Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARÁ
CAMPUS UNIVERSITÁRIO DE BRAGANÇA
INSTITUTO DE ESTUDOS COSTEIROS
LABORATÓRIO DE ECOLOGIA DE MANGUEZAL - LAMA
DIEGO NOVAES CARNEIRO DA SILVA
EFEITO DA SALINIDADE E COMPACTAÇÃO DO SOLO NA ESTRUTURA DO
BOSQUE ANÃO DE AVICENNIA GERMINANS (L.) L. NA PENÍNSULA DE
AJURUTEUA, COSTA AMAZÔNICA BRASILEIRA
Bragança – PA
2015

i
DIEGO NOVAES CARNEIRO DA SILVA
EFEITO DA SALINIDADE E COMPACTAÇÃO DO SOLO NA ESTRUTURA DO
BOSQUE ANÃO DE AVICENNIA GERMINANS (L.) L. NA PENÍNSULA DE
AJURUTEUA, COSTA AMAZÔNICA BRASILEIRA
Trabalho de Conclusão de Curso apresentado ao
Colegiado do Curso de Licenciatura Plena em Ciências
Biológicas, da Universidade Federal do Pará, Campus de
Bragança, como requisito parcial para a obtenção do Grau
de Licenciado em Ciências Biológicas.
Orientador: Prof. Dr. Marcus E.F. Barroncas
Bragança – Pará
2015

ii
DIEGO NOVAES CARNEIRO DA SILVA
EFEITO DA SALINIDADE E COMPACTAÇÃO DO SOLO NA ESTRUTURA DO
BOSQUE ANÃO DE AVICENNIA GERMINANS (L.) L. NA PENÍNSULA DE
AJURUTEUA, COSTA AMAZÔNICA BRASILEIRA
Trabalho de Conclusão de Curso apresentado ao Colegiado do Curso de Licenciatura
Plena em Ciências Biológicas, da Universidade Federal do Pará, Campus de Bragança,
como requisito parcial para a obtenção do grau de Licenciado em Ciências Biológicas.
Data de aprovação: ___ de ____________________ de 2015.
__________________________________________________– Orientador
Prof. Dr. Marcus Emanuel Barroncas Fernandes
Universidade Federal do Pará, Campus de Bragança- UFPA
__________________________________________________– Titular
Profa. Dra. Moirah Paula Machado de Menezes
Universidade Federal do Pará, Campus de Bragança- UFPA
__________________________________________________– Titular
Prof. Dr. Nils Edvin Asp Neto
Universidade Federal do Pará, Campus de Bragança- UFPA
______________________________________________- Suplente
Msc. Danilo Cesar Gardunho - Suplente

iii
DEDICATÓRIA
Dedico a meus pais Isabel Novaes e Fernando Carneiro, ao meu irmão Rian Carneiro e a minha amada
Lanna, meus grandes incentivadores.

iv
AGRADECIMENTOS
À Universidade Federal do Pará e ao Curso de Licenciatura Plena em Ciências
Biológicas pela formação acadêmica e oportunidade de desenvolver esta pesquisa.
Ao Laboratório de Ecologia de Manguezal pelo suporte e apoio para realização
deste trabalho.
A Fundação Amazonia – Banco Nacional de Desenvolvimento Econômico e
Social (BNDES) e a VALE/FAPESPA (edital 001/2010, ICAAF 068/2011. VALE
S.A./FAPESPA, pelo apoio financeiro fundamental ao desenvolvimento desta pesquisa.
Ao Prof. Dr. Marcus pela orientação e oportunidade de realizar este trabalho.
Ao Danilo, Gustavo, Alex e e todos os meu amigos do Laboratório de Ecologia
de Manguezal (LAMA) por ajudarem nas coletas e nas analises de dados, e pelos varios
momentos de descontração.
Ao Hederson por ter me ajudado nas varias excursões para coleta de dados.
À Sanae pela paciência e pelo auxílio na confecção dos mapas.
À todos meus amigos da turma Biologia 2011, pela companheirismo durante
estes quatro anos de curso, e em especial ao meu grupo de trabalho e grandes amigos
Leilton, Amanda, Yrlano, Wellington, Nayara e Dani, pessoas estas que levarei, em
memoria, para o resto da minha vida.
A minha querida, companheira, amiga, amada, sol dos meus dias e lua cheia das
minhas noites, Lanna Jamile, por acreditar nas minhas vitorias e mais do que qualquer
um que eu citar aqui, por ter me apoido e ajudado desde o início desta minha jornada.
Aos meus pais, Isabel Novaes e Fernando Carneiro, meu irmão Rian Carneiro,
e minhas tias, Bia e Zélia, e a todos os meus parentes por terem me apoido, de todas
as formas, nas mudança que decidi tomar na minha vida.
E à todos que, em momentos não menos importantes dos citados acima, me
ajudaram de forma muito importante, os meus sinceros agradecimentos.

v
EPÍGRAFE
Toda a nossa ciência, comparada com a realidade, é primitiva e Infantil. E, no entanto, é a coisa mais preciosa que temos.
Albert Einstein (1879-1955)
É um erro capital teorizar antes de ter os dados. Insensivelmente, começa-se a distorcer os fatos para adaptá-los às teorias, em vez de
fazer com que as teorias se adaptem aos fatos. Comentário de Sherlock Holmes, em A Scandal in Bohemia, de
Arthur Conan Doyle (1891)

vi
SUMÁRIO
RESUMO.................................................................................................... vii
ABSTRACT................................................................................................ viii
Introdução................................................................................................. 1
Material e Métodos................................................................................... 3
Área de Trabalho................................................................................... 3
Atributos Estruturais da vegetação........................................................ 4
Compactação do Solo............................................................................ 4
Salinidade da Água Intersticial............................................................... 4
Análise dos dados.................................................................................. 5
Resultados................................................................................................ 7
Atributos Estruturais.............................................................................. 7
Salinidade, Compactação e Profundidade............................................. 9
Atributos Estruturais X Salinidade e Compactação................................ 10
Discussão................................................................................................. 12
Referências............................................................................................... 16
ANEXO....................................................................................................... 20

vii
Resumo As condições hidroedáficas contribuem para o nanismo das árvores de mangue.
Assim, o presente estudo objetiva verificar como a variação na salinidade e compactação
do solo afetam a altura dos bosques anões de Avicennia germinans. Para caracterizar a
estrutura do bosque foram abertas 90 parcelas de 5x5 m, totalizando 0,25 hectare,
mensurando altura total e circunferência do fuste. Foram medidas a salinidade da água
intersticial (SAL) e campactação do solo (PMC), durante a estação seca. Foram
mensuradas 3982 indivíduos, sendo 99,02% de A. germinans. A análise de cluster
mostrou a presença de três grupos com alturas médias significativamente diferentes:
1,17±0,37; 2,43±0,30 e 3,85±0,70 (KruskalWallis=79,04; gl=2; p<0,001). A análise da
regressão logística mostrou que a relação entre a altura e as variáveis ambientais foi
significativa (χ2=37,55; g.l.=3; p<0,0001). Contudo, somente as variáveis SAL e PMC
apresentaram chances menores do que 10 vezes (odds ratio=0,10 e 0,12, respectivamente)
das árvores mais altas relacionarem-se com a maior salinidades e compactação. De fato,
o bosque anão de A. germinans apresentou grande variabilidade estrutural, com as árvores
adultas que assumiram hábito arbustivo fortemente relacionadas à alta salinidade e
compactação do solo, sendo ambas as variáveis a expressão do déficit hídrico e das
condições edáficas do ambiente.
Palavras-chave Avicennia germinans . Condições hidroedáficas . Costa amazônica
brasileira . Manguezal anão

