Embed Size (px)
Citation preview

UNIVERSIDADE DO ESTADO DO RIO GRANDE DO NORTE
FACULDADE DE ENFERMAGEM
PROGRAMA DE PÓS-GRADUAÇÃO EM SAÚDE E SOCIEDADE
MESTRADO ACADÊMICO EM SAÚDE E SOCIEDADE
ANÁLISE DA PLASTICIDADE CELULAR DA MEDULA ESPINAL
NA PRESENÇA DE MEIO CONDICIONADO DO NERVO
ISQUIÁTICO COM ADIÇÃO DO FATOR DE CRESCIMENTO
FIBROBLÁSTICO - 2 EM CULTURA
CLEBER MAHLMANN VIANA BEZERRA
MOSSORÓ/RN
2015

CLEBER MAHLMANN VIANA BEZERRA
ANÁLISE DA PLASTICIDADE CELULAR DA MEDULA ESPINAL
NA PRESENÇA DE MEIO CONDICIONADO DO NERVO
ISQUIÁTICO COM ADIÇÃO DO FATOR DE CRESCIMENTO
FIBROBLÁSTICO - 2 EM CULTURA
Dissertação apresentada ao Programa
de Pós-graduação em Saúde e
Sociedade da Universidade do Estado
do Rio Grande do Norte, como requisito
final para obtenção do grau de Mestre
em Saúde e Sociedade
Orientador: Dr. Fausto Pierdoná Guzen
MOSSORÓ/RN
2015

94
Catalogação da Publicação na Fonte.
Universidade do Estado do Rio Grande do Norte.
Bibliotecária: Jocelania Marinho Maia de Oliveira CRB 15 / 319
Bezerra, Cleber Mahlmann Viana
Análise da plasticidade celular da medula espinal na presença de meio condicionado do nervo isquiático com adição do fator de crescimento fibroblástico - 2 em cultura. / Cleber Mahlmann Viana Bezerra. – Mossoró, RN, 2014.
93 f. Orientador(a): Prof. Dr. Fausto Pierdoná Guzen
Monografia (Mestre em Saúde e Sociedade). Universidade do Estado do Rio Grande do Norte. Programa de Pós-graduação em Saúde e Sociedade.
1. Cultura da medula espinal. 2. Fator de crescimento fibroblástico -2. 3. Meio
condicionado - Plasticidade neuronal. I. Guzen, Fausto Pierdoná. II. Universidade do Estado do
Rio Grande do Norte. III.Título.


DEDICATÓRIA
À minha família, especialmente aos
meus pais José de Arimatéa e
Sandra Viana, dedico a vocês este
trabalho, pelo incentivo a vencer na
vida e ressaltar que a educação é o
melhor caminho.

AGRADECIMENTOS
Primeiramente a Deus, por ser meu guia mostrando que promover o bem é minha missão.
Aos meus pais pelo apoio que sempre me deram ao longo desta jornada da vida e profissional, proporcionando uma estrutura que me possibilitou chegar até aqui. A distância hoje existente entre nós em virtude dos nossos trabalhos serviu para mostrar ainda mais o tamanho do nosso amor e da nossa cumplicidade, obrigado por essa conquista.
As minhas irmãs Vanessa Viana Bezerra e Waleska Viana Bezerra pelo amor, respeito e amizade que temos um pelo outro e pelo apoio que sempre obtive de vocês.
Aos meus irmãos Abrão Bezerra e Luiza Bezerra que mesmo na inocência de uma criança contribuíram me dando força através do amor que só os irmãos têm.
À Libina Edriana, minha esposa que durante está jornada me incentivou e sempre esteve ao meu lado mesmo nas horas mais complicadas apoiando, e dedicando seu amor incondicional.
Ao professores e orientadores, Fausto Pierdoná Guzen e Eudes Euler Souza Lucena, pelo grande apoio constante e incondicional durante toda a realização deste trabalho, pelo incentivo a participar desse fascinante projeto, por mostrar que o que sabemos é apenas uma gota e o que ignoramos é um oceano e por me conduzir na transposição das minhas dificuldades acreditando no nosso trabalho.
Ao Profº. Rodolfo Lopes, que conheci ao longo dessa caminhada e se tornou um grande amigo. Obrigado pelas oportunidades e ajudas proporcionadas. Serei sempre grato.

Ao Prof. Worgelsanger Pereira, pela disponibilização ao acesso no BIOMOL, e por se fazer extremamente solícito.
A todos os professores do Programa de Pós-Graduação em Saúde e Sociedade da UERN.
Ao amigo Salvador Viana, pela parceria na execução da pesquisa.
A todos os professores e profissionais da Universidade Potiguar – UnP do departamento de Fisioterapia pelo incentivo, ajuda e compreensão das minhas ausências.
A todos os profissionais da Universidade do Estado Rio Grande do Norte, especialmente os parceiros de trabalho diário da Faculdade de Ciências da Saúde - FACS.
Aos meus inestimáveis companheiros de pesquisas do laboratório de Neurologia Experimental da UERN.
A todos os meus alunos, por me ensinaram a lecionar a cada dia melhor.
Muito obrigado!

SUMÁRIO
RESUMO
ABSTRACT
1. INTRODUÇÃO........................................................................................ 16
1.1. O Problema.............................................................................................. 16
1.2. Objetivos.................................................................................................. 17
1.2.1. Objetivo Geral.......................................................................................... 17
1.2.2. Objetivo Específico................................................................................... 17
1.3 Justificativa............................................................................................... 17
2. REVISÃO DE LITERATURA................................................................... 18
2.1. Características Anatômicas do Nervo Isquiático.................................... 18
2.2. O ambiente de lesão e regeneração do sistema nervoso....................... 19
2.3. Fator de crescimento fibroblástico-2 (FGF-2).......................................... 23
3. METODOLOGIA....................................................................................... 26
3.1. Desenho experimental.............................................................................. 26
3.2. Extração e cultivo das células da medula espinal (ME)........................... 27
3.3. Extração e cultivo dos explantes de nervo isquiático............................... 29
3.4. Subcultivos das células da medula espinal e os grupos experimentais 31
3.5. Marcação imunocitoquímica..................................................................... 33
3.6. Dosagem de íons...................................................................................... 36
3.7. Análise dos dados.................................................................................... 37
4. RESULTADOS......................................................................................... 37
4.1. Mudanças morfológicas e expansão das células glias............................. 41
4.2. Mudanças morfológicas e expansão das células neuronais.................... 53
4.3. Fenótipo das células glias e neuronais..................................................... 64

4.4. Dosagem dos íons no meio de cultura..................................................... 69
5. DISCUSSÃO............................................................................................. 73
6. CONCLUSÕES E SUGESTÕES.............................................................. 81
7. REFERÊNCIAS........................................................................................ 82
APÊNDICE............................................................................................... 93

LISTA DE TABELAS E QUADROS
Tabelas Tabela 1. Descrição da amostra dos grupos de acordo com número de células
gliais..................................................................................................................44
Tabela 2. Descrição da amostra dos grupos de acordo com área de células
gliais..................................................................................................................47
Tabela 3. Descrição da amostra dos grupos de acordo com perímetro de
células gliais......................................................................................................50
Tabela 4. Descrição da amostra dos grupos de acordo com número de células
neuronais..........................................................................................................55
Tabela 5. Descrição da amostra dos grupos de acordo com área de células
neuronais..........................................................................................................58
Tabela 6. Descrição da amostra dos grupos de acordo com perímetro de
células neuronais..............................................................................................61
Quadros
Quadro 1. Lista de anticorpos primários...........................................................33

LISTA DE FIGURAS
Figuras
Figura 1. Aspectos Anatômicos do nervo isquiático.........................................18
Figura 2. Ambiente de lesão e regeneração do sistema nervoso.....................21
Figura 3. Extração e preparo da medula espinal para suspensão
celular................................................................................................................27
Figura 4. Troca de meio de cultura celular........................................................28
Figura 5. Acesso cirúrgico ao nervo isquiático..................................................29
Figura 6. Nervo Isquiático dissecado.................................................................29
Figura 7. Remoção do epineuro e perineuro dos ramos nervosos....................30
Figura 8. Placas P60 com meio condicionado por explantes de nervo
isquiático............................................................................................................31
Figura 9. Células precipitadas após centrifugação............................................31
Figura 10. Delineamento do estudo: formação dos grupos e observação
celular................................................................................................................32
Figura 11. Campos de observação celular em placas P60...............................35
Figura 12. Identificação dos meios de cultura após 72 horas para dosagem de
íons....................................................................................................................36
Figura 13. Células de morfologia fibroblastóide................................................37
Figura 14. População de células de Schwann..................................................38
Figura 15. Aspectos morfológicos das células gliais e neuronais.....................39
Figura 16. Morfologia das células após formação dos grupos experimentais por
72 horas. Grupo 1: painéis A (24 horas), B (48 horas), C (72 horas); Grupo 2:
Painéis D (24 horas), E (48 horas), F (72 horas); Grupo 3: Painéis G (24 horas),

H (48 horas), I (72 horas); Grupo 4: Painéis J (24 horas), K (48 horas), L (72
horas). Escala 100µm........................................................................................40
Figura 17. Número de células gliais observadas em cada grupo experimental
de acordo com o dia de observação..................................................................45
Figura 18. Número de células gliais observadas nos dias 1,2 e 3 de acordo com
o grupo experimental.........................................................................................46
Figura 19. Área de células gliais observadas em cada grupo experimental de
acordo com o dia de observação.......................................................................48
Figura 20. Área de células gliais observadas nos dias 1,2 e 3 de acordo com o
grupo experimental............................................................................................49
Figura 21. Perímetro de células gliais observadas em cada grupo experimental
de acordo com o dia de observação..................................................................51
Figura 22. Perímetro de células gliais observadas nos dias 1,2 e 3 de acordo
com o grupo experimental.................................................................................52
Figura 23. Número de células neuronais observadas em cada grupo
experimental de acordo com o dia de observação...........................................56
Figura 24. Número de células neuronais observadas nos dias 1,2 e 3 de acordo
com o grupo experimental.................................................................................57
Figura 25. Área de células neuronais observadas em cada grupo experimental
de acordo com o dia de observação..................................................................59
Figura 26. Área de células neuronais observadas nos dias 1,2 e 3 de acordo
com o grupo experimental.................................................................................60
Figura 27. Perímetro de células neuronais observadas nos dias 1,2 e 3 de
acordo com o grupo experimental.....................................................................62
Figura 28. Perímetro de células neuronais observadas em cada grupo
experimental de acordo com o dia de observação...........................................63

Figura 29. Células do G 2 submetidas a imunofluorecência do anticorpo β-
Tubulina (FITC).................................................................................................64
Figura 30. Células do G 2 submetidas a imunofluorecência do anticorpo GFAP
(Alexa Fluor 488)..............................................................................................65
Figura 31. Células do G 2 submetidas a imunofluorecência do anticorpo NF-200
(Alexa Fluor 488)...............................................................................................65
Figura 32. Células do G 3 submetidas a imunofluorecência do anticorpo GFAP
(Alexa Fluor 488)...............................................................................................66
Figura 33. Células do G 3 submetidas a imunofluorecência do anticorpo NeuN
(Alexa Fluor 488)...............................................................................................66
Figura 34. Células do G 3 submetidas a imunofluorecência do anticorpo OX-42
(Alexa Fluor 488)...............................................................................................67
Figura 35. Células do G 4 submetidas a imunofluorecência do anticorpo GFAP
(Alexa Fluor 488)...............................................................................................67
Figura 36. Células do G 4 submetidas a imunofluorecência do anticorpo NeuN
(Alexa Fluor 488)...............................................................................................68
Figura 37. Células do G 4 submetidas a imunofluorecência do anticorpo NF-200
(Alexa Fluor 488)...............................................................................................68
Figura 38. Células do G 4 submetidas a imunofluorecência do anticorpo OX-42
(Alexa Fluor 488)...............................................................................................69
Figura 39. Dosagem de sódio de acordo com cada grupo
experimental......................................................................................................70
Figura 40. Dosagem de cálcio de acordo com cada grupo
experimental......................................................................................................71
Figura 41. Dosagem de potássio de acordo com cada grupo
experimental......................................................................................................72

LISTA DE ABREVIATURAS
AVL: Analisador de eletrólitos.
°c: grau Celsius.
μg: Micrograma.
μl: Microlitro.
μM: Micromolar.
μmol/L: micromol por litro.
μm2: Micrômetro quadrado.
BDNF: Fator neurotrófico derivado do cérebro.
BSA: Albumina de soro de boi.
β – Tubulina: São as proteínas que compõem os microtúbulos.
Ca: Cálcio
CO2: Gás Carbônico
CS: Célula de Schwann
CEEA: Comissão de Ética de experimentação animal
D-10: Meio de cultura
DMEM: Dulbeco´s Modified Eagle´s Medium
EDTA: Ácido etilenodiamino tetra-acético
ELA: Esclerose Lateral Amiotrófica
FGF-2: Fator de crescimento fibroblástico-2
GFAP: Proteína ácida fibrilar glial
K: Potássio

HRTM: Hospital Regional Tarcísio Maia
IGF: Fator de crescimento tipo insulina
IL-1: Interleucina-1
L-15: Meio Leibovitz 15
LIF: Fator inibidor de leucemia
L4: Quarta vertebra lombar
L5: Quinta vertebra lombar
M: Molar
ml: Mililitro
MAP-2: Proteína associada ao Microtúbulo 2
mm: Milímetro
ME: Medula Espinal
MCNI: Meio condicionado de nervo isquiático
Na: Sódio
NeuN: Marcador nuclear neuronal
NF-200: Neurofilamento 200
NGF: Fator de crescimento do nervo
NT-3: Neurotrofina-3
OX-42: Anticorpo primário
PBS: Tampão fosfato
P60: Placas de petri 60 milímetros
rpm: Rotações por minuto
SNC: Sistema nervoso central

SNP: Sistema nervoso periférico
S1: Primeira vertebra sacral
S2: Segunda vertebra sacral
S3: Terceira vertebra sacral
TGF: Fator transformador de crescimento
UERN: Universidade do Estado do Rio Grande do Norte
UFRN: Universidade Federal do Rio Grande do Norte

RESUMO
Eventos traumáticos são cada vez mais frequentes no cotidiano da sociedade,
gerando lesões graves na medula espinal, implicando na sobrevivência das
células nervosas, prejudicando a regeneração e o restabelicimento dos contatos
funcionais, frequentemente associada à perda permanente da função. Sabe-se
que os nervos periféricos são conhecidos como bons substratos, pois possuem
ambiente ideal para induzir o meio regenerativo. Evidências mostram a influência
da matriz extracelular e do fator de crescimento fibroblástico -2 (FGF-2) no
crescimento de fibras nervosas lesadas no Sistema Nervoso Periférico (SNP).
Nessa perspectiva, esse estudo teve como objetivo analisar a plasticidade
celular da medula espinal na presença de meio condicionado do nervo isquiático
de ratos wistar diante da adição do FGF-2. Esta pesquisa foi aprovada sobre o
parecer consubstanciado CEEA/UERN n0 007/13. O crescimento e a morfologia
celular foram avaliados ao longo de 72 horas. Além disso, a avaliação fenotípica
foi feita a partir da imunocitoquímica para GFAP, OX-42, MAP-2, β-tubulina III,
NeuN e NF-200 no terceiro dia de cultivo também foi realizada a dosagem de
íons presente no meio de cultura. As células cultivadas com meio condicionado
sozinho ou combinado com FGF-2 demonstraram características morfológicas
semelhantes a neurônios e células gliais e uma significativa atividade
proliferativa, crescimento da área e perímetro nos grupos 2, 3 e 4 ao longo dos
dias. As células cultivadas com meio condicionado desprovido de tratamento
com FGF-2 adquiriram fenótipo glial demostrando imunorreatividade para GFAP,
β-tubulina III e NF-200. As células cultivadas com meio condicionado com adição
de FGF-2 expressaram GFAP, OX-42, MAP-2, β-tubulina III, NeuN e NF-200. O
estudo possibilitou a plasticidade de células da medula espinal em linhagens
neuronal e glial.
PALAVRAS CHAVE: Cultura da medula espinal. Fator de crescimento
fibroblástico -2. Meio condicionado. Plasticidade neuronal.

