Embed Size (px)
Citation preview

KARINA TACIANA SANTOS SILVA
IDENTIFICAÇÃO DE PROTEÍNAS ASSOCIADAS À CARCINOGÊNESE
INDUZIDA POR DIBENZOTIOFENO E DIBENZOTIOFENO SULFONA EM
RATOS WISTAR
OURO PRETO – MG, ABRIL DE 2015

UNIVERSIDADE FEDERAL DE OURO PRETO
NÚCLEO DE PESQUISAS EM CIÊNCIAS
BIOLÓGICAS – NUPEB
PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
IDENTIFICAÇÃO DE PROTEÍNAS ASSOCIADAS À CARCINOGÊNESE
INDUZIDA POR DIBENZOTIOFENO E DIBENZOTIOFENO SULFONA EM
RATOS WISTAR
Autora: Karina Taciana Santos Silva
Orientador: Prof. Dr. Milton Hércules Guerra de Andrade
Coorientador: Prof. Dr. William de Castro Borges
Ouro Preto – MG, abril de 2015
Tese submetida ao programa de Pós-
Graduação do Núcleo de Pesquisas em
Ciências Biológicas da Universidade Federal
de Ouro Preto, como parte integrante dos
requisitos para obtenção do título de Doutor
em Ciências Biológicas, área de
concentração: Bioquímica Estrutural e
Biologia Molecular.

S381i Silva, Karina Taciana Santos. Identificação de proteínas associadas à carcinogênese induzida por
Dibenzotiofeno e Dibenzotiofeno Sulfona em ratos Wistar [manuscrito] / Karina Taciana Santos Silva. - 2015.
xxii, 172pf.: il.: color; grafs; tabs.
Orientador: Prof. Dr. Milton Hércules Guerra de Andrade. Coorientador: Prof. Dr. William Castro-Borges.
Tese (Doutorado) - Universidade Federal de Ouro Preto. Núcleo de
Pesquisas em Ciências Biológicas. Programa de Pós-graduação em Ciências Biológicas.
Área de Concentração: Bioquímica Estrutural e Biologia Molecular.
1. Câncer. 2. Dibenzotiofeno. 3. Proteômica. I. Andrade, Milton Hércules Guerra de. II. Castro-Borges, William. III. Universidade Federal de Ouro Preto. IV. Titulo.
CDU: 616-006.6:577.122
Catalogação: www.sisbin.ufop.br

Silva, K.T.S. Ata Defesa
i

Silva, K.T.S. Colaboradores
ii
COLABORADORES
Prof. Dr. Wanderson Geraldo de Lima - UFOP
Profª. Drª. Yara Cristina de Paiva Maia - UFU
Prof. Dr. Luiz Ricardo Goulart Filho – UFU
Dr. Fausto Emílio Capparelli - UFU
Msc. Renata Alves de Oliveira e Castro
Msc. Vinícius Corrêa Rodrigues
Aline Dias de Almeida

Silva, K.T.S. Auxílio
iii
Este trabalho foi realizado no LABORATÓRIO DE ENZIMOLOGIA E
PROTEÔMICA – NUPEB/UFOP, com auxílio da Coordenadoria de
aperfeiçoamento de Pessoal de Nível Superior (CAPES) e Universidade
Federal de Ouro Preto (UFOP).

Silva, K.T.S. Dedicatória
iv
Dedico este trabalho à
minha família e ao Allan,
minhas fontes de força, amor
e apoio incondicionais.

Silva, K.T.S. Epígrafe
v
“O homem interior se renova
sempre. A luta enriquece-o de
experiência, a dor aprimora-lhe as
emoções e o sacrifício tempera-lhe
o caráter.”
(Chico Xavier)

Silva, K.T.S. Agradecimentos
vi
AGRADECIMENTOS
Às agências de fomento FAPEMIG e CNPq pelo apoio financeiro.
Ao Prof. Dr. William de Castro Borges, pela disponibilidade e
aprendizado. Obrigada pela coorientação e por compartilhar os problemas,
alegrias e enfrentar comigo as dificuldades encontradas.
Ao Prof. Dr. Milton Hércules Guerra de Andrade, pela orientação.
Ao Técnico José Henrique Braga Fortes, muito obrigada por toda a ajuda,
pela amizade, risadas e por compartilhar as alegrias, angústias e dúvidas
surgidas durante esses anos de convivência.
Ao Prof. Marcos Aurélio de Santana, pelo apoio e colaboração.
Ao Prof. Dr. Wanderson Geraldo de Lima, pelo aprendizado e
colaboração.
Aos laboratórios LIMP, LIP e LBBM por permitirem o uso de equipamentos
e dependências para realização de parte deste trabalho.
Aos professores do NUPEB que contribuíram de maneira construtiva para
a realização deste trabalho.
À Renata, pela colaboração e disponibilidade dedicadas a este trabalho.
Pela amizade e apoio que recebi durante esses anos de estudo.
Aos alunos de iniciação científica Aline, Vinícius e Pablo, que
participaram diretamente da realização deste trabalho, obrigada pela ajuda
inestimável. Sem vocês, a conclusão deste trabalho teria sido impossível.
Aos amigos que fiz no LEP durante esses 6 anos de pós-graduação, muito
obrigada pelas risadas, conversas e pela ajuda, principalmente nos momentos
finais do desenvolvimento deste trabalho.
Aos meus pais, pelo amor, carinho e apoio incondicionais. Obrigada por
acreditarem nos meus sonhos e muitas vezes abrirem mão dos seus para que
essa conquista se tornasse possível.
A minha irmã, pela amizade e companheirismo mesmo à distância.
Ao Allan, por transformar qualquer dia em um dia especial, pelo amor,
carinho e apoio.
Enfim, agradeço a todos que de maneira direta ou indireta contribuíram
para a realização deste trabalho.

Silva, K.T.S. Índice
vii
ÍNDICE
RESUMO .........................................................................................................................x
LISTA DE ABREVIATURAS ....................................................................................xiii
LISTA DE FIGURAS ..................................................................................................xvi
LISTA DE TABELAS ...................................................................................................xx
1. INTRODUÇÃO ...........................................................................................................2
1.1. Epidemiologia ................................................................................................2
1.2. Aspectos Gerais da Oncogênese .....................................................................3
1.3. Trato Gastrointestinal Inferior ........................................................................4
1.3.1. Características Gerais ......................................................................4
1.3.2. Câncer Intestinal ..............................................................................6
1.3.3. Fatores de Risco e Genéticos ...........................................................8
1.4. Visão Geral de Vias de Sinalização Atuantes no CCR ...................................9
1.4.1. Via Wnt/β-catenina ..........................................................................9
1.4.2. Via Notch .......................................................................................11
1.4.3. Via Hedgehog ................................................................................12
1.4.4. Via PI3K/Akt ..................................................................................14
1.5. Carcinogênese Química ................................................................................16
1.5.1. Dibenzotiofeno e Dibenzotiofeno Sulfona .....................................18
1.5.2. 1,2-Dimetilhidrazina ......................................................................21
1.6. Biomarcadores e Câncer ...............................................................................21
1.6.1. Proteômica e Phage Display ..........................................................23
2. JUSTIFICATIVA ......................................................................................................25
3. OBJETIVOS ..............................................................................................................27
3.1. Objetivo Geral ..............................................................................................27
3.1.1. Objetivos Específicos ....................................................................27
4. MATERIAL E MÉTODOS ......................................................................................29
4.1. Material Biológico ........................................................................................29
4.1.1. Indução de Câncer em Ratos Wistar Tratados com DMH, DBT e
DBTO2 ............................................................................................................................29
4.1.2. Necropsia .......................................................................................30
4.2. Análise Histológica .......................................................................................30
4.2.1. Imunohistoquímica: CEA e CD44 .................................................31

Silva, K.T.S. Índice
viii
4.3. Extração de Proteínas e Análise Proteômica .................................................31
4.3.1. Eletroforese Uni e Bidimensional (1D/2D SDS-PAGE) ................32
4.3.2. Digestão in gel ...............................................................................33
4.3.3. Análise de Proteínas por Espectrometria de Massas .......................34
4.3.4. Categorização Funcional e Mapa de Interação Proteica .................34
4.4. Western bloting …………………………………………………………….35
4.5. Biopanning (Phage Display)……………………………………………….35
4.5.1. Extração de Proteínas para Phage Display .....................................35
4.5.2. Ciclos de Seleção de Phage Display ...............................................36
4.5.3. Titulação de Clones Obtidos no Phage Display ............................37
4.5.4. Extração de DNA de Fagos e Sequenciamento .............................37
4.5.5. Análise in silico dos Peptídeos Obtidos .........................................39
4.6. Síntese de Peptídeos ......................................................................................40
4.6.1. Análise de Pureza e Confirmação de Identidade ............................41
4.7. Síntese de Colunas de Afinidade ...................................................................42
4.7.1. Identificação de Proteínas Ligantes ................................................43
4.7.2. Análise in silico da Interação Proteína/Peptídeo ............................44
4.8. Análise Estatística .........................................................................................44
5. RESULTADOS E DISCUSSÃO ...............................................................................46
5.1. Efeitos Biológicos do Tratamento com DMH, DBT e DBTO2 ......................46
5.1.1. Hemogramas e Dosagens Séricas de ALT, AST e Amilase ............47
5.1.2. Avaliações Morfológicas Macro e Microscópicas de Tecidos
Extraídos Durante a Necropsia ........................................................................................48
5.1.3. Marcação Imunohistoquímica: CEA e CD44 .................................54
5.2. Análise Proteômica .......................................................................................57
5.3. Identificação de Peptídeos Ligantes à Tecidos Neoplásicos Intestinais de
Animais Tratados com DMH, DBT e DBTO2 .................................................................71
5.3.1. Biopanning de Tecido Intestinal ....................................................72
5.3.2. Análise in silico de Peptídeos Selecionados ..................................73
5.3.3. Síntese de Peptídeos .......................................................................75
5.3.4. Isolamento de Proteínas por Afinidade aos Peptídeos Selecionados
por Phage Display ...........................................................................................................77
6. CONCLUSÕES .........................................................................................................82
7. PERSPECTIVAS ......................................................................................................84

Silva, K.T.S. Índice
ix
8. REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................86
9. APÊNDICES ..............................................................................................................99
9.1. Apêndice A – Contagens Global e Diferencial de Leucócitos para Todos os
Animais Tratados ............................................................................................................99
9.2. Apêndice B – Dosagens séricas de AST, ALT e Amilase ............................100
9.3. Apêndice C – Análise Estatística Referente a Avaliação Histológica de
Intestinos Delgado e Grosso Entre Grupos Controle .....................................................101
9.4. Apêndice D – Dados Obtidos por Espectrometria de Massas para 2D SDS-
PAGE e Cromatografia de Afinidade ............................................................................102
9.5. Apêndice E – Western bloting .....................................................................155
9.6. Apêndice F – Alinhamento Entre Sequências Peptídicas e Banco de Dados de
Proteínas para Rattus norvegicus ...................................................................................156
9.7. Apêndice G – Artigo Publicado ..................................................................161

Silva, K.T.S. Resumo
x
RESUMO
O alto conteúdo de compostos organossulfurados nos derivados de petróleo compromete
sua qualidade; assim, sua remoção ou conversão a substâncias menos tóxicas são
importantes etapas durante o processo de refino. Os limites de concentração para esses
compostos estão sob rígido controle em diversos países com o objetivo de minimizar seu
impacto negativo sobre a saúde animal e o ambiente. Dentre eles, o dibenzotiofeno (DBT)
é um hidrocarboneto policíclico aromático que contém um átomo de enxofre em
substituição em sua estrutura. Devido à importância dessas moléculas na oncogênese, e a
escassez de investigações toxicológicas relacionadas ao DBT, este trabalho propôs
realizar uma avaliação dos efeitos tóxicos e moleculares promovidos por um tratamento
crônico de ratos Wistar com dose sub-letal (30 mg/kg) de DBT e seu derivado oxidado
DBTO2 (dibenzotiofeno sulfona), ambos administrados durante 10 semanas. Em paralelo,
seus efeitos tóxicos foram comparados com aqueles provocados pelo tratamento com o
agente mutagênico clássico, 1,2-dimetilhidrazina (DMH), administrado na mesma dose
durante o mesmo período. Na 14ª semana após a última dose, os animais foram
sacrificados para uma avaliação inicial da contagem de células sanguíneas e das funções
hepáticas e pancreáticas. Não foram observadas alterações nesses parâmetros. Entretanto,
análises histopatológicas revelaram lesões de caráter pré-neoplásico nos intestinos dos
animais tratados com DBT e DBTO2 que mostraram-se comparáveis em intensidade e
alterações morfológicas com aquelas encontradas no cólon de animais tratados com
DMH. Em concordância com esse achado, os marcadores tumorais CD44 e CEA
(antígeno carcinoembrionário), utilizados em ensaios de imunohistoquímica
demonstraram aumento de expressão de aproximadamente três vezes para os animais dos
grupos Testes em relação aos animais dos grupos Controle. Análises proteômicas
realizadas por eletroforese bidimensional (2D SDS-PAGE) demonstraram padrões de
expressão proteica diferenciados entre os diferentes grupos. Além da análise proteômica,
foi realizado ensaio de seleção de peptídeos ligantes de proteínas especificamente
relacionados às alterações promovidas pelos indutores químicos utilizados, empregando
a técnica de Phage Dislay. Tal metodologia permitiu a pré-seleção de seis peptídeos
específicos e relacionados aos mecanismos de desenvolvimento/manutenção de câncer.
A identificação das proteínas diferencialmente expressas observadas por 2D SDS-PAGE
e, das proteínas alvo que interagem com peptídeos específicos relacionados às alterações
promovidas por esses agentes químicos, auxilia no avanço do entendimento acerca dos

Silva, K.T.S. Resumo
xi
mecanismos envolvidos no estabelecimento das lesões encontradas nos animais tratados
com DBT e DBTO2.

Silva, K.T.S. Abstract
xii
ABSTRACT
The high content of organosulfur compounds in petroleum derivatives compromises their
quality and, therefore, their removal or conversion to less toxic substances are important
steps during oil refining. The concentration limits of such compounds in fuels are under
strict control worldwide with the aim to minimize their negative impact upon animal
health and the environment. Among them, dibenzothiophene (DBT) is a polycyclic
aromatic hydrocarbon containing a sulfur atom replacing a carbon in the main structure.
Given the importance of such molecules during oncogenesis and the scarcity of
toxicological investigations related to DBT, this work proposes an evaluation of the toxic
effects promoted by a chronic treatment of Wistar rats with a sub-lethal dose (30 mg/kg)
of DBT and its oxidized derivative DBTO2 (dibenzothiophene sulfone), both
administered during 10 weeks. In parallel, their toxicological effects were compared to
those inflicted by treatment with the classic mutagenic compound, 1,2-dimethylhydrazine
(DMH), given at 30 mg/kg for the same period. At the 14th week after the last dose, the
animals were sacrificed for the initial evaluation of blood cell counts and assessment of
hepatic and pancreatic functions. We have observed no alterations in either blood cell
parameters or indication of liver and spleen injuries. However, pre-neoplastic lesions in
the small intestine of DBT and DBTO2 treated animals were comparable in intensity and
morphology to those found in the colon of DMH-treated animals. In agreement with this
finding two tumor markers CD44 and CEA (carcinoembrionic antigen), were detected by
immunohistochemistry and demonstrate approximately three fold higher for treated
animals when compared to control groups. Proteomic analyses by 2D electrophoresis
(2D SDS-PAGE) reveal different expression patterns for Test groups when compared to
Control groups. In addition to the proteomic analysis, we also performed a selection of
test specifically protein binding peptides related to changes produced by chemical
inducers used employing the Phage Display technique. This methodology allowed the
pre-selection of six specific peptides related to the mechanisms of
development/maintenance of cancer. The identification of differentially expressed
proteins observed by 2D SDS-PAGE, and of proteins that interact with specific peptides
related to alterations caused by such agents, assists in advancing understanding of
mechanisms involved in the establishment of lesions found in animals treated DBT and
DBTO2.

Silva, K.T.S. Lista de Abreviaturas
xiii
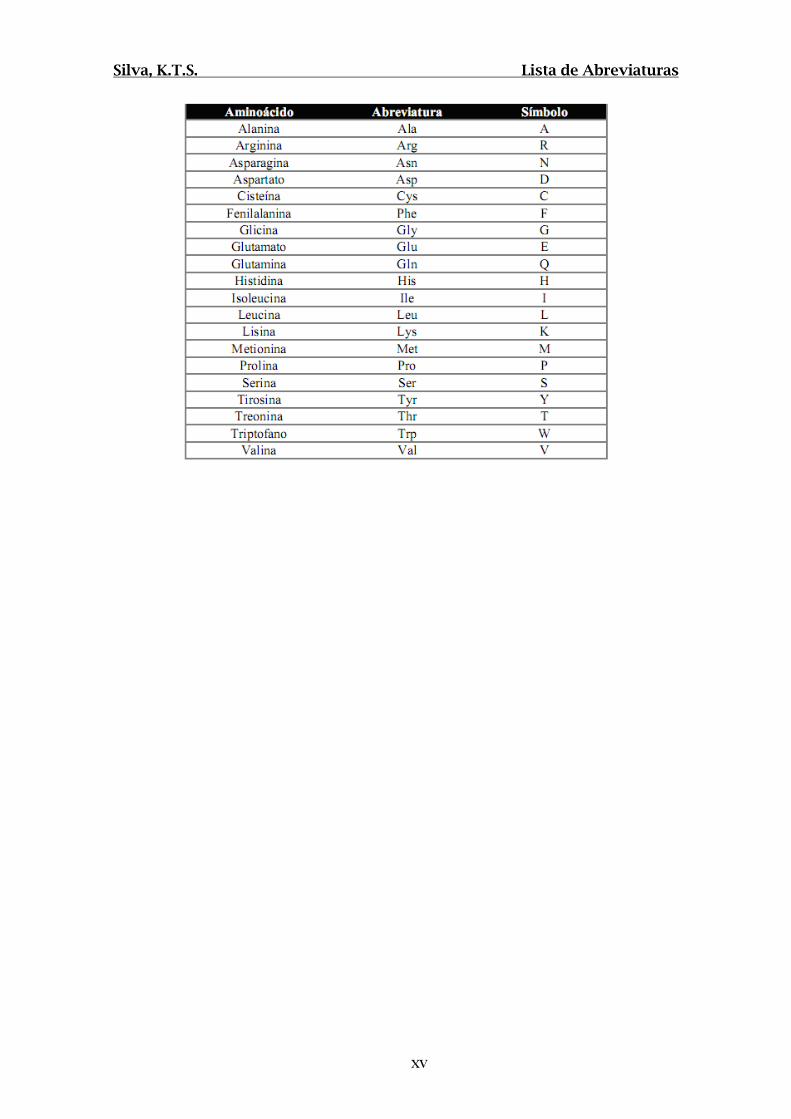
LISTA DE ABREVIATURAS
µg- micro gramas
µL- micro litros
µm- micro metros
2D SDS-PAGE- Eletroforese bidimensional em
gel de poliacrilamida
ACF- Focos de criptas aberrantes
ACN- Acetonitrila
ACTB- Actina subunidade beta
ADH- Álcool dehidrogenase
AHR- Receptor aril hidrocarbono
AKR1B1- Aldose redutase
Akt- Proteína quinase B
ALDOA- Aldolase A
ALT- Alanina amino transferase
ANOVA- Análise de variância
ANXA2- Anexina 2
ANXA5- Anexina 5
AOM- Azoximetanol
APC- Adenomatous polyposis coli
ARHGDIA- Inibidor de dissociação Rho-GDP
AST- Aspartato amino trasnferase
ATIC- Biossíntese de purina bifuncional
BAX- Proteína associada a BCL2
Bcl2- B-cell CLL/lymphoma 2
Bmi- BMI polycomb ring finger oncogene
CA- Carbohydrate antigen
CALR- Calreticulina
CANX- Calnexina
CCA- Centro de Ciência Animal
CCR- Câncer colorretal
CCT6- Proteína complexo T1 sub. zeta
CD44- Cluster of differentiation 44
CEA- Carcinoembrionic antigen
CEUA- Comissão de ética no uso de animais
CFL- Cofilina
c-Jun- Protooncogene jun
CK1- Caseína quinase 1
CKB- Creatina quinase B
CMPK- Citidina monofosfato quinase
c-Myc- Protooncogene myc
CORO1A- Coronina 1A
COX2- Ciclooxigenase
CYP450- Citocromo P450
DBT- Dibenzotiofeno
DBTO2- Dibenzotiofeno sulfona
DCC- Diciclohexilcarbodiimida
DCM- Diclorometano
DL50- Dose Letal 50%
DLD- Dihidrolipoil dehidrogenase
DMF- Dimetilformamida
DMH- 1,2-Dimetilhidrazina
DNA- Ácido desoxiribonucleico
DPP7- Dipeptidil peptidase
Dvl- Dishevelled
Dyrk1-Dual-specificity tyrosine-(Y)-
phosphorylation regulated kinase 1A
E1A- Fator de transcrição E1A
E2F- Fator de transcrição E2F
EEF2- Fator de elongação EEF2
EGFR- Receptor de fator de crescimento
epidermal
ENO1- Enolase alfa
EZR- Ezrina
FAK- Quinase de adesão focal
FDR- False Discovery rate
FITC- Isotiocianato de Fluoresceína
Fz- Frizzled
GAPDH- Gliceraldeído 3-fosfato dehidrogenase
GLI- Fator de transcrição GLI
GLUT- Receptor de glicose
GNB2L1- Proteína ligante de nucleotídeo guanina
GSK3- Glicogênio sintase quinase 3
HBP- 2-Hidróxibifenil
HDAC- Histona deacetilase
HE- Hematoxilina/Eosina
Hh- Hedgehog
HPA- Hidrocarbonetos policíclicos aromáticos
HPAS- Hidrocarbonetos policíclicos aromáticos
sulfurados
HPLC- Cromatografia Líquida de Alta Pressão
HRP- Horseradish peroxidase
HSP70- Heat shock protein 70
HSP90B1- Endoplasmina
HSPA2- Heat shock protein HSPA2
HSPA5- Heat shock protein HSPA5
IARC- Agência internacional para pesquisa em
câncer
IGF-IIR- Receptor de fator de crescimento tipo
insulina 2
IGG2A- Imunoglobulina gama 2A INCA- Instituto Nacional do Câncer
kDa- kilo Dalton
K-ras- Kirsten rat sarcoma viral oncogene
homolog
KRT8- Queratina 8
LDHA- Lactato dehidrogenase A
LEF- Fator potenciador linfóide
LGALS3- Galectina 3

Silva, K.T.S. Lista de Abreviaturas
xiv
LRP6- Lipoprotein receptor related protein 6
LUM- Lumican
M- Molar
MAM- Metil azoximetanol
MCPT1- Protease de mastócitos
mM- mili molar
mm- milímetro
MS- Espectrometria de Massas
mTORC1-
MYL- Miosina cadeia leve
ng- nanogramas
NHS- N-hidroxisuccinimida
NICD- Porção terminal do receptor Notch
NL- Não linear
NuA4- Histona acetiltransferase
OPAS- Organização Pan-Americana de Saúde
p/v- peso/volume
p53- Proteína tumoral p53
PAF- Polipose adenomatosa familiar
PBSS- Tampão Fosfato de Sódio com Salina
PGAM1- Fosfoglicerato mutase 1
PGK1- Fosfoglicerato quinase
pH- potencial hidrogeniônico
PIC- Coquetel de inibidores de protease
PIP- Fosfatidil inositol fosfato
PKM2- Piruvato quinase
PNLIP- Triacilglicerol lipase
ppm- Partes por milhão
PRDX1- Peroxiredoxina 1
Ptch- Proteína Patched
PTEN- Phosphatase and tensin homolog
PVDF- Polyvinylidene fluoride
Rac- Rho-GTPase
Raf- Quinase Raf
Ras- GTPase
RBBP4- Proteína ligante de histona RBBP4
RBP-JK- Recombining binding protein supressor
of hairless
RFESD- Proteína Rieske Fe-S
RNA- Ácido ribonucleico
SAGA- Histona acetiltransferase
SDS- Duodecil sulfato de sódio
SELENBP- Proteína ligante de selênio 1
SOD- Superóxido dismutase
STAGA- Histona acetiltransferase
TAGLN- Transgelina
TBST- Tampão Tris-HCL com Tween 20
TCF- Fator T celular
TF- Transferrina
TFA- Ác. Trifluoroacético
TFTC- TAT-binding protein free TAF-containing
complex
TGFβ- Fator de crescimento transformador
TIMP-1- Tissue inhibitor of metalloproteinases
TIP60- Histona acetiltransferase
TKT- Transcetolase
TNM- Estadiamento tumoral
TPM- Tropomiosina
TRA1- Proteína associada a transcrição 1
UI- Unidades internacionais
UICC- Union for International Cancer Control
v/v- volume/volume
VDAC2- Canal aniônico voltagem dependente
VEGF- Fator de crescimento vascular endotelial
WDR1- Repetição WD
WHO- World Health Organization
YWHAE- 14-3-3 ε
Silva, K.T.S. Lista de Abreviaturas
xv

Silva, K.T.S. Lista de Figuras
xvi
LISTA DE FIGURAS
Figura 01: Distribuição proporcional de óbitos no Brasil por tipo de câncer em 2014 por
sexo....................................................................................................................................2
Figura 02: Corte histológico de seção do duodeno no intestino delgado mostrando as
camadas que compõem o mesmo.......................................................................................5
Figura 03: Corte histológico de seção de intestino grosso mostrando as camadas Mucosa,
Submucosa, Túnica Muscular e Serosa além de componentes característicos..................5
Figura 04: Sequência adenoma-carcinoma em câncer colorretal......................................6
Figura 05: Eventos genéticos envolvidos no desenvolvimento do câncer colorretal.........7
Figura 06: Via Wnt/β-catenina........................................................................................10
Figura 07: Diagrama simplificado da via de sinalização Notch e seus possíveis alvos....12
Figura 08: Via de sinalização Hedgehog em vertebrados................................................13
Figura 09: Via de sinalização PI3K/Akt e vias associadas..............................................15
Figura 10: Vias de sinalização Notch, Wnt, e Hedgehog regulando o destino celular.....16
Figura 11: Estruturas químicas dos principais compostos contendo enxofre encontrados
no petróleo.......................................................................................................................19
Figura 12: “Via de metabolismo 4S para DBT”..............................................................20
Figura 13: Esquema representativo do tempo de tratamento e espera para o
desenvolvimento de alterações patológicas nos animais tratados com DMH, DBT e
DBTO2.............................................................................................................................30

Silva, K.T.S. Lista de Figuras
xvii
Figura 14: Biopanning com a biblioteca de peptídeos Ph.D. Seleção de ligantes
específicos.......................................................................................................................37
Figura 15: Gráfico de acompanhamento de peso dos animais pertencentes aos grupos
controles e testes ao longo de 24 semanas de tratamento/latência.....................................46
Figura 16: A) Fotografia de parte do intestino grosso de animal pertencente ao Grupo III
(Teste DMH 30 mg/kg) onde podem ser observadas as formações de pólipos. B)
Fotografia de parte do intestino delgado de animais pertencentes aos Grupos IV e V
(Testes DBT 30 mg/kg e DBTO2 30 mg/kg respectivamente) onde podem ser observados
pólipos.............................................................................................................................49
Figura 17: Fotomicrografias de cortes histológicos de intestino delgado e grosso corados
pela técnica de HE............................................................................................................53
Figura 18: Fotomicrografias de cortes histológicos de intestino delgado e grosso corados
pela técnica de imunohistoquímica com anti-CEA e anti CD44 e contra corados com
hematoxilina....................................................................................................................56
Figura 19: Géis 2D SDS-PAGE 10% representativos da extração de proteínas solúveis
de intestinos delgados dos A) Grupo Controle Óleo (III) e B) Teste DBT (IV)................58
Figura 20: Géis 2D SDS-PAGE 10% representativos da extração de proteínas solúveis
de intestinos delgados dos A) Grupo Controle Óleo (III) e B) Teste DBTO2 (V)..............58
Figura 21: Géis 2D SDS-PAGE 10% representativos da extração de proteínas solúveis
de intestinos delgados dos A) Grupo Controle Salina (I) e B) Teste DMH (II).................59
Figura 22: Diferenças nos níveis de expressão proteica dos grupos Teste DBT IV (barras
brancas) e Teste DBTO2 V (barras pretas) em relação ao grupo Controle Óleo III...........63
Figura 23: Mapa de interação proteica para proteínas diferencialmente expressas entre
grupos Teste IV e V e Controle III....................................................................................66

Silva, K.T.S. Lista de Figuras
xviii
Figura 24: Diferença nos níveis de expressão proteica entre o grupo Teste DMH (II) e o
grupo Controle Salina (I). Os resultados representam a média ± SEM de triplicatas
biológicas independentes.................................................................................................67
Figura 25: Mapa de interação proteína-metabólito para proteínas diferencialmente
expressas entre grupos Teste II e Controle I.....................................................................70
Figura 26: Perfis eletroforéticos representativos em 1D SDS-PAGE 12% de proteínas de
Intestino Delgado (Controle DBT; Teste DBT (D)) e de Intestino Grosso (Teste DBT (G);
Controle DMH; Teste DMH) para a verificação da qualidade da extração proteica.........71
Figura 27: Fotografias representativas da titulação dos fagos obtidos no último ciclo de
seleção de cada grupo para isolamento das colônias de ER2738 e consequente
amplificação e sequenciamento do DNA.........................................................................73
Figura 28: Alinhamento das sequências peptídicas obtidas após sequenciamento do DNA
dos fagos selecionados randomicamente ao fim do terceiro ciclo de biopanning para os
três grupos avaliados........................................................................................................74
Figura 29: Perfis cromatográficos em sistema HPLC dos peptídeos obtidos em fase
reversa..............................................................................................................................76
Figura 30: Perfis eletroforéticos de 1D SDS-PAGE de extratos proteicos referentes a
grupos Controle e Testes após cromatografia de afinidade...............................................77
Figura 01.A: Contagens global e diferencial de leucócitos provenientes dos grupos
Controles (I e III) e Testes (II, IV e V) ao final do período de tratamento/latência............99
Figura 01.B: Dosagens enzimáticas séricas: A) Razão AST/ALT; B) Amilase
provenientes dos grupos Controles (I e III) e Testes (II, IV e V) ao final do período de
tratamento/latência.........................................................................................................100
Figura 01.E: Validação dos experimentos de imunohistoquímica e análise proteômica.
A) SDS-PAGE 1D de extratos de proteína total de intestino delgado dos animais

Silva, K.T.S. Lista de Figuras
xix
pertencentes aos grupos III, IV e V. B) Western blotting para detecção de CEA, Calnexina
e β-actina........................................................................................................................155

Silva, K.T.S. Lista de Tabelas
xx
LISTA DE TABELAS
Tabela 01: Peptídeos selecionados por Phage display para síntese em fase sólida..........40
Tabela 02: Frequência de lesões observadas no intest. grosso dos animais pertencentes
aos Grupos I-V.................................................................................................................51
Tabela 03: Frequência de lesões observadas no intest. delgado dos animais pertencentes
aos Grupos I-V.................................................................................................................51
Tabela 04: Proteínas identificadas por spot nos géis referentes aos grupos Teste IV e V
que apresentaram alteração na expressão ≥ 1,5 vezes em relação ao grupo Controle III..60
Tabela 05: Proteínas identificadas por spot nos géis referentes ao grupo Teste II que
apresentaram alteração na expressão ≥ 1,5 vezes em relação ao grupo Controle I............61
Tabela 06: Massas moleculares preditas e observadas para cada peptídeo sintetizado....75
Tabela 01.C: Comparação de frequência das lesões observadas no intestino delgado para
os Grupos Controle Óleo e Controle Salina....................................................................101
Tabela 02.C: Comparação de frequência das lesões observadas no intestino grosso para
os Grupos Controle Óleo e Controle Salina....................................................................101
Tabela 01.D: Peptídeos sequenciados por proteína identificada através de espectrometria
de massas para os grupos Teste IV e V em relação ao grupo Controle III no experimento
empregando 2D SDS-PAGE..........................................................................................102
Tabela 02.D: Peptídeos sequenciados por proteína identificada através de espectrometria
de massas para o grupo Teste III em relação ao grupo Controle I no experimento
empregando 2D SDS-PAGE..........................................................................................114

Silva, K.T.S. Lista de Tabelas
xxi
Tabela 03.D: Peptídeos sequenciados por proteína identificada através de espectrometria
de massas para as bandas isoladas por cromatografia de afinidade e selecionadas em 1D
SDS-PAGE....................................................................................................................149
Tabela 01. F: Relação de proteínas que apresentaram região de similaridade com os
peptídeos selecionados por Phage display através do alinhamento de sequências realizado
por BLASTp..................................................................................................................156

Introdução
Silva, K.T.S. Introdução
2
1- INTRODUÇÃO
1.1. Epidemiologia
Apesar dos avanços tecnológicos, a incidência e a mortalidade por câncer têm
aumentado nas últimas décadas. Em 2008, a Agência Internacional para Pesquisa em
Câncer (IARC)/Organização Mundial de Saúde (WHO) estimou que ocorreriam 12,4
milhões de novos casos e 7,6 milhões de óbitos por câncer, doença esta que em 2030 será
responsável por mais de 11 milhões de óbitos em todo o mundo (UICC 2005; WHO 2007,
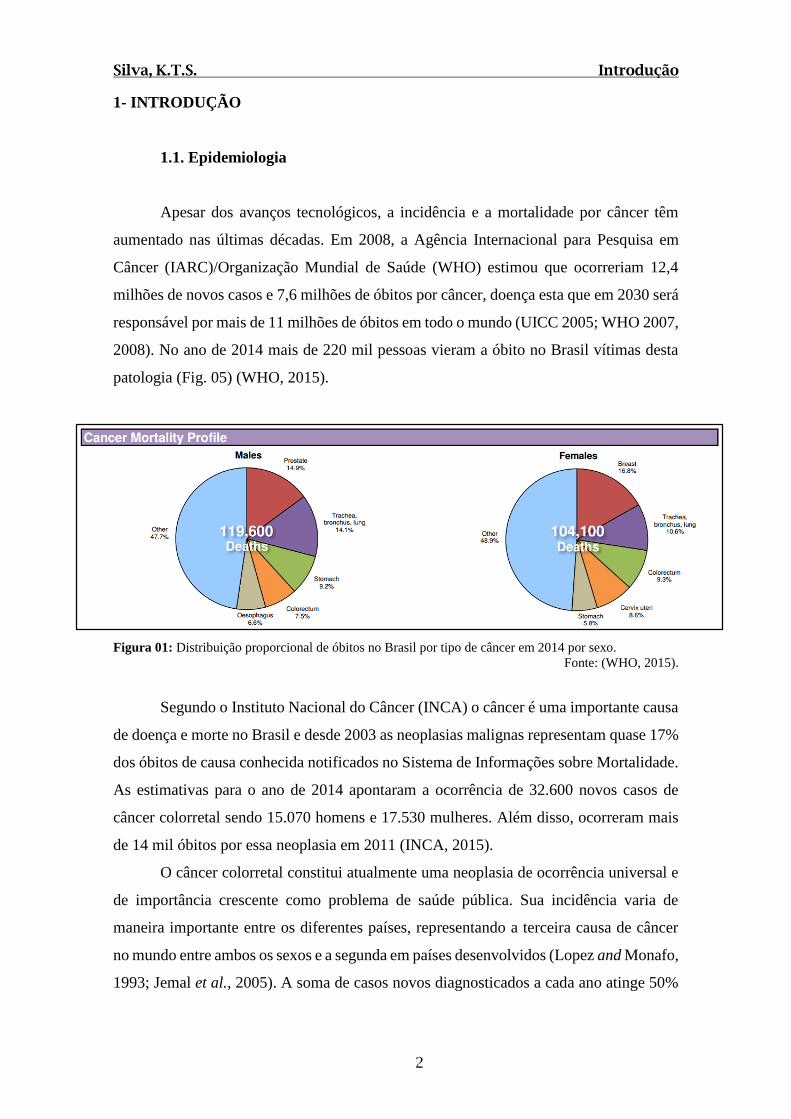
2008). No ano de 2014 mais de 220 mil pessoas vieram a óbito no Brasil vítimas desta
patologia (Fig. 05) (WHO, 2015).
Figura 01: Distribuição proporcional de óbitos no Brasil por tipo de câncer em 2014 por sexo.
Fonte: (WHO, 2015).
Segundo o Instituto Nacional do Câncer (INCA) o câncer é uma importante causa
de doença e morte no Brasil e desde 2003 as neoplasias malignas representam quase 17%
dos óbitos de causa conhecida notificados no Sistema de Informações sobre Mortalidade.
As estimativas para o ano de 2014 apontaram a ocorrência de 32.600 novos casos de
câncer colorretal sendo 15.070 homens e 17.530 mulheres. Além disso, ocorreram mais
de 14 mil óbitos por essa neoplasia em 2011 (INCA, 2015).
O câncer colorretal constitui atualmente uma neoplasia de ocorrência universal e
de importância crescente como problema de saúde pública. Sua incidência varia de
maneira importante entre os diferentes países, representando a terceira causa de câncer
no mundo entre ambos os sexos e a segunda em países desenvolvidos (Lopez and Monafo,
1993; Jemal et al., 2005). A soma de casos novos diagnosticados a cada ano atinge 50%

Silva, K.T.S. Introdução
3
do total observado nos cinco continentes, como registrou em 2002 a Organização Pan-
Americana da Saúde (OPAS) (INCA, 2006).
Diante deste cenário, fica clara a necessidade da continuidade em investimentos
no desenvolvimento de ações para o controle do câncer nos diferentes níveis de atuação,
como na promoção da saúde, na detecção precoce, na assistência aos pacientes, e na
formação de recursos humanos.
1.2. Aspectos Gerais da Oncogênese
Neoplasia (gr. “neo” + “plasis” = neoformação) é o termo utilizado para
caracterizar a proliferação local de clones celulares atípicos, sem causa aparente, de
crescimento excessivo, progressivo, ilimitado, não coordenado, autônomo, irreversível, e
com tendência a perda de diferenciação celular (Robbins and Cotran, 2010). No entanto,
a persistência do crescimento é resultado de alterações genéticas que permitem a
proliferação excessiva e desregulada de forma autônoma; apesar de os tumores
geralmente dependerem do organismo hospedeiro para nutrição e suprimento sanguíneo.
As anomalias genéticas encontradas no câncer afetam tipicamente três classes
gerais de genes (Croce, 2008). Os genes promotores, ou oncogenes (1), os genes
supressores de tumor (2) e, os genes envolvidos no reparo do DNA (3). Quando algum
desses sofre mutação, os controles genéticos pelos quais eles são responsáveis são
perdidos e o processo carcinogênico pode ser iniciado (Vogelstein and Kinzler, 2004; Lea
et al., 2007; Croce, 2008).
Desta forma, o câncer é uma doença complexa e multicausal que ocorre como
consequência do acúmulo progressivo de mutações nos genes onde os produtos protéicos
desempenham importantes papéis no controle da proliferação e diferenciação celular e na
apoptose. Usualmente, diversos fatores promotores atuam anteriormente ao
desenvolvimento neoplásico e com poucas exceções, uma mutação única não é suficiente
para o estabelecimento da doença.
Todos os tumores, benignos ou malignos, apresentam dois componentes básicos:
células neoplásicas clonais que constituem seu parênquima, e estroma reativo, constituído
de tecido conjuntivo, vasos sanguíneos e quantidade variável de macrófagos e linfócitos.
Sabe-se que os tumores malignos induzem resposta inflamatória celular crônica não
relacionada com necrose ou infecção do tumor, e o infiltrado é composto principalmente
de células T e macrófagos. Assim, a defesa imunológica responde a antígenos tumorais

Silva, K.T.S. Introdução
4
presentes na superfície das células malignas expressos diferencialmente daqueles das
células normais e, em alguns casos, pode ser responsável pelo retardo do crescimento
tumoral e de sua progressão (Zbar, 2004).
In vitro, células isoladas do sistema imune têm mostrado serem efetivas contra
tumores através de mecanismos como lise osmótica ou apoptose e citotoxicidade celular.
In vivo, contudo, os tumores parecem desenvolver mecanismos de escape ao sistema
imune. As células neoplásicas podem evadir ao sistema imune por diversas vias, sendo a
capacidade de interferência na função das células imunes o principal motivo de falha no
controle da progressão tumoral (Dunn et al., 2004; Bellati et al., 2009). Além disso, o
papel do microambiente tumoral no reconhecimento e progressão do câncer tem sido cada
vez mais estudado, e as interações entre as células imunes, as células tumorais e os
componentes moleculares do tumor exercem influência significativa na progressão da
doença (Rosenberg et al., 2004).
1.3. Trato Gastrointestinal Inferior
1.3.1. Características Gerais
O trato gastrointestinal inferior está morfologicamente divido em regiões
distintas: intestino delgado (duodeno, jejuno e íleo) e intestino grosso (ceco, cólon, reto,
canal anal e apêndice). É constituído de várias camadas histológicas dispostas
concentricamente e similares em toda a extensão com pequenas modificações e
especificações regionais.
O intestino delgado apresenta modificações estruturais que aumentam a área total
da sua superfície e que promovem a absorção de nutrientes vindos da alimentação. Possui
quatro túnicas que compõem a parede intestinal: (1) mucosa; (2) submucosa; (3) túnica
muscular e (4) serosa, sendo as duas primeiras, os principais sítios de alterações
histológicas observadas em estágios iniciais de transformações neoplásicas (Fig. 01).
Tumores na região do intestino delgado são geralmente raros, correspondendo a cerca de
1 a 6% de todas as neoplasias do trato gastrointestinal (Rangel et al., 2000).
As pregas e vilos encontrados no intestino delgado não são encontrados no
intestino grosso, porém, as mesmas camadas constituem este segmento. As células
epiteliais são mais planas e a mucosa é mais espessa, possuindo longas Glândulas de
Silva, K.T.S. Introdução
5
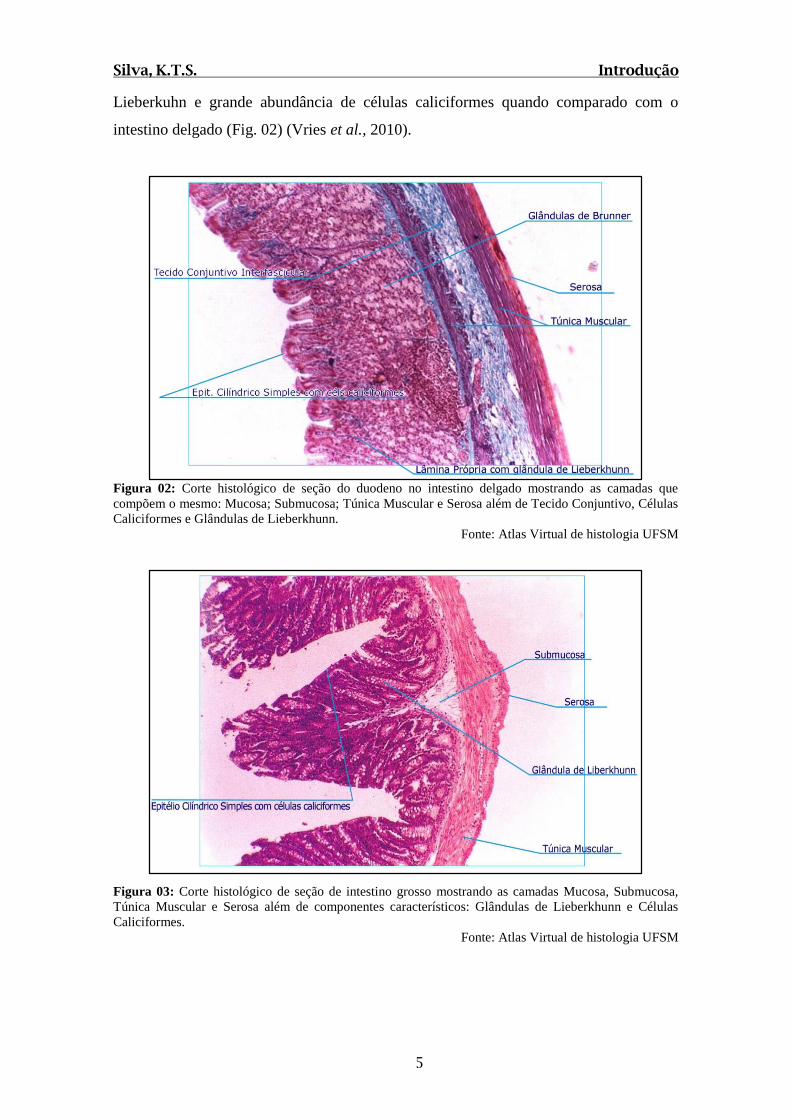
Lieberkuhn e grande abundância de células caliciformes quando comparado com o
intestino delgado (Fig. 02) (Vries et al., 2010).
Figura 02: Corte histológico de seção do duodeno no intestino delgado mostrando as camadas que
compõem o mesmo: Mucosa; Submucosa; Túnica Muscular e Serosa além de Tecido Conjuntivo, Células
Caliciformes e Glândulas de Lieberkhunn.
Fonte: Atlas Virtual de histologia UFSM
Figura 03: Corte histológico de seção de intestino grosso mostrando as camadas Mucosa, Submucosa,
Túnica Muscular e Serosa além de componentes característicos: Glândulas de Lieberkhunn e Células
Caliciformes.
Fonte: Atlas Virtual de histologia UFSM
Silva, K.T.S. Introdução
6
1.3.2. Câncer Intestinal
Os adenomas constituem os tipos de pólipos mais frequentemente encontrados no
intestino delgado, possuindo predileção pelo duodeno distal embora possam ser
encontrados em toda a sua extensão. Histologicamente, eles são similares aos adenomas
encontrados na região colorretal tendendo a apresentar uma arquitetura de vilos mais
pronunciada. Assim como para a região colorretal, o risco de progressão maligna de um
adenoma é maior quanto maior for o pólipo (Brosens et al., 2007).
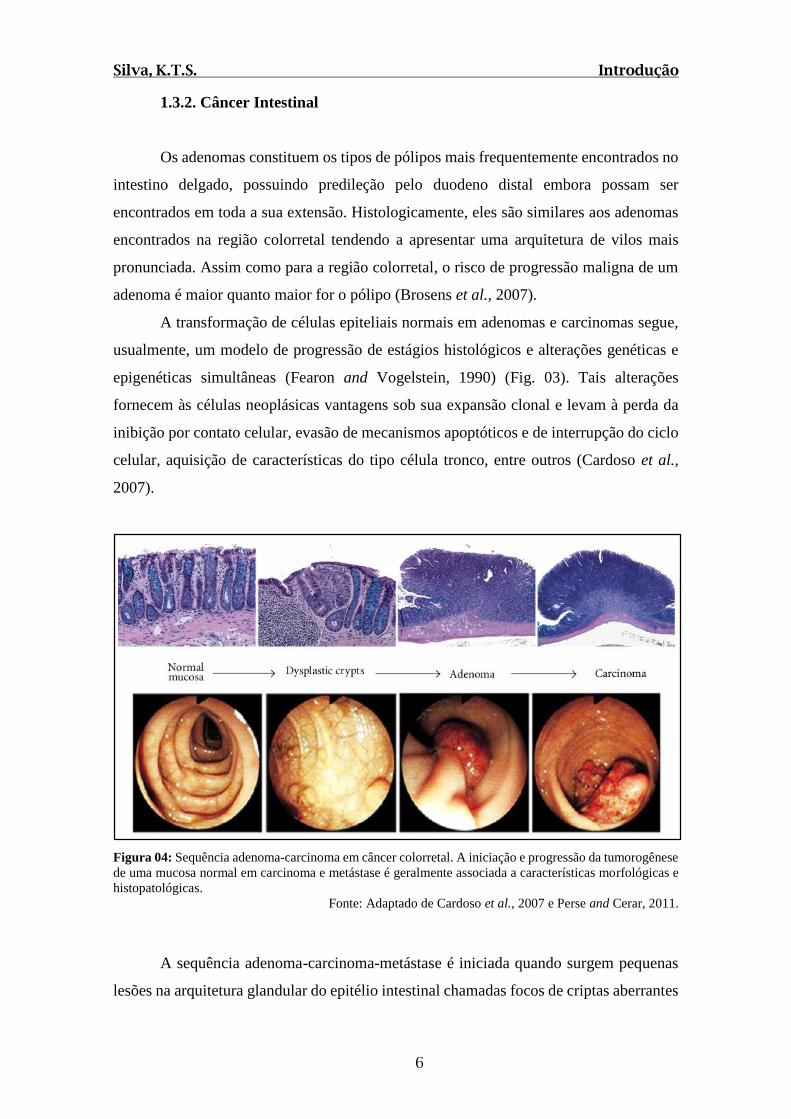
A transformação de células epiteliais normais em adenomas e carcinomas segue,
usualmente, um modelo de progressão de estágios histológicos e alterações genéticas e
epigenéticas simultâneas (Fearon and Vogelstein, 1990) (Fig. 03). Tais alterações
fornecem às células neoplásicas vantagens sob sua expansão clonal e levam à perda da
inibição por contato celular, evasão de mecanismos apoptóticos e de interrupção do ciclo
celular, aquisição de características do tipo célula tronco, entre outros (Cardoso et al.,
2007).
Figura 04: Sequência adenoma-carcinoma em câncer colorretal. A iniciação e progressão da tumorogênese
de uma mucosa normal em carcinoma e metástase é geralmente associada a características morfológicas e
histopatológicas. Fonte: Adaptado de Cardoso et al., 2007 e Perse and Cerar, 2011.
A sequência adenoma-carcinoma-metástase é iniciada quando surgem pequenas
lesões na arquitetura glandular do epitélio intestinal chamadas focos de criptas aberrantes

Silva, K.T.S. Introdução
7
(ACF). Os ACF são agregados de criptas anormais caracterizados por hiperproliferação,
tamanho aumentado, zonas pericrípticas expandidas e fendas alongadas (Rosenberg and
Liu, 1995; Papanikolaou et al., 1998; Papanikolaou et al., 2000).
Algumas mutações estágio-específicas que ocorrem no modelo de progressão
adenoma-carcinoma-metástase são conhecidas, e incluem alterações no gene supressor
de tumor APC (Adenomatous polyposis coli) (Epitélio Normal Adenoma), no proto-
oncogene K-ras (presente em adenomas) e no gene supressor de tumor p53 (na transição
Adenoma Carcinoma) (Nambiar et al., 2010). Além disso, outros genes estão
significativamente alterados em diferentes classes de tumores e envolvidos na
carcinogênese colorretal, no entanto, métodos de classificação e estadiamento do tumor
e realização de prognósticos mais robustos e menos invasivos continuam sendo
necessidades médicas ainda não atendidas (Fig. 04) (Hoops and Traber, 1997; Nambiar
et al., 2010).
Figura 05: Eventos genéticos envolvidos no desenvolvimento do câncer colorretal. A acumulação
gradativa de mutações afeta o controle de crescimento, diferenciação e sobrevivência celular. Para o CCR
os genes cruciais envolvidos no desenvolvimento de pólipos e câncer para cada etapa são mostrados nas
caixas brancas.
Fonte: Adaptado de Simpson and Dorow, 2001.
Os recentes progressos da proteômica têm contribuído para o entendimento da
patofisiologia das neoplasias, propiciando novas perspectivas para diagnósticos e
descoberta de novas drogas anti-câncer. A avaliação sistemática do proteoma, de forma a
comparar a expressão proteica de duas células, tecidos ou organismos em diferentes
condições fisiológicas possibilita a obtenção de informações para identificação,

Silva, K.T.S. Introdução
8
quantificação e caracterização de modificações pós-traducionais presentes em proteínas
e que influenciam diretamente o funcionamento celular (Wilkins et al., 1996; Cho, 2007).
Nesse sentido, a proteômica tem sido utilizada também como nova ferramenta na
busca por biomarcadores moleculares para o câncer colorretal (CCR). As alterações na
expressão proteica envolvidas na transformação maligna podem ser monitoradas de
maneira quantitativa e qualitativa e fornecem valiosas informações que podem ser
utilizadas para a obtenção de diagnósticos e prognósticos mais efetivos. Além disso,
representam um grande impacto na oncologia clínica, especialmente se o biomarcador
pode ser detectado antes dos sintomas clínicos ou se permite um acompanhamento da
resposta ao tratamento (Cho, 2007; Alhquist, 2010).
Embora as alterações moleculares relacionadas ao CCR tenham sido
extensivamente investigadas, poucos pesquisadores têm descrito as aplicações desses
biomarcadores na detecção de estágios primários de diagnóstico e prevenção do câncer.
Considerando que o CCR pode ser prevenido se o adenoma for diagnosticado e removido,
torna-se necessária a identificação de marcadores capazes de diagnosticar de maneira
tecido-específica e precoce as alterações iniciais que ocorrem durante o desenvolvimento
desta doença (Chang et al., 2005).
1.3.3. Fatores de Risco e Genéticos
A explicação para os altos índices encontrados para o CCR está na maior
exposição dos indivíduos a fatores de risco cancerígenos. A redefinição dos padrões de
vida, a partir da uniformização das condições de trabalho, nutrição e consumo
desencadeados pelo processo global de industrialização refletem-se no perfil
epidemiológico das populações (Montesano and Hall, 2001).
Fatores de risco hereditários e ambientais (dietéticos ou não) estão envolvidos no
desenvolvimento do CCR. As alterações genéticas que levam a esta doença podem ser
adquiridas, gerando o chamado câncer esporádico, responsável por 75 a 85% dos casos.
Nele, existe uma ação cumulativa de agentes carcinógenos ambientais e dietéticos sobre
a mucosa, levando a modificações específicas no DNA das células do epitélio intestinal
(Cecconello, 2008). Em outras circunstâncias, o indivíduo já apresenta ao nascimento ou
adquire precocemente mutações específicas que desencadeiam as formas hereditárias do
CCR, representadas principalmente pela polipose adenomatosa familiar (PAF) e pelo
câncer colorretal hereditário não associado à polipose (Hoops and Traber, 1997).

Silva, K.T.S. Introdução
9
Quanto à influência da alimentação, é sabido que a dieta ocidental caracterizada
pelo alto teor de gordura saturada, juntamente com a ingestão excessiva de proteína de
origem animal e baixo teor de fibras, está associada a maior potencial carcinogênico.
Também são considerados nocivos, os fatores relacionados ao estilo de vida como fumo,
sedentarismo, obesidade, consumo excessivo de álcool e ingestão de aminas
heterocíclicas e hidrocarbonetos aromáticos (Marques-Vidal et al., 2006).
1.4. Visão Geral de Vias de Sinalização Atuantes no CCR
No cólon humano, cerca de 10 bilhões de novas células são geradas diariamente
por progenitoras localizadas na base das criptas. O destino desse novo epitélio é
determinado por sinais extracelulares gerados a nível populacional. Tal sinalização
extrínseca é realizada por morfógenos, proteínas solúveis que formam gradientes de
concentração e atuam de maneira dose dependente induzindo respostas celulares distintas
em suas células alvo (Tabata and Takei, 2004).
1.4.1. Via Wnt/β-catenina
A via de sinalização Wnt/β-catenina é altamente conservada em eucariotos e
participa de diversos acontecimentos ligados ao desenvolvimento celular. Ela
desempenha papéis na proliferação, movimento, estabelecimento da polaridade e decisão
do destino celular e manutenção de células tronco. Um dos caminhos mais bem estudados
para essa via é caracterizado pela estabilização e acúmulo de β-catenina no citosol, que
então migra para o núcleo onde participa da ativação da transcrição de proteínas chave na
carcinogênese como c-Myc, VEGF, ciclina D1, entre outras (via canônica) (Krauzova
and Korinek, 2014; Markowska et al., 2014; Klimczak, 2015).
Na ausência de Wnt, a proteína β-catenina é constantemente degradada pela ação
do complexo proteico formado por Axina, APC, CK1 (caseína quinase 1) e GSK3
(glicogênio sintase quinase 3). As proteínas CK1 e GSK3 fosforilam continuamente a
região N terminal de β-catenina que resultam no reconhecimento do produto fosforilado
por β-Trcp, uma subunidade de E3 ubiquitina ligase. A ubiquitinação de β-catenina leva
a sua degradação proteassomal prevenindo assim que esta proteína migre para o núcleo e
ative a transcrição de genes reprimidos pelo complexo TCF/LEF ligado ao DNA (Fator
T celular/Fator Potenciador Linfóide) (Fig. 06, A). A via é ativada quando um ligante

Silva, K.T.S. Introdução
10
Wnt se liga ao receptor transmembrana Fz (Frizzled) e seu co-receptor LRP6 (Lipoprotein
receptor related protein 6). Ocorre a formação de um complexo transmembrana Wnt-Fz-
LRP6 e recrutamento de uma terceira proteína chamada Dvl (Dishevelled), que por
fosforilação resulta no sequestro do complexo de degradação, inibindo-o (Fig. 06, B)
(MacDonald et al., 2009; White et al., 2012).
Figura 06: Via Wnt/β-catenina. A) Na ausência de Wnt, β-catenina é fosforilada pelo complexo formado
por Axina, CK1, GSK3 e APC sendo posteriormente reconhecida e ubiquitinada por β-Trcp e direcionada
para degradação proteassomal. B) Na presença de ligante Wnt, ocorre a formação de um complexo receptor
Fz-LRP6 que sequestra o complexo de degradação inibindo-o e estabilizando a β-catenina presente no
citosol que então migra para o núcleo onde co-ativa TCF levando à transcrição dos regulados por esta
proteína.
Fonte: MacDonald et al., 2009.
Os mamíferos apresentam 19 formas de proteínas Wnt ricas em cisteína e com
peptídeo sinal N-terminal para secreção. Tais proteínas sofrem glicosilação e lipidação
antes da excreção e tais modificações têm papel direto na ativação da via. No CCR cerca
de 90% dos tumores apresentam alguma mutação em fatores regulatórios da via sendo as
mais comuns aquelas que afetam o gene APC ou CTNNB1 (β-catenina). Tais mutações
conferem diferentes níveis de ativação da via e requerem diferentes subconjuntos de
eventos cooperativos (p. ex. silenciamento epigenético, sinais do microambiente ou
alterações em outras vias) para levar à progressão tumoral por diferentes mecanismos
(Brabletz et al., 1998; Vermeulen et al., 2010; White et al., 2012).

Silva, K.T.S. Introdução
11
O crosstalk entre a via Wnt/β-catenina e outras vias de sinalização ligadas ao
desenvolvimento podem modular a via β-catenina no CCR. A proteína de membrana
Notch1 pode se ligar à β-catenina ativa e regular negativamente sua função em células
tronco e tumorais. A via Hedgehog também pode regular a via Wnt/β-catenina. Arimura
e colaboradores demonstraram correlação entre a presença do mediador de Hedgehog,
Smoothened (Smo), e aumento da atividade de Wnt/β-catenina em camundongos
mutantes para APC (Arimura et al., 2009).
Por fim, deve-se ressaltar que embora a via Wnt/β-catenina seja essencial na
reprogramação celular relacionada ao CCR através do estímulo da expressão gênica de
fatores chave, ela possui um efeito transitório e ativo somente nos estágios iniciais da
transformação não sendo necessária para a manutenção celular de pluripotência
(Klimczak, 2015).
1.4.2. Via Notch
A via Notch é essencial na diferenciação de células caliciformes e células
progenitoras, participando da regulação do desenvolvimento e homeostase intestinal. Em
humanos e camundongos os ligantes Notch (Jagged1, Jagged2, DLL1, DLL3 e DLL4)
ativam os receptores Notch (Notch1-4) levando à transcrição de diversos genes alvo (p21,
Hes-1, Dellex, etc) (Fre et al., 2005).
Um ligante Notch presente na membrana de uma célula adjacente liga-se a um
receptor presente na célula receptora. A interação receptor-ligante leva à ativação do
complexo enzimático γ-secretase que cliva a região transmembrana do próprio receptor
Notch (NICD) ativando-o. A porção NICD migra para o núcleo celular onde liga-se a um
regulador transcricional (CBP ou RBP-JK em vertebrados). O complexo formado desloca
co-repressores ligados aos fatores de transcrição e recruta co-ativadores transcricionais
que induzem a expressão gênica (Fig. 07). Diversas vias de sinalização podem ser super
reguladas através da ativação da via Notch, como por exemplo a via PI3K/AKT por
inativação de PTEN (homólogo de fosfatase e tensina deletado no cromossomo 10) e as
vias c-Myc e EGFR (receptor de fator de crescimento epidermal). A ativação da via
parece estar associada com o desenvolvimento primário do CCR indicando que este é um
evento inicial na carcinogênese colorretal (Qiao and Wong, 2009; Miyamoto and
Rosenberg, 2011; Xie et al., 2014).

Silva, K.T.S. Introdução
12
Figura 07: Diagrama simplificado da via de sinalização Notch e seus possíveis alvos. Setas para cima
indicam aumento de expressão regulada por Notch e setas para baixo indicam diminuição de expressão
regulada por Notch.
Fonte: Qiao and Wong, 2009.
1.4.3. Via Hedgehog
Hedgehog são moléculas sinalizadoras secretadas que em mamíferos englobam
três proteínas homólogas com diferentes distribuições temporais e espaciais: Sonic
hedgehog (Shh); Indian hedgehog (Ihh); Desert hedgehog (Dhh) referidas em conjunto
como Hhs (Ingham and McMahon, 2001). Essas proteínas atuam via receptores de
membrana Patched (Ptch), cuja expressão é super regulada pela sinalização de Hh, e
Smoothened (Smo). Na ausência de Hh, Ptch inibe cataliticamente a atividade de Smo
afetando sua localização na membrana celular. Uma molécula endógena que atua como
agonista de Smo é transportada para fora da célula por Ptch prevenindo sua ligação a
Smo. Quando presente Hh liga-se a Patch inativando-a e consequentemente ativando Smo
que traduz o sinal ao citoplasma via GLI (família de fatores de transcrição com motivo
dedo de zinco) (Fig. 08) (Bale and Yu,2001; van den Brink and Hardwick, 2006;
Mazumdar et al., 2011).

Silva, K.T.S. Introdução
13
Figura 08: Via de sinalização Hedgehog em vertebrados. A cascata de Hh é iniciada quando Hh liga-se a
Patch ativando uma segunda proteína transmembrana chamada Smo. Os fatores de transcrição GLI
localizados downstream na via de sinalização ativam a transcrição de genes alvo (GLI A). Na ausência de
Hh Ptch inibe a atividade de Smo afetando sua localização na membrana e levando ao processamento
proteolítico de proteínas GLI reprimindo a transcrição de genes alvo (GLI R).
Fonte: Adaptado de Gupta et al., 2010.
Os fatores GLI existem como três proteínas separadas: GLI1 e GLI2 que atuam
como ativadores transcricionais e GLI3 que atua como repressor transcricional. A
expressão de GLI1 é altamente dependente da ativação da sinalização por Hh. Na
ausência de Hh, Ptch bloqueia a atividade de Smo e as proteínas GLI são
proteoliticamente processadas para gerar o repressor GLIR, largamente derivado de GLI3
que reprime a transcrição. A ligação de Hh a Ptch interrompe a inibição de Smo gerando
o ativador GLIA constituído principalmente por GLI2 e ativando a expressão de genes
alvo (GLI1, Ptch1, proteína de interação com Hh, Ciclina D, Myc, Bmi1, Bcl2, VEGF,
etc). A ativação de GLI é regulada em diferentes níveis via fosforilação por inibidores
como Ren, PKA, GSK3 e ativadores como Dyrk1, Akt e Ras (Gupta et al.,2010).

Silva, K.T.S. Introdução
14
1.4.4. Via PI3K/Akt
A ativação da via PI3K/Akt (fosfatidilinositol-3 quinase) ocorre através da ligação
de fatores de crescimento, citocinas ou insulina a um receptor de membrana tirosina
quinase, levando à autofosforilação de um domínio intracelular do mesmo. Tal
fosforilação permite que ocorram alterações na conformação e subsequente ativação de
PI3K (composta por uma subunidade catalítica – p110 e uma subunidade regulatória –
p85). PI3K promove a transformação de fosfatidilinositol 4,5-bisfosfato (PIP2) em
fosfatidilinositol 3,4,5-trifosfato (PIP3) que, por sua vez, medeia a fosforilação e ativação
de Akt. PIP2 e PIP3 são lipídeos chave responsáveis por mediar respostas celulares ao
estímulo por fatores de crescimento ou citocinas, ligação de integrinas, e sinalização via
integrina quinase ou quinase de adesão focal (FAK) (Markowska et al., 2014).
Akt, também conhecida como proteína quinase B (PKB), pode ser ativada também
em condições de hipóxia, hipoglicemia, ação de radicais livres entre outras. Um dos
principais papéis de Akt é a promoção da proliferação celular e inibição da apoptose por
fosforilação de caspase 9 e interação com o antagonista de morte celular BCL2. A
fosforilação e inativação de GSK3 por Akt leva à inibição da ligação de GSK3 às ciclinas
D e E, e aos fatores de transcrição c-Myc e c-Jun, protegendo-os da degradação
proteolítica e favorecendo a entrada da célula na fase S. Quando ativada, Akt promove a
síntese proteica e crescimento celular por ativação do complexo mTORC1 que ativa a
quinase p70 S6, promovendo aumento na tradução de RNA mensageiro e ativando a
proteína ribossomal S6 e o fator de elongação 2 (EEF2) (Fig. 09). Além disso, Akt
também regula o balanço de carboidratos por aumento de expressão dos transportadores
de glicose GLUT1 e GLUT3, e facilitando a translocação de GLUT4 à membrana celular,
contribuindo para um aumento de influxo de glicose na célula (Brown and Toker, 2015;
Simpson et al., 2015).
Silva, K.T.S. Introdução
15
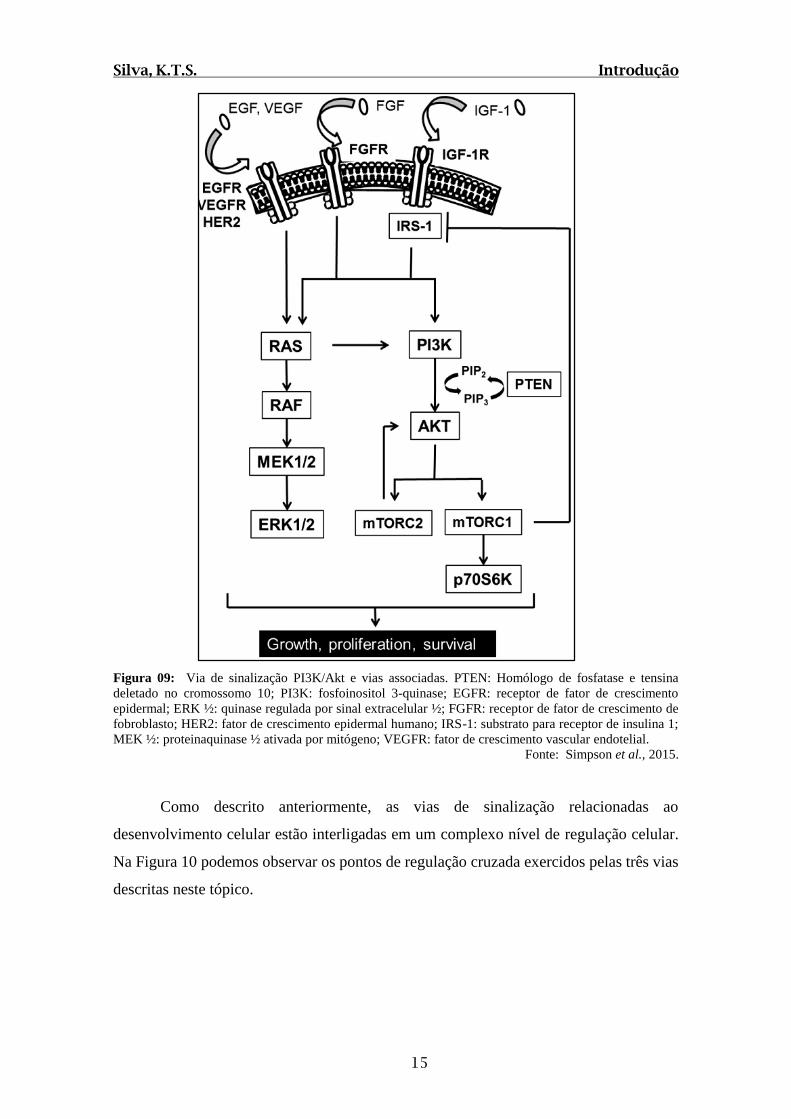
Figura 09: Via de sinalização PI3K/Akt e vias associadas. PTEN: Homólogo de fosfatase e tensina
deletado no cromossomo 10; PI3K: fosfoinositol 3-quinase; EGFR: receptor de fator de crescimento
epidermal; ERK ½: quinase regulada por sinal extracelular ½; FGFR: receptor de fator de crescimento de
fobroblasto; HER2: fator de crescimento epidermal humano; IRS-1: substrato para receptor de insulina 1;
MEK ½: proteinaquinase ½ ativada por mitógeno; VEGFR: fator de crescimento vascular endotelial.
Fonte: Simpson et al., 2015.
Como descrito anteriormente, as vias de sinalização relacionadas ao
desenvolvimento celular estão interligadas em um complexo nível de regulação celular.
Na Figura 10 podemos observar os pontos de regulação cruzada exercidos pelas três vias
descritas neste tópico.

Silva, K.T.S. Introdução
16
Figura 10: Vias de sinalização Notch, Wnt, e Hedgehog regulando o destino celular. A) Via Notch: a
interação ligante-receptor promove a clivagem do domínio NICD por γ-secretase e sua translocação para o
núcleo com consequente indução da transcrição de genes alvo. B) Via Wnt: estabilização e acúmulo nuclear
de β-catenina que se associa com os fatores de transcrição LCF/LEF e co-ativadores (CBP) ativando a
expressão de genes alvo. C) Via Hedgehog: o ligante Hh ativa o receptor Ptch que por sua vez libera o
receptor Smo. Smo então passa a inibir a fosforilação e clivagem de GLI por GSK3, CK1 e PKA
promovendo a formação de GLIA que ativa a transcrição. Notar que a transcrição de genes por Hedgehog
ativa a via Notch e a presença de GSK3 3 CK1 do complexo de degradação de Wnt são também
responsáveis por inibir a via Hedgehog por fosforilação de GLI.
Fonte: Malhotra et al., 2011
1.5. Carcinogênese Química
Os cânceres causados por agentes ambientais frequentemente ocorrem em tecidos
como pulmão, trato gastrointestinal e pele, porque esses órgãos estão expostos ao
ambiente (Shubik and Sice,1956). Recentemente, o estudo da carcinogênese química tem
sido mesclado com estudos de alterações celulares em células cancerígenas,
possibilitando assim a busca por marcadores biológicos e vias metabólicas alteradas.
A carcinogênese química causa alterações genéticas e epigenéticas em células
susceptíveis que podem transmitir vantagem seletiva para o crescimento e sofrer
expansão clonal. Sendo assim, elas se tornam geneticamente instáveis e transformadas

Silva, K.T.S. Introdução
17
em células neoplásicas. O primeiro estágio da carcinogênese, o de iniciação tumoral,
envolve a exposição de células normais a carcinógenos físicos e químicos. Estes
carcinógenos causam danos ao DNA e a outras macromoléculas fornecendo às células
iniciadas responsividade alterada ao microambiente e vantagem proliferativa. No segundo
estágio, a promoção tumoral resulta na proliferação excessiva das células iniciadas e
aumenta a probabilidade de ocorrência de danos genéticos adicionais, incluindo mutações
endógenas que acumulam na população em expansão. No estágio de progressão tumoral
ou terceiro estágio ocorre a aquisição de múltiplas alterações genéticas e multiplicação
descontrolada e irreversível das células alteradas (Loeb and Harris, 2008; Irigaray and
Belpomme, 2010).
O campo de estudos sobre a carcinogênese química provavelmente teve início
com as associações epidemiológicas de ocorrência de tumores com a exposição à fumaça
de tabaco realizadas por Hill e Pott no início do século 18. A partir daí, seguiram-se
observações de tumores no trato urinário de trabalhadores que mantinham contato com
arilaminas nas fábricas européias no final do século 19. Além destes, trabalhos com
animais experimentais envolvendo hidrocarbonetos policíclicos aromáticos (HPA)
derivados de óleos crus serviram desde então como importantes protótipos para o estudo
de HPA (Guengerich,2001).
Os HPA representam uma importante classe de poluentes ambientais os quais têm
recebido atenção especial pelo fato de alguns de seus componentes demonstrarem forte
potencial mutagênico e carcinogênico, além de desempenharem papéis cruciais em três
problemas ambientais importantes: poluição do ar, chuva ácida e mudança climática
global. Esses compostos estão presentes nos combustíveis fósseis e são formados durante
a combustão incompleta e pirólise de matéria orgânica. Eles podem, por exemplo, ser
gerados durante incêndios florestais e erupções vulcânicas além de serem constituintes
naturais dos óleos crus, alguns tipos de alimentos e produtos petroquímicos. Além disso,
podem ser formados pelo gás de cozinha e no tráfego, sendo também uma das diversas
classes de carcinogênicos químicos presentes na fumaça de cigarro (Honer, 2001).
A ampla distribuição dos HPA provenientes de fontes naturais e antropogênicas
juntamente com processos de distribuição e transporte explica sua ocorrência ubíqua. Os
seres humanos são expostos a HPA ambientais, ocupacionais, medicinais e provenientes
da dieta. A absorção através do trato pulmonar, gastrointestinal e da pele ocorre de
maneira dependente do tipo de HPA, do tamanho das partículas onde estão adsorvidos,

Silva, K.T.S. Introdução
18
da composição do adsorvente, e da sua afinidade por gordura (Jacob and Grimmer, 1996;
Angerer et al., 1997; Schoket, 1999; Strickland and Kang, 1999).
O metabolismo desempenha um importante papel na conversão de carcinógenos
químicos em espécies reativas que danificam macromoléculas celulares, interferem com
vias de sinalização e que podem causar câncer. Células de cólon humano sabidamente
metabolizam HPA. Os níveis induzíveis das enzimas do complexo citocromo P450
(CYP450) podem influenciar no desenvolvimento de tumores nos intestinos grosso e
delgado como consequência da ingestão de HPA (Diggs et al., 2011). Em roedores, a
absorção no trato gastrointestinal ocorre rapidamente e picos destes compostos podem ser
identificados no sangue 1-2 horas após a administração (Lipniak et al., 1993).
1.5.1. Dibenzotiofeno e Dibenzotiofeno Sulfona
O petróleo e o carvão são importantes combustíveis fósseis de composição
complexa da qual se distinguem quatro famílias de compostos: hidrocarbonetos alifáticos,
cíclicos, aromáticos e heteromoléculas contendo átomos de nitrogênio (N), enxofre (S)
ou oxigênio (O) na estrutura. Os compostos sulfurados constituem a terceira população
presente nesses combustíveis e, devido à sua difícil biodegradabilidade, são considerados
compostos recalcitrantes podendo servir como marcadores da poluição gerada por óleo.
O dibenzotiofeno (DBT) e seus derivados são os maiores representantes desta classe (Fig.
11) (Kropp et al., 1997; Heilmann et al., 2004).
O conteúdo elevado de hidrocarbonetos aromáticos policíclicos sulfurados
(HPAS) em produtos do petróleo compromete a qualidade dos mesmos e sua remoção ou
conversão em estruturas menos tóxicas é uma importante etapa no processo de refino
(Zhang et al., 2005). Os limites de concentração desses compostos em combustíveis
obedecem às legislações específicas de cada país com o objetivo de minimizar os efeitos
negativos à saúde e ao ambiente. A gasolina e o diesel comercializados na comunidade
Européia e nos EUA devem apresentar, respectivamente, os limites máximos de 30 e 50
ppm de HPAS. A Alemanha limitou o conteúdo de enxofre presente no diesel e na
gasolina a 10 ppm em 2001 (Babich and Moulijn, 2003).

Silva, K.T.S. Introdução
19
Figura 11: Estruturas químicas dos principais compostos contendo enxofre encontrados no petróleo.
Fonte: Adaptado de Speight, 2002.
Visando uma redução da queima e consequente emissão de compostos sulfurados
no ambiente, diversos estudos buscam o desenvolvimento de tecnologias para a
remediação do enxofre de combustíveis. A redução dessas substâncias por parte das
refinarias revela a preocupação do setor em oferecer combustíveis menos tóxicos e menos
poluentes. Neste contexto, o dibenzotiofeno é utilizado na verificação da eficiência de
processos de dessulfurização em derivados do petróleo e, apesar disso, poucos relatos
sobre sua ação tóxica em mamíferos têm sido apresentados (Leighton, 1989; Kobayashi
et al., 2000; Madeira et al., 2008; Scherer et al., 2009).
Trabalhos da década de 80 foram realizados em animais experimentais
empregando doses maciças de DBT e derivados por via oral e subcutânea na expectativa
de demonstrar efeitos tóxicos. Tais trabalhos estabeleceram a dose letal (DL50) de 470
mg/kg e graves lesões no pâncreas e fígado, congestão pulmonar, edema, e hemorragia
intestinal com doses de 335 mg/kg observadas nos ratos que sobreviveram a um tempo
maior que 24 horas após a ingestão do DBT. Outros estudos onde foram administrados o
DBT e outros análogos ativos de petróleo demonstraram, além dos efeitos citados acima,
alterações nos linfonodos mesentéricos e necrose das células do timo (Leighton, 1989).
A toxicidade individual dos HPA de maneira geral está relacionada com a ativação
metabólica e a formação de metabólitos carcinogênicos responsáveis pela alquilação do
DNA e do estágio de iniciação através de mecanismos complexos associados à

Silva, K.T.S. Introdução
20
carcinogênese química (Aas et al., 2000). Os metabólitos ativos formados a partir de HPA
são produtos de reações catalisadas por enzimas CYP450 em particular, pela isoenzima
P4501A1 e, embora os níveis basais dessa isoforma sejam baixos em diversos tecidos,
ocorre uma indução da sua expressão após a exposição a diversos tipos de HPA
(Soontjens et al., 1997; Jones et al., 2000).
Uma das vias de metabolização do DBT inicialmente descrita em espécies de
Rhodococcus sp. IGTS8 é chamada de via sulfóxido-sulfona-sulfonato-sulfato ou “Via
4S”. Durante o metabolismo, o DBT sofre a ação de um sistema multienzimático com
três diferentes atividades, que promovem um ataque oxidativo progressivo no grupo
tiofênico. A primeira enzima da via é uma monooxigenase que oxida o DBT a 5,5’-
dióxido de DBT (DBTO2) em dois passos; a segunda enzima apresenta atividade similar
à uma monoxigenase e converte o 5,5’-dióxido de DBT a 2’- hidroxibifenil-2-sulfinato.
Finalmente, a quebra da ligação C-S transforma o 2’-hidroxibifenil-2-sulfinato em dois
produtos finais, 2-hidroxibifenil (HBP) e sulfito (Fig. 12) (Gray et al., 1996).
Figura 12: “Via 4S”. O enxofre é removido na forma de sulfito, permanecendo intacta a estrutura
hidroxilada.
Fonte: Silva, 2007.

Silva, K.T.S. Introdução
21
Sabe-se que exposições ao DBT presente em ambientes aquáticos podem exercer
efeitos em embriões de zebrafish, como por exemplo, promovendo uma curvatura dorsal
do tronco e cauda, redução do crescimento, e edemas severos do pericárdio e saco
vitelínico. Deformidades similares comumente chamadas de síndrome do saco azul e
reduções no nível de incubação concentração-dependentes também foram observadas
(Wozny et al., 2010).
Como a dieta é uma das principais fontes de exposição humana e animal a HPA,
as células epiteliais intestinais constituem os alvos primários a entrarem em contato com
os contaminantes. O metabolismo de HPA pelos citocromos P450 é bem caracterizado e,
embora exista evidência considerável de que o fígado de mamíferos pode metabolizar tais
substâncias, o metabolismo em tecidos extra-hepáticos como o intestino pode ser de
grande importância (Cavret and Feidt, 2005).
O fato de agentes químicos promoverem alterações randômicas no genoma
justifica o direcionamento de esforços para a quantificação dessas mudanças, diminuição
da exposição a esses agentes e desenvolvimento de alternativas para a quimioprevenção.
1.5.2. 1,2-Dimetilhidrazina
O modelo de indução de CCR utilizando DMH é largamente aplicado por
compartilhar similaridades histológicas, morfológicas e moleculares com o câncer
esporádico observado em humanos (Chen and Huang, 2009; Rosenberg et al., 2009). Tal
substância é metabolizada pelo fígado por diversas reações e intermediários (AOM –
azoxymethanol e MAM – methylazoxumethanol), levando à formação do produto final
altamente reativo: íon metildiazônio. O intermediário MAM pode ser excretado junto à
bile e transportado ao cólon a partir do intestino delgado, ou seguir diretamente às células
epiteliais do cólon via circulação na corrente sanguínea. Este metabólito é responsável
pela metilação de bases do DNA do tecido epitelial resultando em apoptose, aumento na
proliferação e surgimento de mutações nesse tecido (Chang, 1984; Perse and Cerar,
2011).
1.6. Biomarcadores e Câncer
Como importantes indicadores biológicos do estado de progressão tumoral e
condição fisiológica das células, os biomarcadores representam ferramentas poderosas no

Silva, K.T.S. Introdução
22
monitoramento do câncer e aferição da eficácia e segurança de agentes quimioterápicos
(Duffy et al., 2003; Cho, 2007). Tais moléculas podem ser divididas pela habilidade de
auxiliar na prevenção, promover detecção precoce, estabelecer prognóstico e predizer a
resposta do paciente a terapias específicas (McLeod and Murray, 1999; Kulasingam and
Diamandis, 2008). A descoberta de biomarcadores também auxilia no entendimento de
mecanismos biológicos que ocorrem durante o desenvolvimento e progressão da doença.
A ação de circuitos regulatórios e a ocorrência de reações cruzadas entre as vias
envolvidas no processo bioquímico de estabelecimento do câncer dificulta o
entendimento e predição do funcionamento intracelular. Como nas células normais, a
maioria das células tumorais utiliza vias de sinalização redundantes para assegurar a
manutenção e viabilidade das funções críticas à sobrevivência (Cho, 2007).
O estadiamento convencional para o CCR não leva em conta a grande
variabilidade existente entre os pacientes e, alguns estágios de progressão não podem ser
preditos utilizando o critério histopatológico comum (Kahlenberg et al., 2003). Com o
avanço das técnicas de análise molecular e uma melhor compreensão dos mecanismos
envolvidos na tumorogênese, diversos fatores prognósticos moleculares têm sido
identificados.
Entre os marcadores proteicos conhecidos, o antígeno carcinoembrionário (CEA)
é utilizado atualmente na clínica para prognóstico molecular em câncer colorretal. A
concentração sérica desta molécula acima de 5 ng/mL está associada com a recorrência
da doença e metástase. O CEA é uma glicoproteína de membrana de alta massa molecular
pertencente à superfamília das imunoglobulinas. Esta molécula desempenha papéis na
adesão celular, imunidade e apoptose estando implicada na ocorrência de metástase
(Duffy et al., 2003; Kahlenberg et al., 2003).
Embora o CEA seja o biomarcador mais utilizado no CCR, sua detecção somente
torna-se possível em pacientes em estágio de desenvolvimento avançado, o que impede
que ele seja útil na detecção precoce deste tipo de câncer. Outros antígenos tumorais têm
sido estudados como potenciais biomarcadores para o CCR. Entre eles destacam-se o
CA19-9 (antígeno carboidrato 19-9), CA 242, CA-195, CA 50, CA 74-2, CD44 (Cluster
differentiation 44) e o TIMP-1 (inibidor tecidual de metaloproteinase-1). Entretanto,
assim como o CEA, esses marcadores possuem baixa sensibilidade e especificidade não
sendo adequados para propósitos de triagem ou diagnóstico precoce para o CCR (Cho et
al., 1997; Coppola et al., 1998; Li et al., 2001; Qiu et al., 2008).

