Embed Size (px)
Citation preview

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
1
Análise Qualitativa do Modelo de Hodgkin-Huxley
Revisão do Modelo de Hodgkin-Huxley
O modelo de Hodgkin-Huxley para o potencial de ação no axônio gigante de lula é dado
pelo seguinte conjunto de quatro equações diferenciais:
( ) ( ) ( ) injVVNaNaKKm JEVgEVhmgEVngdtdVC +−−−−−−= 34 , (1)
,)()1)(( nVnVdtdn
nn βα −−= (2)
,)()1)(( mVmVdtdm
mm βα −−= (3)
.)()1)(( hVhVdtdh
hh βα −−= (4)
As funções α e β, determinadas por Hodgkin e Huxley a partir do ajuste entre as equações
do modelo e os seus dados experimentais, são:
11010exp
1001,0)(−⎟⎠
⎞⎜⎝
⎛ −−
=VVVnα e ⎟
⎠
⎞⎜⎝
⎛ −=80
exp125,0)( VVnβ , (5)
11025exp
251,0)(−⎟⎠
⎞⎜⎝
⎛ −−
=VVVmα e ⎟
⎠
⎞⎜⎝
⎛ −=18
exp4)( VVmβ , (6)
⎟⎠
⎞⎜⎝
⎛ −=20
exp07,0)( VVhα e 1
1030exp
1)(+⎟⎠
⎞⎜⎝
⎛ −=
VVhβ . (7)
A voltagem V que aparece nestas equações é o potencial de membrana medido em relação
ao potencial de repouso, V = Vm – Vrep, em mV. A densidade de corrente injetada Jinj é
medida em µA/cm2, as condutâncias são medidas em mS/cm2 e a capacitância é medida
em µF/cm2. As outras constantes do modelo (determinadas experimentalmente) são:
3,0 ,36 ,120 === VKNa ggg , (8)
com os respectivos potenciais de Nernst,
6,10 ,12 ,115 =−== VKNa EEE . (9)

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
2
O significado das variáveis n, m e h é o seguinte. Um canal de K+ é composto por quatro
portões de ativação n, idênticos e independentes, de maneira que a probabilidade de que
um canal de K+ esteja aberto é dada por n4. Já um canal de Na+ é formado por três portões
de ativação m, idênticos e independentes, e por um portão de inativação h, independente
dos portões de ativação. A probabilidade de que um canal de Na+ esteja aberto é dada pelo
produto da probabilidade de que seus três portões de ativação estejam abertos, m3, pela
probabilidade de que seu portão de inativação esteja aberto, h: m3h.
A dinâmica das três variáveis, n, m e h, determina o comportamento do sistema quando
uma corrente externa é injetada alterando o valor de V. Para um dado valor de V, essas três
variáveis tendem exponencialmente para os seus respectivos valores estacionários,
)( ),( ),( VhVmVn ∞∞∞ ,
com constantes de tempo iguais a, respectivamente,
)( ),( ),( VVV hmn τττ .
Os gráficos dos valores estacionários e das constantes temporais em função de V são
mostrados abaixo:
Os valores estacionários de n (para o K+) e m (para o Na+) crescem monotonamente à
medida que V aumenta, enquanto que o valor estacionário de h (para o Na+) diminui
monotonamente à medida que V aumenta. É por isso que n e m são chamadas de variáveis
de ativação (do potássio e do sódio, respectivamente) e h é chamada de variável de
inativação (do sódio). Observe que o valor da constante de tempo de m é muito menor do
que os valores das constantes de tempo de n e h.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
3
No equilíbrio, as condutâncias específicas do sódio, gNa, e de vazamento, gv, são baixas e a
condutância específica do potássio, gK, é alta. Desta forma, o termo dominante na equação
(1) é o termo correspondente à corrente de potássio: gK(V − EK). É por isso que o potencial
de equilíbrio (V = 0) é próximo do valor do potencial de Nernst do potássio (EK = − 12). O
potencial de equilíbrio é maior do que EK, o que resulta em um termo de corrente de
potássio (IK) positivo, implicando que a corrente de potássio é para fora da célula numa
tentativa de hiperpolarizar a célula e levar seu potencial de membrana para o potencial de
Nernst do potássio.
Quando uma densidade de corrente despolarizante pequena é injetada, o potencial V cresce
um pouco em relação ao repouso e logo em seguida retorna ao valor de equilíbrio. Durante
este curto período de tempo, a variável que responde mais rapidamente é a variável de
ativação do Na+, m, por causa da sua pequena constante temporal. Isto faz com que m
também se eleve durante este breve período de tempo, tentando atingir o seu respectivo
valor estacionário m∞(V). As demais variáveis, n e h, não respondem de maneira
considerável durante esse curto período de tempo.
Quando uma densidade de corrente despolarizante grande é injetada, ela causa um grande
aumento no valor de V e isto implica que o valor estacionário m∞(V) também será grande.
Se este valor for suficientemente grande (acima do limiar de excitação), então, antes que o
sistema retorne ao repouso, a variável m terá atingido um valor tão grande que o termo da
corrente de sódio na equação (1), m3h(V − ENa), passará a dominá-la. Isso faz com que o
potencial de membrana cresça drasticamente, procurando atingir o potencial de Nernst do
sódio. Enquanto V for menor que ENa (= 115), a corrente de sódio será negativa indicando
que os íons Na+ entram na célula, despolarizando-a ainda mais.
Se nada mais acontecesse, o potencial tenderia a atingir um novo valor de equilíbrio
próximo do potencial de Nernst do sódio ENa. É aqui que as diferenças entre as constantes
temporais desempenham um papel importante.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
4
Quando o potencial de membrana está no seu valor de repouso, a variável de inativação do
sódio, h, é positiva (em torno de 0,6). À medida que o potencial aumenta, o valor
estacionário h∞ diminui e, à medida que isso acontece (h se aproxima de zero), a corrente
de sódio se inativa, pois m3h se aproxima de zero.