viii
Abstract The hydroedaphic conditions contribute to dwarfing of mangrove trees. Thus,
this study aims to verify how the variations in salinity and soil compaction affect the
height of the dwarf Avicennia germinans mangrove. To characterize the structure of forest
were open 90 plots of 5x5 m, totaling 0.25 hectare, measuring total height and stem
circumference. We measured the pore water salinity (SAL) and soil compaction (SC),
during the dry season. A total of 3982 individuals were measured, with 99.02% of A.
germinans. The cluster analysis showed the presence of three groups with significantly
different mean heights: 1.17 ± 0.37; 2.43 ± 0.30 and 3.85 ± 0.70 (Kruskal Wallis = 79.04,
df = 2, p <0.001). The logistic regression analysis showed that the relation between the
height and environmental variables was significant (χ2 = 37.55, df = 3, p <0.0001).
However, only the variables SAL and SC had lower chances than 10 times (odds ratio =
0.10 and 0.12, respectively) of the tallest trees they relate to the highest salinity and
compaction. In fact, the dwarf A. germinans forest showed great structural variability,
with the adult trees that assumed a shrub-like habit strongly related to high salinity and
soil compaction, with both variables the expression of drought and soil conditions of the
environment.
Keywords Avicennia germinans . Brazilian Amazon coast . Dwarf mangrove .
Hydroedaphic conditions

1
Efeito da Salinidade e Compactação do Solo na Estrutura do Bosque Anão de
Avicennia germinans (l.) L. na Península de Ajuruteua, Costa Amazônica Brasileira
Introdução
Os manguezais são formados por espécies herbáceas e arbóreas adaptadas
morfologicamente e fisiologicamente para sobreviver em águas salobras ou salinas, bem
como em solos inconsolidados com baixa concentração de oxigênio (Hogarth 2007).
Essas florestas também apresentam grande variabilidade espacial em resposta a diferentes
fatores ambientais locais, como, por exemplo, a inundação e a salinidade (Naidoo et al.
2002).
A salinidade é um dos principais fatores a influenciar a dinâmica estrutural das
florestas de mangue, como já foi demonstrado em vários estudos prévios (Bunt 1996;
1999; Duke et al. 1998; Matthijs et al. 1999; Joshi e Ghose, 2003). Tal fator pode,
inclusive, conduzir a uma série de disfunções fisiológicas que limitam ou impedem, por
exemplo, o crescimento vegetativo e reprodutivo das árvores (Kozlowski 1997; Sobrado
e Ball 1999). Contudo, embora as espécies arbóreas de mangue apesentem alta adaptação
aos ambientes salinos, a sua capacidade de assimilação de carbono e crescimento é
reduzida de acordo com o aumento da salinidade (Ball e Farquhar 1984; Naidoo e
Chirkoot 2004).
De acordo com Naidoo (2006), as condições hidroedáficas contribuem para aumentar
a salinidade do solo, assim como o estresse hídrico e o desequilíbrio iônico nos tecidos,
incluindo a deficiência de elementos essenciais como o fósforo, o que de forma
sinergética, contribuem para a formação de bosques anões nos manguezais. Essas
condições extremas no manguezal despertaram o interesse sobre a adaptabilidade das
árvores de mangue. A partir desse interesse vários estudos foram realizados na tentativa

2
de explicar as causas do nanismo em bosques de mangue. As discussões envolveram
principalmente os fatores químicos, como salinidade (Ball 1996; Naidoo et al. 2002) e
nutriente (Davis et al. 2001; McKee et al. 2002; Lovelock et al. 2004), bem como aqueles
de origem física, como os ciclos de inundação (Egler 1952) e a compactação do substrato
(Craighead 1971). Segundo Soane e Ouwerkerk (1994), a compactação é o resultado da
expulsão de ar dos poros do substrato, mudando a porosidade, além de aumentar a
densidade e a resistência para penetração, dificultando a permeabilidade e a
disponibilidade de água e nutriente.
Na península de Ajuruteua, localizada na costa amazônica brasileira, os manguezais
ocorrem sob diferentes condições de salinidade e inundação (Medina et al. 2001). As
florestas de mangue ao longo dessa península são formadas principalmente por três
espécies vegetais arbóreas: Rhizophora mangle L. (Rhizophoraceae), Avicennia
germinans (L.) L. (Acanthaceae) e Laguncularia racemosa (L.) Gaertn. f.
(Combretaceae). Dentre as quais A. germinans pode tolerar as maiores taxas de salinidade
e dessecação do solo (Cohen et al. 2004). De acordo com esses autores, ao longo da
península, essas áreas hipersalinas apresentam salinidade da água intersticial de 90 – 100
ppm e são inundadas menos de 28 dias ao ano, sendo os locais onde A. germinans
apresenta tendência para a formação de bosques anões monoespecíficos.
De acordo com Medina et al. (2001), a área de bosque anão existente na península de
Ajuruteua apresenta alta concentração de sódio no sedimento e déficit hídrico no período
seco, aumentando sobremaneira a salinidade da água intersticial. Contudo, ainda são
escassos trabalhos sobre uma abordagem de como as condições edáficas contribuem para
o nanismo das árvores e influenciam a arquitetura dos bosques. Assim, o presente estudo
tem o objetivo de verificar como a variação na salinidade e o déficit hídrico no solo afetam

3
a altura dos bosques anões, bem como determinar as limitações impostas por esses fatores
ao crescimento de A. germinans.
Material e Métodos
Área de estudo
A área de estudo é um bosque de mangue anão com predominância de A. germinans,
com indivíduos de até 5 metros de altura. Essa área está localizada na península de
Ajuruteua, margeada pelo estuário do rio Caeté e do rio Taperaçú, no município de
Bragança-Pará, a 10 km do Furo do Taici (00°53’45,6” S e 046°39’52,1” W), na direção
leste a partir da rodovia PA-458 (Bragança Ajuruteua) (Fig 1).
O clima da região é quente e úmido, com média de precipitação anual de 2508,4 mm,
temperatura média do ar de 25,6 °C (dados da série de 24 anos da estação meteorológica
de Tracuateua-PA, 30 km a sudoeste da área de estudo – INMET, 2014).
Fig. 1 Mapa da península de Ajuruteu, Bragança – Pará. A área hachurada
representa o bosque anão.