Silva, K.T.S. Introdução
23
Sendo assim, o entendimento das bases bioquímicas e alterações moleculares
envolvidas na carcinogênese pode facilitar a detecção de tumores em estágios iniciais e
lesões pré-cancerosas que apresentam risco de sofrer transformações malignas. A
detecção pré-sintomática desses estágios iniciais traz grande benefício na redução dos
níveis de mortalidade (Alquist, 2010).
1.6.1. Proteômica e Phage Display
As aberrações relacionadas aos tumores afetam os níveis genômicos,
transcricionais e pós-transcricionais (Hassaneim et al., 2011). Desde o sequenciamento
do genoma humano, vários esforços na aplicação das análises de expressão gênica na
pesquisa médica, especialmente em câncer, têm sido realizados. Estudos no nível de
expressão gênica resultam em melhor entendimento do comportamento celular tanto nas
células normais quanto nas células tumorais (Wulfkuhle et al., 2003). Entretanto,
geralmente não há relação direta entre alterações na expressão gênica e níveis de
expressão proteica.
As proteínas são os maiores elementos funcionais das células e os estudos de seus
perfis de expressão em processos regulatórios podem auxiliar no entendimento das
alterações moleculares presentes no câncer. Os estudos de interação de proteínas com
outras moléculas, indução e controle de vias metabólicas, proliferação celular,
crescimento, apoptose e senescência, oferecem uma visão única de sistemas biológicos
complexos (Chuthapisith et al., 2007).
Métodos proteômicos utilizados na identificação de marcadores tumorais tem
como objetivo buscar modificações específicas nas proteínas de tecidos tumorais para
permitir a realização de tratamentos individualizados para certos tipos de câncer
(Simpson and Dorow, 2001). Tradicionalmente, a análise do perfil proteômico de
misturas complexas como lisados celulares ou frações sub celulares enriquecidas, envolve
a combinação de diversas técnicas de fracionamento com a espectrometria de massas
(MS) para a identificação das proteínas que compõem a amostra (Cecconi and Zamò,
2011).
Além da identificação e análise dos níveis de expressão proteicos, há um grande
interesse na elucidação das redes de interação de proteínas relacionadas ao câncer. A
habilidade de utilizar bibliotecas sintéticas de peptídeos para obter ligantes específicos de

Silva, K.T.S. Introdução
24
proteínas e entender suas interações permite uma maior compreensão das alterações
relacionadas ao proteoma tumoral (Sidhu and Koide, 2007).
A tecnologia de apresentação por fagos (Phage Display) consiste na fusão gênica
de sequências nucleotídicas que codificam proteínas ou peptídeos ao DNA de um vírus
bacteriófago e resultam na expressão heteróloga dessas sequências na superfície do
capsídeo viral (Smith, 1985). A construção de uma biblioteca peptídica com sequências
aleatórias (geralmente com mais de 1011 sequências) constitui uma população de ligantes
com diferentes capacidades de interação com moléculas alvo provenientes de tecidos ou
células (Scott and Smith,1990; Sidhu et al., 2000).
Técnicas combinatoriais permitem a seleção imparcial de ligantes em um ensaio
funcional sem conhecimento sobre a natureza do alvo na doença. Entretanto, é difícil
isolar ligantes específicos devido à complexidade de alvos expressos simultaneamente
em um tecido ou população celular. Estudos recentes detectaram um grande número de
alvos moleculares super regulados em células tumorais ou associadas a tumores (St Croix
et al., 2000; Ruoslahti, 2001; Vogelstein and Kinzler, 2004).
A identificação de receptores e seus ligantes correspondentes seletivamente
expressos em patologias específicas podem ser úteis no desenvolvimento de sistemas de
entrega de drogas, realização de terapia com o uso de vetores e drogas citotóxicas, o que
aumentaria a janela terapêutica até mesmo para os tratamentos já utilizados (Sergeeva et
al., 2006).

Justificativa

Silva, K.T.S. Justificativa
25
2- JUSTIFICATIVA
A importância dos hidrocarbonetos policíclicos aromáticos e a escassez de estudos
toxicológicos relacionados às consequências da contaminação ambiental pelos agentes
Dibenzotiofeno e Dibenzotiofeno Sulfona, colocam essas substâncias como importantes
objetos de investigação científica. Poucos trabalhos demonstram a caracterização
toxicológica desses compostos em modelos animais, especialmente em mamíferos,
ressaltando a relevância do presente estudo. No que se refere à exposição a substâncias
químicas com baixa solubilidade em água, e presentes em pequenas proporções no
ambiente, a avaliação de tratamentos crônicos utilizando baixas doses é de extrema
importância para uma real avaliação toxicológica. Nesse sentido, esse trabalho tem como
preocupação iniciar uma investigação voltada para a busca de biomarcadores proteicos
associados ao processo de indução crônica de câncer promovida por Dibenzotiofeno e
Dibenzotiofeno Sulfona.

Objetivos

Silva, K.T.S. Objetivos
27
3- OBJETIVOS
3.1- Objetivo Geral
Avaliar as alterações moleculares desenvolvidas após a administração crônica dos
hidrocarbonetos policíclicos aromáticos sulfurados Dibenzotiofeno e Dibenzotiofeno
Sulfona em ratos Wistar.
3.1.1- Objetivos específicos
A) Avaliar possíveis alterações na estrutura tecidual de órgãos relacionados ao
metabolismo de xenobióticos após tratamento com Dibenzotiofeno e
Dibenzotiofeno Sulfona;
B) Avaliar alterações na expressão proteica de tecidos intestinais dos animais
tratados com Dibenzotiofeno e Dibenzotiofeno Sulfona visando a
identificação de marcadores tumorais;
C) Selecionar peptídeos com capacidade de ligação a proteínas com expressão
alterada presentes nos pólipos intestinais resultantes dos tratamentos
empregados;
D) Identificar os alvos de ligação dos peptídeos isolados visando um melhor
entendimento do mecanismo de indução promovido;
E) Realizar estudos histoquímicos utilizando peptídeos sintéticos que
possibilitem a marcação e identificação específica de alterações neoplásicas
intestinais.

Material e Métodos

Silva, K.T.S. Material e Métodos
29
4- MATERIAL E MÉTODOS
Este trabalho foi aprovado pela Comissão de Ética no Uso de Animais (CEUA)
da Universidade Federal de Ouro Preto e catalogado sob o protocolo nº: 2010/14. Para a
realização do mesmo, o número de animais utilizados assim como o tratamento aplicado
restringiu-se ao estabelecido pelos objetivos delineados.
4.1. Material Biológico
4.1.1. Indução de Câncer em Ratos Wistar Tratados com DMH, DBT e
DBTO2
Ratos da linhagem Wistar, machos, com aproximadamente 60 dias de idade,
pesando 200-250 g, foram obtidos no Centro de Ciência Animal (CCA) da Universidade
Federal de Ouro Preto, sendo mantidos à temperatura ambiente com ração e água ad
libitum e ciclo claro/escuro de 12 horas.
Os animais foram divididos em cinco grupos, a saber:
Grupo I: Controle salina (n=5);
Grupo II: Teste DMH 30 mg/kg (n=15);
Grupo III: Controle óleo (n=5);
Grupo IV: Teste DBT 30 mg/kg (n=15);
Grupo V: Teste DBTO2 30 mg/kg (n=10).
Nos animais pertencentes aos grupos I e III administraram-se, respectivamente,
injeções intraperitoneais semanais de salina e óleo vegetal durante 10 semanas. Os
animais dos grupos II, IV e V receberam injeções de soluções de DMH (Sigma-Aldrich)
em salina, DBT e DBTO2 (Sigma-Aldrich) em óleo vegetal na dose de 30 mg/kg no
mesmo regime de tempo dos grupos anteriores. Todos os grupos passaram por um período
de latência de 98 dias (14 semanas) após a última injeção (Fig. 13) para o
desenvolvimento das alterações patológicas, como esperado para a substância de
referência DMH (Newell and Heddle, 2004). Semanalmente, durante os seis meses de

Silva, K.T.S. Material e Métodos
30
execução do experimento, os animais foram pesados e avaliados quanto a aparência
corporal externa.
Figura 13: Esquema representativo do tempo de tratamento e espera para o desenvolvimento de alterações
patológicas nos animais tratados com DMH, DBT e DBTO2.
4.1.2. Necropsia
Decorridos os tempos de injeção e espera, os animais receberam dose letal de
associação quetamina 10% e xilazina 2% através de injeção intraperitoneal.
Posteriormente, realizou-se a coleta do baço, pulmão, fígado, estômago e intestinos para
a realização das análises histológicas, imunohistoquímicas e proteômicas, e do sangue
para análise do hemograma, e dosagens de enzimas hepáticas e pancreáticas.
4.2. Análise Histológica
Os segmentos de tecidos biopsiados foram fixados em formol tamponado e
posteriormente processados e incluídos em parafina. Para a análise histológica, secções
de 4 μm foram coradas com hematoxilina-eosina (HE) e examinadas ao microscópio.
Avaliaram-se qualitativamente os seguintes parâmetros para os intestinos:
presença/ausência de atipias celulares nas camadas mucosa e média, inflamação na
mucosa, presença de necrose, atipias estruturais e hiperplasia de nódulos linfóides. Para
digitalização das imagens utilizou-se o microscópio Leica DM5000B com câmera
acoplada, por meio do programa Leica Application Suite (versão 2.4.0 R1, Leica
Microsystems).

Silva, K.T.S. Material e Métodos
31
4.2.1. Imunohistoquímica: CEA e CD44
A imunohistoquímica foi realizada utilizando-se anticorpos secundários
complexados com biotina seguida de estreptavidina ligada à peroxidase para permitir a
revelação. Para tal, secções histológicas de 4 μm fixadas em formol-parafina foram
desparafinizadas (lavagens sequenciais em xilol, etanol absoluto, etanol 90%, etanol 80%,
etanol 70% e água) e reativadas antigenicamente através do aquecimento das mesmas em
micro-ondas (5 ciclos de 3 minutos cada) em tampão citrato de sódio 0,1 M pH 6,0.
Lavaram-se as lâminas por 3 vezes de 5 min. cada com tampão fosfato de sódio 0,01 M
pH 7,4, NaCl 0,15 M (PBSS). A atividade da peroxidase endógena foi inibida por
incubação com H2O2 3,5% em PBSS por 30 minutos à temperatura ambiente e as lâminas
foram novamente lavadas com PBSS. Bloquearam-se as interações não específicas
utilizando leite em pó 14% em PBSS por 30 minutos em câmara úmida. Os anticorpos
primários anti-CEA e anti-CD44 foram diluídos separadamente na proporção de 1:1.000
sendo utilizados 100 μL por lâmina incubados a 4º C, overnight em câmara úmida.
Após nova lavagem com PBSS os cortes foram cobertos com quantidade
suficiente da solução link (anticorpo secundário-biotina) do kit para imunohistoquímica
LSAB Universal (Dako), e mantidos por 30 minutos à temperatura ambiente em câmara
úmida. Realizou-se a lavagem com PBSS como descrito anteriormente sendo os cortes
cobertos com a solução estreptavidina-peroxidase do kit LSAB, mantidos por mais 30
minutos à temperatura ambiente em câmara úmida.
A formação do precipitado marrom que indica a positividade da reação ocorre
após a adição de solução de cromogêneo DAB 0,05% (3-3’-Diaminobenzidine
Tetrahidrochloride Hidrate – Ambresco) em PBSS acrescido de 0,02% de peróxido de
hidrogênio. As lâminas foram contra coradas por imersão em hematoxilina e montadas
para avaliação.
4.3. Extração de Proteínas e Análise Proteômica
Para extração de proteínas dos tecidos obtidos após a necropsia dos animais foram
utilizados 100 mg de tecido (três amostras de diferentes animais por grupo) em 500 μL
de tampão de rehidratação (uréia 7 M, thio-uréia 2 M, CHAPS 2%, azul de bromofenol
0,002%) adicionados de 10 μL de tampão PIC (Coquetel de Inibidores de Proteases,
Sigma-Aldrich). Homogeneizou-se cada mistura em tubo de vidro com auxílio de motor

Silva, K.T.S. Material e Métodos
32
Potter S (B. Braun Biotech International) por 15 minutos a 1.000 x rpm em banho de gelo,
seguido por três ciclos de sonicação de 20 segundos a 25 W, com intervalos de 45
segundos entre cada ciclo (Sonifier 250, Branson). As suspensões resultantes foram
centrifugadas a 4ºC e 20.000 x g por 1 hora em centrífuga Mikro 200R (Hettich).
Após a centrifugação, separou-se o sobrenadante contendo a fração solúvel para a
realização de eletroforeses uni e bidimensionais e descartou-se os precipitados contendo
o debrit celular.
4.3.1. Eletroforese Uni e Bidimensional (1D/2D SDS-PAGE)
Anteriormente à confecção dos géis uni e bidimensionais, fez-se a dosagem de
proteínas solúveis obtidas conforme o item 4.3 através de densitometria comparativa de
canaletas (30 μg de Proteínas Solúveis) por meio do software Quantity One versão 4.6.9
(Bio-Rad).
Os extratos proteicos foram analisados por meio de eletroforese em gel de
poliacrilamida em condições desnaturantes (10%, 1D SDS-PAGE) como descrito por
Laemmli em 1970. Para o preparo das amostras, cerca de 30 μg de proteína foram
solubilizadas em 10 μL de tampão de amostra (tris-HCl 0,125 M pH 6,8, SDS 4%, glicerol
20%, azul de bromofenol 0,002%) e posteriormente fervidas em banho-maria por 5
minutos. Aplicou-se 20 mA de corrente por gel durante aproximadamente 1,5 horas. Para
coloração, utilizou-se solução de Coomassie Brilhant Blue G250 (Coomassie Brilhant
Blue G 250 0,2%, Etanol 40%, Ácido Acético 7%) por 2 horas e descoloração do
background com solução de Ácido Acético 7% e Etanol 10% overnight. As imagens dos
géis foram capturadas utilizando-se o scanner ImageScanner III (GE Healthcare).
Para a confecção de géis bidimensionais, solubilizaram-se 250 μg de proteína em
tampão de rehidratação contendo dithiotreitol (DTT) 1% e anfólitos pH 3-10 0,8%.
Acondicionou-se as amostras em sarcófagos de porcelana (Strip Holder 13 cm, GE
Healthcare) para incorporação e isoeletrofocalização das proteínas no gel de primeira
dimensão (Immobiline DryStrip Gels 13 cm pH 3-10 NL, GE Healthcare). Após a
focalização isoelétrica, as proteínas presentes nos géis de primeira dimensão foram
submetidas à redução com DTT 1% em solução de equilíbrio (uréia 6 M, tris-HCl 1,5 M
pH 8,8, glicerol 30%, SDS 2%, azul de bromofenol 1%) e, em seguida, alquiladas com
iodoacetamida 4% em solução de equilíbrio.

Silva, K.T.S. Material e Métodos
33
A separação de proteínas por massa molecular (segunda dimensão - 2D SDS-
PAGE) foi realizada em gel de poliacrilamida 10%. A eletroforese foi conduzida a 50
V/gel nos primeiros 10 minutos e, posteriormente a 100 V/gel por aproximadamente 2
horas. Para a coloração, utilizou-se Coomassie Brilhant Blue G 250 Coloidal (Sigma-
Aldrich) (três lavagens de 20 minutos cada com solução de ácido fosfórico 2% e etanol
30%; seguidas de adicionais três lavagens de 10 minutos com solução de ácido fosfórico
2%; e coloração em solução de ácido fosfórico 2%, etanol 18% e sulfato de amônio 15%
contendo Coomassie 0,02%). As imagens foram capturadas com o scanner ImageScanner
III (GE Healthcare) e analisadas através de sobreposição realizada pelo software
SameSpots (TotalLab Ltd. v. 20, UK).
4.3.2. Digestão in gel
Após a obtenção dos géis bidimensionais, os spots de interesse (proteínas
diferencialmente expressas entre os respectivos grupos Controles e Testes) foram
excisados e transferidos para tubos plásticos de 1,5 mL contendo 300 μL de solução
descorante (etanol 20%, ácido acético 7%) e mantidos a 37º C até a completa
descoloração. Retirou-se a solução descorante e lavou-se cada spot com 500 μL de água
mili-Q para remover o excesso da mesma. Em seguida, lavou-se cada spot por 3 vezes de
5 minutos cada com 300 μL de solução NH4HCO3 20 mM em acetonitrila (ACN) 50%.
Os spots foram desidratados em concentrador a vácuo (Speed Vac System –
Iss110, Thermo) durante 30 minutos e rehidratados com 15 μL de solução de tripsina 0,1
μg/μL (Promega) em NH4HCO3 20 mM. Após 20 minutos retirou-se o excesso de solução
de tripsina e recobriu-se os spots com 40 μL de NH4HCO3 20 mM. Os tubos foram
mantidos a 37ºC por 18 horas para digestão das proteínas. Recuperou-se o sobrenadante
da reação contendo peptídeos trípticos e adicionou-se 30 μL de solução de ácido
trifluoroacético (TFA) 2% e ACN 50% aos spots que permaneceram nos tubos para
extrair os fragmentos trípticos que ainda se encontravam no interior dos mesmos.
Decorridos 30 minutos da adição TFA/ACN retirou-se o sobrenadante e combinou-se
com o primeiro sobrenadante obtido para secagem em concentrador a vácuo.
Mantiveram-se as amostras secas a 4ºC para posterior análise por espectrometria de
massas.

Silva, K.T.S. Material e Métodos
34
4.3.3. Análise de Proteínas por Espectrometria de Massas
As amostras obtidas conforme a descrição do item anterior foram ressuspendidas
em 10 μL de ácido fórmico 0,1% para análise por espectrometria de massas. 4,5 μL de
cada preparação de peptídeos foram injetados em cromatógrafo nanoAcquity UPLC
(Waters) sendo direcionado primeiramente à pré-coluna de C18 Trap Symetry (5 μm,
180μm x 20 mm – Waters) e logo após, para a coluna analítica de C18 BEH130 (1,7 μm,
100μm x 100 mm – Waters) condicionada a 35ºC. A cromatografia ocorreu com fluxo
de 0,6 μL/min acoplada com espectrômetro de massas Q-Tof Premier (Waters) com fonte
nanoeletrospray. Foi aplicado um gradiente de 10 minutos com 10-90% de ACN em 0,1%
de ácido fórmico por spot. Os espectros MS e MS/MS foram adquiridos por meio do
software MassLynx v. 4.1.
Os experimentos foram executados utilizando tempo de escaneamento de 1
segundo e razão massa/carga de 100 a 2000 durante o processo cromatográfico. As
aquisições de dados MS/MS foram realizadas utilizando os primeiros cinco íons
precursores com carga +2, +3 ou +4. As buscas foram realizadas por meio do software
Mascot engine v.2.3.02 (Matrix Science Ltd.) utilizando o banco de dados não redundante
para Rattus sp. (NCBInr_012014 - com 61.059 sequências de proteínas).
Os parâmetros de busca utilizados incluíram carbamidometilação de cisteínas
como modificação fixa, oxidação de metionina como modificação variável, permissão
para ocorrência de perda de um sítio de clivagem e tolerância de 0,1 Da para os íons
precursores e fragmentados. A identificação de proteínas foi considerada válida caso a
taxa de falsos positivos fosse ≤ 3% e a porcentagem de cobertura fosse ≥ 8%.
Foram considerados significativos os peptídeos com valor de threshold p ≤ 0,05
sendo identificadas as proteínas cujas entradas mostraram uma porcentagem de cobertura
de sequência ≥ 15%, FDR ≤ 3% e ao menos um peptídeo único identificado.
4.3.4. Categorização Funcional e Mapa de Interação Proteica
Com o objetivo de se obter uma visão integrada das alterações observadas, as
proteínas identificadas por espectrometria de massas foram categorizadas em classes
distintas de acordo com sua função biológica, e submetidas a uma análise de interação
com outras proteínas e moléculas por meio do software STITCH v.4.0 (disponível em
http://stitch.embl.de/).

Silva, K.T.S. Material e Métodos
35
4.4. Western blotting
Realizou-se a técnica de Western blotting para a detecção da expressão das
proteínas ACTB, CANX, e CEA. Foi preparado um extrato total de proteínas solúveis
para os animais pertencentes a todos os grupos conforme descrito no item 4.3.1.
Submeteu-se as amostras de proteínas obtidas a eletroforeses em gel de poliacrilamida
10% e realizou-se a transferência para membranas de PVDF (Invitrogen) a 200 mA em
tampão tris-HCl 25 mM, glicina 192 mM, etanol 20% e SDS 10% sob refrigeração por
duas horas. Em seguida, as membranas foram bloqueadas overnight com tampão tris-HCl
50 mM pH 7,5, Tween-20 0,3% e leite em pó desnatado 5%, sendo posteriormente
lavadas por três vezes com tris-HCl 10 mM pH 7,5. Os anticorpos monoclonais anti-CEA
(1:1000); anti-β-actina (1:2000) e anti-calnexina (1:1000), todos Sigma Aldrich, foram
incubados no tampão de immunoblotting (tris-HCl 50 mM pH 7,5, Tween-20 0,013%,
leite em pó desnatado 5%) à temperatura ambiente por 3 horas e posteriormente
submetidos a detecção com o anticorpo secundário IgG anti-mouse conjugado com
fosfatase alcalina (1:2000) à temperatura ambiente por 2 horas. Após as lavagens com
tampão tris-HCl 10 mM pH 7,5, detectaram-se os sinais com os substratos enzimáticos
nitro-blue tetrazoluim chloride e 5-bromo-4-chloro-31-indolyphosphate p-toluidine salt
(Sigma Aldrich).
4.5. Biopanning (Phage Display)
4.5.1. Extração de Proteínas para Phage Display
A extração de proteínas para a execução da técnica de Phage Display foi realizada
através da maceração de 50 mg de tecido com nitrogênio líquido e suspensão do mesmo
em 1 mL de tampão tris-HCl 20 mM pH 7, sacarose 250 mM e PIC. As amostras foram
sonicadas como descrito no item 4.3, e centrifugadas a 20.000 x g, 4º C por 30 minutos.
A qualidade da extração foi verificada através da visualização das proteínas em 1D SDS-
PAGE corado com solução de Coomassie Brilhant Blue G250 como descrito no item
4.3.1.

Silva, K.T.S. Material e Métodos
36
4.5.2. Ciclos de Seleção de Phage Display
A seleção dos epítopos (expressos nas 5 cópias da proteína III - pIII - do capsídeo
de fagos filamentosos) foi realizada utilizando-se uma biblioteca de peptídeos de 12
aminoácidos (Ph.D.™-12 mer - New England Biolabs). As bibliotecas expressas em
fagos M13 são compostas por peptídeos randômicos seguidos por uma curta sequência
espaçadora Gly-Gly-Gly-Ser fusionada à região N - terminal da proteína capsídica pIII.
Esta biblioteca consiste de aproximadamente 2,0 x 1014 clones com redundância
de 105 clones, os quais representam 2,7 x 109 possíveis combinações entre os resíduos de
aminoácidos. Realizou-se o procedimento de seleção através de três ciclos sequenciais.
Primeiramente sensibilizou-se uma placa de microtitulação (Costar 3590, Corning
Incorporated) com 160 μL de extrato proteico de intestinos provenientes dos Grupos
Teste (1 poço/grupo) na concentração de 100 μg/mL em tampão bicarbonato de sódio 0,1
M pH 8,6 a 4ºC, overnight sob agitação lenta em câmara úmida.
A seguir, secou-se a placa batendo-a contra papel toalha autoclavado. Bloqueou-
se os sítios livres de interação por meio da adição de 250 μL/poço de tampão de bloqueio
(NaHCO3 0,1 M, BSA 5 mg/mL) mantido por 1 hora a 4ºC sob agitação lenta. Após a
incubação, o excesso de tampão de bloqueio foi descartado e a placa foi lavada por 6
vezes com 200 μL/poço de TBS-T (tris-HCl 50 mM pH 7,5, NaCl 150 mM, Tween 20
0,1%) por 1 minuto, sendo seca em seguida com papel toalha.
Realizou-se a diluição de 10 μL (aproximadamente 4 x 1010 fagos) da biblioteca
de fagos em 100 μL de TBS-T e adicionou-se aos poços contendo extrato proteico dos
Grupos Teste (100 μL/poço). Incubou-se a placa por 1 hora sob agitação lenta à
temperatura ambiente. Os fagos não ligantes (F1) foram recolhidos do sobrenadante com
o auxílio de pipeta e descartados. Realizou-se a lavagem com TBS-T 0,1% por 10 vezes
de 1 minuto cada.
A eluição dos fagos ligantes foi realizada com 100 μL de tampão glicina 200 mM
pH 2,2 por 10 minutos sendo os fagos (F2) recolhidos e armazenados em tubo plástico de
0,5 mL. A seguir, adicionou-se 15 μL de tris-HCl 1 M pH 9,1 nos tubos para neutralizar
o pH. Os fagos recuperados (F2) foram amplificados em E. coli ER2738 (OD600nm ≈ 0,3)
em meio Luria-Bertani (LB, triptona 10 g/L, extrato de levedura 5 g/L, NaCl 5 g/L) a
37ºC por 4,5 horas finalizando o primeiro ciclo de seleção (Fig. 14).
Para o segundo e terceiro ciclos de seleção, realizou-se uma pré-seleção
primeiramente utilizando poços contendo proteínas imobilizadas referentes aos grupos

Silva, K.T.S. Material e Métodos
37
Controle. Desta vez, os peptídeos não ligantes (F1) foram recolhidos, amplificados como
descrito anteriormente e então utilizados para a seleção frente aos extratos dos grupos
Testes.
Figura 14: Biopanning com a biblioteca de peptídeos Ph.D. Seleção de ligantes específicos.
4.5.3. Titulação dos Clones Obtidos no Phage Display
A titulação é um procedimento que permite o isolamento dos fagos para posterior
sequenciamento dos mesmos. Para a realização da titulação, 1 μL de solução de fagos foi
diluído em 9 μL de meio LB obtido ao fim do 3º ciclo de seleção e submetido a diluições
seriadas crescentes exponenciais sob log10 em meio LB. Para cada diluição
acrescentaram-se 200 μL de cultura de ER2738 em fase exponencial (OD600nm ≈ 0,5),
agitou-se a solução rapidamente e incubou-se por 5 minutos a temperatura ambiente. As
células bacterianas, agora infectadas, foram transferidas para tubos de cultura contendo 3
mL de Ágar-TOP (LB, MgCl2.6H2O 1 g/L) a 45ºC e espalhadas sobre placas de Petri
contendo meio LB sólido, com IPTG 0,5 mM, Xgal 40 μg/mL e tetraciclina 20 mg/mL.
Para cada diluição foi preparada uma placa. Incubaram-se as placas à 37ºC por 12 horas
para obtenção das colônias isoladas utilizadas no sequenciamento de DNA dos fagos.
4.5.4. Extração de DNA de Fagos e Sequenciamento
Aleatoriamente, 32 colônias referentes a cada grupo foram retiradas das placas de
Petri e transferidas individualmente para poços de placa deepwell contendo 1 mL de meio
LB autoclavado com tetraciclina 100 μL/mL. A placa foi vedada e incubada por 5 horas
a 37ºC sob forte agitação. Após a incubação, centrifugou-se a placa por 40 minutos a
2.500 x g e 800 μL do sobrenadante foram transferidos para outra placa Deepwell estéril.

Silva, K.T.S. Material e Métodos
38
Adicionou-se 200 μL de polietilenoglicol 8000 (PEG)/NaCl (20% PEG, NaCl 2,5 M)
estéril para precipitação dos fagos presentes na solução e incubou-se a temperatura
ambiente por 15 minutos. A placa foi novamente submetida à centrifugação a 2.500 x g
por 40 minutos, sendo o sobrenadante descartado por inversão.
Adicionou-se 100 μl de tampão iodeto (Tris-HCl 10mM pH 8,0, EDTA 1 mM,
NaI 4 M) a cada poço da placa deepwell para se romper as cápsulas dos fagos e precipitar
o DNA. A placa foi vortexada fortemente por 5 minutos. Após a agitação, adicionou-se
250 μL de etanol e manteve-se a placa em repouso absoluto por 10 minutos à temperatura
ambiente. Novamente realizou-se a centrifugação da placa a 2.500 x g por 40 minutos e
descartou-se o sobrenadante por inversão da placa com secagem em papel absorvente.
Lavou-se cada poço com 500 μL de etanol 70% e procedeu-se nova centrifugação com
descarte do sobrenadante por inversão. O DNA purificado foi obtido após a adição de 20
μL de água mili-Q a cada poço. A qualidade do DNA extraído foi avaliada por meio de
eletroforese em gel de agarose 0,8% corado com solução de brometo de etídeo.
Após a observação das bandas eletroforéticas obtidas no gel de agarose, pode-se
concluir que em média, extraiu-se 200-300 ng de DNA/3 μL de solução. Utilizaram-se
aproximadamente 400 ng de DNA molde por reação de sequenciamento acrescidos de 5
pmol de Primer -96 gIII (5’-OHCCC TCA TAG TTA GCG TAA CG-3’ Biolabs), Premix
4 μL (DYEnamic ET Dye Terminator Cycle kit – Amersham Biosciences).
Realizou-se uma reação de 35 ciclos em termociclador de placas (MasterCycler
Eppendorf) nas seguintes condições: (1) Desnaturação – 95ºC por 20 segundos, (2)
Anelamento – 50ºC por 15 segundos, (3) Extensão – 60ºC por 1 minuto. O DNA
amplificado foi precipitado com 1 μL de Acetato de Amônio 7,5 M e 27,5 μL de etanol
absoluto por poço homogeneizando-se a placa com leves batidas sobre a bancada. A placa
foi centrifugada a 2.500 x g por 45 minutos a 4ºC e o sobrenadante foi descartado por
inversão.
Adicionou-se 150 μL de etanol 70% ao precipitado e centrifugou-se a placa a
2.500 x g por 10 minutos. O sobrenadante foi descartado e o etanol remanescente foi
evaporado à temperatura ambiente no escuro. O precipitado resultante foi ressuspendido
em 10 μL de tampão de diluição (DYEnamic ET Dye Terminator Cycle kit – Amersham
Biosciences). Realizou-se o sequenciamento em um sequenciador automático
MegaBACE 1000 (Amersham Biosciences) nas seguintes condições: (1) Voltagem de
Injeção – 2kV, (2) Voltagem de Corrida – 7kV, (3) Tempo de Injeção – 120 segundos,
(4) Tempo de corrida – 200 minutos.

Silva, K.T.S. Material e Métodos
39
A identificação das sequências correspondentes aos peptídeos selecionados foi
realizada manualmente observando-se as sequências Start (ACCTCCACC) e End
(AGAGTGAGA) que indicam respectivamente o início e fim das sequências de
peptídeos de interesse. Selecionou-se as sequências de nucleotídeos obtidas (36
nucleotídeos) e converteu-se as mesmas em sequências de aminoácidos por meio do
software Expasy disponível em http://web.expasy.org/translate/ com frame de leitura 1
3’-5’.
4.5.5. Análise in silico dos Peptídeos Obtidos
Os peptídeos selecionados por Phage Display podem corresponder a um pequeno
fragmento de uma proteína maior que desempenha papel específico em processos
biológicos complexos. A simples seleção de peptídeos marcadores para uma determinada
patologia não fornece informações a respeito desse processo biológico. Sendo assim,
estratégias que utilizam bioinformática podem fornecer informações complementares,
auxiliar na compreensão da doença estudada e relacionar possíveis proteínas envolvidas
aos peptídeos selecionados.
Os métodos aplicados para a obtenção dessas informações foram baseados no
alinhamento das sequências por meio do software ClustalW
(http://www.ebi.ac.uk/tools/msa/clustalw2/), seleção de proteínas alvo por meio do
BlastP do NCBI (http://blast.ncbi.nlm.nih.gov), análise das vias funcionais onde estão
inseridas as proteínas alvo através do software STITCH 4.0 (http://stitch.embl.de).
O BlastP foi selecionado por permitir alinhamento entre proteínas. A base de
dados utilizada foi a RefSeq por apresentar resultados com boa especificidade e
sensibilidade. O organismo selecionado foi Rattus norvegicus devido à espécie utilizada
nesse estudo, sendo os demais parâmetros, ajustados automaticamente pelo aplicativo. As
anotações dos primeiros 40 alvos com melhor score foram analisadas, sendo avaliadas
quais demonstram alguma relação com câncer. Esse número de alvos foi escolhido de
maneira arbitrária até que se obtivesse informação qualitativa significativa.
Devido ao grande número de peptídeos identificados e ao custo e tempo
necessários para que se realizasse a validação da seleção com todos eles, esta estratégia
permitiria selecionar, inicialmente, os peptídeos a serem sintetizados para a realização
dos experimentos subsequentes.

Silva, K.T.S. Material e Métodos
40
4.6. Síntese de Peptídeos
A síntese dos peptídeos propostos (Tab. 01) foi realizada sob a forma solúvel,
utilizando-se um protocolo de síntese em fase sólida estabelecido por Merrifield (1965),
com algumas modificações. Utilizou-se a resina Rink Amide Resin HL (Merck), obtendo-
se um rendimento de aproximadamente 40 µmoles de peptídeo por lote de síntese.
Tabela 01: Peptídeos selecionados por Phage display para síntese em fase sólida.
Grupo Tratamento Peptídeo Selecionado
DMH (Intestino Grosso) QPHKVFFPNLPR
DBT (Intestino Grosso)
EYQLPSRTVPRD
KDPTQFKPMSLW
FDTKAGLTSLVA
DBT (Intestino Delgado) TNTPPSQRVHLS
LAAQQYTSALKS
Para a ativação da resina adotou-se o seguinte procedimento: 40 µmoles de resina
(52 mg) foram adicionados em um tubo de síntese contendo dimetilformamida (DMF)
suficiente para cobrir toda a resina, permanecendo sob agitação constante por três horas
a 37º C. Para a liberação do grupamento Fmoc de proteção, cobriu-se a resina com 3 mL
de 4-metilpiperidina 20% em DMF e lavou-se por 3 vezes de 20 minutos cada, sob
agitação contínua a temperatura de 37º C. Em seguida, a resina foi lavada 3 vezes
alternadamente com metanol e DMF utilizando-se 2 mL de solvente por etapa. Todas as
lavagens foram realizadas com auxílio de uma bomba de vácuo.
Os aminoácidos e demais reagentes auxiliares foram adicionados em
concentrações quatro vezes superiores (160 µmoles) à quantidade de resina para favorecer
o acoplamento. Esses são ligados pelo seu grupamento carboxila ao grupamento amino
da resina, formando uma ligação peptídica. Adicionou-se o primeiro aminoácido ao tubo
de síntese em um volume de 2 mL de DMF, acrescido de 25 µL de di-isopropil
carbodiimida (DIPC) e 23 mg de oxima [ethyl 2-cyano-2-(hydroxyimino)acetate]. Após
2 horas de agitação a 37ºC, retirou-se todo o líquido do tubo de síntese, sendo o peptídeo
crescente submetido à acetilação para prevenção do crescimento de cadeias que não
reagiram com o aminoácido a ser incorporado. Dessa forma, 50 µL de uma solução 1:1
de DIPC e anidrido acético foram adicionados a 1 mL de DMF no tubo de síntese,
permanecendo sob agitação a 37ºC por 30 minutos. Ao fim desta etapa, lavou-se a resina

Silva, K.T.S. Material e Métodos
41
3 vezes alternadamente com metanol e DMF. O grupamento amino desse primeiro
aminoácido acoplado foi então desprotegido, lavando-se a resina com 3 mL de uma
solução de 4-metilpiperidina 20% em DMF, por três vezes de 10 minutos cada, com
agitação contínua a 37°C. Os demais aminoácidos foram processados conforme descrito
anteriormente para o acoplamento do primeiro aminoácido.
Terminados os 12 ciclos de acoplamento, eliminou-se o último grupamento Fmoc
com 4-metilpiperidina 20% em DMF, como descrito previamente, e lavou-se a resina por
4 vezes, durante 5 minutos, com 3 mL de diclorometano (DCM). As cadeias laterais foram
desprotegidas e o peptídeo dissociado da resina pelo uso de 5 mL de uma solução de
clivagem contendo 95% de TFA. O tubo de reação permaneceu sob agitação por quatro
horas. Coletou-se a solução contendo os peptídeos, transferiu-se para tubos de ensaio e
os produtos de síntese foram precipitados com 50 mL de éter etílico, permanecendo em
repouso durante a noite à 4º C. Posteriormente, lavou-se os precipitados com éter etílico
e centrifugou-se por 3 vezes a 1.500 x g por 5 minutos. Na última etapa, desprezou-se o
sobrenadante e ressuspendeu-se o peptídeo em 3 mL de água milli-Q.
Para a obtenção de peptídeos fluorescentes foi utilizado o mesmo protocolo
descrito anteriormente com a adição do fluoróforo 5,6-carboxifluoresceína (Sigma-
Aldrich) na região N-terminal do último aminoácido adicionado. A adição do fluoróforo
foi realizada da mesma maneira que a adição dos aminoácidos constituintes de cada
sequência. Os peptídeos obtidos foram liofilizados, analisados em sistema HPLC (High
Performance Liquid Chromatography) e caracterizados em espectrômetro de massas ESI
LCMS-IT-TOF e através do software BCEPRED (http://www.imtech.res.in).
4.6.1. Análise de Pureza e Confirmação de Identidade
Os peptídeos sintetizados em fase sólida foram submetidos à cromatografia em
fase reversa para a análise de sua pureza. As cromatografias foram realizadas em coluna
C18 (250 mm x 10 mm) (LiChroCART® 250-10 Purospher, Merk), em sistema HPLC
Shimadzu. A coluna foi previamente equilibrada com uma solução de água mili-Q e TFA
0,1%. Inicialmente, eluiram-se alíquotas de todos os peptídeos (20 μL) em um gradiente
de ACN de 20 a 60% e TFA 0,1% durante 80 minutos com um fluxo de 1 mL/minuto.
Monitorou-se a eluição a 280 nm, 260 nm ou 230 nm de maneira dependente da sequência
sintetizada.

Silva, K.T.S. Material e Métodos
42
Após a purificação, os peptídeos foram caracterizados por espectrometria de
massas para confirmação de sua identidade. Para tal, utilizou-se o espectrômetro de
massas LCMS-IT-TOF (Liquid Chromatography Ion Trap Time Of Flight), que opera
por ionização do tipo electrospray (Shimadzu®). O equipamento foi calibrado utilizando
trifluoroacetato de sódio. A voltagem capilar foi de 4500 V no modo positivo de
ionização, com tempo de acumulação de 10 ms. Aplicaram-se as amostras através de
injeções diretas (alíquotas de 5 μL). Utilizou-se a ferramenta ExPASy – Compute PI/Mw
tool (disponível em http://web.expasy.org/compute_pi/) para a predição das massas
moleculares dos peptídeos sintetizados.
4.7. Síntese de Colunas de Afinidade
Para a obtenção de colunas de afinidade com os peptídeos de interesse
imobilizados, realizaram-se as seguintes etapas de ativação/imobilização do suporte
cromatográfico Sepharose 4B (Sigma-Aldrich): ativação com epicloridrina, aminação,
succinilação, ativação com N-hidroxisuccinimida (NHS) e diciclohexilcarbodiimida
(DCC), imobilização dos peptídeos e bloqueio dos grupos carboxílicos livres. As reações
envolvidas nas etapas citadas encontram-se descritas a seguir.
Para a etapa de ativação, lavou-se a resina (Sepharose 4B) em funil de vidro
sinterizado com água destilada e suspendeu-se a mesma em 4 volumes de NaOH 0,4 M
contendo 5% de epicloridrina (Matsumoto, Mizuno et al. 1979). Manteve-se a suspensão
em banho-maria a 40º C por 2 horas sob agitação leve e novamente procedeu-se lavagem
com excesso de água destilada.
A sepharose epóxi-ativada foi aminada utilizando-se 1,5 volumes de solução de
hidróxido de amônio P.A., onde foi suspendida a resina lavada. Manteve-se a suspensão
por 1,5 horas em banho-maria a 40º C sob agitação. Decorrido o período de espera, lavou-
se novamente a resina aminada com água destilada em excesso e NaCl 0,1 M.
Suspendeu-se a resina aminada em 1,5 volumes de NaCl 0,1 M sendo
posteriormente adicionados 0,08 g de anidrido succínico por g de resina para que
ocorresse a succinilação da mesma. Manteve-se o pH da suspensão em 6 através da adição
gradual de NaOH 20%, e, após a estabilização do pH respeitou-se um tempo de espera de
cinco horas à temperatura ambiente. Realizou-se uma nova lavagem com NaOH 0,1 M e
a resina foi novamente suspendida nesta mesma solução por 30 minutos.

Silva, K.T.S. Material e Métodos
43
A sepharose succinilada foi então lavada com 3 volumes de dioxano para tornar a
resina anidra e suspensa em dioxano onde adicionou-se NHS e DCC na concentração de
0,1 M cada para ativação. Esta suspensão foi mantida por 70 minutos sob agitação leve à
temperatura ambiente, sendo então lavada com 8 volumes de dioxano seguidos de 4
volumes de metanol e 3 volumes de dioxano.
Após a ativação, realizou-se uma lavagem rápida com 1 volume de água destilada
sendo a resina rapidamente transferida para um béquer com 3 volumes de solução
NaHCO3 0,01 M, NaCl 0,9% pH 7,5 contendo o peptídeo de interesse na concetração de
1 mg/mL. Após 1 hora à temperatura ambiente seguida de 12 horas sob refrigeração,
ocorreu a imobilização do mesmo.
A etapa final consistiu no bloqueio dos grupos carboxílicos livres com solução de
etanolamina 0,1 M pH 9,0 por 1 hora a temperatura ambiente. Após este procedimento,
procedeu-se nova lavagem com água destilada. Manteve-se o produto a 4º C até o
momento do uso.
4.7.1. Identificação de Proteínas Ligantes
Extratos proteicos de 100 mg de intestino foram preparados em 500 µL de tris-
HCl 50 mM, NaCl 150 mM pH 7,5 e PIC como descrito no item 4.5.1 e submetidos a
cromatografia de afinidade utilizando-se as resinas com peptídeos imobilizados conforme
o item 4.7.
Duas colunas de afinidade por peptídeo, contendo 3 mL de suporte, foram
montadas em seringas plásticas e equilibradas com 30 mL de tampão de equilíbrio tris-
HCl 50 mM, NaCl 150 mM pH 7,5. Amostras relativas aos grupos I e III foram
submetidas à seleção pelo peptídeo QPHKVFFPNLPR e amostras pertencentes aos
grupos II, IV e V foram submetidas à cromatografia com os demais peptídeos
(EYQLPSRTVPRD; KDPTQFKPMSLW; FDTKAGLTSLVA; TNTPPSQRVHLS e
LAAQQYTSALKS).
Os extratos foram expostos por 10 passagens cada um em suas respectivas colunas
de afinidade para assegurar a ligação com os peptídeos imobilizados. As colunas foram
então lavadas com 40 mL de tampão de equilíbrio para remoção das proteínas não ligadas.
A eluição das proteínas especificamente ligadas foi realizada com solução de 2,5 mL de
solução de NaCl 1 M também recromatografada por 10 vezes. As amostras eluídas foram
então dialisadas para remoção do excesso de sal (solução de acetato de amônio 0,01 M

Silva, K.T.S. Material e Métodos
44
pH 7,4 por 24 horas) e concentradas em liofilizador (Benchtop K - Virtis). O precipitado
obtido foi ressuspendido em tampão de amostra e submetido à eletroforese
unidimensional conforme descrito em 4.3.1.
As bandas eletroforéticas observadas para os grupos controle e teste eluídas para
cada peptídeo e coradas com Coomassie G250 foram excisadas para digestão in gel e
posterior identificação por espectrometria de massas como descrito nos itens 4.3.2 e 4.3.3.
4.7.2. Análise in silico da Interação Proteína/Peptídeo
Após a identificação das proteínas alvo ligadas por afinidade aos peptídeos
selecionados por Phage Display, foi realizada uma análise in silico para avaliação da
interação proteína/peptídeo. Para tal, as estruturas tridimensionais referentes às proteínas
disponíveis em RSCB PDB (http://www.rscb.org/pdb) foram utilizadas juntamente com
as estruturas tridimensionais dos peptídeos geradas por meio do software Mobyle@RPBS
(disponível em http://mobyle.rpbs.univ-paris-diderot.fr/cgi-bin/portal.py). A predição de
interação foi realizada por meio do software PatchDock (disponível em
http://bioinfo3d.cs.tau.ac.il/PatchDock/) sem alteração dos parâmetros previamente
setados pelo servidor. As 5 primeiras soluções com melhores scores geométricos, energia
de dessolvatação e área de interface foram avaliadas para observação da região de
interação do peptídeo com a proteína. Tal avaliação poderia fornecer dados sobre o local
de interação dos peptídeos com a estrutura nativa das proteínas ligantes auxiliando no
delineamento de experimentos que utilizassem tais moléculas para a identificação de
alterações relacionadas ao processo carcinogênico.
4.8. Análise Estatística
A análise estatística dos resultados foi realizada por meio do software Graph Pad
Prism (versão 5.0). Utilizou-se ANOVA para testar as diferenças entre os tratamentos,
seguido de pós-teste de Tukey para determinação das diferenças significativas entre os
grupos expostos e os grupos controle no que diz respeito às medições enzimáticas,
hemograma, relações entre os pesos dos animais e diferenças de intensidade entre spots
eletroforéticos. Para análise das diferenças entre os parâmetros histológicos observados
utilizaram-se os testes exato de Fisher e Qui-quadrado. Adotou-se o nível de significância
p < 0,05 para todas as análises.

Resultados e Discussão
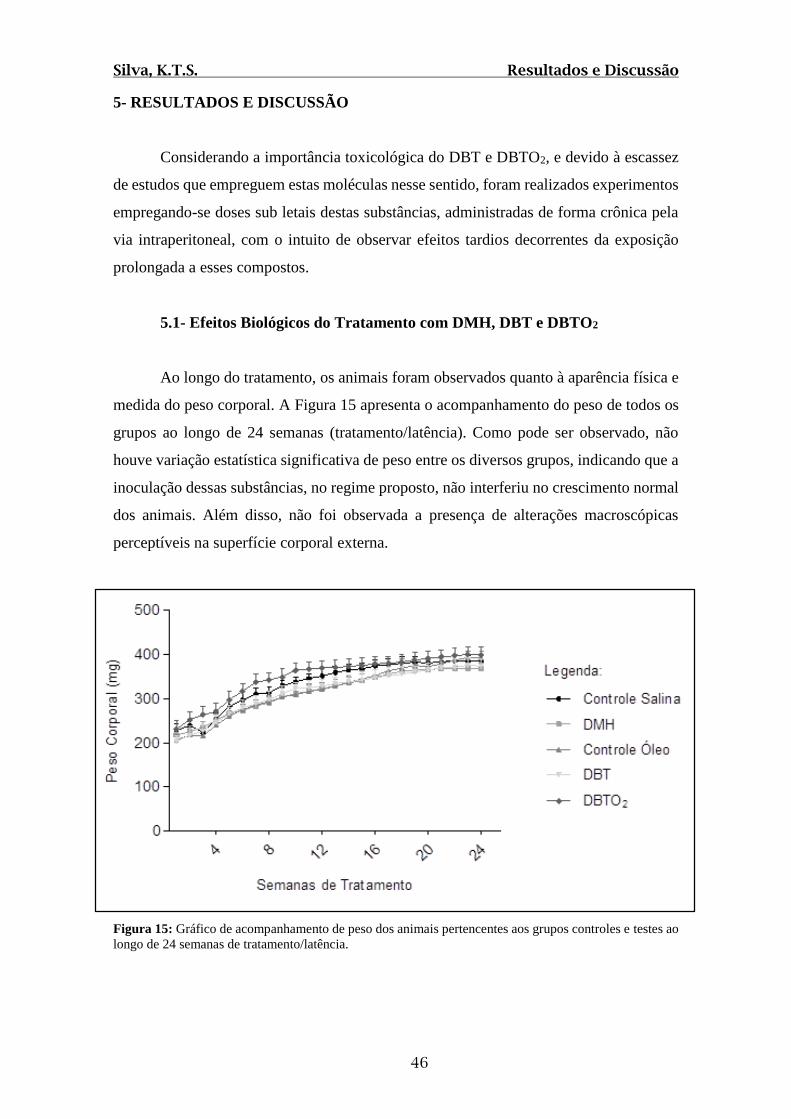
Silva, K.T.S. Resultados e Discussão
46
5- RESULTADOS E DISCUSSÃO
Considerando a importância toxicológica do DBT e DBTO2, e devido à escassez
de estudos que empreguem estas moléculas nesse sentido, foram realizados experimentos
empregando-se doses sub letais destas substâncias, administradas de forma crônica pela
via intraperitoneal, com o intuito de observar efeitos tardios decorrentes da exposição
prolongada a esses compostos.
5.1- Efeitos Biológicos do Tratamento com DMH, DBT e DBTO2
Ao longo do tratamento, os animais foram observados quanto à aparência física e
medida do peso corporal. A Figura 15 apresenta o acompanhamento do peso de todos os
grupos ao longo de 24 semanas (tratamento/latência). Como pode ser observado, não
houve variação estatística significativa de peso entre os diversos grupos, indicando que a
inoculação dessas substâncias, no regime proposto, não interferiu no crescimento normal
dos animais. Além disso, não foi observada a presença de alterações macroscópicas
perceptíveis na superfície corporal externa.
Figura 15: Gráfico de acompanhamento de peso dos animais pertencentes aos grupos controles e testes ao
longo de 24 semanas de tratamento/latência.

Silva, K.T.S. Resultados e Discussão
47
5.1.1- Hemograma e Dosagens Séricas de ALT, AST e Amilase
Os parâmetros hematológicos, dosagens séricas de enzimas pancreáticas e
hepáticas foram avaliadas durante a execução de meu trabalho de mestrado. Os gráficos
relacionados aos resultados obtidos nestes experimentos encontram-se nos Apêndices A
e B respectivamente (Apêndice A – Fig. 01.A e Apêndice B – Fig. 01.B).
Após a necropsia e coleta do sangue realizou-se o hemograma para avaliação de
possíveis alterações das células sanguíneas. O hematócrito foi o primeiro parâmetro
avaliado, não sendo observadas variações estatísticas significativas entre os grupos
controles e testes indicando que o volume globular não é afetado pelos tratamentos
realizados mesmo em doses mais elevadas.
Além do volume globular normal verificado, a observação dos esfregaços de
sangue total não evidenciou alterações morfológicas, presença de inclusões, presença de
hemácias nucleadas ou empilhamento dos glóbulos vermelhos indicando eritropoese
normal e ausência de processos inflamatórios. Tais achados demonstraram que os
tratamentos não interferiram na produção ou sobrevida na circulação das hemácias no
sangue periférico. As contagens globais e diferenciais de leucócitos do sangue periférico
também não demonstraram variações diante da análise estatística aplicada. Além disso,
quando avaliados os resultados individuais das contagens diferenciais de leucócitos não
foi possível detectar diferenças que possam indicar importância diagnóstica.
Determinaram-se também as atividades séricas das enzimas amilase, AST e ALT.
As atividades séricas das enzimas AST e ALT são comumente utilizadas como
marcadores de lesões hepáticas. Em conjunto, as duas enzimas (AST e ALT) são
consideradas os mais importantes parâmetros para detecção de lesões ao fígado
(Anderson et al., 2000).
Escolheu-se medir a atividade das enzimas AST, ALT e amilase por
representarem os principais meios de detecção sérica de alterações das funções hepática
e pancreática respectivamente. Como esses órgãos estão diretamente relacionados ao
metabolismo, as alterações nas funções normais dos mesmos representam informações
importantes a respeito da atividade toxicológica dessas substâncias.
Os resultados encontrados demonstraram que as atividades séricas individuais das
enzimas ALT e AST dos grupos controle salina e controle óleo não apresentaram
diferenças estatísticas significativas. Com relação aos testes DBT e DBTO2, os níveis de
ALT e AST encontraram-se diminuídas estatisticamente em aproximadamente 30%,

Silva, K.T.S. Resultados e Discussão
48
quando comparadas com o grupo controle óleo. Embora a relação AST/ALT tenha se
apresentado ligeiramente maior que 1 para todos os grupos avaliados de forma
indiscriminada entre os grupos controles e testes, não foram observadas lesões hepáticas
no exame histopatológico do fígado.
A atividade sérica de amilase foi avaliada com o objetivo de investigar a
integridade tecidual pancreática. Níveis alterados desta enzima também estão
relacionados à doença da vesícula biliar e alguns distúrbios gastrointestinais. A amilase é
produzida principalmente nas glândulas salivares e no pâncreas. Na pancreatite aguda, os
níveis de amilase podem alcançar valores de quatro a seis vezes o limite superior de
referência, elevando-se de 2 a 12 horas e retornando aos níveis normais após 3 a 4 dias.
A magnitude da elevação da atividade amilásica não se correlaciona com a gravidade da
lesão pancreática. As pancreatites crônicas, assim como o carcinoma pancreático, cursam
com níveis normais ou pouco elevados de amilase (Burtis and Ashwood, 1996).
Entre os grupos estudados não foram detectadas alterações nos níveis séricos de
amilase indicando que não houve a ocorrência de pancreatite aguda. Entretanto, não se
pode concluir que não houve alteração da função pancreática, sendo necessário que esses
dados sejam complementados com outros estudos clínicos para os animais em questão.
5.1.2- Avaliações Morfológicas Macro e Microscópicas de Tecidos Extraídos
Durante a Necropsia
Após avaliações macro e microscópicas do pulmão, fígado, baço e estômago,
observou-se que esses órgãos de todos os grupos estudados apresentaram aspecto e
coloração compatíveis com a normalidade (dados não mostrados). Nos intestinos, o
quadro de aparente normalidade foi observado nos animais dos grupos Controle I e III.
No grupo Teste DMH observou-se a presença de projeções papiliformes na mucosa para
a luz do intestino grosso, de aspecto avermelhado e consistência rígida, sugerindo a
formação de pólipos. Este aspecto não foi observado no intestino delgado desses animais
(Fig. 16 – A). Em contraste, nos animais pertencentes aos grupos Teste DBT e DBTO2
foram visualizadas projeções similares apenas no intestino delgado e em menor
intensidade quando comparadas àquelas visualizadas no intestino grosso dos animais
tratados com DMH (Fig. 16 – B).
O epitélio intestinal contém todas as enzimas envolvidas na ativação e
detoxificação dos HPA, embora essas atividades sejam geralmente menores que no
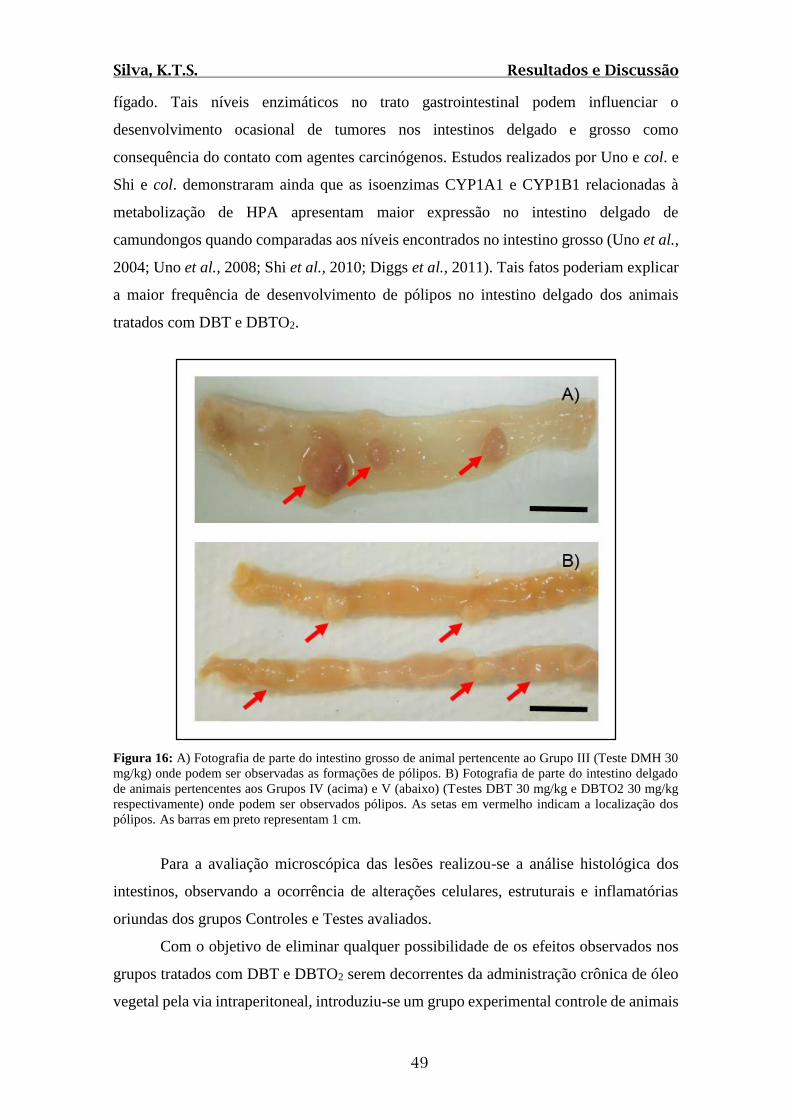
Silva, K.T.S. Resultados e Discussão
49
fígado. Tais níveis enzimáticos no trato gastrointestinal podem influenciar o
desenvolvimento ocasional de tumores nos intestinos delgado e grosso como
consequência do contato com agentes carcinógenos. Estudos realizados por Uno e col. e
Shi e col. demonstraram ainda que as isoenzimas CYP1A1 e CYP1B1 relacionadas à
metabolização de HPA apresentam maior expressão no intestino delgado de
camundongos quando comparadas aos níveis encontrados no intestino grosso (Uno et al.,
2004; Uno et al., 2008; Shi et al., 2010; Diggs et al., 2011). Tais fatos poderiam explicar
a maior frequência de desenvolvimento de pólipos no intestino delgado dos animais
tratados com DBT e DBTO2.
Figura 16: A) Fotografia de parte do intestino grosso de animal pertencente ao Grupo III (Teste DMH 30
mg/kg) onde podem ser observadas as formações de pólipos. B) Fotografia de parte do intestino delgado
de animais pertencentes aos Grupos IV (acima) e V (abaixo) (Testes DBT 30 mg/kg e DBTO2 30 mg/kg
respectivamente) onde podem ser observados pólipos. As setas em vermelho indicam a localização dos
pólipos. As barras em preto representam 1 cm.
Para a avaliação microscópica das lesões realizou-se a análise histológica dos
intestinos, observando a ocorrência de alterações celulares, estruturais e inflamatórias
oriundas dos grupos Controles e Testes avaliados.
Com o objetivo de eliminar qualquer possibilidade de os efeitos observados nos
grupos tratados com DBT e DBTO2 serem decorrentes da administração crônica de óleo
vegetal pela via intraperitoneal, introduziu-se um grupo experimental controle de animais

Silva, K.T.S. Resultados e Discussão
50
tratados com salina para efeito de comparação com animais tratados com óleo. Nestes
grupos, o quadro geral foi compatível com a normalidade histológica onde foi possível a
plena observação das túnicas intestinais bem como de todas as estruturas histológicas do
órgão. No intestino delgado de poucos animais de ambos os grupos Controle observou-
se aumento de células inflamatórias que, quando presentes, se apresentavam de maneira
difusa e de intensidade leve. Além disso, no grupo Controle Óleo foram observadas
reações hiperplásicas dos nódulos linfoides do intestino delgado em apenas 25% dos
animais (Apêndice C – Tab. 01.C). Ao contrário, no intestino grosso de animais
pertencentes ao grupo controle salina foi observado certo grau de inflamação (40%)
(Apêndice C – Tab. 02.C). Porém, considerando o tratamento invasivo ao qual esses
animais foram submetidos, e a diferença estatística encontrada quando analisados esses
dados contra os dados de animais tratados com DMH, DBT e DBTO2, pode-se dizer que
para os animais dos grupos controle o quadro geral observado representa a referência
histológica.
A comparação entre os grupos Controle Salina e Teste DMH nos intestinos grosso
e delgado (Tab. 02 e 03) revelou aumento estatisticamente significativo para todas os
parâmetros avaliadas. As atipias observadas foram principalmente alterações na
morfologia das mucosas, presença de glândulas irregulares e de células com morfologia
irregular de difícil identificação. As áreas de necrose observadas apresentaram-se de
maneira geral focais atingindo principalmente as túnicas mucosas e médias. As
hiperplasias das regiões nodulares eram caracterizadas pelo aumento concêntrico de
linfócitos e macrófagos e apresentavam correlação com as projeções papiliformes
observadas macroscopicamente no intestino grosso (Fig.17).
Uma avaliação semelhante foi realizada com os grupos Controle Óleo, Teste DBT
e DBTO2 (Tab. 02 e 03) revelando também, aumento significativo para todas as alterações
observadas. Apesar de a avaliação macroscópica ter revelado somente a presença de
projeções no intestino grosso para o Grupo Teste DMH, e no intestino delgado para os
Grupos Teste DBT e DBTO2, pode-se observar que quando avaliados
microscopicamente, foram encontradas lesões com características similares em ambos os
intestinos dos 3 grupos de tratamento.
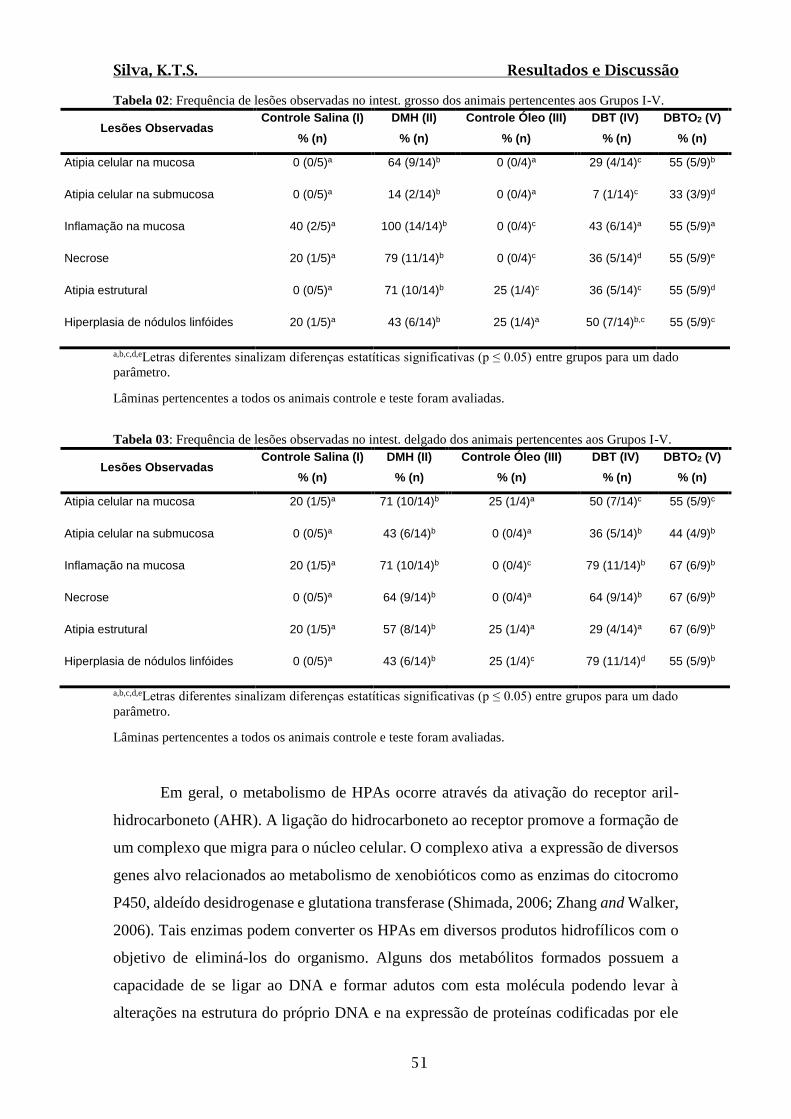
Silva, K.T.S. Resultados e Discussão
51
Tabela 02: Frequência de lesões observadas no intest. grosso dos animais pertencentes aos Grupos I-V.
Lesões Observadas Controle Salina (I)
% (n)
DMH (II)
% (n)
Controle Óleo (III)
% (n)
DBT (IV)
% (n)
DBTO2 (V)
% (n)
Atipia celular na mucosa 0 (0/5)a 64 (9/14)b 0 (0/4)a 29 (4/14)c 55 (5/9)b
Atipia celular na submucosa 0 (0/5)a 14 (2/14)b 0 (0/4)a 7 (1/14)c 33 (3/9)d
Inflamação na mucosa 40 (2/5)a 100 (14/14)b 0 (0/4)c 43 (6/14)a 55 (5/9)a
Necrose 20 (1/5)a 79 (11/14)b 0 (0/4)c 36 (5/14)d 55 (5/9)e
Atipia estrutural 0 (0/5)a 71 (10/14)b 25 (1/4)c 36 (5/14)c 55 (5/9)d
Hiperplasia de nódulos linfóides 20 (1/5)a 43 (6/14)b 25 (1/4)a 50 (7/14)b,c 55 (5/9)c
a,b,c,d,eLetras diferentes sinalizam diferenças estatíticas significativas (p ≤ 0.05) entre grupos para um dado
parâmetro.
Lâminas pertencentes a todos os animais controle e teste foram avaliadas.
Tabela 03: Frequência de lesões observadas no intest. delgado dos animais pertencentes aos Grupos I-V.
Lesões Observadas Controle Salina (I)
% (n)
DMH (II)
% (n)
Controle Óleo (III)
% (n)
DBT (IV)
% (n)
DBTO2 (V)
% (n)
Atipia celular na mucosa 20 (1/5)a 71 (10/14)b 25 (1/4)a 50 (7/14)c 55 (5/9)c
Atipia celular na submucosa 0 (0/5)a 43 (6/14)b 0 (0/4)a 36 (5/14)b 44 (4/9)b
Inflamação na mucosa 20 (1/5)a 71 (10/14)b 0 (0/4)c 79 (11/14)b 67 (6/9)b
Necrose 0 (0/5)a 64 (9/14)b 0 (0/4)a 64 (9/14)b 67 (6/9)b
Atipia estrutural 20 (1/5)a 57 (8/14)b 25 (1/4)a 29 (4/14)a 67 (6/9)b
Hiperplasia de nódulos linfóides 0 (0/5)a 43 (6/14)b 25 (1/4)c 79 (11/14)d 55 (5/9)b
a,b,c,d,eLetras diferentes sinalizam diferenças estatíticas significativas (p ≤ 0.05) entre grupos para um dado
parâmetro.
Lâminas pertencentes a todos os animais controle e teste foram avaliadas.
Em geral, o metabolismo de HPAs ocorre através da ativação do receptor aril-
hidrocarboneto (AHR). A ligação do hidrocarboneto ao receptor promove a formação de
um complexo que migra para o núcleo celular. O complexo ativa a expressão de diversos
genes alvo relacionados ao metabolismo de xenobióticos como as enzimas do citocromo
P450, aldeído desidrogenase e glutationa transferase (Shimada, 2006; Zhang and Walker,
2006). Tais enzimas podem converter os HPAs em diversos produtos hidrofílicos com o
objetivo de eliminá-los do organismo. Alguns dos metabólitos formados possuem a
capacidade de se ligar ao DNA e formar adutos com esta molécula podendo levar à
alterações na estrutura do próprio DNA e na expressão de proteínas codificadas por ele

Silva, K.T.S. Resultados e Discussão
52
(Marinkovic et al., 2013). Em particular, foi demonstrada a citotoxicidade dose
dependente promovida pelo DBT em cultura de células HepG2 e a formação de adutos
de DNA após 24 horas de exposição ao HPAS (Amat et al., 2004).
Diversos trabalhos constataram a presença do metabólito do DBT, o DBTO2, em
modelos animais. Apesar disso, o DBT, assim como outros HPAs de 2-3 anéis, é
considerado um ativador fraco do receptor AHR, consequentemente levando a uma baixa
expressão de enzimas CYP450 (Ciganek et al., 2004; Hecht et al., 2010). Devido a este
fato, tem sido especulado que a toxicidade do DBT ocorre através de outros mecanismos
diferentes da ativação de AHR, mas tais vias ainda permanecem desconhecidas
(Incardona et al., 2004).
A presença de inflamação e necrose aumentadas nos grupos Testes indica o
potencial irritativo e lesivo das drogas utilizadas. Tal observação corrobora com estudos
que demonstraram através da análise de transcritos de zebra fish tratados com DBT, um
aumento de quatro vezes nos níveis da enzima colesterol 25-hidroxilase. Esta molécula
desempenha um conhecido papel no desenvolvimento da resposta inflamatória (Goodale
et al., 2013). Diversos HPAs são conhecidamente capazes de induzir o estresse oxidativo
em culturas de células levando ao aumento da expressão de citocinas inflamatórias
(Suresh et al., 2009; Ovrevik et al., 2010). Além disso, as observações de crescimento
celular aumentado e alterações nas estruturas teciduais demonstram um estágio inicial de
displasia característico de lesões pré-neoplásicas (Fig. 17).
Diante da constatação de indução de lesões de natureza neoplásica nos animais
tratados com DBT e DBTO2 com características semelhantes àquelas provocadas pela
administração crônica de mesma dose de DMH, fica clara a importância desses resultados
como contribuição para a avaliação toxicológica dessas substâncias de irrefutável
relevância ambiental.
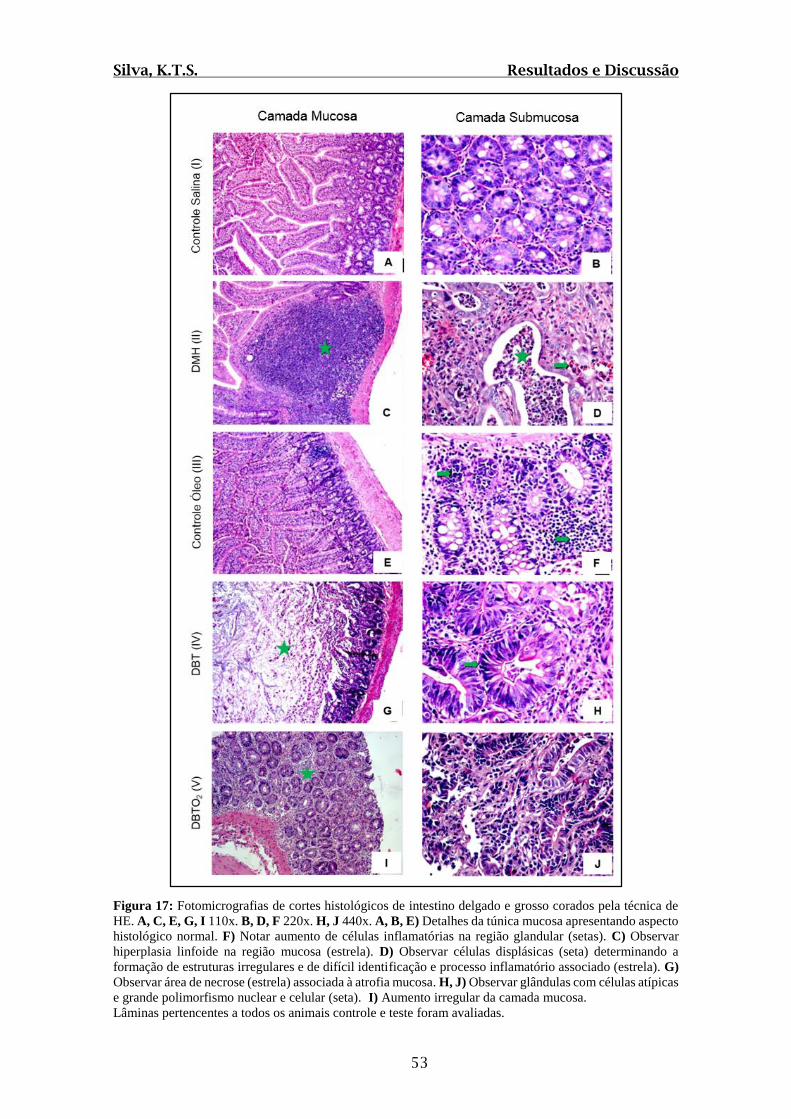
Silva, K.T.S. Resultados e Discussão
53
Figura 17: Fotomicrografias de cortes histológicos de intestino delgado e grosso corados pela técnica de
HE. A, C, E, G, I 110x. B, D, F 220x. H, J 440x. A, B, E) Detalhes da túnica mucosa apresentando aspecto
histológico normal. F) Notar aumento de células inflamatórias na região glandular (setas). C) Observar
hiperplasia linfoide na região mucosa (estrela). D) Observar células displásicas (seta) determinando a
formação de estruturas irregulares e de difícil identificação e processo inflamatório associado (estrela). G)
Observar área de necrose (estrela) associada à atrofia mucosa. H, J) Observar glândulas com células atípicas
e grande polimorfismo nuclear e celular (seta). I) Aumento irregular da camada mucosa.
Lâminas pertencentes a todos os animais controle e teste foram avaliadas.

Silva, K.T.S. Resultados e Discussão
54
5.1.3- Marcação Imunohistoquímica: CEA e CD44
Com o objetivo de confirmar o diagnóstico obtido através da análise histológica
dos intestinos grosso e delgado dos animais pertencentes aos grupos Controles e Testes,
realizou-se a marcação imunohistoquímica com os antincorpos anti-CEA e anti-CD44.
Tais proteínas são conhecidos marcadores tumorais para CCR e outros tipos de câncer
encontrados em diferentes estágios do desenvolvimento tumoral (McLeod and Murray,
1999; Duffy et al., 2003; Wang et al., 2009b).
O Antígeno Carcinoembrionário (CEA) é o marcador proteico mais utilizado na
clínica para avaliação do câncer colorretal. A detecção de altos níveis de expressão desta
proteína no soro ou tecidos está relacionada a um prognóstico ruim da doença (Duffy et
al., 2003). O CEA é um membro da família de imunoglobulinas que atua na adesão
intercelular, função esta, que está intimamente ligada à ocorrência de metástases (Kitadai
et al., 1996).
A avaliação da capacidade celular de polarização do CEA também vem sendo
proposta como método auxiliar no estadiamento do CCR atualmente realizado pela
avaliação das características morfológicas do tumor (TNM). Tal proposta considera as
características morfológicas e funcionais e melhora o entendimento do comportamento
tumoral. Nos tecidos normais ocorre uma polarização da proteína CEA que geralmente
atinge a região apical da célula. Nos tecidos neoplásicos bem polarizados (estágio inicial)
ocorre manutenção parcial da capacidade de polarização, enquanto em estágios mais
avançados ocorre uma distribuição difusa desse antígeno (Priolli et al., 2007).
A proteína CD44 (Cluster of differentiation 44) é um receptor de membrana que
possui uma região ligante para o ácido hialurônico da matriz extracelular através da qual
participa de diversas vias de sinalização celular ligadas à progressão tumoral incluindo
invasão e metástase (Wang et al., 2009a). Essas moléculas de adesão controlam o
comportamento celular através da mediação de contato entre as células ou entre as células
e a matriz extracelular, sendo essenciais para a manutenção da integridade tecidual
(Orian-Rosseau, 2010).
Os membros da família CD44 podem ser encontrados na forma padrão (CD44s),
usualmente associada ao tecido linfóide, ou como isoformas (CD44v) em células
epiteliais embrionárias e em diversos carcinomas humanos (Coppola et al., 1998; de Paula
Carli et al., 2012). No CCR, a isoforma CD44v3 parece estar associada à promoção de
invasão e resistência a apoptose, enquanto a isoforma CD44v6 associa-se com a

Silva, K.T.S. Resultados e Discussão
55
ocorrência de metástase e diminuição na sobrevida do paciente (Kuniyasu et al., 2001;
Kuhn et al., 2007).
A expressão epitelial de CD44 em tecidos intestinais normais também ocorre de
maneira localizada. Rudzki e colaboradores demonstraram por imunohistoquímica que
essa proteína encontra-se limitada à porção interna do epitélio, não sendo observada na
superfície do mesmo. Em tecidos neoplásicos ocorre aumento de expressão das isoformas
CD44v, e esse padrão de distribuição torna-se alterado (Rudzki and Jothy, 1997).
Como pode ser observado na Figura 18, os gráficos 1 e 2 demonstram a contagem
de células intestinais marcadas com os anticorpos anti-CEA e anti-CD44. Em ambos os
gráficos foram encontradas diferenças estatisticamente significativas entre os grupos
Controles e seus respectivos grupos Testes. Além disso, os grupos Controle apresentaram
contagens semelhantes entre si para cada anticorpo utilizado, assim como os grupos
Testes demonstrando similaridade entre os efeitos promovidos pelo DBT, DMH e DBTO2
na alteração dos níveis de expressão desses antígenos. Em ambos os gráficos o aumento
encontrado entre os grupos Controle e Testes foi de aproximadamente 3 vezes,
confirmando o caráter neoplásico das lesões avaliadas histologicamente.
A distribuição celular das proteínas CEA e CD44 também foi avaliada de maneira
qualitativa para auxiliar na caracterização das alterações morfofuncionais observadas nos
animais dos grupos Testes. Pode-se observar que para os animais pertencentes aos grupos
Testes DMH, DBT e DBTO2 a marcação imunohistoquímica com anti-CEA apresentou-
se de maneira relativamente difusa no citoplasma celular quando comparada com a
marcação encontrada nos animais pertencentes aos grupos Controle. Quanto à marcação
obtida com o anticorpo anti-CD44 para os grupos Controle pode-se observar a localização
da expressão proteica no interior da camada submucosa, enquanto para os animais dos
grupos Testes tal marcação foi encontrada por toda a extensão tecidual. Em conjunto, as
duas observações parecem indicar um estágio leve a moderado de progressão tumoral
como descrito anteriormente.
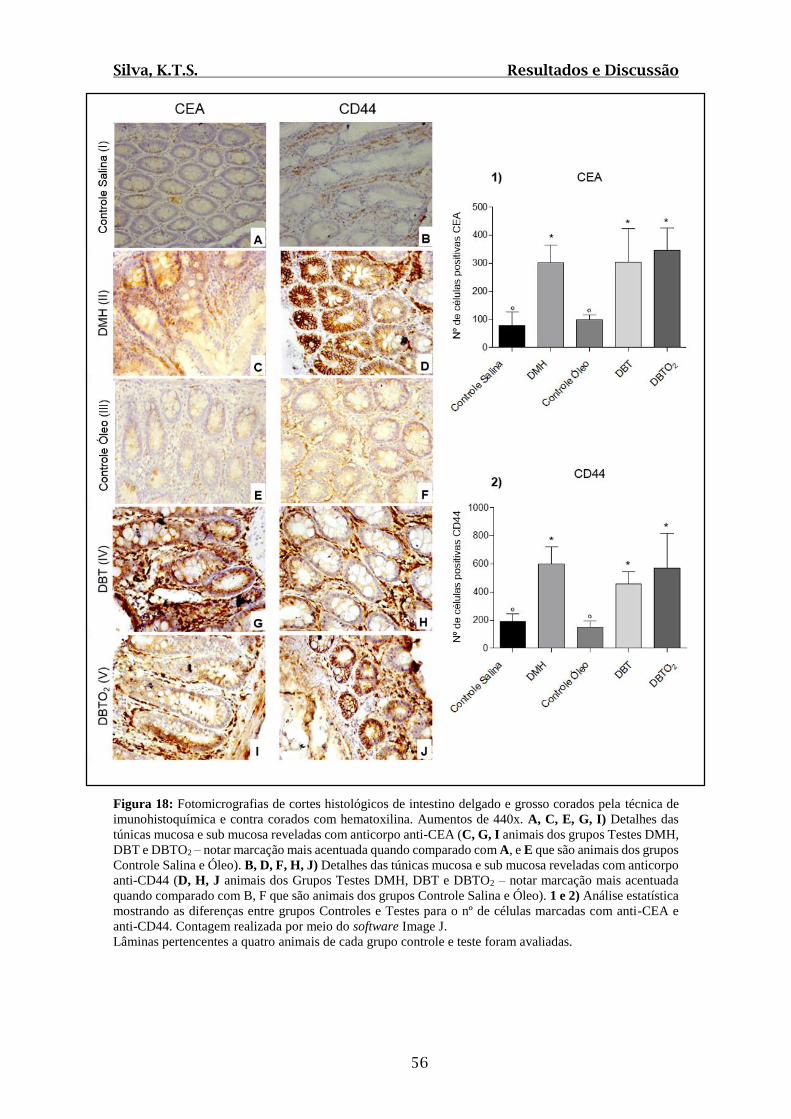
Silva, K.T.S. Resultados e Discussão
56
Figura 18: Fotomicrografias de cortes histológicos de intestino delgado e grosso corados pela técnica de
imunohistoquímica e contra corados com hematoxilina. Aumentos de 440x. A, C, E, G, I) Detalhes das
túnicas mucosa e sub mucosa reveladas com anticorpo anti-CEA (C, G, I animais dos grupos Testes DMH,
DBT e DBTO2 – notar marcação mais acentuada quando comparado com A, e E que são animais dos grupos
Controle Salina e Óleo). B, D, F, H, J) Detalhes das túnicas mucosa e sub mucosa reveladas com anticorpo
anti-CD44 (D, H, J animais dos Grupos Testes DMH, DBT e DBTO2 – notar marcação mais acentuada
quando comparado com B, F que são animais dos grupos Controle Salina e Óleo). 1 e 2) Análise estatística
mostrando as diferenças entre grupos Controles e Testes para o nº de células marcadas com anti-CEA e
anti-CD44. Contagem realizada por meio do software Image J.
Lâminas pertencentes a quatro animais de cada grupo controle e teste foram avaliadas.

