
UNIVERSIDADE ESTADUAL DE SANTA CRUZ
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA E
BIOLOGIA MOLECULAR
RESPOSTAS FISIOLÓGICAS E EXPRESSÃO GÊNICA DE AQUAPO RINA EM
GENÓTIPOS CLONAIS DE CACAU SUBMETIDOS AO SOMBREAMEN TO E AO
ALAGAMENTO DO SOLO
MÁRCIA CHRISTINA DA SILVA BRANCO
ILHÉUS-BAHIA-BRASIL
Agosto de 2010

ii
MÁRCIA CHRISTINA DA SILVA BRANCO
RESPOSTAS FISIOLÓGICAS E EXPRESSÃO GÊNICA DE AQUAP ORINAS EM GENÓTIPOS CLONAIS DE CACAU SUBMETIDOS AO SOMBREAMEN TO E AO
ALAGAMENTO DO SOLO
ILHÉUS – BAHIA – BRASIL
Agosto de 2010
Dissertação apresentada à Universidade Estadual de Santa Cruz, como parte das exigências para obtenção do título de Mestre em Genética e Biologia Molecular.
Área de concentração: Genética e Biologia Molecular
Orientador: Prof. Dr. Alex- Alan Furtado de Almeida

iii
MÁRCIA CHRISTINA DA SILVA BRANCO
RESPOSTAS FISIOLÓGICAS E EXPRESSÃO GÊNICA DE AQUAP ORINAS EM GENÓTIPOS CLONAIS DE CACAU SUBMETIDOS AO SOMBREAMEN TO E AO
ALAGAMENTO DO SOLO
APROVADA: 31 de Agosto de 2010
Prof. Dr. Raul Rene Melendez Valle
CEPEC/CEPLAC
Prof. Dr. Márcio Gilberto Cardoso Costa
UESC
Prof. Dr. Dario Ahnert
CEPEC/CEPLAC
Prof. Dr. Alex -Alan Furtado de Almeida
UESC-Orientador
Dissertação apresentada à Universidade Estadual de Santa Cruz, como parte das exigências para obtenção do título de Mestre em Genética e Biologia Molecular.

iv
DEDICATÓRIA
À minha mãe Edna Lúcia e minha avó Argentina, pelo amor incondicional, dedico.
Ao meu namorado Felipe, por estar sempre ao meu lado, ofereço.

v
AGRADECIMENTOS
À Universidade Estadual de Santa Cruz e ao Programa de pós-graduação em Genética e Biologia Molecular pela oportunidade de realizar este curso.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela concessão da bolsa.
À United States Department of Agriculture (USDA), na pessoa de Virupax C. Baligar, pelo apoio financeiro ao projeto de pesquisa.
Ao Professor Dr. Alex-Alan Furtado de Almeida pela orientação, pela confiança depositada em mim, pelos conhecimentos transmitidos e incentivos.
Ao Professor Dr. Ronan Xavier Corrêa pela co-orientação e pela oportunidade de iniciação na pesquisa científica.
Ao Professor Dr. Fábio Pinto Gomes pela colaboração e sugestões.
Ao Emerson pela paciência e grande colaboração nas análises estatísticas.
Ao Stênio Carvalho pela indispensável ajuda nas análises de Expressão Gênica.
À Renata Silveira pelo auxílio na coleta dos dados e nas análises de teor de carboidratos.
À Seção de Fisiologia Vegetal (SEFIS) do Centro de Pesquisas do Cacau (CEPEC/CEPLAC) pelo suporte nas análises de teor de carboidratos.
À minha mãe, Edna Lúcia, pelo esforço sem medida para que eu chegasse até aqui.
À minha família por apoiarem e por entenderem a minha ausência ou minha presença “relâmpago”.

vi
Ao meu namorado Felipe, companheiro de todas as horas, por todo amor, dedicação, incentivo e por compreender a minha chatice quando estou em fases críticas.
Aos colegas e amigos do Centro de Biotecnologia e Genética pelos momentos de descontração em meio ao trabalho.
Aos colegas do curso de Pós-graduação em Genética e Biologia Molecular pela alegre convivência.
À Bruna, Vânia, Josie e Fabiana pela amizade, companhia e pelas risadas juntas.
Enfim, a todos que colaboraram de alguma maneira para a realização deste trabalho, meus sinceros agradecimentos.

vii
ÍNDICE
EXTRATO .................................................................................................................. ix
ABSTRACT ............................................................................................................... xii
LISTA DE FIGURAS ................................................................................................. xv
LISTA DE TABELAS .............................................................................................. xxvii
1. INTRODUÇÃO ........................................................................................................ 1
2. REVISÃO DE LITERATURA ................................................................................... 5
2.1. Respostas fisiológicas das plantas à variação da irradiância ........................... 5
2.2. Respostas fisiológicas das plantas ao alagamento do solo .............................. 8
2.3. Efeitos isolados da irradiância e do alagamento do solo no cacaueiro ........... 11
2.4. As aquaporinas ............................................................................................... 15
3. MATERIAIS E MÉTODOS..................................................................................... 21
3.1. Material vegetal e condições de cultivo .......................................................... 21
3.2. Parâmetros micrometeorológicos ................................................................... 22
3.3. Medições de trocas gasosas foliares ............................................................. 22
3.4. Medições de fluorescência da clorofila a ........................................................ 22
3.5. Determinação do teor de clorofila a, b, total e carotenóides ........................... 23
3.6. Extravasamento de eletrólitos ......................................................................... 24
3.7. Parâmetros de crescimento ............................................................................ 24
3.8. Teor de carboidratos ....................................................................................... 25
3.8.1. Obtenção do extrato ................................................................................. 25

viii
3.8.2. Quantificação ........................................................................................... 26
3.9. Expressão gênica de aquaporina ................................................................... 26
3.10. Análise estatística ......................................................................................... 27
4. RESULTADOS ...................................................................................................... 28
4.1. Variáveis microclimáticas ................................................................................ 28
4.2. Observações visuais ....................................................................................... 30
4.3. Trocas gasosas foliares .................................................................................. 31
4.4. Emissão de fluorescência da clorofila a .......................................................... 40
4.5. Teores de pigmentos cloroplastídicos ............................................................. 46
4.6. Extravasamento de eletrólitos ......................................................................... 51
4.7. Biomassa seca ............................................................................................... 52
4.8. Teor de carboidratos ....................................................................................... 67
4.8.1. Açúcares solúveis totais ........................................................................... 68
4.8.2. Amido ....................................................................................................... 71
4.9. Expressão gênica da aquaporina PIP1;3 ........................................................ 74
5. DISCUSSÃO ......................................................................................................... 76
5.1. Parâmetros fisiológicos ................................................................................... 76
5.2. Expressão gênica da aquaporina PIP1;3 em plantas submetidas ao
sombreamento e ao alagamento do solo ............................................................... 90
6. CONCLUSÕES ..................................................................................................... 92
7. REFERÊNCIAS ..................................................................................................... 93

ix
EXTRATO
BRANCO, Márcia Christina da Silva, M. S., Universidade Estadual de Santa Cruz,
Ilhéus, agosto de 2010. Respostas fisiológicas e expressão gênica de
aquaporinas em mudas clonais de cacau submetidas ao sombreamento e
alagamento do solo. Orientador: Alex-Alan Furtado de Almeida. Co-orientador:
Ronan Xavier Corrêa. Colaborador: Fábio Pinto Gomes.
O crescimento e desenvolvimento das plantas são freqüentemente limitados por
estresses múltiplos que ocorrem simultaneamente no ambiente. O estudo das
respostas fisiológicas tem-se mostrado como um eficiente meio de elucidar os
efeitos de variáveis ambientais, como a luz e o alagamento do solo, sobre o
crescimento das plantas. O alagamento é um fator estressante, visto que este tipo
de estresse abiótico promove a diminuição do oxigênio disponível no solo para as
raízes e provoca uma série de danos nas plantas, cujas respostas dependem da
espécie, condições fisiológicas e duração do estresse. Por outro lado, a luz é um dos
fatores ambientais que mais influencia o crescimento e a distribuição das espécies
vegetais em diversos ecossistemas. Espécies arbóreas podem diferir grandemente
na maneira com que respondem às variações da irradiância. O presente trabalho
teve como objetivo principal avaliar as características fisiológicas e a expressão da
aquaporina PIP1;3 em dois genótipos clonais de cacau, contrastantes para a
resistência ao alagamento do solo (TSA-792 - resistente e TSH-774 - susceptível),
submetidos a diferentes níveis de sombreamento e a dois regimes hídricos (alagado
e não-alagado), visando elucidar os possíveis mecanismos de sobrevivência ao
sombreamento e a sua interação com o alagamento. As plantas clonais de ambos os

x
genótipos foram crescidas em vasos plásticos com 25 L de solo sob diferentes níveis
de sombreamento [não-alagado (T0), 25% (T25), 50% (T50) e 75% (T75)] em casa
de vegetação por quatro meses. Logo após, parte das plantas foi submetida ao
alagamento do solo, vedando o fundo dos vasos e adicionando água até 2 cm acima
do nível do solo, por um período de 85 dias, e parte mantida com a umidade do solo
próxima à capacidade de campo. Coletaram-se folhas de todos os tratamentos 24 h
após o alagamento do solo para análise de expressão gênica da aquaporina PIP1;3
por meio da qRT-PCR. Fez-se medições simultâneas de fluorescência da clorofila e
de trocas gasosas em nível foliar aos 0, 8, 16, 24, 32, 40 e 48 dias, entre 8 e 12h,
utilizando um medidor portátil de fotossíntese (Licor LI-6400). No final do
experimento, avaliou-se a estabilidade das membranas celulares por meio da
técnica do extravasamento de eletrólitos e determinou-se o teor de pigmentos
cloroplastídicos. Todas as plantas sobreviveram ao período de alagamento, porém
com sintomas visuais de estresse como clorose foliar e lenticelas hipertrofiadas na
base do caule. Os resultados mostraram efeitos significativos (P< 0,05) para
interação dupla (alagamento x sombreamento), em relação à emissão de
fluorescência da clorofila, somente para as variáveis fluorescências inicial e máxima
e eficiência quântica do fotossistema II. Nas trocas gasosas foliares foi observada
interação tripla (genótipo x alagamento x sombreamento) para as variáveis
fotossíntese líquida por unidade de área foliar, condutância estomática ao vapor de
água e eficiência intrínseca do uso de água. Verificaram-se efeitos isolados do
sombreamento nas trocas gasosas foliares das plantas não alagadas. Não se
observou alterações inter e intragenotípicas no extravasamento de eletrólitos para os
tratamentos de alagamento e sombreamento. As estimativas do teor de clorofila
variaram significativamente (p<0,05) entre os níveis de sombreamento e entre os
tratamentos não-alagado e alagado, sem variação intergenotípica. Para as variáveis
de crescimento, não houve interação tripla significativa (p<0,05), contudo observou-
se a interação entre sombreamento e alagamento do solo foi significativa (p<0,05)
para a área foliar total, biomassa seca da raiz, do caule, da folha, total e área foliar
específica. Variações significativas (p<0,05) no teor de açúcares solúveis totais
foram observadas entre os níveis de sombreamento, regime hídrico, órgão e para a
interação nível de sombreamento x regime hídrico, ao passo que, para o teor de
amido, apenas os fatores genótipo, regime hídrico, órgão e a interação regime

xi
hídrico e órgão foram significativos. Verificou-se que o alagamento do solo foi o
principal fator estressor para a maioria das respostas fisiológicas observadas
durante a interação entre sombreamento e alagamento, cujos ajustes fisiológicos
foram distintos entre os genótipos clonais de cacau avaliados. Essas respostas
foram atenuadas para o genótipo clonal TSA-792, que apresentou crescimento e
desenvolvimento superiores em relação ao genótipo clonal TSH-774. A relação
observada entre os níveis de expressão gênica e os parâmetros de trocas gasosas
foliares permitiu afirmar que a expressão do gene PIP1;3 é modulada por anoxia,
principalmente para o genótipo clonal de cacau TSA-792, considerado resistente ao
alagamento do solo.
Palavras-chave: sombreamento, estresse abiótico, Theobroma cacao, aquaporinas,
RT-PCR.

xii
ABSTRACT
BRANCO, Márcia Christina da Silva, M. S., Universidade Estadual de Santa Cruz,
Ilhéus - BA, August, 2010. Physiological responses and gene expression of
aquaporin in cocoa clonal genotypes submitted to th e shading and soil
flooding. Advisor: Alex-Alan Furtado de Almeida. Co-advisor: Ronan Xavier Corrêa.
Collaborator: Fábio Pinto Gomes.
Growth and development of plants frequently are limited by multiple stresses that
occur simultaneously in the environment. Physiological responses have been used
as an efficient tool to elucidate the effect of environment variables, as the light and
the flooding of soil, on the growth in plants. Flooding is a stressing factor because
promotes the reduction of available oxygen from the soil to the roots and provokes a
series of damages to which depends on the species, plants physiological conditions
and duration of the stress. On the other hand, the light is one of the environment
factors that more influence the growth and the distribution of the plant species in
ecosystems. Woody plants can differ greatly in the way with that they answer to the
variations of the irradiance. The present work had as objective to evaluate the
physiological characteristics and the gene expression of aquaporin PIP1;3 in two
clonal genotypes of cacao, contrasting the resistance to soil flooding (TSA-792 -
resistant and TSH-774 - susceptible), submitted to different levels of shading and
water regime, flooded and not flooded, to elucidate the mechanisms of survival to
shading and its interaction with flooding. The clonal plants were grown in plastic
vases each with 25 liters of soil under different levels of shading [(T0), 25% (T25),
50% (T50) e 75% (T75)], in greenhouse, during 85 days. After, part of the plants
were submitted to flooding adding water up to 2 cm above of the level of the ground,
for a period of 85 days. In the other part of the plants, the soil humidity was

xiii
maintained next to the field capacity. Leaves were harvested of all treatments after
24 h of flooding for the gene expression analysis by Real Time qRT-PCR. Gas
exchange and fluorescence chlorophyll at leaf level measurements were made at 0,
8, 16, 24, 32, 40 and 48 days, between 8h and 12h a.m. using a photosynthesis
portatil measurer (Li-Cor 6400). At the end of the experiment, the plasmatic
membrane stability was assessed by electrolytes leakage technique and the
chloroplastidic pigments contents. All the plants survived the period of flooding
however, with stress visual symptoms like leaf chlorosis and lenticels hypertrophied
on the stem base. The results show significant effects (p<0,05) for the double
interaction (flooding x shading) regarding to fluorescence emission only to the
variables minimal and maximal fluorescence and maximum quantum efficiency of
PSII. For the leaf gas exchange was observed triple interaction (genotype x shading
x flooding) for the net photosynthesis per leaf area unit, stomatic conductance to
water vapour, intrinsic water use efficiency. Isolated effects of shading acting on the
leaf gas exchange were observed. No inter- and intragenotypics changes were
observed in the leakage electrolytes in the flooded and shading treatments (p<0, 05).
There was variation in the contents of chlorophylls (p<0,05) among the shading
levels and between the flooded and not flooded treatments but, there was not
variation intragenotypics. Significant double interaction between shading level and
flooded of the soil was observed for the following variables: total and specific leaf
area; root, stem, leaf and total dry biomass. Difference (p<0,05) among the shading
level, water regime, organ and for the interaction shading level x flooding were
observed in the contents of total soluble sugars while in starch, only the factors
genotype, water regime, organ and the interaction between water regime and organ
was significant (p<0,05). Flooding was the main stressor factor for the most
physiological responses observed during the interaction between shading and
flooded soil whose physiological adjustments were different on the clonal cocoa
genotypes assessed. These responses were attenuated for the clonal genotype TSA-
792 that showed the highest grown and development regarding the clonal genotypes
TSH-774. The relationship observed among the levels of gene expression and leaf
gas exchange allowed concluding that the gene expression of PIP1;3 is modulated
by anoxic stress mainly to the clonal genotype of cocoa TSA-792, assumed resistant
to soil flooding.

xiv
Keywords: shade, abiotic stress, Theobroma cacao, aquaporin, Real Time qRT-PCR.

xv
LISTA DE FIGURAS
Figura 1. Efeitos do sombreamento e de fertilizantes sobre o rendimento
do cacaueiro “Amelonado”em Gana (AKENORAH et al.,
1974).........................................................................................
13
Figura 2. Estrutura das aquaporinas, mostrando as seis α-hélices e as
regiões de duplicação gênica (primeira e segunda metade).
NH2 = região aminoterminal; COOH = região carboxiterminal;
A, C, D = loops hidrofílicos; B e E = loops hidrofóbicos; NPA =
Box asparigina–prolina-alanina; SH = local de inibição por
mercúrio (Hg2+); Ser256 = serina 256, local de fosforilação
por proteínas quinases A (PKA) (VERA-ESTRELLA et al.,
2005)...........................................................................................
17
Figura 3. Variáveis microclimáticas medidas no interior da casa de
vegetação. (A) Temperatura (°C) do ar; (B) Umidade relativa
(%) do ar; (C) Valores médios diários (7h – 17h) de radiação
fotossinteticamente ativa (RFA) em cada nível de
sombreamento (NS, %); (▲) T0; (○) T25; (����)T50 e
(����)T75........................................................................................
29
Figura 4. Morfologia externa de plantas dos genótipos clonais de cacau
TSA-792 e TSH-774 submetidos ao sombreamento por quatro
meses e ao sombreamento + alagamento do solo por um
período de 85 dias. (A) Clorose foliar aos oito dias (TSH-774)

xvi
e (B) aos 85 dias após o alagamento do solo (TSA-792); (C)
Lenticelas hipertrofiadas na base do caule oito dias após o
alagamento do
solo.............................................................................................
30
Figura 5. Condutância estomática ao vapor de água (gs) em nível foliar
em genótipos clonais de cacau submetidos ao sombreamento
por quatro meses e ao sombreamento + alagamento do solo
por um período de 48 dias. (A) TSA-792 não-alagado
(y=1,097 – 0,307T - 0,106NS – 0,0875T2 – 0,0047NS2 –
0,0685TNS / R2 = 0,16); alagado (y=1,173 – 1,314***T –
0,3531NS + 0,385*T2 + 0,2806*NS2 + 0,2123TNS/ R2= 0,86);
(B) TSH-774 não-alagado (y= 1,151 – 0,64*T + 0,695NS –
0,239T2 – 0,173NS2 – 0,182TNS / R2 = 0,36); alagado (y=
0,586 – 0,931***T + 0,552*NS + 0,421**T2 – 0,156NS2 –
0,171TNS / R2 = 0,77..................................................................
32
Figura 6. Taxa fotossintética líquida por unidade de área foliar (A) em
genótipos clonais de cacau submetidos ao sombreamento por
quatro meses e ao sombreamento + alagamento do solo por
um período de 48 dias. (A) TSA-792 não-alagado (y=9,95 –
0,172 T + 0,012NS – 0,04T2 – 0,0001NS2 – 0,0015TNS / R2 =
0,80); alagado (y=12,11 – 2,394***T – 0,05*NS + 0,135*T2 +
0,0006*NS2 + 0,0053 TNS/ R2= 0,97); (B) TSH-774 não-
alagado (y= 1,22 – 0,35*T + 0,087NS – 0,037T2 – 0,175NS2 –
0,195TNS/ R2 = 0,70); alagado (y= 0,70 – 0,842*T + 0,63*NS –
0,22T2 – 0,24NS2 – 0,092TNS / R2 =
0,82)............................................................................................
34
Figura 7. Taxa transpiratória foliar (E) em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 48
dias. (A) TSA-792 não-alagado (y= 1,104 – 0,606*T +

xvii
0,082NS – 0,3108T2 – 0,1765NS2 – 0,166TNS / R2 = 0,14);
alagado (y=1,192 – 1,446***T – 0,374NS + 0,505**T2 +
0,394*NS2 + 0,1365TNS/ R2= 0,86); (B) TSH-774 não-alagado
(y= 1,058 – 0,567*T + 0,4177NS – 0,276T2 – 0,397*NS2 –
0,139TNS/ R2 = 0,29); alagado (y= 0,832 – 1,278***T +
0,4621*NS – 0,552*T2 – 0,223NS2 – 0,020TNS / R2 = 0,
80)...............................................................................................
35
Figura 8. Eficiência intrínseca do uso da água (A/gs) em genótipos
clonais de cacau submetidos ao sombreamento por quatro
meses e ao sombreamento + alagamento do solo por um
período de 48 dias. (A) TSA-792 não-alagado (y= 0,4465 –
0,252T + 0,949**NS – 0,283T2 – 0,6273**NS2 – 0,16TNS / R2
= 0,60); alagado (y=0,8542 – 0,053T – 0,4074NS - 0,1648T2 +
0,19NS2 + 0,147TNS/ R2= 0,16); (B) TSH-774 não alagado (y=
0,906 – 0,1374T - 0,842**NS – 0,289*T2 + 0,5532**NS2 –
0,163TNS/ R2 = 0,53); alagado (y= 0,6112 – 0,382T + 0,635NS
– 0,523*T2 – 0,6904*NS2 – 0,1255TNS / R2 =
0,51)............................................................................................
37
Figura 9. Eficiência instantânea do uso da água (A/E) em genótipos
clonais de cacau submetidos ao sombreamento por quatro
meses e ao sombreamento + alagamento do solo por um
período de 48 dias. (A) TSA-792 não-alagado (y= 0,707 –
0,0617T + 1,024**NS – 0,1764T2 – 0,808**NS2 – 0,0075TNS /
R2 = 0,60); alagado (y=0,355 – 0,0862T – 1,121**NS -
0,1145T2 - 0,83**NS2 - 0,1806TNS/ R2= 0,50); (B) TSH-774
não-alagado (y= 0,3822 + 0,0466*T + 0,751**NS – 0,396*T2 -
0,609**NS2 – 0,1455TNS/ R2 = 0,56); alagado (y= 0,6217 +
0,219T + 0,536NS – 0,455T2 – 0,56NS2 – 0,1717TNS / R2 =
0,46.............................................................................................
38
Figura 10. Fluorescência mínima (Fo) foliar em genótipos clonais de

xviii
cacau submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 48
dias. (A) TSA-792 não-alagado (y= 264,04 – 10,19T +
1,63**NS + 1,017T2 + 0,013*NS2 – 0,039TNS / R2 = 0,42);
alagado (y= 241,37 + 15,40T – 1,80*NS – 1,123 T2 +
0,015*NS2 - 0,006TNS/ R2= 0,50); (B) TSH-774 não-alagado
(y= 235,81 + 7,55T - 0,624NS – 1,114T2 + 0,003NS2 –
0,036TNS/ R2 = 0,50); alagado (y= 251,16 + 28,04T –
2,56**NS – 2,10T2 + 0,027**NS2 – 0,141TNS / R2 = 0,63).........
41
Figura 11. Figura 11. Fluorescência máxima (Fm) em nível foliar de
genótipos clonais de cacau submetidos ao sombreamento
por quatro meses e ao sombreamento + alagamento do solo
por um período de 48 dias. (A) TSA-792 não-alagado (y=
1070,18 + 45,04**T + 1,47NS - 2,94T2 - 0,012NS2 –
0,170TNS / R2 = 0,67); alagado (y= 1106,48 + 38,1T +
2,54NS – 6,41*T2 - 0,041*NS2 + 0,335TNS/ R2= 0,46); (B)
TSH-774 não-alagado (y= 1097,71 + 33,84*T + 2,081*NS –
1,77T2 - 0,0155NS2 – 0,308**TNS/ R2 = 0,54); alagado (y=
1120,7 – 10,16T + 3,99*NS + 1,014T2 – 0,05**NS2 +
0,192TNS / R2 =
0,42)..........................................................................................
42
Figura 12. Rendimento quântico máximo de PS2 (Fv/Fm) em folhas de
genótipos clonais de cacau submetidos ao sombreamento
por quatro meses e ao sombreamento + alagamento do solo
por um período de 48 dias. (A) TSA-792 não-alagado (y=
0,7548 + 0,017T + 0,0016**NS – 0,0015T2 - 0,00001*NS2 –
0,0007TNS / R2 = 0,48); alagado (y= 0,782 – 0,008T +
0,002**NS + 0,00005T2 - 0,00002**NS2 + 0,00006TNS/ R2=
0,65); (B) TSH-774 não-alagado (y= 0,7860 – 0,0006T +
0,00085NS + 0,0006T2 - 0,00001NS2 – 0,00002TNS/ R2 =
0,44); alagado (y= 0,7753 – 0,032T + 0,0033**NS + 0,003T2

xix
– 0,00004**NS2 + 0,0001TNS / R2 = 0,63)............................... 44
Figura 13. Teor de clorofila a em genótipos clonais de cacau submetidos
ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 48 dias. (A)
Tratamento não-alagado; (B) Tratamento alagado; (�)
genótipo TSA-792; (�) genótipo TSH-774. Médias intra e
intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste
de Tukey (p<0,05).......................................................................
46
Figura 14. Teor de clorofila b foliar em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 48
dias. (A) Tratamento não-alagado; (B) Tratamento alagado;
(�) genótipo TSA-792; (�) genótipo TSH-774. Médias intra e
intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste
de Tukey (p<0,05). .....................................................................
47
Figura 15. Teor de clorofila total (a + b) foliar em genótipos clonais de
cacau submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 48
dias. (A) Tratamento não-alagado; (B) Tratamento alagado;
(�) genótipo TSA-792; (�) genótipo TSH-774. Médias intra e
intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste
de Tukey (p<0,05). .....................................................................
48
Figura 16. Teor de carotenóides foliar em plantas de genótipos clonais
de cacau submetidos ao sombreamento por quatro meses e
ao sombreamento + alagamento do solo por um período de 48
dias. (A) Tratamento não-alagado; (B) Tratamento alagado;

xx
(�) genótipo TSA-792; (�) genótipo TSH-774. Médias intra e
intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste
de Tukey (p<0,05). .....................................................................
49
Figura 17. Razão clorofila a/b em folhas de genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 48
dias. (A) Tratamento não-alagado; (B) Tratamento alagado;
(�) genótipo TSA-792; (�) genótipo TSH-774. Médias intra e
intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste
de Tukey (p<0,05). .....................................................................
50
Figura 18. Altura da planta em genótipos clonais de cacau submetidos
ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 85 dias. (●) TSA-792
– não-alagado (y = 79,85 – 0,29*NS / R²= 0.94); (○) TSA-792
– alagado (y = 60,95 + 0,443NS + 0,0078NS2/ R²= 0.93); (▼)
TSH-774 – não-alagado (y = 80,43 + 0,252NS -0.006NS² /
R²= 0.76); (∆) TSH-774- alagado (y = 75,90+ 0,111NS –
0,0026NS² / R²= 0.55). ...........................................................
53
Figura 19. Diâmetro do coleto em plantas em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 18,18 + 0,0825NS –
0,0007NS² / R²= 0,98); (○) TSA-792 – alagado (y = 15,16 +
0,0393NS - 0,0011NS² / R²= 0,42); (▼) TSH-774 – não-
alagado (y = 22.28 + 0,1112NS – 0,0008NS² / R²= 0,90); (∆)
TSH-774 alagado (y = 18,34 + 0,2187*NS - 0,0025*NS² / R²=
0,99). ........................................................................................
54

xxi
Figura 20. Número de folhas em plantas de genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 100,5**e-0,009(NS) / R²=
0,99); (○) TSA-792 – alagado (y = 50,3**e-0,009*(NS) / R²=
0,96); (▼) TSH-774 – não-alagado (y = 90,9e-0,008(NS)/ R²=
0,90); (∆) TSH-774- alagado (y = 24,3 + 26,93*e-0,102*(NS) /
R²= 0,99). .................................................................................
55
Figura 21. Biomassa seca da raiz em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 21,73e-0,018*(NS) / R²=
0,95); (○) TSA-792 – alagado (y = 8,85 - 0,065*NS / R²=
0,95); (▼) TSH-774 – não-alagado (y = 26,80*** e-
0,015***(NS)/ R²= 0,99); (∆) TSH-774- alagado (y = 10,76 –
0,07ºNS / R²=
0,85).......................................................................................
56
Figura 22. Biomassa seca do caule em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 32,94 + 0,5954*NS –
0,0044NS²/ R²= 0,99); (○) TSA-792 – alagado (y = 24,31+
0,0441NS – 0,0025*NS²/ R²= 0,99); (▼) TSH-774 – não-
alagado (y = 42,37 + 0,6568*NS – 0,0044*NS² / R²= 0,99);
(∆) TSH-774- alagado (y = 31,09 + 0,3727NS - 0,0024NS² /
R²= 0,97).................................................................................
57
Figura 23. Biomassa seca da folha em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao

xxii
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 38,70e-0,014*(NS)/ R²=
0,97); (○) TSA-792 – alagado (y = 18,5 – 0,14*NS / R²=
0,97); (▼) TSH-774 – não-alagado (y = 37,30 – 0,32*NS /
R²= 0,88); (∆) TSH-774 alagado (y = 22,63 + 0,5741*NS -
0,0053*NS² / R²= 0,99)...........................................................
58
Figura 24. Biomassa seca total em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 92,4***e-0,015*(NS) /
R²= 0,97); (○) TSA-792 – alagado (y = 50,71+ 0,0564NS –
0,0039*NS² / R²= 0,99); (▼) TSH-774 – não-alagado (y =
106,46 e-0,013*(NS)/ R²= 0,99); (∆) TSH-774- alagado (y =
65,11 + 1,0914*NS + 0,0087NS² / R²=
0,99).......................................................................................
59
Figura 25. Proporção de biomassa seca de raiz, caule e folha em
relação à biomassa seca total em genótipos clonais de
cacau submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. Tratamento não-alagado: 0C, 25C, 50C e 75C;
Tratamento alagado: 0A, 25A, 50A e 75A..............................
60
Figura 26. Área folia total em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 85 dias. (●) TSA-
792 – não-alagado (y = 7272,25**e-0,012*(NS) / R²= 0,98); (○)
TSA-792 – alagado (y = 3073,40 + 0,5971NS – 0,3067NS² /
R²= 0,98); (▼) TSH-774 – não-alagado (y = 6033,8e-
0,01*(NS) / R²= 0,80); (∆) TSH-774 alagado (y = 2509,70 +

xxiii
36,179NS + 0,3207NS² / R²= 0,94)........................................ 61
Figura 27. Área foliar específica em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 2,08*e0,01*(NS) / R²=
0,95); (○) TSA-792 – alagado (y = 3,6 e0,011***(NS)/ R²=
0,99); (▼) TSH-774 – não-alagado (y = 1,53 + 0,0654NS -
0.0005NS²/ R²= 0,81); (∆) TSH-774 alagado (y = 2.72+
0,2054NS - 0,0019*NS² / R²= 0,99)......................................
62
Figura 28. Razão de área foliar em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 75,42 + 0,8279NS -
0,009 NS² / R²= 0,85); (○) TSA-792 – alagado (y = 59,85 +
0,1823NS – 0,0032NS²/ R²= 0,50); (▼) TSH-774 – não-
alagado (y = 54,58 + 0,7597NS – 0,0088NS²/ R²= 0,80); (∆)
TSH-774 - alagado (y = 40,867 + 0,0095NS + 0,0016NS² /
R²= 0,75)............................................................
63
Figura 29. Razão de massa foliar em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 0,41 + 0,002NS -
0,0003 NS² / R²= 0,99); (○) TSA-792 – alagado (y = 0,34 –
0,006NS - 0,0006NS² / R²= 0,99); (▼) TSH-774 – não-
alagado (y = 0,36 + 0,0011NS + 0,00002NS² / R²= 0,15);
(∆) TSH-774 alagado (y = 0,34 - 0,0048NS + 0,00005NS² /
R²=0,97)..................................................................................
64
Figura 30. Razão raiz/parte aérea (R/PA) em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao

xxiv
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 0,32 - 0,003NS –
2,965NS² / R²= 0,92); (○) TSA-792 – alagado (y = 0,20 -
0,0002NS + 3,19NS²/ R²= 0,50); (▼) TSH-774 – não-
alagado (y = 0,33 - 0,0011NS + 7,74NS²/ R²= 0,35); (∆)
TSH-774 alagado (y = 0,21 + 0,0021NS - 2,30NS² / R²=
0,43).......................................................................................
65
Figura 31. Teor de açúcares solúveis totais na raiz de genótipos clonais
de cacau submetidos ao sombreamento por quatro meses e
ao sombreamento + alagamento do solo por um período de
85 dias. (●) TSA-792 – não-alagado (y = 56,50 – 1,20NS +
0,012NS2/ R2= 0,99); (○) TSA-792 – alagado (y = 43,5 +
0,39***NS / R2= 0,99); (▼) TSH-774 – não-alagado (y =
44,77 – 0,47NS + 0,003 NS2/ R2= 0,60); (∆) TSH-774
alagado (y = 54,5 + 0,33 NS – 0,004 NS2/ R2= 0,83).............
67
Figura 32. Teor de açúcares solúveis totais no caule de plantas de
genótipos clonais de cacau submetidos ao sombreamento
por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado
(y = 74,93 e-0,02*(NS)/ R2= 0,96); (○) TSA-792 – alagado (y =
78,25 + 0,81NS – 0,014NS2/ R2= 0,97); (▼) TSH-774 –
não-alagado (y = 48,85e- 0,013(NS)/ R2= 0,76); (∆) TSH-774
alagado (y = 75,03 + 0,243NS – 0,006NS2/ R2= 0,99)...........
68
Figura 33. Teor de açúcares solúveis totais em folhas de plantas de
genótipos clonais de cacau submetidos ao sombreamento
por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado
(y = 74,93 e-0,02*(NS)/ R2= 0,96); (○) TSA-792 – alagado (y =

xxv
78,25 + 0,81NS – 0,014NS2/ R2= 0,97); (▼) TSH-774 –
não-alagado (y = 48,85e- 0,013(NS)/ R2= 0,76); (∆) TSH-774
alagado (y = 75,03 + 0,243NS – 0,006NS2/ R2= 0,83)...........
69
Figura 34. Teor de amido na raiz de genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 12,41 – 0,0914NS +
0,0008 NS2/ R2= 0,95). (∆) TSH-774- alagado (y = 10,55 +
0,125NS – 0,002 NS2/ R2= 0,95)............................................
70
Figura 35. Teor de amido no caule de plantas de genótipos clonais de
cacau submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 55,30 – 1,40 NS +
0,0165 NS2/ R2= 0,80); (○) TSA-792 – alagado (y =
128,70e-0,0055*(NS)/ R2= 0,93); (▼) TSH-774 – não-alagado
(y = 46,00 – 0,796 NS + 0,0094 NS2/ R2= 0,96); (∆) TSH-
774 alagado (y = 91,08 + 1,06 NS – 0,018 NS2/ R2= 0,65)....
71
Figura 36. Teor de amido nas folhas de plantas de genótipos clonais de
cacau submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85
dias. (●) TSA-792 – não-alagado (y = 15,25 + 0,0561NS –
0,016 NS2/ R2= 0,70); (○) TSA-792 – alagado (12,36 – 0,21
NS + 0,0033 NS2/ R2= 0,90); (▼) TSH-774 – não-alagado (y
= 15,09 – 0,094 NS + 0,0007NS2/ R2= 0,85); (∆) TSH-774
alagado (y = 16,77 – 0,053 NS + 0,0013 NS2/ R2= 0,40).......
72
Figura 37 . Perfil da expressão do gene PIP1;3 em folhas de plantas de
dois genótipos clonais de T. cacao crescidas em quatro
níveis de sombreamento (T0, T25, T50 e T75) por um
período de quatro meses e submetidas ao alagamento por 24

xxvi
h. ( ) TSA-792 não-alagado; ( ) TSA-792 alagado; ( )
TSH-774 não-alagado; ( ) TSH-774 alagado. Os valores
expressos são relativos ao tratamento não-alagado no
interior da casa de vegetação (T0). Colunas representam os
valores médios ± erro padrão de três réplicas
biológicas..................................................................................
74

xxvii
LISTA DE TABELAS
Tabela 1. Equações utilizadas na determinação das concentrações de
clorofilas a, b e total e de
carotenóides.............................................................................
24
Tabela 2. Valores de probabilidade da ANOVA para os efeitos de
genótipo (G), nível de sombreamento (NS), regime hídrico
(RH), tempo (T) e da interação entre GxNSxRHxT nas trocas
gasosas foliares. (*) p<0,05; (**) p<0,01; (***) p<0,001; (NS)
não significativo..........................................................................
31
Tabela 3. Valores de probabilidade da ANOVA para os efeitos de
genótipo (G), nível de sombreamento (NS), regime hídrico
(RH), tempo (T) e das interações NSxRH e GxNSxRHxT na
emissão de fluorescência da clorofila a. (*) p<0,05 de
probabilidade; (**) p<0,01; (***) p<0,001; (NS) não
significativo.................................................................................
39
Tabela 4. Valores de probabilidade da ANOVA para os efeitos de
genótipo (G), nível de sombreamento (NS), regime hídrico
(RH), tempo (T) e das interações entre NSxRH e GxNSxRS
no teor de pigmentos cloroplastídicos. (*) p<0,05, (**) p<0,01;
(***) p<0,001; (NS) não significativo. .......................................
45
Tabela 5. Valores de probabilidade da ANOVA para os efeitos de

xxviii
genótipo (G), nível de sombreamento (NS), regime hídrico
(RH), tempo (T) e das interações NSxRH e GxNSxRH no
extravasamento de eletrólitos. (*) p<0,05; (**) p<0,01; (***)
p<0,001; (NS) não significativo.................................................
51
Tabela 6. Valores de probabilidade da ANOVA para os efeitos de
genótipo (G), nível de sombreamento (NS), regime hídrico
(RH) e das interações NSxRH e GxNSxRH no
extravasamento de eletrólitos. (*) p<0,05; (**) p<0,01; (***)
p<0,001; (NS) não significativo.................................................
52
Tabela 7. Valores de probabilidade da ANOVA para os efeitos de
genótipo (G), nível de sombreamento (NS), regime hídrico
(RH), órgão e das interações NSxRH, RHxÓrgão,
NSxRHxÓrgão e GxNSxRHxÓrgão no teor de carboidratos.
AST= açúcares solúveis totais (*) p<0,05; (**) p<0,01; (***)
p<0,001; (NS) não significativo...................................................
72

1
1. INTRODUÇÃO
O cacau (Theobroma cacao L.), espécie lenhosa típica de clima tropical,
diplóide (2n = 20), preferencialmente alógama, perene, nativa da região de floresta
úmida da América. Pertencia anteriormente à família Sterculiaceae, mas foi
reclassificado e inserido na família Malvaceae (ALVERSON et al., 1999). A família
Malvaceae Juss. contém cerca de 75 gêneros, com aproximadamente 1.500
espécies, possui distribuição cosmopolita, predominando nos trópicos
(CRONQUIST, 1981), e compreende plantas de grande importância econômica.
Dentre as 22 espécies do gênero Theobroma, apenas T. cacao e Theobroma
grandiflorum (cupuaçu) são exploradas comercialmente em larga escala. O gênero
Theobroma é considerado como de origem exclusivamente neotropical com
dispersão natural em florestas úmidas, estendendo-se da bacia amazônica até o sul
do México, entre as latitudes 18° N e 15° S (CUATRECASAS, 1964).
Do seu provável centro de origem, nas nascentes do rio Amazonas e Orinoco
(ICCO, 2005), espalhou-se em duas principais direções, o que resultou nos dois
principais grupos raciais: o Criollo, cultivado em Venezuela, Colômbia, Equador,
norte da América Central e México; e o Forastero, no norte do Brasil e nas Guianas.
Um terceiro grupo, denominado Trinitário, também é apresentado por alguns autores
como originário de um cruzamento natural entre Criollo e Forastero (BARTLEY,
2005; ALMEIDA; VALLE, 2007). A maior parte (85%) da produção mundial de cacau
provém do grupo Forastero, sendo este predominante também nas plantações
brasileiras. A espécie T. cacao foi introduzida na Bahia em 1746, procedente do
estado do Pará, passando a ser plantado em vários municípios do sul do estado,
onde esses materiais genéticos receberam a denominação de “cacau comum da
Bahia” (LEAL, 2004).

2
A cacauicultura é uma atividade de grande importância econômica. O cacau é
uma importante commodity agrícola de exportação no mundo. Muitos países estão
envolvidos na sua produção, comercialização e consumo. A produção mundial
média, estimada em 3,3 milhões de toneladas (ICCO, 2007) é produzida por mais de
20 países, nas regiões tropicais da África, Américas do Sul e Central e Ásia.
No Brasil, o sul da Bahia é a principal região produtora de cacau onde,
praticamente, 100 municípios têm suas fontes de receitas baseadas no cacau, o
qual é cultivado em vinte e nove mil propriedades, em área superior a 700 mil
hectares (SOUZA; DIAS, 2001). Com a chegada do fungo Moniliophtora perniciosa,
causador da vassoura-de-bruxa, nas lavouras da Bahia em 1989, houve uma grande
mudança no cenário social da população local com perdas de 100% na produção em
algumas fazendas (PEREIRA et al., 1989). Mesmo adotando os métodos tradicionais
de controle houve pouca eficiência do processo, devido às condições climáticas
favoráveis para a disseminação rápida da doença fazendo com que muitos
agricultores abandonassem suas fazendas, provocando demissões em massa e
migrações de pessoas da zona rural para a zona urbana (TREVIZAN, 1996).
Em 1995, a Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC)
iniciou a distribuição de variedades produtivas e resistentes à vassoura-de-bruxa
com a liberação da variedade híbrida Theobahia. Em 1997 foi iniciado um programa
de liberação de variedades clonais que começou com cinco clones, sendo que, até
a presente data, já foram liberadas e, ou recomendadas cerca de 40 variedades
clonais. Entre essas variedades estão os clones, CCN–51, TSA–792, TSH–774 e
TSH–1188, tradicionais fontes de resistência.
Embora o cacau seja uma espécie tolerante a sombra, os efeitos do
sombreamento, além do decréscimo da irradiância, envolve, dentre outros fatores, o
decréscimo da temperatura e do movimento do ar no interior da plantação, afetando
também a umidade relativa do ar e o conteúdo de água no solo (WESSEL, 1989).
Plantas lenhosas são aeróbias e dependem, sobretudo, de um suprimento de
oxigênio molecular do ambiente para sustentar a respiração e várias outras reações
de oxidação e oxigenação. O alagamento afeta (i) o crescimento da parte aérea; (ii)
a expansão de folhas e entrenós, causando a senescência e abscisão prematura
das folhas; (iii) promove clorose, epinastia e necrose foliar; (iv) inibe a fotossíntese e
o transporte de carboidratos; (v) suprime o metabolismo da raiz; (vi) induz o

3
fechamento estomático e a redução da condutância estomática e (vii) promove a
redução do volume das raízes (KOZLOWSKI, 1997) (viii) altera a anatomia e (ix)
induz mortalidade da planta. Apesar disso, muito pouco se conhece a respeito do
comportamento fisiológico desses clones em condições de alagamento associadas
a diferentes níveis de sombra.
As proteínas denominadas aquaporinas (AQPs) controlam o transporte
específico de água através das membranas celulares em todos os organismos vivos
e estão envolvidas na absorção de água pela raiz (HENZLER et al., 1999;
TYERMAN et al., 2002). Algumas AQPs desenvolvem outras funções como
transporte de amônia e outras substâncias, incluindo CO2 no mesofilo da folha
durante a fotossíntese (UEHLEIN et al., 2003; HANBA et al., 2004; FLEXAS et al.,
2006). Apesar disso, pouco se conhece sobre o papel fisiológico das aquaporinas
sob condições desfavoráveis.
Geralmente, o conceito de estresse ambiental em plantas é associado
diretamente com a produção de injúrias e, portanto com perdas econômicas.
Quando se analisam os efeitos de um determinado fator de estresse numa
determinada espécie, seria razoável observar o aparecimento de injúrias que
implicam em perdas mais ou menos importantes tanto na quantidade como na
qualidade do produto final. Porém, nos últimos anos houve um aumento crescente
na quantidade de trabalhos de pesquisa envolvendo possíveis aspectos benéficos
de quantidades moderadas de estresse sobre diferentes tecidos vegetais
(GIAVENO; OLIVEIRA, 2003).
De acordo com Giaveno e Oliveira (2003), um organismo pode ser
considerado como suscetível a um determinado estresse quando sofrer alterações
graves no seu metabolismo sob a forma de injúrias mais ou menos importantes. Já
Morgan (1990) sugeriu que plantas suscetíveis não possuem a capacidade de
detectar a presença do fator estressor ou de reagir ao mesmo por meio de
alterações no sistema hormonal. Por outro lado, se o organismo não apresenta
sintomas de injúrias, por estresse, deve ser considerado como resistente
(GIAVENO; OLIVEIRA, 2003).
A disponibilidade de radiação associada à saturação hídrica frequentemente
observada em solos hidromórficos podem ser fatores limitantes para o
estabelecimento e adaptação às condições que o ambiente oferece. Neste sentido,

4
o presente trabalho teve como objetivo avaliar as características fisiológicas e a
expressão de gene de aquaporina em dois clones de T. cacao, contrastantes para a
resistência ao alagamento do solo (TSA-792 - resistente e TSH-774 - suscetível),
submetidos a diferentes níveis de sombreamento e a dois regimes hídricos (alagado
e não-alagado), visando elucidar os possíveis mecanismos de sobrevivência ao
sombreamento e a sua interação com a anoxia.

5
2. REVISÃO DE LITERATURA
2.1. Respostas fisiológicas das plantas à variação da irradiância
No interior de florestas úmidas, a luz é o recurso limitante para o crescimento
e desenvolvimento das plantas (WHITMORE, 1996). Em geral, apenas 1-2% da
irradiância que atinge o dossel alcança o interior da floresta e pode chegar a 25%
quando uma clareira é formada por abertura no dossel. Neste caso, a luz aumenta
normalmente por um período de meses ou anos, mas decai gradualmente com o
restabelecimento da vegetação. Espécies arbóreas podem diferir extensivamente na
maneira com que respondem a essas variações de irradiância (THOMPSON et al.,
1992). Assim, o conhecimento sobre os requerimentos de luz de espécies arbóreas
tropicais é importante tanto para a recomposição de florestas como para o
desenvolvimento de culturas de espécies economicamente importantes, além de
esclarecer o padrão ótimo de desenvolvimento inicial de cada espécie.
Quando plântulas são expostas às alterações na irradiância, a maioria delas é
capaz de aclimatar-se à mudança ocorrida. A aclimatação envolve mudanças em
uma série de processos metabólicos em resposta à presença do estresse, que
ocorrem num período curto de tempo. Uma planta aclimatada tem sua tolerância ao
estresse aumentada. Em nível foliar, a aclimatação é expressa através de ajustes
fisiológicos das folhas existentes ou por meio de lançamentos foliares apresentando
fisiologia e estrutura anatômica alterada. (WYKA et al., 2007). A aclimatação pode
ser distinguida da adaptação, que se refere a um nível de resistência determinado
geneticamente e adquirido por processos de seleção ao longo de muitas gerações.
A capacidade de algumas espécies em manter populações de plântulas no
sub-bosque de florestas está associada à capacidade das mesmas de aumentar a

6
sobrevivência sob baixas irradiâncias, ou seja, tolerar o sombreamento
(WALTERS;REICH, 2000). A tolerância à sombra é um conceito ecológico que se
refere à capacidade de uma dada espécie de planta de tolerar baixos níveis de
irradiância. Do ponto de vista fisiológico, é definida como o mínimo de irradiância na
qual a planta pode sobreviver. Contudo, a tolerância à sombra é associada com
características como fisiologia, bioquímica, anatomia e morfologia foliar além
daquelas em nível de planta inteira e, embora muitas espécies possam tolerar
condições de baixa luminosidade, somente uma fração delas é capaz de se
reproduzir sob tal condição. Assim, uma definição biológica de tolerância à sombra
deve considerar todo o ciclo de vida da planta. Além disso, a duração da estação de
crescimento e a ocorrência de múltiplos estresses podem alterar a capacidade da
planta de tolerar baixas irradiâncias. De acordo com Niinemets e Valladares (2006),
a tolerância à sombra é inversamente correlacionada com outros fatores limitantes
como déficit hídrico e alagamento.
O crescimento de plântulas arbóreas e a capacidade de adaptar-se às
alterações em determinados ambientes depende de interações entre as
características morfofisiológicas. As características morfológicas incluem área foliar,
espessura e arranjo das folhas, produção de biomassa da raiz, do caule e da folha e
os parâmetros derivados dessas variáveis como área foliar específica, razão
raiz/parte aérea e razão de área e de massa foliar. Para as características
fisiológicas freqüentemente avalia-se a taxa de assimilação de CO2, condutância
estomática, transpiração e a eficiência do uso da água.
Diversas variáveis de crescimento como altura e diâmetro do caule, utilizadas
com maior freqüência, têm sido usadas na avaliação da resposta das plantas à
diferentes níveis de irradiância. Moraes Neto et al. (2000) estudando o
comportamento de espécies arbóreas que ocorrem na mata Atlântica verificaram
diferenças significativas na altura e no diâmetro do caule entre as espécies
estudadas em função dos níveis de irradiância. A produção de biomassa seca, a
área foliar total e específica e as relações entre a biomassa seca da parte aérea e
radicular são também muito utilizadas na avaliação do crescimento de espécies
arbóreas submetidas à variações na irradiância (FARIAS et al., 1997). Verifica-se
uma tendência ao direcionamento da biomassa seca para as raízes em plantas
submetidas a pleno sol. No geral, plântulas crescidas em baixa irradiância alocam

7
biomassa preferencialmente para o caule e folhas do que para as raízes (KITAJIMA,
1994). Scalon et al., (2001) constataram maior distribuição de biomassa na parte
aérea de plantas de pitangueira (Eugenia uniflora L.) submetidas a pleno sol.
Também verificaram que as plantas submetidas a pleno sol apresentaram maior
altura, diâmetro do caule e maior área foliar.
Ao se estudar a capacidade adaptativa de uma determinada espécie a certos
níveis de luminosidade, as variáveis de trocas gasosas foliares são muito úteis. A
taxa fotossintética exerce um importante papel, pois as diferenças de tolerância à
sombra entre espécies estão relacionadas, principalmente, com a capacidade de
adaptação de seu aparelho fotossintético à intensidade de radiação do ambiente
(KRAMER; KOZLOWSKI, 1979). Folhas de plantas aclimatadas ou adaptadas à
altos níveis de irradiância geralmente apresentam taxas fotossintéticas maiores do
que folhas aclimatadas ou adaptadas à baixos níveis de irradiância (BJORKMAN et
al., 1972; BOARDMAN, 1977; BJORKMAN, 1981) Análises feitas por Farquhar e
Sharkey (1982) indicaram que as taxas fotossintéticas superiores apresentadas por
folhas à pleno sol são resultado da alta condutância estomática e da capacidade
fotossintética do mesofilo que apresenta grande quantidade de enzimas
fotossintéticas, inclusive Rubisco.
A adaptação/aclimatação do aparelho fotossintético envolve reduções nos
níveis de enzimas que participam do ciclo de redução do carbono, nos componentes
da cadeia de transporte de elétrons e nas proteínas e pigmentos associados com a
captação da energia luminosa. Geralmente, diferentes graus de luminosidade
causam alterações fisiológicas e morfológicas na planta e o grau de adaptação é
ditado por características genéticas da planta em interação com o seu meio
ambiente (MORAES NETO et al., 2000).
A intensidade de luz também afeta a estrutura e a atividade dos cloroplastos.
Os cloroplastos de plantas de sombra contêm mais tilacóides do que plantas de sol.
Nos tilacóides de plantas sombreadas observa-se um decréscimo na razão clorofila
a/b e na razão proteínas solúveis/clorofila (ANDERSON; OSMOND, 1987). O baixo
conteúdo de proteínas solúveis em plantas sombreadas é refletido também na baixa
atividade da Rubisco (BJORKMAN, 1968). Observações paralelas da atividade de
carboxilase da Rubisco e da taxa de assimilação de CO2 sugerem que a baixa
quantidade da enzima é parcialmente responsável pelo baixo ponto de saturação

8
luminosa observado em plantas sombreadas (BOARDMAN, 1977). Inversamente, as
altas taxas fotossintéticas em plantas de sol devem-se a altas quantidades de
Rubisco e outras enzimas fotossintéticas.
2.2. Respostas fisiológicas das plantas ao alagamen to do solo
O alagamento, um tipo de estresse abiótico, é comum em solos
hidromórficos, em solos de pouca drenagem, em regiões com elevado índice
pluviométrico e em fazendas com larga escala de irrigação (PEZESHKI, 1994;
KOZLOWSKI, 1997). Em condições normais de drenagem, o solo contém poros
preenchidos com ar que apresenta conteúdo de oxigênio similar ao da atmosfera
(cerca de 20%) (PEZESHKI, 1994). A água em excesso substitui o ar presente
nesses poros, restringindo o fluxo de oxigênio no interior do solo, gerando uma
condição de hipoxia (deficiência de oxigênio) ou a anoxia (total ausência de
oxigênio) (PENG et al., 2005). O solo tornando-se hipóxico ou anóxico leva as
raízes a uma situação de estresse fazendo com que as plantas respondam com
maior ou menor eficiência, permitindo a distinção entre espécies tolerantes e
intolerantes à baixa disponibilidade de O2 (ARMSTRONG et al., 1994). A hipoxia é a
forma mais comum de estresse em solos e ocorre durante alagamento a curto prazo
quando as raízes são submersas, porém, a parte aérea permanece em contato com
a atmosfera. A anoxia ocorre em solos que sofrem alagamento por longos períodos
e em plantas completamente submersas em água (PONNAMPERUMA, 1972).
A maioria das respostas de crescimento em raízes hipóxicas e parte aérea
ocorrem em resposta ao etileno. O etileno é sintetizado via rota dependente de O2 e
sua concentração endógena é determinada predominantemente pela taxa de
produção e difusão, ambos afetados pelo alagamento. Os genes que codificam a
ácido1-carboxílico-1-aminociclopropano (ACC) sintase (ACS) e a ACC oxidase
(ACO) são induzidos pelo alagamento (COHEN; KENDE, 1987) enquanto a difusão
do etileno para o ambiente externo é dificultada. O precussor do etileno é o ACC o
qual é sintetizado normalmente nas raízes (BRADFORD; YANG, 1980). Como a
conversão do ACC em etileno necessita de O2, a reação é bloqueada nas células
radiculares em anaerobiose e o ACC é translocado para a região aeróbica da raiz

9
ou para a parte aérea. Como conseqüência, a quantidade de ACC na parte aérea
aumenta.
O etileno inicia e regula muitas respostas morfológicas, químicas e
moleculares, que permitem a planta evitar a anaerobiose nas raízes sob
alagamento, como o desenvolvimento de aerênquima. Aerênquimas são tecidos
com grandes espaços intercelulares que fornecem uma rota de baixa resistência às
trocas gasosas entre a raiz anaeróbica e a parte aérea aeróbica (JACKSON;
ARMSTRONG, 1999). A síntese de etileno é reforçada em raízes sob hipoxia
coincidindo com o desenvolvimento de aerênquima (VISSER, 2003).
O O2 participa da respiração aeróbica como aceptor final de elétrons na
fosforilação oxidativa, na geração de ATP e regenaração de NAD+, e em várias
rotas biossintéticas cruciais como a síntese de clorofila, ácidos graxos e esteróis
(DENNIS et al., 2000). As conseqüências da deficiência de O2 variam desde
aumento da biomassa do caule em relação à raiz até perda das plântulas devido ao
alagamento sazonal (KENNEDDY et al., 1992).
A redução da atividade fotossintética (iniciando a partir de um ou dois dias de
exposição das raízes a esse estresse) pode ocorrer devido à baixa concentração
interna de CO2 nas folhas (ASHRAFF, 2003). Para períodos relativamente longos
de inundação, as limitações da fotossíntese que não ocorrem por razões
estomáticas podem ser atribuídas a diversos fatores como: alteração nas enzimas
do ciclo de Calvin; degradação dos pigmentos fotossintéticos (PEZHESKI, 1994);
potencial hídrico reduzido; deterioração no transporte de fotoassimilados devido à
baixa atividade de absorção (HUANG et al.,1994; LIAO; LIN, 1994). Alguns autores
sugerem que o fechamento estomático pode estar associado com um decréscimo
da condutividade hidráulica das raízes (DAVIES; FLORE, 1986; KOZLOWSKI,
1997), como também na transmissão de sinais hormonais das raízes para a parte
aérea. Dentre esses hormônios estão o ABA e a citocinina (ELSE et al., 1996).
A troca da respiração aeróbica pela anaeróbica afeta severamente a
disponibilidade de energia, requerendo grande quantidade da reserva de
carboidratos existentes na planta, gerando produtos finais potencialmente tóxicos e
causando acidez citoplasmática. Os carboidratos são fornecidos pela respiração
anaeróbica a partir de fontes de reserva ou por meio da translocação dos
fotoassimilados provenientes das folhas ativas fotossinteticamente. A translocação é

10
sensível às condições de anaerobiose e dependendo da planta, não consegue
sustentar a alta demanda de carboidratos exigidos pelas raízes durante o
alagamento, com exceção das espécies tolerantes que possuem aerênquimas.
Como conseqüência, a capacidade em armazenar grande quantidade de
carboidratos em raízes ou rizomas pode conferir à espécie tolerância prolongada à
condição de anoxia (SILVA, 2006).
O fechamento estomático aumenta a possibilidade de sobrevivência da
plântula, pois reduz a demanda por água e nutrientes para a raiz e a absorção de
nutrientes tóxicos (JACKSON, 1994). Porém, em algumas espécies adaptadas a
solos alagados o fechamento estomático não ocorre (JACKSON; DREW, 1984),
como é o caso do Pinus silvestris (ZAERR, 1983) onde não se observaram
diferenças significativas na taxa de fotossíntese entre as plantas alagadas e
controle.
A fase fotoquímica nas folhas também é afetada pelo alagamento do solo.
Nesse caso, ocorre uma redução das reações a nível cloroplastídico e
conseqüentemente uma redução no fornecimento de energia gerada pelo
fotossistema II (ISHIDA, 2004).
O metabolismo anaeróbico leva a geração de energia sem o oxigênio como
aceptor final de elétrons. A nível molecular, a deficiência de O2 induz a produção de
um conjunto de aproximadamente 20 polipeptídios, as proteínas do estresse
anaeróbico (ASP’s – anaerobic stress proteins) sendo, a maioria, enzimas
envolvidas na rota de fermentação (BLOM; VOESENEK, 1996). Os principais
processos envolvidos no metabolismo anaeróbico - fermentação lática e etanólica -
podem exceder o mesmo levando a um acúmulo de diversos produtos de
fermentação tais como etanol, lactato, alanina (derivados do piruvato, produto final
da glicólise), Gaba (ácido alfa-aminobutírico), succinato e malato (RICARD et al.,
1994; DREW, 1991; ). Nesse caso, a respiração mitocondrial é interrompida devido
à ausência de um aceptor final de elétrons. Como resultado a produção de ATP
diminui de 36 para 2 moles por mole de glicose metabolizada (MORARD;
SILVESTRE, 1996), afetando diversos aspectos do metabolismo celular (FUKAO;
BALEY-SERRES, 2004)
Quando organismos aeróbicos são privados de O2, a fosforilação oxidativa na
mitocôndria é inibida, enquanto glicólise e fermentação são promovidas. O acúmulo

11
de piruvato resulta de um declínio na taxa de oxidação e aumento na taxa de
síntese, através da descarboxilação do ácido oxaloacético (AOA) e aumento da
atividade da ácido-oxaloacetato descarboxilase (AOA-DC) sob hipoxia (STREETER;
THOMPSON, 1972).
Abordagens recentes utilizando a técnica de macro e microarranjo e análises
proteômicas ajudam no entendimento da função de genes e proteínas envolvidas na
adaptação ao alagamento e a anaerobiose (DOLFERUS et al., 2003). Com o auxílio
das técnicas de genética molecular, Toojinda et al. (2003), caracterizaram a região
no cromossomo 9 de Oriza sativa como a responsável pela tolerância ao
alagamento, retardamento da senescência e insensibilidade ao etileno.
As espécies não-tolerantes desenvolvem sintomas característicos, os quais
resultam de distúrbios causados pela hipoxia ou anoxia nas raízes. Os mais comuns
são abscisão de folhas, flores e frutos, clorose, redução no comprimento das raízes,
redução no crescimento em altura, inibição da formação dos primórdios foliares,
redução na expansão, podendo tais efeitos levar a morte do vegetal (MEDRI et al.,
1998; PEREIRA et al, 2000; ARMSTRONG et al., 1994; KOZLOWSKI, 1997).
Espécies tolerantes ao alagamento normalmente formam aerênquimas, lenticelas e
raízes adventícias (KOZLOWSKI, 1997). As lenticelas participam da captação e
difusão de O2 para o sistema radicular e na liberação de produtos voláteis
potencialmente tóxicos como etanol, acetaldeído e etileno (MEDRI, 1998).
Alemanno et al (2003), verificaram em condições “in vitro” que o T. cacao em
resposta a uma condição de estresse, pode sintetizar novos compostos fenólicos,
como forma de proteção dos seus tecidos contra a detoxicação.
2.3. Efeitos isolados da irradiância e do alagament o do solo no cacaueiro
O cacau, como espécie nativa da Amazônia, é tolerante ao sombreamento.
Nos sistemas de produção de cacau, o sombreamento destaca-se como uma das
mais importantes técnicas culturais capazes de manter condições ecológicas
indispensáveis ao crescimento, desenvolvimento e produção das plantas
(CUNNINGHAM; BURRIDGE, 1960). Diante dessa característica, a cultura é
normalmente associada a outras espécies, cuja finalidade é a de sombreá-la desde
a fase de implantação até a fase produtiva.

