Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE MINAS GERAIS ESCOLA DE VETERINÁRIA
Colegiado dos cursos de Pós-graduação
ESTUDO DA INTERAÇÃO GENÓTIPO-AMBIENTE
EM CODORNAS DE CORTE
VIVIAN PAULA SILVA FELIPE
BELO HORIZONTE ESCOLA DE VETERINÁRIA - UFMG
2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

1
VIVIAN PAULA SILVA FELIPE
ESTUDO DA INTERAÇÃO GENÓTIPO-AMBIENTE EM CODORNAS DE CORTE
Dissertação apresentada à Escola de Veterinária da Universidade Federal de Minas Gerais, como requisito parcial para obtenção do grau de Mestre em Zootecnia. Área de Concentração: Genética e Melhoramento Animal Orientador: Martinho de Almeida e Silva
Belo Horizonte
UFMG – Escola de Veterinária 2010

2

3
Comissão examinadora:
_________________________________________ Prof. Martinho de Almeida e Silva
(Orientador)
_________________________________________ Prof. Dalton de Oliveira Fontes
_________________________________________ Prof. Robledo de Almeida Torres

4
“All models are wrong, but some are useful” (George E. P. Box)

5
Aos meus pais, Roberto e Luci Mary
Aos meus irmãos, Júnior e Naiara
Aos meus companheirinhos, Nina e Bonifácio
Com todo carinho, DEDICO

6
Agradeço... A Deus em primeiro lugar. Aos meus pais por todo apoio, compreensão e carinho recebido. Aos meus irmãos, que em certa época da vida me ensinaram o sentido literal da palavra lutar, me ensinam diariamente o sentido real da palavra amar. Ao Prof. Martinho pela orientação muito além do acadêmico. Aos professores da banca, Prof. Robledo e Prof. Dalton, pela contribuição ao meu trabalho. Aos meus amiguinhos de estimação Nisinha, Boninho, Sofia, Belito & Belita e Charlinho pelo amor incondicional e pelas horas de alegria e distração. Aos meus amigos pelos momentos de descontração e companheirismo. Aos meus colegas de estudo (e também grandes amigos) Rocha, Luciana, Bruno, Natascha, Fernanda, André’s, Gerusa, Daniel, Parra, Glaucyana e Frederico pelas agradáveis momentos de convivência e ajuda prestada. Ao pessoal do grupo de estudos e às meninas de iniciação científica Nathália, Lissandra e Winnie por toda ajuda e amizade. A todos os essenciais funcionários da UFMG que tornaram tudo mais possível: João, Marciano, Evaristo, Heloísa, Marcos, Nilda, pessoal dos serviços gerais e Xerox, funcionários do LAMA e da fazenda de Igarapé. Às codornas!!! À CAPES pela bolsa concedida. E a todos que, mesmo que não citados, foram importantes pela passagem desta importante etapa da minha vida!

7
SUMÁRIO
1. INTRODUÇÃO ....................................................................................................................................... 10
2. REVISÃO DE LITERATURA.............................................................................................................. 10
2.1 Proteína Ideal............................................................................................................................................. 10
2.2 Interação genótipo-ambiente...................................................................................................................... 11
2.3 Modelos de regressão aleatória.................................................................................................................. 14
3. REFERÊNCIAS BIBLIOGRÁFICAS................................................................................................... 16
CAPÍTULO 1
Efeito da interação genótipo x níveis de metionina+cistina total da dieta sobre o desempenho de codornas de corte.................................................................................................................................. 22
RESUMO................................................................................................................................................... 22
ABSTRACT............................................................................................................................................... 22
1. INTRODUÇÃO......................................................................................................................................... 23
2. MATERIAL E MÉTODOS....................................................................................................................... 24
3. RESULTADOS E DISCUSSÃO............................................................................................................... 27
4. CONCLUSÕES......................................................................................................................................... 31
5. REFERÊNCIAS BIBLIOGRÁFICAS...................................................................................................... 32
CAPÍTULO 2
Efeito da interação nível de metionina+cistina total da dieta x sexo x grupo genético sobre características de carcaça de codornas de corte................................................................................... 34
RESUMO.................................................................................................................................................. 34
ABSTRACT............................................................................................................................................... 34
1. INTRODUÇÃO......................................................................................................................................... 35
2. MATERIAL E MÉTODOS....................................................................................................................... 35
3. RESULTADOS E DISCUSSÃO............................................................................................................... 36
4. CONCLUSÕES......................................................................................................................................... 45
5. REFERÊNCIAS BIBLIOGRÁFICAS...................................................................................................... 45
CAPÍTULO 3
Estudo da sensibilidade dos valores genéticos de peso corporal de codornas de corte à mudança de gradiente nutricional por meio de normas de reação considerando heterocedasticidade residual...................................................................................................................................................... 47
RESUMO................................................................................................................................................... 47
ABSTRACT............................................................................................................................................... 47
1. INTRODUÇÃO......................................................................................................................................... 48
2. MATERIAL E MÉTODOS....................................................................................................................... 49
3. RESULTADOS E DISCUSSÃO............................................................................................................... 52
4. CONCLUSÕES......................................................................................................................................... 57
5. REFERÊNCIAS BIBLIOGRÁFICAS...................................................................................................... 57

8
LISTA DE TABELAS
CAPÍTULO 1
TABELA 1 - Composição percentual da dieta basal formulada para codornas de corte................................ 25
TABELA 2 - Composição percentual da dieta basal formulada para codornas de corte................................ 26
TABELA 3 - Peso final, ganho de peso, consumo de dieta e conversão alimentar de codornas de corte de sete a 21 dias de idade, segundo o nível de metionina + cistina total da dieta.......................... 27
TABELA 4 - Peso final, ganho de peso, consumo de ração e conversão alimentar de codornas de corte de sete a 42 dias de idade, segundo o nível de metionina mais cistina da dieta............................. 29
TABELA 5 - Peso final, ganho de peso, consumo de dieta e conversão alimentar de duas linhagens de codornas de corte de 22 a 42 dias de idade, segundo o nível de metionina+cistina da dieta............................................................................................................................................ 31
CAPÍTULO 2
TABELA 1 - Rendimento de moela de machos e fêmeas de dois grupos genéticos de codornas de corte alimentadas com dietas contendo seis níveis de metionina+cistina.......................................... 37
TABELA 2 - Efeito de sexo sobre o desempenho e características de carcaça de codornas de corte de dois grupos genéticos........................................................................................................................ 37
TABELA 3 - Efeito do grupo genético sobre o rendimento de peito de codornas de corte............................ 37
TABELA 4 - Características de desempenho e cortes nobres de codornas de corte de dois grupos genéticos alimentadas com diferentes níveis de metionina+cistina (Experimento 1)............... 38
TABELA 5 - Características de carcaça (vísceras) de codornas de corte de dois grupos genéticos alimentadas com diferentes níveis de metionina+cistina (Experimento 1)............................... 39
TABELA 6 - Interação nível nutricional de metionina+cistina x sexo x grupo genético sobre o rendimento de carcaça de codornas de corte com base na exigência por fase de crescimento............................................................................................................................... 40
TABELA 7 - Efeito da interação nível de metionina+cistina x grupo genético sobre o peso do coração de codornas de corte ao 42º dia de idade no Experimento 2.......................................................... 41
TABELA 8 - Efeito do nível nutricional de metionina+cistina sobre características de carcaça de codornas de corte (Experimento 2)........................................................................................................... 42
TABELA 9 - Características de desempenho e carcaça de codornas de corte alimentadas com diferentes níveis de metionina+cistina (Experimento 2)............................................................................ 44
TABELA 10 -
Características de carcaça (vísceras) de codornas de corte de dois grupos genéticos alimentadas com diferentes níveis de metionina+cistina (Experimento 2)............................... 45
CAPÍTULO 3
TABELA 1 - Composição das dietas experimentais para codornas de corte.................................................. 49
TABELA 2 - Descrição dos arquivos utilizados nas análises contendo pesos de codornas de corte EV1 e EV2 no 21º e 35º dia de idade................................................................................................... 50
TABELA 3 - Comparação de modelos com heterogeneidade e homogeneidade de variância residual pelo Critério de informação Bayesiano (BIC) .................................................................................. 52
TABELA 4 - Estimativas das correlações genéticas entre os pesos nos diferentes níveis de proteína bruta considerando homogeneidade de variância residual (acima da diagonal) e heterogeneidade de variância residual (abaixo da diagonal)................................................................................ 53
TABELA 5 - Componentes de variância atribuídos aos coeficientes de regressão do intercepto (b0) e linear (b1) do efeito genético aditivo direto em função dos níveis protéicos e componentes de variâncias residuais para cada classe estimados para as linhagens EV1 e EV2 no 21º e 35º dia de idade.......................................................................................................................... 54

9
LISTA DE FIGURAS
CAPÍTULO 1
FIGURA 1 - Regressão do peso médio ao 21º dia de idade (a), ganho de peso do 7º ao 21º dia de idade (b) do 7º ao 21º dia de idade de codornas de corte em relação ao nível de metionina+cistina da dieta...................................................................................................... 28
FIGURA 2 - Regressão do consumo de dieta do 7º ao 21º dia de idade (c) de codornas de corte em relação ao nível de metionina+cistina da dieta....................................................................... 28
FIGURA 3 - Regressão do peso médio ao 42º dia de idade (a), ganho de peso do 7º ao 42º dia de idade (b), consumo de dieta do 7º ao 42º dia de idade para EV1 (c) e para EV2 (d) e conversão alimentar do 7º ao 42º dia de idade do grupo EV1 (e) de codornas de corte em relação ao nível de metionina+cistina total da dieta................................................................................ 30
CAPÍTULO 2
FIGURA 1 - Regressão do rendimento de carcaça de fêmeas EV2 de codornas de corte em relação ao nível de metionina+cistina da dieta........................................................................................ 40
FIGURA 2 - Regressão do peso do coração do grupo genético EV1 (a) e EV2 (b) de codornas de corte em relação ao nível de metionina+cistina da dieta................................................................. 41
FIGURA 3 - Regressão do rendimento de peito (a), rendimento de coração (b), peso das asas (c), rendimento das asas (d), peso do fígado (e) e rendimento de fígado (f) de codornas de corte em relação ao nível de metionina+cistina da dieta........................................................ 43
CAPÍTULO 3
FIGURA 1 - Estrutura de covariância (gr²) do efeito genético aditivo direto em função do nível de proteína bruta da dieta estimada no 21º e 35º dia de idade para os grupos genéticos EV1 e EV2 (a:EV1-21 dias; b:EV1-35dias; c:EV2-21 dias e d:EV2-35 dias).................................. 55
FIGURA 2 - Herdabilidades diretas estimadas dos pesos ao 21º e 35º dias de idade de codornas de corte das linhagens EV1 e EV2 em função do nível de proteína bruta da dieta (a:EV1-21 dias; b:EV1-35dias; c:EV2-21 dias e d:EV2-35 dias)............................................................ 56
FIGURA 3 - Normas de reação de valores genéticos aditivos de 25 indivíduos amostrados arbitrariamente da população avaliada no 21º e 35º dia de idade dos grupos genéticos EV1 e EV2 (a:EV1-21 dias; b:EV1-35dias; c:EV2-21 dias e d:EV2-35 dias)............................... 57

10
1. INTRODUÇÃO
Existem vários fatores que têm contribuído para a criação de codornas no país, entre eles se destacam o rápido crescimento, precocidade na produção, maturidade sexual, alta produtividade, pequenos espaços para implementação da criação, baixo investimento e, consequentemente, rápido retorno financeiro (Pinto et al., 2002). Apesar de no Brasil ainda predominar a exploração de codornas para produção de ovos, há também tanto escassez de conhecimento sobre nutrição, manejo, desempenho e melhoramento genético da espécie para produção de carne (Coturnix coturnix coturnix) quanto falta de disponibilidade no mercado brasileiro de linhagens de corte melhoradas geneticamente. Muitas vezes, a carne de codorna disponível para venda comercial é proveniente de abate de codornas japonesas no final da vida produtiva tendo, portanto, pior qualidade de carcaça se comparada à das codornas de corte criadas especificamente para abate. Em outros países, a importância da carne de aves diferentes do frango de corte é muito maior, cita-se, por exemplo, a França em que 50% da carne de ave consumida é originada de aves exóticas (faisão, codorna, perdiz) em conseqüência da alta qualidade e palatabilidade deste tipo de carne e possibilidade de maior agregação de valor de venda.
Portanto, estudos relacionados à nutrição, fator de maior impacto econômico, e melhoramento genético de codornas tipo carne são essenciais para o estabelecimento da atividade econômica no país. Dentro do melhoramento genético o conhecimento da interação genótipo-ambiente tem se tornado cada vez mais importante. A presença da interação pode causar mudança no ordenamento dos valores genéticos das aves, de forma que aquela que apresenta melhor desempenho em um ambiente pode não ser a melhor em outro ambiente, ou
pode também alterar a variância genética aditiva o que causa maior semelhança entre valores genéticos em um ambiente do que em outro por menor ou maior dispersão desses valores, respectivamente.
Diante deste cenário, os objetivos deste estudo foram avaliar o efeito da interação entre níveis de metionina+cistina total e grupos genéticos de codornas de corte sobre características de desempenho e de carcaça nas diferentes fases do período de crescimento e avaliar a existência de heterogeneidade de sensibilidade dos valores genéticos de linhagens de codornas de corte submetidas a dietas contendo diferentes níveis de proteína bruta no 21º e 35º dias de idade utilizando modelos de regressão aleatória com estrutura heterogênea de variância residual.
2. REVISÃO DE LITERATURA
2.1. Proteína ideal
A “proteína ideal” foi baseada no conceito desenvolvido por H. H. Mitchell e H. M. Scott na Universidade de Illinois no início da década de 60 e tem como objetivo fazer combinação perfeita de aminoácidos indispensáveis que supram todas as necessidades dos animais para crescimento e mantença (EMMERT e BAKER, 1994). Este conceito foi relembrado pelo Agricultural Research Council (ARC, 1981) que publicou pela primeira vez a estimativa de proteína ideal para suínos em fase de crescimento.
O conceito de proteína ideal tem sido aplicado na estimativa de exigências nutricionais de várias espécies pela diminuição no teor de proteína bruta da dieta e suplementação por meio de aminoácidos sintéticos e é definida como o balanço exato de aminoácidos essenciais na dieta, sem deficiências nem excessos de forma que nenhum aminoácido seja

11
utilizado para obtenção de energia. Como vantagens deste conceito citam-se a maximização da utilização dos nutrientes ingeridos pelo animal, maior facilidade de ajuste na formulação de dieta, diminuição do custo da dieta e diminuição na emissão de poluentes pela menor excreção de nitrogênio, preocupação cada vez mais frequente, sendo este responsável por poluição de água (nitritos e nitratos), eutrofização, volatilização de amônia e causador de chuva ácida.
O estudo baseado na proteína ideal usa a lisina como aminoácido referência sendo o requerimento de outros aminoácidos expresso como porcentagem em relação à ela. Segundo Baker (1996), este aminoácido foi escolhido por vários motivos: 1) para frangos de corte é o segundo aminoácido limitante, seguindo os aminoácidos sulfurados, e sua suplementação é economicamente viável, 2) a lisina dietética é utilizada somente para deposição de proteína e mantença (não precursor), 3) variedade de dados a respeito de exigência de lisina em várias circunstâncias ambientais, dietéticas e de composição corporal e 4) a análise deste aminoácido na ração é feita de forma direta.
A possibilidade de utilização de aminoácidos sintéticos, como lisina e metionina, na suplementação de dietas para frangos de corte tem sido avaliada positivamente pelo seu efeito sobre o rendimento de cortes nobres (HICKLING, 1990; MORAN JR e BILGILI, 1990; SCHEUERMANN, 1995), diminuição na excreção de nitrogênio (ALETOR et al., 2000), sem, contudo, prejudicar o desempenho das aves pelo menor conteúdo de proteína bruta dessas dietas (WALDROUP et al., 1976).
A metionina serve como doador de metil para reações de metilação, envolvendo a biossíntese de lipídios, creatina, colina e carnitina entre outros, tendo importante
papel no crescimento e empenamentos das aves (KIM et al., 2006). Autores também relatam a importância deste aminoácido no desenvolvimento do sistema imunológico (TSIAGBE et al., 1987 e D’MELLO, 2003).
Em codornas de corte a metionina também é considerada o primeiro aminoácido limitante e foi demonstrado que a sua suplementação em dietas com baixo teor de proteína bruta permite a redução de proteína da dieta, o custo e a excreção de nitrogênio, controlando a poluição ambiental nas regiões de alta densidade de criação da espécie (SILVA et al., 2006). O que confirma a afirmação de Shirisvastav e Panda (1999) de que a qualidade da proteína é mais importante que seu nível na dieta.
Vários autores têm estudado a exigência de metionina para codornas de postura (MURAKAMI, 1994; STRINGHINI et al., 1998; BELO et al., 2000; PINTO et al., 2003) e de corte (CORRÊA et al., 2006a; VALENTE et al., 2006) e encontraram efeito significativo do nível deste aminoácido sobre características produtivas das aves mostrando que são necessários estudos de requerimento nutricional de metionina+cistina visando ao atendimento da exigência da aves para melhorar o rendimento produtivo, principalmente da espécie destinada para o corte (Coturnix
coturnix coturnix) em razão da escassez de informações científicas na área.
2.2. Interação genótipo-ambiente
O valor fenotípico de determinada característica do indivíduo é função de seu valor genético e da influência de ambiente. Deste modo, o valor fenotípico poderia ser representado por:
P = G + E,

12
em que P é o valor fenotípico, G é o valor genético e E é o desvio de ambiente.
Neste modelo, considera-se que determinadas mudanças no efeito de ambiente causam sempre a mesma influência no fenótipo, seja qual for o genótipo. Quando isto não é verdadeiro, existe interação entre genótipo e ambiente (Falconer e Mackay, 1996).
Deste modo, uma modificação específica de ambiente poderia causar efeito mais intenso em um genótipo do que em outro, podendo existir até mudança na ordem de classificação (ranking) dos melhores valores genéticos quando estes valores são mensurados em diferentes ambientes (Falconer e Mackay, 1996).
Quando ocorre interação entre genótipo e ambiente (GxE) há necessidade de incluir um termo adicional de interação à expressão matemática apresentada anteriormente.
P = G + E + IGxE
É possível que entre dois ambientes distintos sejam notadas modificações apenas na diferença entre expressões fenotípicas de diferentes genótipos, mantendo a ordem da classificação dos melhores valores genéticos. Neste caso, apenas a variância genética se altera. Ambientes menos favoráveis apresentam menor variância fenotípica e, consequentemente, genética. A utilização de material genético de um reprodutor em um ambiente onde a variância genética é menor do que o local onde ele ou seus parentes foram avaliados limita o desempenho de seus filhos e o retorno econômico que estes podem oferecer (Blake et. al., 1988, Stanton et al., 1991). Por outro lado, é possível que a variância genética seja a mesma em dois ambientes distintos, mas que a ordem da classificação dos melhores genótipos seja diferente. Quando ocorre alguma das duas situações citadas
(ou ambas, com alteração de ordem de classificação e variância), existe GxE para a característica considerada (Bowman, 1974, citado por Pereira, 2004).
Desta maneira, se existe interação genótipo-ambiente em determinada característica, a diferença entre fenótipos de dois genótipos pode não ser a mesma em dois ambientes diferentes (Kolmodin, 2003). Sendo assim, filhos de codornas de corte avaliados em dietas com determinado nível protéico não teriam o mesmo desempenho em relação à média da população se alimentados com dietas contendo nível protéico diferente, podendo ocorrer até mesmo mudança na ordem de classificação dos valores genéticos estimados para diferentes níveis protéicos. Neste caso, a predição do valor genético do animal deve ser feita no nível nutricional semelhante ao utilizado em criação comercial. No sentido estatístico, a interação G X E pode ser vista como a necessidade de, por exemplo, dietas com nível protéico específico para grupos genéticos diferentes caso ela seja detectada.
Dentre as formas de se avaliar a interação entre o genótipo e o ambiente, citam-se estudos envolvendo delineamentos em arranjo fatorial, correlação genética da mesma característica em ambientes diferentes e normas de reação.
A análise de variância em experimentos fatoriais inclui todas as combinações de vários conjuntos de níveis e fatores (YATES, 1949) e, portanto, permitem o estudo da interação entre as causas de variação listadas que no caso de estudos de G x E, os genótipos podem ser representados por raças, linhagens, grupos genéticos ou mesmo populações distintas de uma mesma espécie e o ambiente como o tratamento avaliado (temperatura, dietas, densidade populacional, ou qualquer outra variável ambiental que possa ser controlada). Na interpretação dos resultados, a presença de significância do