Silva, K.T.S. Resultados e Discussão
57
5.2- Análise Proteômica
A relação entre biomarcadores tumorais e o CCR tem sido extensivamente
investigada. Proteínas envolvidas em diversos aspectos da progressão da doença
incluindo invasão, metástase, regulação do ciclo celular, fatores de crescimento e
apoptose têm sido descritas como potenciais marcadores (O’Dwyer et al., 2011). As vias
biológicas que podem formar a base para novos agentes terapêuticos têm sido
identificadas para diversos tipos de câncer. Embora algumas mutações de alta frequência
sejam alvos atrativos para o desenvolvimento de drogas, vias de sinalização comuns,
relacionadas a essas mutações, também podem ser alvos para terapia (Markowitz and
Bertagnolli, 2009).
Sendo assim, torna-se interessante um estudo molecular das lesões desenvolvidas
pelos animais tratados com os agentes carcinogênicos, considerando a similaridade entre
as alterações macro e microscópicas dos tecidos intestinais dos ratos tratados com DBT
e DBTO2, e aquelas encontradas nos animais tratados com DMH. Tal abordagem, poderia
auxiliar na verificação das possíveis relações entre as vias biológicas envolvidas no
estabelecimento do processo carcinogênico promovido por ambos os tratamentos. Sendo
assim, inicialmente foram extraídas proteínas solúveis de tecidos intestinais provenientes
de todos os grupos citados para identificação e análise da expressão diferencial das
mesmas.
As figuras 19, 20 e 21 apresentam os perfis eletroforéticos representativos de
triplicatas referentes aos extratos proteicos de fração solúvel obtidos por 2D SDS-PAGE
10% para os grupos III/IV, III/V e I/II, respectivamente. Mediante a análise da
sobreposição e densitometria das imagens realizada através do software SameSpots
(TotalLab), foi possível observar a expressão diferenciada de diversas proteínas entre os
grupos Controle e seus respectivos grupos Testes (indicada pelas setas numeradas).
Foram consideradas quantitativamente significativas, as diferenças de expressão para
cima ou para baixo, com valores de pelo menos 1,5 vez na triplicata biológica realizada,
com um desvio padrão máximo aceitável de 10%.
A qualidade das preparações pode ser atestada pela observação da cobertura de
toda a extensão da faixa de pH provida pela strip durante a isoeletrofocalização (pH3-
10), e também pela presença de spots representando diversas massas moleculares
indicando a manutenção da integridade proteica.

Silva, K.T.S. Resultados e Discussão
58
Figura 19: Géis 2D SDS-PAGE 10% representativos da extração de proteínas solúveis de intestinos delgados dos
A) Grupo Controle Óleo (III) e B) Teste DBT (IV). Nas setas numeradas estão marcados os spots identificados por
espectrometria de massas que apresentaram diferença de 1,5 vez na expressão. Coloração realizada com solução de
Coomassie Coloidal. Para cada experimento foi realizada uma triplicata biológica independente.
Figura 20: Géis 2D SDS-PAGE 10% representativos da extração de proteínas solúveis de intestinos delgados dos
A) Grupo Controle Óleo (III) e B) Teste DBTO2 (V). Nas setas numeradas estão marcados os spots identificados
por espectrometria de massas que apresentaram diferença de 1,5 vez na expressão. Coloração realizada com solução
de Coomassie Coloidal. Para cada experimento foi realizada uma triplicata biológica independente.

Silva, K.T.S. Resultados e Discussão
59
Figura 21: Géis 2D SDS-PAGE 10% representativos da extração de proteínas solúveis de intestinos delgados dos
A) Grupo Controle Salina (I) e B) Teste DMH (II). Nas setas numeradas estão marcados os spots identificados por
espectrometria de massas que apresentaram diferença de 1,5 vez na expressão. Coloração realizada com solução de
Coomassie Coloidal. Para cada experimento foi realizada uma triplicata biológica independente.
Em média, 1700 spots foram detectados por par de gel controle/teste analisado. A
grande maioria destas proteínas mostrou níveis equivalentes de expressão entre os grupos
avaliados. Entretanto, foi possível detectar alterações em 84 spots pertencentes aos grupos
Controle e Teste DBT e DBTO2. Destes, 16 spots relativos ao grupo IV e 20 relativos ao
grupo V demonstraram diminuição na expressão dos testes em relação ao controle e 68 e
64 spots revelaram aumento na expressão de proteínas relativas aos mesmos grupos (não
mostrado nas figuras), respectivamente. Quanto à avaliação dos géis preparados para os
grupos Controle Salina e Teste DMH foram detectados 88 spots com alteração na
expressão proteica. Destes, 19 mostraram diminuição e 69, aumento na densidade dos
spots avaliados.
Todos os spots que demonstraram algum tipo de alteração de expressão foram
excisados e digeridos com tripsina para identificação por espectrometria de massas. Após
o sequenciamento e aplicação dos critérios de seleção descritos no item 4.3.3, 22 proteínas
isoladas nos géis relativos aos grupos III, IV e V e 38 proteínas isoladas nos géis relativos
aos grupos I e II foram identificadas e agrupadas de acordo com as funções moleculares
por elas desempenhadas nas células (Tab. 04 e 05). Dados referentes à identificação dos
peptídeos trípticos encontram-se no Apêndice D ao fim deste trabalho (Apêndice D – Tab.
01.D e Tab. 02.D).
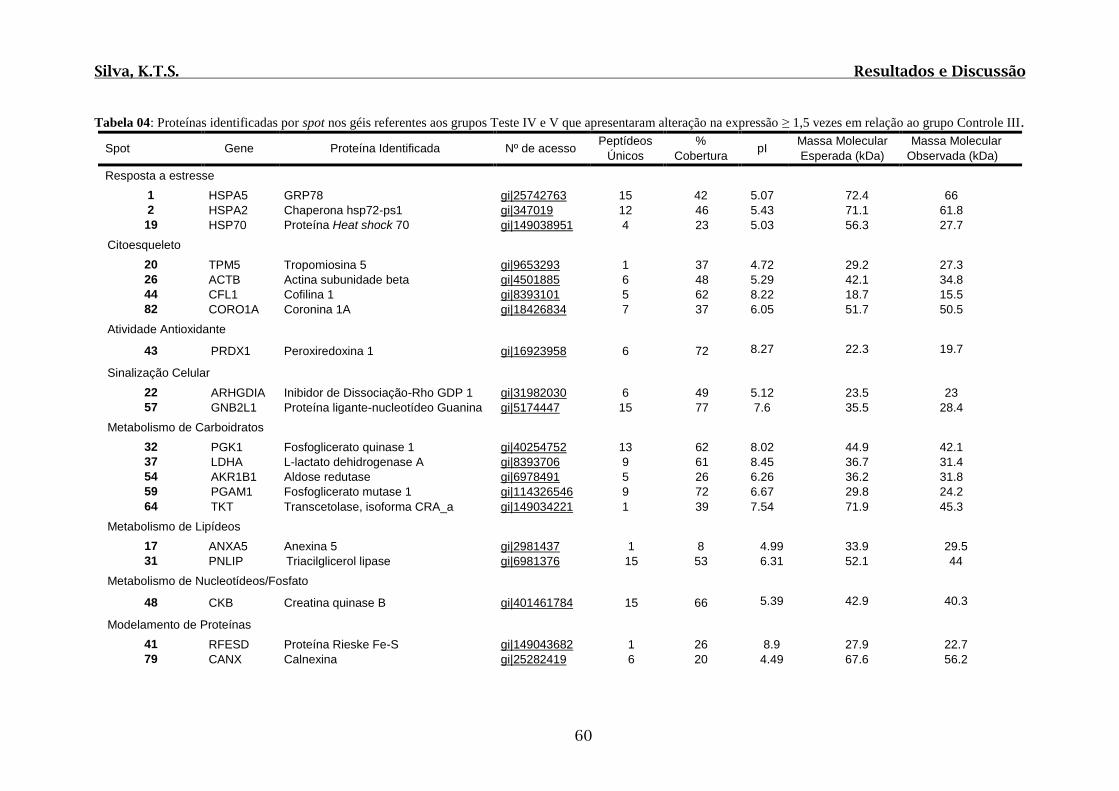
Silva, K.T.S. Resultados e Discussão
60
Tabela 04: Proteínas identificadas por spot nos géis referentes aos grupos Teste IV e V que apresentaram alteração na expressão ≥ 1,5 vezes em relação ao grupo Controle III.
Spot Gene Proteína Identificada Nº de acesso Peptídeos
Únicos
%
Cobertura pI
Massa Molecular
Esperada (kDa)
Massa Molecular
Observada (kDa)
Resposta a estresse
1
2
19
HSPA5
HSPA2
HSP70
GRP78
Chaperona hsp72-ps1
Proteína Heat shock 70
gi|25742763
gi|347019
gi|149038951
15
12
4
42
46
23
5.07
5.43
5.03
72.4
71.1
56.3
66
61.8
27.7
Citoesqueleto
20
26
44
82
TPM5
ACTB
CFL1
CORO1A
Tropomiosina 5
Actina subunidade beta
Cofilina 1
Coronina 1A
gi|9653293
gi|4501885
gi|8393101
gi|18426834
1
6
5
7
37
48
62
37
4.72
5.29
8.22
6.05
29.2
42.1
18.7
51.7
27.3
34.8
15.5
50.5
Atividade Antioxidante
43 PRDX1 Peroxiredoxina 1 gi|16923958 6 72 8.27 22.3 19.7
Sinalização Celular
22
57
ARHGDIA
GNB2L1
Inibidor de Dissociação-Rho GDP 1
Proteína ligante-nucleotídeo Guanina
gi|31982030
gi|5174447
6
15
49
77
5.12
7.6
23.5
35.5
23
28.4
Metabolismo de Carboidratos
32
37
54
59
64
PGK1
LDHA
AKR1B1
PGAM1
TKT
Fosfoglicerato quinase 1
L-lactato dehidrogenase A
Aldose redutase
Fosfoglicerato mutase 1
Transcetolase, isoforma CRA_a
gi|40254752
gi|8393706
gi|6978491
gi|114326546
gi|149034221
13
9
5
9
1
62
61
26
72
39
8.02
8.45
6.26
6.67
7.54
44.9
36.7
36.2
29.8
71.9
42.1
31.4
31.8
24.2
45.3
Metabolismo de Lipídeos
17
31
ANXA5
PNLIP
Anexina 5
Triacilglicerol lipase
gi|2981437
gi|6981376
1
15
8
53
4.99
6.31
33.9
52.1
29.5
44
Metabolismo de Nucleotídeos/Fosfato
48 CKB Creatina quinase B gi|401461784 15 66 5.39 42.9 40.3
Modelamento de Proteínas
41
79
RFESD
CANX
Proteína Rieske Fe-S
Calnexina
gi|149043682
gi|25282419
1
6
26
20
8.9
4.49
27.9
67.6
22.7
56.2

Silva, K.T.S. Resultados e Discussão
61
Outros Processos
23
62
63
FCGBP
TF
TF
Fragmento Fc de IgG
Transferrina
Transferrina
gi|257467629
gi|1854476
gi|1854476
10
16
7
9
33
24
4.92
6.94
6.94
28.7
78.5
78.5
93
69.3
63.2
Tabela 05: Proteínas identificadas por spot nos géis referentes ao grupo Teste II que apresentaram alteração na expressão ≥ 1,5 vezes em relação ao grupo Controle I.
Spot Gene Proteína Identificada Nº de acesso Peptídeos
Únicos % Cobertura pI
Massa Molecular
Esperada (kDa)
Massa Molecular
Observada (kDa)
Síntese/Degradação de Proteínas
25
57
EEF2
MCPT1
Fator de elongação 2
Protease de mastócitos 1
gi|8393296
gi|8393764
19
11
38
68
6,41
6,55
96,2
27,6
86,1
20,5
Citoesqueleto
1
5/6
12
45
70
75
76
TPM1
ACTB
TAGLN
TPM5
MYL
KRT8
EZR
Tropomiosina 1, alfa
Actina subunidade beta
Transgelina
Tropomyosina 5
Miosina, cadeia leve
Citoqueratina 8
Ezrina
gi|149028892
gi|4501883
gi|13928744
gi|9653293
gi|71037403
gi|203734
gi|40804381
13
13
11
4
9
24
9
58
56
53
51
66
63
29
4.69
5,23
8,87
4,72
4,67
5,49
5,83
32,7
42,4
22,6
29,1
19,9
52,7
69,4
32,9
36,7
15,1
24,9
15,9
46,7
70,5
Atividade Antioxidante
36
38
44
PRDX1
ADH
SOD
Peroxiredoxina 1
Álcool Dehidrogenase
Superóxido Dismutase [Mn]
gi|16923958
gi|202727
gi|8394331
8
1
9
77
69
42
8,27
8,52
8,96
22.3
40,5
24,9
40,3
35,9
17,9
Sinalização Celular
60ª
60b
61a
61b
63
72
88
DPP7
TRA1
LUM
RBBP4
YWHAE
ARHGDIA
LGALS3
Dipeptidil peptidase 2
Proteína associada a transcrição 1
Lumican
Proteína ligante de histona RBBP4
14-3-3 proteína epsilon
Inibidor de Dissociação-Rho GDP 1
Galectina 3
gi|14010871
gi|51858886
gi|13591983
gi|5032027
gi|5803225
gi|31982030
gi|13929190
5
10
5
9
14
8
8
22
23
21
30
58
57
37
4,87
5,02
6
4,74
4,63
5.12
8,93
55,6
74,3
38,6
47,9
29,3
23,4
27,3
46,7
46,7
44,4
44,4
24,3
21,8
22,3
Metabolismo de Lipídeos
42 ANXA2 Anexina 2 gi|9845234 24 75 7,55 38,9 32,1
Silva, K.T.S. Resultados e Discussão
62
Metabolismo de Carboidratos
30
31
33
39
41
54
55
83
TKT
PKM2
DLD
ALDOA
GAPDH
GAPDH
ENO1
PGK1
Transcetolase
Piruvato quinase M2
Dihidrolipoil dehidrogenase
Aldolase A
Glicerald.-3-fosf. Dehidrogenase
Glicerald.-3-fosf. Dehidrogenase
Enolase alfa
Fosfoglicerato quinase 1
gi|12018252 gi|206205
gi|40786469
gi|202837
gi|8393418
gi|56188
gi|38649320
gi|40254752
1
25
10
1
1
6
5
15
39
61
28
60
62
21
23
64
7,54
7,15
7,96
8,31
8,14
8,43
6,7
8
71,9
58,3
54,6
39,7
36,1
36,1
51,7
44,9
60,7
52
50,6
34,3
31,5
30,1
43,3
38,4
Metabolismo de Nucleotídeos/Fosfato
29
80
ATIC
CMPK
Biossíntese de purina bifunc. PURH
Citidina monofosfato quinase
gi|48675845
gi|150383503
16
10
41
62
6,69
5,66
64,6
22,4
53,5
17,5
Modelamento de Proteínas
18
20
29
CALR
HSP90B1
CCT6
Calreticulina
Endoplasmina
Proteína Complexo-T 1 sub. zeta
gi|11693172
gi|210032365
gi|76253725
15
19
14
52
30
48
4,33
4,72
6,63
48,2
92,9
58,4
50,6
88,9
53,5
Outros Processos
7
27
43
50
SELENBP
WDR1
VDAC2
IGG2A
Proteína ligante de selênio 1
Repetição WD contendo proteína 1
Canal seleção ânion voltagem dep.2
Imunogl. Gama-2a cadeia pesada
gi|18266692
gi|62078997
gi|13786202
gi|1220486
25
17
11
10
58
32
51
30
6,1
6,15
7,44
8,15
53,1
66,8
32,4
52,2
47,9
58,7
25,4
43,3
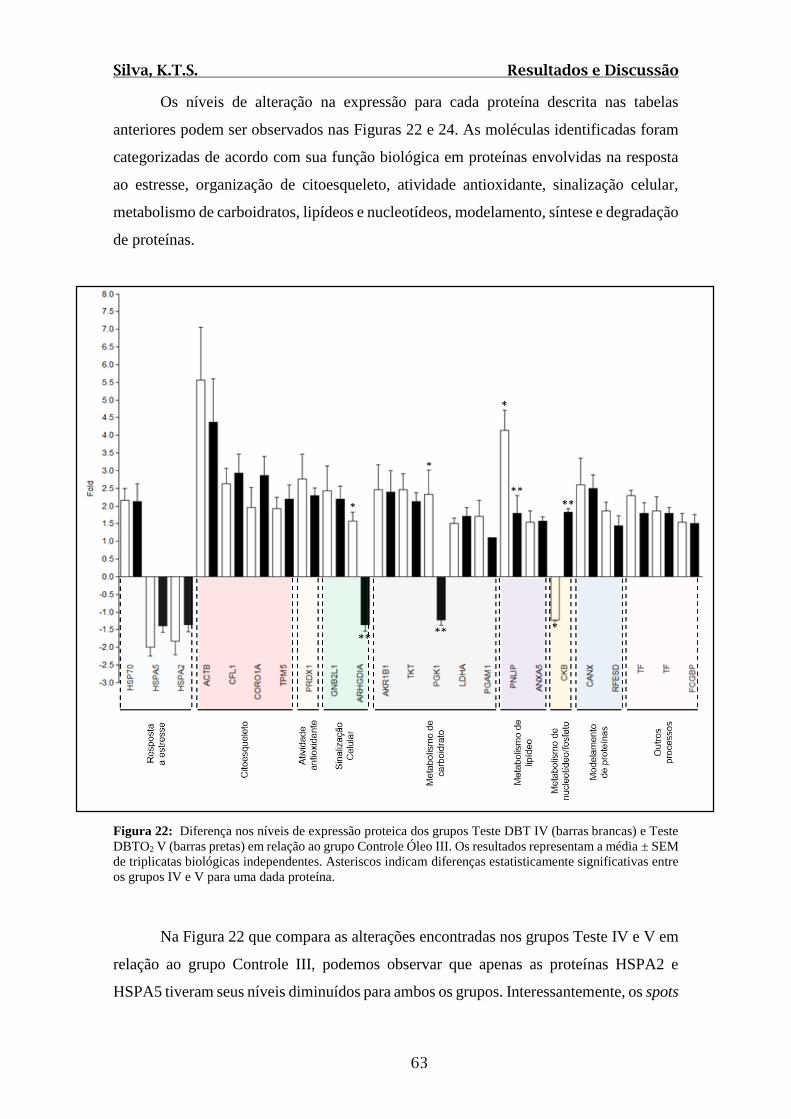
Silva, K.T.S. Resultados e Discussão
63
Os níveis de alteração na expressão para cada proteína descrita nas tabelas
anteriores podem ser observados nas Figuras 22 e 24. As moléculas identificadas foram
categorizadas de acordo com sua função biológica em proteínas envolvidas na resposta
ao estresse, organização de citoesqueleto, atividade antioxidante, sinalização celular,
metabolismo de carboidratos, lipídeos e nucleotídeos, modelamento, síntese e degradação
de proteínas.
Figura 22: Diferença nos níveis de expressão proteica dos grupos Teste DBT IV (barras brancas) e Teste
DBTO2 V (barras pretas) em relação ao grupo Controle Óleo III. Os resultados representam a média ± SEM
de triplicatas biológicas independentes. Asteriscos indicam diferenças estatisticamente significativas entre
os grupos IV e V para uma dada proteína.
Na Figura 22 que compara as alterações encontradas nos grupos Teste IV e V em
relação ao grupo Controle III, podemos observar que apenas as proteínas HSPA2 e
HSPA5 tiveram seus níveis diminuídos para ambos os grupos. Interessantemente, os spots

Silva, K.T.S. Resultados e Discussão
64
identificados como ARHGDIA, PGK1 e CKB demonstraram níveis de expressão opostos
entre os dois grupos Testes em questão. As proteínas ACTB e CANX tiveram níveis
aumentados de expressão e foram utilizadas na validação dos resultados obtidos por
análise proteômica via 2D SDS-PAGE através de Western blotting (Apêndice E – Fig.
01.E).
Moléculas envolvidas na organização do citoesqueleto como cofilina 1, coronina
1, actina e tropomiosina 5, merecem atenção por estarem super expressas. Nesse contexto,
diversos trabalhos já demonstraram aumento nos níveis de expressão da proteína coronina
1 em tecidos tumorais de mama onde ela parece contribuir para invasão e migração das
células tumorais. A super expressão de CORO1A parece estar relacionada à translocação
de Rac1 para a membrana plasmática. Rac1 GTPase, por sua vez, controla diversos
processo gerais como a dinâmica do citoesqueleto, regulação de tamanho celular e entrada
no ciclo celular além da transcrição gênica. Para tal, Rac1 interage com efetores como
proteínas quinases, enzimas, proteínas de modelamento e reguladores diretos do
crescimento, estabilização e ramificação do filamento de actina. A atividade de Rac1 é
controlada através da transição do estado inativo, ligado a GDP, ao estado ativo, ligado a
GTP. Fatores de troca GDP/GTP, proteínas ativadoras de GTPases e inibidores de
dissociação de Rho GDP (RhoGDI) realizam esse controle. CORO1A parece estar
envolvida também na modulação da resposta imune desempenhando papel na
sobrevivência, migração, ativação e regulação de Ca2+ em células T além de suprimir a
ativação de NF-κB inibindo a degradação de IκBα e a translocação nuclear de p65 (Kim
et al., 2009; Castro-Castro et al., 2011; Kaminski et al., 2011; Liu et al., 2014).
Níveis aumentados de cofilina 1 parecem estar associados com um prognóstico
ruim em pacientes com câncer de pulmão. Esta proteína desempenha um papel central no
controle da polimerização de filamentos de actina. Sua atividade pode ser regulada por
fosforilação/defosforilação em resíduos de serina após a estimulação de receptores
ligados à proteína G regulados predominantemente por via dependente de Rac2 e GSK3.
PI3K pode promover ativação da fosfatase SSH1 induzida por insulina e defosforilação
de CFL1 em células de câncer de mama (Peng et al., 2011; Bravo-Cordero et al.,2013).
A diminuição da expressão da tropomiosina 5 em células de melanoma de
camundongos levou ao comprometimento e diminuição da motilidade, contribuindo para
a redução do potencial metastático dessas células, demonstrando o papel desempenhado
por essa proteína no estabelecimento e migração tumoral (Miyado et al., 1996). A
expressão alterada de isoformas celulares e nucleares de actina tem sido demonstrada

Silva, K.T.S. Resultados e Discussão
65
durante a tumorogênese como resultado de sinalização epigenética e perfil genético
alterado para as sequências específicas relacionadas com tais genes (Rao et al., 1997).
Interessantemente, a proteína ARHGDIA (Rho-GDI), que também apresentou
níveis aumentados de expressão celular, encontra-se relacionada à YWHAE atuando em
vias de sinalização relacionadas. ARHGDIA regula a troca GDP/GTP em proteínas
GTPases relacionadas a Ras (subfamília de proteínas Rho - CDC42, RAC1 e RHOA)
impedindo a dissociação de GDP e mantendo-as em estado inativado, protegendo-as de
degradação. Níveis elevados de Rho-GDI podem resultar em migração celular aumentada
e prevenção de apoptose contribuindo para a sobrevivência e migração tumoral como
descrito para câncer de pulmão e cólon, além de serem associados com quimioresistência
em diversas linhagens celulares. A força da interação Rac1/RhoGDI é modulada
negativamente por diversos sinais incluindo proteína quinase C (Adra et al., 1997; Faure
and Dagher, 2001; Wei et al., 2002; Dovas and Couchman, 2005; Zhao et al., 2008).
Enzimas envolvidas no metabolismo de lipídeos e carboidratos também
apresentaram aumento significativo nos tecidos displásicos avaliados. Um aumento
significativo na expressão da enzima triacilglicerol lipase aponta para um papel chave
desempenhado por essa proteína para sustentar a alta demanda energética requerida pelas
células proliferativas. Em paralelo, a super expressão de enzimas envolvidas no
metabolismo de carboidratos, particularmente aquelas relacionadas à via glicolítica,
podem permitir a utilização do glicerol proveniente do metabolismo lipídico aumentado.
O mapa de interação proteica observado na Figura 23, ilustra estas relações. Mais
de 50% das proteínas identificadas demonstram algum grau de interação intracelular. De
relevância particular, os altos níveis de triacilglicerol lipase e de enzimas envolvidas no
processamento de glicerol/carboidrato, fornecem um indício primário sobre o
metabolismo dos tecidos displásicos induzidos pela exposição ao DBT e DBTO2.
Ainda em concordância com o exposto anteriormente, os níveis aumentados
observados para peroxiredoxina 1 podem explicar seu papel no metabolismo de radicais
livres, particularmente peróxidos, gerados por β-oxidação de ácidos graxos. Um papel
adicional para o aumento de expressão para proteínas relacionadas ao estresse oxidativo,
incluindo HSP70, seria controlar o efeito tóxico promovido pelo intenso infiltrado
inflamatório observado. Membros da família da HSP70 relacionam-se com o
modelamento de proteínas contribuindo significativamente para a homeostase proteica.
A isoforma HSP70-1 é expressa em altos níveis em tecidos normais e tumorais
sendo simplesmente referida como HSP70 (Murphy, 2013). Em contraste, a expressão de

Silva, K.T.S. Resultados e Discussão
66
HSP70-2, conhecida como HSPA2, mostra níveis diminuídos em diversos tecidos, com
exceção descrita em literatura para testículos de ratos (Scieglinska et al., 2011).
Concordando com os resultados obtidos neste trabalho, Scieglinska e colaboradores
reportaram níveis indetectáveis de HSPA2 em três diferentes linhagens de células
tumorais incluindo HCT116 de CCR (Scienglinska et al., 2008). A proteína HSPA5
também demonstrou baixos níveis de expressão nos grupos Teste IV e V, mas seu papel
na progressão de neoplasias em intestino delgado continua obscuro.
Figura 23: Mapa de interação proteica para proteínas diferencialmente expressas entre grupos Teste IV e
V e Controle III. Linhas azuis indicam interações proteína-proteína. A espessura das linhas indica a força
das associações. Mapa montado utilizando o software STITCH v. 4.0.
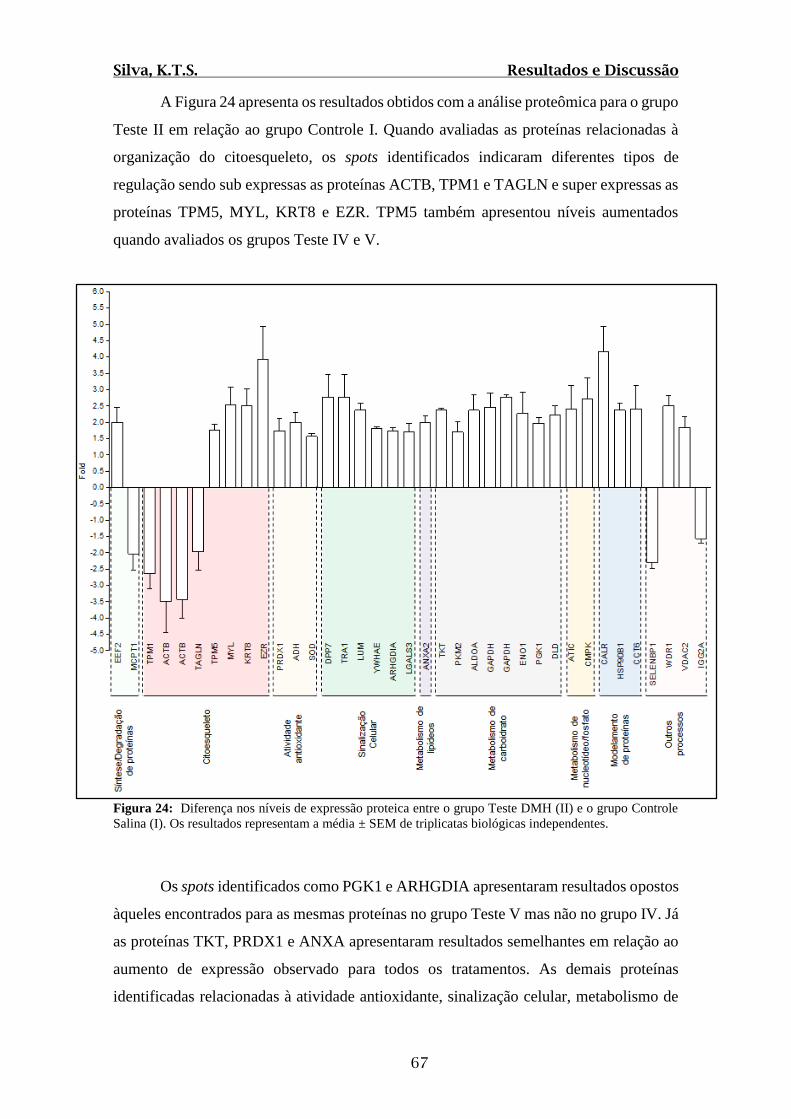
Silva, K.T.S. Resultados e Discussão
67
A Figura 24 apresenta os resultados obtidos com a análise proteômica para o grupo
Teste II em relação ao grupo Controle I. Quando avaliadas as proteínas relacionadas à
organização do citoesqueleto, os spots identificados indicaram diferentes tipos de
regulação sendo sub expressas as proteínas ACTB, TPM1 e TAGLN e super expressas as
proteínas TPM5, MYL, KRT8 e EZR. TPM5 também apresentou níveis aumentados
quando avaliados os grupos Teste IV e V.
Figura 24: Diferença nos níveis de expressão proteica entre o grupo Teste DMH (II) e o grupo Controle
Salina (I). Os resultados representam a média ± SEM de triplicatas biológicas independentes.
Os spots identificados como PGK1 e ARHGDIA apresentaram resultados opostos
àqueles encontrados para as mesmas proteínas no grupo Teste V mas não no grupo IV. Já
as proteínas TKT, PRDX1 e ANXA apresentaram resultados semelhantes em relação ao
aumento de expressão observado para todos os tratamentos. As demais proteínas
identificadas relacionadas à atividade antioxidante, sinalização celular, metabolismo de

Silva, K.T.S. Resultados e Discussão
68
carboidrato, lipídeos, de nucleotídeo/fosfato e modelamento de proteínas apresentaram
níveis aumentados de expressão celular.
Assim como encontrado para os grupos Teste IV e V, diversas proteínas
relacionadas ao metabolismo de carboidratos (TKT, PKM2, ALDOA, GAPDH, ENO1,
PGK1, DLD) e a atividade antioxidante (PRDX1, ADH e SOD) mostraram aumento nos
níveis de expressão.
A proteína TAGLN está relacionada à manutenção da organização dos filamentos
de actina in vivo e a diminuição de sua expressão por inativação via Ras pode ser
observada em linhagens celulares tumorais como mama e cólon, sendo considerada um
marcador no início da transformação tumoral (Zhang et al., 2010; Thompson et al., 2012).
Em concordância com o achado de Mlakar e colaboradores observamos a sub expressão
da isoforma TPM1 em nossas amostras de CCR (Mlakar et al., 2009). Alterações no nível
de expressão da proteína EZR estão relacionados com a ocorrência de metástase e
prognóstico ruim em diversos tipos de câncer. Tal proteína interage com diversas
moléculas de sinalização intracelular envolvidas na progressão tumoral como receptor
Met de fator de crescimento de hepatócito (HGF), integrina β4 e membros da família de
quinases Src (Ghaffari et al., 2014).
Em relação à síntese e degradação de proteínas observou-se um aumento nos
níveis de EEF2 que realiza a translocação da cadeia polipeptídica crescente do sítio A ao
sítio P do ribossomo. A super expressão dessa proteína foi relatada em tumores gástricos,
intestinais, entre outros e parece desempenhar também um papel regulatório na
progressão do ciclo celular nas fases G2/M (Oji et al., 2014). MCPT1 é uma protease
envolvida na degradação de matriz extracelular e favorecimento da angiogênese. Alguns
trabalhos vêm relacionando os níveis de expressão desta e de outras proteases secretadas
com o comportamento tumoral. Foi observado que a presença de mastócitos MCPT1
negativos atuam inibindo o crescimento tumoral e a angiogênese por meio da liberação
de diferentes estímulos (Dyduch et al., 2012).
É importante ressaltar o aumento observado na expressão de diversas proteínas de
sinalização celular. Entre elas, TRA1 se destaca por ser um componente integral dos
complexos histona acetiltransferase STAGA, TFTC e PCAF, equivalentes encontrados
em mamíferos para SAGA, e TIP60, o equivalente para NuA4 de leveduras. TRA1 é um
membro da família de proteínas quinase fosfatidilinositol 3-quinase (PI3K), responsáveis
por regular diversas vias de sinalização por fosforilação de serina/treoninas além de ser
alvo de fatores de transcrição oncogênicos que incluem c-myc, E2F e E1a (Esseghir et

Silva, K.T.S. Resultados e Discussão
69
al., 2007; Helmlinger et al., 2011; Knutson and Hahn, 2011). A proteína de matriz
extracelular LUM está relacionada com a modulação do comportamento celular incluindo
migração e proliferação através da reorganização do citoesqueleto de actina e
desestabilização de complexos de adesão celular. Diversos estudos relatam a super
expressão desta proteína em tumores no útero, pâncreas e cólon intestinal, aparentemente
relacionada a estágios iniciais do desenvolvimento (Lu et al., 2012; Seya et al., 2006;
Pietraszek et al., 2013; Nikitovic et al., 2014).
A família de proteínas regulatórias 14-3-3 (YWHAE) possui mais de 100 ligantes
descritos incluindo moléculas de sinalização celular como a proteína quinase C (PKC),
PI3K e fosfatase cdc25 de maneira dependente de fosforilação. Sua ligação à Raf é
indispensável para a via de sinalização Ras/MAPK, enquanto a ligação à Bcl-2 está
relacionada à inibição de apoptose. A isoforma ε liga-se a p190RhoGEF através de
interação independente de fosforilação e modula a formação do citoesqueleto via FAK.
YWHAE demonstra super expressão nos cânceres de mama, pulmão, tireóide e cólon
sendo que a presença da isoforma ε parece proteger as células tumorais e endoteliais de
cólon de apoptose induzida por estresse oxidativo (Zhai et al., 2003; Satoh et al., 2004;
Liu et al., 2013).
A proteína Galectina-3 (LGALS3) atua como fator de splicing de pré-mRNAs no
núcleo além de estar envolvida na resposta inflamatória aguda incluindo ativação e adesão
de neutrófilos, quimioatração de monócitos e ativação de mastócitos. Parece participar da
regulação de diversas vias de sinalização celular: (1) envolvimento na translocação
nuclear de β-catenina juntamente com as proteínas importina-α/β, nucleoporinas e
FoxM1, na fosforilação de GSK3 e ativação de TCF na via Wnt/β-catenina; (2)
envolvimento na via de sinalização Notch através da interação com a proteína de
membrana Tumor-derived jagged-1 (Laubli et al, 2014; Nakajima et al., 2014; Shim et
al., 2015).
Como pode ser observado na Figura 25, mais de 85% das proteínas que
apresentaram algum grau de alteração na expressão encontram-se relacionadas
intracelularmente, reforçando a ideia de que elas participam de vias de sinalização ligadas
ao desenvolvimento tumoral nos animais estudados.
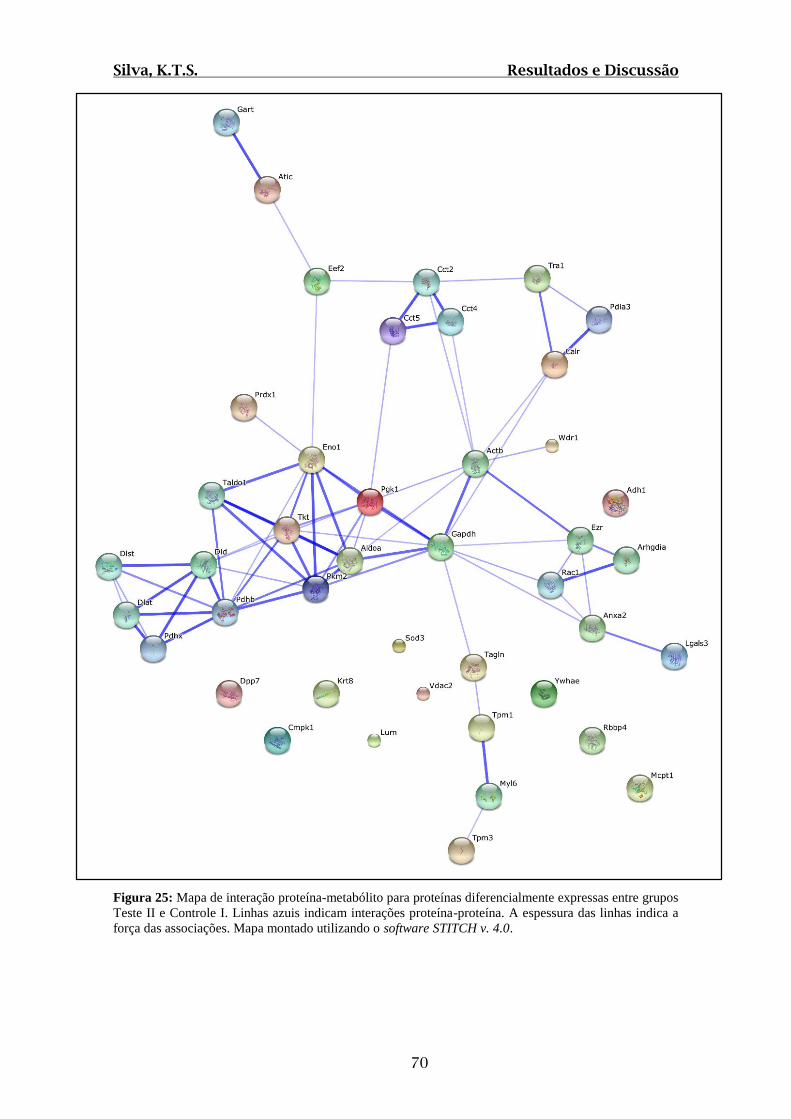
Silva, K.T.S. Resultados e Discussão
70
Figura 25: Mapa de interação proteína-metabólito para proteínas diferencialmente expressas entre grupos
Teste II e Controle I. Linhas azuis indicam interações proteína-proteína. A espessura das linhas indica a
força das associações. Mapa montado utilizando o software STITCH v. 4.0.
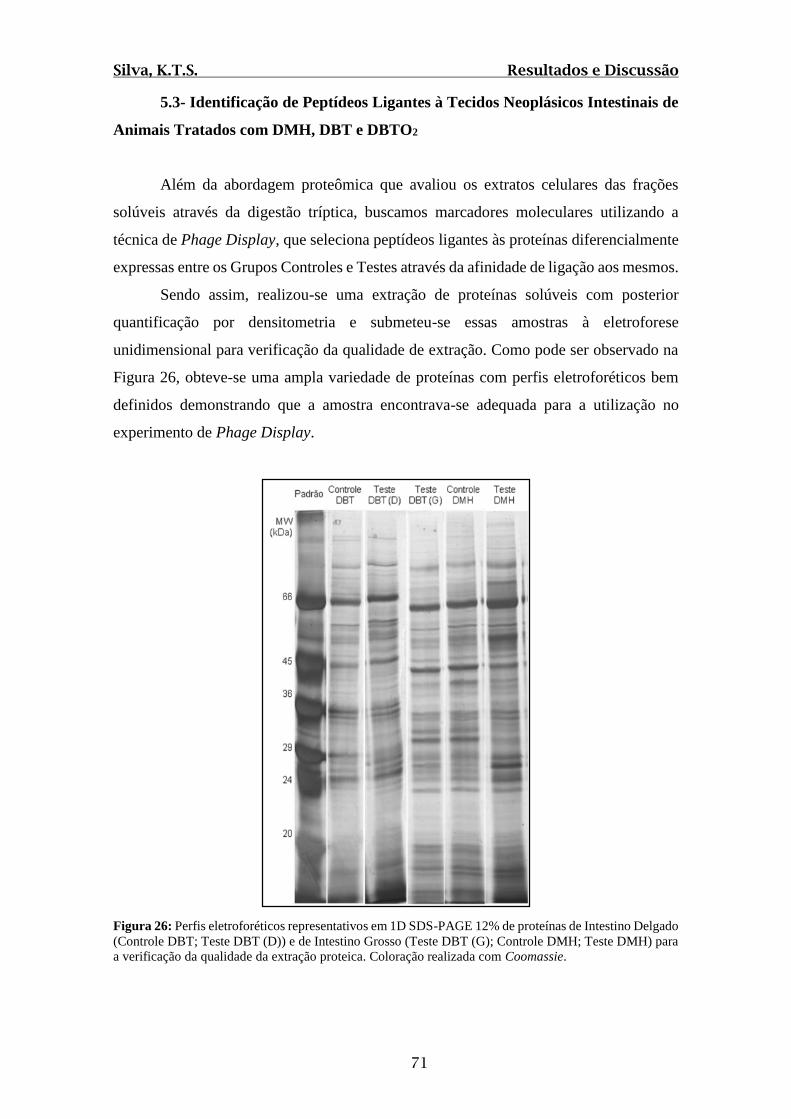
Silva, K.T.S. Resultados e Discussão
71
5.3- Identificação de Peptídeos Ligantes à Tecidos Neoplásicos Intestinais de
Animais Tratados com DMH, DBT e DBTO2
Além da abordagem proteômica que avaliou os extratos celulares das frações
solúveis através da digestão tríptica, buscamos marcadores moleculares utilizando a
técnica de Phage Display, que seleciona peptídeos ligantes às proteínas diferencialmente
expressas entre os Grupos Controles e Testes através da afinidade de ligação aos mesmos.
Sendo assim, realizou-se uma extração de proteínas solúveis com posterior
quantificação por densitometria e submeteu-se essas amostras à eletroforese
unidimensional para verificação da qualidade de extração. Como pode ser observado na
Figura 26, obteve-se uma ampla variedade de proteínas com perfis eletroforéticos bem
definidos demonstrando que a amostra encontrava-se adequada para a utilização no
experimento de Phage Display.
Figura 26: Perfis eletroforéticos representativos em 1D SDS-PAGE 12% de proteínas de Intestino Delgado
(Controle DBT; Teste DBT (D)) e de Intestino Grosso (Teste DBT (G); Controle DMH; Teste DMH) para
a verificação da qualidade da extração proteica. Coloração realizada com Coomassie.

Silva, K.T.S. Resultados e Discussão
72
5.3.1- Biopanning de Tecido Intestinal
A seleção de peptídeos foi realizada utilizando-se uma biblioteca expressa na
superfície de fagos filamentosos M13. Para tal, utilizaram-se 3 etapas de subtração
empregando amostras proteicas de tecidos intestinais de grupos Controle e Testes para
DMH e DBT. Como as alterações apresentadas pelo grupo Teste DBTO2 não
apresentaram diferenças substanciais do grupo Teste DBT diante da abordagem
proteômica, e, presumivelmente este tratamento promove alterações moleculares através
de vias semelhantes, o primeiro não foi utilizado para seleção de peptídeos por essa
técnica a fim de se simplificar o processo.
Finalizados os 3 ciclos de seleção para extratos provenientes do intestino grosso
dos grupos Controle/Teste DMH, intestino grosso Controle/Teste DBT e intestino
delgado Controle/Teste DBT, as colônias de ER2738 contendo os fagos relativos ao
último ciclo de seleção foram plaqueadas e tituladas para que se pudesse obter fagos
isolados para extração e sequenciamento do DNA. Como pode ser observado na Figura
27, não houve aparecimento de colônias brancas indicando ausência de contaminação das
bactérias por fagos do tipo selvagem, demonstrando que a infecção das bactérias ER2738
foi adequada (colônias azuis indicando a presença de fagos M13 que carregam o gene da
β-galactosidase).
Selecionaram-se de maneira randômica 32 colônias isoladas pertencentes a cada
grupo avaliado para nova amplificação e extração e sequenciamento do DNA a fim de se
obter as sequências de peptídeos específicas ligantes para os grupos testes. O DNA
extraído foi submetido à eletroforese em gel de agarose 0,8% para verificar sua qualidade
e estimar sua concentração. Realizou-se então a reação de PCR para amplificação do
DNA e sequenciamento utilizando-se o sequenciador automático MegaBACE 1000.
Foram obtidas ao todo 27 sequências para o grupo DMH sendo 16 delas sequências únicas
(não repetidas), 25 sequências para o grupo DBT intestino grosso, sendo 10 delas únicas
e 29 sequências para o grupo DBT intestino delgado sendo 11 únicas.
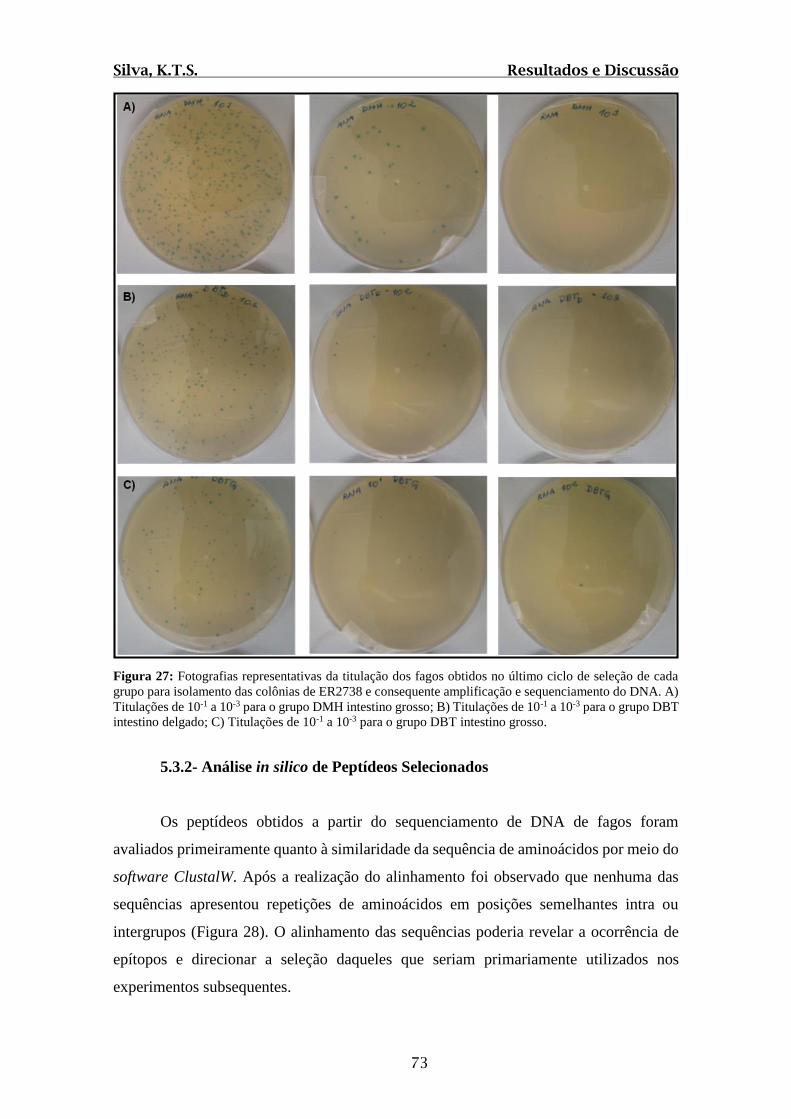
Silva, K.T.S. Resultados e Discussão
73
Figura 27: Fotografias representativas da titulação dos fagos obtidos no último ciclo de seleção de cada
grupo para isolamento das colônias de ER2738 e consequente amplificação e sequenciamento do DNA. A)
Titulações de 10-1 a 10-3 para o grupo DMH intestino grosso; B) Titulações de 10-1 a 10-3 para o grupo DBT
intestino delgado; C) Titulações de 10-1 a 10-3 para o grupo DBT intestino grosso.
5.3.2- Análise in silico de Peptídeos Selecionados
Os peptídeos obtidos a partir do sequenciamento de DNA de fagos foram
avaliados primeiramente quanto à similaridade da sequência de aminoácidos por meio do
software ClustalW. Após a realização do alinhamento foi observado que nenhuma das
sequências apresentou repetições de aminoácidos em posições semelhantes intra ou
intergrupos (Figura 28). O alinhamento das sequências poderia revelar a ocorrência de
epítopos e direcionar a seleção daqueles que seriam primariamente utilizados nos
experimentos subsequentes.

Silva, K.T.S. Resultados e Discussão
74
Figura 28: Alinhamento das sequências peptídicas obtidas após sequenciamento do DNA dos fagos
selecionados randomicamente ao fim do terceiro ciclo de biopanning para os três grupos avaliados.
Utilizado o software ClustalW. As cores iguais indicam a similaridade entre os resíduos alinhados.
Escolheram-se as sequências para a realização dos testes posteriores com base na
frequência de repetição das mesmas dentro de cada grupo e também na relação que cada
peptídeo apresentou com proteínas relacionadas ao câncer quando sua sequência foi
alinhada com sequências de proteínas através do software BLASTp e base de dados
RefSeq. Através de busca na literatura utilizando o banco de dados NCBI pode-se avaliar
a relação entre as proteínas que apresentaram regiões de similaridade com as sequências
peptídicas e diversos tipos de câncer. Considerando que os peptídeos selecionados com
base na frequência de repetição apresentaram de maneira geral, alinhamento com
proteínas que desempenham importantes papéis no desenvolvimento tumoral já relatado
por diversos pesquisadores (Apêndice F – Tab. 03.F), julgamos que a escolha de tais
peptídeos para a síntese e realização de testes posteriores, ainda que primariamente, é
adequada à realização dos objetivos propostos neste trabalho.
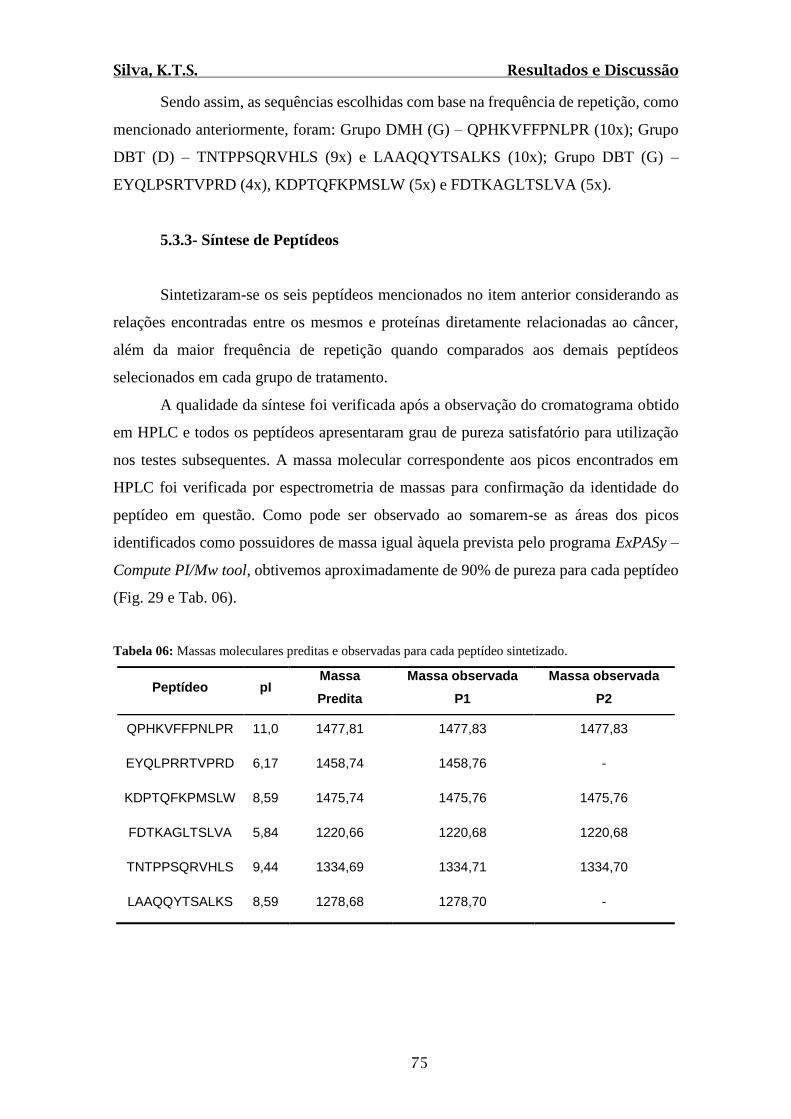
Silva, K.T.S. Resultados e Discussão
75
Sendo assim, as sequências escolhidas com base na frequência de repetição, como
mencionado anteriormente, foram: Grupo DMH (G) – QPHKVFFPNLPR (10x); Grupo
DBT (D) – TNTPPSQRVHLS (9x) e LAAQQYTSALKS (10x); Grupo DBT (G) –
EYQLPSRTVPRD (4x), KDPTQFKPMSLW (5x) e FDTKAGLTSLVA (5x).
5.3.3- Síntese de Peptídeos
Sintetizaram-se os seis peptídeos mencionados no item anterior considerando as
relações encontradas entre os mesmos e proteínas diretamente relacionadas ao câncer,
além da maior frequência de repetição quando comparados aos demais peptídeos
selecionados em cada grupo de tratamento.
A qualidade da síntese foi verificada após a observação do cromatograma obtido
em HPLC e todos os peptídeos apresentaram grau de pureza satisfatório para utilização
nos testes subsequentes. A massa molecular correspondente aos picos encontrados em
HPLC foi verificada por espectrometria de massas para confirmação da identidade do
peptídeo em questão. Como pode ser observado ao somarem-se as áreas dos picos
identificados como possuidores de massa igual àquela prevista pelo programa ExPASy –
Compute PI/Mw tool, obtivemos aproximadamente de 90% de pureza para cada peptídeo
(Fig. 29 e Tab. 06).
Tabela 06: Massas moleculares preditas e observadas para cada peptídeo sintetizado.
Peptídeo pI Massa
Predita
Massa observada
P1
Massa observada
P2
QPHKVFFPNLPR 11,0 1477,81 1477,83 1477,83
EYQLPRRTVPRD 6,17 1458,74 1458,76 -
KDPTQFKPMSLW 8,59 1475,74 1475,76 1475,76
FDTKAGLTSLVA 5,84 1220,66 1220,68 1220,68
TNTPPSQRVHLS 9,44 1334,69 1334,71 1334,70
LAAQQYTSALKS 8,59 1278,68 1278,70 -
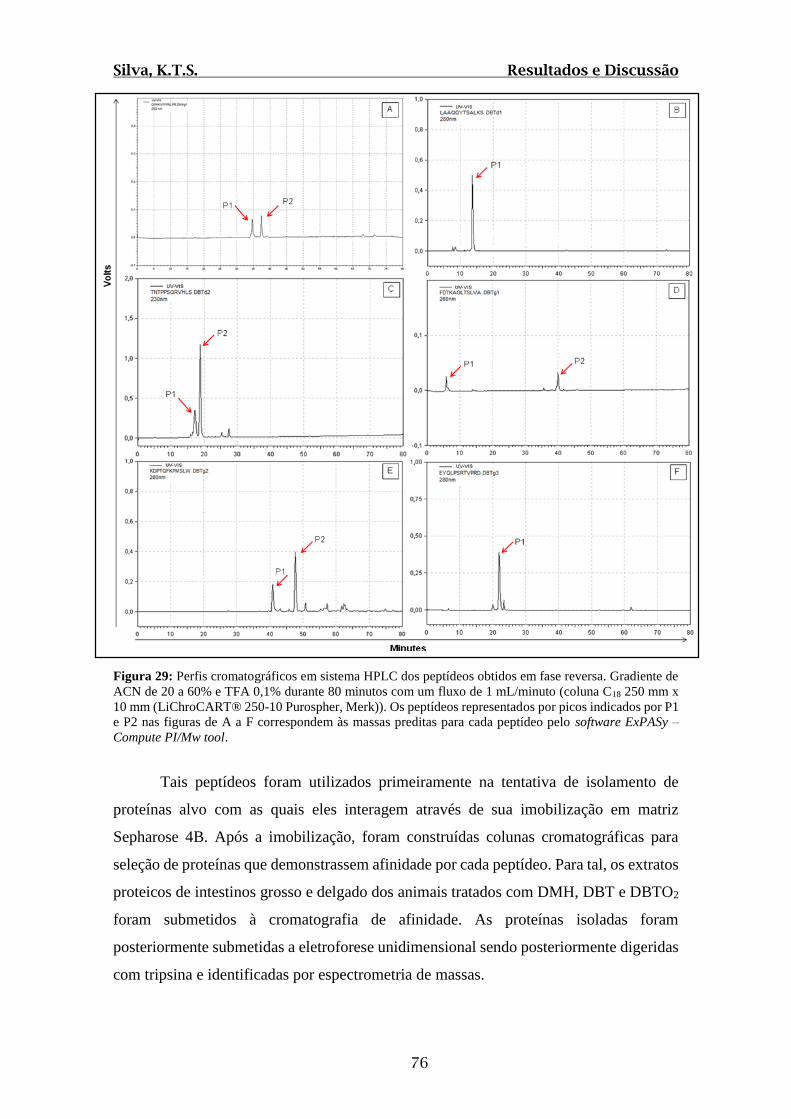
Silva, K.T.S. Resultados e Discussão
76
Figura 29: Perfis cromatográficos em sistema HPLC dos peptídeos obtidos em fase reversa. Gradiente de
ACN de 20 a 60% e TFA 0,1% durante 80 minutos com um fluxo de 1 mL/minuto (coluna C18 250 mm x
10 mm (LiChroCART® 250-10 Purospher, Merk)). Os peptídeos representados por picos indicados por P1
e P2 nas figuras de A a F correspondem às massas preditas para cada peptídeo pelo software ExPASy –
Compute PI/Mw tool.
Tais peptídeos foram utilizados primeiramente na tentativa de isolamento de
proteínas alvo com as quais eles interagem através de sua imobilização em matriz
Sepharose 4B. Após a imobilização, foram construídas colunas cromatográficas para
seleção de proteínas que demonstrassem afinidade por cada peptídeo. Para tal, os extratos
proteicos de intestinos grosso e delgado dos animais tratados com DMH, DBT e DBTO2
foram submetidos à cromatografia de afinidade. As proteínas isoladas foram
posteriormente submetidas a eletroforese unidimensional sendo posteriormente digeridas
com tripsina e identificadas por espectrometria de massas.
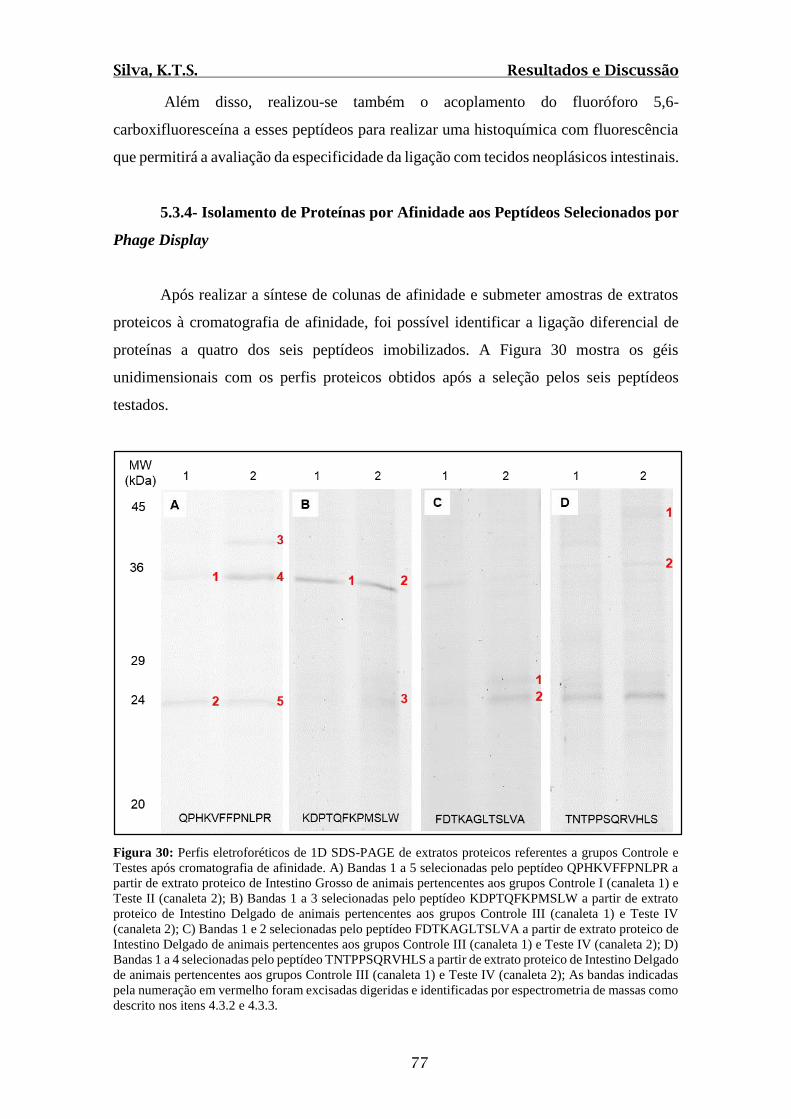
Silva, K.T.S. Resultados e Discussão
77
Além disso, realizou-se também o acoplamento do fluoróforo 5,6-
carboxifluoresceína a esses peptídeos para realizar uma histoquímica com fluorescência
que permitirá a avaliação da especificidade da ligação com tecidos neoplásicos intestinais.
5.3.4- Isolamento de Proteínas por Afinidade aos Peptídeos Selecionados por
Phage Display
Após realizar a síntese de colunas de afinidade e submeter amostras de extratos
proteicos à cromatografia de afinidade, foi possível identificar a ligação diferencial de
proteínas a quatro dos seis peptídeos imobilizados. A Figura 30 mostra os géis
unidimensionais com os perfis proteicos obtidos após a seleção pelos seis peptídeos
testados.
Figura 30: Perfis eletroforéticos de 1D SDS-PAGE de extratos proteicos referentes a grupos Controle e
Testes após cromatografia de afinidade. A) Bandas 1 a 5 selecionadas pelo peptídeo QPHKVFFPNLPR a
partir de extrato proteico de Intestino Grosso de animais pertencentes aos grupos Controle I (canaleta 1) e
Teste II (canaleta 2); B) Bandas 1 a 3 selecionadas pelo peptídeo KDPTQFKPMSLW a partir de extrato
proteico de Intestino Delgado de animais pertencentes aos grupos Controle III (canaleta 1) e Teste IV
(canaleta 2); C) Bandas 1 e 2 selecionadas pelo peptídeo FDTKAGLTSLVA a partir de extrato proteico de
Intestino Delgado de animais pertencentes aos grupos Controle III (canaleta 1) e Teste IV (canaleta 2); D)
Bandas 1 a 4 selecionadas pelo peptídeo TNTPPSQRVHLS a partir de extrato proteico de Intestino Delgado
de animais pertencentes aos grupos Controle III (canaleta 1) e Teste IV (canaleta 2); As bandas indicadas
pela numeração em vermelho foram excisadas digeridas e identificadas por espectrometria de massas como
descrito nos itens 4.3.2 e 4.3.3.

Silva, K.T.S. Resultados e Discussão
78
Na Figura 30 – A podemos observar cinco bandas de proteínas isoladas a partir de
cromatografia de afinidade utilizando o peptídeo QPHKVFFPNLPR e extratos proteicos
provenientes de intestino grosso dos animais Controle I e Teste II. A identificação das
proteínas por espectrometria de massas, revelou que as bandas 1 e 4 são compostas
principalmente por tropomiosina e as bandas 2 e 6 pela proteína ribossomal L10A. A
banda 3 visível somente na canaleta 2 e referente ao grupo Teste II é composta pela
proteína SET. Esta proteína pertence à classe das metiltransferases que estão envolvidas
na biossíntese de lipídeos, inativação hormonal, diferenciação tecidual e no controle
epigenético (Lehman et al., 2008; Albert and Helin, 2010; Eom et al., 2011). Uma parcela
significante de nossas proteínas (208 proteínas humanas) compreende as enzimas que
catalisam a transferência do grupo metil do cofator S-adenosil-L-metionina para um
substrato. Destas, aproximadamente 30% estão associadas a doenças sendo mais
frequentemente ligadas a desordens do sistema nervoso central e câncer (Martin and
McMillan, 2002; Schubert et al., 2003; Petrossian and Clarke, 2011).
Com base em seus arranjos estruturais, as metiltransferases podem ser agrupadas
em duas classes: membros que possuem o domínio SET e membros que possuem o
domínio DOT1 (Jones and Gelbart, 1993; Tschiersch et al., 1994; Stassen et al., 1995).
A função regulatória da metilação de proteínas não é restrita ao código de histonas, sendo
vinculada a diversos processos celulares. Esta modificação pode ocorrer também em
proteínas citosólicas e nucleares, sendo que os efeitos destas metilações ainda pouco
caracterizados (Egorova et al., 2010; Zhang et al., 2012).
Uma das modificações mais estudadas é a metilação do fator supressor de tumor
p53. Esta proteína possui papel central no controle do ciclo celular apoptose e reparo de
DNA em resposta a vários tipos de estresse (Brooks and Gu, 2010). A metilação dos
resíduos de lisina presentes no domínio regulatório C-terminal regula positiva e
negativamente sua função. A metilação realizada por SET7/9 em Lys372 promove
aumento de expressão dos genes alvo de p53. Em contraste, a metilação de Lys382 por
SET8 e Lys370 por SYMD2 suprime a atividade transcricional (Chuikov et al., 2004;
Huang et al., 2006; Shi et al., 2007).
A banda 3, referente à Figura 30 – B, apresentou ligação diferencial entre controle
e teste sendo identificada como histona H4 e H2. A unidade básica da cromatina
eucariótica, o nucleossomo, consiste em uma volta de DNA sobre um octâmero de
histonas (dímeros H2A/H2B e tetrâmero H3/H4). Modificações covalentes nestas
proteínas podem alterar o grau de compactação do DNA, inativar a eucromatina ou ativar

Silva, K.T.S. Resultados e Discussão
79
a heterocromatina. Essas modificações ocorrem na região N-terminal das histonas, são
reversíveis e incluem acetilação, metilação, fosforilação e ubiquitinação. Diversas
famílias de proteínas podem realizar tais modificações (acetiltransferases, deacetilases,
metiltransferases, entre outras) (Wong et al., 2007).
Geralmente a estrutura ativa da cromatina que corresponde a atividade
transcricional aumentada está associada a aumento na acetilação de histonas. Um padrão
de alterações nas histonas H4 caracterizado pela perda de monoacetilação em Lys16 e de
trimetilação em Lys20 tem sido proposto como marcador universal de transformação
maligna. Além disso, as histonas podem ser substituídas por variantes com pequenas
variações de sequência que levam a alterações no padrão de expressão como por exemplo
a substituição de H2A por H2A.Z (Goel and Boland, 2012; Jia and Guo, 2013).
Ambos os extratos utilizados para cromatografia com o peptídeo
KDPTQFKPMSLW mostraram interação entre o peptídeo e a proteína dermicidina
(canaleta 1 e 2 – B). Tal proteína possui propriedades antimicrobianas e é
constitutivamente expressa em humanos pelas glândulas sudoríparas, leucócitos
periféricos, células neuronais e placenta. Ela pode estar também relacionada a outras
doenças humanas como câncer e aterosclerose (Schittek, 2012; Wang, 2014).
A cromatografia de afinidade com o peptídeo FDTKAGLTSLVA isolou as
proteínas ribossomais 60S P0 e L10A (Fig. 30 – C, 1 e 2). P0 é uma fosfoproteína
ribossomal que forma heterodímeros com as fosfoproteínas acídicas P1 e P2 localizadas
na parte ativa do ribossomo. O complexo formado atua interagindo com mRNAs, tRNAs
e fatores de tradução (EF2) sendo, a presença de P0, mas não de P1 e P2, essencial para
esta função. O aumento na expressão de P0 foi relatado em células tumorais de cólon e
fígado sugerindo que esta proteína exerça outras funções celulares além da tradução de
proteínas (Barnard et al., 1992; Kondoh et al., 1999). Chang e colaboradores
demonstraram que P0 pode interagir com a proteína GCIP (Grap2 and Cyclin D
Interacting Protein) levando ao aumento de proliferação celular por inibição da redução
de fosforilação de pRb (Retinoblastome protein) e aumentando a expressão de ciclina D1
(Chang et al., 2008).
Por último, após espectrometria de massas, foi possível identificar as proteínas
asporina e caseína quinase II (Fig. 30 – D, 1 e 2) isoladas por cromatografia utilizando o
peptídeo TNTPPSQRVHLS como ligante. A asporina é um proteoglicano de matriz
extracelular rica em leucinas. O aumento de expressão dessa proteína e de outros
proteoglicanos como decorina e biglicano foi encontrado em diversos pacientes com

Silva, K.T.S. Resultados e Discussão
80
câncer gástrico, pancreático, de mama e próstata. Quadros de inflamação também
demonstram elevação na expressão dessa molécula. A asporina parece coordenar invasão
de células cancerosas no tecido normal adjacente por meio da ativação de Rac1 via
interação com CD44 (Nakajima et al., 2007; Turtoi et al., 2011; Ding et al., 2015;
Satoyoshi et al., 2015).
Níveis elevados da proteína caseína quinase 2 (CK2) foram encontrados em
tumores de mama, próstata, rins, coloretal, cabeça e pescoço e pulmão. Sabe-se que essa
proteína está envolvida em diversas vias de sinalização que regulam a proliferação
celular, diferenciação e apoptose. A CK2 parece regular a via Wnt em múltiplos níveis
sendo capaz de fosforilar o co-fator β-catenina, Dvl, APC e também o fator de transcrição
TCF/LEF. Além disso, associações funcionais sugerem que CK2, NF-κB e a via Wnt
podem atuar em conjunto na promoção da tumorogênese na glândula mamária. CK2
promove degradação constitutiva de IκB e ativação de NF-κB, atuando também em
pontos posteriores da via (Dominguez et al., 2009; Montenarh, 2014).

Conclusões

Silva, K.T.S. Conclusões
82
6- CONCLUSÕES
O tratamento crônico a que os animais foram submetidos permitiu as seguintes
conclusões:
- Os perfis de células sanguíneas e de enzimas utilizadas na avaliação da função
hepática e pancreática não foram alterados, corroborando com a análise histológica e
observação de esfregaços sanguíneos;
- A dose de 30 mg/Kg utilizada para as substâncias DMH, DBT e DBTO2 foi
suficiente para induzir alterações de caráter neoplásico nos intestinos dos animais
tratados, porém, não provocou alterações hepáticas ou nos demais tecidos avaliados
histologicamente (estômago, pulmão e pâncreas);
- As lesões identificadas pelas análises histopatológicas dos intestinos grosso e
delgado indicaram a ocorrência de processo neoplásico em fase inicial de
desenvolvimento tanto para os animais tratados com DBT e DBTO2 quanto para os
animais tratados com DMH com características e intensidade similares. Tal fato situa os
agentes DBT e DBTO2 como indutores de câncer de expressão toxicológica;
- A análise comparativa dos perfis eletroforéticos dos extratos proteicos totais dos
grupos Controles e Testes revelou diferenças na expressão de algumas proteínas, levando
a identificação de marcadores proteicos para as alterações celulares consistentes com
alterações neoplásicas, decorrentes da exposição a esses agentes;
- A identificação de proteínas ligantes a peptídeos selecionados por Phage Display
por cromatografia de afinidade, juntamente com a identificação de proteínas
diferencialmente expressas selecionadas por 2D SDS-PAGE, proporcionou um avanço
no entendimento acerca das alterações bioquímicas e possíveis mecanismos envolvidos
no estabelecimento do processo neoplásico promovido pelo DBT e DBTO2. Foi possível
observar alterações em vias de sinalização gerais de maneira comum aos 3 grupos Teste
avaliados.

Perspectivas

Silva, K.T.S. Perspectivas
84
7- PERSPECTIVAS
Considerando a limitação no diagnóstico de lesões tumorais decorrentes de
indução química com os agentes estudados, e a importância ambiental dos mesmos, torna-
se atraente a busca por novos marcadores proteicos e o desenvolvimento de métodos
diagnósticos para a detecção dos mesmos. Sendo assim, pretende-se avançar com os
experimentos utilizando os peptídeos selecionados por Phage Display através da
realização de histoquímica com peptídeos fluorescentes. Pretende-se também empregar
tais peptídeos na avaliação de uma possível atividade antitumoral utilizando cultura de
células tumorais.

Referências Bibliográficas

Silva, K.T.S. Referências Bibliográficas
86
8- REFERÊNCIAS BIBLIOGRÁFICAS
Aas, E., Baussant, T., Balk, L., Liewenborg, B. and Andersen, O.K., 2000. PAH
metabolites in bile, cytochrome P4501A and DNA adducts as environmental risk
parameters for chronic oil exposure: a laboratory experiment with Atlantic cod.
Aquatic toxicology, 51(2): 241-58.
Adra, C.N. et al., 1997. RhoGDIgamma: a GDP-dissociation inhibitor for Rho proteins
with preferential expression in brain and pancreas. Proceedings of the National
Academy of Sciences of the United States of America, 94(9): 4279-84.
Ahlquist, D.A., 2010. Molecular detection of colorectal neoplasia. Gastroenterology,
138(6): 2127-39.
Albert, M. and Helin, K., 2010. Histone methyltransferases in cancer. Seminars in cell &
developmental biology, 21(2): 209-20.
Anderson, F.H., Zeng, L., Rock, N.R. and Yoshida, E.M., 2000. An assessment of the
clinical utility of serum ALT and AST in chronic hepatitis C. Hepatology research
: the official journal of the Japan Society of Hepatology, 18(1): 63-71.
Amat, A., Pfohl-Leszkowicz, A., Castegnaro, M., 2004. Genotoxic activity of thiophenes
on liver human cell line (HepG2). Polycyclic Aromatic Compounds, 24: 733−742.
Angerer, J., Mannschreck, C. and Gundel, J., 1997. Biological monitoring and
biochemical effect monitoring of exposure to polycyclic aromatic hydrocarbons.
International archives of occupational and environmental health, 70(6): 365-77.
Arimura, S. et al., 2009. Reduced level of smoothened suppresses intestinal tumorigenesis
by down-regulation of Wnt signaling. Gastroenterology, 137(2): 629-38.
Babich, I.V. and Moulijn, J.A., 2003. Science and technology of novel processes for deep
desulfurization of oil refinery streams: a review. Fuel, 82(6): 607-631.
Bale, A.E. and Yu, K.P., 2001. The hedgehog pathway and basal cell carcinomas. Human
molecular genetics, 10(7): 757-62.
Barnard, G.F. et al., 1992. Increased expression of human ribosomal phosphoprotein P0
messenger RNA in hepatocellular carcinoma and colon carcinoma. Cancer
research, 52(11): 3067-72.
Bellati, F. et al., 2009. Immunology of gynecologic neoplasms: analysis of the prognostic
significance of the immune status. Current cancer drug targets, 9(4): 541-65.
Brabletz, T. et al., 1998. Nuclear overexpression of the oncoprotein beta-catenin in
colorectal cancer is localized predominantly at the invasion front. Pathology,
research and practice, 194(10): 701-4.
Bravo-Cordero, J.J., Magalhaes, M.A., Eddy, R.J., Hodgson, L. and Condeelis, J., 2013.
Functions of cofilin in cell locomotion and invasion. Nature reviews. Molecular
cell biology, 14(7): 405-15.
Brooks, C.L. and Gu, W., 2010. New insights into p53 activation. Cell research, 20(6):
614-21.
Brosens, L.A. et al., 2007. Gastrointestinal polyposis syndromes. Current molecular
medicine, 7(1): 29-46.

Silva, K.T.S. Referências Bibliográficas
87
Brown, K.K. and Toker, A., 2015. The phosphoinositide 3-kinase pathway and therapy
resistance in cancer. F1000prime reports, 7: 13.
Burtis, C.A. and Ashwood, E.R., 1996. Tietz fundamentals of clinical chemistry.
Philadelphia [etc.], W. B. Saunders Company.
Cardoso, J., Boer, J., Morreau, H. and Fodde, R., 2007. Expression and genomic profiling
of colorectal cancer. Biochimica et biophysica acta, 1775(1): 103-37.
Castro-Castro, A. et al., 2011. Coronin 1A promotes a cytoskeletal-based feedback loop
that facilitates Rac1 translocation and activation. The EMBO journal, 30(19):
3913-27.
Cavret, S. and Feidt, C., 2005. Intestinal metabolism of PAH: in vitro demonstration and
study of its impact on PAH transfer through the intestinal epithelium.
Environmental research, 98(1): 22-32.
Cecconello, I., 2008. Atualização em cirurgia do aparelho digestivo e em coloproctologia.
São Paulo, Departamento de gastroenterologia da Faculdade de Medicina da
Universidade de São Paulo.
Cecconi, D. and Zamò, A., 2011. Proteomics of human cancer tissues and cells. TrAC
Trends in Analytical Chemistry, 30(2): 346-359.
Chang, H. et al., 2005. Using a combination of cytochrome P450 1B1 and beta-catenin
for early diagnosis and prevention of colorectal cancer. Cancer detection and
prevention, 29(6): 562-9.
Chang, T.W. et al., 2008. Ribosomal phosphoprotein P0 interacts with GCIP and
overexpression of P0 is associated with cellular proliferation in breast and liver
carcinoma cells. Oncogene, 27(3): 332-8.
Chang, W.W., 1984. Histogenesis of colon cancer in experimental animals. Scandinavian
journal of gastroenterology. Supplement, 104: 27-43.
Chen, J. and Huang, X.F., 2009. The signal pathways in azoxymethane-induced colon
cancer and preventive implications. Cancer biology & therapy, 8(14): 1313-7.
Cho, M. et al., 1997. Mucins secreted by cell lines derived from colorectal mucinous
carcinoma and adenocarcinoma. European journal of cancer, 33(6): 931-41.
Cho, W.C., 2007. Contribution of oncoproteomics to cancer biomarker discovery.
Molecular cancer, 6: 25.
Chuikov, S. et al., 2004. Regulation of p53 activity through lysine methylation. Nature,
432(7015): 353-60.
Chuthapisith, S., Layfield, R., Kerr, I.D. and Eremin, O., 2007. Principles of proteomics
and its applications in cancer. The surgeon : journal of the Royal Colleges of
Surgeons of Edinburgh and Ireland, 5(1): 14-22.
Ciganek, M., Neca, J, Adamec, V., Janosek, J., Machala, M., 2004. A combined chemical
and bioassay analysis of traffic-emitted polycyclic aromatic hydrocarbons. Sci.
Total Environ., 334−335, 141−148.
Coppola, D. et al., 1998. CD44V6 expression in human colorectal carcinoma. Human
pathology, 29(6): 627-35.
Croce, C.M., 2008. Oncogenes and cancer. The New England journal of medicine,
358(5): 502-11.

Silva, K.T.S. Referências Bibliográficas
88
da Silva Madeira, L., Ferreira-Leitão, V.S., et al., 2008. Dibenzothiophene oxidation by
horseradish peroxidase in organic media: Effect of the DBT:H2O2 molar ratio and
H2O2 addition mode. Chemosphere, 71(1): 189-194.
de Paula Carli, A. et al., 2012. Bowman-Birk inhibitors, proteasome peptidase activities
and colorectal pre neoplasias induced by 1,2-dimethylhydrazine in Swiss mice.
Food and chemical toxicology : an international journal published for the British
Industrial Biological Research Association, 50(5): 1405-12.
Diggs, D.L. et al., 2011. Polycyclic aromatic hydrocarbons and digestive tract cancers: a
perspective. Journal of environmental science and health. Part C, Environmental
carcinogenesis & ecotoxicology reviews, 29(4): 324-57.
Ding, Q., Zhang, M. and Liu, C., 2015. Asporin participates in gastric cancer cell growth
and migration by influencing EGF receptor signaling. Oncology reports, 33(4):
1783-90.
Dominguez, I., Sonenshein, G.E. and Seldin, D.C., 2009. Protein kinase CK2 in health
and disease: CK2 and its role in Wnt and NF-kappaB signaling: linking
development and cancer. Cellular and molecular life sciences : CMLS, 66(11-12):
1850-7.
Dovas, A. and Couchman, J.R., 2005. RhoGDI: multiple functions in the regulation of
Rho family GTPase activities. The Biochemical journal, 390(Pt 1): 1-9.
Duffy, M.J. et al., 2003. Clinical utility of biochemical markers in colorectal cancer:
European Group on Tumour Markers (EGTM) guidelines. European journal of
cancer, 39(6): 718-27.
Dunn, G.P., Old, L.J. and Schreiber, R.D., 2004. The immunobiology of cancer
immunosurveillance and immunoediting. Immunity, 21(2): 137-48.
Dyduch, G., Kaczmarczyk, K. and Okon, K., 2012. Mast cells and cancer: enemies or
allies? Polish journal of pathology : official journal of the Polish Society of
Pathologists, 63(1): 1-7.
Egorova, K.S., Olenkina, O.M. and Olenina, L.V., 2010. Lysine methylation of
nonhistone proteins is a way to regulate their stability and function. Biochemistry.
Biokhimiia, 75(5): 535-48.
Eom, G.H. et al., 2011. Histone methyltransferase SETD3 regulates muscle
differentiation. The Journal of biological chemistry, 286(40): 34733-42.
Esseghir, S. et al., 2007. Identification of NTN4, TRA1, and STC2 as prognostic markers
in breast cancer in a screen for signal sequence encoding proteins. Clinical cancer
research : an official journal of the American Association for Cancer Research,
13(11): 3164-73.
Faure, J. and Dagher, M.C., 2001. Interactions between Rho GTPases and Rho GDP
dissociation inhibitor (Rho-GDI). Biochimie, 83(5): 409-14.
Fearon, E.R. and Vogelstein, B., 1990. A genetic model for colorectal tumorigenesis.
Cell, 61(5): 759-67.
Fre, S. et al., 2005. Notch signals control the fate of immature progenitor cells in the
intestine. Nature, 435(7044): 964-8.
Ghaffari, A. et al., 2014. A novel role for ezrin in breast cancer angio/lymphangiogenesis.
Breast cancer research : BCR, 16(5): 438.