Entretanto, como a constante temporal τh(V) é muito maior que τm(V), existe um atraso
considerável entre o momento em que a corrente de sódio é ativada pelo aumento de m e o
momento em que ela é inativada pela diminuição de h. O efeito líquido dessa diferença de
escalas temporais para as variáveis m e h é que a corrente de sódio inicialmente sofre um
aumento brusco fazendo com que o potencial também cresça rapidamente e, depois, ela cai
em direção a zero arrastando consigo o valor do potencial.
Mais ou menos no mesmo momento em que a corrente de sódio é inativada, a corrente de
potássio para fora da célula é ativada. Isto ocorre por causa da similaridade entre as escalas
de tempo das variáveis h e n, τh(V) e τn(V). A inativação da corrente de sódio e a ativação
da corrente de potássio fazem com que, mais uma vez, o termo da corrente de potássio,
gK(V − EK), domine a equação (1). Isto faz com que o potencial V tenda a atingir o
potencial de Nernst do potássio e é por isso que V, em sua queda, chega a ultrapassar o
valor de repouso e assume valores negativos.
A partir do momento em que V torna-se negativo, o valor de n diminui consideravelmente
e h volta a crescer. As variações desses dois parâmetros, porém, ocorrem lentamente por
causa das suas grandes constantes temporais. É por isso que o potencial de membrana leva
muito tempo (mais ou menos uma dezena de milisegundos) para retornar ao seu valor de
repouso após ele se tornar negativo.
A figura a seguir ilustra o comportamento temporal do potencial de membrana, das
variáveis n, m e h e das condutâncias e correntes de sódio e potássio do modelo de
Hodgkin-Huxley em resposta a dois pulsos de corrente injetada: um supralimiar, isto é,
com amplitude acima do limiar capaz de gerar um potencial de ação e o outro sublimiar.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
5
Adaptada da Figura 2.15 do livro de Izhikevich, E.M., Dynamical Systems in Neuroscience. MIT Press, Cambridge, MA, 2007.
Existem quatro fases diferentes durante um potencial de ação: fase de subida brusca; fase excitada;
fase refratária; e fase de recuperação (ou pós-hiperpolarização). A fase refratária ocorre logo após a
fase excitada e é um período durante o qual um novo estímulo não consegue provocar qualquer
resposta, mesmo que o potencial esteja próximo do seu valor de repouso. A impossibilidade de
resposta é devida ao fato de que nessa fase a variável h ainda é pequena, fazendo com que os canais
de sódio estejam inativados. À medida que h retorna gradualmente ao seu valor de repouso um
potencial de ação torna-se possível (daí o nome período refratário relativo).

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
6
Redução da Dimensionalidade do Modelo de Hodgkin-Huxley: o plano de fase rápido
No início da década de 1960, o biofísico norte-americano Richard FitzHugh (1922-2007)
desenvolveu uma elegante análise qualitativa do modelo de Hodgkin-Huxley que permite
um melhor entendimento do seu comportamento e que influenciou um grande número de
trabalhos posteriores. A abordagem de FitzHugh se baseia no fato de que algumas
variáveis do modelo variam de maneira rápida enquanto que outras variam de maneira
lenta: a variável de ativação do sódio, m, e o potencial de membrana, V, variam
rapidamente; por outro lado, a variável de ativação do potássio, n, e a variável de
inativação do sódio, h, variam lentamente.
Dessa forma, durante os estágios iniciais de um potencial de ação as variáveis n e h
permanecem essencialmente constantes enquanto que m e V variam. Essa constatação
permite que o sistema de quatro equações diferenciais possa ser aproximado por um
sistema de apenas duas variáveis rápidas, com as duas variáveis lentas sendo tratadas
como constantes. Essa descrição é válida apenas para os estágios iniciais de um potencial
de ação, mas mesmo assim ela permite um bom entendimento do processo de excitação.
Segundo essa aproximação, os valores das variáveis n e h são fixados nos seus valores de
repouso, que chamaremos de n0 e h0 respectivamente, e apenas os valores de m e V
poderão variar no tempo. Sendo assim, as equações do modelo de Hodgkin-Huxley se
reduzem ao sistema de duas equações:
( ) ( ) ( ) injVVNaNaKKm JEVgEVhmgEVngdtdVC +−−−−−−= 0
340 , (10)
.)()1)(( mVmVdtdm
mm βα −−= (11)
Esta última equação é equivalente a
,mmdtdm
m −= ∞τ (12)
onde
.)()(
1 e )()(
)(VVVV
Vm
mmm
mm
m
βατ
βαα
+=
+=∞ (13)

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
7
A representação da dinâmica da variável m na forma definida pelas equações (12) e (13)
será mais interessante para os propósitos do estudo a ser feito aqui.
O sistema bi-dimensional definido pelas equações (10) e (12) pode ser estudado no espaço
de fase (m, V), em que a variável m é colocada no eixo das ordenadas (vertical) e a
variável V é colocada no eixo das abscissas (horizontal). O tempo t faz o papel de um
parâmetro: dada uma condição inicial, V(0), m(0), a integração das equações (10) e (12)
resultará em soluções V(t) e m(t) de maneira que para cada valor de t haverá um ponto
correspondente no gráfico de m versus V. A união desses pontos corresponderá a certa
trajetória m = m(V) no espaço de fase. O espaço de fase das variáveis (m, V) é chamado de
espaço de fase rápido, pois essas duas variáveis são rápidas. Da mesma forma,
chamaremos o sistema de equações definido por (10) e (12) de sistema rápido.