13
termo de interação indica que cada grupo genético apresenta um ambiente para expressão máxima da característica e, por isso, qualquer consideração feita a respeito do ambiente deve ser feita especificando-se o genótipo nele avaliado. Por outro lado, a ausência de interação indica que a variável ambiental imposta apresenta o mesmo efeito para os grupos genéticos avaliados. Como resposta nestes ensaios é possível obter a definição do melhor ambiente para cada “genótipo” quando o efeito de ambiente não se manifesta igualmente para todos os grupos genéticos. A principal desvantagem desta metodologia está na pressuposição da ANOVA que diz respeito a homocedasticidade, o que na realidade não ocorre devido à tendência de maior variação nos melhores ambientes (Burdon, 1977). Em codornas de corte, estudos com este tipo de análise foram feitos por Corrêa et al. (2006b), Valente et al. (2006) e Dionello et al. (2006) que, ao envolverem a avaliação de grupos genéticos submetidos a diferentes planos nutricionais, mostraram que o máximo desempenho é obtido quando as linhas são alimentadas com o nível adequado da dieta, mostrando que a estimativa da exigência de uma linhagem não deve ser extrapolada para outra. Ahmad et al. (2003) apresentaram resultados semelhantes estudando o efeito de G x E sobre a viabilidade de codornas japonesas e encontraram efeito de interação significativo entre estação-grupo genético e nível protéico da dieta-grupo genético sobre a porcentagem de mortalidade das aves. Por outro lado, Felipe et al. (2007) não encontraram efeito de interação nível de metionina+cistina total da dieta e grupos genéticos de fêmeas de codornas de corte concluindo que a mesma dieta poderia ser utilizada para qualquer um dos grupos tendo em vista o máximo ganho de peso.
A utilização da correlação genética para estudo da interação genótipo-ambiente é uma ferramenta útil para o conhecimento do quanto do melhoramento feito em um
ambiente é “carregado” para o outro ambiente e mostra o quanto pode-se esperar ganhar ou perder fazendo a seleção em um ambiente distinto do qual a população melhorada necessita viver (FALCONER e MACKAY, 1996). Para aplicação desta solução considera-se a mesma característica expressa em dois ambientes como duas características distintas e a correlação entre elas representa o quanto se espera de associação da expressão genética nos dois ambientes. Uma alta correlação genética significa que todos os genótipos reagem aproximadamente da mesma maneira às mudanças ambientais e a baixa correlação genética indica que os genótipos reagem de maneira distinta à variação do ambiente. A correlação genética abaixo de 0,80 indica a presença de interação genótipo-ambiente, segundo Robertson (1959), já para Falconer (1987) a correlação abaixo de 1 já é suficiente para estabelecer a existência de G X E. Vários pesquisadores têm utilizado a correlação genética como forma de avaliação da interação genótipo-ambiente e seu impacto sobre como deve ser feita a seleção entre diferentes ambientes/países em gado de corte (DE MATTOS et al., 2000; LEE e BERTRAND, 2002; SIMONELLI et al., 2004; FRIDRICH et al.,2008) e gado de leite (CARABAÑO et al., 1990; RAFFRENATO et al., 2003; HAYES et al., 2003).
A norma de reação é descrita como a mudança sistemática na expressão média de uma característica fenotípica que ocorre em resposta a mudança sistemática de uma variável ambiental (JONG, 1990), isto é, o fenótipo expresso a partir de um genótipo é função do ambiente. Para a predição da norma de reação são utilizadas funções de covariância que modelam as características longitudinais que se modificam ao longo de um gradiente ambiental por meio de modelos de regressão aleatória. E a variação nos coeficientes polinomiais indica G x E, ou seja, variação genética atribuída à sensibilidade ambiental. Mudanças na

14
sensibilidade podem ser resultado da seleção agindo diretamente nos coeficientes da norma de reação ou uma resposta correlacionada à seleção sobre valores fenotípicos dentro dos diferentes ambientes (VIA et al, 1995). A vantagem deste modelo é que o valor genético é predito ao longo do gradiente ambiental de forma contínua e é possível avaliar a sensibilidade da característica às mudanças de ambiente (JONG e BIJMA, 2002). O modelo foi utilizado pela primeira vez para identificar GxE por Kolmodin et al. (2002) que estudaram a sensibilidade ambiental em bovinos de leite Nórdicos em quatro países (Finlândia, Suíça, Dinamarca e Noruega) e encontraram reordenamento dos reprodutores entre os ambiente extremos. Calus et al. (2003) utilizaram modelos de normas de reação para estimar parâmetros de variância genética e sensibilidade ambiental para características de produção de leite e concluíram que a heterogeneidade de variância genética encontrada era considerável porém com variação limitada da herdabilidade e pouco reordenamento ocorreu baseado no índice de seleção sugerindo ausência da interação G X E. Em codornas de corte, Santos et al. (2008) utilizaram normas de reação para descrever a sensibilidade de peso em duas idades de codornas de corte de dois grupos genéticos alimentadas com dietas contendo diferentes níveis protéicos e mostraram a variação nos componentes de (co)variância e herdabilidade com a mudança de gradiente ambiental, usando normas de reação para caracterizar a presença ou ausência de interação genótipo-ambiente.
Diferenças nas estimativas de herdabilidade da mesma característica, medidas sob diferentes condições ambientais, podem ser indicativas da presença de interação genótipo-ambiente (VAN VLECK, 1963; VENCOVSKY e PACKER, 1976). Isto ocorrerá sempre que diferenças entre os ambientes sejam suficientes para influir sobre a expressão da variância genética
aditiva. Entretanto, poderá ocorrer mudança também na variância total apresentada pela característica, imposta por modificações da variância residual, constituindo causa das diferenças observadas nas estimativas da herdabilidade.
2.3. Modelos de regressão aleatória
Características que podem ser medidas várias vezes ao longo da vida são conhecidas como longitudinais. Exemplos disso são peso corporal, produção de leite, produção de ovos, deposição de gordura, eficiência alimentar, entre outras. Os modelos de regressão aleatória, também conhecidos como modelos de dimensão infinita, podem ser aplicados para avaliação genética resultando em maior acurácia com relação ao modelo multicaracterístico e repetibilidade (Meyer, 2004), além disso, são utilizados para predição de normas de reação, curvas de crescimento, curvas de lactação ou curvas de produção de ovos. Em essência as funções de covariância são um caso especial de modelos de regressão aleatória e descrevem a covariância entre medidas tomadas em certo nível da covariável controle como função dessa mesma covariável controle. A principal vantagem deste modelo é que, além de fazer melhor uso das informações disponíveis, inclui uma função aninhada dentro de um efeito aleatório, o que permite que os componentes de variância se modifiquem ao longo de uma trajetória (Kolmodin, 2003). Na abordagem usual do modelo multicaracterístico a medida de escala contínua é dividida em intervalos arbitrários e faz-se o ajuste para medidas tomadas fora do período/idade estipulado e é estimada apenas uma curva para todos os animais avaliados. Já no modelo repetibilidade assume-se que as variâncias genéticas são as mesmas para todos os níveis da covariável controle (idade, ambiente, dia do controle).

15
Modelos de regressão aleatória têm sido extensivamente aplicados no melhoramento animal para modelar registros diários de produção de leite durante a lactação de bovinos leiteiros, assim como no crescimento corporal em bovinos de corte (MEYER, 2000; ALBUQUERQUE E MEYER, 2001; SAKAGUTI et al., 2002; NOBRE et al., 2003), na avaliação da trajetória de crescimento de codornas de corte (Dionello, 2008) e no estudo da interação genótipo x ambiente em características de produção de leite (KOLMODIN et al., 2003) e de desempenho de codornas de corte (SANTOS et al. 2008).
Modelos de regressão aleatória permitem ajustar a trajetória aleatória para cada indivíduo como desvios de uma trajetória média da população, descrevendo os desvios genéticos a partir de regressões fixas, permitindo que cada animal tenha uma forma diferente da trajetória de seus desempenhos em termos genéticos (MERCADANTE et al., 2002).
Em notação matricial os MRA podem ser representados por:
∑ ∑−
=
−
=
+++=1
0
1
0
** )()(A Rk
m
k
m
iijmimijmimij ttFy εφγφα
, em que:
F refere-se ao efeito fixo;
)( *1
0ijmim
k
m
tA
φα∑−
=
é o termo referente ao efeito
aleatório genético aditivo para a covariável tij;
∑−
=
1
0
* )(Rk
m
ijmim tφγ é o termo referente ao efeito
de ambiente permanente; e
iε refere-se ao componente residual.
Quanto às pressuposições de variância e covariância dos modelos de regressão aleatória tem-se:
⊗
⊗
=
D
IK
AK
V pe
a
200
00
00
εσε
γ
α
, em
que:
Ka: matriz de (co)variâncias dos coeficientes de regressão aleatório genéticos aditivos;
Kpe: matriz de (co)variâncias dos coeficientes de regressão aleatório de ambiente permanente;
A: Matriz dos numeradores dos coeficientes de parentesco de Wright;
D: Matriz diagonal;
2εσ : variância dos erros de medida.
E a partir das covariâncias entre os coeficientes de regressão aleatória é possível obter a covariâncias entre produções entre outras covariáveis controle quaisquer.
Os modelos de regressão aleatória, além de permitirem a modelagem de efeitos aleatórios utilizando funções polinomiais, também possibilitam que se considere estrutura de variância residual heterogênea por meio de formação de classes de resíduo ou ajuste de funções polinomiais residuais. A importância disto reside no fato de que, ao se considerar a heterocedasticidade residual, pode-se melhorar a partição da variância total levando ao aumento da acurácia na estimativa de parâmetros e valores genéticos maximizando, assim, a predição da resposta à seleção. Garrick e Van Vleck (1987) mostram perda de 3,3% na taxa de ganho genético quando a heterogeneidade de variância não é considerada na avaliação de bovinos de leite em dados simulados. Outros autores

16
têm demonstrado a necessidade de se utilizarem modelos de regressão aleatória que consideram variância residual heterogênea para estudo de curvas de crescimento (LEWIS E BROTHERSTONE, 2002; SOUSA et al., 2008; BONAFÉ, 2008), produção de leite no dia do controle (EL FARO E ALBUQUERQUE, 2003). Strandberg (2006) afirma que para o estudo de interação genótipo-ambiente utilizando modelos de regressão aleatória não é adequado que a variância residual seja considerada constante ao longo de todo gradiente ambiental, pois para muitas características e escalas ambientais isso pode não ser verdadeiro. Pode-se ter uma noção da necessidade de se considerar heterocedasticidade do resíduo dividindo-se a escala da covariável em classes e depois calculando o valor da variância residual dentro delas, caso a heterogeneidade de variância seja verificada, a sua inclusão no modelo de avaliação deve ser considerada (MEYER,2001).
3. REFERÊNCIAS
BIBLIOGRÁFICAS
AHMAD, M., RAJMANE, B.V., NARAYANKHEDKAR, S.G.. Effect of GxE interaction on livability in quails. Indian Veterinary Journal. V.80, n.9, p.874-876, 2003.
ALBUQUERQUE, L.G. e MEYER, K. Estimates of covariance functions for growth from birth to 630 days of age in Nelore cattle. Journal of Animal Science. v.79, p. 2776-2789, 2001.
ALETOR, V.A., HAMID, I.I., NIEB, E. e PFEFFER, E. Low-protein amino acid-supplemented diets in broiler chickens: effects on performance, carcass
characteristics, whole body composition and efficiencies of nutrient utilisation. Journal of the Science of Food and Agriculture. v.80, p.547-554, 2000.
ARC. 1981. The Nutrient Requirements of Pigs. Commonwealth Agricultural Bureaux, Slough, U.K.
BAKER,D.H. Advances in amino acid nutrition and metabolism of swine and poultry. In: KORNEGAY, E.T. (Ed). Nutrient management of food animals to enhance and protect the environment. Danvers, EUA: CRC Press, 1996. p.41-54.
BELO, M.T.S., COTTA, J.T.B., OLIVEIRA, A.I.G.. Níveis de metionina em rações de codornas japonesas (Coturnix
coturnix japônica) na fase inicial de postura. Agrotec., Lavras, v.24. n.4, p.1068-1078, 2000.
BLAKE, R. W.; HOLMANN, F. J.; GUTIERREZ, J.; CEVALLOS, G. F.; Comparative profitability of United States Holstein artificial insemination on sires in Mexico. Journal of Dairy Science, v. 71, n. 5, p. 1378, 1988.
BONAFÉ, C.M. Avaliação do crescimento de codornas de corte utilizando modelos de regressão aleatória. 2008. 58f. Dissertação (Mestrado em Genética e Melhoramento) – Universidade Federal de Viçosa, Viçosa. 2008.
BURDON, R. Genetic correlation as a concept for studying genotype-environment interaction in forest tree breeding. Silvae Genetica. v.26, n.5–6,p.168–175, 1977.

17
CALUS, M. P. L. AND VEERKAMP, R. F.. Estimation of environmental sensitivity of genetic merit for milk production traits using a random regression model. Journal of Dairy Science. v.86, p.3756–3764, 2003.
CARABAÑO, M.J., WADE, K.M., VAN VLECK, L.D.. Genotype by environment interactions for milk fat production across regions of the united states. Journal of Dairy Science. v.73, p.173-180, 1990.
CORRÊA, G.S.S.; SILVA, M.A.; CORRÊA, A.B. et al. Exigência de metionina + cistina total para codornas de corte em crescimento. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.58, n.3, p.414-420, 2006a.
CORRÊA, G.S.S.; SILVA, M.A.; DIONELLO, N.J.L. Genotype by environment interaction for production traits of meat type quail during the growing phase. In: World Congress on Genetics Aplied to Livestock Production, 2006, Belo Horizonte. Proceedings... 8th World Congress on Genetics Aplied to Livestock Production, 2006b. CD-ROM.
D’MELLO, J.P.F. Amino acid in animal forms. Wallingford: CABI, 2003. 430p.
DE MATTOS, D., BERTRAND, J.K., MISZTAL,I.. Investigation of genotype x environment interactions for weaning weight for Herefords in three countries. Journal of Animal Science. v.78, p.2121-2126, 2000.
DIONELLO, N.J.L.; CORRÊA, G.S.S.; SILVA, M.A. Interação genótipo x ambiente em características produtivas de codornas de corte no período de
crescimento. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 43., 2006, João Pessoa. Anais... João Pessoa, PB: SBZ. CD-ROM.
DIONELLO, N.J.L., CORRÊA, G.S.S., SILVA, M.A., CORRÊA, A.B., SANTOS, G.G.. Estimativas da trajetória genética de crescimento de codornas de corte utilizando modelos de regressão aleatória. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. v.60, n.2, p.454-460, 2008.
EL FARO, L.; ALBUQUERQUE, L.G. Utilização de modelos de regressão aleatória para produção de leite no dia do controle, com diferentes estruturas de variâncias residuais. Revista Brasileira de Zootecnia, v.32, p.1104-1113, 2003.
EMMERT, M.W.; BAKER, D.H. Use of the ideal protein concept for precision formulation of amino acid levels in broilers diets. Journal of Applied Poultry Research, v.6, p.462-470, 1997.
Falconer, D.S.. Introdução à genética quantitativa. Editora UFV, Viçosa, 279p. 1987.
FALCONER, D.S.; MACKAY, T.F.C. Introduction to quantitative genetics. 4.ed. London: Longman. 1996.
FELIPE, V.P.S.; CORRÊA, G.S.S.; SILVA, M.A. Interação genótipo-ambiente em características de desempenho e de carcaça de fêmeas híbridas de codornas de corte na fase de crescimento. In: Congresso Internacional de Zootecnia, 9., 2007, Londrina. Anais... Londrina: UEL, 2007. p. 1-5.

18
FRIDRICH, A.B., SILVA, M.A., VALENTE, B.D., SOUSA, J.E.R et al. Interação genótipo x ambiente e estimativas de parâmetros genéticos dos pesos aos 205 e 365 dias de idade de bovinos Nelore. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. v.60, n.4, p.917-925, 2008.
GARRICK, D. J., Van VLECK, L. D. 1987. Aspects of selection for performance in several environments with heterogeneous variances. Journal Animal Science., v.65, p. 409 – 421, 1987.
HAYES, B.J., CARRICK, M., BOWMAN, P., GODDARD, M.E.. Genotype x Environment interaction for milk production of daughters of Australian dairy sires from test-day records. Journal of Dairy Science. v.86, p.3736-3744, 2003.
HICKLING, D., GUENTER, W., JACKSON, M.E. The effects of dietary methionine and lysine on broiler chicken performance and breast meat yield. Canadian Journal of Animal Science,v.70, p.673-678. 1990.
JONG, G. e BIJMA, P.. Selection and phenotypic plasticity in evolutionary biology and animal breeding. Livestock Production Science. v.78, p.195-214, 2002.
JONG, G. Quantitative genetics of reaction norms. Journal of evolutionary biology, v.3, p. 447-468, 1990.
KIM, W.K.; FROELICH JR., C.A.; PATTERSON, P.H. The potential to reduce poultry nitrogen emissions with dietary methionine or methionine analogues
supplementation. World’s Poult. Sci. Journ., v.62, p.338-353, 2006.
KOLMODIN, R.; STRAMBERG, E.; MADSEN, P. et al. Genotype by environment interaction in Nordic dairy cattle studied using reaction norms. Acta Agriculture Scandinavia, Sect. A, Animal Science, Oslo, Norway, v.52, p.11-24, 2002.
KOLMODIN, R. Reaction norms for the study of genotype by environment interaction in animal breeding. Uppsala: Swedish University of Agricultural Sciences, 2003. 35p. Doctoral thesis. Department of Animal Breeding and Genetics, 2003.
LEE, D.H. E BERTRAND,J.K.. Investigation of genotype x country interaction for growth traits in beef cattle. Journal of Animal Science, v.80, p.330-337, 2002.
LEWIS, R.M.; BROTHERSTONE, S. A genetic evaluation of growth in sheep using random regression techniques. Journal of Animal Science, v.74, p.63-70, 2002.
MERCADANTE, M. E. Z., PACKER, I. U., RAZOOK, A. G. et al. Dias ao parto de fêmeas Nelore de um experimento de seleção para crescimento. II – Modelo de regressão aleatória. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.31, n.4, p. 1726 – 1733, 2002.
MEYER, K. Random regressions to model phenotypic variation in monthly weights of Australian beef cows. Livestock Production Science, v.65, p. 19 – 38, 2000.