4
Atributos estruturais da vegetação
Para caracterizar a estrutura do bosque anão foram demarcadas 90 parcelas de 5x5 m,
totalizando 0,25 hectare (= 2.250 m2; Fig 1). Em cada parcela, todos os indivíduos foram
mensurados quanto à altura total (ALT) e circunferência do fuste. A circunferência das
árvores foi medida com uma fita métrica à altura do peito (CAP; cm) e dos arbustos à 30
cm do nível do chão, próximo à porção basal da planta (CAB; cm). Este último
procedimento foi adotado mesmo quando a primeira bifurcação ou rebrota ocorreu abaixo
de 30 cm (Felfili e Fagg 2007). A partir das medidas de circunferência (CAP e CAB)
foram estimados o diâmetro do fuste (D; cm) e estimados os seguintes atributos
estruturais: Frequência (F), Densidade (De; ind.m-2), Dominância (área basal – Do; m2.ha-
1) e Valor de Importância (VI) para as espécies identificadas (Cintrón e Schaeffer-Novelli
1974).
Compactação do solo
A textura do solo foi medida com um analisador de compactação penetroLOG da
FALKER Automação Agrícola Ltda. Foram mensuradas i) a Pressão Máxima de
Compactação (PMC; kgf.cm-2) e ii) a profundidade (PROF; cm) em que essa pressão
ocorreu. Os dados foram acessados com o auxílio do programa para visualização e análise
de dados de compactação do solo FALKER v.1.41.
Salinidade da água intersticial
A salinidade (SAL) da água intersticial foi obtida com auxílio de um trado de 50 cm de
diâmetro a um metro de profundidade. As amostras de água foram retiradas com o auxílio
de uma seringa de 50 ml e uma mangueira flexível de um metro fixada na seringa. Para

5
aferição da salinidade foi utilizado um refratômetro graduado de 0 a 100, com precisão
de 01 ups (American Optical modelo A366ATC).
As coletas, de salinidade e compactação do solo, foram realizadas pontualmente entre
os meses de setembro e outubro de 2014, na estação seca.
Análise dos dados
Os dados fitossociológicos e ambientais (salinidade, compactação e profundidade da
compactação do solo) foram testados quanto aos pressupostos da normalidade (Teste de
Shapiro-Wilk) e homocedasticidade das variâncias (Teste de Levene). A análise de
variância de Kruskal-Wallis e o teste a posteriori de Dunn foram utilizados quando esses
pressupostos foram violados ao nível de significância de p<0,05. Essas análises foram
realizadas no programa BioEstat 5.0 (Ayres et al. 2007) e foram utilizadas para comparar
os atributos estruturais apenas dos espécimes de A. germinans das 90 parcelas e dos
grupos formados após a análise de Cluster. Essa análise foi utilizada para identificar a
formação de grupos (clusters), com base na altura das árvores e arbustos mensurados
através do índice de Bray-Curtis, no pacote estatístico Primer v.6 (Clarke e Gorley 2006).
O efeito hidroedáfico sobre o bosque de mangue foi investigado através de modelos
de regressão logística (Logit) (Hosmer e Lemeshow 2000). A altura é a variável
dependente (resposta), onde plantas menores e maiores do que o valor estimado da
mediana da altura vão ser associadas às variáveis independentes (preditoras) i) SAL
(baixa e alta), ii) PMC (baixa e alta) e iii) PROF (baixa e alta) (Tabela 1).
Os valores médios das variáveis quantitativas, dependente e independente, foram
transformados em variáveis binárias tomando como base a mediana; salinidade Baixa=0
(0,49-0,67 ppm) e Alta=1 (0,68-0,99 ppm), PMC Baixa=0 (kgf.cm2) e Alta=1 (kgf.cm2) e
profundidade da PMC Baixa=0 (cm) e Alta=1 (cm).

6
Tabela 1: Valores da mediana e o código binário de
sucesso e insucesso da variável resposta Altura (ALT)
e das variáveis preditoras Salinidade (SAL), Pressão
Máxima de Compactação (PMC; kgf.cm-2) e
Profundidade (PROF; cm).
Variável Mediana Insucesso
baixo (0)
Sucesso
alto (1)
ALT 2,55 0,39 – 2,54 2,56 – 5,66
SAL 0,68 0,49 – 0,67 0,68 – 0,99
PMC 5,99 3,47 – 5,83 5,99 – 14,35
PROF 50,0 40,0 – 49,0 50,0 – 60,0
A associação entre essas variáveis é feita a partir do modelo logístico que é o odds
ratio (OR = eβ1), ou seja, é a chance da variável resposta (𝑌) acontecer em relação à
variável preditora (𝑥).
O modelo calcula a probabilidade da associação acontecer, com base nos dados
amostrais obtidos pelo método da máxima verossimilhança (maximiza a probabilidade de
obter o grupo observado de dados), através da seguinte fórmula:
𝑃(𝑌) =1
1 + 𝑒−(𝛼+𝛽1𝑥1+…+𝛽𝑘𝑥𝑘)
Onde: α e β representam parâmetros desconhecidos que serão estimados. Os cálculos
foram realizados no programa BioEstat 5.0 (Ayres et al. 2007).
O método utilizado para a análise da distribuição das variável resposta (ALT) e
preditoras (SAL e PCM) foi através da ferramenta Interpolate to Raster do módulo
Spatial Analyst (opção Inverse Distance Weighted - IDW) no programa ArcGIS 10
(2013). Posteriormente, mapas foram gerados com a representação gráfica dos diferentes
níveis percentuais das variáveis medidas.

