajoso contemplar a variação das réplicas (S) no cálculo da
intensidade do efeito. A diferença entre duas médias portanto, é padronizada pelo
desvio conjunto de ambos os tratamentos. O desvio padrão agrupado é calculado
conforme a expressão:
)2())(1())(1( 22
−+
−+−=
CT
CCTT
nnSDnSDnS
Onde n é o número de réplicas e SD o desvio padrão dos tratamentos. J é calculado
como:
1)2(431
−−+−=
CT nnJ
Desta forma para cada semana (exceto a 1 semana) o tamanho do efeito da presença
de peixes (Dpeixes) foi calculado tanto para a situação ambiente, onde XT foi o
tratamento PEIXE e Xc o CTRL, quanto para a situação de maior trofia, onde XT foi o
tratamento PEIXE+NUT e Xc o tratamento NUT. O mesmo procedimento foi adotado
para o cálculo dos efeitos dos nutrientes (Dnutrientes), onde os efeitos dos nutrientes
foram calculados na ausência de peixe (XT tratamento NUT e Xc o CTRL) e na
presença de peixes (XT tratamento PEIXE+NUT e Xc tratamento PEIXE). A magnitude
do efeito cumulativo de cada fator sobre a biomassa total e dos diferentes grupos
planctônicos foi calculada a partir da integração das respectivas magnitudes
calculadas em cada semana através do programa Metawin (versão 2.0 para Windows)
de acordo com Rosenberg et al. (2000). Em seguida para saber se a variação na força
do efeito dos nutrientes e dos peixes sobre a biomassa de cada grupo foi dependente
da presença do outro fator, (isto é, se o efeito do peixe muda dependendo da condição
trófica ou se o efeito dos nutrientes muda dependendo da presença ou ausência de
peixes) as magnitudes dos efeitos de cada fator foram comparadas nas duas
situações.

46
Para traçar uma comparação entre a intensidade de cascata trófica promovida
por peixes onívoros e zooplanctívoros, confrontamos os dados de intensidade do
efeito de peixe onívoros calculados neste experimento com os dados de 11
experimentos que examinaram a intensidade do efeito de peixes zooplanctívoros. A
compilação destes 11 experimentos foi apresentado em uma meta-análise de Brett &
Goldman (1996), que utilizou a mesma métrica de cálculo de intensidade do efeito
descrita para este estudo (Gurevitch & Hedges 1993), tornando portanto os dados
comparáveis. Estudos prévios sugerem que peixes onívoros e zooplanctívoros
desencadeiam cascata trófica por diferentes vias (Drenner et al. 1996), mas a
comparação da intensidade de seus efeitos é bastante escassa (Lazzaro et al. 1992;
Williams & Moss 2003; Boveri & Quiros 2007). Embora nosso experimento não tenha
sido desenhado para testar no campo a diferença entre diferentes estratégias de
alimentação de peixes, a proposta de comparação da intensidade do efeito de cascata
trófica utilizando dados obtidos aqui e na literatura, permite que nossa comparação
seja bastante abrangente.

47
Resultados
Eficiência da manipulação experimental
As variáveis abióticas coletadas na fase pré-tratamento foram avaliadas através de
análises de variância (ANOVA) simples que demonstraram não haver diferenças
significativas nas condições iniciais dos 16 mesocosmos. A semana 1, anterior à adição de
nutrientes e peixes, apresentou temperatura média máxima da água de 23,36°C [(±0,42),
ANOVA F = 0,29; P = 0,83], concentração de oxigênio máxima de 3,52 mg.L-1 [(±0,47),
ANOVA F = 0,63; P = 0,59], NID de 6,31mmol. L-1 [(±2,27), ANOVA F = 0,26; P = 0,85),
ortofosfato de 0.11 mmol. L-1 [(±0,02), ANOVA F = 0,55; P = 0,65] e razão de
NID:ortofosfato de 57,07 [(±18,77), ANOVA F = 0,26; P = 0,84] (Tabela 1).
As condições iniciais da biomassa e da composição das comunidades
zooplanctônicas e fitoplanctônica também não foram significativamente diferentes nos 4
tratamentos [Fig. 4A (F = 0,02, P = 0,99); 4B (F = 2,63, P = 0,09); 4C (F = 0,85, P = 0,49);
4D (F = 0,85, P = 0,49), 4E (F = 1,35, P = 0,30) e Fig. 5A (F = 0,26, P = 0,84); 5B (F = 0,99,
P = 0,42); 5C (F = 1,24, P = 0,33); 5D(F = 0,71, P = 0,33); 5E (F = 0,17, P = 0,91); 5F (F =
0,35, P = 0,78)] fato que garantiu que a manipulação experimental fosse comparável.
Variáveis Abióticas
A análise de variância repetida (RM-ANOVA) sobre as concentrações de nutrientes
dissolvidos (NID e ortofosfato) demonstrou um aumento significativo e consistente com o
enriquecimento de nutrientes a partir da segunda semana. Com a adição (correção)
semanal sistemática de nutrientes, estes valores persistiram inalterados até a décima
primeira semana, tanto no tratamento NUT quanto PEIXE+NUT. A razão NID:ortofosfato
também permaneceu constante e próxima do valor desejado (5:1) nos tratamentos onde
foram adicionados nutrientes. As concentrações de nutrientes no CTRL e no tratamento
PEIXE permaneceram estáveis e semelhantes às concentrações do ambiente no início do
experimento (Tabela 1). Desta forma, concluímos que a manipulação experimental dos
nutrientes foi bem sucedida.

48
A temperatura variou entre as três profundidades mensuradas mas não sofreu
nenhuma alteração significativa no curso do experimento entre os diferentes tratamentos
(RM-ANOVA; Tabela 1). No entanto, as concentrações de oxigênio na superfície sofreram
um significativo aumento nos tratamentos com nutrientes (particularmente o tratamento
PEIXE+NUT), como resultado provável de maior atividade fotossintética pelo aumento da
biomassa fitoplanctônica (RM-ANOVA; Tabela 1).
Zooplâncton
Tanto a presença de peixes como a adição de nutrientes e a interação entre ambos
os fatores afetaram de forma global e significativa a biomassa da comunidade
zooplanctônica (RM-MANOVA, Tabela 2A).
A presença dos peixes como fator principal diminuiu significativamente a biomassa
dos cladóceros, dos copépodes e da soma de todos os organismos da comunidade
zooplanctônica, e por outro lado, aumentou significativamente a biomassa dos rotíferos
(RM-ANOVA, Tabela 3, Fig. 4A, B, C, E). A redução da biomassa dos cladóceros foi maior
no tratamento PEIXE do que no tratamento PEIXE+NUT (Análise de Contraste Pareada,
Fig. 4B). O aumento da biomassa dos rotíferos foi maior no tratamento PEIXE+NUT do que
no tratamento PEIXE (Análise de Contraste Pareada, Fig. 4A). O efeito da presença de
peixes foi dependente do tempo, mostrando um efeito interativo significativo para a
dinâmica da biomassa dos rotíferos, copépodes, náuplios e da biomassa total (Tabela 3,
Fig. 4).
O efeito interativo da adição de nutrientes e da presença de peixes foi significativo
para a biomassa de rotíferos e cladóceros (Tabela 3). Para os rotíferos, o tratamento
PEIXE+NUT produziu um efeito global de elevação de biomassa (Análise de Contraste
Pareada, Fig. 4A). Para os cladóceros, a direção oposta dos fatores principais produziu
efeito de compensação que aproximou o resultado do tratamento PEIXE+NUT ao
CONTROLE (Análise de Contraste Pareada, Fig. 4B). A interação da adição de nutrientes e

49
da presença de peixes foi dependente do tempo e influenciou significativamente a dinâmica
da biomassa dos rotíferos, cladóceros e da biomassa zooplanctônica total (Tabela 3, Fig. 4).
A adição de nutrientes como fator principal aumentou significativamente a biomassa
dos cladóceros, dos náuplios e da soma de todos os organismos da comunidade
zooplanctônica (RM-ANOVA, Tabela 3, Fig. 4B, D, E). Entretanto, o tratamento NUT não foi
diferente do tratamento PEIXE+NUT para nenhum dos componentes da comunidade
zooplanctônica (Análise de Contraste Pareada, Fig. 4). A interação dos nutrientes com o
tempo também produziu aumento significativo da biomassa total do zooplâncton, bem como
da biomassa de rotíferos e náuplios (Tabela 3).
O tempo foi um fator importante que independente e interativamente com os fatores
manipulados, afetou significativamente a dinâmica da comunidade zooplanctônica de forma
transiente (Tabela 3, Fig. 4).
A análise da contribuição proporcional dos grupos zooplanctônicos também dá
indícios do efeito dos peixes e dos nutrientes na composição da comunidade (Fig. 6). Nos
tratamentos onde os peixes foram incluídos, é clara a redução da contribuição de
cladóceros e o aumento dos rotíferos (Fig. 6B e D). Por outro lado, nos tratamentos onde os
peixes foram excluídos houve um aumento significativo de cladóceros e uma drástica
redução de rotíferos a partir da quarta semana (Fig. 6A e C), ocorrência que ficou ainda
mais evidente quando combinada a adição de nutrientes (Fig. 6C). De modo geral, os
copépodes, representados principalmente por calanóides, foram os organismos com maior
contribuição na biomassa total em todos os tratamentos (Fig. 6). No entanto, no tratamento
PEIXE+NUT, os copépodes ciclopóides co-dominaram com os calanóides (Fig. 6D). Os
copépodes foram um grupo susceptível à predação por peixes tendo uma significativa
redução de sua biomassa (Tabela 3), no entanto, sua contribuição proporcional foi sempre
constante (Fig. 6).

50
Fitoplâncton
Os efeitos independentes e interativos da adição de nutrientes e da presença de
peixes influenciaram significativamente o resultado global do biovolume da comunidade
fitoplanctônica (RM-MANOVA, Tabela 2B).
A presença de peixes como fator principal aumentou significativamente o biovolume
de Chlorophyceae, Cryptophyceae, Euglenophyceae e o biovolume total da comunidade
fitoplanctônica (RM-ANOVA, Tabela 4, Fig. 5B, C, E, F). O efeito interativo do tempo com a
presença de peixes também explicou o aumento significativo do biovolume total, bem como
dos grupos Chlorophyceae, Cryptophyceae e Cyanophyceae (RM-ANOVA, Tabela 4, Fig.
5). A comparação planejada entre tratamentos com a presença de peixes demonstrou um
aumento significativo do biovolume no tratamento PEIXE+NUT, com exceção da classe
Euglenophyceae, para a qual não se detectou diferença entre os tratamentos PEIXE e
PEIXE+NUT (Análise de Contraste Pareada, Fig. 5).
A adição de nutrientes como fator principal aumentou significativamente o biovolume
dos grupos Chlorophyceae, Cryptophyceae, Cyanophyceae e o biovolume total da
comunidade fitoplanctônica (RM-ANOVA, Tabela 4, Fig. 5B, C, D, F). A comparação
planejada entre tratamentos onde foram adicionados nutrientes, demonstrou um aumento
significativo do biovolume total e de Chlorophyceae no tratamento PEIXE+NUT (Análise de
Contraste Pareada, Fig. 5B, F). A análise do efeito interativo do tempo com a adição de
nutrientes novamente indicou aumento do biovolume total, de Chlorophyceae e
Cyanophyceae (Tabela 4). Adicionalmente, este efeito interativo também resultou em
aumento de biovolume de Bacillariophyceae e Euglenophyceae, embora para estas, não
tenha havido diferença entre os tratamentos NUT e PEIXE+NUT (Análise de Contraste
Pareada, Fig. 5A, E).
O efeito interativo da adição de nutrientes e da presença de peixes com o tempo foi
significativo na explicação da dinâmica do biovolume total da comunidade fitoplanctônica,
de Chlorophyceae e Cyanophyceae (RM-ANOVA, Tabela 4, Fig. 5). Com isso, podemos
afirmar que o tempo foi um fator que de forma independente ou interativa com os fatores

51
manipulados influenciou a dinâmica de todos os grupos da comunidade fitoplanctônica (RM-
ANOVA, Tabela 4, Fig. 5).
Das 9 classes da comunidade fitoplanctônica registradas durante o experimento, 5
tiveram uma contribuição relevante para o biovolume total e responderam de forma
significativa aos fatores manipulados (Fig. 7). A partir de uma comunidade quase
inteiramente dominada por Euglenophyceae na primeira semana pré-tratamento, foram
registrados aumentos na contribuição proporcional do biovolume de Bacillariophyceae,
Chlorophyceae, Cryptophyceae e Cyanophyceae (Fig. 7). De maneira geral, nos
tratamentos onde foram incluídos os peixes e conseqüentemente houve uma redução de
herbivoria, as classes Chlorophyceae e Cryptophyceae sofreram um aumento de biovolume
proporcional (Fig. 7B e D). A adição dos nutrientes teve um efeito significativo no aumento
do biovolume proporcional do grupo Cyanophyceae (Fig. 7C e D), que se tornou dominante
no tratamento NUT ao fim do experimento (Fig. 7C). A adição dos nutrientes aliada à
redução da herbivoria do zooplâncton elevou não só o biovolume do grupo Chlorophyceae,
como também sua contribuição proporcional que chegou a 89% no tratamento PEIXE+NUT
no final do experimento (Fig. 7D).
Controle Ascendente e Descente
A regressão temporal entre a biomassa das comunidades fitoplanctônica e
zooplanctônica, levando-se em consideração a média semanal de todos os tratamentos,
demonstra não haver relação significativa entre estas comunidades (Fig. 10A). Este
resultado foi ditado pelos tratamentos sem a presença de peixes, onde o aumento da
biomassa do zooplâncton controlou o crescimento do fitoplâncton, mesmo quando
nutrientes foram adicionados (Fig. 10B). Nos tratamentos onde os peixes estavam
presentes, a pressão de predação sobre o zooplâncton reduziu a herbivoria e
conseqüentemente o fitoplâncton demonstrou um crescimento significativo, que foi ainda
maior no tratamento PEIXE+NUT (Fig. 10C). A regressão linear positiva e significativa entre
a biomassa do zooplâncton e o biovolume do fitoplâncton é resultado do crescimento do

52
fitoplâncton num gradiente de condição trófica (com e sem nutriente) (Fig. 10), que
sustentou um aumento da biomassa do zooplâncton mesmo sob pressão de predação do
peixe.
Intensidade de Cascata Trófica “Effect Sizes”
A intensidade e a direção do efeito dos peixes na biomassa do zooplâncton
dependeram do grupo taxonômico e da condição trófica (Fig. 8A). Para os rotíferos, a
presença dos peixes teve um efeito de elevação da biomassa, que foi ligeiramente mais
intenso com o enriquecimento de nutrientes, embora não significativamente diferente da
condição sem nutrientes. Para os demais componentes do zooplâncton e para biomassa
total da comunidade, a predação por peixes teve um efeito de depleção de biomassa, que
foi mais intenso quando os nutrientes não foram adicionados. A força de predação dos
peixes na biomassa total, de cladóceros e de copépodes foi significativa e menos variável
(menores intervalos de confiança) na condição sem nutrientes, enquanto na condição com
nutrientes foi mais variável (maiores intervalos de confiança) e significativa apenas para
cladóceros (Fig. 8A).
De maneira geral, a adição de nutrientes teve efeito positivo na biomassa do
zooplâncton. No entanto, apenas na presença de peixes, este efeito foi consistente e
significativo para todos os componentes da comunidade (Fig. 8B), sugerindo que o efeito da
adição de nutrientes sobre a biomassa zooplanctônica foi intensificado pelo controle
descendente dos peixes.
Houve uma tendência geral de efeito positivo dos nutrientes e dos peixes no
biovolume do fitoplâncton (Fig. 9). A intensidade do efeito dos peixes (Dpeixe) no biovolume
total e das diferentes classes do fitoplâncton variou entre 0,21 e 0,91 na condição sem
nutrientes, e entre 0,14 a 1,15 na condição com nutrientes (Fig. 9A). O aumento do
biovolume do fitoplâncton pela supressão da herbivoria do zooplâncton pelos peixes
(controle descendente) nas duas condições (com e sem nutrientes) confirma a ocorrência
de cascata trófica. A intensidade do efeito dos nutrientes (Dnutriente) no biovolume total e das

53
diferentes classes do fitoplâncton variou entre -0,06 e 2,03 na ausência de peixes, e entre
0,17 e 2,39 na presença de peixes (Fig. 9B). A comparação da intensidade do efeito dos
peixes e dos nutrientes demonstra que os nutrientes tiveram maior efeito sobre o
fitoplâncton (Fig. 9A e B), embora os amplos intervalos de confiança de Dnutriente
demonstrem grande variabilidade de resposta. Desta forma, peixes e nutrientes foram ao
mesmo tempo efetivos na resposta do fitoplâncton, porém a regulação ascendente dos
nutrientes foi mais importante.
A confrontação do efeito médio do H. bifasciatus e de diferentes espécies de peixes
nas comunidades fito e zooplanctônicas estabeleceu uma medida comparativa da
ocorrência e da intensidade de cascata trófica entre peixes zooplanctívoros estritos e o
peixe onívoro não filtrador utilizado neste estudo (Fig. 11). A maioria dos tratamentos com
peixes resultaram em evidente redução da biomassa zooplanctônica (Fig. 11A) e aumento
da biomassa fitoplanctônica (Fig. 11B). Este resultado dá suporte inequívoco à existência
de cascata trófica, independente do hábito trófico do peixe estudado. A comparação dos
resultados do ponto de vista da intensidade do efeito mostra que o hábito onívoro do H.
bifasciatus pode ter menor efeito de atenuação sobre a comunidade zooplanctônica em
condições naturais (comparação com modelo misto de Brett e Goldman, 1996). Mas é
importante ressaltar uma redução na intensidade deste efeito, na condição onde os
nutrientes foram adicionados. Por outro lado, a intensidade da cascata trófica sobre a
biomassa do fitoplâncton foi enfraquecida pelo hábito onívoro não-filtrador de H. bifasciatus
(comparação com modelo misto de Brett e Goldman, 1996) e neste caso, não houve
diferença na resposta de acordo com a disponibilidade de nutrientes.

54
Tabela 1 - Comparações gerais das variáveis abióticas da coluna d’água entre tratamentos e a condição inicial (CI) do experimento. Os resultados são a média (± DP) ao longo das semanas (segunda a décima primeira, n = 28). Resultados marcados com letras diferentes são estatisticamente distintos (RM-ANOVA, Análise de Contraste como teste post hoc). CI Controle Peixe Nutriente Peixe+Nut
a. Temperatura (oC) Prof. (m) 0,10 23,36 (±0,42) 22,57 (±1,09)a 22,74 (±1,22)a 22,86 (±1,17)a 22,94 (±1,27)a 0,90 22,53 (±0,11) 21,47 (±0,83)a 21,48 (±0,84)a 21,44 (±0,84)a 21,46 (±0,86)a 1,80 22,40 (±0,06) 21,29 (±0,81)a 21,31 (±0,82)a 21,29 (±0,81)a 21,28 (±0,80)a
b. Oxigênio (mg.L-1) Prof. (m) 0,10 3,52 (±0,47) 4,41 (±0,68)a 4,61 (±1,12)ab 6,37 (±3,72)b 9,23 (±3,64)c 0,90 3,03 (±0,48) 3,95 (±0,73)a 4,04 (±0,98)a 4,07 (±2,29)a 5,38 (±1,91)a 1,80 2,88 (±0,47) 3,22 (±0,86)a 3,18 (±0,95)a 2,82 (±1,61)a 3,52 (±1,49)a
c. NID (µmol.L-1)
6,31 (±2,27) 11,86 (±9,64)a 5,88 (±4,10)a 128,8 (±47,11)b 135,8 (±56,56)b
d. PID (µmol.L-1) 0,11 (±0,02) 0,21 (±0,38)a 0,11 (±0,15)a 28,73 (±3,99)b 32,01 (±6,70)b
e. Razão NID:PID 57,07 (±18,77) 231,4 (±370,5)a 240,0 (±371,1)a 4,44 (±1,40)b 4,27 (±1,54)b CI - condição inicial
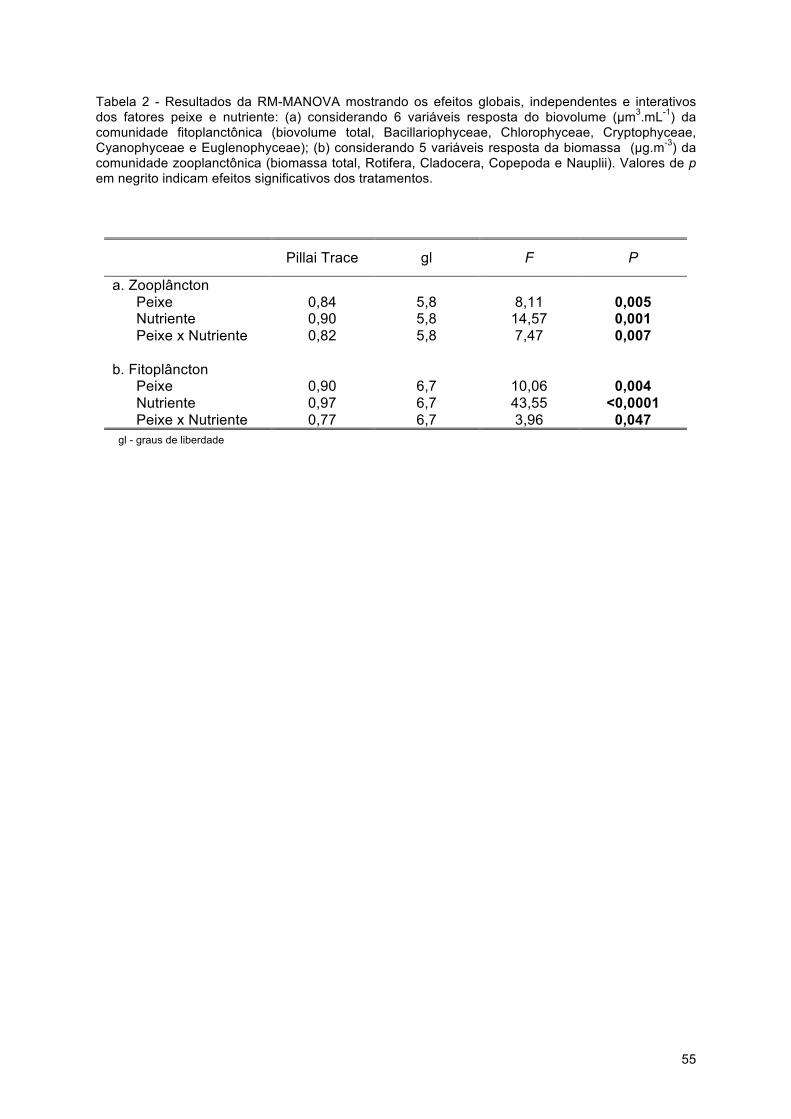
55
Tabela 2 - Resultados da RM-MANOVA mostrando os efeitos globais, independentes e interativos dos fatores peixe e nutriente: (a) considerando 6 variáveis resposta do biovolume (µm3.mL-1) da comunidade fitoplanctônica (biovolume total, Bacillariophyceae, Chlorophyceae, Cryptophyceae, Cyanophyceae e Euglenophyceae); (b) considerando 5 variáveis resposta da biomassa (µg.m-3) da comunidade zooplanctônica (biomassa total, Rotifera, Cladocera, Copepoda e Nauplii). Valores de p em negrito indicam efeitos significativos dos tratamentos.
Pillai Trace gl F P
a. Zooplâncton Peixe 0,84 5,8 8,11 0,005 Nutriente 0,90 5,8 14,57 0,001 Peixe x Nutriente 0,82 5,8 7,47 0,007
b. Fitoplâncton Peixe 0,90 6,7 10,06 0,004 Nutriente 0,97 6,7 43,55 <0,0001 Peixe x Nutriente 0,77 6,7 3,96 0,047
gl - graus de liberdade

