Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
MAURÍCIO E CASTRO CABRAL DA SILVA
“A GLIA DE MÜLLER COMO FONTE DE
FATORES TRÓFICOS PARA NEURÔNIOS
CENTRAIS E PERIFÉRICOS”
TESE SUBMETIDA À UNIVERSIDADE FEDERAL DO
RIO DE JANEIRO VISANDO A OBTENÇÃO DO GRAU
DE DOUTOR EM CIÊNCIAS
Universidade Federal do Rio de Janeiro
Centro de Ciências da Saúde Instituto de Biofísica Carlos Chagas Filho
RIO DE JANEIRO, 11 DE ABRIL DE 2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
CABRAL-DA-SILVA, Maurício e Castro. A Glia de Müller Como Fonte de Fatores Tróficos Para Neurônios Centrais e Periféricos Orientador: Ricardo Augusto de Melo Reis Rio de Janeiro, UFRJ/IBCCF, 2007. X, 159 52f. 2t. Tese: Doutorado em Ciências Biológicas (Biofísica). Universidade do Brasil - UFRJ/IBCCF - Programa de Neurobiologia, Laboratório de neuroquímica. 1. Glia de Müller; 2. Fatores Tróficos; 3. Neurônios Simpáticos de embrião de pinto; 4. Células Ganglionares da Retina; 5. Sobrevivência Neuronal In vitro.

iii
Esta dissertação foi elaborada durante a vigência de auxílio financeiro
do Conselho Nacional de Pesquisa e Desenvolvimento (CNPq), da Fundação de Amparo a Pesquisa do Estado do Rio de Janeiro (FAPERJ), da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) e do Wellcome Trust (Reino Unido).

iv
DEDICATÓRIA Ao Leandro, Meu irmão, amigo e compadre.
6 de Abril de 1978 - †18 de Janeiro de 2007 Você faz falta...

v
AGRADECIMENTOS A todos que de alguma forma contribuíram com a conclusão desta dissertação... A minha esposa que foi capaz de compreender que, durante o período em trabalhei nesta dissertação, ficou em segundo plano. A minha esposa novamente por, mesmo em segundo plano, ter me ajudado a formatar a dissertação … ...Obrigado!

vi
EPÍGRAFE “É tão estranho, os bons morrem jovens. Assim parece ser quando me lembro de você. Que acabou indo embora, cedo demais... ... Eu continuo aqui, meu trabalho e meus amigos E me lembro de você em dias assim dia de chuva, dia de sol E o que sinto eu não sei dizer... ...Só que este ano o verão acabou Cedo demais...”
Renato Russo Love in the afternoon
Álbum: O Descobrimento do Brasil, 1993

vii
SUMÁRIO
Resumo ix Abstract x Lista de figuras e tabelas xi Lista de abreviaturas xiii 1.Introdução 14
1.1 Fatores Neurotróficos 16 1.1.2 Receptores para fatores tróficos 22 1.2 Tipos de morte celular_________________________ ______________ 26 1.2.1 Regulação da apoptose 27 1.2.2 Apoptose em neurônios simpáticos_ 32 1.3 Estrutura da retina_______________________ ________________ 35
1.3.1 Células ganglionares da retina _______________________________ 39 1.3.2 Fatores tróficos retinianos_____ ______________________ _______ 40 2. Objetivos_______ __________________________________________________ 42 3. Materiais e métodos____ _____________________________ ______________ 45 3.1 Materiais e reagentes_____________________ _____________________ 46 3.2 Soluções______ _________________________________________________ 47 3.3 Preparação das placas de cultura_____________________ _________ 48 3.4 Obtenção e cultura dos neurônios periféricos____ _______________ 48 3.5 Contagem de células___________________________________ 52 3.6 Meio condicionado por explantes de retina____________ ________ 53 3.7 Meio condicionado por glia de Müller de camundongo neonato 54 3.8 Meio condicionado por glia de Müller de embriões de galinha___ 56 3.9 Meio condicionado radioativo______________________________ 57 3.10 Imunopurificação de CGR de camundongo___________________ 59 3.11 Fracionamento do meio condicionado por glia de Müller em coluna de heparina
62
3.12 Ensaio de regeneração in vivo________________ 66 3.13 Imunocitoquímica________ 69 3.14 Imunodetecção de proteínas (western blot) 70 3.15 Análise estatística____________ 71 4. Resultados_______________________________________________ 72 4.1 RCM tem efeito trófico em neurônios simpáticos 73 4.1.2 A secreção do fator retiniano não é dependente do desenvolvimento nem de excitabilidade elétrica do tecido
75
4.1.3 O efeito trófico promovido por RCM não é devido à ação individual de neurotrofinas
78
4.1.4 Atividade trófica do RCM é mantida após diálise, é retida em coluna de heparina e é sensível a temperatura
82
4.1.5 RCM induz sobrevivência sinalizando pelas vias da Pi3K e JAK2 mas não pela via das ERKs
84
4.1.6 A liberação de fator retiniano depende de eventos mediados neurotrofinas
86

viii
4.1.7 A atividade trófica promovida por RCM não é bloqueada com anticorpos bloqueadores anti-TGF-β, anti-GDNF nem anti-CNTF
86
4.2 Cultura de células de Müller purificadas de camundongos 88 4.2.1 MMG ativa as vias da pERK, pAKT e pSTAT3 simultaneamente em neurônios do gânglio superior cervical de ratos
90
4.2.2 MMG possui componente ativo que se liga a heparina, potente ação trófica em neurônios simpáticos de ave que não foi abolida por anticorpos bloqueadores anti-NT3, anti-GDNF ou anti-CNTF
92
4.2.3 Fatores tróficos da glia de Müller de camundongos induzem neuritogênese e sobrevivência das CGRS
93
4.2.4 Monocamada de glia de Müller é um excelente substrato para a sobrevida das CGRs__ ___
99
4.2.5 Fatores tróficos secretados por células de Müller induzem proliferação de glia de Müller de camundongos
101
4.2.6 Meio condicionado radioativo 103 4.3 Culturas de células de Müller purificadas de pinto 105 4.3.1 AMG apresenta atividade trófica tanto na fração que se liga à heparina quanto na fração livre______
106
4.3.2 Fracionamento do AGM em coluna de heparina 108 4.3.3 AMG induz migração de células de Schwann a partir de explantes de nervo ciático de ratos adultos
113
4.3.4 AMG induz regeneração de neurônios periféricos e centrais 115 5. Discussão________________________ 120 6. Conclusões________________________________ 134 Referências bibliográficas______________________ 135 Anexo_________________________ 155

ix
RESUMO
A sobrevivência de neurônios do Sistema Nervoso Periférico (SNP) e
Central (SNC) depende de fatores tróficos sintetizados e secretados por alvos biológicos. Por outro lado, células de glia de Müller influenciam e são influenciadas por atividade neuronal na retina. Nós investigamos a atividade trófica de meios condicionados (MC) obtidos de culturas purificadas de glia de Müller de embrião pinto e camundongo pós-natal em culturas purificadas de neurônios simpáticos de pinto e rato, e em células ganglionares da retina (CGRs) de camundongos pós-natal. Também estudamos as propriedades regenerativas desses fatores num modelo de transecção do nervo ciático de rato. As culturas purificadas de glia de Müller de camundongo foram positivamente marcadas com anticorpos anti-vimentina, anti-GFAP e anti-S-100, mas foram negativas para anticorpos contra marcadores neuronais.
Meio condicionado de retina (RCM, obtido de explantes de retinas de 11 dias mantido por 4 dias) manteve 90% de neurônios simpáticos (E10) em cultura por 48h, comparado com < 5% do controle (meio DMEM +10% soro fetal bovino). Meio condicionado de glia de Müller de aves (AMG) manteve 100% de neurônios simpáticos viáveis por 48 horas in vitro seu o efeito trófico não foi bloqueado por anticorpos anti-NGF e anti-NT3 ou com o uso de receptores TrkA, B ou C solúveis usados independentemente. Além disso, BDNF ou NT4 (0.1-50ng/ml) não apresentaram efeitos tróficos sobre neurônios simpáticos. O efeito trófico não foi bloqueado por anticorpos anti-GDNF, anti-TGFβ, e anti-CNTF e não foi mimetizado por FGFb (0.1-10nM). LY294002, um inibidor da Pi3Cinase (50μM), mas não PD098059 (inibidor das MAPCinases), bloqueou a sobrevida de neurônios simpáticos induzido por RCM ou AMG.
Meio condicionado de glia de Müller de camundongos (MMG) também manteve a viabilidade de 100% de neurônios simpáticos de pinto, e de rato após 48 horas em cultura. MMG estimulou as vias de AKT, ERK e STAT3 em neurônios simpáticos, efeito não bloqueado por anticorpos anti-NGF ou anti-CNTF. MMG promoveu crescimento neurítico em explantes de retinas P4 de camundongos, e foi tão efetivo quanto BDNF + CNTF + forscolina (BCF, 45% sobrevida em culturas após 72h, comparado com a condição controle + forscolina). As CGRs plaqueadas sobre a monocamada de células de Müller apresentaram uma sobrevida maior (>80%, 72h) que a observada quando estas eram plaqueadas sobre laminina e poli-L-lisina.
AMG induziu sobrevida e regeneração de células do gânglio da raiz dorsal (GRD) e motoneurônios no segmento L4/5 de ratos adultos cujos nervos ciáticos foram seccionados, porém não teve efeito nem na sobrevida nem na neuritogênese de CGRs de camundongo em cultura.
Em conclusão, nossos resultados mostram que a atividade trófica de moléculas solúveis produzidas por células de Müller em cultura é capaz de manter a viabilidade de diversas populações de neurônios periféricos e centrais in vitro e in vivo, além de promover neuritogênese.

x
ABSTRACT
Neuronal survival in the vertebrate peripheral or central nervous system
depends on neurotrophic factors available from target tissues. Müller glia cells influence and are influenced by neuronal activity throughout the retina. We have examined the trophic effects of conditioned media obtained from purified chick or mice Müller glia cells on purified sympathetic chick embryos neurons and retinal ganglion cells (RGCs) from postnatal mice. We also studied regenerative properties of these factors on a sciatic nerve transection model on adult rats. Purified chick and mice Müller glia cultures were obtained from whole retina cultures. Mice Müller glia were positive for vimentin, GFAP or S100, but negative for neuronal markers. Retina conditioned media (RCM) from explants of embryonic day 11 retinas maintained for 4 days in vitro supported 90% of E10 chick sympathetic neurons after 48 hours. Conditioned medium from purified Avian Müller Glial (AMG) cells supported nearly 100% of E10 chick sympathetic neurons. Anti-NGF (1μg/ml) blocked the survival effect of NGF, but did not block the trophic effect of RCM or AMG. Neither BDNF nor NT4 (0.1-50ng/ml) supported E10 sympathetic neuron survival. Incubation of chimeric immunoglobulin-receptors TrkA, TrkB or TrkC had no effect on RCM or AMG induced sympathetic neuron survival. The survival effects were not blocked by anti-GDNF, anti-TGFβ, and anti-CNTF and were not mimicked by FGFb (0.1-10nM). LY294002 at 50μM (a Pi3K pathway blocker), but not PD098059 at 50μM (a MAPKs pathway blocker) blocked sympathetic survival induced by RCM. Mice Müller Glial conditioned medium (MMG) also supported 100% of the survival of chick or rat sympathetic neurons after 48h compared to < 5% in controls. MMG stimulated AKT, ERK and STAT3 in sympathetic neurons, and this effect was not blocked by anti-NGF or anti-CNTF blocking antibodies. MMG induced neurite outgrowth in P4 mice retina explants and was as effective as BDNF + CNTF + forskolin (45% survival, 72h compared to 8% control). RGCs plated on top of Müller glia cells had a much better survival rate (>80%, after 96h) compared to laminin + poly-L-lysine substrates. Avian Müller Glial conditioned medium (AMG) induced survival and regeneration of L4/5 DRG and motoneurons in vivo, but had no effect on mice RGCs survival or neuritogenesis in vitro.
In conclusion, we show that purified Müller glia culture secrete trophic molecules that support in vitro and in vivo peripheral and central neuronal survival and neuritogenesis.

xi
LISTA DE FIGURAS E TABELAS Figuras Figura 1. ________________________________________________________________18 Figura 2. ________________________________________________________________24 Figura 3. ________________________________________________________________30 Figura 4. ________________________________________________________________31 Figura 5. ________________________________________________________________37 Figura 6. ________________________________________________________________49 Figura 7. ________________________________________________________________51 Figura 8. ________________________________________________________________53 Figura 9. ________________________________________________________________55 Figura 10. _______________________________________________________________58 Figura 11. _______________________________________________________________61 Figura 12. _______________________________________________________________62 Figura 13. _______________________________________________________________64 Figura 14. _______________________________________________________________65 Figura 15. _______________________________________________________________67 Figura 16. _______________________________________________________________68 Figura 17. _______________________________________________________________74 Figura 18. _______________________________________________________________76 Figura 19. _______________________________________________________________77 Figura 20. _______________________________________________________________78 Figura 21. _______________________________________________________________79 Figura 22. _______________________________________________________________80 Figura 23. _______________________________________________________________81 Figura 24. _______________________________________________________________82 Figura 25. _______________________________________________________________83 Figura 26. _______________________________________________________________85 Figura 27. _______________________________________________________________86 Figura 28. _______________________________________________________________88 Figura 29. _______________________________________________________________89 Figura 30. _______________________________________________________________91 Figura 31. _______________________________________________________________92 Figura 32. _______________________________________________________________94 Figura 33. _______________________________________________________________95 Figura 34. _______________________________________________________________97 Figura 35. _______________________________________________________________98 Figura 36. ______________________________________________________________100 Figura 37. ______________________________________________________________101 Figura 38. ______________________________________________________________102 Figura 39. ______________________________________________________________104 Figura 40. ______________________________________________________________105 Figura 41. ______________________________________________________________107 Figura 42. ______________________________________________________________107 Figura 43. ______________________________________________________________109

xii
Figura 44. ______________________________________________________________110 Figura 45. ______________________________________________________________111 Figura 46. ______________________________________________________________112 Figura 47. ______________________________________________________________112 Figura 48. ______________________________________________________________114 Figura 49. ______________________________________________________________116 Figura 50. ______________________________________________________________118 Figura 51. ______________________________________________________________119 Figura 52. ______________________________________________________________133 Tabelas Tabela 1.________________________________________________________________21 Tabela 2._______________________________________________________________100

xiii
LISTA DE ABREVIATURAS AA - Ácido ascórbico. AKT/PKB - Proteína cinase B. AMG - (do inglês, Avian Müller Glia) Meio condicionado por culturas de glia de Müller de embrião de pinto. B27 - Suplemento de fatores para meio de cultura neurobasal. BAD - Proteína BH3 pro-apoptótica. BCF - BDNF + CNTF + Forscolina. Bcl-2 - Proteína BH4 antiapoptótica. BDNF - Fator neurotrófico derivado de cérebro. BFC – Neurônios colinérgicos do prosencéfalo basal BME - Meio basal de Eagle. Caspase - Cisteína aspartato protease CGRs - Células ganglionares da retina. Chat - Colina acetil transferase. CMF - Solução de Hanks sem cálcio e magnésio. CNQX - Inibidor competitivo do receptor não-NMDA. CNTF - Fator neurotrófico do gânglio ciliar. DFA - Diacetato de Fluoresceína DMEM - Meio de Eagle modificado por Dulbecco. e.p.m. - erro padrão da média ERK - Cinase regulada por sinalização extracelular. F - Forscolina FGFb - Fator básico de crescimento derivado de fibroblastos. FHKR - Fator de transcrição forkhead. FPLC - Cromatografia rápida de proteínas. GRD - Gânglio da raiz dorsal. GCS - Gânglio Cervical Superior GSK - Cinase da sintase do glucagon. IRS - Substrato do Receptor da Insulina JAK – Cinase Ativada por Janus. LIF - Fator de Inibição da Leucemia. LY294002 - Inibidor da Pi3K. MAPK - Cinase ativada por fatores mitogênicos. MC - Meio condicionado. MCP - Morte celular programada. MK 801 - Inibidor não competitivo do receptor NMDA. MMG – (do inglês, Mice Müller Glia) Meio condicionado por culturas de glia de Müller de camundongo neonato (P1). NB - Meio de cultura Neurobasal. NGF - Fator de crescimento do nervo. NT3 - Neurotrofina 3. NT4/5 - Neurotrofina 4/5. NT6 - Neurotrofina 6. NT7 - Neurotrofina 7. P75 - Subunidade de baixa afinidade para neurotrofinas.

xiv
PACAP - Polipeptídeo pituitário ativador da adenilato ciclase. PBS - Solução salina tamponada de fosfato. PCR - Reação da polimerase em cadeia. PD098059 - Inibidor da MAPK. Pi3C - Fosfoinositideo 3-OH Cinase. PIP2 - Fosfatidilinositol-4,5-bifosfato PIP3 - Fosfatidilinositol-3,4,5-trifosfato RCM - (do inglês, Retina Conditioned Media) Meio condicionado por explantes de retina de embrião de aves. SFB - Soro fetal bovino. SNC - Sistema nervoso central. SNP - Sistema Nervoso Periférico. STAT - Transdutor de sinal e ativador de transcrição. TNF - Fator de Necrose Tumoral. Trk A - Receptor tirosina cinase de alta afinidade para neurotrofinas ativado por NGF. Trk B - Receptor tirosina cinase de alta afinidade para neurotrofinas ativado por BDNF e/ou NT 4. Trk C - Receptor tirosina cinase de alta afinidade para neurotrofinas ativado por NT 3. TTBS – Solução tamponada de Tris + 0,1% Tween TTX - Tetrodotoxina. VIP- Peptídeo vaso-ativo intestinal

15
1. INTRODUÇÃO

16
1.1 FATORES NEUROTRÓFICOS
A sobrevivência de neurônios no Sistema Nervoso Periférico (SNP) e
Central (SNC) depende de quantidades limitadas de fatores neurotróficos
secretados por tecidos alvos (DAVIES, 1994; GOLDBERG e BARRES, 2000).
Entretanto a capacidade de regeneração do SNP, mas não de neurônios
centrais, é uma característica marcante dos vertebrados superiores
(GOLDBERG e BARRES, 2000). Um dos maiores desafios da neurobiologia
moderna encontra-se exatamente em se descobrir como funciona o
mecanismo que define a vida ou a morte de diferentes tipos neuronais. Se
pudermos identificar o porquê da irreversibilidade de lesões das vias nervosas
centrais após um trauma talvez possamos reverter e recuperar a função
dessas vias.
Trabalhos pioneiros referentes à manutenção de populações de
neurônios sensoriais e simpáticos por quantidades mínimas de fatores solúveis
foram realizados em meados do século passado por Rita Levi-Montalcini e
Victor Hamburger (LEVI-MONTALCINI e HAMBURGER, 1951, 1953).
Como descrito por Cowan (2001), a colaboração entre Montalcini e
Hamburger que deveria ser de apenas alguns meses, no máximo um ano,
durou décadas e determinou o futuro da neurobiologia do desenvolvimento.
No final de 1951, a italiana Levi-Montalcini, em colaboração com a
Doutora Hertha Meyer, veio ao Brasil e passou alguns meses em seu
laboratório na Universidade do Brasil, na época situada na Praia Vermelha.
Desta colaboração resultou uma descoberta que muito acrescentou à

17
neurobiologia moderna, o papel de fatores neurotróficos solúveis na
manutenção da viabilidade neuronal.
A descoberta do Fator de Crescimento do Nervo (NGF, do inglês Nerve
Growth Factor) representou um marco numa área da pesquisa hoje muito
bem estabelecida, que é a área das neurotrofinas e fatores neurotróficos.
Nas primeiras observações de Levi-Montalcini no Rio de Janeiro, a
presença de um explante de sarcoma, um tecido tumoral, em culturas de
explantes de gânglios sensoriais, provocava extensa neuritogênese no tecido
nervoso logo nas primeiras 24 horas. O que foi observado foi um
impressionante aumento do número e tamanho dos neuritos que projetavam
do explante de gânglio da raiz dorsal (GRD) (figura1) (Purves e Lichman,
1985). Com base nestas observações Rita Levi-Montalcini concluiu que o
agente de tal efeito era, possivelmente, uma molécula solúvel que se
difundia no meio e provocava tais alterações nos explantes de tecido
nervoso (LEVI-MONTALCINI e HAMBURGER, 1953). O fenômeno ilustrado por
seus desenhos ganhou a capa de publicações importantes e livros textos em
neurobiologia. Alguns anos mais tarde, com a chegada do bioquímico
Stanley Cohen ao laboratório de Victor Hamburger (em Saint Louis, Missouri),
deu-se o isolamento e a purificação de frações ativas do sarcoma e
também de veneno de cobra e glândula submandibular de rato. A
purificação do fator revelou uma estrutura pentamérica polipeptídica mais
tarde denominada de NGF. Nesta molécula, o dímero das subunidades β é o
responsável pela atividade trófica do NGF, a subunidade γ possui atividade
serina proteinase capaz de processar a forma precursora do β-NGF

18
enquanto a subunidade α-NGF é uma serina proteinase inativa (McDONALD
et al., 1995) portanto ainda desconhecidas quanto à função (PURVES e
LICHMAN, 1985).
Figura 1. A descoberta do primeiro membro da família das neurotrofinas, o NGF, por Rita Levi-montalcini. Observando projeções neuríticas que emanavam de um explante de GRD na presença de um explante de sarcoma concluiu que este liberava fatores solúveis que induziam neuritogênese em explantes de gânglios periféricos (compare a diferença entre os explantes na presença do sarcoma e na ausência, em vermelho). Posteriormente, o bioquímico Stanley Cohen se juntou ao grupo e seqüenciou a estrutura primária das subunidades da proteína NGF.
Além do NGF, fazem parte da família das neurotrofinas o fator
neurotrófico derivado de cérebro (BDNF, do inglês Brain Derived
Neurotrophic Factor), que foi isolado inicialmente a partir de grande
quantidade de cérebro de porcos (BARDE et al., 1982), e teve
posteriormente seu gene clonado com a técnica de DNA recombinante
(LEIBROCK et al., 1989). A clonagem e a expressão do gene do BDNF permitiu

19
novos avanços na área dos fatores neurotróficos. A comparação entre as
estruturas do NGF e do BDNF revelou uma homologia de aproximadamente
50% na cadeia peptídica dos homodímeros básicos (Leibrock et al., 1989). O
uso da técnica de reação da polimerase em cadeia (PCR, do inglês
Polimerase Chain Reaction) com seqüências iniciadoras (primers) para
regiões conservadas das neurotrofinas permitiu a identificação de membros
adicionais desta família. Assim, a neurotrofina-3 (NT-3, HOHN et al., 1990;
MAISONPIERRE et al., 1990; JONES e REICHARDT, 1990; ROSENTHAL et al.,
1990), a neurotrofina 4/5 (NT-4/5, BERKEMEIER et al., 1991; HALLBÖÖK et al.,
1991; IP et al., 1992), mais recentemente a neurotrofina 6 (NT-6; GOTZ et al.,
1996) e a neurotrofina 7 (NT-7; NILSSON et al., 1998) foram obtidas através de
clonagem e seqüenciadas sem a purificação protéica prévia (JOHNSON,
1999). Estas moléculas tróficas caracterizam-se por manter populações
distintas de neurônios no sistema nervoso periférico e central (Tabela 1). O
NGF, por exemplo, tem efeito sobre a sobrevivência e diferenciação em
neurônios simpáticos e sensoriais além de neurônios colinérgicos do
prosencéfalo basal. Estes últimos parecem ser afetados no mal de Alzheimer,
doença neurodegenerativa que afeta a cognição geralmente de pessoas
idosas.
Nos últimos anos, ficou evidente que muitos fatores de crescimento
preenchem uma definição funcional de fator neurotrófico podendo regular
a sobrevivência e a diferenciação de células nervosas. Assim, vários fatores
tróficos que não são estruturalmente relacionados com as neurotrofinas
também mantêm neurônios em cultura. Entre estes fatores temos, o fator

20
neurotrófico derivado do gânglio ciliar (CNTF, do inglês Ciliary Neurotrophic
Factor; SENDTNER et al., 1994), o fator de crescimento de fibroblastos tipo 2
(FGF-2 , do inglês Fibroblast Growth Factor type 2; UNSICKER et al., 1987;
ECKENSTEIN, 1994), o fator inibidor de leucemia (LIF, do inglês Leukemia
inhibitory Factor; MURPHY et al., 1993), o fator neurotrófico derivado de
linhagem glial (GDNF, do inglês Glial Derived Neurotrophic Factor; UNSICKER,
1999), e os fatores análogos à insulina 1 e 2 (IGF, do inglês Insulin-like Growth
Factor; ISHII et al., 1993). A interação da insulina, ou de IGFs com seus
respectivos receptores leva ao recrutamento e a fosforilação do substrato do
receptor da insulina (IRS, do inglês Insulin Receptor Substrate), e ao
subseqüente recrutamento da enzima Pi3Cinase (Pi3C). Pi3C se liga a IRS
convertendo fosfatidilinositol-4,5-bifosfato (PIP2) na membrana celular a
fosfatidilinositol-3,4,5-trifosfato (PIP3). O acúmulo de PIP3 é antagonizado pela
fosfatase homóloga da tensina (PTEN, do inglês Phosphatase and Tensin
Homolog). PIP3 ainda participa da interação com proteína cinase
dependente de fosfoinositídeo (PDK1, do inglês Phosphoinositide-dependent
Protein Kinase 1) e proteína cinase B (PKB, do inglês Protein Kinase B; também
chamada AKT) na membrana, resultando na fosforilação de PKB por PDK1.
Hoje o cenário relacionado com os fatores tróficos encontra-se
pautado nas famílias gênicas como as do CNTF (VERGARA e RAMIREZ, 2004)
e outros membros da mesma família protéica como por exemplo LIF,
Interleucina-6 (IL-6 , do inglês Interleukin-6), IL-11, oncostatina M e
cardiotrofina.

21
Adaptado de Tuszynski e Kordower, 1999
Tabela1: Diversos fatores com conhecida atividade trófica no sistema nervoso de mamíferos.
Neurônios sensoriais, do tronco cerebral.
Fatores Neurotróficos e Alguns de seus Alvos no Sistema Nervoso de MamíferosNome do Fator de Crescimento Alvo
Fator de Crescimento do Nervo (NGF) Neurônios do estriato, nociceptivos, sensoriais, simpáticos e BFC.
Fator Trófico Derivado do Cérebro (BDNF) Neurônios corticais, mecanoreceptores, sensoriais, motores (α), vestibulares, auditivos, CGRs, BFC, HPC.
Fator de Crescimento de Fibroblastos-1 ( FGFα)
Fator de Crescimento Transformante β (TGF β 1-3)
Activina Neurônios motores (α).
Fator Neurotrófico Derivado de Linhagem Glial (GDNF)
Neurturina
Artemina
Fator Neurotrófico Ciliar (CNTF)
Fator inibidor de Leucemia (LIF) ou Fator de diferenciação Colinérgica (CDF)
Cardiotrofina-1
Interleucina-6
Neurotrofina 4/5 (NT 4/5)
Neurotrofina 3 (NT-3) Neurônios corticais, proprioceptores, sensoriais, do fuso muscular, auditivos, BFC, HPC, oligodendrócitos.
Neurotrofina 6 e 7 (NT-6 e NT-7) Não descritas em mamíferos.
Neurônios motores (α), CGRs.
Fator de Crescimento de Fibroblastos-2 ( FGFβ)
Fator de Crescimento de Fibroblastos 3-17
Vários tipos neuronais da medula e tronco cerebral, neurônios corticais. Neurônios dopaminérgicos, neurônios corticais, CGRs, BFC, células tronco.
Diversos tipos neuronais.
Neurônios motores (α), sensoriais.
Neurônios dopaminérgicos, motores (α e corticais), sensoriais, simpáticos.
Persefina
Neurônios motores (α e corticais), corticais, simpáticos,glias.
Fatores Morfogenéticos Ósseos (BMPs)
Neuregulinas Maturação glial e de sinapses.
Fator de Crescimento do Hepatócito (HGF)
Imunofilinas
Midcina
Eritropoetina BFC.
Fator de Crescimento do tipo Insulina-I (IGF-1)
Fator de Crescimento do tipo Insulina-2 (IGF-2)
Fator de Crescimento Epidérmico (EGF)
CNS, formador de padrão durante o desenvolvimento do SNP.
Neurônios motores (α).
Neurônios sensoriais, simpáticos.
Fator de Crescimento Transformante α (TGF α) Neurônios dopaminérgicos, HPC, corticais (in vitro).
Neurônios motores (α), sensoriais, HPC, células de Schwann.
Fotoreceptores.
Neurônios dopaminérgicos, motores (α e corticais), sensoriais.
Neurônios dopaminérgicos, motores (α e corticais).
Neurônios dopaminérgicos, corticais, HPC, células tronco.
Neurônios motores (α), sensoriais, diversos tipos neuronais do cérebro, oligodendrócitos .
Neurônios motores (α e corticais), do estriato, parassimpáticos, sensoriais, simpáticos, BFC, HPC.
Neurônios simpáticos.
Neurônios diversos centrais e periféricos (inclui simpático.
Neurônios sensoriais, do tronco cerebral.
Fatores Neurotróficos e Alguns de seus Alvos no Sistema Nervoso de MamíferosNome do Fator de Crescimento Alvo
Fator de Crescimento do Nervo (NGF) Neurônios do estriato, nociceptivos, sensoriais, simpáticos e BFC.
Fator Trófico Derivado do Cérebro (BDNF) Neurônios corticais, mecanoreceptores, sensoriais, motores (α), vestibulares, auditivos, CGRs, BFC, HPC.
Fator de Crescimento de Fibroblastos-1 ( FGFα)
Fator de Crescimento Transformante β (TGF β 1-3)
Activina Neurônios motores (α).
Fator Neurotrófico Derivado de Linhagem Glial (GDNF)
Neurturina
Artemina
Fator Neurotrófico Ciliar (CNTF)
Fator inibidor de Leucemia (LIF) ou Fator de diferenciação Colinérgica (CDF)
Cardiotrofina-1
Interleucina-6
Neurotrofina 4/5 (NT 4/5)
Neurotrofina 3 (NT-3) Neurônios corticais, proprioceptores, sensoriais, do fuso muscular, auditivos, BFC, HPC, oligodendrócitos.
Neurotrofina 6 e 7 (NT-6 e NT-7) Não descritas em mamíferos.
Neurônios motores (α), CGRs.
Fator de Crescimento de Fibroblastos-2 ( FGFβ)
Fator de Crescimento de Fibroblastos 3-17
Vários tipos neuronais da medula e tronco cerebral, neurônios corticais. Neurônios dopaminérgicos, neurônios corticais, CGRs, BFC, células tronco.
Diversos tipos neuronais.
Neurônios motores (α), sensoriais.
Neurônios dopaminérgicos, motores (α e corticais), sensoriais, simpáticos.
Persefina
Neurônios motores (α e corticais), corticais, simpáticos,glias.
Fatores Morfogenéticos Ósseos (BMPs)
Neuregulinas Maturação glial e de sinapses.
Fator de Crescimento do Hepatócito (HGF)
Imunofilinas
Midcina
Eritropoetina BFC.
Fator de Crescimento do tipo Insulina-I (IGF-1)
Fator de Crescimento do tipo Insulina-2 (IGF-2)
Fator de Crescimento Epidérmico (EGF)
CNS, formador de padrão durante o desenvolvimento do SNP.
Neurônios motores (α).
Neurônios sensoriais, simpáticos.
Fator de Crescimento Transformante α (TGF α) Neurônios dopaminérgicos, HPC, corticais (in vitro).
Neurônios motores (α), sensoriais, HPC, células de Schwann.
Fotoreceptores.
Neurônios dopaminérgicos, motores (α e corticais), sensoriais.
Neurônios dopaminérgicos, motores (α e corticais).
Neurônios dopaminérgicos, corticais, HPC, células tronco.
Neurônios motores (α), sensoriais, diversos tipos neuronais do cérebro, oligodendrócitos .
Neurônios motores (α e corticais), do estriato, parassimpáticos, sensoriais, simpáticos, BFC, HPC.
Neurônios simpáticos.
Neurônios diversos centrais e periféricos (inclui simpático.

22
A proteína FGFb é um outro fator trófico identificado inicialmente em
neurônios por imunocitoquímica (PETTMAN et al., 1986) e que mantém a
sobrevida de neurônios do gânglio ciliar, mas em contraste com o CNTF, não
induz a sobrevida de neurônios do GRD nem de neurônios simpáticos
(UNSICKER et al., 1987). Até o momento já foram descritos e isolados cerca
de 23 membros da família do FGF, com enfoque particular ao FGFb (FGF
básico), que tem função neurotrófica (ABE e SAITO, 2001); para a família do
fator transformante do crescimento beta (TGFβ, do inglês Transforming
Growth Factor) que no SNC tem uma subfamília liderada pelo GDNF que
hoje apresenta 3 novos membros, persefina, artemina e neurturina (BALOH et
al., 2000).
1.1.2 RECEPTORES PARA FATORES TRÓFICOS
É fato que a maioria, se não todos os fatores neurotróficos, ativam seus
respectivos neurônios através de receptores compostos por conjuntos
protéicos multiméricos (complexos de subunidades protéicas) como os
exemplificados na figura 2. Nesta figura podemos observar a ilustração
esquemática de quatro receptores multiméricos de famílias protéicas
diferentes. O primeiro receptor da esquerda para a direita é um modelo de
receptor para ligantes da família das neurotrofinas. Este receptor possui dois
conjuntos de subunidades, uma de alta afinidade chamada receptor de
cinases relacionados à tropomiosina (Trk, do inglês Tropomyosin-related
Kinase; KLEIN et al., 1991; BARBACID, 1991) que tem descritas três isoformas

23
TrkA, TrkB e TrkC com especificidade para diferentes neurotrofinas e outra
subunidade de menor afinidade chamada p75 (CHAO, 1994). Existe pelo
menos uma subunidade de alta afinidade para cada neurotrofina, onde
TrkA tem maior afinidade e eficiência para NGF, TrkB para BDNF e NT4/5,
enquanto TrkC tem grande especificidade para NT-3. Todas as subunidades
Trks contém 3 domínios ricos em leucina, dois concentrados em cisteína, dois
domínios do tipo imunoglobulina na região extracelular do receptor além do
domínio tirosina cinase no lado citoplasmático (JOHNSON, 1999), entretanto
é importante ressaltar o grande número de isoformas existentes para esses
receptores, truncadas ou não, que desempenham variados efeitos
fisiológicos durante o desenvolvimento (NINKINA et al., 1996). Da mesma
maneira as neurotrofinas também são encontradas em diferentes isoformas.
A subunidade p75, por sua vez, membro da família do Fator de
Necrose Tumoral (TNF, do inglês Tumor Necrosis Factor), apresenta
características estruturais com domínios envolvidos em morte celular,
(Domínios de morte, Death Domains, DR1-DR4; KHURSIGARA et. al., 2001). Esta
subunidade apresenta várias funções, desde a potencialização da resposta
ao NGF durante o desenvolvimento (HORTON et al., 1997), até o
envolvimento na resposta de morte celular programada, dependendo do
tecido envolvido e da fase do desenvolvimento. Isto foi demonstrado em
particular na retina de aves por Frade e colaboradores, (1996).