7
Resultados
Atributos estruturais
A área do bosque anão de mangue analisado foram mensuradas 3982 indivíduos, dos
quais 3943 (99,02%) pertenciam a A. germinans. As demais espécies R. mangle e L.
racemosa corresponderam a menos de 1% da amostragem geral. Indivíduos esparsos de
espécies típicas do estrato herbáceo também foram registrados, como: Spartina
alterniflora Loisel, Salicornia gaudichaudiana Moq., Batis marítima L., Sesuvium
portulacastrum (L.) L. e Blutaparon verniculare (L.) Mears., porém somente as espécies
do estrato arbustivo e arbóreo foram considerados para a análise dos atributos estruturais
do bosque anão.
Foi registrada a ocorrência de indivíduos de R. mangle e L. racemosa, em apenas 7
(FR=6,5%) e 11 (FR=10,2%), respectivamente, das 90 parcelas mensuradas (Tabela 1).
Ao contrário, A. germinans ocorreu em todas as parcelas, apresentando o maior diâmetro
do fuste (D=6,36±3,87 cm) e altura das árvores (ALT=2,51±1,19 m). A. germinans foi a
espécie que apresentou os maiores percentuais em todos os atributos [FR (83,3%), DeR
(99,02%), DoR (94,2%)], portanto a mais importante (VI=276,6) no bosque anão (Tabela
2).
A análise de variância entre as espécies de mangue mostrou que há diferença
significativa para todos os atributos estruturais (ALT - H=16,42; gl=2; p<0,001; D - H=
8,82; gl= 2; p<0,001; DeA - H= 177,46; gl= 2; p<0,001 e DoA - H= 18,14; gl= 2;
p<0,001) (Tabela 2).
8
Tabela 2: Atributo estrutural com valores médios (±Desvio Padrão) para a área de estudo na
península de Ajuruteua, Bragança-PA. Avg=Avicennia germinans; Rhm=Rhizophora
mangle; Lar=Laguncularia racemosa; D=Diâmetro do fuste, FR= Frequência Relativa,
DeA=Densidade Absoluta, DeR=Densidade Relativa, DoA=Dominância Absoluta,
DoR=Dominância Relativa e VI=Valor de Importância. Ind.=Indivíduo; n.a.=não aplicável;
p=valor de significância em 0,05.
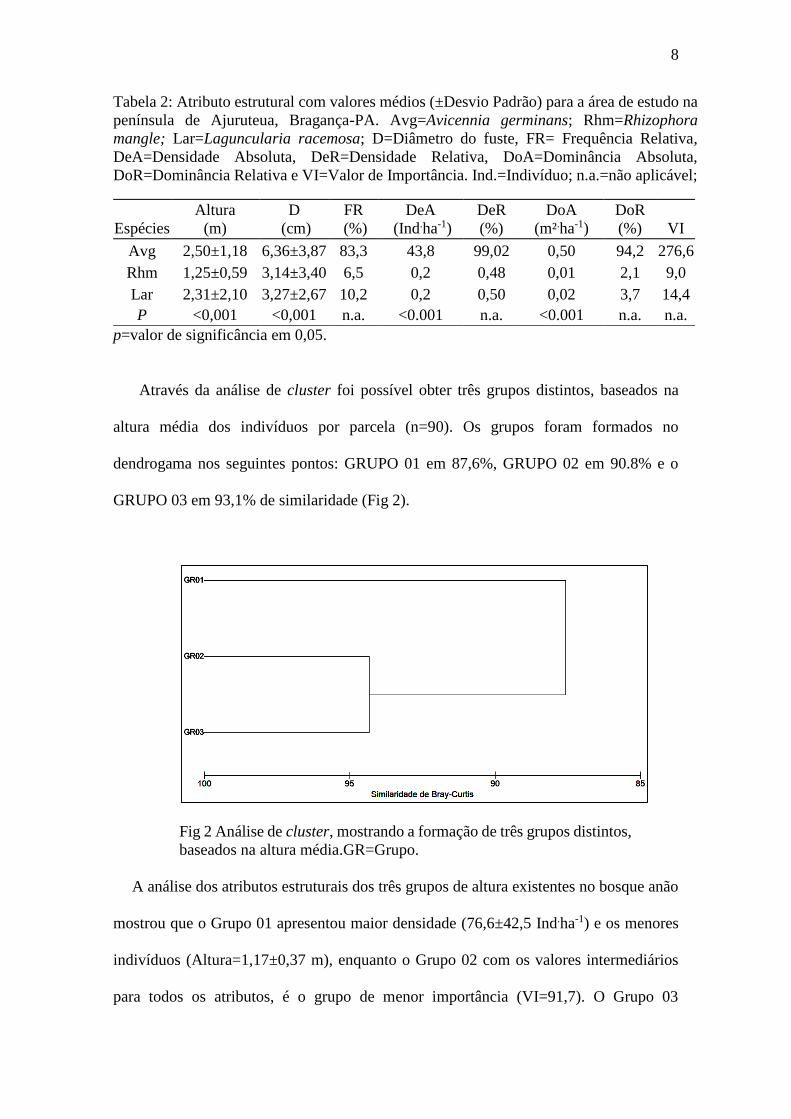
Através da análise de cluster foi possível obter três grupos distintos, baseados na
altura média dos indivíduos por parcela (n=90). Os grupos foram formados no
dendrogama nos seguintes pontos: GRUPO 01 em 87,6%, GRUPO 02 em 90.8% e o
GRUPO 03 em 93,1% de similaridade (Fig 2).
Fig 2 Análise de cluster, mostrando a formação de três grupos distintos,
baseados na altura média.GR=Grupo.
A análise dos atributos estruturais dos três grupos de altura existentes no bosque anão
mostrou que o Grupo 01 apresentou maior densidade (76,6±42,5 Ind.ha-1) e os menores
indivíduos (Altura=1,17±0,37 m), enquanto o Grupo 02 com os valores intermediários
para todos os atributos, é o grupo de menor importância (VI=91,7). O Grupo 03
Espécies
Altura
(m)
D
(cm)
FR
(%)
DeA
(Ind.ha-1)
DeR
(%)
DoA
(m².ha-1)
DoR
(%) VI
Avg 2,50±1,18 6,36±3,87 83,3 43,8 99,02 0,50 94,2 276,6
Rhm 1,25±0,59 3,14±3,40 6,5 0,2 0,48 0,01 2,1 9,0
Lar 2,31±2,10 3,27±2,67 10,2 0,2 0,50 0,02 3,7 14,4
P <0,001 <0,001 n.a. <0.001 n.a. <0.001 n.a. n.a.
9
apresentou valores mais altos em quase todos os atributos e também é o grupo de
vegetação mais importante (VI=113,8) do bosque anão (Tabela 3).
A análise de variância mostrou que todos os atributos estruturais apresentam diferença
significativa entre os grupos de vegetação (ALT - H=79,04; gl=2; p<0,001; D - H=67,15;
gl= 2; p<0,001; DeA - H=47,8; gl= 2; p<0,001 e DoA - H=32,75; gl= 2; p<0,001) (Tabela
3).
Tabela 3. Valores médios (±Desvio Padrão) dos atributos estruturais dos grupos de
Avicennia germinans gerados através da análise de cluster, para a península de Ajuruteua,
Bragança-PA. D=Diâmetro do fuste, Sal=Salinidade, PMC=Pressão Máxima de
Compactação, FR=Frequência Relativa, DeA=Densidade Absoluta, DeR=Densidade
Relativa, DoA=Dominância Absoluta, DoR=Dominância Relativa e VI=Valor de
Importância. n.a.= não aplicável; p=valor de significância em 0,05.
Salinidade, compactação e profundidade
Os valores médios dos fatores abióticos apresentaram variação significativa entre os três
agrupamentos do bosque anão. Apenas a variável “profundidade da pressão máxima”
(PROF) não é capaz de explicar a composição estrutural do bosque, sendo necessária a
correlação com as variáveis “salinidade” (SAL) e “pressão máxima de compactação”
(PMC) (Tabela 4). O Grupo 03, formado por indivíduos com maior média de estatura
(Tabela 3), apresentou as menores médias de atributos abióticos (SAL=0,61±0,09;
Altura
(m)
D
(cm)
DeA
(Ind.ha-1)
DeR
(%)
DoA
(m².ha-1)
DoR
(%) VI
GRUPO 01 1,17±0,37 2,83±0,76 76,6±42,5 57
0,001±0,000
7 6,1 94,5
GRUPO 02 2,43±0,30 5,99±2,23 39,6±20,4 30 0,004±0,003 26,6 91,7
GRUPO 03 3,85±0,70 10,05±3,71 17,6±6,13 13 0,011±0,010 67,3 113,8
P <0.001 <0.001 <0.001 n.a. <0.001 n.a. n.a.