19
MEYER, K.. Scope for a random regression model in genetic evaluation of beef cattle for growth. Livestock Production Science,v. 86, p.69–83, 2004.
MEYER, K. Estimating genetic covariance functions assuming a parametric correlation structure for environmental effects. Genetic Selection Evolution, v.33, p.557-585, 2001.
MORAN Jr, E.T., BILGILI, S.F. Processing losses, carcass quality and meat yields for broiler chicken, receiving diets marginally deficient to adequate in lysine prior to marketing. Poultry Science, v.69, p.702-710. 1990.
MURAKAMI, A.E.; FURLAN, A.C.; TATEISHI, A. et al. Exigência de metionina para codornas japonesas (Coturnix coturnix japonica) em postura. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 31., 1994, Maringá. Anais... Maringá: Sociedade Brasileira de Zootecnia, 1994. p.64-67.
NOBRE, P. R. C., I. MISZTAL, S. TSURUTA, J. K. BERTRAND, L. O. C. SILVA, AND P. S. LOPES. Analysis of growth curves of Nellore cattle by multiple-trait and random regression models. Journal of Animal Science, v.81, p.918–926, 2003.
PEREIRA, J. C. P. Melhoramento Genético Aplicado à Produção Animal, Belo Horizonte, FEPMVZ Editora, 2004, 609 p.
PINTO, R.; FERREIRA, A.S.; ALBINO, L.F.T.; GOMES,P.C., VARGAS JÚNIOR, J.G.. Níveis de Proteína e Energia para
Codornas Japonesas em Postura. Revista Brasileira de Zootecnia, v.31, n.4, p.1761-1770, 2002.
PINTO, R., FERREIRA, A.S., DONZELE, J.L. et al. Exigência de metionina mais cistina para codornas japonesas em crescimento. Revista Brasileira de Zootecnia, v.32, n.5, p.1174-1181, 2003.
RAFFRENATO, E., BLAKE, R.W., OLTENACU, P.A. et al. Genotype by environment interaction for yield and somatic cell score with alternative environmental definitions. Journal of Dairy Science, v.86, p.2470-2479, 2003.
ROBERTSON, A.. The sampling variance of the genetic correlation coefficient. Biometrics, v.15, p.469–485, 1959.
SAKAGUTI, E.S.; SILVA, M.A.; MARTINS, E.N. et al.Trajetória de crescimento e efeito da idade da vaca nos modelos de regressão aleatória de bovinos jovens da raça Tabapuã. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.54, n.4, p.414-423, 2002.
SANTOS, G.G., CORRÊA, G.S.S., VALENTE, B.D., SILVA, M.A. et al. Sensibilidade de valores genéticos de codornas de corte em crescimento às modificações de níveis de proteína das dietas. Arquivo Brasileiro de Medicina Veterinária e Zootecnia ,v.60, n.5, p.1188-1196, 2008.
SCHEUERMANN, G.N.; MAIER, J.C.; BELLAVER, C. et al. Metionina e lisina no desenvolvimento de frangos de corte. Revista Brasileira de Agrociência, v.1, n.2, p. 75-86, Mai-Ago, 1995.

20
SHRISVASTAV, A.K.; PANDA, B.A. A review of quail nutrition research in India. World’s Poultry Science Journal, v.55, n.3, p.73-81, 1999.
SILVA, E.L.; SILVA, J.H.V.; JORDÃO FILHO, J. Redução dos níveis protéicos e suplementação com metionina e lisina em rações para poedeiras leves. Revista Brasileira de Zootecnia, v.35, n.2, p.491-496, 2006.
SIMONELLI, S.M., MARTINS, E.N., SAKAGUTI, E.S. et al. Interação genótipo x ambiente para pesos à desmama, ano e sobreano na raça Nelore. In: V Simpósio da Sociedade Brasileira de Melhoramento Animal, 5º, 2004, Pirassununga. Anais...
Pirassununga, SP: SBMA. CD-ROM.
SOUSA, J.E.R., SILVA, M.A., SARMENTO, J.L.R. et al. Homogeneidade e heterogeneidade de variância residual em modelos de regressão aleatória sobre o crescimento de caprinos Anglo-Nubianos. Pesquisa Agropecuária Brasileira, v.43, n.12, p.1725-1732, 2008.
STANTON, T. L.; BLAKE, R. W.; QUAAS, R. L.; VAN VLECK, L. D.; CARABAÑO, M. J. Genotype by Environment interaction for Holstein Milk Yield in Colombia, Mexico and Puerto Rico. Journal of Dairy Science, v. 74, n. 5, p. 1700 – 1714, 1991.
STRANDBERG, E.; Analysis of genotype by environment interaction using random regression models, 8th World Congress on Genetics Aplied to Livestock Production, 2006, Belo Horizonte, Procedings... 25-05. STRINGHINI, J.H., MOGYCA, N.S., CAFÉ, M.B. et al. Níveis de energia
metabolizável e de metionina em rações para codornas (Coturnix coturnix japônica) em postura. Acta Scientiarium, v.20, n.3, p.407-411, 1998.
TSIAGBE, V.K., COOK, M.E., HARPER, A.E., SUNDAE, M.L.. Enhanced immune responses in broiler chicks fed methionine-supplemented diets. Poultry Science. v.66, n.7, p.1147-1154. 1987.
VALENTE, B.D.; CORRÊA, G.S.S.; DIONELLO, N.J.L. Interaction between methionine of diet and genotype in meat type quail. In: World Congress on Genetics Aplied to Livestock Production, 2006, Belo Horizonte. Proceedings... 8th World Congress on Genetics Aplied to Livestock Production, 2006. CD-ROM.
VAN VLECK, L.D.. Genotype and environment in sire evolution. Journal of Dairy Science, v.46, n.9, p.983-987, 1963.
VENCOVSKY, R. e PACKER, I.U.. Interação genótipo ambiente no melhoramento de bovinos de corte leiteiro. In: Simpósio sobre melhoramento genético de bovinos. Anais... Jaboticabal. UNESP, p.91-103, 1976.
VIA, S., GOMULKIEWICZ, R., DEJONG, G..Adaptive phenotypic plasticity – consensus and controversy. Trends Ecol. Evol. 10: 212–217.1995
WALDROUP, P.W.; MITCHEL, R.J.; PAYNE, J.R. et al. Performance of chicks fed diets formulated to minimize excess levels of essential amino acids. Poultry Science, v.55, p.243-253, 1976.

21
YATES, F.. Sampling method for censuses and surveys. Londres. Charles Griffin, 278p. 1949.

22
CAPÍTULO 1
Efeito da interação genótipo x níveis de metionina+cistina total da dieta sobre o desempenho de codornas de corte
Genotype by total methionine+cystine level of diet interaction for performance traits of
meat type quails
RESUMO
Estudou-se o efeito da interação grupo genético x nível nutricional de metionina+cistina total sobre o desempenho de codornas de corte durante as fases inicial (7 ao 21º dia de idade), final (22º ao 42º dia de idade) e total (7º ao 42º dia de idade) do período de crescimento. Foram realizados dois experimentos, no primeiro foram utilizadas 468 codornas da linhagem EV1 e 390 codornas da linhagem EV2 para estimar o efeito da interação genótipo-ambiente nas fases inicial e total do período de crescimento; no segundo foram utilizadas 288 codornas de cada uma das linhagens avaliadas para estimar o efeito da interação genótipo-ambiente na fase final do período de crescimento. No período inicial de crescimento, não houve interação significativa dos níveis do aminoácido testado x grupos genéticos para as características avaliadas e a exigência nesta fase, com base no ganho de peso, foi estimada em 0,94% de met+cist. No período final de crescimento, houve efeito significativo interação genótipo x nível de metionina+cistina apenas para o consumo de dieta e, em razão da não significância do efeito do nível de metionina+cistina total sobre o desempenho das aves, dietas contendo 0,63% de metionina+cistina total podem ser adotadas nesta fase. No período total de crescimento, compreendido entre o 7º e 42º dia de idade, houve efeito de interação genótipo-nível nutricional de met+cist sobre o consumo de dieta e conversão alimentar e a exigência nutricional, objetivando-se o máximo ganho de peso, foi estimada em 0,98% de met+cist. Palavras-chave: interação genótipo-ambiente, codornas de corte, requerimento nutricional, proteína ideal, metionina
ABSTRACT
The effects of genetic group x methionine + cystine level of diet interaction on performance traits of meat type quails during three phases of the growth period, initial (7-21 days of age), finishing (22-42 days of age) and total ( 7-42 days of age) were evaluated in two experiments. In the first experiment a total of 468 quails of EV1 strain and 390 of EV2 were used to estimate the effect of the genotype x environment interaction during the initial and total phases of the growth period and the second experiment to a total of 288 quails were used to evaluate the effect of the interaction during the finishing phase of the growth period. No interaction effect was observed during the initial phase and the methionine + cystine requirement was estimated as .94%. In the finishing phase a significant effect of the interaction was observed only for feed intake. Due to the no significant effect of the methionine + cystine of the analyzed traits the requirement for this amino acid was estimated as .63%. During the total period (7-42 days of

23
age) significant effect of genotype by methionine + cystine on feed intake, feed: weight gain ratio, and the requirement in this amino acid for maximum weight gain was estimated as .98%.
Keywords: genotype by environment interaction, meat type quails, nutritional requirement,
ideal protein, methionine
1. INTRODUÇÃO
A coturnicultura é atividade alternativa para a produção de proteína de qualidade para o consumo humano, uma vez que requer baixo investimento inicial, utiliza pequenas áreas e promove rápido retorno do capital. Além disso, a carne de codorna é altamente palatável e tem valor diferenciado dentro do mercado cada vez mais exigente por qualidade. Porém, existem poucos estudos científicos sobre codornas tipo corte nas áreas de nutrição, melhoramento genético e produção nas condições de exploração brasileiras o que muitas vezes pode inviabilizar a expansão desta atividade no Brasil.
A dieta de codornas de corte é formulada com base nas exigências nutricionais contidas no NRC (Nutrient... 1994), ou em extrapolações das exigências nutricionais de frangos de corte ou codornas de postura, o que pode causar impacto negativo na atividade seja por superalimentação, pelo aumento no custo de produção e contaminação ambiental, ou pela subestimação das exigências nutricionais de codornas de corte o que gera aproveitamento inadequado do potencial zootécnico da espécie.
A nutrição adequada é o primeiro passo para otimizar o crescimento e o rendimento de carcaça, e reduzir os impactos negativos do excesso de excreção de nitrogênio no ambiente.Uma das formas recomendadas para reduzir a excreção e emissão é diminuir o nível de proteína bruta da dieta por meio da suplementação aminoacídica (KIM et al., 2006). Com a possibilidade do uso de aminoácidos sintéticos nas dietas,
pode-se evitar a perda energética da ração resultante do desbalanceamento de aminoácidos como também diminuir o custo na formulação de dietas utilizando níveis menores de proteína bruta. A metionina é considerada o primeiro aminoácido limitante para aves, e sua deficiência ou excesso pode prejudicar características de carcaça e de desempenho. Este aminoácido, na forma S-adenosilmetionina, é o mais importante doador do radical metil no organismo, sendo exigido para a biossíntese de muitas substâncias importantes envolvidas no crescimento, como creatina, carnitina, poliaminas, epinefrina, colina e melatonina (BAKER, 1991).
Além do ambiente (nutrição, temperatura, umidade, estresse, densidade, entre outros), a genética é fator crucial para expressão das características produtivas das codornas. Em muitos casos, a interação entre estes dois fatores é responsável por mudança na resposta de diferentes genótipos submetidos às variações ambientais, que pode gerar alterações na ordenação dos valores genéticos de acordo com o ambiente no qual estão sendo criadas (Falconer & Mackay, 1996) caracterizando a interação genótipo-ambiente. E os melhores desempenhos são obtidos com a utilização da dieta mais adequada para cada grupo genético.
O objetivo deste estudo foi avaliar os efeitos dos níveis de metionina + cistina, grupos genéticos e a interação entre estes fatores sobre características de desempenho avaliadas em codornas de corte nos períodos inicial (do 7º e 21º dia de idade), final (do 22º e 42º dia de idade) e total de

24
crescimento (do 7 e 42º dia de idade) e estimar as exigências nutricionais de cada grupo genético.
2. MATERIAL E MÉTODOS
O primeiro experimento foi conduzido no aviário experimental do Setor de coturnicultura da Escola de Veterinária da Universidade Federal de Minas Gerais para avaliar o efeito da interação genótipo x níveis de metionina+cistina total da dieta sobre características avaliadas no período inicial e total de crescimento. As codornas com peso médio inicial de 25,05g foram alojadas em baterias de arame galvanizado com dimensões de 0,82m de largura x 0,41m de profundidade x 0,27cm de altura por unidade experimental, equipadas com bebedouro copo e comedouro tipo calha.
O programa de luz adotado foi contínuo (24 horas de luz artificial) durante todo o período experimental e as dietas foram fornecidas à vontade. Diariamente era feita a limpeza dos bebedouros e gaiolas. Foram avaliadas características de desempenho (peso final, ganho de peso, consumo de dieta e conversão alimentar) nas fases inicial (7º ao 21º dia) e total (7º ao 42º dia) de crescimento por meio de pesagens semanais e controle da quantidade de dieta oferecida.
Foram utilizadas 468 codornas da linhagem EV1 e 390 codornas da linhagem EV2 de ambos os sexos do 7º ao 42º dia de idade. O delineamento experimental adotado foi inteiramente ao acaso com seis níveis de metionina+cistina (0,73 a 1,03%), dois grupos genéticos, sendo 5 repetições para o grupo EV1 e 6 repetições para o grupo EV2 contendo 13 aves por unidade experimental.Nos primeiros 7 dias de vida, as aves foram criadas em boxes no chão com dietas contendo 28% de proteína bruta e 1,05% de metionina+cistina, adotada como padrão na Programa de
Melhoramento Genético de Codornas de Corte da UFMG, devido à dificuldade de alojamento dos pintinhos nas baterias de gaiola nos primeiros dias. A partir do 7º dia de vida as aves foram alimentadas com dieta experimental formulada à base de milho e farelo de soja, contendo 23,9% de proteína bruta (PB) e 2900 Kcal de EM/kg de dieta. A dieta basal (tabela 1) foi suplementada com DL-metionina (99%), em substituição ao amido de milho, o que corresponde aos níveis de 0,73% (dieta basal sem suplementação); 0,79; 0,85; 0,91; 0,97 e 1,03% de metionina mais cistina total, permanecendo as dietas isocalóricas e isoprotéicas.
As dietas foram formuladas com base nas exigências nutricionais contidas no NRC (Nutrient... 1994) exceto para metionina + cistina e na composição dos ingredientes segundo Rostagno et al. (2000).
No modelo de avaliação, além do efeito de grupo genético e do nível aminoacídico, foi inserido o efeito da interação entre grupo genético e nível de metionina+cistina e as exigências para este aminoácido foram estimadas por derivação das equações de regressão polinomiais, respeitando-se a interpretação biológica das variáveis. As análises estatísticas foram realizadas por meio do programa SAEG (Sistema... 2004) de acordo com o seguinte modelo:
yijk = µ + Gi + Mj + GxMij + eijk , em que:
yijk = observação k, do grupo genético i, do nível de metionina+cistina total j;
µ = média geral;
Gi = efeito do grupo genético, i = 1 e 2;
Mi = efeito do nível de metionina+cistina total, j = 1, 2...6;
GxMij = interação do grupo genético vs. nível de metionina + cistina total da dieta;
eijk = erro associado a cada observação yijk;

25
Tabela 1. Composição percentual da dieta basal formulada para codornas de corte (Experimento 1)
Ingrediente % Milho 53,5 Farelo de soja 41,9 Calcário 1,07 Óleo de soja 1,02 Fosfato bicálcico 0,96 Amido 0,50 Suplemento mineral e vitamínico¹ 0,50 Sal comum 0,26 L-treonina 0,26 Composição calculada Proteína Bruta (%) 23,89 Energia Metabolizável (Kcal/kg) 2,90 Cálcio (%) 0,80 Fósforo disponível (%) 0,30 Sódio (%) 0,15 Aminoácido total % Lisina 1,30 Metionina+cistina 0,73 Treonina 1,19 Triptofano 0,30 Metionina 0,46 Arginina 1,61 Isoleucina 1,04 Valina 1,11
¹ Composição por kilo de produto: Vit. A – 2.000.000UI; vit. D3 – 375.000 UI; vit E – 3.750mg; vit. K3 – 500mg; vit.B1 - 250mg; vit. B2 – 750mg; vit. B6 – 500mg; vit. B12 – 3.750mcg; niacina – 6.250mg; ác. Pantotênico – 2.500mg; biotina – 10mg; ác. fólico – 125mg; colina – 75.000mg; metionina – 250.000mg; selênio – 45mg; iodo – 175mg; ferro – 12.525mg; cobre – 2.500mg; manganês – 19.500mg; zinco – 13.750mg; avilamicina – 15.000mg; narasin – 12.250mg; B.H.T. – 500mg; vit. C – 12.500mg.
O segundo experimento foi conduzido para avaliar o efeito da interação genótipo x nível de metionina+cistina total sobre características na fase final do período de crescimento das codornas de corte. Foram utilizadas 576 codornas, sendo 288 da linhagem EV1 e 288 da EV2 de ambos os sexos do 21º ao 42º dia de idade. As codornas com peso médio ao 21º dia de idade de 139,95g do grupo genético EV1 e 141,93g do EV2 foram alojadas em baterias de arame galvanizado com dimensões de 0,82m de largura x 0,41m de profundidade x 0,27cm de altura por unidade experimental, equipadas com bebedouro copo e comedouro tipo calha. Durante a fase inicial (1º ao 21º dia idade) as aves foram criadas em boxes no chão em
condições de produção comercial e alimentadas com dieta contendo 0,94% de metionina+cistina, previamente estimada como exigência nesta fase (Experimento 1). O delineamento experimental adotado foi inteiramente ao acaso com seis níveis de metionina+cistina total (0,63; 0,73; 0,83; 0,93; 1,03 e 1,13%), sendo o menor nível mais baixo que o testado no Experimento 1 com o objetivo de se determinar a exigência no período final da fase de crescimento caso ela fosse menor que 0,73%, e 4 repetições, com 12 aves por unidade experimental. As aves foram submetidas às dietas experimentais formuladas à base de milho e farelo de soja, contendo 20,4% de proteína bruta (PB) e 2900 Kcal de EM/kg de dieta. A dieta basal (tabela 2) foi