56
Tabela 3 - Resultados da RM-ANOVA mostrando os efeitos independentes e interativos do tempo, presença de peixe e adição de nutriente sobre a biomassa (µg.m-3) total e dos grupos zooplanctônicos. Valores de P em negrito indicam efeitos significativos dos tratamentos. Setas ao lado dos valores de P indicam a direção do efeito.
Rotifera Cladocera Copepoda Nauplii Biomassa total Fatores gl SQ F P SQ F P SQ F P SQ F P SQ F P FIXOS Peixe (P) 1 18,0 52,5 <10-4 30,4 33,6 <10-4 2343,9 12,8 0,004 1639,4 1,8 0,204 3,1 13,8 0,002 Nutriente (N) 1 0,2 0,7 0,432 13,6 15,1 0,002 5,6 0,8 0,386 0,4 8,5 0,013 1,3 6,0 0,031 P Х N 1 2,8 8,3 0,014 4,3 4,8 0,048 0,3 0,3 0,567 2,0 0,9 0,361 0,4 1,7 0,212 Resíduo 12 4,1 10,8 0,2 0,2 2,7 REPETIDOS (valores de P ajustados pela correção de Huynh-Feldt) Tempo (T) 6 24,5 17,3 <10-4 22,3 10,2 <10-4 26,7 39,4 <10-4 20,0 35,2 <10-4 15,4 36,9 <10-4 T Х P 6 9,5 6,7 <10-4 4,6 2,1 0,065 3,6 5,4 10-4 6,0 10,6 <10-4 2,6 6,2 <10-4 T Х N 6 5,1 3,6 0,004 4,3 2,0 0,084 1,2 1,8 0,115 1,6 2,8 0,016 1,4 3,4 0,005 T Х P Х N 6 11,5 8,1 <10-4 6,1 2,8 0,016 1,3 1,9 0,089 1,1 2,0 0,078 1,1 2,7 0,021 Resíduo 72 17,0 26,2 8,1 6,8 5,0 gl - graus de liberdade SQ - soma dos quadrados

57
Tabela 4 - Resultados da RM-ANOVA mostrando os efeitos independentes e interativos do tempo, presença de peixe e adição de nutriente sobre o biovolume (µm3.mL-1) das classes dominantes e da comunidade fitoplanctônica total. Valores de P em negrito indicam efeitos significativos dos tratamentos. Setas ao lado dos valores de P indicam a direção do efeito.
Bacillariophyceae Chlorophyceae Cryptophyceae Cyanophyceae Euglenophyceae Biovolume total Fatores gl SQ F P SQ F P SQ F P SQ F P SQ F P SQ F P FIXOS Peixe (P) 1 6,9 2,5 0,139 11,4 18,2 0,001 5,0 8,2 0,014 0,3 0,1 0,673 19,0 10,0 0,008 5,1 17,1 0,001 Nutriente (N) 1 1,3 0,4 0,509 27,8 44,4 <10-4 21,8 35,8 10-4 60,6 42,4 <10-4 6,0 3,1 0,101 24,9 83,8 <10-4 P Х N 1 0,2 0,1 0,782 2,2 3,6 0,083 0,4 0,6 0,447 1,0 0,7 0,408 6,1 3,2 0,097 0,1 0,3 0,567 Resíduo 12 33,0 7,5 7,3 17,1 22,8 3,6 REPETIDOS (valores de P ajustados pela correção de Huynh-Feldt) Tempo (T) 6 61,8 4,1 0,001 10,3 1,7 <10-4 4,2 1,4 0,193 61,0 23,3 <10-4 80,1 4,8 0,001 2,8 4,0 0,002 T Х P 6 24,6 1,6 0,153 5,6 0,9 <10-4 15,9 5,6 10-4 14,8 5,6 10-4 12,8 0,8 0,591 3,1 4,4 0,001 T Х N 6 81,2 5,4 10-4 6,3 1,0 <10-4 4,8 1,6 0,138 27,6 10,5 <10-4 42,7 2,6 0,025 6,2 9,0 <10-4 T Х P Х N 6 6,7 0,4 0,848 4,3 0,7 <10-4 5,5 1,9 0,084 15,1 5,7 10-4 13,5 0,8 0,559 2,3 3,3 0,006 Resíduo 72 181,8 7,7 0,1 34,1 31,4 198,1 8,3 gl - graus de liberdade SQ - Soma dos quadrados

58
Figura 4 - Resposta temporal da biomassa (µg.m-3) dos principais grupos taxonômicos (A - C), nauplii (D) e da biomassa total da comunidade zooplanctônica (E) aos tratamentos peixe e nutriente. A semana 1 refere-se às amostras coletadas antes da adição de peixes e nutrientes aos tratamentos e não foram incluídas na analise. Os valores representam a média (n = 4 ± EP). Os asteriscos indicam os níveis de significância dados pela RM-ANOVA (*P < 0,05, **P < 0,01, ***P < 0,001) para os efeitos do peixe (P), nutriente (N), tempo (T) e das interações entre eles. Letras iguais ao lado das legendas que identificam os tratamentos indicam inexistência de diferença estatística entre eles (Análise de contraste pareado com correção de Bonferroni seqüencial). Note diferenças entre as escalas das figuras.
59
Figura 5 - Resposta temporal do biovolume (µm3.mL-1) dos principais grupos taxonômicos (A - E) e do biovolume total da comunidade fitoplanctônica (F) aos tratamentos peixe e nutriente. A semana 1 refere-se às amostras coletadas antes da adição de peixes e nutrientes e não foram incluídas na analise. Os valores representam a média (n = 4 ± EP). Os asteriscos indicam os níveis de significância dados pela RM-ANOVA (*P < 0,05, **P < 0,01, ***P < 0,001) para os efeitos do peixe (P), nutriente (N), tempo (T) e da interação entre eles. Letras iguais ao lado das legendas que identificam os tratamentos indicam inexistência de diferença estatística entre eles (Análise de contraste pareado com correção de Bonferroni seqüencial). Note diferenças entre as escalas das figuras.

60
Figura 6 - Contribuição proporcional dos grupos zooplanctônicos à biomassa total da comunidade, nos diferentes tratamentos ao longo das semanas: (A) CTRL; (B) PEIXE; (C) NUT; (D) PEIXE+NUT.

61
Figura 7 - Contribuição proporcional das classes de algas ao biovolume total da comunidade fitoplanctônica, nos diferentes tratamentos ao longo das semanas: (A) CTRL; (B) PEIXE; (C) NUT; (D) PEIXE+NUT.

62
Figura 8 - Magnitude do efeito (± 95% “bootstrapped” intervalos de confiança) da: (A) predação de peixes sobre a biomassa total e dos diferentes grupos da comunidade zooplanctônica, com e sem adição de nutrientes; (B) adição de nutrientes sobre a biomassa total e dos diferentes grupos da comunidade zooplanctônica, na presença e na ausência de peixes. Efeitos são significativos quando as barras de intervalo de confiança não sobrepõem a linha pontilhada.
Figura 9 - Magnitude do efeito (±95% “bootstrapped” intervalos de confiança) da: (A) presença de peixes sobre o biovolume total e dos diferentes grupos da comunidade fitoplanctônica, com e sem adição de nutrientes; (B) adição de nutrientes sobre o biovolume total e dos diferentes grupos da comunidade fitoplanctônica, na presença e na ausência de peixes. Efeitos são significativos quando as barras de intervalo de confiança não sobrepõem a linha pontilhada.

63
Figura 10 – Regressões lineares entre o biovolume total do fitoplâncton e a biomassa total do zooplâncton (a) ao longo de todos os tratamentos, (b) na ausência de peixe e (c) na presença de peixe. Dados são médias semanais entre as quatro réplicas de cada tratamento.

64
Figura 11 - Comparação dos valores de magnitude do efeito (Hedges’ d) da presença de peixes sobre o zooplâncton (A) e o fitoplâncton (B) registrados no presente estudo (barras escuras) com valores apresentados em uma meta-análise de 11 experimentos (Brett e Goldmand, 1996).

65
Discussão
Os resultados desta pesquisa sugerem que a predação por peixes onívoros não-
filtradores, a disponibilidade de nutrientes, e a interação de ambos os fatores tiveram efeito
significativo na biomassa do fitoplâncton e do zooplâncton. A presença do peixe reduziu
significativamente a biomassa total do zooplâncton e aumentou a biomassa total do
fitoplâncton, produzindo um resultado que suporta a ocorrência de cascata trófica. No
entanto, a magnitude e a direção do efeito dos peixes foi variável nos diferentes grupos
taxonômicos de fitoplâncton e zooplâncton e foi significativamente influenciado pela adição
de nutrientes.
Adequação Experimental
Este experimento foi desenhado de acordo com recomendações de Frost et al.
(2001) que dizem que muitas das importantes interações que acontecem no plâncton estão
bem representadas em experimentos de mesocosmos de mais 1 m3, com um tempo de
duração de pelo menos uma semana. Nosso experimento conduzido no campo incluiu
elementos que representam importantes componentes dos ecossistemas aquáticos, como o
padrão de circulação térmica, o contato com o sedimento e interações com a comunidade
perifítica. As manipulações dos peixes e dos nutrientes foram eficientes, i.e., a densidade
de peixes foi estável durante as 11 semanas experimentais, e a concentração de N e P
dissolvido foi mantida maior nos tratamentos enriquecidos do que nos tratamentos com
concentração ambiente. Sendo assim, estamos certos que as condições realísticas
adotadas neste experimento revelaram padrões confiáveis na distribuição de biomassa dos
níveis tróficos em resposta a pressão de predação e o efeito de nutrientes.

66
Dinâmica Temporal das Comunidades Planctônicas em Resposta à Manipulação
Experimental.
Os resultados obtidos no presente estudo indicam que o efeito das variáveis
categóricas manipuladas (peixe e nutriente), seja na biomassa do zooplâncton ou no
biovolume do fitoplâncton, foi dependente do tempo experimental (Tabela. 3 e 4, Fig. 4 e 5).
A expressão dos efeitos foi mais evidente após algumas semanas do início do experimento,
provavelmente devido ao atraso na resposta das comunidades aos fatores manipulados. Os
resultados globais foram consistentes com padrões de cascata trófica. Diferentemente das
comunidades planctônicas instituídas logo após a montagem das unidades experimentais in
situ, as comunidades bentônica e perifítica levaram um tempo maior para colonizar as
unidades. Sendo assim, foi fundamental considerar uma duração experimental compatível
com o desenvolvimento das interações tróficas em cadeias múltiplas do
Hyphesobrycon bifasciatus.
Vários estudos demonstraram a importância de levar em consideração o tempo em
experimentos de perturbação permanente (“press pertubation”), sobretudo porque a
intensidade e a direção das interações ecológicas são altamente variáveis com o tempo
(Paine 1988; Yodzis 1988; Menge et al. 1994; Power et al. 1996; Sarnelle 1997; Attayde &
Hansson 2001b; Bell et al. 2003). É esperado que o aumento da duração experimental
produza resultados mais previsíveis, uma vez transposta a fase transiente do processo
sucessional pós-perturbação (Hastings 1996; Attayde & Hansson 2001b). A maior parte dos
estudos demonstra que experimentos de longa duração produzem cascatas tróficas de
grande intensidade, mas estes efeitos podem se estabilizar, aumentar ou mesmo decrescer
após algumas gerações do produtor primário (Polis et al. 1996a; Persson 1999; Polis 1999;
Bell et al. 2003). Esta variação de resposta é conseqüência de que nem todas as espécies
de um mesmo nível trófico (i.e. fitoplâncton ou zooplâncton) são igualmente palatáveis e
vulneráveis ao consumo. A heterogeneidade específica intra-níveis tróficos tem
considerável influência na dinâmica temporal das interações entre espécies (Polis et al.
1996b; Attayde & Hansson 2001b; Bell et al. 2003; Hansson et al. 2004) e pode influenciar

67
também o quanto as forças ascendentes e descendentes irão controlar um nível trófico em
particular (Henry et al. 2006).
Experimentos em mesocosmos feitos in situ têm a vantagem de incorporar a
diversidade natural de formas, tamanho e palatabilidade das espécies. Os resultados deste
experimento demonstraram que a heterogeneidade intra-níveis tróficos teve um efeito
substancial na resposta das comunidades fitoplanctônicas e zooplanctônicas aos fatores
manipulados, na dinâmica predador-presa e conseqüentemente na ocorrência de cascata
trófica.
Nos tratamentos onde os peixes não foram incluídos (CTRL e NUT), o biovolume
total do fitoplâncton aumentou em resposta à adição de nutrientes (Fig. 5, Tabela 4). No
entanto, a partir da 3a semana, o aumento da biomassa total do zooplâncton no tratamento
NUT (Fig. 4) gerou um controle descendente eficaz sobre o fitoplâncton (Fig. 10B),
demonstrado pela tendência de queda do biovolume total até a 7a semana (Fig. 5). Os
grupos zooplanctônicos dominantes no tratamento NUT foram cladóceros e copépodes
calanóides (Fig. 6C) que são reconhecidos por apresentarem altas taxas de filtração e
potencial para o controle de florações de algas (Hansson 1992; Sarnelle 1993). Como
resultado da intensa herbivoria entre a 3a e a 7a semana no tratamento NUT, foi observado
um decréscimo significativo no biovolume de Cryptophyceae e Chlorophyceae, que são
classes de algas consideradas respectivamente de alta e média qualidade nutricional e
palatabilidade para o zooplâncton (Brett et al. 2000). Simultaneamente ao decréscimo de
biovolume do fitoplâncton no tratamento NUT, foi registrado um aumento da contribuição
relativa do biovolume de Cyanophyceae, que chegou a dominar inteiramente o tratamento
na 11a semana (Fig. 7C). A sucessão na comunidade fitoplanctônica deu-se pela
substituição de algas de crescimento rápido e palatáveis como Cryptophyceae e
Chlorophyceae, por algas de crescimento lento e reconhecida resistência à herbivoria,
como são as Cyanophyceae. Esta substituição por algas impalatáveis pode ter enfraquecido
o controle imposto pela herbivoria do zooplâncton sobre o fitoplâncton. E as conseqüências
desta mudança na qualidade do alimento aparentemente propagaram-se à comunidade

68
zooplanctônica, que apresentou redução da biomassa de crustáceos entre a 6a e a 11a
semana, quando os rotíferos mostraram significativo restabelecimento em sua biomassa.
Nossos resultados dão suporte a uma previsão secundária pouco explorada da
teoria de cascatas tróficas aquáticas, que prevê atenuação do controle descendente em
comunidades heterogêneas onde algas resistentes à herbivoria atingem altas biomassas
(Agrawal 1998; Bell 2002). Segundo esta previsão, algas de menor tamanho, com
crescimento mais rápido e mais palatáveis dominam em sistemas onde a herbivoria for
mínima; por outro lado, algas resistentes que crescem mais lentamente como resultado do
investimento em defesa, devem se desenvolver e dominar sob intensa herbivoria. Existe um
balanço entre habilidade competitiva e palatabilidade na comunidade de algas, que é
fundamental em mediar a interação entre os níveis tróficos (Agrawal 1998; Hansson et al.
1998; Bell 2002).
Quando os peixes foram incluídos (PEIXE e PEIXE+NUT), a biomassa do
zooplâncton foi reduzida em relação aos tratamentos sem peixe (Fig. 4E, Tabela 3),
enquanto o biovolume total e o biovolume da maioria das classes do fitoplâncton tiveram um
aumento significativo (Fig. 5, Tabela 4). Isto sugere que a presença de peixes foi um
importante fator na redução da pressão de herbivoria do zooplâncton sobre o fitoplâncton,
induzindo a ocorrência de cascata trófica. Apesar dos efeitos positivos globais dos peixes
sobre o fitoplâncton, o aumento do biovolume desta comunidade foi ainda maior no
tratamento PEIXE+NUT, quando houve a combinação de efeitos descendentes e
ascendentes (Fig. 5F e 10C, Tabela 4). Embora neste experimento mecanismos
relacionados à interação trófica entre o fitoplâncton e o zooplâncton não tenham sido
controlados, dois mecanismos não mutuamente exclusivos parecem ter contribuído para o
resultado acentuado desta combinação de fatores.
Primeiro, H. bifasciatus demonstrou ser um peixe de hábito seletivo na predação de
organismos zooplanctônicos de maior tamanho, produzindo um efeito global de redução da
biomassa de cladóceros e copépodes (Tabela 4), que são considerados herbívoros de
maior eficiência (Gilbert 1985, 1988; Hansson 1992; Sarnelle 1997). Especificamente no

69
tratamento PEIXE+NUT, a redução da biomassa de copépodes não foi significativa (Fig.
4C, Tabela 4). O aumento da contribuição relativa de copépodes da ordem Cyclopoida (Fig.
6D), neste tratamento, provavelmente compensou a perda total do grupo por predação. Tal
fato pode estar relacionado ao maior poder de evasibilidade dos copépodes ciclopóides à
predação, comportamento muitas vezes descrito para este grupo (Drenner & McComas
1980; Zaret 1980). A redução da biomassa do macrozooplâncton pela predação dos peixes
também produziu um efeito indireto de aumento da biomassa de rotíferos, pelo relaxamento
das pressões competitivas inter-específicas. Diversos trabalhos experimentais demonstram
a vantagem competitiva de organismos zooplanctônicos de maior tamanho (i.e.
macrozooplâncton) na coleta de partículas em suspensão (Gilbert 1988; Jack & Gilbert
1994; Sarnelle 1997; Hansson et al. 1998). Esta vantagem, no entanto, é suprimida pela
presença de peixes que exibem predação seletiva por tamanho e favorecem o crescimento
de organismos menores como rotíferos, ou com maior capacidade de fuga (Brooks &
Dodson 1965; Grygierek et al. 1966; Hall et al. 1970; Lynch 1979; Hurlbert & Mulla 1981;
Langeland 1982; Langeland & Reinertsen 1982). O favorecimento do microzooplâncton
mediado pela presença de peixes foi significativamente maior no tratamento PEIXE+NUT
(Fig. 4A, Tabela 3). De forma geral, a presença de peixes combinada à adição de nutrientes
produziu um aumento de biomassa total do zooplâncton (Fig. 10C). Este aumento deu-se
principalmente pelo crescimento do microzooplâncton, poder de evasibilidade dos
copépodes ciclopóides à predação, e redução não tão drástica da biomassa de cladóceros
por predação (i.e. as perdas podem ter sido compensadas pelo aumento de recursos
disponíveis) (Fig. 4B). De modo que, embora com uma biomassa zooplanctônica total
maior, o tratamento PEIXE+NUT sofreu uma redução funcional de herbivoria, uma vez que
as espécies presentes eram pouco eficientes no controle do fitoplâncton, o que resultou em
aumento considerável do biovolume das algas (Fig. 5 e 10C, Tabela 4).
Segundo, é conhecido que a distribuição de tamanho da comunidade zooplanctônica
pode afetar também a distribuição de tamanho das algas de várias maneiras (Bogdan et al.
1980; DeMott 1985). Em geral, grande abundância de microzooplâncton deve reduzir o

70
biovolume de algas pequenas, liberando o grupo de algas maiores e/ou impalatáveis (e.g.
Cyanophyceae) da competição com as espécies pequenas e de crescimento rápido (e.g.
Chlorophyceae e Cryptophyceae) (McCauley & Briand 1979; Reynolds 1984; Kerfoot et al.
1988; Leibold 1989; Gliwicz 1990; Sarnelle 1993; Agrawal 1998). Embora este efeito do
microzooplâncton sobre algas tenha presumivelmente ocorrido neste experimento, uma vez
que registramos um aumento simultâneo de náuplios, rotíferos e Cyanophyceae, este não
parece ter sido o principal mecanismo responsável pelo aumento adicional de biovolume
total do fitoplâncton no tratamento PEIXE+NUT. Algumas vezes o crescimento rápido de
algas pequenas pode ser por si só um mecanismo de defesa contra herbivoria (Coley et al.
1985). Em condições eutróficas, algas de crescimento rápido podem suplantar a pressão de
herbivoria do microzooplâncton, e exibir um crescimento líquido maior do que em situação
de limitação por nutrientes. Assim, o grande aumento de biovolume de algas palatáveis
como Chlorophyceae e Cryptophyceae no tratamento PEIXE+NUT, mesmo sob pressão de
herbivoria do microzooplâncton, pode ter sido função da dinâmica de nutrientes que as
favoreceram. Há também a possibilidade de classes consideradas palatáveis, como
Chlorophyceae, conterem espécies de tamanho grande para o consumo do
microzooplâncton, e com isso terem se tornado abundantes no tratamento PEIXE+NUT.
Segundo Agrawal (1998), palatabilidade e resistência à herbivoria são características
relativas à espécie de herbívoro que se estuda, e portanto, é difícil fazer classificações
rigorosas. Assim sendo, o aumento de biovolume de algas impalatáveis, somado ao
aumento do biovolume de algas de crescimento rápido que foram estimuladas pela adição
de nutrientes, parecem ter gerado o padrão de cascata trófica, com a adição de um terceiro
nível trófico (Fig. 10C). Efeitos não mutuamente exclusivos da adição de nutrientes e da
predação por peixes modificaram a composição das comunidades fitoplanctônica e
zooplanctônicas alterando a força de interação trófica entre estas comunidades e
provocando o padrão de aumento linear observado entre a biomassa zooplanctônica e
fitoplanctônica nos tratamentos com a presença de peixes em diferentes condições tróficas
(Fig. 10C).

71
O modelo tradicional de cadeias tróficas presa-dependente (prey-dependent model)
e suas modificações (Hairston et al. 1960; Fretwell 1977; Oksanen et al. 1981; Scheffer
1991) enfatizam o controle dos produtores primários pelos consumidores, e prevêem que
em sistemas de dois níveis tróficos (fitoplâncton e zooplâncton), os efeitos do
enriquecimento por nutrientes são convertidos em aumento de biomassa apenas para os
herbívoros; mas que em sistemas de três níveis tróficos (fitoplâncton, zooplâncton e peixes)
o aumento de biomassa é previsto apenas para os organismos posicionados nos níveis
tróficos ímpares. Nossos resultados para sistemas de dois níveis tróficos (CONTROLE e
NUT, Fig. 10B) são consistentes com o modelo presa-dependente. No entanto, em
contraste a este modelo, a maior biomassa de fitoplâncton e zooplâncton observada nos
tratamentos com peixes (PEIXE e PEIXE+NUT, Fig. 10C), parece corroborar o modelo de
resposta razão-dependente (ratio-dependent model), o qual prevê que o aumento na
disponibilidade de recursos propaga-se em um constante aumento da biomassa dos níveis
tróficos superiores (Arditi & Ginzburg 1989). Vários estudos de campo e laboratório em teias
tróficas aquáticas demonstraram resultado similar de aumento de biomassa simultâneo em
sistemas de dois e três níveis tróficos (Mazumder 1994; Leibold et al. 1997; Persson et al.
2001; Vakkilainen et al. 2004). De forma geral, a adição de nutrientes pareceu determinar a
biomassa total de organismos em cada nível trófico, enquanto a herbivoria e a predação
pelos peixes pareceu regular tanto a biomassa total quanto relativa dos diferentes grupos
funcionais (i.e. algas palatáveis e impalatáveis; macro e microzooplâncton). Isto enfatiza a
importância da heterogeneidade trófica entre os níveis tróficos e dentro deles nas respostas
aos fatores manipulados; e revela a limitação dos modelos teóricos de capturar toda a
variabilidade natural dos sistemas (Leibold et al. 1997; Hulot et al. 2000).
Resultados semelhantes a estes já foram descritos por vários autores, que avaliaram
o efeito de peixes zooplanctívoros e mesmo onívoros filtradores em cadeias tróficas
pelágicas (Leibold 1989; Mittelbach et al. 1995; Vanni & Layne 1997; Ives et al. 1999;
Vakkilainen et al. 2004; Okun et al. 2008; Rondel et al. 2008). No entanto, o efeito potencial
de peixes onívoros multi-cadeia e não filtradores nas comunidades planctônicas era incerto.