Silva, K.T.S. Referências Bibliográficas
89
Goel, A. and Boland, C.R., 2012. Epigenetics of colorectal cancer. Gastroenterology,
143(6): 1442-1460 e1.
Goodale, B.C. et al., 2013. Structurally distinct polycyclic aromatic hydrocarbons induce
differential transcriptional responses in developing zebrafish. Toxicology and
applied pharmacology, 272(3): 656-70.
Gray, K.A. et al., 1996. Molecular mechanisms of biocatalytic desulfurization of fossil
fuels. Nature biotechnology, 14(13): 1705-9.
Guengerich, F.P., 2001. Forging the links between metabolism and carcinogenesis.
Mutation research, 488(3): 195-209.
Gupta, S., Takebe, N. and Lorusso, P., 2010. Targeting the Hedgehog pathway in cancer.
Therapeutic advances in medical oncology, 2(4): 237-50.
Hassanein, M., Rahman, J.S., Chaurand, P. and Massion, P.P., 2011. Advances in
proteomic strategies toward the early detection of lung cancer. Proceedings of the
American Thoracic Society, 8(2): 183-8.
Hecht, S.S., Carmella, S.G., Villalta, P.W. and Hochalter, J.B., 2010. Analysis of
phenanthrene and benzo[a]pyrene tetraol enantiomers in human urine: relevance
to the bay region diol epoxide hypothesis of benzo[a]pyrene carcinogenesis and
to biomarker studies. Chemical research in toxicology, 23(5): 900-8.
Heilmann, J., Boulyga, S.F. and Heumann, K.G., 2004. Accurate determination of sulfur
in gasoline and related fuel samples using isotope dilution ICP-MS with direct
sample injection and microwave-assisted digestion. Analytical and bioanalytical
chemistry, 380(2): 190-7.
Helmlinger, D. et al., 2011. Tra1 has specific regulatory roles, rather than global
functions, within the SAGA co-activator complex. The EMBO journal, 30(14):
2843-52.
Höner, A. 2001. Chapter 4 Polycyclic aromatic hydrocarbon (PAH) metabolites.
Handbook of Analytical Separations. K. Wolfgang, Elsevier Science B.V, 3: 99-
121.
Hoops, T.C. and Traber, P.G., 1997. Molecular pathogenesis of colorectal cancer.
Hematology/oncology clinics of North America, 11(4): 609-33.
Huang, J. et al., 2006. Repression of p53 activity by Smyd2-mediated methylation.
Nature, 444(7119): 629-32.
IARC Publications, 2007. The World Health Organization’s Fight Against Cancer:
Strategies That Prevent, Cure and Care. WHO Library. Geneva, Switzerland.
IARC Publications, 2008. World Cancer Report 2008. Boyle, P.; Levin, B. WHO Library.
WHO Library. Geneva, Switzerland.
Incardona, J.P., Collier, T.K. and Scholz, N.L., 2004. Defects in cardiac function precede
morphological abnormalities in fish embryos exposed to polycyclic aromatic
hydrocarbons. Toxicology and applied pharmacology, 196(2): 191-205.
Ingham, P.W. and McMahon, A.P., 2001. Hedgehog signaling in animal development:
paradigms and principles. Genes & development, 15(23): 3059-87.
International Union Against Cancer, 2005. Introduction UICC Global Cancer Control.
Geneve, Switzerland, UICC.

Silva, K.T.S. Referências Bibliográficas
90
Irigaray, P. and Belpomme, D., 2010. Basic properties and molecular mechanisms of
exogenous chemical carcinogens. Carcinogenesis, 31(2): 135-48.
Jacob, J. and Grimmer, G., 1996. Metabolism and excretion of polycyclic aromatic
hydrocarbons in rat and in human. Central European journal of public health, 4
Suppl: 33-9.
Jemal, A. et al., 2005. Cancer statistics, 2005. CA: a cancer journal for clinicians, 55(1):
10-30.
Jia, Y. and Guo, M., 2013. Epigenetic changes in colorectal cancer. Chinese journal of
cancer, 32(1): 21-30.
Jones, J.M., Anderson, J.W. and Tukey, R.H., 2000. Using the metabolism of PAHs in a
human cell line to characterize environmental samples. Environmental toxicology
and pharmacology, 8(2): 119-126.
Jones, R.S. and Gelbart, W.M., 1993. The Drosophila Polycomb-group gene Enhancer of
zeste contains a region with sequence similarity to trithorax. Molecular and
cellular biology, 13(10): 6357-66.
Kahlenberg, M.S., Sullivan, J.M., Witmer, D.D. and Petrelli, N.J., 2003. Molecular
prognostics in colorectal cancer. Surgical oncology, 12(3): 173-86.
Kaminski, S. et al., 2011. Coronin 1A is an essential regulator of the TGFbeta
receptor/SMAD3 signaling pathway in Th17 CD4(+) T cells. Journal of
autoimmunity, 37(3): 198-208.
Kim, D.H. et al., 2009. Proteomic analysis of breast cancer tissue reveals upregulation of
actin-remodeling proteins and its relevance to cancer invasiveness. Proteomics.
Clinical applications, 3(1): 30-40.
Kitadai, Y. et al., 1996. Regulation of carcinoembryonic antigen expression in human
colon carcinoma cells by the organ microenvironment. The American journal of
pathology, 149(4): 1157-66.
Klimczak, M., 2015. Oncogenesis and induced pluripotency - commonalities of signalling
pathways. Contemporary oncology, 19(1A): A16-21.
Knutson, B.A. and Hahn, S., 2011. Domains of Tra1 important for activator recruitment
and transcription coactivator functions of SAGA and NuA4 complexes. Molecular
and cellular biology, 31(4): 818-31.
Kobayashi, M. et al., 2000. Desulfurization of alkylated forms of both dibenzothiophene
and benzothiophene by a single bacterial strain. FEMS microbiology letters,
187(2): 123-6.
Kondoh, N. et al., 1999. Identification and characterization of genes associated with
human hepatocellular carcinogenesis. Cancer research, 59(19): 4990-6.
Krausova, M. and Korinek, V., 2014. Wnt signaling in adult intestinal stem cells and
cancer. Cellular signalling, 26(3): 570-9.
Kropp, K.G., Andersson, J.T. and Fedorak, P.M., 1997. Bacterial transformations of
1,2,3,4-tetrahydrodibenzothiophene and dibenzothiophene. Applied and
environmental microbiology, 63(8): 3032-42.
Kuhn, S. et al., 2007. A complex of EpCAM, claudin-7, CD44 variant isoforms, and
tetraspanins promotes colorectal cancer progression. Molecular cancer research :
MCR, 5(6): 553-67.

Silva, K.T.S. Referências Bibliográficas
91
Kulasingam, V. and Diamandis, E.P., 2008. Strategies for discovering novel cancer
biomarkers through utilization of emerging technologies. Nature clinical practice.
Oncology, 5(10): 588-99.
Kuniyasu, H., Oue, N., Tsutsumi, M., Tahara, E. and Yasui, W., 2001. Heparan sulfate
enhances invasion by human colon carcinoma cell lines through expression of
CD44 variant exon 3. Clinical cancer research : an official journal of the American
Association for Cancer Research, 7(12): 4067-72.
Laubli, H. et al., 2014. Lectin galactoside-binding soluble 3 binding protein (LGALS3BP)
is a tumor-associated immunomodulatory ligand for CD33-related Siglecs. The
Journal of biological chemistry, 289(48): 33481-91.
Lea, I.A. et al., 2007. Genetic pathways and mutation profiles of human cancers: site- and
exposure-specific patterns. Carcinogenesis, 28(9): 1851-8.
Lehmann, L., Jiang, L. and Wagner, J., 2008. Soy isoflavones decrease the catechol-O-
methyltransferase-mediated inactivation of 4-hydroxyestradiol in cultured MCF-
7 cells. Carcinogenesis, 29(2): 363-70.
Leighton, F.A., 1989. Acute oral toxicity of dibenzothiophene for male CD-1 mice:
LD50, lesions, and the effect of preinduction of mixed-function oxidases.
Fundamental and applied toxicology : official journal of the Society of
Toxicology, 12(4): 787-92.
Li, A. et al., 2001. Comparative study for histology, proliferative activity, glycoproteins,
and p53 protein between old and recent colorectal adenomas in Japan. Cancer
letters, 170(1): 45-52.
Lipniak, M., Brandys, J. and Stepniewska, B., 1993. [Determination of polycyclic
aromatic hydrocarbons in grass]. Roczniki Panstwowego Zakladu Higieny, 44(2-
3): 205-12.
Liu, C. et al., 2014. Porcine coronin 1A contributes to nuclear factor-kappa B (NF-
kappaB) inactivation during Haemophilus parasuis infection. PloS one, 9(8):
e103904.
Liu, T.A. et al., 2013. 14-3-3epsilon overexpression contributes to epithelial-
mesenchymal transition of hepatocellular carcinoma. PloS one, 8(3): e57968.
Loeb, L.A. and Harris, C.C., 2008. Advances in chemical carcinogenesis: a historical
review and prospective. Cancer research, 68(17): 6863-72.
Lopez, M.J. and Monafo, W.W., 1993. Role of extended resection in the initial treatment
of locally advanced colorectal carcinoma. Surgery, 113(4): 365-72.
Lu, Y.P. et al., 2002. Expression of lumican in human colorectal cancer cells. Pathology
international, 52(8): 519-26.
MacDonald, B.T., Tamai, K. and He, X., 2009. Wnt/beta-catenin signaling: components,
mechanisms, and diseases. Developmental cell, 17(1): 9-26.
Malhotra, G.K., Zhao, X., Band, H. and Band, V., 2011. Shared signaling pathways in
normal and breast cancer stem cells. Journal of carcinogenesis, 10: 38.
Marinkovic, N., Pasalic, D. and Potocki, S., 2013. Polymorphisms of genes involved in
polycyclic aromatic hydrocarbons' biotransformation and atherosclerosis.
Biochemia medica, 23(3): 255-65.

Silva, K.T.S. Referências Bibliográficas
92
Markowitz, S.D. and Bertagnolli, M.M., 2009. Molecular origins of cancer: Molecular
basis of colorectal cancer. The New England journal of medicine, 361(25): 2449-
60.
Markowska, A., Pawalowska, M., Lubin, J. and Markowska, J., 2014. Signalling
pathways in endometrial cancer. Contemporary oncology, 18(3): 143-8.
Marques-Vidal, P., Ravasco, P. and Ermelinda Camilo, M., 2006. Foodstuffs and
colorectal cancer risk: a review. Clinical nutrition, 25(1): 14-36.
Martin, J.L. and McMillan, F.M., 2002. SAM (dependent) I AM: the S-
adenosylmethionine-dependent methyltransferase fold. Current opinion in
structural biology, 12(6): 783-93.
Mazumdar, T. et al., 2011. Hedgehog signaling drives cellular survival in human colon
carcinoma cells. Cancer research, 71(3): 1092-102.
McLeod, H.L. and Murray, G.I., 1999. Tumour markers of prognosis in colorectal cancer.
British journal of cancer, 79(2): 191-203.
Merrifield, R.B., 1965. Solid-Phase Peptide Syntheses. Endeavour, 24: 3-7.
Miyado, K., Kimura, M. and Taniguchi, S., 1996. Decreased expression of a single
tropomyosin isoform, TM5/TM30nm, results in reduction in motility of highly
metastatic B16-F10 mouse melanoma cells. Biochemical and biophysical research
communications, 225(2): 427-35.
Miyamoto, S. and Rosenberg, D.W., 2011. Role of Notch signaling in colon homeostasis
and carcinogenesis. Cancer science, 102(11): 1938-42.
Mlakar, V. et al., 2009. Presence of activating KRAS mutations correlates significantly
with expression of tumour suppressor genes DCN and TPM1 in colorectal cancer.
BMC cancer, 9: 282.
Montenarh, M., 2014. Protein kinase CK2 and angiogenesis. Advances in clinical and
experimental medicine : official organ Wroclaw Medical University, 23(2): 153-
8.
Montesano, R. and Hall, J., 2001. Environmental causes of human cancers. European
journal of cancer, 37 Suppl 8: S67-87.
Murphy, M.E., 2013. The HSP70 family and cancer. Carcinogenesis, 34(6): 1181-8.
Nakajima, K. et al., 2014. Galectin-3 inhibits osteoblast differentiation through notch
signaling. Neoplasia, 16(11): 939-49.
Nakajima, M. et al., 2007. Mechanisms for asporin function and regulation in articular
cartilage. The Journal of biological chemistry, 282(44): 32185-92.
Nambiar, P.R., Gupta, R.R. and Misra, V., 2010. An "Omics" based survey of human
colon cancer. Mutation research, 693(1-2): 3-18.
Newell, L.E. and Heddle, J.A., 2004. The potent colon carcinogen, 1,2-dimethylhydrazine
induces mutations primarily in the colon. Mutation research, 564(1): 1-7.
Nikitovic, D., Papoutsidakis, A., Karamanos, N.K. and Tzanakakis, G.N., 2014. Lumican
affects tumor cell functions, tumor-ECM interactions, angiogenesis and
inflammatory response. Matrix biology : journal of the International Society for
Matrix Biology, 35: 206-14.

Silva, K.T.S. Referências Bibliográficas
93
O'Dwyer, D., Ralton, L.D., O'Shea, A. and Murray, G.I., 2011. The proteomics of
colorectal cancer: identification of a protein signature associated with prognosis.
PloS one, 6(11): e27718.
Oji, Y. et al., 2014. The translation elongation factor eEF2 is a novel tumorassociated
antigen overexpressed in various types of cancers. International journal of
oncology, 44(5): 1461-9.
Orian-Rousseau, V., 2010. CD44, a therapeutic target for metastasising tumours.
European journal of cancer, 46(7): 1271-7.
Ovrevik, J. et al., 2010. Differential effects of nitro-PAHs and amino-PAHs on cytokine
and chemokine responses in human bronchial epithelial BEAS-2B cells.
Toxicology and applied pharmacology, 242(3): 270-80.
Papanikolaou, A., Wang, Q.S., Delker, D.A. and Rosenberg, D.W., 1998. Azoxymethane-
induced colon tumors and aberrant crypt foci in mice of different genetic
susceptibility. Cancer letters, 130(1-2): 29-34.
Papanikolaou, A., Wang, Q.S., Papanikolaou, D., Whiteley, H.E. and Rosenberg, D.W.,
2000. Sequential and morphological analyses of aberrant crypt foci formation in
mice of differing susceptibility to azoxymethane-induced colon carcinogenesis.
Carcinogenesis, 21(8): 1567-72.
Peng, X.C. et al., 2011. Comparative proteomic approach identifies PKM2 and cofilin-1
as potential diagnostic, prognostic and therapeutic targets for pulmonary
adenocarcinoma. PloS one, 6(11): e27309.
Perse, M. and Cerar, A., 2011. Morphological and molecular alterations in 1,2
dimethylhydrazine and azoxymethane induced colon carcinogenesis in rats.
Journal of biomedicine & biotechnology, 2011: 473964.
Petrossian, T.C. and Clarke, S.G., 2011. Uncovering the human methyltransferasome.
Molecular & cellular proteomics : MCP, 10(1): M110 000976.
Pietraszek, K. et al., 2013. Lumican - derived peptides inhibit melanoma cell growth and
migration. PloS one, 8(10): e76232.
Priolli, D.G., Cardinalli, I.A., et al., 2007. Proposta para estadiamento do câncer colorretal
baseada em critérios morfofuncionais: correlação com níveis séricos do antígeno
carcinoembrionário. Revista Brasileira de Coloproctologia, 27: 374-383.
Qiao, L. and Wong, B.C., 2009. Role of Notch signaling in colorectal cancer.
Carcinogenesis, 30(12): 1979-86.
Qiu, Y. et al., 2008. Plasma glycoprotein profiling for colorectal cancer biomarker
identification by lectin glycoarray and lectin blot. Journal of proteome research,
7(4): 1693-703.
Rangel, M.F., Silva, M.V.M., et al., 2000. Tumores malignos do intestino delgado.
Revista do Colégio Brasileiro de Cirurgiões 27: 385-388.
Rao, J.Y. et al., 1997. Quantitative changes in cytoskeletal and nuclear actins during
cellular transformation. International journal of cancer. Journal international du
cancer, 70(4): 423-9.
Robbins, S.L., V. Kumar, et al., 2010. Robbins and Cotran pathologic basis of disease.
Philadelphia, PA, Saunders/Elsevier.

Silva, K.T.S. Referências Bibliográficas
94
Rosenberg, D.W., Giardina, C. and Tanaka, T., 2009. Mouse models for the study of
colon carcinogenesis. Carcinogenesis, 30(2): 183-96.
Rosenberg, D.W. and Liu, Y., 1995. Induction of aberrant crypts in murine colon with
varying sensitivity to colon carcinogenesis. Cancer letters, 92(2): 209-14.
Rosenberg, S.A., Yang, J.C. and Restifo, N.P., 2004. Cancer immunotherapy: moving
beyond current vaccines. Nature medicine, 10(9): 909-15.
Rudzki, Z. and Jothy, S., 1997. CD44 and the adhesion of neoplastic cells. Molecular
pathology : MP, 50(2): 57-71.
Ruoslahti, E., 2002. Specialization of tumour vasculature. Nature reviews. Cancer, 2(2):
83-90.
Satoh, J., Yamamura, T. and Arima, K., 2004. The 14-3-3 protein epsilon isoform
expressed in reactive astrocytes in demyelinating lesions of multiple sclerosis
binds to vimentin and glial fibrillary acidic protein in cultured human astrocytes.
The American journal of pathology, 165(2): 577-92.
Satoyoshi, R., Kuriyama, S., Aiba, N., Yashiro, M. and Tanaka, M., 2015. Asporin
activates coordinated invasion of scirrhous gastric cancer and cancer-associated
fibroblasts. Oncogene, 34(5): 650-60.
Scherer, R.P., Malvesti, Á.L. et al., 2009. Estudo de adsorção de compostos sulfurados
utilizando um diesel comercial dopado com benzotiofeno e dibenzotiofeno.
Química Nova, 32: 34-37.
Schittek, B., 2012. The multiple facets of dermcidin in cell survival and host defense.
Journal of innate immunity, 4(4): 349-60.
Schoket, B., 1999. DNA damage in humans exposed to environmental and dietary
polycyclic aromatic hydrocarbons. Mutation research, 424(1-2): 143-53.
Schubert, H.L., Blumenthal, R.M. and Cheng, X., 2003. Many paths to methyltransfer: a
chronicle of convergence. Trends in biochemical sciences, 28(6): 329-35.
Scieglinska, D., Piglowski, W., Chekan, M., Mazurek, A. and Krawczyk, Z., 2011.
Differential expression of HSPA1 and HSPA2 proteins in human tissues; tissue
microarray-based immunohistochemical study. Histochemistry and cell biology,
135(4): 337-50.
Scieglinska, D. et al., 2008. The HspA2 protein localizes in nucleoli and centrosomes of
heat shocked cancer cells. Journal of cellular biochemistry, 104(6): 2193-206.
Scott, J.K. and Smith, G.P., 1990. Searching for peptide ligands with an epitope library.
Science, 249(4967): 386-90.
Sergeeva, A., Kolonin, M.G., Molldrem, J.J., Pasqualini, R. and Arap, W., 2006. Display
technologies: application for the discovery of drug and gene delivery agents.
Advanced drug delivery reviews, 58(15): 1622-54.
Seya, T. et al., 2006. Lumican expression in advanced colorectal cancer with nodal
metastasis correlates with poor prognosis. Oncology reports, 16(6): 1225-30.
Shi, X. et al., 2007. Modulation of p53 function by SET8-mediated methylation at lysine
382. Molecular cell, 27(4): 636-46.
Shi, Z. et al., 2010. Organ-specific roles of CYP1A1 during detoxication of dietary
benzo[a]pyrene. Molecular pharmacology, 78(1): 46-57.

Silva, K.T.S. Referências Bibliográficas
95
Shim, A.R., Dong, G.Z., Lee, H.J. and Ryu, J.H., 2015. Atractylochromene Is a Repressor
of Wnt/beta-Catenin Signaling in Colon Cancer Cells. Biomolecules &
therapeutics, 23(1): 26-30.
Shimada, T., 2006. Xenobiotic-metabolizing enzymes involved in activation and
detoxification of carcinogenic polycyclic aromatic hydrocarbons. Drug
metabolism and pharmacokinetics, 21(4): 257-76.
Shubik, P. and Sice, J., 1956. Chemical carcinogenesis as a chronic toxicity test; a review.
Cancer research, 16(8): 728-42.
Sidhu, S.S. and Koide, S., 2007. Phage display for engineering and analyzing protein
interaction interfaces. Current opinion in structural biology, 17(4): 481-7.
Sidhu, S.S., Lowman, H.B., Cunningham, B.C. and Wells, J.A., 2000. Phage display for
selection of novel binding peptides. Methods in enzymology, 328: 333-63.
Simpson, D.R., Mell, L.K. and Cohen, E.E., 2015. Targeting the PI3K/AKT/mTOR
pathway in squamous cell carcinoma of the head and neck. Oral oncology, 51(4):
291-298.
Simpson, R.J. and Dorow, D.S., 2001. Cancer proteomics: from signaling networks to
tumor markers. Trends in biotechnology, 19(10 Suppl): S40-8.
Smith, G.P., 1985. Filamentous fusion phage: novel expression vectors that display
cloned antigens on the virion surface. Science, 228(4705): 1315-7.
Soontjens, C.D., Holmberg, K., et al., 1997. Characterization of polycyclic aromatic
compounds in diesel exhaust particulate extract responsible for aryl hydrocarbon
receptor activity. Atmospheric Environment, 31(2): 219-225.
Speight, J.G., 2002. Handbook of petroleum product analysis. from
http://www.knovel.com/knovel2/Toc.jsp?BookID=1108.
St Croix, B. et al., 2000. Genes expressed in human tumor endothelium. Science,
289(5482): 1197-202.
Stassen, M.J., Bailey, D., Nelson, S., Chinwalla, V. and Harte, P.J., 1995. The Drosophila
trithorax proteins contain a novel variant of the nuclear receptor type DNA
binding domain and an ancient conserved motif found in other chromosomal
proteins. Mechanisms of development, 52(2-3): 209-23.
Strickland, P. and Kang, D., 1999. Urinary 1-hydroxypyrene and other PAH metabolites
as biomarkers of exposure to environmental PAH in air particulate matter.
Toxicology letters, 108(2-3): 191-9.
Suresh, R. et al., 2009. Assessment of association of exposure to polycyclic aromatic
hydrocarbons with bronchial asthma and oxidative stress in children: A case
control study. Indian journal of occupational and environmental medicine, 13(1):
33-7.
Tabata, T. and Takei, Y., 2004. Morphogens, their identification and regulation.
Development, 131(4): 703-12.
Thompson, O., Moghraby, J.S., Ayscough, K.R. and Winder, S.J., 2012. Depletion of the
actin bundling protein SM22/transgelin increases actin dynamics and enhances
the tumourigenic phenotypes of cells. BMC cell biology, 13: 1.

Silva, K.T.S. Referências Bibliográficas
96
Tschiersch, B. et al., 1994. The protein encoded by the Drosophila position-effect
variegation suppressor gene Su(var)3-9 combines domains of antagonistic
regulators of homeotic gene complexes. The EMBO journal, 13(16): 3822-31.
Turtoi, A. et al., 2011. Identification of novel accessible proteins bearing diagnostic and
therapeutic potential in human pancreatic ductal adenocarcinoma. Journal of
proteome research, 10(9): 4302-13.
Uno, S. et al., 2004. Oral exposure to benzo[a]pyrene in the mouse: detoxication by
inducible cytochrome P450 is more important than metabolic activation.
Molecular pharmacology, 65(5): 1225-37.
Uno, S. et al., 2008. Basal and inducible CYP1 mRNA quantitation and protein
localization throughout the mouse gastrointestinal tract. Free radical biology &
medicine, 44(4): 570-83.
van den Brink, G.R. and Hardwick, J.C., 2006. Hedgehog Wnteraction in colorectal
cancer. Gut, 55(7): 912-4.
Vermeulen, L. et al., 2010. Wnt activity defines colon cancer stem cells and is regulated
by the microenvironment. Nature cell biology, 12(5): 468-76.
Vogelstein, B. and Kinzler, K.W., 2004. Cancer genes and the pathways they control.
Nature medicine, 10(8): 789-99.
Vries, R.G., Huch, M. and Clevers, H., 2010. Stem cells and cancer of the stomach and
intestine. Molecular oncology, 4(5): 373-84.
Wang, G., 2014. Human antimicrobial peptides and proteins. Pharmaceuticals, 7(5): 545-
94.
Wang, S.J., Wong, G., de Heer, A.M., Xia, W. and Bourguignon, L.Y., 2009a. CD44
variant isoforms in head and neck squamous cell carcinoma progression. The
Laryngoscope, 119(8): 1518-30.
Wang, Y., Qu, L. and Hao, Y., 2009b. [The pitfall of transbronchial lung biopsy in the
small cell carcinoma--attach 64 cases immunohistochemistry research.].
Zhongguo fei ai za zhi = Chinese journal of lung cancer, 12(8): 901-4.
Wei, L. et al., 2002. Inhibition of Rho family GTPases by Rho GDP dissociation inhibitor
disrupts cardiac morphogenesis and inhibits cardiomyocyte proliferation.
Development, 129(7): 1705-14.
White, B.D., Chien, A.J. and Dawson, D.W., 2012. Dysregulation of Wnt/beta-catenin
signaling in gastrointestinal cancers. Gastroenterology, 142(2): 219-32.
Wilkins, M.R. et al., 1996. Progress with proteome projects: why all proteins expressed
by a genome should be identified and how to do it. Biotechnology & genetic
engineering reviews, 13: 19-50.
Wong, J.J., Hawkins, N.J. and Ward, R.L., 2007. Colorectal cancer: a model for
epigenetic tumorigenesis. Gut, 56(1): 140-8.
Wozny, M. et al., 2010. CYP1A expression in liver and gills of rainbow trout
(Oncorhynchus mykiss) after short-term exposure to dibenzothiophene (DBT).
Chemosphere, 79(1): 110-2.
Wulfkuhle, J.D., Liotta, L.A. and Petricoin, E.F., 2003. Proteomic applications for the
early detection of cancer. Nature reviews. Cancer, 3(4): 267-75.

Silva, K.T.S. Referências Bibliográficas
97
Xie, Z.Y., Gong, L.B., Zhang, W.T. and Lv, X.P., 2014. Expression of Notch-signaling
pathway in familial adenomatous polyposis and its clinical significance. European
review for medical and pharmacological sciences, 18(21): 3183-8.
Zbar, A.P., 2004. The immunology of colorectal cancer. Surgical oncology, 13(2-3): 45-
53.
Zhai, J. et al., 2003. Direct interaction of focal adhesion kinase with p190RhoGEF. The
Journal of biological chemistry, 278(27): 24865-73.
Zhang, N. and Walker, M.K., 2007. Crosstalk between the aryl hydrocarbon receptor and
hypoxia on the constitutive expression of cytochrome P4501A1 mRNA.
Cardiovascular toxicology, 7(4): 282-90.
Zhang, X., Wen, H. and Shi, X., 2012. Lysine methylation: beyond histones. Acta
biochimica et biophysica Sinica, 44(1): 14-27.
Zhang, Z.W., Yang, Z.M., Zheng, Y.C. and Chen, Z.D., 2010. Transgelin induces
apoptosis of human prostate LNCaP cells through its interaction with p53. Asian
journal of andrology, 12(2): 186-95.
Zhang, Z.Z. et al., 2005. [Comparative study on the genotoxicity of gasoline-fueled
vehicle exhaust and methanol-fueled vehicle exhaust]. Sichuan da xue xue bao.
Yi xue ban = Journal of Sichuan University. Medical science edition, 36(2): 249-
52.
Zhao, L., Wang, H., Li, J., Liu, Y. and Ding, Y., 2008. Overexpression of Rho GDP-
dissociation inhibitor alpha is associated with tumor progression and poor
prognosis of colorectal cancer. Journal of proteome research, 7(9): 3994-4003.

Apêndice
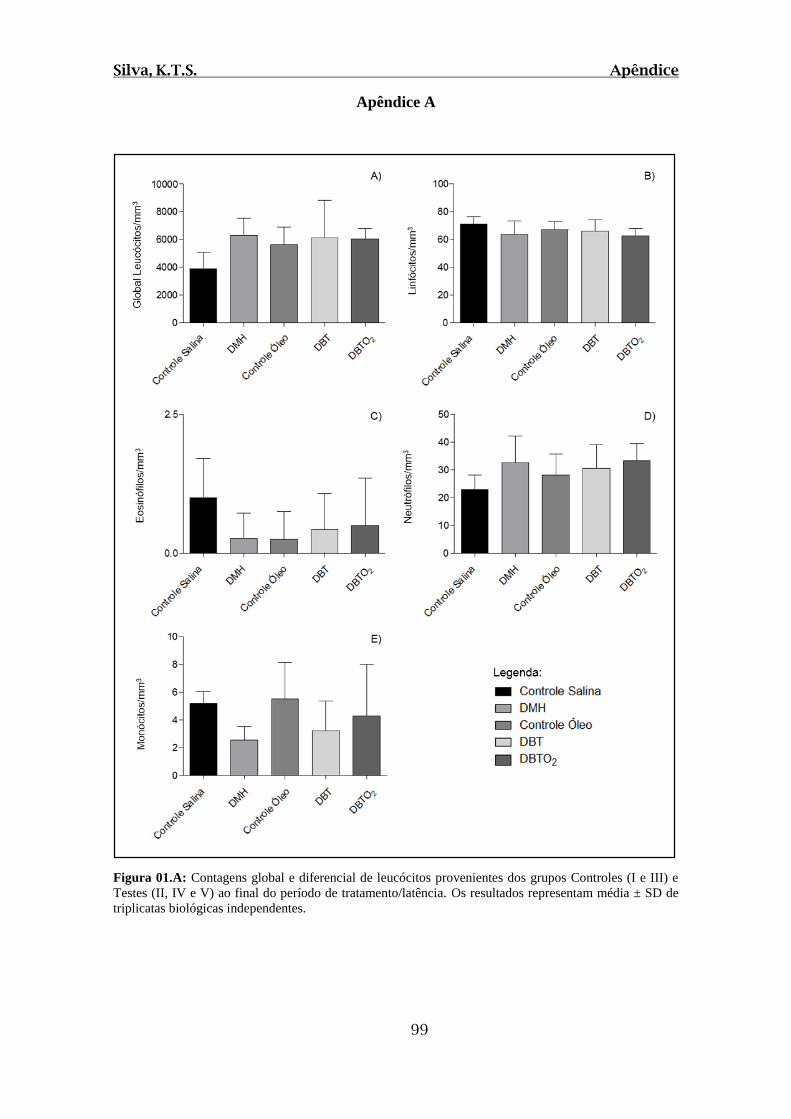
Silva, K.T.S. Apêndice
99
Apêndice A
Figura 01.A: Contagens global e diferencial de leucócitos provenientes dos grupos Controles (I e III) e
Testes (II, IV e V) ao final do período de tratamento/latência. Os resultados representam média ± SD de
triplicatas biológicas independentes.

Silva, K.T.S. Apêndice
100
Apêndice B
Figura 01.B: Dosagens enzimáticas séricas: A) Razão AST/ALT; B) Amilase provenientes dos grupos
Controles (I e III) e Testes (II, IV e V) ao final do período de tratamento/latência. Os resultados representam
média ± SD de triplicatas biológicas independentes.
Silva, K.T.S. Apêndice
101
Apêndice C
Tabela 01.C: Comparação de frequência das lesões observadas no intestino delgado para os grupos
Controle Óleo e Controle Salina.
Lesões Observadas Controle Óleo
% (n)
Controle Salina
% (n) p
Atipias Celulares na Mucosa 25 (1/4) 20 (1/5) 0,4985
Atipias Celulares na Submucosa 0 (0/4) 0 (0/5) -
Inflamação na Mucosa 0 (0/4) 20 (1/5) < 0,0001
Necrose 0 (0/4) 0 (0/5) -
Atipias Estruturais 25 (1/4) 20 (1/5) 0,4985
Hiperplasia de Nódulos Linfóides 25 (1/4) 0 (0/5) < 0,0001
Tabela 02.C: Comparação de frequência das lesões observadas no intestino grosso para os grupos Controle
Óleo e Controle Salina.
Lesões Observadas Controle Óleo
% (n)
Controle Salina
% (n) p
Atipias Celulares na Mucosa 0 (0/4) 0 (0/5) -
Atipias Celulares na Submucosa 0 (0/4) 0 (0/5) -
Inflamação na Mucosa 0 (0/4) 40 (2/5) < 0,0001
Necrose 0 (0/4) 20 (1/5) < 0,0001
Atipias Estruturais 25 (1/4) 0 (0/5) < 0,0001
Hiperplasia de Nódulos Linfóides 25 (1/4) 20 (1/5) 0,4985
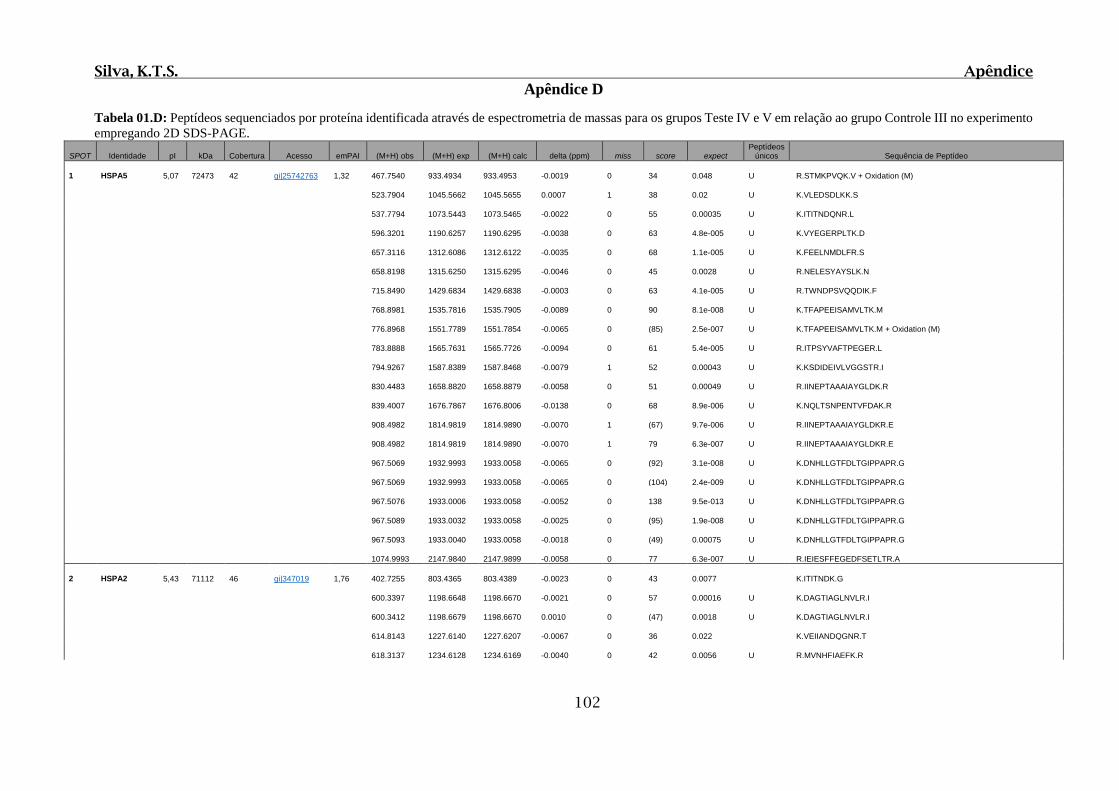
Silva, K.T.S. Apêndice
102
Apêndice D
Tabela 01.D: Peptídeos sequenciados por proteína identificada através de espectrometria de massas para os grupos Teste IV e V em relação ao grupo Controle III no experimento
empregando 2D SDS-PAGE.
SPOT Identidade pI kDa Cobertura Acesso emPAI (M+H) obs (M+H) exp (M+H) calc delta (ppm) miss score expect Peptídeos
únicos Sequência de Peptídeo
1 HSPA5 5,07 72473 42
gi|25742763 1,32 467.7540 933.4934 933.4953 -0.0019 0 34 0.048 U R.STMKPVQK.V + Oxidation (M)
523.7904 1045.5662 1045.5655 0.0007 1 38 0.02 U K.VLEDSDLKK.S
537.7794 1073.5443 1073.5465 -0.0022 0 55 0.00035 U K.ITITNDQNR.L
596.3201 1190.6257 1190.6295 -0.0038 0 63 4.8e-005 U K.VYEGERPLTK.D
657.3116 1312.6086 1312.6122 -0.0035 0 68 1.1e-005 U K.FEELNMDLFR.S
658.8198 1315.6250 1315.6295 -0.0046 0 45 0.0028 U R.NELESYAYSLK.N
715.8490 1429.6834 1429.6838 -0.0003 0 63 4.1e-005 U R.TWNDPSVQQDIK.F
768.8981 1535.7816 1535.7905 -0.0089 0 90 8.1e-008 U K.TFAPEEISAMVLTK.M
776.8968 1551.7789 1551.7854 -0.0065 0 (85) 2.5e-007 U K.TFAPEEISAMVLTK.M + Oxidation (M)
783.8888 1565.7631 1565.7726 -0.0094 0 61 5.4e-005 U R.ITPSYVAFTPEGER.L
794.9267 1587.8389 1587.8468 -0.0079 1 52 0.00043 U K.KSDIDEIVLVGGSTR.I
830.4483 1658.8820 1658.8879 -0.0058 0 51 0.00049 U R.IINEPTAAAIAYGLDK.R
839.4007 1676.7867 1676.8006 -0.0138 0 68 8.9e-006 U K.NQLTSNPENTVFDAK.R
908.4982 1814.9819 1814.9890 -0.0070 1 (67) 9.7e-006 U R.IINEPTAAAIAYGLDKR.E
908.4982 1814.9819 1814.9890 -0.0070 1 79 6.3e-007 U R.IINEPTAAAIAYGLDKR.E
967.5069 1932.9993 1933.0058 -0.0065 0 (92) 3.1e-008 U K.DNHLLGTFDLTGIPPAPR.G
967.5069 1932.9993 1933.0058 -0.0065 0 (104) 2.4e-009 U K.DNHLLGTFDLTGIPPAPR.G
967.5076 1933.0006 1933.0058 -0.0052 0 138 9.5e-013 U K.DNHLLGTFDLTGIPPAPR.G
967.5089 1933.0032 1933.0058 -0.0025 0 (95) 1.9e-008 U K.DNHLLGTFDLTGIPPAPR.G
967.5093 1933.0040 1933.0058 -0.0018 0 (49) 0.00075 U K.DNHLLGTFDLTGIPPAPR.G
1074.9993 2147.9840 2147.9899 -0.0058 0 77 6.3e-007 U R.IEIESFFEGEDFSETLTR.A
2 HSPA2 5,43 71112 46
gi|347019 1,76 402.7255 803.4365 803.4389 -0.0023 0 43 0.0077 K.ITITNDK.G
600.3397 1198.6648 1198.6670 -0.0021 0 57 0.00016 U K.DAGTIAGLNVLR.I
600.3412 1198.6679 1198.6670 0.0010 0 (47) 0.0018 U K.DAGTIAGLNVLR.I
614.8143 1227.6140 1227.6207 -0.0067 0 36 0.022 K.VEIIANDQGNR.T
618.3137 1234.6128 1234.6169 -0.0040 0 42 0.0056 U R.MVNHFIAEFK.R
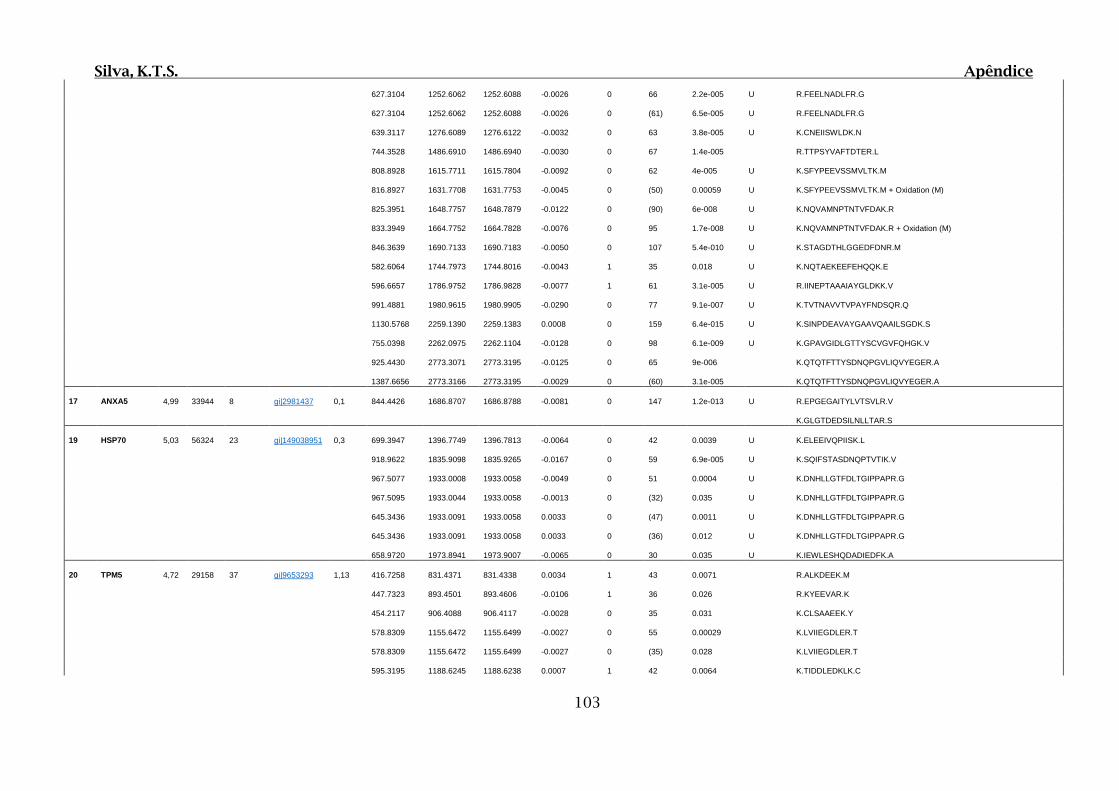
Silva, K.T.S. Apêndice
103
627.3104 1252.6062 1252.6088 -0.0026 0 66 2.2e-005 U R.FEELNADLFR.G
627.3104 1252.6062 1252.6088 -0.0026 0 (61) 6.5e-005 U R.FEELNADLFR.G
639.3117 1276.6089 1276.6122 -0.0032 0 63 3.8e-005 U K.CNEIISWLDK.N
744.3528 1486.6910 1486.6940 -0.0030 0 67 1.4e-005 R.TTPSYVAFTDTER.L
808.8928 1615.7711 1615.7804 -0.0092 0 62 4e-005 U K.SFYPEEVSSMVLTK.M
816.8927 1631.7708 1631.7753 -0.0045 0 (50) 0.00059 U K.SFYPEEVSSMVLTK.M + Oxidation (M)
825.3951 1648.7757 1648.7879 -0.0122 0 (90) 6e-008 U K.NQVAMNPTNTVFDAK.R
833.3949 1664.7752 1664.7828 -0.0076 0 95 1.7e-008 U K.NQVAMNPTNTVFDAK.R + Oxidation (M)
846.3639 1690.7133 1690.7183 -0.0050 0 107 5.4e-010 U K.STAGDTHLGGEDFDNR.M
582.6064 1744.7973 1744.8016 -0.0043 1 35 0.018 U K.NQTAEKEEFEHQQK.E
596.6657 1786.9752 1786.9828 -0.0077 1 61 3.1e-005 U R.IINEPTAAAIAYGLDKK.V
991.4881 1980.9615 1980.9905 -0.0290 0 77 9.1e-007 U K.TVTNAVVTVPAYFNDSQR.Q
1130.5768 2259.1390 2259.1383 0.0008 0 159 6.4e-015 U K.SINPDEAVAYGAAVQAAILSGDK.S
755.0398 2262.0975 2262.1104 -0.0128 0 98 6.1e-009 U K.GPAVGIDLGTTYSCVGVFQHGK.V
925.4430 2773.3071 2773.3195 -0.0125 0 65 9e-006 K.QTQTFTTYSDNQPGVLIQVYEGER.A
1387.6656 2773.3166 2773.3195 -0.0029 0 (60) 3.1e-005 K.QTQTFTTYSDNQPGVLIQVYEGER.A
17 ANXA5 4,99 33944 8
gi|2981437 0,1 844.4426 1686.8707 1686.8788 -0.0081 0 147 1.2e-013 U R.EPGEGAITYLVTSVLR.V
K.GLGTDEDSILNLLTAR.S
19 HSP70 5,03 56324 23
gi|149038951 0,3 699.3947 1396.7749 1396.7813 -0.0064 0 42 0.0039 U K.ELEEIVQPIISK.L
918.9622 1835.9098 1835.9265 -0.0167 0 59 6.9e-005 U K.SQIFSTASDNQPTVTIK.V
967.5077 1933.0008 1933.0058 -0.0049 0 51 0.0004 U K.DNHLLGTFDLTGIPPAPR.G
967.5095 1933.0044 1933.0058 -0.0013 0 (32) 0.035 U K.DNHLLGTFDLTGIPPAPR.G
645.3436 1933.0091 1933.0058 0.0033 0 (47) 0.0011 U K.DNHLLGTFDLTGIPPAPR.G
645.3436 1933.0091 1933.0058 0.0033 0 (36) 0.012 U K.DNHLLGTFDLTGIPPAPR.G
658.9720 1973.8941 1973.9007 -0.0065 0 30 0.035 U K.IEWLESHQDADIEDFK.A
20 TPM5 4,72 29158 37
gi|9653293 1,13 416.7258 831.4371 831.4338 0.0034 1 43 0.0071 R.ALKDEEK.M
447.7323 893.4501 893.4606 -0.0106 1 36 0.026 R.KYEEVAR.K
454.2117 906.4088 906.4117 -0.0028 0 35 0.031 K.CLSAAEEK.Y
578.8309 1155.6472 1155.6499 -0.0027 0 55 0.00029 K.LVIIEGDLER.T
578.8309 1155.6472 1155.6499 -0.0027 0 (35) 0.028 K.LVIIEGDLER.T
595.3195 1188.6245 1188.6238 0.0007 1 42 0.0064 K.TIDDLEDKLK.C
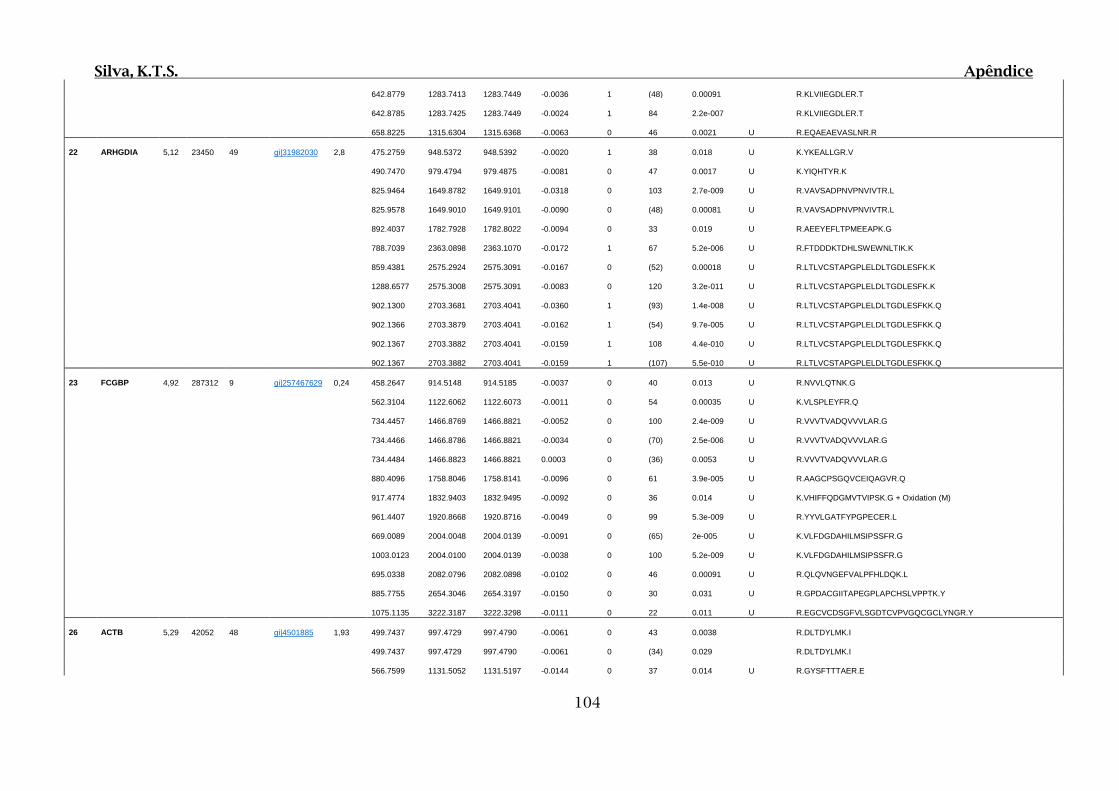
Silva, K.T.S. Apêndice
104
642.8779 1283.7413 1283.7449 -0.0036 1 (48) 0.00091 R.KLVIIEGDLER.T
642.8785 1283.7425 1283.7449 -0.0024 1 84 2.2e-007 R.KLVIIEGDLER.T
658.8225 1315.6304 1315.6368 -0.0063 0 46 0.0021 U R.EQAEAEVASLNR.R
22 ARHGDIA 5,12 23450 49
gi|31982030 2,8 475.2759 948.5372 948.5392 -0.0020 1 38 0.018 U K.YKEALLGR.V
490.7470 979.4794 979.4875 -0.0081 0 47 0.0017 U K.YIQHTYR.K
825.9464 1649.8782 1649.9101 -0.0318 0 103 2.7e-009 U R.VAVSADPNVPNVIVTR.L
825.9578 1649.9010 1649.9101 -0.0090 0 (48) 0.00081 U R.VAVSADPNVPNVIVTR.L
892.4037 1782.7928 1782.8022 -0.0094 0 33 0.019 U R.AEEYEFLTPMEEAPK.G
788.7039 2363.0898 2363.1070 -0.0172 1 67 5.2e-006 U R.FTDDDKTDHLSWEWNLTIK.K
859.4381 2575.2924 2575.3091 -0.0167 0 (52) 0.00018 U R.LTLVCSTAPGPLELDLTGDLESFK.K
1288.6577 2575.3008 2575.3091 -0.0083 0 120 3.2e-011 U R.LTLVCSTAPGPLELDLTGDLESFK.K
902.1300 2703.3681 2703.4041 -0.0360 1 (93) 1.4e-008 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1366 2703.3879 2703.4041 -0.0162 1 (54) 9.7e-005 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1367 2703.3882 2703.4041 -0.0159 1 108 4.4e-010 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1367 2703.3882 2703.4041 -0.0159 1 (107) 5.5e-010 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
23 FCGBP 4,92 287312 9
gi|257467629 0,24 458.2647 914.5148 914.5185 -0.0037 0 40 0.013 U R.NVVLQTNK.G
562.3104 1122.6062 1122.6073 -0.0011 0 54 0.00035 U K.VLSPLEYFR.Q
734.4457 1466.8769 1466.8821 -0.0052 0 100 2.4e-009 U R.VVVTVADQVVVLAR.G
734.4466 1466.8786 1466.8821 -0.0034 0 (70) 2.5e-006 U R.VVVTVADQVVVLAR.G
734.4484 1466.8823 1466.8821 0.0003 0 (36) 0.0053 U R.VVVTVADQVVVLAR.G
880.4096 1758.8046 1758.8141 -0.0096 0 61 3.9e-005 U R.AAGCPSGQVCEIQAGVR.Q
917.4774 1832.9403 1832.9495 -0.0092 0 36 0.014 U K.VHIFFQDGMVTVIPSK.G + Oxidation (M)
961.4407 1920.8668 1920.8716 -0.0049 0 99 5.3e-009 U R.YYVLGATFYPGPECER.L
669.0089 2004.0048 2004.0139 -0.0091 0 (65) 2e-005 U K.VLFDGDAHILMSIPSSFR.G
1003.0123 2004.0100 2004.0139 -0.0038 0 100 5.2e-009 U K.VLFDGDAHILMSIPSSFR.G
695.0338 2082.0796 2082.0898 -0.0102 0 46 0.00091 U R.QLQVNGEFVALPFHLDQK.L
885.7755 2654.3046 2654.3197 -0.0150 0 30 0.031 U R.GPDACGIITAPEGPLAPCHSLVPPTK.Y
1075.1135 3222.3187 3222.3298 -0.0111 0 22 0.011 U R.EGCVCDSGFVLSGDTCVPVGQCGCLYNGR.Y
26 ACTB 5,29 42052 48
gi|4501885 1,93 499.7437 997.4729 997.4790 -0.0061 0 43 0.0038 R.DLTDYLMK.I
499.7437 997.4729 997.4790 -0.0061 0 (34) 0.029 R.DLTDYLMK.I
566.7599 1131.5052 1131.5197 -0.0144 0 37 0.014 U R.GYSFTTTAER.E
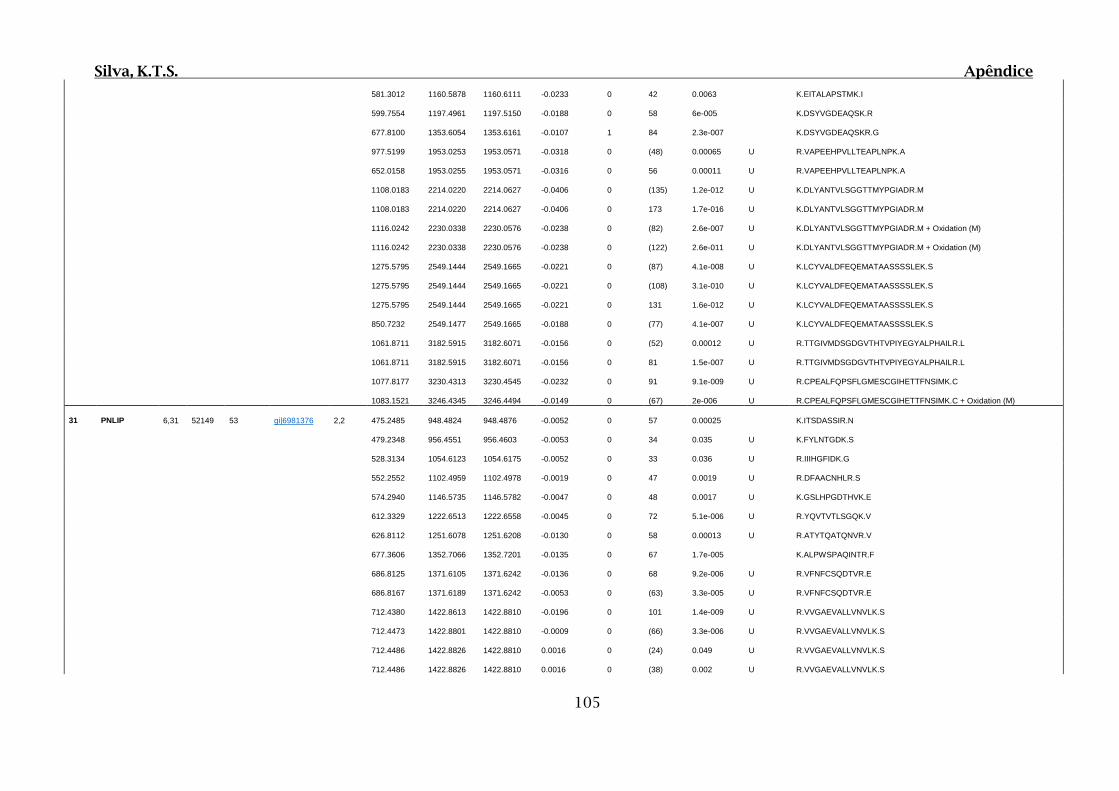
Silva, K.T.S. Apêndice
105
581.3012 1160.5878 1160.6111 -0.0233 0 42 0.0063 K.EITALAPSTMK.I
599.7554 1197.4961 1197.5150 -0.0188 0 58 6e-005 K.DSYVGDEAQSK.R
677.8100 1353.6054 1353.6161 -0.0107 1 84 2.3e-007 K.DSYVGDEAQSKR.G
977.5199 1953.0253 1953.0571 -0.0318 0 (48) 0.00065 U R.VAPEEHPVLLTEAPLNPK.A
652.0158 1953.0255 1953.0571 -0.0316 0 56 0.00011 U R.VAPEEHPVLLTEAPLNPK.A
1108.0183 2214.0220 2214.0627 -0.0406 0 (135) 1.2e-012 U K.DLYANTVLSGGTTMYPGIADR.M
1108.0183 2214.0220 2214.0627 -0.0406 0 173 1.7e-016 U K.DLYANTVLSGGTTMYPGIADR.M
1116.0242 2230.0338 2230.0576 -0.0238 0 (82) 2.6e-007 U K.DLYANTVLSGGTTMYPGIADR.M + Oxidation (M)
1116.0242 2230.0338 2230.0576 -0.0238 0 (122) 2.6e-011 U K.DLYANTVLSGGTTMYPGIADR.M + Oxidation (M)
1275.5795 2549.1444 2549.1665 -0.0221 0 (87) 4.1e-008 U K.LCYVALDFEQEMATAASSSSLEK.S
1275.5795 2549.1444 2549.1665 -0.0221 0 (108) 3.1e-010 U K.LCYVALDFEQEMATAASSSSLEK.S
1275.5795 2549.1444 2549.1665 -0.0221 0 131 1.6e-012 U K.LCYVALDFEQEMATAASSSSLEK.S
850.7232 2549.1477 2549.1665 -0.0188 0 (77) 4.1e-007 U K.LCYVALDFEQEMATAASSSSLEK.S
1061.8711 3182.5915 3182.6071 -0.0156 0 (52) 0.00012 U R.TTGIVMDSGDGVTHTVPIYEGYALPHAILR.L
1061.8711 3182.5915 3182.6071 -0.0156 0 81 1.5e-007 U R.TTGIVMDSGDGVTHTVPIYEGYALPHAILR.L
1077.8177 3230.4313 3230.4545 -0.0232 0 91 9.1e-009 U R.CPEALFQPSFLGMESCGIHETTFNSIMK.C
1083.1521 3246.4345 3246.4494 -0.0149 0 (67) 2e-006 U R.CPEALFQPSFLGMESCGIHETTFNSIMK.C + Oxidation (M)
31 PNLIP 6,31 52149 53
gi|6981376 2,2 475.2485 948.4824 948.4876 -0.0052 0 57 0.00025 K.ITSDASSIR.N
479.2348 956.4551 956.4603 -0.0053 0 34 0.035 U K.FYLNTGDK.S
528.3134 1054.6123 1054.6175 -0.0052 0 33 0.036 U R.IIIHGFIDK.G
552.2552 1102.4959 1102.4978 -0.0019 0 47 0.0019 U R.DFAACNHLR.S
574.2940 1146.5735 1146.5782 -0.0047 0 48 0.0017 U K.GSLHPGDTHVK.E
612.3329 1222.6513 1222.6558 -0.0045 0 72 5.1e-006 U R.YQVTVTLSGQK.V
626.8112 1251.6078 1251.6208 -0.0130 0 58 0.00013 U R.ATYTQATQNVR.V
677.3606 1352.7066 1352.7201 -0.0135 0 67 1.7e-005 K.ALPWSPAQINTR.F
686.8125 1371.6105 1371.6242 -0.0136 0 68 9.2e-006 U R.VFNFCSQDTVR.E
686.8167 1371.6189 1371.6242 -0.0053 0 (63) 3.3e-005 U R.VFNFCSQDTVR.E
712.4380 1422.8613 1422.8810 -0.0196 0 101 1.4e-009 U R.VVGAEVALLVNVLK.S
712.4473 1422.8801 1422.8810 -0.0009 0 (66) 3.3e-006 U R.VVGAEVALLVNVLK.S
712.4486 1422.8826 1422.8810 0.0016 0 (24) 0.049 U R.VVGAEVALLVNVLK.S
712.4486 1422.8826 1422.8810 0.0016 0 (38) 0.002 U R.VVGAEVALLVNVLK.S
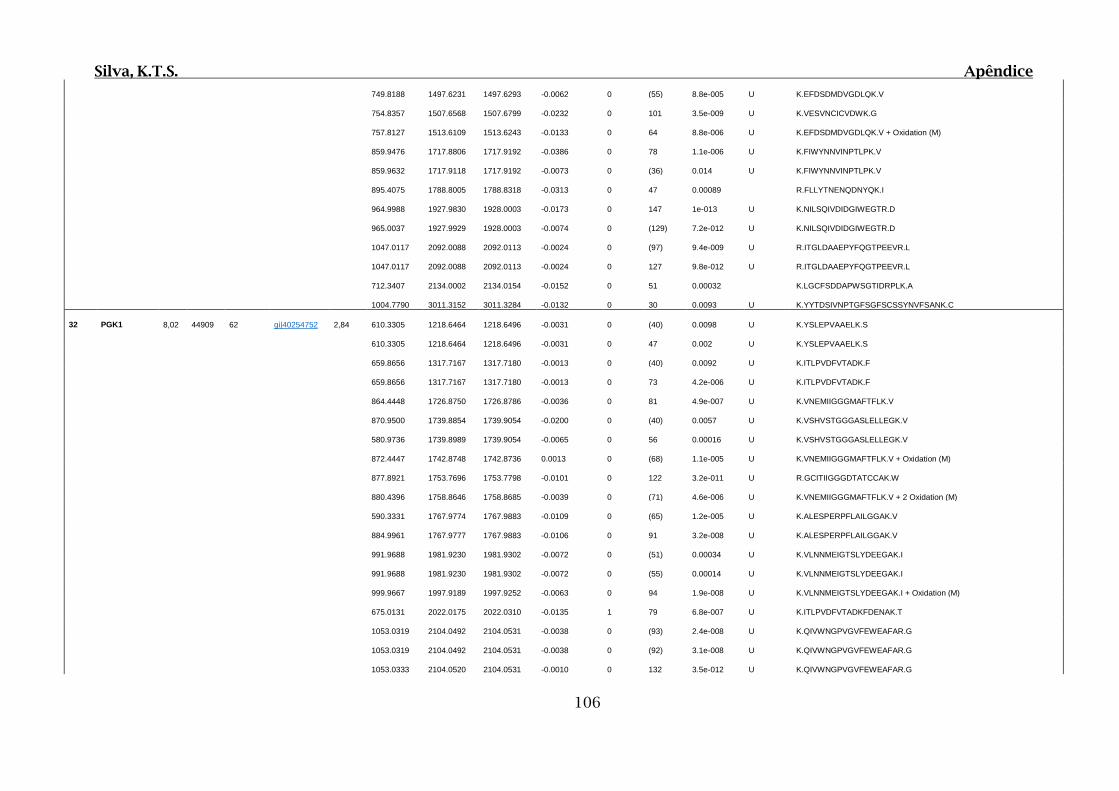
Silva, K.T.S. Apêndice
106
749.8188 1497.6231 1497.6293 -0.0062 0 (55) 8.8e-005 U K.EFDSDMDVGDLQK.V
754.8357 1507.6568 1507.6799 -0.0232 0 101 3.5e-009 U K.VESVNCICVDWK.G
757.8127 1513.6109 1513.6243 -0.0133 0 64 8.8e-006 U K.EFDSDMDVGDLQK.V + Oxidation (M)
859.9476 1717.8806 1717.9192 -0.0386 0 78 1.1e-006 U K.FIWYNNVINPTLPK.V
859.9632 1717.9118 1717.9192 -0.0073 0 (36) 0.014 U K.FIWYNNVINPTLPK.V
895.4075 1788.8005 1788.8318 -0.0313 0 47 0.00089 R.FLLYTNENQDNYQK.I
964.9988 1927.9830 1928.0003 -0.0173 0 147 1e-013 U K.NILSQIVDIDGIWEGTR.D
965.0037 1927.9929 1928.0003 -0.0074 0 (129) 7.2e-012 U K.NILSQIVDIDGIWEGTR.D
1047.0117 2092.0088 2092.0113 -0.0024 0 (97) 9.4e-009 U R.ITGLDAAEPYFQGTPEEVR.L
1047.0117 2092.0088 2092.0113 -0.0024 0 127 9.8e-012 U R.ITGLDAAEPYFQGTPEEVR.L
712.3407 2134.0002 2134.0154 -0.0152 0 51 0.00032 K.LGCFSDDAPWSGTIDRPLK.A
1004.7790 3011.3152 3011.3284 -0.0132 0 30 0.0093 U K.YYTDSIVNPTGFSGFSCSSYNVFSANK.C
32 PGK1 8,02 44909 62
gi|40254752 2,84 610.3305 1218.6464 1218.6496 -0.0031 0 (40) 0.0098 U K.YSLEPVAAELK.S
610.3305 1218.6464 1218.6496 -0.0031 0 47 0.002 U K.YSLEPVAAELK.S
659.8656 1317.7167 1317.7180 -0.0013 0 (40) 0.0092 U K.ITLPVDFVTADK.F
659.8656 1317.7167 1317.7180 -0.0013 0 73 4.2e-006 U K.ITLPVDFVTADK.F
864.4448 1726.8750 1726.8786 -0.0036 0 81 4.9e-007 U K.VNEMIIGGGMAFTFLK.V
870.9500 1739.8854 1739.9054 -0.0200 0 (40) 0.0057 U K.VSHVSTGGGASLELLEGK.V
580.9736 1739.8989 1739.9054 -0.0065 0 56 0.00016 U K.VSHVSTGGGASLELLEGK.V
872.4447 1742.8748 1742.8736 0.0013 0 (68) 1.1e-005 U K.VNEMIIGGGMAFTFLK.V + Oxidation (M)
877.8921 1753.7696 1753.7798 -0.0101 0 122 3.2e-011 U R.GCITIIGGGDTATCCAK.W
880.4396 1758.8646 1758.8685 -0.0039 0 (71) 4.6e-006 U K.VNEMIIGGGMAFTFLK.V + 2 Oxidation (M)
590.3331 1767.9774 1767.9883 -0.0109 0 (65) 1.2e-005 U K.ALESPERPFLAILGGAK.V
884.9961 1767.9777 1767.9883 -0.0106 0 91 3.2e-008 U K.ALESPERPFLAILGGAK.V
991.9688 1981.9230 1981.9302 -0.0072 0 (51) 0.00034 U K.VLNNMEIGTSLYDEEGAK.I
991.9688 1981.9230 1981.9302 -0.0072 0 (55) 0.00014 U K.VLNNMEIGTSLYDEEGAK.I
999.9667 1997.9189 1997.9252 -0.0063 0 94 1.9e-008 U K.VLNNMEIGTSLYDEEGAK.I + Oxidation (M)
675.0131 2022.0175 2022.0310 -0.0135 1 79 6.8e-007 U K.ITLPVDFVTADKFDENAK.T
1053.0319 2104.0492 2104.0531 -0.0038 0 (93) 2.4e-008 U K.QIVWNGPVGVFEWEAFAR.G
1053.0319 2104.0492 2104.0531 -0.0038 0 (92) 3.1e-008 U K.QIVWNGPVGVFEWEAFAR.G
1053.0333 2104.0520 2104.0531 -0.0010 0 132 3.5e-012 U K.QIVWNGPVGVFEWEAFAR.G
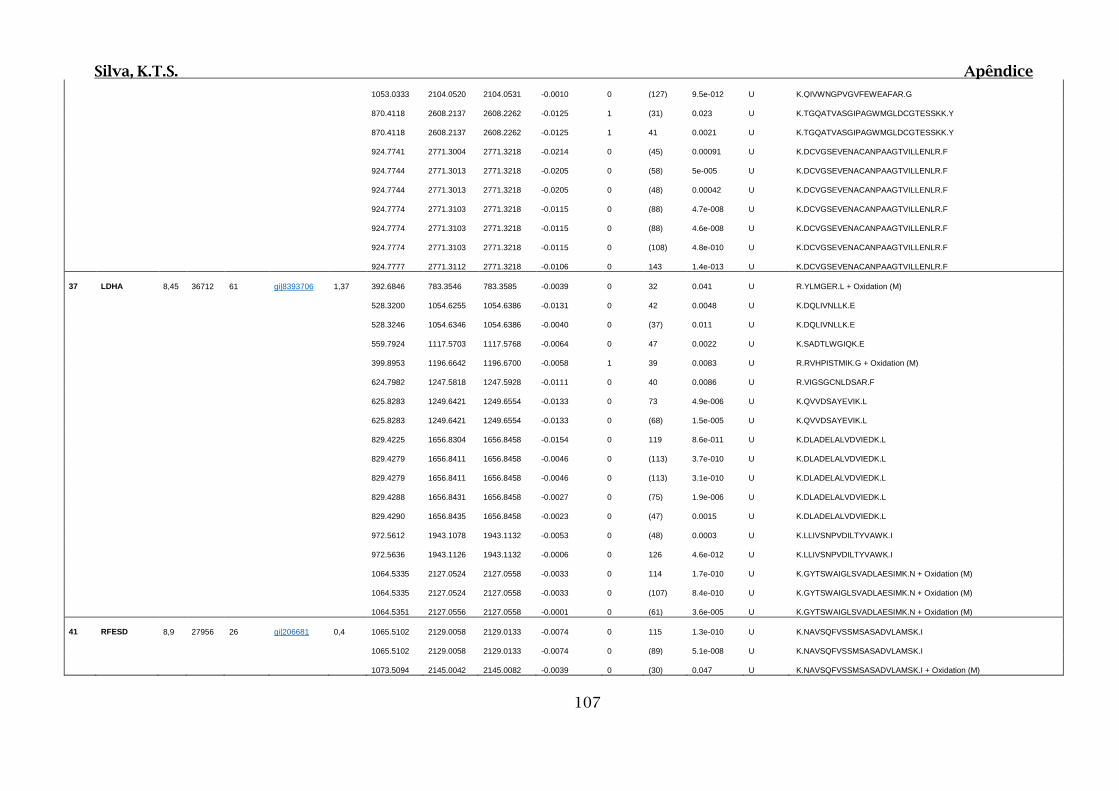
Silva, K.T.S. Apêndice
107
1053.0333 2104.0520 2104.0531 -0.0010 0 (127) 9.5e-012 U K.QIVWNGPVGVFEWEAFAR.G
870.4118 2608.2137 2608.2262 -0.0125 1 (31) 0.023 U K.TGQATVASGIPAGWMGLDCGTESSKK.Y
870.4118 2608.2137 2608.2262 -0.0125 1 41 0.0021 U K.TGQATVASGIPAGWMGLDCGTESSKK.Y
924.7741 2771.3004 2771.3218 -0.0214 0 (45) 0.00091 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7744 2771.3013 2771.3218 -0.0205 0 (58) 5e-005 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7744 2771.3013 2771.3218 -0.0205 0 (48) 0.00042 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7774 2771.3103 2771.3218 -0.0115 0 (88) 4.7e-008 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7774 2771.3103 2771.3218 -0.0115 0 (88) 4.6e-008 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7774 2771.3103 2771.3218 -0.0115 0 (108) 4.8e-010 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7777 2771.3112 2771.3218 -0.0106 0 143 1.4e-013 U K.DCVGSEVENACANPAAGTVILLENLR.F
37 LDHA 8,45 36712 61
gi|8393706 1,37 392.6846 783.3546 783.3585 -0.0039 0 32 0.041 U R.YLMGER.L + Oxidation (M)
528.3200 1054.6255 1054.6386 -0.0131 0 42 0.0048 U K.DQLIVNLLK.E
528.3246 1054.6346 1054.6386 -0.0040 0 (37) 0.011 U K.DQLIVNLLK.E
559.7924 1117.5703 1117.5768 -0.0064 0 47 0.0022 U K.SADTLWGIQK.E
399.8953 1196.6642 1196.6700 -0.0058 1 39 0.0083 U R.RVHPISTMIK.G + Oxidation (M)
624.7982 1247.5818 1247.5928 -0.0111 0 40 0.0086 U R.VIGSGCNLDSAR.F
625.8283 1249.6421 1249.6554 -0.0133 0 73 4.9e-006 U K.QVVDSAYEVIK.L
625.8283 1249.6421 1249.6554 -0.0133 0 (68) 1.5e-005 U K.QVVDSAYEVIK.L
829.4225 1656.8304 1656.8458 -0.0154 0 119 8.6e-011 U K.DLADELALVDVIEDK.L
829.4279 1656.8411 1656.8458 -0.0046 0 (113) 3.7e-010 U K.DLADELALVDVIEDK.L
829.4279 1656.8411 1656.8458 -0.0046 0 (113) 3.1e-010 U K.DLADELALVDVIEDK.L
829.4288 1656.8431 1656.8458 -0.0027 0 (75) 1.9e-006 U K.DLADELALVDVIEDK.L
829.4290 1656.8435 1656.8458 -0.0023 0 (47) 0.0015 U K.DLADELALVDVIEDK.L
972.5612 1943.1078 1943.1132 -0.0053 0 (48) 0.0003 U K.LLIVSNPVDILTYVAWK.I
972.5636 1943.1126 1943.1132 -0.0006 0 126 4.6e-012 U K.LLIVSNPVDILTYVAWK.I
1064.5335 2127.0524 2127.0558 -0.0033 0 114 1.7e-010 U K.GYTSWAIGLSVADLAESIMK.N + Oxidation (M)
1064.5335 2127.0524 2127.0558 -0.0033 0 (107) 8.4e-010 U K.GYTSWAIGLSVADLAESIMK.N + Oxidation (M)
1064.5351 2127.0556 2127.0558 -0.0001 0 (61) 3.6e-005 U K.GYTSWAIGLSVADLAESIMK.N + Oxidation (M)
41 RFESD 8,9 27956 26
gi|206681 0,4 1065.5102 2129.0058 2129.0133 -0.0074 0 115 1.3e-010 U K.NAVSQFVSSMSASADVLAMSK.I
1065.5102 2129.0058 2129.0133 -0.0074 0 (89) 5.1e-008 U K.NAVSQFVSSMSASADVLAMSK.I
1073.5094 2145.0042 2145.0082 -0.0039 0 (30) 0.047 U K.NAVSQFVSSMSASADVLAMSK.I + Oxidation (M)
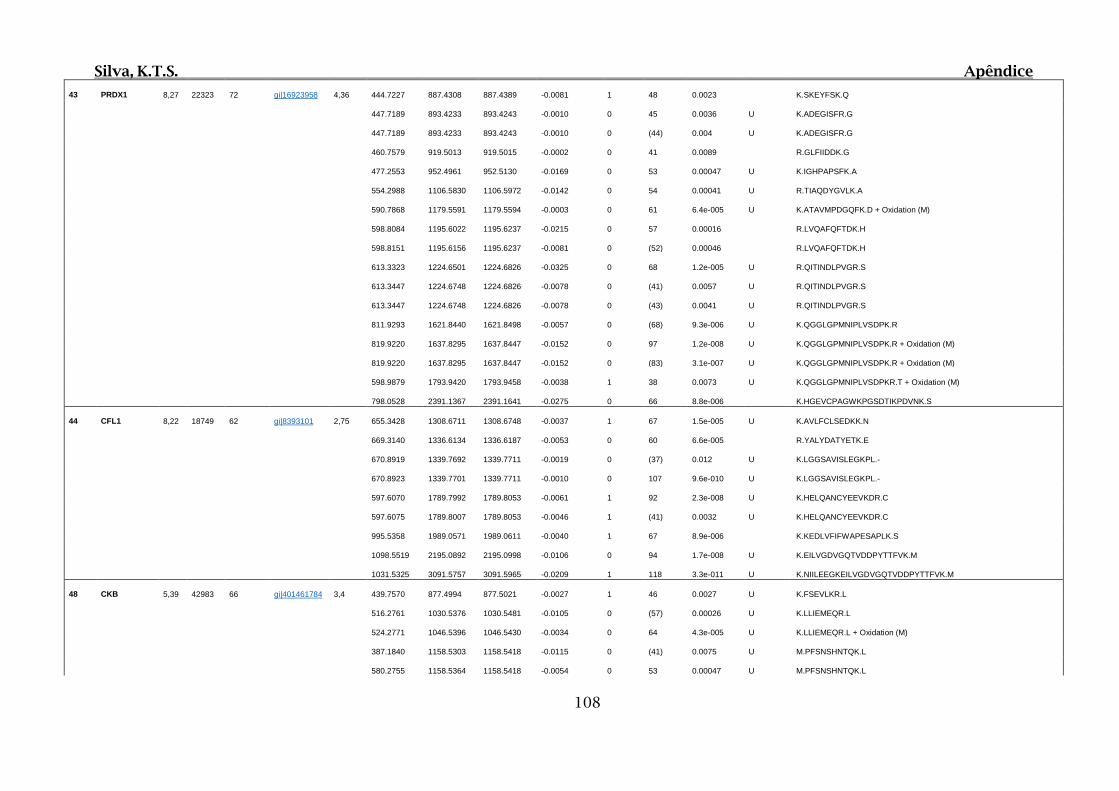
Silva, K.T.S. Apêndice
108
43 PRDX1 8,27 22323 72
gi|16923958 4,36 444.7227 887.4308 887.4389 -0.0081 1 48 0.0023 K.SKEYFSK.Q
447.7189 893.4233 893.4243 -0.0010 0 45 0.0036 U K.ADEGISFR.G
447.7189 893.4233 893.4243 -0.0010 0 (44) 0.004 U K.ADEGISFR.G
460.7579 919.5013 919.5015 -0.0002 0 41 0.0089 R.GLFIIDDK.G
477.2553 952.4961 952.5130 -0.0169 0 53 0.00047 U K.IGHPAPSFK.A
554.2988 1106.5830 1106.5972 -0.0142 0 54 0.00041 U R.TIAQDYGVLK.A
590.7868 1179.5591 1179.5594 -0.0003 0 61 6.4e-005 U K.ATAVMPDGQFK.D + Oxidation (M)
598.8084 1195.6022 1195.6237 -0.0215 0 57 0.00016 R.LVQAFQFTDK.H
598.8151 1195.6156 1195.6237 -0.0081 0 (52) 0.00046 R.LVQAFQFTDK.H
613.3323 1224.6501 1224.6826 -0.0325 0 68 1.2e-005 U R.QITINDLPVGR.S
613.3447 1224.6748 1224.6826 -0.0078 0 (41) 0.0057 U R.QITINDLPVGR.S
613.3447 1224.6748 1224.6826 -0.0078 0 (43) 0.0041 U R.QITINDLPVGR.S
811.9293 1621.8440 1621.8498 -0.0057 0 (68) 9.3e-006 U K.QGGLGPMNIPLVSDPK.R
819.9220 1637.8295 1637.8447 -0.0152 0 97 1.2e-008 U K.QGGLGPMNIPLVSDPK.R + Oxidation (M)
819.9220 1637.8295 1637.8447 -0.0152 0 (83) 3.1e-007 U K.QGGLGPMNIPLVSDPK.R + Oxidation (M)
598.9879 1793.9420 1793.9458 -0.0038 1 38 0.0073 U K.QGGLGPMNIPLVSDPKR.T + Oxidation (M)
798.0528 2391.1367 2391.1641 -0.0275 0 66 8.8e-006 K.HGEVCPAGWKPGSDTIKPDVNK.S
44 CFL1 8,22 18749 62
gi|8393101 2,75 655.3428 1308.6711 1308.6748 -0.0037 1 67 1.5e-005 U K.AVLFCLSEDKK.N
669.3140 1336.6134 1336.6187 -0.0053 0 60 6.6e-005 R.YALYDATYETK.E
670.8919 1339.7692 1339.7711 -0.0019 0 (37) 0.012 U K.LGGSAVISLEGKPL.-
670.8923 1339.7701 1339.7711 -0.0010 0 107 9.6e-010 U K.LGGSAVISLEGKPL.-
597.6070 1789.7992 1789.8053 -0.0061 1 92 2.3e-008 U K.HELQANCYEEVKDR.C
597.6075 1789.8007 1789.8053 -0.0046 1 (41) 0.0032 U K.HELQANCYEEVKDR.C
995.5358 1989.0571 1989.0611 -0.0040 1 67 8.9e-006 K.KEDLVFIFWAPESAPLK.S
1098.5519 2195.0892 2195.0998 -0.0106 0 94 1.7e-008 U K.EILVGDVGQTVDDPYTTFVK.M
1031.5325 3091.5757 3091.5965 -0.0209 1 118 3.3e-011 U K.NIILEEGKEILVGDVGQTVDDPYTTFVK.M
48 CKB 5,39 42983 66
gi|401461784 3,4 439.7570 877.4994 877.5021 -0.0027 1 46 0.0027 U K.FSEVLKR.L
516.2761 1030.5376 1030.5481 -0.0105 0 (57) 0.00026 U K.LLIEMEQR.L
524.2771 1046.5396 1046.5430 -0.0034 0 64 4.3e-005 U K.LLIEMEQR.L + Oxidation (M)
387.1840 1158.5303 1158.5418 -0.0115 0 (41) 0.0075 U M.PFSNSHNTQK.L
580.2755 1158.5364 1158.5418 -0.0054 0 53 0.00047 U M.PFSNSHNTQK.L
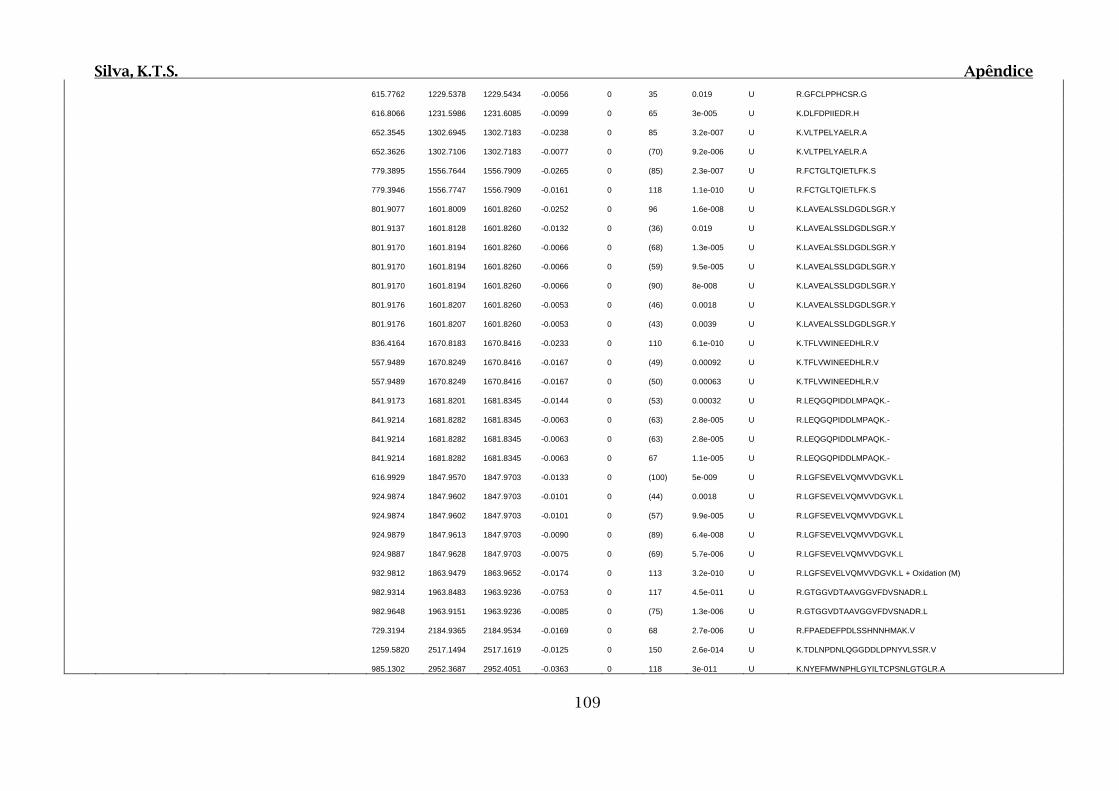
Silva, K.T.S. Apêndice
109
615.7762 1229.5378 1229.5434 -0.0056 0 35 0.019 U R.GFCLPPHCSR.G
616.8066 1231.5986 1231.6085 -0.0099 0 65 3e-005 U K.DLFDPIIEDR.H
652.3545 1302.6945 1302.7183 -0.0238 0 85 3.2e-007 U K.VLTPELYAELR.A
652.3626 1302.7106 1302.7183 -0.0077 0 (70) 9.2e-006 U K.VLTPELYAELR.A
779.3895 1556.7644 1556.7909 -0.0265 0 (85) 2.3e-007 U R.FCTGLTQIETLFK.S
779.3946 1556.7747 1556.7909 -0.0161 0 118 1.1e-010 U R.FCTGLTQIETLFK.S
801.9077 1601.8009 1601.8260 -0.0252 0 96 1.6e-008 U K.LAVEALSSLDGDLSGR.Y
801.9137 1601.8128 1601.8260 -0.0132 0 (36) 0.019 U K.LAVEALSSLDGDLSGR.Y
801.9170 1601.8194 1601.8260 -0.0066 0 (68) 1.3e-005 U K.LAVEALSSLDGDLSGR.Y
801.9170 1601.8194 1601.8260 -0.0066 0 (59) 9.5e-005 U K.LAVEALSSLDGDLSGR.Y
801.9170 1601.8194 1601.8260 -0.0066 0 (90) 8e-008 U K.LAVEALSSLDGDLSGR.Y
801.9176 1601.8207 1601.8260 -0.0053 0 (46) 0.0018 U K.LAVEALSSLDGDLSGR.Y
801.9176 1601.8207 1601.8260 -0.0053 0 (43) 0.0039 U K.LAVEALSSLDGDLSGR.Y
836.4164 1670.8183 1670.8416 -0.0233 0 110 6.1e-010 U K.TFLVWINEEDHLR.V
557.9489 1670.8249 1670.8416 -0.0167 0 (49) 0.00092 U K.TFLVWINEEDHLR.V
557.9489 1670.8249 1670.8416 -0.0167 0 (50) 0.00063 U K.TFLVWINEEDHLR.V
841.9173 1681.8201 1681.8345 -0.0144 0 (53) 0.00032 U R.LEQGQPIDDLMPAQK.-
841.9214 1681.8282 1681.8345 -0.0063 0 (63) 2.8e-005 U R.LEQGQPIDDLMPAQK.-
841.9214 1681.8282 1681.8345 -0.0063 0 (63) 2.8e-005 U R.LEQGQPIDDLMPAQK.-
841.9214 1681.8282 1681.8345 -0.0063 0 67 1.1e-005 U R.LEQGQPIDDLMPAQK.-
616.9929 1847.9570 1847.9703 -0.0133 0 (100) 5e-009 U R.LGFSEVELVQMVVDGVK.L
924.9874 1847.9602 1847.9703 -0.0101 0 (44) 0.0018 U R.LGFSEVELVQMVVDGVK.L
924.9874 1847.9602 1847.9703 -0.0101 0 (57) 9.9e-005 U R.LGFSEVELVQMVVDGVK.L
924.9879 1847.9613 1847.9703 -0.0090 0 (89) 6.4e-008 U R.LGFSEVELVQMVVDGVK.L
924.9887 1847.9628 1847.9703 -0.0075 0 (69) 5.7e-006 U R.LGFSEVELVQMVVDGVK.L
932.9812 1863.9479 1863.9652 -0.0174 0 113 3.2e-010 U R.LGFSEVELVQMVVDGVK.L + Oxidation (M)
982.9314 1963.8483 1963.9236 -0.0753 0 117 4.5e-011 U R.GTGGVDTAAVGGVFDVSNADR.L
982.9648 1963.9151 1963.9236 -0.0085 0 (75) 1.3e-006 U R.GTGGVDTAAVGGVFDVSNADR.L
729.3194 2184.9365 2184.9534 -0.0169 0 68 2.7e-006 U R.FPAEDEFPDLSSHNNHMAK.V
1259.5820 2517.1494 2517.1619 -0.0125 0 150 2.6e-014 U K.TDLNPDNLQGGDDLDPNYVLSSR.V
985.1302 2952.3687 2952.4051 -0.0363 0 118 3e-011 U K.NYEFMWNPHLGYILTCPSNLGTGLR.A
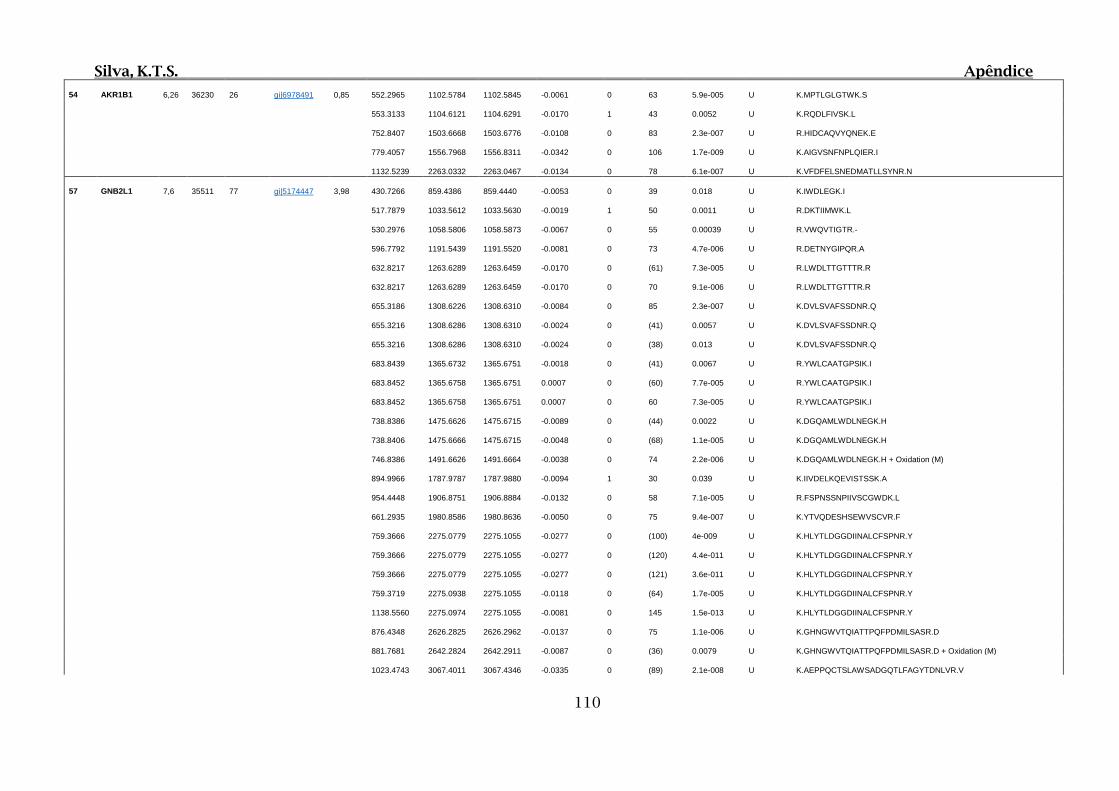
Silva, K.T.S. Apêndice
110
54 AKR1B1 6,26 36230 26
gi|6978491 0,85 552.2965 1102.5784 1102.5845 -0.0061 0 63 5.9e-005 U K.MPTLGLGTWK.S
553.3133 1104.6121 1104.6291 -0.0170 1 43 0.0052 U K.RQDLFIVSK.L
752.8407 1503.6668 1503.6776 -0.0108 0 83 2.3e-007 U R.HIDCAQVYQNEK.E
779.4057 1556.7968 1556.8311 -0.0342 0 106 1.7e-009 U K.AIGVSNFNPLQIER.I
1132.5239 2263.0332 2263.0467 -0.0134 0 78 6.1e-007 U K.VFDFELSNEDMATLLSYNR.N
57 GNB2L1 7,6 35511 77
gi|5174447 3,98 430.7266 859.4386 859.4440 -0.0053 0 39 0.018 U K.IWDLEGK.I
517.7879 1033.5612 1033.5630 -0.0019 1 50 0.0011 U R.DKTIIMWK.L
530.2976 1058.5806 1058.5873 -0.0067 0 55 0.00039 U R.VWQVTIGTR.-
596.7792 1191.5439 1191.5520 -0.0081 0 73 4.7e-006 U R.DETNYGIPQR.A
632.8217 1263.6289 1263.6459 -0.0170 0 (61) 7.3e-005 U R.LWDLTTGTTTR.R
632.8217 1263.6289 1263.6459 -0.0170 0 70 9.1e-006 U R.LWDLTTGTTTR.R
655.3186 1308.6226 1308.6310 -0.0084 0 85 2.3e-007 U K.DVLSVAFSSDNR.Q
655.3216 1308.6286 1308.6310 -0.0024 0 (41) 0.0057 U K.DVLSVAFSSDNR.Q
655.3216 1308.6286 1308.6310 -0.0024 0 (38) 0.013 U K.DVLSVAFSSDNR.Q
683.8439 1365.6732 1365.6751 -0.0018 0 (41) 0.0067 U R.YWLCAATGPSIK.I
683.8452 1365.6758 1365.6751 0.0007 0 (60) 7.7e-005 U R.YWLCAATGPSIK.I
683.8452 1365.6758 1365.6751 0.0007 0 60 7.3e-005 U R.YWLCAATGPSIK.I
738.8386 1475.6626 1475.6715 -0.0089 0 (44) 0.0022 U K.DGQAMLWDLNEGK.H
738.8406 1475.6666 1475.6715 -0.0048 0 (68) 1.1e-005 U K.DGQAMLWDLNEGK.H
746.8386 1491.6626 1491.6664 -0.0038 0 74 2.2e-006 U K.DGQAMLWDLNEGK.H + Oxidation (M)
894.9966 1787.9787 1787.9880 -0.0094 1 30 0.039 U K.IIVDELKQEVISTSSK.A
954.4448 1906.8751 1906.8884 -0.0132 0 58 7.1e-005 U R.FSPNSSNPIIVSCGWDK.L
661.2935 1980.8586 1980.8636 -0.0050 0 75 9.4e-007 U K.YTVQDESHSEWVSCVR.F
759.3666 2275.0779 2275.1055 -0.0277 0 (100) 4e-009 U K.HLYTLDGGDIINALCFSPNR.Y
759.3666 2275.0779 2275.1055 -0.0277 0 (120) 4.4e-011 U K.HLYTLDGGDIINALCFSPNR.Y
759.3666 2275.0779 2275.1055 -0.0277 0 (121) 3.6e-011 U K.HLYTLDGGDIINALCFSPNR.Y
759.3719 2275.0938 2275.1055 -0.0118 0 (64) 1.7e-005 U K.HLYTLDGGDIINALCFSPNR.Y
1138.5560 2275.0974 2275.1055 -0.0081 0 145 1.5e-013 U K.HLYTLDGGDIINALCFSPNR.Y
876.4348 2626.2825 2626.2962 -0.0137 0 75 1.1e-006 U K.GHNGWVTQIATTPQFPDMILSASR.D
881.7681 2642.2824 2642.2911 -0.0087 0 (36) 0.0079 U K.GHNGWVTQIATTPQFPDMILSASR.D + Oxidation (M)
1023.4743 3067.4011 3067.4346 -0.0335 0 (89) 2.1e-008 U K.AEPPQCTSLAWSADGQTLFAGYTDNLVR.V
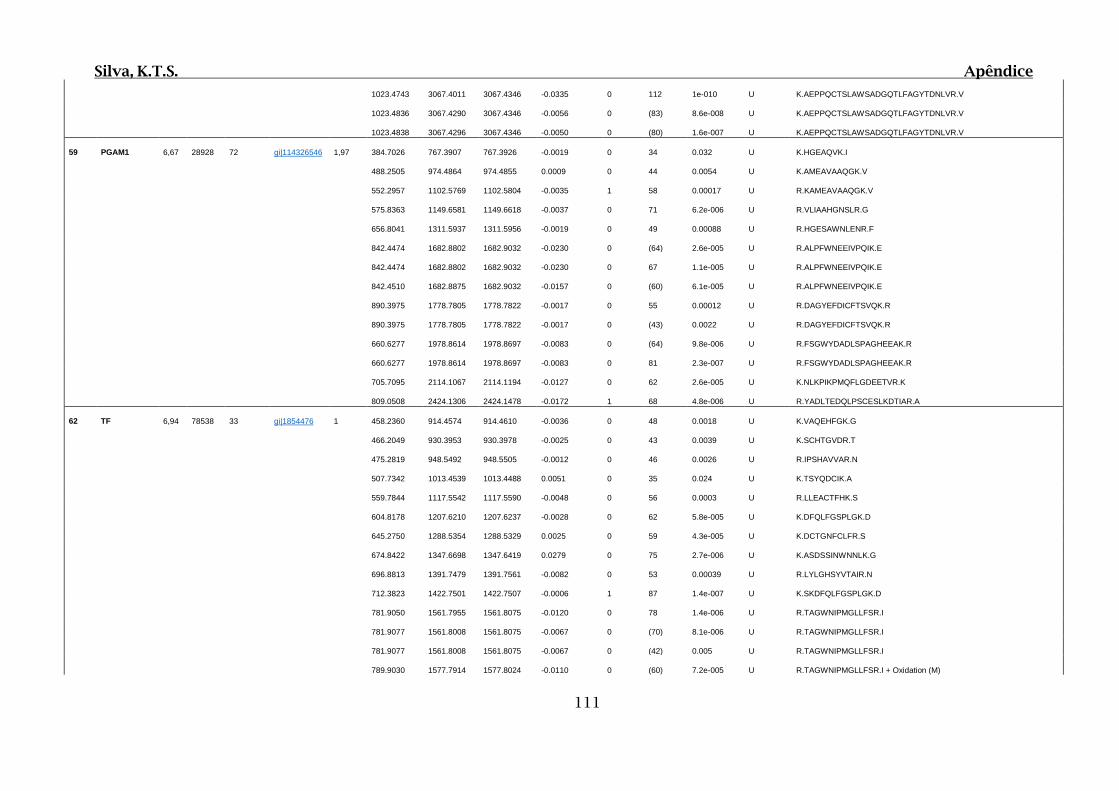
Silva, K.T.S. Apêndice
111
1023.4743 3067.4011 3067.4346 -0.0335 0 112 1e-010 U K.AEPPQCTSLAWSADGQTLFAGYTDNLVR.V
1023.4836 3067.4290 3067.4346 -0.0056 0 (83) 8.6e-008 U K.AEPPQCTSLAWSADGQTLFAGYTDNLVR.V
1023.4838 3067.4296 3067.4346 -0.0050 0 (80) 1.6e-007 U K.AEPPQCTSLAWSADGQTLFAGYTDNLVR.V
59 PGAM1 6,67 28928 72
gi|114326546 1,97 384.7026 767.3907 767.3926 -0.0019 0 34 0.032 U K.HGEAQVK.I
488.2505 974.4864 974.4855 0.0009 0 44 0.0054 U K.AMEAVAAQGK.V
552.2957 1102.5769 1102.5804 -0.0035 1 58 0.00017 U R.KAMEAVAAQGK.V
575.8363 1149.6581 1149.6618 -0.0037 0 71 6.2e-006 U R.VLIAAHGNSLR.G
656.8041 1311.5937 1311.5956 -0.0019 0 49 0.00088 U R.HGESAWNLENR.F
842.4474 1682.8802 1682.9032 -0.0230 0 (64) 2.6e-005 U R.ALPFWNEEIVPQIK.E
842.4474 1682.8802 1682.9032 -0.0230 0 67 1.1e-005 U R.ALPFWNEEIVPQIK.E
842.4510 1682.8875 1682.9032 -0.0157 0 (60) 6.1e-005 U R.ALPFWNEEIVPQIK.E
890.3975 1778.7805 1778.7822 -0.0017 0 55 0.00012 U R.DAGYEFDICFTSVQK.R
890.3975 1778.7805 1778.7822 -0.0017 0 (43) 0.0022 U R.DAGYEFDICFTSVQK.R
660.6277 1978.8614 1978.8697 -0.0083 0 (64) 9.8e-006 U R.FSGWYDADLSPAGHEEAK.R
660.6277 1978.8614 1978.8697 -0.0083 0 81 2.3e-007 U R.FSGWYDADLSPAGHEEAK.R
705.7095 2114.1067 2114.1194 -0.0127 0 62 2.6e-005 U K.NLKPIKPMQFLGDEETVR.K
809.0508 2424.1306 2424.1478 -0.0172 1 68 4.8e-006 U R.YADLTEDQLPSCESLKDTIAR.A
62 TF 6,94 78538 33
gi|1854476 1 458.2360 914.4574 914.4610 -0.0036 0 48 0.0018 U K.VAQEHFGK.G
466.2049 930.3953 930.3978 -0.0025 0 43 0.0039 U K.SCHTGVDR.T
475.2819 948.5492 948.5505 -0.0012 0 46 0.0026 U R.IPSHAVVAR.N
507.7342 1013.4539 1013.4488 0.0051 0 35 0.024 U K.TSYQDCIK.A
559.7844 1117.5542 1117.5590 -0.0048 0 56 0.0003 U R.LLEACTFHK.S
604.8178 1207.6210 1207.6237 -0.0028 0 62 5.8e-005 U K.DFQLFGSPLGK.D
645.2750 1288.5354 1288.5329 0.0025 0 59 4.3e-005 U K.DCTGNFCLFR.S
674.8422 1347.6698 1347.6419 0.0279 0 75 2.7e-006 U K.ASDSSINWNNLK.G
696.8813 1391.7479 1391.7561 -0.0082 0 53 0.00039 U R.LYLGHSYVTAIR.N
712.3823 1422.7501 1422.7507 -0.0006 1 87 1.4e-007 U K.SKDFQLFGSPLGK.D
781.9050 1561.7955 1561.8075 -0.0120 0 78 1.4e-006 U R.TAGWNIPMGLLFSR.I
781.9077 1561.8008 1561.8075 -0.0067 0 (70) 8.1e-006 U R.TAGWNIPMGLLFSR.I
781.9077 1561.8008 1561.8075 -0.0067 0 (42) 0.005 U R.TAGWNIPMGLLFSR.I
789.9030 1577.7914 1577.8024 -0.0110 0 (60) 7.2e-005 U R.TAGWNIPMGLLFSR.I + Oxidation (M)
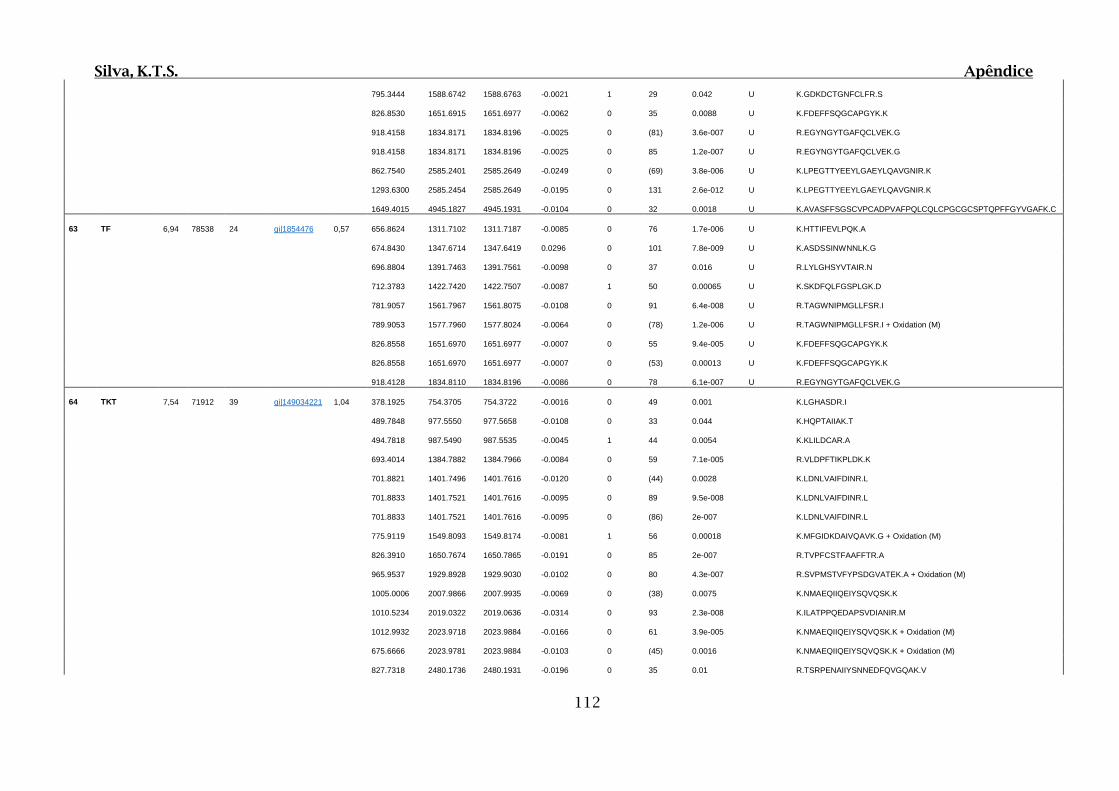
Silva, K.T.S. Apêndice
112
795.3444 1588.6742 1588.6763 -0.0021 1 29 0.042 U K.GDKDCTGNFCLFR.S
826.8530 1651.6915 1651.6977 -0.0062 0 35 0.0088 U K.FDEFFSQGCAPGYK.K
918.4158 1834.8171 1834.8196 -0.0025 0 (81) 3.6e-007 U R.EGYNGYTGAFQCLVEK.G
918.4158 1834.8171 1834.8196 -0.0025 0 85 1.2e-007 U R.EGYNGYTGAFQCLVEK.G
862.7540 2585.2401 2585.2649 -0.0249 0 (69) 3.8e-006 U K.LPEGTTYEEYLGAEYLQAVGNIR.K
1293.6300 2585.2454 2585.2649 -0.0195 0 131 2.6e-012 U K.LPEGTTYEEYLGAEYLQAVGNIR.K
1649.4015 4945.1827 4945.1931 -0.0104 0 32 0.0018 U K.AVASFFSGSCVPCADPVAFPQLCQLCPGCGCSPTQPFFGYVGAFK.C
63 TF 6,94 78538 24
gi|1854476 0,57 656.8624 1311.7102 1311.7187 -0.0085 0 76 1.7e-006 U K.HTTIFEVLPQK.A
674.8430 1347.6714 1347.6419 0.0296 0 101 7.8e-009 U K.ASDSSINWNNLK.G
696.8804 1391.7463 1391.7561 -0.0098 0 37 0.016 U R.LYLGHSYVTAIR.N
712.3783 1422.7420 1422.7507 -0.0087 1 50 0.00065 U K.SKDFQLFGSPLGK.D
781.9057 1561.7967 1561.8075 -0.0108 0 91 6.4e-008 U R.TAGWNIPMGLLFSR.I
789.9053 1577.7960 1577.8024 -0.0064 0 (78) 1.2e-006 U R.TAGWNIPMGLLFSR.I + Oxidation (M)
826.8558 1651.6970 1651.6977 -0.0007 0 55 9.4e-005 U K.FDEFFSQGCAPGYK.K
826.8558 1651.6970 1651.6977 -0.0007 0 (53) 0.00013 U K.FDEFFSQGCAPGYK.K
918.4128 1834.8110 1834.8196 -0.0086 0 78 6.1e-007 U R.EGYNGYTGAFQCLVEK.G
64 TKT 7,54 71912 39
gi|149034221 1,04 378.1925 754.3705 754.3722 -0.0016 0 49 0.001 K.LGHASDR.I
489.7848 977.5550 977.5658 -0.0108 0 33 0.044 K.HQPTAIIAK.T
494.7818 987.5490 987.5535 -0.0045 1 44 0.0054 K.KLILDCAR.A
693.4014 1384.7882 1384.7966 -0.0084 0 59 7.1e-005 R.VLDPFTIKPLDK.K
701.8821 1401.7496 1401.7616 -0.0120 0 (44) 0.0028 K.LDNLVAIFDINR.L
701.8833 1401.7521 1401.7616 -0.0095 0 89 9.5e-008 K.LDNLVAIFDINR.L
701.8833 1401.7521 1401.7616 -0.0095 0 (86) 2e-007 K.LDNLVAIFDINR.L
775.9119 1549.8093 1549.8174 -0.0081 1 56 0.00018 K.MFGIDKDAIVQAVK.G + Oxidation (M)
826.3910 1650.7674 1650.7865 -0.0191 0 85 2e-007 R.TVPFCSTFAAFFTR.A
965.9537 1929.8928 1929.9030 -0.0102 0 80 4.3e-007 R.SVPMSTVFYPSDGVATEK.A + Oxidation (M)
1005.0006 2007.9866 2007.9935 -0.0069 0 (38) 0.0075 K.NMAEQIIQEIYSQVQSK.K
1010.5234 2019.0322 2019.0636 -0.0314 0 93 2.3e-008 K.ILATPPQEDAPSVDIANIR.M
1012.9932 2023.9718 2023.9884 -0.0166 0 61 3.9e-005 K.NMAEQIIQEIYSQVQSK.K + Oxidation (M)
675.6666 2023.9781 2023.9884 -0.0103 0 (45) 0.0016 K.NMAEQIIQEIYSQVQSK.K + Oxidation (M)
827.7318 2480.1736 2480.1931 -0.0196 0 35 0.01 R.TSRPENAIIYSNNEDFQVGQAK.V
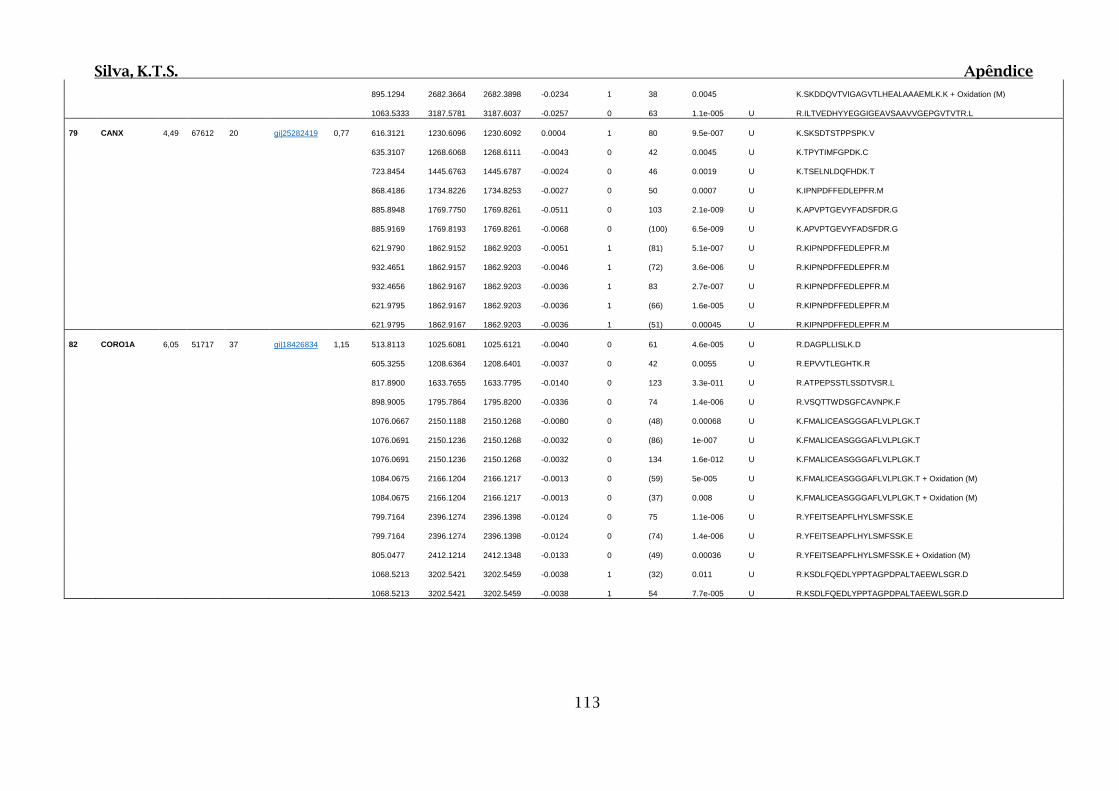
Silva, K.T.S. Apêndice
113
895.1294 2682.3664 2682.3898 -0.0234 1 38 0.0045 K.SKDDQVTVIGAGVTLHEALAAAEMLK.K + Oxidation (M)
1063.5333 3187.5781 3187.6037 -0.0257 0 63 1.1e-005 U R.ILTVEDHYYEGGIGEAVSAAVVGEPGVTVTR.L
79 CANX 4,49 67612 20
gi|25282419 0,77 616.3121 1230.6096 1230.6092 0.0004 1 80 9.5e-007 U K.SKSDTSTPPSPK.V
635.3107 1268.6068 1268.6111 -0.0043 0 42 0.0045 U K.TPYTIMFGPDK.C
723.8454 1445.6763 1445.6787 -0.0024 0 46 0.0019 U K.TSELNLDQFHDK.T
868.4186 1734.8226 1734.8253 -0.0027 0 50 0.0007 U K.IPNPDFFEDLEPFR.M
885.8948 1769.7750 1769.8261 -0.0511 0 103 2.1e-009 U K.APVPTGEVYFADSFDR.G
885.9169 1769.8193 1769.8261 -0.0068 0 (100) 6.5e-009 U K.APVPTGEVYFADSFDR.G
621.9790 1862.9152 1862.9203 -0.0051 1 (81) 5.1e-007 U R.KIPNPDFFEDLEPFR.M
932.4651 1862.9157 1862.9203 -0.0046 1 (72) 3.6e-006 U R.KIPNPDFFEDLEPFR.M
932.4656 1862.9167 1862.9203 -0.0036 1 83 2.7e-007 U R.KIPNPDFFEDLEPFR.M
621.9795 1862.9167 1862.9203 -0.0036 1 (66) 1.6e-005 U R.KIPNPDFFEDLEPFR.M
621.9795 1862.9167 1862.9203 -0.0036 1 (51) 0.00045 U R.KIPNPDFFEDLEPFR.M
82 CORO1A 6,05 51717 37
gi|18426834
1,15 513.8113 1025.6081 1025.6121 -0.0040 0 61 4.6e-005 U R.DAGPLLISLK.D
605.3255 1208.6364 1208.6401 -0.0037 0 42 0.0055 U R.EPVVTLEGHTK.R
817.8900 1633.7655 1633.7795 -0.0140 0 123 3.3e-011 U R.ATPEPSSTLSSDTVSR.L
898.9005 1795.7864 1795.8200 -0.0336 0 74 1.4e-006 U R.VSQTTWDSGFCAVNPK.F
1076.0667 2150.1188 2150.1268 -0.0080 0 (48) 0.00068 U K.FMALICEASGGGAFLVLPLGK.T
1076.0691 2150.1236 2150.1268 -0.0032 0 (86) 1e-007 U K.FMALICEASGGGAFLVLPLGK.T
1076.0691 2150.1236 2150.1268 -0.0032 0 134 1.6e-012 U K.FMALICEASGGGAFLVLPLGK.T
1084.0675 2166.1204 2166.1217 -0.0013 0 (59) 5e-005 U K.FMALICEASGGGAFLVLPLGK.T + Oxidation (M)
1084.0675 2166.1204 2166.1217 -0.0013 0 (37) 0.008 U K.FMALICEASGGGAFLVLPLGK.T + Oxidation (M)
799.7164 2396.1274 2396.1398 -0.0124 0 75 1.1e-006 U R.YFEITSEAPFLHYLSMFSSK.E
799.7164 2396.1274 2396.1398 -0.0124 0 (74) 1.4e-006 U R.YFEITSEAPFLHYLSMFSSK.E
805.0477 2412.1214 2412.1348 -0.0133 0 (49) 0.00036 U R.YFEITSEAPFLHYLSMFSSK.E + Oxidation (M)
1068.5213 3202.5421 3202.5459 -0.0038 1 (32) 0.011 U R.KSDLFQEDLYPPTAGPDPALTAEEWLSGR.D
1068.5213 3202.5421 3202.5459 -0.0038 1 54 7.7e-005 U R.KSDLFQEDLYPPTAGPDPALTAEEWLSGR.D
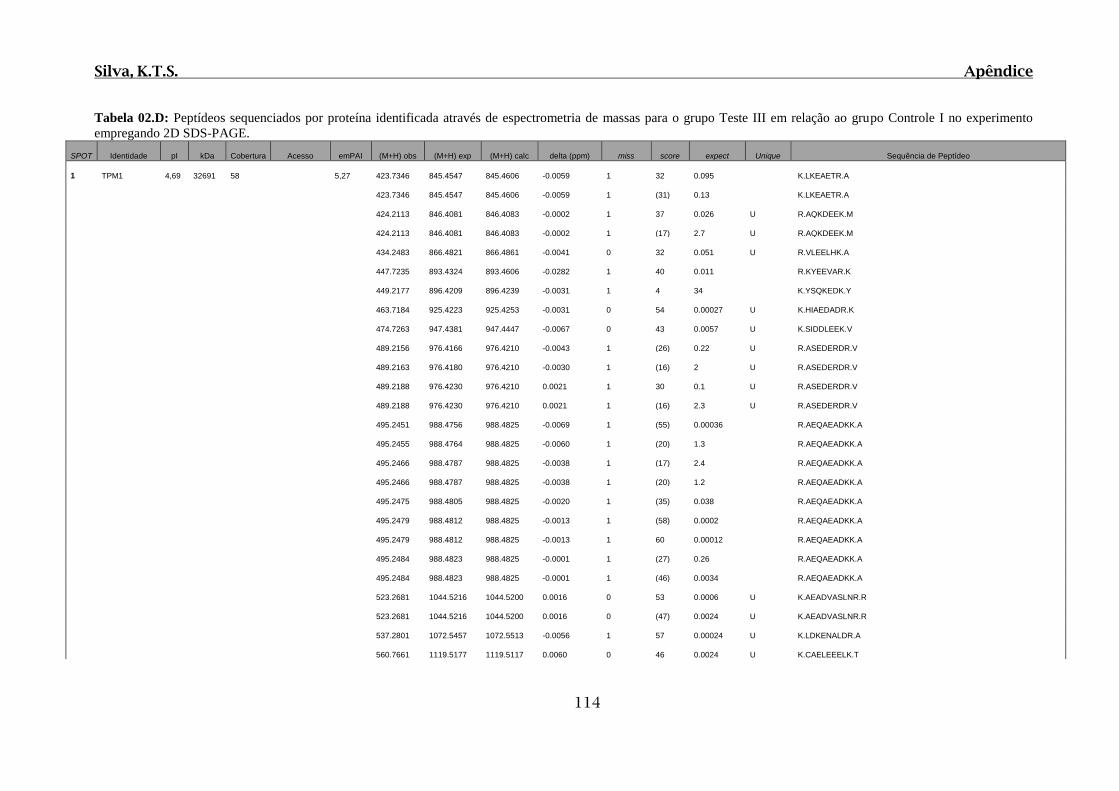
Silva, K.T.S. Apêndice
114
Tabela 02.D: Peptídeos sequenciados por proteína identificada através de espectrometria de massas para o grupo Teste III em relação ao grupo Controle I no experimento
empregando 2D SDS-PAGE.
SPOT Identidade pI kDa Cobertura Acesso emPAI (M+H) obs (M+H) exp (M+H) calc delta (ppm) miss score expect Unique Sequência de Peptídeo
1 TPM1 4,69 32691 58
gi|149028892 5,27 423.7346 845.4547 845.4606 -0.0059 1 32 0.095 K.LKEAETR.A
423.7346 845.4547 845.4606 -0.0059 1 (31) 0.13 K.LKEAETR.A
424.2113 846.4081 846.4083 -0.0002 1 37 0.026 U R.AQKDEEK.M
424.2113 846.4081 846.4083 -0.0002 1 (17) 2.7 U R.AQKDEEK.M
434.2483 866.4821 866.4861 -0.0041 0 32 0.051 U R.VLEELHK.A
447.7235 893.4324 893.4606 -0.0282 1 40 0.011 R.KYEEVAR.K
449.2177 896.4209 896.4239 -0.0031 1 4 34 K.YSQKEDK.Y
463.7184 925.4223 925.4253 -0.0031 0 54 0.00027 U K.HIAEDADR.K
474.7263 947.4381 947.4447 -0.0067 0 43 0.0057 U K.SIDDLEEK.V
489.2156 976.4166 976.4210 -0.0043 1 (26) 0.22 U R.ASEDERDR.V
489.2163 976.4180 976.4210 -0.0030 1 (16) 2 U R.ASEDERDR.V
489.2188 976.4230 976.4210 0.0021 1 30 0.1 U R.ASEDERDR.V
489.2188 976.4230 976.4210 0.0021 1 (16) 2.3 U R.ASEDERDR.V
495.2451 988.4756 988.4825 -0.0069 1 (55) 0.00036 R.AEQAEADKK.A
495.2455 988.4764 988.4825 -0.0060 1 (20) 1.3 R.AEQAEADKK.A
495.2466 988.4787 988.4825 -0.0038 1 (17) 2.4 R.AEQAEADKK.A
495.2466 988.4787 988.4825 -0.0038 1 (20) 1.2 R.AEQAEADKK.A
495.2475 988.4805 988.4825 -0.0020 1 (35) 0.038 R.AEQAEADKK.A
495.2479 988.4812 988.4825 -0.0013 1 (58) 0.0002 R.AEQAEADKK.A
495.2479 988.4812 988.4825 -0.0013 1 60 0.00012 R.AEQAEADKK.A
495.2484 988.4823 988.4825 -0.0001 1 (27) 0.26 R.AEQAEADKK.A
495.2484 988.4823 988.4825 -0.0001 1 (46) 0.0034 R.AEQAEADKK.A
523.2681 1044.5216 1044.5200 0.0016 0 53 0.0006 U K.AEADVASLNR.R
523.2681 1044.5216 1044.5200 0.0016 0 (47) 0.0024 U K.AEADVASLNR.R
537.2801 1072.5457 1072.5513 -0.0056 1 57 0.00024 U K.LDKENALDR.A
560.7661 1119.5177 1119.5117 0.0060 0 46 0.0024 U K.CAELEEELK.T
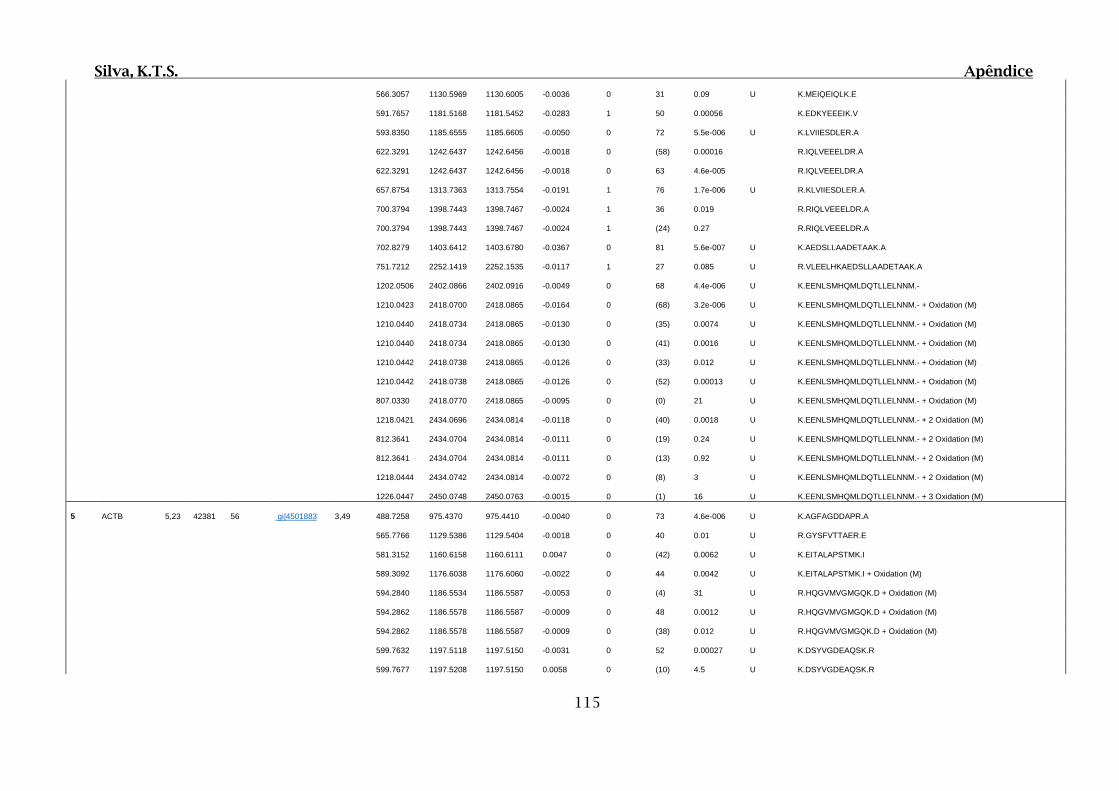
Silva, K.T.S. Apêndice
115
566.3057 1130.5969 1130.6005 -0.0036 0 31 0.09 U K.MEIQEIQLK.E
591.7657 1181.5168 1181.5452 -0.0283 1 50 0.00056 K.EDKYEEEIK.V
593.8350 1185.6555 1185.6605 -0.0050 0 72 5.5e-006 U K.LVIIESDLER.A
622.3291 1242.6437 1242.6456 -0.0018 0 (58) 0.00016 R.IQLVEEELDR.A
622.3291 1242.6437 1242.6456 -0.0018 0 63 4.6e-005 R.IQLVEEELDR.A
657.8754 1313.7363 1313.7554 -0.0191 1 76 1.7e-006 U R.KLVIIESDLER.A
700.3794 1398.7443 1398.7467 -0.0024 1 36 0.019 R.RIQLVEEELDR.A
700.3794 1398.7443 1398.7467 -0.0024 1 (24) 0.27 R.RIQLVEEELDR.A
702.8279 1403.6412 1403.6780 -0.0367 0 81 5.6e-007 U K.AEDSLLAADETAAK.A
751.7212 2252.1419 2252.1535 -0.0117 1 27 0.085 U R.VLEELHKAEDSLLAADETAAK.A
1202.0506 2402.0866 2402.0916 -0.0049 0 68 4.4e-006 U K.EENLSMHQMLDQTLLELNNM.-
1210.0423 2418.0700 2418.0865 -0.0164 0 (68) 3.2e-006 U K.EENLSMHQMLDQTLLELNNM.- + Oxidation (M)
1210.0440 2418.0734 2418.0865 -0.0130 0 (35) 0.0074 U K.EENLSMHQMLDQTLLELNNM.- + Oxidation (M)
1210.0440 2418.0734 2418.0865 -0.0130 0 (41) 0.0016 U K.EENLSMHQMLDQTLLELNNM.- + Oxidation (M)
1210.0442 2418.0738 2418.0865 -0.0126 0 (33) 0.012 U K.EENLSMHQMLDQTLLELNNM.- + Oxidation (M)
1210.0442 2418.0738 2418.0865 -0.0126 0 (52) 0.00013 U K.EENLSMHQMLDQTLLELNNM.- + Oxidation (M)
807.0330 2418.0770 2418.0865 -0.0095 0 (0) 21 U K.EENLSMHQMLDQTLLELNNM.- + Oxidation (M)
1218.0421 2434.0696 2434.0814 -0.0118 0 (40) 0.0018 U K.EENLSMHQMLDQTLLELNNM.- + 2 Oxidation (M)
812.3641 2434.0704 2434.0814 -0.0111 0 (19) 0.24 U K.EENLSMHQMLDQTLLELNNM.- + 2 Oxidation (M)
812.3641 2434.0704 2434.0814 -0.0111 0 (13) 0.92 U K.EENLSMHQMLDQTLLELNNM.- + 2 Oxidation (M)
1218.0444 2434.0742 2434.0814 -0.0072 0 (8) 3 U K.EENLSMHQMLDQTLLELNNM.- + 2 Oxidation (M)
1226.0447 2450.0748 2450.0763 -0.0015 0 (1) 16 U K.EENLSMHQMLDQTLLELNNM.- + 3 Oxidation (M)
5 ACTB 5,23 42381 56 gi|4501883 3,49 488.7258 975.4370 975.4410 -0.0040 0 73 4.6e-006 U K.AGFAGDDAPR.A
565.7766 1129.5386 1129.5404 -0.0018 0 40 0.01 U R.GYSFVTTAER.E
581.3152 1160.6158 1160.6111 0.0047 0 (42) 0.0062 U K.EITALAPSTMK.I
589.3092 1176.6038 1176.6060 -0.0022 0 44 0.0042 U K.EITALAPSTMK.I + Oxidation (M)
594.2840 1186.5534 1186.5587 -0.0053 0 (4) 31 U R.HQGVMVGMGQK.D + Oxidation (M)
594.2862 1186.5578 1186.5587 -0.0009 0 48 0.0012 U R.HQGVMVGMGQK.D + Oxidation (M)
594.2862 1186.5578 1186.5587 -0.0009 0 (38) 0.012 U R.HQGVMVGMGQK.D + Oxidation (M)
599.7632 1197.5118 1197.5150 -0.0031 0 52 0.00027 U K.DSYVGDEAQSK.R
599.7677 1197.5208 1197.5150 0.0058 0 (10) 4.5 U K.DSYVGDEAQSK.R