10
PMC=5,47±1,78; PROF=0,51±0,06), enquanto o Grupo 01 exibiu os maiores valores
médio de SAL, PMC (Tabela 4). PMC (kgf.cm-2)
Tabela 4 Comparação dos valores médios (±Desvio padrão) dos fatores
ambientais entre os grupos de vegetação formados no bosque anão, na
península de Ajuruteua, costa amazônica brasileira. SAL=Salinidade,
PMC=pressão máxima de compactação, PROF=profundidade, p=valor
de significância em 0,05.
SAL PMC (kgf.cm-2) PROF
GRUPO 01 0,85±0,19 8,15±2,95 44,0±15,0
GRUPO 02 0,72±0,14 6,25±1,72 43,0±11,0
GRUPO 03 0,61±0,09 5,47±1,78 51,0±6,0
p <0,001 <0,001 <0,05
Atributos estruturais x salinidade e compactação
As variáveis preditoras salinidade (SAL), pressão máxima de compactação do solo
(PMC) e profundidade (PROF) foram correlacionados, através da Regressão Logística
(Logit), com a variável resposta altura (ALT). A relação entre a variável resposta e as
variáveis preditoras, através do Teste da Razão da Verossimilhança (TRV=43,60), foi
bastante significativa (χ2=37,55; g.l.=3; p<0,0001). Contudo, de acordo com os valores
do teste de Wald, somente a variável PROF não apresentou diferença significativa em
relação à altura (z=-0,5832; p=0,5595) (Tabela 5). O modelo também calcula a razão de
chances (odds ratio) para cada variável preditora em relação à variável resposta (ALT),
ou seja, as relações de sucesso de uma com a outra (Tabela 5). As variáveis SAL e PMC
apresentaram chances muito baixas, menor cerca de 10 vezes (OR=0,10 e 0,12,
respectivamente) dos bosques mais altos estarem relacionados com as mais altas
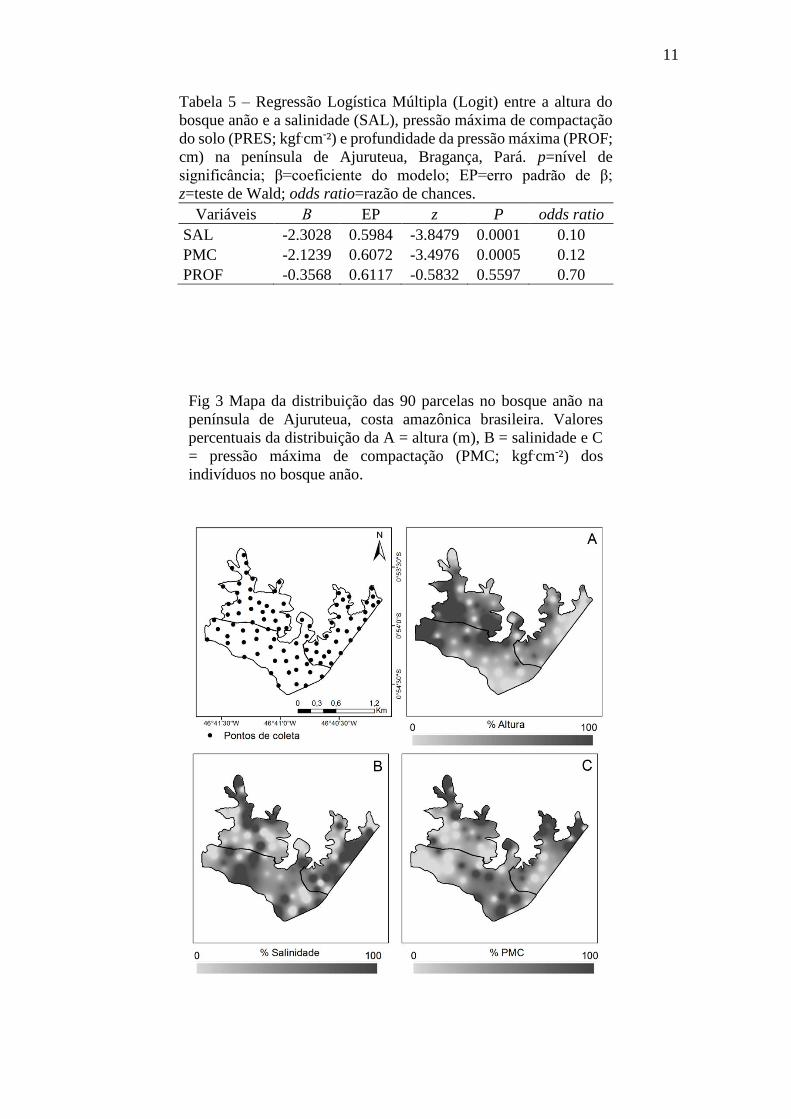
salinidades e maiores valores de compactação (Tabela 5; Fig 3).
11
Tabela 5 – Regressão Logística Múltipla (Logit) entre a altura do
bosque anão e a salinidade (SAL), pressão máxima de compactação
do solo (PRES; kgf.cm-²) e profundidade da pressão máxima (PROF;
cm) na península de Ajuruteua, Bragança, Pará. p=nível de
significância; β=coeficiente do modelo; EP=erro padrão de β;
z=teste de Wald; odds ratio=razão de chances.
Variáveis Β EP z P odds ratio
SAL -2.3028 0.5984 -3.8479 0.0001 0.10
PMC -2.1239 0.6072 -3.4976 0.0005 0.12
PROF -0.3568 0.6117 -0.5832 0.5597 0.70
Fig 3 Mapa da distribuição das 90 parcelas no bosque anão na
península de Ajuruteua, costa amazônica brasileira. Valores
percentuais da distribuição da A = altura (m), B = salinidade e C
= pressão máxima de compactação (PMC; kgf.cm-²) dos
indivíduos no bosque anão.