26
suplementada com DL-metionina (99%), em substituição ao amido de milho.
As dietas foram formuladas com base nas exigências nutricionais contidas no NRC (Nutrient... 1994) exceto para metionina + cistina e na composição dos ingredientes segundo Rostagno et al. (2000). As
exigências para este aminoácido foram estimadas por derivação das equações de regressão polinomiais, respeitando-se a interpretação biológica das variáveis. As análises estatísticas foram realizadas por meio do programa SAEG (Sistema... 2004) de acordo com o mesmo modelo já apresentado no Experimento 1.
Tabela 2. Composição percentual da dieta basal formulada para codornas de corte (Experimento 2)
Ingrediente %
Milho 58,4
Farelo de soja 33,2
Calcário 1,12
Óleo de soja 1,50
Fosfato bicálcico 1,05
Amido 0,50
Suplemento mineral e vitamínico¹ 0,50
Sal comum 0,33
L-treonina 0,25
L-lisina HCL 0,31
Composição calculada
Proteína Bruta (%) 20,40
Energia Metabolizável (Kcal/kg) 2,90
Cálcio (%) 0,80
Fósforo disponível (%) 0,30
Sódio (%) 0,15
Aminoácido total %
Lisina 1,30
Metionina+cistina 0,63
Treonina 1,02
Triptofano 0,25
Arginina 1,33
Isoleucina 0,98
¹ Composição por kilo de produto: Vit. A – 2.000.000UI; vit. D3 – 375.000 UI; vit E – 3.750mg; vit. K3 – 500mg; vit.B1 - 250mg; vit. B2 – 750mg; vit. B6 – 500mg; vit. B12 – 3.750mcg; niacina – 6.250mg; ác. Pantotênico – 2.500mg; biotina – 10mg; ác. fólico – 125mg; colina – 75.000mg; metionina – 250.000mg; selênio – 45mg; iodo – 175mg; ferro – 12.525mg; cobre – 2.500mg; manganês – 19.500mg; zinco – 13.750mg; avilamicina – 15.000mg; narasin – 12.250mg; B.H.T. – 500mg; vit. C – 12.500mg.
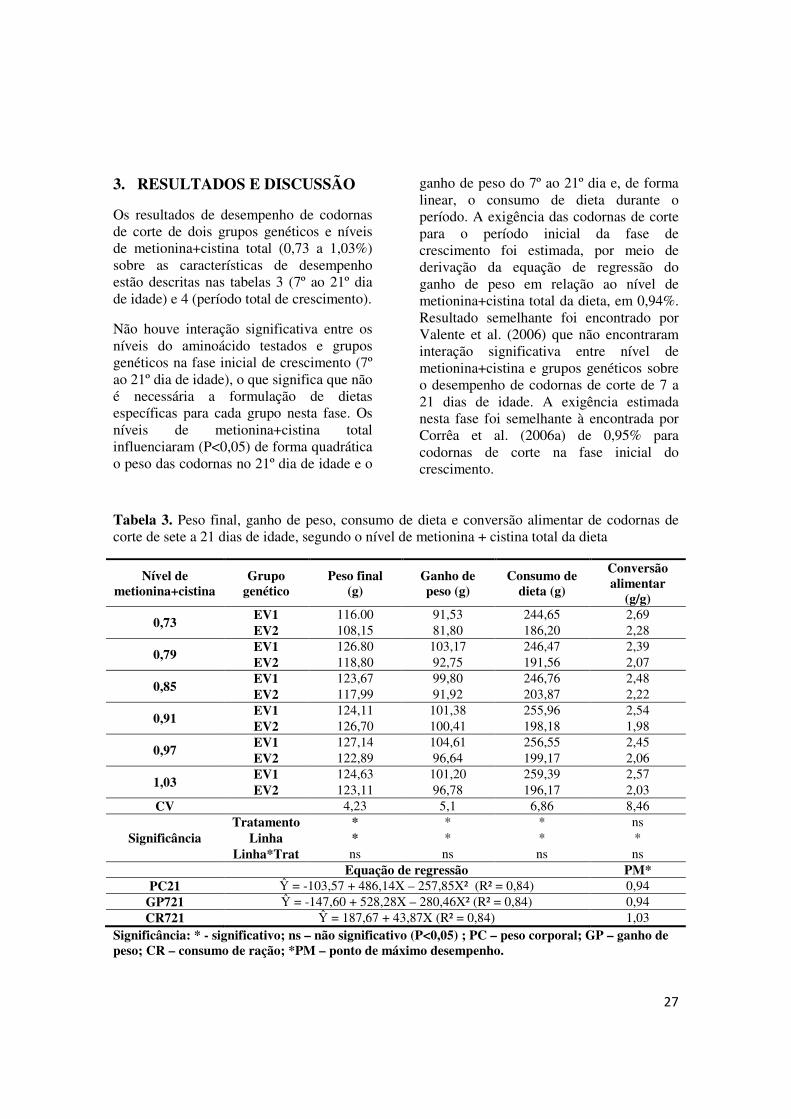
27
3. RESULTADOS E DISCUSSÃO
Os resultados de desempenho de codornas de corte de dois grupos genéticos e níveis de metionina+cistina total (0,73 a 1,03%) sobre as características de desempenho estão descritas nas tabelas 3 (7º ao 21º dia de idade) e 4 (período total de crescimento).
Não houve interação significativa entre os níveis do aminoácido testados e grupos genéticos na fase inicial de crescimento (7º ao 21º dia de idade), o que significa que não é necessária a formulação de dietas específicas para cada grupo nesta fase. Os níveis de metionina+cistina total influenciaram (P<0,05) de forma quadrática o peso das codornas no 21º dia de idade e o
ganho de peso do 7º ao 21º dia e, de forma linear, o consumo de dieta durante o período. A exigência das codornas de corte para o período inicial da fase de crescimento foi estimada, por meio de derivação da equação de regressão do ganho de peso em relação ao nível de metionina+cistina total da dieta, em 0,94%. Resultado semelhante foi encontrado por Valente et al. (2006) que não encontraram interação significativa entre nível de metionina+cistina e grupos genéticos sobre o desempenho de codornas de corte de 7 a 21 dias de idade. A exigência estimada nesta fase foi semelhante à encontrada por Corrêa et al. (2006a) de 0,95% para codornas de corte na fase inicial do crescimento.
Tabela 3. Peso final, ganho de peso, consumo de dieta e conversão alimentar de codornas de corte de sete a 21 dias de idade, segundo o nível de metionina + cistina total da dieta
Nível de metionina+cistina
Grupo genético
Peso final (g)
Ganho de peso (g)
Consumo de dieta (g)
Conversão alimentar
(g/g)
0,73 EV1 116.00 91,53 244,65 2,69 EV2 108,15 81,80 186,20 2,28
0,79 EV1 126.80 103,17 246,47 2,39 EV2 118,80 92,75 191,56 2,07
0,85 EV1 123,67 99,80 246,76 2,48 EV2 117,99 91,92 203,87 2,22
0,91 EV1 124,11 101,38 255,96 2,54 EV2 126,70 100,41 198,18 1,98
0,97 EV1 127,14 104,61 256,55 2,45 EV2 122,89 96,64 199,17 2,06
1,03 EV1 124,63 101,20 259,39 2,57 EV2 123,11 96,78 196,17 2,03
CV 4,23 5,1 6,86 8,46
Significância Tratamento * * * ns
Linha * * * * Linha*Trat ns ns ns ns
Equação de regressão PM* PC21 Ŷ = -103,57 + 486,14X – 257,85X² (R² = 0,84) 0,94
GP721 Ŷ = -147,60 + 528,28X – 280,46X² (R² = 0,84) 0,94 CR721 Ŷ = 187,67 + 43,87X (R² = 0,84) 1,03
Significância: * - significativo; ns – não significativo (P<0,05) ; PC – peso corporal; GP – ganho de peso; CR – consumo de ração; *PM – ponto de máximo desempenho.
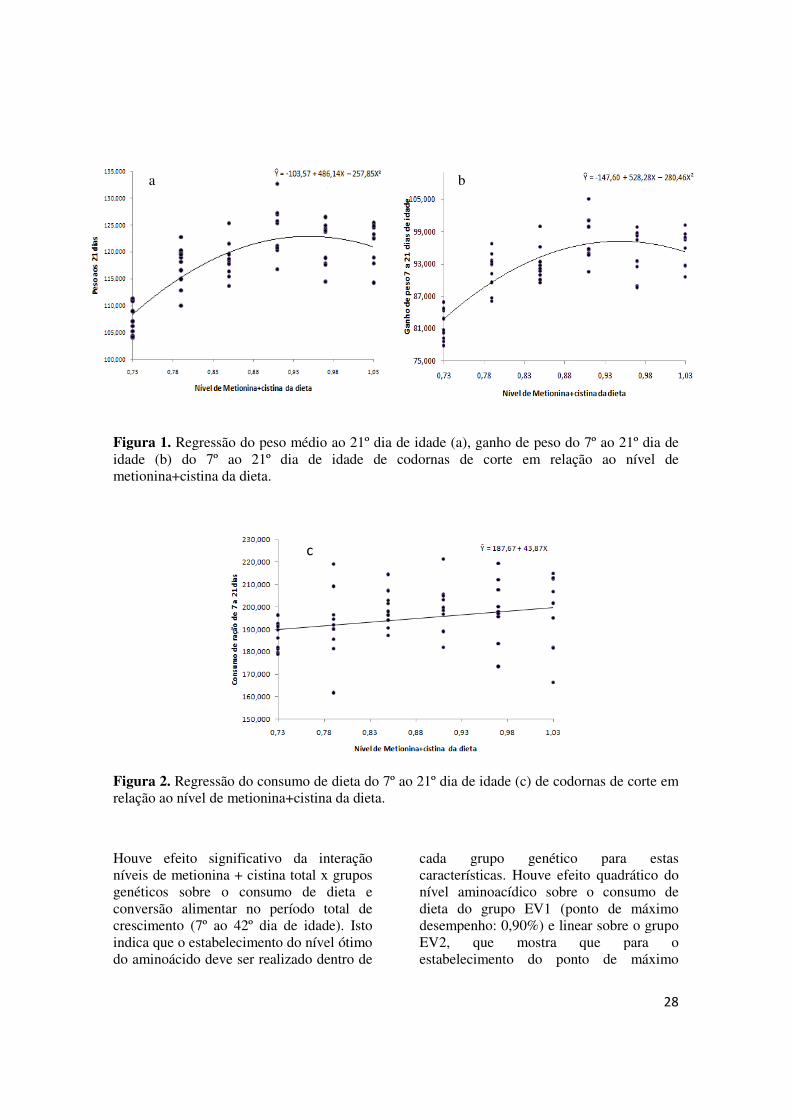
Figura 1. Regressão do peso médio ao 21º dia de idade (a), ganho de peso do 7º ao 21º dia de idade (b) do 7º ao 21º dia de idade de codornas de corte em relação ao nível de metionina+cistina da dieta.
Figura 2. Regressão do consumo de dieta do 7º ao 21º dia de idade (c) de codornas de corte em relação ao nível de metionina+cistina da dieta.
Houve efeito significativo da interação níveis de metionina + cistina total x grupos genéticos sobre o consumo de dieta e conversão alimentar no período total de crescimento (7º ao 42º dia de idade). Isto indica que o estabelecimento do nível ótimo do aminoácido deve ser realizado dentro de
a
Regressão do peso médio ao 21º dia de idade (a), ganho de peso do 7º ao 21º dia de idade (b) do 7º ao 21º dia de idade de codornas de corte em relação ao nível de
Regressão do consumo de dieta do 7º ao 21º dia de idade (c) de codornas de corte em relação ao nível de metionina+cistina da dieta.
Houve efeito significativo da interação níveis de metionina + cistina total x grupos
bre o consumo de dieta e conversão alimentar no período total de crescimento (7º ao 42º dia de idade). Isto indica que o estabelecimento do nível ótimo do aminoácido deve ser realizado dentro de
cada grupo genético para estas características. Houve efeito nível aminoacídico sobre o consumo de dieta do grupo EV1 (ponto de máximo desempenho: 0,90%) e linear sobre o grupo EV2, que mostra que para o estabelecimento do ponto de máximo
c
b
28
Regressão do peso médio ao 21º dia de idade (a), ganho de peso do 7º ao 21º dia de idade (b) do 7º ao 21º dia de idade de codornas de corte em relação ao nível de
Regressão do consumo de dieta do 7º ao 21º dia de idade (c) de codornas de corte em
cada grupo genético para estas características. Houve efeito quadrático do nível aminoacídico sobre o consumo de dieta do grupo EV1 (ponto de máximo desempenho: 0,90%) e linear sobre o grupo EV2, que mostra que para o estabelecimento do ponto de máximo

29
desempenho deste último grupo deveriam ser testados níveis mais altos de metionina + cistina. Por outro lado, houve efeito quadrático do nível de metionina + cistina total sobre a conversão alimentar no período total de crescimento do grupo genético EV1, sendo que o ponto de máximo desempenho (mínima conversão) estimado foi de 0,85%. Não foi verificada influência dos níveis deste aminoácido sobre a conversão alimentar do grupo EV2. Estas diferenças mostram que é importante que sejam avaliados os requerimentos nutricionais dentro de cada grupo para maximização do desempenho das codornas. Resultados semelhantes foram encontrados por Valente et al. (2006) para o período total de crescimento de codornas de corte, os autores encontraram efeito da interação grupo genético x nível de metionina sobre
peso final no 42º dia de idade, consumo de dieta e conversão alimentar. Vários autores como Almeida et al. (2002), Dionello et al.(2006) , Felipe et al. (2007), Wenceslau et al. (2007) e Corrêa et al. (2006b) sugerem a existência de interação genótipo x ambiente entre componentes nutricionais da dieta de codornas de corte em diferentes fases da sua vida produtiva o que mostra a relevância deste tema.
Para as demais características avaliadas, como ganho de peso e peso final, durante o período total de crescimento foi verificado efeito quadrático do nível de metionina + cistina total. A exigência dos dois grupos genéticos, estabelecida por meio da derivação da equação de regressão do ganho de peso em relação ao nível de metionina+cistina, foi de 0,98%.
Tabela 4. Peso final, ganho de peso, consumo de ração e conversão alimentar de codornas de corte de sete a 42 dias de idade, segundo o nível de metionina mais cistina da dieta
Nível de metionina +cistina
Grupo genético Peso final
(g) Ganho de peso (g)
Consumo de ração (g)
Conversão alimentar (g/g)
0,73 EV1 222,43 197,97 707,71 3,58 EV2 240,02 213,67 736,48 3,45
0,79 EV1 238,97 215,33 776,50 3,61 EV2 243,68 217,63 768,98 3,54
0,85 EV1 235,60 211,80 782,89 3,70 EV2 242,19 216,12 767,47 3,55
0,91 EV1 241,26 218,53 786,30 3,60 EV2 253,93 227,65 777,12 3,42
0,97 EV1 240,83 217,80 788,54 3,62 EV2 250,18 223,93 767,69 3,43
1,03 EV1 243,12 219,52 749,05 3,41 EV2 249,93 223,60 805,49 3,60
CV 3,11 3,43 3,91 4,28
Significância Tratamento * * * ns
Linha * * ns * Linha*Trat ns ns * *
Equação de regressão PM* PC42 Ŷ = 27,67 + 443,88X – 225,77X² (R² = 0,72) 0,98
GP742 Ŷ = -17,11 + 488,25X – 249,56X² (R² =0,84) 0,98
CR742 EV1 Ŷ = -1452,39 + 4988,71X – 2767,88X² (R² = 0,92) 0,90 EV2 Ŷ = 623,52 + 167,07X (R² =0,72) -
CA742 EV1 Ŷ = -1,588 + 12,37X – 7,28X² (R² =0,92) 0,85 EV2 - -
Significância: * - significativo; ns – não significativo (P<0,05); PC – peso corporal; GP – ganho de peso; CR – consumo de ração; *PM – ponto de máximo desempenho.

Figura 3. Regressão do peso médio ao 42º dia de idade (a), ganho de peso do 7º ao 42º dia de idade (b), consumo de dieta do 7º ao 42º dia de idade para EV1 (c) e para EV2 (d) e conversão alimentar do 7º ao 42º dia de idade de metionina+cistina total da dieta.
Os resultados para o desempenho das diferentes linhagens de codornas de corte na fase final de crescimento (2
a
c
. Regressão do peso médio ao 42º dia de idade (a), ganho de peso do 7º ao 42º dia de
idade (b), consumo de dieta do 7º ao 42º dia de idade para EV1 (c) e para EV2 (d) e conversão alimentar do 7º ao 42º dia de idade do grupo EV1 (e) de codornas de corte em relação ao nível de metionina+cistina total da dieta.
Os resultados para o desempenho das diferentes linhagens de codornas de corte na fase final de crescimento (22 a 42 dias –
Experimento 2) tratadas com dietas contendo seis níveis diferentes de metionina+cistina constam na tabela 5.
b
d
e
30
. Regressão do peso médio ao 42º dia de idade (a), ganho de peso do 7º ao 42º dia de idade (b), consumo de dieta do 7º ao 42º dia de idade para EV1 (c) e para EV2 (d) e conversão
do grupo EV1 (e) de codornas de corte em relação ao nível
Experimento 2) tratadas com dietas contendo seis níveis diferentes de metionina+cistina constam na tabela 5.

31
Tabela 5. Peso final, ganho de peso, consumo de dieta e conversão alimentar de duas linhagens de codornas de corte de 22 a 42 dias de idade, segundo o nível de metionina+cistina da dieta
Nível de metionina+cistina
Grupo genético
Peso final (g)
Ganho de peso (g)
Consumo de ração (g)
Conversão alimentar
(g/g)
0,63 EV1 276,94 137,65 640,02 4,66 EV2 289,38 147,08 652,69 4,44
0,73 EV1 287,83 148,65 641,60 4,32 EV2 284,52 143,52 643,22 4,51
0,83 EV1 281,25 142,23 626,81 4,41 EV2 290,08 149,01 653,38 4,39
0,93 EV1 289,56 148,50 644,85 4,35 EV2 290,87 148,56 641,54 4,32
1,03 EV1 289,10 148,25 654,67 4,42 EV2 279,90 138,44 614,08 4,45
1,13 EV1 287,53 147,26 630,49 4,29 EV2 284,71 141,27 614,42 4,37
CV 3,39 6,28 3,05 4,82
Significância Tratamento ns ns ns ns
Linha ns ns ns ns Linha*Trat ns ns * ns
Significância: * - significativo a 5%; ns - Não significativo, pelo teste Fisher
Verifica-se que não houve efeito significativo dos tratamentos sobre as características de desempenho avaliadas, o que indica que níveis de metionina+cistina total na dieta de codornas de corte na fase final do crescimento podem ser mantidos mais baixos o que diminuiria o custo da dieta. Portanto, é um período em que as codornas são menos exigentes para estes níveis de aminoácido, contrastando com as exigências encontradas por Felipe et al. (2008), que na fase inicial de crescimento encontraram influência significativa para níveis de metionina+cistina no peso final, ganho de peso e consumo de dieta.
Encontrou-se efeito significativo da interação níveis de metionina+cistina total da dieta x grupo genético sobre o consumo de dieta, o que significa que considerações feitas a respeito do consumo de dieta destas codornas de corte deverão ser feitas considerando a linhagem, isto é, cada linhagem apresenta resposta diferente aos níveis de metionina+cistina. Não houve,
para nenhuma das linhagens, efeito dos níveis de metionina+cistina total testados sobre o consumo de dieta.
4. CONCLUSÕES
Dietas para máximo ganho de peso de codornas de corte durante a fase inicial do crescimento (7 a 21 dias de idade) devem conter 0,94% de metionina+cistina para os dois grupos genéticos e uma relação de 0,72 metionina+cistina/lisina . Para o período total de crescimento (7 a 42 dias) as dietas devem conter 0,98% de metionina+cistina e uma relação de 0,75 metionina+cistina/lisina, para ambos os grupos genéticos.
Codornas de corte durante a fase final do crescimento (22 a 42 dias) são menos exigentes quanto ao nível de metionina+cistina total para características de desempenho e podem ser alimentadas

32
com dietas contendo 0,63% de metionina+cistina dando uma relação de 0,49 metionina+cistina/lisina independente da linhagem sem que haja perdas no desempenho.
Para sistemas de produção que adotam dietas por fase de crescimento, as dietas devem ser formuladas para conter 0,94% de metionina+cistina total na primeira fase e 0,63% de metionina+cistina total para a última fase para ambos os grupos genéticos. Sistemas de criação de codornas que adotem dieta única para todo o período de crescimento devem utilizar dietas contendo 0,98% de metionina+cistina total para ambos os grupos genéticos.
5. REFERÊNCIAS BIBLIOGRÁFICAS
ALMEIDA M.I.M.; OLIVEIRA, E.G.; RAMOS, P.R.R et al. Desempenho produtivo para corte de machos de codornas (Coturnix sp.) de duas linhagens, submetidos a dois ambientes nutricionais. In: SOCIEDADE BRASILEIRA DE MELHORAMENTO ANIMAL, 4., 2002. Anais... Disponível em: http://www.sbmaonline.org.br/anais/iv/trabalhos/aves/ivt02av.pdf
BAKER, D.H. Partitioning of nutrients for growth and other metabolic functions. Poultry Science, v.70, p.1797-1805, 1991.
CORRÊA, G.S.S.; SILVA, M.A.; CORRÊA, A.B. et al. Exigência de metionina + cistina total para codornas de corte em crescimento. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.58, n.3, p.414-420, 2006a.
CORRÊA, G.S.S.; SILVA, M.A.; DIONELLO, N.J.L. Genotype by environment interaction for production traits of meat type quail during the growing
phase. In: World Congress on Genetics Aplied to Livestock Production, 2006, Belo Horizonte. Proceedings... 8th World Congress on Genetics Aplied to Livestock Production, 2006b. CD-ROM.
DIONELLO, N.J.L.; CORRÊA, G.S.S.; SILVA, M.A. Interação genótipo x ambiente em características produtivas de codornas de corte no período de crescimento. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 43., 2006, João Pessoa. Anais... João Pessoa, PB: SBZ. CD-ROM.
FALCONER, D.S.; MACKAY, T.F.C. Introduction to quantitative genetics. 4.ed. London: Longman. 1996.
FELIPE, V.P.S., CORRÊA, G.S.S., CORRÊA, A.B.. Interação entre nível de metionina+cistina total da dieta e dois grupos genéticos de codornas de corte durante o período de crescimento. In: Reunião Anual da Sociedade Brasileira de Zootecnia, 45ª, 2008, Lavras. Anais... Lavras, MG: SBZ. CD-ROM.
FELIPE, V.P.S.; CORRÊA, G.S.S.; SILVA, M.A. Interação genótipo-ambiente em características de desempenho e de carcaça de fêmeas híbridas de codornas de corte na fase de crescimento. In: Congresso Internacional de Zootecnia, 9., 2007, Londrina. Anais... Londrina: UEL, 2007. p. 1-5.
KIM, W.K.; FROELICH JR., C.A.; PATTERSON, P.H. The potential to reduce poultry nitrogen emissions with dietary methionine or methionine analogues supplementation. World’s Poult. Sci. Journ., v.62, p.338-353, 2006.
NUTRIENT requirements of poultry. Washington: National Academy of Sciences, 1994. p.44-45.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. et al. Composição de

33
alimentos e exigências nutricionais. Tabelas Brasileiras. Viçosa, MG: Universidade Federal de Viçosa, 2000. 141p.
SISTEMA de análises estatísticas e genéticas – SAEG. Versão 9.0. Viçosa, MG: UFV, 1999.
VALENTE, B.D.; CORRÊA, G.S.S.; DIONELLO, N.J.L. Interaction between methionine of diet and genotype in meat type quail. In: World Congress on Genetics
Aplied to Livestock Production, 2006, Belo Horizonte. Proceedings... 8th World Congress on Genetics Aplied to Livestock Production, 2006. CD-ROM.
WENCESLAU, R.R.; CORRÊA, G.S.S.; SILVA, M.A. Interação genótipo ambiente para características de desempenho em codornas de corte durante o período inicial da fase de crescimento. . In: Congresso Internacional de Zootecnia, 9., 2007, Londrina. Anais... Londrina: UEL, 2007. p. 1-6.