12
Nos trópicos úmidos (precipitação anual excede 2000 mm), a exemplo do sul
da Bahia, Brasil, o cacau é cultivado em sistema de ‘Cabruca’, onde o cultivo é
realizado sob sombra de Mata Atlântica raleada (LOBÃO et al., 2007), sob sombra
de árvores como Erythrina fusca ou em intercultivo com palmáceas para diversificar
a produção e para explorar os benefícios da interação árvore-cultivo. As árvores têm
várias funções incluindo sombreamento da cultura para evitar a evapotranspiração,
controle da erosão e ciclagem de nutrientes (BEER, 1997; YOUNG, 1997;
HARTEMINK, 2005). Dependendo da espécie, as árvores utilizadas podem ser
regularmente podadas para melhoria do solo (ex: Erytrina spp.) ou deixá-las crescer
para produção de lenha e madeira, como Cordia spp sendo que vários sistemas
agroflorestais utilizam mais do que três espécies (MÜLLER; GAMA RODRIGUES,
2007). O sombreamento deve permitir a passagem de 50 a 60% de luz, para
cacaueiros adultos (GRAMACHO et al. 1992; ALVIM, 1977). De acordo com
Enríquez (1986), deve-se sombrear o cacaueiro durante os três primeiros anos e,
após, mantê-lo em torno de 50% de sombra, garantindo, assim, um bom
desenvolvimento das plantas.
Apesar de o cacaueiro ser planta típica de sombra, as vantagens do uso do
sombreamento não estão propriamente relacionadas com os níveis de intensidade
luminosa mais favoráveis ao crescimento e produção do cacaueiro (ALVIM, 1977).
Seus principais benefícios consistem na (i) conservação do solo e dos recursos
hídricos, (ii) na maior longevidade das plantações, (iii) na maior estabilidade da
produção, (iv) na menor incidência de pragas e doenças, na redução da velocidade
do vento, importante para plantas jovens de cacau que são sensíveis à dessecação
(LEITE, 1981); (v) na redução de plantas invasoras, (vi) na diversificação da
exploração agrícola com o uso de plantas de interesse econômico e (vii) na redução
do desequilíbrio nutricional (BEER, 1987).
O cacau, em condições de sombreamento, é influenciado por diversos fatores
ambientais, pois, além da redução da intensidade de radiação luminosa, o
sombreamento promove variações na temperatura, velocidade do vento e umidade
relativa do ar e do solo (DIAS, 2001; ANIM-KWAPONG, 2003). No entanto, diversos
estudos têm demonstrado que a produção aumenta com o aumento da irradiância
na área de cultivo (ZUIDEMA et al., 2005). Vários autores (AKENORAH et al., 1974;
CABALA-ROSAND et al., 1972; BYRNE, 1972), estudando o rendimento de

13
cacaueiros fertilizados e não sombreados, relataram que o rendimento mais alto foi
obtido com cacaueiros fertilizados e sem sombreamento. Entretanto, os tratamentos
sem sombra apresentaram maior variabilidade na produção e a mesma não era
mantida por mais de dez anos (Figura 1 ).
Figura 1. Efeitos do sombreamento e de fertilizantes sobre o rendimento do
cacaueiro “Amelonado” em Gana (AKENORAH et al, 1974).
Doenças como a podridão-parda (Phytphora palmivora) do cacaueiro são
favorecidas pelo aumento da umidade devido ao aumento no sombreamento
(DAKWA, 1980; SMITH, 1979). Meléndez (1993) comparando as condições
microclimáticas, a liberação de esporos e a incidência da doença (Moniliophthora
roreri) em cacaueiros crescendo sob o sombreamento de Erythrina poeppigiana,
Gliricidia sepium e Inga edulis não encontrou diferenças significativas, exceto para
liberação de esporos que foi maior sob o sombreamento de E. poeppigiana.
Somadas à incidência de pragas e doenças, a produção e a competição por água
são questões fundamentais na controvérsia sobre o uso de árvores para fins de
sombreamento na cultura do cacau.
O sol, assim como a sombra, traz vantagens e desvantagens para o cultivo do
cacau. Os problemas causados a uma plantação a pleno sol são superexposição e
desequilíbrio nutricional. Tal plantação é também afetada pelo estresse hídrico,

14
insetos e doenças (CUBA, 1972). Em solos férteis, livre de insetos, pragas e
doenças, com disponibilidade hídrica e nutrientes, o cacau apresenta maior
produção à pleno sol do que sob sombra (ALVIM, 1958). Um exemplo é na região
de Mucugê (semi-árido baiano), onde o cacau recentemente está sendo cultivado a
pleno sol com sistema de quimirrigação por gotejamento e com produção elevada
(ALMEIDA; VALLE, 2007).
A capacidade de produção das plantas é função direta da área foliar. Altas
produções não podem ser esperadas de plantas pequenas com reduzida área foliar.
O uso da irradiância deve ser o mais eficiente possível e para isso, boa parte das
folhas deve receber luz solar. A fotossíntese das folhas localizadas no interior do
dossel é reduzida, o que faz com que algumas folhas funcionem mais como dreno
do que como fonte de fotoassimilados (KHAN et al., 1987; DIAS, 2001) reduzindo
também a produção de sementes (ZUIDEMA et al., 2005) .No entanto, sob cultivo a
pleno sol, sua longevidade não ultrapassa os seis meses. Muller e Biehl (1993)
encontraram que a longevidade média das folhas de cacaueiro, medida em dias
após o lançamento, de plântulas desenvolvidas em baixa e alta luminosidade foi de
450 e 250 dias, respectivamente.
Augusto (1997), estudando trocas gasosas em cacaueiros submetidos à
irrigação complementar no Espírito Santo, verificou que cacaueiros irrigados
apresentaram valores crescentes de taxa fotossintética líquida, em intensidade
luminosa de até 600 µmol fótons m-2 s-1, com tendência da fotossíntese se
estabilizar em valores próximos de 5,6 µmol CO2 m-2 s-1. Por outro lado, cacaueiros
não irrigados apresentam valores crescentes de fotossíntese, até intensidade
luminosa de 250 µmol fótons m-2 s-1, estabilizando em torno de 3,4 µmol CO2 m-2 s-1.
Ao considerar um dia típico de sol de verão das regiões tropicais onde se cultiva
cacau, pode-se atingir uma luminosidade em torno de 2000 µmol fótons m-2 s-1 e o
cacaueiro, por ser uma planta C3, parece apresentar seu ponto de saturação de luz
em torno de 400 µmol fótons m-2 s-1 (MIELKE et al., 2005b).
O cacau é uma espécie sensível à escassez de água e também ao
alagamento do solo. Por isso, necessita de solos com uma boa drenagem de água.
O alagamento é uma importante barreira para o crescimento inicial e o
estabelecimento do cacaueiro em locais sujeitos a alagamentos periódicos como no
Brasil, Gana, Nigéria e Costa do Marfim onde a precipitação total excede a

15
evapotranspiração e o solo torna-se hipóxico (SENA GOMES; KOZLOWSKY, 1986).
Rehem (2009) avaliando as respostas fisiológicas de mudas clonais de cacau ao
alagamento do substrato observou a ocorrência de decréscimo no crescimento, na
área foliar, na condutância estomática e nas taxas de assimilação de CO2; epinastia
e queda das folhas; hipertrofia de lenticelas e formação de raízes adventícias na
parte submersa do caule. Tais alterações, morfológicas e fisiológicas, são
dependentes do genótipo avaliado. Sena Gomes e Kozlowsky (1986) relataram
resultados similares para plântulas de cacau do genótipo Catongo destacando o
fechamento dos estômatos, que teve início 2h após o alagamento do solo. Apesar
disso, muito pouco se conhece à respeito do comportamento fisiológico do
cacaueiro em condições de alagamento.
2.4. As aquaporinas
A absorção de água e seu movimento através da planta são necessários para
um grande número de processos celulares e fisiológicos tais como: aumento celular,
movimento estomatal, fotossíntese, transporte no floema e transpiração. A água é
absorvida pela raiz dentro do apoplasto, mas, para atravessar a barreira hidrofóbica
das estrias de Caspary (espessamento de suberina presente nas células da
endoderme), as moléculas de águas são transferidas para o simplasto via caminho
transcelular, o qual é proposto ser regulado pelas aquaporinas (AMODEO et al.,
1999; JOHANSSON et al., 2000). As aquaporinas (AQP) são proteínas de 26-30
kDa, pertencentes a superfamília das proteínas integrais da membrana (MIP). Essas
proteínas especificamente facilitam o fluxo passivo de moléculas de água através
das membranas celulares, parecem regular a rota transcelular de água (AGRE et
al., 1993; MAUREL, 1997) e realizam um papel vital no transporte de grandes
volumes de água com gasto mínimo de energia. Algumas aquaporinas realizam
outras funções tais como transporte de amônia e outras substâncias (LUU;
MAUREL, 2005; FLEXAS et al., 2006). Desde a descoberta da primeira aquaporina
em eritrócitos humanos e túbulos renais (AGRE, MIPs têm sido identificadas em
outros organismos incluindo plantas (MAUREL et al., 1997), insetos (BEURON et
al., 1995), leveduras (CARBREY et al., 2001), bactérias (CALAMITA et al., 1995),
protozoários (MITRA et al., 2000) e Archae (KOZONO et al., 2003).

16
Proteínas integrais de membrana individuais variam no tamanho desde 195
(animais) a > 500 aminoácidos (fungos) (ZARDOYA, 2005). Elas são conhecidas
por funcionar como um tetrâmero, composto por unidades monoméricas individuais.
Cada monômero de MIP tem uma estrutura altamente conservada e tipicamente
exibe (1) seis alfa-hélices hidrofóbicas transmembrana (H1 – H6); (2) cinco loops
inter- hélices (LA – LE) sendo que os loops A, C e D são hidrofílicos com localização
citoplasmática ou extracelular, enquanto os loops B e E são hidrofóbicos e estão
parcialmente embebidos na membrana; (3) uma sequência AEF (Ala-Glu-Phe) ou
AEFXXT no domínio N-terminal freqüentemente observado (ZARDOYA; VILLALBA,
2001); (4) duas sequências NPA (asparigina-prolina-alanina) ou “box NPA”
altamente conservadas nos loops B e E (REIZER et al., 1993). A proteína é
estruturada em duas metades, cada uma com três hélices transmembranas (H1 –
H3 ou H4 – H6) e um pequeno loop (LB ou LE) contendo o box NPA. Com base na
similaridade das seqüências as aquaporinas identificadas em plantas nos últimos 10
anos podem ser classificadas dentro de quatro principais subfamílias: as proteínas
integrais da membrana plasmática (PIPs), as proteínas integrais do tonoplasto
(TIPs), as proteínas integrais da membrana similares à nodulina-26 (NIPs) e as
pequenas proteínas integrais básicas da membrana (SIPs) (JOHANSON et al.,
2001; ZARDOYA, 2005).

17
Figura 2. Estrutura das aquaporinas, mostrando as seis α-hélices e as regiões de
duplicação gênica (primeira e segunda metade). NH2 = região
aminoterminal; COOH = região carboxiterminal; A, C, D = loops
hidrofílicos; B e E = loops hidrofóbicos; NPA = Box asparigina–prolina-
alanina; SH = local de inibição por mercúrio (Hg2+); Ser256 = serina 256,
local de fosforilação por proteínas quinases A (PKA) (VERA-ESTRELLA
et al., 2005).
Desde o primeiro relato de aquaporina em plantas por Maurel et al., (1993),
um aumento no número de genes de aquaporinas em plantas têm sido relatados e
alteraram o conhecimento de como as plantas podem regular o fluxo de água em
diferentes condições fisiológicas (TYERMAN et al.,1999,2002; JOHANSSON et al.,
2000; JAVOT; MAUREL, 2002; MAUREL et al., 2002). Diferente dos animais, as
plantas expressam um grande número de aquaporinas. No genoma de Arabdopsis
thaliana 35 genes de aquaporinas foram identificados (JOHANSON et al.,2001;
QUIGLEY et al.,2002), em milho (Zea mays), 36 genes foram identificados

18
(CHAUMONT et al.,2001) enquanto que em vertebrados somente 11 -13 tipos
diferentes de aquaporinas existem (AQP1-AQP11 ou 13).
As proteínas integrais da membrana plasmática (PIP) constituem a maior
subfamília com 13 isoformas em Arabdopsis e 14 em Zea mays (JOHANSON et al.,
2001; CHAUMONT et al., 2001). A maioria das PIPs está localizada na membrana
plasmática e subdividem-se em dois grupos filogenéticos denominados PIP1 e PIP2
(ZARDOYA, 2005). As isoformas de PIP1 possuem 90% de identidade entre as
sequências, entretanto sua permeabilidade e função divergem. Expressões
heterólogas em ovócitos de Xenopus de isorfomas de PIP demonstraram uma baixa
ou nenhuma atividade de transporte de água nessas proteínas (CHAUMONT et al.,
2000; FETTER et al., 2004), porém, a permeabilidade a pequenos solutos e gases,
como glicerol e CO2 mostrou-se aumentada (UEHLEIN et al., 2003; BIELA et al.,
1999). Fetter et al. (2004), investigando a influência da heteromerização em
aquaporinas na permeabilidade a água mostrou que a co-expressão de duas
isoformas de PIPs em milho aumentou a permeabilidade à água em relação a
avaliações das proteínas individualmente. Nesse caso, a heteromerização é
necessária para as PIPS atuarem como canal funcional.
As aquaporinas pertencentes à subfamília PIP2 são mais eficientes como
canais de água do que os membros da subfamília PIP1. Geralmente, PIP2s
apresentam uma região N-terminal mais curta e uma região C-terminal mais longa
além de uma sequência adicional de 4-10 aminoácidos no loop A. Essas proteínas
podem estar envolvidas no transporte celular de água nas raízes, folhas (LOPEZ et
al., 2003; MARTRE et al., 2003), órgãos reprodutivos (BOTS et al., 2005) e na
germinação da semente (SCHUURMANS et al., 2003). Além disso, Hanba et al.,
(2004) superexpressando a HvPIP2;1 (Hordeum vulgare) confirmou a hipótese de
que essa proteína facilita o transporte de CO2.
Uma das primeiras proteínas com função de aquaporina em plantas foi
identificada na membrana do vacúolo de Arabidopsis thaliana. Essa proteína,
chamada TIP1;1 e inicialmente denominada γ-TIP (MAUREL et al., 1993), foi
altamente seletiva a água, quando expressa em ovócitos de Xenopus. No genoma
de Arabdopsis, foram identificadas 10 TIPs divididas em cinco grupos filogenéticos
(TIP1, TIP2, TIP3, TIP4 e TIP5). Medidas isoladas da permeabilidade a água de
vacúolos isolados e membranas plasmáticas mostrou uma permeabilidade 100

19
vezes mais alta do tonoplasto (MAUREL et al., 1997) evidenciando o papel do
tonoplasto na regulação do fluxo de água na célula vegetal. Boursiac et al. (2005)
avaliando a expressão de TIPs em Arabdopsis após estresse por salinidade
observou uma redução de 75% na abundância de transcritos após exposição ao
tratamento. Também, como demonstrado por Gerbeau et al. (2009), algumas TIPs
podem equilibrar as concentrações de uréia entre o vacúolo e o citoplasma.
Segundo Holm et al. (2005) e Loque et al. (2005), o transporte de amônio (NH4+) e
sua base conjugada gasosa, amônia (NH3), são mediadas pelas proteínas integrais
do tonoplasto.
A nodulina 26 foi descrita como as maiores proteínas integrais da membrana
do simbiossomo correspondendo a 10% das proteínas totais da membrana. No
genoma de Arabidopsis, nove proteínas integrais similares à nodulina 26,
distribuídas em dois grupos (NIP1 e NIP2) foram identificadas. Expressão
heteróloga em ovócitos de Xenopus demonstrou que nodulinas 26 são permeáveis
à glicerol (RIVERS et al., 1997) e amônia (NIEMIETZ et al., 2000). Em não
leguminosas, as proteínas integrais similares a nodulina 26 desempenham um
importante papel nas relações hídricas, sendo expressas em tecidos vegetativos e
reprodutivos além de transportar ácido bórico (TAKANO et al., 2006) e ácido lático
(CHOI; ROBERTS, 2007). Quando comparadas a outras aquaporinas, NIPs
possuem uma baixa atividade de permeabilidade a água.
As pequenas proteínas integrais da membrana (SIPs) foram identificadas por
Johansson et al. (2002), porém, informações acerca dessas aquaporinas ainda são
escassas. Sabe-se que são pequenas em tamanho e diferem das outras MIPs por
serem altamente básicas. O seu pequeno tamanho deve-se à região N-terminal que
é mais curta quando comparada as outras MIPs, mas similar a AqpZ (aquaporina Z)
de E.coli. Ishikawa et al., (2005) estudando a localização celular de SIPs em
Arabidopsis, verificou que SIPs eram mais abundantes no retículo endosplamático.
Apesar disso, o relacionamento entre o papel das aquaporinas na regulação
do estado hídrico na planta e a regulação da expressão dos seus genes é confuso.
Algumas aquaporinas são constitutivamente expressas (JOHANSSON et al., 1996),
enquanto a expressão de outras é regulada por diferentes estímulos tais como
estágio de desenvolvimento, hormônios ou por condições ambientais adversas
(VERA-ESTRELLA et al., 2004). A expressão dos genes das AQPs aumenta,

20
diminui ou permanece inalterada sob condições de estresse hídrico a curto prazo
(ALEXANDERSSON et al., 2005; AHARON et al., 2003). Além disso, embora
respostas da expressão gênica a mudanças ambientais possam ocorrer dentro de
horas, o padrão de expressão das AQPs após estresse hídrico a longo prazo,
envolvendo mecanismos de aclimatação, continuam a serem elucidados.

21
3. MATERIAIS E MÉTODOS
3.1. Material vegetal e condições de cultivo
O experimento foi conduzido em condições de casa de vegetação da
Universidade Estadual de Santa cruz (UESC), Ilhéus, BA, Brasil. Foram utilizados
dois clones de T. cacao contrastantes para resistência ao alagamento do solo (TSA-
792- resistente e TSH-774 – suscetível). Esses clones foram fornecidos pelo Instituto
Biofábrica do Cacau (IBC) com aproximadamente 4 a 6 meses de idade após o
enraizamento das estacas obtidas das extremidades de ramos plagiotrópicos de
plantas matriz com 5 a 10 anos de idade.
Posteriormente, as plantas clonais foram transplantadas e cultivadas em
vasos plásticos com capacidade de 25 L contendo solo como substrato. Quarenta
plantas de cada clone foram submetidas aos tratamentos de sombra por um período
de quatro meses. Tais tratamentos foram denominados como T0 (no interior da casa
de vegetação), T25 (sombrite de 25%, no interior da casa de vegetação), T50
(sombrite de 50%, no interior da casa de vegetação) e T75 (sombrite de 75%, no
interior da casa de vegetação), sendo 10 plantas de cada clone por tratamento
perfazendo um total de 80 plantas clonais.
Após o quarto mês de sombreamento, submeteram-se as plantas ao
alagamento do solo por um período de 85 dias (sombreamento + alagamento). O
tratamento alagado foi obtido vedando os furos de drenagem localizados no fundo
dos vasos e adicionando água, até 20 mm acima do nível do solo, à metade do
número de vasos. No tratamento não-alagado, vinte plantas de cada clone foram
mantidas em vasos de mesma capacidade, mas com fundos perfurados para
escoamento do excesso de água da irrigação.

22
3.2. Parâmetros micrometeorológicos
Os parâmetros micrometereológicos avaliados, durante o período de 85 dias,
da interação sombreamento x alagamento do solo na casa de vegetação, foram
umidade relativa e temperatura do ar. Os dados foram coletados por meio de uma
estação climatológica Hobo Micro Station Data Logger (OnSet, USA), com sensores
internos que registram em ambiente Windows. A coleta dos dados
micrometeorológicos foi automatizada com o armazenamento das leituras e posterior
transferência, através da porta serial, para um microcomputador. A radiação
fotossinteticamente ativa (RAF) foi determinada utilizando-se sensores quânticos S-
LIA-M003 (faixa espectral: 400 – 700 nm), acoplados à referida estação
climatológica. Posteriormente, foi realizada a análise calculando as médias diárias.
3.3. Medições de trocas gasosas foliares
As medições de trocas gasosas foliares foram realizadas ao longo do período
de interação sombreamento x alagamento do solo utilizando o medidor portátil LI-
6400 (Li-Cor Inc., Nebrasca, USA). As medições pontuais foram realizadas entre 8
e12 h. A irradiância foi mantida em 800 µmol m-2 s-1, acima da irradiância de
saturação de luz das plantas não alagadas. As taxas de fotossíntese líquida por
unidade de área foliar (A), a condutância estomática ao vapor de água (gs), a taxa
transpiratória foliar (E) foram estimadas a partir dos valores da variação de CO2 e
da umidade do ar no interior da câmara, determinados pelo analisador de gases a
infravermelho do referido aparelho. O tempo mínimo, pré-estabelecido para a
estabilização das leituras foi de 60 s e o máximo, para salvar cada leitura, de 120 s.
O coeficiente de variação (CV) máximo admitido para salvar cada leitura será de
0,8%. Além da radiação fotossinteticamente ativa (RFA), foram mantidos constantes
a temperatura do bloco a 26°C e o CO2 atmosférico no interior da câmara foliar. A
concentração de CO2 intercelular (Ci) foi calculada pelo equipamento a partir dos
valores de A, gs e E (VON CAEMMERER; FARQUHAR, 1981). Foram estimados
também a eficiência intrínseca (A/gs) e a instantânea do uso da água (WUE).
3.4. Medições de fluorescência da clorofila a

23
As medições de fluorescência da clorofila a foram efetuadas nas mesmas
épocas e folhas das plantas usadas para as medições de trocas gasosas usando
também um medidor portátil de fotossíntese (LI-COR, Inc. Lincoln, Nebraska, USA),
modelo Li-6400, com câmara Li-6400-40 para medições simultâneas de emissão de
fluorescência e de trocas gasosas foliares. Em cada medição, um clip foliar foi
colocado na folha por 30 minutos para a reflexão da radiação solar, o decréscimo da
temperatura foliar e a oxidação de todo o sistema de transporte fotossintético de
elétrons. Os sinais de fluorescência foram registrados no sistema de aquisição de
dados do LI-6400 que calculou automaticamente as fluorescências mínima (Fo),
máxima (Fm), e o rendimento quântico potencial máximo (Fv/Fm).
3.5. Determinação do teor de clorofila a, b, total e carotenóides
O teor de clorofila por área foi determinado em extratos dimetilsulfóxido
(DMSO) de discos foliares segundo Hiscox e Israelstam (1979), com algumas
modificações. Após incubação de três discos foliares (0,5 cm2) com 1 mL de DMSO
saturado com CaCO3 por 24 h a 65 ºC, a absorbância dos extratos foi lida no
espectrofotômetro de microplacas (VERSAmax) nos comprimentos de onda 645 nm,
663 nm e 480 nm, para a determinação das concentrações das clorofilas a, b e total
e de carotenóides, respectivamente (Tabela 1). A concentração de clorofila total foi
estimada pela soma dos teores de clorofila a e b.

24
Tabela 1. Equações utilizadas na determinação das concentrações de clorofilas a, b
e total e de carotenóides.
Comprimento de onda (nm) Pigmento
480 Carotenóides
645 Clorofila b
663 Clorofila a
Quantidade de pigmento ( µg/cm 2)
12,7 x A663 - 2,69 x A645
22,9 x A645 - 4,68 x A663
(A480 + 0,114 x A663 - 0,668 x A645) x V x 103/112,5 x 0,63702
3.6. Extravasamento de eletrólitos
Para acessar a estabilidade das membranas foi utilizada a técnica do
extravasamento de eletrólitos (BAJJI et al., 2001). Os discos foliares (3) com ½ cm
de diâmetro foram lavados em água destilada para a retirada do conteúdo das
células rompidas durante a remoção e de outros eletrólitos aderidos. Após a
lavagem, os discos foram secos em papel absorvente e colocados em frascos
contendo 10 mL de água destilada a 25ºC por 6 h sob agitação constante. Em
seguida, a condutividade elétrica foi medida (C1) e os frascos com os discos foram
colocados a 90 ºC por 2 h. Depois do equilíbrio da temperatura, a condutividade
elétrica máxima foi medida (C2) e o extravasamento de eletrólitos calculado por meio
da seguinte fórmula: (C1/C2) x 100.
3.7. Parâmetros de crescimento
No final do período experimental, mediram-se (i) a altura e o diâmetro do
coleto das plantas, usando régua e paquímetro respectivamente; e (ii) a área foliar
total por planta, usando um medidor de área LI-3100 (LI-COR, Inc. Lincoln,
Nebraska, USA) e contou o número de folhas por planta. Em seguida, para
obtenção da biomassa seca, as plantas dos genótipos clonais foram divididas em
partes (raiz, caule e folha), armazenadas em sacos de papel e secas em estufa de
ventilação forçada de ar a 75 °C até massa constante. A partir das variáveis de
crescimento e da biomassa seca determinaram-se: (i) razão de área foliar (RAF=

25
área foliar total / biomassa seca total); (ii) área foliar específica (AFE = área foliar /
biomassa seca foliar); (iv) razão de massa foliar (RMF = biomassa seca foliar /
biomassa seca total); e (v) razão raiz / parte aérea (R/PA = biomassa seca da raiz /
biomassa seca da parte aérea).
3.8. Teor de carboidratos
3.8.1. Obtenção do extrato
A) Açúcares solúveis totais - Inicialmente, 0,1g de cada amostra seca foi
submetida a uma extração com 5 mL de etanol 80% a quente, homogeneizado e
deixado em repouso durante 5 minutos. Em seguida acrescentou-se mais 5 mL de
etanol e as amostras foram centrifugadas por 10 minutos a 5.000 rpm sendo o
sobrenadante transferido para balão volumétrico (50 mL). Esta etapa foi repetida
mais três vezes, os sobrenadantes foram combinados e o volume completado para
50 mL com água destilada (extrato etanólico I).
As amostras foram clarificadas pela adição de 100 mg de carvão vegetal
ativado e filtração em papel de filtro Whatiman nº 1 ou similar (extrato etanólico II).
Do extrato etanólico II, tomaram-se alíquotas de 5 mL que foram
acondicionadas em béquer de 25 mL e evaporadas em chapa de aquecimento a
60°C até a secura. O resíduo resultante foi dissolvido em 5 mL de água destilada
(extrato etanólico III) e acondicionado em frascos de vidro tipo penicilina a -20°C.
B) Amido - Ao resíduo das extrações alcoólicas adicionaram-se 2,5 mL de
água destilada e 3,25 mL de ácido perclórico a 52%. As amostras foram
homogeneizadas, deixadas em repouso por 20 min., e novamente homogeneizadas.
Após, acrescentou-se 2,5 mL de água destilada e as amostras foram centrifugadas
por 10 minutos a 5.000 rpm sendo o sobrenadante transferido para balão
volumétrico de 50 mL. Esta etapa foi repetida, porém o tempo de repouso das
amostras foi de 30 minutos. Os sobrenadantes foram combinados e o volume
completado para 50 mL com água destilada. Alíquotas de 5 mL foram
acondicionadas em frascos de vidro tipo penicilina a -20 °C.

26
3.8.2. Quantificação
Para a quantificação dos açúcares solúveis totais e amidos foi utilizado o
método calorimétrico descrito por Laurentin e Edwards (2003). A concentração de
açúcares solúveis totais foi calculada tomando-se por base os dados da curva
padrão obtida com D-glicose. Para isso uma solução de glicose (1000 µg/mL) foi
preparada diluindo-se 0,1g de glicose em 100 mL de água destilada. Alíquotas de 10
mL (100 µg /mL), 7,5 mL (70 µg /mL), 5,0 mL (50 µg /mL) e 2,5 mL (25 µg /mL)
foram transferidas para balões volumétricos e o volume completado para 100 mL.
3.9. Expressão gênica de aquaporina
Para as análises de expressão gênica de aquaporina via Real Time
qRT-PCR foram realizadas coletas de folhas maduras de plantas não alagadas e
alagadas pertencentes aos genótipos TSA-792 e TSH-774 no tempo de 24 h após a
aplicação do tratamento alagado. As plantas não alagadas já se encontravam
aclimatadas aos níveis de irradiância T0, T25, T50 e T75. As amostras foram
armazenadas a –80°C após fixação em nitrogênio líquido e em seguida foram
liofilizadas. Foram realizadas extrações de RNA das folhas do tratamento não-
alagado e do tratamento alagado utilizando o RNAqueous Kit (Ambion – Applied
Biosystems). Em seguida, as amostras foram submetidas à eletroforese em gel de
agarose a 1% para avaliar se o RNA estava livre de DNA e íntegro para ser
submetido à reação de síntese de primeira fita do cDNA.
A síntese de cDNA foi realizada utilizando 1 µl de RNA total como molde, 20
unidades do RiboLock™ RNase Inhibitor (Fermentas), 10 mmol L-1 de dNTPs Mix
(Fermentas), 0,5 µg de primer Oligo d(T)18 e 200 unidades de RevertAidTM H-Minus
M-MuLV (Fermentas). A reação foi incubada a 42ºC por 60 min finalizando com 70ºC
durante 10 min. para inativação da enzima.
O RT-PCR quantitativo foi realizado em um termociclador Real Time PCR
(Applied Biosystems, modelo 7500), usando o kit The Maxima™ SYBR Green/ROX
qPCR Master Mix (Fermentas). A abundância de transcritos da proteína integral da
membrana plasmática PIP1;3 foi analisada por meio do seguinte primer: Forward-5’-
GACTTCCATCTCCTCTCTCTT-3’ / Reverse-5’-ATGGGCAGGGAAGGATAAAAG-3’

27
(Tm = 55ºC) (BAIGES et al., 2001). O mix para a reação foi composto por 500 ηg de
cDNA como molde, 5 pM de cada primer e uma quantidade suficiente do fluoróforo
SYBR Green I (Fermentas) para um volume final da reação de 20 µL. As condições
da reação foram 50ºC por 2 min., 95ºC por 10 min., 50 repetições de 95ºC por 15 s e
55ºC por 1 min..
Posteriormente, as leituras de fluorescência foram analisadas pelo Sequence
Detection Software (SDS, Applied Biosystems). Todas as reações foram submetidas
as mesmas condições de análise e normalizadas pelo sinal do corante de referência
passiva ROX para correção de flutuações na leitura decorrentes de variação no
volume e evaporação ao longo da reação. Os valores para o Ciclo Threshold (CT)
foram determinados e os números da expressão relativa dos genes foram calculados
como uma porcentagem da amostra não alagada do tratamento T0 utilizando o
método 2–∆∆Ct (LIVAK; SCHMITTGEN, 2001) com o gene da subunidade ribossomal
18S como controle endógeno.
3.10. Análise estatística
O experimento foi conduzido em condições de casa de vegetação em
delineamento experimental inteiramente casualizado em fatorial 2x4x2, referentes
aos 2 genótipos clonais (TSH-774 e TSA–792), 4 níveis de sombreamento (T0, T25,
T50 e T75) e 2 regimes hídricos (alagado e não-alagado), com cinco repetições,
perfazendo um total de 80 mudas clonais. Os dados foram submetidos à ANOVA,
teste de Tukey (p < 0,05) e análises de regressão.

28
4. RESULTADOS
4.1. Variáveis microclimáticas
Durante o período de 16/04/09 a 09/07/09 foram monitoradas a temperatura
(°C) e a umidade relativa do ar (UR, %), no interior da casa de vegetação, e de
16/04/09 a 22/05/09 a radiação fotossinteticamente ativa (RFA, µmol fótons m-2 s-1)
nos ambientes de cultivo T0, T25, T50 e T75 (Figura 1). A temperatura máxima
observada durante o período da interação sombreamento x alagamento do solo foi
28°C e a mínima 21°C. A umidade relativa mínima observada durante o período
experimental foi de 73% e a máxima de 95%. A partir dos dados de RFA coletados
no interior dos ambientes de cultivo foi determinado que 1) no tratamento T0, sem
tela sombrite, o valor médio de RFA foi de 194,6 µmol fótons m-2 s-1; 2) no
tratamento T25, com tela sombrite de 25% de atenuação da radiação solar total
incidente, o valor médio de RFA foi de 93,3 µmol fótons m-2 s-1, que corresponde a
47% de RFA do tratamento T0; 3) no tratamento T50, com sombrite de 50%, o valor
médio de RFA foi de 61 µmol fótons m-2 s-1, correspondendo a 31,5% de RFA do
tratamento T0; 4) no tratamento T75, com sombrite de atenuação de 75%, o valor
médio de RFA foi de 44,1 µmol fótons m-2 s-1; equivalente a 22% de RFA do
tratamento T0 a pleno sol.

29
Figura 3. Variáveis microclimáticas medidas no interior da casa de vegetação. (A)
Temperatura (°C) do ar; (B) Umidade relativa (%) do ar; (C) Valores
médios diários (7h – 17h) de radiação fotossinteticamente ativa (RFA) em
cada nível de sombreamento (NS, %); (▲) T0; (○) T25; (����)T50 e (����)T75.
(A)
(B)
(C)

30
4.2. Observações visuais
Todas as plântulas sobreviveram ao período de alagamento, porém
com sintomas de estresse como clorose nas folhas (Figura 4). Esta condição foi
mais evidente em plantas alagadas a pleno sol do que em plantas alagadas sob
sombreamento. Além disso, lenticelas hipertrofiadas na região submersa do caule,
sinais de decomposição nas raízes inundadas, ausência de raízes adventícias e
ausência de sintomas de injúria característicos de plantas intolerantes como necrose
e senescência foliar foram observadas em ambos os genótipos. Esses sintomas
ocorreram em todos os níveis de luminosidade.
Figura 4. Morfologia externa de plantas dos genótipos clonais de cacau TSA-792 e
TSH-774 submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85 dias. (A)
Clorose foliar aos oito dias (TSH-774) e (B) aos 85 dias após o
alagamento do solo (TSA-792); (C) Lenticelas hipertrofiadas na base do
caule oito dias após o alagamento do solo.