ABSTRACT
Traumatic events are more frequent in everyday society, causing severe spinal
cord injuries, resulting in the survival of nerve cells, which impair regeneration
and reestablishment of functional contacts, often associated with permanent loss
of function. It is known that peripheral nerves are known as good substrates
because they have ideal environment to induce the regeneration medium. It is
known that peripheral nerves are known as good substrates because they have
ideal environment to induce the regeneration medium. Evidence shows the
influence of extracellular matrix and fibroblast growth factor -2 (FGF-2) in the
growth of injured nerve fibers in the peripheral nervous system (PNS). From this
perspective, this study aimed to analyze the cellular plasticity of the spinal cord
in the presence of conditioned media of the sciatic nerve of Wistar rats before the
addition of FGF-2. This research was approved on the opinion embodied
CEEA/UERN n0 007/13. Growth and cell morphology were assessed over 72
hours. In addition, phenotypic analysis was performed from
immunocytochemistry GFAP, OX-42, MAP-2, β-tubulin III, NeuN, NF 200 and
third day of cultivation in the dosage of ions present were also carried out in the
culture medium . Cells cultured with conditioned medium alone or with FGF-2
showed morphological features similar to neurons and glial cells and a significant
proliferative activity of the growth area and perimeter in groups 2, 3 and 4
throughout the day. Cells cultured with conditioned medium devoid of FGF-2
treatment with glial phenotype demonstrating acquired immunoreactivity for
GFAP, β-tubulina III e NF-200. Cells cultured with conditioned medium with FGF-
2 expressed addition GFAP, OX-42, MAP-2, β-tubulina III, NeuN e NF-200. The
study enabled the plasticity of spinal cord cells in neuronal and glial lineages.
KEY WORKS: Spinal cord culture. fibroblast growth factor -2. Conditioned medium. Neuronal plasticity.

15
I INTRODUÇÃO
O Sistema Nervoso é responsável pelas funções orgânicas e pela
integração do homem e dos demais animais ao meio ambiente, uma vez que ele
controla e coordena todas as funções dos demais sistemas corporais, recebendo
e interpretando estímulos oriundos do meio ambiente, desencadeando respostas
específicas a cada estímulo recebido, sejam elas voluntárias ou involuntárias. É
descrito que o Sistema Nervoso divide-se anatomicamente em Sistema Nervoso
Central (SNC) e Sistema Nervoso Periférico (SNP), onde o primeiro tem como
função receber, interpretar e dar respostas aos estímulos conduzidos pelo SNP,
o qual transporta informações de todas as áreas do organismo. Embora tenham
funções e localizações diferentes, o SNC e o SNP são interdependentes (Lundy-
Ekman, 2008).
Pela sua disposição anatômica os nervos periféricos apresentam
possibilidades de lesões frequentes e com mecanismos de injurias variados com
eventos clínicos e traumáticos, cada vez mais comum no cotidiano da sociedade,
como por exemplo: os acidentes automobilísticos, responsáveis por problemas
graves, como dor e sequelas muitas vezes permanentes. Estes danos diminuem
a qualidade de vida das pessoas acometidas, pois geram a incapacitação física
e a perda parcial ou total de suas atividades produtivas, o que origina importantes
consequências econômicas, além do altíssimo custo social gerado pelo aumento
nas despesas da saúde pública (Noble et al., 1998; Santos et al., 2012).
Uma parcela significativa da população Brasileira afetada por lesões em
nervos periféricos encontram-se na plenitude da idade produtiva e
independência funcional, posteriormente ao evento passam para um estágio
onde necessitam da ajuda de terceiros, redução da autoestima e aumento do
custo de vida. Esta afecção vem a constitui-se então, um dos desafios para os
profissionais e pesquisadores da saúde, devido possuir uma alta incidência com
a inexistência de tratamento satisfatório (Cristante et al., 2010; Brito et al., 2011).

16
Estima-se que a incidência das lesões traumáticas em alguns países seja
superior a 500.000 novos casos anuais, dos quais 2,8% dos pacientes adquirem
incapacidades vitalícias em virtude da dificuldade regenerativa, o qual o tempo
é um fator crucial para este evento. No Brasil são poucos os trabalhos publicados
a respeito desta epidemiologia, porém estima-se que ocorram mais de 10.000
novos casos por ano (Noble et al., 1998; Novak e Mackinnon, 2005).
Na busca do modelo mais adequado para pesquisa experimental de
substâncias sistêmicas que atuem na regeneração pós-traumática do SNC,
encontram-se nas literaturas diversos métodos, com variações nas espécies
animais, sítio e método de lesão, tempo de evolução, e meios de análise
funcional e histológica. Diversos estudos publicados demonstram as influências
de inúmeros fatores na recuperação, abordando técnica cirúrgica, enxertos de
fregmentos de nervos e células, administração de substâncias neurotróficas,
laser e até exposição a pulsos eletromagnéticos (Shacklock, 1995; Choi e Dunn,
2001; Boyd et al., 2005; Monte-Raso, 2005; Guo e Dong, 2009; Lukas, 2011;
Souza et al., 2011).
1.1 O PROBLEMA
O crescente número de pessoas no Brasil e no mundo estão sendo
acometidas por doenças neurodegenerativas como a Esclerose Lateral
Amiotrófica, Esclerose Múltipla, Hanseníase, entre outras e por acidentes
traumáticos com mecanismos de injúrias variados, entre eles destacam-se os
acidentes por armas brancas, armas de fogo, acidentes automobilísticos e os
acidentes com motocicletas. Estes acidentes e as doenças neurodegenerativas,
promovem lesões neurológicas de ordem central e ou periférica gerando uma
dependência funcional e passam para um estágio onde necessitam da ajuda de
terceiros, redução da autoestima e aumento do custo social. Esta afecção vem
a constitui-se então, um dos desafios para os profissionais e pesquisadores da
saúde em virtude da inexistência de tratamentos satisfatórios.

17
1.2 OBJETIVOS 1.2.1 OBJETIVO GERAL
Analisar a plasticidade celular da medula espinal na presença de meio
condicionado, na presença e ausência do FGF-2. 1.2.2 OBJETIVOS ESPECÍFICOS
Avaliar a plasticidade das células da medula espinal promovida pelo meio
condicionado do nervo isquiático e pelo fator FGF-2 a partir dos seguintes
parâmetros:
• Evolução de número, área, perímetro e morfologia celular por 72 horas;
• A imunorreatividades das proteínas gliais: proteína ácida fibrilar glial
(GFAP), marcador microglial OX-42 através da imunocitoquímica das
células gliais da medula espinal;
• A imunorreatividades da proteína associada ao microtúbulo-2 (MAP-2), β-
tubulina III, das proteínas neuronais de neurofilamentos 200 (NF-200) e
da proteína nuclear neuronal (NeuN) através da imunocitoquímica das
células da medula espinal;
• A dosagem dos íons Ca, K e Na no meio de cultura utilizado.
1.3 JUSTIFICATIVA
A pesquisa irá contribuir de forma somatória as discussões sobre o tema,
fomentando a relevância das intervenções mais adequadas, visando à melhoria
da qualidade de vida e a promoção da saúde.
Nesse contexto, visando suprimir as lacunas existentes, a terapia com uso
de meio condicionado do nervo isquiático (MCNI) e a utilização do FGF-2 por ter
se mostrado bastante promissor, possibilitando um alto nível de proliferação
celular, um bom controle de sua atividade proliferativa, plasticidade fenotípica e
regeneração nervosa.

18
II REVISÃO DE LITERATURA
2.1 CARACTERÍSTICAS ANATÔMICAS DO NERVO ISQUIÁTICO
Considerado o maior nervo do corpo, o nervo isquiático faz parte do SNP,
pertencendo em carnívoros tanto ao plexo sacral quanto ao lombossacral
(Schwarze e Schröder, 1970; Ellenberger e Baum, 1985; Konig e Liebich 2004).
É um nervo motor que envia, primariamente, a mensagem ao músculo
para que haja contração. Isto ocorre porque o neurônio motor recebe um impulso
nervoso que passa para o corpo celular do neurônio. Esse impulso segue para
o axônio, local onde haverá a despolarização e gerará um potencial de ação na
célula (Konig e Liebich 2004).
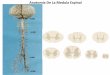
Morfologicamente o ramo anastomótico de L4 se une ao L5 constituíndo
o tronco lombossacral, esta constituição se une com S1 e depois
sucessivamente ao S2, S3 e S4 (Figura 1) atravessando o forame isquiático
maior e na sequência emite seus ramos colaterais, chegando ao ramo terminal
chamado de nervo isquiático, por isso denominado como um nervo periférico
(Sobotta e Becher, 2013).
Fonte: Atlas de Anatomia Humana Petra Kopf-Maier 6aed.
Figura 1. Aspectos anatômicos do nervo isquiático.

19
Caudalmente, passa pela coxa entre o trocânter maior do fêmur e a
tuberosidade isquiática, ao longo da superfície lateral do músculo
semimembranáceo, emitindo ramificações para o músculo glúteo médio,
semimembranáceo, semitendíneo, bíceps femoral, gêmeos, quadrado femoral e
adutores. Na região medial da coxa, o nervo isquiático dobra-se distalmente e
se bifurca em nervo fibular comum (L4, L5, S1 e S2) e nervo tibial formado por
L4, L5, S1, S2 e S3. O nervo fibular comum já na fossa poplítea dirige-se
obliquamente para baixo e lateralmente se bifurcando em nervos fibulares
superficial e profundo (Monte-Raso, 2006; Aversi-Ferreira, 2011).
Para os músculos da região glútea vão os nervos glúteo superior (L4, L5
e S1) e glúteo inferior (L5, S1 e S2). Um ramo sensitivo importante é o nervo
cutâneo posterior da coxa, formado por S1, S2 e S3. Para o períneo, temos o
nervo pudendo formado a partir de S2, S3 e S4. Da região glútea em direção à
face posterior da coxa, onde se situa sobre o músculo adutor magno e é cruzado
posteriormente, pela cabeça longa do músculo bíceps femoral, o nervo também
supre ramos articulares para o quadril e ramos musculares para os músculos do
jarrete (Sobotta e Becher, 2013).
2.2 O AMBIENTE DE LESÃO E REGENERAÇÃO DO SISTEMA NERVOSO
O SNP é composto pelas fibras nervosas, formadas por um ou mais
axônios, envoltos pelas células de Schwann (CSs), mantidas pelo endoneuro,
mais material amorfo da matriz extracelular, capilares, fibroblastos e mastócitos,
assim como o epineuro e o perineuro (Birdi e Antia, 2003). Dentro da bainha
perineural, os axônios e as CSs são envolvidos pela lâmina basal e composta
por várias moléculas secretadas por múltiplas células, dentre elas, as de
Schwann (Alberts et al., 2010). A lâmina basal, através de interações, com
receptores da membrana celular, participa no metabolismo celular, na
organização das proteínas das membranas plasmáticas, na migração celular,
durante a embriogênese e na diferenciação celular. Além disso, influencia a
regeneração axonal, servindo de guia das fibras nervosas e possui ainda funções
estruturais e sinalizadoras (Stoll e Muller, 1999).
Nos anos 80, os experimentos de Aguayo et al.(1981) demonstraram o
potencial de crescimento de fibras de neurônios no modelo de transecção de

20
nervo óptico de ratos adultos, que aloja as fibras nervosas que ligam a retina aos
núcleos subcorticais responsáveis por algumas das funções visuais. Após a
transecção, segmento de nervo isquiático foi interposto entre o coto proximal do
nervo óptico seccionado e o colículo superior, no mesencéfalo. Todo o trajeto do
nervo interposto se fez do lado de fora do crânio. Após alguns meses, a
regeneração foi constatada pela obtenção de registros elétricos no mesencéfalo
após estimulação visual.
O ambiente de lesão dos axônios do SNC difere muito daquele do SNP.
Ao contrário dos nervos, os axônios do SNC não são estruturalmente separados
por bainhas perineurais e neurilemais, estruturas que fornecem um substrato
anatômico para o crescimento da fibra lesada (Junqueira e Carneiro, 2012).
Como os neurônios dos mamíferos geralmente não se dividem, a destruição de
um neurônio representa perda permanente. Seus prolongamentos, no entanto,
dentro de certos limites podem regenerar-se devido à atividade sintética dos
respectivos corpos celulares. Por isso, as fibras se regeneram, embora com
dificuldade. Quando uma célula nervosa é destruída, as que a ela se ligam nada
sofrem, exceto nos raros casos em que um neurônio recebe impulsos
exclusivamente de outro. Neste caso, o neurônio que fica completamente
privado de impulsos nervosos, pela destruição do outro, sofre a chamada
degeneração transneuronal (Purves et al., 2010).
Ao contrário dos elementos nervosos, as células da glia do SNC, e as do
SNP (CSs e células satélites dos gânglios), são dotadas de grande capacidade
de proliferação. Os espaços deixados pelas células e fibras nervosas do SNC
destruído por acidentes ou doença são preenchidos por células da neuroglia
(Alberts et al., 2010). Quando um nervo é seccionado, ocorrem alterações
degenerativas, seguidas de uma fase de reparação. No nervo lesado deve-se
distinguir a parte da fibra que, pela lesão, desligou-se do seu neurônio (parte
distal) e a parte que continua unida ao neurônio (parte proximal). O segmento
proximal, por manter contato com o corpo celular, que é o centro trófico,
frequentemente é regenerado, enquanto o segmento distal degenera totalmente
e acaba por ser reabsorvido (Machado e Haertel, 2014).
No coto distal, tanto o axônio, agora separado de seu centro trófico, como
a bainha de mielina degeneram totalmente. Enquanto se processam essas
alterações, as CSs proliferam, formando colunas celulares compactas. Essas

21
colunas servirão de guia para os axônios que vão crescer durante a fase de
regeneração. Por outro lado, o segmento proximal do axônio cresce e se
ramifica, formando vários filamentos que progridem em direção às colunas de
CSs. Todavia, somente as fibras que penetram nessas colunas têm possibilidade
de alcançar um órgão efetor (Figura 2). Quando o segmento distal do nervo é
perdida, como ocorre na amputação de um membro, as fibras nervosas crescem
ao acaso (Junqueira e Carneiro, 2013).
Fonte: http://new.lakeforest.edu
Figura 2. Ambiente de lesão e regeneração do sistema nervoso.
A eficiência funcional da regeneração depende das fibras ocuparem as
colunas de CSs destinadas aos locais corretos. Num nervo misto, por exemplo,
se as fibras sensitivas regeneradas ocuparem colunas destinadas às placas
motoras de um músculo estriado, a função do músculo não será restabelecida.
A possibilidade de recuperação funcional é aumentada pelo fato de cada fibra
em regeneração dar origem a vários prolongamentos e cada coluna receber
prolongamentos de várias fibras (Purves et al., 2010).