34
CAPÍTULO 2
Efeito da interação nível de metionina+cistina total da dieta x sexo x grupo genético sobre características de carcaça de codornas de corte
Genotype by total methionine+cystine level and sex interaction for carcass traits of meat type
quails
RESUMO
Estudou-se o efeito da interação nível de metionina+cistina total da dieta x grupos genéticos x sexo de codornas de corte sobre características de carcaça em sistemas de alimentação por fase de crescimento (inicial e final) e alimentação com dieta única. Foram realizados dois experimentos, distintos pelo sistema de criação, e ao final de cada um deles, dois machos e duas fêmeas foram abatidos de cada unidade experimental, totalizando 192 aves sendo 96 de cada grupo genético. Para sistemas de dieta única houve efeito da interação tripla apenas para rendimento de moela e não houve efeito dos níveis de met+cist testados sobre características de carcaça e cortes nobres avaliados. Em sistemas de alimentação por fase do período de crescimento houve efeito da interação tripla sobre o rendimento de carcaça e, após o desdobramento dos efeitos, houve efeito significativo do tratamento somente para fêmeas do grupo EV2 com mínimo desempenho em dietas contendo 0,88% de met+cist total; houve efeito linear significativo do nível de met+cist sobre o rendimento de peito, peso e rendimento de asas, peso e rendimento do fígado e rendimento do coração.
Palavras-chave: interação genótipo-ambiente, codornas de corte, rendimento de carcaça, metionina+cistina
ABSTRACT
The effect of methionine + cystine level of diet x genetic group x sex interaction on carcass
traits of meat type in two feeding systems: diets by period of the growth phase (initial and
finishing diets), and common diet for both periods. Two experiments were carried out and at the
end of both experiments two males and two females of each experimental unit were slaughtered,
totaling 192 quails, 96 of each genetic group. In the common diet experiment for both periods a
triple significant interaction effect was observed only for gizzard yield but no effects of
methionine + cystine were observed for carcass traits and main carcass cuts. In diet by phase of
the growth period system triple interaction significant effect was observed for carcass yield.
Significant effect of methionine + cystine level of diet on traits of quails of strain EV2 suggested
minimum performance for quails fed .88% diets. Linear effect of met + cyst level on breast
yield, weight and wing yield, weight and liver yield and heart yield.
Keywords: genotype by environment interaction, meat type quails, carcass yield, methionine
plus cystine

35
1. INTRODUÇÃO
A genética e o ambiente são fatores importantes no controle de expressão de genes responsáveis pelas características produtivas das aves. Em muitos casos, a interação entre estes fatores é responsável por mudança na resposta de diferentes genótipos submetidos às variações ambientais, e pode gerar alterações na classificação das aves de acordo com o ambiente no qual estão sendo criadas.
O progresso genético em termos de melhoria na taxa de crescimento, conversão alimentar e rendimento de carne é desafio constante do ponto de vista nutricional (SCHEUERMANN et al., 1995). Os aminoácidos metionina+cistina são de grande importância por se qualificarem como essenciais, sendo a metionina considerada o primeiro aminoácido limitante para o empenamento e crescimento de aves. Além disso, dietas com melhor balanço de aminoácidos e níveis mais baixos de proteína podem favorecer o desempenho e também facilitam o ajuste na formulação das dietas. O excesso no aporte de aminoácidos essenciais pode resultar em incremento calórico desnecessário, além de causar aumento na excreção de ácido úrico, sobrecarga de rins e fígado, aumento do requerimento energético e do custo da dieta.
Em frangos de corte, estudos mostram que dietas deficientes em metionina podem aumentar o consumo das aves o que causa incremento energético e gera menor teor de proteína na carcaça e maior deposição de gordura corporal e, consequentemente, menor rendimento de abate (Moran, 1994), enquanto que níveis mais elevados de metionina reduzem a quantidade de gordura abdominal (Albino et al., 1999) e aumentam o rendimento de peito (Atêncio et al., 2004; Garcia Neto et al., 2000).
O objetivo deste estudo foi verificar o efeito da interação níveis de metionina+cistina total da dieta x grupos genéticos x sexo de codornas de corte sobre as características de carcaça avaliadas ao 42º dia de idade em sistemas que adotem dieta única no período total de criação ou dieta por fase de crescimento.
2. MATERIAL E MÉTODOS
Para avaliação das características de carcaça, no 42o dia, quatro codornas, sendo dois machos e duas fêmeas (96 aves para cada grupo genético ou 192 codornas por grupo genético e nível de metionina+cistina total), de cada unidade experimental foram amostradas, pesadas, abatidas, depenadas, evisceradas e avaliadas quanto aos pesos e rendimentos de carcaças em relação ao peso vivo e rendimentos dos cortes no final dos Experimentos 1 e 2. O primeiro avaliou o efeito da interação genótipo x nível nutricional de metionina+cistina total em sistemas de dieta única e o segundo para sistema de criação que adota dieta por fase do período de crescimento (inicial e final). O modelo de análise utilizado foi:
hijkhijij
hjhijihhijk
eGMcSMcS
GSGMcSMcGy
+++
+++++= µ
em que:
=hijky observações das características de
carcaça; =µ média geral;
=hG efeito do grupo genético h = 1,2;
=iMc efeito do nível de metionina+cistina
total i = 1,...,6; =jS efeito do sexo j = 1,2;

36
=hiGMc interação dos efeitos de grupo
genético x nível de metionina+cistina da dieta;
=hjGS interação grupo genético e sexo;
=ijMcS interação nível de
metionina+cistina total x sexo; =hijGMcS interação grupo genético x nível
de metionina+cistina total x sexo; =hijke erro aleatório associado a cada
observação hijky .
As variáveis analisadas foram peso ao abate, peso da carcaça, rendimento de carcaça, peso do peito, rendimento de peito, peso das coxas, rendimento de coxa, peso das asas, rendimento de asa, pesos de fígado, moela, coração e seus respectivos rendimentos, peso e rendimento de gordura abdominal.
As análises estatísticas foram realizadas por meio do programa SAEG (2004). Foram realizadas análises de variância para avaliarem os efeitos dos níveis de grupo genético, metionina+cistina, sexo e suas interações sobre as características de carcaça. O efeito do nível de metionina+cistina total sobre características de carcaça foi avaliado por equação de regressão respeitando-se todas as interações por ventura significativas e a interpretação biológica da característica.
3. RESULTADOS E DISCUSSÃO
Os efeitos do sexo, grupo genético e dos níveis de metionina+cistina total em sistemas de alimentação com dieta única sobre as características analisadas são apresentadas nas tabelas 1, 2, 3, 4 e 5. Houve efeito de interação grupo genético x nível de metionina+cistina x sexo apenas para a característica rendimento de moela
(tabela 1). Não houve efeito das interações duplas do modelo sobre nenhuma das características avaliadas. Por outro lado, houve efeito significativo de sexo sobre o peso vivo, peso de carcaça, rendimento de carcaça, peso das coxas, peso do peito, peso e rendimento de gordura abdominal, peso e rendimento do fígado e peso do coração (tabela 2). Fêmeas apresentaram maior peso corporal e consequentemente de carcaça, maior peso de peito e coxas, como também maior peso e rendimento de fígado e gordura abdominal e maior peso do coração em relação aos machos. Apesar do maior peso das fêmeas em relação ao dos machos, estes apresentam maior rendimento de carcaça o que seria vantagem em razão do maior rendimento de abate, o que pode ser economicamente interessante. Estes resultados estão de acordo com os descritos na literatura que mostram que codornas fêmeas tendem a ser mais pesadas que machos (CORRÊA et al., 2008, SANTOS et al., 2008, OLIVEIRA et al., 2002),o que pode ser justificado pelo maior peso de vísceras do aparelho reprodutor (oviduto), fígado e maior proporção de gordura abdominal. Ressalta-se também que as codornas apresentam acentuado dimorfismo sexual (já a partir da 3º semana) sendo que fêmeas apresentam desenvolvimento mais precoce em relação aos machos. Em frangos de corte observa-se o comportamento inverso, machos são, em média, mais pesados enquanto que fêmeas apresentam maior rendimento de carcaça.
Não houve efeito de tratamento sobre nenhuma das variáveis relacionadas à carcaça, cortes nobres e vísceras comestíveis avaliadas. Isto indica que dietas contendo 0,98% de metionina+cistina total, nível estimado como exigência para o período total de crescimento (7-42 dias), não causariam efeito negativo sobre as características de carcaça e cortes nobres avaliadas.

37
Tabela 1. Rendimento de moela de machos e fêmeas de dois grupos genéticos de codornas de corte alimentadas com dietas contendo seis níveis de metionina+cistina
GG Sexo Rendimento
de moela CV
Significância (Tratamento)
EV1 M 2,65 10,17 ns F 2,78 12,90 ns
EV2 M 2,58 7,6 * F 2,74 11,8 *
Equação de regressão Mínimo
desempenho
EV2 M Ŷ = 9,37-15,8X+9,12X² 0,86 F Ŷ = 15,2-27,5X+14,9X² 0,92
Tabela 2. Efeito de sexo sobre o desempenho e características de carcaça de codornas de corte de dois grupos genéticos
Sexo Variáveis CV Macho Fêmea Peso vivo (g) 10,18 214,22B 248,33A Peso de carcaça (g) 10,49 155,20B 169,48A Rendimento de carcaça (%) 5,18 72,47A 68,39B Peso coxas (g) 12,93 39,62B 42,55A Peso do peito (g) 12,75 61,20B 67,65A Peso gordura abdominal (g) 45,80 2,33A 3,09B Rendimento gordura abdominal (%) 44,15 1,48A 1,81B Peso Fígado (g) 19,87 3,79B 5,96A Rendimento fígado (%) 20,35 2,44B 3,54A Peso Coração (g) 15,22 2,23B 2,41A
Médias seguidas de letras distintas nas linhas diferem entre si pelo teste Fisher (P<0,05).
Houve efeito de linhagem apenas sobre o rendimento de peito (tabela 3), o que mostra que a linhagem EV1 apresenta maior proporção de peito em relação à linhagem
EV2 decorrente de algum tipo de seleção diferenciado na formação destes dois grupos genéticos.
Tabela 3. Efeito do grupo genético sobre o rendimento de peito de codornas de corte
Variáveis GG Níveis de metionina+cistina
0,73 0,79 0,85 0,91 0,97 1,03 Média Rend. de peito (%)
EV1 40,91 39,22 40,83 41,09 39,87 40,17 40,35 A EV2 40,80 39,57 38,24 38,88 36,99 39,47 38,99 B
Médias seguidas de letras distintas nas colunas diferem entre si pelo teste Fisher (P<0,05).
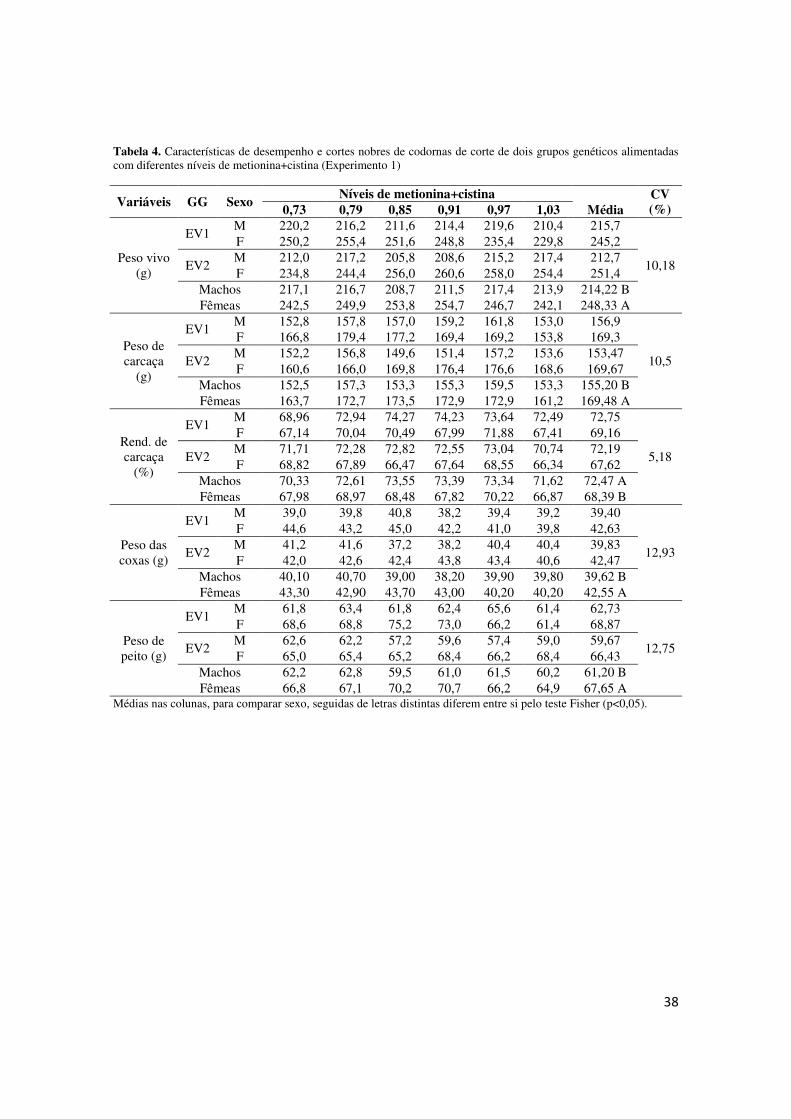
38
Tabela 4. Características de desempenho e cortes nobres de codornas de corte de dois grupos genéticos alimentadas com diferentes níveis de metionina+cistina (Experimento 1)
Variáveis GG Sexo Níveis de metionina+cistina CV
(%) 0,73 0,79 0,85 0,91 0,97 1,03 Média
Peso vivo (g)
EV1 M 220,2 216,2 211,6 214,4 219,6 210,4 215,7
10,18
F 250,2 255,4 251,6 248,8 235,4 229,8 245,2
EV2 M 212,0 217,2 205,8 208,6 215,2 217,4 212,7 F 234,8 244,4 256,0 260,6 258,0 254,4 251,4
Machos 217,1 216,7 208,7 211,5 217,4 213,9 214,22 B Fêmeas 242,5 249,9 253,8 254,7 246,7 242,1 248,33 A
Peso de carcaça
(g)
EV1 M 152,8 157,8 157,0 159,2 161,8 153,0 156,9
10,5
F 166,8 179,4 177,2 169,4 169,2 153,8 169,3
EV2 M 152,2 156,8 149,6 151,4 157,2 153,6 153,47 F 160,6 166,0 169,8 176,4 176,6 168,6 169,67
Machos 152,5 157,3 153,3 155,3 159,5 153,3 155,20 B Fêmeas 163,7 172,7 173,5 172,9 172,9 161,2 169,48 A
Rend. de carcaça
(%)
EV1 M 68,96 72,94 74,27 74,23 73,64 72,49 72,75
5,18
F 67,14 70,04 70,49 67,99 71,88 67,41 69,16
EV2 M 71,71 72,28 72,82 72,55 73,04 70,74 72,19 F 68,82 67,89 66,47 67,64 68,55 66,34 67,62
Machos 70,33 72,61 73,55 73,39 73,34 71,62 72,47 A Fêmeas 67,98 68,97 68,48 67,82 70,22 66,87 68,39 B
Peso das coxas (g)
EV1 M 39,0 39,8 40,8 38,2 39,4 39,2 39,40
12,93
F 44,6 43,2 45,0 42,2 41,0 39,8 42,63
EV2 M 41,2 41,6 37,2 38,2 40,4 40,4 39,83 F 42,0 42,6 42,4 43,8 43,4 40,6 42,47
Machos 40,10 40,70 39,00 38,20 39,90 39,80 39,62 B Fêmeas 43,30 42,90 43,70 43,00 40,20 40,20 42,55 A
Peso de peito (g)
EV1 M 61,8 63,4 61,8 62,4 65,6 61,4 62,73
12,75
F 68,6 68,8 75,2 73,0 66,2 61,4 68,87
EV2 M 62,6 62,2 57,2 59,6 57,4 59,0 59,67 F 65,0 65,4 65,2 68,4 66,2 68,4 66,43
Machos 62,2 62,8 59,5 61,0 61,5 60,2 61,20 B Fêmeas 66,8 67,1 70,2 70,7 66,2 64,9 67,65 A
Médias nas colunas, para comparar sexo, seguidas de letras distintas diferem entre si pelo teste Fisher (p<0,05).