31
4.3. Trocas gasosas foliares
As superfícies de resposta ilustram as diferenças entre os tratamentos
(alagado e não-alagado) para ambos os clones TSA-792 e TSH-774, e as diferenças
apresentadas na interação genótipo x nível de sombra x alagamento x época.
A análise de variância indicou que as váriáveis apresentaram diferenças
intergenotípicas significativas para tratamentos, nível de sombreamento, época e
para a interação entre ambos (Tabela 2).
Tabela 2. Valores de probabilidade da ANOVA para os efeitos de genótipo (G), nível
de sombreamento (NS), regime hídrico (RH), tempo (T) e da interação
entre GxNSxRHxT nas trocas gasosas foliares. (*) p<0,05; (**) p<0,01;
(***) p<0,001; (NS) não significativo.
Para a condutância estomática (gs) foi verificado que houve interação entre
genótipo, nível de sombreamento, regime hídrico e duração do tratamento (Tabela
2). Nota-se, nas plantas não alagadas do genótipo TSA-792, que não houve efeito
do sombreamento (Figura 5A). No tratamento alagado, os valores de gs foram
similares aos das plantas não alagadas. Entretanto, com o tempo de alagamento,
verificou-se uma redução em gs a partir da primeira medição. As plantas não
alagadas do genótipo TSH-774 sofreram influência do sombreamento com tendência
à redução de gs com o aumento do sombreamento e com o tempo de exposição ao
G NS RH T
A 0,0000 *** 0,0000 *** 0,0000 *** 0,0000 *** 0,0000 ***
gs 0,0001 *** 0,0000 *** 0,0000 *** 0,0000 *** 0,0096 **
E 0,0009 *** 0,0000 *** 0,0000 *** 0,0000 *** 0,2356 NS
A/gs 0,5585 NS 0,0352 * 0,0336 * 0,0000 *** 0,2064 NS
A/E 0,2551 NS 0,3494 NS 0,0004 *** 0,0000 *** 0,0000 ***(µmol CO2 mmol H2O-1)
Variável
(µmol CO2 mol H2O)
GxNSxRHxT
(mmol H2O m-2 s-1)
(µmol CO2 m-2 s-1)
(mol m-2 s-1)

32
alagamento (Figura 5B). O valor de gs nas plantas alagadas foi consideravelmente
menor quando comparada com as do genótipo TSA-792 no tratamento a pleno sol,
porém observou-se um aumento significativo com o aumento do nível de sombra,
alcançando valores próximos aos das plantas não alagadas.
Comparando os coeficientes de determinação das equações de regressão do
tratamento não-alagado em ambos os genótipos, verificou-se que apenas 16% da
variação em gs para o genótipo TSA-792 e 36 % para o genótipo TSH-774 deveu-se
ao nível de sombreamento e o tempo, o que pode ser considerado pouco
significativo.

Figura 5. Condutância estomática ao vapor de água (
sombreamento por quatro meses e ao
não-alagado (y=1,097 – 0,307T - 0,
1,314***T – 0,3531NS + 0,385*T2 + 0,
0,695NS – 0,239T2 – 0,173NS2 –
0,156NS2 –
(A)
33
Condutância estomática ao vapor de água (gs) em nível foliar em genótipos clonais de cacau
e ao sombreamento + alagamento do solo por um período de
0,106NS – 0,0875T2 – 0,0047NS2 – 0,0685TNS / R2 = 0,
+ 0,2806*NS2 + 0,2123TNS/ R2= 0,86); (B) TSH-774 não-alagado
0,182TNS / R2 = 0,36); alagado (y= 0,586 – 0,931***T
0,171TNS / R2
(B)
) em nível foliar em genótipos clonais de cacau submetidos ao
alagamento do solo por um período de 48 dias. (A) TSA-792
= 0,16); alagado (y=1,173 –
alagado (y= 1,151 – 0,64*T +
+ 0,552*NS + 0,421**T2 –
= 0,77

34
A taxa fotossintética líquida por unidade de área foliar (A) nas plantas não
alagadas, em ambos os genótipos, manteve-se constante em relação ao nível de
sombreamento (Figura 6), porém decrescendo ao longo do período dos tratamentos
(tempo). Entretanto, observaram-se diferenças intergenotípicas significativas para as
plantas alagadas. Para o clone TSA-792, a taxa fotossintética em relação ao nível de
sombreamento apresentou valores médios similares aos das plantas não alagadas
(Figura 6A). Em contrapartida, o clone TSH-774 apresentou valores inferiores de A
no tratamento a pleno sol (6 µmol m2 s-1 ) quando comparado ao clone TSA-792 (11
µmol m2 s-1), alcançando valores próximos aos das plantas não alagadas com o
aumento do nível de sombreamento (Figura 6B). Para ambos os genótipos, os
valores de A foram reduzidos significativamente com o tempo de alagamento, com
uma média de 3,0 µmol m2 s-1 para o clone TSA-792 e de 2,0 µmol m2 s-1 para o
clone TSH-774 aos 48 dias após a aplicação do tratamento alagado.
Seguindo o mesmo padrão de respostas de gs e A, a transpiração foliar (E) foi
significativamente reduzida nas plantas submetidas ao alagamento do solo desde o
início do tratamento e manteve-se durante todo o experimento (Figura 7). Entretanto,
para a variável E, não houve interação significativa entre genótipos, nível de
sombreamento, regime hídrico e tempo (p<0,05) (Tabela 2). Verificou-se que a
queda nos valores de E com o tempo de alagamento foi mais acentuada para o
genótipo TSH-774 (Figura 7A), sendo que o genótipo TSA-792 (Figura 7B)
apresentou valor de E superior ao do genótipo TSH-774 ao final do experimento.

Figura 6. Taxa fotossintética líquida por unidade de área foliar
por quatro meses e ao sombreamento +
(y=9,95 – 0,172 T + 0,012NS – 0,04T
+ 0,135*T2 + 0,0006*NS2 + 0,0053 TNS/ R
0,175NS2 – 0,195TNS/ R2 = 0,70); alagado (
(A)
35
Taxa fotossintética líquida por unidade de área foliar (A) em genótipos clonais de cacau submetidos ao sombreamento
sombreamento + alagamento do solo por um período de 48 dias.
0,04T2 – 0,0001NS2 – 0,0015TNS / R2 = 0,80); alagado (y=12,11
+ 0,0053 TNS/ R2= 0,97); (B) TSH-774 não-alagado (y= 1,22 – 0,35*
; alagado (y= 0,70 – 0,842*T + 0,63*NS – 0,22T2 – 0,24NS2
(B)
submetidos ao sombreamento
(A) TSA-792 não-alagado
); alagado (y=12,11 – 2,394***T – 0,05*NS
35*T + 0,087NS – 0,037T2 –
– 0,092TNS / R2 = 0,82).

Figura 7. Taxa transpiratória foliar (E) em genótipos clonais de cacau
sombreamento + alagamento do solo por um período de
0,082NS – 0,3108T2 – 0,1765NS2
0,394*NS2 + 0,1365TNS/ R2= 0,86);
0,139TNS/ R2 = 0,29); alagado (y=
(A)
36
) em genótipos clonais de cacau submetidos ao sombreamento
alagamento do solo por um período de 48 dias. (A) TSA-792 não-alagado
– 0,166TNS / R2 = 0,14); alagado (y=1,192 – 1,446***T
= 0,86); (B) TSH-774 não-alagado (y= 1,058 – 0,567*T + 0,4177NS
0,832 – 1,278***T + 0,4621*NS – 0,552*T2 – 0,223NS2
(B)
submetidos ao sombreamento por quatro meses e ao
alagado (y= 1,104 – 0,606*T +
***T – 0,374NS + 0,505**T2 +
NS – 0,276T2 – 0,397*NS2 – 2 – 0,020TNS / R2 = 0, 80).

37
Para a eficiência intrínseca do uso da água (A/gs) (Figura 8), a ANOVA não
foi significativa para o fator genótipo e para a interação entre genótipos, nível de
sombreamento, regime hídrico e tempo (p<0,05). Entretanto, observaram-se
diferenças significativas entre plantas não alagadas e alagadas em condições de
sombreamento e com o tempo de exposição ao tratamento. Por outro lado, a
eficiência instantânea do uso de água (A/E) aumentou com o nível de
sombreamento, com reduções significativas em relação ao tempo de exposição aos
diferentes níveis de sombreamento nas plantas não alagadas do genótipo TSA-792
(Figura 8A). Nas plantas alagadas, houve uma redução pouco significativa da A/E
em relação ao nível de sombreamento e um incremento com a duração do mesmo
(Figura 8A). Comportamento inverso pôde ser verificado nas plantas não alagadas
pertencentes ao genótipo TSH-774 (Figura 8B), que reduziu a A/E com o aumento
do nível de sombreamento e aumentou com o tempo de exposição. Entretanto, nas
plantas alagadas, houve um leve aumento nos valores de A/E, seguido por uma
redução significativa com o nível de sombreamento, sendo que o mesmo
comportamento foi observado com o tempo de alagamento.
Nas plantas alagadas pertencentes ao genótipo TSA-792, o valor de A/E
apresentou comportamento similar ao das plantas não alagadas, entretanto, em um
patamar inferior (Figura 9A). Os valores de A/E foram superiores nos tratamentos de
sombreamento T25 e T50, tanto para o tratamento não-alagado quanto para o
alagado. Pequenos decréscimos foram observados com o tempo de alagamento.
Para o genótipo TSH-774 (Figura 9B), o tratamento não-alagado apresentou
comportamento similar ao não-alagado do genótipo TSA-792, porém com valores
inferiores de A/E. Nas plantas alagadas, A/E foi reduzido a partir do tratamento T25
de sombreamento e também com o tempo de alagamento. Mesmo sob alagamento,
as plantas do genótipo TSA-792 apresentaram A/gs similar ao das plantas não
alagadas do genótipo TSH-774.

Figura 8. Eficiência intrínseca do uso da água (
meses e ao sombreamento + alagamento do solo por um período de
0,252T + 0,949**NS – 0,283T2 – 0,
0,1648T2 + 0,19NS2 + 0,147TNS/ R
0,5532**NS2 – 0,163TNS/ R2 = 0,53
R2 = 0,51).
(A)
38
Eficiência intrínseca do uso da água (A/gs) em genótipos clonais de cacau submetidos ao sombreamento
alagamento do solo por um período de 48 dias. (A) TSA-792
0,6273**NS2 – 0,16TNS / R2 = 0,60); alagado (y=0,8542
0,147TNS/ R2= 0,16); (B) TSH-774 não-alagado (y= 0,906 – 0,1374T
= 0,53); alagado (y= 0,6112 – 0,382T + 0,635NS – 0,523*T2 –
(B)
submetidos ao sombreamento por quatro
792 não-alagado (y= 0,4465 –
0,8542 – 0,053T – 0,4074NS -
T - 0,842**NS – 0,289*T2 +
0,6904*NS2 – 0,1255TNS /

Figura 9. Eficiência instantânea do uso da água (
meses e ao sombreamento + alagamento do solo por um período de
0,0617T + 1,024**NS – 0,1764T2 –
0,1145T2 - 0,83**NS2 - 0,1806TNS/ R
- 0,609**NS2 – 0,1455TNS/ R2 = 0,56
0,46.
(A)
39
Eficiência instantânea do uso da água (A/E) em genótipos clonais de cacau submetidos ao sombreamento
alagamento do solo por um período de 48 dias. (A) TSA-792
– 0,808**NS2 – 0,0075TNS / R2 = 0,60); alagado (y=0,355
0,1806TNS/ R2= 0,50); (B) TSH-774 não-alagado (y= 0,3822 + 0,0466*
= 0,56); alagado (y= 0,6217 + 0,219T + 0,536NS – 0,455T2 – 0,
(B)
submetidos ao sombreamento por quatro
792 não-alagado (y= 0,707 –
0,355 – 0,0862T – 1,121**NS -
0466*T + 0,751**NS – 0,396*T2
0,56NS2 – 0,1717TNS / R2 =

40
4.4. Emissão de fluorescência da clorofila a
A análise de variância (ANOVA) para a emissão de fluorescência da clorofila
a foi significativa para regime hídrico e nível de sombreamento. Para os genótipos
avaliados somente não foi significativa a variável fluorescência máxima (Fm). A
ANOVA também não foi significativa para a interação genótipo x nível de
sombreamento x regime hídrico (Tabela 3).
Tabela 3. Valores de probabilidade da ANOVA para os efeitos de genótipo (G), nível
de sombreamento (NS), regime hídrico (RH), tempo (T) e das interações
NSxRH e GxNSxRHxT na emissão de fluorescência da clorofila a. (*)
p<0,05; (**) p<0,01; (***) p<0,001; (NS) não significativo.
Observa-se que nas plantas não alagadas do genótipo TSA-792, a
fluorescência mínima (Fo) não sofreu variações com a duração do tratamento
(Figura 10). Entretanto, Fo foi bastante reduzido com o aumento do nível de
sombreamento (Figura 10A). Essa redução foi observada também nas plantas
alagadas. Houve um leve aumento de Fo com o período de alagamento para o
genótipo TSA-792 (Figura 10A). Em contrapartida, para o genótipo TSH-774, não se
observou alterações nos valores de Fo nas plantas não alagadas, para o nível de
sombreamento (Figura 10B). Já para as plantas alagadas, verificou-se uma
diminuição nos valores de Fo nos tratamentos T25 e T50, com um aumento pouco
significativo para o tratamento T75. Embora os dois genótipos tenham apresentado
aumento de Fo com o tempo de exposição ao alagamento, evidenciou-se para o
genótipo TSA-792, resistente ao alagamento, esse aumento foi gradual, enquanto
G NS RH T NS x RH GxNSxRHxTFo 0,0095 ** 0,0000 *** 0,0000 *** 0,0356 * 0,020596 * 0,2681 NS
Fm 0,2943 NS 0,0000 *** 0,0254 * 0,0000 *** 0,000022 *** 0,4188 NS
Fv/Fm 0,0059 ** 0,0000 *** 0,0000 *** 0,0347 * 0,000068 *** 0,1230 NS
Variável

41
para o genótipo TSH-774, suscetível ao alagamento, o aumento pôde ser observado
a partir dos primeiros dias de alagamento do solo. As plantas não alagadas do
genótipo TSA-792 apresentaram valores maiores de Fo no tratamento a pleno sol do
que as plantas não alagadas do genótipo TSH-774 (300 e 250, respectivamente) e
valores menores no tratamento T75 (200 e 250, respectivamente.
Para a variável fluorescência máxima (Fm), não houve diferença significativa
em relação ao nível de sombreamento para as plantas não alagadas do genótipo
TSA-792 (Figura 11A). Nas plantas alagadas, os valores de Fm aumentaram nos
tratamentos T25 e T50 e diminuíram no T75 em comparação com o tratamento T0.
Por outro lado, houve um aumento significativo (p<0,05) de Fm em relação à
duração do período de alagamento. Nas plantas não alagadas do genótipo TSH-774
(Figura 11B), verificou-se um comportamento similar àquelas do genótipo TSA-792,
cujos níveis de sombreamento influenciaram pouco nos valores de Fm. No
tratamento alagado, os valores de Fm foram similares para os tratamentos T0 e T75
e os maiores valores foram encontrados nos tratamentos T25 e T50. Entretanto,
para o genótipo TSH-774, o valor de Fm se manteve constante durante todo o
período de alagamento (Figura 11B).

Figura 10. Fluorescência mínima (Fo) foliar em genótipos clonais de cacau
sombreamento + alagamento do solo por um período de
1,63**NS + 1,017T2 + 0,013*NS2
0,015*NS2 - 0,006TNS/ R2= 0,50);
0,036TNS/ R2 = 0,50); alagado (y=
(A)
42
) foliar em genótipos clonais de cacau submetidos ao sombreamento
alagamento do solo por um período de 48 dias. (A) TSA-792 não-alagado
– 0,039TNS / R2 = 0,42); alagado (y= 241,37 + 15,40
= 0,50); (B) TSH-774 não-alagado (y= 235,81 + 7,55T - 0,624NS
251,16 + 28,04T – 2,56**NS – 2,10T2 + 0,027**NS2 – 0,141TNS
(B)
submetidos ao sombreamento por quatro meses e ao
alagado (y= 264,04 – 10,19T +
15,40T – 1,80*NS – 1,123 T2 +
NS – 1,114T2 + 0,003NS2 –
141TNS / R2 = 0,63).
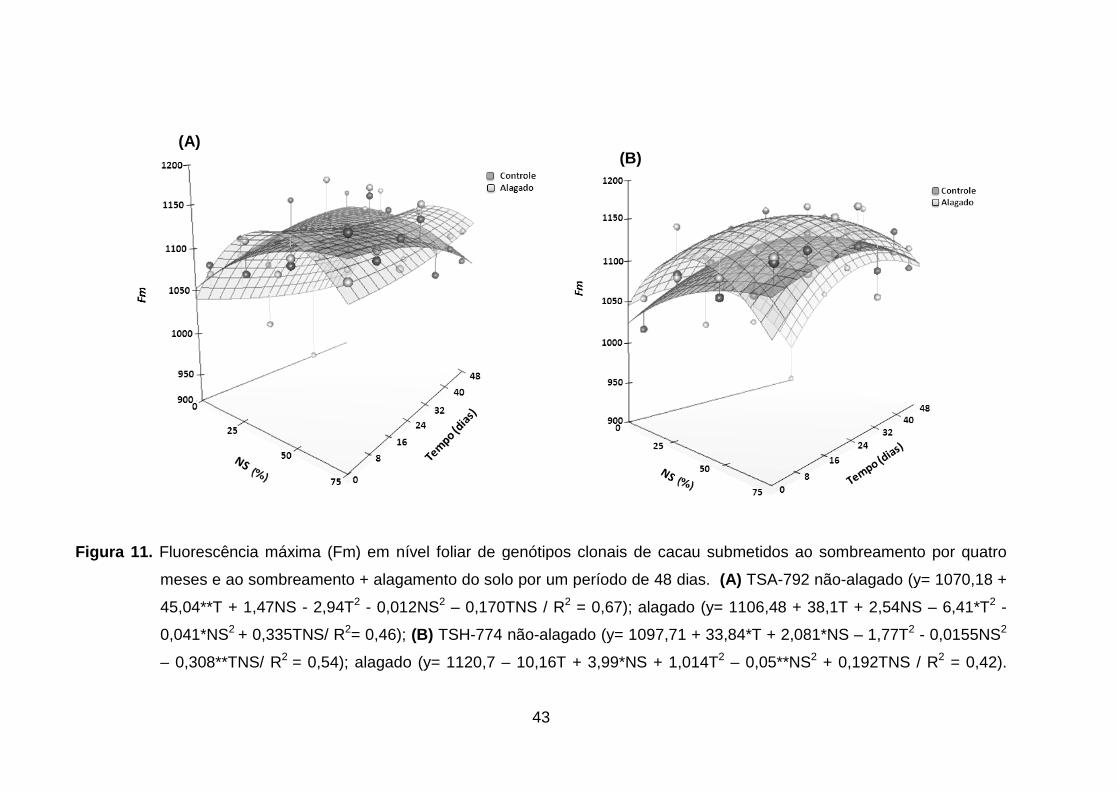
Figura 11. Fluorescência máxima (Fm) em nível foliar de genótipos clonais de cacau
meses e ao sombreamento + alagamento do solo por um período de
45,04**T + 1,47NS - 2,94T2 - 0,012
0,041*NS2 + 0,335TNS/ R2= 0,46);
– 0,308**TNS/ R2 = 0,54); alagado (
(A)
43
) em nível foliar de genótipos clonais de cacau submetidos ao sombreamento
alagamento do solo por um período de 48 dias. (A) TSA-792
0,012NS2 – 0,170TNS / R2 = 0,67); alagado (y= 1106,48 + 38,1
= 0,46); (B) TSH-774 não-alagado (y= 1097,71 + 33,84*T + 2,081*
; alagado (y= 1120,7 – 10,16T + 3,99*NS + 1,014T2 – 0,05**NS
(B)
submetidos ao sombreamento por quatro
792 não-alagado (y= 1070,18 +
38,1T + 2,54NS – 6,41*T2 -
2,081*NS – 1,77T2 - 0,0155NS2
NS2 + 0,192TNS / R2 = 0,42).

44
Verificou-se um incremento significativo no rendimento quântico do
fotossistema II com o aumento do nível de sombreamento nas plantas não alagadas
do genótipo TSA-792 (Figura 12A). Nesse caso, as plantas submetidas ao
tratamento a pleno sol apresentaram valores para Fv/Fm de 0,600 e as plantas
submetidas ao tratamento T75 de sombreamento apresentaram valores de 0,800.
Não houve variação no tempo de exposição ao sombreamento. Já nas plantas
alagadas esse aumento foi bastante expressivo, sendo que as plantas submetidas a
pleno sol apresentaram valores equivalentes às plantas não alagadas. Contudo, nos
tratamento T25 e T75 os valores de Fv/Fm alcançaram 0,9 e não sofreram
alterações com a duração do período de alagamento.
Nas plantas não alagadas do genótipo TSH-774, observou-se um aumento
pouco significativo nos valores de Fv/Fm entre os níveis de sombreamento (Figura
12B), que variaram em torno de 0,800 e 0,900 para os tratamento T0 e T75,
respectivamente. Não foram observadas alterações significativas (p<0,05) com o
tempo de exposição aos tratamentos de sombra. Verificou-se um comportamento
similar das plantas alagadas deste genótipo com àquelas do genótipo TSA-792 em
relação ao nível de sombreamento. Entretanto, com o tempo de alagamento, as
plantas do genótipo TSH-774 exibiram um decréscimo acentuado de Fv/Fm, quando
comparadas as do genótipo TSA-792, a partir dos primeiros dias após a aplicação
do tratamento alagado.

Figura 12. Rendimento quântico máximo de PS2 (
por quatro meses e ao sombreamento +
0,7548 + 0,017T + 0,0016**NS –
0,002**NS + 0,00005T2 - 0,00002**
0,00085NS + 0,0006T2 - 0,00001NS
– 0,00004**NS2
(A)
45
Rendimento quântico máximo de PS2 (Fv/Fm) em folhas de genótipos clonais de cacau submetidos ao sombreamento
sombreamento + alagamento do solo por um período de 48 dias. (A)
0,0015T2 - 0,00001*NS2 – 0,0007TNS / R2 = 0,48); alagado (y=
00002**NS2 + 0,00006TNS/ R2= 0,65); (B) TSH-774 não-alagado
NS2 – 0,00002TNS/ R2 = 0,44); alagado (y= 0,7753 – 0,032T
+ 0,0001TNS / R2
(B)
submetidos ao sombreamento
(A) TSA-792 não-alagado (y=
); alagado (y= 0,782 – 0,008T +
alagado (y= 0,7860 – 0,0006T +
0,032T + 0,0033**NS + 0,003T2
= 0,63).

46
4.5. Teores de pigmentos cloroplastídicos
Não houve interação intergenotípíca significativa (p<0,05) para nível de
sombreamento e regime hídrico (Tabela 4). Verificou-se que os teores de pigmentos
cloroplastídicos foram significativamente maiores nas plantas não alagadas em
relação às plantas alagadas.
Tabela 4. Valores de probabilidade da ANOVA para os efeitos de genótipo (G), nível
de sombreamento (NS), regime hídrico (RH), tempo (T) e das interações
entre NSxRH e GxNSxRS no teor de pigmentos cloroplastídicos. (*)
p<0,05, (**) p<0,01; (***) p<0,001; (NS) não significativo.
O teor de clorofila a foi significativo para o nível de sombreamento, regime
hídrico e para a interação entre ambos (p< 0,05) (Tabela 4). Não foram observadas
diferenças significativas em relação ao teor de clorofila a para as plantas não
alagadas do genótipo TSA-792 entre os níveis de sombreamento, enquanto que
para genótipo TSH-774 houve diferença entre os tratamentos T0 e T75 (Figura 13A).
Nas plantas submetidas ao tratamento alagado, o teor de clorofila a aumentou com a
elevação do nível de sombreamento, sendo a diferença mais significativa observada
nas plantas crescidas sob o tratamento T75 (Figura 13B). Neste tratamento, o teor
de clorofila a não diferiu entre os dois genótipos estudados.
G NS RH NS x RH G x NS x RH
Clorofila a (µg cm-2) 0,6791 NS 0,0000***
0,0000***
0,0425*
0,9376 NS
Clorofila b (µg cm-2) 0,1413 NS 0,1995 NS 0,0059 ** 0,2503 NS 0,2805 NS
Clorofila total (µg cm-2) 0,1913 NS 0,0001 *** 0,0000 *** 0,0441 * 0,3713 NS
Carotenóides (µg cm-2) 0,4216 NS 0,0008 *** 0,0001 *** 0,5390 NS 0,9710 NS
Clorila a/b 0,0033 ** 0,0017 ** 0,0001 *** 0,0394 * 0,4226 NS
Variável

47
Figura 13. Teor de clorofila a em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 48 dias. (A) Tratamento não-alagado; (B)
Tratamento alagado; (�) genótipo TSA-792; (�) genótipo TSH-774.
Médias intra e intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste de Tukey
(p<0,05).
O teor de clorofila b apresentou um padrão de resposta similar ao de clorofila
a, porém em concentrações significativamente menores (Figura 14). Nas plantas não
alagadas, não houve diferenças significativas entre os níveis de sombreamento para
ambos os genótipos (Figura 14A). Nas plantas alagadas, o teor de clorofila b foi
bastante reduzido, ao passo que nas plantas submetidas à baixa irradiância os
valores se aproximaram aos das plantas não alagadas. Entretanto, em T75 os
valores duplicaram em relação aos das plantas cultivadas a pleno sol (Figura 14B).
Diferenças intergenotípicas não foram observadas para as plantas não alagadas,
entretanto o genótipo TSH-774 tendendo a apresentar um teor de clorofila b maior
do que o TSA-792.
(A) (B)

48
Figura 14. Teor de clorofila b foliar em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 48 dias. (A) Tratamento não-alagado; (B)
Tratamento alagado; (�) genótipo TSA-792; (�) genótipo TSH-774.
Médias intra e intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste de Tukey
(p<0,05).
O teor de clorofila total (a + b) não apresentou diferenças intragenotípicas
significativas (p<0,05) entre as médias dos quatro tratamentos de sombreamento
(Figura 15). Verificou-se uma tendência de as plantas não alagadas do genótipo
TSH-774 apresentarem uma maior concentração de clorofila total nos tratamento
T25, T50, T75. Nas plantas alagadas, o teor de clorofila total foi bastante reduzido,
ao passo que as plantas submetidas ao tratamento T75 apresentaram uma
concentração de clorofila duas vezes maior em relação às plantas cultivadas a pleno
sol. Pôde-se observar também que tanto para o teor de clorofila a e b (Figura 13B e
13B), quanto para o teor de clorofila total (Figura 15B), o genótipo TSA-792 foi
superior ao TSH-774 nos tratamento alagado dos níveis T0 e T25 de sombreamento.
(A) (B)

49
Figura 15. Teor de clorofila total (a + b) foliar em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 48 dias. (A) Tratamento não-
alagado; (B) Tratamento alagado; (�) genótipo TSA-792; (�) genótipo
TSH-774. Médias intra e intergenotípica com as mesmas letras
minúsculas e maiúsculas, respectivamente, não diferem entre si pelo teste
de Tukey (p<0,05).
O teor de carotenóides não apresentou diferenças intragenotípicas
significativas (p<0,05) entre as plantas não alagadas nos tratamentos de
sombreamento T0 e T25, porém nos níveis de sombreamento T50 e T75, o genótipo
TSH-774 foi superior ao genótipo TSA-792 em relação à concentração de
carotenóides (Figura 16A). Para as plantas alagadas, o genótipo TSA-792
apresentou teor de carotenóides, nos tratamentos T0 e T25, superiores ao genótipo
TSH-774 (Figura 16B).
(A) (B)

50
Figura 16. Teor de carotenóides foliar em plantas de genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 48 dias. (A) Tratamento não-
alagado; (B) Tratamento alagado; (�) genótipo TSA-792; (�) genótipo
TSH-774. Médias intra e intergenotípica com as mesmas letras
minúsculas e maiúsculas, respectivamente, não diferem entre si pelo teste
de Tukey (p<0,05).
Para a razão clorofila a/b, não houve diferenças intragenotípicas nos quatro
níveis de sombreamento (Figura 17). Entre os genótipos, houve uma tendência do
clone TSA-792 não-alagado apresentar uma razão clorofila a/b um pouco maior do
que o clone TSH-774 nos tratamentos T25, T50 e T75 (Figura 17A). Nas plantas
alagadas, verificou-se, para o genótipo TSA-792, que a razão clorofila a/b foi maior
nos tratamentos T0, T25 e T50, quando comparado ao genótipo TSH-774 (Figura
17B).
(A) (B)

51
Figura 17. Razão clorofila a/b em folhas de genótipos clonais de cacau submetidos
ao sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 48 dias. (A) Tratamento não-alagado; (B)
Tratamento alagado; (�) genótipo TSA-792; (�) genótipo TSH-774.
Médias intra e intergenotípica com as mesmas letras minúsculas e
maiúsculas, respectivamente, não diferem entre si pelo teste de Tukey
(p<0,05).
4.6. Extravasamento de eletrólitos
De acordo com a análise de variância, o extravasamento de eletrólitos não foi
significativo (p<0,05) para genótipo, nível de sombreamento, regime hídrico e a
interação entre ambos (Tabela 5). Entretanto, observou-se, nas plantas alagadas,
uma tendência de acréscimo na porcentagem de eletrólitos nos tratamentos T50 e
T75 para o genótipo TSA-792 (Figura 16A) e nos tratamentos T0 e T25 para o
genótipo TSH-774 (Figura 16B).
(A) (B)

52
Tabela 5. Valores de probabilidade da ANOVA para os efeitos de genótipo (G), nível
de sombreamento (NS), regime hídrico (RH), tempo (T) e das interações
NSxRH e GxNSxRH no extravasamento de eletrólitos. (*) p<0,05; (**)
p<0,01; (***) p<0,001; (NS) não significativo.
4.7. Biomassa seca
Não houve efeitos significativos da interação tripla genótipo x nível de
sombreamento x regime hídrico para as variáveis de biomassa seca analisadas
(p<0,05, Tabela 6). A interação dupla entre nível de sombreamento x regime hídrico
foi significativa para área foliar total, biomassa seca de raiz, caule, folha e total e a
área foliar específica (p<0,05).
G NS RH NSxRH GxNSxRH
0,0694 NS 0,1999 NS 0,3296 NS 0,9802 NS 0,6067 NS
Variável
Extravasamento de eletrólitos (%)

53
Tabela 6. Valores de probabilidade da ANOVA para os efeitos de genótipo (G), nível
de sombreamento (NS), regime hídrico (RH) e das interações GxRH e
GxNSxRH no extravasamento de eletrólitos. (*) p<0,05; (**) p<0,01; (***)
p<0,001; (NS) não significativo.
Variável G NS RH NSxRH
Altura 0,0000 *** 0,0000 *** 0,0064 ** 0,1989 NS 0,5121 NS
Diâmetro do coleto 0,0000 *** 0,0024 ** 0,0000 *** 0,4754 NS 0,1964 NS
Nº de folhas 0,0600 NS 0,0000 *** 0,0000 *** 0,0590 NS 0,7495 NS
Área foliar total 0,0222 * 0,0000 *** 0,0000 *** 0,0018 *** 0,3671 NS
Biomassa seca
Raiz 0,0013 ** 0,0000 *** 0,0000 *** 0,0000 *** 0,7762 NS
Caule (g) 0,0000 *** 0,0000 *** 0,0710 NS 0,0070 ** 0,6720 NS
Folha (g) 0,9039NS
0,0000 *** 0,0000 *** 0,0082 ** 0,2890 NS
Total 0,0052 ** 0,0000 *** 0,0000 *** 0,0001 *** 0,3305 NS
RAF (x 10-2 m2 g-1) 0,0000 *** 0,0719 * 0,0000 *** 0,2598 NS 0,6134 NS
AFE 0,9559NS
0,0000 *** 0,0000 *** 0,0210 * 0,3222 NS
RMF (x 10-2 m2 g-1) 0,0016 ** 0,1944 NS 0,0000 *** 0,0836 NS 0,7218 NS
R/PA 0,0248 * 0,8451 NS 0,0002 *** 0,3973 NS 0,7299 NS
GxNSxRH
(cm)
(mm)
(g)
(x 10-2 m2)
(g)
A altura das plantas não foi influenciada pelas interações entre tratamentos
(Tabela 6), porém observaram-se efeitos significativos (p<0,001) isolados do nível de
sombreamento e do regime hídrico, com tendência à diminuição da altura com o
aumento do nível de sombreamento. No tratamento a pleno sol e T25, as plantas
apresentaram altura média similar, exceto para o tratamento alagado do genótipo
TSA-792 que apresentou uma redução de 25% na altura em relação ao tratamento
não-alagado do mesmo genótipo e aos tratamentos não-alagado e alagado do
genótipo TSH-774 (Figura 18).
No tratamento T50 e T75 de sombreamento, as médias de altura das plantas
não alagadas de ambos os genótipos foram similares às das plantas alagadas. No
geral, as plantas alagadas e alagadas do genótipo TSH-774 tenderam a apresentar
altura superior ao das plantas do genótipo TSA-792.