Um sistema dinâmico bi-dimensional como o do nosso caso pode ser descrito de forma
genérica pelas equações,
onde f e g são funções conhecidas das variáveis x e y. Para cada ponto (x, y) no espaço de
fase (x no eixo das abscissas, y no eixo das ordenadas), as duas equações acima definem
um vetor no ponto cujas componentes são ( !x, !y) . Esse vetor, chamado vetor velocidade,
define a direção instantânea que a trajetória y(x) tem no ponto. Como cada ponto no
espaço de fase pode ser a condição inicial para o sistema acima, todo o espaço de fase está
preenchido por vetores velocidade (ele forma um campo vetorial).
O conhecimento desse campo vetorial é muito útil para estudar um sistema dinâmico como
o acima, pois mesmo que não se saiba como resolver analiticamente as equações (como é
o caso, em geral) ele permite que se determine o chamado retrato de fase do sistema. O
retrato de fase nos dá um entendimento qualitativo das soluções do sistema, isto é, das
trajetórias descritas pelas soluções y(x) no espaço de fase. Para ver exemplos de retratos de
fase, consulte, por exemplo, a página da Wikipédia em inglês sobre phase portraits:
https://en.wikipedia.org/wiki/Phase_portrait).

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
8
Quando se estuda um sistema dinâmico (x, y) no espaço de fase, é muito importante para o
entendimento das suas trajetórias a determinação das chamadas isóclinas. Uma isóclina é
uma curva no plano (x, y) ao longo da qual a derivada temporal de uma das duas variáveis,
!x = dx dt ou !y = dy dt , é constante. Em particular, uma isóclina nula é uma curva nesse
plano ao longo da qual uma dessas duas derivadas temporais é zero.
No caso do espaço de fase (m, V), as duas isóclinas nulas são determinadas por:
⇒= 0dtdVCm
( ) ( ) ( ) ⇒=−−+−+−⇒ 0034
0 injVVNaNaKK JEVgEVhmgEVng
.0
340
034
0
VNaK
injVVNaNaKK
ghmgngJEgEhmgEng
V++
+++=⇒ (14)
e
⇒= 0dtdm
mτ
. )()(
)(VV
Vmmmm
m
βαα+
==⇒ ∞ (15)
O gráfico abaixo (A) mostra essas isóclinas nulas para o caso em que Jinj = 0 traçadas no
plano de fase m × V. O detalhe do cruzamento entre as duas curvas no canto inferior
esquerdo do gráfico está mostrado no zoom ao lado (B).
Exercício: reproduza o gráfico acima.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
9
Observe que as duas isóclinas nulas se intersectam em três pontos, indicados no gráfico
por Vr, Vs e Ve. Esses três pontos são os pontos fixos do sistema rápido, em que as duas
derivadas, dV/dt e dm/dt, se igualam a zero simultaneamente. Se o estado do sistema
rápido começar exatamente em um desses três pontos, ele permanecerá nele para sempre.
Por essa razão, chamamos a esses três pontos de pontos de equilíbrio do sistema rápido.
Note que esses três pontos não são pontos de equilíbrio do sistema completo de quatro
equações do modelo de Hodgkin-Huxley. Eles são apenas os pontos de equilíbrio do
sistema rápido e, para sermos rigorosos, deveríamos chamá-los de pontos de pseudo-
equilíbrio. No entanto, no contexto do estudo do sistema rápido que estamos fazendo,
podemos continuar a chamá-los de pontos de equilíbrio.
Os pontos denominados de Vr e Ve são pontos de equilíbrio estável e o ponto denominado
de Vs é um ponto de sela. Os índices r, e e s indicam “repouso”, “excitado” e “sela”,
respectivamente.
Como saber se um ponto fixo de um sistema dinâmico é um ponto de equilíbrio estável.
Instável ou um ponto de sela? A maneira usual de se fazer isso é linearizando as equações
do sistema em torno de seus pontos fixos. Você fará isso na Lista 3 tendo como exemplo o
sistema de equações para o modelo de neurônio de FitzHugh-Nagumo.
Outra maneira de analisar a estabilidade de um ponto fixo é fazendo um estudo qualitativo
(geométrico) a partir do esboço do retrato de fase do sistema nas vizinhanças de seus
pontos fixos.
Para entender como isso pode ser feito, consideremos, como exemplo ilustrativo, o
seguinte sistema dinâmico:
As isóclinas nulas das variáveis x e y são dadas por:

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
10
• Isóclina nula de x (dx/dt = 0):
• Isóclina nula de y (dy/dt = 0):
Note que ao longo da isóclina nula de x os vetores velocidade são sempre verticais e que
ao longo da isóclina nula de y os vetores velocidade são sempre horizontais (veja a figura
abaixo). Um ponto fixo ocorre quando as isóclinas nulas se cruzam. No caso do exemplo,
o sistema tem um ponto fixo em (x = ½, y = ½). Enquanto percorremos uma isóclina nula
antes de chegar a um ponto fixo, a direção do campo vetorial sobre ela não muda; mas
quando se cruza um ponto fixo a direção muda. As isóclinas nulas dividem o espaço de
fase em torno de um ponto fixo em quatro regiões. Em cada uma dessas regiões, o campo
vetorial aponta em uma mesma direção dada por uma das quatro combinações possíveis:
(i) dx/dt > 0, dy/dt > 0; (ii) dx/dt > 0, dy/dt < 0; (iii) dx/dt < 0, dy/dt > 0; (iv) dx/dt < 0,
dy/dt < 0.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
11
Para determinar a direção do vetor velocidade ao longo de uma isóclina nula, escolhe-se
um ponto sobre a isóclina nula e calcula-se o vetor velocidade naquele ponto. A direção
desse vetor será a direção do vetor velocidade sobre todo o segmento da isóclina nula entre
os dois pontos fixos que o delimitam.