72
Desta forma, nossos resultados indicam que peixes onívoros não-filtradores como
H. bifasciatus, a exemplo dos demais peixes onívoros filtradores e zooplanctívoros, também
produzem cascata trófica através da predação seletiva do zooplâncton. Observamos que
substanciais desdobramentos na dinâmica das comunidades se deram em função da
duração experimental, dos grupos taxonômicos avaliados e da disponibilidade de recursos
no sistema.
Efeito da manipulação experimental na intensidade de cascata trófica
A onivoria tem sido apontada como um mecanismo que tanto pode enfraquecer
quanto intensificar os efeitos dos consumidores sobre os níveis tróficos inferiores. Esta
controvérsia tornou o comportamento onívoro um tema muito abordado na Ecologia
recente, onde se postula que o controle descendente dos onívoros não se expressa com
intensidade similar em todos os ambientes (Pace et al. 1999; Persson 1999; Polis et al.
2000; Power 2000; Schmitz et al. 2000). Tem se discutido que a comum ocorrência de
fortes cascatas tróficas em ambientes aquáticos se deve à linearidade das cadeias e à
homogeneidade dos níveis tróficos, quando comparados a sistemas terrestres (Hairston &
Hairston 1993; Polis et al. 2000). A generalização de que cadeias tróficas de ambientes
aquáticos são sempre simples e favorecem cascata trófica de grande intensidade tem sido
desafiada empírica (Chase 2000; Power 2000) (Capítulo 1) e teoricamente (Hastings 1997;
Scheffer 1998; Vadeboncoeur et al. 2005). A maioria dos estudos empíricos realizados foi
basicamente a respeito de peixes onívoros filtradores, que exploram produtores primários
de cadeias pelágicas (Vanni & Findlay 1990; Fernando 1994; Vanni & Layne 1997; Vanni et
al. 1997). Nestes trabalhos, a adição de interações onívoras em cadeias alimentares
conferiu maior reticularidade às interações tróficas, fato que gerou o enfraquecimento da
cascata trófica (Fagan 1997; McCann & Hastings 1997). No entanto, pouco se sabe
empiricamente sobre os efeitos diretos e indiretos da onivoria multi-cadeia na intensidade
do controle de biomassa dos níveis tróficos em lagos; e como estes efeitos são
influenciados pelo estado trófico.

73
Neste experimento, os resultados observados com relação à intensidade de controle
trófico tanto para o zooplâncton quanto para o fitoplâncton refletiram o que foi observado na
dinâmica destas comunidades, onde a intensidade e a direção dos efeitos foram variáveis
dependendo do nível trófico e do grupo taxonômico avaliado (Fig. 8 e 9). De maneira geral,
o peixe onívoro teve um efeito negativo na biomassa total do zooplâncton de maior
intensidade na ausência dos nutrientes (Fig. 8A). Três mecanismos podem ter contribuído
para o efeito mais intenso do peixe sobre o zooplâncton na condição sem adição de
nutrientes (Fig 8B, painel superior) e menos consistente na condição com adição de
nutrientes (Fig. 8B, painel inferior). (1) As perdas por controle de predação na condição
oligotrófica não foram compensadas por um estímulo ascendente. Este mecanismo é
corroborado pelo fato do zooplâncton ter aumentado sua biomassa pelo estímulo de
nutrientes (Fig. 8B), o que demonstra que o zooplâncton era limitado indiretamente pelos
nutrientes. (2) Na condição eutrófica, o peixe pode ter saciado sua fome ao acessar os
herbívoros zooplanctônicos, que embora sofressem com o impacto da predação, tinham
uma taxa de renovação rápida pela alta disponibilidade de alimento; somado a isso, a
adição de nutrientes favoreceu o aumento de espécies menos vulneráveis à predação (i.e.
rotíferos e copépodes ciclopóides). (3) Na condição eutrófica, os peixes predaram
significativamente mais invertebrados bentônicos (Guariento 2008). Esta troca no alvo
alimentar pode ter relaxado a pressão de predação sobre o zooplâncton e permitido que a
comunidade atingisse valores de biomassa mais elevados.
A diferença de intensidade do efeito do peixe onívoro no zooplâncton em diferentes
condições tróficas não se propagou em efeito positivo significativamente diferente no
biovolume total do fitoplâncton (Fig. 9A). A menor intensidade de predação do zooplâncton
pelos peixes na condição eutrófica (Fig. 8A) não se traduziu em um efeito de cascata trófica
enfraquecido (Fig. 9A). Nesta condição, os organismos zooplanctônicos que sobreviveram à
predação eram herbívoros menos eficientes (rotíferos, náuplios, copépodes ciclopóides).
Somado a isto, o estímulo ascendente produzido pela adição de nutrientes ao biovolume do
fitoplâncton (Fig. 9B), favoreceu o crescimento de espécies resistentes à herbivoria, bem

74
como acelerou o crescimento de outras, mesmo que vulneráveis à herbivoria.
Vários autores prevêem que a predação por peixes planctívoros atua sinergicamente
com o estado trófico do sistema, alterando a intensidade da cascata trófica, no entanto, os
resultados são controversos. A hipótese proposta por McQueen et al. (1986) prevê que o
aumento da biomassa do fitoplâncton pelo efeito indireto de peixes planctívoros seria mais
forte em sistemas oligotróficos, onde a supressão do zooplâncton herbívoro por predação
liberaria as populações de algas para crescer, mesmo que limitadas por nutrientes. Por
outro lado, em um ambiente eutrófico, a introdução de peixes planctívoros não implicaria em
aumento de biomassa fitoplanctônica, uma vez que as algas dominantes destes sistemas
são comumente mais resistentes à herbivoria (e.g. cianobactérias). Segundo alguns
autores, esta hipótese seria apenas aplicável a peixes zooplanctívoros visuais que
ativamente selecionam o macrozooplâncton e indiretamente aumentam a biomassa do
fitoplâncton pela supressão eficiente da herbivoria (Lazzaro 1987; Northcote 1988; Vanni &
Findlay 1990). Para planctívoros facultativos como peixes onívoros filtradores, a hipótese
proposta por Drenner et al. (1996) prevê intensificação da cascata trófica com o aumento da
eutrofização. O comportamento alimentar de onívoros filtradores não apenas suprime o
macrozooplâncton, mas também age como “bomba de nutrientes” enquanto os peixes
consomem nutrientes em forma de detrito no sedimento e excretam nutrientes dissolvidos
na coluna d’água (Lamarra 1975; Brabrand et al. 1990; Vanni 2002). Este efeito dos peixes
onívoros na reciclagem dos nutrientes tende a ser mais forte em sistemas mais eutróficos,
onde o maior acúmulo de nutrientes no sedimento potencializa a ação do peixe como
“bomba de nutrientes”. Desta forma, em condições eutróficas, tanto a supressão da
herbivoria quanto a reciclagem de nutrientes mediada pelos onívoros filtradores estimulam o
crescimento das algas, intensificando o efeito de cascata trófica (Drenner et al. 1996; Vanni
et al. 2006).
Baseados no presente experimento, observamos que o comportamento de peixes
onívoros não filtradores, como o H. bifasciatus, não interagiu sinergicamente com o estado
trófico para modificar a intensidade de cascata trófica (Fig. 9A). Testes experimentais desta

75
natureza se limitaram durante vários anos a avaliar o efeito dos zooplanctívoros visuais em
sua maioria (Post & McQueen 1987; McQueen et al. 1989; Lancaster & Drenner 1990;
Lafontaine & McQueen 1991; DeMelo et al. 1992) e de onívoros filtradores mais raramente
(Drenner et al. 1996; Okun et al. 2008). O comportamento alimentar de peixes, a
produtividade do sistema, o tempo de substituição das espécies e a palatabilidade das
algas são fatores interligados que afetam a distribuição de biomassa em uma rede trófica
(Cebrian 1999; Persson 1999; Polis 1999; Borer et al. 2005). Portanto, as controvérsias de
resultados aqui demonstrados indicam que mais pesquisas sobre estas relações são
necessárias. A inclusão dos efeitos de peixes onívoros não filtradores multi-cadeia nos
modelos atuais pode significativamente contribuir para o entendimento da dinâmica de
redes tróficas (Polis et al. 1996b; Vadeboncoeur et al. 2005) sobretudo em regiões tropicais
onde a diversidade e abundância destes peixes é maior (Flecker & Hu 1992; Fernando
1994; Pough et al. 2005).
O experimento com H. bifsaciatus não corroborou a hipótese de que o estado trófico
altera a intensidade de cascata trófica sobre o fitoplâncton. No entanto, nossos resultados
são congruentes com vários estudos teóricos que prevêem o enfraquecimento da
intensidade da cascata trófica pela presença de ligações tróficas difusas e mais fracas
resultantes do comportamento onívoro dos predadores (Fagan 1997; Leibold et al. 1997;
Agrawal 1998; Borer et al. 2005). As comparações entre a intensidade de cascata trófica
promovida pelo peixe onívoro no presente estudo e espécies de peixes zooplanctívoros
estritos (retirados da meta-análise de (Brett & Goldman 1996), demonstraram que H.
bifasciatus promoveu enfraquecimento significativo da cascata trófica ao nível do
fitoplâncton (Fig. 11B). Este resultado indica que peixes onívoros não filtradores multi-
cadeia têm efeito de enfraquecimento semelhante ao previsto para onívoros filtradores, no
entanto, este efeito não se dá por mecanismos diretos de consumo do fitoplâncton. Os
mecanismos indiretos pelos quais os peixes onívoros multi-cadeia podem interferir na
intensidade de cascata trófica devem estar relacionados ao forrageamento adaptativo
destes peixes, que modificam sua interação com os herbívoros (i.e. tanto na distribuição de

76
biomassa quanto a composição de espécies) e reverberam seus efeitos até a comunidade
fitoplanctônica (Vadeboncoeur et al. 2005). Resultados apresentados por Guariento (2008)
mostraram que H. bifasciatus alimentou-se também no compartimento bentônico durante
todo o experimento, com preferência por algas perifíticas na condição sem nutrientes e por
macroinvertebrados bentônicos na condição com nutrientes. É provável que este
comportamento alimentar de H. bifasciatus tenha gerado uma interação trófica mais fraca
com o zooplâncton, que liberado de uma pressão de predação intensa manteve o
fitoplâncton sob relativo controle descendente.
Entretanto é importante ressaltar ainda que a intensidade do efeito de cascata trófica
promovido pelo H. bifasciatus pode ter sido superestimado devido a artefatos experimentais
relacionados à modificação do habitat da espécie. As condições experimentais dos
mesocosmos reduziram a complexidade estrutural existente naturalmente no habitat
litorâneo em virtude da ausência da vegetação aquática, que é um importante fator
ambiental mediador da força de interação predador-presa em comunidades naturais. Em
lagos rasos não estratificados, ou mesmo nas regiões litorâneas, a complexidade estrutural
imposta pela vegetação desempenha um importante papel na redução da eficiência de
forrageamento de diversas espécies de peixes (Timms & Moss 1984; Jeppesen et al. 1998).
A disponibilidade de refúgios pode alterar a força da interação trófica entre peixes onívoros
e organismos zooplanctônicos, dificultando o acesso dos peixes a seu item alimentar
preferencial (herbívoros) e aumentando a expressão do comportamento onívoro (via maior
consumo de produtores primários e detritos). Sendo assim, numa condição de maior
complexidade estrutural, conforme a existente no habitat natural da espécie estudada, seria
esperado uma cascata trófica ainda mais fraca do peixe sobre a comunidade
fitoplanctônica.
Os resultados deste trabalho evidenciaram que onívoros não filtradores podem
significativamente afetar a ocorrência e a força de cascatas tróficas. Estudos futuros,
sobretudo na região tropical onde vivem mais de 50% das espécies de peixes sendo a sua
maioria de hábitos onívoros, devem enfatizar o papel de onívoros multi-cadeia sobre a

77
distribuição da biomassa em cadeias tróficas planctônicas. Do ponto de vista prático da
gestão dos recursos hídricos, o melhor entendimento dos mecanismos de controle trófico
desencadeados por estes peixes pode subsidiar estratégias de biomanipulação mais
adequadas para a recuperação de sistemas degradados tropicais. Do ponto de vista teórico,
o aprofundamento sobre o conhecimento dos efeitos indiretos de peixes onívoros multi-
cadeia irão significativamente contribuir para o melhor entendimento dos efeitos da onivoria
na dinâmica de cadeias tróficas aquáticas.

78
Capítulo III
Efeitos Diretos e Indiretos da Predação de um Peixe Onívoro na
Estabilidade Temporal da Biomassa e Composição de Espécies de
Comunidades Planctônicas

79
Introdução
A relação entre complexidade (maior número de interações tróficas) e estabilidade
de comunidades ecológicas tem historicamente recebido considerável atenção no âmbito da
ecologia e há muito é considerada como um tópico de intensa controvérsia (Pimm 1980;
Lawton & Warren 1989; Diehl 1995; McCann et al. 1998a; Montoya et al. 2003; Montoya et
al. 2006). O entendimento da relação entre complexidade e estabilidade pode ajudar a
elucidar como cadeias tróficas respondem a perturbações, incluindo a extinção de espécies
causada por atividades humanas. O conceito de estabilidade de ecossistemas abrange
tanto a resistência ou sensibilidade (o nível pelo qual um sistema é afetado por uma
perturbação) quanto a resiliência (a taxa na qual um sistema retorna à sua condição pré-
perturbação).
Os primeiros estudos teóricos a respeito da relação entre complexidade e
estabilidade indicaram que teias tróficas simples eram mais estáveis do que teias tróficas
mais reticuladas e complexas (May 1973) e que onívoros (espécies que ocupam mais de
um nível trófico) aumentavam a complexidade e por isso desestabilizavam cadeias tróficas
naturais (Pimm & Lawton 1978). Entretanto, comunidades naturais são complexas, com
predominância de onívoros (Polis et al. 1989; Winemiller 1990; Polis 1991; Diehl 1993; Coll
& Guershon 2002; Denno & Fagan 2003; Diehl 2003; Arim & Marquet 2004), e persistem ao
longo do tempo, o que contraria a noção de que sistemas complexos são instáveis (Polis
1991, 1998).
Vários estudos teóricos têm recentemente re-analisado a relação entre onivoria e
estabilidade assumindo entretanto pressupostos mais reais. Os resultados continuam
entretanto, apontando para divergências, com alguns modelos prevendo que a onivoria
estabiliza dinâmicas tróficas (McCann & Hastings 1997; McCann et al. 1998a; Kuijper et al.
2003; Emmerson & Yearsley 2004; Vadeboncoeur et al. 2005), enquanto outros sugerem
um efeito desestabilizador (Morin & Lawler 1996; Tanabe & Namba 2005) ou que a relação
entre onivoria e estabilidade depende de características particulares da cadeia trófica (e.g.
topologia das interações) (Vandermeer 2006). A aparente discrepância entre estudos

80
teóricos pode derivar de diferentes abordagens de modelagem, diferentes critérios de
estabilidade e/ou tipo de onivoria explorada [e.g. onivoria intra-cadeia ou onivoria de
cadeias múltiplas (Vadeboncoeur et al. 2005; Montoya et al. 2006; Ives & Carpenter 2007)].
Além disso, os poucos estudos empíricos designados a avaliar o papel da onivoria na
estabilidade, também apresentaram resultados conflitantes. Experimentos de microcosmos
em laboratório demonstraram que a variação temporal na população de presas (bactérias e
algas) não apresentou resposta diferencial à presença de consumidores onívoros e
predadores de topo (Holyoak & Sachdev 1998). Por outro lado, artrópodes onívoros
conferiram estabilidade populacional a afídeos herbívoros submetidos à ação de pesticidas
(Fagan 1997). A discrepância destes resultados gerou também críticas pelo fato de que os
sistemas experimentais utilizados não compreendiam em suas abordagens características
biológicas e físicas dinamicamente ativas que, entre outros aspectos, são responsáveis por
determinar a ocorrência e a força da expressão do forrageamento adaptativo característico
do hábito onívoro (Rosenheim & Corbett 2003; Singer & Bernays 2003). Desta forma, é
evidente que perspectivas teóricas a respeito da relação entre onivoria e estabilidade
carecem não só de um maior número de abordagens empíricas, mas também que estas
abordagens sejam conduzidas em sistemas ecológicos que conservem parte da
complexidade de teias tróficas naturais (Neutel et al. 2002; Neutel et al. 2007).
Experimentos envolvendo os efeitos de peixes onívoros na distribuição da biomassa
de suas presas, notadamente experimentos a respeito de cascatas tróficas envolvendo
comunidades zooplanctônicas e fitoplanctônicas, são relativamente comuns na literatura
ecológica (Drenner et al. 1984a; Drenner et al. 1996; Drenner et al. 1998; Schaus & Vanni
2000; Chumchal & Drenner 2004; Chumchal et al. 2005; Figueredo & Giani 2005; Vanni et
al. 2006; Okun et al. 2008). Entretanto, surpreendentemente, nenhum estudo até o
momento destinou-se a testar empiricamente os efeitos de peixes onívoros sobre a
estabilidade de níveis tróficos inferiores. Nas abordagens teóricas que envolveram esta
questão, o principal mecanismo proposto pelo qual peixes onívoros estabilizariam variações
temporais de comunidades planctônicas seria o fato de que na presença de sua presa

81
preferencial (e.g. zooplâncton) o peixe mantém sua taxa de forrageamento sobre este item,
mas em caso de redução dos estoques desta presa o peixe muda o seu forrageamento
para o fitoplâncton. Este desacoplamento adaptativo em relação a um tipo específico de
presa/alimento resulta em interações fracas e gera independência trófica do consumidor
onívoro a uma determinada presa, o que minimiza as clássicas flutuações temporais denso-
dependentes entre predador e presa responsáveis pelo aumento da probabilidade
estocástica de extinções (McCann et al. 1998a; Vadeboncoeur et al. 2005).
Este tipo de mecanismo assume interações tróficas diretas entre o peixe onívoro e
suas presas, e portanto, é limitado a explicar situações onde o peixe onívoro alimenta-se
tanto do fitoplâncton quanto do zooplâncton. Entretanto, interações tróficas indiretas são
onipresentes em teias tróficas naturais (Strauss 1991) e em muitos casos mais importantes
em determinar padrões de topologia (Miller & Travis 1996) e fluxo de energia do que
interações tróficas diretas (Reisewitz et al. 2006), sobretudo em ecossistemas tropicais,
onde a maior biodiversidade favorece o aumento de interações indiretas (Flecker & Hu
1992).
Embora existam alguns exemplos que avaliaram a importância de interações
indiretas mediadas por onívoros sobre presas (Diehl 1993, 1995), nenhum deles é
destinado a testar efeitos diretos e indiretos de peixes onívoros sobre a estabilidade de
comunidades planctônicas. Uma série de mecanismos diretos e indiretos mediados por
peixes onívoros foram ainda pouco estudados. Por exemplo, a dieta mais diversificada de
peixes onívoros leva a uma redução da força de interação/predação sobre determinada
presa. A ação direta deste mecanismo pode ser de intensidade intermediária de forma a ser
fraco o bastante para não provocar drásticas reduções na população de presas, mas forte o
suficiente para relaxar interações competitivas, estabilizando a comunidade de presas
(Paine 1966; Chase et al. 2002; HilleRisLambers & Dieckmann 2003). Além disso, conforme
tem sido proposto na literatura, a pressão de predação intermediária (e.g. onivoria) aumenta
a diversidade de presas, o que pode maximizar a coexistência de espécies (Worm et al.
2002). A maior diversidade de espécies em teias tróficas sujeitas à predação por peixes

82
onívoros poderia indiretamente aumentar a estabilidade da comunidade através da relação
entre diversidade vs. estabilidade, a qual postula por uma série de mecanismos que
comunidades mais diversas são mais estáveis (McNaughton 1978; Tilman 1996; Hughes &
Roughgarden 2000; McCann 2000; Kiessling 2005; Tilman et al. 2006; Ives & Carpenter
2007). Além disso, pouco se sabe se tais mecanismos estabilizadores diretos e indiretos da
predação de peixes onívoros sobre níveis tróficos adjacentes (e.g. zooplâncton) reverberam
de forma descendente na cadeia trófica.
Neste estudo, foram avaliados os efeitos de um peixe onívoro não filtrador
(Hyphessobrycon bifasciatus), sobre a estabilidade das comunidades zooplanctônicas e
fitoplanctônicas. A variabilidade temporal da biomassa e composição destas comunidades
foi utilizadas como medida de estabilidade. Embora a estabilidade da biomassa das
comunidades seja uma medida comumente usada, a estabilidade da composição de
espécies tem sido largamente negligenciada (Frank & McNaughton 1991; Sankaran &
McNaughton 1999; Forrest & Arnott 2006). Além disso, pelo fato da força de interação entre
predador e presa bem como os efeitos indiretos descendentes sobre a estrutura das
comunidades poderem variar em função do estado trófico (Rosenzweig 1971; Worm et al.
2002; Hillebrand et al. 2007), foi verificado como os efeitos do peixe onívoro sobre a
estabilidade das comunidades interagem com a produtividade do ambiente, manipulada
aqui pela adição de nutrientes. Para elucidar possíveis mecanismos indiretos pelos quais os
efeitos da onivoria e estado trófico pudessem afetar a estabilidade dos parâmetros e
propriedades das comunidades avaliadas, foram analisados também como a presença do
peixe onívoro e a adição de nutrientes modificam estas variáveis. Desta forma duas
hipóteses gerais foram testadas através de um experimento de campo onde a presença de
um peixe onívoro e o estado trófico foram manipulados.
(1) Efeitos diretos e indiretos da predação do peixe onívoro aumentam a
estabilidade temporal da biomassa e da composição das comunidades
fitoplanctônica e zooplanctônica.

83
(2) A adição de nutrientes enfraquece os efeitos estabilizadores da predação por
peixes onívoros sobre a biomassa e composição das comunidades fito e
zooplanctônicas.
Os resultados deste experimento foram utilizados para explorar relações de duas
classes de mecanismos pelas quais a predação por peixes onívoros e a adição de
nutrientes podem afetar a estabilidade das comunidades planctônicas: (1) mediando
interações competitivas e (2) modificando a diversidade de espécies.