24
Figura 2. Variedade de receptores multiméricos para fatores neurotróficos. Da esquerda para a direita temos: o receptor de alta afinidade para neurotrofinas TrkA, TrkB e TrkC; o receptor de baixa afinidade para neurotrofinas, p75; o receptor para FGF com a porção heparan-sulfato que compõe o complexo ativo do receptor; o receptor para GDNF com a sua subunidade α e a proteína acessória (ret) visto que este receptor não possui domínio catalítico e o receptor trimérico para CNTF com sua subunidade gp130 que compõe a tríade do receptor para todos os membros descritos da família do CNTF.
O segundo receptor ilustrado na figura 2 recebe ligantes da família do
FGF. Este receptor tem duas subunidades, uma de alta afinidade com
domínio intracelular tirosina cinase, e uma subunidade essencial contendo
proteoglicanos com caudas de heparan sulfato importantes na resposta
funcional dos fatores tróficos desta família (SIMON, 2000). O terceiro receptor
da figura 2 representa um receptor que tem especificidade por ligantes da
família do GDNF. Estes receptores que começaram a ser identificados em
meados da década de 90 por técnicas de biologia molecular (JING et al.,
1996, TREANNOR et al., 1996) são compostos por pelo menos uma

25
subunidade de grande afinidade para cada membro até então identificado
(GRFα1-4). Onde GRFα1 responde preferencialmente para GDNF, GRFα2
para neurturina, GRFα3 para artemina e GRFα4 para persefina (BALOH et al.,
2000; SAARMA, 2000). Estes estudos foram confirmados por expressão destas
subunidades associadas a fosforilação de proteínas sinalizadoras e sobrevida
de neurônios centrais e periféricos em cultura e in vivo. É válido mencionar
que GFRα4 só foi identificado em aves (ENOKIDO et al., 1998). Os receptores
GRFα1-4 se ligam à membrana plasmática com caudas de
glicofosfatidilinositol e se associam a uma proteína chamada ret, que possui
um domínio tirosina cinase, responsável pela fosforilação de proteínas alvo.
A proteína ret já era conhecida pelo seu papel em diversos tumores do
SNC e dos rins tendo seu gene clonado desde o final da década de 80
(TAKAHASHI et al., 1988), mas até então permanecera como um receptor
órfão.
O CNTF foi o terceiro fator neurotrófico a ser purificado. Diferente do
NGF e demais neurotrofinas, o CNTF é membro de uma ampla família de
protéica denominada de família das citocinas neuropoiéticas (PATTERSON,
1991; JOHNSON, 1999). Os receptores da família do CNTF (figura 2) são
compostos por 3 proteínas que formam o complexo funcional. A
composição do trímero varia de acordo com o do membro que ativa o seu
respectivo receptor (STAHL e YANCOPOULOS, 1996; IP e YANCOPOULOS,
1996). Quando CNTF é o ligante, os receptores são formados por uma
proteína receptora do CNTF (CNTFRα, solúvel ou não), por uma proteína
receptora do LIF (LIFR) e por uma proteína chamada gp130. A subunidade

26
CNTFR também se associa à membrana por cauda de glicofosfatidilinositol. É
válido notar, que diferente das outras três famílias de fatores tróficos, estas
subunidades não apresentam domínios intracelulares tirosina cinase
necessitando, desta forma, recrutar e se associar à proteínas adaptadoras
que possuem função de fosforilar demais proteínas em resíduos tirosina
(TOUW et al., 2000). Estas proteínas adaptadoras são da família das Janus
cinases (JAK, do inglês, Janus Activated Kinase) e geram respostas
fosforilando fatores de transcrição chamados Transdutores de Sinal e
Ativadores de Transcrição (STAT, do inglês, Signal Transduction Activated
transcription) que irão regular a transcrição de diversos genes.
1.2 TIPOS DE MORTE CELULAR
A morte neuronal é parte de um processo na biologia do sistema
nervoso presente tanto no desenvolvimento quanto em desordens
degenerativas. Vários tipos de morte celular têm sido descritos de acordo
com características morfológicas e bioquímicas. Entre os diferentes tipos de
morte celular, o processo apoptótico tem sido o mais estudado. Na
apoptose, a transcrição de genes é ativa, há síntese de proteínas, a
atividade de cisteína aspartato protease (caspases) é aumentada, ocorre
diminuição do volume celular, picnose nuclear, fragmentação do DNA
nuclear e a manutenção da membrana plasmática (SUBRAMANIAM e
UNSICKER, 2006). Em contraste, na necrose não ocorre síntese protéica nem
atividade de caspases, e o núcleo assim como a estrutura do DNA são

27
pouco afetados. No entanto, existe um aumento no volume celular e a
membrana plasmática sofre danos. Autofagia é um outro processo de morte
celular que vem ganhando maior atenção nos últimos anos, e refere-se à
autodestruição da célula caracterizada pelo seqüestro do conteúdo
citoplasmático por vacúolos chamados de autofagosomas, sendo
transportado aos lisossomas para degradação (BROKER et al., 2005). No
processo autofágico, a participação de genes e proteínas foi inicialmente
observada em leveduras e alguns ortólogos de mamíferos. As células que
morrem por autofagia sofrem o processo de forma independente de
caspases (GOZUACIK e KIMCHI, 2004).
1.2.1 REGULAÇÃO DA APOPTOSE
Nos últimos dez anos, a grande quantidade de publicações sobre os
mecanismos moleculares envolvidos na apoptose levou a um grande
avanço no entendimento deste fenômeno. O conceito de apoptose surgiu
na década de 70, quando Kerr e colaboradores (1972) mostraram que este
tipo de fenômeno de Morte Celular Programada (MCP) é conservado na
evolução e tem implicações abrangentes na biologia celular ocorrendo
durante o desenvolvimento e processos patológicos nos mais diversos
sistemas de invertebrados e vertebrados. Neste processo, observa-se a
redução citoplasmática, condensação da cromatina, fragmentação do
DNA genômico e formação de circunvoluções na superfície celular, as quais
dão origem aos corpos apoptóticos. Acredita-se que a ativação

28
descontrolada da MCP no sistema nervoso esteja envolvida com a evolução
de doenças neurodegenerativas como Alzheimer e Parkinson (DESHMUKH e
JOHNSON, 1997). Já foram descritas vias bioquímicas distintas que levam a
MCP envolvendo ou não a mitocôndria além de vias de sinalização ativadas
por receptores com domínio de morte, ambas culminando com a ativação
de caspases (DANIAL e KORSMEYER, 2004). A morte apoptótica requer a
síntese de proteínas e a transcrição de genes apropriados para o fenômeno
apoptótico (MARTIN et al., 1988; ESTUS et al., 1994; FREEMAN et al., 1994) e se
opõe a necrótica, visto que esta última está envolvida com extravasamento
citoplasmático, ruptura de organelas e inflamação tecidual.
As primeiras observações genéticas a respeito dos mecanismos
envolvidos na apoptose foram realizados no nematódeo Caenorhabditis
elegans (C. elegans) durante a década de 80 pelo grupo de Robert Horvitz,
em Boston, EUA (ELLIS e HORVITZ, 1986). Este nematódeo tem um número fixo
de 1090 células no seu organismo. Destas, 131 sofrem apoptose durante o
desenvolvimento. Utilizando mutantes e técnicas bioquímicas, genéticas e
moleculares, três genes críticos nesse processo foram identificados: ced-3
(que codifica uma caspase), ced-4 que em humanos tem seu homólogo,
Fator Ativador de Proteases Apoptóticas 1 (APAF1, do inglês Apoptosis
Protease Activating Factor-1) que ativa caspases induzindo apoptose e ced-
9 que é homólogo ao proto-oncogene humano Bcl-2 (do inglês B-cell
Lymphoma/Leukemia-2), que impede a apoptose quando expressa em altas
quantidades em diversos tipos celulares.

29
Assim como no C. elegans, as células dos vertebrados também
apresentam na sua estrutura proteínas pró-apoptóticas (homólogas às
proteínas sintetizadas pelos genes ced-3 e ced-4) e anti-apoptóticas
(homólogas a ced-9). A principal família de proteínas envolvida neste
processo é a da bcl-2 que é amplamente estudada quanto a sua estrutura e
função. Os membros da família bcl-2 foram sendo elucidados na década de
90, em grande parte pelos trabalhos de Stanley Korsmeyer (Harvard Medical
School, Boston, EUA). O gene da proteína bcl-2 foi descrito em 1984 em
células do sistema imune induzindo um linfoma folicular a uma proliferação
desenfreada quando tal proteína era expressa em altos níveis. A família
possui até 4 domínios de homologia a bcl-2 (BH), correspondendo a regiões
de hélice e que podem ser classificadas em 3 categorias. As proteínas bcl-2,
bcl-xl, bcl-w, bfl-1, Mcl-1 e A1 e alguns outros membros são anti-apoptóticos
e compartilham 4 domínios BH semelhantes. O segundo grupo consiste de
proteínas pro-apoptóticas (Bax, Bak e Bok) que compartilham os domínios
BH1-3 (DONOVAN e COTTER, 2004), dos quais Bax e Bak são essenciais para
inúmeros estímulos de morte (WEI et al., 2001). O terceiro grupo, a subfamília
BH3, que contém apenas esse domínio, consiste de Bim, Bid, Bad, Puma,
Noxa, Hrk/DP5, Bik e Bmf que agem como sentinelas dos sinais de morte
celular e heterodimerizam com membros anti-apoptóticos do grupo Bcl-2
levando a ativação de Bax/Bak e liberação de fatores mitocondriais (WILLIS
e ADAMS, 2005). No entanto, este grupo não pode iniciar a permeabilidade
mitocondrial diretamente sem Bax ou Bak (ZONG et al., 2001).

30
A mitocôndria emergiu como o local essencial na efetivação da MCP
devido a múltiplos sinais bioquímicos, convergindo na liberação de
citocromo C que interagindo com APAF-1 e caspase-9 formam uma estrutura
chamada de apoptosoma. Assim, membros pró-apoptóticos da família bcl-2
podem induzir um aumento na permeabilidade mitocondrial levando a
liberação de citocromo C do espaço mitocondrial para o citoplasma,
induzindo morte por apoptose (ZORNIG et al., 2001). Membros anti-
apoptóticos preservam a integridade mitocondrial e portanto bloqueiam a
liberação de proteínas solúveis inter-membranares (figura 3).
Adaptado de Chao, 1994.
Figura 3. Esquema ilustrativo da complexa regulação do processo apoptótico. Fatores tróficos atuam extracelularmente ativando receptores que vão ativar vias de sinalização intracelulares impedindo a liberação de citocromo C pela mitocôndria. Proteínas pró- e anti-apoptóticas controlam a permeabilidade da membrana mitocondrial. Durante a apoptose, citocromo C liberado interage com APAF-1 e caspase 9, levando a formação do complexo apoptosoma. A apoptose se caracteriza por fragmentação de DNA, fragmentação do DNA e de organelas intracelulares entre outras características.

31
Dois mecanismos são atualmente propostos para isso. No primeiro,
proteínas pró-apoptóticas formariam poros na membrana permitindo a
liberação de citocromo C (idéia defendida por Jean Claude Martinou e
colaboradores, Geneva, Suíça e Richard Youle e colaboradores, Boston,
EUA) e/ou proteínas pró-apoptóticas interagiriam com as anti-apoptóticas
numa estequiometria que impediria que as proteínas pró-apoptóticas fiquem
livres no citoplasma e ativem cascatas de caspases (defendido por
KORSMEYER et al., 1999). Outras proteínas como a 14-3-3, que se situa no
citoplasma, também teriam um papel de tamponamento funcional, por
exemplo se ligando a proteína bad fosforilada, impedindo de exercer seu
efeito apoptótico (figura 4) (DATTA et al., 2000). A fosforilação da proteína
bad e de outras moléculas pela ativação de receptores por fatores tróficos
causa uma mudança nas vias de sinalização intracelulares que impede os
elementos das vias apoptóticas de iniciar a apoptose por mecanismos
diversos.
Figura 4. Esquema ilustrando a inativação de Bad fosforilada pela proteína 14-3-3. Bad fosforilada se desloca da membrana para o citosol onde se liga a 14-3-3 tornando-se inativa.
Adaptado de Datta et al., 2000

32
Como mencionado anteriormente, alguns processos apoptóticos não
envolvem diretamente a mitocôndria, como por exemplo, a apoptose
disparada pela ativação da caspase 12, uma protease localizada no retículo
endoplasmático (NAKAGAWA et al., 2000), e caspase 8, associada com
receptores que possuem domínios de morte, como Fas (van EIJK et al., 2001).
1.2.2 APOPTOSE EM NEURÔNIOS SIMPÁTICOS
Neurônios simpáticos dependentes de NGF são amplamente usados
como modelo no estudo da MCP neuronal. A maioria das populações
neuronais sofre MCP durante o período em que é estabelecida a inervação
de seus alvos. Esta morte se dá porque a sobrevivência destes neurônios
durante o desenvolvimento é dependente de fatores tróficos secretados por
tecidos alvo e/ou células vizinhas em quantidades limitadas. Esta é a
chamada “hipótese neurotrófica” (PURVES e LITCHMAN., 1985). Em ratos os
neurônios simpáticos dependem de NGF de forma aguda para a sua
sobrevivência a partir da idade embrionária de 16 dias (E16) até a primeira
semana pós-natal (HENDRY e CAMPBELL, 1976) e em aves a partir de E8 até o
nascimento. Evidências para essas observações vêm de experimentos com a
administração de NGF exógeno aumentando a sobrevivência dos neurônios
simpáticos (HENDRY e CAMPBELL, 1976), ou pela neutralização do NGF com
o uso de anticorpos específicos (LEVI-MONTALCINI e BOOKER, 1960) e
conseqüentemente diminuindo a população dos neurônios simpáticos. Mais
recentemente, a deleção do gene do NGF (CROWLEY et al., 1994) ou do

33
receptor TrkA (SMEYNE et al., 1994) em camundongos por recombinação
homóloga resultou em vasta morte de neurônios simpáticos. De acordo com
os trabalhos de Davies et al. (1988) o estágio E10 é o mais apropriado para a
maioria dos ensaios envolvendo apoptose e fatores neurotróficos em
embriões de galinha e será o estágio embrionário usado neste trabalho para
culturas de neurônios do gânglio simpático e GRD.
Neurônios simpáticos do gânglio cervical superior (GSC) de
camundongos ou ratos P1 (1 dia pós-natal) e do gânglio tóraco-lombar de
embriões de pinto E10, purificados e mantidos em cultura, dependem de
NGF para manutenção da viabilidade celular e projeção neurítica. A
remoção do NGF, ou a ausência desse fator inicia a MCP e promove a morte
apoptótica destes neurônios em cultura em sua totalidade em
aproximadamente 48 horas após a privação do fator.
Uma das grandes vantagens desse modelo é a relativa
homogeneidade da população dos neurônios simpáticos que sofrem
apoptose, de forma sincrônica e reprodutível (DESHMUKH e JOHNSON, 1997).
Durante as primeiras 8 horas de privação de NGF, os neurônios simpáticos
não exibem qualquer alteração morfológica. Após esse período os neuritos
começam a degenerar, a membrana plasmática perde a sua aparência
lisa, e o corpo celular fica irregular (DESHMUKH e JOHNSON, 1997). Mais de
95% dos neurônios estão mortos por apoptose após 48 horas em cultura
privados de NGF.
Além do NGF, outras moléculas são capazes de inibir a MCP em
neurônios simpáticos. Entre estas moléculas estão análogos de adenosina

34
mono-fosfato cíclico (AMPc) ou ativação direta da proteína cinase A (PKA)
(RYDEL e GREENE, 1988; EDWARDS et al., 1991), despolarização com KCl em
gânglio da raiz dorsal (SCOTT e FISHER, 1970, EDWARDS et al., 1991) e
peptídeos ativos, como o polipeptídeo pituitário que ativador da adenilato
ciclase (PACAP, do inglês Pituitary Adenylate Cyclase-Activating
Polypeptide; PRZYWARA et al., 1998) além da aplicação de ciclohexemida
na cultura visto que esta morte é dependente de síntese protéica (MARTIN,
1988; SCOTT e DAVIES, 1990). Entretanto, ocorrem algumas diferenças de
resposta de sobrevivência em neurônios simpáticos entre as diferentes
espécies. O AMPc está envolvido na manutenção da sobrevivência de
neurônios simpáticos de roedores o que não acontece em neurônios
simpáticos embrionários de pinto (WAKADE et al., 1990). Se NGF, AMPc/PKA e
KCl impedem a morte apoptótica através de vias convergentes ou pelos
mesmos mediadores é algo ainda sem resposta, porém os níveis de Ca2+
intracelulares parecem ser importantes (KOIKE et al., 1989). Quando uma
célula é exposta a um fator trófico, a interação vai levar à fosforilação do
domínio tirosina cinase intracelular desse receptor ou ao recrutamento de
proteínas acessórias que ativam vias enzimáticas fundamentais na
manutenção da sobrevivência e neuritogênese. Assim, o NGF induz tanto
crescimento neurítico quanto sobrevivência em neurônios simpáticos,
eventos disparados pela interação do ligante com seu receptor específico
(TrkA) e sua conseqüente auto-fosforilação. A ativação de TrkA resulta na
ativação de várias vias de sinalização, incluindo a da Ras, das cinases
ativadas por fatores mitogênicos (MAPKs, do inglês, Mitogen Activated

35
Protein Kinases) e Pi3C (KAPLAN e MILLER, 2000). A via Pi3C ativa a PKB, que
por sua vez fosforila diversos alvos responsáveis por manter a integridade, a
diferenciação, a neuritogênese e a proliferação celular (DATTA et al. 1999).
Subseqüentemente à ativação da MAPK por NGF, há a fosforilação das
cinases reguladas por fatores extracelulares (ERKs, do inglês Extracellular
Regulated Kinases) ERK1 (p42) e ERK2 (p44), que também fosforilam alvos
intracelulares (figura 3). Entretanto, essa via, diferentemente da AKT, não
parece ser essencial para a sobrevivência neuronal (VIRDEE e TOLKOVSKY,
1996; CROWDER e FREEMAN, 1998).
1.3 ESTRUTURA DA RETINA
A retina, devido às suas características anatômicas e histológicas, tem
sido amplamente utilizada nos estudos sobre o desenvolvimento e
funcionamento do SNC. Anatomicamente a retina é um tecido de fácil
acesso enquanto, histologicamente a retina é um tecido composto por tipos
celulares organizados de forma complexa constituindo micro-ambientes
caracterizados pela sinalização por diferentes neurotransmissores, ademais, a
retina é constituída por tipos celulares sobre os quais existe amplo
conhecimento morfológico e funcional, em todas as espécies. Essas
propriedades da retina fazem deste tecido um modelo rico para o estudo de
várias propriedades do SNC. Especialmente em embriões de pinto, a retina é
uma estrutura de fácil dissecação durante todo o desenvolvimento
embrionário.

36
A retina é formada durante o desenvolvimento a partir da
invaginação do prosencéfalo, que forma a vesícula óptica, posteriormente
dando origem ao cálice óptico. O cálice óptico apresenta em sua camada
mais externa o epitélio pigmentado e na parte mais interna o tecido neural.
O tecido retiniano é formado pela proliferação e diferenciação de
uma população de células denominadas neuroblastos. Estas células sofrem
várias divisões mitóticas até a diferenciação, dando origem aos diferentes
tipos celulares existentes na retina.
Em retina de pinto as primeiras células a se diferenciarem são as
ganglionares entre o 3o e o 7o dia embrionário. Ocorre em seguida a
diferenciação de células amácrinas, horizontais e fotorreceptores, sendo a
glia de Müller e as células bipolares as últimas a se diferenciarem (KANH,
1974; MISHIMA e FUJITA, 1978; PRADA et al., 1991; SNOW e ROBSON, 1994).
Após a etapa de saída do ciclo mitótico ocorre a migração radial formando
uma estrutura em camadas que é observada no tecido adulto. A retina é
organizada anatomicamente em cinco camadas (figura 5), dentre as quais
três são formadas por corpos celulares: nuclear externa (ONL, do inglês Outer
Nuclear Layer), nuclear interna (INL, do inglês Inner Nuclear Layer) e camada
de células ganglionares (GCL, do inglês Ganglion Cells Layer) e duas
camadas de plexos: plexiforme externa (OPL, do inglês Outer Plexiform Layer)
e plexiforme interna (IPL, do inglês Inner Plexiform Layer), formadas pelos
prolongamentos celulares, onde ocorrem contatos sinápticos.
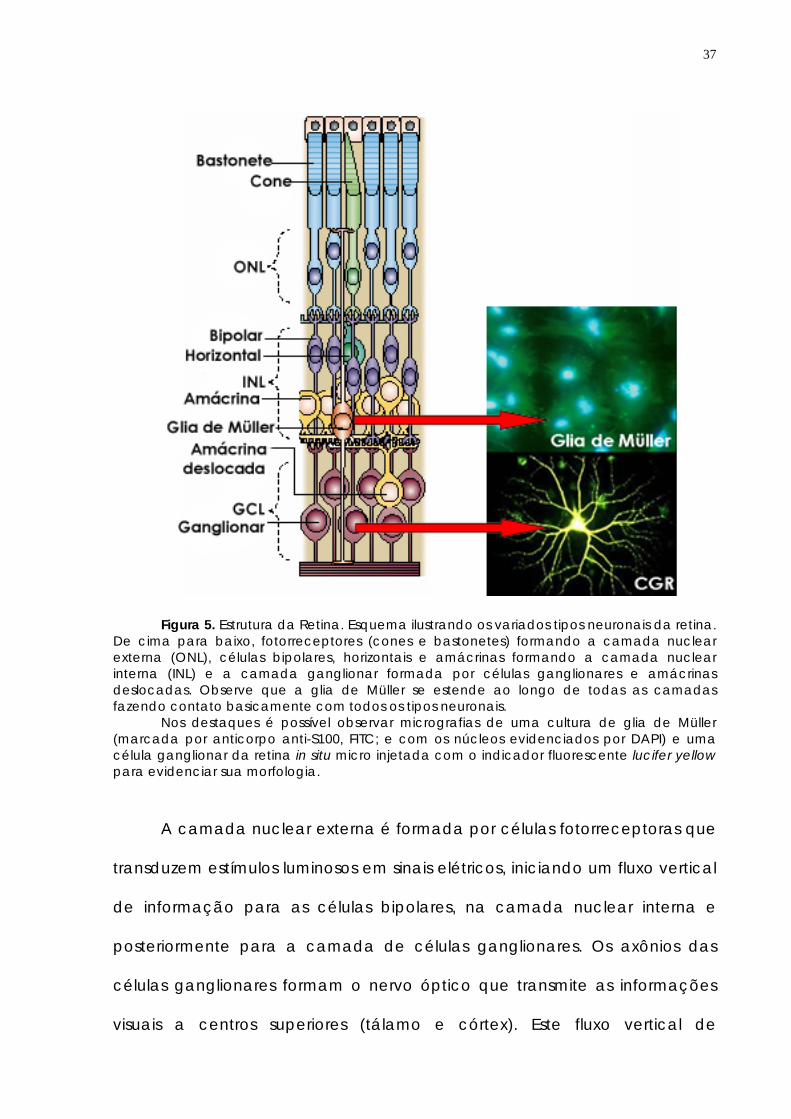
37
Figura 5. Estrutura da Retina. Esquema ilustrando os variados tipos neuronais da retina. De cima para baixo, fotorreceptores (cones e bastonetes) formando a camada nuclear externa (ONL), células bipolares, horizontais e amácrinas formando a camada nuclear interna (INL) e a camada ganglionar formada por células ganglionares e amácrinas deslocadas. Observe que a glia de Müller se estende ao longo de todas as camadas fazendo contato basicamente com todos os tipos neuronais.
Nos destaques é possível observar micrografias de uma cultura de glia de Müller (marcada por anticorpo anti-S100, FITC; e com os núcleos evidenciados por DAPI) e uma célula ganglionar da retina in situ micro injetada com o indicador fluorescente lucifer yellow para evidenciar sua morfologia.
A camada nuclear externa é formada por células fotorreceptoras que
transduzem estímulos luminosos em sinais elétricos, iniciando um fluxo vertical
de informação para as células bipolares, na camada nuclear interna e
posteriormente para a camada de células ganglionares. Os axônios das
células ganglionares formam o nervo óptico que transmite as informações
visuais a centros superiores (tálamo e córtex). Este fluxo vertical de

38
informação sofre processamento lateral, nas camadas plexiformes externa e
interna, por interneurônios que são respectivamente as células horizontais e
amácrinas. A glia de Müller se estende verticalmente na retina
estabelecendo contato com todas as camadas da retina.
A partir do 13o dia embrionário são estabelecidos os primeiros contatos
sinápticos na camada plexiforme interna e no 15o dia na camada plexiforme
externa (COULOMBRE, 1955). Estudos sobre o comprometimento fenotípico
dos precursores celulares retinianos têm tentado esclarecer os princípios
fundamentais que definem o “destino” final dessas células, estabelecendo
um modelo que combina componentes extrínsecos e intrínsecos no controle
da determinação fenotípica. No centro dessa proposta, encontram-se
progenitores capazes de dar origem um subconjunto limitado de tipos
celulares a partir de sinais extrínsecos (LIVESEY e CEPKO, 2001). Durante o
desenvolvimento da retina, seis tipos de neurônios e um tipo de célula glial
são gerados numa ordem geralmente conservada através de diversas
espécies de vertebrados estudadas – inicialmente são geradas as células
ganglionares da retina, seguidas pelos bastonestes e bipolares, enquanto a
glia de Müller esta entre os últimos tipos celulares a surgir na retina (CEPKO et
al., 1996). Uma série de estudos clássicos levou ao estabelecimento de
propriedades chave desse processo, como a multipotencialidade dos
progenitores e ao fato de que a determinação dos progenitores não é
restrita a um ou dois tipos celulares (TURNER et al., 1990; TURNER e CEPKO et
al., 1987).

39
1.3.1 CÉLULAS GANGLIONARES DA RETINA
A axotomia do nervo óptico tem sido usado na última década para
investigar os mecanismos da apoptose em CGRs (QUIGLEY et al., 1995). O
curso temporal da morte das CGRs é bem estabelecido in vivo em ratos
adultos (BERKELAAR et al., 1994; CORDEIRO et al., 2004). Trabalhos recentes
têm mostrado um curso apoptótico semelhante para culturas de explantes
de retina de camundongo adulto (McKERNAN et al., 2006)
As CGRs surgem na retina de camundongos entre os dias embrionários
E10 e E18, com uma taxa máxima de proliferação em E15 (ROHRER et al.,
2001). A morte das CGRs, por outro lado, ocorre em duas fases distintas. Uma
primeira ainda em fase embrionária, com um pico em E15.5 (FRADE e BARDE,
1999). Esse processo apoptótico é mediado por NGF que interagindo com
receptores p75 de baixa afinidade, expresso em altas quantidades na
camada de CGRs em desenvolvimento, leva a uma extensa degeneração
dessas células. Uma segunda onda de apoptose atinge as CGRs no período
neonatal, com um pico em P2 (YOUNG et al., 1984). Esse fenômeno se
correlaciona temporalmente com as projeções das CGRs para as áreas alvo
localizadas no colículo superior, com a elaboração dos contatos sinápticos e
a inervação apropriada. Acredita-se que quantidades limitantes de fatores
tróficos secretados por essas células alvo no colículo controlem essa fase
apoptótica. Quando BDNF ou NT4 é injetado no colículo superior de ratos ou
hamster observa-se uma redução na taxa de apoptose de CGRs durante
essa fase (CUI e HARVEY, 1995; MA et al., 1998). CGRs de animais adultos

40
também são reguladas por neurotrofinas, pois a apoptose induzida por
axotomia é retardada em ratos ou camundongos adultos tratados com
BDNF (MEY e THANOS, 1993; MANSOUR-ROBAEY et al., 1994). BDNF é também
um fator trófico para CGRs mantidas in vitro (MEYER-FRANKE et al., 1995), é
expresso nas camadas retino-recipientes do colículo superior (MA et al., 1998)
e é transportado retrógradamente (von BARTHELD et al., 1996).
1.3.2 FATORES TRÓFICOS RETINIANOS
Tem sido relatado que a retina secreta vários fatores tróficos que
influenciam a sobrevivência e a proliferação de células do sistema visual. A
retina de aves e mamíferos em desenvolvimento expressa neurotrofinas e
seus receptores em camadas e células específicas (LLAMOSAS et al., 1997;
BENNETT et al., 1999). Um trabalho pioneiro realizado por Araújo e Linden
(1993) mostrou que o meio condicionado (MC) por tecido retiniano de rato
apresentava atividade trófica que mantinha a viabilidade de CGRs de ratos
em cultura. Mais tarde, Ary-Pires e colaboradores (1997) mostraram que MC
de explantes e agregados retinianos de embriões de aves promovem
também a sobrevivência de CGRs de ratos dissociadas em cultura.
Trabalhos prévios haviam relatado que retinas de embriões de pinto
secretavam um fator capaz de manter neurônios colinérgicos em cultura.
Esse fator era capaz de induzir a atividade da enzima colina acetiltransferase
(ChAT), além de aumentar a expressão de receptores muscarínicos (De
MELLO et al., 1990). Este mesmo fator estava relacionado com a transcrição

41
de um receptor muscarínico (SKOURUPA e KLEIN, 1993) por células maduras
em cultura, mas não por culturas imaturas. Esse fator seria sensível à protease,
possui cerca de 30-50KDa, é regulado por desenvolvimento e induz a
transcrição seletiva do RNA para o receptor M2 muscarínico colinérgico além
do aumento da respectiva proteína (McKINNON et al., 1998). Além de
neurotrofinas, vários outros fatores com ação trófica no sistema nervoso têm
sido relatados na retina, tais como FGF, TGFβ, GPA, GDNF, IGF e neurocinas,
entre outras moléculas. Como cada um destes fatores tem diversos efeitos no
SNC e SNP, nosso objetivo a princípio foi caracterizar componentes do MC
de explantes de retina com atividade trófica em populações neuronais.
Com base nos dados acima introduzidos buscamos caracterizar o
efeito trófico do meio condicionado por tecido retiniano, identificar as
células responsáveis pela produção dos fatores responsáveis pela ação
trófica do MC, fracionar e, se possível, identificar fatores retinianos com
atividade trófica em populações neuronais diversas centrais e periféricas.

42
2. OBJETIVOS

43
1. Desenvolver culturas de explantes (simpático e GRD) ou purificadas de
neurônios periféricos (simpático) como biosensores da atividade trófica
dos meios condicionados (MCs) retinianos.
2. Caracterizar o efeito trófico de MCs sobre neurônios simpáticos de
pinto isolados em cultura identificando algumas das vias de sinalização
ativadas pelo conjunto de moléculas secretadas pelo tecido retiniano.
3. Purificar glia de Müller a partir de culturas de retina total (neurônios +
glia) de embriões de pinto e camundongo e investigar o efeito trófico
dos MCs por culturas purificadas de glia de Müller em neurônios
periféricos (simpático e GRD) e centrais (CGRs).
4. Purificar uma população de neurônios centrais, as CGRs, de
camundongo pelo método denominado de imunopurificação
(immunopanning - Barres et al., 1988) e investigar o efeito trófico do
MC por glia de Müller sobre a sobrevivência e a neuritogênese destas
células isoladas em cultura em diferentes substratos de adesão.
5. Fracionar e concentrar o MC por glia de Müller no intuito de isolar
frações que induzem sobrevida e/ou neuritogênese em neurônios
periféricos e centrais.

44
6. Estudar as propriedades regenerativas do MC por glia de Müller de
embrião de pinto em neurônios periféricas (GRD) e centrais
(motoneurônios) de ratos adultos num modelo de transecção do nervo
ciático (in vivo).

45
3. MATERIAIS E MÉTODOS

46
3.1 MATERIAIS E REAGENTES
Ácido ascórbico, fatores tróficos, anticorpos, Hoesch 33328, merosina,
poli-L-lisina, Dnase tipo I e II, Albumina bovina sérica (BSA, do inglês, Bovine
Serum Albumin; Sigma), laminina (Invitrogen, SP), tripsina, papaína e solução
de ovomucóide (Wortington Biochemical Corporation), Soro Fetal Bovino
(SFB) GIBCO ou do Biocampo. Fator de crescimento do nervo (NGF 2.5S;
Invitrogen, SP). Meio de cultura celular, meio basal de Eagle (BME do inglês,
Basal Medium Eagle) e Meio basal de Eagle modificado por Dulbecco
(DMEM do inglês, Dulbecco’s modified Eagle’s medium; Invitrogen, SP),
Cloridrato de Xilazina (Rumpom, Bayer), Cloridrato de Quetamina (Vetaset,
Fort Dodge Laboratories), bis-acrilamida, SDS, Tris, metanol. Placas de cultura
(24 poços, 4 poços, 35mm, 60mm ou 90mm Nunc ou Corning
preferencialmente). Filtros Centricon ® (Millipore Corporation). Solução de
Matrigel ® (Collaborative Biomedical Products). Para os experimentos de
imunopurificação utilizamos placas sem tratamento, seguindo a orientação
do protocolo descrito pelo Dr. Ben Barres (Universidade de Stanford, EUA). Os
demais sais utilizados na parte experimental desta tese, seja no Laboratório
de Neuroquímica, Instituto de Biofísica Carlos Chagas Filho (IBCCF), Instituto
de Bioquímica Medica (BioqMed) ou no Departamento de Anatomia
Humana e Genética, em Oxford, foram de pureza Analítica.

47
3.2 SOLUÇÕES
PBS completo (136 mM de cloreto de sódio, 2.7 mM de cloreto de
potássio, 8 mM de fosfato dibásico de sódio, 1.47 mM de fosfato monobásico
de potássio, 0.68 mM de Cloreto de Cálcio e 0.49 mM de Sulfato de
Magnésio, pH 7.4).
Solução de Ponceau S (0.1% ponceau, 5% ácido acético).
Solução salina livre de Ca2+ e Mg2+, CMF (131mM de cloreto de sódio,
4.09 mM de cloreto de potássio, 0.92 mM de fosfato dibásico de sódio, 0.45
mM de fosfato monobásico de potássio, 9.4 mM de bicarbonato de sódio e
12.2 mM de glicose).
TBS (Para 1L: 6,1g Tris, 9g NaCl, pH 7.6).
TTBS (TBS adicionado de 0,1% Tween-20).
Tampão de carregamento SDS 4x (0.25M Tris-HCl pH 6.8, 8% SDS, 30%
glicerol, 0.02% azul de bromofenol, 10% B-ME ou 0.3M DTT).
Tampão de corrida SDS (Para 1L: 3,03g de Tris base, 14,4g de glicina, 1g
de SDS).
Tampão de lise (50mM Tris-HCl Ph 7,4, 1mM EDTA, 60mM pirofosfato de
Na+).
Tampão de transferência (Para 1L: 25mM Tris pH8.5, 0,2M glicina, 20%
metanol. Aferir volume com água destilada).

48
3.3 PREPARAÇÃO DAS PLACAS DE CULTURA
Para melhor adesão dos neurônios ao plástico das placas de cultura, a
superfície das placas foi incubada com solução de poli-L-lisina 10µg/ml por
aproximadamente 12 horas (overnight). Após o período de incubação, as
placas foram lavadas por três vezes com água Milli-Q estéril e
completamente secas dentro da capela do fluxo laminar por 30 minutos . As
placas então receberam solução de laminina (20 µg/mL) em meio DMEM por
pelo menos 2 horas em estufa úmida com 5% CO2. A partir deste ponto as
placas estavam prontas para receber as células. O excesso de solução de
laminina foi aspirado pouco antes do plaqueamento das células.
3.4 OBTENÇÃO E CULTURA DOS NEURÔNIOS PERIFÉRICOS
Ovos embrionados de galinha White Leghorn estagiados segundo
Hamburger e Hamilton (1951) foram obtidos de uma granja local (Tolomei) e
mantidos em estufa à 38ºC em atmosfera úmida. Embriões de 10 dias (E10)
foram sacrificados por decapitação para dissecação do GRD, ou da porção
lombar para-vertebral da cadeia ganglionar do simpático. Para evidenciar a
cadeia ganglionar do simpático o embrião foi aberto ventralmente e
eviscerado tomando-se cuidado para não danificar as estruturas mais
profundas onde se encontram os gânglios (figura 6). A dissecação dos
gânglios foi feita com agulhas de tungstênio afinadas por eletrólise (Davies,
1989). A dissecação foi inteiramente realizada sob lupa em placa de Petri

49
com o embrião imerso em solução salina livre de Ca2+ e Mg2+ (CMF). Os
gânglios isolados e livres de tecidos contaminantes foram postos em CMF
adicionado de tripsina para uma concentração final de 0,05% e incubados à
37ºC por 12 minutos para posterior dissociação das células. Após
tripsinização, os gânglios foram lavados uma vez com DMEM + 10% SFB e três
vezes com CMF, centrifugados e dissociados mecanicamente em
aproximadamente 1,5mL de CMF com pipeta Pasteur de ponta fina.
Figura 6. Esquema ilustrativo da localização anatômica da cadeia ganglionar do
simpático (em vermelho) e do gânglio da raiz dorsal (bulbos para-vertebrais) em embrião de pinto.
A suspensão de células foi cuidadosamente aplicada em uma coluna
de sedimentação (figura 7) preenchida com 60ml de DMEM + 10% SFB
previamente preparada e mantida à 4ºC por pelo menos 12 horas. As células

50
permaneceram na coluna por 1 hora. O propósito desta coluna é separar
por sedimentação diferencial, baseado na gravidade e complexidade dos
corpos celulares, neurônios íntegros de células não neuronais e restos
celulares gerados no processo de dissociação mecânica (pedaços de
membrana), além de pedaços de tecido mal dissociados. A viscosidade do
DMEM + 10% SFB mantido em baixa temperatura permite que os neurônios,
mais pesados que as células satélites e restos celulares, atravessem um
caminho mais longo na coluna e sejam encontrados em frações mais baixas
que as células menores após uma hora. Da coluna foram coletadas 12
frações de aproximadamente 5mL, que foram analisadas sob microscopia
ótica para confirmação da eficiência da separação das células e
identificação das alíquotas enriquecidas em neurônios (figura 7). As alíquotas
selecionadas foram misturadas e, se necessário, diluídas para uma
concentração de 600 a 1000 células/mL. As células foram imediatamente
plaqueadas nas placas previamente preparadas para cada condição
experimental. Os MCs foram utilizados na proporção de 1:2 em relação ao
DMEM (meio controle). A adição de NGF 2.5S na concentração de 10-20
ng/mL à cultura manteve a viabilidade de aproximadamente 90-95% dos
neurônios viáveis após 48 horas e foi usado como controle positivo neste tipo
de cultura. Culturas de neurônios mantidas apenas com DMEM + 10% SFB
apresentaram menos que 5% de sobrevivência neuronal após 48 horas sendo
esta condição utilizada como nosso controle negativo. Pelo menos quatro
experimentos independentes em duplicata foram feitos para cada condição
experimental.

51
Figura 7. Ilustração da coluna de sedimentação e exemplo fotográfico das frações obtidas após 1 hora de sedimentação da suspensão de células. As micrografias ilustram, no alto, restos celulares e pequenas partículas; no meio frações de interesse onde os neurônios estão contidos (observar a existência de prolongamento em alguns corpos celulares) e em baixo, pedaços de tecido mal dissociado.
Os gânglios utilizados no desenvolvimento desta tese ou foram
utilizados na forma dissociada em células ou na forma de explantes, que
foram que foram cultivados sobre os mesmo substratos de adesão utilizados
para células isoladas (poli-L-lisina e laminina) para os ensaios de
neuritogênese apresentados neste trabalho.

52
Retinas de camundongo também foram utilizadas na forma de
explantes para ensaios de neuritogênese. Para tanto camundongos P1-P8
foram mortos por decapitação, tiveram os olhos removidos e enucleados
para remoção da retina. A retina destes animais foi cortada em explantes de
aproximadamente 1mm2 e cultivadas sobre o mesmo substrato de adesão
utilizado para células isoladas.
3.5 CONTAGEM DE CÉLULAS
A quantificação da sobrevivência neuronal foi feita por contagem
visual de neurônios aderidos numa placa quadriculada, com soma íntegro e
liso e neuritos projetados (pelo menos 2 vezes maior que o soma) e aderidos
em uma cultura de neurônios de baixa densidade celular (600-1000
células/placa de 35mm dispersos numa área de 12x12 mm2). Foram feitas
duas contagens por placa, a primeira feita cerca de 4h após plaqueamento,
quando os neurônios já estavam aderidos (é importante mencionar que na
primeira contagem ainda não era possível observar projeção neurítica
devido à ação da tripsina na dissociação do tecido). Esta primeira
contagem foi a nossa referência do total de neurônios aderidos (100%). A
segunda contagem foi feita 48 horas após o plaqueamento e revelou a
percentagem de sobrevivência dos neurônios plaqueados. Os resultados
foram expressos em percentagem do total de células plaqueado.