Por exemplo, seja o ponto (x = 1/3, y = 1) sobre a isóclina nula de y. Neste ponto:
Portanto, para todos os pontos ao longo da isóclina nula de y (linha vermelha no gráfico
acima) entre o eixo-y e o ponto fixo o vetor velocidade é horizontal e aponta para a
esquerda (por causa do sinal negativo de dx/dt). Por outro lado, para os ponto ao longo da
isóclina nula de y entre o ponto fixo e o eixo-x o vetor velocidade é horizontal e aponta
para a direita.
Esses vetores velocidade nos dão uma informação a mais: note que como dx/dt = 0 ao
longo da isóclina nula de x (linha azul no gráfico acima), a componente x do vetor
velocidade tem o mesmo sinal em todos os pontos acima dessa isóclina nula e o sinal
oposto em todos os pontos abaixo dessa isóclina nula. Portanto, basta determinar o sinal de
dx/dt em um ponto acima (ou abaixo) da isóclina nula de x para se conhecer como a
componente ao longo da direção x do campo vetorial se comporta em todos os pontos
acima e abaixo da isóclina nula de x. O ponto (x = 1/3, y = 1) está acima da isóclina nula
de x e o campo vetorial nele aponta para a esquerda. Portanto, para todos os pontos acima
da isóclina nula de x a componente x do campo vetorial aponta para a esquerda e para
todos os pontos abaixo da isóclina nula de x a componente x do campo vetorial aponta para
a direita (veja o inset da direita no gráfico acima).
Um raciocínio análogo vale para os pontos à esquerda e à direita da isóclina nula de y. No
caso, escolheu-se o ponto (x = 2/3, y = 1/3) sobre a isóclina nula de x. Neste ponto:
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
12
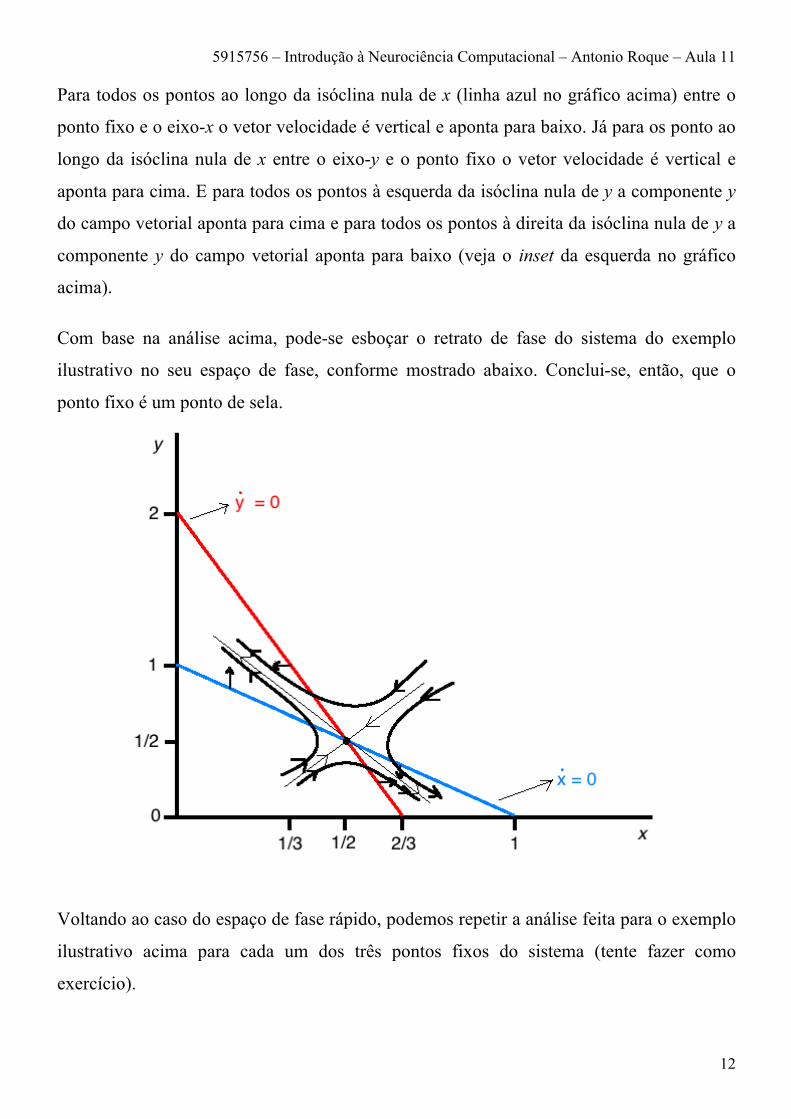
Para todos os pontos ao longo da isóclina nula de x (linha azul no gráfico acima) entre o
ponto fixo e o eixo-x o vetor velocidade é vertical e aponta para baixo. Já para os ponto ao
longo da isóclina nula de x entre o eixo-y e o ponto fixo o vetor velocidade é vertical e
aponta para cima. E para todos os pontos à esquerda da isóclina nula de y a componente y
do campo vetorial aponta para cima e para todos os pontos à direita da isóclina nula de y a
componente y do campo vetorial aponta para baixo (veja o inset da esquerda no gráfico
acima).
Com base na análise acima, pode-se esboçar o retrato de fase do sistema do exemplo
ilustrativo no seu espaço de fase, conforme mostrado abaixo. Conclui-se, então, que o
ponto fixo é um ponto de sela.
Voltando ao caso do espaço de fase rápido, podemos repetir a análise feita para o exemplo
ilustrativo acima para cada um dos três pontos fixos do sistema (tente fazer como
exercício).

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
13
Para o primeiro ponto fixo, que foi chamado de Vr, essa análise nos leva ao seguinte
esboço do retrato de fase (o gráfico está fora de escala e é apenas uma ilustração):
Note que os vetores apontam para o ponto fixo de todos os quatro quadrantes do espaço.
Portanto, o ponto fixo Vr é um ponto de equilíbrio estável.