54
Figura 18. Altura da planta em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y = 79,85 –
0,29*NS / R²= 0.94); (○) TSA-792 – alagado (y = 60,95 + 0,443NS +
0,0078NS2/ R²= 0.93); (▼) TSH-774 – não-alagado (y = 80,43 +
0,252NS - 0,006NS² / R²= 0.76); (∆) TSH-774 alagado (y = 75,90 +
0,111NS – 0,0026NS² / R²= 0.55).
O diâmetro do coleto foi pouco afetado pelo sombreamento nas plantas não
alagadas (Figura 19). Nesse caso, observou-se tendência à diminuição com o
aumento do nível de sombreamento, nas plantas não alagadas de ambos os
genótipos avaliados e no tratamento não-alagado do genótipo TSA-792, sendo que
as plantas não alagadas do genótipo TSH-774 apresentaram valores médios
superiores aos do genótipo TSA-792. No tratamento alagado, o diâmetro do coleto
no genótipo TSH-774 foi maior nas plantas submetidas aos tratamentos T0 e T75 de
sombreamento e menor nas plantas submetidas aos tratamentos T25 e T50. Esse
resultado é válido também na comparação com as plantas alagadas do genótipo
TSA-792.
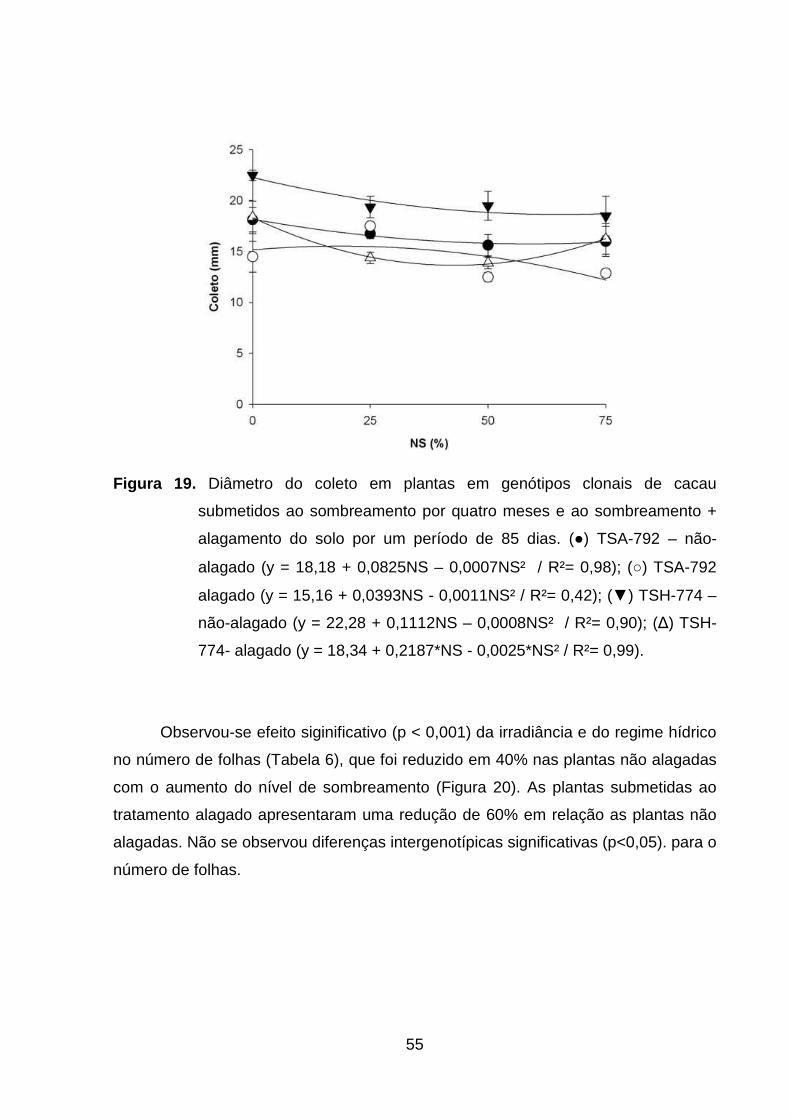
55
Figura 19. Diâmetro do coleto em plantas em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 85 dias. (●) TSA-792 – não-
alagado (y = 18,18 + 0,0825NS – 0,0007NS² / R²= 0,98); (○) TSA-792
alagado (y = 15,16 + 0,0393NS - 0,0011NS² / R²= 0,42); (▼) TSH-774 –
não-alagado (y = 22,28 + 0,1112NS – 0,0008NS² / R²= 0,90); (∆) TSH-
774- alagado (y = 18,34 + 0,2187*NS - 0,0025*NS² / R²= 0,99).
Observou-se efeito siginificativo (p < 0,001) da irradiância e do regime hídrico
no número de folhas (Tabela 6), que foi reduzido em 40% nas plantas não alagadas
com o aumento do nível de sombreamento (Figura 20). As plantas submetidas ao
tratamento alagado apresentaram uma redução de 60% em relação as plantas não
alagadas. Não se observou diferenças intergenotípicas significativas (p<0,05). para o
número de folhas.

56
Figura 20. Número de folhas em plantas de genótipos clonais de cacau submetidos
ao sombreamento por quatro meses e ao sombreamento + alagamento
do solo por um período de 85 dias. (●) TSA-792 – não-alagado (y =
100,5**e-0,009(NS) / R²= 0,99); (○) TSA-792 – alagado (y = 50,3**e-
0,009*(NS) / R²= 0,96); (▼) TSH-774 – não-alagado (y = 90,9e-0,008(NS)/
R²= 0,90); (∆) TSH-774 alagado (y = 24,3 + 26,93*e-0,102*(NS) / R²= 0,99).
A biomassa seca da raiz (BSR) foi significativamente (p<0,001) influenciada
pelo nível de sombreamento e pelo regime hídrico e pela interação entre ambos os
fatores (Tabela 6). Nas plantas não alagadas, observou-se uma redução
exponencial de BSR com o aumento do nível de sombreamento, sendo que as
plantas submetidas ao tratamento T75 de sombreamento apresentaram BSR
equivalente a 33% ao das plantas do tratamento T0 (Figura 21). Já nas plantas
alagadas, a redução de BSR foi linear, porém pouco significativa em relação a BSR
das plantas cultivadas a pleno sol e no sombreamento denso (T75).
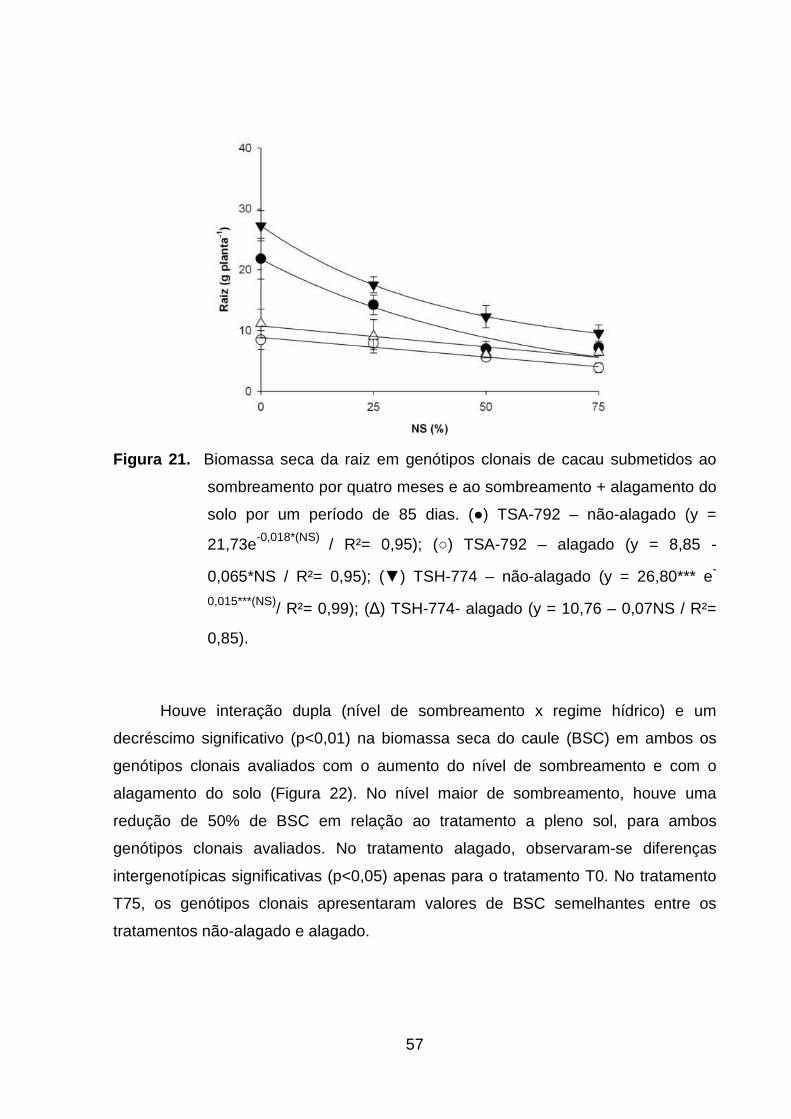
57
Figura 21. Biomassa seca da raiz em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y =
21,73e-0,018*(NS) / R²= 0,95); (○) TSA-792 – alagado (y = 8,85 -
0,065*NS / R²= 0,95); (▼) TSH-774 – não-alagado (y = 26,80*** e-
0,015***(NS)/ R²= 0,99); (∆) TSH-774- alagado (y = 10,76 – 0,07NS / R²=
0,85).
Houve interação dupla (nível de sombreamento x regime hídrico) e um
decréscimo significativo (p<0,01) na biomassa seca do caule (BSC) em ambos os
genótipos clonais avaliados com o aumento do nível de sombreamento e com o
alagamento do solo (Figura 22). No nível maior de sombreamento, houve uma
redução de 50% de BSC em relação ao tratamento a pleno sol, para ambos
genótipos clonais avaliados. No tratamento alagado, observaram-se diferenças
intergenotípicas significativas (p<0,05) apenas para o tratamento T0. No tratamento
T75, os genótipos clonais apresentaram valores de BSC semelhantes entre os
tratamentos não-alagado e alagado.

58
Figura 22. Biomassa seca do caule em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y = 32,94
+ 0,5954*NS – 0,0044NS²/ R²= 0,99); (○) TSA-792 – alagado (y = 24,31
+ 0,0441NS – 0,0025*NS²/ R²= 0,99); (▼) TSH-774 – não-alagado (y =
42,37 + 0,6568*NS – 0,0044*NS² / R²= 0,99); (∆) TSH-774- alagado (y
= 31,09 + 0,3727NS - 0,0024NS² / R²= 0,97).
A biomassa seca de folha (BSF) foi afetada significativamente (p<0,01) pela
interação nível de sombreamento x regime hídrico (Tabela 6). Tanto no tratamento
não-alagado quanto no alagado, os valores de BSF foram reduzidos em 50% com o
aumento do nível de sombreamento (Figura 23). Verificou-se que, no tratamento
não-alagado, não houve diferenças significativas (p<0,05) entre os genótipos clonais
avaliados. No tratamento alagado, o decréscimo de BSF, para o genótipo TSA-792,
apresentou comportamento linear, ao passo que para o genótipo TSH-774, a
resposta foi quadrática, com a redução máxima no tratamento T50 de
sombreamento.

59
Figura 23. Biomassa seca da folha em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y =
38,70e-0,014*(NS)/ R²= 0,97); (○) TSA-792 – alagado (y = 18,5 – 0,14*NS
/ R²= 0,97); (▼) TSH-774 – não-alagado (y = 37,30 – 0,32*NS / R²=
0,88); (∆) TSH-774 alagado (y = 22,63 + 0,5741*NS - 0,0053*NS² / R²=
0,99).
Houve interação dupla significativa (p<0,001), entre nível de sombreamento e
regime hídrico, para biomassa seca total (BST) (Tabela 6). Os valores de BST
sofreram um decréscimo de 60% nas plantas não alagadas dos genótipos clonais
avaliados (Figura 24). Observou-se uma tendência de as plantas não alagadas do
genótipo TSH-774 apresentarem valores de BST superiores ao genótipo TSA-792.
Nas plantas alagadas, respostas em relação a BST foram similares as de BSF.
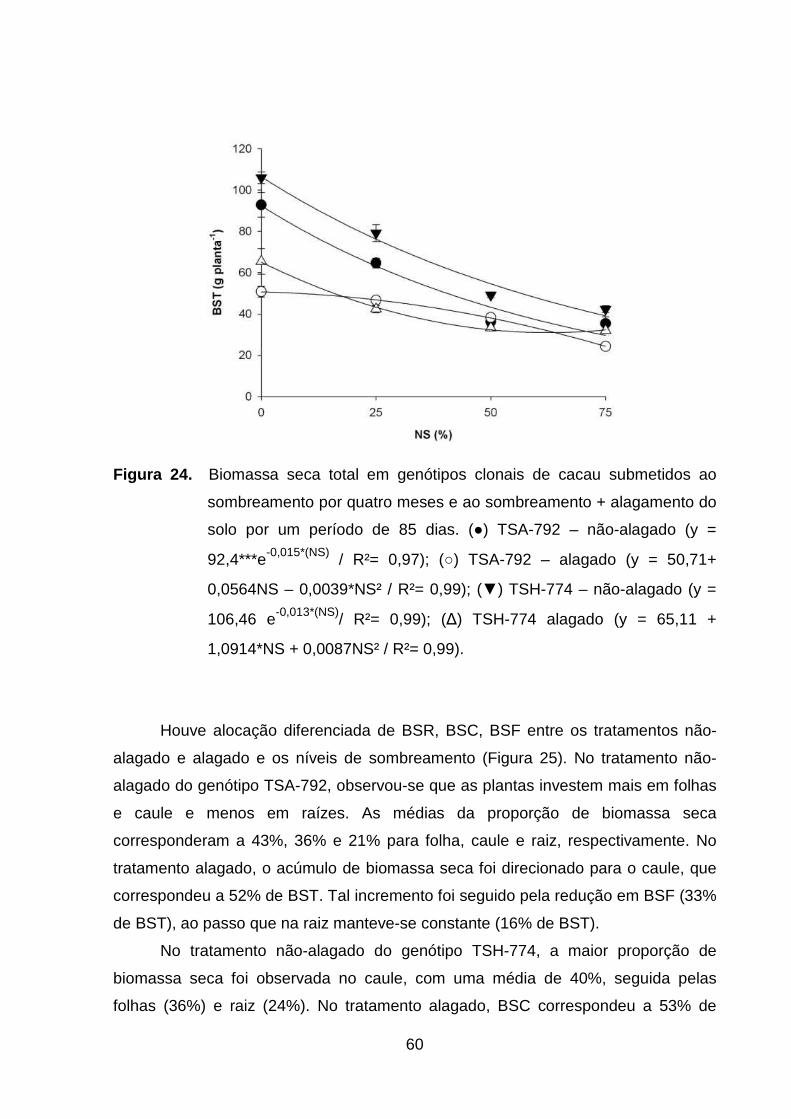
60
Figura 24. Biomassa seca total em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y =
92,4***e-0,015*(NS) / R²= 0,97); (○) TSA-792 – alagado (y = 50,71+
0,0564NS – 0,0039*NS² / R²= 0,99); (▼) TSH-774 – não-alagado (y =
106,46 e-0,013*(NS)/ R²= 0,99); (∆) TSH-774 alagado (y = 65,11 +
1,0914*NS + 0,0087NS² / R²= 0,99).
Houve alocação diferenciada de BSR, BSC, BSF entre os tratamentos não-
alagado e alagado e os níveis de sombreamento (Figura 25). No tratamento não-
alagado do genótipo TSA-792, observou-se que as plantas investem mais em folhas
e caule e menos em raízes. As médias da proporção de biomassa seca
corresponderam a 43%, 36% e 21% para folha, caule e raiz, respectivamente. No
tratamento alagado, o acúmulo de biomassa seca foi direcionado para o caule, que
correspondeu a 52% de BST. Tal incremento foi seguido pela redução em BSF (33%
de BST), ao passo que na raiz manteve-se constante (16% de BST).
No tratamento não-alagado do genótipo TSH-774, a maior proporção de
biomassa seca foi observada no caule, com uma média de 40%, seguida pelas
folhas (36%) e raiz (24%). No tratamento alagado, BSC correspondeu a 53% de

61
BST. Houve um incremento de BSC com o aumento do sombreamento até o
tratamento T50, ao passo no T75, as plantas apresentaram proporção de BSC
equivalente ao de T25.
Figura 25. Proporção de biomassa seca de raiz, caule e folha em relação à
biomassa seca total em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. Tratamento não-alagado: 0C, 25C,
50C e 75C; Tratamento alagado: 0A, 25A, 50A e 75A.
TSA-792
TSH-774

62
A área foliar total (AFT) por planta foi afetada significativamente (p<0,01) pela
interação nível de sombreamento e pelo regime hídrico (Tabela 6). Nos tratamentos
não-alagado, AFT foi inversamente proporcional ao sombreamento disponível para
ambos os genótipos clonais e apresentou um modelo exponencial (Figura 26).
Diferenças intergenotípicas para AFT foram observadas somente no tratamento T0,
sendo o TSA-792 o genótipo clonal com maior AFT. O tratamento alagado reduziu
AFT em 60% tanto para o genótipo TSA-792 quanto para o genótipo TSH-774, com
tendência de TSA-792 apresentar maior AFT do que o TSH-774.
Figura 26. Área folia total em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y =
7272,25**e-0,012*(NS) / R²= 0,98); (○) TSA-792 – alagado (y = 3073,40 +
0,5971NS – 0,3067NS² / R²= 0,98); (▼) TSH-774 – não-alagado (y =
6033,8e-0,01*(NS)/ R²= 0,80); (∆) TSH-774 alagado (y = 2509,70 +
36,179NS + 0,3207NS² / R²= 0,94).

63
Houve interação significativa (p<0,05) entre nível de sombreamento e regime
hídrico para o parâmetro área foliar específica (AFE) (Tabela 6), que variou de 2 a 4
cm2 g-1 no tratamento não-alagado e de 3 a 8 cm2 g-1 no tratamento alagado. As
plantas submetidas aos tratamentos de sombreamento e aos dois regimes hídricos
apresentaram aumento de AFE com o aumento do sombreamento (Figura 27). Esse
acréscimo foi mais pronunciado nas plantas submetidas ao alagamento. Para o
genótipo TSA-792 as respostas de AFE ao nível de sombreamento e ao alagamento
do solo foram exponenciais, enquanto que para o genótipo TSH-774 as respostas
foram quadráticas.
Figura 27. Área foliar específica em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y =
2,08*e0,01*(NS) / R²= 0,95); (○) TSA-792 – alagado (y = 3,6 e0,011***(NS)/
R²= 0,99); (▼) TSH-774 – não-alagado (y = 1,53 + 0,0654NS –
0,0005NS²/ R²= 0,81); (∆) TSH-774 alagado (y = 2,72 + 0,2054NS -
0,0019*NS² / R²= 0,99).

64
A razão de área foliar (RAF) foi significativa (p<0,001) apenas para regime
hídrico e genótipos clonais avaliados (Tabela 6). Entretanto, observou-se uma
tendência de as plantas não alagadas apresentarem aumento de RAF com o
aumento do nível de sombreamento (Figura 28). Os valores de RAF variaram de 50
a 80 cm2 g-1, para as plantas não alagadas, e de 40 a 60 cm2 g-1 para as alagadas.
As plantas do genótipo TSA-792 apresentaram RAF bastante superior ao do
genótipo TSH-774 nos tratamentos não-alagado e alagado até o nível T50 de
sombreamento. No nível T75 de sombreamento, as plantas alagadas dos genótipos
avaliados mostraram valores de RAF similares.
a
Figura 28. Razão de área foliar em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y = 75,42
+ 0,8279NS - 0,009 NS²/ R²= 0,85); (○) TSA-792 – alagado (y = 75.42 +
0,1823NS – 0,0032NS²/ R²= 0,50); (▼) TSH-774 – não-alagado (y =
54,58 + 0,7597NS – 0,0088NS²/ R²= 0,80); (∆) TSH-774 alagado (y =
40,867 + 0,0095NS + 0,0016NS² / R²= 0,75).

65
Para a razão de massa foliar (RMF), verificaram-se diferenças
intergenotípicas significativas (p<0,01) entre os tratamentos não-alagado e alagado
(Tabela 6). Não houve diferenças significativas entre os níveis de sombreamento. As
plantas do genótipo TSA-792 do tratamento não-alagado apresentaram RMF maior
do que as plantas do genótipo TSH-774 submetidas ao mesmo tratamento (Figura
29). Nas plantas alagadas, observou-se uma tendência de decréscimo de RMF com
o aumento do nível de sombreamento. Essa tendência foi mais acentuada no
genótipo TSH-774 até o nível T50 de sombreamento. No nível T75 os dois genótipos
apresentaram valores de RMF equivalentes.
Figura 29. Razão de massa foliar em genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y = 0,41 +
0,002NS - 0,0003 NS²/ R²= 0,99); (○) TSA-792 – alagado (y = 0,34 –
0,006NS - 0,0006NS²/ R²= 0,99); (▼) TSH-774 – não-alagado (y = 0,36
+ 0,0011NS + 0,00002NS²/ R²= 0,15); (∆) TSH-774- alagado (y = 0,34 -
0,0048NS + 0,00005NS² / R²= 0,97).

66
A análise de variância indicou que a razão raiz/parte aérea (R/PA) foi
significativa apenas para o fatores genótipo (p<0,05) e regime hídrico) (p<0,001)
(Tabela 6). No tratamento não-alagado e alagado, R/PA foi maior no genótipo TSH-
774 (Figura 30). Não houve resposta significativa (p<0,05) de R/PA com o aumento
do nível de sombreamento nas plantas alagadas e não alagadas. Entretanto, o
genótipo TSH-774 apresentou tendência de maior acréscimo de R/PA com o
aumento dos níveis de sombreamento.
Figura 30. Razão raiz/parte aérea (R/PA) em genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 85 dias. (●) TSA -792 – não-
alagado (y = 0,32 - 0,003NS – 2,965NS²/ R²= 0,92); (○) TSA-792 –
alagado (y = 0,20 - 0,0002NS + 3,19NS²/ R²= 0,50); (▼) TSH-774 –
não-alagado (y = 0,33 - 0,0011NS + 7,74NS²/ R²= 0,35); (∆) TSH-774-
alagado (y = 0,21 + 0,0021NS - 2,30NS² / R²= 0,43).

67
4.8. Teor de carboidratos
De acordo com a análise de variância (Tabela 7), os fatores nível de
sombreamento, regime hídrico, órgão (raiz, caule e folha) e a interação entre nível
de sombreamento x regime hídrico foram significativos (p<0,05) para o teor de
açúcares solúveis totais (AST) em plantas submetidas ao sombreamento e ao
alagamento do solo. Entretanto, para o teor de amido, houve diferenças
intergenotípicas significativas (p<0,05) em relação ao regime hídrico e órgão e à
interação entre nível de sombreamento e regime hídrico (p<0,05).
Tabela 7. Valores de probabilidade da ANOVA para os efeitos de genótipo (G), nível
de sombreamento (NS), regime hídrico (RH), órgão e das interações
NSxRH, RHxÓrgão, NSxRHxÓrgão e GxNSxRHxÓrgão no teor de
carboidratos. AST= açúcares solúveis totais (*) p<0,05; (**) p<0,01; (***)
p<0,001; (NS) não significativo.
Fator AST Amido
G 0,4886 NS 0,0403 *
NS 0,0018 ** 0,2128 NS
RH 0,0000 *** 0,0000 ***
Órgão 0,0000 *** 0,0000 ***
NS x RH 0,0019 ** 0,2603 NS
RH x Órgão 0,0835 NS 0,0000 ***
NS x RH x Órgão 0,8178 NS 0,0753 NS
G x NS x RH x Órgão 0,6418 NS 0,8104 NS
Carboidrato (mg g-1)

68
4.8.1. Açúcares solúveis totais
Nas raízes dos genótipos clonais de cacau avaliados, houve decréscimo no
teor de AST com os tratamentos de sombreamento e alagado (Figura 31). Por
apresentar teor AST de 50% maior no tratamento T0, a redução nas plantas não
alagadas do genótipo TSA-792 foi mais expressiva do que no genótipo TSH-774. O
tratamento alagado promoveu o aumento linear de AST nas raízes de clones TSA-
792 com o aumento do sombreamento. Esse aumento correspondeu a 75% quando
comparado aos tratamentos T0 e T75. Já para o genótipo TSH-774, não houve
variação do teor de AST em raízes de plantas sombreadas, com uma média de 60
mg g-1 de matéria seca.
Figura 31. Teor de açúcares solúveis totais na raiz de genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 85 dias. (●) TSA-792 – não-
alagado (y = 56,50 – 1,20NS + 0,012NS2/ R2= 0,99); (○) TSA-792 –
alagado (y = 43,5 + 0,39***NS / R2= 0,99); (▼) TSH-774 – não-alagado
(y = 44,77 – 0,47NS + 0,003 NS2/ R2= 0,60); (∆) TSH-774- alagado (y =
54,5 + 0,33 NS – 0,004 NS2/ R2= 0,83).

69
O teor de AST no caule foi reduzido drasticamente nas plantas não alagadas
submetidas aos tratamentos de sombreamento. Para o genótipo TSA-792, o teor de
AST correspondeu a 25% do observado no tratamento T0, ao passo que para o
genótipo TSH-774 foi de 50% (Figura 32). No tratamento alagado, houve tendência
de manutenção do teor de AST entre os níveis de sombreamento para o genótipo
TSH-774. Entretanto, para o genótipo TSA-792, verificou-se uma tendência de
redução no teor de AST no tratamento T75 de sombreamento.
Figura 32. Teor de açúcares solúveis totais no caule de plantas de genótipos
clonais de cacau submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85 dias. (●)
TSA-792 – não-alagado (y = 74,93 e-0,02*(NS)/ R2= 0,96); (○) TSA-792 –
alagado (y = 78,25 + 0,81NS – 0,014NS2/ R2= 0,97); (▼) TSH-774 –
não-alagado (y = 48,85e- 0,013(NS)/ R2= 0,76); (∆) TSH-774- alagado (y =
75,03 + 0,243NS – 0,006NS2/ R2= 0,99).

70
Não houve diferenças significativas (p<0,05) entre o teor AST nas folhas dos
genótipos clonais avaliados no tratamento não-alagado (Figura 33). Observou-se,
para ambos os genótipos, no tratamento não-alagado, a tendência de AST diminuir
com o aumento do sombreamento. Observou-se uma redução no teor de AST entre
os níveis de sombreamento para o genótipo TSA-792 e de aumento para o genótipo
TSH-774.
Figura 33. Teor de açúcares solúveis totais em folhas de plantas de genótipos
clonais de cacau submetidos ao sombreamento por quatro meses e ao
sombreamento + alagamento do solo por um período de 85 dias. (●)
TSA-792 – não-alagado (y = 74,93 e-0,02*(NS)/ R2= 0,96); (○) TSA-792 –
alagado (y = 78,25 + 0,81NS – 0,014NS2/ R2= 0,97); (▼) TSH-774 –
não-alagado (y = 48,85e- 0,013(NS)/ R2= 0,76); (∆) TSH-774- alagado (y =
75,03 + 0,243NS – 0,006NS2/ R2= 0,83).

71
4.8.2. Amido
Não houve ajuste de modelos para os pontos referentes ao tratamento
alagado do genótipo clonal TSA-792 e para o tratamento não-alagado do TSH-774
(Figura 34). Foi observada tendência de redução no teor de amido no sistema
radicular com o aumento do nível de sombreamento para o genótipo TSA-792 não-
alagado e aumento para o TSH-774 alagado.
Figura 34. Teor de amido na raiz de genótipos clonais de cacau submetidos ao
sombreamento por quatro meses e ao sombreamento + alagamento do
solo por um período de 85 dias. (●) TSA-792 – não-alagado (y = 12,41
– 0,0914NS + 0,0008 NS2/ R2= 0,95). (∆) TSH-774- alagado (y = 10,55
+ 0,125NS – 0,002 NS2/ R2= 0,95)
Os tratamentos de sombreamento não influenciaram significativamente
(p<0,05) o teor de amido no caule, tanto para o tratamento não-alagado quanto para
o alagado. Nas plantas não alagadas, houve diferenças intragenotípicas em relação
ao teor de amido no caule. Observou-se, para estas plantas, tendência de maior

72
acúmulo de amido no tratamento T75 de sombreamento (Figura 35). Os maiores
teores de amido foram verificados nas plantas alagadas submetidas aos tratamentos
T0, T25 e T50 de sombreamento, ao passo que no tratamento T75 os valores se
aproximaram ao das plantas não alagadas. Além disso, neste regime hídrico, o
comportamento foi inversamente proporcional aos tratamentos de sombreamento.
Para o genótipo TSA-792, o acúmulo de amido no caule das plantas alagadas do
tratamento T0 foi 140% maior em relação as plantas não alagadas, enquanto que
para o genótipo TSH-774 esse acúmulo foi de 85%.
Figura 35. Teor de amido no caule de plantas de genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 85 dias. (●) TSA-792 – não-
alagado (y = 55,30 – 1,40 NS + 0,0165 NS2/ R2= 0,80); (○) TSA-792 –
alagado (y = 128,70e-0,0055*(NS)/ R2= 0,93); (▼) TSH-774 – não-alagado
(y = 46,00 – 0,796 NS + 0,0094 NS2/ R2= 0,96); (∆) TSH-774- alagado
(y = 91,08 + 1,06 NS – 0,018 NS2/ R2= 0,65).

73
Não houve variação do teor de amido nas folhas das plantas de ambos os
genótipos clonais submetidos aos tratamentos de sombra e alagamento do solo, a
exceção do tratamento T75, onde se observou um aumento do teor de amido nas
folhas das plantas alagadas e uma redução na folhas das plantas não alagadas
(Figura 36).
Figura 36. Teor de amido nas folhas de plantas de genótipos clonais de cacau
submetidos ao sombreamento por quatro meses e ao sombreamento +
alagamento do solo por um período de 85 dias. (●) TSA-792 – não-
alagado (y = 15,25 + 0,0561 NS – 0,016 NS2/ R2= 0,70); (○) TSA-792 –
alagado (12,36 – 0,21 NS + 0,0033 NS2/ R2= 0,90); (▼) TSH-774 –
não-alagado (y = 15,09 – 0,094 NS + 0,0007 NS2/ R2= 0,85); (∆) TSH-
774- alagado (y = 16,77 – 0,053 NS + 0,0013 NS2/ R2= 0,40).

74
4.9. Expressão gênica da aquaporina PIP1;3
No presente estudo, a técnica de PCR em tempo real quantitativa foi utilizada
para determinar o perfil da expressão gênica da proteína integral da membrana
plasmática PIP1;3 em plantas de ambos os genótipos clonais de cacau submetidos
a dois regimes hídricos, não-alagado e alagado, e a quatro níveis de irradiância (T0,
T25, T50 e T75). As amostras coletadas no tratamento T0 foram utilizadas como
calibradoras, para resultados comparativos, já que não receberam tratamentos de
sombreamento no interior da casa de vegetação. Houve variações intergenotípicas
em relação ao perfil da expressão do gene PIP1;3 em nível foliar (Figura 37).
Observou-se, nas plantas não-alagadas dos genótipos clonais TSA 792 e TSH-774,
uma repressão da expressão gênica de PIP1;3 nos níveis T25, T50 e T75 em
comparação ao tratamento T0. Nesse caso, o gene PIP1;3 sofreu maior repressão
nas plantas pertencentes ao genótipo TSA-792. Diferenças intergenotípicas na
abundância de transcritos foram evidenciadas também nas plantas alagadas e
submetidas aos diferentes níveis de sombreamento. O tratamento alagado induziu a
expressão gênica da aquaporina PIP1;3 somente nas plantas do genótipo TSA-792,
ao contrário do genótipo TSH-774, cuja expressão deste gene foi reprimida em
maior intensidade, quando comparada a expressão no tratamento não-alagado.

75
Figura 37 . Perfil da expressão do gene PIP1;3 em folhas de plantas de dois
genótipos clonais de T. cacao crescidas em quatro níveis de
sombreamento (T0, T25, T50 e T75) por um período de quatro meses e
submetidas ao alagamento por 24 h. ( ) TSA-792 não-alagado; ( )
TSA-792 alagado; ( ) TSH-774 não-alagado; ( ) TSH-774 alagado.
Os valores expressos são relativos ao tratamento não-alagado no interior
da casa de vegetação (T0). Colunas representam os valores médios ±
erro padrão de três réplicas biológicas.