84
Materiais e Métodos
Informações relacionadas a características da área de estudo, estrutura dos
mesocosmos, desenho experimental, procedimento experimental podem ser
acessadas no capítulo II.
Amostragem e Análise das Variáveis
Amostragens semanais, conduzidas consecutivamente entre a 1ª e a 7ª semana e
uma última na 11ª semana, foram realizadas totalizando 8 amostragens ao longo do
experimento. Para evitar efeitos relacionados a variações diárias das amostragens foram
sempre conduzidas entre 10 h e 12 h. Como variáveis foram analisados a estrutura (número
de espécies) e o biovolume fitoplanctônico e biomassa zooplanctônica. Para cada
mesocosmo, amostras de água da superfície (0,3 m) e fundo (1,6 m) foram coletadas com
auxílio de garrafa tipo Van Dorn e integradas em um balde de onde foram retiradas
amostras de 100 ml para determinação do biovolume (contagem de células) e identificação
das espécies fitoplanctônicas. O zooplâncton foi coletado através de 1 arrasto vertical (1,6
m) realizado com uma rede de plâncton de 65 µm. O fitoplâncton foi fixado com lugol e o
zooplâncton em solução açucarada de formalina a 4%. Em laboratório células
fitoplanctônicas foram contadas através de microscopia invertida, identificadas ao mais
detalhado nível taxonômico (em geral espécie ou gênero) e medidas de acordo com sua
conformação geométrica para o cálculo do biovolume celular (Hillebrand et al. 1999).
Organismos zooplanctônicos foram identificados ao menor nível taxonômico (exceto
náuplios) e contados através de microscópio estereoscópico. Apenas os náuplios foram
medidos. Os organismos foram agrupados em cladóceros, copépodes (calanóides e
ciclopóides), náuplios e rotíferos. Para determinação da biomassa de microcrustáceos
(copépodes e cladóceros), os primeiros 20 exemplares (somente adultos para copépodes)
contados de cada espécie foram agrupados, secos em estufa a 60oC por 24 h e em seguida
pesados em microbalança Metler modelo UMT2 para a determinação da biomassa por
indivíduo. A biomassa de adultos de microcrustáceos foi calculada multiplicando o peso
médio por indivíduo de cada espécie pela sua respectiva densidade na amostra. Para a

85
determinação da biomassa de náuplios foram utilizadas equações alométricas (peso x
comprimento) segundo Culver et al. (1985). Em seguida o peso médio por indivíduo foi
multiplicado pela densidade de náuplios na amostra. Para a determinação da biomassa de
rotíferos, a densidade de cada espécie foi multiplicada por seus respectivos pesos médios
obtidos da literatura (Wetzel & Likens 1991).
Análises dos Dados
Foram utilizados dois métodos para analisar os efeitos independentes e interativos
da adição de nutrientes e da presença do peixe sobre a diversidade das comunidades
fitoplanctônicas e zooplanctônicas. Curvas de rarefação foram utilizadas como método mais
conservativo e que permitiam a análise global dos efeitos dos fatores sobre a diversidade
planctônica. Curvas de rarefação são constituídas do acúmulo do número de espécies em
função do número ou biomassa de indivíduos amostrados. O número esperado de espécies
é calculado por sucessivas amostragens sobre uma distribuição que assume que indivíduos
de cada espécie são aleatoriamente distribuídos entre as amostragens (Gotelli & Colwell
2001). Desta forma curvas de rarefação permitem calcular a riqueza de espécies de
diferentes comunidades extraindo a interferência relacionada ao esforço de amostragem
(Connor & McCoy 1979) ou a densidade de indivíduos (Gotelli & Colwell 2001). As curvas
de rarefação foram construídas para cada tratamento considerando a riqueza e a biomassa
das espécies por amostra (mesocosmo) incluindo todas as réplicas temporais e espaciais
através do programa EstimateS versão 8.0 para Windows (Colwell 2006). Para a construção
das curvas a rotina utilizada incluiu formulas clássicas para o cálculo de Chao1 e Chao2,
com 50 permutações sem substituição (bootstrap). Para identificar diferenças estatísticas
entre as curvas, foram feitas comparações da sobreposição dos intervalos de confiança de
95% das mesmas até o limite de biomassa (zooplanctônica ou fitoplanctônica) determinado
pela menor curva. Isto permitiu comparar a diversidade de espécies de cada tratamento
descontando o efeito da abundância das comunidades (Gotelli & Colwell 2001). A
equitabilidade do zooplâncton e do fitoplâncton foi calculada semanalmente através da

86
proporção relativa entre o índice de Shannon-Wiever (real das amostras) e o índice de
Shannon-Wiever assumindo proporção relativa máxima entre as espécies. O número de
espécies total calculado da media dos valores semanais observados em cada tratamento
também foi utilizado como medida de diversidade de espécies.
A estabilidade temporal foi estimada para biovolume das classes fitoplanctônicas e
biomassa dos grupos zooplanctônicos, bem como, para o biovolume e biomassa totais
dessas comunidades em cada tratamento, através do índice de variabilidade populacional
(PV) proposto por Heath (2006). Este índice foi escolhido em detrimento de outras medidas
padrões de variabilidade estatística, tal como o coeficiente de variação (CV), pelo fato dele
ser mais robusto a eventos populacionais raros, valores zero e outros aspectos que
infringem a distribuição normal dos dados, uma vez que ele quantifica a variabilidade como
a diferença média percentual originada das comparações realizadas entre todos os valores
de abundância/biomassa observados (Heath 2006). Este índice foi calculado para as
diferentes classes fitoplanctônicas e grupos zooplanctônicos bem como para a biomassa e
biovolume total destas comunidades utilizando os seus valores registrados na série
temporal de um mesmo mesocosmo (excluindo a primeira semana pré-tratamento).
Portanto, cada tratamento conteve quatro réplicas independentes do cálculo de
variabilidade do índice PV. Para todos os cálculos do índice PV foi utilizado o programa
MATLAB versão 6.5 para Windows. Os efeitos globais independentes e interativos dos
peixes e adição de nutrientes sobre a estabilidade temporal do biovolume do fitoplâncton e
da biomassa do zooplâncton foram então primeiramente testados em um modelo
multivariado através de análise de variância múltipla (MANOVA) considerando nutrientes e
peixes como fatores categóricos e o índice PV das comunidades fitoplanctônicas e
zooplanctônicas como variável dependente. Em seguida, análises de variância bi-fatoriais
(two-way ANOVA) foram realizadas para cada variável resposta individualmente para
detectar padrões específicos de resposta da estabilidade temporal das diferentes
comunidades aos fatores manipulados (Scheiner 1993). Para diferenciar, os efeitos
individuais de cada tratamento sobre a estabilidade de cada comunidade, análises de

87
contraste com posterior correção seqüencial de Bonferroni para múltiplas comparações
(Rice 1989) foram realizadas (ver capítulo 2 para detalhes do procedimento). As análises de
variância foram realizadas através do programa STATISTICA 7.0 (StatSoft 2001).
Para elucidar possíveis mecanismos pelos quais a predação pelos peixes onívoros e
a adição de nutrientes poderiam influenciar direta e/ou indiretamente a estabilidade
temporal da biomassa dos níveis tróficos planctônicos analisados, os valores de PV
referentes à biomassa total das comunidades fitoplanctônicas e zooplanctônicas foram
modelados separadamente em função da riqueza de espécies, equitabilidade e biomassa
(Log10) da própria comunidade e da comunidade adjacente de forma a explorar a expressão
intra-trófica e inter-trófica de importantes fatores relacionados a características ecológicas
como redundância funcional, capacidade de compensação, dominância competitiva,
pressão de predação e disponibilidade de recursos, todos considerados mecanismos chave
na determinação da estabilidade de comunidades naturais (MacArthur 1955; Rosenzweig
1971; May 1973; Tilman 1999; Cottingham et al. 2001; Halpern et al. 2005; Polley et al.
2007; Hillebrand et al. 2008). Para isso foi desenvolvida uma série de modelos lineares
generalizados (Generalized Linear Models-GLZ) para observar os efeitos quantitativos dos
atributos das comunidades planctônicas sobre a sua estabilidade. De forma a selecionar o
modelo mais parcimonioso em relação à maior porcentagem de explicabilidade dos dados,
foi utilizado o Akaike’s Information Criterion (AICc, AIC de segunda-ordem, o qual é indicado
para tamanhos amostrais reduzidos), uma abordagem de seleção de modelos baseada na
teoria da informação (Johnson & Omland 2004). Diferenças no AICc (Δi) foram calculadas
entre todos os modelos candidatos da série (Burnham & Anderson 2002). A parcimônia do
modelo é inversamente proporcional ao valor de Δi de forma que valores de Δi conferem um
suporte empírico para um dado modelo (Burnham & Anderson 2002). Além disso, os
valores de Δi foram usados para calcular a medida de força do modelo (Akaike’s weight) ou
(wi) o que fornece evidência de que o modelo é verdadeiramente o mais parcimonioso na
explicação dos resultados. Os valores de wi são padronizados pelas suas somas ao longo

88
de todos os modelos avaliados de forma que seu valor é dependente da série de modelos
utilizados. A vantagem de se usar o critério de seleção de modelos é que o mesmo permite
testar estatisticamente a resposta da variável dependente a múltiplos concomitantes efeitos
ecológicos indiretos, os quais são difíceis de controlar em experimentos de campo onde os
fatores manipulados adicionam considerável complexidade à dinâmica das comunidades
(Johnson & Omland 2004). As análises para a seleção de modelos pelo método de Akaike
foram conduzidas utilizando o programa SAM (Spatial Analysis in Macroecology) versão 3.0
para Windows (Rangel et al. 2006).
Para analisar os efeitos dos fatores manipulados sobre a estabilidade temporal da
composição de espécies das comunidades fitoplanctônicas e zooplanctônicas (náuplios e
copepoditos não inclusos), foi utilizada a análise explanatória (NMDS - Non-metric
Multidimensional Scaling) baseada na dissimilaridade de Bray-Curtis calculada através das
variáveis coletadas entre semanas adjacentes (a partir da segunda semana) para cada
mesocosmo. Ao contrário de algumas outras técnicas de ordenação, a NMDS é adequada
para analisar a composição de espécies e não apresenta restrição à cerca da forma da
relação entre abundância de espécies e gradientes ambientais, o que a torna ideal para a
análise dos dados gerados através de manipulação experimental de múltiplos fatores
(Quinn & Keough 2002). A distância de Bray-Curtis foi calculada, utilizando dados da
biomassa de espécie não transformados. A melhor solução foi encontrada após 400
permutações. Os eixos foram considerados significantes se eles reduziram pelo menos 5%
do estresse total, quando comparado a menor dimensão adjacente e se produziram um
valor de estresse final significativamente menor do que o produzido por 50 permutações
aleatórias de Monte Carlo. A NMDS foi realizada utilizando o programa estatístico PC-Ord
versão 4.17 (McCune & Mefford 1999). O cálculo da distância de Bray-Curtis considera
informações a respeito da abundância de espécies, além efetivamente da presença de
espécies, e peculiaridades relacionadas ao seu cálculo infringem a premissa de
independência das réplicas necessárias para a análise de variância (Anderson 2001). Desta
maneira uma abordagem alternativa não paramétrica foi adotada para a comparação da

89
distância média de Bray-Curtis utilizando os valores entre semanas adjacentes (i.e. 2a - 3a,
3a - 4a, 4a- 5a, etc..) como réplicas do tratamento, o que fornece uma medida de similaridade
média temporal de todo o experimento. Medidas dos intervalos de confiança de 95% ao
redor dos valores médios da distância de Bray-Curtis foram construídas através da técnica
de “bootstrap” com 4999 permutações (Anderson 2001). Diferenças na dissimilaridade de
Bray-Curtis foram consideradas significantes se os intervalos de confiança de 95% em torno
da média de cada tratamento não fossem sobrepostos. Entretanto, considerando que a
similaridade de Bray-Curtis calculada entre semanas adjacentes pode ser variável ao longo
do tempo, ou seja, pode apresentar grandes mudanças na composição entre semanas
adjacentes ao mesmo tempo que se mantém pouco alterada entre outras, foi também
calculada a variabilidade (através do índice de PV) entre as diferenças de Bray-Curtis
semanais. Tal cálculo fornece uma medida de previsibilidade da mudança composicional e
permite inferir se a mudança na composição de espécies entre semanas adjacentes
mantém uma taxa constante (i.e. mais previsível) ou variável (i.e. mais imprevisível) ao
longo do tempo em resposta ao tratamento empregado. Desta forma, foram considerados
como tendo efeitos consistentes no aumento da estabilidade composicional das
comunidades somente os tratamentos onde a redução média na dissimilaridade de Bray-
Curtis entre semanas adjacentes não foi associada à grande variabilidade
(imprevisibilidade) da mudança de composição ao longo do tempo (Ives & Carpenter 2007).

90
Resultados
A presença de peixe e a adição de nutrientes afetaram significativamente porém de
forma independente e antagônica a estabilidade temporal das comunidades fitoplanctônicas
e zooplanctônicas (MANOVA; Tabela 1). A presença de peixes promoveu a redução global
da variabilidade temporal da biomassa (i.e. aumentou a estabilidade) da comunidade
zooplanctônica, enquanto que a adição de nutrientes globalmente contribuiu para o
aumento da variabilidade temporal das comunidades zooplanctônicas e fitoplanctônicas
(MANOVA; Tabela 1A, B). Para ambas as comunidades não foi observado nenhum efeito
interativo multivariado entre os fatores, provavelmente neutralizado pela direção oposta que
o efeito dos peixes e nutrientes demonstraram isoladamente (Tabela 1).
Tabela 1: Resultados da MANOVA mostrando os efeitos globais independentes e interativos da presença de peixes e adição de nutrientes na estabilidade temporal (estimada pelo índice PV) das comunidades: (a) zooplanctônica considerando a biomassa (µg.m-3) de Rotifera, Cladocera, Copepoda, Nauplii e de toda a comunidade zooplanctônica e (b) fitoplanctônica considerando o biovolume (µm3.mL-1) total da comunidade e das classes Bacillariophyceae, Chlorophyceae, Cryptophyceae, Cyanophyceae e Euglenophyceae. Valores de p em negrito indicam efeitos significativos dos tratamentos. Setas ao lado do valor de p indicam se o fator aumentou ou diminuiu a variabilidade temporal da comunidade.
Pillai Trace gl F p (a) Zooplâncton Peixe 0,86 5,8 10,26 0,0025↓ Nutriente 0,89 5,8 14,33 0,0008↑ Peixe × Nutriente 0,67 5,8 3,25 0,0673↑ (b) Fitoplâncton Peixe 0,61 6,7 1,83 0,2220↑ Nutriente 0,89 6,7 10,25 0,0035↑ Peixe × Nutriente 0,51 6,7 1,21 0,3969↑
gl – graus de liberdade
Análises de variância bi-fatoriais (ANOVA two-way) sobre os componentes
individuais das comunidades (grupos e classes) mostraram que para o zooplâncton os
efeitos da predação por peixes e adição de nutrientes foram significativos apenas para os
rotíferos, onde a presença de peixes diminuiu e a adição de nutrientes aumentou a
variabilidade temporal da biomassa deste grupo (Fig. 1A). Para o fitoplâncton, os efeitos
desestabilizadores da adição de nutrientes foram significativos sobre o biovolume das

91
classes Chlorophyceae, Cyanophyceae e sobre o biovolume total da comunidade (Fig. 2B,
D, F). Embora, a presença do peixe como fator principal (independente da condição trófica)
não tenha exercido efeito significativo sobre nenhum dos componentes da comunidade
fitoplanctônica, comparações pareadas entre tratamentos mostraram que a variabilidade
temporal da biomassa total fitoplanctônica no tratamento PEIXE foi significativamente
menor do que os demais tratamentos e o CTRL (Fig. 2F).
As curvas de rarefação demonstraram que a presença do peixe afetou
positivamente a diversidade da comunidade zooplanctônica (Fig. 3A). Os tratamentos
PEIXE e PEIXE+NUT apresentaram os maiores valores de diversidade, embora não
tenham sido diferentes entre si. O tratamento PEIXE foi significativamente diferente do
CTRL e do tratamento NUT, sendo este último o que apresentou a menor diversidade
zooplanctônica (Fig. 3A detalhe). O tratamento PEIXE+NUT foi significativamente diferente
apenas do tratamento NUT (Fig. 3A detalhe). Estes resultados refletem o mesmo padrão
observado para a estabilidade temporal da comunidade zooplanctônica, onde a presença
do peixe e a adição de nutrientes afetaram de forma contrária os padrões de diversidade da
comunidade zooplanctônica, sendo os efeitos dos peixes positivos e dos nutrientes
negativos. Em relação ao fitoplâncton, as curvas de rarefação mostraram que a diversidade
de algas foi significativamente diferente entre todos os tratamentos, sendo maior nos
tratamentos PEIXE, CTRL, NUT e PEIXE+NUT respectivamente (Fig. 3B detalhe). Houve
significativa interação entre os fatores peixe e nutrientes, que quando combinados
contribuíram para gerar os menores valores de diversidade fitoplanctônica.
Pelas análises de regressão linear, apenas a biomassa zooplanctônica foi
significativamente relacionada à variabilidade temporal (PV) do zooplâncton, sendo os
maiores valores de biomassa zooplanctônica associados à maior instabilidade da
comunidade zooplanctônica (Fig. 4A). O critério de seleção de Akaike também selecionou a
biomassa zooplanctônica como o principal preditor da variabilidade temporal do zooplâncton
(Tabela 2). Entretanto, o biovolume e riqueza do fitoplâncton foram selecionados no
segundo modelo mais parcimonioso proposto pelo método Akaike (Tabela 2).
92
Figura 1: Média [n=4 (± EP)] da variabilidade temporal da biomassa de rotíferos (a), cladóceros (b), copépodes (c), náuplios (d) e toda a comunidade zooplanctônica (e) em resposta aos tratamentos experimentais. P (peixe), N (nutriente) denotam casos onde os fatores foram significativos. *** P < 0,001. Letras diferentes sobre as barras indicam diferenças estatísticas entre os tratamentos (análise de contraste para múltiplas comparações com correção seqüencial de Bonferroni). Valores menores de PV indicam maior estabilidade temporal.
Quatro variáveis foram significativamente relacionadas à variabilidade temporal do
biovolume total fitoplanctônico (Fig. 5). A biomassa e a equitabilidade zooplanctônica e o
biovolume fitoplanctônico foram positivamente relacionados à maior variabilidade temporal
da comunidade fitoplanctônica (Fig. 5A, D, E), enquanto a equitabilidade fitoplanctônica
levou a um significativo decréscimo da variabilidade temporal do fitoplâncton (Fig. 5B). O
critério de seleção Akaike, selecionou o modelo contendo o biovolume e a riqueza de

93
espécies fitoplanctônicas como sendo o que explicou a maior variabilidade do fitoplâncton
(Tabela 3). Entretanto, razões entre as medidas de força do melhor modelo em relação aos
4 seguintes modelos (w1/wi) mostraram que a diferença entre estas medidas são muito
pequenas (8,192 – 8,875), o que indica considerável incerteza na seleção de modelos, e
Figura 2: Média [n=4 (± EP)] da variabilidade temporal do biovolume de Bacillariophyceae (a), Chlorophyceae (b), Cryptophyceae (c), Cyanophyceae (d), Euglenophyceae (e) e de toda a comunidade fitoplanctônica (f) em resposta aos tratamentos experimentais. P (peixe), N (nutriente) denotam casos onde os fatores foram significativos. * P < 0,05, ** P < 0,01, *** P < 0,001. Letras diferentes sobre as barras indicam diferenças estatísticas entre os tratamentos (análise de contraste para múltiplas comparações com correção seqüencial de Bonferroni). Valores menores de PV indicam maior estabilidade temporal.

94
portanto todas as variáveis contidas nos demais modelos podem ser importantes na
explicação da variabilidade do fitoplâncton (Tabela 3).
Medidas da distância de Bray-Curtis, evidenciaram que o tratamento PEIXE
estabilizou significativamente a variabilidade temporal da composição de espécies
zooplanctônicas ao longo do tempo (fig. 6a) e a variabilidade (previsibilidade) das
mudanças de composição entre semanas adjacentes não respondeu significativamente a
nenhum tratamento (fig. 6b), indicando a consistência do efeito estabilizador do peixe neste
tratamento. Para o fitoplâncton, foi observado o menor valor médio da distância de Bray-
Curtis no tratamento PEIXE+NUT (fig. 6c). Entretanto, a análise de variabilidade da
mudança composicional entre semanas adjacentes, mostrou que em média o tratamento
PEIXE+NUT foi o mais imprevisível (maior variabilidade) em relação às taxas de mudança
da composição de espécies fitoplanctônicas entre semanas adjacentes (fig. 6d), indicando
que entre certas semanas a magnitude de mudança da composição foi baixa, mas em
outras elevada, caracterizando um sistema pouco previsível.
A análise exploratória (NMDS) da composição das comunidades fitoplanctônica e
zooplanctônica em resposta aos fatores manipulados reforçou o resultado de haver
considerável mudança na composição entre semanas adjacentes (Fig. 7). Para o
zooplâncton, a NMDS mostra que o CTRL e os tratamentos começam a divergir a partir da
segunda semana (Fig. 7A). O CTRL e o tratamento PEIXE mostraram trajetórias temporais
praticamente paralelas (mais semelhantes entre si) em relação à composição de espécies,
mas o tratamento PEIXE, nitidamente apresenta distâncias menores entre os pontos ao
longo da progressão temporal, indicando maior estabilidade da composição. A partir da 3ª
semana, o tratamento NUT mostrou um padrão de sucessão da comunidade não direcional,
indicando instabilidade na trajetória sucessional da comunidade. A trajetória temporal mais
isolada do tratamento PEIXE+NUT indica baixa similaridade composicional deste
tratamento com os demais.
A NMDS para o fitoplâncton mostrou divergência entre os tratamentos a partir da 2ª
semana do experimento (Fig. 7B). A composição de espécies fitoplanctônicas divergiu mais

95
claramente entre os 4 tratamentos do que foi observado para o zooplâncton, mas este
padrão foi mais evidente entre as trajetórias temporais dos tratamentos com nutrientes
(NUT e PEIXE+NUT) em relação aos tratamentos sem nutrientes (CTRL e PEIXE) (Fig. 7B).
As trajetórias dos tratamentos CTRL e PEIXE foram distribuídas ao longo do eixo 2,
enquanto as trajetórias dos tratamentos PEIXE+NUT e NUT distribuídas ao longo do eixo 1.
Entretanto, assim como o observado para o zooplâncton, a distância entre os pontos que
denotam a progressão temporal do experimento foram em geral menores no tratamento
PEIXE (principalmente até a 7ª semana), do que o observado nos outros tratamentos,
indicando também um efeito estabilizador do peixe sobre o fitoplâncton em condições sem
adição de nutrientes.

96
Figura 3: Estimativa da diversidade de espécies zooplanctônicas (a) e fitoplanctônicas (b) aos efeitos individuais e interativos da adição de nutrientes e da presença de peixes estimada por curvas de rarefação. As curvas mostram a riqueza de espécies esperada para uma dada biomassa/biovolume de organismos amostrados. Diferenças estatísticas entre as curvas foram feitas analisando a sobreposição dos intervalos de confiança de 95% (barra de erros) entre as curvas até o limite fixo de biomassa/biovolume indicados pelas setas e representados em detalhe nos gráficos internos. Letras diferentes denotam que os tratamentos diferem significativamente um do outro.

97
Figura 4: Relações entre a variabilidade temporal da biomassa total zooplanctônica em relação à biomassa total (A), equitabilidade (B) e riqueza de espécies (C) do zooplâncton e ao biovolume total (D), equitabilidade (E) e riqueza de espécies (F) do fitoplâncton ao longo dos tratamentos experimentais. Pontos representam a média temporal (2a -11a semana) da variável em cada mesocosmo. A linha tracejada demarca o limite do intervalo de confiança de 95%. Valores menores de PV indicam maior estabilidade.
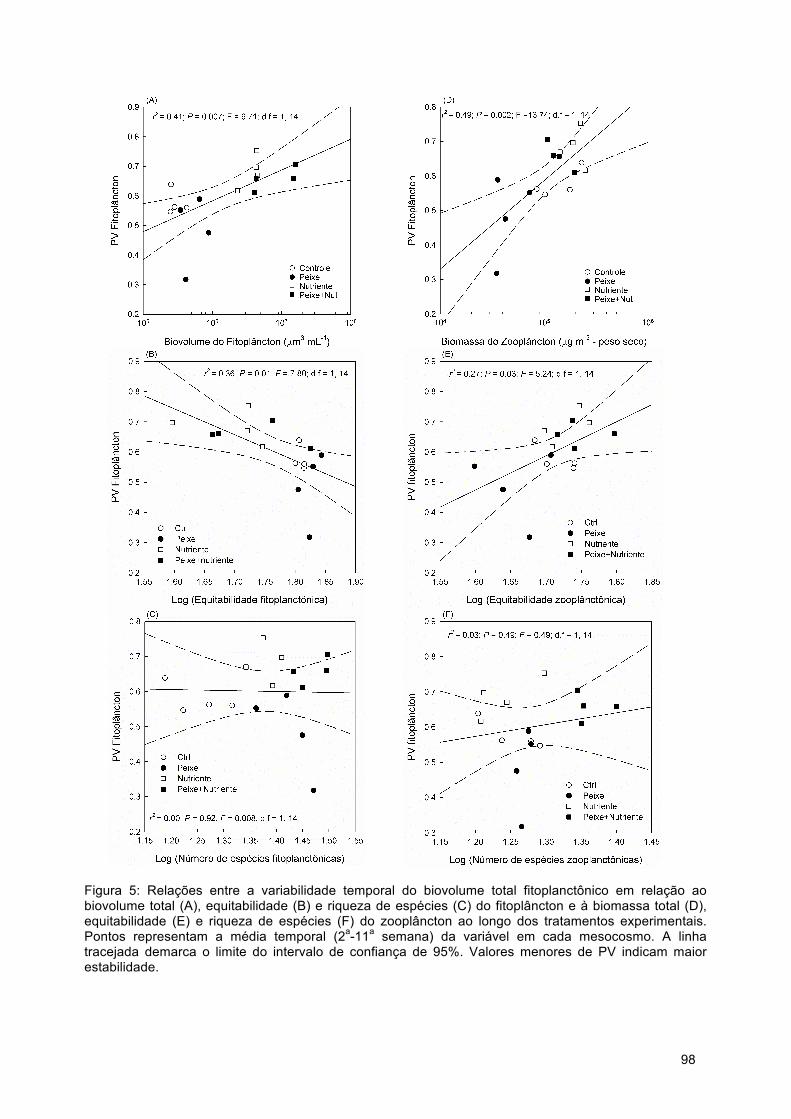
98
Figura 5: Relações entre a variabilidade temporal do biovolume total fitoplanctônico em relação ao biovolume total (A), equitabilidade (B) e riqueza de espécies (C) do fitoplâncton e à biomassa total (D), equitabilidade (E) e riqueza de espécies (F) do zooplâncton ao longo dos tratamentos experimentais. Pontos representam a média temporal (2a-11a semana) da variável em cada mesocosmo. A linha tracejada demarca o limite do intervalo de confiança de 95%. Valores menores de PV indicam maior estabilidade.