22
Após a lesão, axônios fragmentados e restos de bainha de mielina não
são rapidamente e eficientemente removidos do sítio da ferida no SNC, em parte
porque o processo de ativação microglial é inferior ao observado pelos
macrófagos nos nervos. Alguns trabalhos sugerem que células como as de
linhagem glial e os fagócitos são decisivas no processo de reparo e cicatrização,
com interferência no fenômeno de crescimento de fibras nervosas (Bootcov et
al., 1997; Zeev-brann et al., 1998; Chan et al., 2001; Markus et al., 2002b).
Mudanças na expressão local de citocinas ocorrem após a lesão do SNC.
Citocinas como interleucina-1 (IL-1) e fator de crescimento tumoral-α (TNF-α)
são expressas pelas células que invadem o sítio de lesão da medula espinal e
contribuir para o recrutamento de células inflamatórias e modulação da resposta
glial à cicatrização (Ghirnikar et al., 1998). TNF-α e o fator de crescimento de
colônia-1 (CSF-1) aumentam a migração de macrófagos para a região de nervos
lesados (Lotan e Schwartz, 1994), bem como a adesividade destas células,
sugerindo que elas poderiam melhorar a permissividade do crescimento de fibras
(Prewitt et al., 1997).
As lesões completas, com perda de substância, raramente apresentam
recuperação sem intervenção cirúrgica, e as técnicas atuais de reparação
oferecem resultados aleatórios e frequentemente insatisfatórios. O enxerto
autólogo de nervo proporciona os melhores resultados no reparo quando há
transecção de nervo periférico. Este, porém, apresenta limitações, como uma
maior morbidade no local de retirada do enxerto, escassez de sítios doadores de
nervo, diferenças estruturais entre o nervo doador e receptor, além do déficit
sensitivo resultante na área da qual foi retirado (Oliveira et al., 2004; Ichihara et
al., 2008). Nesse contexto, muitos pesquisadores buscam terapias alternativas,
como transplante de células tronco (CTs) autólogas, técnicas de tubulização com
o uso de diferentes materiais e aplicação de fatores tróficos, com o propósito de
otimizar o reparo de nervos periféricos danificados (Oliveira et al., 2004).
Diversos fatores, como a expressão diferencial de genes envolvidos com
o crescimento, os genes relacionados à estrutura do axônio, as moléculas de
adesão celular e outras da matriz extracelular, o suprimento de fatores
neurotróficos, a presença das citocinas inflamatórias e dos fatores inibidores
relacionados à substância branca, em combinações específicas, determinam

23
como o crescimento axonal será sustentado após a lesão nervosa (Bethea e
Dietrich, 2002; Condic e Lemons, 2002; Nguyen et al., 2002; Snider et al., 2002).
2.3 FATOR DE CRESCIMENTO FIBROBLÁSTICO-2 (FGF-2)
Os fatores neurotróficos são polipeptídeos que auxiliam no processo
regenerativo do SNP. Compreendem basicamente um conjunto de famílias de
moléculas e seus receptores responsáveis por manter o crescimento e a
sobrevivência dos axônios e neurônios motores e sensitivos, após danos
teciduais. Diversos fatores tróficos, também conhecidos como fatores de
crescimento, são utilizados e testados in vitro e in vivo na regeneração de nervos
periféricos. Essas proteínas atuam diretamente na proliferação e diferenciação
de diferentes tipos celulares, sendo capazes de promover reparo tecidual e
recuperação funcional (Boyd e Gordon, 2003).
Vários fatores neurotróficos são liberados e atuam conjuntamente após
uma lesão neural periférica a fim de estimular a regeneração neural; incluem o
fator de crescimento do nervo (NGF), o fator neutrófico derivado do cérebro
(BDNF) (Boyd e Gordon, 2003), a neurotrofina-3 (NT-3) (Markus et al., 2002), os
fatores de crescimento tipo insulina I e II (IGF-I e IGF-II) (Perlson et al., 2004), o
FGF-2 (Terenghi, 1999), entre outros.
No que diz respeito às interações tróficas que determinariam o melhor
microambiente à regeneração, uma nova fase se iniciou com a descoberta do
NGF por Rita Levi Montalcini. Foi descrito que a interação entre o neurônio e as
CSs poderia implicar na sinalização recíproca, envolvendo a liberação de
moléculas com atividade trófica (Levi-Montalcini e Angeletti, 1968).
De forma ampla, os fatores neurotróficos influenciam na atividade neural,
incluindo-se a atividade sináptica, o desenvolvimento durante o período
embrionário, a manutenção e sustentação durante a vida adulta e a
sobrevivência dos neurônios após a lesão nervosa. Estes são produzidos pelos
órgãos alvo e transportados retrogradamente ao corpo celular dos neurônios.
Quando ocorre uma lesão nervosa, há a interrupção no fornecimento destes
fatores, podendo levar à morte neuronal e consequentemente à ausência

24
regenerativa (Terenghi, 1999). Em virtude disso, os componentes não neurais,
por exemplo as CSs, passam a produzi-los, de forma a proteger os neurônios e
estimular a regeneração axonal (Lewin et al., 1997). Os fatores neurotróficos são
divididos em duas classes: as neurotrofinas e as neurocitocinas. Exemplos
clássicos de neurotrofinas são o NGF, BDNF, NT-3 e NT-4. Como exemplo de
neurocitocinas, destacam-se o FGF, interleucina (IL-1), o fator de crescimento
transformado (TGF), e o fator inibidor de leucemia (LIF) (Lewin et al., 1997).
Estes fatores neurotróficos são importantes para o desenvolvimento e
sobrevivência dos neurônios sensitivos, motores, simpáticos do SNP e neurônios
colinérgicos presentes no SNC. Fatores como o FGF, promovem a sobrevivência
neural, proliferação das CSs e interação entre as células da glia e os neurônios
(Sendtner et al., 1992; Koop et al., 1997; Wang et al., 2008).
O transplante de populações purificadas das CSs parece ser uma
alternativa viável na tentativa de contornar fatores restritivos relacionados ao
microambiente regenerativo e ao crescimento de fibras nervosas. As CSs
secretam fatores neurotróficos, expressam moléculas de adesão e produzem
numerosas moléculas da matriz extracelular, que sabidamente influenciam o
crescimento das fibras nervosas (Xu et al., 1997). Além disso, a injeção local de
CSs está relacionada à diminuição da cicatriz glial (Martini, 1994). Outra
vantagem do uso dessas células deve-se as características técnicas, já que, o
seu número pode ser rapidamente aumentado in vitro em um curto período de
tempo (Morrissey et al., 1991).
As CSs produzem os fatores neurotróficos NGF, BDNF e outras
neurotrofinas, além de membros da família dos fatores de crescimento de
fibroblastos, como o FGF-2 e membros dos IGFs (Yamamoto et al., 1993;
Springer et al., 1994; Hammarberg et al., 1996; Menei et al., 1998; Sayers et al.,
1998; Russell et al., 2000; Kubo et al., 2002). A expressão gênica de vários
fatores neurotróficos ocorre nas CSs nas porções proximal e distal dos cotos
nervosos após a lesão do nervo isquiático do rato (Cheng et al., 1996; Frostick
et al., 1998; Kirsch et al., 1998). O suprimento aumentado de fatores
neurotróficos por estas células serviria para proteger o axônio de uma maior
degeneração retrógrada e, consequentemente, o corpo celular do neurônio, bem

25
como promover o crescimento da fibra no coto distal do nervo lesado (Raivich e
Kreutzberg, 1993; Henderson et al., 1994).
Sabe-se que o FGF-2 é uma proteína mitogênica capaz de agir em
múltiplos tipos de células tal como fibroblastos, neurônios e células da glia (Davis
e Stroobant, 1990; Baird e Klagsbrun, 1991). A proteína e o RNAm do FGF-2
têm sido identificados no citoplasma de neurônios e no núcleo de astrócitos de
muitas regiões do encéfalo (Matsuyama et al., 1992; Woodward et al., 1992;
Humpel et al., 1993; Fuxe et al., 1996; Chadi e Fuxe, 1998). FGF-2 tem uma
função no desenvolvimento neuronal durante a vida prenatal e também influencia
a sobrevivência e a plasticidade de neurônios no SNC maduro. Por exemplo,
favorece neuroproteção para neurônios e astrócitos do núcleo do nervo
hipoglosso após axotomia deste nervo (Walicke, 1988; Chadi et al., 1993).
No SNC, FGF-2 pode causar remielinização e também proteger diferentes
células neuronais de danos induzidos por morte (Butt e Dinsdale, 2005). O FGF-
2 exógeno uma vez aplicado no sistema adulto nigroestriatal, promove a
sobrevivência e crescimento de neurônios dopaminérgicos e os protege da morte
induzida por neurotoxinas in vivo e in vitro (Grothe e Timmer, 2006).
As atividades mitogênicas e neurotróficas do FGF-1 e 2 incluem sua
capacidade de melhorar a sobrevivência e o crescimento de vários tipos de
células neuronais, como do neocórtex, hipocampo, cerebelo, medula espinal e
neurônios sensoriais isolados do SNC de adultos (Matsuda et al., 1990; Peulve
et al., 1994; Himmelseher et al., 1997; Dono, 2003). Eles também influenciam na
migração e diferenciação das células neuronais (Anderson, 1993; Reuss et al.,
2003). Além disso, ambos FGFs como seus receptores são aumentados após
lesão periférica e central do sistema nervoso (Gomez-Pinilla et al., 1992; Logan
et al., 1992; Mocchetti e Wrathall, 1995; Grothe et al., 2001).
A capacidade dos FGFs para atuarem como potentes fatores
neurotróficos tem sido amplamente demonstrada através da prevenção da morte
induzida por axotomia de neurônios glutamatérgicos (Peterson et al., 1996). In
vitro, o FGF-2 protege os neurônios contra citotoxicidade, diminui a morte celular
apoptótica, forma novos vasos sanguíneos e promove reconstrução da bainha
de mielina, exercendo um efeito neuroprotetor nos neurônios motores após
transecção do nervo isquiático ou da injúria contusa da medula espinal (Mattson

26
et al., 1989; Mattson et al., 1993; Fressinaud, 1994; Baffour et al., 1995; Teng et
al., 1998; Teng et al., 1999; Romero et al., 2001).
Em 2009, Guzen et al. avaliaram o potencial regenerativo do GDNF
adicionado a segmentos de nervos isquiáticos interpostos em medulas espinais
transeccionadas de ratos. O GDNF adicionado favoreceu a recuperação motora,
o crescimento de fibras neuronais locais e a neuroplasticidade. Em 2012, Guzen
et al. utilizaram o mesmo modelo de lesão descrito, bem como o mesmo método
terapêutico. No entanto, foi adicionado FGF-2 no espaço promovido pela
transecção completa da medula espinal avaliando a possibilidade de aumentar
a capacidade do enxerto do nervo isquiático em melhorar a recuperação
sensitivo-motora e tecidual observando o desempenho dos membros posteriores
avaliados semanalmente, durante 8 semanas, usando pontuação do
comportamento motor (BBB) e testes sensoriais ligados ao comportamento da
pontuação combinada (CBS), que indicam o grau de melhoria motora e a
porcentagem de déficit funcional, respectivamente. O tratamento da medula
espinal com nervo isquiático e nervo isquiático na adição do FGF-2 permitiu a
recuperação parcial dos movimentos dos membros posteriores em relação ao
controle, manifestado por escores significativamente mais elevados de
comportamento. O FGF-2 adicionado no enxerto de nervo favoreceu a
recuperação motora parcial e fibras imunorreativas para MAP-2, GAP-43 e NF-
200 onde foram encontrados em maior quantidade no interior do enxerto
adicionado de FGF-2.
III METODOLOGIA 3.1 DESENHO EXPERIMENTAL
Para a realização do experimento foram utilizados 12 animais (ratos da
linhagem Wistar - Rattus novergicus), 06 destes animais, machos com idade
entre 40 a 50 dias e peso aproximado de 250 gramas que permaneceram no
biotério por uma semana antes de serem utilizados, mantidos em gaiolas
plásticas (30 x 16 x 19 cm) coletivas com, no máximo, três animais em cada

27
gaiola e com temperatura média de 22 ± 2ºC, alimentados com ração padrão e
água de torneira fornecida ad libitum. E outros 06 com idade de 02 dias oriundos
do Biotério da Universidade do Estado do Rio Grande do Norte (UERN). O
projeto atendeu as normas para a realização de pesquisa em animais com todos
os procedimentos, passando pela Comissão de Ética no Uso de Animais da
(UERN) sendo aprovado e autorizado através do parecer consubstanciado
CEEA/UERN n0 007/13 (Apêndice 1).
3.2 EXTRAÇÃO E CULTIVO DAS CÉLULAS DA MEDULA ESPINAL (ME)
As células da medula espinal foram coletadas de seis ratos Wistar com 2
dias de vida. Para a extração da (ME) foi utilizado o protocolo modificado de
Cheng e colaboradores (1996). Foi realizado uma eutanásia nos neonatos
utilizando pentobarbital, 60-80mg/kg por via intraperitoneal, posteriormente foi
realizado uma incisão longitudinal paralela a coluna vertebral, retirando toda a
coluna e transferido para tubo falcon de 15mL contendo 4mL de meio Leibovitz-
15 (L-15: GIBCO Invitrogen Corporation). Sob fluxo laminar, teve inicio o
processo de extração da medula, com auxílio de uma tesoura e uma espátula
pequena (Figura 3).
Figura 3. Extração da medula espinal e preparados da medula para a suspensão celular.

28
Novos tubos cônicos facon de 15mL com 4mL de meio Knockout DMEM
low (Dulbecco`s modified Eagle`s medium), suplementado com 10% de soro
bovino fetal e 10U/ml de penicilina G, 10μg/ml de estreptomicina e 25μg/ml de
anfotericina B (D-10), todos obtidos da Cultilab® foram preparados e receberam
a medula extraída para realizar a suspensão celular. A suspensão foi
centrifugada a 3000rpm durante cinco minutos a uma temperatura de 370C, após
o sobrenadante foi desprezado e as células foram ressuspendidas em 1mL de
meio, procedimento repetido por três vezes. Placas para cultura com 60mm
(P60) para o plaqueamento foram preparadas com 1mL de soro bovino fetal,
retirado e desprezado após 30 minutos, na sequência foi adicionado 4,5mL de
D-10 nas P60, em seguida foi realizado o gotejamento das células recém
extraídas, mantidas em estufa úmida a 370C com 5% de CO2 e 95% de ar. A
microscopia de luz invertida com contraste de fases foi utilizada para observação
da adesão celular no fundo das placas. Para possibilitar um suprimento
nutricional adequado, eliminar células hematopoiéticas e desprendidas, bem
como possibilitar uma adequada adesão celular no fundo das placas, o meio de
cultura D-10 foi trocado a cada três dias (Figura 4).
Figura 4. Troca de meio de cultura celular.

29
3.3 EXTRAÇÃO E CULTIVO DOS EXPLANTES DE NERVO ISQUIÁTICO
Sob o fluxo laminar, placas para cultura, P60 foram preparadas com 5ml
do meio L-15. Nos animais que foram submetidos à extração do nervo isquiático,
foi realizada a eutanásia com pentobarbital, 60-80mg/kg por via intraperitoneal e
posteriormente a realização de uma tricotomia da região posterior do dorso e
assepsia local com álcool a 70%. A seguir, foram feitos acessos cirúrgicos na
região posterior da coxa nos dois antímeros, afastando os planos musculares,
resultando na exposição do nervo isquiático e promovendo a retirada do mesmo
(Figura 5).
Figura 5. Acesso cirúrgico ao nervo isquiático.
Cada segmento apresentou aproximadamente 35 mm de comprimento
por 1 mm de diâmetro (Figura 6).
Os nervos isquiáticos foram retirados e colocados nas P60 com meio L-
15, sob técnica cirúrgica asséptica com o auxílio de microinstrumentos (tesoura,
pinça, afastadores). Todo excesso de tecido (músculo, gordura e vasos
sanguíneos) aderido aos nervos foi removido sob magnificação por lupa
estereoscópica SZ61 (Olympus®).

30
Figura 6. Nervo isquiático dissecado.
A seguir, o epineuro e perineuro dos nervos foram removidos sob
magnificação e técnica microcirúrgica (Figura 7).
Figura 7. Remoção do Epineuro e Perineuro do nervo isquiático.
Os nervos dissecados foram segmentados em explantes de 1mm de
comprimento cada. Sob fluxo laminar, os fragmentos dos nervos foram
colocados nas P60 com meio de cultura D-10.
O excesso do meio foi removido de forma que os explantes não ficassem
flutuando, nem tão pouco submersos no meio (Figura 8). Após o quinto dia, o
meio D-10 destas culturas foi trocado 02 vezes por semana, sendo os explantes
transferidos para uma placa nova, com meio novo 01 vez por semana. O meio
trocado foi imediatamente desprezado. Esse procedimento possibilitou um
suprimento nutricional adequado aos explantes, bem como a sua reatividade.
Foram plaqueados 18 explantes de nervo isquiático em cada P60.

31
Figura 8. Placas P60 com meio condicionado por explantes de nervo isquiático.
3.4 SUBCULTIVOS DAS CÉLULAS DA MEDULA ESPINAL E OS GRUPOS EXPERIMENTAIS
Quando as células da ME atingiram 70-90% de confluência no fundo da
placa, o meio básico foi removido e adicionados às placas 2 ml de tripsina/EDTA
(0,25% de tripsina contendo 1 mM de EDTA-Cutilab/Brasil®). A suspensão
celular foi colocada em tubo cônico tipo Falcon com o mesmo volume de meio
DMEM suplementado com 10% de soro bovino fetal por 10 minutos, com o
objetivo de inativar a tripsina. A suspensão foi centrifugada a 1200 rpm durante
dez minutos, após o sobrenadante foi desprezado e as células foram
ressuspendidas em 1 mL de meio (Figura 9).
Figura 9. Células precipitadas após centrifugação.

32
As células foram depositadas em 12 P60, sendo 3 placas para cada grupo
e observadas em três períodos de tempo: 24, 48 e 72 horas (Figura 10), desta
forma foi possível avaliar a aderência e proliferação das células da ME nos
seguintes grupos:
Grupo 1 (G1): ME + Meio D-10;
Grupo 2 (G2): ME + Meio de cultura do nervo isquiático (MCNI);
Grupo 3 (G3): ME + MCNI + FGF-2;
Grupo 4 (G4): ME + FGF-2.
Figura 10. Delineamento do estudo: Formação de grupos e observação celular.
No grupo G1 encontramos células da ME na presença do meio D-10, no
G2 encontramos células da ME na presença de MCNI. Nos grupos de estudo G3
e G4 foi acrescido 1μl de FGF-2 (Sigma®) na concentração 1:10 (Guzen et al.,
2012). Já nos grupos 2 e 3 foi acrescido 1ml de meio (D-10), o qual foi coletado
do meio que estava plaqueado os explantes.
Para observação celular foi utilizado um microscópio invertido com
contraste de fases CKX41 (Olympus®) com câmera digital Moticam 3.0 (Motic®)
acoplada.