53
3.6 MEIO CONDICIONADO POR EXPLANTES DE RETINA
Embriões de pinto de 9 dias (E9) foram decapitados, seus globos
oculares foram removidos, e imediatamente transferidos para uma placa de
Petri com CMF. O cristalino e o humor vítreo foram removidos e descartados
e a retina foi dissecada livre de epitélio pigmentar. A retina foi cortada em
explantes de aproximadamente 1mm2 que eram mantidos em frasco
Erlenmeyer em meio de cultura Meio Basal de Eagle (BME) + 10% de SFB por 4
dias em agitador orbital (70 r.p.m.) à 37ºC em uma atmosfera de 5% de CO2
(figura 8). Após quatro dias de condicionamento o meio foi coletado, filtrado
estéril em filtro Millipore ® 0,22µm, adicionado com SFB (5%), e armazenado
em freezer à -70ºC para posterior utilização.
Figura 8. Esquema do preparo do RCM. A retina é dissecada e cortada em explantes. Os explantes permanecem condicionando o meio por 4 dias em agitador orbital. Após condicionamento o meio é filtrado e armazenado a -70oC para posterior análise.

54
3.7 MEIO CONDICIONADO POR GLIA DE MÜLLER DE CAMUNDONGO NEONATO
Camundongos SV129 do biotério do Departamento de Anatomia
Humana e Genética (Oxford, UK) eram acasalados (1 macho jovem com
pelo menos 8 fêmeas) que forneciam ninhadas semanalmente de 3-12
animais, que eram usados nos estágios determinados.
Camundongos P0-P2 foram sacrificados com dose letal de pentobarbital
injetado intraperitonealmente. Após a morte os animais foram decapitados e
tiveram os olhos removidos e enucleados sob lupa tendo a retina removida
livre de epitélio pigmentado e humor vítreo. Inicialmente, foram feitas
culturas diluídas (baixa densidade celular) de retina total que
permaneceram em estufa úmida a 37oC, até que a glia atingisse
confluência. Após confluência, estas culturas foram submetidas a um
tratamento com 4mM de ácido ascórbico (concentração final) por
aproximadamente 3 horas sendo observadas em intervalos de 30 minutos
para monitoração da morte neuronal. Após lesão dos neurônios, a cultura foi
lavada uma vez com CMF e meio de cultura neurobasal (NB) foi adicionado
com suplemento B27 (Gibco) à cultura. No dia seguinte a cultura foi lavada
mais uma vez com CMF e teve NB + B27 restabelecido. A partir deste ponto o
que temos é uma monocamada de glia de Müller isenta de neurônios (figura
9). O sobrenadante desta cultura passou a ser coletado a cada 48 horas
resultando em no máximo 3 coletas. O meio condicionado por estas culturas
foi coletado, filtrado estéril e armazenado em alíquotas de 50mL em freezer

55
-70oC para posterior utilização. O MC obtido de culturas de glia de Müller de
camundongo foi denominado MMG (do inglês, Mice Müller Glia).
Figura 9. Esquema de preparo do AMG ou MMG. As retinas são dissecadas, tripsinizadas, dissociadas e plaqueadas. Quando a cultura de retina total atinge confluência (A, C7) é tratada com solução de ácido ascórbico 4mM por 3 horas(B, C8). Após lesão dos neurônios e lavagens da cultura temos uma monocada de glia de Müller purificada (C).
Eventualmente algumas culturas apresentavam contaminação por
microglia o que veio a ser evidenciado por marcação com isolectina B4 (um
marcador isolado da planta Griffonia simplicifolia que se liga a
glicoconjugados, que possuem terminal alfa-D-galactose, presentes na
superfície da microglia e macrófagos) conjugada a fluoresceína. A
contaminação veio a ser contornada através de várias lavagens
consecutivas ou replaqueamento isto é, tripsinização e dissociação de uma

56
cultura de monocamada de glia de Müller contaminada com microglia e
plaqueá-las novamente em nova placa.
3.8 MEIO CONDICIONADO POR GLIA DE MÜLLER DE EMBRIÕES DE GALINHA
Inicialmente, foram desenvolvidas culturas primárias de retina total
(neurônios e glia). Para tanto, retinas de embrião de galinha E9 foram
dissecadas livre de epitélio pigmentar em CMF, transferidas para tubo de
ensaio e tripsinizadas por 12 minutos a 37oC, dissociadas mecanicamente e
plaqueadas na proporção de 8 retinas E9 para 20 placas de 100mm (cerca
de 20 milhões de células/placa de 100mm). A cultura foi mantida em estufa
úmida em 5% CO2 em 8ml de DMEM + 10% SFB. A primeira troca de meio foi
feita no dia seguinte ao plaqueamento (E9C1, idade embrionária de 9 dias,
um dia de cultura) para remover as células não aderidas ou mortas, além de
restos celulares oriundos da dissociação mecânica do tecido. A partir deste
ponto foi feita uma troca de meio a cada 48 horas, até a obtenção de
confluência pelas culturas (uma semana em média). Após a confluência, os
neurônios foram eliminados com a adição de 4mM de ácido ascórbico
(concentração final). Após aplicação do ácido, as culturas foram
observadas em intervalos de aproximadamente uma hora para
acompanhamento da morte neuronal. O tratamento foi de 3 horas, tempo
suficiente para lesar os neurônios em sua totalidade. As culturas foram
lavadas uma vez com DMEM, eliminando os resíduos de ácido ascórbico e os
restos de membrana dos neurônios lesados, e incubada com DMEM + 10%

57
SFB por pelo menos 12 horas (overnight) à 37oC em estufa úmida de CO2. No
dia seguinte, a monocamada de glia de Müller foi lavada por duas vezes
com DMEM sem SFB e com a cultura livre de neurônios e soro, restituiu-se 8ml
de DMEM sem SFB à cultura que foi mantida em estufa (figura 9). O MC pela
glia foi coletado a cada intervalo de 48 horas. Este meio foi denominado
“meio condicionado sem soro” (MC-SFB). Foram feitas pelo menos duas
coletas de MC por cultura. O meio foi filtrado estéril, aliquotado e
armazenado em freezer a -70ºC para posterior utilização. Meio condicionado
com soro (MC+SFB) também foi obtido da mesma forma, porém restitui-se
meio com soro à cultura após eliminação dos neurônios com ácido
ascórbico.
3.9 MEIO CONDICIONADO RADIOATIVO
O MC radioativo a partir de culturas purificadas de glia de Müller
obtidas como descrito acima. A cultura sem soro foi incubada com DMEM
livre de metionina (Gibco) adicionado de metionina radioativa (Met-S35). A
cultura foi mantida em estufa a 37oC e o MC radioativo coletado após 48
horas de incubação.
Culturas de neurônios isolados do GSC de rato tratadas com MC
radioativo interagem e internalizam moléculas radioativas que podem nos
auxiliar na identificação das moléculas tróficas produzidas pela glia de
Müller, já que grande parte dos receptores contendo tirosina cinase é
internalizada com seus ligantes (Hendry et al., 1975). Com base nesta
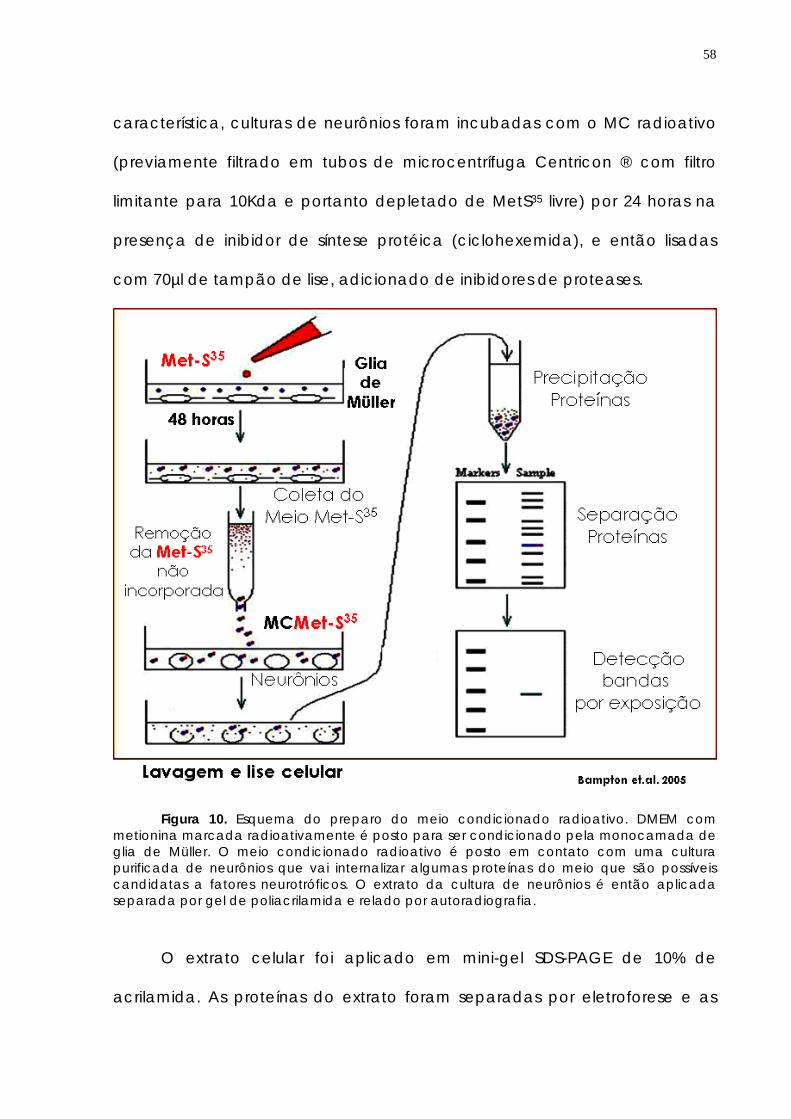
58
característica, culturas de neurônios foram incubadas com o MC radioativo
(previamente filtrado em tubos de microcentrífuga Centricon ® com filtro
limitante para 10Kda e portanto depletado de MetS35 livre) por 24 horas na
presença de inibidor de síntese protéica (ciclohexemida), e então lisadas
com 70µl de tampão de lise, adicionado de inibidores de proteases.
Figura 10. Esquema do preparo do meio condicionado radioativo. DMEM com metionina marcada radioativamente é posto para ser condicionado pela monocamada de glia de Müller. O meio condicionado radioativo é posto em contato com uma cultura purificada de neurônios que vai internalizar algumas proteínas do meio que são possíveis candidatas a fatores neurotróficos. O extrato da cultura de neurônios é então aplicada separada por gel de poliacrilamida e relado por autoradiografia.
O extrato celular foi aplicado em mini-gel SDS-PAGE de 10% de
acrilamida. As proteínas do extrato foram separadas por eletroforese e as

59
bandas radioativas evidenciadas por auto-radiografia do gel. Como não
havia Met-S35 livre no MC apresentado à cultura de neurônios e como estes
se encontravam na presença de um inibidor de síntese protéica, as bandas
radioativas presentes no lisado da cultura de neurônios são provenientes da
glia que incorporou Met-S35 e produziu proteínas marcadas tanto
constitutivas quanto secretadas no MC (figura 10).
3.10 IMUNOPURIFICAÇÃO DE CGR DE CAMUNDONGO
O método de purificação de CGRs foi elaborado originalmente para
imunopurificação de CGRs de rato utilizando-se do anticorpo Thy1.1 por
Barres e colaboradores (1988). Nesse trabalho, adaptamos o método
utilizando o anticorpo Thy1.2 (BD Biosciences Pharmingen, Cat# 553008) que
permite a purificação de CGRs de camundongo. Placas de petri de plástico
foram incubadas com solução do anticorpo primário purificado (anti-IgG de
rato, 10µg/ml) em Tris 50mM pH9.5 por pelo menos 12h (overnight) à 4oC e
então lavadas três vezes com solução salina tamponada de fosfato (PBS) em
pH 7.2. De forma a evitar a interação inespecífica de células ao plástico, as
placas foram incubadas à 37oC em solução de bloqueio (PBS + 0,2%
albumina sérica bovina, BSA) por 30 minutos antes de receber o anticorpo
primário. Após bloqueio, as placas foram novamente lavadas três vezes com
PBS e incubadas com solução de anticorpo monoclonal Thy 1.2 (ou anti-
CD90.2, denominação alternativa) sem azida e baixo nível de endotoxina
(NA/LE do inglês, No Azide and Low Endotoxin; clone 30-H12, isotipo de rato,

60
IgG2b, kappa) por 3 horas a 37oC. Após incubação, a placa foi novamente
lavada três vezes com PBS. Para cada experimento, cerca de 10 retinas de
camundongos P1 (1 dia pós-natal) foram dissecadas, lavadas com CMF e
então incubadas em tubo de microcentrífuga (1,5 mL, eppendorf) com
solução de papaína (20 unidades/mL) por 15 minutos a 37oC. Após
incubação, a solução de papaína foi aspirada e as retinas lavadas em 500µl
de solução de ovomucóide (inibidor da papaína). Após a lavagem, o
sobrenadante foi removido e 250µl de ovomucóide + 0,4% dnase (tipos I e II)
foram adicionados às retinas, que foram imediatamente dissociadas
mecanicamente com micropipeta. A suspensão de células foi centrifugada
a 2000 r.p.m. em microcentrífuga por 5 minutos. O sobrenadante foi
descartado e as células foram novamente suspensas em 100µl de
ovomucóide. Adicionou-se 1400µl de meio de cultura aos 100µl de suspensão
de células em ovomucóide. O PBS da placa de petri previamente tratada
com anticorpos foi removido e transferiu-se a suspensão de células (1500µl)
para a placa. A placa foi mantida por 20-30 minutos a 37oC em estufa úmida
de CO2. Durante o período de incubação das células, a placa foi levemente
agitada a cada 5-6 minutos e observada em microscópio ótico para
visualização das células aderidas (figura 11). A placa com as células foi
lavada de 6 a 8 vezes com meio de cultura até que somente as células
aderidas aos anticorpos, as CGRs, permanecessem na placa (figura 12). As
CGRs foram removidas da placa com tratamento brando de tripsina 0,01%
por 5-8 minutos a 37oC e lavagem vigorosa com meio de cultura. Todo o
meio foi coletado em tubo de 15 mL e centrifugado para recuperar as CGRs

61
isoladas. A partir deste ponto as CGRs (cerca de 5000) foram plaqueadas no
substrato de adesão apropriado (laminina, merosina, lamina basal ou
monocamada de glia de Müller).
A sobrevida das CGRs foi quantificada através de contagem visual de
culturas de baixa densidade após 48, 72 ou 96 horas (dependendo do
experimento) observadas sob microscópio de contraste de fase ou por
imagens de fluorescência. Utilizamos a molécula Hoesch 33328, que tem
afinidade por DNA e evidencia o aspecto nuclear mostrando se a célula
está viável ou apoptótica com base na visualização da fragmentação do
núcleo, assim como a molécula Diacetato de Fluoresceína (DFA), que é um
éster e marca células viáveis, além de anticorpos específicos para
marcadores gliais ou neuronais.
Figura 11. Ilustração esquemática do processo de imuno-seleção de células ganglionares da retina (CGRs). Esta técnica permite isolar um único tipo celular a partir da

62
suspensão de células de um tecido com base no reconhecimento, por meio de anticorpos, de epitopos específicos de superfície de determinados tipos celulares.
Neurobasal (NB) suplementado com B27 foi utilizado como controle
negativo e NB + B27 adicionado dos fatores BDNF (50ng/mL) + CNTF
(50ng/mL)+ forscolina (F - 5µM) (BCF) foi o controle positivo que manteve até
60 % das CGRs viáveis após 48 horas.
As micrografias de contraste de fase e fluorescência foram obtidas
com o sistema Axiophoto da Zeiss.
Figura 12. Imuno-seleção de células ganglionares da retina (CGRs). As micrografias ilustram uma suspensão de células total da retina (A) e CGRs purificadas aderidas à placa após várias lavagens (B). 3.11 FRACIONAMENTO DO MEIO CONDICIONADO POR GLIA DE MÜLLER EM
COLUNA DE HEPARINA
No fracionamento utilizamos lotes de MC sem soro devido à grande
quantidade de proteínas presentes no soro. Cinqüenta mililitros de MC
injetados manualmente com seringa em uma coluna de heparina (Hi-Trap
Amersham-Biosciences, 5mL). O MC que passa através da coluna,

63
denominado fração livre (FF, free fraction), foi coletado para posterior
análise. Antes de começar a eluição da fração ligada à coluna (BF, bound
fraction) foi necessário primeiro remover as moléculas de eluição lenta, além
do indicador vermelho de fenol presente no MC contido na coluna, para
que a leitura à 214nm volte à linha de base equivalente a leitura do tampão
fosfato utilizado como padrão. Para tanto a coluna foi conectada a um
sistema Cromatografia Rápida de Proteínas (FPLC do inglês, Fast Protein
Liquid Chromatography) e lavada por 1 hora com tampão fosfato 50mM, pH
7,2 com fluxo de 2mL/ minuto. O sistema FPLC é um sistema de bombas que
permite gerar gradientes de acordo com a programação desejada e com a
concentração dos tampões ou solventes carregados em cada uma das
bombas. Após lavagem, uma alíquota do tampão que sai da coluna foi
comparada com uma alíquota do mesmo tampão estoque quanto à
absorbância à 214nm. A leitura de ambas as alíquotas após lavagem de 1
hora deve ser a mesma confirmando que a coluna está livre de moléculas
de eluição lenta e somente moléculas aderidas a matriz de heparina
permaneceram na coluna para posterior eluição com gradiente de NaCl.
Com a coluna lavada, foi aplicado um gradiente de NaCl à coluna visando
eluir a BF. Utilizamos tampão fosfato 50mM, pH 7.2 na bomba A e o mesmo
tampão adicionado de 2M de NaCl na bomba B. O sistema foi programado
para estabelecer um gradiente linear de 0 a 1,5 M de NaCl em 40 minutos
com fluxo de 2mL/minuto. Ao atingir 1,5mM, o programa alternou para
bomba B (tampão fosfato + 2M NaCl) e lavou a coluna por 15 minutos. Para
fim de manutenção da coluna após fracionamento do MC a coluna foi

64
lavada com 30 mL de tampão fosfato 50mM, 30 mL de água Milli-Q e 30 mL
de solução de etanol 20% seqüencialmente. A coluna foi mantida em
solução de etanol 20% para uso posterior.
O coletor automático do sistema foi programado para coletar frações
de 2mL. A absorvância de cada alíquota foi medida a 214nm tendo o
tampão fosfato como parâmetro (branco). As alíquotas de interesse foram
dessalinizadas e concentradas em filtro Centricon ® de membrana de 10kDa
e testadas quanto a sua atividade biológica em culturas de neurônios
isolados da cadeia ganglionar do simpático (figura 13). Amostras das
alíquotas com atividade em cultura foram submetidas à eletroforese em mini
gel SDS-PAGE de concentração de 10% de poliacrilamida.
Figura 13. Esquema de fracionamento do AMG. Após lavagem a coluna é eluida com um gradiente de NaCl e as amostras coletadas a cada 1,5mL, dessalinizadas e testadas em culturas purificadas de neurônios para identificar as alíquotas com atividade trófica.
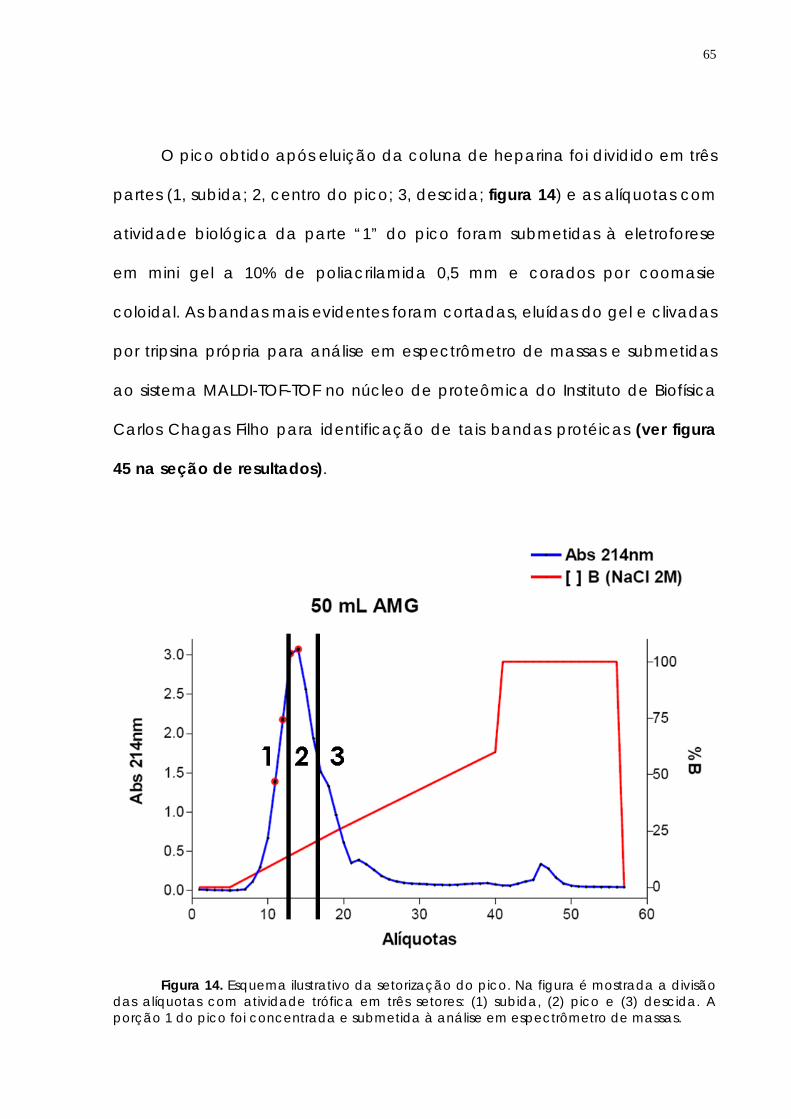
65
O pico obtido após eluição da coluna de heparina foi dividido em três
partes (1, subida; 2, centro do pico; 3, descida; figura 14) e as alíquotas com
atividade biológica da parte “1” do pico foram submetidas à eletroforese
em mini gel a 10% de poliacrilamida 0,5 mm e corados por coomasie
coloidal. As bandas mais evidentes foram cortadas, eluídas do gel e clivadas
por tripsina própria para análise em espectrômetro de massas e submetidas
ao sistema MALDI-TOF-TOF no núcleo de proteômica do Instituto de Biofísica
Carlos Chagas Filho para identificação de tais bandas protéicas (ver figura
45 na seção de resultados).
Figura 14. Esquema ilustrativo da setorização do pico. Na figura é mostrada a divisão das alíquotas com atividade trófica em três setores: (1) subida, (2) pico e (3) descida. A porção 1 do pico foi concentrada e submetida à análise em espectrômetro de massas.

66
3.12 ENSAIO DE REGENERAÇÃO in vivo
Ratos machos adultos (idade em torno de 3 meses) foram utilizados
nos experimentos de transecção e reconexão do nervo ciático. Os animais
foram anestesiados com éter, seguida de profunda anestesia intraperitoneal
com cloridrato de xilazina 0,5% e cloridrato de quetamina 5%, utilizando-se
100μL de cada um dos anestésicos para cada 100g de peso do animal. O
tempo de anestesia variou entre 20 a 30 minutos, não havendo necessidade
de reaplicação.
O nervo ciático foi exposto através de incisão realizada na coxa
direita. O acesso ao nervo foi feito através do afastamento da musculatura
da coxa, atingindo-se então a fossa poplítea, por onde o nervo foi projetado
em direção aos músculos da perna (solear, gastrocnêmio lateral e tibial
anterior). Foi feita a completa transecção deste nervo à 1cm de sua região
de bifurcação. Antes do início do procedimento cirúrgico, um segmento de
tubo de polietileno com 7mm foi preparado de forma que através desse
cilindro tínhamos o cruzamento de dois segmentos de linha de algodão
comum. Antes da transecção, as linhas foram levemente amarradas ao
nervo. Um espaço de aproximadamente 4mm foi deixado entre as amarras e
a transecção foi feita neste espaço com auxílio de um bisturi. Imediatamente
após a transecção, o nervo foi conectado ao tubo de tal maneira que
aproximadamente 2mm do coto proximal e 1mm do coto distal foram
inseridos no tubo. Ao final do procedimento, tínhamos um espaço de
aproximadamente 4mm entre os cotos distal e proximal. Para cada

67
experimento, dois grupos experimentais foram utilizados e cada grupo foi
composto por 3-4 animais.
No grupo controle, o espaço entre os cotos proximal e distal no interior
do tubo de polietileno foi preenchido com solução de Matrigel ® em DMEM.
O volume total injetado por animal foi cerca de 15μL. No segundo grupo de
animais foi injetada solução de Matrigel ® contendo MC total sem soro
(AMG) na proporção de 1:3. As diferentes condições foram injetadas no
interior dos tubos utilizando-se uma microseringa Hamilton com agulha sem
bisel (figura 15). Estabelecidas as condições, os nervos conectados através
do tubo foram acondicionados no interior da fossa poplítea e a incisão
costurada com fio de sutura não absorvível (Clone I Nylon 5-0, Shalon
Suturas).
Figura 15. Esquema ilustrativo da secção do ciático. Após o corte as extremidades do ciático são conectadas a um tubo de polietileno. Dentro do tubo são injetadas, cuidadosamente, as condições experimentais. No nosso caso, AMG, meio controle negativo ou positivo.

68
Logo após o procedimento cirúrgico foi possível observar severo
comprometimento dos movimentos da pata cujo nervo foi seccionado.
Figura 16. Localização do cristal DiI. O cristal de DiI é posto a 4mm do coto distal. Somente os axônios que regenerarem vão transportar partículas de DiI para o soma marcando a célula que regenerou.
Quarenta dias após o procedimento de secção do nervo ciático os
animais foram novamente abertos para receber um cristal de DiI a 2mm do
coto distal (figura 16). Os terminais provenientes da medula espinhal ou do
GRD, cruzaram os 4mm entre os cotos distal e proximal e adentraram no
coto distal atingindo o cristal de DiI, que foi transportado retrogradamente
na forma de partículas para os corpos celulares, marcando as células que
regeneraram após o corte. Após inserção do cristal, os animais foram
novamente suturados e devolvidos ao biotério. Dois dias após inserção do
DiI, os animais foram sacrificados para remoção do ciático, GRDs (na altura
de L4 e L5) e cortes de medula espinhal para análise dos motoneurônios que
foram mantidos em solução de 4% paraformaldeído por pelo menos 24

69
horas. Os tecidos foram incluídos em blocos de parafina para corte em
micrótomo. No micrótomo foram feitos cortes de 12μm que foram montados
em lamínulas pré-tratadas com poli-L-lisina e observados sob microscópio de
fluorescência para identificação e contagem das células positivas para DiI.
3.13 IMUNOCITOQUÍMICA
As culturas submetidas à marcação imunocitoquímica foram
reproduzidas em escala reduzida em placas de quatro poços (10mm cada
poço, volume de 0,5mL).
Foram feitas três lavagens de 5 minutos em cada placa com PBS
completo (todas as lavagens foram de 5 minutos). As culturas foram fixadas
em metanol absoluto por 5 minutos. Para remoção do excesso de metanol
as placas foram novamente lavadas três vezes com PBS. Em cada poço foi
adicionado 200-300 µL de soluções de anticorpos primários preparados com
3% de BSA adicionado de 0,01% Tween 20 em PBS. As placas foram
incubadas na geladeira sob constante agitação por 12 horas. Antes de
incubar as culturas com os anticorpos secundários apropriados produzidos
em camundongo (Ms) ou coelho (Rb) (IgG -FITC ou IgG-TRITC), as culturas
foram lavadas três vezes com PBS e em seguida incubadas com soluções de
anticorpos secundários, preparadas em PBS, por um período de 2 horas em
uma estufa úmida a 37°C, 5% de CO2.

70
Passado o período de incubação as culturas foram novamente
lavadas três vezes com PBS e mantidas em 500µL de PBS para observação
em microscópio de fluorescência.
3.14 IMUNODETECÇÃO DE PROTEÍNAS (WESTERN BLOT)
Neurônios do GSC de ratos Wistar cultivados sob diferentes condições
experimentais (CNTF, 50ng/mL; NGF 20ng/mL ou MMG diluído 1:2) em placas
pré-tratadas com substrato de adesão apropriado (poli-L-lisina + laminina),
ou mantidos em suspensão em tubos eppendorf, foram lisados com tampão
de lise adicionado de inibidores de proteases e fosfatases (NP-40 1%;
Deoxicolato de sódio 0,25%; NaCl 15mM; ; PMSF 1 mM; aprotinina 5µg/mL;
pepstatina 5µg/mL; leupeptina 5µg/mL; NaF 1mM; Na3VO4 1mM) e
coletados. As concentrações de proteína de cada amostra foram
determinadas pelo método de Lowry (BioRad), utilizando albumina sérica
bovina para construção da curva padrão. As amostras de cada condição
foram diluídas em tampão de carregamento para uma concentração final
de 2µg/µL de proteína, fervidas por 3 minutos e submetidas à eletroforese em
gel de poliacrilamida (SDS-PAGE) numa concentração de 10% (p/v)
recebendo 15µL em cada poço. As proteínas do gel foram transferidas para
uma membrana de nitrocelulose eletroforeticamente em cuba de
transferência de imersão (BioRad) por 12 horas a 20V em câmara fria (4oC). A
membrana de nitrocelulose foi incubada em solução de vermelho de
Ponceau para verificar a eficiência da transferência e logo após lavada

71
com água deionizada. A membrana foi então incubada por 12 horas a 4oC
com anticorpos primários monoclonais anti-rato produzidos em camundongo
e cabra específicos contra fosfo-STAT3 (p-STAT3), fosfo-AKT (p-AKT), fosfo-ERKs
(p-ERKs) e contra STAT3 total (controle de carregamento do gel) e
bloqueada por 5 minutos com PBS 1% caseína. A incubação com os
anticorpos secundários foi de uma hora a temperatura ambiente após
lavagem com solução tamponada de Tris adicionada de 0,1% Tween (TTBS
do inglês, Tween-Tris Buffered Saline). Os anticorpos secundários eram
conjugados a fluoróforos (FITC ou TRITC) e o resultado foi analisado sob
detector de quimioluminescência (Sistema Odyssey – Li-Cor Biosciences) de
acordo com instruções do fabricante.
3.15 ANÁLISE ESTATÍSTICA
Desde que não especificado, a maioria dos resultados foram expressos
como média ± erro padrão da média (e.p.m.) de pelo menos três
experimentos independentes realizados em triplicata. A análise estatística foi
feita através do teste t de Student para dois grupos ou análise de variância
(ANOVA) seguido do teste de comparação múltipla de Bonferroni para mais
de três grupos com uma significância estatística de p<0,05.

72
4. RESULTADOS

73
4.1 RCM TEM EFEITO TRÓFICO EM NEURÔNIOS SIMPÁTICOS
É fato que a retina produz fatores diversos com atividade neurotrófica.
Em um estudo pioneiro realizado por Araújo e Linden (1993) foi mostrado que
RCM de rato fornecia suporte trófico à células ganglionares da retina de
ratos. Com base na diversidade de fatores produzida pela retina já descrita
na literatura, investigamos, inicialmente a possibilidade de RCM de embriões
de pinto E9 exercer efeito trófico em neurônios isolados do gânglio simpático
de embriões de pinto E10, neurônios do SNP. Nesta idade embrionária estes
neurônios ainda são neurotrofina dependentes sendo, portanto, um ótimo
modelo para detecção do efeito trófico promovido por RCM. A
sobrevivência induzida por RCM em culturas de neurônios isolados se mostrou
bastante potente sendo equivalente ao controle positivo para este tipo
celular. Cerca de 95% dos neurônios simpáticos entram em apoptose nas
primeiras 48 horas se cultivados na ausência de NGF (figura 17A). Em
contraste, NGF 10ng/mL mantém a viabilidade celular em torno de 80% na
mesma escala de temporal (figura 17B). RCM condicionado na ausência de
soro fetal bovino manteve a sobrevivência de neurônios simpáticos em torno
de 50% (n=5) (figura 17C) enquanto quando condicionado na presença de
5% de SFB e utilizado numa diluição de 1:2 com DMEM (1 parte MC para 2
volume final) manteve a viabilidade celular próxima de 90% (n=30) (figura
17D).
Desta forma, nesta primeira parte do trabalho, todos os experimentos
foram realizados com meio condicionado por retina de embrião de pinto na
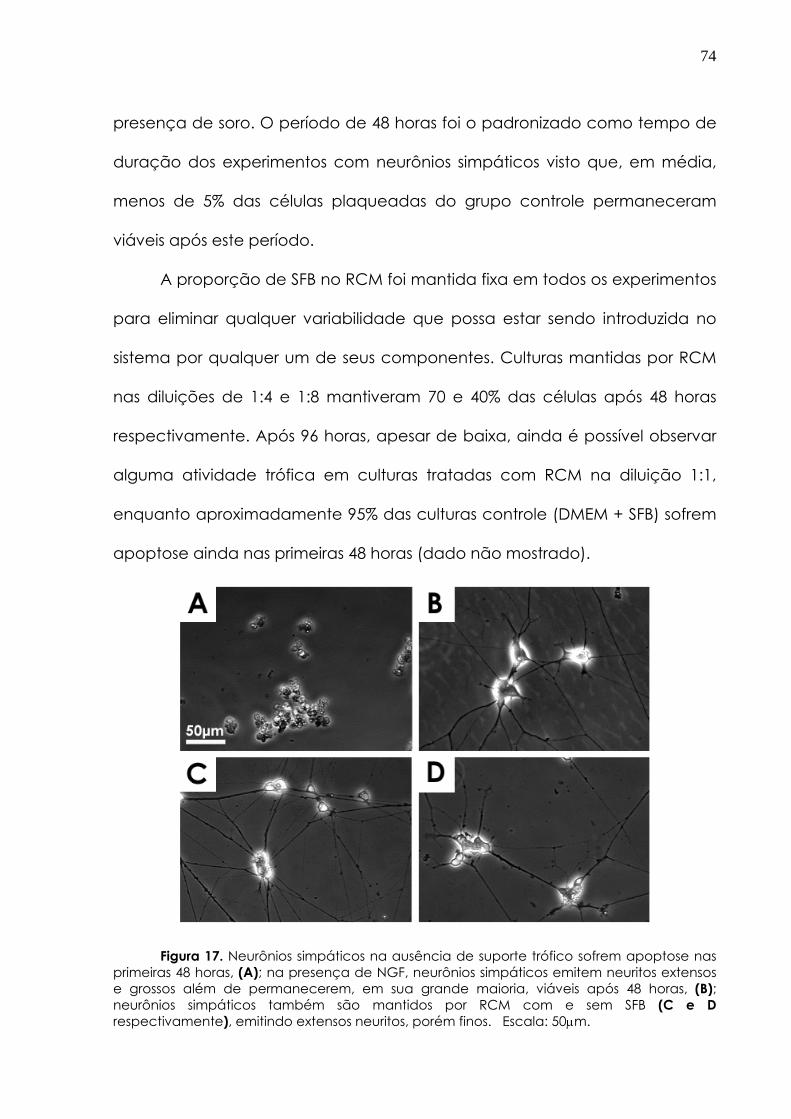
74
presença de soro. O período de 48 horas foi o padronizado como tempo de
duração dos experimentos com neurônios simpáticos visto que, em média,
menos de 5% das células plaqueadas do grupo controle permaneceram
viáveis após este período.
A proporção de SFB no RCM foi mantida fixa em todos os experimentos
para eliminar qualquer variabilidade que possa estar sendo introduzida no
sistema por qualquer um de seus componentes. Culturas mantidas por RCM
nas diluições de 1:4 e 1:8 mantiveram 70 e 40% das células após 48 horas
respectivamente. Após 96 horas, apesar de baixa, ainda é possível observar
alguma atividade trófica em culturas tratadas com RCM na diluição 1:1,
enquanto aproximadamente 95% das culturas controle (DMEM + SFB) sofrem
apoptose ainda nas primeiras 48 horas (dado não mostrado).
Figura 17. Neurônios simpáticos na ausência de suporte trófico sofrem apoptose nas primeiras 48 horas, (A); na presença de NGF, neurônios simpáticos emitem neuritos extensos e grossos além de permanecerem, em sua grande maioria, viáveis após 48 horas, (B); neurônios simpáticos também são mantidos por RCM com e sem SFB (C e D respectivamente), emitindo extensos neuritos, porém finos. Escala: 50μm.

75
Neurônios mantidos por NGF apresentam como característica, um
corpo celular grande além neuritos calibrosos e longos. Já neurônios
simpáticos tratados com RCM apresentam um corpo celular menor, com
neuritos também longos, porém mais finos (figura 17A-D).
A ação combinada de NGF e RCM não apresentou acréscimo
significativo na sobrevivência induzida por NGF sozinho (viabilidade em torno
de 80%).
4.1.2 A SECREÇÃO DO FATOR RETINIANO NÃO É DEPENDENTE DO
DESENVOLVIMENTO NEM DE EXCITABILIDADE ELÉTRICA DO TECIDO.
Sabendo que o perfil proteômico da retina se modifica ao longo do
desenvolvimento questionamos a possibilidade de que RCMs de diferentes
idades (desde embrião a um dia pós-natal) pudessem apresentar diferentes
resultados quanto ao efeito trófico promovido em neurônios simpáticos. Para
responder se o efeito trófico em questão é influenciado pelo
desenvolvimento, RCM de diferentes estágios (E5 a P1) foram coletados e
testados quanto à sua atividade trófica em neurônios simpáticos em cultura.
Não foi observada diferença significativa entre a atividade trófica dos
diferentes RCMs, não sendo portanto regulada por desenvolvimento (figura
18). O efeito neuritogênico, visualmente, também se mostrou equivalente
entre os neurônios tratados com os meio condicionados por diferentes
estágios (dado não mostrado).