Silva, K.T.S. Apêndice
116
599.7677 1197.5208 1197.5150 0.0058 0 (10) 5.5 U K.DSYVGDEAQSK.R
677.8144 1353.6143 1353.6161 -0.0018 1 51 0.00049 U K.DSYVGDEAQSKR.G
750.8570 1499.6994 1499.7005 -0.0011 0 1 51 U K.QEYDEAGPSIVHR.K
751.3669 1500.7192 1500.7262 -0.0070 0 (42) 0.0049 U K.IWHHSFYNELR.V
501.2473 1500.7201 1500.7262 -0.0061 0 53 0.00034 U K.IWHHSFYNELR.V
895.9433 1789.8720 1789.8846 -0.0126 0 (89) 7.4e-008 U K.SYELPDGQVITIGNER.F
895.9471 1789.8797 1789.8846 -0.0050 0 101 5.2e-009 U K.SYELPDGQVITIGNER.F
895.9471 1789.8797 1789.8846 -0.0050 0 (101) 5.3e-009 U K.SYELPDGQVITIGNER.F
978.5157 1955.0167 1955.0364 -0.0196 0 84 2e-007 U R.VAPEEHPTLLTEAPLNPK.A
652.6813 1955.0220 1955.0364 -0.0144 0 (63) 2.3e-005 U R.VAPEEHPTLLTEAPLNPK.A
1114.5319 2227.0492 2227.0579 -0.0087 0 (118) 6e-011 U K.DLYANNVLSGGTTMYPGIADR.M
1122.5256 2243.0366 2243.0528 -0.0162 0 126 8.7e-012 U K.DLYANNVLSGGTTMYPGIADR.M + Oxidation (M)
846.0524 2535.1353 2535.1509 -0.0155 0 (79) 2.5e-007 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5772 2535.1398 2535.1509 -0.0110 0 137 3.6e-013 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5808 2535.1470 2535.1509 -0.0038 0 (33) 0.0096 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5808 2535.1470 2535.1509 -0.0038 0 (31) 0.015 U K.LCYVALDFENEMATAASSSSLEK.S
1276.5815 2551.1484 2551.1458 0.0027 0 (14) 0.75 U K.LCYVALDFENEMATAASSSSLEK.S + Oxidation (M)
1276.5815 2551.1484 2551.1458 0.0027 0 (25) 0.06 U K.LCYVALDFENEMATAASSSSLEK.S + Oxidation (M)
1063.4917 3187.4533 3187.4664 -0.0132 0 87 2.9e-008 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C
1066.2055 3195.5947 3195.6023 -0.0076 0 (64) 8.9e-006 U R.TTGIVLDSGDGVTHNVPIYEGYALPHAIMR.L
1066.2055 3195.5947 3195.6023 -0.0076 0 84 9.2e-008 U R.TTGIVLDSGDGVTHNVPIYEGYALPHAIMR.L
1066.2055 3195.5947 3195.6023 -0.0076 0 (59) 2.9e-005 U R.TTGIVLDSGDGVTHNVPIYEGYALPHAIMR.L
1068.8230 3203.4472 3203.4613 -0.0142 0 (37) 0.0028 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C + Oxidation (M)
1068.8230 3203.4472 3203.4613 -0.0142 0 (22) 0.09 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C + Oxidation (M)
1068.8245 3203.4517 3203.4613 -0.0097 0 (46) 0.0003 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C + Oxidation (M)
6 ACTB 5,31 42249 58 gi|4501889 2,86 488.7269 975.4392 975.4410 -0.0018 0 70 1e-005 U K.AGFAGDDAPR.A
581.3119 1160.6093 1160.6111 -0.0018 0 58 0.00018 U K.EITALAPSTMK.I
589.3089 1176.6033 1176.6060 -0.0027 0 (18) 1.6 U K.EITALAPSTMK.I + Oxidation (M)
594.2862 1186.5578 1186.5587 -0.0009 0 (3) 43 U R.HQGVMVGMGQK.D + Oxidation (M)
594.2873 1186.5601 1186.5587 0.0014 0 62 6e-005 U R.HQGVMVGMGQK.D + Oxidation (M)
599.7651 1197.5156 1197.5150 0.0006 0 51 0.00037 U K.DSYVGDEAQSK.R
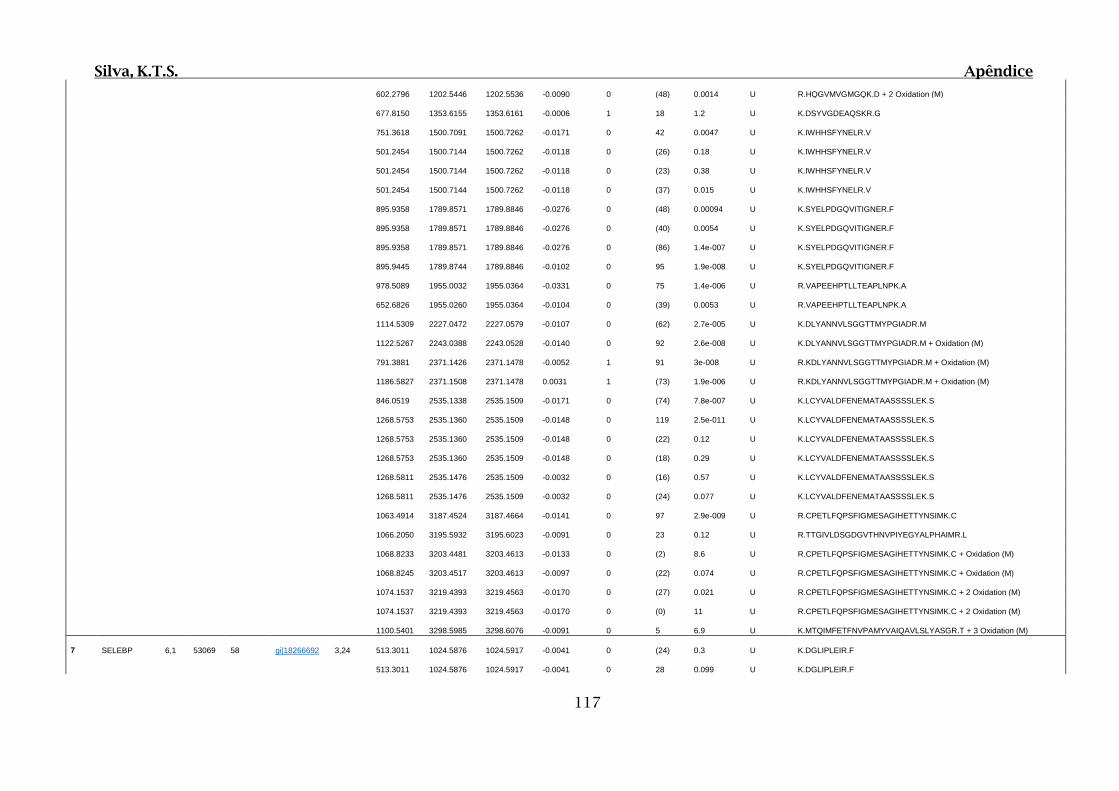
Silva, K.T.S. Apêndice
117
602.2796 1202.5446 1202.5536 -0.0090 0 (48) 0.0014 U R.HQGVMVGMGQK.D + 2 Oxidation (M)
677.8150 1353.6155 1353.6161 -0.0006 1 18 1.2 U K.DSYVGDEAQSKR.G
751.3618 1500.7091 1500.7262 -0.0171 0 42 0.0047 U K.IWHHSFYNELR.V
501.2454 1500.7144 1500.7262 -0.0118 0 (26) 0.18 U K.IWHHSFYNELR.V
501.2454 1500.7144 1500.7262 -0.0118 0 (23) 0.38 U K.IWHHSFYNELR.V
501.2454 1500.7144 1500.7262 -0.0118 0 (37) 0.015 U K.IWHHSFYNELR.V
895.9358 1789.8571 1789.8846 -0.0276 0 (48) 0.00094 U K.SYELPDGQVITIGNER.F
895.9358 1789.8571 1789.8846 -0.0276 0 (40) 0.0054 U K.SYELPDGQVITIGNER.F
895.9358 1789.8571 1789.8846 -0.0276 0 (86) 1.4e-007 U K.SYELPDGQVITIGNER.F
895.9445 1789.8744 1789.8846 -0.0102 0 95 1.9e-008 U K.SYELPDGQVITIGNER.F
978.5089 1955.0032 1955.0364 -0.0331 0 75 1.4e-006 U R.VAPEEHPTLLTEAPLNPK.A
652.6826 1955.0260 1955.0364 -0.0104 0 (39) 0.0053 U R.VAPEEHPTLLTEAPLNPK.A
1114.5309 2227.0472 2227.0579 -0.0107 0 (62) 2.7e-005 U K.DLYANNVLSGGTTMYPGIADR.M
1122.5267 2243.0388 2243.0528 -0.0140 0 92 2.6e-008 U K.DLYANNVLSGGTTMYPGIADR.M + Oxidation (M)
791.3881 2371.1426 2371.1478 -0.0052 1 91 3e-008 U R.KDLYANNVLSGGTTMYPGIADR.M + Oxidation (M)
1186.5827 2371.1508 2371.1478 0.0031 1 (73) 1.9e-006 U R.KDLYANNVLSGGTTMYPGIADR.M + Oxidation (M)
846.0519 2535.1338 2535.1509 -0.0171 0 (74) 7.8e-007 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5753 2535.1360 2535.1509 -0.0148 0 119 2.5e-011 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5753 2535.1360 2535.1509 -0.0148 0 (22) 0.12 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5753 2535.1360 2535.1509 -0.0148 0 (18) 0.29 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5811 2535.1476 2535.1509 -0.0032 0 (16) 0.57 U K.LCYVALDFENEMATAASSSSLEK.S
1268.5811 2535.1476 2535.1509 -0.0032 0 (24) 0.077 U K.LCYVALDFENEMATAASSSSLEK.S
1063.4914 3187.4524 3187.4664 -0.0141 0 97 2.9e-009 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C
1066.2050 3195.5932 3195.6023 -0.0091 0 23 0.12 U R.TTGIVLDSGDGVTHNVPIYEGYALPHAIMR.L
1068.8233 3203.4481 3203.4613 -0.0133 0 (2) 8.6 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C + Oxidation (M)
1068.8245 3203.4517 3203.4613 -0.0097 0 (22) 0.074 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C + Oxidation (M)
1074.1537 3219.4393 3219.4563 -0.0170 0 (27) 0.021 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C + 2 Oxidation (M)
1074.1537 3219.4393 3219.4563 -0.0170 0 (0) 11 U R.CPETLFQPSFIGMESAGIHETTYNSIMK.C + 2 Oxidation (M)
1100.5401 3298.5985 3298.6076 -0.0091 0 5 6.9 U K.MTQIMFETFNVPAMYVAIQAVLSLYASGR.T + 3 Oxidation (M)
7 SELEBP 6,1 53069 58 gi|18266692 3,24 513.3011 1024.5876 1024.5917 -0.0041 0 (24) 0.3 U K.DGLIPLEIR.F
513.3011 1024.5876 1024.5917 -0.0041 0 28 0.099 U K.DGLIPLEIR.F

Silva, K.T.S. Apêndice
118
525.7880 1049.5615 1049.5658 -0.0043 0 44 0.0042 U K.QFYPNLIR.E
549.8442 1097.6738 1097.6808 -0.0070 0 57 9.5e-005 U K.LILPSIISSR.I
553.7579 1105.5013 1105.5040 -0.0027 0 55 0.00026 U K.NEGGTWSVEK.V
575.3137 1148.6129 1148.6189 -0.0060 0 (43) 0.0051 U K.VIEPNEIHAK.C
575.3137 1148.6129 1148.6189 -0.0060 0 51 0.00082 U K.VIEPNEIHAK.C
612.3155 1222.6164 1222.6207 -0.0043 0 (15) 2.7 U K.SPHYSQVIHR.L
612.3171 1222.6197 1222.6207 -0.0010 0 24 0.35 U K.SPHYSQVIHR.L
612.3171 1222.6197 1222.6207 -0.0010 0 (22) 0.53 U K.SPHYSQVIHR.L
617.3253 1232.6361 1232.6401 -0.0040 0 71 7.2e-006 U R.IYVVDVGSEPR.A
632.3331 1262.6516 1262.6659 -0.0144 0 64 3.6e-005 U K.LNPNFLVDFGK.E
632.3331 1262.6516 1262.6659 -0.0144 0 (18) 1.5 U K.LNPNFLVDFGK.E
632.3331 1262.6516 1262.6659 -0.0144 0 (4) 34 U K.LNPNFLVDFGK.E
651.8580 1301.7015 1301.7092 -0.0076 0 41 0.007 U K.EPLGPALAHELR.Y
666.8904 1331.7662 1331.7813 -0.0151 0 84 2.1e-007 U R.LTGQIFLGGSIVK.G
671.4050 1340.7954 1340.8027 -0.0073 1 31 0.036 U R.DKLILPSIISSR.I
685.2892 1368.5639 1368.5656 -0.0017 0 18 0.5 U R.YPGGDCSSDIWI.-
685.2892 1368.5639 1368.5656 -0.0017 0 (18) 0.55 U R.YPGGDCSSDIWI.-
705.8200 1409.6254 1409.6319 -0.0065 0 24 0.2 U K.CGPGYATPLEAMK.G + Oxidation (M)
727.8642 1453.7139 1453.7275 -0.0136 0 48 0.0011 U R.EEIVYLPCIYR.N
755.8888 1509.7630 1509.7715 -0.0086 0 (33) 0.037 U K.GGFVLLDGETFEVK.G
755.8904 1509.7662 1509.7715 -0.0053 0 69 9.3e-006 U K.GGFVLLDGETFEVK.G
755.8918 1509.7691 1509.7715 -0.0024 0 (52) 0.00047 U K.GGFVLLDGETFEVK.G
770.4101 1538.8056 1538.8127 -0.0070 0 (37) 0.014 U R.VPGGPQMIQLSLDGK.R
773.8876 1545.7606 1545.7715 -0.0109 0 90 8.9e-008 U R.LYVTTSLYSAWDK.Q
778.4055 1554.7965 1554.8076 -0.0111 0 70 7.5e-006 U R.VPGGPQMIQLSLDGK.R + Oxidation (M)
834.4166 1666.8186 1666.8256 -0.0070 0 43 0.0034 U R.FLYFSNWLHGDIR.Q
556.6146 1666.8221 1666.8256 -0.0035 0 (23) 0.3 U R.FLYFSNWLHGDIR.Q
848.4609 1694.9072 1694.9138 -0.0065 1 57 0.00011 U K.RVPGGPQMIQLSLDGK.R
571.3087 1710.9042 1710.9087 -0.0045 1 (46) 0.0012 U K.RVPGGPQMIQLSLDGK.R + Oxidation (M)
930.4531 1858.8916 1858.9036 -0.0120 0 2 34 U R.HNIMVSTEWAAPNVFK.D + Oxidation (M)
959.4615 1916.9083 1916.9367 -0.0284 0 90 4.6e-008 U R.NTGIEAPDYLATVDVDPK.S
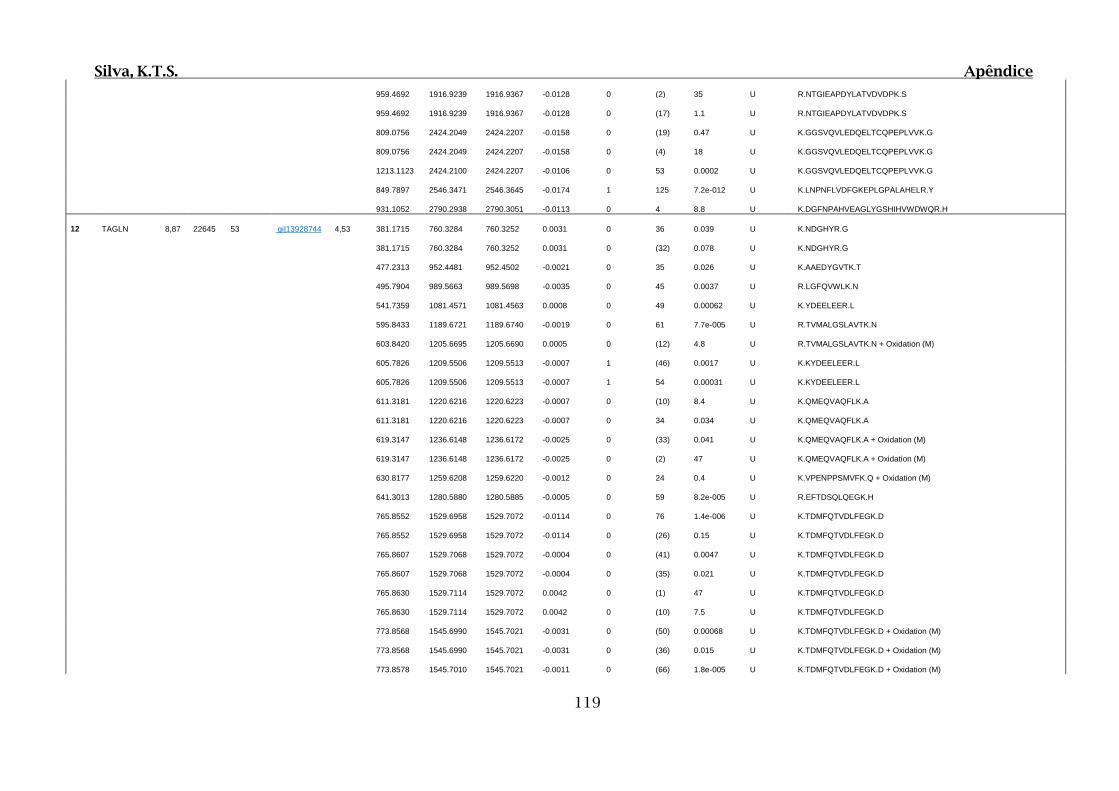
Silva, K.T.S. Apêndice
119
959.4692 1916.9239 1916.9367 -0.0128 0 (2) 35 U R.NTGIEAPDYLATVDVDPK.S
959.4692 1916.9239 1916.9367 -0.0128 0 (17) 1.1 U R.NTGIEAPDYLATVDVDPK.S
809.0756 2424.2049 2424.2207 -0.0158 0 (19) 0.47 U K.GGSVQVLEDQELTCQPEPLVVK.G
809.0756 2424.2049 2424.2207 -0.0158 0 (4) 18 U K.GGSVQVLEDQELTCQPEPLVVK.G
1213.1123 2424.2100 2424.2207 -0.0106 0 53 0.0002 U K.GGSVQVLEDQELTCQPEPLVVK.G
849.7897 2546.3471 2546.3645 -0.0174 1 125 7.2e-012 U K.LNPNFLVDFGKEPLGPALAHELR.Y
931.1052 2790.2938 2790.3051 -0.0113 0 4 8.8 U K.DGFNPAHVEAGLYGSHIHVWDWQR.H
12 TAGLN 8,87 22645 53 gi|13928744 4,53 381.1715 760.3284 760.3252 0.0031 0 36 0.039 U K.NDGHYR.G
381.1715 760.3284 760.3252 0.0031 0 (32) 0.078 U K.NDGHYR.G
477.2313 952.4481 952.4502 -0.0021 0 35 0.026 U K.AAEDYGVTK.T
495.7904 989.5663 989.5698 -0.0035 0 45 0.0037 U R.LGFQVWLK.N
541.7359 1081.4571 1081.4563 0.0008 0 49 0.00062 U K.YDEELEER.L
595.8433 1189.6721 1189.6740 -0.0019 0 61 7.7e-005 U R.TVMALGSLAVTK.N
603.8420 1205.6695 1205.6690 0.0005 0 (12) 4.8 U R.TVMALGSLAVTK.N + Oxidation (M)
605.7826 1209.5506 1209.5513 -0.0007 1 (46) 0.0017 U K.KYDEELEER.L
605.7826 1209.5506 1209.5513 -0.0007 1 54 0.00031 U K.KYDEELEER.L
611.3181 1220.6216 1220.6223 -0.0007 0 (10) 8.4 U K.QMEQVAQFLK.A
611.3181 1220.6216 1220.6223 -0.0007 0 34 0.034 U K.QMEQVAQFLK.A
619.3147 1236.6148 1236.6172 -0.0025 0 (33) 0.041 U K.QMEQVAQFLK.A + Oxidation (M)
619.3147 1236.6148 1236.6172 -0.0025 0 (2) 47 U K.QMEQVAQFLK.A + Oxidation (M)
630.8177 1259.6208 1259.6220 -0.0012 0 24 0.4 U K.VPENPPSMVFK.Q + Oxidation (M)
641.3013 1280.5880 1280.5885 -0.0005 0 59 8.2e-005 U R.EFTDSQLQEGK.H
765.8552 1529.6958 1529.7072 -0.0114 0 76 1.4e-006 U K.TDMFQTVDLFEGK.D
765.8552 1529.6958 1529.7072 -0.0114 0 (26) 0.15 U K.TDMFQTVDLFEGK.D
765.8607 1529.7068 1529.7072 -0.0004 0 (41) 0.0047 U K.TDMFQTVDLFEGK.D
765.8607 1529.7068 1529.7072 -0.0004 0 (35) 0.021 U K.TDMFQTVDLFEGK.D
765.8630 1529.7114 1529.7072 0.0042 0 (1) 47 U K.TDMFQTVDLFEGK.D
765.8630 1529.7114 1529.7072 0.0042 0 (10) 7.5 U K.TDMFQTVDLFEGK.D
773.8568 1545.6990 1545.7021 -0.0031 0 (50) 0.00068 U K.TDMFQTVDLFEGK.D + Oxidation (M)
773.8568 1545.6990 1545.7021 -0.0031 0 (36) 0.015 U K.TDMFQTVDLFEGK.D + Oxidation (M)
773.8578 1545.7010 1545.7021 -0.0011 0 (66) 1.8e-005 U K.TDMFQTVDLFEGK.D + Oxidation (M)

Silva, K.T.S. Apêndice
120
773.8578 1545.7010 1545.7021 -0.0011 0 (71) 4.9e-006 U K.TDMFQTVDLFEGK.D + Oxidation (M)
709.6838 2126.0295 2126.0401 -0.0106 0 (33) 0.024 U R.LVEWIVMQCGPDVGRPDR.G
1064.0252 2126.0358 2126.0401 -0.0043 0 (13) 2.5 U R.LVEWIVMQCGPDVGRPDR.G
715.0156 2142.0251 2142.0350 -0.0100 0 34 0.018 U R.LVEWIVMQCGPDVGRPDR.G + Oxidation (M)
18 CALR 4,33 48137 52 gi|11693172 1,21 369.2126 736.4106 736.4119 -0.0013 0 (12) 7.2 U K.FVLSSGK.F
369.2126 736.4106 736.4119 -0.0013 0 17 2.1 U K.FVLSSGK.F
487.7690 973.5234 973.5233 0.0001 0 22 0.72 U K.LFPGGLDQK.D
488.2467 974.4788 974.4781 0.0007 0 29 0.15 U K.GLQTSQDAR.F
488.2467 974.4788 974.4781 0.0007 0 (15) 3.7 U K.GLQTSQDAR.F
565.2631 1128.5116 1128.5087 0.0028 1 55 0.00022 U K.FYGDQEKDK.G
609.8081 1217.6017 1217.6000 0.0017 1 51 0.0008 U K.DKGLQTSQDAR.F
610.3520 1218.6895 1218.6972 -0.0078 0 60 8.4e-005 U K.GQTLVVQFTVK.H
726.3301 1450.6457 1450.6477 -0.0020 0 77 1.1e-006 U K.EQFLDGDAWTNR.W
738.8263 1475.6380 1475.6463 -0.0083 0 68 6.7e-006 U K.HEQNIDCGGGYVK.L
595.6178 1783.8315 1783.8377 -0.0062 1 69 6.1e-006 U K.IKDPDAAKPEDWDER.A
892.9239 1783.8333 1783.8377 -0.0044 1 (59) 6.5e-005 U K.IKDPDAAKPEDWDER.A
1196.0507 2390.0868 2390.0914 -0.0046 0 121 2.7e-011 U K.IDNSQVESGSLEDDWDFLPPK.K
1196.0520 2390.0894 2390.0914 -0.0020 0 (65) 1.1e-005 U K.IDNSQVESGSLEDDWDFLPPK.K
1196.0520 2390.0894 2390.0914 -0.0020 0 (31) 0.026 U K.IDNSQVESGSLEDDWDFLPPK.K
1196.0544 2390.0942 2390.0914 0.0028 0 (1) 27 U K.IDNSQVESGSLEDDWDFLPPK.K
825.0055 2471.9947 2472.0032 -0.0085 0 38 0.00083 U K.DMHGDSEYNIMFGPDICGPGTK.K + 2 Oxidation (M)
840.3968 2518.1685 2518.1864 -0.0178 1 76 8.5e-007 U K.IDNSQVESGSLEDDWDFLPPKK.I
840.3978 2518.1716 2518.1864 -0.0148 1 (57) 7.3e-005 U K.IDNSQVESGSLEDDWDFLPPKK.I
840.3978 2518.1716 2518.1864 -0.0148 1 (65) 1.2e-005 U K.IDNSQVESGSLEDDWDFLPPKK.I
952.7977 2855.3713 2855.3800 -0.0087 1 1 18 U R.CKDDEFTHLYTLIVRPDNTYEVK.I
1090.1683 3267.4831 3267.4884 -0.0053 0 67 3.1e-006 U K.SGTIFDNFLITNDEAYAEEFGNETWGVTK.A
1379.6700 4135.9882 4135.9843 0.0038 0 13 0.55 U K.GTWIHPEIDNPEYSPDANIYAYDSFAVLGLDLWQVK.S
20 HSP90B1 4,72 92998 30
gi|210032365 0,37 377.2083 752.4020 752.4068 -0.0048 1 28 0.15 U K.KSDYIK.L
502.7785 1003.5424 1003.5451 -0.0027 1 7 22 U K.KTFEINPR.H
541.2781 1080.5417 1080.5352 0.0065 0 9 12 U K.FAFQAEVNR.M
570.2937 1138.5729 1138.5731 -0.0002 0 72 6.6e-006 U K.LGVIEDHSNR.T
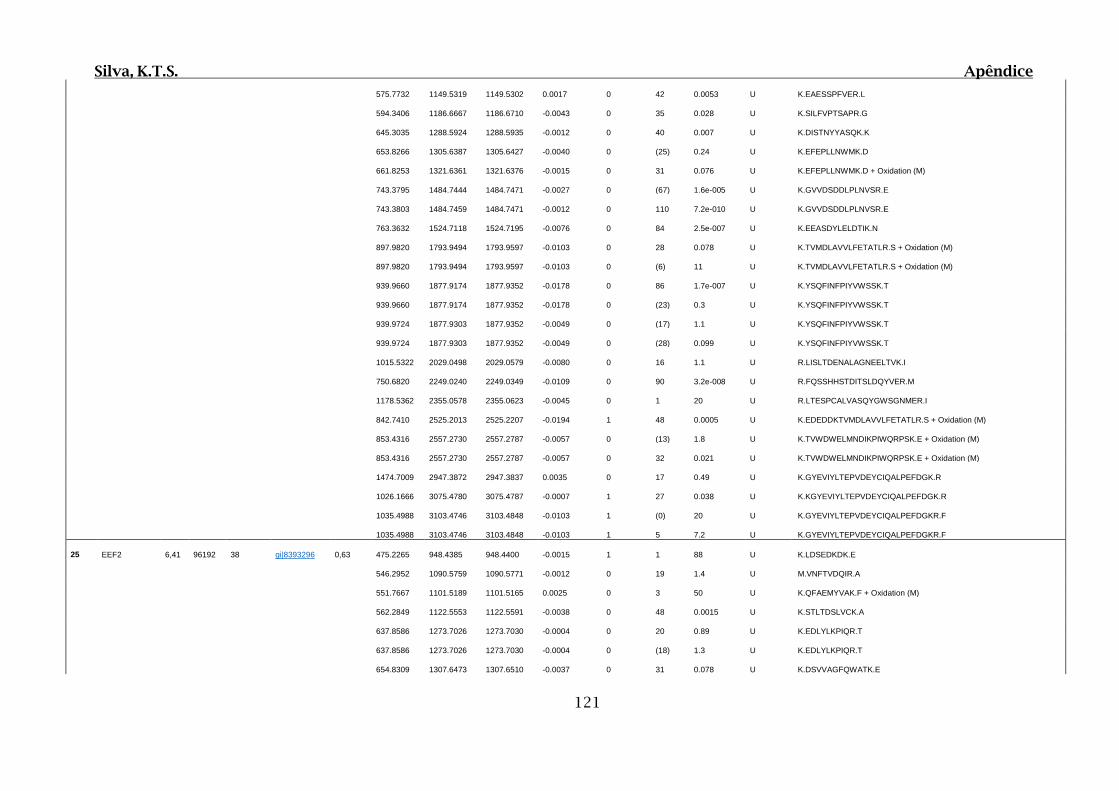
Silva, K.T.S. Apêndice
121
575.7732 1149.5319 1149.5302 0.0017 0 42 0.0053 U K.EAESSPFVER.L
594.3406 1186.6667 1186.6710 -0.0043 0 35 0.028 U K.SILFVPTSAPR.G
645.3035 1288.5924 1288.5935 -0.0012 0 40 0.007 U K.DISTNYYASQK.K
653.8266 1305.6387 1305.6427 -0.0040 0 (25) 0.24 U K.EFEPLLNWMK.D
661.8253 1321.6361 1321.6376 -0.0015 0 31 0.076 U K.EFEPLLNWMK.D + Oxidation (M)
743.3795 1484.7444 1484.7471 -0.0027 0 (67) 1.6e-005 U K.GVVDSDDLPLNVSR.E
743.3803 1484.7459 1484.7471 -0.0012 0 110 7.2e-010 U K.GVVDSDDLPLNVSR.E
763.3632 1524.7118 1524.7195 -0.0076 0 84 2.5e-007 U K.EEASDYLELDTIK.N
897.9820 1793.9494 1793.9597 -0.0103 0 28 0.078 U K.TVMDLAVVLFETATLR.S + Oxidation (M)
897.9820 1793.9494 1793.9597 -0.0103 0 (6) 11 U K.TVMDLAVVLFETATLR.S + Oxidation (M)
939.9660 1877.9174 1877.9352 -0.0178 0 86 1.7e-007 U K.YSQFINFPIYVWSSK.T
939.9660 1877.9174 1877.9352 -0.0178 0 (23) 0.3 U K.YSQFINFPIYVWSSK.T
939.9724 1877.9303 1877.9352 -0.0049 0 (17) 1.1 U K.YSQFINFPIYVWSSK.T
939.9724 1877.9303 1877.9352 -0.0049 0 (28) 0.099 U K.YSQFINFPIYVWSSK.T
1015.5322 2029.0498 2029.0579 -0.0080 0 16 1.1 U R.LISLTDENALAGNEELTVK.I
750.6820 2249.0240 2249.0349 -0.0109 0 90 3.2e-008 U R.FQSSHHSTDITSLDQYVER.M
1178.5362 2355.0578 2355.0623 -0.0045 0 1 20 U R.LTESPCALVASQYGWSGNMER.I
842.7410 2525.2013 2525.2207 -0.0194 1 48 0.0005 U K.EDEDDKTVMDLAVVLFETATLR.S + Oxidation (M)
853.4316 2557.2730 2557.2787 -0.0057 0 (13) 1.8 U K.TVWDWELMNDIKPIWQRPSK.E + Oxidation (M)
853.4316 2557.2730 2557.2787 -0.0057 0 32 0.021 U K.TVWDWELMNDIKPIWQRPSK.E + Oxidation (M)
1474.7009 2947.3872 2947.3837 0.0035 0 17 0.49 U K.GYEVIYLTEPVDEYCIQALPEFDGK.R
1026.1666 3075.4780 3075.4787 -0.0007 1 27 0.038 U K.KGYEVIYLTEPVDEYCIQALPEFDGK.R
1035.4988 3103.4746 3103.4848 -0.0103 1 (0) 20 U K.GYEVIYLTEPVDEYCIQALPEFDGKR.F
1035.4988 3103.4746 3103.4848 -0.0103 1 5 7.2 U K.GYEVIYLTEPVDEYCIQALPEFDGKR.F
25 EEF2 6,41 96192 38 gi|8393296 0,63 475.2265 948.4385 948.4400 -0.0015 1 1 88 U K.LDSEDKDK.E
546.2952 1090.5759 1090.5771 -0.0012 0 19 1.4 U M.VNFTVDQIR.A
551.7667 1101.5189 1101.5165 0.0025 0 3 50 U K.QFAEMYVAK.F + Oxidation (M)
562.2849 1122.5553 1122.5591 -0.0038 0 48 0.0015 U K.STLTDSLVCK.A
637.8586 1273.7026 1273.7030 -0.0004 0 20 0.89 U K.EDLYLKPIQR.T
637.8586 1273.7026 1273.7030 -0.0004 0 (18) 1.3 U K.EDLYLKPIQR.T
654.8309 1307.6473 1307.6510 -0.0037 0 31 0.078 U K.DSVVAGFQWATK.E
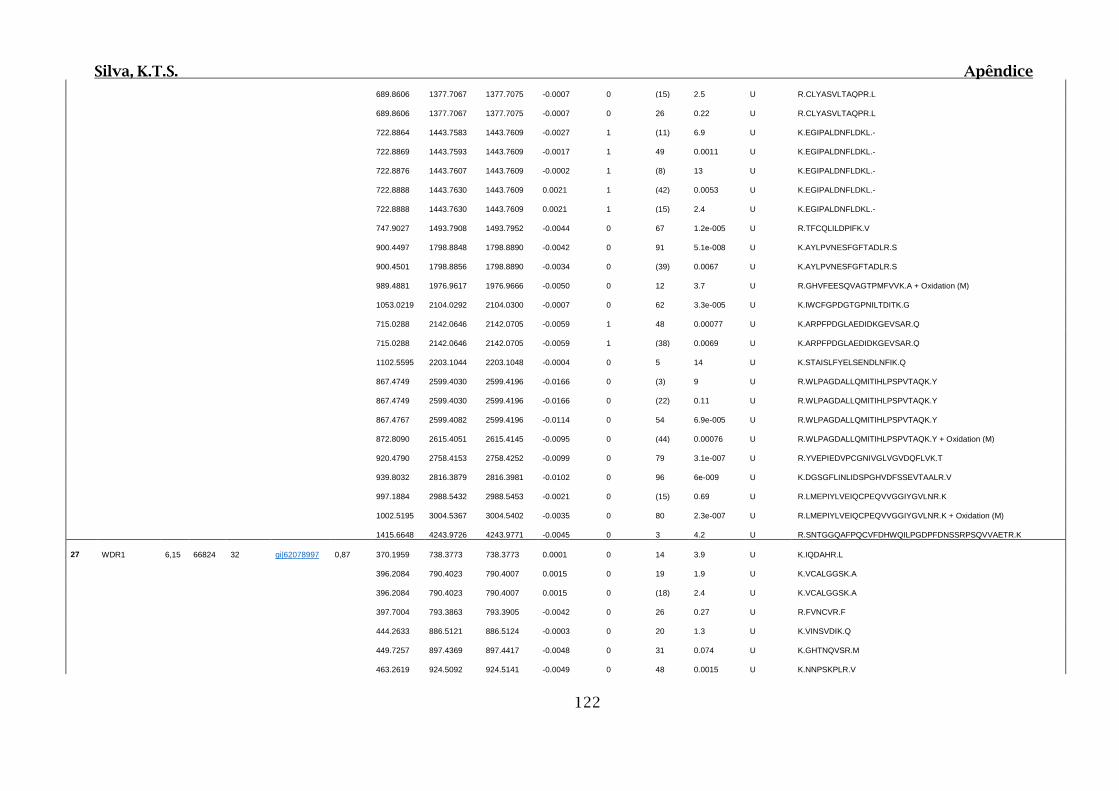
Silva, K.T.S. Apêndice
122
689.8606 1377.7067 1377.7075 -0.0007 0 (15) 2.5 U R.CLYASVLTAQPR.L
689.8606 1377.7067 1377.7075 -0.0007 0 26 0.22 U R.CLYASVLTAQPR.L
722.8864 1443.7583 1443.7609 -0.0027 1 (11) 6.9 U K.EGIPALDNFLDKL.-
722.8869 1443.7593 1443.7609 -0.0017 1 49 0.0011 U K.EGIPALDNFLDKL.-
722.8876 1443.7607 1443.7609 -0.0002 1 (8) 13 U K.EGIPALDNFLDKL.-
722.8888 1443.7630 1443.7609 0.0021 1 (42) 0.0053 U K.EGIPALDNFLDKL.-
722.8888 1443.7630 1443.7609 0.0021 1 (15) 2.4 U K.EGIPALDNFLDKL.-
747.9027 1493.7908 1493.7952 -0.0044 0 67 1.2e-005 U R.TFCQLILDPIFK.V
900.4497 1798.8848 1798.8890 -0.0042 0 91 5.1e-008 U K.AYLPVNESFGFTADLR.S
900.4501 1798.8856 1798.8890 -0.0034 0 (39) 0.0067 U K.AYLPVNESFGFTADLR.S
989.4881 1976.9617 1976.9666 -0.0050 0 12 3.7 U R.GHVFEESQVAGTPMFVVK.A + Oxidation (M)
1053.0219 2104.0292 2104.0300 -0.0007 0 62 3.3e-005 U K.IWCFGPDGTGPNILTDITK.G
715.0288 2142.0646 2142.0705 -0.0059 1 48 0.00077 U K.ARPFPDGLAEDIDKGEVSAR.Q
715.0288 2142.0646 2142.0705 -0.0059 1 (38) 0.0069 U K.ARPFPDGLAEDIDKGEVSAR.Q
1102.5595 2203.1044 2203.1048 -0.0004 0 5 14 U K.STAISLFYELSENDLNFIK.Q
867.4749 2599.4030 2599.4196 -0.0166 0 (3) 9 U R.WLPAGDALLQMITIHLPSPVTAQK.Y
867.4749 2599.4030 2599.4196 -0.0166 0 (22) 0.11 U R.WLPAGDALLQMITIHLPSPVTAQK.Y
867.4767 2599.4082 2599.4196 -0.0114 0 54 6.9e-005 U R.WLPAGDALLQMITIHLPSPVTAQK.Y
872.8090 2615.4051 2615.4145 -0.0095 0 (44) 0.00076 U R.WLPAGDALLQMITIHLPSPVTAQK.Y + Oxidation (M)
920.4790 2758.4153 2758.4252 -0.0099 0 79 3.1e-007 U R.YVEPIEDVPCGNIVGLVGVDQFLVK.T
939.8032 2816.3879 2816.3981 -0.0102 0 96 6e-009 U K.DGSGFLINLIDSPGHVDFSSEVTAALR.V
997.1884 2988.5432 2988.5453 -0.0021 0 (15) 0.69 U R.LMEPIYLVEIQCPEQVVGGIYGVLNR.K
1002.5195 3004.5367 3004.5402 -0.0035 0 80 2.3e-007 U R.LMEPIYLVEIQCPEQVVGGIYGVLNR.K + Oxidation (M)
1415.6648 4243.9726 4243.9771 -0.0045 0 3 4.2 U R.SNTGGQAFPQCVFDHWQILPGDPFDNSSRPSQVVAETR.K
27 WDR1 6,15 66824 32 gi|62078997 0,87 370.1959 738.3773 738.3773 0.0001 0 14 3.9 U K.IQDAHR.L
396.2084 790.4023 790.4007 0.0015 0 19 1.9 U K.VCALGGSK.A
396.2084 790.4023 790.4007 0.0015 0 (18) 2.4 U K.VCALGGSK.A
397.7004 793.3863 793.3905 -0.0042 0 26 0.27 U R.FVNCVR.F
444.2633 886.5121 886.5124 -0.0003 0 20 1.3 U K.VINSVDIK.Q
449.7257 897.4369 897.4417 -0.0048 0 31 0.074 U K.GHTNQVSR.M
463.2619 924.5092 924.5141 -0.0049 0 48 0.0015 U K.NNPSKPLR.V
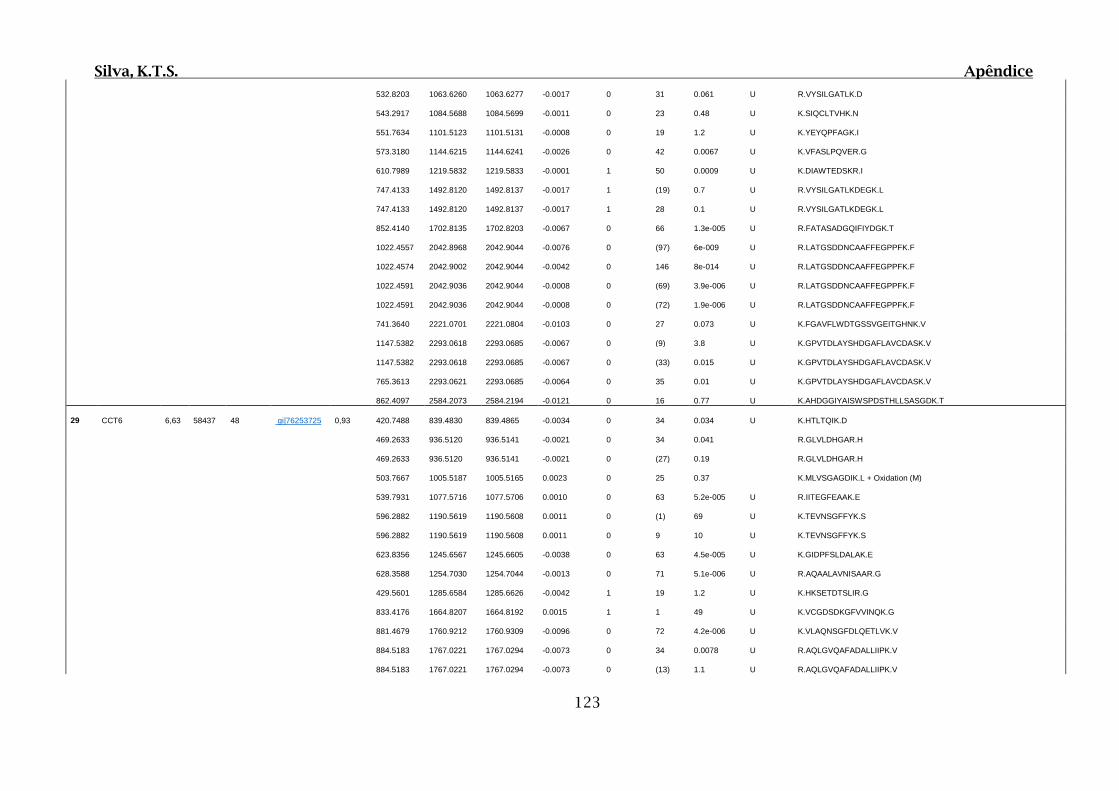
Silva, K.T.S. Apêndice
123
532.8203 1063.6260 1063.6277 -0.0017 0 31 0.061 U R.VYSILGATLK.D
543.2917 1084.5688 1084.5699 -0.0011 0 23 0.48 U K.SIQCLTVHK.N
551.7634 1101.5123 1101.5131 -0.0008 0 19 1.2 U K.YEYQPFAGK.I
573.3180 1144.6215 1144.6241 -0.0026 0 42 0.0067 U K.VFASLPQVER.G
610.7989 1219.5832 1219.5833 -0.0001 1 50 0.0009 U K.DIAWTEDSKR.I
747.4133 1492.8120 1492.8137 -0.0017 1 (19) 0.7 U R.VYSILGATLKDEGK.L
747.4133 1492.8120 1492.8137 -0.0017 1 28 0.1 U R.VYSILGATLKDEGK.L
852.4140 1702.8135 1702.8203 -0.0067 0 66 1.3e-005 U R.FATASADGQIFIYDGK.T
1022.4557 2042.8968 2042.9044 -0.0076 0 (97) 6e-009 U R.LATGSDDNCAAFFEGPPFK.F
1022.4574 2042.9002 2042.9044 -0.0042 0 146 8e-014 U R.LATGSDDNCAAFFEGPPFK.F
1022.4591 2042.9036 2042.9044 -0.0008 0 (69) 3.9e-006 U R.LATGSDDNCAAFFEGPPFK.F
1022.4591 2042.9036 2042.9044 -0.0008 0 (72) 1.9e-006 U R.LATGSDDNCAAFFEGPPFK.F
741.3640 2221.0701 2221.0804 -0.0103 0 27 0.073 U K.FGAVFLWDTGSSVGEITGHNK.V
1147.5382 2293.0618 2293.0685 -0.0067 0 (9) 3.8 U K.GPVTDLAYSHDGAFLAVCDASK.V
1147.5382 2293.0618 2293.0685 -0.0067 0 (33) 0.015 U K.GPVTDLAYSHDGAFLAVCDASK.V
765.3613 2293.0621 2293.0685 -0.0064 0 35 0.01 U K.GPVTDLAYSHDGAFLAVCDASK.V
862.4097 2584.2073 2584.2194 -0.0121 0 16 0.77 U K.AHDGGIYAISWSPDSTHLLSASGDK.T
29 CCT6 6,63 58437 48 gi|76253725 0,93 420.7488 839.4830 839.4865 -0.0034 0 34 0.034 U K.HTLTQIK.D
469.2633 936.5120 936.5141 -0.0021 0 34 0.041 R.GLVLDHGAR.H
469.2633 936.5120 936.5141 -0.0021 0 (27) 0.19 R.GLVLDHGAR.H
503.7667 1005.5187 1005.5165 0.0023 0 25 0.37 K.MLVSGAGDIK.L + Oxidation (M)
539.7931 1077.5716 1077.5706 0.0010 0 63 5.2e-005 U R.IITEGFEAAK.E
596.2882 1190.5619 1190.5608 0.0011 0 (1) 69 U K.TEVNSGFFYK.S
596.2882 1190.5619 1190.5608 0.0011 0 9 10 U K.TEVNSGFFYK.S
623.8356 1245.6567 1245.6605 -0.0038 0 63 4.5e-005 U K.GIDPFSLDALAK.E
628.3588 1254.7030 1254.7044 -0.0013 0 71 5.1e-006 U R.AQAALAVNISAAR.G
429.5601 1285.6584 1285.6626 -0.0042 1 19 1.2 U K.HKSETDTSLIR.G
833.4176 1664.8207 1664.8192 0.0015 1 1 49 U K.VCGDSDKGFVVINQK.G
881.4679 1760.9212 1760.9309 -0.0096 0 72 4.2e-006 U K.VLAQNSGFDLQETLVK.V
884.5183 1767.0221 1767.0294 -0.0073 0 34 0.0078 U R.AQLGVQAFADALLIIPK.V
884.5183 1767.0221 1767.0294 -0.0073 0 (13) 1.1 U R.AQLGVQAFADALLIIPK.V
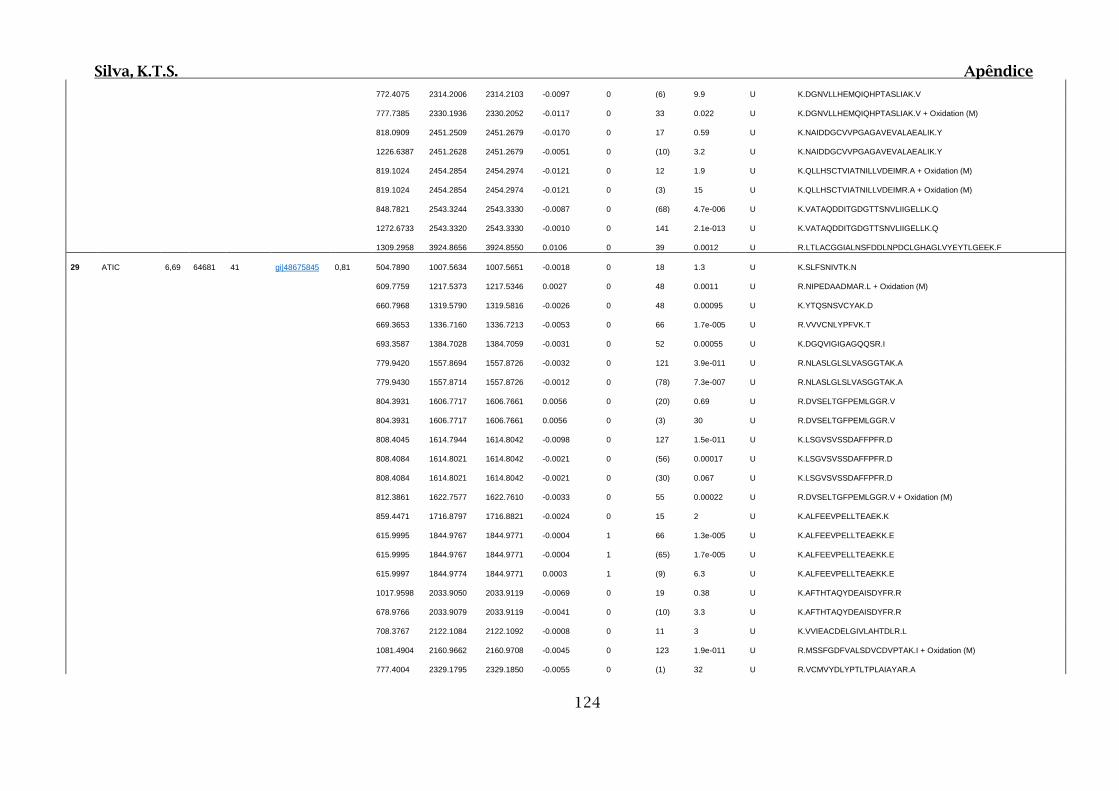
Silva, K.T.S. Apêndice
124
772.4075 2314.2006 2314.2103 -0.0097 0 (6) 9.9 U K.DGNVLLHEMQIQHPTASLIAK.V
777.7385 2330.1936 2330.2052 -0.0117 0 33 0.022 U K.DGNVLLHEMQIQHPTASLIAK.V + Oxidation (M)
818.0909 2451.2509 2451.2679 -0.0170 0 17 0.59 U K.NAIDDGCVVPGAGAVEVALAEALIK.Y
1226.6387 2451.2628 2451.2679 -0.0051 0 (10) 3.2 U K.NAIDDGCVVPGAGAVEVALAEALIK.Y
819.1024 2454.2854 2454.2974 -0.0121 0 12 1.9 U K.QLLHSCTVIATNILLVDEIMR.A + Oxidation (M)
819.1024 2454.2854 2454.2974 -0.0121 0 (3) 15 U K.QLLHSCTVIATNILLVDEIMR.A + Oxidation (M)
848.7821 2543.3244 2543.3330 -0.0087 0 (68) 4.7e-006 U K.VATAQDDITGDGTTSNVLIIGELLK.Q
1272.6733 2543.3320 2543.3330 -0.0010 0 141 2.1e-013 U K.VATAQDDITGDGTTSNVLIIGELLK.Q
1309.2958 3924.8656 3924.8550 0.0106 0 39 0.0012 U R.LTLACGGIALNSFDDLNPDCLGHAGLVYEYTLGEEK.F
29 ATIC 6,69 64681 41 gi|48675845 0,81 504.7890 1007.5634 1007.5651 -0.0018 0 18 1.3 U K.SLFSNIVTK.N
609.7759 1217.5373 1217.5346 0.0027 0 48 0.0011 U R.NIPEDAADMAR.L + Oxidation (M)
660.7968 1319.5790 1319.5816 -0.0026 0 48 0.00095 U K.YTQSNSVCYAK.D
669.3653 1336.7160 1336.7213 -0.0053 0 66 1.7e-005 U R.VVVCNLYPFVK.T
693.3587 1384.7028 1384.7059 -0.0031 0 52 0.00055 U K.DGQVIGIGAGQQSR.I
779.9420 1557.8694 1557.8726 -0.0032 0 121 3.9e-011 U R.NLASLGLSLVASGGTAK.A
779.9430 1557.8714 1557.8726 -0.0012 0 (78) 7.3e-007 U R.NLASLGLSLVASGGTAK.A
804.3931 1606.7717 1606.7661 0.0056 0 (20) 0.69 U R.DVSELTGFPEMLGGR.V
804.3931 1606.7717 1606.7661 0.0056 0 (3) 30 U R.DVSELTGFPEMLGGR.V
808.4045 1614.7944 1614.8042 -0.0098 0 127 1.5e-011 U K.LSGVSVSSDAFFPFR.D
808.4084 1614.8021 1614.8042 -0.0021 0 (56) 0.00017 U K.LSGVSVSSDAFFPFR.D
808.4084 1614.8021 1614.8042 -0.0021 0 (30) 0.067 U K.LSGVSVSSDAFFPFR.D
812.3861 1622.7577 1622.7610 -0.0033 0 55 0.00022 U R.DVSELTGFPEMLGGR.V + Oxidation (M)
859.4471 1716.8797 1716.8821 -0.0024 0 15 2 U K.ALFEEVPELLTEAEK.K
615.9995 1844.9767 1844.9771 -0.0004 1 66 1.3e-005 U K.ALFEEVPELLTEAEKK.E
615.9995 1844.9767 1844.9771 -0.0004 1 (65) 1.7e-005 U K.ALFEEVPELLTEAEKK.E
615.9997 1844.9774 1844.9771 0.0003 1 (9) 6.3 U K.ALFEEVPELLTEAEKK.E
1017.9598 2033.9050 2033.9119 -0.0069 0 19 0.38 U K.AFTHTAQYDEAISDYFR.R
678.9766 2033.9079 2033.9119 -0.0041 0 (10) 3.3 U K.AFTHTAQYDEAISDYFR.R
708.3767 2122.1084 2122.1092 -0.0008 0 11 3 U K.VVIEACDELGIVLAHTDLR.L
1081.4904 2160.9662 2160.9708 -0.0045 0 123 1.9e-011 U R.MSSFGDFVALSDVCDVPTAK.I + Oxidation (M)
777.4004 2329.1795 2329.1850 -0.0055 0 (1) 32 U R.VCMVYDLYPTLTPLAIAYAR.A