33
A contagem das células foi realizada usando microscopia de contraste de
fases em 4 campos não sobrepostos no aumento de 200x. Foram feitas
microfotografias dos 4 grupos em 24 horas, 48 horas e 72 horas, sendo que,
após 72 horas de observação celular, procedeu a imunocitoquímica.
3.5 MARCAÇÃO IMUNOCITOQUÍMICA
No terceiro dia (72 horas), o meio foi retirado e as células lavadas em duas
etapas de cinco minutos com tampão de fosfato (PBS) de sódio, 0,1M, pH 7,4,
fixadas em paraformaldeído (4%) por trinta minutos e, novamente, lavadas em
três banhos de PBS, cinco minutos cada. Em seguida, as células foram tratadas
com Triton à 0,5% (Sigma®) por 10 minutos e lavadas em PBS. Posteriormente,
bloqueios de sítios inespecíficos foram realizados durante 30 minutos em
solução PBS 0,1M contendo 0,2% de Triton e 1% de albumina do soro de boi
(BSA). As células foram, então incubadas com um dos anticorpos primários
descritos abaixo por 2 horas à temperatura ambiente (quadro 1).
Quadro 1. Lista de anticorpos primários.
Anticorpo 1° Animal Concentração Fornecedor
GFAP Mouse 1:400 Sigma
OX 42 Mouse 1:500 Millipore
MAP-2 Mouse 1:2000 Abcam
β-tubulina Rabbit 1:500 Millipore
NeuN Mouse 1:500 Millipore
NF-200 Mouse 1:1200 Abcam
1. Anti-proteína ácida fibrilar glial (GFAP) feito em camundongo
(Sigma) para marcação de astrócitos na concentração de 1:400; Este gene
codifica uma das principais proteínas de filamentos intermediários de astrócitos

34
maduros, sendo utilizado como um marcador para distinguir os astrócitos de
outras células gliais durante o desenvolvimento.
2. Anti-CD11b antibody [OX-42]: Este marcador reconhece a maioria
dos macrófagos, células de Kupffer e também rotula extensões de células
dendríticas, granulócitos e células com a morfologia da microglia no cérebro.
3. Anti-proteína associada ao microtúbulo-2 (MAP-2) feito em
camundongo (Abcam) para marcação de fibras na concentração 1:2000; Este
gene codifica uma proteína que pertence à família das proteínas associadas aos
microtúbulos. Tais proteínas estão envolvidas com a montagem de microtúbulos
no citoesqueleto dos neurônios, determinando e estabilizando a forma dendrítica
durante o desenvolvimento neuronal.
4. Anti-β-tubulina: O anticorpo reconhece todas as cinco isoformas de
β-tubulina (β1-β5) 0,1 reage com o β-Lc e fragmentos β-sc na parte carboxi-
terminal da β-tubulina em imunotransferência. A polimerização de dímeros de
tubulina estão relacionados à formação de microtúbulos. O anticorpo pode ser
usado para localizar β-tubulina em cultura de células ou secções de tecido.
5. Anti-NeuN feito em camundongo (Millipore) para marcação de
corpos neuronais na concentração 1:500; Trata-se de uma proteína nuclear
específica para neurônios que é identificado por imunorreatividade com um
anticorpo monoclonal anti-NeuN. Tem sido amplamente utilizado como uma
ferramenta confiável para detectar tipos de células neuronais pós-mitótica em
neurociência, biologia do desenvolvimento, e diagnóstico histopatológico.
6. Anti-neurofilamento-200 (NF-200) feito em camundongo (Abcam)
para marcação de fibras na concentração 1:1200; Este gene codifica a proteína
de neurofilamento dos axônios, mantendo o calibre e podendo desempenhar um
papel no transporte intracelular. Esta proteína é normalmente utilizada como um
biomarcador de danos neuronais.
Ao término desta etapa, as células foram lavadas em PBS (0,1M; pH 7,4)
por cinco minutos e incubadas por 1 hora com anticorpo secundário anti-mouse
e anti-rabbit produzidos em donkey (Jackson, EUA) conjugados ao fluorofore
AlexaFluor 488 ou AlexaFluor 594 ou FitC ou TritC mantidas em ambiente
refrigerado com ausência de luz. Após a incubação secundária, as células foram
lavadas com PBS por cinco minutos e, imediatamente, examinadas no
microscópio de fluorescência do Laboratório de Neurologia Experimental da

35
UERN (Eclipse E200, Nikon®) e posteriormente no microscópio de fluorescência
do Laboratório de Estudos Neuroquímicos da Universidade Federal do Rio
Grande do Norte / UFRN (Eclipse Ni, Nikon®).
As fotomicrografias foram realizadas com as câmeras digitais Moticam 3.0
e 5.0 (Motic®) nos aumentos de 40x, 100x, 200x em 4 campos numa sequência
definida previamente em cada placa (Figura 11). Na ocorrência de marcações
fluorescentes registrada nas células da ME, existiu o cuidado de examinar o
compartimento subcelular, citoplasmático ou nuclear das imunorreatividades.
Figura 11. Campos de observação celular nas P60.
Dois avaliadores independentes quantificaram as células por campo em
números absolutos, aferindo o perímetro (μm) e a área (μm2) de cada célula em
campos visuais não sobrepostos aleatoriamente, usando cultura de células de
no mínimo 3 experimentos diferentes com aumento de 200x, através de
softwares previamente calibrados (kappa = 0,94). Os softwares utilizados foram
Motic Images Plus 2.0 (Motic®) para observação morfológica, o software Image-
J para contagem celular. Para correção mínima de brilho e contraste das
fotomicrografias, foi utilizado o software Adobe Photshop CS-6.0 (Adobe®).

36
3.6 DOSAGEM DE ÍONS Ao completar 72 horas do subcultivos dos grupos experimentais o meio
de cultura foi retirado das P60 e sob o fluxo laminar colocados em tubos cônicos
tipo falcon de 15mL devidamente identificados (Figura 12) e posteriormente
levados ao setor de análises clínicas do Hospital Regional Tarcísio Maia (HRTM)
para a realização das dosagens dos íons sódio (Na), potássio (K) e cálcio (Ca).
Figura 12. Identificação dos meios de cultura após 72 horas para dosagem de íons.
As dosagens foram realizadas em um analisador (9180 electrolyte
analyzer) AVL automatizado da Roche®. Inicialmente foi realizado uma limpeza
do analisador utilizando o Cleaning solution (Roche®) e com o Sodium electrode
conditioner (solução condicionadora da Roche®), ambas realizadas através da
agulha analisadora de forma automatizada, na sequência solicitou a
autocalibração.
Foram separados tubos de hemólise esterilizados e identificados para que
pudessem receber 100μl de cada meio de cultura que foi analisado. Os tubos de
hemólise já com os meios de cultura foram levados de forma individual ao
analisador e realizado a dosagem dos íons.

37
3.7 ANÁLISES DOS DADOS
O banco de dados da pesquisa foi construído na plataforma do software
SPSS® (Statistical Package for Social Sciences) versão 22.0, com posterior
verificação de consistência da digitação. Após a estruturação final do banco de
dados, foi realizada inicialmente uma análise descritiva de todos os dados
(número, área, perímetro celular e dosagem de íons).
Os dados de expansão celular (número de células, área e perímetro) ao
longo de 72 horas e a dosagem dos íons foram comparados estatisticamente
através da análise de variância (ANOVA) com teste de Bonferone considerando
significante quando p < 0,05.
IV RESULTADOS
Nas placas com explantes de nervo isquiático (seta), constatou-se
inicialmente migração de células de morfologia fibroblastóide (cabeça de seta)
dos fragmentos (Figura 13). Nesta fase, os explantes foram removidos após
cinco dias para uma nova placa de cultura, o que levou à eliminação progressiva
das células que migraram do tecido para o fundo da placa e a possibilidade de
migração de população de CSs (Figura 14)

38
Figura 13. Células de morfologia fibroblastóide.
Figura 14. População de células de Schwann.
Após os procedimentos de subcultivos e formação dos grupos de estudo,
as células ficaram aderidas ao fundo das placas e foram acompanhadas ao longo
de 3 dias para observação do crescimento populacional, área, perímetro,
mudanças morfológicas e fenotípicas.
As culturas de células realizadas, quando cultivadas em MCNI acrescido
de FGF-2, apresentaram alterações mais visíveis quando comparadas com
culturas desprovidas desse tratamento, possibilitando a identificação mais
evidente da morfologia das células gliais e neuronais, resultando desta forma em
efeito plástico (Figura 15).

39
Figura 15. Aspectos Morfológicos das células gliais (cabeça de seta) e células neuronais (seta).
As populações de células da ME utilizadas foram morfologicamente
homogêneas. As culturas de células derivadas da ME, quando cultivadas em
MCNI e FGF-2, apresentaram alterações mais visíveis quando comparadas com
as culturas cultivadas somente em meio D-10, ou somente com FGF-2,
sobretudo no terceiro dia de observação microscópica (Figura 16).

40
Figura 16. Morfologia das células após formação dos grupos experimentais por 72 horas. Grupo 1: painéis
A (24 horas), B (48 horas), C (72 horas); Grupo 2: Painéis D (24 horas), E (48 horas), F (72 horas); Grupo
3: Painéis G (24 horas), H (48 horas), I (72 horas); Grupo 4: Painéis J (24 horas), K (48 horas), L (72 horas).
Escala 100µm.
A B C
D
G
E F
H I
J K L

41
4.1. MUDANÇAS MORFOLÓGICAS E EXPANSÃO DAS CÉLULAS GLIAIS
Com o acompanhamento das células por 72 horas, verificou-se que as
populações se multiplicaram progressivamente desde o primeiro dia até o último
dia de observação em todos os grupos experimentais. No entanto, percebe-se
que no grupo 1 o número de células gliais era maior que o número de células
neuronais. No primeiro dia de registro no grupo 1, alguns campos foram
microfotografados e apresentaram poucas células gliais, especificamente no
primeiro dia de observação. Em contrapartida no segundo dia de observação
foram contabilizadas 116 células no grupo 2 e 106 células no grupo 4.
Contabilizando todas as células ao longo dos três dias de observação em
números absolutos, destacaram-se o grupo 2 (284) e o grupo 4 (239) com maior
número de células, seguido do grupo 3 (146) e do grupo 1 com 96 células gliais
(Tabela 1).
Especificamente, no grupo 1 ocorreu diferença estatística significante
entre o número de células nos 3 dias de observação, onde em 72 horas
(p=0,001) a média das células foi maior que em 24 horas e 48 horas (p=0,004)
maior que 24 horas. No grupo 2, a média de células observadas foi maior no
terceiro dia quando comparado ao dia 1 (p=0,001). Ainda no grupo 2 observa-se
que o número de células foi maior em 48 horas (p=0,001) quando comparado a
24 horas e que em 72 horas (p=0,004) apresentou um número maior quando
comparado a 48 horas. No grupo 3, a média de células contadas em 72 horas
foi superior ao número de células registradas em 24 (p=0,001) e 48 horas
(p=0,007). Porém, 48 horas (p=0,002) foi maior que 24 horas no grupo 3. No
grupo 4, houve diferença estatística significante em 48 horas (p= 0,001) e em 72
horas (p= 0,001) quando comparadas ao número de células registradas em 24
horas. E entre 72 horas (p=0,001) e 48 horas (Figura 17).
Comparando a média do número de células observadas de todos os
grupos experimentais por dia de registro (Dias 1, 2 e 3), verificou-se que não
houver diferença estatística significante no dia 1 e no dia 2. Porém, no dia 3
observa-se uma diferença significante entre os grupos 2 (p= 0,012), 3 (p=0,032)
e 4(0,012) quando comparado com o grupo 1.

42
Considerando a área como aspecto morfológico das células, os grupos 1,
2, 3 e 4 mantiveram ao longo dos três dias de observação um crescimento das
células gliais. Destaque para o grupo 4 por apresentar uma média de
865334184,50 μm2 em 72 horas das células gliais (Tabela 2).
Nos três dias (24 horas, 48 horas e 72 horas) de observação, a área média
das células gliais do grupo 4 foi superior às dos grupos 1, 2 e 3 (Figura 20).
Na observação da área glial do grupo 1 verificou-se que no grupo 1 não
houve diferença estatística significante. No grupo 2, a área glial observada foi
maior no dia 2 (p=0,001) e no dia 3 (p=0,001) quando comparado ao dia 1, ainda
no grupo 2 observou que no dia 3 (p=0,001) a área foi maior que o dia 2. No
grupo 3, a área das células gliais em 72 horas foi superior a área das células em
24 (p=0,001) e 48 horas (p=0,001). O grupo 4, também apresentou diferença
estatística significante na área das células glias no dia 2 (p=0,001) e no dia 3
(p=0,001) quando comparado com dia 1. O dia 3 (p=0,001) também, apresentou
áreas gliais maiores que o dia 2 do grupo 4 (Figura 19).
Observou-se que a área das células gliais de todos os grupos
experimentais por dia de registro (Dias 1, 2 e 3), apresentaram crescimento
maior que as do grupo 1. No dia 1 percebeu que a área celular das glias do grupo
3 (p=0,021) foi maior que o grupo 1 e maior que do grupo 2 (p=0,001). Ainda no
dia 1 o grupo 4 (p=0,001/p=0,001) apresentou área maior que a do grupo 1 e 2
respetivamente. No dia 2 verificou-se que os grupos 2 (p=0,001), 3 (p=0,001) e
4 (p=0,001) apresentaram células gliais com áreas maiores que o grupo 1. Já no
dia 3 foi possível observar que os grupos 2 (p=0,001), 3 (p=0,001) e 4 (p=0,001)
apresentaram áreas celulares maiores que o grupo 1 e que o grupo 3 (p=0,009)
e o grupo 4 (p=0,001) apresentou células com áreas maiores que o grupo 2
(Figura 20).
Após a medição do perímetro das células gliais, observou-se que ao longo
dos três dias de observação dos grupos 1, 2, 3 e 4 as células gliais aumentaram
seu perímetro. No dia 2 de observação (48 horas), o perímetro médio das células
do grupo 2 e 3 foi superior às dos grupos 1 e 4. No dia 3 de aferição, os
perímetros médios das células gliais dos grupos 3 e 4 foram bastante
semelhantes e maiores que dos grupos 1 e 2 (Tabela 3).
Verificou-se que no grupo 1 não apresentou diferença estatística
significante entre os grupos. No grupo 2, o perímetro das células observadas foi

43
maior no terceiro dia quando comparado ao dia 1 (p=0,001) e ao dia 2 (p=0,001).
No grupo 3, o perímetro das células gliais em 48 horas (p=0,001) foi superior ao
registrado em 24 horas, bem como, em 72 horas o perímetro das células gliais
foi superior ao registrado em 24 (p=0,001) e 48 horas (p=0,001). O grupo 4,
houve diferença estatística significativa do perímetro glial no terceiro dia
comparado aos dois dias anteriores (p=0,001/24horas; p=0,001/48horas) (Figura
21).
Comparando a média do perímetro das células gliais observadas de todos
os grupos experimentais ao longo dos três dias (Dias 1, 2 e 3), verificou-se que
em 24 horas, a média do perímetro das células no grupo 2 (p=0,001), grupo 3
(p=0,001) e do grupo 4 (p=0,002) foi superior a perímetro das células gliais do
grupo 1. Em 48 horas de observou-se que a média dos perímetros das células
gliais dos grupos 2, 3 e 4 foram superiores ao grupo 1 (Grupo 2/p=0,001; Grupo
3/p=0,001; Grupo4/p=0,001), que o grupo 3 (p=0,015) foi superior ao grupo 2 e
que o grupo 3 (p=0,065) foi superior ao grupo 4. No último dia de observação
dos perímetros gliais os grupos 2, 3 e 4 foram superiores ao grupo 1 (Grupo
2/p=0,001; Grupo 3/p=0,001; Grupo4/p=0,001) e os grupos 3 (p=0,001) e 4
(p=0,001) foram superior ao grupo 2 (Figura 22).