99
Tabela 2: Modelos a priori sobre as variáveis determinantes da estabilidade temporal da biomassa total zooplanctônica determinado pelo critério de seleção de Akaike. (Akaike’s selection criterion values). Apenas modelos mais parcimoniosos com valores de (Δ ) menores que 7 são mostrados.
Modelo AICc Δ AICc wi w1/wi
B zoo -37.995 0 0.202 B fito, S fito -36.005 1.990 0.075 2.693 B zoo, S zoo -35.272 2.724 0.052 3.885 E zoo -35.250 2.745 0.051 3.961 B zoo, E zoo -35.248 2.748 0.051 3.961 E fito -35.168 2.827 0.049 4.122 S fito, E fito -35.083 2.912 0.047 4.298 B zoo, E fito -35.034 2.961 0.046 4.391 B zoo, S fito -34.719 3.277 0.039 5.179 S fito, S zoo, E fito -34.569 3.426 0.036 5.611 B fito, B zoo -34.362 3.633 0.033 6.121 S fito, E zoo -33.961 4.034 0.027 7.481 S fito -33.628 4.368 0.023 8.783 B fito, S fito, S zoo -33.243 4.752 0.019 10.632 B fito -33.039 4.956 0.017 11.882 S zoo -32.848 5.147 0.015 13.467 B zoo, S fito, S zoo -32.698 5.297 0.014 14.429 E fito, E zoo -32.550 5.445 0.013 15.538 S fito, S zoo -32.524 5.471 0.013 15.538 B fito, S fito, E fito -32.440 5.555 0.013 15.538 B fito, S fito, E zoo -32.157 5.838 0.011 18.364 B zoo, S fito, E fito -32.090 5.905 0.011 18.364 S fito, E fito, E zoo -31.918 6.078 0.010 20.200 B fito, B zoo, S fito -31.892 6.103 0.010 20.200 S zoo, E fito -31.848 6.147 0.009 22.444 B zoo, S fito, E zoo -31.801 6.194 0.009 22.444 B fito, E fito -31.702 6.293 0.009 22.444 S zoo, E zoo -31.685 6.310 0.009 22.444 B fito, E zoo -31.624 6.371 0.008 25.250 B zoo, S zoo, E fito -31.393 6.602 0.007 28.857 B zoo, S zoo, E zoo -31.360 6.635 0.007 28.857 B fito, B zoo, S zoo -31.227 6.768 0.007 28.857 S fito, S zoo, E zoo -31.172 6.823 0.007 28.857 B zoo, E fito, E zoo -31.111 6.884 0.006 33.667 B fito, B zoo, E zoo -31.090 6.905 0.006 33.667 B fito, B zoo, E fito -31.028 6.967 0.006 33.667 B = Biomassa; S = Número de espécies; E = Equitabilidade

100
Tabela 3: Modelos a priori sobre as variáveis determinantes da estabilidade temporal do biovolume total fitoplanctônico determinado pelo critério de seleção de Akaike. (Akaike’s selection criterion values). Apenas modelos mais parcimoniosos com valores de (D ) menores que 7 são mostrados.
Modelo AICc Δ AICc wi w1/wi
B fito, S fito -42.151 0 0.639 B fito, S fito, E zoo -37.954 4.197 0.078 8.192 B fito, B zoo, S fito -37.890 4.261 0.076 8.408 B fito, S fito, S zoo -37.803 4.349 0.073 8.753 B fito, S fito, E fito -37.794 4.357 0.072 8.875 B = Biomassa; S = Número de espécies; E = Equitabilidade

101
Figura 7: Dissimilaridade média (a, c) e variabilidade da dissimilaridade (b, d) entre semanas adjacentes na composição das comunidades zooplanctônica (a, b) e fitoplanctônica (b, d). A dissimilaridade foi estimada a partir da distância de Bray-Curtis. A distância de Bray–Curtis pode variar de 0 (similaridade máxima) até 1,0 (dissimilaridade máxima), portanto valores médios menores denotam uma menor ou mais gradual modificação da composição das comunidades entre semanas adjacentes. O cálculo de Bray-Curtis (a, c) foi aplicado para cada mesocosmo comparando semanas adjacentes. Em seguida foram calculadas médias temporais entre todas as semanas para cada mesocosmo que foram utilizadas para calcular a média geral do tratamento e os intervalos de confiança de 95% (barra de erros) através da técnica de bootstrap com 4000 iterações. A variabilidade temporal da dissimilaridade foi calculada através do índice de variabilidade de PV, entre os valores da distância de Bray-Curtis entre semanas adjacentes calculados para cada mesocosmo. Valores mais elevados da variabilidade do índice de Bray-Curtis significam que a composição da comunidade muda em taxas diferentes entre semanas adjacentes ao longo do experimento (maior instabilidade nas taxas de mudança composicional). Letras diferentes indicam tratamentos onde não houve sobreposição entre os intervalos de confiança (para a, c) e onde os tratamentos foram diferentes entre si com pós-teste de Tukey (b, d) evidenciando diferenças estatísticas significativas entre os mesmos. Símbolo N, denota o fator Nutriente e ** significa P < 0,01. Barras de erros em (b, d) são ±DP.
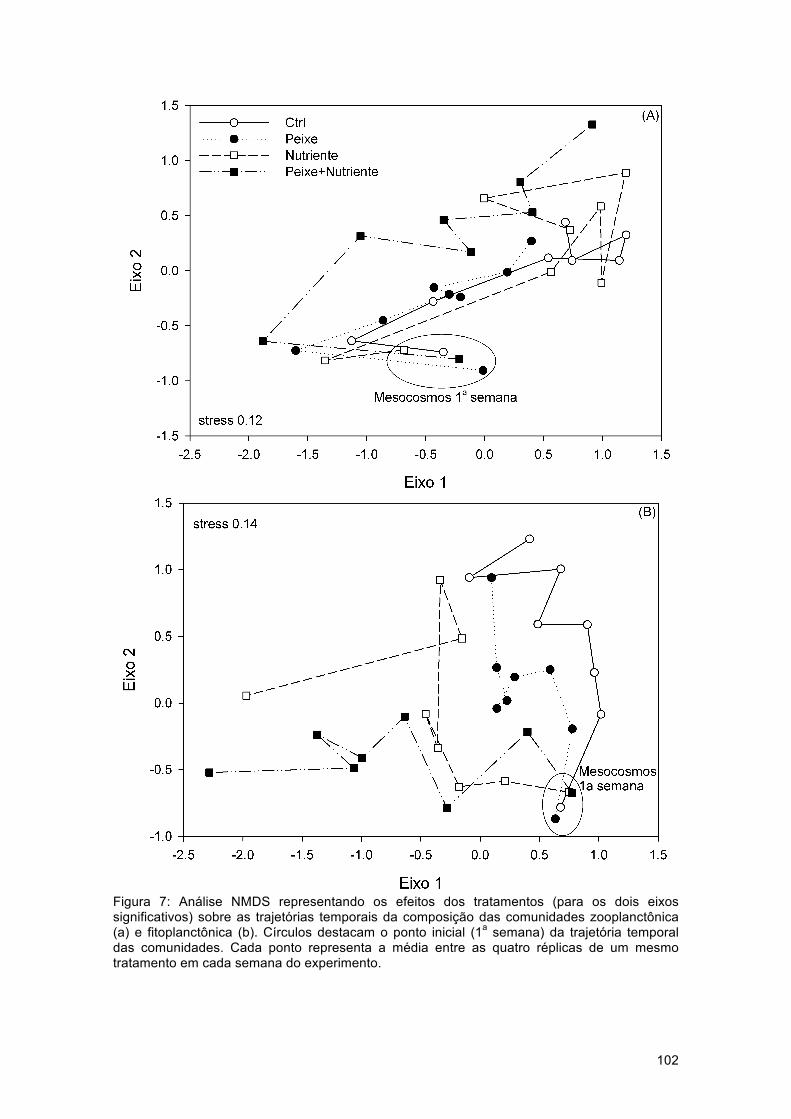
102
Figura 7: Análise NMDS representando os efeitos dos tratamentos (para os dois eixos significativos) sobre as trajetórias temporais da composição das comunidades zooplanctônica (a) e fitoplanctônica (b). Círculos destacam o ponto inicial (1a semana) da trajetória temporal das comunidades. Cada ponto representa a média entre as quatro réplicas de um mesmo tratamento em cada semana do experimento.

103
Discussão
Embora problemas de escala sejam inerentes a todas as pesquisas
experimentais, estudos realizados em mesocosmos in situ têm sido considerados uma
ferramenta realística na representação da dinâmica em larga escala de ecossistemas
naturais (Kemp et al. 2001). De fato, experimentos em mesocosmos já foram usados
com sucesso para elucidar inúmeras relações ecológicas como interação predador-
presa (Luckinbi 1974), diversidade de espécies (Dickerson & Robinson 1986),
estrutura de redes tróficas (Spencer & Warren 1996) e padrões de produtividade
primária (Petersen et al. 1997). Contudo, ainda não foram testes das predições
teóricas da relação onivoria-estabilidade em escala experimental adequada. A
abordagem em mesocosmos adotada neste estudo incluiu ambiente físico e estrutura
das comunidades bastante próximas da condição real do sistema, simulando as
interações tróficas peculiares ao forrageamento adaptativo de onívoros multi-cadeia.
Os resultados deste estudo mostraram que as interações fracas decorrentes da
predação por peixes onívoros aumentaram a estabilidade temporal da biomassa e da
composição das comunidades planctônicas, no entanto, este efeito foi dependente da
condição trófica do sistema. O efeito estabilizador da onivoria foi anulado pelo efeito
antagônico desestabilizador da adição de nutrientes. Estes resultados corroboram as
argumentações teóricas de que o forrageamento adaptativo dos predadores onívoros
estabelece caminhos alternativos para o fluxo de energia, conferindo ao sistema
ecológico maior estabilidade (Polis et al. 1996b; Fagan 1997; Holt & Polis 1997;
McCann & Hastings 1997; McCann et al. 1998a; Kondoh 2003). Adicionalmente, este
estudo forneceu evidências empíricas de que predadores onívoros que acoplam
cadeias tróficas de diferentes compartimentos (i.e. onívoros multi-cadeia) também
agem como estabilizadores de redes tróficas (Polis et al. 1996b; Vadeboncoeur et al.
2005). Além disso, o fato da estabilidade ter sido dependente da trofia do sistema,
sendo as comunidades consistentemente desestabilizadas pelo aumento de
produtividade, corrobora a clássica hipótese do paradoxo do enriquecimento

104
(Rosenzweig 1971; Gilpin 1972; McCauley et al. 1999). Assim, os resultados indicam
que a onivoria multi-cadeia e a produtividade do sistema interagem para determinar
diferentes aspectos da estabilidade das comunidades.
O debate a respeito dos mecanismos que estabilizam as interações
consumidor-recurso em ecossistemas aquáticos sujeitos a onivoria considera em sua
maioria efeitos diretos da alternância na seleção alimentar. Ou seja, a habilidade de
um predador onívoro trocar de um tipo de alimento para outro, quando o primeiro está
em baixa densidade, tende a manter as populações longe da probabilidade estatística
de se tornar extinta, e assim reduzem a amplitude das oscilações de biomassa de
consumidores e recursos, fato que é considerado um mecanismo importante na
manutenção da estabilidade das comunidades naturais. O presente estudo mostra que
não apenas os mecanismos estabilizadores diretos, mas também indiretos
relacionados a mudanças na estrutura e composição das comunidades podem mediar
a estabilidade. Tais mecanismos indiretos envolvem alteração da força de interação
trófica e competitiva entre as espécies e resposta das comunidades a variabilidade
ambiental (Tilman 1999; Yachi & Loreau 1999; Hillebrand et al. 2008).
Estabilidade na biomassa do zooplâncton e no biovolume do fitoplâncton
De acordo com a predição dos modelos teóricos, as interações onívoras em
redes tróficas aquáticas reduziram a variabilidade temporal na biomassa do
zooplâncton e no biovolume do fitoplâncton, no entanto, diferentes mecanismos foram
responsáveis pelos resultados em cada uma das comunidades.
O efeito de um predador na estabilidade temporal de uma comunidade de
presas pode ter dois mecanismos diretos e indiretos de ação. O primeiro é resultado
do consumo direto e direcional do peixe reduzindo a biomassa de microcrustáceos, e
com isso, produzindo relaxamento das pressões competitivas intra e inter-específicas
e prevenindo grandes oscilações de biomassa total (Paine 1966; Chase et al. 2002). O
segundo refere-se à mudança na estrutura da comunidade das presas pela predação

105
seletiva dos competidores superiores, mecanismo que aumenta a coexistência e
diversidade de espécies e indiretamente aumenta a estabilidade local (Chesson 2000;
Chase et al. 2002; Miracle et al. 2007).
No presente experimento, Hyphesobrycon bifasciatus tanto aumentou
significativamente a diversidade da comunidade zooplanctônica (Fig. 3A) quanto
reduziu sua biomassa total (Capítulo 2, Fig. 1E). No entanto, o critério de informação
Akaike apontou a redução de biomassa total como o modelo mais importante para
explicar a maior estabilidade temporal da comunidade zooplanctônica (Tab. 2, Fig.
4A). Ou seja, a redução de biomassa do zooplâncton causada pela predação do peixe
parece ser um dos principais fatores estabilizadores de sua oscilação temporal. Nos
tratamentos com presença de peixe foram registrados os menores valores de
biomassa do zooplâncton, um padrão evidente sobretudo no tratamento PEIXE (Fig.
4A). Esta redução na biomassa total refletiu a predação seletiva do peixe
principalmente sobre cladóceros e copépodes, e esta preferência alimentar mediou
relações inter-especificas na comunidade zooplanctônica, possibilitando o aumento da
biomassa de rotíferos. Vários trabalhos encontraram que os rotíferos, que são
competidores inferiores, se tornaram abundantes no plâncton apenas quando as
populações de microcrustáceos foram suprimidas por peixes (Grygierek et al. 1966;
Gilbert 1985; Sarnelle 1997).
Como fonte de mortalidade, a predação atua como um caso especial de
distúrbio em que o predador geralmente seleciona determinados membros (e.g. por
espécie, classe de tamanho) da comunidade (Huston 1994). Assim, se de alguma
forma a predação causar uma mortalidade diferenciada nos membros dominantes,
liberando espaço e/ou recursos para exploração de outras espécies, é provável que
haja uma maior coexistência de espécies na comunidade (Paine 1966; Leibold 1996;
Roy & Chattopadhyay 2007). Neste estudo, a predação por H. bifasciatus agiu como
um distúrbio, não extinguindo completamente os microcrustáceos e permitindo aos
rotíferos se estabelecerem. O caráter intermediário da pressão de predação deste

106
peixe deu-se pelo seu comportamento adaptativo de trocar de uma presa para outra
por questões energéticas (ver discussão em Pyke et al. 1977). Esta condição ocorreu
quando o predador onívoro passou a consumir tanto o zooplâncton quanto o perifíton,
visando complementar sua dieta (Guariento 2008).
No entanto, o efeito de controle descendente intermediário do peixe parece não
ter sido forte o suficiente para o relaxamento da competição entre os membros da
comunidade zooplanctônica, quando o efeito do peixe foi combinado com a adição de
nutrientes (tratamento PEIXE+NUT) (Fig. 1). Inúmeros trabalhos demonstraram o
efeito desestabilizador do enriquecimento de nutrientes em razão do estímulo ao
rápido crescimento das populações de produtores e consumidores, que se contrapõe
ao controle descendente, e provoca grandes oscilações populacionais (Rosenzweig
1971; Deangelis et al. 1989; PerezFuentetaja et al. 1996; Persson et al. 2001;
Romanuk et al. 2006). Os resultados do presente experimento corroboram o efeito
desestabilizador da adição de nutrientes na comunidade zooplanctônica, e
demonstram que o efeito antagônico de peixes e nutrientes contribuiu para a ausência
de efeito interativo destes fatores na estabilidade global da comunidade (Tabela 1A).
O critério de informação Akaike incluiu em seu segundo melhor modelo
(∆AIC<2, ver Burnham & Anderson 2002) os parâmetros biomassa e riqueza do
fitoplâncton, Bfito e Sfito respectivamente, como importantes variáveis para explicar a
estabilidade temporal da comunidade zooplanctônica (Tabela 2). As relações
observadas experimentalmente foram que (1) a menor estabilidade temporal do
zooplâncton se relacionou aos maiores valores de biomassa do fitoplâncton, que por
sua vez foram registrados nos tratamentos com adição de nutrientes (Fig. 4D); e (2) a
maior estabilidade temporal do zooplâncton se relacionou aos maiores valores de
riqueza do fitoplâncton (Fig. 4F), registrados nos tratamentos com presença de peixe
(principalmente no tratamento PEIXE, Fig. 3B). Portanto, a estabilidade da
comunidade zooplanctônica parece ter sido também influenciada pela comunidade
fitoplanctônica através de mecanismos indiretos relacionados à presença de peixe e

107
adição de nutrientes. O efeito estabilizador da riqueza de espécies de fitoplâncton na
comunidade zooplanctônica em mesocosmos sem nutrientes (tratamento PEIXE), e a
ausência de efeito da riqueza em mesocosmos enriquecidos com nutrientes
(tratamento PEIXE+NUT), sugerem que o aumento do número de espécies
fitoplanctônicas deve contribuir para uma redução na variabilidade da comunidade
zooplanctônica através do mecanismo de complementaridade no uso dos recursos
(Norberg 2000; Loreau & Hector 2001; Cardinale et al. 2002; Romanuk et al. 2006).
Por outro lado, nos mesocosmos com alta proporção de recursos (fitoplâncton)
disponíveis (tratamento NUT e PEIXE+NUT), a biomassa das populações
zooplanctônicas deve flutuar mais que nos mesocosmos onde o uso dos recursos é
completo (tratamentos CTRL e PEIXE) (Rosenzweig 1971; Deangelis et al. 1989;
Romanuk et al. 2006).
O critério de informação AIC selecionou a biomassa e a riqueza do fitoplâncton
como as melhores variáveis para explicar estabilidade temporal do fitoplâncton (Tabela
3). Em geral, o maior biovolume do fitoplâncton foi registrado nos tratamentos com
adição de nutrientes (NUT e PEIXE+NUT), que também apresentaram a maior
instabilidade temporal da comunidade (Fig. 5A). Este resultado corrobora o efeito
global significativo do nutriente de aumentar a variabilidade temporal da comunidade
fitoplanctônica (Tabela 1B). Por outro lado, os menores valores médios de biovolume
do fitoplâncton foram observados nos tratamentos CONTROLE e PEIXE (Fig. 5A) e
embora, neste último, a presença do peixe tenha estimulado um aumento de
biovolume fitoplanctônico significativamente maior do que no controle (Fig. 2F do
Capítulo 2), ainda assim, a maior estabilidade temporal do fitoplâncton foi
significativamente maior no tratamento PEIXE (Fig. 2F). Isso indica que o efeito da
estabilidade do fitoplâncton mediada pelo seu biovolume sofreu influência tanto da
adição de nutrientes quanto da presença de peixes. No entanto, o resultado
ascendente da adição dos nutrientes teve um efeito desestabilizador no biovolume do
fitoplâncton mais forte que o efeito estabilizador da presença do peixe (Tabela 1B).

108
Alguns estudos sugerem assimetria na intensidade relativa dos controles ascendentes
e descendentes na distribuição de biomassa em comunidades pelágicas de água doce
(Brett & Goldman 1997; Borer et al. 2006). É previsto que o controle ascendente deve
ser maior na base das cadeias tróficas, e o controle descendente maior nos níveis
tróficos superiores (McQueen et al. 1986). Porém, este é o primeiro trabalho que
estende a previsão de assimetria no controle ascendente e descendente da biomassa
dos níveis tróficos, para a estabilidade temporal das comunidades. As implicações
práticas deste resultado demonstram que a eutrofização de sistemas aquáticos pode
eliminar o efeito estabilizador previsto para peixes onívoros.
A riqueza de espécies fitoplanctônicas apresentou, segundo o critério AIC, uma
correlação negativa significativa com a variabilidade temporal do biovolume total da
comunidade (r = -0,945), ou seja, quanto maior foi a riqueza, maior foi também a
estabilidade do fitoplâncton. Apesar da análise de regressão não ter apresentado
relação entre estas variáveis (Fig. 5C), o critério AIC selecionou a riqueza como um
bom modelo explicador da variabilidade temporal do fitoplâncton, pois ele leva em
consideração não apenas o ajuste dos dados à regressão, mas também a
complexidade entre as variáveis incluídas no modelo (i.e. como as variáveis interagem
entre si). A análise detalhada dos pontos da regressão (Fig. 5C) bem como das curvas
de rarefação (Fig. 3B) indica que os tratamentos onde os peixes foram incluídos
(PEIXE e PEIXE+NUT) apresentaram os maiores valores de riqueza fitoplanctônica,
mas apenas no tratamento PEIXE os maiores valores de riqueza estiveram
verdadeiramente relacionados à maior estabilidade temporal do fitoplâncton (Fig. 2F).
O aumento da riqueza do fitoplâncton mediada pela presença do peixe pode ter
vias descendentes e ascendentes de atuação. A via descendente diz respeito à
redução da biomassa e aumento da diversidade de espécies do zooplâncton
resultantes da pressão de predação seletiva do H. bifaciatus (i.e. diminuição das
populações de microcrustáceos possibilitando a coexistência com rotíferos). Desta
forma, o equilíbrio entre atenuação de herbivoria e manutenção de espécies de

109
herbívoros com habilidades variadas de consumir algas sem pressioná-las à extinção,
pode ter contribuído para estabelecer uma comunidade fitoplanctônica mais diversa no
tratamento PEIXE (Fig. 3B) (Agrawal 1998; Scheffer et al. 2003; Hillebrand et al.
2007). A via ascendente pela qual o peixe influencia a comunidade fitoplanctônica
pode estar relacionada à translocação de nutrientes do habitat bentônico/litorâneo
para o pelágico via excreção. Neste experimento, o hábito onívoro em cadeias
múltiplas do H. bifasciatus provavelmente atuou translocando nutrientes do perifíton
para a coluna d’água (Guariento 2008). Vários estudos recentes têm demonstrado a
importância da excreção de peixes como uma fração substancial da demanda das
algas por nutrientes (Vanni 2002); sua contribuição relativa parece ser determinante
em ambientes oligotróficos (Gido 2002) estimulando o aumento da produtividade
primária e da diversidade de espécies pela disponibilização de recursos limitantes
(Srivastava & Lawton 1998). Entretanto, na condição eutrófica (tratamento
PEIXE+NUT), a atenuação da herbivoria mediada pelo peixe reduziu a diversidade
provavelmente por favorecer a dominância de espécies (Fig. 3B). Este resultado é
compatível com o modelo que prevê que uma pressão de herbivoria acentuada tem
efeito negativo na diversidade de produtores primários, quando estes estão sob regime
de recursos limitantes, e portanto, a atenuação da herbivoria promove um aumento na
diversidade em ambientes oligotróficos (Worm et al. 2002).
Inúmeras evidências empíricas demonstram que a diversidade de espécies e a
estabilidade estão positivamente correlacionadas (Frank & McNaughton 1991; Worm &
Duffy 2003; Kiessling 2005; Tilman et al. 2006). Os resultados apresentados neste
experimento para a comunidade fitoplanctônica corroboram esta tendência (Tabela 3).
No entanto, os mecanismos por trás desta proposição parecem ser mais complexos do
que o simples aumento do número de espécies e das possibilidades de interação
trófica entre elas (McCann 2000). Os argumentos básicos para esta correlação
positiva entre diversidade e estabilidade dos produtores primários em nível de
comunidade podem ser classificados em duas hipóteses mecanicistas, não

110
mutuamente exclusivas. A primeira prevê que quanto maior a diversidade maior é a
probabilidade de co-ocorrência de espécies que se complementem em suas funções
no ambiente através de efeitos de diferenciação de nicho; este mecanismo deve
supostamente relaxar a competição inter-específica, prevenindo grandes oscilações
populacionais ("complementarity hypothesis", Tilman 1996; Hooper & Vitousek 1998;
Loreau 1998). A segunda hipótese inclui dois pressupostos probabilísticos que podem
ou não considerar interação entre as espécies; uma explicação é que aumentando a
diversidade aumenta a chance de pelo menos algumas espécies responderem
diferencialmente à perturbações e à condições flutuantes do ambiente (Lawton &
Brown 1994; Naeem & Li 1997; Naeem 1998); a explicação seguinte considera que
com o aumento da diversidade aumenta as chances do sistema apresentar
redundância funcional por conter espécies capazes de compensar a perda de
espécies importantes (Lawton & Brown 1994; Naeem & Li 1997; Naeem 1998). A
combinação destes dois pressupostos probabilísticos é a base conceitual da “hipótese
da seguridade” ("insurance hypothesis", Yachi & Loreau 1999) que atualmente tem
sido amplamente aceita como explicação para a relação positiva entre diversidade e
estabilidade. Isolar dentre os vários mecanismos apresentados, aquele que seja o
principal gerador da estabilidade é uma tarefa difícil para experimentos em escala de
redes tróficas, sendo na realidade possível que eles todos estejam ocorrendo
simultaneamente. O presente experimento sugere que a onivoria teve um importante
efeito estabilizador na variabilidade temporal da comunidade fitoplanctônica, e que
este efeito foi indiretamente mediado pela riqueza de espécies.
Estabilidade Temporal da Composição do Zooplâncton e do Fitoplâncton
A estabilidade de redes tróficas tem sido amplamente discutida baseada na
variabilidade agregada das comunidades, ou seja, o foco está nas mudanças em
variáveis que integram múltiplas espécies, como por exemplo abundância total,
produtividade, ciclagem de nutrientes ou biomassa da comunidade (Micheli et al.