76
5. DISCUSSÃO
5.1. Parâmetros fisiológicos
A resposta específica de plantas ao alagamento varia com muitos fatores
incluindo espécie, genótipo, idade e condição da planta, qualidade da água e
duração do período de alagamento (KOZLOWSKI, 1984; KRAMER; KOZLOWSKI,
1979). Como o alagamento elimina o O2 presente no solo, a absorção do mesmo é
favorecida pela produção de lenticelas hipertrofiadas na base do caule. Assim, as
lenticelas observadas nas mudas dos genótipos de cacaueiro avaliados neste
experimento (Figura 4) poderiam ser interpretadas como uma adaptação para
aumentar o transporte de O2 dos tecidos da parte aérea para as raízes. Ferreira et al
(2001), avaliando o crescimento inicial de Piptadenia gonoacantha sob inundação
em diferentes níveis de luz, observaram a protusão de lenticelas desde o primeiro
dia de avaliação. Ainda, compostos potencialmente tóxicos associados com a
anaerobiose (acetaldeído, etanol e etileno) são liberados das plantas pelas lenticelas
(CHIRKOVA; GUTMAN 1972). De acordo com Lobo e Joly (1998), a produção de
lenticelas em plantas inundadas nem sempre tem valor adaptativo, podendo ser
resultantes de alterações hormonais. Mielke et al (2005a, 2003), avaliando as
respostas de Annona glabra e Genipa Americana ao alagamento do solo, relataram
para as duas espécies a presença de lenticelas, leve clorose foliar e ausência de
necrose e senescência foliar.
A condutância estomática (gs) é diretamente proporcional à abertura do poro
estomático. A maior abertura que pode ocorrer no poro estomático, a qual depende
do formato e das propriedades da parede celular, determina o limite máximo da taxa
de entrada de gás (LARCHER, 2000). O decréscimo de gs em folhas de plantas

77
arbóreas tolerantes ou não-tolerantes é uma resposta comum ao alagamento do
solo e está, na maioria dos casos, ligado ao fechamento estomático ocasionado pela
diminuição da difusão de CO2 (KOZLOWSKI, 1997, KOZLOWSKI et al., 2002,
CARVALHO; ISHIDA et al., 2002; OLIVEIRA, 2007). Tanto o genótipo TSA-792
quanto o TSH-774 apresentaram valores de gs variando de 0,020 a 0,120 µmol m-2
s-1 (Figura 5). Estes valores estão de acordo com Nobel (1991), que afirmou que
plantas lenhosas comumente apresentam valores de gs nesta mesma faixa, quando
os estômatos encontram-se abertos.
Bertolde et al. (2010), avaliando os mesmos genótipos, encontraram valores
de 0,08 µmol m-2 s-1 para plantas não alagadas do genótipo TSA-792 e de 0,05 µmol
m-2 s-1 após 45 dias de alagamento. Já para o genótipo TSH-774 os valores de gs
foram 0,06 e 0,02 µmol m-2 s-1, nas plantas não alagadas e alagadas,
respectivamente. Segundo estes autores, os genótipos apresentaram respostas que
permitiram a caracterização de ambos como tolerante (TSA-792) e suscetível (TSH-
774) ao alagamento do solo. Sena Gomes e Kozlowski (1986), avaliando os efeitos
do alagamento do substrato nas relações hídricas e no crescimento de plântulas de
cacau da variedade Catongo relataram que o fechamento dos estômatos foi iniciado
duas horas após a aplicação do tratamento de alagamento. Parolin (2001) e Lopez e
Kursar (2003) demonstraram redução em gs, quando as plantas de floresta
sazonalmente inundada foram submetidas ao alagamento. Entretanto, os valores de
gs em plântulas de Annona glabra, submetidas ao alagamento do solo, foram
reduzidos no quarto dia, mas retornou aos valores controle no 11º, 18º e 56º dia
após a aplicação do tratamento (MIELKE et al., 2003).
A redução acentuada de gs para as plantas alagadas do genótipo TSH-774
(Figura 5B) afetou a assimilação de CO2 nos tratamentos a pleno sol, mas sob
sombreamento os valores de A mantiveram-se elevados. Nesse caso, sugere-se que
sob condições de alagamento, o genótipo TSH-774 parece aclimatar-se sob baixa
incidência de luz, porém, sob alta intensidade de radiação luminosa, a luz torna-se
uma fator de redução da atividade fotossintética. De acordo com DaMatta (2004), a
menor eficiência de assimilação de carbono a pleno sol deve-se principalmente ao
fechamento estomático.
De acordo com Lenssen et al. (1999), há uma hierarquia de efeitos
ambientais, ou seja, qual fator deve ser considerado primeiro como o de maior

78
influência nas respostas das plantas aos efeitos ambientais. Assim, espécies
tolerantes podem ser afetadas independentemente por ambos os fatores, mas, para
espécies intolerantes ao alagamento, os efeitos do sombreamento podem ser
reduzidos porque o alagamento é o fator dominante (LENSSEN et al., 1999).
O melhor desempenho das plantas alagadas do genótipo TSA-792 sob altas
irradiâncias (Figura 5A) deveu-se possivelmente ao aumento da produção de
Rubisco e, ou da capacidade de regeneração de RuBP (ribulose-1,5 bisfosfato –
substrato para a carboxilação) (Long e Hällgren, 1993), a alta atividade de enzimas
fotossintéticas e ao transporte de fotoassimilados mais eficiente do que o genótipo
TSH-774. Entretanto, análises da atividade enzimática e quantificação do teor de
Rubisco nas folhas seriam necessárias para sustentar essa afirmação.
A taxa de assimilação de CO2 nas plantas não alagadas (Figura 6) não foi
influenciada pelo regime de irradiância e só foram observadas variações com o
tempo de exposição aos tratamentos de sombra. Este fato deveu-se à idade
fisiológica das folhas já que pouco ou nenhum lançamento foliar foi observado nas
plantas submetidas aos tratamentos de sombreamento. Quando as folhas crescem,
sua capacidade para fotossintetizar aumenta até elas atingirem a maturidade, a
partir de então, a taxa de assimilação de CO2 começa a decrescer. Yamaguchi e
Friend (1979), trabalhando com plantas de Coffea arabica, demonstraram que as
taxas fotossintéticas são baixas nas folhas em início de expansão (par +1),
alcançam valores máximos nas folhas recém expandidas (par +2) e decrescem
tornando-se constantes com a idade nas folhas fisiologicamente maduras (par+3,
par +4).
Vários estudos demonstram que o alagamento do solo é capaz de diminuir,
de forma significativa, a capacidade fotossintética das plantas sensíveis ao estresse.
Espécies que são sensíveis ao alagamento frequentemente exibem reduções
severas em A e gs (PEZESHKI, 1993; GRAVATT; KIRBY, 1998), sendo estas
variáveis úteis na determinação do grau de tolerância das plantas ao alagamento do
solo (GRAVATT; KIRBY, 1998). De acordo com Pezeshki (1993), um indicador da
tolerância ao alagamento é a inibição inicial das trocas gasosas seguida por uma
recuperação. Bertolde et al. (2010) encontraram valores similares de taxa de
assimilação de CO2 em genótipos de TSA-792 e TSH-774 submetidos ao
alagamento do substrato.

79
A transpiração foliar (E) tem sido pouco estudada em experimentos de
alagamento do solo. Quando E excede a capacidade de reposição da água perdida,
devido a absorção radicular deficiente em condições de alagamento, os estômatos
se fecham parcial ou totalmente com consequente redução em gs. Com essa
restrição às trocas gasosas foliares, a atividade do sistema fotossintético torna-se
reduzida e a taxa de assimilação de CO2 diminui (JACKSON, 2002). Os resultados
do presente trabalho mostram que os efeitos do alagamento causaram redução da
taxa de transpiração tanto para o genótipo TSA-792 (Figura 7A) quanto para o
genótipo TSH-774 (Figura 7B), acompanhada pela diminuição de gs e A, indicando a
existência de mecanismos de adaptação no sentido de diminuir as perdas de água.
Resultados semelhantes foram observados por Rehem et al. (2009) e Bertolde et al.
(2010).
As plantas alagadas do genótipo TSA-792 sobressaíram-se em relação à
eficiência intrínseca do uso da água (A/gs, Figura 8A), quando comparado com as
plantas alagadas do genótipo TSH-774 (Figura8B). A diminuição de gs foi
proporcional ao decréscimo de A, mantendo constante a razão A/gs. Nessas
circunstâncias, o genótipo TSA-792 mostra-se mais tolerante ao estresse, por meio
da economia de água durante as trocas gasosas. As plantas alagadas do genótipo
TSH-774 reduziram a A/gs em relação ao nível de sombreamento e período de
alagamento, devido ao decréscimo mais acentuado de A em relação a gs. A forma
como A relaciona-se com gs tem importância ecológica, pois, se A e gs variam
proporcionalmente, é possível que a concentração interna de CO2 (Ci) no mesofilo
foliar e as eficiências intrínsecas (A/E) e instantâneas (A/gs) do uso de água se
mantenham constantes, no sentido de otimização das trocas gasosas (SCHULZE;
HALL, 1982). A razão A/E, um parâmetro fisiológico capaz de auxiliar a
compreensão das relações hídricas, expressa quantitativamente o comportamento
momentâneo das trocas gasosas em nível floliar (LARCHER, 2000). Segundo Ito
(1978), o aumento de A/E, produzido pela queda de gs e E, é devido à manutenção
da taxa fotossintética das plantas, mesmo com o fechamento parcial dos estômatos.
Os dados de trocas gasosas foliares do presente estudo confirmam os
encontrados por Bertolde et al. (2010) que concluíram que, mesmo apresentando
alterações no comportamento fisiológico, o genótipo TSA-792 apresentou maior
tolerância ao alagamento do que o genótipo TSH-774.

80
A determinação dos teores de clorofila em folhas é extremamente importante,
pois a atividade fotossintética da planta depende da capacidade da folha para
absorver luz (MARENCO; LOPES, 2005). Embora o alagamento cause redução no
teor de clorofila, devido à degradação e, ou decréscimo da sua síntese, as plantas
alagadas dos dois genótipos clonais de cacau avaliados, submetidas ao nível mais
alto de sombreamento, apresentaram teor de clorofila a equivalente ao das plantas
não alagadas a pleno sol (Figura 13). O fato de as plantas, mantidas em tratamentos
mais sombreados, apresentarem maiores teores de clorofilas está relacionado ao
efeito compensatório da espécie, quando submetidas à baixa intensidade de luz
(ALMEIDA et al., 2004) Outros autores sugerem, além disso, redução na sua síntese
em conseqüência do acúmulo de etileno (SENA GOMES; KOZLOWSKI, 1988).
A clorofila b é um dos pigmentos acessórios sintetizado por meio da oxidação
do grupo metil da clorofila a para um grupo aldeído (STREIT et al., 2005). Segundo
Vieira (1996), o estresse provocado na planta pela luz é freqüente sob condições
tropicais, e a concentração de clorofilas e carotenóides são indicadores da
suscetibilidade da planta à variação da intensidade da luz. Nas plantas não-alagadas
Figura 14A), o genótipo TSH-774 tendeu apresentar concentrações de clorofila a e
b maiores do que o genótipo TSA-792 nos tratamentos T25, T50 e T75. Sugere-se
que o genótipo TSH-774 seja mais tolerante à sombra do que o genótipo TSA-792.
Kramer e Kozlowiski (1979) afirmaram que folhas de sombra apresentam uma maior
concentração de clorofilas do que as folhas de sol. Esses resultados estão de
acordo com Rêgo e Possamai (2006), que observaram que níveis de sombreamento
elevado proporcionaram maiores teores de clorofila a e b em jequitibá-rosa; e com
Carvalho (1996), que encontrou maior teor de clorofila b nos níveis mais altos de
sombreamento para as espécies Cabralea canjarana e Centrolobium robustum. O
aumento da clorofila pode ser associado também a uma maior quantidade de grana
em cloroplastos de folhas de sombra em relação às folhas de sol (MEBRAHTU;
HAVOVER, 1991).
A combinação das clorofilas (a + b) e dos pigmentos acessórios, como os
carotenóides, capacita as plantas a captarem a maior parte de energia disponível da
luz solar (MARTINAZZO et al., 2007). Embora o genótipo TSH-774 apresente maior
acúmulo de clorofila total e de carotenóides, sob condições normais de irrigação
(Figura 15A), o genótipo TSA-792 apresentou maior acúmulo de clorofila total e

81
carotenóides nas plantas alagadas do tratamento T0 e T25 (Figura 15B e 16B).
Esses resultados esclarecem, em parte, a maior eficiência fotossintética deste
genótipo (Figura 6A). De acordo com Engel e Poggiani (1991) e Atroch et al.,(2001),
as clorofilas são um dos fatores ligados à eficiência fotossintética de plantas, ao
crescimento e a adaptabilidade a diversos ambientes.
Em geral, o alagamento do solo reduziu o teor de clorofila a,e b e total nos
dois genótipos estudados. Pezeshki et al. (1996) também observaram resultados
semelhantes ao estudarem espécies arbóreas em condição de solo alagado. Em
plântulas de Eryithrina variegata, a diminuição no conteúdo de clorofilas também foi
detectada por Muthuchelian et al (1995), após 10 dias de inundação.
Freqüentemente é observado que a razão clorofila a/b tende a diminuir com o
aumento do nível de sombreamento, devido uma maior proporção de clorofila b em
ambientes sombreados. Este fato ocorre devido à lenta degradação da clorofila b em
plantas de sombra, em relação à clorofila a (ENGEL; POGGIANI, 1991). Nos
resultados obtidos no presente trabalho, essa maior razão clorofila a/b não foi
observada (Figura 17), já que a contribuição da clorofila b foi menor. Estes
resultados estão de acordo com Martinazzo et al. (2007), que demonstraram que os
níveis de sombreamento não afetaram, significamente, a relação clorofila a/b em
mudas de Eugenia uniflora. As plantas não alagadas, em todos os níveis de
sombreamento, apresentaram razões clorofila a/b em torno de 5 ug cm2 e plantas
alagadas de 3,5 a 4,5 ug cm2 com o aumento do nível de sombreamento. Conforme
Seybold e Egle (1970), plantas expostas a elevadas intensidades de luz apresentam
razões clorofila a/b em torno de 3,2 a 4,0 ug cm-2 e plantas crescendo em ambientes
com baixa irradiância possuem razões a/b em torno de 2,5 a 2,9 ug cm-2.
Uma característica comum em plantas submetidas a estresses abióticos é o
aumento da permeabilidade da membrana, expresso pelo aumento na liberação de
íons ou extravasamento de eletrólitos. Alguns trabalhos evidenciaram a utilidade
desta variável, junto com parâmetros fisiológicos, na avaliação das respostas de
espécies vegetais ao estresse e na distinção entre genótipos tolerantes e sensíveis
a um determinado estresse abiótico (BRITO et al., 2009; KARLIDAG, 2009; MARTIN
et al., 1987). Os resultados apresentados neste estudo mostram a suscetibilidade do
genótipo TSH-774 à alta irradiância quando em condições de alagamento (Figura
18). Esse fato pode ser atribuído à indução de respostas ao estresse oxidativo,

82
levando à peroxidação lipídica, permeabilização da membrana e morte celular
(SCANDALIOS, 1993).
O registro da fluorescência em folhas intactas (fluorescência da clorofila in
vivo), especialmente durante a fase de indução pela exposição à radiação após uma
fase de escuro (efeito Kautsky), permite avaliar o estado do fotossistema para a
síntese de ATP, ou seja, avaliar a adaptação à radiação e detecção das mudanças
do aparato fotossintético durante o desenvolvimento da planta ou durante o estresse
(LARCHER, 2005; STRANT; ÖQUIST, 1988; BAKER, 2008).
O valor Fo representa a emissão de luz pelas moléculas de clorofilas a do
complexo antena, antes dos elétrons terem migrado para o centro de reação do
fotossistema 2 (PS2) (MATHIS; PAILLOTIN, 1981) e é um valor referência na
determinação das outras variáveis de fluorescência (HIPKINS; BAKER, 1986).
Segundo Bolhàr-Nordenkampf et al. (1989), o valor de Fo nem sempre é uma
constante e, geralmente, aumenta por dois motivos: os centros de reação de PS2
está comprometido ou a transferência da energia de excitação do complexo antena
para os centros de reação está prejudicada. De acordo com os resultados obtidos
neste experimento, o aumento nos valores de Fo para as plantas alagadas de
ambos os genótipos (Figura 10) pode ter sido causado por alterações estruturais nos
pigmentos fotossintéticos, já que diferenças foram observadas entre plantas não
alagadas e alagadas, quanto ao teor de pigmentos (Figuras 13, 14, 15, 16 e 17).
Estes resultados estão de acordo com Bertolde et al. (2010).
A partir de Fo, a emissão de fluorescência continua a crescer até alcançar a
fluorescência máxima, significando que as moléculas de plastoquinona a receberam
os elétrons do PS 2 e reduziram-se (MATHIS; PAILLOTIN, 1981). Assim, a
fluorescência máxima representa o nível de fluorescência quando toda a quinona
(Qa) é reduzida (BAKER, 2008).
O rendimento quântico potencial máximo (Fv/Fm) (Fm-Fo/Fm) indica a
eficiência máxima na qual a luz absorvida pelo PS 2 é utilizada para a redução da
quinona (BAKER, 2008) e permite, avaliar o rendimento do fotossistema de PS2
após a adaptação ao escuro. A invariabilidade de Fv/Fm nas plantas do genótipo
TSA-792 (Figura 12A), com o período de alagamento, demonstra que a eficiência
fotossintética de PS2 não foi prejudicada e pode ser considerado um indicativo da
resistência deste genótipo à condição de alagamento. Maurenza et al. (2009),

83
avaliando plântulas de Pouteria glomerata (Sapotaceae) parcialmente inundadas,
encontraram valores de Fv/Fm variando entre 0,78 e 0,80 para os tratamentos
alagado e não-alagado, respectivamente, considerando-as com ausência de danos
em PS2. Tais valores foram similares aos encontrados neste trabalho e aqueles
encontrados por Bertolde et al (2010). Valores inferiores a 0,75 indicam situação de
estresse e, portanto, redução do potencial fotossintético da planta (MAXWELL;
JOHNSON, 2000). De acordo com Schreiber et al. (1998), na prática, a razão entre a
fluorescência máxima (Fm) [toda quinona (Qa) reduzida] e a fluorescência mínima
(Fo) (toda Qa oxidada) é aproximadamente 5 a 6 em tecidos fotossintetizantes
saudáveis e adaptados ao escuro.
Os diferentes tratamentos induziram a alocação diferenciada de biomassa nos
diferentes órgãos das plantas de ambos os genótipos clonais. Essas diferenças nos
padrões de biomassa podem ser um fator considerado importante na interpretação
da capacidade adaptativa de cada genótipo ao sombreamento e ao alagamento do
solo. Um maior investimento no crescimento em altura em ambientes sombreados é
uma resposta comum, relatada em vários trabalhos (CANCIAN; CORDEIRO 1998,
POORTER 1999, FERRER; DILLENBURG 2000, JURADO et al., 2006). De acordo
com Ryser e Eek (2000), o aumento no crescimento em altura pode ser vantajoso
como resposta ao sombreamento em curto prazo. No presente estudo, o
crescimento em altura dos genótipos clonais avaliados foi reduzido em condições de
baixa irradiância (Figura 19). Essa resposta poderia ser interpretada como uma
conservação dos recursos por parte da planta, já que um rápido crescimento em
altura estaria associado principalmente a plantas não tolerantes ao sombreamento
(MORGAN; SMITH 1979, BROKAW 1987). Carelli e Fahl (2000) observaram menor
crescimento em altura em mudas de Coffea arabica com excesso de sombreamento
(70% de sombreamento) e não detectaram diferenças significativas entre as plantas
a pleno sol e a 50 % de sombreamento. Pedroso e Varela (1995) e Barbosa (1990)
não encontraram diferenças significativas em crescimento em altura ao testarem
diferentes níveis de sombreamento em mudas de Ceiba pentandra e Jacaranda
copaia, respectivamente.
A redução no crescimento em altura nas plantas alagadas e sombreadas
(Figura 19) poderia ser explicada pelo menor transporte e distribuição de nutrientes
minerais nas plantas (REHEM et al., 2009) e pelo decréscimo na produção de

84
fotoassimilados, evidenciado pela baixa taxa fotossintética liquida apresentadas pelo
genótipo clonal TSH-774 crescido nessa condição. Em solos alagados o crescimento
caulinar pode ser auxiliado pela ação das raízes adventícias que, durante sua
atividade, facilitam a captação de nutrientes. Contudo, não foi relatada presença de
raízes adventícias nos genótipos avaliados. Bertolde et al. (2010) relataram
presença de quantidade média e baixa de raízes adventícias para os genótipos TSA-
792 e TSH-774, respectivamente, submetidos ao alagamento do substrato.
Decréscimo no diâmetro do coleto em plantas não alagadas sob diferentes
níveis de sombreamento foi relatado por Almeida et al. (2005) em mudas de Acacia
mangium submetidas a três condições de sombreamento (pleno sol, 30% e 50%).
Fonseca et al. (2002) observaram que ocorreu decréscimo linear do diâmetro do
coleto em função do aumento do período de permanência sob sombreamento. De
acordo com Fonseca et al. (2002), a redução no diâmetro do coleto é um fator
desfavorável, pois implica na redução da biomassa seca do sistema radicular. Engel
et al. (1991) também relataram que mudas produzidas à pleno sol são mais
adequadas, pois apresentam maior diâmetro do coleto e, provavelmente, um
sistema radicular melhor desenvolvido. No caso de ambos os genótipos clonais
avaliados, a comparação deste parâmetro entre os diferentes níveis de irradiância
testados permite inferir que o sombreamento acentuado (T75) inibiu o crescimento
da planta como um todo (Figura 19), ao passo que o crescimento em altura e em
diâmetro foram favorecidos nos tratamentos T0 e T25 de sombreamento. Em
condições de anoxia, as alterações no diâmetro do coleto estão associadas à
hipertrofia da base do caule e à formação de aerênquima (MIELKE et al., 2005 b).
Apesar da ausência de interação entre níveis de sombreamento e regimes
hídricos, os efeitos do sombreamento e do alagamento foram evidentes, em relação
ao no número de folhas, para os genótipos clonais TSA-792 e TSH-774 (Figura 20).
Nesse caso, o sombreamento acentuado (T75) reduziu o número de folhas em 40%
e o alagamento do solo em 60% em ambos os genótipos clonais avaliados. Nos
tratamentos mais sombreados, os genótipos avaliados não produziram novas folhas,
mas as manteve desde o início do experimento em bom estado fisiológico. Em
contrapartida, o maior número de folhas observadas no tratamento não-alagado
proporcionou uma maior produção de fotoassimilados e partição de biomassa entre
os órgãos vegetativos, favorecendo o seu crescimento e desenvolvimento, exemplo

85
do maior crescimento em altura (Figura 19), diâmetro do coleto (Figura 19) e
produção de biomassa seca total (Figura 24). Ferreira et al. (2001), avaliando o
crescimento de mudas de Piptadenia gonoacantha, submetidas ao alagamento, em
três níveis de irradiância (40%, 70% e 100%) relataram que houve uma redução de
45% no número de folhas nas plantas alagadas, e que não houve interação
significativa entre nível de sombreamento e regime hídrico. Campos e Uchida
(2002), trabalhando com uma espécie nativa da Amazônia (Jacarandá copaia),
encontraram maior número de folhas em ambiente sob 30% de sombreamento.
O decréscimo da biomassa seca de raiz, em função dos tratamentos de
sombreamento nas plantas não alagadas (Figura 21), revela a existência de uma
estreita relação entre a disponibilidade de radiação luminosa e a aquisição de
biomassa pela planta. Os menores valores de BSR observados em plantas mais
sombreadas está relacionado aos menores comprimentos e densidades das raízes
(Lei e Lechowicz, 1998). Segundo Claussen (1996) plantas que recebem altos níveis
de irradiância investem mais em raízes. Assim, por terem uma maior demanda
evapotranspiratória, um maior investimento no crescimento do sistema radicular
assume maior importância nas plantas sob altas irradiâncias do que nas
sombreadas. Nas plantas submetidas ao tratamento alagado, a redução observada
na biomassa seca da raiz (65%) deveu-se à decomposição causada por atividade
fúngica e à susceptibilidade do sistema radicular aos microorganismos presentes em
solos alagados (KOZLOWSKI, 1997). Esses resultados estão de acordo com Mielke
et al. (2003), que observaram uma redução de 49,7% na biomassa da raiz em
plantas de Genipa americana sob estresse por alagamento.
O acúmulo de biomassa seca no caule foi favorecido pelos tratamentos sem
sombreamento (Figura 22). Campos e Uchida (2002) relataram que o nível de 30%
de sombreamento promoveu maiores índices de biomassa seca no caule em mudas
de Ochroma lagopus, enquanto que a do sistema radicular foi reduzida em 50% e
70% de sombreamento. Silva et al. (2007) avaliando plantas de Theobroma
grandiflorum em três níveis de sombreamento (0%, 50% e sombra natural) observou
maior investimento em biomassa seca do caule nas plantas que estavam na
condição de 50% de sombreamento.
O maior acúmulo de biomassa seca em folhas observada no tratamento T0,
com maior nível de irradiância (Figura 23), para ambos os genótipos clonais

86
avaliados, se deveu, em grande parte, ao maior número e espessura das folhas
associado a altas taxas fotossintéticas. Segundo Scalon et al. (2001), o aumento da
espessura do limbo foliar, em condições de alta irradiância, é um dos recursos
utilizados pelas plantas como proteção dos pigmentos fotossintéticos. Almeida et al.
(2005) ao avaliarem o crescimento de Acacia mangium a pleno sol, 30 e 70% de
sombreamento, também verificaram que o acúmulo de biomassa foliar foi
proporcional ao aumento da irradiância. Por outro lado, redução da biomassa seca
foliar no tratamento alagado, principalmente para o genótipo clonal TSH 774, se
deveu principalmente à redução nos lançamentos foliares, além de clorose seguida
de abscisão das folhas.
O decréscimo na área foliar total com o aumento do nível de sombreamento
observado no presente estudo (Figura 26) não está de acordo com o que é
normalmente observado. Segundo Scalon et al. (2001) e Campos e Uchida (2002),
ao avaliarem as respostas das plantas ao sombreamento, há uma necessidade de
ampliação da superfície fotossintetizante para maximizar a captação da baixa
radiação disponível. Kluge (1998) verificou redução da área foliar em Passiflora sp.
sob reduzida intensidade de luz. Entretanto, Santos (2005) observou que a área
foliar foi dez vezes maior em plantas de Cedrela fissilis submetidas à pleno sol. No
tratamento alagado, a diferença na área foliar total, em relação ao tratamento não-
alagado, indica que a produção de folhas novas bem como a expansão de folhas já
existentes são afetadas negativamente pelo alagamento (KOZLOWSKI et al., 1991;
KOZLOWSKI; PALLARDY 1997).
A área foliar específica é um parâmetro que relaciona a área foliar individual e
a biomassa seca da folha (DIAS-FILHO, 1997), ou seja, é a área foliar por unidade
de massa da foliar (KITAJIMA, 1996). Embora não tenha havido diferenças
intergenotípicas entre as plantas não alagadas (Figura 27), houve aumento da área
foliar específica à medida que se reduziu a irradiância disponível. Tal incremento da
área foliar específica é uma resposta comum observadas em plantas sob baixas
irradiâncias (FAHL et al., 1994; STONEMAN; DELL, 1993), pois, é reflexo de
modificações nas dimensões e formas das folhas que tornam-se maiores e mais
delgadas do que folhas produzidas sob altas intensidades de luz. Por outro lado, sob
anoxia, o aumento na área foliar específica, em ambos os genótipos clonais

87
avaliados, demonstra que o cacau tende a reter mais fotoassimilados nas folhas em
situação de estresse.
A razão de área foliar (RAF) (área foliar total/biomassa seca total) é a área
foliar, em cm2, utilizada pela planta para produzir 1 g de biomassa seca
(BENINCASA, 1988). Desse modo, plantas com sombreamento mais intenso tendem
a possuir maior RAF, pois necessitam de maior área foliar para interceptação da
radiação total incidente. A tendência ao aumento de RAF em plantas não alagadas e
mais sombreadas (Figura 28) está de acordo com Kitajima (1996), que verificou que
plantas arbóreas tropicais apresentavam baixo investimento em raízes e elevada
RAF. Os genótipos TSA-792 e TSH-774 apresentaram, no tratamento alagado,
valores de RAF mais baixos em comparação com o tratamento não-alagado. Rehem
et al. (2009), avaliando o efeito do alagamento do substrato no crescimento de
genótipos clonais de cacau, verificaram que os clones CCN-10 e CP-49, tolerantes
ao alagamento, caracterizaram-se por apresentar baixos valores de RAF,
favorecendo a obtenção de altas taxas fotossintéticas.
A razão de massa foliar (RMF) é a razão entre a biomassa seca acumulada
nas folhas e a biomassa seca acumulada na planta toda. Como as folhas (fonte)
produzem biomassa seca a partir da fotossíntese e que os demais órgãos (drenos)
dependem da exportação de fotoassimilados, quanto maior o valor de RMF maior é
a quantidade de material não exportado. No presente trabalho, verificaram-se
diferenças em RMF apenas entre os genótipos e o regime hídrico (Figura 29). No
tratamento não-alagado, o genótipo TSA-792 acumulou mais biomassa seca foliar
do que o genótipo TSH-774, e no alagado essa diferença foi observada somente nos
tratamentos de sombreamento T25 e T50. No tratamento alagado, as plantas dos
genótipos clonais avaliados apresentaram menor RMF em relação as não alagadas,
indicando que mais biomassa seca foi distribuída para raízes e caules do que para
os órgãos fotossintéticos. Esse fato pode ser confirmado na Figura 25 que
demonstra a proporção de biomassa seca total que foi alocada para a raiz, o caule e
folha.
Segundo o mecanismo de alocação de biomassa raiz/parte aérea, sugerido
por Brouwer (1983), as plantas direcionam a biomassa para a parte aérea, se a
assimilação de carbono é limitada por luz e CO2, ou para as raízes se os recursos
limitantes são nutrientes minerais e água. Por isso, espera-se que sob altas

88
irradiâncias a alocação de biomassa para as folhas decresça e aumente para as
raízes, devido à maior demanda evapotranspiratória, sendo o inverso para baixas
irradiâncias. Esse modelo de alocação não se aplica ao observado no presente
estudo, já que, de acordo com a ANOVA (Tabela 7), não houve influência dos
tratamentos de sombreamento na razão raiz/parte aérea.
Plantas alagadas do genótipo clonal TSH-774 apresentaram uma tendência
de acréscimo na razão raiz/parte aérea, enquanto que para o genótipo TSA-792
essa razão se manteve constante (Figura 30). Esse resultado indica que o
crescimento da parte aérea, para o genótipo TSH-774, foi reduzido em relação a
raiz, ao passo que para o genótipo TSA-792 o balanço entre o crescimento de raiz e
parte aérea foi mantido constante. Segundo Kozlowski (1997) a raiz é o órgão mais
diretamente afetado pelo alagamento do solo, sendo, tipicamente, mais reduzido do
que a parte aérea. A supressão na formação e expansão das folhas deve ter
provocado a redução no crescimento da parte aérea em plantas do genótipo TSH-
774. Ferreira et al. (2001), avaliando plântulas de Piptadenia gonoacantha com cinco
meses de idade, submetidas a dois regimes hídricos (não-alagado e alagado) e três
níveis de irradiância, verificaram que plântulas alagadas apresentaram, aos 60 dias
de aplicação dos tratamentos, menor razão raiz/parte aérea.
A classificação das espécies em tolerantes ao alagamento deve ser baseada
na capacidade em manter ou aumentar a biomassa seca da parte aérea durante
períodos de alagamento, segundo critério sugerido por Lobo e Joly (1998). Nesse
caso, plantas alagadas do genótipo clonal TSH-774 mostraram-se pouco tolerantes
ao alagamento do solo, devido à redução da biomassa da parte aérea em relação ao
sistema radicular (Figura 30).
O acúmulo de carboidratos durante o crescimento é essencial para a
sobrevivência das plantas. O armazenamento de carboidratos é importante tanto
para o metabolismo, germinação da semente, sobrevivência e crescimento da
plântula quanto para defesa (KOZLOWSKI, 1992). A redução observada no teor de
AST nas plantas não alagadas, em função do sombreamento, para ambos os
genótipos coincidiu com a redução no crescimento em altura, indicando que os AST
não foram direcionados dos órgãos fonte para o crescimento em altura das plantas
como forma de evitar o sombreamento.