44
Tabela 1. Descrição da amostra dos grupos de acordo com número de células
gliais
Grupo ME + D10
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 6,50 + 2,08 6,50 4,50 – 8,50 4,00 9,00
48 horas 13,75 + 2,21 14,00 11,50 – 15,75 11,00 16,00
72 horas 17,50 + 5,25 16,00 13,50 – 23,00 13,00 25,00
Grupo ME + MCNI
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 8,75 + 1,75 9,00 7,25 – 10,50 6,00 11,00
48 horas 17,75 + 5,06 17,50 13,75 – 22,75 10,00 25,00
72 horas 25,75 + 10,87 25,00 15,25 – 34,75 11,00 43,00
Grupo ME + MCNI + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 10,75 + 3,30 11,00 507, – 13,75 7,00 13,75
48 horas 18,57 + 10,22 17,00 8,00 – 29,00 8,00 34,00
72 horas 20,60 + 5,98 22,00 15,00 – 25,50 12,00 28,00
Grupo ME + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 8,50 + 2,07 8,00 7,25 – 9,00 6,00 13,00
48 horas 20,62 + 9,11 17,50 13,25 – 29,25 13,00 37,00
72 horas 29,00 + 7,98 28,00 21,25 – 36,00 20,00 41,00

45
Figura 17. Número de células gliais observadas em cada grupo experimental
de acordo com o dia de observação. P valores (a= 0,001; b= 0,004; c= 0,001;
d= 0,004; e= 0,001; f= 0,001; g=0,002; h=0,007; i=0,001; j=0,001; k=0,001).
c
e
f g
i j
a b
d
h
k

46
Figura 18. Número de células gliais observadas nos dias 1,2 e 3 de acordo com
o grupo experimental. P valores (a= 0,012; b= 0,032; c=0,012)
c b a

47
Tabela 2. Descrição da amostra dos grupos de acordo com área de células gliais.
Grupo ME + D10
Número celular N Média ± dp Mediana Q 25-75 Mín Máx
24 horas
10 5721,60 + 2165,91 5879,50 4337,00 –
7816,00 1652,0
0 8601,00
48 horas
49
21016,57 + 25336,45 4516,00 2595,50 –
40846,50 1026,0
0 84262,00
72 horas
37
68255,02 + 87138,46 44519,00 36589,00 –
65702,00 22717,
00 476453,0
0
Grupo ME + MCNI
Número celular N Média ± dp Mediana Q 25-75 Mín Máx
24 horas
54
46840556,72 + 17924530,42
44137825,00
33784893,75 – 62845764,25
10629512,00
86345550,00
48 horas
116
368286411,90 + 209321088,20
414975264,00
310676512,50 – 511482261,50
11885136,00
853617150,00
72 horas
114
497461767,20 + 227521356,40
523512596,00
325670366,50 – 655605458,00
12224695,00
999989150,00
Grupo ME + MCNI + FGF2
Número celular N Média ± dp Mediana Q 25-75 Mín Máx
24 horas 30 379012033,20 +
710738308,80 19089846
9,00 128766837,50 – 278886268,80
51955319,00
3999765450,00
48 horas 42 331334737,40 +
411798168,40 31120497
0,00 44840904,00 – 431186594,50
13582997,00
2659654803,00
72 horas
74
695408816,10 + 124179050,70
648775176,00
625773755,80 – 778678484,00
420782039
945638928,00
Grupo ME + FGF2
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas
35
200956213,50 + 183191773,30
212021050,00
54332162,00 – 228592303,00
14465808,00
876634100,00
48 horas 106 436897368,60 +
296110245,20 62101973
8,00 108816907,80 – 664853500,00
10119340,00
837421359,00
72 horas 98 616116568,40 +
416186295,70 86533418
4,50 58406984,50 – 943499614,50
10323103,00
998341156,00

48
Figura19. Área de células gliais observadas em cada grupo experimental de
acordo com o dia de observação. P valores (a= 0,001; b= 0,001; c= 0,001; d=
0,001; e= 0,001; f= 0,001; g= 0,001; h= 0,001).
f g h
d e
a b c

49
Figura 20. Área de células gliais observadas nos dias 1, 2 e 3 de acordo com o
grupo experimental. P valores (a= 0,001; b= 0,021; c= 0,001; d= 0,001; e=
0,001; f= 0,001; g= 0,001; h=0,001; i= 0,001; j= 0,001; k= 0,001; L=0,009).
a b
e
g f
h i
j k
L
c d

50
Tabela 3. Descrição da amostra dos grupos de acordo com perímetro de células gliais.
Grupo ME + D10
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas 10 3626,30 + 1025,68 3379,50 3105,25 –
3993,50 2112,0
0 5770,00
48 horas 49 13853,46 +
16763,11 2839,00 2078,00 – 27739,50
1042,00 81940,00
72 horas 37 89447,48 +
51266,64 124027,00 30421,50 – 133703,00
15798,00
138814,00
Grupo ME + MCNI
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas 54 4110458,70 +
4363676,97 3191872,5
0 2412034,75 – 4214390,50
1146618,00
29449369,00
48 horas 116 4492756,17 +
3117513,20 6138479,0
0 312081,75 – 7200282,75
134379,00
9246509,00
72 horas 114 6879317,72 +
1315599,66 7198521,5
0 6253811,25 – 7413427,75
255190,00
9391048,00
Grupo ME + MCNI + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 30 4489999,40 +
1354535,22 4480039,5
0 3341483,75 – 5389645,25
1364420,00
7383281,00
48 horas 42 5898764,40 +
2236620,32 6240485,5
0 5269889,25 – 7260866,00
178229
9296201,00
72 horas 74 8128709,71 +
1090469,74 8258491,5
0 7276986,75 – 9206516,25
5179175,00
9405780,00
Grupo ME + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 35 4013214,45 +
1570409,09 4157091,0
0 3154367,00 – 5187032,00
1158938,00
8231739,00
48 horas 106 4724517,30 +
2695571,12 5572404,5
0 1357602,50 – 6674496,50
189190,00
8793721,00
72 horas 98 8219812,76 +
1001163,02 8278656,0
0 7311828,75 – 9237308,75
6130492,00
9399978,00

51
Figura 21. Perímetro de células gliais observadas em cada grupo experimental
de acordo com o dia de observação. P valores (a= 0,001; b= 0,001; c= 0,001;
d= 0,001; e= 0,001; f= 0,001; g= 0,001).
g f
c d e
a b

52
Figura 22. Perímetro de células gliais observadas nos dias 1, 2 e 3 de acordo
com o grupo experimental. P valores (a= 0,002; b= 0,001; c= 0,001; d= 0,001;
e= 0,001; f= 0,001; g= 0,015; h= 0,065; i= 0,001; j= 0,001; k= 0,001; L= 0,001;
m= 0,001).
a b
c
d e
f g h
i j
k m L

53
4.2. MUDANÇAS MORFOLÓGICAS E EXPANSÃO DAS CÉLULAS NEURONAIS
Nas observações realizadas das células neurais, destacamos o grupo 2
no dia 2, onde foram contabilizadas 182 células neuronais e 117 células no grupo
4. Quando contabilizado todas as células neuronais ao longo dos três dias em
números absolutos, destacaram-se o grupo 2 (290) e o grupo 4 (196) com maior
número de células, seguidos do grupo 3 (153) e do grupo 1 com 15 células
neuronais (Tabela 4).
Verificou-se que no grupo 1 não apresentou diferença estatística
significante entre o número de células entre 24 horas e 48 horas e 72 horas. No
grupo 2, a média de células observadas foi maior no terceiro dia quando
comparado ao dia 1 (p=0,001) e ao dia 2 (p=0,001). Ainda no grupo 2 foi possível
observar que o dia 2 (p=0,001) apresentou um número maior de células quando
comparado ao dia 1. No grupo 3, a média de células contadas em 72 horas foi
superior ao número de células registradas em 24 (p=0,001) e 48 horas (p=0,001).
No grupo 3 também foi possível observar que a média de células contadas em
48 horas foi superior ao número de células registradas em 24 (p=0,001). O grupo
4, houve diferença estatística significante no número de células vistas no terceiro
dia para os dois dias anteriores (p=0,001/24horas; p=0,006/48horas) e entre as
células observadas no dia 2 e 1 (p=0,001) (Figura 23).
Comparando a média do número de células neuronais observadas de
todos os grupos experimentais por dia de registro (Dias 1, 2 e 3), verificou-se
que em 24 horas, a média de células contadas no grupo 2 foi superior ao número
de células do grupo 1 (p=0,002). Após 48 horas de indução, a média de células
neuronais observadas no grupo 3 foi superior aos demais grupos e a média do
grupo 2 (p=0,001) e do grupo 4 (p=0,001). Foi superior à do grupo 1. No último
dia de contagem os grupos 2 (p=0,001), 3 (p=0,001) e 4 (p=0,001) foram
superiores ao grupo 1. É possível observar também que o grupo 3
(p=0,001/p=0,001) apresentou uma média de células neuronais maior que o
grupo 2 e 4 respectivamente (Figura 24).
Considerando a área como aspecto morfológico das células, observou
que ao longo das 72 horas uma semelhança na média das áreas dos grupos 3 e
4 em relação ao grupo 1 e 2 (Tabela 5).

54
Quando observado a área das células neuronais do grupo 1 foi possível
identificar que não houver diferença estatística significante. No grupo 2 a área
entre 48 horas (p=0,019) e 24 horas apresentou diferença estatística, da mesma
forma quando comparado 72 horas (p=0,001) entre 24 horas e 72 horas (0,001)
entre 48 horas. No grupo 3 observou que entre 48 horas (p=0,024) e 24 horas
apresentou diferença estatística significante, da mesma forma quando
comparado 72 horas (p=0,001) entre 24 horas e 72 horas (0,001) entre 48 horas.
No grupo 4 a área apresentou diferença entre 48 horas (p=0,001) e 24 horas e
entre 72 horas (p=0,001) e 24 horas (Figura 25).
Comparando a média da área das células neuronais observadas de todos
os grupos experimentais ao longo dos três dias (Dias 1, 2 e 3), percebeu que no
dia 1 o grupo 4 (p=0,0047) foi superior ao grupo 1. Porém, não apresentou
diferença estatística significativa para os demais grupos. No dia 2 de observação
os grupos 2 (0,001), grupo 3 (p=0,001) e o grupo 4 (p=0,001) foram superiores
ao grupo 1 e ainda apresentou o grupo 4 (p=0,004) superior ao grupo 2. No dia
3 os grupos 2 (p=0,001), 3 (0,001) e 4 (p=0,003) foram superiores ao grupo 1 e
ainda foi possível observar que o grupo 2 (p=0,000) foi superior ao grupo 3 e
grupo 4.
Na observação do perímetro das células neuronais do grupo 1 não houve
diferença estatística significativa entre os três dias. No grupo 2, o perímetro das
células observadas foi maior em 72 horas (p=0,001) quando comparado ao dia
1 e também, foi maior em 72 horas (p=0,001) quando comparado ao dia 2. No
grupo 3, o perímetro das células neuronais em 48 horas (p=0,001) e em 72horas
(0,001) foi superior ao registrado em 24 horas, bem como, em 72 horas (p=0,001)
o perímetro das células foi superior ao registrado em 48 horas. Já no grupo 4 o
perímetro das células neuronais foi superior em 72 horas ao registrado, quando
comparado aos dois dias anteriores (p=0,001/24horas; p=0,001/48horas) e é
possível observar também que em 48 horas (p=0,000) foi superior a 24horas
(Figura 27).
Quando observado a média do perímetro das células neuronais de todos
os grupos experimentais ao longo dos três dias (Dias 1, 2 e 3), foi possível
perceber que no dia 1, a média do perímetro das células no grupo 2
(p=0,038/p=0,012), foi superior ao perímetro das células do grupo 4 e grupo 1.
Todavia, a média dos perímetros dos grupos 4 e 3 não apresentaram diferença

55
estatística significante quando comparado ao grupo 1. No dia 2 observou-se que
a média dos perímetros das células neuronais dos grupos 2, 3 e 4 foram
superiores ao grupo 1 (Grupo 2/p=0,010; Grupo 3/p=0,001; Grupo4/p=0,001) e
que o grupo 4 (p=0,001) foi superior ao grupo 2. Ainda no segundo dia observou
que o grupo 3 (p=0,001) foi maior que o grupo 2. No último dia de observação
dos perímetros neuronais os grupos 2, 3 e 4 foram superiores ao grupo 1 (Grupo
2/p=0,001; Grupo 3/p=0,001; Grupo4/p=0,001) (Figura 28).
Tabela 4. Descrição da amostra dos grupos de acordo com número de células neuronais
Grupo ME + D10
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 2,75 + 0,95 2,50 2,00-3,75 2,00 4,00
48 horas 6,00 + 1,63 6,00 4,50-7,50 4,00 8,00
72 horas 13,50 + 1,29 13,50 12,25-14,75 12,25 15,00
Grupo ME + MCNI
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 9,12 + 2,47 9,00 8,25-9,75 5,00 14,00
48 horas 22,87 + 5,54 22,00 21,25-25,00 13,00 33,00
72 horas 38,25 + 11,32 37,00 32,00-49,00 20,00 54,00
Grupo ME + MCNI + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 7,00 + 3,38 6,00 5,00-8,75 3,00 14,00
48 horas 35,62 + 8,22 34,00 29,50-40,00 26,00 58,00
72 horas 50,87 + 14,38 48,50 37,75-61,75 32,00 79,00
Grupo ME + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 6,00 + 2,00 5,00 5,00-7,50 4,00 10,00
48 horas 21,00 + 4,98 22,50 17,00-23,75 12,00 28,00
72 horas 33,12 + 10,78 32,50 23,50-39,50 21,00 54,00

56
Figura 23. Número de células neuronais observadas em cada grupo
experimental de acordo com o dia de observação. P valores (a= 0,001; b=
0,001; c= 0,001; d= 0,001; e= 0,001 f= 0,001; g= 0,001; h= 0,006; i= 0,001).
c
a b
e f
d
g h
i

57
Figura 24. Número de células neuronais observadas nos dias 1,2 e 3 de acordo
com o grupo experimental. P valores (a= 0,002; b= 0,001; c= 0,001; d= 0,001;
e= 0,001; f= 0,001; g=0,001; h=0,001; i=0,001; j= 0,001).
c b
a
e
g f h i j
d

58
Tabela 5. Descrição da amostra dos grupos de acordo com área de células neuronais.
Grupo ME + D10
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas 3 6203,00 + 3092,81 5872,00 3289,00 –
5872,00 3289,0
0 9448,00
48 horas 7
6658,65 + 2675,66 7695,00 5641,47 – 8183,12
1210,94 9309,94
72 horas 5 32033,20 +
26075,64 18032,00 16145,50 – 54921,50
15720,00 76953,00
Grupo ME + MCNI
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas 33 27392813,36 +
27423816,07 21250985,
00 2457550,00 – 46276380,00
1145212,00
91324800,00
48 horas 182 48399920,33 +
27656817,45 37125570
0,00 22642650,00 – 72620550,00
13555862,00
13555862,00
72 horas 75 160300768,70 +
63801592,00 13555886
2,00 129446600,00 – 173523012,00
91121700,00
428653400,00
Grupo ME + MCNI + FGF2
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas 21 35036361,10 +
17290108,95 32118650,
00 23205492,50 – 50833884,50
12136412,00
72548610,00
48 horas 45 52612032,82 +
26619898,43 49726012,
00 30261139,00 – 75817468,50
92611881,00
99521825,00
72 horas
87
72735070,51 + 23702981,00
76875362,00
61958400,00 – 92661425,00
4207438,00
98958400,00
Grupo ME + FGF2
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas 25 36670041,52 +
20310508,07 31307500,
00 21338700,00 – 53323313,00
1145327,00
83458450,00
48 horas 117 59146501,26 +
29613594,19 61119025,
00 32517600,00 – 86456375,00
3259100,00
99697431,00
72 horas 54 69697985,26 +
27417198,90 78305190,
00 61213631,50 – 89661138,00
9597800,00
99880179,00

59
Figura25. Área de células neuronais observadas em cada grupo experimental
de acordo com o dia de observação. P valores (a= 0,001; b= 0,019; c= 0,001;
d= 0,001; e= 0,024; f= 0,001; g=0,001; h= 0,001).
g h
a c b
d f
e

60
Figura 26. Área de células neuronais observadas nos dias 1,2 e 3 de acordo
com o grupo experimental. P valores (a= 0,047; b= 0,001; c= 0,001; d= 0,001;
e= 0,004; f= 0,003; g= 0,001; h= 0,001; i= 0,001; j= 0,001).
a
b c
d e
f g
h j i