76
Figura 18. A liberação dos fatores tróficos retinianos que mantém neurônios simpáticos em cultura não é regulada durante o desenvolvimento. RCM preparado a partir de explantes retinianos de embriões (E5 a E16) e pós-natal 1 (P1) exibem semelhante atividade trófica.
Para avaliar o tempo mínimo necessário de condicionamento do
meio, RCMs foram coletados em intervalos de tempo entre 30 minutos e 96
horas. Com este experimento observamos que 30 minutos de
condicionamento do meio por explantes de retina é suficiente para manter
cerca a viabilidade de cerca de 30% de neurônios simpáticos em cultura. O
efeito observado é aumentado para cerca de 60% com 6 horas de
condicionamento e é máximo após 12 horas de condicionamento (figura
19).
A excitabilidade elétrica do tecido nervoso, neste caso a retina, é
outro fator que pode influenciar atenuando ou potencializando os efeitos
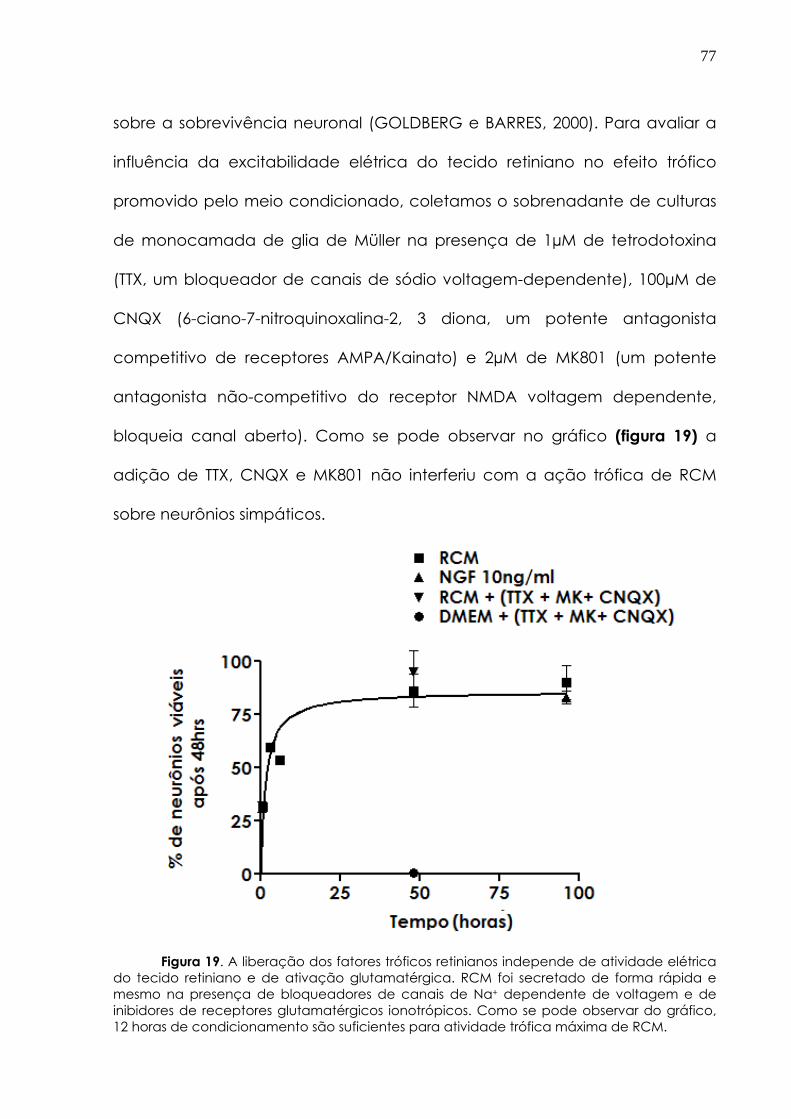
77
sobre a sobrevivência neuronal (GOLDBERG e BARRES, 2000). Para avaliar a
influência da excitabilidade elétrica do tecido retiniano no efeito trófico
promovido pelo meio condicionado, coletamos o sobrenadante de culturas
de monocamada de glia de Müller na presença de 1µM de tetrodotoxina
(TTX, um bloqueador de canais de sódio voltagem-dependente), 100µM de
CNQX (6-ciano-7-nitroquinoxalina-2, 3 diona, um potente antagonista
competitivo de receptores AMPA/Kainato) e 2µM de MK801 (um potente
antagonista não-competitivo do receptor NMDA voltagem dependente,
bloqueia canal aberto). Como se pode observar no gráfico (figura 19) a
adição de TTX, CNQX e MK801 não interferiu com a ação trófica de RCM
sobre neurônios simpáticos.
Figura 19. A liberação dos fatores tróficos retinianos independe de atividade elétrica do tecido retiniano e de ativação glutamatérgica. RCM foi secretado de forma rápida e mesmo na presença de bloqueadores de canais de Na+ dependente de voltagem e de inibidores de receptores glutamatérgicos ionotrópicos. Como se pode observar do gráfico, 12 horas de condicionamento são suficientes para atividade trófica máxima de RCM.

78
4.1.3 EFEITO TRÓFICO PROMOVIDO POR RCM NÃO É DEVIDO À AÇÃO
INDIVIDUAL DE NEUROTROFINAS.
A adição de NGF 10 ng/mL à culturas de neurônios simpáticos isolados
é capaz de promover uma sobrevida de 80 ± 8% após 48 horas. O efeito
trófico de NGF é completamente abolido na presença de 1μg/mL de anti-
NGF (anticorpo monoclonal bloqueador anti-NGF de rato produzido em
camundongo) entretanto a atividade trófica de RCM se manteve quando
incubado na presença de 1µg/mL de anti-NGF (figura 20).
Figura 20. A ação trófica de RCM não é inibida por anticorpo anti-NGF. Neurônios simpáticos na ausência de fatores tróficos morrem quase em sua totalidade nas primeiras 48 horas. O NGF, principal fator trófico para este tipo neuronal foi efetivamente bloqueado pela ação do anticorpo. O anticorpo porem não foi capaz de inibir a atividade trófica de RCM no simpático.

79
Para excluir a possibilidade do anticorpo não estar reconhecendo o
NGF de ave utilizamos também receptores quimera solúveis, constituídos em
parte, pelo domínio extracelular do receptor de alta afinidade do NGF mas
sem a porção intracelular necessária para desencadear a sinalização (TrkA-
IgG, figura 21).
Figura 21. Receptores Trk-IgG. Estes receptores solúveis são usados em abundancia no meio de modo a seqüestrar todo ligante presente e evitar que estes interajam com os receptores de membrana e ativem suas respectivas vias de sinalização.
Esta molécula interage e seqüestra o NGF presente no meio impedindo
que este interaja com os receptores presentes na superfície dos neurônios
simpáticos em cultura e exercer seu papel biológico. Como esperado, 2.4
µg/mL de TrkA solúvel eliminou completamente o efeito de NGF. A mesma

80
concentração de TrkA solúvel não foi capaz de interferir com a ação trófica
promovido por RCM em neurônios simpáticos (figura 22).
Figura 22. A ação trófica de RCM não é inibida pela presença do TrkA solúvel. Como esperado, neurônios simpáticos na ausência de fatores tróficos, em sua grande maioria, morrem nas primeiras 48 horas. O NGF, principal fator trófico para este tipo neuronal foi efetivamente bloqueado pelo receptor solúvel. Entretanto o receptor solúvel não foi capaz de inibir a atividade trófica de RCM no simpático.
Nem BDNF (10 ng/mL), nem NT4 (0.1–10 ng/mL) mantiveram neurônios
simpáticos em cultura de forma significativa. Altas concentrações de NT-4 (50
–100ng/mL) mantiveram cerca de 15-30% dos neurônios viáveis após 48
horas. Entretanto, estes números ainda são muito baixos quando
comparados aos valores obtidos pelo tratamento com NGF ou RCM. TrkB-
IgG, que interage tanto com BDNF quanto com NT-4 não teve efeito algum
na sobrevivência induzida por RCM. Como esperado, TrkB-IgG não bloqueou

81
a atividade trófica do NGF (que é ligante de TrkA) confirmando a
especificidade desta molécula (figura 23).
Figura 23. A ação trófica de RCM não é inibida pela presença do TrkB solúvel e ligantes do receptor TrkB não exercem atividade trófica sobre neurônios simpático. O NGF, principal fator trófico para este tipo neuronal não teve sua ação bloqueada pelo receptor solúvel TrkB, confirmando a especificidade da molécula. Novamente o receptor solúvel não foi capaz de inibir a atividade trófica de RCM no simpático.
Por fim, investigamos o envolvimento do NT-3 na sobrevivência
neuronal induzida por RCM utilizando o receptor quimera TrkC-IgG.
Entretanto, o efeito trófico promovido por RCM nos neurônios simpáticos foi
mantido quando incubado com TrkC-IgG 500 ng/mL, enquanto o efeito
trófico promovido por NT-3 foi completamente abolido quando co-incubado
com TrkC-IgG na mesma concentração acima. É interessante notar que
TrkC-IgG inibiu parcialmente a sobrevivência induzida por NGF (figura 24), o
que sugere uma possível interação do receptor quimérico TrkC com o NGF.

82
Ironicamente, justamente o oposto é descrito na literatura onde artigos
discutem a ativação de TrkA por NGF e também NT-3 (BELLIVEAU et al. 1997).
Figura 24. A ação trófica de RCM não é inibida pela presença do TrkC solúvel. O receptor solúvel inibiu seu respectivo ligante (NT-3). O NGF, sofreu inibição na presença do receptor TrkC solúvel o que sugere uma possível interação entre essas moléculas. Entretanto o receptor solúvel não foi capaz de inibir a atividade trófica de RCM no simpático.
4.1.4 ATIVIDADE TRÓFICA DO RCM É MANTIDA APÓS DIÁLISE, É RETIDA EM
COLUNA DE HEPARINA E É SENSÍVEL A TEMPERATURA.
A atividade trófica do RCM foi mantida mesmo após diálise de 24 e 48
horas, logo, depende de moléculas com mais de 12KDa (figura 25).
A atividade trófica promovida por RCM é bastante inibida quando
este é aquecido por 45 minutos a 65ºC e completamente abolida quando
aquecido por cinco minutos a 95ºC confirmando que o fator é sensível à
temperatura (figura 25).

83
Diversos fatores presentes na retina de embrião de pinto, como por
exemplo o FGFb, se ligam com grande afinidade à heparina (DESIRÉ et al.,
1998). Sendo assim, utilizamos uma coluna de heparina para “capturar”
possíveis fatores tróficos com afinidade à heparina presentes no RCM. O RCM
foi então injetado através de uma coluna de heparina (Hi-Trap) e dividido
em duas frações, uma que se liga (BF, do inglês Bound Fraction) e outra que
não se liga à heparina (FF, do inglês Free Fraction). A fração FF do RCM não
apresentou atividade trófica, enquanto a fração BF, após eluição e
dessalinização, manteve a viabilidade de cerca de 75% dos neurônios
simpáticos em cultura após 48 horas (figura 25).
Figura 25. Fatores secretados pelos explantes retinianos são sensíveis a aquecimento não é dialisável em membrana de 12KDa e tem componente com afinidade a heparina. BF, do inglês Bound Fraction e FF, Free Fraction sendo respectivamente as frações que se ligam e que não se ligam à heparina.
Na faixa de concentração de 0.1 à 50nM o FGFb sozinho não exerce
atividade trófica em neurônios simpáticos em cultura. Neurônios simpáticos

84
quando na presença desta molécula entram em processo apoptótico ainda
nas primeiras 48 horas. A viabilidade neuronal promovida por FGFb ficou
próxima dos valores obtidos nas culturas controle (3-5% de células viáveis
após 48 horas) (dado não mostrado).
4.1.5 RCM INDUZ SOBREVIVÊNCIA SINALIZANDO PELAS VIAS DA Pi3K e JAK2
MAS NÃO PELA VIA DAS ERKs.
Vários trabalhos recentes têm mostrado que fatores tróficos promovem
a sobrevivência de neurônios através de vias dependentes de PI3-K ou
MAPKs. Sendo assim decidimos testar a influência de um inibidor específico
da PI3-K (LY294002) e um inibidor de ERKs (PD098059) na sobrevivência
induzida por RCM em neurônios simpáticos. Enquanto PD098059 até 10 ou
50µM não interferiu na sobrevivência induzida por RCM, como mostrado na
figura 26, LY294002 (50µM) bloqueou cerca de 90% da sobrevivência
neuronal promovida por RCM.
Nós também testamos um inibidor de JAK (especificamente JAK-2)
visto que esta é a via estimulada na ativação por citocinas retinianas (ex:
CNTF). AG490 (50µM) bloqueou completamente a sobrevivência induzida
por RCM como pode ser visto na figura 26.

85
figura 26. A sobrevivência de neurônios simpáticos induzida por RCM depende das
vias PI3-K e JAK2, mas independe da via MAPK. Efeito do PD098059 (10-50μM), um inibidor da MAPK ou de LY294002 (10-50μM), um inibidor da PI3-K na sobrevivência de neurônios simpáticos induzida por RCM. (A). AG490, um inibidor da via JAK2-STAT (50μM), inibe por completo a sobrevivência de neurônios simpáticos induzida por RCM (B).

86
4.1.6 A LIBERAÇÃO DO FATOR RETINIANO DEPENDE DE EVENTOS MEDIADOS
POR NEUROTROFINAS.
Moléculas como NT-3, BDNF e NGF estão presentes na retina
embrionária de pinto (HALBÖÖK et al., 1996; BOVOLENTA et al., 1996;
KARLSSON et al., 1998; FRADE et al., 1999), a presença destas moléculas pode
vir a influenciar a atividade trófica promovida por RCM. Para analisar esta
possibilidade, RCMs foram preparados na presença de TrkA-IgG (2.4 μg/mL),
TrkB-IgG (1.2 μg/mL) ou TrkC-IgG (0.5 μg/mL) prevenindo individualmente a
ação de cada uma das neurotrofinas durante os quatro dias de
condicionamento. RCMs preparados na presença de TrkA-IgG ou TrkB-IgG
apresentaram redução de suas atividades tróficas, mantendo 60 e 40% das
células viáveis após 48 horas enquanto TrkC-IgG não teve efeito algum na
sobrevivência induzida por RCM (figura 28).
Figura 27. A secreção dos fatores tróficos por explantes de retina de embrião de pinto depende de eventos mediados por receptores TrkA e TrkB, mas não por TrkC. RCM preparado por explantes retinianos incubados por 4 dias com TrkA-IgG (2.4 μg/mL), TrkB-IgG (1.2 μg/mL), ou TrkC-IgG (0.5 μg/mL) apresentaram efeitos diversos na sobrevida dos neurônios simpáticos.

87
Esses dados sugerem que a atividade trófica do RCM em neurônios
simpáticos apesar de ser independente da ação de neurotrofinas isoladas
(como visto anteriormente) mostrou-se ser dependente da ativação de seus
receptores (TrkA e TrkB) para a produção ou secreção, pelo tecido retiniano,
de pelo menos alguns dos fatores que compõem o RCM.
4.1.7 A ATIVIDADE TRÓFICA PROMOVIDA POR RCM NÃO É BLOQUEADA COM
ANTICORPOS BLOQUEADORES ANTI-TGF- β, ANTI-GDNF NEM ANTI-CNTF
Em busca da identidade do fator responsável pelo efeito trófico
promovido por RCM, utilizamos anticorpos gerados contra outros fatores com
conhecida ação trófica em neurônios simpáticos, como por exemplo o CNTF
e o GDNF. Utilizamos também anticorpo bloqueador contra TGF-β. Membros
da família do TGF-β (BMPs) estão relacionados com estimulação de
crescimento dendrítico em neurônios simpáticos promovido pela glia (LEIN et
al. 2002). Entretanto, como mostrado no gráfico da figura 28, nenhum dos
três anticorpos interferiu com a sobrevivência e neuritogênese induzida por
RCM.
Como a retina é um tecido complexo com uma variedade de tipos
neuronais e pelo menos um tipo glial predominante, a glia de Müller,
procuramos nos ater a um único tipo celular com o qual pudéssemos levar
adiante os estudos em fatores tróficos retinianos. No nosso caso, vamos
trabalhar com a glia de Müller dada a multi potencialidade desta célula (na
produção de fatores, modulação do micro-ambiente ou no caso de injúria,

88
por exemplo) e também porque meio condicionado de culturas
enriquecidas de neurônios da retina não apresentam atividade trófica
significativa. Encerram aqui, portanto, os resultados gerados com os estudos
com RCM.
Figura 28. A sobrevivência induzida por RCM não é afetada pela presença dos anticorpos anti-TGFb, anti-CNTF nem anti-GDNF, excluindo a possibilidade da ação trófica de RCM ser devido a ação isolada de qualquer um destes fatores. 4.2 CULTURAS DE CÉLULAS DE MÜLLER PURIFICADAS DE CAMUNDONGO
Para obter culturas de monocamada de glia de Müller a partir retinas
de camundongos P1 empregamos o mesmo protocolo utilizado por Reis et
al. (2002) na obtenção de culturas purificadas de glia de Müller de embriões
de pinto. Após a confluência de uma cultura de células retinianas total (em
torno de uma semana) uma grande quantidade de neurônios encontra-se
disposta sobre uma monocamada de células de Müller (figura 29A). Nesse
estágio, as culturas foram tratadas com solução de ácido ascórbico (AA)

89
4mM por um período de 3 horas (figura 29B), lavadas com meio DMEM e
mantidas por 24 horas em DMEM + 10% SFB para recuperação da glia após
tratamento com AA. Após 24 horas, novamente lavadas, as culturas
apresentavam somente uma monocamada de glia de Müller purificada,
isenta de neurônios como pode ser observado na figura 29C. A partir deste
ponto as culturas passaram a ter o meio condicionado coletado a cada 48
horas e substituído por meio fresco. Pelo menos duas coletas foram possíveis
para cada placa de cultura.
Figura 29. Culturas de células retinianas obtidas a partir de camundongo pós-natal P1 As retinas foram dissecadas, dissociadas e as células plaqueadas. Após confluência (A) numerosos neurônios são observados sobre a monocamada da glia de Müller. As culturas foram tratadas com ácido ascórbico 4mM por 3h (B) com a remoção seletiva dos neurônios. Após dois dias as culturas apresentavam apenas células de glia de Müller (C, G, H). Estas culturas foram positivamente marcadas com anticorpos contra GFAP (D), vimentina (E) e S-100 (F), porém foram negativas para HNK-NCAM ou RT-97 (marcadores neuronais).
A glia de camundongos, apesar de seu tamanho, oferece inúmeras
vantagens sendo uma delas a ampla variedade de anticorpos presentes no

90
mercado que interagem eficientemente com proteínas de roedores (e não
de aves, a exemplo do anticorpo contra GFAP). Diversos anticorpos contra
marcadores gliais foram utilizados para atestar a pureza da cultura para glia.
As culturas purificadas foram eficientemente marcadas com anticorpos
contra GFAP (figura 29D), vimentina (figura 29E) e S-100 (figura 29F), 3
marcadores encontrados em células de Müller e em astrócitos. É interessante
observar a marcação dos filamentos do citoesqueleto, em particular a GFAP
(figura 29D), que se manifesta após condições de insulto no caso de gliose
reativa (BRINGMANN et al., 2006). Estas mesmas culturas, porém, foram
negativas para HNK-NCAM ou RT-97 (marcadores neuronais) atestando,
portanto, a pureza destas culturas. A partir destas culturas foi obtido o MMG
utilizado nesta parte do trabalho.
4.2.1 MMG ATIVA AS VIAS DA pERK, pAKT e pSTAT3 SIMULTANEAMENTE EM
NEURÔNIOS DO GÂNGLIO SUPERIOR CERVICAL DE RATOS
O NGF é um fator trófico sem o qual neurônios simpáticos em cultura
entram em processo apoptótico. O NGF tem por característica de ação
estimular as vias da ERK e da AKT que são vias de sinalização relacionadas à
manutenção da sobrevivência em neurônios simpáticos a longo prazo
mantendo estas células em cultura por longo tempo. O CNTF também é um
fator trófico para neurônios simpáticos mas se comparado ao NGF é um fator
que oferece suporte trófico somente para curto prazo (BURNHAM et al.,
1994). O CNTF também ativa a via das ERKs mas apenas de forma transiente.

91
É desconhecido se a ativação de AKT via CNTF ocorre também de forma
transiente. Por outro lado, somente CNTF (e não NGF) ativa a via de JAK-STAT
fosforilando STAT3 (IP e YANCOPOULOS, 1996). Desta forma, decidimos
investigar a possibilidade de envolvimento de tais vias de sinalização para
sobrevivência no modelo simpático do gânglio superior cervical (GSC) de
rato estimulado por MMG. Como mostrado na figura 30 MMG estimula
ambas as vias (NGF e CNTF) promovendo a fosforilação de ERKs, AKT e STAT3
após 90 minutos de incubação. Em paralelo, para descartar a presença de
NGF no MMG, anticorpo monoclonal anti-NGF (1μg/mL) foi incubado com a
cultura tratada com MMG mas não alterou a sobrevivência promovida por
MMG nestas células.
Figura 30. Imuno detecção de proteínas chave fosfoforiladas de duas vias ativadas por fatores tróficos. Nesta figura pode-se observar que apos incubação de 90 minutos, NGF ativa preferencialmente a via da AKT/PKB enquanto CNTF ativa exclusivamente STAT3. MMG ativa ambas as via simultaneamente em culturas de neurônios do GSC de ratos.

92
4.2.2 MMG POSSUI COMPONENTE ATIVO QUE SE LIGA A HEPARINA, POTENTE
AÇÃO TRÓFICA EM NEURÔNIOS SIMPÁTICOS DE AVE QUE NÃO FOI ABOLIDA
POR ANTICORPOS BLOQUEADORES anti-NT3, anti-GDNF OU anti-CNTF.
Culturas de neurônios simpáticos de embrião de pinto E10 quando
incubadas com MMG apresentam 100% de sobrevivência após 48 horas em
cultura (figura 31B) (estado que se mantém por quase 72 horas) este efeito
não foi abolido quando MMG foi incubado com anticorpos bloqueadores
anti-GDNF (31C), anti-NT3 (31D) ou anti-CNTF (31E) (apesar de estimular
fortemente a via de sinalização ativada por CNTF como mostrado no
resultado anterior, figura 30) em concentrações que bloquearam
eficientemente seus respectivos alvos.
Figura 31. A sobrevivência induzida por RCM não é afetada pela presença dos anticorpos anti-GDNF, anti-NT3 ou anti-CNTF o que exclui a possibilidade da ação trófica de MMG ser devido à ação isolada de qualquer um destes fatores. MMG também apresentou componente com forte atividade trófica que se liga a heparina.
MMG ao ser injetado através de coluna de heparina (Hi-Trap –
Amersham) é dividido em fração ligada (BF) que quando eluída e

93
dessalinizada reproduz o efeito do MMG total (figura 31F) e fração livre (FF)
que possui atividade trófica um pouco inferior se comparada ao MMG total
mas equivalente ao controle positivo (NGF (31B), ≈85%)(dado não mostrado).
4.2.3 FATORES TRÓFICOS DA GLIA DE MÜLLER DE CAMUNDONGOS INDUZEM
NEURITOGÊNESE E SOBREVIDA DAS CGRS
Até aqui demonstramos que os fatores secretados pela glia de Müller
de camundongos mantêm neurônios simpáticos (periféricos) em cultura. No
entanto, a glia de Müller é oriunda do SNC. A glia de Müller é encontrada na
retina de vertebrados posicionada de forma a interagir com a maioria,
senão todos os neurônios retinianos, cruzando todas as camadas da retina
(ver figura 2 da introdução) (SARTHY e RIPPS, 2001). Nós nos perguntamos
então sobre a influência que o MMG viria a ter na neuritogênese em
explantes de retina de camundongo in vitro. Como pode ser observado na
figura 32, explantes de retina de camundongos P4 foram mantidos por 72
horas em três diferentes condições, todas na presença de 5μM forscolina (F),
e tiveram suas projeções neuríticas quantificadas. Neurobasal (NB) foi usado
como controle e BDNF + CNTF (B+C, na concentração de 50ng/mL cada) ou
MMG foram as condições experimentais. Poucos neuritos foram projetados
na situação controle (NB), com valores do raio médio (média das distâncias
entre o ponto central do explante e os neuritos mais longos) não maiores que
300μm após 72h (figura 32, NB+F). Entretanto, BCF aumentou
significativamente a extensão neurítica alcançando valores em torno de 400
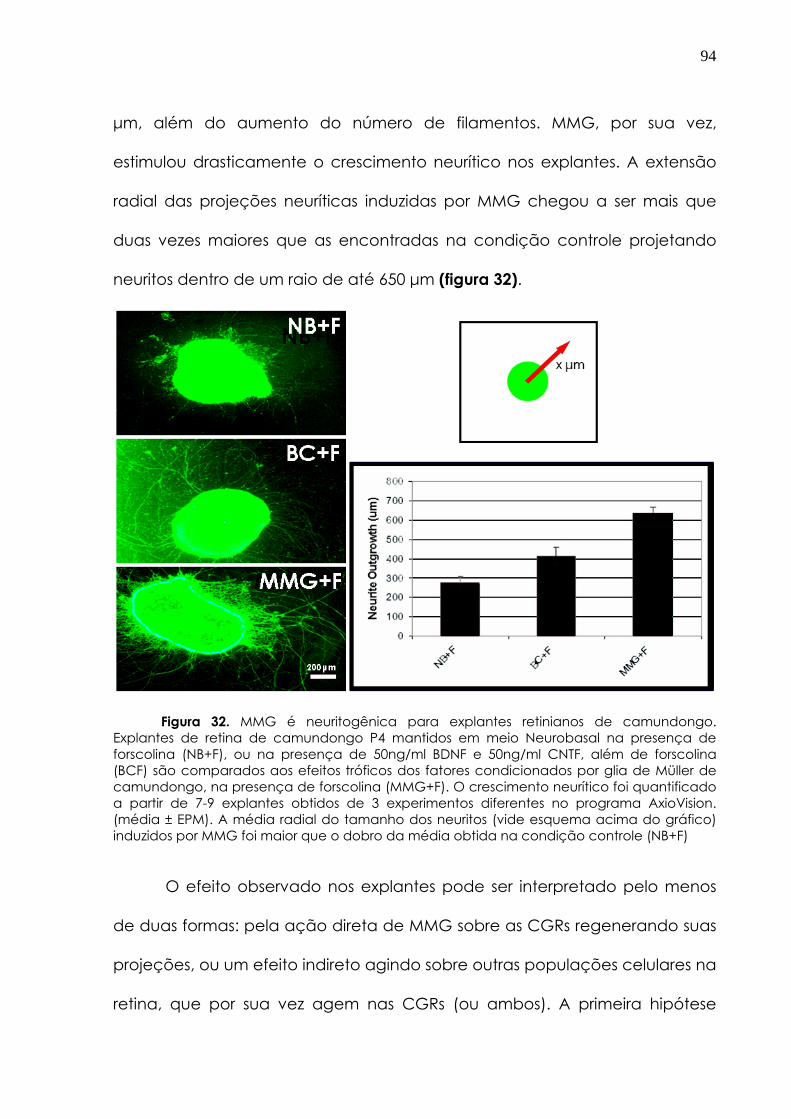
94
μm, além do aumento do número de filamentos. MMG, por sua vez,
estimulou drasticamente o crescimento neurítico nos explantes. A extensão
radial das projeções neuríticas induzidas por MMG chegou a ser mais que
duas vezes maiores que as encontradas na condição controle projetando
neuritos dentro de um raio de até 650 μm (figura 32).
Figura 32. MMG é neuritogênica para explantes retinianos de camundongo. Explantes de retina de camundongo P4 mantidos em meio Neurobasal na presença de forscolina (NB+F), ou na presença de 50ng/ml BDNF e 50ng/ml CNTF, além de forscolina (BCF) são comparados aos efeitos tróficos dos fatores condicionados por glia de Müller de camundongo, na presença de forscolina (MMG+F). O crescimento neurítico foi quantificado a partir de 7-9 explantes obtidos de 3 experimentos diferentes no programa AxioVision. (média ± EPM). A média radial do tamanho dos neuritos (vide esquema acima do gráfico) induzidos por MMG foi maior que o dobro da média obtida na condição controle (NB+F)
O efeito observado nos explantes pode ser interpretado pelo menos
de duas formas: pela ação direta de MMG sobre as CGRs regenerando suas
projeções, ou um efeito indireto agindo sobre outras populações celulares na
retina, que por sua vez agem nas CGRs (ou ambos). A primeira hipótese

95
apresentou-se mais factível de ser investigada devido à existência de um
método eficiente para isolamento de CGRs que se utiliza um antígeno de
superfície especifico para CGRs (Thy1.2/CD90.2). Através do método de
imunopurificação de CGRs isolamos e realizamos experimentos com MMG
em culturas purificadas de CGRs de camundongo. Como visto na figura 33A,
CGRs foram isoladas e purificadas com sucesso das células retinianas de
camundongo utilizando um método adaptado descrito anteriormente para
ratos (BARRES et a., 1988).
Figura 33. CGRs de camundongo foram purificadas pelo método de imunopurificação (immunopanning). (A). CGRs aderidas à placa por meio de anticorpos, selecionadas pelo processo de imunopurificação. Essas células foram soltas da placa mediante tripsina, coletadas e plaqueadas (5000 células) em diferentes substratos e em diferentes condições em meio neurobasal (NB); (B). CGRs de camundongo P8 plaqueados sobre poli-L-lisina e laminina na presença de BCF após 72 horas em cultura; (C). CGRs de camundongo P4 plaqueados sobre poli-L-lisina e laminina na ausência de fatores tróficos, 72 horas; (D). CGRs de camundongo P1 plaqueados sobre glia de Müller, 15 dias; (E). CGRs de camundongos P8 mantidos por MMG em poli-L-lisina e laminina, 72 horas e (F). CGRs de camundongos P8 mantidos por BCF em lamina basal de pinto E9 por 21 dias. Barra de calibração 50μm.

96
As CGRs plaqueadas sobre laminina na presença de BCF apresentam
numerosos neuritos próximos ao soma e corpos celulares brilhantes (figura
33B). Por outro lado, na ausência de fatores tróficos apropriados, poucas
CGRs sobrevivem (figura 33C, 4% após 72hs). Quando plaqueávamos as
CGRs sobre uma monocamada de glia em um sistema de co-cultura as
células se apresentavam saudáveis e emitindo longas projeções após 15 dias
em cultura mostrando que a glia é um excelente substrato para os neurônios
retinianos (figura 33D). MMG (figura 33E), mas não AMG (não mostrado,
CGRs de camundongo não parecem se beneficiar do AMG) manteve as
CGRs após 72hs em cultura promovendo a projeção de longos neuritos. Sob
condições ótimas de cultura (BCF ou MMG+F, plaqueados sobre lamina
basal de pinto E9), é possível manter a viabilidade celular mesmo após 3
semanas em cultura (figura 33F).
A quantificação da sobrevida das CGR isoladas de camundongos P1,
P4 e P8 foi auxiliada pelo uso de um marcador fluorescente que evidencia
células viáveis na cor verde (diacetato de fluoresceína, 10μg/ml FDA), além
de 2 μg/ml Hoesch 23392 (azul), um marcador fluorescente que se intercala
ao DNA, e indica o número de núcleos (e conseqüentemente, corpos
celulares), que neste caso, quando apresentavam somente a cor azul, não
se apresentam mais viáveis. As CGRs que foram plaqueadas em meio NB
eram pouco marcadas com FDA após 72 horas (figura 34A), o que indicava
que na sua maioria estavam em processo de apoptose, sendo marcadas
com Hoesch. A adição de 5μM forscolina (NB+F) aumentou
consideravelmente o número de células viáveis (figura 34B). Na presença de

97
BDNF 50ng/ml + forscolina (B+F), um maior número de CGRs permaneceram
viáveis, com inúmeros neuritos porém pequenos emergindo do corpo celular
(figura 34C). Por outro lado, CNTF 50ng/ml + forscolina (C+F) induziu um
menor número desses neuritos pequenos que emergem do soma
apresentando predominantemente projeções longas quando comparados
a condição anterior (figura 34D). O plaqueamento das CGR na presença de
BDNF, CNTF e forscolina (B+C+F) resultou em um grande número de células
viáveis, com inúmeros neuritos (figura 34E). Por fim, na presença de MMG, o
número de células viáveis foi semelhante ao obtido com BCF, porém a
extensão dos axônios foi muito maior, geralmente acima de 20 vezes o
diametro do soma (figura 34F).
Figura 34. Sobrevida e Neuritogênese de CGR. Aproximadamente 5000 CGR foram plaqueadas sobre 10 μg/ml laminina e 10 μg/ml poli-L-lisina em diferentes condições mantidas em meio NB (sem soro) por 72h. (A). Controle (NB); (B). Adição de 5 μM forscolina; (C). Adição de 50ng/ml BDNF + Forscolina; (D). Adição de 50ng/ml CNTF + Forscolina; (E). Adição de BDNF, CNTF e Forscolina (BCF); (F). Adição de MMG (1:1) + Forscolina. Barra de calibração 50μm.

98
A quantificação da sobrevida das CGRs de retinas de camundongos
P1 em diversas condições descritas acima se encontra na figura 35.
Figura 35. Média de sobrevivência de CGRs de camundongo P1 plaqueadas sobre poli-L-lisina + laminina em diferentes condições após 72 horas em cultura. Observar que a adição de forscolina a qualquer uma das condições aumenta consideravelmente a taxa de sobrevivência neuronal (CGRs). Algumas observações devem ser apontadas:
1. A simples adição de F ao meio NB duplica ou triplica a taxa de sobrevida
das CGRs em 48h ou 72h em cultura.
2. A adição de BDNF ou CNTF as CGRs teve efeitos semelhantes quanto a
sobrevida, e ambas foram potenciadas por forscolina.
3. A sobrevida das CGRs na presença de BCF ou de MMG não foi diferente
significativamente nas idades dos camundongos e nos tempos de cultura
estudados.

99
4.2.4 MONOCAMADA DE GLIA DE MÜLLER É UM EXCELENTE SUBSTRATO PARA A
SOBREVIDA DAS CGR
Como as células gliais de Müller secretam fatores tróficos que
promovem sobrevivência e neuritogênese em CGRs e além disso a troca de
moléculas entre tais células através do contato pode vir a potencializar tal
efeito, decidimos então plaquear CGRs diretamente sobre uma
monocamada de glia na forma de uma co-cultura. A quantificação da
sobrevida das CGRs foi feita através da contagem dos neurônios viáveis
marcados com os anticorpos anti-neurofilamento (clone RT97) (ou anti-HNK-
NCAM 2H3) que reconhecem CGRs, mas não células gliais (que foram
marcadas com anticorpos contra GFAP ou S-100). Como mostrado na figura
36, as CGR plaqueadas sobre a monocamada de glia de Müller se
apresentaram saudáveis e com neuritos projetados, seja na situação
controle, na presença de BCF, ou de MMG após 4 dias em cultura.
Quantificamos o número de CGRs fluorescentes por marcação com
anticorpo anti-neurofilamento (anti-NF) e não vimos diferença na
percentagem de CGRs viáveis após 3 ou 4 dias em cultura (tabela 2).
Quantificamos também o número de CGRs marcadas previamente com
DAPI na concentração de 0,5 μg/mL (fluoróforo que se liga a DNA) após 48 e
72h em cultura, confirmando os dados obtidos com neurônios marcados
para neurofilamento. Assim, o plaqueamento das CGRs sobre as células gliais
resultou em sobrevivência de cerca de 80% dos neurônios após 3 dias em

100
cultura (o dobro do número de neurônios quando comparado aos
plaqueados sobre laminina e poli-L-lisina).
Figura 36. Sobrevida das CGR plaqueadas sobre o tapete de células gliais de Müller.
Os neurônios eram estão marcados em verde com anticorpos específicos contra antígenos neuronais (neurofilamento, HNK-NCAM, RT97, ou 2H3) e as células gliais, em vermelho, com anticorpos anti-GFAP. A quantificação é apresentada na tabela abaixo das micrografias. Quando plaqueadas sobre monocamada de gia de Müller é necessária a adição de fatores tróficos, pois a sobrevida das CGRs foi semelhante na condição controle (Neurobasal + foscolina – NBF) comparada a BCF ou MMG.
Tabela 2. Sobrevida das CGR plaqueadas na ausência ou presença de células gliais de Müller em cultura por 72h. A sobrevida dos neurônios foi o dobro quando plaqueados em tapete de glia comparada à condição sem glia (laminina e poli-L-Lisina).
Na figura 37 podemos comparar o aspecto nuclear (evidenciado por
DAPI) de CGRs plaqueadas sobre a glia de Müller com CGRs plaqueadas
sobre poli-L-lisina + laminina. Na condição em que CGRs foram sem
monocamada de glia de Müller os núcleos dos neurônios se encontram
fragmentados (degenerando, possivelmente comprometido com o
programa de morte celular programada) enquanto a presença da
monocamada de glia de Müller preservou a viabilidade celular refletida pela
integridade nuclear.
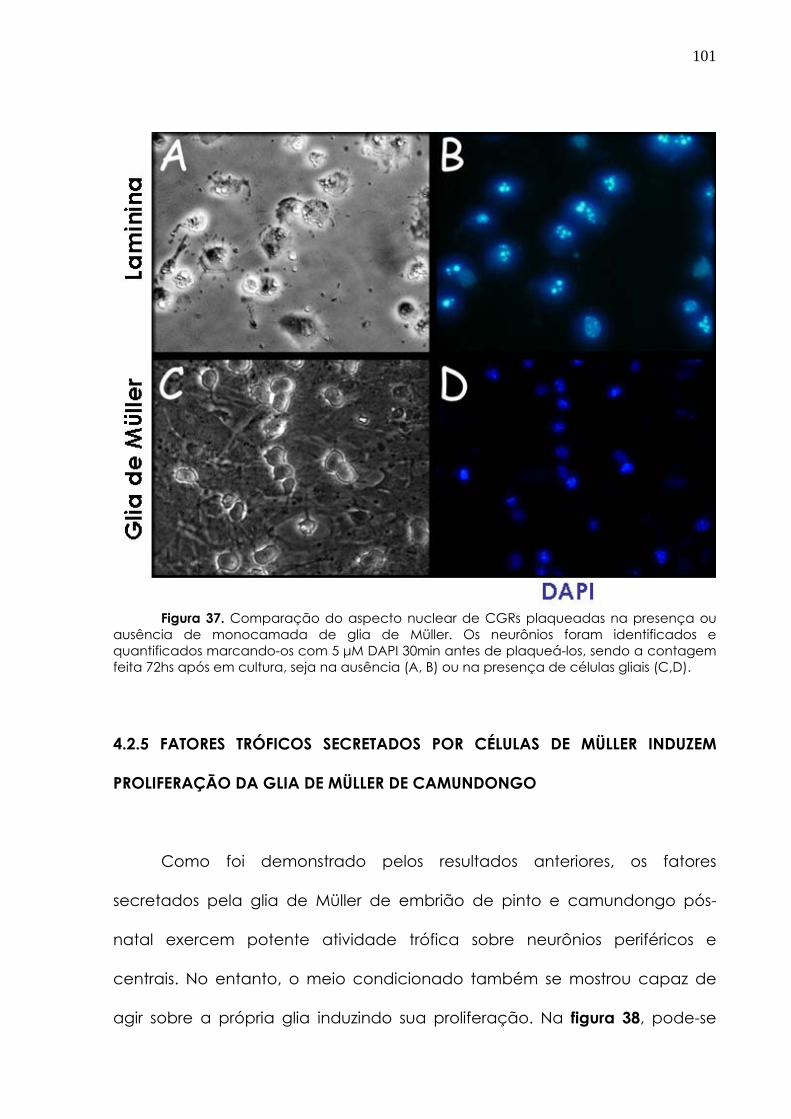
101
Figura 37. Comparação do aspecto nuclear de CGRs plaqueadas na presença ou
ausência de monocamada de glia de Müller. Os neurônios foram identificados e quantificados marcando-os com 5 μM DAPI 30min antes de plaqueá-los, sendo a contagem feita 72hs após em cultura, seja na ausência (A, B) ou na presença de células gliais (C,D).
4.2.5 FATORES TRÓFICOS SECRETADOS POR CÉLULAS DE MÜLLER INDUZEM
PROLIFERAÇÃO DA GLIA DE MÜLLER DE CAMUNDONGO
Como foi demonstrado pelos resultados anteriores, os fatores
secretados pela glia de Müller de embrião de pinto e camundongo pós-
natal exercem potente atividade trófica sobre neurônios periféricos e
centrais. No entanto, o meio condicionado também se mostrou capaz de
agir sobre a própria glia induzindo sua proliferação. Na figura 38, pode-se

102
observar as células remanescentes da imunopurificação de CGRs a partir de
retinas de camundongo P1. Estas células foram plaqueadas sob diferentes
condições, sendo elas: DMEM (controle), forscolina 5μM, AMG ou MMG. Estas
células não tiveram o meio trocado ao longo de alguns dias o que faz com
que os neurônios morram privilegiando as células da glia que, por outro lado
resistem e continuam proliferando.
Figura 38. Proliferação de células de glia de Müller de camundongo por 120 horas em meio DMEM sob diversas condições. 6 retinas de animais P1 foram dissecadas e dissociadas, as CGR foram isoladas e o restante foi plaqueado em lamínulas nas condições acima (Controle, 5μM Forscolina, AMG 1:2 e MMG 1:2). O meio não foi trocado durante os 5 dias em cultura, as lamínulas foram lavadas com PBS, fixadas e reagidas com anticorpo contra S100, um marcador glial e DAPI.
As células gliais foram marcadas com anticorpo contra S-100
marcador específico glial (em vermelho; TRITC). Na presença de forscolina,
houve um aumento do número de células plaqueadas se comparado ao
controle. Na presença de AMG ou de MMG o número de células gliais foi
drasticamente maior.