89
Uma proporção maior de AST ficou alocada nas folhas das plantas dos
genótipos clonais submetidos ao alagamento do solo (Figura 33). Segundo
Kozlowski (1997) e Pezeshki (1994), a translocação de açúcares solúveis das folhas
para as raízes é bastante afetada pelo alagamento do solo. Deste modo, os
açúcares solúveis produzidos na fotossíntese tendem a se acumular nas folhas, e
não são translocados para as raízes, onde são necessários para manter a via
glicolítica em funcionamento. Portanto, a maior disponibilidade de substrato
respiratório, como a glicose, pode ser determinante na sobrevivência dos tecidos
radiculares em ambientes alagados (CRAWFORD; BRAENDLE, 1996; DREW, 1997;
VARTAPETIAN; JACKSON, 1997). Os resultados correspondem aos encontrados
por Carvalho e Ishida (2002), que observaram maior acúmulo de AST na folha e
aumento em raízes de Bactris gasipaes submetidas ao alagamento do solo.
Nas raízes de plantas alagadas do genótipo clonal TSA-792, observaram-se
aumento do teor de AST (Figura 31) em função do nível de sombreamento e
manutenção do teor de amido (Figura 34). Porém, para o genótipo TSH-774 não
houve resposta em relação ao teor de AST, enquanto o teor de amido manteve-se
abaixo daquele observado para o genótipo TSA-792. Possivelmente, plantas do
genótipo TSA-792 sintetizam mais açúcares solúveis como forma de tolerância ao
estresse por alagamento, indicando uma eficiente atividade fotossintética mesmo
sob deficiência de O2 no sistema radicular. Segundo Menezes et al. (2006), o
aumento na concentração de açúcares é conseqüência da hidrólise do amido,
mecanismo pelo qual as plantas adquirem tolerância ao estresse hídrico.
Nas plantas não alagadas de ambos os genótipos clonais de T. cacao, com
ou sem sombreamento, o alto teor de amido no caule (Figura 35), em relação às
raízes e folhas (Figuras 35 e 37), sugerem que o caule parece ser o órgão de
estocagem de amido na planta. Em folhas de plantas alagadas destes genótipos, o
teor de amido foi muito baixo em relação ao caule e similar ao teor de amido nas
raízes. Segundo Liao e Lin (2001), um grande acúmulo de amido nas folhas de
plantas sob estresse abiótico poderia causar inibição da fotossíntese (feedback
inhibition). Entretanto, outros fatores como redução na condutância estomática e no
teor de clorofila total nas plantas alagadas pode ter contribuído para o decréscimo
na atividade fotossintética, observado nos dois genótipos clonais de cacau
avaliados.

90
5.2. Expressão gênica da aquaporina PIP1;3 em plant as submetidas ao
sombreamento e ao alagamento do solo
A descoberta das aquaporinas alterou a visão de como a água é transportada
rapidamente através da membrana e como a planta é capaz de regular o fluxo de
água em diferentes condições fisiológicas. As aquaporinas desempenham também
um importante papel em resposta ao estresse abiótico. Caracterização molecular e
funcional de aquaporinas tem demonstrado sua importância na regulação da rota
transcelular de água, em resposta a fatores abióticos adversos como alagamento,
seca, salinidade e temperatura (ZHAO et al, 2007).
Jang et al. (2004) avaliaram a expressão de 13 genes de aquaporinas em
plântulas de Arabidopsis thaliana submetidas à estresses abióticos e classificaram o
gene PIP1;3 como de expressão intermediária, apresentando 2000 – 5000 cópias
por nanograma de RNA total. Fraysse et al. (2005), utilizando a técnica de
imunolocalização em células de raiz, caule e folha de Spinacia oleracea,
demonstraram que a aquaporina SoPIP1;1, homóloga de AtPIP1;3 em A. thaliana, é
altamente expressa em células-guarda da epiderme foliar. Resultados similares
foram encontrados por Sun et al. (2001) que, por meio da técnica de hibridização in
situ de RNA, afirmaram que células-guarda de Vicia faba apresentam altos níveis de
expressão de aquaporinas da subfamília PIP1 e, provavelmente, estão envolvidas
no controle do movimento estomático. De acordo com Sarda et al. (1997),
aquaporinas pertencentes às subfamílias PIP e TIP são induzidas em células-guarda
de plantas de Helianthus annuus sob estresse hídrico.
A indução da expressão gênica de PIP1;3, observada em plantas clonais do
genótipo TSA-792, submetidas ao alagamento do solo, possivelmente contribuiu
para a regulação do volume e turgor das células-guarda em nível foliar; uma vez que
as relações hídricas nessas células são similares às demais, ou seja, à medida que
o potencial osmótico decresce, como resultado da absorção iônica, a água move-se
para dentro das células-guarda (TAIZ; ZEIGER, 2004) via aquaporinas e a pressão
de turgor aumenta mantendo os ostíolos abertos. Verificou-se que o genótipo TSA-
792, considerado resistente ao alagamento do solo, apresentou taxas fotossintética
(A) e transpiratória (E) e condutância estomática (gs) eficientes aos 48 dias de
interação sombreamento x alagamento do solo, quando comparado ao genótipo

91
TSH-774, cujos valores de A, E e gs se aproximaram de zero e os níveis de
expressão de PIP1;3 foram equivalentes ao tratamento não-alagado.
Sob condições normais de temperatura e umidade relativa do ar, com água e
radiação solar disponível, a fotossíntese é favorecida e os estômatos encontram-se
amplamente abertos. Como os estudos têm demonstrado que as aquaporinas
participam dos mecanismos de abertura e fechamento do poro estomático
(FRAYSSE et al., 2005; SUN et al., 2001; SARDA et al., 1997), sugere-se que a
repressão do gene PIP1;3, observada em plantas não-alagadas de ambos os
genótipos clonais de cacau, é uma resposta esperada; já que, em condições
ambientais ótimas, os genes que apresentam altos níveis de expressão, em
resposta a estresses ambientais diversos, são reprimidos e a síntese protéica é
verificada somente em níveis necessários à manutenção celular.
Apesar de o mecanismo que regula o movimento estomático ser bem
documentado, os sinais que regulam as aquaporinas, nesse processo, ainda são
desconhecidos. Entretanto, os resultados do perfil de expressão do gene de
aquaporina PIP1;3 forneceram novas informações acerca das respostas de
genótipos clonais de cacau aos estresses abióticos estudados, tornando-se ponto de
partida para avaliações futuras do papel de cada aquaporina nas relações hídricas
do cacaueiro.

92
6. CONCLUSÕES
O alagamento do solo foi o principal fator estressor, para a maioria das
respostas fisiológicas observadas, em ambos os genótipos de cacau avaliados,
durante a interação entre sombreamento e alagamento do solo.
O tempo de exposição ao alagamento do solo e a intensidade de radiação
luminosa interferiram nas trocas gasosas e na emissão de fluorescência da clorofila
em nível foliar, cujos ajustes fisiológicos foram distintos entre os genótipos clonais
de cacau avaliados.
As respostas fisiológicas, resultantes da interação entre níveis de
sombreamento e alagamento do solo, foram atenuadas para o genótipo clonal TSA-
792, que apresentou crescimento e desenvolvimento superiores em relação ao
genótipo clonal TSH-774.
A relação observada entre os níveis de expressão gênica e os parâmetros de
trocas gasosas foliares permitiu afirmar que a expressão do gene PIP1;3 é
modulada por anoxia, principalmente para o genótipo clonal de cacau TSA-792,
considerado resistente ao alagamento do solo.

93
7. REFERÊNCIAS
AGRE, P.; SASAKI, S.; CHRISPEELS, M. Aquaporins: a family of water channel
proteins. American Journal of Physiology, v. 261, p. 461, 1993.
AHARON, R.; SHAHAK, Y.; WININGER, S. Overexpression of a plasma membrane
aquaporin in transgenic tobacco improves plant vigor under favorable growth
conditions but not under drought or salt stress. Plant Cell , v. 15, p.439-447, 2003.
AIME, M. C.; MORA, W. P. The causal agents of witches’ broom and frosty pod rot of
cacao (chocolate, Theobroma cacao) from a new lineage Marasmiaceae. Mycologia .
v. 97, n. 5, p. 1012 – 1022, 2005.
AKENKORAH, Y.; AKROFI, G. S.; ADRI, A.K. The end of the first cocoa shade and
manorial experiment at the Cocoa Research Institute of Ghana. Journal of
Horticultural Science , v.49, p. 43 – 51, 1974.
ALEMANNO, L.; RAMOS, T.; GARGADENEC, A.; ANDARY, C.; FERRIERI, N.
Localization and indetification of phenolic compounds in Theobroma cacao L.
somatic embryogenesis. Annals of Botany, v. 92, p. 613-623, 2003.
ALEXANDERSSON, E.; FRAYSSE, L., SJÖVALL-LARSEN, S., GUSTAVSSON, S.,
FELLERT, M., KARLSSON, M., JOHANSON, U., KJELLBOM, P. Whole gene family
expression and drought stress regulation of aquaporins. Plant Molecular Biology , v.
59, p. 469–484, 2005.

94
ALMEIDA, S. M. Z., SOARES, A. M., CASTRO, E. M., VIEIRA, C. V., GAJEGO, E. B.
Alterações morfológicas e alocação de biomassa em plantas jovens de espécies
florestais sob diferentes condições de sombreamento. Ciência Rural , v. 35, n.1, p-
62-68, 2005.
ALMEIDA, S.S.; ARAGÃO, I. L. G.; SILVA, P. J. D. Efeito de clareiras naturais na
estrutura de plântulas de Vochysia guianensis Aubl. (Vochysiaceae) em floresta 50
amazônica de terra firme. Boletim Museu Paraense Emilio Goeldi, Nova Serie
Botânica , v.10, p.91-103, Belém. 1994.
ALMEIDA, L. P. de, ALVARENGA, A. A. de; CASTRO, E. M. de; ZANELA, S.
M.;VIEIRA, C. V. Crescimento inicial de plantas de Cryptocaria aschersoniana Mez.
submetidas a níveis de radiação solar. Ciência Rural , Santa Maria, v. 34, n. 1, p. 83-
88, 2004.
ALMEIDA, A-A. F., VALLE, R. R. Ecophysiology of the cacao tree. Brazilian Journal
of Plant Physiology , v. 19, n. 4, p. 425-448, 2007.
ANDERSON, J.M., Photoregulation of the composition, function, and structure of
thylakoid membranes. Annual Review of Plant Physiology . v. 37, p. 93-136. 1986.
ANDERSON, J.M.; OSMOND, C.B. Shade-sun responses: compromises between
acclimation and photoinhibition. In: KYLE, D.J.; OSMOND C.B.; ARNTZEN, C.J.
(Eds.) Photoinhibition. Amsterdam: Elsevier. p 1-38, 1987.
ALVERSON, W. S.; WHITLOCK, B. A.; NYFFELER, R.; BAYER, C.; BAUM, D. A.
Phylogeny of the core Malvales: evidence from ndhF sequence data. American
Journal of Botany , n. 86, p.1474-1486, 1999.
ALVIM, P. T. El problema del sombreamiento del cacao desde el punto de vista
fisiológico. Agronomia (Peru), n. 25, p. 34 – 42, 1958.

95
ALVIM P. DE T CACAO. In: ALVIM, P. DE T. AND KOZLOWSKI, T.T. (eds)
Ecophysiology of Tropical Crops , p. 279–313, Academic Press, New York, 1977.
AMODEO, G.; DORR, R.; VALLEJO, A.; STUKA, M.; PARISI, M. Radial and axial
water transport in the sugar beet storage root. Journal of Experimental Botany, v.
50, p. 509 – 516, 1999.
ANIM-KWAPONG, G. J. Potential of some neotropical Albizia species as shade trees
when replanting cacao in Ghana. Agroforest Systems , n. 58, p.185-193, 2003.
ARMSTRONG, W.; BRÄNDLE, R.; JACKSON, M. B. Mechanisms of flood tolerance
in plants. Acta Botanica Neerlandica , v.43, p.307-358, 1994.
ARNON, D. I. Cooper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta
vulgaris. Plant Physiology , Bethesda, v. 24, n. 1, p. 1-15, 1949.
ASHRAF, M. Relationships between leaf gas exchange characteristics and growth of
differently adapted populations of Blue panicgrass (Panicum antidotale Retz) under
salinity or waterlogging. Plant Science , v. 165, p. 69-75, 2003.
ATROCH, E. M. A. C.; SOARES, A. M.; ALVARENGA, A. A.; CASTRO, E. M.
Crescimento, teor de clorofilas, distribuição de biomassa e características
anatômicas de plantas jovens de Bauhinia forticata submetidas a diferentes
condições de sombreamento. Ciência e Agrotecnologia , Lavras, v. 25, n. 4, p. 853-
862, 2001.
AUGUSTO, S. G. Irrigação complementar nos diferentes estádios fenológicos do
cacaueiro (Theobroma cacao L.).UFV, Viçosa (Tese de Doutorado), 1997.
BAIGES, I., SCHAFFNER, A.R.; MAS, A. Eight cDNA encoding putative in Vitis
hybrid Richter-110 and their differential expression. Journal of Experimental
Botany v. 52, p. 1949–1951, 2001.

96
BAJJI, M.; KINET, J-M.; LUTTS, S. The use of the electrolyte leakage method for
assessing cell membrane stability as a water stress tolerance test in durum wheat.
Plant Growth Regulation. v.0, p.1-10, 2001.
BARBOSA, A. P. O. O crescimento de mudas de caroba (Jacaranda copaia Aubl. D.
Don. Bignoniaceae) cultivadas sob diferentes níveis de sombreamento e
espaçamento. In: Anais do 6º Congresso Florestal Brasileiro . Campos do Jordão:
Sociedade Brasileira de Silvicultura, p. 526-534, 1990.
BARTLEY, B. G. D. The genetic diversity of cacao and its utilization. CABI
Publishing, Wallingford, UK, 2005.
BAZZAZ, F.A., CARLSON, R.W..Photosynthetic acclimation to variability in light
environment of early and late successional plants. Oecologia , n. 54, p. 313-316,
1982
BAJJI, M.; KINET, J-M.; LUTTS, S. The use of the electrolyte leakage method for
assessing cell membrane stability as a water stress tolerance test in durum wheat.
Plant Growth Regulation. v.0, p.1-10, 2001.
BAILEY-SERRES, J. & VOESENEK, L. A. C. J. Flooding stress: aclimatations and
genetic diversity. Annual Review Plant Biology, n. 59, p. 313-339, 2008.
BAKER, N.R. Chlorophyll Fluorescence: A probe of photosynthesis in vivo. Annual
Review Plant Biology , n. 59, p.89–113, 2008.
BEER, J. Advantages, disadvantages and desirable characteristics of shade trees for
coffee, cacao and tea. Agroforest Systems, v. 5, p.3–13, 1987.
BENINCASA, M. M. P. Análise de crescimento de plantas: noções básicas. Jaboticabal: FUNEP, 1988. 41p.

97
BERTOLDE, F. Z., ALMEIDA, A-A. F., CORRÊA, R. X., GOMES, F. P., GAIOTTO, F.
A., BALIGAR, V. C., LOGUÉRCIO, L., L. Molecular,physiological and morphological
analysis of waterlogging tolerance in clonal genotypes of Theobroma cacao L. Tree
Physiology, v. 30, n.1, p. 56-67, 2010.
BEURON, F.,CAHÉREC, F. L., GUILLAM, M. T., CAVALIERA, G., TASSAN, J. P.,
DELAMAR, C. H. E. C., SCHULTZ, P., MALLOU, H. V., ROLLAND, J. P.,HUBERT,
J. F., GOURANTON, J., THOMAS, D. Structural analysis of a MIP family protein from
the digestive tract of Cicadella viridis. Journal of Biological Chemistry , n. 270, p.
17414–17422, 1995.
BIELA, A.; GROTE, K.; OTTO, B.; HOTH, S.; HEDRICH, R.; KALDENHOFF, R. The
Nicotiana tabacum plasma membrane aquaporin NtAQP1 is mercury insensitive and
permeable for glycerol, Plant Journal , n.18, p.565–570, 1999.
BJORKMAN, O. Carboxydismutase activity in shade-adapted and sun adapted
species of higher plants. Physiologia Plantarum , n.21, p.1-10, 1968.
BJÖRKMAN, O. Responses to different quantum flux densities. In: Encyclopedia of
Plant Physiology . Vol. I. New Series. LANGE, O.L.; NOBEL, P.S.; OSMOND, C.B.;
ZIEGLER, H. (Eds.) Berlin: Springer-Verlag. p. 57-107, 1981.
BJÖRKMAN, O., BOARDMAN, N.K., ANDERSON, J.M., THORNE, S.W.,
GOODCHILD, D.J., PYLIOTIS, N.A. Effect of light intensity during growth of Atriplex
patula on the capacity of photosynthetic reactions, chloroplast components and
structure. Carnegie Institute Washington Year Book . v. 71, p. 115-135, 1972.
BLOM, C.W.P.M. & VOESENEK, L.A.C.J. Flooding: the survival strategies of plants.
Trends Ecology and Evolution , v. 11, p. 290-295, 1996.
BOARDMAN, N. K. Comparative Photosynthesis of sun and shade plants. Annual
Review of Plant Physiology , n. 28, p. 355-377, 1977.

98
BOLHÀR-NORDENKAMPF, H.R.; LONG, S.P., BAKER, N.R., et al. Chlorophyll
fluorescence as probe of the photosynthetic competence of leaves in the field: a
review of current instrument. Functional Ecology , v.3, p.497-514, 1989.
BOTS, M., FERON, R., UEHLEIN, N., WETERINGS, K., KALDENHOFF, R.,
MARIANI, T. PIP1 and PIP2 aquaporins are differentially expressed during tobacco
anther and stigma development, Journal Experimental Botany , n. 56, p. 113–121,
2005.
BOURSIAC Y, CHEN, S., LUU, D. T., SORIEUL, M., VANDENDRIES, N., MAUREL
C. Early effects of salinity on water transport in Arabidopsis roots: molecular and
cellular features of aquaporin expression. Plant Physiology , v. 139, 790–805, 2005.
BRADFORD, K.J.; YANG, S.F. Xylem transport of 1-aminocyclopropano-1-carboxylic
acid an ethylene precursor, in waterlogged tomato plants. Plant Physiology , v. 65,
p. 322-326, 1980.
BRAGA, J. M.; DEFELIPO, B.. Determinação espectrofotométrica de fósforo em
extratos de solos e plantas. Revista Ceres , v. 21, p. 73-85, 1974.
BRITO, G. G., LIMA, M. M. A., CARVALHO, L. P., SOFIATTI, V., OLIVEIRA, F. M,
SILVA, V. N. B. Indicadores fisiológicos como ferramenta em screening para
tolerância ao déficit hídrico em algodão. Congresso brasileiro do algodão. VII
Congresso Brasileiro do Algodão, Foz do Iguaçu, PR, p. 836-845, 2009.
BROKAW, N.V.L. Gap-phase regeneration of three pioneer tree species in a tropical
forest. Journal of Ecology , n. 75, p. 9-19, 1987.
BROUWER, R. Distribution of dry matter in the plant. Netherlands Journal of
Agricultural Science , v.10, p.361-376, 1962.

99
BYRNE, P. N. Cacao shade spacing and fertilizing trial in Papua and New Guinea.
In: International Cocoa Research and Conference , St. Augustine, Tri. p. 275 –
286, 1972.
CABALA, P.; MIRANDA, E. R., SANTANA, C. J. L. Interacción sombra fertilizantes
em cacao tales de Bahia. In: International Cocoa Research and St. Augustine, Tri.
p. 181 – 189, 1972.
CANCIAN, M. A. E.; CORDEIRO, L. Efeito do sombreamento no crescimento inicial
de Lonchocarpus muehlbergianus Hassl. Acta Botanica Brasilica , n. 12, p. 367-
373, 1998.
CALAMITA, G.; BISHAI, W. R.; PRESTON, G. M.; GUGGINO, W. B.; AGRE, P.
Molecular cloning and characterization of AqpZ, a water channel from Escherichia
coli. Journal of Biological Chemistry , n. 270, p. 29063–29066, 1995.
CAMPOS, M.A.S.; UCHIDA, T. Infuência do sombreamento no crescimento de
mudas de três espécies amazônicas. Pesquisa Agropecuária Brasileira , v. 37, n.
3, p. 281-288, 2002.
CARBREY, J. M., BONHIVER, S. M., BOEKE, J. D., AGRE, P. Aquaporins in
Saccharomyces: characterization of a second functional water channel protein. Proc
Natl Acad Sci n. 98, p.1000–1005, 2001.
CARELLI, M.L.C.; FAHL, J.I. Crecimiento y asimilación del carbono y nitrógeno en
plantas jóvenes de Coffea en condiciones de sol y de sombra. In: XIX Simposio
Latinoamericao de Caficultura , Costa Rica. ICAFE. Memoria, p.101-108, 2000.
CARVALHO, C. J. R. de; ISHIDA, F. Y. Respostas de pupunheiras (Bactris gasipaes
Kunth) jovens ao alagamento. Pesquisa Agropecuária Brasileira , v. 37, n. 9, p.
1231-1237, 2002.

100
CARVALHO, P. E. R. Influência da intensidade luminosa e do substrato no
crescimento,no conteúdo de clorofila e na fotossíntese de Cabralea canjerana (Vell.)
MART. Subsp. Canjerana, Calophyllum brasiliense CAMB. e Centrolobium robustum
(Vell) MART. EX Benth., na fase juvenil. 1996. 157 f. Tese (Doutorado) -
Universidade Federal do Paraná, Curitiba.
CHAUMONT, F.; BARRIEU, F.; JUNG, R.; CHRISPEELS, M. J. Plasma membrane
intrinsic proteins from maize cluster in two sequence subgroups with differential
aquaporin activity. Plant Physiology , n.122, p.1025–1034, 2000.
CHIRKOVA, T. V.; T. S. GUTMAN. Physiological role of branch lenticels in willow and
poplarunder conditions of root anaerobiosis. Soviet Pl. Physiol , n. 19, p. 289 – 295,
1972.
CHOI, W. G.; ROBERTS, D. M. Arabidopsis NIP2;1, a major intrinsic protein
transporter of lactic acid induced by anoxic stress. Journal of Biological Chemistry ,
n. 282, p. 24209–18, 2007.
CRAWFORD, R. M. M.; BRAENDLE, R. Oxygen deprivation stress in a changing
environment. Journal of Experimental Botany, v. 47, n. 295, p. 145-159, 1996.
CRONQUIST, A. An integrated system of classification of flowering plants. New York,
Columbia University Press., 1981. 1262p.
COHEN, E.; KENDE, H. In vivo 1-aminocyclopropane-1-carboxylate synthase activity
in internodes of deepwater rice. enhancement by submergence and low oxygen
levels. Plant Physiology , v. 84, p. 282–286, 1987.
CUATRECASAS, J. Cacao and its allies: a taxonomic revision of the genus
Theobroma. Contributions from the United States National Herba rium . v.35, p.
379-614, 1964.

101
CUNNINGHAM, R. K.; BURRIDGE, J. C. The growth of cacao (Theobroma cacao)
with and without shade. Annals of Botany . v. 24, n. 96, p.458 – 462, 1960.
DaMATTA, F. M. Exploring drought tolerance in coffee: a physiological approach with
some insights for plant breeding. Brazilian Journal of Plant Physiology , v. 16, p. 1-
6, 2004.
DAKWA, J. T. The effects of shade and NPK fertilizers on the incidence of cocoa
black pod disease in Ghana. Horticultural Abstracts n. 50, v.3, p. 187, 1980.
DAVIES, F. S; FLORE, J. A. short-term flooding effects on gas exchange and
quantum yield of rabbit eye blueberry (Vaccinium ashei Reade). Plant Physiology .
v. 81, p. 289-292, 1986.
DENNIS, E. S.; DOLFERUS, R.; ELLIS, M.; RAHMAN, M.; WU, Y.; HOEREN, F. U;
GROOVER, A.; ISMOND, K. P.; GOOD, A. G.; PEACOCK, W. J. Molecular
strategies for improving waterlogging tolerance in plants. Journal of Experimental
Botany. v. 51, p. 89-97, 2000.
DIAS, L. A. S. Melhoramento genético do cacaueiro. Viçosa, MG, FUNAPE, UFG. 2001, 501 p. DIAS-FILHO, M.B. Physiologycal response of Solanum crinitum Lam. to contrasting
light environments. Pesquisa Agropececuária Brasileira, v. 32, p. 789-796, 1997.
DOLFERUS, R., KLOK, E. J., DELESSERT, C., WILSON, S., ISMOND, K. P.,
GOOD, A. G., PEACOCK, W. J., DENNIS, E. S. Enhancing the anaerobic response.
Annals of Botany , n. 91, p. 111-117, 2003.
DREW, M. C. Oxygen deficiency in the root environmentand plant mineral nutrition.
In: JACKSON, M. B.; DAVIS, D. D.; LAMBERS, H. (Ed.). Plant life under oxygen

102
deprivation: ecology, physiology and biochemistry . The Hague: SPB Academic,
p. 303-316, 1991.
EHLERINGER, J. Leaf absorptances of Mohave and Sonoran desert plants.
Oecologia , n. 49, p.366-370, 1981.
ELSE, M. A.; TIEKSTRA, A. E.; CROKER, S. J.; DAVIES, W. J.; JACKSON, M. B.
Stomatal closure in flooded tomato plants involves abscisic acid and a chemically
unidentified anti-transpirant in xylem sap. Plant Physiology . v. 112, p. 239-247,
1996.
ENGEL, V. L., POGGIANI, F. Influência do sombreamento sobre o crescimento de
mudas de algumas essencias nativas e suas implicações ecológicas e silviculturais.
IPEF, n. 43, v. 44, p. 1-10, 1990.
ENGEL, V. L.; POGGIANI, F. Estudo da concentração de clorofila nas folhas e seu
espectro de absorção de luz em função do sombreamento em mudas de quatro
espécies florestais nativas. Revista Brasileira de Fisiologia Vegetal . v. 3, n. 1, p.
39 - 45, 1991.
ENRIQUEZ, G. A. Respuesta del cacao hibrido a dos sistemas de sombra em
Turrialba, Costa Rica. In: Proceedings Seminar Advances in Agroforestry
Research. J. W. BEER; H. W. FASSBENDER (Eds.), Turrialba, C. R., CATIE, p. 141
– 154, 1986.
FAHL, J. I.; CARELLI, M. L. C.; VEGA, J.; MAGALHÃES, A. C. Nitrogen and
irradiance levels affecting net photosynthesis and growth of young coffee plants
(Coffea arabica L.). Journal of Horticultural science , v. 69, p. 161-169, 1994.
FARIAS, V.C.C.; COSTA, S. S.; BATALHA, L. F. P. Análise de crescimento de
mudas de cedrorana (Cedrelinga catenae formis (Duke) Deike) cultivadas em
condições de viveiro. Revista Brasileira de Sementes , v. 19, n.2, p. 193-200, 1997.

103
FARQUHAR, G.D.; SHARKEY, T.D. Stomatal conductance and photosynthesis.
Annual Review of Plant Physiology, v. 33, p.317-345, 1982.
FERRER, R. S.; DILLENBURG, L. R. Efeitos da disponibilidade de luz no
crescimento inicial e ecofisiologia de Scutia buxifolia Reissek (Rhamnaceae).
Hoehnea , n. 27, p. 147-157, 2000.
FERREIRA, J. N.; RIBEIRO, J. F.; FONSECA, C. E. L. Crescimento inicial de
Piptadenia gonoacantha (Leguminosae, Mimosoideae) sob inundação em diferentes
nÌveis de luminosidade. Revista Brasileira de Botânica , v. 24, n.4 (suplemento), p.
561-566, 2001.
FETCHER, N.; STRAIN, B.R.; OBERBAUER, S. F. Effect of light regime on the
growth, leaf morphology, and water relations of seedlings of two species of tropical
trees. Oecologia , n. 58, p. 314-319, 1983.
FETTER, K.; VANWILDER, V.; MOSHELION, M.; CHAUMONT, F. Interactions
between plasma membrane aquaporins modulate their water channel activity. Plant
Cell, n.16, p. 215–228, 2004.
FLEXAS, J.; RIBAS-CARBÓ, M.; HANSON, D. T.; BOTA, J.; OTTO, B.; CIFRE, J.;
MCDOWELL, N.; MEDRANO, H.; KALDENHOFF, R. Tobacco aquaporin NtAQP1 is
involved in mesophyll conductanve to CO2 in vivo. Plant Journal .
doi:10.1111/j.1365-313X.2006.02879.x, 2006.
FONSECA, E. P.; VALÉRI, S. V.; MIGLIORANZA, E.; FONSECA, N. A. N.; COUTO
L. Padrão de qualidade de mudas de Trema micrantha (L.) Blume, produzidas sob
diferentes períodos de sombreamento. Revista Árvore , n. 26, v. 4, p. 515-523,
2002.
FRAYSSE,L.; WELLS, B.; MCCANN, M. C.; KJELLBOM, P. Specific plasma
membrane aquaporins of the PIP1 subfamily are expressed in sieve elements and
guard cells. Biology Cell. n. 97, p. 519–34, 2005.

104
FREITAS, R. B., OLIVIERA, L. E. M., FILHO, N. D., SOARES, A. M. Influência de
diferentes níveis de sombreamento no comportamento fisiológico de cultivares de
cafeeiro (Coffea arabica L.). Ciência. Agrotécnica de Lavras , v .27, n.4, p.804-810,
jul./ago., 2003
FUKAO, T.; BAILEY-SERRES, J. Plant response to hypoxia – is survival a balancing
act ?. Trends in Plant Science , v. 9, p. 449-456, 2004.
GERBEAU, P.; GUCLU, J.; RIPOCHE, P.; MAUREL, C. Aquaporin Nt-TIP a can
account for the high permeability of tobacco cell vacuolar membrane to small neutral
solutes. Plant Journal , n. 18, p. 577–587, 1999.
GIAVENO, C. D.; OLIVEIRA, R. F. Apostila Estresse Ambiental: Conceitos Gerais.
Depto de Ciências Biológicas da ESALQ/USP – Lab. Fisiologia de Plantas
Cultivadas sob Condições de Estresse. Piracicaba, SP. 22 p., 2003. Disponível em:
http://www.sbfv.org.br/materialdidatico.php.
GRAMACHO, I. C. P.; MAGNO, A. E. S.; MANDARINO, E. P.; MATOS, A. Cultivo e
beneficiamento do cacau na Bahia. CEPLAC, Ilhéus.1992.
GRAVATT, D. A.; KIRBY, C. J. Patterms of photosynthesis and starch allocation in
seedlings of four bottomland hardwood tree species subjected to flooding. Tree
Physiology , v. 18, p. 411-417, 1998.
HANBA, Y. T., SHIBASAKA, M., HAYASHI, Y., HAYAKAWA, T., KASAMO, K.,
TERASHIMA, I., KATSUHARA, M. Overexpression of the barley aquaporin HvPIP2;1
increases internal CO2 conductance and CO2 assimilation in the leaves of transgenic
rice plants. Plant Cell Physiology , n. 45, p. 521–529, 2004.
HARTEMINK, A. E. Nutrient stocks, nutrient cycling, and soil changes in cocoa
ecosystems: a review. Adv. Agro. n. 86, p. 227–253, 2005.