61
Tabela 6. Descrição da amostra dos grupos de acordo com perímetro de células neuronais.
Grupo ME + D10
Número celular n Média ± dp Mediana Q 25-75 Mín Máx
24 horas 3 7222,00 + 2250,41 8346,00 4631,00 –
8346,00 4631,0
0 8689,00
48 horas 7
6137,44 + 2375,91 7073,10 5245,75 – 7553,00
1137,84 8034,55
72 horas 5 44932,80 +
30006,41 46316,00 14710,50 – 74463,50
11074,00 75531,00
Grupo ME + MCNI
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 33 2249212,57 +
2380104,07 1583960,0
0 481223,50 – 3323205,50
121348,00
9333004,00
48 horas 181 3542497,84 +
3161528,43 2368394,0
0 1611633,00 – 2786498,00
1006773,00
21557498,00
72 horas 75 6639607,30 +
3141382,26 7824220,0
0 4753585,00 – 8914317,00
1051820,00
9958042,00
Grupo ME + MCNI + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 21 1417263,38 +
776067,21 1243945,0
0 988706,00 – 2122440,50
125391,00
3226332,00
48 horas 44 4758732,95 +
2856708,86 5681733,5
0 1292299,75 – 7083893,00
555003,00
9068450,00
72 horas 87 6705048,55 +
2466938,74 7512291,0
0 5512291,00 – 8587652,00
512291,00
9937354,00
Grupo ME + FGF2
Número celular Média ± dp Mediana Q 25-75 Mín Máx
24 horas 25 1071766,64 +
700911,06 1178471,0
0 230296,50 – 1732129,50
121480,00 1898466
48 horas 117 5380350,53 +
2635628,01 6117800,0
0 2573369,50 – 7585009,00
1004938,00
9793721,00
72 horas 54 7387879,59 +
1625457,47 8070309,0
0 6265169,50 – 8265643,00
222611,00
9836400,00

62
Figura 27. Perímetro de células neuronais observadas nos dias 1,2 e 3 de
acordo com o grupo experimental. P valores (a= 0,001; b= 0,001; c= 0,001; d=
0,001; e= 0,001; f= 0,001; g= 0,001; h=0,001).
f g h
d e c
b
a

63
Figura 28. Perímetro de células neuronais observadas em cada grupo
experimental de acordo com o dia de observação. P valores (a=0,012; b=
0,038; c= 0,001; d= 0,001; e= 0,001; f= 0,010; f= 0,001; g= 0,001; h=0,001;
i=0,001; j=0,001).
b
c d e
f
h i
j
a
g

64
4.3. FENÓTIPO DAS CÉLULAS GLIAIS E NEURONAIS
Após 72 horas de observação, procedeu-se a imunocitoquímica dos 4
grupos do experimento. Foi realizado o fechamento dos filtros do microscópio de
fluorescência para validar a marcação. As células do grupo 1 não expressou
nenhuma proteína glial ou neuronal, em contrapartida, as populações do grupo
2 expressaram β- tubulina (Figura 29), GFAP (Figura 30) e NF-200 (Figura 31).
As populações de células do grupo 3 expressaram GFAP (Figura 32), NeuN
(Figura 33) e OX-42 (Figura 34). No grupo 4 as células expressaram GFAP
(Figura 35), NeuN (Figura 36), NF-200 (Figura 37) e OX-42 (Figura 38). No
entanto, as alterações morfológicas das células se fazem novamente presentes
e, agora, mais frequentes no grupo 4.
Figura 29. Células do G 2 submetidas a imunofluorecência do anticorpo β- Tubulina (FITC). Escala
100µm.

65
Figura 30. Células do G 2 submetidas a imunofluorecência do anticorpo GFAP (Alexa Fluor 488). Escala
100µm.
Figura 31. Células do G 2 submetidas a imunofluorecência do anticorpo NF-200 (Alexa Fluor 488). Escala
100µm.

66
Figura 32. Células do G 3 submetidas a imunofluorecência do anticorpo GFAP (Alexa Fluor 488). Escala
100µm.
Figura 33. Células do G 3 submetidas a imunofluorecência do anticorpo NeuN (Alexa Fluor 488). Escala
100µm.

67
Figura 34. Células do G 3 submetidas a imunofluorecência do anticorpo OX-42 (Alexa Fluor 488). Escala
100µm.
Figura 35. Células do G 4 submetidas a imunofluorecência do anticorpo GFAP (Alexa Fluor 488). Escala
100µm.

68
Figura 36. Células do G 4 submetidas a imunofluorecência do anticorpo NeuN (Alexa Fluor 488). Escala
100µm.
Figura 37. Células do G 4 submetidas a imunofluorecência do anticorpo NF-200 (Alexa Fluor 488). Escala
100µm.

69
Figura 38. Células do G 4 submetidas a imunofluorecência do anticorpo OX-42 (Alexa Fluor 488). Escala
100µm.
4.4. DOSAGEM DOS ÍONS NO MEIO DE CULTURA
Em nosso experimento a dosagem do íon de Na apresentou uma
diferença estatística significante em Micromol por litro entre os grupos
experimentais. No grupo 2 (p=0,001), grupo 3 (p=0,001) e grupo 4 (p=0,001)
quando comparado com o grupo 1. Além disso, foi possível observar também
que o grupo 2 (p=0,001) apresentou uma superioridade em relação ao grupo 3,
sendo o grupo 3 (p=0,001) superior ao grupo 4 (Figura 39).
Na dosagem de Ca os grupos 2 (p=0,001), 3 (p=0,001) e 4 (0,001)
apresentaram significância estatística quando comparado com o grupo 1. O
grupo 2 (p=0,001/p=0,001) apresentou superioridade ao grupo 3 e 4
respectivamente. Quando comparado o grupo 3 (p=0,019) com o grupo 4
percebeu uma diferença estatística significante (Figura 40).
Quando observado o Micromol de K por litro entre os grupos
experimentais percebeu que o grupo 1 foi inferior aos grupos 2, 3 e 4 (Grupo
2/p=0,001; grupo 3/p=0,001 e grupo 4/p=0,001). No entanto, o grupo 2 (p=0,001)

70
apresentou Micromol de K por litro maior que no grupo 3 e 4. Quando comparado
o grupo 3 (p=0,001) com o grupo 4 percebe-se uma superioridade estatística
(Figura 41).
Figura 39. Dosagem de sódio de acordo com cada grupo experimental. P
valores (a= 0,001; b= 0,001; c= 0,001; d= 0,001; e= 0,001).
a b
d c
e

71
Figura 40. Dosagem de cálcio de acordo com cada grupo experimental. P
valores (a= 0,001; b= 0,001; c= 0,001; d=0,001; e=0,001 e f=0,019).
a b
c
d
f
e

72
Figura 41. Dosagem de potássio de acordo com cada grupo experimental. P
valores (a= 0,001; b= 0,001; c= 0,001; d= 0.001; e= 0,001; f=0,001).
a b c d e
f

73
V DISCUSSÃO
O presente estudo objetivou a analisar a plasticidade celular da medula
espinal na presença do MCNI, na adição ou ausência do FGF-2 em cultura.
O complexo sistema nervoso de mamíferos nunca alcança um patamar
de organização encontrado antes de lesões ou injúrias. Este pode ser um
mecanismo de proteção fisiológico inerente para prevenir uma excessiva
regeneração desorganizada. Especificamente, o reparo nervoso periférico
requer uma complexa interação entre um guia para crescimento axonal, células
de suporte como as CSs, fatores de crescimento e matriz extracelular (Costa et
al., 2009). O reparo de defeitos periféricos nervosos tem sido um problema para
os cirurgiões em diferentes especialidades.
Várias modalidades de tratamento têm sido investigadas ou
recomendadas. A neurorrafia direta, coaptação dos cotos distal e proximal, é a
escolha de preferência (Di Summa et al., 2011). No entanto, em muitos casos,
isto não é possível devido ao longo espaço (defeito) entre os segmentos
nervosos separados. Enxertos nervosos autólogos, interposições de enxertos
musculares, enxertos venosos e arteriais e o uso de condutos nervosos têm sido
utilizados como alternativas. Muitos modelos de lesão, diferentes combinações
de tipos celulares, o uso de diversos tipos de condutos, bem como os protocolos
de avaliação embasam esse novo campo da ciência (Ghoreishian et al., 2013).
As CSs são células gliais do SNP, que envolvem os axônios e facilitam a
condução do impulso nervoso. Na degeneração axonal Waleriana, as CSs
aliadas aos macrófagos medeiam os passos iniciais para remoção de mielina.
Elas proliferam, migram para formar as bandas de Büngner e secretam fatores
neurotróficos que ajudam a estabelecer um ambiente favorável para inervação
precisa do tecido alvo. No entanto, existem limitações inerentes de uso direto no
reparo nervoso experimental, já que essas células vêm de recursos restritos e
tem disponibilidade limitada (Wei et al., 2010). A aplicação de CSs em nervos
artificiais tem se tornado concreto, no entanto, existem certas limitações como o

74
recurso limitado de células, a necessidade de uma cirurgia secundária e a
rejeição imunológica de enxertos exógenos (Guo e Dong, 2009).
Para que as células funcionem adequadamente, elas devem se organizar
no espaço e interagir mecanicamente com o ambiente ao seu redor. Devem
apresentar uma conformação correta, serem fisicamente robustas e estarem
estruturadas de forma adequada internamente. Muitas células devem também
ser capazes de modificar sua forma e migrar para outros locais. Além disso, toda
célula deve ser capaz de reorganizar seus componentes internos como
decorrência dos processos de crescimento, divisão e/ou adaptação a mudanças
no ambiente (Alberts et al., 2010).
O aspecto mais importante para a sobrevivência celular em um tecido
receptor é o microambiente. Inicialmente, esse aspecto está relacionado à
expressão celular de marcadores de adesão de superfície que interagem com
componentes da matriz extracelular. Adicionado aos efeitos parácrinos dos
fatores de crescimento secretados pelas células adjacentes, o microambiente
possibilita condições para a sobrevivência, migração, invasão tecidual e
diferenciação (Caddick et al., 2006). Correntemente, a diferenciação neuronal
pode ser atingida de 4 maneiras: Uso de fatores neurotróficos ou citocinas,
exposição a indutores químicos, a combinação desses ou com co-culturas de
células nervosas (Yang et al., 2014).
Em nosso trabalho, o meio criado a partir de fragmentos nervosos
simulou uma situação in vivo de injuria nervosa. Dessa maneira, esperava-se
que os explantes se tornassem reativos e secretassem naturalmente fatores que
possibilitassem um ambiente favorável à regeneração nervosa pós-trauma.
O número superior de células contadas nos grupos 2, 3 e 4, bem como
as mudanças morfológicas mais evidentes são teoricamente reflexos do meio
condicionado. O meio D-10, utilizado como um controle foi muito importante para
averiguar se as células da ME adquiriam fenótipos distintos sem nenhuma
indução no decorrer dos dias. Como as propriedades dos fatores neurotróficos
são bastante discutidas, o uso do FGF-2 poderia potencializar os efeitos
fenotípicos das células, bem como induzir uma maior atividade proliferativa e
efeito neuroprotetor.

75
Lucena et al. (2014), investigou a plasticidade de CTs mesenquimais de
medula óssea de ratos na presença de meio condicionado do nervo facial do
FGF – 2, sendo possível perceber que, o uso do FGF-2 potencializou os efeitos
fenotípicos das células,assim como induziu uma maior atividade proliferativa. Em
2008, Lee et al. realizou um modelo transecção completa da ME em que o
segmento da coluna vertebral T8 foi removido para estudar o efeito do tratamento
combinado de enxerto de nervo periférico e aplicação do fator neurotrófico sobre
o meio ambiente da glia e concluiu que a presença do fator neurotrófico
possibilitou uma mudança do ambiente glial possibilitando uma regeneração do
SNC.
Parece plausível que as interações entre FGF-2 e células neurogliais
influenciam o microambiente da lesão, restaurando a homeostase metabólica,
proporcionando a proteção para as células não danificadas e, possivelmente,
resgatando neurônios prejudicados. Uma hipótese provisória é pela diminuição
da degeneração Walleriana e/ou preservação dos oligodendrócitos, preservando
a função de um determinado subconjunto de axônios supraespinais, incluindo os
tratos não piramidais que é importante para modular o desempenho motor
grosseiro (Fehling e Tator, 1995; Yasuda et al., 1995).
O FGF-2 medeia atividade neurotrófica em neurônios dopaminérgicos
mesencefálicos cultivados de ratos embrionários, demonstrando papel
importante no tratamento da doença de Parkinson (Grothe et al., 2000).
Descrição do papel das CSs em neurônios dopaminégicos embrionários em
cultura também é mencionada por Collier e Martin (1993), os quais
demonstraram que essas células promovem sobrevivência e extensão neurítica
em neurônios cultivados. Além disso, ações do FGF-2 e do FGF-20 são descritas
em desordens motoras como a esclerose múltipla, esclerose lateral amiotrófica
e a distrofia muscular (Webster, 1997; Rizvanov et al., 2011).
Trabalho publicado em 2009 utilizando o fator neurotrófico GDNF em
modelo de transecção da ME submetida ao enxerto de nervo periférico
demonstrou que o fator aumenta a resposta sensitivo-motora dos animais
quando comparado aos animais desprovidos da substância (Guzen et al., 2009).
Fazendo uma analogia entre os fatores neurotróficos GDNF e FGF-2 observa-

76
se que os animais que receberam inoculação do nervo periférico na presença do
GDNF cursam com melhora da resposta motora quando comparado aos animais
que receberam inoculação de FGF-2. Animais que recebem GDNF apresentam
no final da análise do BBB pontuação motora em torno de 8, enquanto os animais
que recebem FGF-2 apresentam no final da análise do BBB pontuação em torno
de 5. Além disso, no estudo realizado por Guzen e colaboradores em 2009
demonstra efeito do GDNF na resposta sensitiva analisada através do CBS.
Entretanto, na análise do FGF-2 não observamos efeito na resposta sensitiva em
ambos os testes quando comparado ao grupo que recebeu enxerto de nervo
isquiático na ausência do FGF-2.
Neste estudo, foram utilizados 4 meios de cultura distintos. As células da
ME pertencentes ao grupo 1 foram cultivadas apenas com meio D-10. No grupo
2, o meio D-10 foi condicionado a partir de explantes do nervo isquiático. No
grupo 3 foi usado o MCNI acrescido de FGF-2. Ao meio D-10 foi adicionado FGF-
2 para o cultivo celular do grupo 4. Nossos resultados demonstraram que a
média de células gliais e neuronais contadas nos grupos 2, 3 e 4 aumentaram
após 24 horas. De uma forma geral, a atividade proliferativa encontrada,
sobretudo nos grupos 2, 3 e 4 revela uma importante característica das células
da ME: uma rápida expansão num curto período de tempo.
Este fato deve-se também em virtude do FGF-2 demonstrar um papel
importante, possibilitando o aumento da regulação celular após a injúria do
nervo, promovendo suporte de crescimento neurítico e possibilitando a
sobrevivência de neurônios sensitivos durante a regeneração nervosa (Grothe
et al., 1997, 2006). In vitro, o FGF-2 protege neurônios contra excitotoxicidade,
diminui a morte celular apoptótica e promove reconstrução da bainha de mielina
exercendo um efeito neuroprotetor nos neurônios motores após transecção do
nervo isquiático ou da injúria contusa da medula espinal (Mattson et al., 1989;
Romero et al., 2001).
Outro aspecto importante destacado em nosso estudo foi à área e o
perímetro celular. Pois foram parâmetros elencados primordialmente para

77
descrição morfológica das células ao longo dos dias. Onde apresentaram,
relevância na análise estatística ao longo dos 3 dias de observação. O controle
do tamanho da célula é um aspecto importante do desenvolvimento celular e o
controle da morfologia é essencial para a diferenciação e para o
desenvolvimento de órgãos e tecidos. O tamanho da célula depende do tipo
celular e de seu estágio no ciclo celular (Kharitonova e Vasiliev, 2008). No cultivo,
as alterações do número, tamanho e área celular acontecem durante o
crescimento (Vasiliev, 2004). A forma é controlada por estruturas do
citoesqueleto interno e estruturas de adesão fixando a célula à superfície
(Moizhess e Vasiliev, 2001; Rovensky et al., 2001).
A diferenciação celular é mais bem compreendida considerando que
cada célula é dotada de duas características: diferenciação e potencialidade.
Diferenciação é o grau de especialização da célula, enquanto a potencialidade é
a capacidade que a célula tem de originar outros tipos celulares. As primeiras
células embrionárias (blastômeros) da maioria das espécies podem originar
qualquer tipo celular. Essas células tem o grau de diferenciação zero, e portanto
100% de potencialidade. Dessa maneira, são classificadas como totipotentes.
São consideradas pluripotentes as células derivadas dos folhetos embrionários
e multipotentes as células que derivam progenitores exclusivos de suas
linhagens (Junqueira e Carneiro, 2013).
O controle da forma celular envolve uma organização da membrana
plasmática dentro de uma superfície de contato com o ambiente extracelular
(células vizinhas, matriz extracelular, meio de cultura etc) (Levayer e Lecuit,
2007). As integrinas são versáteis receptores que possibilitam o intercâmbio do
meio celular externo com o meio interno. Muitos processos como morfologia
celular, motilidade, proliferação, diferenciação e morte celular são possíveis,
devido esse trânsito (Docheva et al., 2007). Isso é explicado pelo fenótipo dos
anticorpos neuronais. Claramente vê-se uma tendência que essas células
apresentaram maiores mudanças morfológicas condizentes com a organização
do nosso sistema nervoso: maior número de células com áreas menores e
morfologias distintas (explicado pelo perímetro). Uma completa caracterização
das interações celulares, bioquímicas e moleculares das células da ME com o