111
1999). Pouca atenção tem sido destinada à variabilidade na composição de espécies,
que leva em consideração mudanças relativas na abundância das espécies
componentes (Frank & McNaughton 1991; Sankaran & McNaughton 1999). Até o
momento, nenhum estudo foi feito para testar o papel da onivoria na estabilidade da
composição de espécies. Além disso, dos raros estudos que contemplam
simultaneamente a variabilidade de propriedades agregadas das comunidades e da
composição de espécies, apenas três o fizeram de forma a associar resultados
(Silvertown et al. 1994; Tilman 1996; Sankaran & McNaughton 1999). A proposta de
lidar claramente com estas duas vertentes da variabilidade das comunidades pode
promover o entendimento holístico dos fatores que promovem a estabilidade, além de
ajudar a solucionar um longo caminho de controvérsias na ecologia de comunidades.
Por exemplo, May (1973) sugeriu que comunidades mais ricas em espécies seriam
também menos estáveis, enquanto vários trabalhos empíricos sugerem o oposto
(Tilman & Downing 1994; Tilman 1996). Na verdade, estes dois resultados podem não
ser necessariamente contraditórios porque May (1973) lidou com a variabilidade da
composição, enquanto os estudos empíricos focaram na variabilidade agregada. O
modelo conceitual apresentado por Micheli et al. (1999) demonstrou que medidas
integradas (e.g., biomassa da comunidade) e a composição de espécies podem
responder independentemente a alterações ambientais ou da comunidade. Nosso
estudo é pioneiro no sentido que experimentalmente demonstrou que a onivoria pode
desencadear mecanismos capazes de estabilizar tanto medidas integradas da
comunidade quanto a composição de espécies.
A exemplo do que foi observado para biomassa do zooplâncton (Tabela 1), o
peixe onívoro também estabilizou a composição de espécies do zooplâncton entre
semanas adjacentes, e em contraponto, a adição de nutrientes produziu um efeito
desestabilizador que neutralizou o efeito da presença do peixe (Fig. 6A e 7A).
A literatura tem demonstrado uma dualidade na natureza dos mecanismos que
regem a estabilidade da dinâmica das espécies e das propriedades agregadas que

112
descrevem a comunidade. Um modelo conceitual proposto por Micheli et al. (1999)
apresentou como ambos os componentes da variabilidade da comunidade podem
estar relacionados positiva ou negativamente, produzindo cenários extremos com
quatro possíveis combinações de baixa e alta variabilidade da composição e das
propriedades agregadas. A confrontação deste modelo com os resultados
experimentais da variabilidade do zooplâncton apontou dois cenários de relação
positiva. A presença do peixe onívoro produziu baixa variabilidade temporal da
biomassa e baixa variabilidade da composição de espécies zooplanctônicas, cenário
denominado como estase (“stasis”). O mecanismo proposto por Micheli (1999) prevê
que este cenário ocorre sob condições de equilíbrio competitivo entre as espécies, em
um ambiente relativamente estável. No tratamento PEIXE, o relaxamento de
interações de competição assimétrica entre as espécies do zooplâncton, mediada pela
predação seletiva do peixe sobre o competidor superior (i.e., microcrustáceos), parece
ter sido o principal mecanismo estabilizador. O efeito do peixe na redução da
biomassa total da comunidade, simultânea à manutenção da simetria na intensidade
do efeito de competição per capita de uma espécie na outra, reduziu a variação tanto
na flutuação temporal da biomassa quanto na substituição das espécies. Acreditamos
que o comportamento onívoro do peixe teve um papel importante neste cenário, pois o
forrageamento adaptativo em dois compartimentos (pelágico e bentônico) reduz a
probabilidade de grandes flutuações populacionais das presas (Polis et al. 1996b;
Schindler & Scheuerell 2002; Vadeboncoeur et al. 2005).
Por outro lado, encontramos que a adição de nutrientes desacopla o efeito
estabilizador do peixe onívoro, e em conseqüência, observamos uma alta variabilidade
tanto da biomassa quanto na composição de espécies do zooplâncton, produzindo um
cenário de assincronia (“asynchrony”) no tratamento PEIXE+NUT. A assincronia
ocorre quando as espécies respondem de forma complexa e independente à fatores
abióticos, produzindo flutuações populacionais de grande amplitude, semelhantes aos
observados em ambientes com alta disponibilidade de recursos (Rosenzweig 1971;

113
Deangelis et al. 1989). No tratamento PEIXE+NUT, o enriquecimento teve um papel
crucial em manter espécies competidoras superiores no sistema, que de outra forma
seriam vulneráveis a predação. Isso gerou dinâmicas transientes de substituição de
espécies zooplanctônicas e portanto resultou em maior variabilidade temporal da
biomassa e da composição.
Para a comunidade fitoplanctônica, a presença do peixe onívoro produziu um
efeito de estabilização do biovolume total apenas no tratamento PEIXE (Fig. 2F), e um
efeito de estabilização menos evidente na composição de espécies (Fig. 6C e D, 7A).
A similaridade na composição de espécies fitoplanctônicas entre semanas adjacentes
apontou apenas o tratamento PEIXE+NUT como o mais estável (Fig. 6C), no entanto,
este resultado apresenta um aspecto peculiar que deve ser interpretado à luz da
variabilidade temporal das mudanças registradas na composição (Fig. 6D). Após a
adição de nutrientes, um subconjunto das espécies fitoplanctônicas foi perdida,
permaneceram no sistema aquelas que se tornaram muito abundantes; a dominância
relativa destas espécies aumentou significativamente a variabilidade em resposta aos
nutrientes (Fig. 6D). A dominância de espécies aumenta a importância relativa das
interações competitivas intra-especificas para as espécies dominantes, que vão exibir
dinâmicas de crescimento logístico até o limite da capacidade suporte do sistema,
quando decrescem bruscamente (Hillebrand et al. 2008). Assim, em sistemas de alta
dominância, onde uma ou apenas poucas espécies fazem muita diferença para o
biovolume total e para a composição, dinâmicas de substituição entre a espécies criam
situações de alta similaridade entre semanas adjacentes durante as florações, e por
outro lado, criam situações de alta dissimilaridade nos períodos de transiência
(Cottingham et al. 2001). Isto explica a maior similaridade média na composição do
fitoplâncton entre semanas adjacentes em resposta aos nutrientes (Fig. 6C), no
entanto, este efeito não pode ser considerado estabilidade per se, pois foi acoplado a
menor previsibilidade nas taxas semanais de mudança composicional (Fig. 6D).

114
Sobretudo no tratamento PEIXE+NUT, a presença do peixe agravou a ocorrência de
florações de algas, pois o controle descendente da herbivoria foi reduzido.
O resultado da MNDS mostra a distinção na trajetória dos tratamentos onde
nutrientes foram adicionados (NUT e PEIXE+NUT), além de corroborar graficamente a
alta variabilidade na mudanças da composição de espécies fitoplanctônicas em
resposta ao enriquecimento (i.e., existem trajetórias longas e curtas entre semanas
adjacentes, que representam baixa e alta similaridade, respectivamente) (Fig. 7B). Por
outro lado, o tratamento PEIXE mostrou a menor dispersão de pontos na trajetória
temporal entre as semanas, o que é um forte indicativo de estabilidade na composição
de espécie do fitoplâncton (Fig. 7B). Assim, a presença do peixe onívoro parece
estabilizar tanto a composição do fitoplâncton quanto o biovolume total da comunidade
através de mecanismos ascendentes e descentes, que atuam impedindo que alguma
espécie obtenha monopólio competitivo (Cottingham et al. 2001; Hillebrand et al.
2008). Novamente, o efeito do peixe onívoro na estabilidade temporal da composição
do fitoplâncton parece depender decisivamente do estado trófico do sistema.
Nosso resultados mostram que o peixe onívoro não filtrador tem efeito
estabilizador tanto nas propriedades agregadas (i.e., biomassa e diversidade) quanto
na composição de espécies dos níveis tróficos basais (i.e., zooplâncton e fitoplâncton).
Este resultado é consistente com a idéia de que redes tróficas alimentares com
interações onívoras fracas são mais estáveis e por isso, são comumente encontradas
na natureza. Os mecanismo responsáveis pelo efeito estabilizador da onivoria
envolvem interações diretas (e.g., predação seletiva do zooplâncton e excreção de
nutrientes disponíveis ao fitoplâncton) e indiretas (e.g., alteração das habilidades
competitivas intra e inter-específicas) que modificam atributos das comunidades (e.g.,
riqueza, equitabilidade) capazes de promover estabilidade temporal. Nosso resultados
mostram ainda que o papel estabilizador da onivoria é dependente da integração entre
compartimentos (litorâneo/bentônico) quando os nutrientes são limitantes, no entanto,
esta “via de estabilização” é enfraquecida em resposta ao enriquecimento trófico. Do

115
ponto de vista teórico estes resultados são importantes no que diz respeito à teoria
diversidade-estabilidade, pois mostram que modificações reais da diversidade de
espécies significativamente afetam a estabilidade de propriedades agregadas e
populacionais de comunidades naturais, uma questão que têm sido basicamente
explorada matematicamente ou utilizando comunidades formadas experimentalmente
(Ives & Cardinale 2004). Sob este ponto de vista, os resultados indicam que peixes
onívoros podem desempenhar funções chave em redes tróficas reais. Sob o aspecto
prático, os resultados apontam tendências preocupantes, visto que tanto o
enriquecimento trófico, quanto a redução e extinção de populações e espécies de
peixe apresentam um crescente aumento em regiões tropicais. Considerando a
prevalência do hábito onívoro entre os peixes de água doce nestes sistemas, tais
modificações podem significativamente alterar a forma pela qual as comunidades
resistem e respondem a flutuações ambientais, potencializando a chance de extinções
por eventos estocásticos populacionais e conseqüentemente contribuindo para a
perda de biodiversidade aquática.

116
Discussão e Conclusão Geral
Desde que foi proposta, a teoria de cascata trófica tem estimulado
controvérsias sobre a importância dos fatores ascendentes (nutrientes) e
descendentes (predação) na determinação de estrutura de redes tróficas (Brett &
Goldman 1997; Borer et al. 2006; Gruner et al. 2008). Vários estudos empíricos se
voltaram para esta questão, mas uma crítica veemente do início da década de 90
ponderou que “mesmo uma breve análise da literatura específica indica que, longe de
ser robusta, a teoria de cascata trófica está baseada em meias-verdades e
extrapolação exagerada dos dados”. Desde então, muitos estudos de síntese
buscaram generalizações na literatura sobre a ocorrência de cascata trófica em uma
ampla variedade de ecossistemas aquáticos e terrestres (Shurin et al. 2002; Borer et
al. 2005); vários modelos matemáticos acrescentaram à teoria elementos com
interações onívoras e heterogeneidade dos níveis tróficos (McCann et al. 1998b;
Persson 1999). Mas a união das vertentes teóricas e empíricas permanece como uma
lacuna até os dias atuais. Dentro do escopo restrito de ambientes aquáticos de água
doce, este estudo teve como objetivo reconciliar proposições empíricas e teóricas.
A meta-análise e o experimento apresentados neste estudo promoveram
suporte irrestrito à ocorrência de cascata trófica em ecossistemas aquáticos de água
doce. Tanto peixes onívoros quanto planctívoros tiveram grande impacto na biomassa
do zooplâncton; no entanto, a reverberação deste impacto para o fitoplâncton sofreu
influência da heterogeneidade intra-trófica e da disponibilidade de nutrientes. Apesar
de evidente na maior parte dos experimentos apresentados na meta-análise, a
intensidade da cascata trófica foi significativamente reduzida pela presença de peixes
onívoros, assim como previsto pela teoria (Polis et al. 1996b).
O segundo aspecto do efeito da onivoria muito abordado, porém ainda
controverso, é a estabilidade promovida por ela. MacArthur (1995) pioneiramente
previu que o aumento de “caminhos de fluxo de energia” promovido por interações

117
onívoras preveniria grande flutuações das comunidades de presa, enquanto Pimm e
Lawton (1978) e May (1972 e 1973) demonstraram matematicamente que o aumento
dos possíveis tipos de interação predador-presa aumentaria significativamente a
freqüência de extinções. Seguiram-se, a estas duas vertentes, várias discussões
teóricas, porém a ausência de síntese sobre o assunto mantém a questão em aberto.
Os resultados meta-analíticos demonstraram que, se por um lado, a maior
conectividade entre níveis tróficos originadas pela presença do peixe onívoro afetou
significativamente a intensidade de cascata trófica, por outro lado, não promoveu
maior estabilidade. Em nosso experimento, porém, a onivoria teve efeito estabilizador
tanto nas propriedades agregadas (biomassa do zooplâncton e biovolume do
fitoplâncton), quanto na composição de espécies.
A comparação do resultado meta-analítico e do experimento chama atenção
para outra variável importante para o estudo de redes tróficas: o tipo de onivoria
abordado. A definição de onivoria mais amplamente aceita e estudada é baseada no
hábito do predador de alimentar-se em mais de um nível trófico, porém na mesma
cadeia alimentar. Peixes onívoros desta natureza geralmente são filtradores que tem
impacto direto nas comunidades de fito e zooplâncton. No entanto, a estrutura de
ecossistemas raramente pode ser reduzida a uma simples cadeia (Polis e Strong
1996) e predadores, presas e recursos normalmente atravessam os limites de habitas
e ecossistemas (Polis 1991). Esta diversidade de caminhos tróficos é característica
dos onívoros multi-cadeia, aos quais é atribuído um efeito teórico de conectividade e
estabilidade ainda mais consistentes (Vadeboncoeur, 2005).
A discordância entre os resultados da meta-análise e do experimento sugere
que o tipo de onívoro pode ser realmente uma variável importante de se considerar,
uma vez que a maioria dos peixes incluídos na meta-análise eram onívoros de cadeia
simples, enquanto H. bifasciatus um onívoro multi-cadeia. Neste contexto, é
indispensável que experimentos em cadeias tróficas daqui para frente incluam peixes
onívoros multi-cadeia, para que seja construído um cenário mais amplo sobre os

118
efeitos da onivoria na distribuição de biomassa e estabilidade de propriedades
agregadas e composição de espécies. Além disso, analisar estes efeitos
simultaneamente sobre o maior número possível de comunidades (e.g., planctônicas,
litorâneas e bentônicas) e compartimentos (e.g., pelágico, litorâneo e bentônico) pode
gerar melhores conclusões sobre como o forrageamento adaptativo pode influenciar a
estabilidade ao nível de ecossistema.
Tanto na meta-análise quanto no experimento, o enriquecimento trófico foi um
importante modificador dos efeitos da onivoria, reduzindo seu efeito estabilizador.
Deste modo, a eutrofização antropogênica pode alterar significativamente os caminhos
evolutivos e ecológicos pelos quais a onivoria se expressa em ecossistemas
aquáticos. Esta afirmação é particularmente importante para ecossistemas tropicais,
onde grande parte das espécies de peixes são onívoras, e onde a degradação dos
ecossistemas aquáticos continentais é uma crescente realidade (Schindler 2007).

119
Referências Bibliográficas
Agrawal A.A. (1998). Algal defense, grazers, and their interactions in aquatic trophic cascades. Acta Oecologica-International Journal of Ecology, 19, 331-337.
Agrawal A.A. & Karban R. (1997). Domatia mediate plant-arthropod mutualism. Nature, 387, 562-563.
Anderson M.J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecology, 26, 32-46.
Arditi R. & Ginzburg L.R. (1989). Coupling in predator prey dynamics - Ratio-Dependence. Journal of Theoretical Biology, 139, 311-326.
Arim M. & Marquet P.A. (2004). Intraguild predation: a widespread interaction related to species biology. Ecology Letters, 7, 557-564.
Armsworth P.R. & Roughgarden J.E. (2003). The economic value of ecological stability. Proceedings of the National Academy of Sciences of the United States of America, 100, 7147-7151.
Arnqvist G. & Wooster D. (1995). META-ANALYSIS - SYNTHESIZING RESEARCH FINDINGS IN ECOLOGY AND EVOLUTION. Trends in Ecology & Evolution, 10, 236-240.
Attayde J.L. & Hansson L.A. (2001a). Fish-mediated nutrient recycling and the trophic cascade in lakes. Canadian Journal of Fisheries and Aquatic Sciences, 58, 1924-1931.
Attayde J.L. & Hansson L.A. (2001b). Press perturbation experiments and the indeterminacy of ecological interactions: effects of taxonomic resolution and experimental duration. Oikos, 92, 235-244.
Bell T. (2002). The ecological consequences of unpalatable prey: phytoplankton response to nutrient and predator additions. Oikos, 99, 59-68.
Bell T., Neill W.E. & Schluter D. (2003). The effect of temporal scale on the outcome of trophic cascade experiments. Oecologia, 134, 578-586.
Bogdan K.G., Gilbert J.J. & Starkweather P.L. (1980). In situ clearance rates of planktonic rotifers. Hydrobiologia, 73, 73-77.
Borer E.T., Halpern B.S. & Seabloom E.W. (2006). Asymmetry in community regulation: Effects of predators and productivity. Ecology, 87, 2813-2820.
Borer E.T., Seabloom E.W., Shurin J.B., Anderson K.E., Blanchette C.A., Broitman B., Cooper S.D. & Halpern B.S. (2005). What determines the strength of a trophic cascade? Ecology, 86, 528-537.
Bottrell H., Duncan A., Gliwicz Z., Grygierek E., Herzig A., Hillbricht-Ilkowska A., Kurasawa H., Larsson P. & Weglenska T. (1976). A review of some problems in zooplankton production studies. Norwegian Journal of Zoology, 24, 419-456.
Boveri M.B. & Quiros R. (2007). Cascading trophic effects in pampean shallow lakes: results of a mesocosm experiment using two coexisting fish species with different feeding strategies. Hydrobiologia, 584, 215-222.
Brabrand A., Faafeng B.A. & Nilssen J.P.M. (1990). Relative importance of phosphorus supply to phytoplankton production - Fish excretion versus external loading. Canadian Journal of Fisheries and Aquatic Sciences, 47, 364-372.
Brett M.T. & Goldman C.R. (1996). A meta-analysis of the freshwater trophic cascade. Proceedings of the National Academy of Sciences of the United States of America, 93, 7723-7726.
Brett M.T. & Goldman C.R. (1997). Consumer versus resource control in freshwater pelagic food webs. Science, 275, 384-386.
Brett M.T., Muller-Navarra D.C. & Park S.K. (2000). Empirical analysis of the effect of phosphorus limitation on algal food quality for freshwater zooplankton. Limnology and Oceanography, 45, 1564-1575.
Brooks J. & Dodson S. (1965). Predation, Body Size, and Composition of Plankton. Science, 150, 28-35.
Burnham K.P. & Anderson D.R. (2002). Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach. 2 edn. Springer, New York, USA.
Byrnes J., Stachowicz J.J., Hultgren K.M., Hughes A.R., Olyarnik S.V. & Thornber C.S. (2006). Predator diversity strengthens trophic cascades in kelp forests by modifying herbivore behaviour. Ecology Letters, 9, 61-71.
Caramaschi E.P., Sánchez-Botero J.I., Hollanda-Carvalho P., Brandão C.A.S., Soares C.L., Novaes J.L.C. & Bartolette R. (2004). Peixes das lagoas costeiras do norte fluminense: estudo de caso. In: Pesquisas de Longa Duração na Restinga de Jurubatiba: ecologia,

120
história natural e conservação (eds. Rocha CFD, Esteves FA & Scarano FR). Rima Editora Rio de Janeiro, p. 376.
Cardinale B.J., Palmer M.A. & Collins S.L. (2002). Species diversity enhances ecosystem functioning through interspecific facilitation. Nature, 415, 426-429.
Carpenter S., Kitchell J. & Hodgson J. (1985). Cascading trophic interactions and lake productivity. Bioscience, 35, 634-639.
Carpenter S.R., Brock W.A., Cole J.J., Kitchell J.F. & Pace M.L. (2008). Leading indicators of trophic cascades. Ecology Letters, 11, 128-138.
Carpenter S.R., Cole J.J., Hodgson J.R., Kitchell J.F., Pace M.L., Bade D., Cottingham K.L., Essington T.E., Houser J.N. & Schindler D.E. (2001). Trophic cascades, nutrients, and lake productivity: whole-lake experiments. Ecological Monographs, 71, 163-186.
Castelo-Branco C.W. (1998). Composição e aspectos ecológicos das comunidades zooplanctônicas nas lagoas Imboassica, Cabiúnas e Comprida. In: Ecologia das Lagoas Costeiras do Parque Nacional da Restinga de Jurubatiba e do Município de Macaé (RJ). (ed. Esteves FA). Nupem/UFRJ Rio de Janeiro, p. 464.
Cebrian J. (1999). Patterns in the fate of production in plant communities. American Naturalist, 154, 449-468.
Chase J.M. (2000). Are there real differences among aquatic and terrestrial food webs? Trends in Ecology & Evolution, 15, 408-412.
Chase J.M. (2003). Strong and weak trophic cascades along a productivity gradient. Oikos, 101, 187-195.
Chase J.M., Abrams P.A., Grover J.P., Diehl S., Chesson P., Holt R.D., Richards S.A., Nisbet R.M. & Case T.J. (2002). The interaction between predation and competition: a review and synthesis. Ecology Letters, 5, 302-315.
Chesson P. (2000). Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 31, 343-+.
Chumchal M.M. & Drenner R.W. (2004). Interrelationships between phosphorus loading and common carp in the regulation of phytoplankton biomass. Archiv Fur Hydrobiologie, 161, 147-158.
Chumchal M.M., Nowlin W.H. & Drenner R.W. (2005). Biomass-dependent effects of common carp on water quality in shallow ponds. Hydrobiologia, 545, 271-277.
Coley P.D., Bryant J.P. & Chapin F.S. (1985). Resource availability and plant antiherbivore defense. Science, 230, 895-899.
Colfer R. & Rosenheim J. (2001). Predation on immature parasitoids and its influence of aphid population suppression. Oecologia, 126, 292-304.
Coll M. & Guershon M. (2002). Omnivory in terrestrial arthropods: Mixing plant and prey diets. Annual Review of Entomology, 47, 267-297.
Colwell R.K. (2006). EstimateS: Statistical estimation of species richness and shared species samples. Version 8. <purl.oclc.org/estimates>. URL <purl.oclc.org/estimates>
Confer J.L. & Blades P.I. (1975). OMNIVOROUS ZOOPLANKTON AND PLANKTIVOROUS FISH. Limnology and Oceanography, 20, 571-579.
Connor E.F. & McCoy E.D. (1979). STATISTICS AND BIOLOGY OF THE SPECIES-AREA RELATIONSHIP. American Naturalist, 113, 791-833.
Cottingham K.L., Brown B.L. & Lennon J.T. (2001). Biodiversity may regulate the temporal variability of ecological systems. Ecology Letters, 4, 72-85.
Cottingham K.L., Glaholt S. & Brown A.C. (2004). Zooplankton community structure affects how phytoplankton respond to nutrient pulses. Ecology, 85, 158-171.
de Mello S. & Suzuki M.S. (1998). Variações temporais e espaciais do fitoplâncton das lagoas Imboassica, Cabiúnas e Comprida. In: Ecologia das Lagoas Costeiras do Parque Nacional da Restinga de Jurubatiba e do Município de Macaé (RJ). (ed. Esteves FA). Nupem/UFRJ Rio de Janeiro, p. 464.
Deangelis D.L., Mulholland P.J., Palumbo A.V., Steinman A.D., Huston M.A. & Elwood J.W. (1989). Nutrient Dynamics and Food-Web Stability. Annual Review of Ecology and Systematics, 20, 71-95.
DeMelo R., France R. & McQueen D.J. (1992). Biomanipulation - Hit or Myth? Limnology and Oceanography, 37, 192-207.
DeMott W.R. (1985). Relations between filter mesh-size, feeding mode, and capture efficiency for cladocerans feeding on ultrafine particles. Ergeb. Limnol, 21, 125-l.
Denno R.F. & Fagan W.F. (2003). Might nitrogen limitation promote omnivory among carnivorous arthropods? Ecology, 84, 2522-2531.