39
Tabela 5. Características de carcaça (vísceras) de codornas de corte de dois grupos genéticos alimentadas com diferentes níveis de metionina+cistina (Experimento 1)
Variáveis GG Sexo Níveis de metionina+cistina
Média CV (%) 0,73 0,79 0,85 0,91 0,97 1,03
Peso gord. Abdominal
(g)
EV1 M 2,00 2,40 2,64 2,48 2,40 2,60 2,42
45,8
F 3,46 3,32 3,06 2,60 2,56 3,32 3,05
EV2 M 1,82 2,76 1,74 1,50 3,08 2,64 2,26 F 1,76 4,00 3,36 2,98 3,78 2,88 3,13
Machos 1,91 2,58 2,19 1,99 2,74 2,62 2,33 A Fêmeas 2,61 3,66 3,21 2,79 3,17 3,10 3,09 B
Rend. de gord.
abdominal (%)
EV1 M 1,31 1,51 1,62 1,56 1,49 1,57 1,51
44,1
F 2,06 1,88 1,76 1,50 1,49 2,18 1,81
EV2 M 1,20 1,74 1,12 0,99 1,99 1,68 1,45 F 1,03 2,38 1,96 1,68 2,16 1,71 1,82
Machos 1,25 1,62 1,37 1,27 1,74 1,62 1,48 A Fêmeas 1,54 2,12 1,86 1,59 1,82 1,94 1,81 B
Peso fígado (g)
EV1 M 4,06 4,00 4,00 4,20 3,70 3,84 3,97
19,9
F 6,20 6,34 5,92 6,52 6,06 5,20 6,04
EV2 M 3,90 3,74 3,32 3,34 3,76 3,70 3,63 F 5,06 6,14 5,54 6,14 6,04 6,36 5,88
Machos 3,98 3,87 3,66 3,77 3,73 3,77 3,79 B Fêmeas 5,63 6,24 5,73 6,33 6,05 5,78 5,96 A
Rend. de fígado (%)
EV1 M 2,65 2,54 2,56 2,63 2,28 2,53 2,53
20,3
F 3,70 3,64 3,38 3,94 3,60 3,39 3,61
EV2 M 2,58 2,38 2,20 2,21 2,40 2,39 2,36 F 3,16 3,73 3,25 3,48 3,43 3,80 3,47
Machos 2,61 2,45 2,38 2,42 2,34 2,46 2,44 B Fêmeas 3,42 3,68 3,31 3,71 3,51 3,59 3,54 A
Peso de coração
(g)
EV1 M 2,34 2,14 2,32 2,28 2,22 2,34 2,27
15,3
F 2,58 2,56 2,42 2,34 2,34 2,08 2,39
EV2 M 2,04 2,32 2,08 2,22 2,38 2,18 2,20 F 2,30 2,52 2,72 2,18 2,56 2,42 2,45
Machos 2,19 2,23 2,20 2,25 2,30 2,26 2,23 B Fêmeas 2,44 2,54 2,57 2,26 2,45 2,25 2,41 A
Médias nas colunas, para comparar sexo, seguidas de letras distintas diferem entre si pelo teste Fisher (p<0,05).
Os resultados dos efeitos dos níveis de metionina+cistina, sexo, grupo genético e interações com base na formulação de dietas por fase do período de crescimento (final) estão descritos nas tabelas 6, 7, 8, 9 e 10. Houve interação tripla (nível de metionina+cistina total x grupo genético x sexo) apenas para o rendimento de carcaça, o que sugere que cada sexo dentro de cada grupo genético apresenta exigência diferente de metionina+cistina total para esta característica. Após a realização dos desdobramentos e análises, encontrou-se
efeito significativo do tratamento apenas para o rendimento de carcaça de fêmeas do grupo EV2, em que o mínimo desempenho estimado seria para codornas alimentadas com dietas contendo 0,88% de metionina+cistina total, o que não justifica a separação das dietas por sexo já que não foi estimado o nível de máximo rendimento da carcaça (tabela 6 e figura 1). Portanto, dietas contendo 0,63% de metionina+cistina total podem ser utilizadas tanto para machos quanto para fêmeas de codornas de corte sem danos para o rendimento de
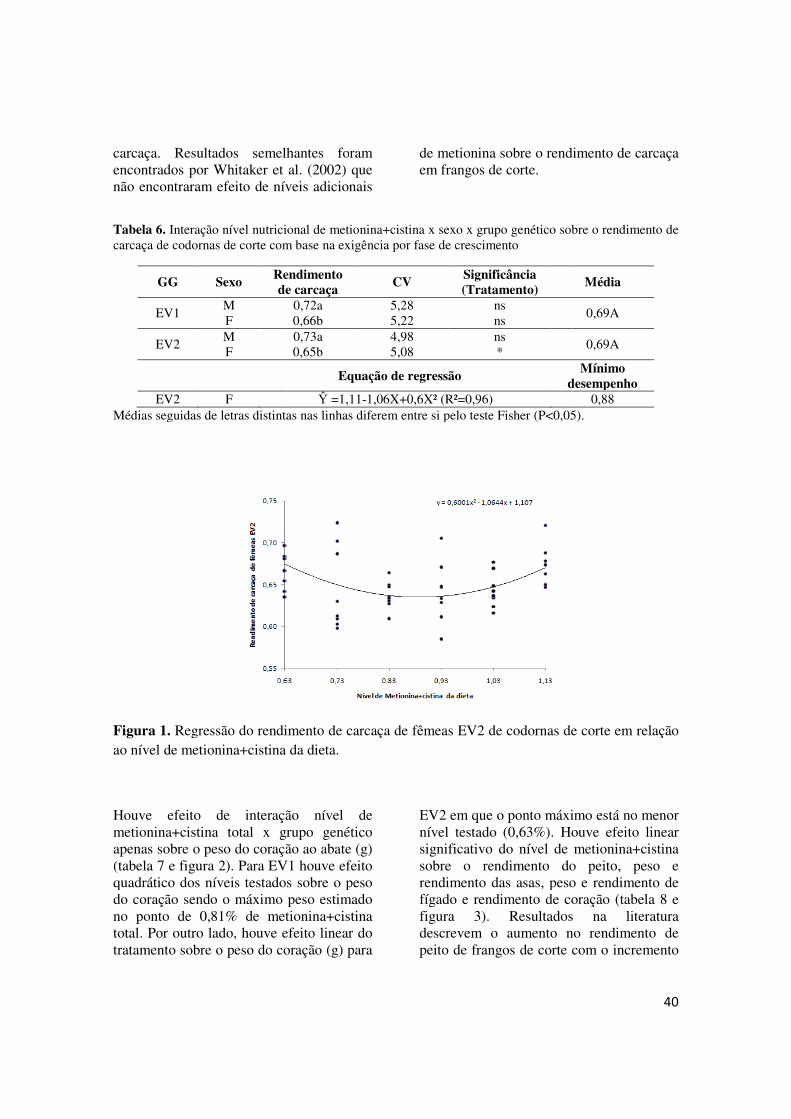
carcaça. Resultados semelhantes foencontrados por Whitaker et al. (2002) que não encontraram efeito de níveis adicionais
Tabela 6. Interação nível nutricional de metionina+cistina x sexo x grupo genético sobre o rendimento de carcaça de codornas de corte com base na exigência por fase de crescimento
GG Sexo Rendimento de carcaça
EV1 M F
EV2 M F
EV2 F Médias seguidas de letras distintas nas linhas diferem entre si pelo teste Fisher
Figura 1. Regressão do rendimento de carcaça de fêmeas EV2 de codornas de corte em relação ao nível de metionina+cistina da dieta.
Houve efeito de interação nível de metionina+cistina total x grupo genético apenas sobre o peso do coração ao abate (g) (tabela 7 e figura 2). Para EV1 houve efeito quadrático dos níveis testados sobre o peso do coração sendo o máximo peso estimado no ponto de 0,81% de metionina+cistina total. Por outro lado, houve efeito linear do tratamento sobre o peso do coração (g) para
carcaça. Resultados semelhantes foram encontrados por Whitaker et al. (2002) que não encontraram efeito de níveis adicionais
de metionina sobre o rendimento de carcaça em frangos de corte.
Interação nível nutricional de metionina+cistina x sexo x grupo genético sobre o rendimento de carcaça de codornas de corte com base na exigência por fase de crescimento
Rendimento de carcaça
CV Significância (Tratamento)
0,72a 5,28 ns 0,66b 5,22 ns 0,73a 4,98 ns 0,65b 5,08 *
Equação de regressão desempenho
Ŷ =1,11-1,06X+0,6X² (R²=0,96) Médias seguidas de letras distintas nas linhas diferem entre si pelo teste Fisher (P<0,05).
Regressão do rendimento de carcaça de fêmeas EV2 de codornas de corte em relação ao nível de metionina+cistina da dieta.
Houve efeito de interação nível de metionina+cistina total x grupo genético apenas sobre o peso do coração ao abate (g) (tabela 7 e figura 2). Para EV1 houve efeito quadrático dos níveis testados sobre o peso do coração sendo o máximo peso estimado
o de 0,81% de metionina+cistina total. Por outro lado, houve efeito linear do tratamento sobre o peso do coração (g) para
EV2 em que o ponto máximo está no menor nível testado (0,63%). Houve efeito linear significativo do nível de metionina+cistina sobre o rendimento do peito, peso e rendimento das asas, peso e rendimento de fígado e rendimento de coração (tabela 8 e figura 3). Resultados na literatura descrevem o aumento no rendimento de peito de frangos de corte com o incremento
40
de metionina sobre o rendimento de carcaça
Interação nível nutricional de metionina+cistina x sexo x grupo genético sobre o rendimento de
Média
0,69A
0,69A
Mínimo desempenho
0,88 (P<0,05).
Regressão do rendimento de carcaça de fêmeas EV2 de codornas de corte em relação
EV2 em que o ponto máximo está no menor nível testado (0,63%). Houve efeito linear significativo do nível de metionina+cistina
rendimento do peito, peso e rendimento das asas, peso e rendimento de fígado e rendimento de coração (tabela 8 e figura 3). Resultados na literatura descrevem o aumento no rendimento de peito de frangos de corte com o incremento

de nível de metionina+cistina total da dieta (Amarante Jr. et al.,2005; Rodrigueiro et al., 2000; Moran et al., 1994), explicado por Sibbald & Wolinetz (1986) pelo fato do requerimento de aminoácidos essenciais, principalmente sulfurosos, para o máximo rendimento de peito estar aciexigência adequada para o crescimento, o que é verificado neste estudo já que a
Tabela 7. Efeito da interação nível de metionina+cistina x grupo genético sobre o peso do coração de codornas de corte ao 42º dia de idade no Experimento 2
Nível de M+C
0,63 0,73 0,83 0,93 1,03 1,13
Médias ANOVA
MC GG
MC x GG CV
EV1 EV2
Médias seguidas de letras minúsculas distintas nas linhas e letras maiúsculas distintas nas colunas diferem entre si pelo teste SNK (p<0,05). PM: ponto de máximo desempenho.
Figura 2. Regressão do peso do coração do grupo genético EV1 (a) e EV2 (b) de codornas de
corte em relação ao nível de metionina+cistina da dieta.
a
ina total da dieta (Amarante Jr. et al.,2005; Rodrigueiro et al., 2000; Moran et al., 1994), explicado por Sibbald & Wolinetz (1986) pelo fato do requerimento de aminoácidos essenciais, principalmente sulfurosos, para o máximo rendimento de peito estar acima da exigência adequada para o crescimento, o que é verificado neste estudo já que a
exigência estimada para a fase final (22 a 42 dias) foi de 0,63% de metionina+cistina enquanto que para se obter máximo rendimento de peito são necessárias dietas contendo nível deste aminoácido acima do valor estimado em decorrência do efeito linear positivo nível de metionina+cistina total sobre o rendimento de peito.
Efeito da interação nível de metionina+cistina x grupo genético sobre o peso do coração de codornas de corte ao 42º dia de idade no Experimento 2
Peso do coração (g) EV1 EV2 Médias2,43a 2,65a 2,54A2,28a 2,48a 2,38AB2,57a 2,34a 2,45AB2,55a 2,46a 2,50A2,21a 2,20a 2,20AB1,88b 2,42a 2,15B2,32 2,43
* * ns ns * *
26,15 16,32 Equação de regressão Ŷ = -1,49+9,91X-6,10X²
Ŷ =2,90-0,54X Médias seguidas de letras minúsculas distintas nas linhas e letras maiúsculas distintas nas colunas diferem entre si pelo teste SNK (p<0,05). PM: ponto de máximo desempenho.
Regressão do peso do coração do grupo genético EV1 (a) e EV2 (b) de codornas de
corte em relação ao nível de metionina+cistina da dieta.
b
41
exigência estimada para a fase final (22 a 42 dias) foi de 0,63% de metionina+cistina enquanto que para se obter máximo rendimento de peito são necessárias dietas
o nível deste aminoácido acima do valor estimado em decorrência do efeito linear positivo nível de metionina+cistina total sobre o rendimento de peito.
Efeito da interação nível de metionina+cistina x grupo genético sobre o peso do
Médias 2,54A
2,38AB 2,45AB 2,50A
2,20AB 2,15B
PM*
0,81 0,63
Médias seguidas de letras minúsculas distintas nas linhas e letras maiúsculas distintas nas colunas diferem
Regressão do peso do coração do grupo genético EV1 (a) e EV2 (b) de codornas de

42
Tabela 8. Efeito do nível nutricional de metionina+cistina sobre características de carcaça de codornas de corte (Experimento 2)
Nível de M+C
Variáveis Rend.
peito (%) Peso asas
(g) Rend. asas
(%) Peso
fígado (g) Rend. fígado
(%)
Rd coração (%)
0,63 40,21 16,93 8,73 6,63 3,35 1,31 0,73 39,79 16,93 8,68 6,34 3,21 1,22 0,83 40,46 16,80 8,46 6,28 3,15 1,24 0,93 39,66 16,62 8,35 6,09 3,04 1,25 1,03 40,14 16,45 8,49 6,18 3,15 1,14 1,13 41,72 15,59 7,92 5,44 2,76 1,10
Médias 40,33 16,52 8,44 6,16 3,11 1,21 ANOVA
MC * * * * * * GG ns * ns ns ns ns
S * * * * * ns MC x S ns ns ns ns ns ns
MC x GG ns ns ns ns ns ns GG x S ns ns ns ns ns ns
MC x GG x S ns ns ns ns ns ns CV 5,68 12,30 10,71 21,83 19,75 18,96
Variável Equação de regressão PM* Rpeito Ŷ = 38,4+2,23X 1,13 Pasas Ŷ = 18,83-2,61X 0,63 Rasas Ŷ = 9,6-1,34X 0,63
Pfígado Ŷ = 7,8-1,90X 0,63 Rfígado Ŷ = 3,9-0,93X 0,63
Rcoração Ŷ = 1,53-0,36X 0,63 Significância: * - significativo a 5%; ns - Não significativo, pelo teste Fisher, PM*: Ponto de máximo desempenho

Figura 3. Regressão do rendimento de peito (a), rendimento de coração (b), peso das asas (c), rendimento das asas (d), peso do fígado (e) e rendimento de fígado (f) de codornas de corte em relação ao nível de metionina+cistina da dieta.
Houve efeito de sexo sobre o peso vivo, peso da carcaça, peso do peito, rendimento de peito, peso das coxas, rendimento de coxas, peso da moela, rendimento de moela, peso das asas, rendimento de asas, peso do fígado e rendimento de fígado (tabela 9 e 10). Resultados semelhantes do efeito do
a
c
e
Regressão do rendimento de peito (a), rendimento de coração (b), peso das asas (c), rendimento das asas (d), peso do fígado (e) e rendimento de fígado (f) de codornas de corte em relação ao nível de metionina+cistina da dieta.
uve efeito de sexo sobre o peso vivo, peso da carcaça, peso do peito, rendimento de peito, peso das coxas, rendimento de coxas, peso da moela, rendimento de moela, peso das asas, rendimento de asas, peso do fígado e rendimento de fígado (tabela 9 e
sultados semelhantes do efeito do
sexo foram já descritos no experimento 1, em todos os casos as fêmeas tenderam a ser mais pesadas que os machos porém apresentam menor rendimento de carcaça em consequência do maior peso de vísceras relacionadas, principalmente, ao aparelho reprodutor feminino.
b
d
f
43
Regressão do rendimento de peito (a), rendimento de coração (b), peso das asas (c), rendimento das asas (d), peso do fígado (e) e rendimento de fígado (f) de codornas de corte em
sexo foram já descritos no experimento 1, em todos os casos as fêmeas tenderam a ser mais pesadas que os machos porém apresentam menor rendimento de carcaça em consequência do maior peso de vísceras
mente, ao aparelho

44
Tabela 9. Características de desempenho e carcaça de codornas de corte alimentadas com diferentes níveis de metionina+cistina (Experimento 2)
Variáveis GG Sexo Níveis de metionina+cistina CV
(%) 0,63 0,73 0,83 0,93 1,03 1,13 Média
Peso vivo (g)
EV1 M 237,8 260,0 261,0 266,9 252,6 244,3
282,0B
9,8
F 312,7 309,8 302,2 317,0 317,9 301,3
EV2 M 269,6 258,8 269,3 281,5 250,8 262,8
290,1A F 303,3 322,0 314,9 320,1 311,6 316,8
Machos 253,7 259,4 265,1 274,2 251,7 253,6 259,6b Fêmeas 308,0 315,9 208,5 318,6 314,7 309,0 312,5a
Peso de carcaça (g)
EV1 M 172,5 179,9 189,3 189,4 185,9 182,0
193,1B
10,0
F 205,5 203,9 203,0 208,5 205,2 192,1
EV2 M 195,2 190,6 201,7 195,0 186,1 193,0
199,4A F 205,4 207,5 200,7 204,7 200,4 212,9
Machos 183,8 185,2 195,5 192,2 186,0 187,5 188,4b Fêmeas 205,5 205,7 201,9 206,6 202,8 202,5 204,2a
Peso do peito
EV1 M 67,9 70,6 74,7 71,2 72,9 73,9
76,5B
12,3
F 84,1 79,4 81,4 81,5 82,5 78,4
EV2 M 78,8 75,1 81,1 80,2 76,7 81,2
82,0A F 82,6 86,6 84,5 83,8 80,6 92,6
Machos 73,4 72,8 77,9 75,7 74,8 77,5 75,4b Fêmeas 83,4 83,0 83,0 82,6 81,6 85,5 83,2a
Rendimento de peito
EV1 M 39,4 39,1 39,4 37,6 39,1 40,7
39,6B
5,6
F 40,9 39,0 40,1 39,1 40,1 40,9
EV2 M 40,3 39,4 40,2 41,1 41,2 42,1
41,1A F 40,3 41,7 42,1 40,9 40,1 43,3
Machos 39,9 39,3 39,8 39,3 40,1 41,4 40,0b Fêmeas 40,6 40,3 41,1 40,0 40,2 42,1 40,7a
Peso das coxas (g)
EV1 M 41,6 42,3 43,7 45,0 43,4 41,3
44,9B
10,1
F 48,3 47,8 47,0 47,5 47,1 43,1
EV2 M 45,7 44,6 46,8 46,4 43,8 44,6
46,4A F 46,5 47,1 47,5 48,4 46,7 48,3
Machos 43,7 43,4 45,3 45,7 43,6 42,9 44,1b Fêmeas 47,4 47,5 47,3 48,0 46,9 45,8 47,1a
Rendimento de coxas (g)
EV1 M 24,1 23,6 23,1 23,8 23,4 22,7
23,3
5,1
F 23,6 23,5 23,1 22,7 23,0 22,5
EV2 M 23,5 23,5 23,2 23,8 23,6 23,1
23,3 F 22,6 22,7 23,7 23,7 23,3 22,7
Machos 23,8 23,5 23,2 23,8 23,5 22,9 23,5a Fêmeas 23,1 23,1 23,4 23,2 23,1 22,6 23,1b
Peso das asas (%)
EV1 M 15,5 15,2 14,8 15,3 15,9 14,3
16,2B
12,3
F 18,5 17,8 17,3 17,1 16,4 16,1
EV2 M 15,7 16,6 17,1 15,8 15,8 15,1
16,9A F 18,1 18,1 18,0 18,2 17,8 16,1
Machos 15,6 15,9 15,9 15,6 15,8 14,7 15,6b Fêmeas 18,3 17,9 17,7 17,7 17,1 16,1 17,5a
Rendimento das asas (g)
EV1 M 8,9 8,5 7,8 8,1 8,6 7,9
8,4
10,7
F 9,0 8,8 8,5 8,3 8,0 8,4
EV2 M 8,1 8,8 8,5 8,1 8,5 7,8
8,5 F 8,9 8,7 9,0 8,9 8,9 7,6
Machos 8,5 8,6 8,2 8,1 8,5 7,8 8,3b Fêmeas 8,9 8,7 8,8 8,6 8,5 8,0 8,6a
Médias seguidas de letras minúsculas distintas nas colunas (sexos), e maiúsculas nas colunas (grupos genéticos) dentro de cada variável diferem entre si pelo teste Fisher (P<0,05).

45
Tabela 10. Características de carcaça (vísceras) de codornas de corte de dois grupos genéticos alimentadas com diferentes níveis de metionina+cistina (Experimento 2)
Variáveis GG Sexo Níveis de metionina+cistina total
Média CV (%) 0,63 0,73 0,83 0,93 1,03 1,13
Peso do fígado (g)
EV1 M 4,3 4,5 5,1 4,4 4,4 4,0
6,0
21,8
F 8,3 7,8 7,6 8,0 7,7 6,3
EV2 M 5,1 4,4 4,5 4,8 4,6 4,4
6,3 F 8,9 8,6 7,9 7,2 8,1 7,1
Machos 4,7 4,5 4,7 4,6 4,5 4,2 4,5b Fêmeas 8,6 8,2 7,8 7,6 7,9 6,7 7,8a
Peso da moela (g)
EV1 M 3,9 3,9 4,0 3,9 4,0 3,5
4,2
15,7
F 4,5 4,6 5,0 4,4 4,5 4,0
EV2 M 4,1 4,0 3,8 3,8 3,8 3,7
4,2 F 4,4 5,1 4,2 4,7 4,4 4,8
Machos 4,0 3,9 3,9 3,9 3,9 3,6 3,9b Fêmeas 4,5 4,9 4,6 4,5 4,5 4,4 4,6a
Médias seguidas de letras minúsculas distintas nas colunas (sexos), e maiúsculas nas colunas (grupos genéticos) dentro de cada variável diferem entre si pelo teste Fisher (P<0,05).
4. CONCLUSÕES
Não foi possível estabelecer a exigência nutricional de metionina+cistina total tanto em sistemas de alimentação que utilizam de dieta única durante a fase de crescimento quanto para sistemas que utilizam dietas por fase do período de crescimento para as características de carcaça.
5. REFERÊNCIAS BIBLIOGRÁFICAS
ALBINO LFT, SILVA SHM, VARGAS JR JGV, ROSTAGNO HS, SILVA MA. Níveis de metionina + cistina para frangos de corte de 1 a 21 e 22 a 42 dias de idade. Revista Brasileira de Zootecnia, v.28, n.3, p.519-525,1999.
AMARANTE JR, V.S., COSTA, F.G.P., BARROS, L.R. et al.. Níveis de metionina+cistina para frangos de corte nos períodos de 22 a 42 e de 43 a 49 dias de idade. Revista Brasileira de Zootecnia, v.34, n.4, p.1195-1201, 2005.
ATÊNCIO, A., ALBINO, L.F.T., ROSTAGNO, H.S. et al.. Exigências de metionina+cistina para frangos de corte machos em diferentes fases de criação. Revista Brasileira de Zootecnia, v.33, n.5, p.1152-1166, 2004.
CORRÊA, G.S.S., SILVA, M.A., CORRÊA, A.B. et al.. Níveis de proteína bruta para codornas de corte durante o período de crescimento. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.60, n.1, p.209-217, 2008.
GARCIA NETO, M., PESTI, G.M., BAKALLI, R.I.. Influence os dietary protein level on the broiler chicken’s response to methionine and betaine supplements. Poutry Science, v.79, p.1478-1484, 2000.
MORAN, E.T.. Response of broiler strains differing in body fat to inadequate methionine: live performance and processing yields. Poultry Science, v.73, n.7, p.1116-1126, 1994.