Para o segundo ponto fixo, que foi chamado de Vs, o esboço do retrato de fase é o seguinte
(novamente, o gráfico abaixo é meramente ilustrativo):
Portanto, o ponto fixo Vs é um ponto de sela.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
14
Finalmente, para o terceiro ponto fixo (chamado de Ve), o esboço ilustrativo do retrato de
fase é:
Portanto, o ponto fixo Ve é estável.
Pergunta: a natureza deste terceiro ponto fixo poderia ser diferente de “estável”?
Para saber mais sobre o campo da matemática por trás do tipo de análise que está sendo
feita aqui, chamado de Sistemas Dinâmicos, recomendo o livro de Strogatz:
Strogatz, S.H., Nonlinear Dynamics and Chaos. Addison-Wesley, Reading, MA, 1994.
Como Vs é um ponto de sela, existe uma linha separatriz no plano de fase passando por Vs
que divide o plano em duas regiões: Qualquer trajetória que comece à esquerda dessa linha
jamais atingirá o ponto de equilíbrio excitado Ve e, com o tempo, atingirá o ponto de
repouso Vr. Por outro lado, qualquer trajetória que comece à direita dessa linha jamais
atingirá o ponto de repouso Vr e acabará atingindo o ponto excitado Ve. A figura a seguir
ilustra o que foi dito.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
15
A linha separatriz, em combinação com os dois pontos de equilíbrio estável Vr e Ve, causa
um fenômeno de tipo limiar: qualquer perturbação em V (por exemplo, causada por uma
corrente injetada Jinj) a partir do estado de repouso que não seja grande o suficiente para
elevar o valor de V acima da linha separatriz irá, com o tempo, retornar ao estado de
repouso. E qualquer perturbação que seja grande o suficiente para elevar o valor de V
acima da separatriz provocará uma longa excursão da voltagem que a levará ao estado
excitado Ve.
Se V e m fossem as únicas variáveis do modelo, então, quando V chegasse a Ve ela
permaneceria neste valor para sempre. Entretanto, como já dito anteriormente, Ve não é um
ponto de equilíbrio do modelo completo. Portanto, para ver o que acontece em uma escala
de tempo mais longa, devemos considerar como as variações lentas em n e h afetam as
propriedades qualitativas do plano de fase rápido.
Primeiramente, note que, como Ve > Vr, h∞(Ve) < h∞(Vr) e n∞(Ve) > n∞(Vr). Portanto,
enquanto V estiver no estado excitado h diminuirá, levando à inativação dos canais de
sódio, e n aumentará, levando à ativação dos canais de potássio.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
16
Note ainda que, embora a isóclina nula de m no plano de fase rápido seja independente de
n e h, a isóclina nula de V não é. Isso implica que, para valores diferentes de n(V) e h(V), a
curva da isóclina nula de V terá formas diferentes no plano de fase rápido (veja abaixo).
O gráfico acima mostra a isóclina nula de V para quatro combinações diferentes de n0 e h0
(com n0 aumentando e h0 diminuindo). Os valores de n0 e h0 usados para plotar essas
quatro isóclinas nulas de V são, respectivamente: (1) n0 = 0,3176 e h0 = 0,596; (2) n0 = 0,5
e h0 = 0,4; (3) n0 = 0,7 e h0 = 0,2; e (4) n0 = 0,8 e h0 = 0,1.
Vemos que, à medida que n aumenta e h diminui, a isóclina nula de V se move para cima e
para a esquerda. Durante esse movimento, os pontos de equilíbrio Vs e Ve vão se
aproximando um do outro e o ponto de equilíbrio Vr se move para a esquerda.
Durante esse processo o estado do sistema está no ponto de equilíbrio estável Ve (estado
excitado), o qual vai lentamente diminuindo enquanto que o ponto de sela Vs vai subindo e
se aproximando cada vez mais de Ve até que, em um dado momento, os dois pontos
coalescem e desaparecem.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
17
Matematicamente, quando o número de pontos fixos ou a natureza deles muda quando um
ou mais parâmetros do sistema passam por certos pontos diz-se que o sistema sofre uma
bifurcação. O(s) parâmetro(s) cuja variação causa(m) a bifurcação é(são) chamado(s) de
parâmetro(s) de bifurcação. No caso em questão, o tipo de bifurcação em que um ponto
fixo estável e um ponto de sela coalescem e desaparecem é chamado de bifurcação sela-
nó. Os parâmetros ds bifurcação são n0 e h0.
Depois que a bifurcação acontece, Vr torna-se o único ponto de equilíbrio do sistema e o
estado do sistema tem, necessariamente, que retornar a ele. Note que, como Vr está muito à
esquerda, ele não é o ponto de equilíbrio do sistema completo. Porém, quando a voltagem
atinge o valor de Vr os valores de n e h mudam novamente e são tais que a isóclina nula de
V muda de forma, levando Vr a crescer lentamente em direção ao valor de repouso inicial,
que é o valor de repouso do sistema completo.
Quando Vr retorna ao valor de repouso do sistema completo, o ciclo correspondente a um
potencial de ação se completa.
A figura acima ilustra as principais etapas de um potencial de ação. A: um estímulo
supralimiar causa um rápido aumento em V, levando-o ao estado excitado Ve. B: V está no
estado excitado Ve e começa a diminuir lentamente à medida que n aumenta e h diminui.
C: Os estados excitado, Ve, e de ponto de sela, Vs, movem-se um em direção ao outro e
desaparecem em uma bifurcação de tipo sela-nó. D: O único estado de equilíbrio restante é
Vr e o potencial de membrana decai em direção a ele. E: As variáveis n e h retornam
lentamente aos seus estados de repouso e, à medida que isso acontece, Vr aumenta
lentamente até atingir o valor de repouso do sistema completo em quatro dimensões.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
18
Redução da Dimensionalidade do Modelo de Hodgkin-Huxley: o plano de fase
rápido-lento
Como o sistema de equações do modelo de Hodgkin-Huxley é tetra-dimensional, há várias
maneiras de combinar suas variáveis em um gráfico bi-dimensional. A maneira vista
acima, com o plano de fase rápido, é apenas uma delas. Usando outras combinações de
duas variáveis pode-se obter um entendimento de outros aspectos do modelo de Hodgkin-
Huxley.