78
seu nicho é necessária para compreender como estas células podem ser
reguladas in vitro.
Nesse universo, a especificidade primorosa dos anticorpos por antígenos
específicos, transformam-nos em valiosos reagentes para a detecção,
purificação e quantificação de antígenos e consequentemente possibilita o uso
de protocolos para denotar diferenciação ou plasticidade celular. Nessa
perspectiva, esse estudo também, utilizou a imunocitoquímica como técnica de
escolha para possível identificação de plasticidade das células da ME diante de
meios de cultivo distintos. Para tanto, foram elencados marcadores de linhagem
neuronal (MAP-2, β-tubulina III, NeuN e NF-200) e marcadores de linhagem glial
(GFAP e OX-42).
Proteínas que se ligam lateralmente aos microtúbulos são coletivamente
chamadas de proteínas de associação a microtúbulos (MAPs). Assim, as MAPs
podem estabilizar os microtúbulos, prevenindo sua dissociação. Células que
superexpressam MAP-2, a qual apresenta longos domínios projetados, formam
feixes de microtúbulos estáveis com um amplo espaçamento. Os microtúbulos
são longos, cilindros, ocos formados pela proteína tubulina. A tubulina ocorre em
todas as células eucarióticas, podendo ser encontrada sob múltiplas isoformas.
As diferentes formas de tubulina são bastante similares e geralmente
copolimerizam em microtúbulos mistos em testes in vitro. No entanto, elas
apresentam posicionamento celular distinto e realizam tarefas relativamente
diferentes (Alberts et al., 2010).
No desenvolvimento neurogênico, β-Tubulina III é expressa em
neurônios em um estágio mais precoce do desenvolvimento, e certas isoformas
de MAP-2 são expressas precocemente durante a diferenciação neuronal de
células precursoras (Huang et al., 2007). β -tubulina III é um confiável marcador
de diferenciação neuronal por ser expresso em neurônios maduros e células
neurais progenitoras (Schwindt et al., 2009).
NeuN é uma proteína nuclear neuronal específica de vertebrados. A
coloração imunocitoquímica é primeiramente vista no núcleo se estendendo até
o citoplasma. Sua imunoreatividade é observada precocemente após mitoses
neuronais. É um excelente marcador de neurônios em culturas primárias. Os

79
filamentos intermediários são fibras compostas por proteínas de filamentos
intermediários, as quais pertencem a uma grande e heterogênea família
(Schwindt et al., 2009).
Os neurofilamentos são encontrados em altas concentrações nos
axônios dos neurônios dos vertebrados. Durante o crescimento do axônio, novas
subunidades de neurofilamentos são incorporadas ao axônio em um processo
dinâmico que envolve tanto a adição de subunidades longitudinalmente ao
comprimento do filamento quanto a adição de subunidades às extremidades do
filamento (Schwindt et al., 2009). A proteína ácida fibrilar glial é um elemento
estrutural de astrócitos fibrilares. GFAP é usado como um marcador astrocítico,
mas também pode ser expresso em células neurais progenitoras imaturas
(Schwindt et al., 2009) e o anticorpo OX-42 marca células dendríticas
extensivamente, granulócitos e células com morfologia de micróglia no cérebro.
Nossos experimentos demonstraram que as células do grupo 2
expressaram GFAP, NF-200 e β– Tubulina III. Já no grupo 4 que apresentava
apenas FGF-2 as células expressaram GFAP, NeuN, NF-200 e OX-42. Além
desses achados, as células que foram cultivas com o meio condicionado
acrescido de FGF-2 expressaram GFAP, NeuN e OX-42. Hipotetizamos que o
fator de crescimento fibroblástico potencializou a plasticidade das células do
grupo 4. A expressão desses anticorpos num curto período de tempo pós
indução (3 dias) pode ser justificado pela evidente reatividade dos explantes
nervosos. A forma histológica com que o nervo isquiático está organizado
possivelmente explica a alta e rápida reatividade bem como uma possível
deposição de fatores neurotróficos no meio condicionado. O microambiente de
lesão periférica simulado pelo meio condicionado a partir de explantes de nervo
isquiático pode justificar a rapidez da plasticidade neuronal e glial encontrada.
Outro achado importante em nosso experimento foi à identificação de que
o nosso meio de cultura apresentou um ambiente rico, nutritivo e com um grande
potencial elétrico, pois através da dosagem dos íons foi observado que os grupo
2, 3 e 4 apresentaram grandes concentrações de Na, K e Ca. A maioria das
células animais apresentam diferenças de potencial elétrico (voltagem), através
de suas membranas plasmáticas. O citoplasma costuma ser eletricamente

80
negativo em relação ao líquido extracelular. Para que haja troca de moléculas e
íons entre a célula e seu meio ambiente, a membrana plasmática possui
proteínas transportadoras e um desses recursos é a bomba de sódio e potássio.
Essa bomba transportadora de íons é fundamental para a sobrevivência dos
seres vivos (Goff,2006).
Para alguns íons, como o Na, os gradientes de concentração e voltagem
atuam na mesma direção criando um gradiente eletroquímico relativamente alto.
O Na é o íon positivamente carregado mais abundante fora da célula, logo, tende
a entrar nas células se tiver oportunidade. Já no íon K os gradientes de
concentração e de voltagem possuem efeitos opostos e o gradiente
eletroquímico é pequeno. O K é um íon positivamente carregado que está
presente em muito maior concentração dentro das células do que fora então, por
causa do efeito oposto, esse íon possui pouco movimento resultante através da
membrana (Jamor et. al., 2012)
O Ca é o principal mineral do esqueleto e um dos cátions mais abundantes
no organismo, representando cerca de 2% do peso corporal. Aproximadamente
99% do Ca corporal está sob a forma inorgânica no esqueleto, principalmente
sob a forma de cristais de hidroxiapatita, o restante (1%) encontra-se no espaço
vascular e intracelular (membrana e retículo endoplasmático). Cerca de 1% do
Ca ósseo é livremente intercambiável com o cálcio do fluido extracelular (Goff,
2006).
Jamor et. al. (2012) descreve que os sais de cálcio têm a função de manter
a integridade da estrutura do esqueleto, e os íons de cálcio estão nos fluidos
intracelular e extracelular, sendo um instrumento de controle de um grande
número de processos bioquímicos. De fato, enquanto os íons de cálcio
intracelular são necessários na atividade de um grande número de enzimas e
também estão envolvidos na transmissão de informações desde a superfície até
o interior da célula, os íons na porção extracelular são necessários para a
excitabilidade neuromuscular, coagulação do sangue e secreção hormonal,
entre muitas outras funções.

81
VI CONCLUSÃO
As células da ME demonstraram uma morfologia típica e rápida expansão,
sobretudo as células que foram expostas ao MCNI. Além disso, a significante
atividade proliferativa nos grupos 3 e 4 em um curto período asseguram seu
potencial de diferenciação in vitro sob a exposição a estímulos adequados. De
uma forma geral, o ambiente condicionado por explantes de nervo isquiático
circundante às células da ME, induziu mudanças morfológicas, bem como
possibilitou plasticidade nervosa.
O FGF-2 adicionado ao meio condicionado potencializou esse efeito, de
forma que as células da ME expressaram proteínas neuronais (MAP-2, β-
tubulina III, NeuN e NF-200) e proteína gliais (GFAP e OX-42). Em média, a área
e perímetro das células dos grupos 3 e 4 foram maiores do que as dos grupos 1
e 2. Essa potencialização e a maior sobrevivência das células se deve ao meio
de cultura utilizado na pesquisa pois apresenta grandes concentrações de Na, K
e Ca possibilitando um ambiente rico, nutritivo e com um grande potencial
elétrico, ideal para a sobrevivência da célula.
Nessa perspectiva construímos um protocolo viável, de fácil execução, em
curto espaço de tempo que possibilitou plasticidade de células da ME numa
linhagem neuronal e glial. O presente estudo melhora nosso conhecimento sobre
a plasticidade de células da ME e facilita a compreensão na busca por melhores
técnicas com terapia celular no uso de doenças e traumas no SNC.
Nesse contexto, vale salientar a importância dos resultados de outras
pesquisas que reproduzam as condições clínicas e a necessidade de utilização
de outros métodos de avaliação.

82
VII REFERÊNCIAS
AGUAYO A. J, DAVID S, BRAY G. M. Influences of the glial environment on the elongation of axons after injury: transplantation studies in adult rodents. J Exp Biol, 95: 231-240, 1981.
ALBERTS B, BRAY O, HOPKIN K, JOHNSON A, LEWIS J, RAFF M, ROBERTS K, WALTER P. Biologia molecular da célula. 5a ed. Porto Alegre: Artmed, 2010.
ANDERSON, D. J. Cell fate determination in the peripheral nervous system: the sympathoadrenal progenitor. J Neurobiol, 24:185-198, 1993.
AVERSI-FERREIRA R. A. G. M. F, MARIN K. A, SILVA F. O. C. & AVERSI-
FERREIRA T. A. Comparative anatomy of the thigh nerves of Cebus libidinosus (Rylands et al., 2000). Pesq. Vet. Bras. 31:261-266, 2011.
BAFFOUR R, ACHANTA K, KAUFMAN J, BERMAN J, GARB J. L, RHEE S, FRIEDMANN P. Synergistic effect of basic fibroblast growth factor and methulprednisolone on neurological function after experimental spinal cord injury. J Neurosurg, 83:105-110, 1995.
BAIRD A, KLAGSBRUN M. The fibroblast growth factor family. Cancer Cells; 3(6): 239-243, 1991
BALLESTEROS PLAZA, V. et al., spinal cord injury: literature update: physiopathology and initial treatment. coluna/columna, v. 11, n. 1, p. 73-76, 2012.
BETHEA J. R, DIETRICH W. D. Targeting the host inflammatory response in traumatic spinal cord injury. Curr Opin Neurol, 15:355-360, 2002.
BIRDI T. J, ANTIA N. H. Mechanisms involved in peripheral nerve damage in leprosy with special reference to insights obtained from in vitro studies and the experimental mouse model. Int J Lepr Other Mycobact Dis, 71:345-354, 2003. BRACKEN, M. B. Steroids for acute spinal cord injury. Cochrane Database Syst Rev, v. 1, p. CD001046, 2012. BRITO, L. M. O. et al., Epidemiological evaluation of victims of spinal cord injury. Rev. Col. Bras. Cir. v. 38, n. 5, p. 304-309, 2011.
BOYD J. G, GORDON T. Neurotrophic factors and their receptors in axonal regeneration and functional recovery after peripheral nerve injury. Mol Neurobiol, 27:277-324, 2003.

83
BOYD B. S, PUTTLITZ C, GAN J, TOPP K. S. Strain and excursion in the rat sciatic nerve during a modified straight leg raise are altered after traumatic nerve injury. J Orthop Res. 23:764-0, 2005.
BOTELHO, R. V. et al. Efetividade da metilprednisolona na fase aguda do trauma raquimedular: revisão sistemática dos ensaios clınicos randomizados Effectiveness of methylprednisolone in the acute phase of spinal cord injuries: a systematic review of randomized controlled trials. Rev Assoc Med Bras, v. 55, n. 6, p. 729-37, 2009.
BOOTCOV M. R, et al., MIC-1, a novel macrophage inhibitory cytokine is a divergent member of the TGF-β superfamily. Immunol Cell Biol, 94:11514-11519, 1997. BUTT A. M, DINSDALE J. Fibroblast growth factor 2 induces loss of adult oligodendrocytes and myelin in vivo. Exp Neurol, 192:125-133, 2005.
CADDICK J et al., Phenotypic and functional characteristics of mesenchymal stem cells differentiated along Schwann cell lineage. Glia, 54:840-840, 2006. CITERO, E. M. M, EDUARDO M, FONTES S. V. Efeitos da fisioterapia com trampolim no traumatismo raquimedular: estudo de caso. Rev. Neurociência, São Paulo, v.. 20, n. 2, p. 222-232, 2012.
CHADI G, FUXE K. Analysis of trophic responses in lesioned brain: Focus on basic fibroblast growth factor mechanisms. Braz J Med Biol Res 31(2): 231-241, 1998.
CHADI G. et al., Protective actions of human recombinant basic fibroblast growth factor on MPTP-lesioned nigrostriatal dopamine neurons after intraventricular infusion. Exp Brain Res 97(1): 145-158, 1993.
CHAN A, MAGNUS T, GOLD R. Phagocytosis of apoptotic inflammatory cells by microglia and modulation by different cytokines. Glia, 33:87-95, 2001. CHENG H. L. et al, Characterization of insulin-like growth factor-I and its receptor and binding proteins in transected nerves and cultured Schwann cells. J Neurochem, 66:525-536,1996.
CHOI D, DUNN L. T. Facial nerve repair and regeneration an overview of basis principles for neurosurgeons. Acta Neurochir (Wien), 143:107-114, 2001. CONDIC M. L, LEMONS M. L. Extracellular matrix in spinal cord regeneration: getting beyond attraction and inhibition. Neuroreport, 13:37-48, 2002.

84
COSTA M. P. et al., Poliglycolic acid tube associated with GM1 in regeneration of peripheral nerves. Acta Ortop Bras, 17:286-290, 2009. CRISTANTE A. F, TARICCO M. A, COLARES G. Estudo experimental da ação da metilprednisolona utilizada antes do traumatismo raquimedular em ratos Wistar. Acta Ortopédica Brasileira, v. 18, n. 1, p. 26-30, 2010.
DAVIS J. B, STROOBANT P. Platelet-derived growth factors and fibroblast growth factors are mitogens for rat Schwann cells. J Cell Biol 110(4): 1353-1360, 1990.
DIMAR, J. R. et al. Predictors of complications after spinal stabilization of thoracolumbar spine injuries. J Trauma, v. 69, n. 6, p. 1497-500, Dec 2010.
DI SUMMA P. G. et al., G. Long-term in vivo regeneration of peripheral nerves through bioengineered nerve grafts. Neuroscience, 181: 278-291, 2011.
DONO R. Fibroblast growth factors as regulators of central nervous system development and function. Am J Physiol Regul Integr Comp Physiol, 284:867-881, 2003.
ELLENBERGER W, BAUM H. Handbuch der vergleichenden Anatomie der Haustiere. Springer, Berlin. 917p, 1985.
FEHLINGS M. G, TATOR C. H. The relationships among the severity of spinal cord injury, residual neurological function, axon counts, and counts of retrogradely labeled neurons after experimental spinal cord injury. Exp
Neurol 1995;132(2): 220-228
FUXE K. et al., Computer-assisted mapping of basic fibroblast growth factor immunoreactive nerve cell populations in the rat brain. J Chem Neuroanat
11(1): 13-35, 1996.
FRESSINAUD C. Basic fibroblast growth factor improves recovery after chemically induced breakdown of myelin-like membranes in pure oligodendrocyte cultures. J Neurosci Res, 38:202-213, 1994.
FROSTICK S. P, YIN Q, KEMP G. J. Schwann cells, neurotrophic factors, and peripheral nerve regeneration. Microsurgery, 18:397-405,1998.
GHIRNIKAR R. S, LEE Y. L, ENG L. F. Inflammation in traumatic brain injury: role of cytokines and chemokines. Neurochem Res, 23:329-340, 1998.