46
OLIVEIRA, E.G., ALMEIDA, M.I.M., MENDES, A.A. et al. Desempenho produtivo de codornas de ambos os sexos para corte alimentadas com dietas com quatro níveis protéicos. Archives of Veterinary Science, v.7, n.2, p.75-80, 2002.
RODRIGUEIRO, R.J.B., ALBINO, L.F.T., ROSTAGNO, H.S. et al. Exigência de metionina+cistina para frangos de corte na fase de crescimento e acabamento. Revista Brasileira de Zootecnia, v.29, n.2, p.507-517, 2000.
SISTEMA de análises estatísticas e genéticas – SAEG. Versão 9.0. Viçosa, MG: UFV, 1999.
SANTOS, G.G., CORRÊA, G.S.S., CORRÊA, A.B. et al. Interação grupo genético x nível de lisina x sexo sobre características de carcaça de codornas de
corte. In: Simpósio brasileiro de Melhoramento Animal, VII, 2008, São Carlos. Anais... SBMA, 2008. CD-ROM.
SCHEUERMANN, G.N.; MAIER, J.C.; BELLAVER, C. et al. Metionina e lisina no desenvolvimento de frangos de corte. Revista Brasileira de Agrociência, v.1, n.2, p. 75-86, Mai-Ago, 1995.
SIBBALD, I.R.; WOLYNETZ, M.S. Effects of dietary lysine and intake on energy utilization and tissue synthesis by broiler chicks. Poultry Science, v.65, p.98-105, 1986.
WHITAKER, H.M.A.; MENDES, A.A.; GARCIA, E.A. et al. Efeito da suplementação de Metionina sobre o desempenho e a avaliação de carcaças de frangos de corte. Revista Brasileira de Ciência Avícola, v.4, n.1, p. 001-009, Jan-Mar 2002.

47
CAPÍTULO 3
Estudo da sensibilidade dos valores genéticos de peso corporal de codornas de corte à mudança de gradiente nutricional por meio de normas de reação
considerando heterocedasticidade residual
Genetic value sensitivity of meat type quails to changes in the nutritional levels of diets
using reaction norms under residual variance heterogeneity assumption
RESUMO
Avaliou-se a sensibilidade dos valores genéticos do peso de codornas de corte, alimentadas com dietas contendo diferentes níveis de proteína bruta, ao 21º e 35º dias de idade provenientes de duas linhagens (EV1 e EV2) por meio de modelos de regressão aleatória considerando heterogeneidade de variância residual por divisão em quatro classes. Modelos de regressão aleatória, para estudo da interação genótipo-ambiente, que consideram a variância residual heterogênea foram melhores do que os que consideraram homogeneidade de variância do resíduo dado pelo menor valor de BIC (Critério de Informação Bayesiano). Os valores dos componentes de variância residual apresentaram variação considerável de acordo com a classe nas quatro análises feitas correspondentes às combinações de grupo genético e idade. Os coeficientes de regressão aleatória do intercepto (b0) e linear (b1) apresentaram correlação positiva em todas as quatro análises, porém a linhagem EV2 apresentou maior magnitude dos valores de correlação em ambas as idades (0,92 e 0,98 ao 21º e 35º dias de idade, respectivamente). Houve heterogeneidade de variância genética aditiva com a mudança no nível de proteína bruta da dieta para ambos os grupos genéticos em todas as idades avaliadas e, por conseqüência, mudança nos valores de herdabilidade em função dos componentes de variância genética aditiva e residual. As normas de reação do grupo EV1 mostram presença de G x E em ambas as idades com alteração do ordenamento das codornas pelo valor genético do peso nas duas idades em função do nível protéico da dieta. Houve interação valor genético x nível de PB da dieta para EV2 em razão do aumento da dispersão dos valores genéticos. Portanto, a avaliação genética de codornas de corte alimentadas com dietas contendo certo nível de proteína bruta não permite a predição de valores genéticos válidos em outros níveis protéicos.
Palavras-chave: normas de reação, regressão aleatória, componentes de (co)variância, avaliação genética, codornas de corte, heterogeneidade de variância
ABSTRACT
The sensitivity of genetic values for body weight of meat type quails fed diets with different
crude protein levels were evaluated at 21 and 35 days of age of quails from EV1 and EV2
strains using random regression model under heterogeneity of residual variance assumption
with four classes of residual variance. Random regression models under residual variance
heterogeneity of residual variance showed better good of fit than model that disregarded the
heterogeneity of residual variance e using the BIC test (Bayesian Information criterion). The

48
residual component of variance showed a remarked variation according to the class of residual
variance considered. The random regression coefficients for intercept (bo) and linear (b1) were
positively correlated in all the four analyses results, but these correlation coefficients were
larger in the EV2 data analyses in both ages (.92 and .98 at 21 and 35 days of age,
respectively). Additive genetic variance heterogeneity with the change in the environment
gradient for both genetic strains and ages and consequently changes in the estimated
heritability’s that are functions of the additive and residual components of variance The
reaction norms for EV1 strain showed a significant interaction for both ages (21 and 35 days)
with remarked changes in the ranking of body weight breeding values in both ages as a result
of changes in the crude protein level of. An interaction breeding value x crude protein level of
diet for EV2 quails was due to an increase in the dispersion of the genetic values according to
protein level of diet. The breeding values for body weight of quails predicted for specific level of
crude protein in the diet do accurately predict breeding values for other level of crude protein
level of diet.
Keywords: reaction norms, random regression, covariance components, genetic evaluation,
meat type quails, variance heterogeneity
1. INTRODUÇÃO
A proteína é o componente mais caro utilizado na formulação de dietas para codornas de corte, portanto tem grande impacto sobre o custo de programas de seleção. Pode-se diminuir este custo pela utilização de dietas com níveis mais baixos de PB para codornas de corte dentro do programa de seleção do que o adotado para codornas destinadas à produção comercial.
As diferenças na sensibilidade dos indivíduos às mudanças de ambiente resultam na interação genótipo-ambiente (GXE) (FALCONER & MACKAY, 1996). Desta maneira, se existe (GXE) para determinada característica, a diferença entre fenótipos de dois genótipos pode não ser a mesma em dois ambientes diferentes (KOLMODIN, 2003). Sendo assim, progênie de codornas de corte avaliada em dietas com determinado nível de PB não teriam o mesmo desempenho em relação à média da população se alimentados com dietas contendo nível diferente de PB, podendo ocorrer até mesmo mudança na ordem de classificação dos valores genéticos estimados para diferentes níveis protéicos. Neste caso, a predição do valor
genético do animal deve ser feita no nível nutricional semelhante ao que é utilizado em criação comercial.
O estudo dessas diferenças na sensibilidade dos valores genéticos pode ser feito utilizando norma de reação que é conceitualmente definida como a mudança sistemática na expressão média de uma característica fenotípica que ocorre em resposta à mudança sistemática de uma variável ambiental (JONG, 1990), isto é, o fenótipo expresso a partir de um genótipo é função do ambiente. Para a predição da mesma são utilizadas funções de covariância (FC) (Kirkpatrick et al.,1990) que modelam as características longitudinais que se modificam ao longo de um gradiente ambiental com a utilização de modelos de regressão aleatória. Além da modelagem de características aleatórias utilizando funções polinomiais, os modelos de regressão aleatória e FC permitem que seja considerada a heterogeneidade de variância residual em cada classe ou mesmo modelar o resíduo por meio de função polinomial. Considerar variâncias residuais heterogêneas pode ter impacto positivo sobre a partição da variação total resultando, assim, em parâmetros como a

49
herdabilidade, por exemplo, mais acurados. Entretanto, modelos que consideram heterogeneidade residual contêm quantidade maior de parâmetros e, por conseqüência, suas matrizes de incidência são mais complexas o que pode dificultar a convergência na estimação dos componentes de variância, principalmente no caso de banco de dados com número elevado de informações.
Portanto, o objetivo deste estudo foi avaliar a existência de interação genótipo-ambiente em codornas de corte EV1 e EV2 em duas idades (21º e 35º dia de idade) alimentadas com dietas contendo diferentes níveis de proteína bruta por meio de modelos de
regressão aleatória considerando a heterogeneidade de variância residual entre classes.
2. MATERIAL E MÉTODOS
As informações utilizadas neste estudo foram provenientes do Setor de Coturnicultura da Escola de Veterinária da UFMG localizado em Igarapé - MG. Progênies de 100 reprodutores e 200 reprodutrizes de cada linhagem EV1 e EV2 foram alimentadas com dietas contendo 24, 26, 28 e 30% de proteína bruta e 2900kcal de energia metabolizável (tabela 1) e pesadas semanalmente.
Tabela 1. Composição das dietas experimentais para codornas de corte
Ingrediente Nível de Proteína Bruta (%)
24 26 28 30 Milho 46,862 40,640 33,812 27,280 Farelo de soja 42,087 47,461 53,112 58,620 Farelo de trigo 4,000 4,000 4,000 4,000 Óleo de soja 3,736 4,715 5,871 6,941 Calcário 1,086 0,998 1,041 1,018 Fosfato bicálcico 0,924 0,921 0,867 0,839 Sup. Min. e vit.¹ 0,500 0,500 0,500 0,500 Sal comum 0,266 0,236 0,256 0,251 DL-metionina 0,138 0,193 0,229 0,275 L-treonina 0,102 0,136 0,162 0,195 Inerte 0,300 0,200 0,150 0,080 Total 100,00 100,00 100,00 100,00 Composição calculada Proteína bruta (%) 24,00 26,00 28,00 30,00 En. metab. (Kcal/kg) 2900 2900 2900 2900 Cálcio (%) 0,80 0,80 0,80 0,80 Fósforo disp. (%) 0,30 0,30 0,30 0,30 Met.+cistina (%) 0,87 0,99 1,05 1,14 Metionina (%) 0,60 0,68 0,74 0,81 Lisina (%) 1,31 1,48 1,59 1,73 Treonina (%) 1,02 1,13 1,24 1,35
¹ Composição por kilo de produto: Vit. A – 2.000.000UI; vit. D3 – 375.000 UI; vit E – 3.750mg; vit. K3 – 500mg; vit.B1 - 250mg; vit. B2 – 750mg; vit. B6 – 500mg; vit. B12 – 3.750mcg; niacina – 6.250mg; ác. Pantotênico – 2.500mg; biotina – 10mg; ác. fólico – 125mg; colina – 75.000mg; metionina – 250.000mg; selênio – 45mg; iodo – 175mg; ferro – 12.525mg; cobre – 2.500mg; manganês – 19.500mg; zinco – 13.750mg; avilamicina – 15.000mg; narasin – 12.250mg; B.H.T. – 500mg; vit. C – 12.500mg.

50
Os dados foram constituídos de 5.256 pesagens registradas de codornas de corte das linhagens EV1 e EV2 nos 21º e 35º dia de idade. Foram retirados registros de aves sem identificação de sexo ou com peso vivo abaixo ou acima de três desvios-padrão em relação à média. O banco de dados continha a identificação do animal, pai, mãe, sexo,
valor do peso à idade de avaliação e nível de proteína da dieta (24, 26, 28 e 30%). O banco de dados foi separado em quatro partes seguindo a combinação de idades (21 e 35 dias) e linhagem (EV1 e EV2) para realização das análises e está descrito na tabela 2.
Tabela 2. Descrição dos arquivos utilizados nas análises contendo pesos de codornas de corte EV1 e EV2 no 21º e 35º dia de idade
Itens Grupo genético/Idade
EV1 EV2 21 dias 35 dias 21 dias 35 dias
Número de observações 1467 1445 1180 1164 Número de observações (24% PB) 600 601 516 518 Número de observações (26% PB) 182 174 91 91 Número de observações (28% PB) 383 388 190 192 Número de observações (30% PB) 302 282 383 363 Animais incluídos na matriz de parentesco 1729 1709 1386 1371
As análises foram realizadas sob modelo animal unicaracterístico por meio de Regressão Aleatória. A regressão fixa para peso médio dentro de sexo assim como as regressões aleatórias genético-aditivas do animal foram modeladas utilizando-se polinômios ortogonais de Legendre. O mesmo modelo de avaliação foi utilizado nos quatro arquivos e foi definido como:
∑ ∑= =
++=1
0
1
0
)()(k k
hijikhkikjkhij epapby φφ
em que:
hijy é a observação do animal h, do sexo j, alimentado com dieta de nível protéico i;
jkb é o coeficiente regressão fixo do polinômio de legendre k atribuído ao sexo j aninhado ao nível protéico;
hka é o coeficiente de regressão aleatória k de efeito genético aditivo direto atribuído ao animal h;
)( ipkφ é o polinômio de Legendre k
atribuído ao nível protéico i, e
hije é o resíduo associado à observação hijy . Na forma matricial este modelo é especificado como:
eZaXby ++= , em que y é o vetor de observações, b é o vetor de coeficientes de regressão fixos relacionado ao sexo, a é o vetor de coeficientes de regressão aleatória de atribuído ao efeito genético aditivo direto, X e Z sendo os vetores de incidência de efeitos em b e a, contendo polinômios de Legendre relativos ao nível da variável controle adotada (PB) utilizado na observação e e o vetor de resíduos. A comparação de modelos que consideravam a variância residual homogênea e heterogênea foi feita pelo