12
Paralelamente, valores probabilísticos (Logot Pi) percentuais mais exatos dessas
relações foram calculados, onde estimou-se que existe alta probabilidade (92,9 e 90,1%)
da porção do bosque anão mais alta ocorrer tanto nas manchas com baixos níveis de SAL,
de PMC e baixa PROF quanto com baixos níveis de SAL, de PMC e alta PROF,
respectivamente (Tabela 6; Fig 3).
Tabela 6 – Probabilidade de variação na altura do bosque
anão em função do estresse salino e hídrico na península de
Ajuruteua, Bragança, Pará. SAL=salinidade;
PRES=pressão máxima de compactação do solo (kgf.cm-²);
PROF=profundidade da pressão máxima (cm); ALT=altura
(m); p=probabilidade de ocorrência.
Interações Logit Pi p (ALT; %)
SAL= 0; PMC= 0; PROF= 0 2,5697 92,9
SAL= 0; PMC= 0; PROF= 1 2,2129 90,1
SAL= 0; PMC= 1; PROF= 0 0,4458 61,0
SAL= 0; PMC= 1; PROF= 1 0,0890 52,2
SAL= 1; PMC= 0; PROF= 0 0,0890 56,6
SAL= 1; PMC= 0; PROF= 1 -0,0899 47,8
SAL= 1; PMC= 1; PROF= 0 -1,8570 13,5
SAL= 1; PMC= 1; PROF= 1 -2,2138 09,9
Discussão
Apesar da ocorrência das espécies R. mangle e L. racemosa na área do bosque anão, suas
contribuições percentuais, no que se refere aos atributos estruturais, não foram
expressivas (Tabela 1). A presença dessas espécies foi registrada em função de algumas
parcelas cobrirem a parte limítrofe do bosque anão, próximo aos canais-de-maré, onde há
maior disponibilidade de água e o terreno é menos salino. De fato a região do bosque
anão é dominada por A. germinans com quase a totalidade (99,02%) de seus indivíduos
distribuídos ao longo da área amostral, reiterando o fato de que essa espécie é mais

13
tolerante às altas taxas de salinidade que caracteriza o ambiente. De acordo com Krauss
e Ball (2013), em geral, espécies de mangue são halófitas facultativas verdadeiras com
necessidade obrigatória à salinidade, ou seja, são capazes de crescer em água doce, mas
respondem com o aumento da salinidade até um crescimento ótimo acima do qual
decresce.
A alta salinidade registrada no bosque anão promove o estresse salino, que por sua
vez, reflete-se na arquitetura do bosque anão. Mesmo em campo, é notória a variação no
porte dos indivíduos considerados anões. Essa observação foi confirmada com a análise
de cluster, gerada a partir da altura média dos indivíduos de cada parcela, formando três
grupos distintos (Fig 2). Esse mesmo bosque anão caracterizado por Medina et al. (2001),
cuja descrição foi de uma floresta monoespecífica de A. germinans com características
arbustivas (shrub-like), composta de indivíduos de até 3 m de altura e diâmetro do fuste
médio de 4,1 cm.
Adicionalmente, a comparação dos valores médios dos atributos estruturais, mostrou
variabilidade significativa entre os grupos, sendo a altura, diâmetro do fuste, densidade e
dominância todos significativamente diferentes (Tabela 3). O GRUPO 03 se destaca, por
ter os maiores valores médios dos atributos altura e diâmetro do fuste (Alt=3,85 m;
D=10,06 cm), sendo também o grupo mais dominância (0,33 m².ha-1) (Tabela 2). Quando
comparamos as medias da salinidade (Sal=0,61 ppm) e da pressão máxima de
compactação (PMC=5,47 kgf.cm2) (Tabela 5), com os dados estruturais do GRUPO 03 e
do GRUPO 01, é possível observar uma relação inversamente proporcional do
desenvolvimento dos atributos bióticos em função dos dados abióticos.
A diferença encontrada na arquitetura do bosque anão, inclusive fazendo com que A.
germinans apresente um porte arbustivo e não arbóreo, é certamente uma resposta aos

14
diferentes fatores ambientais que promovem os distintos níveis de estresse registrados na
área amostral (Tabela 4).
Através da regressão logística foi possível relacionar os três grupos, definidos pela
altura dos indivíduos, com os fatores ambientais de salinidade, compactação do solo e
profundidade da compactação. Os indivíduos do Grupo 01 apresentaram porte mais
arbustivo e estão altamente relacionados com as manchas com maior salinidade da água
intersticial e maior compactação do solo. A profundidade de compactação do solo aparece
como uma variável com pouca correlação com a altura do bosque anão, pois tanto os
valores altos e baixos dessa variável não interferem nas altas probabilidades das outras
variáveis preditoras (salinidade e compactação) se correlacionarem com os indivíduos de
menor altura no bosque (Tabela 5 e 6).
O fato de que a alta salinidade afeta negativamente o porte de indivíduos de A.
germinans, desde há muito tempo já tem sido relatado na literatura científica (Tomlinson
1986; Jiménez e Lugo 2000). Da mesma forma outros efeitos, além da alta salinidade,
agem sobre a estruturação de espécies vegetais. A densidade do solo é enfatizada como
uma das influência que atuam diretamente nas relações bioquímicas e nos aspectos
morfológicos das raízes das plantas (Whalley et al. 2006). De fato, a textura do solo pode
modificar, de forma drástica, a disponibilidade de água no sistema radicular por
resistência mecânica do solo, sendo suficiente para interromper o crescimento radicular,
afetando diretamente a produtividade do bosque. (Bengough et al. 2011).
É importante salientar que a análise de fatores abióticos tem sido usada para avaliar o
crescimento das plantas (Beutler et al. 2006a,b; Beutler et al. 2008; Kaiser et al. 2009).
No caso dos bosques anões, as discussões envolveram principalmente os fatores
químicos, como salinidade (Ball 1996; Naidoo et al. 2002) e nutriente (Davis et al. 2001;

15
McKee et al. 2002; Lovelock et al. 2004), bem como aqueles de origem física, como os
ciclos de inundação (Egler 1952) e a compactação do substrato (Craighead 1971).
De acordo com Naidoo (2006), tanto a hipersalinidade como a alta evapotranspiração
dos bosques com pouca cobertura foliar e maior exposição do solo (como é o caso do
bosque anão do presente estudo) é capaz de promover a maior concentração de íons
inorgânicos, contribuindo para o baixo potencial hídrico do solo. Este, por sua vez, produz
uma série de efeitos semelhantes ao estresse hídrico, sendo então um fator determinante
da produção de biomassa, afetando o porte dos indivíduos. Segundo Krauss e Ball (2013),
o crescimento e o ótimo fisiológico se cruzam em concentrações moderadas de salinidade
nos manguezais. Mas é importante ficar atento para o fato de que as evidências aqui
apresentadas mostram que a grande variação na altura dos indivíduos do bosque anão de
0,14 à 7,80 m, não é dependente apenas da variação da salinidade e da necessidade que a
planta tem de sal no seu organismo, mas também do déficit hídrico distribuído entre as
manchas do bosque anão (Fig 3).
Por fim, o presente estudo mostrou que bosques anões apresentam maior variabilidade
estrutural do que o esperado, mostrando uma significante associação com à variação do
déficit hídrico, expresso pela salinidade e compactação do solo. No entanto, como
sugerido anteriormente por alguns autores (McKee et al. 2002; Lovelock et al. 2004;
Naidoo 2006), a formação de manguezais anões é um fenômeno complexo e influenciado
por uma gama de condições hidroedáficas, cujos estudos sobre o balanço de nutrientes
disponíveis no solo e a topografia local são essenciais para explicar as características
singulares desta região.