85
GOFF, J. P. Macromineral physiology and application to the feeding of the dairy cow for prevention of milk fever and other periparturient mineral disorders. An Feed Sci Tech. v.126,p. 237–257. 2006. GOMEZ-PINILLA F, LEE J. W, COTMAN C. W. Basic FGF in adult rat brain: cellular distribution and response to entorhinal lesion and fimbria-fornix transection. J Neurosci, 12:345-355, 1992.
GROTHE C, MEISINGER C, CLAUS P. In vivo expression and localization of the fibroblast growth factor system in the intact and lesioned rat peripheral nerve and spinal ganglia. J Comp Neurol, 434:342-357, 2001.
GROTHE C, TIMMER M. The physiological and pharmacological role of basic fibroblast growth factor in the dopaminergic nigrostriatal system. Brain Res Rev, 51:293-299, 2006.
GROTHE C. et al., The high molecular weight fibroblast growth factor-2 isoforms (21,000 mol. wt and 23,000 mol. wt) mediate neurotrophic activity on rat embryonic mesencephalic dopaminergic neurons in vitro. Neuroscience 2000;100(1): 73-86.
GUO B. F, DONG M. M. Application of neural stem cells in tissue-engineered artificial nerve. Otolaryngol Head Neck Surg, 140:159-164, 2009.
GUZEN F. P. et al., Glial cell line-derived neurotrophic factor added to a sciatic nerve fragment in a spinal cord gap ameliorates motor impairments in rats and increases local axonal growth. Restor Neurol Neurosci, 27:1-16, 2009.
GUZEN F. P. et al., Sciatic nerve grafting and inoculation of FGF-2 promotes improvement of motor behavior and fiber regrowth in rats with spinal cord transection. Restor Neurol Neurosci, 30:265-275, 2012. HAMMARBERG H. et al., GDNF mRNA in Schwann cells and DRG satellite cells after chronic sciatic nerve injury. Neuroreport, 7:857-860, 1996.
HENDERSON C. E. et al., GDNF: a potent survival factor for motoneurons present in peripheral nerve and muscle. Science, 266:1062-1064, 1994.
HIMMELSEHER S, PFENNINGER E, GEORGIEFF M. Effects of basic fibroblast growth factor on hippocampal neurons after axonal injury. J Trauma, 42:659-664, 1997.
HUANG T, HE D, KLEINER G, KULUZ J. Neuron-like differentiation of adipose-derived stem cells from infant piglets in vitro. J Spinal Cord Med, 30:35-40, 2007. HUMPEL C. et al., Fast and widespread increase of basic fibroblast growth factor messenger RNA and protein in the forebrain after kainate-induced

86
seizures. Neuroscience 57(4): 913-922, 1993.
ICHIHARA S, INADA Y, NAKAMURA T. Artificial nerve tubes and their application for repair of peripheral nerve injury: na update of current concepts. Injury, 39:29-39, 2008.
JAWOR, P. E. et al., Associations of subclinical hypocalcemia at calving with milk yield, and feeding, drinking, and standing behaviors around parturition in Holstein cows. J. Dairy Sci. v.95, p.1240–1248. 2012. JUNQUEIRA L, CARNEIRO J. Biologia celular e molecular. Rio de Janeiro: Guanabara Koogan, 9ª ed, 2012.
JUNQUEIRA L, CARNEIRO J. Histologia Básica. Rio de Janeiro: Guanabara Koogan, 12ª ed, 2013.
KHARITINOVA M. A, VASILEV J. M. Controlling cell length. Semi Cell Dev Biol, 19:480-484, 2008. KIRSCH M, SCHNEIDER T, LEE M. Y, HOFMANN H. D. Lesion-induced changes in the expression of ciliary neurotrophic factor and its receptor in rat optic nerve. Glia, 23:239-248,1998.
KOOP D. M, TRACHTENBERG J. T, THOMPSON W. J. Glial growth factor rescue Schwann cells of mechanoreceptors from denervation-induced apoptosis. The Journal of Neurosc, 17:6697-6706, 1997.
KONIG H.E. & LIEBICH H.G. Anatomia dos Animais Domésticos: texto e atlas
colorido. Artmed, Porto Alegre. 400p, 2004.
KUBO T, YAMASHITA T, YAMAGUCHI A, HOSOKAWA K, TOHYAMA M. Analysis of genes induced in peripheral nerve after axotomy using cDNA microarrays. J Neurochem, 82:1129-1136, 2002.
LEVAYER R, LECUIT T. Breaking down EMT. Nat Cell Biol, 10:757-759, 2008. LEVI-MONTALCINI R, ANGELETTI P. U. Nerve growth factor. Physiol Rev, 48:534-569, 1968.
LEWIN S. L, UTLEY D. S, CHENG E. T, VERITY A. N, TERRIS D. J. Simultaneous treatment with BDNF and CNTF after peripheral nerve transection and repair enhance rates of functional recovery compared with BDNF treatment alone. The Laryngosc, 107:992-999, 1997.
LEE, MENG-JEN et al. Combined treatment using peripheral nerve graft and FGF-1: Changes to the glial environment and differential macrophage reaction in a complete transected spinal cord. Neuroscience Letters 433 (2008) 163–169.

87
LOGAN A, FRAUTSCHY S. A, GONZALEZ A. M, BAIRD A. A time course for the focal elevation of synthesis of basic fibroblast growth factor and one of its high-affinity receptors (flg) following a localized cortical brain injury. J Neurosci, 12:3828-3837, 1992.
LOTAN M, SCHWARTZ M. Cross talk between the immune system and the Nervous system in response to injury: implications for regeneration. FASEB J, 8:1026-1033, 1994. LUCENA, S. E. E et al. Experimental considerations concerning the use of stem cells and tissue engineering for facial nerve regeneration: A systematic reviw. American Association of oral and maxillofacial Surgeons, 1-12, 2013 LUKAS, R. et al. Current role of methylprednisolone in the treatment of acute spinal cord injury. Acta Chir Orthop Traumatol Cech, v. 78, n. 4, p. 305-13, 2011. LUNDY-EKMAN L. Neurociências: fundamentos para reabilitação. 3 ed. Rio de
Janeiro: Elsevier,2008.
MACHADO A, HAERTEL LM. Neuroanatomia funcional. São Paulo: Atheneu,
3ª ed, 2014.
MAKWANA M, RAIVICH G. Molecular mechanisms in successful peripheral regeneration. FEBS J 272(11): 2628-2638, 2005.
MARKUS A, PATEL T. D, SNIDER W. D. Neurotrophic factors and axonal growth. Curr Opin Neurobiol, 12: 523-531, 2002a. MARKUS S, FAILING K, BAUMGARTNER W. Increased expression of pro-inflammatory cytokines and lack of up-regulation of anti-inflammatory cytokines in early distemper CNS lesions. J Neuroimmunol, 125: 30-41, 2002b. MARTINI, R. Expression and functional roles of neural cell surface molecules and extracellular components during development and regeneration of peripheral nerves. J Neurocytol, 23:1-28,1994.
MATTSON M. P, MURRAIN M, GUTHRIE P. B, KATER S. B. Fibroblast growth factor and glutamate: opposing roles in the generation and degeneration of hippocampal neuroarchitecture. J Neurosci, 9: 3728-3740, 1989.
MATTSON M. P, KUMAR K. N, WANG H, CHENG B, MICHAELIS E. K. Basic FGF regulates the expression of a functional 71-kDa NMDA receptor protein that mediates calcium influx and neurotoxicity in hipoccampal neurons. J Neurosci, 13: 4574-4588, 1993.
MATSUDA S, SAITO H, NISHIYAMA N. Effect of basic fibroblast growth factor on neurons cultured from various regions of postnatal rat brain. Brain Res, 520:310-316, 1990.

88
MATSUYAMA A. et al., Localization of basic fibroblast growth factor-like immunoreactivity in the rat brain. Brain Res 587(1): 49-65, 1992.
MENEI P. et al., Schwann cells genetically modified to secrete human BDNF promote enhanced axonal regrowth across transected adult rat spinal cord. Eur J Neurosci, 10:607-621,1998.
MOCCHETTI I, WRATHALL J. R. Neurotrophic factors in central nervous system trauma. J Neurotrauma, 12:853-870, 1995.
MOIZHESS T. G, VASILIEV J. M. Substract-induced polarization of cultured epitheliocytes and fibroblasts: non-reactivity of Ras-transformed cells. Cell Biol Int, 25:931-934, 2001. MORRISSEY T. K, KLEITMAN N, BUNGE, R. P. Isolation and functional characterization of Schwann cells derived from adult peripheral nerve. J Neurosci, 11:2433-42,1991.
MONTE-RASO V. V, BARBIERI C. H, MAZZER N, FASAN V. S. Can therapeutic ultrasound influence the regeneration of peripheral nerves? J Neurosci Methods. 142:185-92, 2005.
MONTE-RASO V. V. Índice funcional do ciático nas lesões por esmagamento do nervo ciático de ratos: avaliação da reprodutibilidade do método entre examinadores. Acta Ortop Bras. 14:133-6, 2006.
NGUYEN M. D, JULIEN J. P, RIVEST S. Innate immunity: the missing link in neuroprotection and neurodegeneration? Nat Rev Neurosci, 3:216-227,2002. NOBLE J, MUNRO C. A, PRASAD V. S, MIDHA R. Analysis of upper and lower extremity peripheral nerve injuries in a population of patients with multiple injuries. J Trauma. 45(1):116-22, 1998.
NOVAK C. B, MACKINNON S. E. Evaluation of nerve injury and nerve compression in the upper quadrant. J Hand Ther. 18:230-40, 2005. OLIVEIRA A, PIERUCCI A, PEREIRA K. Peripheral nerve regeneration through the nerve tubulization technique. Braz J of Morphol Sci, 21: 225-231, 2004. PERLSON E. et al., From snails to sciatic nerve: retrograde injury signaling from axon to soma in lesioned neurons. J Neurobiol, 58: 287-294, 2004.
PURVES D. et al., Neurociências. São Paulo: Artmed, 4ª ed, 2010.
PETERSON D. A, LUCIDI-PHILLIPI C. A, MURPHY D. P, RAY J, GAGE H. F. Fibroblast growth factor-2 protects entorhinal layer II glutamatergic neurons from axotomy-induced death. J Neurosci, 16:886-898, 1996.

89
PEULVE P, LAQUERRIERE A, HEMET J, TADIE M. Comparative effect of α-MSH and b-FGF on neurite extension of fetal rat spinal cord neurons in culture. Brain Res, 654:319-323, 1994.
PREWITT C. M, NIESMAN I. R, KANE C. J, HOULÉ J. D. Activated macrophage/microglial cells can promote the regeneration of sensory axons into the injured spinal cord. Exp Neurol, 148:433-443, 1997. RAIVICH G, KREUTZBERG G. W. Nerve growth factor and regeneration of peripheral nervous system. Clin Neurol Neurosurg, 95:84-88, 1993.
REUSS B, BOHLEN V, HALBACH O. Fibroblast growth factors and their receptors in the central nervous system. Cell Tissue Res, 313:139-157, 2003.
RIZVANOV A. A. et al., Genetically modified human umbilical cord blood cells expressing vascular endothelial growth factor and fibroblast growth factor 2 differentiate into glial cells after transplantation into amyotrophic lateral sclerosis transgenic mice. Exp Biol Med (Maywood) 2011;236(1): 91-98.
ROMERO M. I, RANGAPPA N, GARRY M. G, SMITH G. M. Functional regeneration of chronically injured sensory afferents into adult spinal cord after neurotrophin gene therapy. J Neurosci, 21:8408-8416, 2001.
ROVENSKY Y. U. A, DOMNINA L. V, IVANOVA O. Y. U, VASILIEV J. M. Responses of epitelial and fibroblast-like cells to discontinuos configuration of the culture substrate. Membr Cell Biol, 14:617-627, 2001. RUSSELL F. D, KOISHI K, JIANG Y, MCLENNAN I. S. Anterograde axonal transport of glial cell line-derived neurotrophic factor and its receptors in rat hypoglossal nerve. Neuroscience, 97:575-580, 2000.
SANTOS, T. S. C.; GUIMARÃES, R. M.; BOEIRA, S. F. Epidemiologia do trauma raquimedular em emergências públicas no município de Rio de Janeiro. 2012.
SANTOS, J.E.P. Distúrbios Metabólicos. In: BERCHIELLI, T.T; PIRES, A.V.; OLIVEIRA, S.G. Nutrição de Ruminantes. Jaboticabal:Funep, p. 439 – 461. 2006.
SAYERS S.T. et al., Preparation of brain-derived neurotrophic factor- and neurotrophin-3-secreting Schwann cells by infection with a retroviral vector. J Mol Neurosci, 10:143-160, 1998.
SCHWARZE E. & SCHRÖDER L. Introdución a la Anatomia Veterinária del Aparato Locomotor. Acribia, Zaragoza, 318p, 1970.
SCHWINDT T.T. et al., Effects of FGF-2 and EGF removal on the differentiation of mouse neural precursor cells. An Acad Bras Cienc, 81:443-452. 2009.

90
SENDTNER M, STÖCKLI K. A, THOENEN H. Synthesis and localization of ciliary neurotrophic factor in the sciatic nerve of the adult rat after lesion and during regeneration. The J of Cell Biol, 118:139-148, 1992.
SHACKLOCK M. Neurodynamics. Physiotherapy. 81:9-16, 1995.
SNIDER W. D, ZHOU F. Q, ZHONG J, MARKUS A. Signaling the pathway to regeneration. Neuron, 35:13-16, 2002. SOBOTTA J, BECHER H. Atlas de Anatomia Humana. Rio de Janeiro:
Guanabara Koogan, 23ª ed, 2013.
SOUZA F. I. D, BARROS FILHO T. E. P, CRISTANTE A. F. Evaluation of the use of GM1 after experimental spinal cord injury in rats. Coluna/Columna, v. 10, n. 4, p. 305-308, 2011.
SPRINGER J.E, MU X, BERGMANN L.W, TROJANOWSKI J.Q. Expression of GDNF mRNA in rat and human nervous tissue. Exp Neurol, 127:167-170, 1994.
STOLL G, MULLER H.W. Nerve injury, axonal degeneration and neural regeneration: basic insights. Brain Pathol, 9:313-35, 1999.
TENG YD, MOCCHETTI I, WRATHALL J.R. Basic and acidic fibroblast growth factors protect spinal motor neurones in vivo after experimental spinal cord injury. Eur J Neurosci, 10: 798-802, 1998.
TENG Y.D. et al., Basic fibroblast growth factor increases long-term survival of spinal motor neurons and improves respiratory function after experimental spinal cord injury. J Neurosci, 19:7037-7047, 1999.
TERENGHI G. Peripheral nerve regeneration and neurotrophic factors. J Anat, 194: 1-14, 1999.
VASILIEV J.M. Cytoskeletal mechanisms responsible for invasive migration of neoplastic cells. Int J Dev Biol, 48: 425-439, 2004.
WALICKE P.A. Basic and acidic fibroblast growth factors have trophic effects on neurons from multiple CNS regions. J Neurosci 8(7): 2618-2627, 1988.
WARING III, W. P. et al. 2009 review and revisions of the international standards for the neurological classification of spinal cord injury. The journal of spinal cord medicine, v. 33, n. 4, p. 346, 2010.
WANG D. et al., Bridging small-gap peripheral nerve defects using acellular nerve allograft implanted with autologous bone marrow stromal cells in primates. Brain Res, 1188:44-53, 2008.

91
WEBSTER H.D. Growth factors and myelin regeneration in multiple sclerosis. Mult Scler 1997;3(2): 113-120.
WEI Y. et al., Schwann-like cell differentiation of rat adipose-derived stem cells by indirect co-culture with Schwann cells in vitro. Cell Prolif, 43: 606-616, 2010.
WOODWARD W.R. et al., Nuclear and cytoplasmic localization of basic fibroblast growth factor in astrocytes and CA2 hippocampal neurons. J Neurosci 12(1): 142-152, 1992.
XU X.M, CHEN A,GUENARD V, KLEITMAN N, BUNGE M.B. Bridging Schwann cell transplants promote axonal regeneration from both the rostral and caudal stumps of transected adult rat spinal cord. J Neurocytol, 26:1-16,1997.
YAMAMOTO M, SOBUE G, LI M, ARAKAWA Y, MITSUMA T, KIMATA K. Nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF) and low-affinity nerve growth factor receptor (LNGFR) mRNA levels in cultured rat Schwann cells: differential time- and dose-dependent regulation by cAMP. Neurosci Lett, 152:37-40, 1993.
YANG J.D, CHENG-HUANG, WANG J.C, FENG X.M, LI Y.N, XIAO H.X. The isolation and cultivation of bone marrow stem cells and evaluation of differences for neural-like cells differentiation under the induction with neurotrophic factors. Cytotechnology, in press, 2014.
ZEEV-BRANN A.B, LAZAROV-SPIEGLER O, BRENNER T, SCHWARTZ M. Differential effects of central and peripheral nerves on macrophages and microglia. Glia, 23:181-190, 1998.

92
APÊNDICE

93
APÊNDICE 1
Parecer Consubstanciado do CEEA/UERN N0 007/13