Silva, K.T.S. Apêndice
125
1165.6014 2329.1882 2329.1850 0.0032 0 54 0.00014 U R.VCMVYDLYPTLTPLAIAYAR.A
1204.5963 2407.1780 2407.1755 0.0026 0 18 0.48 U R.AEVSNAIDQYVTGTIGEGEDLVK.W
1204.5963 2407.1780 2407.1755 0.0026 0 (12) 2.1 U R.AEVSNAIDQYVTGTIGEGEDLVK.W
851.7937 2552.3592 2552.3698 -0.0106 0 135 8.5e-013 U K.TVASPDVTVEAAVEQIDIGGVTLLR.A
851.7955 2552.3646 2552.3698 -0.0052 0 (9) 3.1 U K.TVASPDVTVEAAVEQIDIGGVTLLR.A
851.7955 2552.3646 2552.3698 -0.0052 0 (8) 3.5 U K.TVASPDVTVEAAVEQIDIGGVTLLR.A
1277.1926 2552.3706 2552.3698 0.0009 0 (73) 1.2e-006 U K.TVASPDVTVEAAVEQIDIGGVTLLR.A
30 TKT 7,54 71940 39 gi|12018252 1,12 375.2001 748.3857 748.3868 -0.0011 0 23 0.74 R.AFDQIR.M
378.1925 754.3705 754.3722 -0.0016 0 49 0.001 K.LGHASDR.I
482.2322 962.4499 962.4531 -0.0033 0 28 0.18 R.MPTPPNYK.V + Oxidation (M)
489.7848 977.5550 977.5658 -0.0108 0 33 0.044 K.HQPTAIIAK.T
494.7818 987.5490 987.5535 -0.0045 1 44 0.0054 K.KLILDCAR.A
462.6026 1384.7859 1384.7966 -0.0107 0 (15) 1.9 R.VLDPFTIKPLDK.K
693.4014 1384.7882 1384.7966 -0.0084 0 59 7.1e-005 R.VLDPFTIKPLDK.K
701.8821 1401.7496 1401.7616 -0.0120 0 (44) 0.0028 K.LDNLVAIFDINR.L
701.8833 1401.7521 1401.7616 -0.0095 0 89 9.5e-008 K.LDNLVAIFDINR.L
701.8833 1401.7521 1401.7616 -0.0095 0 (86) 2e-007 K.LDNLVAIFDINR.L
767.9130 1533.8114 1533.8225 -0.0111 1 (14) 2.5 K.MFGIDKDAIVQAVK.G
775.9119 1549.8093 1549.8174 -0.0081 1 56 0.00018 K.MFGIDKDAIVQAVK.G + Oxidation (M)
826.3910 1650.7674 1650.7865 -0.0191 0 85 2e-007 R.TVPFCSTFAAFFTR.A
826.3981 1650.7816 1650.7865 -0.0049 0 (15) 2 R.TVPFCSTFAAFFTR.A
826.3981 1650.7816 1650.7865 -0.0049 0 (19) 0.71 R.TVPFCSTFAAFFTR.A
957.9539 1913.8933 1913.9081 -0.0148 0 (17) 0.96 R.SVPMSTVFYPSDGVATEK.A
965.9537 1929.8928 1929.9030 -0.0102 0 80 4.3e-007 R.SVPMSTVFYPSDGVATEK.A + Oxidation (M)
965.9543 1929.8941 1929.9030 -0.0089 0 (23) 0.21 R.SVPMSTVFYPSDGVATEK.A + Oxidation (M)
670.3354 2007.9845 2007.9935 -0.0090 0 (11) 4.2 K.NMAEQIIQEIYSQVQSK.K
1005.0006 2007.9866 2007.9935 -0.0069 0 (38) 0.0075 K.NMAEQIIQEIYSQVQSK.K
1010.5234 2019.0322 2019.0636 -0.0314 0 93 2.3e-008 K.ILATPPQEDAPSVDIANIR.M
1012.9932 2023.9718 2023.9884 -0.0166 0 61 3.9e-005 K.NMAEQIIQEIYSQVQSK.K + Oxidation (M)
675.6646 2023.9721 2023.9884 -0.0163 0 (18) 0.83 K.NMAEQIIQEIYSQVQSK.K + Oxidation (M)
675.6666 2023.9781 2023.9884 -0.0103 0 (45) 0.0016 K.NMAEQIIQEIYSQVQSK.K + Oxidation (M)
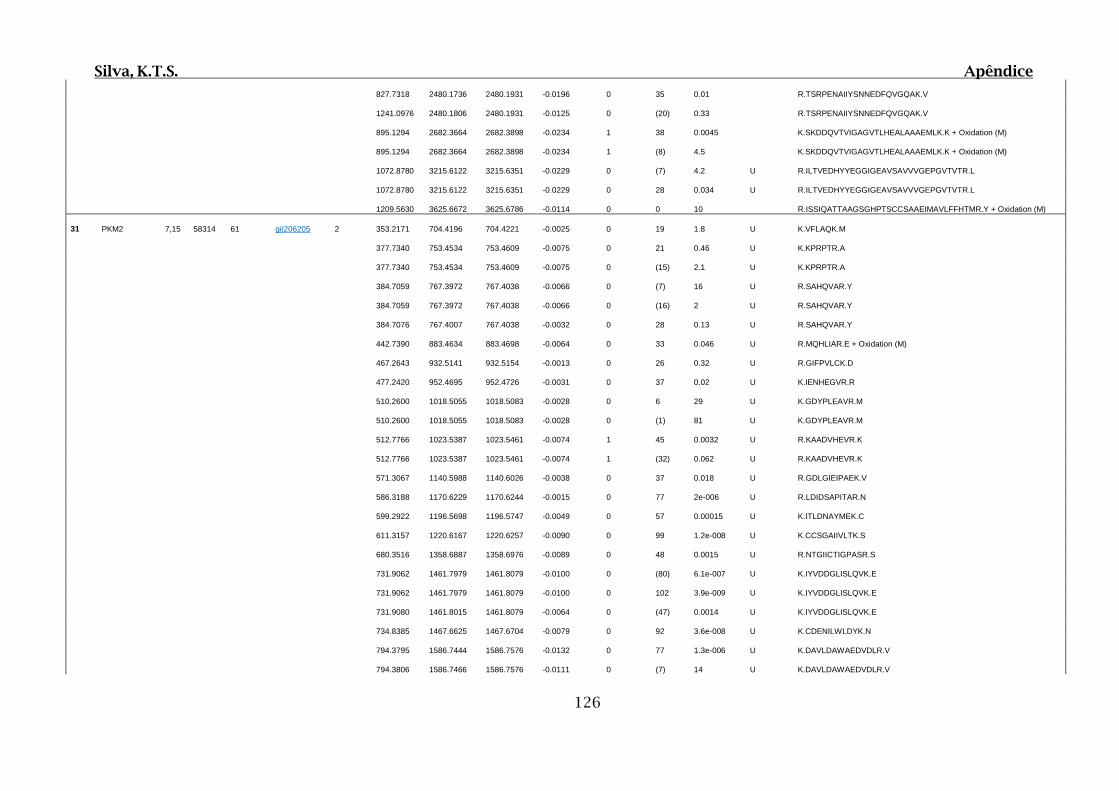
Silva, K.T.S. Apêndice
126
827.7318 2480.1736 2480.1931 -0.0196 0 35 0.01 R.TSRPENAIIYSNNEDFQVGQAK.V
1241.0976 2480.1806 2480.1931 -0.0125 0 (20) 0.33 R.TSRPENAIIYSNNEDFQVGQAK.V
895.1294 2682.3664 2682.3898 -0.0234 1 38 0.0045 K.SKDDQVTVIGAGVTLHEALAAAEMLK.K + Oxidation (M)
895.1294 2682.3664 2682.3898 -0.0234 1 (8) 4.5 K.SKDDQVTVIGAGVTLHEALAAAEMLK.K + Oxidation (M)
1072.8780 3215.6122 3215.6351 -0.0229 0 (7) 4.2 U R.ILTVEDHYYEGGIGEAVSAVVVGEPGVTVTR.L
1072.8780 3215.6122 3215.6351 -0.0229 0 28 0.034 U R.ILTVEDHYYEGGIGEAVSAVVVGEPGVTVTR.L
1209.5630 3625.6672 3625.6786 -0.0114 0 0 10 R.ISSIQATTAAGSGHPTSCCSAAEIMAVLFFHTMR.Y + Oxidation (M)
31 PKM2 7,15 58314 61 gi|206205 2 353.2171 704.4196 704.4221 -0.0025 0 19 1.8 U K.VFLAQK.M
377.7340 753.4534 753.4609 -0.0075 0 21 0.46 U K.KPRPTR.A
377.7340 753.4534 753.4609 -0.0075 0 (15) 2.1 U K.KPRPTR.A
384.7059 767.3972 767.4038 -0.0066 0 (7) 16 U R.SAHQVAR.Y
384.7059 767.3972 767.4038 -0.0066 0 (16) 2 U R.SAHQVAR.Y
384.7076 767.4007 767.4038 -0.0032 0 28 0.13 U R.SAHQVAR.Y
442.7390 883.4634 883.4698 -0.0064 0 33 0.046 U R.MQHLIAR.E + Oxidation (M)
467.2643 932.5141 932.5154 -0.0013 0 26 0.32 U R.GIFPVLCK.D
477.2420 952.4695 952.4726 -0.0031 0 37 0.02 U K.IENHEGVR.R
510.2600 1018.5055 1018.5083 -0.0028 0 6 29 U K.GDYPLEAVR.M
510.2600 1018.5055 1018.5083 -0.0028 0 (1) 81 U K.GDYPLEAVR.M
512.7766 1023.5387 1023.5461 -0.0074 1 45 0.0032 U R.KAADVHEVR.K
512.7766 1023.5387 1023.5461 -0.0074 1 (32) 0.062 U R.KAADVHEVR.K
571.3067 1140.5988 1140.6026 -0.0038 0 37 0.018 U R.GDLGIEIPAEK.V
586.3188 1170.6229 1170.6244 -0.0015 0 77 2e-006 U R.LDIDSAPITAR.N
599.2922 1196.5698 1196.5747 -0.0049 0 57 0.00015 U K.ITLDNAYMEK.C
611.3157 1220.6167 1220.6257 -0.0090 0 99 1.2e-008 U K.CCSGAIIVLTK.S
680.3516 1358.6887 1358.6976 -0.0089 0 48 0.0015 U R.NTGIICTIGPASR.S
731.9062 1461.7979 1461.8079 -0.0100 0 (80) 6.1e-007 U K.IYVDDGLISLQVK.E
731.9062 1461.7979 1461.8079 -0.0100 0 102 3.9e-009 U K.IYVDDGLISLQVK.E
731.9080 1461.8015 1461.8079 -0.0064 0 (47) 0.0014 U K.IYVDDGLISLQVK.E
734.8385 1467.6625 1467.6704 -0.0079 0 92 3.6e-008 U K.CDENILWLDYK.N
794.3795 1586.7444 1586.7576 -0.0132 0 77 1.3e-006 U K.DAVLDAWAEDVDLR.V
794.3806 1586.7466 1586.7576 -0.0111 0 (7) 14 U K.DAVLDAWAEDVDLR.V
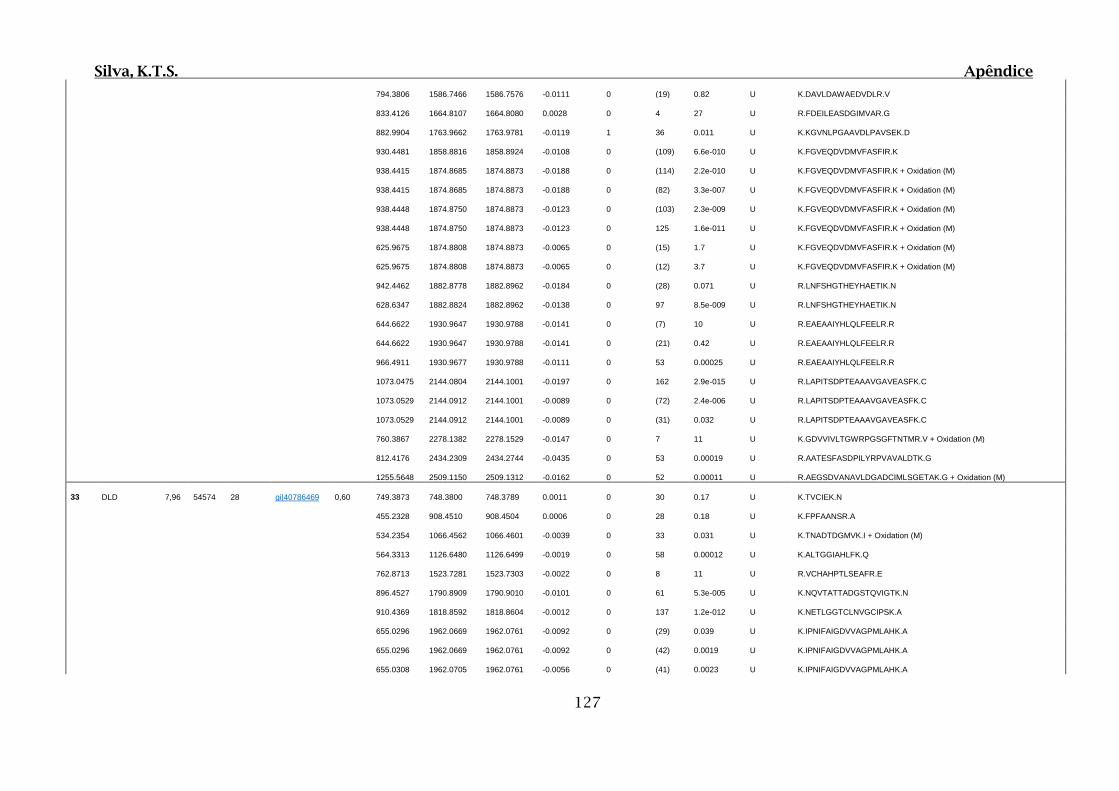
Silva, K.T.S. Apêndice
127
794.3806 1586.7466 1586.7576 -0.0111 0 (19) 0.82 U K.DAVLDAWAEDVDLR.V
833.4126 1664.8107 1664.8080 0.0028 0 4 27 U R.FDEILEASDGIMVAR.G
882.9904 1763.9662 1763.9781 -0.0119 1 36 0.011 U K.KGVNLPGAAVDLPAVSEK.D
930.4481 1858.8816 1858.8924 -0.0108 0 (109) 6.6e-010 U K.FGVEQDVDMVFASFIR.K
938.4415 1874.8685 1874.8873 -0.0188 0 (114) 2.2e-010 U K.FGVEQDVDMVFASFIR.K + Oxidation (M)
938.4415 1874.8685 1874.8873 -0.0188 0 (82) 3.3e-007 U K.FGVEQDVDMVFASFIR.K + Oxidation (M)
938.4448 1874.8750 1874.8873 -0.0123 0 (103) 2.3e-009 U K.FGVEQDVDMVFASFIR.K + Oxidation (M)
938.4448 1874.8750 1874.8873 -0.0123 0 125 1.6e-011 U K.FGVEQDVDMVFASFIR.K + Oxidation (M)
625.9675 1874.8808 1874.8873 -0.0065 0 (15) 1.7 U K.FGVEQDVDMVFASFIR.K + Oxidation (M)
625.9675 1874.8808 1874.8873 -0.0065 0 (12) 3.7 U K.FGVEQDVDMVFASFIR.K + Oxidation (M)
942.4462 1882.8778 1882.8962 -0.0184 0 (28) 0.071 U R.LNFSHGTHEYHAETIK.N
628.6347 1882.8824 1882.8962 -0.0138 0 97 8.5e-009 U R.LNFSHGTHEYHAETIK.N
644.6622 1930.9647 1930.9788 -0.0141 0 (7) 10 U R.EAEAAIYHLQLFEELR.R
644.6622 1930.9647 1930.9788 -0.0141 0 (21) 0.42 U R.EAEAAIYHLQLFEELR.R
966.4911 1930.9677 1930.9788 -0.0111 0 53 0.00025 U R.EAEAAIYHLQLFEELR.R
1073.0475 2144.0804 2144.1001 -0.0197 0 162 2.9e-015 U R.LAPITSDPTEAAAVGAVEASFK.C
1073.0529 2144.0912 2144.1001 -0.0089 0 (72) 2.4e-006 U R.LAPITSDPTEAAAVGAVEASFK.C
1073.0529 2144.0912 2144.1001 -0.0089 0 (31) 0.032 U R.LAPITSDPTEAAAVGAVEASFK.C
760.3867 2278.1382 2278.1529 -0.0147 0 7 11 U K.GDVVIVLTGWRPGSGFTNTMR.V + Oxidation (M)
812.4176 2434.2309 2434.2744 -0.0435 0 53 0.00019 U R.AATESFASDPILYRPVAVALDTK.G
1255.5648 2509.1150 2509.1312 -0.0162 0 52 0.00011 U R.AEGSDVANAVLDGADCIMLSGETAK.G + Oxidation (M)
33 DLD 7,96 54574 28 gi|40786469 0,60 749.3873 748.3800 748.3789 0.0011 0 30 0.17 U K.TVCIEK.N
455.2328 908.4510 908.4504 0.0006 0 28 0.18 U K.FPFAANSR.A
534.2354 1066.4562 1066.4601 -0.0039 0 33 0.031 U K.TNADTDGMVK.I + Oxidation (M)
564.3313 1126.6480 1126.6499 -0.0019 0 58 0.00012 U K.ALTGGIAHLFK.Q
762.8713 1523.7281 1523.7303 -0.0022 0 8 11 U R.VCHAHPTLSEAFR.E
896.4527 1790.8909 1790.9010 -0.0101 0 61 5.3e-005 U K.NQVTATTADGSTQVIGTK.N
910.4369 1818.8592 1818.8604 -0.0012 0 137 1.2e-012 U K.NETLGGTCLNVGCIPSK.A
655.0296 1962.0669 1962.0761 -0.0092 0 (29) 0.039 U K.IPNIFAIGDVVAGPMLAHK.A
655.0296 1962.0669 1962.0761 -0.0092 0 (42) 0.0019 U K.IPNIFAIGDVVAGPMLAHK.A
655.0308 1962.0705 1962.0761 -0.0056 0 (41) 0.0023 U K.IPNIFAIGDVVAGPMLAHK.A
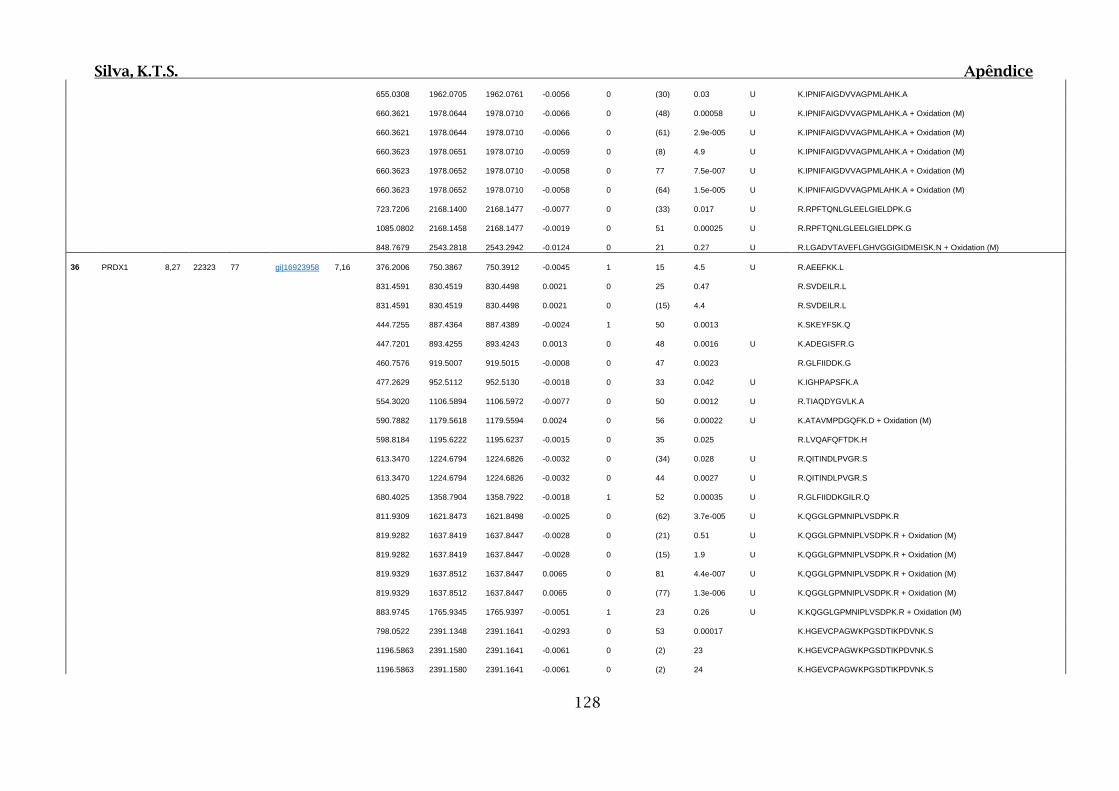
Silva, K.T.S. Apêndice
128
655.0308 1962.0705 1962.0761 -0.0056 0 (30) 0.03 U K.IPNIFAIGDVVAGPMLAHK.A
660.3621 1978.0644 1978.0710 -0.0066 0 (48) 0.00058 U K.IPNIFAIGDVVAGPMLAHK.A + Oxidation (M)
660.3621 1978.0644 1978.0710 -0.0066 0 (61) 2.9e-005 U K.IPNIFAIGDVVAGPMLAHK.A + Oxidation (M)
660.3623 1978.0651 1978.0710 -0.0059 0 (8) 4.9 U K.IPNIFAIGDVVAGPMLAHK.A + Oxidation (M)
660.3623 1978.0652 1978.0710 -0.0058 0 77 7.5e-007 U K.IPNIFAIGDVVAGPMLAHK.A + Oxidation (M)
660.3623 1978.0652 1978.0710 -0.0058 0 (64) 1.5e-005 U K.IPNIFAIGDVVAGPMLAHK.A + Oxidation (M)
723.7206 2168.1400 2168.1477 -0.0077 0 (33) 0.017 U R.RPFTQNLGLEELGIELDPK.G
1085.0802 2168.1458 2168.1477 -0.0019 0 51 0.00025 U R.RPFTQNLGLEELGIELDPK.G
848.7679 2543.2818 2543.2942 -0.0124 0 21 0.27 U R.LGADVTAVEFLGHVGGIGIDMEISK.N + Oxidation (M)
36 PRDX1 8,27 22323 77 gi|16923958 7,16 376.2006 750.3867 750.3912 -0.0045 1 15 4.5 U R.AEEFKK.L
831.4591 830.4519 830.4498 0.0021 0 25 0.47 R.SVDEILR.L
831.4591 830.4519 830.4498 0.0021 0 (15) 4.4 R.SVDEILR.L
444.7255 887.4364 887.4389 -0.0024 1 50 0.0013 K.SKEYFSK.Q
447.7201 893.4255 893.4243 0.0013 0 48 0.0016 U K.ADEGISFR.G
460.7576 919.5007 919.5015 -0.0008 0 47 0.0023 R.GLFIIDDK.G
477.2629 952.5112 952.5130 -0.0018 0 33 0.042 U K.IGHPAPSFK.A
554.3020 1106.5894 1106.5972 -0.0077 0 50 0.0012 U R.TIAQDYGVLK.A
590.7882 1179.5618 1179.5594 0.0024 0 56 0.00022 U K.ATAVMPDGQFK.D + Oxidation (M)
598.8184 1195.6222 1195.6237 -0.0015 0 35 0.025 R.LVQAFQFTDK.H
613.3470 1224.6794 1224.6826 -0.0032 0 (34) 0.028 U R.QITINDLPVGR.S
613.3470 1224.6794 1224.6826 -0.0032 0 44 0.0027 U R.QITINDLPVGR.S
680.4025 1358.7904 1358.7922 -0.0018 1 52 0.00035 U R.GLFIIDDKGILR.Q
811.9309 1621.8473 1621.8498 -0.0025 0 (62) 3.7e-005 U K.QGGLGPMNIPLVSDPK.R
819.9282 1637.8419 1637.8447 -0.0028 0 (21) 0.51 U K.QGGLGPMNIPLVSDPK.R + Oxidation (M)
819.9282 1637.8419 1637.8447 -0.0028 0 (15) 1.9 U K.QGGLGPMNIPLVSDPK.R + Oxidation (M)
819.9329 1637.8512 1637.8447 0.0065 0 81 4.4e-007 U K.QGGLGPMNIPLVSDPK.R + Oxidation (M)
819.9329 1637.8512 1637.8447 0.0065 0 (77) 1.3e-006 U K.QGGLGPMNIPLVSDPK.R + Oxidation (M)
883.9745 1765.9345 1765.9397 -0.0051 1 23 0.26 U K.KQGGLGPMNIPLVSDPK.R + Oxidation (M)
798.0522 2391.1348 2391.1641 -0.0293 0 53 0.00017 K.HGEVCPAGWKPGSDTIKPDVNK.S
1196.5863 2391.1580 2391.1641 -0.0061 0 (2) 23 K.HGEVCPAGWKPGSDTIKPDVNK.S
1196.5863 2391.1580 2391.1641 -0.0061 0 (2) 24 K.HGEVCPAGWKPGSDTIKPDVNK.S
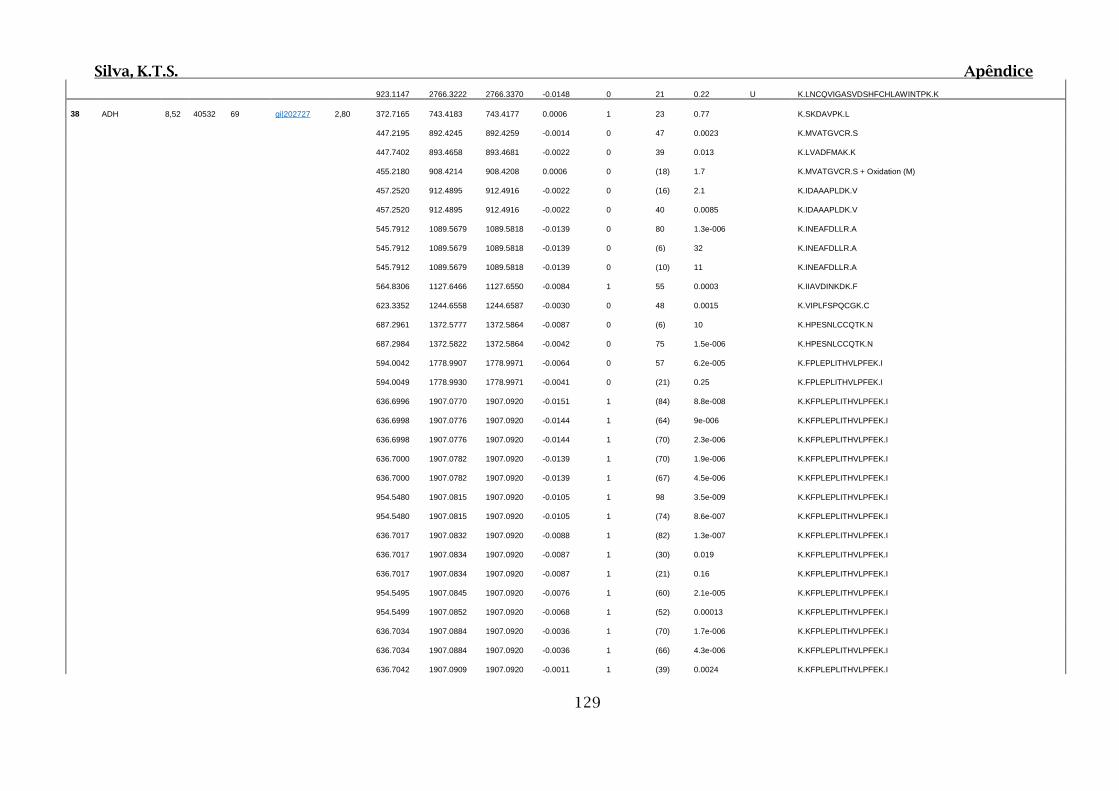
Silva, K.T.S. Apêndice
129
923.1147 2766.3222 2766.3370 -0.0148 0 21 0.22 U K.LNCQVIGASVDSHFCHLAWINTPK.K
38 ADH 8,52 40532 69 gi|202727 2,80 372.7165 743.4183 743.4177 0.0006 1 23 0.77 K.SKDAVPK.L
447.2195 892.4245 892.4259 -0.0014 0 47 0.0023 K.MVATGVCR.S
447.7402 893.4658 893.4681 -0.0022 0 39 0.013 K.LVADFMAK.K
455.2180 908.4214 908.4208 0.0006 0 (18) 1.7 K.MVATGVCR.S + Oxidation (M)
457.2520 912.4895 912.4916 -0.0022 0 (16) 2.1 K.IDAAAPLDK.V
457.2520 912.4895 912.4916 -0.0022 0 40 0.0085 K.IDAAAPLDK.V
545.7912 1089.5679 1089.5818 -0.0139 0 80 1.3e-006 K.INEAFDLLR.A
545.7912 1089.5679 1089.5818 -0.0139 0 (6) 32 K.INEAFDLLR.A
545.7912 1089.5679 1089.5818 -0.0139 0 (10) 11 K.INEAFDLLR.A
564.8306 1127.6466 1127.6550 -0.0084 1 55 0.0003 K.IIAVDINKDK.F
623.3352 1244.6558 1244.6587 -0.0030 0 48 0.0015 K.VIPLFSPQCGK.C
687.2961 1372.5777 1372.5864 -0.0087 0 (6) 10 K.HPESNLCCQTK.N
687.2984 1372.5822 1372.5864 -0.0042 0 75 1.5e-006 K.HPESNLCCQTK.N
594.0042 1778.9907 1778.9971 -0.0064 0 57 6.2e-005 K.FPLEPLITHVLPFEK.I
594.0049 1778.9930 1778.9971 -0.0041 0 (21) 0.25 K.FPLEPLITHVLPFEK.I
636.6996 1907.0770 1907.0920 -0.0151 1 (84) 8.8e-008 K.KFPLEPLITHVLPFEK.I
636.6998 1907.0776 1907.0920 -0.0144 1 (64) 9e-006 K.KFPLEPLITHVLPFEK.I
636.6998 1907.0776 1907.0920 -0.0144 1 (70) 2.3e-006 K.KFPLEPLITHVLPFEK.I
636.7000 1907.0782 1907.0920 -0.0139 1 (70) 1.9e-006 K.KFPLEPLITHVLPFEK.I
636.7000 1907.0782 1907.0920 -0.0139 1 (67) 4.5e-006 K.KFPLEPLITHVLPFEK.I
954.5480 1907.0815 1907.0920 -0.0105 1 98 3.5e-009 K.KFPLEPLITHVLPFEK.I
954.5480 1907.0815 1907.0920 -0.0105 1 (74) 8.6e-007 K.KFPLEPLITHVLPFEK.I
636.7017 1907.0832 1907.0920 -0.0088 1 (82) 1.3e-007 K.KFPLEPLITHVLPFEK.I
636.7017 1907.0834 1907.0920 -0.0087 1 (30) 0.019 K.KFPLEPLITHVLPFEK.I
636.7017 1907.0834 1907.0920 -0.0087 1 (21) 0.16 K.KFPLEPLITHVLPFEK.I
954.5495 1907.0845 1907.0920 -0.0076 1 (60) 2.1e-005 K.KFPLEPLITHVLPFEK.I
954.5499 1907.0852 1907.0920 -0.0068 1 (52) 0.00013 K.KFPLEPLITHVLPFEK.I
636.7034 1907.0884 1907.0920 -0.0036 1 (70) 1.7e-006 K.KFPLEPLITHVLPFEK.I
636.7034 1907.0884 1907.0920 -0.0036 1 (66) 4.3e-006 K.KFPLEPLITHVLPFEK.I
636.7042 1907.0909 1907.0920 -0.0011 1 (39) 0.0024 K.KFPLEPLITHVLPFEK.I

Silva, K.T.S. Apêndice
130
636.7042 1907.0909 1907.0920 -0.0011 1 (65) 5.1e-006 K.KFPLEPLITHVLPFEK.I
1037.5033 2072.9920 2073.0024 -0.0103 0 (146) 1.1e-013 K.VCLIGCGFSTGYGSAVQVAK.V
1037.5033 2072.9920 2073.0024 -0.0103 0 180 4.2e-017 K.VCLIGCGFSTGYGSAVQVAK.V
1037.5061 2072.9976 2073.0024 -0.0047 0 (50) 0.00044 K.VCLIGCGFSTGYGSAVQVAK.V
779.0695 2334.1868 2334.2076 -0.0209 0 (86) 9.2e-008 K.VTPGSTCAVFGLGGVGLSVVIGCK.T
1168.1031 2334.1916 2334.2076 -0.0160 0 136 8.1e-013 K.VTPGSTCAVFGLGGVGLSVVIGCK.T
829.7741 2486.3006 2486.3209 -0.0203 0 61 2.6e-005 K.AAVLWEPHKPFTIEDIEVAPPK.A
1244.1660 2486.3174 2486.3209 -0.0035 0 (13) 1.4 K.AAVLWEPHKPFTIEDIEVAPPK.A
954.4936 2860.4591 2860.4760 -0.0169 0 (17) 0.48 U R.GKPIHHFLSTSTFSQYTVVDDIAVAK.I
954.4945 2860.4616 2860.4760 -0.0144 0 81 2.2e-007 U R.GKPIHHFLSTSTFSQYTVVDDIAVAK.I
1368.3499 4102.0279 4102.0318 -0.0039 0 (23) 0.056 R.SDDHAVSGSLFTPLPAVLGHEGAGIVESIGEGVTCVKPGDK.V
1368.3499 4102.0279 4102.0318 -0.0039 0 24 0.042 R.SDDHAVSGSLFTPLPAVLGHEGAGIVESIGEGVTCVKPGDK.V
1419.7131 4256.1175 4256.1313 -0.0139 0 (5) 2.7 R.LDTMTSALLSCHSACGVSVIVGVPPSAQSLSVNPMSLLLGR.T + 2
Oxidation (M)
1419.7131 4256.1175 4256.1313 -0.0139 0 (6) 2.2 R.LDTMTSALLSCHSACGVSVIVGVPPSAQSLSVNPMSLLLGR.T + 2
Oxidation (M)
1419.7131 4256.1175 4256.1313 -0.0139 0 (8) 1.3 R.LDTMTSALLSCHSACGVSVIVGVPPSAQSLSVNPMSLLLGR.T + 2
Oxidation (M)
1419.7158 4256.1256 4256.1313 -0.0058 0 25 0.024 R.LDTMTSALLSCHSACGVSVIVGVPPSAQSLSVNPMSLLLGR.T + 2
Oxidation (M)
39 ALDOA 8,31 39691 60 gi|202837 2,06 462.7451 923.4756 923.4825 -0.0069 0 45 0.0032 K.ELADIAHR.I
464.2148 926.4150 926.4168 -0.0018 0 2 42 R.CQYVTEK.V
522.7861 1043.5576 1043.5611 -0.0035 0 46 0.0025 R.QLLLTADDR.V
671.8531 1341.6916 1341.7041 -0.0125 0 36 0.025 K.ADDGRPFPQVIK.S
823.9015 1645.7885 1645.8019 -0.0135 1 21 0.53 R.LQSIGTENTEENRR.F
549.6054 1645.7943 1645.8019 -0.0077 1 (2) 40 R.LQSIGTENTEENRR.F
549.6054 1645.7943 1645.8019 -0.0077 1 (0) 70 R.LQSIGTENTEENRR.F
826.9132 1651.8118 1651.8239 -0.0121 0 (100) 7.1e-009 K.FSNEEIAMATVTALR.R
834.9062 1667.7978 1667.8188 -0.0211 0 100 6.2e-009 K.FSNEEIAMATVTALR.R + Oxidation (M)
603.6506 1807.9299 1807.9443 -0.0145 0 44 0.0023 K.CPLLKPWALTFSYGR.A
904.9727 1807.9308 1807.9443 -0.0135 0 (25) 0.21 K.CPLLKPWALTFSYGR.A
696.6955 2087.0646 2087.0874 -0.0228 0 99 6.9e-009 R.VNPCIGGVILFHETLYQK.A
1044.5422 2087.0698 2087.0874 -0.0175 0 (35) 0.015 R.VNPCIGGVILFHETLYQK.A
697.7048 2090.0925 2090.1120 -0.0195 0 88 5.9e-008 U K.IGEHTPSSLAIVENANVLAR.Y
1046.0555 2090.0964 2090.1120 -0.0155 0 (1) 27 U K.IGEHTPSSLAIVENANVLAR.Y
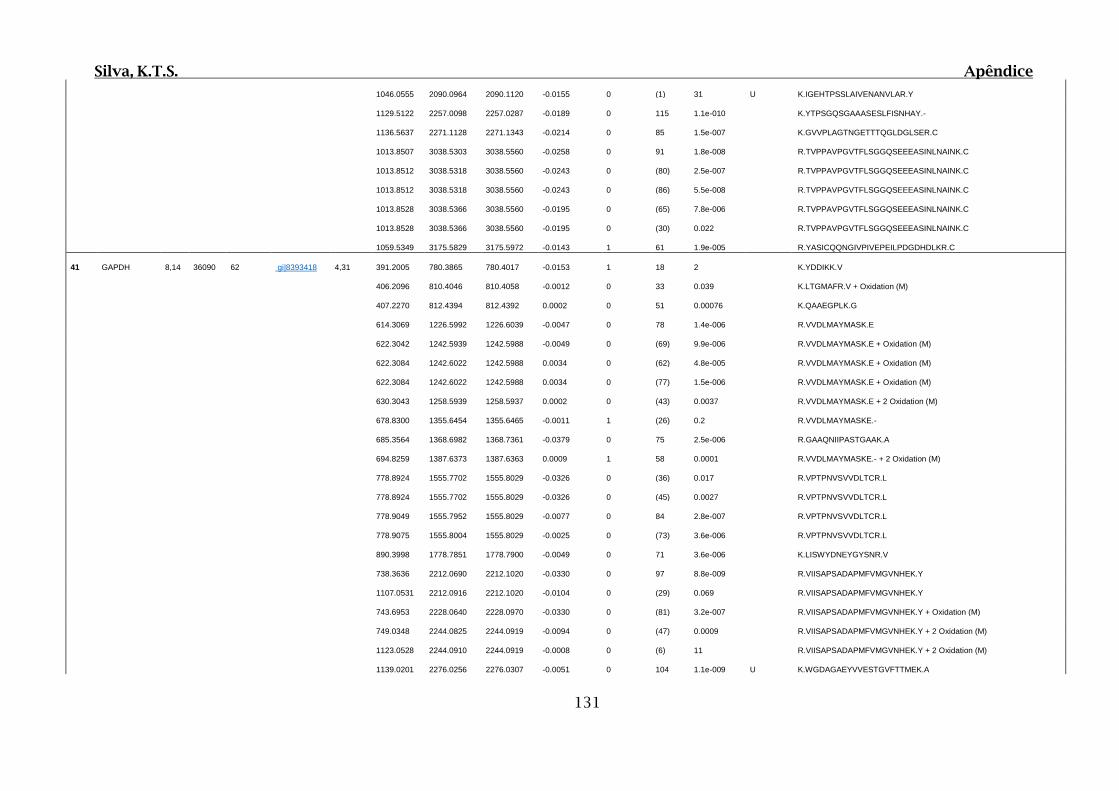
Silva, K.T.S. Apêndice
131
1046.0555 2090.0964 2090.1120 -0.0155 0 (1) 31 U K.IGEHTPSSLAIVENANVLAR.Y
1129.5122 2257.0098 2257.0287 -0.0189 0 115 1.1e-010 K.YTPSGQSGAAASESLFISNHAY.-
1136.5637 2271.1128 2271.1343 -0.0214 0 85 1.5e-007 K.GVVPLAGTNGETTTQGLDGLSER.C
1013.8507 3038.5303 3038.5560 -0.0258 0 91 1.8e-008 R.TVPPAVPGVTFLSGGQSEEEASINLNAINK.C
1013.8512 3038.5318 3038.5560 -0.0243 0 (80) 2.5e-007 R.TVPPAVPGVTFLSGGQSEEEASINLNAINK.C
1013.8512 3038.5318 3038.5560 -0.0243 0 (86) 5.5e-008 R.TVPPAVPGVTFLSGGQSEEEASINLNAINK.C
1013.8528 3038.5366 3038.5560 -0.0195 0 (65) 7.8e-006 R.TVPPAVPGVTFLSGGQSEEEASINLNAINK.C
1013.8528 3038.5366 3038.5560 -0.0195 0 (30) 0.022 R.TVPPAVPGVTFLSGGQSEEEASINLNAINK.C
1059.5349 3175.5829 3175.5972 -0.0143 1 61 1.9e-005 R.YASICQQNGIVPIVEPEILPDGDHDLKR.C
41 GAPDH 8,14 36090 62 gi|8393418 4,31 391.2005 780.3865 780.4017 -0.0153 1 18 2 K.YDDIKK.V
406.2096 810.4046 810.4058 -0.0012 0 33 0.039 K.LTGMAFR.V + Oxidation (M)
407.2270 812.4394 812.4392 0.0002 0 51 0.00076 K.QAAEGPLK.G
614.3069 1226.5992 1226.6039 -0.0047 0 78 1.4e-006 R.VVDLMAYMASK.E
622.3042 1242.5939 1242.5988 -0.0049 0 (69) 9.9e-006 R.VVDLMAYMASK.E + Oxidation (M)
622.3084 1242.6022 1242.5988 0.0034 0 (62) 4.8e-005 R.VVDLMAYMASK.E + Oxidation (M)
622.3084 1242.6022 1242.5988 0.0034 0 (77) 1.5e-006 R.VVDLMAYMASK.E + Oxidation (M)
630.3043 1258.5939 1258.5937 0.0002 0 (43) 0.0037 R.VVDLMAYMASK.E + 2 Oxidation (M)
678.8300 1355.6454 1355.6465 -0.0011 1 (26) 0.2 R.VVDLMAYMASKE.-
685.3564 1368.6982 1368.7361 -0.0379 0 75 2.5e-006 R.GAAQNIIPASTGAAK.A
694.8259 1387.6373 1387.6363 0.0009 1 58 0.0001 R.VVDLMAYMASKE.- + 2 Oxidation (M)
778.8924 1555.7702 1555.8029 -0.0326 0 (36) 0.017 R.VPTPNVSVVDLTCR.L
778.8924 1555.7702 1555.8029 -0.0326 0 (45) 0.0027 R.VPTPNVSVVDLTCR.L
778.9049 1555.7952 1555.8029 -0.0077 0 84 2.8e-007 R.VPTPNVSVVDLTCR.L
778.9075 1555.8004 1555.8029 -0.0025 0 (73) 3.6e-006 R.VPTPNVSVVDLTCR.L
890.3998 1778.7851 1778.7900 -0.0049 0 71 3.6e-006 K.LISWYDNEYGYSNR.V
738.3636 2212.0690 2212.1020 -0.0330 0 97 8.8e-009 R.VIISAPSADAPMFVMGVNHEK.Y
1107.0531 2212.0916 2212.1020 -0.0104 0 (29) 0.069 R.VIISAPSADAPMFVMGVNHEK.Y
743.6953 2228.0640 2228.0970 -0.0330 0 (81) 3.2e-007 R.VIISAPSADAPMFVMGVNHEK.Y + Oxidation (M)
749.0348 2244.0825 2244.0919 -0.0094 0 (47) 0.0009 R.VIISAPSADAPMFVMGVNHEK.Y + 2 Oxidation (M)
1123.0528 2244.0910 2244.0919 -0.0008 0 (6) 11 R.VIISAPSADAPMFVMGVNHEK.Y + 2 Oxidation (M)
1139.0201 2276.0256 2276.0307 -0.0051 0 104 1.1e-009 U K.WGDAGAEYVVESTGVFTTMEK.A
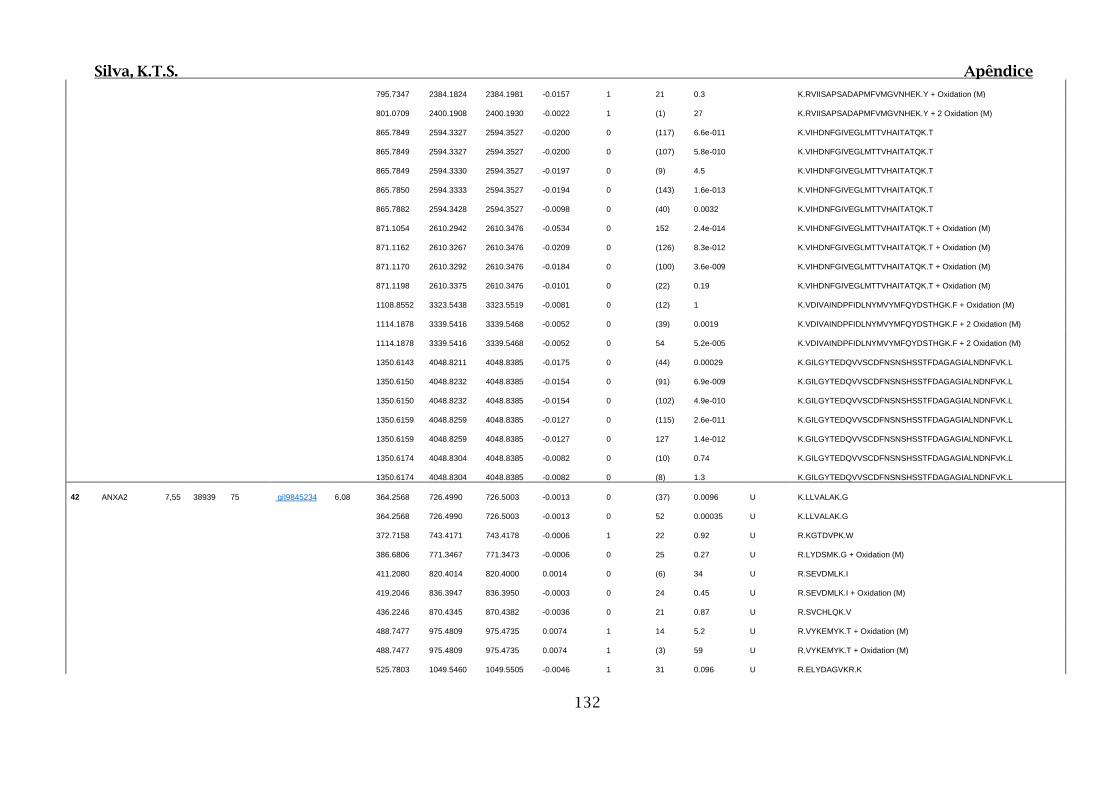
Silva, K.T.S. Apêndice
132
795.7347 2384.1824 2384.1981 -0.0157 1 21 0.3 K.RVIISAPSADAPMFVMGVNHEK.Y + Oxidation (M)
801.0709 2400.1908 2400.1930 -0.0022 1 (1) 27 K.RVIISAPSADAPMFVMGVNHEK.Y + 2 Oxidation (M)
865.7849 2594.3327 2594.3527 -0.0200 0 (117) 6.6e-011 K.VIHDNFGIVEGLMTTVHAITATQK.T
865.7849 2594.3327 2594.3527 -0.0200 0 (107) 5.8e-010 K.VIHDNFGIVEGLMTTVHAITATQK.T
865.7849 2594.3330 2594.3527 -0.0197 0 (9) 4.5 K.VIHDNFGIVEGLMTTVHAITATQK.T
865.7850 2594.3333 2594.3527 -0.0194 0 (143) 1.6e-013 K.VIHDNFGIVEGLMTTVHAITATQK.T
865.7882 2594.3428 2594.3527 -0.0098 0 (40) 0.0032 K.VIHDNFGIVEGLMTTVHAITATQK.T
871.1054 2610.2942 2610.3476 -0.0534 0 152 2.4e-014 K.VIHDNFGIVEGLMTTVHAITATQK.T + Oxidation (M)
871.1162 2610.3267 2610.3476 -0.0209 0 (126) 8.3e-012 K.VIHDNFGIVEGLMTTVHAITATQK.T + Oxidation (M)
871.1170 2610.3292 2610.3476 -0.0184 0 (100) 3.6e-009 K.VIHDNFGIVEGLMTTVHAITATQK.T + Oxidation (M)
871.1198 2610.3375 2610.3476 -0.0101 0 (22) 0.19 K.VIHDNFGIVEGLMTTVHAITATQK.T + Oxidation (M)
1108.8552 3323.5438 3323.5519 -0.0081 0 (12) 1 K.VDIVAINDPFIDLNYMVYMFQYDSTHGK.F + Oxidation (M)
1114.1878 3339.5416 3339.5468 -0.0052 0 (39) 0.0019 K.VDIVAINDPFIDLNYMVYMFQYDSTHGK.F + 2 Oxidation (M)
1114.1878 3339.5416 3339.5468 -0.0052 0 54 5.2e-005 K.VDIVAINDPFIDLNYMVYMFQYDSTHGK.F + 2 Oxidation (M)
1350.6143 4048.8211 4048.8385 -0.0175 0 (44) 0.00029 K.GILGYTEDQVVSCDFNSNSHSSTFDAGAGIALNDNFVK.L
1350.6150 4048.8232 4048.8385 -0.0154 0 (91) 6.9e-009 K.GILGYTEDQVVSCDFNSNSHSSTFDAGAGIALNDNFVK.L
1350.6150 4048.8232 4048.8385 -0.0154 0 (102) 4.9e-010 K.GILGYTEDQVVSCDFNSNSHSSTFDAGAGIALNDNFVK.L
1350.6159 4048.8259 4048.8385 -0.0127 0 (115) 2.6e-011 K.GILGYTEDQVVSCDFNSNSHSSTFDAGAGIALNDNFVK.L
1350.6159 4048.8259 4048.8385 -0.0127 0 127 1.4e-012 K.GILGYTEDQVVSCDFNSNSHSSTFDAGAGIALNDNFVK.L
1350.6174 4048.8304 4048.8385 -0.0082 0 (10) 0.74 K.GILGYTEDQVVSCDFNSNSHSSTFDAGAGIALNDNFVK.L
1350.6174 4048.8304 4048.8385 -0.0082 0 (8) 1.3 K.GILGYTEDQVVSCDFNSNSHSSTFDAGAGIALNDNFVK.L
42 ANXA2 7,55 38939 75 gi|9845234 6,08 364.2568 726.4990 726.5003 -0.0013 0 (37) 0.0096 U K.LLVALAK.G
364.2568 726.4990 726.5003 -0.0013 0 52 0.00035 U K.LLVALAK.G
372.7158 743.4171 743.4178 -0.0006 1 22 0.92 U R.KGTDVPK.W
386.6806 771.3467 771.3473 -0.0006 0 25 0.27 U R.LYDSMK.G + Oxidation (M)
411.2080 820.4014 820.4000 0.0014 0 (6) 34 U R.SEVDMLK.I
419.2046 836.3947 836.3950 -0.0003 0 24 0.45 U R.SEVDMLK.I + Oxidation (M)
436.2246 870.4345 870.4382 -0.0036 0 21 0.87 U R.SVCHLQK.V
488.7477 975.4809 975.4735 0.0074 1 14 5.2 U R.VYKEMYK.T + Oxidation (M)
488.7477 975.4809 975.4735 0.0074 1 (3) 59 U R.VYKEMYK.T + Oxidation (M)
525.7803 1049.5460 1049.5505 -0.0046 1 31 0.096 U R.ELYDAGVKR.K