105
HENZLER, T.; WATERHOUSE, R. N.; SMYTH, A. J.; CARVAJAL, M.; COOKE, D.
T.; SCHÄFFNER, A. R.; STEUDLE, E.; CLARKSON, D. T. Diurnal variations in
hydraulic conductivity and root pressure can be correlated with the expression of
putative aquaporins in the roots of Lotus japonicus. Planta . v. 210, p. 50 – 60, 1999.
HIPKINS, M. F.; BAKER, N. R. Photosynthesis energy transduction a practical
approach. Eds Hipkins, M. F. and Baker, N. R. IRL Press, Oxford, England, 1986.
HISCOX, J.D., ISRAELSTAM, G. F. Different methods of chlorophyll extraction. Can.
J. Bot., 57: 1332-1332, 1979.
HOLM, L. M., JAHN, T. P., MOLLER, A. L., SCHJOERRING, J. K., FERRI, D.,
KLAERKE, D. A., ZEUTHEN, T. NH3 and NH4+ permeability in aquaporin- expressing
Xenopus oocytes, Pflugers Arch , n. 450, p. 415–428, 2005.
HUANG, B.; JOHNSON, J. W.; NESMITH, S., BRIDGE, D.C. Growth, physiological
and anatomical responses of two wheat genotypes to waterlogging and nutrient
supply. Journal of Experimental Botany, v. 45, p.193-202, 1994.
ICCO, INTERNATIONAL COCOA ORGANIZA TION. Quarterly Bulletin of Cocoa
Statistic. Londres, ICCO, Vol XXXI n.2, 2005. 73 p.
ICCO. Annual report, 2007. 43 p.
ISHIDA, F. Y.; OLIVEIRA, L. E. M. de; CARVALHO, C. J. R. de; ALVES, J. D. Efeitos
da inundação parcial e total sobre o crscimento, teor de clorofila e fluorescência de
Setaria anceps e Paspalum repens. Ciência Agrotécnica , v. 26, n. 6, p. 1152-1159,
2002.
ISHIKAWA, F., SUGA, S., UEMURA, T., SATO, M. H., MAESHIMA, M., Novel type
Aquaporin SIPs are mainly localized to the ER membrane and show cell-Specific
expression in Arabidopsis thaliana, FEBS Lett., n. 579, p. 5814–5820, 2005.

106
ITO, O.; ELLA, E.; KAWANO, N;. Physiological basis of submergence tolerante in
rainfed lowland rice ecosystem. Field Crops Research . v. 64, p. 75-90, 1999.
JACKSON, M.L.. Nitrogen determinations for soil and plant tissue. In: Soil chemical
analysis. Engle wood Chiffs Prentice – Hall. 1958.
JACKSON, M.B., W. ARMSTRONG. Formation of aerenchyma and the processes of
plantventilation in relation to soil flooding and submergence. Plant Biology , v. 1, p.
274-287, 1999.
JACKSON, M.B. Root-to-shoot communication in flooded plants: Involvement of
abscisic acid, ethylene and 1-aminocyclopropane-1-carboxilic acid. Agronomic
Journal , Madison, v. 86, p. 775-782, 1994.
JACKSON, MB.. Long distance signalling from roots to shoots assessed: the flooding
story. Journal of Experimental Botany v.53, p.175–181.2002.
JACKSON, M.B.; DREW, M.C. Effects of flooding on growth and metabolism of
herbacious plants. In: KOZLOWSKI, T.T. (Ed.). Flooding and plant growth. Orlando:
Academic Press, p. 47-128, 1984.
JAVOT, H. AND MAUREL,C. The role of aquaporinas in root water uptake. Annals
of .Botany , n. 90, 301–313, 2002.
JOHANSSON, I.; LARSSON, C.; EK, B.; KJELLBOM, P. The major integral proteins
of spinach leaf plasma membranes are putative aquaporins and are phosphorylated
in response to Ca2+ and apoplastic water potential. Plant Cell , v. 8, p. 1181-1191,
1996.
JOHANSSON, I.; KARLSSON, M.; JOHANSSON, U.; LARSSON, C.; KJELLBOM, P.
The role of aquaporins in cellular and whole plant water balance. Biochimica et
Biophysica Acta, v. 146, p.324-342, 2000.

107
JOHANSON, U., KARLSSON, M., JOHANSSON, I., GUSTAVSSON, S., SJOVALL,
S. FRAYSSE, L., WEIG, A.R., KJELLBOM, P. The complete set of genes encoding
major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature
for major intrinsic proteins in plants, Plant Physiology , n. 126, p. 1358–1369, 2001.
JURADO, E; GARCÍA, J., FLORES, J., ESTRADA, E. Leguminous seedling
establishment in Tamaulipan thorn scrub of northeastern Mexico. Forest Ecology
and Management , n. 221, p. 133-139, 2006.
KARLIDAG, H. H., YILDIRIM, E., TURAN, M. Salicylic acid ameliorates the adverse
effect of salt stress on strawberry. Scientia Agricola . (Piracicaba, Braz.), v.66, n.2,
p.180-187, 2009.
KENNEDY, R. A.; RUMPHO, M. E; FOX, T. C. Anaerobic metabolism in plants.
Plant Physiology, v. 100, p.1-6, 1992.
KHAN, M. N.; PATTERSON, G. R.; MATLICK, B. K. Effect of supplemental water
supplied through drip irrigation on cocoa yield at humming bird Hershey LTD., Belize,
C.A. . 10th International Cocoa Research Conference . San Domingo, Republica
Dominicana, p 185-191, 1987.
KITAJIMA, K. Ecophysiology of tropical tree seedlings. In: MUKLEY, S. S.;
CHAZDON, R. L.; SMITH, A. P. (Eds). Tropical Forest plant ecophysiology . New
York, Chapman and Hall, p- 559-596, 1996.
KLUGE, R. A. Maracujazeiro (Passiflora sp.). In: CASTRO, PRC; KLUGE, R. A.
Ecofisiologia de fruteiras tropicais . São Paulo: Nobel, p. 32-47, 1998.
KOZLOWSKI, T. T. Responses of woody plants to flooding and salinity. Tree
Physiology Monograph, v. 1, p. 1 – 29, 1997.

108
KOZLOWSKI, T. T. Physiological-ecological impacts of flooding on riparian forest
ecosystems. Wetlands , v. 22, n. 3, p. 550-561, 2002.
KOZLOWSKY, T. Responses of woody plants to flooding. p. 129 – 163. In: T. T.
KOZLOWSKI (ed.), Flooding and plant growth . Academic Press, Orlando, FL.
KOZLOWSKI, T.T.; PALLARDY, S.G. Physiology of woody plants . Academic Press, San Diego, 1997, 811p.
KRAMER, T.; KOZLOWSKI, T. T. Physiology of woody plants . New York:
Academic Press, 1979. 811 p.
KOZONO, D. ,DING, X. D., IWASAKI, I., MENG, X. Y., KAMAGATA, Y., AGRE, P. ,
KITAGAWA, Y. Functional expression and characterization of an archaeal aquaporin-
AqpM from Methanothermobacter marburgensis. Journal of Biological Chemistry ,
n. 278, p.10649–10656, 2003.
LAISK, A.; LORETO, F. Determining photosynthetic parameters from leaf CO2
exchange and chlorophyll fluorescence. Plant Physiology, v. 110, p. 903-912, 1996.
LARCHER, W. Ecofisiologia vegetal. São Carlos: Rima, 2000. 531 p.
LAURENTIN, A.; EDWARDS, C.A. A microtiter modification of the anthrone-sulfuric
acid colorimetric asay for glucose based carbohydrates. Anal. Biochem ., v. 315, p.
143-145, 2003.
LEAL, J. B. Diversidade genética de cacaueiros (Theobroma cacao L.) resistentes à
vassoura-de-bruxa com base em marcadores RAPD e microssatélites. Dissertação
(Mestrado em Genética e biologia Molecular), Universidade Estadual de Santa Cruz,
Ilhéus, Bahia, 2004. 61p.
LEITE, R. M. DE O., ALVIM, R. AND ALVIM, P. DE T. Effects of wind and solar
radiation on the mechanical rupture of the cacao pulvinus (Abstract), 1981. In:

109
Proceedings of the VII International 161 Cocoa Rese arch Conference ,
Cameroun, 1979, p. 129. Lagos, Nigeria. Cocoa Producers Alliance.
LEI, T.T.; LECHOWICZ, M.J.. Diverse responses ofmaple saplings to forest light
regimes. Annals of Botany, n. 82, p. 9-19, 1998.
LENSSEN, J.P.M., MENTING, F.B.J., VAN DER PUTTEN, W.H., BLOM, C.W.P.M.
Control of plant species richness and zonation of functional groups along a
freshwater flooding gradient. Oikos , n. 80, p. 523–534, 1999.
LIAO, C. T.; LIN, C.-H. Physiological adaptation of crop plants to flooding stress.
Proceedings of the National Science Council, Roc (B), v. 25, p. 148-157, 2001.
LIAO, C. T.; LIN, C.H. Effect of flooding stress on photosynthetic activities of
Momordica charantia. Plant Physiology and Biochemistry, v. 32, p. 479-485, 1994.
LIVAK, K. J.; SCHMITTGEN, T. D. Analysis of relative gene expression data using
real-time quantitative PCR and the 2–∆∆Ct method. Methods , v. 25, p. 402–408,
2001.
LOBÃO, D. E.; SETENTA, W. C.; LOBÃO, E. S. P.; CURVELO, K.; VALLE, R. R.
Cacau cabruca: sistema agrossilvicultural tropical. In: Valle RR (ed), Ciência,
Tecnologia e Manejo do Cacaueiro , pp.290-323, Gráfica e Editora Vital Ltda,
Ilhéus, 2007.
LOBO, P.C., JOLY, C.A. Tolerance to hypoxia andanoxia in neotropical tree species.
In: Ecophysiological strategies of xerophytic and amphi bious plants in the
neotropics. (F.R. SCARANO & A.C. FRANCO, eds.). Series Oecologia Brasiliensis,
PPGE-UFRJ, Rio de Janeiro, v. 4, p.137-1561998 .
LONG, S.P.;HÄLLGREN, J.E. Measurement of CO2 assimilation by plants in the field
and the laboratory. In: HALL, D.O.; SCURLOCK, J.M.O.; BOLHÀR-NORDENKAMPF,
H.R.; LEEGOOD, R.C.; LONG, S.P. (Ed.). Photosynthesis and production in a

110
changing environment: a field and laboratory manual . London, Glasgow, New
York, Tokyo, Melbourne, Madras: Chapman e Hall, 1993. p.129-167.
LOPEZ, F., BOUSSER, A., SISSOEFF, I., GASPAR, M., LACHAISE, B., HOARAU,
J., A. MAHE, Diurnal regulation of water transport and aquaporin gene expression in
maize roots:contribution of PIP2 proteins. Plant Cell Physiology , n. 44, p. 1384–
1395, 2003.
LOPEZ, O. R.; KURSAR, T. A. 2003. Does flood tolerance explain tree species
distribution in tropical seasonally flooded habitats?. Oecologia , n. 136, p.193-204,
2003.
LOQUE, D.; LUDEWIG, U.; YUAN, L.; VONWIREN, N. Tonoplast intrinsic proteins
AtTIP2;1 and AtTIP2;3 facilitate NH3 transport in to the vacuole. Plant Physiology ,
n. 137, p. 671–680, 2005.
LUU, D. T.; MAUREL, C. Aquaporins in a challenging environment: molecular gears
for adjusting plant water status. Plant Cell Environmental, n. 28, p. 85–96, 2005.
MARENCO, R. A.; LOPES, N. F. Fisiologia Vegetal: Fotossíntese, respiração,
relações hídricas e nutrição mineral . Editora UFV. Viçosa, MG. 2005, 451p.
MARTIN, U.; ALLADRU, S. G.; BAHARI, Z. A. Dehydratation tolerance of leaf tissues
of six woody angiosperm species. Plant Physiology , v. 69, p. 182-186, 1987.
MARTRE, P., MORILLON, R., BARRIEU, F., NORTH, G.B., NOBEL, P.S.,
CHRISPEELS, M.J., Plasma membrane aquaporins play a significant role during
recovery from water deficit, Plant Physiology , n. 130, p. 2101–2110, 2003.
MARTINAZZO, E. G; ANESE, S.;WANDSCHEER, A. C. D.; PASTORINI, L. H. Efeito
do Sombreamento sobre o Crescimento Inicial e Teor de Clorofila Foliar de Eugenia
uniflora Linn (Pitanga) – Família Myrtaceae. Revista Brasileira de Biociências ,
Porto Alegre, v. 5, supl. 2, p. 162-164, jul. 2007.

111
MATHIS, P. AND PAILLOTIN, G. Primary processes of photosynthesis. In: HATCH,
M. D. AND BOARDMAN, N. K. (eds). The Biochemistry of Plants , Academic Press,
New York, v. 8, p. 97-161, 1981.
MAUREL, C. Aquaporins and water permeability of membranes. Annual Review
Plant Physiology, v. 48, p. 399 – 429, 1997.
MAUREL, C., TACNET, F., GUCLU, J., GUERN, J., RIPOCHE, P., Purified vesicles
of tobacco cell vacuolar and plasma membranes exhibit dramatically different water
permeability and water channel activity. Proc. Natl. Acad Sci. USA, n. 94, p. 7103–
7108, 1997.
MAUREL, C., JAVOT, H., LAUVERGEAT, V., GERBEAU, P., TOURNAIRE, C.,
SANTONI, V. AND HEYES, J. Molecular physiology of aquaporinas in plants. Int.
Rev. Cytol. , n. 215, p. 105–148, 2002.
MAURENZA, D.; MARENCO, R. A.; PIEDADE, M. T. F. Efeito da inundação de longa
duração sob o crescimento de Pouteria glomerata (Sapotaceae), uma arbórea da
várzea da Amazônia Central. Acta Amazonica [online], v. 39, n. 3, 2009.
MAXWELL, K., JOHNSON, G.N., Chlorophyll fluorescence - a practical guide.
Journal of Experimental Botany , v. 51, p. 659-668, 2000.
MEBRAHTU, T.; HAVOVER, J. W. Leaf age effects on photosynthesis and stomatal
conductance of black locust seedlings. Photosynthetica , Prague, v. 25, n. 4,p. 537-
544, 1991.
MEDRI, M. E. Aspectos morfo-anatômicos e fisiológicos de Peltophorum dubium
(Spr.) taub. submetida ao alagamento e à aplicação de etrel. Revista Brasileira de
Botânica, v. 21, p. 261-267, 1998.
MELÉNDEZ, L. Microambiente, cantidad de esporas en el aire e incidencia del
hongo Moniliophtora roreri (Cif e Par) Evans et al. bajo tres sistemas de manejo de

112
sombra leguminosa en cacao (Theobroma cacao). M.Sc.thesis , CATIE, Turrialba,
Costa Rica, 1993.
MIELKE, M.S.; MATOS, E.M.; COUTO, V.B.; ALMEIDA, A.F.; GOMES, F.B. &
MANGABEIRA, P.A.O. Some photosynthetic and growth responses of Annona
glabra L. seedlings to sol flooding. Acta Botanica Brasilica, n.19, p. 905-911,
2005a.
MIELKE MS, ALMEIDA A-AF, GOMES FP. Photosynthetic traits of five neotropical
rainforest tree species: interactions between light response curves and leaf-to-air
vapour pressure deficit. Brazilian Archives of Biology andTechnology , n.48,
p.815-824, 2005b.
MIELKE,M.S.; ALMEIDA, A.F.; GOMES,F.P.; AGUILAR,M.A., MANGABEIRA, P.A.;
Leaf gas exchange, chlorophyll fluorescence and growth responses of Genipa
americana seedlings to soil flooding. Environmental and Experimental Botany n.50 p.
221-231, 2003.
MITRA, B. N., YOSHINO, R., MORIO, T., YOKOYAMA, M, MAEDA M,
URUSHIHARA H, TANAKA, Y. Loss of a member of the aquaporin gene family, aqpA
affects spore dormancy in Dictyostelium. Gene, n. 27, p. 131–139, 2000.
MORAES NETO, S. P.; GONÇALVES, J. L. de M.; TAKAKI, M.; CENCI, S.;
GONÇALVES, J. C. Crescimento de mudas de algumas espécies arbóreas que
ocorrem na mata Atlântica em função do nível de luminosidade. Revista Árvore , v.
24, n.1, p. 35-45, 2000.
MORARD, P., SILVESTRE, J. Plant injury due to oxygen deficiency in the root
environment of soilless culture: a review. Plant and Soil, v.184, p. 243-254, 1996.
MORGAN, D. C.; SMITH, H. A systematic relationship between phytochrome-
controlled development and species habitat, for plants grown in simulated natural
irradiation. Planta, n. 145, p. 534-536, 1979.

113
MORGAN, P. W. Effects of abiotic stresses on plant hormone systems. In Stress
responses in plants: adaptation and acclimation mec hanisms . Willey-Liss Ed.
Chapter 6., p. 113-146, 1990.
MÜLLER, M. W.; BIEHL, B. Mudanças na capacidade fotossintética de folhas de
cacaueiro (Theobroma cacao L.) influenciadas pela intensidade de luz, durante o
período de vida. In: International Cocoa Research Conference, 11, Procee dings .
Cocoa Producers’ Alliance, London, p. 597-605, 1993.
MÜLLER, M. W., GAMA-RODRIGUES, A. C. Sistemas agroflorestais com cacaueiro.
In: VALLE, R. R. (ed), Ciência, Tecnologia e Manejo do Cacaueiro , p .246-271.
Gráfica e Editora Vital Ltda, Ilhéus, 2007.
MUTHUCHELIAN, K.; MURUGAN, C.; HARIGOVIDAN, R.; NEDUNCHEZHIAN, N.;
KULANDAIVELU, G. Effects of triacontanol in flooded Erythrina variegata seedlings.
1. Changes in growth, photosynthetic pigments and biomass productivity.
Photosynthetica , The Hague, v. 31, n. 2, p. 269-275, 1995.
NIINEMETS, Ü; VALLADARES, F. Shade tolerance, a key plant feature of complex
nature and consequences. Annual Review of Ecology, Evolution, and
Systematics , v. 39, p. 237-257, 2006.
NOBEL, P. S. Leaves and fluxes. In: NOBEL, P. S. Physicochemical and
environmental plant physiology . San Diego: Academic Press, p.393-472, 1991.
OKALI, D.H.; OWUSU, J.K. Growth analysis and photosyntetic rates of cocoa
(Theobroma cacao L.) seedlings in relation to varying shade and nutrient regimens.
Ghana Journal Agricultural Science. v. 8, p. 51-69, 1975.
OLIVEIRA,V.C.; Efeito da saturação hídrica do solo na taxa de assimilação de CO2
e no desenvolvimento de Caliphyllum brasiliense Camb. (Clusiaceae). 2007. 93f.
Dissertação (Mestrado – Programa de Pós- Graduação em Biologia Vegetal)
Universidade Estadual de Campinas, SP, 2007.

114
PAROLIN, P. Morphological and physiological adjustments to waterlogging and
droutht in seedlings of Amazonian flooplain trees. Oecologia , n. 128, p. 326-335,
2001.
PEDROSO, S.G., VARELA, V.P. Efeito do sombreamento no crescimento de mudas
de sumaúna (Ceiba pentandra (L.) Gaertn). Revista Brasileira de Sementes, n. 17,
v. 1: 47-51.
PENG, H.; LIN, T.; WANG, N.; SHIH, M. Differential expression of genes encoding 1-
aminocyclopropane-1-carboxylate synthase in Arabidopsis during hypoxia. Plant
Molecular Biology . v. 58, p. 15-25, 2005.
PEREIRA, J. L. M.; RAM, A.; FIGUEIREDO, J. M.; ALMEIDA, L. C. C. Primeira
ocorrência de vassoura-de-bruxa na principal região produtora de cacau do Brasil.
Agrotrópica, v.1, p.79-81. 1989.
PEREIRA, J. N.; RIBEIRO, J. F.; FONSECA, C. E. L. Crescimento inicial de
Piptadenia gonoacantha (Legiminosae, Mimosoideae) sob inundação em diferentes
níveis de luminosidade. Revista Brasileira de Botânica, v. 24, n. 4, p. 561-566,
2001.
PEZESHKI, S. R., PARDUE, J. H.; DeLAUNE, R. D. Leaf gas exchange and growth
of flood-tolerant and flood-sensitive tree species under low soil redox conditions.
Tree Physiology , Victoria, v. 16, p. 453-458, 1996.
PEZESHKI, S. R. Differences in patterns of photosynthetic responses to hypoxia in
flood-tolerant and flood-sensitive tree species. Phtosynthetica . v. 28, p. 423-430,
1993.
PONNAMPERUMA, F.N. The chemistry of submerged soils. Advance in Agronomy ,
v. 24, p. 29-96, 1972.

115
POORTER, L. Growth responses of 15 rain-forest tree species to a light gradient: the
relative importance of morphological and physiological traits. Functional Ecology , n.
13, p.396-410, 1999.
QUIGLEY, F., ROSENBERG, J. M., SHACHAR-HILL, Y. AND BOHNERT, H. J. From
genome to function: the Arabidopsis aquaporins. Genome Biology, v. 31, n. , p. 1-
17, 2001.
REGO, G. da M., POSSAMAI, E. Efeito do Sombreamento sobre o Teor de Clorofila
e Crescimento Inicial do Jequitibá-rosa. Boletim de Pesquisa Florestal, Embrapa
Florestas, n. 53, p.179-194, 2006.
REIZER, J.; REIZER, A.; SAIER, M. H. J. The MIP family of integral membrane
channel proteins: sequence comparisons, evolutionary relationships, reconstructed
pathway of evolution, and proposed functional differentiation of the two repeated
halves of the proteins. Critical Review Biochemistry and Molecular Biology, v. 28,
p. 235 – 257, 1993.
REHEM, B. C., ALMEIDA, A-A. F., MIELKE, M. S., GOMES, F. P. Efeitos do
alagamento do substrato no crescimento e na composição química de genótipos
clonais de Theobroma cacao L. Revista. Brasileira de fruticultura , v. 31, n. 3,
p.805-815, 2009
RICARD B.; COUÉE I.; RAYMOND P.; SAGLIO P.H.; SAINT-GES V.; PRADET A.
Plant metabolism under hypoxia and anoxia. Plant Physiology and Biochemistry,
v. 32, p. 1-10, 1994.
ROHÁCEK, K. Chlorophyll fluorescence parameters: the definitions, photosynthetic
meaning and mutual relationships. Photosynthetica , v.40, p.13-29, 2002.
RICARD B.; COUÉE I.; RAYMOND P.; SAGLIO P.H.; SAINT-GES V.; PRADET A.
Plant metabolism under hypoxia and anoxia. Plant Physiology and Biochemistry,
v. 32, p. 1-10, 1994.

116
RYSER P., EEK L. Consequences of phenotypic plasticity vs. interspecific variation
in leaf and root traits for acquisition of above ground and below ground resources.
American Journal of Botany , v. 87, p. 402-411, 2000.
SANTOS, D. L., TAKAKI, M. Fenologia de Cedrella fissilis Vell (Meliaceae) na região
rural de Itirapina, São Paulo, Brasil. Acta Botanica Brasileira , v. 19, n. 3, p. 625-
632, 2005.
SARDA,X.,TOUSCH, D., FERRARE, K., LEGRAND, E., DUPUIS, J. M. et al.
TwoTIP-like genes encoding aquaporins are expressed in sunflower guard cells.
Plant Journal , n. 12, p. 1103–11, 1997.
SCANDALIOS, J. G. Oxygen stress and superoxide dismutases. Plant
Physiolology , v. 101, p. 7-12, 1993.
SCALON, S.P.Q. et al. Germinação e crescimento de mudas de pitangueira
(Eugenia uniflora L.) sob condições de sombreamento. Revista Brasileira de
Fruticultura, v. 23, n. 3, p. 652-655, 2001.
SCHREIBER, U.; BILGER, W.; HORMANN, H.; NEUBAUER, C. In: Photosynthesis:
a comprehensive treatise. (ed.) Raghavendra, A.S.. Cambridge University Press,
Cambridge, 1998.
SCHULZE, E.D., ROBICHAUX, R.H., GRACE, J., RUNDEL, P.W, EHLERINGER, J.
R. Plant water balance. BioScience , n. 37, p. 30-37, 1987.
SCHULZE, E.-D. & HALL, A.E. Stomatal responses, water loss and CO2 assimilation
rates of plants in contrasting environments. In: LANGE, O.L.; NOBEL, P.S.;
OSMOND, C.B. & ZIEGLER, H. eds. Physiological Plant Ecology. II. Water
relations and carbon assimilation . Berlin, Spring-Verlag, p.181-229, 1982.

117
SCHUURMANS, J. A., VANDONGEN, J. T., RUTJENS, B. P., BOONMAN, A.,
PIETERSE, C. M., BORSTLAP, A. C. Members of the aquaporin family in the
developing pea seed coat include representatives of the PIP, TIP, and NIP
subfamilies, Plant Molecular Biology , n. 53, p. 633–645, 2003.
SENA GOMES, A. R., KOZLOWSKI, T. T. The effects of flooding on water relations
and growth of Theobroma cacao var. catongo seedlings. Journal of Horticultural
Science , n. 61, p. 265-276, 1986.
SENA-GOMES, A.R.; KOZLOWSKI, T.T. Physiological and growth responses to
flooding of seedling of Hevea brasiliensis. Biotropica, n. 20, p. 286-293, 1988.
SEYBOLD, A.; EGLE, K. Planta , n. 26, p. 491. 1970.
SILVA, R. R.; FREITAS, G. A.; SIEBENEICHLER, S. C.; DA MATA, J. F.; CHAGAS,
J. R. Desenvolvimento inicial de plântulas de Theobroma grandiflorum (Willd. ex
Spreng.) Schum. sob influência de sombreamento. Acta Amazonica . v.37, n.3,
2007.
SMITH, E. S. C. The interrelationships between shade types and cocoa pest and
disease problems in Papua New Guinea. In: Proceedings of the VII International
Cocoa ResearchConference , Cameroun, 1979, p. 37–43. Cocoa Producers
Alliance, Lagos, Nigeria.
STONEMAN, G.L.; DELL, B. Growth of Eucalyptus marginata (Jarrah) seedlings in a
greenhouse in response to shade and soil temperature. Tree Physiology , v. 13, p.
239-252, 1993.
STRANT, M.; ÖQUIST, G. Effects of frost hardening, dehardening and freezing
stress on in vivo chlorophyll fluorescence of scots pine seedlings (Pynus sylvestris
L.). Plant Cell Environment, Oxford, v.11, p.231-238, 1988.

118
STREIT, N. M., CANTERLE, L. P., CANTO, M. W. DO, HYCHECKI HECKTHEUER,
L. H. H. As Clorofilas. Ciência Rural , v.35, n.3, 2005.
STREETER, J.G.; THOMPSON, J.F. In vivo and in vitro studies on γ-aminobutyric
acid metabolism with the radish plant (Raphanus sativus L.). Plant Physiology, v.
49, p. 579-584, 1972.
SOUZA, C. A. S.; DIAS, L. A. S. Melhoramento ambiental e sócio-economia. In:
DIAS, L. A. S. (Ed) Melhoramento Genético do Cacaueiro . Editora Folha de Viçosa
Ltda, Viçosa. p. 1-47, 2001.
SUN, M-H; XU, WEN; ZHU, Y-F; SU, W-A; TANG, Z-C. A simple method for in situ
hybridization to RNA in guard cells of Vicia faba L.: the expression of aquaporins in
guard cells. Plant Molecular Biology Reporter , n. 19, p. 129 135, 2001.
TAIZ, L.; ZEIGER, E., 2004. Fisiologia vegetal. 3. ed. Piracicaba, Artmed. 720p.
TAKANO, J. ,WADA, M.,LUDEWIG, U.,SCHAAF, G. ,VON WIREN, N. ,FUJIWARA,
T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron
uptake and plant development under boron limitation. Plant Cell , n. 18, p. 1498–509,
2003.
TERASHIMA, I., HIKOSAKA, K. Comparative ecophysiology of leaf and canopy
photosynthesis. Plant, Cell and Environment . v. 18, p. 1111-1128, 1995.
TOOJINDA, T., SIANGLIW, M. TRAGOORANG, S., VANAVICHIT, A. Molecular
genetics of submergence tolerance in rice: QTL analysis of key traits. Annals of
Botany , n. 91, p. 243-253, 2003.
TOURNAIRE-ROUX, C., SUTKA, M.N., JAVOT, H., GOUT, E., GERBEAU, P., LUU,
D.T., BLIGNY, R.; MAUREL, C. Cytosolic pH regulates root water transport during
anoxic stress through gating of aquaporins. Nature, n. 425, p. 393-397, 2003.

119
TREVIZAN, S. D. P. Mudanças no sul da Bahia associada a vassoura-de-bruxa do
cacau. In: International Cocoa Research Conference , 12, Salvador. 1996.
Proceeding, Lagos: Cocoa Producers Aliance, p. 1109 – 1116, 1996.
TYERMAN, S. D., BOHNERT, H. J., MAUREL, C., STEUDLE, E., SMITH, J. A. C.
Plant aquaporins: their molecular biology, biophysics and significance for plant water
relations. Journal Experimental Botany , n. 50, p. 1055–1071, 1999.
TYERMAN S. D., NIEMIETZ, C.M., BRAMLEY, H. Plant aquaporins: multifunctional
water and solute channels with expanding roles. Plant Cell Environmental , n. 25,
p.173–194, 2002.
TYREE, M.T.; HAMMEL, M. The measurements of turgor pressure and water
relations of plants by the pressure bomb techniques. Journal of Experimental
Botany, v.23, p.267-283, 1982.
UEHLEIN, N., LOVISOLO, C., SIEFRITZ, F., KALDENHOFF, R., The tobacco
aquaporin NtAQP1 is a membrane CO2 pore with physiological functions, Nature, n.
425, p. 734–737, 2003.
VARTAPETIAN, B. B.; JACKSON, M. B. Plant adaptations to anaerobic stress.
Annals of Botany , v. 79, p. 3-20, 1997. Supplement A.
VERA-ESTRELLA, R.; BARKLA, B. J.; BOHNERT, H. J.; PANTOJA, O. Novel
regulation of aquaporins during osmotic stress. Plant Physiology, v. 135, p. 2318-
2329, 2004.
VERA-ESTRELLA R., BARKLA, B.J., PANTOJA, O. La biologia y la fisiologia de las
acuaporinas de plantas. Biojournal.net , n. 1, 2005.
VIEIRA, G. Gap dynamics in managed Amazonian forest:Structural and
ecophysiological aspects. 1996. 162f. Tese(Doutorado em Ecologia Tropical) -
University of Oxford, Grã-Bretanha.

120
VISSER, E. J. W., VOESENEK, L. A. C. J., VARTAPETIAN, B. B., M. B. JACKSON.
Flooding and plant growth. Annals of Botany , v. 91, n. 2, p. 107-109, 2003.
VON CAEMMERER S., FARQUHAR G.D. Some relationships between the
biochemistry of photosynthesis and the gas exchange of leaves. Planta. v. 153, p.
376-387, 1981.
WALTERS, M. B., REICH, P. B. Seed size, nitrogen supply and growth rate affect tree seedling survival in deep shade. Ecology , n. 81, p. 1887-1901, 2000.
WELLBURN, A.R. The spectral determination of chlorophylls a and b, as well as total
carotenoids, using various solvents with spectrophotometers of different resolution.
Journal of Plant Physiology, v.144, p.307-314, 1994.
WESSEL, M. Shade and nutrition in cocoa. In: Wood GAR, Lass RA (eds) Cocoa .
Longmans Scientific & Technical Publication, 167p., 1989.
WHITMORE, T. C. A review of some aspects of tropical rain forest seedling ecology
with suggestion for further enquiry. In: Swaine, M. D. (ed.). Ecology of Tropical
Forest Tree Seedlings , UNESCO/Parthenon, Paris/Carnforth. p. 3-39, 1996.
WYKA, T.; ROBAKOWSKY, P.; YTKOWIAK, R. Acclimation of leaves to contrasting
irradiance in juvenile trees differing in shade tolerance. Tree Physiology , v. 27, p.
1293 – 1306, 2007.
YAMAGUCHI, T.; FRIEND, D.J.C. Effects of leaf age and irradiance on
photosynthesis of Coffea arabica. Photosynthetica , Prague, v.13, p.271-278, 1979.
YOUNG, A. Agroforestry for Soil Management, 2. ed. CAB International, Wallingford,
UK, 1997, 320p.

121
ZAERR, J. B. Short term flooding and net photosynthesis in seedling of three
conifers. Forest Science , v. 29, p. 71-78, 1983.
ZARDOYA, R.; VILLALBA, S. A. Phylogenetic framework for the aquaporin family in
eukaryotes. Journal of Molecular Evolution, v.52, p. 391 – 404, 2001.
ZARDOYA, R. Phylogeny and evolution of the major intrinsic protein family. Biology
Cell , v. 97, p. 397-414, 2005.
ZHAO, C-X; SHAO, H-B;CHU, L-Y. Aquaporin structure–function relationships: Water
flow through plant living cells. Colloides and surfaces B: Biointerfaces , v.62, n.2,
p. 163-172, 2007
ZUIDEMA, P. A.; LEFFELAAR, P. A.; GERRISTMA, W.; MOMMER, L.; ANTEN, N.
P. R. A physiological production model for cocoa (Theobroma cacao): model
presentation, validation and application. Agricultural Systems . n. 84, p. 95 – 225,
2005.
Recommended

