121
Dickerson J.E. & Robinson J.V. (1986). The controlled assembly of microcosmic communities - The selective extinction hypothesis. Oecologia, 71, 12-17.
Diehl S. (1992). Fish Predation and Benthic Community Structure - the Role of Omnivory and Habitat Complexity. Ecology, 73, 1646-1661.
Diehl S. (1993). RELATIVE CONSUMER SIZES AND THE STRENGTHS OF DIRECT AND INDIRECT INTERACTIONS IN OMNIVOROUS FEEDING RELATIONSHIPS. Oikos, 68, 151-157.
Diehl S. (1995). Direct and Indirect Effects of Omnivory in a Littoral Lake Community. Ecology, 76, 1727-1740.
Diehl S. (2003). The evolution and maintenance of omnivory: Dynamic constraints and the role of food quality. Ecology, 84, 2557-2567.
Diehl S. & Feissel M. (2001). Intraguild prey suffer from enrichment of their resources: A microcosm experiment with ciliates. Ecology, 82, 2977-2983.
Drenner R.W., Denoyelles F. & Kettle D. (1982). SELECTIVE IMPACT OF FILTER-FEEDING GIZZARD SHAD ON ZOOPLANKTON COMMUNITY STRUCTURE. Limnology and Oceanography, 27, 965-968.
Drenner R.W., Gallo K.L., Baca R.M. & Smith J.D. (1998). Synergistic effects of nutrient loading and omnivorous fish on phytoplankton biomass. Canadian Journal of Fisheries and Aquatic Sciences, 55, 2087-2096.
Drenner R.W., Hambright K.D., Vinyard G.L., Gophen M. & Pollingher U. (1987). EXPERIMENTAL-STUDY OF SIZE-SELECTIVE PHYTOPLANKTON GRAZING BY A FILTER-FEEDING CICHLID AND THE CICHLIDS EFFECTS ON PLANKTON COMMUNITY STRUCTURE. Limnology and Oceanography, 32, 1138-1144.
Drenner R.W. & McComas S.R. (1980). The roles of zooplankter escape ability and fish size selectivity in the selective feeding and impact of planktivorous fish. In: Evolution and ecology of zooplankton communities (ed. Kerfoot WC). University Press of New England Hanover, pp. 587–593.
Drenner R.W., Mummert J.R., Denoyelles F. & Kettle D. (1984a). SELECTIVE PARTICLE INGESTION BY A FILTER-FEEDING FISH AND ITS IMPACT ON PHYTOPLANKTON COMMUNITY STRUCTURE. Limnology and Oceanography, 29, 941-948.
Drenner R.W., Smith J.D. & Threlkeld S.T. (1996). Lake trophic state and the limnological effects of omnivorous fish. Hydrobiologia, 319, 213-223.
Drenner R.W., Taylor S.B., Lazzaro X. & Kettle D. (1984b). PARTICLE-GRAZING AND PLANKTON COMMUNITY IMPACT OF AN OMNIVOROUS CICHLID. Transactions of the American Fisheries Society, 113, 397-402.
Drenner R.W., Threlkeld S.T. & McCracken M.D. (1986). EXPERIMENTAL-ANALYSIS OF THE DIRECT AND INDIRECT EFFECTS OF AN OMNIVOROUS FILTER-FEEDING CLUPEID ON PLANKTON COMMUNITY STRUCTURE. Canadian Journal of Fisheries and Aquatic Sciences, 43, 1935-1945.
Emmerson M. & Yearsley J.M. (2004). Weak interactions, omnivory and emergent food-web properties. Proceedings of the Royal Society of London Series B-Biological Sciences, 271, 397-405.
Englund G., Sarnelle O. & Cooper S. (1999). THE IMPORTANCE OF DATA-SELECTION CRITERIA: META-ANALYSES OF STREAM PREDATION EXPERIMENTS. Ecology, 80, 1132-1141.
Enrich-Prast A., Bozelli R.L., Esteves F.A. & Meirelles F.P. (2004). Lagoas costeiras da restinga de jurubatiba: descrição de suas variáveis limnológicas. In: Pesquisas de Longa Duração na Restinga de Jurubatiba: ecologia, história natural e conservação (eds. Rocha CFD, Esteves FA & Scarano FR). Rima Editora Rio de Janeiro, p. 376.
Fagan W.F. (1997). Omnivory as a stabilizing feature of natural communities. American Naturalist, 150, 554-567.
Fernando C.H. (1994). Zooplankton, fish and fisheries in tropical fresh-waters. Hydrobiologia, 272, 105-123.
Figueredo C.C. & Giani A. (2005). Ecological interactions between Nile tilapia (Oreochromis niloticus, L.) and the phytoplanktonic community of the Furnas Reservoir (Brazil). Freshwater Biology, 50, 1391-1403.
Finke D.L. & Denno R.F. (2002). Intraguild predation diminished in complex-structured vegetation: Implications for prey suppression. Ecology, 83, 643-652.
Finke D.L. & Denno R.F. (2004). Predator diversity dampens trophic cascades. Nature, 429, 407-410.

122
Finke D.L. & Denno R.F. (2006). Spatial refuge from intraguild predation: implications for prey suppression and trophic cascades. Oecologia, 149, 265-275.
Flecker A.S. (1997). Habitat modification by tropical fishes: Environmental heterogeneity and the variability of interaction strength. Journal of the North American Benthological Society, 16, 286-295.
Flecker A.S. & Hu (1992). Fish trophic guilds and the structure of a tropical stream - weak direct vs. strong indirect effects. Ecology, 73, 927-940.
Forrest J. & Arnott S.E. (2006). Immigration and zooplankton community responses to nutrient enrichment: a mesocosm experiment. Oecologia, 150, 119-131.
Frank D.A. & McNaughton S.J. (1991). Stability Increases with Diversity in Plant-Communities - Empirical-Evidence from the 1988 Yellowstone Drought. Oikos, 62, 360-362.
Fretwell S.D. (1977). Regulation of plant communities by food-chains exploiting them. Perspectives in Biology and Medicine, 20, 169-185.
Frost T.M., Ulanowicz R.E., Blumenshine S.C., Allen T.F.H., Taub F. & Rodgers J.H. (2001). Scaling issues in experimental ecology: freshwater ecosystems. Scaling relations in experimental ecology. Columbia University Press, New York, 253–280.
Gido K.B. (2002). Interspecific comparisons and the potential importance of nutrient excretion by benthic fishes in a large reservoir. Transactions of the American Fisheries Society, 131, 260-270.
Gilbert J.J. (1985). Competition between rotifers and Daphnia. Ecology, 66, 1943-1950. Gilbert J.J. (1988). Supression of rotifer populations by Daphnia - A review of the evidence, the
mechanisms, and the effects on zooplankton community structure. Limnology and Oceanography, 33, 1286-1303.
Gilpin M. & Rosenzweig M. (1972). Enriched Predator-Prey Systems: Theoretical Stability. In, pp. 902-904.
Gilpin M.E. (1972). Enriched predator-prey systems - Theoretical stability. Science, 177, 902-&. Gliwicz Z.M. (1990). Why do cladocerans fail to control algal blooms. Hydrobiologia, 200, 83-97. Golterman H.L., Clymo R.S. & Ohnstad M.A.M. (1978). Methods for physical and chemical
analysis of freshwater. Blackwell Scientific Publication, Oxford. Gotelli N.J. & Colwell R.K. (2001). Quantifying biodiversity: procedures and pitfalls in the
measurement and comparison of species richness. Ecology Letters, 4, 379-391. Grabowski J.H. (2004). Habitat complexity disrupts predator-prey interactions but not the trophic
cascade on oyster reefs. Ecology, 85, 995-1004. Gruner D.S., Smith J.E., Seabloom E.W., Sandin S.A., Ngai J.T., Hillebrand H., Harpole W.S.,
Elser J.J., Cleland E.E., Bracken M.E.S., Borer E.T. & Bolker B.M. (2008). A cross-system synthesis of consumer and nutrient resource control on producer biomass. Ecology Letters, 11, 740-755.
Grygierek E., Hillbrkxt-Ilkowska A. & Spodniewska I. (1966). The effect of fish on plankton community in ponds. Int. Ver. Theor. Angew. Limnol. Verh, 16, 1359-1366.
Guariento R.D. (2008). Efeito da adição de recursos e da configuração da teia trófica na biomassa e estequiometria (C:N:P) da comunidade perifítica de uma lagoa costeira tropical, Lagoa Cabiúnas – Macaé, RJ. In: Departamento de Ecologia. Universidade Ferderal do Rio de Janeiro Rio de Janeiro, p. 43.
Gurevitch J. & Hedges L.V. (1993). Meta-analysis: combining the results of independent experiments. In: Design and Analysis of Ecological Experiments (eds. Scheiner SM & Gurevitch J). Chapmann and Hall, New York, pp. 378–398.
Gurevitch J. & Hedges L.V. (1999). Statistical issues in ecological meta-analyses. Ecology, 80, 1142-1149.
Hairston N.G. & Hairston N.G. (1993). Cause-effect relationships in energy-flow, trophic structure, and interspecific interactions. American Naturalist, 142, 379-411.
Hairston N.G., Smith F.E. & Slobodkin L.B. (1960). Community Structure, Population Control, and Competition. The American Naturalist, 94, 421.
Halaj J. & Wise D.H. (2001). Terrestrial trophic cascades: How much do they trickle? American Naturalist, 157, 262-281.
Hall D.J., Cooper W.E. & Werner E.E. (1970). Experimental approach to production dynamics and structure of freshwater animal communities. Limnology and Oceanography, 15, 839-&.
Hall S.R., Shurin J.B., Diehl S. & Nisbet R.M. (2007). Food quality, nutrient limitation of secondary production, and the strength of trophic cascades. Oikos, 116, 1128-1143.

123
Halpern B.S., Borer E.T., Seabloom E.W. & Shurin J.B. (2005). Predator effects on herbivore and plant stability. Ecology Letters, 8, 189-194.
Hansson L.A. (1992). The role of food-chain composition and nutrient availability in shaping algal biomass development. Ecology, 73, 241-247.
Hansson L.A., Bergman E. & Cronberg G. (1998). Size structure and succession in phytoplankton communities: the impact of interactions between herbivory and predation. Oikos, 81, 337-345.
Hansson L.A., Gyllstrom M., Stahl-Delbanco A. & Svensson M. (2004). Responses to fish predation and nutrients by plankton at different levels of taxonomic resolution. Freshwater Biology, 49, 1538-1550.
Hart D.R. (2002). Intraguild predation, invertebrate predators, and trophic cascades in lake food webs. Journal of Theoretical Biology, 218, 111-128.
Hastings A. (1996). What equilibrium behavior of Lotka-Volterra models does not tell us about food webs. Food Webs: Integration of Patterns and Dynamics, 211–217.
Hastings A. (1997). Population Biology: Concepts and Models. Springer-Verlag, New York. Heath J.P. (2006). Quantifying temporal variability in population abundances. Oikos, 115, 573-
581. Hedges L.V., Gurevitch J. & Curtis P.S. (1999). The meta-analysis of response ratios in
experimental ecology. Ecology, 80, 1150-1156. Henry M., Stevens M.H.H. & Steiner C.E. (2006). Effects of predation and nutrient enrichment
on a food web with edible and inedible prey. Freshwater Biology, 51, 666-671. Hillebrand H., Bennett D.M. & Cadotte M.W. (2008). Consequences of dominance: A review of
evenness effects on local and regional ecosystem processes. Ecology, 89, 1510-1520. Hillebrand H., Durselen C.D., Kirschtel D., Pollingher U. & Zohary T. (1999). Biovolume
calculation for pelagic and benthic microalgae. Journal of Phycology, 35, 403-424. Hillebrand H., Gruner D.S., Borer E.T., Bracken M.E.S., Cleland E.E., Elser J.J., Harpole W.S.,
Ngai J.T., Seabloom E.W., Shurin J.B. & Smith J.E. (2007). Consumer versus resource control of producer diversity depends on ecosystem type and producer community structure. Proceedings of the National Academy of Sciences of the United States of America, 104, 10904-10909.
HilleRisLambers R. & Dieckmann U. (2003). Competition and predation in simple food webs: intermediately strong trade-offs maximize coexistence. Proceedings of the Royal Society of London Series B-Biological Sciences, 270, 2591-2598.
HilleRisLambers R., van de Koppel J. & Herman P.M.J. (2006). Persistence despite omnivory: benthic communities and the discrepancy between theory and observation. Oikos, 113, 23-32.
Holt R.D. & Polis G.A. (1997). A theoretical framework for intraguild predation. American Naturalist, 149, 745-764.
Holyoak M. & Sachdev S. (1998). Omnivory and the stability of simple food webs. Oecologia, 117, 413-419.
Hooper D.U. & Vitousek P.M. (1998). Effects of plant composition and diversity on nutrient cycling. Ecological Monographs, 68, 121-149.
Hrbacek J., Dvorakova M., Korinek V. & Prochazkova L. (1961). Demonstration of the effect of the fish stock on the species composition of zooplankton and the intensity of metabolism of the whole plankton association. Ver. Limnol, 14, 192-195.
Hughes J.B. & Roughgarden J. (2000). Species diversity and biomass stability. American Naturalist, 155, 618-627.
Hulot F.D., Lacroix G., Lescher-Moutoue F.O. & Loreau M. (2000). Functional diversity governs ecosystem response to nutrient enrichment. Nature, 405, 340-344.
Hurlbert S.H. & Mulla M.S. (1981). Impacts of mosquitofish (Gambusia affinis) predation on plankton communities. Hydrobiologia, 83, 125-151.
Huston M. (1994). Biological Diversity: The Coexistence of Species on Changing Landscapes. Cambridge University Press.
Huxel G.R., McCann K. & Polis G.A. (2002). Effects of partitioning allochthonous and autochthonous resources on food web stability. Ecological Research, 17, 419-432.
Ives A.R. & Cardinale B.J. (2004). Food-web interactions govern the resistance of communities after non-random extinctions. Nature, 429, 174-177.
Ives A.R. & Carpenter S.R. (2007). Stability and diversity of ecosystems. Science, 317, 58-62. Ives A.R., Carpenter S.R. & Dennis B. (1999). Community interaction webs and zooplankton
responses to planktivory manipulations. Ecology, 80, 1405-1421.

124
Ives A.R., Klug J.L. & Gross K. (2000). Stability and species richness in complex communities. Ecology Letters, 3, 399-411.
Jack J.D. & Gilbert J.J. (1994). EFFECTS OF DAPHNIA ON MICROZOOPLANKTON COMMUNITIES. Journal of Plankton Research, 16, 1499-1512.
Jeppesen E., Lauridsen T.L., Kairesalo T. & Perrow M.R. (1998). Impact of submerged macrophytes on fish-zooplankton interactions in lakes (Chapter 5). Jeppesen, E., M. Søndergaard & Christoffersen (eds), The Structuring Role of Submerged Macrophytes in Lakes. Ecological Studies, 131, 91-114.
Jeppessen E., Sondergaard M., Jensen J., Havens K., Aanneville O., Carvalho L., Coveney M., Deneke R., Dokulil M. & Foy B. (2005). Lake responses to reduced nutrient loading-an analysis of contemporary long-term data from 35 case studies. Freshwater Biology, 50, 1747-1771.
Johnson J.B. & Omland K.S. (2004). Model selection in ecology and evolution. Trends in Ecology & Evolution, 19, 101-108.
Kemp W.M., Petersen J.E. & Gardner R.H. (2001). Scale-dependence and the problem of extrapolation. In: Scaling relations in experimental ecology. (eds. Gardner RH, Kemp WM, Kennedy VS & Petersen JE). Columbia University Press, New York, pp. 3–57.
Kerfoot W.C., Levitan C. & Demott W.R. (1988). Daphnia-phytoplankton interactions - Density-dependent shifts in resource quality. Ecology, 69, 1806-1825.
Kiessling W. (2005). Long-term relationships between ecological stability and biodiversity in Phanerozoic reefs. Nature, 433, 410-413.
Komarkova J. (1998). Fish stock as a variable modifying trophic pattern of phytoplankton. Hydrobiologia, 370, 139-152.
Kondoh M. (2003). Foraging adaptation and the relationship between food-web complexity and stability. Science, 299, 1388-1391.
Koroleff F. (1978). Determination of ammonia. In: Methods of sea water analysis (eds. Grasshoff K, Ehrhardt M & Kremling K). Verlag Chemie Republic of Germany, pp. 151-157.
Kuijper L.D.J., Kooi B.W., Zonneveld C. & Kooijman S. (2003). Omnivory and food web dynamics. Ecological Modelling, 163, 19-32.
Lafontaine N. & McQueen D.J. (1991). Contraing trophic level interactions in lake St. George and Haynes Lake (Ontario, Canada). Canadian Journal of Fisheries and Aquatic Sciences, 48, 356-363.
Lamarra V.A. (1975). Digestive activities of carp as a major contributor to the nutrient loading of lakes. Verh. int. Ver. Limnol, 19, 2461-2468.
Lancaster H.F. & Drenner R.W. (1990). Experimental mesocosm study of the separate and interaction effects of phosphorus and mosquitofish (Gambusia affinis) on plankton community structure. Canadian Journal of Fisheries and Aquatic Sciences, 47, 471-479.
Lancaster J., Bradley D.C., Hogan A. & Waldron S. (2005). Intraguild omnivory in predatory stream insects. Journal of Animal Ecology, 74, 619-629.
Lande R. (1993). Risks of Population Extinction from Demographic and Environmental Stochasticity and Random Catastrophes. American Naturalist, 142, 911.
Langeland A. (1982). Interactions between zooplanckton and fish in a fertilized lake. Holarctic Ecology, 5, 273-310.
Langeland A. & Reinertsen H. (1982). Interactions between phytoplankton and zooplankton in a fertilized lake. Holarctic Ecology, 5, 253-272.
Lawton J.H. & Brown V.K. (1994). Redundancy in ecosystems. In: Biodiversity and ecosystem function (eds. Schulze ED & Mooney HA). Springer-Verlag Berlin, pp. Pages 255-270.
Lawton J.H. & Warren P.H. (1989). Patterns in Food Webs - Reply. Trends in Ecology & Evolution, 4, 50-50.
Lazzaro X. (1987). A review of planktivorous fishes - Their evolution, feeding behaviors, selectivities, and impacts. Hydrobiologia, 146, 97-167.
Lazzaro X., Bouvy M., Ribeiro R.A., Oliviera V.S., Sales L.T., Vasconcelos A.R.M. & Mata M.R. (2003). Do fish regulate phytoplankton in shallow eutrophic Northeast Brazilian reservoirs? Freshwater Biology, 48, 649-668.
Lazzaro X., Drenner R.W., Stein R.A. & Smith J.D. (1992). PLANKTIVORES AND PLANKTON DYNAMICS - EFFECTS OF FISH BIOMASS AND PLANKTIVORE TYPE. Canadian Journal of Fisheries and Aquatic Sciences, 49, 1466-1473.
Leibold M.A. (1989). Resource edibility and the effects of predators and productivity on the outcome of trophic interactions. American Naturalist, 134, 922-949.

125
Leibold M.A. (1996). A graphical model of keystone predators in food webs: Trophic regulation of abundance, incidence, and diversity patterns in communities. American Naturalist, 147, 784-812.
Leibold M.A., Chase J.M., Shurin J.B. & Downing A.L. (1997). Species turnover and the regulation of trophic structure. Annual Review of Ecology and Systematics, 28, 467-494.
Loreau M. (1998). Biodiversity and ecosystem functioning: A mechanistic model. Proceedings of the National Academy of Sciences of the United States of America, 95, 5632-5636.
Loreau M. & Hector A. (2001). Partitioning selection and complementarity in biodiversity experiments. Nature, 412, 72-76.
Luckinbi L.S. (1974). Effects of space and enrichment on a predator-prey system. Ecology, 55, 1142-1147.
Lynch M. (1979). Predation, competition, and zooplankton community structure - Experimental study. Limnology and Oceanography, 24, 253-272.
Lynch M. & Shapiro J. (1981). Predation, enrichment, and phytoplankton community structure. Limnol. Oceanogr, 26, 86-102.
MacArthur R.H. (1955). Fluctuations of animal populations and a measure of community stability. Ecology, 36, 533-536.
MacRae V. & Croft B. (1996). Differential impact of Zetzellia ma/i (Acari: Stigmaeidae) on Metaseiulus occidentalis and Typh/odromus pyri (Acari: Phytoseiidae). Exp. AppI. Acarol, 20, 143-1.
Maxwell S.E. & Delaney H.D. (2004). Designing Experiments and Analyzing Data: A Model Comparison Perspective. Lawrence Erlbaum Associates.
May R.M. (1973). Stability and complexity in model ecosystems. Princeton University Press, Princeton.
Mazumder A. (1994). Patterns of algal biomass in dominant odd-link vs. even-link lake ecosystems. Ecology, 75, 1141-1149.
McCann K. & Hastings A. (1997). Re-evaluating the omnivory-stability relationship in food webs. Proceedings of the Royal Society of London Series B-Biological Sciences, 264, 1249-1254.
McCann K., Hastings A. & Huxel G.R. (1998a). Weak trophic interactions and the balance of nature. Nature, 395, 794-798.
McCann K.S. (2000). The diversity-stability debate. Nature, 405, 228-233. McCann K.S., Hastings A. & Strong D.R. (1998b). Trophic cascades and trophic trickles in
pelagic food webs. Proceedings of the Royal Society of London Series B-Biological Sciences, 265, 205-209.
McCann K.S., Rasmussen J.B. & Umbanhowar J. (2005). The dynamics of spatially coupled food webs. Ecology Letters, 8, 513-523.
McCauley E. & Briand F. (1979). Zooplankton grazing and phytoplankton species richness - Field tests of the predation hypothesis. Limnology and Oceanography, 24, 243-252.
McCauley E., Nisbet R.M., Murdoch W.W., de Roos A.M. & Gurney W.S.C. (1999). Large-amplitude cycles of Daphnia and its algal prey in enriched environments. Nature, 402, 653-656.
McCune B. & Mefford M.J. (1999). Multivriate analysis of ecological data, version 4.17. MjM Software, Gleneden Beach, OR.
McIntyre P.B., Flecker A.S., Vanni M.J., Hood J.M., Taylor B.W. & Thomas S.A. (2008). Fish distributions and nutrient cycling in streams: Can fish create biogeochemical hotspots? Ecology, 89, 2335-2346.
McIntyre P.B., Jones L.E., Flecker A.S. & Vanni M.J. (2007). Fish extinctions alter nutrient recycling in tropical freshwaters. Proceedings of the National Academy of Sciences of the United States of America, 104, 4461-4466.
McNaughton S.J. (1978). STABILITY AND DIVERSITY OF ECOLOGICAL COMMUNITIES. Nature, 274, 251-253.
McQueen D.J., Johannes M.R.S., Post J.R., Stewart T.J. & Lean D.R.S. (1989). Bottom-up and Top-Down Impacts on Fresh-Water Pelagic Community Structure. Ecological Monographs, 59, 289-309.
McQueen D.J., Post J.R. & Mills E.L. (1986). Trophic relationships in fresh-water pelagic ecosystems. Canadian Journal of Fisheries and Aquatic Sciences, 43, 1571-1581.
Menge B.A., Berlow E.L., Blanchette C.A., Navarrete S.A. & Yamada S.B. (1994). The Keystone Species Concept - Variation in Interaction Strength in a Rocky Intertidal Habitat. Ecological Monographs, 64, 249-286.