Uma dessas combinações, também estudada por FitzHugh na década de 1960, mostrou-se
bastante útil, não só para o entendimento do modelo de Hodgkin-Huxley, mas também
para a construção de outros modelos para neurônios. Essa combinação envolve uma
variável rápida e uma variável lenta, levando ao que se chama de plano de fase rápido-
lento.
Para construí-lo, FitzHugh raciocinou da seguinte maneira: observando as variações
temporais das variáveis rápidas, V e m, vemos que m varia no tempo de maneira ainda
mais rápida que V. Desta maneira, pode-se aproximar o valor de m pelo seu valor de
estado estacionário m∞, como se m atingisse esse valor instantaneamente e permanecesse
nele por todo o tempo. Por outro lado, durante um potencial de ação as variáveis h e n se
comportam de maneira aproximadamente simétrica (por reflexão especular). De fato, um
gráfico de h versus m mostra que elas estão aproximadamente relacionadas por h ≈ 0,8 − n
(você deve ter verificado isso no item b da primeira questão do Projeto Computacional 3).
Assumindo esta relação funcional, isto é, impondo que h = 0,8 − n por todo o tempo, pode-
se eliminar uma das variáveis lentas do sistema de equações de Hodgkin-Huxley, por
exemplo, a variável h.
Combinando as duas aproximações feitas (m = m∞ e h = 0,8 − n) , o modelo de Hodgkin-
Huxley pode ser reduzido ao seguinte sistema de duas equações, uma para a variável
rápida V e outra para a variável lenta n:

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
19
( ) ( )( ) ( ) ,8,0)(34injVVNaNaKKm JEVgEVnVmgEVng
dtdVC +−−−−−−−= ∞ (16)
e
.nndtdn
n −= ∞τ (17)
Assim como no caso do plano de fase rápido, podemos plotar as isóclinas nulas desse
sistema bidimensional no plano de fase rápido-lento. Isto está feito abaixo, para quatro
valores diferentes de Jinj (Jinj será o parâmetro de bifurcação aqui). Os gráficos também
mostram (em vermelho) as trajetórias do sistema a partir do repouso (V = 0, n = 0,3176).
Nesses gráficos, os valores de Jinj são mantidos constantes nos valores indicados de t = 0 a
t = 50 ms.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
20
Os gráficos abaixo mostram as curvas de V versus t para os quatro casos do gráfico
anterior. Vemos que no primeiro caso (J = 2 µA/cm2) ocorre apenas uma flutuação que
decai rapidamente; no segundo caso (J = 5 µA/cm2) ocorre um único evento tipo potencial
de ação; e nos dois últimos casos (J = 15 µA/cm2 e J = 30 µA/cm2) ocorrem eventos
repetidos (como trens regulares de potenciais de ação) que se mantêm enquanto durar o
estímulo.
Exercício: Escreva um programa em MATLAB que reproduza os dois gráficos acima.
Vamos agora interpretar qualitativamente os resultados desses dois gráficos. Para facilitar,
vamos nos referir ao primeiro gráfico (o dos espaços de fase) como gráfico 1 e ao segundo
gráfico (de V versus t) como gráfico 2.
A isóclina nula de n no gráfico 1 (indicada por dn/dt = 0) é a curva n∞(V). Ela aumenta
monotonicamente com V. Já a isóclina nula de V (indicada por dV/dt = 0) tem um formato
cúbico.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
21
Existe uma única intersecção entre as duas isóclinas nulas (pelo menos para os casos
mostrados), indicando que há apenas um ponto de equilíbrio. Nos quatro casos mostrados,
J é constante e diferente de zero, de maneira que os quatro pontos de equilíbrio são
diferentes do ponto de repouso de onde parte o sistema (o ponto de repouso só coincide
com o ponto de equilíbrio quando J = 0). Por causa disso, vemos que enquanto a isóclina
nula de n é a mesma para os quatro casos (pois não depende de J), a isóclina nula de V
desloca-se para cima com o aumento de J. Isto implica que o ponto de intersecção entre as
duas isóclinas nulas (e, portanto, o ponto de equilíbrio) também move-se para cima com o
aumento de J.
Mais do que isso, note que o ponto de equilíbrio se desloca para a direita do ponto onde a
isóclina nula de V para de diminuir e começa a crescer (o “joelho” da isóclina nula de V).
Nos dois diagramas da parte de baixo do gráfico 1 (que correspondem aos trens de disparo
no gráfico 2) o ponto de equilíbrio do sistema está claramente à direita do joelho da
isóclina nula de V.
O fato de que a isóclina nula de V é cúbica sugere que uma perturbação a partir do repouso
que mantenha o ponto de equilíbrio à esquerda do joelho da isóclina nula de V não
conseguirá excitar o sistema e este retornará rapidamente ao repouso (como na figura
ilustrativa da esquerda abaixo). Já se a perturbação for suficientemente forte para mover o
ponto de equilíbrio para a direita do joelho da isóclina nula de V, então a trajetória do
sistema apresentará um grande aumento em V, correspondendo a um potencial de ação,
antes de retornar ao repouso. Se a perturbação for tão forte a ponto de mover o ponto de
equilíbrio bem para a direita do joelho da isóclina nula de V, a trajetória poderá não
convergir mais para o repouso e o sistema entra em o que se chama de ciclo limite e passa
a girar (no sentido anti-horário) em torno do ponto de repouso enquanto durar a
perturbação (como na figura ilustrativa da direita abaixo).