103
Com os estudos que fizemos não é possível concluir se os fatores que
induzem proliferação glial são os mesmos que induzem sobrevida e
neuritogênese dos neurônios simpáticos e de CGRs.
4.2.6 MEIO CONDICIONADO RADIOATIVO
O propósito deste experimento foi fazer com que as culturas até então
utilizadas como sensores do efeito biológico dos MCs (culturas purificadas de
neurônios simpáticos) venham a funcionar como etapa de purificação de
possíveis fatores tróficos. Desta forma, culturas de glia de Müller de
camundongo em monocamada foram obtidas utilizando o mesmo
protocolo utilizado para o preparo do MMG. A partir do ponto de
monocamada purificada de glia de Müller a cultura foi lavada e recebeu
DMEM isento de metionina. Ao sobrenadante foram adicionados 200μCi de
metionina marcada com enxofre radiativo (Met-S35) e a cultura foi mantida
em estufa para condicionamento do meio por 24 horas. O meio
condicionado radioativo obtido foi depletado de Met-S35 livre através de
centrifugação em filtros Centricon ® de corte baixo (10Kda) e utilizado para
estimular culturas purificadas de neurônios simpáticos do GSC de ratos. Após
exposição dos neurônios ao meio radioativo era esperado que estes viessem
a interagir com as proteínas marcadas produzidas pela glia e que,
conseqüentemente algumas destas proteínas marcadas fossem identificadas
no extrato protéico da cultura de neurônios simpáticos. A evidência de
interação e possível internalização destas proteínas gliais faz destas molécula

104
possíveis candidatas a estarem mediando o efeito trófico promovido por
MMG. O extrato protéico da cultura de neurônios e da monocamada de
glia de Müller além do próprio MMG marcado foram separados
eletroforeticamente em gel SDS-PAGE 10% de poliacrilamida. O gel foi
revelado por auto radiografia. Pelo menos quatro bandas radioativas foram
visualizadas no extrato protéico da cultura de neurônios. Várias bandas
radioativas foram visualisadas no MMG e, obviamente, um “rastro” radioativo
foi visualizado no extrato protéico da monocamada de glia de Müller (figura
39).
Figura 39. Auto-radiografia do gel com AMG marcado radiativamente. Foram detectadas pelo menos quatro bandas radiativas no extrato de neurônios simpáticos do GSC que foram expostos ao meio condicionado radiativo.

105
4.3 CULTURAS DE CÉLULAS DE MÜLLER PURIFICADAS DE PINTO.
A glia de Müller é o principal tipo glial encontrado nas retinas de
vertebrados e tem um papel fundamental na manutenção da sobrevida das
células ganglionares da retina.
Em 2002 desenvolvemos um método de cultivo celular que permitiu a
obtenção de monocamada de glia de Müller livre de neurônios a partir de
culturas de retina total (glia + neurônios). Na figura 40 é possível observar três
diferentes estágios da purificação de glia de Müller de ave. Após a
confluência (neurônios + glia figura 40A). Tratamento com solução de ácido
ascórbico (AA) 4mM por 3 horas (figura 40B), e monocamada de glia de
Müller purificada após lavagens com meio DMEM e mantida por 24 horas em
DMEM + 10% SFB para recuperação da glia após tratamento com AA (figura
40C). A partir deste ponto as culturas passaram a receber DMEM sem soro e
ter o meio condicionado coletado a cada 48 horas e substituído por meio
fresco. Pelo menos duas coletas foram possíveis para cada placa de cultura.
Figura 40. Passos da purificação de glia de Müller de embrião de pinto para obtenção de AMG. (A), cultura de células da retina (neurônios + glia); (B), lesão de neurônios por tratamento agudo com 4mM de AA por 3 horas; e (C), monocamada de glia de Müller purificada após lavagens e recuperação da glia.

106
4.3.1 AMG APRESENTA ATIVIDADE TRÓFICA TANTO NA FRAÇÃO QUE SE LIGA À
HEPARINA QUANTO NA FRAÇÃO LIVRE.
O AMG é mais efetivo na manutenção de neurônios simpáticos em
cultura se comparado ao RCM, mantendo a maioria (≈ 100%) das células
viáveis após 48 horas (figura 41) (contra ≈ 90% mantidas por RCM).
Assim, como foi demonstrado para o RCM, o AMG é também sensível
à temperatura e tem sua atividade trófica mantida após diálise (dado não
mostrado). A atividade trófica do AMG também é mantida na presença de
receptores quimera de imunoglobulinas que mimetizam receptores Trk (TrkA-
IgG, TrkB-IgG ou TrkC-IgG), não sendo também inibida na presença de
anticorpos anti-GDNF, anti-TGFβ ou anti-CNTF. Por outro lado, diferentemente
do RCM, o AMG apresentou atividade trófica tanto na fração que se liga à
heparina (BF) quanto na fração livre (FF) (figura 41). Esse último dado sugere
a glia de Müller está secretando pelo menos dois fatores (ou grupos de
fatores) capazes de promover a viabilidade de neurônios simpáticos em
cultura considerando que a coluna não está saturada. Desta forma as
moléculas presentes no AMG, podem ser, através de uma coluna de
heparina dividido em duas frações ativas (BF e FF) com diferentes afinidades
pela matriz de heparina.
A partir deste ponto passamos a trabalhar somente com BF por se
tratar de uma fração ativa com atividade trófica comparável ao AMG total
e por possuir menos proteína por volume quando comparada a FF, isto é, BF
é a fração mais indicada para as análises bioquímicas posteriores.

107
Figura 41. Caracterização do AMG. O AMG possui dois componentes quanto a afinidade a heparina. Ambos exercem forte atividade trófica no simpático de embriões de pinto E10. A presença de receptores solúveis e de anticorpos contra fatores com conhecida atividade trófica no simpático embrionário de pintos ano interferiu com a atividade trófica de AMG.
Figura 42. Efeito de AMG em diferentes populações neuronais periféricas (simpático e GRD). AMG exerce forte efeito neuritogênico sobre explantes de gânglios periféricos como pode ser observado da figura acima.
Na figura 42 pode-se observar que o AMG dá suporte trófico a mais de
uma população neuronal periférica como é o caso do gânglio da raiz dorsal

108
de embriões de pinto E10. O efeito de AMG sobre GRD, talvez, seja menos
expressivo que sobre o simpático visto que a população neuronal do
simpático e mais homogênea enquanto o GRD é composto por três
diferentes tipos neuronais responsivos a diferentes fatores tróficos durante o
desenvolvimento.
4.3.2 FRACIONAMENTO DO AMG EM COLUNA DE HEPARINA
Com o intuito de fracionar o MC para possível isolamento e
identificação de fatores com atividade trófica em neurônios simpáticos in
vitro, alíquotas de 50 mL de AMG sem soro foram separadas em fração
ligada (BF, bound fraction) e fração livre (FF, free fraction) com auxílio de
uma coluna de 5 mL de heparina (matriz de sefarose de alta performance –
Hi-Trap, Amersham) acoplada a um sistema FPLC.
A coluna, transparente, enquanto carregada apresentava cor
vermelha devido ao indicador de pH (vermelho de fenol) presente no meio
de cultura condicionado pela glia. Como a cor do indicador poderia
interferir na leitura das alíquotas obtidas do FPLC a coluna foi lavada até
retornar a cor branca original. Outro intuito da lavagem foi a exclusão de
moléculas de eluição lenta portanto a coluna permaneceu sendo lavada
por aproximadamente 1 hora mesmo após eluição do vermelho. Quando a
leitura a 214nm da alíquota obtida após 1 hora de lavagem se igualou a do
tampão fosfato 50mM a coluna estava pronta para ser submetida ao
programa de eluição por gradiente de NaCl do sistema FPLC.

109
O perfil de eluição da coluna pode ser observado na figura 43. O
gráfico foi construído a partir do gradiente linear de NaCl (linha vermelha)
medido em percentagem de B (tampão fosfato adicionado de NaCl 2M;
sendo NaCl 2M 100% de B) e da leitura da absorbância a 214nm das
alíquotas obtidas da eluição da coluna (linha azul) em unidade arbitrária. O
coletor de alíquotas foi programado para coletar alíquotas de 1,5mL.
Figura 43. Perfil de eluição do AMG. Uma fração ativa do AMG pode sr eluída da coluna de heparina com um gradiente linear de tampão fostato (50mM) adicionado de 2M de NaCl (em vermelho, [ ] de B). Após eluição do pico o FPLC alterna para 100% de B para lavagem da coluna. Em azul, temos a leitura a 214nm da absorbância das alíquotas (pontos no traço azul).
Cada alíquota foi dessalinizada e testada quanto a sua atividade
biológica em culturas de neurônios simpáticos (ver figura 13 da seção de
materiais e métodos). As alíquotas com maior atividade trófica no simpático

110
foram as que se encontravam compreendidas dentro do pico de eluição da
coluna como pode ser observado na figura 44.
Figura 44. Micrografias ilustrando a localização, no pico, da atividade trófica em neurônios simpáticos. As setas vermelhas indicam pontos sem atividade biológica no simpático enquanto as setas pretas apontam alíquotas com potente ação trófica em neurônios simpáticos em cultura. O efeito trófico está compreendido entre as alíquotas 5 e 17.
Alíquotas, com atividade biológica, compreendidas no pico de
eluição da coluna foram concentradas 40x em filtros Centricon ® e
submetidas à separação em mini gel SDS-PAGE 10 % poliacrilamida. O gel foi
corado pelo método de impregnação por prata e a fração que apresentou
menos bandas foi submetida à análise em espectrômetro de massas (Maldi-
TOF-TOF) figura 45 no Núcleo de Proteômica do Instituto de Biofísica Carlos
Chagas Filho chefiado pelo Dr. Paulo Bisch. O resultado da análise porém,

111
apenas nos revelou que moléculas do soro ainda estavam presentes em
abundância em tal fração o que pôde ser contornado com adicionais
lavagens da cultura de glia para remoção de resíduos de soro.
Figura 45. Análise das bandas mais expressivas de fração do AMG em espectrômetro de massas. A análise revelou proteínas que apesar de estarem descritas na literatura como indutoras ou auxiliares no efeito trófico em neurônios simpáticos (em vermelho) foram identificadas como proteínas bovinas. As demais proteínas são claramente proteínas oriundas do soro fetal bovino. As bandas de número 2 e 6 não foram identificadas.
O uso da coluna de heparina como único instrumento de
fracionamento do meio condicionado se mostrou ineficiente na busca do
isolamento dos possíveis fatores tróficos presentes no AMG pois, na tentativa
de fracionar o pico obtido por gradiente linear e conseqüentemente obter
uma fração mais purificada submeti 50mL de AMG ao FPLC programado
para executar um gradiente em degraus, o que dividiu o pico anterior em
três frações (figura 46) As duas primeiras frações apresentaram potente
efeito trófico em cultura quando utilizados na diluição de 1:10 ou 1:20 (figura

112
47) sendo a terceira fração menos potente na indução de sobrevivência e
neuritogênese in vitro (dado não mostrado).
Figura 46. Fracionamento em degraus do primeiro pico de eluição. O primeiro pico foi dividido pico em três frações através da eluição da coluna por gradiente em degraus. Os picos foram dessalinizadas e testados em cultura quanto à atividade trófica. Os picos 1 e 2 demonstraram potente atividade trófica em neurônios simpáticos em cultura.
Figura 47. Micrografias da atividade trófica promovida pelos picos 1 e 2 em culturas de neurônios simpáticos de embrião de pinto E10. A esta figura foi adicionado um controle adicional, em (D) e possível visualizar o efeito trófico de uma cultura total de retina embrionária de pinto mostrando que o efeito trófico de AMG (e possivelmente também aplicável ao MMG) não se deve a ações indiretas do ácido ascórbico.

113
Nenhum dos três picos foi submetido à análise em espectrômetro de
massas. É fato, porém, que o efeito trófico promovido pelas alíquotas dos
dois primeiros picos em culturas de neurônios do simpático se equivale ao
promovido pelo AMG total (figura 47).
4.3.3 AMG INDUZ MIGRAÇÃO DE CÉLULAS DE SCHWANN A PARTIR DE
EXPLANTES DE NERVO CIÁTICO DE RATOS ADULTOS.
O AMG é composto por uma variedade de fatores secretados pela
glia de Müller de embrião de pinto e possui, além de um componente com
potente atividade trófica sobre neurônios centrais e periféricos, um
componente que atua sobre tipos gliais centrais promovendo proliferação (o
mesmo efeito observado em MMG sobre a proliferação glial, figura 38) e
periféricos promovendo migração como demonstrado na figura 48.
Com o objetivo de estimular a migração de células de Schwann para
fora do explante de nervo ciático de rato adulto, explantes deste nervo
foram plaqueados sobre poli-L-lisina e laminina e cultivados sob diferentes
condições experimentais. Meio de cultura DMEM foi usado como controle e
NGF (50ng/mL), BDNF (50ng/mL) isoladamente ou combinados (NGF + BDNF)
e AMG foram usados como condições experimentais.
Na condição controle (DMEM) não há migração glial para fora do
explante. Entretanto, NGF e BDNF (isolados ou combinados) resultaram em
modesta migração glial quando comparada à migração promovida por
AMG.

114
Meio condicionado por células mesenquimais de rato (MC-MO, meio
condicionado de medula óssea), que são as células aderentes de uma
cultura de medula óssea. Tais células são descritas na literatura como
produtoras de uma série de fatores neurotróficos, entre eles NGF, BDNF, NT-3
e GDNF (CHEN et al., 2005; YE et al., 2005). Este MC que é objeto de estudo
do Laboratório de Neurobiologia Celular e Molecular da Doutora Rosália
Mendez-Otero também foi usado como condição experimental mas não
apresentou efeito significativo na migração glial quando comparado a
migração promovida por AMG. A migração promovida por MC-MO foi
equivalente à promovida por NGF e/ou BDNF.
Figura 48. Perfil de migração de células de Schwann sob diferentes condições. A migração inexiste na condição controle e é equivalente entre as demais condições com exceção do explante tratado com AMG onde é possível observar uma abundante quantidade de células migrando para fora do explante.
Não é possível definir a partir dos dados gerados se a ação do AMG
sobre a glia central ou periférica provém do mesmo conjunto de moléculas
que agem com atividade trófica sobre neurônios.

115
4.3.4 AMG INDUZ REGENERAÇÃO DE NEURÔNIOS PERIFÉRICOS E CENTRAIS
Os resultados mostrados até agora evidenciaram a potente atividade
trófica promovida por AMG em populações de neurônios centrais (CGRs) e
periféricos (simpático) in vitro. Neste sentido, tivemos o interesse de investigar
se o conjunto de moléculas solúveis presentes no AMG era capaz de exercer
alguma atividade trófica em um modelo in vivo.
Em colaboração com o laboratório de Neurobiologia Celular e
Molecular do Instituto de Biofísica, chefiado pela Doutora Rosália Mendez-
Otero e em colaboração com o aluno de doutorado Vitor Resende,
testamos alíquotas sem soro de AMG num modelo de secção de nervo
ciático por ser uma técnica de domínio e uso rotineiro em tal laboratório e
também pelo motivo do nervo ciático ter constituição mista, ou seja, por ser
composto por projeções de neurônios motores (cujos corpos celulares
encontram-se na medula espinhal) e por neurônios sensoriais do gânglio da
raiz dorsal (GRD) no nível dos segmentos da vértebra lombar 4 (L4) e da
vértebra lombar 5 (L5) da coluna vertebral de ratos adultos. A lesão foi
efetuada em animais profundamente anestesiados com o auxílio de um
bisturi. Seccionamos por completo o nervo e inserimos as duas extremidades
em um tubo de polietileno de 7mm de comprimento. No interior deste tubo,
em cada animal, componentes das diferentes condições experimentais
foram injetados. O corte foi suturado e os animais devolvidos a uma caixa de
biotério para recuperação. Após 40 dias da cirurgia, foram implantados
cristais de DiI no coto distal do nervo (figura 49) para que partículas destes

116
cristais fossem transportadas retrógradamente e marcasse os corpos
celulares cujos axônios regeneraram sob influência ou não do AMG
possibilitando assim, a quantificação da atividade trófica do AMG na
regeneração deste nervo. Através da análise dos cortes foi evidente a
distinção entre células positivas e negativas para DiI. É importante mencionar
que todos os tipos neuronais do GRD (corpos pequenos, médios e grandes,
uma característica deste gânglio) foram efetivamente representados como
positivos para DiI, indicando a regeneração de todos os tipos neuronais que
compõem o GRD. Como os diferentes tipos neuronais do GRD respondem de
forma distinta após lesão do ciático (TANDRUP et al., 2000) nós não podemos
afirmar que todos os tipos neuronais marcados regeneraram devido a ação
trófica de AMG. Na figura 49 é também possível observar a distinção entre
células positivas e negativas para DiI no GRD.
Figura 49. Inserção do cristal de DiI no coto distal do nervo ciático (A) e células
positivas e negativas para DiI (lado a lado) confirmando que não há efeito de difusão.

117
Após as seis semanas do tratamento os animais foram sacrificados e os
gânglios, o nervo ciático e a medula espinhal dissecados e fixados para
corte. Quando comparamos os cortes do GRD no nível da vértebra L4 entre
os grupos experimentais, os animais tratados com AMG revelaram um
aumento significativo do número de células DiI positivas (AMG, 207,7 ± 7,5
p<0,001) quando comparados aos tratados com meio de cultura (DMEM,
159,8 ± 4,3) ou com o controle positivo (BDNF + NGF, 162,6 ± 4,7) (figura 50).
Analisando os cortes do GRD no nível da vértebra L5 tratados com MC
(AMG, 198.0 ± 8,9) com os tratados com meio de cultura (DMEM, 151,3 ± 6,7)
observamos um aumento significativo na regeneração deste nervo (refletido
pelo numero de células DiI positivas nos cortes). Nessa população de
neurônios, o MC se mostrou um pouco inferior ao controle positivo (BDNF +
NGF, 223,5 ± 10,9) (figura 50).
A análise dos cortes de medula (motoneurônios) localizados na mesma
altura de segmentos da coluna vertebral revelou uma equivalência na
captação de partículas de DiI entre os animais tratados com MC (AMG, 22,6
± 0,9) quando comparado com o controle positivo (BDNF + NGF, 22,2 ± 1.2),
porém foi evidente que a regeneração promovida pelo tratamento com MC
foi significativamente superior à regeneração observada nos animais
tratados somente com meio de cultura (DMEM, 15,3 ± 0,6) como pode ser
observado na figura 51.
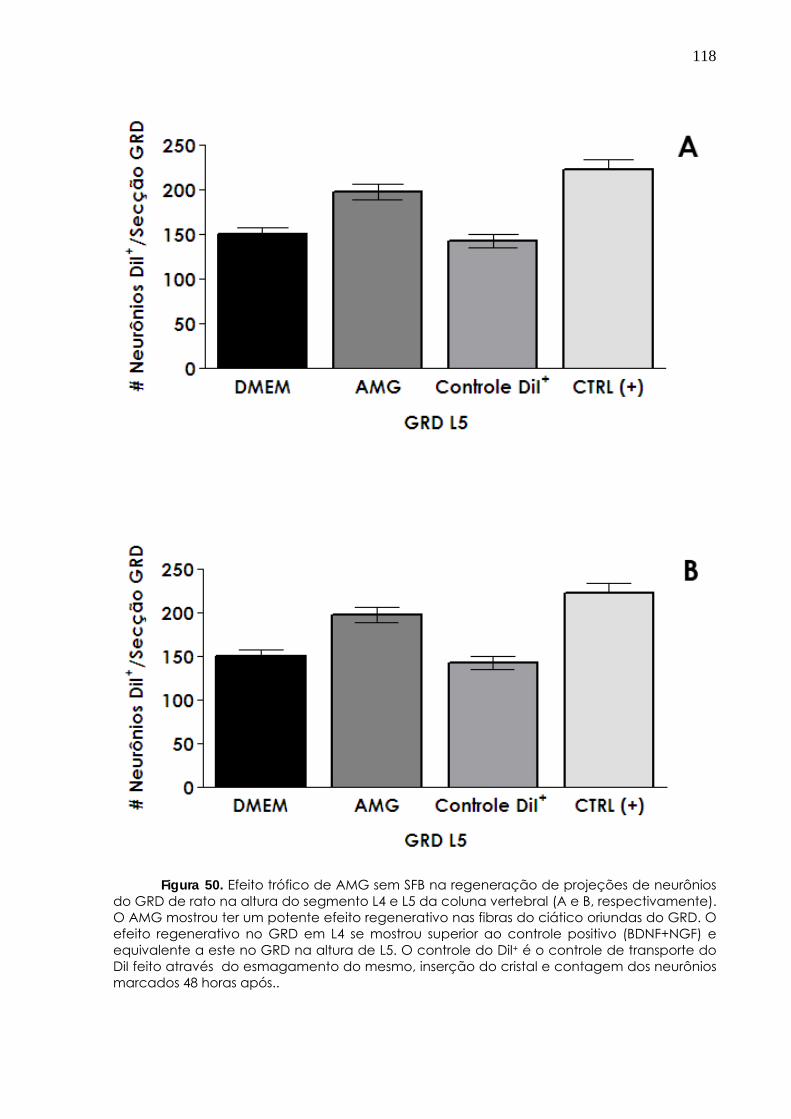
118
Figura 50. Efeito trófico de AMG sem SFB na regeneração de projeções de neurônios do GRD de rato na altura do segmento L4 e L5 da coluna vertebral (A e B, respectivamente). O AMG mostrou ter um potente efeito regenerativo nas fibras do ciático oriundas do GRD. O efeito regenerativo no GRD em L4 se mostrou superior ao controle positivo (BDNF+NGF) e equivalente a este no GRD na altura de L5. O controle do DiI+ é o controle de transporte do DiI feito através do esmagamento do mesmo, inserção do cristal e contagem dos neurônios marcados 48 horas após..

119
Figura 51. Efeito trófico de AMG sem SFB na regeneração de projeções de motoneurônios da medula de rato adulto na altura do segmento L4 e L5 da coluna vertebral. O AMG mostrou ter um potente efeito regenerativo nos axônios dos motoneurônios que compõem o ciático sendo equivalente os controle positivo do experimento.

120
5. DISCUSSÃO

121
DISCUSSÃO
Neste trabalho investigamos a natureza e o efeito trófico promovido
por moléculas secretadas por explantes de retina de embriões de pinto e de
culturas de glia de Müller purificadas, de ave e de camundongo.
Descrevemos inúmeros efeitos tróficos em neurônios periféricos e centrais,
analisando vias de ativação intracelulares pelas quais o MC sinaliza, e
descartando a ação isolada de possíveis fatores conhecidos responsáveis
por efeitos semelhantes.
Como foi demonstrado em estudos anteriores, tanto o MC de retina de
rato pós-natal (ARAÚJO e LINDEN, 1993) quanto o MC de retina de embrião
de pinto (ARY-PIRES et al., 1997) promovem a sobrevivência de CGRs em
cultura, de forma dependente do desenvolvimento. Os fatores que
compõem o MC ainda não foram esclarecidos, mas acredita-se que
contenha uma variedade de substâncias tróficas dentre outros fatores. A
retina com seus múltiplos tipos e subtipos celulares, é um ambiente propício
para interações bastante complexas como as de efeitos secundários, onde
fatores produzidos por uma célula agem em uma segunda promovendo
uma resposta possível de influenciar uma terceira célula.
Ao longo do desenvolvimento do nosso trabalho, fizemos uma série de
experimentos in vitro utilizando uma população relativamente pura de
células secretoras, as células gliais de Müller, em busca de neurotrofinas e
fatores tróficos específicos. Investigamos efeitos na sobrevida e
neuritogênese de populações purificadas de neurônios, periféricos e centrais,

122
tentando interferir em pontos em comum das cascatas de sinalização tendo
como alvo algumas já descritas, como por exemplo as das MAPK e da Pi3C
ativadas por NGF, para o modelo simpático em cultura.
Neste trabalho mostramos que MC por explantes de retina ou por glia
de Müller de ave ou de camundongo purificadas aumentam a
sobrevivência de neurônios simpáticos de embrião de pinto em cultura de
maneira dependente da concentração. Embora estudos prévios tenham
mostrado que NGF, BDNF e NT-3 são expressos na retina de ave durante o
desenvolvimento (HALBÖÖK et al., 1996; FRADE et al., 1999; DAS et al., 1997),
a atividade trófica observada não é devido a nenhuma neurotrofina
clássica. A sobrevivência induzida tanto por RCM quanto por AMG não foi
bloqueada nem por anti-NGF nem por receptores quimera de
imunoglobulinas específicos para neurotrofinas, que impediriam neurotrofinas
de se ligar aos seus respectivos receptores, TrkA, TrkB, ou TrkC.
A glia de Müller é o tipo macroglial majoritário presente na retina e
muitas funções são atribuídas a essas células, o que inclui papéis estruturais e
de nutrição a neurônios (SARTHY e RIPPS, 2001), sinalização por Ca2+ quando
estimulada por ligantes (METEA e NEWMAN, 2006), remoção
(captação/tamponamento) de íons e neurotransmissores do meio
extracelular, além do controle do pH no meio extracelular (NEWMAN e
REICHENBACH, 1996) e controle do movimento de água do espaço
extracelular (WURM et al., 2006). Após lesões agudas, a glia de Müller é uma
fonte potencial para a regeneração neural, reentrando no ciclo celular e
gerando novos neurônios e células gliais (REH e FISCHER, 2006). Uma das

123
propriedades mais interessantes da glia de Müller é a de neuroproteção,
através da liberação de fatores tróficos e de agentes anti-oxidantes, como a
glutationa (GARCÍA e VECINO, 2003). É sabido que alguns fatores tróficos são
secretados pela glia de Müller, sendo capazes de manter alguns tipos
neuronais, como CGRs de ratos em desenvolvimento (RAJU e BENNETT, 1986;
MEYER-FRANKE et al., 1995), CGRs de porcos adultos (GARCÍA et al., 2002),
neurônios simpáticos de embrião de pinto (REIS et al., 2002), entre outros.
Um dos fatores presentes em abundância na retina de galinha e que
tem afinidade por heparina é o FGFb (DESIRÉ et al., 1998; CHAI e MORRIS,
1999), e questionando este seria o fator ou um dos fatores envolvidos na
sobrevivência neuronal simpática. A aplicação do RCM através de uma
coluna de heparina (Hi-Trap) revelou que a fração do RCM que se liga à
heparina possui efeito trófico em neurônios simpáticos, enquanto a fração
livre não apresentou efeito algum. Entretanto, o FGFb (0.1-50nM) não foi
capaz de induzir sobrevivência em culturas de neurônios simpáticos.
Testamos também anticorpos anti-TGFβ, anti-GDNF e anti-CNTF tanto no RCM
como no AMG. Esses anticorpos não tiveram efeito algum na sobrevivência
promovida por esses meios condicionados, mas bloquearam
completamente o efeito dos seus respectivos fatores tróficos. É importante
ressaltar que pelo fato de termos apenas uma pequena alíquota de cada
anticorpo que bloqueia os fatores tróficos, em momentos diferentes da tese,
não pudemos realizar experimentos com combinações de anticorpos em
uma única condição experimental. Seria ideal realizar tal experimento e
observar influência do bloqueio múltiplo sobre a sobrevida dos neurônios

124
simpáticos e das CGRs talvez respondendo o quanto determinado conjunto
de moléculas contribui com o efeito trófico observado revelando a ação
múltipla ou não destes fatores. Esse experimento será realizado quando
tivermos a disposição estes bloqueadores.
Ao investigar que vias intracelulares estavam sendo ativadas por RCM
e MMG, observamos que a enzima Pi3-cinase está envolvida na
sobrevivência neuronal promovida pelos fatores tróficos retinianos e da glia
de Müller. LY294002 (um inibidor da via da Pi3C) em uma concentração de
50μM bloqueou totalmente o efeito trófico em neurônios simpáticos mantidos
por RCM. Muitos trabalhos têm descrito que fatores tróficos como NGF e
fatores de crescimento do tipo insulina (IGF) promovem a sobrevivência de
neurônios através de vias dependentes de Pi3C (SEGAL e GREENBERG, 1996;
CROWDER e FREEMAN, 1998). A capacidade dos fatores tróficos de
promover a sobrevivência tem sido atribuída, pelo menos em parte, à via de
sinalização da Pi3C/c-AKT cinase. Vários pontos desta via de sinalização
(PI3C/c-AKT) foram identificados e podem esclarecer parte dessa cascata
regulatória na indução de sobrevivência. Entre esses pontos estão inclusos
dois componentes envolvidos em morte celular, como as proteínas Bad e
caspase 9, fatores de transcrição da família forkhead, e a cinase IKK, que
regula o fator de transcrição NF-κβ (DATTA et al., 1999).
Estudamos ainda o efeito da tirfostina (AG490), um inibidor específico
da tirosina cinase JAK-2 envolvida na liberação de fatores de crescimento
mediada por estresse. Citocinas da família da interleucina-6 (IL-6) ativam JAK
e efetores acessórios desta via como STAT (PETERSON et al., 1999). AG490

125
(50μM) bloqueou os efeitos de sobrevivência promovidos por RCM, sugerindo
algum envolvimento da via JAK/STAT na mediação do efeito trófico.
A via das MAPK/ERK representa uma distinta via de sinalização
intracelular que tem mostrado ser ativada por neurotrofinas e estímulos
mediados por estresse (SEGAL e GREENBERG, 1996). O PD098059, um inibidor
seletivo de MAPK, quando utilizado em concentrações até 50μM, não teve
efeito algum na sobrevivência de neurônios simpáticos induzida por RCM,
mostrando que o efeito trófico do RCM se dá de forma independente desta
via.
Tendo excluído a ação isolada de neurotrofinas conhecidas como
sendo responsáveis pelos efeitos de sobrevivência promovidos tanto por
RCM quanto por AMG, algumas considerações podem ser feitas a partir das
propriedades ativas desses fatores tróficos.
1. A morfologia dos neurônios simpáticos mantidos por RCM é diferente
daquela vista na presença de NGF. Os neurônios têm corpo pequeno e
refringente com neuritos finos e longos quando mantidos por RCM. Por outro
lado, neurônios tratados com NGF retêm características típicas como um
grande corpo celular refringente, núcleo facilmente identificável, além de
neuritos longos e grossos. Em culturas de células do gânglio da raiz dorsal
adulto foi demonstrado que diferentes neurotrofinas promovem mudanças
morfológicas em subclasses definidas de neurônios sensoriais (GAVAZZI et al.,
1999). Não está claro como cada efeito pode ocorrer, mas há clara
evidência de remodelamento dendrítico local induzido por neurotrofinas
específicas (McALLISTER et al., 1997). A recente demonstração da possível

126
ligação entre o receptor de baixa afinidade p75 e a GTPase Rho permite
estabelecer uma relação entre a ação das neurotrofinas, a regulação de
receptores e o rearranjo do citoesqueleto que podem resultar em cada uma
das mudanças na morfologia celular (YAMASHITA et al., 1999).
2. McKinnon et al. (1998) mostraram que MC de culturas envelhecidas
de retina contém um fator não identificado que induz expressão seletiva do
gene cm2 do receptor muscarínico, porém não induzindo a expressão de
cm3 e nem de cm4. Incubando células da retina com 14 diferentes fatores
tróficos conhecidos, não houve aumento da expressão de cm2, sugerindo
então a possibilidade de um novo fator retiniano, regulado pelo
desenvolvimento. Recentemente, Belmonte et al. (2000) relataram que esses
efeitos eram induzidos por um fator de peso molecular maior que 30 kD,
sensível à clivagem proteolítica e secretado por células de Müller. Nós não
examinamos que efeitos RCM, AMG ou MMG podem ter na expressão
gênica em neurônios simpáticos. Entretanto, para o aspecto que
observamos (sobrevivência e neuritogênese) o fator não apresenta
regulação pelo desenvolvimento.
3. Ary-Pires et al. (1997) mostrou que o MC de retina de embrião de
pinto que promove a sobrevivência de CGRs é sensível à tripsina, sugerindo
que um efeito similar pode se esperado para o fator que promove a
sobrevivência de neurônios simpáticos.
4. O AMG manteve a sobrevivência de neurônios simpáticos em cerca
de 97%, e o efeito durou pelo menos 5 dias em cultura. MMG foi ainda mais
potente, mantendo 100% dos neurônios retinianos por mais de 5 dias em

127
cultura. Em contraste, RCM mostrou uma resposta de sobrevivência inferior,
de cerca de 80%, e por somente 2 dias in vitro (por volta de 72 horas a
maioria das células estava morta). Com estas observações temos algumas
evidências das diferenças, em termos funcionais, da atividade entre o
conjunto de fatores tróficos que compõem o RCM, o AMG e o MMG. Os
testes dos efeitos de neurotrofinas na secreção do fator retiniano (RCM)
revelaram a existência de um efeito indireto de ambos NGF e BDNF no efeito
trófico final do MC. É possível que existam outras complexas interações
regulando a secreção que podem estar ausentes em culturas purificadas de
glias de Müller. Essa diferença pode refletir na perda celular nos explantes de
retina na ausência de neurotrofinas adicionais ou a supressão de vias de
secreção das células de Müller por outras células da retina. Assim, pode ser
que estejamos observando efeitos de mais de um fator, como observado no
AMG no qual tanto a fração que se liga como a que não se liga à heparina
promovem sobrevivência neuronal. Ou como mostrado com MMG no
modelo simpático (GSC) de rato, onde múltiplas vias intracelulares foram
ativadas. Além do mais, fatores retinianos isolados de colículo superior de
roedor têm sido descritos como sendo um proteoglicano condroitina-
sulfatado, que provavelmente contém fatores de sobrevivência ativos
ligados formando um complexo (SCHULZ et al., 1990; HUXLIN et al., 1995).
Em relação as CGRs, estes são neurônios de projeção que ligam a
retina aos centros cerebrais superiores essenciais para a visão. As CGRs
constituem apenas uma pequena percentagem da população total de
células retinianas (≈ 0,5%). Sendo assim, é difícil detectar alterações

128
específicas nas CGRs no lisado total de retina. O uso da técnica de
imunopurificação das CGRs foi neste sentido apropriada para observar
mudanças na neuritogênese e na sobrevida desta população livre de
influências de outras células. Adaptamos o procedimento para
camundongos pós-natais utilizando anticorpo (Thy1.2/CD90.2) que
reconhecem moléculas de superfície das CGRs desta espécie, e que nos
permitiu isolar com sucesso CGRs entre P1 e P8. A segunda fase de morte
celular (que ocorre por volta de P2 nos camundongos) está correlacionada
temporalmente com a chegada dos axônios retinianos no colículo superior,
alvo das CGRs (YOUNG, 1984). Frade e Barde (1999) mostraram que,
enquanto a primeira fase de morte das CGRs está relacionada com a
interação do NGF com a subunidade de baixa afinidade p75, a segunda
fase por outro lado, está associada à competição por pequenas
quantidades de fatores tróficos disponíveis nos tecidos alvo. Alguns achados
sustentam tal afirmação, como a aplicação de BDNF diretamente no
colículo superior de hamster, reduzindo a taxa de apoptose de CGRs (MA et
al., 1998). De forma semelhante, a aplicação de injeções de BDNF ou de NT4
no colículo superior resgata CGRs neonatais que normalmente morreriam por
apoptose (CUI e HARVEY, 1995).
Após lesão no nervo óptico, as CGRs não se regeneram, e assim como
a maioria dos neurônios centrais, morrem por apoptose. Em ratos adultos, por
exemplo, cerca de 85% das CGRs morrem no período de 2 semanas após a
transecção do nervo óptico (VILLEGAS-PÉREZ et al., 1988), devido à privação
dos fatores tróficos dos alvos distantes e das células não neurais que mantém