Silva, K.T.S. Apêndice
133
526.2664 1050.5183 1050.5168 0.0015 0 22 0.63 U K.WISIMTER.S + Oxidation (M)
544.3036 1086.5927 1086.5921 0.0006 0 53 0.00059 U R.DALNIETAIK.T
544.3036 1086.5927 1086.5921 0.0006 0 (39) 0.015 U R.DALNIETAIK.T
556.2791 1110.5436 1110.5458 -0.0022 0 66 2.2e-005 U R.QDIAFAYQR.R
556.2791 1110.5436 1110.5458 -0.0022 0 (54) 0.00031 U R.QDIAFAYQR.R
611.7996 1221.5846 1221.5877 -0.0031 0 (55) 0.00027 U K.TPAQYDASELK.A
611.8002 1221.5859 1221.5877 -0.0018 0 (55) 0.00026 U K.TPAQYDASELK.A
611.8002 1221.5859 1221.5877 -0.0018 0 73 4.6e-006 U K.TPAQYDASELK.A
622.8040 1243.5934 1243.6156 -0.0222 0 64 3.5e-005 U R.TNQELQEINR.V
622.8144 1243.6143 1243.6156 -0.0014 0 (26) 0.21 U R.TNQELQEINR.V
684.3417 1366.6689 1366.6729 -0.0040 1 42 0.0049 U K.DIISDTSGEFRK.L
703.3467 1404.6788 1404.6925 -0.0137 0 (54) 0.00032 U K.SLYYFIQQDTK.G
703.3508 1404.6870 1404.6925 -0.0055 0 63 4.1e-005 U K.SLYYFIQQDTK.G
730.8267 1459.6389 1459.6653 -0.0263 0 (24) 0.21 U K.SYSPYDMLESIR.K
730.8267 1459.6389 1459.6653 -0.0263 0 62 3e-005 U K.SYSPYDMLESIR.K
730.8383 1459.6620 1459.6653 -0.0033 0 (41) 0.0052 U K.SYSPYDMLESIR.K
738.8346 1475.6546 1475.6602 -0.0056 0 (24) 0.24 U K.SYSPYDMLESIR.K + Oxidation (M)
771.9227 1541.8309 1541.8413 -0.0104 0 110 5.9e-010 U K.GVDEVTIVNILTNR.S
771.9230 1541.8314 1541.8413 -0.0099 0 (75) 2.3e-006 U K.GVDEVTIVNILTNR.S
771.9257 1541.8368 1541.8413 -0.0046 0 (54) 0.00025 U K.GVDEVTIVNILTNR.S
834.9610 1667.9075 1667.9280 -0.0205 0 (104) 2e-009 U K.SALSGHLETVMLGLLK.T
556.9800 1667.9180 1667.9280 -0.0100 0 (68) 6.7e-006 U K.SALSGHLETVMLGLLK.T
834.9669 1667.9192 1667.9280 -0.0088 0 (124) 1.6e-011 U K.SALSGHLETVMLGLLK.T
834.9669 1667.9192 1667.9280 -0.0088 0 126 1e-011 U K.SALSGHLETVMLGLLK.T
834.9680 1667.9215 1667.9280 -0.0065 0 (52) 0.00026 U K.SALSGHLETVMLGLLK.T
834.9680 1667.9215 1667.9280 -0.0065 0 (44) 0.0017 U K.SALSGHLETVMLGLLK.T
842.9589 1683.9033 1683.9229 -0.0196 0 (85) 1.8e-007 U K.SALSGHLETVMLGLLK.T + Oxidation (M)
562.3127 1683.9164 1683.9229 -0.0066 0 (57) 0.0001 U K.SALSGHLETVMLGLLK.T + Oxidation (M)
562.3127 1683.9164 1683.9229 -0.0066 0 (19) 0.61 U K.SALSGHLETVMLGLLK.T + Oxidation (M)
842.9660 1683.9175 1683.9229 -0.0054 0 (3) 26 U K.SALSGHLETVMLGLLK.T + Oxidation (M)
842.9660 1683.9175 1683.9229 -0.0054 0 (12) 3.1 U K.SALSGHLETVMLGLLK.T + Oxidation (M)
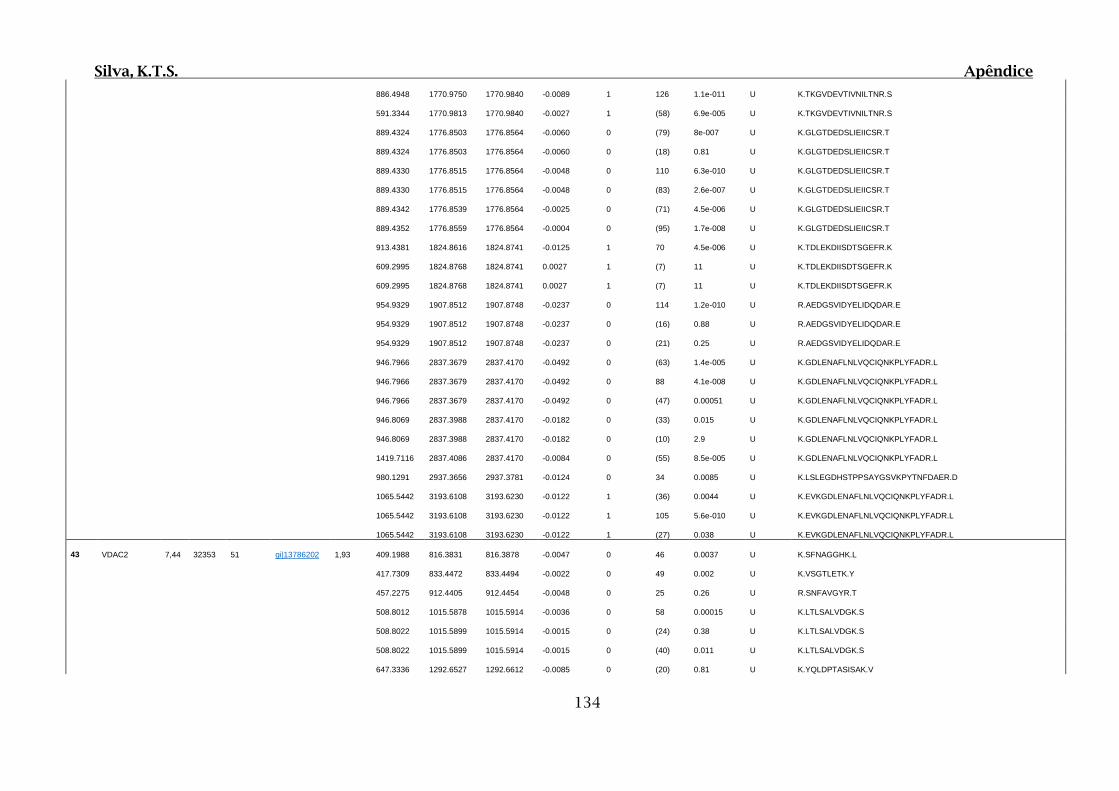
Silva, K.T.S. Apêndice
134
886.4948 1770.9750 1770.9840 -0.0089 1 126 1.1e-011 U K.TKGVDEVTIVNILTNR.S
591.3344 1770.9813 1770.9840 -0.0027 1 (58) 6.9e-005 U K.TKGVDEVTIVNILTNR.S
889.4324 1776.8503 1776.8564 -0.0060 0 (79) 8e-007 U K.GLGTDEDSLIEIICSR.T
889.4324 1776.8503 1776.8564 -0.0060 0 (18) 0.81 U K.GLGTDEDSLIEIICSR.T
889.4330 1776.8515 1776.8564 -0.0048 0 110 6.3e-010 U K.GLGTDEDSLIEIICSR.T
889.4330 1776.8515 1776.8564 -0.0048 0 (83) 2.6e-007 U K.GLGTDEDSLIEIICSR.T
889.4342 1776.8539 1776.8564 -0.0025 0 (71) 4.5e-006 U K.GLGTDEDSLIEIICSR.T
889.4352 1776.8559 1776.8564 -0.0004 0 (95) 1.7e-008 U K.GLGTDEDSLIEIICSR.T
913.4381 1824.8616 1824.8741 -0.0125 1 70 4.5e-006 U K.TDLEKDIISDTSGEFR.K
609.2995 1824.8768 1824.8741 0.0027 1 (7) 11 U K.TDLEKDIISDTSGEFR.K
609.2995 1824.8768 1824.8741 0.0027 1 (7) 11 U K.TDLEKDIISDTSGEFR.K
954.9329 1907.8512 1907.8748 -0.0237 0 114 1.2e-010 U R.AEDGSVIDYELIDQDAR.E
954.9329 1907.8512 1907.8748 -0.0237 0 (16) 0.88 U R.AEDGSVIDYELIDQDAR.E
954.9329 1907.8512 1907.8748 -0.0237 0 (21) 0.25 U R.AEDGSVIDYELIDQDAR.E
946.7966 2837.3679 2837.4170 -0.0492 0 (63) 1.4e-005 U K.GDLENAFLNLVQCIQNKPLYFADR.L
946.7966 2837.3679 2837.4170 -0.0492 0 88 4.1e-008 U K.GDLENAFLNLVQCIQNKPLYFADR.L
946.7966 2837.3679 2837.4170 -0.0492 0 (47) 0.00051 U K.GDLENAFLNLVQCIQNKPLYFADR.L
946.8069 2837.3988 2837.4170 -0.0182 0 (33) 0.015 U K.GDLENAFLNLVQCIQNKPLYFADR.L
946.8069 2837.3988 2837.4170 -0.0182 0 (10) 2.9 U K.GDLENAFLNLVQCIQNKPLYFADR.L
1419.7116 2837.4086 2837.4170 -0.0084 0 (55) 8.5e-005 U K.GDLENAFLNLVQCIQNKPLYFADR.L
980.1291 2937.3656 2937.3781 -0.0124 0 34 0.0085 U K.LSLEGDHSTPPSAYGSVKPYTNFDAER.D
1065.5442 3193.6108 3193.6230 -0.0122 1 (36) 0.0044 U K.EVKGDLENAFLNLVQCIQNKPLYFADR.L
1065.5442 3193.6108 3193.6230 -0.0122 1 105 5.6e-010 U K.EVKGDLENAFLNLVQCIQNKPLYFADR.L
1065.5442 3193.6108 3193.6230 -0.0122 1 (27) 0.038 U K.EVKGDLENAFLNLVQCIQNKPLYFADR.L
43 VDAC2 7,44 32353 51 gi|13786202 1,93 409.1988 816.3831 816.3878 -0.0047 0 46 0.0037 U K.SFNAGGHK.L
417.7309 833.4472 833.4494 -0.0022 0 49 0.002 U K.VSGTLETK.Y
457.2275 912.4405 912.4454 -0.0048 0 25 0.26 U R.SNFAVGYR.T
508.8012 1015.5878 1015.5914 -0.0036 0 58 0.00015 U K.LTLSALVDGK.S
508.8022 1015.5899 1015.5914 -0.0015 0 (24) 0.38 U K.LTLSALVDGK.S
508.8022 1015.5899 1015.5914 -0.0015 0 (40) 0.011 U K.LTLSALVDGK.S
647.3336 1292.6527 1292.6612 -0.0085 0 (20) 0.81 U K.YQLDPTASISAK.V
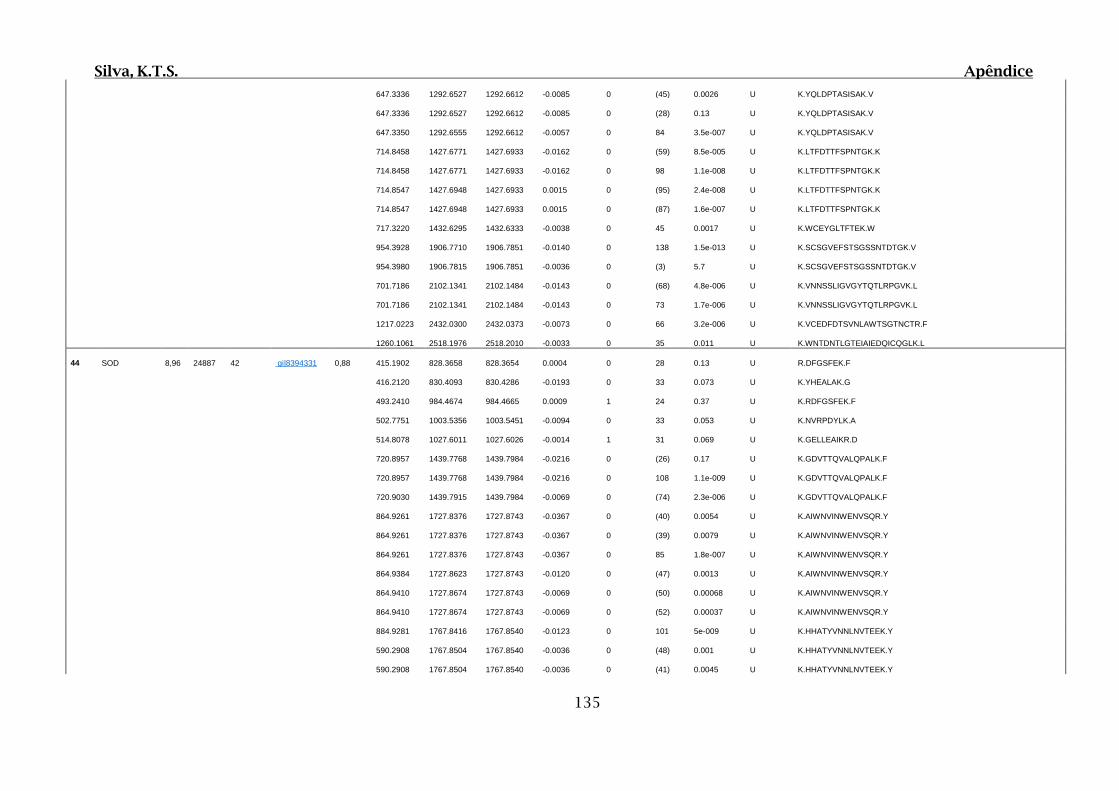
Silva, K.T.S. Apêndice
135
647.3336 1292.6527 1292.6612 -0.0085 0 (45) 0.0026 U K.YQLDPTASISAK.V
647.3336 1292.6527 1292.6612 -0.0085 0 (28) 0.13 U K.YQLDPTASISAK.V
647.3350 1292.6555 1292.6612 -0.0057 0 84 3.5e-007 U K.YQLDPTASISAK.V
714.8458 1427.6771 1427.6933 -0.0162 0 (59) 8.5e-005 U K.LTFDTTFSPNTGK.K
714.8458 1427.6771 1427.6933 -0.0162 0 98 1.1e-008 U K.LTFDTTFSPNTGK.K
714.8547 1427.6948 1427.6933 0.0015 0 (95) 2.4e-008 U K.LTFDTTFSPNTGK.K
714.8547 1427.6948 1427.6933 0.0015 0 (87) 1.6e-007 U K.LTFDTTFSPNTGK.K
717.3220 1432.6295 1432.6333 -0.0038 0 45 0.0017 U K.WCEYGLTFTEK.W
954.3928 1906.7710 1906.7851 -0.0140 0 138 1.5e-013 U K.SCSGVEFSTSGSSNTDTGK.V
954.3980 1906.7815 1906.7851 -0.0036 0 (3) 5.7 U K.SCSGVEFSTSGSSNTDTGK.V
701.7186 2102.1341 2102.1484 -0.0143 0 (68) 4.8e-006 U K.VNNSSLIGVGYTQTLRPGVK.L
701.7186 2102.1341 2102.1484 -0.0143 0 73 1.7e-006 U K.VNNSSLIGVGYTQTLRPGVK.L
1217.0223 2432.0300 2432.0373 -0.0073 0 66 3.2e-006 U K.VCEDFDTSVNLAWTSGTNCTR.F
1260.1061 2518.1976 2518.2010 -0.0033 0 35 0.011 U K.WNTDNTLGTEIAIEDQICQGLK.L
44 SOD 8,96 24887 42 gi|8394331 0,88 415.1902 828.3658 828.3654 0.0004 0 28 0.13 U R.DFGSFEK.F
416.2120 830.4093 830.4286 -0.0193 0 33 0.073 U K.YHEALAK.G
493.2410 984.4674 984.4665 0.0009 1 24 0.37 U K.RDFGSFEK.F
502.7751 1003.5356 1003.5451 -0.0094 0 33 0.053 U K.NVRPDYLK.A
514.8078 1027.6011 1027.6026 -0.0014 1 31 0.069 U K.GELLEAIKR.D
720.8957 1439.7768 1439.7984 -0.0216 0 (26) 0.17 U K.GDVTTQVALQPALK.F
720.8957 1439.7768 1439.7984 -0.0216 0 108 1.1e-009 U K.GDVTTQVALQPALK.F
720.9030 1439.7915 1439.7984 -0.0069 0 (74) 2.3e-006 U K.GDVTTQVALQPALK.F
864.9261 1727.8376 1727.8743 -0.0367 0 (40) 0.0054 U K.AIWNVINWENVSQR.Y
864.9261 1727.8376 1727.8743 -0.0367 0 (39) 0.0079 U K.AIWNVINWENVSQR.Y
864.9261 1727.8376 1727.8743 -0.0367 0 85 1.8e-007 U K.AIWNVINWENVSQR.Y
864.9384 1727.8623 1727.8743 -0.0120 0 (47) 0.0013 U K.AIWNVINWENVSQR.Y
864.9410 1727.8674 1727.8743 -0.0069 0 (50) 0.00068 U K.AIWNVINWENVSQR.Y
864.9410 1727.8674 1727.8743 -0.0069 0 (52) 0.00037 U K.AIWNVINWENVSQR.Y
884.9281 1767.8416 1767.8540 -0.0123 0 101 5e-009 U K.HHATYVNNLNVTEEK.Y
590.2908 1767.8504 1767.8540 -0.0036 0 (48) 0.001 U K.HHATYVNNLNVTEEK.Y
590.2908 1767.8504 1767.8540 -0.0036 0 (41) 0.0045 U K.HHATYVNNLNVTEEK.Y
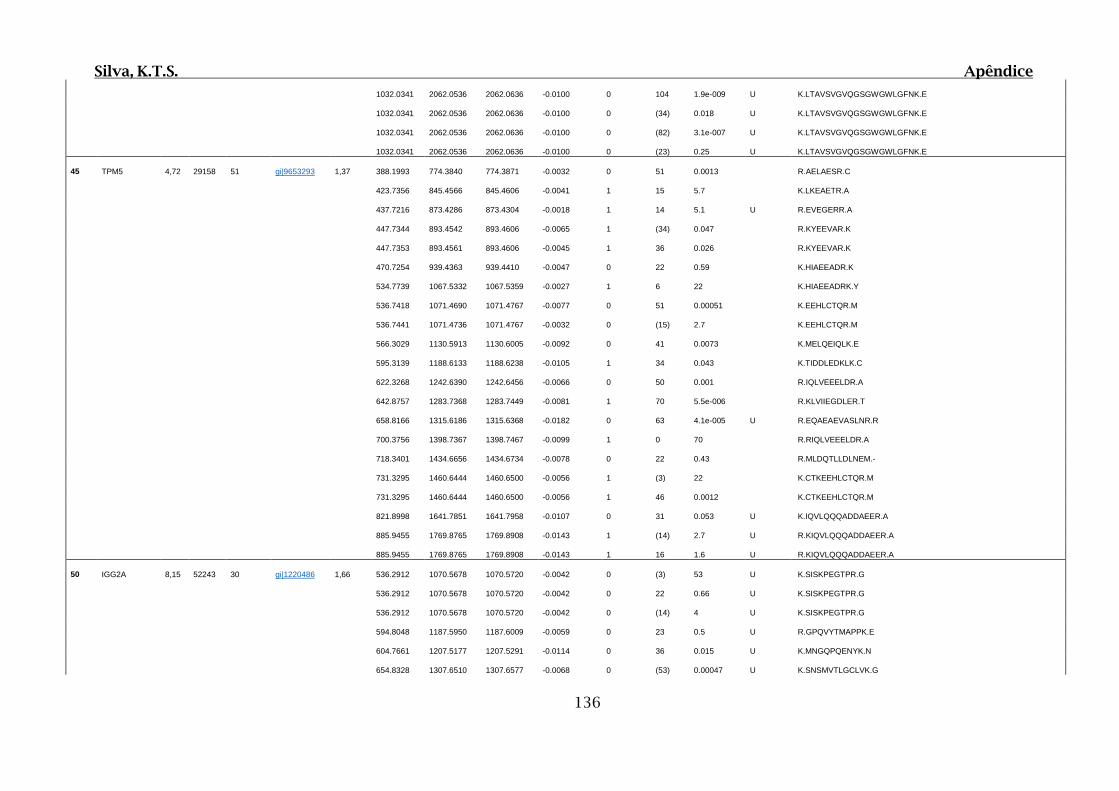
Silva, K.T.S. Apêndice
136
1032.0341 2062.0536 2062.0636 -0.0100 0 104 1.9e-009 U K.LTAVSVGVQGSGWGWLGFNK.E
1032.0341 2062.0536 2062.0636 -0.0100 0 (34) 0.018 U K.LTAVSVGVQGSGWGWLGFNK.E
1032.0341 2062.0536 2062.0636 -0.0100 0 (82) 3.1e-007 U K.LTAVSVGVQGSGWGWLGFNK.E
1032.0341 2062.0536 2062.0636 -0.0100 0 (23) 0.25 U K.LTAVSVGVQGSGWGWLGFNK.E
45 TPM5 4,72 29158 51 gi|9653293 1,37 388.1993 774.3840 774.3871 -0.0032 0 51 0.0013 R.AELAESR.C
423.7356 845.4566 845.4606 -0.0041 1 15 5.7 K.LKEAETR.A
437.7216 873.4286 873.4304 -0.0018 1 14 5.1 U R.EVEGERR.A
447.7344 893.4542 893.4606 -0.0065 1 (34) 0.047 R.KYEEVAR.K
447.7353 893.4561 893.4606 -0.0045 1 36 0.026 R.KYEEVAR.K
470.7254 939.4363 939.4410 -0.0047 0 22 0.59 K.HIAEEADR.K
534.7739 1067.5332 1067.5359 -0.0027 1 6 22 K.HIAEEADRK.Y
536.7418 1071.4690 1071.4767 -0.0077 0 51 0.00051 K.EEHLCTQR.M
536.7441 1071.4736 1071.4767 -0.0032 0 (15) 2.7 K.EEHLCTQR.M
566.3029 1130.5913 1130.6005 -0.0092 0 41 0.0073 K.MELQEIQLK.E
595.3139 1188.6133 1188.6238 -0.0105 1 34 0.043 K.TIDDLEDKLK.C
622.3268 1242.6390 1242.6456 -0.0066 0 50 0.001 R.IQLVEEELDR.A
642.8757 1283.7368 1283.7449 -0.0081 1 70 5.5e-006 R.KLVIIEGDLER.T
658.8166 1315.6186 1315.6368 -0.0182 0 63 4.1e-005 U R.EQAEAEVASLNR.R
700.3756 1398.7367 1398.7467 -0.0099 1 0 70 R.RIQLVEEELDR.A
718.3401 1434.6656 1434.6734 -0.0078 0 22 0.43 R.MLDQTLLDLNEM.-
731.3295 1460.6444 1460.6500 -0.0056 1 (3) 22 K.CTKEEHLCTQR.M
731.3295 1460.6444 1460.6500 -0.0056 1 46 0.0012 K.CTKEEHLCTQR.M
821.8998 1641.7851 1641.7958 -0.0107 0 31 0.053 U K.IQVLQQQADDAEER.A
885.9455 1769.8765 1769.8908 -0.0143 1 (14) 2.7 U R.KIQVLQQQADDAEER.A
885.9455 1769.8765 1769.8908 -0.0143 1 16 1.6 U R.KIQVLQQQADDAEER.A
50 IGG2A 8,15 52243 30 gi|1220486 1,66 536.2912 1070.5678 1070.5720 -0.0042 0 (3) 53 U K.SISKPEGTPR.G
536.2912 1070.5678 1070.5720 -0.0042 0 22 0.66 U K.SISKPEGTPR.G
536.2912 1070.5678 1070.5720 -0.0042 0 (14) 4 U K.SISKPEGTPR.G
594.8048 1187.5950 1187.6009 -0.0059 0 23 0.5 U R.GPQVYTMAPPK.E
604.7661 1207.5177 1207.5291 -0.0114 0 36 0.015 U K.MNGQPQENYK.N
654.8328 1307.6510 1307.6577 -0.0068 0 (53) 0.00047 U K.SNSMVTLGCLVK.G
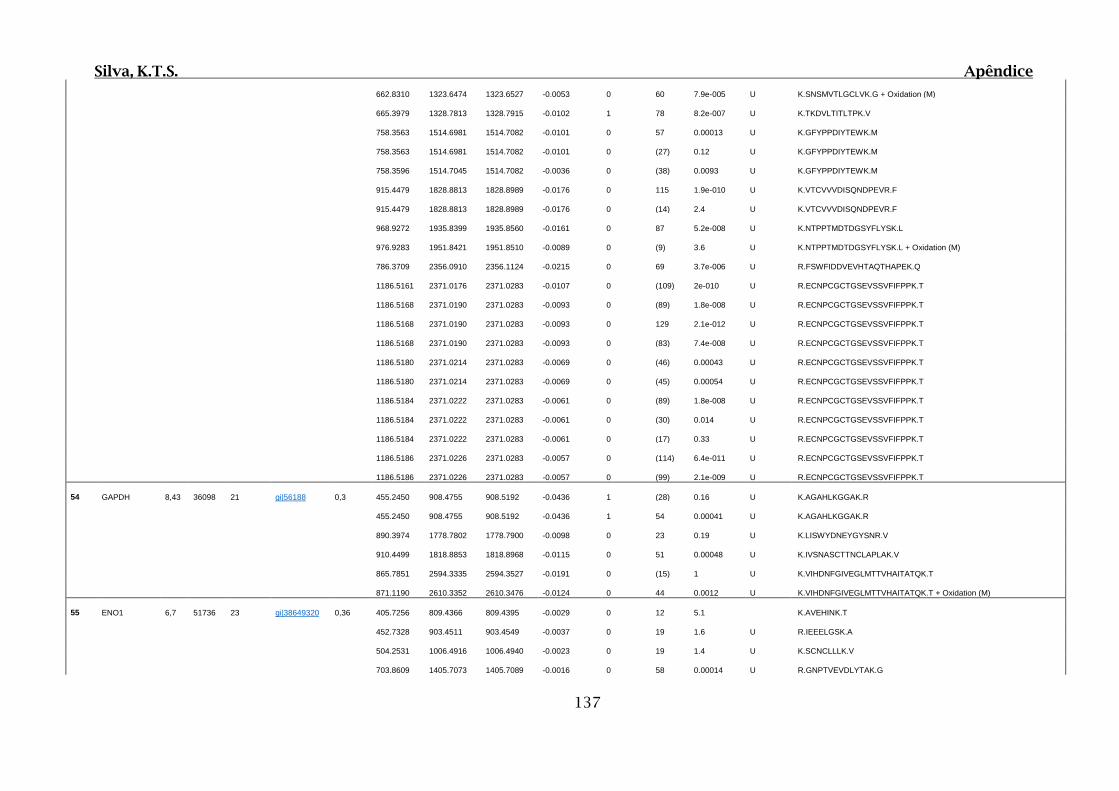
Silva, K.T.S. Apêndice
137
662.8310 1323.6474 1323.6527 -0.0053 0 60 7.9e-005 U K.SNSMVTLGCLVK.G + Oxidation (M)
665.3979 1328.7813 1328.7915 -0.0102 1 78 8.2e-007 U K.TKDVLTITLTPK.V
758.3563 1514.6981 1514.7082 -0.0101 0 57 0.00013 U K.GFYPPDIYTEWK.M
758.3563 1514.6981 1514.7082 -0.0101 0 (27) 0.12 U K.GFYPPDIYTEWK.M
758.3596 1514.7045 1514.7082 -0.0036 0 (38) 0.0093 U K.GFYPPDIYTEWK.M
915.4479 1828.8813 1828.8989 -0.0176 0 115 1.9e-010 U K.VTCVVVDISQNDPEVR.F
915.4479 1828.8813 1828.8989 -0.0176 0 (14) 2.4 U K.VTCVVVDISQNDPEVR.F
968.9272 1935.8399 1935.8560 -0.0161 0 87 5.2e-008 U K.NTPPTMDTDGSYFLYSK.L
976.9283 1951.8421 1951.8510 -0.0089 0 (9) 3.6 U K.NTPPTMDTDGSYFLYSK.L + Oxidation (M)
786.3709 2356.0910 2356.1124 -0.0215 0 69 3.7e-006 U R.FSWFIDDVEVHTAQTHAPEK.Q
1186.5161 2371.0176 2371.0283 -0.0107 0 (109) 2e-010 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5168 2371.0190 2371.0283 -0.0093 0 (89) 1.8e-008 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5168 2371.0190 2371.0283 -0.0093 0 129 2.1e-012 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5168 2371.0190 2371.0283 -0.0093 0 (83) 7.4e-008 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5180 2371.0214 2371.0283 -0.0069 0 (46) 0.00043 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5180 2371.0214 2371.0283 -0.0069 0 (45) 0.00054 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5184 2371.0222 2371.0283 -0.0061 0 (89) 1.8e-008 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5184 2371.0222 2371.0283 -0.0061 0 (30) 0.014 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5184 2371.0222 2371.0283 -0.0061 0 (17) 0.33 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5186 2371.0226 2371.0283 -0.0057 0 (114) 6.4e-011 U R.ECNPCGCTGSEVSSVFIFPPK.T
1186.5186 2371.0226 2371.0283 -0.0057 0 (99) 2.1e-009 U R.ECNPCGCTGSEVSSVFIFPPK.T
54 GAPDH 8,43 36098 21 gi|56188 0,3 455.2450 908.4755 908.5192 -0.0436 1 (28) 0.16 U K.AGAHLKGGAK.R
455.2450 908.4755 908.5192 -0.0436 1 54 0.00041 U K.AGAHLKGGAK.R
890.3974 1778.7802 1778.7900 -0.0098 0 23 0.19 U K.LISWYDNEYGYSNR.V
910.4499 1818.8853 1818.8968 -0.0115 0 51 0.00048 U K.IVSNASCTTNCLAPLAK.V
865.7851 2594.3335 2594.3527 -0.0191 0 (15) 1 U K.VIHDNFGIVEGLMTTVHAITATQK.T
871.1190 2610.3352 2610.3476 -0.0124 0 44 0.0012 U K.VIHDNFGIVEGLMTTVHAITATQK.T + Oxidation (M)
55 ENO1 6,7 51736 23 gi|38649320 0,36 405.7256 809.4366 809.4395 -0.0029 0 12 5.1 K.AVEHINK.T
452.7328 903.4511 903.4549 -0.0037 0 19 1.6 U R.IEEELGSK.A
504.2531 1006.4916 1006.4940 -0.0023 0 19 1.4 U K.SCNCLLLK.V
703.8609 1405.7073 1405.7089 -0.0016 0 58 0.00014 U R.GNPTVEVDLYTAK.G
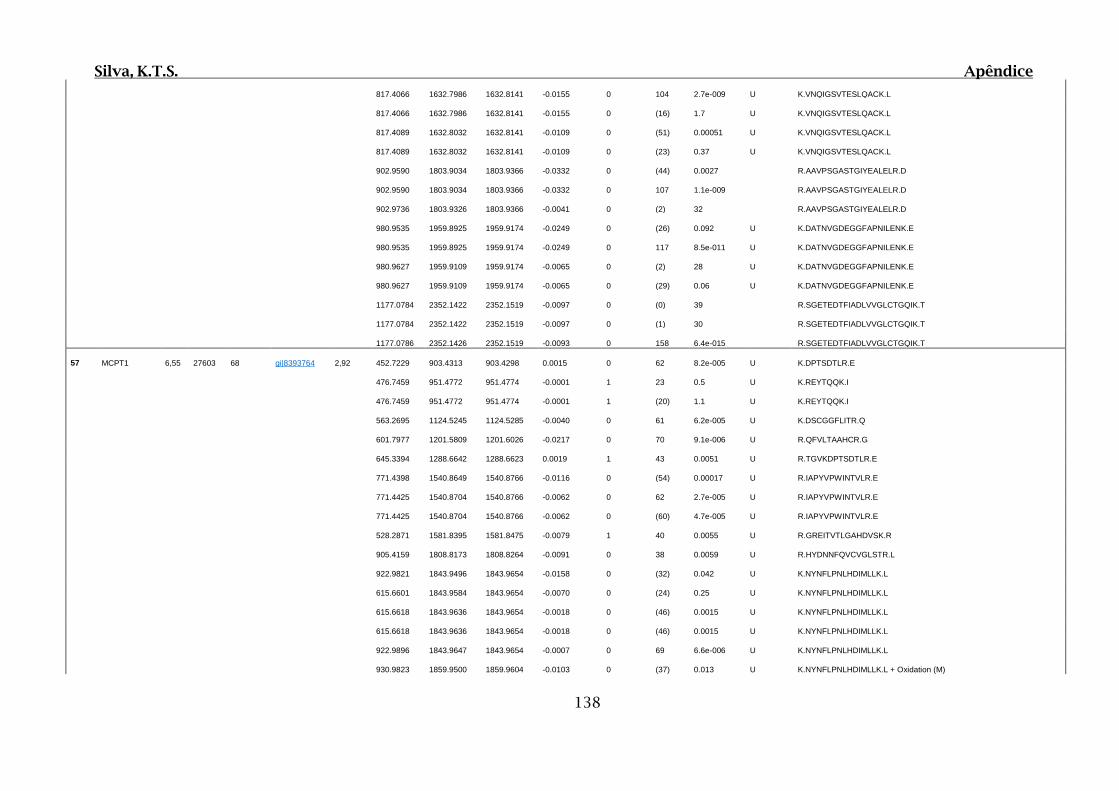
Silva, K.T.S. Apêndice
138
817.4066 1632.7986 1632.8141 -0.0155 0 104 2.7e-009 U K.VNQIGSVTESLQACK.L
817.4066 1632.7986 1632.8141 -0.0155 0 (16) 1.7 U K.VNQIGSVTESLQACK.L
817.4089 1632.8032 1632.8141 -0.0109 0 (51) 0.00051 U K.VNQIGSVTESLQACK.L
817.4089 1632.8032 1632.8141 -0.0109 0 (23) 0.37 U K.VNQIGSVTESLQACK.L
902.9590 1803.9034 1803.9366 -0.0332 0 (44) 0.0027 R.AAVPSGASTGIYEALELR.D
902.9590 1803.9034 1803.9366 -0.0332 0 107 1.1e-009 R.AAVPSGASTGIYEALELR.D
902.9736 1803.9326 1803.9366 -0.0041 0 (2) 32 R.AAVPSGASTGIYEALELR.D
980.9535 1959.8925 1959.9174 -0.0249 0 (26) 0.092 U K.DATNVGDEGGFAPNILENK.E
980.9535 1959.8925 1959.9174 -0.0249 0 117 8.5e-011 U K.DATNVGDEGGFAPNILENK.E
980.9627 1959.9109 1959.9174 -0.0065 0 (2) 28 U K.DATNVGDEGGFAPNILENK.E
980.9627 1959.9109 1959.9174 -0.0065 0 (29) 0.06 U K.DATNVGDEGGFAPNILENK.E
1177.0784 2352.1422 2352.1519 -0.0097 0 (0) 39 R.SGETEDTFIADLVVGLCTGQIK.T
1177.0784 2352.1422 2352.1519 -0.0097 0 (1) 30 R.SGETEDTFIADLVVGLCTGQIK.T
1177.0786 2352.1426 2352.1519 -0.0093 0 158 6.4e-015 R.SGETEDTFIADLVVGLCTGQIK.T
57 MCPT1 6,55 27603 68 gi|8393764 2,92 452.7229 903.4313 903.4298 0.0015 0 62 8.2e-005 U K.DPTSDTLR.E
476.7459 951.4772 951.4774 -0.0001 1 23 0.5 U K.REYTQQK.I
476.7459 951.4772 951.4774 -0.0001 1 (20) 1.1 U K.REYTQQK.I
563.2695 1124.5245 1124.5285 -0.0040 0 61 6.2e-005 U K.DSCGGFLITR.Q
601.7977 1201.5809 1201.6026 -0.0217 0 70 9.1e-006 U R.QFVLTAAHCR.G
645.3394 1288.6642 1288.6623 0.0019 1 43 0.0051 U R.TGVKDPTSDTLR.E
771.4398 1540.8649 1540.8766 -0.0116 0 (54) 0.00017 U R.IAPYVPWINTVLR.E
771.4425 1540.8704 1540.8766 -0.0062 0 62 2.7e-005 U R.IAPYVPWINTVLR.E
771.4425 1540.8704 1540.8766 -0.0062 0 (60) 4.7e-005 U R.IAPYVPWINTVLR.E
528.2871 1581.8395 1581.8475 -0.0079 1 40 0.0055 U R.GREITVTLGAHDVSK.R
905.4159 1808.8173 1808.8264 -0.0091 0 38 0.0059 U R.HYDNNFQVCVGLSTR.L
922.9821 1843.9496 1843.9654 -0.0158 0 (32) 0.042 U K.NYNFLPNLHDIMLLK.L
615.6601 1843.9584 1843.9654 -0.0070 0 (24) 0.25 U K.NYNFLPNLHDIMLLK.L
615.6618 1843.9636 1843.9654 -0.0018 0 (46) 0.0015 U K.NYNFLPNLHDIMLLK.L
615.6618 1843.9636 1843.9654 -0.0018 0 (46) 0.0015 U K.NYNFLPNLHDIMLLK.L
922.9896 1843.9647 1843.9654 -0.0007 0 69 6.6e-006 U K.NYNFLPNLHDIMLLK.L
930.9823 1859.9500 1859.9604 -0.0103 0 (37) 0.013 U K.NYNFLPNLHDIMLLK.L + Oxidation (M)
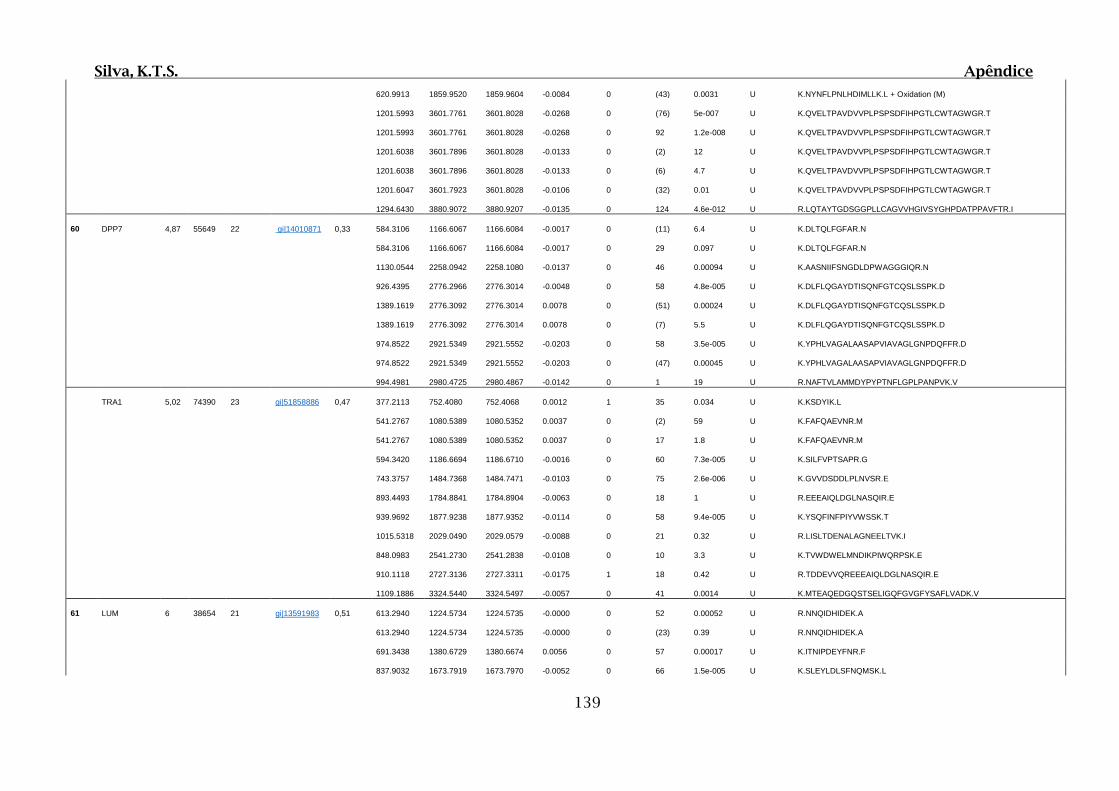
Silva, K.T.S. Apêndice
139
620.9913 1859.9520 1859.9604 -0.0084 0 (43) 0.0031 U K.NYNFLPNLHDIMLLK.L + Oxidation (M)
1201.5993 3601.7761 3601.8028 -0.0268 0 (76) 5e-007 U K.QVELTPAVDVVPLPSPSDFIHPGTLCWTAGWGR.T
1201.5993 3601.7761 3601.8028 -0.0268 0 92 1.2e-008 U K.QVELTPAVDVVPLPSPSDFIHPGTLCWTAGWGR.T
1201.6038 3601.7896 3601.8028 -0.0133 0 (2) 12 U K.QVELTPAVDVVPLPSPSDFIHPGTLCWTAGWGR.T
1201.6038 3601.7896 3601.8028 -0.0133 0 (6) 4.7 U K.QVELTPAVDVVPLPSPSDFIHPGTLCWTAGWGR.T
1201.6047 3601.7923 3601.8028 -0.0106 0 (32) 0.01 U K.QVELTPAVDVVPLPSPSDFIHPGTLCWTAGWGR.T
1294.6430 3880.9072 3880.9207 -0.0135 0 124 4.6e-012 U R.LQTAYTGDSGGPLLCAGVVHGIVSYGHPDATPPAVFTR.I
60 DPP7 4,87 55649 22 gi|14010871 0,33 584.3106 1166.6067 1166.6084 -0.0017 0 (11) 6.4 U K.DLTQLFGFAR.N
584.3106 1166.6067 1166.6084 -0.0017 0 29 0.097 U K.DLTQLFGFAR.N
1130.0544 2258.0942 2258.1080 -0.0137 0 46 0.00094 U K.AASNIIFSNGDLDPWAGGGIQR.N
926.4395 2776.2966 2776.3014 -0.0048 0 58 4.8e-005 U K.DLFLQGAYDTISQNFGTCQSLSSPK.D
1389.1619 2776.3092 2776.3014 0.0078 0 (51) 0.00024 U K.DLFLQGAYDTISQNFGTCQSLSSPK.D
1389.1619 2776.3092 2776.3014 0.0078 0 (7) 5.5 U K.DLFLQGAYDTISQNFGTCQSLSSPK.D
974.8522 2921.5349 2921.5552 -0.0203 0 58 3.5e-005 U K.YPHLVAGALAASAPVIAVAGLGNPDQFFR.D
974.8522 2921.5349 2921.5552 -0.0203 0 (47) 0.00045 U K.YPHLVAGALAASAPVIAVAGLGNPDQFFR.D
994.4981 2980.4725 2980.4867 -0.0142 0 1 19 U R.NAFTVLAMMDYPYPTNFLGPLPANPVK.V
TRA1 5,02 74390 23 gi|51858886 0,47 377.2113 752.4080 752.4068 0.0012 1 35 0.034 U K.KSDYIK.L
541.2767 1080.5389 1080.5352 0.0037 0 (2) 59 U K.FAFQAEVNR.M
541.2767 1080.5389 1080.5352 0.0037 0 17 1.8 U K.FAFQAEVNR.M
594.3420 1186.6694 1186.6710 -0.0016 0 60 7.3e-005 U K.SILFVPTSAPR.G
743.3757 1484.7368 1484.7471 -0.0103 0 75 2.6e-006 U K.GVVDSDDLPLNVSR.E
893.4493 1784.8841 1784.8904 -0.0063 0 18 1 U R.EEEAIQLDGLNASQIR.E
939.9692 1877.9238 1877.9352 -0.0114 0 58 9.4e-005 U K.YSQFINFPIYVWSSK.T
1015.5318 2029.0490 2029.0579 -0.0088 0 21 0.32 U R.LISLTDENALAGNEELTVK.I
848.0983 2541.2730 2541.2838 -0.0108 0 10 3.3 U K.TVWDWELMNDIKPIWQRPSK.E
910.1118 2727.3136 2727.3311 -0.0175 1 18 0.42 U R.TDDEVVQREEEAIQLDGLNASQIR.E
1109.1886 3324.5440 3324.5497 -0.0057 0 41 0.0014 U K.MTEAQEDGQSTSELIGQFGVGFYSAFLVADK.V
61 LUM 6 38654 21 gi|13591983 0,51 613.2940 1224.5734 1224.5735 -0.0000 0 52 0.00052 U R.NNQIDHIDEK.A
613.2940 1224.5734 1224.5735 -0.0000 0 (23) 0.39 U R.NNQIDHIDEK.A
691.3438 1380.6729 1380.6674 0.0056 0 57 0.00017 U K.ITNIPDEYFNR.F
837.9032 1673.7919 1673.7970 -0.0052 0 66 1.5e-005 U K.SLEYLDLSFNQMSK.L
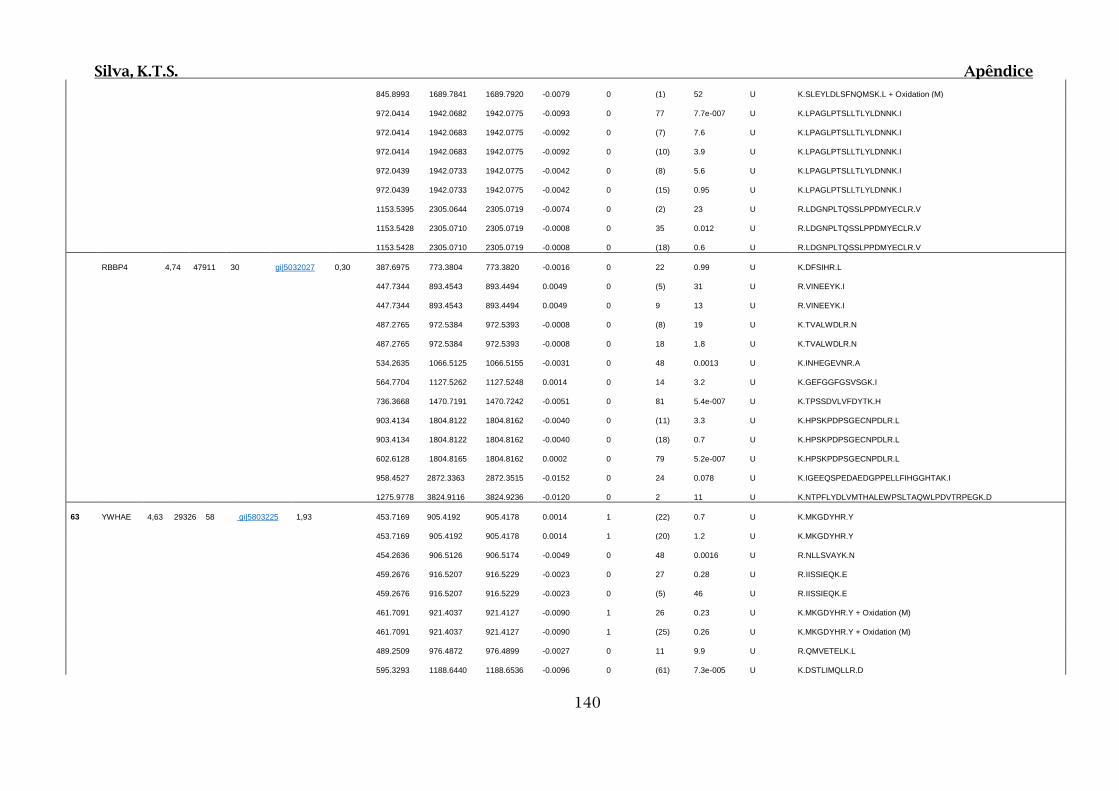
Silva, K.T.S. Apêndice
140
845.8993 1689.7841 1689.7920 -0.0079 0 (1) 52 U K.SLEYLDLSFNQMSK.L + Oxidation (M)
972.0414 1942.0682 1942.0775 -0.0093 0 77 7.7e-007 U K.LPAGLPTSLLTLYLDNNK.I
972.0414 1942.0683 1942.0775 -0.0092 0 (7) 7.6 U K.LPAGLPTSLLTLYLDNNK.I
972.0414 1942.0683 1942.0775 -0.0092 0 (10) 3.9 U K.LPAGLPTSLLTLYLDNNK.I
972.0439 1942.0733 1942.0775 -0.0042 0 (8) 5.6 U K.LPAGLPTSLLTLYLDNNK.I
972.0439 1942.0733 1942.0775 -0.0042 0 (15) 0.95 U K.LPAGLPTSLLTLYLDNNK.I
1153.5395 2305.0644 2305.0719 -0.0074 0 (2) 23 U R.LDGNPLTQSSLPPDMYECLR.V
1153.5428 2305.0710 2305.0719 -0.0008 0 35 0.012 U R.LDGNPLTQSSLPPDMYECLR.V
1153.5428 2305.0710 2305.0719 -0.0008 0 (18) 0.6 U R.LDGNPLTQSSLPPDMYECLR.V
RBBP4 4,74 47911 30 gi|5032027 0,30 387.6975 773.3804 773.3820 -0.0016 0 22 0.99 U K.DFSIHR.L
447.7344 893.4543 893.4494 0.0049 0 (5) 31 U R.VINEEYK.I
447.7344 893.4543 893.4494 0.0049 0 9 13 U R.VINEEYK.I
487.2765 972.5384 972.5393 -0.0008 0 (8) 19 U K.TVALWDLR.N
487.2765 972.5384 972.5393 -0.0008 0 18 1.8 U K.TVALWDLR.N
534.2635 1066.5125 1066.5155 -0.0031 0 48 0.0013 U K.INHEGEVNR.A
564.7704 1127.5262 1127.5248 0.0014 0 14 3.2 U K.GEFGGFGSVSGK.I
736.3668 1470.7191 1470.7242 -0.0051 0 81 5.4e-007 U K.TPSSDVLVFDYTK.H
903.4134 1804.8122 1804.8162 -0.0040 0 (11) 3.3 U K.HPSKPDPSGECNPDLR.L
903.4134 1804.8122 1804.8162 -0.0040 0 (18) 0.7 U K.HPSKPDPSGECNPDLR.L
602.6128 1804.8165 1804.8162 0.0002 0 79 5.2e-007 U K.HPSKPDPSGECNPDLR.L
958.4527 2872.3363 2872.3515 -0.0152 0 24 0.078 U K.IGEEQSPEDAEDGPPELLFIHGGHTAK.I
1275.9778 3824.9116 3824.9236 -0.0120 0 2 11 U K.NTPFLYDLVMTHALEWPSLTAQWLPDVTRPEGK.D
63 YWHAE 4,63 29326 58 gi|5803225 1,93 453.7169 905.4192 905.4178 0.0014 1 (22) 0.7 U K.MKGDYHR.Y
453.7169 905.4192 905.4178 0.0014 1 (20) 1.2 U K.MKGDYHR.Y
454.2636 906.5126 906.5174 -0.0049 0 48 0.0016 U R.NLLSVAYK.N
459.2676 916.5207 916.5229 -0.0023 0 27 0.28 U R.IISSIEQK.E
459.2676 916.5207 916.5229 -0.0023 0 (5) 46 U R.IISSIEQK.E
461.7091 921.4037 921.4127 -0.0090 1 26 0.23 U K.MKGDYHR.Y + Oxidation (M)
461.7091 921.4037 921.4127 -0.0090 1 (25) 0.26 U K.MKGDYHR.Y + Oxidation (M)
489.2509 976.4872 976.4899 -0.0027 0 11 9.9 U R.QMVETELK.L
595.3293 1188.6440 1188.6536 -0.0096 0 (61) 7.3e-005 U K.DSTLIMQLLR.D
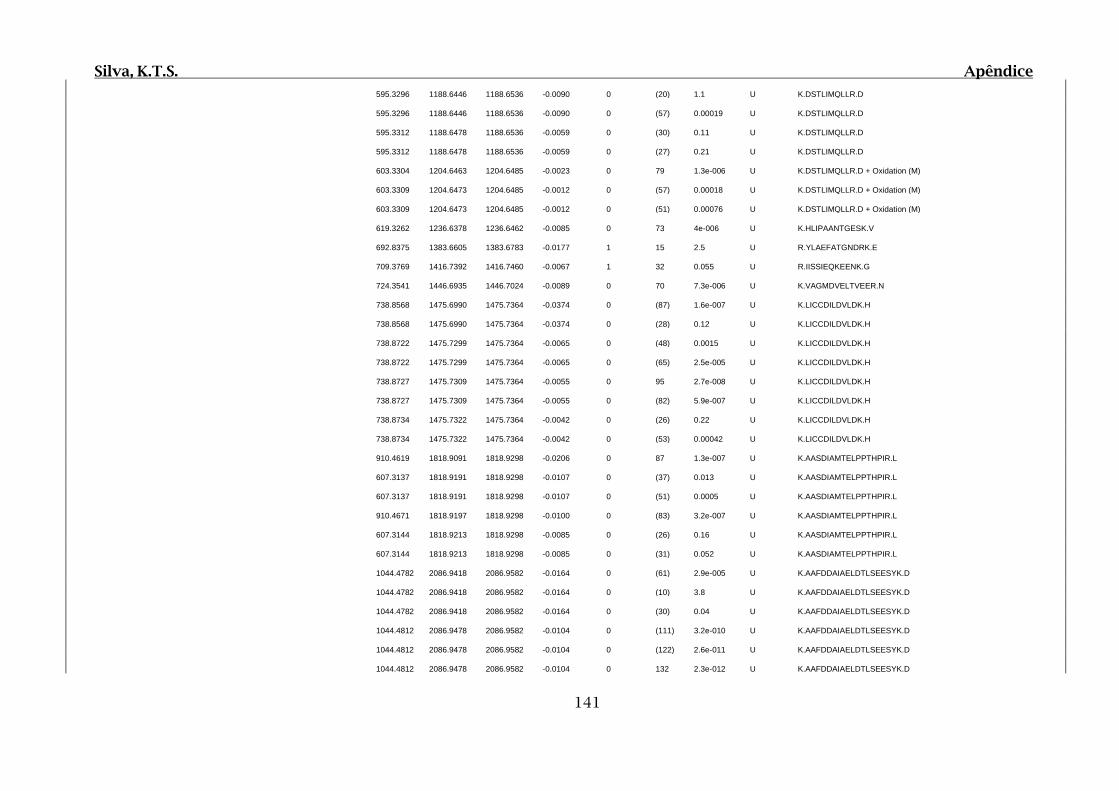
Silva, K.T.S. Apêndice
141
595.3296 1188.6446 1188.6536 -0.0090 0 (20) 1.1 U K.DSTLIMQLLR.D
595.3296 1188.6446 1188.6536 -0.0090 0 (57) 0.00019 U K.DSTLIMQLLR.D
595.3312 1188.6478 1188.6536 -0.0059 0 (30) 0.11 U K.DSTLIMQLLR.D
595.3312 1188.6478 1188.6536 -0.0059 0 (27) 0.21 U K.DSTLIMQLLR.D
603.3304 1204.6463 1204.6485 -0.0023 0 79 1.3e-006 U K.DSTLIMQLLR.D + Oxidation (M)
603.3309 1204.6473 1204.6485 -0.0012 0 (57) 0.00018 U K.DSTLIMQLLR.D + Oxidation (M)
603.3309 1204.6473 1204.6485 -0.0012 0 (51) 0.00076 U K.DSTLIMQLLR.D + Oxidation (M)
619.3262 1236.6378 1236.6462 -0.0085 0 73 4e-006 U K.HLIPAANTGESK.V
692.8375 1383.6605 1383.6783 -0.0177 1 15 2.5 U R.YLAEFATGNDRK.E
709.3769 1416.7392 1416.7460 -0.0067 1 32 0.055 U R.IISSIEQKEENK.G
724.3541 1446.6935 1446.7024 -0.0089 0 70 7.3e-006 U K.VAGMDVELTVEER.N
738.8568 1475.6990 1475.7364 -0.0374 0 (87) 1.6e-007 U K.LICCDILDVLDK.H
738.8568 1475.6990 1475.7364 -0.0374 0 (28) 0.12 U K.LICCDILDVLDK.H
738.8722 1475.7299 1475.7364 -0.0065 0 (48) 0.0015 U K.LICCDILDVLDK.H
738.8722 1475.7299 1475.7364 -0.0065 0 (65) 2.5e-005 U K.LICCDILDVLDK.H
738.8727 1475.7309 1475.7364 -0.0055 0 95 2.7e-008 U K.LICCDILDVLDK.H
738.8727 1475.7309 1475.7364 -0.0055 0 (82) 5.9e-007 U K.LICCDILDVLDK.H
738.8734 1475.7322 1475.7364 -0.0042 0 (26) 0.22 U K.LICCDILDVLDK.H
738.8734 1475.7322 1475.7364 -0.0042 0 (53) 0.00042 U K.LICCDILDVLDK.H
910.4619 1818.9091 1818.9298 -0.0206 0 87 1.3e-007 U K.AASDIAMTELPPTHPIR.L
607.3137 1818.9191 1818.9298 -0.0107 0 (37) 0.013 U K.AASDIAMTELPPTHPIR.L
607.3137 1818.9191 1818.9298 -0.0107 0 (51) 0.0005 U K.AASDIAMTELPPTHPIR.L
910.4671 1818.9197 1818.9298 -0.0100 0 (83) 3.2e-007 U K.AASDIAMTELPPTHPIR.L
607.3144 1818.9213 1818.9298 -0.0085 0 (26) 0.16 U K.AASDIAMTELPPTHPIR.L
607.3144 1818.9213 1818.9298 -0.0085 0 (31) 0.052 U K.AASDIAMTELPPTHPIR.L
1044.4782 2086.9418 2086.9582 -0.0164 0 (61) 2.9e-005 U K.AAFDDAIAELDTLSEESYK.D
1044.4782 2086.9418 2086.9582 -0.0164 0 (10) 3.8 U K.AAFDDAIAELDTLSEESYK.D
1044.4782 2086.9418 2086.9582 -0.0164 0 (30) 0.04 U K.AAFDDAIAELDTLSEESYK.D
1044.4812 2086.9478 2086.9582 -0.0104 0 (111) 3.2e-010 U K.AAFDDAIAELDTLSEESYK.D
1044.4812 2086.9478 2086.9582 -0.0104 0 (122) 2.6e-011 U K.AAFDDAIAELDTLSEESYK.D
1044.4812 2086.9478 2086.9582 -0.0104 0 132 2.3e-012 U K.AAFDDAIAELDTLSEESYK.D
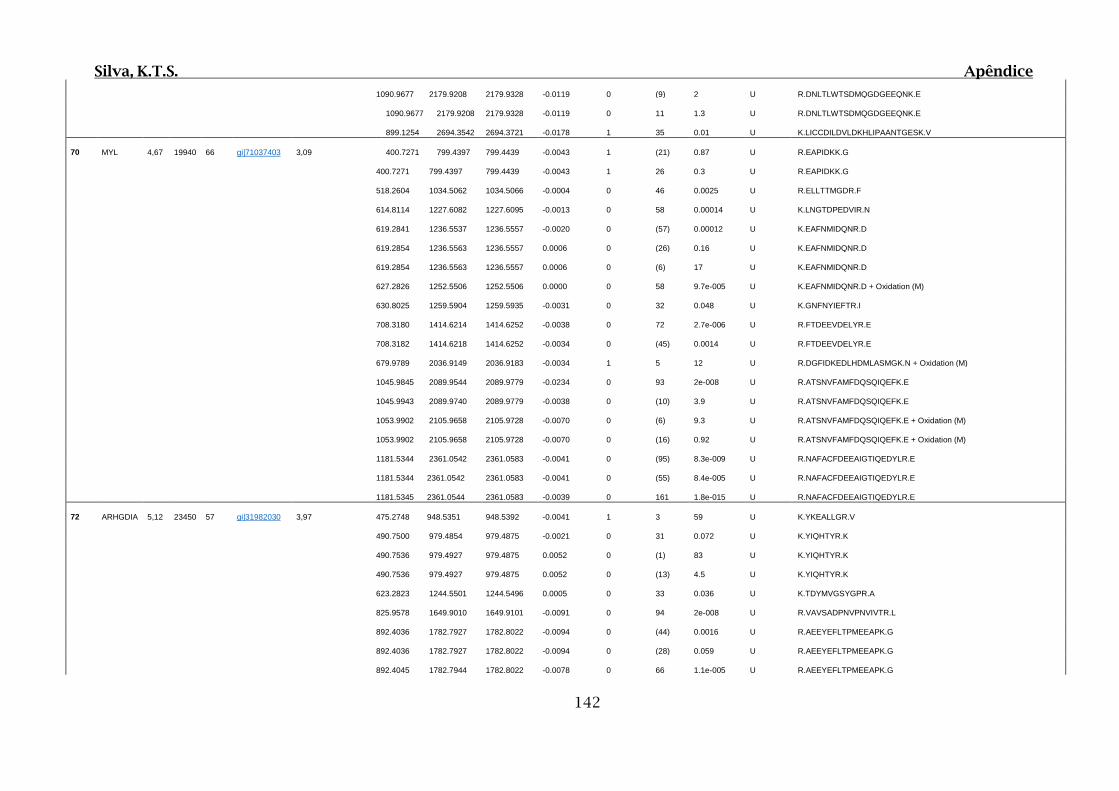
Silva, K.T.S. Apêndice
142
1090.9677 2179.9208 2179.9328 -0.0119 0 (9) 2 U R.DNLTLWTSDMQGDGEEQNK.E
1090.9677 2179.9208 2179.9328 -0.0119 0 11 1.3 U R.DNLTLWTSDMQGDGEEQNK.E
899.1254 2694.3542 2694.3721 -0.0178 1 35 0.01 U K.LICCDILDVLDKHLIPAANTGESK.V
70 MYL 4,67 19940 66 gi|71037403 3,09 400.7271 799.4397 799.4439 -0.0043 1 (21) 0.87 U R.EAPIDKK.G
400.7271 799.4397 799.4439 -0.0043 1 26 0.3 U R.EAPIDKK.G
518.2604 1034.5062 1034.5066 -0.0004 0 46 0.0025 U R.ELLTTMGDR.F
614.8114 1227.6082 1227.6095 -0.0013 0 58 0.00014 U K.LNGTDPEDVIR.N
619.2841 1236.5537 1236.5557 -0.0020 0 (57) 0.00012 U K.EAFNMIDQNR.D
619.2854 1236.5563 1236.5557 0.0006 0 (26) 0.16 U K.EAFNMIDQNR.D
619.2854 1236.5563 1236.5557 0.0006 0 (6) 17 U K.EAFNMIDQNR.D
627.2826 1252.5506 1252.5506 0.0000 0 58 9.7e-005 U K.EAFNMIDQNR.D + Oxidation (M)
630.8025 1259.5904 1259.5935 -0.0031 0 32 0.048 U K.GNFNYIEFTR.I
708.3180 1414.6214 1414.6252 -0.0038 0 72 2.7e-006 U R.FTDEEVDELYR.E
708.3182 1414.6218 1414.6252 -0.0034 0 (45) 0.0014 U R.FTDEEVDELYR.E
679.9789 2036.9149 2036.9183 -0.0034 1 5 12 U R.DGFIDKEDLHDMLASMGK.N + Oxidation (M)
1045.9845 2089.9544 2089.9779 -0.0234 0 93 2e-008 U R.ATSNVFAMFDQSQIQEFK.E
1045.9943 2089.9740 2089.9779 -0.0038 0 (10) 3.9 U R.ATSNVFAMFDQSQIQEFK.E
1053.9902 2105.9658 2105.9728 -0.0070 0 (6) 9.3 U R.ATSNVFAMFDQSQIQEFK.E + Oxidation (M)
1053.9902 2105.9658 2105.9728 -0.0070 0 (16) 0.92 U R.ATSNVFAMFDQSQIQEFK.E + Oxidation (M)
1181.5344 2361.0542 2361.0583 -0.0041 0 (95) 8.3e-009 U R.NAFACFDEEAIGTIQEDYLR.E
1181.5344 2361.0542 2361.0583 -0.0041 0 (55) 8.4e-005 U R.NAFACFDEEAIGTIQEDYLR.E
1181.5345 2361.0544 2361.0583 -0.0039 0 161 1.8e-015 U R.NAFACFDEEAIGTIQEDYLR.E
72 ARHGDIA 5,12 23450 57 gi|31982030 3,97 475.2748 948.5351 948.5392 -0.0041 1 3 59 U K.YKEALLGR.V
490.7500 979.4854 979.4875 -0.0021 0 31 0.072 U K.YIQHTYR.K
490.7536 979.4927 979.4875 0.0052 0 (1) 83 U K.YIQHTYR.K
490.7536 979.4927 979.4875 0.0052 0 (13) 4.5 U K.YIQHTYR.K
623.2823 1244.5501 1244.5496 0.0005 0 33 0.036 U K.TDYMVGSYGPR.A
825.9578 1649.9010 1649.9101 -0.0091 0 94 2e-008 U R.VAVSADPNVPNVIVTR.L
892.4036 1782.7927 1782.8022 -0.0094 0 (44) 0.0016 U R.AEEYEFLTPMEEAPK.G
892.4036 1782.7927 1782.8022 -0.0094 0 (28) 0.059 U R.AEEYEFLTPMEEAPK.G
892.4045 1782.7944 1782.8022 -0.0078 0 66 1.1e-005 U R.AEEYEFLTPMEEAPK.G
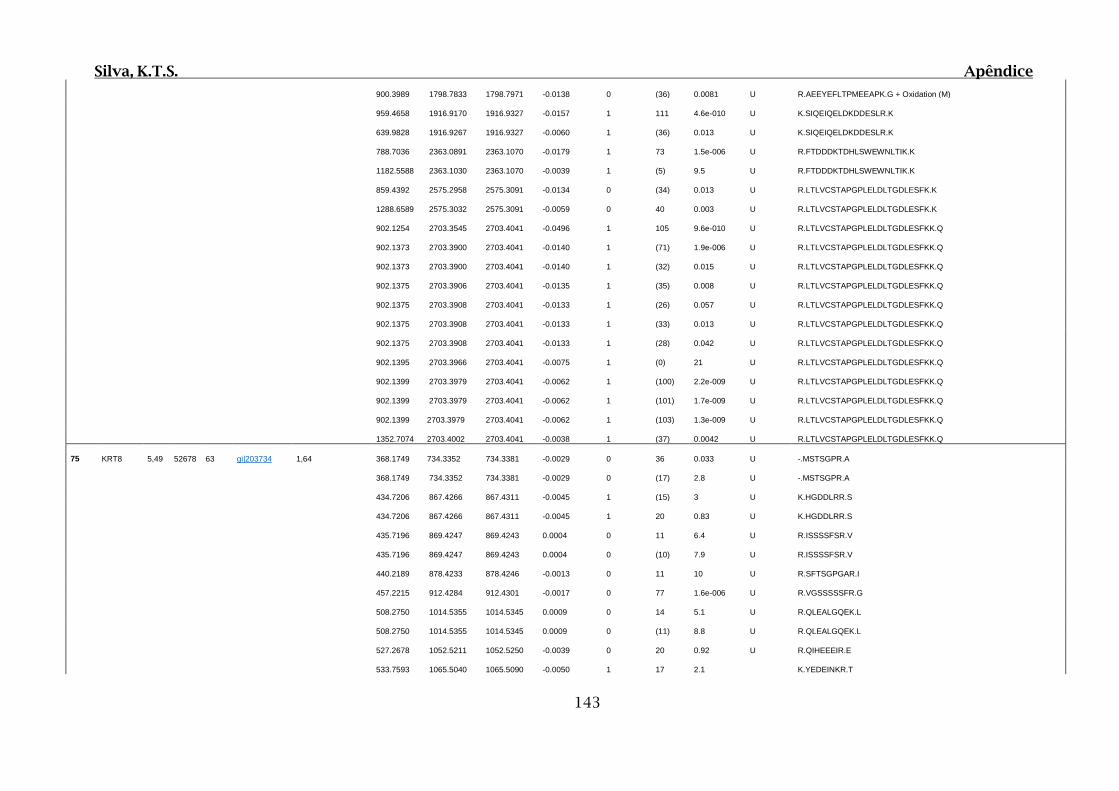
Silva, K.T.S. Apêndice
143
900.3989 1798.7833 1798.7971 -0.0138 0 (36) 0.0081 U R.AEEYEFLTPMEEAPK.G + Oxidation (M)
959.4658 1916.9170 1916.9327 -0.0157 1 111 4.6e-010 U K.SIQEIQELDKDDESLR.K
639.9828 1916.9267 1916.9327 -0.0060 1 (36) 0.013 U K.SIQEIQELDKDDESLR.K
788.7036 2363.0891 2363.1070 -0.0179 1 73 1.5e-006 U R.FTDDDKTDHLSWEWNLTIK.K
1182.5588 2363.1030 2363.1070 -0.0039 1 (5) 9.5 U R.FTDDDKTDHLSWEWNLTIK.K
859.4392 2575.2958 2575.3091 -0.0134 0 (34) 0.013 U R.LTLVCSTAPGPLELDLTGDLESFK.K
1288.6589 2575.3032 2575.3091 -0.0059 0 40 0.003 U R.LTLVCSTAPGPLELDLTGDLESFK.K
902.1254 2703.3545 2703.4041 -0.0496 1 105 9.6e-010 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1373 2703.3900 2703.4041 -0.0140 1 (71) 1.9e-006 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1373 2703.3900 2703.4041 -0.0140 1 (32) 0.015 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1375 2703.3906 2703.4041 -0.0135 1 (35) 0.008 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1375 2703.3908 2703.4041 -0.0133 1 (26) 0.057 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1375 2703.3908 2703.4041 -0.0133 1 (33) 0.013 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1375 2703.3908 2703.4041 -0.0133 1 (28) 0.042 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1395 2703.3966 2703.4041 -0.0075 1 (0) 21 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1399 2703.3979 2703.4041 -0.0062 1 (100) 2.2e-009 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1399 2703.3979 2703.4041 -0.0062 1 (101) 1.7e-009 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
902.1399 2703.3979 2703.4041 -0.0062 1 (103) 1.3e-009 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
1352.7074 2703.4002 2703.4041 -0.0038 1 (37) 0.0042 U R.LTLVCSTAPGPLELDLTGDLESFKK.Q
75 KRT8 5,49 52678 63 gi|203734 1,64 368.1749 734.3352 734.3381 -0.0029 0 36 0.033 U -.MSTSGPR.A
368.1749 734.3352 734.3381 -0.0029 0 (17) 2.8 U -.MSTSGPR.A
434.7206 867.4266 867.4311 -0.0045 1 (15) 3 U K.HGDDLRR.S
434.7206 867.4266 867.4311 -0.0045 1 20 0.83 U K.HGDDLRR.S
435.7196 869.4247 869.4243 0.0004 0 11 6.4 U R.ISSSSFSR.V
435.7196 869.4247 869.4243 0.0004 0 (10) 7.9 U R.ISSSSFSR.V
440.2189 878.4233 878.4246 -0.0013 0 11 10 U R.SFTSGPGAR.I
457.2215 912.4284 912.4301 -0.0017 0 77 1.6e-006 U R.VGSSSSSFR.G
508.2750 1014.5355 1014.5345 0.0009 0 14 5.1 U R.QLEALGQEK.L
508.2750 1014.5355 1014.5345 0.0009 0 (11) 8.8 U R.QLEALGQEK.L
527.2678 1052.5211 1052.5250 -0.0039 0 20 0.92 U R.QIHEEEIR.E
533.7593 1065.5040 1065.5090 -0.0050 1 17 2.1 K.YEDEINKR.T