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
22
A inclinação da curva que descreve uma trajetória no espaço de fase é determinada por
dn/dV (= (dn/dt)/(dV/dt)). Durante um potencial de ação, V varia muito mais rapidamente
que n. Então, a inclinação dn/dV passa a maior parte do tempo tendo valores baixos
(indicando muita variação em V para pouca variação em n). Isto implica que as trajetórias
no espaço de fase são praticamente horizontais, com exceção das regiões em que elas estão
próximas da isóclina nula de V (em que dV/dt ≈ 0).
A isóclina nula de V é chamada tecnicamente de “variedade lenta”. O conceito matemático
de variedade está além dos objetivos deste curso, portanto vamos chamar a isóclina nula
de V de “curva lenta”. O motivo para isso é que quando a trajetória do sistema está sobre a
curva lenta, dV/dt ≈ 0 e o seu movimento é governado pela variável lenta n. Os
movimentos do sistema quando sua trajetória está sobre a curva lenta (os trechos com
componentes verticais nas figuras ilustrativas acima) são lentos e os movimentos do
sistema quando sua trajetória está longe da curva lenta são rápidos e predominantemente
horizontais.
Vamos nos referir à curva lenta como sendo composta de três ramos: o ramo da esquerda,
o ramo do meio e o ramo da direita (veja a figura a seguir).

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
23
Quando a trajetória do sistema está sobre a curva lenta, o sentido do movimento do
sistema é determinado pelo sinal de dn/dt. Na região do espaço de fase à direita da isóclina
nula de n essa derivada é positiva, implicando que quando o sistema está sobre o ramo da
direita da curva lenta ele se move para cima (sentido de aumento de n). Por outro lado, na
região do espaço de fase à esquerda da isóclina nula de n, dn/dt < 0 e o sentido do
movimento do sistema é para baixo. A conseqüência disso é que o sistema se move no
sentido anti-horário (veja a ilustração abaixo).
Observando a trajetória do sistema, vemos que quando ela está sobre o ramo do meio da
curva lenta na sua parte baixa, o sistema se move em direção ao ramo da direita. Já quando
a trajetória do sistema está sobre a parte alta do ramo do meio, ele se move em direção ao
ramo da esquerda. Em qualquer caso, o sistema sempre irá se afastar do ramo do meio. Por
este motivo, o ramo do meio da curva lenta é chamado também de ramo instável da curva
lenta.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
24
O ramo instável da curva lenta atua como um limiar (na realidade, não uma curva limiar
bem definida como a separatriz do exemplo anterior, mas um continuum). Suponha que a
densidade de corrente injetada seja tão pequena que não consiga fazer o sistema, a partir
do seu estado de repouso, chegar à região do ramo instável. Em tal caso, a trajetória do
sistema move-se só um pouquinho para a direita, mas logo em seguida curva-se para a
esquerda e vai para o ponto de equilíbrio. Esta situação corresponde ao caso em que J = 2
µA/cm2 nos gráficos 1 e 2.
Suponha, entretanto, que a densidade de corrente injetada seja suficientemente grande para
levar o sistema a penetrar na região do ramo instável. Neste caso, a trajetória
obrigatoriamente deve se mover para a direita até alcançar o ramo da direita, que
corresponde ao estado excitado (note que V torna-se muito alto). Uma vez sobre o ramo da
direita, o sistema move-se lentamente para cima e para a esquerda (portanto, reduzindo V)
até atingir novamente o ramo instável. Nesse ponto n não pode mais crescer, pois o ramo
da direita deixa de existir e o sistema move-se rapidamente em direção ao ramo da
esquerda. Após atingir o ramo da esquerda, o sistema move-se para baixo e um pouco para
a direita, (aumentando V lentamente, correspondendo à fase de repolarização do potencial
de membrana após um potencial de ação) até atingir o ponto de equilíbrio e completar o
potencial de ação. Esta situação corresponde ao caso em que J = 5 µA/cm2 nos gráficos 1 e
2.
Por outro lado, quando o sistema é estimulado por uma densidade de corrente constante
ainda maior ele entra em um regime em que sua trajetória torna-se cíclica, repetindo-se
indefinidamente sobre o mesmo caminho enquanto durar o estímulo. Exemplos desse caso
são dados pelos dois últimos casos dos gráficos 1 e 2, para J = 15 µA/cm2 e J = 20
µA/cm2. Esse regime corresponde à emissão de um trem estacionário de potenciais de
ação.
Observando os dois gráficos da parte de baixo do gráfico 1, vemos que a trajetória cíclica
e repetitiva do sistema passa abaixo do ponto de equilíbrio (o cruzamento entre as duas
isóclinas nulas).

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
25
O motivo pelo qual a trajetória do sistema evita o ponto de equilíbrio nesses dois casos é
que agora o ponto de equilíbrio tornou-se instável. Já vimos que à medida que a densidade
de corrente injetada J aumenta, o cruzamento entre as duas isóclinas nulas é desviado para
cima e para a direita do joelho da isóclina nula de V. Ele, portanto, passa a cair sobre o
ramo do meio da isóclina nula de V e, por causa disso, torna-se instável.
Em outras palavras, o cruzamento entre as isóclinas nulas de V e n determina o ponto de
equilíbrio do sistema. Abaixo de um certo valor de J, esse ponto de equilíbrio é estável e
as trajetórias do sistema convergem para ele. Os dois primeiros casos do gráfico 1 são
exemplos desse comportamento. Porém, acima de um certo valor de J o ponto de
equilíbrio torna-se instável (ele cai sobre o ramo do meio da isóclina nula de V). Isso faz
com que a trajetória do sistema se afaste do ponto de equilíbrio neste caso, passando
abaixo dele e continuando em direção ao ramo da direita da curva lenta.