129
a harmonia do micro ambiente que envolve os axônios retinianos. A morte
decorrente da axotomia das CGRs de ratos adultos foi caracterizada como
apoptótica (BERKELAAR et al., 1994; GARCIA-VALENZUELA et al., 1994;
ISENMANN et al., 1997). O que tem se tentado para evitar a degeneração
das CGRs após lesão do nervo óptico é a eventual aplicação de fatores
tróficos diretamente sobre as CGRs, como uma estratégia terapêutica para
resgatar neurônios que foram submetidos a axotomia. Nesse sentido, uma
única injeção de BDNF no vítreo resgata por completo as CGRs de ratos
adultos 1 semana após a axotomia (MANSOUR-ROBAEY et al., 1994). Da
mesma forma, o aumento da expressão de CNTF induzida através da
aplicação de adenovírus contendo o gene do CNTF (Ad-CNTF) no vítreo
também se revelou como uma estratégia muito eficiente para o resgate de
CGRs de ratos adultos, aumentando em 155% o número destas células após
14 dias da axotomia, comparado com o animal injetado apenas com o
adenovírus controle(WEISE et al., 2000).
Nossos resultados mostraram que a atividade trófica da glia de Müller
de camundongo, mas não de aves, induz sobrevida e neuritogênese de
CGRs, e que a glia de Müller de camundongo é um excelente substrato para
a manutenção e para o desenvolvimento de CGRs de camundongos pós-
natais. Trabalhos anteriores mostraram que durante o desenvolvimento, o
contato celular entre a glia de Müller e as CGRs é um passo necessário para
o crescimento neurítico desses neurônios de projeção, porém os fatores
solúveis liberados por essas células não eram suficientes para o processo
(RAJU e BENNETT, 1986). Mais recentemente, foi mostrado que a glia de

130
Müller aumenta a sobrevida e induz neuritogênese em CGRs de retinas de
porcos adultos (GARCÍA et al., 2002; GARCIA e VECINO, 2003),
provavelmente por meio de moléculas secretadas, e não apenas por
mecanismos envolvendo contato entre as células. É sabido que a glia de
Müller de diferentes espécies secreta diversas moléculas tróficas, como BDNF
(CAMINOS et al., 1999; SEKI et al., 2005), FGFb (GAO et al., 1992; HARADA et
al., 2000), GDNF (HARADA et al., 2003), que entre outros compostos,
poderiam participar da modulação dos processos de neuritogênese e
sobrevivência das CGRs. Nossos dados confirmam que a glia de Müller
apresenta atividade trófica capaz de manter neurônios periféricos
(simpáticos) e centrais (CGRs), além de induzir atividade regenerativa em
duas outras populações de neurônios (GRD e motoneurônios). Alguns
trabalhos prévios mostraram que a manipulação de tipos celulares como
fibroblastos e astrócitos com a introdução de genes responsáveis pela síntese
de fatores tróficos resgata neurônios de lesões. O mesmo foi observado
quando a glia de Müller foi introduzida com adenovirus contendo o gene do
BDNF, resgatando CGRs após axotomia (Di POLO et al, 1998). Nossos
resultados sugerem que a manipulação da glia de Müller de aves e de
camundongos possa no futuro ser usada para recuperar e regenerar
populações específicas neuronais lesadas.
Embora não tenha sido possível o isolamento e a identificação dos
fatores gliais, nossos resultados sugerem que um dos compostos secretados
seja um dos membros da família do CNTF. O composto AG490, que bloqueia
a via de sinalização (JAK-STAT) ativada por receptores dessa família, inibiu

131
por completo a sobrevida dos neurônios mantidos com RCM. MMG estimulou
a fosforilação de STAT3, como demonstrado em neurônios simpáticos de
rato. E um anticorpo que bloqueia receptores que apresentam a subunidade
gp130 (presente no receptor trimérico característico de membros da família
do CNTF, como LIF e Cardiotrofina), bloqueou a expressão do peptídeo VIP
por neurônios simpáticos embrionários de aves (H. Rohrer, comunicação
pessoal; figura 52. Além disso, nas CGRs plaqueadas na presença de MMG, o
tamanho do axônio, que geralmente era longo e fino, assemelhava-se em
tamanho com os tratados com CNTF, mas não BDNF. De fato, encontramos
base na literatura para afirmar que CNTF (e LIF) é um fator axogênico para as
CGRs, que os difere de BDNF, por exemplo (JO et al., 1999).
No nosso laboratório, já estamos de posse de uma alíquota fracionada
capaz de induzir todos esses efeitos descritos para o MC total em neurônios
periféricos, além de outros (proliferação e migração de células de Schwann).
Após separação das proteínas da alíquota em gel de poliacrilamida a
análise de algumas bandas protéicas em espectrômetro de massas no
revelou que um dos componentes secretados pela glia de aves é a
fibronectina, um componente da matriz extracelular responsável por adesão
e sinalização neuronal. Um outro componente de interesse encontrado no
AMG foi o fator derivado do epitélio pigmentar (PEDF), um fator trófico
capaz de manter inúmeras populações neuronais, além de proteger
neurônios de insultos excitotóxicos e radicais livres (TOMBRAN-TINK e
BARNSTABLE, 2002). Porém, é necessário ter cautela na interpretação dos
resultados, pois a análise das bandas pelo programa mascot, revelou que a

132
fonte de PEDF não era de aves (como foi a de fibronectina), mas sim bovina.
Portanto, uma possibilidade que temos que ressaltar é que nesta primeira
análise as frações obtidas ainda continham diversas proteínas
contaminantes do soro. Mesmo após sucessivas lavagens o soro fetal bovino
ainda se manteve presente com traços significativos após concentração da
fração para análise, a ponto de ter vários dos seus componentes presentes
no AMG. Sabe-se que PEDF é produzido pela glia de Müller (EICHLER et al.,
2004) e está envolvido com a regulação de angiogênese,
contrabalançando os efeitos de um outro fator, o fator de crescimento do
endotélio vascular (VEGF). Mas se PEDF é responsável por parte dos efeitos
descritos neste trabalho, não temos ainda evidências concretas para
qualquer afirmação.
O AMG também mostrou ter um componente ativo na proliferação da
própria glia de Müller (central) e na proliferação e/ou migração de células
de Schwann (periférico). É sabido que a glia de Müller produz FGFb e que
esta molécula é capaz de induzir proliferação glial porém nossas
observações não nos permitem concluir exatamente que moléculas são
responsáveis pela migração de células de Schwann para fora do explante
de ciático visto que a migração promovida por AMG foi superior à induzida
por BDNF e NGF.
O que sabemos é que a atividade trófica retiniana é capaz de
promover regeneração do nervo ciático de ratos adultos, resgatando os
neurônios do GRD e motoneurônios com uma eficiência comparada ao uso
combinado de BDNF e NGF, fatores usados como controle positivo do

133
experimento. Observamos que neurônios com soma pequeno (responsáveis
pela codificação de dor e temperatura), médio (táteis) e grande
(propriocepção) foram igualmente resgatados após 40 dias de tratamento.
Os motoneurônios presentes na seção L4/L5 da medula também apresentam
uma melhora significativa se comparados aos dos animais controle. Assim, o
interesse em descobrir que moléculas são responsáveis por efeitos tão
diversos e importantes continua, e num futuro em breve, esperamos ter mais
resultados que mostrem quem são os componentes que estão por trás destes
efeitos de tamanha importância fisiológica.
Figura 52. Indução de expressão de peptídeo vaso-ativo intestinal (VIP) em neurônios simpáticos tratados com CNTF ou RCM ou AMG. Nesta figura é possível observar que CNTF, RCM e AMG são capazes de induzir a expressão de VIP, mesmo que em diferentes proporções, em neurônios embrionários do simpático de aves. AMG se destaca neste gráfico como um potente indutor da expressão de VIP pro neurônios simpáticos. Um anticorpo contra a subunidade gp130 (uma das proteínas que compõem o trímero que forma o receptor para proteínas do CNTF) foi capaz de inibir a ação indutora da expressão de VIP tanto por CNTF quanto por AMG o que nos leva a crer que pelo menos um membro desta família protéica (principalmente se levarmos em conta o resultado da figura 30) seja um dos componentes do AMG e esteja envolvido com a ação trófica produzido por este em neurônios simpáticos.

134
6. CONCLUSÕES

135
1- Fatores tróficos retinianos (RCM) mantém neurônios periféricos em cultura.
2- O efeito trófico de RCM não é devido à ação isolada de fatores de neurotróficos clássicos.
3- Componentes da fração ativa dos MCs por glia de Müller tem afinidade por heparina.
4- RCM exerce seu efeito trófico sinalizando pelas vias da PI3K e JAK2.
5- A produção ou secreção dos componentes do RCM é mediada por neurotrofinas.
6- A glia de Müller é a grande responsável pela produção dos fatores tróficos retinianos.
7- MMG ativa as vias da ERKs, AKT/PKB e STAT3 em neurônios simpáticos (GSC)de rato.
8- MMG exerce atividade trófica sobre células ganglionares da retina (CGRs) isoladas em cultura.
9- Glia de Müller se mostrou um ótimo substrato para cultivo de CGRs no sistema de co-cultura mantendo alta sobrevivência e extensão neurítica.
10- Moléculas do MMG (oriundas da glia de Müller) são internalizadas por neurônios simpáticos.
11- MCs por glia de Müller possui componente que induz proliferação e migração da glia periférica (Schwann) e central (Müller).
12- E experimentos in vivo, AMG induz regeneração de neurônios centrais (motoneurônios) e periféricos (gânglio da raiz dorsal) em modelo de secção do nervo ciático.

136
REFERÊNCIAS BIBLIOGRÁFICAS

137
REFERÊNCIAS BIBLIOGRÁFICAS ABE, K.; SAITO, H. Effects of Basic Fibroblast Growth Factor on Central Nervous
System Functions. Pharmacol. Res. 43:307-312. (2001) ARAÚJO, E.G.; LINDEN, R. Trophic Factors Produced by Retinal Cells Increase
the Survival of Retinal Ganglion Cells in vitro. Eur. J. Neurosci. 5:1181-1189. (1993)
ARY-PIRES, R.; NAKATANI, M.; REHEN, S.K.; LINDEN, R. Developmentally
Regulated Release of Intraretinal Neurotrophic Factors in vitro. Int. J. Dev. Neurosci. 15:239-255. (1997)
BALOH, R.H.; ENOMOTO, H.; JOHNSON, J.R.E.M.; MILBRANT, J. The GDNF Family
Ligands and Receptors – Implications for Neural Development. Curr. Op. Neurobiol. 10:103-110. (2000)
BARBACID, M.; LAMBALLE, F.; PULIDO, D.; KLEIN, R. The Trk Family of Tyrosine
Protein Kinase Receptors. Biochim Biophys Acta 1072:115-127. (1991) BARDE, Y.-A.; EDGAR D.; THOENEN, H. Purification of a New Neurotrophic
Factor from Mammalian Brain. EMBO J. 1:549-553. (1982) BARRES, B.A.; SILVERSTEIN, B.E.; COREY, D.P.; CHUN, L.L. Immunological,
Morphological, and Electrophysiological Variation among Retinal Ganglion Cells Purified by Panning. Neuron 1:791-803. (1988)
BAX, B.; BLUNDELL, T.L.; MURRAY-RUST, J.; McDONALD, N.Q. Structure of Mouse
7S NGF: a Complex of Nerve Growth Factor with Four Binding Proteins. Structure 5:1275-1285. (1997)
BELLIVEAU, D.J.; KRIVKO, I.; KOHN, J.; LACHANCE, C.; POZNIAK, C.; RUSAKOV,
D.; KAPLAN, D.; MILLER, F.D. NGF and Neurotrophin-3 both Activate Trka on Sympathetic Neurons but Differentially Regulate Survival and Neuritogenesis. J. Cell Biol. 136:375-388. (1997)

138
BELMONTE, K.E.; McKINNON, L.A.; NATHANSON, N.M. Developmental Expression of Muscarinic Acetylcholine Receptors in Chick Retina: Selective Induction of m2 Muscarinic Receptor Expression in ovo by a Factor Secreted by Müller Glial Cells. J. Neurosci. 20:8417-8425. (2000)
BENNETT, J.L.; ZEILER, S.R.; JONES, K.R. Patterned Expression of BDNF and NT-3 in
the Retina and Anterior Segment of the Developing Mammalian Eye. Invest. Ophthalmol. Vis. Sci. 40:2996-3005 (1999)
BERKELAAR, M.; CLARKE, D.B.; WANG, Y.C.; BRAY, G.M.; AGUAYO, A.J.
Axotomy Results in Delayed Death and Apoptosis of Retinal Ganglion Cells in Adult Rats. J. Neurosci. 14:4368-4374. (1994)
BERKEMEIER, L.R.; WINSLOW, J.W.; KAPLAN, D.R.; NIKOLICS, K.; GOEDDEL, D.V.;
ROSENTHAL, A. Neurotrophin-5: a Novel Neurotrophic Factor that Activates Trk and Trkb. Neuron 7:857-866. (1991)
BOVOLENTA, P.; FRADE, J.M.; MARTI, E.; RODRIGUEZ-PENA, M.A.; BARDE, Y.A.;
RODRIGUEZ-TEBAR, A. Neurotrophin-3 Antibodies Disrupt the Normal Development of the Chick Retina. J. Neurosci. 16:4402-4410. (1996)
BRINGMANN, A.; PANNICKE, T.; GROSCHE, J.; FRANCKE, M.; WIEDEMANN, P.;
SKATCHKOV, S.N.; OSBORNE, N.N.; REICHENBACH, A. Müller Cells in the Healthy and Diseased Retina. Progress in Retinal and Eye Research 25:397–424. (2006)
BROKER, L.E.; KRUYT, F.A.; GIACCONE, G. Cell Death Independent of
Caspases: A Review. Clin. Cancer Res. 11:3155–3162. (2005) BURNHAM, P.; LOUIS, J.C.; MAGAL, E.; VARON, S. Effects of Ciliary Neurotrophic
Factor on the Survival and Response to Nerve Growth Factor of Cultured Rat Sympathetic Neurons. Dev. Biol. 161:96-106. (1994)
CAMINOS, E.; BECKER, E.; MARTÍN-ZANCA, D.; VECINO, E.. Neurotrophins and
their Receptors in the Tench Retina during Optic Nerve Regeneration. J. Comp. Neurol. 404:321-331. (1999)

139
CASACCIA-BONNEFIL, P.; KONG, H.; CHAO, M.V. Neurotrophins: the biological paradox of survival factors eliciting apoptosis. Cell Death Differ. 5:357-364. (1998)
CEPKO, C.L.; AUSTIN, C.P.; YANG, X.; ALEXIADES, M.; EZZEDDINE, D. Cell Fate
Determination in the Vertebrate Retina. Proc. Natl. Acad. Sci. USA 93:589-595. (1996)
CHAI, L.; MORRIS, J.E. Heparan Sulfate in the Inner Limiting Membrane of
Embryonic Chicken Retina Binds Basic Fibroblast Growth Factor to Promote Axonal Outgrowth. Exp. Neurol. 160:175-185. (1999)
CHAO, M.V. The p75 Neurotrophin Receptor. J. Neurobiol. 25:1373-1385. (1994) CHEN, Q.; LONG, Y.; YUAN, X.; ZOU, L.; SUN, J.; CHEN, S.; PEREZ-POLO, J.R.;
YANG, K.J. Protective Effects of Bone Marrow Stromal Cell Transplantation in Injured Rodent Brain: Synthesis of Neurotrophic Factors. Neurosci. Res. 80:611-619. (2005)
CORDEIRO, M.F.; GUO, L.; LUONG, V.; HARDING, G.; WANG, W.; JONES, H.E.;
MOSS, S.E.; SILLITO, A.M.; FITZKE, F.W. Real-Time Imaging of Single Nerve Cell Apoptosis in Retinal Neurodegeneration. Proc. Natl. Acad. Sci. USA 101:13352-13356. (2004)
COULOMBRE, A.J. Correlations of structural and biochemical changes in the
developing retina of the chick. Am. J. Anat. 96:153-89. (1955) COWAN, W.M. Viktor Hamburger and Rita Levi-Montalcini: the Path to the
Discovery of Nerve Growth Factor. Annu. Rev. Neurosci. 24:551-600. (2001)
CROWDER, R.J.; FREEMAN, R.S. Phosphatidylinositol 3-Kinase and AKT Protein
Kinase are Necessary and Sufficient for the Survival of Nerve Growth Factor-Dependent Sympathetic Neurons. J. Neurosci. 18:2933-2943. (1998)

140
CROWLEY, C.; SPENCER, S.D.; NISHIMURA, M.C.; CHENKS; PITTS-MEEK, S.; ARMANINI, M.P.; LING, L.H.; MACMAHON, S.B.; SHELTON, D.L.; LEVINSON, A.D.; PHILLIPS, H.S. Mice Lacking Nerve-Growth Factor Display Perinatal Loss of Sensory and Sympathetic Neurons yet Develop Basal Forebrain Cholinergic Neurons. Cell 76:1001-1011. (1994)
CUI, O.; HARVEY, A.R. At Least Two Mechanisms are Involved in the Death of
Retinal Ganglion Cells Following Target Ablation in Neonatal Rats. J. Neurosci. 15:8143-8155. (1995).
DANIAL, N.N.; KORSMEYER, S.J. Cell death: critical control points. Cell 116:205-
219. (2004) DAS, I.; HEMPSTEAD, B.L.; MACLEISH, P.R.; SPARROW, J.R.Immunohistochemical
Analysis of the Neurotrophins BDNF and NT-3 and their Receptors Trk B, Trk C, and p75 in the Developing Chick Retina. Vis. Neurosci. 14:835-842. (1997)
DATTA, S.R.; BRUNET, A.; GREENBERG, M.E. Cellular Survival: a Play in Three
AKTs. Genes Dev. 13:2905-2927. (1999) DATTA, S.R.; KATSOV, A.; HU, L.; PETROS, A.; FESIK, S.W.; YAFFE, M.B.;
GREENBERG, M.E. 14-3-3 Proteins and Survival Kinases Cooperate to Inactivate BAD by BH3 Domain Phosphorylation. Mol. Cell. 6:41-51. (2000)
DAVIES, A.M. Neurotrophic Factor Bioassay Using Dissociated Neurons. In
Nerve Growth Factors Ed. R. Rush. J. Wiley&Sons, Chichester, pp95-109, Ch. 5. (1988)
DAVIES, A.M. Role of Neurotrophins in the Developing Nervous System. J.
Neurobiol. 25:1334-1348. (1994) De MELLO, F.G.; DE MELLO, M.C.F.; HUDSON, R.; KLEIN, W.L. Selective
Expression of Factors Preventing Cholinergic Dedifferentiation. J. Neurochem. 54:886-892. (1990)

141
DESHMUKH, M.; JOHNSON JR, E.M. Programmed Cell Death in Neurons: Focus on the Pathway of Nerve Growth Factor Deprivation-Induced Death of Sympathetic Neurons. Molecular Pharmacology 51:897-906. (1997)
DESIRÉ, L.; HEAD, M.W.; FAYEIN, N.A.; COURTOIS, Y.; JEANNY, J.C. Suppression
of Fibroblast Growth Factor 2 Expression by Antisense Oligonucleotides Inhibits Embryonic Chick Neural Retina Cell Differentiation and Survival in vivo. Dev. Dyn. 212:63–74. (1998)
Di POLO, A.; AIGNER, L.J.; DUNN, R.J.; BRAY, G.M.; AGUAYO, A.J. Prolonged
Delivery of Brain-Derived Neurotrophic Factor by Adenovirus-Infected Müller Cells Temporarily Rescues Injured Retinal Ganglion Cells. Proc. Natl. Acad. Sci. USA 95:3978-3983. (1998)
DONOVAN, M.; COTTER, T.G. Control of Mitochondrial Integrity by Bcl-2 Family
Members and Caspase-Independent Cell Death. Biochim Biophys Acta 1644:133-147. (2004)
ECKENSTEIN, F.P. Fibroblast Growth Factors in the Nervous System. J. Neurobiol.
25:1467-1480. (1994) EDWARDS, S.N.; BUCKMASTER, A.E.; TOLKOVSKY, A.M. The Death Programme in
Cultured Sympathetic Neurons Can Be Suppressed at the Posttranslational Level by Nerve Growth Factor, Cyclic AMP and Depolarization. J. Neurochem. 57:2140-2143. (1991)
EICHLER, W.; YAFAI, Y.; KELLER, T.; WIEDEMANN, P.; REICHENBACH, A. PEDF
Derived from Glial Müller Cells: a Possible Regulator of Retinal Angiogenesis. Exp. Cell Res. 299:68-78. (2004)
ELLIS, H.M.; HORVITZ, H.R. Genetic Control of Programmed Cell Death in the
Nematode C. Elegans. Cell 44:817-829. (1986) ENOKIDO, Y.; DE SAUVAGE, F.; HONGO, J.A.; NINKINA, N.; ROSENTHAL, A.;
BUCHMAN, V.L.; DAVIES, A.M. Grfα4 and the Tyrosine Kianse Ret Form a Functional Receptor Complex for Persephin. Curr. Biol. 8:1019-1022. (1998)

142
ESTUS, S.; ZAKS, W.J.; FREEMAN, R.S.; GRUDA, M.; BRAVO, R.; JOHNSON JR., E. Altered Gene Expression in Neurons During Programmed Cell Death: Identification of C-Jun as Necessary for Neuronal Apoptosis. J. Cell Biol. 127:1717-1727. (1994)
FRADE, J.M.; RODRIGUEZ-TEBAR, A.; BARDE, Y.A. Induction of Cell Death by
Endogenous Nerve Growth Factor through Its p75 Receptor. Nature 383:166-168. (1996)
FRADE, J.M.; BARDE, Y.A. Genetic Evidence for Cell Death Mediated by Nerve
Growth Factor and the Neurotrophin Receptor p75 in the Developing Mouse Retina and Spinal Cord. Development 126:683-690. (1999)
FREEMAN, R.S.; ESTUS, S.; JOHNSON JR., E.M. Analysis of Cell Cycle-Related
Gene Expression in Postmitotic Neurons: Selective Induction of Cyclin D1 during Programmed Cell Death. Neuron 12:343-355. (1994)
GAO, H.; HOLLYFIELD, J.G. Basic Fibroblast Growth Factor (bFGF)
Immunolocalization in The Rodent Outer Retina Demonstrated With an Anti-Rodent bFGF Antibody. Brain Res. 585:355-360. (1992)
GARCIA, M.; FORSTER, V.; HICKS, D.; VECINO, E. Effects of Müller Glia on Cell
Survival and Neuritogenesis in Adult Porcine Retina in vitro. Invest. Ophthalmol. Vis. Sci. 43:3735-3743. (2002)
GARCIA, M.; VECINO, E. Role of Müller Glia in Neuroprotection and
Regeneration in the Retina. Histol. Histopathol. 18:1205-1218. (2003) GARCÍA-VALENZUELA, E.; GORCZYCA, W.; DARZYNKIEWICZ, Z.; SHARMA, S.C.
Apoptosis in Adult Retinal Ganglion Cells after Axotomy. J. Neurobiol. 25:431-438. (1994)
GAVAZZI, I.; KUMAR, R.D.; McMAHON, S.B.; COHEN, J. Growth Responses of
Different Subpopulations of Adult Sensory Neurons to Neurotrophic Factors in vitro. Eur. J. Neurosci. 11:3405-3414. (1999)
GOLDBERG, J.L.; BARRES, B.A. The Relationship between Neuronal Survival and
Regeneration. Ann. Rev. Neurosci. 23:579-612. (2000)

143
GOTZ, R.; KOSTER, R.; WINKLER, C.; RAULF, F.; LOTTSPEICH, F.; SCHARTL, M.;
THOENEN, H. Neurotrophin-6 Is a New Member of the Nerve Growth Factor Family. Nature 372:266-269. (1996)
GOZUACIK, D.; KIMCHI, A. Autophagy as a Cell Death and Tumor Suppressor
Mechanism. Oncogene 23:2891–2906. (2004) HALLBÖÖK, F.; IBANEZ, C.F.; PERSSON, H. Evolutionary Studies of the Nerve
Growth Factor Family Reveal a Novel Member Abundantly Expressed in Xenopus Ovary. Neuron 6:845-858. (1991)
HALLBÖÖK, F.; BACKSTROM, A.; KULLANDER, K.; EBENDAL, T.; CARRI, N.G.
Expression of Neurotrophins and Trk Receptors in the Avian Retina. J. Comp. Neurol. 364:664-676. (1996)
HAMBURGER, V.; HAMILTON, H.L. A Serial of Normal Stages in the
Development of the Chick Embryo. J. Morphol. 88:49-92 (1951). HENDRY, I.A.; CAMPBELL, J. Morphometric Analysis of Rat Superior Cervical
Ganglion after Axotomy and Nerve Growth Factor Treatment. J. Neurocytol. 5:351-360. (1976)
HARADA, C.; HARADA, T.; QUAH, H.M.; MAEKAWA, F.; YOSHIDA, K.; OHNO, S.;
WADA, K.; PARADA, L.F.; TANAKA, K. Potential Role of Glial Cell Line-Derived Neurotrophic Factor Receptors in Müller Glial Cells during Light-Induced Retinal Degeneration. Neuroscience 122:229-235. (2003)
HOHN, A.; LEIBROCK, J.; BAILEY, K.; BARDE, Y.A. Identification and
Characterization of a Novel Member of the Nerve Growth Factor/Brain-Derived Neurotrophic Factor Family. Nature 344:339-341. (1990)
HORTON, A.; LARAMEE, G.; WYATT, S.; SHIT, A.; WINSLOW, J.; DAVIES, A.M. NGF
Binding to p75 Enhances the Sensitivity of Sensory and Sympathetic Neurons to NGF at Different Stages of Development. Mol. Cell Neurosci. 10:162-172. (1997)

144
HUXLIN, K.R.; DREHER, B.; SCHULZ, M.; SEFTON, A.J.; BENNETT, M.R. Effect of Collicular Proteoglycan on the Survival of Adult Rat Retinal Ganglion Cells Following Axotomy. Eur. J. Neurosci. 7:96-107. (1995)
IP, N.Y.; IBANEZ, C.F.; NYE, S.H.; McCLAIN, J.; JONES, P.F.; GIES, D.R.; BELLUSCIO,
L.; LE BEAU, M.M.; ESPINOSA, R. 3rd, SQUINTO, S.P. Mammalian Neurotrophin-4: Structure, Chromosomal Localization, Tissue Distribution, and Receptor Specificity. Proc. Natl. Acad. Sci. USA 89:3060-3064. (1992)
IP, N.Y.; YANCOPOULOS, G. The neurotrophins and CNTF: Two families of
collaborative neurotrophic factors. Ann. Rev. Neurosci. 19:491-515. (1996). ISENMANN, S.; WAHL, C.; KRAJEWSKI, S.; REED, J.C.; BAHR, M. Up-Regulation of
Bax Protein in Degenerating Retinal Ganglion Cells Precedes Apoptotic Cell Death after Optic Nerve Lesion in the Rat. Eur. J. Neurosci. 9:1763-1772. (1997)
ISHII, D.N.; MARSH, D.J. On the Therapeutic Potential for Insulin-Like Growth
Factor Use in Motor Neuron Disease. Exp. Neurol. 124:96-99. (1993) JING, S.; WEN, D.; YU, Y.; HOLST, P.L.; LUO, Y.; FANG, M.; TAMIR, R.; ANTONIO, L.;
HU, Z.; CUPPLES, R.; LOUIS, J.C.; HU, S.; ALTROCK, B.W.; FOX, G.M. GDNF-Induced Activation of the Ret Protein Tyrosine Kinase Is Mediated by GDNFR-Alpha, a Novel Receptor for GDNF. Cell 85:1113-1124. (1996)
JO, S.A.; WANG, E.; BENOWITZ, L.I. Ciliary Neurotrophic Factor Is and
Axogenesis Factor for Retinal Ganglion Cells. Neuroscience 89:579-591. (1999)
JOHNSON, J.E. Neurotrophic Factors. In Fundamental Neuroscience. Eds. MJ
Zigmond, FE Bloom, SC Landis, JL Roberts and LR Squire. Academic Press. pp 611-635. Ch. 21. (1999)
JONES, K.R.; REICHARDT, L. Molecular Cloning of a Human Gene that Is a
Member of the Nerve Growth Factor Family. Proc. Natl. Acad. Sci. USA 87:8060-8064. (1990)

145
KAHN, A.J. An Autoradiographic Analysis of the Time of Appearance of Neurons in the Developing Chick Neural Retina. Dev. Biol. 38:30-40. (1974)
KAPLAN, D.R.; MILLER, F.D. Neurotrophin signal transduction in the nervous
system. Curr Opin Neurobiol. 10:381-391. (2000) KARLSSON, M.; CLARY, D.O.; LEFCORT, F.B.; REICHARDT, L.F.; KARTEN, H.J.;
HALLBOOK, F. Nerve Growth Factor Receptor Trka Is Expressed by Horizontal and Amacrine Cells during Chicken Retinal Development. J. Comp. Neurol. 400:408-416. (1998)
KERR, J.F.R.; WYLLIE, A.H.; CURRIE, A.R. Apoptosis: a Basic Biological
Phenomenon with Wide-Ranging Implications in Tissue Kinetics. Br. J. Cancer 26:239-257. (1972)
KHURSIGARA, G.; BERTIN, J.; YANO, H.; MOFFETT, H.; DISTEFANO, P.S.; CHAO,
MV. A Prosurvival Function for the p75 Receptor Death Domain Mediated Via the Caspase Recruitment Domain Receptor-Interacting Protein 2. J. Neurosci. 21:5854-5863. (2001)
KLEIN, R.; JING, S.Q.; NANDURI, V.; O'ROURKE, E.; BARBACID, M. The Trk Proto-
Oncogene Encodes a Receptor for Nerve Growth Factor. Cell 65:189-197. (1991)
KOIKE, T.; MARTIN, D.P.; JOHNSON JR, E.M. Role Of Ca2+ Channels in the Ability
of Membrane Depolarization to Prevent Neuronal Death Induced by Trophic-Factor Deprivation: Evidence that Levels of Internal Ca2+ Determine Nerve Growth Factor Dependence of Sympathetic Ganglion Cells. Proc. Natl. Acad. Sci. USA. 86:6421-6425. (1989)
KORSMEYER, S.J.; WALDMANN, T.A. Immunoglobulin Genes: Rearrangement
and Translocation in Human Lymphoid Malignancy. J. Clin. Immunol. 4:1-11. (1984)
KORSMEYER, S.J.; GROSS, A.; HARADA, H.; ZHA, J.; WANG, K.; YIN, X.M.; WEI, M.;
ZINKEL, S. Death and Survival Signals Determine Active/Inactive Conformations of Pro-Apoptotic BAX, BAD and BID Molecules. Cold Spring Harb. Symp. Quant. Biol. 64:343-350. (1999)

146
LEIBROCK, J.; LOTTSPEICH, F.; HOHN, A.; HOFER, M.; GENGERER, B.; MAIASKOWSKI, P.; THOENEN, H.; BARDE, Y. Molecular Cloning and Expression of Brain-Derived Neurotrophic Factor. Nature 341:149-152. (1989)
LEIN, P.J.; BECK, H.N.; CHANDRASEKARAN, V.; GALLAGHER, P.J.; CHEN, H.L.;
LIN, Y.; GUO, X.; KAPLAN, P.L.; TIEDGE, H.; HIGGINS, D. Glia Induce Dendritic Growth in Cultured Sympathetic Neurons by Modulating the Balance between Bone Morphogenetic Proteins (Bmps) and BMP Antagonists. J. Neurosci. 22:10377–10387. (2002)
LEVI-MONTALCINI, R.; HAMBURGER, V. Selective Growth-Stimulating Effects of
Mouse Sarcoma on the Sensory and Sympathetic Neurvous System of the Chick Embryo. J. Exp. Zool. 116:321-361. (1951)
LEVI-MONTALCINI, R.; HAMBURGER, V. A Diffusible Agent of Mouse Sarcome,
Producing Hyperplasia of Sympathetic Ganglia and Hyperneurotization of Viscera in the Chick Embryo. J. Exp. Zool. 123:233-287. (1953)
LEVI-MONTALCINI, R.; BOOKER, B. Destruction of the Sympathetic Ganglion in
Mammals by an Anti-Serum to the Nerve-Growth Protein. Proc. Natl. Acad. Sci. USA 46:384-391. (1960)
LIVESEY, R.; CEPKO, C. Vertebrate Neural Cell Fate Determination – Lessons
from the Retina. Nature Neuroscience Reviews 2:109-118. (2001) LLAMOSAS, M.M.; CERNUDA-CERNUDA, R.; HUERTA, J.J.; VEGA, J.A.; GARCIA-
FERNANDEZ, J.M. Neurotrophin Receptors Expression in the Developing Mouse Retina: an Immunohistochemical Study. Anat Embryol (Berl). 195:337-344. (1997)
MA, Y.T.; HSIEH, T.; FORBES, M.E.; JOHNSON, J.E.; FROST, D.O. BDNF Injected into
the Superior Colliculus Reduces Developmental Retinal Ganglion Cell Death. J. Neurosci. 18:2097-2107. (1998)
MAISONPIERRE, P.C.; BELLUSCIO, L.; SQUINTO, S.; IP, N.Y.; FURTH, M.E.; LINDSAY,
R.M.; YANCOPOULOS, G.D. Neurotrophin-3: a Neurotrophic Factor Related to NGF and BDNF. Science 247:1443-1451. (1990)

147
MANSOUR-ROBAEY, S.; CLARKE, D.B.; WANG, Y.C.; BRAY, G.M.; AGUAYO, A.J. Effects of Ocular Injury and Administration of Brain-Derived Neurotrophic Factor on Survival and Regrowth of Axotomized Retinal Ganglion Cells. Proc. Natl. Acad. Sci. USA. 91:1632-1636. (1994)
MARTIN, D.P.; SCHMIDT, R.E., DISTEFANO, O.S.; LOWRY, O.H.; CARTER, J.G.;
JOHNSON JR, E.M. Inhibitors of Protein Synthesis Prevent Neuronal Death Caused by Nerve-Growth Factor Deprivation. J. Cell Biol. 106:829-844. (1988)
MARTINOU, J.C.; GREEN, D.R. Breaking the Mitochondrial Barrier. Nat. Rev.
Mol. Cell Biol. 2:63-67. (2001) MARTINOU, J.C.; YOULE, R.J. Which Came First, the Cytochrome C Release or
the Mitochondrial Fission? Cell Death Differ. 13:1291-1295. (2006) McALLISTER, A.K.; KATZ, L.C.; LO, D.C. Opposing Roles for Endogenous BDNF
and NT-3 in Regulating Cortical Dendritic Growth. Neuron. 18:767-778. (1997)
McDONALD, N.Q.; CHAO, M.V. Structural Determinants of Neurotrophin
Action. J. Biol. Chem. 270:19669-19672. (1995) McKERNAN, D.P.; CAPLIS, C.; DONOVAN, M.; O'BRIEN, C.J.; COTTER, T.G. Age-
Dependent Susceptibility of the Retinal Ganglion Cell Layer to Cell Death. Invest. Ophthalmol. Vis. Sci. 47:807-814. (2006)
McKINNON, L.A.; GUNTHER, E.C.; NATHANSON, N.M. Developmental
Regulation of the Cm2 Muscarinic Acetylcholine Receptor Gene: Selective Induction by a Secreted Factor Produced by Embryonic Chick Retinal Cells. J. Neurosci. 18:59-69. (1998)
METEA, M.R.; NEWMAN, E.A. Calcium Signaling in Specialized Glial Cells. Glia
54:650–655. (2006) MEY, J.; THANOS, S. Intravitreal Injections of Neurotrophic Factors Support the
Survival of Axotomized Retinal Ganglion Cells in Adult Rats in vivo. Brain Res. 602:304-317. (1993)

148
MEYER-FRANKE, A.; KAPLAN, M.R.; PFRIEGER, F.W.; BARRES, B.A. Characterization of the Signalling Interactions that Promote the Survival and Growth of Developing Retinal Ganglion Cells in Culture. Neuron 15:805-819. (1995)
MISHIMA, H.; FUJITA, H. Studies on the cytodifferentiation of the neuroblasts
and visual cells in the chick embryo retina, using the electron-microscopic autoradiography of 3H-thymidine. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 206:1-10. (1978)
MURPHY, M.; REID, K.; BROWN, M.A.; BARTLETT, P.F. Involvement of Leukemia
Inhibitory Factor and Nerve Growth Factor in the Development of Dorsal Root Ganglion Neurons. Development 117:1173-1182. (1993)
NAKAGAWA, T.; ZHU, H.; MORISHIMA, N.; LI, E.; XU, J.; YANKNER, B.A.; YUAN, J.
Caspase-12 Mediates Endoplasmic-Reticulum-Specific Apoptosis and Cytotoxicity by Amyloid-Beta. Nature 403:98-103. (2000)
NEWMAN, E.; REICHENBACH, A. The Müller Cell: a Functional Element of the
Retina. Trends Neurosci. 19:307–312. (1996) NILSSON, A.S.; FAINZILBER, M.; FALCK, P.; IBANEZ, C.F. Neurotrophin-7: a Novel
Member of the Neurotrophin Family from the Zebrafish. FEBS Lett. 424:285-290. (1998)
NINKINA, N.; ADU, J.; FISCHER, A.; PINON, L.G.; BUCHMAN, V.L.; DAVIES, A.M.
Expression and Function of Trkb Variants in Developing Sensory Neurons. EMBO J. 15:6385-6393. (1996)
PATTERSON, P. The Emerging Neuropoietic Cytokine Family. Curr. Op.
Neurobiol. 2:94-97. (1991) PETERSON, W.M.; WANG, Q.; TZEKOVA, R.; WIEGAND, S.J. Ciliary Neurotrophic
Factor and Stress Stimuli Activate the Jak-STAT Pathway in Retinal Neurons and Glia. J. Neurosci. 20:4081-4090. (2000)