16
Referências
Ayres M, Ayres MJ, Ayres DL, Santos AA. (2007) BIOESTAT 5.0 – Aplicações
estatísticas nas áreas das ciências biomédicas. Instituto de Desenvolvimento
Sustentável Mamirauá.
Ball MC (1996) Comparative ecophysiology of mangrove forest and tropical lowland
moist rainforest. In: Mulkey SS, Chazdon RL, Smith AP (eds.) Tropical forest
ecophysiology. New York: Chapman and Hall, pp. 461–496.
Ball MC, Farquhar GD (1984) Photosynthetic and stomatal responses of two mangroves,
Aegiceras corniculatum and Avicennia marina, to long-term salinity and humidity
conditions. Plant Physiology 74:1–6.
Bengough AG, McKenzie BM, Hallett PD, Valentine TA (2011) Root elongation, water
stress, and mechanical impedance: A review of limiting stresses and beneûcial root
tip traits. Journal of Experimental Botany 62:59-68.
Beutler AN, Centurion JF, Silva AP, Barbosa JC (2006a) Intervalo hídrico ótimo e
produtividade de cultivares de soja. Revista Brasileira de engenharia Agrícola e
florestal 10:639-645.
Beutler AN, Centurion JF, Centurion MAPC, Silva AP (2006b) Efeito da compactação
na produtividade de cultivares de soja em Latossolo Vermelho. Revista Brasileira de
Ciência do Solo 30:787-794.
Beutler AN, Centurion JF, Silva AP, Centurion MAPC, Leonel CL, Freddi OS (2008)
Soil compaction by machine traffic and least limiting water range related to soybean
yield. Pesquisa Agropecuária Brasileira 43:1591-1600.
Bunt JS (1999) Overlap in mangrove species zonal patterns: some methods of analysis.
Mangroves and Salt Marshes 3:155-164.

17
Bunt JS (1996) Mangrove zonation: an examination of data from seventeen riverine
estuaries in tropical Australia. Annals of Botany 78:333-341.
Cintrón GM, Schaeffer-Novelli Y (1984) Methods for studying mangrove structure. In:
The mangrove Ecosystem: research methods. Monogr. Oceanogr. Methodology
8:91-113.
Clarke KR, Gorley RN (2006) PRIMER v6: User Manual/Tutorial. PRIMER-E,
Plymouth.
Cohen MCL, Lara RJ, Szlafsztein C, Dittmar T (2004) Mangrove inundation and nutrient
dynamics from a GIS perspective. Wetlands Ecology and Management 12:81-86.
Craighead FC (1971) The trees of south Florida. Vol. 1. The natural environments and
their succession. Miami, FL: University of Miami Press.
Davies SE, Childers DL, Day JW, Rudnick DT, Sklar FH (2001) Wetland-water column
exchanges of carbon, nitrogen, and phosphorus in a southern Everglades dwarf
mangrove. Estuaries 24:610–622.
Duke NC, Ball MC, Ellison JC (1998). Factors influencing in mangroves biodiversity and
distributional gradients. Global Ecology and Biogeography Letters 7:27-47.
Egler FE (1952) Southeast saline Everglades vegetation, Florida, and its management.
Felfili JM, Fagg CW (2007) Floristic composition, diversity and structure of the
“cerrado”sensu stricto on rocky soils in northern Goiás and southern Tocantins,
Brazil. Revista Brasileira de Botânica 30:375-385.
Hogarth PJ (2007) The Biology of Mangroves and Seagrasses. Oxford University Press.
Hosmer DWJr, Lemeshow S (2000) Applied Logistic Regression. 2nd ed. New York:
Wiley.
INMET (2014) Instituto Nacional de Meteorologia. http://www.inmet.gov.br/. Acessado
em 19 de Dezembro de 2014.

18
Joshi H, Ghose M (2003) Forest structure and species distribution along soil salinity and
pH gradient in mangrove swamps of the Sundarbans. Tropical Ecology 44:197-206.
Jiménez JA, Lugo AE (2000) Avicennia germinans (L.) L. Southern Forest Experiment
Station. New Orleans, LA: U.S. Department of Agriculture, Forest Service.
Kaiser DR, Reinert DJ, Reichert JM, Collares GL, Kunz M (2009) Intervalo hídrico ótimo
no perfil explorado pelas raízes de feijoeiro em um Latossolo sob diferentes níveis
de compactação. Revista Brasileira de Ciências do Solo 33:845-855.
Kozlowski TT (1997) Responses of woody plants to flooding and salinity. Tree
Physiology, Monography 1:1-29.
Krauss KW, Ball MC (2013). On the halophytic nature of mangroves. Trees 27:7-11.
Lovelock CE, Feller IC, McKee KL, Engelbrecht BMJ, Ball MC (2004) The effect of
nutrient enrichment on growth, photosynthesis and hydraulic conductance of dwarf
mangroves in Panama. Functional Ecology 18:25–33.
Matthijs S, Tack J, Speybroeck D, Koedam N (1999) Mangrove species zonation and soil
redox state, sulphide concentration and salinity in Gazi Bay (Kenya), a preliminary
study. Mangrove and Salt Marshes 3:243-249.
McKee KL, Feller IC, Popp M, Wanek W (2002) Mangrove isotopic fractionation (δ18 N
and δ13 C) across a nitrogen versus phosphorus limitation gradient. Ecology 83:1065–
1075.
Medina E, Giarrizzo T, Menezes MP, Carvalho ML, Carvalho EA, Peres A, Silva AB,
Viilhena R, Reise A, Braga CE (2001) Mangal communities of the “Salgado
Paraense”: Ecological heterogeneity along the Bragança península assessed through
soil and leaf analyses. Amazoniana 16:397-416.

19
Naidoo G, Tuffers AV, von Willert DJ (2002) Changes in gas exchange and chlorophyll
fluorescence characteristics of two mangroves and a mangrove associate in response
to salinity in the natural environment. Trees 16:140–146.
Naidoo G, Chirkoot D (2004) The effects of coal dust on photosynthetic performance of
the mangrove Avicennia marina in Richards Bay, South Africa. Environmental
Pollution 127:359–366.
Naidoo G (2006) Factors Contributing to Dwarfing in the Mangrove Avicennia marina.
Annals of Botany 97:1095–1101.
Soane BD, Ouwerkerk, C (1994) Soil compaction in croproduction. Amsterdam, Elsevier.
Sobrado MC, Ball MC (1999) Light use in relation to carbon gain in the mangrove,
Avicennia marina, under hypersaline conditions. Australian Journal of Plant
Physiology 26:245–251.
Tomlinson PB (1986) The Botany of Mangroves. Cambridge, Cambridge University
Press.
Whalley W, Clark L, Gowing D, Cope R, Lodge R, Leeds-Harrison P (2006) Does soil
strength play a role in wheat yield losses caused by soil drying?. Plant Soil 280:279-
290.

20
ANEXO
WETLANDS - OFFICIAL SCHOLARLY JOURNAL OF THE SOCIETY OF WETLAND SCIENTISTS
MANUSCRIPT SUBMISSION
Submission of a manuscript implies: that the work described has not been published before; that it is not under consideration for publication anywhere else; that its publication has been approved by all co-authors, if any, as well as by the responsible authorities – tacitly or explicitly – at the institute where the work has been carried out. The publisher will not be held legally responsible should there be any claims for compensation.
Permissions
Authors wishing to include figures, tables, or text passages that have already been published elsewhere are required to obtain permission from the copyright owner(s) for both the print and online format and to include evidence that such permission has been granted when submitting their papers. Any material received without such evidence will be assumed to originate from the authors.
Online Submission
Authors should submit their manuscripts online. Electronic submission substantially reduces the editorial processing and reviewing times and shortens overall publication times. Please follow the hyperlink “Submit online” on the right and upload all of your manuscript files following the instructions given on the screen.
Cover letter: mandatory
All submissions should be accompanied by a cover letter which should include: What is the scientific question you are addressing? What are the key findings that answer this question? Why is this work important and timely? Why is it of interest to the international readership of WETLANDS?
Article Types
The word count should include title, abstract, keywords, body of the text, figures, and tables but excluding authors affiliations, references and on-line supplementary material.
Original research (5,000-6,000): Articles reporting original research about wetlands, natural or constructed, including, but not limited to mechanisms underlying ecosystem processes, the values of wetlands to society, their management, quality assessment and restoration.
Short communications (2,000-3,000): Short articles in the same format as ‘Original Research’ articles reporting on new findings that are deemed to be

21
potentially very important or timely, but do not require a full-size article. Short communications should not be used for reporting preliminary data.
Review article (6,000): Articles summarizing past and recent research on a subject relevant to wetland science and management, often to synthesize over-arching patterns or opinions. The subject and content should be discussed with the Editor-in-Chief ([email protected]) prior to submission.
Invited reviews (8,000): The Mark Brinson Review series was initiated in 2013 and aims to bring together in-depth, over-arching reviews in the fields of wetland science and management, in the widest sense of those terms. These reviews may synthesize observations over long periods of time or may pull together information from various studies to form an opinion. They are either invited, may be regular reviews that are recognized by the Editorial Board of the journal as important enough to be part of the series, or intending authors may contact the Editor-in-Chief.
In exceptional cases longer articles may be submitted, but only after consultation with the Editor in Chief.
Title page
The title page should include: The name(s) of the author(s) A concise and informative title The affiliation(s) and address(es) of the author(s) The e-mail address, telephone and fax numbers of the corresponding author Abstract Please provide an abstract of maximum 200 words. The abstract should not
contain any undefined abbreviations or unspecified references. Keywords Please provide 4 to 6 keywords which can be used for indexing purposes.
Scientific style
Please always use internationally accepted signs and symbols for units (SI units).
Text
• Use a normal, plain font (e.g., 12-point Times Roman) for text. • The text section should be double spaced. • Use italics for emphasis. • Use the automatic line number function to the manuscript. • Use the automatic page numbering function to number the pages. • Do not use field functions. • Use tab stops or other commands for indents, not the space bar. • Use the table function, not spreadsheets, to make tables. • Use the equation editor or MathType for equations. Note: If you use Word 2007, do not create the equations with the default
equation editor but use the Microsoft equation editor or MathType instead. • Save your file in doc format. Do not submit docx files. LaTeX

22
Manuscripts with mathematical content can also be submitted in LaTeX. Headings Please use no more than three levels of displayed headings. Abbreviations Abbreviations should be defined at first mention and used consistently
thereafter. Footnotes Footnotes on the title page are not given reference symbols. Footnotes to the
text are numbered consecutively; those to tables should be indicated by superscript lower-case letters (or asterisks for significance values and other
statistical data). Footnotes can be used to give additional information, which may include the
citation of a reference included in the reference list. They should not consist solely of a reference citation, and they should never include the bibliographic
details of a reference. They should also not contain any figures or tables. Footnotes to the text are numbered consecutively; those to tables should be
indicated by superscript lower-case letters (or asterisks for significance values and other statistical data). Footnotes to the title or the authors of the article are not given reference symbols.
Always use footnotes instead of endnotes. Acknowledgments Acknowledgments of people, grants, funds, etc. should be placed in a separate
section before the reference list. The names of funding organizations should be written in full.
References
Citations within the text, they should be cited chronologically (oldest to newest),
The list of references should only include works that are cited in the text and that have been published or accepted for publication. Personal communications and unpublished works should only be mentioned in the text.
Do not use footnotes or endnotes as a substitute for a reference list. Reference list entries should be alphabetized by the last names of the first
author of each work. All journal names must be written out in full. • Journal article Gamelin FX, Baquet G, Berthoin S, Thevenet D, Nourry C, Nottin S, Bosquet
L (2009) Effect of high intensity intermittent training on heart rate variability in prepubescent children. European Journal of Applied
Physiology 105:731-738. doi: 10.1007/s00421-008-0955-8 Ideally, the names of all authors should be provided, but the usage of “et al” in
long author lists will also be accepted: Smith J, Jones M Jr, Houghton L et al (1999) Future of health insurance. New
England Journal of Medicine 965:325–329 • Article by DOI Slifka MK, Whitton JL (2000) Clinical implications of dysregulated cytokine
production. Journal of Molecular Medicine. Doi:10.1007/s001090000086 • Book South J, Blass B (2001) The future of modern genomics. Blackwell, London • Book chapter

23
Brown B, Aaron M (2001) The politics of nature. In: Smith J (ed) The rise of modern genomics, 3rd edn. Wiley, New York, pp 230-257
• Online document Doe J (1999) Title of subordinate document. In: The dictionary of substances
and their effects. Royal Society of Chemistry. Available via DIALOG. http://www.rsc.org/dose/title of subordinate document.
Accessed 15 Jan 1999 • Dissertation Trent JW (1975) Experimental acute renal failure. Dissertation, University of
California
Tables
All tables are to be numbered using Arabic numerals. Tables should always be cited in text in consecutive numerical order. For each table, please supply a table caption (title) explaining the
components of the table. Identify any previously published material by giving the original source in
the form of a reference at the end of the table caption. Footnotes to tables should be indicated by superscript lower-case letters
(or asterisks for significance values and other statistical data) and included beneath the table body.
Figure Captions
Each figure should have a concise caption describing accurately what the figure depicts. Include the captions in the text file of the manuscript, not in the figure file.
Figure captions begin with the term Fig. in bold type, followed by the figure number, also in bold type.
No punctuation is to be included after the number, nor is any punctuation to be placed at the end of the caption.
Identify all elements found in the figure in the figure caption; and use boxes, circles, etc., as coordinate points in graphs.
Identify previously published material by giving the original source in the form of a reference citation at the end of the figure caption.





