51
critério de informação bayesiano (BIC), proposto por Schwarz (1978) para cada arquivo formado. Este teste de comparação permite a escolha de modelos não-aninhados mais parcimoniosos, pois aplica fator de “penalização” proporcional ao número de parâmetros do modelo como forma de regularização:
)ln(ln2 npLBIC +−= em que, L refere-se ao valor maximizado da função de verossimilhança do modelo estimado, p ao número de parâmetros e n ao número de observações ou tamanho da amostra. Considerou-se a existência de heterogeneidade de variância residual por divisão dos níveis da variável controle (nível de proteína bruta da dieta) em quatro classes de resíduo (24 a 25% de PB, 26 a 27% de PB, 28 a 29% de PB, 30 a 31% de PB) em todas as análises. Assumindo-se que:
=
0
0
Xb
e
a
y
E
= 2
2
110
100
bbb
bbbG
σσ
σσ
RZGAZyV +⊗= ')()(
⊗=
R
AG
e
aV
0
0, onde
=
2
2
2
2
4
3
2
1
000
000
000
000
e
e
e
e
R
σ
σ
σ
σ
e,
2
0bσ e 2
1bσ são os componentes de variância
dos coeficientes de regressão do polinômio de Legendre relacionados ao intercepto e à inclinação da reta atribuída ao efeito genético aditivo direto, respectivamente,
10bbσ é o componente de covariância entre
estes coeficientes, R é a componente de variância residual heterogêneo dividido em quatro classes e A é a matriz de numeradores dos coeficientes de Wright. No modelo que considera homocedasticidade residual 2
eIR σ= .
Para obtenção da covariância genética aditiva entre dois níveis protéicos i e j utilizou-se a seguinte fórmula:
'jia GZZ
ij=σ , em que:
)]()([ 10 iiZ i φφ=
)]()([ 10 jjZ j φφ=
Também foi feita a estimativa da herdabilidade direta em função do nível de proteína bruta da dieta e classe residual da seguinte maneira:
2'
'2
j
i
eii
ii
dGZZ
GZZh
σ+= , em que representa o
nível protéico da dieta e j refere-se à classe residual a qual este nível faz parte. Os componentes de covariância atribuídos a cada efeito aleatório foram estimados por meio do programa Wombat (2006) que utiliza a Máxima Verossimilhança Restrita (REML). Funções de covariância foram utilizadas para estimar a estrutura de covariância genética aditiva e a herdabilidade direta em função dos níveis de proteína bruta da dieta e classe residual. O critério de convergência adotado, definido pelo valor do quadrado das diferenças entre estimativas, foi 1x10-10.
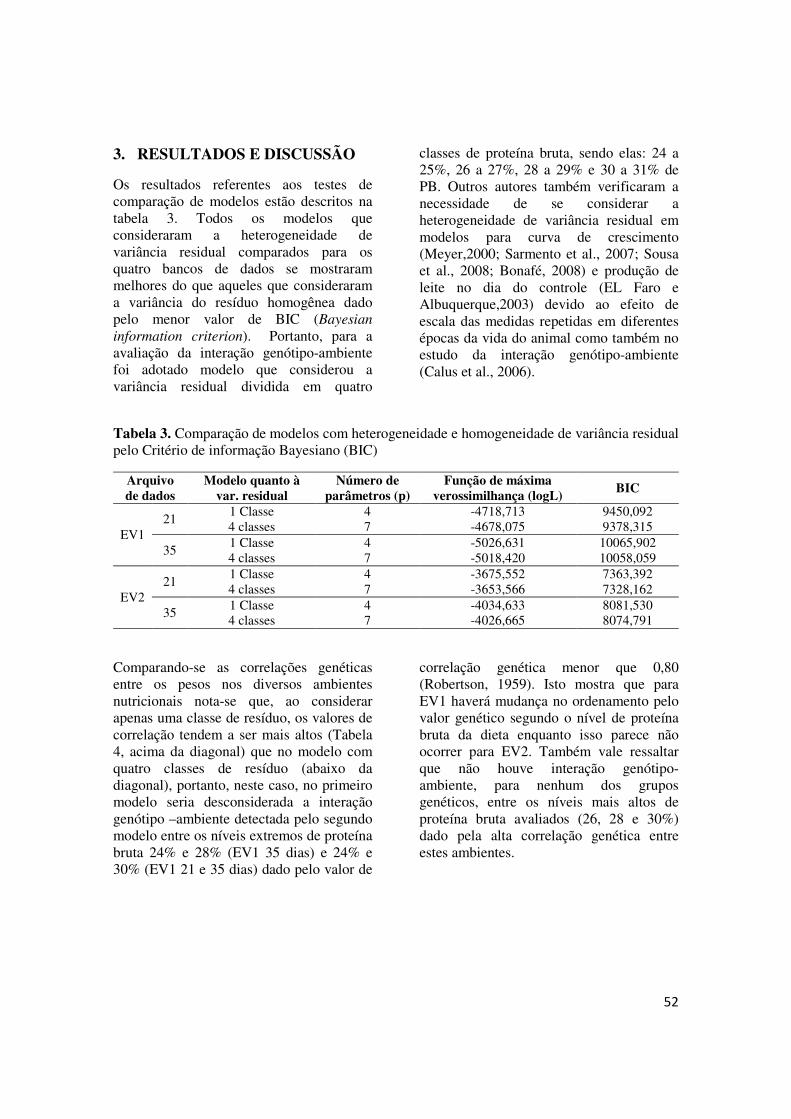
52
3. RESULTADOS E DISCUSSÃO
Os resultados referentes aos testes de comparação de modelos estão descritos na tabela 3. Todos os modelos que consideraram a heterogeneidade de variância residual comparados para os quatro bancos de dados se mostraram melhores do que aqueles que consideraram a variância do resíduo homogênea dado pelo menor valor de BIC (Bayesian
information criterion). Portanto, para a avaliação da interação genótipo-ambiente foi adotado modelo que considerou a variância residual dividida em quatro
classes de proteína bruta, sendo elas: 24 a 25%, 26 a 27%, 28 a 29% e 30 a 31% de PB. Outros autores também verificaram a necessidade de se considerar a heterogeneidade de variância residual em modelos para curva de crescimento (Meyer,2000; Sarmento et al., 2007; Sousa et al., 2008; Bonafé, 2008) e produção de leite no dia do controle (EL Faro e Albuquerque,2003) devido ao efeito de escala das medidas repetidas em diferentes épocas da vida do animal como também no estudo da interação genótipo-ambiente (Calus et al., 2006).
Tabela 3. Comparação de modelos com heterogeneidade e homogeneidade de variância residual pelo Critério de informação Bayesiano (BIC)
Arquivo de dados
Modelo quanto à var. residual
Número de parâmetros (p)
Função de máxima verossimilhança (logL)
BIC
EV1 21
1 Classe 4 -4718,713 9450,092 4 classes 7 -4678,075 9378,315
35 1 Classe 4 -5026,631 10065,902 4 classes 7 -5018,420 10058,059
EV2 21
1 Classe 4 -3675,552 7363,392 4 classes 7 -3653,566 7328,162
35 1 Classe 4 -4034,633 8081,530 4 classes 7 -4026,665 8074,791
Comparando-se as correlações genéticas entre os pesos nos diversos ambientes nutricionais nota-se que, ao considerar apenas uma classe de resíduo, os valores de correlação tendem a ser mais altos (Tabela 4, acima da diagonal) que no modelo com quatro classes de resíduo (abaixo da diagonal), portanto, neste caso, no primeiro modelo seria desconsiderada a interação genótipo –ambiente detectada pelo segundo modelo entre os níveis extremos de proteína bruta 24% e 28% (EV1 35 dias) e 24% e 30% (EV1 21 e 35 dias) dado pelo valor de
correlação genética menor que 0,80 (Robertson, 1959). Isto mostra que para EV1 haverá mudança no ordenamento pelo valor genético segundo o nível de proteína bruta da dieta enquanto isso parece não ocorrer para EV2. Também vale ressaltar que não houve interação genótipo-ambiente, para nenhum dos grupos genéticos, entre os níveis mais altos de proteína bruta avaliados (26, 28 e 30%) dado pela alta correlação genética entre estes ambientes.
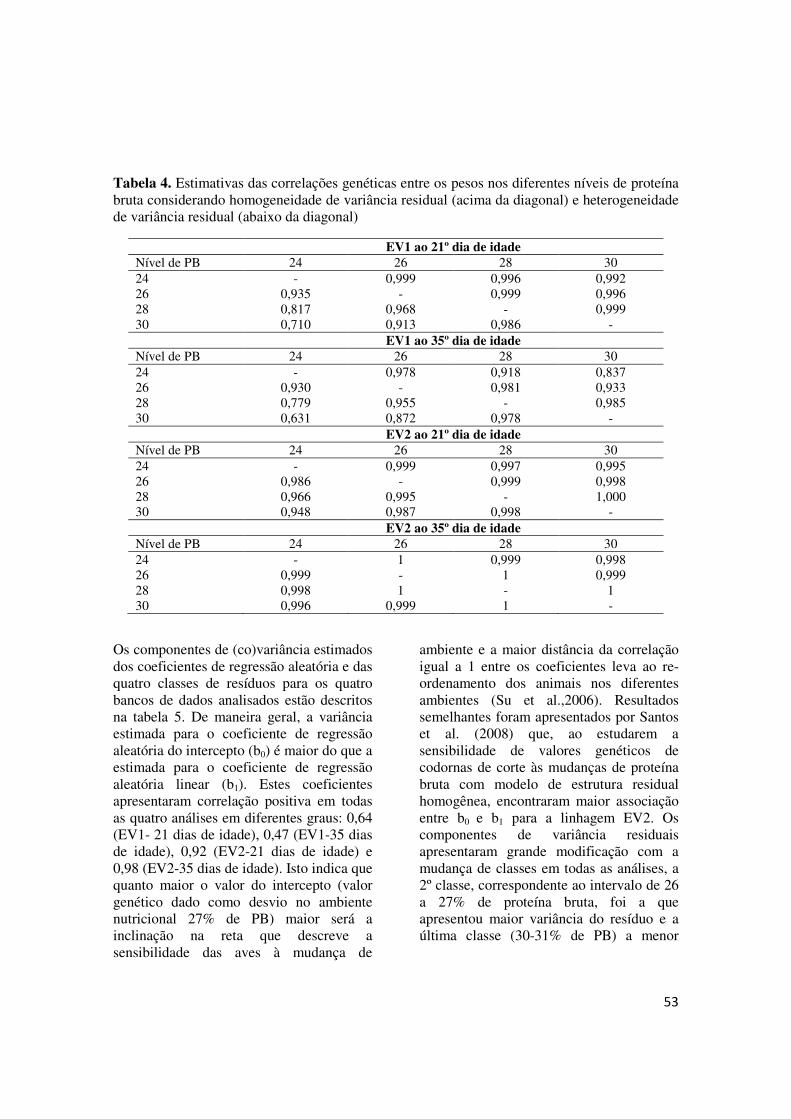
53
Tabela 4. Estimativas das correlações genéticas entre os pesos nos diferentes níveis de proteína bruta considerando homogeneidade de variância residual (acima da diagonal) e heterogeneidade de variância residual (abaixo da diagonal)
EV1 ao 21º dia de idade Nível de PB 24 26 28 30 24 - 0,999 0,996 0,992 26 0,935 - 0,999 0,996 28 0,817 0,968 - 0,999 30 0,710 0,913 0,986 - EV1 ao 35º dia de idade Nível de PB 24 26 28 30 24 - 0,978 0,918 0,837 26 0,930 - 0,981 0,933 28 0,779 0,955 - 0,985 30 0,631 0,872 0,978 - EV2 ao 21º dia de idade Nível de PB 24 26 28 30 24 - 0,999 0,997 0,995 26 0,986 - 0,999 0,998 28 0,966 0,995 - 1,000 30 0,948 0,987 0,998 - EV2 ao 35º dia de idade Nível de PB 24 26 28 30 24 - 1 0,999 0,998 26 0,999 - 1 0,999 28 0,998 1 - 1 30 0,996 0,999 1 -
Os componentes de (co)variância estimados dos coeficientes de regressão aleatória e das quatro classes de resíduos para os quatro bancos de dados analisados estão descritos na tabela 5. De maneira geral, a variância estimada para o coeficiente de regressão aleatória do intercepto (b0) é maior do que a estimada para o coeficiente de regressão aleatória linear (b1). Estes coeficientes apresentaram correlação positiva em todas as quatro análises em diferentes graus: 0,64 (EV1- 21 dias de idade), 0,47 (EV1-35 dias de idade), 0,92 (EV2-21 dias de idade) e 0,98 (EV2-35 dias de idade). Isto indica que quanto maior o valor do intercepto (valor genético dado como desvio no ambiente nutricional 27% de PB) maior será a inclinação na reta que descreve a sensibilidade das aves à mudança de
ambiente e a maior distância da correlação igual a 1 entre os coeficientes leva ao re-ordenamento dos animais nos diferentes ambientes (Su et al.,2006). Resultados semelhantes foram apresentados por Santos et al. (2008) que, ao estudarem a sensibilidade de valores genéticos de codornas de corte às mudanças de proteína bruta com modelo de estrutura residual homogênea, encontraram maior associação entre b0 e b1 para a linhagem EV2. Os componentes de variância residuais apresentaram grande modificação com a mudança de classes em todas as análises, a 2º classe, correspondente ao intervalo de 26 a 27% de proteína bruta, foi a que apresentou maior variância do resíduo e a última classe (30-31% de PB) a menor

54
variância de resíduo indicando que a divisão do resíduo em classes realmente
pode levar à melhor partição da variação do componente residual.
Tabela 5. Componentes de variância atribuídos aos coeficientes de regressão do intercepto (b0) e linear (b1) do efeito genético aditivo direto em função dos níveis protéicos e componentes de variâncias residuais para cada classe estimados para as linhagens EV1 e EV2 no 21º e 35º dia de idade
EV1 Idade
21 35 b0 b1 b0 b1
b0 99,084 17,799 294,71 41,234 b1 7,8899 25,913
Residual CL1 CL2 CL3 CL4 CL1 CL2 CL3 CL4
177,82 353,69 210,20 60,628 264,73 375,32 279,68 135,46 EV2
Idade 21 35
b0 b1 b0 b1 b0 106,77 21,970 279,35 34,958 b1 5,2513 4,5392
Residual CL1 CL2 CL3 CL4 CL1 CL2 CL3 CL4
137,03 332,23 166,26 89,760 299,09 397,32 315,26 169,80
As estruturas de covariância genéticas aditivas diretas em função do nível de PB bruta da dieta estimadas para os dois grupos genéticos em ambas as idades de avaliação estão apresentadas na figura 1. Observa-se que a estrutura de covariância genética aditiva direta apresenta inclinação em direção aos maiores valores de proteína bruta considerados indicando presença de maior variação nos níveis de PB mais altos testados, além disso, verifica-se também maior covariância entre os maiores níveis de proteína bruta dado pela maior inclinação da superfície em direção aos eixos x e y. A forma da superfície de representação da estrutura de (co)variância apresentou-se semelhante nas diferentes idades dentro de cada linhagem, desconsiderando-se a questão da escala. Foi observada mudança nos valores de variância e covariância do efeito genético aditivo em todas as combinações grupo genético e idade com a mudança do nível
protéico da dieta o que mostra a utilidade das funções de covariância para este tipo de estudo.
As estimativas de herdabilidade para os grupos genéticos EV1 e EV2 nas diferentes idades estão apresentadas na figura 2. Houve mudança no valor de herdabilidade para todas as idades/linhagens com a mudança no nível de proteína bruta, em razão da utilização de modelo que considera classes residuais é possível identificar visualmente a divisão das mesmas pela “quebra” no gráfico. Isto ocorre em decorrência do valor diferenciado do componente de variância residual, que entra como denominador no cálculo do valor de h², e, como consequência, os menores valores de herdabilidade, em todas as figuras (a, b, c e d), estão compreendidos no intervalo de 26 a 27% de PB onde o valor de variância residual foi maior. Considerando os valores de herdabilidade

55
dentro de cada intervalo de classe de resíduo, a linha de tendência de descrição deste parâmetro é crescente, isto é, aumenta gradualmente, em maior ou menor escala dependendo do grau de inclinação da estrutura de (co)variância já apresentada, com o aumento do nível protéico. Para ambos os grupos genéticos o valor da herdabilidade tendeu a ser mais alto no 35º dia de idade do que no 21º dia de idade considerando-se as diferentes classes residuais. Portanto, a utilização do valor genético ao 35º dia de idade no critério de seleção é mais indicada com base na maior intensidade de resposta à seleção em
relação à variância fenotípica. Houve também forte variação nos valores de herdabilidade em função dos níveis de proteína da dieta, ou seja, aumento de h² com o aumento de PB da dieta, o que também causaria maior resposta à seleção em relação à variância fenotípica. Santos et al. (2008) chegaram a conclusões semelhantes acerca da herdabilidade porém considerando homogeneidade de variância residual, sendo a mudança em h² consequência apenas na mudança na variância genética aditiva nos diferentes níveis protéicos.
Figura 1. Estrutura de covariância (gr²) do efeito genético aditivo direto em função do nível de proteína bruta da dieta estimada no 21º e 35º dia de idade para os grupos genéticos EV1 e EV2 (a:EV1-21 dias; b:EV1-35dias; c:EV2-21 dias e d:EV2-35 dias).

56
Figura 2. Herdabilidades diretas estimadas dos pesos ao 21º e 35º dias de idade de codornas de corte das linhagens EV1 e EV2 em função do nível de proteína bruta da dieta (a:EV1-21 dias; b:EV1-35dias; c:EV2-21 dias e d:EV2-35 dias).
Foram traçadas as normas de reação de 25 codornas amostrados aleatoriamente em cada uma das quatro análises que são apresentadas na figura 3. Houve heterogeneidade de sensibilidade dos valores genéticos submetidos à mudança no gradiente nutricional em todas as análises combinando idade e grupo genético. A linhagem EV1 apresentou mudança no ordenamento das aves segundo o nível de proteína bruta da dieta tanto ao 21º quanto ao 35º dia de idade, o que já era esperado
em decorrência dos valores de correlação genética e dos coeficientes de regressão aleatória. As normas de reação do grupo genético EV2 apresentam apenas aumento na dispersão dos valores genéticos com o aumento do gradiente protéico dietético não havendo re-ordenamento de aves pelo valor genético. Portanto, detectou-se a presença de interação genótipo-ambiente para ambos os grupos genéticos seja por re-ordenamento dos valores genéticos ou pelo aumento na dispersão dos mesmos.
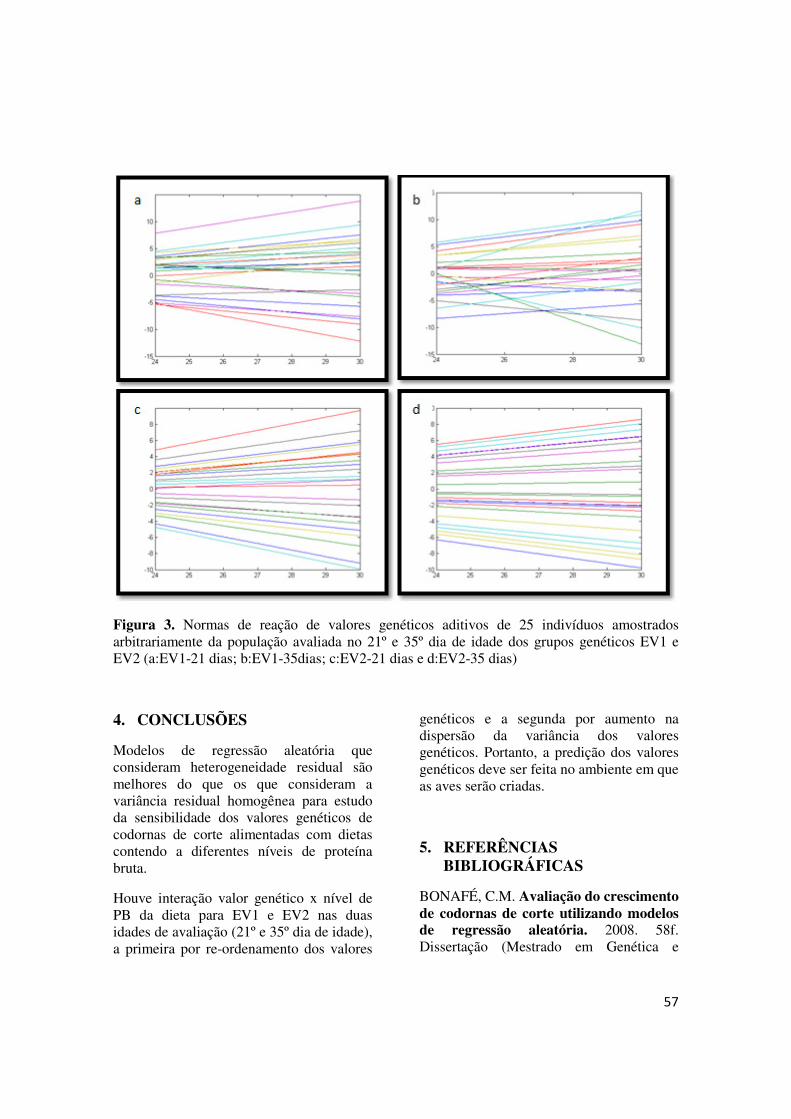
57
Figura 3. Normas de reação de valores genéticos aditivos de 25 indivíduos amostrados arbitrariamente da população avaliada no 21º e 35º dia de idade dos grupos genéticos EV1 e EV2 (a:EV1-21 dias; b:EV1-35dias; c:EV2-21 dias e d:EV2-35 dias)
4. CONCLUSÕES
Modelos de regressão aleatória que consideram heterogeneidade residual são melhores do que os que consideram a variância residual homogênea para estudo da sensibilidade dos valores genéticos de codornas de corte alimentadas com dietas contendo a diferentes níveis de proteína bruta.
Houve interação valor genético x nível de PB da dieta para EV1 e EV2 nas duas idades de avaliação (21º e 35º dia de idade), a primeira por re-ordenamento dos valores
genéticos e a segunda por aumento na dispersão da variância dos valores genéticos. Portanto, a predição dos valores genéticos deve ser feita no ambiente em que as aves serão criadas.
5. REFERÊNCIAS BIBLIOGRÁFICAS
BONAFÉ, C.M. Avaliação do crescimento de codornas de corte utilizando modelos de regressão aleatória. 2008. 58f. Dissertação (Mestrado em Genética e

58
Melhoramento) – Universidade Federal de Viçosa, Viçosa. 2008.
CALUS, M.P.L; POOL, M.H.; VEERKAMP, R.F. Heterogeneous variances and genotype x environment interaction in a random regression test-day model. In: World Congress on Genetics Aplied to Livestock Production, 2006, Belo Horizonte. Proceedings... 8th World Congress on Genetics Aplied to Livestock Production, 2006. CD-ROM.
EL FARO, L.; ALBUQUERQUE, L.G. Utilização de modelos de regressão aleatória para produção de leite no dia do controle, com diferentes estruturas de variâncias residuais. Revista Brasileira de Zootecnia, v.32, p.1104-1113, 2003.
FALCONER, D.S.; MACKAY, T.F.C. Introduction to quantitative genetics. 4.ed. London: Longman. 1996.
JONG, G. Quantitative genetics of reaction norms. Journal of evolutionary biology, v.3, p. 447-468, 1990.
KIRKPATRICK, M.; LOFSVOLD, D.; BULMER, M.; Analysis of the inheritance, selection and volution of growth trajectories. Genetics, v.124, p.979-993, 1990.
KOLMODIN, R. Reaction norms for the study of genotype by environment interaction in animal breeding. Uppsala: Swedish University of Agricultural Sciences, 2003. 35p. Doctoral thesis. Department of Animal Breeding and Genetics, 2003.
MEYER, K. Random regressions to model phenotypic variation in monthly weights of Australian beef cows. Livestock Production Science, v.65, p.19-38, 2000.
MEYER, K. “WOMBAT” - Digging deep for quantitative genetic analyses by restricted maximum likelihood. In: WORLD CONGRESS ON GENETIC APPLIED TO LIVESTOCK PRODUCTION, 8, 2006, Belo Horizonte. Proceedings... Belo Horizonte, 2006, CD-ROM.
ROBERTSON, A.. The sampling variance of the genetic correlation coefficient. Biometrics, v.15, p.469–485, 1959.
SANTOS, G.G., CORRÊA, G.S.S., VALENTE, B.D., SILVA, M.A. et al. Sensibilidade de valores genéticos de codornas de corte em crescimento às modificações de níveis de proteína das dietas. Arquivo Brasileiro de Medicina Veterinária e Zootecnia ,v.60, n.5, p.1188-1196, 2008.
SARMENTO, J.L.R. Modelos de regressão aleatória para avaliação genética da curva de crescimento de ovinos da raça Santa Inês. Tese (Doutorado em genética e melhoramento), Universidade Federal de Viçosa, Viçosa, MG, 2007. 101p.
SCHWARZ, G. Estimeting the dimension of a models. The Annals of Statistics, v. 6, n.2, p.461-464, 1978.
SOUSA, J.E.R., SILVA, M.A., SARMENTO, J.L.R. et al. Homogeneidade e heterogeneidade de variância residual em modelos de regressão aleatória sobre o crescimento de caprinos Anglo-Nubianos. Pesquisa Agropecuária Brasileira, v.43, n.12, p.1725-1732, 2008.
SU, G.; MADSEN P.; LUND M. S. et al. Bayesian analysis of the linear reaction norm model with unknown covariates. J. Anim. Sci., v.84, p.1651-1657, 2006.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