149
PETTMANN, B.; LABOURDETTE, G.; WEIBEL, M.; SENSENBRENNER, M. The Brain Fibroblast Growth Factor (FGF) Is Localized in Neurons. Neurosci. Lett. 68:175-180. (1986)
PRADA, C.; PUGA, J.; PÉREZ-MÉNDEZ, L.; LÓPEZ, R.; RAMÍREZ, G. Spatial and
Temporal Patterns of Neurogenesis in the Chick Retina. Eur. J. Neurosci. 3:559-569. (1991)
PRZYWARA, D.A.; KULKARNI, J.S.; WAKADE, T.D.; LEONTIEV, D.V.; WAKADE, A.R.
Pituitary Adenylyl Cyclase-Activating Polypeptide and Nerve Growth Factor Use the Proteasome to Rescue Nerve Growth Factor-Deprived Sympathetic Neurons Cultured from Chick Embryos. J. Neurochem. 71:1889-1897. (1998)
PURVES, D.; LICHTMAN, J.W. Trophic Effects of Targets on Neurons, In Principles
of Neural Development. Sinauer, Ch. 7, 155-178. (1985) QUIGLEY, H.A.; NICKELLS, R.W.; KERRIGAN, L.A.; PEASE, M.E.; THIBAULT, D.J.;
ZACK, D.J. Retinal Ganglion Cell Death in Experimental Glaucoma and after Axotomy Occurs by Apoptosis. Invest. Ophthalmol. Vis. Sci. 36:774-786. (1995)
RAJU, T.R.; BENNETT, M.R. Retinal Ganglion Cell Survival Requirements: a Major
but Transient Dependence on Müller Glia during Development. Brain Research 383:165-176. (1986)
REH, T.A.; FISCHER, A.J. Retinal Stem Cells. Methods Enzymol. 419:52-73. (2006) REIS, R.A.; CABRAL DA SILVA, M.C.; LOUREIRO DOS SANTOS, N.E.; BAMPTON, E.;
TAYLOR, J.S.; DE MELLO, F.G.; LINDEN, R. Sympathetic Neuronal Survival Induced by Retinal Trophic Factors. J. Neurobiology 50:13-23. (2002)
ROHRER, B.; LAVAIL, M.M.; JONES, K.R.; REICHARDT, L.F. Neurotrophin Receptor
Trkb Activation Is not Required for the Postnatal Survival of the Retinal Ganglion Cells in vivo. Experimental Neurology 172:81-91. (2001)

150
ROSENTHAL, A.; GOEDDEL, D.V.; NGUYEN, T.; LEWIS, M.; SHIH, A.; LARAMEE, G.R.; NIKOLICS, K.; WINSLOW, J.W. Primary Structure and Biological Activity of a Novel Human Neurotrophic Factor. Neuron 4:767-773. (1990)
RYDEL, R.E.; GREENE, L.A. cAMP Analogs Promote Survival and Neurite
Outgrowth in Cultures of Rat Sympathetic and Sensory Neurons Independently of Nerve Growth Factor. Proc. Natl. Acad. Sci. USA 85:1257-1261. (1988)
SAARMA, M. GDNF - a Stranger in the TGF-β Superfamily? Eur. J. Bioch.
267:6968-6971. (2000) SAPORITO, M.S.; HUDKINS, R.L.; MARONEY, A.C. Discovery of CEP-1347/KT-
7515, an Inhibitor of the JNK/SAPK Pathway for the Treatment of Neurodegenerative Diseases. Prog. Med. Chem. 40:23-62. (2002)
SARTHY, V.; RIPPS, H. Structural Organization of Retinal Glia. In the Retinal
Müller Cell: Structure and Function. Perspectives in Vision Research, Kluwer Academic, pp 1-33. (2001)
SCHOBER, A.; HERTEL, R.; ARUMAE, U.; FARKAS, L.; JASZAI, J.; KRIEGLSTEIN, K.;
SAARMA, M.; UNSICKER, K. Glial Cell Line-Derived Neurotrophic Factor Rescues Target-Deprived Sympathetic Spinal Cord Neurons but Requires Transforming Growth Factor-Beta as Cofactor in vivo. J. Neurosci. 19:2008-2015. (1999)
SCHULZ, M.; RAJU, T.; RALSTON, G.; BENNETT, M.R. A Retinal Ganglion Cell
Neurotrophic Factor Purified from the Superior Colliculus. J. Neurochem. 55:832-841. (1990)
SCOTT, S.A.; DAVIES, A.M. Inhibition of Protein Synthesis Prevents Cell Death in
Sensory and Parasympathetic Neurons Deprived of Neurotrophic Factor in vitro. J. Neurobiol. 21:630-638. (1990)
SCOTT, B.S.; FISHER, K.C. Potassium Concentration and Number of Neurons in
Cultures of Dissociated Ganglia. Exp. Neurol. 27:16-22. (1970)

151
SEGAL, R.A.; GREENBERG, M.E. Intracellular Signaling Pathways Activated by Neurotrophic Factors. Annu. Rev. Neurosci. 19:463-89. (1996)
SEKI, M.; TANAKA, T.; SAKAI, Y.; FUKUCHI, T.; ABE, H.; NAWA, H.; TAKEI, N. Müller
Cells as a Source of Brain-Derived Neurotrophic Factor in the Retina: Noradrenaline Upregulates Brain-Derived Neurotrophic Factor Levels in Cultured Rat Müller Cells. Neurochem. Res. 30:1163-1170. (2005)
SENDTNER, M.; CARROLL, P.; HOLTMANN, B.; HUGHES, R.A.; THOENEN, H. Ciliary
Neurotrophic Factor. J. Neurobiol. 25:1436–1453. (1994) SIMON, M.A. Receptor Tyrosine Kinases: Specific Outcomes from General
Signals. Cell 103:5-13. (2000) SKORUPA, A.F.; KLEIN, W.L. Developmentally Regulated Secreted Factor
Controls Expression of Muscarinic Subtypes in Embryonic Chick Retina. J. Neurochem. 60:2087-2097. (1993)
SMEYNE, R.J.; KLEIN, R.; SCHNAPP, A.; LONG, L.K.; BRYANT, S.; LEWIN, A.; LIRA,
A.S.; BARBACID, M. Severe Sensory and Sympathetic Neuropathies in Mice Carrying a Disrupted Trk/NGF Receptor Gene. Nature 368:246-249. (1994)
SNOW, R.L.; ROBSON, J.A. Ganglion Cell Neurogenesis, Migration and Early
Differentiation in the Chick Retina. Neuroscience 58:399-409. (1994) STAHL, N.; YANCOPOULOS, G. The Tripartite CNTF Receptor Complex:
Activation and Signalling Involves Components Shared with other Citokines. J. Neurobiol 25:1454-1466. (1994)
SUBRAMANIAM, S.; UNSICKER, K. Extracellular Signal-Regulated Kinase as an
Inducer of Non-Apoptotic Neuronal Death. Neuroscience 138:1055-1065 (2006)
TAKAHASHI, M.; BUMA, Y.; IWAMOTO, T.; INAGUMA, Y.; IKEDA, H.; HIAI, H.
Cloning and Expression of the Ret Proto-Oncogene Encoding a Tyrosine Kinase with Two Potential Transmembrane Domains. Oncogene 3:571-578. (1988)

152
TANDRUP T.; WOOLF C.J.; COGGESHALL R.E. Delayed Loss of Small Dorsal Root Ganglion Cells after Transection of the Rat Sciatic Nerve. J Comp Neurol. 422:172-80. (2000)
TOMBRAN-TINK, J.; BARNSTABLE, C.J. PEDF: a Multifaceted Neurotrophic Factor.
Nat. Rev. Neurosci. 4:628-636. (2003) TOUW, I.P.; DE KONING, J.P.; WARD, A.C.; HERMANS, M.H. Signaling
Mechanisms of Cytokine Receptors and their Perturbances in Disease. Mol. Cell Endocrinol. 160:1-9. (2000)
TREANOR, J.J.; GOODMAN, L.; DE SAUVAGE, F.; STONE, D.M.; POULSEN, K.T.;
BECK, C.D.; GRAY, C.; ARMANINI, M.P.; POLLOCK, R.A.; HEFTI, F.; PHILLIPS, H.S.; GODDARD, A.; MOORE, M.W.; BUJ-BELLO, A.; DAVIES, A.M.; ASAI, N.; TAKAHASHI, M.; VANDLEN, R.; HENDERSON, C.E.; ROSENTHAL, A. Characterization of a Multicomponent Receptor for GDNF. Nature 382:80-83. (1996)
TURNER, D. L.; CEPKO, C.L. A Common Progenitor for Neurons and Glia Persists
in Rat Retina Late in Development. Nature 328:131-136. (1987) TURNER, D.L.; SNYDER, E.Y.; CEPKO, C.L. Lineage Independent Determination of
Cell Type in the Embryonic Mouse Retina. Neuron 4:833–845. (1990) TUSZYNSKI, M.H.; KORDOWER, J.A. CNS Regeneration: Basic Science and
Clinical Advances. Academic Press, San Diego. Tabela p.55 (1999) TUSZYNSKI, M.H.; THAL, L.; U, H.S.; PAY, M.M.; BLESCH, A.; CONNER, J.;
VAHLSING, H.L. Nerve Growth Factor Gene Therapy for Alzheimer's Disease. J. Mol. Neurosci. 19:207. (2002)
UNSICKER, K.; REICHERT-PREIBSCH, H.; SCHMIDT, R.; PETTMAN, B.; LABOURDETTE,
G.; SENSENBRENNER, M. Astroglial and Fibroblast Growth Factors Have Neurotrophic Functions for Cultured Peripheral and Central Nervous System Neurons. Proc. Natl. Acad. Sci. USA 84:5459–5463. (1987)

153
UNSICKER, K. Glial Cell Line-Derived Neurotrophic Factor Rescues Target-Deprived Sympathetic Spinal Cord Neurons but Requires Transforming Growth Factor-Beta as Cofactor in vivo. J. Neurosci. 19:2008-2015. (1999)
van EIJK, M.; DEFRANCE, T.; HENNINO, A.; DE GROOT, C. Death-Receptor
Contribution To The Germinal-Center Reaction. Trends Immunol. 22:677-682. (2001)
VILLEGAS-PÉREZ, M.P.; VIDAL-SANZ, M.; BRAY, G.M.; AGUAYO, A.J. Influences
of Peripheral Nerve Grafts on the Survival and Regrowth of Axotomized Retinal Ganglion Cells in Adult Rats. J. Neurosci. 8:265-280. (1988)
VERGARA, C.; RAMIREZ, B. CNTF, a Pleiotropic Cytokine: Emphasis on Its
Myotrophic Role. Brain Res. Rev. 47:161-173. (2004) VIRDEE, K.; TOLKOVSKY, A.M. Inhibition of p42 and p44 Mitogen Activated
Protein Kinase Activity by PD98059 Does not Suppress Nerve Growth Factor-Induced Survival of Sympathetic Neurons. J. Neurochem. 67:1801-1805. (1996)
von BARTHELD, C.S.; WILLIAMS, R.; LEFCORT, F.; CLARY, D.O.; REICHARDT, L.F.;
BOTHWELL, M. Retrograde Transport of Neurotrophins from the Eye to the Brain in Chick Embryos: Roles of the p75ntr and Trkb Receptors. J. Neurosci. 16:2995-3008. (1996)
WAKADE, A.R.; BHAVE, S.V.; MALHOTRA, R.K.; WAKADE, T.D. Forskolin Mediates
the Survival of Nerve Growth Factor-Dependent Sympathetic Neurons of Chick Embryo by a Cyclic AMP-Independent Mechanism. J. Neurochem. 54:1281-1287. (1990)
WEI, M.C.; ZONG, W.X.; CHENG, E.H.; LINDSTEN, T.; PANOUTSAKOPOULOU, V.;
ROSS, A.J.; ROTH, K.A.; MACGREGOR, G.R.; THOMPSON, C.B.; KORSMEYER, S.J. Proapoptotic BAX and BAK: a Requisite Gateway to Mitochondrial Dysfunction and Death. Science 292:727-730. (2001)
WEISE, J.; ISENMANN, S.; KLOCKER, N;, KUGLER, S.; HIRSCH, S.; GRAVEL, C.;
BAHR, M. Adenovirus-Mediated Expression of Ciliary Neurotrophic Factor (CNTF) Rescues Axotomized Rat Retinal Ganglion Cells but Does not Support Axonal Regeneration in vivo. Neurobiol. Dis. 7:212-223. (2000)

154
WILLIS, S.N.; ADAMS, J.M. Life in the Balance: How BH3-Only Proteins Induce
Apoptosis. Curr. Opin. Cell. Biol. 17:617-625. (2005) WURM, A.; PANNICKE, T.; IANDIEV, I.; WIEDEMANN, P.; REICHENBACH, A.;
BRINGMANN, A. The Developmental Expression of K1 Channels in Retinal Glial Cells Is Associated with a Decrease of Osmotic Cell Swelling. Glia 54:411–423. (2006)
YAMASHITA, T.; TUCKER, K.L.; BARDE, Y.A. Neurotrophin Binding to the p75
Receptor Modulates Rho Activity and Axonal Outgrowth. Neuron 24:585-593. (1999)
YE, M.; CHEN, S.; WANG, X.; QI, C.; LU, G.; LIANG, L.; XU, J. Glial Cell Line-
Derived Neurotrophic Factor in Bone Marrow Stromal Cells of Rat. Neuroreport 16:581-584. (2005)
YOUNG, R.W. Cell Death during Differentiation of the Retina in the Mouse. J.
Comp. Neurol. 229:362-373. (1984). ZONG, W.X.; LINDSTEN, T.; ROSS, A.J.; MACGREGOR, G.R.; THOMPSON, C.B.
BH3-Only Proteins that Bind Pro-Survival Bcl-2 Family Members Fail to Induce Apoptosis in the Absence of Bax and Bak. Genes Dev. 15:1481-1486. (2001)
ZORNIG, M.; HUEBER, A.O.; BAUM, W.; EVAN, G. Apoptosis Regulators and their
Role in Tumorigenesis. Bioch. Biophys. Acta 1551:1-37. (2001)

155
ANEXO-1

Herbimycin A induces sympathetic neuron survivaland protects against hypoxia
M. C. Cabral-da-Silva,1 N. E. Loureiro dos Santos,1 F. G. de Mello,1 J. S. H. Taylor,2 A. M. Tolkovsky,3
R. Linden1 and R. A. M. Reis1,2,3,CA
1Programa de Neurobiologia, IBCCF,UFRJ, Rio de Janeiro, Brazil; 2Department of Human Anatomy and Genetics,Oxford University, South Parks Road,Oxford OX13QX; 3Department of Biochemistry,Cambridge University,Tennis Court Road,Cambridge,UK
CA,2Corresponding Author and Address: [email protected]
Received1July 2003; accepted 21September 2003
DOI:10.1097/01.wnr.0000099603.19426.c8
We have examined how herbimycin a¡ects the survival and neur-itogenesis of avian sympathetic neurons. Herbimycin promotedsympathetic neuron survival and neuritogenesis. At higher concen-trations (Z100ng/ml), herbimycin still enhanced neuron survivalbut blocked neuritogenesis. Addition of herbimycin (10^30 ng/ml)to neurons cultured in the presence of NGF or retinal conditionedmedium altered neuronal morphology, with an increase in the
number of neurites. Addition of NGF during hypoxia rescued52% of the neurons compared to14% survival in control conditions.Herbimycin alone rescued about 50% of the neurons. In the pre-sence of NGF and100ng/ml herbimycin, 81% of the neurons survivedhypoxia. Our results show that herbimycin promotes survival ofchick sympathetic neurons and potentiates the e¡ects of NGF.NeuroReport14:2397^2401�c 2003 LippincottWilliams &Wilkins.
Key words: Apoptosis; Cell death; Chick; Neurite outgrowth; NGF;Tyrosine kinase inhibitor
INTRODUCTIONNeuronal survival is mediated by neurotrophic factors,which bind to specific receptors and activate downstreamsignaling pathways that suppress apoptosis [1]. Interferencewith the apoptotic pathways, either by suppressing thefunctions of pro-apoptotic proteins or by preventing theactivation of apoptotic pathways following neurotrophinwithdrawal, rescues neurons [2]. There is therefore muchinterest in identifying drugs that might mimic the neuro-trophic effect.Herbimycin A is an ansamycin antibiotic reported to
inhibit the tyrosine kinase (TK) p60c-src and platelet-derived growth factor (PDGF)-induced phospholipase Dactivation, but not phosphokinase A or phosphokinase C.TK inhibitors such as herbimycin or genistein have beenshown to prevent internucleosomal DNA fragmentation, ahallmark of apoptosis [3]. TK inhibitors have also beenshown to enhance neuronal survival of sensory neurons inthe presence of nerve growth factor (NGF) [4] and toincrease neurite outgrowth in PC12 cells [5]. Radicicol, anantifungal antibiotic that also blocks TK p60c-src, enhancesthe survival and neurite outgrowth of neurons fromembryonic chick dorsal root ganglia (DRG) and sympatheticganglia, potentiating the effects of neurotrophins [6]. Incontrast there are also reports showing that TK inhibitorsstimulate apoptosis in cells such as fibroblasts [7] or gliomacells [8].
We showed recently that avian sympathetic neurons inculture are maintained by trophic factors contained inmedium conditioned by retinal tissue (retinal conditionedmedium; RCM). These are not classic neurotrophic factors,as the effects are not blocked by chimera antibodies thatinterfere with neurotrophic factor binding through theirreceptors [9]. We have therefore examined other factors andpathways that might mediate these survival- and regenera-tion-promoting effects. Here we have tested whetherherbimycin would affect survival and neuritogenesis underconditions of NGF withdrawal or hypoxia.
MATERIALS AND METHODSWhite Leghorn chicken eggs, obtained from a local hatchery,were incubated at 381C in a forced-draft incubator. Allexperiments were conducted in accordance with the rulesand guidance governing the use of animals in experimentswithin the UK and Brazil. The lumbar sympathetic chainwas dissected from E10 embryos in DMEM. The isolatedsympathetic ganglia were incubated in trypsin (0.05%,10min, 371C), then centrifuged, washed with 10ml DMEMplus 10% fetal bovine serum (FCS) and triturated to a singlecell suspension. Non-neuronal cells were removed bydifferential sedimentation in a column containing FCS.Neurons were plated at a density of 600–1000/well on poly-L-lysine (10 mg/ml) and laminin (20 mg/ml). The growth
0959-4965�c LippincottWilliams &Wilkins Vol 14 No 18 19 December 2003 2397
MOLECULARNEUROSCIENCES NEUROREPORT
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
medium consisted of DMEM, 10% FCS, gentamicin (10 mg/ml), and survival factors or conditioned medium added justbefore the neurons were plated.Neurons to be grown under hypoxic conditions were
prepared from E10 embryos. After plating, the cultures weretransferred immediately to a hypoxia chamber and main-tained at 371C for up to 36 h. The hypoxia chamber wasfilled with a gas mixture of 85% nitrogen, 10% hydrogenand 5% carbon dioxide. The oxygen concentration in thechamber was monitored with an O2 analyser and keptbelow 1%.Neuronal survival was assessed by two different meth-
ods: counting the number of attached neurons 4 h afterplating and again after 48 h in a 12� 12mm grid in low-density, glial-free cultures [9]; and counting the number offragmented (apoptotic) or healthy nuclei using Hoechst33342 (bisbenzamide) under a fluorescence microscope(Zeiss Axioscope). Neurite outgrowth from sympatheticneuronal cultures was quantified from 5–10 individualimages and was measured using the Zeiss KS400 Imageanalyser (Version 3).At least three independent experiments were performed
in duplicate for each experimental paradigm. Means werecompared with Student’s unpaired t-test assuming equalvariances.
RESULTSE10 sympathetic neurons cultured in the presence of 10 ng/ml NGF had a low level of cell death (10%) while 95% of theneurons were dead after 48 h in cultures grown in theabsence of NGF (Fig. 1a). These dead cells showed classicalhallmarks of apoptosis, such as fragmented nuclei whenstained with Hoechst (not shown). Herbimycin (1 ng/ml)added to neurons deprived of NGF at (Fig. 1b) had no effect,but at 10 ng/ml (Fig. 1c) or 30 ng/ml (Fig. 1d), neuronscultured with herbimycin displayed phase-bright cell bodiesand short neurites. Herbimycin reduced apoptosis from 95%to 30% after 48 h with an EC50 of B3 ng/ml (Fig. 1e).Chick sympathetic neurons grown in the absence of
trophic factors underwent apoptosis and had no neuriteoutgrowth (Fig. 1a, Fig. 2a) while neurons grown in thepresence of 10 ng/ml NGF had phase-bright cell bodies andmany neurites (Fig. 2b). Herbimycin (1 ng/ml) did notincrease the number of neurites when compared to NGF(Fig. 2c). However, addition of increasing concentrations ofherbimycin (10 ng/ml, Fig. 2d; 30 ng/ml, Fig. 2e) in thepresence of NGF altered neuronal morphology, with anincrease in the number of small neurites and in thecomplexity of neurite branching. Total neurite outgrowthwas increased significantly over NGF controls (po 0.05; Fig.2e). When a high concentration of herbimycin (100 ng/ml)was added to NGF-treated neurons, neurite outgrowth wasblocked (Fig. 2f).In the presence of RCM, neurons were phase bright and
showed extensive neurite outgrowth (Fig. 2h). Addition ofherbimycin at 1 ng/ml showed little effect when comparedwith the RCM alone (Fig. 2i). However, at 10 ng/ml (Fig. 2j)and 30 ng/ml (Fig. 2k) herbimycin, neurons were large andphase-bright and the number and extent of neurites wasincreased significantly (po 0.05) over RCM controls. Cellaggregation was pronounced and a web-like array of fine
neurites extended between the islands of neurons. Thebranching pattern increased with the increase in herbimycindose. At 100 ng/ml, however, herbimycin blocked neurito-genesis, but maintained cell survival (Fig. 2l). A summaryfigure that quantifies the total neurite outgrowth under thevarious treatments is shown in Fig. 2m.Sympathetic neurons cultured in medium containing
10 ng/ml NGF, herbimycin at varying doses, or both NGFand herbimycin were placed in the hypoxia chamber with aconcentration of O2o 1% for 24 h.In the absence of trophic factors, 86% of the neurons were
dead at 24 h (Fig. 3a), compared to 42% death undernormoxic conditions after the same period (not shown).NGF (10 ng/ml) reduced the level of death with 527 6% ofcells surviving compared to 897 5% survival undernormoxic conditions after 24 h (Fig. 3b, quantified inFig. 3e). The presence of herbimycin at 10 ng/ml (457 5%survival) or 100 ng/ml (577 7% survival) in mediumduring hypoxia did not have a significant effect on cellsurvival (Fig. 3c,e) compared to the effects of NGF alone.
100
75
50
25
0
% N
euro
nal s
urvi
val
af
ter
48h
1 10 100 1000[Herbimycin A] ng/ml
(e)
(a) (b)
(c) (d)
Fig. 1. The e¡ects of herbimycin on E10 sympathetic neurons culturedfor 48h. (a) Without addition of factors, neurons are apoptotic. (b) With1ng/ml herbimycin there is no e¡ect and the neurons die. (c) With herbi-mycin at 10 ng/ml and (d) 30 ng/ml cells are phase bright and show goodneurite outgrowth. (e) Quanti¢cation of survival e¡ects after 48 h: herbi-mycin (squares) induces a dose-dependent increase in neuronal survivalalmost as robust as survival induced by 10ng/ml NGF (circle). In the ab-sence of NGF neurons die (triangle). The data are from three indepen-dently performed experiments of duplicate assays (mean7s.e., n¼3).Bar¼20mm.
239 8 Vol 14 No 18 19 December 2003
NEUROREPORT M.C.CABRAL-DA-SILVA ETAL.
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

Cell survival with 10 ng/ml herbimycin and NGF (587 7%)was marginally higher than with NGF alone (Fig. 3e), butthis was not statistically significant. However, treatmentwith NGF and 100 ng/ml herbimycin significantly reducedthe effects of hypoxia, with 817 7% (po 0.05) of neuronssurviving after 24 h (Fig. 3d,e).After 24 h hypoxia in the presence of 10 ng/ml NGF,
some neurites had degenerated consistent with the death ofsome of the cell bodies (Fig. 3b). This neurite degeneration
was also observed in neurons treated with 10 ng/mlherbimycin (Fig. 3c). However, herbimycin added incombination with NGF showed clear ameliorative effectson some of the neurites (Fig. 3d), although some signsof neurite degeneration were also observed and thelevel and complexity of branching were reduced whencompared to control normoxic cultures. As with normoxia,herbimycin caused some aggregation of neuronal cell bodies(Fig. 3d).
Tota
l neu
rite
leng
th
afte
r 48
h (A
.U.)
NGF 10 n
g
NGF + H
erb1
ng
NGF + H
erb1
0 ng
NGF + H
erb3
0 ng
RCM + H
erb1
ng
RCM + H
erb1
0 ng
RCM + H
erb3
0 ng
RCM + H
erb1
00 ng
NGF + H
erb1
00 ng
RCM
(a) (b)
(d) (e) (f)
(g) (h) (i)
(j) (k) (l)
(c)
(m)
* *
* * * *
Fig. 2. Herbimycin augments theneuritogenic e¡ects of NGF and RCM. (a)With noNGF, neurons die. (b) NGF (10ng/ml) induces sympathetic neuronalsurvival and neurite outgrowth. (c^e) NGF (10ng/ml) together with herbimycin at 1ng/ml (c), 10 ng/ml (d), or 30ng/ml. (e) Note the increased level andcomplexity of neurite outgrowth. (f ) 100 ng/ml herbimycin inhibits neurite outgrowth. (g,h) RCM induces survival and neuritogenesis compared to con-trol lacking NGF. (i) Addition of herbimycin at 1ng/ml has no e¡ect on neuritogenesis but at 10 ng/ml (j) and 30ng/ml (k) it increases the extent andcomplexity of neurite outgrowth. (l) At 100ng/ml, herbimycin blocks neurite outgrowth. Bar¼20mm. (m) Total neurite length quanti¢ed from 5^10 in-dividual images of sympathetic neuronal cultures grown under the above conditions (mean7 s.e., n¼3. *po 0.05 vs 10ng/ml NGF; **po 0.05 vs RCM,unpaired Student’s t-test assuming equal variance.
Vol 14 No 18 19 December 2003 239 9
HERBIMYCINAND SYMPATHETICNEURONAL SURVIVAL NEUROREPORT
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

DISCUSSIONSympathetic neurons have been used extensively as a modelfor the study of neuronal apoptosis induced by NGFwithdrawal [10]. Neural cell death has been shown to bemediated by an active process that requires changes in theactivation of many internal cell-signalling systems [11]. It isperhaps not surprising that disruption of these processescan modulate the complex pathways involved in apoptosis.For example, blocking some cell-signalling pathways hasbeen shown to reduce the requirement of cells forneurotrophins. The inhibitors, herbimycin, genistein, tyr-phostin, or radicicol, all potentiate the effects of low levels ofNGF on neuron survival [4,6]. In Bartlett’s study of chickDRGs, herbimycin was much more effective than other TKinhibitors in promoting cell survival and neurite outgrowth.In the present study we showed that herbimycin aloneinduced chick sympathetic neuronal survival, as well aspotentiating the effects of NGF. Herbimycin was a far bettertrophic agent for sympathetic neurons than genistein (datanot shown). This result can be explained by the differentmechanism of action of herbimycin, which can affect the SHdomain of the Src family non-receptor-type TKs whilegenistein directly inhibits catalytic activity.Our results showed an increase in neurite outgrowth
and in complexity of neurites, including branching, aftertreatment with herbimycin. Neurotrophins have beenshown to differentially affect process outgrowth inneurons [12], an effect that is thought to be regulated byGTPases associated with actin assembly and cytoskeletalre-arrangement [13]. Over the last few years there has been a
substantial amount of work exploring the multiple connec-tions between integrins, cytoskeletal components, and TKcascades [14]. Initial studies showed that herbimycin,among several other TK inhibitors, stimulated neuriteoutgrowth of cerebellar granule cells in culture [15]. It hasalso been shown that herbimycin enhances glial-cell-line-derived neurotrophic factor (GDNF)-induced neurite for-mation in a human neuroblastoma cell line [16]. The focaladhesion kinase pp125FAK (FAK) may be an indirect targetfor the neuritogenic effects of herbimycin. FAK is a non-receptor tyrosine kinase that is dependent upon TK activityof herbimycin-sensitive kinases such as c-Src for its function.FAK is also capable of binding to a number of othersignalling and structural proteins, including PI-3-kinase,GRAF (a RhoGAP), paxillin, talin, and p130Cas, which playimportant roles in the regulation of cell motility and in thecontrol of apoptosis [14,17]. FAK is localised to filopodiaand it interacts with b1 integrin, which has been shown tobe involved in the growth of Xenopus retinal ganglion cellaxons [18]. Therefore, it is reasonable to suggest thatherbimycin could be inducing a rearrangement of thecytoskeleton in the presence of trophic factors throughinteraction with TKs such as FAK.The effects of herbimycin in protecting sympathetic
neurons from hypoxia are striking. Herbimycin at highdoses (100 ng/ml) greatly potentiated the survival effects ofNGF on sympathetic neurons. The cell bodies and neuritesof these neurons were strong and healthy and the number ofapoptotic bodies seen with the DNA stain Hoescht 33342was reduced (not shown).
100
80
60
40
20
0
% N
euro
nal S
urvi
val
aft
er 4
8 h
Control
NGF
Herbim
ycin1
0
Herbim
ycin1
00
NGF+H10
NGF+H10
0
[5]
[5] [6] [4] [6] [6]
*(e)
(a) (b) (c) (d)
Fig. 3. Herbimycin protects neurons from hypoxia. (a) 24 h hypoxia results in neuronal death. (b) In the presence of 10 ng/ml NGF 50% of the cellssurvive. (c)With10 or100ng/ml herbimycin survival is similar to that inducedbyNGF. (d) The combined e¡ects of NGF and100ng/ml herbimycin rescuessigni¢cantlymore cells (81%).Bar¼20mm. (e) Graph showing cell survival after 24h hypoxiawith thevarious treatments.Number of repeats shown insidebrackets. *po 0.05, unpaired Student’s t-test assuming equal variances
24 0 0 Vol 14 No 18 19 December 2003
NEUROREPORT M.C.CABRAL-DA-SILVA ETAL.
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

Hypoxia occurs during acute and chronic vasculardisease, pulmonary disease and cancer [19]. Hypoxiaactivates apoptotic pathways, resulting in neuronal death[20]. One way that cells respond to reduced oxygen levels isthrough increased expression of hypoxia-inducible tran-scription factor 1 (HIF-1). Trophic factors such as EGF, IGF-1or PDGF, which are known to activate tyrosine kinasereceptors, increase expression of HIF1a [21]. HIF-1 is linkedto an up-regulation of VEGF in ischemic neural tissue [22]and VEGF has known survival and neuritogenic properties[23]. Herbimycin has been recently shown to elevate calciumlevels and to induce an increase in mRNA for HSP70 in anepidermal cell line [24]. This result provides a possibleexplanation for the anti-apoptotic effects of herbimycin inneurons, where up-regulation of HSP-70 has been shown toprotect neurons from hypoxia [25].In conclusion, our data indicate that herbimycin supports
the survival of chick sympathetic neurons in culture at lowdoses, potentiates the effects of trophic factors, and protectsneurons against hypoxia. It will be interesting to identifywhich of the TKs that regulate survival and neuritogenesisare sensitive to herbimycin A.
REFERENCES1. Miller FD and Kaplan DR. Neurotrophin signalling pathways regulating
neuronal apoptosis. Cell Mol Life Sci 58, 1045–1053 (2001).
2. Sofroniew MV, Howe CL and Mobley WC. Nerve growth factor signaling,
neuroprotection, and neural repair. Annu Rev Neurosci 24, 1217–1281 (2001).
3. Ji L, Zhang G and Hirabayashi Y. Inhibition of tumor necrosis factor alpha-
and ceramide-induced internucleosomal DNA fragmentation by herbimycin
A in U937 cells. Biochem Biophys Res Commun 212, 640–647 (1995).
4. Bartlett SE. Protein tyrosine kinase inhibitors synergize with nerve
growth factor in embryonic chick sensory neuronal cell survival. NeurosciLett 227, 87–90 (1997).
5. Miller DR, Lee GM and Manness PF. Increased neurite outgrowth
induced by inhibition of protein tyrosine kinase activity in PC12
pheochromocytoma cells. J Neurochem 60, 2134–2144 (1993).
6. Sano M, Yoshida M, Fukui S and Kitajima S. Radicicol potentiates
neurotrophin-mediated neurite outgrowth and survival of cultured
sensory neurons from chick embryo. J Neurochem 72, 2256–2263 (1999).
7. Hiwasa T, Arase Y, Chen Z, Kita K, Umezawa K, Ito H and Suzuki N.
Stimulation of ultraviolet-induced apoptosis of human fibroblast Uvr-1
cells by tyrosine kinase inhibitors. FEBS Lett 444, 173–179 (1999).
8. Chin LS, Murray SF, Zitnay KM and Rami B. K252a inhibits proliferation
of glioma cells by blocking platelet-derived growth factor signal
transduction. Clin Cancer Res 3, 771–776 (1997).
9. Reis RAM, Silva MCC, Loureiro dos Santos NE, Bampton E, Taylor JSH,
de Mello FG and Linden R. Sympathetic neuronal survival induced by
retinal trophic factors. J Neurobiol 50, 13–23 (2002).
10. Deshmukh M and Johnson EM Jr. Programmed cell death in neurons:
focus on the pathway of nerve growth factor deprivation-induced death
of sympathetic neurons. Mol Pharmacol 51, 897–906 (1997).
11. Miller FD and Kaplan DR. Neurotrophin signalling pathways regulating
neuronal apoptosis. Cell Mol Life Sci 58, 1045–1053 (2001).
12. McAllister AK. Neurotrophins and neuronal differentiation in the central
nervous system. Cell Mol Life Sci 58, 1054–1060 (2001).
13. Negishi M and Katoh H. Rho family GTPases as key regulators for
neuronal network formation. J Biochem (Tokyo) 132, 157–166 (2002).
14. Juliano RL. Signal transduction by cell adhesion receptors and the
cytoskeleton: functions of integrins, cadherins, selectins, and
immunoglobulin-superfamily members. Annu Rev Pharmacol Toxicol 42,283–323 (2002).
15. Doherty P, Furness J, Williams EJ and Walsh FS. Neurite outgrowth
stimulated by the tyrosine kinase inhibitor Herbimycin A requires
activation of tyrosine kinases and protein kinase C. J Neurochem 62, 2124–
2131 (1994).
16. Hiwasa T, Kondo K, Hishiki T, Koshizawa S, Umezawa K and
Nakagawara A. GDNF-induced neurite formation was stimulated by
protein kinase inhibitors and suppressed by Ras inhibitors. Neurosci Lett238, 115–118 (1997).
17. Abbi S and Guan JL. Focal adhesion kinase: protein interactions and
cellular functions. Histol Histopathol 17, 1163–1671 (2002).
18. Worley TL and Holt CE. Expression and Herbimycin A-sensitive
localization of pp125FAK in retinal growth cones. Neuroreport 7,
1133–1137 (1996).
19. Harris AL. Hypoxia: a key regulatory factor in tumour growth. NatureRev Cancer 2, 38–47 (2002).
20. Plesnila N, Zinkel S, Le DA, Amin-Hanjani S, Wu Y and Qiu J et al.BID mediates neuronal cell death after oxygen/glucose deprivation
and focal cerebral ischemia. Proc Natl Acad Sci USA 98, 15318–15323
(2001).
21. Zelzer E, Levy Y, Kahana C, Shilo BZ, Rubinstein M and Cohen B. Insulin
induces transcription of target genes through the hypoxia-inducible
factor HIF-1alpha/ARNT. EMBO J 17, 5085–94 (1998).
22. Jin KL, Mao XO and Greenberg DA. Vascular endothelial growth factor:
direct neuroprotective effect in in vitro ischemia. Proc Natl Acad Sci USA97, 10242–10247 (2000).
23. Bocker-Meffert S, Rosenstiel P, Rohl C, Warneke N, Held-Feindt J,
Sievers J and Lucius R. Erythropoietin and VEGF promote neural
outgrowth from retinal explants in postnatal rats. Invest Ophthalmol VisSci 43, 2021–2026 (2002).
24. Kiang JG. Genistein inhibits herbimycin A-induced over-expression
of inducible heat shock protein 70 kDa. Mol Cell Biochem 245, 191–199
(2003).
25. Lee JE, Yenari MA, Sun GH, Xu L, Emond MR, Cheng D et al. Differential
neuroprotection from human heat shock protein 70 overexpression in
in vitro and in vivo models of ischemia and ischemia-like conditions. ExpNeurol 170, 129–139 (2001).
Acknowledgements:The authors thank Ann Stanmore for helping with the Zeiss KS400 Image analyser.This work was supportedbygrants fromCAPES, FAPERJ,CNPq,PRONEXand theWellcomeTrust.M.C.C.-da-S. holds a PhD fellowship fromCNPq.R.A.M.R.
holds an International Biomedical Fellowship from theWellcomeTrust.
Vol 14 No 18 19 December 2003 24 01
HERBIMYCINAND SYMPATHETICNEURONAL SURVIVAL NEUROREPORT
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