126
Micheli F., Cottingham K.L., Bascompte J., Bjornstad O.N., Eckert G.L., Fischer J.M., Keitt T.H., Kendall B.E., Klug J.L. & Rusak J.A. (1999). The dual nature of community variability. Oikos, 85, 161-169.
Miller T.E. & Travis J. (1996). The evolutionary role of indirect effects in communities. Ecology, 77, 1329-1335.
Miracle M.R., Alfonso M.T. & Vicente E. (2007). Fish and nutrient enrichment effects on rotifers in a Mediterranean shallow lake: a mesocosm experiment. Hydrobiologia, 593, 77-94.
Mittelbach G.G., Turner A.M., Hall D.J., Rettig J.E. & Osenberg C.W. (1995). Perturbation and resilience - A long-term, whole-lake study of predator extinction and reintroduction. Ecology, 76, 2347-2360.
Montoya J.M., Pimm S.L. & Sole R.V. (2006). Ecological networks and their fragility. Nature, 442, 259-264.
Montoya J.M., Rodriguez M.A. & Hawkins B.A. (2003). Food web complexity and higher-level ecosystem services. Ecology Letters, 6, 587-593.
Morin P.J. & Lawler S.P. (1996). Effects of food chain length and omnivory on population dynamics in experimental food webs. In: Food Webs: Integration of Patterns and Dynamics (eds. Polis GA & Winemiller KO). Chapman and Hall, New York, pp. 218–230.
Moss B., Stephen D., Balayla D.M., Becares E., Collings S.E., Fernandez-Alaez C., Fernandez-Alaez M., Ferriol C., Garcia P., Goma J., Gyllstrom M., Hansson L.A., Hietala J., Kairesalo T., Miracle M.R., Romo S., Rueda J., Russell V., Stahl-Delbanco A., Svensson M., Vakkilainen K., Valentin M., Van de Bund W.J., Van Donk E., Vicente E. & Villena M.J. (2004). Continental-scale patterns of nutrient and fish effects on shallow lakes: synthesis of a pan-European mesocosm experiment. Freshwater Biology, 49, 1633-1649.
Murdoch W. & Bence J. (1987). General predators and unstable prey populations. Predation: Direct and Indirect Impacts on Aquatic Communities, 17–30.
Naeem S. (1998). Species redundancy and ecosystem reliability. Conservation Biology, 12, 39-45.
Naeem S. & Li S.B. (1997). Biodiversity enhances ecosystem reliability. Nature, 390, 507-509. Nakagawa S. & Cuthill I.C. (2007). Effect size, confidence interval and statistical significance: a
practical guide for biologists. Biological Reviews, 82, 591-605. Neutel A.M., Heesterbeek J.A.P. & de Ruiter P.C. (2002). Stability in real food webs: Weak links
in long loops. Science, 296, 1120-1123. Neutel A.M., Heesterbeek J.A.P., van de Koppel J., Hoenderboom G., Vos A., Kaldeway C.,
Berendse F. & de Ruiter P.C. (2007). Reconciling complexity with stability in naturally assembling food webs. Nature, 449, 599-U11.
Norberg J. (2000). Resource-niche complementarity and autotrophic compensation determines ecosystem-level responses to increased cladoceran species richness. Oecologia, 122, 264-272.
Northcote T.G. (1988). Fish in the sctructure and function of fresh-water ecosystems - A top-down view. Canadian Journal of Fisheries and Aquatic Sciences, 45, 361-379.
Oksanen L. (1991). Trophic Levels and Trophic Dynamics - a Consensus Emerging. Trends in Ecology & Evolution, 6, 58-60.
Oksanen L., Fretwell S.D., Arruda J. & Niemela P. (1981). Exploitation ecosystems in gradients of primary productivity. American Naturalist, 118, 240-261.
Okun N., Brasil J., Attayde J.L. & Costa I.A.S. (2008). Omnivory does not prevent trophic cascades in pelagic food webs. Freshwater Biology, 53, 129-138.
Osenberg C., Sarnelle O. & Cooper S. (1997). Effect Size in Ecological Experiments: The Application of Biological Models in Meta-Analysis. The American Naturalist, 150, 798-812.
Otto S.B., Berlow E.L., Rank N.E., Smiley J. & Brose U. (2008). Predator diversity and identity drive interaction strength and trophic cascades in a food web. Ecology, 89, 134-144.
Pace M.L., Cole J.J., Carpenter S.R. & Kitchell J.F. (1999). Trophic cascades revealed in diverse ecosystems. Trends in Ecology & Evolution, 14, 483-488.
Paine R.T. (1966). FOOD WEB COMPLEXITY AND SPECIES DIVERSITY. American Naturalist, 100, 65-&.
Paine R.T. (1988). Food Webs - Road maps of interactions or grist for theoretical development. Ecology, 69, 1648-1654.

127
Panosso R.F., Attayde J.L. & Muehe D. (1998). Morfometria das lagoas Imboassica, Cabiúnas, Comprida e Carapebus: implicações para seu funcionamento e manejo. In: Ecologia das Lagoas Costeiras do Parque Nacional da Restinga de Jurubatiba e do Município de Macaé (RJ). (ed. Esteves FA). Nupem/UFRJ Rio de Janeiro, p. 464.
Peacor S.D. & Werner E.E. (1997). Trait-mediated indirect interactions in a simple aquatic food web. Ecology, 78, 1146-1156.
PerezFuentetaja A., McQueen D.J. & Demers E. (1996). Stability of oligotrophic and eutrophic planktonic communities after disturbance by fish. Oikos, 75, 98-110.
Persson A., Hansson L.A., Bronmark C., Lundberg P., Pettersson L.B., Greenberg L., Nilsson P.A., Nystrom P., Romare P. & Tranvik L. (2001). Effects of enrichment on simple aquatic food webs. American Naturalist, 157, 654-669.
Persson L. (1999). Trophic cascades: abiding heterogeneity and the trophic level concept at the end of the road. Oikos, 85, 385-397.
Petersen J.E., Chen C.C. & Kemp W.M. (1997). Scaling aquatic primary productivity: Experiments under nutrient- and light-limited conditions. Ecology, 78, 2326-2338.
Petrucio M.M. (1998). Caracterização das lagoas Imboassica, Cabiúnas, Comprida e Carapebus a partir da temperatura, salinidade, condutividade, alcalinidade, O2 dissolvido, pH, transparência e material em suspensão. In: Ecologia das Lagoas Costeiras do Parque Nacional da Restinga de Jurubatiba e do Município de Macaé (RJ). (ed. Esteves FA). Nupem/UFRJ Rio de Janeiro, p. 464.
Pimm S.L. (1980). Properties of Food Webs. Ecology, 61, 219-225. Pimm S.L. & Lawton J.H. (1978). FEEDING ON MORE THAN ONE TROPHIC LEVEL. Nature,
275, 542-544. Pimm S.L., Lawton J.H. & Cohen J.E. (1991). Food Web Patterns and Their Consequences.
Nature, 350, 669-674. Polis G.A. (1991). Complex Trophic Interactions in Deserts - an Empirical Critique of Food-Web
Theory. American Naturalist, 138, 123-155. Polis G.A. (1998). Ecology - Stability is woven by complex webs. Nature, 395, 744-745. Polis G.A. (1999). Why are parts of the world green? Multiple factors control productivity and the
distribution of biomass. Oikos, 86, 3-15. Polis G.A., Holt R.D., Menge B.A. & Winemiller K.O. (1996a). Time, space, and life history:
influences on food webs. Pages. 435–460. Food Webs: Integration of Patterns and Dynamics. New York: Chapman and Hall.
Polis G.A., Myers C.A. & Holt R.D. (1989). THE ECOLOGY AND EVOLUTION OF INTRAGUILD PREDATION - POTENTIAL COMPETITORS THAT EAT EACH OTHER. Annual Review of Ecology and Systematics, 20, 297-330.
Polis G.A., Sears A.L.W., Huxel G.R., Strong D.R. & Maron J. (2000). When is a trophic cascade a trophic cascade? Trends in Ecology & Evolution, 15, 473-475.
Polis G.A., Strong D.R. & Uk (1996b). Food web complexity and community dynamics. American Naturalist, 147, 813-846.
Polley H.W., Wilsey B.J. & Derner J.D. (2007). Dominant species constrain effects of species diversity on temporal variability in biomass production of tallgrass prairie. Oikos, 116, 2044-2052.
Post J.R. & McQueen D.J. (1987). The impact of planktivorous fish on the structure of a plankton community. Freshwater Biology, 17, 79-89.
Pough F.H., Janis C.M. & Heiser J.B. (2005). Vertebrate Life. In. New Jersey: Pearson Education, Inc.
Power M.E. (1990). Effects of Fish in River Food Webs. Science, 250, 811-814. Power M.E. (2000). What enables trophic cascades? Commentary on Polis et al. Trends in
Ecology & Evolution, 15, 443-444. Power M.E., Tilman D., Estes J.A., Menge B.A., Bond W.J., Mills L.S., Daily G., Castilla J.C.,
Lubchenco J., Paine R.T. & Vd (1996). Challenges in the quest for keystones. Bioscience, 46, 609-620.
Pyke G.H., Pulliam H.R. & Charnov E.L. (1977). Optimal Foraging - Selective review of theory and test. Quarterly Review of Biology, 52, 137-154.
Quinn G.P. & Keough M.J. (2002). Experimental Design and Data Analysis for Biologists. Cambridge University Press.
Rangel T., Diniz-Filho J.A.F. & Bini L.M. (2006). Towards an integrated computational tool for spatial analysis in macroecology and biogeography. Global Ecology and Biogeography, 15, 321-327.

128
Reisewitz S.E., Estes J.A. & Simenstad C.A. (2006). Indirect food web interactions: sea otters and kelp forest fishes in the Aleutian archipelago. Oecologia, 146, 623-631.
Reynolds C.S. (1984). The Ecology of Freshwater Phytoplankton. Cambridge University Press. Rice W.R. (1989). Analyzing tables of statistical tests. Evolution, 43, 223-225. Ricklefs R. (1990). Ecology. WH Freeman, New York. Romanuk T.N., Vogt R.J. & Kolasa J. (2006). Nutrient enrichment weakens the stabilizing effect
of species richness. Oikos, 114, 291-302. Rondel C., Arfi R., Corbin D., Le Bihan F., Ndour E.H. & Lazzaro X. (2008). A cyanobacterial
bloom prevents fish trophic cascades. Freshwater Biology, 53, 637-651. Rosenberg M.S., Adams D.C. & Gurevitch J. (2000). MetaWin: statistical software for meta-
analysis. Version 2.0. Sinauer Associates, Sunderland, Massachusetts. Rosenheim J.A. & Corbett A. (2003). Omnivory and the indeterminacy of predator function: Can
a knowledge of foraging behavior help? Ecology, 84, 2538-2548. Rosenzweig M.I. (1971). PARADOX OF ENRICHMENT - DESTABILIZATION OF
EXPLOITATION ECOSYSTEMS IN ECOLOGICAL TIME. Science, 171, 385-&. Roy S. & Chattopadhyay J. (2007). Towards a resolution of ‘the paradox of the plankton’: A brief
overview of the proposed mechanisms. Ecological Complexity, 4, 26-33. Sánchez-Botero R.I. (2005). Efeitos de distúrbios naturais e antrópicos sobre a fauna ictíica em
lagoas costeiras na região de Macaé. In: Programa de pós-graduação em Ecologia (PPGE). Universidade Federal do Rio de Janeiro Rio de Janeiro, p. 70.
Sankaran M. & McNaughton S.J. (1999). Determinants of biodiversity regulate compositional stability of communities. Nature, 401, 691-693.
Sarnelle O. (1993). Herbivore effects on phytoplankton succession in a eutrophic lake. Ecological Monographs, 63, 129-149.
Sarnelle O. (1997). Daphnia effects on microzooplankton: Comparisons of enclosure and whole-lake responses. Ecology, 78, 913-928.
Schaus M.H. & Vanni M.J. (2000). Effects of gizzard shad on phytoplankton and nutrient dynamics: Role of sediment feeding and fish size. Ecology, 81, 1701-1719.
Schaus M.H., Vanni M.J. & Wissing T.E. (2002). Biomass-dependent diet shifts in omnivorous gizzard shad: Implications for growth, food web, and ecosystem effects. Transactions of the American Fisheries Society, 131, 40-54.
Schaus M.H., Vanni M.J., Wissing T.E., Bremigan M.T., Garvey J.E. & Stein R.A. (1997). Nitrogen and phosphorus excretion by detritivorous gizzard shad in a reservoir ecosystem. Limnology and Oceanography, 42, 1386-1397.
Scheffer M. (1991). Fish and nutrients interplay determines algal biomass - A minimal model. Oikos, 62, 271-282.
Scheffer M. (1998). Ecology of shallow lakes. Kluwer Academic Publishers, Dordrecht, The Netherlands.
Scheffer M., Rinaldi S., Huisman J. & Weissing F.J. (2003). Why plankton communities have no equilibrium: solutions to the paradox. Hydrobiologia, 491, 9-18.
Scheffer M., Rinaldi S. & Kuznetsov Y.A. (2000). Effects of fish on plankton dynamics: a theoretical analysis. Canadian Journal of Fisheries and Aquatic Sciences, 57, 1208-1219.
Scheiner S.M. (1993). MANOVA: multiple response variables and multispecies interactions. p 94–112. In: Design and analysis of ecological experiments. Chapman and Hall, New York (eds. Scheiner SM & Gurevitch J). Oxford University Press, New York, pp. 445–112.
Scheiner S.M. (2001). Multiple response variables and multispecies interactions. In: Design and Analysis of Ecological Experiments (eds. Scheiner SM & Gurevitch J). Oxford University Press, New York, pp. 94–112.
Schindler D.E. (2007). Fish extinctions and ecosystem functioning in tropical ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 104, 5707-5708.
Schindler D.E., Carpenter S.R., Cole J.J., Kitchell J.F. & Pace M.L. (1997). Influence of food web structure on carbon exchange between lakes and the atmosphere. Science, 277, 248-251.
Schindler D.E. & Scheuerell M.D. (2002). Habitat coupling in lake ecosystems. Oikos, 98, 177-189.

129
Schmitz O.J., Hamback P.A. & Beckerman A.P. (2000). Trophic cascades in terrestrial systems: A review of the effects of carnivore removals on plants. American Naturalist, 155, 141-153.
Shurin J.B., Borer E.T., Seabloom E.W., Anderson K., Blanchette C.A., Broitman B., Cooper S.D. & Halpern B.S. (2002). A cross-ecosystem comparison of the strength of trophic cascades. Ecology Letters, 5, 785-791.
Shurin J.B., Gruner D.S. & Hillebrand H. (2006). All wet or dried up? Real differences between aquatic and terrestrial food webs. Proceedings of the Royal Society B-Biological Sciences, 273, 1-9.
Shurin J.B. & Seabloom E.W. (2005). The strength of trophic cascades across ecosystems: predictions from allometry and energetics. Journal of Animal Ecology, 74, 1029-1038.
Sih A. (1993). Effects of Ecological Interactions on Forager Diets: Competition, Predation Risk, Parasitism and Prey Behaviour. Diet Selection: An Interdisciplinary Approach to Foraging Behaviour.
Silvertown J., Dodd M.E., McConway K., Potts J. & Crawley M. (1994). Rainfall, biomass variation, and community composition in the park grass experiment. Ecology, 75, 2430-2437.
Singer M.S. & Bernays E.A. (2003). Understanding omnivory needs a behavioral perspective. Ecology, 84, 2532-2537.
Smith D.W. (1989). THE FEEDING SELECTIVITY OF SILVER CARP, HYPOPHTHALMICHTHYS-MOLITRIX VAL. Journal of Fish Biology, 34, 819-828.
Snyder W.E. & Ives A.R. (2001). Generalist predators disrupt biological control by a specialist parasitoid. Ecology, 82, 705-716.
Soffiati A. (2000). Aspectos históricos das restingas da eco-região Norte do Estado do Rio de Janeiro - Brasil. In: Ecologia de Restingas e Lagoas Costeiras (eds. Esteves FA & Lacerda LD). Núcleo de Pesquisas Ecológicas de Macaé (NUPEM/UFRJ) Rio de Janeiro, p. 446.
Sondergaard M., Jensen J.P. & Jeppesen E. (2003). Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia, 506, 135-145.
Spencer M. & Warren P.H. (1996). The effects of energy input, immigration and habitat size on food web structure: A microcosm experiment. Oecologia, 108, 764-770.
Srivastava D.S. & Lawton J.H. (1998). Why more productive sites have more species: An experimental test of theory using tree-hole communities. American Naturalist, 152, 510-529.
StatSoft (2001). Statistica for Windows version 6.0. Statsoft Inc., Tulsa, OK, USA. Stephen D., Balayla D.M., Becares E., Collings S.E., Fernandez-Alaez C., Fernandez-Alaez M.,
Ferriol C., Garcia P., Goma J., Gyllstrom M., Hansson L.A., Hietala J., Kairesalo T., Miracle M.R., Romo S., Rueda J., Stahl-Delbanco A., Svensson M., Vakkilainen K., Valentin M., Van de Bund W.J., Van Donk E., Vicente E., Villena M.J. & Moss B. (2004). Continental-scale patterns of nutrient and fish effects on shallow lakes: introduction to a pan-European mesocosm experiment. Freshwater Biology, 49, 1517-1524.
Stief P. & Holker F. (2006). Trait-mediated indirect effects of predatory fish on microbial mineralization in aquatic sediments. Ecology, 87, 3152-3159.
Straskraba M. (1965). The effect of fish on the number of invertebrates in ponds and streams. Mitt. int. Ver. Limnol, 13, 106-127.
Strauss S.Y. (1991). Indirect Effects in Community Ecology - Their Definition, Study and Importance. Trends in Ecology & Evolution, 6, 206-210.
Strong D.R. (1992). Are Trophic Cascades All Wet - Differentiation and Donor-Control in Speciose Ecosystems. Ecology, 73, 747-754.
Tanabe K. & Namba T. (2005). Omnivory creates chaos in simple food web models. Ecology, 86, 3411-3414.
Thompson C.M. & Gese E.M. (2007). Food webs and intraguild predation: Community interactions of a native mesocarnivore. Ecology, 88, 334-346.
Tilman D. (1996). Biodiversity: Population versus ecosystem stability. Ecology, 77, 350-363. Tilman D. (1999). The ecological consequences of changes in biodiversity: A search for general
principles. Ecology, 80, 1455-1474. Tilman D. & Downing J.A. (1994). Biodiversity and Stability in Grasslands. Nature, 367, 363-
365. Tilman D., Reich P.B. & Knops J.M.H. (2006). Biodiversity and ecosystem stability in a decade-
long grassland experiment. Nature, 441, 629-632.

130
Timms R.M. & Moss B. (1984). Prevention of growth of potentially dense phytoplankton populations by zooplankton grazing, in the presence of zooplanktivorous fish, in a shallow wetland ecosystem. Limnology and Oceanography, 29, 472-486.
Vadeboncoeur Y., McCann K.S., VanderZanden M.J. & Rasmussen J.B. (2005). Effects of multi-chain omnivory on the strength of trophic control in lakes. Ecosystems, 8, 682-693.
Vadeboncoeur Y., Vander Zanden M.J. & Lodge D.M. (2002). Putting the lake back together: Reintegrating benthic pathways into lake food web models. Bioscience, 52, 44-54.
Vakkilainen K., Kairesalo T., Hietala J., Balayla D.M., Becares E., Van de Bund W.J., Van Donk E., Fernandez-Alaez M., Gyllstrom M., Hansson L.A., Miracle M.R., Moss B., Romo S., Rueda J. & Stephen D. (2004). Response of zooplankton to nutrient enrichment and fish in shallow lakes: a pan-European mesocosm experiment. Freshwater Biology, 49, 1619-1632.
Van de Bund W.J., Romo S., Villena M.J., Valentin M., Van Donk E., Vicente E., Vakkilainen K., Svensson M., Stephen D., Stahl-Delbanco A., Rueda J., Moss B., Miracle M.R., Kairesalo T., Hansson L.A., Hietala J., Gyllstrom M., Goma J., Garcia P., Fernandez-Alaez M., Fernandez-Alaez C., Ferriol C., Collings S.E., Becares E., Balayla D.M. & Alfonso T. (2004). Responses of phytoplankton to fish predation and nutrient loading in shallow lakes: a pan-European mesocosm experiment. Freshwater Biology, 49, 1608-1618.
Van De Bund W.J. & Van Donk E. (2004). Effects of fish and nutrient additions on food-web stability in a charophyte-dominated lake. Freshwater Biology, 49, 1565-1573.
Vander Zanden M.J., Essington T.E. & Vadeboncoeur Y. (2005). Is pelagic top-down control in lakes augmented by benthic energy pathways? Canadian Journal of Fisheries and Aquatic Sciences, 62, 1422-1431.
Vander Zanden M.J. & Vadeboncoeur Y. (2002). Fishes as integrators of benthic and pelagic food webs in lakes. Ecology, 83, 2152-2161.
Vandermeer J. (2006). Omnivory and the stability of food webs. Journal of Theoretical Biology, 238, 497-504.
Vanni M.J. (1986a). COMPETITION IN ZOOPLANKTON COMMUNITIES - SUPPRESSION OF SMALL SPECIES BY DAPHNIA-PULEX. Limnology and Oceanography, 31, 1039-1056.
Vanni M.J. (1986b). FISH PREDATION AND ZOOPLANKTON DEMOGRAPHY - INDIRECT EFFECTS. Ecology, 67, 337-354.
Vanni M.J. (1987). EFFECTS OF NUTRIENTS AND ZOOPLANKTON SIZE ON THE STRUCTURE OF A PHYTOPLANKTON COMMUNITY. Ecology, 68, 624-635.
Vanni M.J. (2002). Nutrient cycling by animals in freshwater ecosystems. Annual Review of Ecology and Systematics, 33, 341-370.
Vanni M.J., Arend K.K., Bremigan M.T., Bunnell D.B., Garvey J.E., Gonzalez M.J., Renwick W.H., Soranno P.A. & Stein R.A. (2005). Linking landscapes and food webs: Effects of omnivorous fish and watersheds on reservoir ecosystems. Bioscience, 55, 155-167.
Vanni M.J., Bowling A.M., Dickman E.M., Hale R.S., Higgins K.A., Horgan M.J., Knoll L.B., Renwick W.H. & Stein R.A. (2006). Nutrient cycling by fish supports relatively more primary production as lake productivity increases. Ecology, 87, 1696-1709.
Vanni M.J. & Findlay D.L. (1990). Trophic Cascades and Phytoplankton Community Structure. Ecology, 71, 921-937.
Vanni M.J. & Layne C.D. (1997). Nutrient recycling and herbivory as mechanisms in the ''top-down'' effect of fish on algae in lakes. Ecology, 78, 21-40.
Vanni M.J., Layne C.D. & Arnott S.E. (1997). ''Top-down'' trophic interactions in lakes: Effects of fish on nutrient dynamics. Ecology, 78, 1-20.
Vanni M.J., Luecke C., Kitchell J.F., Allen Y., Temte J. & Magnuson J.J. (1990). EFFECTS ON LOWER TROPHIC LEVELS OF MASSIVE FISH MORTALITY. Nature, 344, 333-335.
Vinyard G.L., Drenner R.W., Gophen M., Pollingher U., Winkelman D.L. & Hambright K.D. (1988). AN EXPERIMENTAL-STUDY OF THE PLANKTON COMMUNITY IMPACTS OF 2 OMNIVOROUS FILTER-FEEDING CICHLIDS, TILAPIA-GALILAEA AND TILAPIA-AUREA. Canadian Journal of Fisheries and Aquatic Sciences, 45, 685-690.
Werner E.E. & Peacor S.D. (2003). A review of trait-mediated indirect interactions in ecological communities. Ecology, 84, 1083-1100.
Wetzel R.G. & Likens G.E. (1991). Limnological Analysis. Springer Verlag, New York.

131
Williams A.E. & Moss B. (2003). Effects of different fish species and biomass on plankton interactions in a shallow lake. Hydrobiologia, 491, 331-346.
Winemiller K.O. (1990). Spatial and Temporal Variation in Tropical Fish Trophic Networks. Ecological Monographs, 60, 331-367.
Wojdak J.M. & Luttbeg B. (2005). Relative strengths of trait-mediated and density-mediated indirect effects of a predator vary with resource levels in a freshwater food chain. Oikos, 111, 592-598.
Worm B. & Duffy J.E. (2003). Biodiversity, productivity and stability in real food webs. Trends in Ecology & Evolution, 18, 628-632.
Worm B., Lotze H.K., Hillebrand H. & Sommer U. (2002). Consumer versus resource control of species diversity and ecosystem functioning. Nature, 417, 848-851.
Yachi S. & Loreau M. (1999). Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proceedings of the National Academy of Sciences of the United States of America, 96, 1463-1468.
Yodzis P. (1984). How Rare is Omnivory? Ecology, 65, 321-323. Yodzis P. (1988). The indeterminacy of ecological interactions as perceived through
perturbation experiments Ecology, 69, 508-515. Zaret T.M. (1980). The effect of prey motion on planktivore choice. In: Evolution and Ecology of
Zooplankton Communities (ed. Kerfoot WC). University Press of New England Hanover, pp. 594-603.





