Silva, K.T.S. Apêndice
144
547.2652 1092.5159 1092.5199 -0.0040 0 44 0.0046 U R.AQYEEIANR.S
558.7981 1115.5817 1115.5822 -0.0005 1 21 0.88 U K.DANAKLEDLK.N
592.2887 1182.5628 1182.5590 0.0037 0 6 19 U R.AEAETMYQIK.Y
598.2950 1194.5754 1194.5802 -0.0048 0 80 8.2e-007 U K.LVSESSDIMSK.-
627.3431 1252.6717 1252.6775 -0.0058 0 43 0.0042 U K.LEVDPNIQAVR.T
639.3552 1276.6959 1276.7027 -0.0067 0 79 1.1e-006 K.LALDIEIATYR.K
674.8364 1347.6583 1347.6704 -0.0121 0 88 1.5e-007 U R.SLDMDSIIAEVR.A
676.8372 1351.6598 1351.6693 -0.0095 0 (81) 6.4e-007 U R.TEMENEFVLIK.K
676.8372 1351.6598 1351.6693 -0.0095 0 89 1e-007 U R.TEMENEFVLIK.K
676.8397 1351.6647 1351.6693 -0.0046 0 (3) 46 U R.TEMENEFVLIK.K
679.8431 1357.6716 1357.6837 -0.0121 0 105 2.7e-009 U R.ATLEAAIADAEQR.G
682.8386 1363.6627 1363.6653 -0.0026 0 (13) 3.9 U R.SLDMDSIIAEVR.A + Oxidation (M)
710.3687 1418.7229 1418.7405 -0.0176 0 90 7.2e-008 U R.LEGLTDEINFLR.Q
710.3687 1418.7229 1418.7405 -0.0176 0 (69) 9.4e-006 U R.LEGLTDEINFLR.Q
710.3687 1418.7229 1418.7405 -0.0176 0 (72) 4.9e-006 U R.LEGLTDEINFLR.Q
710.3687 1418.7229 1418.7405 -0.0176 0 (51) 0.00059 U R.LEGLTDEINFLR.Q
713.8530 1425.6914 1425.6922 -0.0008 1 18 1.1 U R.SRAEAETMYQIK.Y
910.9514 1819.8883 1819.9026 -0.0143 0 145 1.8e-013 U K.LEVELGNMQGLVEDFK.N
924.3975 1846.7804 1846.7978 -0.0174 0 89 2.5e-008 U R.SNMDNMFESYINNLR.R
940.3966 1878.7786 1878.7876 -0.0090 0 (78) 3e-007 U R.SNMDNMFESYINNLR.R + 2 Oxidation (M)
668.6363 2002.8872 2002.8989 -0.0117 1 35 0.01 U R.SNMDNMFESYINNLRR.Q
688.0287 2061.0644 2061.0816 -0.0172 1 20 0.5 U K.LKLEVELGNMQGLVEDFK.N
1055.0050 2107.9954 2108.0056 -0.0101 0 22 0.29 U R.ELQSQISDTSVVLSMDNSR.S
862.1333 2583.3781 2583.4021 -0.0240 0 119 2.3e-011 U R.GSLGGFGGAGVGGITAVTVNQSLLNPLK.L
1292.7013 2583.3880 2583.4021 -0.0140 0 (61) 1.2e-005 U R.GSLGGFGGAGVGGITAVTVNQSLLNPLK.L
1273.6896 3818.0470 3818.0691 -0.0221 1 2 3.2 U R.GSLGGFGGAGVGGITAVTVNQSLLNPLKLEVDPNIQAVR.T
1403.9512 4208.8318 4208.8546 -0.0228 0 17 0.077 U K.TTSGYAGGLSSSYGGLTSPGFSYGMSSFQPGFGSVGGSSTYSR.T
+ Oxidation (M)
1403.9512 4208.8318 4208.8546 -0.0228 0 (10) 0.37 U K.TTSGYAGGLSSSYGGLTSPGFSYGMSSFQPGFGSVGGSSTYSR.T
+ Oxidation (M)
76 EZR 5,83 69477 29 gi|40804381 0,59 545.2825 1088.5505 1088.5462 0.0043 1 6 33 U R.ITEAEKNER.V
552.7913 1103.5680 1103.5764 -0.0084 0 56 0.00026 K.IGFPWSEIR.N
552.7948 1103.5751 1103.5764 -0.0012 0 (35) 0.033 K.IGFPWSEIR.N
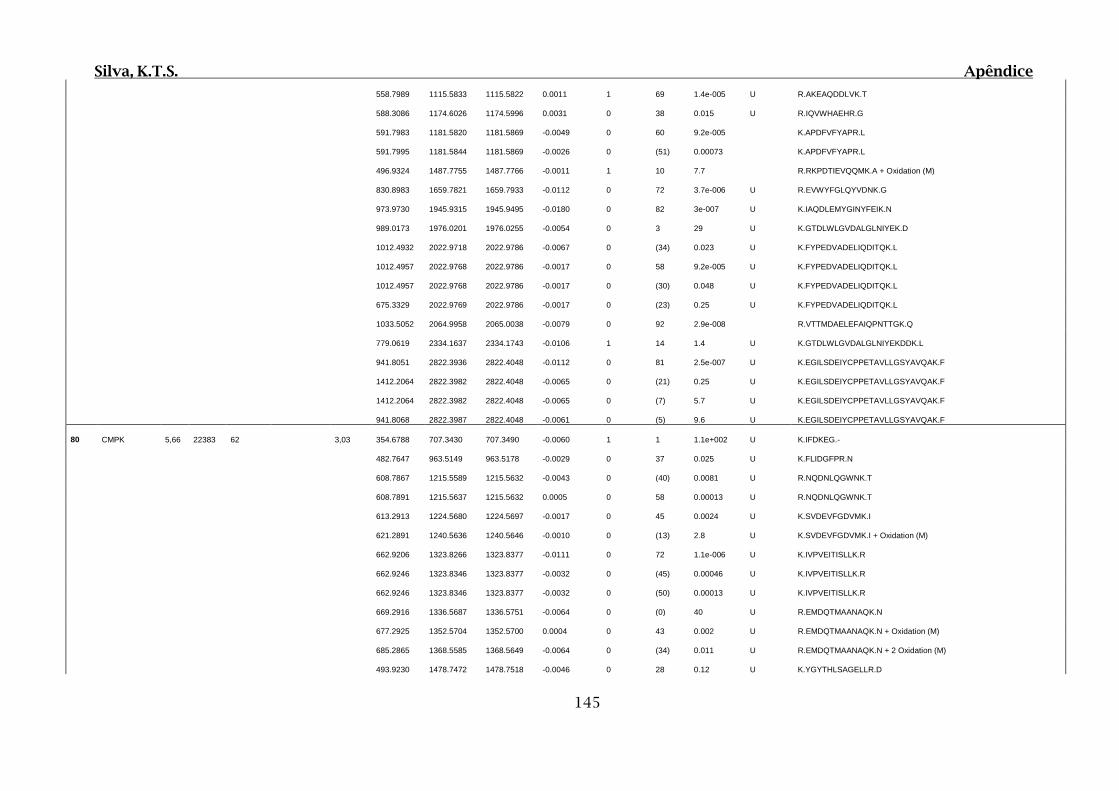
Silva, K.T.S. Apêndice
145
558.7989 1115.5833 1115.5822 0.0011 1 69 1.4e-005 U R.AKEAQDDLVK.T
588.3086 1174.6026 1174.5996 0.0031 0 38 0.015 U R.IQVWHAEHR.G
591.7983 1181.5820 1181.5869 -0.0049 0 60 9.2e-005 K.APDFVFYAPR.L
591.7995 1181.5844 1181.5869 -0.0026 0 (51) 0.00073 K.APDFVFYAPR.L
496.9324 1487.7755 1487.7766 -0.0011 1 10 7.7 R.RKPDTIEVQQMK.A + Oxidation (M)
830.8983 1659.7821 1659.7933 -0.0112 0 72 3.7e-006 U R.EVWYFGLQYVDNK.G
973.9730 1945.9315 1945.9495 -0.0180 0 82 3e-007 U K.IAQDLEMYGINYFEIK.N
989.0173 1976.0201 1976.0255 -0.0054 0 3 29 U K.GTDLWLGVDALGLNIYEK.D
1012.4932 2022.9718 2022.9786 -0.0067 0 (34) 0.023 U K.FYPEDVADELIQDITQK.L
1012.4957 2022.9768 2022.9786 -0.0017 0 58 9.2e-005 U K.FYPEDVADELIQDITQK.L
1012.4957 2022.9768 2022.9786 -0.0017 0 (30) 0.048 U K.FYPEDVADELIQDITQK.L
675.3329 2022.9769 2022.9786 -0.0017 0 (23) 0.25 U K.FYPEDVADELIQDITQK.L
1033.5052 2064.9958 2065.0038 -0.0079 0 92 2.9e-008 R.VTTMDAELEFAIQPNTTGK.Q
779.0619 2334.1637 2334.1743 -0.0106 1 14 1.4 U K.GTDLWLGVDALGLNIYEKDDK.L
941.8051 2822.3936 2822.4048 -0.0112 0 81 2.5e-007 U K.EGILSDEIYCPPETAVLLGSYAVQAK.F
1412.2064 2822.3982 2822.4048 -0.0065 0 (21) 0.25 U K.EGILSDEIYCPPETAVLLGSYAVQAK.F
1412.2064 2822.3982 2822.4048 -0.0065 0 (7) 5.7 U K.EGILSDEIYCPPETAVLLGSYAVQAK.F
941.8068 2822.3987 2822.4048 -0.0061 0 (5) 9.6 U K.EGILSDEIYCPPETAVLLGSYAVQAK.F
80 CMPK 5,66 22383 62
gi|150383503 3,03 354.6788 707.3430 707.3490 -0.0060 1 1 1.1e+002 U K.IFDKEG.-
482.7647 963.5149 963.5178 -0.0029 0 37 0.025 U K.FLIDGFPR.N
608.7867 1215.5589 1215.5632 -0.0043 0 (40) 0.0081 U R.NQDNLQGWNK.T
608.7891 1215.5637 1215.5632 0.0005 0 58 0.00013 U R.NQDNLQGWNK.T
613.2913 1224.5680 1224.5697 -0.0017 0 45 0.0024 U K.SVDEVFGDVMK.I
621.2891 1240.5636 1240.5646 -0.0010 0 (13) 2.8 U K.SVDEVFGDVMK.I + Oxidation (M)
662.9206 1323.8266 1323.8377 -0.0111 0 72 1.1e-006 U K.IVPVEITISLLK.R
662.9246 1323.8346 1323.8377 -0.0032 0 (45) 0.00046 U K.IVPVEITISLLK.R
662.9246 1323.8346 1323.8377 -0.0032 0 (50) 0.00013 U K.IVPVEITISLLK.R
669.2916 1336.5687 1336.5751 -0.0064 0 (0) 40 U R.EMDQTMAANAQK.N
677.2925 1352.5704 1352.5700 0.0004 0 43 0.002 U R.EMDQTMAANAQK.N + Oxidation (M)
685.2865 1368.5585 1368.5649 -0.0064 0 (34) 0.011 U R.EMDQTMAANAQK.N + 2 Oxidation (M)
493.9230 1478.7472 1478.7518 -0.0046 0 28 0.12 U K.YGYTHLSAGELLR.D
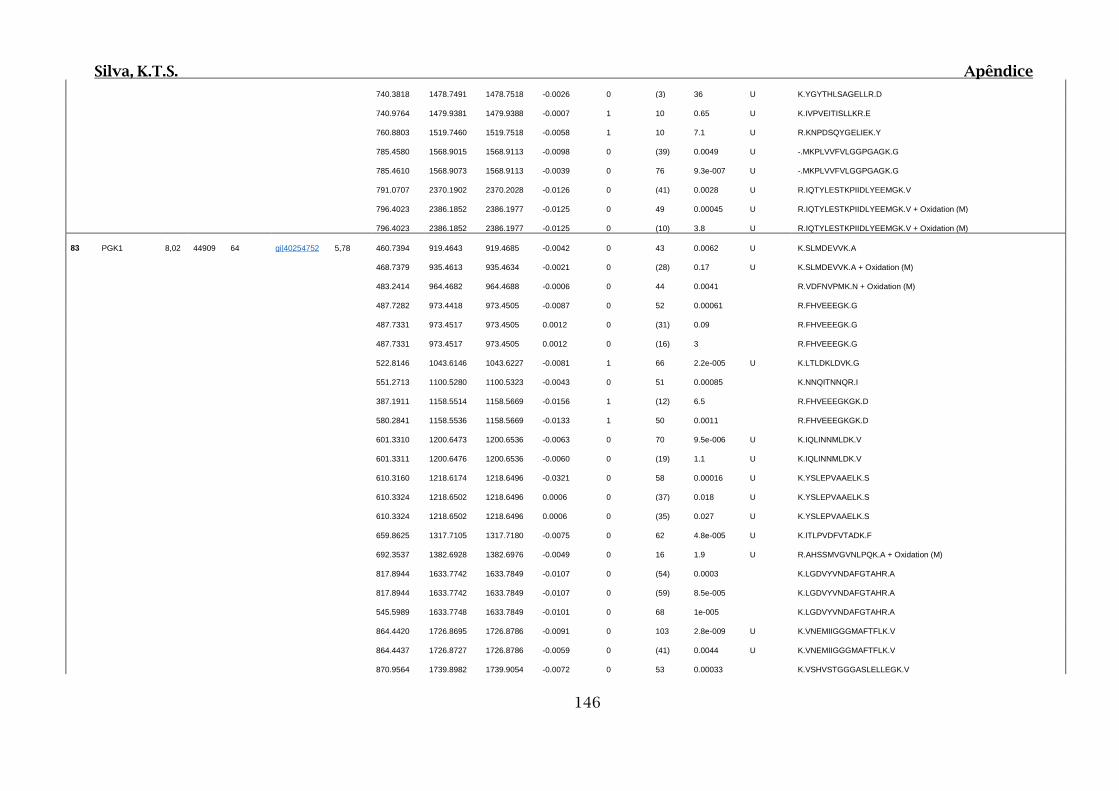
Silva, K.T.S. Apêndice
146
740.3818 1478.7491 1478.7518 -0.0026 0 (3) 36 U K.YGYTHLSAGELLR.D
740.9764 1479.9381 1479.9388 -0.0007 1 10 0.65 U K.IVPVEITISLLKR.E
760.8803 1519.7460 1519.7518 -0.0058 1 10 7.1 U R.KNPDSQYGELIEK.Y
785.4580 1568.9015 1568.9113 -0.0098 0 (39) 0.0049 U -.MKPLVVFVLGGPGAGK.G
785.4610 1568.9073 1568.9113 -0.0039 0 76 9.3e-007 U -.MKPLVVFVLGGPGAGK.G
791.0707 2370.1902 2370.2028 -0.0126 0 (41) 0.0028 U R.IQTYLESTKPIIDLYEEMGK.V
796.4023 2386.1852 2386.1977 -0.0125 0 49 0.00045 U R.IQTYLESTKPIIDLYEEMGK.V + Oxidation (M)
796.4023 2386.1852 2386.1977 -0.0125 0 (10) 3.8 U R.IQTYLESTKPIIDLYEEMGK.V + Oxidation (M)
83 PGK1 8,02 44909 64 gi|40254752 5,78 460.7394 919.4643 919.4685 -0.0042 0 43 0.0062 U K.SLMDEVVK.A
468.7379 935.4613 935.4634 -0.0021 0 (28) 0.17 U K.SLMDEVVK.A + Oxidation (M)
483.2414 964.4682 964.4688 -0.0006 0 44 0.0041 R.VDFNVPMK.N + Oxidation (M)
487.7282 973.4418 973.4505 -0.0087 0 52 0.00061 R.FHVEEEGK.G
487.7331 973.4517 973.4505 0.0012 0 (31) 0.09 R.FHVEEEGK.G
487.7331 973.4517 973.4505 0.0012 0 (16) 3 R.FHVEEEGK.G
522.8146 1043.6146 1043.6227 -0.0081 1 66 2.2e-005 U K.LTLDKLDVK.G
551.2713 1100.5280 1100.5323 -0.0043 0 51 0.00085 K.NNQITNNQR.I
387.1911 1158.5514 1158.5669 -0.0156 1 (12) 6.5 R.FHVEEEGKGK.D
580.2841 1158.5536 1158.5669 -0.0133 1 50 0.0011 R.FHVEEEGKGK.D
601.3310 1200.6473 1200.6536 -0.0063 0 70 9.5e-006 U K.IQLINNMLDK.V
601.3311 1200.6476 1200.6536 -0.0060 0 (19) 1.1 U K.IQLINNMLDK.V
610.3160 1218.6174 1218.6496 -0.0321 0 58 0.00016 U K.YSLEPVAAELK.S
610.3324 1218.6502 1218.6496 0.0006 0 (37) 0.018 U K.YSLEPVAAELK.S
610.3324 1218.6502 1218.6496 0.0006 0 (35) 0.027 U K.YSLEPVAAELK.S
659.8625 1317.7105 1317.7180 -0.0075 0 62 4.8e-005 U K.ITLPVDFVTADK.F
692.3537 1382.6928 1382.6976 -0.0049 0 16 1.9 U R.AHSSMVGVNLPQK.A + Oxidation (M)
817.8944 1633.7742 1633.7849 -0.0107 0 (54) 0.0003 K.LGDVYVNDAFGTAHR.A
817.8944 1633.7742 1633.7849 -0.0107 0 (59) 8.5e-005 K.LGDVYVNDAFGTAHR.A
545.5989 1633.7748 1633.7849 -0.0101 0 68 1e-005 K.LGDVYVNDAFGTAHR.A
864.4420 1726.8695 1726.8786 -0.0091 0 103 2.8e-009 U K.VNEMIIGGGMAFTFLK.V
864.4437 1726.8727 1726.8786 -0.0059 0 (41) 0.0044 U K.VNEMIIGGGMAFTFLK.V
870.9564 1739.8982 1739.9054 -0.0072 0 53 0.00033 K.VSHVSTGGGASLELLEGK.V
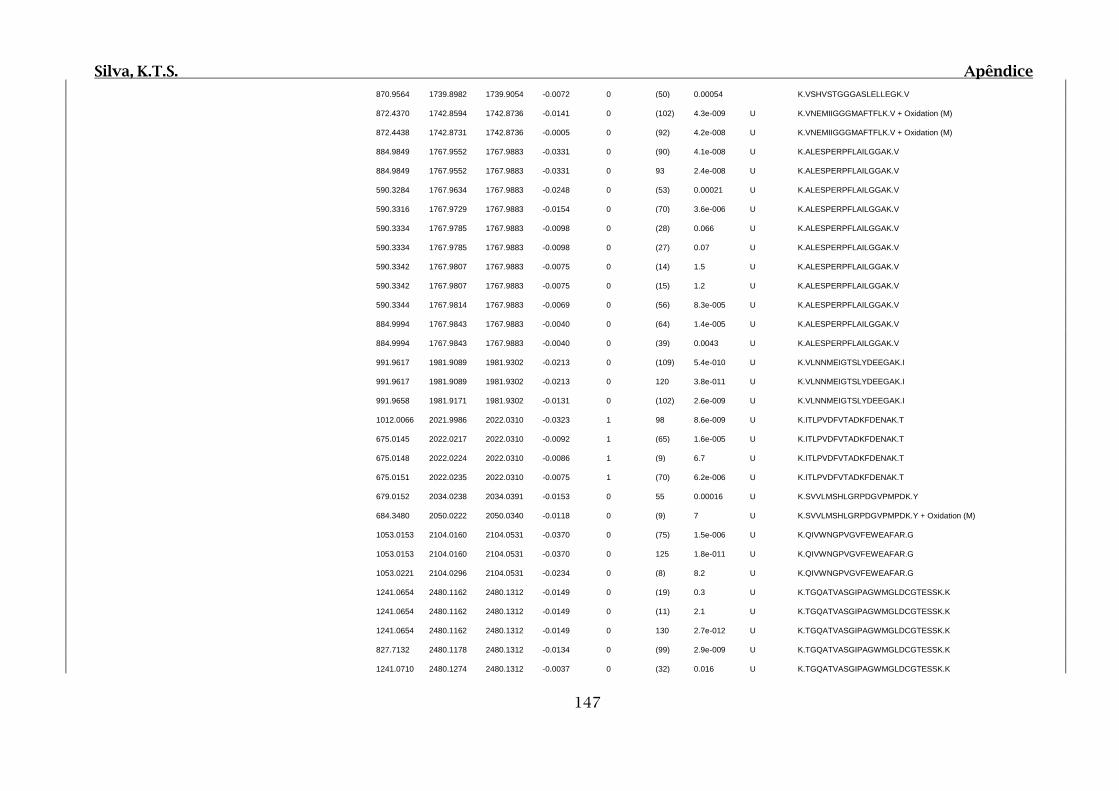
Silva, K.T.S. Apêndice
147
870.9564 1739.8982 1739.9054 -0.0072 0 (50) 0.00054 K.VSHVSTGGGASLELLEGK.V
872.4370 1742.8594 1742.8736 -0.0141 0 (102) 4.3e-009 U K.VNEMIIGGGMAFTFLK.V + Oxidation (M)
872.4438 1742.8731 1742.8736 -0.0005 0 (92) 4.2e-008 U K.VNEMIIGGGMAFTFLK.V + Oxidation (M)
884.9849 1767.9552 1767.9883 -0.0331 0 (90) 4.1e-008 U K.ALESPERPFLAILGGAK.V
884.9849 1767.9552 1767.9883 -0.0331 0 93 2.4e-008 U K.ALESPERPFLAILGGAK.V
590.3284 1767.9634 1767.9883 -0.0248 0 (53) 0.00021 U K.ALESPERPFLAILGGAK.V
590.3316 1767.9729 1767.9883 -0.0154 0 (70) 3.6e-006 U K.ALESPERPFLAILGGAK.V
590.3334 1767.9785 1767.9883 -0.0098 0 (28) 0.066 U K.ALESPERPFLAILGGAK.V
590.3334 1767.9785 1767.9883 -0.0098 0 (27) 0.07 U K.ALESPERPFLAILGGAK.V
590.3342 1767.9807 1767.9883 -0.0075 0 (14) 1.5 U K.ALESPERPFLAILGGAK.V
590.3342 1767.9807 1767.9883 -0.0075 0 (15) 1.2 U K.ALESPERPFLAILGGAK.V
590.3344 1767.9814 1767.9883 -0.0069 0 (56) 8.3e-005 U K.ALESPERPFLAILGGAK.V
884.9994 1767.9843 1767.9883 -0.0040 0 (64) 1.4e-005 U K.ALESPERPFLAILGGAK.V
884.9994 1767.9843 1767.9883 -0.0040 0 (39) 0.0043 U K.ALESPERPFLAILGGAK.V
991.9617 1981.9089 1981.9302 -0.0213 0 (109) 5.4e-010 U K.VLNNMEIGTSLYDEEGAK.I
991.9617 1981.9089 1981.9302 -0.0213 0 120 3.8e-011 U K.VLNNMEIGTSLYDEEGAK.I
991.9658 1981.9171 1981.9302 -0.0131 0 (102) 2.6e-009 U K.VLNNMEIGTSLYDEEGAK.I
1012.0066 2021.9986 2022.0310 -0.0323 1 98 8.6e-009 U K.ITLPVDFVTADKFDENAK.T
675.0145 2022.0217 2022.0310 -0.0092 1 (65) 1.6e-005 U K.ITLPVDFVTADKFDENAK.T
675.0148 2022.0224 2022.0310 -0.0086 1 (9) 6.7 U K.ITLPVDFVTADKFDENAK.T
675.0151 2022.0235 2022.0310 -0.0075 1 (70) 6.2e-006 U K.ITLPVDFVTADKFDENAK.T
679.0152 2034.0238 2034.0391 -0.0153 0 55 0.00016 U K.SVVLMSHLGRPDGVPMPDK.Y
684.3480 2050.0222 2050.0340 -0.0118 0 (9) 7 U K.SVVLMSHLGRPDGVPMPDK.Y + Oxidation (M)
1053.0153 2104.0160 2104.0531 -0.0370 0 (75) 1.5e-006 U K.QIVWNGPVGVFEWEAFAR.G
1053.0153 2104.0160 2104.0531 -0.0370 0 125 1.8e-011 U K.QIVWNGPVGVFEWEAFAR.G
1053.0221 2104.0296 2104.0531 -0.0234 0 (8) 8.2 U K.QIVWNGPVGVFEWEAFAR.G
1241.0654 2480.1162 2480.1312 -0.0149 0 (19) 0.3 U K.TGQATVASGIPAGWMGLDCGTESSK.K
1241.0654 2480.1162 2480.1312 -0.0149 0 (11) 2.1 U K.TGQATVASGIPAGWMGLDCGTESSK.K
1241.0654 2480.1162 2480.1312 -0.0149 0 130 2.7e-012 U K.TGQATVASGIPAGWMGLDCGTESSK.K
827.7132 2480.1178 2480.1312 -0.0134 0 (99) 2.9e-009 U K.TGQATVASGIPAGWMGLDCGTESSK.K
1241.0710 2480.1274 2480.1312 -0.0037 0 (32) 0.016 U K.TGQATVASGIPAGWMGLDCGTESSK.K
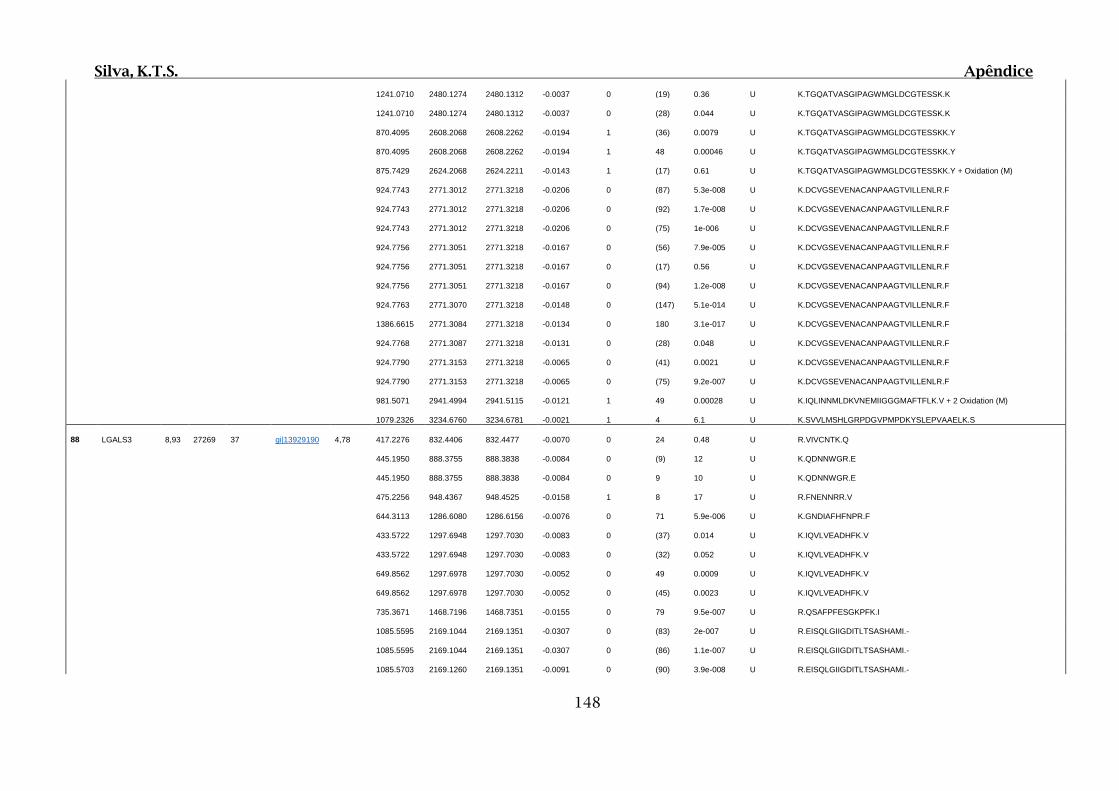
Silva, K.T.S. Apêndice
148
1241.0710 2480.1274 2480.1312 -0.0037 0 (19) 0.36 U K.TGQATVASGIPAGWMGLDCGTESSK.K
1241.0710 2480.1274 2480.1312 -0.0037 0 (28) 0.044 U K.TGQATVASGIPAGWMGLDCGTESSK.K
870.4095 2608.2068 2608.2262 -0.0194 1 (36) 0.0079 U K.TGQATVASGIPAGWMGLDCGTESSKK.Y
870.4095 2608.2068 2608.2262 -0.0194 1 48 0.00046 U K.TGQATVASGIPAGWMGLDCGTESSKK.Y
875.7429 2624.2068 2624.2211 -0.0143 1 (17) 0.61 U K.TGQATVASGIPAGWMGLDCGTESSKK.Y + Oxidation (M)
924.7743 2771.3012 2771.3218 -0.0206 0 (87) 5.3e-008 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7743 2771.3012 2771.3218 -0.0206 0 (92) 1.7e-008 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7743 2771.3012 2771.3218 -0.0206 0 (75) 1e-006 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7756 2771.3051 2771.3218 -0.0167 0 (56) 7.9e-005 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7756 2771.3051 2771.3218 -0.0167 0 (17) 0.56 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7756 2771.3051 2771.3218 -0.0167 0 (94) 1.2e-008 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7763 2771.3070 2771.3218 -0.0148 0 (147) 5.1e-014 U K.DCVGSEVENACANPAAGTVILLENLR.F
1386.6615 2771.3084 2771.3218 -0.0134 0 180 3.1e-017 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7768 2771.3087 2771.3218 -0.0131 0 (28) 0.048 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7790 2771.3153 2771.3218 -0.0065 0 (41) 0.0021 U K.DCVGSEVENACANPAAGTVILLENLR.F
924.7790 2771.3153 2771.3218 -0.0065 0 (75) 9.2e-007 U K.DCVGSEVENACANPAAGTVILLENLR.F
981.5071 2941.4994 2941.5115 -0.0121 1 49 0.00028 U K.IQLINNMLDKVNEMIIGGGMAFTFLK.V + 2 Oxidation (M)
1079.2326 3234.6760 3234.6781 -0.0021 1 4 6.1 U K.SVVLMSHLGRPDGVPMPDKYSLEPVAAELK.S
88 LGALS3 8,93 27269 37 gi|13929190 4,78 417.2276 832.4406 832.4477 -0.0070 0 24 0.48 U R.VIVCNTK.Q
445.1950 888.3755 888.3838 -0.0084 0 (9) 12 U K.QDNNWGR.E
445.1950 888.3755 888.3838 -0.0084 0 9 10 U K.QDNNWGR.E
475.2256 948.4367 948.4525 -0.0158 1 8 17 U R.FNENNRR.V
644.3113 1286.6080 1286.6156 -0.0076 0 71 5.9e-006 U K.GNDIAFHFNPR.F
433.5722 1297.6948 1297.7030 -0.0083 0 (37) 0.014 U K.IQVLVEADHFK.V
433.5722 1297.6948 1297.7030 -0.0083 0 (32) 0.052 U K.IQVLVEADHFK.V
649.8562 1297.6978 1297.7030 -0.0052 0 49 0.0009 U K.IQVLVEADHFK.V
649.8562 1297.6978 1297.7030 -0.0052 0 (45) 0.0023 U K.IQVLVEADHFK.V
735.3671 1468.7196 1468.7351 -0.0155 0 79 9.5e-007 U R.QSAFPFESGKPFK.I
1085.5595 2169.1044 2169.1351 -0.0307 0 (83) 2e-007 U R.EISQLGIIGDITLTSASHAMI.-
1085.5595 2169.1044 2169.1351 -0.0307 0 (86) 1.1e-007 U R.EISQLGIIGDITLTSASHAMI.-
1085.5703 2169.1260 2169.1351 -0.0091 0 (90) 3.9e-008 U R.EISQLGIIGDITLTSASHAMI.-
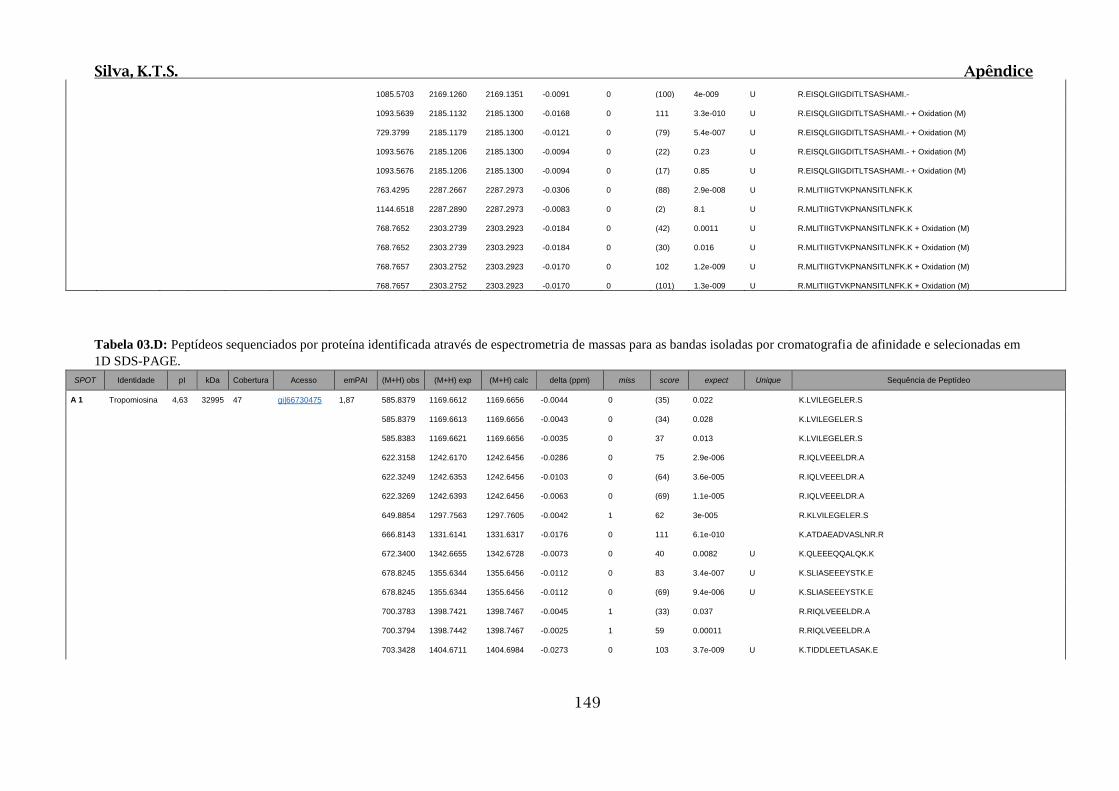
Silva, K.T.S. Apêndice
149
1085.5703 2169.1260 2169.1351 -0.0091 0 (100) 4e-009 U R.EISQLGIIGDITLTSASHAMI.-
1093.5639 2185.1132 2185.1300 -0.0168 0 111 3.3e-010 U R.EISQLGIIGDITLTSASHAMI.- + Oxidation (M)
729.3799 2185.1179 2185.1300 -0.0121 0 (79) 5.4e-007 U R.EISQLGIIGDITLTSASHAMI.- + Oxidation (M)
1093.5676 2185.1206 2185.1300 -0.0094 0 (22) 0.23 U R.EISQLGIIGDITLTSASHAMI.- + Oxidation (M)
1093.5676 2185.1206 2185.1300 -0.0094 0 (17) 0.85 U R.EISQLGIIGDITLTSASHAMI.- + Oxidation (M)
763.4295 2287.2667 2287.2973 -0.0306 0 (88) 2.9e-008 U R.MLITIIGTVKPNANSITLNFK.K
1144.6518 2287.2890 2287.2973 -0.0083 0 (2) 8.1 U R.MLITIIGTVKPNANSITLNFK.K
768.7652 2303.2739 2303.2923 -0.0184 0 (42) 0.0011 U R.MLITIIGTVKPNANSITLNFK.K + Oxidation (M)
768.7652 2303.2739 2303.2923 -0.0184 0 (30) 0.016 U R.MLITIIGTVKPNANSITLNFK.K + Oxidation (M)
768.7657 2303.2752 2303.2923 -0.0170 0 102 1.2e-009 U R.MLITIIGTVKPNANSITLNFK.K + Oxidation (M)
768.7657 2303.2752 2303.2923 -0.0170 0 (101) 1.3e-009 U R.MLITIIGTVKPNANSITLNFK.K + Oxidation (M)
Tabela 03.D: Peptídeos sequenciados por proteína identificada através de espectrometria de massas para as bandas isoladas por cromatografia de afinidade e selecionadas em
1D SDS-PAGE. SPOT Identidade pI kDa Cobertura Acesso emPAI (M+H) obs (M+H) exp (M+H) calc delta (ppm) miss score expect Unique Sequência de Peptídeo
A 1 Tropomiosina 4,63 32995 47 gi|66730475 1,87 585.8379 1169.6612 1169.6656 -0.0044 0 (35) 0.022 K.LVILEGELER.S
585.8379 1169.6613 1169.6656 -0.0043 0 (34) 0.028 K.LVILEGELER.S
585.8383 1169.6621 1169.6656 -0.0035 0 37 0.013 K.LVILEGELER.S
622.3158 1242.6170 1242.6456 -0.0286 0 75 2.9e-006 R.IQLVEEELDR.A
622.3249 1242.6353 1242.6456 -0.0103 0 (64) 3.6e-005 R.IQLVEEELDR.A
622.3269 1242.6393 1242.6456 -0.0063 0 (69) 1.1e-005 R.IQLVEEELDR.A
649.8854 1297.7563 1297.7605 -0.0042 1 62 3e-005 R.KLVILEGELER.S
666.8143 1331.6141 1331.6317 -0.0176 0 111 6.1e-010 K.ATDAEADVASLNR.R
672.3400 1342.6655 1342.6728 -0.0073 0 40 0.0082 U K.QLEEEQQALQK.K
678.8245 1355.6344 1355.6456 -0.0112 0 83 3.4e-007 U K.SLIASEEEYSTK.E
678.8245 1355.6344 1355.6456 -0.0112 0 (69) 9.4e-006 U K.SLIASEEEYSTK.E
700.3783 1398.7421 1398.7467 -0.0045 1 (33) 0.037 R.RIQLVEEELDR.A
700.3794 1398.7442 1398.7467 -0.0025 1 59 0.00011 R.RIQLVEEELDR.A
703.3428 1404.6711 1404.6984 -0.0273 0 103 3.7e-009 U K.TIDDLEETLASAK.E
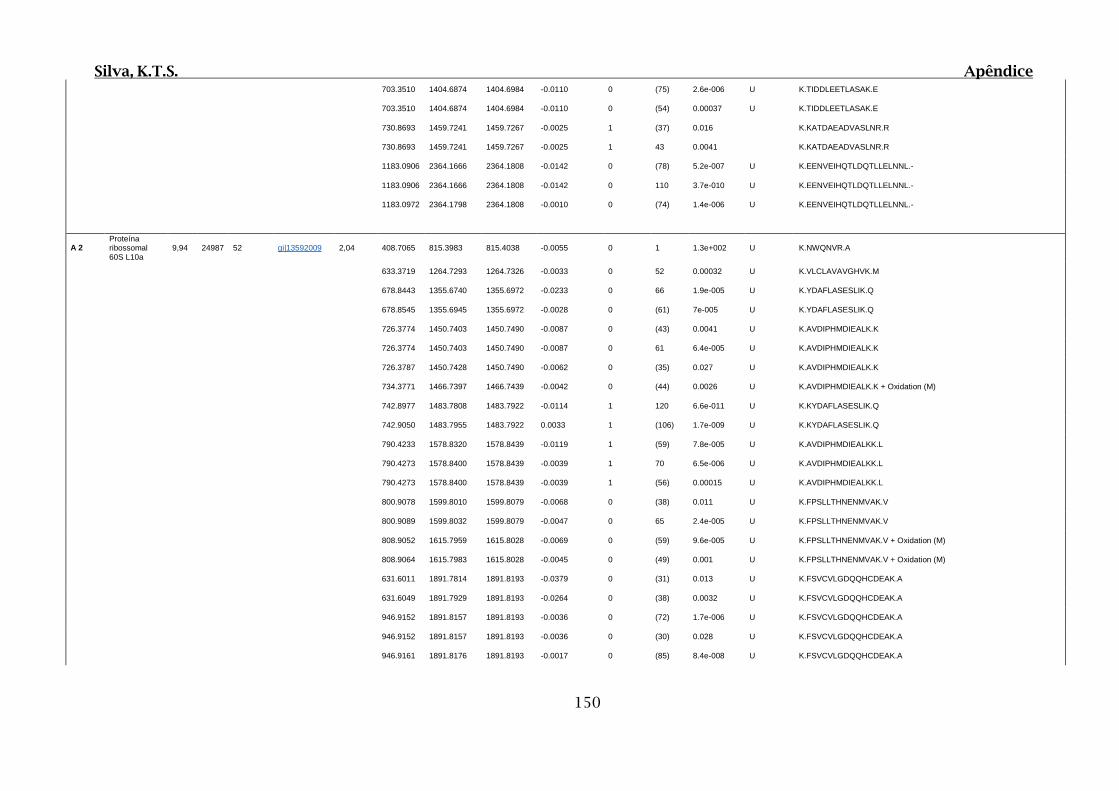
Silva, K.T.S. Apêndice
150
703.3510 1404.6874 1404.6984 -0.0110 0 (75) 2.6e-006 U K.TIDDLEETLASAK.E
703.3510 1404.6874 1404.6984 -0.0110 0 (54) 0.00037 U K.TIDDLEETLASAK.E
730.8693 1459.7241 1459.7267 -0.0025 1 (37) 0.016 K.KATDAEADVASLNR.R
730.8693 1459.7241 1459.7267 -0.0025 1 43 0.0041 K.KATDAEADVASLNR.R
1183.0906 2364.1666 2364.1808 -0.0142 0 (78) 5.2e-007 U K.EENVEIHQTLDQTLLELNNL.-
1183.0906 2364.1666 2364.1808 -0.0142 0 110 3.7e-010 U K.EENVEIHQTLDQTLLELNNL.-
1183.0972 2364.1798 2364.1808 -0.0010 0 (74) 1.4e-006 U K.EENVEIHQTLDQTLLELNNL.-
A 2 Proteína ribossomal 60S L10a
9,94 24987 52 gi|13592009 2,04 408.7065 815.3983 815.4038 -0.0055 0 1 1.3e+002 U K.NWQNVR.A
633.3719 1264.7293 1264.7326 -0.0033 0 52 0.00032 U K.VLCLAVAVGHVK.M
678.8443 1355.6740 1355.6972 -0.0233 0 66 1.9e-005 U K.YDAFLASESLIK.Q
678.8545 1355.6945 1355.6972 -0.0028 0 (61) 7e-005 U K.YDAFLASESLIK.Q
726.3774 1450.7403 1450.7490 -0.0087 0 (43) 0.0041 U K.AVDIPHMDIEALK.K
726.3774 1450.7403 1450.7490 -0.0087 0 61 6.4e-005 U K.AVDIPHMDIEALK.K
726.3787 1450.7428 1450.7490 -0.0062 0 (35) 0.027 U K.AVDIPHMDIEALK.K
734.3771 1466.7397 1466.7439 -0.0042 0 (44) 0.0026 U K.AVDIPHMDIEALK.K + Oxidation (M)
742.8977 1483.7808 1483.7922 -0.0114 1 120 6.6e-011 U K.KYDAFLASESLIK.Q
742.9050 1483.7955 1483.7922 0.0033 1 (106) 1.7e-009 U K.KYDAFLASESLIK.Q
790.4233 1578.8320 1578.8439 -0.0119 1 (59) 7.8e-005 U K.AVDIPHMDIEALKK.L
790.4273 1578.8400 1578.8439 -0.0039 1 70 6.5e-006 U K.AVDIPHMDIEALKK.L
790.4273 1578.8400 1578.8439 -0.0039 1 (56) 0.00015 U K.AVDIPHMDIEALKK.L
800.9078 1599.8010 1599.8079 -0.0068 0 (38) 0.011 U K.FPSLLTHNENMVAK.V
800.9089 1599.8032 1599.8079 -0.0047 0 65 2.4e-005 U K.FPSLLTHNENMVAK.V
808.9052 1615.7959 1615.8028 -0.0069 0 (59) 9.6e-005 U K.FPSLLTHNENMVAK.V + Oxidation (M)
808.9064 1615.7983 1615.8028 -0.0045 0 (49) 0.001 U K.FPSLLTHNENMVAK.V + Oxidation (M)
631.6011 1891.7814 1891.8193 -0.0379 0 (31) 0.013 U K.FSVCVLGDQQHCDEAK.A
631.6049 1891.7929 1891.8193 -0.0264 0 (38) 0.0032 U K.FSVCVLGDQQHCDEAK.A
946.9152 1891.8157 1891.8193 -0.0036 0 (72) 1.7e-006 U K.FSVCVLGDQQHCDEAK.A
946.9152 1891.8157 1891.8193 -0.0036 0 (30) 0.028 U K.FSVCVLGDQQHCDEAK.A
946.9161 1891.8176 1891.8193 -0.0017 0 (85) 8.4e-008 U K.FSVCVLGDQQHCDEAK.A
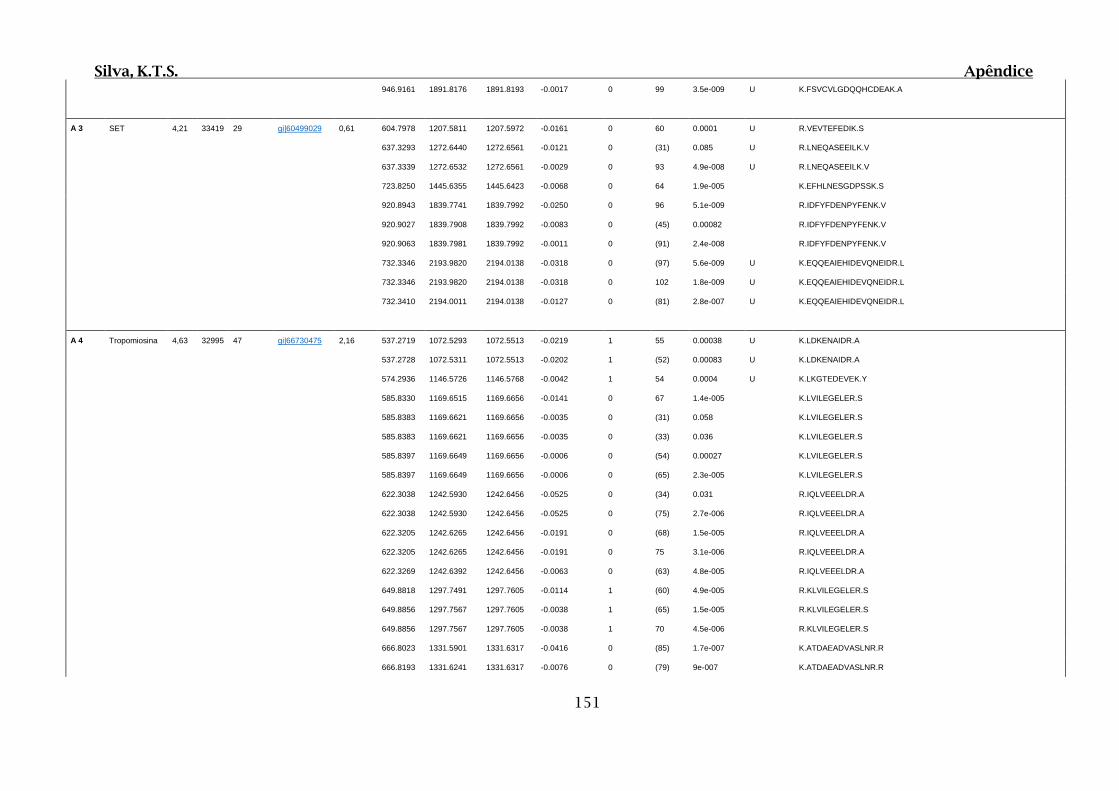
Silva, K.T.S. Apêndice
151
946.9161 1891.8176 1891.8193 -0.0017 0 99 3.5e-009 U K.FSVCVLGDQQHCDEAK.A
A 3 SET 4,21 33419 29 gi|60499029 0,61 604.7978 1207.5811 1207.5972 -0.0161 0 60 0.0001 U R.VEVTEFEDIK.S
637.3293 1272.6440 1272.6561 -0.0121 0 (31) 0.085 U R.LNEQASEEILK.V
637.3339 1272.6532 1272.6561 -0.0029 0 93 4.9e-008 U R.LNEQASEEILK.V
723.8250 1445.6355 1445.6423 -0.0068 0 64 1.9e-005 K.EFHLNESGDPSSK.S
920.8943 1839.7741 1839.7992 -0.0250 0 96 5.1e-009 R.IDFYFDENPYFENK.V
920.9027 1839.7908 1839.7992 -0.0083 0 (45) 0.00082 R.IDFYFDENPYFENK.V
920.9063 1839.7981 1839.7992 -0.0011 0 (91) 2.4e-008 R.IDFYFDENPYFENK.V
732.3346 2193.9820 2194.0138 -0.0318 0 (97) 5.6e-009 U K.EQQEAIEHIDEVQNEIDR.L
732.3346 2193.9820 2194.0138 -0.0318 0 102 1.8e-009 U K.EQQEAIEHIDEVQNEIDR.L
732.3410 2194.0011 2194.0138 -0.0127 0 (81) 2.8e-007 U K.EQQEAIEHIDEVQNEIDR.L
A 4 Tropomiosina 4,63 32995 47 gi|66730475 2,16 537.2719 1072.5293 1072.5513 -0.0219 1 55 0.00038 U K.LDKENAIDR.A
537.2728 1072.5311 1072.5513 -0.0202 1 (52) 0.00083 U K.LDKENAIDR.A
574.2936 1146.5726 1146.5768 -0.0042 1 54 0.0004 U K.LKGTEDEVEK.Y
585.8330 1169.6515 1169.6656 -0.0141 0 67 1.4e-005 K.LVILEGELER.S
585.8383 1169.6621 1169.6656 -0.0035 0 (31) 0.058 K.LVILEGELER.S
585.8383 1169.6621 1169.6656 -0.0035 0 (33) 0.036 K.LVILEGELER.S
585.8397 1169.6649 1169.6656 -0.0006 0 (54) 0.00027 K.LVILEGELER.S
585.8397 1169.6649 1169.6656 -0.0006 0 (65) 2.3e-005 K.LVILEGELER.S
622.3038 1242.5930 1242.6456 -0.0525 0 (34) 0.031 R.IQLVEEELDR.A
622.3038 1242.5930 1242.6456 -0.0525 0 (75) 2.7e-006 R.IQLVEEELDR.A
622.3205 1242.6265 1242.6456 -0.0191 0 (68) 1.5e-005 R.IQLVEEELDR.A
622.3205 1242.6265 1242.6456 -0.0191 0 75 3.1e-006 R.IQLVEEELDR.A
622.3269 1242.6392 1242.6456 -0.0063 0 (63) 4.8e-005 R.IQLVEEELDR.A
649.8818 1297.7491 1297.7605 -0.0114 1 (60) 4.9e-005 R.KLVILEGELER.S
649.8856 1297.7567 1297.7605 -0.0038 1 (65) 1.5e-005 R.KLVILEGELER.S
649.8856 1297.7567 1297.7605 -0.0038 1 70 4.5e-006 R.KLVILEGELER.S
666.8023 1331.5901 1331.6317 -0.0416 0 (85) 1.7e-007 K.ATDAEADVASLNR.R
666.8193 1331.6241 1331.6317 -0.0076 0 (79) 9e-007 K.ATDAEADVASLNR.R
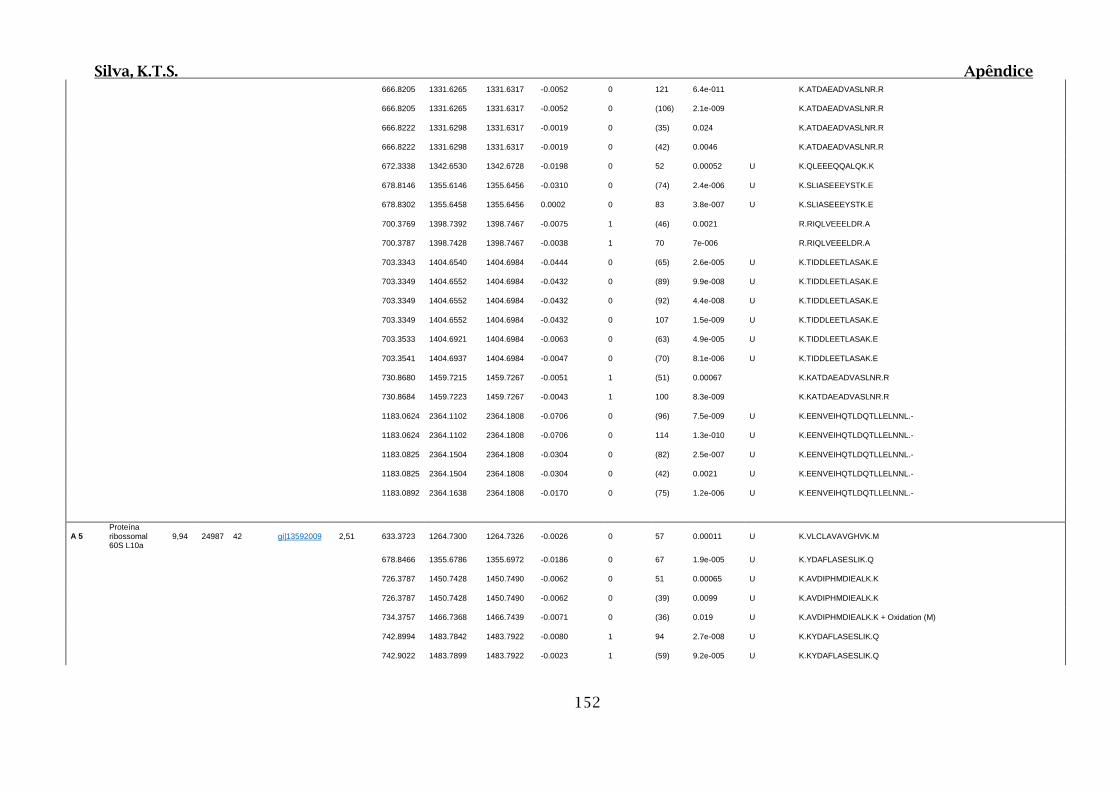
Silva, K.T.S. Apêndice
152
666.8205 1331.6265 1331.6317 -0.0052 0 121 6.4e-011 K.ATDAEADVASLNR.R
666.8205 1331.6265 1331.6317 -0.0052 0 (106) 2.1e-009 K.ATDAEADVASLNR.R
666.8222 1331.6298 1331.6317 -0.0019 0 (35) 0.024 K.ATDAEADVASLNR.R
666.8222 1331.6298 1331.6317 -0.0019 0 (42) 0.0046 K.ATDAEADVASLNR.R
672.3338 1342.6530 1342.6728 -0.0198 0 52 0.00052 U K.QLEEEQQALQK.K
678.8146 1355.6146 1355.6456 -0.0310 0 (74) 2.4e-006 U K.SLIASEEEYSTK.E
678.8302 1355.6458 1355.6456 0.0002 0 83 3.8e-007 U K.SLIASEEEYSTK.E
700.3769 1398.7392 1398.7467 -0.0075 1 (46) 0.0021 R.RIQLVEEELDR.A
700.3787 1398.7428 1398.7467 -0.0038 1 70 7e-006 R.RIQLVEEELDR.A
703.3343 1404.6540 1404.6984 -0.0444 0 (65) 2.6e-005 U K.TIDDLEETLASAK.E
703.3349 1404.6552 1404.6984 -0.0432 0 (89) 9.9e-008 U K.TIDDLEETLASAK.E
703.3349 1404.6552 1404.6984 -0.0432 0 (92) 4.4e-008 U K.TIDDLEETLASAK.E
703.3349 1404.6552 1404.6984 -0.0432 0 107 1.5e-009 U K.TIDDLEETLASAK.E
703.3533 1404.6921 1404.6984 -0.0063 0 (63) 4.9e-005 U K.TIDDLEETLASAK.E
703.3541 1404.6937 1404.6984 -0.0047 0 (70) 8.1e-006 U K.TIDDLEETLASAK.E
730.8680 1459.7215 1459.7267 -0.0051 1 (51) 0.00067 K.KATDAEADVASLNR.R
730.8684 1459.7223 1459.7267 -0.0043 1 100 8.3e-009 K.KATDAEADVASLNR.R
1183.0624 2364.1102 2364.1808 -0.0706 0 (96) 7.5e-009 U K.EENVEIHQTLDQTLLELNNL.-
1183.0624 2364.1102 2364.1808 -0.0706 0 114 1.3e-010 U K.EENVEIHQTLDQTLLELNNL.-
1183.0825 2364.1504 2364.1808 -0.0304 0 (82) 2.5e-007 U K.EENVEIHQTLDQTLLELNNL.-
1183.0825 2364.1504 2364.1808 -0.0304 0 (42) 0.0021 U K.EENVEIHQTLDQTLLELNNL.-
1183.0892 2364.1638 2364.1808 -0.0170 0 (75) 1.2e-006 U K.EENVEIHQTLDQTLLELNNL.-
A 5 Proteína ribossomal 60S L10a
9,94 24987 42 gi|13592009 2,51 633.3723 1264.7300 1264.7326 -0.0026 0 57 0.00011 U K.VLCLAVAVGHVK.M
678.8466 1355.6786 1355.6972 -0.0186 0 67 1.9e-005 U K.YDAFLASESLIK.Q
726.3787 1450.7428 1450.7490 -0.0062 0 51 0.00065 U K.AVDIPHMDIEALK.K
726.3787 1450.7428 1450.7490 -0.0062 0 (39) 0.0099 U K.AVDIPHMDIEALK.K
734.3757 1466.7368 1466.7439 -0.0071 0 (36) 0.019 U K.AVDIPHMDIEALK.K + Oxidation (M)
742.8994 1483.7842 1483.7922 -0.0080 1 94 2.7e-008 U K.KYDAFLASESLIK.Q
742.9022 1483.7899 1483.7922 -0.0023 1 (59) 9.2e-005 U K.KYDAFLASESLIK.Q
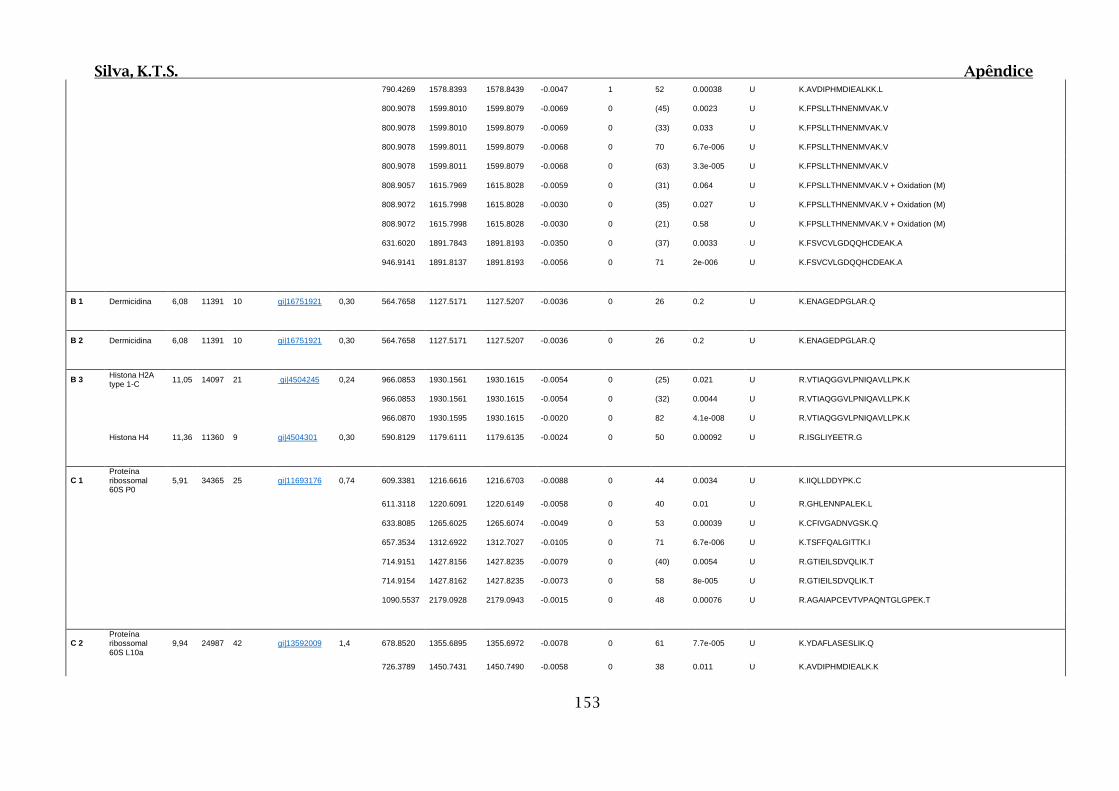
Silva, K.T.S. Apêndice
153
790.4269 1578.8393 1578.8439 -0.0047 1 52 0.00038 U K.AVDIPHMDIEALKK.L
800.9078 1599.8010 1599.8079 -0.0069 0 (45) 0.0023 U K.FPSLLTHNENMVAK.V
800.9078 1599.8010 1599.8079 -0.0069 0 (33) 0.033 U K.FPSLLTHNENMVAK.V
800.9078 1599.8011 1599.8079 -0.0068 0 70 6.7e-006 U K.FPSLLTHNENMVAK.V
800.9078 1599.8011 1599.8079 -0.0068 0 (63) 3.3e-005 U K.FPSLLTHNENMVAK.V
808.9057 1615.7969 1615.8028 -0.0059 0 (31) 0.064 U K.FPSLLTHNENMVAK.V + Oxidation (M)
808.9072 1615.7998 1615.8028 -0.0030 0 (35) 0.027 U K.FPSLLTHNENMVAK.V + Oxidation (M)
808.9072 1615.7998 1615.8028 -0.0030 0 (21) 0.58 U K.FPSLLTHNENMVAK.V + Oxidation (M)
631.6020 1891.7843 1891.8193 -0.0350 0 (37) 0.0033 U K.FSVCVLGDQQHCDEAK.A
946.9141 1891.8137 1891.8193 -0.0056 0 71 2e-006 U K.FSVCVLGDQQHCDEAK.A
B 1 Dermicidina 6,08 11391 10 gi|16751921 0,30 564.7658 1127.5171 1127.5207 -0.0036 0 26 0.2 U K.ENAGEDPGLAR.Q
B 2 Dermicidina 6,08 11391 10 gi|16751921 0,30 564.7658 1127.5171 1127.5207 -0.0036 0 26 0.2 U K.ENAGEDPGLAR.Q
B 3 Histona H2A type 1-C
11,05 14097 21 gi|4504245 0,24 966.0853 1930.1561 1930.1615 -0.0054 0 (25) 0.021 U R.VTIAQGGVLPNIQAVLLPK.K
966.0853 1930.1561 1930.1615 -0.0054 0 (32) 0.0044 U R.VTIAQGGVLPNIQAVLLPK.K
966.0870 1930.1595 1930.1615 -0.0020 0 82 4.1e-008 U R.VTIAQGGVLPNIQAVLLPK.K
Histona H4 11,36 11360 9 gi|4504301 0,30 590.8129 1179.6111 1179.6135 -0.0024 0 50 0.00092 U R.ISGLIYEETR.G
C 1 Proteína ribossomal 60S P0
5,91 34365 25 gi|11693176 0,74 609.3381 1216.6616 1216.6703 -0.0088 0 44 0.0034 U K.IIQLLDDYPK.C
611.3118 1220.6091 1220.6149 -0.0058 0 40 0.01 U R.GHLENNPALEK.L
633.8085 1265.6025 1265.6074 -0.0049 0 53 0.00039 U K.CFIVGADNVGSK.Q
657.3534 1312.6922 1312.7027 -0.0105 0 71 6.7e-006 U K.TSFFQALGITTK.I
714.9151 1427.8156 1427.8235 -0.0079 0 (40) 0.0054 U R.GTIEILSDVQLIK.T
714.9154 1427.8162 1427.8235 -0.0073 0 58 8e-005 U R.GTIEILSDVQLIK.T
1090.5537 2179.0928 2179.0943 -0.0015 0 48 0.00076 U R.AGAIAPCEVTVPAQNTGLGPEK.T
C 2 Proteína ribossomal 60S L10a
9,94 24987 42 gi|13592009 1,4 678.8520 1355.6895 1355.6972 -0.0078 0 61 7.7e-005 U K.YDAFLASESLIK.Q
726.3789 1450.7431 1450.7490 -0.0058 0 38 0.011 U K.AVDIPHMDIEALK.K
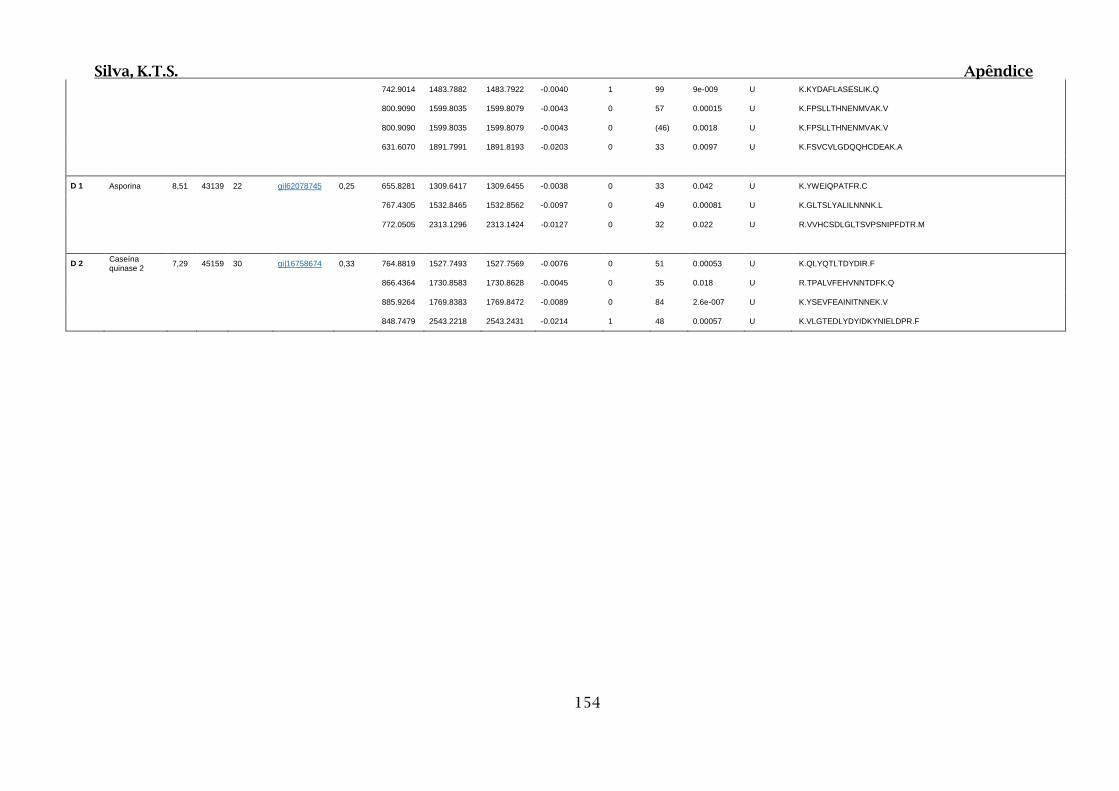
Silva, K.T.S. Apêndice
154
742.9014 1483.7882 1483.7922 -0.0040 1 99 9e-009 U K.KYDAFLASESLIK.Q
800.9090 1599.8035 1599.8079 -0.0043 0 57 0.00015 U K.FPSLLTHNENMVAK.V
800.9090 1599.8035 1599.8079 -0.0043 0 (46) 0.0018 U K.FPSLLTHNENMVAK.V
631.6070 1891.7991 1891.8193 -0.0203 0 33 0.0097 U K.FSVCVLGDQQHCDEAK.A
D 1 Asporina 8,51 43139 22 gi|62078745 0,25 655.8281 1309.6417 1309.6455 -0.0038 0 33 0.042 U K.YWEIQPATFR.C
767.4305 1532.8465 1532.8562 -0.0097 0 49 0.00081 U K.GLTSLYALILNNNK.L
772.0505 2313.1296 2313.1424 -0.0127 0 32 0.022 U R.VVHCSDLGLTSVPSNIPFDTR.M
D 2 Caseína quinase 2
7,29 45159 30 gi|16758674 0,33 764.8819 1527.7493 1527.7569 -0.0076 0 51 0.00053 U K.QLYQTLTDYDIR.F
866.4364 1730.8583 1730.8628 -0.0045 0 35 0.018 U R.TPALVFEHVNNTDFK.Q
885.9264 1769.8383 1769.8472 -0.0089 0 84 2.6e-007 U K.YSEVFEAINITNNEK.V
848.7479 2543.2218 2543.2431 -0.0214 1 48 0.00057 U K.VLGTEDLYDYIDKYNIELDPR.F
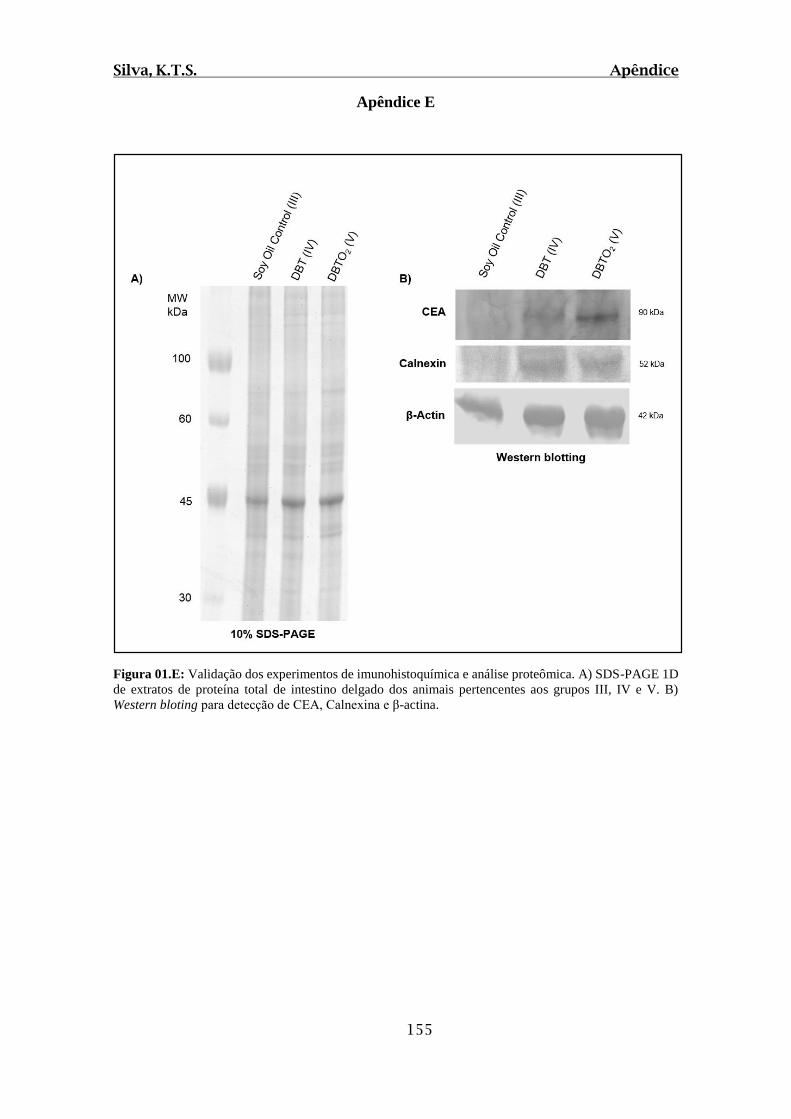
Silva, K.T.S. Apêndice
155
Apêndice E
Figura 01.E: Validação dos experimentos de imunohistoquímica e análise proteômica. A) SDS-PAGE 1D
de extratos de proteína total de intestino delgado dos animais pertencentes aos grupos III, IV e V. B)
Western bloting para detecção de CEA, Calnexina e β-actina.
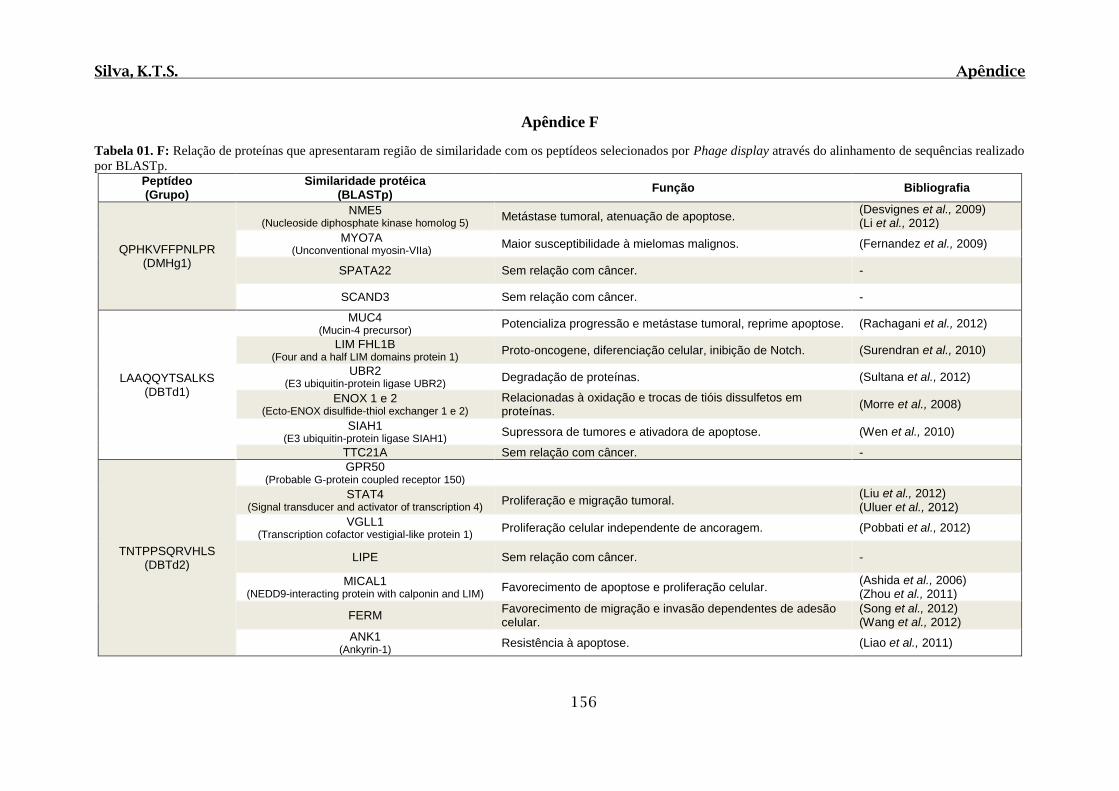
Silva, K.T.S. Apêndice
156
Apêndice F
Tabela 01. F: Relação de proteínas que apresentaram região de similaridade com os peptídeos selecionados por Phage display através do alinhamento de sequências realizado
por BLASTp. Peptídeo (Grupo)
Similaridade protéica (BLASTp)
Função Bibliografia
QPHKVFFPNLPR (DMHg1)
NME5 (Nucleoside diphosphate kinase homolog 5)
Metástase tumoral, atenuação de apoptose. (Desvignes et al., 2009) (Li et al., 2012)
MYO7A (Unconventional myosin-VIIa)
Maior susceptibilidade à mielomas malignos. (Fernandez et al., 2009)
SPATA22 Sem relação com câncer. -
SCAND3 Sem relação com câncer. -
LAAQQYTSALKS (DBTd1)
MUC4 (Mucin-4 precursor)
Potencializa progressão e metástase tumoral, reprime apoptose. (Rachagani et al., 2012)
LIM FHL1B (Four and a half LIM domains protein 1)
Proto-oncogene, diferenciação celular, inibição de Notch. (Surendran et al., 2010)
UBR2 (E3 ubiquitin-protein ligase UBR2)
Degradação de proteínas. (Sultana et al., 2012)
ENOX 1 e 2 (Ecto-ENOX disulfide-thiol exchanger 1 e 2)
Relacionadas à oxidação e trocas de tióis dissulfetos em proteínas.
(Morre et al., 2008)
SIAH1 (E3 ubiquitin-protein ligase SIAH1)
Supressora de tumores e ativadora de apoptose. (Wen et al., 2010)
TTC21A Sem relação com câncer. -
TNTPPSQRVHLS (DBTd2)
GPR50 (Probable G-protein coupled receptor 150)
STAT4 (Signal transducer and activator of transcription 4)
Proliferação e migração tumoral. (Liu et al., 2012) (Uluer et al., 2012)
VGLL1 (Transcription cofactor vestigial-like protein 1)
Proliferação celular independente de ancoragem. (Pobbati et al., 2012)
LIPE Sem relação com câncer. -
MICAL1 (NEDD9-interacting protein with calponin and LIM)
Favorecimento de apoptose e proliferação celular. (Ashida et al., 2006) (Zhou et al., 2011)
FERM Favorecimento de migração e invasão dependentes de adesão celular.
(Song et al., 2012) (Wang et al., 2012)
ANK1 (Ankyrin-1)
Resistência à apoptose. (Liao et al., 2011)
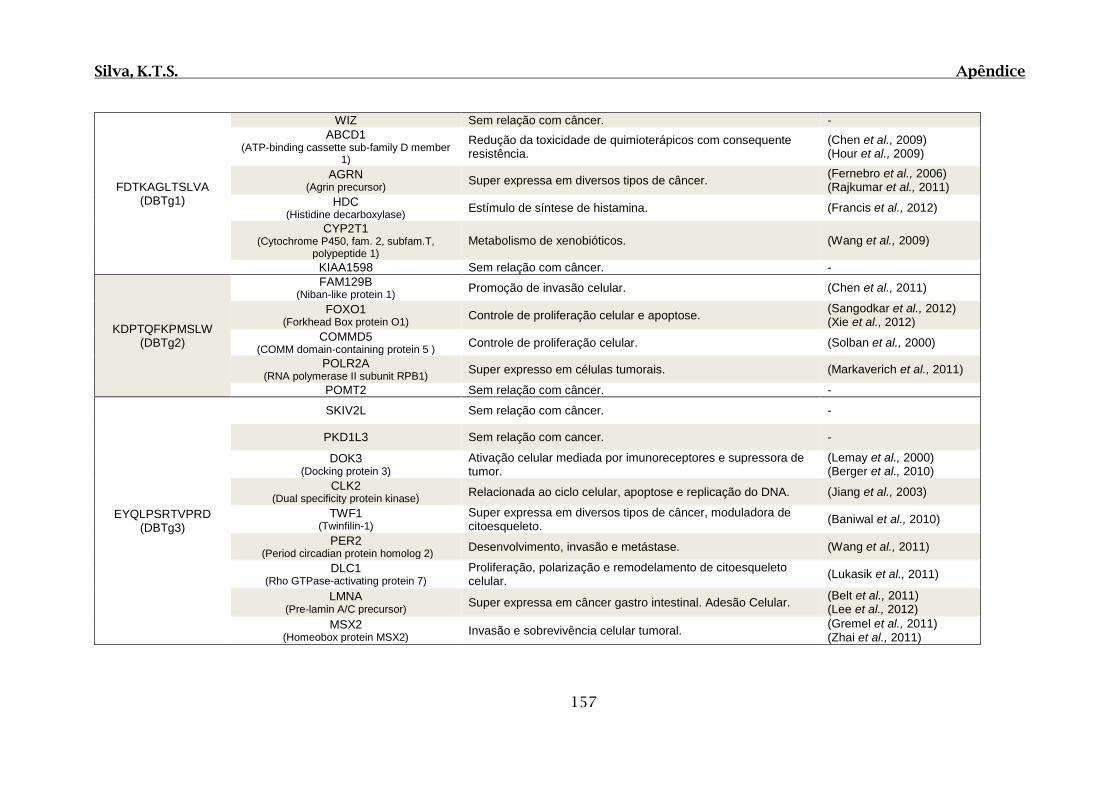
Silva, K.T.S. Apêndice
157
FDTKAGLTSLVA (DBTg1)
WIZ Sem relação com câncer. -
ABCD1 (ATP-binding cassette sub-family D member
1)
Redução da toxicidade de quimioterápicos com consequente resistência.
(Chen et al., 2009) (Hour et al., 2009)
AGRN (Agrin precursor)
Super expressa em diversos tipos de câncer. (Fernebro et al., 2006) (Rajkumar et al., 2011)
HDC (Histidine decarboxylase)
Estímulo de síntese de histamina. (Francis et al., 2012)
CYP2T1 (Cytochrome P450, fam. 2, subfam.T,
polypeptide 1) Metabolismo de xenobióticos. (Wang et al., 2009)
KIAA1598 Sem relação com câncer. -
KDPTQFKPMSLW (DBTg2)
FAM129B (Niban-like protein 1)
Promoção de invasão celular. (Chen et al., 2011)
FOXO1 (Forkhead Box protein O1)
Controle de proliferação celular e apoptose. (Sangodkar et al., 2012) (Xie et al., 2012)
COMMD5 (COMM domain-containing protein 5 )
Controle de proliferação celular. (Solban et al., 2000)
POLR2A (RNA polymerase II subunit RPB1)
Super expresso em células tumorais. (Markaverich et al., 2011)
POMT2 Sem relação com câncer. -
EYQLPSRTVPRD (DBTg3)
SKIV2L Sem relação com câncer. -
PKD1L3 Sem relação com cancer. -
DOK3 (Docking protein 3)
Ativação celular mediada por imunoreceptores e supressora de tumor.
(Lemay et al., 2000) (Berger et al., 2010)
CLK2 (Dual specificity protein kinase)
Relacionada ao ciclo celular, apoptose e replicação do DNA. (Jiang et al., 2003)
TWF1 (Twinfilin-1)
Super expressa em diversos tipos de câncer, moduladora de citoesqueleto.
(Baniwal et al., 2010)
PER2 (Period circadian protein homolog 2)
Desenvolvimento, invasão e metástase. (Wang et al., 2011)
DLC1 (Rho GTPase-activating protein 7)
Proliferação, polarização e remodelamento de citoesqueleto celular.
(Lukasik et al., 2011)
LMNA (Pre-lamin A/C precursor)
Super expressa em câncer gastro intestinal. Adesão Celular. (Belt et al., 2011) (Lee et al., 2012)
MSX2 (Homeobox protein MSX2)
Invasão e sobrevivência celular tumoral. (Gremel et al., 2011) (Zhai et al., 2011)

Silva, K.T.S. Apêndice
158
Referências bibliográficas – Apêndice F
Ashida, S. et al., 2006. Expression of novel molecules, MICAL2-PV (MICAL2 prostate
cancer variants), increases with high Gleason score and prostate cancer
progression. Clinical cancer research : an official journal of the American
Association for Cancer Research, 12(9): 2767-73.
Baniwal, S.K. et al., 2010. Runx2 transcriptome of prostate cancer cells: insights into
invasiveness and bone metastasis. Molecular cancer, 9: 258.
Belt, E.J. et al., 2011. Loss of lamin A/C expression in stage II and III colon cancer is
associated with disease recurrence. European journal of cancer, 47(12): 1837-45.
Berger, A.H. et al., 2010. Identification of DOK genes as lung tumor suppressors. Nature
genetics, 42(3): 216-23.
Chen, K.G., Valencia, J.C., Gillet, J.P., Hearing, V.J. and Gottesman, M.M., 2009.
Involvement of ABC transporters in melanogenesis and the development of
multidrug resistance of melanoma. Pigment cell & melanoma research, 22(6):
740-9.
Chen, S., Evans, H.G. and Evans, D.R., 2011. FAM129B/MINERVA, a novel adherens
junction-associated protein, suppresses apoptosis in HeLa cells. The Journal of
biological chemistry, 286(12): 10201-9.
Desvignes, T., Pontarotti, P., Fauvel, C. and Bobe, J., 2009. Nme protein family
evolutionary history, a vertebrate perspective. BMC evolutionary biology, 9: 256.
Fernebro, J. et al., 2006. Gene expression profiles relate to SS18/SSX fusion type in
synovial sarcoma. International journal of cancer. Journal international du cancer,
118(5): 1165-72.
Francis, H. et al., 2012. Inhibition of histidine decarboxylase ablates the autocrine
tumorigenic effects of histamine in human cholangiocarcinoma. Gut, 61(5): 753-
64.
Gremel, G. et al., 2011. Functional and prognostic relevance of the homeobox protein
MSX2 in malignant melanoma. British journal of cancer, 105(4): 565-74.
Hour, T.C. et al., 2009. Downregulation of ABCD1 in human renal cell carcinoma. The
International journal of biological markers, 24(3): 171-8.
Jiang, N., Benard, C.Y., Kebir, H., Shoubridge, E.A. and Hekimi, S., 2003. Human CLK2
links cell cycle progression, apoptosis, and telomere length regulation. The
Journal of biological chemistry, 278(24): 21678-84.
Lee, W.S. et al., 2012. Methylation status of lamin A/C in gastric cancer cell lines.
Hepato-gastroenterology, 59(116): 1313-8.
Lemay, S., Davidson, D., Latour, S. and Veillette, A., 2000. Dok-3, a novel adapter
molecule involved in the negative regulation of immunoreceptor signaling.
Molecular and cellular biology, 20(8): 2743-54.
Li, F. et al., 2012. Identification of NME5 as a contributor to innate resistance to
gemcitabine in pancreatic cancer cells. The FEBS journal, 279(7): 1261-73.

Silva, K.T.S. Apêndice
159
Liao, Y.H., Hsu, S.M., Yang, H.L., Tsai, M.S. and Huang, P.H., 2011. Upregulated
ankyrin repeat-rich membrane spanning protein contributes to tumour progression
in cutaneous melanoma. British journal of cancer, 104(6): 982-8.
Liu, S. et al., 2012. The oncoprotein HBXIP uses two pathways to up-regulate S100A4
in promotion of growth and migration of breast cancer cells. The Journal of
biological chemistry, 287(36): 30228-39.
Lukasik, D., Wilczek, E., Wasiutynski, A. and Gornicka, B., 2011. Deleted in liver cancer
protein family in human malignancies (Review). Oncology letters, 2(5): 763-768.
Markaverich, B.M., Shoulars, K. and Rodriguez, M.A., 2011. Luteolin Regulation of
Estrogen Signaling and Cell Cycle Pathway Genes in MCF-7 Human Breast
Cancer Cells. International journal of biomedical science : IJBS, 7(2): 101-111.
Morre, D.J., Hostetler, B., Weston, N., Kim, C. and Morre, D.M., 2008. Cancer type-
specific tNOX isoforms: A putative family of redox protein splice variants with
cancer diagnostic and prognostic potential. BioFactors, 34(3): 201-7.
Pobbati, A.V., Chan, S.W., Lee, I., Song, H. and Hong, W., 2012. Structural and
functional similarity between the Vgll1-TEAD and the YAP-TEAD complexes.
Structure, 20(7): 1135-40.
Rachagani, S. et al., 2012. MUC4 potentiates invasion and metastasis of pancreatic cancer
cells through stabilization of fibroblast growth factor receptor 1. Carcinogenesis,
33(10): 1953-64.
Rajkumar, T. et al., 2011. Identification and validation of genes involved in cervical
tumourigenesis. BMC cancer, 11: 80.
Sangodkar, J. et al., 2012. Targeting the FOXO1/KLF6 axis regulates EGFR signaling
and treatment response. The Journal of clinical investigation, 122(7): 2637-51.
Solban, N. et al., 2000. HCaRG, a novel calcium-regulated gene coding for a nuclear
protein, is potentially involved in the regulation of cell proliferation. The Journal
of biological chemistry, 275(41): 32234-43.
Song, X. et al., 2012. A novel membrane-dependent on/off switch mechanism of talin
FERM domain at sites of cell adhesion. Cell research, 22(11): 1533-45.
Sultana, R., Theodoraki, M.A. and Caplan, A.J., 2012. UBR1 promotes protein kinase
quality control and sensitizes cells to Hsp90 inhibition. Experimental cell
research, 318(1): 53-60.
Surendran, K., Selassie, M., Liapis, H., Krigman, H. and Kopan, R., 2010. Reduced Notch
signaling leads to renal cysts and papillary microadenomas. Journal of the
American Society of Nephrology : JASN, 21(5): 819-32.
Uluer, E.T., Aydemir, I., Inan, S., Ozbilgin, K. and Vatansever, H.S., 2012. Effects of 5-
fluorouracil and gemcitabine on a breast cancer cell line (MCF-7) via the
JAK/STAT pathway. Acta histochemica, 114(7): 641-6.
Wang, S.J., Wong, G., de Heer, A.M., Xia, W. and Bourguignon, L.Y., 2009. CD44
variant isoforms in head and neck squamous cell carcinoma progression. The
Laryngoscope, 119(8): 1518-30.
Wang, T. et al., 2012. FERM-containing protein FRMD5 is a p120-catenin interacting
protein that regulates tumor progression. FEBS letters, 586(19): 3044-50.

Silva, K.T.S. Apêndice
160
Wang, Y., Hua, L., Lu, C. and Chen, Z., 2011. Expression of circadian clock gene human
Period2 (hPer2) in human colorectal carcinoma. World journal of surgical
oncology, 9: 166.
Wen, Y.Y. et al., 2010. SIAH1 induced apoptosis by activation of the JNK pathway and
inhibited invasion by inactivation of the ERK pathway in breast cancer cells.
Cancer science, 101(1): 73-9.
Xie, L. et al., 2012. FOXO1 is a tumor suppressor in classical Hodgkin lymphoma. Blood,
119(15): 3503-11.
Zhai, Y. et al., 2011. MSX2 is an oncogenic downstream target of activated WNT
signaling in ovarian endometrioid adenocarcinoma. Oncogene, 30(40): 4152-62.
Zhou, Y. et al., 2011. MICAL-1 is a negative regulator of MST-NDR kinase signaling
and apoptosis. Molecular and cellular biology, 31(17): 3603-15.

Silva, K.T.S. Apêndice
161
Apêndice G
Artigo publicado – Revista: Journal of Proteome Research
Fator de Impacto: 5,001. Qualis CAPES Área CBIOL I: A1

Silva, K.T.S. Apêndice
162

Silva, K.T.S. Apêndice
163
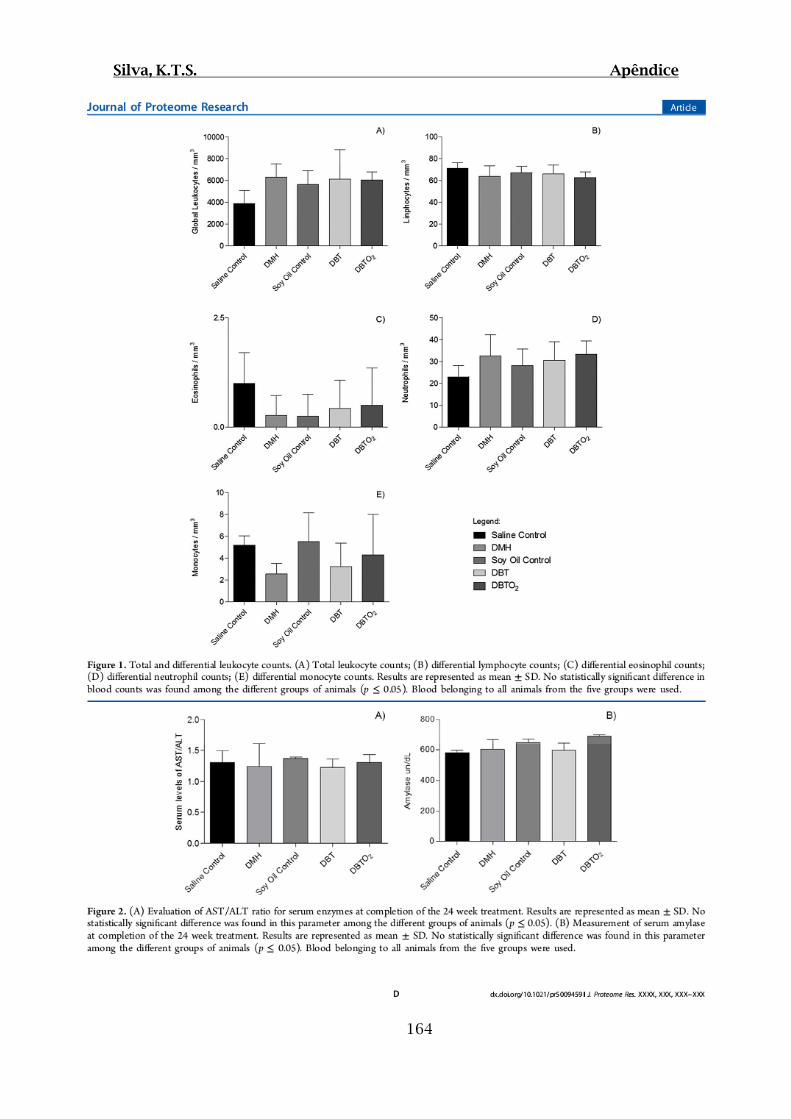
Silva, K.T.S. Apêndice
164
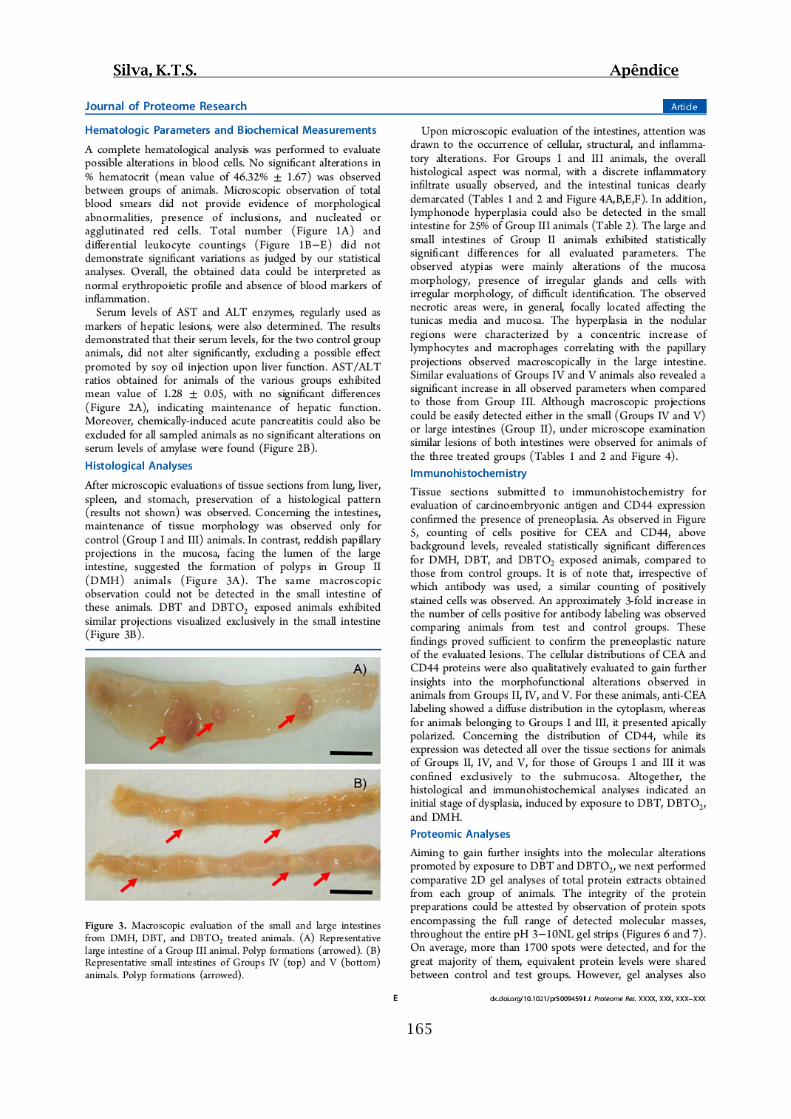
Silva, K.T.S. Apêndice
165
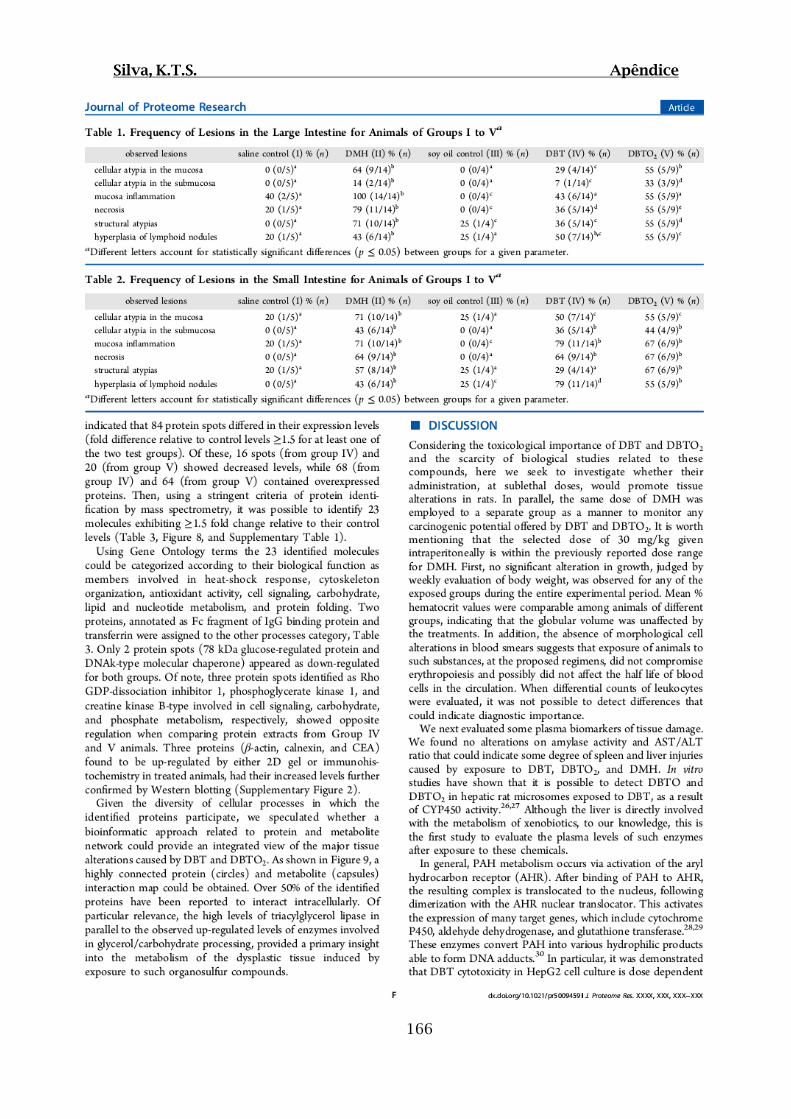
Silva, K.T.S. Apêndice
166
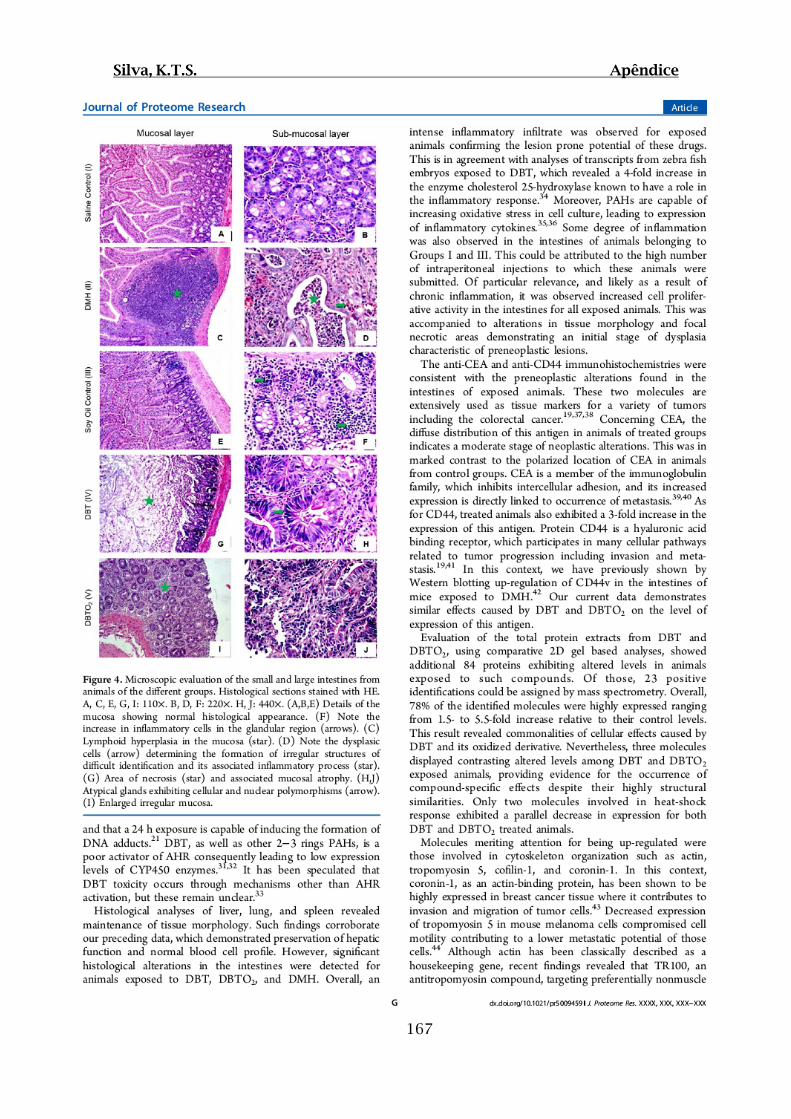
Silva, K.T.S. Apêndice
167
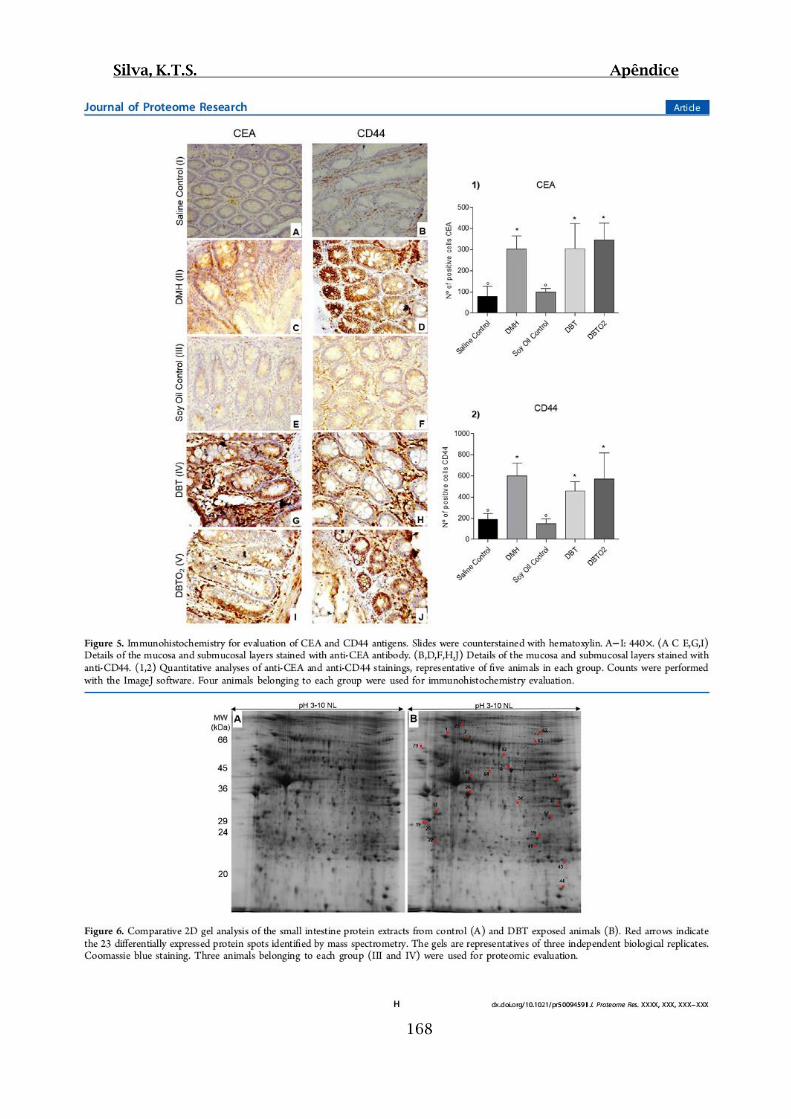
Silva, K.T.S. Apêndice
168
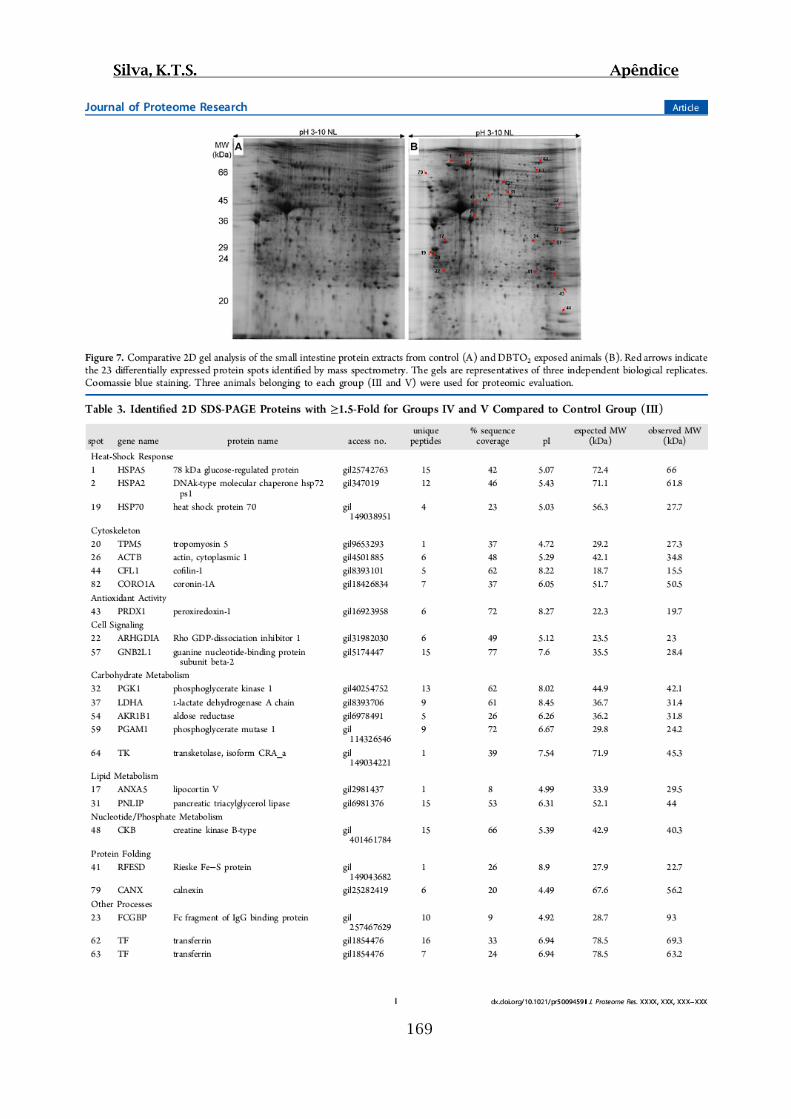
Silva, K.T.S. Apêndice
169
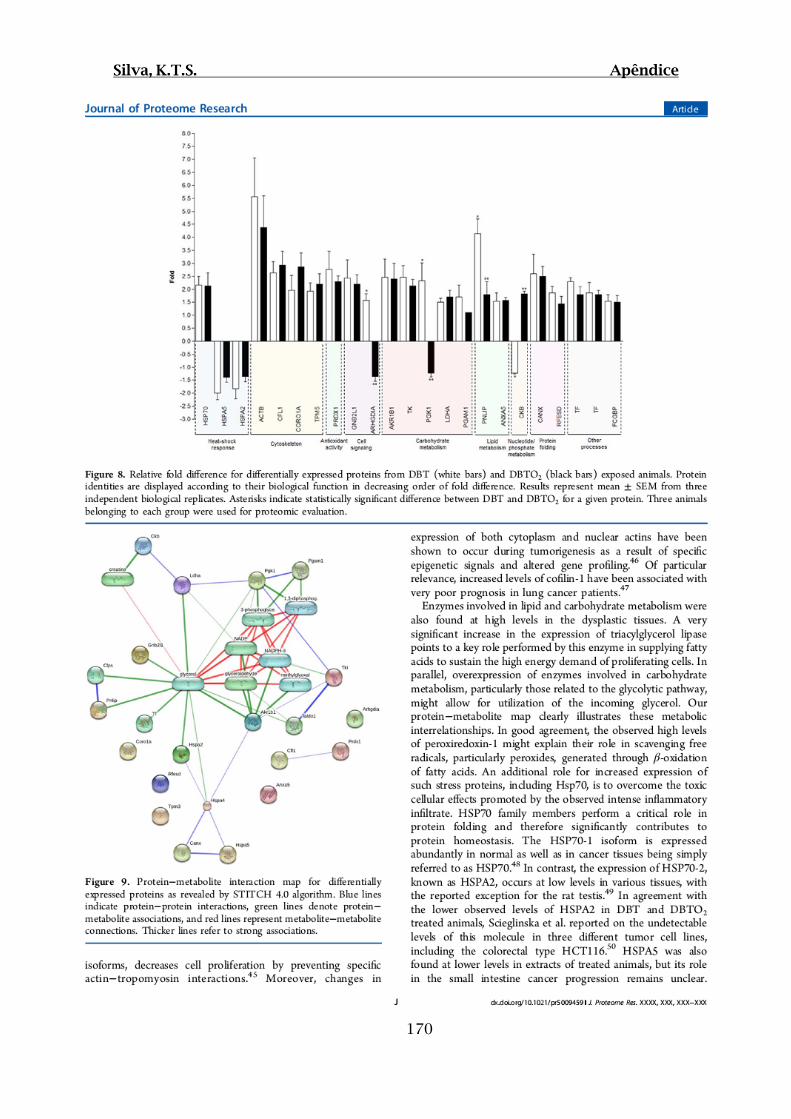
Silva, K.T.S. Apêndice
170

Silva, K.T.S. Apêndice
171

Silva, K.T.S. Apêndice
172





