Esse comportamento cíclico e estável do sistema é chamado na literatura de sistemas
dinâmicos de ciclo limite estável. Os possíveis comportamentos do sistema em tal caso são
os seguintes: se o estado inicial do sistema estiver dentro da região do espaço de fase
delimitada pelo ciclo limite, sua trajetória se espiralará para fora até se encontrar com o
ciclo limite, permanecendo sobre ele daí por diante. Da mesma forma, se o estado inicial
do sistema estiver fora da região delimitada pelo ciclo limite sua trajetória convergirá de
maneira espiral em direção a ele. Portanto, a bacia de atração do ciclo limite é o plano de
fase inteiro (com exceção do ponto de equilíbrio).
O ciclo limite (isto é, para onde as trajetórias tendem) é chamado de estável porque
pequenas perturbações que desviem o sistema de sua trajetória cíclica são rapidamente
suprimidas e o sistema retorna para o ciclo limite.
Na literatura de sistemas dinâmicos, o tipo de comportamento em que oscilações estáveis e
de grande amplitude aparecem abruptamente quando um parâmetro de bifurcação (a
densidade de corrente J neste caso) varia é chamado de bifurcação de Hopf subcrítica.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
26
Existe um outro fenômeno, conhecido como bifurcação de Hopf supercrítica, em que as
oscilações abruptas começam com amplitudes arbitrariamente pequenas que vão
aumentando à medida que o parâmetro de bifurcação aumenta, mas este não é o tipo de
bifurcação que governa o aparecimento dos disparos repetidos no modelo de Hodgkin-
Huxley.
Em problemas de engenharia, bifurcações de Hopf subcríticas são indesejáveis pois
produzem oscilações abruptas cujas amplitudes podem ser bem grandes. No entanto, no
caso de neurônios (pelo menos nos da classe do modelo de Hodgkin-Huxley) ela é um
fenômeno crucial. O que caracteriza uma bifurcação de Hopf subcrítica é a mudança de
um ponto de equilíbrio estável de tipo espiral em um ponto de equilíbrio instável, também
de tipo espiral, circundado por um ciclo limite estável. Para saber mais sobre esses tipos de
pontos de equilíbrio, veja os livros de Strogatz (1994) e de Kaplan e Glass (1995)
indicados na Bibliografia.
Uma das características de uma bifurcação de Hopf é que as oscilações que aparecem
abruptamente já começam com uma frequência finita (isto é, a frequência inicial não pode
ser arbitrariamente pequena). Este é o motivo pelo qual o modelo de Hodgkin-Huxley
corresponde a um neurônio de tipo 2 (veja a aula 2).
Estudos qualitativos como o feito aqui, mas baseados em outros modelos de neurônios que
conseguem reproduzir as propriedades de neurônios de tipo 1 (cujas frequências iniciais de
disparo podem ser arbitrariamente pequenas) indicam que o tipo de bifurcação que
acontece no caso dos neurônios de tipo 1 é o de sela-nó (na realidade, a bifurcação é
chamada de “sela-nó sobre um ciclo limite”). Para maiores detalhes sobre tais estudos,
sugere-se a leitura da bibliografia indicada no fim deste texto.
Baseado na análise que FitzHugh fez do modelo de Hodgkin-Huxley no plano de fase
rápido-lento, ele propôs um modelo simplificado de neurônio (com apenas duas variáveis)
que possui as mesmas características qualitativas do modelo de Hodgkin-Huxley no plano
de fase rápido-lento.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
27
As equações do modelo (que você estudará na Lista 3) podem ser escritas de várias
formas. Uma das mais populares é a seguinte:
(18)
onde 0 < a < 1, ϵ > 0 e γ ≥ 0. A variável V representa o potencial de membrana do
neurônio e a variável w é chamada de variável de recuperação, pois é a responsável pelo
lento retorno de V ao valor de repouso após um potencial de ação (como a variável de
ativação do potássio, n, no modelo de Hodgkin-Huxley). Para que a dinâmica de w seja
mais lenta que a de V, o parâmetro ϵ deve ser pequeno.
Note que a isóclina nula de V acima é uma função cúbica como a do modelo de Hodgkin-
Huxley no espaço de fase rápido-lento e a isóclina nula de ω é uma reta, simplificando a
curva sigmoidal do modelo de Hodgkin-Huxley no espaço de fase rápido-lento, mas
mantendo o mesmo comportamento qualitativo: as isóclinas nulas de V e w se cruzam em
um único ponto fixo que se torna instável via uma bifurcação de Hopf quando I é
suficientemente forte, produzindo um ciclo limite que corresponde a disparos periódicos
do modelo.
Mais ou menos na mesma época em que FitzHugh propôs este modelo, o engenheiro
japonês Jin-Ichi Nagumo (1926-1999) e seus colaboradores desenvolveram um modelo
parecido e, por causa disso, atualmente este modelo é conhecido como modelo de
FitzHugh-Nagumo.
Para saber mais sobre o modelo de FitzHugh-Nagumo e ver algumas animações mostrando
seu comportamento dinâmico, recomendo o artigo da Scholarpédia sobre ele:
http://www.scholarpedia.org/article/FitzHugh-Nagumo_model

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 11
28
Bibliografia:
- FitzHugh, R., Thresholds and plateaus in the Hodgkin-Huxley nerve equations. Journal
of General Physiology, 43: 867-896, 1960;
- FitzHugh, R., Impulses and physiological states in theoretical models of nerve
membrane. Biophysical Journal, 1: 445-466, 1961;
- Kaplan, D. and Glass, L., Understanding Nonlinear Dynamics. Springer, New York,
1995;
- Rinzel, J. and Ermentrout, B., Analysis of neural excitability and oscillations. In: Koch,
C. and Segev, I., Methods in Neuronal Modeling: from ions to networks. (2nd Ed.),
MIT Press, Cambridge, MA, 1998. Chapter 7, pp. 251-291.
- Strogatz, S.H., Nonlinear Dynamics and Chaos. Addison-Wesley, Reading, MA, 1994.