161
ANEXO-2

In vitro toxicity induced by methylmercury on sympathetic neurons
is reverted by L-cysteine or glutathione
Ricardo Augusto de Melo Reis a, Anderson Manoel Herculano b,*,Maurıcio Castro Cabral da Silva a, Ronald Marques dos Santos c,
Jose Luis Martins do Nascimento b
a Laboratory Neurochemistry, Program in Neurobiology IBCCF, UFRJ, Rio de Janeiro, Brazilb Instituto de Biologia, UFPA, Belem, Brazil
c Departamento de Fisiologia, UFF, Niteroi, Brazil
Received 11 January 2007; accepted 26 March 2007
Abstract
Methylmercury (MeHg) is related to several deleterious effects on the vertebrate nervous system and part of these effects are through interaction
with sulfhydryl (–SH) group found in cellular proteins. We decided to characterize the dose-dependent effect of MeHg on the neurotoxicity and the
neurite outgrowth induced effects on chick sympathetic neurons dissociated and purified in culture. In this model, MeHg inhibited neurite
outgrowth (1–10 mM) and induced cell death (1–10 mM) after 48 h in culture. Since metal toxicity often generates reactive oxygen species, we
tested if antioxidant compounds such as glutathione (GSH) or L-cysteine (L-cys) could block the deleterious effects promoted by MeHg.
L-Methionine, another sulfur-containing amino acid, but without a SH group, was also used in this work. We show that GSH (10–100 mM) and
L-cys (100 mM), but not L-methionine (100 mM), fully blocked neurite degeneration and sympathetic neuron cell death in culture.
# 2007 Published by Elsevier Ireland Ltd and the Japan Neuroscience Society.
Keywords: Ngf; Sympathetic neurons; Methylmercury; Glutathione; Apoptosis
1. Introduction
Mercury is an environmental harmful chemical that
bioaccumulates through the food chain increasing human risk
of poisoning (Debes et al., 2006). Within the environment,
mercury can exist as elemental mercury vapor, inorganic
mercury salts, and organic mercury (Clarkson and Magos,
2006). Methylmercury (MeHg) is the most prevalent form of
mercury found in the environment, it is highly lipophilic and it
causes selective cell death in certain neuronal populations
(reviewed in Aschner and Walker, 2002).
The initial studies regarding MeHg intoxication were linked
to the poisoning of the Minamata Bay (Eto, 2000) what caused
the death of a large number of patients by neurological
complications. However, the mechanisms of MeHg neurotoxi-
city are not fully understood.
Due to the importance of this matter, several primary neuronal
cultures have been used to study the toxic effects of MeHg such
as sympathetic (Soderstrom and Ebendal, 1995), sensorial
(Wilke et al., 2003), cortical (Zhang et al., 2003), cerebellar
(Edwards et al., 2005), hippocampal (Szucs et al., 1997) and
retinal cells (Herculano et al., 2006), as well as astrocytes
(Mullaney et al., 1994), neuronal-astrocyte co-cultures (Morken
et al., 2005) and human cell lines (Crespo-Lopez et al., 2007).
Sympathetic and sensorial neurons in culture are supported
by nerve growth factor (NGF) and represent an important
model of a population that is maintained by trophic factors
released by its targets. Previous studies have demonstrated that
MeHg intoxication induces sympathetic ganglia toxicity and
neurite outgrowth inhibition (Soderstrom and Ebendal, 1995).
Although the toxic effect of the MeHg in the ganglion neurons
has been previously characterized, additional studies were
performed lately related to agents that could promote cellular
protection against mercury toxicity in neurons
www.elsevier.com/locate/neures
Neuroscience Research xxx (2007) xxx–xxx
* Corresponding author at: Departamento de Fisiologia, Centro de Ciencias
Biologicas, Universidade Federal do Para, Rua Augusto Correa No. 1, Campus
do Guama, 66075-110 Belem, PA, Brazil. Tel.: +55 91 31831202.
E-mail address: [email protected] (A.M. Herculano).
+ Models
NSR-2645; No of Pages 7
0168-0102/$ – see front matter # 2007 Published by Elsevier Ireland Ltd and the Japan Neuroscience Society.
doi:10.1016/j.neures.2007.03.010
Please cite this article in press as: de Melo Reis, R.A. et al., In vitro toxicity induced by methylmercury on sympathetic neurons is reverted by
L-cysteine or glutathione, Neurosci. Res. (2007), doi:10.1016/j.neures.2007.03.010

Several reports showed that sulfhydryl (–SH) groups
represent the main chemical target of mercury conjugation
in biological system (Hughes, 1957; Mullaney et al., 1994) and
since SH groups are ubiquitous within cells, many different
functions are altered. This phenomenon is strongly associated
to protein inactivation and consequently cell death in the CNS
(Crespo-Lopez et al., 2007). In this context, compounds that
possess SH group could promote reduction of mercury toxicity
in neuronal cells.
MeHg neurotoxicity has been correlated to the production
of reactive oxygen species (ROS) (Sarafian and Verity, 1991).
Many works demonstrated that L-cysteine (L-cys) and reduced
gluthathione (GSH) represent important antioxidants agents in
the cellular environment and L-cys has been shown to increase
the levels of GSH. Previous reports showed that L-cys and GSH
have important functions as antioxidant and on detoxification
of xenobiotica like mercury (Monks et al., 1999). In this sense,
we decided to evaluate the protective role of GSH and L-cys
against MeHg toxicity using avian sympathetic neurons in
culture. We also used L-methionine, another sulfur-containing
amino acid, which not display –SH group, in order to
investigate the importance of SH-reactive compounds against
MeHg toxicity.
2. Materials and methods
2.1. Materials
Dulbecco’s modified Eagle’s medium (DMEM) and fetal calf serum (FCS),
laminin, mouse NGF (2.5 S) and gentamicin reagent solution was obtained from
Gibco. Tissue culture dishes were from Nalge Nunc International (Naperville,
IL, USA). All other reagents, as well as rabbit anti-neurofilament 200 (N4142)
were obtained from Sigma–Aldrich, Inc., Tritc goat anti-rabbit IgG antibody
was from molecular probes.
2.2. Neuron cultures
White leghorn chicken eggs obtained in a local hatchery were incubated at
38 8C in a forced-draft incubator. The lumbar sympathetic chain was dissected
in DMEM and cleaned with electrolytically sharpened tungsten needles as
described before (Reis et al., 2002). Embryos were staged according to
Hamburger and Hamilton (1951). E10 embryo sympathetic ganglia were
incubated at 37 8C with 0.05% trypsin (Worthington Biochemical Corp.,
Freehold, NJ) for 10 min in Ca2+- and Mg2+-free solution (CMF). After
Fig. 1. Temporal effect of MeHg on the avian sympathetic neuron culture survival. Embryonic chick sympathetic neurons were cultured for 48 h and then submitted
to the following treatments: control, NGF 20 ng/ml, NGF 20 ng/ml + MeHg 0.1 mM, NGF 20 ng/ml + MeHg 1 mM or NGF 20 ng/ml + MeHg 10 mM. The toxicity of
MeHg disrupts sympathetic neuronal neurite outgrowth and survival at concentrations equal or above 1 mM, scale 50 mm.
R.A. de Melo Reis et al. / Neuroscience Research xxx (2007) xxx–xxx2
+ Models
NSR-2645; No of Pages 7
Please cite this article in press as: de Melo Reis, R.A. et al., In vitro toxicity induced by methylmercury on sympathetic neurons is reverted by
L-cysteine or glutathione, Neurosci. Res. (2007), doi:10.1016/j.neures.2007.03.010
centrifugation and removal of the trypsin solution, the ganglia were washed with
10 ml of DMEM plus 10% fetal calf serum (FCS) and triturated mechanically
with a fire-polished, siliconized Pasteur pipette to give a single cell suspension.
Non-neuronal cells were removed by differential sedimentation (Davies, 1989).
Neuronal cell suspensions were plated at a density of 600–1000 neurons in a 35-
mm plastic tissue culture dishes with a 2 mm grid (Nalge Nunc Intl.) that had
been precoated with poly-L-lysine (10 mg/ml, overnight) and laminin (20 mg/ml
for at least 4 h).
The neurons were incubated at 37 8C in a humidified 5% CO2 incubator.
DMEM was suplemented with gentamicin (10 mg/ml). NGF 2.5 S (10 ng/ml)
was used as a positive control for the sympathetic neurons, keeping survival
near 90% at 48 h after cell plating. At least three independent experiments were
done in duplicate for each experimental paradigm.
2.3. MeHg incubation protocol
MeHg was prepared in a stock solution (5 mM) in deionised H2O and on the
day of the experiment it was added to the plated neurons of E10 chick embryos
in the presence of 20 ng/ml NGF for 48 h (Figs. 1–3). Alternatively, MeHg was
added to E10C2 sympathetic neuronal cultures maintained by 20 ng/ml NGF
(48 h neurons in culture) in the presence or not of the following agents GSH
(10–100 mM), L-cys (10–100 mM) or methionine (100 mM) for 24 h (Figs. 4
and 5). Neurite outgrowth from sympathetic neuronal cultures was quantified as
described before (Cabral-da-Silva et al., 2003) from four individual images for
each condition and was measured using the ImagePro analyser. Neuronal
survival was assessed by counting the number of attached neurons in a low
density, glial free sympathetic neuron culture isolated from lumbar paraver-
tebral sympathetic chain in a 12-mm � 12-mm grid in the center of each 35-mm
culture dish essentially as described before (Reis et al., 2002; Cabral-da-Silva
et al., 2003). This selected area was counted 3 h (not less than 150 neurons/dish)
after plating and again after 24 or 48 h to assess survival. At least two
independent experiments were performed in duplicate for each experimental
paradigm. Means were compared with an analysis of variance (ANOVA) with
Bonferroni’s multiple comparison test.
2.4. Immunocytochemistry procedure and neurite outgrowth
quantification
In order to visualize the neurite mesh, sympathetic neuronal cultures from
E10C2 treated or not with MeHg were washed three times with phosphate buffer
saline (PBS) and fixed by immersion in absolute methanol for 5 min. The tissue
was then rinsed with PBS and then pre-incubated with bovine serum albumin
(BSA) 5% for 30 min, followed by incubation (24 h) with primary antibody
against neurofilament 200 (N-4142; Sigma–Aldrich, Inc.), a polyclonal rabbit
antiserum (1:1000). After several rinses in PBS, the neuronal culture was
incubated with Tritc goat anti-rabbit IgG (molecular probes) diluted 1:500 in
PBS for 1.5 h. Further washes in PBS were made and the cultures were incubated
in PBS for 2 h. Immunohistochemical control sections were incubated with PBS
or BSA in the absence of primary antibody and no immunoreactivity was detected.
Fig. 2. Sympathetic neuronal survival quantification for the in vitro toxicity
induced MeHg. There was no statistical difference between NGF-treated
neurons and the ones submitted to NGF 20 ng/ml + MeHg 0.1 mM. However,
MeHg added at 1 or 10 mM on NGF-treated neurons induced 55% or 100% cell
death, respectively, 24 h after plating. The data are from triplicate assays
(means � S.E.M.) from one of the three independently performed experiments,
each with similar results.
Fig. 3. Neurite outgrowth on NGF-treated sympathetic neurons treated or not with MeHg. While control neurons (without NGF) had a very few neurites (A), 20 ng/ml
NGF showed a huge neurite network between neurons 48 h after plating (B), addition of 1 mM (D) or 10 mM (E), but not 0.1 mM MeHg (C) induced disruption of the
neurite mesh between sympathetic neurons. The neurite network was revealed after an immuno-detection using a anti-neurofilament 200 antibody attached to a Tritc
fluorochrome. (F) Sympathetic neuronal neurite outgrowth quantification, scale 50 mm.
R.A. de Melo Reis et al. / Neuroscience Research xxx (2007) xxx–xxx 3
+ Models
NSR-2645; No of Pages 7
Please cite this article in press as: de Melo Reis, R.A. et al., In vitro toxicity induced by methylmercury on sympathetic neurons is reverted by
L-cysteine or glutathione, Neurosci. Res. (2007), doi:10.1016/j.neures.2007.03.010

3. Results
In order to study the effects of MeHg intoxication on
dissociated sympathetic neurons, we separated neurons from
glia and plated them on a poly-lysine and laminin substrate.
Sympathetic neurons started to make long neurites within the
first 2 h after plating. Withdrawal of NGF from cultures of E10
sympathetic neurons induced more than 95% of neuronal death
within 48 h (Reis et al., 2002). Indeed, up to 50% sympathetic
neurons in the absence of NGF undergo apoptosis 24 h after
plating (Fig. 1). As shown in Fig. 2, less than 5% of the
sympathetic neurons plated without NGF are found with bright
cell bodies and neurites after 48 h in culture. In contrast, 20 ng/
ml NGF induced neurites a few hours after plating and
maintained bright cell bodies and extensive projections in the
majority of sympathetic neuron cultures 48 h after plating
(Fig. 1). NGF maintained more than 90% of the neurons alive
(Fig. 2). Added 0.1 mM MeHg to NGF-treated neurons had no
effect on the survival rate of the neurons. However, there were
several blebs and ruptures on the neurite outgrowth at 1 mM
MeHg plus 20 ng/ml NGF a few hours after plating (Fig. 1) and
60% of the sympathetic neurons died after 48 h (Fig. 2). The
presence of 10 mM MeHg on NGF-treated neurons induced a
rapid and massive degeneration on the neurite network a few
minutes after the addition, detaching all cells from the plastic
dishes (Fig. 1). There were no viable cells 24 h after plating.
The neurite network induced by sympathetic neurons was
also evaluated using an anti-NF200 antibody attached to a
fluorochome. Neurite outgrowth of control neurons was less
than 5% of the neuritic mesh found on NGF-treated neurons
48 h after plating (Fig. 3A). NGF 20 ng/ml induces a complex
Fig. 4. L-Cys or GSH, but not L-methionine protect in vitro toxicity induced by 1 mM MeHg on E10C2 sympathetic neurons (obtained from an E10 embryo and
maintained for 2 days in culture) treated for 24 h. Sympathetic neurons treated with NGF 20 ng/ml show extensive neurites and bright cell bodies (A), addition of
100 mM GSH (B), 100 mM L-cys (C) or 100 mM methionine (D) had no effect on NGF-treated neurons. Addition of MeHg 1 mM induced a massive cell death 24 h
after treatment (E). However, addition of 100 mM GSH (G) or 100 mM L-cys (I), but not 100 mM methionine (J) fully protected NGF sympathetic neurons poisoned
with 1 mM MeHg for 24 h. While 10 mM GSH (F) partially protected NGF sympathetic neurons treated with MeHg, 10 mM L-cys (H) or 100 mM methionine had no
protective effects on MeHg intoxication (J).
Fig. 5. Quantification of the in vitro toxicity effects on sympathetic neuronal
survival maintained by NGF and treated with MeHg 1 mM in the presence or
not of L-cys, GSH, or L-methionine for 24 h. GSH (10 and 100 mM) and
Cys (100 mM) protected sympathetic neurons treated with MeHg 1 mM
( p < 0.001), but 10 mM Cys or 100 mM methionine had not protective effect.
An ANOVA with Bonferroni’s multiple comparison test were performed (n = 3
experiments in duplicate).
R.A. de Melo Reis et al. / Neuroscience Research xxx (2007) xxx–xxx4
+ Models
NSR-2645; No of Pages 7
Please cite this article in press as: de Melo Reis, R.A. et al., In vitro toxicity induced by methylmercury on sympathetic neurons is reverted by
L-cysteine or glutathione, Neurosci. Res. (2007), doi:10.1016/j.neures.2007.03.010

mesh of neurites 48 h after plating the cells (Fig. 3B). Addition
of 0.1 mM MeHg to 20 ng/ml NGF-treated neurons had no
effect on the neuritic density of sympathetic cells (Fig. 3C), but
1 mM MeHg plus 20 ng/ml NGF had a profound effect,
destroying most of the contacts between cells 48 h after plating
(Fig. 3D). As expected for neuronal survival, the presence of
10 mM MeHg plus 20 ng/ml NGF eliminated all contacts
within a few minutes after addition of the contaminant
(Fig. 3E). Quantification of neurite outgrowth in response to
these treatments is shown in Fig. 3F.
Since L-cys and reduced GSH have been shown to act as
antioxidants, we evaluated the protective role of GSH and L-cys
against MeHg toxicity. As shown in Fig. 4, GSH and L-cys
(100 mM) fully protected NGF-treated sympathetic neurons
after 24 h in culture ( p < 0.001) when cells were pre-incubated
with antioxidants during plating. The same effect was not
observed for methionine (100 mM), and sympathetic neurons
were entirely destroyed upon MeHg treatment (Fig. 4J). The
quantification of neuronal survival following these treatments is
shown in Fig. 5. The number of neurons counted in a 144 mm2
grid showed that 100 mM L-cys and 100 mM GSH fully
protected and maintained the same level of survival of NGF-
treated neurons when MeHg 1 mM was added (nearly 100%
survival); the same was not true for NGF + 100 mM methio-
nine, since only 13 � 4% sympathetic neurons survived 24 h
after plating. As also shown, while GSH 10 mM treatment
produced survival (61 � 10%) of sympathetic neurons
( p < 0.01), 10 mM L-cys (26 � 10%) treatment had no effect
on the survival of sympathetic neurons treated with MeHg. In
conclusion, our results show that GSH (10–100 mM) and
100 mM L-cys are effective compounds in blocking cell death
induced by MeHg.
4. Discussion
Neuronal survival in the vertebrate peripheral nervous
system depends on neurotrophic factors available from target
tissues (Deshmukh and Johnson, 1997). Developing chick
sympathetic neurons within the paravertebral chains are
generated in excess and virtually all sympathetic neurons
depend on NGF during development (Levi-Montalcini and
Booker, 1960). NGF withdrawal leads to apoptotic sympathetic
neuron cell death (Reis et al., 2002) that otherwise would
promote survival of neurons through phosphoinositide 3-
kinase- or mitogen activated protein kinase-dependent path-
ways. We used a purified sympathetic neuronal free from
satellite glia (Davies, 1989) to study the toxic effects of MeHg
and we asked if SH physiological compounds such as cysteine
and GSH could protect these neurons in culture. Glia cells
secrete trophic factors that could interfere with the end result of
neuronal survival since several reports refer to different
mechanisms related to MeHg intoxication related to glia
(astrocytes) and neurons. In addition, astrocytes appear to
increase neuronal resistance, indicating a possible protective
role for astrocytes in MeHg neurotoxicity (Morken et al., 2005).
In the present study we show that MeHg exposure induces
cellular toxicity in neurons cultured from sympathetic chick
ganglion and these results are in agreement with previous works
that demonstrated that chick sympathetic and sensory dorsal
root ganglia are very sensitive to MeHg injury (Nakada et al.,
1981; Soderstrom and Ebendal, 1995; Miura et al., 2000). In
fact, many reports have shown that adult and embryonic
peripheral nervous system represents an important target of
MeHg toxicity (Sanfeliu et al., 2003; Crespo-Lopez et al.,
2005).
In this context, cultures of sympathetic neurons can
represent an interesting experimental model in which to study
the mechanism of action of several neurotoxic agents. Although
the toxicicity induced by MeHg in sympathetic neurons is well
characterized, the cellular mechanism involved in this process
is not completely understood. Soderstrom and Ebendal (1995)
reported that MeHg induces a dose-dependent intoxication, at
low micromolar concentrations, on purified sympathetic
neurons in culture. Some adverse effects were reported on
NGF responses in ganglion cultured neurons and change levels
of NGF production in fibroblasts.
Studies performed by Baxter and Smith (1998) also
demonstrated that mouse sensory neurons showed changes in
the pattern of neuropeptide expression after MeHg exposure.
Several works support that the main mechanism of mercury
toxicity in biological systems is related to the production of
ROS and high conjugation with –SH groups in amino acids and
proteins (Philbert et al., 2000; Allen et al., 2002).
In the present work we also demonstrated by immunocyto-
chemical analysis that MeHg intoxication promotes alterations
on the normal pattern of neurofilament organization, suggesting
that MeHg exposure can promote disruption on the axonal
development and consequently disruption of the normal axonal
transport. Previous reports have shown that abnormal assembly
of neurofilaments in motor neuron cell bodies and axons is
strongly associated with neurodegenerative diseases (Liu et al.,
2004). In this context, the evaluation of cellular mechanism of
MeHg toxicity in the nervous system is very important to
optimization of alternative therapies against mercurial toxicity.
To our knowledge, this is the first demonstration that GSH
and cysteine treatment promotes a complete protection against
MeHg toxicity in peripheral ganglion cultured neurons. This
protective action could be related with two distinct processes,
one is scavenge of MeHg by GSH and L-cys and another is
related to antioxidant potential exerted by those substances.
Previous reports also related that modulation of GSH and L-cys
contents influences the sensitivity of astrocytes and neurons to
toxicity induced by MeHg in the central nervous system (Jain
et al., 1991; Graff et al., 1993; Allen et al., 2002; Fonnum and
Lock, 2004; Kaur et al., 2006). As the association of MeHg-
GSH and MeHg-cysteine, as well as, anti-oxidant activity of
GSH and L-cys is mediated by –SH groups, in this study we
demonstrated that L-methionine, an amino acid similar to
cysteine, but without –SH group, did not protect ganglion
neurons from MeHg toxicity confirming the importance of –SH
contents to protection against MeHg intoxication.
The cellular or molecular mechanisms which contribute to
the differential neurotoxic effects of MeHg on peripheral or
central neurons are, as yet, unknown. Several different factors
R.A. de Melo Reis et al. / Neuroscience Research xxx (2007) xxx–xxx 5
+ Models
NSR-2645; No of Pages 7
Please cite this article in press as: de Melo Reis, R.A. et al., In vitro toxicity induced by methylmercury on sympathetic neurons is reverted by
L-cysteine or glutathione, Neurosci. Res. (2007), doi:10.1016/j.neures.2007.03.010

have been related to MeHg intoxication such as production of
nitric oxide, disruption of Ca2+ homeostasis, impairment of
synaptic transmission, glutamate receptor subtype expression,
and ROS generation (Komulainen and Bondy, 1987; Marty and
Atchison, 1997; Sarafian and Verity, 1991; Limke and Atchison,
2002). One of the hallmarks of cellular degeneration is related to
microtubule since much of the necessary neuronal traffic and
support is generated by the microtubule transport system
(Goldstein and Yang, 2000). A single, 2-h exposure to
submicromolar MeHg caused a transient depolymerization of
neuronal microtubules and reduced the number of developing
axons of E8 chick forebrain neurons, without causing significant
cell death for at least 2 days after treatment (Heidemann et al.,
2001).
In the present work we conclude that primary cultures of
neurons from sympathetic ganglion represent an efficient
experimental model to evaluate the protective effect of agents
against neurotoxicants such as mercury. In summary, MeHg
induces severe alterations on the neurotubulin organization and
that GSH and L-cys could represent important therapeutic agents
against MeHg toxicity in neurons of peripheral nervous system.
Moreover, knowledge originated from this work opens an
important perspective for prevention strategies on local
populations as found in the Amazon region where MeHg
exposure is endemic (Pinheiro et al., 2006, 2007; Bastos et al.,
2007). Mercury from different sources is available for
methylation processes contaminating the fish resources, which
constitute a dietary support for the large population living along
the riverbanks (Lebel et al., 1997; Dolbec et al., 2001). This
approach is sustained by the report performed by Demirkol et al.
(2004) which demonstrated that several fruits and vegetables
such as papaya, mango, orange, asparagus, avocado and
cucumber presented high levels of GSH and L-cys. Our study
validates a previous report that fruit consumption could possibly
interfere against mercury exposure and toxicity (Passos et al.,
2003). Also, a recent publication from the same group show that
fruit consumption may provide a protective effect for Hg
exposure in Amazonian riparian communities (Passos et al.,
2007). Therefore, prevention based on high GSH or L-cys food
compounds could be an effective prophylactic strategy to reduce
Hg exposure in fish eating communities.
Acknowledgements
This work was supported by grants from PADCT/CNPq
and FAPERJ. RAM Reis and JLM do Nascimento are supported
by a research fellowship from the Conselho Nacional de
Desenvolvimento Cientifico e Tecnologico (CNPq).
References
Allen, J.W., Shanker, G., Tan, K.H., Aschner, M., 2002. The consequences of
methylmercury exposure on interactive functions between astrocytes and
neurons. Neurotoxicology 23, 755–759.
Aschner, M., Walker, S.J., 2002. The neuropathogenesis of mercury toxicity.
Mol. Psychiatry 7, S40–S44.
Bastos, W.R., de Almeida, R., Dorea, J.G., Barbosa, A.C., 2007. Annual
flooding and fish-mercury bioaccumulation in the environmentally
impacted Rio Madeira (Amazon). Ecotoxicology 16, 341–346.
Baxter, G.J., Smith, R.A., 1998. Changes in neuropeptide immunoreactivity in
cultured adult mouse sensory neurons following methylmercury chloride
treatments. Neurosci. Lett. 246, 13–16.
Cabral-da-Silva, M.C., Loureiro dos Santos, N.E., de Mello, F.G., Taylor,
J.S.H., Tolkovsky, A.M., Linden, R., Reis, R.A.M., 2003. Herbimycin a
induces sympathetic neuron survival and protects against hypoxia. Neu-
roreport 14, 2397–2401.
Clarkson, T.W., Magos, L., 2006. The toxicology of mercury and its chemical
compounds. Crit. Rev. Toxicol. 36, 609–662.
Crespo-Lopez, M.E., Herculano, A.M., Corvelo, T.C., Do Nascimento, J.L.,
2005. Mercury and neurotoxicity. Rev. Neurol. 40, 441–447.
Crespo-Lopez, M.E., Lima de As, A., Herculano, A.M., Rodriguez Burbano, R.,
Do Nascimento, J.L., 2007. Methylmercury genotoxicity: a novel effect
in human cell lines of the central nervous system. Environ. Int. 33, 141–
146.
Davies, A.M., 1989. Neurotrophic factor bioassay using dissociated neurons. In:
Rush, R. (Ed.), Nerve Growth Factors. John Wiley & Sons, Chichester, pp.
95–109.
Debes, F., Budtz-Jørgensen, E., Weihe, P., White, R.F., Grandjean, P., 2006.
Impact of prenatal methylmercury exposure on neurobehavioral function at
age 14 years. Neurotoxicol. Teratol. 28, 536–547.
Demirkol, O., Adams, C., Ercal, N., 2004. Biologically important thiols in
various vegetables and fruits. J. Agric. Food Chem. 52, 8151–8154.
Deshmukh, M., Johnson Jr., E.M., 1997. Programmed cell death in neurons:
focus on the pathway of nerve growth factor deprivation-induced death of
sympathetic neurons. Mol. Pharmacol. 51, 897–906.
Dolbec, J., Mergler, D., Larribe, F., Roulet, M., Lebel, J., Lucotte, M., 2001.
Sequential analysis of hair mercury levels in relation to fish diet of an
Amazonian population. Braz. Sci. Total Environ. 271, 87–97.
Edwards, J.R., Marty, M.S., Atchison, W.D., 2005. Comparative sensitivity of
rat cerebellar neurons to dysregulation of divalent cation homeostasis and
cytotoxicity caused by methylmercury. Toxicol. Appl. Pharmacol. 208,
222–232.
Eto, K., 2000. Minamata disease. Neuropathology 20, S14–S19.
Fonnum, F., Lock, E.A., 2004. The contributions of excitotoxicity, glutathione
depletion and DNA repair in chemically induced injury to neurones:
exemplified with toxic effects on cerebellar granule cells. J. Neurochem.
88, 513–531.
Goldstein, L.S.B., Yang, Z., 2000. Microtubule-based transport systems
in neurons: the roles of kinesins and dyneins. Ann. Rev. Neurosci. 23,
39–71.
Graff, R.D., Philbert, M.A., Lowndes, H.E., Reuhl, K.R., 1993. The effect of
glutathione depletion on methyl mercury-induced microtubule disassembly
in cultured embryonal carcinoma cells. Toxicol. Appl. Pharmacol. 120,
20–28.
Hamburger, V., Hamilton, H.L., 1951. A series of normal stages in the
development of the chicken embryo. J. Morphol. 88, 49–92.
Heidemann, S.R., Lamoureux, P., Atchison, W.D., 2001. Inhibition of axonal
morphogenesis by nonlethal, submicromolar concentrations of methylmer-
cury. Toxicol. Appl. Pharmacol. 174, 49–59.
Herculano, A.M., Crespo-Lopez, M.E., Lima, S.M., Picanco-Diniz, D.L., Do
Nascimento, J.L., 2006. Methylmercury intoxication activates nitric
oxide synthase in chick retinal cell culture. Braz. J. Med. Biol. Res. 39,
415–418.
Hughes, W.H., 1957. A physicochemical rationale for the biological activity of
mercury and its compounds. Ann. NY Acad. Sci. 65, 454–460.
Jain, A., Martensson, J., Stole, E., Auld, P.A., Meister, A., 1991. Glutathione
deficiency leads to mitochondrial damage in brain. Proc. Natl. Acad. Sci.
U.S.A. 88, 1913–1917.
Kaur, P., Aschner, M., Syversen, T., 2006. Glutathione modulation influences
methyl mercury induced neurotoxicity in primary cell cultures of neurons
and astrocytes. Neurotoxicology 27, 492–500.
Komulainen, H., Bondy, S.C., 1987. Increased free intrasynaptosomal Ca2+ by
neurotoxic organometals: distinctive mechanisms. Toxicol. Appl. Pharma-
col. 88, 77–86.
Lebel, J., Roulet, M., Mergler, D., Lucotte, M., Larribe, F., 1997. Fish diet and
mercury exposure in a riparian Amazonian population. Water Air Soil
Pollut. 97, 31–44.
R.A. de Melo Reis et al. / Neuroscience Research xxx (2007) xxx–xxx6
+ Models
NSR-2645; No of Pages 7
Please cite this article in press as: de Melo Reis, R.A. et al., In vitro toxicity induced by methylmercury on sympathetic neurons is reverted by
L-cysteine or glutathione, Neurosci. Res. (2007), doi:10.1016/j.neures.2007.03.010

Levi-Montalcini, R., Booker, B., 1960. Destruction of the sympathetic ganglia
in mammals by an antiserum to a nerve growth protein. Proc. Natl. Acad.
Sci. U.S.A. 46, 384–391.
Limke, T.L., Atchison, W.D., 2002. Acute exposure to methylmercury opens the
mitochondrial permeability transition pore in rat cerebellar granule cells.
Toxicol. Appl. Pharmacol. 178, 52–61.
Liu, Q., Xie, F., Siedlak, S.L., Nunomura, A., Honda, K., Moreira, P.I., Zhua, X.,
Smith, A., Perry, G., 2004. Neurofilament proteins in neurodegenerative
diseases. Cell Mol. Life Sci. 61, 3057–3075.
Marty, M.S., Atchison, W.D., 1997. Pathways mediating Ca2+ entry in rat
cerebellar granule cells following in vitro exposure to methyl mercury.
Toxicol. Appl. Pharmacol. 147, 319–330.
Miura, K., Himeno, S., Koide, N., Imura, N., 2000. Effects of methylmercury
and inorganic mercury on the growth of nerve fibers in cultured chick dorsal
root ganglia. Tohoku J. Exp. Med. 192, 195–210.
Monks, T.J., Ghersi-Egea, J.F., Philbert, M., Cooper, A.J., Lock, E.A., 1999.
Symposium overview: the role of glutathione in neuroprotection and
neurotoxicity 51. Toxicol. Sci. 161–177.
Morken, T.S., Sonnewald, U., Aschner, M., Syversen, T., 2005. Effects of
methylmercury on primary brain cells in mono- and co-culture. Toxicol. Sci.
87, 169–175.
Mullaney, K.J., Fehm, M.N., Vitarella, D., Wagoner, D.E., Aschner, M., 1994. The
role of –SH groups in methyl mercuric chloride-induced D-aspartate and
rubidium release from rat primary astrocyte cultures. Brain Res. 641, 1–9.
Nakada, S., Saito, H., Imura, N., 1981. Effect of methylmercury and inorganic
mercury on the nerve growth factor-induced neurite outgrowth in chick
embryonic sensory ganglia. Toxicol. Lett. 8, 23–28.
Passos, C.J., Mergler, D., Gaspar, E., Morais, S., Lucotte, M., Larribe, F.,
Davidson, R., de Grosbois, S., 2003. Eating tropical fruit reduces mercury
exposure from fish consumption in the Brazilian Amazon. Environ. Res. 93,
123–130.
Passos, C.J., Mergler, D., Fillion, M., Lemire, M., Mertens, F., Guimaraes,
J.R.D., Philibert, A., 2007. Epidemiologic confirmation that fruit consump-
tion influences mercury exposure in riparian communities in the Brazilian
Amazon. Environm. Res., doi:10.1016/j.envres.2007.01.012, in press.
Philbert, M.A., Billingsley, M.L., Reuhl, K.R., 2000. Mechanisms of injury in
the central nervous system. Toxicol. Pathol. 28, 43–53.
Pinheiro, M.C., Oikawa, T., Vieira, J.L., Gomes, M.S., Guimaraes, G.A.,
Crespo-Lopez, M.E., Muller, R.C., Amoras, W.W., Ribeiro, D.R., Rodri-
gues, A.R., Cortes, M.I., Silveira, L.C., 2006. Comparative study of human
exposure to mercury in riverside communities in the Amazon region. Braz.
J. Med. Biol. Res. 39, 411–414.
Pinheiro, M.C.N., Crespo-Lopez, M.E., Vieira, J.L.F., Oikawa, T., Guimaraes,
G.A., Araujo, C.C., Amoras, W.W., Ribeiro, D.R., Herculano, A.M., do
Nascimento, J.L.M., Silveira, L.C.L., 2007. Mercury pollution and child-
hood in Amazon riverside villages. Environ. Int. 33, 56–61.
Reis, R.A.M., Silva, M.C.C., Loureiro dos Santos, N.E., Bampton, E., Taylor,
J.S.H., de Mello, F.G., Linden, R., 2002. Sympathetic neuronal survival
induced by retinal trophic factors. J. Neurobiol. 50, 13–23.
Sarafian, T., Verity, M.A., 1991. Oxidative mechanisms underlying methylmer-
cury neurotoxicity. Int. J. Dev. Neurosci. 9, 147–153.
Sanfeliu, C., Sebastia, J., Cristofol, R., Rodriguez-Farre, E., 2003. Neurotoxi-
city of organomercurial compounds. Neurotox. Res. 5, 283–305.
Soderstrom, S., Ebendal, T., 1995. In vitro toxicity of methyl mercury: effects
on nerve growth factor (NGF)-responsive neurons and on NGF synthesis in
fibroblasts. Toxicol. Lett. 75, 133–144.
Szucs, A., Angiello, C., Salanki, J., Carpenter, D.O., 1997. Effects of inorganic
mercury and methylmercury on the ionic currents of cultured rat hippo-
campal neurons. Cell Mol. Neurobiol. 17, 273–288.
Wilke, R.A., Kolbert, C.P., Rahimi, R.A., Windebank, A.J., 2003. Methylmer-
cury induces apoptosis in cultured rat dorsal root ganglion neurons.
Neurotoxicology 24, 369–378.
Zhang, J., Miyamoto, K., Hashioka, S., Hao, H.P., Murao, K., Saido, T.C.,
Nakanishi, H., 2003. Activation of m-calpain in developing cortical
neurons following methylmercury treatment. Dev. Brain Res. 142,
105–110.
R.A. de Melo Reis et al. / Neuroscience Research xxx (2007) xxx–xxx 7
+ Models
NSR-2645; No of Pages 7
Please cite this article in press as: de Melo Reis, R.A. et al., In vitro toxicity induced by methylmercury on sympathetic neurons is reverted by
L-cysteine or glutathione, Neurosci. Res. (2007), doi:10.1016/j.neures.2007.03.010

169
ANEXO-3

Elsevier Editorial System(tm) for Brain Research Manuscript Draft Manuscript Number: BRES-D-07-00619 Title: MULLER GLIA FACTORS INDUCE SURVIVAL AND NEURITOGENESIS OF PERIPHERAL AND CENTRAL NEURONS Article Type: Research Report Section/Category: Nervous System Development, Regeneration and Aging Keywords: Muller glia; cell death; sympathetic; RGC; apoptosis; neurotrophin Corresponding Author: Dr. Ricardo AM Reis, PhD Corresponding Author's Institution: Lab. Neurochemistry, Program in Neurobiology IBCCF, UFRJ, Rio de Janeiro, Brazil First Author: Ricardo AM Reis, PhD Order of Authors: Ricardo AM Reis, PhD; Mauricio C Cabral-da-Silva, PhD; Fernando G de Mello, PhD; Jeremy S Taylor, PhD Manuscript Region of Origin: BRAZIL Abstract: We have examined the trophic effects of conditioned media obtained from purified mice Müller glia cells on purified sympathetic chick and rat neuronal cells and on Retinal Ganglion Cells (RGC) from postnatal mice. Purified mice Müller glia cultures stained for vimentin, GFAP or S100, but were negative for neuronal markers. Mice Müller Glial conditioned medium (MMG) was concentrated and at 1:1 dilution supported 100% survival of chick or rat sympathetic neurons after 48h compared to < 5% in controls. Partial purification of the MMG using Centriprep concentrators showed that trophic activity is from molecules above 10kDa. MMG stimulated AKT, ERK and pStat3 in sympathetic neurons, and this effect was not blocked by NGF or CNTF blocking antibodies. MMG induced neurite outgrowth in P4 mice retina explants, and on

2
MULLER GLIA FACTORS INDUCE SURVIVAL AND NEURITOGENESIS OF
PERIPHERAL AND CENTRAL NEURONS
Ricardo Augusto de Melo Reis1, Mauricio e Castro Cabral-da-Silva1, Fernando Garcia de
Mello1, Jeremy SH Taylor3
Lab Neurochemistry1 IBCCF, UFRJ, Rio de Janeiro, 21949-900 Brazil and Department
Physiology, Anatomy and Genetics3, Oxford University, OX13QX Oxford UK
Correspondence Address: Dr. Ricardo AM Reis, Lab Neuroquímica, IBCCF C1-031,
CCS, UFRJ, Ilha do Fundão, Rio de Janeiro-RJ 21949-900 Brasil
Fax number: 00 55 21 22808193
Email: [email protected]
Running title: Muller glia factors and neuronal survival
Number of pages: 34
Number of figures: 06
Number of tables: 02

3
ABSTRACT
We have examined the trophic effects of conditioned media obtained from
purified mice Müller glia cells on purified sympathetic chick and rat neuronal cells and
on Retinal Ganglion Cells (RGC) from postnatal mice. Purified mice Müller glia cultures
stained for vimentin, GFAP or S100, but were negative for neuronal markers. Mice
Müller Glial conditioned medium (MMG) was concentrated and at 1:1 dilution supported
100% survival of chick or rat sympathetic neurons after 48h compared to < 5% in
controls. Partial purification of the MMG using Centriprep concentrators showed that
trophic activity is from molecules above 10kDa. MMG stimulated AKT, ERK and pStat3
in sympathetic neurons, and this effect was not blocked by NGF or CNTF blocking
antibodies. MMG induced neurite outgrowth in P4 mice retina explants, and on isolated
RGC and it was as effective as BDNF + CNTF + forskolin in maintaining survival of
isolated panned RGCs (45% survival, 72h compared to 8% control). RGCs plated on top
of Muller glia cells had a much better survival rate (>80%, 96h) compared to laminin +
poly-L-lysine substrates. In conclusion, we show that purified Muller glia culture secrete
trophic molecules that support peripheral and central neuronal survival and
neuritogenesis.
Section: Nervous System Development, Regeneration and Aging
Key words: Muller glia, cell death, sympathetic, RGC, apoptosis, neurotrophin

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo